Область техники
Изобретение относится к микробной биотехнологии, микробиологической промышленности и касается штамма микроорганизма Streptomyces rochei МР21, продуцирующего кирромицин - поликетидный антибиотик полиеновой структуры.
Изобретение может быть использовано для получения антибиотика кирромицина, обладающего особой селективной активностью в отношении ограниченного круга бактерий, в том числе представителей родов Streptococcus sp. и Mycoplasma sp. Его практическое применение возможно также при разработке лечебных кормов и ветеринарных препаратов для сельскохозяйственных животных.
Уровень техники
Кирромицин (моцимицин, дельвомицин, антибиотик MYC8003 и т.д.) является ингибитором синтеза белка у прокариотических организмов, принадлежащим по механизму действия к группе так называемых «эльфамицинов»: веществ, блокирующих активность термонестабильного фактора элонгации EF-Tu - необходимого функционального звена, обеспечивающего жизненно важную клеточную функцию синтеза белка. У прокариот основной функцией EF-Tu является транспортировка заряженной соответствующей аминокислотой тРНК (аа-тРНК) к А-сайту рибосомы для удлинения синтезируемой полипептидной цепи. EF-Tu представляет собой G-белок, обладающий внутренней способностью гидролизовать ГТФ до ГДФ (гуанозинтрифосфат до гуанозиндифосфата). EF-Tu образует комплекс с ГТФ и аа-тРНК и способствует общей точности трансляции.
Если с кодоном мРНК связывается соответствующий ему антикодон тРНК, то рибосома изменяет конфигурацию и изменяет геометрию ГТФазного домена EF-Tu, что приводит к гидролизу ГТФ в ГДФ. При гидролизе ГТФ конформация EF-Tu резко изменяется, и он диссоциирует от комплекса аа-тРНК и рибосомы. Затем аа-тРНК полностью попадает в А-сайт, где ее аминокислота приближается к полипептиду Р-сайта, а рибосома катализирует ковалентный перенос полипептида на аминокислоту - так растет полипептидная цепь.
Кирромицин препятствует высвобождению EF-Tu из рибосомы после гидролиза ГТФ [Wolf, Chinali, Parmeggiani, 1974] и, таким образом, нарушает процесс удлинения пептидной цепи. Такой механизм активности проявляется как в отношении грамположительных, так и грамотрицательных бактерий [Jotte, 2016].
Наряду с рибосомой, EF-Tu является одной из наиболее важных мишеней для антибиотик-опосредованного ингибирования трансляции [Krab, Parmeggiani, 2002], поскольку не способствует формированию перекрестной устойчивости к веществам, непосредственно связывающимся с рибосомой и блокирующим ее работу, многие из которых широко используются в клинической практике. Природные ингибиторы факторов трансляции в настоящее время вызывают особый интерес у исследователей [Dmitriev, Vladimirov, Lashkevich, 2020], так как на их основе могут быть получены комбинированные антибактериальные препараты из нескольких активных субстанций, действующих синергично.
В 1972 Вольфом и Захнером при скрининге природных антибактериальных субстанций с узким спектром действия в культуральной жидкости Streptomyces collinus Tu 365 было обнаружено и выделено вещество, названное кирромицином [Wolf, Zahner, 1972]. Примерно в то же время, Boc и Вервель выделили вещество моцимицин, которое было идентично кирромицину (дельвомицин, диметиллауродокс) [Maehr et al., 1973; Vos, Verwiel, 1973]. Также были описаны и другие вещества микробного происхождения близкой к кирромицину структуры: ауродокс [Berger et al., 1973], эфротомицин [Wax et al., 1976], хенейкомицин [Zimmerman et al., 1979], кирротрицин [Thein-Schranner et al., 1982], ганефромицин [Carter et al., 1988].
Вскоре после открытия была установлена и редкая биологическая мишень, на которую воздействуют кирромицин и его аналоги: они осуществляют специфическое взаимодействие с фактором элонгации у прокариот (EF-Tu) [Parmeggiani, Swart, 1985; Wolf, Chinali, Parmeggiani, 1974; Wolf, Zahner, 1972]. Благодаря общей мишени эти антибиотики были объединены в группу так называемых эльфамицинов (термин, образованный от сокращения словосочетания elongation factor).
Совместная кристаллизация ауродокса (N-метил-производного кирромицина) с очищенным фактором элонгации, полученным из Thermus aquaticus, выявила, что вещество связывается с доменами I и II фактора элонгации EF-Tu [Vogeley et al., 2001]. Эта связь приводит к изменению конформации данного белкового фактора, повышая его сродство к рибосоме, и тем самым препятствуя его освобождению, что приводит к остановке рибосомы и в конечном счете нарушению процесса синтеза белка. Подобный механизм действия делает кирромицин и его аналоги важными средствами для изучения процессов регуляции синтеза белка и поиска веществ, ценных для фармакологии [Weber et al., 2008].
В патенте US 3927211 описан антибиотик кирромицин (моцимицин, МУС 8003), полученный в микроорганизме Streptomyces ramocissimus. В экспериментах in vitro было установлено, что МУС 8003 обладает селективным ингибирующим действием на ряд микоплазм, возбудителей пневмоний, особенно в отношении Mycoplasma gallisepticum, вызывающего заболевания респираторного тракта у домашней птицы.
Было показано также, что кирромицин обладает низкой минимальной действующей концентрацией (МИК) в отношении Bacillus cereus АТСС 9139, Actinobacillus equuli subsp. equuli T9 и ряда других бактерий [патент US 3927211], особенно выраженная антагонистическая активность отмечена против представителей родов Streptococcus, Enterococcus, Neisseria gonorrhoeae и Haemophilus influenzae [Tavecchia et al., 1996], в то время как белок EF-Tu, выделенный из Staphylococcus aureus оказался устойчивым к присутствию кирромицина [Hall, Watkins, Georgopapadakou, 1989].
Хотя не сообщалось о токсическом действии кирромицина на эукариотические клетки [Schmid, Anke, Wolf, 1978], кирромицин оказался активным против малярийного паразита Plasmodium falciparum в культурах крови, вызывая у плазмодия нарушение синтеза белка в органеллах [Clough et al., 1999].
Благодаря узкому спектру действия кирромицин может рассматриваться как селективный, специфичный агент, ориентированный на ограниченную группу чувствительных микроорганизмов. Кроме того, данное вещество может быть использовано как стимулятор роста животных: было показано, что дозы 2,5-20 мг/кг массы вызывают положительный эффект на прирост биомассы кур [патент US 3927211].
Существуют две причины того, что микроорганизмы могут быть устойчивы к кирромицину: во-первых, они могут иметь мутации в гене EF-Tu, приводящие к изменению сайта связывания с кирромицином и делающие фактор элонгации способным выполнять свои функции в присутствии антибиотика [Mesters et al., 1994]; во-вторых, можно предположить, что у некоторых микроорганизмов кирромицин не может проникнуть внутрь клетки, либо выводится из нее прежде, чем произойдет взаимодействие с EF-Tu [Weber et al., 2008].
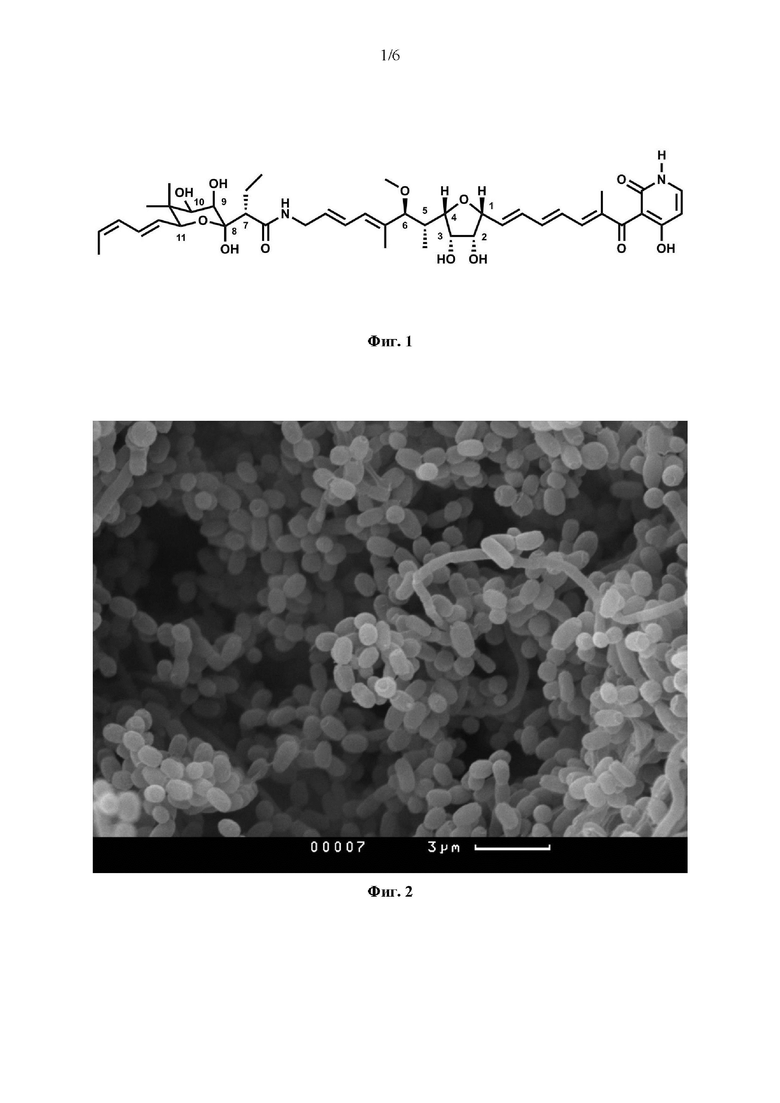
Молекула кирромицина линейный поликетид, содержащий три циклические структуры: пиридоновое кольцо, центральную тетрагидрофурановую структуру и сахароподобную структуру, так называемую голдиноновую кислоту (Фиг. 1). В 2008 году был описан биосинтетический кластер генов, участвующих в синтезе кирромицина, и показано, что поликетидный остов кирромицина синтезируется очень необычным способом с помощью гибридной нерибосомальной пептид с интетазы (NRPS)/поликетидсинтазы (PKS), а также прояснен путь биосинтеза пиридонового кольца [Weber et al., 2008]. Дальнейшие исследования генов, кодирующих «сшивочные ферменты» в биосинтетическом кластере позволили установить их роль на поздних стадиях формирования конечной структуры кирромицина [Robertsen et al., 2018].
Известны несколько продуцентов кирромицина (моцимицина), принадлежащие к роду Streptomyces vi Actinoplanes.
Из работы Рюкерта с соавторами известен нетиповой штамм Streptomyces collinus Tti 365 (DSMZ 40733) - продуцент кирромицина; в результате полногеномного секвенирования выявлены гены биотехнологического кластера, ответственного за синтез кирромицина [Rückert et al., 2013]. Продуктивность этого штамма составляет порядка 6-7 мг/л кирромицина [Weber et al., 2008].
Из статьи Олсторн-Тилеман соавторами и патента US 3927211 известен штамм S. ramocissimus CBS 190-63 [Olsthoorn-Tieleman et al., 2007], описанный как представитель нового вида, однако, не включенный в базу данных LPSN, признанную в настоящее время основой таксономической номенклатуры прокариот (https://lpsn.dsmz.de/). Продуктивность данного штамма составляет 16,1 мг/л кирромицина [патент US 3927211]
Из работы Беретты с соавторами известен штамм Actinoplanes sp. А8924 - продуцент кирромицина [Beretta et al., 1993]. Гастальдо с соавторами путем направленных мутаций и подбора условий улучшили производительность этого штамма с 30-50 мг/л до 350 мг/л кирромицина [Gastaldo et al., 1996]. Однако, в настоящее время судьба данного мутантного штамма Actinoplanes sp. A8924/F неизвестна, поскольку за последующие 28 лет публикаций, упоминающих этот штамм, не обнаружено. Так же нам не удалось обнаружить сам этот штамм в российских и зарубежных коллекциях микроорганизмов.
К серьезным недостаткам других упомянутых штаммов - продуцентов кирромицина, можно отнести малую продуктивность при выработке целевого антибиотика.
Технической задачей, на решение которой направлено настоящее изобретение, является решение как минимум одной из вышеуказанных в уровне техники проблем.
Сущность изобретения
Технической решением является использование описанного признаками в пунктах формулы изобретения.
Одной из возможных технических задач, на решение которой может быть направлено настоящее изобретение, являлось расширение арсенала технических средств, предназначенных для получения антибиотика кирромицина, в частности получение бактериального штамма, способного продуцировать ферментационную жидкость с высоким содержанием антибиотика кирромицина - не менее 200 мг/л.
Задача может решаться тем, что выделен и охарактеризован штамм Streptomyces rochei МР21.
Штамм Streptomyces rochei МР21 был выделен при поверхностном посеве на минеральный агар 1 [Гаузе et al., 1983] из образца, представляющего собой крупную фракцию (диаметром 2-3 мм) морских отложений. Образцы были собраны на городском пляже Федеральной территории Сириус (43.39774 с.ш. 39.96303 в.д) в августе 2021 года.
Штамм депонирован во Всероссийской Коллекции Микроорганизмов ФГБУ «ФИЦ «Пущинский Научный Центр Биологических Исследований Российской Академии Наук» (Россия, 142290, Московская обл., Пущино, пр. Науки, 5, ИБФМ) под регистрационным номером ВКМ Ac-2939D.
Видовая идентификация штамма МР21 основывалась на методологии полифазной таксономии, сочетающей анализ данных секвенирования нуклеотидных последовательностей гена 16s рРНК и сопоставления этих данных с последовательностями, депонированными в базе GenBank [https://www.ncbi.nlm.nih.gov/genbank]. наряду с изучением фенотипических признаков: культуральных, морфологических и физиологических. Данный подход в настоящее время считается абсолютно надежным для идентификации мицелиальных актинобактерий на уровне рода и в большинстве случаев достаточным для определения видовой принадлежности.
Штамм Streptomyces rochei МР21 характеризуется культурально-морфологическими и физиолого-биохимическими признаками, согласующимися с таковыми, приведенными для представителей рода Streptomyces [Goodfellow, 2015].
Культурально-морфологические признаки
Культуральные характеристики штамма Streptomyces rochei МР21 ВКМ Ac-2939D описаны на спектре твердых питательных сред (Фиг. 8), рекомендованных для фенотипического описания актиномицетов в рамках Международного проекта по изучению стрептомицетов - International Streptomyces Project [Shirling, Gottlieb, 1966].
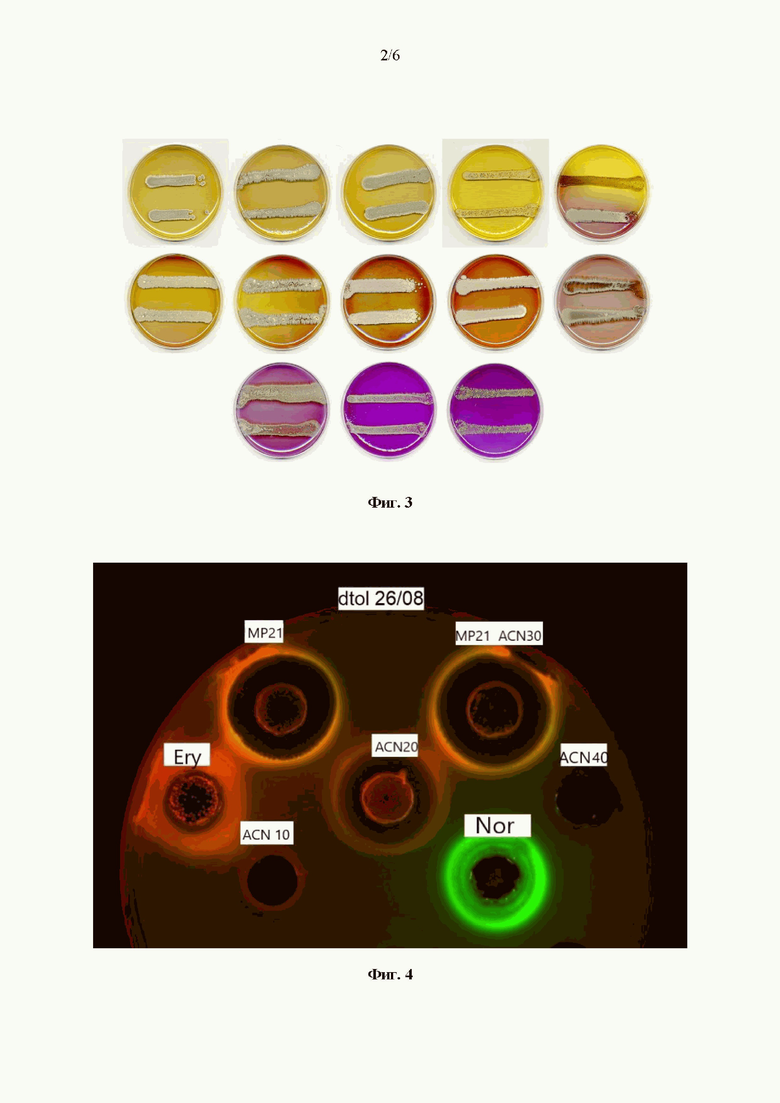
Изучение морфологии Streptomyces rochei MP21 на 14 сутки роста на овсяном агаре с помощью сканирующего электронного микроскопа (Фиг. 2) позволило установить, что штамм образует хорошо развитый разветвленный воздушный мицелий (0,43-0,45 мкм), несущий короткие прямые цепочки гладких овальных спор (1,25-1,27×0,75-0,81 мкм).
Физиолого-биохимические признаки
Штамм растет в аэробных условиях, в диапазоне температур от +20 до +45°С, с оптимумом роста при 37±2°С.
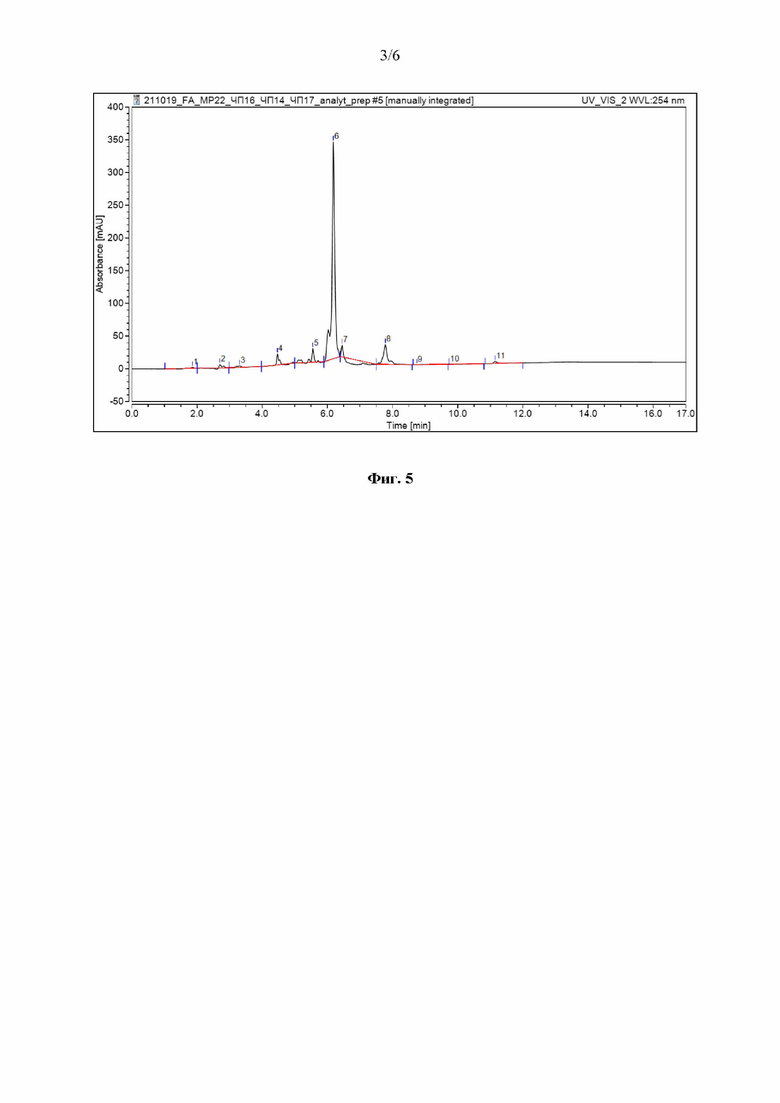
Способен образовывать кислоту при росте на арабинозе, фруктозе, глюкозе, инозитоле, сахарозе, маннозе, рамнозе; слабее: на галактозе, лактозе, мальтозе, ксилозе, маннитоле. Не использует сорбитол, сахарозу (Фиг. 3).
Способен к гидролизу казеина, крахмала, карбоксиметилцеллюлозы.
В качестве источника азота использует нитратные, аммонийные соединения, гидролизаты белков, аминокислоты.
Чувствительность к антибиотикам, определенная диско-диффузионным методом составляет: для эритромицина (5 мкг/диск), левофлоксацина (5 мкг/диск), ванкомицина (5 мкг/диск) и амоксициллина (20 мкг/диск).
Образует антибиотик кирромицин, характеризующийся выраженной антибиотической активностью в отношении ряда патогенных анаэробных бактерий, а также представителей родов Neisseria sp. и Streptococcus sp. [Gastaldo et al., 1996].
Штамм Streptomyces rochei MP21 BKM Ac-2939D не патогенен.
Хранение и поддержание жизнеспособности штамма
Штамм можно хранить при +4°С и поддерживать путем пересева раз в 3-4 недели в пробирки со скошенным овсяным агаром.
Длительное хранение штамма при температуре от -20 до -80°С может быть обеспечено культивированием в жидкой питательной среде, содержащей глюкозу, гидролизат белков, дрожжевой экстракт, минеральные соли в течение 3-5 суток, и последующим разливом культуральной жидкости в специальные емкости для хранения при низких температурах с добавлением веществ-криопротекторов и соблюдением условий стерильности.
Изобретение иллюстрируется следующими графическими материалами:
На Фиг. 1 изображена структурная формула молекулы кирромицина.
На Фиг. 2 представлена электронная микрофотография мицелия и спор штамма Streptomyces rochei МР21 ВКМ Ac-2939D, сделанная на 14-е сутки роста культуры на овсяном агаре при 28°С (Cambridge Instruments CamScan S2).
На Фиг. 3 отражена способность штамма Streptomyces rochei МР21 ВКМ Ac-2939D образовывать кислоту при росте на среде с единственным источником углерода: верхний ряд слева направо: арабиноза, фруктоза, манноза, рамноза, глюкоза; средний ряд: инозитол, ксилоза, мальтоза, галактоза, лактоза; нижний ряд: сахароза, сорбитол, минеральная основа без источника углерода.
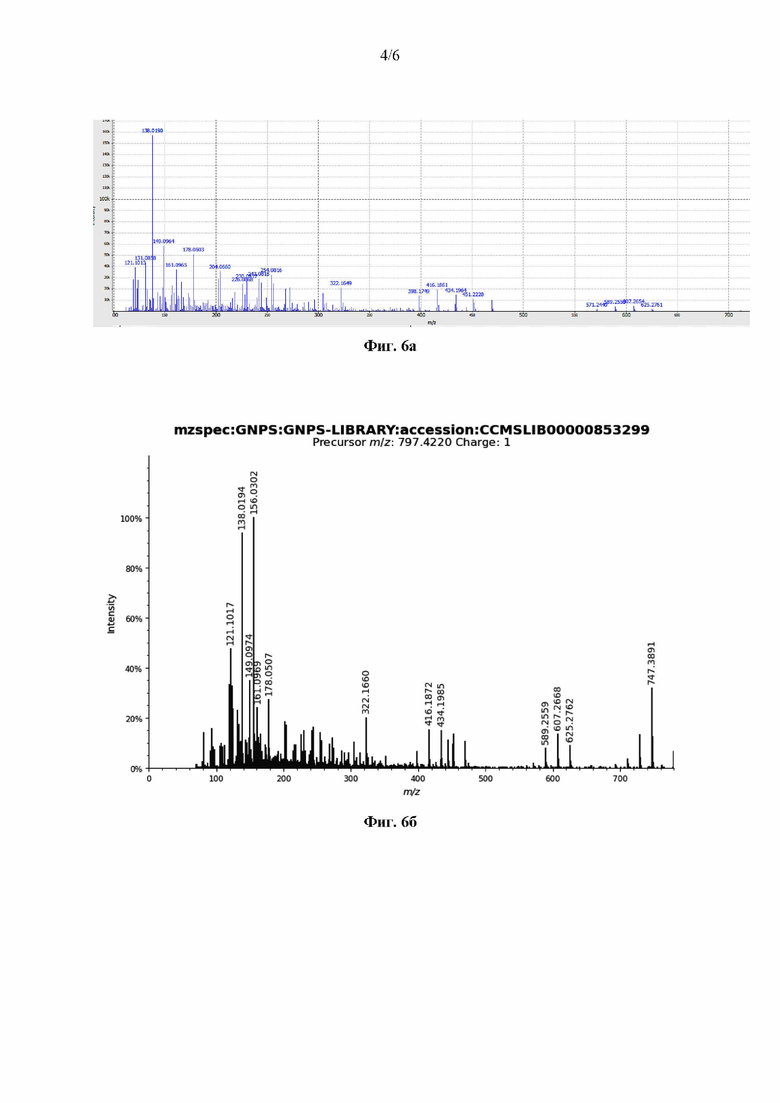
На Фиг. 4 представлены результаты тестирования LPS-фракций на репортерном тест-организме Е.coli dtolC JW5503: МР21 - исходная культуральная жидкость, МР21 (ACN10-ACN50) - фракции культуральной жидкости штамма МР21, элюированные растворами ацетонитрила различных концентраций, нанесены референсные антибиотики эритромицин (Ery, 2 мкг) и норфлоксацин (Nor, 0,04 мкг).
На Фиг. 5 представлены результаты ВЭЖХ-анализа активной фракции, извлеченной 30% раствором ацетонитрила в воде из культуральной жидкости Streptomyces rochei МР21 ВКМ Ac-2939D.
На Фиг. 6а представлены результаты масс-спектрометрического обнаружения кирромицина в культуральной жидкости Streptomyces rochei МР21 ВКМ Ac-2939D (фрагментация иона [М+Н]+, m/z 797.421 Da): спектр фрагментации совпадает с литературными данными (Фиг. 6б).
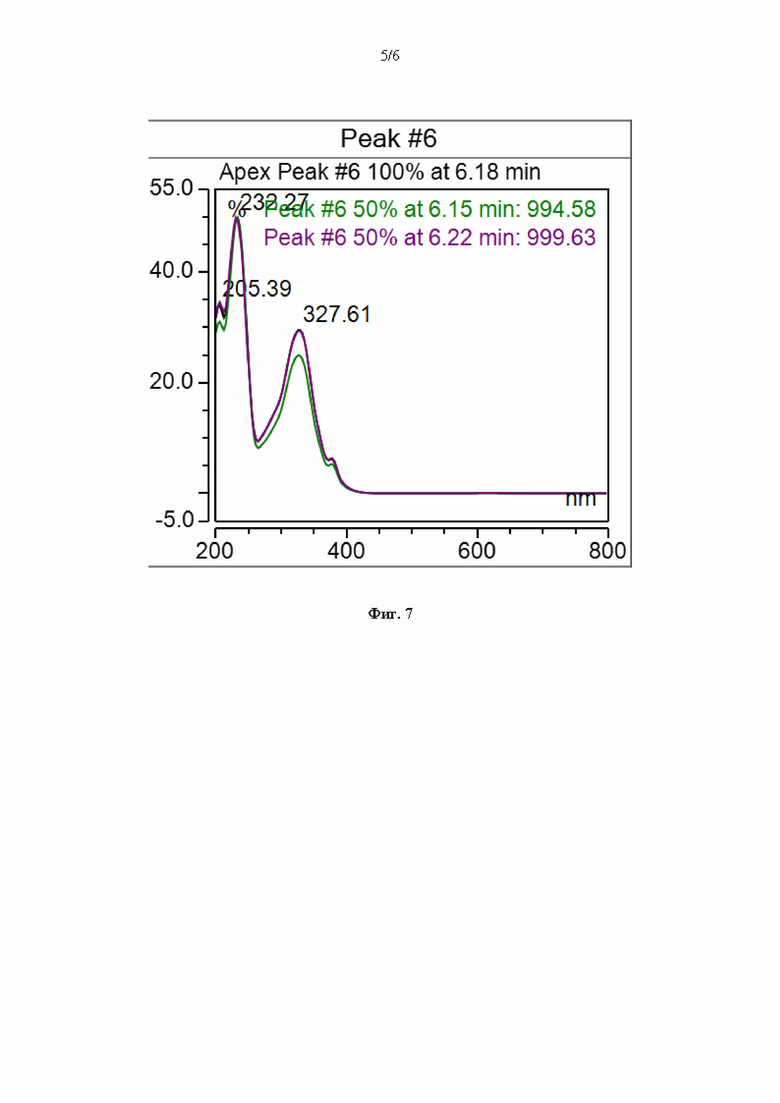
На Фиг. 7 представлен спектр оптического поглощения активного компонента в ультрафиолетовой и видимой областях.
На Фиг. 8 представлена таблица культуральных характеристик штамма Streptomyces rochei МР21 ВКМ Ac-2939D.
Примеры осуществления изобретения
Сущность и практическая применимость настоящего изобретения поясняется следующими примерами:
Пример 1. Подготовка штамма к длительному низкотемпературному хранению.
Штамм Streptomyces rochei МР21 ВКМ Ac-2939D культивируют в жидкой питательной среде следующего состава: глюкоза - 10 г, пептон - 10 г, гидролизат казеина 2 г, дрожжевой экстракт 2 г, NaCl 6 г, вода 1 л, при 28°С. Затем в 3-5 суточную культуральную жидкость в качестве криопротектора асептически вводят стерильный раствор 50% глицерола в объемном соотношении 1:1. Полученную смесь раскапывают по криопробиркам и помещают на хранение при температуре -80°С.
Пример 2. Выделение и культивирование Streptomyces rochei МР21 ВКМ Ac-2939D
Для исследования отбирали образцы морских отложений с территории пляжа ФТ «Сириус» (43.39774 с.ш. 39.96303 в.д.) в стерильные контейнеры, транспортировали в лабораторию и сразу производили посев из серии десятикратных разведений, приготовленных с использованием стерильной морской воды. Использовали питательную среду следующего состава (г/л): крахмал растворимый - 20,0; K2HPO4 - 0,5; MgSO4×7H2O - 0,5; KNO3 - 1,0; NaCl - 0,5; FeSO4×7H2O - 0,01; агар - 20 [Гаузе et al., 1983]. Для ограничения роста грибов и грамотрицательных бактерий в среды перед разливом добавляли нистатин (250 мкг/мл) и налидиксовую кислоту (10 мкг/мл) соответственно. Засеянные чашки Петри инкубировали при 28°С в течение двух недель.
Выросшие колонии мицелиальных прокариот выделяли в отдельные изоляты, среди которых проводили скрининг на антагонистическую активность с помощью репортерной системы Е. coli JW5503 ΔtolC-pDualrep2 [Osterman et al., 2016].
Разработанная репортерная конструкция представляет собой штамм грамотрицательной бактерии Е. coli JW5503 с инактивированным геном tolC, который кодирует один из компонентов AcrAB-TolC системы, ответственной за откачку из клетки различных ксенобиотиков. Модифицированные таким образом клетки лишены способности к синтезу белка TolC, а значит, являются значительно более чувствительными ко многим антибиотикам, в том числе к тем, которые подавляют рост только грамположительных бактерий. Кроме того, данный штамм трансформирован плазмидой pDualrep2, несущей гены для синтеза двух флюоресцентных белков TurboRFP и Katushka2S, которые позволяют определять два из возможных механизмов ингибирующего действия исследуемого агента на репортерные клетки. В случае попадания в клетку веществ, вызывающих повреждения ДНК, в ней активируется синтез белков, необходимых для репарации ДНК (так называемый SOS-ответ), однако, в репортерной системе в этом случае происходит увеличение экспрессии флюоресцентного белка TurboRFP, который можно детектировать при 553/574 нм.
Если сублетальные концентрации тестируемого вещества нарушают процесс синтеза белка в клетках репортерного штамма, то под действием генетически измененного аттенюатора триптофанового оперона в них вырабатывается другой флюоресцентный белок - Katushka2S (максимум поглощения/испускания составляет 588/633 нм).
Пример 3. Оценка ингибирующей активности и определение возможного механизма действия соединения, синтезируемого штаммом Streptomyces rochei МР21 ВКМ Ac-2939D, в отношении репортерного штамма Е. coli JW5503 ΔtolC-pDualrep2.
Ночную культуру штамма E.coli JW5503 ΔtolC, трансформированного плазмидой pDualrep2, разбавляли в десять раз свежей средой LB и выливали на чашки с агаризованной средой LB и канамицином (50 мкг/мл). После высыхания жидкости на поверхности агара, в среде проделывали лунки диаметром 8 мм, в которые вносили по 100 мкл исследуемого образца: культуральную жидкость штамма, полученную способом, описанным в Примере 4.1. (Фиг. 4), фракции, полученные путем твердофазной экстракции посредством элюирования водными растворами ацетонитрила 10%, 20%, 30% и 40%, описанными в Примере 4.3 (Фиг. 4). Также на поверхность агара наносили антибиотики, для верификации отклика репортерного штамма: эритромицин (Ery, 2 мкг), нарушающий процесс трансляции, и норфлоксацин (Nor, 0,04 мкг), вызывающий повреждение ДНК (Фиг. 4).
После культивировании в течении 18 часов при температуре 37°С полученные чашки сканировали при помощи системы для визуализации ChemiDoc® (Bio-Rad) в каналах Су3 (для детекции TurboRFP) и Су5 (для детекции Katushka2S).
Вокруг лунок с культуральной жидкостью и экстрактами штамма Streptomyces rochei МР21 ВКМ Ac-2939D наблюдалась зона отсутствия роста репортерного штамма, а также активная экспрессия белка Katushka2S в зоне сублетальных концентраций (Фиг. 4). Таким образом, было выявлено, что штамм Streptomyces rochei МР21 ВКМ Ac-2939D, при культивировании в условиях, указанных в примере 4.1., вырабатывает вещество, вызывающее повреждение синтеза белка у репортерного штамма E.coli JW5503 ΔtolC-pDualrep2.
Пример 4. Получение кирромицина с использованием нового штамма-продуцента
Пример 4.1. Культивирование штамма-продуцента. Клетки Streptomyces rochei МР21 ВКМ Ac-2939D выращивали в 100 мл жидкой питательной среды следующего состава (г/л): глюкоза - 10, пептон - 10, дрожжевой экстракт - 2, гидролизат казеина - 2, NaCl - 6,0, вода - остальное (рН после автоклавирования при 0,5 атм составляет 7,0-7,2). Культивирование производили в 250 мл конических колбах при 28°С и перемешивании на орбитальном шейкере в течение 3 суток. Полученную культуральную жидкость с клетками продуцента в концентрации 106 кл/мл использовали как посевной материал, засевая им в объеме внесения 5-10% конические колбы общим объемом 750 мл, содержащие 100-150 мл жидкой питательной среды вышеприведенного состава. Засеянные колбы в течение 5 суток инкубировали на шейкере при 28°С.
К концу ферментации содержание кирромицина в культуральной жидкости составляло не менее 270 мг/л.
По окончании срока ферментации культуральную жидкость с клетками микроорганизмов сливали из колб и центрифугировали 10 минут при скорости 4500 об/мин, чтобы отделить кондиционированную культуральную жидкость от клеточной биомассы.
Пример 4.2. Подготовка хроматографической колонки. В чистую и сухую колонку для твердофазной экстракции помещали 1 мл (0,3 г) сухого сорбента LPS-500H (сополимер дивинилбензола/ гидрофильного мономера), заливали 15 мл 50% (v/v) раствора ацетонитрила в воде, после полного просачивания через сорбент заливали 15 мл 10% v/v раствора ацетонитрила в воде и в заключение трижды промывали колонку дистиллированной водой. Данная процедура применяется для гарантированной очистки сорбента от посторонних соединений.
Пример 4.3. Идентификация и выделение кирромицина из культуральной жидкости.
Освобожденную от биомассы с помощью центрифугирования кондиционированную культуральную жидкость Streptomyces rochei МР21 ВКМ Ac-2939D наносили на хроматографическую колонку с подготовленным как описано в Примере 4.2. сорбентом. После нанесения всей культуральной жидкости на колонку, ее промывали дистиллированной водой, а затем элюировали 30% (v/v) раствором ацетонитрила в воде.
Фракция, экстрагированная 30% раствором ацетонитрила, продемонстрировала наибольшую антагонистическую активность при подавлении роста репортерного штамма E.coli dtolC JW5503, трансформированного плазмидой pDualrep2, скрининг проводили способом, описанным в Примере 2. [Osterman et al., 2016] (Фиг. 4).
Полученный элюат разделяли при помощи высокоэффективной хроматографии высокого давления (Фиг. 5) на ВЭЖХ-системе Vanquish Flex UHPLC System, используя детектор Diode Array (ThermoFisher Scientific), оснащенный колонкой Luna® 5 μm C18(2) 100 Å, 250 x 4.6 mm (Phenomenex)). Все полученные пробы тестировали на репортерном штамме E.coli dtolC JW5503 как описано в Примере 2, определяли активную фракцию и концентрировали ее при помощи роторного испарителя.
Идентификацию активного вещества проводили методом хромато-масс-спектрометрии высокого разрешения, с использованием масс-спектрометра Bruker maXis II 4G ETD, хроматографа UltiMate 3000, оснащенного колонкой Acclaim RSLC 120 C18 2.2um 2.1x100mm. Масс-спектры обрабатывали с использованием программ OpenChrom Lablicate Edition (1.4.0.202201211106), TOPPView v. 2.6.0 [Kohlbacher et al., 2007]. Поиск природных соединений-кандидатов проводили с помощью баз данных GNPS [Wang et al., 2016], NP Atlas [Santen van et al., 2019; Santen van et al., 2022], Dictionary of Natural Products.
Брутто-формула соединения C43H60N2O12, содержащегося в активной фракции, устанавливали на основании данных масс-спектрометрии высокого разрешения, отклонение массы составило 0.5 ppm. В режиме регистрации положительных ионов наблюдали ион [М+Н]+ с m/z 797.422 (расчетная масса [М+Н]+ для состава C43H60N2O12 - 797.4224554). Приоритизацию кандидатов проводили на основании данных о времени удерживания, происхождении и механизме действия активного соединения (ингибитор биосинтеза белка), выявляли кирромицин как удовлетворяющий всем заданным условиям.
Подтверждение структуры проводили на основе совпадения зарегистрированного УФ-спектра с максимумом на 327-328 нм с литературными данными [Maehr et al., 1973; Vos, Verwiel, 1973; Vost, Verwiel, 1973] в пределах погрешности. Совпадение структуры дополнительно подтверждали сравнением основных ионов в спектре фрагментации CID выделенного соединения со спектром фрагментации кирромицина из базы данных масс-спектров GNPS (номер спектра CCMSLIB00000853299), в котором наблюдались идентичные фрагментные ионы в области m/z 550-650 (589.3, 607.3, 625.3); в области m/z 400-450 (416.2, 434.2), а также фрагментные ионы с m/z 322.2, 178.1, 138.0, 121.1 и др. (Фиг. 6а, 6б). Различия в интенсивностях фрагментных ионах обусловлены разными условиями съемки и энергиями ионизации, приводящими к изменению глубины фрагментации молекулы; совпадение более чем 6 фрагментных ионов и точной массы однозначно указывает на идентичности структуры выделенного соединения кирромицину.
Таким образом, установлено, что фракция, полученная экстракцией 30% v/v раствором ацетонитрила из культуральной жидкости Streptomyces rochei МР21 ВКМ Ас-2939D, нарушает синтез белка у репортерного штамма JW5503 ΔtolC -pDualrep2 и представлена антибиотиком кирромицином (моцимицином, дельвомицином).
Заявитель просит рассмотреть представленные материалы заявки «Штамм Streptomyces rochei МР21 - продуцент антибиотика кирромицина» на предмет выдачи патента на изобретение.
Литература
1. Гаузе Г.Ф. и др. Определитель актиномицетов. Роды Streptomyces, Streptoverticillium, Chainia. Москва: Наука, 1983. 248 с.
2. Beretta G. et al. A novel producer of the antibiotic kirromycin belonging to the genus Actinoplanes // J. Antibiot. 1993. T. 46. №7. C. 1175-1177.
3. Berger J. et al. A new antibiotic X-5108 OF Streptomyces origin. I. Production, isolation and properties. // J Antibiot (Tokyo). 1973. T. 26. №1. C. 15-22.
4. Carter G.T. et al. LL-E19020 alpha and beta, novel growth promoting agents: isolation, characterization and structures. // J Antibiot (Tokyo). 1988. T. 41. №10. C. 1511-1514.
5. Clough B. et al. Antibiotic inhibitors of organellar protein synthesis in Plasmodium falciparum. // Protist. 1999. T. 150. №2. C. 189-195.
6. Dmitriev S.E., Vladimirov D.O., Lashkevich K.A. A Quick Guide to Small-Molecule Inhibitors of Eukaryotic Protein Synthesis // Biochemistry (Moscow). 2020. T. 85. №11. C. 1389-1421.
7. Gastaldo L. et al. Improvement of the kirromycin fermentation by resin addition // Journal of Industrial Microbiology and Biotechnology. 1996. T. 16. №5. C. 305 308.
8. Goodfellow M. Actinobacteria // Bergey's Manual of Systematics of Archaea and Bacteria.: John Wiley & Sons, Ltd, 2015. C. 1-2.
9. Hall С.C., Watkins J.D., Georgopapadakou N.H. Effects of elfamycins on elongation factor Tu from Escherichia coli and Staphylococcus aureus. // Antimicrob Agents Chemother. 1989. T. 33. №3. C. 322-325.
10. Jotte M. Kirromycin Enacyloxin Ha report for BioOrganic Chemistry., 2016.
11. Kohlbacher О et al. TOPP--the OpenMS proteomics pipeline. // Bioinformatics. 2007. T. 23. №2. C. e191-197.
12. Krab I.M., Parmeggiani A. Mechanisms of EF-Tu, a pioneer GTPase // Progress in Nucleic Acid Research and Molecular Biology.: Academic Press, 2002. C. 513-551.
13. Maehr H. et al. Antibiotic X-5108. V. Structures of antibiotic X-5108 and mocimycin. // J Am Chem Soc. 1973. T. 95. №25. C. 8449 8450.
14. Mesters J. R. et al. The structural and functional basis for the kirromycin resistance of mutant EF-Tu species in Escherichia coli. // EMBO J. 1994. T. 13. №20. C. 4877-4885.
15. Olsthoorn-Tieleman L.N. et al. Elongation factor Tu3 (EF-Tu3) from the kirromycin producer Streptomyces ramocissimus Is resistant to three classes of EF-Tu-specific inhibitors. // J Bacterid. 2007. T. 189. №9. C. 3581-3590.
16. Osterman I.A. et al. Sorting Out Antibiotics' Mechanisms of Action: a Double Fluorescent Protein Reporter for High-Throughput Screening of Ribosome and DNA Biosynthesis Inhibitors // Antimicrob Agents Chemother. 2016. T. 60. №12. C. 7481-7489.
17. Parmeggiani A., Swart G.W. Mechanism of action of kirromycin-like antibiotics. // Annu Rev Microbiol. 1985. T. 39. C. 557-577.
18. Robertsen H.L. et al. Filling the Gaps in the Kirromycin Biosynthesis: Deciphering the Role of Genes Involved in Ethylmalonyl-CoA Supply and Tailoring Reactions. // Sci Rep. 2018. T. 8. №1. C. 3230.
19. Rtickert C. et al. Complete genome sequence of the kirromycin producer Streptomyces collinus Tti 365 consisting of a linear chromosome and two linear plasmids. // J Biotechnol. 2013. T. 168. №4. C. 739-740.
20. Santen J.A. van et al. The Natural Products Atlas: An Open Access Knowledge Base for Microbial Natural Products Discovery. // ACS Cent Sci. 2019. T. 5. №11. C. 1824-1833.
21. Santen J. A. van et al. The Natural Products Atlas 2.0: a database of microbially-derived natural products. // Nucleic Acids Res. 2022. T. 50. №D1. C. D1317-D1323.
22. Schmid В., Anke Т., Wolf H. Action of pulvomycin and kirromycin on eukaryotic cells. // FEBS Lett. 1978. T. 96. №1. C. 189 191.
23. Shirling E.В., Gottlieb D. Methods for characterization of Streptomyces species // International Journal of Systematic Bacteriology. 1966. T. 16. №3. C. 313-340.
24. Tavecchia P. et al. Synthesis and biological evaluation of new fragments from kirromycin antibiotic. // J Antibiot (Tokyo). 1996. T. 49. №12. C. 1249-1257.
25. Thein-Schranner I. et al. Metabolic products of microorganisms. 209 Kirrothricin, anew member of the kirromycin-group.// J Antibiot (Tokyo). 1982. T. 35. №8. C. 948-956.
26. Vogeley L. et al. Conformational change of elongation factor Tu (EF-Tu) induced by antibiotic binding. Crystal structure of the complex between EF-Tu.GDP and aurodox. // J Biol Chem. 2001. T. 276. №20. C. 17149-17155.
27. Vos C., Verwiel P.E. J. Structure of the new antibiotic mocimycin (myc 8003): chromophore and furopyranone fragment // Tetrahedron Letters. 1973. T. 14. №30. C. 2823-2826.
28. Vost C., Verwiel P. E. J. The total structure of the novel antibiotic mocimycin (MYC 8003)//Tetrahedron Letters. 1973. T. 14. C. 5173-5176.
29. Wang M. et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. // Nat Biotechnol. 2016. T. 34. №8. C. 828-837.
30. Wax R. et al. Efrotomycin, a new antibiotic from Streptomyces lactamdurans. // J Antibiot (Tokyo). 1976. T. 29. №6. C. 670-673.
31. Weber Т. et al. Molecular analysis of the kirromycin biosynthetic gene cluster revealed beta-alanine as precursor of the pyridone moiety. // Chem Biol. 2008. T. 15. №2. C. 175-188.
32. Wolf H., Chinali G, Parmeggiani A. Kirromycin, an inhibitor of protein biosynthesis that acts on elongation factor Tu. // Proc Natl Acad Sci USA. 1974. T. 71. №12. C. 4910-4914.
33. Wolf H., Zahner H. [Metabolic products of microorganisms. 99. Kirromycin]. // Arch Mikrobiol. 1972. T. 83. №2. C. 147-154.
34. Zimmerman S.B. et al. Heneicomycin, anew antibiotic (A21A): fermentation, isolation, and antibacterial spectrum. // J Antibiot (Tokyo). 1979. T. 32. №6. C. 665-666.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм Streptomyces mirabilis KB13 - продуцент дутомицина | 2022 |
|
RU2798195C1 |
| Штамм Streptomyces sp. YVZ013 - продуцент антибиотика поликетомицина | 2023 |
|
RU2813975C1 |
| Штамм Streptomyces iakyrus Pe6 ВКПМ Ас-2084 - продуцент антибиотика нибомицина | 2018 |
|
RU2696029C1 |
| Штамм Amycolatopsis rifamycinica - продуцент антибиотика тетраценомицина Х | 2019 |
|
RU2724537C1 |
| Штамм Streptomyces sp. YVZ014 - продуцент антибиотика лизолипина X | 2023 |
|
RU2817695C1 |
| СПОСОБ ПОЛУЧЕНИЯ НИБОМИЦИНА | 2018 |
|
RU2718802C1 |
| Штамм Streptomyces sp. YVZ012 - продуцент суругамида А | 2023 |
|
RU2821907C1 |
| Штамм Streptomyces sp., выделенный из байкальских эндемичных амфипод - продуцент антибиотика актифенола | 2018 |
|
RU2704862C1 |
| Штамм Streptomyces sp. YVZ016 ВКМ Ас-2999D - продуцент веществ, способствующих защите сельскохозяйственных культур от заболеваний, вызываемых бактериями Clavibacter michiganensis | 2024 |
|
RU2840022C1 |
| Штамм Streptomyces sp. YVZ020 ВКМ Ас-3001D - продуцент метаболитов, обладающих фитостимулирующим и фитозащитным действием против возбудителя бактериоза Clavibacter michiganensis | 2024 |
|
RU2840021C1 |

Изобретение относится к области биотехнологии и представляет собой штамм Streptomyces rochei MP21, депонированный во Всероссийской Коллекции Микроорганизмов ФГБУ «ФИЦ «Пущинский Научный Центр Биологических Исследований Российской Академии Наук» под регистрационным номером ВКМ Ас 2939D, - продуцент антибиотика кирромицина. Изобретение позволяет расширить арсенал технических средств, предназначенных для получения антибиотика кирромицина, в частности получить бактериальный штамм, способный продуцировать ферментационную жидкость с высоким содержанием антибиотика кирромицина - не менее 200 мг/л. 8 ил., 4 пр.
Штамм Streptomyces rochei MP21, депонированный во Всероссийской Коллекции Микроорганизмов ФГБУ «ФИЦ «Пущинский Научный Центр Биологических Исследований Российской Академии Наук» под регистрационным номером ВКМ Ас-2939D, – продуцент антибиотика кирромицина.
| US 3927211 А1, 16.12.1975 | |||
| BERETTA G | |||
| et al | |||
| A novel producer of the antibiotic kirromycin belonging to the genus Actinoplanes, J | |||
| Antibiot | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| т | |||
| Способ изготовления звездочек для французской бороны-катка | 1922 |
|
SU46A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| c | |||
| Музыкальный инструмент | 1924 |
|
SU1175A1 |
| ГАУЗЕ Г.Ф | |||
| и др | |||
| Определитель актиномицетов | |||
| Роды Streptomyces, Streptoverticillium, Chainia | |||
| Москва: Наука, 1983 | |||
| Деревянная повозка с кузовом, устанавливаемым на упругих дрожинах | 1920 |
|
SU248A1 |
