[Область техники]
Настоящая заявка относится к варианту супероксиддисмутазы 1 и способу получения глутатиона или его производных с применением указанного варианта. [Уровень техники]
Глутатион (GSH) представляет собой сераорганическое соединение, наиболее часто встречающееся в клетках, и он существует в форме трипептида, в котором конъюгированы три аминокислоты (то есть глицин, глутамат и цистеин).
В организме глутатион существует в двух формах: восстановленный глутатион (GSH) и окисленный глутатион (GSSG). Восстановленный глутатион в общем случае присутствует в относительно высоком процентном отношении. Он в основном распределен в клетках печени и кожи человеческого организма и играет важную роль в антиоксидантных функциях, таких как разложение и удаление свободных кислородных радикалов, детоксикационных функциях, таких как удаление чужеродных соединений (то есть токсичных веществ), и осветляющих функциях, которые ингибируют выработку меланина, и т.д.
Благодаря указанным разнообразным функциям глутатион привлек большое внимание в качестве вещества в различных областях, таких как фармацевтика, оздоровительные функциональные пищевые продукты, косметика и т.д., а также его применяют для получения ароматизирующих ингредиентов, пищевых продуктов и кормовых добавок. Известно, что глутатион обладает значительным эффектом усиления вкусоароматических свойств сырья и сохранения насыщенных вкусоароматических свойств и может быть применен отдельно в виде усилителя вкусоароматических свойств кокуми (kokumi) или в комбинации с другими веществами. Как правило, вещество кокуми обладает более насыщенными вкусоароматическими свойствами, чем вещество умами iumami), такое как нуклеиновая кислота и глутамат натрия (MSG), и известно, что оно образуется посредством разложения и старения белков. Кроме того, глутатион также может быть превращен в другие гамма-глутамилпептиды (Sofyanovich, О. A. et al., (2019) Multiple pathways for the formation of the γ-glutamyl peptides γ-glutamyl-valine and γ-glutamyl-valyl glycine in Saccharomyces cerevisiae. PLoS ONE 14 (5): e0216622). гамма-Глутамилпептид представляет собой низкомолекулярное соединение, содержащее гамма-карбоксильную группу глутаминовой кислоты на N-конце молекулы, и известно, что его широко применяют в качестве усилителя вкусоароматических свойств кокуми.
Однако, несмотря на возросший спрос на глутатион и его производные, которые могут быть применены в различных областях, как описано выше, процесс синтеза ферментов до сих пор не коммерциализирован из-за высоких производственных затрат, и, соответственно, рынок существенно не активизировался из-за значительных затрат, необходимых для промышленного производства глутатиона и его производных.
[Описание]
[Техническая проблема]
Для решения указанных проблем в настоящей заявке предложены вариант супероксиддисмутазы 1 и способ получения глутатиона или его производных с применением указанного варианта. [Техническое решение]
Одной из задач настоящей заявки является обеспечение варианта супероксиддисмутазы 1, где аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту.
Другой задачей настоящей заявки является обеспечение полинуклеотида, кодирующего указанный вариант, и вектора, содержащего указанный полинуклеотид.
Еще одной задачей настоящей заявки является обеспечение микроорганизма, содержащего любое одно или более из указанного варианта и полинуклеотида, кодирующего указанный вариант.
Еще одной задачей настоящей заявки является обеспечение способа получения глутатиона или его производных, включающего культивирование указанного микроорганизма.
[Предпочтительные эффекты]
Новый вариант супероксиддисмутазы 1 согласно настоящей заявке может быть эффективно применен для получения глутатиона или его производных в больших количествах.
[Краткое описание графических материалов]
На Фиг. 1 и 2 представлены результаты подтверждения уровня получения глутатиона посредством введения указанного варианта супероксиддисмутазы 1 в штамм Saccharomyces cerevisiae.
[Подробное описание предпочтительных воплощений]
Настоящая заявка будет подробно описана следующим образом. При этом каждое описание и воплощение, раскрытое в настоящем документе, может быть применено к другим описаниям и воплощениям, соответственно. То есть, все комбинации различных элементов, раскрытых в настоящем документе, входят в объем настоящей заявки. Кроме того, объем настоящей заявки не ограничен конкретным описанием, приведенным ниже.
Кроме того, средние специалисты в данной области техники могут распознать или подтвердить с применением только традиционных экспериментов многие эквиваленты конкретных аспектов настоящего изобретения, описанного в настоящем документе. Кроме того, также предполагается, что указанные эквиваленты включены в настоящую заявку.
Кроме того, на протяжении настоящего описания был указан посредством ссылок и упомянут ряд научных статей и патентных документов. Содержание упомянутых научных статей и патентных документов полностью включено в настоящее описание посредством ссылок, и уровень области техники, к которой относится настоящее изобретение, и содержание настоящего изобретения будут описаны более ясно.
В одном из аспектов настоящей заявки предложен вариант супероксиддисмутазы 1, содержащий аминокислотную замену в белке, обладающем активностью супероксиддисмутазы 1 (SOD1), где указанная замена включает замену аминокислоты, соответствующей положению 37 от N-конца SEQ ID NO: 1, на другую аминокислоту.
Указанный вариант может представлять собой вариант белка, где аминокислота, соответствующая положению 37 от N-конца в аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту.
«Супероксиддисмутаза 1» согласно настоящей заявке представляет собой фермент, также называемый «SOD1» или «[Cu-Zn] супероксиддисмутазой». Известно, что указанная супероксиддисмутаза катализирует реакцию по следующей схеме:
2О2-+2Н+→H2O2+O2
В контексте настоящего описания аминокислотная последовательность супероксиддисмутазы 1 представляет собой аминокислотную последовательность, кодируемую геном sod1, и может быть названа «белком SOD1». Аминокислота, входящая в состав супероксиддисмутазы 1 согласно настоящей заявке, может быть получена из известной базы данных GenBank от Национального центра биотехнологической информации США (NCBI). Указанная супероксиддисмутаза 1 может представлять собой белок, содержащий аминокислотную последовательность SEQ ID NO: 1, но не ограничивается им. В другом примере указанная супероксиддисмутаза 1 может быть получена из Saccharomyces cerevisiae. В еще одном примере она может представлять собой белок, где аминокислота, соответствующая положению 37 в аминокислотной последовательности SEQ ID NO: 1, представляет собой глицин, но не ограничивается им, и любая последовательность, обладающая такой же активностью супероксиддисмутазы 1, как указанная выше аминокислотная последовательность, может быть включена без ограничения.
В конкретном примере супероксиддисмутаза 1 согласно настоящей заявке может представлять собой белок, содержащий аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, имеющую гомологию или идентичность с ней, составляющую 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более. Кроме того, очевидно, что любой белок, содержащий аминокислотную последовательность, где часть указанной аминокислотной последовательности удалена, модифицирована, заменена или добавлена, также может входить в объем белка, являющегося целью модификации согласно настоящей заявке при условии, что указанная аминокислотная последовательность представляет собой аминокислотную последовательность, имеющую такую гомологию или идентичность и проявляющую эффект, соответствующий эффекту указанного выше белка.
Кроме того, хотя в настоящей заявке указанная супероксиддисмутаза 1 в одном из примеров определяется как белок, содержащий аминокислотную последовательность SEQ ID NO: 1, это не исключает мутации, которая может возникнуть в результате добавления «бессмысленной» последовательности выше или ниже по ходу транскрипции от аминокислотной последовательности SEQ ID NO: 1, или которая может встречаться в природе, или его молчащей мутации, и специалистам в данной области техники будет очевидно, что любой белок, обладающий такой же активностью или активностью, соответствующей активности белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, может относиться к супероксиддисмутазе 1 согласно настоящей заявке.
То есть, хотя в настоящей заявке она выражается как «белок или полипептид, имеющий аминокислотную последовательность согласно конкретному SEQ ID NO», или «белок или полипептид, содержащий аминокислотную последовательность согласно конкретному SEQ ID NO», очевидно, что любой белок, содержащий аминокислотную последовательность, где часть указанной аминокислотной последовательности удалена, модифицирована, заменена или добавлена, также может быть применен в настоящей заявке, при условии, что указанный белок обладает такой же активностью или активностью, соответствующей активности полипептида, состоящего из аминокислотной последовательности согласно соответствующему SEQ ID NO.
В контексте настоящего описания термин «вариант» или «модифицированный полипептид» относится к белку содержащему одну или более аминокислот, отличающихся от аминокислот указанной последовательности в результате консервативных замен и/или модификаций, так что функции или свойства указанного белка сохраняются.
Указанные варианты отличаются от идентифицированных последовательностей в результате замены, удаления или добавления нескольких аминокислот. Такие варианты в общем случае могут быть идентифицированы посредством модификации одной или более из указанных выше аминокислотных последовательностей белка и оценки свойств модифицированного белка. То есть, способность указанных вариантов может быть повышена, не изменена или снижена по сравнению с нативным белком. Кроме того, некоторые варианты могут включать модифицированные полипептиды, в которых удален один или более участков, таких как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, в которых участок удален из N- и/или С-конца зрелого белка. Термин «вариант» или «модифицированный полипептид» может быть применен взаимозаменяемо с такими терминами, как модификация, модифицированный белок, мутант, мутеин, дивергент, вариант и т.д., при условии, что указанные термины применяют для обозначения вариации, но термины не ограничиваются ими.
В контексте настоящего описания термин «консервативная замена» относится к замене аминокислоты на другую аминокислоту, обладающую сходными структурными и/или химическими свойствами. Указанный вариант может иметь, например, одну или более консервативных замен с сохранением при этом одной или более биологических активностей. Такая аминокислотная замена в общем случае может происходить на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатка.
Например, среди аминокислот, содержащих электрически заряженную боковую цепь, положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; отрицательно заряженные (кислые) аминокислоты включают глутаминовую кислоту и аспарагиновую кислоту; и аминокислоты, содержащие незаряженную боковую цепь, включают глицин, аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан, пролин, серии, треонин, цистеин, тирозин, аспарагин и глутамин.
Кроме того, указанный вариант также может включать удаление или добавление аминокислот, которые оказывают минимальное влияние на свойства и вторичную структуру полипептида. Например, указанный полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью на N-конце, участвующей в котрансляционном или посттрансляционном переносе белков. Кроме того, указанный полипептид также может быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза указанного полипептида.
В одном из воплощений вариант согласно настоящей заявке может представлять собой вариант супероксиддисмутазы 1, где аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, или модифицированный полипептид, обладающий активностью супероксиддисмутазы 1, наряду с супероксиддисмутазой 1, описанной выше. Вариант согласно настоящей заявке, то есть «вариант супероксиддисмутазы 1», может быть выражен как «(модифицированный) полипептид, обладающий активностью супероксиддисмутазы 1», или «вариант SOD1» или может представлять собой полипептид, который может увеличивать получение глутатиона по сравнению с природным полипептидом дикого типа или немодифицированным полипептидом, но не ограничивается им.
В любом из описанных выше воплощений указанный вариант может представлять собой вариант супероксиддисмутазы 1, где аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту.
В настоящей заявке «замена на другую аминокислоту» не ограничивается при условии, что она представляет собой аминокислоту, отличающуюся от аминокислоты до замены. При этом, когда в настоящей заявке указано, что «конкретная аминокислота была заменена», очевидно, что указанная аминокислота заменена на аминокислоту, отличающуюся от аминокислоты до замены, даже если специально не указано, что указанная аминокислота была заменена на другую аминокислоту.
В любом из описанных выше воплощений указанная «другая аминокислота» может представлять собой аминокислоту за исключением глицина. В частности, указанная другая аминокислота может представлять собой аминокислоту, выбранную из аланина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, цистеина, тирозина, аспарагина, глутамина, аспарагиновой кислоты, глутаминовой кислоты, лизина, аргинина и гистидина, но не ограничивается ей.
В контексте настоящего описания «соответствующее положение» относится к аминокислотному остатку в положении, указанном в белке или пептиде, или аминокислотному остатку, который подобен, идентичен или гомологичен остатку, указанному в белке или пептиде. В контексте настоящего описания «соответствующий участок» в общем случае относится к сходному или соответствующему положению в родственном белке или эталонном белке.
В настоящей заявке может быть применена определенная нумерация положений аминокислотных остатков в белке, применяемом в настоящем документе. Например, возможно изменение нумерации положений аминокислотных остатков белка согласно настоящей заявке на соответствующие положения посредством выравнивания пептидной последовательности белка согласно настоящей заявке с белком-мишенью, подлежащим сравнению.
Указанный вариант супероксиддисмутазы 1, в котором аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, может представлять собой белок, в котором аминокислота, соответствующая положению 37 SEQ ID NO: 1, заменена на другую аминокислоту в аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, имеющей гомологию или идентичность с ней, составляющую 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более.
Такой вариант может представлять собой вариант, имеющий гомологию или идентичность с SEQ ID NO: 1, составляющую 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более, и имеющий гомологию или идентичность с SEQ ID NO: 1, составляющую менее 100%, но не ограничивается им.
Указанный вариант супероксиддисмутазы 1, в котором аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, может содержать любую из аминокислотных последовательностей SEQ ID NO: 12-30. В частности, указанный вариант может по существу состоять из любой из аминокислотных последовательностей SEQ ID NO: 12-30 и более конкретно состоять из любой из аминокислотных последовательностей SEQ ID NO: 12-30.
Кроме того, указанный вариант может содержать любую из аминокислотных последовательностей SEQ ID NO: 12-30 или может содержать аминокислотную последовательность, в которой аминокислота в положении 37 в указанной аминокислотной последовательности является фиксированной (то есть, в аминокислотной последовательности указанного варианта аминокислота, соответствующая положению 37 в аминокислотной последовательности SEQ ID NO: 12-30, идентична аминокислоте в положении 37 в аминокислотной последовательности SEQ ID NO: 12-30), и которая имеет гомологию или идентичность с ней, составляющую 80% или более, но не ограничивается ими.
В частности, вариант согласно настоящей заявке может содержать любую из аминокислотных последовательностей SEQ ID NO: 12-30 и может иметь гомологию или идентичность аминокислотных последовательностей с любой из аминокислотных последовательностей SEQ ID NO: 12-30, составляющую по меньшей мере 80%, 90%, 95%, 96%, 97%, 98% или 99, где аминокислота, соответствующая положению 37 в аминокислотной последовательности любой из SEQ ID NO: 12-30, может включать полипептид, который представляет собой аминокислоту, отличную от глицина. Кроме того, очевидно, что любой белок, содержащий аминокислотную последовательность, в котором часть указанной аминокислотной последовательности удалена, модифицирована, заменена или добавлена, может входить в объем настоящей заявки при условии, что указанная аминокислотная последовательность имеет такую гомологию или идентичность и демонстрирует эффективность, соответствующую эффективности указанного варианта.
В контексте настоящего описания термин «гомология» или «идентичность» относится к степени родства между двумя указанными аминокислотными последовательностями или нуклеотидными последовательностями и может быть выражен в процентах. Термины «гомология» и «идентичность» часто могут использоваться взаимозаменяемо друг с другом.
Гомология или идентичность последовательностей консервативных полинуклеотидов или полипептидов может быть определена с помощью стандартных алгоритмов выравнивания и может быть применена со штрафом за гэп по умолчанию, установленным применяемой программой. Гомологичные или идентичные последовательности по существу могут гибридизоваться в умеренно или очень жестких условиях, так что вся длина последовательности или по меньшей мере примерно 50%, 60%, 70%, 80% или 90% или более от полной длины может гибридизоваться. Очевидно, что гибридизация с полинуклеотидами, содержащими общие кодоны или вырожденные кодоны, также включена.
Обладают ли какие-либо две полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, может быть определено, например, с помощью известного компьютерного алгоритма, такого как программа «FASTA» (Pearson et al, (1988) Proc. Natl. Acad. Sci. USA 85:2444), с применением параметров по умолчанию. В качестве альтернативы это может быть определено с помощью алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), который выполняют с применением программы Needleman из пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al, 2000, Trends Genet. 16:276-277) (предпочтительно версии 5.0.0 или более поздней) (пакета программ GCG (Devereux, J. et al, Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F. et al, J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994 и CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, гомология, сходство или идентичность могут быть определены с применением программы BLAST или ClustalW от Национального центра биотехнологической информации США (NCBI).
Гомология, сходство или идентичность полинуклеотидов или полипептидов могут быть определены, например, посредством сравнения информации о последовательностях с применением, например, компьютерной программы GAP, такой как в Needleman et al. (1970), J Mol Biol. 48:443, как раскрыто в Smith and Waterman, Adv. Appl. Math (1981) 2:482. Вкратце программа GAP определяет гомологию, сходство или идентичность как значение, полученное посредством деления числа одинаково выровненных символов (то есть, нуклеотидов или аминокислот) на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать (1) бинарную матрицу сравнения (содержащую значение 1 для идентичностей и 0 для не идентично стей) и взвешенную матрицу сравнения согласно Gribskov et al. (1986), Nucl. Acids Res. \A\61A5, как раскрыто в Schwartz and Dayhoff, eds., Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или матрицу замен EDNAFULL (NUC4.4 в версии EMBOSS от NCBI)); (2) штраф за каждый гэп, составляющий 3,0, и дополнительный штраф за каждый символ в каждом гэпе, составляющий 0,10 (или штраф за открытие гэпа, составляющий 10, и штраф за продолжение гэпа, составляющий 0,5); и (3) отсутствие штрафа за концевые гэпы.
Кроме того, обладают ли какие-либо две полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью друг с другом, может быть обнаружено посредством сравнения указанных последовательностей в эксперименте по Саузерн-гибридизации в жестких условиях согласно определению, и соответствующие определенные условия гибридизации находятся в компетенции специалиста в данной области техники и могут быть определены способом, хорошо известным специалистам в данной области техники (например, см. J. Sambrook et al, Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F. M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
В другом аспекте настоящей заявки может быть предложен полинуклеотид, кодирующий указанный вариант.
В контексте настоящего описания термин «полинуклеотид», который представляет собой полимер нуклеотидов, состоящий из нуклеотидных мономеров, соединенных в длинную цепь ковалентной связью, представляет собой нить ДНК или РНК, имеющую по меньшей мере некоторую длину.
Ген, кодирующий супероксиддисмутазу 1 согласно настоящей заявке, может представлять собой ген sodl.
Указанный ген может быть получен из дрожжей. В частности, он может быть получен из дрожжей рода Saccharomyces, более конкретно из Saccharomyces cerevisiae. В частности, может быть включен любой ген без ограничения при условии, что он кодирует полипептид, обладающий активностью супероксиддисмутазы 1, полученной из Saccharomyces cerevisiae. В одном из воплощений он может представлять собой ген, кодирующий аминокислотную последовательность SEQ ID NO: 1, и в другом воплощении он может включать нуклеотидную последовательность SEQ ID NO: 2 или SEQ ID NO: 3, но не ограничивается ими.
Полинуклеотид, кодирующий вариант супероксиддисмутазы 1 согласно настоящей заявке, может включать любой полинуклеотид, кодирующий указанный вариант супероксиддисмутазы 1 или полипептид, обладающий активностью, соответствующей его активности, без ограничения.
Супероксиддисмутаза 1 согласно настоящей заявке или полинуклеотид, кодирующий ее вариант, могут подвергаться различным модификациям в кодирующем участке в пределах, которые не меняют аминокислотной последовательности указанного полипептида, из-за вырождения кодонов или с учетом кодонов, предпочтительных для организма, в котором должен экспрессироваться указанный полипептид.
В частности, полинуклеотид, кодирующий вариант супероксиддисмутазы 1 согласно настоящей заявке, может включать любую полинуклеотидную последовательность без ограничения при условии, что она кодирует вариант белка, в котором аминокислота, соответствующая положению 37 в аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту.
Например, полинуклеотид, кодирующий вариант белка согласно настоящей заявке, может представлять собой вариант белка согласно настоящей заявке, в частности полинуклеотидную последовательность, кодирующую любую из аминокислотных последовательностей SEQ ID NO: 12-30 или полипептид, имеющий гомологию или идентичность с ней, и последовательность, кодирующая аминокислоту, соответствующую положению 37 в любой из аминокислотных последовательностей SEQ ID NO: 12-30, может представлять собой полинуклеотидную последовательность, кодирующую аминокислоту за исключением глицина, но не ограничивается ей. Указанные гомология и идентичность являются такими, как описано выше.
Кроме того, полинуклеотид, кодирующий вариант белка согласно настоящей заявке, может включать зонд, который может быть получен из известной последовательности гена, например, любой последовательности, кодирующей вариант белка, где аминокислота, соответствующая положению 37 в аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, который может гибридизоваться с последовательностью, комплементарной всей или части указанной нуклеотидной последовательности, в жестких условиях без ограничения.
Термин «жесткие условия» относится к условиям, в которых обеспечивается возможность специфической гибридизации между полинуклеотидами. Такие условия конкретно описаны в источниках (например, J. Sambrook et al.). Например, указанные жесткие условия могут включать условия, в которых гены, имеющие высокую гомологию или идентичность, составляющую 40% или более, в частности 90% или более, более конкретно 95% или более, 96% или более, 97% или более, 98% или более или еще более конкретно 99% или более, гибридизуются друг с другом, и гены, имеющие гомологию или идентичность более низкую, чем указанные выше гомологии или идентичности, не гибридизуются друг с другом, или условия промывания при Саузерн-гибридизации, то есть промывание один раз, в частности два или три раза, при концентрации соли и температуре, соответствующих 60°С, 1×SSC (стандартный солевой раствор), 0,1% SDS (додецилсульфат натрия), в частности 60°С, 0,1×, 0,1% SDS и более конкретно 68°С, 0,1×SSC, 0,1% SDS.
Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несоответствия между основаниями в зависимости от жесткости гибридизации. Термин «комплементарный» применяют для описания взаимосвязи между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в отношении ДНК аденин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, полинуклеотид согласно настоящей заявке может включать выделенные фрагменты нуклеиновой кислоты, комплементарные всей последовательности, а также последовательности нуклеиновой кислоты, по существу сходные с ними.
В частности, полинуклеотиды, имеющие гомологию или идентичность, могут быть обнаружены с применением условий гибридизация, включающих стадию гибридизации при значении Tm, составляющем 55°С, в описанных выше условиях. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, но не ограничивается ими, и может быть подходящим образом отрегулировано специалистами в данной области техники в зависимости от их цели.
Соответствующая жесткость гибридизации указанных полинуклеотидов зависит от длины указанных полинуклеотидов и степени комплементарности, и указанные параметры хорошо известны в данной области техники (например, см. приведенный выше Sambrook et al, 9.50-9.51, 11.7-11.8).
В еще одном аспекте настоящей заявки может быть предложен вектор, содержащий полинуклеотид, кодирующий указанный вариант белка.
В контексте настоящего описания термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего белок-мишень, функционально связанную с подходящим участком, регулирующим экспрессию (или последовательностью, регулирующей экспрессию), чтобы иметь возможность экспрессировать целевой полипептид в подходящей клетке-хозяине. Указанный участок, регулирующий экспрессию, может содержать промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции указанной транскрипции, последовательность, кодирующую подходящий мРНК-сайт связывания рибосомы, и последовательность для регуляции терминации транскрипции и трансляции. После трансформации в подходящую клетку-хозяина указанный вектор может реплицироваться или функционировать независимо от генома хозяина или может интегрироваться в его геном.
В одном из примеров полинуклеотид, кодирующий белок-мишень, в хромосоме может быть заменен на модифицированный полинуклеотид посредством вектора для внутриклеточной хромосомной вставки. Вставка указанного полинуклеотида в хромосому может быть выполнена любым способом, известным в данной области техники, например, посредством гомологичной рекомбинации, но не ограничивается ей. Указанный вектор может дополнительно содержать селективный маркер для подтверждения указанной вставки в хромосому. Указанный селективный маркер предназначен для отбора клеток, трансформированных указанным вектором, то есть, для подтверждения того, была ли осуществлена вставка целевой молекулы нуклеиновой кислоты, и могут быть применены маркеры, которые обеспечивают селектируемые фенотипы, такие как устойчивость к лекарственным средствам, ауксотрофия, устойчивость к клеточным токсичным агентам или экспрессия поверхностных белков. Только клетки, экспрессирующие указанный селективный маркер, способны выживать или демонстрировать различные фенотипы в среде, обработанной селективным агентом, и, таким образом, указанные трансформированные клетки могут быть отобраны.
Вектор, применяемый в настоящей заявке, не имеет конкретных ограничений, и может быть применен любой вектор, известный в данной области техники. Дрожжевой вектор экспрессии может включать как дрожжевую интегративную плазмиду (YIp), так и внехромосомный плазмидный вектор. Указанный внехромосомный плазмидный вектор может включать дрожжевую эписомальную плазмиду (Yep), дрожжевые репликативные плазмиды (YRp) и дрожжевую центромерную плазмиду (YCp). Кроме того, дрожжевые искусственные хромосомы (YACs) также могут быть применены в качестве вектора согласно настоящей заявке. Конкретные примеры вектора, который может быть применен, могут включать pESCHIS, pESC-LEU, pESC-TRP, pESC-URA, Gateway pYES-DEST52, pA0815, pGAPZ A, pGAPZ B, pGAPZ C, pGAPα A, pGAPα B, pGAPα C, pPIC3.5K, pPIC6 A, pPIC6 В, pPIC6 C, pPIC6α A, pPIC6α B, pPIC6α C, pPIC9K, pYC2/CT, вектор дрожжевого дисплея pYD1, pYES2, pYES2/CT, pYES2/NT A, pYES2/NT B, pYES2/NT C, pYES2/CT, pYES2.1, pYES-DEST52, pTEF1/Zeo, pFLD1, PichiaPinkTM, p427-TEF, p417-CYC, pGAL-MF, p427-TEF, p417-CYC, PTEF-MF, pBY011, pSGP47, pSGP46, pSGP36, pSGP40, ZM552, pAG303GAL-ccdB, pAG414GAL-ccdB, pAS404, pBridge, pGAD-GH, pGAD T7, pGBK T7, pHIS-2, pOBD2, pRS408, pRS410, pRS418, pRS420, pRS428, дрожжевую микронную плазмиду формы A (yeast micron A form), pRS403, pRS404, pRS405, pRS406, pYJ403, pYJ404, pYJ405 и pYJ406, но не ограничиваются ими.
В контексте настоящего описания, термин «трансформация» относится к введению вектора, содержащего полинуклеотид, кодирующий белок-мишень, в клетку-хозяина, так что белок, кодируемый указанным полинуклеотидом, может экспрессироваться в указанной клетке-хозяине. При условии, что трансформированный полинуклеотид может экспрессироваться в клетке-хозяине, не имеет значения, интегрирован ли указанный трансформированный полинуклеотид в хромосому указанной клетки-хозяина и расположен в ней или расположен внехромосомно, и оба случая могут быть включены. Кроме того, указанный полинуклеотид может включать ДНК и РНК, кодирующие указанный белок-мишень. Указанный полинуклеотид может быть введен в любой форме при условии, что он может быть введен в указанную клетку-хозяина и экспрессироваться в ней. Например, указанный полинуклеотид может быть введен в указанную клетку-хозяина в форме кассеты экспрессии, которая представляет собой генную конструкцию, содержащую все элементы, необходимые для ее автономной экспрессии. Указанная кассета экспрессии обычно может содержать промотор, функционально связанный с указанным полинуклеотидом, терминатор транскрипции, сайт связывания рибосомы или терминатор трансляции. Указанная кассета экспрессии может находиться в форме самореплицируемого вектора экспрессии. Кроме того, указанный полинуклеотид может быть введен в указанную клетку-хозяина в том виде, в каком он есть, и функционально связан с последовательностями, необходимыми для экспрессии в указанной клетке-хозяине, но не ограничивается этим.
Кроме того, в контексте настоящего описания термин «функционально связанный» означает, что последовательность гена функционально связана с последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой полипептид согласно настоящей заявке.
Способ трансформации вектора согласно настоящей заявке может включать любой способ, который может обеспечивать введение нуклеиновой кислоты в клетку, и указанная трансформация может быть выполнена посредством выбора соответствующего стандартного метода, известного в данной области техники, в соответствии с клеткой-хозяином. Например, указанный способ может включать электропорацию, осаждение фосфатом кальция (CaPO4,), осаждение хлоридом кальция (CaCl2), микроинъекцию, способ с применением полиэтиленгликоля (ПЭГ), метод с применением диэтиламиноэтилдекстрана (DEAE-декстрана), способ с применением катионных липосом и способ с применением ацетата лития-диметилсульфоксида (ДМСО) и т.д., но не ограничивается ими.
В настоящей заявке может быть предложен микроорганизм, содержащий любое одно или более из супероксиддисмутазы 1 и полинуклеотида, кодирующего указанный вариант.
В контексте настоящего описания термин «микроорганизм» включает все микроорганизмы дикого типа или генетически модифицированные микроорганизмы естественным или искусственным способом и представляет собой комплексное понятие, включающее все микроорганизмы, у которых конкретный механизм ослаблен или усилен из-за вставки чужеродного гена или усиления или ослабления активности эндогенного гена.
В одном из воплощений указанный микроорганизм может представлять собой микроорганизм, который обеспечивает получение глутатиона. В контексте настоящего описания термин «микроорганизм, который обеспечивает получение глутатиона» может быть применен взаимозаменяемо с такими терминами, как «микроорганизм, обеспечивающий получение глутатиона», «микроорганизм, обладающий способностью обеспечивать получение глутатиона», «штамм, обеспечивающий получение глутатиона», «штамм, обладающий способностью обеспечивать получение глутатиона» и т.д.
В любом из описанных выше воплощений указанный микроорганизм может представлять собой микроорганизм, который обеспечивает получение производного глутатиона.
В любом из описанных выше воплощений микроорганизм согласно настоящей заявке может содержать вектор, содержащий полинуклеотид, кодирующий супероксиддисмутазу 1.
Термин «глутатион» согласно настоящей заявке может быть применен взаимозаменяемо с термином «глутатион» или «GSH» и относится к трипептиду, состоящему из трех аминокислот (то есть, глутамата, цистеина, глицина). Глутатион может быть применен в качестве сырья для фармацевтических средств, оздоровительных функциональных пищевых продуктов, ароматизирующих веществ, пищевых продуктов, кормовых добавок, косметических средств и т.д., но не ограничивается этим.
«Производное глутатиона» согласно настоящей заявке может включать любое вещество, которое может быть подвергнуто превращению с применением глутатиона в качестве исходного вещества без ограничения.
В настоящей заявке глутатион или его производное в совокупности могут быть названы «гамма-глутамилпептидом». гамма-Глутамилпептид относится к низкомолекулярному соединению, содержащему гамма-карбоксильную группу глутаминовой кислоты на N-конце указанной молекулы.
В частности, указанный гамма-глутамилпептид может представлять собой соединение, представленное общей формулой 1 ниже:
[Общая формула 1]
γ-Glu X Y
В общей формуле выше X представляет собой цистеин, глутаминовую кислоту, тирозин, метионин, валин, триптофан, аспарагиновую кислоту, изолейцин, лейцин, аспарагин, глутамин, гистидин, лизин, орнитин, аргинин, S-метилцистеин или S-этилцистеин;
Y представляет собой глицин, аланин, лейцин, изолейцин, серии, треонин, метионин, цистеин, аспарагиновую кислоту, аспарагин, глутамин, лизин, орнитин, аргинин, фенилаланин, тирозин, пролин, гидроксипролин, альфа-аминомасляную кислоту или отсутствует.
В одном из воплощений указанный гамма-глутамил пептид может быть выбран из γ-Glu-Cys-Gly (глутатиона), γ-Glu-Glu (γ-EF), γ-Glu-Tyr (γ-EY), γ-Glu-Met (γ-EM), γ-Glu-Val (γ-EV), γ-Glu-Trp (γ-EW), γ-Glu-Asp (γ-ED), γ-Glu-Ile (γ-EI), γ-Glu-Leu (γ-EL), γ-Glu-Cys-Gly (γ-ECG), γ-Glu-Val-Gly (γ-EVG), γ-глутамил-S-метилцистеина (γ-Е-S-метил-С), γ-глутамил-S-этенилцистеина (γ-Е-S-этенил-С) и γ-глутамилцистеинил-β-аланина (γ-Е-С-β-А).
В любом из описанных выше воплощений указанный гамма-глутамилпептид может представлять собой глутатион, но не ограничивается им.
Микроорганизм согласно настоящей заявке может относиться к микроорганизму, способному обеспечивать получение избытка желаемого глутатиона, содержащему супероксиддисмутазу 1, по сравнению с микроорганизмом дикого типа или немодифицированным микроорганизмом. Указанный микроорганизм не имеет конкретных ограничений по своему типу при условии, что он может обеспечивать получение глутатиона, но может представлять собой микроорганизм рода Saccharomyces, в частности Saccharomyces cerevisiae. Однако указанный микроорганизм не ограничивается им.
Родительский штамм микроорганизма, обеспечивающего получение глутатиона, содержащего указанный вариант, не имеет конкретных ограничений при условии, что он способен обеспечивать получение глутатиона. Указанный микроорганизм может дополнительно включать мутации, такие как усиление путей биосинтеза для повышения способности получения глутатиона, снятие ингибирования по принципу обратной связи или инактивация генов, которая ослабляет пути деградации или биосинтетические пути, и указанные мутации не исключают мутаций, которые встречаются в природе.
В контексте настоящего описания термин «усиление» активности полипептида означает, что активность полипептида увеличивается по сравнению с его эндогенной активностью. Указанный термин «усиление» может быть применен взаимозаменяемо с такими терминами, как «активация», «повышающая регуляция», «сверхэкспрессия», «увеличение» и т.д. В частности, указанные активация, усиление, повышающая регуляция, сверхэкспрессия и увеличение могут включать оба случая: когда проявляется активность, которой изначально не было, или когда активность усиливается по сравнению с эндогенной активностью или активностью до модификации. Термин «эндогенная активность» относится к активности конкретного полипептида, которой первоначально обладал родительский штамм до трансформации или немодифицированный микроорганизм, когда признак изменяется посредством генетической модификации, вызванной естественными или искусственными факторами, и может быть применен взаимозаменяемо с термином «активность до модификации». «Усиление», «повышающая регуляция», «сверхэкспрессия» или «увеличение» активности полипептида по сравнению с его эндогенной активностью означает, что активность и/или концентрация (уровень экспрессии) указанного полипептида повышается по сравнению с активностью и/или концентрацией (уровнем экспрессии) конкретного полипептида, которой первоначально обладал родительский штамм до трансформации или немодифицированный микроорганизм.
Указанное усиление может быть достигнуто посредством введения чужеродного полипептида или посредством повышения активности и/или концентрации (уровня экспрессии) эндогенного полипептида. Усиление активности полипептида может быть подтверждено посредством увеличения уровня активности указанного полипептида, уровня экспрессии или количества продукта, выделяемого из указанного полипептида.
Усиление активности полипептида может быть применено различными способами, хорошо известными в данной области техники, и не ограничивается при условии, что они могут усиливать активность целевого полипептида по сравнению с указанной активностью микроорганизма до модификации. В частности, может быть применена генная инженерия и/или белковая инженерия, хорошо известная специалистам в данной области техники, которая представляет собой распространенный метод молекулярной биологии, но указанный способ не ограничивается ей (например, см. Sitnicka et al. Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol.2. 1-16, Sambrook et al. Molecular Cloning 2012 и т.д.).
В частности, указанное усиление полипептида согласно настоящей заявке может быть достигнуто посредством:
1) увеличения числа внутриклеточных копий полинуклеотида, кодирующего указанный полипептид;
2) замены регулирующего экспрессию участка гена, кодирующего указанный полипептид, на хромосоме на последовательность, обладающую сильной активностью;
3) модификации нуклеотидной последовательности, кодирующей инициаторный кодон или 5'-нетранслируемую область (5'-UTR) транскрипта гена, кодирующего указанный полипептид;
4) модификации аминокислотной последовательности указанного полипептида таким образом, что активность указанного полипептида усиливается;
5) модификации полинуклеотидной последовательности, кодирующей указанный полипептид, таким образом, что активность указанного полипептида усиливается (например, модификации полинуклеотидной последовательности гена указанного полипептида для кодирования полипептида, который был модифицирован для усиления активности указанного полипептида);
6) введения чужеродного полипептида, проявляющего активность указанного полипептида, или чужеродного полинуклеотида, кодирующего его;
7) оптимизации кодонов полинуклеотида, кодирующего указанный полипептид;
8) анализа третичной структуры указанного полипептида и вследствие этого отбора и модификации подверженного воздействию сайта или его химической модификации; или
9) комбинации двух или более способов, выбранных из указанных выше (1)-(8), но не ограничивается конкретно ими.
Более конкретно
способ 1) увеличения числа внутриклеточных копий полинуклеотида, кодирующего указанный полипептид, может быть достигнут посредством введения вектора, который функционально связан с указанным полинуклеотидом, кодирующим указанный полипептид, и способен реплицироваться и функционировать независимо от клетки-хозяина, в указанную клетку-хозяина. В качестве альтернативы указанный способ может быть достигнут посредством введения одной копии или двух копий полинуклеотидов, кодирующих указанный полипептид, в хромосому клетки-хозяина. Указанное введение в хромосому может быть выполнено посредством введения вектора, который способен вставлять указанный полинуклеотид в хромосому клетки-хозяина, в указанную клетку-хозяина, но не ограничивается этим. Указанный вектор является таким, как описано выше.
Способ 2) замены регулирующего экспрессию участка (или регулирующей экспрессию последовательности) гена, кодирующего указанный полипептид, на хромосоме на последовательность, обладающую сильной активностью, может быть достигнут посредством индуцирования модификации в указанной последовательности посредством удаления, вставки, неконсервативной или консервативной замены или их комбинации для дополнительного усиления активности указанного участка, регулирующего экспрессию, или посредством замены указанной последовательности на последовательность, обладающую более сильной активностью. Указанный участок, регулирующий экспрессию, может содержать, но не ограничивается конкретно ими, промотор, операторную последовательность, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, и т.д. В одном из примеров указанный способ может включать замену исходного промотора на сильный промотор, но не ограничивается этим.
Примеры промоторов, известных для эукариот, включают промоторы для фактора элонгации трансляции 1 (TEF1), глицерол-3-фосфатдегидрогеназы 1 (GPD1), 3-фосфоглицераткиназы или других гликолитических ферментов, например енолазы, глицеральдегид-3-фосфатдегидрогеназы, гексокиназы, пируватдекарбоксилазы, фосфофруктокиназы, глюкозо-6-фосфатизомеразы, 3-фосфоглицератмутазы,
пируваткиназы, триозофосфатизомеразы, фосфоглюкозоизомеразы и глюкокиназы; примеры других дрожжевых промоторов, которые являются индуцируемыми промоторами с дополнительным преимуществом транскрипции, контролируемой условиями роста, включают промоторы для алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, деструктивных ферментов, связанных с азотистым обменом, металлотионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, ответственных за утилизацию мальтозы и галактозы; промоторы, которые могут быть применены, когда указанная клетка-хозяин представляет собой дрожжи, могут включать промотор TEF1, промотор TEF2, промотор GAL10, промотор GAL1, промотор ADH1, промотор ADH2, промотор PHO5, промотор GAL1-10, промотор TDH3 (промотор GPD), промотор TDH2, промотор TDH1, промотор PGK1, промотор PYK2, промотор ENO1, промотор ENO2 и промотор TPI1; и подходящие векторы и промоторы для применения для дрожжевой экспрессии дополнительно описаны в европейском патенте №073657, но промоторы не ограничиваются ими. Кроме того, дрожжевые энхансеры также могут быть успешно применены в комбинации с дрожжевыми промоторами, но не ограничиваются этим.
Способ 3) модификации нуклеотидной последовательности, кодирующей инициаторный кодон или 5'-UTR транскрипта гена, кодирующего указанный полипептид, может быть достигнут, например, посредством замены указанной нуклеотидной последовательности на нуклеотидную последовательность, кодирующую другой инициаторный кодон, имеющий более высокую частоту экспрессии указанного полипептида по сравнению с эндогенным инициаторный кодоном, но не ограничивается этим.
Способы 4) и 5) модификации указанной аминокислотной последовательности или указанной полинуклеотидной последовательности могут быть достигнуты посредством индуцирования модификации в указанной последовательности посредством удаления, вставки, неконсервативной или консервативной замены в аминокислотной последовательности указанного полипептида или полинуклеотидной последовательности, кодирующей указанный полипептид, или их комбинации для усиления активности указанного полипептида или посредством замены указанной последовательности на аминокислотную последовательность или полинуклеотидную последовательность, модифицированную так, чтобы обладать более сильной активностью, или аминокислотную последовательность или полинуклеотидную последовательность, модифицированную так, чтобы усиливать указанную активность, но не ограничиваются этим. Указанная замена, в частности, может быть выполнена посредством вставки указанного полинуклеотида в хромосому посредством гомологичной рекомбинации, но не ограничивается ей. Вектор, применяемый в настоящем документе, может дополнительно содержать селективный маркер для подтверждения указанной вставки в хромосому. Указанный селективный маркер является таким, как описано выше.
Способ 6) введения чужеродного полинуклеотида, обеспечивающего активность указанного полипептида, может быть достигнут посредством введения в клетку-хозяина чужеродного полинуклеотида, кодирующего полипептид, который проявляет активность, совпадающую/сходную с активностью указанного полипептида. Указанный чужеродный полинуклеотид может быть применен без ограничения независимо от его происхождения или последовательности при условии, что он обеспечивает активность, совпадающую/сходную с активностью указанного полипептида. Указанное введение может быть выполнено средними специалистами в данной области техники посредством выбора подходящим образом способа трансформации, известного в данной области техники, и экспрессия введенного полинуклеотида в указанной клетке-хозяине обеспечивает получение указанного полипептида с увеличением тем самым его активности.
Способ 7) оптимизации кодонов полинуклеотида, кодирующего указанный полипептид, может быть достигнут посредством оптимизации кодонов эндогенного полинуклеотида для увеличения транскрипции или трансляции внутри клетки-хозяина или посредством оптимизации его кодонов таким образом, чтобы могли быть достигнуты оптимизированные транскрипция и трансляция чужеродного полинуклеотида внутри указанной клетки-хозяина.
Способ 8) анализа третичной структуры указанного полипептида и вследствие этого отбора и модификации подверженного воздействию сайта или его химической модификации может быть достигнут, например, посредством сравнения информации о последовательности полипептида, подлежащего анализу, с базой данных, в которой хранится информация о последовательностях известных белков, для определения кандидатов в матричные белки в соответствии со степенью сходства последовательностей и, таким образом, подтверждения указанной структуры на основе указанной информации с отбором и трансформацией или модификацией вследствие этого подверженного воздействию сайта, подлежащего модификации или химической модификации.
Такое усиление активности указанного полипептида может означать, что активность или концентрация соответствующего полипептида увеличивается по сравнению с активностью или концентрацией полипептида, экспрессированного в микроорганизме дикого типа или микроорганизме до модификации, или что увеличивается количество продукта, полученного из указанного полипептида, но не ограничивается этим.
Модификация части или всего указанного полинуклеотида в микроорганизме согласно настоящей заявке может быть достигнута посредством (а) гомологичной рекомбинации с применением вектора для хромосомной вставки в указанном микроорганизме или редактирования генома с применением сконструированной нуклеазы (например, CRISPR-Cas9) и/или (b) может быть индуцирована светом, таким как ультрафиолетовые лучи и радиация и т.д., и/или химической обработкой, но не ограничивается этим. Способ модификации части или всего указанного гена может включать способ с применением технологии рекомбинации ДНК. Например, часть или весь указанный ген может быть удален посредством инъекции нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, гомологичную целевому гену, в микроорганизм для индуцирования гомологичной рекомбинации. Инъецированные нуклеотидная последовательность или вектор могут содержать доминантный селективный маркер, но не ограничиваются этим.
В контексте настоящего описания термин «ослабление» полипептида представляет собой комплексное понятие, включающее как пониженную активность, так и отсутствие активности по сравнению с его эндогенной активностью. Термин «ослабление» может быть применен взаимозаменяемо с такими терминами, как «инактивация», «недостаточность», «понижающая регуляция», «уменьшение», «снижение», «аттенюация» и т.д.
Указанное ослабление также может включать случай, когда активность полипептида сама по себе уменьшается или устраняется по сравнению с активность указанного полипептида, которой первоначально обладал микроорганизм, из-за мутации полинуклеотида, кодирующего указанный полипептид; случай, когда общий уровень внутриклеточной активности и/или концентрации (уровень экспрессии) полипептида снижается по сравнению с природным штаммом из-за ингибирования экспрессии гена полинуклеотида, кодирующего указанный полипептид, или ингибирования трансляции в указанный полипептид и т.д.; случай, когда указанный полинуклеотид вообще не экспрессируется; и/или случай, когда отмечается отсутствие активности полипептида, даже когда указанный полинуклеотид экспрессируется. В контексте настоящего описания термин «эндогенная активность» относится к активности конкретного полипептида, которой первоначально обладал родительский штамм до трансформации, микроорганизм дикого типа или немодифицированный микроорганизм, когда признак изменяется посредством генетической модификации, вызванной естественными или искусственными факторами, и может быть применен взаимозаменяемо с термином «активность до модификации». Выражение о том, что активность полипептида «инактивируется, является недостаточной, уменьшается, понижающе регулируется, снижается или ослабляется» по сравнению с его эндогенной активностью означает, что активность указанного полипептида уменьшается по сравнению с активностью конкретного полипептида, которой первоначально обладал родительский штамм до трансформации или немодифицированный микроорганизм.
Ослабление активности полипептида может быть выполнено любым способом, известным в данной области техники, но указанный способ не ограничивается этим, и может быть достигнуто посредством применения различных способов, хорошо известных в данной области техники (например, см. Nakashima N. et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014;15(2):2773-2793, Sambrook et al. Molecular Cloning 2012 и т.д.).
В частности, ослабление активности полипептида согласно настоящей заявке может быть достигнуто посредством:
1) удаления части или всего гена, кодирующего указанный полипептид;
2) модификации участка, регулирующего экспрессию (последовательности, регулирующей экспрессию), таким образом, что экспрессия гена, кодирующего указанный полипептид, уменьшается;
3) модификации аминокислотной последовательности, составляющей указанный полипептид, таким образом, что активность указанного полипептида устраняется или ослабляется (например, удаления/замены/добавления одной или более аминокислот в указанной аминокислотной последовательности);
4) модификации последовательности гена, кодирующей указанный полипептид, таким образом, что активность указанного полипептида устраняется или ослабляется (например, удаления/замены/добавления одного или более нуклеотидов в нуклеотидной последовательности гена указанного полипептида для кодирования полипептида, который был модифицирован для устранения или ослабления активности указанного полипептида);
5) модификации нуклеотидной последовательности, кодирующей инициаторный кодон или 5'-UTR транскрипта гена, кодирующего указанный полипептид;
6) введения антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующим указанный полипептид;
7) добавления последовательности, комплементарной последовательности Шайна Дальгарно (ШД), на переднем конце последовательности SD гена, кодирующего указанный полипептид, для образования вторичной структуры с ингибированием тем самым прикрепления рибосомы;
8) инженерии с применением обратной транскрипции (reverse transcription engineering, RTE), которая добавляет промотор, подлежащий обратной транскрипции, на 3'-конце открытой рамки считывания (ORF) последовательности гена, кодирующей указанный полипептид; или
9) комбинации двух или более способов, выбранных из указанных выше способов (1)-(8), но не ограничивается конкретно ими.
Например,
способ 1) удаления части или всего гена, кодирующего указанный полипептид, может быть достигнут посредством удаления всего полинуклеотида, кодирующего эндогенный целевой полипептид, внутри хромосомы или посредством замены указанного полинуклеотида на полинуклеотид, имеющий частично удаленную нуклеотидную последовательность, или на маркерный ген.
Способ 2) модификации участка, регулирующего экспрессию (последовательности, регулирующей экспрессию), может быть достигнут посредством индуцирования модификации в указанном участке, регулирующем экспрессию (указанной последовательности, регулирующей экспрессию), посредством удаления, вставки, неконсервативной замены или консервативной замены или их комбинации или посредством замены указанной последовательности на последовательность, обладающую более слабой активностью. Указанный участок, регулирующий экспрессию, может содержать промотор, операторную последовательность, последовательность, кодирующую сайт связывания рибосомы, и последовательность для регуляции терминации транскрипции и трансляции, но не ограничивается ими.
Способ 5) модификации нуклеотидной последовательности, кодирующей инициаторный кодон или 5'-UTR транскрипта гена, кодирующего указанный полипептид, может быть достигнут, например, посредством замены указанной нуклеотидной последовательности на нуклеотидную последовательность, кодирующую другой инициаторный кодон, имеющий более низкую частоту экспрессии полипептида, чем эндогенный инициаторный кодон, но не ограничивается этим.
Способы 3) и 4) модификации указанной аминокислотной последовательности или указанной полинуклеотидной последовательности могут быть достигнуты посредством индуцирования модификации в указанной последовательности посредством удаления, вставки, неконсервативной или консервативной замены в аминокислотной последовательности указанного полипептида или полинуклеотидной последовательности, кодирующей указанный полипептид, или их комбинации для ослабления активности указанного полипептида или посредством замены указанной последовательности на аминокислотную последовательность или полинуклеотидную последовательность, модифицированную так, чтобы обладать более слабой активностью, или аминокислотную последовательность или полинуклеотидную последовательность, модифицированную так, чтобы не обладать активностью, но не ограничиваются этим. Например, экспрессия указанного гена может быть ингибирована или ослаблена посредством введения мутации в указанную полинуклеотидную последовательность с образованием терминирующего кодона, но не ограничивается этим.
Способ 6) введения антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующим указанный полипептид, можно найти в источниках (Weintraub, Н. et al., Antisense-RNA as a molecular tool for genetic analysis, Reviews - Trends in Genetics, Vol.1(1) 1986).
Способ 7) добавления последовательности, комплементарной последовательности Шайна Дальгарно (ШД), на переднем конце последовательности SD гена, кодирующего указанный полипептид, для образования вторичной структуры с ингибированием тем самым прикрепления рибосомы может быть достигнут посредством ингибирования трансляции мРНК или снижения ее скорости.
8) Инженерия с применением обратной транскрипции (reverse transcription engineering, RTE), которая добавляет промотор, подлежащий обратной транскрипции, на 3'-конце открытой рамки считывания (ORF) последовательности гена, кодирующей указанный полипептид, может быть достигнута посредством образования антисмыслового нуклеотида, комплементарного транскрипту гена, кодирующему указанный полипептид, для ослабления указанной активности.
В любом из описанных выше воплощений микроорганизм согласно настоящей заявке может содержать мутацию в регулирующем экспрессию участке глутаматцистеинлигазы (GSH1) для увеличения способности получения глутатиона. Указанная мутация может представлять собой любую одну или более мутаций, выбранных из -250(С→Т), -252(G→A), 398(А→Т), -399(А→С), -407(Т→С), и 409(Т→С) выше по ходу транскрипции от GSH1 ORF.
В любом из описанных выше воплощений указанный микроорганизм может содержать мутацию, где 86-я аминокислота и/или 653-я аминокислота глутаматцистеинлигазы (GSH1) заменены на другие аминокислоты. В частности, указанный микроорганизм может содержать мутацию, где 86-я аминокислота заменена на аргинин, и 653-я аминокислота заменена на метионин, но указанная мутация не ограничивается этим.
В любом из описанных выше воплощений указанный микроорганизм может содержать мутацию, при которой активность глутаматцистеинлигазы (GSH1) усиливается, но не ограничивается этим.
Микроорганизм, содержащий любое одно или более из варианта супероксиддисмутазы 1 согласно настоящей заявке и полинуклеотида, кодирующего указанный вариант, может представлять собой «микроорганизм, экспрессирующий супероксиддисмутазу 1, где аминокислота, соответствующая положению 37 в аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту», но не ограничивается им.
Указанные супероксиддисмутаза 1 и ее варианты являются такими, как описано выше.
В контексте настоящего описания термин «экспрессироваться/экспрессирующийся» в отношении белка относится к состоянию, в котором белок-мишень вводят в микроорганизм, или белок-мишень модифицируют так, чтобы он экспрессировался в микроорганизме.
Микроорганизм, экспрессирующий вариант белка согласно настоящей заявке, может представлять собой микроорганизм, который был модифицирован так, чтобы экспрессировать вариант белка согласно настоящей заявке, и, соответственно, в еще одном аспекте настоящей заявки предложен способ получения микроорганизма, экспрессирующего вариант белка согласно настоящей заявке.
В контексте настоящего описания термин «штамм до модификации» или «микроорганизм до модификации» не исключает штамма, содержащего мутацию, которая может возникнуть в микроорганизме естественным путем, и может относиться к штамму дикого типа или природному штамму самому по себе или штамму до того, как признак изменяется вследствие генетической модификации, вызванной естественными или искусственными факторами. Термин «штамм до модификации» или «микроорганизм до модификации» может быть применен взаимозаменяемо с терминами «немутантный штамм», «немодифицированный штамм», «немутантный микроорганизм», «немодифицированный микроорганизм» или «эталонный микроорганизм».
В настоящей заявке микроорганизм, содержащий вариант супероксиддисмутазы 1 согласно настоящей заявке, полинуклеотид, кодирующий указанный вариант, или вектор, содержащий указанный полинуклеотид, может представлять собой рекомбинантный микроорганизм, и рекомбинация может быть достигнута посредством генетической модификации, такой как трансформация.
Например, указанный микроорганизм может представлять собой рекомбинантный микроорганизм, полученный посредством трансформации с применением вектора, содержащего указанный полинуклеотид, но не ограничивается им. Указанный рекомбинантный микроорганизм может представлять собой дрожжи, например микроорганизм рода Saccharomyces, в частности дрожжи Saccharomyces cerevisiae, но не ограничивается ими.
В еще одном аспекте настоящей заявки предложен способ получения глутатиона или его производных, включающий культивирование указанного микроорганизма. Указанные микроорганизм, глутатион или его производные являются такими, как описано выше.
Среда и другие условия культивирования, применяемые для культивирования штамма согласно настоящей заявке, могут представлять собой любую среду, применяемую для традиционного культивирования микроорганизмов рода Saccharomyces, без какого-либо конкретного ограничения. В частности, штамм согласно настоящей заявке может быть культивирован в аэробных или анаэробных условиях в традиционной среде, содержащей соответствующий источник углерода, источник азота, источник фосфора, неорганическое соединение, аминокислоту и/или витамин, при регулировании температуры, рН и т.д.
В настоящей заявке указанный источник углерода может включать углеводы, такие как глюкоза, фруктоза, сахароза, мальтоза и т.д.; сахарные спирты, такие как маннит, сорбит и т.д.; органические кислоты, такие как пировиноградная кислота, молочная кислота, лимонная кислота и т.д.; аминокислоты, такие как глутамат, метионин, лизин и т.д., но не ограничивается ими. Кроме того, указанный источник углерода может включать природные органические питательные вещества, такие как гидролизат крахмала, меласса, сырая меласса, рисовые отруби, маниок, меласса из сахарного тростника и жидкий кукурузный экстракт и т.д., и могут быть применены углеводы, такие как глюкоза и стерилизованная предварительно обработанная меласса (то есть, меласса, превращенная в восстанавливающий сахар). Кроме того, различные другие источники углерода в соответствующем количестве могут быть применены без ограничения. Указанные источники углерода могут быть применены по отдельности или в комбинации из двух или более видов, но не ограничиваются этим.
Указанный источник азота может включать неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и т.д.; и органические источники азота, такие как аминокислоты, пептон, NZ-амин (NZ-amine), мясной экстракт, дрожжевой экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыба или продукт ее разложения, обезжиренный соевый жмых или продукт его разложения и т.д. Указанные источники азота могут быть применены по отдельности или в комбинации из двух или более видов, но не ограничиваются этим.
Указанный источник фосфора может включать дигидрофосфат калия, гидрофосфат калия или соответствующие натрийсодержащие соли и т.д. Примеры указанного неорганического соединения могут включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и т.д.
Кроме того, в указанную среду могут быть включены аминокислоты, витамины и/или соответствующие предшественники. В частности, к культуральной среде для указанного штамма могут быть добавлены L-аминокислоты и т.д. В частности, могут быть добавлены глицин, глутамат и/или цистеин, и при необходимости могут быть дополнительно добавлены L-аминокислоты, такие как лизин, но настоящая заявка не ограничивается этим.
Указанные среда или предшественник могут быть добавлены к культуральному продукту периодически или непрерывно, но не ограничиваются этим.
В настоящей заявке рН культурального продукта может быть отрегулирован в процессе культивирования указанного штамма посредством добавления такого соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота, серная кислота и т.д., к указанному культуральному продукту соответствующим образом. Кроме того, в процессе указанного культивирования для предупреждения образования пены может быть добавлен антивспенивающий агент, такой как сложный полигликолевый эфир жирной кислоты. Кроме того, газообразный кислород или кислородсодержащий газ может быть инъецирован в указанный культуральный продукт, чтобы поддерживать аэробное состояние указанного культурального продукта; или может быть инъецирован газообразный азот, водород или диоксид углерода, или газ можно не инъецировать, чтобы поддерживать анаэробное или микроаэробное состояние указанной среды.
Температура указанного культурального продукта может составлять от 25°С до 40°С, и в частности от 28°С до 37°С, но не ограничивается указанными значениями. Указанное культивирование может быть продолжено до тех пор, пока не будет получено желаемое количество подходящих для применения веществ, в частности в течение 1-100 часов, но не ограничивается указанными значениями.
Указанный способ получения может дополнительно включать дополнительный процесс после стадии культивирования. Указанный дополнительный процесс может быть подходящим образом выбран в зависимости от применения глутатиона или его производных.
В частности, указанный способ получения глутатиона или его производных может включать стадию выделения глутатиона или его производных, накопленных в клетках посредством стадии культивирования, например указанный способ может включать стадию выделения глутатиона или его производных из по меньшей мере одного вещества, выбранного из указанных штаммов, их высушенного продукта, их экстракта, их культурального продукта и их лизата, после стадии культивирования. В другом примере указанный способ получения производных глутатиона может включать стадию превращения глутатиона в производное.
Указанный способ может дополнительно включать стадию лизиса указанного штамма перед указанной стадией выделения или одновременно с ней. Лизис указанного штамма может быть выполнен способом, обычно применяемым в области техники, к которой относится настоящая заявка, например, с помощью буферного раствора для лизиса, ультразвукового аппарата, термообработки и френч-пресса и т.д. Кроме того, указанная стадия лизиса может включать, но не ограничивается ими, ферментативные реакции, такие как реакции под действием лиаз клеточных стенок, лиаз нуклеиновых кислот, трансфераз нуклеиновых кислот и протеаз и т.д.
Сухие дрожжи, дрожжевой экстракт, порошковая смесь дрожжевого экстракта, чистый глутатион или его производные, имеющие высокое содержание глутатиона или его производных, могут быть получены посредством способа получения согласно настоящей заявке, но не ограничивается этим, и могут быть подходящим образом получены в соответствии с желаемым продуктом.
В настоящей заявке термин «сухие дрожжи» может быть применен взаимозаменяемо с таким термином, как «высушенный продукт указанного штамма». Указанные сухие дрожжи могут быть получены посредством сушки дрожжевых клеток, в которых накоплен глутатион, и, в частности, могут быть включены в композицию для корма, композицию для пищевого продукта и т.д., но не ограничиваются этим.
В контексте настоящего описания термин «дрожжевой экстракт» может быть применен взаимозаменяемо с таким термином, как «экстракт указанного штамма». Указанный экстракт указанного штамма может означать вещество, остающееся после отделения клеточной стенки от клеток указанного штамма. В частности, он может означать остальные компоненты за исключением клеточной стенки среди компонентов, полученных посредством лизиса указанных клеток. Указанный экстракт указанного штамма содержит глутатион или его производные, и другие компоненты могут включать один или более компонентов, выбранных из белков, углеводов, нуклеиновых кислот и волокон, но не ограниваются ими.
На стадии выделения целевые вещества, то есть глутатион или его производные, могут быть выделены подходящим способом, известным в данной области техники.
Указанная стадия выделения может включать процесс очистки. Указанный процесс очистки может представлять собой очистку посредством отделения от указанного штамма только глутатиона или его производных. В результате указанного процесса очистки может быть получен чистый глутатион.
При необходимости указанный способ получения глутатиона может дополнительно включать стадию смешивания вещества, выбранного из указанных штаммов, полученных после стадии культивирования, их высушенного продукта, их экстракта, их культурального продукта, их лизата и выделенного из них глутатиона, с эксципиентом. Посредством указанной стадии смешивания может быть получена порошковая смесь дрожжевого экстракта. То же самое может быть применено к производным глутатиона.
Указанный эксципиент может быть подходящим образом выбран и применен согласно предполагаемому применению или форме, например, он может быть выбран из крахмала, глюкозы, целлюлозы, лактозы, гликогена, D-маннита, сорбита, лактита, мальтодекстрина, карбоната кальция, синтетического силиката алюминия, гидроортофосфата кальция, сульфата кальция, хлорида натрия, гидрокарбоната натрия, очищенного ланолина, декстрина, альгината натрия, метилцеллюлозы, коллоидного силикагеля, гидроксипропилкрахмала, гидроксипропилметилцеллюлозы,
пропиленгликоля, казеина, лактата кальция, Primojel® и гуммиарабика, в частности одного или более компонентов, выбранных из крахмала, глюкозы, целлюлозы, лактозы, декстрина, гликогена, D-маннита и мальтодекстрина, но не ограничивается ими.
Указанный эксципиент может включать, например, консервант, смачивающий агент, диспергирующий агент, суспендирующий агент, буфер, стабилизирующий агент или изотонический агент, но не ограничивается ими.
[Способ осуществления изобретения]
Настоящая заявка будет подробно описана посредством примеров и экспериментальных примеров. Однако указанные примеры и экспериментальные примеры предоставлены исключительно для иллюстративных целей, и объем настоящей заявки не предназначен для ограничения указанными примерами и экспериментальными примерами.
Пример 1: Улучшение штамма, обеспечивающего получение глутатиона
Для дополнительного улучшения способности обеспечивать получение глутатиона у штамма CJ-5 (KR 10-2222210 В1), который представляет собой обеспечивающий получение глутатиона штамм, депонированный под регистрационным номером КССМ12568Р, индуцировали мутации следующим образом.
Указанный штамм CJ-5 культивировали на твердой среде, а затем инокулировали в бульон с получением культурального раствора, и облучали клетки ультрафиолетовым изучением (УФ) с применением УФ-лампы. После этого посредством выделения получали только мутантные штаммы, которые образовывали колонии после размазывания УФ-облученного культурального раствора по среде для чашек Петри, и выделяли штамм с наибольшим улучшением способности получения глутатиона.
По результатам подтверждения генома выделенного штамма возникли мутации -250(С→Т), -252(G→A), -398(А→Т), -399(А→С), -407(Т→С) и -409(Т→С) выше по ходу транскрипции от GSH1 ORF, и было подтверждено, что цистеин, аминокислота в положении 86 белка GSH1, кодируемого геном gshl, был заменен на аргинин, и аминокислота в положении 653 (глицин) была заменена на метионин.
Указанный выше штамм был назван СС02-2816 и депонирован в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM) согласно Будапештскому договору 8 декабря 2020 года под регистрационным номером КССМ12891Р.
Пример 2: Дополнительный эксперимент по улучшению штамма
Для дополнительного улучшения способности обеспечивать получение глутатиона у штамма СС02-2816 согласно примеру 1 индуцировали мутацию следующим образом.
Указанный штамм СС02-2816 культивировали на твердой среде, а затем инокулировали в бульон с получением культурального раствора, и клетки облучали УФ-светом с применением УФ-лампы. После этого посредством выделения получали только мутантные штаммы, которые образовывали колонии после размазывания УФ-облученного культурального раствора по среде для чашек Петри, и анализировали нуклеотидные последовательности.
По результатам эксперимента было подтверждено, что аминокислота в положении 37 (глицин) SOD1 (SEQ ID NO: 1), которая представляет собой белок, кодируемый sodl, кодирующим супероксиддисмутазу 1 штамма, имеющего улучшенное на 26% содержание глутатиона, была заменена на аргинин.
Пример 3: Эксперимент по мутации 37-го остатка белка SOD1
По результатам примера 2 было установлено, что 37-е положение белка SOD1 будет иметь большое значение для получения глутатиона, и штамм S. cerevisiae дикого типа CEN.PK2-1D и штамм СС02-2816 согласно примеру 1 конструировали для экспрессии мутированного белка, где 37-я аминокислота белка SOD1 была заменена на другую аминокислоту, с определением тем самым, увеличилось ли получение глутатиона.
Для получения штамма, где 37-я аминокислота белка SOD1 из дрожжей S. cerevisiae была заменена на аргинин, применяли плазмиды pWAL100 и pWBR100 со ссылкой на содержание, раскрытое в источнике Lee Т. Н. et al. {J. Microbiol. Biotechnol. (2006), 16(6), 979-982).
В частности, ПЦР проводили следующим образом с применением геномной ДНК штамма CJ-5 в качестве матрицы. ПЦР проводят с применением праймеров SEQ ID NO: 4 и SEQ ID NO: 6 с получением частичной последовательности N-конца SOD1, содержащей N-концевую BamHI-фланкирующую последовательность, инициаторный кодон SOD1 ORF и кодирующую последовательность мутации G37R, и с получением частичной последовательности С-конца SOD1, содержащей С-концевую XhoI-фланкирующую последовательность, терминирующий кодон SOD1 ORF и кодирующую последовательность мутации G37R, с применением праймеров SEQ ID NO: 5 и SEQ ID NO: 7. После этого в результате проведения перекрывающейся ПЦР с применением SEQ ID NO: 4 и SEQ ID NO: 7 на основе указанных двух последовательностей в качестве матрицы получали фрагменты SOD1 ORF, содержащие кодирующую последовательность мутантного белка SOD1, в которой 37-я аминокислота была заменена на аргинин, N-концевой фермент рестрикции BamHI и С-концевой фермент рестрикции XhoI. После обработки с применением BamHI и Xhol указанные фрагменты ORF клонировали в вектор pWALlOO, обработанный тем же ферментом, и получали вектор pWAL100-SODl(G37R)).
Кроме того, проводили ПЦР с применением SEQ ID NO: 8 и SEQ ID NO: 9 на основе геномной ДНК штамма CJ-5 в качестве матрицы с получением фрагмента размером 500 п. о. (пар оснований, bp) ниже по ходу транскрипции от терминирующего кодона SOD1 ORF, содержащего N-концевую последовательность фермента рестрикции SpeI и С-концевую последовательность фермента рестрикции NcoI, и указанный фрагмент обрабатывали ферментами рестрикции SpeI и NcoI. После клонирования указанного фрагмента в pWBR100, обработанный тем же ферментом рестрикции, получали вектор pWBR100-SOD1I.
Для получения фрагмента ДНК, в конечном итоге подлежащего введению в дрожжи, продукт ПЦР, содержащий кодирующую последовательность мутации, содержащей аргинин, и часть K1URA3, получали на основе ранее полученного вектора pWAL100-SOD1 (G37R) в качестве матрицы с применением праймеров SEQ ID NO: 4 и SEQ ID NO: 10, и продукт ПЦР, содержащий часть K1URA3 и фрагмент размером 500 п. о. ниже по ходу транскрипции от терминирующего кодона SOD1, получали на основе вектора pWBR100-SOD1 в качестве матрицы с применением праймеров SEQ ID NO: 11 и SEQ ID NO: 9. Каждый продукт ПЦР трансформировали в S. cerevisiae CEN.PK2-1D и S. cerevisiae СС02-2816 в одном и том же молярном соотношении. ПЦР проводили в условиях денатурации при 95°С в течение 5 минут, отжига при 53°С в течение 1 минуты и полимеризации при 72°С в течение 1 минуты на 1 т.п.о. (тысяч пар оснований, kb). Для трансформации дрожжей применяли метод ацетата лития, модифицированный по сравнению с источником авторства Geitz (Nucleic Acid Research, 20(6), 1425). В частности, после промывания дрожжевых клеток с оптической плотностью (OD), составлявшей от 0,7 до 1,2, дважды с применением буфера, содержавшего ацетат лития/ТЕ, продукты ПЦР и одноцепочечную ДНК (Sigma D-7656) смешивали и подвергали стационарному культивированию в буфере, содержавшем ацетат лития/ТЕ/40% ПЭГ, при 30°С в течение 30 минут и при 42°С в течение 15 минут. После этого указанные клетки культивировали на чашке с синтетической полной (SC) агаровой средой (2% глюкозы) без содержания урацила до тех пор, пока не были отмечены колонии, с получением штамма, в который были введены кодирующая последовательность мутации G37R SOD1 и ген K1URA3. Впоследствии, чтобы удалить K1URA3, каждый штамм культивировали в 2 мл YPD в течение ночи, разбавляли 1/100 и высевали на чашке с SC агаровой средой (2% глюкозы), содержавшей 0,1% 5-фтороротовой кислоты (5-FOA), для конструирования SOD1 G37R-мутантного штамма S. cerevisiae CEN.PK2-1D и SOD G37R-мутанта ого штамма S. cerevisiae СС02-2816, из которых был удален урациловый маркер. Штаммы, способные экспрессировать мутантный белок SOD1 с заменой на аминокислоту, отличающуюся от аргинина, также получали таким же образом за исключением того, что применяли пару праймеров, где последовательность, кодирующая 37-ю аминокислоту, представлявшую собой аргинин, в последовательностях праймеров SEQ ID NO: 5 и SEQ ID NO: 6, была заменена на последовательность, кодирующую другую аминокислоту.
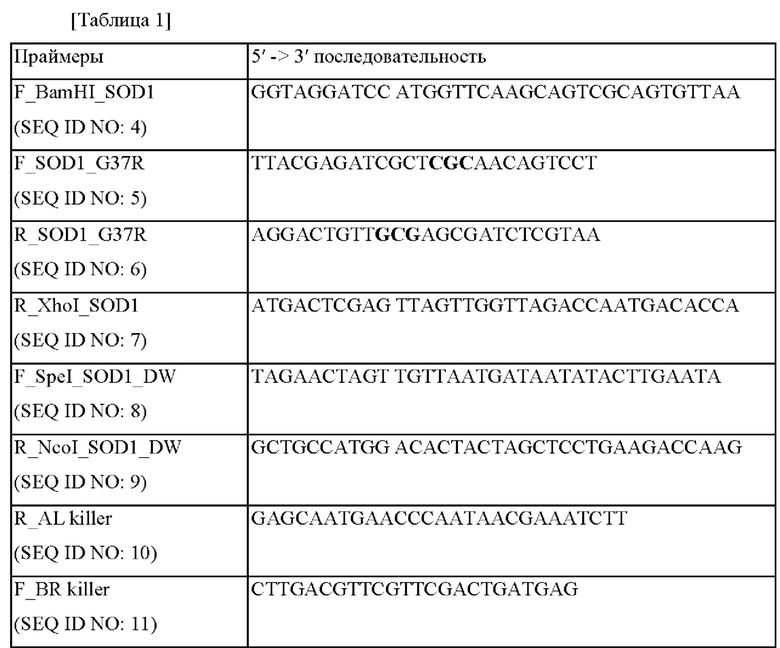
Результаты измерения концентрации и содержания глутатиона (GSH), полученного посредством культивирования каждого штамма, полученного выше, в течение 26 часов, представлены в таблицах 2 и 3 ниже и на фиг. 1 и 2.
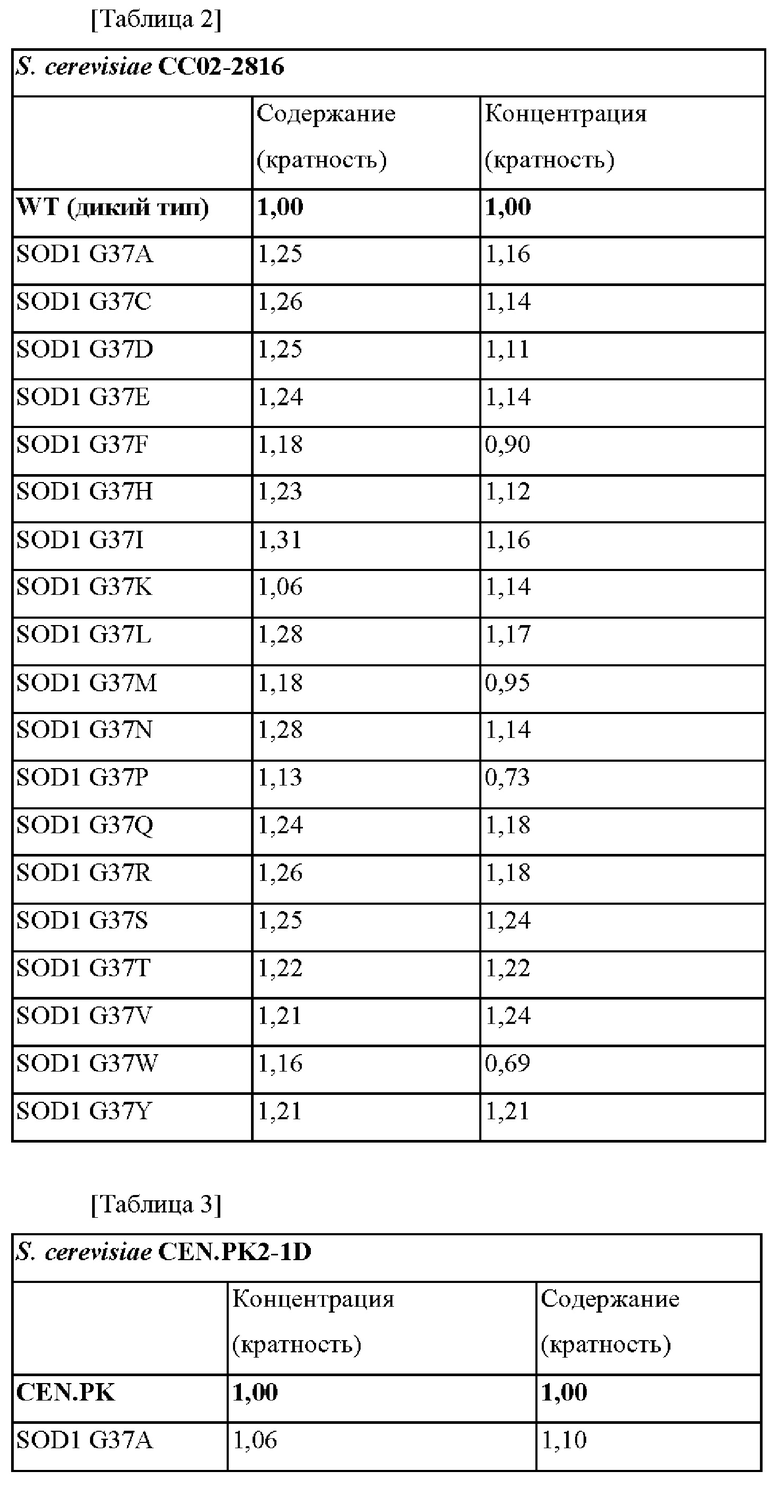
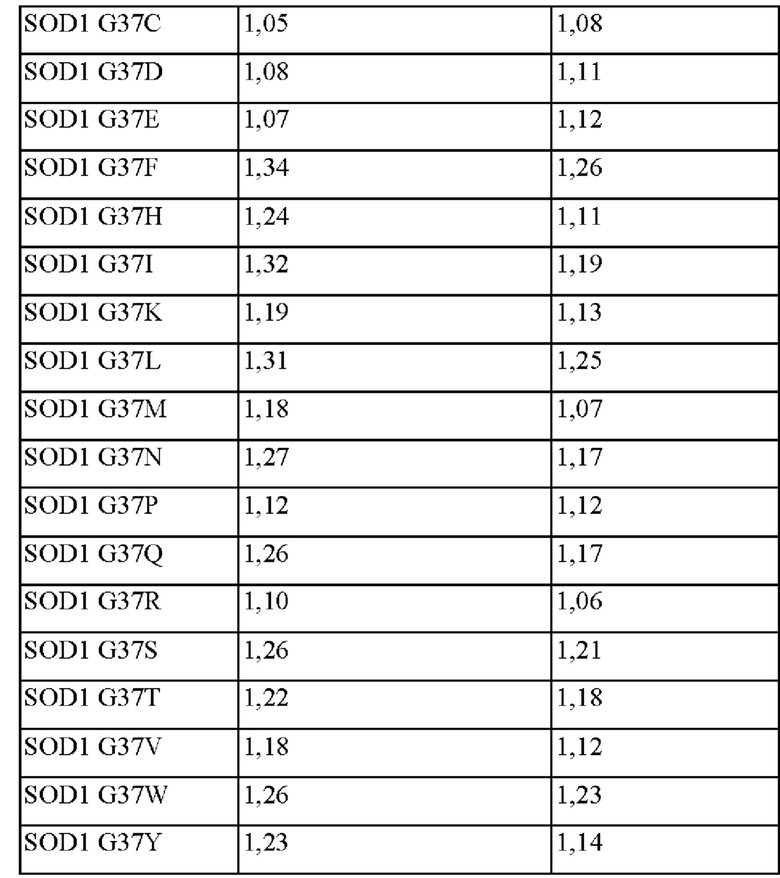
На основании результатов было подтверждено, что способность обеспечивать получение глутатиона может быть увеличена посредством замены аминокислоты в положении 37 на другие аминокислоты, и, соответственно, может подразумеваться, что вариант белка SOD1, предложенный в настоящей заявке, может быть эффективно применен для получения галша-глутамилпептидов.
Из вышеизложенного специалист в области техники, к которой относится настоящая заявка, сможет понять, что настоящая заявка может быть воплощена в других конкретных формах без изменения технических понятий или существенных характеристик настоящей заявки. В этой связи иллюстративные воплощения, раскрытые в настоящем документе, предназначены исключительно для иллюстративных целей и не должны рассматриваться как ограничивающие объем настоящей заявки. Таким образом, объем настоящей заявки определяется прилагаемой формулой настоящего изобретения, а не вышеизложенным описанием. Все изменения, которые подпадают под смысл и диапазон эквивалентности формулы настоящего изобретения, должны быть включены в объем настоящей заявки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> Варинат супероксиддисмутазы 1 и способ получения глутатиона
или его производного с применением указанного варианта
<130> OPA22037
<150> KR 10-2021-0075689
<151> 2021-06-10
<160> 30
<170> KoPatentIn 3.0
<210> 1
<211> 154
<212> PRT
<213> Saccharomyces cerevisiae
<400> 1
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Gly Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 2
<211> 465
<212> DNA
<213> Saccharomyces cerevisiae
<400> 2
atggttcaag cagtcgcagt gttaaagggt gatgccggtg tctctggtgt tgtcaagttc 60
gaacaggctt ccgaatccga gccaaccact gtctcttacg agatcgctgg taacagtcct 120
aacgcagaac gtgggttcca cattcatgag tttggagatg ccaccaatgg ttgtgtctct 180
gctggtcctc acttcaatcc tttcaagaag acacatggtg ctccaactga cgaagtcaga 240
catgtcggtg acatgggtaa cgtaaagacg gacgaaaatg gtgtggccaa gggctccttc 300
aaggactctt tgatcaagct tatcggtcct acctccgttg taggcagaag cgtcgttatc 360
cacgccggcc aagatgactt aggtaagggt gacactgaag aatctttgaa gactggtaat 420
gccggtccaa gaccagcctg tggtgtcatt ggtctaacca actaa 465
<210> 3
<211> 465
<212> DNA
<213> Unknown
<220>
<223> Saccharomyces cerevisiae_CC02-2816_sod1
<400> 3
atggttcaag cagtcgcagt gttaaagggt gatgccggtg tctctggtgt tgtcaagttc 60
gaacaggctt ccgaatccga gccaaccact gtctcttacg agatcgctgg taacagtcct 120
aacgcagaac gtgggttcca tattcatgag tttggagatg ccaccaatgg ttgtgtctct 180
gctggtcctc acttcaatcc tttcaagaag acacacggtg ctccaactga cgaagtcaga 240
catgtcggtg acatgggtaa cgtaaagacg gacgaaaatg gtgtggccaa gggctccttc 300
aaggactctt tgatcaagct tatcggtcct acctccgttg taggcagaag cgtcgttatc 360
cacgccggcc aagatgactt aggtaagggt gacactgaag aatctttgaa gactggtaat 420
gccggtccaa gaccagcctg tggtgtcatt ggtctaacca actaa 465
<210> 4
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> F_BamHI_SOD1
<400> 4
ggtaggatcc atggttcaag cagtcgcagt gttaa 35
<210> 5
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> F_SOD1_G37R
<400> 5
ttacgagatc gctcgcaaca gtcct 25
<210> 6
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> R_SOD1_G37R
<400> 6
aggactgttg cgagcgatct cgtaa 25
<210> 7
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> R_XhoI_SOD1
<400> 7
atgactcgag ttagttggtt agaccaatga cacca 35
<210> 8
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> F_SpeI_SOD1_DW
<400> 8
tagaactagt tgttaatgat aatatacttg aata 34
<210> 9
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> R_NcoI_SOD1_DW
<400> 9
gctgccatgg acactactag ctcctgaaga ccaag 35
<210> 10
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> R_AL killer
<400> 10
gagcaatgaa cccaataacg aaatctt 27
<210> 11
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> F_BR killer
<400> 11
cttgacgttc gttcgactga tgag 24
<210> 12
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37A
<400> 12
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Ala Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 13
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37C
<400> 13
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Cys Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 14
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37D
<400> 14
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Asp Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 15
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37E
<400> 15
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Glu Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 16
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37F
<400> 16
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Phe Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 17
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37H
<400> 17
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala His Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 18
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37I
<400> 18
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Ile Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 19
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37K
<400> 19
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Lys Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 20
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37L
<400> 20
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Leu Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 21
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37M
<400> 21
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Met Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 22
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37N
<400> 22
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Asn Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 23
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37P
<400> 23
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Pro Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 24
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37Q
<400> 24
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Gln Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 25
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37R
<400> 25
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Arg Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 26
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37S
<400> 26
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Ser Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 27
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37T
<400> 27
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Thr Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 28
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37V
<400> 28
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Val Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 29
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37W
<400> 29
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Trp Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<210> 30
<211> 154
<212> PRT
<213> Artificial Sequence
<220>
<223> SOD1 G37Y
<400> 30
Met Val Gln Ala Val Ala Val Leu Lys Gly Asp Ala Gly Val Ser Gly
1 5 10 15
Val Val Lys Phe Glu Gln Ala Ser Glu Ser Glu Pro Thr Thr Val Ser
20 25 30
Tyr Glu Ile Ala Tyr Asn Ser Pro Asn Ala Glu Arg Gly Phe His Ile
35 40 45
His Glu Phe Gly Asp Ala Thr Asn Gly Cys Val Ser Ala Gly Pro His
50 55 60
Phe Asn Pro Phe Lys Lys Thr His Gly Ala Pro Thr Asp Glu Val Arg
65 70 75 80
His Val Gly Asp Met Gly Asn Val Lys Thr Asp Glu Asn Gly Val Ala
85 90 95
Lys Gly Ser Phe Lys Asp Ser Leu Ile Lys Leu Ile Gly Pro Thr Ser
100 105 110
Val Val Gly Arg Ser Val Val Ile His Ala Gly Gln Asp Asp Leu Gly
115 120 125
Lys Gly Asp Thr Glu Glu Ser Leu Lys Thr Gly Asn Ala Gly Pro Arg
130 135 140
Pro Ala Cys Gly Val Ile Gly Leu Thr Asn
145 150
<---
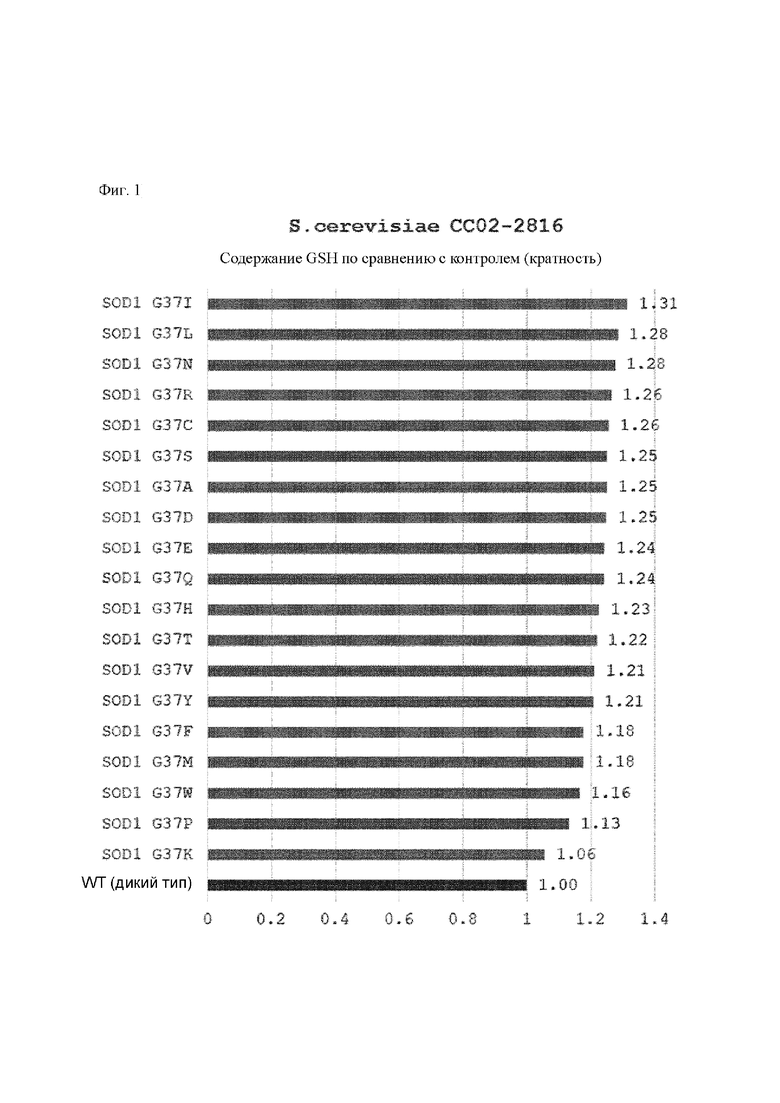
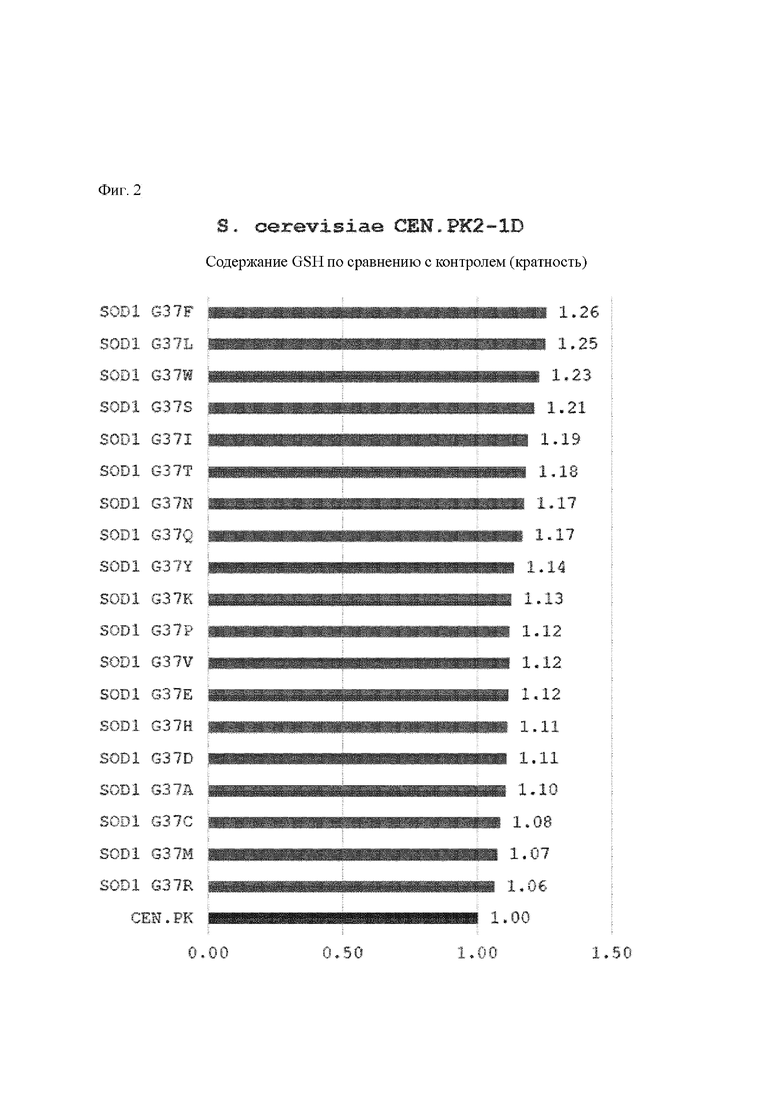
Изобретение относится к биотехнологии. Предложен полипептид, участвующий в продуцировании глутатиона, где аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту. Также предложены полинуклеотид, кодирующий указанный полипептид, микроорганизм, который продуцирует глутатион и содержит указанный полипептид или полинуклеотид, кодирующий указанный полипептид, и способ продуцирования глутатиона. Изобретение позволяет увеличить выход глутатиона. 4 н. и 6 з.п. ф-лы, 2 ил., 3 табл., 3 пр.
1. Полипептид, участвующий в продуцировании глутатиона, где аминокислота, соответствующая положению 37 от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту.
2. Полипептид по п. 1, где указанная другая аминокислота выбрана из аланина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, цистеина, тирозина, аспарагина, глутамина, аспарагиновой кислоты, глутаминовой кислоты, лизина, аргинина и гистидина.
3. Полипептид по п. 1, содержащий любую из аминокислотных последовательностей SEQ ID NO: 12-30.
4. Полинуклеотид, кодирующий полипептид по любому из пп. 1-3.
5. Микроорганизм для продуцирования глутатиона, содержащий любое одно или более из полипептида по любому из пп. 1-3 и полинуклеотида, кодирующего указанный полипептид.
6. Микроорганизм по п. 5, где микроорганизм продуцирует глутатион.
7. Микроорганизм по п. 5, где микроорганизм относится к роду Saccharomyces.
8. Микроорганизм по п. 7, представляющий собой Saccharomyces cerevisiae.
9. Способ получения глутатиона, включающий культивирование микроорганизма, содержащего любое одно или более из полипептида по любому из пп. 1-3 и полинуклеотида, кодирующего указанный полипептид, в среде.
10. Способ по п. 9, дополнительно включающий выделение глутатиона из по меньшей мере одного вещества, выбранного из культивируемого микроорганизма, его высушенного продукта, его экстракта, его культурального продукта и его лизата.
| Центробежный регулятор скорости, действующий торможением | 1927 |
|
SU12638A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| AI Y., et al | |||
| Inhibition effect of expression of Cu/Zn superoxide dismutase from rice on synthesis of Glutathione | |||

