Группа изобретений относится к области биотехнологии, иммунологии и вирусологии, а именно к моноклональным антителам и их антигенсвязывающим фрагментам, которые связываются с рецептор-связывающим доменом (RBD) Spike-белка вируса SARS-CoV-2, нейтрализующие вирус SARS-CoV-2, к фрагментам ДНК, кодирующим указанные антитела.
За три года с начала пандемии, вызванной бета-коронавирусом SARS-CoV-2, была зарегистрирована гибель почти 7 млн человек. Использование ряда средств профилактики и терапии коронавирусной инфекции, вызванной SARS-CoV-2 (также называемой в данном документе ковид-19), позволяет снизить вероятность заражения, госпитализации и летального исхода. К таким средствам относятся вакцинопрофилактика, использование вирус-специфических малых молекул, препаратов плазмы реконвалесцентов, рекомбинантных Spike-специфичных белков (в том числе антител) и их производных. Несмотря на повсеместное проведение кампаний по иммунизации населения и идентификации нейтрализующих моноклональных антител против Spike-белка вируса SARS-CoV-2, существует значительная потребность в получении новых вариантов таких антител для расширения арсенала диагностических, профилактических и терапевтических средств особенно с учетом появления новых вариантов вируса. Кроме того, существуют группы населения, которым вакцинация либо не показана по медицинским соображениям, либо у которых не может быть выработан протективный иммунный ответ. Таким образом, проведение пассивной иммунизации - введение препаратов моноклональных антител в качестве пред- или пост-экспозиционной профилактики или терапии позволяет защитить таких людей, обеспечить менее длительное и тяжелое течение болезни, в том числе предотвратить развитие пост-ковидных осложнений, и снизить вероятность дальнейшей передачи вируса.
Используемый в настоящей заявке термин «антитело» предназначен для обозначения молекулы иммуноглобулина, составленной из четырех полипептидных цепей, при этом две тяжелые (Н) цепи и две легкие (L) цепи связываются друг с другом дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно называемую в настоящем документе VH) и константную область тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов: CH1, СН2 и СН3. Каждая легкая цепь содержит вариабельную область легкой цепи (сокращенно называемую в настоящем документе VL) и константную область легкой цепи. Константная область легкой цепи состоит из одного домена: CL.
Области VH и VL могут далее подразделяться на гипервариабельные районы, которые называются определяющими комплементарность областями (CDR), перемежаемые областями с более высоким уровнем консервативности, называемыми каркасными областями (FR). Каждая область VH и VL образована тремя CDR и четырьмя FR, расположенными от амино-терминального конца к карбокси-терминальному концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
Четыре каркасных участка формируют конформацию типа бета-складчатого слоя. Участки CDR расположены в близком соседстве друг с другом благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDR и каркасные участки антител могут быть определены путем ссылки на нумерационную систему Кабата (Kabat numberings ystem, Kabat et al., 1987, "Sequences of Proteins of Immunological Interest", US Dept. Of Health and Human Services, US Government Printing Office) в сочетании с данными рентгеноструктурного анализа, как указано в заявке WO 91/09967. Участки CDR и каркасные участки антител могут также быть определены по номенклатуре Международной информационной системы по иммуногенетике (IMGT, International Immunogenetics Information System, www.imgt.org) (Lefranc, M.P. and Lefranc, G. Methods Mol Biol. 2552: 3-59 (2023)).
Если не указано иное, любая представленная в описании полипептидная цепь имеет аминокислотную последовательность, которая начинается на N-конце и заканчивается на С-конце. Если антигенсвязывающий центр содержит и VH-, и VL-области, то они могут быть расположены на одной и той же полипептидной молекуле или предпочтительно каждая область может располагаться на разных цепях, VH-область может быть частью тяжелой цепи иммуноглобулина или ее фрагмента, a VL-область - частью легкой цепи иммуноглобулина или ее фрагмента.
В настоящем изобретении термин «антитело, обладающее вируснейтрализующей активностью» означает молекулу, которая связывается с SARS-CoV-2, образует стабильный комплекс и препятствует прикреплению и/или проникновению вируса SARS-CoV-2 в клетку. Способность антитела к связыванию с антигеном может быть определена специалистом в данной области с использованием методов, включающих, но не ограничивающихся, методом иммуноферментного анализа (ИФА), равновесным диализом, с использованием плазмон-поверхностного резонанса и биослойной интерферометрии (BLI). Методы определения аффинности хорошо известны специалисту в данной области техники, подробно описаны Janeway и соавт. (Immunobiology: The Immune System in Health and Disease (Garland Publishing Company, 1996)).
Используемый в настоящем изобретении термин «антигенсвязывающий фрагмент» антитела обозначает фрагмент антитела, который сохраняет способность к специфическому связыванию с антигеном. К примерам таких фрагментов, включенных в термин «антигенсвязывающий фрагмент» антитела, относятся: (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1; (ii) фрагмент F(ab')2, бивалентный фрагмент, состоящий из двух фрагментов Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH отдельной области антитела; (v) фрагмент dAb (Ward, E.S. et al. Nature. 241, 544-546, (1989)), состоящий из домена VH; и (vi) отдельная определяющая комплементарность область (CDR). Более того, несмотря на то, что два домена во фрагменте Fv, VL и VH кодируются различными генами, их можно объединить, используя рекомбинантные технологии, посредством синтетического линкера, который позволяет построить из них единую белковую цепочку, в которой области VL и VH связываются с образованием моновалентных молекул (известных как одноцепочечные антитела или scFv; см., например, Bird, R.E. et al. Science. 24, 423-426 (1988); Huston, J.S. et al. Proc. Natl. Acad. Sci. USA 85: 5879-5883 (1988)). Такие одноцепочечные антитела также включаются в сферу охвата термина «антигенсвязывающий фрагмент» антитела.
Специалисту в данной области известно, что за взаимодействие антитела с антигеном наибольший вклад вносит тяжелая цепь (Н). Однако известны случаи, когда в распознавании антигена участвует только одна цепь (или Н, или L), а вторую цепь можно заменить без потери антигенсвязывающих свойств антителом (Shiraiwa, Н. et al. Engineering a bispecific antibody with a common light chain: Identification and optimization of an anti-CD3 epsilon and anti-GPC3 bispecific antibody, ERY974. Methods. 154: 10-20 (2009); Senn, B.M. et al. Combinatorial immunoglobulin light chain variability creates sufficient В cell diversity to mount protective antibody responses against pathogen infections. Eur J Immunol. 33(4): 950-61 (2003)). Таким образом, в широком понимании в сферу охвата термина «антигенсвязывающий фрагмент» антитела можно включить любые белковые молекулы, содержащие хотя бы один из VH и VL доменов заданного антитела.
Известно, что из трех поверхностных белков SARS-CoV-2 лишь один, Spike, вызывает образование вируснейтрализующих антител. Также было показано, что подавляющее число полученных к настоящему моменту Spike-специфических моноклональных антител, обладающих способностью нейтрализовать вирус, направлены против RBD-домена Spike, ответственного за связывание с основным входным рецептором вируса, белком АСЕ2.
Идентификация SARS-CoV-2-специфических нейтрализующих моноклональных антител, в основном, осуществляется методом секвенирования генов иммуноглобулинов из единичных В-клеток людей, перенесших ковид-19 (Pinto, D. et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 583, 290-295 (2020)).
В настоящий момент уже созданы сотни различных SARS-CoV-2-специфических, в том числе и вируснейтрализующих антител, обладающих различными профилями потентности и широты нейтрализации. Однако разработанные моноклональные антитела могут быть неактивными в отношении появившихся новых вариантов SARS-CoV-2. Так, например, моноклональное антитело бамланивимаб не способно нейтрализовать дельта-вариант SARS-CoV-2 (Planas, D. et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 596, 276-280 (2021)). В отношении бета-варианта бамланивимаб и этесивимаб полностью неактивны, а активность касиривимаба значительно снижена (Wang, P. Et al. Antibody resistance of SARS-CoV-2 variants B. 1.351 and B.1.1.7. Nature 593, 130-135 (2021)).
Антитела REGN10987 и REGN10933 (имдевимаб и касиривимаб) на 2-3 порядка хуже нейтрализуют вирус штаммов Омикрон ВА.1 и ВА.2, а сотровимаб и антитело S2X259 - чувствительны к этим же штаммам, но в меньшей степени (Iketani, S. et al., Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 604, 553-556. (2022)).
Таким образом, идентификация и всесторонняя характеризация новых SARS-CoV-2-специфических антител является актуальной задачей и позволяет создавать композиции антител, против новых вирусных вариантов SARS-CoV-2 и потенциально против других, в том числе, неописанных коронавирусов.
Существует множество публикаций, описывающих получение и характеризацию RBD-специфических моноклональных антител человека, в том числе и обладающих вируснейтрализующей активностью.
Например, известны человеческие RBD-специфические нейтрализующие антитела бамланивимаб (LY-CoV555) (Jones, В. et al. The neutralizing antibody, LY-CoV555, protects against SARS-CoV-2 infection in nonhuman primates. Sci. Transl. Med. 13(593), 1-17 (2021) и этесивимаб (LY-C0V016/JS016/CB6) (Shi, R. et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 584, 120-124 (2020)), последовательности которых известны. Однако существенным недостатком данных антител является их неспособность нейтрализовать ряд циркулирующих вирусных вариантов SARS-CoV-2 (https://covdb.stanford.edu/page/susceptibility-data/), что ограничивает их профилактическое и терапевтическое использование.
Известна панель моноклональных антител человека iB1-iB26, которая нейтрализует Wu-1 и Delta варианты SARS-CoV-2 (патент RU 2800649 С2, опубл. 25.07.2023). Недостатком данного решения является отсутствие информации о нейтрализации вариантов вируса более поздних вариантов.
Известна общедоступная база данных CoV-AbDab (http://opig.stats.ox.ac.uk/webapps/covabdab/), содержащая информацию о последовательностях и свойствах сотен RBD-специфических моноклональных антител различного происхождения (в том числе и человеческих), многие из которых обладают вируснейтрализующей активностью в отношении SARS-CoV-2.
Недостатками большинства таких антител является их неполная характеризация в плане устойчивости к циркулирующим вариантам SARS-CoV-2, поскольку такие варианты появились позже, чем были получены антитела, а также в плане эпитопной специфичности, широты нейтрализации, наличия или отсутствия ауто- и полиреактивности и т.д. Таким образом, создание новых детально охарактеризованных RBD-специфических моноклональных антител, как нейтрализующих так и ненейтрализующих, является крайне актуальной задачей, поскольку может обеспечить рациональный дизайн композиций антител, предотвращающих мутационное ускользание вируса, а также более точную диагностику, эффективную терапию и профилактику коронавирусной инфекции.
Наиболее близкими к заявляемой группе изобретений - прототипом, являются моноклональное антитело мыши IgM-изотипа и его антигенсвязывающий фрагмент, селективно взаимодействующие с RBD фрагментом в составе S белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью к SARS-CoV-2. Моноклональное антитело получено путем иммунизации мышей Balb/C человеческим рекомбинантным RBD фрагментом S-белка вируса SARS-CoV-2, экспрессированным в E. coli и полноразмерным S-белком вируса SARS-CoV-2 в составе клеток линии НЕК293, получения и селекции линий гибридом, продуцирующих моноклональные антитела к рекомбинантному RBD фрагменту в составе S белка вируса SARS-CoV-2, анализа аффинности и специфичности отобранного МАт, определения нуклеотидной последовательности его вариабельных доменов. Специфичность моноклонального антитела к RBD фрагменту в составе S белка вируса SARS-CoV-2 подтверждена методами иммуноферментного анализа, иммуноблоттинга, иммуноцитохимического и иммуногистохимического окрашивания. Установлена нуклеотидная последовательность вариабельных доменов моноклонального антитела (заявка WO 2021211775 А1, опубл. 21.10.2021).
Недостатками данного мышиного антитела и его антигенсвязывающего фрагмента для целей терапии и профилактики коронавирусной инфекции, вызванной SARS-CoV-2, являются его чужеродность для организма человека, вследствие чего введение такого антитела пациентам может быть иммуногенным и сопровождаться нежелательными побочными реакциями. Также не известен уровень (ауто)реактивности данного антитела по отношению к клеткам человека, поскольку оно было получено в иммунизированных мышах и в процессе созревания В-клетки проходило негативную селекцию против антигенов мыши, а не человека.
Также недостатком данного мышиного антитела является его принадлежность к IgM классу: (i) это снижает время его полужизни, а, значит, будет требовать дополнительных инъекций антитела пациенту; (ii) производство антител IgM изотипа является крайне трудоемким и сопровождается низким выходом, а сохранение всех заявленных свойств данного антитела при его возможной конверсии в IgG1-формат для преодоления данного ограничения является сомнительным, поскольку будет связано, как минимум, со снижением его авидности.
Также недостатком данного изобретения является то, что нейтрализующая активность мышиного антитела была показана только in vitro на клетках Vero Е6 и с использованием раннего штамма SARS-CoV-2 Wuhan-Hu-1. Таким образом, уровень и механизм нейтрализующей активности данного антитела в отношении SARS-CoV-2, а также детали эпитопной специфичности (следовательно, и узнавание им вирусных вариантов, среди новых циркулирующих изолятов) - неизвестны.
Задачей заявляемой группы изобретений является получение новых моноклональных антител человека IgG1 подкласса, способных к связыванию с RBD Spike-белка вируса SARS-CoV-2, а также антигенсвязывающих фрагментов таких антител, обладающих вируснейтрализующей активностью широкого спектра, а также получение ДНК фрагментов, кодирующих антитело и его антигенсвязывающий фрагмент.
Техническим результатом группы изобретений является создание нового моноклонального антитела человека iC2, отличающегося по аминокислотной последовательности вариабельных доменов легкой и тяжелой цепей от известных моноклональных антител к RBD фрагменту в составе Spike-белка вируса SARS-CoV-2 и его антигенсвязывающего фрагмента, способного к связыванию с RBD Spike-белка вируса SARS-CoV-2 и нейтрализации различных вариантов вируса штамма Омикрон (ВА.1-ВА.3).
Также техническим результатом является повышение аффинности созданного антитела, что позволяет снизить его дозировку при применении, что в значительной степени снижает его стоимость при производстве, а значит - повышает привлекательность для коммерциализации.
Также техническим результатом является способность созданного антитела нейтрализовать различные варианты вируса SARS-CoV-2 штамма Омикрон, что позволяет использовать его для профилактики и терапии заболеваний, вызываемых вирусом SARS-CoV-2 этого штамма.
Поставленная задача решается созданием моноклонального антитела человека iC2, селективно связывающего RBD фрагмент Spike-белка вируса SARS-CoV-2, состоящего из тяжелой (Н) и легкой (L) цепей, имеющих аминокислотные последовательности SEQ ID NO: 1 и 2, а также созданием кодирующих их фрагментов ДНК, имеющих нуклеотидные последовательности SEQ ID NO: 5 и 6, соответственно.
В аминокислотных последовательностях цепей антитела iC2 можно выделить вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), имеющих аминокислотные последовательности SEQ ID NO: 3 и 4, кодируемые нуклеотидными последовательностями SEQ ID NO: 7 и 8, соответственно.
Вариабельный домен тяжелой цепи (VH) содержит последовательности гипервариабельных районов: CDRH1, имеющего аминокислотную последовательность SEQ ID NO: 9 (GFTFEDYA), CDRH2, имеющего аминокислотную последовательность SEQ ID NO: 10 (ISWNGGII), и CDRH3, имеющего аминокислотную последовательность SEQ ID NO: 11 (TKVLYHYDSSGYYPGPFDY). Вариабельный домен легких цепей (VL) содержит последовательности гипервариабельных районов: CDRL1, имеющего аминокислотную последовательность SEQ ID NO: 12 (QSVSSY), CDRL2, имеющего аминокислотную последовательность SEQ ID NO: 13 (DAS) и CDRL3, имеющего аминокислотную последовательность SEQ ID NO: 14 (QQRSDWPPYT). Участки CDR в VH и VL были определены по номенклатуре IMGT (Lefranc, М.Р. and Lefranc, G. Methods Mol Biol. 2552: 3-59 (2023)).
Как хорошо известно, минорные изменения аминокислотной последовательности, такие как делеция, добавление или замена одной, небольшого количества или даже нескольких аминокислот, могут приводить к получению аллельной формы исходного белка, которая обладает практически идентичными свойствами. Также известен способ «перевивки» CDR-петель одного антитела в контекст каркасных последовательностей другого антитела, также с сохранением исходных свойств, что в частности позволяет производить гуманизацию антител, полученных в других млекопитающих. Таким образом, антитело изобретения включает те варианты, аминокислотная последовательность которых вне CDR петель отличается от приведенных в настоящей заявке последовательностей, но которые сохранили ту же антигенсвязывающую активность.
Антитело iC2 было получено способом, включающим следующие стадии: (i) выделение В-лимфоцитов периферической крови доноров, перенесших вакцинацию и коронавирусную инфекцию ковид-19 и сформировавших высокие нейтрализующие титры антител в сыворотке крови; (ii) окрашивание В-лимфоцитов флуоресцентно меченными препаратами RBD SARS-CoV-1; (iii) сортировка единичных окрашенных RBD+ В-лимфоцитов; (iv) проведение обратной транскрипции и ПЦР с наборами вырожденных праймеров, комплементарных лидерным и константным последовательностям генов иммуноглобулинов; (v) секвенирование полученных ПЦР продуктов с целью выяснения нуклеотидной и аминокислотной последовательностей, экспрессируемых такими В-клетками антител; (vi) клонирование нуклеотидных последовательностей таких антител в экспрессионные векторы для наработки в культуре клеток млекопитающих; (vii) очистка, анализ специфичности и активности рекомбинантных моноклональных антител человека. В последующем также предусмотрено определение аминокислотной последовательности вариабельных доменов таких антител, измерение аффинности и специфичности, нейтрализующих свойств in vitro и in vivo.
Использованный метод является общепринятым, однако был модифицирован таким образом, чтобы отбираемые моноклональные антитела человека были обогащены антителами, способными распознавать консервативные участки RBD Spike-белка SARS-CoV-2. Достигается это тем, что окрашивание В-лимфоцитов (пункт ii) проводят флуоресцентно меченными препаратами RBD SARS-CoV-1, а не SARS-CoV-2, как это делается обычно. Данная модификация метода серьезно влияет на специфичность получаемых антител. Суть замены меченного препарата заключается в том, что тем самым производится отбор антител, распознающих SARS-CoV-1, но из крови реконвалесцентов, которые были вакцинированы прививкой на основе RBD SARS-CoV-2 и переболели ковид-19, а значит обладают антителами, распознающими SARS-CoV-2. Таким образом, происходит отбор только тех антител, которые распознают эпитопы, присутствующие в Spike-белках обоих вирусов (SARS-CoV-1 и SARS-CoV-2). Учитывая, что данные белки идентичны только на 76% по аминокислотной последовательности, отбираемые антитела с высокой долей вероятности будут распознавать эпитопы, которые являются консервативными, то есть не будут меняться в ходе дальнейшего мутагенеза SARS-CoV-2.
Также настоящим изобретением создан антигенсвязывающий фрагмент моноклонального антитела iC2, селективно связывающий RBD Spike-белка вируса SARS-CoV-2, обладающий вируснейтрализующей активностью, имеющий аминокислотные последовательности вариабельной области тяжелой цепи (VH) - SEQ ID NO: 3, и вариабельной области легкой цепи (VL) - SEQ ID NO: 4, кодируемые нуклеотидными последовательностями SEQ ID NO: 7 и 8, соответственно.
У белков, созданных в рамках заявляемого технического решения, CDR участки являются идентичными, то есть CDRH1 участок антитела iC2 совпадает с CDRH1 участком антигенсвязывающего фрагмента моноклонального антитела iC2, и т.д.
Антитело iC2 человека селективно связывается с RBD Spike-белка SARS-CoV-2, содержит приведенные выше последовательности вариабельных областей и может содержать консервативные участки обеих цепей, необходимые для функционирования указанного антитела. Консервативные участки тяжелых цепей иммуноглобулинов классов IgG, IgM, IgA, IgD или IgE человека и других млекопитающих могут быть использованы в качестве консервативных участков для тяжелых цепей согласно настоящему изобретению. Например, антитело или его антигенсвязывающий фрагмент, согласно изобретению, могут содержать консервативные участки тяжелых цепей, принадлежащие к изотипу IgG1, IgG2, IgG3 или IgG4. Консервативные участки легкой цепи иммуноглобулинов изотипов каппа или лямбда человека и других млекопитающих могут быть использованы в качестве консервативных участков для легких цепей антител согласно настоящему изобретению.
Антитело настоящего изобретения является человеческим антителом. В частном случае антитело может быть гуманизованным антителом, мышиным антителом, химерным антителом или другим генно-инженерным антителом.
В ходе решения поставленной задачи были установлены последовательности нуклеиновых кислот, кодирующие тяжелую (Н) цепь антитела iC2 (нуклеотидная последовательность SEQ ID NO: 5), легкую (L) цепь антитела iC2 (нуклеотидная последовательность SEQ ID NO: 6), вариабельный VH домен антитела iC2 (нуклеотидная последовательность SEQ ID NO: 7) и вариабельный VL домен антитела iC2 (нуклеотидная последовательность SEQ ID NO: 8).
Специалистам в данной области техники известно, что ввиду вырожденности генетического кода могут быть различия в последовательности ДНК кодирующей одну и ту же аминокислотную последовательность (Льюин Б. Гены. Москва: Мир, 1987). Поэтому последовательности нуклеиновых кислот, согласно настоящему изобретению, могут отличаться, при условии, что они кодируют участки цепей антитела iC2, содержащего последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 3, и последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 4.
Вышеназванные аминокислотные последовательности антитела iC2 (SEQ ID NO: 1-4), кодируемые нуклеотидными последовательностями (SEQ ID NO: 5-8), а также аминокислотные последовательности антигенсвязывающего фрагмента (SEQ ID NO: 3-4), кодируемые нуклеотидными последовательностями (SEQ ID NO: 7-8) представлены в перечне последовательностей.
Для целей терапии RBD-специфические моноклональные антитела могут быть использованы при введении человеку или другому млекопитающему в короткий срок после появления первых симптомов заболевания, вызванного SARS-CoV-2. Пути введения антител могут включать внутривенную и внутримышечную инъекцию, доставку аэрозолем в дыхательные пути или подкожное введение. Антитело или его антигенсвязывающий фрагмент изобретения могут быть введены системно, назально или путем прямой доставки кодирующих его нуклеиновых кислот в организм млекопитающих. Помимо терапевтического применения антитела могут быть введены заранее в качестве меры профилактики. В таком случае целесообразно использование антител для лиц, имеющих высокий риск заражения вирусом SARS-CoV-2, тяжелого течения заболевания и не подлежащих вакцинации по медицинским или иным причинам. При профилактическом использовании необходимо учитывать период полужизни антител в кровотоке, который в среднем для антител изотипа IgG1 составляет 21 день, но может быть значительно увеличен за счет модификации последовательности аминокислотных остатков в Fc-части.
Краткое описание чертежей и таблиц, иллюстрирующих изобретение
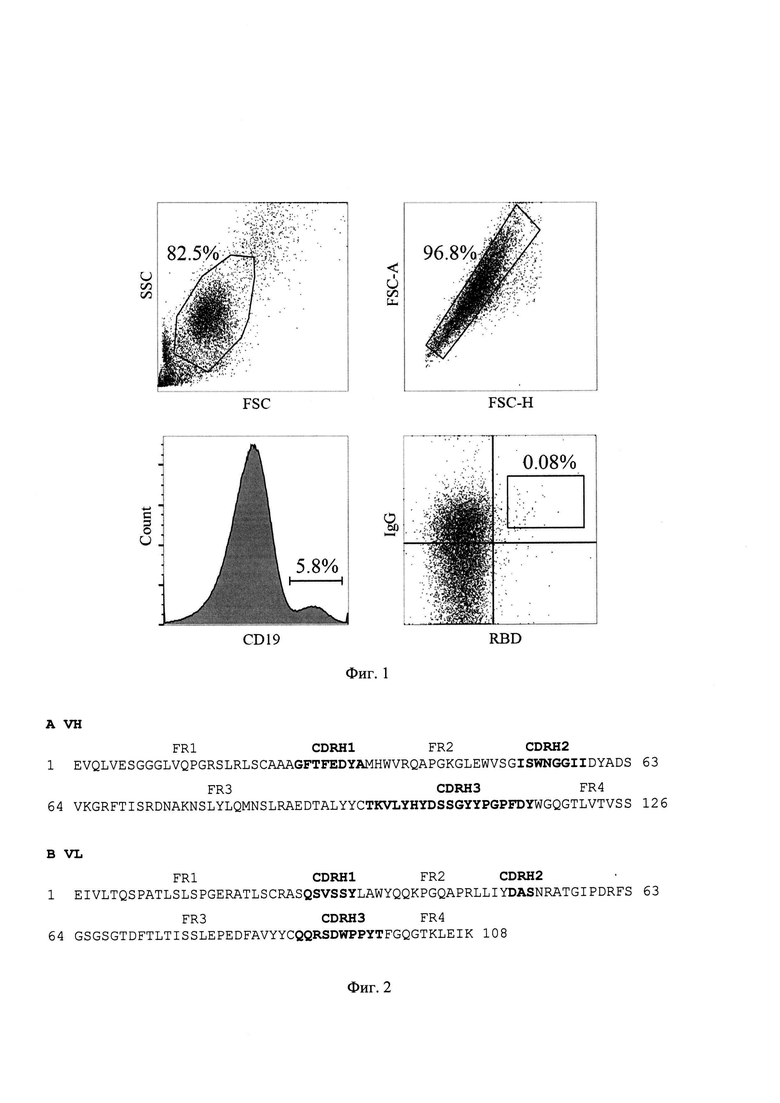
На Фиг. 1 показана стратегия гейтирования и результаты FACS-окрашивания В-лимфоцитов донора-реконвалесцента.
На Фиг. 2 представлены аминокислотные последовательности вариабельных доменов тяжелой VH (А) и легкой VL (В) цепей моноклонального антитела человека iC2. CDR-районы отмечены жирным шрифтом.
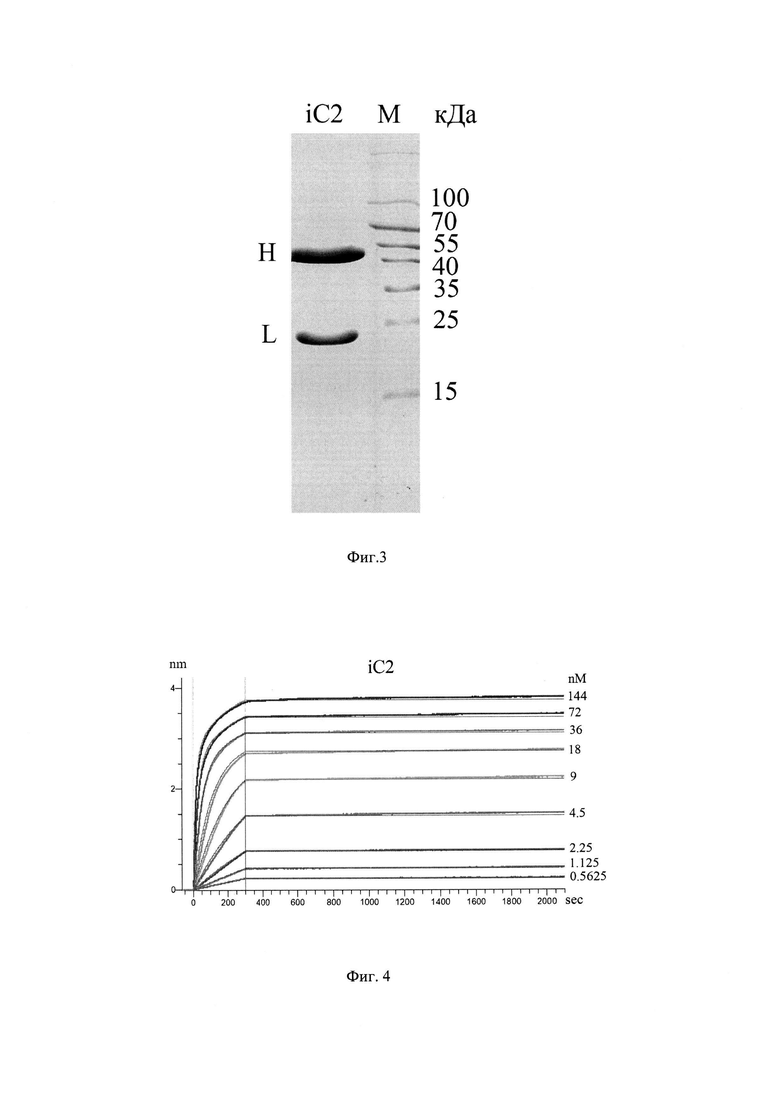
На Фиг. 3 показаны результаты разделения очищенного рекомбинантного антитела iC2 в SDS-полиакриламидном геле, окрашенном Кумасси, где М - маркер молекулярного веса, L - легкая цепь антитела, Н - тяжелая цепь антитела.
На Фиг. 4 приведена серия сенсограмм, полученная при инжекции разных концентраций аналита методом биослойной интерферометрии (BLI). Полученные данные позволяют количественно рассчитать аффиность взаимодействия мноклонального антитела iC2 с рекомбинантным RBD SARS-CoV-2. Концентрация аналита (антитела iC2) указана справа от каждой из сенсограмм.
Последующие примеры приведены исключительно для целей объяснения и не ограничивают каким-либо образом объем настоящего изобретения.
Пример 1. Выделение В-лимфоцитов человека, специфических к RBD Spike-белка вируса SARS-CoV-1.
У доноров был произведены забор периферической венозной крови и выделение живых CD19+IgG+В-лимфоцитов (окрашивание DAPI, конъюгатами anti-human CD19-PE (ООО «Сорбент»), anti-human IgG-FITC (ООО «Сорбент»), 5 мкл/106 мононуклеарных клеток крови). При отборе доноров выбирали только тех, кто был вакцинирован и ревакцинирован (вакциной на основе Wu-1 штамма), а также переболел ковид-19 хотя бы один раз после начала циркуляции штаммов Дельта или Омикрон. В-лимфоциты были дополнительно окрашены биотинилированным рекомбинантным RBD SARS-CoV-1 (2 мкг/мл) с последующей детекцией при помощи конъюгата АРС-стрептавидин (Thermo Fisher Scientific). Поверхностный фенотип целевой популяции В-лимфоцитов для сортировки единичных клеток в индивидуальные ПЦР-пробирки, таким образом, был RBD+CD19+IgG+DAPI- (Фиг. 1). Окрашенные аналогичным образом В-лимфоциты здоровых доноров были использованы в качестве негативного контроля.
Пример 2. Получение моноклонального антитела человека против RBD Spike-белка вируса SARS-CoV-1.
Процедуры синтеза кДНК с единичных В-лимфоцитов и ПЦР с обратной транскрипцией проводили, как описано в работе Guselnikov и соавт. (Guselnikov, S.V. et al. A simple way to increase recovery of the expressed VH and VL genes in single-sorted human В cells. BioTechniques 67, 184-187 (2019)), за исключением того, что для амплификации последовательностей, кодирующих γ - и λ - цепи антител, был использован протокол для μ-цепей. В частности, для амплификации последовательностей генов VHγ, Vκ и Vλ был использован набор вырожденных праймеров из работы Tiller и соавт. (Tiller, Т. et al. Efficient generation of monoclonal antibodies from single human В cells by single cell RT-PCR and expression vector cloning. J. Immunol. Methods 329, 112-124 (2008)). Парные образцы ПЦР-ампликонов (VHγ+Vκ или Vλ) секвенировали по Сэнгеру и клонировали в векторы для эукариотической экспрессии, кодирующие константные районы y1, к или Х-цепей иммуноглобулинов человека.
В результате была расшифрована пара нуклеотидных последовательностей VH (SEQ ID NO: 7) и VL (SEQ ID NO: 8) генов из одиночного В-лимфоцита. С учетом генетического кода были определены аминокислотные последовательности VH (SEQ ID NO: 3) и VL (SEQ ID NO: 4) участков моноклонального антитела iC2, специфичного к RBD Spike-белка SARS-CoV-2. Анализ последовательности VL, а также ассоциированного с ним CL, показал принадлежность легкой цепи iC2 антитела к каппа типу (κ). Участки CDRs VH (CDRH1 - SEQ ID NO: 9; CDRH2 - SEQ ID NO: 10; CDRH3 - SEQ ID NO: 11) и VL (CDRL1 - SEQ ID NO: 12; CDRL2 - SEQ ID NO: 13; CDRL3 - SEQ ID NO: 14) были определены по номенклатуре IMGT (Lefranc M.P., Lefranc G. MethodsMolBiol. 2552: 3-59 (2023)) (Фиг. 2).
Проведенный биоинформатический анализ последовательности iC2 антитела против известных последовательностей антител к SARS-CoV-2 не выявил антител, на 100% совпадающих по последовательностям легкой и тяжелой цепи с антителом изобретения.
В дальнейшем нуклеотидная последовательность VH была клонирована в вектор для эукариотической экспрессии в одной рамке с последовательностью, кодирующей константную область тяжелой цепи иммуноглобулина человека (Fc(IgG1)), в результате чего была получена нуклеотидная последовательность (SEQ ID NO: 5), кодирующая аминокислотную последовательность (SEQ ID NO: 1). Аналогичным образом, VL последовательность была клонирована в одной рамке с константной областью лямбда легкой цепи иммуноглобулина человека (Сκ), в результате чего была получена нуклеотидная последовательность (SEQ ID NO: 6), кодирующая аминокислотную последовательность (SEQ ID NO: 2).
Смесь плазмидных ДНК экспрессионных конструкций, кодирующих легкую или тяжелую цепь антитела iC2, была доставлена в клетки линии HEK293T, используя метод кальций-фосфатной трансфекции (Chenuet, S. et al. Calcium phosphate transfection generates mammalian recombinant cell lines with higher specific productivity than polyfection. Biotechnol. Bioeng. 101, 937-945 (2008)). Как правило, за 12 часов до проведения трансфекции 107 клеток рассаживали в культуральные флаконы площадью 150 см2 в среде IMDM (Gibco, Thermo Fisher Scientific) с добавлением 10% FBS (Gibco, Thermo Fisher Scientific) и смеси пенициллина/стрептомицина (Gibco, Thermo Fisher Scientific). Плазмидные ДНК смешивали в молярном соотношении 1:1, к полученной смеси добавляли 2 М СаС12 до финальной концентрации 25 мМ и доводили суммарный объем смеси стерильной водой до 3 мл. Затем смесь медленно добавляли к 3 мл раствора 2×HBS (50 мМ HEPES, 1,5 мМ Na2HPO4, 280 мМ NaCl, 10 мМ KCl, 12 мМ сахароза, рН=7.11) при постоянном интенсивном перемешивании. Полученный раствор добавляли по каплям к клеткам. Спустя 8 часов, культуральную среду заменяли на бессывороточную среду ЕХ-CELL® 293 Serum-Free Medium (Sigma). Трансфицированные клетки росли в течение 6 дней в CO2-инкубаторе при 37°С, 5% СО2. Супернатантны, очищенные от клеточного дебриса при помощи центрифугирования (4000×g, 10 мин), фильтровали через 0.22 мкм PES-фильтр (ТРР). Антитела очищали на колонке с Protein А агарозой (McLab, USA, #РРА-503) в соответствии с инструкциями производителя. Вкратце, после загрузки колонку промывали дважды PBS. Антитело элюировали раствором 0,1 М глицина, рН 2,7, 150 мМ NaCl в нейтрализующий буфер состава 1 М Tris, рН 8,0, в соотношении 1:10. Затем проводили диализ в течение ночи в PBS и концентрировали с использованием колонок Ultra-15 Ultracel-100K (Amicon) до финальной концентрации 2 мг/мл. Чистоту и выход препарата антитела контролировали посредством разделения в 15% SDS-полиакриламидном геле (Фиг. 3).
Пример 3. Определение аффинности связывания моноклонального антитела iC2 человека и разных вариантов рекомбинантого RBD SARS-CoV-2.
Для измерения кинетики взаимодействия антитела iC2 с RBD вариантом дикого типа (Wu-1) была использована биослойная интерферометрия на приборе Octet (ForteBio). С этой целью была вначале проведена иммобилизация антитела iC2 на поверхности биосенсора АНС (45 мкг/мл). На референсном сенсоре в качестве нерелевантного антитела было использовано рекомбинантное моноклональное антитело человека glVRC01 (Georgiev, I.S. et al. Antibodies VRC01 and 10E8 neutralize HIV-1 with high breadth and potency even with Ig-framework regions substantiaily reverted to germline. J Immunol. 192(3), 1100-1106 (2014)). Характеризация связывания была проведена в диапазоне девяти концентраций RBD (0.5625-144 нМ) (Фиг. 4). Для обработки данных были использованы параметры полного глобального фиттинга, бивалентная модель, R^2>0,9, Х^2<3 и последовательный переход между режимами: Baseline 60 с, Loading 120 с, Association 300 с, Dissociation 1800 с, Regeneration 3 раза по 5 с. В Табл. 1 представлены результаты измерения аффинности связывания рекомбинантного RBD с антителом iC2 методом биослойной интерферометрии, где KD - равновесная константа диссоциации, ka - константа скорости ассоциации, a kdis - константа скорости диссоциации.

Из таблицы 1 видно, что KD имеет высокое значение, что позволяет использовать iC2 для детекции вируса, например, в образцах слюны человека.
Пример 4. Определение вируснейтрализующей активности моноклонального антитела iC2 человека in vitro.
Для оценки уровня вируснейтрализующей активности моноклонального антитела iC2 использовали лентивирусные частицы, псевдотипированные разными вариантами S-белка SARS-CoV-2. Детекцию уровня заражения клеток HEK293T проводили с использованием люминометрического теста (Табл. 2).
Для получения лентивирусных частиц, псевдотипированных Spike-белком SARS-CoV-2, проводили кальций-фосфатную трансфекцию клеток линии НЕК293Т смесью плазмид psPAX2, pLV-Luc и pCAGGS-SpikeΔ19 в молярном соотношении 4:6:3. Плазмида pCAGGS-SpikeΔ19 кодировала укороченный с С-конца на 19 аминокислотных остатков Spike-белок, соответствующий вариантам Spike SARS-CoV-2 (SΔ19 штамма Омикрон. Через восемь часов после трансфекции среду заменяли на Opti-Mem (Gibco, Thermo Fisher Scientific) с добавлением 2,5% FBS (Gibco, Thermo Fisher Scientific). Супернатанты собирали два дня спустя, центрифугировали при 4000 об/мин в течение 5 минут, после чего лентивирусные частицы концентрировали путем центрифугирования при 20000×g, 4°С в течение 90 минут. Для проведения нейтрализационного теста в качестве мишеней использовали клетки линии АСЕ2-HEK293T со стабильной поверхностной экспрессией белка АСЕ2, рассаженные с плотностью 30000 клеток/лунку в 96-луночном штативе в день проведения теста. Антитело iC2 серийно двукратно разводили в среде Opti-MEM с добавлением 2,5% FBS в диапазоне от 0,5 нг/мл до 1 мкг/мл и ко-инкубировали с 20000 лентивирусных частиц, псевдотипированных Spike-белком SARS-CoV-2 в течение 30 мин при 37°С в объеме 100 мкл. Полученную смесь антител и лентивирусных частиц добавляли к клеткам АСЕ2-HEK293T и возвращали в CO2-инкубатор. Спустя 48 часов после трансдукции измеряли уровень люминесценции клеточного лизата с использованием люминометра. Значение концентрации полумаксимального ингибирования IC50 определяли, используя нелинейную регрессию, как концентрацию антитела, обеспечивавшую нейтрализацию 50% псевдотипированных лентивирусных частиц.
В Табл. 2 продемонстрирована способность антитела iC2 нейтрализовать лентивирусы, псевдотипированные разными вариантами Spike-белка SARS-CoV-2 штамма Омикрон с помощью люминометрического теста.

Таким образом, было установлено, что моноклональное антитело iC2 является высокопотентным нейтрализатором различных вариантов (ВА.1-ВА.3) штамма Омикрон SARS-CoV-2 (IC50<100 нг/мл).
Высокая аффинность заявляемого антитела позволит снизить его дозировку при диагностическом, профилактическом и терапевтическом применении, а способность нейтрализовать различные варианты штамма Омикрон SARS-CoV-2 говорят о его широком спектре действия и наличии потенциала для нейтрализации новых вариантов SARS-CoV-2.
--->
Перечень последовательностей
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Перечень
последовательностей iC1.xml" softwareName="WIPO Sequence"
softwareVersion="2.0.0" productionDate="2022-06-08">
<ApplicantFileReference>1</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение науки Институт молекулярной и клеточной биологии
Сибирского отделения Российской академии наук (ИМКБ СО
РАН)</ApplicantName>
<ApplicantNameLatin>Institute of Molecular and Cellular Biology,
Siberian Branch of Russian Academy of Sciences</ApplicantNameLatin>
<InventionTitle languageCode="ru">Моноклональное антитело iC2,
селективно связывающее рецептор-связывающий домен Spike-белка вируса
SARS-CoV-2 и его антигенсвязывающий фрагмент, обладающие
вируснейтрализующей активностью</InventionTitle>
<SequenceTotalQuantity>14</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>456</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..456</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>EVQLVESGGGLVQPGRSLRLSCAAAGFTFEDYAMHWVRQAPGKGLEWVS
GISWNGGIIDYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCTKVLYHYDSSGYYPGPFDYWGQG
TLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLY
SLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDT
LMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYK
CKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENN
YKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK</INSDSeq_seq
uence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>215</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..215</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIY
DASNRATGIPDRFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSDWPPYTFGQGTKLEIKRTVAAPSVFIF
PPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEK
HKVYACEVTHQGLSSPVTKSFNRGEC</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>126</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..126</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>EVQLVESGGGLVQPGRSLRLSCAAAGFTFEDYAMHWVRQAPGKGLEWVSGIS
WNGGIIDYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCTKVLYHYDSSGYYPGPFDYWGQGTLV
TVSS</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>108</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..108</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDAS
NRATGIPDRFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSDWPPYTFGQGTKLEIK</INSDSeq_sequ
ence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>1371</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1371</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCC
TGAGACTCTCCTGTGCAGCCGCTGGATTCACCTTTGAAGATTATGCCATGCACTGGGTCCGGCAAGCTCC
AGGGAAGGGCCTGGAGTGGGTCTCAGGCATTAGTTGGAATGGTGGTATCATAGACTATGCGGACTCTGTG
AAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTCTATCTGCAAATGAACAGTCTGAGAG
CTGAGGACACGGCCTTGTATTACTGTACAAAAGTATTGTATCACTATGATAGTAGTGGTTATTACCCTGG
CCCCTTTGACTATTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCGTCGACCAAGGGCCCATCGGTC
TTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACT
ACTTCCCCGAACCTGTGACGGTCTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGC
TGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACC
CAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAAT
CTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCT
CTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGAC
GTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGA
CAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGA
CTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACC
ATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGA
CCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGA
GAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTC
CTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGC
ATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA</INSDSeq_s
equence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>648</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..648</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAA
GAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGG
CCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGACAGGTTCAGTGGC
AGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACT
GTCAGCAGCGTAGCGACTGGCCTCCGTACACTTTTGGCCAGGGGACCAAGCTGGAGATCAAACGTACGGT
GGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTG
TGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGG
GTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGAC
GCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCG
CCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTGA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="7">
<INSDSeq>
<INSDSeq_length>378</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..378</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCC
TGAGACTCTCCTGTGCAGCCGCTGGATTCACCTTTGAAGATTATGCCATGCACTGGGTCCGGCAAGCTCC
AGGGAAGGGCCTGGAGTGGGTCTCAGGCATTAGTTGGAATGGTGGTATCATAGACTATGCGGACTCTGTG
AAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTCTATCTGCAAATGAACAGTCTGAGAG
CTGAGGACACGGCCTTGTATTACTGTACAAAAGTATTGTATCACTATGATAGTAGTGGTTATTACCCTGG
CCCCTTTGACTATTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="8">
<INSDSeq>
<INSDSeq_length>324</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..324</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAA
GAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGG
CCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGACAGGTTCAGTGGC
AGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACT
GTCAGCAGCGTAGCGACTGGCCTCCGTACACTTTTGGCCAGGGGACCAAGCTGGAGATCAAA</INSDSe
q_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="9">
<INSDSeq>
<INSDSeq_length>8</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>GFTFEDYA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="10">
<INSDSeq>
<INSDSeq_length>8</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ISWNGGII</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="11">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
/INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TKVLYHYDSSGYYPGPFDY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="12">
<INSDSeq>
<INSDSeq_length>6</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..6</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>QSVSSY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="13">
<INSDSeq>
<INSDSeq_length>3</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..3</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>DAS</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="14">
<INSDSeq>
<INSDSeq_length>10</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..10</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>QQRSDWPPYT</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ РЕЦЕПТОР-СВЯЗЫВАЮЩЕГО ДОМЕНА SPIKE-БЕЛКА ВИРУСА SARS-COV-2 И ИХ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ, КОДИРУЮЩИЕ ИХ НУКЛЕИНОВЫЕ КИСЛОТЫ, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ | 2021 |
|
RU2800649C2 |
| Однодоменное антитело ламы Н5 и его производное H5-Fc, специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2022 |
|
RU2793967C1 |
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |
| Универсальный интеграционный вектор SB7G_HIGH и рекомбинантная плазмида SB7G_HIGH_DEV_57, обеспечивающая синтез и секрецию рекомбинантного человеческого антитела к рецептор-связывающему домену (RBD) коронавируса SARS-CoV-2 в клетках млекопитающих и полученная с использованием универсального вектора SB7G_HIGH, и рекомбинантное моноклональное антитело DEV_K57, обладающее вируснейтрализующей активностью в отношении SARS-CoV-2 | 2023 |
|
RU2829359C1 |
| Широко нейтрализующее антитело против SARS-CoV-2 | 2022 |
|
RU2810476C1 |
| БИСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ SARS-COV-2 | 2022 |
|
RU2791749C1 |
| Тяжелоцепочечные моноклональные антитела, специфически связывающиеся с S белком вируса SARS-CoV-2, и способ их применения для терапии заболеваний, вызываемых различными вариантами вируса SARS-CoV-2 | 2024 |
|
RU2836313C1 |
| Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2 | 2021 |
|
RU2794141C2 |
| ПОЛИВАЛЕНТНАЯ ВЕКТОРНАЯ ВАКЦИНА HVT | 2020 |
|
RU2832110C1 |
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен (RBD) Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью. В одном из вариантов реализации изобретения моноклональное антитело или его антигенсвязывающий фрагмент включают вариабельную область тяжелой цепи (VH), содержащую последовательность аминокислот SEQ ID NO: 3, и вариабельную область легкой цепи (VL), содержащую последовательность аминокислот SEQ ID NO: 4. Изобретение направлено на разработку нового моноклонального антитела, связывающегося с RBD Spike-белка вируса SARS-CoV-2, имеющего повышенную аффинность и способного нейтрализовать различные варианты вируса SARS-CoV-2 штамма Омикрон. 2 н. и 5 з.п. ф-лы, 4 ил., 2 табл., 4 пр.
1. Моноклональное антитело iC2, селективно связывающее рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающее вируснейтрализующей активностью, состоящее из тяжелой (Н) цепи, имеющей последовательность аминокислот SEQ ID NO: 1, и легкой (L) цепи, имеющей последовательность аминокислот SEQ ID NO: 2, включающее вариабельную область тяжелой цепи (VH) указанного антитела, содержащую последовательность аминокислот SEQ ID NO: 3, и вариабельную область легкой цепи (VL), содержащую последовательность аминокислот SEQ ID NO: 4, кодируемые нуклеотидными последовательностями SEQ ID NO: 5-8 соответственно.
2. Антигенсвязывающий фрагмент моноклонального антитела iC2, селективно связывающий рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающий вируснейтрализующей активностью, включающий вариабельную область тяжелой цепи (VH) указанного антитела, содержащую последовательность аминокислот SEQ ID NO: 3, и включающий вариабельную область легкой цепи (VL), содержащую последовательность аминокислот SEQ ID NO: 4, кодируемые нуклеотидными последовательностями SEQ ID NO: 7 и 8 соответственно.
3. Моноклональное антитело iC2 или его антигенсвязывающий фрагмент по пп. 1, 2, имеющие определяющие комплементарность области (CDR) со следующими аминокислотными последовательностями:
(i) CDRH1 - SEQ ID NO: 9;
(ii) CDRH2 - SEQ ID NO: 10;
(iii) CDRH3 - SEQ ID NO: 11;
(iv) CDRL1 - SEQ ID NO: 12;
(v) CDRL2 - SEQ ID NO: 13;
(vi) CDRL3 -SEQ ID NO: 14.
4. Моноклональное антитело iC2 или его антигенсвязывающий фрагмент по пп. 1, 2, содержащее вариабельную область тяжелой цепи (VH), содержащую последовательность аминокислот, не менее чем на 90% сходную с SEQ ID NO: 3, и вариабельную область легкой цепи (VL), содержащую последовательность аминокислот, не менее чем на 90% сходную с SEQ ID NO: 4.
5. Моноклональное антитело iC2 или его антигенсвязывающий фрагмент по пп. 1, 2, где антитело или его антигенсвязывающий фрагмент содержат консервативные участки тяжелых цепей, относящиеся к классу IgG, IgM, IgA, IgD или IgE.
6. Моноклональное антитело iC2 или его антигенсвязывающий фрагмент по пп. 1, 2, где антитело или его антигенсвязывающий фрагмент содержат консервативные участки тяжелых цепей, принадлежащие к изотипу IgG1, IgG2, IgG3 или IgG4.
7. Антигенсвязывающий фрагмент по п. 2, где антигенсвязывающий фрагмент представляет собой Fv, scFv, Fab, F(ab')2, Fab' или любой иной белок, содержащий VH и/или VL последовательности антитела iC2.
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |
| Гуманизированное моноклональное антитело, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, средство и способ для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2765731C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ РЕЦЕПТОР-СВЯЗЫВАЮЩЕГО ДОМЕНА SPIKE-БЕЛКА ВИРУСА SARS-COV-2 И ИХ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ, КОДИРУЮЩИЕ ИХ НУКЛЕИНОВЫЕ КИСЛОТЫ, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ | 2021 |
|
RU2800649C2 |
| WO 2022065445 A1, 31.03.2022 | |||
| US 11370828 B2, 28.06.2022 | |||
| WO 2021045836 A1, 11.03.2021 | |||
| Bordoloi D | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

