Область техники
Настоящее изобретение относится к новому промотору, вектору, включающему его, микроорганизму, включающему его, и способу получения глутатиона с его использованием.
Предшествующий уровень техники
Глутатион (GSH) как органическое соединение серы, обычно присутствующее в большинстве клеток, представляет собой трипептид, состоящий из трех аминокислот: глицин, глутамат и цистеин.
Глутатион присутствует в живом организме в восстановленной форме глутатиона (GSH) и в окисленной форме глутатиона (GSSG). Восстановленная форма глутатиона (GSH), которая присутствует в относительно высокой пропорции в нормальных условиях, в основном распределяется в клетках печени и кожи в организме человека и играет важные роли антиоксидантной функции при разложении и удалении реакционноспособного кислорода, детоксикационной функции при удалении ксенобиотических соединений, таких как токсичные вещества, и отбеливающей функции при ингибировании выработки меланина (Sipes IG et at., "The role of glutathione in the toxicity of xenobiotic compounds: metabolic activation of 1,2-dibromoethane by glutathione", Adv Exp Med Biol., 1986;197:457-67).
Поскольку выработка глутатиона постепенно уменьшается по мере прогрессирования процесса старения, а снижение выработки глутатиона, играющего важную роль в антиоксидантной и дезинтоксикационной функциях, способствует накоплению реакционноспособного кислорода, что является основной причиной старения, необходимо поставлять глутатион извне.
Имея различные функции, как описано выше, глутатион привлек внимание как вещество в различных областях, таких как фармацевтика, полезные для здоровья пищевые продукты и косметика, а также используется для производства вкусовых ингредиентов и пищевых и кормовых добавок. Известно, что глутатион оказывает сильное влияние на обогащение вкуса сырых ингредиентов и сохранение богатого аромата, и что его можно использовать отдельно или в сочетании с другими веществами в качестве усилителя вкуса кокуми (kokumi). В целом известно, что вещества кокуми имеют более насыщенный вкус, чем вещества умами (umami), такие как известные нуклеиновые кислоты и глутамат натрия (MSG), и известно, что они образуются в результате разложения белка во время созревания.
Хотя существует растущий спрос на глутатион для применения в различных областях, рынок для него не сформирован из-за высоких затрат на промышленное производство глутатиона, так как процессы синтеза ферментов не получили промышленной реализации из-за высокой стоимости, а способы извлечения глутатиона из микроорганизмов дают низкие выходы.
Описание изобретения
Техническая задача
В настоящем изобретении предложены новый промотор, вектор, включающий его, микроорганизм, включающий его, и способ получения глутатиона с его использованием.
Техническое решение
В настоящем изобретении предложен полинуклеотид, обладающий промоторной активностью и где по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид.
В настоящем изобретении предложен вектор, включающий полинуклеотид, обладающий промоторной активностью.
В настоящем изобретении предложен микроорганизм, принадлежащий к роду Saccharomyces sp., включающий один или более из: полинуклеотида, обладающего промоторной активностью, и гена, кодирующего целевой белок; и вектора, включающего его.
В настоящем изобретении предложен способ получения глутатиона, включающий культивирование микроорганизма в культуральной среде.
Полезные эффекты
Последовательность нового промотора по настоящему изобретению значительно увеличивает продукцию глутатиона и, таким образом, может быть использована для получения глутатиона с высоким выходом.
Наилучший способ осуществления изобретения
Ниже настоящее изобретение будет описано подробно. При этом каждое описание и воплощение, раскрытое в настоящем изобретении, может быть применено к описанию других описаний и воплощений. Другими словами, все комбинации различных компонентов, раскрытых в настоящем описании изобретения, включены в объем настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать подробным описанием, представленным ниже.
Специалисты в данной области техники будут понимать или способны определить, используя только рутинные эксперименты, множество воплощений, эквивалентных конкретным воплощениям настоящего изобретения. Подразумевается, что объем настоящего изобретения охватывает такие эквивалетные воплощения.
В одном аспекте настоящего изобретения предложен полинуклеотид, обладающий промоторной активностью, где по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид.
Используемый здесь термин «полинуклеотид» относится к нити ДНК, имеющей определенную минимальную длину как полимер нуклеотидов, в котором мономеры нуклеотидов связаны друг с другом в виде длинной цепи ковалентными связями.
Используемый здесь термин «полинуклеотид, обладающий промоторной активностью» относится к области ДНК, расположенной в непосредственной близости к сайту, где транскрипция гена, подлежащего экспрессии, т.е. целевого гена, инициируется, причем эта область включает сайт, с которым РНК-полимер аз а, энхансер и т.д., связываются для экспрессии целевого гена.
Полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может быть использован как промотор общего применения для усиления.
Например, полинуклеотид может быть использован в качестве промотора, способного усиливать экспрессию полипептида, обладающего активностью глутамат-цистеинлигазы. Кроме того, полинуклеотид может представлять собой полинуклеотид, используемый для увеличения продукции или выхода глутатиона. Полинуклеотид по настоящему изобретению может включать любую полинуклеотидную последовательность, обладающую промоторной активностью.
В настоящем изобретении полинуклеотидная последовательность SEQ ID NO: 1 или 2 может представлять собой последовательность, способную функционировать в качестве промотора глутамат-цистеинлигазы.
Однако, полинуклеотидная последовательность SEQ ID NO: 1 или 2 является репрезентативной полинуклеотид ной последовательностью, указывающей положение модификации, и любая полинуклеотидная последовательность, соответствующая ей и обладающая промоторной активностью, также может быть включена в последовательности, допускающие введение модификации. Например, любая полинуклеотидная последовательность, способная функционировать в качестве промотора глутамат-цистеинлигазы или полипептида, обладающего эквивалентной ей активностью, может быть включена в объем последовательности, в которую введена модификация по настоящему изобретению, без ограничения. С такими последовательностями, когда по меньшей мере один из нуклеотидов, соответствующих нуклеотидам по положениям 92, 94, 102, 103, 249 и 251 SEQ ID NO: 1 или 2, заменен на другой нуклеотид, может быть предложен промотор, обладающий более высокой активностью по сравнению с незамещенной (немодифицированной) промоторной последовательностью.
Нуклеотидная последовательность SEQ ID NO: 1 или 2 может быть получена из известной базы данных NCBI GenBank. Последовательности, соответствующие SEQ ID NO: 1 или 2 и служащие в качестве промотора глутамат-цистеинлигазы, могут быть получены из микроорганизма, принадлежащего к роду Saccharomyces sp., в частности Saccharomyces cerevisiae, но не ограничиваясь им, и могут включать любую последовательность, обладающую активностью, эквивалентной активности полинуклеотид а, без ограничения.
В настоящем изобретении полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотидную последовательность SEQ ID NO: 1 или 2 или полинуклеотидную последовательность, имеющую по меньшей мере 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологии или идентичности с SEQ ID NO: 1 или 2, в которых по меньшей мере один нуклеотид заменен на другой нуклеотид в указанных конкретных положениях или положениях, соответствующих им. Полинуклеотидная последовательность, имеющая гомологию или идентичность, может исключать последовательность, имеющую идентичность 100%, или может представлять собой последовательность, имеющую идентичность менее 100%.
В то же время, хотя в настоящем описании используют термины «полинуклеотид, имеющий полинуклеотидную последовательность заданной SEQ ID NO» и «полинуклеотид, включающий нуклеотидную последовательность заданной SEQ ID NO», очевидно, что в настоящем описании также можно использовать любой полинуклеотид, имеющий полинуклеотидную последовательность, включающую делецию, модификацию, замену или добавление одного или нескольких нуклеотидов, при условии что этот полинуклеотид обладает активностью, идентичной или эквивалентной полинуклеотиду, состоящему из полинуклеотидной последовательности заданной SEQ ID NO.
Например, очевидно, что любой полинуклеотид, включающий добавление бессмысленной последовательности внутри или на конце полинуклеотидной последовательности заданной SEQ ID NO или делецию части полинуклеотидной последовательности заданной SEQ ID NO внутри или на ее конце, также может входить в объем настоящего изобретения, при условии что этот полинуклеотид обладает активностью, идентичной или эквивалентной активности полинуклеотид а по настоящему изобретению.
Гомология и идентичность относятся к степени родства между двумя данными нуклеотидными последовательностями и могут быть выражены в процентах.
Термины «гомология» и «идентичность» часто могут быть использованы взаимозаменяемо.
Гомология или идентичность последовательности консервативных полинуклеотидов может быть определена с помощью стандартного алгоритма выравнивания и установленных программой штрафов на введение гэпа по умолчанию, которые можно использовать в комбинации. По существу гомологичные или идентичные последовательности могут гибридизоваться друг с другом на по меньшей мере примерно 50%, 60%, 70%, 80% или 90% от всей последовательности или по всей длине последовательности в условиях умеренной или высокой жесткости. Что касается гибридизуемых полинуклеотидов, можно также рассматривать полинуклеотиды, включающие вместо кодона вырожденный кодон.
Гомология, подобие или идентичность между двумя данными полинуклеотидными последовательностями могут быть определены с использованием известного компьютерного алгоритма, такого как программа FASTA, используя параметры по умолчанию, как описано в работе Pearson et al. (1988, Proc. Natl. Acad. Sci. USA 85:2444). Альтернативно, их можно определить, используя алгоритм Нидлмана-Вунша (1970, J. МЫ. Biol. 48:443-453) выполняемый программой Нидлмана в пакете программ от The European Molecular Biology Open Software Suite (EMBOSS) (Rice et al, 2000, Trends Genet. 16:276-277) (версия 5.0.0 или более поздняя) (включая пакет программ GCG (Devereux, J. et al, Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F. et al, J Molec Biol 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego,1994; and Carillo et al. (1988) SIAMJApplied Math 48:1073). Например, гомологию, подобие или идентичность можно определить с помощью программ BLAST от Национального центра биотехнологической информации или ClustalW.
Гомологию, подобие или идентичность полинуклеотидов можно определять путем сравнения информации о последовательности с помощью компьютерной программы GAP, такой как в Needleman et al. (1970), J Mol Biol 48: 443, как описано в работе Smith and Waterman, Adv. Appl. Math (1981) 2:482. Вкратце, программа GAP определяет подобие как результат деления количества выровненных символов (т.е. нуклеотидов или аминокислот), являющихся подобными, на общее количество символов в более короткой из двух последовательностей. Параметры программы GAP по умолчанию могут включать: (1) однокомпонентную матрицу сравнений (содержащую значение 1 для идентичности и 0 для отсутствия идентичности) и взвешенную матрицу сравнений или матрицу замен Gribskov et al. (1986) Nucl. Acids Res. 14:6745, раскрытую в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или EDNAFULL (версия EMBOSS NCBI NUC4.4)); (2) штраф 3,0 за каждый гэп и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф на открытие гэпа 10, штраф на удлинение гэпа 0,5); и (3) отсутствие штрафа для концевых гэпов. Таким образом, используемый здесь термин «гомология» или «идентичность» отражает родство между последовательностями.
Кроме того, полинуклеотид может включать любой зонд, полученный из любых известных последовательностей генов, например, полинуклеотидную последовательность, гибридизованную с последовательностью, полностью или частично комплементарную вышеописанной полинуклеотидной последовательности, в жестких условиях и обладающую такой же активностью, без ограничения. Термин «жесткие условия» относится к условиям, в которых обеспечивается специфическая гибридизация между полинуклеотидами. Такие условия подробно раскрыты в известных документах (например, см. J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F. M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York). Например, жесткие условия могут включать осуществление гибридизации генов, имеющих высокую гомологию или идентичность, например гомологию или идентичность 40% или конкретно 70% или более, 80% или более, 85% или более, 90% или более, более конкретно 95% или более, еще более конкретно 97% или более, и наиболее конкретно 99% или более, без осуществления гибридизации генов, имеющих гомологию или идентичность более низкую, чем указанные выше гомологии или идентичности, или выполнение отмывки один раз, конкретно два или три раза, в обычных условиях отмывки для Саузерн-гибридизации с концентрацией соли и температурой 60°С, 1×SSC и 0,1% SDS, конкретно 60°С, 0,1×SSC и 0,1% SDS, и более конкретно 68°С, 0,1×SSC и 0,1% SDS.
Для гибридизации требуется, чтобы два нуклеотида имели комплементарные последовательности, хотя возможно ошибочное спаривание оснований в зависимости от степени жесткости гибридизации. Термин «комплементарный» используют для описания взаимоотношения между нуклеотидными основаниями, способными гибридизоваться друг с другом. Например, что касается ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, настоящее изобретение может включать не только по существу подобную нуклеиновокислотную последовательность, но также фрагмент нуклеиновой кислоты, выделенный, но комплементарный целой последовательности.
В частности, полинуклеотид, имеющий гомологию или идентичность, может быть обнаружен с использованием описанных выше условий гибридизации, включающих стадию гибридизации при значении Tm 55°С. Также, значение Tm может представлять собой 60°С, 63°С или 65°С, без ограничения ими, и может подходящим образом регулироваться специалистом в данной области техники в соответствии с задачами.
Степень жесткости условий для гибридизации полинуклеотида может зависеть от длины полинуклеотида и степени комплементарности, и переменные хорошо известны в данной области техники (Sambrook et al., см. выше, 9.50-9.51, 11.7-11.8).
Полинуклеотид, обладающий промоторной активностью, предложенный в настоящем изобретении, может иметь повышенную промоторную активность благодаря замене нуклеотида в конкретном положении описанной выше полинуклеотидной последовательности, обладающей промоторной активностью.
В одном воплощении полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может включать полинуклеотид, обладающий промоторной активностью, в котором по меньшей мере один нуклеотид нуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид. В частности, полинуклеотид может состоять из полинуклеотида, обладающего промоторной активностью, в котором по меньшей мере один нуклеотид нуклеотидной последовательности SEQ ID NO: 1 или 2 заменен на другой нуклеотид. Полинуклеотид, обладающий промоторной активностью, может быть использован взаимозаменяемо в настоящем описании с «мутантным промотором». Мутантный промотор может представлять собой промотор, в котором нуклеотид заменен на другой нуклеотид в одном или более положениях, двух или более положениях, трех или более положениях, четырех или более положениях, пяти или более положениях или во всех шести положениях, или положениях, соответствующих им.
В одном воплощении полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотид, включающий полинуклеотидную последовательность SEQ ID NO: 1 или 2, в которой по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251, заменен на другой нуклеотид и который обладает промоторной активностью.
«Другой нуклеотид» конкретно не ограничен, при условии что он представляет собой нуклеотид, отличный от нуклеотида до замены. Например, когда нуклеотид в положении 92 включает тимин (Т) в полинуклеотидной последовательности SEQ ID NO: 1, выражение «нуклеотид в положении 92 SEQ ID NO: 1 заменен на другой нуклеотид», означает, что тимин (Т) заменен на цитозин (С), аденин (А) или гуанин (G), отличные от тимина (Т). Кроме того, в настоящем изобретении, если не указано иное, описание нуклеотида как «замененного» означает, что нуклеотид заменен на другой нуклеотид, отличный от нуклеотида до замены.
В то же время, специалисты в данной области могут определить нуклеотиды по положениям, соответствующим положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2 по настоящему изобретению, в любой полинуклеотидной последовательности посредством выравнивания последовательностей, известного в данной области техники, и очевидно, что выражение «нуклеотид по определенному положению определенной SEQ ID NO» включает «нуклеотид по положению, соответствующему ему» в любой полинуклеотидной последовательности, если в настоящем изобретении не указано иное. Таким образом, любая полинуклеотидная последовательность полинуклеотида, обладающего промоторной активностью, где по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов, соответствующих нуклеотидам по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид, включена в объем настоящего изобретения.
«Положения 92, 94, 102, 103, 249 и 251 SEQ ID NO: 1 или 2» соответствуют 409 нуклеотиду (нт), 407 нт, 399 нт, 398 нт, 252 нт и 250 нт выше А в качестве точки отсчета (0) в ATG, который является кодоном инициации, в полинуклеотидах, обладающих промоторной активностью, имеющих происхождение из штаммов Saccharomyces cerevisiae CEN KSD-Yc, YJM1450, YJM1401, YJM1307 и штамма, депонированного в соответствии с Будапештским договором в Корейском центре культур микроорганизмов (KCCM) с регистрационным номером KCCM12568P, т.е. в SEQ ID NO: 2, и, таким образом, положения могут быть указаны как 409-е, 407-е, 399-е, 398-е, 252-е и 250-е положения, соответственно, выше ORF в соответствии со способом, обычно используемым в данной области техники.
В то же время, в отношении полинуклеотида, обладающего промоторной активностью, имеющего происхождение из Saccharomyces cerevisiae CEN.PK1-D, т.е. SEQ ID NO: 1, в которой нуклеотид по положению 74 выше ORF (т.е. 74-е положение) удален в промоторной последовательности Saccharomyces cerevisiae CEN KSD-Yc, «положения 92, 94, 102, 103, 249 и 251 SEQ ID NO: 1 или 2» соответствуют положениям 408, 406, 398, 397, 251 и 249 выше ORF, соответственно.
В одном воплощении полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может представлять собой полинуклеотид, обладающий промоторной активностью, в котором по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид.
В частности, в настоящем изобретении полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотид, в котором два или более нуклеотидов, выбранных из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменены на другие нуклеотиды.
В частности, в настоящем изобретении полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотид, в котором четыре или более нуклеотидов, выбранных из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменены на другие нуклеотиды.
В частности, полинуклеотид может представлять собой полинуклеотид, в котором все шесть нуклеотидов, выбранных из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменены на другие нуклеотиды.
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотид, в котором нуклеотиды по положениям 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменены на другие нуклеотиды.
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотид, в котором нуклеотиды по положениям 92, 94, 102 и 103 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменены на другие нуклеотиды.
Однако, настоящее изобретение не ограничено ими.
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может включать:
замену тимина (Т) по положению 92 на гуанин (G), цитозин (С) или аденин (А);
замену тимина (Т) по положению 94 на гуанин (G), цитозин (С) или аденин (А);
замену аденина (А) по положению 102 на гуанин (G), цитозин (С) или тимин (Т);
замену аденина (А) по положению 103 на гуанин (G), цитозин (С) или тимин (Т);
замену гуанина (G) по положению 249 на тимин (Т), цитозин (С) или аденин (А);
замену цитозина (С) по положению 251 на тимин (Т), гуанин (G) или аденин (А);
или любую их комбинацию в полинуклеотидной последовательности SEQ ID NO: 1 или 2.
В одном воплощении тимин (Т) по положению 92 может быть заменен на цитозин (С). Это может быть также записано как 92 (Т→С) или 409 (Т→С), а в соответствии с референсной последовательностью как -408 (Т→С).
В одном воплощении тимин (Т) по положению 94 может быть заменен на цитозин (С). Это может быть также записано как 94 (Т→С) или 407 (Т→С), а в соответствии с референсной последовательностью как -406 (Т→С).
В одном воплощении аденин (А) по положению 102 может быть заменен на цитозин (С). Это может быть также записано как 102 (А→С) или -399 (А→С), а в соответствии с референсной последовательностью как -398 (А→-С).
В одном воплощении аденин (А) по положению 103 может быть заменен на тимин (Т). Это может быть также записано как 103 (А→Т) или -398 (А→Т), а в соответствии с референсной последовательностью как 397 (А→Т).
В одном воплощении гуанин (G) по положению 249 может быть заменен на аденин (А). Это может быть также записано как 249 (G→А) или -252 (G→А), а в соответствии с референсной последовательностью как -251 (G→А).
В одном воплощении цитозин (С) по положению 251 может быть заменен на тимин (Т). Это может быть также записано как 251 (С→Т) или -250 (С→Т), а в соответствии с референсной последовательностью как -249 (С→Т).
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может включать по меньшей мере одну из замен 92 (Т→С), 94 (Т→С), 102 (А→С), 103 (А→Т), 249 (G→A) и 251 (С→Т).
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может включать замены 249 (G→А) и 251 (С→Т).
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может включать замены 92 (Т→С), 94 (Т→С), 102 (А→С) и 103 (А→Т).
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может включать все замены из 92 (Т→С), 94 (Т→С), 102 (А→С), 103 (А→Т), 249 (G^A) и 251 (С→Т).
В одном воплощении настоящего изобретения полинуклеотид, обладающий промоторной активностью, может включать по меньшей мере одну полинуклеотидную последовательность, выбранную из SEQ ID NO: с 3 по 32. В частности, полинуклеотид может состоять из одной полинуклеотидной последовательности, выбранной из SEQ ID NO: с 3 по 32, без ограничения ими.
Как описано выше, хотя выражения «полинуклеотид, имеющий нуклеотидную последовательность определенной SEQ ID NO:» или «полинуклеотид, включающий нуклеотидную последовательность определенной SEQ ID NO:» используют в настоящем изобретении, очевидно, что любой полинуклеотид, имеющий нуклеотидную последовательность, включающую делецию, модификацию, замену или добавление одного или нескольких нуклеотидов, также может быть использован в настоящем изобретении, при условии что полинуклеотид имеет активность, идентичную или эквивалентную активности полинуклеотида, состоящего из нуклеотидной последовательности конкретной SEQ ID NO.
Кроме того, настоящее изобретение не ограничено вышеупомянутыми воплощениями, и полинуклеотидная последовательность может включать различные модификации в диапазоне, который не оказывает существенного влияния на активность промотора.
Полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может быть использован в качестве промотора.
Промотор может быть расположен в 5'-области сайта инициации транскрипции мРНК.
Промотор может иметь повышенную промоторную активность по сравнению с обычными промоторами. То есть, промотор может повышать не только экспрессию целевого гена в клетках-хозяевах, но также экспрессию и/или активность белка, кодируемого данным целевым геном. С учетом задач настоящего изобретения, целевой ген для усиления экспрессии может быть модифицирован в соответствии с получаемым продуктом, а промотор может быть использован в качестве промотора общего назначения для усиления целевого гена.
«Целевой ген» относится к гену, экспрессия которого должна регулироваться промоторной последовательностью по настоящему изобретению с учетом задач по настоящему изобретению. Белок, кодируемый целевым геном, может быть назван «целевым белком», а ген, кодирующий «целевой белок», может быть назван «целевым геном».
Кроме того, полинуклеотид, кодирующий целевой белок, может иметь различные модификации, выполненные в кодирующей области, при условии что они не изменяют аминокислотную последовательность белка, экспрессируемого из кодирующей области, вследствие вырожденности кодонов или с учетом кодонов, предпочтительных для живого организма, в котором этот полинуклеотид экспрессируется. Описания полинуклеотидной последовательности являются такими как приведено выше.
В одном воплощении целевой белок может представлять собой полипептид, обладающий активностью глутамат-цистеинлигазы. То есть, целевым геном промотора может быть ген, кодирующий полипептид, имеющий глутамат-цистеинлигазу.
В настоящем изобретении «глутамат- цисте инлигаза» представляет собой фермент, также называемый «фермент, связывающий глутамат-цистеин», «гамма-глутамилцистеинсинтетаза (GCS)» или «белок GSH1».
Известно, что глутамат-цистеинлигаза катализирует следующую реакцию:
L-глутамат + L-цистеин + АТФ ↔ гамма-глутамилцистеин + АДФ + Pi
Кроме того, реакция, катализируемая глутамат-цистеинлигазой, известна как первая стадия синтеза глутатиона.
Аминокислотная последовательность, составляющая глутамат-цисте инлигазу, может быть получена из известной базы данных NCBI GenBank. Например, глутамат-цистеинлигаза может иметь происхождение из Saccharomyces cerevisiae. Например, глутамат-цистеинлигаза может представлять собой белок, включающий аминокислотную последовательность SEQ ID NO: 33, но может включать любую последовательность, обладающую такой же активностью, что и указанная аминокислотная последовательность, без ограничения.
Кроме того, «полипептид, обладающий активностью глутамат-цистеинлигазы» по настоящему изобретению может включать не только немодифицированную или встречающуюся в природе форму глутамат-цистеинлигазы дикого типа, но также варианты, имеющие такую же или усиленную активность глутамат-цистеинлигазы.
В настоящем изобретении термин «модифицированный полипептид», имеющий то же значение, что и «вариант», может относиться к белку, полученному консервативной заменой и/или модификацией по меньшей мере одной аминокислоты, отличной от той, которая указана в данной последовательности, при сохранении функций или свойств белка. Например, модифицированный полипептид может представлять собой вариант, в котором аминокислота по положению 86 OTN-конца SEQ ID NO: 33 заменена на другой аминокислотный остаток, отличный от цистеина, в описанной выше глутамат-цистеинлигазе.
Вариант отличается от идентифицированной последовательности заменой, делецией или добавлением нескольких аминокислот. Такие варианты могут быть получены путем модификации одной или более аминокислот в указанной выше аминокислотной последовательности белка и идентифицированы путем оценки свойств модифицированного белка. То есть, способность варианта может быть усилена по сравнению с нативным белком. Кроме того, некоторые варианты могут включать варианты, из которых удалена по меньшей мере одна часть, такая как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, в которых часть удалена с N- и/или С-конца зрелого белка.
Термин «вариант» также может быть использован взаимозаменяемо с другими терминами, такими как модификация, модифицированный белок, модифицированный полипептид, мутант, мутеин и дивергент, и любые термины, используемые для обозначения вариации, также могут быть использованы без ограничения. С учетом задач настоящего изобретения вариант может иметь повышенную активность по сравнению с белками дикого типа или ^модифицированными белками, не ограничиваясь этим.
Используемый здесь термин «консервативная замена» относится к замене одной аминокислоты на другую аминокислоту, имеющую сходные структурные и/или химические свойства. Вариант может иметь по меньшей мере одну консервативную замену при сохранении по меньшей мере одной биологической активности. Такая аминокислотная замена обычно может происходить на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатка.
Варианты также могут включать делецию или добавление аминокислот, оказывающих минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью на N-конце белка, которая котрансляционно или посттрансляционно направляет перенос белка. Полипептид также может быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
В настоящем изобретении «замена на другую аминокислоту» конкретно не ограничена, если аминокислота после замены отличается от аминокислоты до замены. То есть, замена цистеина, который является 86-й аминокислотой от N-конца аминокислотной последовательности SEQ ID NO: 33, на аминокислоту, отличную от цистеина, также может быть выражена как «замена аминокислоты по положению 86 на другую аминокислоту». В то же время, в настоящем изобретении очевидно, что выражение «определенная аминокислота заменена» означает, что аминокислота после замены отличается от аминокислоты до замены, если только не используется выражение «заменена на другую аминокислоту».
«Вариант глутамат-цистеинлигазы» по настоящему изобретению может быть также обозначен как «(модифицированный) полипептид, обладающий активностью глутамат-цистеинлигазы» или «вариант GSH1», который может усиливать продукцию глутатиона по сравнению с белком до модификации, полипептидом дикого типа или немодифицированным полипептидом, не ограничиваясь этим.
В данном варианте по меньшей мере одна аминокислота аминокислотной последовательности SEQ ID NO: 33 может быть заменена на другую аминокислоту. В частности, вариант может включать замену аминокислоты, соответствующей положению 86 аминокислотной последовательности SEQ ID NO: 33, на другую аминокислоту. Другая аминокислота может быть выбрана из глицина, аланина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, тирозина, аспарагина, глутамата, глутамина, аспартата, лизина, аргинина и гистидина.
В одном воплощении аминокислота, соответствующая положению 86 аминокислотной последовательности SEQ ID NO: 33, может быть заменена на аргинин, не ограничиваясь этим.
В настоящем изобретении очевидно, что «вариант, в котором 86-я аминокислота от N-конца аминокислотной последовательности SEQ ID NO: 33 заменена на другую аминокислоту», включает вариант, в котором аминокислота, соответствующая положению 86 аминокислотной последовательности SEQ ID NO: 33, заменена на другую аминокислоту, хотя аминокислота находится в положении, отличном от 86-го, вследствие делеции/вставки/добавления или т.п. аминокислоты на N- или С-конце или в середине аминокислотной последовательности SEQ ID NO: 33. Кроме того, хотя вариант, в котором 86-я аминокислота с N-конца аминокислотной последовательности SEQ ID NO: 33 заменена на другую аминокислоту, раскрыт в качестве примера варианта глутамат-цистеинлигазы по настоящему изобретения, очевидно, что вариант глутамат-цистеинлигазы по настоящему изобретению не ограничивается этим вариантом аминокислотной последовательности SEQ ID NO: 33, и вариант, в котором аминокислота, соответствующая положению 86 аминокислотной последовательности SEQ ID NO: 33, заменена на другую аминокислоту в любой аминокислотной последовательности, обладающей активностью глутамат-цистеинлигазы, также входит в объем варианта глутамат-цистеинлигазы по настоящему изобретению. В любой аминокислотной последовательности «аминокислота, соответствующая положению 86 аминокислотной последовательности SEQ ID NO: 33», может быть идентифицирована различными способами выравнивания последовательностей, хорошо известными в данной области техники.
Вариант глутамат-цистеинлигазы по настоящему изобретению, в котором аминокислота, соответствующая положению 86 от N-конца аминокислотной последовательности SEQ ID NO: 33, заменена на другую аминокислоту, может представлять собой белок, включающий аминокислотную последовательность SEQ ID NO: 33 или аминокислотную последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологии или идентичности с ней. Кроме того, очевидно, что любой белок, имеющий аминокислотную последовательность, включающую делецию, модификацию, замену или добавление одной или нескольких аминокислот по положению, отличному от 86-го, входит в объем настоящего изобретения, при условии что белок сохраняет вышеописанную гомологию или идентичность и эффект, эквивалентный эффекту варианта. Гомология и идентичность являются такими, как описано выше.
Ген, кодирующий полипептид, обладающий активностью глутамат-цистеинлигазы, по настоящему изобретению может быть назван «геном GSH1».
Данный ген может иметь происхождение из дрожжей. В частности, ген может иметь происхождение из микроорганизма, принадлежащего к роду Saccharomyces, более конкретно Saccharomyces cerevisiae. В частности, ген может представлять собой ген, кодирующий аминокислотную последовательность SEQ ID NO: 33, но не ограничиваясь этим.
В настоящем изобретении «ген GSH1», т.е. полинуклеотид, кодирующий полипептид, обладающий активностью глутамат-цистеинлигазы, может иметь различные модификации, выполненные в кодирующей области, при условии что аминокислотная последовательность полипептида, экспрессируемого из кодирующей области, не изменяется вследствие вырожденности кодонов или с учетом кодонов, предпочтительных для живого организма, в котором данный полинуклеотид экспрессируется.
Поскольку полипептид, обладающий активностью глутамат-цистеинлигазы по настоящему изобретению, также включает последовательности вариантов, любые полинуклеотидные последовательности, кодирующие варианты белка, в которых аминокислота по положению 86 аминокислотной последовательности SEQ ID NO: 33 по настоящему изобретению заменена на другую аминокислоту, также могут быть включены без ограничения.
Кроме того, полинуклеотид может включать полинуклеотидную последовательность, которая гибридизуется с зондом, сконструированным с использованием известной генной последовательности, например полинуклеотидной последовательности, полностью или частично комплементарной полинуклеотидной последовательности, в жестких условиях для кодирования варианта белка, в котором аминокислота, соответствующая положению 86 аминокислотной последовательности SEQ ID NO: 33, заменена на другую аминокислоту, без ограничения.
В другом аспекте настоящего изобретения предложена композиция для экспрессии гена, включающего полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
Композиция для экспрессии гена относится к композиции, способной экспрессировать ген, использующий полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
Например, композиция для экспрессии гена включает полинуклеотид, обладающий промоторной активностью, по настоящему изобретению, и может дополнительно включать любые другие компоненты, способные обеспечить функционирование полинуклеотида в качестве промотора.
В композиции для экспрессии гена по настоящему изобретению полинуклеотид может быть в форме, включенной в вектор, способный к экспрессии гена, функционально связанного с ним, в клетке-хозяине.
В другом аспекте настоящего изобретения предложен вектор, включающий либо полинуклеотид, обладающий промоторной активностью, либо полинуклеотид и ген, кодирующий целевой белок.
В одном воплощении целевой белок может представлять собой полипептид, обладающий активностью глутамат-цистенилигазы.
Используемый здесь термин «вектор» относится к конструкции ДНК, включающей полинуклеотидную последовательность, кодирующую целевой белок, которая функционально связана с соответствующей регуляторной последовательностью для экспрессии целевого белка в подходящей клетке-хозяине.
С учетом задач настоящего изобретения регуляторная последовательность может включать полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
В то же время, регуляторная последовательность может включать промотор для инициации транскрипции, последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания мРНК с рибосомой, и последовательность, регулирующую терминацию транскрипции и трансляции. После введения вектора в подходящую клетку-хозяина он может реплицироваться или функционировать независимо от генома хозяина и может интегрироваться в геном.
Вектор, используемый в настоящем изобретении, конкретно не ограничен, и можно использовать любой вектор, известный в данной области техники. В качестве вектора, экспрессируемого в дрожжах, можно использовать как интегративные дрожжевые плазмиды (Yip), так и внехромосомные плазмидные векторы.
Внехромосомный плазмидный вектор может включать эписомальные дрожжевые плазмиды (YEp), репликативные дрожжевые плазмиды (YRp) и дрожжевые центромерные плазмиды (YCp).
Кроме того, в качестве вектора по настоящему изобретению можно также использовать хромосомы искусственных дрожжей (YAC).
В качестве конкретного примера, доступные векторы могут включать pESCHIS, pESC-LEU, pESC-TRP, pESC-URA, Gateway pYES-DEST52, pAO815, pGAPZ A, pGAPZ B, pGAPZ C, pGAPα A, pGAPα B, pGAPα С, pPIC3.5K, pPIC6 A, pPIC6 В, pPIC6 C, pPIC6α A, pPIC6α B, pPIC6α C, pPIC9K, pYC2/CT, pYD1 дисплейный вектор дрожжей, pYES2, pYES2/CT, pYES2/NT A, pYES2/NT B, pYES2/NT C, pYES2/CT, pYES2.1, pYES-DEST52, pTEF1/Zeo, pFLD1, PichiaPinkTM, p427-TEF, p417-CYC, pGAL-MF, p427-TEF, p417-CYC, PTEF-MF, pBY011, pSGP47, pSGP46, pSGP36, pSGP40, ZM552, PAG303GAL-ccdB, PAG414GAL-ccdB, pAS404, pBridge, pGAD-GH, pGAD T7, pGBK T7, pHIS-2, pOBD2, pRS408, pRS410, pRS418, pRS420, pRS428, форму микрон А дрожжей, pRS403, pRS404, pRS405, pRS406, pYJ403, pYJ404, pYJ405 и pYJ406, не ограничиваясь ими.
Встраивание полинуклеотида в хромосому можно осуществить любым способом, известным в данной области техники, например, гомологичной рекомбинацией, не ограничиваясь этим. Маркер селекции может быть дополнительно включен для подтверждения хромосомной вставки. Маркер селекции используют для селекции клеток, которые трансформируют вектором, то есть для идентификации вставки желаемой молекулы нуклеиновой кислоты, и примеры маркера селекции могут включать маркеры, обеспечивающие селектируемые фенотипы, такие как устойчивость к лекарственным средствам, потребность в питательных веществах, устойчивость к цитотоксическим агентам, или поверхностную экспрессию вариантного полипептида. Только клетки, экспрессирующие маркер селекции, способны выживать или проявлять разные фенотипы в среде, обработанной селективным агентом, и, таким образом, могут быть отобраны трансформированные клетки. Например, полинуклеотид дикого типа может быть заменен на мутантный полинуклеотид с использованием вектора для хромосомной вставки в клетки.
Используемый здесь термин «трансформация» относится к процессу встраивания вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяин, для обеспечения возможности экспрессировать целевой белок в клетке-хозяине.
Трансформированный полинуклеотид может быть либо в форме, встроенной в хромосому клетки-хозяина, либо в форме, расположенной вне хромосомы, при условии что белок экспрессируется в клетке-хозяине. Кроме того, полинуклеотид, кодирующий целевой белок, может включать ДНК и РНК, кодирующие целевой белок. Полинуклеотид, кодирующий целевой белок, может быть введен в клетку-хозяин в любой форме, при условии что полинуклеотид вводится в клетку-хозяин и белок экспрессируется в ней. Например, полинуклеотид, кодирующий целевой белок, может быть введен в клетку-хозяин в виде экспрессионной кассеты, которая представляет собой генную конструкцию, включающую все основные элементы, необходимые для саморепликации.
Экспрессионная кассета обычно может включать промотор, функционально связанный с полинуклеотидом, кодирующим целевой белок, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции. Экспрессионная кассета может быть в форме самореплицирующегося вектора экспрессии. Кроме того, полинуклеотид, кодирующий целевой белок, может быть введен в клетку-хозяина в его исходной форме и функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине, не ограничиваясь этим.
Кроме того, используемый здесь термин «функционально связанный» означает, что полинуклеотидная последовательность, кодирующая целевой белок по настоящему изобретению, функционально связана с последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотидной последовательности.
С учетом задач настоящего изобретения промотор может представлять собой полинуклеотид, обладающий промоторной активностью по настоящему изобретению.
Способы трансформации согласно настоящему изобретению включают любые способы, позволяющие вводить вектор в клетку-хозяина, и могут быть осуществлены с помощью подходящих стандартных методик, хорошо известных в данной области техники, выбираемых в соответствии с клеткой-хозяином. Например, могут быть использованы электропорация, осаждение фосфатом кальция (СаРО4), осаждение хлоридом кальция (CaCl2), микроинъекция, метод полиэтиленгликоля (PEG), метод DEAE-декстрана, метод катионных липосом и метод ацетата лития-DMSO, но настоящее изобретение не ограничивается этим.
В другом аспекте настоящего изобретения предложен микроорганизм, включающий полинуклеотид, обладающий промоторной активностью, по настоящему изобретению, полинуклеотид, включающий описанный выше полинуклеотид и ген, кодирующий целевой белок, или вектор, содержащий его.
Целевой белок может представлять собой полипептид, обладающий активностью глутамат-цистеинлигазы. Полинуклеотид, обладающий промоторной активностью, по настоящему изобретению, целевой белок, полипептид, обладающий активностью глутамат-цистеинлигазы, и вектор являются такими, как описано выше.
Микроорганизм может представлять собой дрожжи, в частности, микроорганизм, принадлежащий к роду Saccharomyces sp., и более конкретно Saccharomyces cerevisiae.
Микроорганизм может представлять собой микроорганизм, экспрессирующий глутамат-цистеинлигазу, микроорганизм, экспрессирующий полипептид, обладающий активностью глутамат-цистеинлигазы, или микроорганизм, в который введен полипептид, обладающий активностью глутамат-цистеинлигазы, не ограничиваясь этим.
В одном воплощении микроорганизм может представлять собой микроорганизм, обладающий способностью продуцировать глутатион, микроорганизм, полученный путем усиления способности продуцировать глутатион у исходного штамма, имеющего от природы низкую способность продуцировать глутатион, или микроорганизм, полученный путем обеспечения способности продуцировать глутатион для родительского штамма, неспособного продуцировать глутатион. В одном воплощении микроорганизм может представлять собой микроорганизм, экспрессирующий вариант глутамат-цистеинлигазы, включающий по меньшей мере одну аминокислотную вариацию в аминокислотной последовательности SEQ ID NO: 33, и аминокислотная вариация может включать замену 86-й аминокислоты с N-конца SEQ ID NO: 33 на другую аминокислоту. Однако настоящее изобретение не ограничивается этим. Вариант глутамат-цистеинлигазы является таким, как описано выше.
Используемый в настоящем документе термин «белок, подлежащий экспрессии/экспрессированный» означает состояние, при котором целевой белок, например глутамат-цистеинлигаза или ее вариант, вводится или модифицируется для экспрессии в микроорганизме. В случае, когда белок присутствует в микроорганизме, активность белка повышается по сравнению с активностью его эндогенного белка или активностью до модификации.
В частности, термин «введение белка» относится к приданию активности конкретного белка микроорганизму, который не обладает этим белком, или усилению активности белка по сравнению с собственной активностью белка или активностью до модификации. Например, введение белка может относиться к введению полинуклеотида, кодирующего белок, в хромосому или к введению вектора, включающего полинуклеотид, кодирующий конкретный белок, в микроорганизм, чтобы тем самым проявить активность белка.
Кроме того, «усиление активности» может означать, что активность конкретного белка микроорганизма повышается по сравнению с собственной активностью или активностью до модификации. Термин «внутренняя активность» может относиться к активности конкретного белка, которым обладает родительский штамм до трансформации, когда микроорганизм трансформируется путем естественной или искусственной генетической вариации.
С учетом задач настоящего изобретения усиление активности может быть достигнуто путем использования полинуклеотидной последовательности, обладающей промоторной активностью, по настоящему изобретению в качестве регуляторной последовательности экспрессии целевого белка. Целевой белок может представлять собой белок дикого типа или вариант, как описано выше, регуляторная последовательность экспрессии может представлять собой регуляторную последовательность экспрессии гена, кодирующего вариант белка, или регуляторную последовательность экспрессии хромосомного гена, кодирующего белок дикого типа.
Кроме того, могут быть использованы любые другие методы повышения активности или комбинированный метод. Например, в дополнение к способу использования полинуклеотидной последовательности, обладающей промоторной активностью, по настоящему изобретению в качестве регуляторной последовательности экспрессии целевого белка, может быть использован по меньшей мере один способ, выбранный из группы, состоящей из способа увеличения числа копий гена, кодирующего целевой белок в клетках, способа замены хромосомного гена, кодирующего белок дикого типа, на ген, кодирующий вариант белка, способа дополнительного введения мутации в ген, кодирующий ген, кодирующий белок, для усиления активности варианта белка и способа введения варианта белка в микроорганизм, не ограничиваясь этим.
Активность целевого белка можно повысить, используя полинуклеотид, обладающий промоторной активностью, по настоящему изобретению в качестве регуляторной последовательности экспрессии целевого белка в микроорганизме.
Например, активность или концентрация белка обычно может быть увеличена минимум на 1%, 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500%, максимум до 1000% или 2000%, по сравнению с активностью или концентрацией штамма микроорганизма дикого типа или немодифицированного микроорганизма, не ограничиваясь этим.
Используемый здесь термин «немодифицированный микроорганизм» не исключает штаммы, имеющие мутацию, которая может встречаться в природе в микроорганизмах, и может представлять собой штамм дикого типа, микроорганизм, не включающий полинуклеотид, обладающий промоторной активностью, по настоящему изобретению или микроорганизм, не трансформированный вектором, включающий полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
Микроорганизм по настоящему изобретению может представлять собой микроорганизм, продуцирующий глутатион.
Используемый здесь термин «глутатион» может быть использован взаимозаменяемо с «GSH» и относится к трипептидному соединению, состоящему из трех аминокислот: глутамат, цистеин и глицин. Глутатион может быть использован в качестве сырья для фармацевтических препаратов, полезных для здоровья пищевых продуктов, вкусовых ингредиентов, пищевых и кормовых добавок, косметики и т.п., но не ограничивается этим.
Используемый здесь термин «микроорганизм, продуцирующий глутатион» включает микроорганизмы, в которых происходит естественная или искусственная генетическая модификация, и может относиться к микроорганизму, в котором конкретный механизм ослаблен или усилен путем введения экзогенного гена или усиления или инактивации эндогенного гена путем генетической модификации с целью получения глутатиона. С учетом задач настоящего изобретения, микроорганизм, продуцирующий глутатион, может относиться к микроорганизму, включающему полинуклеотид, обладающий промоторной активностью, по настоящему изобретению и способному продуцировать большее количество целевого глутатиона по сравнению с микроорганизмами дикого типа или ^модифицированными микроорганизмами.
Термин «микроорганизм, продуцирующий глутатион» может быть использован взаимозаменяемо с терминами «глутатион-продуцирующий микроорганизм», «микроорганизм, обладающий способностью продуцировать глутатион», «штамм, продуцирующий глутатион», «штамм, обладающий способностью продуцировать глутатион» и т.п.
Микроорганизм, продуцирующий глутатион, может представлять собой рекомбинантный микроорганизм. Рекомбинантный микроорганизм является таким, как описано выше. Микроорганизм может включать мутацию для усиления пути биосинтеза для увеличения способности продуцировать глутатион, высвобождения ингибирования по принципу обратной связи или инактивации генов, которые ослабляют путь деградации или путь биосинтеза, и такая мутация может быть вызвана искусственным методом, например УФ-облучением, и не исключает естественной мутации.
Другой аспект настоящего изобретения относится к способу получения глутатиона, включающему культивирование микроорганизма в культуральной среде. Микроорганизм и глутатион являются такими, как описано выше. Глутатион может накапливаться в микроорганизме при культивировании штамма.
Что касается питательной среды, используемой для культивирования штамма по настоящему изобретению, или других условий культивирования, то без ограничений можно использовать любую питательную среду, обычно используемую для культивирования микроорганизмов, принадлежащих к роду Saccharomyces, и, в частности, штамм по настоящему изобретению можно культивировать в обычной среде, содержащей подходящие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины в аэробных или анаэробных условиях при регулировании температуры, рН и т.п.
В настоящем изобретении в качестве источников углерода могут быть использованы углеводы, такие как глюкоза, фруктоза, сахароза и мальтоза; сахарные спирты, такие как маннит и сорбит; органические кислоты, такие как пировиноградная кислота, молочная кислота и лимонная кислота; и аминокислоты, такие как глутамат, метионин и лизин, не ограничиваясь ими. Кроме того, можно использовать натуральные органические питательные вещества, такие как гидролизаты крахмала, меласса, патока, рисовые отруби, маниока, багасса сахарного тростника и кукурузный экстракт, а также углеводы, такие как глюкоза и стерильная предварительно обработанная меласса (т.е. меласса, превращенная в восстановленные сахара), и подходящие количества любых других источников углерода также могут быть использованы без ограничений. Эти источники углерода можно использовать по отдельности или в комбинации по меньшей мере двух из них.
В качестве источников азота можно использовать неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; и источники органического азота, такие как аминокислоты, пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, кукурузный экстракт, гидролизат казеина, рыба или продукты ее разложения, а также обезжиренный соевый жмых или продукты его разложения. Эти источники азота можно использовать по отдельности или в комбинации по меньшей мере двух из них.
В качестве источников фосфора можно использовать монофосфат калия, дифосфат калия или соответствующие им натрийсодержащие соли. В качестве неорганических соединений могут быть использованы хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и т.п.
Культуральная среда может дополнительно включать аминокислоты, витамины и/или подходящие предшественники. В частности, L-аминокислоты и т.п.могут быть добавлены в культуральную среду для штамма. В частности, в культуральную среду могут быть добавлены глицин, глутамат и/или цистеин, и L-аминокислоты, такие как лизин, могут быть дополнительно добавлены при необходимости, но настоящее изобретение не ограничивается этим.
Культуральную среду и предшественники можно добавлять к культурам в периодическом или непрерывном процессе, не ограничиваясь этим.
В настоящем изобретении в процессе культивирования штамма такие соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, могут быть надлежащим образом добавлены к культурам для корректировки рН культур. Кроме того, может быть добавлен пеногаситель, такой как полигликолевый эфир жирной кислоты, для подавления пенообразования во время культивирования. Кроме того, в культуры можно вводить кислород или кислородсодержащий газ, чтобы поддерживать культуры в аэробных условиях, а азот, водород или углекислый газ можно вводить в культуры, чтобы поддерживать культуры в анаэробных и микроаэробных условиях, без подачи каких-либо других газов для этого.
Температуру культур можно поддерживать на уровне от 25°С до 40°С, более конкретно от 28°С до 37°С, не ограничиваясь этим. Культивирование можно продолжать до тех пор, пока не будет получено желаемое количество продукта, в частности, в течение от 1 до 100 часов, не ограничиваясь этим.
Способ получения глутатиона может дополнительно включать дополнительный процесс после стадии культивирования. Дополнительный процесс может быть соответствующим образом выбран в соответствии с целью использования глутатиона.
В частности, способ получения глутатиона может дополнительно включать выделение глутатиона из по меньшей мере одного выбранного из культивируемого микроорганизма, высушенного продукта микроорганизма, экстракта микроорганизма, культурального продукта микроорганизма и лизата микроорганизма после стадии культивирования.
Способ может дополнительно включать лизис микроорганизма (штамма) до или одновременно со стадией выделения. Лизис штамма может быть осуществлен любым способом, обычно используемым в данной области техники, к которому относится настоящее изобретение, например, путем тепловой обработки или с использованием буферного раствора для лизиса, ультразвукового аппарата и френч-пресса. Кроме того, стадия лизиса может включать ферментативную реакцию с использованием литического фермента клеточной стенки, нуклеазы, транснуклеотидазы, протеазы и т.п., не ограничиваясь этим.
С учетом задач настоящего изобретения, в соответствии со способом получения глутатиона могут быть приготовлены сухие дрожжи, дрожжевой экстракт и порошок смеси дрожжевого экстракта, каждый из которых имеет высокое содержание глутатиона, и чистый глутатион. Однако настоящее изобретение не ограничивается этим, и эти продукты могут быть приготовлены соответствующим образом в соответствии с желаемыми продуктами.
В настоящем изобретении сухие дрожжи могут быть использованы взаимозаменяемо с терминами «сухой продукт микроорганизма», «сухой продукт штамма» и т.п. Сухие дрожжи могут быть получены путем высушивания штамма дрожжей, в котором накоплен глутатион, и, в частности, они могут быть включены в кормовую композицию, пищевую композицию и т.п., не ограничиваясь этим.
В настоящем изобретении экстракт дрожжей можно использовать взаимозаменяемо с такими терминами, как «экстракт микроорганизма», «экстракт штамма» и т.п.Экстракт штамма может относиться к веществам, оставшимся после отделения клеточных стенок от штамма. В частности, экстракт штамма может относиться к компонентам, оставшимся после удаления клеточных стенок из компонентов, полученных путем лизиса клеток. Экстракт штамма включает глутатион и один или более других компонентов, выбранных из белков, углеводов, нуклеиновых кислот и волокон, в дополнение к глутатиону, но не ограничиваясь этим.
Стадия выделения может быть выполнена с использованием любого подходящего способа, хорошо известного в данной области техники, и глутатион может быть выделен в качестве целевого вещества.
Стадия выделения может включать процесс очистки. Процесс очистки может быть осуществлен путем выделения из штамма только глутатиона. В процессе очистки можно получить чистый глутатион.
При необходимости способ получения глутатиона может дополнительно включать смешивание эксципиента с одним выбранным из штамма, высушенного продукта, экстракта, культурального продукта и его лизата, и выделенного из них глутатиона. Путем смешивания можно приготовить порошок смеси дрожжевого экстракта.
Эксципиент может быть соответствующим образом выбран и использован в соответствии с предназначенными применением или формой и может быть, например, выбран из крахмала, глюкозы, целлюлозы, лактозы, гликогена, D-маннита, сорбита, лактита, мальтодекстрина, карбоната кальция, синтетического силиката алюминия, моногидрофосфата кальция, сульфата кальция, хлорида натрия, гидрокарбоната натрия, очищенного ланолина, декстрина, альгината натрия, метил целлюлозы, коллоидного силикагеля, гидроксипропилкрахмала, гидроксипропилметилцеллюлозы, пропиленгликоля, казеина, лактата кальция, приможеля и аравийской камеди, и в частности, он может включать по меньшей мере один компонент, выбранный из крахмала, глюкозы, целлюлозы, лактозы, декстрина, гликогена, D-маннита и мальтодекстрина, не ограничиваясь этим.
Эксципиент может включать, например, консервант, увлажнитель, диспергирующий агент, суспендирующий агент, буфер, стабилизатор или изотонический агент, не ограничиваясь этим.
Другой аспект настоящего изобретения относится к применению полинуклеотида, в котором по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид, в качестве промотора.
Полинуклеотид является таким, как описано выше.
Осуществление изобретения
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие примеры и экспериментальные примеры. Однако следующие примеры и экспериментальные примеры представлены исключительно для иллюстрации настоящего изобретения, и объем настоящего изобретения ими не ограничивается.
Пример 1: Получение штамма CJ-5, обладающего способностью продуцировать глутатион
Штаммы получали из блоков дрожжей, содержащих различные штаммы, и их характеристики улучшали для отбора штаммов, обладающих способностью продуцировать глутатион.
В частности, образцы зерна, такие как рис, ячмень, бобы мунг и овес, собирали в 20 районах, таких как Хвасон, Пхентхэк, Йонъин и т.д. в Кенгидо, Республика Корея, затем измельчали, смешивали, завертывали в ткань, плотно сжатую для придания формы, скручивали для ферментации в течение 10 суток и медленно сушили для приготовления блоков дрожжей.
Для выделения различных штаммов из приготовленных блоков дрожжей был проведен следующий эксперимент. К 5 г блоков дрожжей добавляли 45 мл солевого раствора и измельчали с помощью миксера. Чтобы полностью выделить штамм дрожжей, полученную смесь разбавляли серийными разведениями, распределяли на агаре YPD (10 г/л дрожжевого экстракта, 20 г/л бактопептона и 20 г/л глюкозы на 1 л дистиллированной воды) и культивировали при 30°С в течение 48 часов. Затем, в соответствии с морфологией колоний и микроскопической проверкой, колонии дрожжей высевали штрихами на агар YPD. 25 мл бульона YPD высевали в коническую колбу на 250 мл и в нее инокулировали полностью выделенный штамм и культивировали во встряхивающем инкубаторе в течение 48 часов при 30°С и 200 об/мин. Штаммы подвергали скринингу, определяя продукцию глутатиона.
Для улучшения первично выделенных штаммов в этих выделенных штаммах индуцировали случайную мутацию. В частности, из блоков дрожжей был выделен штамм, обладающий способностью продуцировать глутатион, который получил название штамм CJ-37. Этот штамм CJ-37 культивировали в твердой среде и инокулировали в бульон для получения его культурального раствора и этот культуральный раствор подвергали воздействию УФ-света с использованием УФ-лампы. После того как культуральный раствор, подвергнутый воздействию УФ-лучей, высевали на среду для чашек, был выделен только мутантный штамм, который образовал колонии, и была идентифицирована его продукция глутатиона.
В результате, среди мутантных штаммов, штамм, демонстрирующий наибольшую продукцию глутатиона, был выбран в качестве штамма, продуцирующего глутатион, который получил название штамм CJ-5 и был депонирован в Корейском центре культур микроорганизмов (KCCM) в соответствии с Будапештским договором под номером доступа No. KCCM12568P 31 июля 2019 г.
Пример 2: Разработка модифицированной последовательности GSH1 путем эксперимента для дополнительного улучшения штамма CJ-5
Пример 2-1: Индукция мутации и идентификация модифицированной последовательности
Мутацию индуцировали следующим образом для дальнейшего улучшения способности штамма CJ-5 продуцировать глутатион.
Штамм CJ-5 культивировали в твердой среде и инокулировали в бульон для получения культурального раствора и этот культуральный раствор подвергали воздействию УФ-света с использованием УФ-лампы. После того как культуральный раствор, подвергнутый воздействию УФ-света, высевали на среду для чашек, был выделен только мутантный штамм, образовавший колонии. Штамм, демонстрирующий наибольшую продукцию глутатиона, был выделен и назван штаммом СС02-2490, который был депонирован в Корейском центре культур микроорганизмов (KCCM) в соответствии с Будапештским договором под номером доступа KCCM12659P 17 января 2020 г. В результате анализа последовательности оснований гена биосинтеза глутатиона, GSH1, в отношении усиления способности штамма продуцировать глутатион, было подтверждено, что цистеин, который является 86-й аминокислотой белка GSH1, кодируемого геном GSH1, был заменен на аргинин.
Пример 2-2: Эксперимент по замене остатка С86 белка GSH1
Учитывая, что аминокислота по положению 86 белка GSH1 важна для продукции глутатиона на основании результатов Примера 2-1, мутантные штаммы Saccharomyces cerevisiae (S. cerevisiae) CEN.PK2-1D и штамм Saccharomyces cerevisiae (S.cerevisiae) CJ-5 получали для экспрессии вариантов белка, в которых цистеин по положению 86 белка GSH1 был заменен на другую аминокислоту, и было выявлено увеличение продукции глутатиона.
Для получения штаммов, в которых цистеин по положению 86 белка GSH1 Saccharomyces cerevisiae был заменен на аргинин, использовали плазмиды pWAL100 и pWBR100 со ссылкой на публикацию Lee ТН, et al. (J. Microbiol. Biotechnol. (2006), 16(6), 979-982). В частности, полимеразную цепную реакцию (ПЦР) проводили следующим образом, используя в качестве матрицы геномную ДНК штамма CJ-5. Частичную последовательность N-конца белка GSH1, включающую N-концевую фланкирующую последовательность BamHI, кодон инициации ORF GSH1 и последовательность, кодирующую мутацию C86R, получали путем проведения ПЦР с использованием праймеров SEQ ID NO: 34 и 35, а частичную последовательность С-конца белка GSH1, включающую С-концевую фланкирующую последовательность XhoI, кодон терминации ORF GSH1 и последовательность, кодирующую мутацию C86R, получали путем проведения ПЦР с использованием праймеров SEQ ID NO: 36 и 37. Затем, в результате проведения ПЦР с перекрытием с использованием двух последовательностей в качестве матриц с праймерами SEQ ID NO: 34 и 37, был получен фрагмент ORF GSH1, включающий последовательность, кодирующую вариант GSH1, в котором цистеин по положению 86 был заменен на аргинин, и N-концевую BamHI и С-концевую XhoI последовательности рестриктаз. Фрагмент ORF обрабатывали BamHI и XhoI, а затем клонировали в вектор pWAL100, обработанный теми же ферментами, с получением вектора pWAL100-GSHl(C86R).
Кроме того, путем проведения ПЦР с использованием геномной ДНК штамма CJ-5 в качестве матрицы и праймеров SEQ ID NO: 38 и 39 был получен фрагмент 500 п.о. (пар оснований) ниже кодона терминации ORF GSH1, включающий N-концевую SpeI и С-концевую NcoI последовательности рестриктаз, и обработан рестриктазами SpeI и NcoI. Затем полученный продукт клонировали в pWBR100, обработанный теми же ферментами, с получением вектора pWBR100-GSHl.
Перед получением окончательного фрагмента ДНК для введения в дрожжи ПЦР-продукты, включающие последовательность, кодирующую мутацию аргинина, и часть KlURA3, получали с использованием вектора pWAL100-GSH1 (C86R), полученного как описано выше, в качестве матрицы и праймеров SEQ ID NOS: 34 и 40, а ПЦР-продукты, включающие часть KlURA3 и 500 п.о. ниже кодона терминации GSH1, получали с использованием вектора pWBR100-GSH1 в качестве матрицы и праймеров SEQ ID NO: 41 и 39. S. cerevisiae CEN.PK2-1D и S. cerevisiae CJ-5 трансформировали этими ПЦР-продуктами в одинаковом молярном соотношении. ПЦР проводили путем денатурации при 95°С в течение 5 минут, отжига при 53°С в течение 1 минуты и полимеризации в течение 1 минуты на килограмм при 72°С, и трансформацию дрожжей проводили по литий-ацетатному методу, модифицированному из метода, раскрытого в публикации Geitz (Nucleic Acid Research, 20(6), 1425). В частности, клетки дрожжей с О.Д. от 0,7 до 1,2 дважды промывали буфером ацетата лития/ТЕ и смешивали с ПЦР-продуктами и одноцепочечной ДНК (Sigma D-7656). Смесь культивировали в условиях статической культуры в буфере ацетат лития/ТЕ/40% ПЭГ при 30°С в течение 30 минут и при 42°С в течение 15 минут. Затем клетки культивировали в чашке с SC (2% глюкозы) с агаром, не содержащим урацил, до тех пор, пока не становились видны колонии, с получением штамма, в который были введены последовательность, кодирующая мутацию C86R GSH1 и ген KlURA3. Затем для удаления KlURA3 штаммы культивировали в течение ночи в 2 мл YPD, разбавляли в соотношении 1/100, высевали на чашку с агаром SC (2% глюкозы), содержащую 0,1% 5-FOA, с получением мутантного штамма S. cerevisiae CEN PK2-1D GSH1 C86R и мутантного штамма S. cerevisiae CJ-5 GSH1 C86R, из которого был удален урациловый маркер. Штаммы, способные экспрессировать варианты GSH1, в которых цистеин был заменен на 18 типов аминокислот, отличных от аргинина, были получены таким же образом, за исключением пары праймеров SEQ ID NO: 35 и 36, в которой последовательность, кодирующая 86-й аргинин, была заменена на последовательность, кодирующую другую аминокислоту.
После культивирования штаммов, полученных как описано выше, в течение 26 часов измеряли концентрацию продуцированного глутатиона (GSH), как перечислено в Таблицах 2 и 3.
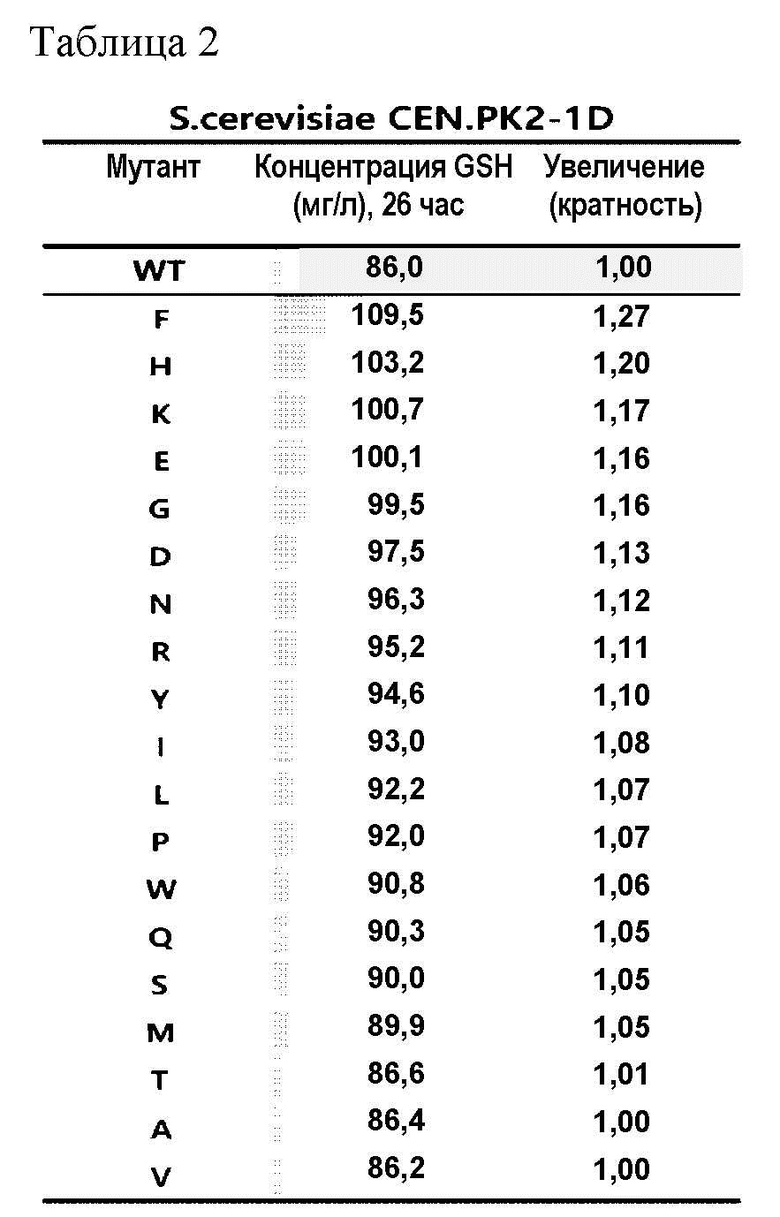
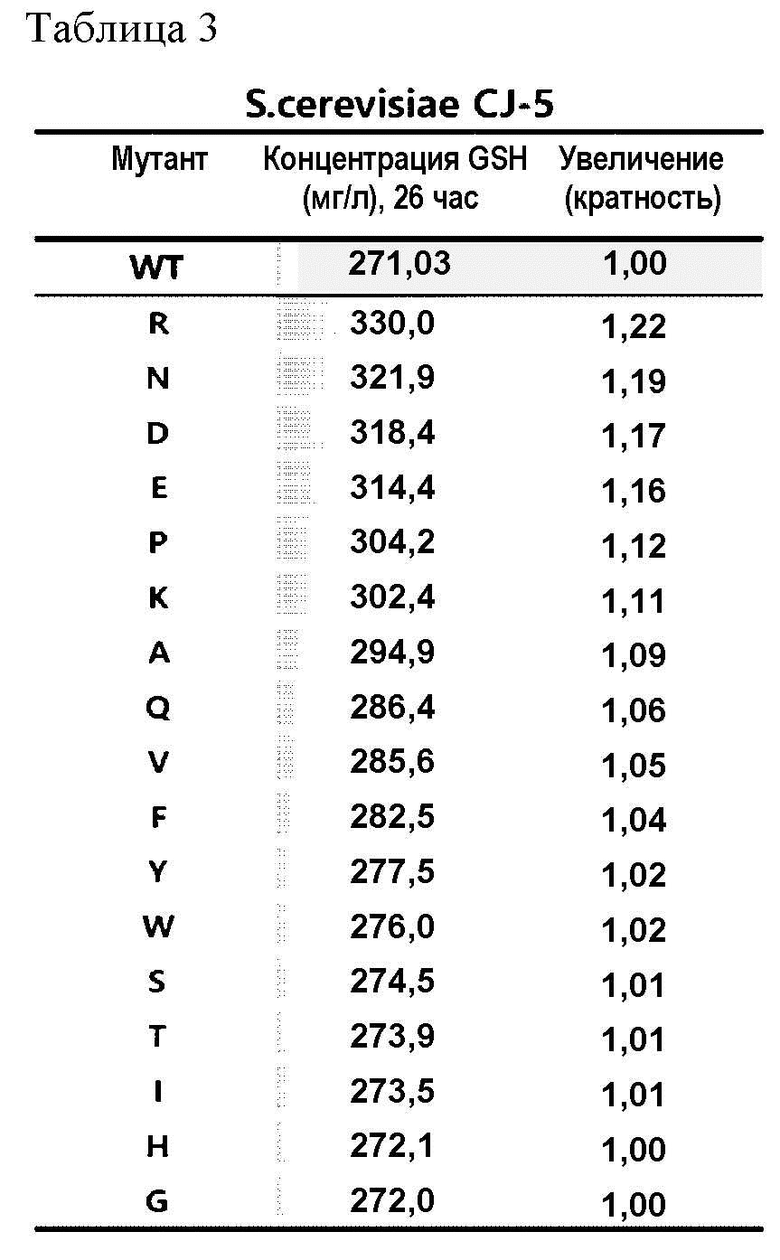
В результате этого эксперимента было подтверждено, что способность продуцировать глутатион, полученная заменой цистеина по положению 86 белка GSH1 на другую аминокислоту, увеличилась вплоть до 27% по сравнению со способностью продуцировать глутатион, полученной белком GSH1 дикого типа.
Пример 2-3: Индукция дополнительной мутации для усиления способности продуцировать глутатион и идентификация модифицированной последовательности
Мутацию индуцировали следующим образом для дальнейшего усиления способности штамма СС02-2490 продуцировать глутатион.
Штамм СС02-2490 культивировали в твердой среде и инокулировали в бульон для получения культурального раствора и этот культуральный раствор подвергали воздействию УФ-света с использованием УФ-лампы. После того как культуральный раствор, подвергнутый воздействию УФ-света, высевали на среду для чашек, был выделен только мутантный штамм, сформировавший колонии, и были проанализированы последовательности GS1-кодирующей области и ее вышерасположенной области в штамме с наиболее усиленной способностью продуцировать глутатион.
В результате было подтверждено, что модификация происходит по положениям -250 (С→Т), -252 (G^A), -398 (А→Т), -399 (А→С), -407 (Т→С) и -409 (Т→С) выше последовательности ORF GSH1. Штамм получил название штамм СС02-2544 и был депонирован в Корейском центре культур микроорганизмов (KCCM) в соответствии с Будапештским договором под номером доступа KCCM12674P 20 февраля 2020 г.
Пример 2-4: Идентификация способности продуцировать глутатион в соответствии с мутацией в промоторе GSH1
Для сравнения способности продуцировать глутатион в присутствии или в отсутствие мутации в данном промоторе измеряли способность продуцировать глутатион для штамма СС02-2490, полученного в Примере 2-1, и штамма СС02-2544, полученного в Примере 2-3. Способность продуцировать глутатион для штамма CEN.PK1-D дикого типа и штамма CJ-5 измеряли в качестве контроля, как показано в Таблице 4 ниже.
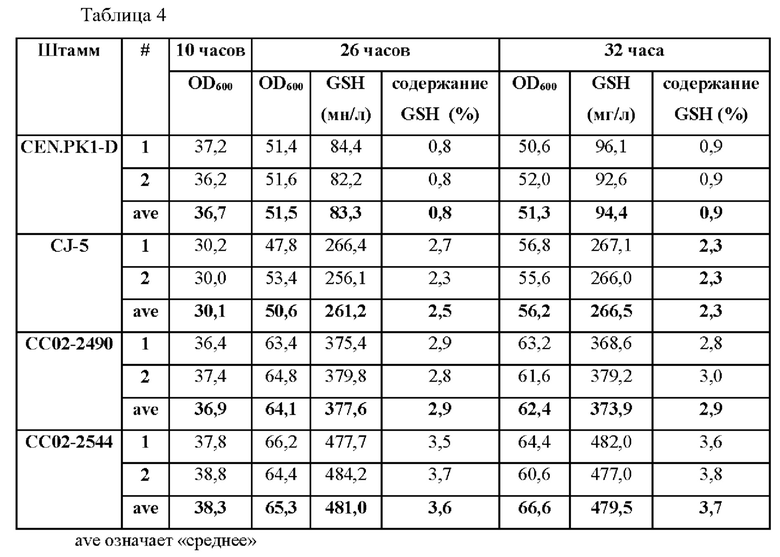
В результате эксперимента было подтверждено, что продукция глутатиона у штамма СС02-2544 значительно увеличилась на 508% по сравнению со штаммом CEN.PK1-D дикого типа.
Кроме того, продукция глутатиона штаммом СС02-2544 увеличилась на 128% и более по сравнению с родительским штаммом СС02-2490.
В результате было подтверждено, что мутация промоторной последовательности гена GSH1 может приводить к увеличению продукции глутатиона.
Пример 3: Эксперимент по введению мутации в промотор GSH1 штамма
Для того чтобы идентифицировать влияние мутации в промоторе на штаммы, мутацию промотора, подтвержденную в Примере 2-3, вводили в штамм дикого типа, штамм CJ-5 и штамм СС02-2490, полученные в Примере 2-1, и оценивали их способности продуцировать глутатион.
Сначала, в результате выравнивания последовательностей для введения мутации и идентификации последовательности выше ORF GSH1 (SEQ ID NO: 2) штамма CJ-5 было подтверждено, что штамм CJ-5 имеет ту же последовательность промотора GSH1, что и штаммы Saccharomyces cerevisiae CEN KSD-Yc, YJM1450, YJM1401 и YJM1307.
Однако, при сравнении с последовательностью выше ORF GSH1 (SEQ ID NO: 1) штамма дикого типа (Saccharomyces cerevisiae CEN.PK1-D) было подтверждено, что по положению 74 промотора штамма CJ-5 имела место вставка аденина.
На основании этого, в последовательности выше ORF GSH1 Saccharomyces cerevisiae CEN.PK1-D положения, соответствующие положениям -250, -252, -398, -399, 407 и 409 последовательности выше ORF GSH1 штамма CJ-5 определили как положения -249, -251, -397, -398, -406 и -408 согласно выравниванию последовательностей.
Затем, вышеописанные шесть типов мутаций вводили в области выше ORF GSH1 каждого из штаммов Saccharomyces cerevisiae CEN.PK1-D, CJ-5 и CC02-2490.
Более конкретно, для получения штаммов, в которых вышеописанные шесть типов мутаций были введены в область выше ORF GSH1, использовали плазмиды pWAL100 и pWBR100 со ссылкой на публикацию Lee ТН, et al. (J. Microbiol. Biotechnol. (2006), 16(6), 979-982).
После синтеза гена фрагмента, включающего область выше ORF GSH1, включающую шесть типов мутации и область ORF GSH1, проводили ПЦР с использованием праймеров SEQ ID NO: 34 и 37, с тем чтобы получить фрагмент области выше ORF GSH1, включающий N-концевую последовательность рестриктазы BamHI, С-концевую последовательность рестриктазы XhoI и шесть типов мутаций. Затем, pWAL100 и этот фрагмент обрабатывали BamHI и XhoI с последующим лигированием для получения плазмиды.
Кроме того, путем проведения ПЦР с использованием геномной ДНК штамма CJ-5 в качестве матрицы и праймеров SEQ ID NO: 38 и 39 был получен фрагмент 500 п. о. ниже кодона терминации ORF GSH1, включающий N-концевую SpeI и С-концевую NcoI последовательности рестриктаз, и обработан рестриктазами SpeI и NcoI. Затем полученный продукт лигировали в pWBR100, обработанный теми же ферментами, для получения плазмиды.
Перед получением окончательного фрагмента ДНК для введения в дрожжи ПЦР-продукты, включающие область выше ORF GSH1, включающую шесть типов мутации, область ORF GSH1 и часть KlURA3, получали с использованием плазмиды pWAL, полученной как описано выше, в качестве матрицы и праймеров SEQ ID NO: 34 и 40, а ПЦР-продукты, включающие часть KlURA3 и 500 п.о. ниже кодона терминации ORF GSH1, получали с использованием плазмиды pWBR в качестве матрицы и праймеров SEQ ID NO: 41 и 39. Штаммы S. cerevisiae CEN.PK2-1D и S. cerevisiae CJ-5 и CC02-02490 трансформировали этими ПЦР-продуктами в одинаковом молярном соотношении. ПЦР проводили путем денатурации при 95°С в течение 5 минут, отжига при 53°С в течение 1 минуты и полимеризации в течение 1 минуты на килограмм при 72°С и трансформацию дрожжей проводили по литий-ацетатному методу, модифицированному из метода, раскрытого в публикации Geitz (Nucleic Acid Research, 20(6), 1425). В частности, клетки дрожжей с О.Д. от 0,7 до 1,2 дважды промывали буфером ацетата лития/ТЕ дважды. Эту ДНК и одноцепочечную ДНК (Sigma D-7656) смешивали и смесь культивировали в условиях статической культуры в буфере ацетат лития/ТЕ/40% ПЭГ при 30°С в течение 30 минут и при 42°С в течение 15 минут. Затем клетки культивировали в чашке с SC (2% глюкозы) с агаром, не содержащим урацил, до тех пор, пока не становились видны колонии, с получением штаммов, в которые были введены указанные шесть типов мутаций и ген KlURA3 был введен в область выше ORF GSH1. Затем для удаления KlURA3 штаммы культивировали в течение ночи в 2 мл YPD, разбавляли в соотношении 1/100, высевали на чашку с агаром SC (2% глюкозы), содержащую 0,1% 5-FOA, с получением имеющих происхождение из S. cerevisiae CEN.PK2-1D мутантных штаммов, имеющих шесть типов мутаций в области выше ORF GSH1, и имеющих происхождение из S. cerevisiae CJ-5 и СС02-2490 мутантных штаммов, имеющих шесть типов мутаций в области выше ORF GSH1.
Продукция GSH указанных штаммов показана в Таблице 5 ниже.
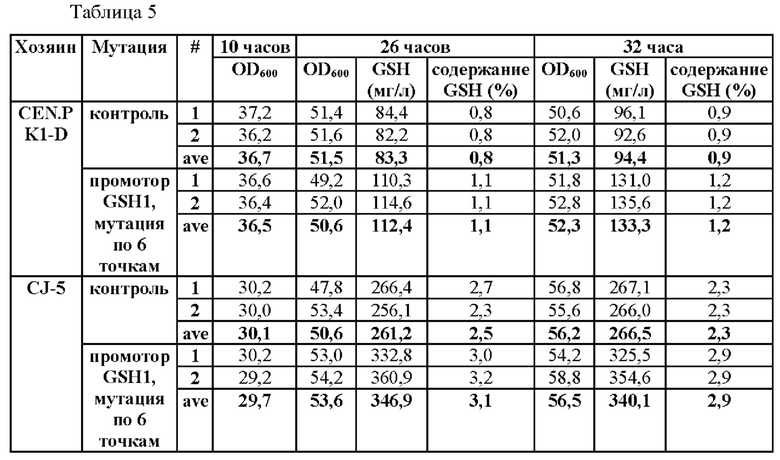
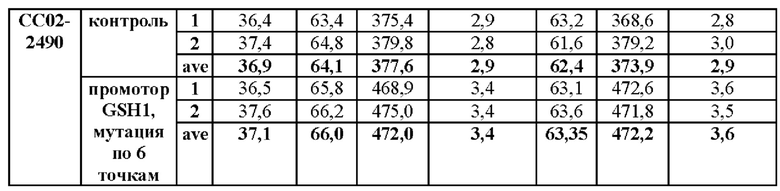
В результате эксперимента было подтверждено увеличение продукции глутатиона вплоть до 141% у того же штамма в соответствии с мутацией в промоторе.
В результате было подтверждено, что мутация промоторной последовательности гена GSH1 может приводить к увеличению продукции глутатиона.
Пример 4: Эксперимент по введению мутации в промотор GSH1 (1)
На основании результатов, подтвержденных в вышеописанных примерах, был проведен эксперимент для определения того, усиливается ли способность продуцировать глутатион путем введения мутации только по некоторым из указанных положений.
В частности, мутация была введена по двум точкам по положениям -250 и -252 выше ORF GSH1 (-250 (С→Т) и -252 (G→А)) и по четырем точкам по положениям -398, -399, -407 и -409 (-398 (А→Т), -399 (А→С), -407 (Т→С) и -409 (Т→С)) в CEN.PK1-D, CJ-5 и СС02-2490, соответственно. (В случае CEN.PK1-D мутация была введена по соответствующим им положениям -249, -251, -397, -398, -406 и -408, которые будут применены при последующем описании). Штаммы получали таким же образом, как в Примере 3, за исключением того, что синтезировали ген фрагмента, включающего область выше ORF GSH1, включающую мутацию по двум точкам или четырем точкам, и область ORF GSH1, а затем проводили ПЦР с использованием праймеров SEQ ID NO: 34 и 37, с тем чтобы получить фрагмент области выше ORF GSH1, включающей N-концевую последовательность рестриктазы BamHI, С-концевую последовательность рестриктазы а XhoI и мутацию по двум точкам или четырем точкам.
Результаты данного эксперимента показаны в Таблицах 6-8.
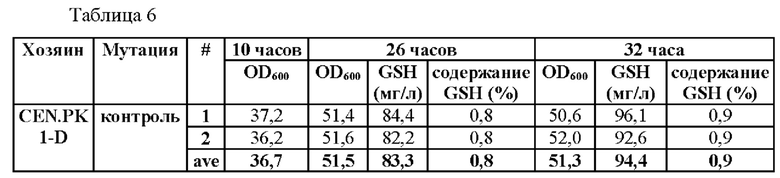
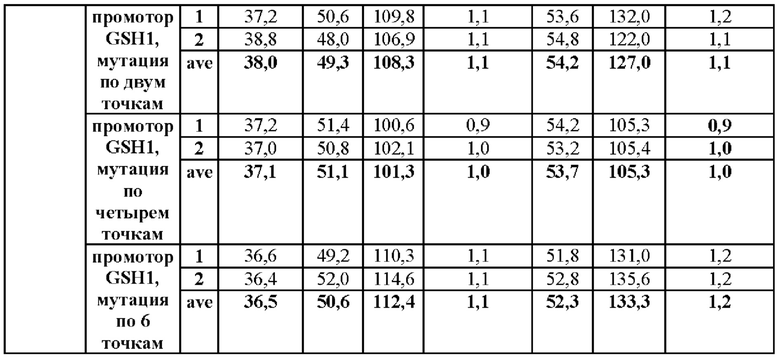
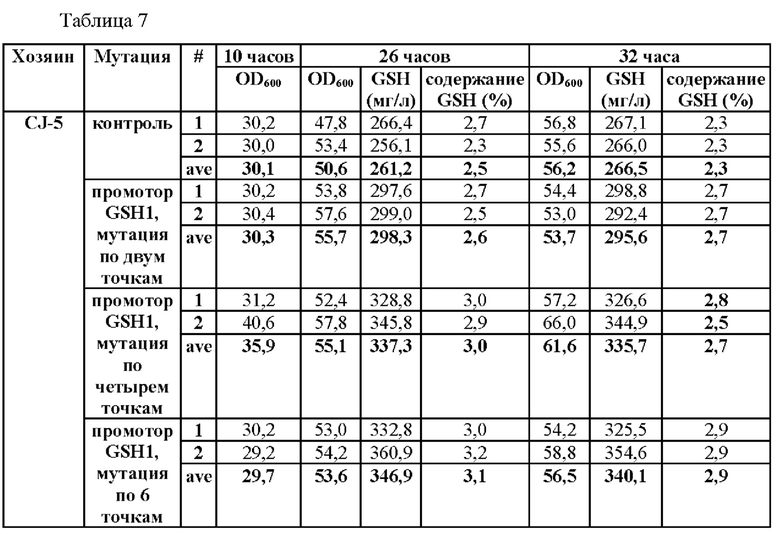
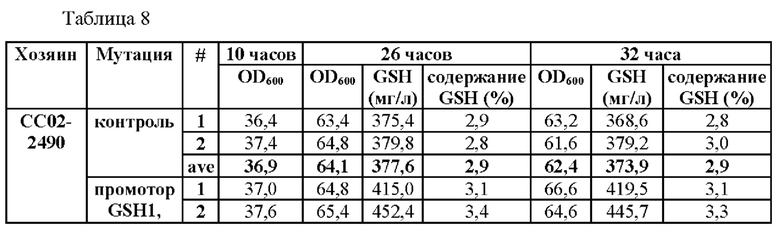
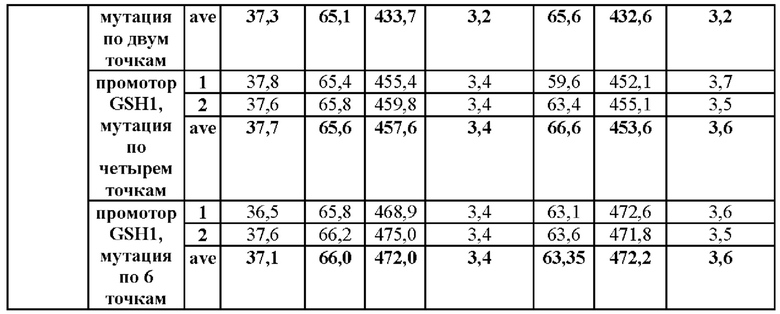
В результате данного эксперимента было подтверждено, что способность продуцировать глутатион усиливалась при введении мутаций по двум точкам (-250 (С→Т) и -252 (G→А)) или по четырем точкам (-398 (А→Т)), -399 (А→С), -407 (Т→С) и -409 (Т→С)) по сравнению со штаммами без мутации в промоторной области, а продукция глутатиона увеличивалась вплоть до 135%.
На основании этого можно подтвердить, что способность продуцировать глутатион значительно усиливается за счет мутации, введенной только в некоторые положения промотора, разработанного в настоящем изобретении, а также во все шесть первоначально подтвержденных положений.
Пример 5: Эксперимент по введению мутации в промотор GSH1 (2)
На основании результатов, подтвержденных в вышеописанных примерах, был проведен эксперимент для определения того, усиливается ли способность продуцировать глутатион путем введения мутации только в некоторые из положений. В частности, мутацию вводили по одной точке по положению -250 (С→Т) (мутация по одной точке (1)) или по положению -252 (G→А) (мутация по одной точке (2)) и по двум точкам по обоим положениям в штаммах CEN.PK1-D и CJ-5, и их способности продуцировать глутатион измеряли, а результаты показаны в Таблицах 9 и 10.
Для этого штаммы получали таким же образом, как в Примере 3, за исключением того, что синтезировали фрагмент гена, включающий область выше ORF GSH1, включающую мутацию по одной точке или двум точкам, и область ORF GSH1, и затем проводили ПЦР с использованием праймеров SEQ ID NO: 34 и 37, чтобы таким образом получить фрагмент области выше ORF GSH1, включающий N-концевую последовательность рестриктазы BamHI, С-концевую последовательность рестриктазы XhoI и мутацию по одной точке или двум точкам.
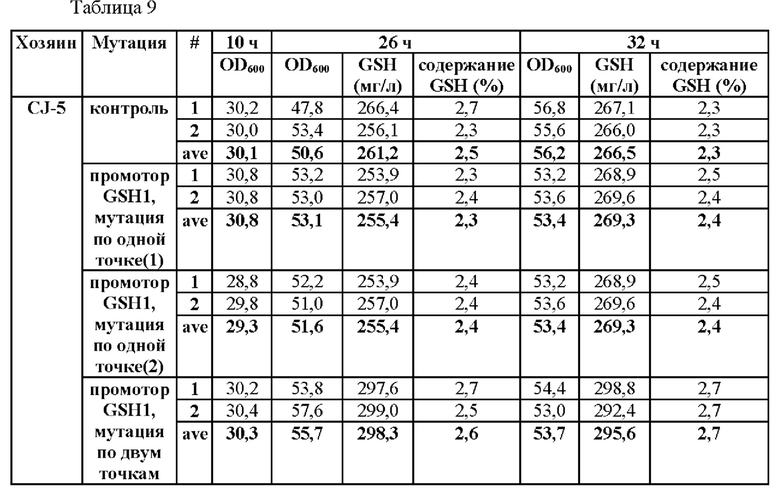
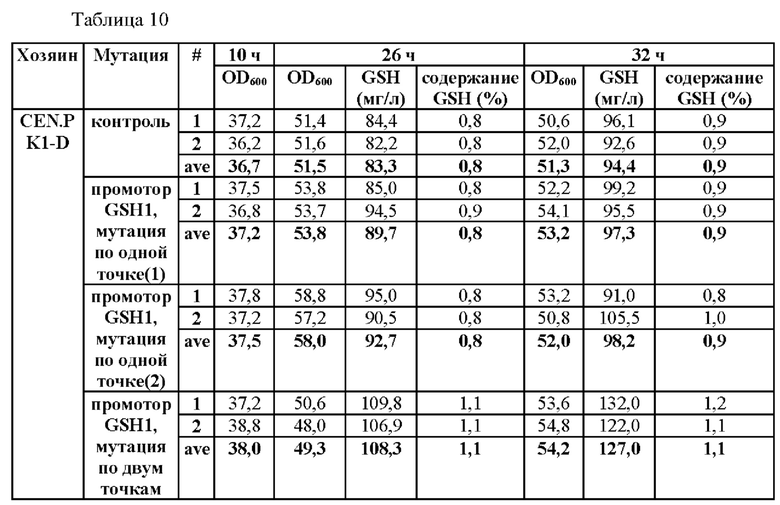
В результате данного эксперимента было подтверждено, что способность продуцировать глутатион усиливалась при введении мутации только в некоторые точки.
На основании этого можно подтвердить, что способность продуцировать глутатион значительно усиливается за счет мутации, введенной только в некоторые положения промотора, разработанного в настоящем изобретении, а также во все шесть первоначально подтвержденных положений.
Пример 6: Эксперимент по введению мутации в промотор GSH1 (3)
На основании результатов, подтвержденных в вышеописанных примерах, был проведен эксперимент для определения того, усиливается ли способность продуцировать глутатион путем введения мутации только в некоторые из указанных положений.
В частности, мутацию вводили по четырем точкам по положениям -398, -399, -407 и -409 (-398 (А→Т), -399 (А→С), -407 (Т→С) и -409 (Т→С)), независимо или в комбинации.
Мутации были следующими:
1) мутация по одной точке (1): GSH1 -398 (А→Т)
2) мутация по одной точке (2): GSH1 -399 (А→С)
3) мутация по одной точке (3): GSH1 -407 (Т→С)
4) мутация по одной точке (4): GSH1 -409 (Т→С)
5) мутация по двум точкам (1): GSH1 -398 (А→Т) и -399 (А→С)
6) мутация по двум точкам (2): GSH1 -398 (А→Т) и -407 (Т→С)
7) мутация по двум точкам (3): GSH1 -398 (А→Т) и -409 (Т→С)
8) мутация по двум точкам (4): GSH1 -399 (А→С) и -407 (Т→С)
9) мутация по двум точкам (5): GSH1 -399 (А→С) и -409 (Т→С)
10) мутация по двум точкам (6): GSH1 -407 (Т→С) и -409 (Т→С)
11) мутация по четырем точкам: GSH1 -398 (А→Т), -399 (А→С), -407 (Т→С) и 409 (Т→С)
Для этого штаммы получали таким же образом, как в Примере 3, за исключением того, что синтезировали фрагмент гена, включающий область выше ORF GSH1, включающую мутацию по одной точке, двум точкам или четырем точкам, и область ORF GSH1, и затем проводили ПЦР с использованием праймеров SEQ ID NO: 34 и 37, с тем чтобы получить фрагмент области выше ORF GSH1, включающий N-концевую последовательность рестриктазы BamHI, С-концевую последовательность рестриктазы XhoI и мутацию по одной точке, двум точкам или четырем точкам.
Определяли способность штаммов продуцировать глутатион, и результаты показаны в Таблицах 11 и 12 ниже.
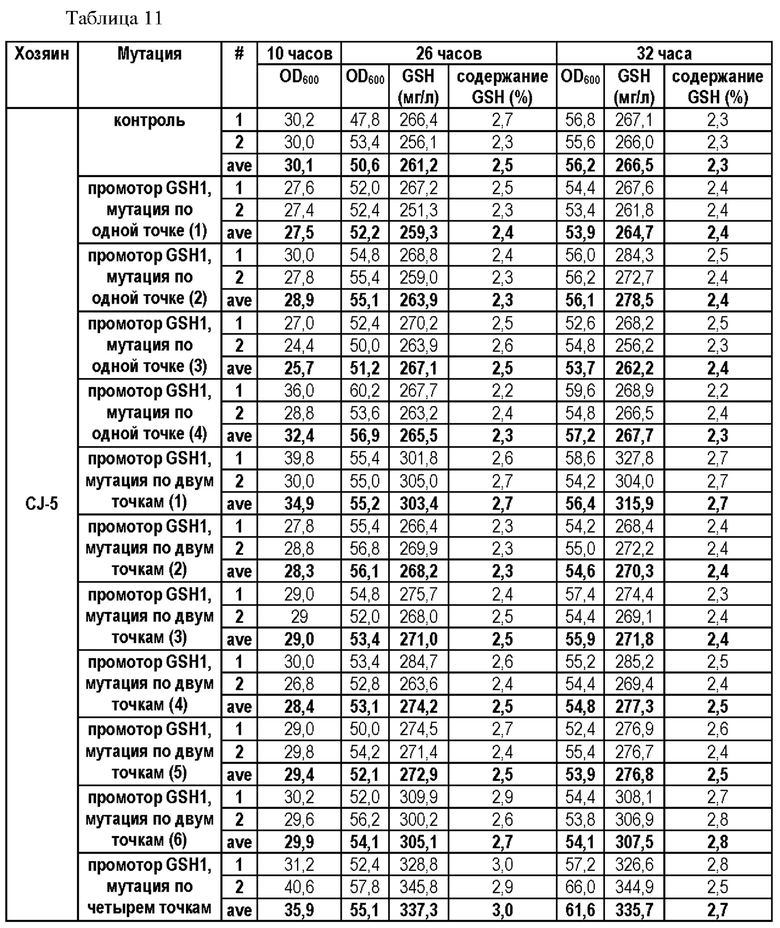
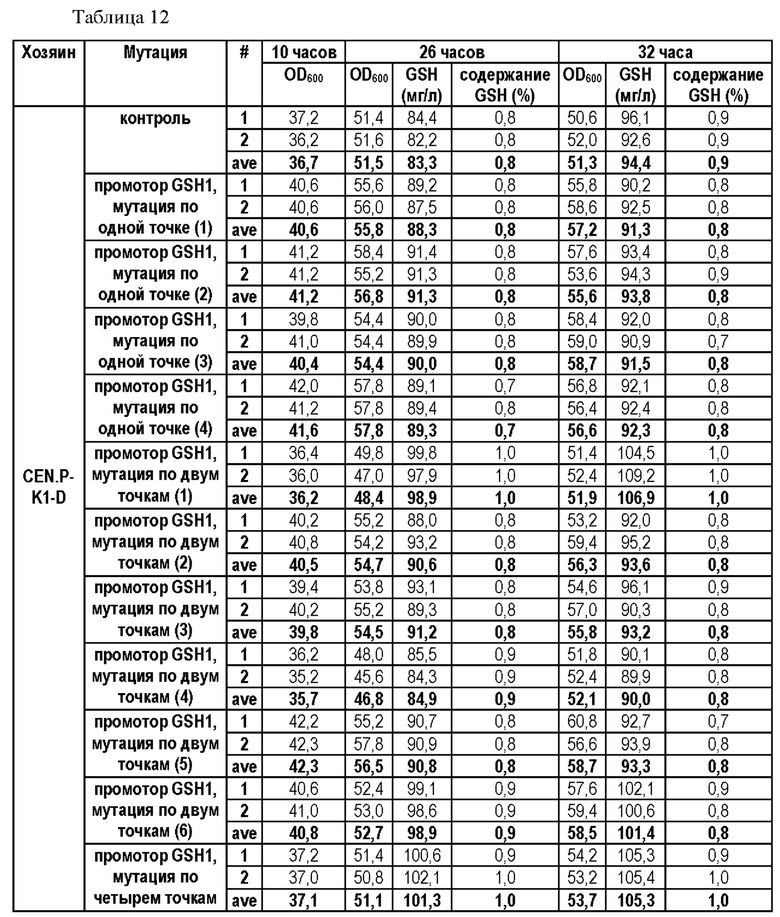
В результате данного эксперимента было подтверждено, что способность продуцировать глутатион усиливалась при введении мутации только в некоторые из четырех точек по сравнению со штаммами без мутации в промоторной области, а также при введении мутации во все четыре точки. В частности, в случаях 5) мутация по двум точкам (1): GSH1 -398 (А→Т) и -399 (А→С) и 10) мутация по двум точкам (6): GSH1 -407 (Т→С) и 409 (Т→С), способность продуцировать глутатион была значительно усилена по сравнению со штаммами без мутации в промоторной области.
На основании этого можно подтвердить, что способность продуцировать глутатион значительно усиливается за счет мутации, введенной только в некоторые положения промотора, разработанного в настоящем изобретении, а также во все шесть первоначально подтвержденных положений.
Вышеприведенное описание настоящего изобретения предоставлено с целью иллюстрации, и специалистам в данной области техники будет понятно, что различные изменения и модификации могут быть сделаны без изменения технической концепции и существенных признаков настоящего изобретения. Таким образом, понятно, что вышеописанные воплощения являются иллюстративными во всех аспектах и не ограничивают настоящее изобретение. Различные воплощения, раскрытые здесь, не предназначены для ограничения, а истинный объем и сущность указаны в следующей формуле изобретения. Настоящее изобретение следует считать ограниченным только признаками прилагаемой формулы изобретения, наряду с полным объемом эквивалентов, на которые распространяется такая формула изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> НОВЫЙ ПРОМОТОР И СПОСОБ ПОЛУЧЕНИЯ ГЛУТАТИОНА С ЕГО ИСПОЛЬЗОВАНИЕМ
<130> OPA21012
<150> KR 10-2020-0041184
<151> 2020-04-03
<160> 41
<170> KoPatentIn 3.0
<210> 1
<211> 499
<212> DNA
<213> Unknown
<220>
<223> Saccharomyces cerevisiae CEN.PK-1D
<400> 1
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 2
<211> 500
<212> DNA
<213> Unknown
<220>
<223> Saccharomyces cerevisiae CJ-5
<400> 2
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 3
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-92 94 102 103 249 251
<400> 3
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccctctgcc cctcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattag tcatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 4
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-92 94 102 103
<400> 4
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccctctgcc cctcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 5
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-249 251
<400> 5
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattag tcatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 6
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-102 103
<400> 6
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc cctcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 7
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-92 102
<400> 7
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccttctgcc ccacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 8
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-92 103
<400> 8
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccttctgcc catcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 9
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-94 102
<400> 9
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcctctgcc ccacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 10
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-94 103
<400> 10
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcctctgcc catcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 11
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-92 94
<400> 11
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccctctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 12
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-249
<400> 12
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattag ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 13
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-251
<400> 13
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg tcatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 14
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-92
<400> 14
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 15
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-94
<400> 15
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcctctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 16
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-102
<400> 16
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc ccacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 17
<211> 499
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CEN.PK-1D промотор-мутация-103
<400> 17
ctcttgaatg gcgacagcct attgccccag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc catcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
gccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcctcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaattga gcagatttag tatagggcta cattgtaggg tggtttagag tatcgaaaat 480
atacatatag aagaataaa 499
<210> 18
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-92 94 102 103 249 251
<400> 18
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccctctgcc cctcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattag tcatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 19
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-92 94 102 103
<400> 19
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccctctgcc cctcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 20
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-249 251
<400> 20
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattag tcatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 21
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-102 103
<400> 21
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc cctcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 22
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-92 102
<400> 22
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccttctgcc ccacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 23
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-92 103
<400> 23
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccttctgcc catcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 24
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-94 102
<400> 24
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcctctgcc ccacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 25
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-94 103
<400> 25
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcctctgcc catcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 26
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-92 94
<400> 26
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccctctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 27
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-249
<400> 27
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattag ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 28
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-251
<400> 28
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg tcatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 29
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-92
<400> 29
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt cccttctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 30
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-94
<400> 30
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcctctgcc caacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 31
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-102
<400> 31
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc ccacgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 32
<211> 500
<212> DNA
<213> Artificial Sequence
<220>
<223> S. cerevisiae CJ-5 промотор-мутация-103
<400> 32
ctcttgaatg gcgacagcct attgcctcag tgttccctca acaaccttgg tagttggagc 60
gcaattagcg tatcctgtac catactaatt ctcttctgcc catcgacggc tgccattagt 120
cagcatggcg cgcacgtgac tacaactgtg gctggaaacc ttttcgtcct ccccggtttt 180
tcagtgagcc gactctacta caatgctttt tcatttttca ctcagaaaaa cctgcaattt 240
tccaaattgg ccatgctctg tgcctccctt gacaaaggac atcttccctg tttataaacg 300
gcggcttacc aaaagttgaa gcttgttctt gcttcttatg agtggagcaa tcgattatat 360
tgaatcgttg tgctggagta gttggatctt tccacgtggt ctcgagtcac ttgtagaagc 420
tgaaaaattg agcaggttta gtatagggct acattgtagg gtggtttaga gtatcgaaaa 480
tatacatata gaagaataaa 500
<210> 33
<211> 678
<212> PRT
<213> Saccharomyces cerevisiae
<400> 33
Met Gly Leu Leu Ala Leu Gly Thr Pro Leu Gln Trp Phe Glu Ser Arg
1 5 10 15
Thr Tyr Asn Glu His Ile Arg Asp Glu Gly Ile Glu Gln Leu Leu Tyr
20 25 30
Ile Phe Gln Ala Ala Gly Lys Arg Asp Asn Asp Pro Leu Phe Trp Gly
35 40 45
Asp Glu Leu Glu Tyr Met Val Val Asp Phe Asp Asp Lys Glu Arg Asn
50 55 60
Ser Met Leu Asp Val Cys His Asp Lys Ile Leu Thr Glu Leu Asn Met
65 70 75 80
Glu Asp Ser Ser Leu Cys Glu Ala Asn Asp Val Ser Phe His Pro Glu
85 90 95
Tyr Gly Arg Tyr Met Leu Glu Ala Thr Pro Ala Ser Pro Tyr Leu Asn
100 105 110
Tyr Val Gly Ser Tyr Val Glu Val Asn Met Gln Lys Arg Arg Ala Ile
115 120 125
Ala Glu Tyr Lys Leu Ser Glu Tyr Ala Arg Gln Asp Ser Lys Asn Asn
130 135 140
Leu His Val Gly Ser Arg Ser Val Pro Leu Thr Leu Thr Val Phe Pro
145 150 155 160
Arg Met Gly Cys Pro Asp Phe Ile Asn Ile Lys Asp Pro Trp Asn His
165 170 175
Lys Asn Ala Ala Ser Arg Ser Leu Phe Leu Pro Asp Glu Val Ile Asn
180 185 190
Arg His Val Arg Phe Pro Asn Leu Thr Ala Ser Ile Arg Thr Arg Arg
195 200 205
Gly Glu Lys Val Cys Met Asn Val Pro Met Tyr Lys Asp Ile Ala Thr
210 215 220
Pro Glu Thr Asp Asp Ser Ile Tyr Asp Arg Asp Trp Phe Leu Pro Glu
225 230 235 240
Asp Lys Glu Ala Lys Leu Ala Ser Lys Pro Gly Phe Ile Tyr Met Asp
245 250 255
Ser Met Gly Phe Gly Met Gly Cys Ser Cys Leu Gln Val Thr Phe Gln
260 265 270
Ala Pro Asn Ile Asn Lys Ala Arg Tyr Leu Tyr Asp Ala Leu Val Asn
275 280 285
Phe Ala Pro Ile Met Leu Ala Phe Ser Ala Ala Ala Pro Ala Phe Lys
290 295 300
Gly Trp Leu Ala Asp Gln Asp Val Arg Trp Asn Val Ile Ser Gly Ala
305 310 315 320
Val Asp Asp Arg Thr Pro Lys Glu Arg Gly Val Ala Pro Leu Leu Pro
325 330 335
Lys Tyr Asn Lys Asn Gly Phe Gly Gly Ile Ala Lys Asp Val Gln Asp
340 345 350
Lys Val Leu Glu Ile Pro Lys Ser Arg Tyr Ser Ser Val Asp Leu Phe
355 360 365
Leu Gly Gly Ser Lys Phe Phe Asn Arg Thr Tyr Asn Asp Thr Asn Val
370 375 380
Pro Ile Asn Glu Lys Val Leu Gly Arg Leu Leu Glu Asn Asp Lys Ala
385 390 395 400
Pro Leu Asp Tyr Asp Leu Ala Lys His Phe Ala His Leu Tyr Ile Arg
405 410 415
Asp Pro Val Ser Thr Phe Glu Glu Leu Leu Asn Gln Asp Asn Lys Thr
420 425 430
Ser Ser Asn His Phe Glu Asn Ile Gln Ser Thr Asn Trp Gln Thr Leu
435 440 445
Arg Phe Lys Pro Pro Thr Gln Gln Ala Thr Pro Asp Lys Lys Asp Ser
450 455 460
Pro Gly Trp Arg Val Glu Phe Arg Pro Phe Glu Val Gln Leu Leu Asp
465 470 475 480
Phe Glu Asn Ala Ala Tyr Ser Val Leu Ile Tyr Leu Ile Val Asp Ser
485 490 495
Ile Leu Thr Phe Ser Asp Asn Ile Asn Ala Tyr Ile His Met Ser Lys
500 505 510
Val Trp Glu Asn Met Lys Ile Ala His His Arg Asp Ala Ile Leu Phe
515 520 525
Glu Lys Phe His Trp Lys Lys Ser Phe Arg Asn Asp Thr Asp Val Glu
530 535 540
Thr Glu Asp Tyr Ser Ile Ser Glu Ile Phe His Asn Pro Glu Asn Gly
545 550 555 560
Ile Phe Pro Gln Phe Val Thr Pro Ile Leu Cys Gln Lys Gly Phe Val
565 570 575
Thr Lys Asp Trp Lys Glu Leu Lys His Ser Ser Lys His Glu Arg Leu
580 585 590
Tyr Tyr Tyr Leu Lys Leu Ile Ser Asp Arg Ala Ser Gly Glu Leu Pro
595 600 605
Thr Thr Ala Lys Phe Phe Arg Asn Phe Val Leu Gln His Pro Asp Tyr
610 615 620
Lys His Asp Ser Lys Ile Ser Lys Ser Ile Asn Tyr Asp Leu Leu Ser
625 630 635 640
Thr Cys Asp Arg Leu Thr His Leu Asp Asp Ser Lys Gly Glu Leu Thr
645 650 655
Ser Phe Leu Gly Ala Glu Ile Ala Glu Tyr Val Lys Lys Asn Lys Pro
660 665 670
Ser Ile Glu Ser Lys Cys
675
<210> 34
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> F_BamHI_GSH1
<400> 34
ggtaggatcc atgggactct tagctttggg cac 33
<210> 35
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> R_GSH1_C86R
<400> 35
ttagcctccc taagggacga atcct 25
<210> 36
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> F_GSH1_C86R
<400> 36
cgtcccttag ggaggctaac gatgt 25
<210> 37
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> R_XhoI_GSH1
<400> 37
atgactcgag ttaacatttg ctttctattg aaggc 35
<210> 38
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> F_SpeI_GSH1_DW
<400> 38
tagaactagt actcctttta tttcggttgt gaa 33
<210> 39
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> R_NcoI_GSH1_DW
<400> 39
gctgccatgg gaatagtgtg aaccgataac tgtgt 35
<210> 40
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> R_AL киллер
<400> 40
gagcaatgaa cccaataacg aaatctt 27
<210> 41
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> F_BR киллер
<400> 41
cttgacgttc gttcgactga tgag 24
<---
Изобретение относится к биотехнологии. Предложены полинуклеотид, обладающий промоторной активностью, композиция для экспрессии гена, содержащая указанный полинуклеотид, вектор экспрессии, содержащий указанный полинуклеотид, и применение указанного полинуклеотида в качестве промотора. Также предложены микроорганизм, принадлежащий к роду Saccharomyces sp., для получения целевого белка, содержащий указанный полинуклеотид, и способ получения глутатиона с использованием микроорганизма. Изобретение обеспечивает получение глутатиона с высоким выходом. 6 н. и 7 з.п. ф-лы, 12 табл., 6 пр.
1. Полинуклеотид, обладающий промоторной активностью, где по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид,
где замена нуклеотида по положению 92 представляет собой замену на цитозин (C);
замена нуклеотида по положению 94 представляет собой замену на цитозин (C);
замена нуклеотида по положению 102 представляет собой замену на цитозин (C);
замена нуклеотида по положению 103 представляет собой замену на тимин (T);
замена нуклеотида по положению 249 представляет собой замену на аденин (A);
и
замена нуклеотида по положению 251 представляет собой замену на тимин (T).
2. Полинуклеотид по п. 1, где нуклеотиды по положениям 249 и 251 полинуклеотида, обладающего промоторной активностью, заменены на другие нуклеотиды.
3. Полинуклеотид по п. 1, где нуклеотиды по положениям 92, 94, 102 и 103 полинуклеотида, обладающего промоторной активностью, заменены на другие нуклеотиды.
4. Полинуклеотид по п. 1, где нуклеотиды по положениям 92, 94, 102, 103, 249 и 251 полинуклеотида, обладающего промоторной активностью, заменены на другие нуклеотиды.
5. Полинуклеотид по п. 1, где полинуклеотид, обладающий промоторной активностью, состоит из одной полинуклеотидной последовательности, выбранной из SEQ ID NO: с 3 по 32.
6. Композиция для экспрессии гена, содержащая полинуклеотид по любому из пп. 1-5 и ген, кодирующий целевой белок.
7. Вектор экспрессии, содержащий полинуклеотид по любому из пп. 1-5 и ген, кодирующий целевой белок.
8. Вектор по п. 7, где целевой белок имеет активность глутамат-цистеинлигазы.
9. Микроорганизм, принадлежащий к роду Saccharomyces sp., для получения целевого белка, содержащий полинуклеотид по любому из пп. 1-5, полинуклеотид, содержащий указанный полинуклеотид и ген, кодирующий целевой белок, или вектор экспрессии, включающий указанный полинуклеотид и ген, кодирующий целевой белок.
10. Микроорганизм по п. 9, где целевой белок представляет собой полипептид, имеющий активность глутамат-цистеинлигазы.
11. Способ получения глутатиона, включающий культивирование микроорганизма по п. 9 в культуральной среде.
12. Способ по п. 11, дополнительно включающий выделение глутатиона из по меньшей мере одного выбранного из культивируемого микроорганизма, высушенного продукта микроорганизма, экстракта микроорганизма, культурального продукта микроорганизма и лизата микроорганизма.
13. Применение полинуклеотида, где по меньшей мере один нуклеотид, выбранный из группы, состоящей из нуклеотидов по положениям 92, 94, 102, 103, 249 и 251 полинуклеотидной последовательности SEQ ID NO: 1 или 2, заменен на другой нуклеотид, в качестве промотора,
где замена нуклеотида по положению 92 представляет собой замену на цитозин (C);
замена нуклеотида по положению 94 представляет собой замену на цитозин (C);
замена нуклеотида по положению 102 представляет собой замену на цитозин (C);
замена нуклеотида по положению 103 представляет собой замену на тимин (T);
замена нуклеотида по положению 249 представляет собой замену на аденин (A);
и
замена нуклеотида по положению 251 представляет собой замену на тимин (T).
| BIOT-PELLETIER D | |||
| ET AL | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| ПОРШНЕВОЙ КОМПРЕССОР СО ЗВЕЗДООБРАЗНО РАСПОЛОЖЕННЫМИ ЦИЛИНДРАМИ | 1935 |
|
SU48991A1 |

