ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к трансформанту, полученному посредством введения гена, кодирующего цис-пренилтрансферазу, и гена, кодирующего рецептор Nogo-B, в хозяина, что позволяет хозяину экспрессировать цис-пренилтрансферазу и рецептор Nogo-B, и к способу получения полиизопреноида с использованием указанного трансформанта.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
В настоящее время натуральный каучук (один пример полиизопреноидов) для применения в промышленных изделиях из каучука получают из растений, продуцирующих изопреноиды, таких как гевея бразильская (Hevea brasiliensis), принадлежащая к семейству Euphorbiaceae, или фикус каучуконосный (Ficus elastica), принадлежащий к семейству Moraceae.
Сейчас Hevea brasiliensis является фактически единственным источником натурального каучука, используемого в промышленных изделиях из каучука. Hevea brasiliensis представляет собой растение, которое можно выращивать только в определенных географических областях, таких как Юго-Восточная Азия и Южная Америка. Более того, деревья Hevea brasiliensis необходимо выращивать на протяжении приблизительно семи лет, начиная с посадки, чтобы вырастить деревья, достаточно зрелые для получения из них каучука, при этом получать из них натуральный каучук можно лишь в течение периода продолжительностью от 20 до 30 лет. Согласно прогнозам, спрос на натуральный каучук в будущем будет расти, особенно в развивающихся странах, но по причинам, обсуждаемым выше, существенно увеличить количество натурального каучука, получаемого из Hevea brasiliensis, затруднительно. Поэтому есть опасение, что источники натурального каучука иссякнут, и существует потребность в разработке стабильных источников натурального каучука, отличных от зрелых деревьев Hevea brasiliensis, и повышении продуктивности при получении натурального каучука из Hevea brasiliensis.
Натуральный каучук имеет цис-1,4-полиизопреновую структуру с изопентенилдифосфатной (IPP) единицей, и природа этой структуры указывает на то, что в биосинтез натурального каучука вовлечена цис-пренилтрансфераза (CPT). Например, несколько CPT обнаружено у Hevea brasiliensis, включая трансферазу 1 каучука Hevea (HRT1) и трансферазу 2 каучука Hevea (HRT2) (см., например, непатентные литературные источники 1 и 2). Также известно, что синтез каучука у вида одуванчика Taraxacum brevicorniculatum может быть ослаблен посредством подавления экспрессии CPT (см., например, непатентный литературный источник 3).
Предшествующие исследования белков, связанных с биосинтезом натурального каучука, были сосредоточены на факторе элонгации каучука (rubber elongation factor, REF) и белке малых частиц каучука (small rubber particle protein, SRPP) (см., например, непатентные литературные источники 4 и 5). Тем не менее, взаимосвязи между этими белками и CPT не вполне понятны.
Также было высказано предположение о том, что рецептор Nogo-B (NgBr) вовлечен в биосинтез долихолов человеческой CPT (см., например, непатентный литературный источник 6).
Список процитированной литературы
Непатентные литературные источники
Непатентный литературный источник 1: Rahaman et al., ВМС Genomics, 2013, vol. 14.
Непатентный литературный источник 2: Asawatreratanakul et al, European Journal of Biochemistry, 2003, vol. 270, pp. 4671-4680.
Непатентный литературный источник 3: Post et al., Plant Physiology, 2012, vol. 158, pp. 1406-1417.
Непатентный литературный источник 4: Hillebrand et al., PLoS ONE, 2012, vol. 7.
Непатентный литературный источник 5: Priya et al., Plant Cell Reports, 2007, vol. 26, pp. 1833-1838.
Непатентный литературный источник 6: K.D. Harrison et al., The EMBO Journal, 2011, vol. 30, pp. 2490-2500.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Техническая проблема
Как обсуждено выше, существует потребность в разработке стабильных источников натурального каучука, отличных от зрелых деревьев Hevea brasiliensis, и повышении продуктивности при получении натурального каучука из Hevea brasiliensis. Тем не менее, в настоящее время механизм биосинтеза натурального каучука и, в частности, механизм его регуляции, остаются в значительной степени неясными, и все еще сохраняется большой потенциал для улучшения и значительного увеличения количества получаемого натурального каучука. В этом контексте одним возможным способом решения этих проблем является стабилизация и усиление активности СРТ при биосинтезе натурального каучука для увеличения количества получаемого натурального каучука.
В свете этих обстоятельств настоящее изобретение направлено на обеспечение трансформанта, полученного посредством введения гена, кодирующего цис-пренилтрансферазу, и гена, кодирующего рецептор Nogo-B, которые считают вовлеченными в биосинтез полиизопреноидов, в хозяина, что позволяет хозяину экспрессировать цис-пренилтрансферазу и рецептор Nogo-B, и способа получения полиизопреноида с использованием данного трансформанта.
Решение проблемы
Настоящее изобретение относится к трансформанту, полученному посредством введения гена, кодирующего цис-пренилтрансферазу, и гена, кодирующего рецептор Nogo-B, в хозяина, что позволяет хозяину экспрессировать цис-пренилтрансферазу и рецептор Nogo-B.
Предпочтительно, по меньшей мере один из гена, кодирующего цис-пренилтрансферазу, или гена, кодирующего рецептор Nogo-B, имеет происхождение из растения.
Предпочтительно, по меньшей мере один из гена, кодирующего цис-пренилтрансферазу, или гена, кодирующего рецептор Nogo-B, имеет происхождение из растения, продуцирующего изопреноиды.
Предпочтительно, по меньшей мере один из гена, кодирующего цис-пренилтрансферазу, или гена, кодирующего рецептор Nogo-B, имеет происхождение из Hevea brasiliensis.
Предпочтительно, ген, кодирующий рецептор Nogo-B, представляет собой любую из следующих ДНК:
[3] ДНК, имеющая нуклеотидную последовательность SEQ ID NO: 5; и
[4] ДНК, способная гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 5, в жестких условиях.
Предпочтительно, ген, кодирующий цис-пренилтрансферазу, представляет собой любую из следующих ДНК:
[1] ДНК, имеющая нуклеотидную последовательность SEQ ID NO: 1 или 3; и
[2] ДНК, способная гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1 или 3, в жестких условиях и кодирующая белок, имеющий ферментативную активность, катализирующий реакцию элонгации цис-цепи изопреноидного соединения.
Предпочтительно, ген, кодирующий цис-пренилтрансферазу, представляет собой любую из следующих ДНК:
[1-1] ДНК, имеющая нуклеотидную последовательность SEQ ID NO: 1; и
[2-1] ДНК, способная гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1, в жестких условиях и кодирующая белок, имеющий ферментативную активность, катализирующий реакцию элонгации цис-цепи изопреноидного соединения.
Хозяин предпочтительно представляет собой растение, продуцирующее изопреноиды.
Настоящее изобретение также относится к способу получения полиизопреноида с использованием описанного выше трансформанта.
Полезные эффекты изобретения
Трансформант по настоящему изобретению получен посредством введения гена, кодирующего цис-пренилтрансферазу (СРТ), и гена, кодирующего рецептор Nogo-B (NgBr), в хозяина, что позволяет хозяину экспрессировать СРТ и NgBr. Поскольку СРТ и NgBr коэкспрессированы в хозяине, ожидают, что активность СРТ будет стабилизирована и усилена. Соответственно, ожидают, что у трансформанта количество продуктов биосинтеза через реакции, которые катализирует СРТ, и, следовательно, количество полученных полиизопреноидов, будет увеличено, и, таким образом, использование такого трансформанта в получении полиизопреноидов может привести к повышению выхода полиизопреноидов.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 представлены фотографии, на которых показаны результаты двугибридного анализа у дрожжей.
ОПИСАНИЕ ВОПЛОЩЕНИЙ ИЗОБРЕТЕНИЯ
Автор настоящего изобретения проводил различные исследования для улучшения продуктивности при получении полиизопреноидов. В этих исследованиях он сосредоточился на цис-пренилтрансферазе (СРТ), поскольку ее считают одним из ферментов, играющих ключевую роль в биосинтезе полиизопреноидов. Согласно одному сообщению (непатентный литературный источник 6), рецептор Nogo-B (NgBr) взаимодействует с СРТ у людей, улучшая стабильность белка СРТ, и посредством этого стабилизирует и усиливает активность биосинтеза долихолов. Это позволяет предполагать возможное вовлечение NgBr в активность СРТ у организмов в целом. В свете описанного выше, автор настоящего изобретения получил трансформант, конструированный для экспрессии СРТ и NgBr. Ожидают, что у такого трансформанта, благодаря сосуществованию СРТ и NgBr, активность СРТ будет стабилизирована и усилена. Соответственно, ожидают, что количество продуктов биосинтеза через реакции, которые катализирует СРТ, и, следовательно, количество полученных полиизопреноидов, будет увеличено, приводя к повышению выхода полиизопреноидов.
Трансформант по настоящему изобретению получен посредством введения гена, кодирующего цис-пренилтрансферазу (СРТ), и гена, кодирующего рецептор Nogo-B (NgBr), в хозяина, что позволяет хозяину экспрессировать СРТ и NgBr.
Ген, кодирующий СРТ, и/или ген, кодирующий NgBr, могут иметь любое происхождение, и предпочтительно имеют происхождение из растения, более предпочтительно растения, продуцирующего изопреноиды. Еще более предпочтительно оба гена имеют происхождение из по меньшей мере одного растения, продуцирующего изопреноиды, выбранного из группы, состоящей из, среди прочего, Hevea brasiliensis, Sonchus oleraceus, Parthenium argentatum и Taraxacum kok-saghyz, особенно предпочтительно Hevea brasiliensis
(Аминокислотная последовательность цис-пренилтрансферазы (СРТ))
Конкретным примером СРТ является следующий белок [1]:
[1] белок, имеющий аминокислотную последовательность SEQ ID NO: 2 или 4.
Известно, что белки имеющие одну или более чем одну аминокислотную замену, делецию, вставку или присоединение относительно исходной аминокислотной последовательности, могут сохранять функцию исходного белка. С учетом этого факта, другим конкретным примером СРТ является следующий белок [2]:
[2] белок, имеющий аминокислотную последовательность, содержащую одну или более чем одну аминокислотную замену, делецию, вставку и/или присоединение относительно аминокислотной последовательности SEQ ID NO: 2 или 4, и имеющий присущую ему функцию.
Функция, присущая белку с SEQ ID NO: 2 или 4, относится здесь к функции, присущей СРТ, то есть ферментативной активности, катализирующей реакцию элонгации цис-цепи изопреноидного соединения.
Для сохранения ферментативной активности аминокислотная последовательность предпочтительно содержит одну или более, более предпочтительно 1-58, еще более предпочтительно 1-44, еще более предпочтительно 1-29, особенно предпочтительно 1-15, наиболее предпочтительно 1-6, наиболее предпочтительно 1-3 аминокислотных замен, делеций, вставок и/или присоединений относительно аминокислотной последовательности SEQ ID NO: 2.
Кроме того, для сохранения ферментативной активности аминокислотная последовательность предпочтительно содержит одну или более, более предпочтительно 1-57, еще более предпочтительно 1-43, еще более предпочтительно 1-29, особенно предпочтительно 1-15, наиболее предпочтительно 1-6, наиболее предпочтительно 1-3 аминокислотных замен, делеций, вставок и/или присоединений относительно аминокислотной последовательности SEQ ID NO: 4.
Среди прочих аминокислотных замен, предпочтительны консервативные замены. Конкретные примеры включают замены в пределах каждой из следующих групп в скобках: (глицин, аланин), (валин, изолейцин, лейцин), (аспарагиновая кислота, глутаминовая кислота), (аспарагин, глутамин), (серии, треонин), (лизин, аргинин), (фенилаланин, тирозин) и тому подобное.
Также известно, что белки с аминокислотными последовательностями, в высокой степени идентичными исходной аминокислотной последовательности, могут также иметь сходную функцию. С учетом этого факта, другим конкретным примером СРТ является следующий белок [3]:
[3] белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 80% идентичности с аминокислотной последовательностью SEQ ID NO: 2 или 4, и имеющий присущую ему функцию.
Для сохранения присущей функции, то есть ферментативной активности, описанной выше, идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 2 или 4 составляет предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95%, особенно предпочтительно по меньшей мере 98%, наиболее предпочтительно по меньшей мере 99%.
Идентичность двух аминокислотных последовательностей или нуклеотидных последовательностей может быть определена с использованием алгоритма BLAST [Pro. Natl. Acad. Sci. USA, 90, 5873 (1993)], разработанного Karlin и Altschul, или FASTA [Methods Enzymol., 183, 63 (1990)].
Наличие или отсутствие у белка описанной выше ферментативной активности может быть определено обычными методиками, такими как экспрессия целевого белка в трансформанте, полученном посредством введения гена, кодирующего целевой белок, в Escherichia coli или другие организмы-хозяева, и определение наличия или отсутствия функции целевого белка соответствующим методом измерения активности.
Предпочтительно, СРТ представляет собой любой из следующих белков:
[1-1] белок, имеющий аминокислотную последовательность SEQ ID NO: 2;
[2-1] белок, имеющий аминокислотную последовательность, содержащую одну или более чем одну аминокислотную замену, делецию, вставку и/или присоединение относительно аминокислотной последовательности SEQ ID NO: 2, и имеющий присущую ему функцию; и
[3-1] белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 90% идентичности с аминокислотной последовательностью SEQ ID NO: 2, и имеющий присущую ему функцию. Автор настоящего изобретения обнаружил, что, когда СРТ представляет собой любой из указанных выше белков, он взаимодействует с NgBr. В этом случае ожидают, что активность СРТ будет дополнительно стабилизирована и усилена.
(Аминокислотная последовательность рецептора Nogo-B (NgBr))
Конкретным примером NgBr является следующий белок [4]:
[4] белок, имеющий аминокислотную последовательность SEQ ID NO: 6.
Как описано выше, известно, что белки имеющие одну или более чем одну аминокислотную замену, делецию, вставку или присоединение относительно исходной аминокислотной последовательности, могут иметь присущую функцию. Таким образом, другим конкретным примером NgBr является следующий белок [5]:
[5] белок, имеющий аминокислотную последовательность, содержащую одну или более чем одну аминокислотную замену, делецию, вставку и/или присоединение относительно аминокислотной последовательности SEQ ID NO: 6, и имеющий присущую ему функцию.
Функция, присущая белку по SEQ ID NO: 6, относится здесь к функции, присущей NgBr, то есть функциям связывания с мембраной через один или более чем один трансмембранный домен на N-концевой стороне белка и взаимодействия с другим белком на его С-концевой стороне. Известно, что в качестве функции рецептора Nogo-B в нейронах NgBr взаимодействует с ингибирующим белком миелина (например, Nogo-A) или сходными с ним белками и, посредством этого, вовлечен в передачу сигналов, приводящих к коллапсу конуса роста нейронов и ингибированию отрастания нейритов. Тем не менее, здесь взаимодействие, вовлеченное в передачу сигналов, описанную выше, не включено в функцию, присущую белку по SEQ ID NO: 6.
Для сохранения присущей функции, то есть функции NgBr, аминокислотная последовательность предпочтительно содержит одну или более, более предпочтительно от 1 до 52, еще более предпочтительно от 1 до 39, еще более предпочтительно от 1 до 26, особенно предпочтительно от 1 до 13, наиболее предпочтительно от 1 до 6, наиболее предпочтительно от 1 до 3 аминокислотных замен, делеций, вставок и/или присоединений относительно аминокислотной последовательности SEQ ID NO: 6.
Аналогично описанному выше, среди прочих аминокислотных замен, предпочтительны консервативные замены. Конкретные примеры включают замены в пределах каждой из следующих групп в скобках: (глицин, аланин), (валин, изолейцин, лейцин), (аспарагиновая кислота, глутаминовая кислота), (аспарагин, глутамин), (серии, треонин), (лизин, аргинин), (фенилаланин, тирозин) и тому подобное.
Как описано выше, также известно, что белки с аминокислотными последовательностями, в высокой степени идентичными исходной аминокислотной последовательности, могут также иметь сходную функцию. Таким образом, другим конкретным примером NgBr является следующий белок [6]:
[6] белок, имеющий аминокислотную последовательность имеющую, по меньшей мере 80% идентичности с аминокислотной последовательностью SEQ ID NO: 6, и имеющий присущую ему функцию.
Для сохранения присущей функции, то есть функции NgBr, идентичность последовательности с аминокислотной последовательностью SEQ ID NO: 6 составляет предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95%, особенно предпочтительно по меньшей мере 98%, наиболее предпочтительно по меньшей мере 99%.
Наличие или отсутствие у белка структуры NgBr может быть определено обычными методиками, такими как экспрессия целевого белка в трансформанте, полученном посредством введения гена, кодирующего целевой белок, в Escherichia coli или другие организмы-хозяева, разрушение трансформанта с последующим разделением на фракции посредством центрифугирования и наблюдением после этого сильной экспрессии в мембранной фракции посредством вестерн-блот-анализа с использованием коммерческого антитела против рецептора Nogo (например, Millipore, GeneTex).
(ДНК, кодирующая цис-пренилтрансферазу (СРТ))
ДНК, кодирующая СРТ, может представлять собой любую из следующих ДНК:
[1] ДНК, имеющая нуклеотидную последовательность SEQ ID NO: 1 или 3; и
[2] ДНК, способная гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1 или 3, в жестких условиях и кодирующая белок, имеющий присущую ему функцию.
Функция, присущая белку, кодируемому ДНК, имеющей нуклеотидную последовательность SEQ ID NO: 1 или 3, представляет собой здесь то же, что функция, присущая белку по SEQ ID NO: 2 или 4.
При использовании здесь термин «гибридизация» обозначает процесс, при котором ДНК гибридизуется с ДНК, имеющей определенную нуклеотидную последовательность, или частью этой ДНК. Соответственно, ДНК, имеющая определенную нуклеотидную последовательность, или часть этой ДНК могут иметь нуклеотидную последовательность, длина которой достаточна для использования в качестве зонда в нозерн- или Саузерн-блот-анализе или в качестве олигонуклеотидного праймера в анализе посредством полимеразной цепной реакции (ПЦР). ДНК, используемая в качестве зонда, может иметь длину по меньшей мере 100 оснований, предпочтительно по меньшей мере 200 оснований, более предпочтительно по меньшей мере 500 оснований, хотя она может представлять собой ДНК длиной по меньшей мере 10 оснований, предпочтительно по меньшей мере 15 оснований.
Методики проведения экспериментов по гибридизации ДНК хорошо известны. Условия гибридизации, в которых проводят эксперименты, могут быть определены согласно, например, Molecular Cloning, 2nd ed. and 3rd ed. (2001), Methods for General and Molecular Bacteriology, ASM Press (1994), Immunology methods manual, Academic press (Molecular), и по многим другим стандартным руководствам.
Жесткие условия могут включать, например, инкубацию фильтра с иммобилизованной ДНК и ДНК-зондом в течение ночи при 42°C в растворе, содержащем 50% формамида, 5х SSC (750 мМ хлорида натрия, 75 мМ цитрата натрия), 50 мМ фосфата натрия (рН 7,6), 5х раствор Денхардта, 10% декстрансульфата и 20 мкг/л денатурированной ДНК молок лосося, с последующей промывкой фильтра, например, в растворе 0,2х SSC при приблизительно 65°C. Также могут быть использованы менее жесткие условия. Жесткость можно менять, изменяя концентрацию формамида (уменьшение процентного содержания формамида приводит к меньшей жесткости), концентрации солей или температуру. Например, условия низкой жесткости включают инкубацию в течение ночи при 37°C в растворе, содержащем 6х SSCE (20х SSCE: 3 моль/л хлорида натрия, 0.2 моль/л д и гидрофосфата натрия, 0,02 моль/л EDTA (этилендиаминтетрауксусная кислота), pH 7,4), 0,5% SDS (додецилсульфат натрия), 30% формамида и 100 мкг/л денатурированной ДНК молок лосося, с последующей промывкой в растворе 1х SSC, содержащем 0,1% SDS, при 50°C. Кроме того, для еще большего снижения жесткости, промывки, проводимые после гибридизации, можно проводить при более высоких концентрациях солей (например, 5х SSC) в указанных выше условиях низкой жесткости.
Возможны изменения различных указанных выше условий посредством включения или замены блокирующих реагентов, используемых для подавления фона в экспериментах по гибридизации. Для включения блокирующих реагентов может быть необходимым модифицировать условия гибридизации для обеспечения совместимости.
ДНК, способная гибридизоваться в жестких условиях, как описано выше, может иметь нуклеотидную последовательность имеющую, по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, еще более предпочтительно по меньшей мере 98%, особенно предпочтительно по меньшей мере 99% идентичности с нуклеотидной последовательностью SEQ ID NO: 1 или 3, согласно расчетам с использованием такой программы, как BLAST или FASTA, с параметрами, указанными выше.
Определить, кодирует ли ДНК, способная гибридизоваться с указанной выше ДНК в жестких условиях, белок, имеющий предопределенную ферментативную активность, можно обычными методиками, такими как экспрессия целевого белка в трансформанте, полученном посредством введения гена, кодирующего целевой белок, в Escherichia coli или другие организмы-хозяева, и определение наличия или отсутствия функции целевого белка соответствующим методом измерения активности.
ДНК, кодирующая СРТ, предпочтительно представляет собой любую из следующих ДНК:
[1-1] ДНК, имеющая нуклеотидную последовательность SEQ ID NO: 1; и
[2-1] ДНК, способная гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1, в жестких условиях, и кодирующая белок, имеющий присущую ему функцию. Автор настоящего изобретения обнаружил, что, когда ДНК, кодирующая СРТ, представляет собой любую из ДНК с описанными выше последовательностями, экспрессируемая СРТ взаимодействует с NgBr. В этом случае ожидают, что активность СРТ будет дополнительно стабилизирована и усилена.
(ДНК, кодирующая рецептор Nogo-B (NgBr))
ДНК, кодирующая NgBr, может представлять собой любую из следующих ДНК:
[3] ДНК, имеющая нуклеотидную последовательность SEQ ID NO: 5; и
[4] ДНК, способная гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 5, в жестких условиях и кодирующая белок, имеющий присущую ему функцию.
Функция, присущая белку, кодируемому ДНК, имеющей нуклеотидную последовательность SEQ ID NO: 5, представляет собой здесь то же, что функция, присущая белку по SEQ ID NO: 6.
Используемый здесь термин «гибридизация» соответствует описанному выше. Жесткие условия также соответствуют описанному выше.
ДНК, способная гибридизоваться в жестких условиях, как описано выше, может иметь нуклеотидную последовательность, имеющую по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, еще более предпочтительно по меньшей мере 98%, особенно предпочтительно по меньшей мере 99% идентичности с нуклеотидной последовательностью SEQ ID NO: 5, согласно расчетам с использованием такой программы, как BLAST или FASTA, с параметрами, указанными выше.
Определить, кодирует ли ДНК, способная гибридизоваться с указанной выше ДНК в жестких условиях, белок, имеющий предопределенную структуру, можно обычными методиками, такими как экспрессия целевого белка в трансформанте, полученном посредством введения гена, кодирующего целевой белок, в Escherichia coli или другие организмы-хозяева, разрушение трансформанта с последующим разделением на фракции посредством центрифугирования и наблюдением после этого сильной экспрессии в мембранной фракции посредством вестерн-блот-анализа с использованием коммерческого антитела против рецептора Nogo (например, Millipore, GeneTex).
Для определения аминокислотной последовательности или нуклеотидной последовательности белков могут быть применены обычные методики. Например, из растущего растения выделяют общую РНК, возможно очищают мРНК (матричная РНК), и посредством реакции обратной транскрипции синтезируют кДНК (комплементарная ДНК). Затем на основе аминокислотной последовательности известного белка, соответствующего целевому белку, разрабатывают вырожденные праймеры, проводят частичную амплификацию фрагмента ДНК посредством ОТ-ПЦР (полимеразная цепная реакция с обратной транскрипцией) и частичное определение последовательности. Затем применяют метод RACE или похожий метод для определения полноразмерной нуклеотидной последовательности или аминокислотной последовательности. Метод RACE (метод быстрой амплификации концов кДНК) относится к методу, в котором, когда нуклеотидная последовательность кДНК частично известна, ПЦР проводят, исходя из информации о нуклеотидной последовательности такой известной области, для клонирования неизвестной области, распространяющейся до конца кДНК, и этот метод позволяет клонировать полноразмерную кДНК посредством ПЦР без получения библиотеки кДНК.
Каждый из вырожденных праймеров может предпочтительно быть получен из последовательности растительного происхождения, часть которой имеет высокое сходство с целевым белком.
Если нуклеотидная последовательность, кодирующая белок, известна, полноразмерную нуклеотидную последовательность или аминокислотную последовательность можно определить, разработав праймер, содержащий инициирующий кодон, и праймер, содержащий терминирующий кодон, используя известную нуклеотидную последовательность, с последующим проведением ОТ-ПЦР с использованием синтезированной кДНК в качестве матрицы.
(Трансформант)
Ген, кодирующий СРТ, и ген, кодирующий NgBr, вводят в хозяина с получением организма (трансформанта), трансформированного для экспрессии СРТ и NgBr. Поскольку СРТ и NgBr коэкспрессированы в трансформанте, ожидают, что активность СРТ будет стабилизирована и усилена. Соответственно, ожидают, что у трансформанта количество продуктов биосинтеза через реакции, которые катализирует СРТ, и, следовательно, количество полученных полиизопреноидов будет увеличено, подходящим образом приводя к повышению выхода полиизопреноидов.
Далее кратко описано, как получить организм (трансформант), трансформированный для экспрессии СРТ и NgBr. Такой трансформант может быть получен обычными известными способами.
Конкретно, например, трансформант может быть получен следующим образом. ДНК, содержащую нуклеотидную последовательность SEQ ID NO: 1 или 3, и ДНК, содержащую нуклеотидную последовательность SEQ ID NO: 5, вводят ниже промотора подходящего вектора экспрессии с использованием подходящих рестриктаз и тому подобного, получая рекомбинантную ДНК. Эта рекомбинантная ДНК может затем быть введена в клетки-хозяева, совместимые с вектором экспрессии, с получением трансформированных клеток. Альтернативно, вектор экспрессии, где ДНК, содержащая нуклеотидную последовательность SEQ ID NO: 1 или 3, введена ниже промотора с использованием подходящих рестриктаз и тому подобного, и вектор экспрессии, где ДНК, содержащая нуклеотидную последовательность SEQ ID NO: 5, введена ниже промотора с использованием подходящих рестриктаз и тому подобного, используют для получения рекомбинантных ДНК, и эти рекомбинантные ДНК могут затем быть введены в клетки-хозяева, совместимые с векторами экспрессии, с получением трансформированных клеток.
Может быть использован любой хозяин (клетки-хозяева), способный экспрессировать интересующие гены, такой как микроорганизмы, дрожжи, клетки животных, клетки насекомых, клетки растений и другие организмы, предпочтительно эукариоты. Поскольку улучшенную продуктивность при получении полиизопреноидов и повышенный выход полиизопреноидов можно ожидать, в частности, при экспрессии СРТ и NgBr у организмов, способных к биосинтезу полиизопреноидов, хозяин предпочтительно представляет собой растение, более предпочтительно, среди прочего, растение, продуцирующее изопреноиды, и клетки-хозяева предпочтительно представляют собой клетки растения, более предпочтительно клетки растения, продуцирующего изопреноиды. Таким образом, в другом подходящем воплощении настоящего изобретения хозяин представляет собой растение, продуцирующее изопреноиды.
Каких-либо конкретных ограничений относительно растения, продуцирующего изопреноиды, нет, при условии, что оно способно продуцировать изопреноид. Примеры включают растения рода Hevea, такие как Hevea brasiliensis; растения рода Sonchus, такие как Sonchus oleraceus, Sonchus asper и Sonchus brachyotus; растения рода Solidago, такие как Solidago altissima, Solidago virgaurea подвид asiatica, Solidago virgaurea подвид leipcarpa, Solidago virgaurea подвид leipcarpa f. paludosa, Solidago virgaurea подвид gigantea и Solidago gigantea Ait. var. leiophylla Fernald; растения рода Helianthus, такие как Helianthus annuus, Helianthus argophyllus, Helianthus atrorubens, Helianthus debilis, Helianthus decapetalus и Helianthus giganteus; растения рода Taraxacum, такие как одуванчики (Taraxacum), Taraxacum venustum H. Koidz, Taraxacum hondoense Nakai, Taraxacum platycarpum Dahlst, Taraxacum japonicum, Taraxacum officinale Weber и Taraxacum kok-saghyz; растения рода Ficus, такие как Ficus carica, Ficus elastica, Ficus pumila L, Ficus erecta Thumb., Ficus ampelas Burm. f., Ficus benguetensis Merr., Ficus irisana Elm., Ficus microcarpa L.f., Ficus septica Burm. f. и Ficus benghalensis; растения рода Parthenium, тате как Parthenium argentatum, Parthenium hysterophorus и Ambrosia artemisiifolia (Parthenium hysterophorus); латук (Lactuca sativa) (Lactuca serriola) и Ficus benghalensis. Растение, продуцирующее изопреноиды, предпочтительно представляет собой по меньшей мере одно, выбранное из группы, состоящей из растений родов Hevea, Sonchus, Taraxacum и Parhenium, более предпочтительно по меньшей мере одно, выбранное из группы, состоящей, среди прочего, из Hevea brasiliensis, Sonchus oleraceus, Parthenium argentatum и Taraxacum kok-saghyz.
Вектор экспрессии может представлять собой вектор, способный к автономной репликации в клетках-хозяевах или включению в их хромосомы и содержащий промотор в положении, позволяющем проводить транскрипцию рекомбинантной ДНК.
В случае, когда в качестве клеток-хозяев используют клетки растений, в качестве вектора экспрессии могут быть использованы, например, вектор рВI, вектор pUC, вектор на основе Ti-плазмиды или вируса табачной мозаики.
Может быть использован любой промотор, функционирующий в клетках растений. Примеры включают промотор 35S вируса мозаики цветной капусты (CaMV), промотор рисового актина-1, промотор гена нопалинсинтазы, промотор 35S вируса табачной мозаики и промотор гена рисового актина.
Предпочтительны векторы экспрессии с промоторами, специфично экспрессируемыми в тканях, в которых происходит биосинтез изопреноидных соединений, таких как млечники. Замедление роста растений и другие нежелательные эффекты можно уменьшить посредством специфичной экспрессии в ткани, в которой происходит биосинтез изопреноида.
Рекомбинантный вектор может быть введен любым способом, позволяющим вводить ДНК в клетки-хозяева. Примеры включают способы с использованием агробактерий (JP S59-140885 A, JP S60-70080 А, WO 94/00977), электропорацию (JP S60-251887 А) и способы с использованием биобаллистических пушек (генных пушек) (JP 2606856 В, JP 2517813 В).
Трансформант (трансгенные растительные клетки) может быть получен описанными выше или другими способами.
Согласно настоящему изобретению также предложено растение, продуцирующее изопреноиды, в которое были введены ген, кодирующий СРТ, и ген, кодирующий NgBr. Каких-либо конкретных ограничений относительно растения, продуцирующего изопреноиды, нет, при условии, что оно представляет собой растение, продуцирующее изопреноиды, содержащее трансгенные растительные клетки. Растение, продуцирующее изопреноиды, концептуально включает не только трансгенные растительные клетки, полученные описанными выше способами, но также, например, все их потомство или клоны и даже потомство из растений, полученных посредством пассирования этих клеток. После получения трансгенных растительных клеток, в геном которых были введены ДНК или вектор, из трансгенных растительных клеток могут быть получены потомство или клоны посредством полового или бесполого размножения, тканевой культуры, клеточной культуры, слияния клеток или другими методиками. Кроме того, трансгенные растительные клетки или их потомство или клоны могут быть использованы для получения репродуктивных материалов (например, семян, плодов, черенков, корневищ, клубней, ростков, придаточных почек, придаточных зародышей, каллусов, протопластов), которые могут затем быть использованы для крупномасштабного получения растения, продуцирующего изопреноиды.
Методики регенерации растений из трансгенных растительных клеток уже известны; например, в Doi et al. раскрыты методики для эвкалипта (JP Н11-127025 А), в Fujimura et al. раскрыты методики для риса (Fujimura et al., (1995), Plant Tissue Culture Lett., vol. 2: p. 74-), в Shillito et al. раскрыты методики для кукурузы (Shillito et al., (1989), Bio/Technology, vol. 7: p. 581-), в Visser et al. раскрыты методики для картофеля (Visser et al., (1989), Theor. Appl. Genet., vol. 78: p. 589-), и в Akama et al. раскрыты методики для Arabidopsis thaliana (Akama et al., (1992), Plant Cell Rep., vol. 12: p. 7-). Специалисты в данной области могут осуществить регенерацию растений из трансгенных растительных клеток, следуя этим документам.
Наличие или отсутствие экспрессии гена целевого белка в регенерированном растении может быть определено хорошо известными методами. Например, для оценки экспрессии целевого белка может быть применен вестерн-блот-анализ.
Семена могут быть получены от трансгенного растения, например, следующим образом: трансгенное растение укореняется в подходящей среде, его переносят на водосодержащую почву в вегетационном сосуде и выращивают при подходящих условиях культивирования с получением, в конечном счете, семян, которые затем собирают. Далее, растения можно выращивать из семян, например, следующим образом: семена, полученные от трансгенного растения, как описано выше, высевают в водосодержащую почву и выращивают при подходящих условиях культивирования с получением растений.
Согласно настоящему изобретению ожидают, что растение, продуцирующее изопреноиды, в которое были введены ген, кодирующий СРТ, и ген, кодирующий NgBr, может быть использовано в получении полиизопреноида с улучшенной продуктивностью. Конкретно, получение полиизопреноида можно осуществлять путем культивирования трансгенных растительных клеток, полученных, как описано выше, каллусов, полученных из таких трансгенных растительных клеток, клеток, редифференцированных из таких каллусов, или тому подобного в подходящей среде или путем выращивания трансгенных растений, регенерированных из трансгенных растительных клеток, растений, выращенных из семян, собранных с таких трансгенных растений, или тому подобного при подходящих условиях культивирования. Ожидают, что трансформант по настоящему изобретению будет иметь стабилизированную и усиленную активность СРТ, благодаря введенным в него белкам. Соответственно, ожидают, что количество продуктов биосинтеза через реакции, которые катализирует СРТ, и, следовательно, количество полученного полиизопреноида, будет увеличено, приводя к повышению выхода полиизопреноида.
При использовании здесь термин «полиизопреноид» является общим термином, используемым для обозначения полимеров, имеющих изопреновые (C5H8) единицы. Примеры полиизопреноидов включают такие полимеры, как монотерпены (C10), сесквитерпены (C15), дитерпены (C20), сестертерпены (C25), тритерпены (C30), тетратерпены (C40) и натуральный каучук. При использовании здесь термин «изопреноид» относится к соединению, имеющему изопреновые (C5H8) единицы, и концептуально включает полиизопреноиды.
Как описано выше, поскольку в настоящем изобретении происходит одновременная экспрессия СРТ и NgBr, ожидают, что активность СРТ будет стабилизирована и усилена. Соответственно, ожидают, что у трансформанта количество продуктов биосинтеза через реакции, которые катализирует СРТ, и, следовательно, количество полученного полиизопреноида, будет увеличено, приводя к повышению выхода полиизопреноида. Таким образом, другой аспект настоящего изобретения относится к способу получения полиизопреноида с использованием трансформанта, полученного посредством введения гена, кодирующего СРТ, и гена, кодирующего NgBr, в хозяина, что позволяет хозяину экспрессировать СРТ и NgBr.
Возможные способы повышения выхода полиизопреноида в присутствии СРТ и NgBr включают, в дополнение к способу применения трансформанта, конструированного для экспрессии как СРТ, так и NgBr in vivo, как описано выше, способ, включающий присутствие как СРТ, так и NgBr in vitro, например, при выделении неочищенных ферментов из клеток, очистки СРТ и NgBr и помещения их вместе в пробирку.
В способе, включающем присутствие как СРТ, так и NgBr in vitro, СРТ и NgBr могут быть получены посредством введения гена, кодирующего СРТ, и/или гена, кодирующего NgBr, в подходящий вектор и использование трансформированных Escherichia coli, дрожжей или растений, бесклеточных систем экспрессии белка или других средств.
Каких-либо конкретных ограничений относительно источника СРТ, используемой в способе, включающем присутствие как СРТ, так и NgBr in vitro, нет, и СРТ предпочтительно имеет происхождение из эукариота, более предпочтительно растения, еще более предпочтительно растения, продуцирующего изопреноиды. СРТ может также быть модифицирована любым способом, таким как добавление модифицирующей группы к ферменту (например, фосфорилирование, метилирование, ацетилирование, пальмитоилирование, миристоилирование, фарнезилирование, добавление сахарной цепи или убиквитинирование), или окисление/восстановление дисульфидой группы, или структурная модификация протеазой.
Каких-либо конкретных ограничений относительно источника NgBr, используемого в способе, включающем присутствие как СРТ, так и NgBr in vitro, нет, и NgBr предпочтительно имеет происхождение из эукариота, более предпочтительно растения, еще более предпочтительно растения, продуцирующего изопреноиды. NgBr может также быть модифицирован любым способом, таким как добавление модифицирующей группы к ферменту (например, фосфорилирование, метилирование, ацетилирование, пальмитоилирование, миристоилирование, фарнезилирование, добавление сахарной цепи или убиквитинирование), или окисление/восстановление дисульфидой группы, или структурная модификация протеазой.
Как описано выше, автор настоящего изобретения обнаружил, что СРТ, кодируемая ДНК, имеющей нуклеотидную последовательность SEQ ID NO: 1, или ДНК, способной гибридизоваться с ДНК, имеющей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1, в жестких условиях и кодирующей белок, имеющий присущую ему функцию, взаимодействует с NgBr. Считают, что, в дополнение к СРТ, NgBr также взаимодействует с третьим белком. Примеры вариантов такого взаимодействия включают следующие два варианта.
(1) СРТ взаимодействует с третьим белком через NgBr, или, иными словами, NgBr, СРТ и третий белок одновременно взаимодействуют друг с другом.
(2) Взаимодействие между NgBr и СРТ и взаимодействие между NgBr и третьим белком происходят по отдельности.
Неограничивающие примеры третьего белка включают фактор элонгации каучука (REF), белок малых частиц каучука (SRPP) и фарнезилдифосфатсинтазу (FPP-синтазу).
В способе, включающем присутствие как СРТ, так и NgBr in vitro, в дополнение к СРТ и NgBr, может дополнительно быть добавлен другой фермент, для совместного присутствия с ними. Примеры фермента включают ферменты, о которых известно, что они присутствуют в латексе, такие как фактор элонгации каучука (REF), белок малых частиц каучука (SRPP), малый GTP(гуанозинтрифосфат)-связывающий белок, гевеин, β-1,3-глюканаза, фарнезилдифосфатсинтаза (FPP-синтаза) и белки-ингибиторы протеаз. Среди них особенно предпочтительны ферменты, взаимодействующие с NgBr, такие как REF.
В способе, включающем присутствие как СРТ, так и NgBr in vitro, дополнительные компоненты, которые могут быть добавлены для совместного присутствия в дополнение к СРТ и NgBr, не ограничены ферментами и могут включать мембраны, используемые для включения в них продуктов взаимодействия. Каких-либо конкретных ограничений относительно типа мембраны нет, и она может представлять собой естественную мембрану, такую как клеточная мембрана или малая частица каучука, или искусственную мембрану, такую как липосома.
Липид, образующий мембрану, может представлять собой липид, способный образовывать двухслойную липидную мембрану, и примеры включают известные глицерогликолипиды, сфингогликолипиды, холестерин и фосфолипиды.
Примеры глицерогликолипидов включают сульфоксирибозилглицериды, дигликозилдиглицериды, дигалактозилдиглицериды, галактозилдиглицериды и гликозилдиглицериды. Примеры сфингогликолипидов включают галактозилцереброзиды, лактозилцереброзиды и ганглиозиды.
Примеры фосфолипидов включают природные или синтетические фосфолипиды, такие как фосфатидилхолины, фосфатидилэтаноламины, фосфатидилсерины, фосфатидные кислоты, фосфатидилдиглицерины, фосфатидилинозиты, лизофосфатидилхолины, сфингомиелины, лецитин яичного желтка, соевый лецитин и гидрогенизированные фосфолипиды.
Примеры фосфатидилхолинов включают соевый фосфатидилхолин, фосфатидилхолин яичного желтка, дилауроилфосфатидилхолин, димиристоилфосфатидилхолин, дипальмитоилфосфатидилхолин, диолеоилфосфатидилхолин и дистеароилфосфатидилхолин.
Примеры фосфатидилэтаноламинов включают диолеоилфосфатидилэтаноламин, дилауроилфосфатидилэтаноламин, димиристоилфосфатидилэтаноламин, дипальмитоилфосфатидилэтаноламин и дистеароилфосфатидилэтаноламин.
Примеры фосфатидилсеринов включают дилауроилфосфатидилсерин, димиристоилфосфатидилсерин, дипальмитоилфосфатидилсерин и дистеароилфосфатидилсерин.
Примеры фосфатидилдиглицеринов включают дилауроилфосфатидилдиглицерин, димиристоилфосфатидилдиглицерин, дипальмитоилфосфатидилдиглицерин и дистеароилфосфатидилдиглицерин.
Примеры фосфатидилинозитов включают дилауроилфосфатидилинозит, димиристоилфосфатидилинозит, дипальмитоилфосфатидилинозит и дистеароилфосфатидил инозит.
ПРИМЕРЫ
Настоящее изобретение конкретно объяснено со ссылкой на примеры, но настоящее изобретение не ограничено этими примерами.
[Клонирование]
(Определение аминокислотной последовательности и нуклеотидной последовательности целевого белка)
Из латекса Hevea brasiliensis выделяли общую РНК методом с использованием горячего фенола. К 6 мл латекса добавляли 6 мл буфера со 100 мМ ацетата натрия, 1 мл 10%-го раствора SDS и затем 12 мл насыщенного водой фенола, предварительно разогретого при 65°C. Смесь инкубировали 5 минут при 65°C, перемешивали с использованием вихревой мешалки и центрифугировали в течение 10 минут при комнатной температуре при 7000 об/мин. После центрифугирования супернатант переносили в новую пробирку, добавляли 12 мл раствора фенола : хлороформа (1:1) и смесь перемешивали посредством встряхивания в течение 2 минут. После перемешивания полученную смесь центрифугировали еще раз в течение 10 минут при комнатной температуре при 7000 об/мин, супернатант переносили в новую пробирку, добавляли 12 мл раствора хлороформа : изоамилового спирта (24:1) и смесь перемешивали посредством встряхивания в течение 2 минут. После перемешивания полученную смесь центрифугировали еще раз в течение 10 минут при комнатной температуре при 7000 об/мин, супернатант переносили в новую пробирку, добавляли 1,2 мл 3 М раствора ацетата натрия и 13 мл изопропанола и смесь перемешивали с использованием вихревой мешалки. Полученную смесь инкубировали в течение 30 минут при -20°C для осаждения общей РНК. Инкубированную смесь центрифугировали в течение 10 минут при 4°C и 15000 об/мин и удаляли супернатант, получая осажденную общую РНК. Полученную общую РНК промывали дважды 70%-м этанолом и затем растворяли в воде, свободной от РНКаз.
Из полученной общей РНК синтезировали кДНК. Синтез кДНК проводили с использованием набора для синтеза первой цепи кДНК PrimeScript II (Takara), следуя инструкции.
Гены СРТ и NgBr получали, используя полученную первую цепь кДНК в качестве матрицы. ПЦР проводили с использованием KOD-plus-Neo (Toyobo Co., Ltd.), следуя инструкции. ПЦР-реакция включала 35 циклов, каждый из которых состоял из 10 секунд при 98°C, 30 секунд при 58°C и 1 минуты при 68°C.
Ген СРТ был получен с использованием следующих праймеров:
праймер 1: 5'-tttggccattacggccatggaattatacaacggtgagagg-3';
праймер 2: 5'-tttggccgaggcggccttattttaagtattccttatgtttc-3'.
Ген NgBr был получен с использованием следующих праймеров:
праймер 3: 5'-tttggccattacggccatggatttgaaacctggag-3';
праймер 4: 5'-tttggccgaggcggcctcatgtaccataattttgctgcac-3'.
Два типа генов СРТ (HRT1 и HRT2) и один тип гена NgBr (NgBr) были получены, как описано выше. Эти три типа генов секвенировали для определения полноразмерной нуклеотидной последовательности и аминокислотной последовательности. Нуклеотидная последовательность HRT1 приведена в SEQ ID NO: 1, нуклеотидная последовательность HRT2 приведена в SEQ ID NO: 3, и нуклеотидная последовательность NgBr приведена в SEQ ID NO: 5. Аминокислотная последовательность HRT1 приведена в SEQ ID NO: 2, аминокислотная последовательность HRT2 приведена в SEQ ID NO: 4, и аминокислотная последовательность NgBr приведена в SEQ ID NO: 6.
(Конструирование векторов)
Полученные амплифицированные ДНК-фрагменты трех типов подвергали добавлению dA и затем вводили в векторы pGEM-T Easy с использованием pGEM-T Easy Vector System (Promega), получая pGEM-HRT1, pGEM-HRT2 и pGEM-NgBr.
(Трансформация E. coli)
Полученными векторами трансформировали Escherichia coli DH5a, трансформантов культивировали на среде с LB-агаром, содержащей ампициллин и X-gal, и клетки Е. coli, несущие введенные целевые гены, отбирали посредством скрининга синей/белой окраски.
Клетки Е. coli, трансформированные плазмидами с целевыми генами, культивировали в течение ночи при 37°C в жидкой среде LB. После культивирования клетки и плазмиды собирали. Для сбора плазмид использовали набор FastGene Plasmid mini kit (Nippon Genetics Co., Ltd.).
Посредством секвенирования было подтверждено, что в нуклеотидных последовательностях генов, введенных в собранные плазмиды, не было мутаций.
[Получение дрожжей, трансформированных для экспрессии гена, кодирующего СРТ, и гена, кодирующего NgBr]
Гены HRT1, HRT2 и NgBr для введения в дрожжевые векторы экспрессии были получены посредством ПЦР с использованием векторов, полученных в приведенном выше разделе «Конструирование векторов», в качестве матриц.
Гены HRT1 и HRT2 были получены с использованием следующих праймеров:
праймер 5: 5'-gacgcccgggaggccatgaa-3';
праймер 6: 5'-cagcttcctcccgggctttg-3'.
Ген NgBr был получен с использованием следующих праймеров:
праймер 7: 5'-tttctcgagatggatttgaaacctggagctg-3';
праймер 8: 5'-tttctcgagtgtaccataattttgctgcac-3'.
Полученные ДНК-фрагменты подвергали добавлению dA и затем вводили в векторы pGEM-T Easy с использованием pGEM-T Easy Vector System (Promega), получая pGEM-HRT1 YE, pGEM-HRT2 YE и pGEM-NgBr YE.
Трансформацию E. coli, сбор плазмид и подтверждение нуклеотидной последовательности проводили, как описано выше в разделе «Трансформация Е. coli», но с использованием полученных векторов. pGEM-HRT1 YE и pGEM-HRT2 YE, для которых было подтверждено отсутствие мутаций нуклеотидной последовательности, обрабатывали рестриктазой SmaI и вводили в pGK426, обработанный сходным образом с использованием SmaI, получая pGK-HRT1 и pGK-HRT2.
Сходным образом, pGEM-NgBrYE, для которой было подтверждено отсутствие мутаций нуклеотидной последовательности, обрабатывали рестриктазой XbaI и вводили в pGK425, обработанный сходным образом с использованием XbaI, получая pGK-NgBr.
Полученными плазмидами трансформировали дрожжевой штамм SNH23-7D. Для двойной трансформации дрожжей применяли метод с использованием PEG (полиэтиленгликоль). Для скрининга трансформированные клетки культивировали при 23°C в течение трех суток на среде SD без Leu и Ura. При получении двойных трансформантов дрожжей были использованы следующие пары.
(1) SNH23-7D/pGK426, pGK425 (без введения генов).
(2) SNH23-7D/pGK-HRT1, pGK425 (штамм, экспрессирующий только HRT1).
(3) SNH23-7D/pGK-HRT2, pGK425 (штамм, экспрессирующий только HRT2).
(4) SNH23-7D/pGK-HRT1, pGK-NgBr (штамм, экспрессирующий HRT1 и NgBr одновременно).
(5) SNH23-7D/pGK-HRT2, pGK-NgBr (штамм, экспрессирующий HRT2 и NgBr одновременно).
Трансформантов SNH23-7D/pGK426, pGK-NgBr (штаммов, экспрессирующих только NgBr) не получали.
[Дрожжевой двугибридный скрининг]
Целевые гены, введенные в плазмиды, полученные в приведенном выше разделе «Клонирование», расщепляли, обрабатывая их рестриктазой SfiI, для введения целевых генов в плазмиды для дрожжевого двугибридного скрининга.
(Конструирование векторов)
Расщепленные гены вводили в дрожжевые двугибридные плазмиды с использованием Ligation high ver. 2 (Toyobo). Использованная дрожжевая двугибридная плазмида-«приманка» представляла собой вектор-«приманку» рВТ3-SUC, и использованная дрожжевая двугибридная плазмида-«добыча» представляла собой вектор-«добычу» pPR3-N. HRT1 или HRT2 вводили в вектор-«добычу» (pPR-HRT1, pPR-HRT2) и HRT1, HRT2 или NgBr вводили в вектор-«приманку» (pBT-HRT1, pBT-HRT2, pBT-NgBr).
(Трансформация Е. coli)
Дрожжевыми двугибридными векторами, полученными, как описано выше, трансформировали Escherichia coli DH5α. Трансформированные клетки культивировали на среде с LB-агаром, содержащей ампициллин, и отбирали клетки Е. coli, несущие введенные целевые гены.
Клетки Е. coli, трансформированные плазмидами с целевыми генами, культивировали в течение ночи при 37°C в жидкой среде LB. После культивирования клетки и плазмиды собирали. Для сбора плазмид использовали набор FastGene Plasmid mini kit (Nippon Genetics Co., Ltd.).
(Двойная трансформация дрожжей)
Собранными плазмидами трансформировали дрожжевой штамм NMY-51. Для двойной трансформации дрожжей применяли PEG-метод. Для скрининга трансформированные клетки культивировали при 30°C в течение трех суток на среде SD без Trp и Leu. Пары, использованные при получении двойных трансформантов дрожжей, показаны в Таблице 1.
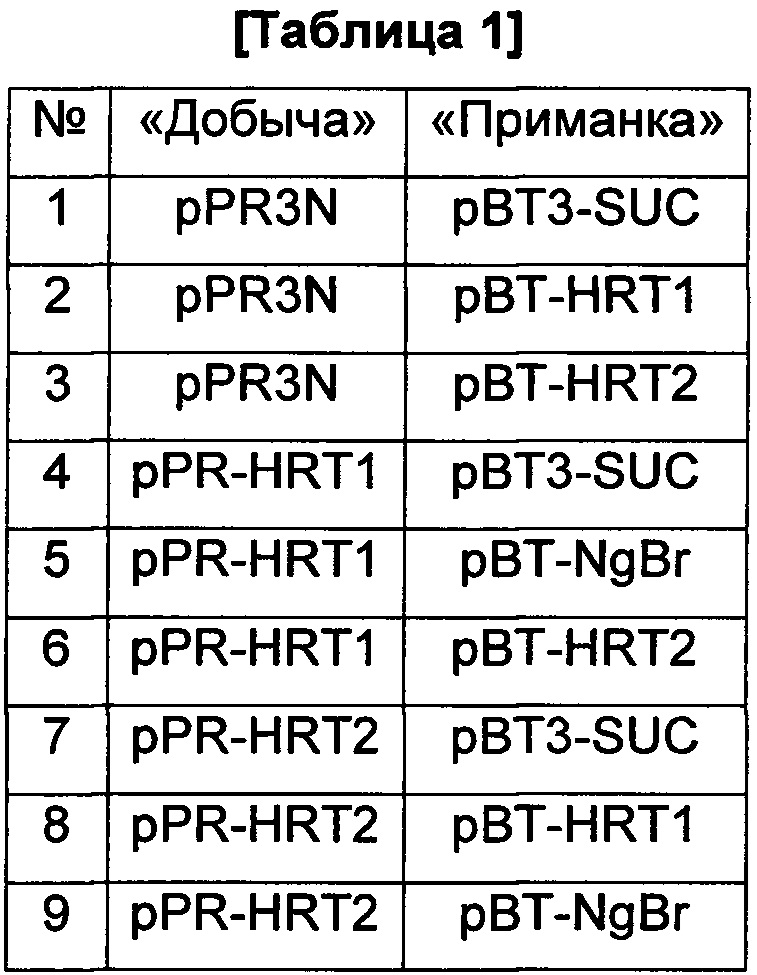
Двойных трансформантов дрожжей наблюдали на предмет взаимодействий между ферментами посредством дрожжевого двугибридного скрининга, культивируя их на среде SD без Trp, Leu, Ade и His в течение трех суток при 30°C. В этом эксперименте дрожжи могут расти на селективной среде только тогда, когда белок, кодируемый геном, введенным в плазмиду-«приманку», взаимодействует с белком, кодируемым геном, введенным в плазмиду-«добычу».
Результаты анализа посредством дрожжевого двугибридного скрининга показаны на Фиг. 1. На Фиг. 1(a) показаны результаты, демонстрирующие наличие или отсутствие взаимодействия HRT1 с HRT2 или NgBr. На Фиг. 1(b) показаны результаты, демонстрирующие наличие или отсутствие взаимодействия HRT2 с HRT1 или NgBr. На Фиг. 1(a) сектора, расположенные по часовой стрелке, начиная с верхнего правого сектора, соответствуют результатам, полученным для пар №№1, 4, 5, 6 и 3, соответственно, в Таблице 1. На Фиг. 1(b) сектора, расположенные по часовой стрелке, начиная с верхнего правого сектора, соответствуют результатам, полученным для пар №№1, 7, 8, 9 и 2, соответственно, в Таблице 1.
Как показано на Фиг. 1, только результаты, полученные для пары №5 в Таблице 1, то есть пары HRT1 и NgBr, на Фиг. 1(a) продемонстрировали взаимодействие. Это показывает, что NgBr взаимодействует с HRT1, являющимся одним типом СРТ, присутствующим в латексе, но не взаимодействует с HRT2. Таким образом, продемонстрировано, что NgBr взаимодействует не со всеми типами СРТ, но взаимодействует с определенной СРТ.
На основании этих результатов можно полагать, что активность HRT1 может быть дополнительно стабилизирована и усилена при комбинировании HRT1 с NgBr.
Перечень условных обозначений
1: пара плазмиды-«добычи» pPR3N и плазмиды-«приманки» рВТ3-SUC.
2: пара плазмиды-«добычи» pPR3N и плазмиды-«приманки» pBT-HRT1.
3: пара плазмиды-«добычи» pPR3N и плазмиды-«приманки» pBT-HRT2.
4: пара плазмиды-«добычи» pPR-HRT1 и плазмиды-«приманки» рВТ3-SUC.
5: пара плазмиды-«добычи» pPR-HRT1 и плазмиды-«приманки» pBT-NgBr.
6: пара плазмиды-«добычи» pPR-HRT1 и плазмиды-«приманки» PBT-HRT2.
7: пара плазмиды-«добычи» pPR-HRT2 и плазмиды-«приманки» рВТ3-SUC.
8: пара плазмиды-«добычи» pPR-HRT2 и плазмиды-«приманки» pBT-HRTL
9: пара плазмиды-«добычи» pPR-HRT2 и плазмиды-«приманки» pBT-NgBr.
(Текстовый перечень последовательностей)
SEQ ID NO: 1: нуклеотидная последовательность гена, кодирующего HRT1 Hevea brasiliensis.
SEQ ID NO: 2: аминокислотная последовательность HRT1 Hevea brasiliensis.
SEQ ID NO: 3: нуклеотидная последовательность гена, кодирующего HRT2 Hevea brasiliensis.
SEQ ID NO: 4: аминокислотная последовательность HRT2 Hevea brasiliensis.
SEQ ID NO: 5: нуклеотидная последовательность гена, кодирующего NgBr Hevea brasiliensis.
SEQ ID NO: 6: аминокислотная последовательность NgBr Hevea brasiliensis.
SEQ ID NO: 7: праймер 1.
SEQ ID NO: 8: праймер 2.
SEQ ID NO: 9: праймер 3.
SEQ ID NO: 10: праймер 4.
SEQ ID NO: 11: праймер 5.
SEQ ID NO: 12: праймер 6.
SEQ ID NO: 13: праймер 7.
SEQ ID NO: 14: праймер 8.







Изобретение относится к области биотехнологии, в частности к трансформированной клетке для экспрессии цис-пренилтрансферазы и рецептора Nogo-B, полученной посредством введения в клетку кодирующего гена, кодирующего цис-пренилтрансферазу, и гена, кодирующего рецептор Nogo-B. Также раскрыт способ получения полиизопреноида, включающий культивирование вышеуказанной трансформированной клетки. Изобретение обеспечивает получение полиизопреноидов с высокой эффективностью. 2 н. и 2 з.п. ф-лы, 1 ил., 1 табл.
1. Трансформированная клетка для экспрессии цис-пренилтрансферазы и рецептора Nogo-B, полученная посредством введения гена, кодирующего цис-пренилтрансферазу, и гена, кодирующего рецептор Nogo-B, в клетку,
где ген, кодирующий рецептор Nogo-B, представляет собой любую из следующих ДНК:
ДНК, содержащая нуклеотидную последовательность SEQ ID NO: 5; и
ДНК, способная гибридизоваться с ДНК, содержащей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 5, в жестких условиях; и
где ген, кодирующий цис-пренилтрансферазу, представляет собой любую из следующих ДНК:
ДНК, содержащая нуклеотидную последовательность SEQ ID NO: 1 или 3; и
ДНК, способная гибридизоваться с ДНК, содержащей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1 или 3, в жестких условиях.
2. Трансформированная клетка по п. 1, где ген, кодирующий цис-пренилтрансферазу, представляет собой любую из следующих ДНК:
ДНК, содержащая нуклеотидную последовательность SEQ ID NO: 1; и
ДНК, способная гибридизоваться с ДНК, содержащей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO: 1, в жестких условиях, и кодирующая белок, имеющий ферментативную активность, катализирующую реакцию элонгации цис-цепи изопреноидного соединения.
3. Трансформированная клетка по п. 1 или 2, где клетка представляет собой клетку растения, продуцирующего изопреноиды.
4. Способ получения полиизопреноида, включающий культивирование трансформированной клетки по любому из пп. 1-3 в подходящей среде.
| HARRISON K.D | |||
| et al.: "Nogo-B receptor is necessary for cellular dolichol biosynthesis and protein N-glycosylation", THE EMBO JOURNAL, vol | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Пропеллер с наружным обедом и поворотными лопастями | 1924 |
|
SU2490A1 |
| TAKAHASHI S | |||
| et al.: "Characterization of cis-prenyltransferases from the rubber producing plant Hevea brasiliensis heterologously expressed in yeast and plant cells", PLANT BIOTECHNOLOGY, vol | |||
| Солесос | 1922 |
|
SU29A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Способ укрепления электродов в катодных лампах | 1923 |
|
SU411A1 |
| SURMACZ L | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Способ укрепления лопаток цепного движителя | 1922 |
|
SU1841A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Русская печь | 1919 |
|
SU240A1 |
| POST J | |||
| et al.: "Laticifer-specific cis-prenyltransferase silencing affects the rubber, triterpene, and inulin content of Taraxacum brevicorniculatum", PLANT PHYSIOLOGY, vol | |||
| Система механической тяги | 1919 |
|
SU158A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Электролитический счетчик ампер-часов | 1925 |
|
SU1406A1 |
| ЛУТОВА Л.А., Биотехнология высших растений, Издательство С.-Петербургского университета, 2010, стр | |||
| Способ крашения тканей | 1922 |
|
SU62A1 |

