Настоящее изобретение относится к антисмысловым олигонуклеотидам, их применению в качестве ингибиторов сигнального пути TGF-R, фармацевтическим композициям, содержащим такие антисмысловые олигонуклеотиды, и применению для профилактики и лечения неврологических, нейродегенеративных и гиперпролиферативных заболеваний, в том числе онкологических заболеваний.
В организме человека TGF-β находится в трех известных подтипах, TGF-β1, TGF-β2 и TGF-β3. Они подвергаются повышающей регуляции при нейродегенеративных заболеваниях, таких как ALS, и также при некоторых злокачественных заболеваниях человека, и также было показано, что повышенная экспрессия этого фактора роста имеет место при таких патологических состояниях, как нейродегенеративные заболевания, острая травма и нейровоспаление, и при старении. Полагают, что изоформы трансформирующего фактора роста бета (TGF-β1) также участвуют в патогенезе преэклампсии.
Активированные TGF-β оказывают свои эффекты на клетки-мишени через три различных класса рецепторов: тип I (TGFRI), также называемый активин-подобными киназами (ALK; 53 кДа), тип II (TGFRII; 70-100 кДа) и тип III (TGFRIII; 200-400 кДа). Рецепторы TGF-β представляют собой однопроходные рецепторы с серин-треонин-киназной активностью. В то время, как рецептор типа II киназы является конститутивно активным, рецептор типа I должен активироваться. Данный процесс инициируется посредством связывания лиганда с TGFRII; это «запускает» образование временного комплекса, который включает лиганд и рецептор типа I и II. Принимая во внимание димерный состав лиганда, рецепторный комплекс, скорее всего, состоит из тетрамерной структуры, образованной двумя парами рецепторов каждого типа.
Трансдукция сигналов TGF-β происходит через его рецепторы и ниже через белки Smad. Smad-зависимая трансдукция клеточных сигналов, инициируемая связыванием изоформы TGF-β с парой специфичных рецепторов TGFRI/II, приводит к фосфорилированию внутриклеточных Smads и затем транслокации активированного комплекса Smad в ядро, чтобы оказать влияние на экспрессию конкретного гена-мишени. Дивергенция сигналов на другие пути и конвергенция с соседних сигнальных путей генерирует очень сложную сеть. В зависимости от окружающей и клеточной среды сигнальный путь TGF-β приводит к различным клеточным реакциям, таким как пролиферация, дифференцировка, подвижность клеток и апоптоз опухолевых клеток. При раке TGF-β может непосредственно влиять на рост опухоли (упоминаемый как внутренний эффект сигнального пути TGF-β) или опосредованно (упоминаемый как внешний эффект) посредством стимуляции роста опухоли, индукцией эпителиально-мезенхимального перехода (EMT), блокированием противоопухолевых иммунных ответов, повышением опухолеассоциированного фиброза, модулированием внеклеточного матрикса (ECN) и миграцией клеток, и, наконец, усилением ангиогенеза. Факторы (например, концентрация, время, локальное воздействие), определяющие, имеет сигнальный путь TGF-β промоторную или супрессорную функцию для опухоли, являются предметом интенсивных исследований и дискуссий. В настоящее время считается, что супрессорная функция сигнального пути TGF-β для опухолей отсутствует на ранних стадиях рака аналогично рецессивным мутациям с потерей функции у других опухолевых супрессоров. Следовательно, существует несколько фармакологических подходов к лечению различных видов рака блокированием сигнальных путей TGF-β, таких как исследование галуницертиба и TEW-7197, которые оба являются низкомолекулярными ингибиторами TGFRI и находятся на стадии клинических испытаний, и LY3022859, антитело против TGFRII.
Сигналы, обеспечиваемые белками семейства трансформирующего фактора роста (TGF-β), представляют собой систему посредством которой нейрональные стволовые клетки регулируются в физиологических условиях, но по аналогии с другими типами клеток освобождаются от этого контроля после трансформации в раковые стволовые клетки. TGF-β представляет собой многофункциональный цитокин, участвующий в различных физиологических и патофизиологических процессах в головном мозге. Он индуцируется в мозге взрослого человека после травмы или гипоксии и во время нейродегенерации, когда он модулирует и гасит воспалительные реакции. После травмы, несмотря на то, что TGF-β в общем, является нейропротективным, он ограничивает самовосстановление головного мозга ингибированием пролиферации нервных стволовых клеток и индукцией фиброза/глиоза для образования рубца. Подобно своему действию на нейрональные стволовые клетки, TGF-β оказывает антипролиферативный контроль в отношении большинства типов клеток; однако, как это ни парадоксально, многие опухоли избегают контроля TGF-β. Более того, в таких опухолях развиваются механизмы, которые изменяют антипролиферативное влияние TGF-β и превращают его в онкогенные сигналы, в основном, организуя многочисленные TGF-β-опосредованные эффекты на матрикс, миграцию и инвазию, ангиогенез, и, самое главное, механизмы избежания иммунного надзора. Таким образом, TGF-β участвует в прогрессировании опухолей (смотри фиг. 3).
Следовательно, рецептор II TGF (рецептор II трансформирующего фактора роста бета; аналогично используемые символы: рецептор типа II TGF-бета, TGFBR2; AAT3; FAA3; LDS1B; LDS2; LDS2B; MFS2; RIIC; TAAD2; TGFR-2; TGFbeta-RII, TGF-RII, TGF-RII), и, в частности, его ингибирование, был признан в качестве мишени для лечения нейродегенеративных заболеваний, таких как ALS, и гиперпролиферативных заболеваний, таких как рак и фиброзные болезни.
Таким образом, целью настоящей заявки является обеспечение фармацевтически активных соединений, способных ингибировать экспрессию рецептора II TGF (TGF-RII), и, следовательно, уменьшать количество рецептора II TGF (TGF-RII) и снижать активность сигнального пути ниже по отношению TGF-β.
Цель настоящего изобретения достигается с помощью принципов независимых пунктов формулы изобретения. Другие преимущественные признаки, аспекты и детали изобретения станут очевидными из зависимых пунктов формулы изобретения, описания изобретения, фигур и примеров настоящей заявки.
С удивлением было установлено, что тысячи соединений-кандидатов, таких как белоково-нуклеотидные комплексы, миРНК, микроРНК (миРНК), рибозимы, аптамеры, CpG-олигонуклеотиды, ДНК-зимы, рибопереключатели, липиды, пептиды, небольшие молекулы, модификаторы raft или кавеолина, модификаторы аппарата Гольджи, антитела и их производные, в частности, химеры, Fab-фрагменты и Fc-фрагменты, антисмысловые олигонуклеотиды, содержащие LNA (LNA®: «замкнутые» нуклеиновые кислоты), являются наиболее перспективными кандидатами для применений, раскрытых здесь.
Таким образом, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4) или последовательность CCCTAAACAC (SEQ. ID NO: 5), или ACTACCAAAT последовательность (SEQ. ID NO: 6), или последовательность GGACGCGTAT (SEQ. ID NO: 7), или последовательность GTCTATGACG (SEQ. ID NO: 8), или последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, способную к гибридизации с указанной последовательностью TGGTCCATTC (SEQ. ID NO: 4) или последовательностью CCCTAAACAC (SEQ. ID NO: 5), или последовательностью ACTACCAAAT (SEQ. ID NO:6), или последовательностью GGACGCGTAT (SEQ. ID NO: 7), или последовательностью GTCTATGACG (SEQ. ID NO: 8), или последовательностью TTATTAATGC (SEQ. ID NO: 9) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4) или последовательность CCCTAAACAC (SEQ. ID NO: 5) или ACTACCAAAT последовательность (SEQ. ID NO: 6), или последовательность GGACGCGTAT (SEQ. ID NO: 7), или последовательность GTCTATGACG (SEQ. ID NO: 8), или последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, способную к гибридизации с указанной последовательностью TGGTCCATTC (SEQ. ID NO: 4) или последовательностью CCCTAAACAC (SEQ. ID NO: 5), или последовательностью ACTACCAAAT (SEQ. ID NO:6), или последовательностью GGACGCGTAT (SEQ. ID NO: 7), или последовательностью GTCTATGACG (SEQ. ID NO: 8), или последовательностью TTATTAATGC (SEQ. ID NO: 9) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4) или последовательность CCCTAAACAC (SEQ. ID NO: 5), или ACTACCAAAT последовательность (SEQ. ID NO: 6), или последовательность GGACGCGTAT (SEQ. ID NO: 7), или последовательность GTCTATGACG (SEQ. ID NO: 8), или последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TGGTCCATTC (SEQ. ID NO: 4) или последовательности CCCTAAACAC (SEQ. ID NO: 5), или последовательности ACTACCAAAT (SEQ. ID NO:6), или последовательности GGACGCGTAT (SEQ. ID NO: 7), или последовательности GTCTATGACG (SEQ. ID NO: 8), или последовательности TTATTAATGC (SEQ. ID NO: 9) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID No. 4) или последовательность CCCTAAACAC (SEQ. ID NO: 5) или последовательность ACTACCAAAT (SEQ. ID NO: 6), или последовательность GGACGCGTAT (SEQ. ID NO: 7), или последовательность GTCTATGACG (SEQ. ID NO: 8), или последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TGGTCCATTC (SEQ. ID NO: 4) или последовательности CCCTAAACAC (SEQ. ID NO: 5) или последовательности ACTACCAAAT (SEQ. ID NO: 6) или последовательности GGACGCGTAT (SEQ. ID NO: 7) или последовательности GTCTATGACG (SEQ. ID NO: 8) или последовательности TTATTAATGC (SEQ. ID NO: 9) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TGGTCCATTC (SEQ. ID NO: 4), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TGGTCCATTC (SEQ. ID NO: 4), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TGGTCCATTC (SEQ. ID NO: 4), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4), и антисмысловой олигонуклеотид содержит последовательность, комплементарную указанной последовательности TGGTCCATTC (SEQ. ID NO: 4), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CCCTAAACAC (SEQ. ID NO: 5), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CCCTAAACAC (SEQ. ID NO: 5), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CCCTAAACAC (SEQ. ID NO: 5), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CCCTAAACAC (SEQ. ID NO: 5), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CCCTAAACAC (SEQ. ID NO: 5), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CCCTAAACAC (SEQ. ID NO: 5), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CCCTAAACAC (SEQ. ID NO: 5), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CCCTAAACAC (SEQ. ID NO: 5), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTACCAAAT (SEQ. ID NO: 6), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью ACTACCAAAT (SEQ. ID NO: 6), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTACCAAAT (SEQ. ID NO: 6), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью ACTACCAAAT (SEQ. ID NO: 6), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTACCAAAT (SEQ. ID NO: 6), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности ACTACCAAAT (SEQ. ID NO: 6), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTACCAAAT (SEQ. ID NO: 6), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности ACTACCAAAT (SEQ. ID NO: 6), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GGACGCGTAT (SEQ. ID NO: 7), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью GGACGCGTAT (SEQ. ID NO: 7), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GGACGCGTAT (SEQ. ID NO: 7), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью GGACGCGTAT (SEQ. ID NO: 7), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GGACGCGTAT (SEQ. ID NO: 7), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности GGACGCGTAT (SEQ. ID NO: 7), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GGACGCGTAT (SEQ. ID NO: 7), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности GGACGCGTAT (SEQ. ID NO: 7), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GTCTATGACG (SEQ. ID NO: 8), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью GTCTATGACG (SEQ. ID NO: 8), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GTCTATGACG (SEQ. ID NO: 8), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью GTCTATGACG (SEQ. ID NO: 8), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GTCTATGACG (SEQ. ID NO: 8), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности GTCTATGACG (SEQ. ID NO: 8), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GTCTATGACG (SEQ. ID NO: 8), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности GTCTATGACG (SEQ. ID NO: 8), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TTATTAATGC (SEQ. ID NO: 9), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TTATTAATGC (SEQ. ID NO: 9), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TTATTAATGC (SEQ. ID NO: 9), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком открытой рамки считывания гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок открытой рамки считывания гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TTATTAATGC (SEQ. ID NO: 9), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TTATTAATGC (SEQ. ID NO: 9), и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды по настоящему изобретению предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют собой звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и, более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце и/или на 5'терминальном конце.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, которые содержат 3-10 звеньев LNA и которые, в частности, содержат 1-5 звеньев LNA на терминальном 5'-конце и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида, и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные. Антисмысловые олигонуклеотиды по настоящему изобретению также могут содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида.
Таким образом, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297), CTGGTCCATTCA (SEQ. ID NO: 298), TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300), TCCCTAAACACT (SEQ. ID NO: 301), CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO: 303), CACTACCAAATA (SEQ. ID NO: 304), TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306), TGGACGCGTATC (SEQ. ID NO: 307), GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309), GGTCTATGACGA (SEQ. ID NO: 310), TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297), CTGGTCCATTCA (SEQ. ID NO: 298), TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300), TCCCTAAACACT (SEQ. ID NO: 301), CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO:303), CACTACCAAATA (SEQ. ID NO: 304), TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306), TGGACGCGTATC (SEQ. ID NO: 307), GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309), GGTCTATGACGA (SEQ. ID NO: 310), TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313 соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297), CTGGTCCATTCA (SEQ. ID NO: 298), TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300), TCCCTAAACACT (SEQ. ID NO: 301), CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO:303), CACTACCAAATA (SEQ. ID NO: 304), TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306), TGGACGCGTATC (SEQ. ID NO: 307), GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309), GGTCTATGACGA (SEQ. ID NO: 310), TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297), CTGGTCCATTCA (SEQ. ID NO: 298), TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300), TCCCTAAACACT (SEQ. ID NO: 301), CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO:303), CACTACCAAATA (SEQ. ID NO: 304), TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306), TGGACGCGTATC (SEQ. ID NO: 307), GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309), GGTCTATGACGA (SEQ. ID NO: 310), TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313 соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297) или CTGGTCCATTCA (SEQ. ID NO: 298), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297) или CTGGTCCATTCA (SEQ. ID NO: 298) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297) или CTGGTCCATTCA (SEQ. ID NO: 298), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CTGGTCCATTC (SEQ. ID NO: 296), TGGTCCATTCA (SEQ. ID NO: 297) или CTGGTCCATTCA (SEQ. ID NO: 298) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300) и TCCCTAAACACT (SEQ. ID NO: 301), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300) и TCCCTAAACACT (SEQ. ID NO: 301) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300) или TCCCTAAACACT (SEQ. ID NO: 301), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TCCCTAAACAC (SEQ. ID NO: 299), CCCTAAACACT (SEQ. ID NO: 300) или TCCCTAAACACT (SEQ. ID NO: 301) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO:303) или CACTACCAAATA (SEQ. ID NO: 304), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO: 303) или CACTACCAAATA (SEQ. ID NO: 304) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO: 303) или CACTACCAAATA (SEQ. ID NO: 304), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CACTACCAAAT (SEQ. ID NO: 302), ACTACCAAATA (SEQ. ID NO: 303) или CACTACCAAATA (SEQ. ID NO: 304) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306) или TGGACGCGTATC (SEQ. ID NO: 307), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306) или TGGACGCGTATC (SEQ. ID NO: 307) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306) или TGGACGCGTATC (SEQ. ID NO: 307), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TGGACGCGTAT (SEQ. ID NO: 305), GGACGCGTATC (SEQ. ID NO: 306) или TGGACGCGTATC (SEQ. ID NO: 307) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309) или GGTCTATGACGA (SEQ. ID NO: 310), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309) или GGTCTATGACGA (SEQ. ID NO: 310) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309) или GGTCTATGACGA (SEQ. ID NO: 310), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности GGTCTATGACG (SEQ. ID NO: 308), GTCTATGACGA (SEQ. ID NO: 309) или GGTCTATGACGA (SEQ. ID NO: 310) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности TTTATTAATGC (SEQ. ID NO: 311), TTATTAATGCC (SEQ. ID NO: 312) или TTTATTAATGCC (SEQ. ID NO: 313 соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды по настоящему изобретению предпочтительно содержат 3-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют собой звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и, более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце и/или на 5'терминальном конце.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, которые содержат 3-10 звеньев LNA и которые, в частности, содержат 1-5 звеньев LNA на терминальном 5'-конце и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида, и, между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида.
Таким образом, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317), ACTGGTCCATTCAT (SEQ. ID NO: 318), CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322), CTCCCTAAACACTA (SEQ. ID NO: 323), ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327), ACACTACCAAATAG (SEQ. ID NO: 328), GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332), GTGGACGCGTATCG (SEQ. ID NO: 333), CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337), CGGTCTATGACGAG (SEQ. ID NO: 338), CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317), ACTGGTCCATTCAT (SEQ. ID NO: 318), CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322), CTCCCTAAACACTA (SEQ. ID NO: 323), ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327), ACACTACCAAATAG (SEQ. ID NO: 328), GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332), GTGGACGCGTATCG (SEQ. ID NO: 333), CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337), CGGTCTATGACGAG (SEQ. ID NO: 338), CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Альтернативно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317), ACTGGTCCATTCAT (SEQ. ID NO: 318), CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322), CTCCCTAAACACTA (SEQ. ID NO: 323), ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327), ACACTACCAAATAG (SEQ. ID NO: 328), GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332), GTGGACGCGTATCG (SEQ. ID NO: 333), CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337), CGGTCTATGACGAG (SEQ. ID NO: 338), CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317), ACTGGTCCATTCAT (SEQ. ID NO: 318), CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322), CTCCCTAAACACTA (SEQ. ID NO: 323), ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327), ACACTACCAAATAG (SEQ. ID NO: 328), GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332), GTGGACGCGTATCG (SEQ. ID NO: 333), CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337), CGGTCTATGACGAG (SEQ. ID NO: 338), CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317), ACTGGTCCATTCAT (SEQ. ID NO: 318), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317), ACTGGTCCATTCAT (SEQ. ID NO: 318) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317) или ACTGGTCCATTCAT (SEQ. ID NO: 318), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности ACTGGTCCATTC (SEQ. ID NO: 314), TGGTCCATTCAT (SEQ. ID NO: 315), CTGGTCCATTCAT (SEQ. ID NO: 316), ACTGGTCCATTCA (SEQ. ID NO: 317) или ACTGGTCCATTCAT (SEQ. ID NO: 318) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322) или CTCCCTAAACACTA (SEQ. ID NO: 323), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322) или CTCCCTAAACACTA (SEQ. ID NO: 323) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322) или CTCCCTAAACACTA (SEQ. ID NO: 323), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CTCCCTAAACAC (SEQ. ID NO: 319), CCCTAAACACTA (SEQ. ID NO: 320), TCCCTAAACACTA (SEQ. ID NO: 321), CTCCCTAAACACT (SEQ. ID NO: 322) или CTCCCTAAACACTA (SEQ. ID NO: 323) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327) или ACACTACCAAATAG (SEQ. ID NO: 328), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327) или ACACTACCAAATAG (SEQ. ID NO: 328) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327) или ACACTACCAAATAG (SEQ. ID NO: 328), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности ACACTACCAAAT (SEQ. ID NO: 324), ACTACCAAATAG (SEQ. ID NO: 325), CACTACCAAATAG (SEQ. ID NO: 326), ACACTACCAAATA (SEQ. ID NO: 327) или ACACTACCAAATAG (SEQ. ID NO: 328) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332) или GTGGACGCGTATCG (SEQ. ID NO: 333), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332) или GTGGACGCGTATCG (SEQ. ID NO: 333) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332) или GTGGACGCGTATCG (SEQ. ID NO: 333), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности GTGGACGCGTAT (SEQ. ID NO: 329), GGACGCGTATCG (SEQ. ID NO: 330), TGGACGCGTATCG (SEQ. ID NO: 331), GTGGACGCGTATC (SEQ. ID NO: 332) или GTGGACGCGTATCG (SEQ. ID NO: 333) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337) или CGGTCTATGACGAG (SEQ. ID NO: 338), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337) или CGGTCTATGACGAG (SEQ. ID NO: 338) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CGGTCTATGACG (SEQ. ID NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337) или CGGTCTATGACGAG (SEQ. ID NO: 338), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CGGTCTATGACG (SEQ NO: 334), GTCTATGACGAG (SEQ. ID NO: 335), GGTCTATGACGAG (SEQ. ID NO: 336), CGGTCTATGACGA (SEQ. ID NO: 337) или CGGTCTATGACGAG (SEQ. ID NO: 338) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Предпочтительно настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343), и антисмысловой олигонуклеотид содержит последовательность, способную гибридизоваться с указанной последовательностью CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Другими словами, настоящее изобретение также относится к антисмысловому олигонуклеотиду(ам), состоящему из 14-20, более предпочтительно 14-18 нуклеотидов, и, по меньшей мере, четыре из 14-20, более предпочтительно 14-18 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343), и антисмысловой олигонуклеотид содержит последовательность, комплементарную последовательности CTTTATTAATGC (SEQ. ID NO: 339), TTATTAATGCCT (SEQ. ID NO: 340), TTTATTAATGCCT (SEQ. ID NO: 341), CTTTATTAATGCC (SEQ. ID NO: 342) или CTTTATTAATGCCT (SEQ. ID NO: 343) соответственно, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды по настоящему изобретению предпочтительно содержат 4-11 звеньев LNA, более предпочтительно 4-10 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют собой звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и, более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце и/или на 5'терминальном конце.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, которые содержат 3-10 звеньев LNA и которые, в частности, содержат 1-5 звеньев LNA на терминальном 5'-конце и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида, и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида.
Таким образом, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N1-GTCATAGA-N2-3' (SEQ. ID NO: 12) или 5'-N3-ACGCGTCC-N4-3' (SEQ. ID NO: 98), или 5'-N11-TGTTTAGG-N12-3'(SEQ. ID NO: 10), или 5'-N5-TTTGGTAG-N6-3' (SEQ. ID NO: 11), или 5'-N7-AATGGACC-N8-3' (SEQ. ID NO: 100), или 5'-N9-ATTAATAA-N10-3' (SEQ. ID NO: 101), где
N1 представляет: CATGGCAGACCCCGCTGCTC-, ATGGCAGACCCCGCTGCTC-, TGGCAGACCCCGCTGCTC-, GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA, -CCGAGCCCCCAGCGCAG, -CCGAGCCCCCAGCGCAGC, -CCGAGCCCCCAGCGCAGCG или -CCGAGCCCCCAGCGCAGCGG;
N3 представляет: GGTGGGATCGTGCTGGCGAT-, GTGGGATCGTGCTGGCGAT-, TGGGATCGTGCTGGCGAT-, GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT,- TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACGATGTGCAGCGGC, -ACAGGACGATGTGCAGCGG, -ACAGGACGATGTGCAGCG, -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC, или -A;
N5 представляет: GCCCAGCCTGCCCCAGAAGAGCTA-, CCCAGCCTGCCCCAGAAGAGCTA-, CCAGCCTGCCCCAGAAGAGCTA-, CAGCCTGCCCCAGAAGAGCTA-, AGCCTGCCCCAGAAGAGCTA-, GCCTGCCCCAGAAGAGCTA-, CCTGCCCCAGAAGAGCTA-, CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, ТА- или A-;
N6 представляет: -TGTTTAGGGAGCCGTCTTCAGGAA, -TGTTTAGGGAGCCGTCTTCAGGA, -TGTTTAGGGAGCCGTCTTCAGG, -TGTTTAGGGAGCCGTCTTCAG, -TGTTTAGGGAGCCGTCTTCA, -TGTTTAGGGAGCCGTCTTC, -TGTTTAGGGAGCCGTCTT, -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -Т;
N7 представляет: TGAATCTTGAATATCTCATG-, GAATCTTGAATATCTCATG-, AATCTTGAATATCTCATG-, ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-;
N8 представляет: -AGTATTCTAGAAACTCACCA, -AGTATTCTAGAAACTCACC, -AGTATTCTAGAAACTCAC, -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A;
N9 представляет: ATTCATATTTATATACAGGC-, TTCATATTTATATACAGGC-, TCATATTTATATACAGGC-, CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или С-;
N10 представляет: -AGTGCAAATGTTATTGGCTA, -AGTGCAAATGTTATTGGCT, -AGTGCAAATGTTATTGGC, -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A;
N11 представляет: TGCCCCAGAAGAGCTATTTGGTAG-, GCCCCAGAAGAGCTATTTGGTAG-, CCCCAGAAGAGCTATTTGGTAG-, CCCAGAAGAGCTATTTGGTAG-, CCAGAAGAGCTATTTGGTAG-, CAGAAGAGCTATTTGGTAG-, AGAAGAGCTATTTGGTAG-, GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-,
N12 представляет: -GAGCCGTCTTCAGGAATCTTCTCC, -GAGCCGTCTTCAGGAATCTTCTC, -GAGCCGTCTTCAGGAATCTTCT, -GAGCCGTCTTCAGGAATCTTC, -GAGCCGTCTTCAGGAATCTT, -GAGCCGTCTTCAGGAATCT, -GAGCCGTCTTCAGGAATC, -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -Ga или -G;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Таким образом, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N1-GTCATAGA-N2-3' (SEQ. ID NO: 12) или 5'-N3-ACGCGTCC-N4-3' (SEQ. ID NO: 98), или 5'-N11-TGTTTAGG-N12-3'(SEQ. ID NO: 10), или 5'-N5-TTTGGTAG-N6-3' (SEQ. ID NO: 11), или 5'-N7-AATGGACC-N8-3' (SEQ. ID NO: 100), или 5'-N9-ATTAATAA-N10-3' (SEQ. ID NO: 101), где остатки N1-N12 имеют значения, в частности, дополнительно ограниченные значениями, раскрытыми здесь, и солям и оптическим изомерам указанного антисмыслового олигонуклеотида.
Кроме того, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N1-GTCATAGA-N2-3' (SEQ. ID NO: 12), где:
N1 представляет: CATGGCAGACCCCGCTGCTC-, ATGGCAGACCCCGCTGCTC-, TGGCAGACCCCGCTGCTC-, GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA, -CCGAGCCCCCAGCGCAG, -CCGAGCCCCCAGCGCAGC, -CCGAGCCCCCAGCGCAGCG или -CCGAGCCCCCAGCGCAGCGG;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды формулы S1 (SEQ ID NO:12) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и, более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце и/или на терминальном 5'-конце.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые, в частности, содержат 1-5 звеньев LNA на терминальном 5'-конце и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида, и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце и 2-4 звена LNA на терминальном 3'-конце антисмыслового олигонуклеотида, и еще более предпочтительно содержат 3-4 звена LNA на терминальном 5'-конце и 3-4 звена LNA на терминальном 3'-конце антисмыслового олигонуклеотида, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S1):
где:
N1 представляет: CATGGCAGACCCCGCTGCTC-, ATGGCAGACCCCGCTGCTC-, TGGCAGACCCCGCTGCTC-, GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 выбран из: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA, -CCGAGCCCCCAGCGCAG, -CCGAGCCCCCAGCGCAGC, -CCGAGCCCCCAGCGCAGCG или -CCGAGCCCCCAGCGCAGCGG.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S1) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S1) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S1) содержит 12-20 нуклеотидов, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S1):
5'-N1-GTCATAGA-N2-3'
где:
N1 представляет: TGGCAGACCCCGCTGCTC-, GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-; и
N2 выбран из: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA, -CCGAGCCCCCAGCGCAG или -CCGAGCCCCCAGCGCAGC.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S1):
5'-N1-GTCATAGA-N2-3'
где:
N1 представляет: GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-; и
N2 выбран из: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG или -CCGAGCCCCCAGC.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S1):
5'-N1-GTCATAGA-N2-3'
где:
N1 представляет: CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-; и
N2 выбран из: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC или -CCGAGCCC.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N1A представляет: CATGGCAGACCCCGCTGCT-, ATGGCAGACCCCGCTGCT-, TGGCAGACCCCGCTGCT-, GGCAGACCCCGCTGCT-, GCAGACCCCGCTGCT-, CAGACCCCGCTGCT-, AGACCCCGCTGCT-, GACCCCGCTGCT-, ACCCCGCTGCT-, CCCCGCTGCT-, CCCGCTGCT-, CCGCTGCT-, CGCTGCT-, GCTGCT-, CTGCT-, TGCT-, GCT-, CT- или T-;
N2A представляет: -C, -CG, -CGA, -CGAG, -CGAGC, -CGAGCC, -CGAGCCC, -CGAGCCCC, -CGAGCCCCC, -CGAGCCCCCA, -CGAGCCCCCAG, -CGAGCCCCCAGC, -CGAGCCCCCAGCG, -CGAGCCCCCAGCGC, -CGAGCCCCCAGCGCA, -CGAGCCCCCAGCGCAG, -CGAGCCCCCAGCGCAGC, -CGAGCCCCCAGCGCAGCG или -CGAGCCCCCAGCGCAGCGG;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Предпочтительно N1A представляет: TGGCAGACCCCGCTGCT-, GGCAGACCCCGCTGCT-, GCAGACCCCGCTGCT-, CAGACCCCGCTGCT-, AGACCCCGCTGCT-, GACCCCGCTGCT-, ACCCCGCTGCT-, CCCCGCTGCT-, CCCGCTGCT-, CCGCTGCT-, CGCTGCT-, GCTGCT-, CTGCT-, TGCT-, GCT-, CT- или T-; и
N2A представляет: -C, -CG, -CGA, -CGAG, -CGAGC, -CGAGCC, -CGAGCCC, -CGAGCCCC, -CGAGCCCCC, -CGAGCCCCCA, -CGAGCCCCCAG, -CGAGCCCCCAGC, -CGAGCCCCCAGCG, -CGAGCCCCCAGCGC, -CGAGCCCCCAGCGCA, -CGAGCCCCCAGCGCAG или -CGAGCCCCCAGCGCAGC.
Более предпочтительно N1A представляет: GACCCCGCTGCT-, ACCCCGCTGCT-, CCCCGCTGCT-, CCCGCTGCT-, CCGCTGCT-, CGCTGCT-, GCTGCT-, CTGCT-, TGCT-, GCT-, CT- или T-; и
N2A представляет: -C, -CG, -CGA, -CGAG, -CGAGC, -CGAGCC, -CGAGCCC, -CGAGCCCC, -CGAGCCCCC, -CGAGCCCCCA, -CGAGCCCCCAG или -CGAGCCCCCAGC.
Еще более предпочтительно N1A представляет: CGCTGCT-, GCTGCT-, CTGCT-, TGCT-, GCT-, CT- или T-; и
N2A представляет: -C, -CG, -CGA, -CGAG, -CGAGC, -CGAGCC или -CGAGCCC.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S1A/SEQ. ID NO: 69) содержит 12-24 нуклеотида и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S1A) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S1A) содержит 12-20 нуклеотидов, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Кроме того, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N3-ACGCGTCC-N4-3' (SEQ. ID NO: 98), где:
N3 представляет: GGTGGGATCGTGCTGGCGAT-, GTGGGATCGTGCTGGCGAT-, TGGGATCGTGCTGGCGAT-, GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACGATGTGCAGCGGC, -ACAGGACGATGTGCAGCGG, -ACAGGACGATGTGCAGCG, -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды общей формулы S2 (SEQ. ID NO: 98) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые предпочтительно содержат 1-5 звеньев LNA на терминальном 5'-конце, и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце, и 2-4 звена LNA на терминальном 3'-конце, и еще более предпочтительные содержат 3-4 звена LNA на терминальном 5'-конце, и 3-4 звена LNA на терминальном 3'-конце, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S2):
где:
N3 представляет: GGTGGGATCGTGCTGGCGAT-, GTGGGATCGTGCTGGCGAT-, TGGGATCGTGCTGGCGAT-, GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACGATGTGCAGCGGC, -ACAGGACGATGTGCAGCGG, -ACAGGACGATGTGCAGCG, -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S2) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S2) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S2) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S2):
5'-N3-ACGCGTCC-N4-3'
где:
N3 представляет: TGGGATCGTGCTGGCGAT-, GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-; и
N4 представляет: -ACAGGACGATGTGCAGCG, -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S2):
5'-N3-ACGCGTCC-N4-3'
где:
N3 представляет: TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S2):
5'-N3-ACGCGTCC-N4-3'
где:
N3 представляет: CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N3A представляет: GGTGGGATCGTGCTGGCGA-, GTGGGATCGTGCTGGCGA-, TGGGATCGTGCTGGCGA-, GGGATCGTGCTGGCGA-, GGATCGTGCTGGCGA-, GATCGTGCTGGCGA-, ATCGTGCTGGCGA-, TCGTGCTGGCGA-, CGTGCTGGCGA-, GTGCTGGCGA-, TGCTGGCGA-, GCTGGCGA-, CTGGCGA-, TGGCGA-, GGCGA-, GCGA-, CGA-, GA- или A-;
N4A представляет: -CAGGACGATGTGCAGCGGC, -CAGGACGATGTGCAGCGG, -CAGGACGATGTGCAGCG, -CAGGACGATGTGCAGC, -CAGGACGATGTGCAG, -CAGGACGATGTGCA, -CAGGACGATGTGC, -CAGGACGATGTG, -CAGGACGATGT, -CAGGACGATG, -CAGGACGAT, -CAGGACGA, -CAGGACG, -CAGGAC, -CAGGA, -CAGG, -CAG, -CA или -C;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Предпочтительно N3A представляет: TGGGATCGTGCTGGCGA-, GGGATCGTGCTGGCGA-, GGATCGTGCTGGCGA-, GATCGTGCTGGCGA-, ATCGTGCTGGCGA-, TCGTGCTGGCGA-, CGTGCTGGCGA-, GTGCTGGCGA-, TGCTGGCGA-, GCTGGCGA-, CTGGCGA-, TGGCGA-, GGCGA-, GCGA-, CGA-, GA- или A-; и
N4A представляет: -CAGGACGATGTGCAGCG, -CAGGACGATGTGCAGC, -CAGGACGATGTGCAG, -CAGGACGATGTGCA, -CAGGACGATGTGC, -CAGGACGATGTG, -CAGGACGATGT, -CAGGACGATG, -CAGGACGAT, -CAGGACGA, -CAGGACG, -CAGGAC, -CAGGA, -CAGG, -CAG, -CA или -C.
Более предпочтительно N3A представляет: TCGTGCTGGCGA-, CGTGCTGGCGA-, GTGCTGGCGA-, TGCTGGCGA-, GCTGGCGA-, CTGGCGA-, TGGCGA-, GGCGA-, GCGA-, CGA-, GA- или A-; и
N4A представляет: -CAGGACGATGTG, -CAGGACGATGT, -CAGGACGATG, -CAGGACGAT, -CAGGACGA, -CAGGACG, -CAGGAC, -CAGGA, -CAGG, -CAG, -CA или -C.
Еще более предпочтительно N3A представляет: CTGGCGA-, TGGCGA-, GGCGA-, GCGA-, -CAG, -CA или -C; и
N4A представляет: -CAGGACG, -CAGGAC, -CAGGA, -CAGG, -CAG, -CA или -C.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S2A/SEQ. ID NO: 70) содержит 12-24 нуклеотида и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S2A) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S2A) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Кроме того, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N11-TGTTTAGG-N12-3' (SEQ. ID NO: 10), где:
N11 представляет: TGCCCCAGAAGAGCTATTTGGTAG-, GCCCCAGAAGAGCTATTTGGTAG-, CCCCAGAAGAGCTATTTGGTAG-, CCCAGAAGAGCTATTTGGTAG-, CCAGAAGAGCTATTTGGTAG-, CAGAAGAGCTATTTGGTAG-, AGAAGAGCTATTTGGTAG-, GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-;
N12 представляет: -GAGCCGTCTTCAGGAATCTTCTCC, -GAGCCGTCTTCAGGAATCTTCTC, -GAGCCGTCTTCAGGAATCTTCT, -GAGCCGTCTTCAGGAATCTTC, -GAGCCGTCTTCAGGAATCTT, -GAGCCGTCTTCAGGAATCT, -GAGCCGTCTTCAGGAATC, -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды общей формулы S3 (SEQ. ID NO: 10) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце и терминальном 5'-конце. Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые предпочтительно содержат 1-5 звеньев LNA на терминальном 5'-конце, и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце, и 2-4 звена LNA на терминальном 3'-конце, и еще более предпочтительно содержат 3-4 звена LNA на терминальном 5'-конце, и 3-4 звена LNA на терминальном 3'-конце, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S3):
где:
N11 представляет: TGCCCCAGAAGAGCTATTTGGTAG-, GCCCCAGAAGAGCTATTTGGTAG-, CCCCAGAAGAGCTATTTGGTAG-, CCCAGAAGAGCTATTTGGTAG-, CCAGAAGAGCTATTTGGTAG-, CAGAAGAGCTATTTGGTAG-, AGAAGAGCTATTTGGTAG-, GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTCTTCAGGAATCTTCTCC, -GAGCCGTCTTCAGGAATCTTCTC, -GAGCCGTCTTCAGGAATCTTCT, -GAGCCGTCTTCAGGAATCTTC, -GAGCCGTCTTCAGGAATCTT, -GAGCCGTCTTCAGGAATCT, -GAGCCGTCTTCAGGAATC, -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S3) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S3) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S3) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S3):
5'-N11-TGTTTAGG-N12-3'
где:
N11 представляет: AGAAGAGCTATTTGGTAG-, GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTCTTCAGGAATC, -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S3):
5'-N11-TGTTTAGG-N12-3'
где:
N11 представляет: AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S3):
5'-N11-TGTTTAGG-N12-3'
где:
N11 представляет: TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N11A-GTGTTTAGGG-N12A-3' (SEQ. ID NO: 71), где:
N11A представляет: TGCCCCAGAAGAGCTATTTGGTA-, GCCCCAGAAGAGCTATTTGGTA-, CCCCAGAAGAGCTATTTGGTA-, CCCAGAAGAGCTATTTGGTA-, CCAGAAGAGCTATTTGGTA-, CAGAAGAGCTATTTGGTA- AGAAGAGCTATTTGGTA-, GAAGAGCTATTTGGTA-, AAGAGCTATTTGGTA-, AGAGCTATTTGGTA-, GAGCTATTTGGTA-, AGCTATTTGGTA-, GCTATTTGGTA-, CTATTTGGTA-, TATTTGGTA-, ATTTGGTA-, TTTGGTA-, TTGGTA-, TGGTA-, GGTA-, GTA-, TA- или A-;
N12A представляет: -AGCCGTCTTCAGGAATCTTCTCC, -AGCCGTCTTCAGGAATCTTCTC, -AGCCGTCTTCAGGAATCTTCT, -AGCCGTCTTCAGGAATCTTC, -AGCCGTCTTCAGGAATCTT, -AGCCGTCTTCAGGAATCT, -AGCCGTCTTCAGGAATC, -AGCCGTCTTCAGGAAT, -AGCCGTCTTCAGGAA, -AGCCGTCTTCAGGA, -AGCCGTCTTCAGG, -AGCCGTCTTCAG, -AGCCGTCTTCA, -AGCCGTCTTC, -AGCCGTCTT, -AGCCGTCT, -AGCCGTC, -AGCCGT, -AGCCG, -AGCC, -AGC, -AG или -A;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Предпочтительно N11A представляет: AGAAGAGCTATTTGGTA-, GAAGAGCTATTTGGTA-, AAGAGCTATTTGGTA-, AGAGCTATTTGGTA-, GAGCTATTTGGTA-, AGCTATTTGGTA-, GCTATTTGGTA-, CTATTTGGTA-, TATTTGGTA-, ATTTGGTA-, TTTGGTA-, TTGGTA-, TGGTA-, GGTA-, GTA-, TA- или A-; и
N12A представляет: -AGCCGTCTTCAGGAATC, -AGCCGTCTTCAGGAAT, -AGCCGTCTTCAGGAA, -AGCCGTCTTCAGGA, -AGCCGTCTTCAGG, -AGCCGTCTTCAG, -AGCCGTCTTCA, -AGCCGTCTTC, -AGCCGTCTT, -AGCCGTCT, -AGCCGTC, -AGCCGT, -AGCCG, -AGCC, -AGC, -AG или -A.
Более предпочтительно N11A представляет: AGCTATTTGGTA-, GCTATTTGGTA-, CTATTTGGTA-, TATTTGGTA-, ATTTGGTA-, TTTGGTA-, TTGGTA-, TGGTA-, GGTA-, GTA-, TA- или A-; и
N12A представляет: -AGCCGTCTTCAG, -AGCCGTCTTCA, -AGCCGTCTTC, -AGCCGTCTT, -AGCCGTCT, -AGCCGTC, -AGCCGT, -AGCCG, -AGCC, -AGC, -AG или -A.
Еще более предпочтительно N11A представляет: TTTGGTA-, TTGGTA-, TGGTA-, GGTA-, GTA-, TA- или A-; и
N12A представляет: -AGCCGTC, -AGCCGT, -AGCCG, -AGCC, -AGC, -AG или -A.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S3A/SEQ. ID NO: 71) содержит 12-24 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S3A) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S3A) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Кроме того, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N5-TTTGGTAG-N6-3' (SEQ. ID NO: 11), где:
N5 представляет: GCCCAGCCTGCCCCAGAAGAGCTA-, CCCAGCCTGCCCCAGAAGAGCTA-, CCAGCCTGCCCCAGAAGAGCTA-, CAGCCTGCCCCAGAAGAGCTA-, AGCCTGCCCCAGAAGAGCTA-, GCCTGCCCCAGAAGAGCTA-, CCTGCCCCAGAAGAGCTA-, CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-;
N6 представляет: -TGTTTAGGGAGCCGTCTTCAGGAA, -TGTTTAGGGAGCCGTCTTCAGGA, -TGTTTAGGGAGCCGTCTTCAGG, -TGTTTAGGGAGCCGTCTTCAG, -TGTTTAGGGAGCCGTCTTCA, -TGTTTAGGGAGCCGTCTTC, -TGTTTAGGGAGCCGTCTT, -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды общей формулы S4 (SEQ. ID NO: 11) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и более предпочтительно, по меньшей мере, два звена LNA находятся на терминальном 3'-конце и терминальном 5'-конце. Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые предпочтительно содержат 1-5 звеньев LNA на терминальном 5'-конце, и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце, и 2-4 звена LNA на терминальном 3'-конце, и еще более предпочтительные содержат 3-4 звена LNA на терминальном 5'-конце и 3-4 звена LNA на терминальном 3'-конце, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S4):
где:
N5 представляет: GCCCAGCCTGCCCCAGAAGAGCTA-, CCCAGCCTGCCCCAGAAGAGCTA-, CCAGCCTGCCCCAGAAGAGCTA-, CAGCCTGCCCCAGAAGAGCTA-, AGCCTGCCCCAGAAGAGCTA-, GCCTGCCCCAGAAGAGCTA-, CCTGCCCCAGAAGAGCTA-, CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 выбран из: -TGTTTAGGGAGCCGTCTTCAGGAA, -TGTTTAGGGAGCCGTCTTCAGGA, -TGTTTAGGGAGCCGTCTTCAGG, -TGTTTAGGGAGCCGTCTTCAG, -TGTTTAGGGAGCCGTCTTCA, -TGTTTAGGGAGCCGTCTTC, -TGTTTAGGGAGCCGTCTT, -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S4) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S4) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S4) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S4):
5'-N5-TTTGGTAG-N6-3'
где:
N5 представляет: CCTGCCCCAGAAGAGCTA-, CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 выбран из: -TGTTTAGGGAGCCGTCTT, -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S4):
5'-N5-TTTGGTAG-N6-3'
где:
N5 представляет: CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 выбран из: -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S4):
5'-N5-TTTGGTAG-N6-3'
где:
N5 представляет: AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 выбран из: -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N5A-ATTTGGTAGT-N6A-3' (SEQ. ID NO: 72), где:
N5A представляет: GCCCAGCCTGCCCCAGAAGAGCT-, CCCAGCCTGCCCCAGAAGAGCT-, CCAGCCTGCCCCAGAAGAGCT-, CAGCCTGCCCCAGAAGAGCT-, AGCCTGCCCCAGAAGAGCT-, GCCTGCCCCAGAAGAGCT-, CCTGCCCCAGAAGAGCT-, CTGCCCCAGAAGAGCT-, TGCCCCAGAAGAGCT-, GCCCCAGAAGAGCT-, CCCCAGAAGAGCT-, CCCAGAAGAGCT-, CCAGAAGAGCT-, CAGAAGAGCT-, AGAAGAGCT-, GAAGAGCT-, AAGAGCT-, AGAGCT-, GAGCT-, AGCT-, GCT-, CT- или T-; и
N6A представляет: -GTTTAGGGAGCCGTCTTCAGGAA, -GTTTAGGGAGCCGTCTTCAGGA, -GTTTAGGGAGCCGTCTTCAGG, -GTTTAGGGAGCCGTCTTCAG, -GTTTAGGGAGCCGTCTTCA, -GTTTAGGGAGCCGTCTTC, -GTTTAGGGAGCCGTCTT, -GTTTAGGGAGCCGTCT, -GTTTAGGGAGCCGTC, -GTTTAGGGAGCCGT, -GTTTAGGGAGCCG, -GTTTAGGGAGCC, -GTTTAGGGAGC, -GTTTAGGGAG, -GTTTAGGGA, -GTTTAGGG, -GTTTAGG, -GTTTAG, -GTTTA, -GTTT, -GTT, -GT или -G;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Предпочтительно N5A представляет: CCTGCCCCAGAAGAGCT-, CTGCCCCAGAAGAGCT-, TGCCCCAGAAGAGCT-, GCCCCAGAAGAGCT-, CCCCAGAAGAGCT-, CCCAGAAGAGCT-, CCAGAAGAGCT-, CAGAAGAGCT-, AGAAGAGCT-, GAAGAGCT-, AAGAGCT-, AGAGCT-, GAGCT-, AGCT-, GCT-, CT- или T-; и
N6A представляет: -GTTTAGGGAGCCGTCTT, -GTTTAGGGAGCCGTCT, -GTTTAGGGAGCCGTC, -GTTTAGGGAGCCGT, -GTTTAGGGAGCCG, -GTTTAGGGAGCC, -GTTTAGGGAGC, -GTTTAGGGAG, -GTTTAGGGA, -GTTTAGGG, -GTTTAGG, -GTTTAG, -GTTTA, -GTTT, -GTT, -GT или -G.
Более предпочтительно N5A представляет: CCCAGAAGAGCT-, CCAGAAGAGCT-, CAGAAGAGCT-, AGAAGAGCT-, GAAGAGCT-, AAGAGCT-, AGAGCT-, GAGCT-, AGCT-, GCT-, CT- или T-; и
N6A представляет: -GTTTAGGGAGCC, -GTTTAGGGAGC, -GTTTAGGGAG, -GTTTAGGGA, -GTTTAGGG, -GTTTAGG, -GTTTAG, -GTTTA, -GTTT, -GTT, -GT или -G.
Еще более предпочтительно N5A представляет: AAGAGCT-, AGAGCT-, GAGCT-, AGCT-, GCT-, CT- или T-; и
N6A представляет: -GTTTAGG, -GTTTAG, -GTTTA, -GTTT, -GTT, -GT или -G.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S4/SEQ. ID NO: 72) содержит 12-24 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S4A) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S4) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Кроме того, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N7-AATGGACC-N8-3' (SEQ. ID NO: 100), где:
N7 представляет: TGAATCTTGAATATCTCATG-, GAATCTTGAATATCTCATG-, AATCTTGAATATCTCATG-, ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-;
N8 представляет: -AGTATTCTAGAAACTCACCA, -AGTATTCTAGAAACTCACC, -AGTATTCTAGAAACTCAC, -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды общей формулы S6 (SEQ. ID NO: 100) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и более предпочтительно, по меньшей мере, две звена LNA находятся на терминальном 3'-конце и терминальном 5'-конце. Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые предпочтительно содержат 1-5 звеньев LNA на терминальном 5'-конце, и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце, и 2-4 звена LNA на терминальном 3'-конце, и еще более предпочтительно содержат 3-4 звена LNA на терминальном 5'-конце, и 3-4 звена LNA на терминальном 3'-конце, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S6):
где:
N7 представляет: TGAATCTTGAATATCTCATG-, GAATCTTGAATATCTCATG-, AATCTTGAATATCTCATG-, ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 выбран из: -AGTATTCTAGAAACTCACCA, -AGTATTCTAGAAACTCACC, --AGTATTCTAGAAACTCAC, -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S6) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S6) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S6) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S6):
5'-N7-AATGGACC-N8-3'
где:
N7 представляет: AATCTTGAATATCTCATG-, ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 выбран из: -AGTATTCTAGAAACTCAC, -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S6):
5'-N7-AATGGACC-N8-3'
где:
N7 представляет: TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 выбран из: -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S6):
5'-N7-AATGGACC-N8-3'
где:
N7 представляет: ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 выбран из: -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью
где:
N7A представляет: TGAATCTTGAATATCTCAT-, GAATCTTGAATATCTCAT-, AATCTTGAATATCTCAT-, ATCTTGAATATCTCAT-, TCTTGAATATCTCAT-, CTTGAATATCTCAT-, TTGAATATCTCAT-, TGAATATCTCAT-, GAATATCTCAT-, AATATCTCAT-, ATATCTCAT-, TATCTCAT-, ATCTCAT-, TCTCAT-, CTCAT-, TCAT-, CAT-, AT- или T-;
N8A представляет: -GTATTCTAGAAACTCACCA, -GTATTCTAGAAACTCACC, -GTATTCTAGAAACTCAC, -GTATTCTAGAAACTCA, -GTATTCTAGAAACTC, -GTATTCTAGAAACT, -GTATTCTAGAAAC, -GTATTCTAGAAA, -GTATTCTAGAA, -GTATTCTAGA, -GTATTCTAG, -GTATTCTA, -GTATTCT, -GTATTC, -GTATT, -GTAT, -GTA, -GT или -G;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Предпочтительно N7A представляет: AATCTTGAATATCTCAT-, ATCTTGAATATCTCAT-, TCTTGAATATCTCAT-, CTTGAATATCTCAT-, TTGAATATCTCAT-, TGAATATCTCAT-, GAATATCTCAT-, AATATCTCAT-, ATATCTCAT-, TATCTCAT-, ATCTCAT-, TCTCAT-, CTCAT-, TCAT-, CAT-, AT- или T-; и
N8A представляет: -GTATTCTAGAAACTCAC, -GTATTCTAGAAACTCA, -GTATTCTAGAAACTC, -GTATTCTAGAAACT, -GTATTCTAGAAAC, -GTATTCTAGAAA, -GTATTCTAGAA, -GTATTCTAGA, -GTATTCTAG, -GTATTCTA, -GTATTCT, -GTATTC, -GTATT, -GTAT, -GTA, -GT или -G.
Более предпочтительно N7A представляет: TGAATATCTCAT-, GAATATCTCAT-, AATATCTCAT-, ATATCTCAT-, TATCTCAT-, ATCTCAT-, TCTCAT-, CTCAT-, TCAT-, CAT-, AT- или T-; и
N8A представляет: -GTATTCTAGAAA, -GTATTCTAGAA, -GTATTCTAGA, -GTATTCTAG, -GTATTCTA, -GTATTCT, -GTATTC, -GTATT, -GTAT, -GTA, -GT или -G.
Еще более предпочтительно N7A представляет: ATCTCAT-, TCTCAT-, CTCAT-, TCAT-, CAT-, AT- или T-; и
N8A представляет: -GTATTCT, -GTATTC, -GTATT, -GTAT, -GTA, -GT или -G.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S6А/SEQ. ID NO: 73) содержит 12-24 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S6A) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S6A) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Кроме того, настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N9-ATTAATAA-N10-3' (SEQ. ID NO: 101), где:
N9 представляет: ATTCATATTTATATACAGGC-, TTCATATTTATATACAGGC-, TCATATTTATATACAGGC-, CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-;
N10 представляет: -AGTGCAAATGTTATTGGCTA, -AGTGCAAATGTTATTGGCT, -AGTGCAAATGTTATTGGC, -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды общей формулы S7 (SEQ. ID NO: 101) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA и наиболее предпочтительно 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и более предпочтительно, по меньшей мере, две звена LNA находятся на терминальном 3'-конце и терминальном 5'-конце. Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые предпочтительно содержат 1-5 звеньев LNA на терминальном 5'-конце, и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце, и 2-4 звена LNA на терминальном 3'-конце, и еще более предпочтительно содержат 3-4 звена LNA на терминальном 5'-конце, и 3-4 звена LNA на терминальном 3'-конце, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S7):
где:
N9 представляет: ATTCATATTTATATACAGGC-, TTCATATTTATATACAGGC-, TCATATTTATATACAGGC-, CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 выбран из: -AGTGCAAATGTTATTGGCTA, -AGTGCAAATGTTATTGGCT, -AGTGCAAATGTTATTGGC, -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S7) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S7) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S7) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S7):
5'-N9-ATTAATAA-N10-3'
где:
N 9 представляет: TCATATTTATATACAGGC-, CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 выбран из: -AGTGCAAATGTTATTGGC, -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S7):
5'-N9-ATTAATAA-N10-3'
где:
N9 представляет: TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 выбран из: -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S7):
5'-N9-ATTAATAA-N10-3'
где:
N9 представляет:: ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 выбран из: -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A.
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 12-24 нуклеотидов, и, по меньшей мере, три из 12-24 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью
где:
N9A представляет: ATTCATATTTATATACAGG-, TTCATATTTATATACAGG-, TCATATTTATATACAGG-, CATATTTATATACAGG-, ATATTTATATACAGG-, TATTTATATACAGG-, ATTTATATACAGG-, TTTATATACAGG-, TTATATACAGG-, TATATACAGG-, ATATACAGG-, TATACAGG-, ATACAGG-, TACAGG-, ACAGG-, CAGG-, AGG-, GG- или G-;
N10A представляет: -GTGCAAATGTTATTGGCTA, -GTGCAAATGTTATTGGCT, -GTGCAAATGTTATTGGC, -GTGCAAATGTTATTGG, -GTGCAAATGTTATTG, -GTGCAAATGTTATT, -GTGCAAATGTTAT, -GTGCAAATGTTA, -GTGCAAATGTT, -GTGCAAATGT, -GTGCAAATG, -GTGCAAAT, -GTGCAAA, -GTGCAA, -GTGCA, -GTGC, -GTG, -GT или -G;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Предпочтительно N9A представляет: TCATATTTATATACAGG-, CATATTTATATACAGG-, ATATTTATATACAGG-, TATTTATATACAGG-, ATTTATATACAGG-, TTTATATACAGG-, TTATATACAGG-, TATATACAGG-, ATATACAGG-, TATACAGG-, ATACAGG-, TACAGG-, ACAGG-, CAGG-, AGG-, GG- или G-; и
N10A представляет: -GTGCAAATGTTATTGGCTA, -GTGCAAATGTTATTGGCT, -GTGCAAATGTTATTGGC, -GTGCAAATGTTATTGG, -GTGCAAATGTTATTG, -GTGCAAATGTTATT, -GTGCAAATGTTAT, -GTGCAAATGTTA, -GTGCAAATGTT, -GTGCAAATGT, -GTGCAAATG, -GTGCAAAT, -GTGCAAA, -GTGCAA, -GTGCA, -GTGC, -GTG, -GT или -G;
Более предпочтительно N9A представляет: TTTATATACAGG-, TTATATACAGG-, TATATACAGG-, ATATACAGG-, TATACAGG-, ATACAGG-, TACAGG-, ACAGG-, CAGG-, AGG-, GG- или G-; и
N10A представляет: -GTGCAAATGTTA, -GTGCAAATGTT, -GTGCAAATGT, -GTGCAAATG, -GTGCAAAT, -GTGCAAA, -GTGCAA, -GTGCA, -GTGC, -GTG, -GT или -G.
Еще более предпочтительно N9A представляет: ATACAGG-, TACAGG-, ACAGG-, CAGG-, AGG-, GG- или G-; и
N10A представляет: -GTGCAAA, -GTGCAA, -GTGCA, -GTGC, -GTG, -GT или -G.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S7A/SEQ. ID NO: 74) содержит 12-24 нуклеотида и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S7A) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S7А) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду(ам), состоящему из 8-18, предпочтительно 10-28 нуклеотидов, и, по меньшей мере, два из 8-28, предпочтительно 10-28 нуклеотидов, представляют LNA, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N13 представляет: CCCAGCCTGCCCCAGAAGAGCTATTTG-, CCAGCCTGCCCCAGAAGAGCTATTTG-, CAGCCTGCCCCAGAAGAGCTATTTG-, AGCCTGCCCCAGAAGAGCTATTTG-, GCCTGCCCCAGAAGAGCTATTTG-, CCTGCCCCAGAAGAGCTATTTG-, CTGCCCCAGAAGAGCTATTTG-, TGCCCCAGAAGAGCTATTTG-, GCCCCAGAAGAGCTATTTG-, CCCCAGAAGAGCTATTTG-, CCCAGAAGAGCTATTTG-, CCAGAAGAGCTATTTG-, CAGAAGAGCTATTTG-, AGAAGAGCTATTTG-, GAAGAGCTATTTG-, AAGAGCTATTTG-, AGAGCTATTTG-, GAGCTATTTG-, AGCTATTTG-, GCTATTTG-, CTATTTG-, TATTTG-, ATTTG-, TTTG-, TTG-, TG- или G-;
N14 выбран из: -TAGGGAGCCGTCTTCAGGAATCTTCTC, -TAGGGAGCCGTCTTCAGGAATCTTCT, -TAGGGAGCCGTCTTCAGGAATCTTC, -TAGGGAGCCGTCTTCAGGAATCTT, -TAGGGAGCCGTCTTCAGGAATCT, -TAGGGAGCCGTCTTCAGGAATC, -TAGGGAGCCGTCTTCAGGAAT, -TAGGGAGCCGTCTTCAGGAA, -TAGGGAGCCGTCTTCAGGA, -TAGGGAGCCGTCTTCAGG, -TAGGGAGCCGTCTTCAG, -TAGGGAGCCGTCTTCA, -TAGGGAGCCGTCTTC, -TAGGGAGCCGTCTT, -TAGGGAGCCGTCT, -TAGGGAGCCGTC, -TAGGGAGCCGT, -TAGGGAGCCG, -TAGGGAGCC, -TAGGGAGC, -TAGGGAG, -TAGGGA, -TAGGG, -TAGG, -TAG, -TA или -T;
m равно 0 или 1;
n равно 0 или 1;
и n+m=1 или 2;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Антисмысловые олигонуклеотиды общей формулы S5 (SEQ. ID NO: 99) предпочтительно содержат 2-10 звеньев LNA, более предпочтительно 3-9 звеньев LNA и еще более предпочтительно 4-8 звеньев LNA и также предпочтительно, по меньшей мере, 6 звеньев, которые не являются LNA, более предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно 8 звеньев, которые не являются LNA. Звенья, которые не являются LNA, предпочтительно представляют звенья ДНК. Звенья LNA предпочтительно расположены на терминальном 3'-конце (также называемом 3'-концом) и терминальном 5'-конце (также называемом 5'-концом). Предпочтительно, по меньшей мере, одно и более предпочтительно, по меньшей мере, две звена LNA находятся на терминальном 3'-конце и терминальном 5'-конце. Таким образом, предпочтительными являются антисмысловые олигонуклеотиды по настоящему изобретению, обозначенные как гапмеры, которые содержат 2-10 звеньев LNA и которые предпочтительно содержат 1-5 звеньев LNA на терминальном 5'-конце, и 1-5 звеньев LNA на терминальном 3'-конце антисмыслового олигонуклеотида и между звеньями LNA, по меньшей мере, 7 и более предпочтительно, по меньшей мере, 8 звеньев ДНК. Более предпочтительно антисмысловые олигонуклеотиды содержат 2-4 звена LNA на терминальном 5'-конце, и 2-4 звена LNA на терминальном 3'-конце, и еще более предпочтительно содержат 3-4 звена LNA на терминальном 5'-конце, и 3-4 звена LNA на терминальном 3'-конце, и содержат предпочтительно, по меньшей мере, 7 звеньев, которые не являются LNA, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA.
Кроме того, антисмысловые олигонуклеотиды могут содержать обычные азотистые основания, такие как аденин, гуанин, цитозин, тимин и урацил, а также их обычные производные, такие как 5-метилцитозин или 2-аминоаденин. Антисмысловые олигонуклеотиды по настоящему изобретению могут также содержать модифицированные межнуклеотидные мостики, такие как фосфоротиоат или фосфородитиоат вместо фосфатных мостиков. Такие модификации могут присутствовать только в сегментах LNA или только в сегменте, который не является LNA, антисмыслового олигонуклеотида. В качестве звеньев LNA, в частности, остатки b1-b9, раскрытые здесь, являются особенно предпочтительными.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды формулы (S5):
5'-(N13)m-GTAGTGTT-(N14)n-3'
где:
N13 представляет: GCCTGCCCCAGAAGAGCTATTTG-, CCTGCCCCAGAAGAGCTATTTG-, CTGCCCCAGAAGAGCTATTTG-, TGCCCCAGAAGAGCTATTTG-, GCCCCAGAAGAGCTATTTG-, CCCCAGAAGAGCTATTTG-, CCCAGAAGAGCTATTTG-, CCAGAAGAGCTATTTG-, CAGAAGAGCTATTTG-, AGAAGAGCTATTTG-, GAAGAGCTATTTG-, AAGAGCTATTTG-, AGAGCTATTTG-, GAGCTATTTG-, AGCTATTTG-, GCTATTTG-, CTATTTG-, TATTTG-, ATTTG-, TTTG-, TTG-, TG- или G-; и
N14 выбран из: -TAGGGAGCCGTCTTC, -TAGGGAGCCGTCTT, -TAGGGAGCCGTCT, -TAGGGAGCCGTC, -TAGGGAGCCGT, -TAGGGAGCCG, -TAGGGAGCC, -TAGGGAGC, -TAGGGAG, -TAGGGA, -TAGGG, -TAGG, -TAG, -TA или -T; и
m равно 0 или 1; n равно 0 или 1; и n+m=1 или 2.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S5) содержит 10-28 нуклеотидов и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S5) содержит 11-24 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце. Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S5) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S5):
5'-(N13)m-GTAGTGTT-(N14)n-3'
где:
N13 представляет: CCCCAGAAGAGCTATTTG-, CCCAGAAGAGCTATTTG-, CCAGAAGAGCTATTTG-, CAGAAGAGCTATTTG-, AGAAGAGCTATTTG-, GAAGAGCTATTTG-, AAGAGCTATTTG-, AGAGCTATTTG-, GAGCTATTTG-, AGCTATTTG-, GCTATTTG-, CTATTTG-, TATTTG-, ATTTG-, TTTG-, TTG-, TG- или G-; и
N14 выбран из: -TAGGGAGCCG, -TAGGGAGCC, -TAGGGAGC, -TAGGGAG, -TAGGGA, -TAGGG, -TAGG, -TAG, -TA или -T; и
m равно 0 или 1; n равно 0 или 1; и n+m=1 или 2.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S5):
5'-(N13)m-GTAGTGTT-(N14)n-3'
где:
N13 представляет: GAAGAGCTATTTG-, AAGAGCTATTTG-, AGAGCTATTTG-, GAGCTATTTG-, AGCTATTTG-, GCTATTTG-, CTATTTG-, TATTTG-, ATTTG-, TTTG-, TTG-, TG- или G-; и
N14 выбран из: -TAGGG, -TAGG, -TAG, -TA или -T; и
m равно 0 или 1; n равно 0 или 1; и n+m=1 или 2.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S5):
5'-(N13)m-GTAGTGTT-(N14)n-3'
где:
N13 представляет: CAGAAGAGCTATTTG-, AGAAGAGCTATTTG-, GAAGAGCTATTTG-, AAGAGCTATTTG-, AGAGCTATTTG-, GAGCTATTTG-, AGCTATTTG-, GCTATTTG-, CTATTTG-, TATTTG-, ATTTG-, TTTG-, TTG-, TG- или G-; и
N14 выбран из: -TAGGGAGCCGTCTTCAGGAATCT, -TAGGGAGCCGTCTTCAGGAATC, -TAGGGAGCCGTCTTCAGGAAT, -TAGGGAGCCGTCTTCAGGAA, -TAGGGAGCCGTCTTCAGGA, -TAGGGAGCCGTCTTCAGG, -TAGGGAGCCGTCTTCAG, -TAGGGAGCCGTCTTCA, -TAGGGAGCCGTCTTC, -TAGGGAGCCGTCTT, -TAGGGAGCCGTCT, -TAGGGAGCCGTC, -TAGGGAGCCGT, -TAGGGAGCCG, -TAGGGAGCC, -TAGGGAGC, -TAGGGAG, -TAGGGA, -TAGGG, -TAGG, -TAG, -TA или -T; и
m равно 0 или 1; n равно 0 или 1; и n+m=1 или 2.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S5):
5'-(N13)m-GTAGTGTT-(N14)n-3'
где:
N13 представляет: GAGCTATTTG-, AGCTATTTG-, GCTATTTG-, CTATTTG-, TATTTG-, ATTTG-, TTTG-, TTG-, TG- или G-; и
N14 выбран из: -TAGGGAGCCGTCTTCAGG, -TAGGGAGCCGTCTTCAG, -TAGGGAGCCGTCTTCA, -TAGGGAGCCGTCTTC, -TAGGGAGCCGTCTT, -TAGGGAGCCGTCT, -TAGGGAGCCGTC, -TAGGGAGCCGT, -TAGGGAGCCG, -TAGGGAGCC, -TAGGGAGC, -TAGGGAG, -TAGGGA, -TAGGG, -TAGG, -TAG, -TA или -T; и
m равно 0 или 1; n равно 0 или 1; и n+m=1 или 2.
Также предпочтительными являются антисмысловые олигонуклеотиды формулы (S5):
5'-(N13)m-GTAGTGTT-(N14)n-3'
где:
N13 представляет: ATTTG-, TTTG-, TTG-, TG- или G-; и
N14 выбран из: -TAGGGAGCCGTCT, -TAGGGAGCCGTC, -TAGGGAGCCGT, -TAGGGAGCCG, -TAGGGAGCC, -TAGGGAGC, -TAGGGAG, -TAGGGA, -TAGGG, -TAGG, -TAG, -TA или -T; и
m равно 0 или 1; n равно 0 или 1; и n+m=1 или 2.
Предпочтительно антисмысловой олигонуклеотид общей формулы (S5/SEQ. ID NO: 99) содержит 12-24 нуклеотида и, по меньшей мере, один нуклеотид LNA на 3'-конце и, по меньшей мере, один нуклеотид LNA на 5'-конце. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)».
Более предпочтительно антисмысловой олигонуклеотид общей формулы (S5) содержит 12-22 нуклеотида и, по меньшей мере, два нуклеотида LNA на 3'-конце и, по меньшей мере, два нуклеотида LNA на 5'-конце.
Еще более предпочтительно антисмысловой олигонуклеотид общей формулы (S5) содержит 12-20, более предпочтительно 13-19 и еще более предпочтительно 14-18 нуклеотидов и, 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце. Предпочтительно антисмысловые олигонуклеотиды представляют гапмеры формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. Предпочтительно антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания».
Еще один аспект настоящего изобретения относится к антисмысловому олигонуклеотиду(ам), имеющему длину из 10-28 нуклеотидов, предпочтительно 10-24 нуклеотидов, более предпочтительно 11-22 нуклеотидов, или 12-20 нуклеотидов, еще более предпочтительно 13-19 нуклеотидов и наиболее предпочтительно 14-18 нуклеотидов, где, по меньшей мере, два из нуклеотидов, предпочтительно, по меньшей мере, три из нуклеотидов, и более предпочтительно, по меньшей мере, четыре из нуклеотидов представляют LNA, и последовательность антисмыслового олигонуклеотида из 10-28 нуклеотидов, предпочтительно 10-24 нуклеотидов, более предпочтительно 11-22 нуклеотидов, или 12-20 нуклеотидов, еще более предпочтительно 13-19 нуклеотидов и наиболее предпочтительно 14-18 нуклеотидов выбрана из группы последовательностей из 10-28 нуклеотидов, предпочтительно 10-24 нуклеотидов, более предпочтительно 11-22 нуклеотидов, или 12-20 нуклеотидов, еще более предпочтительно 13-19 нуклеотидов и наиболее предпочтительно 14-18 нуклеотидов, входящих в состав последовательности, выбранной из следующей группы:
GAATCTTGAATATCTCATGAATGGACCAGTATTCTAGAAAC
(SEQ. ID NO. 75: 383-423 в последовательности SEQ. ID NO: 1),
TTCATATTTATATACAGGCATTAATAAAGTGCAAATGTTAT
(SEQ. ID NO. 77: 2245-2285 в последовательности SEQ. ID NO: 1),
TGAGGAAGTGCTAACACAGCTTATCCTATGACAATGTCAAAG
(SEQ. ID NO. 78: 2315-2356 в последовательности SEQ. ID NO: 1),
GCCTGCCCCAGAAGAGCTATTTGGTAGTGTTTAGGGAGCCGTCTTCAGG
(SEQ. ID NO. 79: 2528-2576 в последовательности SEQ. ID NO: 1),
CGCAGGTCCTCCCAGCTGATGACATGCCGCGTCAGGTACTCCTGTAGGT
(SEQ. ID NO. 81: 3205-3253 в последовательности SEQ. ID NO: 1),
ATGTCGTTATTAACCGACTTCTGAACGTGCGGTGGGATCGTGCTGGCGATACGCGTCCACAGGA
CGATGTGCAGCGGC
(SEQ. ID NO. 83: 4141-4218 в последовательности SEQ. ID NO: 1),
GGCCACAGGCCCCTGAGCAGCCCCCGACCCATGGCAGACCCCGCTGCTCGTCATAGACCGAGC
CCCCAGCGCAG
(SEQ. ID NO. 84: 4216-4289 в последовательности SEQ. ID NO: 1),
ATGTCGTTATTAACCGACTTCTGAACGTGCGGTGGGATCGTGCTGGCGATACGCGTCCACAGGA
CGATGTGCAGCGGCCACAGGCCCCTGAGCAGCCCCCGACCCATGGCAGACCCCGCTGCTCGTC
ATAGACCGAGCCCCCAGCGCAG
(SEQ. ID NO. 86: 4141-4289 в последовательности SEQ. ID NO: 1),
TTGAATATCTCATGAATGGACCAGTATTCTA
(SEQ. ID NO. 87: 388-418 в последовательности SEQ. ID NO: 1),
CAAGTGGAATTTCTAGGCGCCTCTATGCTACTG
(SEQ. ID NO. 88: 483-515 в последовательности SEQ. ID NO: 1),
ATTTATATACAGGCATTAATAAAGTGCAAAT
(SEQ. ID NO. 89: 2250-2280 в последовательности SEQ. ID NO: 1),
AAGTGCTAACACAGCTTATCCTATGACAATGT
(SEQ. ID NO. 90: 2320-2351 в последовательности SEQ. ID NO: 1),
CCCCAGAAGAGCTATTTGGTAGTGTTTAGGGAGCCGTCT
(SEQ. ID NO. 91: 2533-2571 в последовательности SEQ. ID NO: 1),
CTGGTCGCCCTCGATCTCTCAACACGTTGTCCTTCATGCTTTCGACACAGGGGTGCTCCCGCAC
CTTGGAACCAAATG
(SEQ. ID NO. 92: 2753-2830 в последовательности SEQ. ID NO: 1),
GTCCTCCCAGCTGATGACATGCCGCGTCAGGTACTCCTG
(SEQ. ID NO. 93: 3210-3248 в последовательности SEQ. ID NO: 1),
CTCAGCTTCTGCTGCCGGTTAACGCGGTAGCAGTAGAAGA
(SEQ. ID NO. 94: 3655-3694 в последовательности SEQ. ID NO: 1),
GTTATTAACCGACTTCTGAACGTGCGGTGGGATCGTGCTGGCGATACGCGTCCACAGGACGATGTGCA
(SEQ. ID NO. 95: 4146-4213 в последовательности SEQ. ID NO: 1),
CAGGCCCCTGAGCAGCCCCCGACCCATGGCAGACCCCGCTGCTCGTCATAGACCGAGCCCCCAG
(SEQ. ID NO. 96: 4221-4284 в последовательности SEQ. ID NO: 1),
CACGCGCGGGGGTGTCGTCGCTCCGTGCGCGCGAGTGACTCACTCAACTTCA
(SEQ. ID NO. 97: 4495-4546 в последовательности SEQ. ID NO: 1),
где антисмысловой олигонуклеотид способен избирательно гибридизоваться, по отношению ко всему транскриптому человека, только с геном, кодирующим TGF-RII, или с мРНК, кодирующей TGF-RII, и солям и оптическим изомерам антисмыслового олигонуклеотида.
Указанный антисмысловой олигонуклеотид, имеющий последовательность, входящую в состав последовательностей № 75, 77, 78, 79, 81, 83, 84, 86-97, содержит 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 3'-конце, и 2-5, предпочтительно 3-5 и более предпочтительно 3-4 звена LNA на терминальном 5'-конце, и предпочтительно имеют структуру гапмеров формы LNA-сегмент A-сегмент ДНК-LNA-сегмент B. В качестве нуклеотидов LNA (звеньев LNA) подходящими, в частности, являются таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно в разделе «Предпочтительные LNA», и в качестве межнуклеотидных мостиков подходящими являются, в частности, раскрытые в разделе «Межнуклеотидные связи (IL)». Предпочтительно указанные антисмысловые олигонуклеотиды содержат, по меньшей мере, 6, более предпочтительно, по меньшей мере, 7, и наиболее предпочтительно, по меньшей мере, 8 звеньев, которые не являются LNA, таких как звенья ДНК, между двумя сегментами LNA. Подходящие азотистые основания для звеньев, которые не являются LNA, и звеньев LNA описаны в разделе «Азотистые основания». Подходящие примеры указанных антисмысловых олигонуклеотидов представлены формулами (S1)-(S7), (S1A)-(S4A), (S6A) и (S7A).
SEQ. ID NO: 1 представляет антисмысловую цепь кДНК (кДНК) (5'-3' антисмысловую последовательность) рецептора II трансформирующего фактора роста бета человека (TGF-RII), вариант транскрипта 2.
SEQ. ID NO: 2 представляет смысловую цепь кДНК (кДНК) (5'-3' смысловую последовательность) рецептора II трансформирующего фактора роста бета человека (70/80 кДа) (TGF-RII), вариант транскрипта 2. Альтернативно также можно рассматривать последовательность SEQ. ID NO: 2, как представляющую последовательность мРНК рецептора II трансформирующего фактора роста бета человека (TGF-RII), вариант транскрипта 2 (SEQ. ID NO: 3), но представленной в коде ДНК, т.е. представленной в коде G, C, A, T, а не в коде РНК.
SEQ. ID NO: 3 представляет мРНК (5'-3' смысловую последовательность) рецептора II трансформирующего фактора роста бета человека (TGF-RII), вариант транскрипта 2. Очевидно, что мРНК, показанная в SEQ. ID NO: 3, представлена в коде РНК, т.е. представлена в коде G, C, A, U.
Очевидно, понятно, что «кодирующая цепь ДНК», как здесь используется, относится к цепи ДНК, которая идентична мРНК (за исключением того, что представлена в коде ДНК), и которая включает кодоны, используемые для трансляции белка. Она не используется в качестве матрицы для транскрипции в мРНК. Таким образом, термины «кодирующая цепь ДНК», «смысловая цепь ДНК» и «нематричная цепь ДНК» могут быть использованы взаимозаменяемо. Кроме того, как здесь используется, термин «некодирующая цепь ДНК» относится к цепи ДНК, комплементарной «кодирующей цепи ДНК», и служит в качестве матрицы для транскрипции мРНК. Таким образом, термины «некодирующая цепь ДНК», «антисмысловая цепь ДНК» и «матричная цепь ДНК» могут быть использованы взаимозаменяемо.
Термин «антисмысловой олигонуклеотид» относится к олигомеру или полимеру рибонуклеиновой кислоты (РНК) или дезоксирибонуклеиновой кислоты (ДНК) или их миметикам, или вариантам, таким как антисмысловые олигонуклеотиды, имеющие измененную межнуклеотидную связь, такую как фосфортиоатная связь, и/или одно или более модифицированных азотистых оснований, таких как 5-метилцитозин, и/или одну или более модифицированных нуклеотидных звеньев, таких как LNA, например, β-D-окси-LNA. Термин «антисмысловой олигонуклеотид» включает олигонуклеотиды, состоящие из встречающихся в природе азотистых оснований, сахаров и ковалентных межнуклеотидных (остовных) связей, а также олигонуклеотиды, имеющие не встречающиеся в природе фрагменты, которые функционируют аналогично. Такие модифицированные или замещенные олигонуклеотиды часто являются предпочтительными по сравнению с нативными формами за счет наличия желаемых свойств, например, таких как повышенное поглощения клетками, повышенная аффинность для нуклеиновой кислоты-мишени и повышенная стабильность в присутствии нуклеаз. Антисмысловые олигонуклеотиды представляют короткие каталитические РНК или каталитические олигонуклеотиды, которые гибридизуются с нуклеиновой кислотой-мишенью и ингибируют ее экспрессию.
Термин «нуклеозид» хорошо известен специалистам в данной области и относится к пентозной сахарной группе, такой как рибоза, дезоксирибоза или модифицированная или замкнутая рибоза или модифицированная или замкнутая дезоксирибоза, такая как LNA, которые подробно описаны ниже. Азотистое основание связано с гликозидным атомом углерода (положение 1' пентозы), и межнуклеотидная связь образуется между атомом кислорода или серы в положении 3', и предпочтительно атомом кислорода в положении 3' нуклеозида и атомом кислорода или серы в положении 5' и предпочтительно атомом кислорода в положении 5' соседнего нуклеозида, в то время как межнуклеотидная связь отсутствует в нуклеозиде (смотри фиг. 2).
Термин «нуклеотид» хорошо известен специалистам в данной области и относится к пентозной сахарной группе, такой как рибоза, дезоксирибоза или модифицированная или замкнутая рибоза или модифицированная или замкнутая дезоксирибоза, такая как LNA, которые подробно описаны ниже. Азотистое основание связано с гликозидным атомом углерода (положение 1' пентозы), и межнуклеотидная связь образуется между атомом кислорода или серы в положении 3', и предпочтительно атомом кислорода в положении 3' нуклеотида и атомом кислорода или серы в положении 5' и предпочтительно атомом кислорода в положении 5' соседнего нуклеотида, в то время как межнуклеотидная связь является частью нуклеотида (смотри фиг. 2).
Азотистые основания
Термин «азотистое основание», сокращенно здесь как «В», и относится к пяти обычным азотистым основаниям аденину (А), тимину (Т), гуанину (G), цитозину (С) и урацилу (U), а также к их модификациям или аналогам, или аналогам, способным образовывать пары оснований согласно модели Уотсона-Крика с основаниями в комплементарной цепи. Модифицированные азотистые основания включают другие синтетические и природные азотистые основания, такие как 5-метилцитозин (С*), 5-гидроксиметилцитозин, N4-метилцитозин, ксантин, гипоксантин, 7-деазаксантин, 2-аминоаденин, 6-метиладенин, 6-метилгуанин, 6-этиладенин, 6-этилгуанин, 2-пропиладенин, 2-пропилгуанин, 6-карбоксиурацил, 5-галогенурацил, 5,6-дигидроурацил, 5-галогенцитозин, 5-пропинилурацил, 5-пропинилцитозин, 6-азаурацил, 6-азацитозин, 6-азатимин, 5-урацил (псевдоурацил), 4-тиоурацил, 8-фтораденин, 8-хлораденин, 8-бромаденин, 8-иодаденин, 8-аминоаденин, 8-тиоладенин, 8-тиоалкиладенин, 8-гидроксиаденин, 8-фторгуанин, 8-хлоргуанин, 8-бромгуанин, 8-иодгуанин, 8-аминогуанин, 8-тиолгуанин, 8-тиолалкилгуанин, 8-гидроксигуанин, 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-иодурацил, 5-трифторметилурацил, 5-фторцитозин, 5-бромцитозин, 5-хлорцитозин, 5-иодцитозин, 5-трифторметилцитозин, 7-метилгуанин, 7-метиладенин, 8-азагуанин, 8-азааденин, 7-деазагуанин, 7-деазааденин, 7-деаза-8-азааденин, 3-деазагуанин, 3-деазааденин, 2-тиоурацил, 2-тиотимин и 2 тиоцитозин и т.д., где 5-метилцитозин и/или 2-аминоаденин являются предпочтительными, поскольку эти модификации, как было показано, повышают стабильность дуплекса нуклеиновой кислоты.
Предпочтительные антисмысловые олигонуклеотиды по настоящему изобретению могут содержать аналоги азотистых оснований. Азотистое основание только одного нуклеотидного звена антисмыслового олигонуклеотида может быть замещено аналогом азотистого основания, или два, три, четыре, пять или даже все азотистые основания в антисмысловом олигонуклеотиде могут быть замещены аналогами азотистых оснований, такими как 5-метилцитозин или N6-метиладенин, или 2-аминоаденин. Предпочтительно звенья LNA могут быть связаны с аналогами азотистых оснований, такими как 5-метилцитозин.
Следует иметь ввиду, что при обращении к последовательности нуклеотидов или мономеров, то, что называют, представляет собой последовательность оснований, таких как А, Т, G, C или U. Однако за исключением конкретных примеров, приведенных в таблицах 3-8, при представлении антисмысловых олигонуклеотидов буквенным кодом A, T, G, C и U следует понимать, что указанный антисмысловой олигонуклеотид может содержать любые азотистые основания, раскрытые здесь, любую 3'-концевую группу, раскрытую здесь, любую 5'-концевую группу, раскрытую здесь, и любую межнуклеотидную связь (также называемую межнуклеотидным мостиком), описанную здесь. Нуклеотиды А, Т, G, С и U также следует понимать как нуклеотиды LNA или нуклеотиды, которые не являются LNA, предпочтительно такие как нуклеотиды ДНК. Только в отношении конкретных примеров, приведенных в таблицах 4-9, азотистые основания, звенья LNA, звенья, которые не являются LNA, межнуклеотидные связи и концевые группы дополнительно определены, как указано в разделе «Список условных обозначений» перед таблицей 2.
Было доказано, что антисмысловые олигонуклеотиды, а также соли антисмысловых олигонуклеотидов, описанные здесь, являются комплементарными к мишени, которая представляет ген, кодирующий TGF-RII, или мРНК, кодирующую TGF-RII, т.е. гибридизуются достаточно эффективно и с достаточной специфичностью, и особенно избирательностью с получением желаемого ингибирующего эффекта.
Термин «соль» относится к физиологически и/или фармацевтически приемлемым солям антисмысловых олигонуклеотидов по настоящему изобретению. Антисмысловые олигонуклеотиды содержат азотистые основания, такие как аденин, гуанин, тимин, цитозин или их производные, которые являются основными соединениями и которые образуют соли, такие как хлорид или мезилат. Межнуклеотидная связь предпочтительно содержит отрицательно заряженный атом кислорода или серы, которые образуют соли, такие как соли натрия, лития или калия. Таким образом, фармацевтически приемлемые основно-аддитивные соли образованы неорганическими основаниями или органическими основаниями. Примеры подходящих органических и неорганических оснований представляют основания, полученные из ионов металлов, например, алюминия, ионов щелочных металлов, таких как натрий или калий, ионов щелочноземельных металлов, таких как кальций или магний, или иона аминной соли, или гидроксидов щелочных или щелочноземельных металлов, карбонатов или бикарбонатов. Примеры включают водный LiOH, NaOH, KOH, NH4OH, карбонат калия, аммиак и бикарбонат натрия, соли аммония, первичные, вторичные и третичные амины, например, такие как гидроксид тетраалкиламмония, низшие алкиламины, такие как метиламин, трет-бутиламин, прокаин, этаноламин, арилалкиламины, такие как дибензиламин и N,N-дибензилэтилендиамин, низшие алкилпиперидины, такие как N-этилпиперидин, циклоалкиламины, такие как циклогексиламин или дициклогексиламин, морфолин, глюкамин, N-метил- и N,N-диметилглюкамин, 1-адамантиламин, бензатин или соли, полученные из аминокислот, таких как аргинин, лизин, орнитин, или амиды исходно нейтральных или кислых аминокислот, хлорпрокаин, холин, прокаин и тому подобное.
Поскольку антисмысловые олигонуклеотиды являются основными соединениями, то они образуют фармацевтически приемлемые соли с органическими и неорганическими кислотами. Примерами пригодных кислот для образования такой кислотно-аддитивной соли являются соляная кислота, бромистоводородная кислота, серная кислота, фосфорная кислота, уксусная кислота, лимонная кислота, щавелевая кислота, малоновая кислота, салициловая кислота, п-аминосалициловая кислота, яблочная кислота, фумаровая кислота, янтарная кислота, аскорбиновая кислота, малеиновая кислота, сульфоновая кислота, фосфоновая кислота, перхлорная кислота, азотная кислота, муравьиная кислота, пропионовая кислота, глюконовая кислота, молочная кислота, винная кислота, оксималеиновая кислота, пировиноградная кислота, фенилуксусная кислота, бензойная кислота, п-аминобензойная кислота, п-оксибензойная кислота, метансульфоновая кислота, этансульфоновая кислота, азотистая кислота, оксиэтансульфоновая кислота, этиленсульфоновая кислота, п-толуолсульфоновая кислота, нафтилсульфоновая кислота, сульфаниловая кислота, камфорсульфоновая кислота, хиновая кислота, миндальная кислота, о-метилминдальная кислота, гидроксибензолсульфоновая кислота, пикриновая кислота, адипиновая кислота, D-o-толилвинная кислота, тартроновая кислота, α-толуиловая кислота, (о,м,п)-толуиловая кислота, нафтиламинсульфоновая кислота, и другие минеральные или карбоновые кислоты, хорошо известные специалистам в данной области. Соли получают взаимодействием свободного основания с достаточным количеством требуемой кислоты с получением соли обычным способом.
В контексте настоящего изобретения термин «гибридизация» означает гибридизацию нуклеиновых кислот, где одноцепочечная нуклеиновая кислота (ДНК или РНК) взаимодействует с другой одноцепочечной нуклеиновой кислотой, имеющей очень сходную или даже комплементарную последовательность. Таким образом, взаимодействие происходит за счет образования водородных связей между определенными азотистыми основаниями (спаривания оснований).
Как здесь используется, термин «комплементарность» (комплементарность пары оснований ДНК и РНК) относится к способности к точному спариванию между двумя нуклеиновыми кислотами. Нуклеотиды в паре оснований комплементарны друг другу, когда их форма позволяет им связываться друг с другом водородными связями. В результате чего образуется пара аденина и тимидина (или урацила) посредством образования двух водородных связей, и образуется пара цитозин-гуанин посредством образования трех водородных связей. Как здесь используется, термин «комплементарные последовательности» означает последовательности ДНК или РНК, которые, когда они выравниваются антипараллельно друг к другу, то азотистые основания в каждом положении в последовательностях будут комплементарны, как если смотреть в зеркало и видеть оборотную сторону предметов.
Как здесь используется, термин «специфически гибридизуемый» указывает на достаточную степень комплементарности или точного спаривания оснований антисмыслового олигонуклеотида с последовательностью-мишенью таким образом, что происходит стабильное и специфическое связывание между антисмысловым олигонуклеотидом и ДНК- или РНК-мишенью. Последовательность антисмыслового олигонуклеотида по изобретению не должна быть на 100% комплементарной по отношению к его нуклеиновой кислоте-мишени, чтобы быть специфически гибридизуемой, хотя, 100% комплементарность является предпочтительной. При этом «100% комплементарность» означает, что антисмысловой олигонуклеотид гибридизуется с мишенью по ее всей или полной длине без ошибочного спаривания. Другими словами, в настоящем изобретении определено, что антисмысловое соединение является специфически гибридизуемым, когда связывание соединения с молекулой-мишенью ДНК или РНК происходит в физиологических или патологических условиях, а неспецифическое связывание антисмыслового олигонуклеотида с немишеневыми последовательностями весьма маловероятно или даже невозможно.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловым олигонуклеотидам, где антисмысловые олигонуклеотиды связываются со 100% комплементарностью с мРНК, кодирующей TGF RII, и не связываются с каким-либо другим участком в полном транскриптоме человека. Кроме того, предпочтительно настоящее изобретение относится к антисмысловым олигонуклеотидам, где антисмысловые олигонуклеотиды обладают 100% комплементарностью по их полной длине с мРНК, кодирующей TGF RII, и не обладают эффектами вне мишени. Альтернативно настоящее изобретение предпочтительно относится к антисмысловым олигонуклеотидам, обладающим 100% комплементарностью по отношению к мРНК, кодирующей TGF RII, но не обладает комплементарностью к другой мРНК транскриптома человека. Таким образом, термин «транскриптом человека» относится к полному набору транскриптов в человеческом организме, что означает транскрипты всех типов клеток и условий окружающей среды (в любой данный момент времени).
Специфичность
Антисмысловые олигонуклеотиды по настоящему изобретению имеют общее свойство, заключающееся в том, что они специфичны в отношении участка, где они связываются с геном или с мРНК, кодирующей TGF-RII. По настоящему изобретению предпочтительно, чтобы в человеческом транскриптоме антисмысловые олигонуклеотиды обладали 100% комплементарностью по их полной длине только с мРНК, кодирующей TGF-RII. Кроме того, целью настоящего изобретения было найти антисмысловые олигонуклеотиды без перекрестной реактивности в пределах транскриптома млекопитающего, иного, чем обезьяны; в частности, антисмысловые олигонуклеотиды обладают только перекрестной реактивностью с транскриптомом человекообразных обезьян. Это позволит избежать проявления побочных эффектов. Таким образом, антисмысловые олигонуклеотиды по настоящему изобретению обладают высокой специфичностью в отношении гибридизации с геном или с мРНК, кодирующей TGF-RII. Антисмысловые олигонуклеотиды по изобретению предпочтительно связываются по их полной длине со 100% комплементарностью, специфичной для гена, кодирующего TGF-RII, или мРНК, кодирующей TGF-RII, и не связываются с каким-либо другим участком во всем транскриптоме человека. Это означает, что антисмысловые олигонуклеотиды по настоящему изобретению гибридизуются с мишенью (мРНК TGF-RII) без ошибочного спаривания.
Как здесь используется, термин «мРНК может включать мРНК, содержащую интроны (также относится к пре-мРНК), и также мРНК, которая не содержит интронов.
Антисмысловые олигонуклеотиды по настоящему изобретению способны связываться или гибридизоваться с пре-мРНК и/или с мРНК. Это означает, что антисмысловые олигонуклеотиды могут связываться или гибридизоваться в области интрона или внутри области интрона пре-мРНК или могут связываться или гибридизоваться в перекрывающейся области интрон-экзон пре-мРНК или могут связываться или гибридизоваться в области экзона или внутри области экзона пре-мРНК и области экзона мРНК (см. фиг. 1). Предпочтительными являются антисмысловые олигонуклеотиды, которые способны связываться или гибридизоваться с пре-мРНК и мРНК. Связывание или гибридизация антисмысловых олигонуклеотидов (ASO) с пре-мРНК ингибирует образование 5'-кэппа, ингибирует сплайсинг пре-мРНК для образования мРНК, и активирует РНКазу H, которая расщепляет пре-мРНК. Связывание или гибридизация антисмысловых олигонуклеотидов (ASO) с мРНК активирует РНКазу H, которая расщепляет РНК и ингибирует связывание рибосомных субъединиц.
Антисмысловые олигонуклеотиды по настоящему изобретению содержат, по меньшей мере, 10 и не более 28, предпочтительно не более чем 24 и более предпочтительно не более чем 20 нуклеотидов и, следовательно, содержат 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов, предпочтительно 11-20 или 11-19, или 12-19, или 13-19, или 13-18 нуклеотидов и более предпочтительно 14- 18 нуклеотидов, где, по меньшей мере, два, предпочтительно три из этих нуклеотидов представляют замкнутые нуклеиновые кислоты (LNA). Также возможны более короткие антисмысловые олигонуклеотиды, т.е. антисмысловые олигонуклеотиды, содержащие менее 10 нуклеотидов, но чем короче антисмысловые олигонуклеотиды, тем выше риск, что гибридизация будет не достаточно сильной, и селективность будет снижаться или будет вовсе отсутствовать. Неселективные антисмысловые олигонуклеотиды несут риск связываться с нежелательными участками в человеческом транскриптоме и с нежелательными мРНК, кодирующими другие белки, чем TGF-RII, тем самым вызывая нежелательные побочные эффекты. Также возможны более длинные антисмысловые олигонуклеотиды, содержащие более чем 20 нуклеотидов, но дальнейшее увеличение длины делает синтез таких антисмысловых олигонуклеотидов еще более сложным и дорогостоящим, без какой-либо дополнительной пользы в отношении повышения селективности или силы гибридизации, или лучшей стабильности к деградации.
Таким образом, настоящее изобретение относится к антисмысловым олигонуклеотидам, состоящим из 10-20 нуклеотидов, где, по меньшей мере, два нуклеотида, и предпочтительно 3'- и 5'-концевых нуклеотида представляют LNA. Таким образом, предпочтительно, чтобы, по меньшей мере, 3'-концевой нуклеотид представлял LNA, и также, по меньшей мере, 5'-концевой нуклеотид представлял LNA. В том случае, если присутствует более чем 2 LNA, то предпочтительно, чтобы дополнительные LNA были связаны с 3'-или 5'-концевой LNA, как это имеет место в случае гапмеров, описанных здесь.
Один нуклеотидный строительный блок, присутствующий в антисмысловом олигонуклеотиде по настоящему изобретению, может быть представлен следующей общей формулой (B1) и (B2):
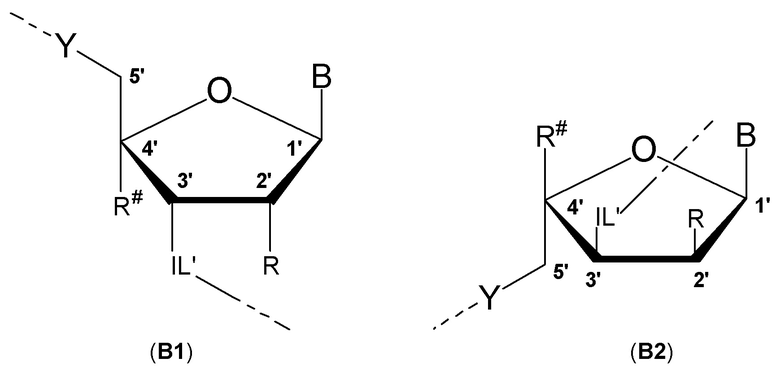
где:
В представляет азотистое основание;
IL' представляет -X''-Р(=Х')(X-);
R представляет -Н, -F, -OH, -NH2, -ОСН3, -OCH2CH2OCH3, и R# представляет -Н;
или R и R#, взятые вместе, образуют мостик -R#-R-, который выбран из -CH2-O-, -CH2-S-, -CH2-NH-, -CH2-N(CH3)-, -CH2-N(C2H5)-, -CH2-CH2-O-, -CH2-CH2-S-, -CH2-CH2-NH-, -CH2-CH2-N(CH3)- или -CH2-CH2-N(C2H5)-;
X' представляет =O или =S;
Х представляет -O-, -OH, -ORH, -NHRH, -N(RH)2, -OCH2CH2ORH, -OCH2CH2SRH, -BH3-, -RH, -SH, -SRH или -S-;
Х'' представляет -O-, -NH-, -NRH-, -CH2-, или -S-;
Y представляет -О-, -NH-, -NRH-, -CH2- или -S-;
RH выбран из атома водорода и С1-4-алкила и предпочтительно -СН3 или -С2Н5 и наиболее предпочтительно -CH3.
Предпочтительно Х- представляет -O-, -OH, -OCH3, -NH(CH3), -N (CH3)2, -OCH2CH2OCH3, -OCH2CH2SCH3, -BH3-, -СН3, -SH, -SCH3 или -S-; и более предпочтительно -O-, -OH, -OCH3, -N(CH3)2, -OCH2CH2OCH3, -BH3-, -SH, -SCH3 или -S-.
IL' предпочтительно представляет -O-P(O)(O-)-, -OP(O)(S-)-, -O-P(S)(S-)-, -S-P(O)(O-)-, -S-P(O)(S-)-, -S-Р(S)(S-)-, -O-P(O)O-)-, -O-P(O)(S-)-, -S-Р(O)(O-)-, -O-P(O)(RH)-, -O-P(O)(ORH)-, -O-P(O) (NHRH)-, -O-P(O)[N(RH)2]-, -O-P(O) (BH3-)-, -O-P(O)(OCH2CH2ORH)-, -O-P(O)(OCH2CH2SRH)-, -O-Р(O)(O-)-, -NRH-P(O)(O-)-, где RH выбран из атома водорода и С1-4-алкила.
Группа -O-P(O)(RH)-O- предпочтительно представляет -O-P(O)(CH3)-O- или -O-P(O)(C2H5)-O-, и наиболее предпочтительно -O-P(O)(CH3)-O-.
Группа -O-P(O)(ORH)-O- предпочтительно представляет -O-P(O)(OCH3)-O- или -O-P(O)(OC2H5)-O-, и наиболее предпочтительно -O-P(O)(OCH3)-O-.
Группа -O-Р(O)(NHRH)-O- предпочтительно представляет -O-P(O)(NHCH3)-O- или -O-P(O)(NHC2H5)-O- и наиболее предпочтительно -O-P(O)(NHCH3)-О-.
Группа -O-Р(O)[N(RH)2]-O- предпочтительно представляет -O-P(O)[N(CH3)2]-O- или -O-P(O)[N(C2H5)2]-О-, и наиболее предпочтительно -O-P(O)[N(CH3)2]-O-.
Группа -O-Р(O)(OCH2CH2ORH)-O- предпочтительно представляет -O-P(O)(OCH2CH2OCH3)-O- или -O-P(O)(OCH2CH2OC2H5)-O- и наиболее предпочтительно -O-P(O)(OCH2CH2OCH3)-О-.
Группа -O-Р(O)(OCH2CH2SRH)-O-, предпочтительно представляет -O-P(O)(OCH2CH2SCH3)-O- или -O-P(O)(OCH2CH2SC2H5)-O-, и наиболее предпочтительно -O-P(O)(OCH2CH2SCH3)-О-.
Группа -O-Р(O)(O-)-NRH- предпочтительно представляет -O-P(O)(O-)-NH- или -O-P(O)(O-)-N(CH3)- и наиболее предпочтительно -O-P(O)(O-)-NH-.
Группа -NRH-P(O)(O-)-O- предпочтительно представляет -NH-P(O)(O-)-O- или -N(CH3)-P(O)(O-)-O- и наиболее предпочтительно -NH-P(O)(O-)-O-.
Еще более предпочтительно IL' представляет -O-P(O)(O-)-, -O-P(O)(S-)-, -O-P(S)(S-)-, -O-Р(O)(NHRH)- или -O-P(O)[N(RH)2] -, и еще более предпочтительно IL' представляет -O-P(O)(O-)-, -O-P(O) (S-)- или -O-P(S)(S-)-, и наиболее предпочтительно IL' представляет -O-P(O)(S-)- или -O-P(S)(S-)-.
Предпочтительно Y представляет -О-.
Предпочтительно В представляет обычное азотистое основание, выбранное из A, T, G, C, U.
Предпочтительно IL представляет -O-P(=O)(S-) - или -O-P(=S) (S-)-.
Приведенные выше определения B, Y и IL' применимы также к формулам b1-b9.
Таким образом, следующие общие формулы (B3)-(B6) являются предпочтительными:
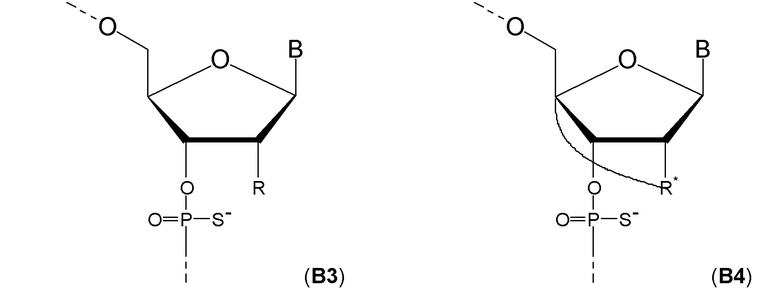
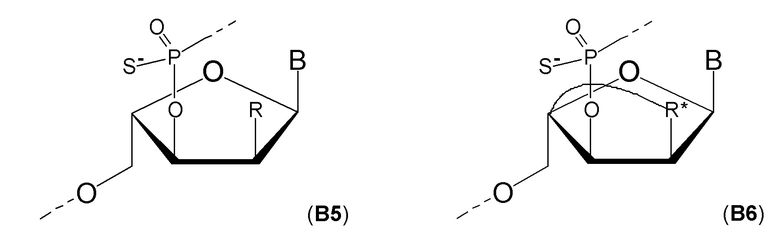
где:
В представляет азотистое основание и предпочтительно А, Т, G, C, U;
R представляет -H, -F, -OH, -NH2, -N(CH3)2, -OCH3, -OCH2CH2OCH3, -OCH2CH2CH2OH, -OCH2CH2CH2NH2 и предпочтительно -H;
R* представляет группу -R#-R-, как это определено ниже, и она, например, предпочтительно выбрана из -C(RaRb)-O-, -C(RaRb)-NRc-, -C(RaRb)-S- и -C(RaRb)-C(RaRb)-O-, где заместители Ra, Rb и Rc имеют значения, определенные здесь. Более предпочтительно R* выбран из -CH2-O-, -CH2-S-, -CH2-NH-, -CH2-N(CH3)-, -CH2-CH2-O- или -CH2-CH2-S-, более предпочтительно из -CH2-O-, -CH2-S-, -CH2-CH2-O- или -CH2-CH2-S-, и еще более предпочтительно из -CH2-O-, -CH2-S- или -CH2-CH2-O-, и еще более предпочтительно из -СН2-О- или -СН2-S-, и наиболее предпочтительно представляет -СН2-О-.
Примерами предпочтительных нуклеотидов, которые не представляют звенья LNA, являются следующие:
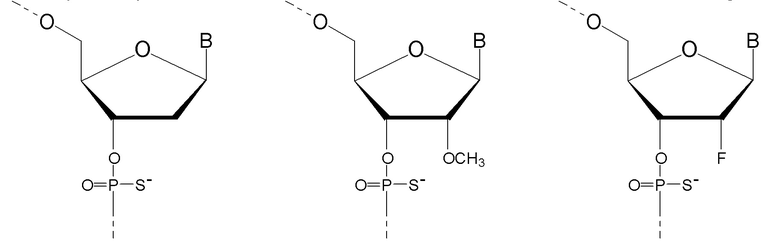

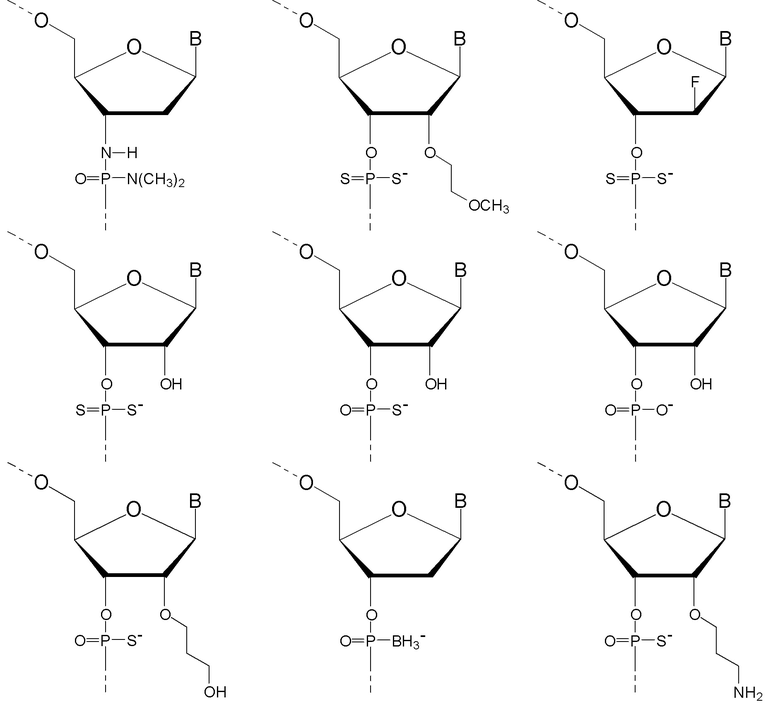
Межнуклеотидные связи (IL)
Мономеры антисмысловых олигонуклеотидов, описанных здесь, соединены друг с другом посредством межнуклеотидной связи. Соответственно, каждый мономер связан со смежным 3'-мономером через межнуклеотидную связь. Специалисту с обычной квалификацией в данной области техники понятно, что, в контексте настоящего изобретения 5'-мономер в конце олигомера не содержит 5'-межнуклеотидную связь, хотя, он может содержать концевую 5'-группу или нет. Термин «межнуклеотидная связь» означает группу, способную ковалентно связывать два нуклеотида, два нуклеотидных аналога, таких как две LNA, и нуклеотид и аналог нуклеотида, такой как LNA. Конкретные и предпочтительные примеры включают фосфатные группы и фосфортиоатные группы.
Нуклеотиды антисмысловых олигонуклеотидов по настоящему изобретению или соседние нуклеотидные последовательности соединены друг с другом посредством межнуклеотидных связей. Соответственно каждый нуклеотид связан через 5'-положение с соседним 3'-нуклеотидом через межнуклеотидную связь.
Антисмысловые олигонуклеотиды можно модифицировать различными способами. Возможны модификации в остове, и они относятся к антисмысловым олигонуклеотидам, где фосфатные группы (также называемые фосфодиэфирными группами) в их межнуклеотидном остове частично или полностью заменены другими группами. Предпочтительные модифицированные остовы антисмысловых олигонуклеотидов включают, например, фосфоротиоаты, хиральные фосфоротиоаты, фосфородитиоаты, фосфотриэфир, аминоалкилфосфотриэфиры, метил-, этил- и С3-С10-алкилфосфонаты, включая 3'-алкиленфосфонаты и хиральные фосфонаты, фосфинаты, фосфороамидаты, включая 3'-аминофосфорамидат и аминоалкилфосфорамидаты, тионофосфорамидаты, тионоалкил фос фонаты, тионоалкилфосфотриэфиры и боранофосфаты, содержащие 3'-5' связи, их 2'-5' связанные аналоги, и таковые, которые имеют инвертированную полярность, где смежные пары нуклеотидных звеньев связаны 3'-5' с 5'-3' или 2'-5' с 5'-2'. Различные соли, смешанные соли и их свободные кислотные формы также включаются и раскрываются более подробно.
Подходящие межнуклеотидные связи включают таковые, которые описаны в международной заявке WO 2007/031091, например межнуклеотидные связи, приведенные в первом абзаце на странице 34 в WO 2007/031091 (включена в данное описание в качестве ссылки). В некоторых вариантах осуществления предпочтительно модифицировать межнуклеотидную связь по сравнению с обычной фосфодиэфирной связью, которая более устойчива к атаке нуклеаз, такую как фосфоротиоат или боранофосфат - эти две, принятые как опосредованное РНКазой Н расщепление, также представляют путь ингибирования антисмысловым олигонуклеотидом, приводящий к снижению экспрессии гена-мишени.
Межнуклеотидная связь состоит из группы IL', которая является группой, связанной с 3'-атомом углерода рибозного фрагмента и группой Y, которая является группой, связанной с 5'-углеродным атомом соседнего рибозного фрагмента, как показано в формуле (IL'Y) ниже:
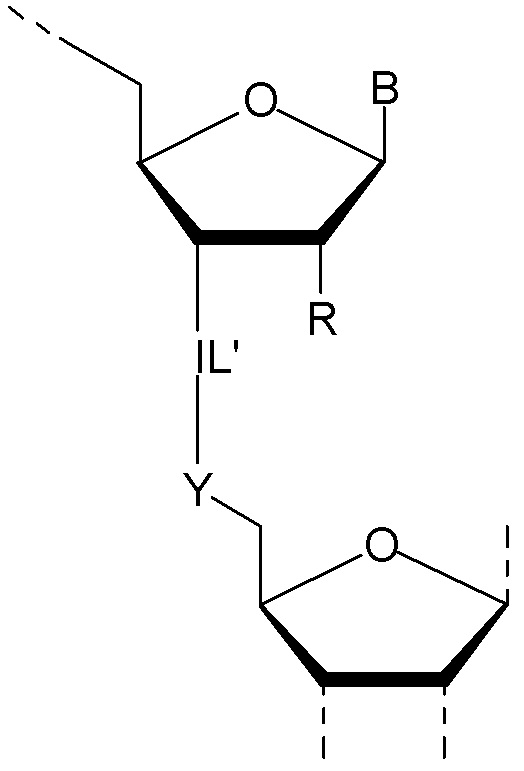
Межнуклеотидная связь IL представлена -IL'-Y-. IL' представляет -X''-P(=X')(Х-)- так, что IL представляет -X''-P(=X')(X-)-Y-, где заместители Х-, Х', Х'' и Y имеют значения, определенные здесь.
Межнуклеотидная связь IL=-X''-P(=X')(X-)-Y- предпочтительно выбрана из группы, состоящей из:
-O-P(O)(O-)-О-, -OP(O)(S-)-О-, -O-P(S)(S-)-О-, -S-P(O)(O-)-О-, -S-P(O)(S-)-О-, -S-Р(S)(S-)-О-, -O-P(O)(O-)-S-, -O-P(O-)(S)-S-, -S-Р(O)(O-)-S-, -O-P(O)(RH)-О-, -O-P(O)(ORH)-О-, -O-P(O)(NHRH)-О-, -O-P(O)[N(RH)2]-О-, -O-P(O)(BH3-)-О-, -O-P(O)(OCH2CH2ORH)-О-, -O-P(O)(OCH2CH2SRH)-O-, -O-Р(O)(O-)-NRH-, -NRH-P(O)(O-)-O-, где RH выбран из атома водорода и С1-4-алкила.
Группа -O-P(O)(RH)-O- предпочтительно представляет -O-P(O)(CH3)-O- или -O-P(O)(C2H5)-O-, и наиболее предпочтительно -O-P(O)(CH3)-O-.
Группа -O-P(O)(ORH)-O- предпочтительно представляет -O-P(O)(OCH3)-O- или -O-P(O)(OC2H5)-O-, и наиболее предпочтительно -O-P(O)(OCH3)-O-.
Группа -O-Р(O)(NHRH)-O- предпочтительно представляет -O-P(O)(NHCH3)-O- или -O-P(O)(NHC2H5)-O- и наиболее предпочтительно -O-P(O)(NHCH3)-О-.
Группа -O-Р(O)[N(RH)2]-O- предпочтительно представляет -O-P(O)[N(CH3)2]-O- или -O-P(O)[N(C2H5)2]-О-, и наиболее предпочтительно -O-P(O)[N(CH3)2]-O-.
Группа -O-Р(O)(OCH2CH2ORH)-O- предпочтительно представляет -O-P(O)(OCH2CH2OCH3)-O- или -O-P(O)(OCH2CH2OC2H5)-O- и наиболее предпочтительно -O-P(O)(OCH2CH2OCH3)-О-.
Группа -O-Р(O)(OCH2CH2SRH)-O- предпочтительно представляет -O-P(O)(OCH2CH2SCH3)-O- или -O-P(O)(OCH2CH2SC2H5)-O-, и наиболее предпочтительно -O-P(O)(OCH2CH2SCH3)-О-.
Группа -O-Р(O)(O-)-NRH- предпочтительно представляет -O-P(O)(O-)-NH- или -O-P(O)(O-)-N(CH3)-, и наиболее предпочтительно -O-P(O)(O-)-NH-.
Группа -NRH-P(O)(O-)-O- предпочтительно представляет -NH-P(O)(O-)-O- или -N(CH3)-P(O)(O-)-O- и наиболее предпочтительно -NH-P(O)(O-)-O-.
Еще более предпочтительно IL представляет -O-P(O)(O-)-О-, -O-P(O)(S-)-О-, -O-P(S)(S-)-О-, -O-Р(O)(NHRH)-О- или -O-P(O)[N(RH)2]-О -, и еще более предпочтительно IL представляет -O-P(O)(O-)-О-, -O-P(O)(S-)-О- или -O-P(S)(S-)-О-, и наиболее предпочтительно IL представляет -O-P(O)(S-)-О- или -O-P(O)(O-)-O-.
Таким образом, IL предпочтительно представляет фосфатную группу (-O-P(O)(O-)-O-), фосфоротиоатную группу (-O-P(O)(S-)-О-) или фосфородитиоатную группу (-O-Р(S)(S-)-О-).
Нуклеотидные звенья или нуклеозиды антисмысловых олигонуклеотидов соединены друг с другом посредством межнуклеотидных связей, таким образом, что в одном антисмысловом олигонуклеотиде могут присутствовать различные межнуклеотидные связи. Звенья LNA предпочтительно связаны межнуклеотидными связями, которые не являются фосфатными группами. Звенья LNA соединены друг с другом группой IL, которая предпочтительно выбрана из -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -O-P(O)(NHRH)-O- и -O- (O)[N(RH)2]-О- и более предпочтительно -O-P(O)(S-)-O- и -O-Р(S) (S-)-О-.
Звенья, которые не являются LNA, связаны друг с другом группой IL, которая предпочтительно выбрана из -O-P(O)(O-)-O-, -O-P(O)(S-)-О-, -О-P (S)(S-)-O-, -O-P(O)(NHRH)-O- и -O-P(O)[N(RH)2]-О- и более предпочтительно -О-P(O)(O-)-O-, -O-P(O)(S-)-O- и -O-Р(S)(S-)-О-.
Звенья, которые не являются LNA, связаны со звеном LNA группой IL, которая предпочтительно выбрана из -O-P(O)(S-)-О-, -О-Р(S)(S-)-O-, -O-P(O)(NHRH)-O- и -O-P(O)[N(RH)2]-О- и более предпочтительно -O-P (O)(S-)-O- и -O -P(S)(S-)-О-.
Как здесь используется, термин «звено LNA» относится к нуклеотиду, который замкнут, т.е. нуклеотиду, который имеет бициклическую структуру, и особенно бициклическую рибозную структуру и особенно бициклическую рибозную структуру, показанную в общей формуле (II). Мост «замыкает» рибозу в 3'-эндо (север) конформации. Рибозный фрагмент нуклеотида LNA модифицируется дополнительным мостиком, соединяющим 2'-кислород и 4'-углерод. Альтернативно используемые термины для LNA представляют бициклические нуклеотиды или связанные мостиком нуклеотиды, таким образом, альтернативный термин для звена LNA представляет бициклическое нуклеотидное звено или связанное мостиком нуклеотидное звено.
Как здесь используется, термин «звено, которое не является LNA» относится к нуклеотиду, который не является замкнутым, т.е. нуклеотиду, который не содержит бициклическую сахарную группу, и особенно не содержит бициклическую рибозную структуру и особенно бициклическую рибозную структуру, показанную в общей формуле (II). Звенья, которые не являются LNA, наиболее предпочтительно представляют звенья ДНК.
Как здесь используется, термин «звено ДНК» относится к нуклеотиду, содержащему 2-дезоксирибозу в качестве сахара. Таким образом, нуклеотид состоит из азотистого основания и 2-дезоксирибозы.
Как здесь используется, термин «звено» относится к части или фрагменту, или группе антисмыслового олигонуклеотида по настоящему изобретению. Таким образом, «звено» не является полной молекулой, оно является частью или его фрагментом, или группой антисмыслового олигонуклеотида, которое имеет, по меньшей мере, одно положение для ковалентного связывания с другой частью или фрагментом, или группой антисмыслового олигонуклеотида. Например, общие структуры (B1)-(B6) представляют звенья, поскольку они могут быть ковалентно связаны через группу Y и IL' или -O- и -O-P(O)(S-)- соответственно. Предпочтительно звено представляет фрагмент, состоящий из пентозной структуры, азотистого основания, соединенного с пентозной структурой 5'-радикальной группой, и радикальной группой IL'.
Как здесь используется, термин «строительный блок» или «мономер» относится к молекуле, и, в частности, к нуклеозиду, который используется в синтезе антисмыслового олигонуклеотида по настоящему изобретению. Примерами являются молекулы LNA общей формулы (I), где Y представляет 5'-концевую группу, и IL' представляет 3'-концевую группу.
Кроме того, предпочтительными являются чистые диастереомерные антисмысловые олигонуклеотиды. Предпочтительными являются Sp- и Rp-диастереомеры, как показано справа:
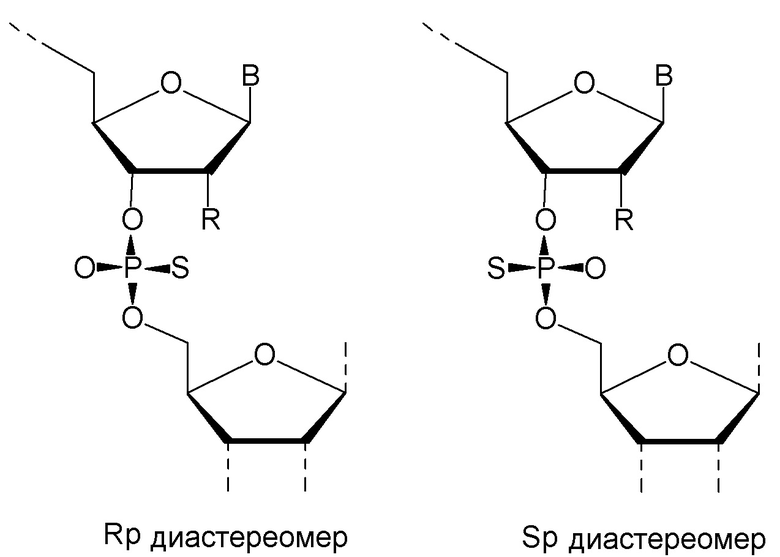
Соответствующие серусодержащие (S) межнуклеотидные связи, как представлено здесь, являются предпочтительными.
Предпочтительными являются фосфоротиоатные группы в остове, где, по меньшей мере, 50% межнуклеотидных связей представляют фосфортиоатные группы. Также предпочтительным является то, чтобы звена LNA, если они присутствуют, были связаны между собой через фосфоротиоаты в качестве межнуклеотидных связей. Наиболее предпочтительным является полностью фосфоротиоатный остов, т.е. наиболее предпочтительным является, когда все нуклеотидные звенья, а также звенья LNA (если они присутствуют) были связаны друг с другом через фосфортиоатные группы, которые определяются следующим образом: -O-P(O)(S-)-O-, что является аналогичным -O-P(O, S)-O- или -O-P(O-)(S)-O-.
В случае, если антисмысловой олигонуклеотид является гапмером, то предпочтительно, чтобы участки LNA имели межнуклеотидные связи, выбранные из -O-P(O)(S-)-O- и -O-P(S)(S-)-O- и чтобы участки, которые не являются LNA, в средней части имели межнуклеотидные связи, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-, и чтобы участки LNA были связаны с участками, которые не являются LNA, через межнуклеотидные связи, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O- и -O-Р(S)(S-)-О-.
Еще более предпочтительно, если все межнуклеотидные связи, которые представляют 9 в 10-мерных и 19 в 20-мерных олигонуклеотидах, были выбраны из -О-P(O)(S-)-O- и -O-P(S)(S-)-О-.Еще более предпочтительным является то, чтобы все межнуклеотидные связи представляли фосфортиоатные группы (-О-Р(О)(S-)-О-) или представляли фосфородитиоатные группы (-О-Р(S)(S-)-О-).
Замкнутые нуклеиновые кислоты (LNA ® )
Особенно предпочтительно, чтобы некоторые из нуклеотидов общей формулы (В1) или (В2) в антисмысловых олигонуклеотидах были замещены так называемыми LNA (замкнутыми нуклеиновыми кислотами). Сокращенное обозначение LNA является зарегистрированным торговым названием, но здесь термин «LNA» используется исключительно в описательной форме. Предпочтительно, чтобы концевые нуклеотиды замещены LNA и более предпочтительно последние 1-4 нуклеотида на 3'-конце и/или последние 1-4 нуклеотида 5'-конце были замещены LNA. Кроме того, предпочтительно, чтобы, по меньшей мере, концевой нуклеотид на 3'-конце и 5'-конце каждый был замещен LNA.
Как здесь используется, термин «LNA» относится к бициклическому нуклеотидному аналогу, известному как «замкнутая нуклеиновая кислота». Он может относиться к мономеру LNA, или, при использовании в контексте «антисмыслового олигонуклеотида с LNA» или «антисмысловому олигонуклеотиду, содержащему LNA», LNA относится к олигонуклеотиду, содержащему один или более таких бициклических нуклеотидных аналогов. Нуклеотиды LNA характеризуются наличием линкерной группы (такой как мостик) между C2' и C4' рибозного сахарного кольца - например, как показано в виде бирадикала R#-R, описанного ниже. LNA, используемая в антисмысловых олигонуклеотидах по настоящему изобретению, предпочтительно имеет структуру общей формулы (I):
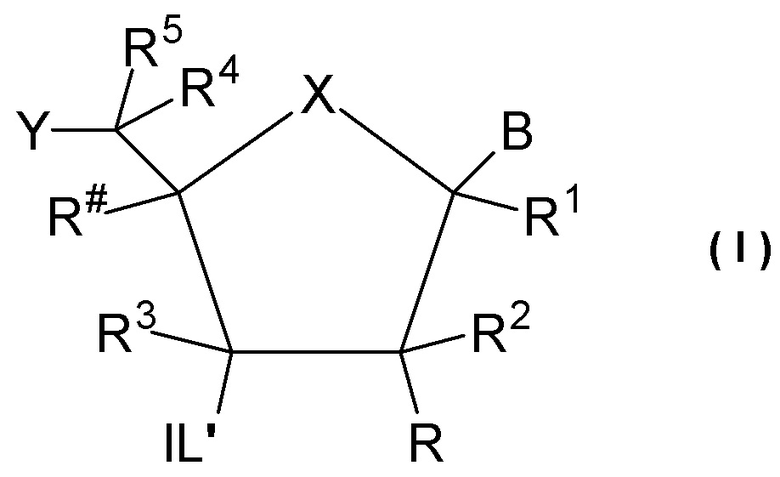
где для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации;
где Х выбран из -О-, -S-, -N(RN)-, -C(R6R7)-, и предпочтительно Х представляет -O-;
В выбран из атома водорода, необязательно замещенного С1-4-алкокси, необязательно замещенного С1-4-алкила,, необязательно замещенного С1-4-ацилокси, азотистых оснований и аналогов азотистых оснований, и предпочтительно В представляет азотистое основание или аналог азотистого основания, и наиболее предпочтительно представляет обычное азотистое основание;
Y представляет часть межнуклеотидной связи с соседним нуклеотидом в случае, если фрагмент общей формулы (I) представляет звено LNA антисмыслового олигонуклеотида по настоящему изобретению, или 5'-концевую группу в случае, если фрагмент общей формулы (I) представляет мономер или строительный блок для синтеза антисмыслового олигонуклеотида по настоящему изобретению. 5'-атом углерода необязательно содержит заместитель R4 и R5;
IL' представляет часть межнуклеотидной связи с соседним нуклеотидом в случае, если фрагмент общей формулы (I) представляет звено LNA антисмыслового олигонуклеотида по настоящему изобретению, или 5'-концевую группу в случае, если фрагмент общей формулы (I) представляет мономер или строительный блок для синтеза антисмыслового олигонуклеотида по настоящему изобретению.
R# и R, взятые вместе, представляют двухвалентную линкерную группу, состоящую из 1-4 групп или атомов, выбранных из -C(RaRb)-, -C(Ra)=C(Rb)-, -C(Ra)=N-, -O-, -Si(Ra)2-, -S-, -SO2-, -N(Rc)- и > C=Z, где Z выбран из -O-, -S- и -N(Ra)-, и Ra, Rb и Rc независимо друг от друга выбраны из атома водорода, необязательно замещенного C1-12-алкила, необязательно замещенного C2-6-алкенила, необязательно замещенного C2-6-алкинила, гидрокси, необязательно замещенного C1-12-алкокси, С1-6-алкокси-С1-6-алкила, С2-6-алкенилокси, карбокси, С1-12-алкоксикарбонила, С1-12-алкилкарбонила, формила, арила, арилоксикарбонила, арилокси, арилкарбонила, гетероарила, гетероарилоксикарбонила, гетероарилокси, гетероарилкарбонила, амино, моно- и ди(С1-6- алкил)амино, карбамоила, моно- и ди(C1-6-алкил)аминокарбонила, амино-C1-6-алкилениламинокарбонила, моно- и ди(С1-6-алкил)амино-С 1-6-алкилениламинокарбонила, C1-6-алкилкарбониламино, карбамидо, С1-6-алканоилокси, сульфоно, С1-6-алкилсульфонилокси, нитро, азидо, сульфанила, С1-6-алкилтио, атома галогена, где арил и гетероарил могут быть необязательно замещены, и где два геминальных заместителя Ra и Rb, взятые вместе, могут представлять необязательно замещенный метилен (=СН2), где для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации; и
каждый из заместителей R1, R2, R3, R4, R5, R6 и R7, которые присутствуют, независимо выбраны из атома водорода, необязательно замещенного C1-12-алкила, необязательно замещенного C2-6-алкенила, необязательно замещенного C2-6-алкинила, гидрокси, C1-12-алкокси, С1-6-алкокси-С1-6-алкила, С2-6-алкенилокси, карбокси, С1-12-алкоксикарбонила, С1-12-алкилкарбонила, формила, арила, арилоксикарбонила, арилокси, арилкарбонила, гетероарила, гетероарилоксикарбонила, гетероарилокси, гетероарилкарбонила, амино, моно- и ди(С1-6-алкил)амино, карбамоила, моно- и ди(C1-6-алкил)аминокарбонила, амино-C1-6-алкиламинокарбонила, моно- и ди(С1-6-алкил)амино-С1-6-алкиламинокарбонила, C1-6-алкилкарбониламино, карбамидо, С1-6-алканоилокси, сульфоно, С1-6-алкилсульфонилокси, нитро, азидо, сульфанила, С1-6-алкилтио, атома галогена, где арил и гетероарил могут быть необязательно замещены, и где два геминальных заместителя, взятые вместе, могут представлять оксо, тиооксо, имино или необязательно замещенный метилен;
где RN выбран из атома водорода и С1-4-алкила, и где два соседних (негеминальных) заместителя могут обозначать дополнительную связь, приводя к образованию двойной связи; и RN, если он присутствует и не входит в состав бирадикала, выбран из атома водорода и С1-4-алкила; и их основно- и кислотно-аддитивные соли. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации.
В предпочтительных вариантах осуществления R# и R, взятые вместе, представляют бирадикал, состоящий из групп, выбранных из группы, состоящей из -C(RaRb)-C(RaRb)-, -C(RaRb)-O-, -C(RaRb)-NRC-, -C(RaRb)-S- и -C(RaRb)-C(RaRb)-O-, где каждый из Ra, Rb и Rc может быть необязательно, независимо выбран.
В некоторых вариантах осуществления Ra и Rb могут быть необязательно, независимо выбраны из группы, состоящей из атома водорода и C1-6-алкила, такого как метил, и предпочтительно является атомом водорода.
В предпочтительных вариантах осуществления R1, R2, R3, R4 и R5 независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного С1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила или замещенного С2-6-алкинила, С1-6-алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, C1-6-аминоалкила или замещенного C1-6-аминоалкила. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации.
В предпочтительных вариантах осуществления R1, R2, R3, R4 и R5 представляют атом водорода.
В некоторых вариантах осуществления R1, R2 и R3 независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного С1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила или замещенного С2-6-алкинила, С1-6-алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, C1-6-аминоалкила или замещенного C1-6-аминоалкила. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации. В предпочтительных вариантах осуществления R1, R2 и R3 представляют атом водорода.
В предпочтительных вариантах осуществления R4 и R5 каждый независимо выбран из группы, состоящей из -H, -CH3, -CH2-CH3, -CH2-O-CH3 и -CH=CH2. Соответственно, в некоторых вариантах осуществления один из R4 или R5 представляет атом водорода, тогда как другой (R4 или R5 соответственно) выбран из группы, состоящей из C1-6-алкила, С2-6-алкенила, С2-6-алкинила, замещенного C1-6-алкила, замещенного С2-6-алкенила, замещенного С2-6-алкинила или замещенного ацила (-C(=O)-); где каждая замещенная группа моно- или полизамещена заместителями, независимо выбранными из атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, -OJ1, -SJ1, -NJ1J2, -N3, -COOJ1, -CN, -О-С(=O)NJ1J2, -N(Н)С(=NH)NJ1J2 или -N(H)С(=Х)N(H)J2, где Х представляет О или S; и каждый J1 и J2 независимо представляет -Н, C1-6-алкил, замещенный C1-6-алкил, С2-6-алкенил, замещенный С2-6-алкенил, С2-6-алкинил, замещенный С2-6-алкинил, С1-6-аминоалкил, замещенный С1-6-аминоалкил или защитную группу. В некоторых вариантах осуществления один из R4 или R5 представляет замещенный C1-6-алкил. В некоторых вариантах осуществления один из R4 или R5 представляет замещенный метилен, где предпочтительные заместители включают одну или более групп, независимо выбранных из -F, -NJ1J2, -N3, -CN, -OJ1, -SJ1, -О-С(=O)NJ1J2, -N(Н)С(=NH)NJ1J2 или -N(H)С(=O)N(H)J2. В некоторых вариантах осуществления каждый J1 и J2 независимо представляет -H или C1-6-алкил. В некоторых вариантах осуществления один из R4 или R5 представляет метил, этил или метоксиметил. В некоторых вариантах осуществления один из R4 или R5 представляет метил. В дополнительном варианте осуществления один из R4 или R5 представляет этиленил. В некоторых вариантах осуществления один из R4 или R5 представляет замещенный ацил. В некоторых вариантах осуществления один из R4 или R5 представляет -О-С(=O)NJ1J2. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации. Такие 5'-модифицированные бициклические нуклеотиды раскрыты в публикации международной заявки WO 2007/134181 А, которая включена здесь в качестве ссылки во всей ее полноте.
В некоторых вариантах осуществления В представляет азотистое основание, в том числе аналоги азотистого основания и встречающиеся в природе азотистые основания, такие как пурин или пиримидин, или замещенный пурин или замещенный пиримидин, такие как азотистое основание в настоящем описании, например, азотистое основание, выбранное из группы, состоящей из аденина, цитозина, тимина, аденина, урацила и/или модифицированного или замещенного азотистого основания, такого как 5-тиазолоурацил, 2-тиоурацил, 5-пропинилурацил, 2'-тиотимин, 5-метилцитозин, 5-тиазолоцитозин, 5-пропинилцитозин и 2,6-диаминопурин.
В предпочтительных вариантах осуществления R# и R, взятые вместе, представляют бирадикал, выбранный из -С(RaRb)-O-, -C(RaRb) -C(RcRd)-O-, -C(RaRb)-C(RcRd)-C(ReRf)-O-, -C(RaRb)-O-С(RdRe)-, -C (RaRb)-O-С(RdRe)-O-, -C (RaRb)-C(RdRe)-, -C(RaRb)-C(RcRd)-C(ReRf)-, -C(Ra)=C(Rb)-C(RdRe)-, -C(RaRb)-N(Rc)-, -C(RaRb)-С(RdRe)-N(Rc)-, -C (RaRb)-N(Rc)-O-, -C(RaRb)-S- и -C(RaRb)-C(RdRe)-S-, где Ra, Rb, Rc, Rd, Re и Rf каждый независимо выбран из атома водорода, необязательно замещенного C1-12-алкила, необязательно замещенного C2-6-алкенила, необязательно замещенного C2-6-алкинила, гидрокси, необязательно замещенного C1-12-алкокси, С1-6-алкокси-С1-6-алкила, С2-6-алкенилокси, карбокси, С1-12-алкоксикарбонила, С1-12-алкилкарбонила, формила, арила, арилоксикарбонила, арилокси, арилкарбонила, гетероарила, гетероарилоксикарбонила, гетероарилокси, гетероарилкарбонила, амино, моно- и ди(С1-6- алкил)амино, карбамоила, моно- и ди(C1-6-алкил)аминокарбонила, амино-C1-6-алкиламинокарбонила, моно- и ди(С1-6-алкил)амино-С1-6-алкиламинокарбонила, C1-6-алкилкарбониламино, карбамидо, С1-6-алканоилокси, сульфоно, С1-6-алкилсульфонилокси, нитро, азидо, сульфанила, С1-6-алкилтио, атома галогена, где арил и гетероарил могут быть необязательно замещены, и где два геминальных заместителя Ra и Rb, взятые вместе, могут представлять необязательно замещенный метилен (=СН2). Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации.
В дополнительном варианте осуществления R# и R, взятые вместе, представляют бирадикал (двухвалентную группу), выбранный из -СН2-O-, -СН2-S-, -СН2-NH-, -СН2-N(CH3)-, -СН2-СН2-О-, -СН2-СН(СН3)-, -СН2-СН2-S-, -СН2-СН2-NH-, -СН2-СН2-СН2-, -СН2-СН2-СН2-О-, -СН2-СН2-СН(СН3)-, -СН=СН-СН2-, -СН2-О-СН2-О-, -СН2-NH-О-, -СН2-N(CH3)-O-, -CH2-O-CH2-, -CH(CH3)-O-, -CH(CH2-O-CH3)-O-, -СН2-СН2-, и -СН=СН-. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации.
В некоторых вариантах осуществления R# и R, взятые вместе, представляют бирадикал -C(RaRb)-N(Rc)-O-, где Ra и Rb независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, С1-6- алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, С1-6-аминоалкила или замещенного C1-6-аминоалкила, такой как атом водорода; где Rc выбран из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, С1-6-алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, С1-6-аминоалкила или замещенного C1-6-аминоалкила, и предпочтительно представляет атом водорода.
В предпочтительных вариантах осуществления R# и R, взятые вместе, представляют бирадикал -C(RaRb)-O-С(RdRe)-O-, где Ra, Rb, Rd и Re независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, С1-6-алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, С1-6-аминоалкила или замещенного C1-6-аминоалкила, и предпочтительно представляют атом водорода.
В предпочтительных вариантах осуществления R# и R образуют бирадикал -CH(Z)-O-, где Z выбран из группы, состоящей из C1-6-алкила, С2-6-алкенила, С2-6-алкинила, замещенного C1-6-алкила, замещенного С2-6-алкенила, замещенного С2-6-алкинила, ацила, замещенного ацила, замещенного амида, тиола или замещенного тио; и где каждая из замещенных групп независимо моно- или полизамещена необязательно защищенными заместителями, независимо выбранными из атома галогена, оксо, гидроксила, -OJ1, -NJ1J2, -SJ1, -N3, -OC(=X)J1, -OC(=X)NJ1J2, -NJ3C(=Х)NJ1J2 и -CN, где каждый J1, J2 и J3, независимо представляет -Н или C1-6-алкил, и Х представляет О, S или NJ1. В предпочтительных вариантах осуществления Z представляет C1-6-алкил или замещенный С1-6-алкил. В других предпочтительных вариантах осуществления Z представляет метил. В предпочтительных вариантах осуществления Z представляет замещенный С1-6-алкил. В предпочтительных вариантах осуществления указанный заместитель представляет собой С1-6-алкокси. В некоторых вариантах осуществления Z представляет CH3OCH2-. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации. Такие бициклические нуклеотиды раскрыты в патенте США № 7399845, который включен здесь в качестве ссылки во всей его полноте. В предпочтительных вариантах осуществления R1, R2, R3, R4 и R5 представляют атом водород. В предпочтительных вариантах осуществления R1, R2 и R3 представляют водород, и один или оба из R4, R5 могут быть другими, чем атом водород, как указано выше, и в международной заявке WO 2007/134181.
В предпочтительных вариантах осуществления R# и R, взятые вместе, представляют бирадикал, который содержит замещенную аминогруппу в мостике, такой как бирадикал -СН2-N(Rc)-, где Rc представляет С1-12-алкилокси. В предпочтительных вариантах осуществления R# и R, взятые вместе, представляют бирадикал Cq3q4-NOR-, где q3 и q4 независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, С1-6-алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, С1-6-аминоалкила или замещенного C1-6-аминоалкила; где каждая замещенная группа независимо моно- или полизамещена заместителями, независимо выбранными из атома галогена, -OJ1, -SJ1, -NJ1J2, -COOJ1, -CN, -OC(=O)NJ1J2, -NH-C(=NH)NJ1J2 или -NH-С(=Х)NHJ2, где Х представляет О или S; и каждый из J1 и J2 независимо представляет -Н, C1-6-алкил, С2-6-алкенил, С2-6-алкинил, С1-6-аминоалкил или защитную группу. Для всех хиральных центров, асимметричные группы могут находиться в любой R- или S- ориентации. Такие бициклические нуклеотиды раскрыты в международной заявке WO2008/150729, которая включена здесь в качестве ссылки во всей ее полноте. В предпочтительных вариантах осуществления R1, R2, R3, R4 и R5 независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, С1-6-алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, С1-6-аминоалкила или замещенного C1-6-аминоалкила. В предпочтительных вариантах осуществления R1, R2, R3, R4 и R5 представляют атом водорода. В предпочтительных вариантах осуществления R1, R2 и R3 представляют атом водорода, и один или оба из R4, R5 могут быть иными, чем атом водорода, как указано выше, и в международной заявке WO 2007/134181.
В предпочтительных вариантах осуществления R# и R, взятые вместе, представляют бирадикал (двухвалентную группу) -C(RaRb)-O-, где Ra и Rb каждый независимо представляет атом галогена, C1-12-алкил, замещенный C1-12-алкил, С2-6-алкенил, замещенный С2-6-алкенил, С2-6-алкинил, замещенный С2-6-алкинил, С1-12-алкокси, замещенный С1-12-алкокси, -OJ1, -SJ1, -S(O)J1, -SO2-J1, -NJ1J2, -N3, -CN, -C(=O)OJ1, -C(=O)NJ1J2, -C(=O)J1, -OC(=O)NJ1J2, -NH-C(=NH) NJ1J2, -NH-С(=O)NJ1J2 или -NH-C(=S)NJ1J2; или Ra и Rb, взятые вместе, представляют =C(q3)(q4); q3 и q4 каждый независимо представляет -Н, атом галогена, С1-12-алкил или замещенный С1-12-алкил; каждая замещенная группа независимо моно- или полизамещена заместителями, независимо выбранными из атома галогена, С1-6-алкила, замещенного С1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, -OJ1, -SJ1, -NJ1J2, -N3, -CN, -C(=O)OJ1, -C(=O)NJ1J2, -C(=O)J1, -OC(=O)NJ1J2, -NH-C(=O)NJ1J2 или -NH-C(=S)NJ1J2; и каждый из J1 и J2 независимо представляет -H, С1-6-алкил, замещенный С1-6-алкил, С2-6-алкенил, замещенный С2-6-алкенил, С2-6-алкинил, замещенный С2-6-алкинил, С1-6-аминоалкил, замещенный С1-6-аминоалкил или защитную группу. Такие соединения описаны в международной заявке WO2009006478A, которая включена здесь в качестве ссылки во всей ее полноте.
В предпочтительных вариантах осуществления R# и R образуют бирадикал -Q-, где Q представляет -C(q1)(g2)C(q3)(q4)-, -C(q1)=С(q3)-, -C[=C(q1)(q2)]-С(q3)(q4)- или -C(q1)(q2)-C[=С(q3) (q4)]-;
q1, q2, q3, q4 каждый независимо друг от друга представляет -Н, атом галогена, C1-12-алкил, замещенный C1-12-алкил, С2-6-алкенил, замещенный С2-6-алкенил, С2-6-алкинил, замещенный С2-6-алкинил, С1-12-алкокси, замещенный С1-12-алкокси, -OJ1, -SJ1, -S(O)J1, -SO2-J1, -NJ1J2, -N3, -CN, -C(=O)OJ1, -C(=O)NJ1J2, -C(=O)J1, -OC(=O)NJ1J2, -NH-C(=NH) NJ1J2, -NH-С(=O)NJ1J2 или -NH-C(=S)NJ1J2; каждый из J1 и J2 независимо друг от друга представляет -H, С1-6-алкил, С2-6-алкенил, С2-6-алкинил, С1-6-аминоалкил или защитную группу; и необязательно, когда Q представляет -C(q1)(q2)C(q3)(q4), и один из q3 или q4 представляет -СН3, то, по меньшей мере, один из q3 или q4 или один из q1 и q2 является иным, чем -Н. В предпочтительных вариантах осуществления R1, R2, R3, R4 и R5 представляют атом водород. Для всех хиральных центров асимметричные группы могут находиться в любой R- или S-ориентации. Такие бициклические нуклеотиды раскрыты в международной заявке WO2008/154401, которая включена здесь в качестве ссылки во всей ее полноте. В предпочтительных вариантах осуществления изобретения R1, R2, R3, R4 и R5 независимо выбраны из группы, состоящей из атома водорода, атома галогена, C1-6-алкила, замещенного C1-6-алкила, С2-6-алкенила, замещенного С2-6-алкенила, С2-6-алкинила, замещенного С2-6-алкинила, С1-6- алкоксила, замещенного С1-6-алкоксила, ацила, замещенного ацила, С1-6-аминоалкила или замещенного C1-6-аминоалкила. В предпочтительных вариантах R1, R2, R3, R4 и R5 представляют атом водорода. В предпочтительных вариантах осуществления R1, R2 и R3 представляют атом водорода, и один или оба из R4, R5 могут быть иными, чем атом водорода, как указано выше и в международных заявках WO 2007/134181 или WO 2009/067647 (альфа-L-бициклические аналоги нуклеиновых кислот).
Как здесь используется, термин «С1-6-алкил» относится к -CH3, -C2H5, -C3H7, -CH(CH3)2, -C4H9, -CH2-CH(CH3)2, -CH(СН3)-C2H5, - C(CH3)3, -C5H11, -CH(СН3)-C3H7, -СН2-СН(СН3)-С2Н5, -СН(СН3)-СН(СН3)2, -C(CH3)2-С2Н5, -СН2-C(CH3)3, -СН(С2Н5)2, -C2H4-CH(CH3)2, -C6H13, -C3H6-CH(CH3)2, -C2H4-CH(CH3)-C2H5, -СН(СН3)-C4H9, -СН2-СН(СН3)-C3H7, -СН (СН3)-СН2-СН(СН3)2, -СН(СН3)-СН(СН3)-С2Н5, -СН2-СН(СН3)-СН(СН3)2, -СН2-С(СН3)2-С2Н5, -С(СН3)2-С3Н7, -С(СН3)2-СН(СН3)2, -C2H4-C(CH3)3, -CH2-CH(С2Н5)2 и -CH(CH3)-C(CH3)3. Термин «С1-С6-алкил» также включает «С1-С6-циклоалкил» такой, как цикло-C3H5, цикло-C4H7, цикло-C5H9 и цикло-С6Н11.
Предпочтительными являются -CH3, -C2H5, -C3H7, -CH(CH3)2, -C4H9, -CH2-CH(CH3)2, -CH(СН3)-C2H5, -C(CH3)3 и -C5H11. Особенно предпочтительными являются -CH3, -C2H5, -C3H7 и -CH(CH3)2.
Термин «C1-C6-алкил» также включает «C1-C6-циклоалкил» такой, как цикло-C3H5, цикло-C4H7, цикло-C5H9, и цикло-С6Н11.
Как здесь используется, термин «C1-C12-алкил» относится к C1-C6-алкилу, -C7H15, -C8H17, -C9H19, -C10H21, -C11H23, -C12H25.
Как здесь используется, термин «С2-С6-алкиленил» относится к -CH2-, -C2H4-, -CH(CH3)-, -C3H6-, -CH2-CH(CH3)-, -CH(CH3)-CH2-, -C(CH3)2-, -C4H8-, -CH2-C(CH3)2-, -C(CH3)2-CH2-, -C2H4-CH(CH3)-, -CH(CH3)-C2H4-, -CH2-CH(CH3)-CH2-, -CH(CH3)-CH(CH3)-, -C5H10-, -CH(CH3)-C3H6-, -CH2-CH(CH3)-C2H4-, -C2H4-CH(CH3)-CH2-, -C3H6-CH(CH3)-, -C2H4-C(CH3)2-, -C(CH3)2-C2H4-, -CH2-C(CH3)2-CH2-, -CH2-CH(CH3)-CH(CH3)-, -CH(CH3)-CH2-CH(CH3)-, -CH(CH3)-CH(CH3)-CH2-, -CH(CH3)-CH(CH3)-CH(CH3)-, -C(CH3)2-C3H6-, -CH2-C(CH3)2-C2H4-, -C2H4-C(CH3)2-CH2-, -C3H6-C(CH3)2-, -CH(CH3)-C4H8-, -C6H12-, -CH2-CH(CH3)-C3H6-, -C2H4-CH(CH3)-C2H4-, -C3H6-CH(CH3)-CH2-, -C4H8-CH(CH3)-, -C2H4-CH(CH3)-CH(CH3)-, -CH2-CH(CH3)-CH(CH3)-CH2-, -CH2-CH(CH3)-CH2-CH(CH3)-, -CH(CH3)-C2H4-CH(CH3)-, -CH(CH3)-CH2-CH(CH3)-CH2- и -CH(CH3)-CH(CH3)-C2H4-.
Как здесь используется, термин «С2-С6-алкенил» относится к -CH=CH2, -CH2-CH=CH2, -C(CH3)=CH2, -CH=CH-CH3, -C2H4-CH=CH2, -CH2-CH=CH-CH3, -CH=CH-C2H5, -CH2-C(CH3)=CH2, -CH(CH3)-CH=CH, -CH=C(CH3)2, -C(CH3)=CH-CH3, -CH=CH-CH=CH2, -C3H6-CH=CH2, -C2H4-CH=CH-CH3, -CH2-CH=CH-C2H5, -CH=CH-C3H7, -CH2-CH=CH-CH=CH2, -CH=CH-CH=CH-CH3, -CH=CH-CH2-CH=CH2, -C(CH3)=CH-CH=CH2, -CH=C(CH3)-CH=CH2, -CH=CH-C(CH3)=CH2, -C2H4-C(CH3)=CH2, -CH2-CH(CH3)-CH=CH2, -CH(CH3)-CH2-CH=CH2, -CH2-CH=C(CH3)2, -CH2-C(CH3)=CH-CH3, -CH(CH3)-CH=CH-CH3, -CH=CH-CH(CH3)2, -CH=C(CH3)-C2H5, -C(CH3)=CH-C2H5, -C(CH3)=C(CH3)2, -C(CH3)2-CH=CH2, -CH(CH3)-C(CH3)=CH2, -C(CH3)=CH-CH=CH2, -CH=C(CH3)-CH=CH2, -CH=CH-C(CH3)=CH2, -C4H8-CH=CH2, -C3H6-CH=CH-CH3, -C2H4-CH=CH-C2H5, -CH2-CH=CH-C3H7, -CH=CH-C4H9, -C3H6-C(CH3)=CH2, -C2H4-CH(CH3)-CH=CH2, -CH2-CH(CH3)-CH2-CH=CH2, -CH(CH3)-C2H4-CH=CH2, -C2H4-CH=C(CH3)2, -C2H4-C(CH3)=CH-CH3, -CH2-CH(CH3)-CH=CH-CH3, -CH(CH3)-CH2-CH=CH-CH3, -CH2-CH=CH-CH(CH3)2, -CH2-CH=C(CH3)-C2H5, -CH2-C(CH3)=CH-C2H5, -CH(CH3)-CH=CH-C2H5, -CH=CH-CH2-CH(CH3)2, -CH=CH-CH(CH3)-C2H5, -CH=C(CH3)-C3H7, -C(CH3)=CH-C3H7, -CH2-CH(CH3)-C(CH3)=CH2, -CH(CH3)-CH2-C(CH3)=CH2, -CH(CH3)-CH(CH3)-CH=CH2, -CH2-C(CH3)2-CH=CH2, -C(CH3)2-CH2-CH=CH2, -CH2-C(CH3)=C(CH3)2, -CH(CH3)-CH=C(CH3)2, -C(CH3)2-CH=CH-CH3, -CH(CH3)-C(CH3)=CH-CH3, -CH=C(CH3)-CH(CH3)2, -C(CH3)=CH-CH(CH3)2, -C(CH3)=C(CH3)-C2H5, -CH=CH-C(CH3)3, -C(CH3)2-C(CH3)=CH2, -CH(C2H5)-C(CH3)=CH2, -C(CH3)(C2H5)-CH=CH2, -CH(CH3)-C(C2H5)=CH2, -CH2-C(C3H7)=CH2, -CH2-C(C2H5)=CH-CH3, -CH(C2H5)-CH=CH-CH3, -C(C4H9)=CH2, -C(C3H7)=CH-CH3, -C(C2H5)=CH-C2H5, -C(C2H5)=C(CH3)2, -C[C(CH3)3]=CH2, -C[CH(CH3)(C2H5)]=CH2, -C[CH2-CH(CH3)2]=CH2, -C2H4-CH=CH-CH=CH2, -CH2-CH=CH-CH2-CH=CH2, -CH=CH-C2H4-CH=CH2, -CH2-CH=CH-CH=CH-CH3, -CH=CH-CH2-CH=CH-CH3, -CH=CH-CH=CH-C2H5, -CH2-CH=CH-C(CH3)=CH2, -CH2-CH=C(CH3)-CH=CH2, -CH2-C(CH3)=CH-CH=CH2, -CH(CH3)-CH=CH-CH=CH2, -CH=CH-CH2-C(CH3)=CH2, -CH=CH-CH(CH3)-CH=CH2, -CH=C(CH3)-CH2-CH=CH2, -C(CH3)=CH-CH2-CH=CH2, -CH=CH-CH=C(CH3)2, -CH=CH-C(CH3)=CH-CH3, -CH=C(CH3)-CH=CH-CH3, -C(CH3)=CH-CH=CH-CH3, -CH=C(CH3)-C(CH3)=CH2, -C(CH3)=CH-C(CH3)=CH2, -C(CH3)=C(CH3)-CH=CH2 и -CH=CH-CH=CH-CH=CH2.
Предпочтительными являются -CH=CH2, -CH2-CH=CH2, -C(CH3)=CH2, -CH=CH-CH3, -C2H4-CH=CH2, -CH2-CH=CH-CH3. Особенно предпочтительными являются -CH=CH2, -CH2-CH=CH2 и -CH=CH-CH3.
Как здесь используется, термин «С2-С6-алкинил» относится к -C≡CH, -C≡C-CH3, -CH2 C≡CH, -C2H4-C≡CH, -CH2-C≡C-CH3, -C≡C-C2H5, -C3H6-C≡CH, -C2H4-C≡C-CH3, -CH2-C≡C-C2H5, -C≡C-C3H7, -CH(CH3)-C≡CH, -CH2-CH(CH3)-C≡CH, -CH(CH3)-CH2-C≡CH, -CH(CH3)-C≡C-CH3, -C4H8-C≡CH, -C3H6-C≡C-CH3, -C2H4-C≡C-C2H5, -CH2-C≡C-C3H7, -C≡C-C4H9, -C2H4-CH(CH3)-C≡CH, -CH2-CH(CH3)-CH2-C≡CH, -CH(CH3)-C2H4-C≡CH, -CH2-CH(CH3)-C≡C-CH3, -CH(CH3)-CH2-C≡C-CH3, -CH(CH3)-C≡C-C2H5, -CH2-C≡C-CH(CH3)2, -C≡C-CH(CH3)-C2H5, -C≡C-CH2-CH(CH3)2, -C≡C-C(CH3)3, -CH(C2H5)-C≡C-CH3, -C(CH3)2-C≡C-CH3, -CH(C2H5)-CH2-C≡CH, -CH2-CH(C2H5)-C≡CH, -C(CH3)2-CH2-C≡CH, -CH2-C(CH3)2-C≡CH, -CH(CH3)-CH(CH3)-C≡CH, -CH(C3H7)-C≡CH, -C(CH3)(C2H5)-C≡CH, -C≡C-C≡CH, -CH2-C≡C-C≡CH, -C≡C-C≡C-CH3, -CH(C≡CH)2, -C2H4-C≡C-C≡CH, -CH2-C≡C-CH2-C≡CH, -C≡C-C2H4-C≡CH, -CH2-C≡C-C≡C-CH3, -C≡C-CH2-C≡C-CH3, -C≡C-C≡C-C2H5, -C≡C-CH(CH3)-C≡CH, -CH(CH3)-C≡C-C≡CH, -CH(C≡CH)-CH2-C≡CH, -C(C≡CH)2-CH3, -CH2-CH(C≡CH)2, -CH(C≡CH)-C≡C-CH. Предпочтительными являются -С≡СН и -С≡С-СН3.
Термин «С1-6-алкоксил» относится к группе «C1-C6-алкил-O-».
Термин «С1-12-алкоксил» относится к группе «C1-C12-алкил-O-».
Термин «С1-6-аминоалкил» относится к группе «H2N-C1-C6-алкил-».
Термин «С2-С6-алкенилокси» относится к группе «С2-С6-алкенил-О-».
Термин «С1-6-алкилкарбонил» относится к группе «C1-C6-алкил-СО-». Также упоминается как «ацил».
Термин «С1-12-алкилкарбонил» относится к «С1-С12-алкил-СО-». Также упоминается как «ацил».
Термин «С1-6-алкоксикарбонил» относится к группе «С1-С6-алкил-О-СО-».
Термин «С1-12-алкоксикарбонил» относится к группе «С1-С12-алкил-СО-О-».
Термин «С1-С6-алаканоилокси» относится к группе «С1-С6-алкил-СО-О-».
Термин «С1-6-алкилтио» относится к группе «С1-С6-алкил-S-».
Термин «С1-6-алкилсульфонилокси» относится к группе «С1-С6-алкил-SO2-O-».
Термин «С1-6-алкилкарбониламино» относится к группе «С1-С6-алкил-СО-NH-».
Термин «С1-6-алкиламино» относится к группе «С1-С6-алкил-NH-».
Термин «(C1-6)2алкиламино» относится к диалкиламиногруппе, такой как «[С1-С6-алкил][С1-С6-алкил]N-».
Термин «С1-6-алкиламинокарбонил» относится к группе «С1-С6-алкил-NH-CO-».
Термин «(C1-6)-алкиламинокарбонил» относится к группе диалкиламинокарбонил, такой как «[C1-C6-алкил][C1-C6-алкил]-N-CO-».
Термин «амино-C1-6-алкиламинокарбонил» относится к группе «H2N-[С1-С6-алкиленил]-HN-СО-».
Термин «C1-6-алкиламино-С1-6-алкиламинокарбонил» относится к группе «C1-С6-алкил-HN-[C1-C6-алкиленил]-NH-СО-».
Термин «(C1-6)2-алкиламино-С1-6-алкиламинокарбонил» относится к группе «[C1-C6-алкил][C1-C6-алкил]-N-[C1-C6-алкиленил]-NH-СО-».
Термин «арил» относится к фенилу, толуилу, замещенному фенилу и замещенному толуилу.
Термин «арилокси» относится к группе «арил-О-».
Термин «арилкарбонил» относится к группе «арил-СО-».
Термин «арилоксикарбонил» относится к группе «арил-O-СО-».
Термин «гетероарил» относится к замещенным или незамещенным гетероароматическиам группам, которые содержат 4-9 атомов в кольце, 1-4 из которых выбраны из О, N и/или S. Предпочтительные «гетероарильные» группы содержат 1 или 2 гетероатома в 5- или 6-членном ароматическом кольце. Включаются моно- и бициклические кольцевые системы. Типичными «гетероарильными» группами являются пиридил, фурил, тиенил, пирролил, оксазолил, тиазолил, имидазолил, изоксазолил, оксадиазолил, пиридазинил, пиримидил, пиразинил, 1,3,5-триазинил, 1,2,3-триазолил, 1,3,4-тиадиазолил, индолизинил, индолил, изоиндолил, бензо[b]фурил, бензо[b]тиенил, индазолил, бензимидазолил, бензтиазолил, пуринил, хинолизинил, хинолил, изохинолил, хиназолинил, хиноксалинил, 1,8-нафтиридинил, тетрагидрохинолил, бензооксазолил, хром-2-онил, индазолил и тому подобное.
Термин «гетероарилокси» относится к группе «гетероарил-О-».
Термин «гетероарилкарбонил» относится к группе «гетероарил-СО-».
Термин «гетероарилоксикарбонил» относится к группе «гетероарил-O-СО- ».
Термин «замещенный» относится к группам, в которых один или более атомов водорода замещены одним или более из следующих заместителей: -OH, -OCH3, -OC2H5, -OC3H7, -O-цикло-C3H5, -OCH(CH3)2, -OCH2Ph, -F, -Cl, -COCH3, -COC2H5, -COC3H7, -CO-цикло-C3H5, -COCH(CH3)2, -COOH, -CONH2, -NH2, -NHCH3, -NHC2H5, -NHC3H7, -NH-цикло-C3H5, -NHCH(CH3)2, -N(CH3)2, -N(C2H5)2, -N(C3H7)2, -N(цикло-C3H5)2, -N[CH(CH3)2]2, -SO3H, -OCF3, -OC2F5, цикло-C3H5, -CH3, -C2H5, -C3H7, -CH(CH3)2, -CH=CH2, -CH2-CH=CH2, -C≡CH и/или -C≡C-CH3.
В случае, когда общая формула (I) представляет мономеры или строительные блоки для синтеза антисмысловых олигонуклеотидов по настоящему изобретению, то концевые группы Y и IL' выбраны независимо друг от друга из атома водорода, азидо, атома галогена, циано, нитро, гидрокси, PG-O-, AG-O-, меркапто, PG-S-, AG-S, С1-6-алкилтио, амино, PG-N(RH)-, AG-N(RH)-, моно- или ди(С1 -6-алкил)амино, необязательно замещенного С1-6-алкокси, необязательно замещенного С1-6-алкила, необязательно замещенного С2-6-алкенила, необязательно замещенного С2-6-алкенилокси, необязательно замещенного С1-6-алкенила, необязательно замещенного С2-6-алкенилокси, необязательно замещенного С2-6-алкинила, монофосфата, монотиофосфата, дифосфата, дитиофосфата, трифосфата, тритиофосфата, карбокси, сульфоно, гидроксиметила, PG-O-CH2, AG-O-CH2-, аминометила, PG-N(RH)-CH2, AG-N(RH)-CH2, карбоксиметила, сульфонометила, где PG представляет собой защитную группу для -ОН, -SH и NH(RH) соответственно, AG является активирующей группой для -ОН, -SH и NH(RH) соответственно, и RH выбран из атома водорода и С1-6-алкила.
Защитные группы PG в гидрокси-заместителях включают замещенный тритил, такой как 4,4'-диметокситритил (DMT), 4-монометокситритил (MMT), необязательно замещенный 9-(9-фенил)ксантенил (пиксил), необязательно замещенный метокситетрагидропиранил (mthp), силил, такой как триметилсилил (™S), триизопропилсилил (TIPS), трет-бутилдиметилсилил (TBDMS), триэтилсилил и фенилдиметилсилил, трет-бутиловые эфиры, ацетали (включающие две гидроксильные группы), ацил, такой как ацетил или, замещенный галогеном ацетил, например, хлорацетил или фторацетил, изобутирил, пивалоил, бензоил и замещенные бензоилы, метоксиметил (MOM), бензиловые эфиры или замещенные бензиловые эфиры, такие как 2,6-дихлорбензил (2,6-Cl2Bzl). Альтернативно, когда Y или IL' представляют гидроксил, то они могут быть защищены присоединением к твердому носителю, необязательно через линкер.
Когда Y или IL' представляет аминогруппу, то иллюстративными примерами защитных групп для аминогруппы являются флуоренилметоксикарбонил (Fmoc), трет-бутилоксикарбонил (ВОС), трифторацетил, аллилоксикарбонил (alloc или АОС), бензилоксикарбонил (Z или Cbz), замещенные бензилоксикарбонилы, такие как 2-хлорбензилоксикарбонил (2-ClZ), монометокситритил (ММТ), диметокситритил (DMT), фталоил и 9-(9-фенил)ксантенил (пиксил).
Act представляет активирующую группу для -ОН, -SH, -NH(RH) соответственно. Такие активирующие группы, например, выбраны из необязательно замещенного O-фосфорамидита, необязательно замещенного O-фосфотриэфира, необязательно замещенного O-фосфодиэфира, необязательно замещенного H-фосфоната и необязательно замещенного O-фосфоната.
В данном контексте термин «фосфорамидит» означает группу формулы -P(ORx)N(Ry)2, где Rx представляет необязательно замещенную алкильную группу, например, метил, 2-цианоэтил или бензил, и каждый из Ry представляет необязательно замещенные алкильные группы, например, этил или изопропил, или группа N(Ry)2 образует морфолиногруппу (N(CH2CH2)2O). Rx предпочтительно обозначает 2-цианоэтил, и два Ry предпочтительно являются одинаковыми и обозначают изопропил. Таким образом, особенно подходящий фосфорамидит представляет N,N-диизопропил-O-(2-цианоэтил)фосфорамидит.
Мономеры LNA или строительные блоки LNA
Мономеры LNA или строительные блоки LNA, используемые в качестве исходных веществ в синтезе антисмысловых олигонуклеотидов по настоящему изобретению, предпочтительно представляют собой нуклеозиды LNA следующих общих формул:
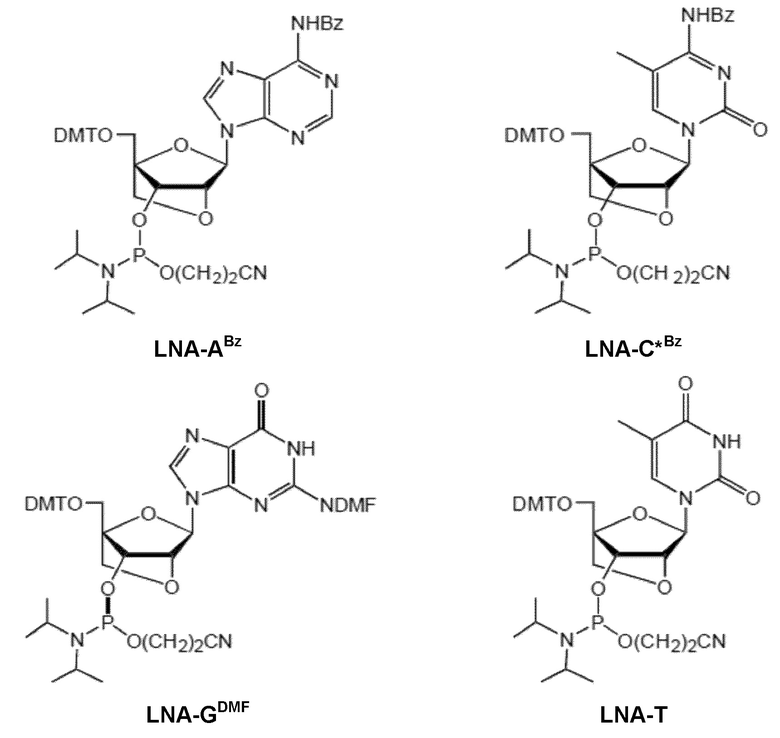
Строительные блоки LNA, как правило, обеспечиваются в виде фосфорамидитов LNA с четырьмя различными азотистыми основаниями: аденином (А), гуанином (G), 5-метилцитозином (С*) и тимином (Т). Антисмысловые олигонуклеотиды по настоящему изобретению, содержащие звенья LNA, синтезируют с использованием стандартной фосфорамидатной химии. В строительных блоках LNA азотистые основания защищены. Предпочтительной защитной группой для аминогруппы пуринового основания является бензоильная группа (BZ), обозначенная как ABZ. Предпочтительной защитной группой для аминогруппы 5-метилпиримидинового основания является бензоильная группа (BZ), обозначенная как C*Bz. Предпочтительной защитной группой для аминогруппы пуринового основания является диметилформамидин (DMF), диэтилформамидин (DEF), дипропилформамидин (DPF), дибутилформамидин (DBF) или изо-бутирил(-СОСН(СН3)2), обозначенные соответственно как GDMF, GDEF, GDPF, GDBF или GiBu. Таким образом, группа -NDMF относится к -N=CH-N(CH3)2. DMT относится к 4,4'-диметокситритилу.
Таким образом, LNA-Т относится к LNA 5'-O-(4,4'-диметокситритил)-3'-O(2-цианоэтил-N,N-диизопропил)фосфорамитид- тимидину. LNA С*Bz относится к LNA 5'-O-(4,4'-диметокситритил)-3'-O-(2-цианоэтил-N,N-диизопропил)фосфорамидит-4-N-бензоил-5-метил-2'-цитидину. LNA-ABZ относится к LNA 5'-O-(4,4'-диметокситритил)-3'-O-(2-цианоэтил-N,N-диизопропил)фосфорамидит-6-N-бензоил-2'-аденозину. LNA-GDMF относится к LNA 5'-O-(4,4'-диметокситритил)-3'-O-(2-цианоэтил-N,N-диизопропил)фосфорамидит-2-N-диметилформамидин-2'-гуанозину. LNA-GiBu относится к LNA 5'-O-(4,4'-диметокситритил)-3'-O-(2-цианоэтил-N,N-диизопропил)фосфор амидит-2-N-бутирил-2'-гуанозину.
Концевые группы
В случае, если Y представляет 5'-концевую группу антисмыслового олигонуклеотида по настоящему изобретению, то остаток Y, также обозначенный как Y5', представляет:
-OH, -O-C1-6-алкил, -S-C1-6-алкил, -O-C6-9-фенил, -O-C7-10-бензил, -NH-C1-6-алкил, -N(C1-6-алкил)2, -O-C2-6-алкенил, -S-C2-6-алкенил, -NH-C2-6-алкенил, -N(C2-6-алкенил)2, -O-C2-6-алкинил, -S-C2-6-алкинил, -NH-C2-6-алкинил, -N(C2-6-алкинил)2, -O-C1-6-алкилeнил-O-C1-6-алкил, -O-[C1-6-алкилeнил-O]m-C1-6-алкил, -O-CO-C1-6-алкил, -O-CO-C2-6-алкенил, -O-CO-C2-6-алкинил, -O-S(O)-C1-6-алкил, -O-SO2-C1-6-алкил, -O-SO2-O-C1-6-алкил, -O-P(O)(O-)2, -O-P(O)(O-)(O-C1-6-алкил), -O-P(O)(O-C1-6-алкил)2, -O-P(O)(S-)2, -O-P(O)(S-C1-6-алкил)2, -O-P(O)(S-)(O-C1-6-алкил), -O-P(O)(O-)(NH-C1-6-алкил), -O-P(O)(O-C1-6-алкил)(NH-C1-6-алкил), -O-P(O)(O-)[N(C1-6-алкил)2], -O-P(O)(O-C1-6-алкил)[N(C1-6-алкил)2], -O-P(O)(O-)(BH3-), -O-P(O)(O-C1-6-алкил)(BH3-), -O-P(O)(O-)(O-C1-6-алкилeнил-O-C1-6-алкил), -O-P(O)(O-C1-6-алкилeнил-O-C1-6-алкил)2, -O-P(O)(O-)(O-C1-6-алкилeнил-S-C1-6-алкил), -O-P(O)(O-C1-6-алкилeнил-S-C1-6-алкил)2, -O-P(O)(O-)(OCH2CH2O-C1-6-алкил), -O-P(O)(OCH2CH2O-C1-6-алкил)2, -O-P(O)(O-)(OCH2CH2S-C1-6-алкил), -O-P(O)(OCH2CH2S-C1-6-алкил)2, -O-P(O)(O-)OC3H6OH, -O-P(O)(S-)OC3H6OH, -O-P(S)(S-)OC3H6OH, где С1-6-алкил, С2-6-алкенил, С2-6-алкинил, -O-С6-9-фенил или -O-С7-10-бензил могут быть дополнительно замещены -F, -ОН, C1-4-алкилом, С2-4-алкенилом и/или С2-4-алкинилом, где m выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
Более предпочтительными являются: -OCH3, -OC2H5, -OC3H7, -O-цикло-C3H5, -OCH(CH3)2, -OC(CH3)3, -OC4H9, -OPh, -OCH2-Ph, -O-COCH3, -O-COC2H5, -O-COC3H7, -O-CO-цикло-C3H5, -O-COCH(CH3)2, -OCF3, -O-S(O)CH3, -O-S(O)C2H5, -O-S(O)C3H7, -O-S(O)-цикло-C3H5, -O-SO2CH3, -O-SO2C2H5, -O-SO2C3H7, -O-SO2-цикло-C3H5, -O-SO2-OCH3, -O-SO2-OC2H5, -O-SO2-OC3H7, -O-SO2-O-цикло-C3H5, -O(CH2)nN[(CH2)nOH], -O(CH2)nN[(CH2)n-H], -O-P(O)(O-)OC3H6OH, -O-P(O)(S-)OC3H6OH,
еще более предпочтительными являются:
-OCH3, -OC2H5, -OCH2CH2OCH3 (также известный как MOE), -OCH2CH2-N(CH3)2 (также известный как DMAOE), -O[(CH2)nO]mCH3, -O(CH2)nOCH3, -O(CH2)nNH2, -O(CH2)nN(CH3)2, -O-P(O)(O-)OC3H6OH, -O-P(O)(S-)OC3H6OH,
где n выбран из 1, 2, 3, 4, 5 или 6; и
где m выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
В случае, когда IL' представляет 3'-концевую группу антисмыслового олигонуклеотида по настоящему изобретению, то остаток IL', также обозначенный как IL3', представляет:
-OH, -O-C1-6-алкил, -S-C1-6-алкил, -O-C6-9-фенил, -O-C7-10-бензил, -NH-C1-6-алкил, -N(C1-6-алкил)2, -O-C2-6-алкенил, -S-C2-6-алкенил, -NH-C2-6-алкенил, -N(C2-6-алкенил)2, -O-C2-6-алкинил, -S-C2-6-алкинил, -NH-C2-6-алкинил, -N(C2-6-алкинил)2, -O-C1-6-алкилeнил-O-C1-6-алкил, -O-[C1-6-алкилeнил-O]m-C1-6-алкил, -O-CO-C1-6-алкил, -O-CO-C2-6-алкенил, -O-CO-C2-6-алкинил, -O-S(O)-C1-6-алкил, -O-SO2-C1-6-алкил, -O-SO2-O-C1-6-алкил, -O-P(O)(O-)2, -O-P(O)(O-)(O-C1-6-алкил), -O-P(O)(O-C1-6-алкил)2, -O-P(O)(S-)2, -O-P(O)(S-C1-6-алкил)2, -O-P(O)(S-)(O-C1-6-алкил), -O-P(O)(O-)(NH-C1-6-алкил), -O-P(O)(O-C1-6-алкил)(NH-C1-6-алкил), -O-P(O)(O-)[N(C1-6-алкил)2], -O-P(O)(O-C1-6-алкил)[N(C1-6-алкил)2], -O-P(O)(O-)(BH3-), -O-P(O)(O-C1-6-алкил)(BH3-), -O-P(O)(O-)(O-C1-6-алкилeнил-O-C1-6-алкил), -O-P(O)(O-C1-6-алкилeнил-O-C1-6-алкил)2, -O-P(O)(O-)(O-C1-6-алкилeнил-S-C1-6-алкил), -O-P(O)(O-C1-6-алкилeнил-S-C1-6-алкил)2, -O-P(O)(O-)(OCH2CH2O-C1-6-алкил), -O-P(O)(OCH2CH2O-C1-6-алкил)2, -O-P(O)(O-)(OCH2CH2S-C1-6-алкил), -O-P(O)(OCH2CH2S-C1-6-алкил)2, -O-P(O)(O-)OC3H6OH, -O-P(O)(S-)OC3H6OH, -O-P(S)(S-)OC3H6OH,
где С1-6-алкил, С2-6-алкенил, С2-6-алкинил, -O-С6-9-фенил или -O-С7-10-бензил могут быть дополнительно замещены -F, -ОН, C1-4-алкилом, С2-4-алкенилом и/или С2-4-алкинилом, где m выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
Более предпочтительными являются: -OCH3, -OC2H5, -OC3H7, -O-цикло-C3H5, -OCH(CH3)2, -OC(CH3)3, -OC4H9, -OPh, -OCH2-Ph, -O-COCH3, -O-COC2H5, -O-COC3H7, -O-CO-цикло-C3H5, -O-COCH(CH3)2, -OCF3, -O-S(O)CH3, -O-S(O)C2H5, -O-S(O)C3H7, -O-S(O)-цикло-C3H5, -O-SO2CH3, -O-SO2C2H5, -O-SO2C3H7, -O-SO2-цикло-C3H5, -O-SO2-OCH3, -O-SO2-OC2H5, -O-SO2-OC3H7, -O-SO2-O-цикло-C3H5, -O(CH2)nN[(CH2)nOH], -O(CH2)nN[(CH2)n-H], -O-P(O)(O-)OC3H6OH, -O-P(O)(S-)OC3H6OH,
еще более предпочтительными являются:
-OCH3, -OC2H5, -OCH2CH2OCH3 (также известный как MOE), -OCH2CH2-N(CH3)2 (также известный как DMAOE), -O[(CH2)nO]mCH3, -O(CH2)nOCH3, -O(CH2)nNH2, -O(CH2)nN(CH3)2, -O-P(O)(O-)OC3H6OH, -O-P(O)(S-)OC3H6OH,
где n выбран из 1, 2, 3, 4, 5 или 6; и
где m выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10.
Предпочтительные LNA
В предпочтительных вариантах осуществления звенья LNA, используемые в антисмысловых олигонуклеотидах по настоящему изобретению, предпочтительно, имеют структуру общей формулы (II):
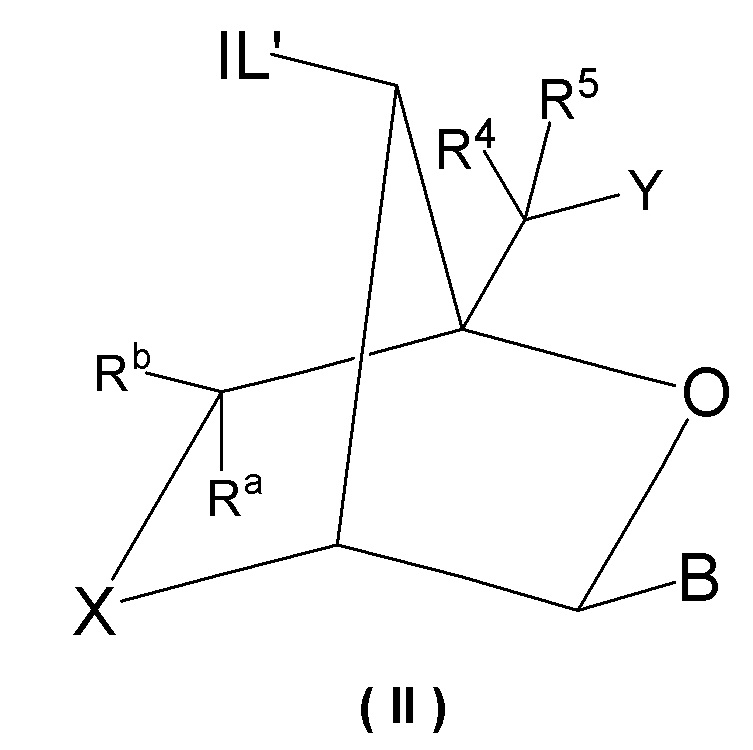
Группа -C(RaRb)-X- предпочтительно представляет -C(RaRb)-O-, -C(RaRb)-NRc-, -C(RaRb)-S- и -C(RaRb)-C(RaRb)-O-, где заместители Ra, Rb и Rc имеют значения, определенные здесь, и предпочтительно представляют C1-6-алкил и более предпочтительно С1-4-алкил. Более предпочтительно -C(RaRb)-X- выбран из -CH2-O-, -CH2-S-, -CH2-NH-, -CH2-N(CH3)-, -CH2-CH2-O- или -CH2-CH2-S-, и более предпочтительно из -CH2-O-, -CH2-S-, -CH2-CH2-O- или -CH2-CH2-S-, и еще более предпочтительно из -CH2-O-, -CH2-S- или -CH2-CH2-O-, и еще более предпочтительно из -CH2-O- или -CH2-S-, и наиболее предпочтительно представляет -CH2-O-.
Все хиральные центры и асимметричные заместители (если таковые имеются) могут находиться в R- или в S-ориентации. Например, два иллюстративных стереохимических изомера представляют бета-D- и альфа-L-изоформы, как показано ниже:
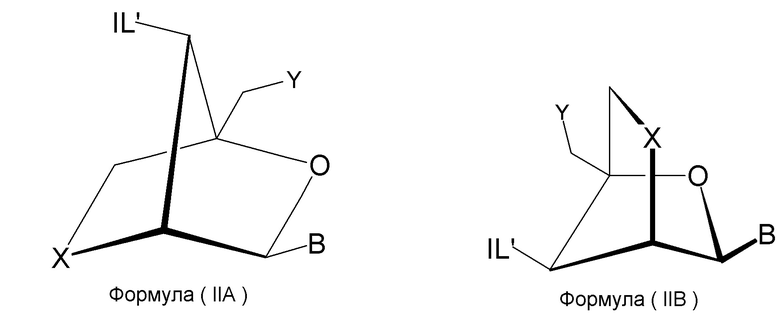
Предпочтительные звенья LNA выбраны из общих формул (b1)-(b9):
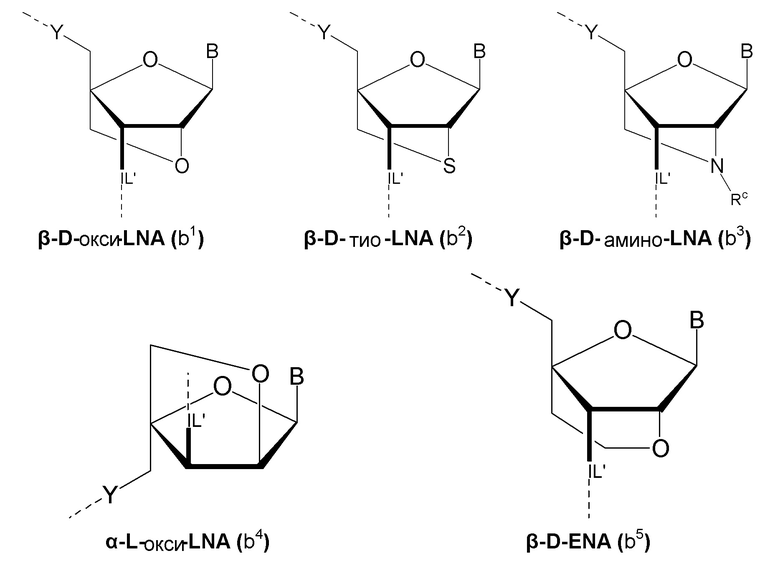
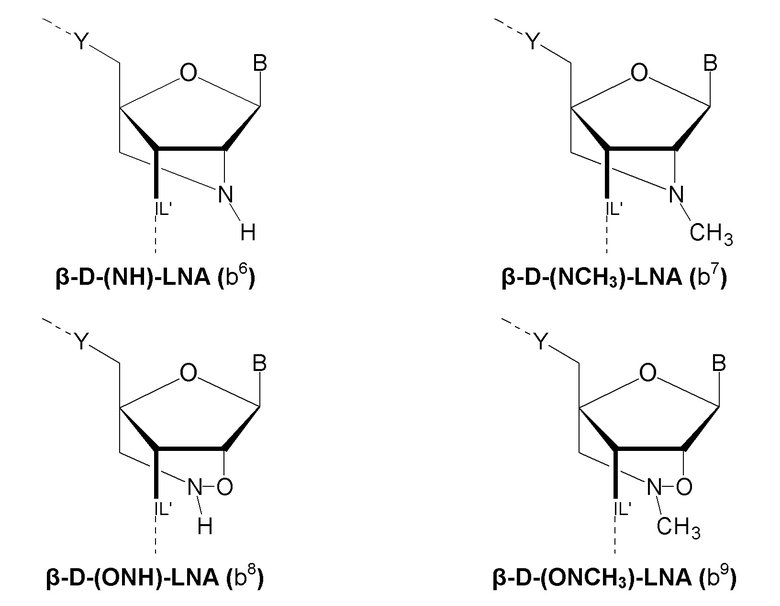
Термин «тио-LNA» включает замкнутый нуклеотид, где Х в общей формуле (II) выбран из -S- или -СН2-S-. Тио-LNA может находиться в бета-D- и альфа-L-конфигурации.
Термин «амино-LNA» включает замкнутый нуклеотид, где Х в общей формуле (II) выбран из -NH-, -N(R)-, -СН2-NH- и -СН2-N(R)-, где R выбран из атома водорода и С1-4-алкила. Амино-LNA может находиться в бета-D- и альфа-L-конфигурации.
Термин «окси-LNA» включает замкнутый нуклеотид, где Х в общей формуле (II) представляет -О-. «Окси-LNA» может находиться в бета-D- и альфа-L-конфигурации.
Термин «ЕNA» включает замкнутый нуклеотид, где Х в общей формуле (II) представляет -CH2-O- (где атом кислорода -СН2-O- присоединен к 2'-положению по отношению к основанию B). Ra и Rb независимо представляют атом водорода или метил.
В предпочтительных примерных вариантах осуществления LNA выбран из бета-D-окси-LNA, альфа-L-окси-LNA, бета-D-амино-LNA и бета-D-тио-LNA, в частности, бета-D-окси-LNA.
Еще более предпочтительными являются следующие антисмысловые олигонуклеотиды (таблица 1):
SP - стартовое положение или стартовый нуклеотид в SEQ ID NO:2
L - длина последовательности.
Антисмысловые олигонуклеотиды, раскрытые здесь, такие как антисмысловые олигонуклеотиды, представленные в таблицах 1-3, и в частности, антисмысловые олигонуклеотиды, представленные в таблицах 4-9, состоят из нуклеотидов, предпочтительно нуклеотидов ДНК, представляющих звенья, которые не являются LNA (также называемые здесь как нуклеотиды, которые не являются LNA), а также звеньев LNA (также называемых здесь как нуклеотиды LNA). Несмотря на то, что точно не указано, антисмысловые олигонуклеотиды с последовательностями SEQ ID NO: 102а-218а из таблицы 1 содержат 2-4 нуклеотида LNA (звена LNA) на 3'-конце и 2-4 нуклеотида LNA (звена LNA) на 5'-конце. Несмотря на то, что точно не указано, «С» в таблице 2, которые относятся к звеньям LNA, предпочтительно содержат 5-метилцитозин (С*) в качестве азотистого основания.
Это означает, несмотря на то, что точно не указано, антисмысловые олигонуклеотиды по настоящему изобретению или, как здесь раскрыто буквенным кодом А, С, G, Т и U, могут содержать любую межнуклеотидную связь, любую концевую группу и любое азотистое основание, описанные здесь. Кроме того, антисмысловые олигонуклеотиды по настоящему изобретению или раскрытые здесь, представляют гапмеры любой гапмерной структуры, как здесь описано, по меньшей мере, с одним звеном LNA на 3'-конце и, по меньшей мере, с одним звеном LNA на 5'-конце. Кроме того, любое звено LNA, как здесь раскрыто, может быть использовано в антисмысловых олигонуклеотидах по настоящему изобретению или раскрытых здесь. Так, например, антисмысловой олигонуклеотид GCTCGTCATAGACCGA (SEQ ID NO: 13) или CGATACGCGTCCACAG (SEQ ID NO: 14) или GTAGTGTTTAGGGAGC (SEQ ID NO: 15) или GCTATTTGGTAGTGTT (SEQ ID NO: 16) или CATGAATGGACCAGTA (SEQ ID NO: 17) или AGGCATTAATAAAGTG (SEQ ID NO: 18) содержит, по меньшей мере, одно звено LNA на 5'-конце и, по меньшей мере, одно звено LNA на 3'-конце, любое азотистое основание, любую 3'-концевую группу, любую 5'-концевую группу, любую гапмерную структуру и любую межнуклеотидную связь, как здесь описано, и также включает соли и оптические изомеры этого антисмыслового олигонуклеотида.
Использование звеньев LNA является предпочтительным, особенно на терминальном 3'-конце и терминальном 5'-конце. Таким образом, предпочтительно, если последние 1-5 нуклеотидов на терминальном 3'-конце и также последние 1-5 нуклеотидов на терминальном 5'-конце, в частности, последовательностей, раскрытых здесь, и в частности, последовательностей SEQ ID NO: 102а-218а в таблице 1, представляют звенья LNA (также называемые нуклеотидами LNA), в то время как между 1-5 звеньями LNA на 3'- и 5'-конце, находится 2-14, предпочтительно 3-12, более предпочтительно 4-10, более предпочтительно 5-9, еще более предпочтительно 6-8 звеньев, которые не являются LNA (также называющиеся нуклеотидами, которые не являются LNA). Такой вид антисмысловых олигонуклеотидов называют гапмерами, и они более подробно описаны ниже. Более предпочтительными являются 2-5 нуклеотидов LNA на 3'-конце и 2-5 нуклеотидов LNA на 5'-конце или 1-4 нуклеотида LNA на 3'-конце и 1-4 нуклеотида LNA на 5'-конце и еще более предпочтительно 2-4 нуклеотида LNA на 3'-конце и 2-4 нуклеотида LNA на 5'-конце антисмысловых олигонуклеотидов с числом предпочтительно 4-10, более предпочтительно от 5-9, еще более предпочтительно 6-8 звеньев, которые не являются LNA, между звеньями LNA на 3'- и 5'-конце.
Кроме того, в качестве межнуклеотидных связей между звеньями LNA и между звеньями LNA и звеньями, которые не являются LNA, предпочтительным является использование фосфоротиоатов или фосфородитиоатов и предпочтительно фосфоротиоатов.
Таким образом, предпочтительными являются антисмысловые олигонуклеотиды, где более чем 50%, предпочтительно более чем 60%, более предпочтительно более чем 70%, еще более предпочтительно более чем 80%, и наиболее предпочтительно более чем 90% межнуклеотидных связей представляют фосфоротиоатные или фосфатные, и более предпочтительно фосфоротиоатные связи, и где последние 1-4 или 2-5 нуклеотидов на 3'-конце представляют звенья LNA, и последние 1-4 или 2-5 нуклеотидов на 5'-конце представляют звенья LNA, и между звеньями LNA на концах последовательности находится 6-14 нуклеотидов, предпочтительно 7-12, предпочтительно 8-11, более предпочтительно 8-10 звеньев, которые не являются LNA, предпочтительно звенья ДНК. Кроме того, предпочтительно, чтобы эти антисмысловые олигонуклеотиды в форме гапмеров состояли в общей сложности из 12-20, предпочтительно 12-18 нуклеотидов.
Гапмеры
Антисмысловые олигонуклеотиды по изобретению могут состоять из нуклеотидных последовательностей, которые содержат нуклеотиды ДНК, представляющие звенья, которые не являются LNA, а также нуклеотиды LNA, и они могут располагаться в виде гапмера.
Таким образом, антисмысловые олигонуклеотиды по настоящему изобретению, предпочтительно представляют собой гапмеры. Гапмер состоит из средней части нуклеотидных звеньев ДНК, которые не являются замкнутыми, и которые, таким образом, не являются звеньями LNA. Нуклеотиды ДНК данной средней части могут быть связаны друг с другом межнуклеотидными связями (IL), описанными здесь, которые предпочтительно представляют фосфатные группы, фосфортиоатные группы или фосфородитиоатные группы, и которые могут содержать аналоги азотистых оснований, такие как 5-пропинилцитозин, 7-метилгуанин, 7-метиладенин, 2-аминоаденин, 2-тиотимин, 2-тиоцитозин или 5-метилцитозин. Такие звенья ДНК или нуклеотиды ДНК не являются бициклическими пентозными структурами. Средняя часть звеньев, которые не являются LNA, фланкирована на 3'-конце и 5'-конце последовательностями, состоящими из звеньев LNA. Таким образом, гапмеры имеют общую формулу:
LNA-последовательность 1 - последовательность, которая не является LNA - LNA-последовательность 2
или
участок A - участок B - участок C
Средняя часть антисмыслового олигонуклеотида, которая состоит из нуклеотидных звеньев ДНК, которые не являются звеньями LNA, когда образует дуплекс с комплементарной РНК-мишенью, способна к рекрутингу РНКазы. 3'- и 5'-концевые нуклеотидные звенья представляют звенья LNA, которые предпочтительно находятся в альфа-L-конфигурации, в частности, предпочтительными являются бета-D-окси-LNA и альфа-L-окси-LNA.
Таким образом, гапмер представляет собой антисмысловой олигонуклеотид, который содержит непрерывный участок нуклеотидов ДНК, который способен к рекрутингу РНКазы, такой как РНКаза H, такой как участок, по меньшей мере, из 6 или 7 нуклеотидов ДНК, которые не являются LNA, называемые здесь средней частью или участком B, где участок В фланкирован на 3'- и 5'-концах областями нуклеотидных аналогов с повышенной аффинностью, которые представляют звенья LNA, например, 1-6 звеньями LNA 5' и 3', к непрерывному участку нуклеотидов ДНК, способному к рекрутингу РНКазы - эти фланкирующие области относятся к участкам А и С соответственно.
Предпочтительно гапмер содержит (поли)нуклеотидную последовательность формулы (5'к 3') A-B-C или необязательно A-B-C-D или D-A-B-C, где; участок А (5'-область) состоит, по меньшей мере, из одного нуклеотидного аналога, например, по меньшей мере, одного звена LNA, например, 1-6 звеньев LNA, и участок В состоит, по меньшей мере, из пяти последовательных нуклеотидов ДНК, представляющих звенья, которые не являются LNA, и которые способны к рекрутингу РНКазы (при образовании дуплекса с комплементарной молекулой РНК, такой как мРНК-мишень), и участок С (3'-область) состоит, по меньшей мере, из одного нуклеотидного аналога, например, по меньшей мере, одного звена LNA, например, 1-6 звеньев LNA, и область D, если она присутствует, состоит из 1, 2 или 3 нуклеотидных звеньев ДНК, представляющих звенья, которые не являются LNA.
В некоторых вариантах осуществления участок А состоит из 1, 2, 3, 4, 5 или 6 звеньев LNA, например, 2-5 звеньев LNA, например, 3 или 4 звеньев LNA; и/или участок С состоит из 1, 2, 3, 4, 5 или 6 звеньев LNA, например, 2-5 звеньев LNA, например, 3 или 4 звеньев LNA.
В некоторых вариантах осуществления участок В состоит из 5, 6, 7, 8, 9, 10, 11 или 12 последовательных нуклеотидов ДНК, которые способны к рекрутингу РНКазы, или 6-10, или 7-9, например, 8 последовательных нуклеотидов, которые способны к рекрутингу РНКазы. В некоторых вариантах осуществления участок В состоит, по меньшей мере, из одного нуклеотидного звена ДНК, например, 1-12 нуклеотидных звеньев ДНК, предпочтительно 4-12 нуклеотидных звеньев ДНК, более предпочтительно 6-10 нуклеотидных звеньев ДНК, еще более предпочтительно, например, 7-10 нуклеотидных звеньев ДНК и наиболее предпочтительно, 8, 9 или 10 нуклеотидных звеньев ДНК, представляющих звенья, которые не являются LNA.
В некоторых вариантах осуществления участок А состоит из 3 или 4 LNA, участок В состоит из 7, 8, 9 или 10 нуклеотидных звеньев ДНК, и участок С состоит из 3 или 4 звеньев LNA. Такие конструкции включают (A-B-C): 1-7-2, 2-7-1, 2-7-2, 3-7-1, 3-7-2, 1-7-3, 2-7-3, 3-7-3, 2-7-4, 3-7-4, 4-7-2, 4-7-3, 4-7-4, 1-8-1, 1-8-2, 2-8-1, 2-8-2, 1-8-3, 3-8-1, 3-8-3, 2-8-3, 3-8-2, 4-8-1, 4-8-2, 1-8-4, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 1-9-1, 1-9-2, 2-9-1, 2-9-2, 2-9-3, 3-9-2, 3-9-3, 1-9-3, 3-9-1, 4-9-1, 1-9-4, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 1-10-1, 1-10-2, 2-10-1, 2-10-2, 1-10-3, 3-10-1, 2-10-2, 2-10-3, 3-10-2, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 1-11-1, 1-11-2, 2-11-1, 2-11-2, 1-11-3, 3-11-1, 2-11-2, 2-11-3, 3-11-2, 3-11-3, 2-11-4, 4-11-2, 3-11-4, 4-11-3, 4-11-4, и могут дополнительно включать участок D, который может содержать 1 или 2 нуклеотидных звена ДНК, которые не являются звеньями LNA.
Другие гапмерные конструкции раскрыты в международной заявке WO2004/046160A, которая включена здесь в качестве ссылки. Предварительная заявка на патент США 60/977409, включенная здесь в качестве ссылки, относится к короткомерным гапмерным антисмысловым олигонуклеотидам, которые также подходят для настоящего изобретения.
В некоторых вариантах осуществления антисмысловой олигонуклеотид состоит из непрерывной нуклеотидной последовательности в общей сложности из 10, 11, 12, 13 или 14 нуклеотидных звеньев (звеньев LNA и звеньев, которые не являются LNA вместе), где эта непрерывная нуклеотидная последовательность имеет формулу (5'-3'), А-В-С, или необязательно A-B-C-D или D-A-B-C, где участок А состоит из 1, 2 или 3 звеньев LNA, и участок В состоит из 7, 8 или 9 непрерывных нуклеотидных звеньев ДНК, которые не являются звеньями LNA, и которые способны к рекрутингу РНКазы при образовании дуплекса с комплементарной молекулой РНК (например, мРНК-мишенью), и участок С состоит из 1, 2 или 3 звеньев LNA. Когда он присутствует, то участок D состоит из одного нуклеотидного звена ДНК, которое не является LNA.
В некоторых вариантах осуществления участок А состоит из 1 звена LNA. В некоторых вариантах осуществления состоит участок А из 2 звеньев LNA. В некоторых вариантах осуществления участок С состоит из 3 звеньев LNA. В некоторых вариантах осуществления участок С состоит из 2 звеньев LNA. В некоторых вариантах осуществления участок С состоит из 3 звеньев LNA. В некоторых вариантах осуществления участок В состоит из 7 нуклеотидных звеньев ДНК, которые не являются LNA. В некоторых вариантах осуществления участок В состоит из 8 нуклеотидных звеньев ДНК. В некоторых вариантах осуществления участок В состоит из 9 нуклеотидных звеньев ДНК. В некоторых вариантах осуществления участок В состоит из 1-9 нуклеотидных звеньев ДНК, например, 2, 3, 4, 5, 6, 7 или 8 нуклеотидных звеньев ДНК. Нуклеотидные звенья ДНК всегда представляют звенья, которые не являются LNA. В некоторых вариантах осуществления участок В содержит 1, 2 или 3 звена LNA, которые предпочтительно находятся в альфа-L-конфигурации и которые более предпочтительно представляют звенья альфа-окси-L-LNA. В некоторых вариантах осуществления число нуклеотидов, присутствующих в A-B-C, выбрано из группы, состоящей из (звенья LNA-участок В-звенья LNA и более предпочтительно звенья альфа-L-окси-LNA (участок А)-участок В-(участок С) звенья альфа-L-окси-LNA): 1-8-1, 1-8-2, 2-8-1, 2-8-2, 1-8-3, 3-8-1, 3-8-3, 2-8-3, 3-8-2, 4-8-1, 4-8-2, 1-8-4, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 1-9-1, 1-9-2, 2-9-1, 2-9-2, 2-9-3, 3-9-2, 3-9-3, 1-9-3, 3-9-1, 4-9-1, 1-9-4, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 1-10-1, 1-10-2, 2-10-1, 2-10-2, 1-10-3, 3-10-1, 2-10-2, 2-10-3, 3-10-2, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 1-11-1, 1-11-2, 2-11-1, 2-11-2, 1-11-3, 3-11-1, 2-11-2, 2-11-3, 3-11-2, 3-11-3, 2-11-4, 4-11-2, 3-11-4, 4-11-3, 4-11-4. В других предпочтительных вариантах осуществления число нуклеотидов в A-B-C выбрано из группы, состоящей из: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3, и еще более предпочтительно представляет: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
Фосфоротиоатные, фосфатные или фосфородитиоатные и особенно фосфортиоатные межнуклеотидные связи также являются предпочтительными, в частности, для участка B гапмера. Фосфоротиоатные, фосфатные или фосфородитиоатные связи и особенно фосфоротиоатные межнуклеотидные связи также могут быть использованы для фланкирующих участков (А и С, и для связывания А или С с D, и в участке D, если он присутствует).
Однако участки A, В и С могут содержать межнуклеотидные связи, иные, чем фосфоротиоатные или фосфородитиоатные, такие как фосфодиэфирные связи, в частности, например, когда применение нуклеотидных аналогов защищает межнуклеотидные связи в участках А и С от разрушения под воздействием эндонуклеазы, когда участки А и С состоят из звеньев LNA.
Межнуклеотидные связи в антисмысловом олигонуклеотиде могут быть фосфодиэфирными, фосфоротиоатными, фосфордитиоатными или боранофосфатными таким образом, чтобы обеспечить расщепление РНК-мишени РНКазой Н. Фосфоротиоат или фосфородитиоат являются предпочтительными для повышения устойчивости к воздействию нуклеаз и по другим причинам, таким простота производства. В одном аспекте олигомера по изобретению, звенья LNA и/или звенья, которые не являются LNA, связаны друг с другом фосфоротиоатными группами.
Следует понимать, что включение фосфодиэфирных связей, например, одной или двух связей, иначе в антисмысловой олигонуклеотид с фосфортиоатными связями, в частности, между или рядом с звеньями LNA (как правило, в участке А и/или С) может изменить биодоступность и/или биологическое распределение антисмыслового олигонуклеотида (см. WO2008/053314A, которая включена здесь в качестве ссылки).
В некоторых вариантах осуществления, например, в последовательностях антисмысловых олигонуклеотидов, раскрытых здесь, и где это подходит и не указано конкретно, все остальные группы межнуклеотидных связей представляют фосфодиэфирные группы или фосфортиоатные группы, или их комбинации.
В некоторых вариантах осуществления все группы межнуклеотидных связей представляют фосфоротиоатные группы. При обращении к специфическим гапмерным последовательностям антисмысловых олигонуклеотидов, описанных здесь, следует понимать, что в различных вариантах осуществления, когда связи представляют фосфоротиоатные связи, то можно использовать альтернативные связи, такие как здесь описаны, например, фосфатные связи (также называемые фосфодиэфирными связями), в частности, для связей между нуклеотидными аналогами, такими как звенья LNA. Аналогично, при обращении к специфическим гапмерным последовательностям антисмысловых олигонуклеотидов, описанных здесь, когда остатки C представляют 5'-метил-модифицированный цитозин, в различных вариантах осуществления, то один или более C, присутствующих в олигомере, могут представлять немодифицированные остатки C.
Список условных обозначений
Как здесь используется, аббревиатуры b, d, s, ss имеют следующее значение:
нуклеотиды, выделенные жирным шрифтом, представляют нуклеотиды LNA
нуклеотиды, не выделенные жирным шрифтом, представляют нуклеотиды, которые не являются LNA.
Гапмерные последовательности
Следующие антисмысловые олигонуклеотиды в форме гапмеров, приведенные в таблицах 2-9 и более предпочтительно в таблицах 4-9, являются особенно предпочтительными.
Предпочтительные антисмысловые олигонуклеотиды
Далее раскрываются предпочтительные антисмысловые олигонуклеотиды по настоящему изобретению.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20, и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и на терминальном 3'-конце последовательности находится, по меньшей мере, 6, предпочтительно, 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N1-GTCATAGA-N2-3' (SEQ. ID NO: 12) или 5'-N3-ACGCGTCC-N4-3' (SEQ. ID NO: 98) или 5'-N11-TGTTTAGG-N12-3' (SEQ. ID NO: 10) или 5'-N5-TTTGGTAG-N6-3' (SEQ. ID NO: 11) или 5'-N7-AATGGACC-N8-3' (SEQ. ID NO: 100) или 5'-N9-ATTAATAA-N10-3' (SEQ. ID NO: 101), где:
N1 представляет: CATGGCAGACCCCGCTGCTC-, ATGGCAGACCCCGCTGCTC-, TGGCAGACCCCGCTGCTC-, GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA, -CCGAGCCCCCAGCGCAG, -CCGAGCCCCCAGCGCAGC, -CCGAGCCCCCAGCGCAGCG или -CCGAGCCCCCAGCGCAGCGG;
N3 представляет: GGTGGGATCGTGCTGGCGAT-, GTGGGATCGTGCTGGCGAT-, TGGGATCGTGCTGGCGAT-, GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACGATGTGCAGCGGC, -ACAGGACGATGTGCAGCGG, -ACAGGACGATGTGCAGCG, -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A;
N5 представляет: GCCCAGCCTGCCCCAGAAGAGCTA-, CCCAGCCTGCCCCAGAAGAGCTA-, CCAGCCTGCCCCAGAAGAGCTA-, CAGCCTGCCCCAGAAGAGCTA-, AGCCTGCCCCAGAAGAGCTA-, GCCTGCCCCAGAAGAGCTA-, CCTGCCCCAGAAGAGCTA-, CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-;
N6 представляет: -TGTTTAGGGAGCCGTCTTCAGGAA, -TGTTTAGGGAGCCGTCTTCAGGA, -TGTTTAGGGAGCCGTCTTCAGG, -TGTTTAGGGAGCCGTCTTCAG, -TGTTTAGGGAGCCGTCTTCA, -TGTTTAGGGAGCCGTCTTC, -TGTTTAGGGAGCCGTCTT, -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T;
N7 представляет: TGAATCTTGAATATCTCATG-, GAATCTTGAATATCTCATG-, AATCTTGAATATCTCATG-, ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-;
N8 представляет: -AGTATTCTAGAAACTCACCA, -AGTATTCTAGAAACTCACC, -AGTATTCTAGAAACTCAC, -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A;
N9 представляет: ATTCATATTTATATACAGGC-, TTCATATTTATATACAGGC-, TCATATTTATATACAGGC-, CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-;
N10 представляет: -AGTGCAAATGTTATTGGCTA, -AGTGCAAATGTTATTGGCT, -AGTGCAAATGTTATTGGC, -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A;
N11 представляет: TGCCCCAGAAGAGCTATTTGGTAG-, GCCCCAGAAGAGCTATTTGGTAG-, CCCCAGAAGAGCTATTTGGTAG-, CCCAGAAGAGCTATTTGGTAG-, CCAGAAGAGCTATTTGGTAG-, CAGAAGAGCTATTTGGTAG-, AGAAGAGCTATTTGGTAG-, GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-;
N12 представляет: -GAGCCGTCTTCAGGAATCTTCTCC,
-GAGCCGTCTTCAGGAATCTTCTC, -GAGCCGTCTTCAGGAATCTTCT, -GAGCCGTCTTCAGGAATCTTC, -GAGCCGTCTTCAGGAATCTT, -GAGCCGTCTTCAGGAATCT, -GAGCCGTCTTCAGGAATC, -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G;
или где N1-N12 представляют любой из ограниченных списков остатков, раскрытых здесь,
и солям и оптическим изомерам антисмыслового олигонуклеотида.
Кроме того, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20, и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и на терминальном 3'-конце последовательности находится, по меньшей мере, 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N1-GTCATAGA-N2-3' (SEQ. ID NO: 12), где:
N1 представляет: GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA или -CCGAGCCCCCAGCGCAG;
и солям и опти ческим изомерам антисмыслового олигонуклеотида.
N1 и/или N2 также могут представлять любой из дополнительных ограниченных списков 3'- и 5'-остатков, раскрытых здесь.
Особенно предпочтительными гапмерными антисмысловыми олигонуклеотидами, подпадающими под общую формулу S1:
являются следующие:
Антисмысловые олигонуклеотиды формулы S1 в форме гапмеров (LNA-сегмент 1-сегмент ДНК -LNA-сегмент 2) содержат сегмент LNA на терминальном 5'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и содержат сегмент LNA на терминальном 3'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и между двумя сегментами LNA один сегмент ДНК, состоящий из 6-14, предпочтительно 7-12 и более предпочтительно 8-11 звеньев ДНК.
Антисмысловые олигонуклеотиды формулы S1 содержат нуклеотиды LNA (звенья LNA), описанные здесь, в частности, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и предпочтительно раскрытые в разделе «Предпочтительные LNA». Звенья LNA и звенья ДНК могут содержать стандартные азотистые основания, такие как аденин (А), цитозин (С), гуанин (G), тимин (Т) и урацил (U), но могут также содержать модифицированные азотистые основания, описанные в разделе «Азотистые основания». Антисмысловые олигонуклеотиды формулы S1 или сегменты LNA и сегмент ДНК антисмыслового олигонуклеотида могут содержать любую межнуклеотидную связь, описанную здесь, и в частности, таковые, раскрытые в разделе «Межнуклеотидные связи (IL)». Антисмысловые олигонуклеотиды формулы S1 необязательно могут также содержать концевые группы в терминальном 3'-конце и/или терминальном 5'-конце и в частности, таковые, раскрытые в разделе «Концевые группы».
Результаты экспериментов показали, что модифицированные азотистые основания незначительно увеличивают или изменяют активность антисмысловых олигонуклеотидов по изобретению в отношении тестированных неврологических и онкологических заболеваний. Было показано, что модифицированные азотистые основания 5-метилцитозин или 2-аминоаденин дополнительно повышают активность антисмысловых олигонуклеотидов формулы S1, особенно если 5-метилцитозин используется только в нуклеотидах LNA или в нуклеотидах LNA и в нуклеотидах ДНК, и/или, если 2-аминоаденин используется в нуклеотидах ДНК и не в нуклеотидах LNA.
Предпочтительная гапмерная структура антисмысловых олигонуклеотидов формулы S1 является следующей: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3 и еще более предпочтительной: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
В качестве звеньев LNA для антисмысловых олигонуклеотидов формулы S1, в частности, β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9) являются предпочтительными. Результаты экспериментов показали, что все эти звенья LNA b1, b2, b4, b5, b6, b7, b8 и b9 можно синтезировать с требуемым усилием и обеспечить антисмысловые олигонуклеотиды с сопоставимой стабильностью и активностью. Однако, основываясь на результатах экспериментов, звенья LNA b1, b2, b4, b5, b6 и b7 являются более предпочтительными. Еще более предпочтительными являются звенья LNA b1, b2, b4, b6 и b7, и еще более предпочтительными являются звенья LNA b1 и b4, и наиболее предпочтительным также с учетом сложности химического синтеза является β-D-окси-LNA (b1).
К настоящему времени не было найдено никакой конкретной 3'-концевой группы или 5'-концевой группы, которая заметно изменяла или повышала стабильность или активность в отношении онкологических или неврологических заболеваний, таким образом, 3'- и 5'-концевые группы возможны, но явно не являются предпочтительными.
Возможны различные межнуклеотидные мостики или межнуклеотидные связи. В приведенных здесь формулах межнуклеотидная связь IL представлена -IL'-Y-. Таким образом, IL=-IL'-Y-=-X''-P(=X')(X-)-Y-, где IL предпочтительно выбрана из группы, состоящей из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-. Предпочтительными являются межнуклеотидные связи IL, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(OCH2CH2OCH3)-O-, и более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, и еще более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, и наиболее предпочтительные, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на 5'-конце и 1-5 нуклеотидов на 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью
где:
N1 представляет: GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-,CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA или -CCGAGCCCCCAGCGCAG; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9); и предпочтительно из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и
межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-;
и предпочтительно из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Более предпочтительно N1 представляет: CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-; и
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC, -CCGAGCCCCC, -CCGAGCCCCCA, -CCGAGCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG или -CCGAGCCCCCAGCGC.
Еще более предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и более предпочтительно 13-19 или 14-18 нуклеотидов, и 2-5 из этих нуклеотидов на терминальном 5'-конце и 2-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 7, предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N1 представляет: ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-; предпочтительно N1 представляет: CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-; и
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC или -CCGAGCCCCC, -CCGAGCCCCCA или -CCGAGCCCCCAG; предпочтительно N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGC, -CCGAGCC, -CCGAGCCC, -CCGAGCCCC или -CCGAGCCCCC;
и нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-; и предпочтительно выбраны из фосфата, фосфоротиоата и фосфородитиоата;
и солям и опти ческим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды последовательностей SEQ ID NO: 19-SEQ ID NO: 53, содержащие сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 3'-конце, и сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 5'-конце, и сегмент, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 звеньев ДНК между двумя сегментами звеньев LNA, где звенья LNA выбраны из: β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из фосфата, фосфоротиоата и фосфородитиоата. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин в звеньях LNA, предпочтительно всех звеньях LNA и/или 2-аминоаденин в некоторых или всех звеньях ДНК, и/или 5-метилцитозин в некоторых или всех звеньях ДНК.
Также особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды из таблицы 4 (SEQ ID NO: 232a-244b).
Кроме того, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью
где:
N3 представляет: GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-;
N4 представляет: -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A;
и солям и оптически изомерам антисмыслового олигонуклеотида.
N3 и/или N4 могут также представлять любой из дополнительных ограниченных списков 3'- и 5'-остатков, раскрытых здесь.
Особенно предпочтительными гапмерными антисмысловыми олигонуклеотидами, подпадающими под общую формулу S2:
являются следующие:
Антисмысловые олигонуклеотиды формулы S2 в форме гапмеров (LNA-сегмент 1-сегмент ДНК-LNA-сегмент 2) содержат сегмент LNA на терминальном 5'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и содержат сегмент LNA на терминальном 3'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и между двумя сегментами LNA один сегмент ДНК, состоящий из 6-14, предпочтительно 7-12 и более предпочтительно 8-11 звеньев ДНК.
Антисмысловые олигонуклеотиды формулы S2 содержат нуклеотиды LNA (звенья LNA), описанные здесь, в частности, таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и, предпочтительно таковые, раскрытые в разделе «Предпочтительные LNA». Звенья LNA и звенья ДНК могут содержать обычные азотистые основания, такие как аденин (А), цитозин (С), гуанин (G), тимин (Т) и урацил (U), но могут также содержать модифицированные азотистые основания, как описано в разделе «Азотистые основания». Антисмысловые олигонуклеотиды формулы S2 или сегменты LNA и сегмент ДНК антисмыслового олигонуклеотида могут содержать любую межнуклеотидную связь, описанную здесь, и в частности, таковые, раскрытые в разделе «Межнуклеотидные связи (IL)». Антисмысловые олигонуклеотиды формулы S2 необязательно могут также содержать концевые группы в терминальном 3'-конце и/или терминальном 5'-конце и в частности, таковые, раскрытые в разделе «Концевые группы».
Результаты экспериментов показали, что модифицированные азотистые основания незначительно увеличивают или изменяют активность антисмысловых олигонуклеотидов по изобретению в отношении тестированных неврологических и онкологических заболеваний. Было показано, что модифицированные азотистые основания 5-метилцитозин или 2-аминоаденин дополнительно повышают активность антисмысловых олигонуклеотидов формулы S1, особенно если 5-метилцитозин используется только в нуклеотидах LNA или в нуклеотидах LNA и в нуклеотидах ДНК и/или, если 2-аминоаденин используется в нуклеотидах ДНК и не в нуклеотидах LNA.
Предпочтительная гапмерная структура антисмысловых олигонуклеотидов формулы S2 является следующей: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3 и еще более предпочтительной: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
В качестве звеньев LNA для антисмысловых олигонуклеотидов формулы S2, в частности, β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9) являются предпочтительными. Результаты экспериментов показали, что все эти звенья LNA b1, b2, b4, b5, b6, b7, b8 и b9 можно синтезировать с требуемым усилием и обеспечить антисмысловые олигонуклеотиды с сопоставимой стабильностью и активностью. Однако, основываясь на экспериментах, звенья LNA b1, b2, b4, b5, b6 и b7 являются более предпочтительными. Еще более предпочтительными являются звенья LNA b1, b2, b4, b6 и b7, и еще более предпочтительными являются звенья LNA b1 и b4, и наиболее предпочтительным также в отношении сложности химического синтеза является β-D-окси-LNA (b1).
К настоящему времени не было найдено никакой конкретной 3'-концевой группы или 5'-концевой группы, которая заметно изменяла или повышала стабильность или активность в отношении онкологических или неврологических заболеваний, но явно не являются предпочтительными.
Возможны различные межнуклеотидные мостики или межнуклеотидные связи. В приведенных здесь формулах межнуклеотидная связь IL представлена -IL'-Y-. Таким образом, IL=-IL'-Y-=-X''-P(=X')(X-)-Y-, где IL предпочтительно выбрана из группы, состоящей из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-. Предпочтительными являются межнуклеотидные связи IL, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(OCH2CH2OCH3)-O-, и более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, и еще более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, и наиболее предпочтительные, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N3 представляет: GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-; и
N4 представляет: -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A, и
и нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9); и предпочтительно из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и
межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-;
и предпочтительно из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Более предпочтительно N3 представляет: GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-; и
N4 представляет: -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A.
Еще более предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и более предпочтительно 13-19 или 14-18 нуклеотидов, и 2-5 из этих нуклеотидов на терминальном 5'-конце и 2-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 7, предпочтительно, по меньшей мере, 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N3-ACGCGTCC-N4-3' (SEQ ID NO: 98), где
N3 представляет: CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-; предпочтительно N3 представляет: TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или T-; и
N4 представляет: -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A; предпочтительно N4 представляет: -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A;
и нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-; и предпочтительно выбраны из фосфата, фосфоротиоата и фосфородитиоата;
и солям и опти ческим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды последовательностей SEQ ID NO: 54-SEQ ID NO: 68 и SEQ ID NO: 349-SEQ ID NO: 368, содержащие сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 3'-конце, и сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 5'-конце, и сегмент, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 звеньев ДНК между двумя сегментами из звеньев LNA, где звенья LNA выбраны из: β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из фосфата, фосфоротиоата и фосфородитиоата. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин в звеньях LNA, предпочтительно всех звеньях LNA и/или 2-аминоаденин в некоторых или всех звеньях ДНК, и/или 5-метилцитозин в некоторых или всех звеньях ДНК.
Также особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды из таблицы 5 (SEQ ID NO: 245a-257b).
Кроме того, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью 5'-N11-TGTTTAGG-N12-3' (SEQ ID NO: 10), где:
N11 представляет: GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-;
N12 представляет: -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
N11 и/или N12 могут также представлять любой из дополнительных ограниченных списков 3'- и 5'-остатков, как здесь раскрыто.
Особенно предпочтительными гапмерными антисмысловыми олигонуклеотидами, подпадающими под общую формулу S3:
являются следующие:
Антисмысловые олигонуклеотиды формулы S3 в форме гапмеров (LNA-сегмент 1-сегмент ДНК-LNA-сегмент 2) содержат сегмент LNA на терминальном 5'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и содержат сегмент LNA на терминальном 3'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и между двумя сегментами LNA один сегмент ДНК, состоящий из 6-14, предпочтительно 7-12 и более предпочтительно 8-11 звеньев ДНК.
Антисмысловые олигонуклеотиды формулы S3 содержат нуклеотиды LNA (звенья LNA), описанные здесь, в частности, таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и, предпочтительно таковые, раскрытые в разделе «Предпочтительные LNA». Звенья LNA и звенья ДНК могут содержать обычные азотистые основания, такие как аденин (А), цитозин (С), гуанин (G), тимин (Т) и урацил (U), но могут также содержать модифицированные азотистые основания, как описано в разделе «Азотистые основания». Антисмысловые олигонуклеотиды формулы S3 или сегменты LNA и сегмент ДНК антисмыслового олигонуклеотида могут содержать любую межнуклеотидную связь, описанную здесь, и в частности, таковые, раскрытые в разделе «Межнуклеотидные связи (IL)». Антисмысловые олигонуклеотиды формулы S3 необязательно могут также содержать концевые группы на терминальном 3'-конце и/или терминальном 5'-конце и в частности, таковые, раскрытые в разделе «Концевые группы».
Результаты экспериментов показали, что модифицированные азотистые основания незначительно увеличивают или изменяют активность антисмысловых олигонуклеотидов по изобретению в отношении тестированных неврологических и онкологических заболеваний. Было показано, что модифицированные азотистые основания 5-метилцитозин или 2-аминоаденин дополнительно повышают активность антисмысловых олигонуклеотидов формулы S3, особенно если 5-метилцитозин используется только в нуклеотидах LNA или в нуклеотидах LNA и в нуклеотидах ДНК и/или, если 2-аминоаденин используется в нуклеотидах ДНК и не в нуклеотидах LNA.
Предпочтительная гапмерная структура антисмысловых олигонуклеотидов формулы S2 является следующей: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3, и еще более предпочтительная: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
В качестве звеньев LNA для антисмысловых олигонуклеотидов формулы S3, в частности, β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9) являются предпочтительными. Результаты экспериментов показали, что все эти звенья LNA b1, b2, b4, b5, b6, b7, b8 и b9 можно синтезировать с требуемым усилием и обеспечить антисмысловые олигонуклеотиды с сопоставимой стабильностью и активностью. Однако, основываясь на экспериментах, звенья LNA b1, b2, b4, b5, b6 и b7 являются более предпочтительными. Еще более предпочтительными являются звенья LNA b1, b2, b4, b6 и b7, и еще более предпочтительными являются звенья LNA b1 и b4, и наиболее предпочтительным также с учетом сложности химического синтеза является β-D-окси-LNA (b1).
К настоящему времени не было найдено никакой конкретной 3'-концевой группы или 5'-концевой группы, которая заметно изменяла или повышала стабильность или активность в отношении онкологических или неврологических заболеваний, таким образом, 3'- и 5'-концевые группы возможны, но явно не являются предпочтительными.
Возможны различные межнуклеотидные мостики или межнуклеотидные связи. В формулах, раскрытых здесь, межнуклеотидная связь IL представлена -IL'-Y-. Таким образом, IL=-IL'-Y-=-X''-P(=X')(X-)-Y-, где IL предпочтительно выбрана из группы, состоящей из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-. Предпочтительными являются межнуклеотидные связи IL, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(OCH2CH2OCH3)-O-, и более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, и еще более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, и наиболее предпочтительные, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N11 представляет: GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9); и предпочтительно из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и
межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-;
и предпочтительно из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Более предпочтительно N11 представляет: AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTCTTCAGGA, -GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G.
Еще более предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и более предпочтительно 13-19 или 14-18 нуклеотидов, и 2-5 из этих нуклеотидов на терминальном 5'-конце и 2-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 7, предпочтительно, по меньшей мере, 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N11 представляет: GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; предпочтительно N11 представляет: TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-; и
N12 представляет: -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G; предпочтительно N12 представляет: -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT, -GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-; и предпочтительно выбраны из фосфата, фосфоротиоата и фосфородитиоата;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды последовательностей SEQ ID NO: 369-SEQ ID NO: 403, содержащие сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 3'-конце, и сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 5'-конце, и сегмент, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 звеньев ДНК между двумя сегментами звеньев LNA, где звенья LNA выбраны из: β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из фосфата, фосфоротиоата и фосфородитиоата. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин в звеньях LNA, предпочтительно всех звеньях LNA и/или 2-аминоаденин в некоторых или всех звеньях ДНК, и/или 5-метилцитозин в некоторых или всех звеньях ДНК.
Также особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды из таблицы 6 (SEQ ID NO: 258a-270b).
Кроме того, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N5 представляет: CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-,CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-;
N6 представляет: -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
N5 и/или N6 могут также представлять любой из дополнительных ограниченных списков 3'- и 5'-остатков, как здесь раскрыто.
Особенно предпочтительными гапмерными антисмысловыми олигонуклеотидами, подпадающими под общую формулу S4:
являются следующие:
Антисмысловые олигонуклеотиды формулы S4 в форме гапмеров (LNA-сегмент 1-сегмент ДНК-LNA-сегмент 2) содержат сегмент LNA на терминальном 5'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и содержат сегмент LNA на терминальном 3'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и между двумя сегментами LNA один сегмент ДНК, состоящий из 6-14, предпочтительно 7-12 и более предпочтительно 8-11 звеньев ДНК.
Антисмысловые олигонуклеотиды формулы S4 содержат нуклеотиды LNA (звенья LNA), как здесь описано, в частности, таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и, предпочтительно таковые, раскрытые в разделе «Предпочтительные LNA». Звенья LNA и звенья ДНК могут содержать обычные азотистые основания, такие как аденин (А), цитозин (С), гуанин (G), тимин (Т) и урацил (U), но могут также содержать модифицированные азотистые основания, как описано в разделе «Азотистые основания». Антисмысловые олигонуклеотиды формулы S4 или сегменты LNA и сегмент ДНК антисмыслового олигонуклеотида могут содержать любую межнуклеотидную связь, как здесь описано, и в частности, таковые, раскрытые в разделе «Межнуклеотидные связи (IL)». Антисмысловые олигонуклеотиды формулы S4 необязательно могут также содержать концевые группы на терминальном 3'-конце и/или терминальном 5'-конце и в частности, таковые, раскрытые в разделе «Концевые группы».
Результаты экспериментов показали, что модифицированные азотистые основания незначительно увеличивают или изменяют активность антисмысловых олигонуклеотидов по изобретению в отношении тестированных неврологических и онкологических заболеваний. Было показано, что модифицированные азотистые основания 5-метилцитозин или 2-аминоаденин дополнительно повышают активность антисмысловых олигонуклеотидов формулы S4, особенно если 5-метилцитозин используется только в нуклеотидах LNA или в нуклеотидах LNA и в нуклеотидах ДНК и/или, если 2-аминоаденин используется в нуклеотидах ДНК и не в нуклеотидах LNA.
Предпочтительная гапмерная структура антисмысловых олигонуклеотидов формулы S4 является следующей: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3 и еще более предпочтительной: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
В качестве звеньев LNA для антисмысловых олигонуклеотидов формулы S4, в частности, β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9) являются предпочтительными. Результаты экспериментов показали, что все эти звенья LNA b1, b2, b4, b5, b6, b7, b8 и b9 можно синтезировать с требуемым усилием и обеспечить антисмысловые олигонуклеотиды с сопоставимой стабильностью и активностью. Однако, основываясь на экспериментах, звенья LNA b1, b2, b4, b5, b6 и b7 являются более предпочтительными. Еще более предпочтительными являются звенья LNA b1, b2, b4, b6 и b7, и еще более предпочтительными являются звенья LNA b1 и b4, и наиболее предпочтительным также с учетом сложности химического синтеза является β-D-окси-LNA (b1).
К настоящему времени не было найдено никакой конкретной 3'-концевой группы или 5'-концевой группы, которая заметно изменяла или повышала стабильность или активность в отношении онкологических или неврологических заболеваний, таким образом, 3'- и 5'-концевые группы возможны, но явно не являются предпочтительными.
Возможны различные межнуклеотидные мостики или межнуклеотидные связи. В приведенных здесь формулах межнуклеотидная связь IL представлена -IL'-Y-. Таким образом, IL=-IL'-Y-=-X''-P(=X')(X-)-Y-, где IL предпочтительно выбрана из группы, состоящей из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-. Предпочтительными являются межнуклеотидные связи IL, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(OCH2CH2OCH3)-O-, и более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, и еще более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, и наиболее предпочтительные, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N5 представляет: CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 представляет: -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9); и предпочтительно из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и
межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-;
и предпочтительно из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Более предпочтительно N5 представляет: GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 представляет: -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T.
Еще более предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и более предпочтительно 13-19 или 14-18 нуклеотидов, и 2-5 из этих нуклеотидов на терминальном 5'-конце и 2-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 7, предпочтительно, по меньшей мере, 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N5 представляет: CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; предпочтительно N5 представляет: AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-; и
N6 представляет: -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG, or -T; предпочтительно N6 представляет: -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-; и предпочтительно выбраны из фосфата, фосфоротиоата и фосфородитиоата;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды последовательностей SEQ ID NO: 404-SEQ ID NO: 438, содержащие сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 3'-конце, и сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 5'-конце, и сегмент, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 звеньев ДНК между двумя сегментами звеньев LNA, где звенья LNA выбраны из: β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из фосфата, фосфоротиоата и фосфородитиоата. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин в звеньях LNA, предпочтительно всех звеньях LNA и/или 2-аминоаденин в некоторых или всех звеньях ДНК, и/или 5-метилцитозин в некоторых или всех звеньях ДНК.
Также особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды из таблицы 7 (SEQ ID NO: 271a-283b).
Кроме того, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N7 представляет: ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-,AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-;
N8 представляет: -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
N7 и/или N8 могут также представлять любой из дополнительных ограниченных списков 3'- и 5'-остатков, как здесь раскрыто.
Особенно предпочтительными гапмерными антисмысловыми олигонуклеотидами, подпадающими под общую формулу S6:
являются следующие:
Антисмысловые олигонуклеотиды формулы S6 в форме гапмеров (LNA-сегмент 1-сегмент ДНК-LNA-сегмент 2) содержат сегмент LNA на терминальном 5'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и содержат сегмент LNA на терминальном 3'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и между двумя сегментами LNA один сегмент ДНК, состоящий из 6-14, предпочтительно 7-12 и более предпочтительно 8-11 звеньев ДНК.
Антисмысловые олигонуклеотиды формулы S6 содержат нуклеотиды LNA (звенья LNA), как здесь описано, в частности, таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и, предпочтительно таковые, раскрытые в разделе «Предпочтительные LNA». Звенья LNA и звенья ДНК могут содержать обычные азотистые основания, такие как аденин (А), цитозин (С), гуанин (G), тимин (Т) и урацил (U), но могут также содержать модифицированные азотистые основания, как описано в разделе «Азотистые основания». Антисмысловые олигонуклеотиды формулы S6 или сегменты LNA и сегмент ДНК антисмыслового олигонуклеотида могут содержать любую межнуклеотидную связь, как здесь описано, и в частности, таковые, раскрытые в разделе «Межнуклеотидные связи (IL)». Антисмысловые олигонуклеотиды формулы S6 необязательно могут также содержать концевые группы на терминальном 3'-конце и/или терминальном 5'-конце и в частности, таковые, раскрытые в разделе «Концевые группы».
Результаты экспериментов показали, что модифицированные азотистые основания незначительно увеличивают или изменяют активность антисмысловых олигонуклеотидов по изобретению в отношении тестированных неврологических и онкологических заболеваний. Было показано, что модифицированные азотистые основания 5-метилцитозин или 2-аминоаденин дополнительно повышают активность антисмысловых олигонуклеотидов формулы S6, особенно если 5-метилцитозин используется только в нуклеотидах LNA или в нуклеотидах LNA и в нуклеотидах ДНК и/или, если 2-аминоаденин используется в нуклеотидах ДНК и не в нуклеотидах LNA.
Предпочтительная гапмерная структура антисмысловых олигонуклеотидов формулы S6 является следующей: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3 и еще более предпочтительной: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
В качестве звеньев LNA для антисмысловых олигонуклеотидов формулы S6, в частности, β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9) являются предпочтительными. Результаты экспериментов показали, что все эти звенья LNA b1, b2, b4, b5, b6, b7, b8 и b9 можно синтезировать с требуемым усилием и обеспечить антисмысловые олигонуклеотиды с сопоставимой стабильностью и активностью. Однако, основываясь на экспериментах, звенья LNA b1, b2, b4, b5, b6 и b7 являются более предпочтительными. Еще более предпочтительными являются звенья LNA b1, b2, b4, b6 и b7, и еще более предпочтительными являются звенья LNA b1 и b4, и наиболее предпочтительным также в отношении сложности химического синтеза является β-D-окси-LNA (b1).
К настоящему времени не было найдено никакой конкретной 3'-концевой группы или 5'-концевой группы, которая заметно изменяла или повышала стабильность или активность в отношении онкологических или неврологических заболеваний, таким образом, 3'- и 5'-концевые группы возможны, но явно не являются предпочтительными.
Возможны различные межнуклеотидные мостики или межнуклеотидные связи. В приведенных здесь формулах межнуклеотидная связь IL представлена -IL'-Y-. Таким образом, IL=-IL'-Y-=-X''-P(=X')(X-)-Y-, где IL предпочтительно выбрана из группы, состоящей из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-. Предпочтительными являются межнуклеотидные связи IL, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(OCH2CH2OCH3)-O-, и более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, и еще более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, и наиболее предпочтительные, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N7 представляет: ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-,AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 представляет: -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A; и
и нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9); и предпочтительно из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и
межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-;
и предпочтительно из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Более предпочтительно N7 представляет: CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 представляет: -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A.
Еще более предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и более предпочтительно 13-19 или 14-18 нуклеотидов, и 2-5 из этих нуклеотидов на терминальном 5'-конце и 2-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 7, предпочтительно, по меньшей мере, 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N7 представляет: GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; предпочтительно N7 представляет: ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 представляет: -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A; предпочтительно N8 представляет: -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-; и предпочтительно выбраны из фосфата, фосфоротиоата и фосфородитиоата;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды последовательностей SEQ ID NO: 439-SEQ ID NO: 473, содержащие сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 3'-конце, и сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 5'-конце, и сегмент, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 звеньев ДНК между двумя сегментами звеньев LNA, где звенья LNA выбраны из: β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из фосфата, фосфоротиоата и фосфородитиоата. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин в звеньях LNA, предпочтительно всех звеньях LNA и/или 2-аминоаденин в некоторых или всех звеньях ДНК, и/или 5-метилцитозин в некоторых или всех звеньях ДНК.
Также особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды из таблицы 8 (SEQ ID NO: 219a-231b).
Кроме того, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N9 представляет: CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-,TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-;
N10 представляет: -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A;
и солям и оптическим изомерам антисмыслового олигонуклеотида.
N9 и/или N10 могут также представлять любой из дополнительных ограниченных списков 3'- и 5'-остатков, как здесь раскрыто.
Особенно предпочтительными гапмерными антисмысловыми олигонуклеотидами, подпадающими под общую формулу S7:
являются следующие:
Антисмысловые олигонуклеотиды формулы S7 в форме гапмеров (LNA-сегмент 1-сегмент ДНК-LNA-сегмент 2) содержат сегмент LNA на терминальном 5'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и содержат сегмент LNA на терминальном 3'-конце, состоящий из 2-5, предпочтительно 2-4 звеньев LNA, и между двумя сегментами LNA один сегмент ДНК, состоящий из 6-14, предпочтительно 7-12 и более предпочтительно 8-11 звеньев ДНК.
Антисмысловые олигонуклеотиды формулы S7 содержат нуклеотиды LNA (звенья LNA), как здесь описано, в частности, таковые, раскрытые в разделе «Замкнутые нуклеиновые кислоты» (LNA®) и, предпочтительно таковые, раскрытые в разделе «Предпочтительные LNA». Звенья LNA и звенья ДНК могут содержать обычные азотистые основания, такие как аденин (А), цитозин (С), гуанин (G), тимин (Т) и урацил (U), но могут также содержать модифицированные азотистые основания, как описано в разделе «Азотистые основания». Антисмысловые олигонуклеотиды формулы S7 или сегменты LNA и сегмент ДНК антисмыслового олигонуклеотида могут содержать любую межнуклеотидную связь, как описано здесь, и в частности, таковые, раскрытые в разделе «Межнуклеотидные связи (IL)». Антисмысловые олигонуклеотиды формулы S7 необязательно могут также содержать концевые группы на терминальном 3'-конце и/или терминальном 5'-конце и в частности, таковые, раскрытые в разделе «Концевые группы».
Результаты экспериментов показали, что модифицированные азотистые основания незначительно увеличивают или изменяют активность антисмысловых олигонуклеотидов по изобретению в отношении тестированных неврологических и онкологических заболеваний. Было показано, что модифицированные азотистые основания 5-метилцитозин или 2-аминоаденин дополнительно повышают активность антисмысловых олигонуклеотидов формулы S7, особенно если 5-метилцитозин используется только в нуклеотидах LNA или в нуклеотидах LNA и в нуклеотидах ДНК и/или, если 2-аминоаденин используется в нуклеотидах ДНК и не в нуклеотидах LNA.
Предпочтительная гапмерная структура антисмысловых олигонуклеотидов формулы S7 является следующей: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3 и еще более предпочтительной: 3-8-3, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 3-10-4, 4-10-3, 4-10-4, 3-11-4 и 4-11-3.
В качестве звеньев LNA для антисмысловых олигонуклеотидов формулы S7, в частности, β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9) являются предпочтительными. Результаты экспериментов показали, что все эти звенья LNA b1, b2, b4, b5, b6, b7, b8 и b9 можно синтезировать с требуемым усилием и обеспечить антисмысловые олигонуклеотиды с сопоставимой стабильностью и активностью. Однако, основываясь на экспериментах, звенья LNA b1, b2, b4, b5, b6 и b7 являются более предпочтительными. Еще более предпочтительными являются звенья LNA b1, b2, b4, b6 и b7, и еще более предпочтительными являются звенья LNA b1 и b4, и наиболее предпочтительным также в отношении сложности химического синтеза является β-D-окси-LNA (b1).
К настоящему времени не было найдено никакой конкретной 3'-концевой группы или 5'-концевой группы, которая заметно изменяла или повышала стабильность или активность в отношении онкологических или неврологических заболеваний, таким образом, 3'- и 5'-концевые группы возможны, но явно не являются предпочтительными.
Возможны различные межнуклеотидные мостики или межнуклеотидные связи. В приведенных здесь формулах межнуклеотидная связь IL представлена -IL'-Y-. Таким образом, IL=-IL'-Y-=-X''-P(=X')(X-)-Y-, где IL предпочтительно выбрана из группы, состоящей из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-. Предпочтительными являются межнуклеотидные связи IL, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(OCH2CH2OCH3)-O-, и более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, и еще более предпочтительные, выбранные из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, и наиболее предпочтительные, выбранные из -O-P(O)(O-)-O- и -O-P(O)(S-)-O-.
Таким образом, настоящее изобретение предпочтительно относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 10-28 нуклеотидов, предпочтительно 11-24 нуклеотидов, более предпочтительно 12-20 и еще более предпочтительно 13-19 или 14-18 нуклеотидов, и 1-5 из этих нуклеотидов на терминальном 5'-конце и 1-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N9 представляет: CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-,TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 представляет: -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A, и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-ENA (b5), β-D-(NH)-LNA (b6), β-D-(NCH3)-LNA (b7), β-D-(ONH)-LNA (b8) и β-D-(ONCH3)-LNA (b9); и предпочтительно из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и
межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-, -O-P(O)(CH3)-O-, -O-P(O)(OCH3)-O-, -O-P(O)(NH(CH3))-O-, -O-P(O)[N(CH3)2]-O-, -O-P(O)(BH3-)-O-, -O-P(O)(OCH2CH2OCH3)-O-, -O-P(O)(OCH2CH2SCH3)-O-, -O-P(O)(O-)-N(CH3)-, -N(CH3)-P(O)(O-)-O-;
и предпочтительно из -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Более предпочтительно N9 представляет: TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 представляет: -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A.
Еще более предпочтительно настоящее изобретение относится к антисмысловому олигонуклеотиду в форме гапмера, состоящему из 11-24 нуклеотидов, более предпочтительно 12-20 нуклеотидов и более предпочтительно 13-19 или 14-18 нуклеотидов, и 2-5 из этих нуклеотидов на терминальном 5'-конце и 2-5 нуклеотидов на терминальном 3'-конце антисмыслового олигонуклеотида представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и терминальном 3'-конце находится последовательность, по меньшей мере, из 7, предпочтительно, по меньшей мере, 8 нуклеотидов ДНК, и антисмысловой олигонуклеотид способен гибридизоваться с участком гена, кодирующего TGF-RII, или с участком мРНК, кодирующей TGF-RII, где антисмысловой олигонуклеотид представлен следующей последовательностью:
где:
N9 представляет: TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; предпочтительно N9 представляет: ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-; и
N10 представляет: -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A; предпочтительно N10 представляет: -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A; и
нуклеотиды LNA выбраны из β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из: -O-P(O)(O-)-O-, -O-P(O)(S-)-O-, -O-P(S)(S-)-O-, -S-P(O)(O-)-O-, -S-P(O)(S-)-O-, -O-P(O)(O-)-S-, -O-P(O)(S-)-S-, -S-P(O)(O-)-S-; и предпочтительно выбраны из фосфата, фосфоротиоата и фосфородитиоата;
и солям и оптическим изомерам антисмыслового олигонуклеотида. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин и/или 2-аминоаденин.
Особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды последовательностей SEQ ID NO: 474-SEQ ID NO: 508, содержащие сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 3'-конце, и сегмент из 2-5, предпочтительно 2-4 и более предпочтительно 3-4 звеньев LNA на 5'-конце, и сегмент, по меньшей мере, из 6, предпочтительно 7 и более предпочтительно 8 звеньев ДНК между двумя сегментами звеньев LNA, где звенья LNA выбраны из: β-D-окси-LNA (b1), β-D-тио-LNA (b2), α-L-окси-LNA (b4), β-D-(NH)-LNA (b6) и β-D-(NCH3)-LNA (b7); и межнуклеотидные связи выбраны из фосфата, фосфоротиоата и фосфородитиоата. Такие предпочтительные антисмысловые олигонуклеотиды могут не содержать какого-либо модифицированного терминального 3'- и 5'-конца или могут не содержать какой-либо 3'- и 5'-концевой группы, и могут в качестве модифицированного азотистого основания содержать 5-метилцитозин в звеньях LNA, предпочтительно всех звеньях LNA и/или 2-аминоаденин в некоторых или всех звеньях ДНК, и/или 5-метилцитозин в некоторых или всех звеньях ДНК.
Также особенно предпочтительными являются гапмерные антисмысловые олигонуклеотиды из таблицы 9 (SEQ ID NO: 284a-236b).
Фармацевтические композиции
Антисмысловые олигонуклеотиды по настоящему изобретению предпочтительно вводят в форме их фармацевтически активных солей, необязательно используя по существу нетоксичные фармацевтически приемлемые носители, эксципиенты, адъюванты, растворители или разбавители. Лекарственные средства по настоящему изобретению формулируют в обычном твердом или жидком носителе или разбавителе и обычном фармацевтически приготовленном адъюванте на подходящем уровне дозирования известным способом. Предпочтительные препараты и композиции находятся в пригодной для введения форме, которая пригодна для инфузии или инъекции (интратекальной, интрацеребровентрикулярной, внутричерепной, внутривенной, интрапаренхимальной, внутриопухолевой, внутриглазной или экстраглазной, внутрибрюшинной, внутримышечной, подкожной), местного введения в мозг, ингаляции, местного введения в солидную опухоль или перорального применения. Однако возможны также другие формы применения, такие как абсорбция через выстилающие эпителиальные или слизисто-кожные оболочки (слизистую оболочку ротовой полости, ректальную и вагинальную эпителиальные оболочки, слизистую оболочку носоглотки, слизистую оболочку кишечника), ректально, трансдермально, местно, внутрикожно, внутрижелудочно, внутримышечно, внутрикожно, подкожно, подъязычно или любыми другими способами, доступными в области фармации.
Вводимые препараты включают, например, инъекционные жидкие композиции, задерживающие композиции, порошки, в частности, для ингаляции, пилюли, таблетки, таблетки с пленочным покрытием, покрытые оболочкой таблетки, диспергируемые гранулы, драже, гели, сиропы, взвеси, суспензии, эмульсии, капсулы и депо-препараты. Возможны и другие способы введения галеновых препаратов, например непрерывная инъекция через имплантированный насос или катетер в головной мозг.
Как здесь используется, термин «фармацевтически приемлемый» относится к любому носителю, который не оказывает отрицательного влияния на эффективность биологической активности антисмысловых олигонуклеотидов, используемых в качестве активного ингредиента в композиции, и не токсичен для хозяина, которому он вводится. Примеры подходящих фармацевтических носителей хорошо известны в данной области и включают забуференные фосфатом физиологические растворы, воду, эмульсии, такие как эмульсии масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Такие носители можно формулировать обычными методами, и активное соединение может вводиться субъекту в эффективной дозе. «Эффективная доза» относится к количеству антисмыслового олигонуклеотида, используемого в качестве активного ингредиента, которое достаточно для воздействия на течение и тяжесть заболевания, приводя к снижению или ремиссии такой патологии. «Эффективная доза», пригодная для лечения и/или профилактики этих заболеваний или расстройств, может быть определена с использованием методов, известных специалистам в данной области. Кроме того, антисмысловые олигонуклеотиды по настоящему изобретению можно смешать и вводить вместе с липосомами, комплексообразующими агентами, молекулами, нацеленными на рецепторы, растворителями, консервантами и/или разбавителями.
Предпочтительными являются фармацевтические препараты в форме инфузионных растворов или твердых матриц для непрерывного высвобождения активного ингредиента, особенно для непрерывной инфузии для интратекального введения, интрацеребровентрикулярного введения или внутричерепного введения, по меньшей мере, одного антисмыслового олигонуклеотида по настоящему изобретению. Также предпочтительными являются фармацевтические препараты в виде растворов или твердых матриц, пригодных для местного введения в головной мозг. Для фиброзных болезней легких особенно предпочтительны композиции для ингаляционного введения.
Готовый к употреблению стерильный раствор содержит, например, по меньшей мере, один антисмысловой олигонуклеотид в концентрации от 1 до 10 мг/мл, предпочтительно от 5 до 10 мг/мл, и изотонический агент, выбранный, например, из сахаров, таких как сахароза, лактоза, маннит или сорбит. Также может быть включен подходящий буферный агент для контроля рН раствора до 6-8 (предпочтительно 7-8). Другим необязательным ингредиентом композиции может быть неионогенное поверхностно-активное вещество, такое как твин 20 или твин 80.
Стерильный лиофилизированный порошок, который восстанавливают для применения, содержит, по меньшей мере, один антисмысловой олигонуклеотид и необязательно объемообразующий агент (например, маннит, трегалозу, сорбит, глицин) и/или криопротектор (например, трегалозу, маннит). Растворителем для восстановления может быть вода для инъекционных препаратов, с буферной солью или без нее для регуляции рН до 6-8.
Аэрозольные препараты, подходящие для ингаляции, могут содержать растворы и твердые вещества в виде порошка, которые могут находиться в комбинации с фармацевтически приемлемым носителем, таким как инертный сжатый газ, например, азот.
Особенно предпочтительной фармацевтической композицией является лиофилизированный (высушенный сублимационной сушкой) препарат (лиофилизат), подходящий для введения ингаляцией или для внутривенного введения. Для получения предпочтительного лиофилизированного препарата, по меньшей мере, один антисмысловой олигонуклеотид по изобретению растворяют в 4-5% (мас./об.) растворе маннита и затем раствор лиофилизуют. Раствор маннита можно также приготовить в подходящем буферном растворе, как описано выше.
Дополнительные примеры подходящих крио/лиопротекторов (иначе относится к объемообразующему агенту или стабилизаторам) включают не содержащий тиоловых групп альбумин, иммуноглобулины, полиалкиленоксиды (например, ПЭГ, полипропиленгликоли), трегалозу, глюкозу, сахарозу, сорбит, декстран, мальтозу, рафинозу, стахиозу и другие сахариды (смотри, например, международную заявку WO 97/29782), в то время как маннит используется предпочтительно. Они могут быть использованы в обычных количествах в обычных способах лиофилизации. Способы лиофилизации хорошо известны в области приготовления фармацевтических композиций.
Для введения ингаляцией диаметр частиц лиофилизированного препарата предпочтительно составляет от 2 до 5 мкм, более предпочтительно от 3 до 4 мкм. Лиофилизованный препарат особенно подходит для введения с использованием ингалятора, например ингалятора OPTINEB® или VENTA NEB® (NEBU TEC, Elsenfeld, Германия). Лиофилизованный продукт можно регидратировать в стерильной дистиллированной воде или любой другой жидкости, подходящей для ингаляционного введения. Альтернативно, для внутривенного введения лиофилизированный продукт может быть регидратирован в стерильной дистиллированной воде или любой другой жидкости, подходящей для внутривенного введения.
После регидратации для введения в стерильной дистиллированной воде или другой подходящей жидкости лиофилизированный препарат должен иметь примерно физиологическую осмоляльность ткани-мишени для регидратированного пептидного препарата, то есть крови при внутривеннои введении или легочной ткани при ингаляционном введении. Таким образом, предпочтительно, чтобы регидратированный препарат был, по существу, изотоническим.
Предпочтительная концентрация для внутривенного, перорального или ингаляционного введения составляет от 10 до 2000 мкмоль/мл и более предпочтительно составляет от 200 до 800 мкмоль/мл.
Для перорального введения в форме таблеток или капсул, по меньшей мере, один антисмысловой олигонуклеотид может быть объединен с любым нетоксичным фармацевтически приемлемым инертным носителем, подходящим для перорального введения, таким как лактоза, крахмал, сахароза, целлюлоза, стеарат магния, дикальций фосфат, сульфат кальция, тальк, маннит, этиловый спирт (жидкие формы) и тому подобное. Кроме того, когда это желательно или необходимо, то также в смесь могут быть включены подходящие связующие, смазывающие вещества, разрыхлители и красители. Порошки и таблетки могут содержать примерно от 5 до 95% композиции по изобретению.
Подходящие связующие вещества включают крахмал, желатин, природные сахара, кукурузные подсластители, природные и синтетические камеди, такие как гуммиарабик, альгинат натрия, карбоксиметилцеллюлоза, полиэтиленгликоль и воски. Среди смазывающих веществ, которые могут быть упомянуты для применения в данных лекарственных формах, находятся борная кислота, бензоат натрия, ацетат натрия, хлорид натрия и тому подобное. Разрыхлители включают крахмал, метилцеллюлозу, гуаровую камедь и тому подобное.
Кроме того, композиции по настоящему изобретению могут быть составлены в форме с замедленным высвобождением для обеспечения контролируемой скорости высвобождения, по меньшей мере, одного антисмыслового олигонуклеотида для оптимизации терапевтических эффектов. Подходящие лекарственные формы для замедленного высвобождения включают имплантируемые биодеградируемые матрицы для замедленного высвобождения, содержащие, по меньшей мере, один антисмысловой олигонуклеотид, многослойные таблетки, содержащие слои с различной скоростью распадаемости, или полимерные матрицы с контролируемым высвобождением, пропитанные, по меньшей мере, одним антисмысловым олигонуклеотидом.
Препараты в жидкой форме включают растворы, суспензии и эмульсии. В качестве примера можно упомянуть водные или водные-пропиленгликолевые растворы для парентеральных инъекций или добавки подсластителей и замутнителей для пероральных растворов, суспензий и эмульсий.
Подходящими разбавителями являются вещества, которые обычно составляют основную часть композиции или лекарственной формы. Подходящие разбавители включают сахара, такие как лактоза, сахароза, маннит и сорбит, крахмалы, полученные из пшеницы, кукурузы, риса и картофеля, и целлюлозы, такие как микрокристаллическая целлюлоза. Количество разбавителей в композиции может составлять примерно от 5% до примерно 95% по отношению к общей массе композиции, предпочтительно примерно от 25% до примерно 75% по отношению к общей массе композиции.
Термин разрыхлители относится к веществам, добавленным к композиции для обеспечения ее разрушения (распадаемости) и высвобождения активных веществ. Подходящие разрыхлители включают крахмалы, «растворимые в холодной воде» модифицированные крахмалы, такие как натрия карбоксиметилкрахмал, натуральные и синтетические камеди, такие как камедь рожкового дерева, карая, гуаровая камедь, трагакант и агар, производные целлюлозы, такие как метилцеллюлоза и натрий карбоксиметилцеллюлоза, микрокристаллические целлюлозы и сшитые микрокристаллические целлюлозы, такие как кроскармеллоза натрия, альгинаты, такие как альгиновая кислота и альгинат натрия, глины, такие как бентониты, и шипучие смеси. Количество разрыхлителя в композиции может составлять примерно от 1 до примерно 40% по отношению к массе композиции, предпочтительно примерно от 2 до 30% по отношению к массе композиции, более предпочтительно примерно от 3 до 20% по отношению к массе композиции, и наиболее предпочтительно примерно от 5 до примерно 10% по отношению к массе композиции.
Связующие вещества представляют вещества, которые связывают или «склеивают» порошки вместе и делают их когезионными, образуя гранулы, таким образом, функционируя в качестве «адгезива» в композиции. Связующие вещества дополнительно повышают прочность сцепления, уже обеспеченную разбавителем или объемообразующим агентом. Подходящие связующие вещества включают сахара, такие как сахароза, крахмалы, полученные из пшеницы, кукурузы, риса и картофеля; природные камеди, такие как гуммиарабик, желатин и трагакант; производные бурых морских водорослей, такие как альгиновая кислота, альгинат натрия и альгинат аммония, альгинат кальция; целлюлозные материалы, такие как метилцеллюлоза и натрий карбоксиметилцеллюлоза и гидроксипропилметилцеллюлоза; поливинилпирролидон; и неорганические вещества, такие как алюмосиликат магния. Количество связующего вещества в композиции может составлять примерно от 1 до примерно 30% по отношению к массе композиции, предпочтительно примерно от 2 до 20% по отношению к массе композиции, более предпочтительно примерно от 3 до 10% по отношению к массе композиции, и еще более предпочтительно примерно от 3 до примерно 6% по отношению к массе композиции.
Смазочное вещество относится к веществу, добавленному в лекарственную форму для того, чтобы обеспечить таблетке, гранулам и т.д. после их прессования, извлечение из прессовальной формы или матрицы, или истирание в результате трения или износа. Подходящие смазывающие вещества включают стеараты металлов, такие как стеарат магния, стеарат кальция или стеарат калия, стеариновую кислоту; высокоплавкие воски; и водорастворимые смазывающие вещества, такие как хлорид натрия, бензоат натрия, ацетат натрия, олеат натрия, полиэтиленгликоли и D,L-лейцин.
Смазывающие вещества обычно добавляют на самой последней стадии перед прессованием, так как они должны присутствовать на поверхностях гранул и между ними и частями таблеточного пресса. Количество смазывающего вещества в композиции может составлять примерно от 0,05 до примерно 15% по отношению к массе композиции, предпочтительно примерно от 0,2 до примерно 5% по отношению к массе композиции, более предпочтительно примерно от 0,3 до примерно 3% по отношению к массе композиции, и наиболее предпочтительно примерно от 0,3 до примерно 1,5% по отношению к массе композиции.
Скользящие вещества представляют вещества, которые предотвращают комкование и улучшают текучесть гранулятов, для того, чтобы поток был гладким и однородным. Подходящие скользящие вещества включают диоксид кремния и тальк. Количество скользящего вещества в композиции может варьировать примерно от 0,01 до примерно 10% по отношению к массе композиции, предпочтительно примерно от 0,1 до примерно 7% по отношению к массе композиции, более предпочтительно примерно от 0,2 до примерно 5% по отношению к массе композиции, и наиболее предпочтительно примерно от 0,5 до примерно 2% по отношению к массе композиции.
В фармацевтические композиции, раскрытые здесь, антисмысловые олигонуклеотиды включают предпочтительно в форме их солей и, необязательно вместе с другими компонентами, которые повышают стабильность антисмысловых олигонуклеотидов, увеличивают рекрутинг РНКазы Н, повышают способность нахождения мишени, усиливают поглощение клетками и тому подобное. Для достижения этих целей антисмысловые олигонуклеотиды можно модифицировать химически вместо или в дополнении к использованию дополнительных компонентов, пригодных для достижения этих целей. Таким образом, антисмысловые олигонуклеотиды по изобретению можно химически связать с группами или компонентами, которые усиливают активность, клеточное распределение или поглощение клетками антисмысловых олигонуклеотидов и т.д.. Такие фрагменты включают липидные группы, такие как холестериловая группа, холевую кислоту, тиоэфир, гексил-S-тритилтиол, тиохолестерин, алифатическую цепь, например, остатки додекандиола или ундецила, фосфолипид, такой как дигексадецил-rac-глицерин или 1,2-ди-O-гексадецил-rac-глицеро-3-H-фосфонат триэтиламмония, цепь полиамина или полиэтиленгликоля, или адамантан-уксусную кислоту, пальмитиловую группу или группу октадециламина или гексиламинокарбонилоксихолестерина. Настоящее изобретение также включает антисмысловые олигонуклеотиды, которые являются химерными соединениями. «Химерные» антисмысловые олигонуклеотиды в контексте настоящего изобретения представляют собой антисмысловые олигонуклеотиды, которые содержат две или более химически различных областей, одна из которых представляет последовательность олигонуклеотида, как здесь раскрыто, которая связана с группой или компонентом, предназначенных для повышения поглощения клетками, повышения устойчивости к деградации под воздействием нуклеаз, повышения аффинности связывания с нуклеиновой кислотой-мишенью, увеличения рекрутинга РНКазы Н и так далее. Например, дополнительная область или группа или компонент антисмыслового олигонуклеотида может служить в качестве субстрата для ферментов, способных расщеплять гибриды РНК:ДНК или молекулы РНК:РНК. В качестве примера: РНКаза Н является клеточной эндорибонуклеазой, которая отщепляет цепь РНК из дуплекса РНК:ДНК. Таким образом, активация РНКазы Н приводит к отщеплению РНК-мишени, которая в данном случае представляет собой мРНК, кодирующую TGF-RII, тем самым существенно повышая эффективность ингибирования экспрессии гена антисмысловым олигонуклеотидом. Следовательно, сопоставимые результаты часто можно получить с более короткими олигонуклеотидами при использовании химерных олигонуклеотидов.
Показания
Настоящее изобретение относится к применению антисмысловых олигонуклеотидов, раскрытых здесь, для профилактики и лечения нейродегенеративных заболеваний, нейротравмы, нейрососудистых и нейровоспалительных заболеваний, включая постинфекционные и воспалительные заболевания центральной нервной системы (ЦНС).
Антисмысловые олигонуклеотиды по настоящему изобретению особенно пригодны для стимуляции регенерации и функционального восстановления поврежденных нервных путей и/или для лечения и компенсации вызванного возрастом снижения обновления нейрональных стволовых клеток.
Таким образом, еще один аспект настоящего изобретения относится к применению антисмыслового олигонуклеотида, раскрытого здесь, для стимуляции регенерации нейрональной ткани посредством реактивации нейрогенеза, обеспечения дифференцировки и миграции нейронов и индукции интеграции новых нейронов в анатомические и функциональные нейрональные цепи.
Дополнительный аспект настоящего изобретения относится к применению антисмыслового олигонуклеотида, раскрытого здесь, для стимуляции регенерации и клинического (структурного) восстановления у пациентов с повреждением нервной системы или повреждением других систем органов, вызванных фиброзом, или потерей оборота стволовых клеток.
Кроме того, антисмысловые олигонуклеотиды пригодны для компенсации и лечения снижения обновления нейрональных стволовых клеток, вызванного возрастом, воспалением или дефектом гена.
Антисмысловые олигонуклеотиды по настоящему изобретению ингибируют экспрессию TGF-RII и, следовательно, используются для лечения заболеваний, ассоциированных с повышающей регуляцией или повышенным уровнем TGF-RII и/или TGF-RII.
Таким образом, еще один аспект настоящего изобретения относится к применению антисмысловых олигонуклеотидов в профилактике и лечении нейродегенеративных заболеваний, нейровоспалительных расстройств, травматических или посттравматических расстройств, сосудистых или точнее нейроваскулярных расстройств, гипоксических расстройств, постинфекционных расстройств центральной нервной системы, фиброзных болезней, гиперпролиферативных заболеваний, рака, опухолей, пресвиакуза и пресбиопии.
Термин «нейродегенеративное заболевание» или «неврологическое заболевание», или «нейровоспалительное расстройство» относится к любому заболеванию, расстройству или состоянию, поражающему центральную или периферическую нервную систему, включая СПИД, неврологические осложнения СПИДа, отсутствие septum pellucidum, приобретенную эпилептиформную афазию, острый диссеминированный энцефаломиелит, адренолейкодистрофию, агнезию мозолистого тела, агнозию, синдром Айкарди, болезнь Александера, болезнь Альперса, альтернирующую гемиплегию, болезнь Альцгеймера, боковой амиотрофический склероз (БАС), анэнцефалию, аневризму, синдром Ангельмана, ангиоматоз, аноксию, афазию, апраксию, арахноидальные кисты, арахноидит, мальформацию Арнольда-Киари, артериовенозную мальформацию, аспартам, синдром Аспергера, атаксию телеангиэктазию, атаксию, синдром дефицита внимания с гиперактивностью, аутизм, вегетативную дисфункцию, боли в спине, синдром Барта, болезнь Баттена, болезнь Бехчета, паралич Белла, доброкачественный эссенциальный блефароспазм, доброкачественную фокальную амиотрофию, доброкачественную внутричерепную гипертензию, синдром Бернарда-Рота, болезнь Бинсвангера, блефароспазм, синдром Блоха-Сульцбергера, родовые травматические повреждения плечевого сплетения, травмы плечевого сплетения, синдром Брэдбери-Эгглстона, аневризму головного мозга, черепно-мозговую травму, опухоли головного и спинного мозга, синдром Броуна-Секара, бульбоспинальную мышечную атрофию, болезнь Канавана, синдром запястного туннеля, каузалгию, каверномы, кавернозную ангиому, кавернозную мальформацию, пирамидный паралич, синдром центрального паралича, центральный болевой синдром, головные расстройства, мозжечковую дегенерацию, гипоплазию мозжечка, церебральную аневризму, церебральный атеросклероз, церебральную атрофию, церебральную форму бери-бери, церебральный гигантизм, церебральную гипоксию, церебральный паралич, церебро-оукло-фасцио-скелетный синдрома, болезнь Шарко-Мари-Тута, ишемический синдром, хорею, хореоакантоцитоз, хроническую воспалительную демиелинизирующую полинейропатию (CIDP), хроническую ортостатическую непереносимость, хроническую боль, синдром Коккейна типа II, синдром Коффина-Лоури, кому, включая стойкое вегетативное состояние, сложный региональный болевой синдром, врожденную лицевую диплегию, врожденную миастению, врожденную миопатию, врожденные пороки развития кавернозных сосудов, кортикобазальную дегенерацию, краниальный артериит, краниосиностоз, болезнь Крейтцфельдта-Якоба, кумулятивные травматические расстройства, синдром Кушинга, цитомегалическую болезнь телец включения (CIBD), цитомегаловирусную инфекцию, синдром танцующих глаз-танцующих ног, синдром Дэнди-Уокера, болезнь Доусона, синдром Де Морсье, паралич Дежерин-Клюмпке, мультиинфарктную деменцию, подкорковую деменцию, деменция с телами Леви, дерматомиозит, диспраксию развития, синдром Девика, диабетическую невропатию, рассеянный склероз, синдром Драве, вегетативную дистонию, дисграфию, дислексию, дисфагию, диспраксию, дистонию, раннюю младенческую эпилептическую энцефалопатию, синдром пустого турецкого седла, летаргический энцефалит, энцефалит и менингит, энцефалоцеле, энцефалопатию, энцефалотригеминальный ангиоматоз, эпилепсию, болезнь Эрба, параличи Эрба-Дюшенна и Дежерина-Клюмпке, болезнь Фабри, синдром Фара, обмороки, семейную дисаутономию, семейную гемангиому, семейную идиопатическую кальцификацию базальных ганглиев, семейный спастический паралич, фебрильные судороги (например, ГЭФС и ГЭФС плюс), синдром Фишера, синдром «вялого ребенка», атаксию Фридрейха, болезнь Гоше, синдром Герстмана, болезнь Герстмана-Штреуслера-Шейнкера, гигантоклеточный артериит, гигантоклеточную болезнь с включениями, глобоидноклеточную лейкодистрофию, глоссофарингеальную невралгию, синдром Гийена-Барре, связанную с HTLV-1 миелопатию, болезнь Галлервордена-Шпатца, черепно-мозговую травму, головную боль, гемикранию континуа, гемифациальный спазм, глазодвигательную гемиплегию, наследственные невропатии, наследственную спастическую параплегию, наследственную полиневропатическую атаксию, опоясывающий лишай уха, опоясывающий лишай, синдром Хираямы, голопрозэнцефалию, болезнь Гентингтона, гидроанэнцефалию, гидроцефалию нормального давления, гидроцефалию (в частности, TGFβ-индуцированную гидроцефалию), гидромиелию, гиперкортицизм, гиперсомнию, гипертонию, гипотонию, гипоксию, иммунно-опосредованный энцефаломиелит, миозит с тельцами включения, недержание пигмента, детскую гипотонию, инфальтивную форму болезни накопления фитановой кислоты, инфантильную болезнь рефсум, инфантильные спазмы, воспалительные миопатии, кишечную липодистрофию, внутричерепные кисты, внутричерепную гипертензию, синдром Исаакса, синдром Жубера, синдром Кернса-Сейра, болезнь Кеннеди, синдром Кинсбурна, синдром Клейна-Левина, синдром Клиппеля-Фейла, синдром Клиппеля-Треноне (KTS), синдром Клювера-Бюси, амнестический синдром Корсакова, болезнь Краббе, болезнь Кугельберга-Веландера, куру, миастенический синдром Ламберта-Итона, синдром Ландау-Клеффнера, компрессию латерального кожного нерва, латеральный синдром продолговатого мозга, необучаемость, болезнь Ли, синдром Леннокса-Гасто, синдром Леша-Нихана, лейкодистрофию, синдром Левина-Критчли, деменцию с тельцами Леви, лиссенцефалию, синдром «запертого человека», болезнь Лу-Герига, неврологические осложнения волчанки, болезнь Лайма-неврологические осложнения, болезнь Мачадо-Джозефа, макренцефалию, мегаленцефалию, синдром Мелькерссона-Розенталя, менингит, болезнь Менкеса, невралгию латерального кожного нерва бедра, метахроматическую лейкодистрофию, микроцефалию, мигрень, синдром Миллера-Фишера, микроинсульт, митохондриальные миопатии, синдром Мебиуса, мономелическую амиотрофию, болезни двигательного нейрона, болезнь Моямоя, муколипидоз, мукополисахаридозы, мультиинфарктную деменцию, мультифокальную двигательную нейропатию, рассеянный склероз (MS), множественную системную атрофию (MSA-C и MSA-P), множественную системную атрофию с ортостатической гипотензией, мышечную дистрофию, врожденную миастению, миастению гравис, диффузный миелинокластический склероз, миоклоническую энцефалопатию младенцев, миоклонус, врожденную миопатию, тиреотоксическую миопатию, миопатию, врожденную миотонию, миотонию, нарколепсию, нейроакантоцитоз, нейродегенерацию с накоплением железа в головном мозге, нейрофиброматоз, злокачественный нейролептический синдром, неврологические осложнения СПИДа, неврологические проявления болезни Помпе, невромиелит оптического нерва, нейромиотонию, нейрональный цероид-липофусциноз, нейрональные расстройства движения, наследственные невропатии, нейросаркоидоз, нейротоксичность, невус кавернозный, болезнь Нимана-Пика, синдром О'Салливана-Маклеода, затылочную невралгию, скрытую дизрафию спинного мозга, синдром Охтахара, оливомостомозжечковую атрофию, синдром опсоклонуса-миоклонуса, ортостатическую гипотензию, синдром Оверуса, хроническую боль, паранеопластические синдромы, парестезии, болезнь Паркинсона, врожденную пармиопатию, пароксизмальный хореоатетоз, пароксизмальную гемикранию, синдром Парри Ромберга, болезнь Пелицеуса-Мерцбахера, синдром Пена-Шокейра, периневральные кисты, периодические параличи, периферическую нейропатию, перивентрикулярную лейкомаляцию, стойкое вегетативное состояние, первазивные расстройства развития, болезнь накопления фитановой кислоты, болезнь Пика, синдром грушевидной мышцы, опухоли гипофиза, полимиозит, болезнь Помпе, поренцефалию, постполиомиелитический синдром, постгерпетическую невралгию, постинфекционный энцефаломиелит, постуральную гипотензию, синдром постуральной ортостатической тахикардии, синдром постуральной тахикардии, первичный боковой склероз, прионные заболевания, прогрессирующую гемифациальную атрофию, прогрессирующую опорно-двигательную атаксию, прогрессирующую мультифокальную лейкоэнцефалопатию, прогрессирующую склерозирующую полиодистрофию, прогрессирующий надъядерный паралич, ложную опухоль головного мозга, пиридоксин-зависимые и пиридоксин-реагирующие судорожные расстройства, синдром Рамсея-Ханта типа I, синдром Рамсея-Ханта типа II, энцефалит Расмуссена и другие аутоиммунные эпилепсии, синдром рефлекторной симпатической дистрофии, детскую форму болезни Рефсума, болезнь Рефсума, повторяющиеся нарушения движения, повторяющиеся стрессовые травмы, синдром беспокойных ног, ретровирус-ассоциированную миелопатию, синдром Ретта, синдром Рейе, синдром Райли-Дея, головную боль SUNCT, кисты крестцовых нервных корешков, «пляска святого Вита», болезнь слюнной железы, болезнь Сандхоффа, болезнь Шильдера, шизенцефалию, судорожные припадки, септооптическую дисплазию, тяжелую миоклоническую эпилепсию младенцев (SMEI), синдром «встряхнутого ребенка», опоясывающий лишай, синдром Шая-Дрейджера, синдром Шегрена, апноэ во сне, сонную болезнь, синдром Сото, спастичность, расщелину позвоночника, инфаркт спинного мозга, повреждение спинного мозга, опухоли спинного мозга, спинальную мышечную атрофию, спиноцеребеллярную атрофию, синдром Стила-Ричардсона-Ольшевского, синдром ригидного человека, стрионигральную дегенерацию, инсульт, синдром Стерджа-Вебера, подострый склерозирующий панэнцефалит, субкортикальную атеросклеротическую энцефалопатию, расстройства глотания, хорею Сиденгама, обмороки, сифилитический спинальный склероз, сирингогидромиелию, сирингомиелию, системную красную волчанку, табес, позднюю дискинезию, кисты Тарлова, болезнь Тея-Сакса, височный артериит, синдром «привязанного» спинного мозга, болезнь Томсена, синдром грудного выхода, тиреотоксическую миопатию, невралгию тройничного нерва, паралич Тодда, синдром Туретта, транзиторную ишемическую атаку, трансмиссивные губчатые энцефалопатии, поперечный миелит, черепно-мозговую травму, тремор, невралгию тройничного нерва, тропический спастический парапарез, клубневой склероз, сосудистую эректильную опухоль, васкулит, включая височный артериит, болезнь фон Экономо, болезнь Гиппеля-Линдау (VHL), болезнь фон Реклингхаузена, синдром Валленберга, болезнь Верднига-Хоффинана, синдром Вернике-Корсакова, западный синдром, болезнь Уиппла, синдром Уильямса, болезнь Вильсона, X-сцепленную и спинальную бульбарную мышечную атрофию и синдром Зеллвегера.
Предпочтительные примеры нейродегенеративных заболеваний и нейровоспалительных расстройств выбраны из группы, состоящей или включающей: болезнь Альцгеймера, болезнь Паркинсона, болезнь Крейтцфельдта-Якоба (CJD), новый вариант болезни Крейтцфельдта-Якоба (nvCJD), болезнь Галлервордена-Шпатца, болезнь Гентингтона, мультисистемную атрофию, деменцию, лобно-височную деменцию, расстройства двигательного нейрона с множественным спонтанным или генетическим фоном, боковой амиотрофический склероз (АЛС), спинномозговую мышечную атрофию, спиноцеребеллярные атрофии (SCA), шизофрению, аффективные расстройства, большое депрессивное расстройство, менингоэнцефалит, бактериальный менингоэнцефалит, вирусный менингоэнцефалит, аутоиммунные расстройства ЦНС, рассеянный склероз (MS), острые ишемические/гипоксические поражения, травму спинного мозга, травму головы и позвоночника, черепно-мозговые травмы, артериосклероз, атеросклероз, микроангиопатическую деменцию, болезнь Бинсвангера (лейкоареоз), дегенерацию сетчатки, улитковую дегенерацию, дегенерацию желтого пятна, улитковую глухоту, деменцию, связанную со СПИДом, пигментный ретинит, Х-сцепленный рецессивный синдром тремора/атаксии (FXTAS), прогрессирующий супрануклеарный паралич (PSP), стриатонигральную дегенерацию (SND), оливопонтомозжечковую дегенерацию (OPCD), синдром Шая-Дрейджера, зависящий от возраста дефицит памяти, неврологические расстройства развития, связанные с деменцией, синдром Дауна, синуклеинопатии, мутации супероксиддисмутазы, расстройства повторов тринуклеотида, такие как болезнь Гентингтона, травму, гипоксию, сосудистые заболевания, сосудистые воспаления, старение ЦНС. Также можно упомянуть возрастное снижение обновления стволовых клеток.
Особо предпочтительные упомянутые примеры нейродегенеративных заболеваний и нейровоспалительных расстройств выбраны из группы, состоящей или включающей: болезнь Альцгеймера, болезнь Паркинсона, болезнь Гентингтона, боковой амиотрофический склероз (БАС), гидроцефалию (в частности, TGF-β-индуцированную гидроцефалию), травму ЦНС и спинного мозга, такую как повреждение спинного мозга, травму головы и позвоночника, травматические повреждения головного мозга, дегенерацию сетчатки, дегенерацию желтого пятна, улитковую глухоту, деменцию, связанную со СПИДом, расстройства повторов тринуклеотида, такие как болезнь Гентингтона, и старение ЦНС.
Антисмысловые олигонуклеотиды также пригодны для профилактики и лечения фиброзных болезней. Фиброз или фиброзная болезнь представляет образование избыточной фиброзной соединительной ткани в органе или ткани при репаративном или реактивном процессе. Это может быть реактивное, доброкачественное или патологическое состояние. В ответ на травму это называется рубцеванием, и если фиброз возникает из одной клеточной линии, то он называется фибромой. Физиологически это действует на отложение внеклеточного матрикса, который может облитерировать архитектонику и функцию основного органа или ткани. Также термин фиброз можно использовать для описания патологического состояния избыточного отложения фиброзной ткани, а также процесса отложения соединительной ткани при заживлении. Фиброз представляет процесс, включающий стимулированные клетки для образования соединительной ткани, включая коллаген и гликозаминогликаны. Затем макрофаги и поврежденная ткань между интерстицием высвобождает TGF-β. TGF-β стимулирует пролиферацию и активацию фибробластов, которые откладывают соединительную ткань. Снижение уровней TGF-β предотвращает и уменьшает образование соединительной ткани и таким образом предотвращает и лечит фиброз.
Примерами фиброзных болезней являются:
Таким образом, еще один аспект настоящего изобретения относится к применению антисмыслового олигонуклеотида для профилактики и/или лечения или к применению антисмыслового олигонуклеотида для приготовления фармацевтической композиции для профилактики и/или лечения фиброза легких, кистозного фиброза, цирроза печени, эндомиокардиального фиброза, «старого» инфаркта миокарда, фиброза предсердий, фиброза средостения, миелофиброза, забрюшинного фиброза, прогрессирующего массивного фиброза, нефрогенного системного фиброза, глаукомы, такой как первичная открытоугольная глаукома, болезни Крона, келоида, системного склероза, артрофиброза, болезни Пейрони, контрактуры Дюпюитрена и остаточных явлений после красной волчанки.
Еще один аспект настоящего изобретения относится к применению антисмыслового олигонуклеотида для профилактики и/или лечения гиперпролиферативных заболеваний, рака, опухолей и их метастазов или к применению антисмыслового олигонуклеотида для приготовления фармацевтической композиции для профилактики и/или лечения гиперпролиферативных заболеваний, рака, опухолей и их метастазов.
Примеры гиперпролиферативных заболеваний, рака, опухолей выбраны из группы, состоящей из или включающей: аденокарциному, меланому, острый лейкоз, акустическую невриному, ампулярную карциному, карциному анального отверстия, астроцитому, базально-клеточную карциному, рак поджелудочной железы, десмоидную опухоль, рак мочевого пузыря, бронхиальную карциному, немелкоклеточный рак легких (NSCLC), рак молочной железы, лимфому Беркитта, рак тела матки, CUP-синдром (неизвестная первичная опухоль), колоректальный рак, рак тонкого кишечника, опухоли тонкого кишечника, рак яичника, рак эндометрия, эпендимому, рак эпителиальных типов, опухоли Юинга, опухоли желудочно-кишечного тракта, рак желудка, рак желчного пузыря, карциномы желчного пузыря, рак матки, рак шейки матки, шейки матки, глиобластомы, гинекологические опухоли, опухоли уха, носа и горла, гематологические неоплазии, волосатоклеточный лейкоз, рак уретры, рак кожи, рак кожи яичка, опухоли головного мозга (глиомы, например астроцитомы, олигодендроглиомы, медуллобластомы, PNET, смешанные глиомы), метастазы в мозг, рак яичка, опухоль гипофиза, карциноиды, саркому Капоши, рак гортани, зародышевоклеточную опухоль, рак кости, колоректальную карциному, опухоли головы и шеи (опухоли в области уха, носа и горла), карциному ободочной кишки, краниофарингиомы, рак ротовой полости (рак в области рта и на губах), рак центральной нервной системы, рак печени, метастазы в печень, лейкоз, опухоль века, рак легких, рак лимфоузлов (ходжкина/неходжкина), лимфомы, рак желудка, злокачественную меланому, злокачественную неоплазию, злокачественные опухоли желудочно-кишечного тракта, карциному молочной железы, рак прямой кишки, медуллобластомы, меланому, менингиомы, болезнь Ходжкина, грибовидный микоз, рак носа, невриному, нейробластому, рак почки, почечноклеточные карциномы, неходжкинские лимфомы, олигодендроглиому, рак пищевода, остеолитические карциномы и остеопластические карциномы, остеосаркомы, карциному яичников, карциному поджелудочной железы, рак полового члена, плазмоцитому, плоскоклеточный рак головы и шеи (SCCHN), рак предстательной железы, рак глотки, карциному прямой кишки, ретинобластому, рак влагалища, рак щитовидной железы, болезнь Шнеебергера, рак пищевода, спиналиомы, Т-клеточную лимфому (грибовидный микоз), тимому, трубчатую карциному, опухоли глаза/окулярные опухоли, рак уретры, урологические опухоли, уротелиальную карциному, рак вульвы, появление бородавок, опухоли мягких тканей, саркому мягких тканей, опухоль Вильмса, карциному шейки матки и рак языка.
Термин «рак» предпочтительно относится к раку, выбранному из группы, состоящей или включающей рак легких, такой как карцинома легких, рак печени, такой как гепатоцеллюлярная карцинома, меланому или злокачественную меланому, рак поджелудочной железы, такой как эпителиоидная карцинома поджелудочной железы или аденокарцинома поджелудочной железы, рак ободочной кишки, такой как колоректальная аденокарцинома, рак желудка или карциному желудка, карциному молочной железы, злокачественную астроцитому, рак предстательной железы, лейкоз, такой как острый миелоидный лейкоз, хронический миелоидный лейкоз, моноцитарный лейкоз, промиелоцитарный лейкоз, лимфоцитарный лейкоз, острый лимфобластный лейкоз, лимфоцитарный лейкоз и острый лимфобластный лейкоз, и лимфому, такую как гистиоцитарная лимфома.
Для лечения гиперпролиферативных заболеваний, рака, опухолей и их метастазов антисмысловые олигонуклеотиды можно вводить с регулярными интервалами (интервалами доз DI) от 3 дней до двух недель, например, 4, 5, 6, 7, 8, 9, 10, 11, 12 или 13 дней, например, примерно 1 неделя, например, 6, 7 или 8 дней. Целесообразно, чтобы, по меньшей мере, две дозы имели период DI между двумя дозами, например, 3, 4, 5, 6, 7, 8, 9 или 10 доз, каждая с интервалом доз (DI) между каждой дозой антисмыслового олигонуклеотида. Период DI между каждой дозой может быть одинаковым, например, составлять 3 дня и две недели, например, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 дней, например, примерно 1 неделю, например, 6, 7 или 8 дней.
Предпочтительно каждая доза антисмыслового олигонуклеотида может составлять примерно от 0,25 до примерно 10 мг/кг, например, примерно 0,5 мг/кг, примерно 1 мг/кг, примерно 2 мг/кг, примерно 3 мг/кг, примерно 4 мг/кг, примерно 5 мг/кг, примерно 6 мг/кг, примерно 7 мг/кг, примерно 8 мг/кг, примерно 9 мг/кг. В некоторых вариантах осуществления каждая доза антисмыслового олигонуклеотида может составлять примерно от 2 до примерно 6 мг/кг или примерно от 4 до примерно 5 мг/кг. В некоторых вариантах осуществления каждая доза антисмыслового олигонуклеотида составляет, по меньшей мере, 2 мг/кг, например, 2, 3, 4, 5, 6, 7 или 8 мг/кг, например, 6 мг/кг. В некоторых вариантах осуществления режим дозирования антисмыслового олигонуклеотида может быть повторен после начального режима дозирования, например, после периода отдыха, когда антисмысловой олигонуклеотид не вводится. Например, период отдыха может длиться более 2 недель, например, примерно 3 недели или примерно 4 недели, или примерно 5 недель или примерно 6 недель. В некоторых вариантах осуществления схема дозирования антисмыслового олигонуклеотида представляет одну дозу в неделю, повторяемую три, четыре или пять раз. Такой режим дозирования затем можно повторить после периода отдыха, например, примерно через 3-5 недели, например, примерно через 4 недели. В некоторых вариантах осуществления антисмысловой олигонуклеотид вводят во время первого режима дозирования с регулярными интервалами дозирования (DI) от 4 до 13 дней между 2-10 введениями.
Введение антисмыслового олигонуклеотида обычно проводят парентеральным введением, таким как подкожное, внутримышечное, внутривенное или внутрибрюшинное введение.
Краткое описание фигур
На фиг. 1 показано ингибирующее действие антисмысловых олигонуклеотидов (ASO). ДНК транскрибируется в пре-мРНК, с которой в ядре клетки антисмысловые олигонуклеотиды (ASO) могут связываться или гибридизоваться с комплементарной последовательностью в экзоне (как представлено первым ASO справа и первым ASO слева) или внутри интрона (как представлено вторым ASO справа) или при расположении, состоящем из области экзона и области смежного интрона (как представлено вторым ASO слева). Посредством посттранскрипционной модификации, т.е. сплайсинга, формируется мРНК, с которой ASO может связываться или гибридизоваться в цитоплазме клетки, чтобы ингибировать трансляцию мРНК в последовательность белка. Таким образом, ASO подвергает нокдауну ген-мишень и экспрессию белка избирательно.
На фиг. 2 показано нуклеозидное звено (без межнуклеотидной связи) или нуклеотидное звено (с межнуклеотидной связью), которые не являются звеньями LNA и которые могут входить в состав антисмысловых олигонуклеотидов по настоящему изобретению, особенно в области B, в том случае, если антисмысловой олигонуклеотид по настоящему изобретению представляет гапмер.
На фиг. 3 показан TGF-бета и его эффекты на нейрональные стволовые клетки, раковые стволовые клетки и опухоли. TGF-бета ингибирует пролиферацию нейрональных стволовых клеток. Он может оказывать влияние на переход клетки в раковую стволовую клетку, которая может выйти из-под контроля роста с участием TGF-бета. Позднее при прогрессировании опухоли TGF-бета функционирует как онкоген; он также способствует росту опухоли за счет стимуляции ангиогенеза и подавления иммунной системы. Кроме того, он способствует миграции клеток, тем самым направляя клетки в метастазы.
На фиг. 4 показан антисмысловой олигонуклеотид SEQ ID NO: 218b в форме гапмера, состоящий из 16 нуклеотидов с 3 звеньями LNA (C*b1 и Ab1 и Tb1) на терминальном 5'-конце и 4 звеньями LNA (Ab1 и Gb1 и Tb1 и Ab1) на терминальном 3'-конце и 9 нуклеотидов ДНК (dG, dA, dA, dT, dG, dG, dA, dC и dC) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) в первом звене LNA от терминального 5'-конца.
На фиг. 5: обработка ASO (SEQ ID NO: 218b) приводит к снижению внутриклеточного уровня белка pSmad2. Мечение антителом к pSmad2 (левый столбец, красный) в клетках A549 (фиг. 5A) и ReNcell CX® (фиг. 5B) после трансфекции гимнозисом ASO SEQ ID NO: 218b в течение 72 ч или 96 ч соответственно. Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b.
На фиг. 6: обработка ASO (SEQ ID NO: 218с) приводит к снижению внутриклеточного уровня белка pSmad2. Мечение антителом к pSmad2 (левый столбец, красный) в клетках A549 (фиг. 6A) и ReNcell CX® (фиг. 6B) после трансфекции гимнозом ASO SEQ ID NO: 218с в течение 72 ч или 96 ч соответственно. Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218с.
На фиг. 7: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218b) приводит к понижающей регуляции мРНК TGF-RII. Сильная понижающая регуляция мРНК TGF-RII после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 7A) и ReNcell CX® (фиг. 7B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанным контрольным образцам. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
На фиг. 8: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218с) приводит к понижающей регуляции мРНК TGF-RII. Сильная понижающая регуляция мРНК TGF-RII после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 8A) и ReNcell CX® (фиг. 8B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанным контрольным образцам. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218с, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
На фиг. 9 показан антисмысловой олигонуклеотид SEQ ID NO: 209y в форме гапмера, состоящий из 16 нуклеотидов с 2 звеньями LNA (Gb1 и Tb1) на терминальном 5'-конце и 3 звеньями LNA (Ab1 и Gb1 и C*b1) на терминальном 3'-конце и 11 нуклеотидов ДНК (dA, dG, dT, dG, dT, dT, dT, dA, dG, dG и dG) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) в последнем звене LNA от терминального 5'-конца.
На фиг. 10 показан антисмысловой олигонуклеотид SEQ ID NO: 210q в форме гапмера, состоящий из 16 нуклеотидов с 4 звеньями LNA (Gb1 и C*b1 и Tb1 и Ab1) на терминальном 5'-конце и 3 звеньями LNA (Gb1 и Tb1 и Tb1) на терминальном 3'-конце и 9 нуклеотидов ДНК (dT, dT, dT, dG, dG, dT, dA, dG и dTs) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) во втором звене LNA от терминального 5'-конца.
На фиг. 11: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218b) приводит к понижающей регуляции мРНК CTGF. Сильная понижающая регуляция мРНК CTGF после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 11A) и ReNcell CX® (фиг. 11B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
На фиг. 12: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218b) приводит к понижающей регуляции мРНК CTGF. Сильная понижающая регуляция мРНК CTGF после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 12A) и ReNcell CX® (фиг. 12B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Клетки метили антителом к CTGF (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 13: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218b) приводит к снижению внутриклеточного уровня белка pSmad2. Экспрессия белка pSmad2 снижалась после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 13A) и ReNcell CX® (фиг. 13B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Клетки метили антителом к pSmad2 (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 14: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218с) приводит к понижающей регуляции мРНК CTGF. Сильная понижающая регуляция мРНК CTGF после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 14A) и ReNcell CX® (фиг. 14B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218с, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
На фиг. 15: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218с) приводит к снижению внутриклеточного уровня белка CTGF. Экспрессия белка CTGF снижалась после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549. ASO инкубировали в течение 72 ч в присутствии TGF-β1. Клетки метили антителом к CTGF (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218с, E=TGF-β1.
На фиг. 16: в присутствии TGF-β1 обработка ASO (SEQ ID NO: 218с) приводит к снижению внутриклеточного уровня белка pSmad2. Экспрессия белка pSmad2 снижалась после трансфекции гимнозом TGF-RII-специфического ASO в предварительно инкубированных с TGF-β1 клетках (48 ч) A549 (фиг. 16A) и ReNcell CX® (фиг. 16B). ASO инкубировали в течение 72 ч или 96 ч в присутствии TGF-β1 соответственно. Клетки метили антителом к pSmad2 (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218с, E=TGF-β1.
На фиг. 17: предварительная обработка ASO (SEQ ID NO: 218b) и последующее совместное воздействие TGF-β1 приводит к снижению уровня мембранного белка TGF-RII. Уровень белка TGF-RII снижался после трансфекции гимнозисом TGF-RII-специфического ASO с последующим совместным воздействием с TGF-β1 в клетках (48 ч) A549 (фиг. 17A) и ReNcell CX® (фиг. 17B). ASO инкубировали в течение 72 ч или 96 ч соответственно перед совместным воздействием с TGF-β1 в течение 48 ч. Клетки метили антителом к TGF-RII (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 18: предварительная обработка ASO (SEQ ID NO: 218b) и последующее совместное воздействие с TGF-β1 приводит к снижению внутриклеточного уровня белка pSmad3. Экспрессия белка pSmad3 снижалась после трансфекции гимнозисом TGF-RII-специфического ASO с последующим совместным воздействием с TGF-β1 в клетках (48 ч) A549 (фиг. 18A) и ReNcell CX® (фиг. 18B). ASO инкубировали в течение 72 ч или 96 ч соответственно перед совместным воздействием с TGF-β1 в течение 48 ч. Клетки метили антителом к pSmad3 (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 19: ASO (SEQ ID NO: 218b) усиливает и TGF-β1 снижает нейрогенез в клетках-предшественниках нейронов человека ReNcell CX®. мРНК маркера нейрогенеза DCX подвергается повышающей регуляции в клетках ReNcell CX® после повторной трансфекции гимнозисом (2×96 ч) ASO по изобретению. Сильное снижение экспрессии мРНК DCX было обнаружено после 8-суточного воздействия TGF-β1. Уровни мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05 по отношению к 2,5 мкМ С, #p<0,05 по отношению к 10 мкМ С.
На фиг. 20: ASO (SEQ ID NO: 218b) усиливает и TGF-β1 снижает пролиферацию в клетках-предшественниках нейронов человека ReNcell CX®. Экспрессия белка-маркера пролиферации Ki67 повышается в клетках ReNcell CX® после повторной трансфекции гимнозисом (2×96 ч) ASO по изобретению. Пониженную экспрессию белка Ki67 обнаруживали после 8-суточного воздействия TGF-β1. Клетки метили антителом к Ki67 (левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 21: независимо от условий пролиферации ASO (SEQ ID NO: 218b) усиливает дифференцировку клеток-предшественников нейронов человека ReNcell CX®. Отмечено присутствие нейрональных маркеров NeuN (фиг. 23A, левый столбец, красный) и βIII-тубулина (фиг. 23B, левый столбец, красный) в клетках ReNcell CX®. Обработку ASO проводили в течение первых 4 суток в условиях пролиферации и затем еще в течение 4 суток в условиях пролиферации (+EGF/FGF) или дифференцировки (-EGF/FGF). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, +EGF/FGF=пролиферация, -EGF/FGF=дифференцировка.
На фиг. 22: ASO-опосредованное (SEQ ID NO: 218b) спасение нейрональных стволовых клеток от остановки пролиферации, индуцированной TGF-β. Пролиферацию клеток-предшественников нейронов человека ReNcell CX® наблюдали с или без воздействия TGF-β1 в течение 7 суток с последующей обработкой ASO в течение 8 суток. Повышающая регуляция мРНК GFAP (фиг. 24А), Ki67 (фиг. 24В) и DCX (фиг. 24С) через 7 суток после предварительной инкубации с TGF-β1 указывает на восстановление пролиферации стволовых клеток. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05 по отношению к А. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
На фиг. 23: ASO снижает пролиферацию клеток рака легких человека (A549). Экспрессия белка-маркера пролиферации Ki67 снижается в клетках A549 после трансфекции гимнозисом (72 ч) ASO по изобретению. Было обнаружно снижение экспрессии белка Ki67 (левый столбец, зеленый). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 26: обработка ASO опосредует антифибротические эффекты опухоли и ослабляет клеточный стресс. Клетки A549 наблюдали после обработки TGF-β1 или после трансфекции гимозом ASO по изобретению (72 ч). Клетки метили антителом к FN (фиг. 30А, левый столбец, зеленый), фаллоидином (актиновый цитоскелет, фиг. 30В, левый столбец, красный). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 27: обработка ASO опосредует антифибротические эффекты опухоли. Клетки рака легких человека A549 исследовали после предварительной инкубации с TGF-β1 (48 ч) с последующей трансфекцией гимнозисом ASO по изобретению и совместным воздействием с обработкой TGF-β1 (72 ч). Клетки метили антителом к CTGF (фиг. 31А, левый столбец, красный) и FN (фиг. 31В, левый столбец, зеленый). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализированы с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1.
На фиг. 28: обработка ASO опосредует антифибротические эффекты опухоли. Клетки рака легких человека A549 наблюдали после предварительной инкубации с TGF-β1 (48 ч) с последующей трансфекцией гимнозисом ASO по изобретению и совместным воздействием с обработкой TGF-β1 (72 ч). Клетки метили антителом к CTGF (фиг. 32А, левый столбец, красный) и FN (фиг. 32В, левый столбец, зеленый). Ядерную ДНК окрашивали DAPI (средний столбец, синий). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с помощью программного обеспечения Image J и программного обеспечения CorelDRAW® X7. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218с, E=TGF-β1.
На фиг. 29 показан антисмысловой олигонуклеотид SEQ ID NO: 209x в форме гапмера, состоящий из 16 нуклеотидов с 2 звеньями LNA (Gb1 и Tb1) на терминальном 5'-конце и 3 звеньями LNA (Ab1 и Gb1 и C*b1) на терминальном 3'-конце и 11 нуклеотидов ДНК (dA, dG, dT, dG, dT, dT, dT, dA, dG, dG и dG) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) в терминальных концевых группах на терминальном 5'-конце и на терминальном 3'-конце.
На фиг. 30 показан антисмысловой олигонуклеотид SEQ ID NO: 152h в форме гапмера, состоящий из 15 нуклеотидов с 4 звеньями LNA (С*b1 и Gb1 и Ab1 и Tb1) на терминальном 5'-конце и 3 звеньями LNA (Ab1 и C*b1 и Ab1) на терминальном 3'-конце и 8 нуклеотидов ДНК (dA, dC, dG, dC, dG, dT, dC и dC) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) в первом и втором последнем звене LNA от терминального 5'-конца.
На фиг. 31 показан антисмысловой олигонуклеотид SEQ ID NO: 143h в форме гапмера, состоящий из 14 нуклеотидов с 2 звеньями LNA (С*b1 и Tb1s) на терминальном 5'-конце и 3 звеньями LNA (C*b1 и C*b1 и Gb1) на терминальном 3'-конце и 9 нуклеотидов ДНК (dC, dG, dT, dC, dA, dT, dA, dG и dA) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) в первом, третьем от последнего и втором звене LNA от терминального 5'-конца.
На фиг. 32 показан антисмысловой олигонуклеотид SEQ ID NO: 213k в форме гапмера, состоящий из 17 нуклеотидов с 3 звеньями LNA (С*b1 и Ab1 и Gb1) на терминальном 5'-конце и 3 звеньями LNA (Gb1 и Tb1 и Gb1) на терминальном 3'-конце и 11 нуклеотидов ДНК (dG, dC, dA, dT, dT, dA, dA, dT, dA, dA и dA) между сегментами LNA с фосфоротиоатными межнуклеотидными связями и азотистым основанием 5-метилцитозином (C*) в первом звене LNA от терминального 5'-конца.
Примеры
Материалы и методы
Большинство антисмысловых олигонуклеотидов, а также контрольных или стандартных олигонуклеотидов, используемых здесь, синтезировали EXIQON в качестве заказных олигонуклеотидов в соответствии с потребностями изобретателей/заявителя. Олигонуклеотиды, имеющие следующие последовательности, использовали в качестве стандартов:
Стандартный олигонуклеотид 0=dCsdAsdGsdCsdCsdCsdCsdCsdGsdAsdCsdCsdCsdAsdTsdG (SEQ ID NO: 147c);
Стандартный олигонуклеотид 1=Ab1sAb1sC*b1sdAsdCsdGsdTsdCsdTsdAsdTsdAsC*b1sGb1sC*b1 (SEQ ID NO: 76);
Стандартный олигонуклеотид 2=C*b1sAb1sGb1sdCsdCsdCsdCsdCsdGsdAsdCsdCsdCsAb1sTb1sGb1 (SEQ ID NO: 147m);
Стандартный олигонуклеотид 3=TTGAATATCTCATGAATGGA; содержащий 2'-MOE-группы (5 звеньев 5'и 3') и фосфоротиоатные связи (SEQ ID NO: 80);
Стандартный олигонуклеотид 4=CAGAAGAGCTATTTGGTAGT; содержащий 2'-MOE-группы (5 звеньев 5'и 3') и фосфоротиоатные связи (SEQ ID NO: 82);
Стандартный олигонуклеотид 5=TGGTAGTGTTTAGGGAGCCG (SEQ ID NO: 85)
Стандартный олигонуклеотид 6=GTGCAGGGGAAAGATGAAAA (SEQ ID NO: 344)
Стандартный олигонуклеотид 7=GAGCTCTTGAGGTCCCTGTG (SEQ ID NO: 345)
Стандартный олигонуклеотид 8=AGCCTCTTTCCTCATGCAAA (SEQ ID NO: 346)
Стандартный олигонуклеотид 9=CCTTCTCTGCTTGGTTCTGG (SEQ ID NO: 347) и
Стандартный олигонуклеотид 10=GCCATGGAGTAGACATCGGT (SEQ ID NO: 348).
Протоколы стандартных процедур:
Культивирование клеток:
карцинома
карцинома
Материал:
FCS (ATCC #30-2020)
Пируват натрия (Sigma #S8636)
Бикарбонат натрия (Sigma #S8761-100ML)
Трансферрин (Sigma #T8158-100MG)
Селенит натрия (Sigma #S5261-10G)
Пенициллин/стрептомицин (P/S) (Sigma-Aldrich #P4458)
Незаменимые аминокислоты (AS) 100× (Sigma #M7145)
Антибиотик/антимикотик (Sigma #A5955)
MEM раствор витаминов (Sigma #M6895)
PBS (Sigma #D8537)
Основный фактор роста фибробластов человека (Millipore #GF003)
Эпидермальный фактор роста человека (Millipore #GF144)
Добавка N-2 (Life Technologies #17502048)
Среда для поддержания нейрональных стволовых клеток ReNcell (Millipore #SCM005)
Культивирование и диссеминирование клеток:
После удаления среды клетки промывали PBS и инкубировали с аккутазой (Sigma-Aldrich #P4458) (5 мин, комнатная температура). После инкубации клетки подвергали разделению и добавляли полную среду (3 мл, компания-производитель: смотри таблицу 10 для соответствующих клеточных линий). Затем клетки переносили в пробирку Эппендорфа емкостью 5 мл и центрифугировали (5 мин, 1000 об/мин, комнатная температура). Осадок из флакона 1 T75 (Sarstedt #833.910.302) ресуспендировали в 2,5 мл свежей среды. Количество клеток в клеточной суспензии определяли с помощью автоматического счетчика клеток Luna-FL™ (Biozym #872040) окрашиванием с использованием набора для анализа жизнеспособности клеток окрашиванием красителем акридиновый оранжевый/пропидиум иодид (Biozym #872045). Для адгезии клеток ReNcell CX® необходимо покрытие планшетов ламинином (Millipore #CC095) перед посевом клеток для экспериментов из расчета 2 мкг/см2. Раствор ламинина-PBS вносили в соответствующем количестве непосредственно в лунки и колбы и инкубировали в течение 1,5 ч при 37°С. Для экспериментов клетки высевали и собирали, как описано в методическом подразделе соответствующего экспериментального раздела. После инкубирования в течение ночи клеток при 37°С и 5% СО2 клетки обрабатывали, как описано в соответствующем описании эксперимента. 500 мкл оставшейся клеточной суспензии помещали в новый флакон Т75, заполненный 10 мл свежей полной среды для культивирования клеток.
Анализ РНК
Полную фракцию РНК для синтеза кДНК выделяли с использованием мининабора innuPREP® (Analytik Jena #845-KS-2040250), следуя инструкциям производителя. Для синтеза кДНК определяли общее содержание РНК с использованием фотометра (Eppendorf, BioPhotometer D30 #6133000907), при разбавлении водой, не содержащей нуклеазы. Затем получали первую цепь кДНК с использованием набора для синтеза кДНК iScript™ (BioRad #170-8891), следуя рекомендациям производителя. Для анализа мРНК проводили ОТ-ПЦР в режиме реального времени с использованием системы детектирования PCR Detection PCR96 Real Time (BioRad #185-5196).
Все пары праймеров, готовые к применению и стандартизованные, смешивали с соответствующим готовым к использованию раствором Mastermix (SsoAdvanced™ Universe SYBR® Green Supermix (BioRad #172-5271), следуя инструкциям производителя (BioRad Prime PCR Quick Guide). Пары праймеров для экспериментов in vivo были адаптированы соответственно отдельным видам.
В качестве матрицы использовали 1 мкл соответствующей кДНК. РНК, которая не подвергалась обратной транскрипции, служила в качестве отрицательного контроля для ОТ-ПЦР в режиме реального времени. Для относительного количественного анализа использовали ген «домашнего хозяйства» бета-субъединицы-2-подобной 1 гуанин-нуклеотид-связывающего белка (GNB2L1). ОТ-ПЦР в режиме реального времени проводили по следующему протоколу:
(градиент 0,5°С)
Затем BioRad CFX Manager 3.1 использовали для количественного определения уровня соответствующей мРНК относительно мРНК GNB2L1 и затем нормализовали к необработанному контролю.
Вестерн-блоттинг:
Для анализа белков клетки/ткани лизировали с использованием реагента для экстракции белков млекопитающих M-PER®/реагента для экстракции белков из тканей T-PER® (Thermo Scientific, № 78501/#78510), следуя инструкциям производителя. SDS-акриламидные гели (10%) готовили с использованием набора TGX Stain Free™ Fast Cast™ Acrylamide (BioRad #161-0183), следуя инструкциям производителя. Образцы белка (20 мкл) разбавляли 1:5 буфером Лэммли (6,5 мкл, Roti®-Load1, Roth #K929.1), инкубировали при 60°С в течение 30 мин и весь объем белкового раствора наносили на гель. Разделение белков проводили электрофорезом с использованием Power Power Supply (Biorad #164-5050SP) и камеры для электрофореза Mini-PROTEAN® Tetra (BioRad #165-8001-SP) (200 В, 45 мин). После электрофореза белки гибридизовали с использованием системы переноса белков с гелей на мембраны Turbo Trans-Blot® (BioRad #170-4155SP). Все материалы для постановки вестерн-блоттинга входили в набор Trans-Blot® Turbo RTA PVDF-Midi (BioRad #170-4273).
PVDF-мембрану для блоттинга активировали в метаноле (Merck #1.06009.2511) и уравновешивали в 1× буфере для переноса. После блоттинга (25 В, 1 А, 30 мин) мембраны промывали (3×, 10 мин, комнатная температура) 1× TBS (Roth #10.60.1), содержащим 0,5 мл твина-20 (Roth #9127.1). После этого мембраны блокировали 5% BSA (не содержащий альбумин IgG, Roth #3737.3), разбавляли TBS-T в течение 1 ч при комнатной температуре, добавляли первичные антитела (разведенные в 0,5% BSA в TBS-T, таблица 13) и инкубировали при 4°С в течение 2 суток. Антитела для экспериментов in vivo выбирали соответственно видовой специфичности.
На следующей стадии мембраны промывали TBS-T (3× 10 мин, комнатная температура) и инкубировали со вторичным антителом (1 ч, комнатная температура, таблица 13). После инкубации блоты промывали TBS-T, проявляли с использованием субстрата Luminata™Forte Western HRP (Millipore #WBLUF0500) и полосы детектировали с помощью люминесцентного анализатора изображений (ImageQuant™ LAS 4000, GE Healthcare). Затем блоты промывали TBS-T (3× 10 мин, комнатная температура) и блокировали 5% BSA, разведенным TBS-T (1 ч, комнатная температура). Для сравнения с геном «домашнего хозяйства» мембраны инкубировали с HRP-конъюгированным анти-альфа-тубулин-антителом (1:2000 в 0,5% BSA, 4°С, в течение ночи). На следующий день блоты проявляли с использованием субстрата Luminata™Forte Western HRP (Millipore #WBLUF0500) и полосы детектировали с помощью люминесцентного анализатора изображений. Наконец, блоты промывали TBS-T (3× 5 мин) и окрашивали с использованием 1х раствора Roti®-Blue (Roth #A152.2) и высушили при комнатной температуре.
Иммуноцитохимия
Клетки обрабатывали и собирали, как описано выше. После фиксации клеток с использованием Roti®-Histofix 4% (Roth #P087.4) в 8-луночных слайд-планшетах для культивирования клеток (6 мин, комнатная температура) их трижды промывали PBS. После блокирования клеток в течение 1 ч при комнатной температуре с помощью блокирующего раствора (Zytomed #ZUC007-100) клетки инкубировали с соответствующими первичными антителами, приведенными в таблице 14, и инкубировали при 4°С в течение ночи.
Затем клеточные культуральные слайды трижды промывали PBS после инкубации со вторичным антителом (1 ч, комнатная температура). Все разведения антител готовили с помощью разбавителя антител (Zytomed #ZUC025-100).
После инкубации со вторичным антителом клетки трижды промывали PBS, покровные стекла снимали с планшетов для культивирования клеток и монтировали препараты с использованием среды для монтировки VECTASHIELD® HardSet™ с DAPI (Biozol #VEC-H-1500). Препараты высушивали в течение ночи при 4°С перед флуоресцентной микроскопией (Zeiss, Axio® Observer.Z1). Изображения анализировали с использованием программного обеспечения Image J и программного обеспечения CorelDRAW® X7.
Эксперименты in vivo
Тест с мононуклеарными клетками периферической крови
РВМС отделяли от лейкоцитарной пленки, соответствующей трансфузионным единицам 500 мл полной крови. Каждую единицу получали от здоровых добровольцев, и цитрат глюкозы использовали в качестве антикоагулянта. Готовили лейкоцитаную пленку крови, и она была доставлена Банком крови Suhl Института трансфузионной медицины, Германия. Кровь от каждого донора проверяли на антитела к ВИЧ, антитела к HCV, антиген HBs, TPHA, ВИЧ-РНК и SPGT (ALAT). Только образцы крови с отрицательным результатом при тестировании на наличие инфекционных агентов и с нормальным значением SPGT использовали для разделения лейкоцитов и эритроцитов низкоскоростным центрифугированием. Выделение РВМС проводили в течение 40 ч с момента отбора у донора градиентным центрифугированием с использованием Ficoll-Histopague® 1077 (Heraeus™ Multifuge™ 3 SR). Для анализа IFN-α PBMC высевали из расчета 100000 клеток/лунку 96-луночного планшета в 100 мкл полной среды плюс добавки (RPMI1640+L-Glu,+10% FCS+PHA-P (5 мкг/мл)+IL-3 (10 мкг/мл)) и добавляли тестируемые соединения (5 мкл) для прямой инкубации (24 ч, 37°С, 5% CO2). Для анализа TNF-α РВМС высевали из расчета 100000 клеток/лунку 96-луночного планшета в 100 мкл полной среды без добавок (RPMI1640+L-Glu+10% FCS) и вносили тестируемые соединения (5 мкл) для прямой инкубации (24 ч, 37°С, 5% СО2). ELISA (измерения в двух повторностях из объединенных супернатантов, 20 мкл) проводили для анализа huIFNα (eBioscience, #BMS216INSTCE), следуя инструкциям производителя. ELISA (измерения в двух повторностях из объединенных супернатантов, 20 мкл) проводили для анализа huTNFα (eBioscience, #BMS223INSTCE), следуя инструкциям производителя.
Анализ bДНК
Уровни мРНК TGF-RII определяли в лизате печени, почек и легких анализом bДНК, следуя инструкциям производителя (набор QuantiGene®, Panomics/Affimetrix).
Иммунофлуоресценция
Из заключенной в парафин ткань спинного мозга и головного мозга готовили срезы толщиной 5 мкм (3-4 среза на предметное стекло). Парафиновые срезы депарафинизировали и демаскировали нагреванием в цитратном буфере (10 мМ, 40 мин) в микроволновой печи. Затем депарафинизированные срезы инкубировали с 0,3% H2O2 (30 мин, комнатная температура), промывали PBS (10 мин, комнатная температура) и блокировали блокирующим раствором (Zytomed #ZUC007-100) в течение 30 мин. После блокирования в течение 1 ч при комнатной температуре с помощью блокирующего раствора (Zytomed) срезы инкубировали с 150 мкл соответствующих первичных антител и инкубировали при 4°С в течение ночи. После промывки PBS (три раза, 5 мин, комнатная температура) срезы инкубировали с вторичным антителом в течение 1 ч при комнатной температуре. Все разведения антител готовили с использованием разбавителя для антител (Zytomed #ZUC025-100). Затем срезы снова промывали PBS (три раза, 5 мин, комнатная температура) и монтировали с использованием среды для монтировки VECTASHIELD® с DAPI (Vector). Антитела для иммунофлюоресценции были сопоставимы с таковыми в экспериментах по культивированию клеток и адаптированы для каждого вида.
Электрохемилюминесценция
Для выявления иммунологических и гематологических изменений использовали метод электрохемилюминесценции (MesoScale Discovery®, Мэриленд, США). Для каждого анализа использовали 25 мкл образцов белка, крови и спинномозговой жидкости, и процедуру выполняли, следуя инструкциям производителя.
Анализ BrdU
Мечение делящихся клеток проводили внутрибрюшинной инъекцией аналога тимидина BrdU (Sigma, Steinheim, Германия) в дозе 50 мг/кг массы тела с использованием стерильного раствора BrdU с концентрацией 10 мг/мл, растворенного в 0,9% (мас./об.) растворе NaCl. Инъекции BrdU проводили ежедневно на последней неделе эксперимента.
Хирургия
Для длительной центральной инфузии животных подвергали хирургической операции для имплантации icv канюли, присоединенной к осмотическому мининаносу Alzet® (мыши, крысы, скорость инфузии 0,25 мкл/ч, Alzet®, модель 2004, Купертино, США) или газовому насосу давления (обезьяны рода Cynomolgus, скорость инфузии 0,25 мл/ч, Tricumed®, Model IP 2000V, Германия). Канюлю и насос стереотаксически имплантировали под анестезией кетамином/ксилацином (Baxter, GmbH, Германия) в полустерильных условиях. Каждый осмотический мининасос/газовый насос давления имплантировали подкожно со стороны брюшной области посредством разреза кожи длиной 1 см на шее животных и соединяли с icv канюлей силиконовыми трубками. Животных помещали в стереотаксическую рамку, и icv канюлю погружали в правый боковой желудочек. Канюлю фиксировали двумя винтами из нержавеющей стали с использованием зубного цемента (Kallocryl, Speiko® Dr. Speier GmbH, Мюнстер, Германия). Кожу на шее закрывали швами. Во время операции температуру тела поддерживали грелкой. Для профилактики послеоперационных инфекций мышей местно обрабатывали препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 0,1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия). Трубку заполняли соответствующим раствором. Для анализа отбирали образцы крови, спинномозговой жидкости и тканей. Гистологическое подтверждение мест icv имплантации проводили на срезах мозга толщиной 40 мкм, окрашенных крезил-фиолетовым.
Показатели клинического исхода и функциональный анализ
Начало симптоматического заболевания, начало первого пареза и выживаемость использовались в качестве конечных точек в опытах in vivo. Начало симптоматического течения заболевания определяли по отсутствию вытягивания конечностей в тесте подвешивания за хвост. Временная точка, на которой были обнаружены нарушения походки (например, качающаяся походка или прихрамывание), принимали за начало первого пареза. Эти показатели определяли ежедневно, начиная с 40 суток.
Для наблюдения за прогрессированием заболевания проводили тест «беличье колесо» (LMTB, Берлин, Германия). Животных помещали в клетки по отдельности с доступом к «беличьему колесу», начиная с 33-дневного возраста. Моторная активность непосредственно коррелировала с количеством оборотов в минуту, проделанных каждым животным на «беличьем колесе». Каждый полный оборот колеса производил два электромагнитных сигнала, напрямую подаваемых в компьютер, подключенный максимум к 120 колесам. Данные по тесту «беличье колесо» записывались и анализировались с помощью программного обеспечения «Maus Vital» (Laser- und Medizin-Technologie, Берлин, Германия). Время оценки составляло 12 ч с 18:00 до 6:00.
Пространственное обучение (водный бассейн Морриса)
Тестирование поведенческих реакций проводили с 8:00 до 13:00. Крыс обучали в черном круглом бассейне (диаметром 1,4 м, высотой 50 см, заполненном теплой водой 20°C до высоты 30 см), для поиска видимой белой мишени (10 см в диаметре, поднятой над поверхностью воды примерно на 1 см), которую располагали в течение всего исследования в центре того же воображаемого квадрата (проксимальный сигнал). Каждое животное обучалось ориентироваться на платформе в 3 последовательных раундах с 12 испытаниями/раунд, один раунд в день и с интервалом между испытаниями 10-20 с.
Микробиологический анализ
Образцы антисмысловых олигонуклеотидов подвергали микробиологическому анализу согласно статье Европейской фармакопеи 2.6.12, статье Фармакопеи США 30 <61> в отношении общего количества аэробных микроорганизмов (TAMC) и общего комбинированного количества дрожжей и форм (TYMC).
Анионобменная высокоэффективная жидкостная хроматография (AEX-HPLC)
Целостность и стабильность образцов антисмысловых олигонуклеотидов (ASO) определяли с помощью AEX-HPLC с использованием системы ÄKTAexplorer™ (GE Healthcare, Фрайбург, Германия). Очищенные образцы ASO обессоливали осаждением этанолом. Идентичность ASO подтверждали методом масс-спектрометрии с ионизацией электрораспылением (ESI-MS) и чистоту определяли AEX-HPLC на колонке Dionex DNAPac™ 200 (4×250 мм).
Пример 1: определение ингибирующей активности антисмысловых олигонуклеотидов по изобретению на уровень мРНК
1.1. Трансфекция антисмысловых олигонуклеотидов
Ингибирующую активность нескольких антисмысловых олигонуклеотидов, направленных против TGF-RII, тестировали на человеческих эпителиальных клетках рака легких (A549). мРНК TGF-RII определяли количественным анализом с использованием метода разветвленной ДНК в общей мРНК, выделенной из клеток, инкубированных с TGF-RII-специфическими олигонуклеотидами.
Описание метода:
Клетки получали и культивировали, как описано выше. Трансфекцию антисмысловых олигонуклеотидов проводили непосредственно после посева 10000 клеток A549/лунку в 96-луночном планшете и проводили с использованием Lipofectamine® 2000 (Invitrogen GmbH, Карлсруэ, Германия, номер по каталогу 11668-019), как описано производителем. В двух независимых экспериментах с одной концентрацией, проведенных в четырех повторностях, олигонуклеотиды трансфектировали при концентрации 20 нМ. После трансфекции клетки инкубировали в течение 24 ч при 37°С и 5% CO2 в увлажненном термостате (Heraeus GmbH, Ханау, Германия). Для измерения мРНК TGF-RII клетки собирали и лизировали при 53°С, следуя процедурам, рекомендованным производителем набора QuantiGene® Explore (Panomics, Fremont, CA, USA, номер по каталогу QG0004) для выделения разветвленной ДНК (bДНК). Для количественного определения содержания мРНК гена «домашнего хозяйства» глицеральдегид-3-фосфатдегидрогеназы (GAPDH) использовали набор QuantiGene® Explore, в то время как количественное определение мРНК TGF-RII проводили с использованием набора QuantiGene® 2.0 (произведено на заказ для Axolabs GmbH, Kulmbach, Германия). После инкубации и лизиса 10 мкл лизатов инкубировали с наборами зондов, специфичных к человеческому TGF-RII и человеческой GAPDH. Оба типа реакции проводили, следуя протоколу производителя, для соответствующего набора QuantiGene®. Хемолюминесценцию измеряли на счетчике Victor2™ multilabel counter (Perkin Elmer, Wiesbaden, Германия) в виде RLU (относительныесветовые единицы), и значения, полученные с наборами зондов к TGF-RII, нормализовали к соответствующим значениям GAPDH для каждой лунки, и затем нормализовали к соответствующему показанию мРНК из ложнообработанных клеток.
Результаты
Результаты свидетельствуют о выраженной подавляющей регуляции TGF-RII несколькими ASO после трансфекции клеток A549. Понижающая регуляция после трансфекции стандартных олигонуклеотидов 6-10 не была столь эффективной и приводила к подавлению не более чем на 60%.
Таблица 15: понижающая регуляция мРНК TGF-RII. Трансфекция TGF-RII-специфическими антисмысловыми олигонуклеотидами (ASO) клеток эпителиальной карциномы легких человека (A549). Количественное определение уровней экспрессии мРНК проводили по отношению к гену «домашнего хозяйства» GAPDH с использованием набора QuantiGene®. Затем зонды нормализовали к соответствующему показанию для мРНК из ложнообработанных клеток.
Заключение
ASO по изобретению были эффективно нацелены на мРНК TGF-RII. Указанные ASO обеспечивали эффективную понижающую регуляцию мРНК-мишени после трансфекции клеток A549.
1.2. Поглощение гимнозисом антисмысловых олигонуклеотидов
1.2.1a Сравнение нокдаунов мишени для ASO по изобретению и последовательностям предшествующего уровня трансфекцией гимнозисом в клетках A549 и Panc-1
Понижающую регуляторную активность нескольких антисмысловых олигонуклеотидов, нацеленных на TGF-RII, тестировали в человеческих эпителиальных клетках рака легких (A549) путем прямого поглощения без использования трансфекционных реагентов («поглощение гимнозисом»). мРНК TGF-RII определяли количественным анализом с использованием метода разветвленной ДНК в общей мРНК, выделенной из клеток, инкубированных с TGF-RII-специфическими олигонуклеотидами.
Описание метода:
Клетки получали и культивировали, как описано в общих методах. Трансфекцию гимнозисом антисмысловых олигонуклеотидов осуществляли приготовлением 96-луночного планшета с соответствующими антисмысловыми олигонуклеотидами и последующим посевом из расчета 10000 клеток (Panc-1) или 8000 клеток (A549)/ лунку. Эксперименты проводили в четырех повторностях, олигонуклеотиды использовали в конечных концентрациях 5 мкМ (Panc-1) и 7,5 мкМ (A549). Клетки инкубировали в течение 72 ч при 37°С и 5% CO2 в увлажненном термостате (Heraeus GmbH, Hanau, Германия). Для измерения мРНК TGF-RII клетки собирали и лизировали при 53°С, следуя процедурам, рекомендованным производителем набора QuantiGene® Explore (Panomics, Fremont, CA, США, номер по каталогу QG0004) для разветвленной ДНК (bДНК). Для количественного определения мРНК гена «домашнего хозяйства» GAPDH использовали набор QuantiGene® Explore, в то время как количественное определение мРНК TGF-RII проводили с использованием набора QuantiGene® 2.0 (произведен по заказу для Axolabs GmbH, Kulmbach, Германия). После инкубации и лизиса 10 мкл лизатов инкубировали с наборами зондов, специфичными к человеческому TGF-RII и человеческому GAPDH. Оба типа реакции проводили, следуя протоколу производителя для соответствующего набора QuantiGene®. Хемолюминесценцию измеряли на счетчике Victor2™ multilabel counter (Perkin Elmer, Wiesbaden, Германия) в виде RLU (относительные световые единицы), и значения, полученные с наборами зондов TGF-RII нормализовали к соответствующим значениям GAPDH для каждой лунки, и затем нормализовали к соответствующему показанию мРНК из обработанных PBS клеток.
Выбранные результаты представлены в таблице 16a. Дальнейшие модифицированные последовательности SEQ ID NO: 209ay, SEQ ID NO: 209аx и SEQ ID NO: 209y, а именно ASO, представленные в таблице 6 описания, показали сравнимые значения с этими тремя антисмысловыми олигонуклеотидами. Кроме того, модификации SEQ ID NO: 152h, а именно ASO, представленный в таблице 5 описания, показали сравнимые значения с этим антисмысловым олигонуклеотидом. Модификации SEQ ID NO: 218b, а именно ASO, представленный в таблице 8 описания, показали сравнимые значения с антисмысло-олигонуклеотидом SEQ ID NO: 218b. Модификации SEQ ID NO: 213k, а именно ASO, представленные в таблице 9 описания, показали сопоставимые значения с антисмысловыми олигонуклеотидом SEQ ID NO: 213k. Также модификации SEQ ID NO: 210q, а именно ASO, представленный в таблице 7 описания, показали сопоставимые значения с антисмысловым олигонуклеотидом SEQ ID NO: 210q. Наконец, модификации SEQ ID NO: 143h, а именно ASO, представленный в таблице 4 описания, показали сопоставимые значения с антисмысловым олигонуклеотидом SEQ ID NO: 143h. Трансфекция антисмысловых олигонуклеотидов, представленных в таблицах 4-9, приводила к более сильной понижающей регуляции мРНК-мишени TGF-RII по сравнению с трансфекцией тестированных стандартных последовательностей (клетки A549: подавление <0,5, клетки Panc1: подавление <0,4).
Таблица 16а: эффективность понижающей регуляции мРНК-мишени посредством трансфекции гимнозисом. Оставшаяся мРНК TGF-RII после поглощения гимнозисом выбранных TGF-RII-специфических ASO в клетках A549 и Panc-1. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» глицеральдегид-3-фосфатдегидрогеназы (GAPDH) и сравнивали с клетками, обработанными PBS, которые использовали в качестве контроля (=1) с использованием набора QuantiGene®.
Заключение:
Трансфекция гимнозисом ASOs по изобретению приводит к неизменно более сильной понижающей регуляции мРНК-мишени TGF-RII, чем трансфекция тестированными контрольными последовательностями. Антисмысловые олигонуклеотиды по изобретению превосходили все тестированные последовательности, известные из предшествующего уровня техники, независимо от выбранной клеточной линии человека. Тем не менее, в целом антисмысловые олигонуклеотиды, имеющие длину 12-20 нуклеотидов, приводят к более эффективной понижающей регуляции мРНК-мишени TGF-RII, чем более короткие или более длинные антисмысловые олигонуклеотиды. Такой эффект был еще более заметным для антисмысловых олигонуклеотидов, имеющих длину 14-18 нуклеотидов, которые, как правило, проявляли наиболее сильные эффекты.
1.2.1b Анализ трансфекции гимнозисом клеток A549 с использованием метода разветвленной ДНК
Наиболее эффективные антисмысловые олигонуклеотиды против TGF-RII по данным скрининга трансфекции были дополнительно охарактеризованы поглощением гимнозисом в клетки A549. мРНК TGF-RII количественно определяли методом разветвленной ДНК в общей мРНК, выделенной из клеток, инкубированных с TGF-RII-специфическими антисмысловыми олигонуклеотидами.
Описание метода:
Клетки A549 культивировали, как описано ранее в стандартных условиях. Для экспериментов с одной концентрацией и исследования зависимости концентрация-эффект 80000 клеток A549/лунку высевали в 6-луночный культуральный планшет и инкубировали непосредственно с олигонуклеотидами в концентрации 7,5 мкМ. Для измерения мРНК TGF-RII клетки собирали, лизировали при 53°С и анализировали методом разветвленной ДНК, следуя процедурам, рекомендованным производителем набора QuantiGene® Explore (Panomics, Fremont, CA, USA, номер по каталогу QG0004), как описано выше (смотри 1.1).
Результаты:
Приведенные ASO в таблице 16b показали снижение уровня мРНК-мишени TGF-RII по сравнению с геном «домашнего хозяйства» GAPDH в клетках A549. Для десяти наиболее эффективных ASO также определяли 50% ингибирующую концентрацию (IC50). Все SEQ ID NO: 209t, SEQ ID NO: 218b, SEQ ID NO: 218с и SEQ ID NO: 209y приводили к наиболее специфическому нокдауну TGF-RII в низких концентрациях.
Таблица 16b: понижающая регуляция мРНК TGF-RII после поглощения гимнозисом TGF-RII-специфических ASO в клетках A549. Уровни мРНК определяли относительно гена «домашнего хозяйства» GAPDH с использованием набора QuantiGene®. IC50=ингибирующая концентрация, вызывающая 50% понижающую регуляцию, Pos. Ctrl: LNA, нацеленные на aha-1=активатор белка теплового шока 90 кДа гомолога ATPазы 1 (Aha1), который использовали в качестве положительного контроля, стандартный образец 1=скремблированный контроль.
n=4
n=4
n=4
Заключение:
Понижающая регуляция мишени наиболее эффективными ASO по изобретению вновь была высокой без использования трансфекционных реагентов. Таким образом, трансфекция гимнозисом может быть осуществимым и предпочтительным методом для дальнейшей разработки лекарственного средства.
1.2.2. Анализ поглощения гимнозисом клетками A549 и ReNcell CX ®
Ингибирующую активность антисмысловых олигонуклеотидов (ASO) в отношении мРНК-мишени определяли в клетках-предшественниках нейронов из кортикальной области мозга человека (клетки ReNcell CX®, Millipore #SCM007). Вопросы относительно взрослого нейрогенеза в качестве терапевтической мишени оценивали в опытах по трансфекции гимнозисом с наиболее эффективными ASO. В качестве контрольной клеточной линии использовали клетки A549.
Описание метода:
Клетки A549 и ReNcell CX® культивировали, как описано выше. Для проведения исследований клетки высевали в 24-луночный культуральный планшет (Sarstedt #83.1836.300) (50000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для обработки клеток A549 и ReNcell CX® среду удаляли и заменяли свежей полной средой (0,5 мл для 24-луночного планшета). Затем стандартный олигонуклеотид 1, ASO с SEQ ID NO: 218b и ASO с SEQ ID NO: 218c добавляли в среду в концентрациях 2,5 и 10 мкМ для анализа понижающей регуляции мишени на различные временные точки (клетки A549: 18 ч, 72 ч, 6 суток, клетки ReNcell CX®: 18 ч, 96 ч, 8 суток) при 37°С и 5% СО2. Для сбора клетки дважды промывали PBS и замораживали при -20°C. Для анализа мРНК с использованием ОТ-ПЦР в режиме реального времени клетки обрабатывали, как описано выше. Использовали готовые к применению и стандартизованные пары праймеров для ОТ-ПЦР в режиме реального времени (смотри таблицу 11) и смешивали с соответствующим готовым к применению раствором Mastermix (SsoAdvanced™ Universe SYBR® Green Supermix (BioRad #172-5271), следуя инструкциям производителя (BioRad Prime PCR Quick Guide). Зонды анализировали в трех повторностях и данные количественно оценивали по отношению к мРНК GNB2L1, используя BioRad CFX Manager™ 3.1, и затем нормализовали к необработанному контролю. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Результаты:
Результаты показали, что трансфекция гимнозисом SEQ ID NO: 218b и 218c приводит к специфической понижающей регуляции мРНК TGF-RII в клетках A549 и ReNcell CX® в зависимости от концентрации и времени (таблица 17). Уровень мРНК-мишени в клетках A549 был достоверно снижен через 18 ч, и через 72 ч и 6 суток был еще более эффективно снижен. Через 18 ч в клетках ReNcell CX® наблюдали только незначительное снижение мРНК TGF-RII после поглощения гимнозисом 10 мкМ, но понижающая регуляция мишени была достоверной через 72 ч для обеих тестируемых концентраций и была стабильной до 8 суток.
Таблица 17: зависимая от концентрации и времени понижающая регуляция мРНК TGF-RII после трансфекции гимнозисом TGF-RII-специфического ASO в клетках A549 и ReNcell CX ®. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, D=SEQ ID NO: 218c, ±=SEM, *p<0,05, **p<0,01 по отношению к A, +p<0,05, ++p<0,01 по отношению к B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
18 ч, n=3
72 ч, n=3
6 суток, n=3
Временная точка
18 ч, n=3
96 ч, n=3
8 суток, n=3
Заключение:
Эффективная и стабильная понижающая регуляция мРНК-мишени посредством поглощения гимнозисом ASO достигается даже при длительном применении. Следовательно, клетки ReNcell CX® можно использовать, например, для экспериментов, направленных на восстановление взрослого нейрогенеза в качестве терапевтического варианта для пациентов. То же самое относится к другим показаниям, как показано результатами экспериментов с клетками А549.
В совокупности, эффективная понижающая регуляция TGF-RII подходит независимо от способа трансфекции и типа клеток. Поглощение гимнозисом ASO является предпочтительным методом трансфекции, поскольку в клинических применениях отсутствие дополнительных трансфекционных агентов обеспечивает высокую безопасность для пациентов.
Пример 2: определение ингибирующей активности антисмысловых олигонуклеотидов, нацеленных на TGF-RII, на уровень белка-мишени
Анализ вестерн-блоттингом и иммуноцитохимией проводили для определения того, приводит ли снижение уровня мРНК TGF-RII, опосредованное антисмысловыми олигонуклеотидами (ASO) по изобретению, в клетках рака легких человека (A549) и клетках-предшественниках нейронов человека (ReNcell CX®) к снижению уровня белка-мишени.
Описание метода:
Клетки культивировали, как описано выше. Для обработки клетки высевали в 6-луночный культуральный планшет (Sarstedt #83.3920.300, 80000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #946140802, 10000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для трансфекции гимнозисом клеток A549 и ReNcell CX® клеточную среду удаляли и заменяли свежей полной средой (1 мл на 6-луночного и 0,5 мл на 8-луночного планшетов). Затем стандартный олигонуклеотид 1 (скремблированный контроль), соответствующий ASO по изобретению добавляли в среду в концентрациях 2,5 и 10 мкМ для анализа понижающей регуляции белка-мишени через 72 ч в клетках A549 и 96 ч в клетках ReNcell CX®. Клетки лизировали и исследовали вестерн-блоттингом, как описано в общем методическом разделе. Первичное антитело к TGF-RII разводили 0,5% BSA в TBS-T и инкубировали при 4°C в течение 2 суток. После этого мембраны инкубировали со вторичным антителом к кроличьему IgG, конъюгированному с HRP, разведенным 0,5% BSA в TBS-T (1 ч, комнатная температура). После инкубации блоты промывали TBS-T, визуализировали с использованием субстрата Luminata™Forte Western HRP (Millipore #WBLUF0500) и полосы детектировали с помощью люминесцентного анализатора изображений (ImageQuant™ LAS 4000, GE Healthcare). Для сравнения с геном «домашнего хозяйства» мембраны инкубировали с HRP-конъюгированным анти-GAPDH (1:1000 в 0,5% Blotto, 4°С, в течение ночи). Денситометрический количественный анализ проводили относительно GAPDH и затем нормализовали к необработанному контролю с помощью программного обеспечения Image Studio™ Lite.
Процедуру иммуноцитохимии проводили, как описано в стандартном протоколе. Для подтверждения понижающей регуляции мишени анти-TGF-RII разбавляли и инкубировали в течение ночи при 4°С. В качестве вторичного антитела использовали Cy3-конъюгированное козье антикроличье антитело. Все разведения антител готовили с разбавителем для антител (Zytomed® #ZUC025-100). Исследование клеток проводили с помощью флуоресцентной микроскопии (Zeiss Axio® Observer.Z1). Изображения анализировали с использованием программного обеспечения Image J и программного обеспечения CorelDRAW® X7.
Результаты после трансфекции гимнозисом:
Анализ вестерн-блоттингом и иммуноцитохимией использовали для подтверждения снижения уровня белка TGF-RII. Через 72 ч после трансфекции гимнозисом уровень белка TGF-RII был достоверно снижен при использовании высокой концентрации различных ASO по изобретению по сравнению с необработанным контролем в клетках A549 (таблица 18). Сниженные уровни TGF-RII также наблюдали в клетках ReNcell CX® (таблица 18). Для обеих клеточных линий снижение уровня белка TGF-RII было продемонстрировано анализом вестерн-блоттингом. Результаты иммуноцитохимии свидетельствовали о выраженом, зависимом от концентрации снижении белка TGF-RII в обеих клеточных линиях по сравнению с необработанными клетками и обработанными скремблированным контролем клетками.
Таблица 18: денситометрический анализ после вестерн-блоттинга TGF-RII. Наблюдали снижение белка TGF-RII после трансфекции гимнозисом TGF-RII-специфических ASO в клетках A549 и ReNcell CX® через 72 ч или 96 ч соответственно. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием програмного обеспечения Image Studio™ Lite и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, D=SEQ ID NO: 218с, F=SEQ ID NO: 210q, G=SEQ ID NO: 213k, H=SEQ ID NO: 143h, I=SEQ ID NO: 152h, J=SEQ ID NO: 209az, К=SEQ ID NO: 209y, ±=SEM, *p<0,05 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
72 ч, n=3
96 ч, n=2
Заключение:
В дополнении к понижающей регуляции мРНК-мишени трансфекция гимнозисом SEQ ID NO: 218b, SEQ ID NO: 218с, SEQ ID NO: 210q, SEQ ID NO: 213k, SEQ ID NO: 143h, SEQ ID NO: 152h, SEQ ID NO: 209az и SEQ ID NO: 209y приводит к выраженному снижению уровня белка-мишени в клетках A549 и ReNcell CX®. Окрашивание на TGF-RII выявило дозозависимое снижение уровня белка TGF-RII после обработки данными ASO в обеих клеточных линиях.
Результаты после трансфекции гимнозисом других дополнительных ASO:
Анализ белка показал пониженное количество TGF-RII в клетках A549 и клетках ReNcell CX® после трансфекции гимнозисом тестированных ASO (10 мкМ, таблица 19). Это также было подтверждено результатами иммуноцитохимии. Для обеих клеточных линий можно было выявить снижение уровня белка TGF-RII под действием трансфекции гимнозисом тестированных ASO по сравнению с необработанными клетками и обработанными скремблированным контролем клетками.
Таблица 19: денситометрический анализ после вестерн-блоттинга TGF-RII. Наблюдали снижение уровня белка TGF-RII после трансфекции гимнозисом TGF-RII-специфических ASO в клетках A549 и ReNcell CX® через 72 ч или 96 ч соответственно. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием программного обеспечения Image Studio™ Lite и затем нормализовали к необработанному контролю. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета. Снижение уровня белка для ASO 1-25 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению с SEQ ID NO: 143h; B=более чем на 10%, но менее чем на 20% ниже по сравнению с SEQ ID NO: 143h; C=более чем на 20%, но менее чем на 30% ниже по сравнению с SEQ ID NO: 143h. Снижение уровня белка для ASO 26-48 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению с SEQ ID NO: 152h; B=более чем на 10%, но менее чем на 20% ниже по сравнению с SEQ ID NO: 152h; C=более чем на 20%, но менее чем на 30% ниже по сравнению с SEQ ID NO: 152h. Снижение уровня белка для ASO 49-74 указано с помощью следующего ключа: A=менее чем на 10% ниже среднего значения, полученного с SEQ ID NO: 209az и SEQ ID NO: 209y; B=более чем на 10%, но менее чем 20% ниже среднего значения, полученного с SEQ ID NO: 209az и SEQ ID NO: 209y; C=более 20%, но менее чем на 30% ниже среднего значения, полученного с SEQ ID NO: 209az и SEQ ID NO: 209y. Снижение уровня белка для ASO 75-100 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению с SEQ ID NO: 210q; B=более чем на 10%, но менее чем на 20% ниже по сравнению с SEQ ID NO: 210q; C=более чем на 20%, но менее чем на 30% ниже по сравнению с SEQ ID NO: 210q. Снижение уровня белка для ASO 101-124 указано с помощью следующего ключа: A=менее чем на 10% ниже среднего значения, полученного с SEQ ID NO: 218b и SEQ ID NO: 218с; B=более чем на 10%, но менее чем на 20% ниже среднего значения, полученного с SEQ ID NO: 218b и SEQ ID NO: 218с; C=более чем на 20%, но менее чем на 30% ниже среднего значения, полученного с SEQ ID NO: 218b и SEQ ID NO: 218с. Снижение уровня белка для ASO 125-152 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению SEQ ID NO: 213k; B=более чем на 10%, но менее чем на 20% ниже по сравнению SEQ ID NO: 213k; C=более чем на 20%, но менее чем на 30% ниже по сравнению SEQ ID NO: 213k.
Заключение:
В совокупности, дозозависимая понижающая регуляция мРНК TGF-RII в результате трансфекции гимнозисом в клетках A549 и ReNcell CX® приводила к дозозависимому снижению уровней белка-мишени. ASO по изобретению являются эффективными в понижающей регуляции белка-мишени, как показано в клетках A549 и ReNcell CX®.
Пример 3: анализ влияния антисмысловых олигонуклеотидов на сигнальный путь ниже к TGF-RII.
Функциональные анализы проводили на клетках рака легких человека (A549) и клетках-предшественниках нейронов человека (ReNcell CX®). Сигнальный путь ниже к TGF-β анализировали, основываясь на эффективной понижающей регуляции мРНК TGF-RII и снижения уровней белка после трансфекции гимнозисом ASO по изобретению. Следовательно, оценивали уровни мРНК и белка фактора роста соединительной ткани (CTGF), известного как медиатор ниже к TGF-β. Кроме того, исследовали фосфорилирование Smad2 (матери против гомолога декапентаплегии 2). Фосфорилирование Smad2 является маркером активного пути TGF-β, с последующей повышающей регуляцией гена-мишени CTGF ниже.
Описание метода:
Клетки культивировали, как описано ранее. Для обработки клетки высевали в 6-луночный культуральный планшет (Sarstedt #83.3920.300) (80000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°С и 5% CO2. Для трансфекции гимнозисом культуральную среду клеток A549 и ReNcell CX® удаляли и заменяли свежей полной средой (1 мл на 6- луночный и 0,5 мл на 8-луночный планшет). Затем в среду добавляли стандартный олигонуклеотид 1 (скремблированный контроль), ASO с последовательностью SEQ ID NO: 218b (SEQ ID NO: 218b), SEQ ID NO: 218c (SEQ ID NO: 218c) в концентрации 2,5 и 10 мкМ и через 72 ч в клетках A549 и 96 ч в клетках ReNcell CX® проводили соответствующий анализ. Для оценки эффектов на уровень мРНК CTGF проводили ОТ-ПЦР в режиме реального времени, как описано ранее. Пара праймеров для анализа CTGF была готова к применению и стандартизирована. Для определения уровней белка CTGF и pSmad2 использовали вестерн-блоттинг и иммуноцитохимию, как описано ранее. Тип и использованные разведения антител для соответствующего метода приведены в таблицах 13 и 14.
3.1. Результаты для SEQ ID NO: 218b
3.1.1. Влияние на уровень мРНК и белка CTGF
Уровень мРНК CTGF достоверно и дозозависимо снижался после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549 (72 ч) и ReNcell CX® (96 ч). Уровень медиатора ниже к TGF-β уменьшался до 52±0,02% в клетках ReNcell CX® и до 39±0,03% в клетках A549 после трансфекции гимнозисом 10 мкМ SEQ ID NO: 218b (таблица 20). В совокупности с этими пониженными уровнями мРНК CTGF наблюдали сильное снижение экспрессии белка CTGF в клетках A549 (таблица 21).
Таблица 20: дозозависимая и достоверная понижающая регуляция мРНК CTGF после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549 и ReNcell CX®. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. ±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
72 ч, n=3
96 ч, n=3
Таблица 21: денситометрический анализ после вестерн-блоттинга CTGF. Наблюдали понижающую регуляцию белка CTGF после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549 через 72 ч. Уровни белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Studio™ Lite и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b.
Временная точка
72 ч, n=1
Заключение:
Функциональное ингибирование сигнального пути TGF-β было достигнуто трансфекцией гимнозисом SEQ ID NO: 218b, что было показано наличием понижающей регуляции мРНК-мишени CTGF и снижением уровней белка CTGF в клетках A549 и ReNcell CX®.
3.1.2. Влияние на уровень белка pSmad2
Уровни белка pSmad2 анализировали, чтобы подтвердить отрицательную регуляцию CTGF, в качестве специфического результата ASO-опосредованного ингибирования сигнального пути TGF-β.
Окрашивание на pSmad2 после трансфекции гимнозисом ASO SEQ ID NO: 218b через 72 ч в клетках A549 и 96 ч в клетках ReNcell CX® показало дозозависимое ингибирование фосфорилирования Smad2 (фиг. 5). Кроме того, снижение уровней экспрессии pSmad2 под действием ASO SEQ ID NO:218b подтверждали анализом вестерн-блоттингом в клетках А549 (таблица 22).
Таблица 22: денситометрический анализ после вестерн-блоттинга pSmad2. Наблюдали понижающую регуляцию белка pSmad2 после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549 через 72 ч. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием программного обеспечения Studio™ Lite и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b.
Временная точка
72 ч, n=1
Заключение:
Трансфекция гимнозисом SEQ ID NO: 218b клеток A549 и ReNcell CX® приводила к дозозависимому ингибированию медиаторов сигнального пути ниже к TGF-β. Уровни CTGF и фосфорилирование Smad2 снижались под действием ASO SEQ ID NO: 218b, оба факта указывают на ингибирование пути TGF-β.
3.2 Результаты для SEQ ID NO: 218c
3.2.1. Влияние на уровень мРНК CTRF и белка pSmad2
Трансфекция гимнозисом ASO SEQ ID NO: 218c приводила к понижающей регуляции мРНК CTGF в клетках A549 и ReNcell CX® (таблица 23). Данные иммуноцитохимии для pSmad2 подтверждали ингибирование сигнального пути TGF-β (фиг. 6). Следовательно, понижающая регуляция мРНК CTGF является прямым эффектом подавления сигнального пути TGF-β.
Таблица 23: установлена достоверная понижающая регуляция мРНК CTGF в клетках A549 и ReNcell CX ® . Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218с. ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
линия
Временная точка
72 ч, n=4
96 ч, n=3
Заключение:
ASO SEQ ID NO: 218c был эффективен в ингибировании сигнального пути TGF-β после понижающей регуляции мРНК-мишени TGF-RII. Это исследовали по определению понижающей регуляции мРНК CTGF и пониженному уровню белка pSmad2 в качестве маркера сигнального пути TGF-β.
В совокупности, ASO по изобретению эффективны в опосредовании функционального ингибирования сигнального пути TGF-β посредством понижающей регуляции TGF-RII. Таким образом, ASO будут полезны для медицинских показаний, в которых участвуют повышенные уровни TGF-β, например, неврологических расстройств, фиброза и прогрессирования опухолей.
Пример 4: ингибирующая активность ASO по изобретению на уровни мРНК-мишени в обработанных TGF-β1 клетках
4.1. Поглощение гимнозисом ASO клетками A549 и ReNcell CX ® после предварительной обработки TGF-β1
Для анализа ингибирующей активности антисмысловых олигонуклеотидов (ASO) в клетках-предшественниках нейронов человека из кортикальной области мозга (ReNcell CX®) при патологических состояниях клетки предварительно обрабатывали трансформирующим фактором роста-1 (TGF-β1). По результатам предыдущих исследований известно, что TGF-β1 обнаруживается в высоких концентрациях в цереброспинальной жидкости (CSF) при всех неврологических расстройствах, например, ALS. Таким образом, ингибирующее воздействие ASO на TGF-β-зависимый сигнальный путь было исследовано после предварительной обработки и в присутствии TGF-β1. В качестве стандартной клеточной линии использовали клетки A549.
Описание метода:
A549 и ReNcell CX® культивировали, как описано выше. Для проведения исследований клетки высевали в 24-луночный культуральный планшет (Sarstedt #83.1836.300) (50000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для обработки клеток A549 и ReNcell CX® среду удаляли и заменяли свежей полной средой (0,5 мл для 24-луночного планшете). После экспозиции TGF-β1 (10 нг/ мл, PromoCell #C-63499) в течение 48 ч среду заменяли, повторную обработку TGF-β1 проводили в комбинации со стандартным олигонуклеотидом 1 (скремблированный контроль, 10 мкМ), ASO SEQ ID NO: 218b (10 мкМ) или ASO SEQ ID NO: 218c (10 мкМ) в среде. Клетки A549 инкубировали еще 72 ч, тогда как клетки ReNcell CX® собирали через 96 ч. Соответственно клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты), как описано выше. Использованные пары праймеров для ОТ-ПЦР в режиме реального времени приведены в таблице 11.
4.1.1 Результаты для SEQ ID NO: 218b
На эффективность понижающей регуляции мРНК TGF-RII под действием ASO SEQ ID NO: 218b предварительная инкубация с TGF-β1 не оказывала вияния в клетках A549 и ReNcell CX® (таблица 24, фиг. 7). Уровень мРНК-мишени в клетках A549 был достоверно снижен после обработки одним ASO (оставшаяся мРНК: 15±0,05%), а также после обработки в присутствии TGF-β1 вслед за предварительной обработкой (оставшаяся мРНК: 7±0,01%). В клетках ReNcell CX® ASO SEQ ID NO: 218b показал сходную эффективность ингибирования мРНК TGF-RII в отсутствии TGF-β1 (25±0,01%) или в присутствии TGF-β1 после предварительной обработки TGF-β1 (17±0,02%).
Таблица 24: в присутствии TGF-β1 ASO SEQ ID NO: 218b приводит к сильной понижающей регуляции мРНК TGF-β1 после трансфекции гимнозисом в клетках A549 и ReNcell CX®. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, С=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
48 ч TGF-β1 → 72 ч/96 ч TGF-β1+ASO/обработка одним ASO
n=4
n=3
Заключение:
мРНК-мишень эффективно подверглась понижающей регуляции примерно до уровня 20% в результате трансфекции гимнозисом ASO по изобретению в присутствии TGF-β1 после предварительной инкубации в обеих тестированных клеточных линиях.
4.1.2 Результаты для SEQ ID NO: 218c
Понижающая регуляция мРНК TGF-RII под действием ASO SEQ ID NO: 218c была эффективной в присутствии TGF-β1 в клетках A549 и ReNcell CX® (таблица 25, фиг. 8). Уровень мРНК-мишени в обеих тестированных клеточных линиях был достоверно снижен независимо от обработки одним ASO SEQ ID NO: 218c или в присутствии TGF-β1.
Таблица 25: в присутствии TGF-β1 ASO SEQ ID NO: 218b приводит к сильной понижающей регуляции мРНК TGF-β1 после трансфекции гимнозисом в клетках A549 и ReNcell CX®. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218с, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
48 ч TGF-β1 → 72 ч/96 ч TGF-β1+ASO/обработка одним ASO
n=2
n=2
Заключение:
В совокупности, ASO по изобретению были эффективны в понижающей регуляции мРНК TGF-RII в присутствии TGF-β1, указывая на то, что ASO функционируют в условиях патологических состояний.
Пример 5: ингибирующая активность ASO по изобретению на уровни белка-мишени в обработанных TGF-β1 клетках
Для анализа ингибирующей активности антисмысловых олигонуклеотидов (ASO) в клетках-предшественниках нейронов человека из кортикальной области мозга (ReNcell CX®) при патологических состояниях клетки предварительно обрабатывали трансформирующим фактором роста-1 (TGF-β1). По результатам предыдущих исследований известно, что TGF-β1 обнаруживается в высоких концентрациях в цереброспинальной жидкости (CSF) при всех неврологических расстройствах, например, ALS. Таким образом, ингибирующая эффективность ASO на TGF-β-сигнальный путь была исследована после предварительной обработки и в присутствии TGF-β1. В качестве стандартной клеточной линии использовали клетки A549.
Описание метода:
Клетки культивировали, как описано ранее в стандартном протоколе. Для обработки клетки высевали в 6-луночный культуральный планшет (Sarstedt #83.3920.300) (80000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°С и 5% CO2. Для исследования эффектов трансфекции гимнозисом (клетки A549 и ReNcell CX®) после предварительной инкубации с TGF-β1 (Promocell #C-63499) среду удаляли и заменяли свежей полной средой (1 мл для 6-луночных планшетов и 8-луночных планшетов). После экспозиции TGF-β1 (10 нг/мл, 48 ч) среду заменяли, к клеткам добавляли TGF-β1 (10 нг/мл), стандартный олигонуклеотид 1 (скремблированный контроль, 10 мкM) и ASO по изобретению (10 мкM) в комбинации и при самостоятельной обработке. Клетки A549 инкубировали еще 72 ч, тогда как клетки ReNcell CX® собирали через 96 ч. Соответственно, клетки дважды промывали PBS и затем использовали для выделения белка (6-луночные планшеты) с последующим анализом вестерн-блоттингом или иммуноцитохимическим исследованием клеток (8-луночные слайд-планшеты для культивирования клеток). Процедуры для использованных методик проводили, как описано ранее. Использованные антитела и разведения для соответствующих методов приведены в таблицах 13 и 14.
Результаты после трансфекции гимнозисом
Вестерн-блоттинг и иммуноцитохимический анализ клеток A549 показали, что ASO, имеющие последовательности SEQ ID NO: 218b, SEQ ID NO: 218с, SEQ ID NO: 210q, SEQ ID NO: 213k, SEQ ID NO: 143h, SEQ ID NO: 152h, SEQ ID NO: 209az, SEQ ID NO: 209y генерируют сильную понижающую регуляцию мишени в присутствии TGF-β1 (таблица 26). Окрашивание на TGF-RII на фиксированных клетках ReNcell CX® подтвердило результаты, наблюдаемые в клетках A549. Тестированные ASO проявили сильную понижающую регуляцию мишени после самостоятельной обработки, а также в присутствии TGF-β1.
Таблица 26: денситометрический анализ после вестерн-блоттинга TGF-RII. Наблюдали снижение уровня белка TGF-RII после предварительной инкубации с TGF-RII с последующей трансфекцией гимнозисом различных ASO в клетках A549. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием программного обеспечения Studio™ Lite и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, D=SEQ ID NO: 218с, F=SEQ ID NO: 210q, G=SEQ ID NO: 213k, H=SEQ ID NO: 143h, I=SEQ ID NO: 152h, J=SEQ ID NO: 209az, К=SEQ ID NO: 209y, E=TGF-β1.
Временная точка
48 ч TGF-β1 → 72 ч TGF-β1
+ ASO/обработка одним ASO
n=1
Заключение:
Предварительная инкубация с TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO: 218b, SEQ ID NO: 218с, SEQ ID NO: 210q, SEQ ID NO: 213k, SEQ ID NO: 143h, SEQ ID NO: 152h, SEQ ID NO: 209az и SEQ ID NO: 209y приводила помимо понижающей регуляции мРНК-мишени, к снижению уровня белка-мишени в клетках A549 и ReNcell CX®.
Результаты после трансфекции гимнозисом других дополнительных ASO:
Анализ вестерн-блоттингом показал пониженное количество белка TGF-RII в клетках A549 (таблица 27) после трансфекции гимнозисом в течение 72 ч по сравнению с необработанными клетками и клетками, обработанными скремблированным контролем. Предварительная инкубация с TGF-β1 с последующей трансфекцией гимнозисом тестированных ASO вызывала снижение по сравнению с клетками, которые были предварительно обработаны TGF-β1, с последующей трансфекцией гимнозисом скремблированного контроля. Иммуноцитохимическое исследование клеток A549 и ReNcell CX® после окрашивания на TGF-RII показало, что тестированные ASO опосредовали выраженное снижение уровня белка-мишени после трансфекции гимнозисом с предварительной обработкой TGF-β1 или без нее.
Таблица 27: денситометрический анализ после вестерн-блоттинга TGF-RII. Наблюдали снижение белка TGF-RII после предварительной инкубации с TGF-RII с последующей трансфекцией гимнозисом TGF-RII-специфических ASO в клетках A549. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием программного обеспечения Studio™ Lite и затем нормализовали к необработанному контролю. Снижение уровня белка для ASO 1-25 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению с SEQ ID NO: 143h; B=более чем на 10%, но менее чем на 20% ниже по сравнению с SEQ ID NO: 143h; C=более чем на 20%, но менее чем на 30% ниже по сравнению с SEQ ID NO: 143h. Снижение уровня белка для ASO 26-48 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению с SEQ ID NO: 152h; B=более чем на 10%, но менее чем на 20% ниже по сравнению с SEQ ID NO: 152h; C=более чем на 20%, но менее чем на 30% ниже по сравнению с SEQ ID NO: 152h. Снижение уровня белка для ASO 49-74 указано с помощью следующего ключа: A=менее чем на 10% ниже среднего значения, полученного с SEQ ID NO: 209az и SEQ ID NO: 209y; B=более чем на 10%, но менее чем 20% ниже среднего значения, полученного с SEQ ID NO: 209az и SEQ ID NO: 209y; C=более чем на 20%, но менее чем на 30% ниже среднего значения, полученного с SEQ ID NO: 209az и SEQ ID NO: 209y. Снижение уровня белка для ASO 75-100 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению с SEQ ID NO: 210q; B=более чем на 10%, но менее чем на 20% ниже по сравнению с SEQ ID NO: 210q; C=более чем на 20%, но менее чем на 30% ниже по сравнению с SEQ ID NO: 210q. Снижение уровня белка для ASO 101-124 указано с помощью следующего ключа: A=менее чем на 10% ниже среднего значения, полученного с SEQ ID NO: 218b и SEQ ID NO: 218с; B=более чем на 10%, но менее чем на 20% ниже среднего значения, полученного с SEQ ID NO: 218b и SEQ ID NO: 218с; C=более чем на 20%, но менее чем на 30% ниже среднего значения, полученного с SEQ ID NO: 218b и SEQ ID NO: 218с. Снижение уровня белка для ASO 125-152 указано с помощью следующего ключа: A=менее чем на 10% ниже по сравнению SEQ ID NO: 213k; B=более чем на 10%, но менее чем на 20% ниже по сравнению SEQ ID NO: 213k; C=более чем на 20%, но менее чем на 30% ниже по сравнению SEQ ID NO: 213k.
в опыте
Заключение:
Даже после предварительной инкубации с TGF-β1 трансфекция гимнозисом ASO по изобретению приводит к снижению уровня белка TGF-RII в клетках A549 и ReNcell CX®.
Пример 6: анализ влияния ASO по изобретению на сигнальный путь ниже к TGF-RII после предварительной инкубации с TGF-β1
Функциональные анализы проводили в клетках рака легких человека (A549) и клетках-предшественниках нейронов человека (ReNcell CX®). Анализировали сигнальный путь TGF-β ниже по отношеню к нему, основываясь на эффективной понижающей регуляции мРНК TGF-RII и снижении уровней белка после трансфекцит гимнозисом ASO по изобретению в присутствии TGF-β1. Следовательно, оценивали уровни мРНК и белка фактора роста соединительной ткани (CTGF), известного как медиатор ниже TGF-β. Кроме того, исследовали фосфорилирование Smad2 (матери против гомолога декапентаплегии 2). Фосфорилирование Smad2 является маркером активного пути TGF-β, с последующей повышающей регуляцией даунстрим гена-мишени CTGF.
Описание метода:
Клетки культивировали, как описано ранее. Для обработки клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (50000 клеток/лунку), 6-луночные культуральные планшеты (Sarstedt #83.3920.300) (80000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°С и 5% CO2. Для исследования эффектов трансфекции гимнозисом (клетки A549 и ReNcell CX®) после предварительной инкубации с TGF-RII культуральную среду удаляли и заменяли свежей полной средой (1 мл на 6-луночных и 8-луночных планшетов). После экспозиции TGF-RII (10 нг/мл, 48 ч) среду заменяли, к клеткам добавляли TGF-β1 (10 нг/мл), стандартный олигонуклеотид 1 (скремблированный контроль, 10 мкМ), ASO с последовательностью SEQ ID NO: 218b (10 мкМ) и ASO с последовательностью SEQ ID NO: 218c (10 мкМ) в комбинации и в виде самостоятельной обработки. Клетки А549 инкубировали еще в течение 72 ч, в то время как клетки ReNcell CX® собирали через 96 ч. Соответственно, клетки дважды промывали PBS и затем использовали для определения РНК (24-луночные планшеты) и выделения белка (6-луночные планшеты) или иммуноцитохимического исследования клеток (8-луночные слайд-планшеты для культивирования клеток). Оценку влияния на уровень мРНК CTGF проводили ОТ-ПЦР в режиме реального времени, как описано ранее. Пара праймеров для анализа CTGF была готова к применению и стандартизирована. Для анализа уровней белка CTGF и pSmad2 использовали вестерн-блоттинг и иммуноцитохимию, как описано ранее. Тип и использованные разведения антител для соответствующего метода приведены в таблицах 13 и 14.
6.1. Результаты для SEQ ID NO: 218b
6.1.1. Влияние на уровни мРНК и белка CTGF
мРНК CTGF подверглась понижающей регуляции после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549 (72 ч, 0,52±0,05) и ReNcell CX® (96 ч, 0,70±0,25), в то время как инкубация с TGF-β1 в течение 5 суток (А549: 48 ч+72 ч, 6,92±2,32) или 6 суток (ReNcell CX®: 48 ч+96 ч, 1,60±015) соответственно, вызывала достоверную повышающую регуляцию мРНК CTGF. ASO SEQ ID NO:218b был достаточно эффективным, чтобы вызвать понижающую регуляцию мРНК CTGF, блокируя эффекты TGF-β1 в присутствии TGF-β1 (таблица 28, фиг. 11). Согласно данным по уровням мРНК иммунохимическое окрашивание на CTGF также подтверждало эти наблюдения для уровней белка (фиг. 12).
Таблица 28: понижающая регуляция мРНК CTGF в присутствии TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO: 218b в клетках A549 и ReNcell CX®. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
Временная точка
48 ч TGF-β1 → 72 ч/96 ч TGF-β1
+ ASO/обработка одним ASO
n=5
n=3
Заключение:
В присутствии TGF-β1 последующая обработка ASO SEQ ID NO: 218b приводила, во-первых, к понижающей регуляция мРНК TGF-RII и, во вторых, к снижению уровней мРНК и белка CTGF в клетках A549 и ReNcell CX®. Данный факт означает, что ASO SEQ ID NO: 218b является достаточно сильным, чтобы быть активным в условиях патологических состояний с высоким уровнем TGF-β1 и способен спасти клетки от последствий эффектов, опосредуемых TGF-β1.
6.1.2. Влияние на уровень белка pSmad2
Для проверки того, является ли понижающая регуляция CTGF следствием специфического ингибирования TGF-β-сигнального пути, опосредованного ASO SEQ ID NO: 218b в присутствии TGF-β1, определяли уровни белка pSmad2.
Окрашивание на pSmad2 после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом ASO SEQ ID NO: 218b с параллельным воздействием TGF-β1 приводит к ингибированию фосфорилирования Smad2 в обеих тестированных клеточных линиях (фиг. 13). Кроме того, снижение уровня белка pSmad2 подтверждали анализом вестерн-блоттингом в клетках A549 и ReNcell CX® (таблица 29).
Таблица 29: денситометрический анализ после вестерн-блоттинга pSmad2. Наблюдали понижающую регуляцию белка pSmad2 после трансфекции гимнозисом ASO SEQ ID NO: 218b. Также была установлена отмена TGF-β1-опосредованных эффектов с помощью ASO по изобретению при сравнении комбинированных обработок. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием программного обеспечения Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
48 ч TGF-β1 → 72 ч/96 ч TGF-β1
+ ASO/обработка одним ASO
n=5
n=3
Заключение:
ASO SEQ ID NO: 218b приводит к функциональному ингибированию сигнального пути TGF-β в клетках A549 и ReNcell CX® в присутствии TGF-β1, что подтверждалось снижением фосфорилирования Smad2.
6.2 Результаты для SEQ ID NO: 218c
6.2.1. Влияние на мРНК и уровень белка CTGF
Полученные данные показывают понижающую регуляцию мРНК CTGF после комбинированной обработки ASO SEQ ID NO: 218c и TGF-β1 (A549: 0,86, ReNcell CX®: 0,23) по сравнению с комбинированной обработкой с использованием скремблированного контроля и TGF-β1 (A549: 5,89, ReNcell CX®: 1,25) (таблица 30 и фиг. 14). В дополнение к этим наблюдениям иммунохимическое окрашивание на CTGF подтверждало наличие отмены TGF-β1-опосредованных эффектов на уровень белка под действием ASO SEQ ID NO: 218с (фиг. 15).
Таблица 30: уровни мРНК CTGF после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO:218c и параллельной обработкой TGF-β1 в клетках A549 и ReNcell CX®. Данные подтвердили эффективное предупреждение эффектов TGF-β1 на уровни мРНК CTGF под воздействием ASO SEQ ID NO: 218с. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218c, E=TGF-β1, ±=SEM, **p<0,01 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
Временная точка
48 ч TGF-β1 → 72 ч/96 ч TGF-β1
+ ASO/обработка одним ASO
n=3
n=2
Заключение:
Данные подтвердили эффективное предупреждение индуцированных TGF-β1 эффектов на уровни мРНК и белка CTGF под воздействием ASO SEQ ID NO: 218с.
6.2.2. Влияние на уровень белка pSmad2
Для проверки того, является ли понижающая регуляция CTGF (6.2.1) следствием ингибирования сигнального пути TGF-β1, опосредованного ASO SEQ ID NO: 218c, даже при наличии предварительной инкубации с TGF-β1, анализировали уровни белка pSmad2.
Фосфорилирование Smad2 индуцировали инкубацией с TGF-β1 (1,52±0,19), в то время как трансфекция гимнозисом ASO обеспечивала снижение pSmad2 в клетках A549 (0,89±0,05). Предварительная инкубация с TGF-β1 с последующей комбинированной обработкой приводит к подавлению эффектов TGF-β1 на фосфорилирование Smad2 (анализ вестерн-блоттингом, таблица 31). Результаты иммуноцитохимии подтверждали данные, полученные анализом вестерн-блоттингом (фиг. 16).
Таблица 31: денситометрический анализ после вестерн-блоттинга pSmad2. Определяли понижающую регуляцию белка pSmad2 после трансфекции гимнозисом с ASO SEQ ID NO: 218c. Показано ингибирование опосредованных TGF-β1 эффектов с помощью ASO по изобретению, когда сравнивались комбинированные обработки. Уровни белка определяли по отношению к гену «домашнего хозяйства» GAPDH с использованием программного обеспечения Studio™ Lite и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218c, E=TGF-β1. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
Временная точка
48 h TGF-β1 → 72 h TGF-β1
+ ASO/обработка одним ASO
n= 2
Заключение:
ASO SEQ ID NO: 218c эффективно ингибирует сигнальный путь TGF-β после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом ASO. Это было показано определением уровней даунстрим белка pSmad2.
В совокупности, ASO по изобретению высокоактивны в опосредовании функционального ингибирования сигнального пути TGF-β в присутствии патологических, высоких уровней TGF-β1 посредством эффективной понижающей регуляций мРНК TGF-RII. Таким образом, ASO по изобретению будут полезны при медицинских показаниях, при которых имеют место повышенные уровни TGF-β, например, неврологических расстройствах, фиброзе, прогрессировании опухолей и других.
Пример 7: определение профилактической активности антисмысловых олигонуклеотидов на уровне мРНК (последующая обработка TGF-β1)
Для анализа профилактической активности антисмысловых олигонуклеотидов (ASO) в клетках-предшественниках нейронов человека из кортикальной области головного мозга (ReNcell CX®), ASO трансфицировали в клетки посредством поглощения гимнозисом с последующей обработкой трансформирующим фактором роста-1 (TGF-β1).
Описание метода:
Клетки A549 и ReNcell CX® культивировали, как описано выше. Для оценки профилактической обработки клетки высевали в 24-луночный культуральный планшет (Sarstedt #83.1836.300) (50000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Затем в среду добавляли стандартный олигонуклеотид 1 (скремблированный контроль, 10 мкМ) или ASO с SEQ ID NO: 218b (10 мкМ) на 72 ч (А549) или 96 ч (ReNcell CX®). Вслед за инкубацией после трансфекции гимнозисом к клеткам добавляли TGF-β1 (10 нг/мл, Promocell #C-63499) без замещения среды на 48 ч. Для сбора клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты) с последующим анализом мРНК ОТ-ПЦР в режиме реального времени. Использовали готовые к применению и стандартизированные пары праймеров для ОТ-ПЦР в режиме реального времени и смешивали с соответствующим готовым к применению раствором Mastermix (SsoAdvanced™ Universial SYBR® Green Supermix (BioRad #172-5271), следуя инструкциям производителя (BioRad Prime PCR Quick Guide). Методы проводили, как описано выше.
7.1 Результаты для SEQ ID NO: 218b
На эффективность понижающей регуляции мРНК TGF-RII под действием ASO SEQ ID NO: 218b последующая инкубация с TGF-β1 не оказывала влияния в клетках A549 и ReNcell CX® (таблица 32). Достоверное снижение уровня мРНК-мишени в клетках ReNcell CX® было показано после обработки одним ASO с SEQ ID NO: 218b (0,33*±0,11). Трансфекция гимнозисом ASO с последующей обработкой TGF-β1 эффективно снижала уровень мРНК-мишени TGF-RII. В клетках A549 SEQ ID NO: 218b показал аналогичную эффективность в ингибировании мРНК TGF-RII при самостоятельной (0,25±0,07) или комбинированной обработке с последующей инкубацией TGF-β1 (0,24±0,06).
Таблица 32: понижающая регуляция мРНК TGF-RII после трансфекции гимнозисом с последующей обработкой TGF-β1 ASO в клетках A549 и ReNcell CX®. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, С=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, **p<0,05 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
Временная точка
72 ч/96 ч ASO → 48 ч TGF-β1
n=3
n=3
Заключение:
Поглощение гимнозисом ASO SEQ ID NO: 218b с последующей инкубацией с TGF-β1 было эффективным в понижающей регуляции мРНК-мишени TGF-RII, указывая, что ASO SEQ ID NO:218b можно использовать для профилактического лечения по медицинским показаниям.
Пример 8: определение ингибирующей активности ASO по изобретению на уровень белка с последующей обработкой TGF-β1
Для того, чтобы оценить профилактическую активность ASO по изобретению в клетках-предшественниках нейронов человека из кортикальной области мозга (ReNcell CX®) ASO трансфицировали в клетки путем трансфекции гимнозисом с последующей обработкой TGF-β1.
Описание метода:
Клетки культивировали, как описано ранее в стандартном протоколе. Для обработки клетки высевали в 8-луночные слайд- планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Затем в среду добавляли стандартный олигонуклеотид 1 (скремблированный контроль, 10 мкМ) или ASO с последовательностью SEQ ID NO: 218b (SEQ ID NO: 218b, 10 мкМ) на 72 ч (A549) или 96 ч (ReNcell CX®). После трансфекции гимнозисом добавляли TGF-β1 (10 нг/мл, Promocell #C-63499) на 48 ч без замещения среды. Для сбора клетки дважды промывали PBS и затем использовали для иммуноцитохимического анализа. Процедуру проводили, как описано выше. Используемые антитела и разведения для соответствующих методов приведены в таблицах 13 и 14.
8.1. Результаты по снижению белка TGF-RII после трансфекции гимнозисом SEQ ID NO: 218b с последующей обработкой TGF-β1
Иммуноцитохимический анализ на TGF-RII для клеток A549 и ReNcell CX® показал, что ASO SEQ ID NO:218b генерирует сильную понижающую регуляцию мРНК-мишени TGF-RII после последующей обработки TGF-β1 (фиг. 17).
Заключение:
Трансфекция гимнозисом ASO SEQ ID NO: 218b с последующей обработкой TGF-β1 приводила к понижающей регуляции мРНК-мишени, а также к выраженному снижению уровня белка TGF-RII в клетках A549 и ReNcell CX®.
В совокупности, эффективность понижающей регуляции белка TGF-RII, опосредуемой ASO SEQ ID NO: 218b, в комбинации с последующей обработкой TGF-β1 по-прежнему проявлялась, что дает основание полагать, что ASO по изобретению эффективны для профилактических применений.
Пример 9: влияние обработки ASO на даунстрим сигнальный путь TGF-RII с последующей обработкой TGF-β1
Эффективность ASO по изобретению в опосредовании ингибирования сигнального пути TGF-β оценивали для обработки TGF-β1 с последующей трансфекцией гимнозисом в клетках рака легких человека (A549) и клетках-предшественниках нейронов человека (ReNcell CX®). Следовательно, анализировали даунстрим молекулы на сигнальном пути TGF-β, Smad3 (матери против гомолога декапентаплегии 2) и фактор роста соединительной ткани (CTGF).
Описание метода:
Клетки культивировали, как описано ранее в стандартном протоколе. Для обработки клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (50000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Затем в среду добавляли стандартный олигонуклеотид 1 (скремблированный контроль, 10 мкМ) или ASO SEQ ID NO: 218b (10 мкМ) на 72 ч (А549) или 96 ч (ReNcell CX®). После трансфекции гимнозисом добавляли TGF-β1 (10 нг/мл, Promocell #C-63499) без замещения среды на последующие 48 ч. Для сбора клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты) или иммуноцитохимического исследования клеток (8-луночные слайд-планшеты для культивирования клеток). Для оценки влияния на уровень мРНК CTGF проводили ОТ-ПЦР в режиме реального времени, как описано ранее. Пара праймеров для анализа CTGF была готова к использованию и стандартизирована. Для определения уровней белка pSmad3 использовали иммуноцитохимию, как описано ранее. Тип и использованные разведения антител для соответствующего метода приводятся в таблицах 13 и 14.
9.1. Результаты для SEQ ID NO: 218b
9.1.1. Влияние на уровень мРНК CTGF и белка pSmad3
Уровень мРНК CTGF снижался после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549 (5 суток: 0,67±0,02) и ReNcell CX® (6 суток: 0,70±0,02). При добавлении TGF-β1 через 72 ч или 96 ч соответственно, клетки реагировали с повышением мРНК CTGF, но по сравнению с трансфекцией гимнозисом скремблированного контроля после обработки TGF-β1 индукция мРНК CTGF была сильно снижена (таблица 33). Для подтверждения того, является ли понижающая регуляция мРНК CTGF следствием ингибирования сигнального пути TGF-β, опосредованной ASO SEQ ID NO: 218b, также после обработки TGF-β1 определяли уровни белка pSmad3. На фиг. 18 показано, что сигнальный путь TGF-β фактически был блокирован трансфекцией гимнозисом ASO SEQ ID NO: 218b в клетках A549 (фиг. 18A) и ReNcell CX® (фиг. 18B). Данный эффект имел место также после трансфекции гимнозисом тестированного ASO с последующей обработкой TGF-β1.
Таблица 33: понижающая регуляция мРНК CTGF после трансфекции гимнозисом ASO SEQ ID NO: 218b с последующей обработкой TGF-β1 в клетках A549 и ReNcell CX®. Уровни экспрессии мРНК определяли количественно по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, *p<0,05, **p<0,01 по отношению к A, +p<0,05, ++p <0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
Временная точка
72 ч/96 ч ASO → +/- 48 ч TGF-β1
n=3
n=3
Заключение:
Трансфекция гимнозисом ASO SEQ ID NO: 218b приводила к понижающей регуляции мРНК и белка TGF-RII, а также к снижению уровней мРНК CTGF и белка pSmad3 в клетках A549 и ReNcell CX® независимо от обработки TGF-β1.
Это означает, что ASO SEQ ID NO: 218b является достаточно эффективным, чтобы проявлять активность в профилактических условиях для отмены или снижения имеющихся TGF-β1-опосредованных эффектов.
Пример 10: анализ потенциальных провоспалительных и токсических эффектов антисмысловых олигонуклеотидов
10.1. Тест с моноядерными клетками периферической крови (РВМС)
Для оценки антисмысловых олигонуклеотидов (ASO) на иммуностимулирующие свойства мононуклеарные клетки периферической крови (РВМС) инкубировали с контрольными ASO и тестируемыми соединениями с последующей постановкой ELISA для анализа IFNα и TGFα.
Описание метода:
Тест с мононуклеарными клетками периферической крови (PBMC)
РВМС отделяли от лейкоцитарной пленки, соответствующей трансфузионным единицам 500 мл полной крови. Каждую единицу получали от здоровых добровольцев, и цитрат глюкозы использовали в качестве антикоагулянта. Готовили лейкоцитаную пленку крови, и она была доставлена Банком крови Suhl Института трансфузионной медицины, Германия. Кровь от каждого донора проверяли на антитела к ВИЧ, антитела к HCV, антиген HBs, TPHA, ВИЧ-РНК и SPGT (ALAT). Только образцы крови с отрицательным результатом при тестировании на наличие инфекционных агентов и с нормальным значением SPGT использовали для разделения лейкоцитов и эритроцитов низкоскоростным центрифугированием. Выделение РВМС проводили в течение 40 ч с момента отбора у донора градиентным центрифугированием с использованием Ficoll-Histopague® 1077 (Heraeus™ Multifuge™ 3 SR). Для анализа IFN-α PBMC высевали из расчета 100000 клеток/лунку 96-луночного планшета в 100 мкл полной среды плюс добавки (RPMI1640+L-Glu,+10% FCS+PHA-P (5 мкг/мл)+IL-3 (10 мкг/мл)) и добавляли тестируемые соединения (5 мкл) для прямой инкубации (24 ч, 37°С, 5% CO2). Для анализа TNF-α РВМС высевали из расчета 100000 клеток/лунку 96-луночного планшета в 100 мкл полной среды без добавок (RPMI1640+L-Glu+10% FCS) и вносили тестируемые соединения (5 мкл) для прямой инкубации (24 ч, 37°С, 5% СО2). ELISA (измерения в двух повторностях из объединенных супернатантов, 20 мкл) проводили для анализа huIFNα (eBioscience, #BMS216INSTCE), следуя инструкциям производителя. ELISA (измерения в двух повторностях из объединенных супернатантов, 20 мкл) проводили для анализа huTNFα (eBioscience, #BMS223INSTCE), следуя инструкциям производителя.
Результаты:
Отсутствовал иммуностимулирующий эффект обработки ASO на РВМС, о чем свидетельствовало отсутствие детектируемой секреции IFNα (таблица 34) и TNFα (таблица 35) при инкубации с ASO. Функциональность анализа подтверждали наличием иммуностимулирующего эффекта иммуностимулирующей, конъюгированной с холестерином siРНК (XD-01024, IFNα) и полиинозиновой:полицитидиловой кислоты (поли I:C; TNFα; InvivoGen #tlrl-pic), которая является синтетическим аналогом двухцепочечной РНК, и связывается с TLR3 и стимулирует иммунную систему.
Таблица 34: ответная реакция IFNα на воздействие ASO по изобретению показывает ответную выработку IFNα РВМС при инкубации с ASO. Количественную оценку уровней экспрессии проводили с положительным контролем (ODN2216 [олигонуклеотид CpG класса A, распознаваемый TLR9 и приводящий к сильным иммуностимулирующим эффектам; InvivoGen tlrl-2216], поли I:C, XD-01024) с использованием анализа ELISA.
Таблица 35: ответная реакция TNFα на воздействие ASO по изобретению. Проводили количественную оценку уровней экспрессии по отношению к контрольным кандидатам (ODN2216, поли I:C, XD-01024) с использованием анализа ELISA.
10.2 Токсикология антисмысловых олигонуклеотидов по изобретению в условиях in vivo
Для анализа токсических свойств антисмысловых олигонуклеотидов (ASO) мышам C57/Bl6N проводили три внутривенных инъекции ASO, и после эвтаназии определяли активность трансаминаз в сыворотке крови, печени и почках.
Описание метода:
Самок мышей C57/Bl6N в возрасте 6 недель обрабатывали тестируемыми соединениями (SEQ ID NO: 218b, SEQ ID NO: 218c) в течение семи суток. ASO (200 мкл, 15 мг/кг/массы тела) вводили внутривенно на сутки 1, 2 и 3 периода обработки. Наблюдали за динамикой привесов (SEQ ID NO: 218c) через день, и на сутки 4 отбирали пробы крови из vena fascicularis и готовили сыворотку. На сутки 8 животных подвергали эвтаназии (CO2) и отбирали пробы крови из полой вены и готовили сыворотку, отбирали пробы печени (кусочки ≤ 50 мг), почек и легких для количественного определения мРНК и трансаминаз. Уровни мРНК TGF-RII определяли в лизате печени, почек и легких методом bDNA (набор QuantiGene®, Panomics/Affimetrix). Активность аспартаттрансаминазы (ASP) и аланинтрансаминазы (ALT) определяли на Cobas Integra® 400 в разведенной сыворотке в соотношении 1:10.
Таблица 36: уровни экспрессии сывороточной аланинтрансаминазы и аспартаттрансаминазы у мышей C57/Bl6N после повторных внутривенных инъекций ASO. Количественную оценку уровней экспрессии проводили сравнением с уровнями экспрессии у животных, обработанных физиологическим раствором. ±=SEM.
Таблица 37: уровни экспрессии TGF-RII в ткани печени, почек и легких у мышей C57/Bl6N после повторных внутривенных инъекций ASO. Количественную оценку уровней экспрессии проводили сравнением с уровнями экспрессии у животных, обработанных физиологическим раствором. ±= SEM.
соединение
Таблица 38: уровни экспрессии аланинтрансаминазы и аспартаттрансаминазы в сыворотке мышей C57/Bl6N после повторных внутривенных инъекций ASO. Количественную оценку уровней экспрессии проводили сравнением с уровнями экспрессии у животных, обработанных физиологическим раствором. ±= SEM.
Таблица 39: уровни экспрессии TGF-RII в ткани печени, почек и легких у мышей C57/Bl6N после повторных внутривенных инъекций ASO. Количественную оценку уровней экспрессии проводили сравнением с уровнями экспрессии у животных, обработанных физиологическим раствором. ±= SEM.
Таблица 40: динамика прироста массы тела в течение 7-суточной схемы введения ASO. Прирост массы тела определяли количественно по сравнению с массой тела на сутки 0, которую принимали за 100%.
Заключение: не наблюдали провоспалительных или токсических эффектов соответствующих ASO по изобретению на РВМС или мышах C57/Bl6N. Таким образом, обработка ASO, нацеленных на TGF-RII, отражает безопасный метод лечения различных связанных с TGF-β расстройств.
Пример 11: оценка интрацеребровентрикулярной инфузии ASO по изобретению на индуцированное TGF-β ингибирование пролиферации нейрональных стволовых клеток и клеток-предшественников нейронов in vivo
Целью настоящего исследования была оценка потенциальной способности ASO по изобретению против TGF-RII i) профилактировать и ii) лечить индуцированные TGF-β1 эффекты на пролиферацию нейрональных стволовых клеток и клеток-предшественников in vivo.
Описание метода:
11.1. Профилактика связанного с TGF-β1 ингибирования нейрогенеза
Двухмесячным самкам крыс Fischer-344 (n=32) проводили интрацеребровентрикулярные инфузии через осмотические мининасосы (модель 2002, Alzet), соединенные с канюлями из нержавеющей стали. Хирургическую имплантацию мининасосов выполняли под глубокой анестезией с использованием внутримышечных инъекций. Животным инфузировали ASO по изобретению (концентрация 1,64 мМ в насосе), контрольный ASO (концентрация 1,64 мМ в насосе) или аCSF (искусственная спинномозговая жидкость) в течение 7 суток. На сутки 8 меняли насосы, и животным вводили i) аCSF, ii) TGF-β1 (500 нг/мл в насосе), iii) TGF-β1 (500 нг/мл в насосе) плюс контрольный ASO (концентрация 1,64 мМ в насосе) или iv) TGF-β1 (500 нг/мл в насосе) плюс ASO по изобретению (концентрация 1,64 мМ в насосе) в течение 14 суток. В конце инфузионного периода всем животным транскардиально перфузировали 4% параформальдегид. Головной мозг исследовали на локализацию пути канюли, и животные с неправильным размещением канюли исключались из анализа. В течение последних 24 ч периода инфузии животным проводили внутрибрюшинную инъекцию бромдезоксиуридина (BrdU) в дозе 200 мг/кг.
Ткань обрабатывали для хромогенного иммунодетектирования BrdU-позитивных клеток в сагитальных срезах тощиной 40 мкм. Подсчитывали количество позитивных на BrdU клеток в трех рамках подсчета размером 50 мкм ×50 мкм на срез, расположенных в самой нижней, средней и верхней частях субвентрикулярной зоны. Положительные профили, пересекавшие самую верхнюю фокальную плоскость (плоскость исключения) или боковые границы рамки для подсчета, не учитывались. Для исследования гиппокампа определяли объем гиппокампа и подсчитывали все положительные клетки внутри и рядом с его границами. Общее количество положительных профилей умножали на отношение эталонного объема к объему выборки для получения установленного количества BrdU-положительных клеток для каждой структуры. Все экстраполяции выполняли для одного полушария головного мозга и затем удваивали для представления суммарных значений для всего мозга. Данные представлены как средние значения±стандартные отклонения (SD). Статистический анализ проводили с использованием непарного двустороннего t-теста сравнения - тест Стьюдента с t-критерием для сравнения обработанной TGF-β1 и контрольной группы (программное обеспечение GraphPad Prism 4, США). Уровень значимости определяли при р<0,05.
11.2. Лечение ассоциированного с TGF-β1 ингибирования нейрогенеза
Животные получали аCSF или рекомбинантный человеческий TGF-β1 (500 нг/мл, в насосе) со скоростью введения 0,5 мкл в час в течение 14 суток. Через 14 суток насосы меняли и животным вводили i) аCSF, ii) рекомбинантный TGF-β1 человека (500 нг/мл в насосе) или совместно с инфузией iii) ASO по изобретению (концентрация 1,64 мМ в насосе) плюс рекомбинантный TGF-β1 человека (500 нг/мл в насосе) или iv) контрольный ASO (концентрация 1,64 мМ в насосе) плюс рекомбинантный TGF-β1 человека (500 нг/мл в насосе). В конце инфузионного периода всем животным транскардиально перфузировали 4% параформальдегид. Головной мозг исследовали на локализацию пути канюли, и животные с неправильным размещением канюли исключались из анализа. В течение последних 24 ч периода инфузии животные получали внутрибрюшинную инъекцию бромдезоксиуридина (BrdU) в дозе 200 мг/кг.
Гистологический анализ проводили, как описано выше (11.1).
Результаты:
Обработка ASO SEQ ID NO: 143aj, SEQ ID NO: 143h и SEQ ID NO: 210q специфически и частично ослабляла эффект TGF-β1 на пролиферацию клеток в гиппокампе и в стенке желудочка. Обработка ASO по изобретению специфически и частично отменяла ингибирующий эффект TGF-β1 на нейрогенез.
Заключение: ASO по настоящему изобретению, демонстрирующие перекрестную реактивность с грызунами, индуцировали нейрогенез в данном эксперименте in vivo. ASO по настоящему изобретению, демонстрирующие отсутствие перекрестной реактивности, оказывали в целом даже более сильные эффекты в экспериментах in vitro. В результате предполагается, что данные ASO по изобретению будут также более эффективными в испытаниях in vivo у людей и приматов, отличных от человека, и поэтому будут функционировать как высокоэффективное лекарственное средство для профилактики или лечения индуцированного TGF-β1 ингибирования пролиферации нейрональных стволовых клеток и клеток-предшественников нейронов.
Пример 12: анализ влияния антисмысловых олигонуклеотидов по изобретению на пролиферацию и специфические маркеры клеток-предшественников нейронов человека
Боковой амиотрофический склероз (ALS) является нейродегенеративным летальным заболеванием без наличия эффективного лечения к настоящему времени. Проводимые в настоящее время молекулярно-генетические исследования все в большей степени раскрывают молекулярный патогенез данного фатального заболевания; по результатам предыдущих исследований известно, что TGF-β обнаруживается в высоких концентрациях в цереброспинальной жидкости (CSF) пациентов с ALS. Известно, что эти высокие уровни циркулирующего TGF-β способствуют вхождению стволовых клеток в состояние покоя и, как следствие, вызывают торможение взрослого нейрогенеза в субвентрикулярной зоне (SVZ) головного мозга. Таким образом, регенерация дегенерирующих нейронов, по-видимому, предотвращается при усилении сигнального пути TGF-β. Для выяснения того, насколько селективное ингибирование сигнального пути TGF-β, опосредованное антисмысловыми олигонуклеотидами по изобретению, может реактивировать взрослый нейрогенез, необходимо вначале подтвердить факт опосредованной TGF-β остановки клеточного цикла.
Описание методов:
Исследования по остановке клеточного цикла. Клетки культивировали, как описано выше в стандартном протоколе. Для экспериментов клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (30000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для определения опосредованного TGF-β1 влияния на клеточный цикл в условиях пролиферации (+EGF/FGF) (Millipore: EGF #GF144, bFGF #GF003) или дифференцировки (-EGF/FGF) клетки обрабатывали в течение 4 суток TGF-β1 (PromoCell #C-63499, 10 или 50 нг/мл) после удаления и замены соответствующей среды. На сутки 4 среду обновляли и проводили повторную обработку TGF-β1 до суток 7. На сутки 7 клетки собирали промыванием дважды PBS и затем использовали для выделения РНК (24-луночные планшеты), как описано выше. Для оценки влияния TGF-β1 на клеточный цикл с использованием ОТ-ПЦР в режиме реального времени анализировали мРНК маркера пролиферации Ki67, гена-супрессора опухолей р53, ингибитора циклинзависимой киназы 1 (p21) и маркера нейрогенеза даблкортина (DCX). Соответствующие пары праймеров приведены в таблице 11.
Анализ мРНК на эффекты ASO SEQ ID NO: 218b на клетках-предшественниках нейронов человека: клетки культивировали, как описано ранее в стандартном протоколе. Для экспериментов клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (30000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для настоящих экспериментов клеточную среду заменяли и к клеткам добавляли на 96 ч стандартный олигонуклеотид 1 (скремблированный контроль, 2,5 и 10 мкМ), ASO с SEQ ID NO: 218b (2,5 и 10 мкМ) или TGF-β1 (10 нг/мл, Promocell #C-63499). По истечении времени инкубации среду снова заменяли и проводили дальнейшую обработку еще 96 ч. Через 8 суток обработки клетки собирали. Клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты). Для оценки влияния на клетки-предшественники определяли уровни мРНК нестина (ранний нейрональный маркер), Sox2 (ранний нейрональный маркер), DCX (маркер нейрогенеза) и Ki67 (маркер пролиферации) с использованием ОТ-ПЦР в режиме реального времени, как описано ранее. Соответствующие пары праймеров приведены в таблице 11.
Пролиферативные и дифференцирующие эффекты TGFRII-специфических ASO трансфекцией гимнозисом клеток ReNcell CX ® . Следующей целью было исследование того, оказывает ли влияние TGF-RII-специфический ASO на пролиферацию клеток ReNcell CX®. Соответственно, клетки культивировали, как описано ранее, и высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (30000 клеток/лунку) или 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°С и 5% СО2. Для построения кривой пролиферации клетки обрабатывали после замены среды в течение 72 ч стандартным олигонуклеотидом 1 (скремблированный контроль, 2,5 и 10 мкМ) и ASO SEQ ID NO: 218b (2,5 и 10 мкМ). После времени инкубации среду заменяли и обработку повторяли два раза. После сбора супернатанта оставшиеся клетки собирали из 24-луночных планшетов для определения количества клеток. Для этой цели оставшиеся клетки промывали PBS (2×), обрабатывали аккутазой (500 мкл/лунку) и инкубировали в течение 5 мин при 37°C. Затем добавляли 500 мкл среды и определяли количество клеток с использованием автоматического флуоресцентного счетчика клеток Luna FL® и устройства для светлопольного освещения (Biozym, #872040), следуя инструкциям производителя. Вкратце, 18 мкл клеточной суспензии добавляли к 2 мкл красителя акридиновый оранжевый/пропидиум иодида из набора для оценки жизнеспособности клеток (Biozym #872045). Через 1 мин отстаивания 10 мкл наносили на стекло для подсчета клеток (Biozym #872011), подсчитывали количество клеток и выражали в виде общего числа клеток/мл и процента живых клеток по сравнению с мертвыми клетками. После трансфекции гимнозисом стандартного олигонуклеотида 1 (10 мкМ), SEQ ID NO: 218b (10 мкМ) и соответствующей обработки TGF-β1 (10 нг/мл) в течение 8 суток, клетки на 8-луночных слайд-планшетах для культивирования клеток фиксировали и окрашивали антителом к Ki67. Для оценки способности клеток ReNcell CX®к дифференцировке после трансфекции гимнозисом другие 8-луночные слайд-планшеты для культивирования клеток обрабатывали стандартным олигонуклеотидом 1 (10 мкМ), SEQ ID NO: 218b (10 мкМ) и TGF-β1 (10 нг/мл) соответственно в течение 96 ч в условиях пролиферации (+EGF/FGF). Затем одну часть клеток обрабатывали еще в течение 96 ч в условиях пролиферации, тогда как другую часть клеток обрабатывали и выдерживали в условиях дифференцировки (-EGF/FGF). После окрашивания клеток определяли уровни экспрессии нейрофиламента N (NeuN) и βIII-тубулина флуоресцентной микроскопией. Протокол сбора, фиксации и окрашивания клеток описан выше, и соответствующие разведения антител приведены в таблице 14.
Анализ мРНК маркеров пролиферации и нейрогенеза после трансфекции гимнозисом после предварительной инкубации с TGF-β1. Клетки культивировали, как описано ранее в стандартном протоколе. Для экспериментов клетки высевали в 24-луночные планшеты для культивирования (Sarstedt #83.1836.300) (30000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для индукции остановки клеточного цикла клетки ReNcell CX® обрабатывали TGF-β1 в течение 4 суток. Затем среду заменяли и вновь добавляли TGF-β1 (10 нг/мл). На сутки 8 среду заменяли еще раз, и проводили трансфекцию гимнозисом в течение 96 ч добавлением стандартного олигонуклеотида 1 (10 мкМ), SEQ ID NO: 218b (10 мкМ) в комбинации с TGF-β1 (10 нг/мл). После инкубации клетки собирали двукратным промыванием PBS. Затем выделяли РНК и проводили анализ мРНК ОТ-ПЦР в режиме реального времени, как описано выше.
12.1.1. Опосредование TGF-β1 остановки клеточного цикла в клетках-предшественниках нейронов человека
Определение маркеров состояния покоя стволовых клеток показало, что TGF-β1 опосредует остановку клеточного цикла через 7 суток после обработки им клеток. Экспрессия мРНК маркера пролифераци Ki67 снижалась в зависимости от концентрации TGF-β1. Также экспрессия мРНК гена-супрессора опухолей р53 подверглась понижающей регуляции, коррелируя с концентрацией TGF-β1. В противоположность, уровень ингибитора циклинзависимой киназы 1 (p21) достоверно повышался под действием TGF-β1. В совокупности, данные результаты указывают на то, что стволовые клетки находились в состоянии покоя, индуцированным TGF-β1. Интересно, что уровень DCX, маркера нейрогенеза, был сильно снижен под действием TGF-β1 (таблица 41).
Таблица 41: экспрессия мРНК Ki67, p27, p21 и DCX через 7 суток после обработки TGF-β1 в клетках ReNcell CX®. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, E=TGF-β1. ±=SEM, *p<0,05 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 7 суток после обработки TGF-β1
n=3
n=3
n=3
n=3
Заключение:
Пролиферация клеток ReNcell CX® блокировалась под действием TGF-β1.
12.1.2. Результаты по оценке влияния антисмысловых олигонуклеотидов на маркеры нейрональных стволовых клеток человека
Для исследования влияния ASO SEQ ID NO: 218b на маркеры стволовых клеток через 8 суток после повторной трансфекции гимнозисом (2×96 ч) в клетках ReNcell CX® анализировали различные маркеры ранних клеток-предшественников нейронов (таблица 42). ASO SEQ ID NO: 218b не оказывал влияния на уровни экспрессии гена нестина и Sox2. Уровень мРНК GFAP несколько повышался после трансфекции гимнозисом 10 мкМ ASO SEQ ID NO: 218b и, напротив, DCX четко повышался после поглощения гимнозисом ASN SEQ ID NO: 218b. Экспрессия всех тестированных маркеров достоверно снижалась после обработки TGF-β1 (8 сутки) (таблица 42, фиг. 19).
Таблица 42: экспрессия мРНК нестина, Sox2, GFAP и DCX через 8 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках ReNcell CX®. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM, +p<0,05 по отношению к 2,5 мкМ C, #p<0,05 по отношению к 10 мкМ C. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 8 суток после трансфекции гимнозисом или обработки TGF-β1
n=4
n=4
n=4
n=4
Заключение:
Результаты анализа мРНК показывают, что ASO SEQ ID NO: 218b направляет клетки ReNcell CX® к еще более высокой клеточной «стволовости» (повышающая регуляция GFAP). Кроме того, индукция DCX указывает на повышенный нейрогенез. Обработка TGF-β1 приводит к противоположному эффекту.
12.1.3. Результаты оценки влияния антисмысловых олигонуклеотидов на пролиферацию нейрональных стволовых клеток человека
Проводили дальнейший анализ для исследования того, насколько трансфекция гимнозисом ASO SEQ ID NO: № 218b действительно влияет на скорость пролиферации подсчетом клеток через 9 суток после повторной трансфекции гимнозисом (3×72 ч) и определением уровней белка Ki67 через 8 суток после трансфекции гимнозисом (2×96 ч).
Результаты:
Количество клеток повышалось после поглощения гимнозисом ASN SEQ ID NO: 218b, согласуясь с повышенной экспрессией белка маркера пролиферации Ki67, выявленной при иммунохимическом окрашивании клеток (таблица 43, фиг. 20). Флуоресцентный анализ иммуноцитохимического окрашивания также свидетельствовал об остановке пролиферации, опосредованной TGF-β1.
Таблица 43: повышение количества клеток через 9 суток после повторной трансфекции гимнозисом (3×72 ч) клеток ReNcell CX®. Количество клеток определяли с использованием автоматического флуоресцентного счетчика клеток Luna FL® и устройства для светлопольного освещения (Biozym, #872040), следуя инструкциям производителя. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM.
Заключение:
Трансфекция гимнозисом ASO SEQ ID NO: 218b клеток ReNcell CX® приводит к увеличению числа клеток параллельно с повышением экспрессии белка Ki67, что указывает на усиление пролиферации предшественников нейронов.
12.1.3. Результаты оценки влияния антисмысловых олигонуклеотидов на способность нейрональных стволовых клеток человека к дифференцировке
Для исключения влияния ASO SEQ ID NO: 218b на способность клеток к дифференцировке ASO SEQ ID NO: 218b трансфицировали в клетки путем поглощения гимнозисом в течение 96 ч в условиях пролиферации (+EGF/FGF). После времени инкубации среду заменяли и к одной части клеток добавляли среду для пролиферации, тогда как к другой части клеток добавляли среду для дифференцировки (-EGF/FGF). Затем проводили еще одну трансфекцию гимнозисом в течение 96 ч. Клетки анализировали определением уровней экспрессии нейрональных маркеров нейрофиламента N (NeuN) и βIII-тубулина.
Результаты:
Иммунохимическое окрашивание на NeuN (фиг. 23A) и βIII-тубулин (фиг. 23B) демонстрирует отсутствие эффектов на способность клеток к дифференцировке после трансфекции гимнозисом ASO в условиях пролиферации, с последующей трансфекцией гимнозисом в условиях дифференцировки. ASO SEQ ID NO: 218b не оказывал влияния на сигнал βIII-тубулина, человеческого нейронспецифического белка, в условиях дифференцировки и был сравним с необработанным контролем. Кроме того, на экспрессию NeuN не оказывала влияния трансфекция гимнозисом в условиях дифференцировки. Таким образом, клетки все еще были способны дифференцироваться в нейроны. Удивительно, что клетки ReNcell CX® экспрессировали нейрональные маркеры NeuN и βIII-тубулин после трансфекции гимнозисом ASO в условиях пролиферации (2×96 ч) во время обоих периодов, указывая на то, что трансфекция гимнозисом ASO может способствовать специфическому сдвигу к дифференцировке нейронов даже в условиях пролиферации. Кроме того, наблюдали повышенные скорости пролиферации клеток-предшественников нейронов (таблица 43, фиг. 20). Также окрашивание на NeuN показало, что клетки, обработанные ASO SEQ ID NO: 218b, выглядят более жизнеспособными по сравнению с клетками, подвергшихся всем другим видам обработки (фиг. 21A). Очевидно, что пролиферативный потенциал клеток, которые были обработаны TGF-β1, был значительно ниже.
Заключение:
ASO SEQ ID NO: 218b по изобретению не оказывал влияния на способность к дифференцировке. Интересно отметить, что клетки ReNcell CX® подвергались дифференцировке в нейроны после трансфекции гимнозисом в условиях пролиферации и дифференцировки. Данный факт с учетом наблюдения о повышении скорости пролиферации указывает на то, что ASO SEQ ID NO: 218b способствует нейрогенезу с тенденцией к усилению дифференцировки в нейроны.
12.1.4. Результаты оценки влияния антисмысловых олигонуклеотидов по изобретению на пролиферацию нейрональных стволовых клеток человека после предварительной инкубации с TGF-β1
Для анализа того, насколько трансфекция гимнозисом ASO SEQ ID NO: 218b является эффективной для отмены TGF-β1-опосредованных эффектов на клетки ReNcell CX®, проводили дополнительные исследования с предварительной инкубацией с TGF-β1 в течение 7 суток с последующей трансфекцией гимнозисом в течение 8 суток (2× 96 ч).
Результаты:
Экспрессия гена GFAP (таблица 44, фиг. 22А) в качестве раннего нейронального маркера, Ki67 (таблица 44, фиг. 22B) в качестве маркера пролиферации и DCX (таблица 44, фиг. 22C) в качестве маркера нейрогенеза была повышена после обработки одним ASO, в то время как TGF-β1 приводил к обратному эффекту. Кроме того, через 7 суток после предварительной инкубации с TGF-β1 обработка ASO по изобретению полностью отменял TGF-β1-индуцированные эффекты. Таким образом, анализ показывает, что ASO SEQ ID NO: 218b является эффективным в восстановлении последствий опосредованных TGF-β1 эффектов на стволовые клетки и маркеры пролиферации.
Таблица 44: экспрессия мРНК GFAP, Ki67 и DCX через 7 суток после предварительной инкубации с TGF-β1 с последующей 2×96 ч трансфекцией гимнозисом SEQ ID NO: 218b в клетках ReNcell CX®. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1, ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 7 суток после предварительной инкубации с TGF-β1
с последующей трансфекцией гимнозисом
2×96 ч
n=2
n=1
n=2
Заключение:
Результаты показывают, что взрослый нейрогенез может быть реактивирован TGF-RII-специфическим ASO-опосредованным блокированием сигнального пути TGF-β.
В совокупности, TGF-RII-специфический ASO SEQ ID NO: 218b «спасал» клетки от опосредованного TGF-β состояния покоя стволовых клеток и стимулировал взрослый нейрогенез, не влияя на дифференцировку. Это делает его идеальным лекарственным препаратом для восстановления головного мозга.
Пример 13: определение терапевтической активности антисмысловых олигонуклеотидов по изобретению при прогрессирующем ALS у мышей SOD1
Для анализа терапевтической потенциальной активности ASO для лечения бокового амиотрофического склероза (ALS) трансгенных мышей самцов и самок SOD1 G93A обрабатывали ASO по изобретению в различных дозах icv введением в боковой желудочек с помощью осмотических мининасосов ALZET®. Кроме того, рилузол использовали в качестве стандарта. Рилузол является лекарственным препаратом, применяемым для лечения бокового амиотрофического склероза, и комерчески доступен от Sanofi Pharmaceuticals. Он задерживает начало зависимости от искусственной вентиляции легких или трахеостомии у отдельных пациентов и может увеличить выживаемость примерно на два-три месяца.
Описание метода:
Для длительной центральной инфузии icv канюлю, присоединенную к осмотическому мининаносу Alzet® (скорость инфузии 0,25 мкл/ч, Alzet®, модель 2004, Купертино, США), стереотаксически имплантировали под анестезией изофлураном (Baxter, GmbH, Германия) в полустерильных условиях. Каждый осмотический мининасос имплантировали подкожно со стороны брюшной области посредством разреза кожи длиной 1 см на шее мыши и соединяли с icv канюлей силиконовыми трубками. Животных помещали в стереотаксическую рамку и icv канюлю (23G, длина 3 мм) погружали в правый боковой желудочек (0,3 мм сзади, 1 мм латеральнее, на глубину 3 мм относительно брегмы). Канюлю фиксировали двумя винтами из нержавеющей стали с использованием зубного цемента (Kallocryl, Speiko® Dr. Speier GmbH, Мюнстер, Германия). Кожу на шее закрывали швами. Во время операции температуру тела поддерживали грелкой. Для профилактики послеоперационных инфекций мышей местно обрабатывали препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 0,1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия). Трубку заполняли соответствующим раствором. Для определения влияния ASO на развитие и прогрессирование ALS, начало проявления симптомов, парез и выживаемость использовали в качестве конечных точек in vivo. В возрасте девяти недель мышей подвергали эвтаназии и извлекали головной мозг для нейропатологического анализа. Гистологическое подтверждение мест icv имплантации проводили на срезах мозга толщиной 40 мкм, окрашенных крезил-фиолетовым.
ASO по изобретению оказывали потенциальные эффекты в экспериментах in vitro. Согласуясь с этим, перекрестно реагирующие на грызунах ASO по изобретению с последовательностями SEQ ID NO: 143aj, SEQ ID NO: 143h и SEQ ID NO: 210q, были также эффективны в вышеописанных экспериментах, результаты которых доказывают эффект в лечении животных моделей ALS. ASO по настоящему изобретению, демонстрирующие отсутствие перекрестной реактивности, оказывали даже более сильные эффекты в экспериментах in vitro. В результате предполагается, что данные ASO по изобретению будут также более эффективными в испытаниях in vivo у приматов, отличных от человека и людей, и поэтому будут функционировать как высокоэффективное лекарственное средство для профилактики или лечения индуцированного TGF-β1 ингибирования пролиферации нейрональных стволовых клеток и клеток-предшественников нейронов, и тем самым лечения ALS и других нейродегенеративных расстройств.
Пример 14: определение терапевтической активности антисмысловых олигонуклеотидов по изобретению, нацеленных на TGF-RII, на развитие и прогрессирование болезни Гентингтона у мышей R6/2
Для анализа терапевтической активности ASO для лечения болезни Гентингтона (HD) трансгенным мышам самцам и самкам R6/2 вводили различные дозы TGF-RII-специфического ASO по изобретению icv введением в боковой желудочек с помощью осмотических мининасосов.
Описание метода:
Для длительной центральной инфузии мышей подвергали хирургической операции для имплантации icv канюли, присоединенной к осмотическому мининаносу Alzet® (скорость инфузии 0,25 мкл/ч, Alzet®, модель 2004, Купертино, США) в возрасте 5 недель. Канюлю и насос стереотаксически имплантировали под анестезией кетамином/ксилацином (Baxter, GmbH, Германия) в полустерильных условиях. Каждый осмотический мининасос имплантировали подкожно со стороны брюшной области посредством разреза кожи длиной 1 см на шее мыши и соединяли с icv канюлей силиконовыми трубками. Животных помещали в стереотаксическую рамку и icv канюлю (23G, длина 3 мм) погружали в правый боковой желудочек (0,3 мм сзади, 1 мм латеральнее, на глубину 3 мм относительно брегмы). Канюлю фиксировали двумя винтами из нержавеющей стали с использованием зубного цемента (Kallocryl, Speiko® Dr. Speier GmbH, Мюнстер, Германия). Кожу на шее закрывали швами. Во время операции температуру тела поддерживали грелкой. Для профилактики послеоперационных инфекций мышей местно обрабатывали препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 0,1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия). Трубку заполняли соответствующим раствором. Для определения влияния ASO на развитие и прогрессирование HD, начало проявления симптомов, силу схвата, общую моторику и выживаемость использовали в качестве конечных точек in vivo. В возрасте девяти недель мышей подвергали эвтаназии и извлекали головной мозг для нейропатологического анализа. Гистологическое подтверждение мест icv имплантации проводили на срезах мозга толщиной 40 мкм, окрашенных крезил-фиолетовым.
ASO по изобретению оказывали потенциальные эффекты в экспериментах in vitro. Согласуясь с этим, перекрестно реагирующие на грызунах ASO по изобретению с последовательностями SEQ ID NO: 143aj, SEQ ID NO: 143h и SEQ ID NO: 210q, были также эффективны в вышеописанных экспериментах, результаты которых доказывают эффект в лечении животных моделей болезни Гентингтона. ASO по настоящему изобретению, демонстрирующие отсутствие перекрестной реактивности, оказывали даже более сильные эффекты в экспериментах in vitro. В результате предполагается, что данные ASO по изобретению будут также более эффективными в испытаниях in vivo у приматов, отличных от человека и людей, и поэтому будут функционировать как высокоэффективное лекарственное средство для профилактики или лечения индуцированного TGF-β1 ингибирования пролиферации нейрональных стволовых клеток и клеток-предшественников нейронов, и тем самым лечения HD и других нейродегенеративных расстройств.
Пример 15: определение терапевтической активности ASO по изобретению при прогрессировании TGF2-индуцированной гидроцефалии и связанного с ней когнитивного дефицита у крыс Fischer-344
Целью настоящего исследования было лечение животных, страдающих TGF-индуцированными эффектами, конкретно на i) пролиферацию и нейрогенез нейрональных стволовых клеток, ii) формирование гидроцефалии и iii) дефицит пространственного обучения путем внутрижелудочковой инфузии ASO по изобретению в зависимости от дозы.
Описание метода: осмотические мининасосы для интрацеребровентрикулярной инфузии имплантировали самкам крыс Fischer-344 массой тела 180-200 г (nобщее=70, nв группе=10). Инфузировали а) искусственную спинномозговую жидкость (aCSF: 148,0 мМ NaCl, 3,0 мМ KCl, 1,4 мМ CaCl2, 0,8 мМ MgCl2, 1,5 мМ Na2HPO4, 0,2 мМ NaH2PO4, 100 мкг/мл крысиного сывороточного альбумина, 50 мкг/мл гентамицина, рН 7,4) в качестве контроля или b) TGF-β1 1 мкг/мл в aCSF с использованием осмотического насоса Alzet® модель 2004 со скоростью введения 0,25 мкл/ч в течение 14 суток. Через 14 суток насосы меняли и использовали осмотические насосы Alzet® модель 2004 (скорость введения 0,25 мкл/ч) для следующих инфузий: aCSF или TGF-β1 (1 мкг/мл) в комбинации с различными концентрациями TGF-RII-специфических ASO (1,1 ммоль/л, 3,28 ммоль/л, 9,84 ммоль/л) или контрольного ASO (3,28 ммоль/л) (2×4 недели). В течение последних четырех дней инфузионного периода животным ежедневно вводили внутрибрюшинно BrdU (50 мг/кг массы тела) для мечения пролиферирующих клеток. Насосы удаляли и через две недели животных обследовали функционально в тесте пространственного обучения (водный бассейн Морриса) в течение 14 суток. Через день животным перфузировали 0,9% NaCl, извлекали головной мозг, ипсилатеральное полушарие фиксировали в 4% параформальдегиде для количественного гистологического анализа PCNA, BrdU, DCX, BrdU/NeuN и BrdU/GFAP, и также для стереологического анализа объема боковых желудочков в качестве показателя гидроцефалии. Далее контралатеральное полушарие рассекали и различные области (стенки желудочков, гиппокамп, кору) обрабатывали для количественной ОТ-ПЦР в целях определения уровней экспрессии TGF-RII. МР-снимки были сделаны на 4 животных 1, 3 и 6 групп на сутки 4 до имплантации насоса, через неделю после имплантации насоса, в день смены первого насоса и затем каждые 2 недели до конца инфузионного периода. Гистологическое подтверждение мест icv имплантации проводили на срезах мозга толщиной 40 мкм с окрашиванием крезил-фиолетовым.
aCSF
недели 3-10: ASO:
3,28 ммоль/л
1 мкг/мл
недели 3-10:
1 мкг/мл
TGF-β1: 1 мкг/мл
недели 3-10:
TGF-β1: 1 мкг/мл
скремблированный ASO: 3,28 ммоль/л
TGF-β1: 1 мкг/мл
недели 3-10:
TGF-β1: 1 мкг/мл
ASO: 1,1 ммоль/л
3,28 ммоль/л
9,84 ммоль/л
ASO по изобретению оказывали потенциальные эффекты в экспериментах in vitro. Согласуясь с этим, перекрестно реагирующие на грызунах ASO по изобретению с последовательностями SEQ ID NO: 143aj, SEQ ID NO: 143h и SEQ ID NO: 210q, были также эффективны в вышеописанных экспериментах, результаты которых доказывают эффект в лечении животных моделей гидроцефалии. ASO по настоящему изобретению, демонстрирующие отсутствие перекрестной реактивности, оказывали даже более сильные эффекты в экспериментах in vitro. В результате предполагается, что данные ASO по изобретению будут также более эффективными в испытаниях in vivo у приматов, отличных от человека и людей, и поэтому будут функционировать как высокоэффективное лекарственное средство для профилактики или лечения индуцированного TGF-β1 ингибирования пролиферации нейрональных стволовых клеток и клеток-предшественников нейронов, и тем самым лечения гидроцефалии и других нейродегенеративных расстройств.
Пример 16: определение терапевтической активности антисмысловых олигонуклеотидов, нацеленных на TGF-RII, для восстановления повреждения спинного мозга у крыс Fischer 344
Для анализа терапевтической активности ASO для лечения повреждения спинного мозга (SCI), самцов и самок крыс Fischer-344 обрабатывали ASO по изобретению в различных дозах icv введением в боковой желудочек с помощью осмотических мини-насосов.
Описание метода:
SCI воспроизводили перерезанием шейного дорсального отдела позвоночника на уровне C3 ножом из вольфрамовой нити. На следующей стадии для длительной центральной инфузии крыс (с массой тела 180-200 г) подвергали хирургической операции для имплантации icv канюли, присоединенной к осмотическому мининаносу Alzet® (скорость инфузии 0,25 мкл/ч, Alzet® модель 2004, Купертино, США). Канюлю и насос стереотаксически имплантировали под анестезией кетамином/ксилацином (Baxter, GmbH, Германия) в полустерильных условиях. Каждый осмотический мининасос имплантировали подкожно со стороны брюшной области посредством разреза кожи длиной 1 см на шее мыши и соединяли с icv канюлей силиконовыми трубками. Животных помещали в стереотаксическую рамку и icv канюлю (23G, длина 3 мм) погружали в правый боковой желудочек (1,0 мм сзади, 1,0 мм латеральнее, на глубину 1,8 мм относительно брегмы). Канюлю фиксировали двумя винтами из нержавеющей стали с использованием зубного цемента (Kallocryl, Speiko® Dr. Speier GmbH, Мюнстер, Германия). Кожу на шее закрывали швами. Во время операции температуру тела поддерживали грелкой. Для профилактики послеоперационных инфекций мышей местно обрабатывали препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 0,1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия). Трубку заполняли соответствующим раствором. Для определения влияния ASO на процесс реабилитации после SCI, через 4 недели после хирургической операции проводили МРТ-визуализацию структуры in vivo (3T MRI, Allegra Siemens, катушка с фазовой решеткой для мелких животных). Через 6 недель животных подвергали эвтаназии, и извлекали спинной мозг для гистологического и иммуногистохимического анализа. Гистологическое подтверждение мест icv имплантации проводили на срезах мозга толщиной 40 мкм, окрашенных крезил-фиолетовым.
ASO по изобретению оказывали потенциальные эффекты в экспериментах in vitro. Согласуясь с этим, перекрестно реагирующие на грызунах ASO по изобретению с последовательностями SEQ ID NO: 143aj, SEQ ID NO: 143h и SEQ ID NO: 210q, были также эффективны в вышеописанных экспериментах, результаты которых доказывают эффект в лечении на крысах Fischer-344, животных моделях параплегии спинного мозга. МРТ-снимки и нейропатологический анализ свидетельствовали о высокой эффективности ASO по изобретению. ASO по настоящему изобретению, демонстрирующие отсутствие перекрестной реактивности, оказывали даже более сильные эффекты в экспериментах in vitro. В результате предполагается, что данные ASO по изобретению будут также более эффективными в испытаниях in vivo у приматов, отличных от человека и людей, и поэтому будут функционировать как высокоэффективное лекарственное средство для профилактики или лечения индуцированного TGF-β1 ингибирования пролиферации нейрональных стволовых клеток и клеток-предшественников нейронов, и тем самым лечения повреждения спинного мозга и других нейродегенеративных расстройств.
Пример 17: опосредованные ASO эффекты на пролиферацию клеточной линии рака легких человека A549
Определяли уровень мРНК Ki67, p53, каспазы 8 (Casp8) и ингибитора ДНК-связывающего белка 2 (ID2) в качестве репрезентативных маркеров пролиферации в нескольких типах опухолевых клетках. По результатам предыдущих исследований известно, что экспрессия гена-супрессора опухолей р53 и ID2 часто резко повышается в опухолевых тканях. Ki67 является маркером пролиферации, и Casp8 представляет индикатор апоптоза. Кроме того, после трансфекции гимнозисом определяли количество клеток.
Описание метода:
Клетки A549 культивировали, как описано выше. Для обработки клеток среду удаляли и заменяли свежей полной средой в 24-луночных культуральных планшетах (Sarstedt #83.1836.300) (30000 клеток/лунку), 6-луночных культуральных планшетах (Sarstedt #83.3920.300) (50000 клеток/лунку) или 8-луночных слайд-планшетах для культивирования клеток (Sarstedt #94.6140.802) (20000 клеток/лунку) (0,5 мл для 24-луночных планшетов и 8-луночных слайд-планшетов для культивирования клеток и 1 мл для 6-луночных планшетов) и инкубировали в течение ночи при 37°С и 5% CO2. Для анализа экспрессии мРНК и влияния на пролиферацию клетки обрабатывали стандартным олигонуклеотидом 1 (скремблированный контроль) и ASO SEQ ID NO: 218b в концентрациях 2,5 мкМ и 10 мкМ и инкубировали в течение 72 ч при 37°С и 5% СО2. Обработку, включающую замену среды, повторяли 3 раза каждые 72 ч (в целом 12 суток). Для иммуноцитохимического анализа пролиферации (Ki67) трансфекцию гимнозисом ASO SEQ ID NO: 218b ограничивали 72 ч. Затем клетки дважды промывали PBS и использовали для выделения белка (6-луночные планшеты), иммуноцитохимии (8-луночные слайд-планшеты для культивирования клеток), построения кривой пролиферации и выделения РНК (24-луночные планшеты). Протоколы для анализа РНК, белка и иммуноцитохимии выполняли, как описано выше. Для построения кривой пролиферации извлекали оставшиеся клетки из 24-луночных планшетов для определения количества клеток. Для этой цели оставшиеся клетки промывали PBS (2×), обрабатывали аккутазой (500 мкл/лунку) и инкубировали в течение 7 мин при 37°C. Затем добавляли 500 мкл среды и определяли количество клеток с использованием автоматического флуоресцентного счетчика клеток Luna FL® (Biozym, #872040), следуя инструкциям производителя. Вкратце, 18 мкл клеточной суспензии добавляли к 2 мкл красителей акридиновый оранжевый/пропидиум иодид из набора для определения жизнеспособности клеток (Biozym #872045). Через 1 мин осаждения 10 мкл наносили на стекло для подсчета клеток (Biozym #872011). Клетки подсчитывали и по отдельности рассчитывали количество живых и мертвых клеток.
17.1 Результаты для ASO SEQ ID NO: 218b
Анализ мРНК показал снижение уровней экспрессии Ki67, p53 и ID2 через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b. В противоположность, экспресия Casp8 повышалась при низких концентрациях ASO SEQ ID NO: 218b (таблица 46). Эти результаты указывают на то, что снижение роста опухоли связано с небольшим увеличением апоптических клеток. Кроме того, анализ вестерн-блоттингом показал снижение уровня белка Ki67 и pAkt через 12 суток после трансфекции гимнозисом ASO по изобретению (таблица 47). Иммунохимическое исследование клеток A549 после трансфекции гимнозисом ASO SEQ ID NO: 218b показало пониженный уровень сигналов Ki67 по сравнению со скремблированным контролем при обеих использованных концентрациях (фиг. 23). Наконец, количество клеток A549 снижалось примерно на 50% через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b (таблица 48).
Таблица 46: экспрессия мРНК Ki67, p53, Casp8 и ID2 через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549. Данные по регуляции тестированных генов демонстрирует снижение скорости пролиферации после трансфекции гимнозисом ASO по изобретению. Снижение уровня мРНК ID2 оказывает положительное действие в подавлении экспансии опухолевых клеток. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=2
n=2
n=2
Таблица 47: денситометрический анализ после вестерн-блоттинга Ki67 и pAkt. Установлена понижающая регуляция белка Ki67 и pAkt через 12 суток после трансфекции гимнозисом TGF-RII-специфического ASO SEQ ID NO: 218b в клетках A549. Уровни белка определяли по отношению к гену «домашнего хозяйства» GNB2L1, используя программное обеспечение Image Studio™ Lite, и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM.
уровни белка через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=1
Таблица 48: количество клеток через 12 суток после повторной трансфекции гимнозисом. Количество клеток определяли через 12 суток после повторных трансфекций гимнозисом (4×72 ч) клеток A549 с использованием автоматического флуоресцентного счетчика клеток Luna FL® и устройства для светлопольного освещения (Biozym, #872040), следуя инструкциям производителя. A=необработанный контроль, C=SEQ ID NO: 218b, ±=SEM.
количество клеток через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
клеток
n=3
n=3
Заключение:
Полученные результаты показывают, что снижение роста опухолей связано с увеличением числа апоптотических клеток. Примечательно, что уровень ID2, который является возможным геном-мишенью для терапии в опухолях, снижается после трансфекции гимнозисом TGF-RII-специфического ASO SEQ ID NO: 218b.
В совокупности, ASO SEQ ID NO: 218b эффективен в минимизации скоростей пролиферации и снижении экспрессии генов, способствующих росту опухолей.
Пример 18: влияние трансфекции гимнозисом ASO на пролиферацию нескольких опухолевых клеточных линий
Сигнальный путь TGF-β является критическим путем в развитии рака. С одной стороны, TGF-β стимулирует факторы, которые подавляют рост опухолей, но с другой стороны, этот ростовый фактор приводит к стимуляции миграции клеток, инвазии клеток, пролиферации клеток, иммунной регуляции и способствует реорганизации среды, способствуя прогрессированию и метастазированию опухолевых клеток. Таким образом, TGF-β является ключевой мишенью в лечении рака. Уровни мРНК и белка маркера пролиферации (Ki67) и количество клеток определяли после поглощения гимнозисом ASO по изобретению, используя первый в качестве маркера скорости пролиферации в опухолевых клетках. Кроме того, определяли уровни мРНК гена-супрессора опухолей р53 и ингибитора ДНК-связывающего белка 2 (ID2).
Описание методов:
A549 культивировали, как описано выше. Для обработки клеток среду удаляли и заменяли свежей полной средой в 24-луночных культуральных планшетах (Sarstedt #83.1836.300) (30000 клеток/лунку), 6-луночных культуральных планшетах (Sarstedt #83.3920.300) (50000 клеток/лунку) или 8-луночных слайд-планшетах для культивирования клеток (Sarstedt #94.6140.802) (20000 клеток/лунку) (0,5 мл для 24-луночных и 1 мл для 6-луночных планшетов) и инкубировали в течение ночи при 37°С и 5% CO2. Для анализа экспрессии мРНК и оценки влияния на пролиферацию клетки обрабатывали стандартным олигонуклеотидом 1 (скремблированный контроль) и ASO SEQ ID NO: 218b в концентрациях 2,5 мкМ и 10 мкМ и инкубировали в течение 72 ч при 37°С и 5% СО2. Обработку, включающую замену среды, повторяли 3 раза каждые 72 ч (в целом 12 суток). Для сбора клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты), выделения белка (6-луночные планшеты) или построения кривой пролиферации. Протоколы для выделения РНК и белка выполняли, как описано выше. Перед подсчетом клеток для построения кривой пролиферации клетки анализирвоали с использованием световой микроскопии (Nicon, TS-100 F LED #MFA33500). Оставшиеся клетки извлекали из 24-луночных планшетов для определения количества клеток. Для этой цели оставшиеся клетки промывали PBS (2×), обрабатывали аккутазой (500 мкл/лунку) и инкубировали в течение 5-7 мин при 37°C. Затем добавляли 500 мкл среды и определяли количество клеток с использованием автоматического флуоресцентного счетчика клеток Luna FL® и устройства для светлопольного освещения (Biozym, #872040), следуя инструкциям производителя. Вкратце, 18 мкл клеточной суспензии добавляли к 2 мкл красителей акридиновый оранжевый/пропидиум иодида из набора для определения жизнеспособности клеток (Biozym #872045). Через 1 мин осаждения 10 мкл наносили на стекло для подсчета клеток (Biozym #872011). Клетки подсчитывали и по отдельности рассчитывали количество живых и мертвых клеток.
18.1 Результаты для SEQ ID NO: 218b
Уровни мРНК Ki67 эффективно снижались независимо (A549, L3.6pl, Panc-1) или в зависимости от использованных концентраций ASO (HT-29, Panc-1, CaCo2) через 12 суток после трансфекции гимнозисом (таблица 40). Уровень экспрессии гена р53 также изменялся в клетках А549, ХТ-29, К562, KG-1, СаСо2 и ТМК-1 (таблица 50). Была показана пониженная экспрессия белка Ki67 в клетках A549, L3.6pl, ™K-1, HT-29 и K562 (таблица 51). Примечательно, что экспрессия мРНК ID2 подверглась последовательной эффективной и дозозависимой понижающей регуляции в клетках A549, HT-29, K562 и ™K-1, опосредуемой ASO SEQ ID NO: 218b (таблица 51). Кроме того, ASO SEQ ID NO: 218b приводил к снижению скорости пролиферации нескольких линий опухолевых клеток (таблица 53). Дозозависимое уменьшение количества клеток было установлено для клеток HPAFII, MCF-7, KG1, K562, U937 и HTZ-19. Клетки злокачественной опухоли легких (A549) показали примерно 50% снижение количества клеток, вызванное ASO SEQ ID NO: 218b. Снижение количества клеток было дополнительно подтверждено световой микроскопией для линий HPAFII, K562, MCF-7, Panc-1 и HTZ-1 через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b (фиг. 24).
Сопоставимые результаты были получены для антисмысловых олигонуклеотидов с последовательностями SEQ ID NO: 141d, 141g, 141i, 143r, 143w, 143af, 143ag, 143ah, 143j, 143p, 143q, 233d, 234d, 235b, 235d, 237b, 237c, 237i, 237m, 238c, 238f, 239e, 240c, 241b, 242a, 246e, 247d, 248b, 248e, 248g, 152k, 152s, 152t, 152u, 152ab, 152ag, 152ah, 152ai, 249c, 249e, 250b, 250g, 251c, 251f, 252e, 253c, 254b, 255a, 259e, 260d, 261b, 261e, 261g, 262d, 262e, 209s, 209v, 209w, 209x, 209ai, 209an, 209at, 209au, 209av, 210o, 210v, 210w, 210x, 210ab, 210ac, 210ad, 210af, 210am, 263b, 263c, 263i, 263m, 264e, 264h, 265e, 266c, 267b, 268a, 272e, 273d, 274a, 274d, 274f, 275g, 275i, 276b, 276c, 276j, 276k, 277d, 277e, 278f, 279c, 280b, 281a, 218ad, 218n, 218t, 218u, 218v, 218ah, 218an, 218ao, 218ap, 220d, 221d, 222b, 222c,222f, 223c, 223f, 224i, 224m, 225c, 225f, 226e, 227c, 213o, 213p, 213q, 213s, 213y, 213z, 213aa, 213af, 228b, 229a, 285d, 286d, 287d, 287e, 287f, 288e, 288i, 289d, 289h, 289o, 289p, 289q, 290c, 290f, 290i, 291c, 292c, 293b и 294a. Большинство вышеуказанных антисмысловых олигонуклеотидов были не более эффективными, чем последовательности SEQ ID NO: 218b и 218c, но, тем не менее, они являются существенно более преимущественными, чем антисмысловые олигонуклеотиды, известные в данной области. Таким образом, антисмысловые олигонуклеотиды по настоящему изобретению очень пригодны для лечения гиперпролиферативных заболеваний, таких как рак и опухоли.
Таблица 49: экспрессия мРНК маркера пролиферации Ki67. Через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b клеток A549, HT-29, L3.6pl, KG1, Panc-1 и CaCo2, уровень мРНК Ki67 снижался во всех клеточных линиях соответственно. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=2
n=2
n=1
n=1
n=1
Таблица 50: экспрессия мРНК супрессора опухолей р53. Через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b клеток A549, HT-29, K562, KG1, CaCo2 и ™K-1, уровень мРНК р53 снижался во всех клеточных линиях соответственно. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, *p<0,05 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=1
n=1
n=1
n=1
n=1
Таблица 51: экспрессия мРНК ID2. Через 12 суток после трансфекции гимнозисом ASO SEQ ID NO: 218b клеток A549, HT-29, K562 и ™K-1, уровень мРНК ID2 снижался в зависимости от концентрации во всех клеточных линиях соответственно. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, *p<0,05 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=1
n=1
n=1
Таблица 52: денситометрический анализ после вестерн-блоттинга Ki67. Установлена понижающая регуляция белка Ki67 после трансфекции гимнозисом ASO SEQ ID NO: 218b. Уровень белка определяли количественно по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Image Studio™ Lite и нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=2
n=2
n=2
n=1
Таблица 53: количество клеток в нескольких линиях раковых клеток через 12 суток после повторной трансфекции гимнозисом (4×72 ч). ASO SEQ ID NO: 218b трансфицировали в несколько линий раковых клеток. Количество клеток определяли с использованием автоматического флуоресцентного счетчика клеток Luna FL® и устройства светопольного освещения (Biozym, #872040), следуя инструкциям производителя. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, a=живые клетки, d=мертвые клетки. ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
B 2,5 мкM против C 10 мкM +p<0,01 C 10 мкM против D 10 мкM #p<0,01
A против C 10 мкM *p<0,01
A против C 10 мкM *p<0,05
B 10 мкM против C 10 мкM +p<0,01
Заключение:
Модуляция мРНК Ki67, p53 и ID2 под действием ASO SEQ ID NO: 218b указывает на положительный эффект в подавлении роста опухолей в нескольких органах и различного происхождения. Известно, что Ki67, ID2 и p53 активируются и способствуют пролиферации клеток в различных типах злокачественных опухолей. Маркеры пролиферации Ki67, p53 и ID2 подвергались эффективной понижающей регуляции. Подсчет клеток и световая микроскопия некоторых типов опухолевых клеток через 12 суток после трансфекции гимнозисом показали, что ASO SEQ ID NO: 218b является высокоэффективным агентом для снижения пролиферации клеток.
В совокупности, TGF-RII-специфический ASO SEQ ID NO: 218b эффективно снижал скорость пролиферации параллельно с установленными модуляциями мРНК Ki67, p53 и ID2. Эти данные дают основание предположить, что ASO по изобретению являются многообещающими кандидатами для применения в качестве лекарственных препаратов в целях снижения прогрессирования опухолевых клеток и метастазирования опухолевых клеток.
Пример 19: анализ влияния антисмысловых олигонуклеотидов на ангиогенез в нескольких линиях опухолевых клеток
Модуляция ангиогенеза необходима для роста и восстановления органов. Нарушения роста кровеносных сосудов вносят свой вклад в развитие различных заболеваний, например, таких как рост опухолей, ишемия, воспалительные и иммунные расстройства. Известно, что TGF-β является проангиогенным фактором. Это может быть наиболее важно для воспалительных и неопластических процессов, когда избыточный ангиогенез ответственен за прогрессирование заболевания. Данные эффекты могут иметь место параллельно с индуцированным TGF-β1 фиброзом. Следовательно, ингибирование сигнального пути TGF-β TGFRII-специфическим ASO может представлять собой адекватный терапевтический подход.
Для проверки этого предположения несколько линий опухолевых клеток трансфицировали данными ASO поглощением гимнозисом. Через 12 суток после многократных трансфекций гимнозисом клеточный супернатант анализировали на уровни белка проангиогенных факторов мультиплексным анализом. Данная технология позволила исследовать несколько проангиогенных белков (VEGF, Tie-2, FLt-1, PIGF и bFGF) с использованием электрохемилюминесценции. Фактор роста эндотелия сосудов (VEGF) является мощным секретируемым опухолями цитокином, который способствует ангиогенезу и тем самым стимулирует, например, пролиферацию опухолей. Tie-2 представляет собой белок, который экспрессируется из активно растущих кровеносных сосудов. Fms-подобная тирозинкиназа 1 (Flt-1), также известная как рецептор 1 фактора роста эндотелия сосудов (VEGFR1), представляет собой трансмембранную рецепторную тирозинкиназу, которая экспрессируется на высоком уровне в сосудистых эндотелиальных клетках, и плацентарный фактор роста (PIGF) функционирует вместе с VEGF и активируется при патологических состояниях, например, при образовании опухоли. Кроме того, основный фактор роста фибробластов (bFGF) является ростовым фактором, который также индуцирует ангиогенез. PAI-1 является геном-мишенью для TGF-β и опосредует образование рубцов и ангиогенные эффекты TGF-β. Таким образом, PAI-1 является также ключевым фактором для инвазии и метастазирования опухолей. Высокая концентрация PAI-1 у пациентов является плохим прогностическим фактором, например, при раке молочной железы, легких, колоректальном раке и раке желудке. Высокие концентрации PAI-1 также являются фактором риска для заболеваний, при которых тромбоз имеет большое значение (например, инфаркт миокарда, инсульт). Таким образом, также исследовали регуляцию мРНК PAI-1 TGF-β-специфическими антисмысловыми олигонуклеотидами.
Описание методов:
Опухолевые клеточные линии культивировали, как описано выше (таблица 10). Для обработки клеток среду удаляли и заменяли свежей полной средой в 24-луночных культуральных планшетах (Sarstedt #83.1836.300) (30000 клеток/лунку), инкубировали в течение ночи при 37°C и 5% CO2. На следующий день к обновленной среде добавляли стандартный олигонуклеотид 1 (скремблированный контроль) и ASO SEQ ID NO: 218b в концентрациях 2,5 и 10 мкМ и инкубировали в течение 72 ч при 37°С и 5% CO2. Обработку, включающую замену среды, повторяли 3 раза каждые 72 ч (в целом 12 суток). После этого клеточный супернатант собирали и анализировали методом MesoScale Discovery® (MSD Discovery), что позволило исследовать несколько проангиогенных белков (VEGF, Tie-2, FLt-1, PIGF и bFGF) с использованием электрохемилюминесценции. Протокол проведения эксперимента и информацию об отдельных ростовых факторах получали из инструкций производителя (MSD MesoScale Discovery®, #K15198G). Результаты были оценены с помощью программного обеспечения GraphPad Prism® 6.0.
Затем клетки дважды промывали PBS и использовали для выделения РНК (24-луночные планшеты) для анализа того, может ли трансфекция гимнозисом ASO регулировать уровни мРНК ингибитора-1 активатора плазминогена (PAI-1) с использованием ОТ-ПЦР в режиме реального времени. Использовали протоколы и праймеры, и они перечислены, как описано выше.
19.1 Результаты для SEQ ID NO: 218b
В таблице 54 показано, что мРНК PAI-1 подвергалась понижающей регуляции дозозависимым образом в клетках нескольких тестированных раковых опухолей (A549: рак легких, HPAFII: аденокарцинома поджелудочной железы, HT-29: колоректальная аденокарцинома, HTZ-19: меланома, ™K-1: карцинома желудка, ТНР-1: моноцитарный лейкоз) после повторной трансфекции гимнозисом ASO SEQ ID NO: 218b. Кроме того, уровни белка VEGF в супернатантах стимулированных клеток также свидетельствовали о дозозависимом снижении в A549, HTZ-19, HPAFII и PC3M (аденокарцинома предстательной железы). Для клеток HPAFII и PC3M понижающая регуляция была статистически достоверной (таблица 55). Влияние ASO SEQ ID NO: 218b на bFGF подтверждалось данными для VEGF, означая, что ASO SEQ ID NO: 218b является эффективным в подавлении ангиогенеза (таблица 56). Результаты для клеток A549 и PC3M также показали достоверное снижение уровня bFGF. Количество белка PIGF в клеточных супернатантах незначительно, но дозозависимо снижалось в клетках A549 и HTZ-19. В клетках PC3M уровень основного эндогенного PIGF был выше, чем во всех других тестированных клетках, и эффект ASO был также очевиден (таблица 57). Наконец, имела место понижающая регуляция белка Flt-1 в клетках HT-29 (таблица 58) и снижение Tie-2 в клетках HTZ-19 (ASO SEQ ID NO: 218b, 2,5 мкМ) и MCF-7 (карцинома молочной железы, 10 мкМ) (таблица 59).
Сопоставимые результаты были получены для антисмысловых олигонуклеотидов с последовательностями SEQ ID NO: 141d, 141g, 141i, 143r, 143w, 143af, 143ag, 143ah, 143j, 143p, 143q, 233d, 234d, 235b, 235d, 237b, 237c, 237i, 237m, 238c, 238f, 239e, 240c, 241b, 242a, 246e, 247d, 248b, 248e, 248g, 152k, 152s, 152t, 152u, 152ab, 152ag, 152ah, 152ai, 249c, 249e, 250b, 250g, 251c, 251f, 252e, 253c, 254b, 255a, 259e, 260d, 261b, 261e, 261g, 262d, 262e, 209s, 209v, 209w, 209x, 209ai, 209an, 209at, 209au, 209av, 210o, 210v, 210w, 210x, 210ab, 210ac, 210ad, 210af, 210am, 263b, 263c, 263i, 263m, 264e, 264h, 265e, 266c, 267b, 268a, 272e, 273d, 274a, 274d, 274f, 275g, 275i, 276b, 276c, 276j, 276k, 277d, 277e, 278f, 279c, 280b, 281a, 218ad, 218n, 218t, 218u, 218v, 218ah, 218an, 218ao, 218ap, 220d, 221d, 222b, 222c,222f, 223c, 223f, 224i, 224m, 225c, 225f, 226e, 227c, 213o, 213p, 213q, 213s, 213y, 213z, 213aa, 213af, 228b, 229a, 285d, 286d, 287d, 287e, 287f, 288e, 288i, 289d, 289h, 289o, 289p, 289q, 290c, 290f, 290i, 291c, 292c, 293b и 294a. Большинство вышеуказанных антисмысловых олигонуклеотидов были не более эффективными, чем последовательности SEQ ID NO: 218b и 218c, но, тем не менее, они являются существенно более преимущественными, чем антисмысловые олигонуклеотиды, известные в данной области. Таким образом, антисмысловые олигонуклеотиды по настоящему изобретению очень пригодны для лечения гиперпролиферативных заболеваний, таких как рак и опухоли.
Таблица 54: экспрессия мРНК PAI-1 через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549, HPAFII, HT-29, HTZ-19, ™K-1 и THP-1. ASO SEQ ID NO: 218b в зависимости от концентрации регулирует экспрессию гена PAI-1 таким образом, что прогноз заболевания улучшается. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=3
n=1
n=2
n=2
n=2
n=2
Таблица 55: уровни белка VEGF в клеточном супернатанте через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549, HPAFII, HTZ-19, PC3M с использованием метода MesoScale Discovery ® (MSD Mesoscale Discovery, #K15198G). Уровни белка определяли анализом электрохемилюминесценции. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, *p<0,05 и **p<0,01 по отношению к A, +p<0,05 и ++p<0,01 по отношению к 2,5 мкМ B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка (пг/мл) через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=2
n=2
n=2
Таблица 56: уровни белка bFGF в клеточном супернатанте через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549 и PC3M с использованием метода MesoScale Discovery ® (MSD Mesoscale Discovery, #K15198G). Уровни белка определяли анализом электрохемилюминесценции. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, *p<0,05 и **p<0,01 по отношению к A, +p<0,05 и ++p<0,01 по отношению к 2,5 мкМ B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка (пг/мл) через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=2
Таблица 57: уровни белка PIGF в клеточном супернатанте через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549, HTZ-19 и PC3M с использованием метода MesoScale Discovery ® (MSD Mesoscale Discovery, #K15198G). Уровни белка определяли анализом электрохемилюминесценции. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка (пг/мл) через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=1
n=2
Таблица 58: уровни белка Flt-1 в клеточном супернатанте через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках HTZ-19 с использованием метода MesoScale Discovery ® (MSD Mesoscale Discovery, #K15198G). Уровни белка определяли анализом электрохемилюминесценции. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка (пг/мл) через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
Таблица 59: уровни белка Tie-1 в клеточном супернатанте через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках HTZ-19 и MCF-7 с использованием метода MesoScale Discovery ® (MSD Mesoscale Discovery, #K15198G). Уровни белка определяли анализом электрохемилюминесценции. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка (пг/мл) через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=1
Заключение:
Все анализированные проангиогенные факторы (VEGF, bFGF, PIGF, Flt-1 и Tie-2) могут регулироваться по действием ASO SEQ ID NO: 218b таким образом, что это обеспечивает подавление прогрессирования опухоли и других патологических механизмов, зависящих от усиленного ангиогенеза. Кроме того, уровень мРНК PAI-1 дозозависимо снижался под действием ASO SEQ ID NO: 218b. Данный фактор, являющийся геном-мишенью для TGF-β и, например, представляет признанный прогностический маркер при раке молочной железы, также подвергался дозозависимой понижающей регуляции.
В совокупности, все тестированные ASO по изобретению были эффективны в подавлении ангиогенных процессов, которые способствуют прогрессированию опухолей, метастазированию, воспалению и тромбозу. Таким образом, ASO по изобретению, нацеленные на TGF-RII, являются эффективным кандидатом для применения в качестве терапевтического средства при различных типах заболеваний, связанных с раком и тромбозом.
Пример 20: Анализ влияния ASO по изобретению на фиброз
TGF-β участвует во многих процессах, таких как пролиферация, миграция клеток, заживление ран, ангиогенез и межклеточные взаимодействия. По результатам нескольких исследований известно, что этот фактор часто повышается во время патогенеза при некоторых заболеваниях, в том числе первичной открытоугольной глаукоме, болезни Альцгеймера, фиброзе легких и диабетической нефропатии. Эти заболевания связаны с патологическими изменениями внеклеточного матрикса (ECM) и актинового цитоскелета. Часто такие наблюдаемые изменения коррелируют с прогрессированием тяжести заболевания и резистентностью к лечению (эпителиально-мезенхимальный переход-EMT-в опухолях). Фактор роста соединительной ткани (CTGF) является медиатором ниже TGF-β и опосредует фибротические эффекты TGF-β. Так, показано, что CTGF опосредует отложение ECM и модулирует реорганизацию актинового цитоскелета. Для исследования того, насколько ASO по изобретению вносят свой вклад в разрешение фибротических процессов посредством ингибирования сигнального пути TGF-β, оценивали уровни CTGF в дополнение к фибронектину (FN) и коллагену IV (ColIV), которые представляют собой два основных компонента ECM в нескольких различных типах раковых клеток. Кроме того, было исследовано влияние ASO на CTGF, FN и актиновый цитоскелет в клетках-предшественниках нейронов (ReNcell CX®) и клетках рака легких человека (A549).
20.1. Фиброз при нейродегенерации
Описание методов:
Клетки культивировали, как описано ранее в стандартном протоколе. Для обработки клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (50000 клеток/лунку), 6-луночные культуральные планшеты (Sarstedt #83.3920.300) (80000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для исследования ответной реакции клеток ReNcell CX® на TGF-β1 их обрабатывали после обновления среды TGF-β1 (2 и 10 нг/мл, PromoCell #C63499) в течение 48 ч с последующим анализом мРНК для CTGF. Для выяснения влияния ASO на CTGF и FN в клетках ReNcell CX® среду удаляли и заменяли свежей полной средой (1 мл на 6-луночный планшет и 0,5 мл на 8-луночный планшет). Затем в среду добавляли стандартный олигонуклеотид 1 (скремблированный контроль), ASO SEQ ID NO: 218b и SEQ ID NO: 218с в концентрациях 2,5 и 10 мкМ и через 96 ч проводили соответствующий анализ (ОТ-ПЦР в режиме реального времени, вестерн-блоттинг и иммуноцитохимию). Для оценки влияния ASO после исследования эффекта предварительной инкубации с TGF-β1 среду удаляли и заменяли свежей полной средой (1 мл для 6-луночных планшетов и 8-луночных слайд-планшетов для культивирования клеток). После экспозиции TGF-β1 (10 нг/мл, 48 ч) среду заменяли, добавляли TGF-β1 (10 нг/мл), стандартный олигонуклеотид 1 (10 мкM), ASO с SEQ ID NO: 218b (10 мкМ) и ASO с SEQ ID NO: 218c (10 мкМ) в комбинации и в виде самостоятельной обработки клеток. Затем клетки ReNcell CX® собирали через 96 ч после трансфекции гимнозисом. Соответственно, клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты) и выделения белка (6-луночные планшеты) или иммуноцитохимического анализа клеток (8-луночные слайд-планшеты для культивирования клеток). Использовали протоколы, антитела, разведения и праймеры, как описано выше.
20.1.1. Результаты оценки эффектов TGF-β1 на клетки-предшественники нейронов (ReNcell CX®)
Данные о реакции клеток ReNcell CX® на TGF-β1 отсутствуют. Следовательно, для выяснения этого вопроса клетки ReNcell CX® обрабатывали в течение 48 ч TGF-β1 в двух разных концентрациях (таблица 60). Оценка ОТ-ПЦР в режиме реального времени выявила дозозависимую индукцию экспрессии генов CTGF и TGF-β1.
Таблица 60: экспрессия мРНК CTGF и TGF-β1 через 48 ч после стимуляции TGF-β1. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, E=TGF-β1. ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК после 48 ч обработки TGF-β1
Временная точка
48 ч
n=3
48 ч
n=3
Заключение:
Клетки ReNcell CX® проявляли ответную реакцию на воздействие TGF-β1, представляющую самоиндукцию TGF-β1, и повышение экспрессии гена-мишени CTGF для TGF-β1. В совокупности, клетки ReNcell CX® идеально подходят для изучения вопросов, касающихся эффектов TGF-β.
20.1.2 Результаты для SEQ ID NO: 218b
20.1.2.1. Эффекты трансфекции гимнозисом
Трансфекция гимнозисом ASO SEQ ID NO: 218b приводит к дозозависимому и статистически достоверному снижению CTGF и FN (таблица 61). Данный эффект ASO SEQ ID NO: 218b подтверждали для уровня белка FN. Уровень белка FN снижался примерно на 70% через 96 ч после трансфекции гимнозисом тестированного ASO, в то время как обработка клеток ReNcell CX® TGF-β1 приводила к 3,4-кратной индукции FN (таблица 62).
Таблица 61: дозозависимая и достоверная понижающая регуляция мРНК CTGF после трансфекции гимнозисом SEQ ID NO: 218b в клетках ReNcell CX ®. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. ±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
уровни мРНК после трансфекции гимнозисом
Временная точка
96 ч, n=3
96 ч, n=3
Таблица 62: денситометрический анализ после вестерн-блоттинга фибронектина. Установлена понижающая регуляция белка FN через 96 ч после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках ReNcell CX®. Уровень белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Image Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b.
уровни белка после трансфекции гимнозисом
Временная точка
96 ч, n=1
Заключение:
ASO SEQ ID NO: 218b был эффективным в понижающей регуляции уровней мРНК CTGF и FN в клетках-предшественниках нейронов человека. Обработка ASO SEQ ID NO: 218b приводила к снижению уровня белка FN через 96 ч после трансфекции гимнозисом. Таким образом, TGF-RII- специфический ASO опосредует блокирование TGF-β-индуцированных фибротических эффектов в клетках ReNcell CX®.
20.1.2.2. Эффекты трансфекции гимнозисом после предварительной инкубации с TGF-β
Для анализа того, насколько ASO SEQ ID NO: 218b также способен подавлять фибротические эффекты, опосредуемые TGF-β при патологических состояниях, клетки ReNcell CX® предварительно инкубировали с TGF-β с последующей трансфекцией гимнозисом в течение 96 ч. Установленные затем уровни мРНК CTGF и FN указывают на выраженное антифибротическое действие ASO SEQ ID NO: 218b также после индукции под действием TGF-β экспрессии гена CTGF и FN (таблица 63). Иммуноцитохимическое окрашивание на CTGF (фиг. 25A) и FN (фиг. 25B) подтвердило данные анализа мРНК. Кроме того, окрашивание фаллоидином для анализа актинового цитоскелета показало индукцию стрессовых волокон после обработки TGF-β, в то время как ASO SEQ ID NO: 218b был эффективен в блокировании индукции стрессовых волокна, опосредуемой TGF-β (фиг. 25С).
Таблица 63: понижающая регуляция мРНК CTGF и FN после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO: 218b в клетках ReNcell CX® (по сравнению со скремблированным контролем). Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β,±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
уровни мРНК через 48 ч TGF-β1 → 96 ч TGF-β1+ASO/обработка одним ASO
Временная точка
96 ч, n=3
96 ч, n=3
Заключение:
ASO SEQ ID NO: 218b демонстрировал выраженные антифибротические эффекты при индуцированных патологических состояниях (предварительной инкубацией с TGF-β1). Помимо понижающей регуляции FN, который является одним из основных компонентов ECM, актиновый цитоскелет также подвергался воздействию ASO по изобретению таким образом, что последний может быть полезен для лучшего исхода при фиброзных болезнях.
20.1.3 Результаты для SEQ ID NO: 218c
20.1.3.1. Эффекты трансфекции гимнозисом
Трансфекция гимнозисом ASO SEQ ID NO: 218c приводит к выраженному и достоверному снижению мРНК CTGF после трансфекции гимнозисом 10 мкМ ASO SEQ ID NO: 218с (таблица 64).
Таблица 64: понижающая регуляция мРНК CTGF после трансфекции гимнозисом SEQ ID NO: 218c в клетках ReNcell CX ® . Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени, и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218с. ±=SEM, *p<0,05 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
уровни мРНК после трансфекции гимнозисом
Временная точка
96 ч, n=3
Заключение:
ASO SEQ ID NO: 218c был эффективным в дозозависимом снижении уровня мРНК CTGF.
20.1.3.2. Эффекты трансфекции гимнозисом после предварительной инкубации с TGF-β
Результаты по трансфекции гимнозисом ASO SEQ ID NO: 218c после предварительной инкубации с TGF-β1 подтверждались эффективным блокированием индуцированных TGF-β1 эффектов на уровень мРНК CTGF (таблица 65). ASO был эффективным в блокировании влияния TGF-β1 на CTGF так, что комбинированная обработка была сравнима с обрабаботкой одним ASO SEQ ID NO: 218c.
Таблица 65: уровень мРНК CTGF после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO: 218c и параллельной обработкой TGF-β1 в клетках ReNcell CX®. Данные подтвердили эффективное блокирование индуцированных TGF-β1 эффектов на уровень мРНК CTRF под действием ASO SEQ ID NO:218c по сравнению с комбинированными обработками. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218c, E=TGF-β1. ±=SEM, *p<0,05 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
уровни мРНК через 48 ч TGF-β1 → 96 ч TGF-β1+ASO/обработка одним ASO
n=3
Заключение:
ASO SEQ ID NO: 218c вызывал выраженную понижающую регуляцию мРНК и белка CTGF даже при искусственных патологических состояниях (предварительной инкубацией с TGF-β1).
В совокупности, помимо сильных антифибротических эффектов, TGF-RII-специфические ASO вызывали модуляцию актинового цитоскелета. Индукция стрессовых волокон может вызывать повышение ригидности и неподвижности клеток, что может играть роль, например, при болезни Альцгеймера и других нейродегенеративных заболеваниях. Отложение ECM также может опосредовать быстрые патологические изменения, например, при первичной открытоугольной глаукоме. Таким образом, уменьшение отложения ECM и подавление формирования стрессовых волокон может быть полезным для улучшения прогноза при неврологических заболеваниях, связанных с фиброзом. Таким образом, TGF-RII-специфические ASO являются эффективными терапевтическими агентами для лечения, например, болезни Альцгеймера и первичной открытоугольной глаукомы.
20.2. Фиброз легких
Описание методов:
Для определения влияния ASO на ECM и актиновый цитоскелет в легких, исследовали клетки рака легких человека (A549) и культивировали, как описано ранее. Для обработки клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (50000 клеток/лунку), 6-луночные культуральные планшеты (Sarstedt #83.3920.300) (80000 клеток/лунку) и 8-луночные слайд-планшеты для культивирования клеток (Sarstedt #94.6140.802) (10000 клеток/лунку) и инкубировали в течение ночи при 37°C и 5% CO2. Для исследования ответной реакции клеток A549 на TGF-β1 после обновления среды их обрабатывали TGF-β1 (2 и 10 нг/мл, PromoCell #C63499) в течение 48 ч с последующим анализом мРНК CTGF. Для оценки влияния ASO на CTGF и FN в клетках A549 среду удаляли и заменяли свежей полной средой (1 мл для 6-луночных и 0,5 мл для 8-луночных планшетов). Затем в среду добавляли стандартный олигонуклеотид 1 (скремлированный контроль), ASO SEQ ID NO: 218b и SEQ ID NO: 218с в концентрациях 2,5 и 10 мкМ и проводили соответствующий анализ (ОТ-ПЦР в режиме реального времени, вестерн-блоттинг и иммуноцитохимию) через 72 ч в клетках ReNcell CX®. Для того, чтобы показать возможное действие ASO после предварительной инкубации с TGF-β1, среду удаляли и заменяли свежей полной средой (1 мл для 6-луночных планшетов и 8-луночных слайд-планшетов для культивирования клеток). После экспозиции TGF-β1 (10 нг/мл, 48 ч) среду заменяли, добавляли TGF-β1 (10 нг/мл), стандартный олигонуклеотид 1 (10 мкM), ASO SEQ ID NO: 218b (10 мкМ) и ASO SEQ ID NO: 218c (10 мкМ) в комбинации и в виде самостоятельной обработки клеток. Затем клетки A549 собирали через 72 ч после трансфекции гимнозисом. Соответственно, клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты) и белка (6-луночные планшеты) или иммуноцитохимического анализа клеток (8-луночные слайд-планшеты для культивирования клеток). Протоколы, использованные антитела, разведения и праймеры были такими, как описано выше.
20.2.1. Результаты оценки эффектов TGF-β1 на клетки рака легких (A549)
Для исследования способности клеток A549 реагировать на экспозицию TGF-β1, их обрабатывали в течение 48 ч TGF-β1 в двух разных концентрациях (таблица 66). Оценка ОТ-ПЦР в режиме реального времени показала, что CTGF и TGF-β1 вызывают дозозависимую индукцию генной экспрессии.
Таблица 66: индуцированная экспрессия мРНК CTGF и TGF-β1 через 48 ч после стимуляции TGF-β1 в клетках A549. Уровни экспрессии мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, E=TGF-β1. ±=SEM, *p <0,05 и **p <0,01 по отношению к A, ++p <0,05 по отношению к 2 нг/мл E. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК после 48 ч обработки TGF-β1
Временная точка
48 ч, n=3
48 ч, n=3
Заключение
Клетки A549 показали дозозависимую и достоверную повышающую регуляцию мРНК CTGF после экспозиции TGF-β1. Кроме того, наблюдали самоиндукцию TGF-β1. В совокупности, клетки A549 являются хорошей моделью для исследования вопросов, касающихся TGF-эффектов в легких и при раке легких.
20.2.2 Результаты для SEQ ID NO: 218b
20.2.2.1 Результаты оценки эффектов трансфекции гимнозисом
Трансфекция гимнозисом ASO SEQ ID NO: 218b вызывает дозозависимое и высокодостоверное снижение экспрессии гена CTGF (таблица 67). Тестированный ASO также оказывал влияние на уровень мРНК FN, но не в зависимости от концентрации. В противоположность, окрашивание на FN выявило дозозависимое снижение уровня FN по сравнению со скремблированным контролем (фиг. 260A). Кроме того, исследовали влияние ASO и TGF-β на актиновый цитоскелет. На фиг. 26B показана индукция актиновых волокон, включая образование стрессовых волокон, после обработки TGF-β1 в клетках A549 в зависимости от концентрации, в то время, как сигнал после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549 был достоверно снижен параллельно с установленной отменой TGF-β1-опосредованных эффектов. В отношение анализа белков, то была показана специфическая понижающая регуляция CTGF параллельно с торможением pErk1/2, посредством которого опосредуются фибротические эффекты CTGF (таблица 68). Кроме того, через 72 ч после трансфекции гимнозисом ASO SEQ ID NO: 218b было отмечено снижение уровней обоих основных компонентов ЕСМ: CollV и FN (таблица 68).
Таблица 67: дозазависимая и достоверная понижающая регуляция мРНК CTGF после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
уровни мРНК после трансфекции гимнозисом
Временная точка
72 ч, n=3
72 ч, n=3
Таблица 68: денситометрический анализ после вестерн-блоттинга CTGF, FN, CollV и pErk11/2. Анализ проводили через 72 ч после трансфекции гимнозисом ASO SEQ ID NO: 218b в клетках A549. Уровень белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Image Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b.
уровни белка после трансфекции гимнозисом
Временная точка
72 ч
n=1
72 ч
n=1
72 ч
n=1
72 ч
n=2
Заключение:
Трансфекция гимнозисом SEQ ID NO: 218b была эффективной в отношении модулирующих факторов, которые участвуют в отложении ECM и реорганизации актинового цитоскелета в клетках легких человека.
20.2.2.2. Результаты оценки эффектов трансфекции гимнозисом после предварительной инкубации с TGF-β1
Результаты трансфекции гимнозисом ASO SEQ ID NO: 218b после предварительной инкубации с TGF-β1 подтверждали эффективное блокирование сильных индуцированных TGF-β1 эффектов на уровни мРНК CTGF и FN (таблица 69). Иммуноцитохимическое окрашивание на CTGF (фиг. 27A) и FN (фиг. 27B) подтвердило обнаружение эффектов на мРНК также на уровне белка.
Таблица 69: уровень мРНК CTGF и FN после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO: 218b и параллельной обработкой с TGF-β1 в клетках A549. Данные подтвердили эффективное блокирование индуцированных TGF-β1 эффектов на уровни мРНК CTGF и FN под действием ASO SEQ ID NO: 218b по сравнению с комбинированными обработками. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, E=TGF-β1. ±=SEM, *p<0,05, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
Временная точка
уровни мРНК через 48 ч TGF-β1 → 72 ч TGF-β1+ASO/обработка одним ASO
n=5
n=3
Заключение:
ASO SEQ ID NO: 218b был эффективным в опосредовании антифибротических эффектов в клетках A549 при искусственных патологических состояниях, имитированных избыточными концентрациями TGF-β1.
20.2.3 Результаты для SEQ ID NO: 218c
20.2.3.1 Результаты оценки эффектов трансфекции гимнозисом
Трансфекция гимнозисом ASO SEQ ID NO: 218c опосредует выраженное дозозависимое и достоверное снижение мРНК CTGF через 72 ч после трансфекции гимнозисом клеток A549 (таблица 70).
Таблица 70: понижающая регуляция мРНК CTGF через 72 ч после трансфекции гимнозисом SEQ ID NO: 218c клеток A549. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218с. ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Даннета.
уровни мРНК после трансфекции гимнозисом
Временная точка
72 ч n=4
Заключение:
Трансфекция гимнозисом ASO SEQ ID NO: 218c была эффективной в снижении уровня мРНК медиатора CTGF даунстрим TGF-β.
20.2.2.2. Результаты оценки эффектов трансфекции гимнозисом после предварительной инкубации с TGF-β
Результаты оценки трансфекции гимнозисом ASO SEQ ID NO: 218c после предварительной инкубации с TGF-β1 подтвердили эффективное блокирование сильных индуцированных TGF-β1 эффектов на уровни мРНК CTRF (таблица 71). Иммуноцитохимическое окрашивание на CTGF подтвердило эти данные на уровне белка (фиг. 28).
Таблица 71: уровни мРНК CTGF после предварительной инкубации с TGF-β1 с последующей трансфекцией гимнозисом SEQ ID NO: 218c и параллельной обработкой TGF-β1 в клетках A549. Данные подтвердили эффективное блокирование индуцированных TGF-β1 эффектов на уровни мРНК CTRF под действием ASO SEQ ID NO: 218c по сравнению с комбинированными обработками. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, D=SEQ ID NO: 218c, E=TGF-β1. ±=SEM, **p<0,01 по отношению к A, ++p<0,01 по отношению к E+B. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
Временная точка
48 ч TGF-β1 → 72 ч TGF-β1+ASO/обработка одним ASO
n=3
Заключение:
ASO SEQ ID NO: 218c был эффективным в опосредовании антифибротических эффектов в клетках A549 при искусственных патологических состояниях, имитированных избыточными концентрациями TGF-β1. В совокупности, ASO SEQ ID NO: 218c является эффективным терапевтическим агентом, поскольку патология фиброза легких может замедляться посредством снижения уровней CTGF, FN и CollV. Кроме того, образование стрессовых волокон может эффективно снижаться под действием TGF-RII-специфических ASO, что делает ASO по изобретению идеальными терапевтическими агентами.
20.3. Влияние на несколько типов раковых клеток
Описание методов:
Для исследования эффектов ASO, направленных против ECM (CTGF, FN, CollV), клетки использовали и культивировали, как описано ранее в стандартном протоколе (таблица 10). Для обработки клетки высевали в 24-луночные культуральные планшеты (Sarstedt #83.1836.300) (30000 клеток/лунку), 6-луночные культуральные планшеты (Sarstedt #83.3920.300) (50000 клеток/лунку) и инкубировали в течение ночи при 37°С и 5% СО2. Для анализа экспрессии и оценки влияния на уровни мРНК и белка CTGF, FN и CollV клетки обрабатывали стандартным олигонуклеотидом 1 (скремлированный контроль) или ASO SEQ ID NO: 218b в концентрациях 2,5 и 10 мкМ и инкубировали в течение 72 ч при 37°С и 5% СО2. Обработку, включая замену среды, повторяли 3 раза каждые 72 ч (в целом 12 суток). Для сбора клетки дважды промывали PBS и затем использовали для выделения РНК (24-луночные планшеты) или выделения белка (6-луночные планшеты). Протоколы выделения РНК и белка, а также использованные антитела и разведения аналогичны описанным выше.
20.3.1 Результаты для SEQ ID NO: 218b
Антифибротические эффекты оценивали анализом уровней мРНК и белка CTGF, FN, CollV. Уровень мРНК CTGF (таблица 72) дозозависимо снижался под действием SEQ ID NO: 218b в клетках HT-29, HTZ-19, MCF-7 и THP-1. Для клеток KG-1 была установлена понижающая регуляция медиатора ниже TGF-β под действием ASO SEQ ID NO: 218b в концентрации 2,5 мкМ. Для клеток A549, Panc-1 и CaCo2 было показано снижение FN (таблица 73), согласуясь с дозозависимым снижением мРНК CollV (таблица 74) в клетках THP-1, HTZ-19 и L3.6pl (таблица 65). Анализ вестерн-блоттингом выявил значительное снижение уровня белка CTGF в клетках HT-29, MCF-7, ™K-1 и L3.6pl. Результат для MCF-7 был статистически значимым (таблица 75). Кроме того, фосфорилирование Erk1/2 в клетках A549 и ™K-1 ингибировалось ASO SEQ ID NO: 218b. PErk1/2 обычно активируется под действием CTGF с индукцией фибротических эффектов, опосредованных TGF-β (таблица 76). Уровни белков FN (A549, MCF-7, HT-29, HTZ-19, HPAFII) и CollV (A549, HTZ-19, HPAFII, PC3M) (таблицы 77 и 78), двух основных компонентов ECM, резко снижались примерно на 50%.
Таблица 72: экспрессия мРНК CTGF через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках HT-29, HTZ-19, KG1, MCF-7 и THP-1. Уровень мРНК CTGF был снижен после трансфекции гимнозисом SEQ ID NO: 218b во всех тестируемых клеточных линиях. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=1
n=1
n=1
n=2
Таблица 73: экспрессия мРНК CTGF через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549, Panc-1 и CaCo2-1. Уровень мРНК FN был снижен после трансфекции гимнозисом SEQ ID NO: 218b во всех тестированных клеточных линиях. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM, **p<0,01 по отношению к A. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=1
n=2
Таблица 74: экспрессия мРНК CollV через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b в клетках A549, HTZ-19, THP-1, L3.6pl, Panc-1 и CaCo2. Уровень мРНК CollV был снижен после трансфекции гимнозисом SEQ ID NO: 218b во всех тестированных клеточных линиях. Уровни мРНК определяли по отношению к гену «домашнего хозяйства» GNB2L1 с использованием количественной ОТ-ПЦР в режиме реального времени и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b, ±=SEM. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни мРНК через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=2
n=2
n=1
n=2
n=1
n=2
Таблица 75: денситометрический анализ после вестерн-блоттинга в клетках HT-29, MCF-7, L3.6pl и ™K-1 через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b. Установлена понижающая регуляция белка CTGF под действием ASO SEQ ID NO: 218b. Уровни белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Image Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка через 12 суток после повторной трансфекции гимнозисом (4× 72 ч)
n=1
n=2
n=1
n=1
Таблица 76: денситометрический анализ после вестерн-блоттинга в клетках A549 и ™K-1 через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b. Установлена понижающая регуляция белка pErk1/2 под действием ASO SEQ ID NO: 218b. Уровни белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечния Image Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=1
Таблица 77: денситометрический анализ после вестерн-блоттинга в клетках A549, MCF-7, HT-29, HTZ-19 и HPAFII через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b. Установлена понижающая регуляция белка FN под действием ASO SEQ ID NO: 218b. Уровни белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Image Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=2
n=1
n=1
n=1
Таблица 78: денситометрический анализ после вестерн-блоттинга в клетках A549, MCF-7, HT-29, HTZ-19 и HPAFII через 12 суток после трансфекции гимнозисом SEQ ID NO: 218b. Установлена понижающая регуляция белка CollV под действием ASO SEQ ID NO: 218b. Уровни белка определяли по отношению к гену «домашнего хозяйства» альфа-тубулина с использованием программного обеспечения Image Studio™ Lite и затем нормализовали к необработанному контролю. A=необработанный контроль, B=стандартный олигонуклеотид 1, C=SEQ ID NO: 218b. Статистический анализ проводили с использованием обычного однофакторного дисперсионного анализа ANOVA с последующими апостериорными сравнениями по методу Тьюки.
уровни белка через 12 суток после повторной трансфекции гимнозисом (4×72 ч)
n=1
n=1
n=1
n=1
Заключение:
Повышенное отложение ECM, опосредуемое TGF-β1, через даунстрим медиатор CTGF, может быть эффективно отменено TGF-RII-специфическими ASO по изобретению в различных линиях опухолевых клеток. Пониженный уровень компонентов ЕСМ может способствовать менее агрессивному течению прогрессирования опухолей. В совокупности, испытуемые ASO могут демонстрировать новую терапевтическую стратегию при различных заболеваниях, связанных с фиброзом.
Пример 21: порог токсичности ASO по изобретению при длительном интрацеребровентрикулярном введении с использованием схемы с возрастающими дозами на обезьянах рода Cynomolgus
Для определения идеального диапазона доз для исследования токсичности по GLP был проведен предварительный опыт с использованием длительного интрацеребровентрикулярного (icv) введения антисмыслового олигонуклеотида(ASO) в возрастающих дозах на обезьянах рода Cynomolgus. Во время схемы введения животным проводили наблюдение за изменениями иммунологических, гематологических и физиологических показателей.
Описание метода:
Для длительной центральной инфузии ASO самцам и самкам обезьян рода Cynomolgus имплантировали подкожно газовый насос давления (0,25 мл/24 ч, Tricumed-IP 2000V®), присоединенный к силиконовому катетеру, направленному на правый боковой желудочек, под анестезией кетамином/ксилацином в полустерильных условиях. Для каждого вида обработки использовали одного самца и одну самку обезьян (SEQ ID NO: 218b, SEQ ID NO: 218c, концентрации приведены в таблице 79). Каждый насос имплантировали подкожно со стороны брюшной области через разрез кожи длиной 10 см на шее обезьяны и соединяли с icv канюлей с помощью силиконового катетера. Животных помещали в стереотаксическую рамку и icv канюлю погружали в правый боковой желудочек. Канюлю фиксировали двумя винтами из нержавеющей стали с использованием зубного цемента (Kallocryl, Speiko®-Dr. Speier GmbH, Мюнстер, Германия). Кожу на шее закрывали швами. Во время операции температуру тела поддерживали грелкой. Для профилактики послеоперационных инфекций обезьян обрабатывали местно препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия). Трубки и соответствующий насос заполняли соответствующим раствором для обработки. Периоды инфузии ASO (1 неделя для каждой дозы) прерывались однонедельным периодом вымывания с введением исключительно 0,9% NaCl. В течение всей схемы введения наблюдали за динамикой прироста массы тела и приемом корма. Кроме того, один раз в неделю отбирали образцы крови и СМЖ для определения гематологических, а также иммунологических изменений, и также концентрации ASO в крови. В последний день опыта животных подвергали эвтаназии и извлекали органы (печень, почки, головной мозг) и анализировали на пролиферацию, апоптоз, нокдаун мРНК и образование опухолей.
Заключение:
Все тестированные ASO по изобретению были, по меньшей мере, нетоксичны в течение недель 1-6, и поэтому их использовали для проведения дальнейших исследований и токсикологических опытов. Однако инфузия антисмысловых олигонуклеотидов SEQ ID NO: 214, SEQ ID NO: 138b, SEQ ID NO: 172b, а также стандартного олигонуклеотида 0 и стандартного олигонуклеотида 5 приводила к проявлению токсических эффектов на ранних стадиях вышеуказанной схемы. Следовательно, данные антисмысловые олигонуклеотиды были признаны, как непригодные в качестве терапевтических агентов, и их не использовали для дальнейших исследований.
Пример 22: определение поведенческих и физиологических нарушений после центрального введения антисмыслового олигонуклеотида
Целью данного опыта было исследование эффектов однократного интрацеребровентрикулярного (icv) введения антисмыслового олигонуклеотида на неврологические и поведенческие показатели у крыс.
Описание метода:
Стереотаксические процедуры проводили под анестезией кетамином/ксилацином в полустерильных условиях. После хирургической операции крысам давали два дня для восстановления.
icv имплантация направляющей канюли
Животных помещали в стереотаксическую рамку и направляющую канюлю (12 мм) имплантировали на 2 мм выше левого бокового желудочка (координаты относительно брегмы: 1,0 мм сзади, 1,6 мм латеральнее к средней линии, 1,8 мм ниже поверхности черепа). Направляющую канюлю закрепляли двумя винтами из нержавеющей стали с использованием зубного акрилового цемента (Kallocryl, Speiko® -Dr. Speier GmbH, Мюнстер, Германия) и закрывали ложной канюлей. Во время операции температуру тела поддерживали грелкой. Для пофилактики послеоперационных инфекций крыс обрабатывали местно препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 0,1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия).
icv инфузия
Легко зафиксированным крысам проводили icv инфузию ASO (2 мкМ/5 мкл, 10 мкМ/5 мкл, 50 мкмоль/5 мкл, 250 мкмоль/5 мкл) или растворителя (5 мкл, 0,9% NaCl, pH 7,4, Braun), используя канюлю размером 27 G, которую вытягивали на 2 мм за направляющей канюлей и оставляли на месте в течение 30 с для обеспечения диффузии. За крысами проводили наблюдение через 15, 30, 60 и 120 мин после icv введения для оценки поведенческих реакций, двигательной активности, возбуждения ЦНС, положения, координации движений, мышечного тонуса, рефлексов и температуры тела.
Подтверждение размещения канюли и микродиализного зонда
После эвтаназии извлекали головной мозг, быстро замораживали и хранили при -80°С до анализа. Гистологическое подтверждение мест icv имплантации проводили на срезах мозга толщиной 40 мкм, окрашенных крезил-фиолетовым.
Настоящие результаты показали, что однократное icv введение ASO (обеих последовательностей SEQ ID NO: 218b, SEQ ID NO: 218c) в различных дозах было безопасным и надежным методом для крыс с учетом отсутствия влияния на неврологические показатели.
Пример 23: определение идеального диапазона доз для исследования токсичности на обезьянах рода Cynomolgus по GLP (предварительный опыт по оценке токсичности на крысах)
Для исследования любых общих токсических эффектов при ежедневном внутривенном (в/в) введении антисмыслового олигонуклеотида (ASO) и для определения идеального диапазона доз для предварительного исследования токсичности по GLP на крысах, проводили предварительный эксперимент по оценке токсичности на крысах.
Описание метода:
Для многократных внутривенных инъекций ASO 20 самцов и 20 самок крыс разделяли на четыре группы по видам обработки: группа с растворителем, группы ASOнизкая доза, ASOсредняя доза и ASOвысокая доза. Данную схему введения использовали для SEQ ID NO: 218b и SEQ ID NO: 218с. Крысам ежедневно вводили внутривенно болюсную инъекцию ASO в течение 15 последовательных суток. Наблюдали за падежом крыс (дважды в день), клиническими симптомами (один раз в день, приростом массы тела (еженедельно)), приемом корма (еженедельно). На 15 сутки схемы эксперимента животных подвергали эвтаназии, извлекали органы (печень, почки, головной мозг) и собирали кровь из туловища. После чего ткани и кровь анализировали на иммунологические и гематологические изменения.
Результаты настоящего исследования показали, что два ASO SEQ ID NO: 218b и SEQ ID NO: 218c являются безопасными лекарственными препаратами для применения при многих различных расстройствах без проявления токсических эффектов при введении в низких и средних дозах.
Пример 24: определение общетоксических эффектов при многократных внутривенных введениях антисмысловых олигонуклеотидов
Целью данного опыта было исследование того, в какой дозе при ежедневном внутривенном (в/в) введении антисмысловой олигонуклеотид (ASO) оказывает какое-либо общетоксическое действие на крыс.
Описание метода:
Для многократных внутривенных инъекций ASO 80 самцов и 80 самок крыс разделяли на четыре группы по видам обработки: группа с растворителем, группы ASOнизкая доза, ASOсредняя доза и ASOвысокая доза. Крысам ежедневно вводили внутривенно болюсную инъекцию ASO в течение 29 последовательных суток. Наблюдали за падежом крыс (дважды в день), клиническими симптомами (один раз в день, приростом массы тела (еженедельно)), приемом корма (еженедельно). На 29 сутки схемы эксперимента животные подвергались эвтаназии, извлекали органы (печень, почки, головной мозг) и собирали кровь из туловища. Также отбирали костный мозг и готовили мазки. Затем ткани и кровь анализировали на иммунологические и гематологические и гистопатологические изменения.
Результаты настоящего исследования показали, что два ASO SEQ ID NO: 218b и SEQ ID NO: 218c являются безопасными лекарственными препаратами для применения при многих различных расстройствах без проявления токсических эффектов при введении в низких и средних дозах.
Пример 25: определение токсических свойств антисмыслового олигонуклеотида при длительном центральном введении на обезьянах рода Cynomolgus
Для определения эффективной и установления токсической дозы самцам и самкам обезьян рода Cynomolgus вводили антисмысловой олигонуклеотид (ASO) по изобретению в различных дозах при длительном интрацеребровентрикулярном введении. Во время схемы введения наблюдали за иммунологическими, гематологическими и физиологическими изменениями у животных.
Описание метода:
Для длительной центральной инфузии ASO самцам и самкам обезьян рода Cynomolgus имплантировали подкожно газовый насос давления (0,25 мл/24 ч, Tricumed-IP 2000V®), присоединенный к силиконовому катетеру, направленному на правый боковой желудочек, под анестезией кетамином/ксилацином в полустерильных условиях. Для каждого вида обработки использовали трех самцов и трех самок обезьян (растворитель, ASOнизкая доза и ASOвысокая доза, концентрации приведены в таблице 79). Кроме того, для определения сроков восстановления двух самцов и двух самок обезьян (растворитель и ASOвысокая доза) подвергали эвтаназии через четыре недели после прекращения введения ASO. Каждый насос имплантировали подкожно со стороны брюшной области через разрез кожи длиной 10 см на шее обезьяны и соединяли с icv канюлей с помощью силиконового катетера. Животных помещали в стереотаксическую рамку и icv канюлю погружали в правый боковой желудочек. Канюлю фиксировали двумя винтами из нержавеющей стали с использованием зубного цемента (Kallocryl, Speiko®-Dr. Speier GmbH, Мюнстер, Германия). Кожу на шее закрывали швами. Во время операции температуру тела поддерживали грелкой. Для профилактики послеоперационных инфекций обезьян обрабатывали местно препаратом бетаизодона® (Mundipharma GmbH, Лимбург, Германия) и вводили 1 мл антибиотиков (п/к, Байтрил® 2,5% Bayer Vital GmbH, Леверкузен, Германия). Трубки заполняли соответствующим раствором для обработки. В течение всей схемы введения наблюдали за динамикой прироста массы тела и приемом корма. Кроме того, один раз в неделю отбирали образцы крови и СМЖ для определения гематологических, а также иммунологических изменений, и также концентрации ASO в крови. В последний день опыта животных подвергали эвтаназии и извлекали органы (печень, почки, головной мозг) и анализировали на пролиферацию, апоптоз, нокдаун мРНК и образование опухолей. Через 57 недель дополнительных животных, используемых для определения сроков восстановления, также подвергали эвтаназии и определяли аналогичные показатели.
Таблица 80: условия обработки и количество животных в каждой группе в 4-недельном исследовании токсичности по GLP и дополнительном 4-недельном периоде восстановления.
Результаты настоящего исследования показали, что при длительном интрацеребровентрикулярном введении ASO является нетоксичным и безопасным лекарственным препаратом, который можно использовать для лечения целого ряда различных заболеваний.
Пример 26: определение стабильности и биологической активности антисмыслового олигонуклеотида в растворах различных носителей
Для исследования того, имеются ли какие-либо эффекты взаимодействия антисмысловых олигонуклеотидов (SEQ ID NO: 218b, SEQ ID NO: 218c) и инфузионного раствора, проводили 29-дневный предварительный опыт. Соответственно, два ASO восстановливали в различных, не содержащих эндотоксины растворах (PBS, вода для инъекций [WFI], 0,9% NaCl) и хранили в разных условиях соответственно. Образцы отбирали один раз в неделю и определяли значение pH, стабильность, содержание и целостность ASO анализом AEX-HPLC. Определяли любые изменения эффективности по эффективности нокдауна мРНК TGF-RII в анализе с клеточной культурой для каждого образца соответственно.
Описание метода:
Лиофилизованные ASO разбавляли раствором соответствующего носителя (вода для инъекций, 0,9% NaCl, PBS) в стерильных условиях (ламинарный поток, BIOWIZARD Golden GL-170 Ergoscience®, режим S1). Маркировали пробирки Эппендорфа емкостью 1,5 мл и заполняли 100 мкл (AEX-HPLC) или 250 мкл (нокдаун мишени) соответствующего раствора ASO (все стадии в ламинарном потоке, BIOWIZARD Golden GL-170 Ergoscience®, режим S1, схему пипетирования/маркировки смотри в таблице 81). На следующей стадии все образцы выдерживали в соответствующих условиях хранения, и раз в неделю отбирали образцы (схему отбора образцов смотри в таблице 82) и хранили при -80°C до анализа.
Таблица 81: схема маркировки в исследовании стабильности в системе ASO-носитель. Схему маркировки выполняли для SEQ ID NO: 218b и SEQ ID NO: 218c (в каждом случае 10 мкМ и 0,24 мМ) и для всех трех носителей WFI, 0,9% NaCl и PBS (=> 12 различных схем).
(WFI, 0,9% NaCL или PBS)
X_-20°C_сутки 6
X_-20°C_сутки 12
X_-20°C_сутки 18
X_-20°C_сутки 24
X_-20°C_сутки 29
X_+4°C_сутки 6
X_+4°C _сутки 12
X_+4°C _сутки 18
X_+4°C _сутки 24
X_+4°C _сутки 29
X_+20°C _сутки 6
X_+20°C _сутки 12
X_+20°C _сутки 18
X_+20°C _сутки 24
X_+20°C _сутки 29
X_+37°C_сутки 6
X_+37°C _сутки 12
X_+37°_сутки 18
X_+37°_сутки 24
X_+37°C _сутки 29
X_40°C_сутки 6
X_40°C _сутки 12
X_40°C _сутки 18
X_40°C _сутки 24
X_40°C _сутки 29
[10 мкM]
X_pH значение_сутки 0
X_значение pH_
сутки 29
[0,24 мM]
X_исходно
[0,24 мM]
X_-20°C_сутки 6
X_-20°C_сутки 12
X_-20°C_сутки 18
X_-20°C_сутки 24
X_-20°C_сутки 29
[0,24 мкM]
X_+4°C_сутки 6
X_+4°C _сутки 12
X_+4°C _сутки 18
X_+4°C _сутки 24
X_+4°C _сутки 29
[0,24 мM]
X_+20°C _сутки 6
X_+20°C _сутки 12
X_+20°C _сутки 18
X_+20°C _сутки 24
X_+20°C _сутки 29
[0,24 мM]
X_+37°C_сутки 6
X_+37°C _сутки 12
X_+37°_сутки 18
X_+37°_сутки 24
X_+37°C _сутки 29
[0,24 мM]
X_40°C_сутки 6
X_40°C _сутки 12
X_40°C _сутки 18
X_40°C _сутки 24
X_40°C _сутки 29
[0,24 мM]
X_значение pH_сутки 0
X_значение pH_
сутки 29
Таблица 82: схема отбора образцов в исследовании стабильности в системе ASO-носитель. Схему отбора образцов выполняли для SEQ ID NO: 218b и SEQ ID NO: 218c (в каждом случае 10 мкМ и 0,24 мМ) и для всех трех носителей WFI, 0,9% NaCl и PBS (=> 12 различных схем).
Поскольку отсутствовали какие-либо отрицательные эффекты для всех растворов носителей в отношении стабильности, содержания и целостности SEQ ID NO: 218b и SEQ ID NO: 218с, то 0,9% NaCl использовали для эксперимента по оценке стабильности ASO при применении.
Пример 27: определение стабильности и биологической активности при применении антисмысловых олигонуклеотидов по изобретению (ASO) в растворе наполнителя
Для исследования того, существуют ли какие-либо эффекты взаимодействия антисмысловых олигонуклеотидов (SEQ ID NO: 218b, SEQ ID NO: 218c) и газового насоса давления или катетера, проводили предварительный 29-дневный опыт. Соответственно, два ASO восстанавливали в 0,9% NaCl, и заполняли насос и катетер, следуя описанию производителя. Образцы отбирали один раз в неделю и определяли значение рН, проводили микробиологический анализ и определяли стабильность, содержание и целостность олигонуклеотидов анализом AEX-HPLC. Любые изменения в эффективности определяли по эффективности нокдауна мРНК TGF-RII в анализе с клеточной культурой с каждым образцом соответственно.
Описание метода:
Лиофилизированные ASO разбавляли 0,9% NaCl в стерильных условиях (ламинарный поток, BIOWIZARD Golden GL-170 Ergoscience®, режим S1). Маркировали пробирки Эппендорфа емкостью 5 мл согласно схеме маркировки (смотри таблицу 83) в стерильных условиях (ламинарный поток, BIOWIZARD Golden GL-170 Ergoscience®, режим S1). Два газовых насоса давления (Tricumed Model IP-2000 V®) и катетер (набор спинальных катетеров 4000) заполняли, следуя описанию производителя, соответствующим раствором ASO (все стадии проводили в ламинарном потоке, BIOWIZARD Golden GL-170 Ergoscience®, режим S1, схему пипетирования/маркировки смотри в таблице 83). На следующей стадии насос, соединенный с катетером, который был соединен с крышкой пробирки Эппендорфа емкостью 5 мл и оставшиеся пробирки хранили в ящике для хранения, где все отверстия были закрыты Parafilm®, для избежания загрязнения. Образцы отбирали раз в неделю, хранили при -80°С до анализа, и катетер, соединенный с крышкой пробирки Эппендорфа емкостью 5 мл, переносили в следующую пробирку для продолжения процедуры отбора проб. Кроме того, один образец отбирали непосредственно из насоса через болюсный порт и хранили при -80°C. В последний день опыта был отобран дополнительный образец для микробиологического анализа.
Таблица 83: схема маркировки в исследовании стабильности ASO при применении. Схему маркировки выполняли для SEQ ID NO: 218b и SEQ ID NO: 218c (в каждом случае 0,24 мМ); PS: (образец из насоса:образец непосредственно из катетера), AS: (дополнительный образец: образец отобран непосредственно из резервуара внутри насоса через болюсный порт, МБ: (микробиологический анализ: 500 мкМ PS и AS).
исходно
PS_сутки 6
PS_сутки 12
PS_сутки 18
PS_сутки 24
PS_сутки 29
AS_сутки 6
AS_сутки 12
AS_сутки 18
AS_сутки 24
AS_сутки 29
MB_сутки 29
значение pH сутки 0
значение pH сутки 29
_+37°C
PS_сутки 6
_+37°C
PS_сутки 12
_+37°C
PS_сутки 18
_+37°C
PS_сутки 24
_+37°C
PS_сутки 29
_+37°C
AS_сутки 6
_+37°C
AS_сутки 12
_+37°C
AS_сутки 18
_+37°C
AS_сутки 24
_+37°C
AS_сутки 29
_+37°C
MB_сутки 29
значение pH сутки 0
значение pH сутки 29
Таблица 84: схема обора образцов в исследовании стабильности ASO при применении. Схему отбора образцов выполняли для SEQ ID NO: 218b и SEQ ID NO: 218c (в каждом случае 0,24 мМ). PS: (образец из насоса:образец непосредственно из катетера), AS: (дополнительный образец: образец отобран непосредственно из резервуара внутри насоса через болюсный порт, МБ: (микробиологический анализ: 500 мкМ PS и AS).
Поскольку отсутствовали какие-либо эффекты насоса и катетера в отношении стабильности, содержания и целостности SEQ ID NO: 218b и SEQ ID NO: 218с, и также отсутствовали заметные проблемы при микробиологческом анализе, то данная схема применения представляет оптимальную методику для интратекального и интрацеребровентрикулярного введения обезьянам рода Cynomolgus и людям.
Химический синтез
Сокращения
Pybop: гексафторфосфат (бензотриазол-1-илокси)трипирролидинфосфония
DCM: дихлорметан
ДМФА: диметилформамид
DMAP: 4-диметиламинопиридин
DMT: 4,4'-диметокситритил
LCAA: длинноцепочечный алкиламино
TRIS: Трис(гидроксиметил)аминометан
TRIS-HCl: гидрохлорид трис(гидроксиметил)аминометана
DEPC: диэтилдикарбонат
Синтез и очистка гапмерных антисмысловых олигонуклеотидов
Антисмысловые олигонуклеотиды в форме гапмеров были собраны на синтезаторе ABI 3900 или ABI 394 или на Expedite™ (Applied Biosystems) в соответствии с фосфорамидитной химией олигомеризации. На синтезаторе ABI3900 твердый носитель нагружали полистиролом UnySupport (производства Glen Research, Sterling, Virginia, США) с получением масштаба синтеза 0,2 мкмоль. На синтезаторе ABI 394 твердым носителем служило стекло с контролируемой пористостью 500 A (CPG), нагруженное Unylinker™ производства Chemgenes (Wilmington, MA, США), с получением масштаба синтеза 3 мкмоль.
Вспомогательные реагенты для синтеза, такие как «Deblock», «Oxidizer», «CapA» и «CapB», а также ДНК фосфорамидиты получали от SAFC Proligo (Гамбург, Германия).
В частности, мономеры дезокситимидина (dT) 5'-O-(4,4'диметокситритил)-2'-O,3'-O-(2-цианоэтил-N,N-диизопропил)фосфорамидит, 4-N-бензоил-2'-дезоксицитидин (dCBz), 6-N-бензоил-2'-дезоксиаденозин (dABz) и 2-N-изобутирил-2'-деоксигуанозин (dGiBu) использовали в качестве строительных блоков ДНК. 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-(2-цианоэтил-N,N-диизопропил)фос форамидит (LNA-GDMF)-O,4'-C-метилентимидин-3'-[(2-цианоэтил)-(N, N-диизопропил)]фосфорамидит (LNA-Tb),5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидит (LNA-ABz), 5'-O-DMT-2'-O-4'-C-метилен-5-метил-N4-бензоилцитидин- -(2-цианоэтил-N,N-диизопропил)фосфорамидит (LNA-C*Bz) использовали в качестве строительных блоков LNA. LNA фосфорамидиты получали от Exiqon (Вебек, Дания).
Как показано на примерах LNA в таблице 85, фосфорамидиты растворяли в сухом ацетонитриле с получением 0,07 М раствора олигонуклеотида, за исключением LNA-C*Bz, который растворяли в смеси ТГФ/ацетонитрил (25/75 (об./об.)).
г/моль
100 мг
25/75 (v/v)
β-D-тио-LNA: 5'-O-DMT-2'-дезокси-2'-меркапто-2'-S, 4'-С-метилен-N6-бензоиладенозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидит, 5'-O-DMT-2'-дезокси-2'-меркапто-2'-S,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидиты, 5'-O-DMT-2'-дезокси-2'-меркапто-2'-S,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидит, 5'-O-DMT-2'-дезокси-2'-меркапто-2'-S,4'-C-метилентимидин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидит и 5'-O-DMT-2'-дезокси-2'-амино-2'-N,4'-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидиты синтезировали, как описано в J. Org. Chem. 1998, 63, 6078-6079.
Синтез β-D-амино-LNA: 5'-O-DMT-2'-дезокси-2'-амино-2'-N,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидитов, 5'-О-DMT-2'-дезокси-2'-амино-2'-N,4'-С-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита, 5'-O-DMT-2'-дезокси-2'-амино-2'-N,4'-C-метилентимидин-3'-[(2-цианоэтил)-(N,N-диизопро пил)]фос форамидита, 5'-O-DMT-2'-дезокси-2'-метиламино-2'-N,4'-метилен-N6-бензоиладенозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита и 5'-O-DMT-2'-дезокси-2'-метиламино-2'-N,4'-С-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидитов, 5'-O-DMT-2'-дезокси-2'-метиламино-2'-N,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил)(N,N-диизопропил)] фосфорамидита и 5'-O-DMT-2'-дезокси-2'-метиламино-2'-N,4'-C-метилентимидин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита проводили, следуя процедуре, описанной в литературе (J. Org Chem., 1998, 63, 6078-6079).
Синтез β-L-окси-LNA: α-L-5'-O-DMT-2'-O,4'-C-метилен-N6- бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита, α-L-5'-O-DMT-2'-O-,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита, α-L-5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита и α-L-5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(2-цианоэтил)-(N,N-диизопропил)] фосфорамидита проводили аналогично процедурам, описанным в литературе (J.Am.Chem.Soc., 2002, 124, 2164-2176; Angew Chem., Int., Ed., 200, 39, 1656-1659).
(β-бензоилмеркапто)этил)пирролидинолтиофосфорамидиты для синтеза олигонуклеотида с фосфоротиоатным скелетом получали по аналогии с протоколом, описанным Caruthers (J. Org. Chem., 1996, 61, 4272-4281).
«Фосфорамидиты-С3»: (3-(4,4'-диметокситритилокси)пропил-1- [(2-цианоэтил)-(N,N-диизопропил)]фосфорамидит и «3'-спейсер C3 CPG» (1-диметокситритилоксипропандиол-3-сукциноил)- длинноцепочечный алкиламино-CPG получали от Glen Research.
Общая процедура
Приготовление конъюгата LNA-твердый носитель:
1) Получение неполного сукцинилового эфира LNA (WO2007/112754)
5'-О-DMT-3'-гидроксинуклеозидный мономер, янтарный ангидрид (1,2 экв) И DMAP (1,2 экв) растворяли в 35 мл дихлорметана (DCM). Реакционную смесь перемешивали при комнатной температуре в течение ночи. После экстракции NaH2PO4 0,1 М pH 5,5 (2×) и насыщенным раствором соли (1×) органический слой дополнительно высушивали над безводным NaSO4, фильтровали и выпаривали. Производное неполного эфира получали с выходом 95% и использовали без дополнительной очистки.
2) Получение конъюгата LNA-носитель (WO2007/112754)
Вышеуказанное полученное производное неполного эфира (90 мкмоль) растворяли в минимальном количестве ДМФА, добавляли DIEA и pyBOP (90 мкмоль) и перемешивали вместе в течение 1 мин. Данную предварительно активированную смесь объединяли с LCAA-CPG (500 Å, размер 80-120 меш, 300 мг) в лабораторном синтезаторе и перемешивали. Через 1,5 ч при комнатной температуре носитель отфильтровывали и промывали ДМФА, DCM и МеОН. После высушивания установленная нагрузка составила 57 мкмоль/г (смотри Tom Brown, Dorcas JS Brown. Modern machine-aided methods of oligodeoxyribonucleotide synthesis. In: F.Eckstein, editor. Oligonucleotides and 35 Analogues A Practical Approach. Oxford: IRL Press, 1991: 13-14).
Удлинение олигонуклеотида (реакция сочетания)
5-Этилтио-1H-тетразол (ЭТТ) в качестве активатора (0,5 М раствор в ацетонитриле) использовали для реакции сочетания фосфорамидитов. Вместо ETT в качестве активатора можно использовать другие реагенты, такие как 4,5-дицианоимидазол (DCI), описанный в международной заявке WO2007/112754, 5- бензилтио-1H-тетразол или сахарин-1-метилимидазол. 0,25 М раствор DCI в ацетонитриле использовали для реакции сочетания с LNA.
Кэпирование
Добавляли 10% уксусный ангидрид (Ac2O) в ТГФ (чистый для ВЭЖХ) и 10% N-метилимидазол (NMI) в смеси ТГФ/пиридин (8:1) (чистый для ВЭЖХ) и давали взаимодействовать.
Окисление
Окисление фосфора (III) до фосфора (V) обычно проводили, например, смесью йод/ТГФ/пиридин/H2O с использованием 0,02 М йода в смеси ТГФ/пиридин/H2O, полученного от Glen Research, или 0,5 М (1S)-(+)-(10-камфорсульфонил)оксазиридина (CSO) в безводном ацетонитриле производства Glen Research.
В случае, когда получали фосфортиоатную межнуклеозидную связь, то стадию тиолирования проводили с использованием 0,05 М раствора 3-((диметиламинометилиден)амино)-3H-1,2,4-дитиазол-3-тиона (DDTT, полученного от Chemgenes (Wilmington, MA, США)) в смеси безводный ацетонитрил/пиридин (1:1 об./об.). В случае использования LNA тиолирование проводили с использованием 0,2 М 3,H-1,2-бензотиол-3-она 1,1-диоксида (реагента Бьюкажа) в безводном ацетонитриле.
В общем тиолирование также можно провести с использованием хлорида ксантана (0,01 М раствор в смеси ацетонитрил/пиридин 10%), как описано в международной заявке WO2007/112754. Альтернативно можно использовать другие реагенты для стадии тиолирования, такие как ксантангидрид (5-имино(1,2,4)дитиазолидин-3-тион), фенилацетилдисульфид (PADS).
В случае синтеза фосфородитиоата полученный тиофосфитный триэфир окисляли до фосфоротиотриэфира добавлением 0,05 М DDTT (3-((диметиламинометилиден)амино)-3Н-1,2,4-дитиазол-3-тиона) в смеси пиридин/ацетонитрил (4:1 об./об.).
Отщепление от твердого носителя и снятие защиты
В конце твердофазного синтеза антисмысловой олигонуклеотид можно отщепить вариантами «DMT-on» или «DMT-off». Вариант «DMT off» означает, что конечную группу 5'-O-(4,4'-диметокситритил) удаляют в синтезаторе с использованием реагента «Deblock», и вариант DMT-on означает, что данная группа остается, в то время как олигонуклеотид отщепляют от твердого носителя. Группы DMT удаляли трихлоруксусной кислотой.
Вариант «DMT-off»
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове. Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и удаляли защитные группы, используя 1-5 мл концентрированного водного раствора аммиака (полученного от Sigma Aldrich) в течение 16 ч при 55°C. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Если олигонуклеотиды содержат фосфородитиоатный триэфир, то с тиоловых групп снимали защиту с помощью смеси тиофенол:триэтиламин:диоксан, 1:1:2, об./об./об., в течение 24 ч, затем олигонуклеотиды отщепляли от твердого носителя с использованием водного аммиака в течение 1-2 ч при комнатной температуре и дополнительно снимают защиту в течение 4 ч при 65°C.
Вариант «DMT-on»
Олигонуклеотиды отщепляли от твердого носителя с использованием водного аммиака в течение 1-2 ч при комнатной температуре и затем снимали защиту в течение 4 ч при 65°C. Олигонуклеотиды очищали обращенно-фазовой ВЭЖХ (ОФ-ВЭЖХ) и затем DMT-группу удаляли обработкой трихлоруксусной кислотой.
Если олигонуклеотиды содержат фосфородитиотный триэфир, то отщепление от твердого носителя и снятие защиты с тиоловой группы проводили добавлением 850 мкл аммиака в концентрированном этаноле (смесь аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч.
Концевые группы
Концевые группы на 5'-конце антисмыслового олигонуклеотида
Олигонуклеотид на твердом носителе обрабатывали 3% трихлоруксусной кислотой в дихлорметане (мас./об.) для полного удаления защитной группы 5'-DMT. Затем соединение превращали с подходящей концевой группой с (цианоэтил-N,N-диизопропил) фосфорамидитной группой. После окисления фосфора (III) до фосфора (V), снятие защиты, отделение от твердого носителя и снятие защиты с последовательности проводили, как описано выше.
Очистка
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял: 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением включения 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в 32 объемах колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом.
Аналитика
Идентичность антисмысловых олигонуклеотидов подтверждали масс-спектрометрией с электрораспылительной ионизацией (ESI-MS) и чистоту определяли аналитическим капиллярным электрофорезом OligoPro (CE).
Очистку дитиоата проводили на приборе Amersham Biosciences P920 FPLC, снабженном колонкой Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Пример 28
Gb1sTb1sdAsdGsdTsdGsdTsdTsdTsdAsdGsdGsdGsAb1sGb1sC*b1 (SEQ ID NO: 209y)
5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоксилцитидин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления 5'-DMT. После нескольких промываний общим количеством 800 мкл ацетонитрила проводили реакцию сочетания с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили в колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать в течение 45 с. В конце данного цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и смеси 448 мкл N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидитов (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-О,4'-метилентимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-О,4'-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове.
Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и затем снимали защиту с использованием 5 мл концентрированного водного аммиака в течение 16 ч при 55°С. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в пределах 32 объемов колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом. Антисмысловой олигонуклеотид получали с чистотой 93,7%. ESI-МС: найдено: 5387,3 Да; вычислено: 5387,80 Да.
Пример 29
Gb1Tb1dAdGdTdGdTdTdTdAdGdGdGAb1Gb1C*b1 (SEQ ID NO: 209u)
LNA связывали с CPG, следуя общей процедуре. Реакцию сочетания и стадию кэпирования также проводили, как описано в примере 28. После стадии кэпирования систему промывали 800 мкл ацетонитрила и 400 мкл 0,02 М раствора йода в смеси ТГФ/пиридин/Н2О наносили на колонку на 45 с. Систему промывали после стадии окисления 24 мкл ацетонитрила. После очистки получали антисмысловой олигонуклеотид с чистотой 95,3%. ESI-МС: найдено: 5146,80 Да; вычислено: 5146,4 Да.
Пример 30
/5SpC3s/Gb1sTb1sdAsdGsdTsdGsdTsdTsdTsdAsdGsdGsdGsAb1sGb1sC*b1 (SEQ ID NO: 209v)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 28. Но за исключением того, что после того, как последний нуклеотид был присоединен к олигонуклеотиду и были проведены стадии окисления и кэпирования добавляли 80 мкл фосфорамидита-C3 (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила. Последующие стадии выполняли, как описано в примере 28. После очистки получали антисмысловой олигонуклеотид с чистотой 97,4%. HRMS (ESI): найдено: 5540,70 Да; вычислено: 5541,4 Да.
Пример 31
Gb1sTb1sdAsdGsdTsdGsdTsdTsdTsdAsdGsdGsdGsAb1sGb1sC*b1/3SpC3s/ (SEQ ID NO: 209w)
3'-Спейсер C3 CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-[(2-цианоэтил-N,N-диизопропил)] фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазола (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать в течение 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT. Последующие реакции проводили, как описано в примере 28. После очистки получали антисмысловой олигонуклеотид с чистотой 92,7%. ESI-sMS: найдено: 5541,70 Да; вычислено: 5541,4 Да.
Пример 32
Gb1ssTb1ssAb1ssdGssdTssdGssdTssdTssdTssdA*ssdGssdGssdGssAb1ssGb1ssC*b1 (SEQ ID NO: 209an)
5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоксицитидин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления 5'-DMT. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(β-бензоилмеркапто)этил]пирролидинолтиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазола(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-[(β-бензоилмеркапто)этил]пирролидинол-тиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Дальнейшее удлинение олигонуклеотида проводили аналогично.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 850 мкл аммиака в концентрированном этаноле (аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч для отделения антисмыслового олигонуклеотида от твердого носителя и снятия защиты с тиоловой группы.
Затем неочищенный антисмысловой олигонуклеотид очищали анионообменной хроматографией с использованием колонки Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Пример 33
Gb1sTb1sAb1sdGsdTsdGsdTsdTsdTsdAsdGsdGsGbsAb1sGb1sC*b1 (SEQ ID NO: 209az)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 28. После очистки получали антисмысловой олигонуклеотид с чистотой 90,5%. ESI-МС: найдено: 5442,9 Да; вычислено: 5443,3 Да.
Пример 34
Gb1sTb1sAb1sGb1sdTsdGsdTsdTsdTsdAsdGsdGsGb1sAb1sGb1sC*b1 (SEQ ID NO: 209ba)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными звеньями ДНК и LNA, как описано в примере 28. После очистки получали антисмысловой олигонуклеотид с чистотой 89,4%. ESI-МС: найдено: 5469,9 Да; вычислено: 5471,3 Да.
Пример 35
Gb1sTb1sAb1sdGsdTsdGsdTsdTsdTsdAsdGsdGsdGsdAsGb1sC*b1 (SEQ ID NO: 209bb)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 28. После очистки получали антисмысловой олигонуклеотид с чистотой 88,4%. ESI-МС: найдено: 5386,5 Да; вычислено: 5387,3 Да.
Пример 36
Gb1Tb1dAsdGsdTsdGsdTsdTsdTsdAsdGsdGsdGsAb1Gb1C*b1 (SEQ ID NO: 209s)
Соединение синтезировали, следуя общей процедуре, как описано в примере 28 и примере 29 с соответствующими строительными блоками ДНК, ДНК-производных и LNA. После очистки получали антисмысловой олигонуклеотид с чистотой 96,8%. ESI-МС: найдено: 5323,30 Да; вычислено: 5323,0 Да.
Пример 37
Gb1sTb1sdA*sdGsdTsdGsdTsdTsdTsdA*sdGsdGsdGsAb1sGb1sC*b1 (SEQ ID NO: 209t)
Соединение синтезировали, следуя общей процедуре, и как описано в примере 28, с соответствующими строительными блоками ДНК и LNA. После очистки получали антисмысловой олигонуклеотид с чистотой 91,4%. ESI-МС: найдено: 5416,30 Да; вычислено: 5417,3 Да.
Пример 38
/5SpC3s/Gb1sTb1sdAsdGsdTsdGsdTsdTsdTsdAsdGsdGsdGsAb1sGb1sC*b1/3SpC3s/ (SEQ ID NO: 209x)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 28, примере 30 и примере 31. После очистки получали антисмысловой олигонуклеотид с чистотой 95,1%. ESI-МС: найдено: 5696,30 Да; вычислено: 5695,5 Да.
Примеры 39-132
Другие олигонуклеотиды из таблицы 6 синтезировали, следуя общей процедуре, и как описано в примерах. Получение антисмыслового олигонуклеотида, содержащего звенья β-D-тио-LNA, α-L-окси-LNA, β-D-(NH)-LNA или β-D-(NCH3)-LNA проводили таким же образом, что и антисмысловые олигонуклеотиды, содержащие звенья β-D-окси-LNA.
Пример 133
Gb1sC*b1sTb1sAb1sdTsdTsdTsdGsdGsdTsdAsdGsdTsGb1sTb1sTb1 (SEQ ID NO: 210q)
5'-O-DMT-2'-O,4'-C-метилентимидин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT. После нескольких промываний общим количеством 800 мкл ацетонитрила проводили реакцию сочетания с 80 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили в колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце данного цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фос форамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фос форамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 M). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол (NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной группы 5'-DMT.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове. Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и удаляли защитные группы, используя 5 мл концентрированного водного раствора аммиака в течение 16 ч при 55°C. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением включения 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в 32 объемах колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом.
Получали антисмысловой олигонуклеотид с чистотой 87,1%. ESI-МС: найдено: 5384,30 Да; вычислено: 5384,3 Да.
Пример 134
Gb1C*b1Tb1Ab1dTdTdTdGdGdTdA*dGdTGb1Tb1Tb1 (SEQ ID NO: 210r)
LNA связывали с CPG, следуя общей процедуре. Реакцию сочетания и стадию кэпирования также проводили, как описано в примере 133. После стадии кэпирования систему промывали 800 мкл ацетонитрила и 400 мкл 0,02 М раствора йода в смеси ТГФ/пиридин/Н2О наносили на колонку на 45 с. Систему промывали после стадии окисления 24 мкл ацетонитрила. После очистки получали антисмысловой олигонуклеотид с чистотой 95,3%.
Пример 135
/5SpC3s/Gb1sC*b1sTb1sAb1sdTsdTsdTsdGsdGsdTsdAsdGsdTsGb1sTb1sTb1 (SEQ ID NO: 210v)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 133. Но за исключением того, что после того, как последний нуклеотид был присоединен к олигонуклеотиду и были проведены стадии окисления и кэпирования добавляли 80 мкл фосфорамидита-C3 (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Сочетание проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила. Последующие стадии выполняли, как описано в примере 133. После очистки получали антисмысловой олигонуклеотид с чистотой 93,9%.
Пример 136
Gb1sC*b1sTb1sAb1sdTsdTsdTsdGsdGsdTsdAsdGsdTsGb1sTb1sTb1/3SpC3s/(SEQ ID NO: 210w)
3'-Спейсер C3 CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать в течение 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. Последующие реакции проводили, как описано в примере 133. После очистки получали антисмысловой олигонуклеотид с чистотой 89,7%.
Пример 137
Gb1C*b1Tb1Ab1dTsdTsdTsdGsdGsdTsdAsdGsdTsGb1Tb1Tb1 (SEQ ID NO: 210o)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 133 и примере 134. После очистки получали антисмысловой олигонуклеотид с чистотой 83,8%. ESI-МС: найдено: 5288,10 Да; вычислено: 5287,9 Да.
Пример 138
Gb1sC*b1sTb1sAb1sdTsdTsdTdGsdGsdTsdA*sdGsdTsGb1sTb1sTb1 (SEQ ID NO: 210p)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 133. После очистки получали антисмысловой олигонуклеотид с чистотой 80,7%. ESI-МС: найдено: 5398,40 Да; вычислено: 5399,3 Да.
Пример 139
Gb1ssC*b1ssTb1ssdAssdTssdTssdTssdGssdGssdTssdA*ssdGssdTssGb1ssTb1ssTb1 (SEQ ID NO: 209af)
5'-O-DMT-2'-O,4'-C-метилентимидин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(β-бензоилмеркап то)этил]пирроли динол тиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(β-бензоилмеркапто)этил]пирролид инол тиофос форамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Дальнейшее удлинение олигонуклеотида проводили аналогично.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 850 мкл аммиака в концентрированном этаноле (аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч для отделения антисмыслового олигонуклеотида от твердого носителя и снятия защиты с тиоловой группы.
Затем неочищенный антисмысловой олигонуклеотид очищали анионообменной хроматографией с использованием колонки Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Примеры 140-233
Другие олигонуклеотиды из таблицы 7 синтезировали, следуя общей процедуре, и как описано в примерах. Получение антисмыслового олигонуклеотида, содержащего единицы β-D-тио-LNA, α-L-окси-LNA, β-D-(NH)-LNA или β-D-(NCH3)-LNA, проводили таким же образом, что и антисмысловые олигонуклеотиды, содержащие звенья β-D-окси-LNA.
Пример 234
C*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdCsdCsAb1sGb1sTb1sAb1 (SEQ ID NO: 210b)
5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-O-сукцинат
5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин (500 мг, 0,73 ммоль), 95 мг янтарного ангидрида (0,95 ммоль, 1,2 экв.) и 116 мг DMAP (0,95 ммоль, 1,2 экв.) растворяли в 35 мл дихлорметана. Реакционную смесь перемешивали при комнатной температуре в течение ночи. Реакционный раствор промывали 2 раза 10 мл NaH2PO4 (0,1 М, рН 5,5) и один раз 10 мл насыщенного раствора соли. Органическую фазу высушивали под безводным NaSO4, фильтровали и концентрировали досуха в вакууме. Производное неполного эфира получали с выходом 95% и использовали без дальнейшей очистки на следующей стадии.
5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-O-сукци ноил-связанный LCAA-CPG
70 мг производного неполного эфира (90 мкмоль) растворяли в 0,3 мл ДМФА, добавляли 11,6 мкл DIEA (90 мкмоль) и pyBOP (90 мкмоль) и перемешивали вместе в течение 1 мин. Эту смесь объединяли с LCAA-CPG (500 Å, размер 80-120 меш, 300 мг) в лабораторном синтезаторе и перемешивали в течение 1,5 ч при комнатной температуре. Носитель отфильтровывали и промывали ДМФА, DCM и МеОН. После высушивания определенная нагрузка равнялась 57 мкмоль/г.
Удлинение
5'-O-DMT-2'-O,4'-C-метилен-N6-бензилоксиаденозин-3'-O-сукци ноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(2-цианоэтил](N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)фос фо рамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл cмеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл cмеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове. Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и удаляли защитные группы, используя 5 мл концентрированного водного раствора аммиака в течение 16 ч при 55°C. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением включения 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в пределах 32 объемов колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом. Получали антисмысловой олигонуклеотид с чистотой 94,8%. ESI-МС: найдено: 5365,80 Да; вычислено: 5365,30 Да.
Пример 235
C*b1Ab1Tb1dGdAdAdTdGdGdAdCdCAb1Gb1Tb1Ab1 (SEQ ID NO: 218r)
LNA связывали с CPG, следуя общей процедуре. Реакцию сочетания и стадию кэпирования также проводили, как описано в примере 234. После стадии кэпирования систему промывали 800 мкл ацетонитрила и 400 мкл 0,02 М раствора йода в смеси ТГФ/пиридин/Н2О наносили на колонку на 45 с. Систему промывали после стадии окисления 24 мкл ацетонитрила. После очистки получали антисмысловой олигонуклеотид с чистотой 97,8%. ESI-МС: найдено: 5125,10 Да; вычислено: 5124,4 Да.
Пример 236
/5SpC3s/C*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdCsdCsAb1sGb1sTb1sAb1 (SEQ ID NO: 218t)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. Но за исключением того, что после того, как последний нуклеотид был присоединен к олигонуклеотиду и были проведены стадии окисления и кэпирования добавляли 80 мкл фосфорамидита-C3 (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Сочетание проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила. Последующие стадии выполняли, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 94,2%. ESI-МС: найдено: 5519,60 Да; вычислено: 5519,4 Да.
Пример 237
C*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdCsdCsAb1sGb1sTb1sAb1s/3SpC3/(SEQ ID NO: 218u)
3'-Спейсер C3 CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать в течение 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. Последующие реакции проводили, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 94,3%. ESI-MS: найдено: 5519,10 Да; вычислено: 5519,4 Да.
Пример 238
C*b1ssAb1ssTb1ssdGssdAssdAssdTssdGssdGssdAssdCssdCssAb1ssGb1ssTb1ssAb1 (SEQ ID NO: 218aa)
5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин- 3'-[(β-бензоилмеркапто)этил]пирролидинолтиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(β-бензоилмеркапто)этил]пирролидинолтиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Дальнейшее удлинение олигонуклеотида проводили аналогично.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 850 мкл аммиака в концентрированном этаноле (аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч для отделения антисмыслового олигонуклеотида от твердого носителя и снятия защиты с тиоловой группы.
Затем неочищенный антисмысловой олигонуклеотид очищали анионообменной хроматографией с использованием колонки Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Пример 239
C*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdC*sdC*sAb1sGb1sTb1sAb1 (SEQ ID NO: 218m)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 93,8%. ESI-МС: найдено: 5394,00 Да; вычислено: 5393,3 Да.
Пример 240
C*b1Ab1Tb1dGsdAsdAsdTsdGsdGsdAsdC*sdC*sAb1Gb1Tb1Ab1 (SEQ ID NO: 218n)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234 и примере 235. После очистки получали антисмысловой олигонуклеотид с чистотой 94,7%. ESI-МС: найдено: 5297,30 Да; вычислено: 5297,0 Да.
Пример 241
C*b1sAb1sTb1sdGsdA*sdA*sdTsdGsdGsdA*sdCsdCsAb1sGb1sTb1sAb1 (SEQ ID NO: 218o)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 92,8%. ESI-МС: найдено: 5410,40 Да; вычислено: 5410,3 Да.
Пример 242
C*b1sAb1sTb1sdGsdA*sdA*sdTsdGsdGsdA*sdC*sdC*sAb1sGb1sTb1sAb1 (SEQ ID NO: 218p)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 95,3%. ESI-МС: найдено: 5437,40 Да; вычислено: 5438,4 Да.
Пример 243
C*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdC*sdCsAbsGb1sTb1sAb1 (SEQ ID NO: 218q)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 93,9%. ESI-МС: найдено: 5378,80 Да; вычислено: 5379,3 Да.
Пример 244
C*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdCsdC*sAb1sGb1sTb1sAb1 (SEQ ID NO: 218с)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 92,9%. ESI-МС: найдено: 5379,10 Да; вычислено: 5379,3 Да.
Пример 245
C*b1sAb1sTb1sdGdAdAdTdGdGdAdC*sdC*sAb1sGb1sTb1sAb1 (SEQ ID NO: 218s)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234. После очистки получали антисмысловой олигонуклеотид с чистотой 94,5%. ESI-МС: найдено: 5152,70 Да; вычислено: 5152,4 Да.
Пример 246
/5SpC3/sC*b1sAb1sTb1sdGsdAsdAsdTsdGsdGsdAsdCsdCsAb1sGb1sTb1sAb1s/3SpC3/ (SEQ ID NO: 218v)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 234, примере 236 и примере 237. После очистки получали антисмысловой олигонуклеотид с чистотой 94,4%. ESI-МС: найдено: 5673,50 Да; вычислено: 5673,5 Да.
Примеры 247-335
Другие олигонуклеотиды из таблицы 8 синтезировали, следуя общей процедуре, и как описано в примерах. Получение антисмыслового олигонуклеотида, содержащего единицы β-D-тио-LNA, α-L-окси-LNA, β-D-(NH)-LNA или β-D-(NCH3)-LNA, проводили таким же образом, что и антисмысловые олигонуклеотиды, содержащие звенья β-D-окси-LNA.
Пример 336
C*b1sGb1sAb1sTb1sdAsdCsdGsdCsdGsdTsdCsdCsAb1sC*b1sAb1 (SEQ ID NO: 152h)
5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-O-сукци но ил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезокситимидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-С-метилентимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метил имидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фос фор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фос фор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове. Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и удаляли защитные группы, используя 5 мл концентрированного водного раствора аммиака в течение 16 ч при 55°C. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением включения 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в 32 объемах колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом.
Пример 337
C*b1Gb1Ab1Tb1dAdCdGdC*dGdTdCdC*Ab1C*b1Ab1 (SEQ ID NO: 152q)
LNA связывали с CPG, следуя общей процедуре. Реакцию сочетания и стадию кэпирования также проводили, как описано в примере 336. После стадии кэпирования систему промывали 800 мкл ацетонитрила и 400 мкл 0,02 М раствора йода в смеси ТГФ/пиридин/Н2О наносили на колонку на 45 с. Систему промывали после стадии окисления 24 мкл ацетонитрила. После очистки получали антисмысловой олигонуклеотид с чистотой 93,1%.
Пример 338
/5SpC3s/C*b1sGb1sAb1sTb1sdAsdC*sdGsdC*sdGsdTsdCsdCsAb1sC*b1sAb1 (SEQ ID NO: 152s)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 336. Но за исключением того, что после того, как последний нуклеотид был присоединен к олигонуклеотиду и были проведены стадии окисления и кэпирования добавляли 80 мкл фосфорамидита-C3 (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Сочетание проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила. Последующие стадии выполняли, как описано в примере 336. После очистки получали антисмысловой олигонуклеотид с чистотой 96,5%.
Пример 339
C*b1sGb1sAb1sTb1sdAsdC*sdGsdCsdGsdTsdCsdC*sAb1sC*b1sAb1/3SpC3s/ (SEQ ID NO: 152t)
3'-Спейсер C3 CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать в течение 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. Последующие реакции проводили, как описано в примере 336. После очистки получали антисмысловой олигонуклеотид с чистотой 92,1%.
Пример 340
C*b1ssGb1ssAb1ssdTssdAssdC*ssdGssdCssdGssdTssdCssdC*ssAb1ssC*b1ssAb1 (SEQ ID NO: 152aa)
5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-O-сукци ноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-[(β-бензоилмеркапто)этил]пирролидинолтио фосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-[(β-бензоилмеркапто)этил]пирролидинолтиофос форамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Дальнейшее удлиненеие олигонуклеотида проводили аналогично.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 850 мкл аммиака в концентрированном этаноле (аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч для отделения антисмыслового олигонуклеотида от твердого носителя и снятия защиты с тиоловой группы.
Затем неочищенный антисмысловой олигонуклеотид очищали анионообменной хроматографией с использованием колонки Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Примеры 341-433
Другие олигонуклеотиды из таблицы 5 синтезировали, следуя общей процедуре, и как описано в примерах. Получение антисмыслового олигонуклеотида, содержащего единицы β-D-тио-LNA, α-L-окси-LNA, β-D-(NH)-LNA или β-D-(NCH3)-LNA, проводили таким же образом, что и антисмысловые олигонуклеотиды, содержащие звенья β-D-окси-LNA.
Пример 434
C*b1sTb1sdCsdGsdTsdCsdAsdTsdAsdGsdAsC*b1sC*b1sGb1 (SEQ ID NO: 143h)
5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодейст вовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-С-метилентимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фос фор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове. Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и удаляли защитные группы, используя 5 мл концентрированного водного раствора аммиака в течение 16 ч при 55°C. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением включения 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в 32 объемах колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом.
Пример 435
C*b1Tb1dC*dGdTdCdAdTdAdGdAC*b1C*b1Gb1 (SEQ ID NO: 143ad)
LNA связывали с CPG, следуя общей процедуре. Реакцию сочетания и стадию кэпирования также проводили, как описано в примере 434. После стадии кэпирования систему промывали 800 мкл ацетонитрила и 400 мкл 0,02 М раствора йода в смеси ТГФ/пиридин/Н2О наносили на колонку на 45 с. Систему промывали после стадии окисления 24 мкл ацетонитрила. После очистки получали антисмысловой олигонуклеотид с чистотой 88,7%.
Пример 436
/5SpC3s/C*b1sTb1sdC*dGdTdC*dA*dTdAdGdA*sC*b1sC*b1sGb1 (SEQ ID NO: 143af)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 434 и примере 435. Но за исключением того, что после того, как последний нуклеотид был присоединен к олигонуклеотиду и были проведены стадии окисления и кэпирования добавляли 80 мкл фосфорамидита-C3 (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Связывание проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила. Последующие стадии выполняли, как описано в примере 434 и примере 435. После очистки получали антисмысловой олигонуклеотид с чистотой 94,4%.
Пример 437
C*b1sTb1sdC*dGdTdC*dA*dTdAdGdA*sC*b1sC*b1sGb1/3SpC3s/ (SEQ ID NO: 143ag)
3'-Спейсер C3 CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фос фор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать в течение 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. Последующие реакции проводили, как описано в примере 434 и примере 435. После очистки получали антисмысловой олигонуклеотид с чистотой 91,6%.
Пример 438
C*b1ssTb1ssC*b1ssdGssdTssdC*ssdAssdTssdAssdGssdAssC*b1ssC*b1ssGb1 (SEQ ID NO: 143t)
5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-[(β-бензоилмеркапто)этил]пирролидинол-тиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-[(β-бензоилмеркапто)этил]пирролидинол тиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Дальнейшее удлинение олигонуклеотида проводили аналогично.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 850 мкл аммиака в концентрированном этаноле (аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч для отделения антисмыслового олигонуклеотида от твердого носителя и снятия защиты с тиоловой группы.
Затем неочищенный антисмысловой олигонуклеотид очищали анионообменной хроматографией с использованием колонки Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Примеры 439-534
Другие олигонуклеотиды из таблицы 4 синтезировали, следуя общей процедуре, и как описано в примерах. Получение антисмыслового олигонуклеотида, содержащего единицы β-D-тио-LNA, α-L-окси-LNA, β-D-(NH)-LNA или β-D-(NCH3)-LNA, проводили таким же образом, что и антисмысловые олигонуклеотиды, содержащие звенья β-D-окси-LNA.
Пример 535
C*b1sAb1sGb1sdGsdCsdAsdTsdTsdAsdAsdTsdAsdAsdAsGb1sTb1sGb1 (SEQ ID NO: 213k)
5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-O-сукциноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин- 3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фос фор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил)(N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил-N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-дезокситимидин-3'-[(2-цианоэтил)(N,N-диизопропил)]фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N6-бензоил-2'-дезоксиаденозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N4-бензоил-2'-дезоксицитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-N2-изобутирил-2'-дезоксигуанозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил-N,N-диизопропил)]фос фор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N6-бензоиладенозин-3'-(2-цианоэтил-N,N-диизопропил)фосфорамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-5-метил-N4-бензоилцитидин-3'-(2-цианоэтил-N,N-диизопропил)фосфор амидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 20% раствором диэтиламина в ацетонитриле (Biosolve BV, Valkenswaard, Нидерланды) в течение 20 мин для удаления цианоэтильных защитных групп на фосфатном остове. Затем антисмысловые олигонуклеотиды отщепляли от твердого носителя и удаляли защитные группы, используя 5 мл концентрированного водного раствора аммиака в течение 16 ч при 55°C. Твердый носитель отделяли от антисмысловых олигонуклеотидов фильтрованием или центрифугированием.
Затем неочищенные антисмысловые олигонуклеотиды очищали анионообменной высокоэффективной жидкостной хроматографией (ВЭЖХ) в системе AKTA Explorer (GE Healthcare, Фрайбург, Германия) и на колонке, заполненной Source Q15 (GE Helthcare). Буфер А представлял 10 мМ перхлората натрия, 20 мМ Трис, 1 мМ ЭДТА, рН 7,4 и содержал 20% ацетонитрила, и буфер В был таким же, как буфер А, за исключением включения 500 мМ перхлората натрия. Использовали градиент от 15% В до 55% В в 32 объемах колонки (CV). В УФ-свете снимали спектр при 280 нм. Соответствующие фракции объединяли и осаждали 3 М NaOAc, рН=5,2 и 70% этанолом. Наконец, осадок промывали 70% этанолом.
Пример 536
C*b1Ab1Gb1dGdC*dAdTdTdAdAdTdAdAdAGb1Tb1Gb1 (SEQ ID NO: 213n)
LNA связывали с CPG, следуя общей процедуре. Реакцию сочетания и стадию кэпирования также проводили, как описано в примере 535. После стадии кэпирования систему промывали 800 мкл ацетонитрила и 400 мкл 0,02 М раствора йода в смеси ТГФ/пиридин/Н2О наносили на колонку на 45 с. Систему промывали после стадии окисления 24 мкл ацетонитрила. После очистки получали антисмысловой олигонуклеотид с чистотой 91,4%.
Пример 537
/5SpC3s/C*b1sAb1sGb1sdGsdC*sdAsdTsdTsdAsdAsdTsdAsdAsdAsGb1sTb1sGb1 (SEQ ID NO: 231o)
Соединение синтезировали, следуя общей процедуре, с соответствующими строительными блоками ДНК и LNA, как описано в примере 535. Но за исключением того, что после того, как последний нуклеотид был присоединен к олигонуклеотиду и были проведены стадии окисления и кэпирования добавляли 80 мкл фосфорамидита-C3 (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Связывание проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила. Последующие стадии выполняли, как описано в примере 535. После очистки получали антисмысловой олигонуклеотид с чистотой 87,1%.
Пример 538
C*b1sAb1sGb1sdGsdC*sdAsdTsdTsdAsdAsdTsdAsdAsdAsGb1sTb1sGb1/3SpC3s/ (SEQ ID NO: 213p)
3'-Спейсер C3 CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 80 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(2-цианоэтил)(N,N-диизо про пил)]фос форамидита (0,07 М) и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 640 мкл реагента Бьюкажа (0,2 М) наносили на колонку на 180 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодейст вовать в течение 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. Последующие реакции проводили, как описано в примере 535. После очистки получали антисмысловой олигонуклеотид с чистотой 95,7%.
Пример 539
C*b1ssAb1ssGb1ssdGssdC*ssdAssdTssdTssdAssdAssdTssdAssdAssAb1ssGb1ssTb1ssGb1 (SEQ ID NO: 213ae)
5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-O-сукци ноил-связанный LCAA-CPG (0,2 мкмоль) обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы. После нескольких промываний общим количеством 800 мкл ацетонитрила реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилентимидин- 3'-[(β-бензоилмеркапто)этил]пирролидинолтиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Реакцию сочетания проводили с 38 мкл 5'-O-DMT-2'-O,4'-C-метилен-N2-диметилформамидингуанозин-3'-[(β-бензоилмеркапто)этил]пирроли динолтиофосфорамидита (0,15 М) в 10% дихлорметане (об./об.) в ацетонитриле и 236 мкл DCI в ацетонитриле (0,25 М). Реакцию сочетания проводили в течение 250 с, и избыток реагентов вымывали 800 мкл ацетонитрила и 900 мкл DDTT (0,05 М в смеси пиридин/ацетонитрил 4:1 об./об.) наносили на колонку на 240 с. Систему промывали 320 мкл ацетонитрила. Для стадии кэпирования добавляли 448 мкл уксусного ангидрида в ТГФ (1:9 об./об.) и 448 мкл смеси N-метилимидазол(NMI)/ТГФ/пиридин (1:8:1) и оставляли взаимодействовать на 45 с. В конце этого цикла систему промывали 480 мкл ацетонитрила. Соединение обрабатывали 1400 мкл 3% трихлоруксусной кислоты в дихлорметане в течение 60 с для полного удаления защитной 5'-DMT-группы.
Дальнейшее удлинение олигонуклеотида проводили аналогично.
По завершении твердофазного синтеза антисмысловые олигонуклеотиды обрабатывали 850 мкл аммиака в концентрированном этаноле (аммиак/этанол 3:1 об./об.) при 55°С в течение 15-16 ч для отделения антисмыслового олигонуклеотида от твердого носителя и снятия защиты с тиоловой группы.
Затем неочищенный антисмысловой олигонуклеотид очищали анионообменной хроматографией с использованием колонки Mono Q 10/100 GL. Буферы готовили с обработанной DEPC водой, и их состав был следующим: буфер А: 25 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0; буфер B: 25 мМ Трис-HCl, 1 мМ ЭДТА, 1 М NaCl, pH 8,0.
Примеры 540-640
Другие олигонуклеотиды из таблицы 9 синтезировали, следуя общей процедуре, и как описано в примерах. Получение антисмыслового олигонуклеотида, содержащего единицы β-D-тио-LNA, α-L-окси-LNA, β-D-(NH)-LNA или β-D-(NCH3)-LNA, проводили таким же образом, что и антисмысловые олигонуклеотиды, содержащие звенья β-D-окси-LNA.
Список последовательностей
SEQ ID NO: 1: рецептор II трансформирующего фактора роста бета (TGFBR2) человека, вариант транскрипта 2 (антисмысловая цепь; код ДНК)
TTTAGCTACT AGGAATGGGA ACAGGAGGCA GGATGCTCAC CTGAGTATTT TGCTTTATTC 60
AATCTAATAA ACATTTTATT TATGTAAAAG ACAAACAATG CATAGAATAA AAATAAGTGC 120
TTGAGACTTT TGATATAAAA AGAGTATATA GCATTCACAT TCCTATTTTA ATACATGAGT 180
ACAGCTGAAG TGTTCCATAA AAGAATAAAA CTTTCCCTTT ATGTATAGTA GTGAAAAAAG 240
TCAGTATTTT TAGGAACTAC AGAATGTTAT TCCTTGGTCT TTTTTCTTGA ATAAGAAAAA 300
AAAACATAAA CAAAACAAGC CACAGTATCC TCTGACACTA CATTCCAGTT TATGCTGATA 360
ACCCAGAAGT GAGAATACTC TTGAATCTTG AATATCTCAT GAATGGACCA GTATTCTAGA 420
AACTCACCAC TAGAGGTCAA TGGGCAACAG CTATTGGGAT GGTATCAGCA TGCCCTACGG 480
TGCAAGTGGA ATTTCTAGGC GCCTCTATGC TACTGCAGCC ACACTGTCTT TAACTCTCAG 540
CCCACCCACA CTGAGGAGGG TGCCTAGAGG TTCTATTTCC AAACCTTTGC ATGTATCTTA 600
AAAATCTCAA TAAAATGAGA CCTTCCACCA TCCAAACAGA GCTGATATTC TCACTACCAG 660
TCCCTCTCTA ATATTCCTAT TTGGCTGAAA ATAAGTAGCT TCAAAAAGTT TTAAAAAAGA 720
GATTACTTGC AGCATTAACA CTTCTTTGTT GATTAACAAG TTTCCTATGG AGTTTTAAAG 780
CTCATACTTT GTTCTTGTCC TTGTGGACAC AAATTTTCTA ACTGCAAATG GGACCTTTGT 840
GTCCCACATT CAAATCCTCT CTAGTAATTT CTGCAAAGGT TGAGAAGGCT GGCATGATGG 900
AGAGAACGGT AACCATGAGG AAAGCTTCTT GGAGTAAAGC ACTCCTCTCT CCAATGCAGA 960
GGGTAAAACT ATTAACATAT AAGCAAAAGA AACTTGGGCT AACTGAGACC CTTAAAGGAG 1020
TTCCCCTTTA GTCCAATAAA AGGCCAACTT CAAATCTTAA CACCAGATAA GGTAGTCAAA 1080
ATCATATTAT ATACCCAGAG AATGACTGCT TGAATGGACA TTTCTTACAA GGGACCTTGG 1140
TTAGGTGCAG ATTTAATTCC TAGACTGGGG TCCAGGTAGG CAGTGGAAAG AGCTAATGTT 1200
TACAGTGAGA AGTGAGGCAG CTTTGTAAGT GTCTCCACAC CTTCACATTT TGTGAACGTG 1260
GACTGGAGAT AACTGAAAAC CATCTGCTAT CCTTACCTGG GGATCCAGAT TTTCCTGCAA 1320
AATCTCCAAA TATTTATAAA GTGGCTTCAC TTTTTGAAAC GCTGTGCTGA CCAAACAAAA 1380
CATATGTTTA GAGTGCCTGA GGTCATAGTC CTGACAATGA TAGTATTGTG TAGTTGAAAT 1440
CCTCTTCATC AGGCCAAACT GTGCTTGAGC AATCAGGAGC CCAGAAAGAT GGAACCCATT 1500
GGTGTTTGTA TAGAAAACTA GAAAATCAAG TCAAGTGTAA TGAAAAAGTA AACACGATAA 1560
AGCCTAGAGT GAGAATTTGC TCCTTTTTAG AAAAGGATGA AGGCTGGGAG CAGAGAATAG 1620
TAACATAAGT GCAGGGGAAA GATGAAAAAA AGAACAATTT TTCATTAGTA GATGGTGGGG 1680
CAATCGCATG GATGGGGACA TCTGTTCTGA TTTTTCTGCA ACCCATGAAG GTAAAAAGTG 1740
GGGTTCAAAA CATTCAAGGT ATTAAAGATG GGGTAGAGTT TCTAAACTAG GTTGAGGGAG 1800
AGTTTCTAAA CTAGCCCCCC AGATTTGGGG CTTGGAGCTT AAATGAAAAG TCCAGGAGAA 1860
ATAAGGGCAC ACAGGAACCC CGGGAACACT GGTCCTCAAA CAGTGCCACT GTACTTAGTT 1920
CCATGGCCAG AAGAGAAGTG CTAGGCAGGG AATGATTATT TTGCAAAAGC AAGTGCAATG 1980
TGGTCATAGC TGGCTGTGAG ACATGGAGCC TCTTTCCTCA TGCAAAGTTC ACTGTTTTAC 2040
AGTCAGAGAA CCACTGCATG TGTGATTGTC AAATGCTAAT GCTGTCATGG GTCCCTTCCT 2100
TCTCTGCTTG GTTCTGGAGT TCTCCAATAA AACCAATTTC CTGGGAATAT TTGATGTTTT 2160
TCCTTGTCTC TTTTCAAGGT ATGGCTATAT ATATAGAGCT ATAGACATAT ATAGATATAT 2220
ATATATATAT ATAAAACATA GCTATTCATA TTTATATACA GGCATTAATA AAGTGCAAAT 2280
GTTATTGGCT ATTGTAAAAA TCAATCTCAT TTCCTGAGGA AGTGCTAACA CAGCTTATCC 2340
TATGACAATG TCAAAGGCAT AGAATGCTCT ATGTCACCCA CTCCCTGCTG CTGTTGTTTC 2400
TGCTTATCCC CACAGCTTAC AGGGAGGGGA GTGACCCCCT TGGTTTTCCA GGAAGCATCA 2460
GTTCAGGGGC AGCTTCCTGC TGCCTCTGTT CTTTGGTGAG AGGGGCAGCC TCTTTGGACA 2520
TGGCCCAGCC TGCCCCAGAA GAGCTATTTG GTAGTGTTTA GGGAGCCGTC TTCAGGAATC 2580
TTCTCCTCCG AGCAGCTCCT CCCCGAGAGC CTGTCCAGAT GCTCCAGCTC ACTGAAGCGT 2640
TCTGCCACAC ACTGGGCTGT GAGACGGGCC TCTGGGTCGT GGTCCCAGCA CTCAGTCAAC 2700
GTCTCACACA CCATCTGGAT GCCCTGGTGG TTGAGCCAGA AGCTGGGAAT TTCTGGTCGC 2760
CCTCGATCTC TCAACACGTT GTCCTTCATG CTTTCGACAC AGGGGTGCTC CCGCACCTTG 2820
GAACCAAATG GAGGCTCATA ATCTTTTACT TCTCCCACTG CATTACAGCG AGATGTCATT 2880
TCCCAGAGCA CCAGAGCCAT GGAGTAGACA TCGGTCTGCT TGAAGGACTC AACATTCTCC 2940
AAATTCATCC TGGATTCTAG GACTTCTGGA GCCATGTATC TTGCAGTTCC CACCTGCCCA 3000
CTGTTAGCCA GGTCATCCAC AGACAGAGTA GGGTCCAGAC GCAGGGAAAG CCCAAAGTCA 3060
CACAGGCAGC AGGTTAGGTC GTTCTTCACG AGGATATTGG AGCTCTTGAG GTCCCTGTGC 3120
ACGATGGGCA TCTTGGGCCT CCCACATGGA GTGTGATCAC TGTGGAGGTG AGCAATCCCC 3180
CGGGCGAGGG AGCTGCCCAG CTTGCGCAGG TCCTCCCAGC TGATGACATG CCGCGTCAGG 3240
TACTCCTGTA GGTTGCCCTT GGCGTGGAAG GCGGTGATCA GCCAGTATTG TTTCCCCAAC 3300
TCCGTCTTCC GCTCCTCAGC CGTCAGGAAC TGGAGTATGT TCTCATGCTT CAGATTGATG 3360
TCTGAGAAGA TGTCCTTCTC TGTCTTCCAA GAGGCATACT CCTCATAGGG AAAGATCTTG 3420
ACTGCCACTG TCTCAAACTG CTCTGAAGTG TTCTGCTTCA GCTTGGCCTT ATAGACCTCA 3480
GCAAAGCGAC CTTTCCCCAC CAGGGTGTCC AGCTCAATGG GCAGCAGCTC TGTGTTGTGG 3540
TTGATGTTGT TGGCACACGT GGAGCTGATG TCAGAGCGGT CATCTTCCAG GATGATGGCA 3600
CAGTGCTCGC TGAACTCCAT GAGCTTCCGC GTCTTGCCGG TTTCCCAGGT TGAACTCAGC 3660
TTCTGCTGCC GGTTAACGCG GTAGCAGTAG AAGATGATGA TGACAGATAT GGCAACTCCC 3720
AGTGGTGGCA GGAGGCTGAT GCCTGTCACT TGAAATATGA CTAGCAACAA GTCAGGATTG 3780
CTGGTGTTAT ATTCTTCTGA GAAGATGATG TTGTCATTGC ACTCATCAGA GCTACAGGAA 3840
CACATGAAGA AAGTCTCACC AGGCTTTTTT TTTTCCTTCA TAATGCACTT TGGAGAAGCA 3900
GCATCTTCCA GAATAAAGTC ATGGTAGGGG AGCTTGGGGT CATGGCAAAC TGTCTCTAGT 3960
GTTATGTTCT CGTCATTCTT TCTCCATACA GCCACACAGA CTTCCTGTGG CTTCTCACAG 4020
ATGGAGGTGA TGCTGCAGTT GCTCATGCAG GATTTCTGGT TGTCACAGGT GGAAAATCTC 4080
ACATCACAAA ATTTACACAG TTGTGGAAAC TTGACTGCAC CGTTGTTGTC AGTGACTATC 4140
ATGTCGTTAT TAACCGACTT CTGAACGTGC GGTGGGATCG TGCTGGCGAT ACGCGTCCAC 4200
AGGACGATGT GCAGCGGCCA CAGGCCCCTG AGCAGCCCCC GACCCATGGC AGACCCCGCT 4260
GCTCGTCATA GACCGAGCCC CCAGCGCAGC GGACGGCGCC TTCCCGGACC CCTGGCTGCG 4320
CCTCCGCGCC GCGCCCTCTC CGGACCCCGC GCCGGGCCGG CAGCGCAGAT GTGCGGGCCA 4380
GATGTGGCGC CCGCTCGCCA GCCAGGAGGG GGCCTGGAGG CCGGCGAGGC GCGGGGAGGC 4440
CCCCGGCGGC CGAGGGAAGC TGCACAGGAG TCCGGCTCCT GTCCCGAGCG GGTGCACGCG 4500
CGGGGGTGTC GTCGCTCCGT GCGCGCGAGT GACTCACTCA ACTTCAACTC AGCGCTGCGG 4560
GGGAAACAGG AAACTCCTCG CCAACAGCTG GGCAGGACCT CTCTCCGCCC GAGAGCCTTC 4620
TCCCTCTCC 4629
SEQ ID NO: 2: рецептор II трансформирующего фактора роста бета (TGFBR2) человека, вариант транскрипта 2, мРНК (смысловая цепь; записано в коде ДНК)
GGAGAGGGAG AAGGCTCTCG GGCGGAGAGA GGTCCTGCCC AGCTGTTGGC GAGGAGTTTC 60
CTGTTTCCCC CGCAGCGCTG AGTTGAAGTT GAGTGAGTCA CTCGCGCGCA CGGAGCGACG 120
ACACCCCCGC GCGTGCACCC GCTCGGGACA GGAGCCGGAC TCCTGTGCAG CTTCCCTCGG 180
CCGCCGGGGG CCTCCCCGCG CCTCGCCGGC CTCCAGGCCC CCTCCTGGCT GGCGAGCGGG 240
CGCCACATCT GGCCCGCACA TCTGCGCTGC CGGCCCGGCG CGGGGTCCGG AGAGGGCGCG 300
GCGCGGAGGC GCAGCCAGGG GTCCGGGAAG GCGCCGTCCG CTGCGCTGGG GGCTCGGTCT 360
ATGACGAGCA GCGGGGTCTG CCATGGGTCG GGGGCTGCTC AGGGGCCTGT GGCCGCTGCA 420
CATCGTCCTG TGGACGCGTA TCGCCAGCAC GATCCCACCG CACGTTCAGA AGTCGGTTAA 480
TAACGACATG ATAGTCACTG ACAACAACGG TGCAGTCAAG TTTCCACAAC TGTGTAAATT 540
TTGTGATGTG AGATTTTCCA CCTGTGACAA CCAGAAATCC TGCATGAGCA ACTGCAGCAT 600
CACCTCCATC TGTGAGAAGC CACAGGAAGT CTGTGTGGCT GTATGGAGAA AGAATGACGA 660
GAACATAACA CTAGAGACAG TTTGCCATGA CCCCAAGCTC CCCTACCATG ACTTTATTCT 720
GGAAGATGCT GCTTCTCCAA AGTGCATTAT GAAGGAAAAA AAAAAGCCTG GTGAGACTTT 780
CTTCATGTGT TCCTGTAGCT CTGATGAGTG CAATGACAAC ATCATCTTCT CAGAAGAATA 840
TAACACCAGC AATCCTGACT TGTTGCTAGT CATATTTCAA GTGACAGGCA TCAGCCTCCT 900
GCCACCACTG GGAGTTGCCA TATCTGTCAT CATCATCTTC TACTGCTACC GCGTTAACCG 960
GCAGCAGAAG CTGAGTTCAA CCTGGGAAAC CGGCAAGACG CGGAAGCTCA TGGAGTTCAG 1020
CGAGCACTGT GCCATCATCC TGGAAGATGA CCGCTCTGAC ATCAGCTCCA CGTGTGCCAA 1080
CAACATCAAC CACAACACAG AGCTGCTGCC CATTGAGCTG GACACCCTGG TGGGGAAAGG 1140
TCGCTTTGCT GAGGTCTATA AGGCCAAGCT GAAGCAGAAC ACTTCAGAGC AGTTTGAGAC 1200
AGTGGCAGTC AAGATCTTTC CCTATGAGGA GTATGCCTCT TGGAAGACAG AGAAGGACAT 1260
CTTCTCAGAC ATCAATCTGA AGCATGAGAA CATACTCCAG TTCCTGACGG CTGAGGAGCG 1320
GAAGACGGAG TTGGGGAAAC AATACTGGCT GATCACCGCC TTCCACGCCA AGGGCAACCT 1380
ACAGGAGTAC CTGACGCGGC ATGTCATCAG CTGGGAGGAC CTGCGCAAGC TGGGCAGCTC 1440
CCTCGCCCGG GGGATTGCTC ACCTCCACAG TGATCACACT CCATGTGGGA GGCCCAAGAT 1500
GCCCATCGTG CACAGGGACC TCAAGAGCTC CAATATCCTC GTGAAGAACG ACCTAACCTG 1560
CTGCCTGTGT GACTTTGGGC TTTCCCTGCG TCTGGACCCT ACTCTGTCTG TGGATGACCT 1620
GGCTAACAGT GGGCAGGTGG GAACTGCAAG ATACATGGCT CCAGAAGTCC TAGAATCCAG 1680
GATGAATTTG GAGAATGTTG AGTCCTTCAA GCAGACCGAT GTCTACTCCA TGGCTCTGGT 1740
GCTCTGGGAA ATGACATCTC GCTGTAATGC AGTGGGAGAA GTAAAAGATT ATGAGCCTCC 1800
ATTTGGTTCC AAGGTGCGGG AGCACCCCTG TGTCGAAAGC ATGAAGGACA ACGTGTTGAG 1860
AGATCGAGGG CGACCAGAAA TTCCCAGCTT CTGGCTCAAC CACCAGGGCA TCCAGATGGT 1920
GTGTGAGACG TTGACTGAGT GCTGGGACCA CGACCCAGAG GCCCGTCTCA CAGCCCAGTG 1980
TGTGGCAGAA CGCTTCAGTG AGCTGGAGCA TCTGGACAGG CTCTCGGGGA GGAGCTGCTC 2040
GGAGGAGAAG ATTCCTGAAG ACGGCTCCCT AAACACTACC AAATAGCTCT TCTGGGGCAG 2100
GCTGGGCCAT GTCCAAAGAG GCTGCCCCTC TCACCAAAGA ACAGAGGCAG CAGGAAGCTG 2160
CCCCTGAACT GATGCTTCCT GGAAAACCAA GGGGGTCACT CCCCTCCCTG TAAGCTGTGG 2220
GGATAAGCAG AAACAACAGC AGCAGGGAGT GGGTGACATA GAGCATTCTA TGCCTTTGAC 2280
ATTGTCATAG GATAAGCTGT GTTAGCACTT CCTCAGGAAA TGAGATTGAT TTTTACAATA 2340
GCCAATAACA TTTGCACTTT ATTAATGCCT GTATATAAAT ATGAATAGCT ATGTTTTATA 2400
TATATATATA TATATCTATA TATGTCTATA GCTCTATATA TATAGCCATA CCTTGAAAAG 2460
AGACAAGGAA AAACATCAAA TATTCCCAGG AAATTGGTTT TATTGGAGAA CTCCAGAACC 2520
AAGCAGAGAA GGAAGGGACC CATGACAGCA TTAGCATTTG ACAATCACAC ATGCAGTGGT 2580
TCTCTGACTG TAAAACAGTG AACTTTGCAT GAGGAAAGAG GCTCCATGTC TCACAGCCAG 2640
CTATGACCAC ATTGCACTTG CTTTTGCAAA ATAATCATTC CCTGCCTAGC ACTTCTCTTC 2700
TGGCCATGGA ACTAAGTACA GTGGCACTGT TTGAGGACCA GTGTTCCCGG GGTTCCTGTG 2760
TGCCCTTATT TCTCCTGGAC TTTTCATTTA AGCTCCAAGC CCCAAATCTG GGGGGCTAGT 2820
TTAGAAACTC TCCCTCAACC TAGTTTAGAA ACTCTACCCC ATCTTTAATA CCTTGAATGT 2880
TTTGAACCCC ACTTTTTACC TTCATGGGTT GCAGAAAAAT CAGAACAGAT GTCCCCATCC 2940
ATGCGATTGC CCCACCATCT ACTAATGAAA AATTGTTCTT TTTTTCATCT TTCCCCTGCA 3000
CTTATGTTAC TATTCTCTGC TCCCAGCCTT CATCCTTTTC TAAAAAGGAG CAAATTCTCA 3060
CTCTAGGCTT TATCGTGTTT ACTTTTTCAT TACACTTGAC TTGATTTTCT AGTTTTCTAT 3120
ACAAACACCA ATGGGTTCCA TCTTTCTGGG CTCCTGATTG CTCAAGCACA GTTTGGCCTG 3180
ATGAAGAGGA TTTCAACTAC ACAATACTAT CATTGTCAGG ACTATGACCT CAGGCACTCT 3240
AAACATATGT TTTGTTTGGT CAGCACAGCG TTTCAAAAAG TGAAGCCACT TTATAAATAT 3300
TTGGAGATTT TGCAGGAAAA TCTGGATCCC CAGGTAAGGA TAGCAGATGG TTTTCAGTTA 3360
TCTCCAGTCC ACGTTCACAA AATGTGAAGG TGTGGAGACA CTTACAAAGC TGCCTCACTT 3420
CTCACTGTAA ACATTAGCTC TTTCCACTGC CTACCTGGAC CCCAGTCTAG GAATTAAATC 3480
TGCACCTAAC CAAGGTCCCT TGTAAGAAAT GTCCATTCAA GCAGTCATTC TCTGGGTATA 3540
TAATATGATT TTGACTACCT TATCTGGTGT TAAGATTTGA AGTTGGCCTT TTATTGGACT 3600
AAAGGGGAAC TCCTTTAAGG GTCTCAGTTA GCCCAAGTTT CTTTTGCTTA TATGTTAATA 3660
GTTTTACCCT CTGCATTGGA GAGAGGAGTG CTTTACTCCA AGAAGCTTTC CTCATGGTTA 3720
CCGTTCTCTC CATCATGCCA GCCTTCTCAA CCTTTGCAGA AATTACTAGA GAGGATTTGA 3780
ATGTGGGACA CAAAGGTCCC ATTTGCAGTT AGAAAATTTG TGTCCACAAG GACAAGAACA 3840
AAGTATGAGC TTTAAAACTC CATAGGAAAC TTGTTAATCA ACAAAGAAGT GTTAATGCTG 3900
CAAGTAATCT CTTTTTTAAA ACTTTTTGAA GCTACTTATT TTCAGCCAAA TAGGAATATT 3960
AGAGAGGGAC TGGTAGTGAG AATATCAGCT CTGTTTGGAT GGTGGAAGGT CTCATTTTAT 4020
TGAGATTTTT AAGATACATG CAAAGGTTTG GAAATAGAAC CTCTAGGCAC CCTCCTCAGT 4080
GTGGGTGGGC TGAGAGTTAA AGACAGTGTG GCTGCAGTAG CATAGAGGCG CCTAGAAATT 4140
CCACTTGCAC CGTAGGGCAT GCTGATACCA TCCCAATAGC TGTTGCCCAT TGACCTCTAG 4200
TGGTGAGTTT CTAGAATACT GGTCCATTCA TGAGATATTC AAGATTCAAG AGTATTCTCA 4260
CTTCTGGGTT ATCAGCATAA ACTGGAATGT AGTGTCAGAG GATACTGTGG CTTGTTTTGT 4320
TTATGTTTTT TTTTCTTATT CAAGAAAAAA GACCAAGGAA TAACATTCTG TAGTTCCTAA 4380
AAATACTGAC TTTTTTCACT ACTATACATA AAGGGAAAGT TTTATTCTTT TATGGAACAC 4440
TTCAGCTGTA CTCATGTATT AAAATAGGAA TGTGAATGCT ATATACTCTT TTTATATCAA 4500
AAGTCTCAAG CACTTATTTT TATTCTATGC ATTGTTTGTC TTTTACATAA ATAAAATGTT 4560
TATTAGATTG AATAAAGCAA AATACTCAGG TGAGCATCCT GCCTCCTGTT CCCATTCCTA 4620
GTAGCTAAA 4629
SEQ ID NO: 3: рецептор II трансформирующего фактора роста бета (TGFBR2) человека, вариант транскрипта 2, мРНК (смысловая цепь; записано в коде РНК)
GGAGAGGGAG AAGGCUCUCG GGCGGAGAGA GGUCCUGCCC AGCUGUUGGC GAGGAGUUUC 60
CUGUUUCCCC CGCAGCGCUG AGUUGAAGUU GAGUGAGUCA CUCGCGCGCA CGGAGCGACG 120
ACACCCCCGC GCGUGCACCC GCUCGGGACA GGAGCCGGAC UCCUGUGCAG CUUCCCUCGG 180
CCGCCGGGGG CCUCCCCGCG CCUCGCCGGC CUCCAGGCCC CCUCCUGGCU GGCGAGCGGG 240
CGCCACAUCU GGCCCGCACA UCUGCGCUGC CGGCCCGGCG CGGGGUCCGG AGAGGGCGCG 300
GCGCGGAGGC GCAGCCAGGG GUCCGGGAAG GCGCCGUCCG CUGCGCUGGG GGCUCGGUCU 360
AUGACGAGCA GCGGGGUCUG CCAUGGGUCG GGGGCUGCUC AGGGGCCUGU GGCCGCUGCA 420
CAUCGUCCUG UGGACGCGUA UCGCCAGCAC GAUCCCACCG CACGUUCAGA AGUCGGUUAA 480
UAACGACAUG AUAGUCACUG ACAACAACGG UGCAGUCAAG UUUCCACAAC UGUGUAAAUU 540
UUGUGAUGUG AGAUUUUCCA CCUGUGACAA CCAGAAAUCC UGCAUGAGCA ACUGCAGCAU 600
CACCUCCAUC UGUGAGAAGC CACAGGAAGU CUGUGUGGCU GUAUGGAGAA AGAAUGACGA 660
GAACAUAACA CUAGAGACAG UUUGCCAUGA CCCCAAGCUC CCCUACCAUG ACUUUAUUCU 720
GGAAGAUGCU GCUUCUCCAA AGUGCAUUAU GAAGGAAAAA AAAAAGCCUG GUGAGACUUU 780
CUUCAUGUGU UCCUGUAGCU CUGAUGAGUG CAAUGACAAC AUCAUCUUCU CAGAAGAAUA 840
UAACACCAGC AAUCCUGACU UGUUGCUAGU CAUAUUUCAA GUGACAGGCA UCAGCCUCCU 900
GCCACCACUG GGAGUUGCCA UAUCUGUCAU CAUCAUCUUC UACUGCUACC GCGUUAACCG 960
GCAGCAGAAG CUGAGUUCAA CCUGGGAAAC CGGCAAGACG CGGAAGCUCA UGGAGUUCAG 1020
CGAGCACUGU GCCAUCAUCC UGGAAGAUGA CCGCUCUGAC AUCAGCUCCA CGUGUGCCAA 1080
CAACAUCAAC CACAACACAG AGCUGCUGCC CAUUGAGCUG GACACCCUGG UGGGGAAAGG 1140
UCGCUUUGCU GAGGUCUAUA AGGCCAAGCU GAAGCAGAAC ACUUCAGAGC AGUUUGAGAC 1200
AGUGGCAGUC AAGAUCUUUC CCUAUGAGGA GUAUGCCUCU UGGAAGACAG AGAAGGACAU 1260
CUUCUCAGAC AUCAAUCUGA AGCAUGAGAA CAUACUCCAG UUCCUGACGG CUGAGGAGCG 1320
GAAGACGGAG UUGGGGAAAC AAUACUGGCU GAUCACCGCC UUCCACGCCA AGGGCAACCU 1380
ACAGGAGUAC CUGACGCGGC AUGUCAUCAG CUGGGAGGAC CUGCGCAAGC UGGGCAGCUC 1440
CCUCGCCCGG GGGAUUGCUC ACCUCCACAG UGAUCACACU CCAUGUGGGA GGCCCAAGAU 1500
GCCCAUCGUG CACAGGGACC UCAAGAGCUC CAAUAUCCUC GUGAAGAACG ACCUAACCUG 1560
CUGCCUGUGU GACUUUGGGC UUUCCCUGCG UCUGGACCCU ACUCUGUCUG UGGAUGACCU 1620
GGCUAACAGU GGGCAGGUGG GAACUGCAAG AUACAUGGCU CCAGAAGUCC UAGAAUCCAG 1680
GAUGAAUUUG GAGAAUGUUG AGUCCUUCAA GCAGACCGAU GUCUACUCCA UGGCUCUGGU 1740
GCUCUGGGAA AUGACAUCUC GCUGUAAUGC AGUGGGAGAA GUAAAAGAUU AUGAGCCUCC 1800
AUUUGGUUCC AAGGUGCGGG AGCACCCCUG UGUCGAAAGC AUGAAGGACA ACGUGUUGAG 1860
AGAUCGAGGG CGACCAGAAA UUCCCAGCUU CUGGCUCAAC CACCAGGGCA UCCAGAUGGU 1920
GUGUGAGACG UUGACUGAGU GCUGGGACCA CGACCCAGAG GCCCGUCUCA CAGCCCAGUG 1980
UGUGGCAGAA CGCUUCAGUG AGCUGGAGCA UCUGGACAGG CUCUCGGGGA GGAGCUGCUC 2040
GGAGGAGAAG AUUCCUGAAG ACGGCUCCCU AAACACUACC AAAUAGCUCU UCUGGGGCAG 2100
GCUGGGCCAU GUCCAAAGAG GCUGCCCCUC UCACCAAAGA ACAGAGGCAG CAGGAAGCUG 2160
CCCCUGAACU GAUGCUUCCU GGAAAACCAA GGGGGUCACU CCCCUCCCUG UAAGCUGUGG 2220
GGAUAAGCAG AAACAACAGC AGCAGGGAGU GGGUGACAUA GAGCAUUCUA UGCCUUUGAC 2280
AUUGUCAUAG GAUAAGCUGU GUUAGCACUU CCUCAGGAAA UGAGAUUGAU UUUUACAAUA 2340
GCCAAUAACA UUUGCACUUU AUUAAUGCCU GUAUAUAAAU AUGAAUAGCU AUGUUUUAUA 2400
UAUAUAUAUA UAUAUCUAUA UAUGUCUAUA GCUCUAUAUA UAUAGCCAUA CCUUGAAAAG 2460
AGACAAGGAA AAACAUCAAA UAUUCCCAGG AAAUUGGUUU UAUUGGAGAA CUCCAGAACC 2520
AAGCAGAGAA GGAAGGGACC CAUGACAGCA UUAGCAUUUG ACAAUCACAC AUGCAGUGGU 2580
UCUCUGACUG UAAAACAGUG AACUUUGCAU GAGGAAAGAG GCUCCAUGUC UCACAGCCAG 2640
CUAUGACCAC AUUGCACUUG CUUUUGCAAA AUAAUCAUUC CCUGCCUAGC ACUUCUCUUC 2700
UGGCCAUGGA ACUAAGUACA GUGGCACUGU UUGAGGACCA GUGUUCCCGG GGUUCCUGUG 2760
UGCCCUUAUU UCUCCUGGAC UUUUCAUUUA AGCUCCAAGC CCCAAAUCUG GGGGGCUAGU 2820
UUAGAAACUC UCCCUCAACC UAGUUUAGAA ACUCUACCCC AUCUUUAAUA CCUUGAAUGU 2880
UUUGAACCCC ACUUUUUACC UUCAUGGGUU GCAGAAAAAU CAGAACAGAU GUCCCCAUCC 2940
AUGCGAUUGC CCCACCAUCU ACUAAUGAAA AAUUGUUCUU UUUUUCAUCU UUCCCCUGCA 3000
CUUAUGUUAC UAUUCUCUGC UCCCAGCCUU CAUCCUUUUC UAAAAAGGAG CAAAUUCUCA 3060
CUCUAGGCUU UAUCGUGUUU ACUUUUUCAU UACACUUGAC UUGAUUUUCU AGUUUUCUAU 3120
ACAAACACCA AUGGGUUCCA UCUUUCUGGG CUCCUGAUUG CUCAAGCACA GUUUGGCCUG 3180
AUGAAGAGGA UUUCAACUAC ACAAUACUAU CAUUGUCAGG ACUAUGACCU CAGGCACUCU 3240
AAACAUAUGU UUUGUUUGGU CAGCACAGCG UUUCAAAAAG UGAAGCCACU UUAUAAAUAU 3300
UUGGAGAUUU UGCAGGAAAA UCUGGAUCCC CAGGUAAGGA UAGCAGAUGG UUUUCAGUUA 3360
UCUCCAGUCC ACGUUCACAA AAUGUGAAGG UGUGGAGACA CUUACAAAGC UGCCUCACUU 3420
CUCACUGUAA ACAUUAGCUC UUUCCACUGC CUACCUGGAC CCCAGUCUAG GAAUUAAAUC 3480
UGCACCUAAC CAAGGUCCCU UGUAAGAAAU GUCCAUUCAA GCAGUCAUUC UCUGGGUAUA 3540
UAAUAUGAUU UUGACUACCU UAUCUGGUGU UAAGAUUUGA AGUUGGCCUU UUAUUGGACU 3600
AAAGGGGAAC UCCUUUAAGG GUCUCAGUUA GCCCAAGUUU CUUUUGCUUA UAUGUUAAUA 3660
GUUUUACCCU CUGCAUUGGA GAGAGGAGUG CUUUACUCCA AGAAGCUUUC CUCAUGGUUA 3720
CCGUUCUCUC CAUCAUGCCA GCCUUCUCAA CCUUUGCAGA AAUUACUAGA GAGGAUUUGA 3780
AUGUGGGACA CAAAGGUCCC AUUUGCAGUU AGAAAAUUUG UGUCCACAAG GACAAGAACA 3840
AAGUAUGAGC UUUAAAACUC CAUAGGAAAC UUGUUAAUCA ACAAAGAAGU GUUAAUGCUG 3900
CAAGUAAUCU CUUUUUUAAA ACUUUUUGAA GCUACUUAUU UUCAGCCAAA UAGGAAUAUU 3960
AGAGAGGGAC UGGUAGUGAG AAUAUCAGCU CUGUUUGGAU GGUGGAAGGU CUCAUUUUAU 4020
UGAGAUUUUU AAGAUACAUG CAAAGGUUUG GAAAUAGAAC CUCUAGGCAC CCUCCUCAGU 4080
GUGGGUGGGC UGAGAGUUAA AGACAGUGUG GCUGCAGUAG CAUAGAGGCG CCUAGAAAUU 4140
CCACUUGCAC CGUAGGGCAU GCUGAUACCA UCCCAAUAGC UGUUGCCCAU UGACCUCUAG 4200
UGGUGAGUUU CUAGAAUACU GGUCCAUUCA UGAGAUAUUC AAGAUUCAAG AGUAUUCUCA 4260
CUUCUGGGUU AUCAGCAUAA ACUGGAAUGU AGUGUCAGAG GAUACUGUGG CUUGUUUUGU 4320
UUAUGUUUUU UUUUCUUAUU CAAGAAAAAA GACCAAGGAA UAACAUUCUG UAGUUCCUAA 4380
AAAUACUGAC UUUUUUCACU ACUAUACAUA AAGGGAAAGU UUUAUUCUUU UAUGGAACAC 4440
UUCAGCUGUA CUCAUGUAUU AAAAUAGGAA UGUGAAUGCU AUAUACUCUU UUUAUAUCAA 4500
AAGUCUCAAG CACUUAUUUU UAUUCUAUGC AUUGUUUGUC UUUUACAUAA AUAAAAUGUU 4560
UAUUAGAUUG AAUAAAGCAA AAUACUCAGG UGAGCAUCCU GCCUCCUGUU CCCAUUCCUA 4620
GUAGCUAAA 4629
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Neurovision Pharma GmbH
<120> Антисмысловые олигонуклеотиды в качестве ингибиторов сигнального пути TGF-R
<130> NVP-P03653WO
<150> EP14193368
<151> 2014-11-16
<160> 508
<170> PatentIn version 3.5
<210> 1
<211> 4629
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Рецептор II трансформирующего фактора роста бета человека
(70/80кДа) (TGFBR2), вариант транскрипта 2, антисмысловая цепь ДНК
<400> 1
tttagctact aggaatggga acaggaggca ggatgctcac ctgagtattt tgctttattc 60
aatctaataa acattttatt tatgtaaaag acaaacaatg catagaataa aaataagtgc 120
ttgagacttt tgatataaaa agagtatata gcattcacat tcctatttta atacatgagt 180
acagctgaag tgttccataa aagaataaaa ctttcccttt atgtatagta gtgaaaaaag 240
tcagtatttt taggaactac agaatgttat tccttggtct tttttcttga ataagaaaaa 300
aaaacataaa caaaacaagc cacagtatcc tctgacacta cattccagtt tatgctgata 360
acccagaagt gagaatactc ttgaatcttg aatatctcat gaatggacca gtattctaga 420
aactcaccac tagaggtcaa tgggcaacag ctattgggat ggtatcagca tgccctacgg 480
tgcaagtgga atttctaggc gcctctatgc tactgcagcc acactgtctt taactctcag 540
cccacccaca ctgaggaggg tgcctagagg ttctatttcc aaacctttgc atgtatctta 600
aaaatctcaa taaaatgaga ccttccacca tccaaacaga gctgatattc tcactaccag 660
tccctctcta atattcctat ttggctgaaa ataagtagct tcaaaaagtt ttaaaaaaga 720
gattacttgc agcattaaca cttctttgtt gattaacaag tttcctatgg agttttaaag 780
ctcatacttt gttcttgtcc ttgtggacac aaattttcta actgcaaatg ggacctttgt 840
gtcccacatt caaatcctct ctagtaattt ctgcaaaggt tgagaaggct ggcatgatgg 900
agagaacggt aaccatgagg aaagcttctt ggagtaaagc actcctctct ccaatgcaga 960
gggtaaaact attaacatat aagcaaaaga aacttgggct aactgagacc cttaaaggag 1020
ttccccttta gtccaataaa aggccaactt caaatcttaa caccagataa ggtagtcaaa 1080
atcatattat atacccagag aatgactgct tgaatggaca tttcttacaa gggaccttgg 1140
ttaggtgcag atttaattcc tagactgggg tccaggtagg cagtggaaag agctaatgtt 1200
tacagtgaga agtgaggcag ctttgtaagt gtctccacac cttcacattt tgtgaacgtg 1260
gactggagat aactgaaaac catctgctat ccttacctgg ggatccagat tttcctgcaa 1320
aatctccaaa tatttataaa gtggcttcac tttttgaaac gctgtgctga ccaaacaaaa 1380
catatgttta gagtgcctga ggtcatagtc ctgacaatga tagtattgtg tagttgaaat 1440
cctcttcatc aggccaaact gtgcttgagc aatcaggagc ccagaaagat ggaacccatt 1500
ggtgtttgta tagaaaacta gaaaatcaag tcaagtgtaa tgaaaaagta aacacgataa 1560
agcctagagt gagaatttgc tcctttttag aaaaggatga aggctgggag cagagaatag 1620
taacataagt gcaggggaaa gatgaaaaaa agaacaattt ttcattagta gatggtgggg 1680
caatcgcatg gatggggaca tctgttctga tttttctgca acccatgaag gtaaaaagtg 1740
gggttcaaaa cattcaaggt attaaagatg gggtagagtt tctaaactag gttgagggag 1800
agtttctaaa ctagcccccc agatttgggg cttggagctt aaatgaaaag tccaggagaa 1860
ataagggcac acaggaaccc cgggaacact ggtcctcaaa cagtgccact gtacttagtt 1920
ccatggccag aagagaagtg ctaggcaggg aatgattatt ttgcaaaagc aagtgcaatg 1980
tggtcatagc tggctgtgag acatggagcc tctttcctca tgcaaagttc actgttttac 2040
agtcagagaa ccactgcatg tgtgattgtc aaatgctaat gctgtcatgg gtcccttcct 2100
tctctgcttg gttctggagt tctccaataa aaccaatttc ctgggaatat ttgatgtttt 2160
tccttgtctc ttttcaaggt atggctatat atatagagct atagacatat atagatatat 2220
atatatatat ataaaacata gctattcata tttatataca ggcattaata aagtgcaaat 2280
gttattggct attgtaaaaa tcaatctcat ttcctgagga agtgctaaca cagcttatcc 2340
tatgacaatg tcaaaggcat agaatgctct atgtcaccca ctccctgctg ctgttgtttc 2400
tgcttatccc cacagcttac agggagggga gtgaccccct tggttttcca ggaagcatca 2460
gttcaggggc agcttcctgc tgcctctgtt ctttggtgag aggggcagcc tctttggaca 2520
tggcccagcc tgccccagaa gagctatttg gtagtgttta gggagccgtc ttcaggaatc 2580
ttctcctccg agcagctcct ccccgagagc ctgtccagat gctccagctc actgaagcgt 2640
tctgccacac actgggctgt gagacgggcc tctgggtcgt ggtcccagca ctcagtcaac 2700
gtctcacaca ccatctggat gccctggtgg ttgagccaga agctgggaat ttctggtcgc 2760
cctcgatctc tcaacacgtt gtccttcatg ctttcgacac aggggtgctc ccgcaccttg 2820
gaaccaaatg gaggctcata atcttttact tctcccactg cattacagcg agatgtcatt 2880
tcccagagca ccagagccat ggagtagaca tcggtctgct tgaaggactc aacattctcc 2940
aaattcatcc tggattctag gacttctgga gccatgtatc ttgcagttcc cacctgccca 3000
ctgttagcca ggtcatccac agacagagta gggtccagac gcagggaaag cccaaagtca 3060
cacaggcagc aggttaggtc gttcttcacg aggatattgg agctcttgag gtccctgtgc 3120
acgatgggca tcttgggcct cccacatgga gtgtgatcac tgtggaggtg agcaatcccc 3180
cgggcgaggg agctgcccag cttgcgcagg tcctcccagc tgatgacatg ccgcgtcagg 3240
tactcctgta ggttgccctt ggcgtggaag gcggtgatca gccagtattg tttccccaac 3300
tccgtcttcc gctcctcagc cgtcaggaac tggagtatgt tctcatgctt cagattgatg 3360
tctgagaaga tgtccttctc tgtcttccaa gaggcatact cctcataggg aaagatcttg 3420
actgccactg tctcaaactg ctctgaagtg ttctgcttca gcttggcctt atagacctca 3480
gcaaagcgac ctttccccac cagggtgtcc agctcaatgg gcagcagctc tgtgttgtgg 3540
ttgatgttgt tggcacacgt ggagctgatg tcagagcggt catcttccag gatgatggca 3600
cagtgctcgc tgaactccat gagcttccgc gtcttgccgg tttcccaggt tgaactcagc 3660
ttctgctgcc ggttaacgcg gtagcagtag aagatgatga tgacagatat ggcaactccc 3720
agtggtggca ggaggctgat gcctgtcact tgaaatatga ctagcaacaa gtcaggattg 3780
ctggtgttat attcttctga gaagatgatg ttgtcattgc actcatcaga gctacaggaa 3840
cacatgaaga aagtctcacc aggctttttt ttttccttca taatgcactt tggagaagca 3900
gcatcttcca gaataaagtc atggtagggg agcttggggt catggcaaac tgtctctagt 3960
gttatgttct cgtcattctt tctccataca gccacacaga cttcctgtgg cttctcacag 4020
atggaggtga tgctgcagtt gctcatgcag gatttctggt tgtcacaggt ggaaaatctc 4080
acatcacaaa atttacacag ttgtggaaac ttgactgcac cgttgttgtc agtgactatc 4140
atgtcgttat taaccgactt ctgaacgtgc ggtgggatcg tgctggcgat acgcgtccac 4200
aggacgatgt gcagcggcca caggcccctg agcagccccc gacccatggc agaccccgct 4260
gctcgtcata gaccgagccc ccagcgcagc ggacggcgcc ttcccggacc cctggctgcg 4320
cctccgcgcc gcgccctctc cggaccccgc gccgggccgg cagcgcagat gtgcgggcca 4380
gatgtggcgc ccgctcgcca gccaggaggg ggcctggagg ccggcgaggc gcggggaggc 4440
ccccggcggc cgagggaagc tgcacaggag tccggctcct gtcccgagcg ggtgcacgcg 4500
cgggggtgtc gtcgctccgt gcgcgcgagt gactcactca acttcaactc agcgctgcgg 4560
gggaaacagg aaactcctcg ccaacagctg ggcaggacct ctctccgccc gagagccttc 4620
tccctctcc 4629
<210> 2
<211> 4629
<212> ДНК
<213> Homo sapiens
<400> 2
ggagagggag aaggctctcg ggcggagaga ggtcctgccc agctgttggc gaggagtttc 60
ctgtttcccc cgcagcgctg agttgaagtt gagtgagtca ctcgcgcgca cggagcgacg 120
acacccccgc gcgtgcaccc gctcgggaca ggagccggac tcctgtgcag cttccctcgg 180
ccgccggggg cctccccgcg cctcgccggc ctccaggccc cctcctggct ggcgagcggg 240
cgccacatct ggcccgcaca tctgcgctgc cggcccggcg cggggtccgg agagggcgcg 300
gcgcggaggc gcagccaggg gtccgggaag gcgccgtccg ctgcgctggg ggctcggtct 360
atgacgagca gcggggtctg ccatgggtcg ggggctgctc aggggcctgt ggccgctgca 420
catcgtcctg tggacgcgta tcgccagcac gatcccaccg cacgttcaga agtcggttaa 480
taacgacatg atagtcactg acaacaacgg tgcagtcaag tttccacaac tgtgtaaatt 540
ttgtgatgtg agattttcca cctgtgacaa ccagaaatcc tgcatgagca actgcagcat 600
cacctccatc tgtgagaagc cacaggaagt ctgtgtggct gtatggagaa agaatgacga 660
gaacataaca ctagagacag tttgccatga ccccaagctc ccctaccatg actttattct 720
ggaagatgct gcttctccaa agtgcattat gaaggaaaaa aaaaagcctg gtgagacttt 780
cttcatgtgt tcctgtagct ctgatgagtg caatgacaac atcatcttct cagaagaata 840
taacaccagc aatcctgact tgttgctagt catatttcaa gtgacaggca tcagcctcct 900
gccaccactg ggagttgcca tatctgtcat catcatcttc tactgctacc gcgttaaccg 960
gcagcagaag ctgagttcaa cctgggaaac cggcaagacg cggaagctca tggagttcag 1020
cgagcactgt gccatcatcc tggaagatga ccgctctgac atcagctcca cgtgtgccaa 1080
caacatcaac cacaacacag agctgctgcc cattgagctg gacaccctgg tggggaaagg 1140
tcgctttgct gaggtctata aggccaagct gaagcagaac acttcagagc agtttgagac 1200
agtggcagtc aagatctttc cctatgagga gtatgcctct tggaagacag agaaggacat 1260
cttctcagac atcaatctga agcatgagaa catactccag ttcctgacgg ctgaggagcg 1320
gaagacggag ttggggaaac aatactggct gatcaccgcc ttccacgcca agggcaacct 1380
acaggagtac ctgacgcggc atgtcatcag ctgggaggac ctgcgcaagc tgggcagctc 1440
cctcgcccgg gggattgctc acctccacag tgatcacact ccatgtggga ggcccaagat 1500
gcccatcgtg cacagggacc tcaagagctc caatatcctc gtgaagaacg acctaacctg 1560
ctgcctgtgt gactttgggc tttccctgcg tctggaccct actctgtctg tggatgacct 1620
ggctaacagt gggcaggtgg gaactgcaag atacatggct ccagaagtcc tagaatccag 1680
gatgaatttg gagaatgttg agtccttcaa gcagaccgat gtctactcca tggctctggt 1740
gctctgggaa atgacatctc gctgtaatgc agtgggagaa gtaaaagatt atgagcctcc 1800
atttggttcc aaggtgcggg agcacccctg tgtcgaaagc atgaaggaca acgtgttgag 1860
agatcgaggg cgaccagaaa ttcccagctt ctggctcaac caccagggca tccagatggt 1920
gtgtgagacg ttgactgagt gctgggacca cgacccagag gcccgtctca cagcccagtg 1980
tgtggcagaa cgcttcagtg agctggagca tctggacagg ctctcgggga ggagctgctc 2040
ggaggagaag attcctgaag acggctccct aaacactacc aaatagctct tctggggcag 2100
gctgggccat gtccaaagag gctgcccctc tcaccaaaga acagaggcag caggaagctg 2160
cccctgaact gatgcttcct ggaaaaccaa gggggtcact cccctccctg taagctgtgg 2220
ggataagcag aaacaacagc agcagggagt gggtgacata gagcattcta tgcctttgac 2280
attgtcatag gataagctgt gttagcactt cctcaggaaa tgagattgat ttttacaata 2340
gccaataaca tttgcacttt attaatgcct gtatataaat atgaatagct atgttttata 2400
tatatatata tatatctata tatgtctata gctctatata tatagccata ccttgaaaag 2460
agacaaggaa aaacatcaaa tattcccagg aaattggttt tattggagaa ctccagaacc 2520
aagcagagaa ggaagggacc catgacagca ttagcatttg acaatcacac atgcagtggt 2580
tctctgactg taaaacagtg aactttgcat gaggaaagag gctccatgtc tcacagccag 2640
ctatgaccac attgcacttg cttttgcaaa ataatcattc cctgcctagc acttctcttc 2700
tggccatgga actaagtaca gtggcactgt ttgaggacca gtgttcccgg ggttcctgtg 2760
tgcccttatt tctcctggac ttttcattta agctccaagc cccaaatctg gggggctagt 2820
ttagaaactc tccctcaacc tagtttagaa actctacccc atctttaata ccttgaatgt 2880
tttgaacccc actttttacc ttcatgggtt gcagaaaaat cagaacagat gtccccatcc 2940
atgcgattgc cccaccatct actaatgaaa aattgttctt tttttcatct ttcccctgca 3000
cttatgttac tattctctgc tcccagcctt catccttttc taaaaaggag caaattctca 3060
ctctaggctt tatcgtgttt actttttcat tacacttgac ttgattttct agttttctat 3120
acaaacacca atgggttcca tctttctggg ctcctgattg ctcaagcaca gtttggcctg 3180
atgaagagga tttcaactac acaatactat cattgtcagg actatgacct caggcactct 3240
aaacatatgt tttgtttggt cagcacagcg tttcaaaaag tgaagccact ttataaatat 3300
ttggagattt tgcaggaaaa tctggatccc caggtaagga tagcagatgg ttttcagtta 3360
tctccagtcc acgttcacaa aatgtgaagg tgtggagaca cttacaaagc tgcctcactt 3420
ctcactgtaa acattagctc tttccactgc ctacctggac cccagtctag gaattaaatc 3480
tgcacctaac caaggtccct tgtaagaaat gtccattcaa gcagtcattc tctgggtata 3540
taatatgatt ttgactacct tatctggtgt taagatttga agttggcctt ttattggact 3600
aaaggggaac tcctttaagg gtctcagtta gcccaagttt cttttgctta tatgttaata 3660
gttttaccct ctgcattgga gagaggagtg ctttactcca agaagctttc ctcatggtta 3720
ccgttctctc catcatgcca gccttctcaa cctttgcaga aattactaga gaggatttga 3780
atgtgggaca caaaggtccc atttgcagtt agaaaatttg tgtccacaag gacaagaaca 3840
aagtatgagc tttaaaactc cataggaaac ttgttaatca acaaagaagt gttaatgctg 3900
caagtaatct cttttttaaa actttttgaa gctacttatt ttcagccaaa taggaatatt 3960
agagagggac tggtagtgag aatatcagct ctgtttggat ggtggaaggt ctcattttat 4020
tgagattttt aagatacatg caaaggtttg gaaatagaac ctctaggcac cctcctcagt 4080
gtgggtgggc tgagagttaa agacagtgtg gctgcagtag catagaggcg cctagaaatt 4140
ccacttgcac cgtagggcat gctgatacca tcccaatagc tgttgcccat tgacctctag 4200
tggtgagttt ctagaatact ggtccattca tgagatattc aagattcaag agtattctca 4260
cttctgggtt atcagcataa actggaatgt agtgtcagag gatactgtgg cttgttttgt 4320
ttatgttttt ttttcttatt caagaaaaaa gaccaaggaa taacattctg tagttcctaa 4380
aaatactgac ttttttcact actatacata aagggaaagt tttattcttt tatggaacac 4440
ttcagctgta ctcatgtatt aaaataggaa tgtgaatgct atatactctt tttatatcaa 4500
aagtctcaag cacttatttt tattctatgc attgtttgtc ttttacataa ataaaatgtt 4560
tattagattg aataaagcaa aatactcagg tgagcatcct gcctcctgtt cccattccta 4620
gtagctaaa 4629
<210> 3
<211> 4629
<212> РНК
<213> Homo sapiens
<400> 3
ggagagggag aaggcucucg ggcggagaga gguccugccc agcuguuggc gaggaguuuc 60
cuguuucccc cgcagcgcug aguugaaguu gagugaguca cucgcgcgca cggagcgacg 120
acacccccgc gcgugcaccc gcucgggaca ggagccggac uccugugcag cuucccucgg 180
ccgccggggg ccuccccgcg ccucgccggc cuccaggccc ccuccuggcu ggcgagcggg 240
cgccacaucu ggcccgcaca ucugcgcugc cggcccggcg cgggguccgg agagggcgcg 300
gcgcggaggc gcagccaggg guccgggaag gcgccguccg cugcgcuggg ggcucggucu 360
augacgagca gcggggucug ccaugggucg ggggcugcuc aggggccugu ggccgcugca 420
caucguccug uggacgcgua ucgccagcac gaucccaccg cacguucaga agucgguuaa 480
uaacgacaug auagucacug acaacaacgg ugcagucaag uuuccacaac uguguaaauu 540
uugugaugug agauuuucca ccugugacaa ccagaaaucc ugcaugagca acugcagcau 600
caccuccauc ugugagaagc cacaggaagu cuguguggcu guauggagaa agaaugacga 660
gaacauaaca cuagagacag uuugccauga ccccaagcuc cccuaccaug acuuuauucu 720
ggaagaugcu gcuucuccaa agugcauuau gaaggaaaaa aaaaagccug gugagacuuu 780
cuucaugugu uccuguagcu cugaugagug caaugacaac aucaucuucu cagaagaaua 840
uaacaccagc aauccugacu uguugcuagu cauauuucaa gugacaggca ucagccuccu 900
gccaccacug ggaguugcca uaucugucau caucaucuuc uacugcuacc gcguuaaccg 960
gcagcagaag cugaguucaa ccugggaaac cggcaagacg cggaagcuca uggaguucag 1020
cgagcacugu gccaucaucc uggaagauga ccgcucugac aucagcucca cgugugccaa 1080
caacaucaac cacaacacag agcugcugcc cauugagcug gacacccugg uggggaaagg 1140
ucgcuuugcu gaggucuaua aggccaagcu gaagcagaac acuucagagc aguuugagac 1200
aguggcaguc aagaucuuuc ccuaugagga guaugccucu uggaagacag agaaggacau 1260
cuucucagac aucaaucuga agcaugagaa cauacuccag uuccugacgg cugaggagcg 1320
gaagacggag uuggggaaac aauacuggcu gaucaccgcc uuccacgcca agggcaaccu 1380
acaggaguac cugacgcggc augucaucag cugggaggac cugcgcaagc ugggcagcuc 1440
ccucgcccgg gggauugcuc accuccacag ugaucacacu ccauguggga ggcccaagau 1500
gcccaucgug cacagggacc ucaagagcuc caauauccuc gugaagaacg accuaaccug 1560
cugccugugu gacuuugggc uuucccugcg ucuggacccu acucugucug uggaugaccu 1620
ggcuaacagu gggcaggugg gaacugcaag auacauggcu ccagaagucc uagaauccag 1680
gaugaauuug gagaauguug aguccuucaa gcagaccgau gucuacucca uggcucuggu 1740
gcucugggaa augacaucuc gcuguaaugc agugggagaa guaaaagauu augagccucc 1800
auuugguucc aaggugcggg agcaccccug ugucgaaagc augaaggaca acguguugag 1860
agaucgaggg cgaccagaaa uucccagcuu cuggcucaac caccagggca uccagauggu 1920
gugugagacg uugacugagu gcugggacca cgacccagag gcccgucuca cagcccagug 1980
uguggcagaa cgcuucagug agcuggagca ucuggacagg cucucgggga ggagcugcuc 2040
ggaggagaag auuccugaag acggcucccu aaacacuacc aaauagcucu ucuggggcag 2100
gcugggccau guccaaagag gcugccccuc ucaccaaaga acagaggcag caggaagcug 2160
ccccugaacu gaugcuuccu ggaaaaccaa gggggucacu ccccucccug uaagcugugg 2220
ggauaagcag aaacaacagc agcagggagu gggugacaua gagcauucua ugccuuugac 2280
auugucauag gauaagcugu guuagcacuu ccucaggaaa ugagauugau uuuuacaaua 2340
gccaauaaca uuugcacuuu auuaaugccu guauauaaau augaauagcu auguuuuaua 2400
uauauauaua uauaucuaua uaugucuaua gcucuauaua uauagccaua ccuugaaaag 2460
agacaaggaa aaacaucaaa uauucccagg aaauugguuu uauuggagaa cuccagaacc 2520
aagcagagaa ggaagggacc caugacagca uuagcauuug acaaucacac augcaguggu 2580
ucucugacug uaaaacagug aacuuugcau gaggaaagag gcuccauguc ucacagccag 2640
cuaugaccac auugcacuug cuuuugcaaa auaaucauuc ccugccuagc acuucucuuc 2700
uggccaugga acuaaguaca guggcacugu uugaggacca guguucccgg gguuccugug 2760
ugcccuuauu ucuccuggac uuuucauuua agcuccaagc cccaaaucug gggggcuagu 2820
uuagaaacuc ucccucaacc uaguuuagaa acucuacccc aucuuuaaua ccuugaaugu 2880
uuugaacccc acuuuuuacc uucauggguu gcagaaaaau cagaacagau guccccaucc 2940
augcgauugc cccaccaucu acuaaugaaa aauuguucuu uuuuucaucu uuccccugca 3000
cuuauguuac uauucucugc ucccagccuu cauccuuuuc uaaaaaggag caaauucuca 3060
cucuaggcuu uaucguguuu acuuuuucau uacacuugac uugauuuucu aguuuucuau 3120
acaaacacca auggguucca ucuuucuggg cuccugauug cucaagcaca guuuggccug 3180
augaagagga uuucaacuac acaauacuau cauugucagg acuaugaccu caggcacucu 3240
aaacauaugu uuuguuuggu cagcacagcg uuucaaaaag ugaagccacu uuauaaauau 3300
uuggagauuu ugcaggaaaa ucuggauccc cagguaagga uagcagaugg uuuucaguua 3360
ucuccagucc acguucacaa aaugugaagg uguggagaca cuuacaaagc ugccucacuu 3420
cucacuguaa acauuagcuc uuuccacugc cuaccuggac cccagucuag gaauuaaauc 3480
ugcaccuaac caaggucccu uguaagaaau guccauucaa gcagucauuc ucuggguaua 3540
uaauaugauu uugacuaccu uaucuggugu uaagauuuga aguuggccuu uuauuggacu 3600
aaaggggaac uccuuuaagg gucucaguua gcccaaguuu cuuuugcuua uauguuaaua 3660
guuuuacccu cugcauugga gagaggagug cuuuacucca agaagcuuuc cucaugguua 3720
ccguucucuc caucaugcca gccuucucaa ccuuugcaga aauuacuaga gaggauuuga 3780
augugggaca caaagguccc auuugcaguu agaaaauuug uguccacaag gacaagaaca 3840
aaguaugagc uuuaaaacuc cauaggaaac uuguuaauca acaaagaagu guuaaugcug 3900
caaguaaucu cuuuuuuaaa acuuuuugaa gcuacuuauu uucagccaaa uaggaauauu 3960
agagagggac ugguagugag aauaucagcu cuguuuggau gguggaaggu cucauuuuau 4020
ugagauuuuu aagauacaug caaagguuug gaaauagaac cucuaggcac ccuccucagu 4080
gugggugggc ugagaguuaa agacagugug gcugcaguag cauagaggcg ccuagaaauu 4140
ccacuugcac cguagggcau gcugauacca ucccaauagc uguugcccau ugaccucuag 4200
uggugaguuu cuagaauacu gguccauuca ugagauauuc aagauucaag aguauucuca 4260
cuucuggguu aucagcauaa acuggaaugu agugucagag gauacugugg cuuguuuugu 4320
uuauguuuuu uuuucuuauu caagaaaaaa gaccaaggaa uaacauucug uaguuccuaa 4380
aaauacugac uuuuuucacu acuauacaua aagggaaagu uuuauucuuu uauggaacac 4440
uucagcugua cucauguauu aaaauaggaa ugugaaugcu auauacucuu uuuauaucaa 4500
aagucucaag cacuuauuuu uauucuaugc auuguuuguc uuuuacauaa auaaaauguu 4560
uauuagauug aauaaagcaa aauacucagg ugagcauccu gccuccuguu cccauuccua 4620
guagcuaaa 4629
<210> 4
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 4
tggtccattc 10
<210> 5
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223 Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 5
ccctaaacac 10
<210> 6
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 6
actaccaaat 10
<210> 7
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 7
ggacgcgtat 10
<210> 8
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 8
gtctatgacg 10
<210> 9
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 9
ttattaatgc 10
<210> 10
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S3
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'TGCCCCAGAAGAGCTATTTGGTAG'3 или последовательности, полученные из 5'TGCCCCAGAAGAGCTATTTGGTAG'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере,G
<220>
<221> misc_feature
<222> (10)..(10)
<223> n может представлять 5'GAGCCGTCTTCAGGAATCTTCTCC'3 или последовательности, полученные из 5'GAGCCGTCTTCAGGAATCTTCTCC'3, где один или более нуклеотидов удалены с 3'-конца, и где n представляет, по меньшей мере,G
<400> 10
ntgtttaggn 10
<210> 11
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S4
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'GCCCAGCCTGCCCCAGAAGAGCTA'3 или последовательности, полученные из 5'GCCCAGCCTGCCCCAGAAGAGCTA'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, A
<220>
<221> misc_feature
<222> (10)..(10)
<223> n может представлять 5'TGTTTAGGGAGCCGTCTTCAGGAA'3 или последовательности, полученные из 5'TGTTTAGGGAGCCGTCTTCAGGAA'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, T
<400> 11
ntttggtagn 10
<210> 12
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S1
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'CATGGCAGACCCCGCTGCTC'3 или последовательности, полученные из 5'CATGGCAGACCCCGCTGCTC'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, C
<220>
<221> misc_feature
<222> (10)..(10)
<223> n может представлять 5'CCGAGCCCCCAGCGCAGCGG'3 или последовательности, полученные из 5'CCGAGCCCCCAGCGCAGCGG'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, C
<400> 12
ngtcatagan 10
<210> 13
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 13
gctcgtcata gaccga 16
<210> 14
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 14
cgatacgcgt ccacag 16
<210> 15
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 15
gtagtgttta gggagc 16
<210> 16
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 16
gctatttggt agtgtt 16
<210> 17
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223 Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 17
catgaatgga ccagta 16
<210> 18
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 18
aggcattaat aaagtg 16
<210> 19
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 19
ccgctgctcg tcatagac 18
<210> 20
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 20
cgctgctcgt catagacc 18
<210> 21
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 21
gctgctcgtc atagaccg 18
<210> 22
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 22
ctgctcgtca tagaccga 18
<210> 23
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 23
tgctcgtcat agaccgag 18
<210> 24
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 24
gctcgtcata gaccgagc 18
<210> 25
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 25
ctcgtcatag accgagcc 18
<210> 26
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 26
tcgtcataga ccgagccc 18
<210> 27
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 27
cgtcatagac cgagcccc 18
<210> 28
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 28
cgctgctcgt catagac 17
<210> 29
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 29
gctgctcgtc atagacc 17
<210> 30
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 30
ctgctcgtca tagaccg 17
<210> 31
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 31
tgctcgtcat agaccga 17
<210> 32
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 32
gctcgtcata gaccgag 17
<210> 33
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 33
ctcgtcatag accgagc 17
<210> 34
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 34
tcgtcataga ccgagcc 17
<210> 35
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 35
cgtcatagac cgagccc 17
<210> 36
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 36
gctgctcgtc atagac 16
<210> 37
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 37
ctgctcgtca tagacc 16
<210> 38
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 38
tgctcgtcat agaccg 16
<210> 39
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 39
gctcgtcata gaccga 16
<210> 40
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 40
ctcgtcatag accgag 16
<210> 41
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 41
tcgtcataga ccgagc 16
<210> 42
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 42
cgtcatagac cgagcc 16
<210> 43
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 43
ctgctcgtca tagac 15
<210> 44
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 44
tgctcgtcat agacc 15
<210> 45
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 45
gctcgtcata gaccg 15
<210> 46
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 46
ctcgtcatag accga 15
<210> 47
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 47
tcgtcataga ccgag 15
<210> 48
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 48
cgtcatagac cgagc 15
<210> 49
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 49
tgctcgtcat agac 14
<210> 50
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 50
gctcgtcata gacc 14
<210> 51
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 51
ctcgtcatag accg 14
<210> 52
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 52
tcgtcataga ccga 14
<210> 53
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 53
cgtcatagac cgag 14
<210> 54
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 54
gctggcgata cgcgtcca 18
<210> 55
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 55
ctggcgatac gcgtccac 18
<210> 56
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 56
tggcgatacg cgtccaca 18
<210> 57
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 57
ggcgatacgc gtccacag 18
<210> 58
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 58
gcgatacgcg tccacagg 18
<210> 59
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 59
cgatacgcgt ccacagga 18
<210> 60
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 60
gatacgcgtc cacaggac 18
<210> 61
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 61
atacgcgtcc acaggacg 18
<210> 62
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 62
tacgcgtcca caggacga 18
<210> 63
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 63
ctggcgatac gcgtcca 17
<210> 64
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 64
tggcgatacg cgtccac 17
<210> 65
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 65
ggcgatacgc gtccaca 17
<210> 66
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 66
gcgatacgcg tccacag 17
<210> 67
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 67
cgatacgcgt ccacagg 17
<210> 68
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223 Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 68
gatacgcgtc cacagga 17
<210> 69
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая последовательность
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'CATGGCAGACCCCGCTGCT'3 или последовательности, полученные из 5'CATGGCAGACCCCGCTGCT'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, T
<220>
<221> misc_feature
<222> (10)..(10)
<223> n может представлять 5'CGAGCCCCCAGCGCAGCGG'3 или последовательности, полученные из 5'CGAGCCCCCAGCGCAGCGG'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, C
<220>
<221> misc_feature
<222> (12)..(12)
<223> n представляет a, c, g, or t
<400> 69
ncgtcataga cn 12
<210> 70
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'GGTGGGATCGTGCTGGCGA'3 или последовательности, полученные из 5'GGTGGGATCGTGCTGGCGA'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, A
<220>
<221> misc_feature
<222> (12)..(12)
<223> n может представлять 5'CAGGACGATGTGCAGCGGC'3 или последовательности, полученные из 5'CAGGACGATGTGCAGCGGC'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, C
<400> 70
ntacgcgtcc an 12
<210> 71
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая последовательность
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'TGCCCCAGAAGAGCTATTTGGTA'3 или последовательности, полученные из 5'TGCCCCAGAAGAGCTATTTGGTA'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, A
<220>
<221> misc_feature
<222> (12)..(12)
<223> n может представлять 5'AGCCGTCTTCAGGAATCTTCTCC'3 или последовательности, полученные из 5'AGCCGTCTTCAGGAATCTTCTCC'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, A
<400> 71
ngtgtttagg gn 12
<210> 72
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая последовательность
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'GCCCAGCCTGCCCCAGAAGAGCT'3 или последовательности, полученные из 5'GCCCAGCCTGCCCCAGAAGAGCT'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, T
<220>
<221> misc_feature
<222> (12)..(12)
<223> n может представлять 5'GTTTAGGGAGCCGTCTTCAGGAA'3 или последовательности, полученные из 5'GTTTAGGGAGCCGTCTTCAGGAA'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, G
<400> 72
natttggtag tn 12
<210> 73
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая последовательность
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'TGAATCTTGAATATCTCAT'3 или последовательности, полученные из 5'TGAATCTTGAATATCTCAT'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, T
<220>
<221> misc_feature
<222> (12)..(12)
<223> n может представлять 5'GTATTCTAGAAACTCACCA'3 или последовательности, полученные из 5'GTATTCTAGAAACTCACCA'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, G
<400> 73
ngaatggacc an 12
<210> 74
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая последовательность
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'ATTCATATTTATATACAGG'3 или последовательности, полученные из 5'ATTCATATTTATATACAGG'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, G
<220>
<221> misc_feature
<222> (12)..(12)
<223> n может представлять 5'GTGCAAATGTTATTGGCTA'3 или последовательности, полученные из 5'GTGCAAATGTTATTGGCTA'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, G
<400> 74
ncattaataa an 12
<210> 75
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 75
gaatcttgaa tatctcatga atggaccagt attctagaaa c 41
<210> 76
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид (скремблированный контроль)
<400> 76
aacacgtcta tacgc 15
<210> 77
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 77
ttcatattta tatacaggca ttaataaagt gcaaatgtta t 41
<210> 78
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 78
tgaggaagtg ctaacacagc ttatcctatg acaatgtcaa ag 42
<210> 79
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 79
gcctgcccca gaagagctat ttggtagtgt ttagggagcc gtcttcagg 49
<210> 80
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 80
ttgaatatct catgaatgga 20
<210> 81
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 81
cgcaggtcct cccagctgat gacatgccgc gtcaggtact cctgtaggt 49
<210> 82
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 82
cagaagagct atttggtagt 20
<210> 83
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 83
atgtcgttat taaccgactt ctgaacgtgc ggtgggatcg tgctggcgat acgcgtccac 60
aggacgatgt gcagcggc 78
<210> 84
<211> 74
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 84
ggccacaggc ccctgagcag cccccgaccc atggcagacc ccgctgctcg tcatagaccg 60
agcccccagc gcag 74
<210> 85
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 85
tggtagtgtt tagggagccg 20
<210> 86
<211> 149
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 86
atgtcgttat taaccgactt ctgaacgtgc ggtgggatcg tgctggcgat acgcgtccac 60
aggacgatgt gcagcggcca caggcccctg agcagccccc gacccatggc agaccccgct 120
gctcgtcata gaccgagccc ccagcgcag 149
<210> 87
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 87
ttgaatatct catgaatgga ccagtattct a 31
<210> 88
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 88
caagtggaat ttctaggcgc ctctatgcta ctg 33
<210> 89
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 89
atttatatac aggcattaat aaagtgcaaa t 31
<210> 90
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 90
aagtgctaac acagcttatc ctatgacaat gt 32
<210> 91
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 91
ccccagaaga gctatttggt agtgtttagg gagccgtct 39
<210> 92
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 92
ctggtcgccc tcgatctctc aacacgttgt ccttcatgct ttcgacacag gggtgctccc 60
gcaccttgga accaaatg 78
<210> 93
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 93
gtcctcccag ctgatgacat gccgcgtcag gtactcctg 39
<210> 94
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 94
ctcagcttct gctgccggtt aacgcggtag cagtagaaga 40
<210> 95
<211> 68
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 95
gttattaacc gacttctgaa cgtgcggtgg gatcgtgctg gcgatacgcg tccacaggac 60
gatgtgca 68
<210> 96
<211> 64
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 96
caggcccctg agcagccccc gacccatggc agaccccgct gctcgtcata gaccgagccc 60
ccag 64
<210> 97
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 97
cacgcgcggg ggtgtcgtcg ctccgtgcgc gcgagtgact cactcaactt ca 52
<210> 98
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S2
<220>
<221> misc_feature
<222> (1)..(1)
<223> w может представлять 5'GGTGGGATCGTGCTGGCGAT'3 или последовательности, полученные из 5'GGTGGGATCGTGCTGGCGAT'3, где один или более нуклеотидов удалены с 5'-конца и где w представляет, по меньшей мере, T
<220>
<221> misc_feature
<222> (10)..(10)
<223> r может представлять 5'ACAGGACGATGTGCAGCGGC'3 или последовательности, полученные из 5'ACAGGACGATGTGCAGCGGC'3, где один или более нуклеотидов удалены с 3'-конца и где r представляет, по меньшей мере, A
<400> 98
nacgcgtccn 10
<210> 99
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S5
<220>
<221> misc_feature
<222> (1)..(1)
<223> w может представлять 5'CCCAGCCTGCCCCAGAAGAGCTATTTG'3 или последовательности, полученные из 5'CCCAGCCTGCCCCAGAAGAGCTATTTG'3, где один или более нуклеотидов удалены с 5'-конца и где w представляет, по меньшей мере, G
<220>
<221> misc_feature
<222> (10)..(10)
<223> r может представлять 5'TAGGGAGCCGTCTTCAGGAATCTTCTC'3 или последовательности, полученные из 5'TAGGGAGCCGTCTTCAGGAATCTTCTC'3, где один или более нуклеотидов удалены с 3'-конца и где r представляет, по меньшей мере, G
<400> 99
ngtagtgttn 10
<210> 100
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S6
<220>
<221> misc_feature
<222> (1)..(1)
<223> w может представлять 5'TGAATCTTGAATATCTCATG'3 или последовательности, полученные из 5'TGAATCTTGAATATCTCATG'3, где один или более нуклеотидов удалены с 5'-конца и где w представляет, по меньшей мере, G
<220>
<221> misc_feature
<222> (1)..(1)
<223> n может представлять 5'TGAATCTTGAATATCTCATG'3 или последовательности, полученные из 5'TGAATCTTGAATATCTCATG'3, где один или более нуклеотидов удалены с 5'-конца и где n представляет, по меньшей мере, G
<220>
<221> misc_feature
<222> (10)..(10)
<223> r может представлять 5'AGTATTCTAGAAACTCACCA'3 или последовательности, полученные из 5'AGTATTCTAGAAACTCACCA'3, где один или более нуклеотидов удалены с 3'-конца и где r представляет, по меньшей мере, A
<400> 100
naatggaccn 10
<210> 101
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Общая формула S7
<220>
<221> misc_feature
<222> (1)..(1)
<223> w может представлять 5'ATTCATATTTATATACAGGC'3 или последовательности, полученные из 5'ATTCATATTTATATACAGGC'3, где один или более нуклеотидов удалены с 5'-конца и где w представляет, по меньшей мере, C
<220>
<221> misc_feature
<222> (10)..(10)
<223> r может представлять 5'AGTGCAAATGTTATTGGCTA'3 или последовательности, полученные из 5'AGTGCAAATGTTATTGGCTA'3, где один или более нуклеотидов удалены с 3'-конца и где r представляет, по меньшей мере, A
<220>
<221> misc_feature
<222> (10)..(10)
<223> n может представлять 5'AGTGCAAATGTTATTGGCTA'3 или последовательности, полученные из 5'AGTGCAAATGTTATTGGCTA'3, где один или более нуклеотидов удалены с 3'-конца и где n представляет, по меньшей мере, A
<400> 101
nattaataan 10
<210> 102
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 102
gcgagtgact cactcaa 17
<210> 103
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 103
cgagtgactc actca 15
<210> 104
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 104
gcgagtgact cactca 16
<210> 105
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 105
cgcgagtgac tcactca 17
<210> 106
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 106
cgagtgactc actc 14
<210> 107
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 107
cgcgagtgac tcactc 16
<210> 108
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 108
gcgcgagtga ctcactc 17
<210> 109
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 109
gcgagtgact cact 14
<210> 110
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 110
gcgcgagtga ctcact 16
<210> 111
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 111
cgcgcgagtg actcact 17
<210> 112
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 112
cgagtgactc ac 12
<210> 113
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 113
gcgagtgact cac 13
<210> 114
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 114
cgcgagtgac tcac 14
<210> 115
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 115
cgcgcgagtg actcac 16
<210> 116
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 116
gcgcgcgagt gactcac 17
<210> 117
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 117
cgcgagtgac tca 13
<210> 118
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 118
gcgcgagtga ctca 14
<210> 119
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 119
cgcgcgagtg actca 15
<210> 120
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 120
gcgcgcgagt gactca 16
<210> 121
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 121
tgcgcgcgag tgactca 17
<210> 122
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 122
cgcgcgagtg actc 14
<210> 123
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 123
tgcgcgcgag tgactc 16
<210> 124
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 124
gtgcgcgcga gtgactc 17
<210> 125
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 125
cgcgcgagtg act 13
<210> 126
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 126
tgcgcgcgag tgac 14
<210> 127
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 127
cgtgcgcgcg agtgac 16
<210> 128
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 128
tgcgcgcgag tga 13
<210> 129
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 129
gtcgtcgctc cgtgcg 16
<210> 130
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 130
gtcgtcgctc cgtgc 15
<210> 131
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 131
gtgtcgtcgc tccgtgc 17
<210> 132
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 132
tcgtcgctcc gtg 13
<210> 133
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 133
tgtcgtcgct ccgtg 15
<210> 134
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 134
tcgtcgctcc gt 12
<210> 135
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 135
gtcgtcgctc cgt 13
<210> 136
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 136
tgtcgtcgct ccgt 14
<210> 137
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 137
gtgtcgtcgc tccgt 15
<210> 138
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 138
ggtgtcgtcg ctccgt 16
<210> 139
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 139
cgtcatagac cgagcc 16
<210> 140
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 140
atagaccgag cc 12
<210> 141
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 141
gctcgtcata gaccga 16
<210> 142
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 142
cgtcatagac cga 13
<210> 143
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 143
ctcgtcatag accg 14
<210> 144
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 144
gctcgtcata gaccg 15
<210> 145
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 145
gctcgtcata gacc 14
<210> 146
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 146
cagcccccga cccatgg 17
<210> 147
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 147
cagcccccga cccatg 16
<210> 148
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 148
agcccccgac ccat 14
<210> 149
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 149
cagcccccga cccaagcccc cgacccat 28
<210> 150
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 150
cgcgtccaca ggacgat 17
<210> 151
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 151
cgcgtccaca ggac 14
<210> 152
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 152
cgatacgcgt ccaca 15
<210> 153
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 153
cgatacgcgt cca 13
<210> 154
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 154
tggcgatacg cgtcca 16
<210> 155
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 155
cgatacgcgt cc 12
<210> 156
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 156
gcgatacgcg tcc 13
<210> 157
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 157
gctggcgata cgcgtcc 17
<210> 158
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 158
ctggcgatac gcgtc 15
<210> 159
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 159
gcgatacgcg tc 12
<210> 160
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 160
gctggcgata cgcgtc 16
<210> 161
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 161
tggcgatacg cgtc 14
<210> 162
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 162
tggcgatacg cgt 13
<210> 163
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 163
ctggcgatac gcgt 14
<210> 164
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 164
ggcgatacgc gt 12
<210> 165
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 165
ctggcgatac gcg 13
<210> 166
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 166
tggcgatacg cg 12
<210> 167
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 167
atcgtgctgg cgatacg 17
<210> 168
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 168
cgtgcggtgg gatcgt 16
<210> 169
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 169
acgtgcggtg ggatcgt 17
<210> 170
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 170
aacgtgcggt gggatcg 17
<210> 171
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 171
aacgtgcggt gggat 15
<210> 172
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 172
tgaacgtgcg gtgggat 17
<210> 173
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 173
cgacttctga acgtgcg 17
<210> 174
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 174
ttaacgcggt agcagta 17
<210> 175
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 175
taacgcggta gcagta 16
<210> 176
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 176
gttaacgcgg tagcagt 17
<210> 177
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 177
ttaacgcggt agcag 15
<210> 178
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 178
taacgcggta gca 13
<210> 179
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 179
taacgcggta gc 12
<210> 180
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 180
ttaacgcggt agc 13
<210> 181
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 181
ttaacgcggt ag 12
<210> 182
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 182
gttaacgcgg tag 13
<210> 183
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 183
cggttaacgc ggtag 15
<210> 184
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 184
ccggttaacg cggtag 16
<210> 185
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 185
cggttaacgc ggta 14
<210> 186
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 186
ggttaacgcg gta 13
<210> 187
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 187
ccggttaacg cggta 15
<210> 188
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 188
gccggttaac gcggta 16
<210> 189
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 189
tgccggttaa cgcggta 17
<210> 190
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 190
cggttaacgc ggt 13
<210> 191
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 191
ccggttaacg cgg 13
<210> 192
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 192
gccggttaac gcgg 14
<210> 193
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 193
tgccggttaa cgcgg 15
<210> 194
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 194
gccggttaac gcg 13
<210> 195
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 195
ctgccggtta acgcg 15
<210> 196
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 196
gctgccggtt aacgcg 16
<210> 197
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 197
atgccgcgtc aggtac 16
<210> 198
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 198
acatgccgcg tca 13
<210> 199
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 199
gatgacatgc cgcgtc 16
<210> 200
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 200
gacatgccgc gt 12
<210> 201
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 201
gatgacatgc cgcgt 15
<210> 202
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 202
atgacatgcc gcg 13
<210> 203
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 203
tcccgcacct tggaacc 17
<210> 204
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 204
cgatctctca acacgt 16
<210> 205
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 205
tcgatctctc aacacgt 17
<210> 206
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 206
cgatctctca acacg 15
<210> 207
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 207
tcgatctctc aacacg 16
<210> 208
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 208
ctcgatctct caacacg 17
<210> 209
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 209
gtagtgttta gggagc 16
<210> 210
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 210
gctatttggt agtgtt 16
<210> 211
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 211
agcttatcct atgac 15
<210> 212
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 212
agcttatcct atga 14
<210> 213
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 213
caggcattaa taaagtg 17
<210> 214
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 214
ctaggcgcct ctatgc 16
<210> 215
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 215
taggcgcctc tatg 14
<210> 216
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 216
ctaggcgcct ctatg 15
<210> 217
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 217
taggcgcctc tat 13
<210> 218
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 218
catgaatgga ccagta 16
<210> 219
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 219
gaatggacca 10
<210> 220
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 220
tgaatggacc ag 12
<210> 221
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 221
tgaatggacc agt 13
<210> 222
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 222
atgaatggac cagt 14
<210> 223
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 223
atgaatggac cagta 15
<210> 224
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 224
catgaatgga ccagtat 17
<210> 225
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 225
tcatgaatgg accagtat 18
<210> 226
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 226
tcatgaatgg accagtatt 19
<210> 227
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 227
ctcatgaatg gaccagtatt 20
<210> 228
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 228
tctcatgaat ggaccagtat tc 22
<210> 229
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 229
atctcatgaa tggaccagta ttct 24
<210> 230
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 230
tatctcatga atggaccagt attcta 26
<210> 231
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 231
atatctcatg aatggaccag tattctag 28
<210> 232
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 232
cgtcatagac 10
<210> 233
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 233
tcgtcataga cc 12
<210> 234
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 234
tcgtcataga ccg 13
<210> 235
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 235
ctcgtcatag accga 15
<210> 236
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 236
atatacaggc attaataaag tgcaaatg 28
<210> 237
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 237
tgctcgtcat agaccga 17
<210> 238
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 238
tgctcgtcat agaccgag 18
<210> 239
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 239
tgctcgtcat agaccgagc 19
<210> 240
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 240
ctgctcgtca tagaccgagc 20
<210> 241
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 241
gctgctcgtc atagaccgag cc 22
<210> 242
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 242
cgctgctcgt catagaccga gccc 24
<210> 243
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 243
ccgctgctcg tcatagaccg agcccc 26
<210> 244
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 244
cccgctgctc gtcatagacc gagccccc 28
<210> 245
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 245
tacgcgtcca 10
<210> 246
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 246
atacgcgtcc ac 12
<210> 247
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 247
gatacgcgtc cac 13
<210> 248
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 248
gatacgcgtc caca 14
<210> 249
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 249
cgatacgcgt ccacag 16
<210> 250
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 250
gcgatacgcg tccacag 17
<210> 251
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 251
gcgatacgcg tccacagg 18
<210> 252
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 252
gcgatacgcg tccacagga 19
<210> 253
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 253
ggcgatacgc gtccacagga 20
<210> 254
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 254
tggcgatacg cgtccacagg ac 22
<210> 255
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 255
ctggcgatac gcgtccacag gacg 24
<210> 256
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 256
gctggcgata cgcgtccaca ggacga 26
<210> 257
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 257
tgctggcgat acgcgtccac aggacgat 28
<210> 258
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 258
gtgtttaggg 10
<210> 259
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 259
agtgtttagg ga 12
<210> 260
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 260
tagtgtttag gga 13
<210> 261
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 261
tagtgtttag ggag 14
<210> 262
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 262
tagtgtttag ggagc 15
<210> 263
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 263
gtagtgttta gggagcc 17
<210> 264
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 264
ggtagtgttt agggagcc 18
<210> 265
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 265
ggtagtgttt agggagccg 19
<210> 266
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 266
tggtagtgtt tagggagccg 20
<210> 267
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 267
ttggtagtgt ttagggagcc gt 22
<210> 268
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 268
tttggtagtg tttagggagc cgtc 24
<210> 269
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 269
atttggtagt gtttagggag ccgtct 26
<210> 270
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 270
tatttggtag tgtttaggga gccgtctt 28
<210> 271
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 271
atttggtagt 10
<210> 272
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 272
tatttggtag tg 12
<210> 273
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 273
tatttggtag tgt 13
<210> 274
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 274
ctatttggta gtgt 14
<210> 275
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 275
ctatttggta gtgtt 15
<210> 276
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 276
gctatttggt agtgttt 17
<210> 277
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 277
agctatttgg tagtgttt 18
<210> 278
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 278
agctatttgg tagtgttta 19
<210> 279
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 279
gagctatttg gtagtgttta 20
<210> 280
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 280
agagctattt ggtagtgttt ag 22
<210> 281
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 281
aagagctatt tggtagtgtt tagg 24
<210> 282
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 282
gaagagctat ttggtagtgt ttaggg 26
<210> 283
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 283
agaagagcta tttggtagtg tttaggga 28
<210> 284
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 284
cattaataaa 10
<210> 285
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 285
gcattaataa ag 12
<210> 286
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 286
gcattaataa agt 13
<210> 287
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 287
ggcattaata aagt 14
<210> 288
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 288
ggcattaata aagtg 15
<210> 289
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 289
aggcattaat aaagtg 16
<210> 290
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 290
caggcattaa taaagtgc 18
<210> 291
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 291
acaggcatta ataaagtgc 19
<210> 292
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 292
acaggcatta ataaagtgca 20
<210> 293
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 293
tacaggcatt aataaagtgc aa 22
<210> 294
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 294
atacaggcat taataaagtg caaa 24
<210> 295
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 295
tatacaggca ttaataaagt gcaaat 26
<210> 296
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 296
ctggtccatt c 11
<210> 297
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 297
tggtccattc a 11
<210> 298
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 298
ctggtccatt ca 12
<210> 299
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 299
tccctaaaca c 11
<210> 300
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 300
ccctaaacac t 11
<210> 301
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 301
tccctaaaca ct 12
<210> 302
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 302
cactaccaaa t 11
<210> 303
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 303
actaccaaat a 11
<210> 304
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 304
cactaccaaa ta 12
<210> 305
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 305
tggacgcgta t 11
<210> 306
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 306
ggacgcgtat c 11
<210> 307
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 307
tggacgcgta tc 12
<210> 308
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 308
ggtctatgac g 11
<210> 309
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 309
gtctatgacg a 11
<210> 310
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 310
ggtctatgac ga 12
<210> 311
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 311
tttattaatg c 11
<210> 312
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 312
ttattaatgc c 11
<210> 313
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 313
tttattaatg cc 12
<210> 314
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 314
actggtccat tc 12
<210> 315
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 315
tggtccattc at 12
<210> 316
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 316
ctggtccatt cat 13
<210> 317
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 317
actggtccat tca 13
<210> 318
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 318
actggtccat tcat 14
<210> 319
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 319
ctccctaaac ac 12
<210> 320
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 320
ccctaaacac ta 12
<210> 321
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 321
tccctaaaca cta 13
<210> 322
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 322
ctccctaaac act 13
<210> 323
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 323
ctccctaaac acta 14
<210> 324
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 324
acactaccaa at 12
<210> 325
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 325
actaccaaat ag 12
<210> 326
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 326
cactaccaaa tag 13
<210> 327
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 327
acactaccaa ata 13
<210> 328
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 328
acactaccaa atag 14
<210> 329
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 329
gtggacgcgt at 12
<210> 330
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 330
ggacgcgtat cg 12
<210> 331
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 331
tggacgcgta tcg 13
<210> 332
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 332
gtggacgcgt atc 13
<210> 333
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 333
gtggacgcgt atcg 14
<210> 334
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 334
cggtctatga cg 12
<210> 335
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 335
gtctatgacg ag 12
<210> 336
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 336
ggtctatgac gag 13
<210> 337
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 337
cggtctatga cga 13
<210> 338
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 338
cggtctatga cgag 14
<210> 339
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 339
ctttattaat gc 12
<210> 340
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 340
ttattaatgc ct 12
<210> 341
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 341
tttattaatg cct 13
<210> 342
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 342
ctttattaat gcc 13
<210> 343
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность-мишень в рецепторе II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2, смысловая цепь
<400> 343
ctttattaat gcct 14
<210> 344
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 344
gtgcagggga aagatgaaaa 20
<210> 345
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 345
gagctcttga ggtccctgtg 20
<210> 346
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 346
agcctctttc ctcatgcaaa 20
<210> 347
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 347
ccttctctgc ttggttctgg 20
<210> 348
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Стандартный олигонуклеотид
<400> 348
gccatggagt agacatcggt 20
<210> 349
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 349
atacgcgtcc acaggac 17
<210> 350
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 350
tacgcgtcca caggacg 17
<210> 351
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 351
tggcgatacg cgtcca 16
<210> 352
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 352
ggcgatacgc gtccac 16
<210> 353
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 353
gcgatacgcg tccaca 16
<210> 354
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 354
cgatacgcgt ccacag 16
<210> 355
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 355
gatacgcgtc cacagg 16
<210> 356
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 356
atacgcgtcc acagga 16
<210> 357
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 357
tacgcgtcca caggac 16
<210> 358
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 358
ggcgatacgc gtcca 15
<210> 359
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 359
gcgatacgcg tccac 15
<210> 360
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 360
cgatacgcgt ccaca 15
<210> 361
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 361
gatacgcgtc cacag 15
<210> 362
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 362
atacgcgtcc acagg 15
<210> 363
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 363
tacgcgtcca cagga 15
<210> 364
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 364
gcgatacgcg tcca 14
<210> 365
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 365
cgatacgcgt ccac 14
<210> 366
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 366
gatacgcgtc caca 14
<210> 367
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 367
atacgcgtcc acag 14
<210> 368
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 368
tacgcgtcca cagg 14
<210> 369
<400> 369
000
<210> 370
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 370
tttggtagtg tttaggga 18
<210> 371
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 371
ttggtagtgt ttagggag 18
<210> 372
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 372
tggtagtgtt tagggagc 18
<210> 373
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 373
ggtagtgttt agggagcc 18
<210> 374
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 374
gtagtgttta gggagccg 18
<210> 375
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 375
tagtgtttag ggagccgt 18
<210> 376
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 376
agtgtttagg gagccgtc 18
<210> 377
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 377
gtgtttaggg agccgtct 18
<210> 378
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 378
tttggtagtg tttaggg 17
<210> 379
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 379
ttggtagtgt ttaggga 17
<210> 380
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 380
tggtagtgtt tagggag 17
<210> 381
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 381
ggtagtgttt agggagc 17
<210> 382
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 382
gtagtgttta gggagcc 17
<210> 383
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 383
tagtgtttag ggagccg 17
<210> 384
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 384
agtgtttagg gagccgt 17
<210> 385
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 385
gtgtttaggg agccgtc 17
<210> 386
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 386
ttggtagtgt ttaggg 16
<210> 387
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 387
tggtagtgtt taggga 16
<210> 388
<400> 388
000
<210> 389
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 389
gtagtgttta gggagc 16
<210> 390
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 390
tagtgtttag ggagcc 16
<210> 391
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 391
agtgtttagg gagccg 16
<210> 392
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 392
gtgtttaggg agccgt 16
<210> 393
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 393
tggtagtgtt taggg 15
<210> 394
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 394
ggtagtgttt aggga 15
<210> 395
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 395
gtagtgttta gggag 15
<210> 396
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 396
tagtgtttag ggagc 15
<210> 397
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 397
agtgtttagg gagcc 15
<210> 398
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 398
gtgtttaggg agccg 15
<210> 399
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 399
ggtagtgttt aggg 14
<210> 400
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 400
gtagtgttta ggga 14
<210> 401
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 401
tagtgtttag ggag 14
<210> 402
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 402
agtgtttagg gagc 14
<210> 403
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 403
gtgtttaggg agcc 14
<210> 404
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 404
gaagagctat ttggtagt 18
<210> 405
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 405
aagagctatt tggtagtg 18
<210> 406
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 406
agagctattt ggtagtgt 18
<210> 407
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 407
gagctatttg gtagtgtt 18
<210> 408
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 408
agctatttgg tagtgttt 18
<210> 409
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 409
gctatttggt agtgttta 18
<210> 410
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 410
ctatttggta gtgtttag 18
<210> 411
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 411
tatttggtag tgtttagg 18
<210> 412
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 412
atttggtagt gtttaggg 18
<210> 413
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 413
aagagctatt tggtagt 17
<210> 414
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 414
agagctattt ggtagtg 17
<210> 415
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 415
gagctatttg gtagtgt 17
<210> 416
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 416
agctatttgg tagtgtt 17
<210> 417
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 417
gctatttggt agtgttt 17
<210> 418
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 418
ctatttggta gtgttta 17
<210> 419
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 419
atttggtagt gtttagg 17
<210> 420
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 420
atttggtagt gtttagg 17
<210> 421
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 421
agagctattt ggtagt 16
<210> 422
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 422
gagctatttg gtagtg 16
<210> 423
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 423
agctatttgg tagtgt 16
<210> 424
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 424
gctatttggt agtgtt 16
<210> 425
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 425
ctatttggta gtgttt 16
<210> 426
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 426
tatttggtag tgttta 16
<210> 427
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 427
atttggtagt gtttag 16
<210> 428
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 428
gagctatttg gtagt 15
<210> 429
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 429
agctatttgg tagtg 15
<210> 430
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 430
gctatttggt agtgt 15
<210> 431
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 431
ctatttggta gtgtt 15
<210> 432
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 432
tatttggtag tgttt 15
<210> 433
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 433
atttggtagt gttta 15
<210> 434
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 434
agctatttgg tagt 14
<210> 435
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 435
gctatttggt agtg 14
<210> 436
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 436
ctatttggta gtgt 14
<210> 437
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 437
tatttggtag tgtt 14
<210> 438
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 438
atttggtagt gttt 14
<210> 439
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 439
tatctcatga atggacca 18
<210> 440
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 440
atctcatgaa tggaccag 18
<210> 441
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 441
tctcatgaat ggaccagt 18
<210> 442
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 442
ctcatgaatg gaccagta 18
<210> 443
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 443
tcatgaatgg accagtat 18
<210> 444
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 444
catgaatgga ccagtatt 18
<210> 445
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 445
atgaatggac cagtattc 18
<210> 446
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 446
tgaatggacc agtattct 18
<210> 447
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 447
gaatggacca gtattcta 18
<210> 448
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 448
atctcatgaa tggacca 17
<210> 449
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 449
tctcatgaat ggaccag 17
<210> 450
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 450
ctcatgaatg gaccagt 17
<210> 451
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 451
tcatgaatgg accagta 17
<210> 452
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 452
catgaatgga ccagtat 17
<210> 453
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 453
atgaatggac cagtatt 17
<210> 454
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 454
tgaatggacc agtattc 17
<210> 455
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 455
gaatggacca gtattct 17
<210> 456
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 456
tctcatgaat ggacca 16
<210> 457
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 457
ctcatgaatg gaccag 16
<210> 458
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 458
tcatgaatgg accagt 16
<210> 459
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 459
catgaatgga ccagta 16
<210> 460
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 460
atgaatggac cagtat 16
<210> 461
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 461
tgaatggacc agtatt 16
<210> 462
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 462
gaatggacca gtattc 16
<210> 463
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 463
ctcatgaatg gacca 15
<210> 464
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 464
tcatgaatgg accag 15
<210> 465
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 465
catgaatgga ccagt 15
<210> 466
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 466
atgaatggac cagta 15
<210> 467
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 467
tgaatggacc agtat 15
<210> 468
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 468
gaatggacca gtatt 15
<210> 469
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 469
tcatgaatgg acca 14
<210> 470
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 470
catgaatgga ccag 14
<210> 471
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 471
atgaatggac cagt 14
<210> 472
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 472
tgaatggacc agta 14
<210> 473
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 473
gaatggacca gtat 14
<210> 474
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 474
tatacaggca ttaataaa 18
<210> 475
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 475
atacaggcat taataaag 18
<210> 476
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 476
tacaggcatt aataaagt 18
<210> 477
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 477
acaggcatta ataaagtg 18
<210> 478
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 478
caggcattaa taaagtgc 18
<210> 479
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 479
aggcattaat aaagtgca 18
<210> 480
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 480
ggcattaata aagtgcaa 18
<210> 481
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 481
gcattaataa agtgcaaa 18
<210> 482
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 482
cattaataaa gtgcaaat 18
<210> 483
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 483
atacaggcat taataaa 17
<210> 484
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 484
tacaggcatt aataaag 17
<210> 485
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 485
acaggcatta ataaagt 17
<210> 486
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 486
caggcattaa taaagtg 17
<210> 487
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 487
aggcattaat aaagtgc 17
<210> 488
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 488
ggcattaata aagtgca 17
<210> 489
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 489
gcattaataa agtgcaa 17
<210> 490
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 490
cattaataaa gtgcaaa 17
<210> 491
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 491
tacaggcatt aataaa 16
<210> 492
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 492
acaggcatta ataaag 16
<210> 493
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 493
caggcattaa taaagt 16
<210> 494
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 494
aggcattaat aaagtg 16
<210> 495
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 495
ggcattaata aagtgc 16
<210> 496
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 496
gcattaataa agtgca 16
<210> 497
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 497
cattaataaa gtgcaa 16
<210> 498
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 498
acaggcatta ataaa 15
<210> 499
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 499
caggcattaa taaag 15
<210> 500
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 500
aggcattaat aaagt 15
<210> 501
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 501
ggcattaata aagtg 15
<210> 502
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 502
gcattaataa agtgc 15
<210> 503
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 503
cattaataaa gtgca 15
<210> 504
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 504
caggcattaa taaa 14
<210> 505
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 505
aggcattaat aaag 14
<210> 506
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 506
ggcattaata aagt 14
<210> 507
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 507
gcattaataa agtg 14
<210> 508
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой олигонуклеотид против рецептора II трансформирующего фактора роста бета человека (70/80кДа) (TGFBR2), вариант транскрипта 2
<400> 508
cattaataaa gtgc 14
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ К АЛЬФА-СИНУКЛЕИНУ И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2773197C2 |
| Специфичные для гена амфирегулина двухцепочечные олигонуклеотиды и содержащие их композиции для профилактики и лечения связанных с фиброзом заболеваний и респираторных заболеваний | 2019 |
|
RU2795179C2 |
| ДВУХЦЕПОЧЕЧНЫЙ ОЛИГОНУКЛЕОТИД, СПЕЦИФИЧНЫЙ К ГЕНУ CTGF, И СОДЕРЖАЩАЯ ЕГО КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ФИБРОЗНЫХ ЗАБОЛЕВАНИЙ И ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ДЫХАТЕЛЬНОЙ СИСТЕМОЙ | 2020 |
|
RU2807108C1 |
| ЛЕЧЕНИЕ СВЯЗАННЫХ С ГЕНОМ-СУПРЕССОРОМ ОПУХОЛЕЙ ЗАБОЛЕВАНИЙ ПОСРЕДСТВОМ ИНГИБИРОВАНИЯ ПРИРОДНОГО ТРАНСКРИПТА В АНТИСМЫСЛОВОЙ ОРИЕНТАЦИИ ОТНОСИТЕЛЬНО ЭТОГО ГЕНА | 2009 |
|
RU2746478C2 |
| ОЛИГОНУКЛЕОТИДЫ ДЛЯ ПОНИЖЕНИЯ ЭКСПРЕССИИ PD-L1 | 2017 |
|
RU2747822C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ | 2022 |
|
RU2840656C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2747734C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2777570C2 |
| ОЛИГОНУКЛЕОТИДЫ ДЛЯ ПОНИЖЕНИЯ ЭКСПРЕССИИ PD-L1 | 2017 |
|
RU2835926C1 |
| КОМПОЗИЦИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С ОЖИРЕНИЕМ, СОДЕРЖАЩАЯ АМФИРЕГУЛИН-СПЕЦИФИЧЕСКУЮ ДВУХЦЕПОЧЕЧНУЮ ОЛИГОНУКЛЕОТИДНУЮ СТРУКТУРУ | 2021 |
|
RU2810514C1 |
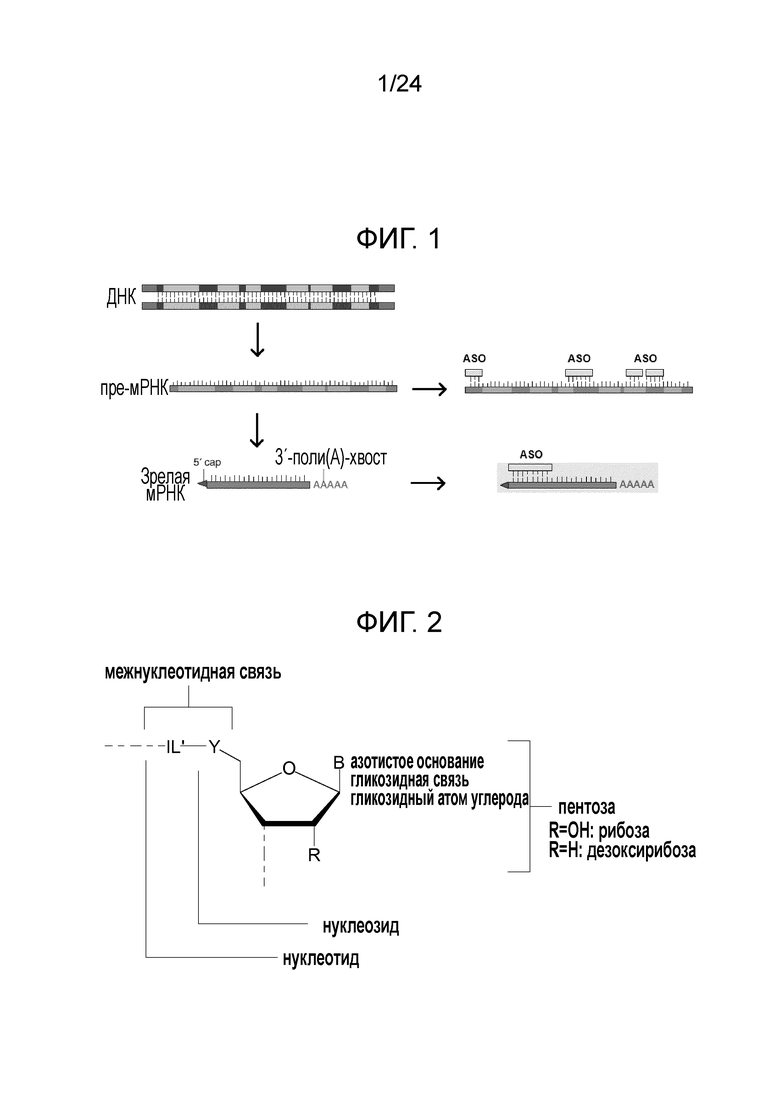
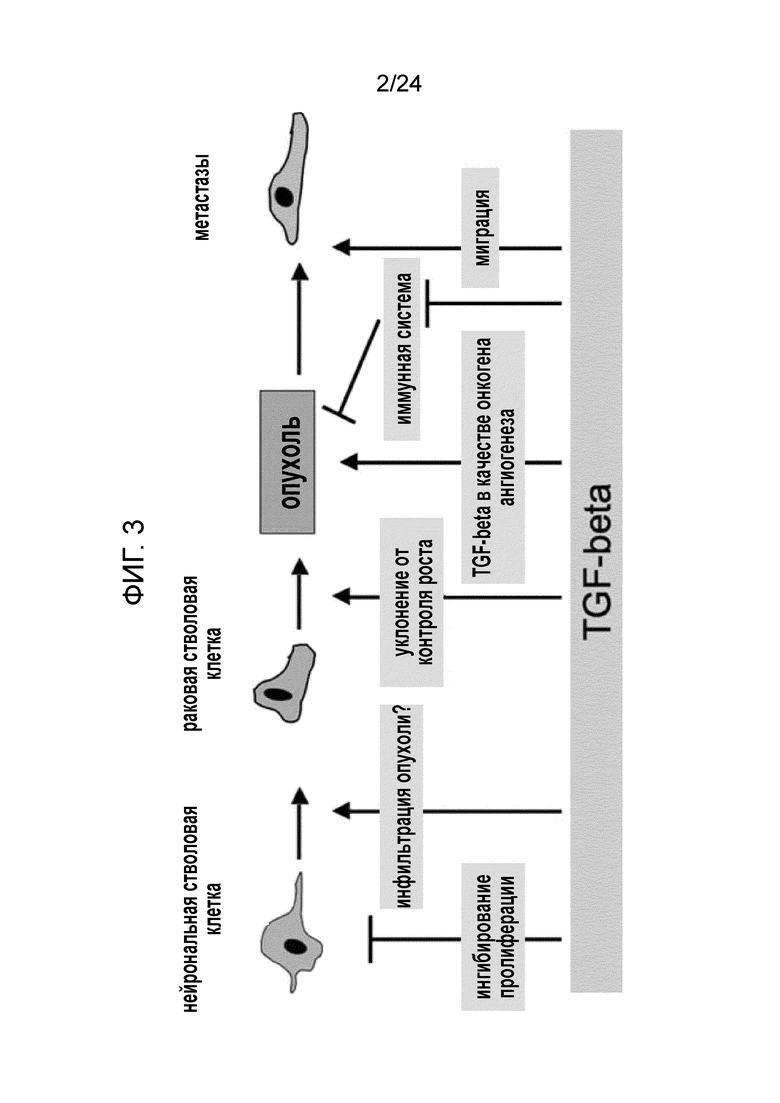
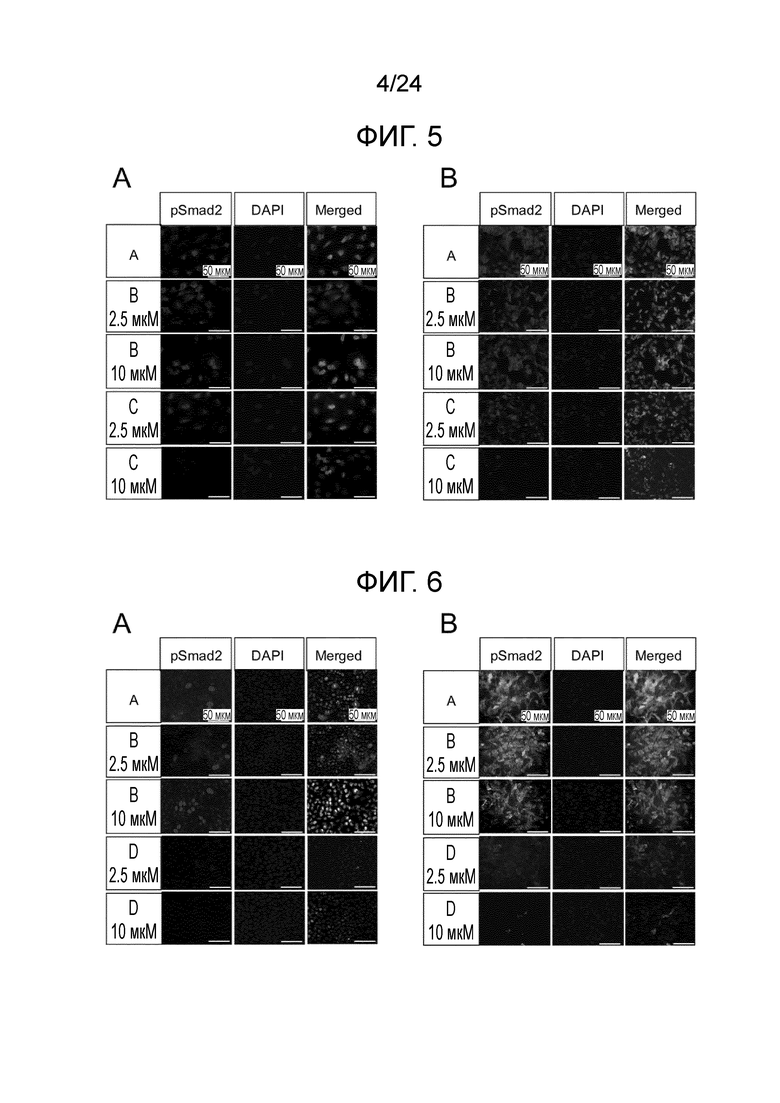
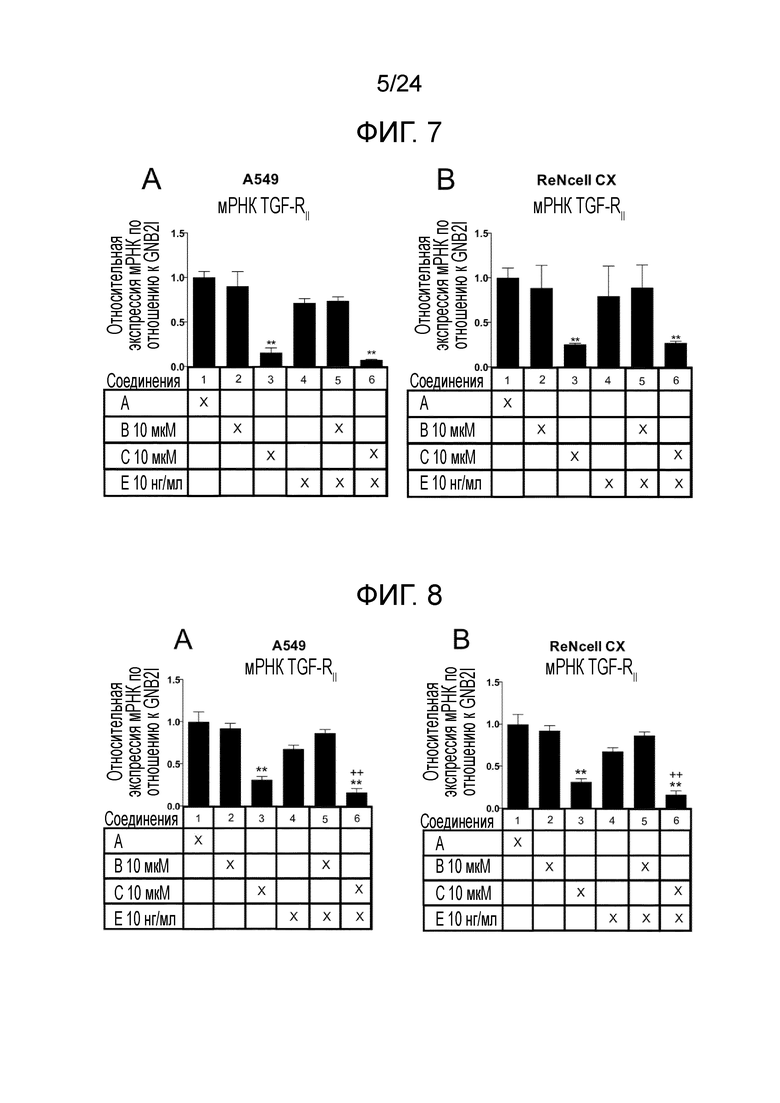
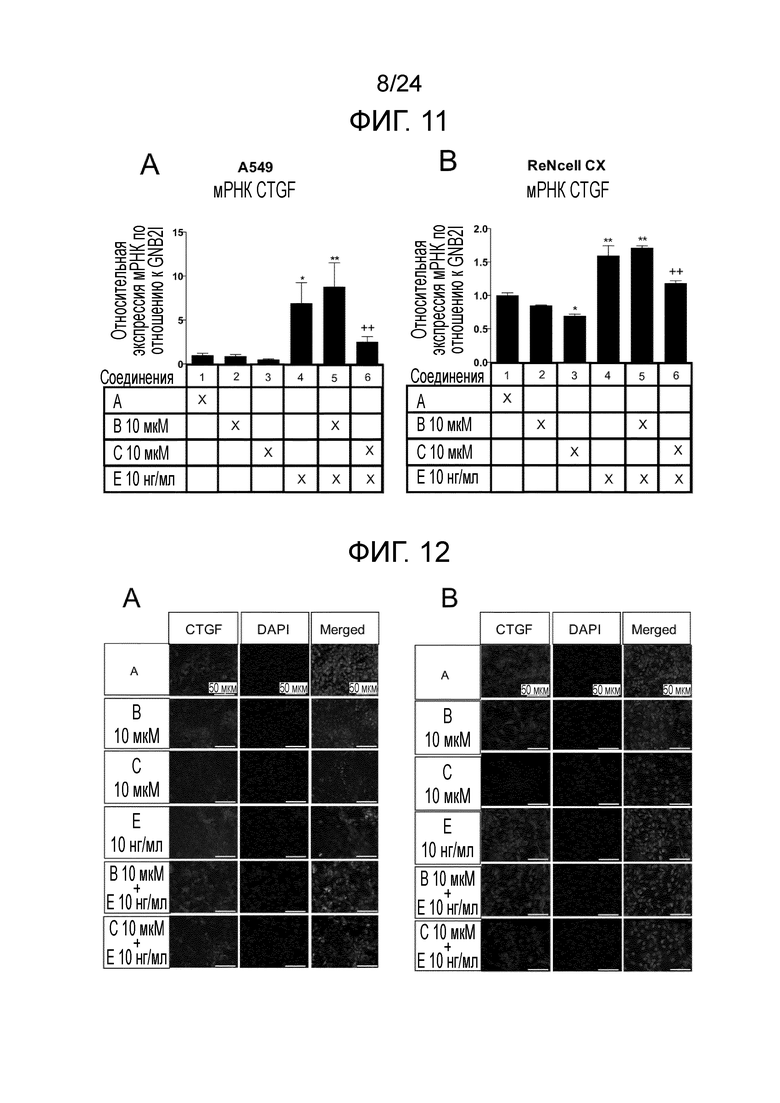
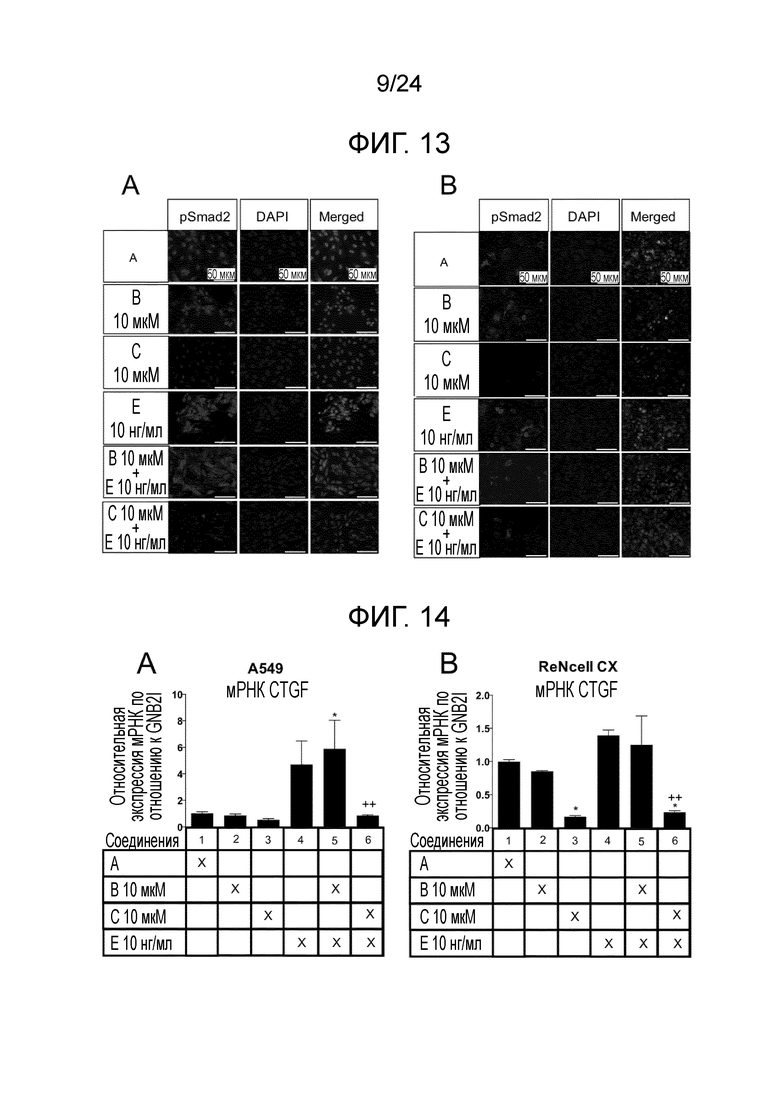

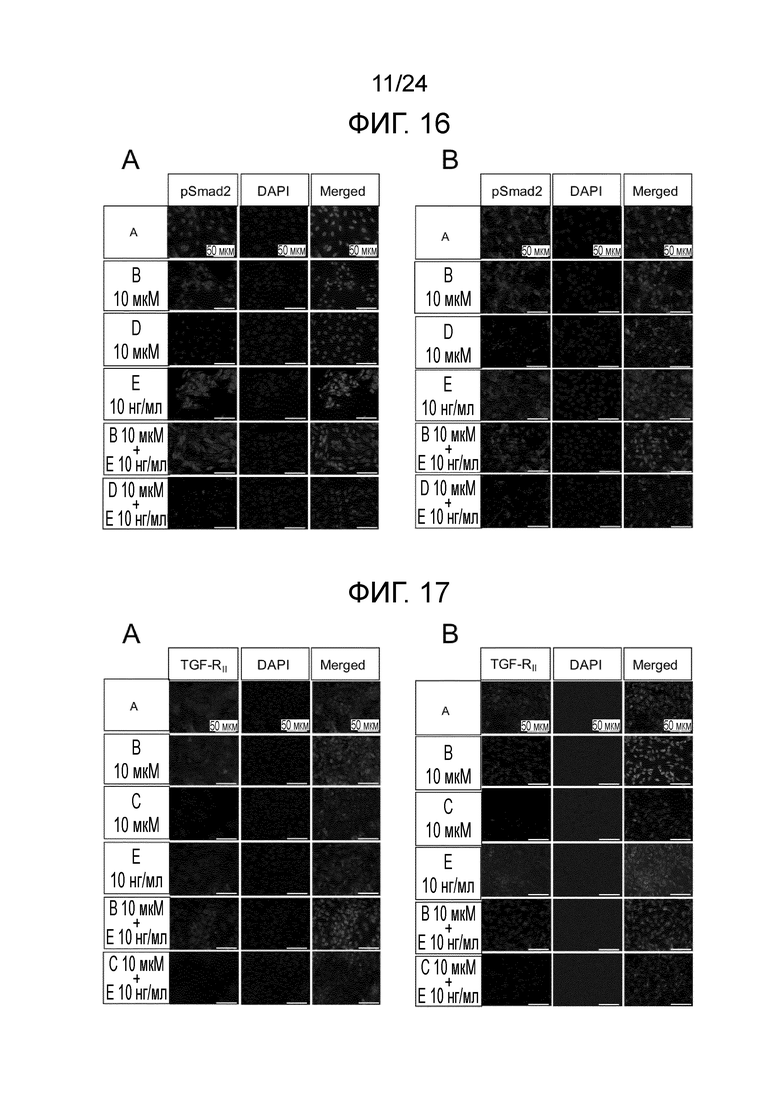
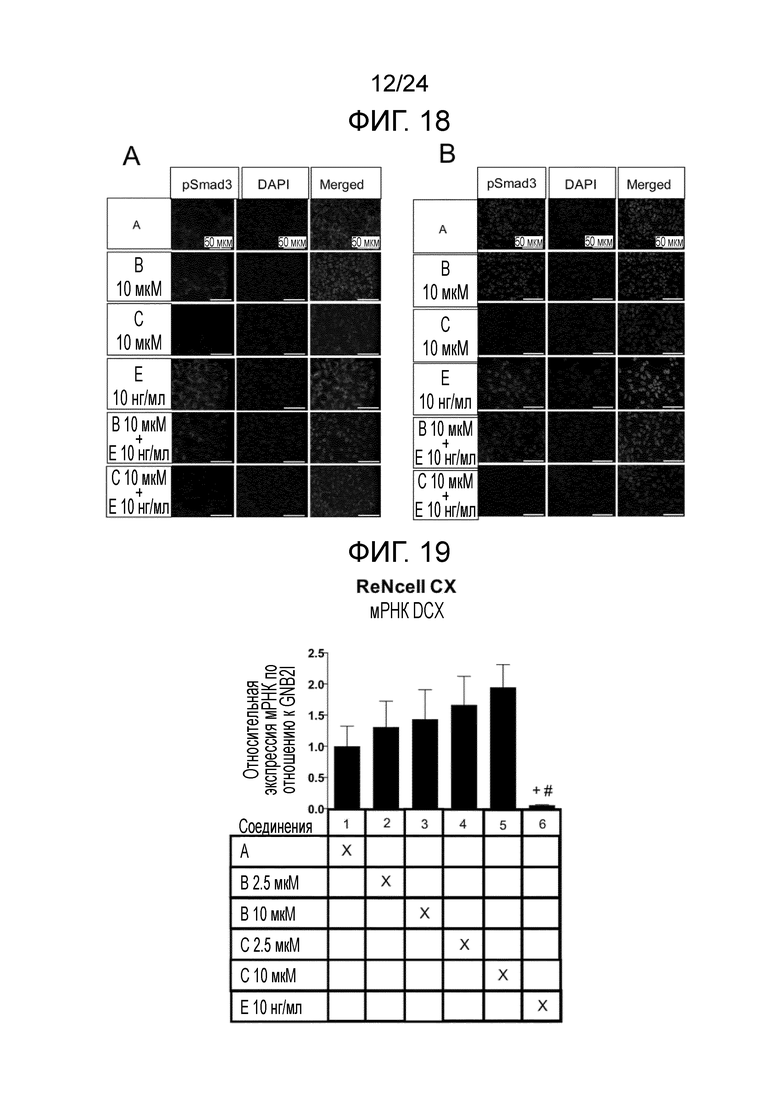
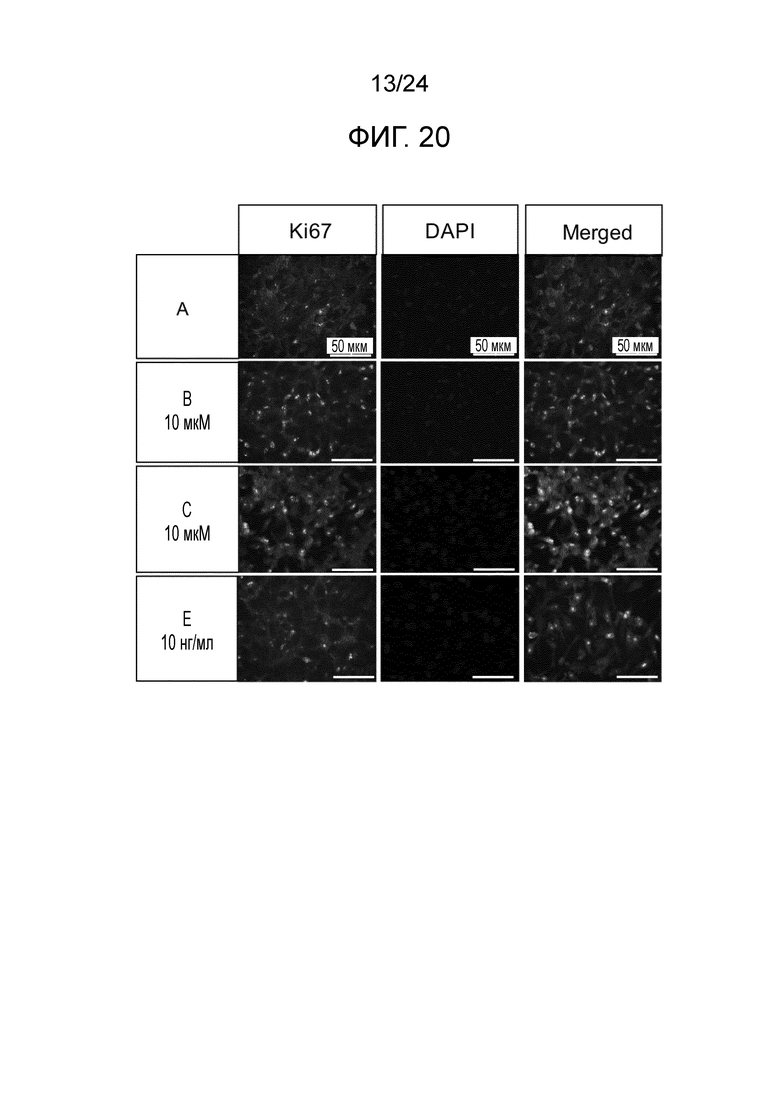

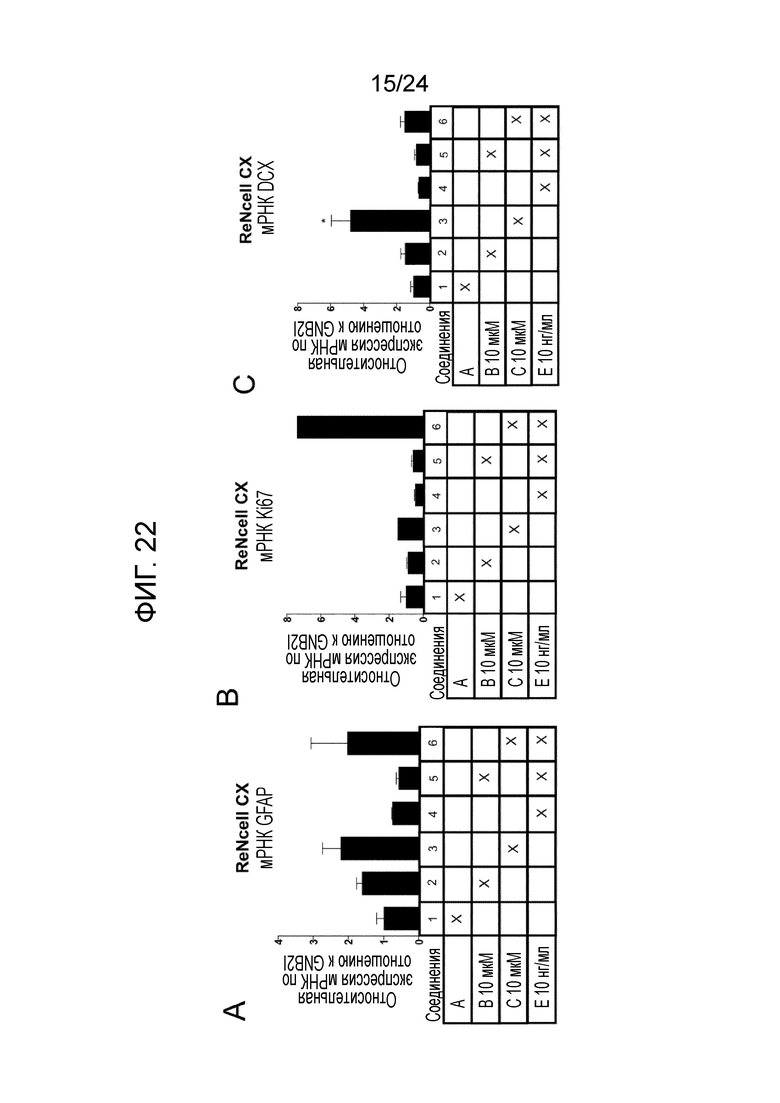
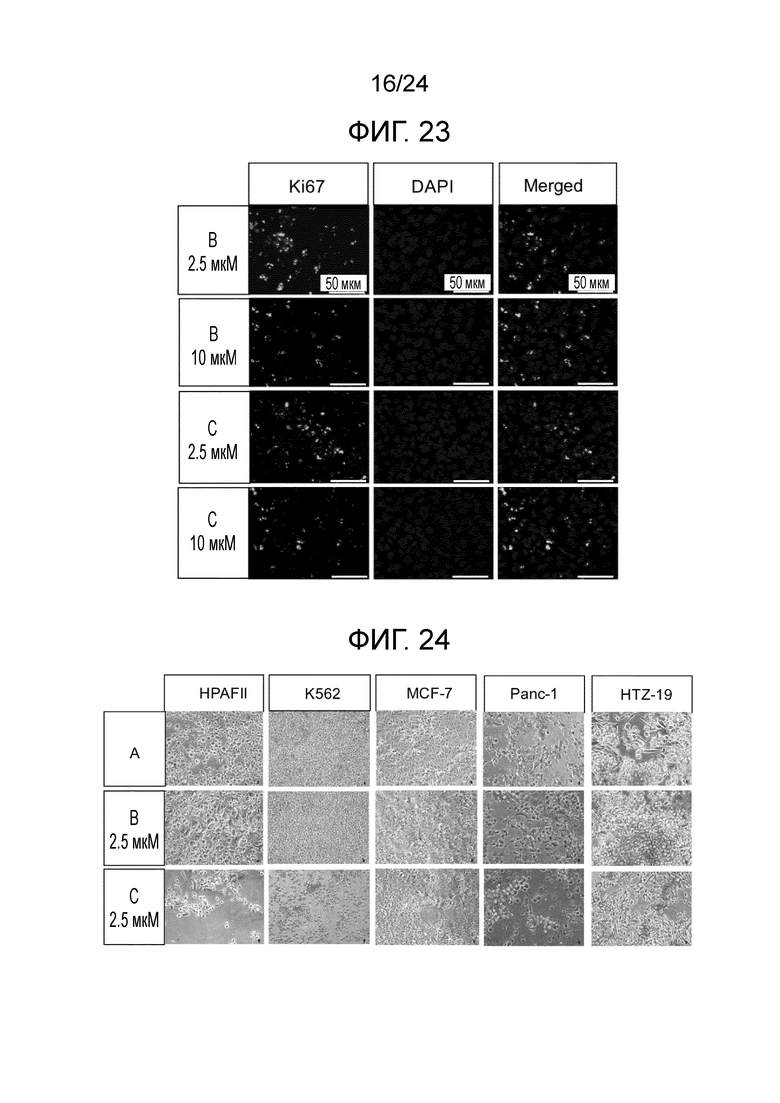
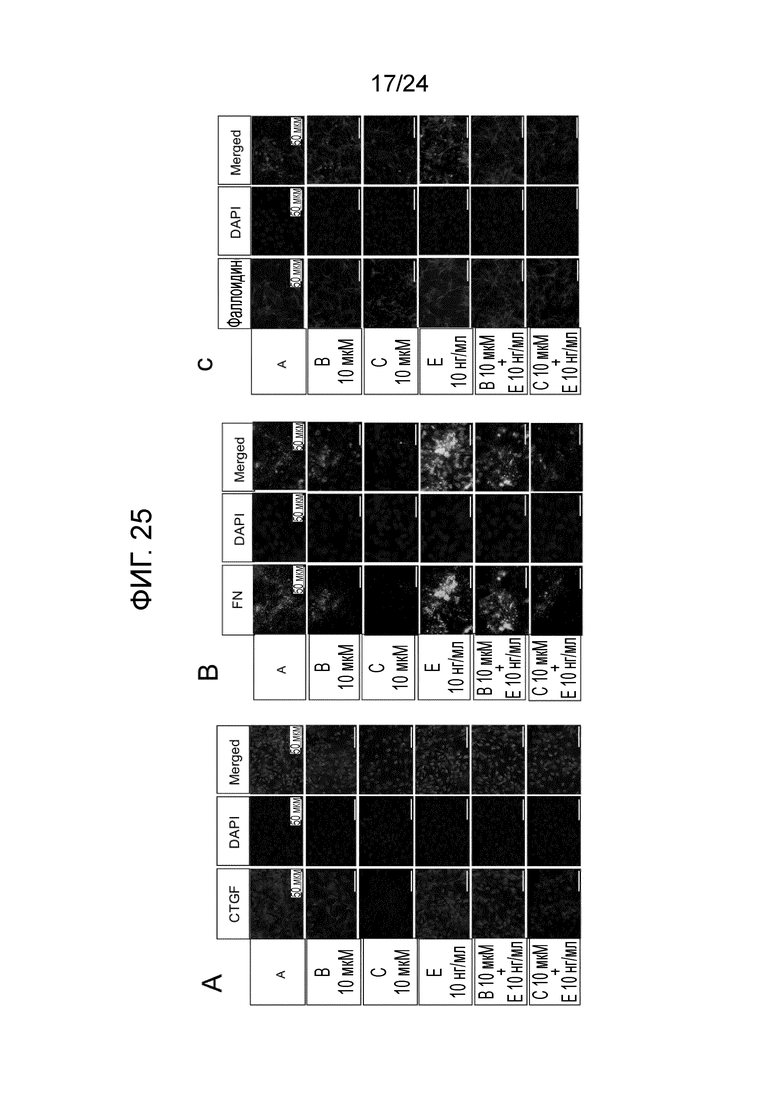



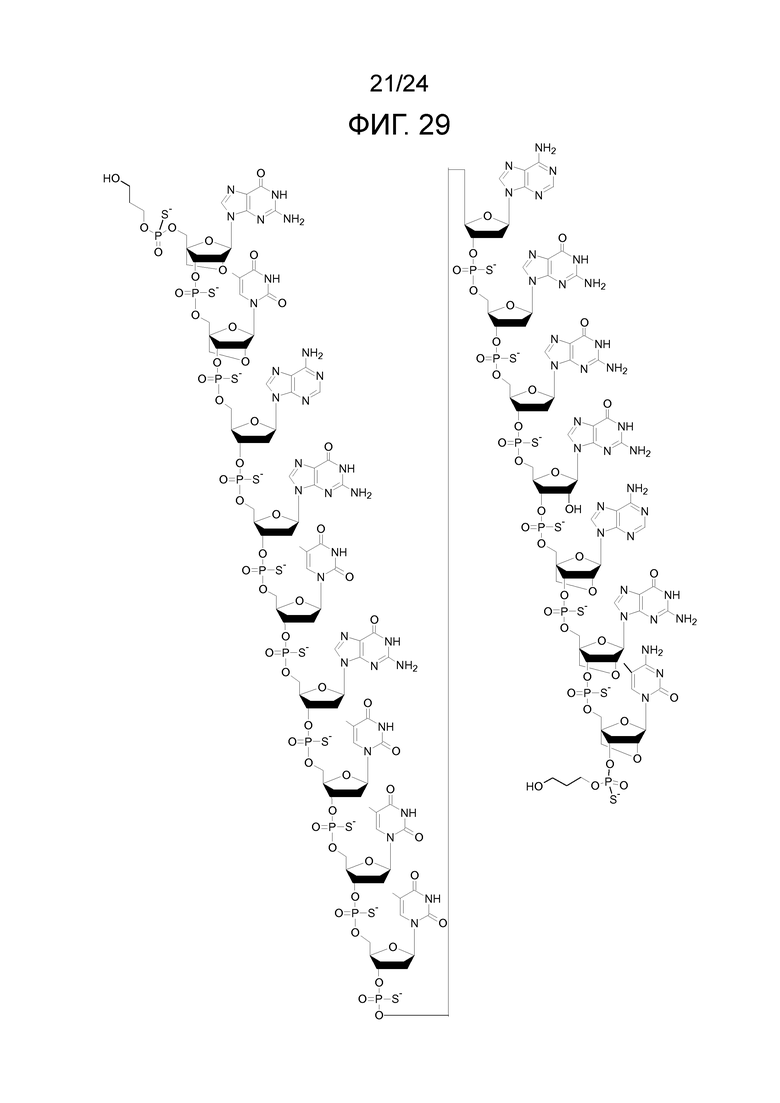
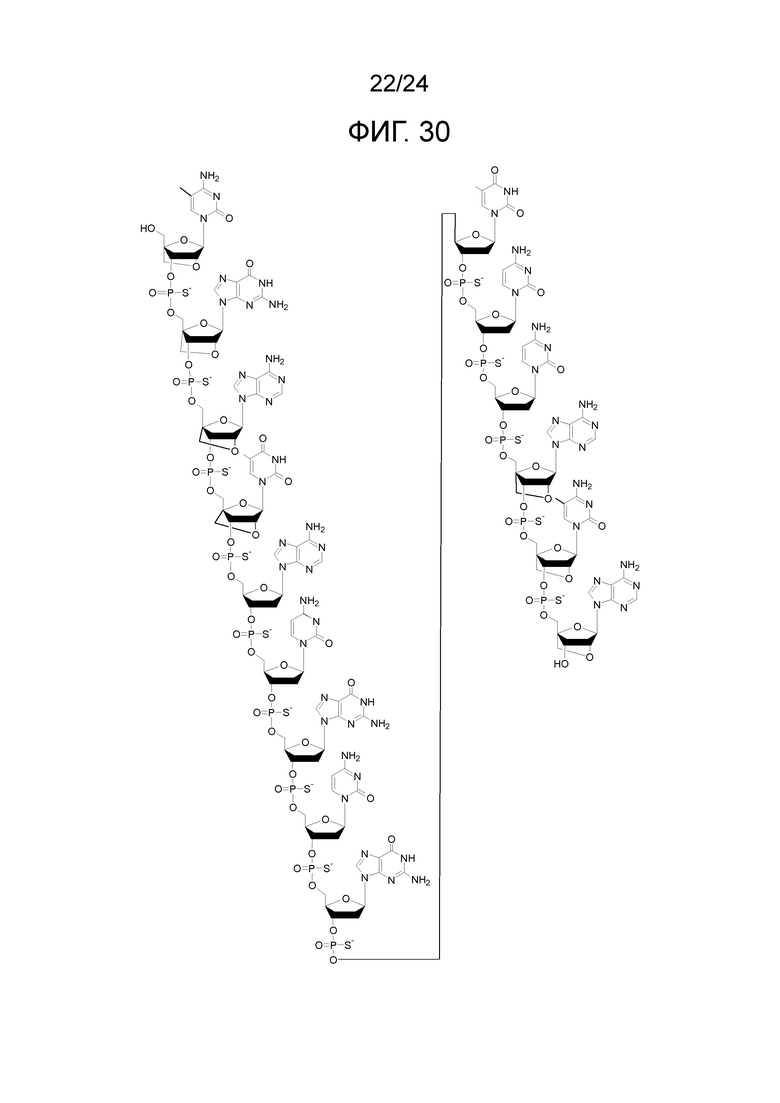
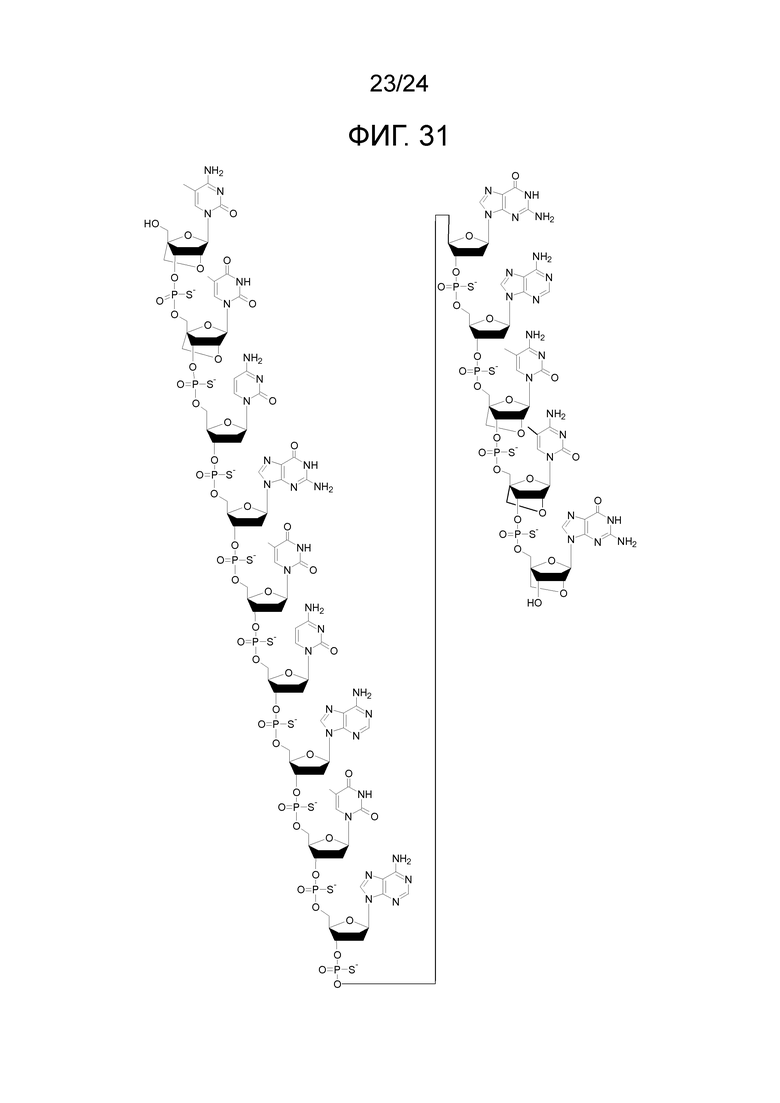

Изобретение относится к биотехнологии. Описаны антисмысловые олигонуклеотиды, состоящие из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид комплементарен участку гена, кодирующего TGF-RII, или участку мРНК, кодирующей TGF-RII. Описана фармацевтическая композиция, содержащая такие антисмысловые олигонуклеотиды. Указанные нуклеотиды применяют для профилактики и лечения неврологических, нейродегенеративных, фиброзных и гиперпролиферативных заболеваний. 3 н. и 11 з.п. ф-лы, 32 ил., 85 табл., 640 пр.
1. Антисмысловой олигонуклеотид, состоящий из 10-28 нуклеотидов, и, по меньшей мере, два из 10-28 нуклеотидов представляют LNA, и антисмысловой олигонуклеотид комплементарен участку гена, кодирующего TGF-RII, или участку мРНК, кодирующей TGF-RII, где участок гена, кодирующего TGF-RII, или участок мРНК, кодирующей TGF-RII, содержит последовательность TGGTCCATTC (SEQ. ID NO: 4)или последовательность CCCTAAACAC (SEQ. ID NO: 5), или последовательность ACTACCAAAT (SEQ. ID NO: 6), или последовательность GGACGCGTAT (SEQ. ID NO: 7), или последовательность GTCTATGACG (SEQ. ID NO: 8), или последовательность TTATTAATGC (SEQ. ID NO: 9),и антисмысловой олигонуклеотид содержит последовательность, комплементарную указанной последовательности TGGTCCATTC (SEQ. ID NO: 4) или последовательности CCCTAAACAC (SEQ. ID NO: 5), или последовательности ACTACCAAAT (SEQ. ID NO: 6), или последовательности GGACGCGTAT (SEQ. ID NO: 7), или последовательности GTCTATGACG (SEQ. ID NO: 8), или последовательности TTATTAATGC (SEQ. ID NO: 9) соответственно, и соли и оптические изомеры указанного антисмыслового олигонуклеотида, где антисмысловой олигонуклеотид ингибирует экспрессию TGF-RII.
2. Антисмысловой олигонуклеотид по п.1, где антисмысловой олигонуклеотид гибридизуется избирательно только с последовательностью TGGTCCATTC (SEQ. ID NO: 4) или последовательностью CCCTAAACAC (SEQ. ID NO: 5), или последовательностью ACTACCAAAT (SEQ. ID NO: 6), или последовательностью GGACGCGTAT (SEQ. ID NO: 7), или последовательностью GTCTATGACG (SEQ. ID NO: 8), или последовательностью TTATTAATGC (SEQ. ID NO: 9) участка гена, кодирующего TGF-RII, или участка мРНК, кодирующей TGF-RII.
3. Антисмысловой олигонуклеотид по п.1 или 2, где антисмысловой олигонуклеотид имеет длину 12-20 нуклеотидов, и/или где антисмысловой олигонуклеотид имеет гапмерную структуру с 1-5 звеньями LNA на терминальном 3'-конце и 1-5 звеньями LNA на терминальном 5'-конце, и/или где антисмысловой олигонуклеотид содержит фосфат, фосфоротиоат и/или фосфородитиоат в качестве межнуклеотидных связей.
4. Антисмысловой олигонуклеотид по любому из пп.1-3, где антисмысловой олигонуклеотид представлен следующей последовательностью:
5'-N1-GTCATAGA-N2-3' (SEQ. ID NO: 12) или
5'-N3-ACGCGTCC-N4-3' (SEQ. ID NO: 98) или
5'-N5-TTTGGTAG-N6-3' (SEQ. ID NO: 11) или
5'-N7-AATGGACC-N8-3' (SEQ. ID NO: 100) или
5'-N9-ATTAATAA-N10-3' (SEQ. ID NO: 101) или
5'-N11-TGTTTAGG-N12-3' (SEQ. ID NO: 10) или
где:
N1 представляет: CATGGCAGACCCCGCTGCTC-, ATGGCAGACCCCGCTGCTC-, TGGCAGACCCCGCTGCTC-, GGCAGACCCCGCTGCTC-, GCAGACCCCGCTGCTC-, CAGACCCCGCTGCTC-, AGACCCCGCTGCTC-, GACCCCGCTGCTC-, ACCCCGCTGCTC-, CCCCGCTGCTC-, CCCGCTGCTC-, CCGCTGCTC-, CGCTGCTC-, GCTGCTC-, CTGCTC-, TGCTC-, GCTC-, CTC-, TC- или C-;
N2 представляет: -C, -CC, -CCG, -CCGA, -CCGAG, -CCGAGCC, -CCGAGCCCCC, -CCGAGCCCCCC, -CCGAGCCCCCCAG, -CCGAGCCCCCAGC, -CCGAGCCCCCAGCG, -CCGAGCCCCCAGCGC, -CCGAGCCCCCAGCGCA, -CCGAGCCCCCAGCGCAG, -CCGAGCCCCCAGCGCAGC, -CCGAGCCCCCAGCGCAGCGG или -CCGAGCCCCCAGCGCAGCGG;
N3 представляет: GGTGGGATCGTGCTGGCGAT-, GTGGGATCGTGCTGGCGAT-, TGGGATCGTGCTGGCGAT-, GGGATCGTGCTGGCGAT-, GGATCGTGCTGGCGAT-, GATCGTGCTGGCGAT-, ATCGTGCTGGCGAT-, TCGTGCTGGCGAT-, CGTGCTGGCGAT-, GTGCTGGCGAT-, TGCTGGCGAT-, GCTGGCGAT-, CTGGCGAT-, TGGCGAT-, GGCGAT-, GCGAT-, CGAT-, GAT-, AT- или Т-;
N4 представляет: -ACAGGACGATGTGCAGCGGC, -ACAGGACGATGTGCAGCGG, -ACAGGACGATGTGCAGCG, -ACAGGACGATGTGCAGC, -ACAGGACGATGTGCAG, -ACAGGACGATGTGCA, -ACAGGACGATGTGC, -ACAGGACGATGTG, -ACAGGACGATGT, -ACAGGACGATG, -ACAGGACGAT, -ACAGGACGA, -ACAGGACG, -ACAGGAC, -ACAGGA, -ACAGG, -ACAG, -ACA, -AC или -A;
N5 представляет: GCCCAGCCTGCCCCAGAAGAGCTA-, CCCAGCCTGCCCCAGAAGAGCTA-, CCAGCCTGCCCCAGAAGAGCTA-, CAGCCTGCCCCAGAAGAGCTA-, AGCCTGCCCCAGAAGAGCTA-, GCCTGCCCCAGAAGAGCTA-, CCTGCCCCAGAAGAGCTA-, CTGCCCCAGAAGAGCTA-, TGCCCCAGAAGAGCTA-, GCCCCAGAAGAGCTA-, CCCCAGAAGAGCTA-, CCCAGAAGAGCTA-, CCAGAAGAGCTA-, CAGAAGAGCTA-, AGAAGAGCTA-, GAAGAGCTA-, AAGAGCTA-, AGAGCTA-, GAGCTA-, AGCTA-, GCTA-, CTA-, TA- или A-;
N6 представляет: -TGTTTAGGGAGCCGTCTTCAGGAA, -TGTTTAGGGAGCCGTCTTCAGGA, -TGTTTAGGGAGCCGTCTTCAGG, -TGTTTAGGGAGCCGTCTTCAG, -TGTTTAGGGAGCCGTCTTCA, -TGTTTAGGGAGCCGTCTTC, -TGTTTAGGGAGCCGTCTT, -TGTTTAGGGAGCCGTCT, -TGTTTAGGGAGCCGTC, -TGTTTAGGGAGCCGT, -TGTTTAGGGAGCCG, -TGTTTAGGGAGCC, -TGTTTAGGGAGC, -TGTTTAGGGAG, -TGTTTAGGGA, -TGTTTAGGG, -TGTTTAGG, -TGTTTAG, -TGTTTA, -TGTTT, -TGTT, -TGT, -TG или -T;
N7 представляет: TGAATCTTGAATATCTCATG-, GAATCTTGAATATCTCATG-, AATCTTGAATATCTCATG-, ATCTTGAATATCTCATG-, TCTTGAATATCTCATG-, CTTGAATATCTCATG-, TTGAATATCTCATG-, TGAATATCTCATG-, GAATATCTCATG-, AATATCTCATG-, ATATCTCATG-, TATCTCATG-, ATCTCATG-, TCTCATG-, CTCATG-, TCATG-, CATG-, ATG-, TG- или G-; и
N8 выбран из: -AGTATTCTAGAAACTCACCA, -AGTATTCTAGAAACTCACC, -AGTATTCTAGAAACTCAC, -AGTATTCTAGAAACTCA, -AGTATTCTAGAAACTC, -AGTATTCTAGAAACT, -AGTATTCTAGAAAC, -AGTATTCTAGAAA, -AGTATTCTAGAA, -AGTATTCTAGA, -AGTATTCTAG, -AGTATTCTA, -AGTATTCT, -AGTATTC, -AGTATT, -AGTAT, -AGTA, -AGT, -AG или -A;
N9 представляет: ATTCATATTTATATACAGGC-, TTCATATTTATATACAGGC-, TCATATTTATATACAGGC-, CATATTTATATACAGGC-, ATATTTATATACAGGC-, TATTTATATACAGGC-, ATTTATATACAGGC-, TTTATATACAGGC-, TTATATACAGGC-, TATATACAGGC-, ATATACAGGC-, TATACAGGC-, ATACAGGC-, TACAGGC-, ACAGGC-, CAGGC-, AGGC-, GGC-, GC- или C-;
N10 представляет: -AGTGCAAATGTTATTGGCTA, -AGTGCAAATGTTATTGGCT, -AGTGCAAATGTTATTGGC, -AGTGCAAATGTTATTGG, -AGTGCAAATGTTATTG, -AGTGCAAATGTTATT, -AGTGCAAATGTTAT, -AGTGCAAATGTTA, -AGTGCAAATGTT, -AGTGCAAATGT, -AGTGCAAATG, -AGTGCAAAT, -AGTGCAAA, -AGTGCAA, -AGTGCA, -AGTGC, -AGTG, -AGT, -AG или -A;
N11 представляет: TGCCCCAGAAGAGCTATTTGGTAG-, GCCCCAGAAGAGCTATTTGGTAG-, CCCCAGAAGAGCTATTTGGTAG-, CCCAGAAGAGCTATTTGGTAG-, CCAGAAGAGCTATTTGGTAG-, CAGAAGAGCTATTTGGTAG-, AGAAGAGCTATTTGGTAG-, GAAGAGCTATTTGGTAG-, AAGAGCTATTTGGTAG-, AGAGCTATTTGGTAG-, GAGCTATTTGGTAG-, AGCTATTTGGTAG-, GCTATTTGGTAG-, CTATTTGGTAG-, TATTTGGTAG-, ATTTGGTAG-, TTTGGTAG-, TTGGTAG-, TGGTAG-, GGTAG-, GTAG-, TAG-, AG- или G-,
N12 представляет: -GAGCCGTCTTCAGGAATCTTCTCC, -GAGCCGTCTTCAGGAATCTTCTC, -GAGCCGTCTTCAGGAATCTTCT, -GAGCCGTCTTCAGGAATCTTC, -GAGCCGTCTTCAGGAATCTT, -GAGCCGTCTTCAGGAATCT, -GAGCCGTCTTCAGGAATC,-GAGCCGTCTTCAGGAAT, -GAGCCGTCTTCAGGAA, -GAGCCGTCTTCAGGA,-GAGCCGTCTTCAGG, -GAGCCGTCTTCAG, -GAGCCGTCTTCA, -GAGCCGTCTTC, -GAGCCGTCTT, -GAGCCGTCT, -GAGCCGTC, -GAGCCGT,-GAGCCG, -GAGCC, -GAGC, -GAG, -GA или -G.
5. Антисмысловой олигонуклеотид по любому из пп.1-4, где последние 2-4 нуклеотида на терминальном 5'-конце представляют нуклеотиды LNA, и последние 2-4 нуклеотида на терминальном 3'-конце представляют нуклеотиды LNA, и между нуклеотидами LNA на терминальном 5'-конце и нуклеотидами LNA на терминальном 3'-конце находится, по меньшей мере, 6 последовательных нуклеотидов, представляющих нуклеотиды, которые не являются LNA, или которые представляют нуклеотиды ДНК.
6. Антисмысловой олигонуклеотид по любому из пп.1-4, где нуклеотиды LNA связаны друг с другом через фосфоротиоатную группу или фосфородитиоатную группу, или где все нуклеотиды связаны друг с другом через фосфатную группу или фосфоротиоатную группу, или фосфородитиоатную группу.
7. Антисмысловой олигонуклеотид по любому из пп.1-4, где нуклеотиды LNA выбраны из следующей группы:
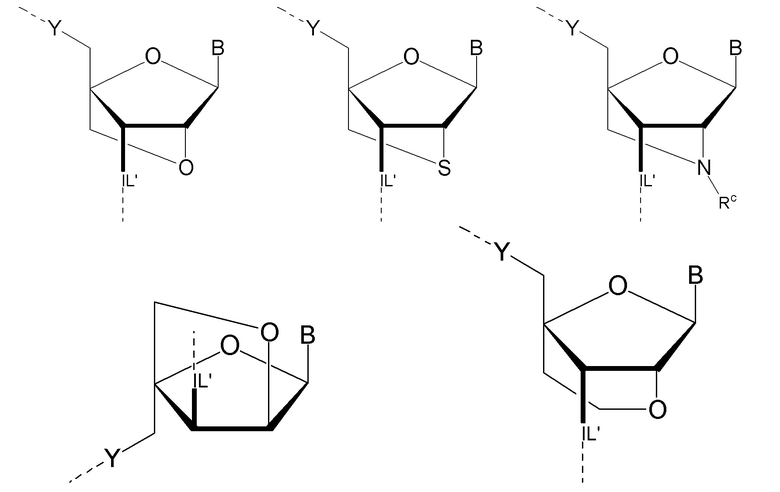
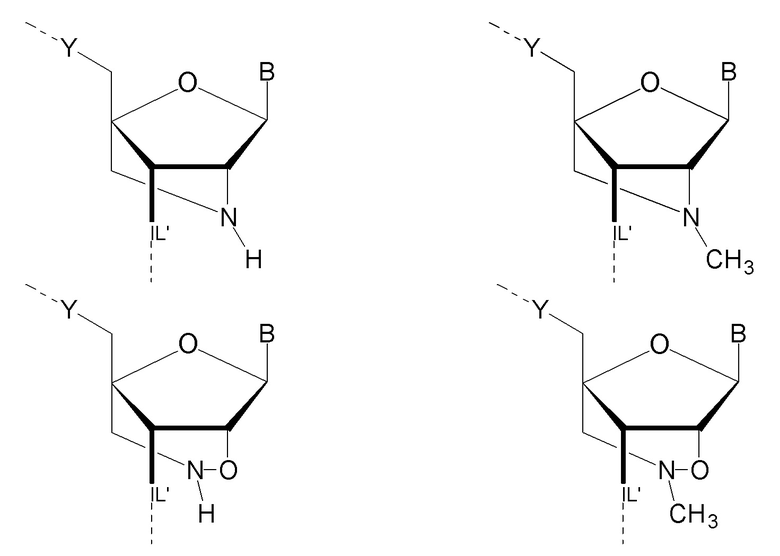
где:
IL' представляет -Xʺ-P(=X')(X-)-;
X' представляет =O или =S;
X- представляет -O-, -OH, -ORH, -NHRH, -N(RH)2, -OCH2CH2ORH, -OCH2CH2SRH, -BH3-, -RH, -SH, -SRH или -S-;
Xʺ представляет -O-, -NH-, -NRH-, -CH2- или -S-;
Y представляет -O-, -NH-, -NRH-, -CH2- или -S-;
RC и RH независимо друг от друг выбраны из атома водорода и C1-4-алкила;
В представляет азотистое основание, выбранное из следующей группы:
аденин, тимин, гуанин, цитозин, урацил, 5-метилцитозин, 5-гидроксиметилцитозин, N4-метилцитозин, ксантин, гипоксантин, 7-деазаксантин, 2-аминоаденин, 6-метиладенин, 6-метилгуанин, 6-этиладенин, 6-этилгуанин, 2-пропиладенин, 2-пропилгуанин, 6-карбоксиурацил, 5,6-дигидроурацил, 5-пропинилурацил, 5-пропинилцитозин, 6-азаурацил, 6-азацитозин, 6-азатимин, 5-урацил, 4-тиоурацил, 8-фтораденин, 8-хлораденин, 8-бромаденин, 8-иодаденин, 8-аминоаденин, 8-тиоладенин, 8-тиоалкиладенин, 8-гидроксиладенин, 8-фторгуанин, 8-хлоргуанин, 8-бромгуанин, 8-иодгуанин, 8-аминогуанин, 8-тиолгуанин, 8-тиоалкилгуанин, 8-гидроксилгуанин, 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-иодурацил, 5-трифторметилурацил, 5-фторцитозин, 5-бромцитозин, 5-хлорцитозин, 5-иодцитозин, 5-трифторметилцитозин, 7-метилгуанин, 7-метиладенин, 8-азагуанин, 8-азааденин, 7-деазагуанин, 7-деазааденин, 7-деаза-8-азааденин, 3-деазагуанин, 3-деазааденин, 2-тиоурацил, 2-тиотимин и 2-тиоцитозин.
8. Антисмысловой олигонуклеотид по любому из пп.1-4, имеющий одну из следующих гапмерных структур: 3-8-3, 4-8-2, 2-8-4, 3-8-4, 4-8-3, 4-8-4, 3-9-3, 4-9-2, 2-9-4, 4-9-3, 3-9-4, 4-9-4, 3-10-3, 2-10-4, 4-10-2, 3-10-4, 4-10-3, 4-10-4, 2-11-4, 4-11-2, 3-11-4, 4-11-3.
9. Антисмысловой олигонуклеотид по любому из пп.1-4, где антисмысловые олигонуклеотиды связываются со 100% комплементарностью с мРНК, кодирующей TGF-RII, и не связываются с каким-либо другим участком в транскриптоме человека.
10. Антисмысловой олигонуклеотид, выбранный из следующей группы:
где аббревиатуры b, d, C*, A*, s, ss имеют следующие значения:
нуклеотиды, выделенные жирным шрифтом, представляют нуклеотиды LNA
нуклеотиды, не выделенные жирным шрифтом, представляют нуклеотиды, которые не являются LNA, и
где антисмысловой олигонуклеотид ингибирует экспрессию TGF-RII.
11. Антисмысловой олигонуклеотид по любому из пп.1-4 для стимуляции регенерации и функционального восстановления поврежденных нервных путей и/или для лечения и компенсации индуцированного возрастом снижения обновления нейрональных стволовых клеток.
12. Антисмысловой олигонуклеотид по любому из пп.1-4 для применения в профилактике и лечении нейродегенеративных заболеваний, нейровоспалительных расстройств, травматических или посттравматических расстройств, нейрососудистых расстройств, гипоксических расстройств, постинфекционных расстройств центральной нервной системы, фиброзных болезней, гиперпролиферативных заболеваний, рака, опухолей, пресбиакузиса и пресбиопии.
13. Антисмысловой олигонуклеотид для применения по п.12, где нейродегенеративные заболевания и нейровоспалительные расстройства выбраны из группы, состоящей из: болезни Альцгеймера, болезни Паркинсона, болезни Крейтцфельдта-Якоба, нового варианта болезни Крейтцфельдта-Якоба, болезни Галлервордена-Шпатца, болезни Гентингтона, мультисистемной атрофии, деменции, лобно-височной деменции, расстройств двигательного нейрона, бокового амиотрофического склероза, спинномозговой мышечной атрофии, спиноцеребеллярных атрофий, шизофрении, аффективных расстройств, большого депрессивного расстройства, менингоэнцефалита, бактериального менингоэнцефалита, вирусного менингоэнцефалита, аутоиммунных расстройств ЦНС, рассеянного склероза, острых ишемических/гипоксических поражений, травмы центральной нервной системы и спинного мозга, травмы головы и позвоночника, черепно-мозговых травм, артериосклероза, атеросклероза, микроангиопатической деменции, болезни Бинсвангера, дегенерации сетчатки, улитковой дегенерации, дегенерации желтого пятна, улитковой глухоты, деменции, связанной со СПИДом, пигментного ретинита, Х-сцепленного рецессивного синдрома тремора/атаксии, прогрессирующего супрануклеарного паралича, стриатонигральной дегенерации, оливопонтомозжечковой дегенерации, синдрома Шая-Дрейджера, зависящего от возраста дефицита памяти, неврологических расстройств развития, связанных с деменцией, синдрома Дауна, синуклеинопатий, мутаций супероксиддисмутазы, расстройств повторов тринуклеотида, травмы, гипоксии, сосудистых заболеваний, сосудистых воспалений и старения ЦНС, и где фиброзные болезни выбраны из группы, состоящей из: фиброза легких, кистозного фиброза, цирроза печени, эндомиокардиального фиброза, «старого» инфаркта миокарда, фиброза предсердий, фиброза средостения, миелофиброза, забрюшинного фиброза, прогрессирующего массивного фиброза, нефрогенного системного фиброза, болезни Крона, келоида, системного склероза, артрофиброза, болезни Пейрони, контрактуры Дюпюитрена и остаточных явлений после красной волчанки.
14. Фармацевтическая композиция, содержащая, по меньшей мере, один антисмысловой олигонуклеотид по любому из пп.1-4 или 10 вместе, по меньшей мере, с одним фармацевтически приемлемым носителем, эксципиентом, адъювантом, растворителем или разбавителем, где антисмысловой олигонуклеотид ингибирует экспрессию TGF-RII.
| ИНГИБИТОРЫ ПЕРЕДАЧИ СИГНАЛА ТРАНСФОРМИРУЮЩИХ ФАКТОРОВ РОСТА (TGF-R) ДЛЯ ЛЕЧЕНИЯ РАССТРОЙСТВ ЦНС | 2005 |
|
RU2385933C2 |
| Дисковый поворотный затвор | 1989 |
|
SU1716235A1 |

