Родственные заявки
По настоящей заявке испрашивается приоритет предварительной заявки США №61/732452, поданной 3-го декабря 2012 года, предварительной заявки США №61/816788, поданной 28-го апреля 2013 года, предварительной заявки США №61/863106, поданной 7-го августа 2013 года, предварительной заявки США №61/881523, поданной 24-го сентября 2013 года, и предварительной заявки США №61/898710, поданной 1-го ноября 2013 года, содержание которых включено в настоящий документ в качестве ссылки в полном объеме.
Область техники, к которой относится изобретение
Изобретение относится к моноклональным и/или моновалентным антителам, которые связывают CD47. Изобретение относится к моноклональным и/или моновалентным антителам, которые связывают CD19. Изобретение также относится к новым биспецифичным моноклональным антителам, имеющим различную специфичность в каждом участке связывания молекулы иммуноглобулина, где один из участков связывания является специфичным к CD47. Изобретение также относится к новым биспецифичным моноклональным антителам, имеющим различную специфичность в каждом участке связывания молекулы иммуноглобулина, где один из участков связывания является специфичным к CD19.
Предпосылки создания изобретения
CD47 или интегрин-связанный белок (IAP) представляет собой экспрессирующийся во всех клетках трансмембранный гликопротеин весом 50 кД, имеющий множество функций в межклеточных взаимодействиях. Он взаимодействует с множеством лигандов, таких как интегрины, SIRPα (сигнал-регуляторный белок альфа), SIRPγ и тромбоспондины (Oldenborg, P.A., CD47: A Cell Surface Glycoprotein Which Regulates Multiple Functions of Hematopoietic Cells in Health and Disease, ISRN Hematol. 2013; 2013: 614619; Soto-Pantoja DR, et al, Therapeutic opportunities for targeting the ubiquitous cell surface receptor CD47 (2012), Expert Opin Ther Targets. 2013 Jan;17(l): 89-103; Sick E, et al., CD47 Update: a multifaced actor in the tumor micro environment of potential therapeutic interest, Br J Pharmacol. 2012 Dec; 167(7): 1415-30). В контексте системы врожденного иммунитета CD47 функционирует в качестве маркера самого себя, передавая ингибирующий сигнал «не убивай меня» через связывание с SIRPα, экспрессируемым миелоидными клетками, такими как макрофаги, нейтрофилы и дендритные клетки. Поэтому, роль повсеместной экспрессии CD47 в физиологической ситуации заключается в защите здоровых клеток от истребления системой врожденного иммунитета (Oldenborg PA, et al., CD47-Signal Regulatory Protein a (Sirpα) Regulates Fcγ and Complement Receptor-Mediated Phagocytosis, J Exp Med. 2001 Apr 2;193(7):855-62; Mattias Olsson, Role of the CD47/SIRPα-interaction in regulation of macrophage phagocytosis, Department of Integrative Medical Biology, Section for Histology and CellBiology, Umeå University, Umeå, Sweden, Thesis; Oldenborg PA., Role of CD47 in erythroid cells and in autoimmunity, Leuk Lymphoma. 2004 Jul; 45(7): 1319-27; Oldenborg PA, et al., Role of CD47 as a Marker of Self on Red Blood Cells., Science. 2000 Jun 16; 288(5473): 2051-4; Brown EJ, Frazier WA., integrin-associated protein (CD47) and its ligands., Trends Cell Biol. 2001 Mar; 11(3): 130-5).
Опухолевые клетки используют этот механизм иммуносупрессии путем сверхэкспрессии CD47, который успешно позволяет им избегать иммунологического надзора и уничтожения клетками врожденной иммунной системы. (Majeti R, Chao et al., CD47 is an adverse prognostic factor and therapeutic antibody target on human acute myeloid leukemia stem cells, Cell. 2009 Jul 23; 138(2): 286-99; S. Jaiswal et al., CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis., Cell. 2009 Jul 23; 138(2): 271-85). Экспрессия CD47 повышена в большинстве раковых опухолей человека (например, неходжскинской лимфоме, остром миелоидном лейкозе, раке грудных желез, ободочной кишки, глиобластоме, раке яичников, мочевого пузыря и простаты), а повышение уровня экспрессии CD47 четко коррелирует с агрессивно развивающимся заболеванием и низкой выживаемостью. (Majeti R, et al., Cell. 2009 Jul 23; 138(2):286-99; S. Jaiswal et al., Cell. 2009 Jul 23; 138(2): 271-85; Willingham SB, et al., The CD47-signal regulatory protein alpha (SIRPα) interaction is a therapeutic target for human solid tumors, Proc Natl Acad Sci U S A. 2012 Apr 24; 109(17): 6662-7; Chao MP, et al., Therapeutic antibody targeting of CD47 eliminates human acute lymphoblastic leukemia., Cancer Res. 2011 Feb 15; 71(4): 1374-84).
Повсеместная экспрессия CD47 в здоровых тканях ставит вопрос безопасности и эффективности лечения: во-первых, направленное воздействие на CD47 с использованием нейтрализующего моноклонального антитела (Mab) может повлиять на здоровые клетки, приводя к тяжелым токсическим эффектам, как было показано в доклинических исследованиях на мышах и яванских макаках (Willingham SB, et al., Proc Natl Acad Sci USA. 2012 Apr 24;109(17): 6662-7; Weiskopf K, et al., Engineered SIRPα Variants as Immunotherapeutic Adjuvants to Anticancer Antibodies, Science. 2013 Jul 5; 341(6141): 88-91). Во-вторых, даже если тяжелые токсические эффекты могли бы быть устранены или ослаблены путем использования альтернативных форматов (Weiskopf K, et al., Science. 2013 Jul 5; 341(6141): 88-91), повсеместная экспрессия CD47 все равно могла бы вызывать быстрое удаление CD47-связывающих молекул посредством мишень-опосредованного распределения лекарственного препарата, приводя к плохой фармакокинетике и снижению эффективности.
Соответственно, существует потребность в антителах и терапевтических средствах, которые обеспечивают направленное воздействие на CD47 и преодолевают эти препятствия.
Сущность изобретения
Изобретение относится к моноклональным антителам, которые связывают CD47. Эти антитела все вместе называются в настоящем документе анти-CD47 моноклональными антителами или анти-CD47 mAb. Предпочтительно, чтобы моноклональные антитела были специфичны по меньшей мере к CD47 человека. В некоторых вариантах осуществления изобретения моноклональные антитела, которые распознают CD47 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD47, не принадлежащим человеку, таким как в качестве неограничивающего примера CD47 приматов, кроме человека, например, CD47 яванского макака, и/или CD47 грызунов. В некоторых вариантах осуществления изобретения эти анти-CD47 моноклональные антитела ингибируют взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения эти анти-CD47 моноклональные антитела ингибируют взаимодействие между CD47 человека и SIRPα человека. Изобретение также включает антитела, которые связываются с тем же эпитопом, что и анти-CD47 моноклональное антитело, раскрытое в настоящем документе, и ингибируют взаимодействие между CD47 и SIRPα, например, между CD47 человека и SIRPα человека.
Изобретение также относится к моновалентным антителам и/или биспецифичным антителам, которые включают по меньшей мере первое плечо, которое специфично к CD47. Предпочтительно, чтобы моновалентные антитела и/или биспецифичные антитела были специфичны по меньшей мере к CD47 человека. В некоторых вариантах осуществления изобретения моновалентные антитела и/или биспецифичные антитела, которые распознают CD47 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD47, не принадлежащим человеку, таким как в качестве неограничивающего примера CD47 приматов, кроме человека, например, CD47 яванского макака, и/или CD47 грызунов. В некоторых вариантах осуществления изобретения эти анти-CD47 моновалентные антитела и/или анти-CD47 биспецифичные антитела ингибируют взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения эти анти-CD47 моновалентные антитела и/или анти-CD47 биспецифичные антитела ингибируют взаимодействие между CD47 человека и SIRPα человека. Изобретение также включает антитела, которые связываются с тем же эпитопом, что и анти-CD47 моновалентное и/или анти-CD47 биспецифичное антитело, раскрытое в настоящем документе, и ингибируют взаимодействие между CD47 и SIRPα, например, между CD47 человека и SIRPα человека.
Изобретение относится к биспецифичным антителам, которые распознают CD47 и вторую мишень. Изобретение обеспечивает идентификацию, продукцию и очистку биспецифичных антител, которые неотличимы по последовательности от стандартных антител, и где один из участков связывания является специфичным к CD47, а второй участок связывания является специфичным к другой мишени, например, опухоль-связанному антигену (ТАА). В некоторых вариантах осуществления изобретения ТАА представляет собой антиген, который экспрессируется на клеточной поверхности раковой клетки. В некоторых вариантах осуществления изобретения раковая клетка выбрана из клетки рака легких, клетки рака бронхов, клетки рака простаты, клетки рака молочной железы, клетки колоректального рака, клетки рака поджелудочной железы, клетки рака яичников, лейкемической клетки, клетки лимфомы, клетки рака пищевода, клетки рака печени, клетки рака мочевыводящих путей и/или мочевого пузыря, клетки рака почек, клетки рака полости рта, клетки рака гортани, клетки рака матки и/или клетки меланомы.
В некоторых вариантах осуществления изобретения подходящие ТАА в качестве неограничивающего примера включают CD20, HER2, HER3, EGFR, IGF1R, c-Met, PDGFR1, CD40, CD40L, CD30, CS1, CD70, глипикан, мезотелин, PSMA, PSCA, MUC1, CA125, CEA, FRA, EpCAM, DR5, HGFR1 и/или 5T4.
CD47 (кластер дифференцировки 47) функционирует в качестве сигнала «не ешь меня» для фагоцитарных клеток и, как известно, сверхэкспрессируется многими опухолями (избегание иммунологического надзора). CD47 взаимодействует с SIRPα, который экспрессируется на фагоцитирующих клетках. CD47 снижает фагоцитарную активность. CD47 ингибирует созревание и активацию дендритных клеток (DC). CD47 также участвует в процессах, таких как, например, апоптоз, выживание, пролиферация, адгезия, миграция и регуляция ангиогенеза, кровяного давления, тканевой перфузии и/или гомеостаза тромбоцитов.
CD47 также участвует в раковых процессах. Например, CD47 сверхэкспрессируется в различных гематологических и солидных опухолях. CD47 является подтвержденным маркером раковых стволовых клеток/инициирующих опухоль клеток. Считается, что сверхэкспрессия CD47 может помогать опухолевым клеткам избегать иммунологического надзора и уничтожения клетками врожденной иммунной системы. Высокий уровень CD47 также ассоциируется с плохим клиническим исходом при раковых заболеваниях, таких как, например, лейкозы, лимфомы, рак молочной железы, рак толстой кишки, рак яичников, рак мочевого пузыря, рак простаты и/или глиома. Поэтому, направленное воздействие на CD47 было бы полезным для лечения, задержки развития или для иного ослабления симптома ракового заболевания.
Поскольку CD47 экспрессируется повсеместно, он представляет собой проблемную мишень для моноклональных антител. Тем не менее, антитела, специфичные к CD47, описанные в настоящем документе, применимы в качестве моноспецифичных антител и могут использоваться для терапевтического вмешательства, или в качестве исследовательского или диагностического реагента. Моноспецифичные антитела по изобретению, которые связывают CD47, а также фрагменты таких моноспецифичных антител, которые иммунологически активны и по-прежнему связывают CD47, включают описанные в настоящем документе иллюстративные антитела, например, антитело 5A3, антитело 5A3M4, антитело 5A3M3, антитело 5A3M5, антитело KE8, антитело KE8-P6H5 (также именуемое в настоящем документе KE8H5), антитело KE8-P3B2 (также именуемое в настоящем документе KE8B2), антитело KE8-P2A2 (также именуемое в настоящем документе KE8A25), антитело KE8F2, антитело KE8G2, антитело KE84G9, антитело KE81G9, антитело KE81A3, антитело KE8E8, антитело KE8G6, антитело KE8H3, антитело KE8C7, антитело KE8A4, антитело KE8A8, антитело KE8G11, антитело KE8B7, антитело KE8F1, антитело KE8C4, антитело KE8A3, антитело KE86G9, антитело KE8H6, антитело KA3, антитело KA3-P5G2 (также именуемое в настоящем документе KA3G2), антитело KA3-P1A3 (также именуемое в настоящем документе KA3A3), антитело KA3-P5C5 (также именуемое в настоящем документе KA3C5), антитело KA3H8, антитело KA3B2, антитело KA3A2, антитело KA3D3, антитело KA3H3, антитело KC4, антитело KC4-P1G11KC4-P4C11, антитело KC4-P6B1KC4-P4F4 и антитело KC4-P2E2 (также именуемое в настоящем документе KC4E2), антитело KC4, антитело KC4F4, антитело KC4A1, антитело KC4C11, антитело KC4E10, антитело KC4B1, антитело KC4C3, антитело KC4A4, антитело KC4G11, антитело KC4G9 и их фрагменты.
Антитела по изобретению, которые связывают CD47, и их фрагменты, служат для модуляции, блокирования, ингибирования, снижения, антагонизма, нейтрализации или иного нарушения функциональной активности CD47. Функциональная активность CD47 включает в качестве неограничивающего примера взаимодействие с SIRPα. Антитела считаются полностью модулирующими, блокирующими, ингибирующими, снижающими, антагонизирующими, нейтрализующими или иным образом нарушающими взаимодействие CD47-SIRPα, когда уровень взаимодействия CD47-SIRPα в присутствии антитела снижается по меньшей мере на 95%, например, на 96%, 97%, 98%, 99% или 100% по сравнению с уровнем взаимодействия CD47-SIRPα в отсутствие связывания с антителом, описанным в настоящем документе. Антитела считаются частично модулирующими, блокирующими, ингибирующими, снижающими, антагонизирующими, нейтрализующими или иным образом нарушающими взаимодействие CD47-SIRPα, когда уровень взаимодействия CD47-SIRPα в присутствии антитела снижается менее чем на 95%, например, на 10%, 20%, 25%, 30%, 40%, 50%, 60%, 75%, 80%, 85% или 90% по сравнению с уровнем взаимодействия CD47-SIRPα в отсутствие связывания с антителом, описанным в настоящем документе.
Изобретение также относится к биспецифичным антителам, в которых по меньшей мере один участок связывания является специфичным к CD47. Биспецифичные антитела по изобретению направлены на CD47 и второй антиген, например, опухоль-связанный антиген (ТАА). В некоторых вариантах осуществления биспецифичные антитела включает функциональный участок Fc. Связывающее ТАА плечо биспецифичного антитела направляет CD47-плечо к опухолевой клетке или раковой стволовой клетке. CD47-плечо блокирует, ингибирует или иным образом снижает взаимодействие между CD47 и SIRPα, тем самым передавая фагоцитам сигнал «съешь меня». В некоторых вариантах осуществления изобретения связывающее ТАА плечо биспецифичного антитела включает последовательность анти-CD19 антитела или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления изобретения биспецифичное антитело проявляет «сбалансированную» аффинность к каждой из двух мишеней. В других вариантах осуществления изобретения биспецифичное антитело проявляет «несбалансированную» аффинность к каждой из двух мишеней. Например, в анти-CD47/CD19 биспецифичном антителе аффинность анти-CD19 плеча увеличена. Например, в анти-CD47/CD19 биспецифичном антителе аффинность анти-CD47 плеча снижена. Например, в анти-CD47/CD19 биспецифичном антителе аффинность анти-CD19 плеча увеличена, а аффинность анти-CD47 плеча снижена. Эти биспецифичные антитела с несбалансированной аффинностью полезны, например, для повышения избирательности к клетке-мишени или группе клеток-мишеней.
В некоторых вариантах осуществления изобретения аффинность анти-CD19 плеча увеличивается по меньшей мере в 100 раз после созревания аффинности. В некоторых вариантах осуществления изобретения аффинность анти-CD47 плеча уменьшается по меньшей мере в 2 раза после снижения аффинности. Например, в некоторых вариантах осуществления изобретения анти-CD47 плечо имеет аффинность к CD47 ниже примерно в 2-100 раз после снижения аффинности.
Биспецифичные антитела по изобретению, которые включают по меньшей мере одно анти-CD47 плечо, служат для модуляции, блокирования, ингибирования, снижения, антагонизма, нейтрализации или иного нарушения функциональной активности CD47. Функциональная активность CD47 включает в качестве неограничивающего примера взаимодействие с SIRPα. Биспецифичные антитела считаются полностью модулирующими, блокирующими, ингибирующими, снижающими, антагонизирующими, нейтрализующими или иным образом нарушающими взаимодействие CD47-SIRPα, когда уровень взаимодействия CD47-SIRPα в присутствии биспецифичного антитела снижается по меньшей мере на 95%, например, на 96%, 97%, 98%, 99% или 100% по сравнению с уровнем взаимодействия CD47-SIRPα в отсутствие связывания с биспецифичным антителом, описанным в настоящем документе. Биспецифичные антитела считаются частично модулирующими, блокирующими, ингибирующими, снижающими, антагонизирующими, нейтрализующими или иным образом нарушающими взаимодействие CD47-SIRPα, когда уровень взаимодействия CD47-SIRPα в присутствии биспецифичного антитела снижается менее чем на 95%, например, на 10%, 20%, 25%, 30%, 40%, 50%, 60%, 75%, 80%, 85% или 90% по сравнению с уровнем взаимодействия CD47-SIRPα в отсутствие связывания с биспецифичным антителом, описанным в настоящем документе.
Анти-CD47 плечи биспецифичных антител по изобретению полезны с рядом плеч, которые связывают другие антигены, например, ТАА. Примеры анти-CD47 плеч, анти-CD47 моновалентных антител и/или биспецифичных антител по изобретению включают антитела, именуемые в настоящем документе как антитело 5A3, антитело 5A3M4, антитело 5A3M3, антитело 5A3M5, антитело KE8, антитело KE8-P6H5 (также именуемое в настоящем документе KE8H5), антитело KE8-P3B2 (также именуемое в настоящем документе KE8B2), антитело KE8-P2A2 (также именуемое в настоящем документе KE8A25), антитело KE8F2, антитело KE8G2, антитело KE84G9, антитело KE81G9, антитело KE81A3, антитело KE8E8, антитело KE8G6, антитело KE8H3, антитело KE8C7, антитело KE8A4, антитело KE8A8, антитело KE8G11, антитело KE8B7, антитело KE8F1, антитело KE8C4, антитело KE8A3, антитело KE86G9, антитело KE8H6, антитело KA3, антитело KA3-P5G2 (также именуемое в настоящем документе KA3G2), антитело KA3-P1A3 (также именуемое в настоящем документе KA3A3), антитело KA3-P5C5 (также именуемое в настоящем документе KA3C5), антитело KA3H8, антитело KA3B2, антитело KA3A2, антитело KA3D3, антитело KA3H3, антитело KC4, антитело KC4-P1G11KC4-P4C11, антитело KC4-P6B1KC4-P4F4 и антитело KC4-P2E2 (также именуемое в настоящем документе KC4E2), антитело KC4, антитело KC4F4, антитело KC4A1, антитело KC4C11, антитело KC4E10, антитело KC4B1, антитело KC4C3, антитело KC4A4, антитело KC4G11, антитело KC4G9 и их фрагменты. В некоторых вариантах осуществления изобретения связывающее ТАА плечо биспецифичного антитела включает последовательность анти-CD19 антитела или его антигенсвязывающий фрагмент.
Изобретение относится к выделенным биспецифичным антителам, имеющим первое плечо, которое включает первую аминокислотную последовательность, связывающую CD47, и второе плечо, которое включает вторую аминокислотную последовательность, не связывающую CD47, причем биспецифичное антитело ингибирует взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения вторая аминокислотная последовательность связывает опухоль-связанный антиген (ТАА). В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека.
В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека на уровне по меньшей мере в десять раз более высоком, чем соответствующий уровень ингибирования взаимодействия CD47 человека/SIRPα человека, проявляемый моновалентным анти-CD47 антителом, которое включает первую аминокислотную последовательность, связывающую CD47, и вторую аминокислотную последовательность, не связывающую белок человека.
В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека на уровне по меньшей мере в 100 раз более высоком, чем соответствующий уровень ингибирования взаимодействия CD47 человека/SIRPα человека, проявляемый моновалентным анти-CD47 антителом, которое включает первую аминокислотную последовательность, связывающую CD47, и вторую аминокислотную последовательность, не связывающую белок человека.
В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека на уровне по меньшей мере в 1000 раз более высоком, чем соответствующий уровень ингибирования взаимодействия CD47 человека/SIRPα человека, проявляемый моновалентным анти-CD47 антителом, которое включает первую аминокислотную последовательность, связывающую CD47, и вторую аминокислотную последовательность, не связывающую белок человека.
В некоторых вариантах осуществления изобретения биспецифичное антитело включает первое плечо, которое ингибирует взаимодействие между CD47 человека на поверхности клеток и растворимым SIRPα человека с IC50 выше 5 нМ в анализе, описанном в примере 4, и в котором моновалентное антитело 5A3M3 имеет IC50 приблизительно 13 нМ.
В некоторых вариантах осуществления изобретения биспецифичное антитело включает первое плечо, которое сохраняется более чем на 80% после инкубации при 37°С в течение 30 минут в цельной крови человека при концентрации 10 мкг/мл, как описано в примере 15.
В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека на уровне по меньшей мере в десять раз, по меньшей мере в 100 раз или по меньшей мере в 1000 раз более высоком, чем соответствующий уровень ингибирования взаимодействия CD47 человека/SIRPα человека, проявляемый моновалентным анти-CD47 антителом, которое включает первую аминокислотную последовательность, связывающую CD47, и вторую аминокислотную последовательность, не связывающую белок человека, и включает первое плечо, которое ингибирует взаимодействие между CD47 человека на поверхности клеток и растворимым SIRPα человека с IC50 выше 5 нМ в анализе, описанном в примере 4, и в котором моновалентное антитело 5A3M3 имеет IC50 приблизительно 13 нМ.
В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека на уровне по меньшей мере в десять раз, по меньшей мере в 100 раз или по меньшей мере в 1000 раз более высоком, чем соответствующий уровень ингибирования взаимодействия CD47 человека/SIRPα человека, проявляемый моновалентным анти-CD47 антителом, которое включает первую аминокислотную последовательность, связывающую CD47, и вторую аминокислотную последовательность, не связывающую белок человека, и включает первое плечо, которое сохраняется более чем на 80% после инкубации при 37°С в течение 30 минут в цельной крови человека при концентрации 10 мкг/мл, как описано в примере 15.
В некоторых вариантах осуществления изобретения ТАА является CD19. В некоторых вариантах осуществления изобретения вторая аминокислотная последовательность не связывает белок человека.
В некоторых вариантах осуществления изобретения первая аминокислотная последовательность включает аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH1) SEQ ID NO: 225, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH2) SEQ ID NO: 226, аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH3) SEQ ID NO: 227, аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области легкой цепи (CDRL1), выбранную из SEQ ID NO: 228-241 и 262-272, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области легкой цепи (CDRL2), выбранную из 242-245 и 273-280, и аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области легкой цепи (CDRH3), выбранную из 246-261 и 281.
В некоторых вариантах осуществления изобретения первая аминокислотная последовательность включает аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO: 114 и аминокислотную последовательность вариабельной области легкой цепи, выбранную из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206.
В некоторых вариантах осуществления изобретения биспецифичное антитело включает две копии полипептида одной тяжелой цепи и первую легкую цепь и вторую легкую цепь, причем первая и вторая легкие цепи являются разными.
В некоторых вариантах осуществления изобретения по меньшей мере участок первой легкой цепи относится к каппа-типу, и по меньшей мере участок второй легкой цепи относится к лямбда-типу. В некоторых вариантах осуществления изобретения первая легкая цепь включает по меньшей мере константную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь включает по меньшей мере константную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь включает константную область каппа-цепи и вариабельную область каппа-цепи, и где вторая легкая цепь включает константную область лямбда-цепи и вариабельную область лямбда-цепи.
В некоторых вариантах осуществления изобретения последовательности каркасных областей константных и вариабельных областей принадлежат человеку.
Изобретение также относится к биспецифичным антителам и/или моновалентным антителам, которые включают по меньшей мере первое плечо, которое ингибирует взаимодействие между CD47 человека на поверхности клеток и растворимым SIRPα человека с IC50 выше 5 нМ в анализе, описанном в примере 4, и в котором моновалентное антитело 5A3M3 имеет IC50 приблизительно 13 нМ.
Изобретение также относится к биспецифичным антителам и/или моновалентным антителам, которые включают по меньшей мере первое плечо, которое сохраняется более чем на 80% после инкубации при 37°С в течение 30 минут в цельной крови человека при концентрации 10 мкг/мл, как описано в примере 15. В некоторых вариантах осуществления изобретения биспецифичное антитело и/или моновалентное антитело ингибирует взаимодействие между CD47 и сигнала-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения биспецифичное антитело и/или моновалентное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека.
Изобретение также относится к выделенным биспецифичным антителам, имеющим первое плечо, которое включает первую аминокислотную последовательность, связывающую CD47, и второе плечо, которое включает вторую аминокислотную последовательность, связывающую CD19, причем биспецифичное антитело ингибирует взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα).
В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека. В некоторых вариантах осуществления изобретения биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека на уровне, который выбран из группы, состоящей из: по меньшей мере десять раз более высоком, по меньшей мере в 100 раз более высоком и по меньшей мере в 1000 раз более высоком, чем соответствующий уровень ингибирования взаимодействия CD47 человека/SIRPα человека, проявляемый моновалентным анти-CD47 антителом, которое включает первую аминокислотную последовательность, связывающую CD47, и вторую аминокислотную последовательность, не связывающую белок человека.
В некоторых вариантах осуществления изобретения первая аминокислотная последовательность включает аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH1) SEQ ID NO: 225, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH2) SEQ ID NO: 226, аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH3) SEQ ID NO: 227, аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области легкой цепи (CDRL1), выбранную из SEQ ID NO: 228-241 и 262-272, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области легкой цепи (CDRL2), выбранную из 242-245 и 273-280, и аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области легкой цепи (CDRH3), выбранную из 246-261 и 281.
В некоторых вариантах осуществления изобретения первая аминокислотная последовательность включает аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO: 114 и аминокислотную последовательность вариабельной области легкой цепи, выбранную из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206.
В некоторых вариантах осуществления изобретения биспецифичное антитело включает две копии полипептида одной тяжелой цепи и первую легкую цепь и вторую легкую цепь, причем первая и вторая легкие цепи являются разными.
В некоторых вариантах осуществления изобретения по меньшей мере участок первой легкой цепи относится к каппа-типу, и по меньшей мере участок второй легкой цепи относится к лямбда-типу. В некоторых вариантах осуществления изобретения первая легкая цепь включает по меньшей мере константную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь включает по меньшей мере константную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь включает константную область каппа-цепи и вариабельную область каппа-цепи, и где вторая легкая цепь включает константную область лямбда-цепи и вариабельную область лямбда-цепи.
В некоторых вариантах осуществления изобретения последовательности каркасных областей константных и вариабельных областей принадлежат человеку.
Изобретение также относится к моновалентным антителам, которые связывают CD47. Эти антитела все вместе именуются в настоящем документе моновалентными анти-CD47 антителами или анти-CD47 моновалентными mAb. Моновалентные антитела по изобретению включают одно плечо, которое специфично распознает CD47, и второе плечо, именуемое в настоящем документе фиктивным плечом. Фиктивное плечо включает аминокислотную последовательность, которая не связывается или иным образом перекрестно реагирует с белком человека. В некоторых вариантах осуществления изобретения фиктивное плечо включает аминокислотную последовательность, которая не связывается или иным образом перекрестно реагирует с белком человека, который присутствует в цельной крови. Специалистам в данной области техники будет понятно, что белки человека, найденные в крови, являются совокупностью, которая представляет все, или практически все, антигены, присутствующие в кровотоке. В некоторых вариантах осуществления изобретения фиктивное плечо включает аминокислотную последовательность, которая не связывается или иным образом перекрестно реагирует с белком человека, который присутствует в солидной ткани. Предпочтительно, чтобы моновалентные антитела были специфичны по меньшей мере к CD47 человека. В некоторых вариантах осуществления изобретения моновалентные антитела, которые распознают CD47 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD47, не принадлежащим человеку, таким как, в качестве неограничивающего примера, CD47 приматов, кроме человека, например, CD47 яванского макака, и/или CD47 грызунов.
Анти-CD47 плечи моновалентных антител по изобретению полезны с любым фиктивным плечом. Примеры анти-CD47 плеч моновалентных антител по изобретению включают антитела, именуемые в настоящем документе как антитело 5A3, антитело 5A3M4, антитело 5A3M3, антитело 5A3M5, антитело KE8, антитело KE8-P6H5 (также именуемое в настоящем документе KE8H5), антитело KE8-P3B2 (также именуемое в настоящем документе KE8B2), антитело KE8-P2A2 (также именуемое в настоящем документе KE8A25), антитело KE8F2, антитело KE8G2, антитело KE84G9, антитело KE81G9, антитело KE81A3, антитело KE8E8, антитело KE8G6, антитело KE8H3, антитело KE8C7, антитело KE8A4, антитело KE8A8, антитело KE8G11, антитело KE8B7, антитело KE8F1, антитело KE8C4, антитело KE8A3, антитело KE86G9, антитело KE8H6, антитело KA3, антитело KA3-P5G2 (также именуемое в настоящем документе KA3G2), антитело KA3-P1A3 (также именуемое в настоящем документе KA3A3), антитело KA3-P5C5 (также именуемое в настоящем документе KA3C5), антитело KA3H8, антитело KA3B2, антитело KA3A2, антитело KA3D3, антитело KA3H3, антитело KC4, антитело KC4-P1G11KC4-P4C11, антитело KC4-P6B1KC4-P4F4 и антитело KC4-P2E2 (также именуемое в настоящем документе KC4E2), антитело KC4, антитело KC4F4, антитело KC4A1, антитело KC4C11, антитело KC4E10, антитело KC4B1, антитело KC4C3, антитело KC4A4, антитело KC4G11, антитело KC4G9 и их фрагменты. В некоторых вариантах осуществления изобретения связывающее ТАА плечо биспецифичного антитела включает последовательность анти-CD19 антитела или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления изобретения моновалентное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека.
В некоторых вариантах осуществления изобретения анти-CD47 плечо моновалентного антитела включает первую аминокислотную последовательность, которая включает аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH1) SEQ ID NO: 225, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH2) SEQ ID NO: 226, аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH3) SEQ ID NO: 227, аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области легкой цепи (CDRL1), выбранную из SEQ ID NO: 228-241 и 262-272, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области легкой цепи (CDRL2), выбранную из 242-245 и 273-280, и аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области легкой цепи (CDRH3), выбранную из 246-261 и 281.
В некоторых вариантах осуществления изобретения анти-CD47 плечо моновалентного антитела включает первую аминокислотную последовательность, которая включает аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO: 114 и аминокислотную последовательность вариабельной области легкой цепи, выбранную из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206.
В некоторых вариантах осуществления изобретения моновалентное антитело включает две копии полипептида одной тяжелой цепи и первую легкую цепь и вторую легкую цепь, причем первая и вторая легкие цепи являются разными.
В некоторых вариантах осуществления изобретения по меньшей мере участок первой легкой цепи относится к каппа-типу, и по меньшей мере участок второй легкой цепи относится к лямбда-типу. В некоторых вариантах осуществления изобретения первая легкая цепь включает по меньшей мере константную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь включает по меньшей мере константную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения вторая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых вариантах осуществления изобретения первая легкая цепь включает константную область каппа-цепи и вариабельную область каппа-цепи, и где вторая легкая цепь включает константную область лямбда-цепи и вариабельную область лямбда-цепи.
В некоторых вариантах осуществления изобретения последовательности каркасных областей константных и вариабельных областей принадлежат человеку.
Биспецифичные антитела по изобретению создают с использованием любых способов, известных в данной области, например, в качестве неограничивающего примера, с использованием сшитых фрагментов, квадром и/или любого из множества рекомбинантных форматов, таких как, в качестве не ограничивающих примеров, связанных фрагментов антител, принудительных гетеродимеров и/или рекомбинантных форматов, основанных на одиночных доменах. Примеры биспецифичных форматов включают, но не ограничиваются ими, биспецифичные IgG на основе обмена Fab-плеч (Gramer et al., 2013 Mabs. 5(6)); CrossMab-формат (Klein C et al., 2012 MAbs 4(6)); множество форматов, основанных на подходах принудительной гетеродимеризации, таких как SEED-технология (Davis JH et al., 2010 Protein Eng Des Sel. 23(4): 195-202), электростатическая адресация (Gunasekaran K et al., J Biol Chem. 2010 285(25): 19637-46.) или подход «узел-впадина» (Ridgway JB et al., Protein Eng. 1996 9(7):617-21.) или другие наборы мутаций, препятствующих образованию гомодимеров (Von Kreudenstein TS et al., 2013 MAbs. 5(5): 646-54.); биспецифичные форматы на основе фрагментов, такие как тандемные scFv (такие как BiTE) (Wolf E et al., 2005 Drug Discov. Today 10(18): 1237-44.); биспецифичные тетравалентные антитела (Pörtner LM et al., 2012 Cancer Immunol Immunother. 61(10): 1869-75.); переориентирующие молекулы с двойной аффинностью (Moore PA et al., 2011 Blood. 117(17): 4542-51), диатела (Kontermann RE et al., Nat Biotechnol. 1997 15(7):629-31).
В некоторых вариантах осуществления изобретения биспецифичные антитела несут различную специфичность в каждом антигенсвязывающем центре и включают две копии полипептида одной тяжелой цепи и первую легкую цепь и вторую легкую цепь, причем первая и вторая легкие цепи являются разными.
В некоторых антителах по меньшей мере первый участок первой легкой цепи относится к каппа-типу, и по меньшей мере участок второй легкой цепи относится к лямбда-типу. В некоторых антителах первая легкая цепь включает по меньшей мере константную область каппа-цепи. В некоторых антителах первая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых антителах первая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых антителах вторая легкая цепь включает по меньшей мере константную область лямбда-цепи. В некоторых антителах вторая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых антителах вторая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых антителах первая легкая цепь включает константную область каппа-цепи и вариабельную область каппа-цепи, и где вторая легкая цепь включает константную область лямбда-цепи и вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения последовательности каркасных областей константных и вариабельных областей принадлежат человеку.
Эти анти-CD47 плечи, моноспецифичные анти-CD47 антитела, моновалентные анти-CD47 антитела и/или биспецифичные антитела, в которых по меньшей мере один участок связывания специфичен к CD47, содержат аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 114, и аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206.
Изобретение относится к моноклональным антителам, которые связывают CD47. Например, данное изобретение относится к моноклональным антителам, которые ингибируют взаимодействие между CD47 человека на поверхности клеток и растворимым SIRPα человека с IC50 выше 0,3 нМ в анализе, описанном в примере 4, и в котором антитело 5A3M3 имеет IC50 приблизительно 0,36 нМ.
Изобретение также относится к моноклональным антителам, которые связывают CD47 и восстанавливаются более чем на 80% после инкубации при 37°С в течение 30 минут в цельной крови человека при концентрации 10 мкл/мл, как описано в примере 15. В некоторых вариантах осуществления изобретения моноклональное антитело ингибирует взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения моноклональное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека.
Изобретение также относится к анти-CD47 моноклональным антителам, которые включают аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH1) SEQ ID NO: 225, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH2) SEQ ID NO: 226, аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области тяжелой цепи (CDRH3) SEQ ID NO: 227, аминокислотную последовательность 1-й определяющей комплементарность области вариабельной области легкой цепи (CDRL1), выбранную из SEQ ID NO: 228-241 и 262-272, аминокислотную последовательность 2-й определяющей комплементарность области вариабельной области легкой цепи (CDRL2), выбранную из 242-245 и 273-280, и аминокислотную последовательность 3-й определяющей комплементарность области вариабельной области легкой цепи (CDRH3), выбранную из 246-261 и 281.
В некоторых вариантах осуществления изобретения анти-CD47 моноклональное антитело включает аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 114. В некоторых вариантах осуществления изобретения анти-CD47 моноклональное антитело включает аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206. В некоторых вариантах осуществления анти-CD47 моноклональное антитело включает аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 114, и аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206.
В некоторых вариантах осуществления изобретения анти-CD47 моноклональное антитело включает аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO: 114 и аминокислотную последовательность вариабельной области легкой цепи, выбранную из SEQ ID NO: 116, 118, 120, 122, 124, 126, 128, 130, 132, 134, 136, 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160, 162, 164, 166, 168, 170, 172, 174, 176, 178, 180, 182, 184, 186, 188, 190, 192, 194, 196, 198, 200, 202, 204 и 206.
В некоторых вариантах осуществления изобретения анти-CD47 антитело включает комбинацию последовательности вариабельной области тяжелой цепи и последовательности вариабельной области легкой цепи, выбранную из группы, состоящей из комбинаций, показанных в 5A3, 5A3M4, 5A3M3, 5A3M5, KE8, KE8F2, KE8G2, KE84G9, KE81G9, KE81A3, KE8E8, KE8G6, KE8H5, KE8A2, KE8H3, KE8C7, KE8B2, KE8A4, KE8A8, KE8G11, KE8B7, KE8F1, KE8C4, KE8A3, KE86G9, KE8H6, KA3, KA3H8, KA3A3, KA3C5, KA3B2, KA3A2, KA3D3, KA3G2, KA3H3, KC4, KC4E2, KC4F4, KC4A1, KC4C11, KC4E10, KC4B1, KC4C3, KC4A4, KC4G11 и KC4G9.
Изобретение относится к моноклональным антителам, которые связывают CD19. Эти антитела все вместе именуются в настоящем документе анти-CD19 моноклональными антителам или анти-CD19 mAb. Предпочтительно, чтобы моноклональные антитела были специфичны по меньшей мере к CD19 человека. В некоторых вариантах осуществления изобретения моноклональные антитела, которые распознают CD19 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD19, не принадлежащим человеку, таким как, в качестве неограничивающего примера, CD19 приматов, кроме человека, например, CD19 яванского макака, и/или CD19 грызунов.
В некоторых вариантах осуществления изобретения анти-CD19 моноклональное антитело включает аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 114. В некоторых вариантах осуществления изобретения анти-CD19 моноклональное антитело включает аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из SEQ ID NO: 208, 210, 212, 214, 216, 218 и 220. В некоторых вариантах осуществления изобретения анти-CD19 моноклональное антитело включает аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности SEQ ID NO: 114, и аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности, выбранной из последовательностей SEQ ID NO: 208, 210, 212, 214, 216, 218 и 220.
В некоторых вариантах осуществления изобретения анти-CD19 моноклональное антитело включает аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO: 114 и аминокислотную последовательность вариабельной области легкой цепи, выбранную из SEQ ID NO: 208, 210, 212, 214, 216, 218 и 220.
Изобретение также относится к моновалентным антителам, которые связывают CD19. Эти антитела все вместе именуются в настоящем документе анти-CD19 моновалентными антителами или анти-CD19 моновалентными mAb. Моновалентные антитела по изобретению включают одно плечо, которое специфично распознает CD19, и второе плечо, именуемое в настоящем документе фиктивным плечом. Фиктивное плечо включает аминокислотную последовательность, которая не связывается или иным образом перекрестно реагирует с белком человека. В некоторых вариантах осуществления изобретения фиктивное плечо включает аминокислотную последовательность, которая не связывается или иным образом перекрестно реагирует с белком человека, который присутствует в цельной крови. В некоторых вариантах осуществления изобретения фиктивное плечо включает аминокислотную последовательность, которая не связывается или иным образом перекрестно реагирует с белком человека, который присутствует в солидной ткани. Предпочтительно, чтобы моновалентные антитела были специфичны по меньшей мере к CD19 человека. В некоторых вариантах осуществления изобретения моновалентные антитела, которые распознают CD19 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD19, не принадлежащим человеку, таким как, в качестве неограничивающего примера, CD19 приматов, кроме человека, например, CD19 яванского макака, и/или CD47 грызунов.
Изобретение также относится к биспецифичным антителам, которые распознают CD19 и вторую мишень. В некоторых вариантах осуществления изобретения вторая мишень представляет собой антиген, который, как известно, связан или иным образом вовлечен в аутоиммунные заболевания и/или воспалительные заболевания, такие как, например, В-клеточные аутоиммунные заболевания и/или воспалительные заболевания, включающие в качестве неограничивающего примера системную красную волчанку (SLE), ревматоидный артрит (RA), идиопатическую тромбоцитопеническую пурпуру (ITP), гипергаммаглобулинемию Вальденстрема, синдром Шегрена, рассеянный склероз (MS) и/или волчаночный нефрит.
В некоторых вариантах осуществления изобретения подходящие вторые мишени включают в качестве неограничивающего примера CD20, CD22, CD40, BAFFR, CD5, CD32b, ICOSL, IL6R и/или IL21R.
Биспецифичные антитела по изобретению, которые распознают CD19 и вторую мишень, создают с использованием любых способов, известных в данной области, например, в качестве неограничивающего примера, с использованием сшитых фрагментов, квадром и/или любого из множества рекомбинантных форматов, таких как, в качестве не ограничивающих примеров, связанных фрагментов антител, принудительных гетеродимеров и/или рекомбинантных форматов, основанных на одиночных доменах. Изобретение обеспечивает идентификацию, продукцию и очистку биспецифичных антител, которые неотличимы по последовательности от стандартных антител, и где один из участков связывания специфичен к CD19, а второй участок связывания специфичен к другой мишени, например, опухоль-связанному антигену (ТАА). Немодифицированная природа антител по изобретению обеспечивает им благоприятные для изготовления и биохимические характеристики, аналогичные стандартным моноклональным антителам.
В некоторых вариантах осуществления изобретения биспецифичные антитела несут различную специфичность в каждом антигенсвязывающем центре и включают две копии полипептида одной тяжелой цепи и первую легкую цепь и вторую легкую цепь, причем первая и вторая легкие цепи являются разными.
В некоторых антителах по меньшей мере первый участок первой легкой цепи относится к каппа-типу, и по меньшей мере участок второй легкой цепи относится к лямбда-типу. В некоторых антителах первая легкая цепь включает по меньшей мере константную область каппа-цепи. В некоторых антителах первая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых антителах первая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых антителах вторая легкая цепь включает по меньшей мере константную область лямбда-цепи. В некоторых антителах вторая легкая цепь дополнительно включает вариабельную область лямбда-цепи. В некоторых антителах вторая легкая цепь дополнительно включает вариабельную область каппа-цепи. В некоторых антителах первая легкая цепь включает константную область каппа-цепи и вариабельную область каппа-цепи, и где вторая легкая цепь включает константную область лямбда-цепи и вариабельную область лямбда-цепи. В некоторых вариантах осуществления изобретения последовательности каркасных областей константных и вариабельных областей принадлежат человеку.
Моноклональные, моновалентные и/или биспецифичные антитела по изобретению могут использоваться для терапевтического вмешательства, или в качестве исследовательского или диагностического реагента. Например, моноклональные, моновалентные и/или биспецифичные антитела по изобретению могут использоваться в способах лечения, профилактики и/или задержки развития патологий, связанных с аномальной экспрессией и/или активностью CD47 и/или CD47-SIRPα или в способах ослабления симптома, связанного с такими патологиями, путем введения антитела по изобретению индивиду, который нуждается в таком лечении или профилактике. Подлежащим лечению индивидом является, например, человек. Моноклональные, моновалентные и/или биспецифичные антитела вводят в количестве, достаточном для лечения, профилактики, задержки развития или для ослабления симптома, связанного с патологией.
В некоторых вариантах осуществления изобретения моноклональные, моновалентные и/или биспецифичные антитела, описанные в настоящем документе, используют в сочетании с одним или несколькими дополнительными средствами или комбинацией дополнительных средств. Подходящие дополнительные средства включают современные фармацевтические и/или хирургические методы лечения предполагаемого состояния, такого как, например, раковые, воспалительные и/или аутоиммунные заболевания. В некоторых вариантах осуществления изобретения моноклональные, моновалентные и/или биспецифичные антитела могут использоваться в сочетании с ритуксимабом.
В некоторых вариантах осуществления изобретения моноклональные, моновалентные и/или биспецифичные антитела и дополнительное средство получают в виде единой терапевтической композиции, и моноклональное, моновалентное и/или биспецифичное антитело и дополнительное средство вводят одновременно. В альтернативном варианте моноклональные, моновалентные и/или биспецифичные антитела и дополнительное средство отделены друг от друга, например, каждый из них получен в виде отдельной терапевтической композиции, и моноклональное, моновалентное и/или биспецифичное антитело и дополнительное средство вводят одновременно, или моноклональные, моновалентные и/или биспецифичные антитела и дополнительное средство вводят в разное время в течение лечения. Например, моноклональное, моновалентное и/или биспецифичное антитело вводят перед введением дополнительного средства, моноклональное, моновалентное и/или биспецифичное антитело вводят после введения дополнительного средства, или моноклональное, моновалентное и/или биспецифичное антитело и дополнительное средство вводят поочередно. Согласно описанию в настоящем документе, моноклональное, моновалентное и/или биспецифичное антитело и дополнительное средство вводят одной дозой или несколькими дозами.
Патологии, подходящие для лечения и/или профилактики с использованием антител по изобретению, включают, например, рак или любое другое заболевание или расстройство, связанное с аномальной экспрессией и/или активностью CD47.
Изобретение также относится к способам получения биспецифичных антител, которые проявляют «несбалансированную» аффинность к каждой из двух мишеней. Например, в некоторых вариантах анти-CD47/CD19 биспецифичного антитела аффинность анти-CD19 плеча увеличена с использованием созревания аффинности. Например, в некоторых вариантах анти-CD47/CD19 биспецифичного антитела аффинность анти-CD47 плеча снижена с использованием снижения аффинности. Например, в некоторых вариантах анти-CD47/CD19 биспецифичного антитела аффинность анти-CD19 плеча увеличена с использованием созревания аффинности, а аффинность анти-CD47 плеча снижена с использованием снижения аффиности. Эти биспецифичные антитела с несбалансированной аффинностью полезны, например, для повышения избирательности к клетке-мишени или группе клеток-мишеней.
Фармацевтические композиции по изобретению могут включать антитело по изобретению и носитель. Эти фармацевтические композиции могут быть включены в наборы, такие как, например, диагностические наборы.
Краткое описание чертежей
Фигура 1 представляет собой иллюстрацию выравнивания последовательностей между последовательностью вариабельной области легкой цепи (VL) анти-CD47 антитела 5A3 (SEQ ID NO: 116) и ближайшей к ней последовательностью зародышевой линии (SEQ ID NO: 282), IGKV1-33 человека, в соответствии с номенклатурой IMGT.
Фигура 2 представляет собой график, изображающий блокирующую способность вариантов антитела, 5A3-М3 и 5A3-М5, по сравнению с родительским антителом 5A3 в анализе связывания CD47/SIRPα.
Фигура 3 представляет собой график, иллюстрирующий специфичность различных анти-CD47 моноклональных антител, показываемую связыванием очищенных анти-CD47 mAb с клетками СНО, трансфицированных CD47 человека, оцениваемую проточной цитометрией (серые столбцы). Анти-CD47 MAb не связываются с нетрансфицированными клетками СНО (черные столбцы).
Фигура 4 представляет собой график, изображающий связывание с нативным CD47 и специфичность анти-CD47 MAb, показанные связыванием очищенных анти-CD47 MAb с клетками НЕК293-Р, которое оценивается проточной цитометрией (серые столбцы). Связывание с клетками НЕК293-Р, стабильно трансфицированными киРНК, специфичной к hCD47, значительно снижено (черные столбцы).
Фигура 5 представляет собой график, изображающий связывание с нативным CD47 человека и перекрестную реактивность с CD47 яванского макака. Оценивали связывание очищенных анти-CD47 MAb с CD4+ Т-клетками из РВМС человека (светло-серые столбцы) и яванского макака (темно-серые столбцы).
Фигура 6 представляет собой график, изображающий способность анти-CD47 Mab блокировать взаимодействие CD47-SIRPα, исследуемую в анализе ингибирования CD47-SIRPα (конкурентное ингибирование связывания рекомбинантного растворимого SIRPα человека с hCD47-экспрессирующими клетками СНО, описанное в примере 4). Показаны величины IC50, полученные в экспериментах доза-ответ. Анти-CD47 Mab сгруппированы по семействам и проиндексированы по блокирующей способности - от наивысшей до самой низкой. Нейтрализующее действие антител по настоящему изобретению сравнивается с коммерчески доступным анти-CD47 антителом B6H12.
Фигура 7 представляет собой иллюстрацию, отражающую гемагглютинирующую активность анти-CD47 антител. О гемагглютинации свидетельствует слипшийся осадок эритроцитов в форме полумесяца по нижнему краю лунки, в то время как неагглютинированные не образуют агрегаты и равномерно распределены по площади поверхности лунки. На фигуре 7 продемонстрировано, что высокоаффинные анти-CD47 моноклональные антитела семейств 5A3, Ke8 и KA3 вызывают гемагглютинацию; в отличие от других трех семейств, антитела семейства Kc4, протестированные в этом эксперименте, не вызывают гемагглютинацию.
Фигура 8 представляет собой график, изображающий кривую «доза-ответ», для FACS-анализа связывания, выполненного на клетках Raji с исходным анти-CD19 клоном 1B7, клоном D11, выявленным после первого раунда созревания аффинности, и конечным клоном L7B7_c2, полученным после второго раунда созревания аффинности.
Фигуры 9A-9C представляют собой серию графиков, демонстрирующих способность анти-CD47xCD19 BsAb (биспецифичных антител) одновременно активировать две мишени на поверхности клеток. Графики на фиг. 9А-9С представляют FACS-профили, полученные с моновалентными и биспецифичными антителами, связывающимися с CD19-отрицательными В-NHL клетками (DS-1) и CD19-положительными клетками лимфомы Беркитта (Raji). Все антитела имели формат IgG1 человека и были протестированы при четырех концентрациях, как указано.
Фигура 10 представляет собой серию графиков, изображающих SIRPα-блокирующую активность моновалентных и биспецифичных антител. На фигуре 10 продемонстрирована совместная активация CD19 и CD47 на поверхности клетки-мишени, путем доказательства того, что нейтрализация взаимодействия CD47-SIRPα с помощью анти-CD47xCD19 BsAb является CD19-зависимой. Эксперименты проводились в квадрупликатах. Показаны средние значения и SEM (стандартная ошибка среднего). Кривые ингибирования доза-ответ были аппроксимированы с помощью программы GraphPad.
На фигурах 11А-11С показаны кривые «доза-ответ» для ADCC, полученные с использованием анти-CD47xCD19 κλ-антител (черная линия) или соответствующих анти-CD47 моновалентных антител (серая линия). ADCC с использованием анти-CD19 Mab, С2, приведены для сравнения (пунктирная серая линия). Анализ ADCC проводили с PBMC из цельной крови человека в качестве эффекторных клеток и окрашенных Calcein АМ клеток Raji (фиг. 11А, 11С) или клеток Ramos (фиг. 11В) в качестве клеток-мишеней (соотношение эффекторных клеток и клеток-мишеней составляло :50). Цитотоксичность рассчитывали по степени высвобождения кальцеина из клеток-мишеней. Показан процент специфичной гибели клеток +/- SD (среднеквадратичное отклонение). Эксперименты проводились в дупликатах. На фигурах 11А-11В продемонстрирована способность анти-CD47xCD19 BsAb убивать CD19-положительные клетки CD19-зависимым образом, тогда как соответствующие анти-CD47 моновалентные антитела были гораздо менее эффективными или вообще неэффективными. На фигуре 11C показано, что эффективность уничтожения клеток Raji с использованием анти-CD47xCD19 антител была сравнима с ритуксимабом и намного выше, чем с использованием анти-CD19 моноклонального антитела, C2.
Фигура 12 представляет собой график, изображающий фагоцитарную активность трех анти-CD47xCD19 κλ-антител по настоящему изобретению (черные линии) по сравнению с соответствующими анти-CD47 моновалентными антителами (серые линии) в ADCP-эксперименте «доза-ответ». Для сравнения показан фагоцитоз с использованием анти-CD47 моноклонального антитела, B6H12 (на основе IgG1 человека, пунктирная черная линия), и с анти-CD19 моноклональным антителом С2 (пунктирная серая линия). Эксперимент по ADCP проводили с макрофагами человека, дифференцированными из моноцитов периферической крови, и клетками Raji в качестве клеток-мишеней (соотношение эффекторных клеток и клеток-мишеней составляло 1:5). Фагоцитоз оценивали с помощью FACS. Показан процент макрофагов, фагоцитировавших по меньшей мере одну клетку-мишень. Анти-CD47xCD19 κλ-антитела способствуют фагоцитозу CD19-положительных клеток CD19-зависимым образом, тогда как соответствующие анти-CD47 моновалентные антитела были гораздо менее эффективными или вообще неэффективными.
Фигура 13 представляет собой график, иллюстрирующий активность различных антител в отношении ксенотрансплантата B-клеточной лимфомы Raji в NOD/SCID-мышах. Лечение антителами начинали после того как размер привитой опухоли достигал примерно 0,1 см3 и заканчивали на 25-й день (D25). Группы лечения (n=5) были, как указано на врезке. Показана эволюция среднего объема опухоли в группе лечения +/- SD. На фигуре 13 показано, что эффективность BsAb аналогична эффективности B6H12 или ритуксимаба, и уничтожение опухоли является CD19-зависимым, поскольку соответствующее моновалентное антитело было менее эффективным.
Фигура 14 представляет собой график, отражающий, что анти-CD47 антитела с высокой и умеренной активностью эффективно сорбируются на эритроцитах. В случае BsAb, это явление ограничено молекулами, имеющими высокую аффинность CD47-плеч, такими как 5A3.
Подробное описание изобретения
Изобретение относится к моноклональным антителам, которые связывают CD47. Эти антитела все вместе называются в настоящем документе анти-CD47 моноклональными антителами или анти-CD47 mAb. Предпочтительно, чтобы моноклональные антитела были специфичны по меньшей мере к CD47 человека. В некоторых вариантах осуществления изобретения моноклональные антитела, которые распознают CD47 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD47, не принадлежащим человеку, таким как, в качестве неограничивающего примера, CD47 приматов, кроме человека, например, CD47 яванского макака, и/или CD47 грызунов. В некоторых вариантах осуществления изобретения эти анти-CD47 моноклональные антитела ингибируют взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения эти анти-CD47 моноклональные антитела ингибируют взаимодействие между CD47 человека и SIRPα человека. Изобретение также включает антитела, которые связываются с тем же эпитопом, что и анти-CD47 моноклональное антитело, раскрытое в настоящем документе, и ингибируют взаимодействие между CD47 и SIRPα, например, между CD47 человека и SIRPα человека.
Изобретение также относится к моновалентным антителам и/или биспецифичным антителам, которые включают по меньшей мере первое плечо, которое является специфичным к CD47. Предпочтительно, чтобы моновалентные антитела и/или биспецифичные антитела были специфичны по меньшей мере к CD47 человека. В некоторых вариантах осуществления изобретения моновалентные антитела и/или биспецифичные антитела, которые распознают CD47 человека, также имеют перекрестную реактивность по меньшей мере с одним другим белком CD47, не принадлежащим человеку, таким как, в качестве неограничивающего примера, CD47 приматов, кроме человека, например, CD47 яванского макака, и/или CD47 грызунов. В некоторых вариантах осуществления изобретения эти анти-CD47 моновалентные антитела и/или анти-CD47 биспецифичные антитела ингибируют взаимодействие между CD47 и сигнал-регуляторным белком альфа (SIRPα). В некоторых вариантах осуществления изобретения эти анти-CD47 моновалентные антитела и/или анти-CD47 биспецифичные антитела ингибируют взаимодействие между CD47 человека и SIRPα человека. Изобретение также включает антитела, которые связываются с тем же эпитопом, что и анти-CD47 моновалентное и/или анти-CD47 биспецифичное антитело, раскрытое в настоящем документе, и ингибируют взаимодействие между CD47 и SIRPα, например, между CD47 человека и SIRPα человека.
Биспецифичные антитела по изобретению позволяю одновременное связывание двумя плечами антитела двух антигенов на поверхности клетки (называемое совместной активацией), что приводит к дополнительному или синергетическому увеличению аффинности за счет механизма авидности. Как следствие, совместная активация придает высокую селективность в отношении клеток, экспрессирующих оба антигена, по сравнению с клетками, которые экспрессируют только один единственный антиген. Дополнительно, аффинность двух плеч биспецифичного антитела к их соответствующим мишеням может быть подобрана таким образом, что связывание с клетками-мишенями в основном обусловлено одним из плеч антитела. В некоторых вариантах осуществления изобретения биспецифичное антитело включает первое плечо, которое связывает CD47, и второе плечо, которое связывает опухоль-связанный антиген (ТАА), причем второе плечо связывается с ТАА с высокой аффинностью, а первое плечо связывается с CD47 с низкой аффинностью, т.е. аффинностью, которой достаточно для ингибирования CD47/SIRPα после связывания с ТАА. Такая конструкция позволяет биспецифичным антителам по изобретению преимущественно ингибировать CD47 в злокачественных, а не в здоровых клетках. В примерах, приведенных в настоящем документе, биспецифичное антитело с первым плечом, которое связывает CD47 с низкой аффинностью, и вторым плечом, которое связывает CD19 с высокой аффинностью (называется анти-CD47xCD19 биспецифичное антитело), позволяет преимущественное ингибирование CD47 в злокачественных, а не в здоровых клетках. Помимо двух антигенсвязывающих плеч анти-CD47xTAA биспецифичному антителу требуется функциональный Fc-участок для привлечения макрофагов и/или других иммунных эффекторных клеток. Формат IgG человека для биспецифичных антител (такой как описанный в настоящем документе формат κλ-антител) хорошо подходит для создания двунаправленных анти-CD47xTAA биспецифичных антител. Как показано в примерах, приведенных в настоящем документе, способность двунаправленных биспецифичных антител совместно связывать CD47 и CD19 приводит к значительному увеличению аффинности связывания с CD19-положительными клетками и CD19-зависимой нейтрализации взаимодействия CD47/SIRPα. Это, в свою очередь, приводит к эффективному и селективному уничтожению злокачественных клеток, опосредованному анти-CD47xCD19 биспецифичными антителами, как было показано в экспериментах по ADCC и ADCP, приведенных в настоящем документе.
Примеры анти-CD47 моноклональных антител, моноспецифичных анти-CD47 антител, анти-CD47 моновалентных антител и/или биспецифичных антител по изобретению, в которых по меньшей мере один участок связывания специфичен к CD47, включают, например, антитело 5A3, антитело 5A3M4, антитело 5A3M3, антитело 5A3M5, антитело KE8, антитело KE8-P6H5 (также именуемое в настоящем документе KE8H5), антитело KE8-P3B2 (также именуемое в настоящем документе KE8B2), антитело KE8-P2A2 (также именуемое в настоящем документе KE8A25), антитело KE8F2, антитело KE8G2, антитело KE84G9, антитело KE81G9, антитело KE81A3, антитело KE8E8, антитело KE8G6, антитело KE8H3, антитело KE8C7, антитело KE8A4, антитело KE8A8, антитело KE8G11, антитело KE8B7, антитело KE8F1, антитело KE8C4, антитело KE8A3, антитело KE86G9, антитело KE8H6, антитело KA3, антитело KA3-P5G2 (также именуемое в настоящем документе KA3G2), антитело KA3-P1A3 (также именуемое в настоящем документе KA3A3), антитело KA3-P5C5 (также именуемое в настоящем документе KA3C5), антитело KA3H8, антитело KA3B2, антитело KA3A2, антитело KA3D3, антитело KA3H3, антитело KC4, антитело KC4-P1G11KC4-P4C11, антитело KC4-P6B1KC4-P4F4 и антитело KC4-P2E2 (также именуемое в настоящем документе KC4E2), антитело KC4, антитело KC4F4, антитело KC4A1, антитело KC4C11, антитело KC4E10, антитело KC4B1, антитело KC4C3, антитело KC4A4, антитело KC4G11 и антитело KC4G9, а также их иммунологически активные и/или антигенсвязывающие фрагменты.
В некоторых вариантах осуществления изобретения примеры анти-CD47 моноклональных антител, моноспецифичных анти-CD47 антител, анти-CD47 моновалентных антител и/или биспецифичных антител по изобретению включают комбинацию определяющих комплементарность областей (CDR) тяжелых и легких цепей, выбранных из последовательностей CDR, представленных в таблицах 1, 2 и 3, где CDR, представленные в таблицах 1, 2 и 3, определяются в соответствии с номенклатурой IMGT.
В некоторых вариантах осуществления изобретения примеры анти-CD47 моноклональных антител, моноспецифичных анти-CD47 антител, анти-CD47 моновалентных антител и/или биспецифичных антител по изобретению включают комбинацию последовательностей CDR тяжелых цепей из таблицы 1 и двух наборов CDR легких цепей, выбранных из последовательностей CDRL1, CDRL2 и CDRL3, представленных в таблицах 2 и 3.
В некоторых вариантах осуществления примеры анти-CD47 моноклональных антител, моноспецифичных анти-CD47 антител, анти-CD47 моновалентных антител и/или биспецифичных антител по изобретению включают комбинацию последовательностей CDR тяжелых цепей из таблицы 1 и первого набора CDR легких цепей, выбранных из последовательностей CDRL1, CDRL2 и CDRL3, представленных в таблице 2, и второго набора CDR легких цепей, выбранных из последовательностей CDRL1, CDRL2 и CDRL3, представленных в таблице 3.
CDR тяжелых цепей анти-CD47 антител
(SEQ ID NO: 225)
(SEQ ID NO: 226)
(SEQ ID NO: 227)
CDR легких каппа-цепей анти-CD47 антител
(SEQ ID NO: 228)
(SEQ ID NO: 242)
(SEQ ID NO: 246)
(SEQ ID NO: 229)
(SEQ ID NO: 243)
(SEQ ID NO: 247)
(SEQ ID NO: 230)
(SEQ ID NO: 244)
(SEQ ID NO: 248)
(SEQ ID NO: 231)
(SEQ ID NO: 245)
(SEQ ID NO: 249)
(SEQ ID NO: 232)
(SEQ ID NO: 250)
(SEQ ID NO: 233)
(SEQ ID NO: 251)
(SEQ ID NO: 234)
(SEQ ID NO: 252)
(SEQ ID NO: 235)
(SEQ ID NO: 253)
(SEQ ID NO: 236)
(SEQ ID NO: 254)
(SEQ ID NO: 237)
(SEQ ID NO: 255)
(SEQ ID NO: 238)
(SEQ ID NO: 256)
(SEQ ID NO: 239)
(SEQ ID NO: 257)
(SEQ ID NO: 240)
(SEQ ID NO: 258)
(SEQ ID NO: 241)
(SEQ ID NO: 259)
(SEQ ID NO: 260)
(SEQ ID NO: 261)
CDR легких лямбда-цепей анти-CD47 антител
(SEQ ID NO: 262)
(SEQ ID NO: 273)
(SEQ ID NO: 281)
(SEQ ID NO: 263)
(SEQ ID NO: 274)
(SEQ ID NO: 264)
(SEQ ID NO: 275)
(SEQ ID NO: 265)
(SEQ ID NO: 276)
(SEQ ID NO: 266)
(SEQ ID NO: 277)
(SEQ ID NO: 267)
(SEQ ID NO: 278)
(SEQ ID NO: 268)
(SEQ ID NO: 279)
(SEQ ID NO: 269)
(SEQ ID NO: 280)
(SEQ ID NO: 270)
(SEQ ID NO: 271)
(SEQ ID NO: 272)
Каждый из примеров анти-CD47, анти-CD19, моновалентных и биспецифичных антител, описанных в настоящем документе, включает общую тяжелую цепь (HC), одну каппа-цепь или одну лямбда-цепь для анти-CD47 и анти-CD19 антител, одну каппа и одну лямбда легкие цепи (LC) для моновалентных и биспецифичных антител, как показано в аминокислотных и соответствующих нуклеотидных последовательностях, приведенных ниже. Каждый из примеров анти-CD47, анти-CD19, моновалентных и биспецифичных антител, описанных ниже, включает общую вариабельную область тяжелой цепи (VH), один вариабельный домен легкой каппа-цепи или один вариабельный домен легкой лямбда-цепи для анти-CD47 и анти-CD19 антител, один вариабельный домен легкой каппа-цепи и один вариабельный домен легкой лямбда-цепи (VL) для моновалентных и биспецифичных антител, как показано в аминокислотных и соответствующих нуклеотидных последовательностях, приведенных ниже.
Хотя указанные ниже последовательности антител приведены в настоящем документе в качестве примеров, следует понимать, что эти последовательности могут быть использованы для создания биспецифичных антител с использованием любой из множества методик, известных в данной области. Примеры биспецифичных форматов включают, но не ограничиваются ими, биспецифичные IgG на основе обмена Fab-плеч (Gramer et al., 2013 Mabs. 5(6)); CrossMab-формат (Klein C et al., 2012 MAbs 4(6)); множество форматов, основанных на подходах принудительной гетеродимеризации, таких как SEED-технология (Davis JH et al., 2010 Protein Eng Des Sel. 23(4): 195-202), электростатическая адресация (Gunasekaran K et al., J Biol Chem. 2010 285(25): 19637-46.) или подход «узел-впадина» (Ridgway JB et al., Protein Eng. 1996 9(7): 617-21.) или другие наборы мутаций, препятствующих образованию гомодимеров (Von Kreudenstein TS et al., 2013 MAbs. 5(5): 646-54.); биспецифичные форматы на основе фрагментов, такие как тандемные scFv (такие asBiTE) (Wolf E et al., 2005 Drug Discov. Today 10(18): 1237-44.); биспецифичные тетравалентные антитела (Pörtner LM et al., 2012 Cancer Immunol Immunother. 61(10): 1869-75.); переориентирующие молекулы с двойной аффинностью (Moore PA et al., 2011 Blood. 117(17): 4542-51), диатела (Kontermann RE et al., Nat Biotechnol. 1997 15(7): 629-31).
Примеры анти-CD47, анти-CD19, моновалентных и биспецифичных антител включают общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1.
Нуклеотидная последовательность общей тяжелой цепи (SEQ ID NO: 1)
GAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGCACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAAGTTATGGTGCTTTTGACTACTGGGGCCAGGGAACCCTGGTCACAGTCTCGAGCGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACAGTCTCGTGGAACTCAGGAGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAGAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTATACCCTGCCCCCATCTCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACTTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAACGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGTCCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTTAA
Аминокислотная последовательность общей тяжелой цепи (SEQ ID NO: 2)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSYGAFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Примеры анти-CD47, анти-CD19, моновалентных и биспецифичных антител включают общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113.
Нуклеотидная последовательность общего вариабельного домена тяжелой цепи (SEQ ID NO: 113)
GAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGCACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAAGTTATGGTGCTTTTGACTACTGGGGCCAGGGAACCCTGGTCACAGTCTCGAGC
Аминокислотная последовательность общего вариабельного домена тяжелой цепи (SEQ ID NO: 114)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSYGAFDYWGQGTLVTVSS
АНТИ-CD47 АНТИТЕЛА
Антитело 5A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 4), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 3.
Нуклеотидная последовательность легкой цепи антитела 5А3 (SEQ ID NO: 3)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGGTGCATCCAGGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела 5А3 (SEQ ID NO: 4)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYGASRLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело 5A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 116), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 115.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела 5А3 (SEQ ID NO: 115)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGGTGCATCCAGGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела 5А3 (SEQ ID NO: 116)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYGASRLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRGPRTFGQGTKVEIK
Антитело 5A3-M4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 6), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 5.
Нуклеотидная последовательность легкой цепи антитела 5A3-M4 (SEQ ID NO: 5)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGGTGCATCCAGGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGAACCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела 5A3-M4 (SEQ ID NO: 6)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYGASRLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRNPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело 5A3-M4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 118), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 117.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела 5А3-М4 (SEQ ID NO: 117)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGGTGCATCCAGGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGAACCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела 5А3-М4 (SEQ ID NO: 118)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYGASRLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRNPRTFGQGTKVEIK
Антитело 5A3-M3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 8), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 7.
Нуклеотидная последовательность легкой цепи антитела 5A3-M3 (SEQ ID NO: 7)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGTCCATTAGTAGTTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGCTGCATCCTCGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела 5A3-M3 (SEQ ID NO: 8)
DIQMTQSPSSLSASVGDRVTITCQASQSISSYLNWYQQKPGKAPKLLIYAASSLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело 5A3-M3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 120), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 119.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела 5А3-М3 (SEQ ID NO: 119)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGTCCATTAGTAGTTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGCTGCATCCTCGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела 5А3-М3 (SEQ ID NO: 120)
DIQMTQSPSSLSASVGDRVTITCQASQSISSYLNWYQQKPGKAPKLLIYAASSLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRGPRTFGQGTKVEIK
Антитело 5A3-M5 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 10), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 9.
Нуклеотидная последовательность легкой цепи антитела 5A3-M5 (SEQ ID NO: 9)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGGTGCATCCAGGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGTACCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела 5A3-M5 (SEQ ID NO: 10)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYGASRLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRYPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело 5A3-M5 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 122), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 121.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела 5А3-М5 (SEQ ID NO: 121)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTACGGTGCATCCAGGTTGGAAACAGGGGTCCCATCAAGGTTCAGTGGAAGTGGATCTGGGACAGATTTTACTTTCACCATCAGCAGCCTGCAGCCTGAAGATATTGCAACATATTACTGTCAGCAGAAGCACCCCCGGTACCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела 5А3-М5 (SEQ ID NO: 122)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYGASRLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQKHPRYPRTFGQGTKVEIK
Антитело Ke8 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 12), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 11.
Нуклеотидная последовательность легкой цепи антитела Ke8 (SEQ ID NO: 11)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCACAAGCGGCGGCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8 (SEQ ID NO: 12)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRRPQTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 124), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 123.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела Ке8 (SEQ ID NO: 123)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCACAAGCGGCGGCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8 (SEQ ID NO: 124)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRRPQTFGQGTKVEIK
Антитело Ke8H5 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 14), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 13.
Нуклеотидная последовательность легкой цепи антитела KE8H5 (SEQ ID NO: 13)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCGAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCATAAGCGTGCGCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8H5 (SEQ ID NO: 14)
DIQMTQSPSSLSASVGDRVTITCRASQSIARYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRAPQTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8H5 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 126), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 125.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8Н5 (SEQ ID NO: 125)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCGAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCATAAGCGTGCGCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8Н5 (SEQ ID NO: 126)
DIQMTQSPSSLSASVGDRVTITCRASQSIARYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRAPQTFGQGTKVEIK
Антитело Ke8B2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 16), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 15.
Нуклеотидная последовательность легкой цепи антитела KE8B2 (SEQ ID NO: 15)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCACCCGCGTGCCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8B2 (SEQ ID NO: 16)
DIQMTQSPSSLSASVGDRVTITCRASQSIGKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRAPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8B2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 128), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 127.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8В2 (SEQ ID NO: 127)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCACCCGCGTGCCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8В2 (SEQ ID NO: 128)
DIQMTQSPSSLSASVGDRVTITCRASQSIGKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRAPRTFGQGTKVEIK
Антитело Ke8A2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 18), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 17.
Нуклеотидная последовательность легкой цепи антитела KE8A2 (SEQ ID NO: 17)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCCCGTGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8A2 (SEQ ID NO: 18)
DIQMTQSPSSLSASVGDRVTITCRASQSIDRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8A2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 130), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 129.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8А2 (SEQ ID NO: 129)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCCCGTGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8А2 (SEQ ID NO: 130)
DIQMTQSPSSLSASVGDRVTITCRASQSIDRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRGPRTFGQGTKVEIK
Антитело Ke8E8 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 20), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 19.
Нуклеотидная последовательность легкой цепи антитела KE8E8 (SEQ ID NO: 19)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCCCGTGGCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8E8 (SEQ ID NO: 20)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8E8 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 132), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 131.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8Е8 (SEQ ID NO: 131)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCCCGTGGCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8Е8 (SEQ ID NO: 132)
DIQMTQSPSSLSASVGDRVTITCQASQDINKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRGPRTFGQGTKVEIK
Антитело Ke8H3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 22), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 21.
Нуклеотидная последовательность легкой цепи антитела KE8H3 (SEQ ID NO: 21)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCGCGTGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8H3 (SEQ ID NO: 22)
DIQMTQSPSSLSASVGDRVTITCRASQSINRYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8H3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 134), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 133.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8Н3 (SEQ ID NO: 133)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCGCGTGGGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8Н3 (SEQ ID NO: 134)
DIQMTQSPSSLSASVGDRVTITCRASQSINRYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRGPRTFGQGTKVEIK
Антитело Ke8G6 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 24), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 23.
Нуклеотидная последовательность легкой цепи антитела KE8G6 (SEQ ID NO: 23)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGCGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8G6 (SEQ ID NO: 24)
DIQMTQSPSSLSASVGDRVTITCRASQSIGRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8G6 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 136), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 135.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE8G6 (SEQ ID NO: 135)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGCGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE8G6 (SEQ ID NO: 136)
DIQMTQSPSSLSASVGDRVTITCRASQSIGRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Ke8A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 26), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 25.
Нуклеотидная последовательность легкой цепи антитела KE8A3 (SEQ ID NO: 25)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGTAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCCCGTGGGCCGAGCACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8A3 (SEQ ID NO: 26)
DIQMTQSPSSLSASVGDRVTITCRVSQSISKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPSTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 138), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 137.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8А3 (SEQ ID NO: 137)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGTAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCCCGTGGGCCGAGCACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8А3 (SEQ ID NO: 138)
DIQMTQSPSSLSASVGDRVTITCRVSQSISKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPSTFGQGTKVEIK
Антитело Ke81A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 28), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 27.
Нуклеотидная последовательность легкой цепи антитела KE81A3 (SEQ ID NO: 27)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCGCGTGCCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE81A3 (SEQ ID NO: 28)
DIQMTQSPSSLSASVGDRVTITCQASQDINRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRAPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke81A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 140), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 139.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ81А3 (SEQ ID NO: 139)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCGCGTGCCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ81А3 (SEQ ID NO: 140)
DIQMTQSPSSLSASVGDRVTITCQASQDINRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRAPRTFGQGTKVEIK
Антитело Ke8A8 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 30), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 29.
Нуклеотидная последовательность легкой цепи антитела KE8A8 (SEQ ID NO: 29)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGCGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8A8 (SEQ ID NO: 30)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYAASTLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8A8 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 142), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 141.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8А8 (SEQ ID NO: 141)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGCGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8А8 (SEQ ID NO: 142)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYAASTLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Ke8C7 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 32), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 31.
Нуклеотидная последовательность легкой цепи антитела KE8C7 (SEQ ID NO: 31)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGCGCCATCCGCGTGGCCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8C7 (SEQ ID NO: 32)
DIQMTQSPSSLSASVGDRVTITCQASQDINRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8C7 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 144), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 143.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8С7 (SEQ ID NO: 143)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTAATAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGCGCCATCCGCGTGGCCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8С7 (SEQ ID NO: 144)
DIQMTQSPSSLSASVGDRVTITCQASQDINRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPRTFGQGTKVEIK
Антитело Ke8G2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 34), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 33.
Нуклеотидная последовательность легкой цепи антитела KE8G2 (SEQ ID NO: 33)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAACAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCCCGTGCGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8G2 (SEQ ID NO: 34)
DIQMTQSPSSLSASVGDRVTITCRASQSIGRYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCQQKHPRAPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8G2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 146), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 145.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE8G2 (SEQ ID NO: 145)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAACAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCCCGTGCGCCGAGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE8G2 (SEQ ID NO: 146)
DIQMTQSPSSLSASVGDRVTITCRASQSIGRYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCQQKHPRAPRTFGQGTKVEIK
Антитело Ke81G9 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 36), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 35.
Нуклеотидная последовательность легкой цепи антитела KE81G9 (SEQ ID NO: 35)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGCGGCATAAGCGTTCCCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE81G9 (SEQ ID NO: 36)
DIQMTQSPSSLSASVGDRVTITCRASQSIDKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHKRSPQTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke81G9 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 148), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 147.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE81G9 (SEQ ID NO: 147)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGCGGCATAAGCGTTCCCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE81G9 (SEQ ID NO: 148)
DIQMTQSPSSLSASVGDRVTITCRASQSIDKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHKRSPQTFGQGTKVEIK
Антитело Ke8F2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 38), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 37.
Нуклеотидная последовательность легкой цепи антитела KE8F2 (SEQ ID NO: 37)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCGCGTGCGCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8F2 (SEQ ID NO: 38)
DIQMTQSPSSLSASVGDRVTITCRASQSIDKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRAPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8F2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 150), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 149.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE8F2 (SEQ ID NO: 149)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCGCGTGCGCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE8F2 (SEQ ID NO: 150)
DIQMTQSPSSLSASVGDRVTITCRASQSIDKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRAPRTFGQGTKVEIK
Антитело Ke8B7 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 40), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 39.
Нуклеотидная последовательность легкой цепи антитела KE8B7 (SEQ ID NO: 39)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGGAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTAGCCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8B7 (SEQ ID NO: 40)
DIQMTQSPSSLSASVGDRVTITCRASQSIGKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8B7 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 152), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 151.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8В7 (SEQ ID NO: 151)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGGAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTAGCCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8В7 (SEQ ID NO: 152)
DIQMTQSPSSLSASVGDRVTITCRASQSIGKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIK
Антитело Ke8C4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 42), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 41.
Нуклеотидная последовательность легкой цепи антитела KE8C4 (SEQ ID NO: 41)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAATTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGGGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8C4 (SEQ ID NO: 42)
DIQMTQSPSSLSASVGDRVTITCRASQSISRYLNWYQQKPGKAPKLLIYAASNLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRGPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8C4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 154), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 153.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8С4 (SEQ ID NO: 153)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAATTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGGGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8С4 (SEQ ID NO: 154)
DIQMTQSPSSLSASVGDRVTITCRASQSISRYLNWYQQKPGKAPKLLIYAASNLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRGPKTFGQGTKVEIK
Антитело Ke8F1 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 44), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 43.
Нуклеотидная последовательность легкой цепи антитела KE8F1 (SEQ ID NO: 43)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCTTCTTATGTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCGGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCATAAGCGTCGGCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8F1 (SEQ ID NO: 44)
DIQMTQSPSSLSASVGDRVTITCRASQSIASYVNWYQQKPGKAPKLLIYAASGLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRRPQTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8F1 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 156), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 155.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE8F1 (SEQ ID NO: 155)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCTTCTTATGTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCGGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCATAAGCGTCGGCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE8F1 (SEQ ID NO: 156)
DIQMTQSPSSLSASVGDRVTITCRASQSIASYVNWYQQKPGKAPKLLIYAASGLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRRPQTFGQGTKVEIK
Антитело Ke8G11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 46), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 45.
Нуклеотидная последовательность легкой цепи антитела KE8G11 (SEQ ID NO: 45)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGGAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGGGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8G11 (SEQ ID NO: 46)
DIQMTQSPSSLSASVGDRVTITCRASQSIGRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRGPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8G11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 158), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 157.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE8G11 (SEQ ID NO: 157)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGGAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCGCGTGGGCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE8G11 (SEQ ID NO: 158)
DIQMTQSPSSLSASVGDRVTITCRASQSIGRYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRGPKTFGQGTKVEIK
Антитело Ke8H6 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 48), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 47.
Нуклеотидная последовательность легкой цепи антитела KE8H6 (SEQ ID NO: 47)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAATGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCGCGTGGGCCGCGCACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8H6 (SEQ ID NO: 48)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYNASTLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8H6 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 160), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 159.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8Н6 (SEQ ID NO: 159)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAATGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCGCGTGGGCCGCGCACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8Н6 (SEQ ID NO: 160)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYNASTLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPRTFGQGTKVEIK
Антитело Ke84G9 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 50), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 49.
Нуклеотидная последовательность легкой цепи антитела KE84G9 (SEQ ID NO: 49)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCGCGTAGCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE84G9 (SEQ ID NO: 50)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRSPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke84G9 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 162), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 161.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE84G9 (SEQ ID NO: 161)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAAGCATCCGCGTAGCCCGCGGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE84G9 (SEQ ID NO: 162)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQKHPRSPRTFGQGTKVEIK
Антитело Ke8A4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 52), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 51.
Нуклеотидная последовательность легкой цепи антитела KE8A4 (SEQ ID NO: 51)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCATAAGCGTAGCCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE8A4 (SEQ ID NO: 52)
DIQMTQSPSSLSASVGDRVTITCRASQSIAKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRSPQTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke8A4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 164), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 163.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КЕ8А4 (SEQ ID NO: 163)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGTTCCATAAGCGTAGCCCGCAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КЕ8А4 (SEQ ID NO: 164)
DIQMTQSPSSLSASVGDRVTITCRASQSIAKYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFHKRSPQTFGQGTKVEIK
Антитело Ke86G9 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 54), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 53.
Нуклеотидная последовательность легкой цепи антитела KE86G9 (SEQ ID NO: 53)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAATGCATCCAATTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCGCGTGGGCCGACCACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KE86G9 (SEQ ID NO: 54)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYNASNLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPTTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ke86G9 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 166), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 165.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KE86G9 (SEQ ID NO: 165)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAATGCATCCAATTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGAGGCATCCGCGTGGGCCGACCACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KE86G9 (SEQ ID NO: 166)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYNASNLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQRHPRGPTTFGQGTKVEIK
Антитело Ka3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 56), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 55.
Нуклеотидная последовательность легкой цепи антитела КА3 (SEQ ID NO: 55)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCACCCGCGCGCCCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3 (SEQ ID NO: 56)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 168), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 167.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3 (SEQ ID NO: 167)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGCAGCTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCACCCGCGCGCCCCGAAGACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3 (SEQ ID NO: 168)
DIQMTQSPSSLSASVGDRVTITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Ka3A2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 58), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 57.
Нуклеотидная последовательность легкой цепи антитела КА3А2 (SEQ ID NO: 57)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCTCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3A2 (SEQ ID NO: 58)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3A2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 170), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 169.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3А2 (SEQ ID NO: 169)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCTCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3А2 (SEQ ID NO: 170)
DIQMTQSPSSLSASVGDRVTITCRASQSISKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIK
Антитело Ka3H3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 60), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 59.
Нуклеотидная последовательность легкой цепи антитела КА3Н3 (SEQ ID NO: 59)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTGCTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCGCTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCTCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3H3 (SEQ ID NO: 60)
DIQMTQSPSSLSASVGDRVTITCQASQDIAKYLNWYQQKPGKAPKLLIYAASALQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3H3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 172), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 171.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3Н3 (SEQ ID NO: 171)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCAGGCGAGTCAGGACATTGCTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCGCTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCTCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3Н3 (SEQ ID NO: 172)
DIQMTQSPSSLSASVGDRVTITCQASQDIAKYLNWYQQKPGKAPKLLIYAASALQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIK
Антитело Ka3A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 62), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 61.
Нуклеотидная последовательность легкой цепи антитела КА3А3 (SEQ ID NO: 61)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCTAGTTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3A3 (SEQ ID NO: 62)
DIQMTQSPSSLSASVGDRVTITCRASQSIASYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 174), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 173.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3А3 (SEQ ID NO: 173)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCTAGTTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3А3 (SEQ ID NO: 174)
DIQMTQSPSSLSASVGDRVTITCRASQSIASYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Ka3H8 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 64), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 63.
Нуклеотидная последовательность легкой цепи антитела КА3Н8 (SEQ ID NO: 63)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCGAGTTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCTCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3H8 (SEQ ID NO: 64)
DIQMTQSPSSLSASVGDRVTITCRASQSIASYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3H8 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 176), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 175.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3Н8 (SEQ ID NO: 175)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGCGAGTTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCGGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCTCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3Н8 (SEQ ID NO: 176)
DIQMTQSPSSLSASVGDRVTITCRASQSIASYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRSPKTFGQGTKVEIK
Антитело Ka3B2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 66), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 65.
Нуклеотидная последовательность легкой цепи антитела КА3В2 (SEQ ID NO: 65)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAACATTGGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAGTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3B2 (SEQ ID NO: 66)
DIQMTQSPSSLSASVGDRVTITCRASQNIGKYLNWYQQKPGKAPKLLIYSASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3B2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 178), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 177.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3В2 (SEQ ID NO: 177)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAACATTGGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATAGTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3В2 (SEQ ID NO: 178)
DIQMTQSPSSLSASVGDRVTITCRASQNIGKYLNWYQQKPGKAPKLLIYSASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Ka3C5 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 68), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 67.
Нуклеотидная последовательность легкой цепи антитела КА3С5 (SEQ ID NO: 67)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATTCTGCATCCTCTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCCCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3C5 (SEQ ID NO: 68)
DIQMTQSPSSLSASVGDRVTITCRASQSISRYLNWYQQKPGKAPKLLIYSASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3C5 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 180), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 179.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КА3С5 (SEQ ID NO: 179)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAGTAGGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATTCTGCATCCTCTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCCCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КА3С5 (SEQ ID NO: 180)
DIQMTQSPSSLSASVGDRVTITCRASQSISRYLNWYQQKPGKAPKLLIYSASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Ka3G2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 70), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 69.
Нуклеотидная последовательность легкой цепи антитела KA3G2 (SEQ ID NO: 69)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGGGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3G2 (SEQ ID NO: 70)
DIQMTQSPSSLSASVGDRVTITCRASQSIDKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRGPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3G2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 182), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 181.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KA3G2 (SEQ ID NO: 181)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGATAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGGGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KA3G2 (SEQ ID NO: 182)
DIQMTQSPSSLSASVGDRVTITCRASQSIDKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRGPKTFGQGTKVEIK
Антитело Ka3D3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую каппа-цепь (SEQ ID NO: 72), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 71.
Нуклеотидная последовательность легкой цепи антитела KA3D3 (SEQ ID NO: 71)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность легкой цепи антитела KA3D3 (SEQ ID NO: 72)
DIQMTQSPSSLSASVGDRVTITCRASQSIGKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Антитело Ka3D3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой каппа-цепи (SEQ ID NO: 184), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 183.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KA3D3 (SEQ ID NO: 183)
GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTGGTAAGTATTTAAATTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGGTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGTCTGCAACCTGAAGATTTTGCAACTTACTACTGTCAGCAGATGCATCCTCGCGCGCCGAAAACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KA3D3 (SEQ ID NO: 184)
DIQMTQSPSSLSASVGDRVTITCRASQSIGKYLNWYQQKPGKAPKLLIYAASRLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQMHPRAPKTFGQGTKVEIK
Антитело Kc4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 74), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 73.
Нуклеотидная последовательность легкой цепи антитела КС4 (SEQ ID NO: 73)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGTGGTTATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGGTCAGTAATCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4 (SEQ ID NO: 74)
QSALTQPASVSGSPGQSITISCTGTSSDVGGYNYVSWYQQHPGKAPKLMIYEVSNRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 186), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 185.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4 (SEQ ID NO: 185)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGTGGTTATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGGTCAGTAATCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4 (SEQ ID NO: 186)
QSALTQPASVSGSPGQSITISCTGTSSDVGGYNYVSWYQQHPGKAPKLMIYEVSNRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4G11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 76), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 75.
Нуклеотидная последовательность легкой цепи антитела KC4G11 (SEQ ID NO: 75)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGGAAGGCGAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGGATAGTGATCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4G11 (SEQ ID NO: 76)
QSALTQPASVSGSPGQSITISCTGTSSDVGKANYVSWYQQHPGKAPKLMIYKDSDRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4G11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 188), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 187.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KC4G11 (SEQ ID NO: 187)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGGAAGGCGAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGGATAGTGATCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KC4G11 (SEQ ID NO: 188)
QSALTQPASVSGSPGQSITISCTGTSSDVGKANYVSWYQQHPGKAPKLMIYKDSDRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4C11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 78), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 77.
Нуклеотидная последовательность легкой цепи антитела КС4С11 (SEQ ID NO: 77)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGGGGAATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAATAGTAAGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4C11 (SEQ ID NO: 78)
QSALTQPASVSGSPGQSITISCTGTSSDVRGNNYVSWYQQHPGKAPKLMIYENSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4C11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 190), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 189.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4С11 (SEQ ID NO: 189)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGGGGAATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAATAGTAAGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4С11 (SEQ ID NO: 190)
QSALTQPASVSGSPGQSITISCTGTSSDVRGNNYVSWYQQHPGKAPKLMIYENSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4A1 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 80), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 79.
Нуклеотидная последовательность легкой цепи антитела КС4А1 (SEQ ID NO: 79)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGTGCGAGGAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAGTAGTAAGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4A1 (SEQ ID NO: 80)
QSALTQPASVSGSPGQSITISCTGTSSDVSARNYVSWYQQHPGKAPKLMIYESSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4A1 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 192), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 191.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4А1 (SEQ ID NO: 191)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGTGCGAGGAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAGTAGTAAGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4А1 (SEQ ID NO: 192)
QSALTQPASVSGSPGQSITISCTGTSSDVSARNYVSWYQQHPGKAPKLMIYESSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4A4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 82), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 81.
Нуклеотидная последовательность легкой цепи антитела КС4А4 (SEQ ID NO: 81)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTAGAACCAGCAGTGACGTTAATAATACTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGACTAGTGGTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4A4 (SEQ ID NO: 82)
QSALTQPASVSGSPGQSITISCTRTSSDVNNTNYVSWYQQHPGKAPKLMIYKTSGRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4A4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 194), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 193.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4А4 (SEQ ID NO: 193)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTAGAACCAGCAGTGACGTTAATAATACTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGACTAGTGGTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4А4 (SEQ ID NO: 194)
QSALTQPASVSGSPGQSITISCTRTSSDVNNTNYVSWYQQHPGKAPKLMIYKTSGRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4E10 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 84), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 83.
Нуклеотидная последовательность легкой цепи антитела КС4Е10 (SEQ ID NO: 83)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAATTCTGCTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGAGTAGTAGTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4E10 (SEQ ID NO: 84)
QSALTQPASVSGSPGQSITISCTGTSSDVNSANYVSWYQQHPGKAPKLMIYKSSSRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4E10 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 196), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 195.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4Е10 (SEQ ID NO: 195)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAATTCTGCTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGAGTAGTAGTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4Е10 (SEQ ID NO: 196)
QSALTQPASVSGSPGQSITISCTGTSSDVNSANYVSWYQQHPGKAPKLMIYKSSSRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4G9 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 86), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 85.
Нуклеотидная последовательность легкой цепи антитела KC4G9 (SEQ ID NO: 85)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCCGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGAGAGGAAGAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGAATAGTACTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4G9 (SEQ ID NO: 86)
QSALTQPASVSGSPGQSITISCTGTSSDVERKNYVSWYQQHPGKAPKLMIYKNSTRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4G9 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 198), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 197.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KC4G9 (SEQ ID NO: 197)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCCGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGAGAGGAAGAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGAATAGTACTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KC4G9 (SEQ ID NO: 198)
QSALTQPASVSGSPGQSITISCTGTSSDVERKNYVSWYQQHPGKAPKLMIYKNSTRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4C3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 88), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 87.
Нуклеотидная последовательность легкой цепи антитела КС4С3 (SEQ ID NO: 87)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGGCGGCTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGAATAGTACTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4C3 (SEQ ID NO: 88)
QSALTQPASVSGSPGQSITISCTGTSSDVRAANYVSWYQQHPGKAPKLMIYKNSTRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4C3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 200), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 199.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4С3 (SEQ ID NO: 199)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGGCGGCTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATAAGAATAGTACTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4С3 (SEQ ID NO: 200)
QSALTQPASVSGSPGQSITISCTGTSSDVRAANYVSWYQQHPGKAPKLMIYKNSTRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4F4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 90), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 89.
Нуклеотидная последовательность легкой цепи антитела KC4F4 (SEQ ID NO: 89)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGAGGGCTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATCAGGATAGTAGTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4F4 (SEQ ID NO: 90)
QSALTQPASVSGSPGQSITISCTGTSSDVRRANYVSWYQQHPGKAPKLMIYQDSSRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4F4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 202), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 201.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела KC4F4 (SEQ ID NO: 201)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGAGGGCTAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATCAGGATAGTAGTCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела KC4F4 (SEQ ID NO: 202)
QSALTQPASVSGSPGQSITISCTGTSSDVRRANYVSWYQQHPGKAPKLMIYQDSSRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4B1 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 92), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 91.
Нуклеотидная последовательность легкой цепи антитела КС4В1 (SEQ ID NO: 91)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGGCTAATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAGTAGTGCGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4B1 (SEQ ID NO: 92)
QSALTQPASVSGSPGQSITISCTGTSSDVRANNYVSWYQQHPGKAPKLMIYESSARPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4B1 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 204), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 203.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4В1 (SEQ ID NO: 203)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTAGGGCTAATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAGTAGTGCGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4В1 (SEQ ID NO: 204)
QSALTQPASVSGSPGQSITISCTGTSSDVRANNYVSWYQQHPGKAPKLMIYESSARPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
Антитело Kc4E2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 94), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 93.
Нуклеотидная последовательность легкой цепи антитела КС4Е2 (SEQ ID NO: 93)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTTATTATAATAAGTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAGTAGTAAGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела KC4E2 (SEQ ID NO: 94)
QSALTQPASVSGSPGQSITISCTGTSSDVYYNKYVSWYQQHPGKAPKLMIYESSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело Kc4E2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 206), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 205.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела КС4Е2 (SEQ ID NO: 205)
CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTTATTATAATAAGTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGAGTAGTAAGCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCAGCTCATATGATTGGTGGTTCCGCCCCAAGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела КС4Е2 (SEQ ID NO: 206)
QSALTQPASVSGSPGQSITISCTGTSSDVYYNKYVSWYQQHPGKAPKLMIYESSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYDWWFRPKVFGGGTKLTVL
АНТИ-CD19 АНТИТЕЛА
Антитело C2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95.
Нуклеотидная последовательность легкой цепи антитела С2 (SEQ ID NO: 95)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGAAGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACCAGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела С2 (SEQ ID NO: 96)
NFMLTQPHSVSESPGKTVTISCTRSSGSIEDKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDQSLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело C2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела С2 (SEQ ID NO: 207)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGAAGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACCAGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела С2 (SEQ ID NO: 208)
NFMLTQPHSVSESPGKTVTISCTRSSGSIEDKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDQSLYGWVFGGGTKLTVL
Антитело A6 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 98), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 97.
Нуклеотидная последовательность легкой цепи антитела А6 (SEQ ID NO: 97)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGGTGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACGTACGACGAGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела А6 (SEQ ID NO: 98)
NFMLTQPHSVSESPGKTVTISCTRSSGSIGDKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDESLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело A6 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 210), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 209.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела А6 (SEQ ID NO: 209)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGGTGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACGTACGACGAGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела А6 (SEQ ID NO: 210)
NFMLTQPHSVSESPGKTVTISCTRSSGSIGDKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDESLYGWVFGGGTKLTVL
Антитело C6 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 100), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 99.
Нуклеотидная последовательность легкой цепи антитела С6 (SEQ ID NO: 99)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCAATGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTTTGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACACCAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела С6 (SEQ ID NO: 100)
NFMLTQPHSVSESPGKTVTISCTRSSGSINDKYVQWYQQRPGSSPTIVIYFDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDTSLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело C6 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 212), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 211.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела С6 (SEQ ID NO: 211)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCAATGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTTTGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACACCAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела С6 (SEQ ID NO: 212)
NFMLTQPHSVSESPGKTVTISCTRSSGSINDKYVQWYQQRPGSSPTIVIYFDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDTSLYGWVFGGGTKLTVL
Антитело C9 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 102), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 101.
Нуклеотидная последовательность легкой цепи антитела С9 (SEQ ID NO: 101)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGCTGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACGAGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела С9 (SEQ ID NO: 102)
NFMLTQPHSVSESPGKTVTISCTRSSGSIADKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDESLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело C9 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 214), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 213.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела С9 (SEQ ID NO: 213)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGCTGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACGAGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела С9 (SEQ ID NO: 214)
NFMLTQPHSVSESPGKTVTISCTRSSGSIADKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDESLYGWVFGGGTKLTVL
Антитело B11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 104), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 103.
Нуклеотидная последовательность легкой цепи антитела В11 (SEQ ID NO: 103)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGAAGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACAACAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела В11 (SEQ ID NO: 104)
NFMLTQPHSVSESPGKTVTISCTRSSGSIEDKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDNSLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело B11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 216), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 215.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела В11 (SEQ ID NO: 215)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGAAGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCATTGTGATCTATTATGATAACGAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGACCTACGACAACAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела В11 (SEQ ID NO: 216)
NFMLTQPHSVSESPGKTVTISCTRSSGSIEDKYVQWYQQRPGSSPTIVIYYDNERPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQTYDNSLYGWVFGGGTKLTVL
Антитело D11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105.
Нуклеотидная последовательность легкой цепи антитела D11 (SEQ ID NO: 105)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCAGCATCGATGATAAGTTTGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCACTGTGATCTATTATGATAACATTAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGTCCTATGACGCGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела D11 (SEQ ID NO: 106)
NFMLTQPHSVSESPGKTVTISCTRSSGSIDDKFVQWYQQRPGSSPTTVIYYDNIRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDASLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело D11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела D11 (SEQ ID NO: 217)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCAGCATCGATGATAAGTTTGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCACTGTGATCTATTATGATAACATTAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGTCCTATGACGCGAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела D11 (SEQ ID NO: 218)
NFMLTQPHSVSESPGKTVTISCTRSSGSIDDKFVQWYQQRPGSSPTTVIYYDNIRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDASLYGWVFGGGTKLTVL
Антитело B7 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, и включает легкую лямбда-цепь (SEQ ID NO: 108), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 107.
Нуклеотидная последовательность легкой цепи антитела В7 (SEQ ID NO: 107)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGCGGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCACTGTGATCTATGAGGATAACCAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGTCCTATGACAGCAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность легкой цепи антитела В7 (SEQ ID NO: 108)
NFMLTQPHSVSESPGKTVTISCTRSSGSIADKYVQWYQQRPGSSPTTVIYEDNQRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDSSLYGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Антитело B7 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, и включает вариабельный домен легкой лямбда-цепи (SEQ ID NO: 220), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 219.
Нуклеотидная последовательность вариабельного домена легкой цепи антитела В7 (SEQ ID NO: 219)
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCTCTATCGCGGATAAGTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCACTGTGATCTATGAGGATAACCAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGTCCTATGACAGCAGCCTGTATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена легкой цепи антитела В7 (SEQ ID NO: 220)
NFMLTQPHSVSESPGKTVTISCTRSSGSIADKYVQWYQQRPGSSPTTVIYEDNQRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDSSLYGWVFGGGTKLTVL
ФИКТИВНЫЕ ЛЕГКИЕ ЦЕПИ
Фиктивная легкая цепь 1 (SEQ ID NO: 110) кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 109.
Нуклеотидная последовательность фиктивной легкой цепи 1 (SEQ ID NO: 109)
CAGTCTGTGTTGACGCAGCCGCCCTCAGTGTCTGCGGCCCCAGGACAGAAGGTCACCATCTCCTGCTCTGGAAGCAGCTCCAATATTGAGACTGGTTCTGTATCCTGGTACCAGCAGCTCCCAGGAACAGCCCCCAAACTCCTCATTTATGACAATAATAAGCGACCCTCAGGGATTCCTGACCGATTCTCTGGCTCCAAGTCTGGCACGTCAGCCACCCTGGGCATCACCGGACTCCAGACTGGGGACGAGGCCGATTATTACTGCGGAACATGGGATGACAGCCTGCCTGGATGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCTTGGAAAGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATAA
Аминокислотная последовательность фиктивной легкой цепи 1 (SEQ ID NO: 110)
QSVLTQPPSVSAAPGQKVTISCSGSSSNIETGSVSWYQQLPGTAPKLLIYDNNKRPSGIPDRFSGSKSGTSATLGITGLQTGDEADYYCGTWDDSLPGWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Вариабельный домен фиктивной легкой цепи 1 (SEQ ID NO: 222) кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
Нуклеотидная последовательность вариабельного домена фиктивной легкой цепи 1 (SEQ ID NO: 221)
CAGTCTGTGTTGACGCAGCCGCCCTCAGTGTCTGCGGCCCCAGGACAGAAGGTCACCATCTCCTGCTCTGGAAGCAGCTCCAATATTGAGACTGGTTCTGTATCCTGGTACCAGCAGCTCCCAGGAACAGCCCCCAAACTCCTCATTTATGACAATAATAAGCGACCCTCAGGGATTCCTGACCGATTCTCTGGCTCCAAGTCTGGCACGTCAGCCACCCTGGGCATCACCGGACTCCAGACTGGGGACGAGGCCGATTATTACTGCGGAACATGGGATGACAGCCTGCCTGGATGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
Аминокислотная последовательность вариабельного домена фиктивной легкой цепи 1 (SEQ ID NO: 222)
QSVLTQPPSVSAAPGQKVTISCSGSSSNIETGSVSWYQQLPGTAPKLLIYDNNKRPSGIPDRFSGSKSGTSATLGITGLQTGDEADYYCGTWDDSLPGWVFGGGTKLTVL
Фиктивная легкая цепь 2 (SEQ ID NO: 112) кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 111.
Нуклеотидная последовательность фиктивной легкой цепи 2 (SEQ ID NO: 111)
GAAATAGTGATGACGCAGTCTCCAGCCACCCTGTCTGTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGACGGTTAAGAATAATTTAGCCTGGTACCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCATCCACCAGGGCCACTGGTATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGAGTTCACTCTCACCATCAGCAGCCTGCAGTCTGAAGATTTTGCAGTTTATTACTGTCAGCAGTATAACAACTGGTTGCCCATCAACCCCTATACCTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA
Аминокислотная последовательность фиктивной легкой цепи 2 (SEQ ID NO: 112)
EIVMTQSPATLSVSPGERATLSCRASQTVKNNLAWYQQKPGQAPRLLIYGASTRATGIPARFSGSGSGTEFTLTISSLQSEDFAVYYCQQYNNWLPINPYTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Вариабельный домен фиктивной легкой цепи 2 (SEQ ID NO: 224) кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 223.
Нуклеотидная последовательность вариабельного домена фиктивной легкой цепи 2 (SEQ ID NO: 223)
GAAATAGTGATGACGCAGTCTCCAGCCACCCTGTCTGTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGACGGTTAAGAATAATTTAGCCTGGTACCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCATCCACCAGGGCCACTGGTATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGAGTTCACTCTCACCATCAGCAGCCTGCAGTCTGAAGATTTTGCAGTTTATTACTGTCAGCAGTATAACAACTGGTTGCCCATCAACCCCTATACCTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотная последовательность вариабельного домена фиктивной легкой цепи 2 (SEQ ID NO: 224)
EIVMTQSPATLSVSPGERATLSCRASQTVKNNLAWYQQKPGQAPRLLIYGASTRATGIPARFSGSGSGTEFTLTISSLQSEDFAVYYCQQYNNWLPINPYTFGQGTKVEIK
МОНОВАЛЕНТНЫЕ АНТИТЕЛА
В некоторых вариантах осуществления изобретения моновалентное антитело 5A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 4), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 3, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело 5A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 116), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 115, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело 5A3-M3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 8), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 7, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело 5A3-M3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 120), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 119, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело 5A3-M5 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 10), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 9, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело 5A3-M5 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 122), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 121, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ke8 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 12), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 11, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ke8 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 124), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 123, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ke8A2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 18), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 17, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ke8A2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 130), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 129, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ke8B2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 16), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 15, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ke8B2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 128), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 127, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ke8G11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 46), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 45, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ke8G11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 158), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 157, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ke8C4 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 42), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 41, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ke8C4 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 154), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 153, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ke8A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 26), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 25, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ke8A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 138), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 137, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ka3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 56), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 55, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ka3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 168), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 167, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ka3A3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 62), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 61, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ka3A3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 174), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 173, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ka3G2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 70), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 69, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ka3G2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 182), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 181, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело Ka3H3 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 60), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 59, и фиктивную легкую лямбда-цепь 1 (SEQ ID NO: 110), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 109. В некоторых вариантах осуществления изобретения моновалентное антитело Ka3H3 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 172), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 171, и вариабельный домен фиктивной легкой лямбда-цепи 1 (SEQ ID NO: 222), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 221.
В некоторых вариантах осуществления изобретения моновалентное антитело C2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, фиктивную легкую каппа-цепь 2 (SEQ ID NO: 112), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 111, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения моновалентное антитело C2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен фиктивной легкой каппа-цепи 2 (SEQ ID NO: 224), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 223, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
БИСПЕЦИФИЧНЫЕ АНТИТЕЛА
В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 4), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 3, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 116), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 115, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M3xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 8), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 7, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M3xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 120), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 119, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M3xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 8), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 7, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M3xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 120), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 119, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M5xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 10), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 9, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M5xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 122), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 121, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M5xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 10), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 9, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело 5A3-M5xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 122), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 121, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 12), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 11, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 124), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 123, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 12), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 11, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 124), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 123, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8A2xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 18), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 17, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8A2xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 130), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 129, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8B2xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 16), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 15, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8B2xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 128), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 127, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8G11xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 46), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 45, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8G11xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 158), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 157, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8C4xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 42), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 41, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8C4xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 154), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 153, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8C4xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 42), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 41, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8C4xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 154), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 153, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8A3xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 26), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 25, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8A3xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 138), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 137, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8A3xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 26), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 25, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело Ke8A3xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 138), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 137, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 56), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 55, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 168), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 167 и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 56), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 55, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 168), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 167, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3A3xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 62), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 61, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3A3xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 174), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 173, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3G2xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 70), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 69, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3G2xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 182), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 181, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3G2xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 70), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 69, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3G2xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 182), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 181, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3H3xD11 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 60), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 59, и легкую лямбда-цепь (SEQ ID NO: 106), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 105. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3H3xD11 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 172), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 171, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 218), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 217.
В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3H3xC2 включает общую тяжелую цепь (SEQ ID NO: 2), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 1, легкую каппа-цепь (SEQ ID NO: 60), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 59, и легкую лямбда-цепь (SEQ ID NO: 96), кодируемую нуклеотидной последовательностью, представленной в SEQ ID NO: 95. В некоторых вариантах осуществления изобретения биспецифичное антитело Ka3H3xC2 включает общий вариабельный домен тяжелой цепи (SEQ ID NO: 114), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 113, вариабельный домен легкой каппа-цепи (SEQ ID NO: 172), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 171, и вариабельный домен легкой лямбда-цепи (SEQ ID NO: 208), кодируемый нуклеотидной последовательностью, представленной в SEQ ID NO: 207.
Определения:
Если не указано иное, научные и технические термины, используемые в настоящем изобретении, должны иметь значения, которые обычно понятны специалистам в данной области. Кроме того, если иное не предусмотрено контекстом, термины в единственном числе должны включать множественное число, а термины во множественном числе должны включать единственное число. Обычно методы и используемая с ними номенклатура клеточной и тканевой культуры, молекулярной биологии, а также белковой и олиго- или полинуклеотидной химии и гибридизации, описанные в настоящем документе, хорошо известны и обычно используются в данной области. Стандартные методы используются для рекомбинантной ДНК, синтеза олигонуклеотидов, а также для тканевой культуры и трансформации (например, электропорации, липофекции). Ферментативные реакции и методы очистки осуществляют в соответствии с инструкциями производителя, или как обычно они осуществляются в данной области, или как описано в настоящем документе. Вышеизложенные методы и процедуры, как правило, выполняют в соответствии с обычными способами, хорошо известными в данной области и как описано в различных общих и более конкретных ссылках, которые цитируются и обсуждаются в настоящем описании. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)). Лабораторные процедуры и методы аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, используемые в настоящем документе, и номенклатуры, используемые в связи с ними, хорошо известны и обычно используются в данной области. Стандартные методы используются для химических синтезов, химических анализов, получения, составления рецептур и доставки фармацевтических препаратов, и для лечения пациентов.
Следует понимать, что при использовании в контексте настоящего изобретения следующие термины, если не указано иное, имеют указанные ниже значения.
Используемый в настоящем документе термин «антитело» относится к молекулам иммуноглобулинов и к иммунологически активным участкам молекул иммуноглобулинов (Ig), т.е. к молекулам, которые содержат антигенсвязывающий участок, который специфичным образом связывается (вступает в иммунологическую реакцию) с антигеном. Под «специфичным связыванием» или «иммунологическим взаимодействием», или «иммуноспецифичным связыванием», подразумевается, что антитело взаимодействует с одной или несколькими антигенными детерминантами целевого антигена и не взаимодействует (т.е. не связывается) с другими полипептидами или связывается с другими полипептидами с гораздо более низкой аффинностью (Kd>10-6). Указанные антитела включают, но не ограничиваются ими, поликлональные, моноклональные, химерные, dAb (доменные антитела), одноцепочечные, Fab-, Fab'- и F(ab')2-фрагменты, scFv и Fab-экспрессирующую библиотеку.
Известно, что основная структурная единица антитела содержит тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, причем каждая пара имеет одну «легкую» (примерно 25 кД) и одну «тяжелую» (примерно 50-70 кД) цепь. Аминоконцевой участок каждой цепи включает вариабельную область из примерно 100-110 или большего числа аминокислот, в основном отвечающих за распознавание антигена. Карбоксиконцевой участок каждой цепи определяет константную область, в основном отвечающую за эффекторную функцию. В общем, молекулы антител человека относятся к любому из классов IgG, IgM, IgA, IgE и IgD, которые отличаются друг от друга по характеру тяжелой цепи, присутствующей в молекуле. Некоторые классы разделяются на подклассы, например, такие, как IgG1, IgG2 и др. Кроме того, у людей легкая цепь может быть представлена каппа- или лямбда-цепью.
Термин «моноклональное антитело» (MAb) или «композиция моноклонального антитела» в контексте настоящего изобретения относится к популяции молекул антител, которые содержат только один тип молекул антител, состоящий из единственного генного продукта легкой цепи и единственного генного продукта тяжелой цепи. В частности, определяющие комплементарность области (CDR) моноклонального антитела одинаковы во всех молекулах в популяции. MAb содержат антигенсвязывающий участок, способный вступать в иммунную реакцию с конкретным эпитопом антигена, отличительной особенностью которого является уникальная аффинность связывания с ним.
Термин «антигенсвязывающий участок» или «участок связывания» относится к части молекулы иммуноглобулина, которая принимает участие в связывании антигена. Антигенсвязывающий участок образован аминокислотными остатками N-концевых вариабельных («V») областей тяжелых («Н») и легких («L») цепей. Три высоковариабельных участка в V-областях тяжелых и легких цепей называются «гипервариабельными областями», которые включены между более консервативными фланкирующими участками, известными как «каркасные области» или «FR». Поэтому, термин «FR» относится к аминокислотным последовательностям, которые в природе находятся между гипервариабельными областями или прилегают к ним в иммуноглобулинах. В молекуле антитела, три гипервариабельные области легкой цепи и три гипервариабельные области тяжелой цепи расположены относительно друг друга в пространстве, образуя антигенсвязывающую поверхность. Антигенсвязывающая поверхность является комплементарной поверхности связываемого антигена, и три гипервариабельные области каждой из тяжелой и легкой цепей называются «определяющими комплементарность областями» или «CDR». Аминокислоты относят к каждому домену в соответствии с определениями Kabat Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987 and 1991)) или Chothia & Lesk J. Mol. Biol. 196:901-917 (1987), Chothia et al. Nature 342: 878-883 (1989).
Используемый в настоящем документе термин «эпитоп» включает любую белковую детерминанту, способную специфично связываться с иммуноглобулином, scFv или Т-клеточным рецептором. Термин «эпитоп» включает любую белковую детерминанту, способную специфично связываться с иммуноглобулином или Т-клеточным рецептором. Эпитопные детерминанты обычно состоят из химически активных поверхностных групп молекул, таких как боковые цепи аминокислот или сахаров, и обычно имеют определенные пространственные характеристики, а также определенные зарядные характеристики. Например, антитела могут быть получены против N-концевых или C-концевых пептидов полипептида. Антитело специфично связывает антиген, если константа диссоциации составляет ≤1 мкМ, например, ≤100 нМ, предпочтительно, ≤10 нМ и, более предпочтительно, ≤1 нМ.
Используемые в настоящем документе термины «иммунологическое связывание» и «свойства иммунологического связывания» относятся к нековалентным взаимодействиям такого типа, какие происходят между молекулой иммуноглобулина и антигеном, к которому специфичен иммуноглобулин. Сила или аффинность взаимодействий при иммунологическом связывании может быть выражена через константу диссоциации (Kd) взаимодействия, где меньшая Kd означает большую аффинность. Свойства иммунологического связывания избранных полипептидов можно количественно определить с помощью хорошо известных способов. Один из таких способов включает измерение скоростей образования и диссоциации комплекса антигенсвязывающего участка и антигена, где указанные скорости зависят от концентраций участников комплекса, аффинности взаимодействия и геометрических параметров, которые одинаково влияют на скорость в обоих направлениях. Таким образом, как «константа скорости ассоциации» (Kon), так и «константа скорости диссоциации» (Koff) могут быть определены путем вычисления концентраций и фактических скоростей ассоциации и диссоциации (см. Nature 361: 186-87 (1993)). Отношение Koff/Kon позволяет сократить все параметры, не связанные с аффинностью, и оно равно константе диссоциации Kd (см. в общем Davies et al. (1990) Annual Rev Biochem 59:439-473). Антитело по настоящему изобретению специфично связывается со своей мишенью, если константа равновесия связывания (Kd) составляет ≤1 мкМ, например, ≤100 нМ, предпочтительно ≤10 нМ и, более предпочтительно, ≤1 нМ, при измерении с помощью методов анализа, таких как анализ радиолигандного связывания или аналогичные методы анализа, известные специалистам в данной области.
Используемый в настоящем документе термин «выделенный полинуклеотид» означает полинуклеотид геномного происхождения, полученный из кДНК, или синтетического происхождения, или какую-либо их комбинацию, причем благодаря своему происхождению «выделенный полинуклеотид» (1) не ассоциирован со всем полинуклеотидом или с его участком, с которым «выделенный полинуклеотид» находится в природе, (2) функционально связан с полинуклеотидом, который не связан с ним в природе, или (3) не встречается в природе как часть более длинной последовательности. Полинуклеотиды по изобретению включают молекулы нуклеиновых кислот, кодирующие молекулы тяжелых цепей иммуноглобулинов, и молекулы нуклеиновых кислот, кодирующие молекулы легких цепей иммуноглобулинов, описанные в настоящем документе.
Используемый в настоящем описании термин «выделенный белок» означает белок, полученный из кДНК, рекомбинантной РНК или синтетически, или с использованием некоторой их комбинации, причем благодаря своему происхождению или источнику получения «выделенный белок» (1) не ассоциирован с белками, с которыми он находится в природе, (2) свободен от других белков из того же источника, например, от морских белков, (3) экспрессируется клеткой, принадлежащей другому виду, или (4) не встречается в природе.
Термин «полипептид» используется в настоящем документе в качестве общего термина для обозначения нативного белка, фрагментов или аналогов полипептидной последовательности. Следовательно, фрагменты и аналоги нативного белка относятся к разновидностям, входящим в понятие «полипептид». Предпочтительные полипептиды по изобретению включают молекулы тяжелых цепей иммуноглобулинов и молекулы легких цепей иммуноглобулинов, описанные в настоящем документе, а также молекулы антител, образованные комбинациями, включающими молекулы тяжелых цепей иммуноглобулинов с молекулами легких цепей иммуноглобулинов, такими как молекулы легких каппа-цепей иммуноглобулиновов, и наоборот, а также их фрагменты и аналоги.
Используемый в настоящем документе термин «встречающийся в природе», применяемый к объекту, относится к тому факту, что объект может встречаться в природе. Например, полипептидная или полинуклеотидная последовательность, которая присутствует в организме (включая вирусы), которая может быть выделена из источника в природе, и которая не была преднамеренно модифицирована человеком в лаборатории или иным образом, является встречающейся в природе.
Используемый в настоящем документе термин «функционально связанный» относится к расположению описанных таким образом компонентов, которые находятся в отношениях, позволяющих им функционировать предназначенным им образом. Регуляторная последовательность, «функционально связанная» с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается при условиях, совместимых с регулирующими последовательностями.
Используемый в настоящем документе термин «регуляторная последовательность» относится к полинуклеотидным последовательностям, которые необходимы для осуществления экспрессии и процессинга кодирующих последовательностей, с которыми они лигированы. Природа таких регуляторных последовательностей различается в зависимости от организма хозяина, у прокариот такие регуляторные последовательности обычно включают промотор, участок связывания рибосомы и последовательность терминации транскрипции, у эукариот обычно такие регуляторные последовательности включают промоторы и последовательность терминации транскрипции. Предполагается, что термин «регуляторная последовательность», включает как минимум все компоненты, присутствие которых существенно для экспрессии и процессинга, а также может включать дополнительные компоненты, присутствие которых является предпочтительным, например, лидерные последовательности и последовательности слитого партнера. Указанный в настоящем документе термин «полинуклеотид» означает полимерный ряд нуклеотидов в длину по меньшей мере из 10 оснований рибонуклеотидов или дезоксирибонуклеотидов, или же модифицированной формы каждого типа нуклеотидов. Термин включает одноцепочечные или двухцепочечные формы ДНК.
Используемые в настоящем документе двадцать обычных аминокислот и их аббревиатуры соответствуют общепринятому применению. См. Immunology - A Synthesis (2nd Edition, E. S. Golub and D. R. Gren, Eds., Sinauer Associates, Sunderland, Mass. (1991)). Стереоизомеры (например, D-аминокислоты) двадцати обычных аминокислот, неприродные аминокислоты, такие как α,α-двухзамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие необычные аминокислоты также могут быть подходящими компонентами для полипептидов по настоящему изобретению. Примеры необычных аминокислот включают: 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, О-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие аналогичные аминокислоты и иминокислоты (например, 4-гидроксипролин). В используемой в настоящем документе записи полипептида левостороннее направление представляет собой N-концевое направление, а правостороннее направление представляет собой С-концевое направление в соответствии со стандартным применением и традицией.
В отношении полипептидов термин «существенная идентичность» означает, что две пептидные последовательности при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT с использованием значений штрафа за разрыв по умолчанию имеют по меньшей мере 80% идентичности по последовательности, предпочтительно, по меньшей мере 90% идентичности по последовательности, более предпочтительно, по меньшей мере 95% идентичности по последовательности и, наиболее предпочтительно, по меньшей мере 99% идентичности по последовательности.
Предпочтительно, чтобы позиции остатков, не являющиеся идентичными, отличались консервативными аминокислотными заменами.
Консервативные аминокислотные замены относятся к взаимозаменяемости остатков, имеющих аналогичные боковые цепи. Например, группу аминокислот, имеющих алифатические боковые цепи, представляют глицин, аланин, валин, лейцин и изолейцин; группу аминокислот, имеющих алифатические-гидроксильные боковые цепи, представляют серин и треонин; группу аминокислот, имеющих содержащие амидогруппу боковые цепи, представляют аспарагин и глутамин; группу аминокислот, имеющих ароматические боковые цепи, представляют фенилаланин, тирозин и триптофан; группу аминокислот, имеющих основные боковые цепи, представляют лизин, аргинин и гистидин; а группу аминокислот, имеющих серосодержащие боковые цепи, представляют цистеин и метионин. Предпочтительными группами консервативных замен аминокислот являются валин-лейцин-изолейцин,индивид фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутаминовая кислота-аспарагиновая кислота и аспарагин-глутамин.
Как обсуждается в настоящем документе, предполагается, что минорные вариации в аминокислотных последовательностях молекул антител или иммуноглобулинов относятся к настоящему изобретению, при условии того, что вариации в аминокислотной последовательности сохраняют по меньшей мере 75%, более предпочтительно, по меньшей мере 80, 90, 95% и, наиболее предпочтительно, 99%. В частности, предполагаются консервативные аминокислотные замены. Консервативными заменами являются замены, которые происходят в пределах семейства аминокислот, объединенных по свойствам своих боковых цепей. Генетически кодируемые аминокислоты, в общем, подразделяются на следующие семейства: (1) кислые аминокислоты, включающие аспартат и глутамат; (2) основные аминокислоты, включающие лизин, аргинин, гистидин; (3) неполярные аминокислоты, включающие аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан и (4) незаряженные полярные аминокислоты, включающие глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Гидрофильные аминокислоты включают аргинин, аспарагин, аспартат, глутамин, глутамат, гистидин, лизин, серин и треонин. Гидрофобные аминокислоты включают аланин, цистеин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан, тирозин и валин. Другие семейства аминокислот включают (i) серин и треонин, которые представляют собой семейство алифатических гидроксисодержащих аминокислот; (ii) аспарагин и глутамин, которые представляют собой содержащее амидогруппу семейство; (iii) аланин, валин, лейцин и изолейцин, которые представляют собой алифатическое семейство; и (iv) фенилаланин, триптофан и тирозин, которые представляют собой ароматическое семейство. Например, имеются основания ожидать, что отдельная замена лейцина на изолейцин или валин, аспартата на глутамат, треонина на серин или аналогичная замена аминокислоты структурно-родственной аминокислотой не будет иметь большого влияния на связывание или свойства полученной в результате молекулы, особенно если замена не затрагивает аминокислоту в каркасном участке. Будет ли аминокислотная замена давать функциональный пептид, можно легко определять путем анализа удельной активности полипептидного производного. В настоящем документе подробно описаны методы анализа. Фрагменты или аналоги молекул антител или иммуноглобулинов могут быть легко получены специалистами в данной области. Предпочтительные N- или С-концы фрагментов или аналогов находятся вблизи границ функциональных доменов. Структурные и функциональные домены могут быть идентифицированы путем сравнения данных по нуклеотидной и/или аминокислотной последовательности с общедоступными или коммерческими базами данных последовательностей. Предпочтительно использовать компьютерные способы сравнения для идентификации мотивов последовательностей или предсказанных конформационных доменов белков, которые присутствуют в других белках с известной структурой и/или функцией. Известны способы идентификации белковых последовательностей, которые сворачиваются в известные трехмерные структуры. Bowie et al. Science 253: 164 (1991). Таким образом, нижеследующие примеры демонстрируют, что специалист в данной области может выявить мотивы последовательности и структурные конформации, которые могут использоваться для определения структурных и функциональных доменов по изобретению.
Предпочтительные аминокислотные замены представляют собой те, которые: (1) снижают чувствительность к протеолизу, (2) снижают чувствительность к окислению, (3) изменяют аффинность связывания при образовании белковых комплексов, (4) изменяют аффинность связывания и (5) создают или модифицируют физикохимические или функциональные свойства таких аналогов. Аналоги могут включать различные мутированные варианты с последовательностью, отличающейся от природной пептидной последовательности. Например, единичные или множественные аминокислотные замены (предпочтительно, консервативные аминокислотные замены) могут быть сделаны в природной последовательности (предпочтительно, в участке полипептида вне домена(ов), образующих межмолекулярные контакты). Консервативная аминокислотная замена не должна по существу изменять структурных характеристик родительской последовательности (например, замещающая аминокислота не должна приводить к разрушению спирали, которая присутствует в родительской последовательности, или не должна разрушать другие типы вторичной структуры, которой характеризуется родительская последовательность). Примеры известных в данной области полипептидных вторичных и третичных структур описаны в Proteins, Structures and Molecular Principles (Creighton, Ed., W.H. Freeman and Company, New York (1984)); Introduction to Protein Structure (C. Branden and J. Tooze, eds., Garland Publishing, New York, N. Y. (1991)); и Thornton et al. Nature 354:105(1991).
Используемые в настоящем документе термины «метка» или «меченный» относится к введению детектируемого маркера, например, путем введения радиоактивной аминокислоты или присоединения к полипептиду биотиновых групп, которые могут быть выявлены меченным авидином (например, стрептавидином, содержащим флуоресцентный маркер или ферментативную активность, которая может выявляться оптическими или колориметрическими методами). В некоторых ситуациях метка или маркер также могут быть терапевтическими. В данной области известны и могут использоваться различные способы мечения полипептидов и гликопротеинов. Примеры меток для полипептидов включают, но не ограничиваются ими, следующие метки: радиоизотопы или радионуклиды (например, 3Н, 14С, 15N, 35S, 90Y, 99Tc, 111In, 125I, 131I), флуоресцентные метки (например, FITC, родамин, лантаниды фосфора), ферментативные метки (например, пероксидазу хрена, β-галактозидазу, люциферазу, щелочную фосфатазу), хемилюминесцентные, биотиновые группы, заданные полипептидные эпитопы, распознаваемые вторичным репортером (например, последовательности пары лейциновых молний, участки связывания вторичных антител, связывающие металл домены, эпитопные теги). В некоторых вариантах осуществления изобретения метки присоединяют с помощью спейсерных плеч различной длины для снижения потенциальных стерических затруднений. Используемый в настоящем документе термин «фармацевтическое средство или лекарственное средство» относится к химическому соединению или композиции, способному вызывать требуемый терапевтический эффект при надлежащем введении пациенту.
Другие химические термины в настоящем документе используются в соответствии с общепринятым применением в данной области, например, проиллюстрированном в The McGraw-Hill Dictionary of Chemical Terms (Parker, S., Ed., McGraw-Hill, San Francisco (1985)).
Используемый в настоящем документе термин «по существу, чистый» означает, что указанный вид является преобладающим присутствующим видом (т.е. на молярной основе он наиболее распространен, чем любой другой индивидуальный вид в композиции), и, предпочтительно, по существу очищенная фракция представляет собой композицию, в которой указанный вид составляет по меньшей мере примерно 50% (на молярной основе) от всех присутствующих видов макромолекул.
В общем, по существу, чистая композиция будет содержать более чем примерно 80% от всех видов макромолекул, присутствующих в композиции, более предпочтительно, более чем примерно 85, 90, 95 и 99%. Наиболее предпочтительно, чтобы указанный вид был очищен до практической гомогенности (когда примеси не могут детектироваться в композиции с помощью стандартных способов детекции), при этом композиция состоит по существу из одного вида макромолекул.
Термин «пациент» включает собой людей и ветеринарных индивидов.
Антитела
Различные способы, известные в данной области, могут использоваться для получения моноклональных антител, направленных против заданной мишени или против ее производных, фрагментов, аналогов, гомологов или ортологов. (См., например, Antibodies: A Laboratory Manual, Harlow E, and Lane D, 1988, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, включенную в настоящий документ в качестве ссылки).
Антитела очищают с помощью хорошо известных методов, таких как хроматография с использованием белка А или белка G, которые обеспечивают в основном IgG-фракцию иммунной сыворотки. После чего (или в альтернативном варианте) специфичный антиген, который является мишенью целевого иммуноглобулина, или его эпитоп, может быть иммобилизован на колонке для очистки иммуноспецифичного антитела путем иммуноаффинной хроматографии. Очистка иммуноглобулинов обсуждается, например, D. Wilkinson (The Scientist, опубликовано The Scientist, Inc., Philadelphia PA, Vol. 14, No. 8 (April 17, 2000), pp. 25-28).
В некоторых вариантах осуществления изобретения антитела по изобретению являются моноклональными антителами. Моноклональные антитела получают, например, с помощью процедур, изложенных в примерах, представленных в настоящем документе. Антитела также получают, например, путем иммунизации мышей BALB/с комбинаций клеточных трансфектантов, экспрессирующих высокий уровень заданной мишени на своей поверхности. Гибридомы, полученные в результате слияния миеломных и В-клеток затем подвергают скринингу на реактивность к выбранной мишени.
Моноклональные антитела получают, например, с помощью гибридомных способов, таких как те, которые описаны Kohler and Milstein, Nature, 256: 495 (1975). В гибридомном способе мышь, хомяка или другое соответствующее животное-хозяина, как правило, иммунизируют иммунизирующим средством, чтобы вызвать появление лимфоцитов, продуцирующих или способных продуцировать антитела, которые будут специфически связываться с иммунизирующим средством. В альтернативном варианте, лимфоциты могут быть иммунизированы in vitro.
Иммунизирующее средство будет, как правило, включать белковый антиген или его фрагмент, или слитый с ним белок. В общем, используют или лимфоциты периферической крови, если желательны клетки человека, или клетки селезенки или клетки лимфоузлов, если желательны клетки млекопитающих кроме человека. Затем лимфоциты сливают с иммортализованной линией клеток с использованием подходящего вызывающего слияние клеток средства, такого как полиэтиленгликоль, для получения гибридомной клетки (Goding, Monoclonal antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103). Иммортализованные клеточные линии обычно представляют собой трансформированные клетки млекопитающих, в частности, клетки миеломы грызунов, бычьей миеломы и миеломы человека. Обычно используют линии клеток миеломы крысы или мыши. Клетки гибридомы можно культивировать в подходящей культуральной среде, предпочтительно, содержащей одно или несколько веществ, подавляющих рост или выживание неслитых иммортализованных клеток. Например, если в родительских клетках отсутствует фермент гипоксантин-гуанин-фосфорибозилтрансфераза (HGPRT или HPRT), культуральная среда для гибридомы, как правило, будет включать гипоксантин, аминоптерин и тимидин («среда HAT») - вещества, предотвращающие рост HGPRT-дефицитных клеток.
Предпочтительными иммортализованными линиями клеток являются клетки, которые эффективно сливаются, поддерживают высокий уровень экспрессии антитела выбранными продуцирующими антитело клетками и являются чувствительными к среде, такой как среда HAT. Более предпочтительными иммортализованными линиями клеток являются линии миеломы мышей, которые можно получить, например, из Salk Institute Cell Distribution Center, San Diego, California и American Type Culture Collection, Manassas, Virginia. Также описана продукция моноклональных антител в линиях клеток миеломы человека и гетеромиеломы мыши и человека. (См. Kozbor, J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal antibody Production Techniques и Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63).
Затем, культуральную среду, в которой культивировали клетки гибридомы, можно проанализировать на присутствие моноклональных антител, направленных против антигена. Предпочтительно, чтобы специфичность связывания моноклонального антитела, продуцируемого клетками гибридомы, была определена иммунопреципитацией или анализом связывания in vitro, таким как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Такие способы и анализы известны в данной области. Аффинность связывания моноклонального антитела можно, например, определять анализом Скэтчарда по Munson and Pollard, Anal. Biochem., 107: 220 (1980). Более того, при терапевтическом применении моноклональных антител важно идентифицировать антитела, имеющие высокую степень специфичности и высокую аффинность связывания к антигену-мишени.
После идентификации желательных клеток гибридомы, клоны можно субклонировать методом лимитирующих разведений и выращивать общепринятыми способами. (См. Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103). Подходящая для данной цели культуральная среда включает, например, модифицированную Дульбекко среду Игла и среду RPMI-1640. В альтернативном варианте, клетки гибридомы можно выращивать in vivo в виде асцитов у млекопитающего.
Моноклональные антитела, секретируемые субклонами, можно выделять или очищать из культуральной среды или асцитной жидкости общепринятыми способами очистки иммуноглобулинов, такими как, например, очистка на белок A-сефарозе, хроматография на гидроксиапатите, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела можно получить также способами рекомбинантных ДНК, такими, как описаны в патенте США № 4816567. ДНК, кодирующую моноклональные антитела по изобретению, можно легко выделить и секвенировать с применением общепринятых процедур (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Клетки гибридомы по изобретению служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно поместить в экспрессирующие векторы, которыми затем трансфецируют клетки-хозяева, такие как обезьяньи клетки COS, клетки яичников китайского хомячка (CHO) или клетки миеломы, которые иначе не продуцируют иммуноглобулиновый белок, для синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК также можно модифицировать, например, путем замены кодирующими последовательностями константных доменов тяжелых и легких цепей человека гомологичных мышиных последовательностей (патент США №4816567; Morrison, Nature 368, 812-13 (1994)) или путем ковалентного присоединения к последовательности, кодирующей иммуноглобулин, всей или части последовательности, кодирующей неиммуноглобулиновый полипептид. Таким неиммуноглобулиновым полипептидом можно заменить константные домены антитела по изобретению или можно заменить вариабельные домены одного антигенсвязывающего центра антитела по изобретению для получения химерного бивалентного антитела.
Моноклональные антитела по изобретению включают гуманизированные антитела или антитела человека. Эти антитела подходят для введения человеку, не вызывая иммунного ответа у человека против вводимого иммуноглобулина. Гуманизированные формы антител представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие субпоследовательности антител), которые в основном состоят из последовательности иммуноглобулина человека и содержат минимальную последовательность, полученную из иммуноглобулина, не принадлежащего человеку. Гуманизацию проводят в целом, следуя способу Winter и сотрудников (Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332: 323-327 (1988); Verhoeyen et al., Science 239: 1534-1536 (1988)), путем замены CDR грызунов соответствующих последовательностей CDR из антитела человека. (См. также патент США №5225539). В некоторых случаях остатки из каркасных участков вариабельной области иммуноглобулина человека заменены соответствующими остатками, не принадлежащими человеку. Гуманизированные антитела также содержат, например, остатки, которые не встречаются ни в реципиентном антителе, ни в импортируемых последовательностях CDR или каркасных участков. В общем, гуманизированное антитело включает по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все CDR соответствуют таковым в иммуноглобулине, не принадлежащем человеку, а все или практически все каркасные области представляют собой таковые из консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также включает по меньшей мере участок константной области иммуноглобулина (Fc), как правило, иммуноглобулина человека (Jones et al, 1986; Riechmann et al, 1988; and Presta, Curr. Op. Struct. Biol, 2: 593-596 (1992)).
Полностью человеческие антитела представляют собой молекулы антител, в которых вся последовательность как легкой цепи, так и тяжелой цепи, включая CDR, происходит из генов человека. В настоящем документе такие антитела называют «антителами человека» или «полностью человеческими антителами». Моноклональные антитела могут быть получены с использованием триомной методики, гибридомной методики с В-клетками человека (см. Kozbor, et al., 1983 Immunol Today 4: 72) и EBV-гибридомной методики для получения моноклональных антител (см. Cole, et al., 1985 In: MONOCLONAL ANTIBODIES AND CANCER THERAPY, Alan R. Liss, Inc., pp. 77-96). Моноклональные антитела могут использоваться и могут быть получены с использованием гибридом человека (см. Cote, et al., 1983. Proc Natl Acad Sci USA 80: 2026-2030) или же путем трансформации В-клеток человека вирусом EBV in vitro (см. Cole, et al., 1985 In: MONOCLONAL ANTIBODIES AND CANCER THERAPY, Alan R. Liss, Inc., pp. 77-96).
Дополнительно антитела человека можно получать с применением других методик, включая библиотеки фагового дисплея (Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991)). Аналогичным образом антитела человека можно создать введением локусов иммуноглобулинов человека в трансгенных животных, например, мышей, у которых эндогенные гены иммуноглобулинов частично или полностью инактивированы. После антигенной стимуляции наблюдается продукция человеческих антител, близко сходная с наблюдаемой у человека продукцией во всех отношениях, включая реаранжировку генов, сборку и репертуар антител. Данный подход описан, например, в патентах США №№5545807, 5545806, 5569825, 5625126, 5633425, 5661016 и в Marks et al., Bio/Technology 10, 779-783 (1992); Lonberg et al., Nature 368 856-859 (1994); Morrison, Nature 368, 812-13 (1994); Fishwild et al., Nature Biotechnology 14, 845-51 (1996); Neuberger, Nature Biotechnology 14, 826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol. 13 65-93 (1995).
Антитела человека можно дополнительно получить с использованием трансгенных животных, которые модифицированы для получения полностью человеческих антител, а не эндогенных антител этого животного, в ответ на антигенную стимуляцию. (См. публикацию РСТ WO 94/02602). Эндогенные гены, кодирующие тяжелые и легкие цепи иммуноглобулинов хозяина (не человека) инактивируют, и вводят в его геном активные локусы, кодирующие легкие и тяжелые цепи иммуноглобулинов человека. Гены человека встраивают, например, с помощью искусственных хромосом дрожжей, содержащих необходимые сегменты ДНК человека. Затем путем перекрестного скрещивания промежуточных трансгенных животных, несущих неполный набор модификаций, в потомстве получают животное, которое обеспечивает все желаемые модификации. Примером таких животных является мышь, которая называется XENOMOUSE™ и описана в публикациях РСТ WO 96/33735 и WO 96/34096. Это животное продуцирует В-клетки, которые секретируют полностью человеческие иммуноглобулины. Антитела можно получать непосредственно из животного после иммунизации представляющим интерес иммуногеном в форме, например, препарата поликлональных антител или, в качестве альтернативы, из иммортализованных В-клеток, полученных из организма животного, таких как гибридомы, продуцирующие моноклональные антитела. Дополнительно можно выделить и экспрессировать гены, кодирующие иммуноглобулины с вариабельными областями человека, для непосредственного получения антител или для дальнейшей их модификации с целью получения аналогов антител, таких как, например, молекулы одноцепочечных Fv (scFv).
Пример способа получения хозяина (не человека) на примере мыши, лишенной экспрессии тяжелых цепей эндогенных иммуноглобулинов, описана в патенте США №5939598. Эта мышь может быть получена способом, который включает удаление генов J-сегмента из по меньшей мере одного эндогенного локуса тяжелых цепей в эмбриональной стволовой клетке, чтобы предотвратить реаранжировку локуса и чтобы предотвратить образование транскрипта реаранжированного локуса тяжелых цепей иммуноглобулинов, причем удаление выполняют с помощью направленного вектора, содержащего ген, кодирующий селектируемый маркер, и получение из эмбриональных стволовых клеток трансгенных мышей, чьи соматические и зародышевые клетки содержат ген, кодирующий селектируемый маркер.
Один способ получения интересующего антитела, такого как антитело человека, раскрыт в патенте США №5916771. Этот способ включает введение вектора экспрессии, который содержит нуклеотидную последовательность, кодирующую тяжелую цепь, в одну клетку млекопитающего-хозяина в культуре, введение вектора экспрессии, содержащего нуклеотидную последовательность, кодирующую легкую цепь, в другую клетку млекопитающего-хозяина в культуре, и слияние двух клеток с образованием гибридной клетки. Гибридная клетка экспрессирует антитело, содержащее и тяжелую цепь, и легкую цепь.
Дальнейшее усовершенствование этой процедуры - способ идентификации клинически значимого эпитопа на иммуногене и соответственный способ отбора антитела, которое специфично связывается со значимым эпитопом, раскрыт в публикации РСТ WO 99/53049.
Антитело можно экспрессировать с помощью вектора, содержащего ДНК-сегмент, кодирующий одноцепочечное антитело, описанное выше.
Они могут включать векторы, липосомы, неконъюгированную ДНК, генную пушку, катетеры и т.д. Векторы включают химические конъюгаты, такие как описанный в WO 93/64701, который имеет направляющую молекулу (например, лиганд рецептора клеточной поверхности) и молекулу, связывающую нуклеиновую кислоту (например, полилизин), вирусный вектор (например, вектор на основе ДНК- или РНК-вирусов), слитые белки, такие как описанный в PCT/US 95/02140 (WO 95/22618), который представляет собой слитый белок, содержащий направляющую молекулу (например, антитело, специфичное в отношении клетки-мишени) и молекулу, связывающую нуклеиновую кислоту (например, протамин), плазмиды, фаги и т.д. Векторы могут быть хромосомными, нехромосомными или синтетическими.
Предпочтительные векторы включают вирусные векторы, слитые белки и химические конъюгаты. Ретровирусные векторы включают вирус лейкоза мышей Молони. Предпочтительными являются вирусные векторы, основанные на ДНК. Эти векторы включают векторы на основе вирусов оспы, такие как векторы на основе ортопоксвирусов или авипоксвирусов, герпесвирусные векторы, такие как вектор на основе вируса простого герпеса I (HSV) (см. Geller, A.I. et al., J. Neurochem., 64: 487 (1995); Lim, F., et al., in DNA Cloning: Mammalian Systems, D. Glover, Ed. (Oxford Univ. Press, Oxford England) (1995); Geller, A.I. et al., Proc. Natl. Acad. Sci.: U.S.A., 90: 7603 (1993); Geller, A.I., et al., Proc Natl. Acad. Sci. USA, 87: 1149 (1990)), аденовирусные векторы (см. LeGal LaSalle et al., Science, 259: 988 (1993); Davidson, et al., Nat. Genet. 3: 219 (1993); Yang, et al., J. Virol. 69: 2004 (1995)) и векторы на основе аденосвязанных вирусов (Kaplitt, M. G. et al., Nat. Genet. 8: 148 (1994)).
Векторы на основе поксвирусов вводят ген в цитоплазму клетки. Векторы на основе авипоксвирусов приводят лишь к кратковременной экспрессии нуклеиновой кислоты. Аденовирусные векторы, векторы на основе аденосвязанных вирусов и векторы на основе вируса простого герпеса (HSV) являются предпочтительными для введения нуклеиновой кислоты в нервные клетки. Аденовирусный вектор приводит к более краткосрочной экспрессии (примерно 2 месяца), чем аденосвязанный вирус (примерно 4 месяца), который, в свою очередь, действует в течение более короткого времени, чем векторы на основе HSV. Выбор конкретного вектора зависит от клетки-мишени и подлежащего лечению состояния. Введение может осуществляться стандартными методами, например, путем инфекции, трансфекции, трансдукции или трансформации. Примеры способов переноса генов включают, например, неконъюгированную ДНК, преципитацию с CaPO4, DEAE-декстран, электропорацию, слияние протопластов, липофекцию, микроинъекцию в клетки и вирусные векторы.
Вектор может использоваться для направленного введения по существу в любую клетку-мишень. Например, для направленного введения векторов (например, аденовирусного, HSV) в требуемое место может использоваться стереотаксическая инъекция. Дополнительно частицы могут доставляться путем интрацеребровентрикулярной (icv) инфузии с использованием инфузионной системы с мининасосом, такой как инфузионная система SynchroMed. Способ, основанный на объемном потоке, называемый конвекцией, также доказал свою эффективность по доставке крупных молекул в удаленные области головного мозга и может использоваться при доставке вектора к клетке-мишени. (См. Bobo et al., Proc. Natl. Acad. Sci. USA 91: 2076-2080 (1994); Morrison et al., Am. J. Physiol. 266: 292-305 (1994)). Другие способы, которые могут использоваться, включают катетеры, внутривенную, парентеральную, внутрибрюшинную и подкожную инъекцию, а также пероральный и другие пути введения.
Биспецифичные антитела представляют собой антитела, которые обладают специфичностью связывания по меньшей мере с двумя различными антигенами. В данном случае одна из специфичностей связывания относится к мишени, такой как CD47 или любой его фрагмент. Второй мишенью связывания является другой антиген, предпочтительно, белок клеточной поверхности или рецептор, или субъединица рецептора.
Способы получения биспецифичных антител известны в данной области. Обычно рекомбинантное получение биспецифичных антител основано на совместной экспрессии двух пар тяжелых и легких цепей иммуноглобулинов, где две тяжелые цепи имеют разную специфичность (Milstein and Cuello, Nature, 305: 537 (1983)). Вследствие случайной перестановки иммуноглобулиновых тяжелых и легких цепей такие гибридомы (квадромы) продуцируют потенциальную смесь из 10 различных молекул антител, из которых только одна имеет правильную биспецифичную структуру. Очистку правильной молекулы обычно проводят с помощью поэтапной аффинной хроматографии. Аналогичные методы раскрыты в WO 93/08829, опубликованной 13-го мая, 1993 г. и в Traunecker et al., EMBO J., 10: 3655 (1991).
Биспецифичные и/или моновалентные антитела по изобретению могут быть получены с использованием любого из множества методов, известных в данной области техники, в том числе тех, которые раскрыты в одновременно рассматриваемой заявке WO 2012/023053, поданной 16-го августа 2011 г., содержание которой включено в настоящее описание в качестве ссылки в полном объеме. Способы, описанные в WO 2012/023053, обеспечивают биспецифичные антитела, идентичные по структуре иммуноглобулину человека. Этот тип молекулы состоит из двух копий полипептида уникальной тяжелой цепи, вариабельной области первой легкой цепи, слитой с константной области каппа-цепи, и вариабельной области второй легкой цепи, слитой с константной областью лямбда-цепи. Каждый центр связывания проявляет различную антигенную специфичность, в которую вносят вклад как тяжелая, так и легкая цепь. Вариабельные области легкой цепи могут принадлежать лямбда- или каппа-семейству и, предпочтительно, быть слитыми с лямбда- и каппа-константными доменами, соответственно. Это является предпочтительным для того, чтобы избежать создания неприродных полипептидных соединений. Однако также возможно получение биспецифичных антител по изобретению путем слияния вариабельной области легкой каппа-цепи с константным доменом лямбда-цепи для первой специфичности и слияния вариабельной области легкой лямбда-цепи с константным доменом каппа-цепи для второй специфичности. Полученные биспецифичные антитела, описанные в WO 2012/023053, называются IgGκλ-антителами или «κλ-антителами», новым форматом полностью человеческих биспецифичных IgG. Этот формат κλ-антител позволяет аффинную очистку биспецифичного антитела, которое невозможно отличить от стандартной молекулы IgG с характеристиками, которые неотличимы от стандартного моноклонального антитела, и, следовательно, является предпочтительным по сравнению с предшествующими форматами.
Необходимой стадией способа является идентификация двух Fv-областей антитела (каждой, состоящей из вариабельного домена легкой цепи и вариабельного домена тяжелой цепи), имеющих различную антигенную специфичность, которые включают одинаковый вариабельный домен тяжелой цепи. Для создания моноклональных антител и их фрагментов описаны многочисленные способы. (См., например, Antibodies: A Laboratory Manual, Harlow E, and Lane D, 1988, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, incorporated herein by reference). Полностью человеческие антитела представляют собой молекулы антител, в которых последовательность как легкой цепи, так и тяжелой цепи, в том числе CDR 1 и 2, происходят из генов человека. CDR3 может принадлежать человеку или может быть создана синтетически. Такие антитела в настоящем документе называют «антителами человека» или «полностью человеческими антителами». Моноклональные антитела человека могут быть получены с использованием триомной методики, гибридомной методики с В-клетками человека (см. Kozbor, et al., 1983 Immunol Today 4: 72) и EBV-гибридомной методики для получения моноклональных антител (см. Cole, et al., 1985 In: MONOCLONAL ANTIBODIES AND CANCER THERAPY, Alan R. Liss, Inc., pp. 77-96). Моноклональные антитела человека могут использоваться и могут быть получены с использованием человеческих гибридом (см. Cote, et al., 1983. Proc Natl Acad Sci USA 80: 2026-2030) или же путем трансформации В-клеток человека вирусом EBV in vitro (см. Cole, et al., 1985 In: MONOCLONAL ANTIBODIES AND CANCER THERAPY, Alan R. Liss, Inc., pp. 77-96).
Моноклональные антитела получают, например, путем иммунизации животного антигеном-мишенью или его иммуногенным фрагментом, производным или вариантом. В альтернативном варианте животное иммунизируют клетками, трансфицированными вектором, содержащим молекулу нуклеиновой кислоты, кодирующую антиген-мишень таким образом, чтобы целевой антиген экспрессировался и был связан с поверхностью трансфицированных клеток. В данной области хорошо известно множество методов получения ксеногенных животных. Например, см. патенты США №6075181 и №6150584, которые включены в настоящий документ в качестве ссылки в полном объеме.
В качестве альтернативы, антитела получают путем скрининга библиотеки, который содержит последовательности антител или их антигенсвязывающих доменов, на связывание с антигеном-мишенью. Эту библиотеку получают, например, в бактериофагах в виде белков или пептидов, слитых с белком оболочки бактериофага, который экспрессируется на поверхности собранных фаговых частиц, а кодирующие последовательности ДНК содержатся в фаговых частицах (т.е. «библиотека фагового дисплея»).
Гибридомы, полученные в результате слияния миеломных и В-клеток, затем подвергают скринингу на реактивность к антигену-мишени. Моноклональные антитела получают, например, с помощью гибридомных способов, таких как те, которые описаны Kohler and Milstein, Nature, 256: 495 (1975). В гибридомном способе мышь, хомяка или другое соответствующее животное-хозяина, как правило, иммунизируют иммунизирующим средством, чтобы вызвать появление лимфоцитов, продуцирующих или способных продуцировать антитела, которые будут специфически связываться с иммунизирующим средством. В альтернативном варианте, лимфоциты могут быть иммунизированы in vitro.
Хотя это и не является совсем невозможным, случайная идентификация различных антител, имеющих один и тот же вариабельный домен тяжелой цепи, но направленных против различных антигенов, является крайне маловероятной. Действительно, в большинстве случаев тяжелая цепь вносит основной вклад в поверхность связывания антигена, а также является наиболее вариабельной по последовательности. В частности, CDR3 тяжелой цепи является наиболее разнообразной CDR по последовательности, длине и структуре. Поэтому, два антитела, специфичные к различным антигенам, почти всегда имеют разные вариабельные домены тяжелой цепи.
Способы, раскрытые в одновременно рассматриваемой заявке WO 2012/023053, преодолевают это ограничение и значительно облегчают выделение антител, имеющих одинаковый вариабельный домен тяжелой цепи, путем использования библиотек антител, в которых вариабельный домен тяжелой цепи является одинаковым для всех членов библиотеки и, таким образом, разнообразие ограничивается вариабельным доменом легкой цепи. Такие библиотеки описаны, например, в одновременно рассматриваемых заявках WO 2010/135558 и WO 2011/084255, каждая из которых включена в настоящее описание в качестве ссылки в полном объеме. Однако поскольку вариабельный домен легкой цепи экспрессируется совместно с вариабельным доменом тяжелой цепи, оба домена могут вносить свой вклад в связывание антигена. Чтобы еще больше облегчить процесс, библиотеки антител, содержащих один и тот же вариабельный домен тяжелой цепи и панель либо вариабельных доменов легких лямбда-цепей, либо вариабельных доменов легких каппа-цепей, могут использоваться параллельно для in vitro отбора антител против различных антигенов. Этот подход позволяет идентифицировать два антитела, имеющих общую тяжелую цепь, но одно несет вариабельный домен легкой лямбда-цепи, а другое - вариабельный домен легкой каппа-цепи, которые могут использоваться в качестве строительных блоков для создания биспецифичных антител в формате полноразмерного иммуноглобулина по изобретению. Полученные биспецифичные антитела по изобретению могут относится к различным изотипам, а их Fc-участок может быть модифицирован для изменения свойств связывания различных Fc-рецепторов и, таким образом, для изменения эффекторных функций антитела, а также фармакокинетических свойств. Многочисленные способы модификации Fc-участка описаны и применимы к антителам по изобретению. (См., например, Strohl, WR Curr Opin Biotechnol 2009 (6):685-91; патент США №6528624; заявку PCT/US 2009/0191199, поданную 9-го января 2009 г.). Способы по настоящему изобретению также могут использоваться для создания биспецифичных антител и смеси антител в F(ab')2-формате, у которого отсутствует Fc-участок.
Общая тяжелая цепь и две различные легкие цепи совместно экспрессируются в одной клетке, что позволяет сборку биспецифичного антитела по изобретению. Если все полипептиды экспрессируются на одном уровне и собираются одинаково хорошо с образованием молекулы иммуноглобулина, то соотношение моноспецифичных (одинаковые легкие цепи) и биспецифичных (две различные легкие цепи) должно составлять 50%. Однако вполне вероятно, что различные легкие цепи экспрессируются на различном уровне и/или не собираются с одинаковой эффективностью. Поэтому, используют подходы для модуляции относительной экспрессии различных полипептидов с целью компенсации их характеристик экспрессии или их различной предрасположенности к сборке с общей тяжелой цепью. Эта модуляция может быть осуществлена посредством силы промотора, использования внутренних сайтов связывания рибосом (IRES), имеющих различную эффективность, или других типов регуляторных элементов, которые могут действовать на транскрипционном или трансляционном уровнях, а также воздействовать на стабильность мРНК. Различные промоторы с различной силой могут включать CMV (очень ранний промотор цитомегаловируса); EF1-1α (промотор 1α-субъединицы фактора элонгации транляции у человека); Ubc (промотор убиквитина С); SV40 (промотр вируса 40 обезьян). Также были описаны различные IRES из млекопитающих и вирусного происхождения. (См., например, Hellen CU and Sarnow P. Genes Dev 2001 15: 1593-612). Эти IRES могут сильно различаться по своей длине и по эффективности привлечения рибосом. Кроме того, возможна дополнительная регуляция активности путем введения множества копий IRES (Stephen et al. 2000 Proc Natl Acad Sci USA 97: 1536-1541). Модуляция экспрессии также может быть осуществлена путем множественных последовательных трансфекций клеток для увеличения копийности отдельных генов, экспрессирующих одну или другую легкую цепь, и, таким образом, для изменения их относительной экспрессии. В примерах, приведенных в настоящем документе, продемонстрировано, что регуляция относительной экспрессии различных цепей принципиальна для максимизации сборки и общего выхода биспецифичного антитела.
Совместная экспрессия тяжелой цепи и двух легких цепей дает смесь трех различных антител в клеточном культуральном супернатанте: двух моноспецифичных бивалентных антител и одного биспецифичного бивалентного антитела. Последнее необходимо очистить из смеси, чтобы получить целевую молекулу. Описанный в настоящем документе способ значительно облегчает процедуру такой очистки за счет использования аффинного хроматографического носителя, который специфично взаимодействует с константными доменами легких каппа- или лямбда-цепей, такого как аффинные носители CaptureSelect Fab Kappa и CaptureSelect Fab Lambda (BAC BV, Holland). Этот подход многостадийной аффинной хроматографической очистки эффективен и обычно применим к антителам по изобретению. Он резко контрастирует со специфичными способами очистки, разработанными и оптимизированными для каждого биспецифичного антитела, полученного из квадром или других клеточных линий, экспрессирующих смеси антител. Действительно, если биохимические характеристики различных антител в смеси являются похожими, то их разделение с использованием стандартной хроматографической методики, такой как ионообменная хроматография, может быть проблематичным или даже совсем невозможным.
Другие подходящие способы очистки включают те, которые раскрыты в одновременно рассматриваемой заявке PCT/IB 2012/003028, поданной 19-го октября 2012 г., опубликованной как WO 2013/088259, содержание которой таким образом включено в качестве ссылки в полном объеме.
В других вариантах получения биспецифичных антител, вариабельные домены антитела с желательными специфичностями связывания (центрами связывания антитела с антигеном) могут быть слиты с последовательностями константных доменов иммуноглобулинов. Слияние предпочтительно проводят с константным доменом тяжелой цепи иммуноглобулина, содержащим по меньшей мере часть шарнирной, CH2- и CH3-областей. Предпочтительно, чтобы по меньшей мере в одном из слитых белков присутствовала первая константная область (CH1) тяжелой цепи, содержащая участок, необходимый для связывания легкой цепи. ДНК, кодирующую слитые тяжелые цепи иммуноглобулина и, если требуется, легкую цепь иммуноглобулина, вводят в отдельные векторы экспрессии и совместно трансфицируют в подходящий организм-хозяин. Дополнительные подробности создания биспецифичных антител см., например, в Suresh et al., Methods in Enzymology, 121: 210 (1986).
Согласно другому подходу, описанному в WO 96/27011, границу раздела между парой молекул антител можно искусственно сконструировать так, чтобы максимально увеличить процентное содержание гетеродимеров, которые выделяются из культуры рекомбинантных клеток. Предпочтительная граница раздела включает по меньшей мере часть CH3-области константного домена антитела. В этом способе одну или несколько некрупных боковых цепей аминокислот из границы раздела первой молекулы антитела заменяют более крупными боковыми цепями (например, тирозина или триптофана). На границе раздела второй молекулы антитела создаются компенсаторные «полости» идентичного или схожего с крупной боковой цепью(ми) размера путем замены крупных боковых цепей аминокислот на меньшие (например, аланина или треонина). Это обеспечивает механизм увеличения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Методы создания биспецифичных антител из фрагментов антител также описаны в литературе. Например, биспецифичные антитела можно получать с помощью химической сшивки. Полученные биспецифичные антитела можно использовать в качестве средств для избирательной иммобилизации ферментов.
Также описаны различные методы получения и выделения фрагментов биспецифичных антител напрямую из культуры рекомбинантных клеток. Например, биспецифичные антитела получали с помощью лейциновых молний. Kostelny et al., J. Immunol, 148(5): 1547-1553 (1992). Пептиды лейциновой молнии из белков Fos и Jun присоединяли к Fab'-участкам двух различных антител путем слияния генов. Гомодимеры антител восстанавливали в шарнирной области для образования мономеров и затем вновь окисляли для образования гетеродимеров антител. Такой способ также можно использовать для получения гомодимеров антител. Технология «диател», описанная Hollinger et al., Proc. Natl Acad. Sci. USA, 90:6444-6448 (1993), обеспечивает альтернативный механизм получения фрагментов биспецифичных антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) с помощью линкера, который слишком короток, чтобы позволить образование пар между двумя доменами одной и той же цепи. Соответственно, VH- и VL-домены одного фрагмента вынужденно образуют пары с комплементарными VL и VH-доменами другого фрагмента, создавая, таким образом, два антигенсвязывающих участка. Также опубликована другая стратегия получения фрагментов биспецифичных антител путем использования димеров одноцепочечных Fv (sFv). См. Gruber et al., J. Immunol, 152: 5368 (1994).
Предусмотрены антитела с более чем двумя валентностями. Например, можно получать триспецифичные антитела. Tutt et al., J. Immunol. 147: 60 (1991).
Типичные биспецифичные антитела могут связываться с двумя различными эпитопами, по меньшей мере один из которых берет свое происхождение от белкового антигена по изобретению. В альтернативном варианте анти-антигенное плечо молекулы иммуноглобулина может быть объединено с плечом, которое связывается с триггерной молекулой на лейкоците, такой как молекула Т-клеточного рецептора (например, CD2, CD3, CD28 или В7) или Fc-рецепторы для IgG (Fc-R), такие как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16), для того чтобы сфокусировать клеточные защитные механизмы на клетке, экспрессирующей определенный антиген. Биспецифичные антитела могут также использоваться для введения цитотоксических средств в клетки, которые экспрессируют определенный антиген. Эти антитела имеют антигенсвязывающее плечо и плечо, которое связывается с цитотоксическим средством или радионуклидным хелатором, таким как EOTUBE, DPTA, DOTA или ТЕТА. Другие биспецифичные антитела, представляющие интерес, связываются с белковым антигеном, описанным в настоящем документе, и дополнительно связывают тканевый фактор (TF).
Гетероконъюгатные антитела также входят в объем настоящего изобретения. Гетероконъюгатные антитела состоят из двух ковалентно сшитых антител. Такие антитела, например, были предложены для нацеливания клеток иммунной системы на нежелательные клетки (см. патент США №4676980) и для лечения ВИЧ-инфекции (см. WO 91/00360, WO 92/00373, EP 03089). Предполагается, что гетероконъюгатные антитела можно получать in vitro с помощью известных способов синтетической белковой химии, в том числе способов, включающих перекрестно-сшивающие средства. Например, иммунотоксины могут быть сконструированы с использованием реакции дисульфидного обмена или путем образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат и те, которые описаны, например, в патенте США №4676980.
Может быть желательно модифицировать эффекторную функцию антитела по изобретению для того, чтобы повысить, например, эффективность антитела при лечении злокачественной опухоли и/или других заболеваний и расстройств, связанных с аберрантной экспрессией и/или активностью CD47. Например, в Fc-область можно ввести остаток(ки) цистеина, тем самым позволяя образование дисульфидной связи в данной области. Гомодимерное антитело, полученное таким образом, может отличаться улучшенной способностью к интернализации и/или повышенной опосредованной комплементом гибелью клеток и антителозависимой клеточной цитотоксичностью (ADCC). (См. Caron et al., J. Exp Med., 176: 1191-1195 (1992) и Shopes, J. Immunol., 148: 2918-2922 (1992)). В альтернативном варианте может быть сконструировано антитело, которое имеет двойные Fc-области, и за счет этого имеет повышенную способность к опосредованной комплементом гибели клеток и антителозависимой клеточной цитотоксичности (ADCC). (См. Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989)).
Изобретение также относится к иммуноконъюгатам, содержащим антитело, конъюгированное с цитотоксическим средством, таким как токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения, или его фрагмент), или радиоактивный изотоп (т.е. радиоактивный конъюгат).
Ферментативно активные токсины и их фрагменты, которые могут использоваться, включают А-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модекцина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII, и PAP-S), ингибитор из Momordica charantia, курцин, кротин, ингибитор из Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Для получения радиоконъюгированных антител доступны различные радионуклиды. Примеры включают 212Bi, 131I, 131In, 90Y и 86Re.
Конъюгаты антитела и цитотоксического средства получают с использованием различных бифункциональных средств для связывания белков, таких как N-сукцинимидил-3-(2-пиридилтиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат-HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидо-соединения (такие как бис-(п-азидобензил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-соединения активного фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, рициновый иммунотоксин может быть получен, как описано в Vitetta et al., Science 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминопентауксусная кислота (MXDTPA) представляет собой типовое хелатирующее средство для конъюгации радионуклида с антителом. (См. WO 94/11026).
Обычным специалистам в данной области будет понятно, что множество разнообразных молекул может быть присоединено к полученным в результате антителам по изобретению. (См., например, "Conjugate Vaccines", Contributions to Microbiology and Immunology, J.M. Cruse and R.E. Lewis, Jr (eds), Carger Press, New York, (1989), содержание которой включено в настоящее изобретение в качестве ссылки в полном объеме).
Связывание может быть выполнено с помощью любой химической реакции, которая будет связывать две молекулы, при условии что антитело и другая молекула сохраняют соответствующую им активность. Данное связывание может включать многие химические механизмы, например, ковалентное связывание, аффинное связывание, интеркаляцию, координатное связывание и комплексообразование. Предпочтительное связывание, однако, является ковалентным. Ковалентное связывание достигается путем прямой конденсации существующих боковых групп или путем введения внешних молекул-мостиков. Многие бивалентные или поливалентные связывающие средства могут использоваться в связывании белковых молекул, таких как антитела по настоящему изобретению, с другими молекулами. Например, типичные связывающие средства могут включать органические соединения, такие как тиоэфиры, карбодиимиды, сукцинимидные сложные эфиры, диизоцианаты, глутаровый альдегид, диазобензолы и гексаметилендиамины. Данный список не предназначен для полного перечисления различных классов связывающих средств, известных в данной области, а скорее является примером наиболее распространенных связывающих средств. (См. Killen and Lindstrom, Jour. Immun. 133: 1335-2549 (1984); Jansen et al., Immunological Reviews 62: 185-216 (1982); и Vitetta et al., Science 238: 1098 (1987)).
Предпочтительные линкеры описаны в литературе. (См., например, Ramakrishnan, S. et al., Cancer Res. 44: 201-208 (1984), где описано применение MBS (M-малеимидобензоил-N-гидроксисукцинимидного эфира). См. также патент США №5030719, в котором описано применение галогенированного производного ацетилгидразида, связанного с антителом посредством олигопептидного линкера. Особенно предпочтительные линкеры включают: (i) EDC (1-этил-3-(3-диметиламинопропил)карбодиимида гидрохлорид; (ii) SMPT (4-сукцинимидилокарбонил-альфа-метил-альфа-(2-пиридилдитио)толуол (Pierce Chem. Co., кат. №21558G); (iii) SPDP (сукцинимидил-6[3-(2-пиридилдитио)пропионамидо]гексанат (Pierce Chem. Co., кат. №21651G); (iv) сульфо-LC-SPDP (сульфосукцинимидил-6-[3-(2-пиридилдитио)пропионамид]гексаноат (Pierce Chem. Co., кат. №2165-G); и (v) сульфо-NHS (N-гидроксисульфосукцинимид: Pierce Chem. Co., кат. №24510), конъюгированный с EDC.
Описанные выше линкеры содержат компоненты, которые имеют различные признаки, что приводит к образованию конъюгатов с отличающимися физико-химическими свойствами. Например, сульфо-NHS-эфиры алкилкарбоксилатов более стабильны, чем сульфо-NHS-эфиры ароматических карбоксилатов. Содержащие NHS-эфиры линкеры менее растворимы, чем сульфо-NHS-эфиры. Кроме того, SMPT-линкер содержит стерически блокированную дисульфидную связь и может образовывать конъюгаты с повышенной стабильностью. Дисульфидные связи в общем менее стабильны, чем другие связи, поскольку дисульфидная связь расщепляется in vitro, что приводит к меньшей доступности конъюгата. Сульфо-NHS, в частности, может усиливать стабильность карбодиимидных связей. Карбодиимидные связи (такие как EDC) при использовании совместно с сульфо-NHS образуют сложные эфиры, которые более устойчивы к гидролизу, чем сама по себе карбодиимидная связь.
Раскрытые в настоящем документе антитела могут быть также представлены в виде иммунолипосом. Содержащие антитело липосомы получают известными в данной области способами, такими как способы, описанные в Epstein et al., Proc. Natl. Acad. Sci. USA 82: 3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA 77: 4030 (1980); и в патентах США №№4485045 и 4544545. Липосомы с повышенным временем нахождения в кровотоке описаны в патенте США №5013556.
Особенно подходящие липосомы могут быть получены способом обращенно-фазового выпаривания с использованием липидной композиции, содержащей фосфатидилхолин, холестерин и -модифицированный фосфатидилэтаноламин (PEG-PE). Для получения липосом с требуемым диаметром проводят экструзию липосом через фильтры с определенным размером пор. Fab'-фрагменты антитела по изобретению могут быть конъюгированы с липосомами, как описано в Martin et al., J. Biol. Chem. 257: 286-288 (1982), посредством реакции дисульфидного обмена.
Применение анти-CD47 антител
Следует понимать, что введение терапевтических молекул в соответствии с настоящим изобретением может осуществляться с подходящими носителями, наполнителями и другими средствами, которые включают в композиции, чтобы обеспечить улучшенную передачу, доставку, толерантность и т.п. Множество подходящих составов можно найти в фармакологическом справочнике, известном всем фармацевтам: Remington's Pharmaceutical Sciences (15th ed, Mack Publishing Company, Easton, PA (1975)), в особенности в его 87-й главе авторов Blaug, Seymour. Эти составы включают, например, порошки, пасты, мази, желе, воски, масла, липиды, липидсодержащие (катионные или анионные) везикулы (например, Lipofectin™), конъюгаты ДНК, безводные абсорбентные пасты, эмульсии масло-в-воде и вода-в-масле, эмульсии с CARBOWAX (полиэтиленгликолями с различной молекулярной массой), полутвердые гели и полутвердые смеси, содержащие CARBOWAX. Любая из вышеуказанных смесей может подходить для лечения и терапевтических методов в соответствии с настоящим изобретением, при условии, что действующий ингредиент в составе не инактивируется составом, а состав является физиологически совместимым и переносимым при данном пути введения. Для дополнительной информации, связанной с составами, вспомогательными веществами и носителями, хорошо известными фармацевтам, см. также Baldrick P. "Pharmaceutical excipient development: the need for preclinical guidance." Regul. Toxicol Pharmacol. 32(2):210-8 (2000), Wang W. "Lyophilization and development of solid protein pharmaceuticals." Int. J. Pharm. 203(1-2): 1-60 (2000), Charman WN "Lipids, lipophilic drugs, and oral drug delivery-some emerging concepts." J Pharm Sci. 89(8):967-78 (2000), Powell et al. "Compendium of excipients for parenteral formulations" PDA J Pharm Sci Technol. 52:238-311 (1998) и приведенные в них ссылки.
Терапевтические составы по изобретению, которые включают антитело по изобретению, используются для лечения или облегчения симптомов, связанных с раком, например, в качестве не ограничивающего примера, лейкозами, лимфомами, раком молочных желез, раком толстой кишки, раком яичников, раком мочевого пузыря, раком простаты, глиомой, раком легких и бронхов, колоректальным раком, раком поджелудочной железы, раком пищевода, раком печени, раком мочевого пузыря, раком почек и почечной лоханки, раком полости рта и гортани, раком тела матки и/или меланомой. Настоящее изобретение также относится к способам лечения или облегчения симптомов, связанных с раком. Терапевтическая схема осуществляется путем идентификации индивида, например, пациента-человека, страдающего от (или имеющего риск развития) рака, с использованием стандартных способов.
Эффективность лечения определяется в сочетании с каким-либо известным способом диагностики или лечения конкретного иммунного расстройства. Ослабление одного или нескольких симптомов иммунного расстройства указывает, что антитело дает положительный клинический эффект.
Способы скрининга антител, которые обладают требуемой специфичностью, включают, но не ограничиваются ими, твердофазный иммуноферментный анализ (ELISA) и другие иммунологически опосредованные методы, известные в данной области техники.
Антитела, направленные против мишени, такой как CD47, опухоль-связанного антигена или другого антигена (или его фрагмента) могут быть использованы в способах, известных в данной области техники, относящихся к локализации и/или количественному определению этих мишеней, например, могут использоваться для измерения уровня этих мишеней в соответствующих физиологических образцах, в диагностических способах, для визуализации белка, и т.п. В данном варианте осуществления изобретения антитела, специфичные к любой из этих целей, или их производное, фрагмент, аналог или гомолог, которые содержат антигенсвязывающий домен антитела, используются в качестве фармакологически активных соединений (далее именуются как «терапевтические средства»).
Антитело по изобретению может использоваться для выделения конкретной мишени с помощью стандартных методов, таких как иммунноаффинные методы, хроматография или иммунопреципитация. Антитела по изобретению (или их фрагмент) могут использоваться в диагностике для контроля уровня белка в ткани как часть процедуры медицинского исследования, например, для определения эффективности данной схемы лечения. Детекция может быть облегчена путем связывания (т.е., физического соединения) антитела с детектируемым веществом. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы и радиоактивные материалы. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазыу или ацетилхолинэстеразу; примеры подходящих простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных материалов включают умбеллиферон, флуоресцеин, изотиоцианат флуоресцеина, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; пример люминесцентного материала включает люминол; примеры биолюминесцентных материалов включают люциферазу, люциферин и экворин; и примеры подходящего радиоактивного материала включают 125I, 131I, 35S или 3H.
Антитела по изобретению, включая поликлональные, моноклональные, гуманизированные и полностью человеческие антитела, могут быть использованы в качестве терапевтических средств. Такие средства обычно будут использоваться для лечения или профилактики заболевания или патологии, связанных с аберрантной экспрессией или активацией данной мишени у индивида. Препарат антитела, предпочтительно имеющий высокую специфичность и высокую аффинность к своему антигену-мишени, вводят индивиду, и он обычно действует за счет своего связывания с мишенью. Введение антитела может блокировать (или ингибировать или препятствовать) сигнальную функцию мишени. Введение антитела может блокировать (или ингибировать или препятствовать) связывание мишени с эндогенным лигандом, с которым она связывается в природе. Например, антитело связывается с мишенью и нейтрализует или иным образом ингибирует взаимодействие между CD47 и SIRPα.
Терапевтически эффективное количество антитела по изобретению обычно относится к количеству, необходимому для достижения терапевтической цели. Как отмечалось выше, это может быть взаимодействие между антителом и его антигеном-мишенью, что в некоторых случаях, препятствует функционированию мишени. Количество, необходимое для введения, будет, кроме того, зависеть от аффинности связывания антитела с его конкретным антигеном, а также будет зависеть от скорости, с которой антитело истощается из свободного объема индивида, которому его вводят. Общие диапазоны терапевтически эффективных доз антитела или фрагмента антитела по изобретению могут составлять, в качестве не ограничивающего примера, от примерно 0,1 мг/кг массы тела до примерно 50 мг/кг массы тела. Общая частота введения может варьировать, например, от двух раз в день до одного раза в неделю.
Антитела или их фрагмент по изобретению могут быть введены для лечения различных заболеваний и расстройств в виде фармацевтических композиций. Принципы и факторы, учитываемые при получении таких композиций, а также руководство по выбору компонентов, предоставлены, например, в Remington: The Science And Practice Of Pharmacy 19th ed. (Alfonso R. Gennaro, et al, editors) Mack Pub. Co., Easton, Pa.: 1995; Drug Absorption Enhancement: Concepts, Possibilities, Limitations, And Trends, Harwood Academic Publishers, Langhorne, Pa., 1994; и Peptide And Protein Drug Delivery (Advances In Parenteral Sciences, Vol. 4), 1991, M. Dekker, New York.
При использовании фрагментов антител, предпочтителен наименьший ингибирующий фрагмент, который специфично связывается со связывающим доменом белка-мишени. Например, на основе последовательностей вариабельных областей антитела могут быть разработаны пептидные молекулы, которые сохраняют способность связывать последовательность белка-мишени. Такие пептиды могут быть синтезированы химически и/или могут быть получены с помощью технологии рекомбинантной ДНК. (См., например, Marasco et al. Proc. Natl. Acad. Sci. USA, 90: 7889-7893 (1993)). Состав также может содержать более одного действующего соединения, необходимого для конкретного показания, которое подлежит лечению, предпочтительно, с комплементарной активностью, которые не оказывают отрицательного влияния друг на друга. В качестве альтернативы или в дополнение композиция может содержать средство, который усиливает ее функцию, такой как, например, цитотоксическое средство, цитокин, химиотерапевтическое средство или ингибирующее рост средство. Такие молекулы обычно присутствуют в комбинации в количествах, эффективных для предполагаемого назначения.
Действующие ингредиенты могут быть также заключены в микрокапсулы, полученные, например, способами коацервации или путем межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии.
Составы для применения in vivo должны быть стерильными. Это легко достигается путем фильтрации через стерильные фильтрационные мембраны.
Могут быть получены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, где матрицы представляют собой формованные изделия, например, пленки или микрокапсулы. Примеры матриц с замедленным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат) или поливиниловый спирт), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (микросферы для инъекций, состоящие из сополимера молочной кислоты и гликолевой кислоты и лейпролида ацетата), и поли-D-(-)-3-гидроксимасляную кислоту. Тогда как полимеры, такие как этиленвинилацетат и сополимеры молочной кислоты и гликолевой кислоты способны высвобождать молекулы в течение более 100 дней, некоторые гидрогели высвобождают белки за более короткие периоды времени.
Антитело по изобретению может быть использовано в качестве средства для детекции присутствия данной мишени (или ее белкового фрагмента) в образце. В некоторых вариантах осуществления изобретения антитело содержит детектируемую метку. Антитела являются поликлональными или, более предпочтительно, моноклональными. Используется интактное антитело или его фрагмент (например, Fab, scFv или F(ab)2). Предполагается, что термин «меченый» в отношении зонда или антитела охватывает прямое мечение зонда или антитела путем связывания (т.е. физического соединения) детектируемого вещества с зондом или антителом, а также непрямое мечение зонда или антитела за счет способности вступать в реакцию с другим напрямую меченным реагентом. Примеры непрямого мечения включают детекцию первичного антитела с использованием флуоресцентно-меченого вторичного антитела и мечение конца ДНК-зонда биотином таким образом, что его можно детектировать с использованием флуоресцентно-меченого стрептавидина. Предполагается, что термин «биологический образец» включает ткани, клетки и биологические жидкости, выделенные из индивида, а также ткани, клетки и жидкости, присутствующие в индивиде. Поэтому в термин «биологический образец» включена кровь, а также фракция или компонент крови, в том числе сыворотка крови, плазма крови или лимфа. То есть способ детекции по изобретению может использоваться для обнаружения анализируемых мРНК, белка или геномной ДНК в биологическом образце in vitro, а также in vivo. Например, методы in vitro для обнаружения анализируемой мРНК включают Нозерн-гибридизацию и гибридизации in situ. Методы in vitro для детекции анализируемого белка включают твердофазный иммуноферментный анализ (ELISA), Вестерн-блот, иммунопреципитацию и иммунофлюоресценцию. Методы in vitro для детекции анализируемой геномной ДНК включают Саузерн-гибридизацию. Процедуры для проведения иммунологических анализов описаны, например, в "ELISA: Theory and Practice: Methods in Molecular Biology", Vol. 42, J.R. Crowther (Ed.) Human Press, Totowa, NJ, 1995; "Immunoassay", E. Diamandis and T. Christopoulus, Academic Press, Inc., San Diego, CA, 1996; и "Practice and Theory of Enzyme Immunoassays", P. Tijssen, Elsevier Science Publishers, Amsterdam, 1985. Кроме того, in vivo методы для детекции анализируемого белка включают введение индивиду меченого антитела против анализируемого белка. Например, антитело может быть помечено радиоактивным маркером, присутствие и расположение которого у индивида могут детектироваться с помощью стандартных методов визуализации.
Фармацевтические композиции
Антитела по изобретению (также именуемые в настоящем документе «действующими соединениями») и их производные, фрагменты, аналоги и гомологи могут быть включены в фармацевтические композиции, подходящие для введения. Такие композиции, как правило, содержат антитело и фармацевтически приемлемый носитель. Предполагается, что используемый в настоящем документе термин «фармацевтически приемлемый носитель» включает любые и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средство, изотонические и замедляющие всасывание средство и т.п., совместимые с фармацевтическим введением. Подходящие носители описаны в последнем издании Remington's Pharmaceutical Sciences, стандартном справочнике в данной области, который включен в настоящее описание в качестве ссылки. Предпочтительные примеры таких носителей или разбавителей включают, но не ограничиваются ими, воду, физиологический раствор, растворы Рингера, раствор декстрозы и 5%-й сывороточный альбумин человека. Также можно использовать липосомы и неводные носители, такие как нелетучие масла. Применение таких сред и средств для фармацевтически активных веществ хорошо известно в данной области. За исключением случаев, когда любая стандартные среда или средство несовместимы с действующим соединением, предусмотрено их применение в композициях. В композиции также могут быть включены дополнительные действующие соединения.
Фармацевтическая композиция по изобретению составлена таким образом, чтобы быть совместимой с его предполагаемым путем введения. Примеры путей введения включают парентеральное, например, внутривенное, внутрикожное, подкожное, пероральное (например, путем ингаляции), трансдермальное (т.е. местное), введение через слизистую и ректальное введение. Растворы или суспензии, используемые для парентерального, внутрикожного или подкожного применения могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные средства, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатирующие средства, такие как этилендиаминтетрауксусная кислота (ЭДТА); буферы, такие как ацетаты, цитраты или фосфаты; и средства для регулирования тоничности, такие как хлорид натрия или декстроза. рН можно регулировать с помощью кислот или оснований, таких как соляная кислота или гидроксид натрия. Препарат для парентерального введения может быть заключен в ампулы, одноразовые шприцы или многодозовые флаконы, изготовленные из стекла или пластика.
Фармацевтические композиции, подходящие для инъекций, включают стерильные водные растворы (в случае водорастворимых композиций) или дисперсии и стерильные порошки для экстемпорального приготовления стерильных инъекционных растворов или дисперсий. Для внутривенного введения подходящие носители включают физиологический раствор, бактериостатическую воду, Cremophor EL™ (фирма BASF, Parsippany, NJ) или фосфатно-солевой буфер (PBS). Во всех случаях композиция должна быть стерильной и должна быть жидкой до такой степени, чтобы могла легко выдавливаться из шприца. Она должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Носитель может быть растворителем или дисперсионной средой, содержащей, например, воду, этанол, многоатомный спирт (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Надлежащую текучесть можно поддерживать, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем использования поверхностно-активных веществ. Предотвращение действия микроорганизмов может быть достигнуто с помощью различных антибактериальных и противогрибковых средств, например, парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимеросала и т.п. Во многих случаях будет предпочтительно включать в композиции изотонические средства, например, сахара, многоатомные спирты, такие как маннит, сорбит, и хлорид натрия. Пролонгированная абсорбция инъекционных композиций может быть осуществлена путем включения в композицию средства, который задерживает абсорбцию, например, моностеарата алюминия и желатина.
Стерильные инъекционные растворы могут быть получены введением действующего соединения в требуемом количестве в соответствующий растворитель с одним из перечисленных выше ингредиентов или их комбинацией (если требуется) с последующей стерилизацией фильтрованием. В общем, дисперсии получают включением действующего соединения в стерильный носитель, который содержит основную дисперсионную среду и требуемые другие ингредиенты из тех, которые перечислены выше. В случае стерильных порошков для приготовления стерильных инъекционных растворов, способами получения являются вакуумная сушка и сушка вымораживанием, которая дает порошок действующего ингредиента плюс любой дополнительный желательный ингредиент из ранее стерилизованного фильтрацией раствора.
Пероральные композиции обычно включают инертный разбавитель или съедобный носитель. Они могут быть заключены в желатиновые капсулы или спрессованы в таблетки. Для целей перорального терапевтического введения действующее соединение может быть включено с вспомогательными веществами и может использоваться в виде таблеток, пастилок или капсул. Пероральные композиции также могут быть получены с использованием жидкого носителя для использования в виде жидкости для полоскания рта, где соединение в жидком носителе принимается перорально и полоскается и выплевывается или проглатывается. Фармацевтически совместимые связующие средства и/или адъюванты могут быть включены в качестве части композиции. Таблетки, пилюли, капсулы, пастилки и т.п. могут содержать любые из следующих ингредиентов или соединений аналогичной природы: связующее вещество, такое как микроцеллюлоза, камедь трагаканта или желатин; вспомогательное вещество, такое как крахмал или лактоза; вещество, способствующее распадаемости, такое как альгиновая кислота, Primogel или кукурузный крахмал; смазывающее вещество, такое как стеарат магния или Sterotes; вещество, способствующее скольжению, такое как коллоидный диоксид кремния; подсластитель, такой как сахароза или сахарин; или вкусо-ароматическое средство, такое как мята перечная, метилсалицилат или апельсиновый ароматизатор.
Для введения путем ингаляции соединения вводятся в виде аэрозольного спрея из находящегося под давлением контейнера или дозатора, который содержит подходящий пропеллент, например, газ, такой как диоксид углерода, или небулайзера.
Системное введение также может осуществляться через слизистую или через кожу. Для введения через слизистую или кожу в составе используются вещества, проникающие через соответствующий барьер. Такие проникающие вещества обычно известны в данной области и включают, например, для введения через слизистую, поверхностно-активные вещества, соли желчных кислот и производные фузидовой кислоты. Введение через слизистую может осуществляться путем использования назальных спреев или суппозиториев. Для введения через кожу действующие соединения вводят в состав мазей, бальзамов, гелей или кремов, как в общем известно в данной области.
Соединения также могут быть получены в виде суппозиториев (например, с традиционными основами для суппозиториев, такими как масло какао и другие глицериды) или удерживающих клизм для ректальной доставки.
В одном из вариантов осуществления изобретения действующие соединения изготавливают с носителями, которые будут защищать соединение от быстрого выведения из организма, например, в виде состава с контролируемым высвобождением, включая имплантаты и микроинкапсулированные системы доставки. Могут использоваться биоразлагаемые, биосовместимые полимеры, например, этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Способы получения таких составов будут очевидны специалистам в данной области. Материалы также могут быть получены коммерчески от Alza Corporation и Nova Pharmaceuticals, Inc. В качестве фармацевтически приемлемых носителей также могут использоваться липосомальные суспензии (включая липосомы, ориентированные на инфицированные клетки с моноклональными антителами к вирусным антигенам). Они могут быть получены в соответствии со способами, известными специалистам в данной области техники, например, как описано в патенте США №4522811.
Особенно предпочтительно получение пероральных или парентеральных композиций в стандартной лекарственной форме для простоты введения и однородности дозировки. Стандартная лекарственная форма в контексте настоящего изобретения относится к физически дискретным единицам, подходящим в качестве единичных доз для индивида, подлежащего лечению, причем каждая единица содержит заданное количество действующего соединения, рассчитанное на получение желаемого терапевтического эффекта в сочетании с требуемым фармацевтическим носителем. Спецификация для стандартных лекарственных форм по изобретению диктуется и непосредственно зависит от уникальных характеристик действующего соединения и конкретного терапевтического эффекта, который должен быть достигнут, а также от ограничений, накладываемых в области рецептур на такое действующее соединение для лечения индивидуумов.
Фармацевтические композиции могут быть включены в контейнер, пакет или дозатор вместе с инструкциями по введению.
Далее изобретение будет описано в следующих ниже примерах, которые не ограничивают объем изобретения, описанный в формуле изобретения.
ПРИМЕРЫ
ПРИМЕР 1: клонирование, экспрессия и очистка анти-CD47 человека
Клонирование. Последовательность, соответствующую внеклеточному домену CD47 человека (hCD47), амплифицировали на кДНК человека с помощью полимеразной цепной реакции (ПЦР) с использованием специфичных олигонуклеотидов. Амплифицированный продукт очищали в геле и клонировали в вектор для экспрессии в клетках млекопитающих, pEAK8 (EDGE Biosystems, Gaithersburg, MD). Вектор дополнительно модифицировали, чтобы ввести Avitag™ (Avidity, Denver CO) и гекса-гистидиновый тег на С-конце, позволяющие биотинилирование белка по одному сайту и очистку с помощью IMAC (аффинной хроматографии на иммобилизованных ионах металла). Конструкции подтверждали секвенированием ДНК.
Экспрессия. Плазмиду затем трансфицировали в клетки млекопитающих с использованием трансфекционного реагента на основе липосом, такого как TransIT-LT1 (Minis, Madison, WI). На стадии трансфекции требуется только небольшое количество ДНК и клеток, как правило, 2×105 клеток и 2 мкг плазмидной ДНК на лунку, а трансфекцию проводили в 6-луночном планшете. Хотя в приведенных ниже примерах могут использоваться различные клеточные линии млекопитающих, проводили трансфекцию трансформированных монослойных эпителиальных клеток почки эмбриона человека (клеток РЕАК). Эти клетки, стабильно экспрессируют ген EBNA-1, дополнительно способствующий процессу эписомальной репликации, являются полуприкрепленными и могут выращиваться при стандартных условиях культивирования в клеточном инкубаторе (при 5% СО2 и 37°С в среде DMEM, дополненной 10%-й фетальной телячьей сывороткой). Через 24 часа клетки помещали в селективные условия путем добавления среды, содержащей 0,5-2 мкг/мл пуромицина, поскольку клетки, несущие эписомальный вектор, устойчивы к этому антибиотику.
Через две-три недели после трансфекции, клетки использовали для посева в Т-флаконы или одноразовые биореакторы CELLine для стадии продукции. CELLine представляет собой двухкамерный биореактор, который может использоваться в стандартном инкубаторе для клеточных культур. Меньшая камера (15 мл) содержит клетки и отделена от большей камеры, содержащей среду (1 л), с помощью полупроницаемой мембраны с размером отсечки 10 кДа (Bruce et al. 2002, McDonald et al. 2005). Эта система позволяет диффузию питательных веществ, газов и продуктов обмена веществ, сохраняя клетки и секретируемые белки в меньшем отсеке. Культуру поддерживают в течение 7-10 дней до сбора супернатанта. Поскольку среда содержит сыворотку, клетки сохраняют хорошую жизнеспособность, и можно провести несколько циклов пролучения с использованием одних и тех же клеток и контейнеров.
Очистка. После сбора клеточные культуральные супернатанты осветляли центрифугированием и фильтровали через мембрану 0,22 мкм. Супернатант из T-флаконов концентрировали в 20-40 раз с помощью концентратора, такого как SartoFlow 200 (Sartorius) с мембраной, имеющей соответствующий размер отсечки, чтобы сохранить представляющий интерес белок. Эта стадия не требуется при использовании биореактора CELLline вследствие малого объема, извлекаемого из клеточной камеры. Кроме того, стадия концентрирования увеличивает концентрацию как представляющего интерес белка, так и высокомолекулярных примесей, таких как бычий альбумин или иммуноглобулины сыворотки. В противоположность этому, супернатант, извлекаемый из клеточной камеры биореактора CELLine, содержит концентрированный рекомбинантный белок и пониженный уровень примесей, поскольку они не могут пересекать мембрану 10 кДа, разделяющую две камеры реактора. Это увеличенное соотношение рекомбинантного белка и примесей значительно повышает эффективность очистки с помощью IMAC. В концентрированный супернатант затем добавляли 100 мМ имидазол и наносили на Ni-NTA-смолу для аффинной хроматографии (Qiagen). Относительно высокая концентрация имидазола минимизирует связывание примесей со смолой. После промывки колонки, белки элюируют при скорости потока 2 мл/мин с использованием 30-миллилитрового градиента имидазола (20-400 мМ имидазол) на хроматографической системе ÄKTA Prime (Amersham Pharmacia Biotech). Элюирование градиентом дополнительно улучшает чистоту рекомбинантного белка, но может быть заменено ступенчатой элюцией, если не доступна система хроматографии. Элюированные фракции могут быть проанализированы с помощью электрофореза в ДСН-ПААГ или ELISA, чтобы определить содержание в них рекомбинантного белка. Фракции, представляющие интерес, объединяли и обессоливали на колонке PD-10 (GE Healthcare), уравновешенной фосфатно-солевым буфером или другим соответствующим буфером. Обессоленные белки затем можно оценить количественно с использованием различных методов, а их чистоту проанализировать с помощью электрофореза в ДСН-ПААГ. Рекомбинантный CD47 биотинилировали in vitro, используя биотин-лигазу (Avidity, Denver CO) в соответствии с инструкциями изготовителя. После обессоливания уровень биотинилирования оценивали с помощью соосаждения на магнитных гранулах со стрептавидином и электрофореза в ДСН-ПААГ.
ПРИМЕР 2: клонирование, экспрессия и очистка CD19 человека
Клонирование. Последовательность, соответствующую внеклеточному домену CD19 человека (hCD19), амплифицировали на кДНК человека с помощью полимеразной цепной реакции (ПЦР) с использованием специфичных олигонуклеотидов. Амплифицированный продукт очищали в геле и клонировали в вектор для экспрессии в клетках млекопитающих, pEAK8 (EDGE Biosystems, Gaithersburg, MD). Вектор дополнительно модифицировали, чтобы ввести Avitag™ (Avidity, Denver CO) и гекса-гистидиновый тег на С-конце, позволяющие биотинилирование белка по одному сайту и очистку с помощью IMAC (аффинной хроматографии на иммобилизованных ионах металла). Конструкции подтверждали секвенированием ДНК.
Экспрессия и очистка. Экспрессию, очистку и биотинилирование растворимого hCD19 проводили, как описано в примере 1.
ПРИМЕР 3: отбор методом фагового дисплея с использованием библиотек scFv человека, содержащих фиксированную вариабельную область тяжелой цепи
Общие процедуры конструирования и работы с библиотекам scFv человека, представленными на бактериофаге M13, описаны в Vaughan et al., (Nat. Biotech. 1996, 14:309-314), включенной в настоящий документ в качестве ссылки в полном объеме. Библиотеки, используемые для отбора и скрининга, кодируют scFv, которые все имеют один и тот же VH-домен и отличаются только своими VL-доменами. Способы создания библиотек с фиксированным VH и их применение для идентификации и сборки биспецифичных антител описаны в US 2012/0184716 и WO 2012/023053, которые включены в настоящее описание в качестве ссылки в полном объеме. Процедуры для идентификации scFv, связывающихся с hCD19 или hCD47, описаны ниже.
Отбор в жидкой фазе. Аликвоты scFv-фаговых библиотек (1012 БОЕ) блокировали PBS, содержащим 3% (масс./объем) обезжиренного молока в течение одного часа при комнатной температуре на ротационном смесителе. Заблокированные фаги затем истощали на стрептавидиновых магнитных шариках (Dynal М-280) в течение одного часа при комнатной температуре на ротационном смесителе. Истощенные фаги затем инкубировали с биотинилированными in vitro hCD19 или hCD47 (100 нМ) в течение двух часов при комнатной температуре на ротационном смесителе. Шарики собирали с помощью магнитной стойки с последующими четырьмя промывками PBS/0,1%-м Tween 20 и 3 промывками PBS. Шарики затем напрямую добавляли к 10 мл экспоненциально растущих клеток TG1 и инкубировали в течение одного часа при 37°С с медленным встряхиванием (100 об./мин.). Аликвоту инфицированных TG1 разбавляли серийными разведения для определения выхода селекции. Остальные инфицированные TG1 центрифугировали при 3000 об./мин. в течение 15 минут и ресуспендировали в 0,5 мл 2xTY-AG (среде 2xTY, содержащей 100 мкг/мл ампициллина и 2% глюкозы) и высевали на чашки Bioassay с 2xTYAG-агаром. После инкубации в течение ночи при 30°С в чашки добавляли по 10 мл 2xTYAG и клетки соскабливали с поверхности и переносили в 50-мл полипропиленовую пробирку. К суспензии клеток добавляли 2xTYAG, содержащую 50% глицерина, чтобы получить конечную концентрацию глицерина 17%. Аликвоты после раунда отбора хранили при -80°С.
Выделение фагов. 100 мкл клеточной суспензии, полученной из предшествующих раундов отбора, добавляли к 20 мл 2xTYAG и выращивали при 37°С и перемешивании (240 об./мин.) до достижения OD600 от 0,3 до 0,5. Затем культуру суперинфицировали 3,3×1010 частицами хелперного фага MK13K07 и инкубировали в течение одного часа при 37°C (150 об./мин.). Затем заменяли среду путем центрифугирования клеток при 2000 об./мин. в течение 10 минут, удаления среды и ресуспендирования осадка в 20 мл 2xTY-АК (100 мкг/мл ампициллина, 50 мкг/мл канамицина). Затем культуру выращивали в течение ночи при 30°С (240 об./мин.). На следующий день содержащий фаги супернатант использовали для следующего раунда отбора.
Отбор на клеточной поверхности. Содержащие фаги супернатанты блокировали PBS, содержащим 3% (масс./объем) обезжиренного молока в течение одного часа при комнатной температуре на ротационном смесителе. Заблокированные фаги затем истощали в течение одного часа на Т-клетках Jukat, которые не экспрессируют CD19 и которые были ранее блокированы PBS, содержащим 2% (масс./объем) обезжиренного молока. Истощенные фаги затем инкубировали с 2×107 клеток Raji, экспрессирующих CD19, в течение одного часа при комнатной температуре и осторожном встряхивании. Затем клетки осаждали и промывали десять раз PBS. Связанные фаги элюировали, добавляя напрямую 10 мл экспоненциально растущих TG1 во флакон T75 и инкубируя в течение одного часа при 37°С и медленном встряхивании. Аликвоту инфицированных TG1 разбавляли серийными разведения для определения выхода селекции. Инфицированные TG1 центрифугировали при 3000 об./мин. в течение 15 минут и ресуспендировали в 0,5 мл 2xTY-AG (среде 2xTY, содержащей 100 мкг/мл ампициллина и 2% глюкозы) и высевали на чашки Bioassay с 2xTYAG-агаром. После инкубации в течение ночи при 30°С в чашки добавляли по 10 мл 2xTYAG и клетки соскабливали с поверхности и переносили в 50-мл полипропиленовую пробирку. К суспензии клеток добавляли 2xTYAG, содержащую 50% глицерина, чтобы получить конечную концентрацию глицерина 17%. Аликвоты после раунда отбора хранили при -80°С.
Получение scFv в периплазме для связывания и функциональных тестов. Индивидуальные клоны вносили в микротитрационный планшет с глубокими лунками, содержащими 0,9 мл среды 2xTYAG (0,1% глюкозы) на лунку и выращивали при 37°С в течение 5-6 ч. (250 об./мин.). Затем добавляли по 100 мкл на лунку 0,2 мМ IPTG в среде 2xTY до получения конечной концентрации 0,02 мМ IPTG. Затем планшеты инкубировали в течение ночи при 30°С при встряхивании при 250 об./мин. Планшеты с глубокими лунками центрифугировали при 2500 об./мин. в течение 10 мин и осторожно удаляли супернатант. Осадки ресуспендировали в 150 мкл буфера TES (50 мМ Трис/HCl (рН 8), 1 мМ ЭДТА (рН 8), 20% сахарозы, дополненного полным протеазным ингибитором, Roche). Гипотонический шок выполняли добавлением 150 мкл разбавленного буфера TES (разведение TES в воде составляло 1:5) и инкубацией на льду в течение 30 мин. Затем планшеты центрифугировали при 4000 об./мин. в течение 10 минут для удаления клеток и дебриса. Супернатанты осторожно переносили в другой микротитрационный планшет и хранили на льду для немедленного тестирования в функциональных анализах или анализах связывания.
Секвенирование фаговых клонов. Одиночные клоны помещали в микротитрационный планшет, содержащий по 150 мкл среды 2xTYAG (2% глюкозы) на лунку и выращивали при 30°C (120 об./мин.) в течение ночи. На следующий день 5 мкл культуры переносили в другой планшет, содержащий 45 мкл дистиллированной воды и перемешивали. Планшет затем замораживали при -20°С. После оттаивания 1 мкл этой суспензии использовали для ПЦР-амплификации с использованием стандартных протоколов ПЦР с праймером, специфичным для pNDS1: mycseq, 5'-CTCTTCTGAGATGAGTTTTTG-3' (SEQ ID NO: 283) и gene3leader, 5'-TTATTATTCGCAATTCCTTTAGTTGTTCCT-3' (SEQ ID NO: 284). ПЦР-реакции очищали в 96-луночном формате с использованием системы Montage PCRμ96 (Millipore). 5 мкл элюированной ДНК секвенировали с использованием праймеров mycseq и gene3leader.
ПРИМЕР 4: скрининг на связывание scFv с hCD47 и ингибирование scFv взаимодействия с SIRPα
Связывание: скрининг на связывание scFv с hCD47 проводили в гомогенном анализе с использованием технологии FMAT. В каждой лунке 384-луночного оптически прозрачного планшета (Costar) смешивали следующие реагенты : 30 мкл суспензии полистирольных шариков со стрептавидином (Spherotech, 3000 шариков/лунку), покрытых биотинилированным hCD47 или контрольным биотинилированным белком (NusA); 60 мкл периплазматического препарата scFv; 10 мкл детектирующего буфера (PBS, содержащего анти-cmyc антитело в концентрации 5 мкг/мл, конъюгированное с AlexaFluor 647 антитело против мышиной Fc-области разводили 1:200). После перемешивания при 450 об./мин. в течение 5 минут, 384-луночные планшеты инкубировали при комнатной температуре и измеряли через 1 и 3 часа на FMAT 8200 Cellular Detection System (Applied Biosystems). Каждый образец scFv тестировали в дубликатах с hCD47 и NusA. Экспрессирующие scFv клоны, дающие специфичный сигнал на hCD47, а не NusA, отбирали для дальнейшего анализа.
Ингибирование взаимодействия CD47-SIRPα: scFv также скринировали по их способности ингибировать взаимодействие между CD47 и SIRPα в гомогенном анализе на основе шариков с использованием технологии FMAT. Полистирольные шарики (Spherotech) инкубировали с 5 мкг/мл козьих антител, специфичных к Fcγ IgG человека (Jackson lmmunoresearch). После промывки шариков добавляли 5 мкг/мл SIRPα-Fc (R&D Systems) для иммобилизации слитого белка на поверхности шариков. После блокирования PBS, содержащим 2% Tropix I-block (Applied Biosystems), в каждую лунку 384-луночного оптически прозрачного планшета (Costar) добавляли по 30 мкл суспензии покрытых шариков (3000 шариков/лунку). В отдельном 96-луночном планшете 120 мкл периплазматических препаратов scFv смешивали с 24 мкл биотинилированного hCD47 (300 нг/мл) и инкубировали в течение 50 минут при комнатной температуре, так чтобы scFv мог связаться с hCD47. После инкубации к смеси добавляли 24 мкл стрептавидина-Cy5 (1 мкг/мл; Invitrogen) и 70 мкл этой конечной смеси добавляли к 30 мкл шариков в каждой лунке 384-луночного планшета. После 3 часов инкубации при комнатной температуре планшет затем измеряли на FMAT 8200 Cellular Detection System (Applied Biosystems). В каждый планшет включали контрольные лунки, не содержащие scFv или содержащие неспецифичные scFv, не связывающие CD47, так чтобы отобрать для дальнейшего анализа экспрессирующие scFv клоны, которые приводили к уменьшению сигнала SIRPα-CD47, измеренного в контрольных лунках.
В альтернативном варианте для скрининга потенциальных антител также использовали клеточный метод анализа, в котором регистрируется взаимодействие растворимого SIRPα с hCD47, экспрессируемым на поверхности стабильно трансфицированной линии клеток яичников китайского хомячка (СНО). В каждую лунку 384-луночного оптически прозрачного планшета (Costar) добавляли по 20 мкл PBS с 2%-м BSA и 0,1%-м азидом, содержащих 3000 клеток CHO, экспрессирующих hCD47. Затем в каждую лунку добавляли по 50 мкл двухкратного разведения периплазматических препаратов scFv и инкубировали при комнатной температуре в течение 30 минут, чтобы позволить scFv связаться с CD47 на клетках. После инкубации добавляли 30 мкл PBS с 2%-м BSA и 0,1%-м азидом, содержащих 10 нг/мл SIRPα-Fc (R&D systems) и конъюгированное с FMAT Blue анти-hIgG-Fc антитело (разведенное 1:2000) и дополнительно инкубировали в течение 3 часов перед измерением на FMAT 8200 Cellular Detection System (Applied Biosystems).
ПРИМЕР 5: скрининг на связывание scFv с hCD19
Скрининг scFv на связывание с рекомбинантным hCD19 проводили в гомогенном анализе с использованием технологии FMAT, как описано в примере 4.
Скрининг также проводили на клетках Raji на связывание с нативной формой hCD19. В каждую лунку 384-луночного оптически прозрачного планшета (Costar) добавляли по 30 мкл PBS с 2%-м BSA и 0,1%-м азидом, содержащих 3000 клеток Raji (В-клеточной лини человека, экспрессирующей CD19) или клеток Jurkat (T-клеточнй линии человека, которая не экспрессирует CD 19). Затем добавляли 30 мкл двухкратного разведения периплазматических препаратов scFv, 30 мкл PBS-BSA2% и 10 мкл 10X детектирующего буфера (PentaHis AF647 антитело от компании Qiagen, разведенное 1:700 в PBS-BSA2%). После смешивания при 450 об./мин.в течение 5 минут, 384-луночные планшеты инкубировали при комнатной температуре и измеряли через 1 и 3 часа на FMAT 8200 Cellular Detection System (Applied Biosystems). Клоны, экспрессирующие scFv, которые давали специфичный сигнал с клетками Raji, но не с клетками Jurkat, были отобраны для дальнейшего анализа.
ПРИМЕР 6: переформатирование в IgG перспективных фиксированных VH и транзиентная экспрессия в клетках млекопитающих
После скрининга перспективные scFv против hCD19 или hCD47 переформатировали в IgG и экспрессировали с помощью транзиентной трансфекции в клетках РЕАК. Последовательности VH и VL выбранного scFv амплифицировали со специфичными олигонуклеотидами и клонировали в вектор экспрессии, содержащий константные области тяжелой и легкой цепей, и конструкции подтверждали секвенированием. Векторы экспрессии трансфицировали в клетки млекопитающих, используя Fugene 6 Transfection Reagent (Roche, Basel, Switzerland). Кратко, клетки РЕАК культивировали в 6-луночных планшетах с концентрацией 6×105 клеток на лунку в 2 мл культуральной среды, содержащей бычью фетальную сыворотку. Векторы экспрессии, кодирующие последовательности перспективных VH и VL совместно трансфицировали в клетки, используя Fugene 6 Transfection Reagent в соответствии с инструкциями производителя. На следующий день после трансфекции культуральную среду аспирировали, а к клеткам добавляли 3 мл свежей бессывороточной среды и культивировали в течение 3 дней при 37°C. После трехдневного периода культивирования собирали супернатант для очистки IgG на колонках для высокоскоростной жидкостной хроматографии, Protein G-Sepharose 4B (Sigma, St. Louis, MO) в соответствии с инструкциями производителя. Кратко, супернатанты от трансфицированных клеток инкубировали в течение ночи при 4°C со связывающим IgG буфером ImmunoPure (G) (Pierce, Rockford, IL). Образцы затем пропускали через колонки для высокоскоростной жидкостной хроматографии, Protein G-Sepharose 4B, и IgG далее очищали, используя буфер для элюции. Элюированную фракцию IgG затем диализовали против PBS, а содержание IgG количественно оценивали по поглощению при 280 нм. Чистоту и целостность IgG подтверждали электрофорезом в ДСН-ПААГ.
ПРИМЕР 7: модуляция аффинности анти-hCD47 антител
(a) Антитела Ka3, Ke8, Kc4
Три антитела, идентифицированных в ходе скринингового процесса, описанного в приведенных выше примерах, которые, как было показано, специфичны к CD47 человека и способны блокировать взаимодействие между CD47 и SIRPα, были отобраны для созревания аффинности с целью повышения их аффинности и силы действия. Все эти антитела имеют одинаковую вариабельную область тяжелой цепи, но имеют различные вариабельные области легких цепей. Ka3 и Ke8 содержат легкую каппа-цепь (IGVK1-39 согласно номенклатуре IMGT), тогда как Kc4 содержит легкую лямбда-цепь (IGVL2-14). Было создано несколько фаговых библиотек, представляющих варианты scFv, путем введения разнообразия в CDR1, CDR2 и CDR3 вариабельной области легкой цепи, оставляя вариабельную область тяжелой цепи немодифицированной. Одна библиотека из 9×107 трансформантов, покрывающая теоретическое разнообразие 7×105, была создана для Ka3, две библиотеки из 2×108 трансформантов каждая, частично покрывающих теоретическое разнообразие 2,4×109, были созданы для Ke8, и одна библиотека из 3,6×107 трансформантов, покрывающая теоретическое разнообразие 2,6×105, была создана Kc4. Эти библиотеки использовали для отбора методом фагового дисплея, как описано в примере 3, за исключением того, что жесткость отбора была увеличена между раундами путем уменьшения концентрации hCD47 между различными раундами: 10 нМ и 1 нМ hCD47 использовали в первом и втором раунде отбора, соответственно. Отобранные варианты скринировали на способность ингибировать взаимодействие между hCD47 и SIRPα с использованием анализа, описанного в примере 4. Положительные клоны затем были переформатированы в IgG и охарактеризованы, как описано в нижеследующих примерах. Эти попытки созревания аффинности привели к идентификации следующих анти-CD47 антител:
- Ke8H6; Ke86G9; Ke8A3; Ke8C4; Ke8F1; Ke8B7; Ke8G1; Ke8A8; Ke8A4; Ke8B2; Ke8C7; Ke8H3; Ke8A2; Ke8H5; Ke8G6; Ke8E8; Ke81A3; Ke81G9; Ke84G9; Ke8G2; Ke8F2
- Ka3G2; Ka3D3; Ka3A2; Ka3B2; Ka3C5; Ka3A3; Ka3H8; Ka3H3
- Kc4E2; Kc4F4; Kc4A1; Kc4C11; Kc4E10; Kc4B1; Kc4C3; Kc4A4;
Kc4G11; Kc4G9
(b) генно-инженерные модификации антитела 5A3 с целью модулирования аффинности
Последовательность VL анти-CD47 антитела 5A3 была модифицирована для снижения его аффинности к мишени. Последовательность VL антитела 5A3 выравнивали с ее ближайшей зародышевой последовательностью, IGKV1-33 человека, в соответствии с номенклатурой IMGT (фиг. 1). С использованием этого выравнивания были идентифицированы несколько остатков в CDRL1 и CDRL2 из VL антитела 5A3, которые не являлись консервативными относительно зародышевой последовательности. Некоторые из этих аминокислот мутировали с целью изменения аффинности связывания антитела. Остатки из CDRL3 антитела 5А3 также были изменены для модулирования связывающей способности антитела, одновременно сохраняя специфичность к тому же эпитопу на CD47. Эти различные стратегии привели к идентификации перспективных антител 5A3-M3 и 5A3-M5.
Эти варианты были сначала протестированы в анализе связывания CD47/SIRPα для определения их блокирующей способности по сравнению с родительским антителом 5А3 (фиг. 2). Оба антитела, 5A3-M3 и 5A3-M5, хуже ингибировали взаимодействие CD47 и SIRPα чем 5A3, причем 5A3-M5 показывало наиболее слабую способность к ингибированию.
Затем оценивали аффинность этих вариантов к CD47 человека методом поверхностного плазмонного резонанса. KD для антител 5A3, 5A3-M3 и 5A3-M5 составляли примерно 2,36E-08, 5,60E-08 и 2,84E-06 M, соответственно. Эти данные подтвердили, что варианты 5А3 связываются с CD47 с меньшей аффинностью по сравнению с родительским антителом, и что 5A3-M5 имеет наименьшую аффинность к CD47 человека, тогда как 5A3-M3 имеет промежуточную аффинность.
ПРИМЕР 8: характеристика антител против CD47
Связывание антител против CD47 с клетками СНО, трансфицированными huCD47
Специфичность анти-CD47 моноклональных антител (Mab) была показана с помощью проточной цитометрии с использованием клеток СНО, стабильно трансфицированных CD47 человека (клетки CHO-huCD47). В качестве контроля использовали нетрансфицированные клетки СНО. Кратко, очищенные анти-CD47 Mab инкубировали с клетками CHO-huCD47 при конечной концентрации 10 мкг/мл в течение 30 минут. После двух промывок связавшиеся анти-CD47 антитела детектировали с использованием конъюгированного с Cy-5 вторичного антитела к Fc человека (BD Biosciences). На фиг. 3 показано заметное связывание анти-CD47 Mab с трансфицированными hu-CD47 клетками CHO, и отсутствие связывания (или фоновое связывание) с нетрансфицированными клетками СНО, таким образом демонстрирующее специфичность анти-CD47 Mab по настоящему изобретению.
Связывание анти-CD47 антител с клетками HEK293-P
Специфичность анти-CD47 моноклональных антител была дополнительно подтверждена в эксперименте с использованием клеток HEK293-P с опосредованным киРНК нокдауном гена CD47. Клетки HEK293-P (клетки PEAK) были получены из клеток почки эмбриона человека и экспрессируют CD47 на низком или среднем уровне. Был создан дефицитный по CD47 вариант клеток PEAK путем их стабильной трансфекции киРНК, специфичной к гену CD47. Экспрессия антигена CD47 на клеточной поверхности снижена в этих клетках PEAK с нокдауном по CD47 более чем на 85% (данные не показаны). На фиг. 4 показано связывание отобранных анти-CD47 Mab с нетрансфицированными клетками РЕАК и и имеющими нокдаун по CD47 клетками PEAK. Связывание анти-CD47 Mab с клетками РЕАК, трансфицированными киРНК к CD47, значительно снижено, таким образом подтверждая их антигенную специфичность.
Перекрестная реактивность анти-CD47 антител с CD47 яванского макака; связывание анти-CD47 антител с CD4+ T-клетками человека и яванского макака
Способность анти-CD47 моноклональных антител по настоящему изобретению перекрестно реагировать с нативным CD47 яванского макака была проверена с помощью проточной цитометрии. Связывание анти-CD47 антител с CD4-положительными T-лимфоцитами яванского макака, присутствующими среди мононуклеарных клеток периферической крови (PBMC) сравнивали со связыванием с соответствующей популяцией клеток человека. Кратко, мононуклеарные клетки периферической крови (PBMC) яванского макака были получены из Ricerca Biosciences. PBMC человека выделяли из лейкоцитарной пленки, используя СРТ-пробирки с фиколлом (Beckton and Dickinson). Для анализа методом проточной цитометрии PBMC предварительно инкубировали с FcgR Blocking Reagent (Miltenyi Biotech) в течение 20 минут для блокирования рецепторов Fc-гамма перед добавлением анти-CD47 антител (конечная концентрация составляла 0,005 мг/мл). После инкубации в течение 30 минут клетки промывали и добавляли конъюгированное с РЕ антитело к CD4 человека (клон L200, BD Pharmingen, разведенный 1/100) и конъюгированное с FMAT Blue антитело к Fc человека (Jackson Immuno Research, 109-005-098). Затем, с помощью проточной цитометрии определяли MFI (среднюю интенсивность флуоресценции) для связывания CD47 (канал FL4) в CD4-положительной популяции (отобранной в канале FL2). Как показано на фиг. 5, анти-CD47 моноклональные антитела по настоящему изобретению связываются с нативным CD47 человека и перекрестно реагируют с CD47 яванского макака.
SIRPα-блокирующая активность анти-CD47 антител
SIRPα-блокирующую активность анти-CD47 антител определяли в конкурентном анализе связывания CD47-SIRPα. Эксперименты доза-ответ с анти-CD47 Mab позволили определить значения IC50 для каждого из анти-CD47 MAb по настоящему изобретению. Кратко, клетки СНО, трансфицированные CD47 человека, инкубировали с His-меченным растворимым SIRPα человека (конечная концентрация составляла 200 нг/мл) и увеличивающимися концентрациями анти-CD47 Mab (от 3,3 пМ до 330 нМ, в квадрупликатах). Детекцию связанного SIRPα проводили, как описано в примере 4. На фиг. 6 показана способность анти-CD47 Mab блокировать взаимодействие CD47-SIRPα, представленную значениями IC50. Анти-CD47 Mab сгруппированы по семействам и ранжированы, начиная от максимальной и до минимальной блокирующей способности. Их нейтрализующую активность сравнивали с доступным в продаже анти-CD47 антителом, B6H12. Из фиг. 6 видно, что нейтрализующие способности анти-CD47 моноклональных антител по настоящему изобретению варьируют в широком диапазоне.
Гемагглютинирующая активность анти-CD47 антител
На фиг. 7 продемонстрировано, что высокоаффинные анти-CD47 Mab из семейств 5A3, Ke8 и Ka3 вызывают гемагглютинацию, однако в отличие от других трех семейств, антитела из семейства Kc4, протестированные в этом эксперименте, по-видимому, не вызывали гемагглютинации, даже одно антитело, сильно связывающееся с CD47 и сильно ингибирующее взаимодействие CD47-SIRPα.
Анти-CD47 MAb были протестированы на свою способность вызывать гомотипическое слипание эритроцитов (гемагглютинацию). 10 мкл цельной крови человека разбавляли 40 мкл раствора антитела в PBS с различными концентрациями (диапазон конечных концентраций Mab составлял от 0,003 мкг/мл до 50 мкг/мл) в плоскодонных 96-луночных планшетах. Смесь крови и антител инкубировали в течение ночи при 37°C без качания. В конце инкубации планшеты встряхивали вручную, наклоняли примерно на 30°C и оставляли примерно на 10 мин.
О гемагглютинации свидетельствует образование сгустка в форме полумесяца на дне по границе лунки. Все анти-CD47 антитела, кроме антител с наименьшей аффинностью, из семейств 5A3, Ke8 и Ka3 (особенно, 5A3M5, Ke8A3, Ka3A3) вызывали гемагглютинацию. В отличие от этого, анти-CD47 антитела из семейства Kc4 не вызывали гемагглютинацию, даже самые высокоаффинные (Kc4E2, Kc4F4).
ПРИМЕР 9: аффинное созревание анти-CD19 антител
(a) Антитело B7
Из антител, идентифицированных в ходе скринингового процесса, описанного в примерах выше, В7 было выбрано для созревания аффинности с целью повышения аффинности к hCD19. Перспективное антитело В7 содержит легкую лямбда-цепь (IGLV6-57) и было создано несколько фаговых библиотек, представляющих варианты scFv, путем введения разнообразия в CDR1, CDR2 и CDR3 вариабельной области легкой цепи, оставляя вариабельную область тяжелой цепи немодифицированной. Различные стратегии введения разнообразия использовали для создания 20 библиотек, содержащих всего 2×109 трансформантов, частично покрывающих теоретическое разнообразие 4×1012.
(b) Антитело L7B7_D11
Антитело D11 было идентифицировано в ходе созревания аффинности антитела В7, описанного выше, и оно связывается с hCD19 с более высокой аффинностью, чем родительское антитело В7. Это антитело было выбрано для второго раунда созревания аффинности по его легкой цепи. Всего было создано 6 библиотек, содержащих 2,8×109 трансформантов, частично покрывающих теоретическое разнообразие 4×109, и использовано для отбора методом фагового дисплея, как описано выше, за исключением того, что для каждого раунда отбора использовали 1 нМ hCD19. Этот второй раунд созревания аффинности привел к идентификации следующих антител с улучшенным связыванием с CD19: L7B7_C2; L7B7_A6; L7B7_B11; L7B7_C6 и L7B7_C9
ПРИМЕР 10: экспрессия и очистка биспецифичных антител, несущих легкую лямбда и каппа-цепь.
Одновременная экспрессия одной тяжелой цепи и двух легких цепей в одной клетке может привести к сборке трех различных антител. Одновременной экспрессии можно добиться различными путями, такими как трансфекция несколькими векторами, которые экспрессируют по одной цепи из подлежащих совместной экспрессии, или путем использования векторов, которые обеспечивают экспрессию нескольких генов. Ранее был создан вектор pNovi κHλ, позволяющий совместную экспрессию одной тяжелой цепи, одной легкой каппа-цепи и одной легкой лямбда-цепи, и описанный в US 2012/0184716 и WO 2012/023053, содержание которых таким образом включено в настоящее описание в качестве ссылки в полном объеме. Экспрессия этих трех генов находится под контролем промоторов цитомегаловируса человека (hCMV), и вектор также содержит ген глутаминсинтетазы (GS), который обеспечивает возможность отбора и получения стабильных клеточных линий. Гены VH и VL анти-hCD19 IgG или анти-hCD47 IgGK были клонированы в вектор pNovi κHλ для транзиентной экспрессии в клетках млекопитающих. Клетки РEАК культивировали в 6-луночных планшетах с концентрацией 6×105 клеток на лунку в 2 мл культуральной среды, содержащей бычью фетальную сыворотку. 2 мкг плазмидной ДНК трансфицировали в клетки с использованием трансфекционного реагента TransIT-LT1 (Mirus) в соответствии с инструкциями изготовителя. Концентрацию антитела в содержащем сыворотку супернатанте трансфицированных клеток измеряли в нескольких временных точках в ходе продукции, используя технологию Интерферометрии биослоя (BLI). Для количественного анализа использовали инструмент OctetRED96 instrument и биосенсоры на белок А (Pall, Basel, Switzerland). Для определения концентрации IgG использовали 200 мкл супернатанта, биосенсоры подготавливали и регенерировали с использованием 10 мМ глицина, рН 1,7, а для создания стандартной кривой были получены IgG-калибраторы, разведенные в кондиционной среде от клеток PEАК. Концентрации определяли с использованием уравнения 5PL-модели (взвешенный Y) для стандартной кривой доза-ответ и уравнения скорости связывания для начального наклона кривой. В зависимости от концентрации антител, супернатанты собирали через 7-10 дней после трансфекции и осветляли центрифугированием при 1300 g в течение 10 мин. Процесс очистки состоял из трех аффинных стадий. Сначала аффинный носитель CaptureSelect™ IgG-CH1 (Life Technologies, Zug, Switzerland) промывали PBS, а затем добавляли в осветленный супернатант. После инкубации в течение ночи при +4°C супернатанты центрифугировали 10 мин. при 1000 g, проскок сохраняли, а смолу промывали дважды, используя PBS. Затем смолу переносили в центрифугируемые колонки, и для элюции использовали раствор, содержащий 50 мМ глицина, рН 2,7. Полученные несколько элюируемых фракций собирали и обессоливали против PBS, используя центрифугируемые фильтровальные устройства 50 kDa Amicon® Ultra (Merck KGaA, Darmstadt, Germany). Конечный продукт, содержащий тотальные IgG человека из супернатанта, оценивали количественно, используя спектрофотометр Nanodrop (NanoDrop Technologies, Wilmington, DE), и инкубировали 15 мин при комнатной температуре и 20 об./мин. с соответствующим объемом аффинного носителя CaptureSelect™ LC-kappa (Hu) (Life Technologies, Zug, Switzerland). Стадии инкубацию, отделения смолы, элюции и обессоливания проводили, как описано ранее. Последнюю стадию аффинной очистки проводили, используя аффинный носитель CaptureSelect™ LC-lambda (Hu) (Life Technologies, Zug, Switzerland) и применяя тот же способ, что и для двух предшествующих очисток. Конечный продукт оценивали количественно с использованием Nanodrop. Очищенные биспецифичные антитела анализировали электрофорезом в денатурирующих и восстановительных условиях. Использовали Agilent 2100 Bioanalyzer с набором Protein 80, как описано изготовителем (Agilent Technologies, Santa Clara, CA, USA). 4 мкл очищенных образцов смешивали с буфером для образцов, содержим дитиотриэтол (DTT; Sigma Aldrich, St. Louis, MO). Образцы прогревали 5 мин. при 95°C и наносили на чип. Все образцы были исследованы на загрязнение эндотоксином с использованием теста Limulus Amebocyte Lysate (LAL; Charles River Laboratories, Wilmington, MA).
ПРИМЕР 11: характеристика моновалентных и биспецифичных антител.
Направленные на две мишени биспецифичные антитела связываются с двумя различными антигенами на поверхности одной клетки. Одновременное связывание двух плеч антитела с двумя антигенами на поверхности клетки (называемое совместной активацией) приводит к аддитивному или синергетическому увеличению аффинности вследствие механизма авидности. Как следствие, совместная активация придает высокую селективность в отношении клеток, экспрессирующих оба антигена, по сравнению с клетками, которые экспрессируют только один единственный антиген. Дополнительно, аффинность двух плеч биспецифичного антитела к их соответствующим мишеням может быть подобрана таким образом, что связывание с клетками-мишенями будет в основном обусловлено одним из плеч антитела. Например, специфичное к двум мишеням κλ-антитело, состоящее из одного плеча, связывающегося с высокой аффинностью с опухоль-связанным антигеном (ТАА), например, CD19, и второго плеча, связывающегося с CD47 с меньшей аффинностью, но достаточной для ингибирования CD47/SIRPα после совместной активации с TAA, должно обеспечивать возможность преимущественного ингибирования CD47 в раковых, а не здоровых клетках. В экспериментах, описанных ниже (фиг. 9-13), сравнивается аффинность связывания, сила нейтрализации взаимодействия CD47-SIRPα и способность убивать раковые клетки анти-CD47xCD19 биспецифичного κλ-антитела и соответствующего моновалентного антитела, то есть имеющего такое же CD47-связывающее плечо плюс «фиктивное» несвязывающее плечо.
Связывание моновалентных и биспецифичных антител с В-клеточными линиями
Для демонстрации того, что связывание анти-CD47xCD19 κλ-антител с клетками-мишенями является CD19-зависимым, был проведен ряд экспериментов на сортере флуоресцентно активированных клеток (FACS), в которых сравнивали связывание анти-CD47xCD19 κλ-антител с их моновалентными аналогами. Использовали два типа клеток, CD19-положительную линию клеток лимфомы Беркитта, Raji (экспрессирующую примерно 65000 молекул CD47 на клетку), и CD19-отрицательную линию клеток неходжскинской В-лимфомы, DS-1 (экспрессирующую примерно 150000 молекул CD47 на клетку), в качестве контроля. На фигурах 9A-9C продемонстрировано, что анти-CD47xCD19 κλ-антитело совместно активирует две мишени на поверхности клеток Raji. Это показано (i) по повышенной аффинности к клеткам Raji по сравнению с клетками DS-1 и (ii) по повышенной аффинности анти-CD47xCD19 κλ-антитела по сравнению с анти-CD47 моновалентным антителом, наблюдаемой с клетками Raji, но не с клетками DS-1. Сравнение профилей FACS, полученных для связывания анти-CD19 моновалентного антитела, анти-CD47 моновалентного антитела и анти-CD47xCD19 κλ-антитела с клетками Raji, ясно показывает, что связывание анти-CD47xCD19 κλ-антитела с клетками-мишенями преимущественно обусловлено CD19-плечом.
SIRPα-блокирующая активность моновалентных и биспецифичных антител
Другой ряд экспериментов обеспечивает дополнительное доказательство совместной активации CD19 и CD47 на поверхности клетки-мишени, показывая, что нейтрализация взаимодействия CD47-SIRPα анти-CD47xCD19 κλ-антителами является CD19-зависимой. В этом эксперименте активность анти-CD47xCD19 κλ-антител и соответствующих моновалентных антител была протестирована в анализе ингибирования CD47-SIRPα, как описано в примере 4. На фигуре 10 показано, что анти-CD47xCD19 κλ-антитела ингибировали взаимодействие CD47-SIRPα в клетках Raji значительно сильнее, чем соответствующие анти-CD47 моновалентные антитела. Эффективная нейтрализация взаимодействия CD47-SIRPα требовала совместной активации CD19. Величины IC50, полученные с анти-CD47xCD19 BsAb, были в 20-1000 раз меньше величин, полученных с соответствующим анти-CD47 моновалентным антителом (см. таблицу 4).
Величины IC50 для анти-CD47 моновалентных и биспецифичных антител
ПРИМЕР 12: ADCC, опосредованная биспецифичными антителами, является CD19-зависимой.
Способность двунаправленных κλ-антител совместно активировать CD47 и CD19 приводит к значительному увеличению аффинности связывания с CD19-положительными клетками и CD19-зависимой нейтрализации взаимодействия CD47-SIRPα. Это, в свою очередь, переходит в эффективное и селективное уничтожение раковых клеток, опосредованное анти-CD47xCD19 κλ-антителом, как продемонстрировано в ADCC- и ADCP-экспериментах, описанных в этом и следующем примере.
ADCC-анализы проводили с клетками-мишенями нефракционированных PBMC человека и В-клеточных лимфом Raji или Ramos. Эксперименты «доза-ответ», показанные на фиг. 11, демонстрируют, что анти-CD47xCD19 κλ-антитела, предложенные в изобретении, убивают клетки В-клеточной лимфомы более эффективно, чем соответствующие анти-CD47 моновалентные антитела. Эффективная ADCC, таким образом, зависит от совместной активации CD19. На фигуре 11С показано, что эффективность ADCC с использованием анти-CD47xCD19 κλ-антител сравнима с ритуксимабом, и значительно выше чем при использовании анти-CD19 Mab, C2.
ПРИМЕР 13: ADCP, опосредованный биспецифичными антителами, является CD19-зависимым
На фигуре 12 продемонстрировано, что анти-CD47xCD19 BsAb, предложенные в настоящем документе, обеспечивают фагоцитоз CD19-положительных клеток CD19-зависимым образом, тогда как соответствующие анти-CD47 моновалентные антитела являются гораздо менее эффективными (если вообще имеют данный вид активности).
ADCP-эксперименты проводили с макрофагами человека, дифференцированными из моноцитов периферической крови и Raji в качестве клеток-мишеней. Макрофаги совместно инкубировали с CFSE-меченными клетками Raji (соотношение эффекторных клеток и клеток-мишеней составляло 1:5) в течение 2,5 часов при 37°C в присутствии увеличивающихся концентраций биспецифичного или моновалентного антитела. После инкубации добавляли биотинилированное антитело против CD14 человека и Strep-Cy5 для мечения макрофагов. Клетки затем промывали и анализировали на FACS. О фагоцитозе свидетельствовали двойные положительные события.
Эксперименты «доза-ответ», показанные на фиг. 12, демонстрируют, что анти-CD47xCD19 κλ-антитела являются более сильными, чем соответствующие анти-CD47 моновалентные антитела. Эффективная ADCC, таким образом, зависит от совместной активации CD19. Совместная активация CD19 биспецифичным антителом обеспечивает эффективность. Более того, эксперименты, показанные на фиг. 12, подтверждают, что блокирование CD47 необходимо для эффективного ADCP, поскольку анти-CD19 Mab, C2, которе связывает клетки-мишени с высокой аффинностью, не вызывает значительного фагоцитоза.
ПРИМЕР 14: in vivo противоопухолевая активность биспецифичных антител
Противоопухолевую активность анти-CD47xCD19 κλ-антитела оценивали в модели лимфомы на основе клеток Raji. 2×106 клеток Raji имплантировали подкожно мышам NOD/SCID. Объем опухолей измеряли 3 раза в неделю. После того, как привитая опухоль достигала 0,1 см3, мышей случайным образом распределяли по 5 группам (по 5 мышей на группу), и начинали лечение антителом. В этом эксперименте сравнивали эффект анти-CD47xCD19 κλ-антитела, Ka3xD11, с эффектом моновалентного антитела Ka3 и двумя моноклональными антителами в качестве положительного контроля, анти-CD47 Mab, B6H12, и ритуксимабом. Антитела вводили внутрибрюшинной инъекцией три раза в неделю до конца эксперимента (день 25). Ритуксимаб вводили в количестве 200 мкг на мышь на инъекцию. Все другие антитела вводили в количестве 400 мкг на мышь на инъекцию.
Как показано на фиг. 13, эффективность анти-CD47xCD19 κλ-антитела была аналогичной B6H12, которое, как известно, сильно связывается с CD47, блокирует взаимодействие CD47-SIRPα и подавляет рост опухоли в этой модели лимфомы. Более того, эффективность анти-CD47xCD19 κλ-антитела была также сравнима с эффективностью ритуксимаба. Моновалентное анти-CD47 антитело было явно менее эффективно чем биспецифичное анти-CD47xCD19 κλ-антитело, демонстрируя, что удаление опухоли является CD19-зависимым.
ПРИМЕР 15: связывание анти-CD47 антитела с эритроцитами
Присутствуя в количестве более 5 миллиардов клеток на мл крови с 25000 молекул CD47 на клетку эритроциты потенциально представляют основную причину антиген-зависимого клиренса CD47-связывающих антител. Для оценки эффекта сорбции эритроцитами анти-CD47 антитела инкубировали с цельной кровью. После инкубации долю анти-CD47 антител, оставшихся в крови, определяли с помощью ELISA.
Кратко, 200 мкл цельной крови, содержащей антикоагулянт, смешивали с 20 мкл антитела (110 мкл/мл в PBS) и инкубировали 30 минут при 37°C и качании. Плазму затем отделяли от клеток центрифугированием, и концентрацию несвязанного антитела определяли с помощью ELISA. Для каждого протестированного антитела полученные результаты сравнивали с контролем, который представлял собой то же антитело, внесенное напрямую в плазму, и нормализовали по несвязывающим IgG, тестируемым параллельно.
На фиг. 14 показано, что анти-CD47 с высокой и средней аффинностью эффективно сорбировались на эритроцитах. Однако в случае BsAb это явление было ограничено молекулами с высокоаффинными анти-CD47 плечами, такими как 5A3. Это позволяет предположить, что в общем BsAb менее подвержены сорбции на эритроцитах и мишень-опосредованному распределению препарата, чем анти-CD47 Mab.
Другие варианты осуществления изобретения
Хотя изобретение описано в сочетании с подробными примерами, предшествующее описание предназначено для иллюстрации и не ограничивает объем изобретения, который определяется объемом прилагаемой формулы изобретения. Другие аспекты, преимущества и модификации находятся в пределах объема прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТИ-SIRPα АНТИТЕЛА | 2019 |
|
RU2812199C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ CEACAM5 И CD47 | 2019 |
|
RU2798246C2 |
| АНТИТЕЛО К SIRPα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822496C1 |
| МЕЧЕНЫЕ АЛЬДЕГИДОМ ПОЛИПЕПТИДЫ ИММУНОГЛОБУЛИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2606016C2 |
| НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19 | 2018 |
|
RU2788127C2 |
| ПОЛУЧЕНИЕ МУЛЬТИСПЕЦИФИЧЕСКИХ СМЕСЕЙ АНТИТЕЛ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2815826C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧНЫХ И МУЛЬТИВАЛЕНТНЫХ АНТИТЕЛ | 2011 |
|
RU2608640C2 |
| АНТИТЕЛО, НАЦЕЛЕННОЕ НА CD47, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2832773C1 |
| АНТИТЕЛЬНЫЕ АГЕНТЫ, СПЕЦИФИЧНЫЕ К CD19 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773317C2 |
| Моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817697C1 |






















































































































































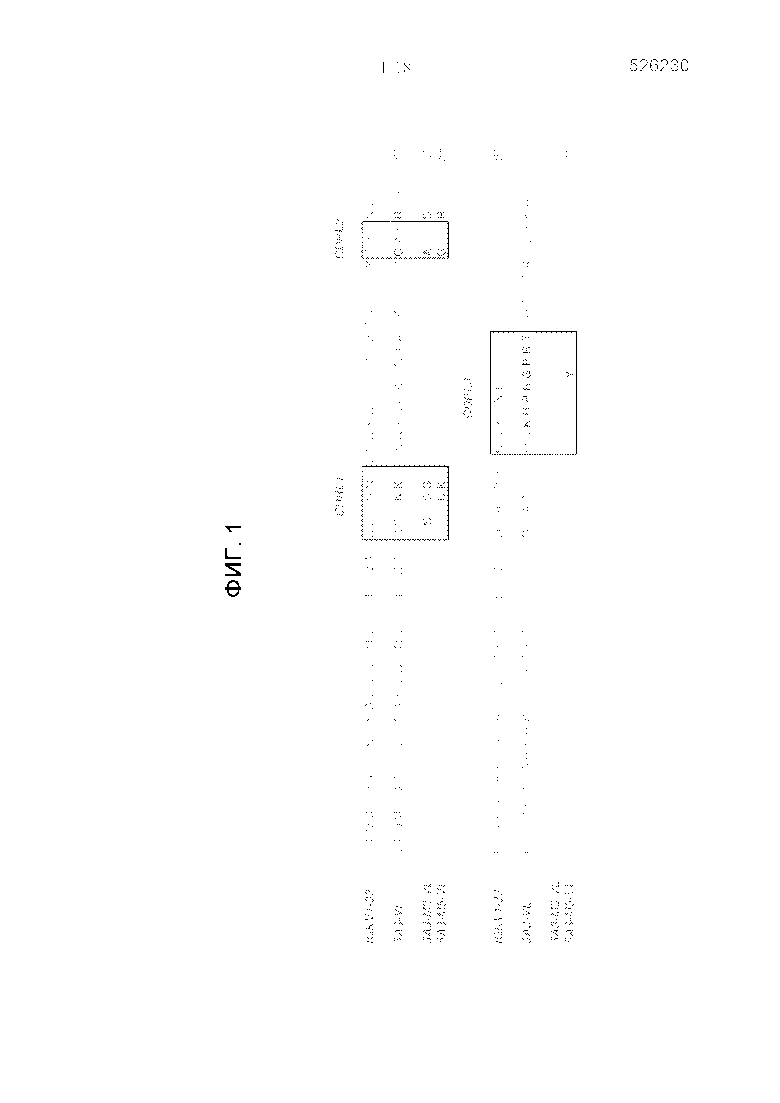





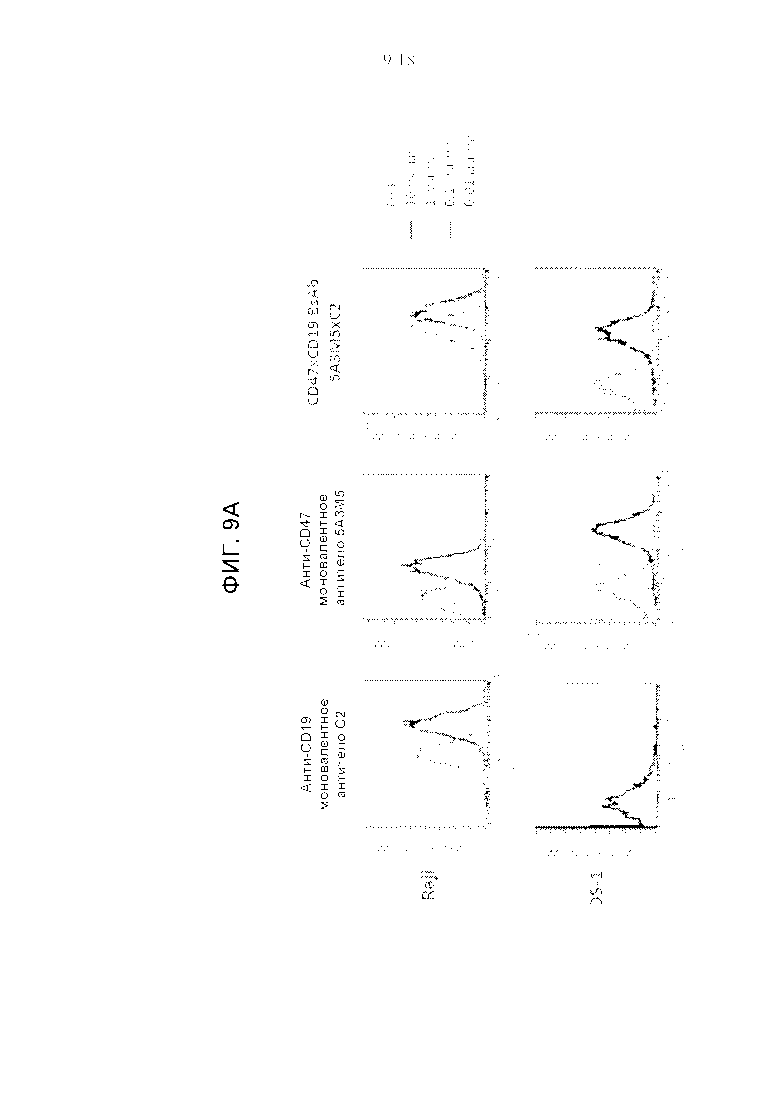

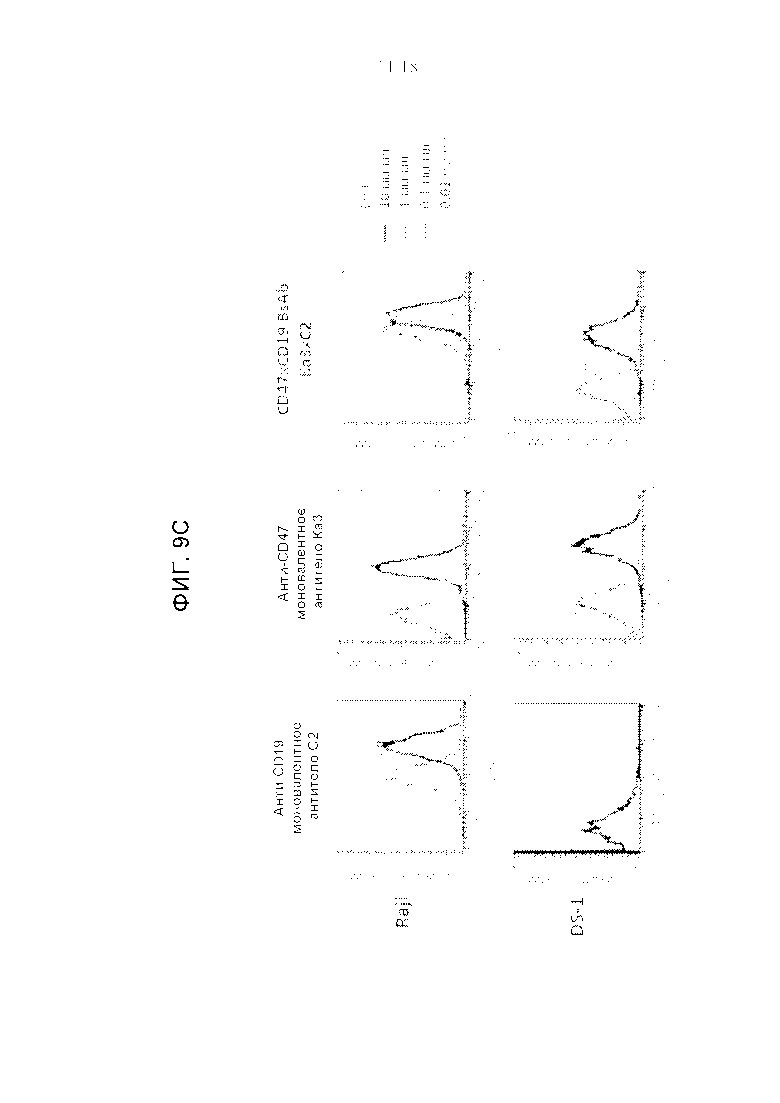


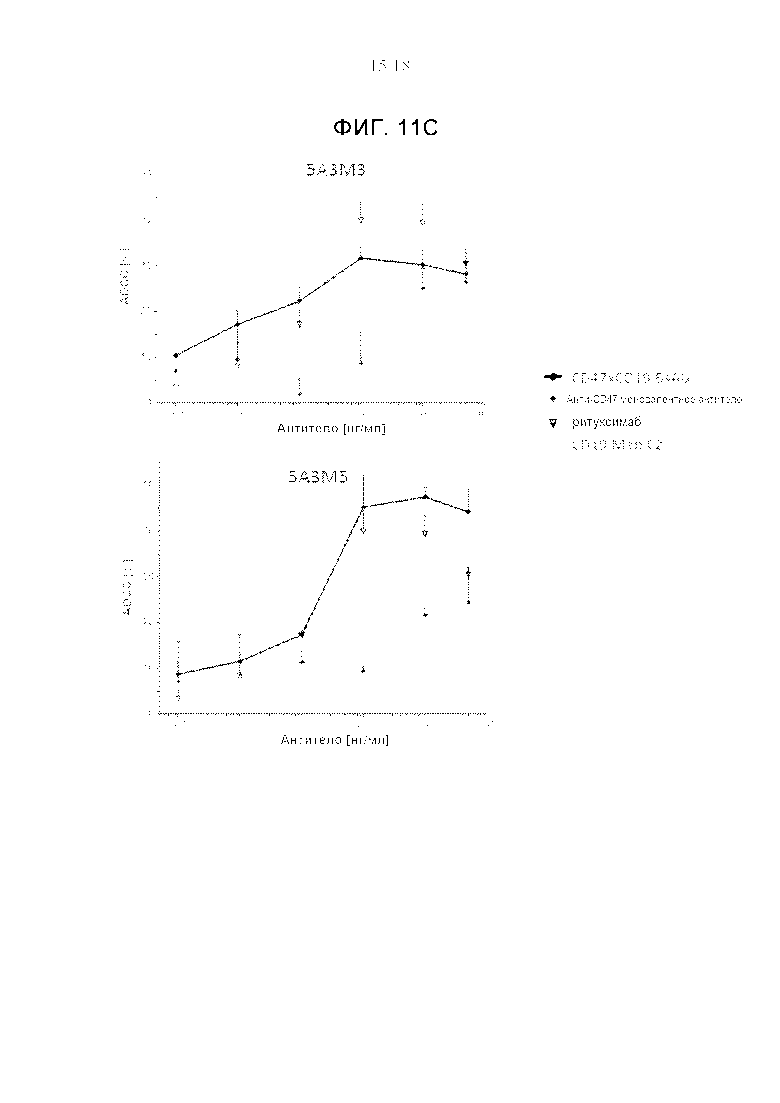
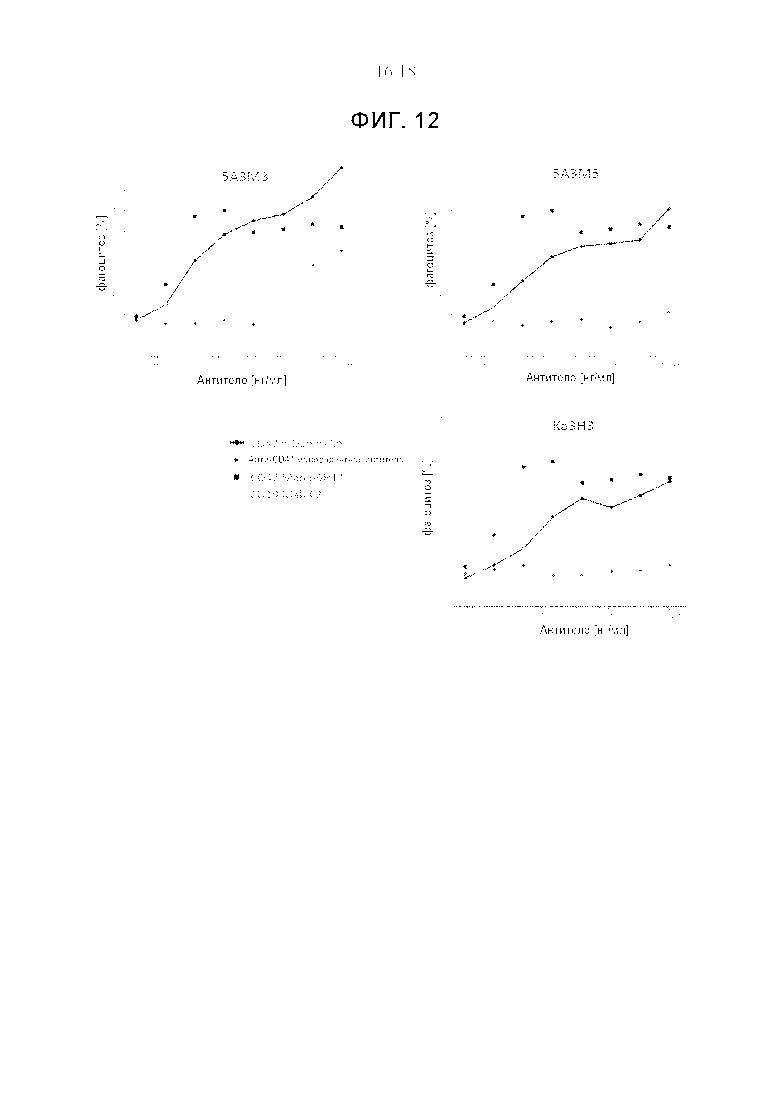


Настоящее изобретение относится к иммунологии. Предложено биспецифическое антитело, которое связывает CD47 и CD19. Данное антитело ингибирует взаимодействие между CD47 и SIRPα и может найти дальнейшее применение в способах лечения различных патологий, связанных с аномальной экспрессией и/или активностью CD47 и CD19, в том числе опухолей. 11 з.п. ф-лы, 14 ил., 4 табл., 15 пр.
1. Биспецифичное антитело, содержащее
(i) две вариабельные области тяжелой цепи, содержащие
определяющую комплементарность область 1 тяжелой цепи (CDRH1), содержащую аминокислотную последовательность SEQ ID NO:225,
определяющую комплементарность область 2 тяжелой цепи (CDRH2), содержащую аминокислотную последовательность SEQ ID NO:226, и
определяющую комплементарность область 3 тяжелой цепи (CDRH3), содержащую аминокислотную последовательность SEQ ID NO:227;
(ii) первую вариабельную область легкой цепи, содержащую
определяющую комплементарность область 1 легкой цепи (CDRL1), содержащую аминокислотную последовательность SEQ ID NO:240,
определяющую комплементарность область 2 легкой цепи (CDRL2), содержащую аминокислотную последовательность SEQ ID NO:242, и
определяющую комплементарность область 3 легкой цепи (CDRL3), содержащую аминокислотную последовательность SEQ ID NO:254; и
(iii) вторую вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:208;
где биспецифическое антитело связывает CD47 и CD19.
2. Выделенное биспецифичное антитело по п.1, где биспецифичное антитело ингибирует взаимодействие между CD47 человека и SIRPα человека.
3. Выделенное биспецифичное антитело по п.1, которое содержит
аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO:114 и
аминокислотную последовательность вариабельной области легкой цепи SEQ ID NO:168.
4. Выделенное биспецифичное антитело по п.1, где по меньшей мере участок первой легкой цепи относится к каппа-типу и по меньшей мере участок второй легкой цепи относится к лямбда-типу.
5. Выделенное биспецифичное антитело по п.4, где первая легкая цепь содержит по меньшей мере константную область каппа-цепи.
6. Выделенное биспецифичное антитело по п.5, где первая легкая цепь содержит вариабельную область каппа-цепи.
7. Выделенное биспецифичное антитело по п.5, где первая легкая цепь содержит вариабельную область лямбда-цепи.
8. Выделенное биспецифичное антитело по п.4, где вторая легкая цепь содержит по меньшей мере константную область лямбда-цепи.
9. Выделенное биспецифичное антитело по п.8, где вторая легкая цепь содержит вариабельную область лямбда-цепи.
10. Выделенное биспецифичное антитело по п.8, где вторая легкая цепь содержит вариабельную область каппа-цепи.
11. Выделенное биспецифичное антитело по п.4,
где первая легкая цепь содержит константную область каппа-цепи и вариабельную область каппа-цепи и
где вторая легкая цепь содержит константную область лямбда-цепи и вариабельную область лямбда-цепи.
12. Выделенное биспецифичное антитело по п.1, где константные и вариабельные последовательности каркасной области принадлежат человеку.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| YU J, LIN MF, "Anti-CD47 monoclonal antibody (B6H12) impairs the maturation and function of human dendritic cells." J Exp Hematol, 2005, 13(2):192-7 | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |

