Предшествующий уровень техники
Область техники, к которой относится изобретение
Настоящее изобретение относится к способу получения вариантов гиалуронидазы PH20, которые включают одну или несколько замен аминокислотных остатков и при необходимости включают делеции некоторых N-концевых и/или С-концевых аминокислотных остатков в гиалуронидазе, в частности, РН20 дикого типа или зрелой PH20 дикого типа, или в аминокислотных последовательностях РН20 дикого типа или зрелой РН20 дикого типа.
Описание предшествующего уровня техники
Гиалуронидазы представляют собой ферменты, расщепляющие гиалуроновую кислоту, присутствующую во внеклеточном матриксе. Гиалуронидазы гидролизуют гиалуроновую кислоту, тем самым снижая вязкость гиалуроновой кислоты во внеклеточном матриксе и увеличивая проникновение в ткани (кожу) (Bookbinder et al., 2006). Гиалуронидазы использовались для улучшения абсорбции жидкостей организма путем подкожной инъекции [Muchmore et al., 2012] или внутримышечной инъекции [Krantz et al., 2016], а также для улучшения диффузии местных анестетиков [Clement et al. et al., 2003]. Препараты, проникновение которых при подкожном введении улучшают таким способом, включают морфин [Thomas et al., 2009], цефтриаксон [Harb et al., 2010], инсулин [Muchmore et al., 2012] и иммуноглобулин [Wasserman et al., 2014]. Гиалуронидазы также используют для улучшения диспергирования жидкостей или лекарств, проникающих в ткани при внутривенной инъекции или диффузии гематомы. Среди гиалуронидаз рекомбинантный человеческий белок PH20, обладающий активностью при нейтральном значении pH, был разработан Halozyme Therapeutics Inc. и продается под торговым названием «Hylenex» (Bookbinder et al., 2006).
Сообщалось, что рекомбинантные белки PH20 человека экспрессируются в дрожжах (P. pastoris), клетках насекомых DS-2 и клетках животных. Рекомбинантные белки РН20, продуцируемые в клетках насекомых и дрожжей, отличаются от РН20 человека характером N-гликозилирования при посттрансляционной модификации, что влияет на их активность и влечет за собой риск возникновения побочных эффектов в организме.
Для разработки биофармацевтических препаратов важными факторами, обеспечивающими гибкость производства, являются однородность продукта и длительный срок хранения. Во время производства возникают различные типы микрогетерогенности из-за различий в размере и заряде, появляющихся в результате ферментативной или спонтанной деградации и модификаций. Каждая из химических и ферментативных модификаций, таких как дезамидирование и сиалилирование, увеличивает суммарный отрицательный заряд и снижает значение pI в антителе, тем самым образуя кислые варианты [Harris RJ et al., 2004]. Кроме того, отщепление С-концевого лизина приводит к потере суммарных положительных зарядов и образованию кислых вариантов. Образование основных вариантов может происходить в результате амидирования С-концевого лизина или глицина, образования сукцинимида, окисления аминокислот или удаления сиаловой кислоты, что устраняет дополнительные положительные или отрицательные заряды; оба типа модификации увеличивают значение pI [Harris RJ et al., 2004].
Гликозилирование представляет собой посттрансляционный процесс у белков в клетках (эукариот) и происходит в эндоплазматическом ретикулуме и аппарате Гольджи. Гликозилирование подразделяется на N-гликозилирование и O-гликозилирование, которые различаются в зависимости от присоединенной функциональной группы. Процесс присоединения сахаров, таких как лактоза или фукоза, к белкам, вырабатываемым в клетках, в широком смысле называется «гликозилированием». Когда гликан соединяется с белком посредством гликозилирования, белок подвергается процессу «фолдинга» с образованием трехмерной структуры. Это придает белку стабильность, так что он может сохраняться в течение длительного времени без высвобождения (развертывания). Этот гликан участвует в важном процессе, который обеспечивает связь и обмен информацией между клетками.
Существует два типа реакций присоединения гликанов к белкам: N-связанное или O-связанное гликозилирование. Эти два процесса гликозилирования различаются по механизму синтеза и присоединения гликанов, и среди них лучше известны механизм и роль N-гликозилирования. Гликаны, добавленные посредством N-гликозилирования, называются «N-гликанами» и образуются в эндоплазматическом ретикулуме. Ряд ферментов ALG (аспарагин-связанного гликозилирования) на долихол-пирофосфате (PP-Dol), присутствующем в мембране эндоплазматического ретикулума, добавляются к N-ацетилглюкозамину (GlcNAc), маннозе (Man), глюкозе (Glc) и т.д. для окончательного синтеза Glc3Man9GlcNAc2-PP-Dol, представляющего собой сложный гликан в форме олигосахарида, связанного с липидом (LLO). Синтезированный LLO переносится на последовательность N-гликозилирования пептида, включающую N-x-S/T, которая напрямую транслируется с рибосомы в эндоплазматический ретикулум посредством механизма ко-трансляционной транслокации с помощью олигосахаридтрансферазы, состоящей из 8 или более субъединиц. N-гликаны, присоединенные к белкам, удаляются один за другим с конца гликана глюкозидазой (α-глюкозидазой I, Gls1p; α-глюкозидазой II, Glsp II), присутствующей в эндоплазматическом ретикулуме. Поскольку вторая и третья глюкоза удаляются медленнее, фолдинг завершается с помощь лектиновых шаперонов, кальнексина и кальретикулина. Когда вся глюкоза отщеплена, фолдинг белка считается завершенным, и присоединенные к белку гликаны Man8GlcNAc2 переносятся в аппарат Гольджи. В связи с этим существует процесс контроля качества, когда еще раз проверяется фолдинг гликопротеинов перед их переносом из эндоплазматического ретикулума в аппарат Гольджи. Если надлежащий фолдинг не достигнут, процесс повторяется, давая возможность завершить фолдинг за счет поступления одной молекулы глюкозы в цикл кальнексина/кальретикулина.
Описанный выше начальный процесс биосинтеза N-гликанов в эндоплазматическом ретикулуме сохраняется почти одинаковым у широкого круга организмов от дрожжей, простых эукариотических микроорганизмов, до животных, т.е. высших организмов. Однако гликаны, перенесенные в аппарат Гольджи, претерпевают различные гликановые модификации, специфичные для каждого вида, что приводит к образованию совершенно разных типов гликанов у дрожжей, насекомых, растений и животных. Однако эти различные гликаны также имеют общий центральный участок, а именно структуру, в которой три маннозы и два GlcNAc связаны с азотом аспарагина, называемую «триманнозильным ядром». Форма, в которой манноза в основном связана с триманнозильным ядром, называется «типом с высоким содержанием маннозы» и часто встречается у дрожжей и плесени. Структура, для которой в аппарате Гольджи последовательно добавляются десятки манноз, также обнаружена у широко известных дрожжей Saccharomyces cerevisiae. С другой стороны, в клетках насекомых обнаружены гликопротеины, имеющие гликаны дефицитного по маннозе типа (олигоманнозного типа). Сначала они обрезаются с образованием Man5GlcNAc2 с помощью маннозидазы IA, IB и IC в аппарате Гольджи, N-ацетилглюкозаминилтрансфераза (GNT) I добавляет один GlcNAc к Man5GlcNAc2, а затем маннозидаза II производит гибридную структуру, в которой один GlcNAc добавлен к триманнозильному ядру. Затем добавленный GlcNAc снова отщепляется, образуя структуру гликана маннозо-дефицитного типа. В клетках животных α(1,6)-фукоза часто добавляется к первому GlcNAc, связанному с аспарагиновым остатком. У животных GNT II действует на структуру, в которой один GlcNAc добавлен к триманнозильному ядру, и добавляет к ним другой GlcNAc, образуя гликан, имеющий двухантенную структуру. Затем GNT IV и V могут обеспечивать формирование четырехантенных структур, а в некоторых случаях GNT VI, IX или VB могут формировать шестиантенные структуры. После добавления GlcNAc для формирования антенного каркаса β-галактозилтрансфераза и α-сиалилтрансфераза, присутствующие в теле Гольджи, формируют сложную гликановую структуру, в которой галактоза и сиаловая кислота добавлены к GlcNAc.
N-гликозилирование может сильно влиять на фолдинг или активность белков, и существует очень высокая вероятность того, что наличие гликозилирования, а также структура или форма гликанов могут варьировать в зависимости от типа клетки-хозяина, метода рекомбинантной манипуляции и условий культивирования (Schilling et al., 2002) при получении белков или их природных вариантов с использованием методов генной инженерии в промышленности. То есть количественные различия в структуре гликана или сахарных компонентах, составляющих гликан, возникают в зависимости от различий в условиях получения в процессе продукции белка. Условия культивирования, влияющие на N-гликозилирование, включают концентрацию глюкозы или глутамина в культуральной среде (Tachibana et al. 1994), концентрацию растворенного кислорода (DO) (Restelli et al. 2006), рН культуральной среды (Borys et al., 1993), концентрацию аммиака в культуральной среде (Borys et al., 1994;), температуру культуры (Clark et al., 2004) и т.п.
В общем, фермент специфически связывается с субстратом с образованием комплекса фермент-субстрат, который действует как катализатор, снижая энергию активации реакции и ускоряя реакцию фермента. Фермент может различать субстрат и молекулы, конкурирующие с субстратом, и способен специфически связываться с субстратом за счет распределения комплементарных зарядов в положении, в котором фермент связывается с субстратом, комплементарной структуры и распределения гидрофильности и гидрофобности. Модель «замок-ключ», в которой фермент и субстрат комплементарны друг другу и имеют геометрическую форму, была предложена для выяснения положения связывания фермента, но не дает удовлетворительного объяснения переходного состояния фермент-субстратного комплекса. Согласно модели индуцированного соответствия, предложенной для преодоления этой проблемы, поскольку фермент и субстрат продолжают взаимодействовать друг с другом в комплексе фермент-субстрат, они изменяют его структуру на основе гибкой структуры ферментного белка. В этом процессе реакции может дополнительно способствовать распределение окружающих электрических зарядов по аминокислотным остаткам или N-гликанам, составляющим активные центры, или распределение гидрофильности/гидрофобности.
Кроме того, взаимодействие зарядов необходимо для реакции гиалуронидазы, которая представляет собой фермент, гидролизующий гиалуроновую кислоту в качестве субстрата. Arming et al. показали, что аргининовый остаток, имеющий положительные заряды в гиалуронидазе PH20, необходим для ферментативной активности для связывания с гиалуроновой кислотой, которая является субстратом, имеющим большое количество распределенных в ней отрицательных зарядов (Arming et al. 1997). Следовательно, можно сделать вывод, что распределение заряда N-гликана также влияет на такую ферментативную активность. Важно доказать, что при связывании с гиалуронидазой гиалуроновой кислоты, которая является субстратом, имеющим большое количество отрицательных зарядов, уровень отрицательно заряженных сахаров, покрывающих сиаловую кислоту в N-гликанах, то есть уровень сиалилирования, влияет на образование фермент-субстратного комплекса или ход ферментативной реакции. Для ограничения уровня сиалилирования следует ограничить перенос сиаловой кислоты на галактозный остаток, провести десиалилирование или ограничить уровень галактозилирования.
Таким образом, на продуктивность и активность рекомбинантной гиалуронидазы PH20 и ее вариантов влияет изменение уровня N-гликанов, поэтому исследования по разработке способа получения гиалуронидазы PH20 или ее варианта, обладающих высокой активностью и продуктивностью, и применимых в промышленности, путем контроля и поддержания уровня N-гликанов, необходимы для эффективного массового производства в данной области техники.
Изложение сущности изобретения
Таким образом, настоящее изобретение было сделано с учетом вышеуказанных проблем, и одной из целей настоящего изобретения является обеспечение способа культивирования клеток-хозяев, которые продуцируют рекомбинантную гиалуронидазу PH20 или ее вариант, и гиалуронидазы PH20 или ее варианта, полученных этим способом, в частности, способа получения гиалуронидазы РН20 или ее варианта, обладающего улучшенной ферментативной активностью и продуктивностью.
Цели настоящего изобретения не ограничиваются описанными выше. Другие цели, не описанные в настоящей заявке, будут понятны специалистам в данной области техники из следующего описания.
В настоящем изобретении, когда клетки-хозяева для продукции рекомбинантной гиалуронидазы PH20 или ее варианта культивируют в определенных условиях культивирования, характеристики N-гликозилирования полученной рекомбинантной гиалуронидазы PH20 или ее варианта и, кроме того, ферментативная активность полученной гиалуронидазы PH20 или ее варианта значительно улучшаются.
В частности, было обнаружено, что можно эффективно повышать активность продуцируемой гиалуронидазы PH20 или ее варианта путем контроля уровней сиалилирования, галактозилирования и/или маннозилирования, в частности, уровня сиалилирования в N-гликанах, и что ферментативное активность и продуктивность гиалуронидазы РН20 или ее варианта по настоящему изобретению могут быть заметно улучшены путем проведения культивирования в течение определенного периода времени после изменения температуры культивирования.
В частности, способ получения гиалуронидазы PH20 или ее варианта по настоящему изобретению включает:
(1) культивирование клеток-хозяев, экспрессирующих рекомбинантную гиалуронидазу PH20 или ее вариант, при температуре культивирования от 35°C до 38°C до интегрального количества жизнеспособных клеток 20×106-120×106 клеток в день/мл; и
(2) снижение температуры культивирования до 28°C-34°C и последующее культивирование клеток-хозяев в течение от 2 до 18 дней при поддержании температуры культивирования в соответствии по меньшей мере с одним способом, выбранным из группы, состоящей из:
(а) культивирования клеток-хозяев при поддержании концентрации остаточной глюкозы в среде от 0,001 г/л до 4,5 г/л в течение периода культивирования; и
(b) культивирование клеток-хозяев при поддержании рН культуральной среды от 6,8 до 7,2.
PH20 или её вариант, произведенный способом получения в соответствии с настоящим изобретением, характеризуется тем, что уровень сиалилирования N-гликана полученной PH20 или ее варианта составляет от 1 до 38%, при этом ферментативная активность заметно повышается, но не ограничивается этим, и значение является экспериментальным значением, имеющим диапазон ошибки 10%. Причина этого заключается в том, что отклонения возникают в зависимости от таких условий, как оборудование, используемое для культивирования, и навыки работы аналитика при установке условий культивирования, поэтому числовые значения, установленные в настоящем изобретении, следует интерпретировать в более широком смысле с учетом отклонений. а не в узком смысле. Клетки-хозяева можно культивировать одним или несколькими способами, выбранными из группы, состоящей из периодического культивирования, повторного периодического культивирования, периодического культивирования с подпиткой, повторного периодического культивирования с подпиткой, непрерывного культивирования и перфузионного культивирования.
Краткое описание чертежей
Вышеупомянутые и другие цели, признаки и другие преимущества настоящего изобретения станут более понятными из следующего подробного описания, взятого вместе с прилагаемыми чертежами, на которых:
фиг. 1 иллюстрирует результат анализа ферментативной активности, результаты изоэлектрофокусирования и определения содержания N-гликанов в отношении гиалуронидазы человека PH20 дикого типа и ее вариантов, где:
фиг. 1A иллюстрирует ферментативную активность гиалуронидазы человека PH20 дикого типа и ее вариантов;
фиг. 1В иллюстрирует результат изоэлектрофокусирования каждого образца; и
фиг. 1C иллюстрирует содержание N-гликанов;
фиг. 2 иллюстрирует результат сравнительного анализа ферментативной активности и профиля изоэлектрофокусирования между очищенными фракциями варианта гиалуронидазы PH20, обработанными PNGase F (пептид-N-гликозидазой F), сиалидазой A и сиалидазой A + галактозидазой, где:
фиг. 2А иллюстрирует результат изоэлектрофокусирования каждого образца;
фиг. 2B иллюстрирует ферментативную активность; и
фиг. 2C иллюстрирует содержание N-гликанов;
фиг. 3 иллюстрирует результат анализа изменений роста клеток, жизнеспособности клеток, pH и концентрации лактата в зависимости от условий культивирования при добавлении N-ацетил-D-маннозамина или галактозы по отношению к клеткам, продуцирующим варианты гиалуронидазы PH20, где:
фиг. 3А иллюстрирует изменение роста клеток;
фиг. 3B иллюстрирует изменение жизнеспособности клеток;
фиг. 3C иллюстрирует изменение рН; а
фиг. 3D иллюстрирует изменение концентрации лактата;
фиг. 4 представляет собой график, показывающий активность собранной клеточной культуральной жидкости для получения вариантов гиалуронидазы РН20 в зависимости от условий культивирования при добавлении N-ацетил-D-маннозамина или галактозы;
фиг. 5 иллюстрирует результат изоэлектрофокусирования собранной клеточной культуральной жидкости для получения вариантов гиалуронидазы PH20 в зависимости от условий культивирования при добавлении N-ацетил-D-маннозамина или галактозы;
фиг. 6 иллюстрирует результат анализа структуры N-гликанов собранной клеточной культуральной жидкости для получения вариантов гиалуронидазы PH20 в зависимости от условий культивирования при добавлении N-ацетил-D-маннозамина или галактозы;
фиг. 7 иллюстрирует изменения роста клеток, жизнеспособности клеток и концентрации аммиака клеток для продукции вариантов гиалуронидазы PH20 в зависимости от количества дней культивирования, где:
фиг. 7А показывает изменение роста клеток;
фиг. 7B показывает изменение жизнеспособности клеток;
фиг. 7C показывает изменение интегральной плотности жизнеспособных клеток; и
фиг. 7D иллюстрирует изменение концентрации аммиака;
фиг. 8 представляет собой график, показывающий удельную активность собранной жидкости от культивирования клеток для получения вариантов гиалуронидазы PH20 в зависимости от дня культивирования клеток;
фиг. 9 иллюстрирует результат изоэлектрофокусирования собранной клеточной культуральной жидкости для получения вариантов гиалуронидазы PH20 в зависимости от дня культивирования клеток;
фиг. 10 иллюстрирует результаты анализа структуры и активности N-гликанов собранной клеточной культуральной жидкости для получения вариантов гиалуронидазы PH20 в зависимости от дня культивирования клеток;
фиг. 11 иллюстрирует изменение роста клеток в зависимости от условий концентрации глюкозы, жизнеспособности клеток, рН, осмоляльности, концентрации глюкозы и концентрации лактата по отношению к клеткам, продуцирующим варианты гиалуронидазы PH20, где:
фиг. 11А иллюстрирует изменение роста клеток;
фиг. 11В иллюстрирует изменение жизнеспособности клеток;
фиг. 11C иллюстрирует изменение pH;
фиг. 11D иллюстрирует изменение осмоляльности;
фиг. 11E иллюстрирует изменение концентрации глюкозы; а
фиг. 11F иллюстрирует изменение концентрации лактата;
фиг. 12 представляет собой график, показывающий удельную активность собранной жидкости от клеточной культуры, продуцирующей варианты гиалуронидазы РН20, в зависимости от концентрации глюкозы;
фиг. 13 иллюстрирует результат изоэлектрофокусирования собранной жидкости от клеточной культуры, продуцирующей варианты гиалуронидазы PH20, в зависимости от концентрации глюкозы;
фиг. 14 иллюстрирует результат анализа структуры N-гликанов собранной жидкости от клеточной культуры для продукции вариантов гиалуронидазы PH20 в зависимости от концентрации глюкозы;
фиг. 15 иллюстрирует результаты анализа изменений роста клеток, жизнеспособности клеток, pH и интегральной плотности жизнеспособных клеток в зависимости от уровней pH по отношению к клеткам, продуцирующим варианты гиалуронидазы PH20, где:
фиг. 15А иллюстрирует изменение роста клеток;
фиг. 15В иллюстрирует изменение жизнеспособности клеток;
фиг. 15C иллюстрирует изменение рН; а
фиг. 15D иллюстрирует изменение интегральной плотности жизнеспособных клеток;
фиг. 16 представляет собой график, показывающий удельную активность собранной культуральной жидкости для клеток, продуцирующих варианты гиалуронидазы PH20, в зависимости от уровней pH;
фиг. 17 иллюстрирует результат изоэлектрофокусирования собранной культуральной жидкости для клеток, продуцирующих варианты гиалуронидазы PH20, в зависимости от уровня pH;
фиг. 18 представляет собой хроматограмму очистки, полученную посредством первичной хроматографии на анионообменной смоле во время очистки варианта PH20 гиалуронидазы;
фиг. 19 представляет собой хроматограмму очистки, полученную с помощью вторичной анионообменной хроматографии во время очистки варианта гиалуронидазы PH20; и
фиг. 20 представляет собой хроматограмму очистки, полученную посредством хроматографии с катионообменной смолой во время очистки варианта гиалуронидазы PH20.
Подробное описание изобретения
Если не указано иное, все технические и научные термины, используемые в настоящей заявке, имеют такие же значения, как их понимают специалисты в области техники, к которой относится настоящее изобретение. В целом, используемая здесь номенклатура хорошо известна и обычно используется в данной области техники.
Настоящее изобретение основано на обнаружении того, что уровни сиалилирования, галактозилирования и/или маннозилирования, в частности, уровень сиалилирования N-гликана, имеют решающее значение для ферментативной активности и продуктивности гиалуронидазы PH20 или ее вариантов при получении гиалуронидазы PH20 или ее вариантов для промышленного применения с использованием метода генной инженерии.
В частности, было обнаружено, что при условиях, когда уровень сиалилирования N-гликана РН20 или ее варианта составляет от 1 до 38%, предпочтительно от 1 до 30%, более предпочтительно от 1,5 до 28% и наиболее предпочтительно от 2 до 25%, ферментативная активность и продуктивность гиалуронидазы PH20 или ее варианта улучшаются до неожиданно высоких уровней.
Более конкретно, было обнаружено, что когда уровень галактозилирования N-гликанов РН20 или ее варианта составляет от 1 до 68%, уровень сиалилирования составляет от 1 до 38%, а уровень маннозилирования составляет от 40 до 63%, предпочтительно, когда уровень галактозилирования составляет от 5 до 60%, уровень сиалилирования составляет от 1 до 30%, а уровень маннозилирования составляет от 42 до 62%, более предпочтительно, когда уровень галактозилирования составляет от 10 до 56%, уровень сиалилирования составляет от 1,5 до 28%, а уровень маннозилирования составляет от 44 до 61%, и наиболее предпочтительно, когда уровень галактозилирования составляет от 15 до 50%, уровень сиалилирования составляет от 2 до 25%, а уровень маннозилирования составляет от 47 до 60%, ферментативная активность и продуктивность гиалуронидазы PH20 или ее варианта повышаются до неожиданно высокого уровня.
Приведенные выше числовые значения получены из экспериментальных результатов примеров, показанных в таблице 1, и представляют собой числовые значения, полученные с доверительным интервалом 95%, и такие числовые значения могут также включать ошибку 10%. Это связано с тем, что измерения уровней сахара (глюкозы) в белках различаются в зависимости от таких условий, как оборудование, используемое для эксперимента, время ферментативной реакции, температура испытания, квалификация аналитика и т.д., поэтому уровень глюкозы, измеренный в настоящем изобретении, следует интерпретировать в более широком смысле с учетом различий между лабораториями, а не в ограниченном смысле.
В настоящем изобретении доля галактозилирования (%) N-гликанов представляет собой сумму доли (%) гликанов, содержащих галактозу на своем конце, таких как G1, G1F, G1F', G2, G2F, A1, A1F, A2 и A2F, среди N-гликанов, доля сиалилирования (%) представляет собой сумму доли (%) N-гликанов, содержащих сиаловую кислоту на своем конце, таких как A1, A1F, A2 и A2F, среди N-гликанов, а доля маннозилирования (%) представляет собой сумму доли N-гликанов, содержащих маннозу на своем конце, таких как M4G0F, M5, M5G0, M6, M7 и M8, среди N-гликанов.
В частности, способ получения гиалуронидазы PH20 или ее варианта с уровнем сиалилирования в N-гликане от 1 до 38% по настоящему изобретению включает:
(1) культивирование клеток-хозяев, экспрессирующих рекомбинантную гиалуронидазу PH20 или ее вариант, при температуре культивирования от 35°C до 38°C до интегральной плотности жизнеспособных клеток от 20×106 до 120×106 клеток в сутки/мл; и
(2) снижение температуры культивирования до 28°C-34°C и последующее культивирование клеток-хозяев в течение от 2 до 18 дней при поддержании температуры культивирования в соответствии по меньшей мере с одним способом, выбранным из группы, состоящей из:
(a) культивирования клеток-хозяев при поддержании концентрации остаточной глюкозы в среде от 0,001 г/л до 4,5 г/л в течение периода времени культивирования; и
(b) культивирования клеток-хозяев при поддержании рН культуральной среды от 6,8 до 7,2.
Приведенное выше значение является экспериментальным значением с погрешностью в 10%. Причина заключается в том, что изменения происходят в зависимости от таких условий, как оборудование, используемое для культивирования, и навыки работы аналитика при установке условий культивирования, поэтому числовые значения, установленные в настоящем изобретении, следует интерпретировать в более широком смысле с учетом таких факторов. разнообразие, а не в узком смысле.
Снижение температуры культуры со стадии (1) на стадию (2) осуществляют, когда интегральная плотность жизнеспособных клеток достигает от 20×106 до 120×106 клеток в сутки/мл, предпочтительно от 40×106 до 100×106 клеток в сутки/мл, более предпочтительно, от 60×106 до 80×106 клеток в сутки/мл, но не ограничивается этим.
Кроме того, период времени культивирования на стадии (2) может составлять от 2 до 18 дней, предпочтительно от 3 до 16 дней, более предпочтительно от 4 до 14 дней, но не ограничивается этим.
Например, в случае использования метода периодического культивирования с подпиткой при производстве РН20 или ее варианта в соответствии с настоящим изобретением можно максимизировать продуктивность РН20 или ее варианта, а ферментативную активность полученной РН20 или ее варианта можно значительно улучшить путем культивирования посевного материала непосредственно перед основной культурой при температуре от 35°C до 38°C посредством перфузионной культуры или периодического культивирования с подпиткой, пока концентрация клеток не достигнет определенного уровня, а затем проведения основной культуры посредством инокуляции при пониженной температуре ниже 35°С в течение от 2 до 18 дней, предпочтительно от 3 до 16 дней, более предпочтительно от 4 до 14 дней.
Концентрация инокулята клеток основной культуры может составлять 1×105 клеток/мл или более, предпочтительно 5×105 клеток/мл или еще более предпочтительно 1×106 клеток/мл или более, но не ограничивается этим.
Выполняя культивирование с использованием комбинации периодического культивирования с подпиткой и перфузионного культивирования при производстве РН20 или ее варианта в соответствии с настоящим изобретением, можно максимизировать продуктивность РН20 или ее варианта, а ферментативная активность полученной РН20 или ее варианта может быть заметно увеличена.
В настоящем изобретении остаточную концентрацию глюкозы в среде во время культивирования поддерживают на уровне от 0,001 до 4,5 г/л, предпочтительно от 0,01 до 4,0 г/л и более предпочтительно от 0,1 до 3,5 г/л, но настоящее изобретение не ограничивается этим.
Используемое в настоящей заявке выражение «остаточную концентрацию глюкозы в среде во время культивирования поддерживают на уровне от 0,001 до 4,5 г/л, предпочтительно от 0,01 до 4,0 г/л и более предпочтительно от 0,1 до 3,5 г/л» означает, что когда остаточная концентрация глюкозы в среде, измеренная в течение периода культивирования с интервалами от 1 до 36 часов, предпочтительно от 3 до 30 часов, более предпочтительно от 6 до 24 часов или в режиме реального времени, ниже установленной контрольной концентрации 0,001 г/л до 4,5 г/л, предпочтительно от 0,01 до 4,0 г/л и более предпочтительно от 0,1 до 3,5 г/л, в среду добавляют базовый раствор глюкозы для получения соответствующей эталонной концентрации при культивировании.
В настоящем изобретении специалистам в данной области техники должно быть понятно, что эталонная концентрация остаточной концентрации глюкозы в среде может быть установлена в пределах от 0,001 г/л до 4,5 г/л, предпочтительно от 0,01 до 4,0 г/л, и более предпочтительно от 0,1 до 3,5 г/л, и эталонная концентрация может соответствующим образом изменяться в течение периода культивирования. Например, эталонная концентрация остаточной глюкозы в среде составляет 2 г/л на 1-й и 2-й дни культивирования, эталонная концентрация снижается до 1,5 г/л на 3-5-й день культивирования, и после этого эталонная концентрация повышается до 2 г/л или устанавливается на уровне ниже 1,5 г/л, например, 1,0 г/л.
Гиалуронидаза РН20 или ее вариант, полученные способом по настоящему изобретению и имеющие заданные уровни сиалилирования, галактозилирования и/или маннозилирования в N-гликановой части, обладают ферментативной гиалуронидазной активностью в культуральной среде 10000 ЕД/мл, предпочтительно 11000 ЕД/мл или выше, более предпочтительно 12000 ЕД/мл или выше, но не ограничиваясь этим.
Кроме того, гиалуронидаза PH20 или ее вариант, полученные способом согласно настоящему изобретению и имеющие заданный уровень сиалилирования, галактозилирования и/или маннозилирования в N-гликане, представляют собой человеческую PH20 дикого типа, полученную обычным способом, и имеют ферментативную активность, повышенную на 10% или более, предпочтительно на 12% или более, более предпочтительно на 15% или более, по сравнению с активностью человеческой PH20 дикого типа, полученной обычным способом, но настоящее изобретение этим не ограничивается.
Предпочтительно, в способе получения гиалуронидазы PH20 или ее варианта по настоящему изобретению культивирование клеток-хозяев на стадии (1) и/или стадии (2) можно проводить при одном или нескольких условиях, выбранных из группы, состоящей из : (i) добавления аммиака в среду или повышения концентрация аммиака в среде до 5 мМ или более; (ii) добавления в среду одного или нескольких веществ, выбранных из группы, состоящей из глутамина, глюкозамина, уридина, глюкозамина и бутирата натрия; и (iii) отсутствия добавления галактозы и manNAc в среду, но способ этим не ограничивается.
Более конкретно, в способе получения гиалуронидазы PH20 или ее варианта по настоящему изобретению культивирование клеток-хозяев на стадии (1) и/или стадии (2) можно проводить путем добавления аммиака в среду, повышения концентрации аммиака до 5 мМ или более, добавления глутамина, отсутствия добавления галактозы и manNAc, добавления уридина или глюкозамина, и добавления бутирата натрия, но способ этим не ограничивается.
Используемый в настоящей заявке термин «гиалуронидаза PH20 или ее вариант» относится к ферменту, расщепляющему гиалуроновую кислоту, находящуюся во внеклеточном матриксе.
Термин «гиалуронидаза PH20» или «PH20», используемый в настоящей заявке, интерпретируется как включающий и PH20 дикого типа, и ее зрелую форму, а термин «вариант гиалуронидазы PH20 или PH20» означает вариант PH20, который включает замены, делеции и вставки одного или нескольких аминокислотных остатков, и при необходимости включает делеции некоторых N-концевых и/или С-концевых аминокислотных остатков в аминокислотной последовательности «гиалуронидазы PH20» или «PH20», но не ограничиваются этим.
Гиалуронидаза по настоящему изобретению относится к гиалуронидазе, полученной из животного или микроорганизма, такого как Streptomyces, и обладающей гиалуронидазной активностью, среди которых «гиалуронидаза PH20» или «PH20» получена из животного или микроорганизма, такого как Streptomyces, и предпочтительно получена от людей, крупного рогатого скота или овец.
«Гиалуронидаза PH20» или «вариант PH20» человеческого происхождения в соответствии с настоящим изобретением проиллюстрированы в международной патентной публикации № 2020/022791 и патенте США № 9,447,401, но не ограничиваются ими, и интерпретируются как включающие любую гиалуронидазу или вариант, который содержит замены, делеции и вставки одного или нескольких аминокислотных остатков, при необходимости включает укорочения одного или нескольких N-концевых и/или С-концевых аминокислотных остатков в аминокислотной последовательности «гиалуронидазы PH20» или «PH20», и обладает ферментативной активностью гиалуронидазы.
Примеры клеток-хозяев, используемых в настоящей заявке для экспрессии белков гиалуронидазы, могут включать клетки животных, дрожжей, актиномицетов и насекомых и т.п., но не ограничиваются ими.
Клетки животных предпочтительно представляют собой клетки млекопитающих, более предпочтительно обычно используемые для культивирования клетки животных, такие как клетки CHO, клетки HEK, клетки COS, клетки 3T3, клетки миеломы, клетки BHK, клетки HeLa и клетки Vero, и особо предпочтительно клетки CHO для массовой экспрессии. Кроме того, для получения необходимых белков, в частности, предпочтительно применяют клетки, подходящие для введения необходимых генов, такие как клетки dhfr-CHO (Proc. Natl. Acad. Sci. USA (1980) 77, 4216-4220) или CHO K-1, которые представляют собой клетки CHO с нокаутом гена DHFR (Proc. Natl. Acad. Sci. USA (1968) 60, 1275), или клетки CHO K-1 (Proc. Natl. Acad. Sci. USA (1968) 60, 1275). Клетки CHO особо предпочтительно представляют собой клеточные линии DG44, DXB-11, K-1 или CHO-S, и введение векторов в клетки-хозяева осуществляют с использованием такого метода, как метод с фосфатом кальция, метод с ДЭАЭ-декстраном, электропорация, или липопротеиновый метод.
Примеры дрожжей включают Saccharomyces sp., Hansenula sp., Kluyveromyces, Pichia sp. и т.п., а актиномицеты, например, включают Streptomyces, но не ограничиваются ими.
В способе получения гиалуронидазы PH20 или ее варианта по настоящему изобретению культивирование клеток-хозяев на стадии (1) и/или стадии (2) можно проводить одним или несколькими способами, выбранными из группы, состоящей из периодического культивирования, повторного периодического культивирования, периодического культивирования с подпиткой, повторного периодического культивирования с подпиткой, непрерывного культивирования и перфузионного культивирования, но не ограничиваясь этим.
Периодическое культивирование представляет собой метод культивирования пролиферирующих клеток без добавления свежей среды или сброса культурального раствора во время культивирования. Непрерывное культивирование — это метод культивирования с непрерывным добавлением и удалением среды во время культивирования. Кроме того, непрерывное культивирование включает перфузионное культивирование. Периодическое культивирование с подпиткой находится между периодическим культивированием и непрерывным культивированием, также называется «полупериодическим культивированием» и включает непрерывное или последовательное добавление среды во время культивирования. Этот метод культивирования предотвращает выброс клеток, даже если культуральный раствор постоянно выгружается. В настоящем изобретении можно использовать любой метод культивирования, причем предпочтительным является периодическое культивирование с подпиткой или непрерывное культивирование, особо предпочтительным является периодическое культивирование с подпиткой.
Как описано выше, в гиалуронидазе РН20 или ее варианте, произведенных способом получения по настоящему изобретению, удельная активность фермента может быть увеличена на 10% или более по сравнению с активностью, полученной обычным способом, в частности, по сравнению с удельной активностью человеческой PH20 дикого типа. Это увеличение ферментативной активности обусловлено изменениями свойств N-гликозилирования и/или модификацией заряда гиалуронидазы PH20 или ее варианта, полученных способом согласно настоящему изобретению.
В частности, ферментативная активность увеличивается в зависимости от изменений сиалилирования, галактозилирования и/или маннозилирования среди паттернов N-гликозилирования. Эти характеристики N-гликозилирования, в частности сиалилирования, галактозилирования и/или маннозилирования, можно регулировать способом получения по настоящему изобретению.
В настоящем изобретении примеры ковалентной связи между глюкозой и белками включают N-гликозидную связь, в которой N-ацетил-D-глюкозамин ковалентно связан с аспарагиновым остатком, составляющим белок (N-гликозидсвязанный гликан), O-гликозидную связь, в которой N-ацетил-D-галактозамин ковалентно связан с сериновым или треониновым остатком (О-гликозид-связанный гликан) и т.п., но нет особого ограничения в отношении типа ковалентной связи между глюкозой и белками в гликопротеине по настоящему изобретению, и гликопротеин, включающий один или оба из N-гликозид-связанного гликана и О-гликозид-связанного гликана, относится к настоящему изобретению.
В этом аспекте настоящее изобретение направлено на гиалуронидазу PH20 или ее вариант, имеющие уровень сиалилирования в содержании N-гликанов от 1 до 38%, предпочтительно от 1 до 30%, более предпочтительно от 1,5 до 28% и наиболее предпочтительно 2%. до 25%.
Более конкретно, настоящее изобретение направлено на гиалуронидазу PH20 или ее вариант, имеющие уровень галактозилирования в содержании N-гликанов от 1 до 68%, уровень сиалилирования в содержании N-гликанов от 1 до 38% и уровень маннозилирования в содержании N-гликанов от 40 до 63%, предпочтительно уровень галактозилирования в содержании N-гликанов от 5 до 60%, уровень сиалилирования в содержании N-гликанов от 1 до 30% и уровень маннозилирования в содержании N-гликанов от 42 до 62%, более предпочтительно уровень галактозилирования в содержании N-гликанов от 10 до 56%, уровень сиалилирования в содержании N-гликанов от 1,5 до 28% и уровень маннозилирования в содержании N-гликанов от 44 до 61%, и наиболее предпочтительно уровень галактозилирования в содержании N-гликанов от 15 до 50%, уровень сиалилирования в содержании N-гликанов от 2 до 25%, и уровень маннозилирования в содержании N-гликанов от 47 до 60%.
Приведенные выше числовые значения выведены из экспериментальных результатов примеров, показанных в Таблице 1, и получены с использованием числового значения, имеющего 95% доверительный интервал, и числовые значения также включают ошибку 10%. Это связано с тем, что при измерении содержания глюкозы в белках существуют различия в зависимости от таких условий, как оборудование, используемое для эксперимента, время ферментативной реакции, температура испытания, квалификация аналитика и т.д., поэтому измеренное содержание глюкозы в настоящем изобретении следует интерпретировать в более широком смысле с учетом различий между лабораториями, а не в ограниченном смысле. Кроме того, модификация заряда PH20 или ее варианта может быть идентифицирована с помощью изоэлектрофокусирования.
Гиалуронидазу РН20 или ее вариант, имеющие специфический уровень сиалилирования и/или галактозилирования и маннозилирования в содержании N-гликанов по настоящему изобретению, предпочтительно производят способом получения по настоящему изобретению, но не ограничиваются этим, но очевидно, что гиалуронидаза PH20 или ее вариант могут быть получены другими способами, которые могут быть модифицированы специалистами в данной области техники.
В настоящем изобретении среда, используемая для культивирования клеток для экспрессии гликопротеинов, предпочтительно, но не ограничиваясь этим, представляет собой бессывороточную среду, и в качестве основной среды можно использовать среду DMEM/F12 (комбинацию сред DMEM и F12). Кроме того, коммерчески доступные бессывороточные среды, например, среда HycellCHO, среда ActiPro (HyClone, США), среда CD OptiCHO™, среда CHO-S-SFM II или среда CD CHO (Gibco, США), среда IS CHO-V™ (Irvine Scientific, США), EX-CELL® Advanced CHO Fed-Batch Medium (Sigma-Aldrich, США) или подобные могут быть использованы в качестве основной среды, но изобретение ими не ограничивается.
В настоящем изобретении питательная среда, используемая при культивировании клеток для экспрессии гликопротеинов, представляет собой бессывороточную среду и, например, Cell Boost™ 1, Cell Boost™ 2, Cell Boost™ 3, Cell Boost™ 4, Cell Boost™ 5, Cell Boost™ 6, Cell Boost™ 7a/7b (HyClone, США), CD CHO EfficientFeed™ A AGT™, CD CHO EfficientFeed™ B AGT™, CD CHO EfficientFeed™ C AGT™, CD CHO EfficientFeed™ A плюс AGT™, CD CHO EfficientFeed™ B плюс AGT™, CD CHO EfficientFeed™ C плюс AGT™ (Gibco, США), BalanCD® CHO Feed 4 (Irvine Scientific, США), EX-CELL® Advanced CHO Feed (Sigma-Aldrich, США), CHO-U Feed Mix U1B7/ CHO-U Feed Mix U2B13 (Kerry, США) можно использовать в качестве питательной среды, но изобретение этим не ограничивается.
Используемые в настоящей заявке термины «питательная среда» и «концентрированная питательная среда» относятся к средам, состоящим из определенного питательного вещества или множества питательных веществ, таких как аминокислоты, витамины, соли, микроэлементы, липиды и глюкоза, и могут быть концентрированными продуктами основной среды. Компоненты и концентрации полученной питательной среды могут варьировать в зависимости от культивируемых клеток. Кроме того, коммерческие питательные среды, например, питательная среда Cell Boost Series (HyClone, США), питательная среда EfficientFeed Supplement, питательная среда GlycanTune (Gibco, США), питательная среда BalanCD CHO (Irvine Scientific, США), питательная среда для культивирования клеток Cellvento® CHO (Merck, США), питательная среда EX-CELL® Advanced CHO (Sigma-Aldrich, США) или подобные могут быть использованы в качестве питательной среды, но изобретение ими не ограничивается.
Используемый в настоящей заявке термин «гидролизат растительного происхождения» относится к продукту, экстрагированному из садового гороха, семян хлопка, пшеничного глютена, соевых бобов или тому подобного, и не содержащему компонентов животного происхождения, который представляет собой добавку, включающую большое количество аминокислот, пептидов, витаминов, углеводов, нуклеотидов, минералов и других ингредиентов, и также может производиться с различными компонентами и концентрациями компонентов в зависимости от культивируемых клеток. Кроме того, коммерческие гидролизаты растительного происхождения, такие как HyPep™ 7404, UltraPep™ Cotton, HyPep™ 7504, HyPep™ 4601N (Kerry, США), Cotton 100, Cotton 200, Phytone™ и Soy 100 (Gibco, США), могут использоваться в качестве добавки, но изобретение не ограничивается этим.
Добавка, используемая для увеличения или уменьшения содержания N-гликанов в гликопротеине, используемом в настоящем изобретении, обычно представляет собой компонент, который, как известно, участвует в гликозилировании белка. В частности, для ограничения уровня сиалилирования следует ограничить доставку сиаловой кислоты к галактозному остатку, провести десиалилирование или ограничить уровень галактозилирования. При культивировании клеток такие добавки, которые добавляют в среду в заданных концентрациях, включают компоненты в качестве предшественников гликозилирования, такие как N-ацетил-D-маннозамин, глюкоза, манноза, глутамин и галактоза, аммиак и масляная кислота.
Большинство культур клеток животных в основном используют среду, содержащую сыворотку. Однако трудно создать среду, пригодную для продукции белка, поскольку среда, содержащая сыворотку, представляет собой сложную композицию, химические компоненты которой не установлены полностью. В основном используют бессывороточную среду или среду, содержащую небольшое количество сыворотки, потому что сыворотка может оказывать негативное влияние на разделение и очистку, а также проблемы, связанные со стоимостью и воспроизводимостью. Поскольку концентрация глюкозы, которая является источником углерода, в бессывороточной среде очень низка, глюкозу дополнительно добавляют в качестве основного источника углерода в среду для культивирования, чтобы поддержать рост клеток и получить высокую концентрацию целевого белка, и глутамин может быть дополнительно добавлен в среду для культивирования. В частности, уровень сиалилирования должен быть повышен для увеличения периода полувыведения in vivo при производстве белковых фармацевтических препаратов. Для этого глюкозу и глутамин в среде следует поддерживать в определенных концентрациях или выше, чтобы они не истощались, а рН культуральной среды также необходимо сохранять на определенном уровне. Концентрации глюкозы и глутамина, измеренные в культуральной среде, относятся к концентрациям остаточной глюкозы и глутамина после потребления клетками. Кроме того, могут быть использованы промоторы для увеличения активности ферментов, связанных с повышением уровня сиалилирования, или ингибиторы для подавления десиалилирования. Кроме того, добавляя предшественники N-гликанов или контролируя промоторы активности родственных ферментов или условия культивирования, можно повысить уровень галактозилирования и, таким образом, также можно повысить уровень сиалилирования.
Однако уровень N-гликанов не может контролироваться вышеописанным способом при производстве гиалуронидазы PH20 или ее варианта, тогда как при использовании способа по настоящему изобретению можно получить гиалуронидазу PH20 или ее вариант, имеющие необходимые уровни гликанов и высокую ферментативную активность.
Способ получения по настоящему изобретению может дополнительно включать выделение и очистку полученной гиалуронидазы PH20 или ее варианта.
Выделение и очистку гиалуронидазы PH20 или ее варианта по настоящему изобретению предпочтительно проводят с использованием не аффинного связывания, а характеристик ионной связи и/или гидрофобного взаимодействия гиалуронидазы PH20 или ее варианта, но изобретение этим не ограничивается.
В частности, разделение и очистку гиалуронидазы РН20 или ее варианта по настоящему изобретению предпочтительно проводят с использованием не аффинной хроматографии, а хроматографии гидрофобного взаимодействия и ионообменной хроматографии, такой как катионообменная хроматография и/или анионообменная хроматография, но изобретение не ограничивается этим.
Кроме того, выделение и очистка гиалуронидазы PH20 или ее варианта по настоящему изобретению направлены на удаление кислой гиалуронидазы PH20 или ее варианта, обладающего низкой ферментативной активностью, и удаление кислой гиалуронидазы PH20 или ее варианта предпочтительно проводят с помощью ионообменной хроматографии.
Необходимо проанализировать скорость каталитической реакции фермента, чтобы определить промышленную применимость фермента. Ферментативную реакцию можно классифицировать как ферментативную реакцию, имеющую активный центр с фиксированной реакционной способностью, и ферментативную реакцию, имеющую множество активных центров, имеющих различную реакционную способность. Известно, что скорость каталитической реакции фермента с одним активным центром с фиксированной реакционной способностью, такого как гиалуронидаза, соответствует кинетическому уравнению Михаэлиса-Ментен.
Ферментативная кинетика Михаэлиса-Ментен основана на предположении, что ферментативная реакция представляет собой двухстадийную реакционную систему, включающую обратимую стадию реакции, на которой образуется комплекс [ES] фермента (Е) и субстрата (S), и необратимую стадию реакции, на которой комплекс ES диссоциирует с образованием продукта (P). В этом случае kf, kr и kcat — константы скорости реакции в каждом направлении (Alan Fersht (1977). Enzyme structure and mechanism).
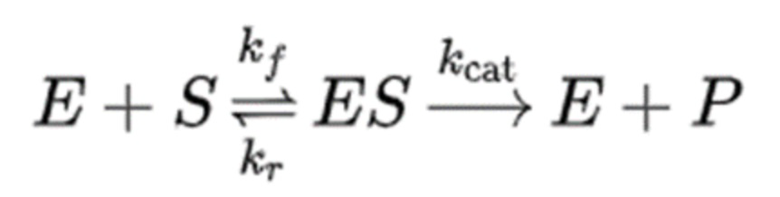
Что касается ферментативной реакции, предполагается, что процесс взаимодействия фермента с субстратом с образованием комплекса ES быстро достигает равновесия или квазистационарного состояния, при условии, что 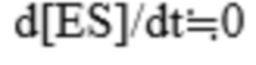 удовлетворяется за счет достаточного снижения концентрации фермента при проведении реакции, поддерживающей достаточно высокую концентрацию субстрата. Поскольку формулы кинетики, предполагающие быстрое равновесие или квазистационарное состояние, выводятся одинаковым образом, в большинстве экспериментов предполагается квазистационарное состояние, в котором концентрация субстрата изначально выше, чем концентрация фермента.
удовлетворяется за счет достаточного снижения концентрации фермента при проведении реакции, поддерживающей достаточно высокую концентрацию субстрата. Поскольку формулы кинетики, предполагающие быстрое равновесие или квазистационарное состояние, выводятся одинаковым образом, в большинстве экспериментов предполагается квазистационарное состояние, в котором концентрация субстрата изначально выше, чем концентрация фермента.
При таких условиях, как «количество фермента постоянно до и после реакции» и на основании предположения «когда химическая реакция достигает химического равновесия, скорость реакции, при которой образуется продукт, равна скорости, при которой продукт снова разлагается», скорость реакции конечного продукта может быть выражена следующим уравнением кинетики Михаэлиса-Ментен. В этом случае, KM = (kr + kcat) / kf, и Vmax = kcat [E]0.
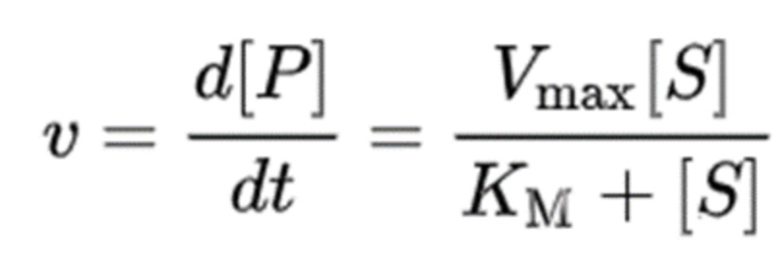
Уравнение Лайнуивера-Берка применяют для экспериментального анализа скорости ферментативной реакции с использованием кинетического уравнения Михаэлиса-Ментен. Это уравнение показывает взаимосвязь между обратной величиной 1/V экспериментально измеренной скорости реакции и обратной величиной 1/[S] данной концентрации субстрата в эксперименте. Статистическая проверка того, что это уравнение представляет собой линейное уравнение, показывает, что ферментативная реакция является реакцией, следующей кинетическому уравнению Михаэлиса-Ментен, и KM и Vmax могут быть рассчитаны с использованием этого уравнения.
Ферменты, катализирующие химическую реакцию, имеют переходное состояние после связывания с субстратом в активном центре, и энергия активации для достижения переходного состояния с высокой энергией снижается из-за множественных связей с субстратом. Константа равновесия для достижения этого переходного состояния пропорциональна kcat/KM. Здесь 1/KM представляет собой показатель, объединяющий степень образования комплекса фермент-субстрат путем связывания фермента с субстратом со степенью, в которой комплекс фермент-субстрат сохраняется без разложения, а kcat представляет собой константу равновесия, при котором продукт образуется из фермент-субстратного комплекса. Следовательно, можно сказать, что kcat/KM является показателем того, сколько продукта может быть получено из субстрата и фермента, то есть показателем каталитической эффективности фермента.
Промышленная пригодность гиалуронидазы пропорциональна ее ферментативной каталитической эффективности. В частности, при подкожном введении фермента вместе с полимерным фармакологически активным веществом, таким как моноклональное антитело, важную роль играет ферментативная каталитическая эффективность гиалуронидазы. В случае, когда вариант по настоящему изобретению имеет более высокое значение kcat/KM, чем PH20 дикого типа, при подкожном введении гиалуронидазы, содержащейся в полимерном фармакологически активном веществе, присутствующая в месте введения гиалуроновая кислота быстро разлагается, и таким образом, можно получить превосходящий эффект быстро диспергирующегося фармакологически активного вещества. Кроме того, когда вариант по настоящему изобретению имеет более высокое значение kcat, чем PH20 дикого типа, максимальная скорость реакции Vmax увеличивается при той же концентрации фермента, тем самым обеспечивая превосходные эффекты разложения большего количества гиалуроновой кислоты за тот же период времени и диспергирования фармакологически активного вещества в более широкой области.
Таким образом, для определения ферментативных свойств варианта РН20 по настоящему изобретению анализировали скорость ферментативной реакции каждого варианта, и Vmax (максимальную скорость ферментативной реакции), Km (концентрацию субстрата в условиях 50% Vmax), kcat (скорость конверсии субстрата) и kcat/Km (эффективность ферментного катализатора) сравнивали в Примере 4. Результаты демонстрируют, что вариант PH20 по настоящему изобретению превосходит PH20 дикого типа.
Примеры
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако специалистам в данной области техники будет очевидно, что эти примеры представлены только для иллюстрации настоящего изобретения и не должны рассматриваться как ограничивающие объем настоящего изобретения.
Пример 1. Взаимосвязь между активностью гиалуронидазы, N-гликаном и характером изоэлектрофокусирования
Результаты анализа активности, характера изоэлектрофокусирования и уровня N-гликанов гиалуронидазы человека PH20 дикого типа и ее варианта HM46 показаны на фиг. 1. Хотя разница в количестве активных единиц двух гиалуронидаз была более чем двукратной, не было большой разницы в диапазоне изоэлектрических точек или уровне N-гликанов.
Был проведен эксперимент для выявления взаимосвязи между N-гликаном, характером изоэлектрофокусирования и активностью гиалуронидазы PH20. В процессе очистки варианта гиалуронидазы PH20 можно было отделить основную фракцию (фракция 1) от кислой фракции (фракция 2), и соответствующие фракции использовали в качестве образцов. Фиг. 2 представляет результаты анализа картины изоэлектрофокусирования и ферментативной активности образца, обработанного PNGase F для удаления всех N-гликанов, образца, обработанного сиалидазой А для удаления терминальной сиаловой кислоты, и образца, обработанного сиалидазой А и галактозидазой для удаления терминальной сиаловой кислоты и галактозы. Две фракции отличались диапазоном изоэлектрической точки, а кислая фракция проявляла пониженную ферментативную активность. После обработки PNGase F две фракции не проявляли активности и имели схожий характер изоэлектрофокусирования. Эти результаты показали, что уровень N-гликанов тесно связан с ферментативной активностью. Кроме того, было обнаружено, что существует взаимосвязь между содержанием терминальной сиаловой кислоты и ферментативной активностью посредством явления, при котором характер изоэлектрофокусирования или ферментативная активность кислой фракции аналогичны активности основной фракции при удалении терминальной сиаловой кислоты. Однако основная фракция с низким содержанием терминальной сиаловой кислоты проявляла улучшенную гиалуронидазную ферментативную активность по сравнению с кислой фракцией с высоким содержанием сиаловой кислоты.
Взаимосвязь между ферментативной активностью и содержанием N-гликанов была дополнительно продемонстрирована на основании результатов оценки гиалуронидаз дикого типа и различных вариантов гиалуронидазы, происходящих из различных клеточных источников культивирования.
Таблица 1
рование
(%)
(%)
Относительная активность: активность, выраженная в процентах отношения (активности образца)/(активности человеческой PH20 дикого типа).
Процент галактозилирования (%): сумма процентного содержания (%) N-гликанов, содержащих терминальную галактозу, таких как G1, G1F, G1F', G2, G2F, A1, A1F, A2 и A2F.
Процент сиалилирования (%): сумма процентного содержания (%) N-гликанов, содержащих терминальную сиаловую кислоту, таких как A1, A1F, A2 и A2F.
Процент маннозилирования (%): сумма процентного содержания (%) N-гликанов, содержащих терминальную маннозу, таких как M4G0F, M5, M5G0, M6, M7, M8 и M9.
Последовательности человеческих PH20 и HM46 дикого типа раскрыты в международной патентной публикации № 2020/022791.
Образцы PH20 овцы и РН20 бонобо готовили способом, показанным в Примере 10, и последовательности показаны в таблице 3.
Как показано в таблице 1, были получены различные типы гиалуронидазы PH20 и вариантных белков, и было исследовано содержание N-гликанов. Для этой цели использовали множество культур с использованием различных источников клеток, таких как временная экспрессионная культура, полученная путем трансфекции клеток ExpiCHO рекомбинантными генами с использованием реагента ExpiFectamine CHO (Gibco, США) каждый раз при проведении культивирования, культура клональной клеточной линии с использованием клонов клеточной линии, полученных с помощью рекомбинантных генов, и культура клеточной линии с использованием скринированных моноклонов, обладающих высокой продуктивностью. В настоящей заявке культура клональных клеточных линий представляет собой культуру с использованием высокопродуктивных клонов клеточных линий, отобранных с использованием селективных маркеров из клонов клеток млекопитающих, трансфицированных рекомбинантными генами, полученную с использованием поликлональной клеточной линии и нацеленную на получение моноклональных клеточных линий. Клеточные линии включают исследовательский банк клеток (RCB), производственный основной банк клеток (MCB) и рабочий банк клеток (WCB). Кроме того, в методе культивирования были опробованы различные комбинации тест-культуры для оптимизации производства, и даже здесь было обнаружено, что существует разница в активности и N-гликане.
Опытная культура клональной клеточной линии № 1 представляет собой периодическую культуру объемом 2 литра, которую выращивают при 37°C без изменения температуры культивирования с использованием среды HycellCHO (HyClone, США). Опытная культура клональной клеточной линии № 2, очищенная фракция № 1, и опытная культура клональной клеточной линии № 2, очищенная фракция № 2, были фракциями, разделенными и очищенными с помощью анионообменной хроматографии из собранной жидкости клеточной культуры, культивируемой при 37°C с использованием периодического метода с подпиткой в среде для культивирования с подпиткой EX-CELL® Advanced CHO (Sigma-Aldrich, США) в партии объемом 8 литров с Cotton 100UF (Gibco), а затем BalanCD® CHO Feed 4 (Irvine Scientific, США). Очищенная фракция № 1 является фракцией элюирования с низким содержанием соли, а очищенная фракция № 2 представляет собой фракцию элюирования с высоким содержанием соли. Опытная культура клеточной линии №1 была получена путем культивирования при температуре культивирования 37°C с использованием периодического метода с подпиткой, с применением среды для подпитки EX-CELL® Advanced CHO (Sigma-Aldrich, США) в 2 литровой загрузке с Cotton 100UF (Gibco), а затем BalanCD® CHO Feed 4 (Irvine Scientific, США), с последующим сдвигом температуры культивирования до 32°C, когда интегральная плотность жизнеспособных клеток достигала заданного уровня. Опытная культура клеточной линии №2 была получена путем культивирования при температуре 37°С с использованием периодического метода с подпиткой, с использованием среды для подпитки EX-CELL® Advanced CHO (Sigma-Aldrich, США) в партии 10 литров с Cotton 100UF (Gibco), а затем CD CHO EfficientFeed™ B плюс AGT™ (Gibco, США), с последующим сдвигом температуры культивирования до 32°C, когда интегральная плотность жизнеспособных клеток достигала заданного уровня, с последующим проведением культивирования в течение 12 дней. Опытная культура клеточной линии №3 была получена путем культивирования при температуре 37°С с применением периодического метода с подпиткой, с использованием среды для подпитки EX-CELL® Advanced CHO (Sigma-Aldrich, США) в партии 50 литров с Cotton 100UF (Gibco, США), а затем CD CHO EfficientFeed™ B плюс AGT™(Gibco, США); с последующим сдвигом температуры культивирования до 32°C, когда интегральная плотность жизнеспособных клеток достигала заданного уровня. Опытная культура клеточной линии № 4 была получена путем культивирования клеток при температуре 37°C с использованием периодического метода с подпиткой, с применением среды для подпитки EX-CELL® Advanced CHO (Sigma-Aldrich, США) в партии 10 литров с Cotton 100UF (Gibco, США), а затем CD CHO EfficientFeed™B плюс AGT™ (Gibco, США) с последующим сдвигом температуры культивирования до 32°С при достижении заданного уровня интегральной плотности жизнеспособных клеток и культивированием клеток в течение 13 суток. Культура клеточной линии № 1, культура клеточной линии № 2, культура клеточной линии № 3 и культура клеточной линии № 4 получены путем культивирования исследовательского банка клеток (RCB), производственного основного банка клеток (MCB) и рабочего банка клеток. (WCB) при температуре культивирования 37°C с использованием периодического метода с подпиткой с применением среды для подпитки EX-CELL® Advanced CHO (Sigma-Aldrich, США) в партии объемом 200 литров с Cotton 100UF (Gibco, США), а затем CD CHO EfficientFeed™ B плюс AGT™ (Gibco, США) с последующим сдвигом температуры культивирования до 32°С при достижении заданного значения интегральной плотности жизнеспособных клеток, и культивированием клеток в течение 12 суток. Основываясь на этих условиях, были проведены эксперименты в условиях культивирования из Примеров 2-5, чтобы установить способ культивирования для поддержания уровней N-гликанов.
Как видно из таблицы 1, человеческая гиалуронидаза PH20 дикого типа и ее вариант, а также гиалуронидаза PH20 млекопитающих должны иметь уровень галактозилирования от 15 до 50%, уровень сиалилирования от 2,5 до 28% и уровень маннозилирования от 47 до 60% в N-гликане, и должны иметь уровень галактозилирования от 1 до 68%, уровень сиалилирования от 1 до 38% и уровень маннозилирования от 40 до 63% в N-гликане при применении 95% доверительного интервала к результаты испытаний, для обеспечения промышленно пригодной гиалуронидазной активности. Такие числовые значения могут также включать погрешность в 10%, поскольку при измерении уровней глюкозы в белках возможны отклонения в зависимости от таких условий, как оборудование, используемое для эксперимента, время ферментативной реакции, температура испытания, квалификация аналитика и т.п., поэтому уровень глюкозы, измеренный в настоящем изобретении, следует интерпретировать в более широком смысле с учетом различий между лабораториями, а не в ограниченном смысле.
Кроме того, в частности, принимая во внимание взаимосвязь между активностью и сиалилированием, можно видеть, что гиалуронидаза, имеющая активность выше активности человеческой PH20 дикого типа и, таким образом, пригодная для промышленного использования, может быть получена только тогда, когда сиалилирование ограничено примерно до 30% или менее. Приведенное выше числовое значение может быть получено в результате экспериментов и может включать погрешность в 10%.
Высококачественную гиалуронидазу, поддерживающую уровень N-гликанов, идентифицировали с помощью временной экспрессионной культуры, а клеточные линии получали путем отбора моноклональных штаммов, обладающих высокой продуктивностью, для стабильного коммерческого производства. Все опытные культуры клеточных линий №2, №3, №4 и культуры клеточных линий №1, №2, №3, №4, представленные в таблице 1, которые получены методами культивирования с использованием результатов Примеров 2, 3, 4 и 5, проявляли уровень экспрессии фермента 10000 единиц/мл или более, что указывает на возможность получения высококачественной гиалуронидазы с высокой эффективностью и низкой стоимостью.
Пример 2. Культура, зависящая от добавки
Вариант гиалуронидазы PH20 инокулировали в концентрации 2×106 клеток/мл в каждую из трех колб Эрленмейера объемом 125 мл, содержащих среду для периодического культивирования с подпиткой EX-CELL® Advanced CHO (Sigma-Aldrich, США) с добавлением или без добавления Cotton 200UF (Gibco, США) и затем 20 мМ N-ацетил-D-маннозамина (NZP, Нидерланды) или 50 мМ галактозы (Pfanstiehl, США), проводили культивирование в периодическом режиме в инкубаторе при 37°C и 8% CO2, а затем периодическое культивирование с подпиткой при пониженной температуре 32°C, когда интегральная плотность жизнеспособных клеток (IVCD) достигла диапазона сдвига. Питательную среду CD CHO EfficientFeed™ B плюс среду AGT™ (Gibco, США) подавали ежедневно в количестве 1,88% от исходного объема культуры в колбе. Образцы клеток собирали ежедневно из культуральных жидкостей и измеряли плотность жизнеспособных клеток, жизнеспособность клеток, рН и уровни лактата. После прекращения культивирования центрифугировали при 2000 об/мин в течение 10 минут для получения надосадочной жидкости культуры. Активность образца, культивируемого в вышеуказанных условиях, определяли с помощью ВЭЖХ и анализа мутности, а белковые характеристики и уровни N-гликанов наблюдали с помощью изоэлектрофокусирования и анализа гликозилирования. Среда с добавлением 50 мМ галактозы показала увеличение галактозилирования на 24% и снижение активности на 2% по сравнению со средой без добавления 50 мМ галактозы, а среда с добавлением 20 мМ N-ацетил-D-маннозамина показала 35% увеличение сиалилирования и снижение активности на 20% по сравнению со средой без добавления 20 мМ N-ацетил-D-маннозамина (фиг. 3, 4, 5 и 6).
Пример 3. Культура в зависимости от периода культивирования
Клетки, сверхэкспрессирующие вариант гиалуронидазы PH20, инокулировали в количестве 2×106 клеток/мл в среду EX-CELL® Advanced CHO Fed-Batch (Sigma-Aldrich, США) в биореакторе Sartorius 200 л. На вторые сутки культивирования проводили периодическое культивирование с использованием Cotton 200UF (Gibco, США) и среды CD CHO EfficientFeed™ B плюс AGT™ (Gibco, США), которая представляет собой концентрированную питательную среду (Gibco, США), в качестве среды подпитки, при рН 7,2±0,4 и DO 40% при скорости 47 об/мин. Исходное культивирование проводили при 37°C, температуру сдвигали до 32°C, когда интегральная плотность жизнеспособных клеток достигала диапазона изменения, и проводили периодическое культивирование с подпиткой. Питательную среду CD CHO EfficientFeedTM B плюс среду AGTTM (Gibco, США) подавали ежедневно в количестве 1,88% от исходного объема культуры в 200-литровом биореакторе. Что касается периода культивирования, культивирование прекращали на 19-й день, когда жизнеспособность клеток падала до 40% или менее. Ежедневно из культуральных жидкостей собирали различные образцы клеток, определяли плотность жизнеспособных клеток, жизнеспособность клеток, интегральную плотность жизнеспособных клеток, измеряли уровень ионов аммония, и культуральный раствор собирали с использованием глубинного фильтра. Активность образцов, культивируемых в вышеуказанных условиях, определяли с помощью ВЭЖХ и анализа мутности, а белковые характеристики и уровни N-гликанов наблюдали с помощью изоэлектрофокусирования и анализа гликозилирования. По мере увеличения количества дней культивирования уровни галактозилирования и сиалилирования снижались, а активность увеличивалась (фиг. 7, 8, 9 и 10).
Пример 4. Культура при контролируемой концентрации глюкозы в питательной среде
Клетки, сверхэкспрессирующие вариант гиалуронидазы PH20, инокулировали в количестве 2×106 клеток/мл в среду EX-CELL® Advanced CHO Fed-Batch (Sigma-Aldrich, США) в биореакторе Sartorius на 2 литра. На вторые сутки культивирования проводили периодическое культивирование с подпиткой, используя Cotton 200UF (Gibco, США) и среду CD CHO EfficientFeed™ B плюс AGT™ (Gibco, США), которая представляет собой концентрированную питательную среду (Gibco, США), при рН 7,2±0,4 и DO 40%, со скоростью 120 об/мин. Исходное культивирование проводили при 37°C, температуру сдвигали до 32°C, когда интегральная плотность жизнеспособных клеток достигала диапазона изменения, и затем проводили периодическое культивирование с подпиткой. Питательную среду CD CHO EfficientFeed™ B плюс среду AGT™ (Gibco, США) подавали ежедневно в количестве 1,88% от исходного объема культуры в 2-литровый биореактор. Ежедневно из культуральных жидкостей собирали различные образцы клеток, и измеряли плотность жизнеспособных клеток, жизнеспособность клеток, pH, осмоляльность и концентрацию глюкозы и лактата.
Концентрацию глюкозы в культуральной среде измеряли ежедневно и контролировали с момента времени, когда концентрация глюкозы, содержащейся в надосадочной жидкости культуры, не превышала стандартную концентрацию 2, 4 или 6 г/л, так как глюкоза потребляется растущими клетками. Когда измеренная концентрация глюкозы составляла 2, 4 или 6 г/л или менее, что является соответствующими стандартными концентрациями, базовый раствор глюкозы 200 г/л добавляли в течение максимального времени 3 часа в количестве, обеспечивающем достижение стандартной концентрации. Когда измеренная концентрация глюкозы составляла 2, 4 или 6 г/л или более, базовый раствор не добавляли для поддержания каждой стандартной концентрации. В целом влияние изменений содержания глюкозы на рост клеток в течение 3 часов при культивировании клеток млекопитающих было незначительным.
В этом случае условие поддержания 2 г/л в качестве стандартной концентрации называлось условием концентрации 1 г/л (±1 г/л), условие поддержания 4 г/л в качестве стандартной концентрации называли условием концентрации 3 г/л (±1 г/л), а условие поддержания 6 г/л в качестве стандартной концентрации называли условием концентрации 5 г/л (±1 г/л). Например, условие поддержания концентрации глюкозы на уровне 1 г/л (±1 г/л) означает, что нижняя граница диапазона контроля концентрации глюкозы установлена на уровне 0 г/л, а верхняя граница диапазона контроля установлена на уровне 2 г/л и означает, что в реальной культуре, когда измеренная концентрация глюкозы в надосадочной жидкости культуры достигает нижнего предела 0 г/л, дополнительно добавляют базовый раствор глюкозы 200 г/л, чтобы максимальная концентрация глюкозы достигала 2 г/л, а когда концентрация глюкозы в надосадочной жидкости культуры достигает верхнего предела 2 г/л, не добавляют базового раствора глюкозы 200 г/л. Следовательно, в двух условиях концентрации, а именно концентрации 1 г/л (±1 г/л) и концентрации 3 г/л (±1 г/л), условие концентрации 2 г/л соответствует верхнему предел по отношению к условию концентрации 1 г/л (±1 г/л) и соответствует нижнему пределу по отношению к условию концентрации 3 г/л (±1 г/л), поэтому эти два условия являются условиями, при которых происходят совершенно разные действия, и поэтому они считаются не перекрывающими друг друга.
После прекращения культивирования надосадочную жидкость культуры получали центрифугированием при 4°С и 10000 об/мин в течение 60 минут. Активность образца, культивируемого в вышеуказанных условиях, определяли с помощью ВЭЖХ и анализа мутности, а белковые характеристики и уровни N-гликанов определяли с помощью изоэлектрофокусирования и анализа гликозилирования. Результаты показали, что по мере увеличения концентрации глюкозы в надосадочной жидкости культуры уровни галактозилирования и сиалилирования возрастали, а активность снижалась (фиг. 11, 12, 13 и 14).
Пример 5. Культура при контролируемом значении рН
Клетки, сверхэкспрессирующие вариант гиалуронидазы PH20, инокулировали в количестве 2×106 клеток/мл в среде EX-CELL® Advanced CHO Fed-Batch (Sigma-Aldrich, США) в биореакторе Sartorius 2 литра. На второй день культивирования проводили периодическое культивирование с подпиткой, используя Cotton 200UF (Gibco, США) и среду CD CHO EfficientFeed™ B плюс AGT™ (Gibco, США), которая представляет собой концентрированную питательную среду (Gibco, США), в качестве подпитки, при DO 40% со скоростью 120 об/мин. Исходную культуру проводили при 37°С, температуру сдвигали до 32°С, когда интегральная плотность жизнеспособных клеток достигла диапазона изменения, и затем проводили периодическое культивирование с подпиткой. Питательную среду CD CHO EfficientFeed™ B плюс AGT™ (Gibco, США) подавали ежедневно в количестве 1,88% от исходного объема культуры в 2-литровый биореактор. Температуру культуры меняли, культуру разделяли и далее культивировали в соответствии с четырьмя условиями pH, а именно pH 6,8±0,1, pH 7,0±0,1, pH 7,2±0,1 и pH 7,4±0,1, чтобы найти улучшенные условия по сравнению с обычным рН 7,2±0,4. Диапазон контроля рН культуры устанавливали в соответствии с этими четырьмя условиями с использованием обычного биореактора, который включает функцию контроля рН. Например, pH 7,0±0,1 означает, что нижний предел диапазона контроля pH составляет pH 6,9, а верхний предел диапазона контроля pH составляет pH 7,1. В реальной культуре, когда рН культуральной среды достигает нижнего предела рН 6,9, добавляют основание для повышения рН, а когда рН культуральной среды достигает верхнего предела рН 7,1, добавляют углекислый газ для снижения рН. Следовательно, в двух условиях рН 6,8±0,1 и рН 7,0±0,1 условию рН 6,9 соответствует верхний предел при рН 6,8±0,1 и нижний предел при рН 7,0±0,1. Поэтому считается, что два условия не перекрывают друг друга. Культивирование проводили до дня, когда жизнеспособность клеток достигала 40% или менее, ежедневно отбирали несколько образцов клеток из культуральных жидкостей, и измеряли плотность жизнеспособных клеток, жизнеспособность клеток, рН и интегральный уровень плотности жизнеспособных клеток. После прекращения культивирования надосадочную жидкость культуры получали центрифугированием при 4°С и 10000 об/мин в течение 60 минут. Активность образца, культивируемого в вышеуказанных условиях, определяли с помощью ВЭЖХ и анализа мутности, а белковые характеристики определяли с помощью изоэлектрофокусирования. Активность менялась в зависимости от pH культуральной среды, а самая высокая активность и самый низкий уровень сиалилирования наблюдались при pH 7,0±0,1, что было улучшенным состоянием по сравнению с обычным состоянием (таблица 2, фиг. 15, 16 и 17).
Таблица 2
(обычные условия)
Пример 6. Очистка гиалуронидазы с использованием надосадочной жидкости культуры клеток животных
Этап 1: Обработка надосадочной жидкости культуры с заменой буфера/сурфактантом
Условия культуральных растворов корректировали до условий уравновешивания первой анионообменной колонки путем контроля рН и электропроводности с помощью УФ/ДФ с использованием мембранного фильтра с пределом отсечения по молекулярной массе 30 кДа. Скорректированный раствор обрабатывали подходящей концентрацией растворителя/поверхностно-активного вещества для инактивации вирусов и проводили реакцию при комнатной температуре в течение примерно 60 минут.
Этап 2: Первичная анионообменная колоночная хроматография (Q Sepharose Fast Flow).
Отфильтрованный белковый раствор пропускали через первичную анионообменную колонку для захвата гиалуронидазы анионообменной смолой и элюировали из колонки при высокой концентрации соли. Перед загрузкой колонку уравновешивали с использованием трометаминового буфера (имеющего рН 8,0 и концентрацию соли 30 мМ). После загрузки колонку промывали тем же буфером (первичное промывание). После первичного промывания колонку вторично промывали тем же буфером (с рН 8,0, но с концентрацией соли 60 мМ, с более высокой проводимостью, чем концентрация соли при первичном промывании). После вторичного промывания проводили элюцию необходимого белка, гиалуронидазы, с использованием соответствующего буфера, имеющего рН 8,0 и концентрацию соли 200 мМ, как показано на фиг. 18.
Этап 3: Вторичная анионообменная колоночная хроматография (Capto Q).
Отфильтрованный белковый раствор пропускали через вторичную анионообменную колонку для удаления кислого варианта гиалуронидазы с помощью анионообменной смолы и элюировали из колонки при высокой концентрации соли. Перед загрузкой колонку уравновешивали, используя буфер Бис-Трис (имеющий pH 6,0 и не содержащий соли). После загрузки колонку промывали тем же буфером Бис-Трис (первичное промывание). После первичного промывания колонку промывали тем же буфером Бис-Трис, который имел концентрацию соли 20 мМ, имеющую более высокую проводимость, чем буфер без соли при первичном промывании). После вторичного промывания проводили элюцию необходимого белка, гиалуронидазы, с использованием буфера Бис-Трис, имеющего концентрацию соли, как показано на фиг. 19.
Этап 4: Катионообменная (Capto MMC) колоночная хроматография
Скорректированный белковый раствор пропускали через катионообменную колонку для удаления кислого варианта гиалуронидазы с помощью катионообменной смолы и элюировали с колонки при высокой концентрации соли. Перед загрузкой колонку уравновешивали, используя цитратный буфер с pH 5,5, имеющий концентрацию соли 80 мМ. После загрузки колонку промывали тем же цитратным буфером (первичное промывание). После первичного промывания колонку промывали соответствующим буфером Бис-Трис с рН 7,5. После вторичного промывания проводили элюцию необходимого белка, гиалуронидазы, с использованием буфера Бис-Трис с рН 8,0, имеющего концентрацию соли 400 мМ, как показано на фиг. 20.
Этап 5: Нанофильтрация/приготовление готового белка
После катионообменной колонки белковый раствор, содержащий необходимую гиалуронидазу, фильтровали через фильтр с размером пор 1 мкм и подвергали нанофильтрации. Нанофильтрованный белковый раствор концентрировали до высокой концентрации 10 мг/мл и доводили посредством УФ/ДФ с использованием мембранного фильтра с MWCO 8 кДа для замены с гистидиновым буфером с рН 7,0, содержащим 145 мМ соли.
Пример 7. Анализ ферментативной активности гиалуронидазы
Ферментативную активность гиалуронидазы PH20 и других гиалуронидаз измеряли с помощью турбидиметрического анализа следующим образом.
Турбидиметрический анализ представляет собой метод измерения осадка, образующегося при смешивании гиалуроновой кислоты с альбумином (БСА), с использованием поглощения. Когда гиалуроновая кислота гидролизуется PH20, поглощение уменьшается при смешивании с альбумином. В целом этот процесс осуществляли следующим образом. Гиалуронидазу PH20 (Sigma) разбавляли до 1; 2; 5; 7,5; 10; 15; 20; 30; 50 или 60 единиц/мл и помещали в соответствующие пробирки. Образец очищенного белка растворяли в буфере для разбавления ферментов (20 мМ Трис·HCl, pH 7,0, 77 мМ NaCl; 0,01% (масс./объем) бычьего сывороточного альбумина) и разбавляли в каждой пробирке в 100, 300, 600, 1200 или 2400 раз. Раствор гиалуроновой кислоты 3 мг/мл разбавляли в 10 раз до 0,3 мг/мл в других пробирках, чтобы довести объем каждой пробирки до 180 мкл. 60 мкл фермента смешивали с разбавленным раствором гиалуроновой кислоты и проводили реакцию при 37°С в течение 45 минут. После реакции в каждую лунку 96-луночного планшета добавляли 50 мкл прореагировавшего фермента и 250 мкл кислого раствора альбумина, встряхивали в течение 10 минут и измеряли поглощение при 600 нм с использованием спектрофотометра. Активность образца в единицах была получена с использованием результатов испытаний стандартного продукта, для которого известна активность в единицах, и результатов испытаний образца.
Пример 8. Анализ гиалуронидазы посредством изоэлектрофокусирования
Изоэлектрофокусирование гиалуронидазы проводили с помощью готового геля (pH 3-7, Invitrogen) и буферного раствора для изоэлектрофокусирования. Образец очищенной гиалуронидазы наносили на готовый гель, и проводили электрофорез с использованием аппарата для электрофореза производства Novex Corporation при 100 В в течение 1 часа, при 200 В в течение 1 часа и при 500 В в течение 30 минут. После завершения анализа гель промывали очищенной водой, белок фиксировали 12% раствором трихлоруксусной кислоты, окрашивали раствором красителя Кумасси голубого R-250 и обесцвечивали раствором уксусной кислоты в метаноле для анализа белковых полос, проявленных на геле.
Пример 9. Анализ уровня N-гликанов гиалуронидазы
Уровень N-гликанов гиалуронидазы анализировали путем маркировки образца N-гликана, разделения путем обработки гиалуронидазы PNGase F (N-гликозидазой F), 2-AB (2-аминобензамидом) с последующим проведением сверхвысокоэффективной жидкостной хроматографии с использованием колонки ACQUITY UPLC Glycan BEH Amide (Waters). Образец очищенной гиалуронидазы обессоливали и подвергали взаимодействию с PNGase F при 37°С в течение 16-18 часов для выделения N-гликана. N-гликан метили 2-АВ и затем проводили реакцию при 65°С в течение 3 часов, после чего из него удаляли избыток 2-АВ. Образец меченого N-гликана разделяли с помощью ВЭЖХ с градиентом ацетонитрила 72%-20%. Разделенный образец выявляли с помощью флуоресцентного детектора (FLD) для анализа уровней N-гликанов. Соответствующие разделенные N-гликаны классифицировали; N-гликаны, содержащие терминальную галактозу (G1, G1F, G1F', G2, G2F, A1, A1F, A2, A2F и т.п.), добавляли для определения процента галактозилирования (%); N-гликаны, содержащие терминальную сиаловую кислоту (A1, A1F, A2, A2F и т.п.), добавляли для определения процента сиалилирования (%); и N-гликаны, содержащие терминальную маннозу (M4G0F, M5, M5G0, M6, M7, M8, M9 и т.п.) добавляли для определения процента маннозилирования (%).
Пример 10. Получение гиалуронидазы животного происхождения и анализ уровня N-гликанов
(1) Производство генов гиалуронидазы РН20 бонобо и овцы
Для исследования активности и уровня N-гликанов гиалуронидазы PH20 животного происхождения от антропоидов, то есть бонобо, и парнокопытных, то есть овец, гиалуронидазу PH20 получали следующим образом. кДНК синтезировали из генов дикого типа и вставляли в сайты рестрикционных ферментов Xho I и Not I вектора pcDNA3.4-TOPO. Для экспрессии в клетках ExpiCHO вместо нативного сигнального пептида PH20 в качестве сигнального пептида использовали один из сигнальных пептидов гормона роста человека, сывороточного альбумина человека или Hyal1 человека. Для очистки белка с использованием колонки HisTrap последовательность ДНК His-метки располагали на 3'-конце кДНК PH20. Каждую последовательность идентифицировали с помощью секвенирования ДНК. В таблице 3 показаны последовательности гиалуронидаз животного происхождения.
Таблица 3


(2) Экспрессия гиалуронидазы бонобо и овечьей PH20
Экспрессию проводили с использованием системы экспрессии ExpiCHO. Когда количество клеток ExpiCHO достигало 6×106 клеток/мл, клетки ExpiCHO трансфицировали с использованием реагента ExpiFectamine CHO с плазмидой, в которой кДНК гиалуронидазы PH20 встраивали в вектор pcDNA3.4-TOPO. Используемая здесь среда для культивирования клеток представляла собой среду для экспрессии ExpiCHO (100~500 мл). После трансфекции клетки ExpiCHO культивировали при встряхивании при 130 об/мин в течение 6 дней. В течение этого периода клетки культивировали при 37°С в течение 1 дня, а затем дополнительно культивировали при пониженной температуре 32°С в течение 5 дней. По завершении культивирования клетки центрифугировали при 10000 об/мин в течение 30 минут для получения клеточной надосадочной жидкости.
(3) Очистка гиалуронидазы PH20 бонобо и овцы
Рекомбинантные белки гиалуронидазы животного происхождения, с His-меткой на С-конце, полученные в клетках ExpiCHO, очищали с помощью двухстадийной колоночной хроматографии с использованием системы AKTA prime (GE Healthcare Systems). PH20 бонобо имеет pI 6, поэтому для нее использовали Q Sepharose для анионообменной хроматографии, а овечья PH20 имеет pI 8 или более, и поэтому была подвергнута одностадийной очистке с использованием колонки Capto S для катионообменной хроматографии. Каждый белок подвергали двухстадийной очистке с использованием колонки HisTrap HP для аффинной хроматографии с His-Tag.
Для очистки белка с использованием колонки Q Sepharose готовили буфер А (20 мМ фосфата натрия, рН 7,5) и буфер В (20 мМ фосфата натрия, рН 7,5; 0,5 М NaCl). Белок связывали с колонкой Q Sepharose, и колонку промывали 5 колоночными объемами буфера А для удаления неспецифически связанных белков, а затем промывали 5 колоночными объемами буфера B при градиенте концентрации от 0 до 100% для элюирования белка.
Для очистки белка с использованием колонки Capto S готовили буфер А (20 мМ фосфат натрия, 15 мМ NaCl, pH 6,0) и буфер B (20 мМ фосфат натрия, 500 мМ NaCl, pH 6,0). Значение рН и проводимость культуральной среды устанавливали такими же, как у буфера А, и культуральную среду фильтровали через мембрану с размером пор 0,22 мкм. Образец белка связывали с колонкой Capto S, и колонку промывали 3 колоночными объемами (CV) буфера А для удаления неспецифически связанных белков. Колонку промывали 4 CV буфера B для элюирования белка.
Для очистки белка с использованием колонки HisTrap HP готовили буфер А (20 мМ фосфат натрия, 500 мМ NaCl, pH 7,5) и буфер B (20 мМ фосфат натрия, 500 мМ NaCl, 500 мМ имидазол, pH 7,5). Образец белка связывали с колонкой HisTrap HP, и колонку промывали 7 CV 7% буфера B для удаления неспецифически связанных белков, а затем промывали 3 CV 40% буфера B для элюирования целевого белка. Элюат колонки подвергали диализу с использованием буфера для диализа (20 мМ фосфата натрия, 100 мМ NaCl, pH 7,0).
(4) Анализ гиалуронидазы PH-20 бонобо и овец.
Анализ активности проводили таким же образом, как в Примере 7, и анализ уровня N-гликанов выполняли таким же образом, как в Примере 9. Результаты представлены в таблице 1.
Пример 11. Анализ ферментативной кинетики вариантов в зависимости от уровня N-гликанов
Для анализа ферментативной кинетики вариантов по настоящему изобретению ферментативную активность измеряли методом Моргана-Элсона (Takahashi, T. et al. (2003) Anal. Biochem. 322:257-263). Метод Моргана-Элсона представляет собой колориметрический метод, который анализирует дающие красный цвет вещества (при 545 нм), возникающие в результате реакции восстановленного терминального N-ацетил-D-глюкозамина (GlcNAc), образующегося при гидролизе гиалуроновой кислоты гиалуронидазой, с пара-диметиламинобензальдегидом (ДМАБ), то есть реактивом Эрлиха. N-ацетил-D-глюкозамин (GlcNAc, Sigma), разбавленный до 0,25; 0,50; 0,75; 1,00 или 1,25 мМ в буферном растворе для разведения (0,1 М NaPi; 0,1 М NaCl; 1,5 мМ 1,4-лактона сахарной кислоты, рН 5,35) восстанавливали обработкой тетраборатом в каждой пробирке, а затем добавляли ДМАБ, чтобы вызвать колориметрическую реакцию. После реакции измеряли оптическую плотность при 545 нм, чтобы создать стандартную кривую реакции для GlcNAc. Гиалуроновую кислоту в качестве субстрата разводили до 0,54; 0,65; 0,87; 1,23 или 2,17 мкМ в буферном растворе для разведения в каждой пробирке, и в каждую пробирку добавляли гиалуронидазу с последующей реакцией при 37°С в течение 5 минут и нагреванием при 100°С в течение 5 минут до завершения ферментативной реакции. Полученное вещество восстанавливали путем обработки тетраборатом и добавляли ДМАБ, чтобы вызвать колориметрическую реакцию. После реакции измеряли поглощение при 545 нм и определяли ферментативную активность, используя стандартную кривую реакции GlcNAc, приведенную выше. С помощью этого метода анализировали ферментативную кинетику PH20 дикого типа с SEQ ID NO: 1 и варианта PH20 по настоящему изобретению. В результате кривая Лайнуивера-Берка оказалась линейной, что означает, что вариант РН20 по настоящему изобретению следует формуле кинетики фермента Михаэлиса-Ментен.
В таблице 4 представлены результаты анализа ферментативной кинетики фракций № 1 и № 2, полученных из опытной культуры клональной клеточной линии № 2, среди образцов в таблице 1 из Примера 1. Результаты показали, что когда уровни галактозилирования и маннозилирования находятся в одинаковых пределах диапазонах и уровень сиалилирования низкий, ферментативная активность рекомбинантной гиалуронидазы повышается за счет высокой каталитической эффективности (kcat/Km) фермента.
Экспериментальный результат доказывает, что на активность фермента с одинаковой аминокислотной структурой может влиять изменение гликозилирования, точнее, изменение уровня сиалилирования. Таким образом, это свидетельствует, что при попытке получения промышленно пригодной гиалуронидазы путем массового производства гиалуронидазы PH20 дикого типа или ее варианта с использованием рекомбинантного метода можно создать фермент, имеющий большую промышленную применимость, путем контроля уровня сиалилирования.
Таблица 4
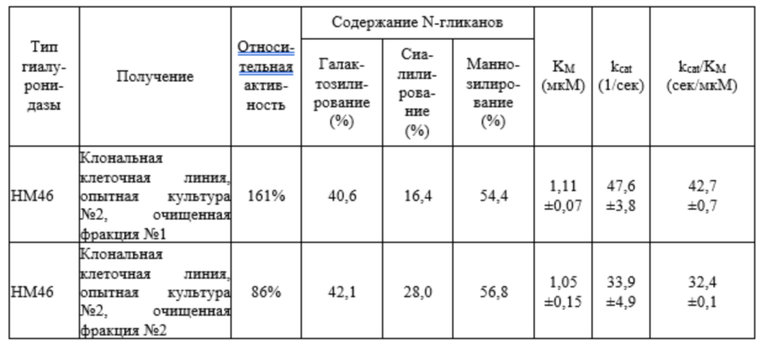
Промышленная применимость
Способ получения рекомбинантного белка гиалуронидазы PH20 или ее варианта в соответствии с настоящим изобретением позволяет получать рекомбинантный белок гиалуронидазы PH20 или ее варианта, обладающий высокой продуктивностью и высокой ферментативной активностью, и, таким образом, реализует массовое производство и поставку рекомбинантного белка гиалуронидазы PH20 или ее варианта.
Хотя были подробно описаны конкретные конфигурации настоящего изобретения, специалистам в данной области техники будет понятно, что это подробное описание представлено в качестве предпочтительных вариантов осуществления только для иллюстративных целей и не должно рассматриваться как ограничивающее объем настоящего изобретения. Таким образом, существенный объем настоящего изобретения определяется прилагаемой формулой изобретения и ее эквивалентами.
Ссылки
1. L. H. Bookbinder, A. Hofer, M. F. Haller, M. L. Zepeda, G-A. Keller, J. E. Lim, T. S. Edgington, H. M. Shepard, J. S. Patton, G. I. Frost. (2006). A recombinant human enzyme for enhanced interstitial transport of therapeutics. J. Control. Release. 114, 230-241.
2. Douglas B. Muchmore, M.D., and Daniel E. Vaughn, Ph.D. (2012). Accelerating and Improving the Consistency of Rapid-Acting Analog Insulin Absorption and Action for Both Subcutaneous Injection and Continuous Subcutaneous Infusion Using Recombinant Human Hyaluronidase. J. Diabetes Sci. and Technol. 6(4): 764-72
3. E. M. Krantz. (1980). Low-dose intramuscular ketamine and hyaluronidase for induction of anaesthesia in non-premedicated children. S. Afr. Med. J. 58(4):161-2.
4. W. A. Clement, S. H. Vyas, J. N. Marshall, J. H. Dempster. (2003). The use of hyaluronidase in nasal infiltration: prospective randomized controlled pilot study. J. Laryngol. Otol. 117(8):614-8.
5. Thomas J. R., Wallace M. S., Yocum R. C., Vaughn D. E., Haller M. F., Flament J. (2009). The INFUSE-Morphine study: use of recombinant human hyaluronidase (rHuPH20) to enhance the absorption of subcutaneously administered morphine in patients with advanced illness. J. Pain and Symptom Manag. 38(5):663-672.
6. George Harb, Francois Lebel, Jean Battikha, Jeffrey W Thackara. (2010). Safety and pharmacokinetics of subcutaneous ceftriaxone administered with or without recombinant human hyaluronidase (rHuPH20) versus intravenous ceftriaxone administration in adult volunteers. Curr. Med. Res. Opin. 26(2):279-88.
7. Richard L. Wasserman. (2014). Overview of recombinant human hyaluronidase-facilitated subcutaneous infusion of IgG in primary immunodeficiencies. Immunotherapy. 6(5):553-67.
8. Harris R. J., Shire S. J., Winter C. (2004). Commercial manufacturing scale formulation and analytical characterization of therapeutic recombinant antibodies. Drug. Dev. Res. 61:137-154.
9. Stephan Schilling, Torsten Hoffmann, Fred Rosche, Susanne Manhart, Claus Wasternack, and Hans-Ulrich Demuth. (2002). Heterologous Expression and Characterization of Human Glutaminyl Cyclase: Evidence for a Disulfide Bond with Importance for Catalytic Activity. Biochemistry, 35, 10849-10857.
10. H. Tachibana 1, K. Taniguchi, Y. Ushio, K. Teruya, K. Osada, H. Murakami. (1994). Changes of monosaccharide availability of human hybridoma lead to alteration of biological properties of human monoclonal antibody. Cytotechnology. 16(3):151-7.
11. Veronica Restelli, Ming-Dong Wang, Norman Huzel, Martin Ethier, Helene Perreault, Michael Butler. (2006). The effect of dissolved oxygen on the production and the glycosylation profile of recombinant human erythropoietin produced from CHO cells. Biotechnol. Bioeng. 94:481-494.
12. M. C. Borys, D. I. Linzer, E. T. Papoutsakis. (1993). Culture pH affects expression rates and glycosylation of recombinant mouse placental lactogen proteins by Chinese hamster ovary (CHO) cells. Biotechnology (NY). 11:720-724.
13. M. C. Borys, D. I. Linzer, E. T. Papoutsakis. (1994). Ammonia affects the glycosylation patterns of recombinant mouse placental lactogen-I by Chinese hamster ovary cells in a pH-dependent manner. Biotechnol. Bioeng. 43:505-514.
14. Clark K. J., Chaplin F. W., Harcum S. W. (2004). Temperature effects on product quality-related enzymes in batch CHO cell cultures producing recombinant tPA. Biotechnol. Prog. 20:1888-1892.
15. Arming S., Strobl B., Wechselberger C., Kreil G. (1997) In-vitro mutagenesis of PH-20 hyaluronidase from human sperm. Eur J. Biochem. 247:810-814
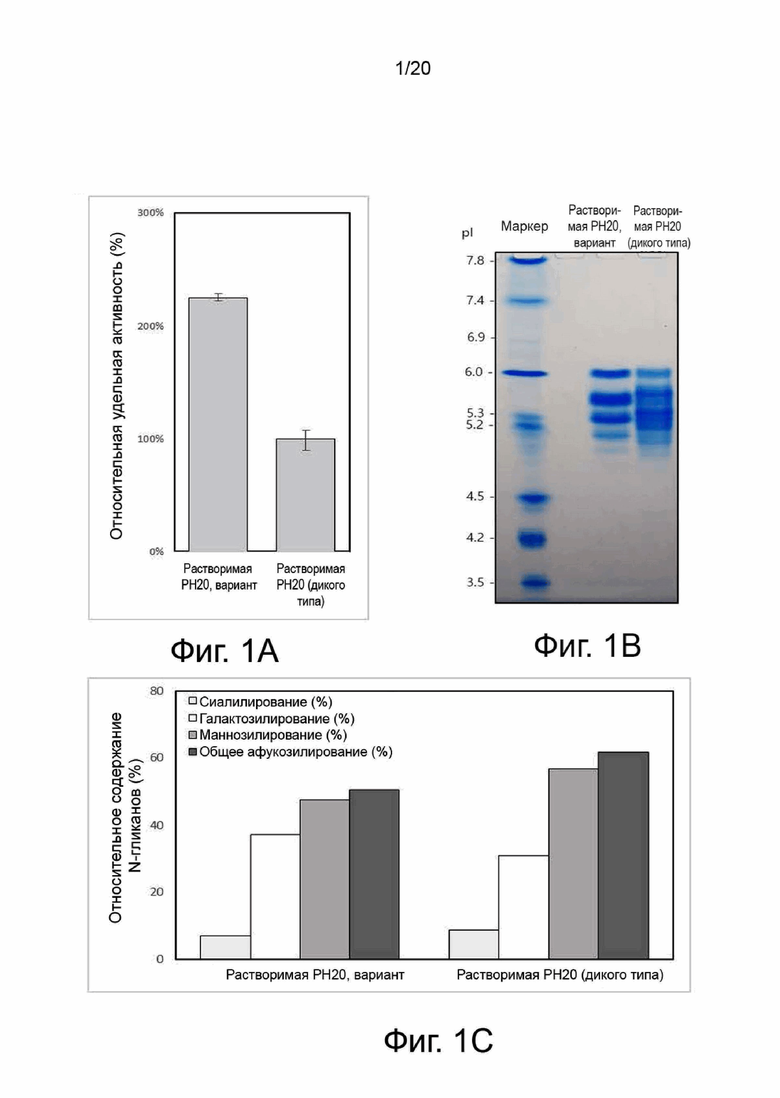
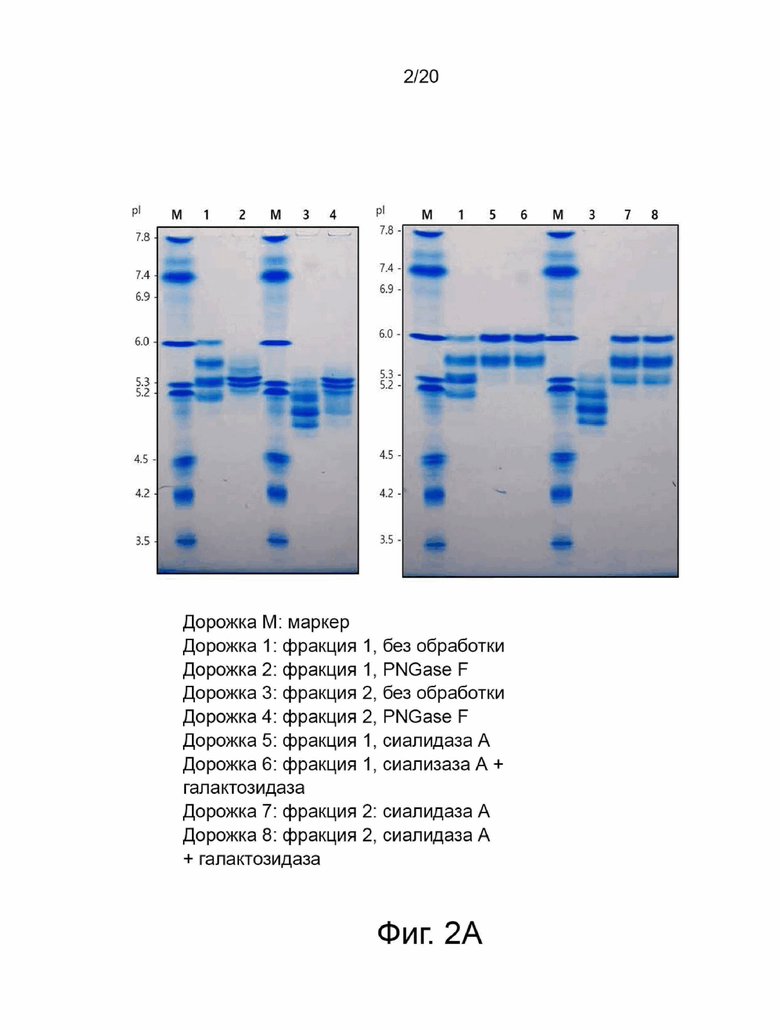
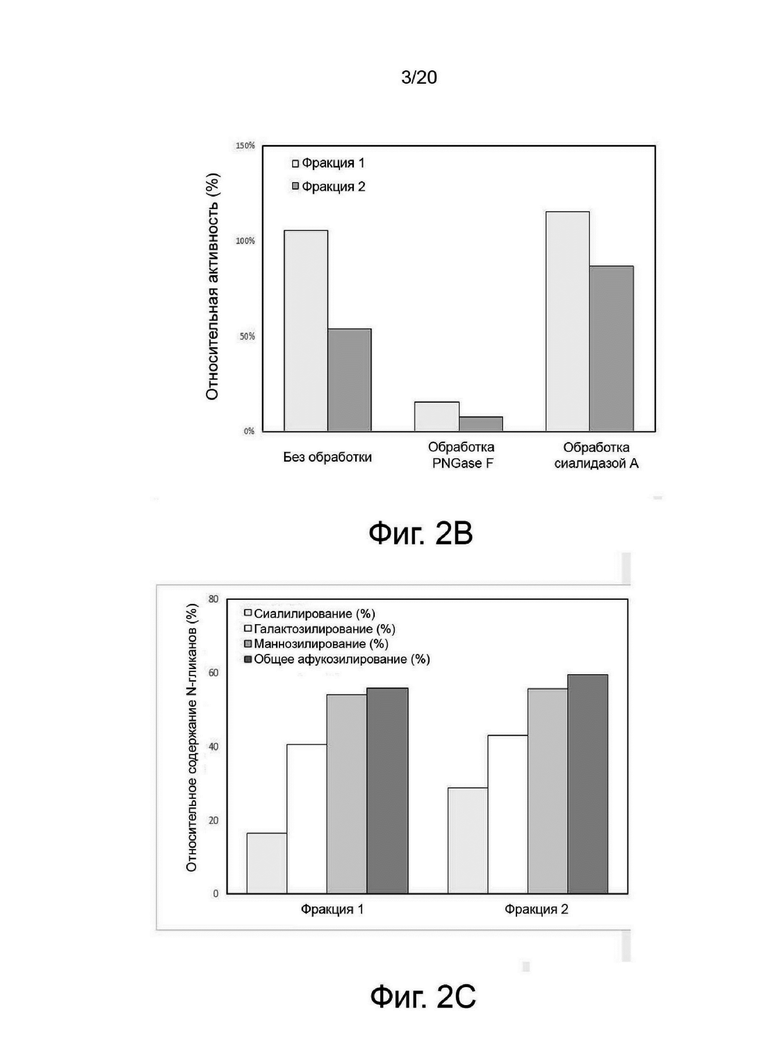
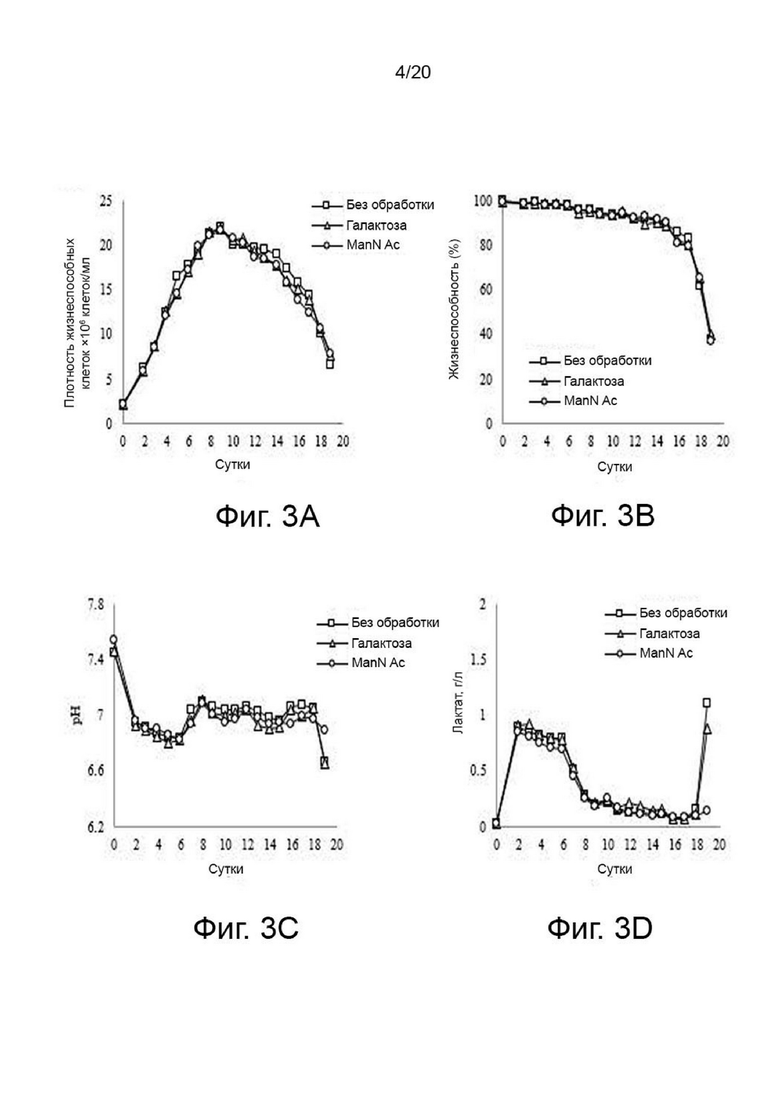
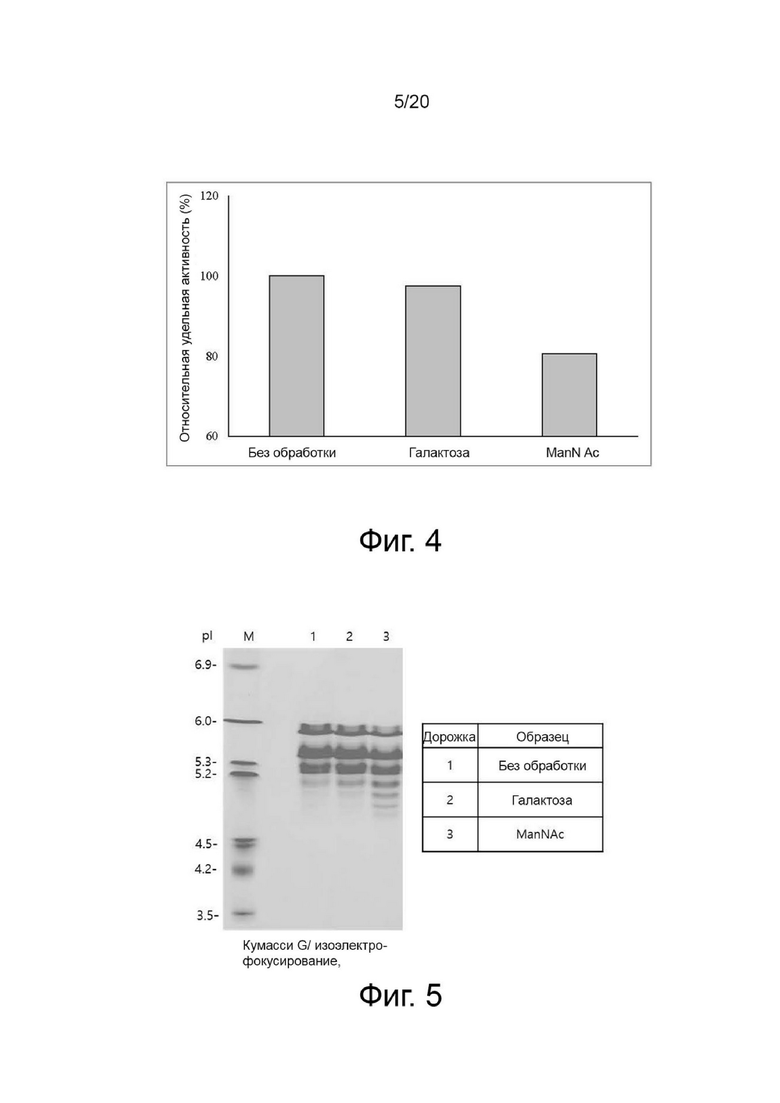
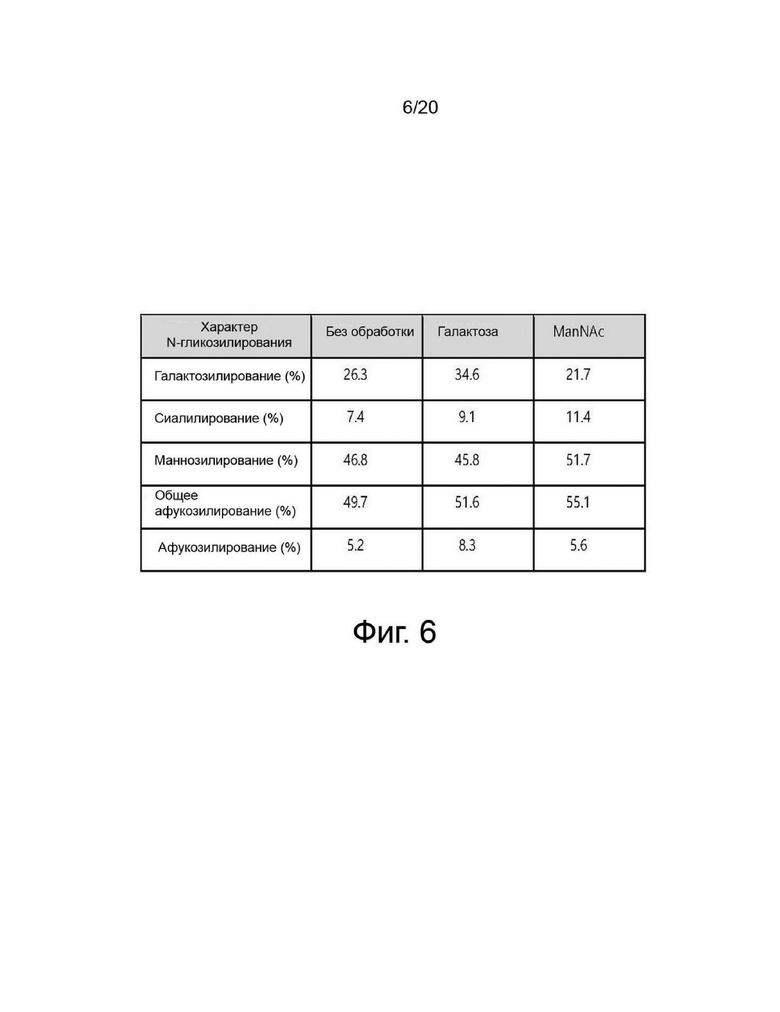
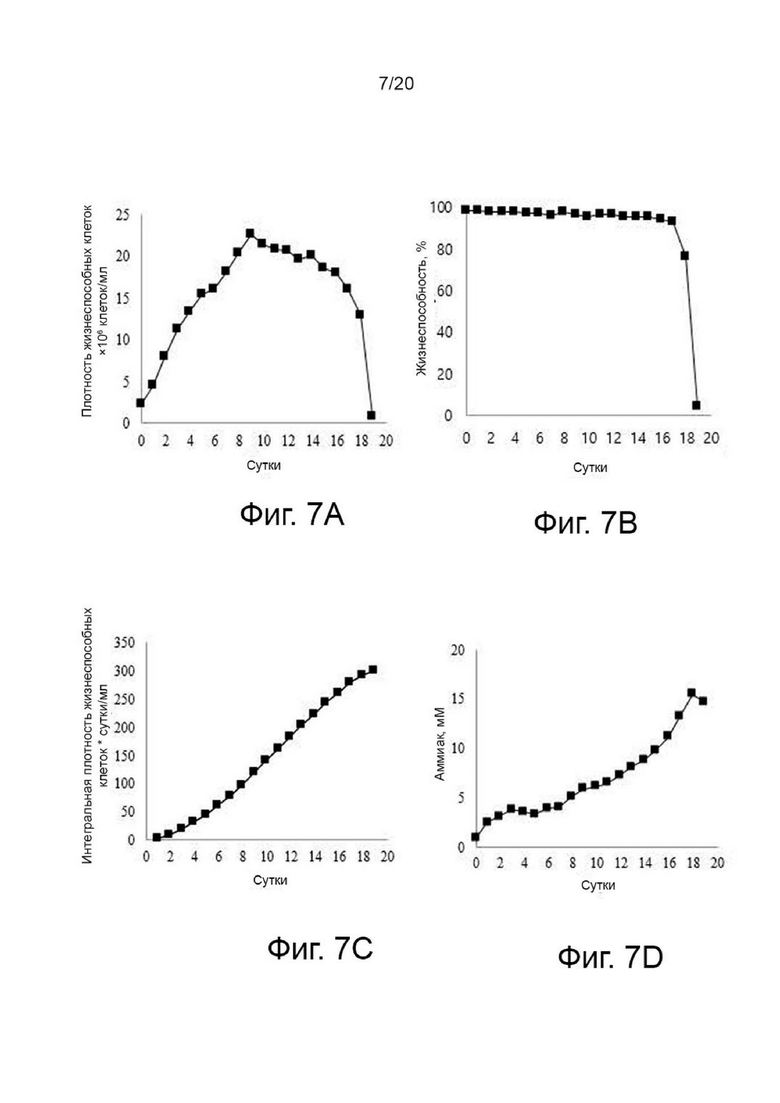
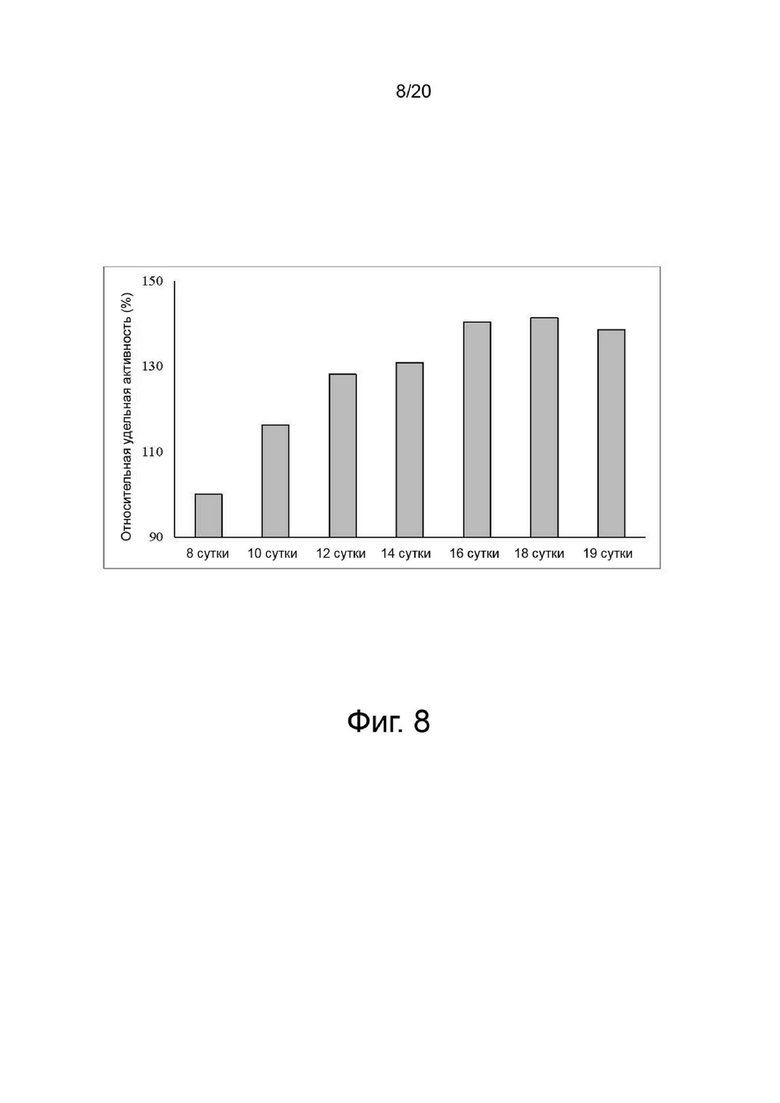
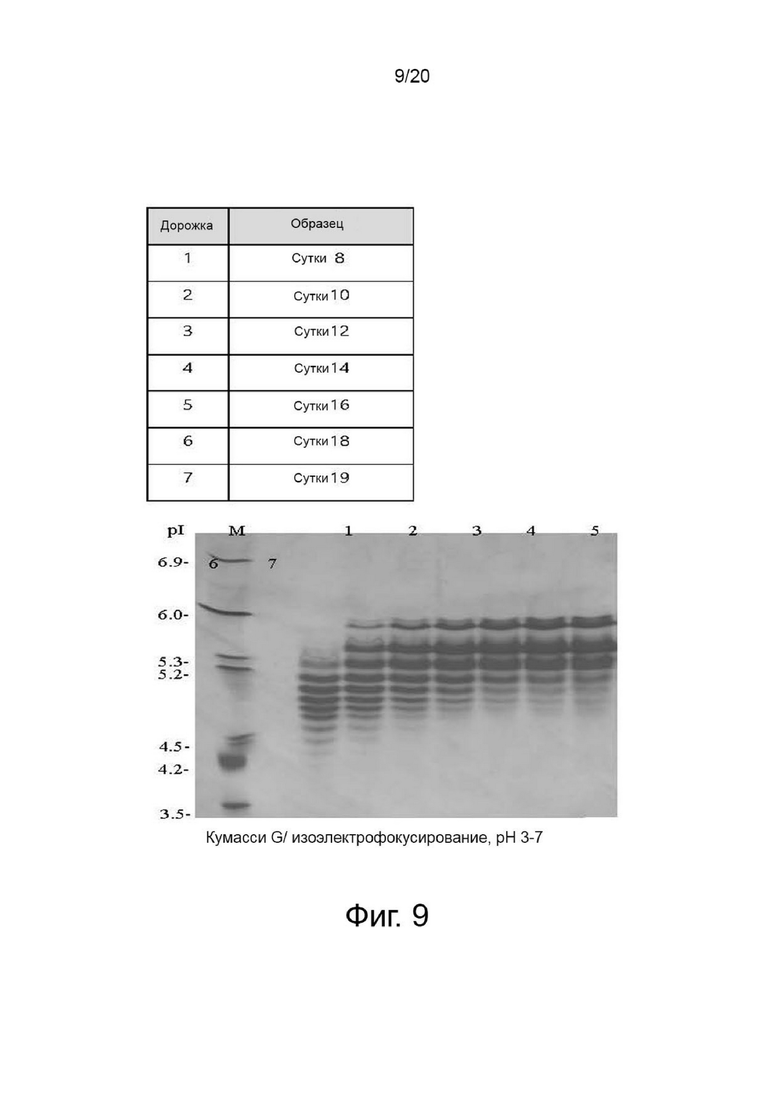
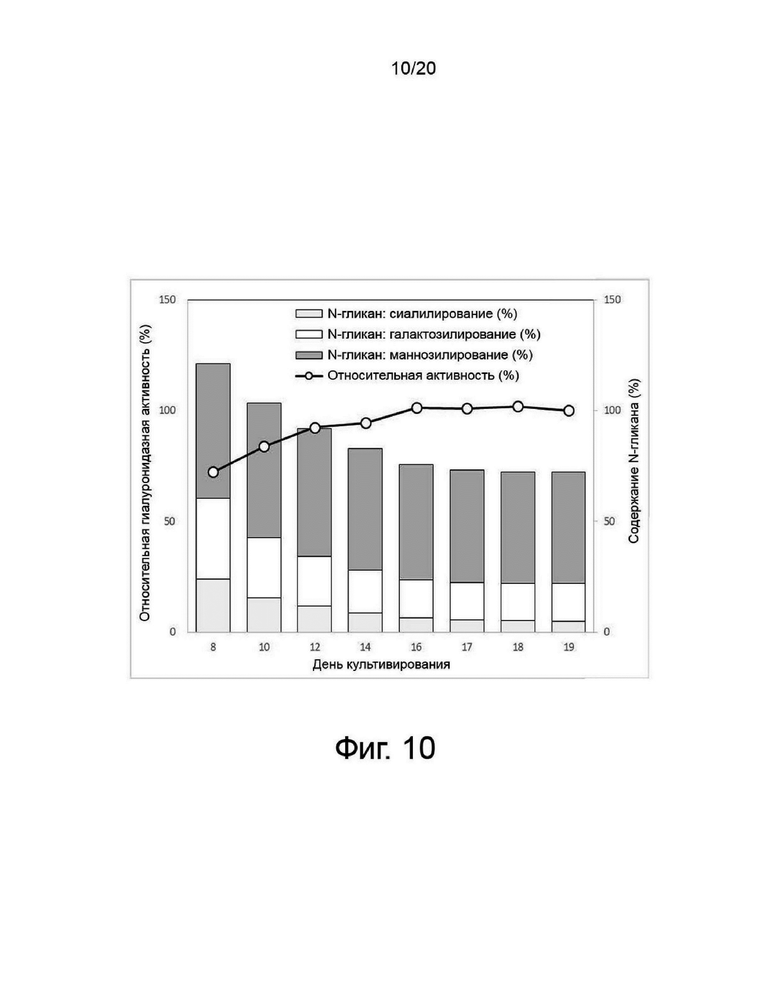
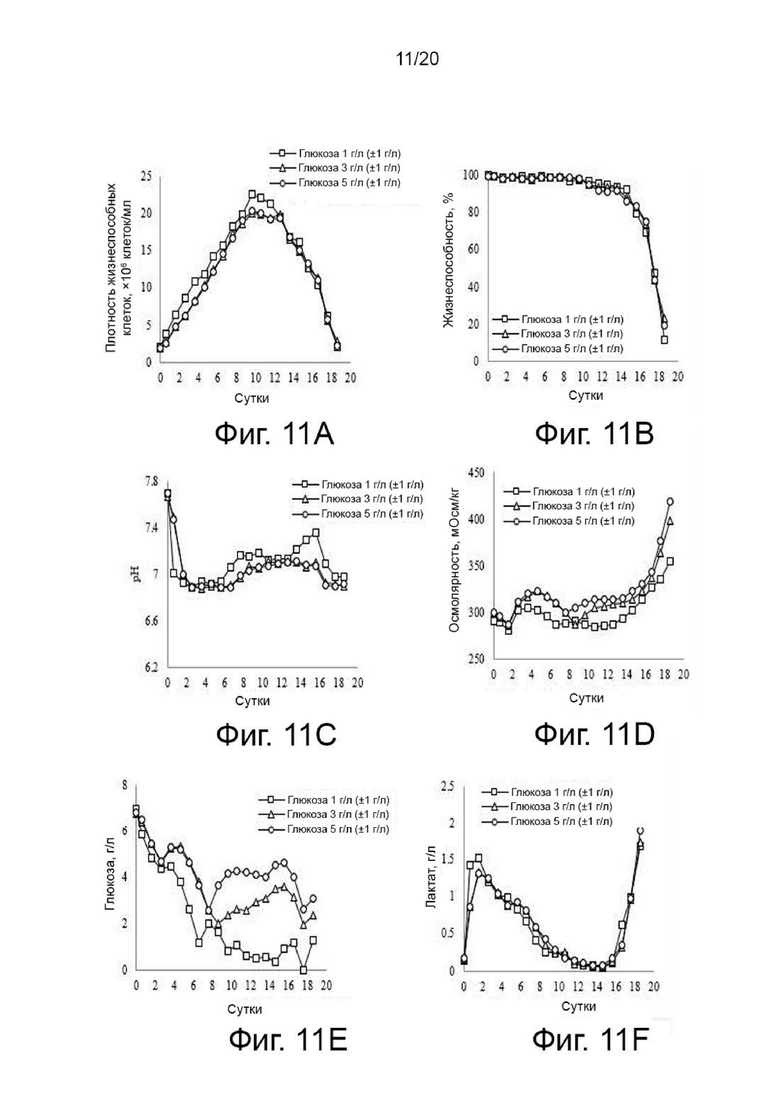
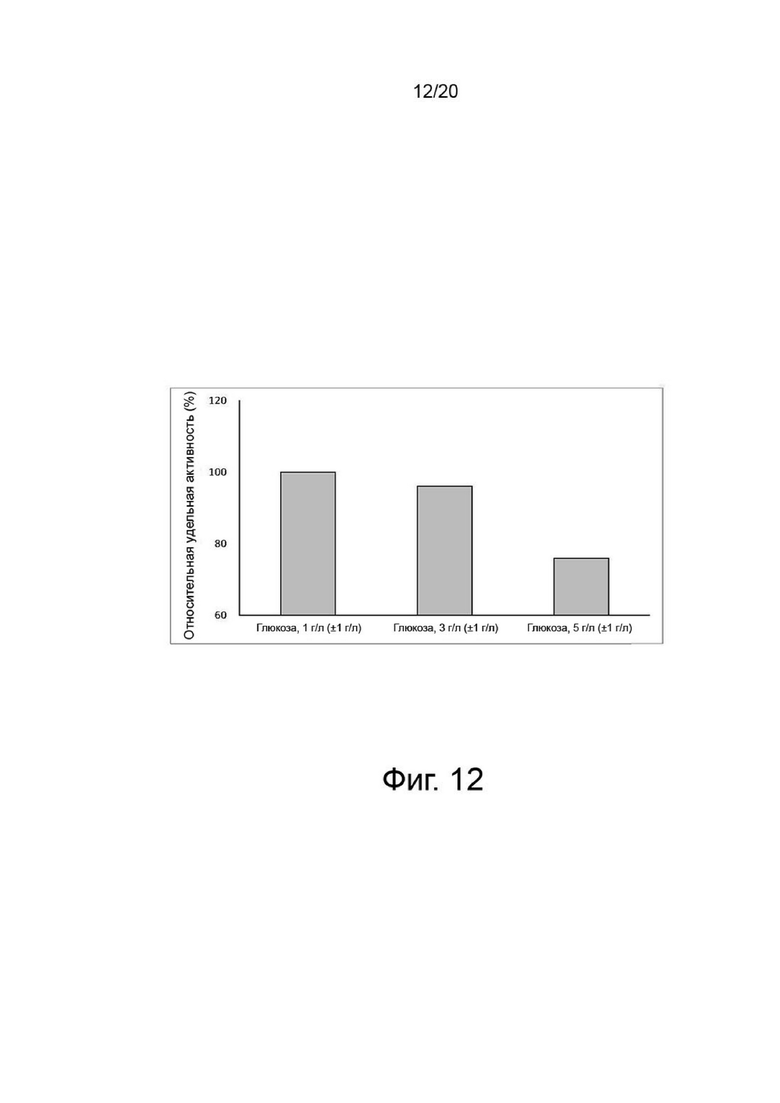

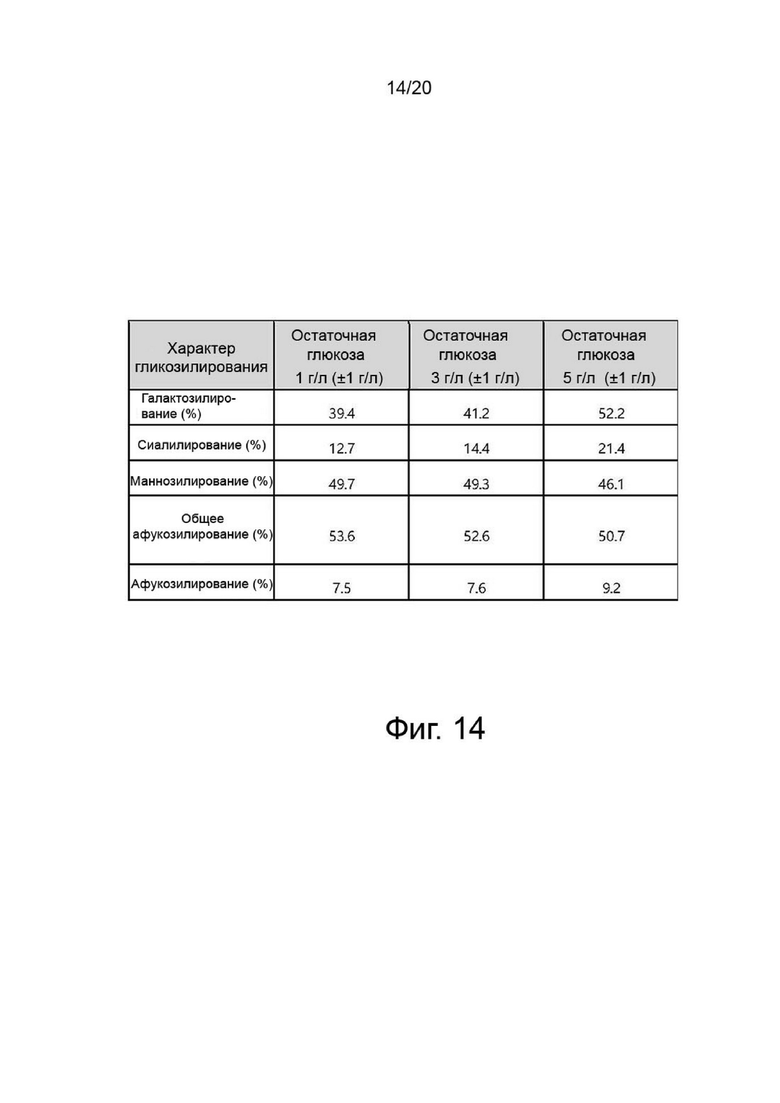
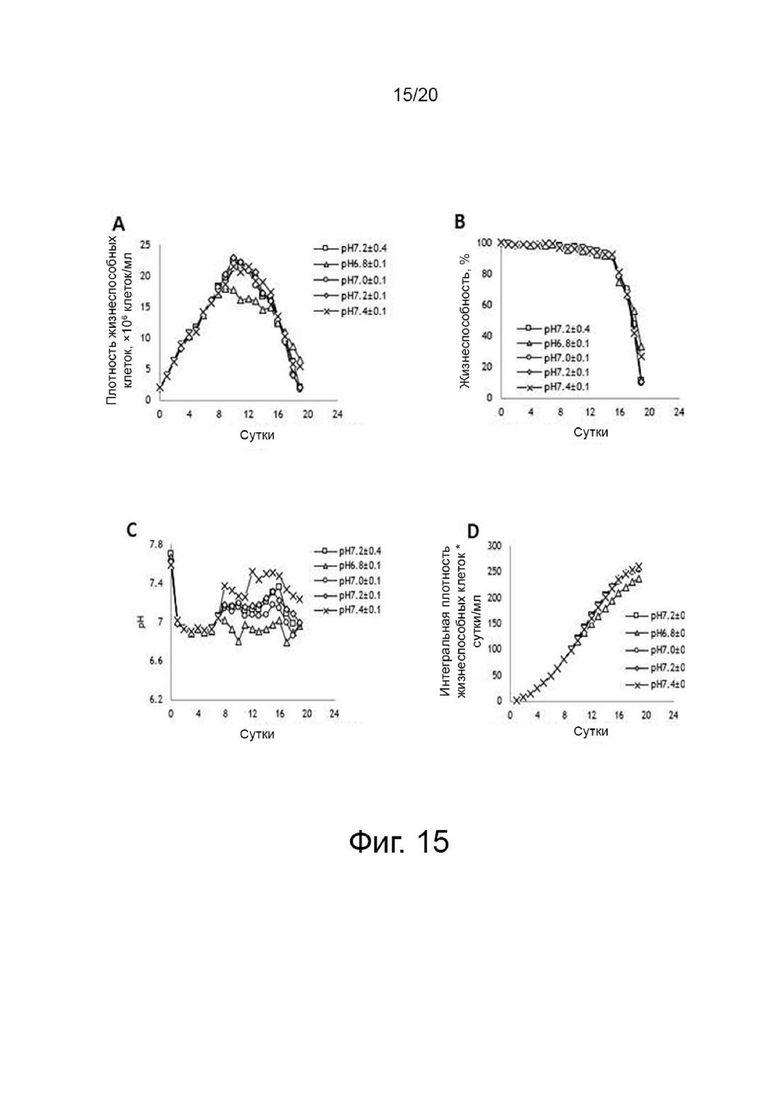
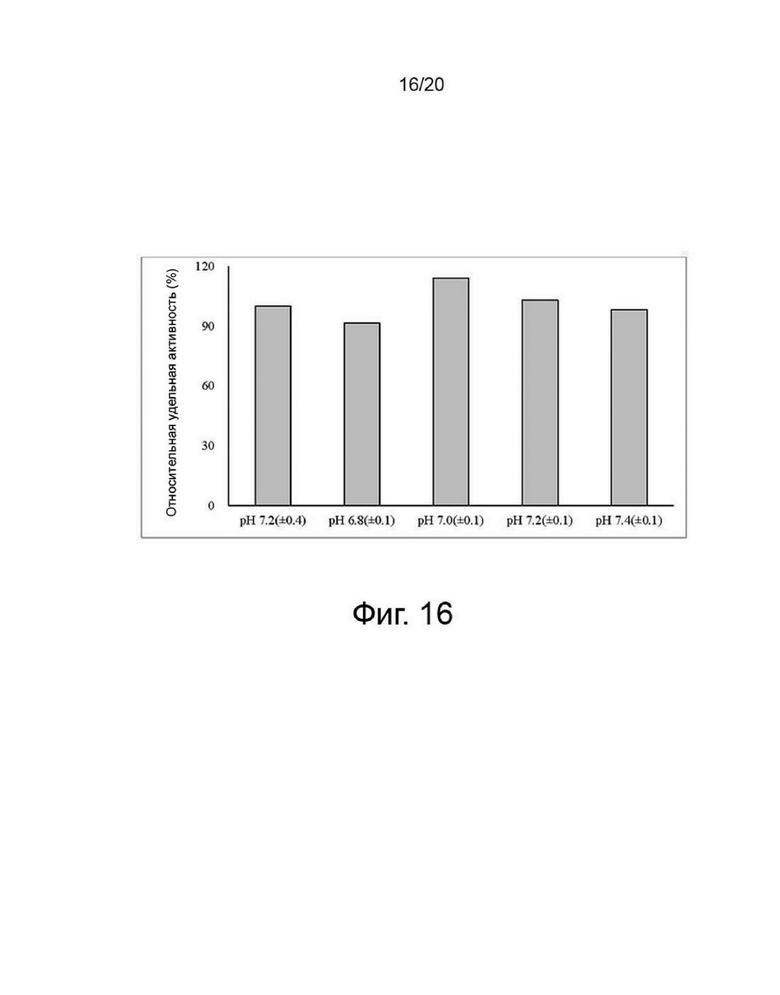
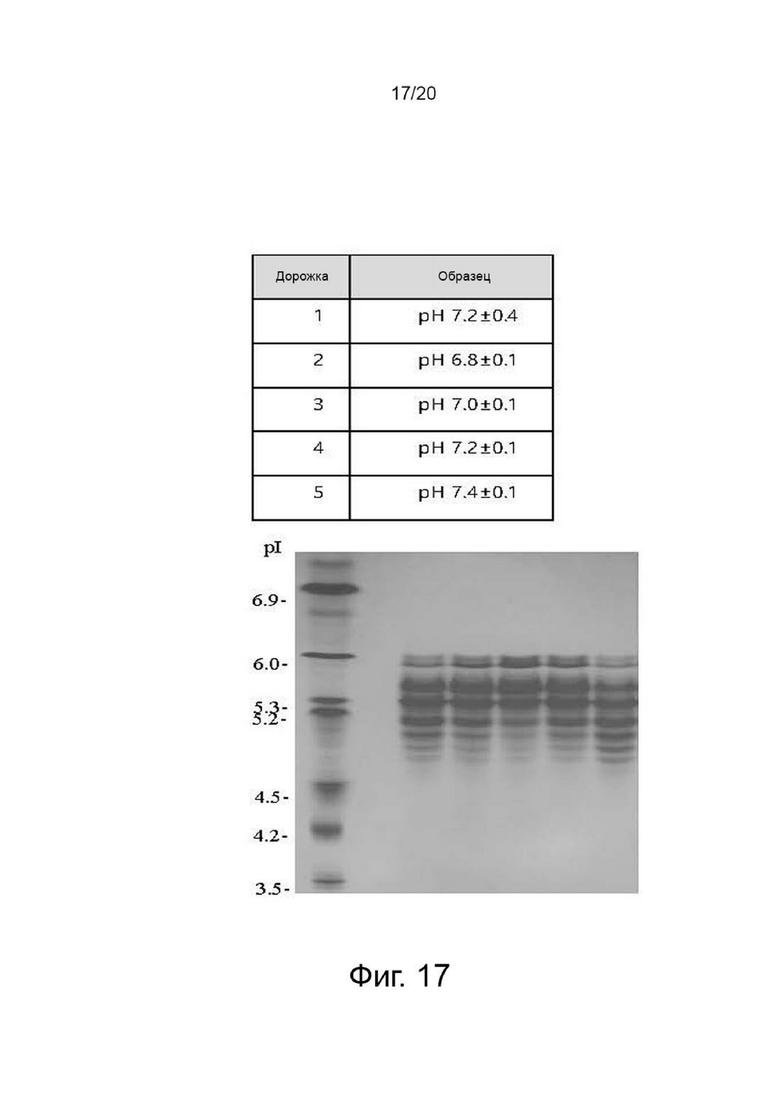
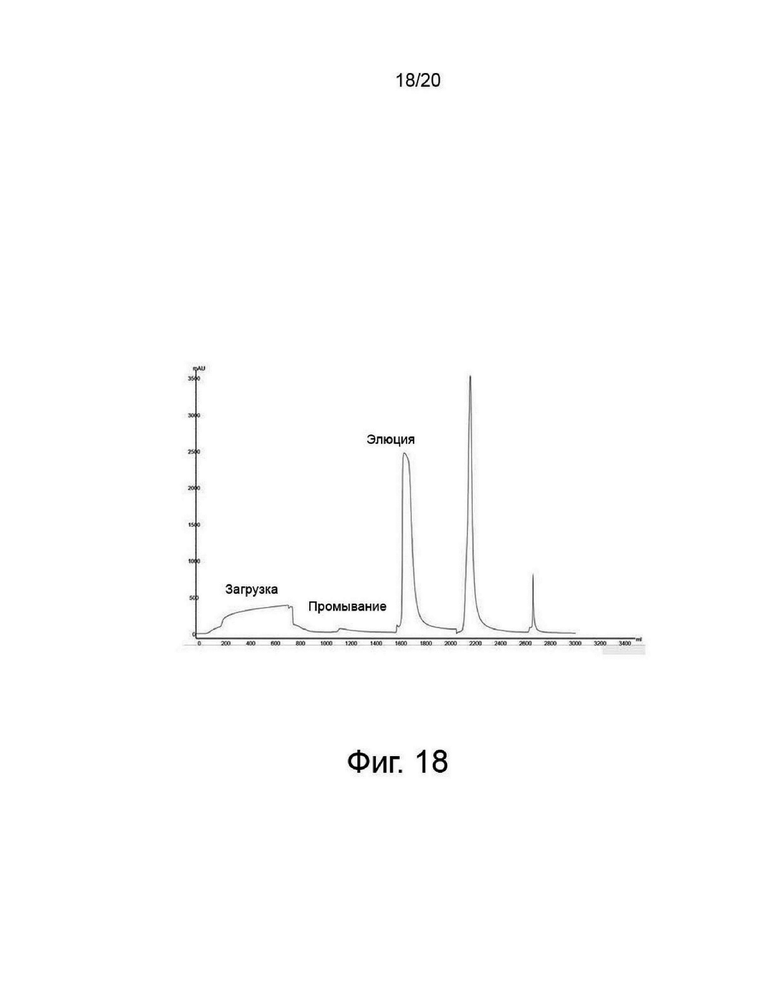
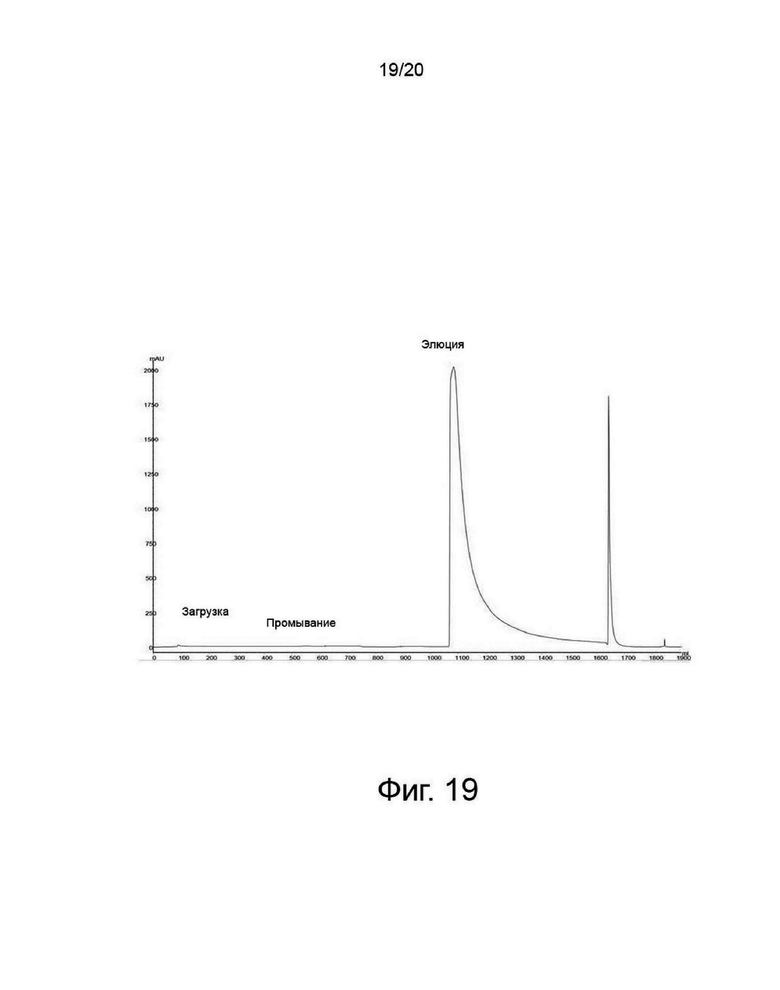
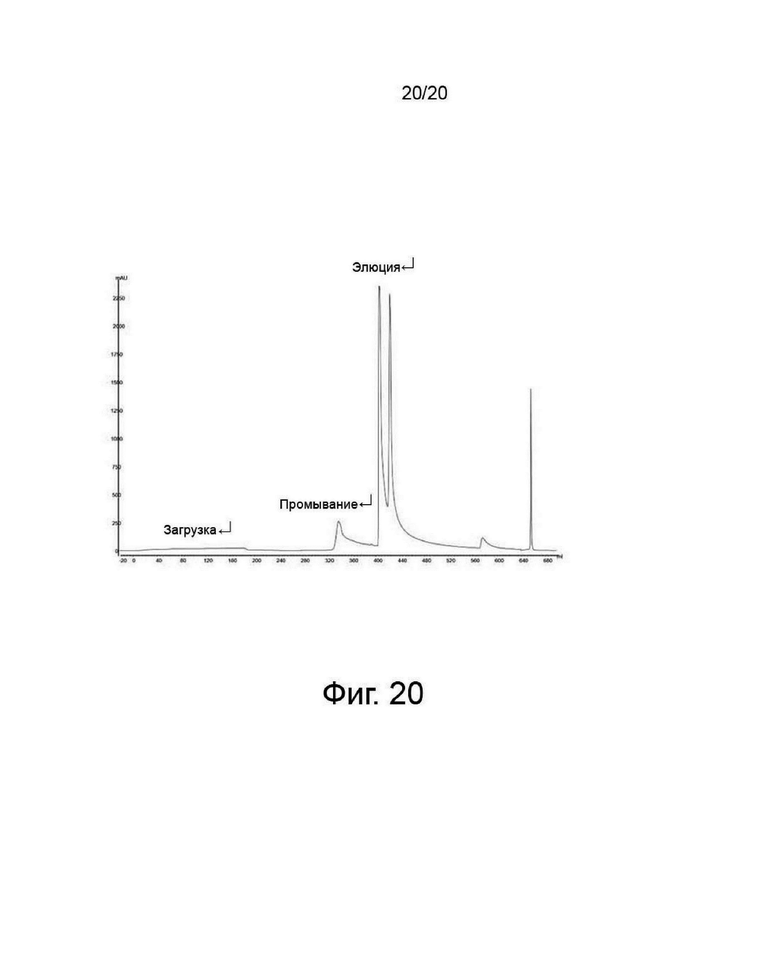
Изобретение относится к биотехнологии. Предложен способ получения рекомбинантной гиалуронидазы PH20 или ее варианта. Способ позволяет изменять уровни N-гликанов в условиях культивирования, включающих контролируемую концентрацию глюкозы в культуральной среде и пониженную температуру культивирования в течение определенного периода времени культивирования. Изобретение обеспечивает получение рекомбинантной гиалуронидазы PH20 или ее варианта с увеличенной удельной активностью и продуктивностью. 12 з.п. ф-лы, 20 ил., 4 табл., 11 пр.
1. Способ получения рекомбинантной гиалуронидазы PH20 или ее варианта, который содержит замены, делеции и инсерции одного или нескольких аминокислотных остатков и, необязательно, содержит делеции некоторых N-концевых и/или С-концевых аминокислотных остатков в аминокислотной последовательности гиалуронидазы РН20, и имеющий активность фермента гиалуронидазы, включающий:
(1) культивирование клеток-хозяев, экспрессирующих рекомбинантную гиалуронидазу PH20 или ее вариант, при температуре культивирования от 35°C до 38°C до тех пор, пока интегральная плотность жизнеспособных клеток клеток-хозяев не достигнет от 20×106 до 120×106 клеток в сутки/мл; и
(2) снижение температуры культивирования до 28-34°С и последующее культивирование клеток-хозяев в течение от 2 до 18 суток; при поддержании температуры культуры:
(а) где остаточную концентрацию глюкозы в среде поддерживают на уровне от 0,001 г/л до 4,5 г/л в течение периода культивирования; и
(b) где pH культуральной среды поддерживают на уровне 6,9~7,1;
при этом уровень сиалилирования N-гликана полученной PH20 или ее варианта составляет от 1 до 21.7%, где доля сиалилирования (%) представляет собой сумму доли (%) N-гликанов, содержащих сиаловую кислоту на своем конце, таких как A1, A1F, A2 и A2F,
при этом доля галактозилирования составляет от 1 до 68%, а доля маннозилирования составляет от до 40 до 63% N-гликанов в полученной PH20, соответственно, где доля галактозилирования (%) представляет собой сумму доли (%) N-гликанов, содержащих галактозу на своем конце, таких как G1, G1F, G1F', G2, G2F, A1, A1F, A2 и A2F, а доля маннозилирования (%) представляет собой сумму доли (%) N-гликанов, содержащих маннозу на своем конце, таких как M4G0F, M5, M5G0, M6, M7, M8, и M9, и
при этом активность гиалуронидазы в культуральной среде составляет 10000 единиц/мл или более.
2. Способ по п. 1, где указанный уровень галактозилирования составляет от 1 до 49,6%, уровень сиалилирования составляет от 1 до 21,7%, а уровень маннозилирования составляет от 40 до 58,3% N-гликанов полученной PH20 соответственно.
3. Способ по п. 1, где указанный уровень галактозилирования составляет от 14,4 до 49,6%, уровень сиалилирования составляет от 2,5 до 21,7%, а уровень маннозилирования составляет от 47,6 до 58,3% N-гликанов полученной PH20.
4. Способ по п. 1, где удельная активность полученной PH20 или ее варианта по меньшей мере на 10% выше, чем удельная активность человеческой PH20 дикого типа.
5. Способ по п. 1, где культивирование клеток-хозяев на стадии (1) и/или стадии (2) осуществляют одним или несколькими способами, выбранными из группы, состоящей из периодического культивирования, повторного периодического культивирования, периодического культивирования с подпиткой, повторного периодического культивирования с подпиткой, непрерывного культивирования и перфузионного культивирования.
6. Способ по п. 1, где культивирование клеток-хозяев на стадии (1) и/или стадии (2) проводят при одном или нескольких условиях, выбранных из группы, состоящей из:
(i) условий, в которых концентрацию аммиака в среде поддерживают на уровне 5 мМ или более;
(ii) условий, в которых в среду добавляют одно или несколько веществ, выбранных из группы, состоящей из глутамина, глюкозамина, уридина, глюкозамина и бутирата натрия; и
(iii) условий, в которых в среду не добавляют галактозу и manNAc.
7. Способ по п. 1, где вариант РН20 включает замену одного или нескольких аминокислотных остатков и при необходимости включает усечение одного или нескольких аминокислотных остатков N-конца и/или С-конца в аминокислотной последовательности РН20 дикого типа.
8. Способ по п. 1, где клетки-хозяева представляют собой клетки животных, дрожжей, актиномицетов или насекомых.
9. Способ по п. 1, дополнительно включающий выделение и очистку полученной PH20 или ее варианта.
10. Способ по п. 9, где выделение и очистку PH20 или ее варианта осуществляют с использованием характеристик ионной связи и/или гидрофобного взаимодействия PH20 или ее варианта, а не аффинного связывания.
11. Способ по п. 10, где выделение и очистку РН20 или ее варианта осуществляют с использованием хроматографии гидрофобного взаимодействия и ионообменной хроматографии, а не аффинной хроматографии.
12. Способ по п. 9, где удаляют кислую РH20 или ее вариант.
13. Способ по п. 12, где удаление кислой гиалуронидазы PH20 или ее варианта проводят с помощью ионообменной хроматографии.
| EP 3037529, 29.06.2016 | |||
| CHEN K.J | |||
| ET AL | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| J Biosci Bioeng | |||
| Токарный резец | 1924 |
|
SU2016A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| FROST GI | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Expert Opin Drug Deliv | |||

