Перекрестная ссылка
Настоящая заявка испрашивает приоритет согласно международной заявке на патент №PCT/US 13/61431, поданной 24 сентября 2013 г., и предварительной заявке на патент США №61/954822, поданной 18 марта 2014 г., каждая из которых полностью включена в настоящую заявку посредством ссылки.
Декларация о правах правительства
Настоящее изобретение было выполнено при поддержке правительства США по грантам R01 СА080192 и R01 HLA078836, присужденных Национальным институтом здоровья. Правительство США имеет определенные права на настоящее изобретение.
Уровень техники
Аденовирусы человека (АВ) классифицируют на шесть типов (от А до F), в настоящее время включающих 51 серотип. Большинство серотипов АВ используют рецептор вируса Коксаки и аденовируса (CAR) в качестве рецептора для первичного прикрепления (Bergelson et al., 1997). Однако это не относится к серотипам аденовирусов типа В. Недавно авторы настоящего изобретения предложили новую классификацию для аденовирусов типа В на основании рецепторов, которые используют эти вирусы (Tuve et al., 2006). Вирусы группы 1 (АВ16, 21, 35, 50) используют в качестве рецептора практически исключительно CD46; вирусы группы 2 (АВ3, АВ7, 14) используют общий неустановленный рецептор(ы), который отличается от CD46 и получил временное название рецептор X; вирусы группы 3 (АВ11) взаимодействуют преимущественно с CD46, но также используют рецептор X, если CD46 блокирован.
Аденовирусы типа В являются распространенными патогенами человека. Начиная с 2005 года, в большинстве военных учебных центров США наблюдали одновременное появление различных серотипов типа В. Эти серотипы включали АВ3, АВ7 и АВ14 (Metzgar et al., 2007). В 2007 году в нескольких точках в США и Азии был обнаружен новый высокопатогенный и, возможно, более вирулентный штамм АВ14, АВ14а (Louie et al., 2008; Tate et al., 2009). Авторы настоящего изобретения недавно показали, что, исходя из типа используемого рецептора, АВ14а относится к группе 2 типа В (Wang et al., 2009). В совокупности, в настоящей заявке все серотипы, использующие рецептор X (АВ3, АВ7, АВ14, АВ14а и АВ11), называются АВВ-2/3.
АВВ-2/3 имеют большую значимость в качестве векторов для переноса генов, особенно в отношении опухолей эпителиального происхождения, представляющих большую часть солидных опухолей (Yamamoto and Curiel, 2010). Эпителиальные клетки сохраняют некоторое количество межклеточных контактов и полярность в направлении от апикальной части клетки к базальной. Основные особенности эпителиальных клеток являются консервативными для эпителиальных типов рака in situ и линий раковых клеток (Turley et al., 2008). CAR и CD46 обычно заключены в пределах плотных и адгезионных контактов эпителиальных раковых клеток и недоступны для аденовирусов, которые используют эти рецепторы для прикрепления (Coyne and Bergelson, 2005; Strauss et al., 2009). Напротив, вирусы АВВ-2/3 эффективно инфицируют эпителиальные раковые клетки, что частично осуществляется за счет индукции процессов, которые напоминают эпителиально-мезенхимальный переход (ЭМП) (Strauss et al., 2009). Еще одной отличительной особенностью АВВ-2/3 является их способность производить во время репликации субвирусные додекаэдрические частицы, состоящие из фибриллы АВ и пентона (Norrby et al., 1967). Полноразмерный основной белок пентона не может образовывать пентон-додекаэдры (PtDd), и для этого требуется его спонтанное протеолитическое укорочение по N-концу между остатками 37 и 38 (Fuschiotti et al., 2006). Этот сайт расщепления является консервативным в АВ3, АВ7, АВ11 и АВ14, но не присутствует в АВ2 и АВ5. В случае АВ3 PtDd образуются при значительном избытке 5,5×106 PtDd на частицу инфекционного вируса (Fender et al., 2005), и было высказано предположение, что PtDd усиливает инфицирующую способность АВ3 путем разрушения межклеточных контактов, тем самым способствуя распространению вируса (Walters et al., 2002).
Краткое описание изобретения
Согласно одному аспекту в настоящем изобретении предложены выделенные полипептиды, содержащие аминокислотную последовательность, приведенную в любой из последовательностей SEQ ID NO: 1-11. Согласно другому аспекту в настоящем изобретении предложены рекомбинантные полипептиды фибриллы АВВ-2/3, содержащие:
a) один или более центральных доменов (shaft-доменов) полипептида фибриллы АВВ-2/3, мотивов центрального домена, или их функциональных эквивалентов;
b) шарообразный домен (knob-домен) полипептида фибриллы АВВ-2/3, функционально связанный и расположенный в направлении С-конца по отношению к одному или более центральным доменам полипептида фибриллы АВВ-2/3 или мотивам центрального домена, причем шарообразный домен полипептида фибриллы АВВ-2/3 содержит полипептид, содержащий любую из последовательностей, приведенных в SEQ ID NO: 1-11; и
с) один или более доменов димеризации, происходящих не из вируса АВВ-2/3, которые функционально связаны и расположены в направлении N-конца по отношению к одному или более центральным доменам полипептида фибриллы АВВ-2/3 или мотивам центрального домена.
Согласно одному варианту реализации настоящего изобретения полипептид фибриллы АВВ-2/3 не содержит хвостового домена полипептида фибриллы АВВ-2/3. Согласно другому варианту реализации настоящего изобретения каждый центральный домен или мотивы центрального домена выбраны из группы, состоящей из центрального домена или мотива центрального домена полипептида фибриллы АВ3, центрального домена или мотива центрального домена полипептида фибриллы АВ7, центрального домена или мотива центрального домена полипептида фибриллы АВ11, центрального домена или мотива центрального домена полипептида фибриллы АВ14, центрального домена полипептида фибриллы АВ14а, или комбинаций мотивов центрального домена, и их функциональных эквивалентов. Согласно другому варианту реализации настоящего изобретения каждый центральный домен или мотив центрального домена содержит аминокислотную последовательность, приведенную в любой из SEQ ID NO: 12-18, SEQ ID NO: 43-48, или их комбинации. Согласно другому варианту реализации настоящего изобретения домен димеризации содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 24 и SEQ ID NO: 25. Согласно другому варианту реализации настоящего изобретения рекомбинантный полипептид фибриллы АВВ-2/3 содержит или состоит из аминокислотной последовательности, приведенной в любой из SEQ ID NO: 28-34. Согласно другому варианту реализации настоящего изобретения полипептид фибриллы АВВ-2/3 мультимеризован, например, димеризован. Согласно другому варианту реализации настоящего изобретения полипептид фибриллы АВВ-2/3 дополнительно содержит одно или более соединений, конъюгированных с рекомбинантным полипептидом фибриллы АВВ-2/3, таких как терапевтические, диагностические и агенты для визуализации.
Согласно другому аспекту в настоящем изобретении предложены выделенные нуклеиновые кислоты, кодирующие выделенный пептид или рекомбинантные полипептиды фибриллы АВВ-2/3 согласно настоящему изобретению, рекомбинантные векторы экспрессии, содержащие выделенные нуклеиновые кислоты, и клетки-хозяева, содержащие рекомбинантные векторы экспрессии.
Согласно другому аспекту в настоящем изобретении предложены фармацевтические композиции, содержащие
(а) мультимер фибриллы АВВ-2/3 в соответствии с любым вариантом реализации или комбинацией вариантов реализации настоящего изобретения; и фармацевтически приемлемый носитель.
Согласно еще одному аспекту в настоящем изобретении предложены способы увеличения эффективности терапевтического лечения или диагностики заболевания, связанного с эпителиальной тканью, и/или визуализации эпителиальных тканей, включающие введение субъекту, нуждающемуся в этом:
a) такого количества одного или более терапевтических агентов, которое достаточно для лечения заболевания, такого количества диагностического агента, которое достаточно для диагностики заболевания, и/или такого количества агента для визуализации, которое достаточно для визуализации эпителиальной ткани; и
b) такого количества мультимера фибриллы АВВ-2/3 или фармацевтической композиции согласно любому из вариантов реализации или комбинации вариантов реализации настоящего изобретения, которое достаточно для повышения эффективности одного или более терапевтических, диагностических и/или агентов для визуализации.
Примеры заболеваний, связанных с эпителиальной тканью, включают солидные опухоли, синдром раздраженного кишечника, воспалительное заболевание кишечника, болезнь Крона, язвенный колит, запор, гастроэзофагеальную рефлюксную болезнь, пищевод Барретта, хроническую обструктивную болезнь легких, астму, бронхит, эмфизему легких, кистозный фиброз, интерстициальную болезнь легких, пневмонию, первичную легочную гипертензию, легочную эмболию, легочной саркоидоз, туберкулез, панкреатит, заболевания протоков поджелудочной железы, непроходимость желчных путей, холецистит, холедохолитиаз, заболевания головного мозга, псориаз, дерматит, гломерулонефрит, гепатит, сахарный диабет, заболевания щитовидной железы, целлюлит, инфекцию, пиелонефрит и камни в желчном пузыре.
Согласно другому аспекту в настоящем изобретении предложены способы лечения заболевания, связанного с эпителиальной тканью, включающие введение субъекту, нуждающемуся в этом, такого количества мультимера фибриллы АВВ-2/3 или фармацевтической композиции в соответствии с любым вариантом или комбинацией вариантов реализации настоящего изобретения, которое достаточно для лечения указанного заболевания. В типичных вариантах реализации настоящего изобретения указанное заболевание может представлять собой вирусную инфекцию или солидную опухоль.
Согласно другому аспекту в настоящем изобретении предложены способы улучшения доставки соединения к эпителиальной ткани, включающие приведение эпителиальной ткани в контакт с одним или более соединениями, которые необходимо доставить к эпителиальной ткани; и с таким количеством мультимера фибриллы АВВ-2/3 или фармацевтической композиции в соответствии с любым вариантом или комбинацией вариантов реализации настоящего изобретения, которое достаточно для улучшения доставки одного или более соединений к эпителиальной ткани. В типичных вариантах реализации настоящего изобретения одно или более соединений могут представлять собой диагностические или агенты для визуализации.
Согласно другому аспекту в настоящем изобретении предложены способы улучшения доставки вещества к ткани, экспрессирующей десмоглеин 2 (DSG2), включающие приведение указанной ткани, экспрессирующей DSG2, в контакт с:
a) одним или более соединениями, которые необходимо доставить к указанной ткани; и
b) таким количеством мультимера фибриллы АВВ-2/3 или фармацевтической композиции в соответствии с любым вариантом или комбинацией вариантов реализации настоящего изобретения, которое достаточно для улучшения доставки одного или более соединений к указанной ткани.
Согласно другому аспекту в настоящем изобретении предложены способы индукции эпителиально-мезенхимального перехода (ЭМП) в ткани, включающие приведение эпителиальной ткани в контакт с таким количеством мультимера фибриллы АВВ-2/3 или фармацевтической композиции в соответствии с любым вариантом или комбинацией вариантов реализации настоящего изобретения, которое достаточно для индукции ЭМП.
Согласно другому аспекту в настоящем изобретении предложены способы выявления соединений-кандидатов для применения в соответствии с одним или более вариантами, выбранными из лечения заболевания, связанного с эпителиальной тканью, улучшения доставки вещества к эпителиальной ткани, улучшения доставки вещества к ткани, экспрессирующей DSG2, индукции ЭМП в ткани, и/или лечения инфекции АВВ-2/3, включающие:
а) приведение мультимера фибриллы АВВ-2/3 в соответствии с любым вариантом или комбинацией вариантов реализации настоящего изобретения в контакт с DSG2 в условиях, которые обеспечивают связывание мультимера с DSG2, причем приведение в контакт осуществляют в присутствии одного или более испытываемых соединений; и
b) выявление положительных испытываемых соединений, которые конкурируют с мультимером фибриллы АВВ-2/3 за связывание с DSG2 по сравнению с контролем;
причем положительные испытываемые соединения представляют собой соединения-кандидаты для применения в соответствии с одним или более вариантами, выбранными из лечения заболевания, связанного с эпителиальной тканью, улучшения доставки вещества к эпителиальной ткани, улучшения доставки вещества к ткани, экспрессирующей DSG2, индукции ЭМП в ткани, и/или лечения инфекции АВВ-2/3.
Описание чертежей
Фигура 1. Остатки, которые, как было установлено, являются критическими для связывания с DSG2. А) Приведены аминокислотные последовательности головной части белка фибриллы АВ3 и АВ14р1. Бета-листы, присутствующие в головной части белка фибриллы АВ3 (номер доступа PDB 1H7Z_A) и АВ14 (PDB: 3F0Y_A) обозначены линиями. Черные стрелки указывают остатки в пределах головной части белка фибриллы АВ3, которые устраняют или уменьшают степень связывания с DSG2 после точечного мутагенеза. По сравнению с исходным штаммом АВ14 (deWit), AB14p1 содержит делецию двух остатков аминокислот в петле FG головной части белка фибриллы (24), показанной с помощью треугольника. В) Схематическое строение димерных мутированных вариантов белка фибриллы АВ3. Головной домен фибриллы и один мотив центральной части белка фибриллы гибридизовали посредством гибкого линкера с гомодимеризующимся доменом К-спирали (41). Белки способны к самопроизвольной димеризации и могут быть очищены с помощью аффинной хроматографии на носителе His-Ni-NTA. C-F) Исследование связывания димерных мутированных вариантов головной части белка фибриллы АВ3 с растворимым DSG2. С и D) Окрашивание Кумасси синим. В дорожку вносили 10 мкг очищенного белка головной части фибриллы АВ3 (не подвергнутого кипячению). Тримерные формы головной части белка фибриллы обозначены стрелкой. Гель содержал ДСН, и буфер для загрузки содержал ДТТ, который, как сообщалось ранее, вызывал разобщение димеров тримерных форм головной части белка фибриллы (41). Е и F) Приведены результаты исследования с помощью вестерн-блоттинга с использованием растворимого рекомбинантного DSG2 в качестве зонда, с последующим внесением моноклонального антитела к DSG2 и IgG-ПХ к иммуноглобулину мыши. Для сравнения приведены результаты, полученные с использованием JO-1 (0,5 мкг/дорожку). Вестерн-блоты сканировали, и количественно оценивали интенсивность сигналов.
Фигура 2. Трехмерная модель головной части белка фибриллы АВ3. Структура основана на последовательности с номером доступа PDB: 1H7Z_A. Верхняя панель: четыре ключевые области, участвующие в связывании с DSG2. Ключевые остатки показаны на изоповерхности тримера головной части белка фибриллы. Вид с верхней стороны (апикальной стороны), обращенной к рецептору. Нижняя панель: Все ключевые остатки в совокупности. Правая сторона: увеличенное изображение борозды после незначительного бокового поворота.
Фигура 3. Конкуренция вируса АВ3 с димеризованными мутированными вариантами головной части белка фибриллы АВ3. А) Относительная величина присоединения Н3-меченого вируса АВ3 в присутствии димерных мутированных вариантов головной части белка фибриллы. 1,8×105 клеток HeLa инкубировали с мутированными вариантами головной части белка фибриллы АВ3 в концентрации 2,5 и 100 мкг/мл на льду в течение 1 часа. Затем на льду добавляли по 400 КОЕ/клетку вируса Н3-АВ3 и дополнительно инкубировали в течение 1 часа. Несвязанные вирусные частицы отмывали. Прикрепление вирусных частиц, инкубированных с фосфатно-солевым буфером (ФСБ), принимали за 100%. N=3. В) Исследование конкуренции с использованием вируса AB3-GFP при инфицировании клеток HeLa. 1,5×105 клеток HeLa высевали в 24-луночные планшеты. Клетки инкубировали с увеличивающимися концентрациями мутированных вариантов головной части белка фибриллы АВ3 в течение одного часа при комнатной температуре. Затем добавляли 100 КОЕ/клетку вируса АВ3-GFP, и исследовали экспрессию GFP через 18 часов с помощью проточной цитометрии. Левая панель: процент GFP-положительных клеток. Правая панель: средняя интенсивность флуоресценции. N=3. Стандартное отклонение составило менее 10%. С) Относительное прикрепление Н3-меченого вируса АВ3 в присутствии димерных мутированных вариантов головной части белка фибриллы, содержащих множественные мутации. Исследование проводили, как описано в В). Стандартное отклонение составило менее 10%. D) Исследование конкуренции с использованием вируса AB3-GFP при инфицировании клеток HeLa. Исследование проводили, как описано в С). Стандартное отклонение составило менее 10%.
Фигура 4. Исследование связывания головной части белка фибриллы АВ3 с растворимым CD46. Варианты головной части белка фибриллы АВ3, содержащие различное количество мотивов центральной части, и головную часть белка фибриллы АВ3 дикого типа (дорожка 1: AB3-S6/Kn, дорожка 2: AB3-S5/Kn, дорожка 3: AB3-S4/Kn, дорожка 4: AB3-S3/Kn, дорожка 5: AB3-S2/Kn, дорожка 6: AB3-S/Kn), JO-1 (дорожка 7) и CD46-связывающаяся головная часть белка фибриллы Ad35 (дорожка 8)), подвергали блоттингу и гибридизовали с растворимым DSG2 (верхняя панель) или растворимым CD46 (нижняя панель). Связывание определяли с помощью моноклонального антитела (мАт) к DSG2 или мАт к CD46.
Фигура 5. Корреляция уменьшенного связывания с DSG2 и способности к размыканию эпителиальных контактов. А) Трансэпителиальное электрическое сопротивление (TEER), измеренное на поляризованных клетках рака толстой кишки Т84. Клетки культивировали в камерах Transwell до получения стабильного сигнала TEER, то есть до образования плотных контактов. После этого в апикальную камеру добавляли в общей сложности 5 мкг димерных головных частей белка фибриллы АВ3 в ФСБ и инкубировали в течение 1 часа. TEER измеряли в указанных временных точках. N=6. Во временных точках 1,5 и 4 часа различие между JO-1 по сравнению с D261N и N186D было статистически значимым (р<0,01). Стрелки указывают на временные точки добавления и удаления головной части белка фибриллы АВ3. В) Повышение эффективности терапии иринотеканом. В общей сложности 4×106 клеток А549 вводили подкожно мышам линии CB17-SCID/beige. После того как опухоль достигала объема ~100 мм3 (15-й день после имплантации), мышам внутривенно вводили 2 мг/кг JO-1, E299V, N186D или ФСБ, с последующей внутривенной инъекцией иринотекана (37,5 мг/кг) через 1 час. Лечение повторяли на 25 день. N=5. Различия между группами «иринотекан» по сравнению с «E299V + иринотекан» или «иринотекан» по сравнению с «N186 + иринотекан» не были статистически достоверными. После 20-го дня различие между группами «иринотекан» по сравнению с «JO-1 + иринотекан» было статистически достоверным (р<0,01).
Фигура 6. Замены аминокислот, которые увеличивают связывание с DSG2. А) Приведена аминокислотная последовательность головной части белка фибриллы АВ3. Бета-листы обозначены линиями. Стрелки указывают на остатки внутри головной части белка фибриллы АВ3, которые после точечного мутагенеза обеспечивают получение более интенсивных сигналов при количественном исследовании колоний с помощью блоттинга, что указывает на прочное связывание с DSG2. В) Изоповерхность трех мономеров головной части белка. Левая панель: вид сверху; правая панель: вид сбоку. V239 и Y250 не экспонируются на поверхности, что с большей вероятностью свидетельствует о структурном изменении в головной части, а не об участии в непосредственном связывании с DSG2. С) Локализация всех мутаций, которые усиливают связывание с DSG2. Остатки показаны пурпурным цветом в двух мономерах головной части белка. Изоповерхность одного мономера показана с помощью полупрозрачной серой заливки.
Фигура 7. Исследование взаимодействий недимеризованной головной части белка фибриллы АВ3 с DSG2 методом поверхностного плазменного резонанса (SPR).
A) DSG2 иммобилизовали на сенсорных чипах, и неспецифический сигнал автоматически вычитали из сигнала контрольной проточной ячейки. Головные части белка фибриллы АВ3 (без домена димеризации: «noDD») в концентрации 2,5 мкг/мл вводили в течение 3 мин, с последующим периодом диссоциации в течение 2,5 мин. В) Сводные данные SPR. Головные части белка вводили в диапазоне концентраций от 2,5 до 10 мкг/мл, и оценивали кинетику и параметры сродства с использованием программного обеспечения BIAeval. Извлеченные данные будут суммированы в таблице. Wt = головная часть белка фибриллы АВ3 без мутаций.
Фигура 8. Результаты исследования с помощью электронной микроскопии и трехмерная структура мутированного варианта головной части белка фибриллы АВ3, JO-2. А-С) Отрицательное окрашивание JO-2 с использованием SST. Можно наблюдать димерные формы, но также присутствуют структуры с более высокой степенью мультимеризации, гетерогенный комплекс размером приблизительно 50 нм показан тонкими стрелками, и меньшая по размеру обычная «додекаэдрическая» частица показана толстыми стрелками. Изображения в увеличенном масштабе приведены на В и С. D-G) кристаллографическая структура недимеризованной формы (мутированный вариант K217E/F224S). D) Кристаллы белка. Е) Головная часть белка АВ3 дикого типа окрашена в серый цвет, петля 217-224 EF выделена другим цветом. Петля EF приобретает неупорядоченную структуру в мутированном варианте. В мутированной структуре нет плотности для этих остатков. F) Мутированный вариант показан в виде схемы. G) Наложение этих двух структур показывает, что в мутированном варианте K217E/F224S петля EF является полностью неупорядоченной. На нижних панелях приведены увеличенные изображения одного мономера. К217 и F224 отображаются в виде палочек.
Фигура 9. Исследование димерных мутированных вариантов головной части белка фибриллы АВ3, имеющих повышенную аффинность в отношении DSG2. А) Конкуренция вирусной инфекции AB3-GFP в клетках HeLa с димерным мутированным вариантом, имеющим повышенную аффинность, Y250F и JO-1 (димерная головная часть белка фибриллы АВ3 дикого типа). Условия проведения экспериментов соответствуют тем, которые описаны для Фиг. 3с. Левая панель: процент GFP-положительных клеток. Правая панель: средняя интенсивность флуоресценции. N=3. Стандартное отклонение составило менее 10%. В) Конкуренция вирусной инфекции AB3-GFP в клетках HeLa с мутированными вариантами головной части белка фибриллы АВ3, проявляющими повышенное связывание с DSG2, но не содержащими домена димеризации. 1,5×105 клеток HeLa высевали в 24-луночные планшеты. Клетки инкубировали с увеличивающимися концентрациями мутированных вариантов головной части белка фибриллы АВ3 в течение одного часа при комнатной температуре. Затем вносили 100 КОЕ/клетку вируса AB3-GFP, и исследовали экспрессию GFP через 18 часов. С) Результаты оценки TEER на клетках рака толстой кишки Т84. Условия экспериментов соответствовали таковым для Фиг. 5а. Приведены результаты оценки TEER через 4 часа. N=3.
Фигура 10. Комбинация варианта JO-1, имеющего повышенную аффинность, с химиотерапией. А) Повышение эффективности терапии иринотеканом (I). Условия экспериментов соответствовали таковым для Фиг. 5b. Различия в группах «JO-1 + I» по сравнению с «JO-2 + иринотекан» и «JO-2 + I» по сравнению с «JO-4 + I» были достоверными после 20-го дня. N=5. В) JO-4 усиливает терапию PLD в модели рака яичников в более низкой дозе, чем JO-1. Опухоли жировой ткани молочной железы были созданы из клеток первичного рака яичников ovc316. Лечение начинали, когда опухоли достигали объема 100 мм3. Мышам вводили внутривенно 2 мг/кг JO-1 или 0,5 мг/кг JO-4, с последующей внутривенной инъекцией пегилированного доксорубицина (PLD) (1 мг/кг) через один час. Лечение повторяли еженедельно. С) JO-4 повышает эффективность терапии в случае тройного негативного рака молочной железы с неблагоприятным прогнозом (TNBC). В общей сложности 4×106 клеток MDA-MB-231 TNBC вводили в жировую ткань молочной железы мышей СВ17 SCID-beige. JO-4 (2 мг/кг) вводили внутривенно за 1 час до применения цетуксимаба (С) (10 мг/кг, внутрибрюшинно) и наб-паклитаксела (nab-P) (5 мг/кг, в/в). Лечение проводили еженедельно. N=10, р<0,01 на 25-й день для наб-паклитаксела + С по сравнению с JO-4 + nab-P + С.
Фигура 11. Фармакокинетика, токсичность и иммуногенность JO-4. А) Клиренс JO-1 и JO-4 из сыворотки крови. Трансгенным мышам hDSG2 с подкожными опухолями TC1-hDSG2 (~600 мм3) внутривенно вводили JO-1 или JO-4 (2 мг/кг), и образцы сыворотки исследовали с помощью твердофазного иммуноферментного анализа (ИФА). N=3. Следует отметить, что ось ординат имеет логарифмическую шкалу. В) Количество лимфоцитов и тромбоцитов у трансгенных мышей hDSG2/TC1-hDSG2 после введения JO-1 или JO-4. N=3. С) Исследования эффективности терапии у иммунокомпетентных трансгенных мышей hDSG2 с опухолями TC1-hDSG2. После того как опухоли достигали объема ~80 мм3, JO-1 или JO-4 (2 мг/кг) или ФСБ вводили внутривенно с последующим введением PLD/Doxil (в дозе 1,5 мг/кг в/в) через 1 ч. Лечение повторяли в моменты времени, показанные стрелками. Опухолям обеспечивали период роста в течение примерно двух недель. Начиная с 15-го дня, антитела к JO-1/JO-4 детектировали в сыворотке крови с помощью ИФА. Два дополнительных цикла лечения проводили на 28-й день и 35-й день. Эффективность JO-1 и JO-4 сохранялась после нескольких циклов лечения, даже в присутствии детектируемых антител. Различие между JO-1/PLD по сравнению с JO-4/PLD является статистически достоверным после 31-го дня. N=10.
Фигура 12. Выравнивание последовательностей головной части белка фибриллы. Показаны остатки, которые устраняют/уменьшают связывание головной части белка фибриллы АВ3 с DSG2.
Фигура 13. Сыворотки, полученные у людей и сверхвакцинированных мышей, не ингибируют активность JO-4. А) Исследование сыворотки крови человека для оценки связывания с JO-4 с помощью ИФА. Для захвата были использованы поликлональные антитела кролика к головной части белка фибриллы АВ3, затем вносили рекомбинантный белок JO-1, сыворотку человека (разведение от 1:20 до 1:1000) и IgG-ПХ к иммуноглобулину человека. В качестве отрицательного контроля использовали коммерческую сыворотку человека, инфицированного АВ, истощенную в отношении IgG (горизонтальная линия). В качестве положительного контроля использовали сыворотку ученого, который постоянно работает с вирусом АВ3. Образцы от Р1 до Р38 представляют собой образцы сыворотки пациентов, страдающих раком яичников, полученные из Тихоокеанского консорциума по изучению рака яичников.
Подробное описание изобретения
Все процитированные литературные источники полностью включены в настоящую заявку посредством ссылки. В тексте настоящей заявки, если не указано иное, использованные методики могут быть найдены в любом из нескольких известных литературных источников, таких как: Molecular Cloning: A Laboratory Manual (Sambrook, et al., 1989, Cold Spring Harbor Laboratory Press), Gene Expression Technology (Methods in Enzymology, Vol. 185, под ред. D. Goeddel, 1991. Academic Press, San Diego, CA), "Guide to Protein Purification" в Methods in Enzymology (под ред. M.P. Deutshcer, (1990) Academic Press, Inc.); PCR Protocols: A Guide to Methods and Applications (Innis, et al. 1990. Academic Press, San Diego, CA), Culture of Animal Cells: A Manual of Basic Technique, 2 изд. (R.I. Freshney. 1987. Liss, Inc. New York, NY), Gene Transfer and Expression Protocols, pp. 109-128, под ред. E.J. Murray, The Humana Press Inc., Clifton, N.J.) и в каталоге Ambion 1998 (Ambion, Остин, Техас, США).
В настоящей заявке формы единственного числа «а», «an» и «the» включают множественное число, если из контекста явно не следует иное. В настоящей заявке «и» используется взаимозаменяемо с «или», если явно не указано иное.
В настоящей заявке аминокислотные остатки сокращены следующим образом: аланин (Ala, А), аспарагин (Asn, N), аспарагиновая кислота (Asp; D), аргинин (Arg; R), цистеин (Cys, С), глутаминовая кислота (Glu, Е), глутамин (Gln, Q), глицин (Gly; G), гистидин (His, Н), изолейцин (Ile; I), лейцин (Leu, L), лизин (Lys; K), метионин (Мет; М), фенилаланин (Phe, F), пролин (Pro, Р), серии (Ser; S), треонин (Thr, Т), триптофан (Tip, W), тирозин (Tyr, Y) и валин (Val; V).
В настоящей заявке аббревиатура «АВ» относится к аденовирусу и, как правило, за ней следует номер, указывающий серотип аденовируса. Например, «АВ3» относится к аденовирусу серотипа 3.
Все варианты реализации любого аспекта настоящего изобретения могут быть использованы в комбинации, если из контекста явно не следует иное.
Согласно первому аспекту в настоящем изобретении предложен выделенный полипептид, содержащий или состоящий из аминокислотной последовательности:

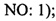 в которой
в которой
Х2 представляет собой Н, L или Р;
Х3 представляет собой K или Е;
Х4 представляет собой Т, F, S или L;
Х5 представляет собой V, D или отсутствует;
Х6 представляет собой Е, G или отсутствует;
Х7 представляет собой Y или F;
Х8 представляет собой Т, K или Е; и
Х9 представляет собой N или S;
причем верным является по меньшей мере одно из следующих условий:
Х2 представляет собой Р;
Х3 представляет собой Е;
Х4 представляет собой S или L;
Х5 представляет собой D;
Х6 представляет собой G;
Х7 представляет собой F;
Х8 представляет собой Е; или
Х9 представляет собой S. Выделенные полипептиды в соответствии с указанным аспектом настоящего изобретения включают мутированные домены головной части белка фибриллы АВВ-2/3, которые можно применять, например, для получения рекомбинантных полипептидов фибриллы АВВ-2/3, которые обеспечивают значительное повышение аффинности в отношении десмоглеина 2 (DSG2) по сравнению с ранее известными DSG2-связывающими полипептидами. Как показано в примерах ниже, рекомбинантные полипептиды фибриллы АВВ-2/3 согласно настоящему изобретению, которые содержат мутированные головные домены согласно первому аспекту настоящего изобретения, как дополнительно установлено, являются терапевтически более эффективными, чем ранее известные DSG2-связывающие полипептиды, для лечения заболевания, связанного с эпителиальной тканью, о чем свидетельствует улучшенная эффективность в нескольких моделях рака. Выделенные пептиды согласно настоящему изобретению также можно применять, например, в качестве антигенов к вирусам АВВ-2/3.
Согласно одному варианту реализации настоящего изобретения выделенные полипептиды в соответствии с первым аспектом настоящего изобретения содержат или состоят из аминокислотной последовательности:
Согласно другому варианту реализации настоящего изобретения выделенные полипептиды в соответствии с первым аспектом настоящего изобретения содержат или состоят из аминокислотной последовательности:

Согласно другому варианту реализации настоящего изобретения выделенные полипептиды в соответствии с первым аспектом настоящего изобретения содержат или состоят из аминокислотной последовательности:

Во всех перечисленных вариантах верным является по меньшей мере одно из следующих условий:
Х2 представляет собой Р;
X3 представляет собой Е;
Х4 представляет собой S или L;
Х5 представляет собой D;
Х6 представляет собой G;
Х7 представляет собой F;
Х8 представляет собой Е; или
Х9 представляет собой S.
В различных вариантах реализации настоящего изобретения верными являются по меньшей мере 2, 3, 4, 5, 6, 7 или все 8 из этих утверждений. Согласно одному типичному варианту реализации настоящего изобретения по меньшей мере Х7 представляет собой F. Согласно другому варианту реализации настоящего изобретения по меньшей мере X3 представляет собой Е, и Х4 представляет собой S. Согласно другому варианту реализации настоящего изобретения по меньшей мере Х9 представляет собой S. Согласно другому варианту реализации настоящего изобретения по меньшей мере Х5 представляет собой D. Согласно другому варианту реализации настоящего изобретения по меньшей мере Х4 представляет собой L. Согласно другому варианту реализации настоящего изобретения по меньшей мере Х2 представляет собой Р, и Х8 представляет собой Е. Согласно другому варианту реализации настоящего изобретения по меньшей мере Х6 представляет собой G, и Х8 представляет собой Е.
В различных других вариантах реализации настоящего изобретения выделенный полипептид содержит или состоит из одного из следующих пептидов:
(а)

(b)

(с)

(d)
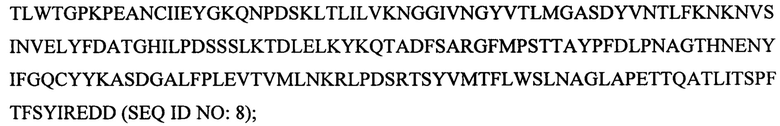
(е)

(f)

(g)
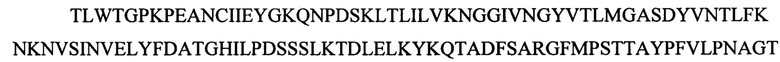
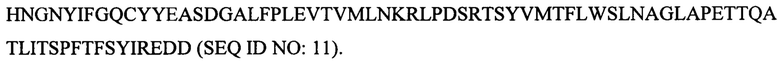
Согласно второму аспекту в настоящем изобретении предложен рекомбинантный полипептид фибриллы АВВ-2/3, содержащий:
a) один или более центральных доменов полипептида фибриллы АВВ-2/3, мотивов центрального домена полипептида фибриллы или их функциональных эквивалентов;
b) головной домен полипептида фибриллы АВВ-2/3, функционально связанный и расположенный в направлении С-конца по отношению к одному или более центральным доменам полипептида фибриллы АВВ-2/3 или мотивам центрального домена полипептида фибриллы АВВ-2/3, причем головной домен полипептида фибриллы АВВ-2/3 содержит полипептид в соответствии с любым вариантом реализации или комбинацией вариантов реализации первого аспекта настоящего изобретения; и
c) один или более доменов димеризации, полученных не из вируса АВВ-2/3, функционально связанных и расположенных в направлении N-конца по отношению к одному или более центральным доменам полипептида фибриллы или мотивам центрального домена фибриллы АВВ-2/3.
В настоящей заявке термин «АВВ-2/3» относится к любому серотипу аденовируса, который использует DSG2 в качестве эпителиального рецептора для связывания вирусной частицы. В настоящее время были выявлены серотипы АВ3, АВ7, АВ11, АВ14 и АВ14а. По мере выявления других серотипов АВ, специалисты в данной области техники могут легко определить те, которые принадлежат к семейству АВВ-2/3 на основании результатов количественных исследований связывания с DSG2, как описано в настоящей заявке. Например, исследования с помощью поверхностного плазмонного резонанса (SPR) с использованием сенсоров, содержащих иммобилизованный рекомбинантный DSG2, в комбинации с исследованиями конкурентного связывания с DSG2, могут быть использованы, чтобы определить, связываются ли новые серотипы АВ с DSG2. Дополнительные примеры исследований, таких как исследования потери и усиления функции, подробно описаны в WO 2011/156761.
Вирион аденовируса представляет собой икосаэдр, для которого характерно наличие фибриллы, расположенной в основании каждой из 12 вершин капсида. Фибрилла на вирионе представляет собой гомотримерную структуру, состоящую из 3-х отдельных полипептидов фибриллы. Каждый полипептид фибриллы аденовируса представляет собой асимметричную структуру, состоящую из N-концевого хвоста, который взаимодействует с пентоновым белком капсида и содержит сигналы, необходимые для переноса белка в ядро клетки; центральной части, которая содержит несколько повторяющихся блоков, содержащих 15 остатков; и С-концевого головного домена, который содержит детерминанты для связывания рецептора (J.S. Hong and J.A. Engler, Journal of Virology 70:7071-7078 (1996)). Все аденовирусы прикрепляются к своим рецепторам с помощью головного домена белка на конце фибриллы. Соответственно, в настоящей заявке термин «полипептид фибриллы» АВВ-2/3 относится к полноразмерному полипептиду фибриллы, который содержит N-концевой хвостовой домен, центральный домен или мотив центрального домена, и С-концевой головной домен. Полипептиды фибриллы самопроизвольно собираются в гомотримеры, называемые «фибриллы», которые расположены на внешней стороне вириона аденовируса в основании каждой из двенадцати вершин капсида.
В предпочтительном варианте реализации настоящего изобретения рекомбинантные полипептиды не содержат хвостовой домен из полипептида фибриллы АВ. Как подробно описано ниже, авторы настоящего изобретения выявили ключевые остатки, мутация которых приводит к получению полипептидов фибриллы со значительно повышенной аффинностью в отношении DSG2, и со значительно повышенной терапевтической эффективностью. Полипептиды согласно этому аспекту настоящего изобретения, следовательно, может применять, например, для образования мультимеров фибриллы АВВ-2/3 для использования в различных способах согласно настоящему изобретению, которые описаны выше. Согласно данному аспекту рекомбинантные полипептиды могут содержать центральные домены или мотивы центрального домена из любого вируса АВВ-2/3, или любые мутированные варианты (содержащие замены, добавления, делеции, химеры и т.д.) таких центральных доменов или мотивов центрального домена, которые сохраняют или улучшают аффинность связывания с DSG2, и способны образовывать мультимеры (например, димеры) с помощью домена димеризации (функциональные эквиваленты). Например, исследования с помощью поверхностного плазмонного резонанса (SPR) с использованием сенсоров, содержащих иммобилизованный рекомбинантный DSG2, в комбинации с исследованиями конкурентного связывания с DSG2, могут быть использованы, чтобы определить, связываются ли оцениваемые рекомбинантные полипептиды с DSG2.
В настоящей заявке термин «полипептид» используется в самом широком смысле для обозначения последовательности остатков аминокислот. Полипептиды согласно настоящему изобретению могут содержать L-аминокислоты, D-аминокислоты (которые являются устойчивыми к действию протеаз, специфичных в отношении L-аминокислот, in vivo) или комбинацию D- и L-аминокислот. Полипептиды, описанные в настоящей заявке, могут быть синтезированы с помощью химических методов или рекомбинантно экспрессированы. Полипептиды могут быть связаны с другими соединениями, чтобы увеличить период полувыведения в условиях in vivo, например, путем присоединения ПЭГ, гидроксиэтилкрахмала, последовательности пролин-аланин-серин, гликозилирования, или могут быть получены в виде гибридного белка с областью Fc или в виде деиммунизированных вариантов. Специалисты в данной области техники поймут, что такая связь может быть ковалентной или нековалентной.
В настоящей заявке термин «функционально связанный» относится к расположению элементов, при котором домены расположены так, что они функционируют как единое целое в соответствии с предполагаемой целью. Термин не подразумевает, что домены расположены в непосредственной близости в полипептиде, поскольку между доменами могут присутствовать спейсерные/линкерные последовательности, длина которых может существенно различаться. Согласно одному неограничивающему варианту длина спейсера между любыми двумя доменами рекомбинантных полипептидов фибриллы АВВ-2/3 может находиться в пределах от приблизительно 0 аминокислот до приблизительно 20 аминокислот. В различных других неограничивающих вариантах длина спейсера может составлять 0-20, 0-19, 0-18, 0-17, 0-16, 0-15, 0-14, 0-13, 0-12, 0-11, 0-10, 0-9, 0-8, 0-7, 0-6, 0-5, 0-4, 0-3, 0-2, 0-1, 1-20, 1-19, 1-18, 1-17, 1-16, 1-15, 1-14, 1-13, 1-12, 1-11, 1-10, 1-9, 1-8, 1- 7, 1-6, 1-5, 1-4, 1-3, 1-2, 2-20, 2-19, 2-18, 2-17, 2-16, 2-15, 2-14, 2-13, 2-12, 2-11, 2-10, 2-9, 2-8, 2-7, 2-6, 2-5, 2-4, 2-3, 3-20, 3-19, 3-18, 3-17, 3-16, 3-15, 3-14, 3-13, 3-12, 3-11, 3-10, 3-9, 3-8, 3-7, 3-6, 3-5, 3-4, 4-20, 4-19, 4-18, 4-17, 4-16, 4-15, 4-14, 4-13, 4-12, 4-11, 4-10, 4-9, 4-8, 4-7, 4-6, 4-5, 5-20, 5-19, 5-18, 5-17, 5-16, 5-15, 5-14, 5-13, 5-12, 5-11, 5-10, 5-9, 5-8, 5-7, 5-6, 6-20, 6-19, 6-18, 6-17, 6-16, 6-15, 6-14, 6-13, 6-12, 6-11, 6-10, 6-9, 6-8, 6-7, 7-20, 7-19, 7-18, 7-17, 7-16, 7-15, 7-14, 7-13, 7-12, 7-11, 7-10, 7-9, 7-8, 8-20, 8-19, 8-18, 8-17, 8-16, 8-15, 8-14, 8-13, 8-12, 8-11, 8-10, 8-9, 9-20, 9-19, 9-18, 9-17, 9-16, 9-15, 9-14, 9-13, 9-12, 9-11, 9-10, 10-20, 10-19, 10-18, 10-17, 10-16, 10-15, 10-14, 10-13, 10-12, 10-11, 11-20, 11-19, 11-18, 11-17, 11-16, 11-15, 11-14, 11-13, 11-12, 12-20, 12-19, 12-18, 12-17, 12-16, 12-15, 12-14, 12-13, 13-20, 13-19, 13-18, 13-17, 13-16, 13-15, 13-14, 14-20, 14-19, 14-18, 14-17, 14-16, 14-15, 15-20, 15-19, 15-18, 15-17, 15-16, 16-20, 16-19, 16-18, 16-17, 17-20, 17-19, 17-18, 18-20, 18-19, 19-20, 20,19, 18,17,16,15,14,13,12,11, 10, 9, 8, 7, 6, 5,4, 3, 2, 1 или 0 аминокислот.
В настоящей заявке термин «рекомбинантный полипептид» означает неприродный белковый продукт, в котором домены рекомбинантного полипептида получены из одного или более других белков или искусственно полученных последовательностей, таких как мутированные полипептиды головного домена согласно настоящему изобретению. Например, каждый центральный домен или мотив центрального домена может быть получен из другого природного белка. Рекомбинантный полипептид может быть сконструирован с помощью различных механизмов, включая, но не ограничиваясь ими, стандартные методики манипулирования ДНК и химическую сборку посредством частей субъединиц рекомбинантного полипептида. Химическая сборка может привести к получению эквивалентной формы, такой как молекулярно-генетическая форма, или альтернативных комплексов с эквивалентной функцией. В предпочтительном варианте реализации настоящего изобретения рекомбинантный полипептид получают с помощью стандартных методик рекомбинантных ДНК. Методики получения рекомбинантных продуктов и выделения рекомбинантных полипептидов согласно настоящему изобретению хорошо известны в пределах уровня компетентности специалиста в данной области техники на основании принципов, изложенных в настоящем изобретении.
Согласно одному варианту реализации настоящего изобретения каждый центральный домен или мотивы центрального домена выбраны из группы, состоящей из центрального домена или мотива центрального домена АВ3, мотива центрального домена АВ5, центрального домена или мотива центрального домена АВ7, центрального домена или мотива центрального домена АВ11, центрального домена или мотива центрального домена АВ14, центрального домена или мотива центрального домена АВ14а, их комбинаций и их функциональных эквивалентов. Центральный домен или мотивы центрального домена требовались для димеризации головной части белка фибриллы, которая необходима для связывания с DSG2 и последующего кратковременного размыкания межклеточных контактов. В настоящей заявке термин «мотив центрального домена» представляет собой любую часть центрального домена, которая обеспечивает димеризацию головной части рекомбинантных полипептидов фибриллы АВВ-2/3 согласно настоящему изобретению. Такие мотивы центральных доменов могут быть легко определены специалистами в данной области техники на основании примеров, представленных ниже. Например, исследования с помощью поверхностного плазмонного резонанса (SPR) с использованием сенсоров, содержащих иммобилизованный рекомбинантный DSG2, в комбинации с исследованиями конкурентного связывания с DSG2, могут быть использованы для определения, связываются ли оцениваемые рекомбинантные полипептиды с DSG2. Дополнительные примеры исследований, таких как исследования потери и усиления функции, подробно описаны в примере 1.
Рекомбинантные полипептиды могут содержать от 1 до 22 центральных доменов или мотивов центрального домена полипептида фибриллы АВВ-2/3. Следовательно, в различных вариантах реализации настоящего изобретения полипептиды содержат 1-22, 1-21, 1-20, 1-19, 1-18, 1-17, 1-16, 1-15, 1-14, 1-13, 1-12, 1-11, 1-10, 1-9, 1-8, 1-7, 1-6, 1-5, 1-4, 1-3, 1-2, 2-22, 2-21, 2-20, 2-19, 2-18, 2-17, 2-16, 2-15, 2-14, 2-13, 2-12, 2-11, 2-10, 2-9, 2-8, 2-7, 2-6, 2-5, 2-4, 2-3, 3-22, 3-21, 3-20, 3-19, 3-18, 3-17, 3-16, 3-15, 3-14, 3-13, 3-12, 3-11, 3-10, 3- 9, 3-8, 3-7, 3-6, 3-5, 3-4, 4-22, 4-21, 4-20, 4-19, 4-18, 4-17, 4-16,4-15, 4-14, 4-13, 4-12,4-11, 4- 10, 4-9, 4-8, 4-7, 4-6, 4-5, 5-22, 5-21, 5-20, 5-19, 5-18, 5-17, 5-16, 5-15, 5-14, 5-13, 5-12, 5-11, 5-10, 5-9, 5-8, 5-7, 5-6, 6-22, 6-21, 6-20, 6-19, 6-18, 6-17, 6-16, 6-15, 6-14, 6-13, 6-12,6-11, 6-10, 6-9, 6-8, 6-7, 7-22, 7-21, 7-20, 7-19, 7-18, 7-17, 7-16, 7-15, 7-14, 7-13, 7-12, 7-11, 7-10, 7-9, 7-8, 8-22, 8-21, 8-20, 8-19, 8-18, 8-17, 8-16, 8-15, 8-14, 8-13, 8-12, 8-11, 8-10, 8-9, 9-22, 9-21, 9-20, 9-19, 9-18, 9-17, 9-16, 9-15, 9-14, 9-13, 9-12, 9-11, 9-10, 10-22, 10-21, 10-20, 10-19, 10-18, 10-17, 10-16, 10-15, 10-14, 10-13, 10-12, 10-11, 11-22, 11-21, 11-20, 11-19, 11-18, 11-17, 11-16, 11-15, 11-14, 11-13, 11-12, 12-22, 12-21, 12-20, 12-19, 12-18, 12-17, 12-16, 12-15, 12-14, 12-13, 13-22, 13-21, 13-20, 13-19, 13-18, 13-17, 13-16, 13-15, 13-14, 14-22, 14-21, 14-20, 14-19, 14-18, 14-17, 14-16, 14-15, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 15-16, 16-22, 16-21, 16-20, 16-19, 16-18, 16-17, 17-22, 17-21, 17-20, 17-19, 17-18, 18-22, 18-21, 18-20, 18-19, 19-22, 19-21, 19-20, 20-22, 20-21, 21-22, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или 22 центральных домена или мотива центрального домена белка фибриллы АВВ-2/3. В том случае, если присутствует более 1 центрального домена или мотива центрального домена белка фибриллы АВВ-2/3, каждый центральный домен или мотив центрального домена может быть идентичен другому, либо одна или более копий центрального домена или мотива центрального домена могут различаться в одном рекомбинантном полипептиде. В предпочтительном варианте реализации настоящего изобретения рекомбинантный полипептид фибриллы АВВ-2/3 содержит один центральный домен или мотив центрального домена.
Согласно другому варианту реализации настоящего изобретения один или более (или все) центральных доменов или мотивов центрального домена в рекомбинантном полипептиде содержат или состоят из аминокислотной последовательности, приведенной в SEQ ID NO: 12:

или SEQ ID NO: 43-48:
мотив центрального домена АВ3: NSIALKNNTL SEQ ID NO: 43
мотив центрального домена АВ7: NSNNICINDNINTL SEQ ID NO: 44
мотив центрального домена АВ5: GAITVGNKNNDKLTL SEQ ID NO: 45
мотав центрального домена АВ11: NSNNICIDDNINTL SEQ ID NO: 46
мотив центрального домена АВ14: NSNNICIDDNINTL SEQ ID NO: 47
мотив центрального домена АВ35: GDICIKDSINTL SEQ ID NO: 48
В указанной последовательности и в других вариабельных последовательностях, приведенных в настоящем описании, вариабельные остатки отмечены в круглых скобках, и «-» указывает на то, что остаток может отсутствовать.
Согласно другому варианту реализации настоящего изобретения один или более (или все) центральных доменов или мотивов центрального домена в рекомбинантном полипептиде содержат или состоят из аминокислотной последовательности, приведенной в SEQ ID NO: 13:
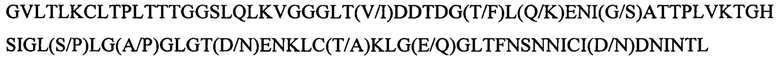
или SEQ ID NO: 43-48.
Согласно другому варианту реализации настоящего изобретения один или более (или все) центральных доменов или мотивов центрального домена в рекомбинантном полипептиде содержат или состоят из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 14 (АВ3), SEQ ID NO: 15 (AB7), SEQ ID NO: 16 (AB11), SEQ ID NO: 17 (AB14), SEQ ID NO: 18 (AB14a) и SEQ ID NO: 43-48.
Головной домен полипептида фибриллы АВВ-2/3 содержит или состоит из любого варианта реализации или комбинации вариантов реализации первого аспекта настоящего изобретения (т.е. содержит любую из последовательностей SEQ ID NO: 1-11); указанные полипептидные домены подробно описаны в первом аспекте настоящего изобретения.
В настоящей заявке термин «домен димеризации» представляет собой пептидную последовательность, которая способствует димеризации в рекомбинантном полипептиде, содержащем ее. Любой подходящий домен димеризации, полученный не из вируса АВВ-2/3, может быть использован в рекомбинантном полипептиде согласно настоящему изобретению, при условии, что он обеспечивает димеризацию рекомбинантного полипептида и, следовательно, связывание с DSG2. Домен димеризации получен не из вируса АВВ-2/3, т.е. он не является природным доменом полипептида фибриллы АВВ-2/3. Неограничивающие примеры многочисленных доменов димеризации, известных специалистам в данной области техники и подходящих для использования согласно настоящему изобретению, включают, но не ограничиваются ими, пептидные спирали, содержащие по меньшей мере одну спираль, или структуру, образованную спиралью, катушку и другую спираль и т.д., суперспиральные структуры, домены димеризации в пределах, например, многих поверхностных сигнальных рецепторов клеток, области Fc или шарнирные области антитела, лейциновые молнии, N-концевой домен белка STAT, РК506-связывающий белок, С-концевой домен белка LexA, ядерные рецепторы, N-концевой домен FkpA, оранжевый каротиноид-протеин из A. maxima, матриксный белок M1 вируса гриппа, нейраминидазу вируса гриппа, L-фукулоза-1-фосфатальдолазу Е. coli; и т.п. (см., например, O'Shea, Science. 254: 539 (1991), Barahmand-Pour et al., Curr. Top. Microbiol. Immunol. 211: 121-128 (1996); Klemm et al., Annu. Rev. Immunol. 16: 569-592 (1998); Klemm et al., Annu. Rev. Immunol. 16: 569-592 (1998); Ho et al., Nature. 382: 822-826 (1996); и Pomeranz et al., Biochem. 37: 965 (1998)). Другие примеры включают остатки от 325 до 410 белка Е2 бычьего папилломавируса (Dostatni, N., et al., EMBO J 7 (1988) 3807-3816; Haugen, Т., et al. EMBO J 7 (1988) 4245-4253; McBride, A., et al., EMBO J 7 (1988) 533-539; McBride, A., et al., Proc Natl Acad Sci USA 86 (1989) 510-514), дейодиназу типа I (Dl): DFLVIYIEEAHASDGW (SEQ ID NO: 19) или ADFL-YI-EAH-DGW (SEQ ID NO: 20); белок капсида ВИЧ-1: QGPKEPFRDYVDRFYKTLRA (SEQ ID NO: 21); мотив димеризации лейциновой молнии дрожжей GCN4: HMKQLDVEELSNYHLNVARLKVGER (SEQ ID NO: 22); лейциновую молнию в белке, предотвращающем терминацию транскрипции, Е. coli; и BglG: GVTQLMREMLQLIKFQFSLNYQEESLSYQRLVT (SEQ ID NO: 23). В предпочтительных вариантах реализации настоящего изобретения домен димеризации содержит одну или более копий EVSALEK (SEQ ID NO: 24) и/или KVSALKE (SEQ ID NO: 25).
Выявление соответствующих пептидных последовательностей, которые могут служить в качестве доменов димеризации, а также их мутированных вариантов, в рекомбинантных полипептидах согласно настоящему изобретению находится в пределах уровня компетентности специалиста в данной области техники. Например, димеризацию рекомбинантных полипептидов фибриллы АВВ-2/3 можно оценить с помощью критериев, включающих седиментацию в градиентах сахарозы, устойчивость к протеолизу трипсином и электрофоретическую подвижность в полиакриламидном геле (Hong and Engler, Journal of Virology 70:7071-7078 (1996)).
Рекомбинантные полипептиды могут содержать один или более доменов димеризации, полученных не из вируса АВВ-2/3. Следовательно, в различных вариантах реализации настоящего изобретения рекомбинантный полипептид содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более доменов димеризации, полученных не из вируса АВВ-2/3. В том случае, если в полипептиде присутствует несколько доменов, предпочтительной является идентичность всех доменов димеризации.
В предпочтительном варианте реализации настоящего изобретения спейсерный пептид расположен между доменом димеризации и одним или более центральными доменами или мотивами центральных доменов. Согласно другому предпочтительному варианту реализации спейсерный пептид представляет собой пептид со структурной гибкостью. Может быть использован практически любой пептид со структурной гибкостью. Например, гибкий пептид может содержать повторы остатков аминокислот, такие как Gly-Gly-Gly-Ser (SEQ ID NO: 26), или любой другой подходящий повтор остатков аминокислот. Согласно другому варианту реализации настоящего изобретения может быть использована шарнирная область антитела. Спейсер может иметь любую подходящую длину, которая поддерживает способность рекомбинантного полипептида к димеризации и поддерживает связывание рекомбинантного полипептида с DSG2.
Согласно одному предпочтительному варианту реализации настоящего изобретения рекомбинантный полипептид АВВ-2/3 содержит один или более центральных доменов, каждый их которых содержит или состоит из центрального домена АВ3 (SEQ ID NO: 14).
Указанный предпочтительный вариант реализации настоящего изобретения может быть использован с любым вариантом реализации или комбинацией вариантов реализации настоящего изобретения, описанными в настоящей заявке. Например, может быть использован любой подходящий головной домен и любой подходящий домен димеризации, включая, но не ограничиваясь ими, одну или более копий EVSALEK (SEQ ID NO: 24) и/или KVSALKE (SEQ ID NO: 25). Аналогичным образом, любые подходящие спейсерные пептиды могут быть использованы между доменом димеризации и центральным доменом или мотивом центрального домена и/или между центральным доменом или мотивом центрального домена и головным доменом. В наиболее предпочтительном варианте реализации настоящего изобретения рекомбинантный полипептид АВВ-2/3 содержит или состоит из JO-1 (SEQ ID NO: 27) или его мультимеров (таких как димер).
Рекомбинантные полипептиды могут содержать дополнительные домены, такие как домен для выделения полипептида и/или домен для детектирования. Домен для выделения может быть добавлен, чтобы облегчить очистку/выделение полипептида после, например, получения рекомбинантного полипептида. Любой подходящий домен для выделения может быть использован, включая, но не ограничиваясь ими, пептидные метки HIS, СВР, CYD (ковалентный, но диссоциируемый пептид NorpD), Strep II, FLAG, НРС (тяжелая цепь белка С), аффинные метки GST и МВР. В настоящей заявке термин «домен детектирования» означает одну или более аминокислотных последовательностей, которые можно детектировать. Любой подходящий домен детектирования может быть использован, включая, но не ограничиваясь ими, природные флуоресцентные белки (например, зеленый флуоресцентный белок и флуоресцентные белки из небиолюминесцентных видов Anthozoa), флуоресцентные или люминесцентные белки, требующие присутствия кофакторов (например, фикобилипротеины или люциферазы), и эпитопы, распознаваемые специфичными антителами, или другие специфичные природные или искусственные связывающие зонды, включая, но не ограничиваясь ими, красители, кофакторы ферментов и сконструированные связывающие молекулы, которые содержат флуоресцентную или люминесцентную метку.
Согласно другим предпочтительным вариантам реализации настоящего изобретения рекомбинантный полипептид фибриллы АВВ-2/3 содержит или состоит из аминокислотной последовательности, приведенной ниже:
(а)(М/-)

(b) (М/-)

(c) (М/-)

(d) (М/-)

(e) (М/-)
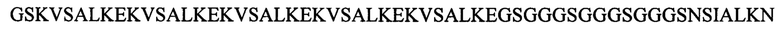

(f) (M/-)

(g) (м/-)

Согласно другому варианту реализации настоящего изобретения рекомбинантные полипептиды находятся в мультимерной форме, такой как димер, тример и т.д. В предпочтительном варианте реализации настоящего изобретения мультимер содержит димер, образованный путем димеризации с помощью доменов димеризации в каждом гомотримере (то есть: полипептид представляет собой гомотример, образованный путем тримеризации домена головной части белка). В мультимерной форме (например, в виде димера) рекомбинантные полипептиды содержат мультимеры фибриллы АВВ-2/3 и могут быть использованы в соответствии с различными способами согласно настоящему изобретению, описанными выше. Специалисты в данной области техники поймут, что такие мультимеры могут содержать мультимеры идентичного рекомбинантного полипептида согласно настоящему изобретению, или могут содержать мультимеры различных рекомбинантных полипептидов согласно настоящему изобретению. Согласно одному варианту реализации настоящего изобретения домены димеризации являются идентичными в каждом рекомбинантном полипептиде, образующем часть мультимера. Согласно другому варианту реализации настоящего изобретения домены димеризации являются различными в каждом рекомбинантном полипептиде, образующем часть мультимера. Согласно другому варианту реализации настоящего изобретения центральные домены и/или головные домены являются идентичными в каждом рекомбинантном полипептиде, образующем часть мультимера. Согласно другому варианту реализации настоящего изобретения центральные домены и/или головные домены являются различными в каждом рекомбинантном полипептиде, образующем часть мультимера.
Степень мультимеризации фибриллы АВВ-2/3 может быть определена в соответствии с методами, хорошо известными специалистам в данной области техники. Например, мультимеризацию рекомбинантных конструкций фибриллы АВВ-2/3 можно оценить с помощью критериев, включающих седиментацию в градиентах сахарозы, устойчивость к протеолизу трипсином и электрофоретическую подвижность в полиакриламидном геле (Hong and Engler, Journal of Virology 70:7071-7078 (1996)). Мультимер фибриллы представляет собой очень устойчивый комплекс в отношении электрофоретической подвижности, и при разделении его молекулярная масса будет соответствовать массе мультимера образца, который не подвергали кипячению перед электрофорезом в ДСН-ПААГ. После кипячения, однако, мультимерная структура нарушается, и при последующем разделении белка его молекулярная масса соответствует таковой для мономера белка.
Рекомбинантные полипептиды или их мультимерные варианты могут храниться в растворе или могут быть заморожены.
Согласно другому варианту реализации настоящего изобретения рекомбинантные полипептиды согласно настоящему изобретению комбинируют (например, конъюгируют) с одним или более терапевтическими агентами для лечения заболевания, связанного с эпителиальной тканью. Такие конъюгаты могут быть использованы, например; в терапевтических способах лечения согласно настоящему изобретению. Способы конъюгации полипептидов согласно настоящему изобретению с терапевтическим агентом, представляющим интерес, например, путем ковалентного связывания или химического сшивания, хорошо известны специалистам в данной области техники. Любой подходящий терапевтический агент может быть использован для образования конъюгата в соответствии с указанным вариантом реализации настоящего изобретения, включая, но не ограничиваясь ими, соединения, разрушающие строму опухоли (такие как релаксин), алкилирующие агенты, ингибиторы ангиогенеза, антитела, антиметаболиты, антимитотики, антипролиферативные средства, ингибиторы Aurora-киназы, ингибиторы промоторов апоптоза (например, Bcl-xL, Bcl-w и Bfl-1), активаторы сигнального пути с участием рецептора гибели, ингибиторы киназы Bcr-Abl, антитела BiTE (биспецифичные антитела, мобилизующие Т-клетки), модификаторы биологического ответа, ингибиторы циклин-зависимой киназы, ингибиторы клеточного цикла, ингибиторы циклооксигеназы-2, ингибиторы факторов роста, ингибиторы белка теплового шока (HSP)-90, деметилирующие агенты, ингибиторы гистондеацетилазы (HDAC), различные виды гормональной терапии, иммунологические агенты, ингибиторы апоптозных белков (IAP), интеркалирующие антибиотики, ингибиторы киназ, ингибиторы мишени рапамицина у млекопитающих, микроРНК-ингибиторы митоген-активируемой протеинкиназы, входящей в состав киназы, регулируемой внеклеточными сигналами (MAP/ERK киназы), поливалентные связывающие белки, нестероидные противовоспалительные препараты (НПВП), ингибиторы полиАДФ (аденозиндифосфат)-рибоза полимеразы (PARP), химиотерапевтические агенты на основе платины, ингибиторы polo-подобной киназы (Plk), ингибиторы протеасомы, аналоги пурина, аналоги пиримидина, ингибиторы рецепторных тирозинкиназ, ретиноидные/дельтоидные растительные алкалоиды, малые ингибирующие рибонуклеиновые кислоты (миРНК), ингибиторы топоизомеразы и т.п.
Типичные терапевтические агенты, входящие в состав перечисленных различных классов, включают, но не ограничиваются ими: доцетаксел, доксорубицин, иринотекан, паклитаксел (таксол®), частицы паклитаксела, связанного с альбумином (абраксан®), липосомальную форму доксорубицина HCl (доксил®), антитела BiTE, такие как адекатумумаб (микромет МТ201), блинатумомаб (микромет МТ103) и т.п., терапевтические агенты на основе миРНК, алкилирующие агенты, включая алтретамин, AMD-473, АР-5280, апазиквон, бендамустин, бросталлицин, бусульфан, карбоквон, кармустин (BCNU), хлорамбуцил, клоретазин® (ларомустин, VNP 40101М), циклофосфамид, дакарбазин, децитабин, 5'-азацитидин, эстрамустин, фотемустин, глюфосфамид, ифосфамид, KW-2170, ломустин (CCNU), мафосфамид, мелфалан, митобронитол, митолактол, нимустин, N-оксид азотистого иприта, ранимустин, темозоломид, тиотепа, треанда® (бендамустин), треосульфан, рофосфамид и т.п.; ингибиторы ангиогенеза, включая ингибиторы специфичных для эндотелия рецепторных тирозинкиназ (Tie-2), ингибиторы рецептора эпидермального фактора роста (EGFR), ингибиторы рецептора инсулиноподобного фактора роста-2 (IGFR-2), ингибиторы матриксной металлопротеиназы-2 (ММР-2), ингибиторы матриксной металлопротеиназы-9 (ММР-9), ингибиторы рецептора тромбоцитарного фактора роста (PDGFR), аналоги тромбоспондина, ингибиторы тирозинкиназы рецепторов фактора роста эндотелия сосудов (VEGFR) и т.п.; антиметаболиты, включая алимта® (динатриевая соль пеметрекседа, LY231514, МТА), 5-азацитидин, кселода® (капецитабин), кармофур, леустат® (кладрибин), клофарабин, цитарабин, окфосфат цитарабина, цитозинарабинозид, децитабин, дефероксамин, доксифлуридин, эфлорнитин, EICAR (5-этинил-1-бета-D-рибофуранозилимидазол-4-карбоксамид), эноцитабин, этнилцитидин, флударабин, 5-фторурацил по отдельности или в комбинации с лейковорином, гемзар® (гемцитабин), гидроксимочевину, алкеран® (мелфалан), меркаптопурин, рибозид 6-меркаптопурина, метотрексат, аналоги метотрексата (например, триметрексат и пралатрексат), микофенолокислоту, неларабин, нолатрексед, окфосфат, пелитрексол, пентостатин, ралтитрексед, рибавирин, триапин, триметрексат, S-1, тиазофурин, тегафур, TS-1, видарабин и т.п.; ингибиторы белка Bcl-2, включая АТ-101 ((-)госсипол), генасенс® (G3139 или облимерсен (антисмысловой олигонуклеотид против Bcl-2)), IPI-194, IPI-565, N-(4-(4-((4'-хлор(1,1'-дифенил)-2-ил)метил)пиперазин-1-ил)бензоил)-4-(((1R)-3-(диметиламино)-1-((фенилсульфанил)метил)пропил)амино)-3-нитробензолсульфонамид) (АВТ-737), N-(4-((2-(4-хлорфенил)-5,5-диметил-1-циклогексен-1-ен-1-ил)метил)пиперазин-1-ил)бензоил)-4-(((1R)-3-(морфолин-4-ил)-1-((фенилсульфанил)метил-)пропил)амино)-3-((трифторметил)сульфонил)бензолсульфонамид (АВТ-263), GX-070 (обатоклакс) и т.п.; ингибиторы киназы BCR-ABL, включая дасатиниб® (BMS-354825), глеевек® (иматиниб) и т.п.; ингибиторы CDK, включая AZD-5438, BMI-1040, BMS-032, BMS-387, CVT-2584, флавопиридол, GPC-286199, MCS-5A, PD0332991, РНА-690509, целициклиб (CYC-202, R-росковитин), ZK-304709 и т.п.; ингибиторы EGFR, включая ABX-EGF, анти-EGFR иммунолипосомы, EGF-вакцину, EMD-7200, эрбитукс® (цетуксимаб), HR3, антитела IgA, пресса® (гефитиниб), тарцева® (эрлотиниб или OSI-774), ТР-38, EGFR-гибридный белок, тайкерб® (лапатиниб) и т.п.; ингибиторы рецептора ErbB2, включая СР-724-714, CI-1033 (канертиниб), герцептин® (трастузумаб), тайкерб® (лапатиниб), омнитарг® (2С4, петузумаб), ТАК-165, GW-572016 (ионафарниб), GW-282974, ЕKВ-569, PI-166, dHER2 (вакцина против HER2), АРС-8024 (вакцина против HER-2), биспецифичное антитело к HER/2neu, B7.her2IgG3, трифункциональные биспецифичные антитела к AS HER2, моноклональные антитела (мАт) AR-209, мАт 2В-1 и т.п.; ингибиторы гистондеацетилазы, включая ромидепсин, LAQ-824, MS-275, трапоксин, субероиланилид гидроксамовой кислоты (SAHA), TSA, вальпроевую кислоту и т.п.; ингибиторы HSP-90, включая AAG-nab, 17-AAG, CNF-101, CNF-1010, CNF-2024, 17-DMAG, гелданамицин, IPI-504, KOS-953, микограб® (человеческое рекомбинантное антитело к HSP-90), NCS-683664, PU24FC1, PU-3, радикикол, SNX-2112, STA-9090 VER49009 и т.п.; активаторы сигнальных путей с участием рецепторов гибели, включая TRAIL, антитела или другие агенты, которые нацелены на TRAIL или рецепторы гибели (например, DR4 и DR5), такие как апомаб, конатумумаб, ETR2-ST01, GDC0145, (лексатумумаб), HGS-1029, LBY-135 PRO-1762 и трастузумаб; химиотерапевтические агенты на основе платины, включая цисплатин, элоксатин® (оксалиплатин) эптаплатин, лобаплатин, недаплатин, параплатин® (карбоплатин), сатраплатин, пикоплатин и т.п.; ингибиторы VEGFR, включая авастин® (бевацизумаб), АВТ-869, АЕЕ-788, акситиниб (AG-13736), AZD-2171, СР-547,632, IM-862, макуген (пегаптамиб), нексавар® (сорафениб, BAY43-9006), пазопаниб (GW-786034), ваталаниб (РТК-787, ZK-222584), сутент® (сунитиниб, SU-11248), VEGF Trap, ZACTIMAThi (вандетаниб, ZD-6474) и т.п.; терапевтические агенты на основе дендритных клеток (сипулейцел-Т, провенг®); ингибиторы топоизомеразы, включая акларубицин, 9-аминокамптотецин, амонафид, амсакрин, бекатекарин, белотекан, BN-80915, камптосар® (гидрохлорид иринотекана), камптотецин, дексразоксин, дифломотекан, эдотекарин, элленс® или фарморубицин® (эпирубицин), этопозид, экзатекан, абраксан, иренотекан, 10-гидроксикамптотецин, гиматекан, луртотекан, митоксантрон, оратецин, пирарубицин, пиксантрон, рубитекан, собузоксан, SN-38, тафлупозид, топотекан и т.п.; антитела, включая авастин® (бевацизумаб), CD40-специфичные антитела, chTNT-1/B, деносумаб, эрбитукс® (цетуксимаб), хумакс-CD4® (занолимумаб), антитела, специфичные к рецептору инсулиноподобного фактора роста I, линтузумаб, панорекс® (эдреколомаб), ренкарекс® (WX G250), ритуксан® (ритуксимаб), тицилимумаб, трастузимаб и т.п.; гормональные терапевтические агенты, включая аримидекс® (анастрозол), аромазин® (экземестан), арзоксифен, касодекс® (бикалутамид), цетротид® (цетрореликс), дегареликс, деслорелин, десопан® (трилостан), дексаметазон, дрогенил® (флутамид), эвиста® (ралоксифен), афема® (фадрозол), фарестон® (торемифен), фаслодекс® (фулвестрант), фемара® (летрозол), форместан, глюкокортикоиды, гекторол® (доксеркальциферол), ренагел® (карбонат севеламера), лазофоксифен, лейпролида ацетат, мегейс® (мегестрол), мифепрекс® (мифепристон), ниландрон® (нилутамид), нолвадекс® (цитрат тамоксифена), пленаксис® (абареликс), преднизолон, пропециа® (финастерид), рилостан, супрефакт® (бусерелин), трелстар® (гормон, высвобождающий лютеинизирующий гормон (LHRH)), вантас® (имплантируемый гистрелин), веторил® (трилостан или модрастан), золадекс® (фосрелин, гозерелин) и т.п.; иммунологические агенты, включая интерферон-альфа, интерферон альфа-2а, интерферон альфа-2b, интерферон-бета, интерферон-гамма 1а, актиммун® (интерферон гамма-1b) или интерферон гамма-n1, их комбинации и т.п. Другие агенты включают альфаферон® (ИФН-альфа), ВАМ-002 (окисленный глутатион), беромун® (тазонермин), бексар® (тозитумомаб), кампат® (алемтузумаб), CTLA4 (антиген 4 цитотоксических лимфоцитов), декарбазин, денилейкин, эпратузумаб, граноцит® (ленограстим), лентинан, лейкоцитарный альфа-интерферон, имиквимод, MDX-010 (анти-CTLA-4), вакцину на основе убитых клеток меланомы, митумомаб, молграмостим, милотарг™ (гемтузумаб озогамицин), нейпоген® (филграстим), онко-VAC-CL, оварекс® (ореговомаб), пемтумомаб (Y-muHMFG1), провенг® (сипулейцел-Т), саргарамостим, сизофилан, тецелейкин, терацис® (бацилла Кальмета-Герена), убенимекс, вирулизин® (иммунотерапевтический агент, Lorus Pharmaceuticals), Z-100 (специфичная субстанция Маруяма (SSM)), WF-10 (тетрахлордекаоксид (TCDO)), пролейкин® (алдеслейкин), задаксин® (тималфазин), зенапакс® (даклизумаб), зевалин® (90Y-ибритумомаб тиуксетан) и т.п.; офатумомаб; модификаторы биологического ответа, включая крестин, лентинан, сизофуран, пицибанил PF-3512676 (CpG-8954), убенимекс и т.п.; аналоги пиримидина, включая цитарабин (ага С или арабинозид С), цитозинарабинозид, доксифлуридин, флудара® (флударабин), 5-ФУ (5-фторурацил), флоксуридин, гемзар® (гемцитабин), томудекс® (ратитрексед), троксатил® (триацетилуридин троксацитабин) и т.п.; аналоги пурина, включая ланвис® (тиогуанин) и пури-нетол® (меркаптопурин); антимитотические агенты, включая батабулин, эпотилон D (KOS-862), N-(2-((4-гидроксифенил)амино)пиридин-3-ил)-4-метоксибензолсульфонамид, иксабепилон (BMS 247550), паклитаксел, таксотер® (доцетаксел), PNU100940 (109881), патупилон, XRP-9881 (ларотаксел), винфлунин, ZK-EPO (синтетический эпотилон) и т.п.; и другие химиотерапевтические агенты, такие как абраксан® (ABI-007), АВТ-100 (ингибитор фарнезилтрансферазы), адвексин® (вакцина Ad5CMV-p53), альтокор® или мевакор® (ловастатин), амплиг® (поли-I:поли-C12U, синтетическая РНК), аптозин® (экзисулинд), аредиа® (памидроновая кислота), арглабин, L-аспарагиназу, атаместан (1-метил-3,17-дион-андроста-1,4-диен), аваг® (тазаротен), AVE-8062 (производное комбрестатина) ВЕС2 (митумомаб), кахектин или кахексии (фактор некроза опухоли), канваксин (вакцина), цеавак® (вакцина против рака), целейк® (целмолейкин), цеплен® (дигидрохлорид гистамина), церварикс® (вакцина против вируса папилломы человека), CHOP® (С: цитоксан® (циклофосфамид); Н: адриамицин® (гидроксидоксорубицин); О: винкристин (онковицин®); Р: преднизолон), CYPAT® (ацетат ципротерона), комбрестатин А4Р, DAB(389)EGF (каталитический и транслокационный домены дифтерийного токсина, гибридизованные посредством линкера His-Ala с эпидермальным фактором роста человека) или TransMID-107R® (дифтерийные токсины), дакарбазин, дактиномицин, 5,6-диметилксантенон-4-уксусная кислота (DMXAA), энилурацил, эвизон™ (лактат скваламина), димерицин® (липосомальный лосьон T4N5), дискодермолид, DX-8951f (мезилат экзатекана), энзастаурин, ЕРО906 (эпитилон В), гардасил® (рекомбинантная вакцина против четырехвалентного папилломавируса человека (типы 6, 11, 16, 18)), гастриммун®, генасенс®, GMK (ганглиозид-конъюгированная вакцина), Джи-Вакс® (вакцина против рака простаты), галофугинон, гистерелин, гидроксикарбамид, ибандроновую кислоту, IGN-101, IL-13-PE38, IL-13-PE38QQR (цинтредекин безудотокс), IL-13-экзотоксин Pseudomonas, интерферон-альфа, интерферон-гамма, юнован® или мепакт® (мифамуртид), лонафарниб, 5,10-метилентетрагидрофолат, мильтефозин (гексадецилфосфохолин), неовастат® (АЕ-941), нейтрексин® (глюкуронат триметрексата), нипент® (пентостатин), онконаз® (рибонуклеазный фермент), онкофаг® (вакцина против меланомы), онковакс® (вакцина на основе IL-2), оратецин® (рубитекан), осидем® (клеточный препарат на основе антител), мАт оварекс® (мышиное моноклональное антитело), паклитаксел, пандимекс® (агликоны сапонинов женьшеня, содержащие 20(S) протопанаксадиол (aPPD) и 20(S) протопанаксатриол (аРРТ)), панитумумаб, панвак®-VF (изучаемая вакцина против рака), пегаспаргаза, ПЭГ-интерферон А, феноксодиол, прокарбазин, ребимастат, ремоваб® (катумаксомаб), ревлимид® (леналидомид), RSR13 (эфапроксирал), соматулин® LA (ланреотид), сориатан® (ацитретин), стауроспорин (Streptomyces staurospores), талабостат (РТ100), таргретин® (бексаротен), таксопрексин® (DHA-паклитаксел), телцита® (канфосфамид, TLK286), темилифен, темодар® (темозоломид), тесмилифен, талидомид, тератоп® (STn-KLH), тимитак (2-амино-3,4-дигидро-6-метил-4-оксо-5-(4-пиридилтио)хиназолина дигидрохлорид), TNFERABE® (аденовектор: носитель ДНК, содержит ген фактора некроза опухоли альфа), траклеер® или завеска® (бозентан), третиноин (ретин-А), тетрандрин, тризенокс®. (триоксид мышьяка), вирулизин®, украин (производное алкалоидов чистотела большого), витаксин (антитело к а5рЗ), эксцитрин® (мотексафин гадолиния), ксинлей® (атразентан), ксиотакс® (паклитаксел полиглумекс), ионделис® (трабектедин), ZD-6126, зинекард® (дексразоксан), зомета® (золедроновая кислота), кризотиниб, зорубицин и т.п.
Согласно другому предпочтительному варианту реализации настоящего изобретения терапевтический агент включает соединение, которое связывается с десмоглеином-2; предпочтительно соединение, которое связывается с DSG2 и размыкает плотные контакты.
Согласно другим вариантам реализации настоящего изобретения терапевтический агент включает радиоактивные частицы/радиотерапевтические лекарственные средства. Любое подходящее радиотерапевтическое лекарственное средство или частица могут быть использованы по усмотрению лечащего врача, включая, но не ограничиваясь ими, кобальт-60, йод-131, иридий-192, стронций-89, самарий-153, рений-186 и свинец-212.
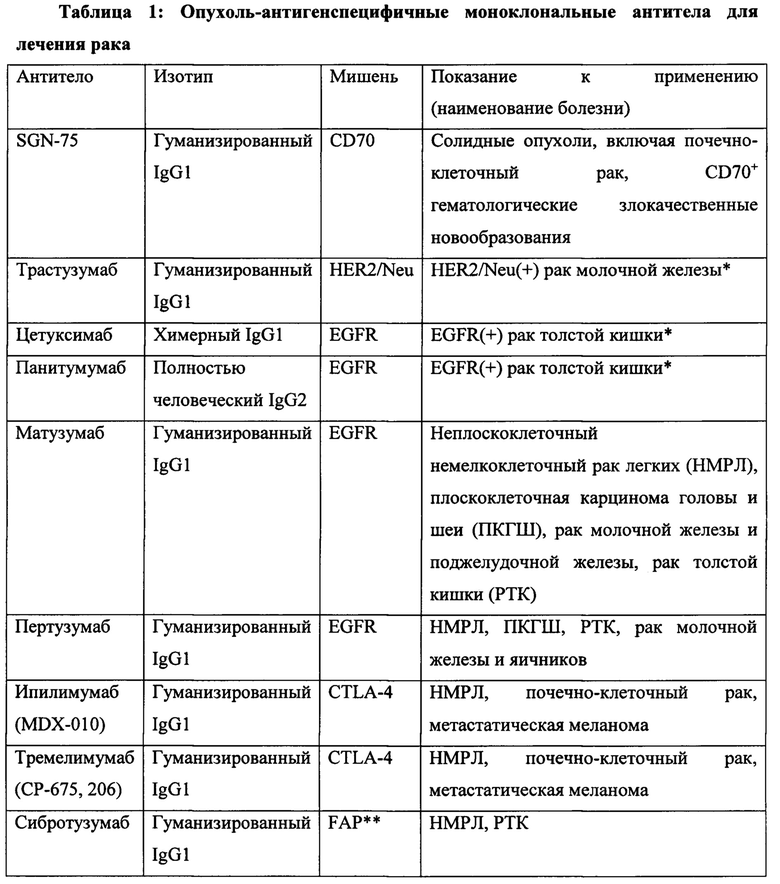
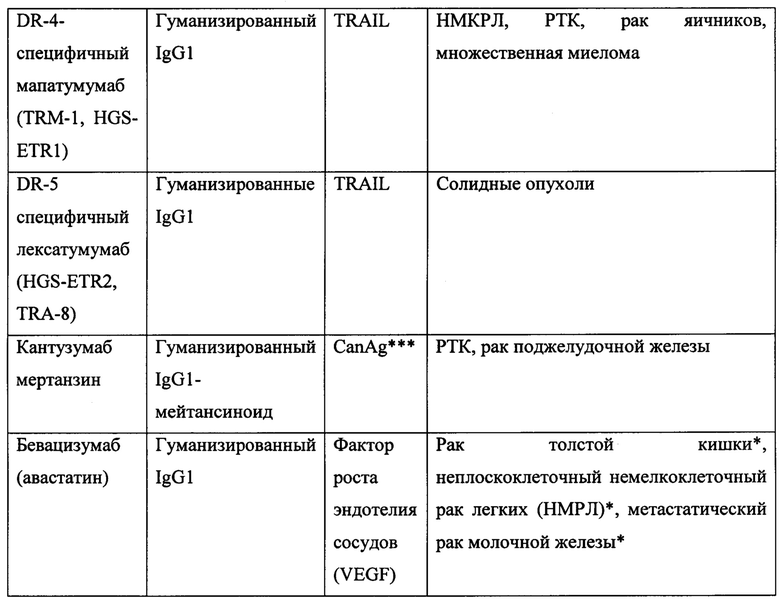
Согласно предпочтительному варианту реализации настоящего изобретения терапевтический агент представляет собой противоопухолевый терапевтический агент и включает химиотерапевтическое или противоопухолевое моноклональное антитело, описанное в настоящей заявке. Согласно другому предпочтительному варианту реализации настоящего изобретения терапевтический агент включает антитело, выбранное из группы, состоящей из трастузумаба, цетумиксимаба, петузумаба, апомаба, конатумумаба, лексатумумаба, бевацизумаба, деносумаба, занолимумаба, линтузумаба, эдреколомаба, ритуксимаба, тицилимумаба, тозитумомаба, алемтузумаба, эпратузумаба, митумомаба, гемтузумаба озогамицина, ореговомаба, пемтумомаба, даклизумаба, панитумумаба, катумаксомаба, офатумумаба и ибритумомаба. Неограничивающие примеры подходящих противоопухолевых мАт и конкретные способы их применения перечислены в таблице 1, и подробно описаны в работе Campoli, М., et al., Principles & Practice of Oncology 23(1&2):1-19 (2009), которая полностью включена в настоящую заявку посредством ссылки.
Согласно другому варианту реализации рекомбинантные полипептиды согласно настоящему изобретению комбинированы (например, конъюгированы) с одним или более диагностическими или агентами для визуализации. Рекомбинантные полипептиды согласно настоящему изобретению и их мультимеры широко применяют для доставки любого диагностического, агента для визуализации или другого соединения к эпителиальной ткани, содержащей межклеточные контакты, в которой доступ к мишени, представляющей интерес, может быть ограничен. В различных неограничивающих вариантах реализации настоящего изобретения агенты для визуализации могут включать любое химическое соединение, которое может производить детектируемый сигнал, прямо или косвенно. Многие из таких агентов для визуализации известны специалистам в данной области техники. Примеры агентов для визуализации, подходящих для применения в соответствии с раскрытыми способами и композициями, включают радиоактивные изотопы, флуоресцентные молекулы, магнитные частицы (включая наночастицы), металлические частицы (включая наночастицы), фосфоресцирующие молекулы, ферменты, антитела, лиганды и их комбинации, тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания эпителиальной ткани, связанное с таким агентом для визуализации. Способы детектирования и измерения сигналов, которые производят агенты для визуализации, также известны специалистам в данной области техники. Например, радиоактивные изотопы можно детектировать с помощью сцинтилляционного счетчика или путем прямой визуализации; флуоресцентные молекулы можно детектировать с помощью флуоресцентных спектрофотометров; фосфоресцентные молекулы можно детектировать с помощью спектрофотометра или путем непосредственной визуализации с помощью камеры; ферменты можно детектировать с помощью обнаружения или визуализации продукта реакции, катализируемой этим ферментом; антитела можно детектировать путем обнаружения вторичной детектируемой метки, связанной с антителом. Согласно одному предпочтительному варианту реализации настоящего изобретения агент для визуализации и/или диагностический агент представляет собой тот, который может быть использован для детектирования опухоли, как путем прямого связывания с опухолью, так и путем присоединения агента для визуализации или диагностического агента к соединению, которое может связываться с опухолью.
В различных вариантах реализации настоящего изобретения агент для визуализации может представлять собой флуоресцентный агент для визуализации, тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания эпителиальной ткани, связанное с флуоресцентным агентом для визуализации. Флуоресцентный агент для визуализации представляет собой любой химический фрагмент, который имеет детектируемый флуоресцентный сигнал. Агент для визуализации может быть использован по отдельности или в комбинации с другими агентами для визуализации. Примеры подходящих флуоресцентных агентов, которые можно использовать в композициях и способах, раскрытых в настоящей заявке, включают, но не ограничиваются ими, флуоресцеин (FITC), 5-карбоксифлуоресцеин-N-гидроксисукцинимидный эфир, 5,6-карбоксиметилкрахмал флуоресцеина, нитробенз-2-окса-1,3-тиадиазол-4-ил (NBD), флуорескамин, ОРА, NDA, индоцианиновый зеленый краситель, цианиновые красители (например, Су3, Су3.5, Су5, Су5.5 и Су7), 4-ацетамидо-4'-изотиоцианатостилбен-2,2-дисульфоновую кислоту, акридин, изотиоцианат акридина, 5-(2'-аминоэтил)аминонафталин-1-сульфоновую кислоту (EDANS), 4-амино-N-[3-винилсульфонил)фенилинафталимид-3,5дисульфонат, N-(4-анилино-1-нафтил)малеимид, антраниламид, BODIPY, бриллиантовый желтый, кумарин, 7-амино-4-метилкумарин (АМС, кумарин 120), 7-амино-4-трифторметилкумарин (кумарин 151), цианозин, 4',6-диаминидино-2-фенилиндол (DAPI), 5',5"-дибромпирогаллолсульфонафталин (бромпирогаллол красный), 7-диэтиламино-3-(4'-изотиоцианатофенил)-4-метилкумариндиэтилентриаминпентаацетат, 4,4'-диизотиоцианатодигидростильбен-2,2'-дисульфоновую кислоту, 4,4'-диизотиоцианатостильбен-2,2'-дисульфоновую кислоту, 5-[диметиламино]нафталин-1-сульфонилхлорид (DNS, дансилхлорид), 4-(4'-диметиламинофенилазо)бензойную кислоту (DABCYL), 4-диметиламинофенилазофенил-4-изотиоцианат (DABITC), эозин, изотиоцианат эозина, эритрозин, эритрозин В, изотиоцианат, бромид этидия, этидий, 5-карбоксифлуоресцеин (FAM), 5-(4,6-дихлортриазин-2-ил)аминофлуоресцеин (DTAF), 2',7'-диметокси-4'5'-дихлор-6-карбоксифлуоресцеин (JOE), изотиоцианат флуоресцеина, IR144, IR1446, малахитовый зеленый изотиоцианат, 4-метилумбеллиферон, орто-крезолфталеин, нитротирозин, парарозанилин, феноловый красный, В-фикоэритрин, орто-фталдиальдегид, пирен, бутират пирена, сукцинимидил-1-пиренбутират, реактивный красный 4 (Cibacron [R] бриллиантовый красный 3В-А), 6-карбокси-Х-родамин (ROX), 6-карбоксиродамин (R6G), лиссамин-родамин В сульфонилхлорид, родамин (Rhod), 5,6-тетраметилродамин, родамин В, родамин 123, родамин X изотиоцианат, сульфородамин В, сульфородамин 101, сульфонилхлоридное производное сульфородамина 101 (техасский красный), N,N,N',N'-тетраметил-6-карбоксиродамин (TAMRA), тетраметилродамин, изотиоцианат тетраметилродамина (TRITC), рибофлавин, кораллин, кумарин-6 и т.п., включая их комбинации. Перечисленные флуоресцентные фрагменты для визуализации могут быть получены из различных коммерческих источников, включая Molecular Probes (Юджин, Орегон, США) и Research Organics (Кливленд, Огайо, США) или могут быть синтезированы специалистам в данной области техники.
В другом примере агенты для визуализации могут содержать агент для магнитно-резонансной томографии (МРТ), тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания эпителиальной ткани, связанное с агентом для МРТ. Агент для МРТ представляет собой любой химический фрагмент, который производит детектируемый сигнал магнитного резонанса или который может повлиять (например, увеличить или сместить) на сигнал магнитного резонанса другого агента. Указанный тип агента для визуализации можно применять по отдельности или в комбинации с другим агентом для визуализации. Согласно другому примеру агент для МРТ на основе гадолиния может служить в качестве агента для визуализации. Примером подходящего агента для МРТ, который может входить в состав агентов для визуализации, раскрытых в настоящем изобретении, является пара-аминобензилдиэтилентриаминопентауксусная кислота (p-NH2-Bz-DTPA, соединение 7), конъюгированная форма диэтилентриаминпентауксусной кислоты (DTPA), которая, как известно, прочно связывает гадолиний и одобрена для клинического применения в качестве контрастного вещества для МРТ. Встраивание агента для МРТ на поверхность большой макромолекулы, такой как дендримерный субстрат, как описано в настоящей заявке, может обеспечить большую релаксацию Т1 (высокий контраст), и включение нескольких копий агента на одну молекулу может увеличить сигнал. При комбинировании агента для визуализации для МРТ и, например, флуоресцентного агента для визуализации, полученный агент можно детектировать, визуализировать и регистрировать его динамику в режиме реального времени с помощью МРТ. Другие агенты для визуализации включают агенты для ПЭТ, которые могут быть получены путем включения 18F или хелатора 64Cu или 68Ga. Кроме того, добавление радионуклида можно использовать для улучшения визуализации методом SPECT или доставки дозы радиации, тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания эпителиальной ткани, связанное с агентом для ПЭТ.
В некоторых вариантах реализации настоящего изобретения диагностический агент представляет собой диагностический агент для визуализации, включая, но не ограничиваясь ими, агенты для позитронно-эмиссионной томографии (ПЭТ), агенты для компьютерной томографии (КТ), агенты для магнитно-резонансной томографии (МРТ), агенты для визуализации для ядерно-магнитного резонанса (ЯМР), агенты для рентгеноскопии и контрастные агенты для ультразвуковых исследований. Перечисленные диагностические агенты включают радиоизотопы таких элементов, как йод (I), включая 123I, 125I, 131I и т.д., барий (Ва), гадолиний (Gd), технеций (Тс), включая 99Тс, фосфор (Р), включая 31Р, железо (Fe), марганец (Mn), таллий (Tl), хром (Cr), включая 51Cr, углерод (С), включая 14С, или т.п., флуоресцентномеченые соединения или их комплексы, хелаты, аддукты и конъюгаты. Любые подходящие агенты для ПЭТ могут быть использованы, включая, но не ограничиваясь ими, углерод-11, азот-13, кислород-15, фтор-18, 11С-метомидат, и аналоги глюкозы, включая, но не ограничиваясь ей, фтордезоксиглюкозу (аналог глюкозы, меченый изотопом фтора-18).
Согласно другим вариантам реализации настоящего изобретения диагностический агент представляет собой маркерный ген, который кодирует белки, которые легко детектируются при экспрессии в клетке (включая, но не ограничиваясь ими, бета-галактозидазу, зеленый флуоресцентный белок, люциферазу и т.п.), и меченые нуклеиновые зонды (например, радиоактивномеченые или флуоресцентномеченые зонды). Согласно некоторым вариантам реализации настоящего изобретения ковалентная конъюгация диагностических или агентов для визуализации с мультимерами АВВ-2/3, предложенными в настоящей заявке, достигается благодаря различным процессам конъюгации. Согласно другим вариантам реализации настоящего изобретения диагностический агент нековалентно связан с мультимерами АВВ-2/3, предложенными в настоящей заявке.
Согласно другому аспекту в настоящем изобретении предложены нуклеиновые кислоты, кодирующие полипептид согласно любому варианту реализации настоящего изобретения. Нуклеиновые кислоты могут содержать РНК или ДНК, а также могут быть получены и выделены с использованием стандартных методик молекулярной биологии, основываясь на принципах, предложенных в настоящей заявке. Нуклеиновые кислоты могут содержать дополнительные домены, которые можно применять для облегчения экспрессии и/или очистки кодируемого белка, включая, но не ограничиваясь ими, последовательности полиА, модифицированные последовательности Козак и последовательности, кодирующие эпитопные метки, сигналы экспорта и сигналы секреции, сигналы ядерной локализации и сигналы локализации на плазматической мембране.
Согласно другому аспекту в настоящем изобретении предложены рекомбинантные векторы экспрессии, содержащие нуклеиновую кислоту согласно любому аспекту настоящего изобретения, функционально связанную с промотором. Термин «рекомбинантный вектор экспрессии» включает векторы, которые функционально связывают нуклеиновую кислоту, кодирующую область или ген, с любым промотором, способным осуществлять экспрессию продукта гена. Последовательность промотора, которая используется для инициации экспрессии раскрытых нуклеиновых кислот в системе экспрессии млекопитающих, может быть конститутивной (инициируется любым из множества промоторов, включая, но не ограничиваясь ими, промотор CMV, SV40, RSV, актина, EF) или индуцибельной (инициируется любым из ряда индуцируемых промоторов, включая, но не ограничиваясь ими, промоторы, реагирующие на тетрациклин, экдизон, стероиды). Конструирование векторов экспрессии для использования в трансфекции прокариотических клеток также хорошо известно в данной области техники, и, следовательно, может быть осуществлено с помощью стандартных методик. (См, например, Sambrook, Fritsch, and Maniatis, в: Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1989; Gene Transfer and Expression Protocols, pp. 109-128, ed. E.J. Murray, The Humana Press Inc., Clifton, N.J.) и каталог Ambion 1998 (Ambion, Остин, Техас, США). Вектор экспрессии должен быть способен к репликации в организмах-хозяевах в виде эписомы либо путем интеграции в хромосомную ДНК хозяина, и может содержать любые другие компоненты, которые буду сочтены целесообразными для данного способа применения, включая, но не ограничиваясь ими, селективные маркеры, такие как ген устойчивости к антибиотику.
Согласно другому аспекту в настоящем изобретении предложены клетки-хозяева, содержащие рекомбинантные векторы экспрессии, раскрытые в настоящей заявке, и их потомство, причем клетки-хозяева могут представлять собой прокариотические или эукариотические клетки. Клетки могут быть кратковременно или стабильно трансфецированы. Трансфекцию векторов экспрессии в прокариотические и эукариотические клетки можно осуществлять с помощью любой методики, известной в данной области техники, включая, но не ограничиваясь ими, стандартные способы трансформации бактерий, соосаждение с фосфатом кальция, электропорацию или трансфекцию, опосредованную липосомами, ДЭАЭ-декстраном, поликатионами или вирусами. (См, например, Molecular Cloning: A Laboratory Manual (Sambrook, et al., 1989, Cold Spring Harbor Laboratory Press; Culture of Animal Cells: A Manual of Basic Technique, 2 изд. (R.I. Freshney. 1987. Liss, Inc. New York, NY). Методики, в которых культивируемые клетки, трансфецированные векторами экспрессии, используют для получения больших количеств полипептидов, хорошо известны в данной области техники.
Согласно другому аспекту в настоящем изобретении предложены фармацевтические композиции, содержащие:
a), мультимер фибриллы АВВ-2/3 согласно настоящему изобретению; и
b). фармацевтически приемлемый носитель.
Мультимер фибриллы АВВ-2/3 может представлять собой любой мультимер, описанный в настоящей заявке, в соответствии с любым аспектом, вариантом реализации или комбинацией вариантов реализации настоящего изобретения, который содержит мутированный полипептид головного домена согласно любому варианту реализации первого аспекта настоящего изобретения (т.е.: SEQ ID NO: 1-11).
Фармацевтическая композиция может дополнительно содержать один или более терапевтических агентов для лечения заболевания, связанного с эпителиальной тканью, включая, но не ограничиваясь ими, те, которые описаны выше. Согласно предпочтительному варианту реализации настоящего изобретения терапевтический агент представляет собой противоопухолевый терапевтический агент и включает химиотерапевтическое или противоопухолевое моноклональное антитело, описанное в настоящей заявке. Согласно другому предпочтительному варианту реализации настоящего изобретения противоопухолевый терапевтический агент включает антитело, выбранное из группы, состоящей из трастузумаба, цетумиксимаба, петузумаба, апомаба, конатумумаба, лексатумумаба, бевацизумаба, деносумаба, занолимумаба, линтузумаба, эдреколомаба, ритуксимаба, тицилимумаба, тозитумомаба, алемтузумаба, эпратузумаба, митумомаба, гемтузумаба озогамицина, ореговомаба, пемтумомаба, даклизумаба, панитумумаба, катумаксомаба, офатумумаба и ибритумомаба.
Фармацевтически приемлемый носитель является нетоксичным, биосовместимым и выбран так, чтобы не оказывать нежелательного влияния на биологическую активность мультимеров (и любых других терапевтических агентов, комбинированных с ним). Примеры фармацевтически приемлемых носителей для пептидов описаны Yamada et al. в патенте США №5211657. Композиции могут быть изготовлены в виде препаратов в твердой, полутвердой, гелевой, жидкой и газообразной формах, таких как таблетки, капсулы, порошки, гранулы, мази, растворы, суппозитории, ингаляционные препараты и инъекции, что позволяет вводить их пероральным, парентеральным или хирургическим путем. Подходящие носители для парентерального пути введения с помощью инъекции, инфузии или орошения и местной доставки включают дистиллированную воду, забуференный фосфатом физиологический раствор, нормальный раствор Рингера или раствор Рингера с добавлением лактата, раствор декстрозы, раствор Хэнка или пропандиол. Помимо этого в качестве растворителя или суспендирующей среды можно применять стерильные нелетучие масла. Любое биосовместимое масло может быть использовано в качестве растворителя, включая синтетические моно- или диглицериды. При изготовлении инъецируемых препаратов также можно использовать жирные кислоты, такие как олеиновая кислота. Носитель и агент могут быть смешаны в виде жидкости, суспензии, полимеризуемого или неполимеризуемого геля, пасты или мази. Носитель также может содержать носитель для доставки, чтобы поддержать (т.е. продлить, задержать или регулировать) доставку агента(ов) или усилить доставку, всасывание, стабильность или фармакокинетику терапевтического агента(ов). Описанный носитель для доставки может содержать, в качестве неограничивающего примера, микрочастицы, микросферы, наносферы или наночастицы, состоящие из белков, липосом, углеводов, синтетических органических соединений, неорганических соединений, полимерных или сополимерных гидрогелей и полимерных мицелл. Подходящие системы доставки на основе гидрогелей и мицелл включают сополимеры РЕО:РНВ:РЕО и комплексы сополимер/циклодекстрин, раскрытые в международной публикации WO 2004/009664 А2, а также комплексы РЕО и РЕО/циклодекстрин, раскрытые в публикации патента США №2002/0019369 А1. Перечисленные гидрогели могут быть введены локально в место предполагаемого действия или подкожно, или внутримышечно, чтобы образовать депо с пролонгированным высвобождением.
В случае интратекального (IT) или интрацеребровентрикулярного (ICV) введения композиций могут быть использованы системы доставки, имеющие надлежащую степень стерильности (например, жидкости; гели, суспензии и т.д.). Для перорального введения непептидергических агентов в качестве носителя композиций может быть использован инертный наполнитель или разбавитель, такой как сахароза, кукурузный крахмал или целлюлоза.
Композиции согласно настоящему изобретению также могут содержать биосовместимые наполнители, такие как диспергирующие или смачивающие агенты, суспендирующие агенты, разбавители, буферы, способствующие проникновению вещества, эмульгаторы, связующие вещества, загустители, ароматизаторы (для перорального введения). Типичные составы можно вводить парентерально в форме инъецируемых доз раствора или суспензии мультимера в физиологически приемлемом разбавителе с фармацевтическим носителем, который может представлять собой стерильную жидкость, такую как вода, масла, физиологический раствор, глицерин или этанол. Помимо этого в композиции, содержащей модифицированные полипептиды, могут присутствовать вспомогательные вещества, такие как смачивающие или эмульгирующие агенты, поверхностно-активные вещества, буферные вещества и т.п. Дополнительные компоненты фармацевтических композиций включают углеводороды (например, животного, растительного или синтетического происхождения), например, соевое масло и минеральное масло. В целом, предпочтительными жидкими носителями для инъекционных растворов являются гликоли, такие как пропиленгликоль или полиэтиленгликоль.
Фармацевтическую композицию также можно вводить в форме депо-инъекций или имплантируемого препарата, который может быть изготовлен так, чтобы обеспечить пролонгированное или пульсирующее высвобождение мультимеров и других терапевтических агентов (если они присутствуют).
Помимо полипептидов согласно настоящему изобретению фармацевтическая композиция может содержать (а) лиопротектор; (b) поверхностно-активное вещество; (с) наполнитель; (d) агент, регулирующий тоничность; (е) стабилизатор; (Е) консервант и/или (g) буфер. Согласно некоторым вариантам реализации настоящего изобретения буфер в фармацевтической композиции представляет собой буфер на основе Триса, гистидина, фосфата, цитрата или ацетата. Фармацевтическая композиция также может содержать лиопротектор, например сахарозу, сорбит или трегалозу. Согласно некоторым вариантам реализации настоящего изобретения фармацевтическая композиция содержит консервант, например, хлорид бензалкония, бензетониум, хлоргексидин, фенол, м-крезол, бензиновый спирт, метилпарабен, пропилпарабен, хлорбутанол, о-крезол, п-крезол, хлоркрезол, нитрат фенилртути, тимеросал, бензойную кислоту, и различные их смеси. Согласно другим вариантам реализации настоящего изобретения фармацевтическая композиция содержит наполнитель, такой как глицин. Согласно другим вариантам реализации настоящего изобретения фармацевтическая композиция содержит поверхностно-активное вещество, например, полисорбат-20, полисорбат-40, полисорбат-60, полисорбат-65, полисорбат-80, полисорбат-85, полоксамер-188, монолаурат сорбита, монопальмитат сорбита, моностеарат сорбита, моноолеат сорбита, трилаурат сорбита, тристеарат сорбита, триолеат сорбита, или их комбинацию. Фармацевтическая композиция также может содержать агент, регулирующий тоничность, например, соединение, которое делает состав по существу изотоничным или изоосмотичным по отношению к крови человека. Типичные агенты, регулирующие тоничность, включают сахарозу, сорбит, глицин, метионин, маннит, декстрозу, инозит, хлорид натрия, аргинин и гидрохлорид аргинина. Согласно другим вариантам реализации настоящего изобретения фармацевтическая композиция дополнительно содержит стабилизатор, например, молекулу, которая в комбинации с представляющим интерес белком по существу предотвращает или снижает химическую и/или физическую нестабильность белка, представляющего интерес, в лиофилизированной или жидкой форме. Примеры стабилизаторов включают сахарозу, сорбит, глицин, инозитол, хлорид натрия, метионин, аргинин и гидрохлорид аргинина.
Фармацевтическая композиция может быть упакована любым подходящим способом. Согласно одному варианту реализации настоящего изобретения фармацевтическая композиция упакована в виде набора, содержащего контейнер (такой как ампула) с мультимером фибриллы АВВ-2/3. Согласно предпочтительному варианту реализации настоящего изобретения набор дополнительно содержит, в этом же или в отдельном контейнере (например, флаконе), терапевтический, диагностический или агент для визуализации для введения субъекту совместно с мультимером фибриллы АВВ-2/3.
Согласно другому аспекту в настоящем изобретении предложены наборы, содержащие (а) один или более рекомбинантных полипептидов/мультимеров фибриллы АВВ-2/3, выделенные нуклеиновые кислоты, рекомбинантные векторы экспрессии и/или клетки-хозяева согласно настоящему изобретению; и (b) инструкции по его/их применению в лечении заболеваний, связанных с эпителиальной тканью. Наборы могут дополнительно содержать терапевтический агент для применения в соответствии со способами согласно настоящему изобретению.
Согласно другому аспекту в настоящем изобретении предложены способы повышения эффективности терапевтического лечения или диагностики заболевания, связанного с эпителиальной тканью, и/или визуализации эпителиальных тканей, включающие введение субъекту, нуждающемуся в этом:
a) одного или более терапевтических агентов в количестве, достаточном для лечения заболевания, диагностического агента в количестве, достаточном для диагностики заболевания, и/или агента для визуализации в количестве, достаточном для визуализации эпителиальной ткани; и
b) такого количества мультимера фибриллы АВВ-2/3 согласно настоящему изобретению или фармацевтической композиции согласно настоящему изобретению, которое достаточно для повышения эффективности одного или более терапевтических, диагностических и/или агентов для визуализации.
Способы в соответствии с указанным аспектом настоящего изобретения могут быть использованы для повышения эффективности терапевтического лечения, диагностики или визуализации заболевания, связанного с эпителиальной тканью, за счет улучшения доступа терапевтического, диагностического и/или агента для визуализации к мишени и его распределения в эпителиальной ткани. Безотносительно к какому-либо конкретному механизму действия, авторы настоящего изобретения полагают, что повышение эффективности происходит за счет дополняющих механизмов: перемещения рецептора-мишени от базолатеральной к апикальной поверхности клетки, обеспечивая тем самым улучшение доступа к мишени в эпителиальной ткани для терапевтических, диагностических и/или агентов для визуализации, которые нацелены на рецептор, таких как мышиные моноклональные антитела, и более эффективного проникновения терапевтического агента посредством разрушения межклеточных контактов. DSG2 является основным высокоаффинным рецептором для АВВ-2/3. DSG2 представляет собой кальций-связывающий трансмембранный гликопротеин, принадлежащий к семейству белков кадгеринов. В эпителиальных клетках DSG2 является компонентом структуры межклеточной адгезии. Его цитоплазматический хвост взаимодействует с несколькими белками, которые непосредственно контактируют с регуляторами клеточной адгезии и межклеточных контактов/морфологии клеток. Было показано, что DSG2 сверхэкспрессируется в ряде эпителиальных злокачественных опухолей, включая рак желудка, плоскоклеточные карциномы, меланому, метастатический рак предстательной железы и рак мочевого пузыря.
Безотносительно к какому-либо конкретному механизму действия, авторы настоящего изобретения полагают, что связывание мультимера фибриллы АВВ-2/3 с DSG2 инициирует кратковременное DSG2-опосредованное размыкание межклеточных контактов, которое улучшает доступ терапевтических, диагностических, агентов для визуализации, или любого другого соединения, представляющего интерес, связывающегося с мишенью в эпителиальных клетках, которое в ином случае было бы задержано, по меньшей мере в некоторой степени, межклеточными контактами. Подробные примеры такого вида активности приведены в настоящей заявке. Способы в соответствии с настоящим изобретением, следовательно, могут быть осуществлены с использованием любого мультимера фибриллы АВВ-2/3 согласно настоящему изобретению, чтобы индуцировать кратковременное DSG2-опосредованное размыкание межклеточных контактов. Типичные мультимеры, содержащие один или более мультимеров фибриллы АВВ-2/3 согласно настоящему изобретению, которые можно применять в соответствии с указанными способами, включают, но не ограничиваются ими, вирионы АВВ-2/3, капсиды АВВ-2/3, додекаэдрические частицы АВВ-2/3 (PtDd) (субвирусные додекаэдрические частицы, которые вырабатываются АВВ-2/3 во время репликации) и рекомбинантные мультимеры фибриллы АВВ-2/3.
Способы согласно настоящему изобретению имеют широкое применение для доставки любого терапевтического, диагностического или агента для визуализации, или другого соединения к эпителиальной ткани, содержащей межклеточные контакты, в которой доступ к представляющей интерес мишени может быть ограничен, поскольку DSG2 широко экспрессируется в эпителиальных клетках. В настоящей заявке термин «заболевание, связанное с эпителиальной тканью» представляет собой любое нарушение, при котором терапевтический, диагностический или агент для визуализации, который вводят в/через эпителиальные клетки/эпителиальную ткань, обеспечивает клиническую пользу для пациента, такую как улучшение эффективности терапии, диагностики и/или визуализации. Такие заболевания включают, но не ограничиваются ими, солидные опухоли (т.е. любую опухоль, содержащую контакты между эпителиальными клетками), заболевания желудочно-кишечного тракта (включая, но не ограничиваясь ими, синдром раздраженного кишечника, воспалительное заболевание кишечника, болезнь Крона, неспецифический язвенный колит, запор, гастроэзофагеальную рефлюксную болезнь, пищевод Барретта и т.д.), кожные заболевания (включая, но не ограничиваясь ими, псориаз и дерматит), заболевания легких (включая, но не ограничиваясь ими, хроническую обструктивную болезнь легких, астму, бронхит, эмфизему легких, кистозный фиброз, интерстициальную болезнь легких, пневмонию, заболевание протоков поджелудочной железы), заболевания головного мозга (то есть любое заболевание головного мозга, при котором пациент может получить клиническую пользу от улучшения транспорта лекарственных препаратов через гематоэнцефалический барьер), первичную легочную гипертензию, легочную эмболию, саркоидоз легких, туберкулез и т.д., заболевания почек (включая, но не ограничиваясь им, гломерулонефрит), заболевания печени (включая, но не ограничиваясь им, гепатит), эндокринные заболевания (включая, но не ограничиваясь ими, диабет и заболевания щитовидной железы), заболевания протоков поджелудочной железы (включая, но не ограничиваясь им, панкреатит), а также заболевания желчных протоков (включая, но не ограничиваясь ими, непроходимость желчных путей, холецистит, холедохолитиаз, камни в желчном пузыре и т.д.), а также инфекции эпителиальных тканей (включая, но не ограничиваясь ими, целлюлит, пневмонию, гепатит и пиелонефрит). Согласно одному предпочтительному варианту реализации настоящего изобретения заболевание, связанное с эпителиальной тканью, представляет собой солидную опухоль, включая, но не ограничиваясь ими, опухоли молочной железы, опухоли легких, опухоли толстой кишки, опухоли прямой кишки, опухоли кожи, эндокринные опухоли, опухоли желудка, опухоли предстательной железы, опухоли яичников, опухоли матки, опухоли шейки матки, опухоли почек, меланомы, опухоли поджелудочной железы, опухоли печени, опухоли головного мозга, опухоли головы и шеи, опухоли носоглотки, опухоли желудка, плоскоклеточные карциномы, аденокарциномы, опухоли мочевого пузыря и опухоли пищевода. Специалисты в данной области техники поймут, что такие опухоли включают первичные опухоли, локально инвазивные опухоли, а также метастазировавшие опухоли.
В настоящей заявке термин «повышение эффективности» означает любое повышение терапевтической, диагностической и/или визуализирующей эффективности по сравнению с той, которую наблюдают при использовании терапевтического, диагностического и/или агента для визуализации по отдельности. Например, количественные показатели терапевтической эффективности будут варьироваться в зависимости от заболевания, подлежащего лечению, но легко выявляются лечащим врачом. Например, повышение эффективности включает, но не ограничивается им, повышение одного или более из следующих условий, по сравнению с лечением с использованием только терапевтического агента: (а) снижение тяжести заболевания; (b) ограничение или предотвращение развития симптомов, характерных для заболевания или заболеваний, подлежащих лечению; (с) ингибирование ухудшения симптомов, характерных для заболевания или заболеваний, которые лечат; (d) ограничение или предотвращение рецидивов заболевания или заболеваний у пациентов, которые ранее имели заболевание или заболевания; и (е) заболевание или предотвращение рецидива симптомов у пациентов, которые ранее имели симптомы заболевания или заболеваний. Согласно одному неограничивающему примеру лечение солидной опухоли обеспечивает возможность индуцировать выход опухолевых рецепторов с базолатеральной стороны эпителиальных клеток, чтобы обеспечить улучшенный доступ и лизис опухоли.
При лечении рака существуют стандарты определения ответа опухоли и стандартные способы измерения ответа. Такие стандарты включают ответ опухоли, который определяется путем мониторинга изменения размера опухоли или уровня сывороточных маркеров заболевания. Частичный ответ определяется как более чем 50% уменьшение размера опухоли, в то время как полный ответ определяется как полное исчезновение опухоли. Методы, используемые для измерения размеров опухолей, хорошо известны врачам и включают физическое обследование, рентгенологическое обследование, такое как КТ, МРТ, ПЭТ, рентгенография, а также определение концентрации сывороточных маркеров, таких как специфичный антиген простаты, который используется для мониторинга рака предстательной железы. Другие показатели терапевтической эффективности лечения рака включают измерение времени до начала прогрессирования, оценку продолжительности выживания без прогрессирования и общей выживаемости.
Улучшенная эффективность диагностики включает любое улучшение эффективности по сравнению с таковой при введении диагностического агента по отдельности, включая, но не ограничиваясь этим, повышение специфичности и/или чувствительности диагностического теста. Улучшение эффективности визуализации включает любое повышение эффективности по сравнению с таковым при введении агента для визуализации по отдельности, включая, но не ограничиваясь ими, повышение специфичности, чувствительности, воспроизводимости, усиление контрастности, детектирование мелких очагов заболевания, более точное установление границ заболевания, например, размера и формы заболеваний, таких как опухоли, абсцессы и т.д.
В различных вариантах реализации настоящего изобретения повышение эффективности в популяции пациентов составляет 5%, 10%, 15%, 20%, 25%, 50%, 75%, 100% или более по сравнению с эффективностью при использовании терапевтического, диагностического и/или агента для визуализации по отдельности.
Любой подходящий субъект может получить лечение с использованием способов согласно настоящему изобретению, предпочтительными субъектами являются люди.
В способах согласно настоящему изобретению может быть использован любой терапевтический, диагностический, агент для визуализации, или другое соединение, которое может направлено воздействовать на эпителиальную ткань и доставка которого к эпителиальной ткани может быть улучшена благодаря кратковременному размыканию межклеточных контактов. Согласно одному варианту реализации настоящего изобретения терапевтический агент выбран из группы, состоящей из антител, иммуноконъюгатов, наночастиц, терапевтических агентов на основе нуклеиновых кислот и их комбинаций, химиотерапевтических агентов, вакцин, терапии с использованием радиоактивных частиц/радиотерапии («облучения»), клеточной иммунотерапии, включая адоптивную Т-клеточную терапию и терапию с использованием дендритных клеток (например, внутриопухолевое проникновение вводимых Т-клеток), ингаляционных терапевтических агентов, конструкций для генной терапии (включая, но не ограничиваясь им, вирус АВВ-2/3 в качестве вектора для генной терапии, а также совместное введение с вектором для генной терапии на основе АВ5), других терапевтических агентов на основе нуклеиновых кислот, а также их комбинаций.
В различных вариантах реализации настоящего изобретения терапевтический агент выбран из группы, состоящей из алкилирующих агентов, ингибиторов ангиогенеза, антител, антиметаболитов, антимитотиков, антипролиферативных средств, ингибиторов Aurora-киназы, ингибиторов промоторов апоптоза (например, Bcl-xL, Bcl-w и Bfl-1), активаторов сигнального пути с участием рецептора гибели, ингибиторов киназы Bcr-Abl, антител BiTE (биспецифичных антител, мобилизующих Т-клетки), модификаторов биологического ответа, ингибиторов циклинзависимой киназы, ингибиторов клеточного цикла, ингибиторов циклооксигеназы-2, ингибиторов фактора роста, ингибиторов белка теплового шока (HSP)-90, деметилирующих агентов, ингибиторов гистондеацетилазы (HDAC), гормональных лекарственных средств, иммунологических лекарственных средств, ингибиторов белков апоптоза (IAP), интеркалирующих антибиотиков, ингибиторов киназ, ингибиторов мишени рапамицина у млекопитающих, микроРНК, ингибирующих митоген-активируемую протеинкиназу, входящую в состав киназы, регулируемой внеклеточными сигналами, поливалентных связывающих белков, нестероидных противовоспалительных препаратов (НПВП), ингибиторов полиАДФ (аденозиндифосфат)-рибоза полимеразы (PARP), химиотерапевтических средств на основе платины, ингибиторов polo-киназы (Plk), ингибиторов протеасомы, аналогов пурина, аналогов пиримидина, ингибиторов рецепторных тирозинкиназ, ретиноидных/дельтоидных растительных алкалоидов, малых ингибирующих рибонуклеиновых кислот (миРНК), ингибиторов топоизомеразы и тому подобного.
Типичные терапевтические агенты, входящие в перечисленные различные классы, включают, но не ограничиваются ими: доцетаксел, доксорубицин, иринотекан, паклитаксел (таксол®), частицы паклитаксела, стабилизированные альбумином (абраксан®), липосомальную форму доксорубицина HCl (доксил®), антитела BiTE, такие как адекатумумаб (микромет МТ201), блинатумомаб (микромет МТ103) и т.п., терапевтические агенты на основе миРНК, алкилирующие агенты, включая алтретамин, AMD-473, AP-5280, апазиквон, бендамустин, бросталлицин, бусульфан, карбоквон, кармустин (BCNU), хлорамбуцил, клоретазин® (ларомустин, VNP 40101М), циклофосфамид, дакарбазин, децитабин, 5'-азацитидин, эстрамустин, фотемустин, глюфосфамид, ифосфамид, KW-2170, ломустин (CCNU), мафосфамид, мелфалан, митобронитол, митолактол, нимустин, N-оксид азотистого иприта, ранимустин, темозоломид, тиотепа, треанда® (бендамустин), треосульфан, рофосфамид и т.п.; ингибиторы ангиогенеза, включая ингибиторы специфичных для эндотелия рецепторных тирозинкиназ (Tie-2), ингибиторы рецептора эпидермального фактора роста (EGFR), ингибиторы рецептора инсулиноподобного фактора роста-2 (IGFR-2), ингибиторы матриксной металлопротеиназы-2 (ММР-2), ингибиторы матриксной металлопротеиназы-9 (ММР-9), ингибиторы рецептора тромбоцитарного фактора роста (PDGFR), аналоги тромбоспондина, ингибиторы тирозинкиназы рецептора фактора роста эндотелия сосудов (VEGFR), и т.п.; антиметаболиты, включая алимта® (динатриевая соль пеметрекседа, LY231514, МТА), 5-азацитидин, кселода® (капецитабин), кармофур, лейстат® (кладрибин), клофарабин, цитарабин, окфосфат цитарабина, цитозинарабинозид, децитабин, дефероксамин, доксифлуридин, эфлорнитин, EICAR (5-этинил-1-бета-D-рибофуранозилимидазол-4-карбоксамид), эноцитабин, этнилцитидин, флударабин, 5-фторурацил по отдельности или в комбинации с лейковорином, гемзар® (гемцитабин), гидроксимочевину, алкеран® (мелфалан), меркаптопурин, 6-меркаптопурина рибозид, метотрексат, аналоги метотрексата (такие как триметрексат и пралатраксат), микофеноловую кислоту, неларабин, нолатрексед, окфосфат, пелитрексол, пентостатин, ралтитрексед, рибавирин, триапин, триметрексат, S-1, тиазофурин, тегафур, TS-1, видарабин и т.п.; ингибиторы белка Bcl-2, включая АТ-101 ((-)госсипол), генасенс® (G3139 или облимерсен (антисмысловой олигонуклеотид, нацеленный против Bcl-2)), IPI-194, IPI-565, N-(4-(4-((4'-хлор(1,1'-дифенил)-2-ил)метил)пиперазин-1-ил)бензоил)-4-(((1R)-3-(диметиламино)-1-((фенилсульфанил)метил)пропил)амино)-3-нитробензенсульфонамид) (АВТ-737), N-(4-(4-((2-(4-хлорфенил)-5,5-диметил-1-циклогексен-1-ен-1-ил)метил)пиперазин-1-ил)бензоил)-4-(((1R)-3-(морфолин-4-ил)-1-((фенилсульфанил)метил-)пропил)амино)-3-((трифторметил)сульфонил)бензолсульфонамид (АВТ-263), GX-070 (обатоклакс) и т.п.; ингибиторы киназы Bcr-Abl, включая дасатиниб® (BMS-354825), глеевек® (иматиниб) и т.п.; ингибиторы CDK, включая AZD-5438, BMI-1040, BMS-032, BMS-387, CVT-2584, флавопиридол, GPC-286199, MCS-5A, PD0332991, РНА-690509, селициклиб (CYC-202, R-росковитин), ZK-304709 и т.п.; ингибиторы EGFR, включая ABX-EGF, анти-EGFR иммунолипосомы, EGF-вакцины, EMD-7200, эрбитукс® (цетуксимаб), HR3, антитела IgA, иресса® (гефитиниб), тарцева® (эрлотиниб или OSI-774), ТР-38, EGFR-гибридный белок, тайкерб® (лапатиниб) и т.п.; ингибиторы рецептора ErbB2, включая СР-724-714, CI-1033 (канертиниб), герцептин® (трастузумаб), тайкерб® (лапатиниб), омнитарг® (2С4, петузумаб), ТАК-165, GW-572016 (ионафарниб), GW-282974, ЕKВ-569, PI-166, dHER2 (вакцина против HER2), АРС-8024 (вакцина против HER-2), биспецифичное антитело к HER/2neu, B7.her2IgG3, трифункциональные биспецифичные антитела AS HER2, мАт AR-209, мАт 2В-1 и т.п.; ингибиторы гистондеацетилазы, включая ромидепсин, LAQ-824, MS-275, трапоксин, субероиланилид гидроксамовую кислоту (SAHA), TSA, вальпроевую кислоту и т.п.; ингибиторы HSP-90, включая 117-AAG-nab, 17-AAG, CNF-101, CNF-1010, CNF-2024, 17-DMAG, гелданамицин, IPI-504, KOS-953, микограб® (человеческое рекомбинантное антитело к HSP-90), NCS-683664, PU24FC1, PU-3, радикикол, SNX-2112, STA-9090 VER49009 и т.п.; активаторы сигнальных путей с участием рецептора гибели, включая TRAIL, антитела или другие агенты, которые нацелены на TRAIL или рецепторы гибели (например, DR4 и DR5), такие как апомаб, конатумумаб, ETR2-ST01, GDC0145, (лексатумумаб), HGS-1029, LBY-135 PRO-1762 и трастузумаб; химиотерапевтические агенты на основе платины, включая цисплатин, элоксатин® (оксалиплатин), эптаплатин, лобаплатин, недаплатин, параплатин® (карбоплатин), сатраплатин, пикоплатин и т.п.; ингибиторы VEGFR, включая авастин® (бевацизумаб), АВТ-869, АЕЕ-788, акситиниб (AG-13736), AZD-2171, СР-547,632, IM-862, макуген (пегаптамиб), нексавар® (сорафениб, BAY43-9006), пазопаниб (GW-786034), ваталаниб (РТК-787, ZK-222584), сутент® (сунитиниб, SU-11248), VEGF Trap, ZACTIMAThi (вандетаниб, ZD-6474) и т.п.; терапевтические агенты на основе дендритных клеток (сипулейцел-Т, провенг®); ингибиторы топоизомеразы, включая акларубицин, 9-аминокамптотецин, амонафид, амсакрин, бекатекарин, белотекан, BN-80915, камптосар® (гидрохлорид иринотекана), камптотецин, дексразоксин, дифломотекан, эдотекарин, элленс® или фарморубицин® (эпирубицин), этопозид, экзатекан, абраксан, иренотекан, 10-гидроксикамптотецин, гиматекан, луртотекан, митоксантрон, оратецин, пирарубицин, пиксантрон, рубитекан, собузоксан, SN-38, тафлупозид, топотекан и т.п.; антитела, включая авастин® (бевацизумаб), CD40-специфичные антитела, chTNT-1/B, деносумаб, эрбитукс® (цетуксимаб), хумакс-CD4® (занолимумаб), антитела, специфичные в отношении рецептора ИФРI, линтузумаб, панорекс® (эдреколомаб), ренкарекс® (WX G250), ритуксан® (ритуксимаб), тицилимумаб, трастузимаб и т.п.; гормональные лекарственные препараты, включая аримидекс® (анастрозол), аромазин® (экземестан), арзоксифен, касодекс® (бикалутамид), цетротид® (цетрореликс), дегареликс, деслорелин, дезопан® (трилостан), дексаметазон, дрогенил® (флутамид), эвиста® (ралоксифен), афема® (фадрозол), фарестон® (торемифен), фаслодекс® (фулвестрант), фемара® (летрозол), форместан, глюкокортикоиды, гекторол® (доксеркальциферол), ренагель® (севеламер карбонат), лазофоксифен, ацетат лейпролида, мегаце® (мегестрол), мифепрекс® (мифепристон), ниландрон® (нилютамид), нолвадекс® (цитрат тамоксифена), пленаксис® (абареликс), преднизолон, пропециа® (финастерид), рилостан, супрефакт® (бусерелин), трелстар® (гормон, высвобождающий лютеинизирующий гормон (LHRH)), вантас® (имплантируемый гистрелин), веторил® (трилостан или нодрастан), золадекс® (фосрелин, гозерелин) и т.п.; иммунологические препараты, включая интерферон-альфа, интерферон альфа-2а, интерферон альфа-2b, интерферон бета, интерферон-гамма-1а, актиммун® (интерферон гамма-1b) или интерферон гамма-n1, их комбинации и т.п. Другие агенты включают альфаферон® (ИФН-альфа), ВАМ-002 (окисленный глутатион), беромун® (тазонермин), бексар® (тозитумомаб), кампат® (алемтузумаб), CTLA4 (антиген 4 цитотоксических лимфоцитов), декарбазин, денилейкин, эпратузумаб, граноцит® (ленограстим), лентинан, лейкоцитарный альфа-интерферон, имиквимод, MDX-010 (анти-CTLA-4), вакцину на основе убитых клеток меланомы, митумомаб, молграмостим, милотарг™ (гемтузумаб озогамицин), нейпоген® (филграстим), онко-VAC-CL, оварекс® (ореговомаб), пемтумомаб (Y-muHMFG1), провенг® (сипулейцел-Т), саргарамостим, сизофилан, тецелейкин, терацис® (бацилла Кальмета-Герена), убенимекс, вирулизин® (иммунотерапевтический агент, Lorus Pharmaceuticals), Z-100 (специфичная субстанция Маруяма (SSM)), WF-10 (тетрахлордекаоксид (TCDO)), пролейкин® (алдеслейкин), задаксин® (тималфазин), зенапакс® (даклизумаб), зевалин® (90Y-ибритумомаб тиуксетан) и т.п.; офатумомаб; модификаторы биологического ответа, включая крестин, лентинан, сизофуран, пицибанил PF-3512676 (CpG-8954), убенимекс и т.п.; аналоги пиримидина, включая цитарабин (ага С или арабинозид С), цитозинарабинозид, доксифлуридин, флудара® (флударабин), 5-ФУ (5-фторурацил), флоксуридин, гемзар® (гемцитабин), томудекс® (ратитрексед), троксатил® (триацетилуридин троксацитабин) и т.п.; аналоги пурина, включая ланвис® (тиогуанин) и пури-нетол® (меркаптопурин); антимитотические агенты, включая батабулин, эпотилон D (KOS-862), N-(2-((4-гидроксифенил)амино)пиридин-3-ил)-4-метоксибензолсульфонамид, иксабепилон (BMS 247550), паклитаксел, таксотер® (доцетаксел), PNU100940 (109881), патупилон, XRP-9881 (ларотаксел), винфлунин, ZK-EPO (синтетический эпотилон) и т.п.; и другие химиотерапевтические агенты, такие как абраксан® (ABI-007), АВТ-100 (ингибитор фарнезилтрансферазы), адвексин® (вакцина Ad5CMV-p53), альтокор® или мевакор® (ловастатин), амплиг® (поли-1:поли-C12U, синтетическая РНК), аптозин® (экзисулинд), аредиа® (памидроновая кислота), арглабин, L-аспарагиназа, атаместан (1-метил-3,17-дион-андроста-1,4-диен), аваг® (тазаротен), AVE-8062 (производное комбрестатина) ВЕС2 (митумомаб), кахектин или кахексии (фактор некроза опухоли), канваксин (вакцина), цеавак® (вакцина против рака), целейк® (целмолейкин), цеплен® (дигидрохлорид гистамина), церварикс® (вакцина против вируса папилломы человека), CHOP® (С: цитоксан® (циклофосфамид); Н: адриамицин® (гидроксидоксорубицин); О: винкристин (онковицин®); Р: преднизолон), CYPAT® (ацетат ципротерона), комбрестатин А4Р, DAB(389)EGF (каталитический и транслокационный домены дифтерийного токсина, гибридизованные посредством линкера His-Ala с эпидермальным фактором роста человека) или TransMID-107R® (дифтерийные токсины), дакарбазин, дактиномицин, 5,6-диметилксантенон-4-уксусную кислоту (DMXAA), энилурацил, эвизон™ (лактат скваламина), димерицин® (липосомальный лосьон T4N5), дискодермолид, DX-8951f (мезилат экзатекана), энзастаурин, ЕРО906 (эпитилон В), гардасил® (четырехвалентная рекомбинантная вакцина против папилломавируса человека (типа 6, 11, 16, 18)), гастриммун®, генасенс®, GMK (ганглиозид-конъюгированная вакцина), Джи-Вакс® (вакцина против рака простаты), галофугинон, гистерелин, гидроксикарбамид, ибандроновую кислоту, IGN-101, IL-13-PE38, IL-13-PE38QQR (цинтредекин безудотокс) IL-13-экзотоксин Pseudomonas, интерферон альфа, интерферон гамма, юнован® или мепакт® (мифамуртид), лонафарниб, 5,10-метилентетрагидрофолат, мильтефозин (гексадецилфосфохолин), неовастат® (АЕ-941), нейтрексин® (глюкуронат триметрексата), нипент® (пентостатин), онконаз® (рибонуклеазный фермент), онкофаг® (вакцина против меланомы), онковакс® (вакцина на основе IL-2), оратецин® (рубитекан), осидем® (клеточный препарат на основе антител), мАт оварекс® (мышиное моноклональное антитело), паклитаксел, пандимекс® (агликоны сапонинов женьшеня, содержащие 20(S) протопанаксадиол (aPPD) и 20(S) протопанаксатриол (аРРТ)), панитумумаб, панвак®-VF (изучаемая вакцина против рака), пегаспаргазу, ПЭГ-интерферон А, феноксодиол, прокарбазин, ребимастат, ремоваб® (катумаксомаб), ревлимид® (леналидомид), RSR13 (эфапроксирал), соматулин® LA (ланреотид), сориатан® (ацитретин), стауроспорин (Streptomyces staurospores), талабостат (РТ100), таргретин® (бексаротен), таксопрексин® (DHA-паклитаксел), телцита® (канфосфамид, TLK286), темилифен, темодар® (темозоломид), тесмилифен, талидомид, тератоп® (STn-KLH), тимитак (2-амино-3,4-дигидро-6-метил-4-оксо-5-(4-пиридилтио)хиназолина дигидрохлорид), TNFERABE® (аденовектор: носитель ДНК, содержит ген фактора некроза опухоли альфа), траклеер® или завеска® (бозентан), третиноин (ретин-А), тетрандрин, тризенокс®. (триоксид мышьяка), вирулизин®, украин (производное алкалоидов чистотела большого), витаксин (антитело к α5β3), эксцитрин® (мотексафин гадолиния), ксинлей® (атразентан), ксиотакс® (паклитаксел полиглумекс), ионделис® (трабектедин), ZD-6126, зинекард® (дексразоксан), зомета® (золедроновая кислота), кризотиниб, зорубицин и т.п.
В другом предпочтительном варианте реализации настоящего изобретения терапевтический агент содержит соединение, которое связывается с десмоглеином-2; предпочтительно соединение, которое связывается с DSG2 и размыкает плотные контакты.
Согласно другим вариантам реализации настоящего изобретения терапевтический агент содержит радиоактивные частицы/радиотерапию. Любой подходящий способ радиоактивной терапии или частица могут быть использованы по усмотрению лечащего врача, включая, но не ограничиваясь ими, кобальт-60, йод-131, иридий-192, стронций-89, самарий-153, рений-186 и свинец-212.
Согласно предпочтительному варианту реализации настоящего изобретения терапевтический агент представляет собой противоопухолевый терапевтический агент и содержит химиотерапевтическое или противоопухолевое моноклональное антитело, описанное в настоящей заявке. Согласно другому предпочтительному варианту реализации настоящего изобретения противоопухолевый терапевтический агент содержит антитело, выбранное из группы, состоящей из трастузумаба, цетумиксимаба, петузумаба, апомаба, конатумумаба, лексатумумаба, бевацизумаба, деносумаба, занолимумаба, линтузумаба, эдреколомаба, ритуксимаба, тицилимумаба, тозитумомаба, алемтузумаба, эпратузумаба, митумомаба, гемтузумаба озогамицина, ореговомаба, пемтумомаба, даклизумаба, панитумумаба, катумаксомаба, офатумумаба и ибритумомаба. Неограничивающие примеры подходящих противоопухолевых мАт и конкретные способы их применения, приведены в таблице 1 выше, и дополнительно описаны в работе Campoli, М., et al., Principles & Practice of Oncology 23(1&2):1-19 (2009), полностью включенной в настоящую заявку посредством ссылки.
Терапевтические моноклональные антитела могут представлять собой любой тип моноклонального антитела, включая, но не ограничиваясь ими, стандартные моноклональные антитела, гуманизированные моноклональные антитела, полностью человеческие антитела, полученные от мышей или из других источников, химерные моноклональные антитела и их фрагменты. Термин «гуманизированные моноклональные антитела» относится к моноклональным антителам, полученным из моноклональных антител видов, не относящихся к человеку, например, из моноклонального антитела мыши. В другом варианте, гуманизированные моноклональные антитела могут быть получены из химерных антител, которые сохраняют или по существу сохраняют антигенсвязывающие свойства исходных нечеловеческих моноклональных антител, но которые при введении человеку проявляют уменьшенную иммуногенность по сравнению с исходным моноклональным антителом. Например, химерные моноклональные антитела могут содержать фрагменты человеческих и мышиных антител, как правило, константные области человека и вариабельные области мыши. Гуманизированные моноклональные антитела могут быть получены с использованием различных способов, известных в данной области техники, включая, но не ограничиваясь ими: (1) прививку гипервариабельных участков моноклонального антитела из вида, не относящегося к человеку, на последовательности каркаса и константных областей человека («гуманизация»), и (2) трансплантацию вариабельных доменов моноклонального антитела из вида, не относящегося к человеку, и их «маскировку» с использованием поверхности, сходной с таковой в человеческих антителах, путем замены поверхностных остатков («рекомбинация поверхностных остатков»). Перечисленные способы описаны, например, в Jones et al., Nature 321:522-525 (1986); Morrison et al., Proc. Natl. Acad. Sci., U.S.A., 81:6851-6855 (1984); Morrison and Oi, Adv. Immunol., 44:65-92 (1988); Verhoeyer et al., Science 239:1534-1536 (1988); Padlan, Molec. Immun. 28:489-498 (1991); Padlan, Molec. Immunol. 31(3):169-217 (1994); и Kettleborough, C.A. et al., Protein Eng. 4(7):773-83 (1991). Моноклональные антитела могут быть фрагментированы с использованием обычных методик, и фрагменты подвергают скринингу для определения возможности их применения с помощью способов, используемых для полноразмерных антител. Например, фрагменты F(ab')2 могут быть получены путем обработки антитела пепсином. Полученный фрагмент F(ab')2 может быть обработан для восстановления дисульфидных мостиков с получением фрагментов Fab'. Фрагменты Fab могут быть получены путем обработки антитела IgG папаином; фрагменты F(ab') могут быть получены путем расщепления антител IgG пепсином. Фрагмент F(ab') также может быть получен путем связывания Fab', как описано ниже, с помощью тиоэфирной связи или дисульфидной связи. Фрагмент Fab' представляет собой фрагмент антитела, полученный путем разрезания дисульфидной связи в шарнирной области F(ab')2. Фрагмент F(ab') может быть получен путем обработки фрагмента F(ab')2 восстанавливающим агентом, таким как дитиотреитол. Пептиды, представляющие собой фрагменты антител, также могут быть получены путем экспрессии нуклеиновых кислот, кодирующих такие пептиды, в рекомбинантных клетках (см, например, Evans et al., J. Immunol. Meth. 184: 123-38 (1995)). Например, химерный ген, кодирующий часть фрагмента F(ab')2 может содержать последовательности ДНК, кодирующие домен СН1 и шарнирную область Н-цепи, после которых расположен трансляционный стоп-кодон, чтобы обеспечить получение молекулы усеченного фрагмента антитела. Неограничивающие примеры фрагментов моноклональных антител включают (i) фрагмент Fab, моновалентный фрагмент, состоящий по существу из доменов VL, VH, CL и CH1; (ii) фрагменты F(ab)2 и F(ab')2, двухвалентные фрагменты, содержащие два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий по существу из доменов VH и CH1; (iv) фрагмент Fv, состоящий по существу из доменов VL и VH одного плеча антитела, (v) фрагмент dAb (Ward et al., (1989) Nature 341:544-546), состоящий по существу из домена VH; и (vi) одну или более выделенных гипервариабельных областей (CDR), или функциональный паратоп.
Согласно одному предпочтительному варианту реализации настоящего изобретения, который можно комбинировать с любым вариантом или комбинацией вариантов реализации настоящего изобретения, заболевание включает HER-2-положительную опухоль, и способ включает совместное введение мультимера фибриллы АВВ-2/3 согласно настоящему изобретению с подходящим терапевтическим моноклональным антителом, по отдельности или в комбинации с химиотерапевтическим агентом, облучением или их комбинацией. Согласно другому предпочтительному варианту реализации настоящего изобретения моноклональное антитело представляет собой трастузумаб. Согласно другому предпочтительному варианту реализации, который можно комбинировать с любым из указанных вариантов реализации настоящего изобретения, HER-2-положительная опухоль выбрана из группы, состоящей из опухоли молочной железы, опухоли желудка, опухоли толстой кишки и опухоли яичников. Согласно другому предпочтительному варианту реализации настоящего изобретения способ осуществляют у пациентов, которые не имеют надлежащего клинического ответа на трастузумаб, например, из-за отсутствия ремиссии опухоли, из-за рецидива опухоли или развития устойчивости к трастузумабу. Способы, раскрытые в указанных вариантах реализации настоящего изобретения, также можно применять для уменьшения дозировки трастузумаба, необходимой для получения терапевтического эффекта, и, следовательно, могут быть использованы для уменьшения побочных эффектов (таких как кардиотоксичность, связанная с приемом трастузумаба).
В другом предпочтительном варианте реализации настоящего изобретения, который можно комбинировать с любым вариантом или комбинацией вариантов реализации настоящего изобретения, заболевание включает EGFR-положительную опухоль, и способ включает совместное введение мультимера фибриллы АВВ-2/3 и подходящего терапевтического моноклонального антитела, по отдельности или в комбинации с химиотерапевтическим агентом, облучением или их комбинацией. Согласно другому предпочтительному варианту реализации настоящего изобретения моноклональное антитело представляет собой цетуксимаб. Согласно другому предпочтительному варианту реализации настоящего изобретения, который можно комбинировать с любым из указанных вариантов реализации настоящего изобретения, EGFR-положительная опухоль выбрана из группы, состоящей из опухоли легкого, опухоли толстой кишки, опухоли молочной железы, опухоли прямой кишки, опухоли головы и шеи и опухоли поджелудочной железы. Согласно другому предпочтительному варианту реализации настоящего изобретения способ осуществляют у пациентов, которые не имеют надлежащего клинического ответа на цетуксимаб, например, из-за отсутствия ремиссии опухоли, из-за рецидива опухоли или развития устойчивости к цетуксимабу. Способы, раскрытые в указанных вариантах реализации настоящего изобретения, также можно применять для уменьшения дозы цетуксимаба, необходимой для получения терапевтического эффекта, и, следовательно, они могут быть использованы для уменьшения побочных эффектов (таких как акне-подобные виды сыпи, которые часто возникают во время терапии цетуксимабом).
Согласно одному предпочтительному варианту реализации настоящего изобретения, который можно комбинировать с любым вариантом или комбинацией вариантов реализации настоящего изобретения, заболевание включает эпителиальную опухоль, и способ включает совместное введение мультимеров фибриллы АВВ-2/3 с ингибитором фактора роста эндотелия сосудов (VEGF), по отдельности или в комбинации с другим химиотерапевтическим агентом, облучением или их комбинацией. Может быть использован любой подходящий ингибитор VEGF, включая, но не ограничиваясь им, бевацизумаб.
Согласно другому варианту реализации настоящего изобретения, который можно комбинировать с любым вариантом или комбинацией вариантов реализации настоящего изобретения, описанными в настоящей заявке, способы, включающие солидные опухоли, дополнительно включают введение соединения, способного разрушать стромальные белки опухоли. Для разрушения стромальных белков опухоли может быть использовано любое подходящее соединение, включая, но не ограничиваясь ими, релаксин, коллагеназу, трипсин, диспазу, ММР (металлопротеиназу)-1 и ММР-8. Доставку указанных соединений можно осуществлять с помощью любого подходящего механизма, включая генную терапию, раздельное введение с мультимером фибриллы АВВ-2/3 и терапевтическим агентом, или введение в виде конъюгата с фибриллой АВВ-2/3 или терапевтическим агентом.
Согласно другому варианту реализации настоящего изобретения, который можно комбинировать с любым вариантом или комбинацией вариантов реализации настоящего изобретения, раскрытыми в настоящей заявке, способы дополнительно включают введение мультимера АВВ-2/3 в комбинации с другими соединениями, размыкающими межклеточные контакты. В настоящей заявке термин «соединение, размыкающее межклеточные контакты» представляет собой соединение, которое способно кратковременно размыкать межклеточные контакты. Могут быть использованы любые подходящие соединения, размыкающие межклеточные контакты. Согласно одному неограничивающему варианту соединение, размыкающее межклеточные контакты, содержит токсин Zona occludens (Zot), токсин, вырабатываемый Vibrio cholerae, который способен обратимо изменять межклеточные контакты эпителиальных клеток кишечника, что позволяет макромолекулам проникать через барьеры слизистых оболочек (Fasano et al. (1991) Proc Natl Acad Sci USA 88: 5242-5246). Был создан гексапептид, полученный из Zot (AT-1001). Согласно другому варианту реализации настоящего изобретения энтеротоксин Clostridium perfringens удаляет клаудины-3 и -4 из плотных контактов, чтобы облегчить проникновение бактерий (Sonoda N, et al. (1999) J Cell Biol 147: 195-204). Согласно другому варианту реализации настоящего изобретения онкобелки, кодируемые АВ, ВПЧ, HTLV-1 человека, могут кратковременно размыкать межклеточные контакты эпителиальных клеток за счет изменения природного расположения белка ZO-1 (Latorre IJ, et al. (2005) J Cell Sci 118: 4283-4293). Согласно другим вариантам реализации настоящего изобретения ряд вирусов человека вовлекает плотные контакты или другие молекулы межклеточных контактов для проникновения в эпителиальные клетки. Такие вирусы включают вирус гепатита С (Evans MJ, et al. (2007) Nature 446: 801-805), реовирус (Barton ES, et al. (2001) Cell 104: 441-451) и вирус простого герпеса (Geraghty RJ, et al. (1998) Science 280: 1618-1620).
Согласно другому варианту реализации настоящего изобретения терапевтический агент представляет собой ингаляционный терапевтический агент. Любой подходящий ингаляционный терапевтический агент может быть использован в соответствии со способами согласно настоящему изобретению. Согласно различным неограничивающим вариантам реализации настоящего изобретения ингаляционный терапевтический агент выбран из группы, состоящей из кортикостероидов, бронходилататоров, бета-агонистов, антихолинергических препаратов, альбутерола (провентил®; венолин®; аккунеб®; проэйр®), левальбутерола (ксопенекс®), пирбутрола (максэйр®), бромида ипратропия (атровент®), беклометазона, будесонида, флунизолида (аэробид®), флутиказона, триамцинолона ацетонида, флутиказона (кортикостероид) и сальметерола (абвэйр®), формоторола (бета-агонист бронходилататор длительного действия) и будесонида (кортикостероид) (симикорт®), альбутерола (бета-агонист) и ипратропиума (комбивент®; холинолитик), будесонида (пулмикорт респулес®) и тиопропиума (спирива®, антихолинергический бронходилататор).
Согласно другому варианту реализации настоящего изобретения соединение содержит диагностический агент или агент для визуализации. Способы согласно настоящему изобретению имеют широкое применение для доставки любого диагностического, агента для визуализации или другого соединения к эпителиальной ткани, содержащей межклеточные контакты, в которой доступ к мишени, представляющей интерес, может быть ограничен. Согласно различным неограничивающим вариантам реализации настоящего изобретения агенты для визуализации могут содержать любое химическое соединение, которое может производить детектируемый сигнал, прямо либо косвенно. Многие такие агенты для визуализации известны специалистам в данной области техники. Примеры агентов для визуализации, подходящих для использования в раскрытых способах и композициях, включают радиоактивные изотопы, флуоресцентные молекулы, магнитные частицы (включая наночастицы), металлические частицы (включая наночастицы), фосфоресцирующие молекулы, ферменты, антитела, лиганды и их комбинации, тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания, ассоциированного с эпителиальной тканью, связанное с таким агентом для визуализации. Способы детектирования и измерения сигналов, которые производят агенты для визуализации, также известны специалистам в данной области техники. Например, радиоактивные изотопы можно детектировать с помощью сцинтилляционного счетчика или прямой визуализации; флуоресцентные молекулы можно детектировать с помощью флуоресцентных спектрофотометров; фосфоресцентные молекулы можно детектировать с помощью спектрофотометра или непосредственной визуализации с использованием камеры; ферменты можно детектировать с помощью обнаружения или визуализации продукта реакции, катализируемой ферментом; антитела можно детектировать путем обнаружения вторичной обнаруживаемой метки, присоединенной к антителу. Согласно одному предпочтительному варианту реализации настоящего изобретения агент для визуализации и/или диагностический агент представляет собой агент, который может быть использован для детектирования опухоли, как путем прямого связывания с опухолью, так и путем соединения агента для визуализации или диагностического агента с соединением, которое может связываться с опухолью.
Согласно одному примеру агенты для визуализации могут содержать флуоресцентный агент для визуализации, тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания, ассоциированного с эпителиальной тканью, связанное с флуоресцентным агентом для визуализации. Флуоресцентный агент для визуализации представляет собой любой химический фрагмент, который производит детектируемый флуоресцентный сигнал. Агент для визуализации можно применять по отдельности или в комбинации с другими агентами для визуализации. Примеры подходящих флуоресцентных агентов, которые можно применять в композициях и способах, раскрытых в настоящей заявке, включают, но не ограничиваются ими, флуоресцеин (FITC), 5-карбоксифлуоресцеин-N-гидроксисукцинимидный эфир, 5,6-карбоксиметилкрахмал флуоресцеина, нитробенз-2-окса-1,3-тиадиазол-4-ил (NBD), флуорескамин, ОРА, NDA, индоцианиновый зеленый краситель, цианиновые красители (например, Су3, Су3.5, Су5, Су5.5 и Су7), 4-ацетамидо-4'-изотиоцианатостилбен-2,2'дисульфоновую кислоту, акридин, изотиоцианат акридина, 5-(2'-аминоэтил)аминонафталин-1-сульфоновую кислоту (EDANS), 4-амино-N-[3-винилсульфонил)фенилинафталимид-3,5дисульфонат, N-(4-анилино-1-нафтил)малеимид, антраниламид, BODIPY, бриллиантовый желтый, кумарин, 7-амино-4-метилкумарин (АМС, кумарин 120), 7-амино-4-трифторметилкумарин (кумарин 151), цианозин, 4',6-диаминидино-2-фенилиндол (DAPI), 5',5"-дибромпирогаллолсульфонафталин (бромпирогаллол красный), 7-диэтиламино-3-(4'-изотиоцианатофенил)-4-метилкумариндиэтилентриаминпентаацетат, 4,4'-диизотиоцианатодигидростильбен-2,2'-дисульфоновую кислоту, 4,4'-диизотиоцианатостильбен-2,2'-дисульфоновую кислоту, 5-[диметиламино]нафталин-1-сульфонилхлорид (DNS, дансилхлорид), 4-(4'-диметиламинофенилазо)бензойную кислоту (DABCYL), 4-диметиламинофенилазофенил-4-изотиоцианат (DABITC), эозин, изотиоцианат эозина, эритрозин В, эритрозин, изотиоцианат, бромид этидия, этидий, 5-карбоксифлуоресцеин (FAM), 5-(4,6-дихлортриазин-2-ил)аминофлуоресцеин (DTAF), 2',7'-диметокси-4'5'-дихлор-6-карбоксифлуоресцеин (JOE), изотиоцианат флуоресцеина, IR144, IR1446, малахитовый зеленый изотиоцианат, 4-метилумбеллиферон, орто-крезолфталеин, нитротирозин, парарозанилин, феноловый красный, В-фикоэритрин, орто-фталдиальдегид, пирен, бутират пирена, сукцинимидил-1-пиренбутират, реактивный красный 4 (Cibacron [R] бриллиантовый красный 3В-А), 6-карбокси-Х-родамин (ROX), 6-карбоксиродамин (R6G), лиссамин-родамин В сульфонилхлорид, родамин (Rhod), 5,6-тетраметилродамин, родамин В, родамин 123, родамин X изотиоцианат, сульфородамин В, сульфородамин 101, сульфонилхлоридное производное сульфородамина 101 (техасский красный), N,N,N',N'-тетраметил-6-карбоксиродамин (TAMRA), тетраметил родамина, изотиоцианат тетраметилродамина (TRITC), рибофлавин, кораллин, кумарин-6 и т.п., включая их комбинации. Перечисленные флуоресцентные фрагменты для визуализации могут быть получены из различных коммерческих источников, включая Molecular Probes (Юджин, Орегон, США) и Research Organics (Кливленд, Огайо, США) или могут быть синтезированы специалистам в данной области техники.
Согласно другому примеру агенты для визуализации могут содержать агент для магнитно-резонансной томографии (МРТ), тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания, ассоциированного с эпителиальной тканью, связанное с агентом для МРТ. Агент для МРТ представляет собой любой химический фрагмент, который производит детектируемый сигнал магнитного резонанса или который может оказать влияние (например, увеличить или сместить) на сигнал магнитного резонанса другого агента. Указанный тип агента для визуализации можно применять по отдельности или в комбинации с другим агентом для визуализации. Согласно другому примеру в качестве агента для визуализации можно применять агент для МРТ на основе гадолиния. Пример подходящего агента для МРТ, который можно включать в состав раскрытых агентов для визуализации, представляет собой пара-аминобензил диэтилентриаминопентауксусную кислоту (p-NH2-Bz-DTPA, соединение 7), конъюгируемую форму диэтилентриаминпентауксусной кислоты (DTPA), которая, как известно, прочно связывает гадолиний и одобрена для клинического применения в качестве контрастного вещества для магнитно-резонансной томографии. Встраивание агента для МРТ в большую макромолекулу, такую как дендримерный субстрат, как раскрыто в настоящей заявке, может обеспечить большую релаксацию Т1 (высокий контраст) и получение нескольких копий агента на одну молекулу, что может увеличить сигнал. При комбинировании агента для МРТ и, например, флуоресцентного агента для визуализации, полученный в результате агент можно детектировать, визуализировать и наблюдать в режиме реального времени с помощью МРТ. Другие агенты для визуализации включают агенты для ПЭТ, которые могут быть получены путем включения F18 или хелатора Cu64 или Ga68. Помимо этого добавление радионуклида можно применять, чтобы облегчить визуализацию с помощью SPECT или доставку дозы радиации, тогда как диагностические агенты могут содержать соединение, которое является диагностическим маркером для конкретного заболевания, ассоциированного с эпителиальной тканью, связанное с агентом для ПЭТ.
Согласно некоторым вариантам реализации настоящего изобретения диагностический агент представляет собой диагностический агент для визуализации, включая, но не ограничиваясь ими, агенты для позитронно-эмиссионной томографии (ПЭТ), агенты для компьютерной томографии (КТ), агенты для магнитно-резонансной томографии (МРТ), агенты для визуализации для ядерно-магнитного резонанса (ЯМР), агенты для рентгеноскопии и контрастные агенты для ультразвуковых исследований. Перечисленные диагностические агенты включают радиоизотопы таких элементов как йод (I), включая 123I, 125I, 131I и т.д., барий (Ва), гадолиний (Gd), технеций (Тс), включая 99Тс, фосфор (Р), включая 31Р, железо (Fe), марганец (Mn), таллий (Tl), хром (Cr), включая 51Cr, углерод (С), включая 14С, или т.п., флуоресцентномеченые соединения, или их комплексы, хелаты, аддукты и конъюгаты. Могут быть использованы любые подходящие агенты для ПЭТ, включая, но не ограничиваясь ими, углерод-11, азот-13, кислород-15, фтор-18, 11С-метомидат, и его глюкозные аналоги, включая, но не ограничиваясь этим, фтордезоксиглюкозу (аналог глюкозы, меченый фтором-18).
Согласно другим вариантам реализации настоящего изобретения диагностический агент представляет собой маркерный ген, кодирующий белки, которые легко детектируются при экспрессии в клетке (включая, но не ограничиваясь ими, бета-галактозидазу, зеленый флуоресцентный белок, люциферазу и т.п.), и меченые нуклеиновые зонды (например, радиоактивномеченые или флуоресцентномеченые зонды). Согласно некоторым вариантам реализации настоящего изобретения ковалентная конъюгация диагностических агентов и мультимеров АВВ-2/3, предложенных в настоящей заявке, достигается в соответствии с различными способами конъюгации. Согласно другим вариантам реализации настоящего изобретения диагностический агент нековалентно связан с мультимерами АВВ-2/3, предложенными в настоящей заявке.
Согласно другому аспекту в настоящем изобретении предложены способы улучшения доставки вещества к эпителиальной ткани, включающие приведение эпителиальной ткани в контакт с (а) одним или более соединениями, которые необходимо доставить к эпителиальной ткани; и (b) таким количеством мультимера фибриллы АВВ-2/3 согласно настоящему изобретению, которое достаточно для улучшения доставки одного или более соединений к эпителиальной ткани. Согласно указанному аспекту соединения могут представлять собой любые подходящие соединения, такие как те, которые подробно описаны выше. Согласно предпочтительному варианту реализации настоящего изобретения одно или более соединений содержат агент для визуализации. Согласно другому предпочтительному варианту реализации настоящего изобретения эпителиальная ткань содержит солидную опухоль, включая любую из опухолей, раскрытых в настоящей заявке. Согласно различным неограничивающим вариантам солидная опухоль выбрана из группы, состоящей из опухолей молочной железы, опухолей легких, опухолей толстой кишки, опухолей прямой кишки, опухолей желудка, опухолей предстательной железы, опухолей яичников, опухолей матки, опухолей кожи, эндокринных опухолей, опухолей шейки матки, опухолей почек, меланом, опухолей поджелудочной железы, опухолей печени, опухолей головного мозга, опухолей головы и шеи, опухолей носоглотки, опухолей желудка, плоскоклеточных карцином, аденокарцином, опухолей мочевого пузыря и опухолей пищевода. Типичные мультимеры, содержащие один или более мультимеров фибриллы АВВ-2/3 согласно настоящему изобретению, которые можно применять в соответствии с указанными способами, включают, но не ограничиваются ими, вирионы АВВ-2/3, капсиды АВВ-2/3, додекаэдрические частицы АВВ-2/3 (PtDd) (субвирусные додекаэдрические частицы, которые вырабатывает АВВ-2/3 во время репликации) и рекомбинантные мультимеры фибриллы АВВ-2/3.
Согласно другому аспекту в настоящем изобретении предложены способы улучшения доставки вещества к клетке или ткани, экспрессирующей десмоглеин 2 (DSG2), включающие приведение клетки или ткани, экспрессирующей DSG2, в контакт с (a) одним или более соединениями, которые необходимо доставить к клетке или ткани; и (b) таким количеством мультимера фибриллы АВВ-2/3 согласно настоящему изобретению, которое достаточно для улучшения доставки одного или более соединений к ткани. Примерные типы тканей, экспрессирующих DSG2, включают, но не ограничиваются ими, эпителиальные клетки/ткани (например, те, которые раскрыты в настоящей заявке), тромбоциты и гранулоциты человека. Как показано в приведенных ниже примерах, DSG2 также выполняет функцию рецептора в неполяризованных клетках. Следовательно, указанные способы можно применять не только в эпителиальных клетках и тканях, но их применение также целесообразно, например, при патогенезе АВВ-2/3 и внутрисосудистом введении векторов АВВ-2/3 для целей генной терапии. Типичные мультимеры, содержащие один или более мультимеров фибриллы АВВ-2/3 согласно настоящему изобретению, которые можно применять в указанных способах, включают, но не ограничиваются ими, вирионы АВВ-2/3, капсиды АВВ-2/3, додекаэдрические частицы АВВ-2/3 (PtDd) (субвирусные додекаэдрические частицы, которые вырабатывает АВВ-2/3 при репликации) и рекомбинантные мультимеры фибриллы АВВ-2/3.
Согласно другому аспекту в настоящем изобретении предложены способы индукции эпителиально-мезенхимального перехода (ЭМП) в ткани, включающие приведение эпителиальной ткани в контакт с количеством мультимера фибриллы АВВ-2/3 согласно настоящему изобретению, достаточным для индукции ЭМП. ЭМП представляет собой клеточную программу трансдифференцировки, при которой эпителиальные клетки теряют такие характеристики как наличие межклеточных соединений и приобретают свойства мезенхимальных клеток. ЭМП характеризуется повышенной экспрессией мезенхимальных маркеров, повышенной экспрессией соединений внеклеточного матрикса, сниженной экспрессией эпителиальных маркеров, измененным расположением факторов транскрипции, и измененной активацией киназ, а также диссоциацией межклеточных контактов. Типичные мультимеры, содержащие один или более мультимеров фибриллы АВВ-2/3 согласно настоящему изобретению, которые можно применять в указанных способах, включают, но не ограничиваются ими, вирионы АВВ-2/3, капсиды АВВ-2/3, додекаэдрические частицы АВВ-2/3 (PtDd) (субвирусные додекаэдрические частицы, которые вырабатывает АВВ-2/3 при репликации) и рекомбинантные мультимеры фибриллы АВВ-2/3.
Во всех аспектах и вариантах реализации способов согласно настоящему изобретению терапевтический, диагностический и/или агент для визуализации может быть введен совместно с мультимером АВВ-2/3 (например, с помощью композиций согласно настоящему изобретению, раскрытых выше), или может быть введен отдельно. Согласно одному варианту реализации настоящего изобретения терапевтический агент соединен с мультимером АВВ-2/3 с помощью любой подходящей ковалентной или нековалентной связи. В одном неограничивающем варианте реализации настоящего изобретения мультимер фибриллы АВВ-2/3 может быть прикреплен к токсину или другому лекарственному препарату, чтобы вызвать гибель клеток солидной опухоли.
Мультимер фибриллы АВВ-2/3 и/или терапевтический агент можно вводить любым способом, который считается подходящим по мнению лечащего врача, в зависимости от того, какой способ введения, локальный или системный, является наиболее подходящим для заболевания, которое лечат. В настоящей заявке термины «системная доставка» и «системное введение» включают, но не ограничиваются ими, пероральные и парентеральные пути введения, включая внутримышечный (в/м), подкожный, внутривенный (в/в), внутриартериальный, ингаляционный, сублингвальный, буккальный, местный, чрескожный, назальный, ректальный, вагинальный и другие пути введения, которые обеспечивают эффективное распространение доставленного агента к одному или нескольким местам предполагаемого терапевтического действия. Предпочтительные пути системного введения композиций согласно настоящему изобретению включают внутривенный, внутримышечный, подкожный и ингаляционный пути введения. В одном предпочтительном варианте реализации настоящего изобретения используется внутривенное введение, например, для лечения диссеминированных опухолей (и для доставки моноклональных антител). Согласно другому варианту реализации настоящего изобретения пероральная доставка может быть предпочтительной, например, для лечения эпителиальных заболеваний желудочно-кишечного тракта (ЖКТ). Согласно другому варианту реализации настоящего изобретения назальная или аэрозольная доставка может быть предпочтительной для доставки в легкие, например, для лечения заболеваний эпителиальной ткани легких.
Мультимер фибриллы АВВ-2/3 может быть введен в комбинации с другой молекулой, такой как липид или липосома, чтобы защитить полипептиды от ферментативного расщепления. Например, ковалентное присоединение полимеров, в частности, полиэтиленгликоля (ПЭГ), было использовано для защиты определенных белков от ферментативного гидролиза в организме, обеспечивая тем самым увеличение периода полувыведения.
Мультимер фибриллы АВВ-2/3 и/или терапевтический агент могут быть введены системно на периодической основе с интервалами, которые, как установлено, поддерживают желаемый уровень терапевтического эффекта. Например, введение путем внутривенной инъекции можно осуществлять один раз в сутки, один раз в неделю, каждые две-четыре недели или с менее частыми интервалами. Схема дозирования будет определена лечащим врачом с учетом различных факторов, которые могут оказывать влияние на действие комбинации агентов. Такие факторы включают степень прогрессирования состояния, подлежащего лечению, возраст, пол и вес пациента, а также другие клинические факторы. Дозировка мультимера фибриллы АВВ-2/3 и/или терапевтического агента будет варьироваться в зависимости от мультимера и/или терапевтического агента, который вводят, а также от наличия и характера любого носителя для доставки лекарственного препарата (например, носитель для доставки с пролонгированным высвобождением). Помимо этого дозировка может быть скорректирована с учетом изменения частоты введения и фармакокинетических показателей доставляемого агента(ов). Дозировки мультимеров фибриллы АВВ-2/3, как правило, находятся в диапазоне от 0,01 до 250 мг/кг, предпочтительно от 0,1 до 10 мг/кг, и более предпочтительно от 0,10 до 0,5 мг/кг. Дозировки одобренных терапевтических агентов могут быть легко определены врачами. Терапевтический агент также можно вводить в сниженной дозе благодаря усиленному проникновению в эпителиальные ткани, например, эпителиальные опухоли.
Мультимер фибриллы АВВ-2/3 можно вводить субъекту перед, одновременно или после введения терапевтического агента. Согласно предпочтительному варианту реализации настоящего изобретения введение терапевтического агента и мультимера фибриллы АВВ-2/3 осуществляют одновременно. Сроки введений терапевтического агента по отношению к мультимеру фибриллы АВВ-2/3 можно варьировать для достижения наибольшего терапевтического эффекта. Предпочтительно терапевтический агент вводят в такой момент времени, чтобы обеспечить его контакт с кратковременным размыканием межклеточных контактов, вызванным связыванием мультимера фибриллы АВВ-2/3 с DSG2. Например, терапевтический агент можно вводить перед, одновременно и после каждого введения мультимера фибриллы АВВ-2/3. В других предпочтительных вариантах реализации настоящего изобретения терапевтический агент можно вводить после введения мультимера фибриллы АВВ-2/3, например, в пределах интервала времени не более 5 минут, 10 минут, 15 минут, 30 минут, 45 минут, 1 часа, 2 часов, 3 часов, 4 часов, 5 часов, 6 часов, 8 часов, 10 часов, 12 часов, 18 часов, 24 часов, 30 часов, 36 часов, 40 часов, 42 часов, 48 часов, 54 часов, 60 часов, 66 часов, 72 часов, 78 часов, 84 часов, 90 часов или даже 96 часов после введения мультимера фибриллы АВВ-2/3.
Согласно другому аспекту в настоящем изобретении предложены способы лечения заболевания, связанного с эпителиальной тканью, включающие введение субъекту, нуждающемуся в этом, такого количества мультимера фибриллы АВВ-2/3 согласно настоящему изобретению, которое достаточно для лечения указанного заболевания. В этом варианте реализации настоящего изобретения не осуществляют введение другого терапевтического агента. В неограничивающих вариантах реализации настоящего изобретения монотерапию используют для лечения заболевания, выбранного из группы, состоящей из вирусной инфекции АВВ-2/3, солидной опухоли или заболевания, которое можно лечить с использованием вектора для доставки гена на основе АВВ-2/3. Например, при лечении солидных опухолей указанный способ включает улучшение доступа клеток иммунной системы к очагу заболевания, например, путем проникновения (например, внутриопухолевое проникновение ранее существовавших природных клеток-киллеров, Т-клеток или дендритных клеток). Указанный способ также может применять для лечения любого из заболеваний, связанных с эпителиальными клетками, рассмотренных выше, в отношении которых улучшенный доступ клеток иммунной системы к эпителиальным клеткам-мишеням обеспечит клиническую пользу. Согласно одному предпочтительному варианту реализации настоящего изобретения заболевание представляет собой солидную опухоль, и способ включает повышение эффективности воздействия иммунной системы на опухоль. Указанный способ можно использовать для любой из солидных опухолей, описанных выше. Все варианты и комбинации вариантов реализации первого аспекта настоящего изобретения могут быть также использованы во втором аспекте, если из контекста явно не следует иное. Типичные мультимеры, содержащие один или более мультимеров фибриллы АВВ-2/3 согласно настоящему изобретению, которые могут быть использованы в соответствии с указанными способами, включают, но не ограничиваются ими, вирионы АВВ-2/3, капсиды АВВ-2/3, додекаэдрические частицы АВВ-2/3 (PtDd) (субвирусные додекаэдрические частицы, которые вырабатываются АВВ-2/3 при репликации) и рекомбинантные мультимеры фибриллы АВВ-2/3.
Согласно другому аспекту в настоящем изобретении предложен способ выявления соединений-кандидатов для одного или более из следующих: лечения заболевания, связанного с эпителиальной тканью, улучшения доставки вещества к эпителиальной ткани, улучшения доставки вещества к ткани, экспрессирующей DSG2, индукции ЭМП в ткани, и/или лечения инфекции АВВ-2/3, включающий (а) приведение мультимера фибриллы АВВ-2/3 согласно настоящему изобретению в контакт с DSG2 в условиях, способствующих связыванию мультимера с DSG2, причем приведение в контакт осуществляют в присутствии одного или более из испытываемых соединений; и (b) выявление положительных испытываемых соединений, которые конкурируют с мультимером фибриллы АВВ-2/3 за связывание с DSG2 по сравнению с контролем; причем положительные испытываемые соединения являются соединениями-кандидатами для одного или более из следующих: лечения заболевания, связанного с эпителиальной тканью, улучшения доставки вещества к эпителиальной ткани, улучшения доставки вещества к ткани, экспрессирующей DSG2, индукции ЭМП в ткани, и/или лечения инфекции АВВ-2/3.
Положительные испытываемые соединения, которые конкурируют с мультимером фибриллы АВВ-2/3 за связывание с DSG2, являются соединениями-кандидатами для кратковременного размыкания межклеточных контактов за счет их взаимодействия с DSG2. Дальнейшие количественные исследования для подтверждения способности соединений кратковременно размыкать межклеточные контакты за счет взаимодействия с DSG2 можно осуществлять с помощью любых подходящих способов, включая, но не ограничиваясь ими, способы исследований, описанные в примерах ниже. Выявленные таким способом соединения для лечения заболевания, связанного с эпителиальной тканью, улучшения доставки вещества к эпителиальной ткани, улучшения доставки вещества к ткани, экспрессирующей DSG2, или индукции ЭМП в ткани, могут быть использованы в качестве замены мультимера АВВ-2/3 в любом из способов согласно настоящему изобретению. Кроме того, АВВ-2/3 является важным возбудителем инфекций дыхательных путей человека (некоторые из которых являются тяжелыми) и аденовирусного конъюнктивита. Следовательно, соединения, обеспечивающие лечение инфекции АВВ-2/3, могут обеспечить клиническую пользу. Как раскрыто в настоящей заявке, DSG2 является основным высокоаффинным рецептором, используемым АВВ-2/3, и, соответственно, соединения, которые могут снизить связывание АВВ-2/3 с DSG2, являются соединениями-кандидатами для лечения или ограничения развития инфекции АВВ-2/3.
Типичные мультимеры, содержащие один или более мультимеров фибриллы АВВ-2/3 согласно настоящему изобретению, которые можно применять в соответствии с указанными способами, включают, но не ограничиваются ими, вирионы АВВ-2/3, капсиды АВВ-2/3, додекаэдрические частицы АВВ-2/3 (PtDd) (субвирусные додекаэдрические частицы, которые вырабатывает АВВ-2/3 при репликации) и рекомбинантные мультимеры фибриллы АВВ-2/3. Может быть использован любой подходящий контроль, включая, но не ограничиваясь этим, связывание мультимера АВВ-2/3 с DSG2 в отсутствие испытываемых соединений.
Согласно одному варианту реализации настоящего изобретения DSG представляет собой рекомбинантный DSG2. В другом варианте реализации в способах согласно настоящему изобретению применяют клетки, экспрессирующие DSG2 (эндогенно или рекомбинантно) на поверхности клеток.
Согласно одному неограничивающему варианту реализации для выявления соединений-кандидатов, которые конкурируют с мультимерами фибриллы АВВ-2/3 за связывание с DSG2, могут быть использованы исследования с помощью поверхностного плазмонного резонанса (SPR) с использованием сенсоров, содержащих иммобилизованный рекомбинантный DSG2, в комбинации с исследованиями конкурентного связывания DSG2. Согласно другому варианту реализации настоящего изобретения выявление соединений включает исследования трансдукции, в которых способность испытываемых соединений ослаблять связывание детектируют как уменьшение способности функциональных вирионов АВВ-2/3 трансдуцировать DSG2-экспрессирующие эпителиальные клетки. Согласно другому варианту реализации настоящего изобретения экстракт DSG2-экспрессирующих клеток подвергают электрофоретическому разделению и исследуют методом вестерн-блоттинга, и меченый мультимер фибриллы АВВ-2/3 используют для зондирования вестерн-блота в присутствии испытываемых соединений. Согласно другому варианту реализации настоящего изобретения могут быть использованы количественные исследования методом дот-блоттинга, такие как те, которые описаны в работе Wang et al., J. Virology (2007) 81:12785-12792; и Wang et al. (2008) 82:10567-10579.
В том случае, если испытываемые соединения содержат полипептидные последовательности, такие полипептиды могут быть синтезированы химически или рекомбинантно экспрессированы. Рекомбинантная экспрессия может быть осуществлена с использованием стандартных способов в данной области техники, раскрытых выше. Векторы для рекомбинантной экспрессии могут содержать бактериальные или вирусные векторы экспрессии, и клетки-хозяева могут представлять собой прокариотические или эукариотические клетки. Синтетические полипептиды, полученные с использованием хорошо известных методик синтеза в твердой фазе или жидкой фазе, или путем конденсации пептидов, или любой их комбинации, могут содержать природные и неприродные аминокислоты. Аминокислоты, используемые для синтеза пептидов, могут представлять собой стандартную смолу с иммобилизованными Boc (Nα-аминозащищенные Nα-трет-бутилоксикарбонилом) аминокислотами и стандартными протоколами для снятия защиты, нейтрализации и отмывки, или стандартные основание-неустойчивые Nα-аминозащищенные 9-фторенилметоксикарбонилом (Fmoc) аминокислоты. Fmoc- и Boc-Nα-аминозащищенные аминокислоты могут быть получены из Sigma, Cambridge Research Biochemical или других химических компаний, известных специалистам в данной области техники. Помимо этого полипептиды могут быть синтезированы с использованием других Na-защитных групп, которые известны специалистам в данной области техники. Твердофазный пептидный синтез можно осуществлять с помощью методик, хорошо известных в данной области техники и предоставляемых на коммерческой основе, например, с использованием автоматических синтезаторов.
В случае если испытываемые соединения содержат антитела, такие антитела могут быть поликлональными или моноклональными. Антитела могут быть гуманизированными, полностью человеческими или мышиными антителами. Такие антитела могут быть получены с помощью хорошо известных способов, таких как те, которые описаны в Harlow and Lane, Antibodies; A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., (1988).
В случае если испытываемые соединения содержат последовательности нуклеиновых кислот, такие нуклеиновые кислоты могут быть синтезированы химически или рекомбинантно экспрессированы. Методы рекомбинантной экспрессии хорошо известны специалистам в данной области техники (см., например, Sambrook, et al., 1989, выше). Нуклеиновые кислоты могут представлять собой ДНК или РНК, и могут быть одноцепочечными или двухцепочечными. Аналогичным образом, нуклеиновые кислоты могут быть химически или ферментативно синтезированы вручную или с применением автоматизированных реакций, используя стандартные методики в данной области техники. Если нуклеиновая кислота синтезирована химическим способом или с помощью ферментативного синтеза в условиях in vitro, она может быть очищена перед введением в клетку. Например, нуклеиновые кислоты могут быть очищены из смеси с помощью экстракции растворителем или смолой, осаждения, электрофореза, хроматографии или их комбинации. В другом варианте нуклеиновые кислоты могут быть использованы без очистки или с минимальной очисткой, чтобы избежать потерь в результате обработки образцов.
В случае если испытываемые соединения содержат, помимо полипептидов, антитела или нуклеиновые кислоты, такие соединения могут быть получены любым из множества способов, известных в данной области техники для проведения органического химического синтеза.
Примеры. Структурные и функциональные исследования взаимодействия головных доменов белка фибриллы аденовирусов и десмоглеина 2.
Реферат
Серотипы аденовирусов человека (АВ) АВ3, АВ7, АВ11, АВ14 и недавно обнаруженный новый штамм АВ14 (АВ14р1) используют десмоглеин 2 (DSG2), белок межклеточных контактов эпителиальных клеток, в качестве рецептора для инфицирования. В отличие от взаимодействия АВ с CAR и CD46, точные структурные параметры связывания АВ с DSG2 до сих пор не ясны. Используя подход, основанный на библиотеках экспрессии Е. coli, содержащих мутированные варианты головного домена белка фибриллы АВ3 и АВ14р1, полученные методом случайного мутагенеза, авторы настоящего изобретения выявили аминокислотные остатки, которые при точечном мутагенезе устраняют или уменьшают связывание головного домена АВ с DSG2. Выявленные остатки образуют три кластера внутри одной борозды на крайнем дистальном конце головной части белка фибриллы. Библиотека, содержащая мутированные варианты головной части белка фибриллы АВ3, также была использована для выявления вариантов с повышенной аффинностью в отношении DSG2. Авторы настоящего изобретения обнаружили ряд мутаций в пределах или вблизи петли EF головной части белка АВ3, которые обеспечивали на несколько порядков большую величину аффинности к DSG2 по сравнению с таковой для головного домена АВ3 дикого типа. Результаты исследования кристаллической структуры одного из мутированных вариантов выявили, что введенные мутации делают петлю EF более гибкой, что могло бы способствовать взаимодействию с DSG2. Выводы авторов настоящего изобретения имеют практическую значимость для терапии рака. Недавно авторы настоящего изобретения сообщили, что головная часть белка фибриллы АВ3, содержащая рекомбинантный белок (JO-1), способна индуцировать размыкание межклеточных контактов между эпителиальными раковыми клетками, что, в свою очередь, значительно улучшило внутриопухолевое проникновение и эффективность терапевтических агентов. В настоящей заявке показано, что варианты JO-1 с улучшенной аффинностью являются терапевтически более эффективными, чем исходный белок в серии моделей рака.
Введение
Недавно авторы настоящего изобретения установили, что DSG2 является основным рецептором для определенной группы аденовирусов типа В, включая аденовирус серотипа 3 (АВ3), широко распространенного серотипа в человеческой популяции (42). Авторы настоящего изобретения обнаружили, что домен(ы) АВ3, взаимодействующий с DSG2, образован несколькими головными частями белка фибриллы (41). Обнаруженный специфичный способ взаимодействия головной части белка фибриллы AB3-DSG2 обеспечивает высокую авидность и является функционально значимым для размыкания контактов между эпителиальными клетками (41, 42). Последнее включает кластеризацию DSG2 и активацию путей, которые напоминают эпителиально-мезенхимальный переход, включая фосфорилирование МАР-киназы и подавление экспрессии белка межклеточных контактов (6, 40, 42). Способность размыкать межклеточные контакты эпителиальных клеток представляется важной для проникновения и распределения АВ3 внутри эпителиальных клеток дыхательных путей (40-42). В недавнем исследовании авторы настоящего изобретения предприняли попытку обнаружить минимальный фрагмент внутри капсида АВ3, который обеспечивает эффективное связывание с DSG2 (41). Был сконструирован небольшой рекомбинантный белок, который содержит головную часть белка фибриллы АВ3 и домен, который обеспечивает самопроизвольную димеризацию тримерных головных частей белка фибриллы АВ3 (JO-1) (41). JO-1 может быть легко получен в Е. coli и очищен с помощью аффинной хроматографии. В культурах поляризованных эпителиальных клеток JO-1 индуцировал размыкание межклеточных контактов, тогда как внутривенная инъекция JO-1 мышам с эпителиальными опухолями обеспечила более эффективное проникновение противоопухолевых препаратов (5, 6).
Первая цель настоящего исследования состояла в том, чтобы дополнительно установить структурные особенности взаимодействия головной части белка фибриллы AB3-DSG2. Цель включала выявление аминокислотных остатков в пределах головной части белка фибриллы АВ3, которые участвуют в связывании с DSG2, и создание мутированных вариантов JO-1 с уменьшенным или устраненным связыванием с DSG2. Вторая цель настоящего исследования, которая имеет трансляционное значение, заключалась в дальнейшем улучшении JO-1 путем усиления его аффинности в отношении DSG2, увеличивая тем самым его терапевтический эффект. Вторая цель была достигнута путем выявления мутированных вариантов с повышенным связыванием с DSG2.
Обе цели были достигнуты с помощью библиотек экспрессии Е. coli, содержащих мутированные варианты головной части белка фибриллы АВ3. Были выявлены остатки в трех различных кластерах внутри головной части белка фибриллы АВ3, которые критически вовлечены в связывание с DSG2. Все остатки локализованы в пределах одной борозды на дистальном конце головной части белка фибриллы, обращенной к рецептору. Затем была проведена оценка влияния идентифицированных мутаций на способность головной части фибриллы размыкать эпителиальные контакты, которую осуществляли путем измерения трансэпителиального электрического сопротивления в поляризованных эпителиальных клетках в условиях in vitro, а также способности повышать эффективность химиотерапевтических препаратов у мышей с ксенотрансплантатами эпителиальных опухолей. Как и следовало ожидать, при введении в JO-1 мутаций, вызывающих снижение аффинности в отношении DSG2, полученные в результате белки обладали меньшей способностью размыкать эпителиальные контакты. С другой стороны, ряд мутаций, которые увеличивали аффинность JO-1 в отношении DSG2, проявляли более сильную активность в размыкании эпителиальных межклеточных контактов. В целом, результаты этих исследований указывают на корреляцию между аффинностью головной части белка фибриллы АВ3 в отношении DSG2 и последующим действием на эпителиальные межклеточные контакты.
Третья цель данного исследования состояла в том, чтобы установить остатки головной части фибриллы, взаимодействующие с DSG2, для другого серотипа, нацеленного на DSG2, недавно открытого штамма АВ14р1 (44), который считается более патогенным/вирулентным, чем исходный штамм (Adl4-deWit) (10, 16, 22). Распределение бета-листов в АВ14р1 отличается от такого в АВ3, что потенциально может привести к различиям в способе связывания с DSG2. В этой связи для того чтобы выявить остатки АВ14р1, взаимодействующие с DSG2, была создана библиотека экспрессии Е. coli, содержащая мутированные варианты головной части белка фибриллы АВ14р1.
Материалы и методы
Белки. Рекомбинантный белок человека DSG2 был получен из Leinco Technologies, Inc. (Сент-Луис, Миссури, США). Головная часть фибриллы АВ3 была получена из вируса АВ3, штамм GB, полученного из АТСС. Головная часть фибриллы АВ14р1 получена из вируса АВ14р1, штамм Portland 2971/2007, предоставленного Центром по контролю и профилактике заболеваний (Атланта, Джорджия, США) (44). Головные части фибриллы с N-концевыми 6-гистидиновыми метками нарабатывали в клетках E.coli, используя вектор экспрессии pQE30 (Qiagen, Валенсия, Калифорния, США), и очищали с помощью хроматографии на Ni-NTA агарозе, как описано в другом месте (43).
Клеточные линии. Клетки HeLa293 и клетки А549 поддерживали в среде DMEM, дополненной 10% ФБС, 100 ед./мл пенициллина, 100 мкг/мл стрептомицина (P/S), 2 мМ глутамина (Glu) и 1× MEM с несущественными аминокислотами (Invitrogen, Карлсбад, Калифорния, США). Клетки рака толстой кишки Т84 (АТСС CCL-248) культивировали в смеси, содержащей среду F12 Хэма и DMEM в соотношении 1:1, а также 10% ФБС, глутамин и P/S. Клетки Ovc316 представляют собой HER/Neu-положительные эпителиальные опухолевые клетки, полученные из биопсии рака яичников (32). Клетки Ovc316 культивировали в MEGM (Lonza, Маплетон, Иллинойс, США), содержащий 3 мкг/л hEGF, 5 мкг/л инсулина, 5 мг/л гидрокортизона, 26 мг/л бычьего гипофизарного экстракта, 25 мг/л амфотерицина В (Lonza) и дополненной 1% ФБС, 100 ME пенициллина, 100 мкг/л стрептомицина, 10 мг/л ципрофлоксацина. Клетки MDA-MB-231, линия тройных негативных клеток рака молочной железы (АТСС-НТВ-26), культивировали в среде L-15 Лейбовица с добавлением 10% ФБС, 100 ME пенициллина, 100 мкг/л стрептомицина. TC1-DSG2 получали из клеток ТС1, линии клеток рака легких C57BL/6, которые экспрессируют HPV16 Е6 и Е7 (36). Клетки ТС1 трансдуцировали VSVG-псевдотипированным лентивирусным вектором, экспрессирующим DSG2 человека (42). Клон, который экспрессировал DSG2 человека на уровне, соответствующем уровню, наблюдаемому в опухолях человека, отбирали для исследований в условиях in vivo.
Аденовирусы. Размножение, включение метки метил-Н3-тимидина, очистку и титрование АВ3 дикого типа проводили, как описано в другом месте (37). AB3-GFP представляет собой вектор на основе АВ3 дикого типа, содержащий кассету экспрессии CMV-GFP, вставленную в область Е3 (42). Концентрации вирусных частиц (ВЧ) определяли спектрофотометрическим способом, измеряя оптическую плотность при 260 нм (OD260). Титры бляшкообразующих единиц (БОЕ) были определены с использованием клеток 293, как описано в другом месте (29). Соотношение ВЧ и БОЕ составило 20:1 для всех вирусных препаратов.
Библиотека головных частей белка фибриллы АВ3. Кодирующую последовательность головной части белка АВ3 (аминокислоты 108-319), содержащую два последних повтора центральной части белка фибриллы, получали с помощью ПЦР из ДНК АВ3 с использованием праймеров Р1: 
 (SEQ ID NO: 35) и Р2:
(SEQ ID NO: 35) и Р2:  (SEQ ID NO: 36), и клонировали в pQE30 (Qiagen, Валенсия, Калифорния, США) для экспрессии в Е. coli. Полученная плазмида была названа pQE-Ad3knob. Случайный мутагенез с помощью ПЦР проводили на основании протокола, опубликованного в другом месте (7, 8). В общих чертах, 20 фмоль ДНК-матрицы pQE-Ad3knob, 30 пмоль (каждого) праймеров для ПЦР (Pmutl:
(SEQ ID NO: 36), и клонировали в pQE30 (Qiagen, Валенсия, Калифорния, США) для экспрессии в Е. coli. Полученная плазмида была названа pQE-Ad3knob. Случайный мутагенез с помощью ПЦР проводили на основании протокола, опубликованного в другом месте (7, 8). В общих чертах, 20 фмоль ДНК-матрицы pQE-Ad3knob, 30 пмоль (каждого) праймеров для ПЦР (Pmutl:  (SEQ ID NO: 37) и Pmut2:
(SEQ ID NO: 37) и Pmut2:  (SEQ ID NO: 38), 2,5 мкл, 3,5 мкл, 5 мкл или 10 мкл 10-кратного буфера для мутагенеза (70 мМ MgCl2, 500 мМ KCl, 100 мМ Трис (рН=8,3 при 25°С), и 0,1% (масс/об.) желатина), 10 мкл 5 мМ MnCl2, 10 мкл смеси дНТФ (2 мМ дГТФ, 2 мМ дАТФ, 10 мМ дЦТФ, 10 мМ дТТФ) и 5 единиц Taq-полимеразы (Promega, Мэдисон, Висконсин, США) смешивали в конечном объеме 100 мкл. Условия проведения ПЦР: 94°С в течение 1 мин, 45°С в течение 1 мин и 72°С в течение 1 мин (30 циклов). Мутированные продукты ПЦР (615 п. н. в длину, содержащие мутации только в рамке считывания в начале головной части белка фибриллы), очищали, расщепляли соответствующими ферментами и клонировали в плазмиду pQE-Ad3knob. Для контроля качества библиотеки, содержащей продукты случайного мутагенеза, клетки E.coli Ml5 трансформировали продуктом лигирования (Qiagen, Валенсия, Калифорния, США), высевали на чашки с канамицином и ампициллином, и случайным образом отбирали 50 колоний для секвенирования.
(SEQ ID NO: 38), 2,5 мкл, 3,5 мкл, 5 мкл или 10 мкл 10-кратного буфера для мутагенеза (70 мМ MgCl2, 500 мМ KCl, 100 мМ Трис (рН=8,3 при 25°С), и 0,1% (масс/об.) желатина), 10 мкл 5 мМ MnCl2, 10 мкл смеси дНТФ (2 мМ дГТФ, 2 мМ дАТФ, 10 мМ дЦТФ, 10 мМ дТТФ) и 5 единиц Taq-полимеразы (Promega, Мэдисон, Висконсин, США) смешивали в конечном объеме 100 мкл. Условия проведения ПЦР: 94°С в течение 1 мин, 45°С в течение 1 мин и 72°С в течение 1 мин (30 циклов). Мутированные продукты ПЦР (615 п. н. в длину, содержащие мутации только в рамке считывания в начале головной части белка фибриллы), очищали, расщепляли соответствующими ферментами и клонировали в плазмиду pQE-Ad3knob. Для контроля качества библиотеки, содержащей продукты случайного мутагенеза, клетки E.coli Ml5 трансформировали продуктом лигирования (Qiagen, Валенсия, Калифорния, США), высевали на чашки с канамицином и ампициллином, и случайным образом отбирали 50 колоний для секвенирования.
Библиотека AB14: Кодирующую последовательность головной части белка АВ14р1 (аминокислоты 108-323), содержащую два последних повтора центральной части белка фибриллы, получали с помощью ПЦР из ДНК АВ14р1 с использованием праймеров Р1: 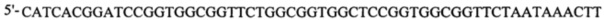
 (SEQ ID NO: 39) и Р2:
(SEQ ID NO: 39) и Р2: 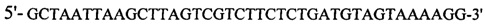 (SEQ ID NO: 40), и клонировали в pQE30 (Qiagen, Валенсия, Калифорния, США) для экспрессии в Е. coli. Полученная плазмида получила название pQE-Adl4plknob. Случайный мутагенез с помощью ПЦР проводили с использованием праймеров для ПЦР (Pmutl:
(SEQ ID NO: 40), и клонировали в pQE30 (Qiagen, Валенсия, Калифорния, США) для экспрессии в Е. coli. Полученная плазмида получила название pQE-Adl4plknob. Случайный мутагенез с помощью ПЦР проводили с использованием праймеров для ПЦР (Pmutl: 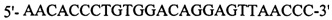 (SEQ ID NO: 41) и Pmut2:
(SEQ ID NO: 41) и Pmut2: 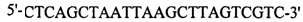 (SEQ ID NO: 42)). Мутированные продукты ПЦР (594 п. н. в длину, содержащие мутации только в рамке считывания в начале головной части белка фибриллы) очищали, расщепляли соответствующими ферментами и клонировали в плазмиду pQE-Adl4plknob. Для контроля качества библиотеки, содержащей продукты случайного мутагенеза, клетки Е. coli M15 трансформировали продуктом лигирования (Qiagen, Валенсия, Калифорния, США), высевали на чашки с канамицином и ампициллином, и случайным образом отбирали 50 колоний для секвенирования.
(SEQ ID NO: 42)). Мутированные продукты ПЦР (594 п. н. в длину, содержащие мутации только в рамке считывания в начале головной части белка фибриллы) очищали, расщепляли соответствующими ферментами и клонировали в плазмиду pQE-Adl4plknob. Для контроля качества библиотеки, содержащей продукты случайного мутагенеза, клетки Е. coli M15 трансформировали продуктом лигирования (Qiagen, Валенсия, Калифорния, США), высевали на чашки с канамицином и ампициллином, и случайным образом отбирали 50 колоний для секвенирования.
Количественные исследования колоний. Плазмидную библиотеку, содержащую мутированные варианты головной части фибриллы АВ3 или АВ14р1, трансформировали в штаммы-хозяева Е. coli XL1 Blue или M15 и высевали на чашки с LB-агаром, содержащие соответствующие антибиотики, т.е. амп. или амп. + кан., соответственно. После культивирования в течение ночи поверх колоний помещали мембранный фильтр с размером пор 0,45 мкм Durapore (Millipore, Биллерика, Массачусетс, США). Мембрану отделяли и осторожно помещали колониями вверх на два листа бумаги толщиной 3 мм, пропитанных средой LB, дополненной антибиотиками и 1 мМ IPTG. Экспрессию белка в колониях индуцировали в течение 6 ч при 30°С, после этого фильтр с колониями помещали на поверхность нитроцеллюлозного фильтра и бумаги Ватман 3 мм, смоченных в нативном буфере для лизиса (20 мМ Трис-Cl (рН=8), 300 мМ NaCl, 50 мМ MgCl2, 0,1 мг/мл лизоцима, 0,75 мг/мл ДНКазы I, половина таблетки, содержащей полный коктейль ингибиторов протеаз, без ЭДТА/10 мл (Roche, Пало-Альто, Калифорния, США)). «Сэндвич-фильтр» инкубировали при комнатной температуре в течение 10 мин, и затем подвергали четырехкратному замораживанию/оттаиванию в течение 10 мин при -80°С и 10 мин при 30°С. Из сэндвича удаляли нитроцеллюлозную мембрану и блокировали 3% БСА в TBST при 4°С в течение ночи. Затем блот инкубировали с 0,1 нг/мл рекомбинантного белка DSG2 (Leinco, Сент-Луис, Миссури, США) в TBST/BSA и затем с мышиными моноклональными антителами к DSG2 (клон 6D8; Serotec Ltd., Оксфорд, Великобритания) и конъюгированным с пероксидазой хрена IgG к иммуноглобулину мыши. Колонии без связывания DSG2 отбирали и культивировали в 3 мл среды LB в течение ночи. Экспрессию белка индуцировали с помощью 1 мМ IPTG в течение 5 часов, затем бактерии осаждали, повторно суспендировали в буфере для загрузки, содержащем ДСН, и подвергали трехкратному замораживанию/оттаиванию. После электрофореза белки переносили на нитроцеллюлозную мембрану и инкубировали с антителами к гексагистидиновой метке (МСА1396, Sertec) для количественной оценки тримеризации головной части фибриллы АВ. Для скрининга мутированных вариантов с более прочным связыванием с DSG2, клетки штамма-хозяина Е. coli Ml5 трансформировали библиотекой, содержащей мутированные варианты головной части фибриллы АВ3. Экспрессию белка в колониях индуцировали в течение не более чем 20 минут при комнатной температуре. Отбирали колонии, которые имели наиболее интенсивный сигнал связывания DSG2.
Вестерн-блоттинг: Для вестерн-блоттинга использовали готовые гели mini-PROTEAN (Bio-Rad, Геркулес, Калифорния, США) с 4-15% градиентом полиакриламида. В дорожку вносили в общей сложности 1 мкг белка, смешанного с двукратным загрузочным буфером (10 мМ Трис-HCl, рН=6,8, 200 мМ ДТТ, 4% ДСН, 20% глицерина, 0,2% бромфенолового синего). Образцы подвергали кипячению (В) в течение 5 мин либо вносили некипячеными (UB). Использовали следующий подвижный буфер: 25 мМ Триса, рН=8,3, 0,192 М глицина, 0,1% ДСН. После электрофореза белки переносили на нитроцеллюлозную мембрану и инкубировали с рекомбинантным белком DSG2 человека и антителами к DSG2, как описано ранее (42). Вестерн-блоты сканировали и количественно оценивали с помощью программного обеспечения Image J 1.32 (Национальный институт здоровья, Бетесда, Мэриленд, США). Интенсивность полосы JO-1 принимали за 100%. Для исследования активности киназы MAP поляризованные культуры Т84 лизировали в буфере, содержащем 20 мМ HEPES (рН=7,5), 2 мМ ЭГТА, 10% глицерина, 1% Тритона X100, 1 мМ PMSF, 200 мкМ Na3VO4 и ингибиторы протеаз, на льду. После обработки ультразвуком образцы осаждали и супернатант, содержащий белок, хранили при -80°С. Для вестерн-блоттинга использовали 15 мкг общего белка и моноклональное антитело к фосфо-р44/42 МАРК (Erk1/2) (Thr202/Tyr204) (Cell Signaling, Дэнверс, Массачусетс, США) или моноклональное антитело к Erk1/2 мыши (Cell Signaling).
Количественные исследования конкурентного связывания. Клетки HeLa отделяли от чашек для культивирования путем инкубации с версеном и промывали ФСБ. В общей сложности по 105 клеток на пробирку ресуспендировали в 50 мкл охлажденного на льду адгезионного буфера (DMEM с добавлением 2 мМ MgCl2, 1% ФБС и 20 мМ HEPES), содержащем различные концентрации белка головной части фибриллы АВ3, и инкубировали на льду в течение 1 часа. Затем в адгезионный буфер вносили Н3-меченый вирус АВ3 дикого типа с показателем множественности инфекции (MOI) 8000 вирусных частиц (ВЧ) на клетку, и конечный объем доводили до 100 мкл. После 1 ч инкубации на льду клетки осаждали и промывали два раза в 0,5 мл охлажденного на льду ФСБ. После последней промывки удаляли супернатант, и связанную с клетками радиоактивность определяли с помощью сцинтилляционного счетчика. Число ВЧ, связанных на клетку, рассчитывали с использованием удельной радиоактивности вириона и количества клеток.
Поверхностный плазменный резонанс: Показания регистрировали на приборе BIAcore 3000. Во всех экспериментах в качестве рабочего буфера использовали HBS-N (GE Healthcare, Питтсбург, Пенсильвания, США) с добавлением 2 мМ CaCl2, при скорости потока 5 мкл/мин. Иммобилизацию на сенсорном чипе СМ4 (BIAcore) проводили с использованием 10 мкг/мл DSG2, разведенного в 10 мМ ацетатном буфере с рН=4,5, который впрыскивали в течение 10 минут на проточную ячейку, активированную этил(диметиламинопропил)карбодиимидом (EDC)/N-гидроксисукцинимидом (NHS). Контрольную проточную ячейку активировали (EDC/NHS) и инактивировали этаноламином. Различные концентрации белков головной части фибриллы АВ3 вводили для связывания в течение 3 минут с последующим периодом диссоциации в течение 2,5 мин, сигнал автоматически вычитали из неспецифичного сигнала проточной ячейки активированной (EDC/NHS) и деактивированной этаноламином. Кинетические константы и параметры аффинности рассчитывали с использованием программного обеспечения BIAeval.
Кристаллография: Условия кристаллизация для wtAB3 и мутированных вариантов головной части белка K217E/F224S создавали с использованием службы лаборатории высокопропускного скрининга при Институте медицинских исследований Гауптман-Вудворда. Для дифракционных исследований wtAB3 и мутированный вариант головной части белка K217E/F224S кристаллизовали с использованием метода висячей капли. Кристаллы выращивали с использованием резервуарного раствора 1,65 MgSO4⋅7H2O в 0,1М буфере TAPS с рН=9,0 и 15 мг/мл раствора белка. Кристаллы замораживали с использованием криопротектора, состоящего из 85% резервуарного буфера и 15% глицерина (об./об.). Сбор данных проводили при 100К на ID 14-4 Европейской установки синхротронного излучения (ESRF) с использованием цепочки процессов технической платформы EDNA (19). Данные индексировали и масштабировали с помощью XDS/XScale (20, 21), и структуру устанавливали путем молекулярного замещения (PDB 1H7Z) с помощью программы PHASER (25). Модель строили и уточняли с использованием Coot (14) и Phenix (2), соответственно (таблица 1а). Запись «Структура мутированного варианта головного домена К217Е и F224S аденовируса 3» получила код доступа RCSB ID - rcsb080687 и код PDB ID - 4LIY.


Статистические данные для самого высокого разрешения детектора приведены в скобках.
Трехмерная структура: Для исследования структуры использовали программное обеспечение PyMOL. Мутации в головном домене АВ3 (PDB 1H7Z) окрашивали с использованием различных цветов на фиолетовой изоповерхности. Мономеры мутированного варианта головной части фибриллы K217E/F224S АВ3 изображали в виде цветных схем с мутациями в палочках и накладывали на серую схему головной части белка фибриллы АВ3 дикого типа.
Электронная микроскопия с негативным контрастированием: Рекомбинантный белок JO-2 визуализировали с помощью электронной микроскопии с негативным контрастированием, чтобы оценить степень его мультимеризации. Стандартный препарат слюда-углерод использовали с белком в концентрации 0,1 мг/мл. Образец окрашивали с использованием 1% (масс./об.) кремневольфрамата натрия (рН=7,0) и визуализировали на электронном микроскопе JEOL-1200 при 100 кВ.
Количественное исследование проницаемости. В общей сложности 5×105 клеток Т84 высевали в 12 мм вставки Transwell (мембраны ПЭТ с размером пор 0,4 мкм (Corning, Нью-Йорк, США) и культивировали в течение >14 дней до достижения стабильных показателей трансэпителиального электрического сопротивления (TEER). Культуральную среду заменяли каждые 2-3 дня. Клетки подвергали воздействию лигандов DSG2 (20 мкг/мл) в адгезионной среде (DMEM, содержащей 1% ФБС, 2 мМ MgCl2, 20 мМ HEPES) в течение 15 мин при комнатной температуре, и TEER измеряли и рассчитывали, как описано в другом месте (39).
Исследования на животных: Все эксперименты с участием животных проводили в соответствии с руководящими принципами организации, установленными Университетом штата Вашингтон. Мышей содержали в помещениях, свободных от специфических патогенов. Иммунодефщитных мышей (СВ17) [название линии: NOD.CB17-Prkdcscid/J] получали из Jackson Laboratory. Мыши, трансгенные по DSG2 человека, содержали локус DSG2 человека размером 90 п. н. и экспрессировали hDSG2 на уровне, который аналогичен уровню экспрессии у человека (40).
Ксенотрансплантаты опухолей А549, MDA-MB-231 и ovc316 создавали путем инъекции соответствующих опухолевых клеток в жировую ткань молочной железы мышей СВ17 (в комбинации с матригелем в соотношении 1:1). Опухоли TC1-DSG2 создавали путем подкожной инъекции клеток TC1-DSG2 DSG2-трансгенным мышам. JO-0, JO-1, JO-2 или JO-4 внутривенно вводили за один час перед применением химиотерапевтических лекарственных препаратов: иринотекана/камптосара™ (Pfizer Inc., Гротон, Коннектикут, США), липосомальной формы пегилированного доксорубицина/липодокса™ (Sun Pharmaceuticals IN, Индия), цетуксимаба/эрбитукса™ (ImClone, Сомервилль, Нью-Джерси, США), наночастиц паклитаксела, конъюгированного с альбумином/абраксана™ (Abraxis Biosciences, Summit, Нью-Джерси, США). Объемы опухолей измеряли три раза в неделю. Каждая группа лечения состояла не менее чем из 5 мышей. Животных умерщвляли и эксперимент прекращали, когда опухоли в одной из групп достигали объема 800 мм3 или имели признаки изъязвления.
Антитела к JO4: концентрации антитела к JO-4 в образцах сыворотки человека измеряли с помощью ИФА. Планшеты покрывали кроличьими поликлональными антителами к белку фибриллы АВ3 (42), затем вносили рекомбинантный JO-4, образцы сыворотки человека (разведение от 1:2 до 1:1000), и IgG-ПХ к иммуноглобулину человека. Образцы сыворотки от пациентов, страдающих раком яичников, были предоставлены Тихоокеанским консорциумом по изучению рака яичников.
Трехмерная структура: Для визуализации трехмерной структуры головного домена фибриллы АВ3 (MMDB ID: 16945, PDB ID: 1H7Z (13)) использовали программное обеспечение PyMOL.
Статистический анализ: Все результаты выражали как среднее значение±SD. Применяли двухфакторный дисперсионный анализ для многократного тестирования. Количество животных и значения Р указаны в подписях к фигурам.
Результаты
Остатки, критические для связывания с DSG2. В первую очередь экспериментальная работа была сосредоточена на АВ3.
Высокоаффинное связывание с DSG2 и последующее размыкание эпителиальных контактов требует присутствия нескольких тримеров головного домена в пространственной констелляции в вирионе, PtDd, или димеризованной (гримерной) головной части фибриллы АВ3 (например, JO-1) (41). Рекомбинантная (тримерная) головная часть фибриллы с двумя мотивами центральной части, но без домена димеризации («мономер головной части фибриллы АВ3»), связывается с DSG2 с аффинностью, величина которой на несколько порядков меньше, чем величина аффинности JO-1, не способна блокировать инфекцию АВ3 и не индуцирует размыкание контактов (37, 41, 42). Однако величина аффинности «мономера головной части фибриллы АВ3» достаточно высока для детектирования связывания с помощью вестерн-блоттинга, при котором в качестве зонда используется растворимый DSG2. В этой связи авторы настоящего изобретения использовали библиотеку экспрессии Е. coli, содержащую мутированные варианты мономера головной части фибриллы АВ3, несущие гексагистидиновую метку, для выявления остатков аминокислот в пределах головной части белка фибриллы АВ3, которые имеют критическое значение для связывания с DSG2. Для создания описанной библиотеки использовали мутагенез с помощью ПЦР (7, 8) в соответствии с протоколом, который в среднем создает от одной до двух замен аминокислот на головную часть фибриллы. Библиотеку белка головной части фибриллы АВ3 в Е. coli XL-1 blue высевали на чашки с агаром, экспрессию головной части белка индуцировали IPTG, и колонии подвергали скринингу для определения связывания с DSG2 с использованием рекомбинантного DSG2 и антител к DSG2. Первый этап скрининга ~10000 колоний для обнаружения вариантов, которые не связываются с DSG2, выявил 240 колоний-кандидатов. При исследовании с помощью вестерн-блоттинга для детектирования гексагистидиновой метки 40 из 240 колоний экспрессировали тримерную головную часть фибриллы, что указывает на отсутствие крупных конформационных изменений. Остальные варианты имели усеченную головную часть белка фибриллы или не образовывали тримеры. Соответствующие 40 плазмид секвенировали. Подавляющее большинство колоний содержали единичные замены аминокислот внутри головной части белка фибриллы. При детектировании нескольких замен аминокислот в головной части белка фибриллы, синтезировали новые геномы головной части белка фибриллы АВ3, содержащие соответствующие мутации по отдельности. Дальнейшие этапы скрининга колоний не выявили другие области, это свидетельствует о том, что были найдены все остатки, взаимодействующие с DSG2. Для последующих исследований впоследствии использовали в общей сложности 8 независимых мутированных вариантов (Фиг. 1а). Для всех последующих исследований создавали самопроизвольно димеризующиеся формы мутированных вариантов фибриллы АВ3 (Фиг. 1b) и очищали их с помощью аффинной хроматографии с использованием колонок Ni-NTA. Очищенные димеризованные мутированные варианты головной части белка исследовали для определения связывания с DSG2 с помощью вестерн-блоттинга (Фиг. 1c-f). Связывание всех мутированных вариантов было значительно слабее, по сравнению с JO-1 (т.е. димеризованной формы, содержащей головную часть белка фибриллы АВ3 дикого типа). Мутированные варианты D261N и F265L практически не связывались с DSG2, в то время как другие мутированные варианты проявляли различные уровни остаточного связывания с DSG2 (3,6% - 20% головной части белка АВ3 дикого типа) (таблица 2, «Вестерн-блот»). Выявленные остатки, критические для связывания головной части белка АВ3 с DSG2, находились в трех различных областях головной части белка фибриллы АВ3 в пределах CD-петли/D-бета-листа (N186D, V189G, S190P), FG-петли/G-бета-листа (D261N, F265L) и Н-бета-листа/HI-петли (L292A, L296R, E299V) (Фиг. 1а). В трехмерной модели головной части белка фибриллы АВ3 (номер доступа PDB: 1H7Z_A) все выявленные остатки были расположены на апикальной стороне головной части белка фибриллы и придерживались одной конкретной борозды в головной части (Фиг. 2). Следует отметить, что кроме F265L, все другие замены привели к изменению заряда в соответствующем остатке. Различия в характере мигрирования в полиакриламидном геле (например, для D261N) показывают, что замены вызвали конформационные изменения. В настоящее время авторы настоящего изобретения предпринимают попытки кристаллизовать полученные мутанты для исследования их трехмерной структуры.
Чтобы создать головную часть белка фибриллы АВ3 и в итоге вирус АВ3 с максимально уменьшенным связыванием с DSG2, множественные мутации были введены в трех определенных областях, в частности, комбинация N186D и D261N, комбинация D261N и L296R, а также комбинация всех трех N186D, D261N и L296R. Как и следовало ожидать, комбинация мутаций во всех трех критических областях привела к самому высокому уровню снижения связывания (таблица 2, «вестерн-блот), Фиг. 1f).
Таблица 2: Исследование мутированных вариантов фибриллы АВ3. Во второй колонке приведены результаты количественного исследования полос вестерн-блота, соответствующих тримерам головной части белка АВ3. Интенсивность для головной части белка фибриллы АВ3 дикого типа была принята за 100%. Данные, приведенные в третьей колонке, отражают способность димерных мутированных вариантов головной части белка фибриллы АВ3 ингибировать инфекцию AB3-GFP (см. Фиг. 3b). Степень ингибирования повышается с увеличением процента. Ингибирование под действием JO-1 (димерный головной домен wtAB3) принимали за 100%. В четвертой колонке приведены соответствующие данные для прикрепления вируса АВ3. N=3. Показаны средние значения. Стандартное отклонение составляло менее 10%.
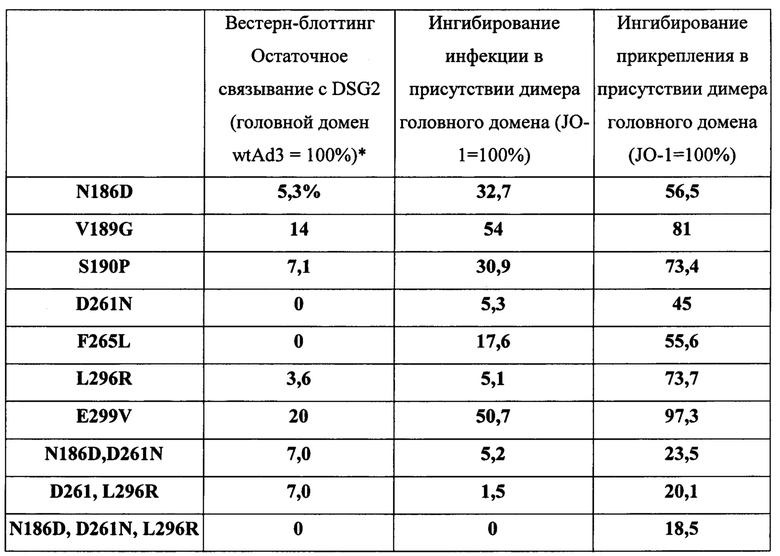
Вследствие значимости АВ14р1 как недавно выявленного патогенна также была создана библиотека («мономерных») мутированных вариантов головной части белка фибриллы АВ14р1. Первый этап скрининга выявил ~300 колоний-кандидатов для вариантов, которые не связываются с DSG2. При исследовании с помощью вестерн-блоттинга для детектирования гексагистидиновой метки 45 из 300 колоний экспрессировали тримерный головной домен фибриллы. Секвенирование этих вариантов выявило 15 независимых мутированных вариантов с ограниченной способностью к связыванию с DSG2 (Фиг. 1а). Интересно отметить, что, несмотря на различное распределение бета-листов, аминокислотные остатки, которые имеют критическое значение для связывания головной части белка АВ14р1, были локализованы в одних и тех же трех областях, которые были выявлены для головной части фибриллы АВ3. Из-за этого сходства дальнейшие исследования были проведены только с выбранными мутированными вариантами головной части белка фибриллы АВ3.
Функциональная проверка. Исследования конкурентного связывания проводили на клетках HeLa, которые экспрессируют DSG2 (42). В первую очередь было исследовано прикрепление Н3-меченого вируса АВ3 после предварительной инкубации клеток с димерными головными частями фибриллы АВ3 (Фиг. 3а). Уменьшение связывания вируса АВ3 сопоставляли со снижением после предварительной инкубацией с JO-1, то есть димерным белком, который содержит головную часть фибриллы АВ3 дикого типа. Ингибирование связывания под действием JO-1 принимали за 100%. Мутированные варианты L296R, D261N и F265L оказывали наименьшее блокирующее действие на связывание вируса АВ3 (5,1, 5,3, и 17,6%), следующими были мутированные варианты S190P, N186D и E299V (уменьшение связывания на 30,9, 32,7 и 50,7%, соответственно) (таблица 2, «Прикрепление»). Аналогичный способ количественного исследования использовали для измерения способности димерных мутированных вариантов головной части белка фибриллы АВ3 блокировать трансдукцию клеток HeLa вектором AB3-GFP. Трансдукцию измеряли на основании экспрессии GFP (Фиг. 3b). Аналогично тому, что наблюдали в исследовании связывания, инфекция AB3-GFP была минимально снижена при предварительной инкубации с мутированными вариантами D261N и F265L, следующими были мутированные варианты N186D, S190P, L296R и V189G. При совокупном анализе данных по связыванию с DSG2 (вестерн-блоттинг), конкурентному связыванию и инфекции был сделан вывод, что область, содержащая остатки с 261 по 265, является наиболее важной областью для связывания с DSG2. Область вокруг остатков 186-190 также вносит вклад в связывание, в то время как область, содержащая остаток 299, по-видимому, лишь незначительно участвует в связывании. Димерные мутированные варианты головной части белка АВ3, содержащие комбинированные мутации, также исследовали для определения их способности конкурировать с вирусом АВ3 за прикрепление (Фиг. 3с) и инфекцию (Фиг. 3d). Мутированный вариант с мутациями во всех трех областях (N185, D261, L296) не блокировал связывание или инфекцию АВ3 даже в концентрациях 200 мкг/мл, это указывает на то, что он практически не устранял связывание с DSG2. Следует отметить, что при использовании sCD46 в качестве зонда для вестерн-блоттинга фибриллы АВ3 дикого типа не наблюдали специфичного связывания (Фиг. 4). Этот результат указывает на то, что АВ3 неэффективно связывается с CD46.
Соотнесение уменьшенного связывания с DSG2 и более слабой способности размыкать эпителиальные контакты. Несложное количественное исследование способности димерных мутированных вариантов головной части белка фибриллы АВ3 размыкать межклеточные контакты основано на измерении трансэпителиального электрического сопротивления (TEER) в клеточных культурах, культивируемых в планшетах Transwell. Эпителиальные раковые клетки культивируют до достижения стабильных показателей TEER, когда образуется большая часть межклеточных контактов. Добавление JO-1 в течение 1 часа к апикальной стороне культур, выращенных в планшетах Transwell, привело к быстрому уменьшению величины TEER, что указывает на размыкание контактов (Фиг. 5а). Инкубация с мутированным вариантом D261N не оказывала влияния на TEER. N186D и E299V оказывали промежуточное действие, которое коррелировало с остаточным связыванием соответствующих головных доменов с DSG2. В предыдущих исследованиях авторы настоящего изобретения также установили, что JO-1 индуцировал изменения в эпителиальных контактах ксенотрансплантатов опухолей и повышал противоопухолевую эффективность высокомолекулярных химиотерапевтических агентов, например иринотекана. (Иринотекан имеет молекулярную массу 586,7 Да и используется для лечения рака толстой кишки и рака легких). Этот эффект был использован для оценки функции димерных мутированных вариантов головной части белка АВ3 в условиях in vivo (Фиг. 5b). Аналогично действию, которое наблюдали в условиях in vitro, JO-1 усиливал эффективность терапии иринотеканом, в то время как мутированные варианты с уменьшенным связыванием с DSG2 не оказывали существенного влияния на эффективность иринотекана.
Димерные головные части белка фибриллы АВ3 с повышенной аффинностью. Как было указано выше, JO-1 имеет практическое значение для терапии рака. Следовательно, очень важно лучше понять структурные особенности его взаимодействия с DSG2 и создать мутированные варианты JO-1 с повышенной аффинностью в отношении DSG2. Повышение аффинности биопрепаратов используют для: i) уменьшения их эффективной дозы, ii) увеличения их периода полувыведения, iii) потенциального увеличения эффективности их терапевтического действия, и iv) устранения нежелательных явлений, вызванных антителами, которые вырабатываются у пациентов в ответ на введение биопрепаратов (например, нейтрализация или изменение фармакокинетических параметров). Для получения аналогов JO-1 с повышенной аффинностью проводили скрининг библиотеки экспрессии Е. coli, содержащей случайные мутации в JO-1, для выявления вариантов с повышенным связыванием с DSG2. Из 10000 высеянных колоний отобрали двадцать колоний с наиболее интенсивными сигналами DSG2, и ДНК плазмиды секвенировали. Было выявлено семь различных мутантов с одной или двумя заменами аминокислот: Y250F, K217E+F224S, N293S, V239D, F224L, E248G+K258E и L277R+N293D. Локализация остатков в первичных и трехмерных структурах головной части белка фибриллы АВ3 показана на Фиг. 6. Следует отметить, что большинство мутаций были локализованы в пределах петли EF, это указывает на то, что эта петля участвует в стабилизации взаимодействия между АВ3 и DSG2. V239 и Y250 не экспонируются на поверхности головной части белка, что скорее указывает на структурные изменения в головном домене, а не непосредственное участие в связывании с DSG2 (Фиг. 6b, правая панель). Рекомбинантные димерные мутированные варианты головной части фибриллы АВ3 затем очищали. Для измерения аффинности мутированных вариантов в отношении DSG2 проводили исследования с помощью поверхностного плазмонного резонанса. Результаты исследований с использованием белков головной части фибриллы, содержащих домен димеризации, были сложными для интерпретации, вероятно, вследствие того, что эти мутированные варианты образовывали многомерные комплексы. Поэтому были проведены исследования с использованием белков головной части фибриллы, не содержащих домена димеризации («noDD»). Величины константы скорости ассоциации (ka или kon) и константы скорости диссоциации (kd или koff), а также KD головной части белка АВ3 дикого типа и всех мутированных вариантов головной части белка приведены на Фиг. 7. В соответствии с данными предыдущих исследований (41), было обнаружено, что головная часть фибриллы АВ3 дикого типа, не содержащая домена димеризации, связывалась с DSG2 только с относительно низкой аффинностью (KD=10 мкМ). За исключением мутированного варианта L227R/noDD+N293D/noDD, все мутированные варианты, выявленные при скрининге колоний с использованием блоттинга, имели более высокую аффинность в отношении DSG2. Следует отметить, что величины аффинности мутированного варианта Y250F/noDD или V239D/noDD были в 885 или 405 раз выше, чем величина аффинности AB3knob/noDD. Высокая аффинность различных мутированных вариантов в основном была обусловлена более быстрым связыванием с DSG2, а не изменением скорости диссоциации. Единственным исключением из этой тенденции был мутированный вариант N293S/noDD, скорость ассоциации которого была самой низкой по сравнению с другими мутированными вариантами. Однако подобная более низкая скорость ассоциации частично компенсировалась более низкой скоростью диссоциации. В совокупности эти результаты указывают на то, что связывание головной части белка фибриллы АВ3 дикого типа (noDD) с DSG2 в основном ограничено более низкой скоростью ассоциации, которая может быть улучшена с помощью панели мутаций. По-видимому, выявленные мутации не изменяют стабильность этого взаимодействия, а смещают баланс ассоциации по отношению к диссоциации, что приводит к повышению аффинности лигандов в отношении рецептора.
Чтобы лучше понять структурные элементы, которые повышают связывание с DSG2, проводили более тщательное исследование с использованием мутированного варианта K217E/F224S. При исследовании с помощью просвечивающей электронной микроскопии мутированного варианта головной части фибриллы K217E/F224S, содержащего домен димеризации, который был окрашен уранилацетатом, были выявлены частицы с 6 головными частями, представляющие димеризованные тримерные фибриллы (Фиг. 8а, толстые стрелки и Фиг. 8b). Интересным фактом является то, что в этих условиях головные части белка фибриллы также образовывали симметричные агрегаты (с диаметром ~30 нм), напоминающие свернутые PtDd (Фиг. 8а, тонкие стрелки и Фиг. 8с). Затем для установления структуры мутированного варианта K217E/F224S на атомном уровне были проведены исследования с помощью рентгеновской кристаллографии (Фиг. 8d-h). Как и ожидалось, мутированный вариант K217E/F224S образовывал монотример из головных частей белка фибриллы (Фиг. 8е). Трехмерную структуру мутированного варианта накладывали на структуру мутированного варианта головной части белка фибриллы АВ3 дикого типа (Фиг. 8f-h). Сопоставление структур выявило, что петля EF в мутированном варианте K217E/F224S имела полностью нарушенную структуру. Указанная петля находится в основании головного домена на стыке с центральным доменом белка фибриллы. Следовательно, мутации K217E/F224S могут обеспечить более легкое связывание за счет увеличения гибкости этой петлевой области.
Соотнесение повышенной аффинности с более сильной способностью размыкать эпителиальные контакты.
Для последующих исследований использовали формы головной части белка фибриллы АВ3, содержащие домен димеризации. Чтобы исследовать отобранные мутированные варианты с высокой аффинностью, проводили исследования конкуренции AB3-GFP и димерных форм мутированных вариантов фибриллы АВ3 с повышенной аффинностью при инфицировании клеток HeLa (Фиг. 9а). На основании экспрессии GFP все димерные мутированные варианты, исключая мутированный вариант L277R+N293D, интибировали инфекцию AB3-GFP в значительно большей степени, чем JO-1. Следует отметить, что недимеризованные формы головной части белка фибриллы АВ3 с повышенной аффинностью в отношении DSG2 не обладали способностью действовать в качестве конкурентов в исследованиях с использованием трансдукции (Фиг. 9b). Более высокая аффинность в отношении DSG2 привела к увеличению способности размыкать эпителиальные контакты в культурах клеток, выращенных в планшетах Transwell (Фиг. 9с). По сравнению с JO-1, величины TEER в культурах, инкубированных с мутированными вариантами V250F, V239D и К217Е + F224S, были значительно выше.
Два варианта JO-1 с повышенной аффинностью, Y293D и V250F, исследовали в условиях in vivo. Указанные мутированные варианты были названы JO-2 и JO-4, соответственно. Первое исследование проводили на ксенотрансплантатной модели, полученной из клеток А549, в условиях, которые были аналогичны таковым в исследовании, описанном для Фиг. 5b. На фигуре 10а показано, что мутированные варианты JO-2 и JO-4 с повышенной аффинностью увеличивали эффективность терапии иринотеканом в значительно большей степени, чем JO-1 (р<0,05, начиная с 27-го дня). Помимо этого JO-4 (KD=11,4 нМ) был значительно более эффективен, чем JO-2 (KD=24,9 нМ), что указывает на корреляцию между аффинностью в отношении DSG2 и терапевтическим действием. Дополнительные исследования проводили на ксенотрансплантатах опухолей, полученных из клеток ovc316 (30, 32). Клетки ovc316 представляют собой HER2/Neu-положительные эпителиальные опухолевые клетки, полученные из биопсии рака яичников. Эти клетки могут подвергаться ЭМП и обратному процессу, мезенхимально-эпителиальному переходу (МЭП), при определенных условиях in vitro и in vivo. Субфракция клеток ovc316, которая является положительной в отношении Nanog, CD133 и Е-кадгерина, обогащена раковыми стволовыми клетками, т.е. самообновляющимися клетками с плюрипотентным потенциалом и способностью образовывать опухоли (30). Следовательно, клетки Ovc316 полностью моделируют гетерогенность и пластичность, наблюдаемую в опухолях in situ. Внутривенная инъекция JO-1 в дозе 2 мг/кг за один час до инъекции пегилированного липосомального доксорубицина (PLD) - препарата, который широко используется для химиотерапии рака яичников, существенно повышала эффективность лечения (Фиг. 10b). Важно отметить, что в дозе 0,5 мг/кг JO-4 оказывал еще больший стимулирующий эффект на терапию PLD. Наконец, JO-4 исследовали в модели тройного отрицательного рака молочной железы (TNBC). TNBC характеризуется отсутствием или минимальной экспрессией рецептора эстрогена (ER), рецептор прогестерона (PR) и отсутствием сверхэкспрессии HER2/Neu. На TNBC приходится 15% всех случаев рака молочной железы. Общая выживаемость является неудовлетворительной по сравнению с таковой у пациентов, которые имеют другие фенотипы. Характерной особенностью TNBC является высокий уровень DSG2 и эпителиальных контактов. Обнадеживающие клинические результаты в лечении TNBC были достигнуты при использовании наночастиц паклитаксела, конъюгированного с альбумином (наб-паклитаксел/абраксан™) по отдельности или в комбинации с мАт к EGFR, цетуксимабом/эрбитуксом™ (38). Результаты исследования авторов настоящего изобретения показали, что JO-4 значительно увеличил эффективность терапии наб-паклитакселом/цетуксимабом в мышиной модели ортотопических опухолей TNBC (Фиг. 10 с). Ввиду его терапевтической значимости, JO-4 также исследовали в соответствующей мышиной опухолевой модели. Поскольку вирус АВ3 и производные головной части белка фибриллы АВ3 не связываются с мышиными клетками и тканями, использовали мышей, трансгенных по гену человека, экспрессирующих человеческий DSG2, характер и уровень экспрессии которого соответствовал таковым у человека (40). Мышам подкожно имплантировали сингенные опухоли TC1-hDSG. Когда опухоли достигали объема ~600 мм3, JO-1 или JO-4 вводили внутривенно для исследований клинической безопасности и эффективности. Концентрации JO-1 и JO-4 в сыворотке снизились более чем на порядок в течение одного часа после инъекции, при этом снижение было значительно более выраженным для JO-4 (р<0,01 в течение 1 ч после инъекции) (Фиг. 11а). Через один час после инъекции концентрации JO-1 и JO-4 достигли плато при ~100 нг/мл. Также были исследованы гематологические параметры после в/в инъекции JO-1 и JO-4. Биохимический анализ крови не выявил патологических изменений. Количество клеток крови соответствовало норме, за исключением количества лимфоцитов и тромбоцитов, которое снизилось вскоре после инъекции (Фиг. 11b). Количество лимфоцитов и тромбоцитов достигло низшей точки через 24 часа после инъекции, причем после инъекции JO-4 уровни лимфоцитов и тромбоцитов были значительно ниже (р<0,01). Интересно отметить, что у мышей, которым вводили JO-4, количество лимфоцитов и тромбоцитов возвращалось к нормальным уровням быстрее, чем у животных, получавших JO-1.
JO-1 и JO-4 представляют собой белки вирусного происхождения и являются иммуногенными. У иммунокомпетентных мышей сывороточные антитела IgG к указанным белкам могут быть детектированы с помощью ИФА через две недели после инъекции (5). Одной из теоретических предпосылок для повышения аффинности терапевтических белков является то, что это позволяет избежать действия нейтрализующих антител в сыворотке крови. Чтобы проверить это предположение, проводили повторные инъекции JO-1 и JO-4 в модели опухоли у иммунокомпетентных мышей hDSG2, несущих опухоли TC1-hDSG2 (Фиг. 11с). Лечение прекращали после двух циклов обработки JO-1 и PLD, и опухолям обеспечивали возможность возобновления роста. 3-й и 4-й циклы лечения начинали на 28-й день и 35-й день, соответственно. Во время 3-го цикла антитела к JO-1 в сыворотке крови детектировали с помощью ИФА. Важно отметить, что в обоих 3-м и 4-м циклах лечения JO-1 и JO-4 усиливали действие терапии PLD, причем усиливающий эффект был значительно сильнее для JO-4.
В совокупности, результаты функциональных исследований с использованием димерных мутированных вариантов фибриллы АВ3 с повышенной аффинностью демонстрируют корреляцию между аффинностью в отношении DSG2 и действием, направленным на размыкание эпителиальных межклеточных контактов, и терапевтическим действием.
Обсуждение
Остатки, вовлеченные в связывание головного домена АВ3 с DSG2. В отличие от взаимодействия АВ с CAR и CD46 (4, 28) структурные параметры, относящиеся к взаимодействию АВ с DSG2, до сих пор не ясны. Несмотря на установление кристаллической структуры головной части белка фибриллы АВ3, для DSG2 доступна лишь трехмерная структура наиболее дистального из четырех внеклеточных доменов (ВКД) (MMDB ID: 59843). Однако результаты предыдущих исследований конкурентного связывания, проведенных авторами настоящего изобретения с использованием моноклональных антител к различным доменам DSG2, показали, что ВКД 3 и 4 участвуют в связывании с АВ3 (42). В данном исследовании авторы настоящего изобретения использовали мутагенез для выявления остатков аминокислот в пределах головной части белка фибриллы АВ3, которые имеют критическое значение для связывания с DSG2. Проведение исследований с использованием мутагенеза DSG2 не представлялось возможным, поскольку при экспрессии в Е. coli белок не связывался с АВ3, это указывает на то, что для создания активных АВ3-связывающих сайтов в белке DSG2 необходимы посттрансляционные модификации (данные не показаны). Выявленные остатки, критические для связывания головной части белка АВ3 с DSG2, находились в трех различных областях головной части белка фибриллы АВ3 и формировали потенциальный карман связывания, локализованный в бороздке на дистальном конце головной части белка фибриллы, обращенной к рецептору. Следует отметить, что связывание других серотипов АВ с CAR или CD46 в первую очередь вовлекает области на боковой или базальной стороне соответствующих головных частей белка фибриллы (27, 43). Данные авторов настоящего изобретения свидетельствуют о том, что АВ3 использует другую стратегию связывания. В настоящее время авторы настоящего изобретения проводят кристаллографические исследования с использованием димерных головных частей фибриллы АВ3 и DSG2. Принимая во внимание, что мультимерные головные части белка фибриллы АВ3 кластеризуют несколько молекул DSG2 (41), следует ожидать, что трехмерная структура этого комплекса будет сложной. Один из вопросов для дальнейшего исследования заключается в установлении того, участвуют ли остатки, критические для связывания головной части белка фибриллы АВ3 с DSG2, в связывании других типов ABB с DSG2. Следует отметить, что, хотя D261, F265 и Е299 сохраняются во всех четырех DSG2-взаимодействующих АВ (АВ3, 7, 11, 14), другие критические остатки (N186, VI89, L296) различаются между этими серотипами (Фиг. 12).
АВ14 является важным объектом исследования из-за недавнего появления нового штамма (АВ14р1). Ранее данный штамм никогда не был задокументирован в Соединенных Штатах Америки, первые сообщения о АВ14р1 появились марте и апреле 2006 года в ходе планового наблюдения в нескольких призывных учебных центрах США (26). В марте - июне следующего года в общей сложности 140 дополнительных случаев подтвержденного респираторного заболевания HABV-B14p1 были зарегистрированы у пациентов в штатах Орегон, Вашингтон и Техас (3). Тридцать восемь процентов этих пациентов были госпитализированы, включая 17% тех, кто был направлен в отделения интенсивной терапии; 5% пациентов умерли. Впоследствии вспышки HABV-B14p1 были обнаружены на других пяти базах и среди гражданского населения в Вашингтоне (1), Орегоне (23), Аляске (15), Висконсине и Пенсильвании (10, 22), а также в Канаде (16), Китае (33) и Южной Корее (34). На данный момент молекулярные основы, лежащие в основе высокой патогенности и/или вирулентности АВ14р1, неясны. Авторы настоящего изобретения предприняли попытку установить структурные компоненты, обуславливающие связывание АВ14р1 с DSG2. Распределение бета-листов в АВ14р1 отличается от такового в АВ3 (Фиг. 1а). Следовательно, аналогично CD46-взаимодействующим серотипам (11, 12), существует возможность того, что DSG2-взаимодействующие АВ различаются по своей стратегии связывания с DSG2, что может обусловить различные области связывания с DSG2. Однако скрининг библиотеки, содержащей мутированные варианты головной части белка фибриллы АВ14р1, не подтвердил эту гипотезу. Области, участвующие в связывании с DSG2, были по существу аналогичны для головной части белка фибриллы АВ3 и АВ14р1. Тем не менее, данные авторов настоящего изобретения имеют значение для лечения вирусемии АВ14р1, в частности, для производства ингибиторов АВ14р1 или высокоаффинных приманок, которые могут индуцировать опсонизацию вируса, присутствующего в кровотоке или дыхательных путях.
Сообщалось, что, помимо DSG2, АВ3 может использовать CD46 как рецептор для инфицирования клеток, если DSG2 отсутствует (35). Ранее авторы настоящего изобретения обнаружили, что в поляризованных нормальных эпителиальных клетках DSG2 заключен внутри плотных контактов и недоступен с апикальной стороны, в то время как CD46 присутствует на обеих сторонах мембраны (42). В этой связи было выдвинуто предположение, что CD46 может играть роль относительно неэффективного рецептора входа для АВ3, в то время как выработанные de novo АВ3 и пентон-додекаэдры АВ3 взаимодействуют с DSG2, размыкают эпителиальные контакты и обеспечивают эффективное латеральное распределение АВ3 или проникновение в более глубокие слои ткани и кровоток. Возможность индивидуального устранения остатков головной части белка АВ3, которые являются критическими для связывания с DSG2 и CD46, соответственно, может позволить доказать выдвинутую гипотезу.
Головные части белка фибриллы с повышенной аффинностью. Большинство мутаций, которые увеличивали аффинность в отношении DSG2, были локализованы в пределах петли EF, это указывает на то, что эта петля участвует в стабилизации взаимодействия между АВ3 и DSG2. Интересно отметить, что, в отличие от АВ7, 11 и 14, головная часть фибриллы АВ3 содержит в этой области два дополнительных остатка (VL), за которыми расположен пролин. Следовательно, эта петля может быть дополнительно продлена, и пролин может ориентировать ее так, чтобы обеспечить лучшее взаимодействие с рецептором. Результаты исследования трехмерной структуры одного из указанных мутированных вариантов на атомном уровне подтверждают этот вывод. Данные этих исследований указывают на то, что введенные мутации делают петлю более гибкой, что может способствовать взаимодействию с DSG2.
Выявление головной части белка АВ3 с более высокой аффинностью, чем у головной части АВ3 дикого типа, имеет практическое применение для АВ3-опосредованной генной терапии. Недавно в клинических испытаниях было показано, что векторы для переноса генов, основанные на АВ3, являются перспективными для терапии рака (18). Теоретически, АВ3-векторы с повышенной аффинностью могут быть использованы в более низких дозах и вытесняют нейтрализующие антитела. В последнее время были предприняты попытки включить высокоаффинные лиганды в вирус кори (17) и векторы на основе АВ5 (3, 9, 45), чтобы повысить эффективность и специфичность инфекции клеток-мишеней в условиях in vivo. На основании результатов, полученных авторами настоящего изобретения в данном исследовании, подобной стратегии теперь можно придерживаться для векторов АВ3.
Помимо улучшения векторов АВ3 варианты JO-1 с повышенной аффинностью имеют значимость для трансляционной медицины. Большинство солидных опухолей имеет эпителиальное происхождение, и, несмотря на то, что злокачественные клетки дедифференцированы, они сохраняют межклеточные контакты, что является ключевой особенностью эпителиальных клеток, как в первичной опухоли, так и в метастатических очагах (5, 31). Межклеточные контакты представляют собой механизм защиты от атак иммунной системы хозяина и создают физические барьеры, которые препятствуют проникновению и распространению противоопухолевых препаратов внутри опухоли, включая моноклональные антитела и химиотерапевтические лекарственные препараты (5, 31). При внутривенном введении мышам с ксенотрансплантатами или сингенными DSG2-трансгенными опухолями JO-1 существенно повышал терапевтический эффект различных химиотерапевтических препаратов, а также моноклональных антител (5, 6). В данном исследовании авторы настоящего изобретения показали, что новые варианты JO-1 с повышенной аффинностью (например, JO-4) увеличивали эффективность противоопухолевых агентов в значительно большей степени, чем JO-1 (иринотекан, наб-паклитаксел, пегилированный липосомальный доксорубицин, цетуксимаб) в четырех моделях опухолей (А549, ovc316, MDA-MB231 и TC1-DSG2). Исследования, проведенные на DSG2-трансгенных мышах с сингенными опухолями, выявили, что сывороточные уровни JO-4 быстро снижаются, вероятнее всего, из-за связывания с DSG2 на поверхности тканей. Предыдущие исследования показали, что, помимо опухолей, лимфоциты и тромбоциты трансгенных мышей hDSG2 экспрессируют hDSG2 (аналогично человеку и обезьянам) (40, 42). В соответствии с этими данными авторы настоящего изобретения обнаружили, что инъекция JO-4 привела к кратковременному уменьшению количества лимфоцитов и тромбоцитов.
Несмотря на тот факт, что примерно одна треть людей имеет нейтрализующие антитела к АВ3 (42), в недавнем исследовании с использованием сыворотки от пациентов, страдающих раком яичников, авторы настоящего изобретения обнаружили детектируемые уровни (связывание) антител к JO-4 лишь у 10% пациентов (N=38) (Фиг. 13). Тем не менее, очевидно, что в организме человека будут развиваться адаптивные иммунные реакции на введенный внутривенно JO-4, в частности, после повторной инъекции. Применительно к этим данным, однако, следует отметить, что антитела к JO-4, которые вырабатываются после инъекции иммунокомпетентным мышам, как оказалось, не оказывают критического ингибиторного действия на функцию JO-4. Данные, приведенные на Фиг. 11с, демонстрируют, что JO-1 и JO-4 остаются эффективными после нескольких циклов лечения, даже в присутствии детектируемых антител. Поскольку терапевтический эффект после повторной инъекции был значительно выше для JO-4, предполагается, что JO-4 является более мощным не только в размыкании межклеточных контактов, но также и в разрушении комплексов между соединением, размыкающим контакты, и сывороточными антителами.
В заключение следует отметить, что исследования авторов настоящего изобретения выявляют важные структурные характеристики связывания головной части белка фибриллы АВ3 и АВ14р1 с DSG2. Также установлена корреляция между аффинностью головной части белка фибриллы АВ3 в отношении DSG2 и последующим действием на эпителиальные контакты. И, наконец, создание рекомбинантных димерных головных частей белка фибриллы АВ3 с повышенной аффинностью имеет практическое значение для терапии рака.
Список литературы
1. 2007. Acute respiratory disease associated with adenovirus serotype 14--four states, 2006-2007. MMWR Morb Mortal Wkly Rep 56:1181-1184.
2. Adams, P.D., P.V. Afonine, G. Bunkoczi, V.B. Chen, I.W. Davis, N. Echols, J.J. Headd, L.W. Hung, G.J. Kapral, R.W. Grosse-Kunstleve, A.J. McCoy, N.W. Moriarty, R. Oeffher, R.J. Read, D.C. Richardson, J.S. Richardson, Т.C. Terwilliger, and P.H. Zwart. 2010. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr 66:213-221.
3. Belousova, N., G. Mikheeva, J. Gelovani, and V. Krasnykh. 2008. Modification of adenovirus capsid with a designed protein ligand yields a gene vector targeted to a major molecular marker of cancer. Journal of virology 82:630-637.
4. Bewley, M.С., K. Springer, Y.B. Zhang, P. Freimuth, and J.M. Flanagan. 1999. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, CAR. Science 286:1579-1583.
5. Beyer, I., H. Cao, J. Persson, H. Song, M. Richter, Q. Feng, R. Yumul, R. van Rensburg, Z. Li, R. Berenson, D. Carter, S. Roffler, C. Drescher, and A. Lieber. 2012. Coadministration of epithelial junction opener JO-1 improves the efficacy and safety of chemotherapeutic drugs. Clin Cancer Res 18:3340-3351.
6. Beyer, I., R. van Rensburg, R. Strauss, Z. Li, H. Wang, J. Persson, R. Yumul, Q. Feng, H. Song, J. Bartek, P. Fender, and A. Lieber. 2011. Epithelial junction opener JO-1 improves monoclonal antibody therapy of cancer. Cancer Res 71:7080-7090.
7. Cadwell, R.C., and G.F. Joyce. 1994. Mutagenic PCR. PCR Methods Appl 3:S136-140.
8. Cadwell, R.C., and G.F. Joyce. 1992. Randomization of genes by PCR mutagenesis. PCR Methods Appl 2:28-33.
9. Campos, S.К., M.B. Parrott, and M.A. Barry. 2004. Avidin-based targeting and purification of a protein IX-modified, metabolically biotinylated adenoviral vector. Mol Ther 9:942-954.
10. Carr, M.J., A.E. Kajon, X. Lu, L. Dunford, P. O'Reilly, P. Holder, C.F. De Gascun, S. Coughlan, J. Connell, D.D. Erdman, and W.W. Hall. 2011. Deaths associated with human adenovirus-14pl infections, Europe, 2009-2010. Emerg Infect Dis 17:1402-1408.
11. Cupelli, K., S. Muller, B. D. Persson, M. Jost, N. Arnberg, and T. Stehle. 2010. Structure of adenovirus type 21 knob in complex with CD46 reveals key differences in receptor contacts among species В adenoviruses. J Virol 84:3189-3200.
12. Cupelli, K., and Т. Stehle. 2011. Viral attachment strategies: the many faces of adenoviruses. Curr Opin Virol 1:84-91.
13. Durmort, С., C. Stehlin, G. Schoehn, A. Mitraki, E. Drouet, S. Cusack, and W.P. Burmeister. 2001. Structure of the fiber head of Ad3, a non-CAR-binding serotype of adenovirus. Virology 285:302-312.
14. Emsley, P., B. Lohkamp, W.G. Scott, and K. Cowtan. 2010. Features and development of Coot. Acta Crystallogr D Biol Crystallogr 66:486-501.
15. Esposito, D.H., T.J. Gardner, E. Schneider, L.J. Stockman, J.E. Tate, C.A. Panozzo, C.L. Robbins, S.A. Jenkerson, L. Thomas, С.M. Watson, A.T. Curns, D.D. Erdman, X. Lu, T. Cromeans, M. Westcott, C. Humphries, J. Ballantyne, G.E. Fischer, J.B. McLaughlin, G. Armstrong, and L. J. Anderson. 2010. Outbreak of pneumonia associated with emergent human adenovirus serotype 14-Southeast Alaska, 2008. J Infect Dis 202:214-222.
16. Girouard, G., R. Garceau, L. Thibault, Y. Oussedik, N. Bastien, and Y. Li. 2013. Adenovirus serotype 14 infection, New Brunswick, Canada, 2011. Emerg Infect Dis 19:119-122.
17. Hasegawa, K., С. Hu, T. Nakamura, J.D. Marks, S.J. Russell, and K.W. Peng. 2007. Affinity thresholds for membrane fusion triggering by viral glycoproteins. Journal of virology 81:13149-13157.
18. Hemminki, О., I. Diaconu, V. Cerullo, S.K. Pesonen, A. Kanerva, T. Joensuu, K. Kairemo, L. Laasonen, K. Partanen, L. Kangasniemi, A. Lieber, S. Pesonen, and A. Hemminki. 2012. Ad3-hTERT-E1A, a Fully Serotype 3 Oncolytic Adenovirus, in Patients With Chemotherapy Refractory Cancer. Mol Ther 20:1821-1830.
19. Incardona, M.F., G.P. Bourenkov, K. Levik, R.A. Pieritz, A.N. Popov, and O. Svensson. 2009. EDNA: a framework for plugin-based applications applied to X-ray experiment online data analysis. J Synchrotron Radiat 16:872-879.
20. Kabsch, W. 2010. Integration, scaling, space-group assignment and post-refinement. Acta Crystallogr D Biol Crystallogr 66:133-144.
21. Kabsch, W. 2010. Xds. Acta Crystallogr D Biol Crystallogr 66:125-132.
22. Kajon, A.E., X. Lu, D.D. Erdman, J. Louie, D. Schnurr, K.S. George, M.P. Koopmans, T. Allibhai, and D. Metzgar. 2010. Molecular epidemiology and brief history of emerging adenovirus 14-associated respiratory disease in the United States. J Infect Dis 202:93-103.
23. Lewis, P.F., M.A. Schmidt, X. Lu, D.D. Erdman, M. Campbell, A. Thomas, P.R. Cieslak, L.D. Grenz, L. Tsaknardis, C. Gleaves, B. Kendall, and D. Gilbert. 2009. A community-based outbreak of severe respiratory illness caused by human adenovirus serotype 14. J Infect Dis 199:1427-1434.
24. Louie, J.K., A.E. Kajon, M. Holodniy, L. Guardia-LaBar, B. Lee, A.M. Petru, J.K. Hacker, and D.P. Schnurr. 2008. Severe pneumonia due to adenovirus serotype 14: a new respiratory threat? Clin Infect Dis 46:421-425.
25. McCoy, A.J., R.W. Grosse-Kunstleve, P.D. Adams, M.D. Winn, L.C. Storoni, and R.J. Read. 2007. Phaser crystallographic software. J Appl Crystallogr 40:658-674.
26. Metzgar, D., M. Osuna, A.E. Kajon, A.W. Hawksworth, M. Irvine, and K.L. Russell. 2007. Abrupt emergence of diverse species В adenoviruses at US military recruit training centers. The Journal of infectious diseases 196:1465-1473.
27. Pache, L., S. Venkataraman, G.R. Nemerow, and V.S. Reddy. 2008. Conservation of fiber structure and CD46 usage by subgroup B2 adenoviruses. Virology 375:573-579.
28. Persson, B.D., D.M. Reiter, M. Marttila, Y.F. Mei, J.M. Casasnovas, N. Arnberg, and T. Stehle. 2007. Adenovirus type 11 binding alters the conformation of its receptor CD46. Nat Struct Mol Biol 14:164-166.
29. Shayakhmetov, D.M., T. Papayannopoulou, G. Stamatoyannopoulos, and A. Lieber. 2000. Efficient gene transfer into human CD34(+) cells by a retargeted adenovirus vector. J Virol 74:2567-2583.
30. Strauss, R., Z.Y. Li, Y. Liu, I. Beyer, J. Persson, P. Sova, T. Moller, S. Pesonen, A. Hemminki, P. Hamerlik, C. Drescher, N. Urban, J. Bartek, and A. Lieber. 2011. Analysis of epithelial and mesenchymal markers in ovarian cancer reveals phenotypic heterogeneity and plasticity. PLoS One 6:e16186.
31. Strauss, R., and A. Lieber. 2009. Anatomical and physical barriers to tumor targeting with oncolytic adenoviruses in vivo. Curr Opin Mol Ther 11:513-522.
32. Strauss, R., P. Sova, Y. Liu, Z.-Y. Li, S. Tuve, D. Pritchard, P. Brinkkoetter, T. Moller, O. Wildner, S. Pesonen, A. Hemminki, N. Urban, C. Drescher, and A. Lieber. 2009. Epithelial phenotype of ovarian cancer mediates resistance to oncolytic adenoviruses. Cancer Research 15:5115-5125.
33. Tang, L., J. An, Z. Xie, S. Dehghan, D. Seto, W. Xu, and Y. Ji. 2013. Genome and Bioinformatic Analysis of a HAdV-B14p1 Virus Isolated from a Baby with Pneumonia in Beijing, China. PLoS One 8:e60345.
34. Trei, J.S., N.M. Johns, J.L. Garner, L.B. Noel, В.V. Ortman, K.L. Ensz, M.C. Johns, M.L. Bunning, and J.C. Gaydos. 2010. Spread of adenovirus to geographically dispersed military installations, May-October 2007. Emerg Infect Dis 16:769-775.
35. Trinh, H.V., G. Lesage, V. Chennamparampil, B. Vollenweider, C.J. Burckhardt, S. Schauer, M. Havenga, U.F. Greber, and S. Hemmi. 2012. Avidity binding of human adenovirus serotypes 3 and 7 to the membrane cofactor CD46 triggers infection. J Virol 86:1623-1637.
36. Tuve, S., В.M. Chen, Y. Liu, T.L. Cheng, P. Toure, P.S. Sow, Q. Feng, N. Kiviat, R. Strauss, S. Ni, Z.Y. Li, S.R. Roffler, and A. Lieber. 2007. Combination of tumor site-located CTL-associated antigen-4 blockade and systemic regulatory T-cell depletion induces tumor-destructive immune responses. Cancer Res 67:5929-5939.
37. Tuve, S., H. Wang, C. Ware, Y. Liu, A. Gaggar, K. Bernt, D. Shayakhmetov, Z. Li, R. Strauss, D. Stone, and A. Lieber. 2006. A new group В adenovirus receptor is expressed at high levels on human stem and tumor cells. J Virol 80:12109-12120.
38. Ueno, N.Т., and D. Zhang. 2011. Targeting EGFR in Triple Negative Breast Cancer. J Cancer 2:324-328.
39. Walters, R.W., P. Freimuth, Т.O. Moninger, I. Ganske, J. Zabner, and M.J. Welsh. 2002. Adenovirus fiber disrupts CAR-mediated intercellular adhesion allowing virus escape. Cell 110:789-799.
40. Wang, H., I. Beyer, J. Persson, H. Song, Z. Li, M. Richter, H. Cao, R. van Rensburg, X. Yao, K. Hudkins, R. Yumul, X.B. Zhang, M. Yu, P. Fender, A. Hemminki, and A. Lieber. 2012. A new human DSG2-transgenic mouse model for studying the tropism and pathology of human adenoviruses. J Virol 86:6286-6302.
41. Wang, H., Z. Li, R. Yumul, S. Lara, A. Hemminki, P. Fender, and A. Lieber. 2011. Multimerization of adenovirus serotype 3 fiber knob domains is required for efficient binding of virus to desmoglein 2 and subsequent opening of epithelial junctions. J Virol 85:6390-6402.
42. Wang, H., Z.Y. Li, Y. Liu, J. Persson, I. Beyer, T. Moller, D. Koyuncu, M.R. Drescher, R. Strauss, X.B. Zhang, J.K. Wahl, 3rd, N. Urban, C. Drescher, A. Hemminki, P. Fender, and A. Lieber. 2011. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat Med 17:96-104.
43. Wang, H., Y.C. Liaw, D. Stone, O. Kalyuzhniy, I. Amiraslanov, S. Tuve, C.L. Verlinde, D. Shayakhmetov, T. Stehle, S. Roffler, and A. Lieber. 2007. Identification of CD46 binding sites within the adenovirus serotype 35 fiber knob. J Virol 81:12785-12792.
44. Wang, H., S. Tuve, D.D. Erdman, and A. Lieber. 2009. Receptor usage of a newly emergent adenovirus type 14. Virology 387:436-441.
45. Zeng, Y., M. Pinard, J. Jaime, L. Bourget, P. Uyen Le, M.D. O'Connor-McCourt, R. Gilbert, and B. Massie. 2008. A ligand-pseudoreceptor system based on de novo designed peptides for the generation of adenoviral vectors with altered tropism. The journal of gene medicine 10:355-367.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТАГОНИСТЫ ALK1 И ИХ ПРИМЕНЕНИЕ В ЛЕЧЕНИИ ПОЧЕЧНО-КЛЕТОЧНОЙ КАРЦИНОМЫ | 2013 |
|
RU2633638C2 |
| АДРЕСНОЕ ВОЗДЕЙСТВИЕ НА УСТОЙЧИВЫЙ К ТРАСТУЗУМАБУ HER2+ РАК МОЛОЧНОЙ ЖЕЛЕЗЫ ПОСРЕДСТВОМ HER3-НАЦЕЛЕННЫХ НАНОЧАСТИЦ | 2015 |
|
RU2692104C2 |
| СПЕЦИФИЧЕСКИЕ К ОПУХОЛИ АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2595403C2 |
| IL-1бета-СВЯЗЫВАЮЩИЕ АНТИТЕЛА И ИХ ФРАГМЕНТЫ | 2006 |
|
RU2518295C2 |
| РЕЦЕПТОР-НАПРАВЛЕННЫЕ КОНСТРУКЦИИ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2682335C2 |
| ШАТТЛ ДЛЯ ГЕМАТОЭНЦЕФАЛИЧЕСКОГО БАРЬЕРА | 2013 |
|
RU2711552C2 |
| ИДЕНТИФИКАЦИЯ, ОЦЕНКА И ЛЕЧЕНИЕ РАКОВЫХ ЗАБОЛЕВАНИЙ С ГЕНЕТИЧЕСКОЙ ИЛИ ПРИОБРЕТЕННОЙ УСТОЙЧИВОСТЬЮ К ИНГИБИТОРАМ ALK | 2011 |
|
RU2589834C2 |
| ИНГИБИТОРЫ АНГИОПОЭТИНПОДОБНОГО БЕЛКА 4, ИХ КОМБИНАЦИИ И ПРИМЕНЕНИЕ | 2005 |
|
RU2392966C2 |
| ШАТТЛ ДЛЯ ГЕМАТОЭНЦЕФАЛИЧЕСКОГО БАРЬЕРА | 2013 |
|
RU2663120C2 |
| ВАРИАНТЫ БЕТА-ЦЕПИ HGF | 2006 |
|
RU2403261C2 |
































































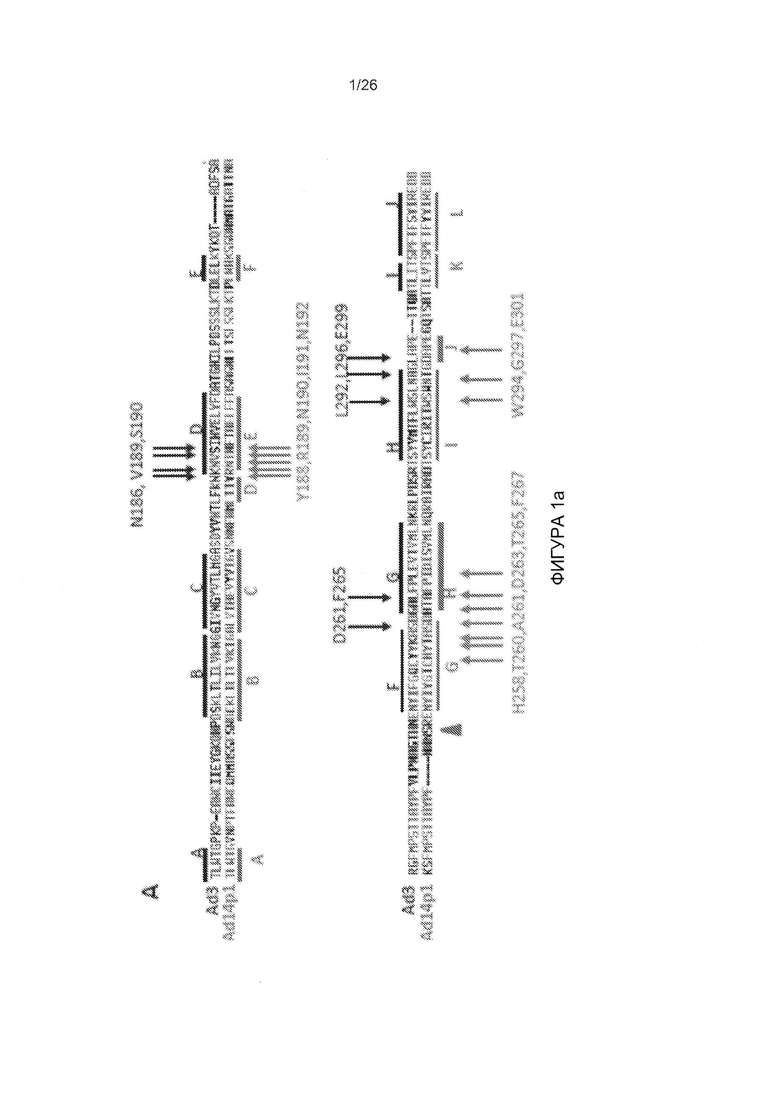
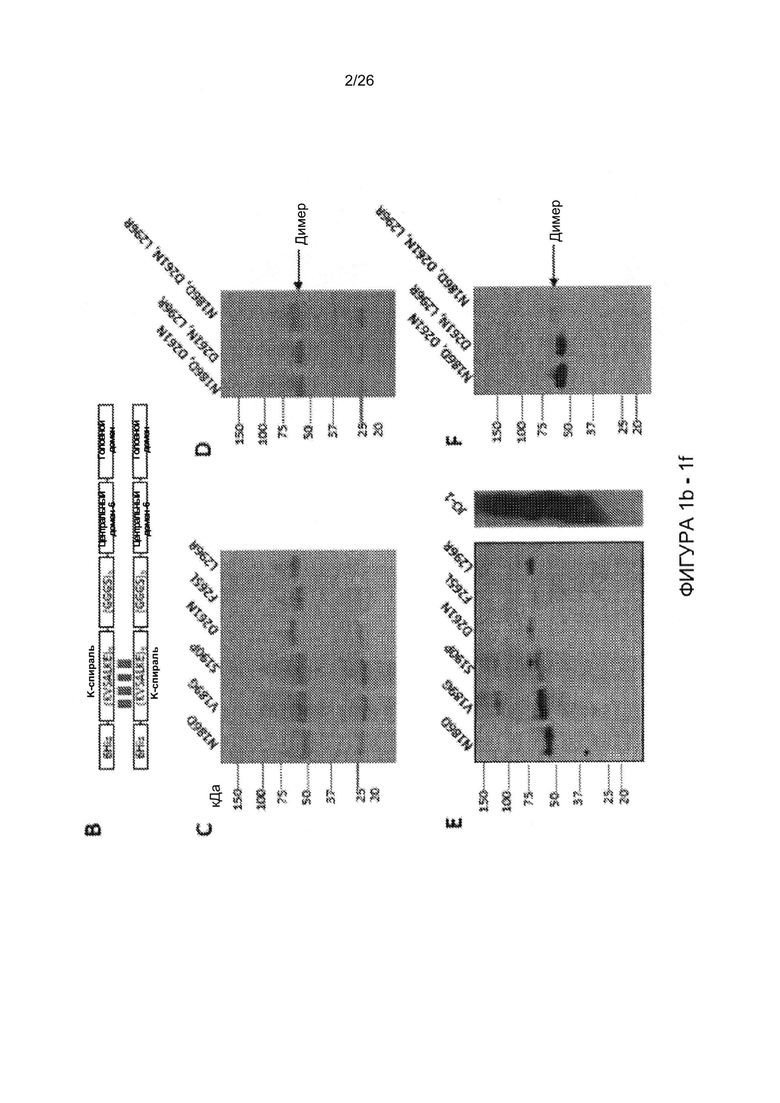
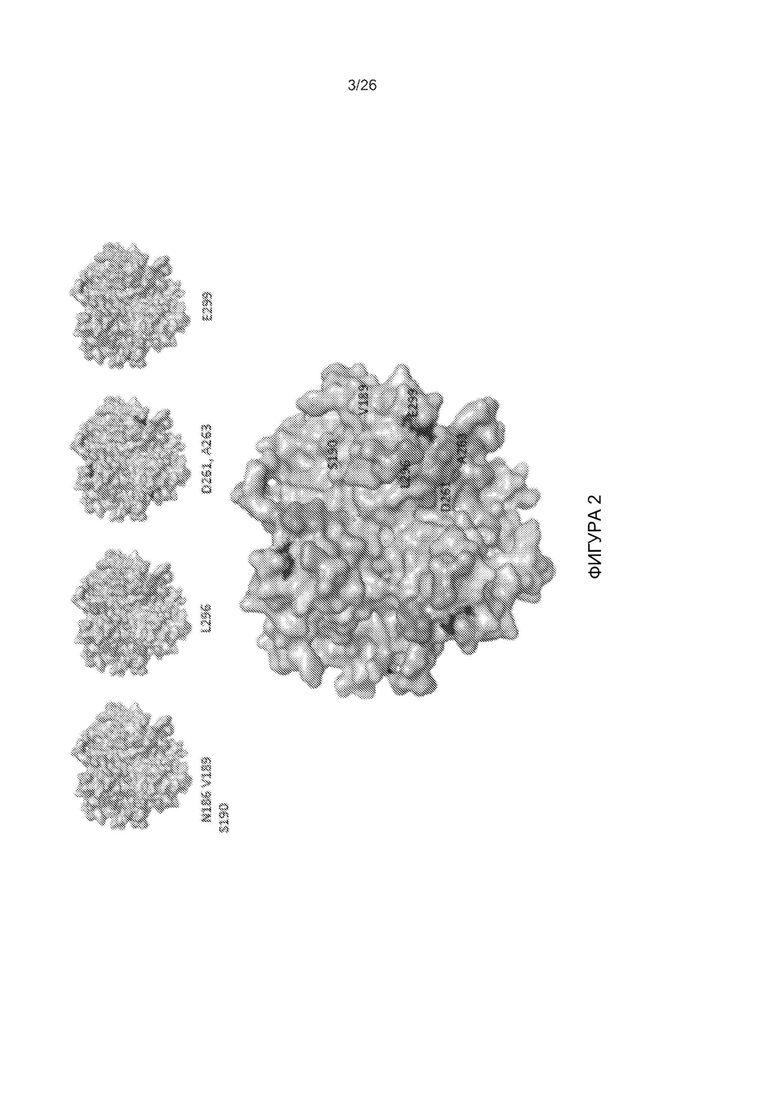
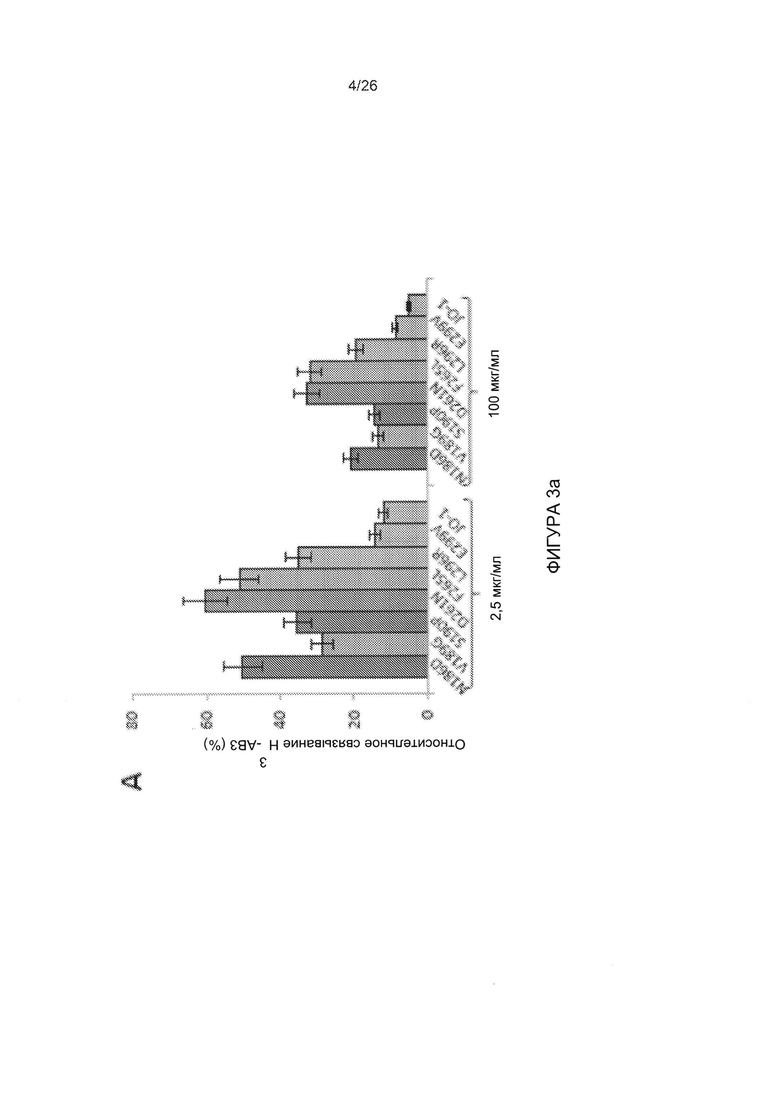
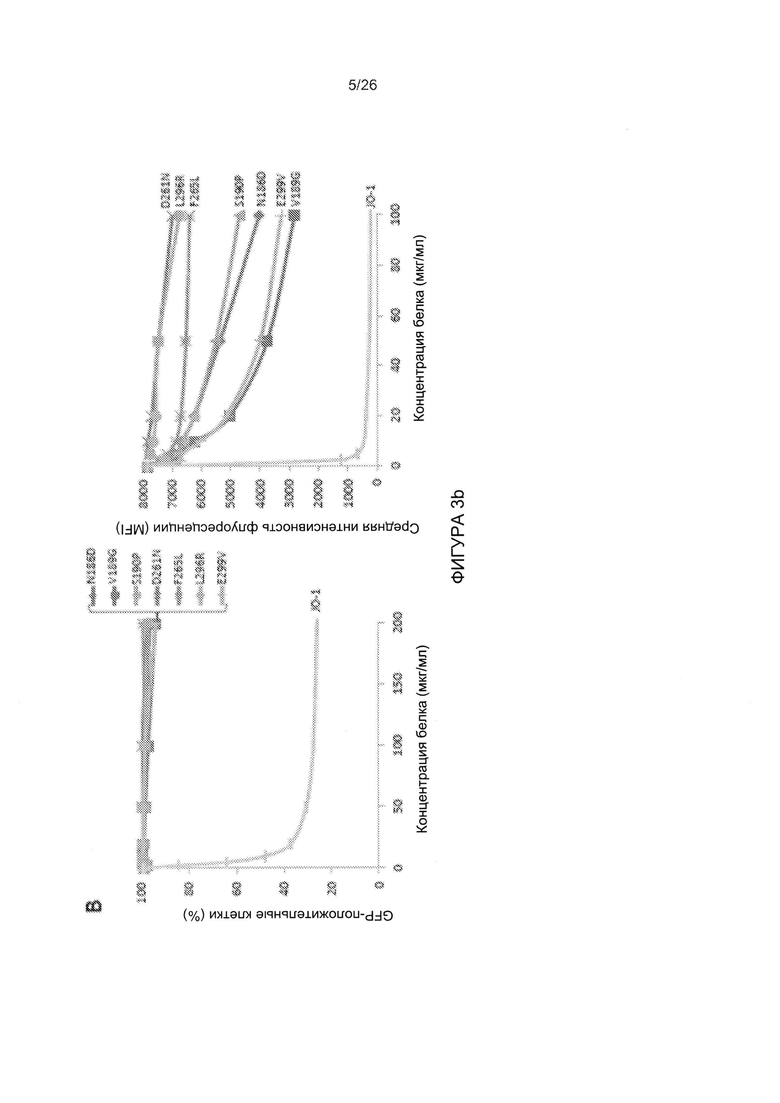
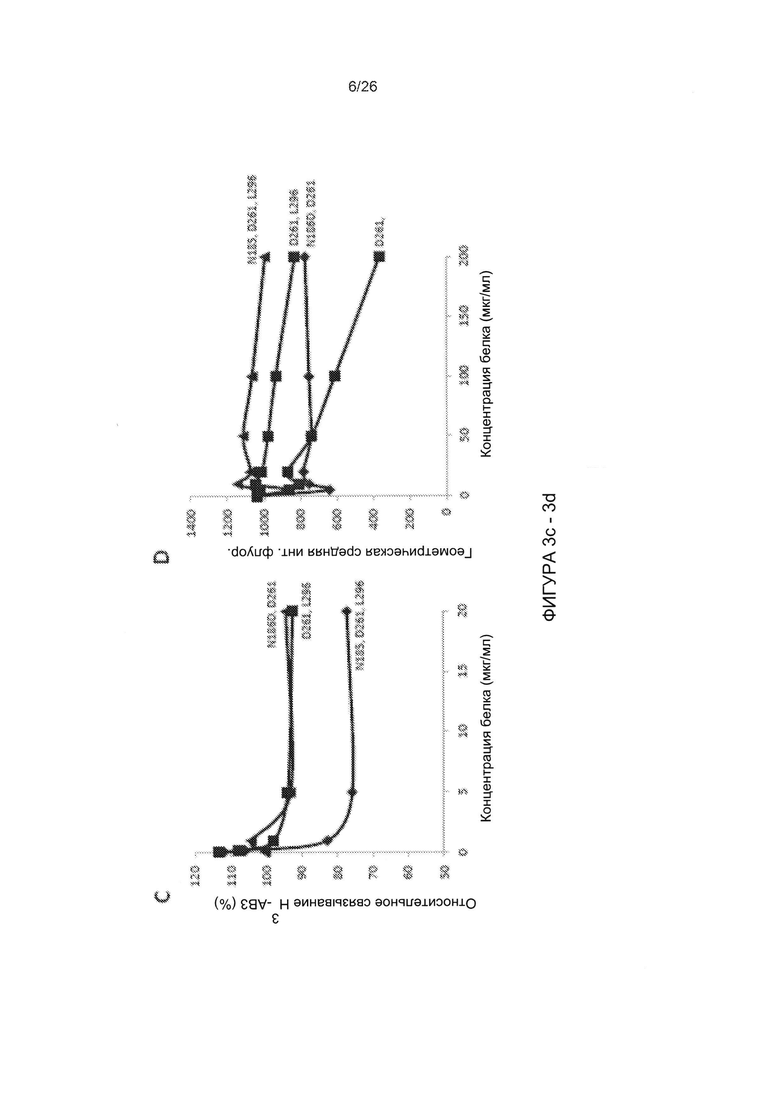
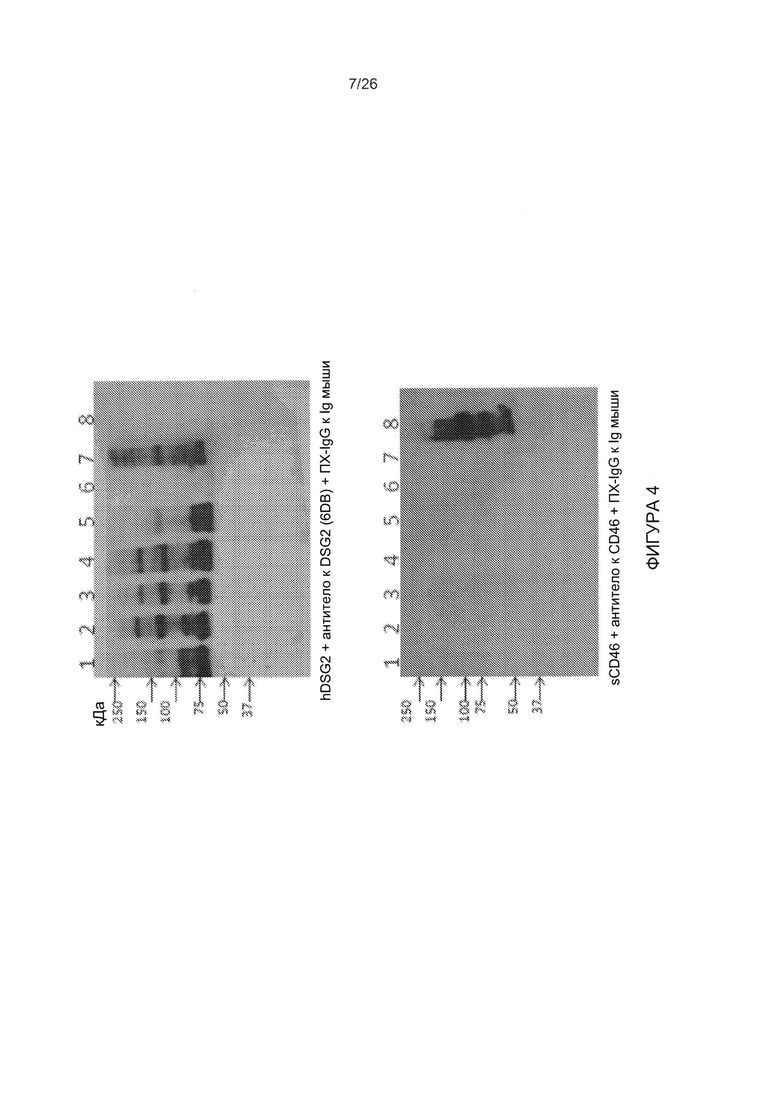
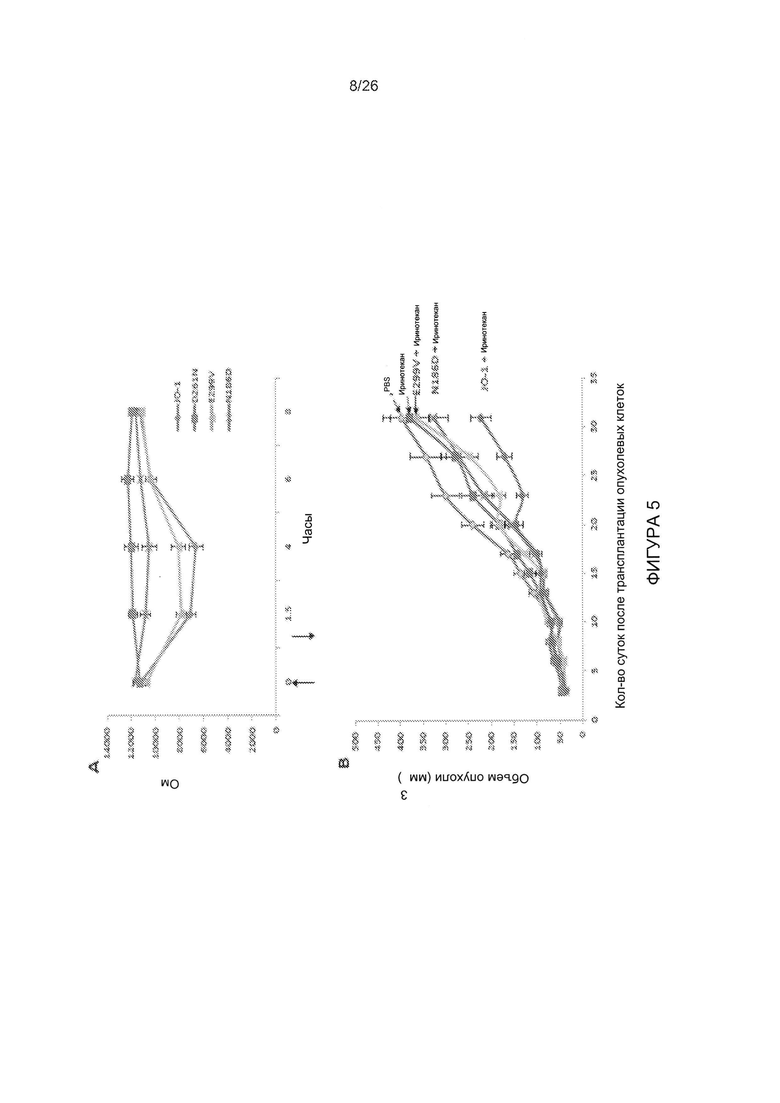
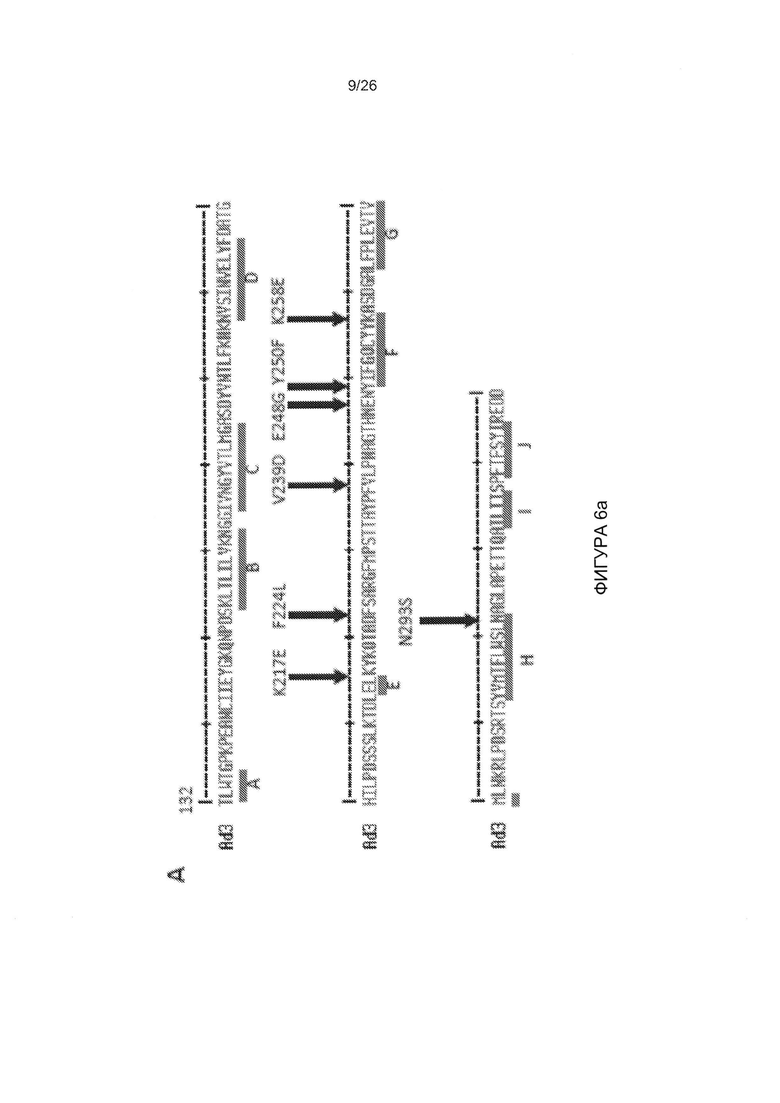

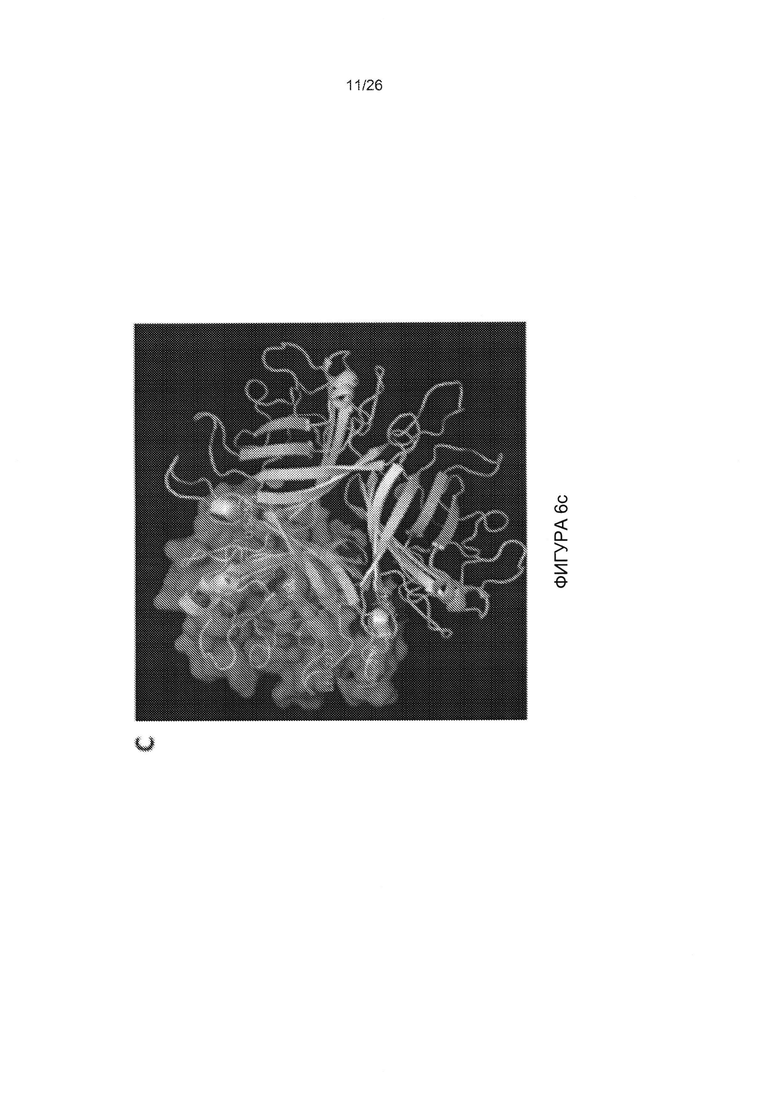
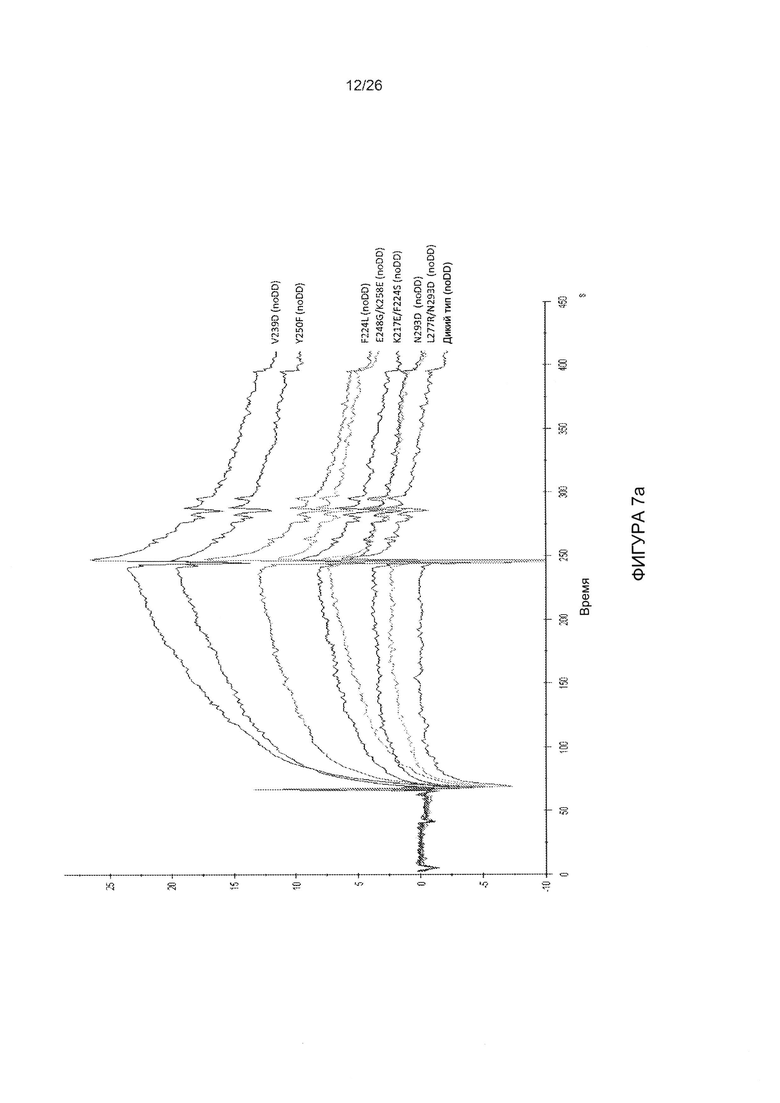
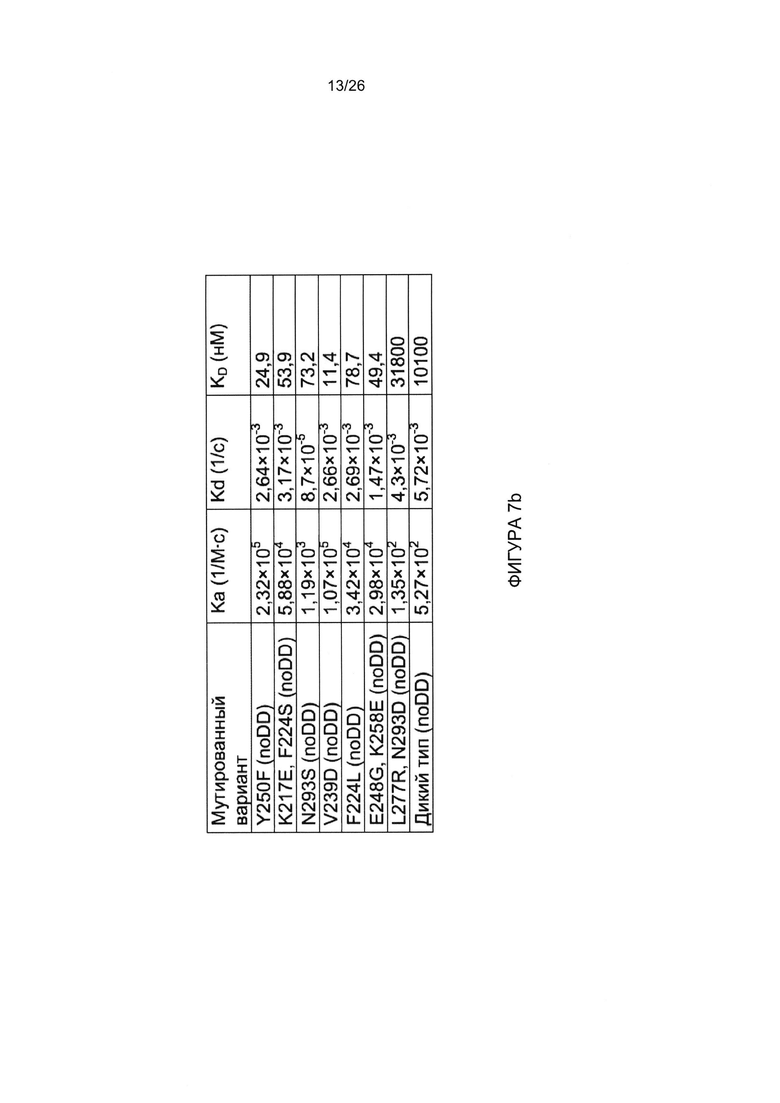
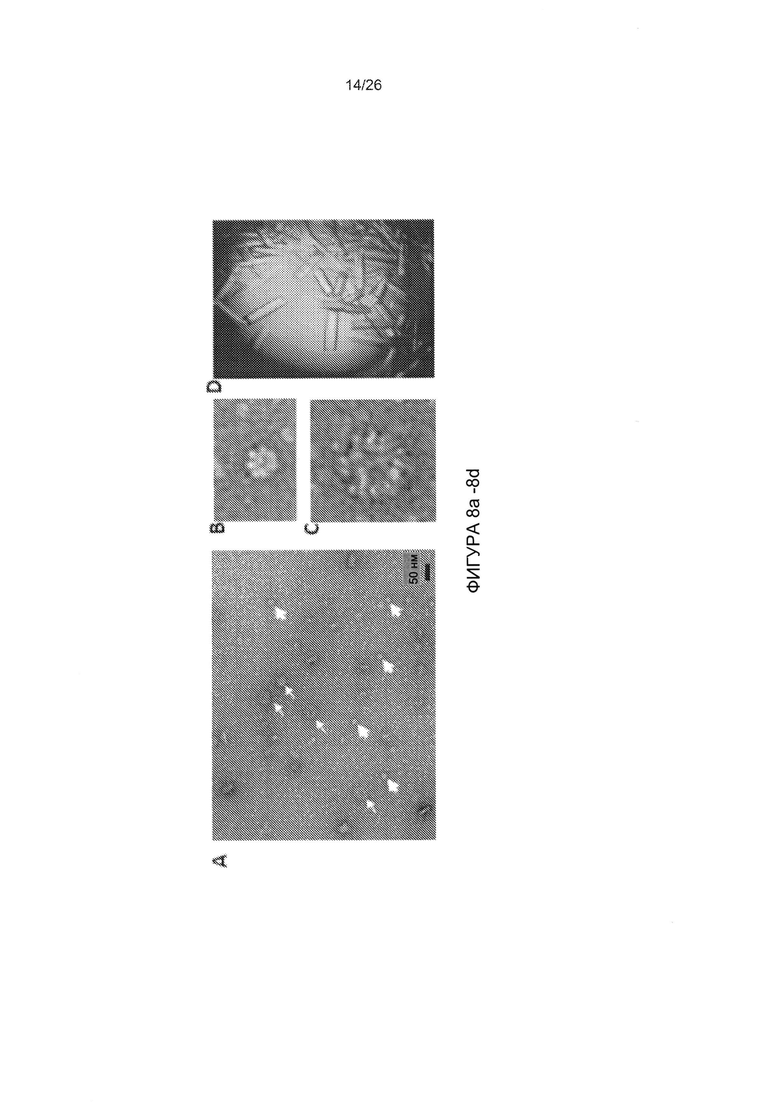
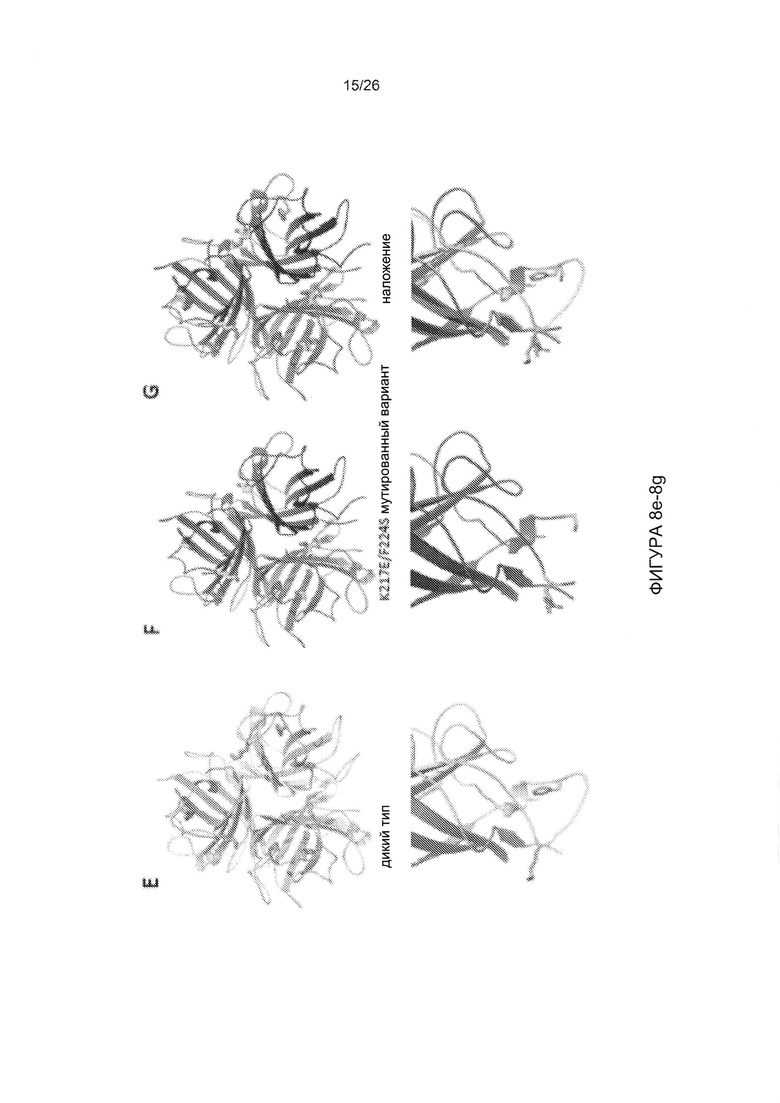
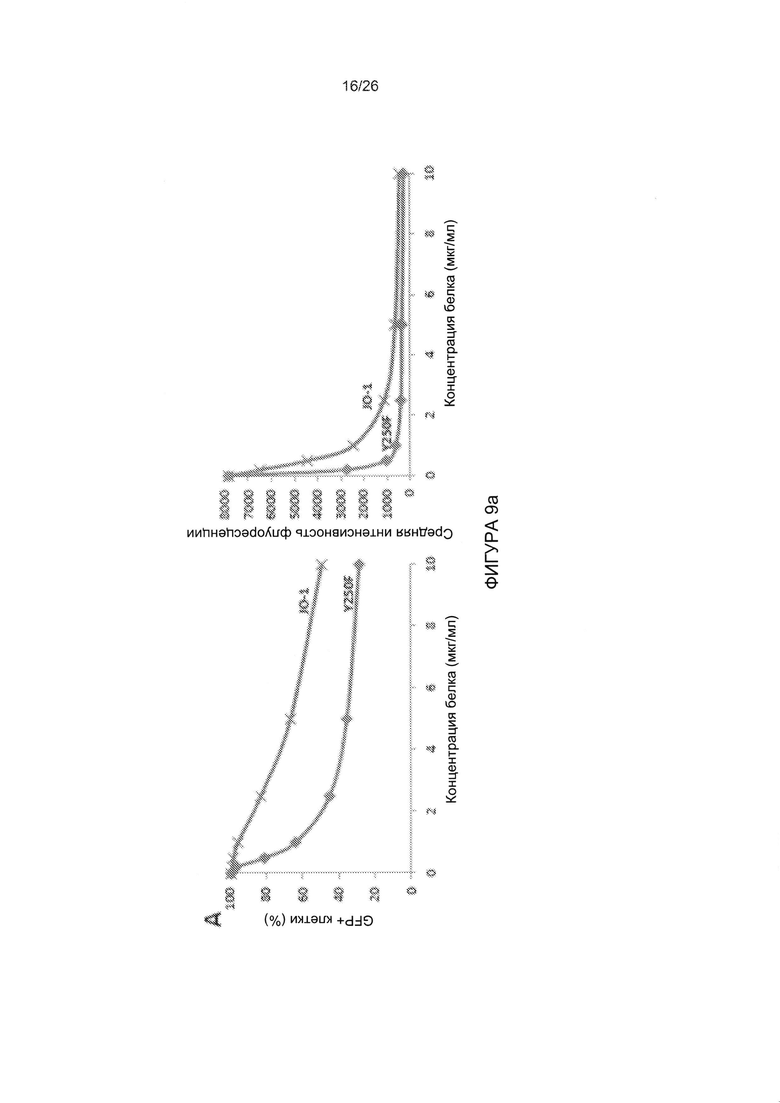
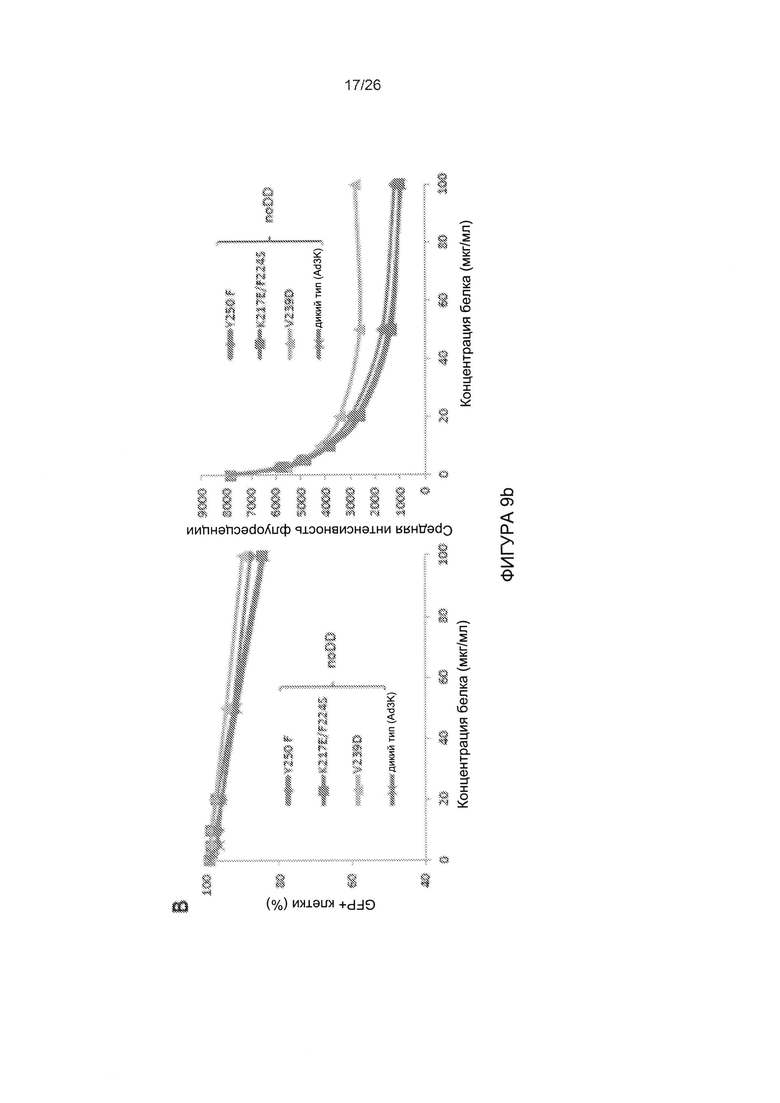
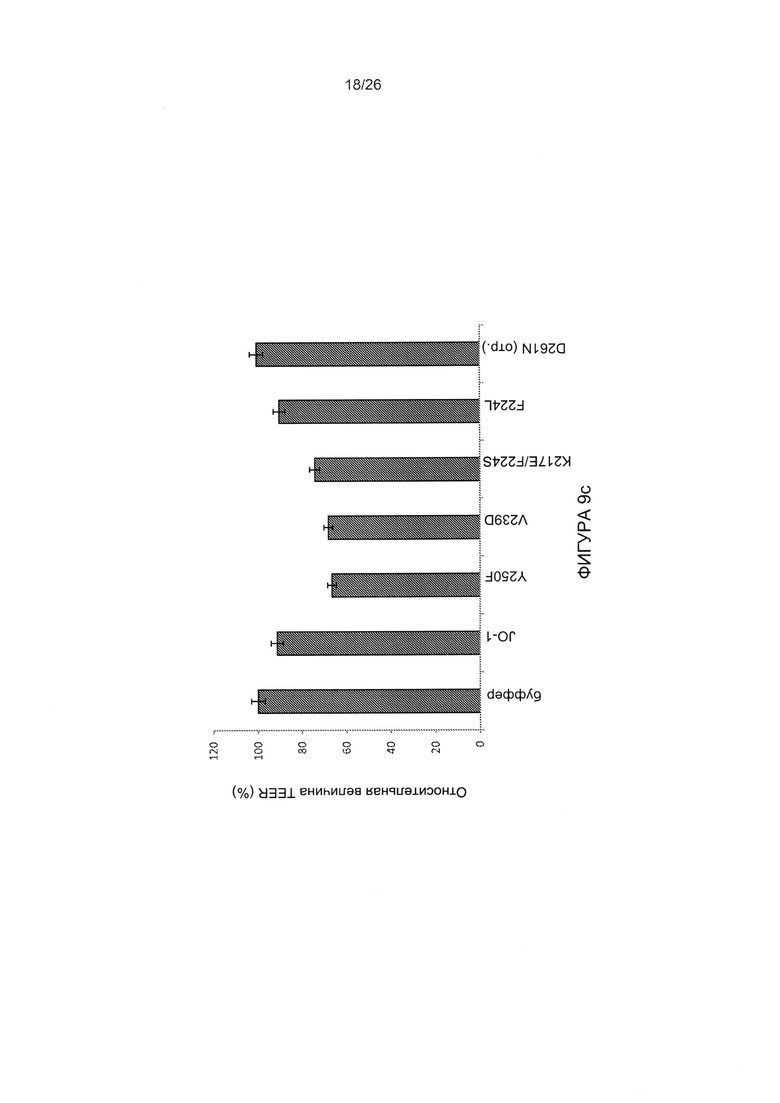

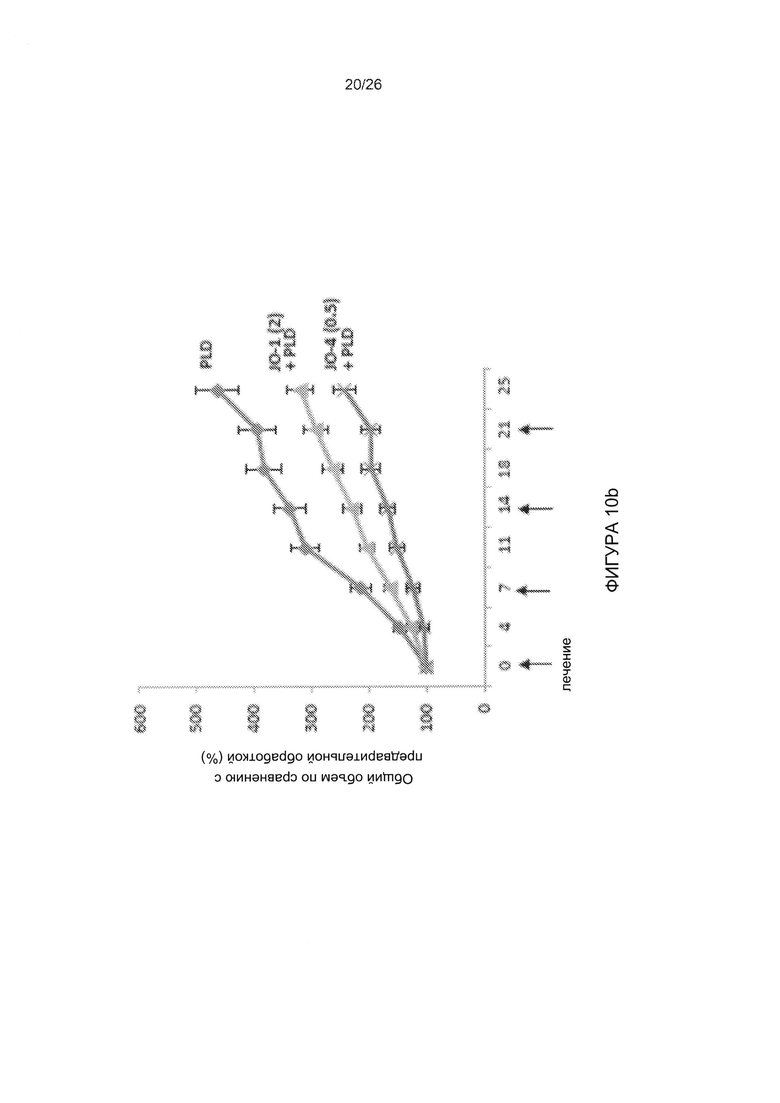
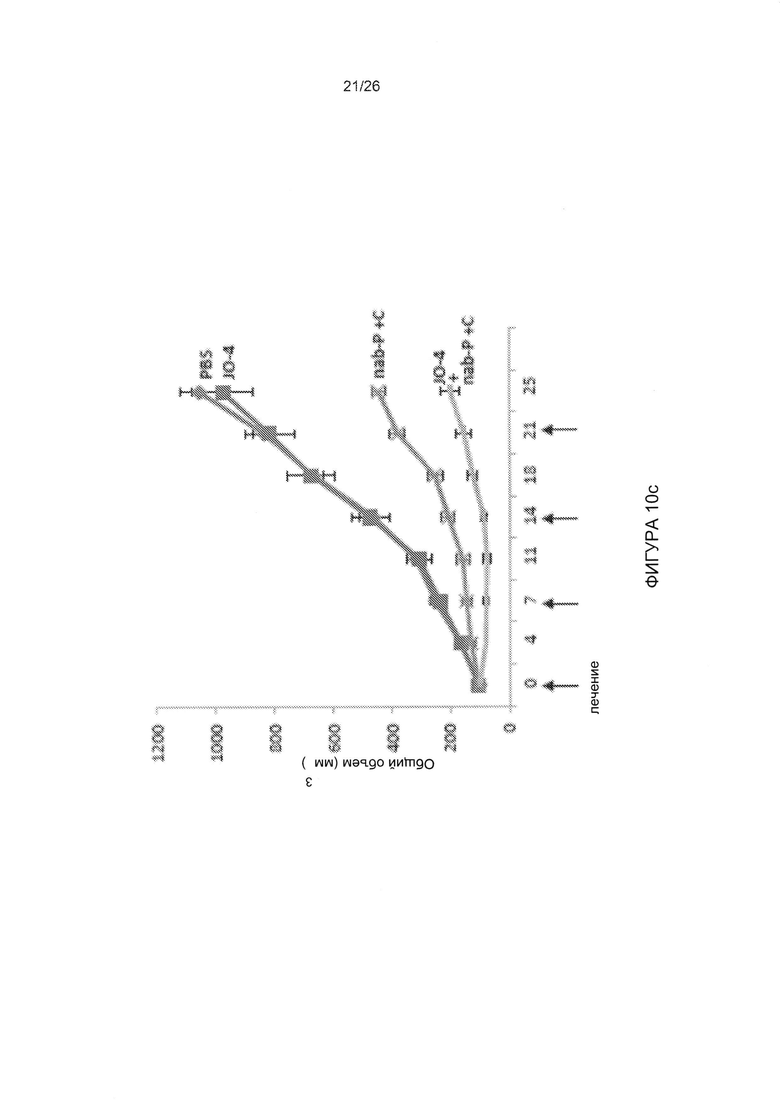
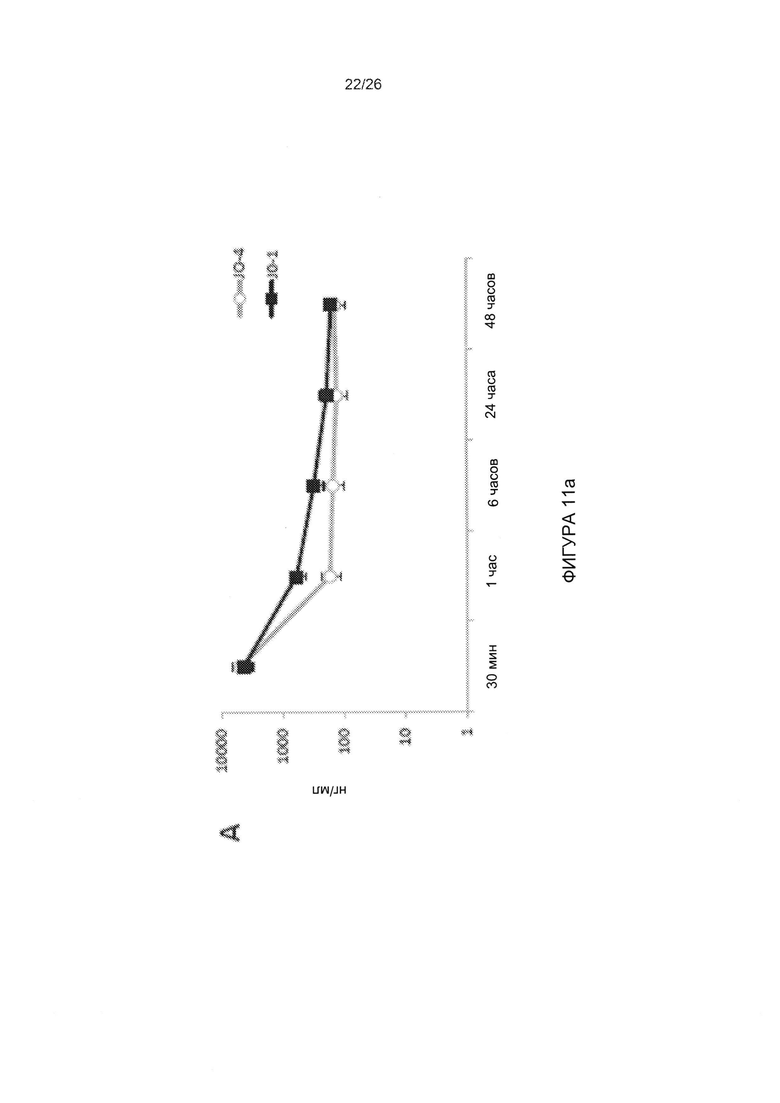
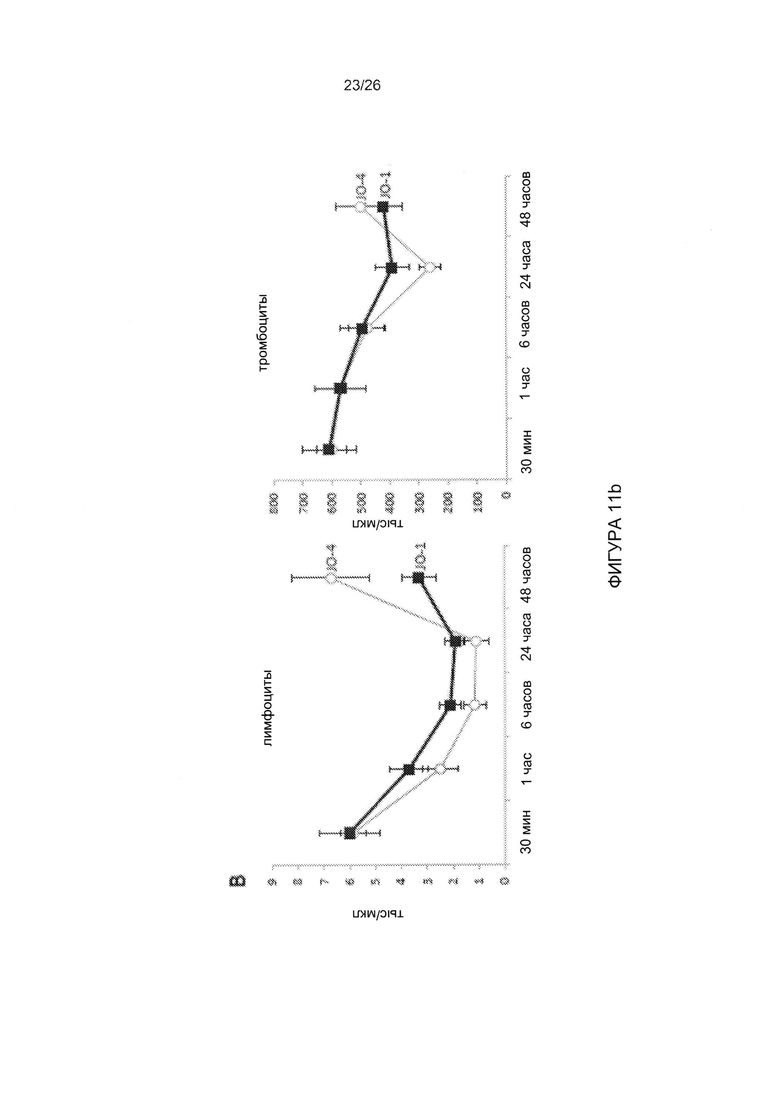
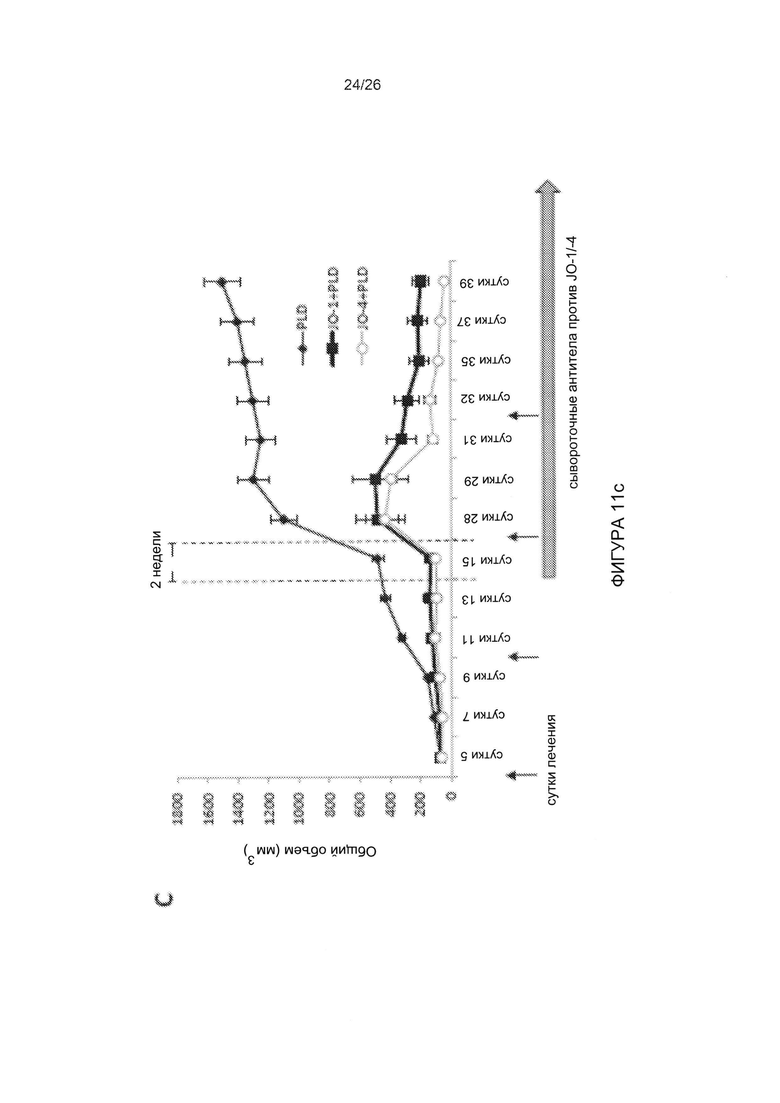
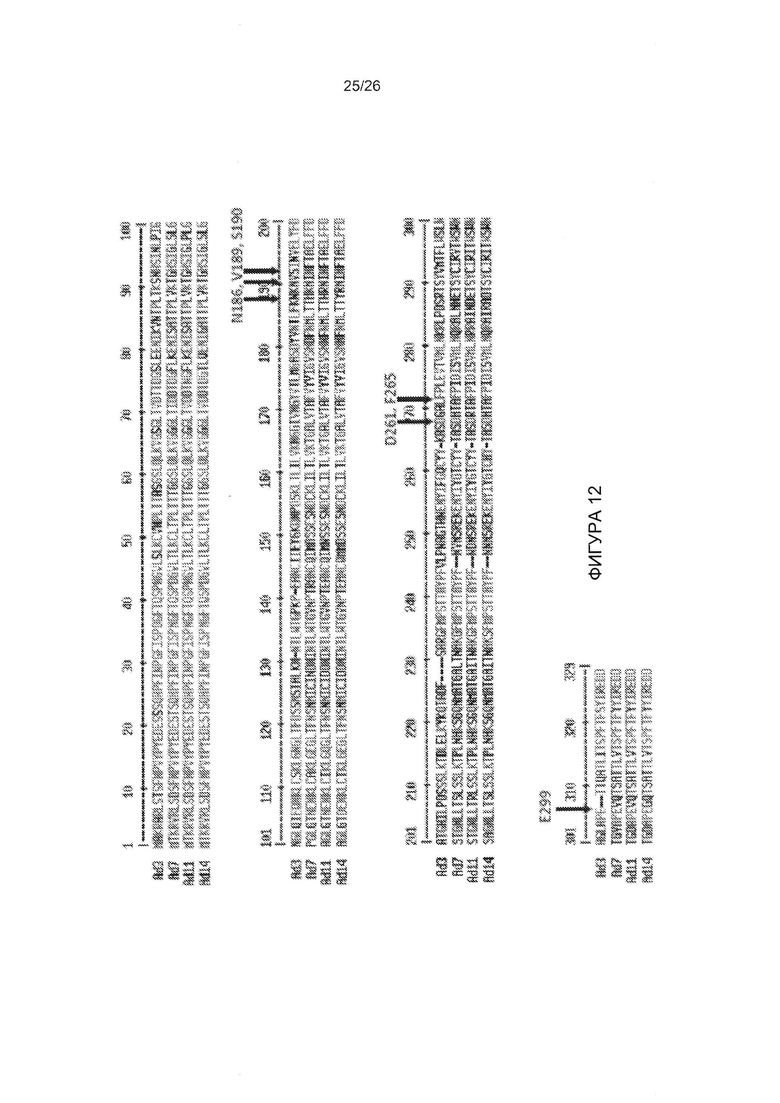
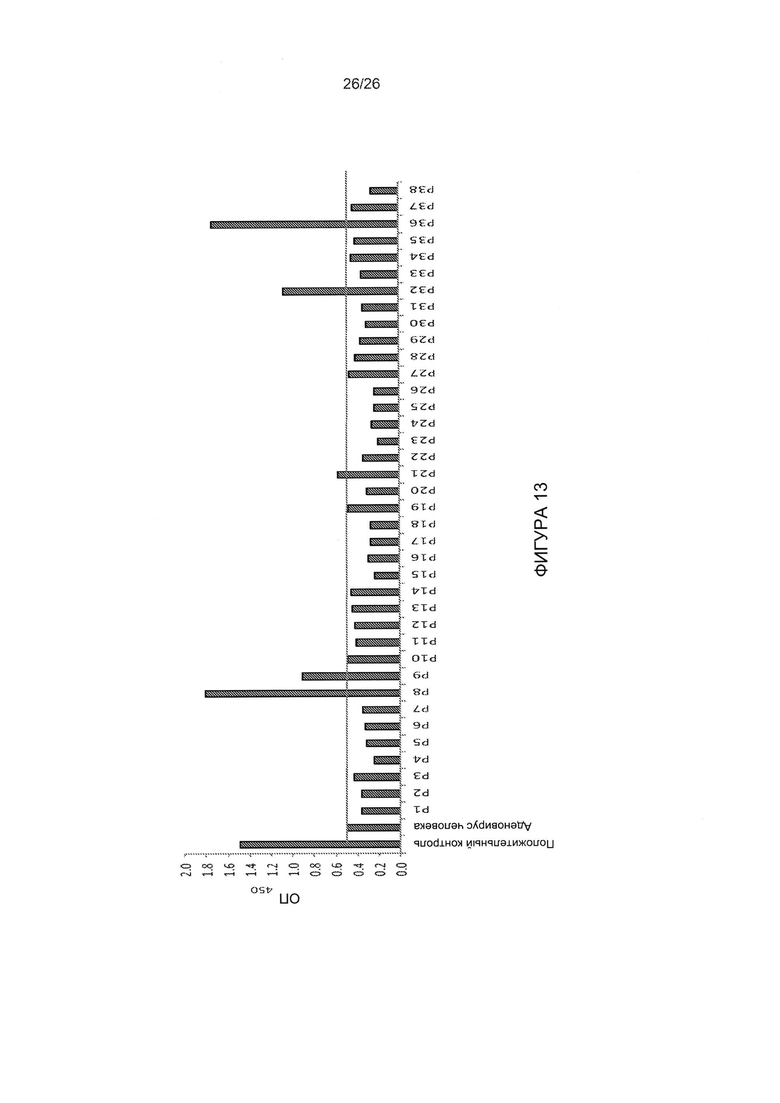
Изобретение относится к области генной инженерии, конкретно к получению рекомбинантных полипептидов аденовируса, и может быть использовано в медицине для увеличения эффективности терапевтического лечения солидной опухоли, экспрессирующей десмоглеин 2 (DSG2). Рекомбинантным путем получают полипептид фибриллы аденовируса типа В 2/3 (АВВ-2/3), содержащий мотив центрального домена полипептида фибриллы АВВ-2/3, головной домен полипептида фибриллы АВВ-2/3, функционально связанный и расположенный в направлении С-конца по отношению к указанному мотиву центрального домена полипептида фибриллы АВВ-2/3, и домен димеризации, полученный не из вируса АВВ-2/3, расположенный в направлении N-конца по отношению к указанному мотиву центрального домена полипептида фибриллы АВВ-2/3. На основе мультимеров указанного полипептида получают конъюгат с цитотоксическим соединением. Изобретение позволяет получить рекомбинантный полипептид с увеличенным связыванием с DSG2 по сравнению с аденовирусом типа В 2/3 дикого типа. 7 н. и 11 з.п. ф-лы, 13 ил., 2 табл., 1 пр.
1. Рекомбинантный полипептид фибриллы аденовируса типа В 2/3 (АВВ-2/3) с увеличенным связыванием с десмоглеином 2 (DSG2) по сравнению с аденовирусом типа В 2/3 дикого типа, содержащий:
a) один или более мотивов центрального домена полипептида фибриллы АВВ-2/3, при этом каждый мотив центрального домена содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 12-18 и 43-48;
b) головной домен полипептида фибриллы АВВ-2/3, функционально связанный и расположенный в направлении С-конца по отношению к указанным одному или более мотивам центрального домена полипептида фибриллы АВВ-2/3, причем указанный головной домен полипептида фибриллы АВВ-2/3 представляет собой пептид, выбранный из любой из последовательностей SEQ ID NO: 5-8 и 10-11; и
c) один или более доменов димеризации, полученных не из вируса АВВ-2/3, расположенных в направлении N-конца по отношению к указанным одному или более мотивам центрального домена полипептида фибриллы АВВ-2/3.
2. Рекомбинантный полипептид фибриллы АВВ-2/3 по п. 1, отличающийся тем, что указанный головной домен полипептида фибриллы АВВ-2/3 содержит аминокислотную последовательность SEQ ID NO: 8.
3. Рекомбинантный полипептид фибриллы АВВ-2/3 по п. 1, отличающийся тем, что каждый мотив центрального домена выбран из группы, состоящей из мотива центрального домена полипептида фибриллы АВ3, мотива центрального домена полипептида фибриллы АВ7, мотива центрального домена полипептида фибриллы АВ11, мотива центрального домена полипептида фибриллы АВ14, мотива центрального домена полипептида фибриллы АВ14а и их комбинаций.
4. Рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-3, отличающийся тем, что указанные один или более мотивов центрального домена содержат 1-22 мотива центрального домена.
5. Рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-3, отличающийся тем, что каждый мотив центрального домена содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 43-48.
6. Рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-3, отличающийся тем, что указанный домен димеризации содержит аминокислотную последовательность, выбранную из группы, состоящей из EVSALEK (SEQ ID NO: 24) и/или KVSALKE (SEQ ID NO: 25).
7. Рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-3, отличающийся тем, что указанные один или более мотивов центрального домена представляют собой мотив центрального домена с последовательностью SEQ ID NO: 43.
8. Рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:28-31 и 33-34.
9. Рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-3, отличающийся тем, что указанный полипептид фибриллы АВВ-2/3 содержит один мотив центрального домена полипептида фибриллы АВВ-2/3.
10. Гомотример рекомбинантного полипептида фибриллы аденовируса типа В 2/3 (АВВ-2/3) с увеличенным связыванием с десмоглеином 2 (DSG2) по сравнению с гомотримером аденовируса типа В 2/3 дикого типа, при этом указанный гомотример полипептида фибриллы АВВ-2/3 содержит три идентичных полипептида, где каждый из индивидуальных полипептидов содержит:
a) один или более мотивов центрального домена полипептида фибриллы АВВ-2/3, при этом каждый мотив центрального домена содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 12-18 и 43-48;
b) головной домен полипептида фибриллы АВВ-2/3, функционально связанный и расположенный в направлении С-конца по отношению к указанным одному или более мотивам центрального домена полипептида фибриллы АВВ-2/3, причем указанный головной домен полипептида фибриллы АВВ-2/3 представляет собой пептид, выбранный из любой из последовательностей SEQ ID NO: 5-8 и 10-11; и
с) один или более доменов димеризации, полученных не из вируса АВВ-2/3, расположенных в направлении N-конца по отношению к указанным одному или более мотивам центрального домена полипептида фибриллы АВВ-2/3.
11. Гомотример рекомбинантного полипептида фибриллы АВВ-2/3 по п. 10, отличающийся тем, что указанный головной домен полипептида фибриллы АВВ-2/3 содержит аминокислотную последовательность SEQ ID NO: 8.
12. Гомотример рекомбинантного полипептида фибриллы АВВ-2/3 по п. 10, отличающийся тем, что каждый индивидуальный полипептид содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 28-31 и 33-34.
13. Гомотример рекомбинантного полипептида фибриллы АВВ-2/3 по п. 10, отличающийся тем, что каждый индивидуальный полипептид содержит аминокислотную последовательность SEQ ID NO: 31.
14. Мультимер, содержащий димер гомотримера рекомбинантного полипептида фибриллы АВВ-2/3 по любому из пп. 10-13, с увеличенным связыванием с десмоглеином 2 (DSG2) по сравнению с мультимером гомотримеров аденовируса типа В 2/3 дикого типа.
15. Конъюгат для увеличения эффективности терапевтического лечения солидной опухоли, экспрессирующей DSG2, содержащий:
a) мультимер по п. 14 и
b) по меньшей мере одно цитотоксическое соединение, конъюгированное с указанным мультимером.
16. Выделенная нуклеиновая кислота, кодирующая указанный рекомбинантный полипептид фибриллы АВВ-2/3 по любому из пп. 1-9.
17. Рекомбинантный вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 16.
18. Клетка-хозяин для экспрессии рекомбинантного полипептида фибриллы АВВ-2/3 по любому из пп. 1-9, содержащая рекомбинантный вектор экспрессии по п. 17.
| WANG HONGJIE et al., Multimerization of Adenovirus Serotype 3 Fiber Knob Domains Is Required for Efficient Binding of Virus to Desmoglein 2 and Subsequent Opening of Epithelial Junctions, JOURNAL OF VIROLOGY, 2011, v | |||
| Устройство для выпрямления опрокинувшихся на бок и затонувших у берега судов | 1922 |
|
SU85A1 |
| Насос | 1917 |
|
SU13A1 |
| ТРАНСПОРТИРУЮЩЕЕ ПРИСПОСОБЛЕНИЕ К СНОПОВЯЗАЛКАМ | 1926 |
|
SU6390A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| BEATTY M.S | |||
| et al., Chapter two--Adenovirus strategies for tissue-specific targeting, Adv | |||
| Cancer Res., 2012, v.115, p.39-67 | |||
| WO 2011156761 A1, 15.12.2011; | |||
| АДЕНОВИРУС С ОБРАЩЕННОЙ ГЕННОЙ ЭКСПРЕССИЕЙ И ЕГО ПРИМЕНЕНИЕ | 2003 |
|
RU2393221C2 |

