Изобретение касается в общем областей молекулярной биологии, биотехнологии и белкового синтеза. Более конкретно, данное изобретение относится к новому способу синтеза флуоресцентно меченого белка в бесклеточной системе белкового синтеза.
Предшествующий уровень техники
Ряд показаний свидетельствуют о том, что насцентные (возникающие) полипептиды приобретают вторичную и третичную структуру на рибосомах. Beckmann и его сотрудники (1990) установили, что Hsp70 взаимодействует с насцентными белками и постулировали ко-трансляционную укладку белка. Cама рибосома может играть важную роль в процессе укладки. Lim и Spirin (1986) высказали гипотезу, что растущий пептид может синтезироваться в виде альфа-спирали. Были рассмотрены применения флуресцентных способов для исследования удлинения и укладки растущих пептидов и белков на рибосомах. Ранее эти способы применялись для демонстрации того, что растущий белок MS2 укладывается, будучи связанным с рибосомами. Было показано, что так называемые "наставники" (факторы, участвующие в обеспечении сборки пространственной структуры белков - прим, переводчика) облегчают укладку многих белков из их денатурированного состояния, но очень мало известно о том, функционируют ли эти белки на рибосомах во время трансляции. Hartl и сотрудники представили данные, свидетельствующие о том, что DnaJ является первым "наставником", который связывается с растущими пептидами люциферазы светляка или хлорамфениколацетилтрансферазы на рибосомах зародышей пшеницы (Hendrick et al., 1993).
В предшествующей области знаний не было эффективных средств синтеза флуоресцентно меченого белка в системе бесклеточного белкового синтеза. Данное изобретение решает эту проблему.
Раскрытие изобретения
В данном изобретении кумарин из кумарин-малеимидил-SAcMet тРНК (CPM-SAcMet-тРНКf) включали в N-конец растущих полипептидов, которые синтезировались на рибосомах в бактериальной бесклеточной сопряженной системе транскрипции/трансляции. Способы с использованием флуоресценции использовали для наблюдения изменений в локальном окружении и подвижности N-концевого зонда и поочередно действий наставников на укладку, активацию и выделение растущих (насцентных) или полноразмерных полипептидов из рибосом. Данное изобретение показывает, что наставники влияют на укладку растущих полипептидов роданезы в то время, когда они связаны в виде пептидил-тРНК на рибосомах. Наставники облегчают их высвобождение из рибосом в виде ферментативно активного белка.
В одном варианте данного изобретения обеспечен способ синтезирования флуоресцентно меченого белка в бесклеточной системе белкового синтеза, предусматривающий следующие стадии: (а) инкубирование пробы рибосом, полученных из бесклеточного экстракта, с плазмидной ДНК, содержащей кодирующую последовательность для целевого белка, причем эту пробу инкубируют в среде для сопряженной транскрипции/трансляции вместе с аминоацил-тРНК, имеющей флуоресцентную метку; (b) частичную очистку флуоресцентно меченого белка путем отделения вновь синтезированного флуоресцентно меченого белка от других флуоресцентных компонентов внутри этой пробы; (с) измерение количества синтезированного белка; (d) определение флуоресценции вновь синтезированного белка и (e) определение биологической активности вновь синтезированного белка.
Другие и дополнительные аспекты, особенности и преимущества данного изобретения будут очевидны из следующего описания предпочтительных в настоящее время вариантов изобретения, даваемых с целью описания.
Краткое описание чертежей
Для более ясного понимания указанных выше особенностей, преимуществ и целей настоящего изобретения представлено более подробное описание изобретения со ссылками на некоторые его варианты, которые иллюстрируются прилагаемыми чертежами. Эти чертежи образуют часть настоящей заявки. Однако следует заметить, что прилагаемые чертежи иллюстрируют предпочтительные варианты изобретения и, следовательно, не должны рассматриваться как ограничивающие изобретение в их объеме.
Фиг.1 показывает анализ свободной и связанной с рибосомами роданезы, либо транслированной в присутствии [14C]-лейцина, либо инициированной СРМ-Sac-[35S]-Met-тРНКf. Роданезу синтезировали посредством сопряженной транскрипции/трансляции либо с [14C] -лейцином (160 Ки/моль), либо с СРМ-Sac-[35S]-Met-тРНКf (4000 Ки/моль) в качестве радиоактивного предшественника. После инкубирования реакционную смесь центрифугировали. Супернатант и ресуспендированные рибосомы анализировали при помощи электрофореза в ПААГ-ДСН и авторадиографии. Показана авторадиограмма (первые 2 части, меченые 14C-лейцином и СРМ [35S] -Met, соответственно). Полосы 1 и 2 - инкубирование с внутренним [14C] -лейцином; полосы 3 и 4- инкубирование с N-концевым СРМ- Sac-[35S]-метионином. Полосы 1 и 3 - 20 мкл супернатанта, полосы 2 и 4 - 20 мкл ресуспендированных рибосом. Третья часть фиг. 1 (меченые антитела против RHO) показывает Вестерн-блот, зондированный антителами против роданезы. Положение пероксидазы хрена, ковалентно связанной со вторым антителом, визуализировали с применением ECL, системы Amersham. Стрелка указывает электрофоретическую подвижность нативной роданезы из бычьей печени.
Фиг. 2 показывает гашение флуоресценции кумарина нитрометаном. Представлены графики Stern - Volmer интенсивности флуоресценции устойчивого состояния для СРМ- роданезы, высвобожденной из рибосом или связанной с рибосомами, в сравнении со свободной СРМ-Sac-Met-тРНК (линия 1) и этой тРНК, связанной с рибосомами (линия 6). Линия 2 - СРМ-роданеза в супернатанте в ферментативно активной форме; линия 3 - СРМ-роданеза, высвобожденная при помощи Dnak линия 4 - СРМ-роданеза, высвобожденная пуромицином; линия 5-СРМ-роданеза, связанная с рибосомами.
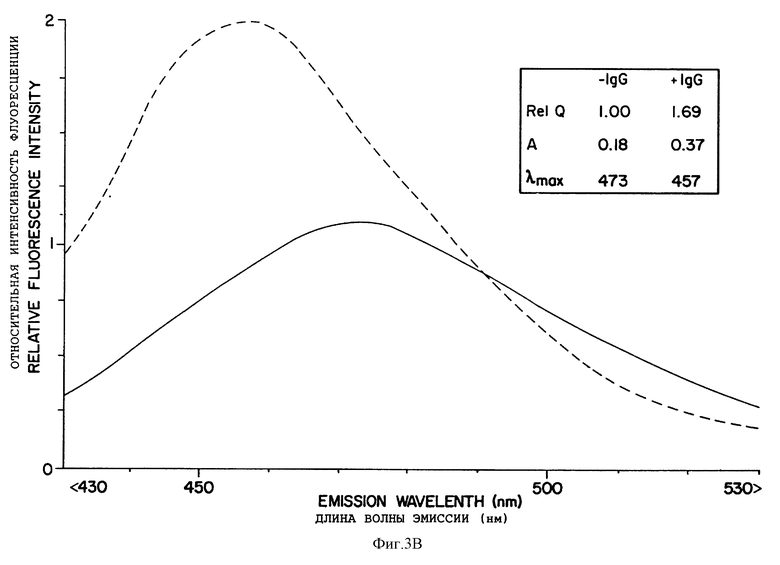
Фиг. 3 показывает спектры испускания флуоресценции СРМ-меченой роданезы перед инкубацией и после инкубации с антителами против кумарина. Анализировали СРМ-роданезу, связанную с рибосомами (A) или выделенную в супернатант (B) (сплошная линия). Затем добавляли IgG против кумарина и снова снимали спектры (пунктирная линия). Вставка дает количественные данные флуоресценции для соответствующих анализированных проб.
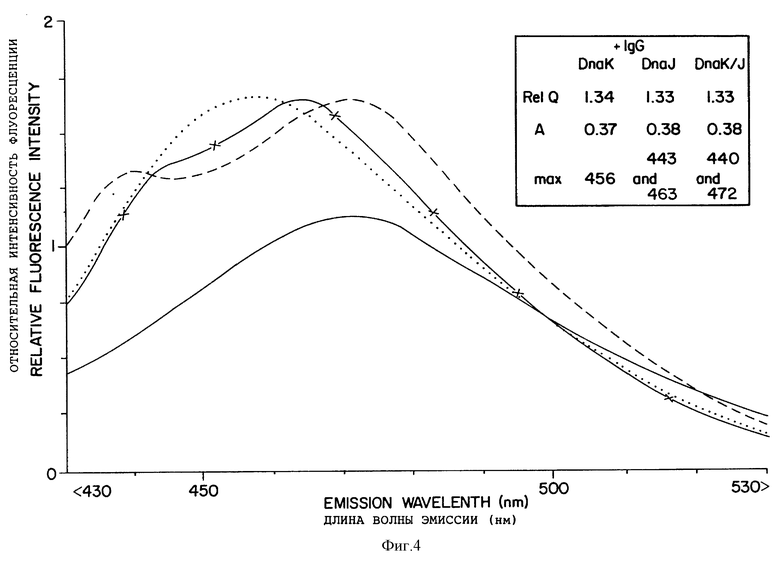
Фиг. 4 показывает, что DnaJ влияет на взаимодействие IgG против кумарина со связанной с рибосомами СРМ-роданезой. Сначала снимали спектр СРМ-роданезы на рибосомах после инкубирования со спарсомицином (сплошная линия). Устанавливали три параллельные пробы (СРМ-роданеза на рибосомах, инкубированная со спарсомицином), к которым добавляли DnaJ или DnaK по отдельности или вместе. Снимали спектры (не показано) и анализировали. Было обнаружено, что эти данные идентичны или очень сходны с данными, представленными в таблице VI. Затем к каждой пробе добавляли IgG против кумарина. После инкубирования в течение 10 минут при комнатной температуре снимали спектры, которые представлены здесь. (x-x)DnaJ+IgG; (...)DnaK+IgG; (- -)DnaJ и DnaK+IgG.
Подробное описание изобретения
В описании данного изобретения используются следующие абревиатуры: RHO-роданеза; СРМ-3-(4-малеимидофенил)-4-метил-7- (диэтиламино)-кумарин; СРМ-роданеза-роданеза, меченая на ее N-концевом метионине СРМ; SDS-PAGE гель-электрофорез в полиакриламидном геле с додецилсульфатом натрия (ПААГ-ДСН); анти-СРМ IqG-кроличьи поликлональные антитела против кумарина; СРМ-SAc-Met-тРНКf-инициирующая тРНК, которая была аминоацилирована метионином и затем помечена на аминогруппе метионина через меркаптоуксусную кислоту при помощи СРМ; IgG-иммуноглобулин; Fab - его генерируемый папаином фрагмент.
Данное изобретение касается также способа синтезирования флуоресцентно меченого белка в бесклеточной системе белкового синтеза, предусматривающий следующие стадии: (a) инкубирование пробы рибосом, полученных из бесклеточного экстракта, с плазмидной ДНК, содержащей кодирующую последовательность для представляющего интерес белка, причем пробу инкубируют в среде для сопряженной транскрипции/трансляции вместе с амино-ацил-тРНК, имеющей флуоресцентную меткую; (b) частичную очистку флуоресцентно меченого белка путем отделения вновь синтезированного флуоресцентно меченого белка от других флуоресцентных компонентов внутри пробы; (с) измерение количества синтезированного белка; (d) определение флуоресценции вновь синтезированного белка и (e) определение биологической активности вновь синтезированного белка.
Обычно, в данном изобретении, флуоресцентная метка находится на N-конце белка. В предпочтительном варианте применяемая в данном способе аминоцил-тРНК представляет собой метионил-тРНК-метионин/f. Предпочтительно, эта флуоресцентная метка ковалентно связана с аминокислотой, инициирующей тРНК.
В способе настоящего изобретения среда для транскрипции/трансляции обычно содержит РНК-полимеразу, нуклеозидтрифосфаты, систему регенерирования энергии и аминокислоты в буферном солевом растворе. В предпочтительном варианте рибосомы выделены из экстракта E.coli, называемого S30. В предпочтительном варианте, плазмидная ДНК является нелинейной. Предпочтительно, инкубирование на стадии (а) проводят приблизительно в течение 30 минут при приблизительно 37oC. Также в способе данного изобретения флуоресцентно меченый белок частично очищают способом, выбранным из группы, состоящей из центрифугирования, ионообменной хроматографии и гель-фильтрации.
Следующие далее примеры даются с целью иллюстрации различных вариантов изобретения и не предназначены для какого-либо ограничения данного изобретения.
Пример 1
Химикалии.
Нуклеозидтрифосфаты и тРНК E.coli были поставлены Boehringer-Mannheim; 3-(4-малеимидофенил)-7-диэтиламино-4-метилкумарин (СРМ) из Molecular Probes Jnc. (bugene, OR). тРНКMetf рифампицин, пуромицин, спарсомицин и все другие реактивы поставлялись Sigma. Смесь рибонуклеазы A и TI ("РНКазный коктейль") был из Ambion (Austin. TX). [14C]-лейцин и [35S-метионин были из NEN-DuPont. Наставники DnaK, DnaJ, GrpE, GroEL и GroES были куплены в Epicentre Technologies (Madison, WI). Исходная содержащая ген роданезы плазмида, роданеза, выделенная из митохондрий бычьей печени, и поликлональные антитела против роданезы были получены от доктора Paul Horowitz (University of Texas Health Science Cenrte, San Antonio, TX). Поликлональные антитела против факторов освобождения (рилизинг-факторов) E. coli 1 и 2 были предоставлены доктором Warren Tate (Universitu of Otago, New Zealand). Вторые антитела (козлиные антитела против кроличьего IgG-пероксидаза хрена и кроличьи антитела против овечьего IgG-пероксидаза хрена) были приобретены из South San Francisco, CA).
Пример 2
In vitro системы
Образование плазмид, выделение РНК-полимеразы SP6 и получение бесклеточного экстракта Е. coli (S30), а также выделение фракции рибосом из S30 выполняли, как описано Kudlicki et al., Anal. Biochem. 206:389 (1992). В основном, плазмидные препараты готовили согласно стандартным процедурам (J. Sambrook et al. , (1989) Molecular Cloning, Cold Spring Harbor Zaboratory Press), за исключением того, что стадию центрифугирования в Cs Cl заменяли Q Cartridge хроматографией (Bio-Rad). РНК-полимеразы SP6 и T7 коммерчески доступны. Применяемую фракцию рибосом отделяли от фракции S30, полученной согласно способу G. Zubay из E. coli K12 (A19). Клетки выращивали при 37oC в LB (Sigma) бульоне, к которому добавляли 20% глюкозу (10 мл на 2 л среды). Клетки отбирали в середине логарифмической фазы роста при 37oC и лизировали в ячейке под давлением. Затем клетки центрифугировали в течение 30 минут при 30000 Xg и получали экстракт S30. К буферу для лизиса добавляли ингибитор протеаз, фенилметилсульфонилфторид, до конечной концентрации 0,5 мМ. Аликвоты S30 (9 мл каждая) центрифугировали при 47000 об/мин, в течение 4 часов в роторе Ti 50 Beckman. Осажденные рибосомы ресуспендировали в 1,0 мл 20 мМ Трис-нCl (pH 7,5), 10 мМ Mg (OAc)2 и 1 мМ ДТТ, получая суспензию с приблизительно 1200 A260-единиц/мл. Эту фракцию рибосом хранили при -70oC в виде небольших аликвот.
Препарат СРМ-SAcMet-тРНКf применяли, как описано ранее, для мечения α-амино-группы Phe-тРНКPheСРМ (Odom et al., 1990). Вкратце, α-амино-группу метионина в [35S]-Met-тРНКf ацилировали дитиодигликолевой кислотой и затем восстанавливали до соответствующего производного тиогликолевой кислоты. Его сульфгидрильная группа реагировала с малеимидом (СРМ). Антитела против СРМ получали в кроликах; фракцию IgG из сыворотки иммунизированного кролика получали, как описано (Picling et al., 1992).
In vitro система для сопряженной транскрипции/трансляции была детально описана (Kudlicki et al., 1992). Вкратце, примененная система для проведения сопряженной транскрипции/трансляции содержала в общем объеме 30 мкл: 50 мМ Трис-ацетата (pH 7,8), 14 мМ Mg (OAc)2, 36 мМ NH4OAc, 72 мМ KOAc, 2 мМ Ca(OAc)2, 0,5 мМ ЭДТА, 2% полиэтиленгликоль-6000, 2 мМ ДТТ (дитиотреитол), 1,2 мМ АТФ, 0,8 мМ каждого из GТР, UТР и СТР (ГТФ, УТФ и ЦТФ), 0,5 мМ цАМФ, 27 мМ фосфоенолпирувата, 0,35 мкг пируваткиназы, 1 мкг фолиновой кислоты, 83 мкМ 14C-лейцин, 330 мкМ каждую из остальных 19 аминокислот, 20 мкг тРНК E. coli (Boehringer) 0,5 мкг рифампицина, 0,3 мМ глюкозо-6-фосфат, 1,2 A260-единиц фракции рибосом Е. coli. 0,5 мкг плазмидной ДНК и 0,5 мкг РНК-полимеразы SP6. Для этого теста 14C-лейцин разбавляли до 40 Ки/моль. Инкубирование проводили в течение 30 минут при 37oC. Для крупномасштабного синтеза СРМ-роданезы объемы реакционной смеси увеличивали до 0,9 мл, и они содержали соли и низкомолекулярные компоненты, перечисленные выше, плюс 5 мМ Na2S2O3. Приблизительно 60 A260-единиц непромытых рибосом инкубировали приблизительно с 10-15 мкг с нелинейной плазмидой (рSP65), содержащей кодирующую роданезу последовательность под контролем промотора SP6, СРМ-SAc[35S]-Met-тРНКf(350 Ки/моль) применяли в качестве радиоактивного предшественника; фолиновую кислоту не включали в смесь. В некоторых контрольных экспериментах использовали [14C] -лейцин (40 Ки/моль) вместе с нерадиоактивной fMet-тРНКf. После инкубирования в течение 30 минут при 37oC пробу наносили на 0,6 мл забуференной сахарозы и центрифугировали в течение 45 минут при 45000 об/мин. Затем собирали и сохраняли супернатант, а слой сахарозы удаляли; осадок рибосом промывали, затем ресуспендировали в 60 мкл 20 мМ Трис-нCl, pH 7,5, 10 мМ Mg (OAc)2, 30 мМ NH4OAc, 1 мМ ДТТ, 5 мМ Na2S2O3 (раствор A). Сразу же после центрифугирования фракцию супернатанта (приблизительно 0,9 мл) обрабатывали рибонуклеазами A и T1 (0,1 мг и 2000 Е/мл, соответственно) в течение 15 минут при 37oC для разрушения оставшейся СРМ-SAc-[35S-Met-тРНКf. Затем полученные реакционные смеси хроматографировали на колонке Sephadex G100 (1х20 см), уравновешенной раствором A, для отделения вновь синтезированной СРМ-меченой роданезы от низкомолекулярных продуктов деградации, содержащих кумарин и [35S]-метионин.
Пример 3
Измерения флуоресценции
Измерения флуоресценции выполняли на считающем фотоны спектрофлуорометре модели 8000C из SLM-Aminco Instruments Inc. (Urbana, IL). Спектры измеряли при интервалах испускания 1 нм при скорости сканирования 0,5 секунд на инкремент (приращение) длины волны; длина волны возбуждения была 390 нм. Спектры и относительные квантовые выходы нормализовали на основе радиоактивности из [35S]-метионина, который присутствовал в виде СРМ-SAc-метионина в пробе. Относительные квантовые выходы флуоресценции определяли из интегрированных эмиссионных спектров. Относительный квантовый выход СРМ-роданезы во фракции супернатанта был принят за 1,00 и все другие квантовые выходы даны по отношению к этой величине. Анизотропию флуоресценции определяли, как описано в Odom et al., (1984), при длине волны эмиссии 480 нм. Для исследований флуоресценции аликвоту ресуспендированных рибосом или фракции обработанного супернатанта инкубировали в кювете в общем объеме 420 мкл, содержащем соли и низкомолекулярные компоненты, описанные выше для сопряженной транскрипции/трансляции, но без аминокислот и без УТФ и ЦТФ (раствор B). После снятия спектра тестируемые компоненты добавляли в минимальном объеме. Кювету инкубировали в течение 10 минут при 37oC и затем снова снимали спектр. Добавки: анти-СРМ-IgG, 0,1 мг; наставники GroES, 1,6 мкг; GroEL, 6 мкг; DnaK, 4 мкг; DnaJ 2 мкг; GrpE:, 3 мкг.
Пример 4
Определения количества роданезы
Осаждение белка трихлоруксусной кислотой (ТХУ) для определения количества полипептидов роданезы, анализ электрофорезом в ПААГ-ДСН и авторадиографию выполняли способами хорошо известными в данной области знаний. Активность роданезы определяли согласно Sorbo (1953) в описании Tsalkova et al. (1993). В основном ферментативную активность роданезы определяли калориметрическим анализом, измеряющим превращение CN- в SCN- ферментом с использованием S2O32- в качестве субстрата. Образовавшийся SCN- детектировали и количественно определяли путем измерения поглощения при 460 нм комплекса между этим продуктом и ионами трехвалентного железа. За одну единицу брали количество фермента, генерирующего 1 мкмоль продукта в минуту при 37oC в этой тест-системе.
Вестерн-блоты из электрофореза в ПААГ-ДСН зондировали бычьими антителами против факторов высвобождения 1 или 2 или кроличьими антителами против роданезы, затем соответствующими вторыми антителами, с которыми была соединена пероксидаза хрена. Пероксидазу детектировали по хемилюминесценции с применением ECL продуктов Amersham.
Пример 5
Укладка роданезы, меченой кумарином на ее N-конце
Для наблюдения действия "наставников" во время укладки и активации насцентной (растущей) роданезы кумарин трансляционно встраивали в N-конце фермента из СРМ-SAc[35S]-Met-тРНКf, образованную ферментативно, дериватизовали химической реакцией и затем использовали для инициации белкового синтеза на рибосомах E.coli, как описано выше. Включение СРМ-SAc[35S]-метионина на N-концы полипептидов, образуемых в системе сопряженной транскрипции/трансляции, наблюдали по радиоактивности после осаждения трихлоруксусной кислотой синтезируемых in vitro полипептидов роданезы. Продукт идентифицировали электрофорезом в ПААГ-ДСН и авторадиографией, как показано на фиг. 1. Анализ проводили после отделения рибосом от фракции супернатанта. Имеется разница в миграции на ДСН-полиакриламидных гелях полноразмерной роданезы в супернатанте и во фракции рибосом, соответственно.
Для дальнейшей идентификации этих полноразмерных форм продукта, синтезированного во время сопряженной транскрипции/трансляции полипептиды из дСН-полиакриламидного геля переносили на PVDF-мембрану и зондировали антителами против роданезы. Поскольку связанный с рибосомой полипептид может быть превращен в ферментативно активный растворимый белок, данное изобретение убедительно показывает, что этот полипептид является предшественником растворимого фермента.
Природа различия между растворимым и связанным с рибосомами полипептидом не известна. Одним из предложений является то, что более медленно мигрирующая форма ковалентно связана с тРНК или произведенным из нее нуклеотидным фрагментом. То, что эта гипотеза неверна, доказывается следующими наблюдениями: на характер электрофоретической миграции не влияет прединкубирование пробы со смесью рибонуклеаз или прединкубирование при pH 10,5 при 100oC в течение 10 минут. Эфирная связь между пептидом и 3'-концевой рибозой тРНК должна была бы гидролизоваться при указанных выше условиях. Кроме того, относительно медленно мигрирующий полипептид не был обнаружен со связанными с рибосомами формами хлорамфеникол-ацетилтрансферазы, рицина или дигидрофолатредуктазы. Следовательно, медленно мигрирующий полипептид относится только к роданезе. Природа медленно мигрирующей формы не была установлена, но наиболее вероятно, что она образуется трансляцией через терминирующий кодон UGA, находящийся на конце кодирующей последовательности роданезы, что приводит к трансляции тринадцати дополнительных кодонов перед наступлением терминации при кодоне UAG в фазе. Однако несколько удивительно, что эта более длинная форма предшественника количественно превращается в растворимую форму во время активации и освобождения полипептида.
Радиоактивно меченый метионин из СРМ-SAc[35S]-Met-тРНКf, добавляемой к реакционной смеси, может находиться только на N-конце роданезы, но не внутри ее. Тиоэфирная связь между СРМ и тиолом метионина, ацилированного меркаптоуксусной кислотой, химически и ферментативно стабильна при применяемых условиях инкубирования. СМР-SAcMet-тРНК не деацилируется метионил-тРНК-синтетазой, и тРНКMetm не может ферментативно аминоацилироваться СМР-SAc[35S] метионином. Кроме того, в реакционную смесь включали большой избыток немеченого метионина (350 мкМ). Это значительно уменьшало удельную радиоактивность свободного [35S]-метионина, который мог присутствовать и обеспечивать включение метионина во внутреннее положение полипептида. Поэтому считали, что молярное количество кумарина прямо пропорционально радиоактивности из [35S] -метионина, и радиоактивность использовали для нормализации интенсивности флуоресценции и спектральных данных таким образом, что величины, приводимые ниже для этих параметров, прямо пропорциональны квантовому выходу флуоресценции и могут сравниваться между различными экспериментами.
Пример 6
Распределение вновь синтезированной СРМ-меченой роданезы
Распределение вновь синтезированной СРМ-меченой роданезы в растворимой фракции и фракции рибосом, соответственно, анализировали после осаждения центрифугированием рибосом через слой раствора сахарозы сразу после ее синтеза. Из таблицы 1 видно, что [14C]-лейцин включался во внутренние положения полипептидов роданезы, N-конец которых не был модифицирован кумарином. В этом случае для инициации белкового синтеза использовали нерадиоактивную fMet-тРНКf. Приблизительно половина этих вновь синтезированных полипептидов (62% в расчете на [35S]-метионин и 43% в расчете на [14C]-лейцин) обнаружена во фракции рибосом. Большая часть оставшегося вновь образованного белка извлекалась из супернатанта. Белковый продукт во фракции супернатанта полностью представлял собой полноразмерную роданезу, как было оценено авторадиографией высушенного полиакриламидного геля, на котором анализировали эти полипептиды (фиг. 1).
Молярное количество роданезы, синтезированной в бесклеточной системе транскрипции/трансляции, определяли из включения [35S] -метионина или [14C-лейцина в полипептиды роданезы. При описанных условиях только одна кодирующая последовательность транскрибировалась из этой плазмиды (pSP65), а именно мРНК для роданезы. Включение [14C]-лейцина в пептиды в реакционной смеси без плазмиды было менее 5% от включения в присутствии плазмиды, и эти данные не учитывали. Цепь полноразмерной роданезы содержит 24 остатка лейцина и имеет молекулярную массу 33000. Таким образом, зная процент полноразмерного продукта (почти 100% для фракции супернатанта), молярное количество полноразмерного белка, которое синтезировалось, можно рассчитать из включения N-концевого метионина или внутреннего лейцина. Эту величину можно использовать для расчета удельной активности фермента, единиц/ мг роданезы. Единицы активности роданезы определяли в молях тиоцианата, образованного в минуту.
Пример 7
Активность роданезы
Удельная ферментативная активность роданезы во фракции супернатанта составляла 744 единиц/мг роданезы относительно включенного [14C]-лейцина. Эта активность сходна с удельной активностью нативной роданезы, выделенной из бычьей печени. Таким образом, почти весь продукт, транслирующийся из рибосом во фракцию супернатанта, укладывался в нативную конформацию. В противоположность этому, растущие полипептиды на рибосомах не обладали ферментативной активностью. Подобная ситуация возникает при образовании кумарин-[35S]-метионином N-конца вновь синтезированной роданезы: роданеза в супернатанте активна, тогда как полипептиды на рибосомах ферментативно неактивны.
Меньшее количество [35S]-метионин-меченой роданезы выделялось из рибосом в ходе белкового синтеза с N-концевым зондом, но ее средняя удельная ферментативная активность (833 единиц/мг роданезы) была примерно на 17% выше, чем активность нативного фермента. Это, по-видимому, обусловлено удалением части N-концевого [35S]-метионина из вновь образованного фермента.
Пример 8
Стабильность вновь синтезированной роданезы
Стабильность СРМ-[35S]-метионина на N-конце вновь синтезированной роданезы испытывали непосредственно в супернатанте и во фракции рибосом, соответственно. Снижение осажденной трихлоруксусной кислотой радиоактивности приблизительно на 10% наблюдали после 60 минут инкубирования при 37oC, тогда как во фракции супернатанта не было потери активности (данные не представлены).
Нативная роданеза не имеет N-концевого метионина. Известны специфические ферменты, удаляющие метионин из N-конца многих белков. В случае in vitro синтезированной роданезы, полипептиды, не имеющие N-концевого кумарин-[35S]-метионина, не детектировались радиоактивностью, что приводило к результату, занижающему количество белка роданезы, присутствующего в этих пробах. Более высокая удельная ферментативная активность фракции супернатанта, рассчитанная по N-концевому метионину (833 единиц/мг по сравнению с 744 ед./мг в расчете на внутренний лейцин, таблица 1), по-видимому, отражает эту недооценку белка роданезы, присутствующего в пробе. Также, небольшая часть молекул роданезы могла инициироваться немеченым эндогенным формилметионином из Met -тРНК, которая не устранялась во время получения фракции S30. Следует заметить, что сразу же после синтеза и центрифугирования супернатант обрабатывали рибонуклеазой, затем хроматографировали на Сефадексе G100 для отделения СРМ-меченой роданезы от низкомолекулярных флуоресцентных соединений. Во всяком случае, таблица 1 указывает на то, что кумарин может включаться в N-конце роданезы, и что эти молекулы ферментативно активны при освобождении из рибосом.
Роданезу синтезировали сопряженной транскрипцией/трансляцией с СРМ-SAc[35S] -Met-тРНК в качестве инициирующей тРНК или с fMet-тРНКf в качестве инициирующей тРНК и [14C]-лейцином в качестве радиоактивной метки. После 30 минут инкубирования при 37oC реакционную смесь центрифугировали, получая фракцию рибосом и супернатант, которые анализировали отдельно на количество синтезированного белка или на ферментативную активность. Количества даны для реакционной смеси объемом 30 мкл. Удельная активность фермента (нативной роданезы) равна приблизительно 750 единиц/мл белка, 1-не рассчитывали из-за присутствия неполной и полноразмерной неактивной роданезы на рибосомах. 2-рассчитывали пмоль фермента в скобках (24 остатка лейцина на полипептид роданезы). 3-эта средняя удельная ферментативная активность постоянно была выше, чем удельная активность нативной роданезы.
Пример 9
Характеристики флуоресценции СРМ-меченой роданезы
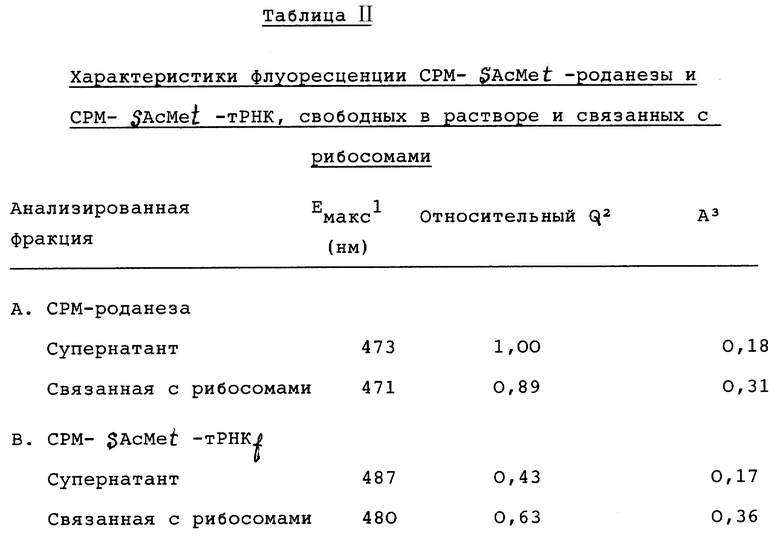
Параметры флуоресценции СРМ-SAc-Met-роданезы, свободной в супернатанте или связанной с рибосомами, суммированы в таблице 11. Соответствующие эмиссионные спектры даны на фиг. 3A и 3B. Соответствующие эмиссионные спектры даны на фиг. 3A и 3B. Для сравнения, в таблицу 11 включены данные для СРМ-SAc-Met-тРНКf, свободной в растворе или связанной с рибосомами. Молярное количество меченой на N-конце СРМ-SAcf-Met-тРНКf или роданезы (в расчете на [35S] -метионин) было примерно одинаково в каждой пробе; затем данные по флуоресценции нормализовали по радиоактивности до общего молярного количества кумарина. Квантовый выход флуоресценции (Q) дан по отношению к квантовому выходу ферментативно активной СРМ-SAcMet-роданезы во фракции супернатанта, для которой относительный квантовый выход был принят за 1,00 (таблица 11). Таким образом, представляется возможность сравнения спектров и относительных квантовых выходов, представленных на чертежах и в таблицах.
Квантовый выход ферментативно активной СРМ-роданезы во фракции супернатанта был несколько выше, чем квантовый выход СРМ-роданезы, связанной с рибосомами (относительный квантовый выход 1,0 против 0,89), но оба были значительно выше, чем флуоресценция из СРМ-SAcMet-тРНКf, как свободной в растворе (Q= 0,43), так и связанной в пептидилтрансферазном центре рибосом (G= 0,63). Последним является случай перед инициированием пептидного синтеза с образованием насцентной (растущей) роданезы с СРМ-SAc-метионином на ее N-конце. Этот квантовый выход отражает местное окружение кумаринового зонда. Для свободного кумарина или СРМ-цистеина квантовый выход увеличивается с батохромным сдвигом в спектре эмиссии с растворителями с понижающейся полярностью. Для сравнения, абсолютный квантовый выход и максимум эмиссии для свободного СРМ-цистеина равны 0,31 и 486 нм, соответственно, в воде, но эти величины равны 0,91 и 470 нм в 95% этаноле, 0,51 и 460 нм в диоксане. Таким образом, спектральные данные таблицы II, по-видимому, указывают на то, что кумариновый зонд СРМ-SAcMet-тРНКf, связанный с рибосомами, встречает более гидрофобное окружение, когда он присутствует на N-конце насцентного пептида, но все еще удерживается в виде пептидил-тРНК на рибосомах. Наблюдалось дальнейшее увеличение квантового выхода по мере освобождения растущей роданезы во фракцию супернатанта в виде ферментативно активной формы. Эти различия, по-видимому, отражают различные стадии в укладке растущей роданезы постепенно в нативную конформацию, как будет рассмотрено ниже. Анизотропия флуоресценции была приблизительно 0,17 для СРМ-Met-тРНКf, свободной в растворе, и приблизительно 0,36 при связанном с рибосомами состоянии (таблица II). Анизотропия для СРМ-Met на N-конце связанной с рибосомами роданезы была 0,31, но падала до 0,18 при освобождении фермента из рибосом в ферментативно активной форме в супернатант. Это различие, по-видимому, отражает затруднение движения флуорофора, создаваемое рибосомой, вследствие стерического препятствия движению самого зонда и/или различием во вращательной релаксации или времени нахождения в растворе свободной роданезы и связанной с рибосомами роданезы, соответственно. Последний параметр отражает различие в массе роданезы и рибосом.
Характеристики флуоресценции для роданезы определяли со сходными или идентичными пробами, применяемыми для определения синтеза и ферментативной активности роданезы, представленных в таблице I. Для сравнения включены данные по флуоресценции для СРМ-SAcMet-тРНК (пункт B. выше). 1Eмакс. - длина волны эмиссионного спектра, 2относ. Q - относительный квантовый выход флуоресценции; его определяли из интегрированного эмиссионного спектра флуоресценции, нормализованного на основе радиоактивности [35S]-метионина и выражали относительно квантового выхода ферментативно активной СРМ-SAcMet-роданезы (сравни таблицу 1) во фракции супернатанта. 3A - анизотропия флуоресценции.
Пример 10
Действие молекулярных "наставников" на связанную с рибосомами СРМ-роданезу
Фермент, освобожденный из рибосомы в присутствии "наставников" был ферментативно активен с удельной активностью, сходной с удельной активностью нативной роданезы, и содержал высокую долю молекул фермента в нативной конформации. В противоположность этому, связанная с рибосомами роданеза была ферментативно неактивной.
Данное изобретение иллюстрирует действие "наставников" (DnaJ, DnaK, GrpE, GroEL, GroES) на флуоресценцию из связанной с рибосомами, ферментативно неактивной растущей СРМ-роданезы и коррелирует наблюдаемые эффекты с высвобождением и удельной ферментативной активностью этого фермента. Последний параметр был взят как мера правильной укладки роданезы в нативную конформацию. Каждый из пяти "наставников" добавляли отдельно, затем все вместе одновременно к реакционной смеси, содержащей рибосомы с полноразмерной роданезой, при нетранслирующих условиях.
Таблица IIIA показывает, что наставники DnaK и GroES (каждый) вызывают значительное освобождение полипептидов роданезы, что отражено уменьшением в анизотропии флуоресценции. В противоположность этому, DnaJ дал небольшое, но значимое увеличение в анизотропии. Наставники GrpE и GroEL оказывали очень небольшое влияние на флуоресценцию. Заметно большее действие как на количество белка, освобожденного из рибосом, так и на флуоресценцию, наблюдали при инкубировании рибосом со всеми наставниками одновременно. Затем реакционные смеси разделяли центрифугированием на фракции рибосом и супернатанта.
Количество в пмолях [35S]-метионина в полипептидах роданезы во фракции супернатанта дано в таблице IIIB. Удельная ферментативная активность роданезы во фракции супернатанта после инкубирования со всеми наставниками вместе приближалась к удельной активности нативной роданезы, 695 единиц/мг по сравнению с 745 для нативного фермента. Квантовый выход, анизотропия и максимум эмиссии освобожденного материала также были очень сходны с этими параметрами для ферментативно активной роданезы (срав. таблицу II). В противоположность этому, роданеза, освобожденная из рибосом в присутствии только DnaK или GroES, была по существу ферментативно неактивна, хотя она и состояла более чем на 90% из полноразмерных полипептидов роданезы (данные не представлены). Также ее квантовый выход и максимум эмиссии отличались от этих параметров ферментативно активной роданезы, что позволяло предположить, что этот материал не был уложен в нативную конформацию. Таким образом, данное изобретение демонстрирует, что молекулярные наставники усиливают эффективное складывание в нативную конформацию только при их совместном действии на связанной с рибосомами растущей роданезе. DnaK или GroES вызывают некоторое высвобождение насцентных цепей из рибосом, но другие наставники являются необходимыми для завершения процесса укладки на рибосомах.
Рибосомы, содержащие 4,5 пмоль СРМ-SAc[35S]-метионин в растущих пептидах роданезы, инкубировали в течение 10 минут при 37oC при нетранслирующих условиях с указанными наставниками. Характеристики флуоресценции реакционных смесей определяли либо сразу (часть A), либо для фракции супернатанта только после центрифугирования (часть B). Ферментативную активность определяли только во фракции супернатанта. 1- сокращения соответствуют тем, что даны в таблице II. *- материал, непригодный для надежного измерения флуоресценции.
Качество коммерческих наставников оценивали электрофорезом в ПААГ-ДСН с последующим окрашиванием Кумасси. Наставники дали единственную полосу ожидаемого размера или ожидаемой электрофоретической подвижности, за исключением GroES, который содержал некоторое количество более высокомолекулярных примесей, и DnaJ, который давал двойную полосу. Все используемые наставники испытывали на присутствие рилизинг-фактора 1 или рилизинг-фактора 2 в виде примеси. Вестерн-блоты из гелей ПААГ-ДСН зондировали соответствующими антителами. Ни один из рилизинг-факторов не был обнаружен ни в одном из препаратов наставников при помощи этого способа (данные не показаны).
Таблица III показывает, что ни один отдельный наставник не был способен превращать неактивную связанную с рибосомами роданезу в природную конформацию. Затем исследовали последовательное добавление одного или двух наставников. Неожиданным аспектом данного изобретения, иллюстрируемым в таблице IV, было то, что, по-видимому, важным является порядок добавления. Ферментативно активную роданезу получали только в том случае, если сначала добавляли DnaJ, а затем DnaK и GrpE и, наконец, GroEL плюс GroES (секция A таблицы IV). Освобождение из рибосом и активация были ассоциированы со снижением в анизотропии и увеличением в относительном квантовом выходе. В противоположность этому, анизотропия оставалась высокой и квантовый выход изменялся очень мало, когда GroEL/ES были первыми наставниками, с которыми инкубировали связанные с рибосомами полипептиды роданезы, а затем добавляли недостающие наставники (секция B в таблице IV). В этой ситуации ферментативная активность была близка к нижнему пределу детектирования. В таблице IV параметры флуоресценции определяли во всей реакционной смеси, тогда как ферментативную активность определяли во фракции супернатанта после центрифугирования, GroEL/ES могут действовать на флуоресцентные молекулы роданезы, все еще связанные с рибосомами, или на молекулы, освобожденные перед добавлением GroEL/ES.
Рибосомы, содержащие 4,5 пмоль СРМ-SAc[35S]-метионина растущих полипептидов роданезы, инкубировали по 10 минут при 37oC с указанными наставниками в порядке, показанном в пунктах A или B. Параметры флуоресценции определяли как описано выше, затем пробы центрифугировали и полученный супернатант анализировали на количество освобожденной из рибосом роданезы и на ферментативную активность, из которой рассчитывали удельную ферментативную активность, 1-сокращения указаны в таблице II. 2-измеряли в супернатанте. После инкубирования при данных условиях и измерений флуоресценции рибосомы отделяли от супернатанта, как описано выше.
Пример 11
Требования для превращения ферментативно неактивной роданезы
Данное изобретение показывает также необходимые условия в отношении наставников и инкубирования для превращения ферментативно неактивной роданезы в ферментативно активную конформацию фермента. Работу, представленную в таблице IV, повторяли до того момента, при котором добавляли DnaK/GrpE (перед добавлением GrpEL+GroES) затем пробу центрифугировали для быстрого отделения освобожденного белка от связанных с рибосомами пептидов роданезы. Супернатант, содержащий освобожденные от рибосом полноразмерные пептиды роданезы, пипетировали обратно в кювету для измерения флуоресценции, определяли параметры флуоресценции (таблица V, секция A, линия 1) и добавляли
GroEL/ES (линия 2). После 10 минут инкубирования снова снимали спектр и рассчитывали анизотропию, как описано выше. Подобным образом анализировали фракцию рибосом, содержащую связанную с рибосомами роданезы, после того как ее ресуспендировали в растворе B и повторно инкубировали в отсутствие или в присутствии GroEL/ES. Затем определяли спектральные свойства этих проб (таблица VB, первые 2 столбца). Наконец, пробы опять центрифугировали, получая супернатант и фракцию рибосом. Количество и удельную ферментативную активность определяли для освобожденной роданезы, которая присутствовала во фракции этого супернатанта (таблица VB, последние 2 столбца). Таблица V показывает, что приблизительно 1,8 пмоль СРМ-роданезы (приблизительно 30% связанной с рибосомами СРМ-роданезы в первоначальной пробе) освобождалось в супернатант после инкубирования с DnaJ, DnaK и GrpE, но без добавления GroEL/ ES. Этот материал имел низкую анизотропию (как и ожидали) и не имел ферментативной активности. Добавление GroEL/ES к этой фракции, содержащей освобожденную СРМ-роданезу, не влияло на анизотропию и оказывало лишь минимальное действие на ферментативную активность. СРМ-роданеза, связанная с рибосомами, имела очень высокую анизотропию перед добавлением GroEL/ES, поскольку СРМ-роданеза, которая освобождалась из рибосом при помощи DnaK во время предшествующего инкубирования, исключалась из пробы центрифугированием. Анизотропия резко падала при добавлении GroEL/ES (таблица VB) и детектировалась ферментативная активность. Удельная ферментативная активность этого освобожденного белка была равна удельной активности нативного фермента.
Таблица V демонстрирует, что GroEL/ES имеет незначительное влияние или не влияет на освобожденные полипептиды роданезы, но, по-видимому, вызывает активацию и высвобождение связанной с рибосомами роданезы после предварительного отдельного инкубирования этих рибосом с DnaK, DnaJ и GrpE. Для сравнения, в таблицу VC включены результаты для полипептидов роданезы, освобожденных из рибосом пуромицином. Ни один из наставников не оказывал детектируемого действия ни на ферментативную активность, ни на флуоресценцию из СРМ-роданезы после инкубирования этих рибосом с пуромицином. Подобным образом, ни один из наставников, отдельно или в комбинации, не оказывал какого-либо действия на ферментативную активность или флуоресценцию ферментативно активной СРМ-роданезы, освобождавшейся из рибосом во время исходной сопряженной транскрипции/трансляции (данные не приведены).
Для таблицы V СРМ-Met-роданезу, связанную с рибосомами, (приблизительно 4,5 пмоль), инкубировали в растворе B с DnaJ, DnaK и GrpE. Затем пробу центрифугировали и анализировали супернатант и фракцию рибосом после повторного суспендирования последней в том же самом объеме раствора B. Снимали спектры и определяли количество и активность роданезы в малой аликвоте. Затем к каждой пробе добавляли GroEL и GroES и измерения повторяли. Что касается фракции рибосом, после добавления GroEL/ES пробу опять центрифугировали в воздушной центрифуге и определяли количество и удельную ферментативную активность.
Для таблицы V, части C, СРМ-роданезу, связанную с рибосомами, инкубировали в течение 10 минут при 37oC с 2 мМ пуромицином, затем эту инкубационную смесь центрифугировали. Анализировали только полученный супернатант после второго инкубирования без наставников или с наставниками. 1-сокращения указаны в таблице II.
Пример 12
Спарсомицин блокирует освобождение связанной с рибосомами СРМ-роданезы
Антибиотик спарсомицин связывается с субъединицами рибосом 50S с Kd приблизительно 0,1 М и ингибирует пептидилтрансферазную реакцию. В микромолярных концентрациях спарсомицин не влиял на флуоресценцию кумарина из рибосом, несущих СРМ-роданезу. Сначала спарсомицин и затем наставники добавляли по отдельности, как описано в таблице III. Таблица VI показывает, что ни один из наставников не оказывал какого-либо детектируемого действия на измеренные параметры флуоресценции. При добавлении наставников в виде комбинаций (не показано) или по отдельности после спарсомицина имелись только очень небольшие изменения в анизотропии по сравнению с данными, представленными в таблице IIIA, что свидетельствует о том, что большая часть полипептидов СРМ- роданезы остается связанной с рибосомами. Это проверяли центрифугированием пробы после выполнения измерения флуоресценции. Менее 10% молекул роданезы освобождалось в супернатант в присутствии спарсомицина и этот освобожденный от рибосом материал был ферментативно неактивен (данные не приведены). Таким образом, спарсомицин предотвращает высвобождение роданезы при помощи DnaK или GroES из рибосом. Поскольку спарсомицин препятствует пептидилтрансферазной реакции, данное изобретение демонстрирует, что СРМ-роданеза удерживается на рибосомах в виде пептидил-тРНК, и что высвобождение при помощи DnaK и/или GroES вовлекает пептидилтрансферазный центр в реакцию, в результате которой гидролизуется связь между тРНК и полипептидом.
Экспериментальные условия были такие же, что и в случае таблицы III, за исключением того, что перед добавлением каждого из наставников по отдельности добавляли 4 мкМ спарсомицин. Ни один из наставников не вызвал более, чем несколько процентов высвобождения СРМ-роданезы в супернатант.
Пример 13
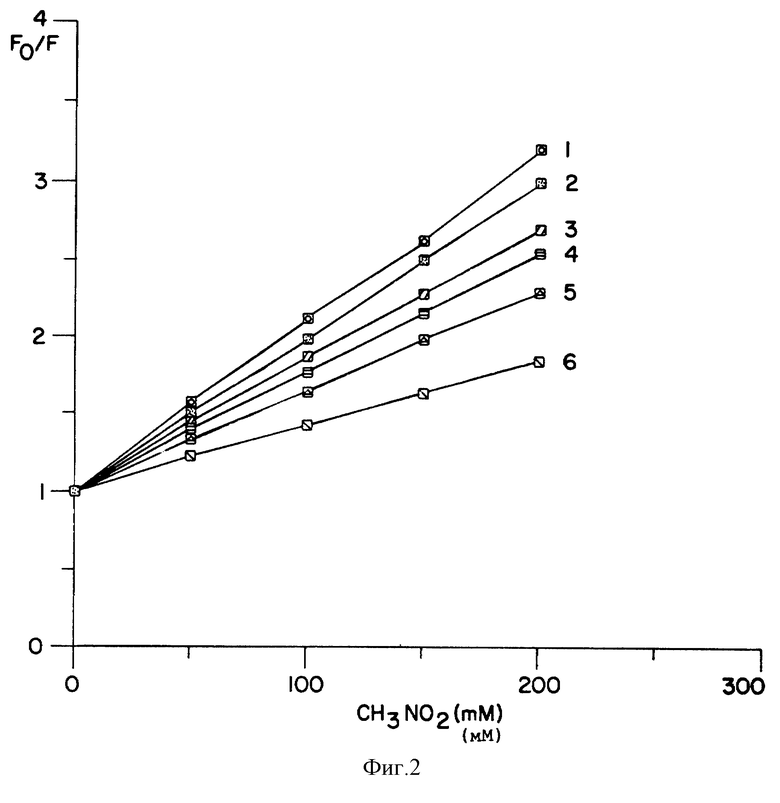
Доступность N-концевого кумарина для нитрометана
Гашение флуоресценции из пептидов СРМ-роданезы нитрометаном оценивали как меру доступности этого зонда в различных ситуациях. Нитрометан представляет собой водорастворимую неионизированную небольшую молекулу, которая эффективно гасит флуоресценцию кумарина в растворе при столкновении с зондом. Кривые Стерна-Вольмера показаны на фиг. 2. СРМ-Met-тРНКf, свободная в растворе и связанная с рибосомами, обнаруживает самое высокое и самое низкое гашение, соответственно. В растворе этот флуорофор относительно доступен, тогда как он в большой мере защищен от нитрометана, когда СРМ-Met-тРНКf связана с рибосомами. СРМ при N-конце ферментативно активной роданезы в растворе был так же относительно доступен. Это согласуется с данными анизотропии для ферментативно активной роданезы (таблица II) и кристаллической структурой роданезы, в которой N-концевой сегмент является открытым. Интересно, что гашение было значительно более низким для роданезы, освобождаемой из рибосом DnaK или пуромицином. Этот материал был ферментативно неактивен, как показано в таблицах III и V. Величины, полученные в обоих случаях, были сходными, но более низкими, чем полученные для свободной, ферментативно активной роданезы. Таким образом, данное изобретение демонстрирует, что N-конец этого освобожденного материала был менее открытым, чем у нативной роданезы и что этот белок не был уложен в его природную конформацию. Еще более низкое гашение в случае связанной с рибосомами роданезы может быть обусловлено отчасти экранированием рибосомами, особенно растущих коротких пептидов, а также неполной укладкой полноразмерных полипептидов.
Пример 14
Промежуточные состояния, детектируемые IgG против кумарина
Поликлональные антитела с высокой специфичностью связывания СРМ-флуорофора использовали для оценки укладки цепей полноразмерной роданезы, связанной с рибосомами, в сравнении с ферментативно активной роданезой, находящейся в свободном виде в растворе. Фиг. 3A и 3B показывают, что этот зонд был доступен для антител против кумарина как на связанной с рибосомами, так и на свободной СРМ-роданезе. Для последней, ферментативно активную роданезу освобождали из рибосом инкубированием со всеми наставниками, затем рибосомы удаляли из реакционной смеси центрифугированием, как описано в связи с таблицей II. Антитела вызывают увеличение в анизотропии и квантовом выходе, который был связан с батохромным сдвигом во флуоресценции как из свободной, так и из связанной с рибосомами СРМ-роданезы. В присутствии IgG против кумарина максимум эмиссии, приблизительно 456 нм, был одинаковым в обоих случаях. Изменение в квантовом выходе и максимуме эмиссии, по-видимому, отражает увеличение в гидрофобности окружения зонда, вызываемое связыванием с иммуноглобулином. Увеличение в квантовом выходе, вызываемое антителами, было меньше в случае связанной с рибосомами роданезы (фиг. 3A), чем в случае ферментативно активной СРМ-роданезы, находящейся в свободном виде в растворе (фиг. 3B). Это различие в квантовом выходе, возможно, обусловлено частичным экранированием более коротких растущих пептидов от антител рибосомами. Этот феномен наблюдали ранее как с растущим на рибосомах полиаланином, так и с растущим белком оболочки MS2. Подобно прежним результатом с СРМ-SAcAla-тРНК, СРМ-SAcMet-тРНКf, связанная в сайте P рибосом, по-видимому, полностью экранирована от IgG, поскольку нет изменения ни в одном из трех параметров при инкубировании с IgG против СРМ (данные не показаны).
Для демонстрации действия наставников на связанную с рибосомами растущую СРМ-роданезу сначала спарсомицин, затем DnaJ или DnaK, отдельно или вместе, добавляли к реакционной смеси, содержащей рибосомы, подобные тем, данные которых отображены на фиг. 3A. В присутствии спарсомицина из рибосом освобождалось очень мало роданезы, как обсуждалось в связи с таблицей VI. После инкубирования добавляли IgG против кумарина. Спектры испускания из полученных смесей показаны на фиг. 4. Для сравнения дается эмиссионный спектр пробы, инкубированной только со спарсомицином (без добавления наставников или IgG против кумарина). GroEL и GroES, добавленные по отдельности или вместе, не дали каких-либо спектральных изменений, подобных изменениям, наблюдаемым с DnaJ без DnaK или с DnaK в присутствии IgG.
Спектры для проб, содержащих DnaK или DnaJ или оба наставника, можно сравнивать непосредственно со спектрами фиг. 3A, полученного для пробы, содержащей связанную с рибосомами роданезу, с IgG или без lqG в отсутствие наставников. Инкубирование с DnaK и затем с IgG имело небольшое действие на полученный эмиссионный спектр, в то время как отчетливо двухвершинная кривая наблюдалась для пробы, инкубированной с DnaJ. Этот эффект был более выраженным для пробы, содержащей как DnaK так и DnaJ. В этом случае отчетливые максимумы видны в спектре при 435 нм и 472 нм. Последний очень близок к максимуму для свободной и связанной с рибосомами роданезы в отсутствие IgG как показано на фиг. 3A и 3B.
Два пика эмиссионного спектра указывают, что в присутствии IgG наблюдается два вида флуресценции в пробах, инкубированных с DnaJ или Dna и DnaK. Это объясняется тем, что антитела против кумарина способны взаимодействовать только с частью СРМ-роданезы, т.к. DnaJ был связан с N-концом некоторых растущих пептидов роданезы. Hartl и сотр. представили данные, свидетельствующие о том, что DnaJ может химически смешиваться со связанными с рибосомами растущими полипептидами. Однако ряд доказательств свидетельствует о том, что DnaJ не связывается с N-концом связанной с рибосомами роданезы (--), по крайней мере, это детектируемо не влияет на N-концевой кумарин. Во-первых, DnaJ не оказывал видимого действия на квантовый выход при его отдельном добавлении, так же как и на анизотропию или эмиссионный спектр, как показано в таблице III. Во-вторых, DnaJ по-видимому, не защищает N-концевой кумарин. Экранирование некоторой части растущей СРМ-роданезы от IgG приводило бы к уменьшению квантового выхода и анизотропии по сравнению с пробой без DnaJ. Такое уменьшение не наблюдалось (сравни вставку фиг. 4 со вставкой фиг. 3A). То, что IgG связывается с кумарином при обоих видах флуоресценции (максимумы приблизительно 440 нм и 470 нм, фиг. 4), доказывается сходным изменением интенсивности флуоресценции, вызываемым IgG обоих областях эмиссионного спектра. Таким образом, фиг. 4 показывает, что DnaJ вызывает изменение в состоянии либо растущего пептида, либо рибосомы, которое влияет на растущий пептид таким образом, что изменяется окружение зонда, когда он связан с IgG. DnaK по-видимому, облегчает это изменение, вызываемое DnaJ.
Пример 15
Другим аспектом настоящего изобретения дрожжевой ТАТА-блок связывающий белок синтезировали в экстракте S30 E.coli посредством сопряженной транскрипции/трансляции из плазмиды, содержащей эту кодирующую последовательность под контролем T7 промотора. В этом случае в инкубационную смесь включали T7 РНК-полимеразу. Инициирующей тРНК была СРМ-SAc[35S]Met-тРНКf.
Из 10 мкл реакционной смеси синтезировали приблизительно 125 нг N (кумарин) белка дрожжей, связывающего ТАТА-блок. Реакционную смесь увеличивали в 62 раза. После инкубирования 600 мкл реакционной смеси, содержащей приблизительно 7,5 мкг меченого белка, наносили на колонку 0,5 мл, содержащую ДЭАЭ-целлюлозу, уравновешенную в 20 мМ Трис-HCl, pH 7,9, 100 мМ KCl, 0,5 мМ этилендиаминтетрауксусную кислоту (ЭДТА), 5 мМ β-меркаптоэтанол и 20% глицерин (буфер A). Приблизительно 0,75 мкг дрожжевого ТАТА-блок связывающего белка не адсорбировались на этом матриксе, приблизительно 1,5 мкг N(кумарин)-белка, связывающего ТАТА-блок элюировали 200 мМ KCl в том же самом буферном растворе. Дополнительные 3,0 мкг кумарин меченого белка элюировали из колонки 350 мМ KCl. Этот материал наносили на гепарин-Сефарозную колонку (объем 1 мл), уравновешенную в буфере А. Приблизительно половина радиоактивно меченого белка адсорбировалась на колонке и элюировалась путем увеличения концентрации соли до 0,7 М KCl в буфере A. Элюированный белок применяли для исследования при помощи флуоресценции связывания дрожжевого ТАТА-блок связывающего белка со специфическими последовательностями ДНК. В качестве контроля использовали мутированную последовательность ДНК. Изменения в квантовом выходе флуоресценции кумарина и анизотропии флуоресценции указывают, что синтезированный in vitro белок, очищенный, как описано выше, специфически взаимодействует с ДНК, содержащей ТАТА-блок.
С применением описанной здесь бесклеточной системы трансляции белковый синтез протекает при относительно высокой линейной скорости приблизительно в течение 20 минут, после чего скорость быстро падает. Ферментативно активная роданеза образуется и высвобождается из рибосом во время периода линейного синтеза, но недостаток этой системы связан с накоплением, по-видимому, полноразмерных полипептидов роданезы в виде пептидил-тРНК в пептидилтрансферазном центре рибосом. О существовании накопленных растущих полипептидов в виде пептидил-тРНК в сайте P рибосом свидетельствует их реакционноспособность с пуромицином и влияние спарсомицина на блокирование последующей терминации и освобождения белка. В применении здесь, термин "терминация" относится только к тем реакциям, которые непосредственно касаются гидролиза эфирной связи между растущим полипептидом и тРНК, тогда как термин "освобождение" относится здесь к отделению белка от рибосомы. Накопленная, связанная с рибосомами насцентная роданеза, была ферментативно неактивна, но освобождалась и превращалась в полностью активный фермент только инкубированием рибосом одновременно с пятью наставниками, DnaK, DnaJ, GrpE, GroEL и GroES. He было детектируемого включения аминокислот в растущий белок во время этого инкубирования при примененных условиях. Роданезу с пониженной ферментативной активностью получали инкубированием рибосом сначала с DnaJ, DnaK и GrpE с последующим вторым инкубированием этих рибосом с GroEL и GrpES. Это, по-видимому, устанавливает порядок этих реакций и подтверждается результатами с DnaJ.
Белок, освобожденный из рибосом реакцией с пуромицином или инкубированием только с DnaK или GroES, был ферментативно неактивным. Этот освобожденный от рибосом белок не активировался инкубированием с любой комбинацией наставников. Это несколько удивительно, поскольку GroEL. и GroES эффективно усиливали повторную укладку в ферментативно активный белок роданезы, которая была денатурирована мочевиной или гуанидин x HCl. Вопрос о том, медиируют ли GroEL и GroES или не медиируют укладку белка на самой рибосоме или они действуют только на освобожденный белок, перед тем как он подвергается вторичным реакциям, которые препятствуют действию GroEL и GroES, требует дополнительных исследований.
Изменения во флуоресценции из кумарина при N-конце растущей роданезы отражают зависимые от наставников реакции, которые имеют место во время активации, терминации и освобождения фермента из рибосомы. Флуоресценция из кумарина очень чувствительна к местному окружению флуорофора, так что изменения в положении зонда в рибосоме, укладка растущего пептида, с которым он связан, или связывание наставника вблизи зонда, по-видимому, отражаются изменениями в квантовом выходе флуоресценции, эмиссионных максимумах и анизотропии. Различия в этих параметрах для полностью ферментативно активной СРМ-роданезы, которая освобождена из рибосом, и связанной с рибосомами СРМ-роданезы очевидны и, по-видимому, отражают различия в конформации белка. Например, можно сравнить относительный квантовый выход ферментативно активной растворимой СРМ-роданезы (таблица II) с выходом ферментативно неактивной растворимой роданезы, которая высвобождалась из рибосом инкубированием с DnaK или GroES (таблица III). Неясно, почему эти явно неродственные белки с АТФ усиливают освобождение роданезы. Их активность в этом отношении напоминает активность рилизинг-фактора 3, RF-3, который имеет ГТФ-азную активность.
То, что роданеза существует в различных конформациях, подтверждается фиг. 2, которая иллюстрирует различия в гашении нитрометаном флуоресценции для белка в различных состояниях. Эти различия, вероятно, отражают различия в экранировании зонда самим белком в различных конформационных состояниях. Однако поразительной особенностью является то, что ни один из наставников, в отдельности или в комбинации, не вызывал резко выраженного изменения во флуоресценции, сравнимого с изменением, наблюдаемым с IgG против кумарина (фиг. 3A и 3B). Это, по-видимому, свидетельствует о том, что ни один из этих наставников не взаимодействует непосредственно с N-концом СРМ-роданезы. Фиг. 4 показывает, что DnaJ усиливает реакцию, которая влияет на связанную с рибосомами растущую роданезу, как это видно из результатов с IgG против кумарина. После реакции с DnaJ N-концевой кумарин существует в двух различных состояниях, которые могут различаться антителами против кумарина. Компонент с максимумом эмиссии вблизи 440 нм образуется одним DnaJ, но усиливается в присутствии DnaK, что позволяет предположить, что последний облегчает реакцию, усиливаемую DnaJ. Возможно, что эти результаты отражают существование растущих цепей разной длины, например, один вид или класс которых взаимодействует с DnaJ или DnaK или обоими, тогда как другой вид, предположительно более короткая форма, может не реагировать с этими наставниками. Эти два (гипотетических) класса разной длины молекул, возможно, представляют собой различно складывающиеся промежуточные продукты, которые существуют при удлинении растущего пептида. Сильно выраженное действие, заметное на фиг. 4, наблюдали только в присутствии спарсомицина. Таким образом, типы флуоресценции с максимумами вблизи 440 и 470 нм отражают промежуточные состояния, генерируемые DnaJ и DnaK, которые улавливаются спарсомицином. Отсюда следует, что они могут предшествовать кодон-направляемому гидролизу пептидил-тРНК в цепи реакций терминации. Применяемые наставники были высокоочищенными и, по-видимому, свободными от рилизинг-факторов. Высокая доля рилизинг-факторов 1 и 2 была связана с рибосомами в клетках Е.Coli.
Неясно, как или почему медиируемые наставниками реакции влияют на медиируемый факторами гидролиз связи пептид-тРНК, терминацию и освобождение растущего белка из рибосомы. Первая реакция блокируется спарсомицином и, как полагают, включает в себя пептидилтрансферазную реакцию. Сходный эффект был отмечен с белками, образующимися на эндоплазматическом ретикулуме. Связь частицы узнавания сигнала с рибосомой ингибирует элонгацию растущего пептида до тех пор, пока комплекс [частица узнавания сигнала-растущий белок-рибосома] не взаимодействует с рецептором или стыкующим белком на мембране эндоплазматического ретикулума.
Возникает вопрос, где в рибосоме происходят реакции терминации, укладки и высвобождения белка. Во время синтеза полипептидов аминоацил-тРНК и короткие пептиды в виде петидил-тРНК удерживаются жестко в гидрофобном сайте, пептидилтрансферазном центре, который расположен у основания центрального бугорка большой субъединицы рибосомы. тРНК связаны со специфическими сайтами на промежуточных поверхностях между двумя субъединицами рибосом с растущим пептидом в виде пептидил-тРНК, расположенным в пептидилтрансферазном центре. Однако электронная микроскопия показывает, что домен выхода, участок, при котором растущий белок выходит из рибосомы, расположен на дистальном конце большой субъединицы на расстоянии до 90  или более от пептидилтрансферазного центра. Длину растущего полиаланинового пептида в остатках аланина применяли для оценки расстояния от пептидилтрансферазного центра до точки, в которой N-концевой зонд пептида становился доступным для IgG или его Fab фрагментов, приблизительно 60 остатков или 90
или более от пептидилтрансферазного центра. Длину растущего полиаланинового пептида в остатках аланина применяли для оценки расстояния от пептидилтрансферазного центра до точки, в которой N-концевой зонд пептида становился доступным для IgG или его Fab фрагментов, приблизительно 60 остатков или 90  (1,5
(1,5  / аминокислотный остаток в спиральной конформации) для IgG и 40 остатков или 60
/ аминокислотный остаток в спиральной конформации) для IgG и 40 остатков или 60  для Fab. Fab фрагмент против кумарина может реагировать с пептидами более короткой средней длины, чем соответствующий IgG, из которого этот фрагмент получен, что позволяет предположить, что его меньший размер позволяет ему проникать дальше в рибосому. Сообщалось о химическом сшивании DnaJ с насцентным пептидом хлорамфениколацетилтрансферазы и люциферазы светляка, таким коротким, как 55 аминокислот в длину, на рибосомах зародышей пшеницы. Эта длина пептида соответствует результатам для полиаланина, приведенным выше.
для Fab. Fab фрагмент против кумарина может реагировать с пептидами более короткой средней длины, чем соответствующий IgG, из которого этот фрагмент получен, что позволяет предположить, что его меньший размер позволяет ему проникать дальше в рибосому. Сообщалось о химическом сшивании DnaJ с насцентным пептидом хлорамфениколацетилтрансферазы и люциферазы светляка, таким коротким, как 55 аминокислот в длину, на рибосомах зародышей пшеницы. Эта длина пептида соответствует результатам для полиаланина, приведенным выше.
Данное изобретение показывает, что амино-конец коротких растущих пептидов экранирован внутри домена рибосомы до тех пор, пока пептиды не удлиняются до длины, при которой их N-конец не будет хорошо удаляться из участка пептидилтрансферазного центра. Этот домен может включать в себя туннель через большую субъединицу рибосомы, как первоначально сообщали Jonath и сотр. Eisenstein et al. (1994) сделали обзор данных по структуре рибосом, показывающий, что в больших субъединицах рибосом существуют один или более туннелей или полостей между пептидилтрансферазным центром и доменом выхода. Crowley и сотр. (1993) представили данные, свидетельствующие о том, что сигнальная последовательность растущих эукариотических мембранных белков перемещается через подобный туннель во время их синтеза. Данное изобретение предполагает, что эта полость внутри большой субъединицы рибосомы обеспечивает убежище, в котором может иметь место, по меньшей мере, часть процесса укладки, в окружении, в котором растущий пептид частично экранирован от протеаз и других разрушающих процессов, которые имеют место в цитоплазме клетки.
Все патенты или публикации, упоминаемые в этой заявке, определяют уровни специалистов в этой области, к которым обращено это изобретение. Эти патенты и публикации включены здесь ссылками, равноценными тому, как если бы каждая отдельная публикация была специально и отдельно указана как включенная в виде ссылки.
Специалисту в данной области будет понятно, что данное изобретение хорошо адаптируется для достижения упомянутых выше целей и получения упомянутых результатов и преимуществ, а также других свойственных ему преимуществ. Представленные примеры вместе со способами, процедурами, обработками, молекулами и характерными соединениями, описанными здесь, представляют собой предпочтительные в настоящее время варианты, являются примерными и не предназначены для ограничений объема изобретения. Изменения в нем и другие применения, которые могут встретиться специалистам в данной области, охватываются существом и объемом изобретения, определяемым объемом формулы изобретения.





Изобретение может быть использовано в биотехнологии. В бесклеточной системе белкового синтеза инкубируют пробы рибосом с плазмидной ДНК, кодирующей необходимый белок. Инкубацию осуществляют в среде для сопряженной транскрипции/трансляции вместе с аминоацил-тРНК, имеющей флуоресцентную метку. Затем полученный белок очищают, измеряют его количество, определяют флуоресценцию и биологическую активность. Нанесение флуоресцентной метки на N-конец синтезированного белка позволяет надежно диагностировать синтезируемые белки, сохраняющие полную ферментативную активность. 8 з.п. ф-лы, 6 табл., 4 ил.
| Analitical Biochemistry, vol | |||
| Гидравлический способ добычи торфа | 1916 |
|
SU206A1 |
| Journal of Biological Chemistry, v | |||
| Аппарат для нагревания окружающей его воды | 1920 |
|
SU257A1 |
| Biochemistry, vol | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Способ получения функционально активных мембранных рецепторных белков | 1990 |
|
SU1723126A1 |

