ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к химерному антигенному рецептору (CAR), содержащему определенный спейсерный домен, который вызывает образование мультимерных молекул CAR на клеточной поверхности. Мультимерная молекула CAR может являться «суперчувствительной» и способна вызывать активацию Т-клеток в ответ на связывание с антигеном, который экспрессируется с низкой плотностью на клетке-мишени.
УРОВЕНЬ ТЕХНИКИ
Химерные антигенные рецепторы (CAR)
Традиционно, антиген-специфичные Т-клетки образуются в результате селективного размножения Т-клеток периферической крови, изначально специфичных к антигену-мишени. Однако трудным и довольно часто невозможным является отбор и размножение большого числа Т-клеток, специфичных к большинству раковых антигенов. Генная терапия с использованием встраивающихся векторов обеспечивает решение этой проблемы, поскольку трансгенная экспрессия химерного антигенного рецептора (CAR) позволяет генерировать большое число Т-клеток, специфичных к любому поверхностному антигену путем ex vivo трансдукции вирусным вектором крупной популяции Т-клеток периферической крови.
Химерные антигенные рецепторы представляют собой белки, которые придают эффекторной функции Т-клеток специфичность антигенсвязывающей молекулы, такой как моноклональное антитело (мАТ). Их обычная форма представляет собой белок с трансмембранным доменом I типа с распознающим антиген аминоконцом, спейсером, трансмембранным доменом, которые все связаны со сложным эндодоменом (внутриклеточным доменом), который передает сигналы пролиферации и активации Т-клеток (см. фиг. 1А).
Наиболее распространенными формами этих молекул являются слитые белки, состоящие из одноцепочечных вариабельных фрагментов (scFv), полученных из моноклональных антител, которые распознают антиген-мишень, слитых через спейсер и трансмембранный домен с сигнальным эндодоменом. Такие молекулы вызывают активацию Т-клеток в ответ на распознавание scFv своей мишени. Когда Т-клетки экспрессируют такой CAR, они распознают и убивают клетки-мишени, которые экспрессируют антиген-мишень. Было разработано несколько CAR к опухоле-ассоциированным антигенам, и подходы адоптивного переноса с использованием таких экспрессирующих CAR Т-клеток в настоящее время проходят клинические испытания для лечения различных видов рака.
CAR часто содержат спейсерный домен, чтобы обеспечить соответствующее расстояние между антигенсвязывающим доменом и клеточной мембраной и для обеспечения подходящей ориентации, достижимости и отделения от фосфатаз при связывании с лигандом.
Стандартно используемыми спейсерами являются Fc из IgG1, стеблевая последовательность из CD8α и CD28, и даже просто шарнир из IgG1 или эктодомен CD247 могут оказаться достаточными в зависимости от антигена (фиг. 2b).
Эти стандартные спейсеры имеют ряд ограничений, поскольку они либо должны содержать целые доменные структуры, чтобы сформировать функциональный спейсер, либо они являются сильно гликозилированными, и изменение длины аминокислотной последовательности может привести к непредсказуемые изменениям размера спейсера. Например, IgG-спейсеры должны содержать целое число иммуноглобулиновых доменов. Это требование целого числа структурных доменов означает, что изменения, которые можно было бы осуществить в спейсерах, ограничены.
Кроме того, перечисленные выше спейсеры, как правило, имеют длинные первичные аминокислотные последовательности, которые сворачиваются с образованием требуемых вторичных и третичных структур. Как таковые, они, как правило, кодируются протяженными нуклеотидными последовательностями. Эта необходимость в наличии длинной нуклеотидной последовательности представляет собой проблему при конструировании векторов, кодирующих CAR.
Классические CAR имеют определенные конструкционные ограничения. Поскольку классический CAR является гомодимером, обычно присутствуют два (одинаковых) связывающих сайта и соотношение 1:1 антигенсвязывающих доменов к внутриклеточным Т-клеточным сигнальным доменам. Это требует определенной стехиометрии и ограничивает гибкость системы в целом.
Таким образом, существует потребность в альтернативных CAR, которые предлагают большую гибкость в плане дизайна.
Проблемы с аффинностью
Связывающие домены CAR, как правило, получены из вариабельной области либо ранее существовавшего антитела, либо антител, отобранных из библиотеки. В результате, большинство отобранных CAR связывают родственный лиганд с наномолярным сродством. В отличие от этого биофизические свойства связывания TCR:пептид:МНС (TCR-pMHC), как правило, составляют 10-1 мкМ (на порядок ниже в аффинности). Хотя взаимодействия с более высокой аффинностью повышают специфичность в отношении лиганда при заданной концентрации рецептора, появились новые данные о том, что TCR эволюционировал до более низкой аффинности для того, чтобы позволить Т-клеткам обнаруживать клетки-мишени, экспрессирующие родственный комплекс пептид:МНС с низкой плотностью.
Сообщалось о том, что Т-клетки могут быть активированы с помощью всего лишь десяти родственных рМНС, и что один комплекс рМНС может вызвать продуктивный сигнал у 200 молекул TCR. Полагают, что это достигается с помощью процесса, известного как последовательная инициация, когда один родственный рМНС, присутствующий на клетке-мишени, может несколько раз пройти через цикл связывания, инициации и последующей диссоциации с TCR, эффективно усиливая сигнал. В результате, только небольшое число родственных рМНС необходимо для передачи продуктивного сигнала.
Более высокая аффинность CAR означает, что молекулярная диссоциация после взаимодействия может занять от несколько минут до нескольких часов, в отличие от TCR, для которого она, как правило, составляет секунды. По этой причине маловероятно, что передача сигнала от CAR дает эффективную стимуляцию ответа, а вместо этого основана на лигировании большего числа рецепторов. Это может ограничить функции CAR взаимодействием только с лигандами, экспрессирующимися на клетках-мишенях с высокой плотностью. Было подсчитано, что для высокоаффинного CAR требуется, чтобы клетка-мишень экспрессировала более 10000 копий молекулы лиганда, чтобы индуцировать эффективный литический ответ. Более конкретно, с использованием первого поколения CAR James и сотр. показали необходимость наличия приблизительно 30000 молекул-мишеней на клетке-мишени (вызывающих эндоцитоз приблизительно 20000 молекул CAR), чтобы вызвать максимальную литическую активность (S. James et al., The Journal of Immunology, vol. 184 (8) 4284-4294, 2010). Модели на животных показывают, что клетки-мишени, которые экспрессируют лиганды в количестве меньшем, чем пороговый уровень для лизиса, могут избежать обнаружения и могут повторно вызвать заболевание (U. Anurathapan et al., Molecular Therapy, vol. 22 (3) 623-633, 2014).
Один способ повышения чувствительности CAR к лигандам с низкой плотностью заключается в использовании низкоаффинного связывающего домена, который, таким образом, может имитировать последовательную инициацию ответа, запускаемую TCR-pMHC. Однако этот подход имеет некоторые ограничения. Применение низкоаффинных CAR в настоящее время не поддается прогнозированию вследствие неизвестного вклада, вносимого костимуляторными молекулами, образованием псеводимеров и адгезионными молекулами в последовательную инициацию ответа, запускаемую TCR-pMHC. В терапии с использованием CAR эти молекулярные взаимодействия варьируют в зависимости от клетки-мишени, и, поэтому, трудно получить надежную последовательную инициацию ответа. Кроме того, способы уменьшения аффинности CAR до одного уровня с TCR, как правило, начинаются с высокоаффинного CAR с последующим мутагенезом CDR. Этот подход является длительным и часто неудачным, что может привести к непредсказуемым результатам и увеличивает риск неспецифичного связывания.
Конструирование CAR, который способен инициировать сигнал при низкой плотности лиганда, позволит использовать этот метод лечения в отношении гораздо большего числа видов рака, а также снизить вероятность того, что раковая опухоль избежит иммунологического контроля.
Таким образом, существует потребность в CAR, которые не имеют проблем, изложенных выше.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ АСПЕКТОВ ИЗОБРЕТЕНИЯ
В первом аспекте настоящее изобретение относится к формирующему химерный антигенный рецептор (CAR) полипептиду, содержащему:
(i) антигенсвязывающий домен;
(ii) суперспиральный спейсерный домен;
(iii) трансмембранный домен; и
(iv) эндодомен.
Настоящее изобретение также относится к вспомогательному полипептиду, содержащему:
(i) суперспиральный спейсер домена;
(iii) трансмембранный домен; и
(iv) эндодомен.
Суперспиральный домен позволяет мультимеризацию множества формирующих CAR полипептидов и/или вспомогательных полипептидов, например, по меньшей мере трех формирующих CAR полипептидов и/или вспомогательных полипептидов с образованием мультимерного CAR.
Суперспиральный домен может быть взят из любого из следующих белков: олигомерного матриксного белка хряща (СОМР), маннозосвязывающего белка А, суперспирального серин-богатого белка 1, полипептид-высвобождающего фактора 2, SNAP-25, SNARE, Lac-репрессора или аполипопротеина Е.
Суперспиральный домен может содержать последовательность, приведенную в SEQ ID NO: 1 или ее фрагмент, или ее вариант, который имеет по меньшей мере 80% идентичности по последовательности.
Эндодомен может содержать по меньшей мере один из эндодомена CD3-дзета, эндодомена CD28, эндодомена 41ВВ и эндодомена OX40.
Эндодомен может содержать последовательность, приведенную в SEQ ID NO: 7, или ее вариант, который имеет по меньшей мере 80% идентичности по последовательности.
Антигенсвязывающий домен может связывать антиген, который экспрессируется с низкой плотностью на клетки-мишени. Так, например, антигенсвязывающий домен может связываться с ROR-1, Typr-1 или ВСМА.
Формирующий CAR полипептид может содержать элемент, способный образовывать мостик с другим CAR.
Например, элемент может быть способен образовывать дисульфидный мостик с другим CAR, который содержит такой элемент.
Второй аспект настоящего изобретения относится к мультимерным CAR, которые образуются в результате взаимодействия между суперспиральными спейсерными доменами или формирующим(и) CAR полипептидом(ами) и/или вспомогательным(и) полипептидом(ами).
В первом варианте осуществления второго аспекта настоящее изобретение относится к мультимерному антигенному химерному рецептору (CAR), содержащему множество формирующих CAR полипептидов, описанных выше.
Во втором варианте осуществления второго аспекта настоящего изобретения предложен мультимерный химерный антигенный рецептор (CAR), содержащий один или несколько формирующих CAR полипептидов и один или несколько вспомогательных полипептидов, описанных выше.
Формирующий(е) CAR полипептид(ы) и/или вспомогательный(е) полипептид(ы) в мультимерном CAR могут содержать различные эндодомены.
Если мультимерный CAR содержит два или несколько формирующих CAR полипептидов, то они могут иметь различные антигенсвязывающие домены, например, антигенсвязывающие домены с различной специфичностью связывания.
Один из эндодоменов формирующего CAR полипептида и вспомогательного полипептида может содержать эндодомен CD3-дзета, а другой эндодомен формирующего CAR полипептида и вспомогательного полипептида может включать в себя эндодомен 41ВВ. Если присутствуют два вспомогательных полипептида, то один может содержать эндодомен 41ВВ, а другой может содержать эндодомен CD28.
Мультимерный CAR может, например, являться димерным, тримерным, тетрамерным, пентамерным, гексамерным или гептамерным.
Пентамерный CAR может включать любую из следующих комбинаций формирующего CAR полипептида и вспомогательных полипептидных цепей:

В случае, когда мультимерный CAR содержит первый и второй формирующие CAR полипептиды по первому аспекту настоящего изобретения, антигенсвязывающий домен первого CAR может связываться с другим эпитопом, чем антигенсвязывающий домен второго CAR.
В этом варианте осуществления антигенсвязывающий домен первого CAR может связываться с другим антигеном, чем антигенсвязывающий домен второго CAR.
Настоящее изобретение также относится к сцепленному комплексу, который содержит по меньшей мере два многомерных CAR по второму аспекту настоящего изобретения, где первый CAR на первом мультимерном CAR образует мостик со вторым CAR на втором мультимерном CAR, таким образом, что первый и второй мультимерные CAR связываются с образованием комплекса.
Мостик может представлять собой дисульфидный мостик или дополнительную суперспиральную структуру.
В третьем аспекте настоящее изобретение относится к сигнальной системе химерного антигенного рецептора (CAR), которая включает:
(i) мультимерный CAR, содержащий формирующий CAR полипептид или вспомогательный полипептид, описанные выше, который включает в себя первым гетеродимеризующийся домен; и
(ii) внутриклеточный сигнальный компонент, содержащий сигнальный домен и второй гетеродимеризующийся домен;
где гетеродимеризация между первыми и вторыми гетеродимеризующимися доменами заставляет мультимерный CAR и сигнальный компонент образовать функциональный CAR-комплекс.
Каждый формирующий(е) CAR полипептид(ы) или вспомогательный(е) полипептид(ы) может содержать множество гетеродимеризующихся доменов, так что один формирующий CAR полипептид или вспомогательный полипептид способен гетеродимеризоваться с множеством сигнальных компонентов.
Сигнальный компонент сигнальной системы CAR может содержать множество сигнальных доменов.
В четвертом аспекте настоящее изобретение относится к нуклеиновой кислоте, которая кодирует формирующий CAR полипептид по первому аспекту настоящего изобретения и/или вспомогательный полипептид, описанные выше.
Пятый аспект настоящего изобретения относится к нуклеотидным конструкциям, которые содержат две или несколько нуклеотидных последовательностей.
В первом варианте осуществления пятого аспекта настоящего изобретения предложена нуклеотидная конструкция, которая кодирует два или несколько формирующих CAR полипептидов по первому аспекту настоящего изобретения.
Во втором варианте осуществления пятого аспекта настоящего изобретения предложена нуклеотидная конструкция, которая кодирует по меньшей мере один формирующий CAR полипептид и по меньшей мере один вспомогательный полипептид, описанные выше.
В первом варианте осуществления пятого аспекта настоящего изобретения предложена нуклеотидная конструкция, которая кодирует:
(i) по меньшей мере один формирующий CAR полипептид по первому аспекту настоящего изобретения, который формирует мультимерный CAR по второму аспекту настоящего изобретения; и
(ii) внутриклеточный сигнальный компонент по пятому аспекту изобретения.
В шестом аспекте настоящее изобретение относится к вектору, который содержит нуклеотидную последовательность по четвертому аспекту настоящего изобретения или нуклеотидную конструкцию по пятому аспекту настоящего изобретения.
Вектор может представлять собой, например, ретровирусный вектор или лентивирусный вектор, или транспозон.
В седьмом аспекте настоящее изобретение относится к клетке, которая экспрессирует формирующий CAR полипептид или вспомогательный полипептид по первому аспекту настоящего изобретения, мультимерный CAR по второму аспекту настоящего изобретения, сигнальную систему CAR по третьему аспекту настоящего изобретения; или сцепленный комплекс, описанный выше.
Клетка может представлять собой Т-клетку или NK-клетку.
В восьмом аспекте настоящее изобретение относится к фармацевтической композиции, которая содержит клетку по седьмому аспекту настоящего изобретения.
В девятом аспекте настоящее изобретение относится к клетке по седьмому аспекту настоящего изобретения для применения в лечении заболевания.
В десятом аспекте настоящее изобретение относится к применению клетки по седьмому аспекту настоящего изобретения в производстве лекарственного средства для лечения заболевания.
В одиннадцатом аспекте настоящее изобретение относится к способу лечения заболевания, который включает стадию введения субъекту клетки по седьмому аспекту настоящего изобретения.
Заболевание может представлять собой рак, например, хронический лимфоцитарный лейкоз (CLL), меланому или миелому.
В двенадцатом аспекте настоящее изобретение относится к набору, который содержит нуклеиновую кислоту по четвертому аспекту настоящего изобретения, нуклеотидную конструкцию по пятому аспекту настоящего изобретения или вектор по шестому аспекту настоящего изобретения.
В тринадцатом аспекте настоящее изобретение относится к набору, который включает клетку по седьмому аспекту настоящего изобретения.
В четырнадцатом аспекте настоящее изобретение относится к способу изготовления клетки по седьмому аспекту настоящего изобретения, который включает стадию введения нуклеиновой кислоты по четвертому аспекту настоящего изобретения, нуклеотидной конструкции по пятому аспекту настоящего изобретения или вектора по шестому аспекту настоящего изобретения.
Клетка может быть выделена из образца, полученного от субъекта.
Использование суперспирального домена в качестве спейсера в CAR обеспечивает ряд преимуществ по сравнению со спейсерами, которые использовались ранее, такими как Fc-домены из IgG.
Например, использование суперспирального домена обеспечивает размеры спейсера, изменяемые с шагом 0,15 нм. Добавление или вычитание отдельных аминокислот или нескольких аминокислот означает, что размер суперспирального спейсера может изменяться ступенчато. В отличие от этого, использование IgG-спейсеров допускает только добавление или удаление целых иммуноглобулиновых доменов. Это означает, что наименьший шаг при изменениях составляет приблизительно 4 нм (т.е. размер уложенного иммуноглобулинового домена).
Суперспиральные домены кодируются меньшим ДНК-фрагментом (например, приблизительно 100 нуклеотидами) по сравнению с Fc-доменом из IgG (приблизительно 700 нуклеотидов). Это позволяет уменьшить размер ДНК-вектора, что является важным для улучшения вирусного титра и эффективности трансдукции.
Использование суперспирального спейсера позволяет выбор из большого числа суперспиральных спейсеров, которые не будут перекрестно гибридизоваться с другими суперспиральными доменами. Это отличает их от других спейсеров, где существует более ограниченное число вариантов спейсеров.
Использование суперспирального спейсера также обеспечивает большую гибкость с точки зрения дизайна CAR, чем классический CAR. Например, он позволяет образовывать гомо- или гетеро-олигомерные комплексы CAR. Гетеро-олигомерные комплексы CAR полезны при конструировании мультицепочечных CAR с эндодоменами CD28/OX40/41ВВ и TCRz для того, чтобы гарантировать, что каждый из эндодоменов находится в оптимальной близости от мембраны и присутствует в желаемых соотношениях.
Авторы настоящего изобретения сконструировали гиперчувствительной CAR без изменения биофизических свойств связывающего домена. Это является желательным, поскольку способы снижения аффинности связывающих фрагментов CAR не поддаются прогнозированию и часто имеют неустановленную специфичность.
Сверхчувствительность CAR обеспечивается за счет увеличения валентности CAR. В частности, использования суперспирального спейсерного домена, который способен взаимодействовать, образуя мультимер, содержащий более двух CAR, что повышает чувствительность к мишеням, экспрессирующих лиганды с низкой плотностью, благодаря усилению ITAM и авидности в олигомерном комплексе CAR.
Чувствительность может быть увеличена путем увеличения соотношения суперспиральный домен-сигнальный домен и scFv-суперспиральный домен, так что каждый scFv соединен с множеством сигнальных элементов (см. фиг. 10с). Чувствительность может также быть повышена путем образования комплексов мультимерных CAR (фиг. 10f).
В мультимерном CAR по изобретению, сигнальные эндодомены находятся в trans-положении близко к мембране, что позволяет точную подстройку комбинаций Т-клеточных сигнальных доменов (фиг. 10а и b); и включено более трех различных внутриклеточных сигнальных доменов (фиг. 10d), так что структура включает в себя больше эндодоменных сигналов, чем третье поколение CAR (фиг. 1d).
Использование отдельной молекулы компонента внутриклеточной сигнализации, которая гетеродимеризуется с CAR внутри клетки, обеспечивает дополнительное усиление благодаря количеству эндодоменов на один антигенсвязывающий домен, давая «супер-CAR» (фиг. 15).
Мультимерный CAR по изобретению может иметь больше одной антигенсвязывающей специфичности, что позволяет направленное взаимодействие с множеством эпитопов или антигенов (фиг. 10е).
Мультимерный CAR, имеющий множество связывающих доменов, будет иметь гораздо большую авидность, чем классический гомодимерный CAR. Это может являться важным, например, для связывания доменов с низкой аффинностью, поскольку накопленная сила множества взаимодействий обеспечивает высокую специфичность связывания. Мультимерный CAR может связывать антиген аналогично IgM, который содержит несколько ковалентно связанных иммуноглобулинов, образуя пентамерную или гексамерную структуру.
ОПИСАНИЕ ФИГУР
Фигура 1 - а) Схематическая диаграмма, иллюстрирующая классический CAR. (b)-(d): Разные поколения и перестановка эндодоменов в CAR: (b) первоначальные конструкции передавали ITAM-сигналы только через эндодомен FcεR1-γ или CD3ζ, тогда как более поздние конструкции передавали дополнительные (с) один или (d) два костимулирующих сигнала в одном сложном эндодомене.
Фигура 2 - а) Схематическая диаграмма, иллюстрирующая классический CAR. b) Схематическая диаграмма, иллюстрирующая стандартные спейсеры в CAR. «S» обозначает дисульфидные связи.
Фигура 3 - Природные димерные, тримерные и тетрамерные суперспиральные структуры (измененное изображение из Andrei N. Lupas and Markus Gruber; Adv Protein Chem. 2005; 70:37-78).
Фигура 4 - Кристаллическая структура пентамерного суперспирального мотива из олигомерного матриксного белка хряща (СОМР) и IgG1 человека. Отдельные цепи изображены в разном цвете. Суперспиральная структура СОМР представлена с N-конца с С-концом, уходящим в страницу, и также представлена сбоку с С-концом слева и N-концом справа. IgG1 человека представлен в боковом изображении с N-концом вверху и С-концом снизу.
Фигура 5 - Суперспиральный спейсер CAR. а) Схематическая диаграмма, иллюстрирующая CAR с пентамерным суперспиральным спейсером, полученным из СОМР. b) Схема конструкции, изображающая ORF для COMP-CAR и контрольного CAR. с) Аминокислотная последовательность ORF из анти-CD33-COMP-CAR и анти-ROR-1-COMP-CAR. d) ДНК-последовательность ORF из анти-CD33-COMP-CAR и анти-ROR-1-COMP-CAR.
Фигура 6 - Уровень экспрессии суперспиральных CAR на поверхности, а) Мышиная Т-клеточная линия была трансдуцирована анти-CD33 СОМР CAR или анти-CD33 IgG1 CAR. Эти клетки затем окрашивали химерным растворимым CD33, слитым с мышиным Fc из IgG2a, перед вторичным окрашиванием конъюгированными с РЕ антителами к мышиным IgG. b) Мышиная Т-клеточная линия была трансдуцирована анти-ROR-l-COMP-CAR или анти-ROR-l-IgG1-CAR. Эти клетки затем окрашивали растворимым His-меченым ROR-1 с последующим вторичным окрашиванием анти-His-биотином и последующим третьим окрашиванием стрептавидином-АРС.
Фигура 7 - Стимуляция анти-ROR-1-COMP-CAR-T-клеток иммобилизованным лигандом. Трансдуцированные мышиные Т-клетки культивировали совместно с анти-His шариками, которые были предварительно покрыты различными концентрациями супернатанта с растворимым His-меченым ROR-1. Количество IL-2 в супернатанте после совместного культивирования анализировали через 16-24 часа с помощью ELISA.
Фигура 8 - Уровень экспрессии ROR-1 на клетках-мишенях. Линия клеток SKW в норме экспрессирует низкий уровень ROR-1. Эти клетки были трансдуцированы ROR-1, чтобы повысить уровень экспрессии. Эти клетки окрашивали анти-ROR-1-APC и сравнивали с неокрашенными клетками.
Фигура 9 - Стимуляция анти-ROR-1-COMP-CAR-T-клеток положительными по ROR-1 клетками SKW. Трансдуцированные мышиные Т-клетки совместно культивировали с клетками-мишенями SKW, которые экспрессируют лиганд ROR-1 с низкой или высокой плотностью. Число Т-клеток поддерживали на постоянном уровне, а количество клеток-мишеней было различным. Количество IL-2 в супернатанте после совместного культивирования анализировали через 16-24 часа с помощью ELISA. Серая область обозначает диапазон стандартной кривой для этого эксперимента. Синяя пунктирная линия представляет собой средние величины секретируемого IL-2 после стимуляции РМА и иономицином. Красная пунктирная линия представляет собой средние величины IL-2, обнаруживаемые в культуре одних Т-клеток (не стимулированных).
Фигура 10 - Конструкции суперспиральных CAR. а) Схематическая диаграмма, иллюстрирующая CAR из формирующего CAR полипептида и вспомогательного полипептида. Формирующий CAR полипептид обеспечивает первый сигнал в Т-клетке и состоит из связывающего элемента scFv на N-конце, за которым следует спейсер СОМР, трансмембранный домен и TCRz. Вспомогательный полипептид обеспечивает третий сигнал в Т-клетке и не имеет N-концевого связывающего лиганда, а начинается со спейсера СОМР с последующими трансмембранным доменом и сигнальньш мотивом 41ВВ; b) схематическая диаграмма, иллюстрирующая другую суперспиральную CAR-систему, которая включает формирующий CAR полипептид и два вспомогательных полипептида. Формирующий CAR полипептид обеспечивает первый сигнал в Т-клетке и состоит из связывающего элемента scFv на N-конце, за которым следует спейсер СОМР, трансмембранный домен и TCRz. Первый вспомогательный полипептид обеспечивает второй сигнал в Т-клетке и не имеет N-концевого связывающего лиганда, а начинается со спейсера СОМР с последующими трансмембранным доменом и сигнальным мотивом CD28. Второй вспомогательный полипептид обеспечивает третий сигнал в Т-клетке и не имеет N-концевого связывающего лиганда, а начинается со спейсера СОМР с последующими трансмембранным доменом и сигнальным мотивом 41ВВ. «S» обозначает дисульфидные связи. Такое расположение позволяет сигнальным эндодоменам находится в trans-положении близко к мембране, что усиливает сигнал; с) схематическая диаграмма, иллюстрирующая другую суперспиральную CAR-систему, в которой соотношение scFv:суперспиральный домен намного меньше соотношения суперспиральный домен:сигнальный домен, так что каждый scFv-фрагмент присоединен к нескольким сигнальным элементам. Эта организация комплекса представляет собой усилительную систему: в результате ограничения числа scFv до одного на комплекс каждый активированный лиганд будет передавать сигнал через 5 TCRz-цепей (два лиганда через 10 TCRz). Это является отличием от традиционных димерных CAR, где два лиганда передают сигнал через 2 цепи TCRz. Эта система будет существенно усиливать инициацию сигнала 5 раз; d) схематическая диаграмма, иллюстрирующая другую суперспиральную CAR-систему, которая содержит больше эндодоменов, чем CAR третьего поколения. Суперспиральный спейсер позволяет введение двух дополнительных сигнальных доменов («EXTRA») относительно традиционного CAR третьего поколения; е) схематическая диаграмма, иллюстрирующая другую суперспиральную CAR-сигнальную систему, которая имеет несколько мишеней (в данном случае показаны участки связывания с двумя мишенями: один участок связывает лиганд А, и другой участок связывает лиганд В). Такой дизайн является альтернативой для архитектуры TanCAR; f) схематическая диаграмма, иллюстрирующая другую суперспиральную CAR-систему, которая содержит элемент, образующий связь, такую как дисульфидный мостик, с другим содержащим суперспиральный спейсер CAR, давая сцепленный комплекс, который дополнительно увеличивает валентность scFv:сигнальный домен. Подобно дизайну, показанному на фиг. 10с, это система является амплифицирующей. Каждый сцепленный комплекс будет передавать сигнал через 10 цепей TCRz. Это является отличием от традиционных димерных CAR, передающих сигнал через цепи 2 TCRz. Эта система будет существенно усиливать инициацию сигнала в 5 раз. Замена шарнирного домена IgG тримерной суперспиральной структурой приведет к увеличению без малого в 8 раз, тогда как тетрамерная биспираль усилила бы инициацию в 10 раз.
Фигура 11 - Укорачивание СОМР-спейсера
a) Схематическая диаграмма, показывающая анти-ROR-l-COMP CAR, где СОМР-спейсер укорочен с N-конца с 45 аминокислот до «X» аминокислот,
b) клетки 293Т, трансфицированные укороченными конструкциями и проанализированные с помощью FACS.
Фигура 12 - Схематическая диаграмма, показывающая мультимерные и классические CAR, протестированные в примере 6:
А) гетеромультимерный CAR, который включает: полипептид, имеющий анти-CD19-антигенсвязывающий домен; суперспиральный спейсерный домен и эндодомен CD3-дзета; и вспомогательный полипептид, имеющий суперспиральный спейсерный домен и эндодомен 41BB. CAR кодируется бицистронной конструкцией, имеющей следующую структуру: aCD19fmc63-COMP-CD28tmZ-2A-COMP-CD28tm-41ВВ. В этой структуре CAR сигнальные мотивы 41ВВ и TCR-дзета расположены параллельно.
B) гомомультимерный CAR из полипептидов, содержащих анти-CD19-антигенсвязывающий домен; суперспиральный спейсерный домен и комбинированный эндодомен 41ВВ/CD3-дзета. CAR кодируется конструкцией, имеющей следующую структуру: aCD19fmc63-COMP-CD8TM-41BB-Z. В этой структуре CAR сигнальные мотивы 41ВВ и TCR-дзета расположены последовательно.
C) классический гомодимерный CAR второго поколения, который состоит из двух полипептидов, имеющих анти-CD19-антигенсвязывающий домен, спейсерный стеблевой домен CD8 и комбинированный эндодомен 41ВВ/CD3-дзета. CAR кодируется бицистронной конструкцией, которая также кодирует ген самоубийства RQR8. Конструкция имеет следующую структуру: RQR8-2A-aCD19fmc63-CD8STK-41BBZ.
Фигура 13 - Уничтожение клеток-мишеней CD19+SupT1 с использованием CAR, показанных на фиг. 12, на 2-й день.
Фигура 14 - Уничтожение клеток-мишеней CD19+SupT1 с использованием CAR, показанных на фиг. 12, на 5-й день.
Фигура 15 - Схематическая диаграмма конструкций суперспиральных супер-CAR, протестированных в примере 7:
aCD19-IgGFc-Z - классический гомодимерный CAR, содержащий на молекулу 2 молекулы TCRz, имеющей fmc63 в качестве CD19-связывающего элемента.
Суперспиральный супер-CAR из пяти полипептидов, каждый из которых содержит четыре отдельных AD1-домена. Поэтому суперспиральный супер-CAR содержит 20 AD1-доменов.
COMP_x4AD1 - суперспиральный cynep-CAR был протестирован в сочетании с сигнальным компонентом, не содержащим копий сигнального домена TCR-дзета. Он использовался в качестве отрицательного контроля.
COMP_x4AD1+Z-DDD1-Z - суперспиральный супер-CAR был протестирован в сочетании с сигнальным компонентом, имеющим 2 копии сигнального домена TCR-дзета. Поскольку DDD1 связывает AD1 со стехиометрией 2:1, эти сигнальные домены дают 80 копий домена TCR-дзета для каждой мишени 5-полипептидного суперспирального CAR.
Фигура 16 - Высвобождение IL-2 после стимуляции клетками-мишенями, экспрессирующими специфичный антиген (CD19), при различных концентрациях: низкой, средней и высокой.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (CAR)
Классические CAR, которые схематически показаны на фиг. 1, представляют собой химерные трансмембранные белки I типа, которые соединяют внеклеточные антиген-распознающие домены (связывающие элементы) с внутриклеточным сигнальным доменом (эндодоменом). Связывающий элемент обычно представляет собой одноцепочечный вариабельный фрагмент (scFv), полученный из моноклональных антител (мАТ), но он может иметь другие форматы, которые содержат антителоподобный или лигандный антигенсвязывающий сайт. Трансмембранный домен удерживает белок в клеточной мембране и соединяет спейсер с эндодоменом.
Ранние конструкции CAR имели эндодомены из внутриклеточных частей гамма-цепи FcεR1 или CD3ζ. Поэтому эти рецепторы первого поколения передают иммунологический сигнал 1, который является достаточным для того, чтобы запустить Т-клеточный лизис распознаваемых клеток-мишеней, но не может полностью активировать пролиферацию и жизнеспособность Т-клеток. Чтобы преодолеть это ограничение, были сконструированы сложные эндодомены: слияние внутриклеточной части Т-клеточной костимуляторной молекулы с внутриклеточной частью из CD3ζ дало рецепторы второго поколения, которые могут передавать активирующий и костимуляторный сигнал одновременно после распознавания антигена. Наиболее часто используемым костимуляторным доменом является домен из CD28. Он обеспечивает самый сильный костимуляторный сигнал, а именно, иммунологический сигнал 2, который запускает пролиферацию Т-клеток. Также были описаны несколько рецепторов, которые включают в себя эндодомены из семейства TNF-рецепторов, такие как близкородственные OX40 и 41ВВ, которые передают сигналы выживаемости. Уже описаны еще более сильные CAR третьего поколения, которые имеют эндодомены, способные передавать сигналы активации, пролиферации и выживаемости.
Кодирующие CAR нуклеиновые кислоты могут быть перенесены в Т-клетки, например, с использованием ретровирусных векторов. Таким образом можно создать большое количество антигенспецифичных Т-клеток для адоптивного переноса клеток. Когда CAR связывает антиген-мишень, это приводит к передаче активирующего сигнала в Т-клетке, на которой он экспрессируется. Таким образом, CAR направляет специфичность и цитотоксичность Т-клеток к клеткам, экспрессирующим антиген-мишень.
CAR по настоящему изобретению содержит антигенсвязывающий домен, суперспиральный спейсерный домен, трансмембранный домен и эндодомен. Суперспиральный спейсерный домен обеспечивает многочисленные преимущества относительно спейсеров, описанных ранее в данной области техники.
СУПЕРСПИРАЛЬНЫЙ ДОМЕН
CAR, как правило, содержат спейсерную последовательность, соединяющую антигенсвязывающий домен с трансмембранным доменом. Спейсер позволяет антигенсвязывающему домену иметь соответствующую ориентацию и быть доступным для антигена. Спейсер также блокирует доступ фосфатазам при взаимодействии с лигандом.
CAR по настоящему изобретению содержит суперспиральный спейсерный домен.
Биспираль представляет собой структурный мотив, в котором от двух до семи альфа-спиралей свернуты вместе, как нити в канате (фиг. 3). Многие эндогенные белки включают суперспиральные домены. Суперспиральный домен может участвовать в сворачивании белка (например, он взаимодействует с несколькими альфа-спиральными мотивами в пределах одной и той же белковой цепи) или может отвечать за белок-белковые взаимодействия. В последнем случае биспираль может инициировать образование гомо- или гетеро-олигомерных структур.
Используемые в настоящем документе термины «мультимер» и «мультимеризация» являются синонимами и взаимозаменяемыми с «олигомером» и «олигомеризацией».
Структура суперспиральных доменов хорошо известна в данной области. Например, она описана Lupas & Gruber (Advances in Protein Chemistry; 2007; 70; 37-38).
Биспирали обычно содержат повторяющийся паттерн, hxxhcxc, из гидрофобных (h) и заряженных (с) аминокислотных остатков, называемый гептадным повтором. Позиции в гептадном повторе обычно помечены ABCDEFG, где and являются гидрофобными позициями, часто занимаемыми изолейцином, лейцином или валином. Сворачивание последовательности с этим повторяющимся паттерном в альфа-спиральную вторичную структуру заставляет гидрофобные остатки выстраиваться в виде «полосы», которая аккуратно закручивается вокруг спирали в левую сторону, образуя амфипатическую структуру. Наиболее выгодным способом нахождения двух таких спиралей в цитоплазме является сворачивание гидрофобных нитей друг против друга между гидрофильными аминокислотами. Таким образом, погружение гидрофобных поверхностей обеспечивает термодинамическую движущую силу олигомеризации. Упаковка в поверхности раздела между биспиралями является исключительно плотной с почти полным ван-дер-Ваальсовым контактом между боковыми цепями остатков a и d.
Альфа-спирали могут быть параллельными или антипараллельными, и обычно имеют левозакрученную биспираль. Хотя они не являются предпочтительными, в природе также было найдено несколько правозакрученных биспиралей, а также они существуют в сконструированных белках.
Суперспиральный домен может являться любым суперспиральным доменом, который способен образовывать суперспиральный мультимер, так что образуется комплекс CAR или вспомогательных полипептидов, содержащих суперспиральные домены.
Зависимость между последовательностью и окончательной свернутой структурой суперспирального домена хорошо известны в данной области (Mahrenholz et al.; Molecular & Cellular Proteomics; 2011; 10 (5):M110.004994). Как таковой, суперспиральный домен может представлять собой искусственно созданный суперспиральный домен.
Примеры белков, которые содержат суперспиральный домен, включают, но не ограничиваются ими, кинезиновый моторный белок, дельта-антиген гепатита D, коровый белок бокса C/D sRNP архей, олигомерный матриксный белок хряща (СОМР), маннозосвязывающий белок А, суперспиральный серин-богатый белок 1, полипептид-высвобождающий фактор 2, SNAP-25, SNARE, Lac-репрессор или аполипопротеин Е.
Последовательность различных суперспиральных доменов показана ниже:
Кинезиновый моторный белок: параллельный гомодимер (SEQ ID NO: 30)
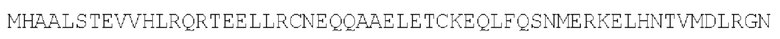
Дельта-антиген гепатита D: параллельный гомодимер (SEQ ID NO: 31)
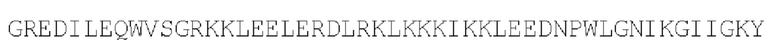
Коровый белок бокса C/D sRNP архей: антипараллельный гетеродимер (SEQ ID NO: 32)
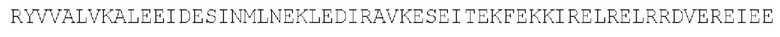
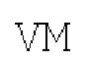
Маннозосвязывающий белок А: параллельный гомотример (SEQ ID NO: 33)
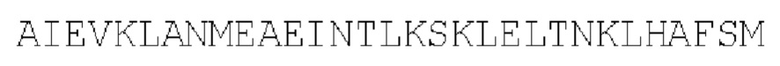
Суперспиральный серин-богатый белок 1: параллельный гомотример (SEQ ID NO: 34)
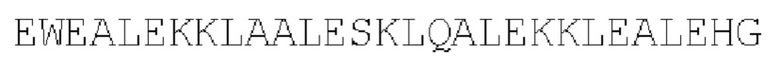
Полипептид-высвобождающий фактор_2: антипараллельный гетеротример
Цепь А:  (SEQ ID NO: 35)
(SEQ ID NO: 35)
Цепь В: 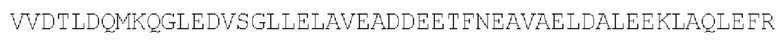 (SEQ ID NO: 36)
(SEQ ID NO: 36)
SNAP-25 и SNARE: параллельный гетеротетрамер
Цепь А: 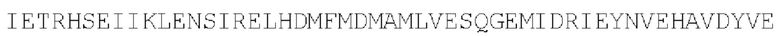 (SEQ ID NO: 37)
(SEQ ID NO: 37)
Цепь В: 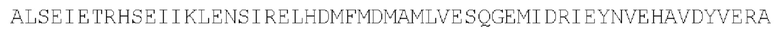
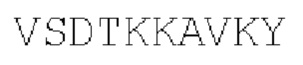 (SEQ ID NO: 38)
(SEQ ID NO: 38)
Цепь С:
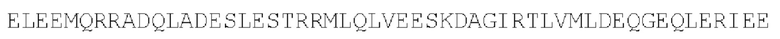
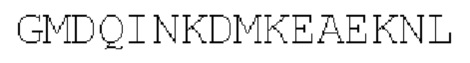 (SEQ ID NO: 39)
(SEQ ID NO: 39)
Цепь D: 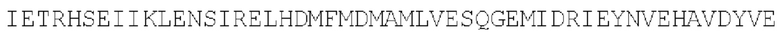 (SEQ ID NO: 40)
(SEQ ID NO: 40)
Lac-репрессор: параллельный гомотетрамер
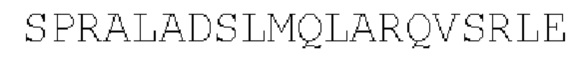 (SEQ ID NO: 41)
(SEQ ID NO: 41)
Аполипопротеин E: антипараллельный гетеротетрамер
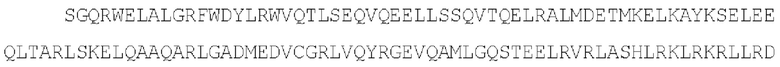
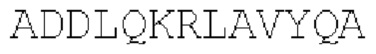 (SEQ ID NO: 42)
(SEQ ID NO: 42)
Суперспиральный домен способен олигомеризоваться. В некоторых вариантах осуществления суперспиральный домен может быть способен образовывать тример, тетрамер, пентамер, гексамер или гептамер.
Суперспиральный домен отличается от лейциновой молнии. Лейциновая молния представляет собой супер-вторичные структуры, которые функционируют в качестве димеризующихся доменов. Их присутствие создает силу притяжения в параллельных альфа-спиралях. Одна лейциновая молния состоит из множества остатков лейцина с интервалами из приблизительно 7 остатков, которые образуют амфипатическую альфа-спираль с гидрофобной областью, проходящей с одной стороны. Этот гидрофобный участок обеспечивает область димеризации, позволяя мотивам соединяться, «застегиваясь на молнию». Лейциновые молнии, как правило, имеют от 20 до 40 аминокислот в длину, например, приблизительно 30 аминокислот.
Лейциновые молнии, как правило, образованы двумя различными последовательностями, например, кислая лейциновая молния гетеродимеризуется с основной лейциновой молнией. Примером лейциновой молнии являются стыковочный домен (DDD1) и якорный домен (AD1), которые описаны более подробно ниже.
Лейциновые молнии образуют димеры, тогда как суперспиральные спейсеры по настоящему изобретению образуют мультимеры (тримеры и мультимеры более высокого порядка). Лейциновые молнии гетеродимеризуются в димеризующемся участке последовательности, тогда как суперспиральные домены гомодимеризуются.
В одном варианте осуществления настоящее изобретение относится к суперчувствительному CAR.
Суперчувствительный CAR обеспечивается за счет увеличения валентности CAR. В частности, использование суперспирального домена, который способен взаимодействовать с образованием мультимера, содержащего более двух суперспиральных доменов, и, следовательно, более двух CAR, повышает чувствительность к мишеням, экспрессирующим лиганды с низкой плотностью, благодаря увеличению количества присутствующих ITAM и авидности олигомерного комплекса CAR.
Таким образом, в одном варианте осуществления настоящий формирующий CAR полипептид содержит суперспиральный спейсерный домен, который позволяет мультимеризацию по меньшей мере трех формирующих CAR полипептидов. Другими словами, CAR содержит суперспиральный домен, которым способен образовывать тример, тетрамер, пентамер, гексамер или гептамер из суперспиральных доменов.
Примеры суперспиральных доменов, которые способны образовывать мультимер, содержащий более двух суперспиральных доменов, включают, но не ограничиваются ими, олигомерный матриксный белок хряща (СОМР), маннозосвязывающий белок А, суперспиральный серин-богатый белок 1, полипептид-высвобождающий фактор 2, SNAP-25, SNARE, Lac-репрессор или аполипопротеин Е (см. SEQ ID NO: 30-42 выше).
Суперспиральный домен может представлять собой суперспиральной домен из СОМР.
СОМР является одним из наиболее стабильных белковых комплексов в природе (стабилен в диапазоне от 0°С-100°С и в широком диапазоне значений рН) и может денатурироваться только в 4-6М гидрохлориде гуанидина. Суперспиральный домен СОМР способен образовывать пентамер. СОМР является также эндогенно экспрессируемым белком, который в природе экспрессируется в межклеточное пространство. Это снижает риск иммуногенности по сравнению с синтетическими спейсерами. Кроме того, решена кристаллическая структура суперспирального мотива СОМР, что дает точную оценку длины спейсера (фиг. 4). Структура СОМР имеет приблизительно 5,6 нм в длину (по сравнению с длиной шарнирного и СН2СН3-доменов IgG человека, которая составляет приблизительно 8,1 нм).
Суперспиральный домен может состоять из или содержать последовательность, показанную в SEQ ID NO: 1, или ее фрагмент.SEQ ID NO: 1

Как показано на фиг. 11, можно укоротить суперспиральный домен СОМР на N-конце и сохранить экспрессию на поверхности. Поэтому суперспиральный домен может включать или состоять из укороченного варианта SEQ ID NO: 1, который укорочен с N-конца. Укороченный СОМР может содержать 5 С-концевых аминокислот SEQ ID NO: 1, то есть последовательности CDACG. Укороченный СОМР может содержать от 5 до 44 аминокислот, например, по меньшей мере 5, 10, 15, 20, 25, 30, 35 или 40 аминокислот. Укороченный СОМР может соответствовать С-концу SEQ ID NO: 1. Например укороченный СОМР, содержащий 20 аминокислот, может содержать последовательность 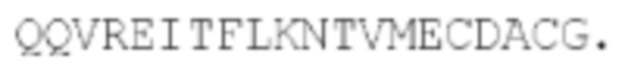 Укороченный СОМР может сохранить остаток(ки) цистеина, участвующие в мультимеризации. Укороченный СОМР может сохранять способность образовывать мультимеры.
Укороченный СОМР может сохранить остаток(ки) цистеина, участвующие в мультимеризации. Укороченный СОМР может сохранять способность образовывать мультимеры.
Известны различные суперспиральные домены, которые образуют гексамеры, такие как gp41 из HIV и искусственно разработанная белковая гексамерная биспираль, описанная N. Zaccai et al. (2011) Nature Chem. Bio., (7) 935-941). Мутантная форма лейциновой молнии GCN4-pl образует гептамерную суперспиральную структуру (J. Liu. et al., (2006) PNAS (103) 154 57-154 62).
Суперспиральный домен может содержать вариант одного из суперспиральных доменов, описанных выше, при условии, что вариант последовательности сохраняет способность образовывать суперспиральный олигомер. Например, суперспиральный домен может содержать вариант последовательности, показанной в SEQ ID NO: 1 или 30-42, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности по последовательности, при условии, что вариант последовательности сохраняет способность образовывать суперспиральный олигомер.
Процент идентичности между двумя полипептидными последовательностями может быть легко определен с помощью таких программ, как BLAST, которая находится в свободном доступе по адресу http://blast.ncbi.nlm.nih.gov.
АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН
Антигенсвязывающий домен представляет собой часть классического CAR, который распознает антиген.
В данной области техники известно множество антигенсвязывающих доменов, включая домены на основе антигенсвязывающего сайта антитела, миметиков антител и Т-клеточных рецепторов. Например, антигенсвязывающий домен может содержать: одноцепочечный вариабельным фрагмент (scFv), полученный из моноклонального антитела; природный лиганд антигена-мишени; пептид с достаточной аффинностью к мишени; однодоменный связывающий элемент, такой как антитело верблюдовых; искусственный связывающий элемент как DARPin; или одноцепочечный связывающий элемент, полученный из Т-клеточного рецептора.
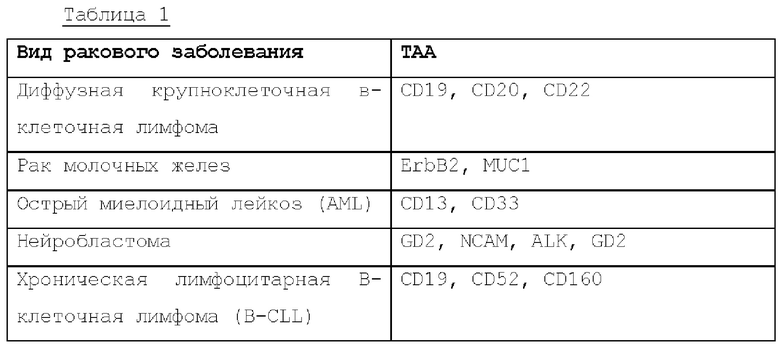
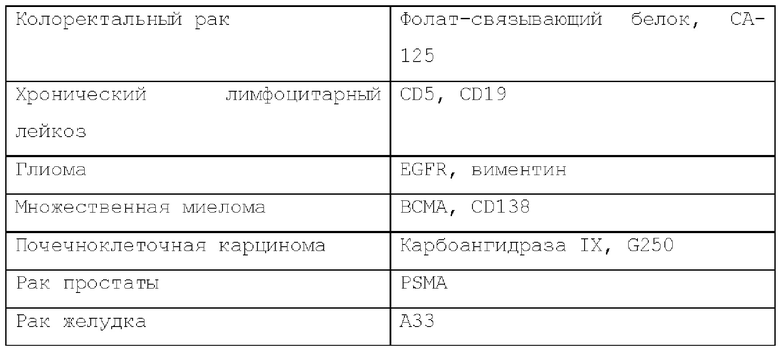
Известны различные опухоле-ассоциированные антигены (ТАА), показанные в нижеследующей таблице 1. Антигенсвязывающий домен, используемый в настоящем изобретении, может представлять собой домен, способный связываться с ТАА, указанной в ней.
В некоторых вариантах осуществления настоящее изобретение относится к суперчувствительному CAR, который способен стимулировать активацию клеток в ответ на антиген, который экспрессируется на клетке-мишени с низкой плотностью.
Антигенсвязывающий домен может связывать ТАА, который экспрессируется на клетке, например, раковой клетке, с низкой плотностью. ТАА, экспрессирующийся с низкой плотностью, может относиться, например, к ТАА, экспрессирующемуся на уровне от 10 до 1000 молекул на клетку.
Примеры ТАА, которые, как известно, экспрессируются с низкой плотностью в некоторых видах рака, включают, но не ограничиваются ими, ROR1 в CLL, Typr-1 в меланоме и ВСМА в миеломе.
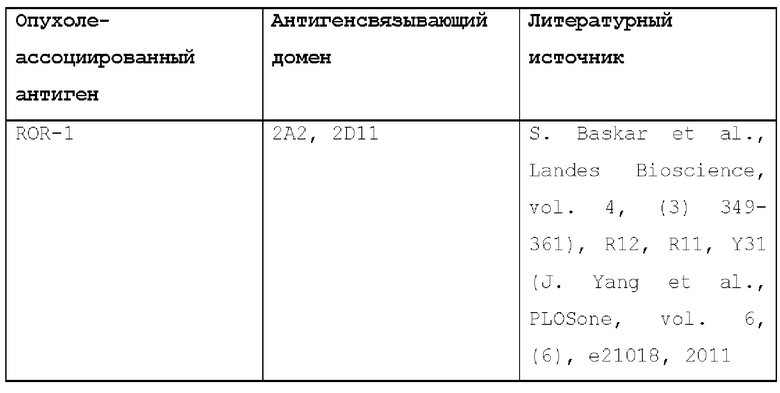
Антигенсвязывающие домены (такие как scFv или мАТ), которые связывают эти ТАА, были описаны ранее, например, приведенные в следующей таблице:

ТРАНСМЕМБРАННЫЙ ДОМЕН
Трансмембранный домен представляет собой последовательность из CAR, которая проходит сквозь мембрану. Он может включать в себя гидрофобную альфа-спираль. Трансмембранный домен может быть получен из CD28, который дает хорошую стабильность рецептора.
СИГНАЛЬНЫЙ ПЕПТИД
Формирующие CAR полипептиды и/или вспомогательные полипептиды по настоящему изобретению могут содержать сигнальный пептид, так что, когда он экспрессируется в клетке, такой, как Т-клетка, синтезируемый белок направляется в эндоплазматический ретикулум, а затем на поверхность клетки, в которой он экспрессируется.
Основа сигнального пептида может содержать длинную цепочку гидрофобных аминокислот, которая имеет тенденцию к образованию одиночной альфа-спирали. Сигнальный пептид может начинаться коротким участком положительно заряженных аминокислот, который помогает обеспечивать правильную топологию полипептида во время переноса белка. На конце сигнального пептида обычно имеется аминокислотный участок, который распознается и расщепляется сигнальной пептидазой. Сигнальная пептидаза может осуществлять расщепление либо во время, либо после завершения переноса белка, давая свободный сигнальный пептид и зрелый белок. Свободные сигнальные пептиды затем расщепляются специфичными протеазами.
Сигнальный пептид может находиться на аминоконце молекулы.
Сигнальный пептид может содержать последовательность, показанную в SEQ ID NO: 2, 3 или 4, или ее вариант, имеющий 5, 4, 3, 2 или 1 мутаций аминокислот (инсерций, замен или дополнений), при условии, что сигнальный пептид все еще функционирует, обеспечивая экспрессию CAR на клеточной поверхности.
SEQ ID NO: 2: 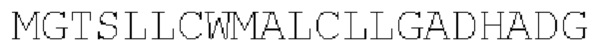
Сигнальный пептид из SEQ ID NO: 2 является компактным и высокоэффективным и получен из бета-цепи TCR. Предполагается, что он обеспечивает примерно 95%-е расщепление после концевого глицина, давая эффективное удаление с помощью сигнальной пептидазы.
SEQ ID NO: 3:
Сигнальный пептид SEQ ID NO: 3 получен из IgG1
SEQ ID NO: 4: 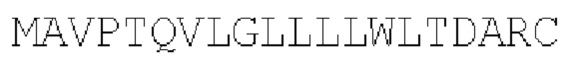
Сигнальный пептид из SEQ ID NO: 4 получен из CD8a.
ЭНДОДОМЕН
Эндодомен является участком классического CAR, который расположен на внутриклеточной стороне мембраны.
Эндодомен представляет собой участок передачи сигнала классического CAR. После распознавания антигена антигенсвязывающим доменом отдельные молекулы CAR образуют кластер, нативные CD45 и CD148 исключаются из синапса, и сигнал передается в клетку.
Эндодомен из суперспирального спейсерного CAR может представлять собой или содержать внутриклеточный сигнальный домен. В альтернативном варианте осуществление эндодомен настоящего CAR может быть способен взаимодействовать с внутриклеточной сигнальной молекулой, которая присутствует в цитоплазме, что приводит к передаче сигнала.
Внутриклеточный сигнальный домен или отдельная внутриклеточная сигнальная молекула может представлять собой или содержать Т-клеточный сигнальный домен.
Наиболее часто используемым сигнальным доменом является эндодомен CD3-дзета, который содержит 3 ITAM. Он передает сигнал активации в Т-клетке после связывания антигена. CD3-дзета не может обеспечить полностью компетентный активационный сигнал, и может быть необходима дополнительная костимуляторная передача сигнала. Например, для передачи сигнала на пролиферацию/выживание совместно с CD3-дзета может быть использован химерный белок CD28 и ОХ40, или все три могут быть использованы совместно (как показано на фиг. 1В).
Настоящий CAR может содержать только эндодомен CD3-дзета, эндодомен CD3-дзета с эндодоменом из CD28 или из ОХ40, либо эндодомен CD28 и эндодомен ОХ40 и CD3-дзета (фиг. 1).
Эндодомен может содержать один или несколько из следующих вариантов: эндодомен ICOS, эндодомен CD27, эндодомен BTLA, эндодомен CD30, эндодомен GITR и эндодомен HVEM.
Эндодомен может содержать последовательность, показанную в SEQ ID NO: 5-13, или ее вариант, имеющий по меньшей мере 80% идентичности по последовательности.
SEQ ID NO: 5 - эндодомен CD3-дзета
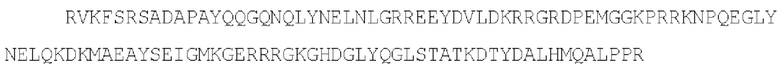
SEQ ID NO: 6 - эндодомены CD28 и CD3-дзета

SEQ ID NO: 7 - эндодомены CD28, ОХ40 и CD3-дзета

SEQ ID NO: 8 - эндодомен ICOS

SEQ ID NO: 9 - эндодомен CD27
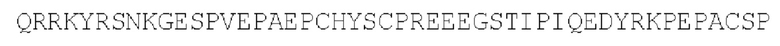
SEQ ID NO: 10 - эндодомен BTLA

SEQ ID NO: 11 - эндодомен CD30

SEQ ID NO: 12 - эндодомен GITR

SEQ ID NO: 13 - эндодомен HVEM
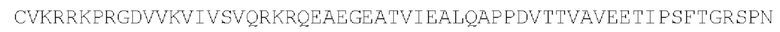

Вариант последовательности может иметь по меньшей мере, 80%, 85%, 90%, 95%, 98% или 99% идентичности по последовательности с SEQ ID NO: 5-13 при условии, что последовательность обеспечивает действующий внутриклеточный сигнальный домен.
ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR)
В одном аспекте настоящее изобретение относится к CAR, содержащему формирующий CAR полипептид по первому аспекту настоящего изобретения и вспомогательный полипептид, который включает (i) суперспиральный спейсерный домен; (ii) трансмембранный домен; и (iii) эндодомен, где суперспиральный спейсерный домен вспомогательного полипептида способен взаимодействовать с суперспиральным доменом формирующего CAR полипептида.
Формирующий CAR полипептид обеспечивает антигенсвязывающий домен и, следовательно, антигенную специфичность.
Вспомогательный полипептид обеспечивает дополнительный эндодомен, который может быть использован для создания желаемого сигнального ответа. Это является преимуществом относительно сложного сигнального домена, поскольку каждый сигнальный домен остается свободным от других сигнальных доменов. Кроме того, это позволяет каждому сигнальному домену находиться в оптимальной близости к мембране для передачи сигнала.
Эндодомен формирующего CAR полипептида может содержать по меньшей мере первый внутриклеточный сигнальный домен; а эндодомен вспомогательного полипептида может содержать по меньшей мере второй внутриклеточный сигнальный домен. Например, один эндодомен формирующего CAR полипептида и вспомогательного полипептида может содержать эндодомен CD3-дзета, а другой эндодомен CAR и вспомогательного полипептида может содержать эндодомен 41 ВВ.
В другом варианте осуществления настоящее изобретение относится к CAR по второму аспекту настоящего изобретения, дополнительно содержащему второй вспомогательный полипептид, включающий: (i) суперспиральный домен; (ii) трансмембранный домен; и (iii) эндодомен; где суперспиральный домен второго вспомогательного полипептида способен взаимодействовать с суперспиральными доменами формирующего CAR полипептида и первого вспомогательного полипептида.
Эндодомен формирующего CAR полипептида может содержать по меньшей мере первый внутриклеточный сигнальный домен, эндодомен первого вспомогательного полипептида может содержать по меньшей мере второй внутриклеточный сигнальный домен, а эндодомен второго вспомогательного полипептида может содержать по меньшей мере третий внутриклеточный сигнальный домен.
Например, эндодомены CAR, первого вспомогательного полипептида и второго вспомогательного полипептида могут включать в себя эндодомен CD3-дзета, эндодомен 41ВВ и эндодомен CD28.
Настоящее изобретение также относится к вспомогательному полипептиду, пригодному для использования в CAR, описанном в настоящем документе.
МУЛЬТИМЕРНЫЙ CAR
Настоящее изобретение относится к мультимерному CAR, который содержит множество формирующих CAR полипептидов по настоящему изобретению и, необязательно, вспомогательный(е) полипептид(ы), которые образуют комплекс в результате взаимодействия между суперспиральными спейсерными доменами.
Мультимерный CAR может быть, например, тримерным, тетрамерным, пентамерным, гексамерным или гептамерным.
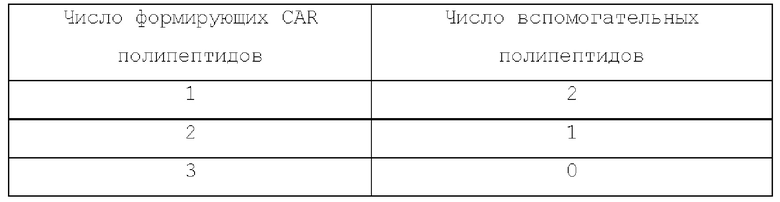
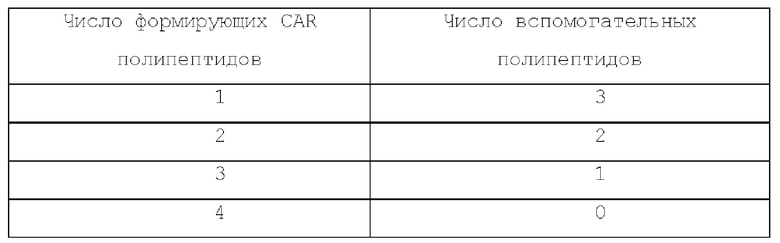
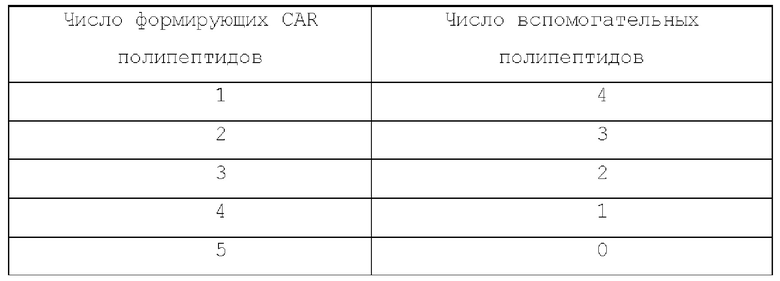
Число формирующих CAR полипептидов относительно вспомогательных белков в каждом типе CAR приведено в таблицах ниже:
Тримерный CAR:
Тетрамерный CAR:
Пентамерный CAR:
Ассоциация формирующих CAR полипептидов и вспомогательных полипептидов в клетке будет случайной, так что параметры, приведенные в таблицах выше, могут относиться к одному мультимерному CAR, в котором число формирующих CAR полипептидов и вспомогательных полипептидов может быть точно определенно, или к среднему числу формирующих CAR полипептидов и вспомогательных полипептидов в мультиплексном CAR, экспрессирующихся на клетке. В системах с высоким соотношением вспомогательного полипептида и формирующего CAR полипептида возможно, что на клеточной поверхности будут экспрессироваться некоторые мультикомплексы только из вспомогательного полипептида. Это не является проблемой при условии, что по меньшей мере некоторые из мультикомплексов, экспрессируемых на поверхности клетки, содержат формирующий CAR полипептид.
Множество формирующих CAR полипептидов и, необязательно, вспомогательного(ых) полипептида (ов) могут содержать один эндодомен.
В альтернативном варианте множество формирующих CAR полипептидов и, необязательно, вспомогательный(е) полипептид(ы) могут содержать различные эндодомены. Таким образом, множество различных эндодоменов могут активироваться одновременно. Это является преимуществом относительно сложного сигнального домена, поскольку каждый сигнальный домен остается свободным от других сигнальных доменов. Кроме того, это позволяет каждому сигнальному домену находиться в оптимальной близости к мембране для передачи сигнала.
В случае, когда мультимерный CAR содержит множество антигенсвязывающих доменов, это приведет к увеличению авидности связывания антигена. Мультимерный CAR может имитировать связывание антигена с IgM, который включает пять или шесть иммуноглобулинов.
СИГНАЛЬНАЯ СИСТЕМА CAR
Настоящее изобретение также относится к сигнальной системе химерного антигенного рецептора (CAR), которая содержит:
(i) мультимерный CAR, содержащий формирующий CAR полипептид или вспомогательный полипептид, описанный выше, который включает в себя первым гетеродимеризующийся домен; и
(ii) внутриклеточный сигнальный компонент, содержащий сигнальный домен и второй гетеродимеризующийся домен;
где гетеродимеризация между первым и вторым гетеродимеризующимися доменами заставляет мультимерный CAR и сигнальный компонент образовывать функциональные комплекс CAR.
Каждый формирующий CAR полипептид или вспомогательный полипептид могут включать в себя множество гетеродимеризующихся доменов, так что один формирующий CAR полипептид или вспомогательный полипептид способен гетеродимеризоваться с множеством сигнальных компонентов. Пример такой системы показан на фиг. 15.
Для дополнительного увеличения соотношения сигнального домена и антигенсвязывающего домена, каждый сигнальный компонент может включать в себя множество сигнальных доменов.
Гетеродимеризация может происходить только в присутствии низкомолекулярного соединения, например, с использованием такой системы, которая описана в WO 2016/030691.
В качестве альтернативы гетеродимеризация может происходить спонтанно. Первый и второй гетеродимеризующиеся домены способны спонтанно димеризоваться друг с другом. Гетеродимеризация происходит только между первым и вторым гетеродимеризующимися доменами без необходимости в какой-либо отдельной молекуле, действующей в качестве «индуктора» димеризации.
Сигнальная система по настоящему изобретению не ограничена положением конкретной пары гетеродимеризующихся доменов. Обеспечивающий направленное действие компонент (т.е. мультимерный CAR) может содержать либо домен из пары гетеродимеризующихся доменов при условии, что сигнальный компонент содержит соответствующий комплементарный домен, который позволит обеспечивающему направленное действие компоненту и сигнальному компонент совместно находиться на клеточной мембране.
Гетеродимеризующиеся домены для использования в данной системе CAR не ограничиваются теми, которые взаимодействуют в соотношении 1:1. Например, гетеродимеризующиеся домены могут взаимодействовать с образованием мультимеров (например, тримеров или тетрамеров). Домены могут взаимодействовать таким образом, который позволяет колокализацию одного первого гетеродимеризующегося домена с несколькими (например, 2 или 3) вторыми гетеродимеризующимися доменами. При этом может быть предпочтительно иметь сигнальный домен, который содержит второй гетеродимеризующийся домен, так что множество сигнальных компонентов могут колокализоваться с одним мультимерный CAR. Например, это может быть предпочтительным, когда требуется высокий уровень сигнала после связывания антигена с мультимерным CAR.
Мультимерный CAR может содержать множество гетеродимеризующихся доменов, так что он будет взаимодействовать с множеством сигнальных компонентов. Например, мультимерный CAR может содержать более двух гетеродимеризующихся доменов, например, от 3 до 10 гетеродимеризующихся доменов. На фиг. 15 показан мультимерный CAR, который содержит 20 гетеродимеризующихся доменов, по четыре на формирующий CAR полипептид/вспомогательный полипептид.
Для удобства, термин гетеродимеризующийся домен используется в настоящем документе для всех доменов, которые опосредуют колокализацию мультимерного CAR и сигнальных компонентов.
В данной области техники известно множество подходящих гетеродимеризующихся доменов, примеры которых приведены в настоящем документе.
Первый и второй гетеродимеризующиеся домены могут представлять собой лейциновые молнии.
Лейциновые молнии хорошо известны в данной области (см., например, Hakoshima; Encyclopedia of Life Sciences; 2005). Лейциновая молния представляет собой супер-вторичную структуру, которая функционирует в качестве димеризующегося домена. Их присутствие создает силу притяжения в параллельных альфа-спиралях. Одна лейциновая молния состоит из множества остатков лейцина с интервалами из приблизительно 7 остатков, которые образуют амфипатическую альфа-спираль с гидрофобной областью, проходящей с одной стороны. Этот гидрофобный участок обеспечивает область димеризации, позволяя мотивам соединяться, «застегиваясь на молнию». Лейциновые молнии, как правило, имеют от 20 до 40 аминокислот в длину, например, приблизительно 30 аминокислот.
Первый и/или второй гетеродимеризующиеся домены могут содержать последовательность, показанную в SEQ ID NO: 43 или 44. Первый гетеродимеризующийся домен может содержать последовательность, показанную в SEQ ID NO: 43, а второй гетеродимеризующийся домен может содержать последовательность, показанную в SEQ ID NO: 44, или наоборот.
SEQ ID NO: 43: 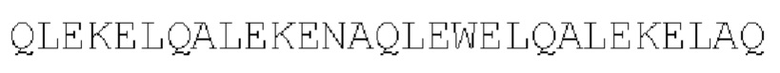
SEQ ID NO: 44: 
В некоторых вариантах осуществления первый и второй гетеродимеризующиеся домены могут быть кислыми (например, SEQ ID NO: 43) или основными (например, SEQ ID NO: 44) лейциновыми молниями. В частности, если первый гетеродимеризующийся домен представляет собой кислую лейциновую молнию, второй гетеродимеризующийся домен является основной лейциновой молнией, и наоборот.
Первый и второй гетеродимеризующиеся домены могут представлять собой димеризующийся и стыковочный домен (DDD1) и якорный домен (AD1). Эти домены и взаимодействия между ними известны в данной области техники (Rossi et al.; PNAS; 2006; 103 (18); 6841-6846).
DDD1 представляет собой короткую альфа-спиральную структуру, полученную из протеинкиназы А (PKA). AD1 представляет собой короткую альфа-спиральную структуру, полученная из А-киназных якорных белков (AKAP).
Домен DDD1 может содержать последовательность, показанную в SEQ ID NO: 45.
SEQ ID NO: 45: 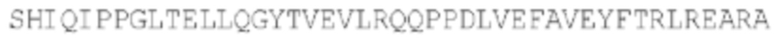
Домен AD1 может содержать последовательность, показанную в SEQ ID NO: 46
SEQ ID NO: 46: 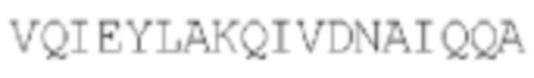
Поскольку взаимодействие DDD1/AD1 является тримерным, домен AD1, присутствующий на эндодомене CAR, будет набирать три сигнальных домена, содержащих домен DDD1. Таким образом, в конкретном варианте осуществления эндодомен CAR содержит домен AD1, а внутриклеточный сигнальный компонент содержит домен DDD1.
Эти гетеродимеризующиеся домены могут быть получены из бактериальной рибонуклеазы (барназы) и пептидов Барнстар.
Барназа представляет собой рибонуклеазный белок из Bacillus amyloliquefaciens. Он состоит на 110 аминокислот. Барнстар функционирует, ингибируя нуклеазную активность барназы и, поэтому, она связывает Барнстар с очень высокой аффинностью (скорость ассоциации 108с-1М-1).
Гетеродимеризующиеся домены могут быть получены из панкреатических РНКаз человека и S-пептида.
Панкреатические РНКазы человека являются пиримидин-специфичными эндонуклеазами. S-пептид является ферментативно неактивным протеолитическим фрагментом РНКазы А, в котором отсутствует сайт связывания РНК.
Настоящее изобретение также охватывает варианты гетеродимеризующихся последовательностей, описанных в настоящем документе, которые сохраняют способность к димеризации с соответствующим гетеродимеризующийся доменом. Гетеродимеризующийся домен может представлять собой вариант, имеющий 5, 4, 3, 2 или 1 аминокислотных мутации (инсерции, замены или дополнения) или по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности по последовательности по сравнению с последовательностью, показанной в SEQ ID NO: 43, 44, 45 или 46 при условии, что они все еще вызывают гетеродимеризацию между CAR и сигнальным компонентом.
НУКЛЕИНОВАЯ КИСЛОТА
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей формирующий CAR полипептид по первому аспекту настоящего изобретения и/или вспомогательный полипептид по первому аспекту настоящего изобретения.
Предполагается, что используемые в данном описании термины «полинуклеотид», «нуклеотид», и «нуклеиновая кислота» являются синонимами друг друга.
Специалисту в данной области будет понятно, что множество различных полинуклеотидов и нуклеиновых кислот могут кодировать один и тот же полипептид в результате вырожденности генетического кода. Кроме того, следует понимать, что опытные специалисты могут, используя обычные методы, создать нуклеотидные замены, которые не влияют на полипептидную последовательность, кодируемую полинуклеотидами, описанными в данном документе, чтобы учесть встречаемость кодонов в любом конкретном организме-хозяине, в котором полипептиды должны экпрессироваться.
Нуклеиновые кислоты по изобретению могут содержать ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Они могут также представлять собой полинуклеотиды, которые включают в себя синтетические или модифицированные нуклеотиды. Ряд различных типов модификаций олигонуклеотидов известен в данной области техники. К ним относятся метилфосфонатные и фосфоротиоатные связи, добавление акридиновых или полилизиновых цепей на 3'- и/или 5'-концы молекулы. В отношении вариантов применения, описанных в данном документе, следует понимать, что полинуклеотиды могут быть модифицированы любым способом, доступным в данной области. Такие модификации могут быть выполнены для того, чтобы повысить активность in vivo или продолжительность жизни полинуклеотидов, представляющих интерес.
Термины «вариант», «гомолог» или «производное» в отношении нуклеотидной последовательности включают в себя любую замену, вариацию, модификацию, замещение, делецию или добавление одной (или большего числа) нуклеиновой кислоты в последовательности.
Настоящее изобретение также относится к последовательности нуклеиновой кислоты, кодирующей вспомогательный полипептид, подходящий для использования в CAR по второму аспекту настоящего изобретения.
НУКЛЕОТИДНЫЕ КОНСТРУКЦИИ
Настоящее изобретение также относится к нуклеотидной конструкции, которая кодирует множество нуклеотидных последовательностей.
Например, нуклеотидная конструкция может кодировать два или несколько формирующих CAR полипептидов по первому аспекту настоящего изобретения.
В этом варианте осуществления нуклеотидная конструкция может содержать по меньшей мере две нуклеотидные последовательности:
(i) первую нуклеотидную последовательность, которая кодирует первый формирующий CAR полипептид; и
(ii) вторую нуклеотидную последовательность, которая кодирует второй формирующий CAR полипептид.
Нуклеотидная конструкция может кодировать по меньшей мере один формирующий CAR полипептид по первому аспекту настоящего изобретения и по меньшей мере один вспомогательный полипептид, описанный выше.
В этом варианте осуществления нуклеотидная конструкция может содержать по меньшей мере две нуклеотидные последовательности:
(i) первую нуклеотидную последовательность, которая кодирует формирующий CAR полипептид; и
(ii) вторую нуклеотидную последовательность, которая кодирует вспомогательный полипептид.
Нуклеотидная конструкция может кодировать:
(i) по меньшей мере, один формирующий CAR полипептид, который образует мультимерный CAR по второму аспекту настоящего изобретения; и
(ii) внутриклеточный сигнальный компонент, описанный выше.
В этом варианте осуществления нуклеотидная конструкция может содержать по меньшей мере две нуклеотидные последовательности:
(i) первую нуклеотидную последовательность, которая кодирует формирующий CAR полипептид; и
(ii) вторую нуклеотидную последовательность, которая кодирует внутриклеточный сигнальный компонент.
Нуклеотидная конструкция может содержать третью нуклеотидную последовательность, которая кодирует вспомогательный полипептид.
Поэтому, нуклеотидная конструкция может продуцировать два или несколько полипептидов, которые соединены сайтом(ами) расщепления. Сайт расщепления может быть саморасщепляющимся, так что при синтезирующийся при трансляции продукт сразу же расщепляется на отдельные полипептиды без необходимости в какой-либо внешней расщепляющей активности.
Сайт расщепления может представлять собой любую последовательность, которая позволяет разделить полипептид, содержащий несколько CAR и/или вспомогательных полипептидов.
Термин «расщепление» используется в данном документе для удобства, но сайт расщепления может обеспечивать разделение пептидов на отдельные объекты с помощью механизма, отличного от классического расщепления. Например, для саморасщепляющегося пептида 2А вируса ящура (FMDV) (см. ниже) были предложены различные модели для объяснения «расщепляющей» активности: протеолиз протеиназой клетки-хозяина, автопротеолиз или трансляционный эффект (Donnelly et al (2001) J. Gen. Virol. 82:102 7-1041). Точный механизм такого «расщепления» не имеет значения для целей настоящего изобретения, до тех пор, пока сайт расщепления, расположенный между нуклеотидными последовательностями, кодирующими белки, приводит к тому, что белки экспрессируются в виде отдельных объектов.
Сайт расщепления может представлять собой сайт расщепления фурина.
Фурин представляет собой фермент, который принадлежит к семейству конвертаз субтилизин-подобных пробелков. Представители этого семейства являются конвертазами пробелков, которые процессируют латентные предшественники белков в их биологически активные продукты. Фурин является кальций-зависимой сериновой эндопротеазой, которая может эффективно расщеплять предшественники белков по их сайтам из парных основных аминокислот. Примеры субстратов фурина включают пропаратироидный гормон, предшественник трансформирующего фактора роста-бета 1, проальбумин, про-бета-секретазу, мембранную матриксную металлопротеиназу 1-го типа, бета-субъединицу профактора роста нервов и фактор фон Виллебранда. Фурин расщепляет белки только после основной аминокислотной последовательности-мишени (каноническая последовательность представляет собой Arg-X-(Arg/Lys)-Arg') и присутствует в основном в аппарате Гольджи.
Сайт расщепления может представлять собой сайт расщепления протеазой из вируса гравировки табака (TEV).
TEV-протеаза представляет собой высокоспецифичную к последовательности цистеиновую протеазу, который является химотрипсин-подобной протеазой. Она является высокоспецифичной к своему сайту расщепления и поэтому часто используется для контролируемого расщепления слитых белков как in vitro, так и in vivo. Консенсусным сайтом расщепления TEV является ENLYFQ\S (где «\» обозначает расщепляемую пептидную связь). Клетки млекопитающих, такие как клетки человека, не экспрессируют TEV-протеазу. Поэтому, в вариантах осуществления, в которых нуклеотидная конструкция по настоящему изобретению содержит сайт расщепления TEV и экспрессируется в клетках млекопитающего, экзогенная TEV-протеаза также должна экспрессироваться в клетках млекопитающего.
Сайт расщепления может кодировать саморасщепляющийся пептид.
«Саморасщепляющийся пептид» относится к пептиду, который функционирует таким образом, что при синтезе продукта, содержащего полипептиды и саморасщепляющийся пептид, он немедленно «расщепляется» или разделяется на отдельные и дискретные первый и второй полипептиды без необходимости в какой-либо внешней расщепляющей активности.
Саморасщепляющийся пептид может представлять собой саморасщепляющийся пептид 2А из афто- или кардиовируса. Первичное расщепление 2А/2В афто- или кардиовирусов опосредуется «расщеплением» 2А по своему собственному С-концу. В афтовирусах, таких как вирус ящура (FMDV) и вирус лошадиного ринита, область 2А представляет собой короткий участок около 18 аминокислот, который вместе с N-концевым остатком белка 2В (консервативным остатком пролина) представляет собой автономный элемент, способный опосредовать «расщепление» на своем собственном С-конце (Donelly et al. (2001), как указано выше).
«2А-подобные» последовательности были обнаружены в других пикорнавирусах, кроме афто- или кардиовирусов, пикорнавирус-подобных вирусах насекомых, ротавирусах типа С и повторяющихся последовательностях из видов Trypanosoma и бактериальной последовательности (Donelly et al (2001), как указано выше). Сайт расщепления может включать в себя одну из этих 2А-подобных последовательностей, таких, как:


Сайт расщепления может включать 2А-подобную последовательность, приведенную в SEQ ID NO: 19 
Настоящее изобретение также относится к набору, содержащему одну или несколько нуклеотидных последовательностей, кодирующих формирующий CAR полипептид по первому аспекту настоящего изобретения и/или вспомогательный полипептид, подходящий для получения CAR по второму аспекту настоящего изобретение.
ВЕКТОР
Настоящее изобретение также относится к вектору или набору векторов, которые содержат одну или несколько нуклеотидных последовательностей или нуклеотидных конструкций, описанных выше. Такой вектор может быть использован для введения нуклеотидной(ых) последовательности(ей) в клетку-хозяина таким образом, что она экспрессирует формирующий CAR полипептид или вспомогательный пептид по первому аспекту настоящего изобретения и/или CAR по второму аспекту настоящего изобретения.
Вектор может, например, представлять собой плазмиду или вирусным вектор, такой как ретровирусный вектор или лентивирусным вектор, либо вектор на основе транспозона или синтетической мРНК.
Вектор может быть способен трансфицировать или трансдуцировать иммунную клетку, такую как Т-клетка или NK-клетка.
КЛЕТКА
Настоящее изобретение также относится к клетке, такой как иммунная клетка, содержащей формирующий CAR полипептид, CAR или сигнальную систему CAR, описанные выше.
Клетка может содержать нуклеиновую кислоту, нуклеотидную конструкцию или вектор по настоящему изобретению.
Клетка может представлять собой иммунную клетку, в частности, цитолитическую иммунную клету, такую как Т-клетка или NK-клетка.
Т-клетки или Т-лимфоциты представляют собой тип лимфоцитов, которые играют центральную роль в клеточном иммунитете. Они отличаются от других лимфоцитов, таких как В-клетки и естественные клетки-киллеры (NK-клетки), присутствием Т-клеточного рецептора (TCR) на клеточной поверхности. Существуют различные типы Т-клеток, описанные ниже.
Хелперные Т-клетки (ТН-клетки) помогают другим бельм клеткам крови в иммунологических процессах, включая созревание В-клеток в плазматические клетки и В-клетки памяти, и активацию цитотоксических Т-клеток и макрофагов. ТН-клетки экспрессируют CD4 на своей поверхности. ТН-клетки активируются при взаимодействии с пептидными антигенами, представляемыми молекулами МНС II класса на поверхности антиген-представляющих клеток (АРС). Эти клетки могут дифференцироваться в один из нескольких подклассов, включающих ТН1, ТН2, ТН3, ТН17, Th9 или TFH, которые секретируют различные цитокины, обуславливающие различные типы иммунных реакций.
Цитолитические Т-клетки (ТС-клетки или CTL) уничтожают инфицированные вирусами клетки и опухолевые клетки, а также участвуют в отторжении трансплантата. CTL экспрессируют CD8 на своей поверхности. Эти клетки распознают свои мишени путем связывания с антигеном, связанного с МНС I класса, который присутствует на поверхности всех клеток, имеющих ядро. С помощью IL-10, аденозина и других молекул, секретируемых регуляторными Т-клетками, CD8+-клетки могут быть инактивированы до энергичного состояния, которое предотвращает аутоиммунные заболевания, такие как экспериментальный аутоиммунный энцефаломиелит.
Т-клетки памяти представляют собой подмножество антиген-специфических Т-клеток, которые сохраняются в течение длительного времени после подавления инфекции. Они быстро размножаются до большого количества эффекторных Т-клеток при повторном воздействии соответствующим антигеном, тем самым обеспечивая иммунную систему «памятью» в отношении перенесенных инфекций. Т-клетки памяти включают три подтипа: Т-клетки центральной памяти (ТСМ-клетки) и два типа Т-клеток эффекторной памяти (ТЕМ-клетки и TEMRA-клетки). Клетки памяти могут быть CD4+или CD8+. Т-клетки памяти, как правило, экспрессируют белок CD45RO на клеточной поверхности.
Регуляторные Т-клетки (Treg-клетки), ранее известные как супрессорные Т-клетки, имеют решающее значение для поддержания иммунологической толерантности. Их главная роль заключается в полной остановке Т-клеточного иммунного ответа и подавлении аутореактивных Т-клеток, которые избежали процесса негативной селекции в тимусе.
Описано два основных класса CD4+ Treg-клеток - природные Treg-клетки и адоптивные Treg-клетки.
Природные Treg-клетки (также известные как CD4+CD25+FoxP3+Treg-клетки) образуются в тимусе и связаны с взаимодействием между развивающимися Т-клеток как с миелоидными (CD11c+), так и с плазмоцитарными (CD123+) дендритными клетками, которые были активированы TSLP. Природные Treg-клетки можно отличить от других Т-клеток по присутствию внутриклеточной молекулы, называющейся FoxP3. Мутации гена FOXP3 могут препятствовать развитию регуляторных Т-клеток, что приводит к смертельному аутоиммунному заболеванию IPEX.
Адоптивные Treg-клетки (также известные как TR1-клетки или Th3-клетки) может образовываться во время нормального иммунного ответа.
Клетка может представлять собой естественную киллерную клетку (или NK-клетку). NK-клетки являются частью врожденной иммунной системы. NK-клетки обеспечивают быструю реакцию на врожденные сигналы от инфицированных вирусом клеток независимые от МНС образом.
NK-клетки (принадлежащие к группе врожденных лимфоидных клеток) определяются как крупные зернистые лимфоциты (LGL) и представляют собой третий вид клеток, которые дифференцируются из общего лимфоидного предшественника, генерирующего В- и Т-лимфоциты. NK-клетки, как известно, дифференцируются и созревают в костном мозге, лимфатических узлах, селезенке, тимусе и в миндалинах, откуда они затем входят в кровоток.
CAR-клетки по настоящему изобретению могут представлять собой любой из типов клеток, перечисленных выше.
Т- или NK-клетки, экспрессирующие CAR по настоящему изобретению или его компоненты, могут быть созданы ex vivo либо из собственной периферической крови больного (1-я сторона), либо из подготовленного трансплантата гемопоэтических стволовых клеток из донорной периферической крови (2-я сторона), либо из периферической крови от не связанного родственно донора (3-я сторона).
В качестве альтернативы, Т или NK-клетки, экспрессирующие CAR по настоящему изобретению или его компоненты, могут быть получены путем ex vivo дифференцировки индуцируемых клеток-предшественников или эмбриональных клеток-предшественников в Т или NK-клетки. В качестве альтернативы может быть использована иммортализованная Т-клеточная линия, которая сохраняет свою литическую функцию и может выступать в качестве терапевтического средства.
Во всех этих вариантах осуществления CAR-клетка создается путем введения ДНК или РНК, кодирующих CAR по изобретению или его компонент, с помощью одного из множества способов, включая трансдукцию вирусным вектором, трансфекцию ДНК или РНК.
CAR-клетки по изобретению могут представлять собой ex vivo Т- или NK-клетки от субъекта. Т- или NK-клетки могут быть получены из образца мононуклеарных клеток периферической крови (РВМС). Т или NK-клетки могут быть активированы и/или размножены до трансдукции нуклеиновой кислотой, кодирующей молекулу, обеспечивающую CAR по изобретению или компоненты CAR, например, путем обработки анти-CD3 моноклональным антителом.
Т- или NK-клетки по изобретению могут быть получены путем:
(i) выделения содержащего Т- или NK-клетки образца из субъекта или других источников, перечисленных выше; и
(ii) трансдукции или трансфекции Т- или NK-клеток одной или несколькими нуклеотидными последовательности или нуклеотидными конструкциями, описанными выше.
Т- или NK-клетки затем могут быть очищены, например, отобраны на основе экспрессии антигенсвязывающего домена антигенсвязывающего полипептида.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Настоящее изобретение также относится к фармацевтической композиции, содержащей множество клеток, экспрессирующих CAR по изобретению или их компоненты.
Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый носитель, разбавитель или вспомогательное вещество. Фармацевтическая композиция, необязательно, может содержать один или несколько дополнительных фармацевтически активных полипептидов и/или соединений. Такая композиция, например, может иметь форму, подходящую для внутривенного вливания.
СПОСОБ ЛЕЧЕНИЯ
Настоящее изобретение относится к способу лечения и/или профилактики заболевания, который включает в себя стадию введения субъекту клеток по настоящему изобретению (например, в фармацевтической композиции, описанной выше).
Способ лечения заболевания относится к терапевтическому применению клеток по настоящему изобретению. При этом клетки можно вводить субъекту, имеющему существующее заболевание или состояние, для того, чтобы уменьшить, снизить или улучшить по меньшей мере один симптом, связанный с заболеванием и/или замедлить, уменьшить или блокировать развитие заболевания.
Способ профилактики заболевания относится к профилактическому использованию клеток по настоящему изобретению. При этом такие клетки могут быть введены субъекту, который еще не заболел, и/или у которого не наблюдается каких-либо симптомов заболевания, чтобы предупредить возникновение или ослабить причину заболевания, или чтобы уменьшить или предотвратить развитие по меньшей мере одного симптома, связанного с заболеванием. Субъект может иметь предрасположенность к заболеванию или может предполагать, что он имеет риск развития заболевания.
Способ может включать следующие стадии:
(i) выделение содержащего Т- или NK-клетки образца;
(ii) трансдукцию или трансфекцию таких клеток нуклеотидной последовательностью, нуклеотидной конструкцией или вектором по изобретению;
(iii) введение субъекту клеток по п. (ii).
Содержащий Т- или NK-клетки образец может быть выделен у субъекта, или из других источников, например, описанных выше. Т-или NK-клетки могут быть выделены из собственной периферической крови субъекта (1-я сторона), либо из подготовленного трансплантата гемопоэтических стволовых клеток из донорной периферической крови (2-я сторона), либо из периферической крови от не связанного родственно донора (3-я сторона).
Настоящее изобретение относится к CAR-клетке по настоящему изобретению для применения при лечении и/или профилактики заболевания.
Изобретение также относится к применению CAR-клетки по настоящему изобретению в производстве лекарственного средства для лечения и/или профилактики заболевания.
Заболевание, подлежащее лечению и/или предупреждению с помощью способов по настоящему изобретению, может представлять собой раковое заболевание, такое как рак мочевого пузыря, рак молочной железы, рак толстой кишки, рак эндометрия, рак почки (почечно-клеточный рак), лейкемия, рак легкого, меланома, неходжкинская лимфома, рак поджелудочной железы, рак предстательной железы и рак щитовидной железы.
CAR-клетки по настоящему изобретению могут быть способны лизировать клетки-мишени, такие как раковые клетки. Клетка-мишень может распознаваться по экспрессии ТАА, например, по экспрессии ТАА, представленного выше в таблице 1.
CAR-клетки по настоящему изобретению могут быть способы лизировать клетки-мишени, такие как раковые клетки, которые экспрессируют ТАА с низкой плотностью. Примеры ТАА, которые, как известно, экспрессируются с низкой плотностью при некоторых типах рака, включают, но не ограничиваются ими, ROR1 в CLL, Typr-1 в меланоме и ВСМА в миеломе.
CAR-клетки и фармацевтические композиции по настоящему изобретению могут подходить для лечения и/или профилактики заболеваний, описанных выше.
CAR-клетки и фармацевтические композиции по настоящему изобретению могут быть пригодны для использования в любом из описанных выше способов.
Изобретение далее дополнительно описано с помощью примеров, которые должны послужить помощью обычным специалистам в данной области техники при осуществлении настоящего изобретения на практике и не предназначены для какого-либо ограничения объема изобретения.
ПРИМЕРЫ
Пример 1 - экспрессия COMP-CAR на поверхности клетки
Мышиная Т-клеточная линия была трансдуцирована анти-CD33-COMP-CAR (аминокислотная последовательность показана на фиг. 5с, а нуклеотидная последовательность показана на фиг. 5d) или анти-CD33-IgG1-CAR.
Эти клетки затем окрашивали химерным растворимым CD33, слитым с мышиным Fc из IgG2a перед вторичным окрашиванием антителами к мышиным IgG, конъюгированными с РЕ (фиг. 6а).
Мышиная Т-клеточная линия была трансдуцирована анти-ROR-l-COMP-CAR (аминокислотная последовательность показана на фиг. 5с, а нуклеотидная последовательность показана на фиг. 5d) или анти-ROR-1-IgG1-CAR.
Эти клетки затем окрашивали растворимым His-меченым ROR-1 с последующим вторичным окрашиванием анти-His-биотином, и последующим третьим окрашиванием стрептавидином-АРС (фиг. 6b).
Все четыре CAR успешно экспрессировались на поверхности клетки. Эти данные также показывают, что связывающий домен в CAR ориентирован так, чтобы не затруднять связывание лиганда, будучи связанным с СОМР-спейсером.
Пример 2 - стимуляция COMP-CAR-T-клеток иммобилизованным лигандом
Т-клетки с шариками, покрытыми иммобилизованным лигандом, были использованы для стимуляции COMP-ROR-l-CAR-T-клеток. Для этого был сконструирован и проэкспрессирован растворимый His-меченый ROR-1. Супернатанты, содержащие эти растворимые лиганды, затем инкубировали при различных концентрациях с определенным количеством анти-His шариков. Затем шарики промывали для удаления несвязанного лиганда, и эти шарики использовали для стимуляции Т-клеток, трансдуцированных либо COMP-CAR, либо эквивалентными CAR с IgG-спейсером.
Трансдуцированные мышиные Т-клетки совместно культивировали с анти-His шариками, которые были предварительно покрыты различными концентрациями супернатанта с растворимые His-меченым ROR-1. Содержание IL-2 в супернатанте после совместного культивирования анализировали с помощью ELISA через 16-24 часа (фиг. 7).
Пример 3 - уровни экспрессии R0R-1 клетками-мишенями
Линия клеток SKW исходно экспрессирует низкий уровень ROR-1. Эти клетки были трансдуцированы ROR-1, чтобы увеличить уровень экспрессии. Эти клетки окрашивали анти-ROR-l-APC и сравнивали с неокрашенными клетками (фиг. 8).
Пример 4 - стимуляция анти-ROR-1-COMP-САР-Т-клеток положительными по ROR-1 клетками SKW
Трансдуцированные мышиные Т-клетки (описанные в примерах 1 и 2) культивировали совместно с клетками-мишенями SKW, которые экспрессируют лиганд ROR-1 с низкой или высокой плотностью. Число Т-клеток поддерживали на постоянном уровне, а число клеток-мишеней было различным. Содержание IL-2 в супернатанте после совместно культивирования анализировали через 16-24 часа с помощью ELISA (фиг. 9).
Более высокие уровни IL-2 были обнаружены при совместном культивировании анти-ROR-l-COMP-CAR-T-клеток с клетками-мишенями SKW, экспрессирующими лиганд ROR-1 с низкой плотностью, по сравнению с анти-ROR-l-IgG1-CAR.
Оба CAR были способны инициировать активирующий ответ у клеток SKW с высокой экспрессией мишени.
Пример 5 - укорачивание СОМР-спейсера
СОМР-спейсер в анти-ROR-l-CAR был укорочен относительно своей первоначальной длины в 45 аминокислот. Этими конструкциями с укороченным СОМР трансфицировали клетки 2 93Т, которые затем окрашивали на поверхностную экспрессию CAR с использованием sROR-1-His с последующим окрашиванием анти-His-биотином и последующим окрашиванием стрептавидином-РЕ.Су7. Эти клетки также окрашивали на маркер RQR8 антителом анти-CD34-FITC. Данные FACS-диаграмм показывают стабильную поверхностную экспрессию различных укороченных форм СОМР-спейсера, отражая возможность варьировать длину суперспирального спейсера всего с помощью одной или нескольких аминокислот (фиг. 11).
Пример 6 - сравнение мультимерных CAR с суперспиралями с классическим димерным CAR
Для того чтобы сравнить функцию CAR с суперспиральными спейсерами по изобретению с обычным CAR, был изготовлен ряд конструкций с одинаковыми антигенсвязывающими доменами и эквивалентными эндодоменами, но с разными спейсерами, что приводит к совершенно иной структуре CAR. Различные CAR показаны на фиг. 12.
Все CAR содержали анти-CD19-антигенсвязывающий домен, основанный на fmc63, и эндодомен «второго поколения», содержащий эндодомены 41ВВ и CD3-дзета. Были протестированы следующие форматы:
a) гетеромультимерный CAR, который содержит: полипептид, имеющий анти-CD19-антигенсвязывающий домен, суперспиральный спейсерный домен и эндодомен CD3-дзета, и вспомогательный полипептид, имеющий суперспиральный спейсерный домен и эндодомен 41 ВВ (фиг. 12А);
b) гомомультимерный CAR из полипептидов, содержащих анти-CD19-антигенсвязывающий домен, суперспиральный спейсерный домен и комбинированный эндодомен 41ВВ/CD3-дзета (фиг. 12В); и
c) классический гомодимерный CAR второго поколения, который содержит два полипептида, имеющих анти-CD19-антигенсвязывающий домен, спейсерный домен из стеблевой области CD8 и комбинированный эндодомен 41ВВ/CD3-дзета (фиг. 12С).
Векторами, кодирующими CAR, показанные на фиг. 12, были
Был получен RD114-псевдотипированный ретровирус, кодирующий различные структуры CAR.
Т-клетки истощали на экспрессирующие CD56 клетки и совместно культивировали с равным количеством клеток SupT1 для достижения соотношения эффектор:мишень 1:1. Перед анализом методом проточной цитометрии равное количество флуоресцентных счетных шариков добавляли к каждой совместной культуре для нормализации числа клеток и для учета каких-либо отличий в анализируемых объемах. CAR-опосредованную цитотоксичность оценивали методом проточной цитометрии следующим образом: Т-клетки отличали от опухолевых клеток путем окрашивания на экспрессию CD3 относительно FCS-параметра, а опухолевые клетки определяли по отсутствию CD3 на них и по более высокому FCS-сигналу. Жизнеспособность оценивали путем окрашивания мертвых клеток красителем 7-AAD, и жизнеспособные клетки определяли как те, которые не поглощали краситель. Жизнеспособные клетки опухолей подсчитывали для каждой совместной культуры, и процент цитотоксичности рассчитывали путем нормализации количества жизнеспособных опухолевых клеток на их количество, получаемое при совместном культивировании с нетрансдуцированными РВМС (100%). Лизис клеток-мишеней оценивали на 2-й и 5-й день.
Результаты показаны на фиг. 13 (день 2) и фиг. 14 (день 5). Через два дня все три структуры CAR лизировали клетки-мишени CD19+ SupT1. Два мультимерных CAR давали лучший лизис относительно эквивалентного классического гомодимерного CAR. Гомомультимерный CAR (Fmc63-COMP-41BBz) показал наибольший лизис. На 5-й день, некоторые остаточные клетки-мишени оставались при использовании классического гомодимерого CAR (Fmc63-CD8STK-41BBz), при этом для обоих мультимерных CAR практически невозможно было обнаружить жизнеспособных клеток-мишеней.
Пример 7 - получение и тестирование суперспиральных супер-CAR
Одной из главных проблем в технологиях CAR является обнаружение антигенов, которые присутствуют с низкой плотностью на клетках-мишенях. Для решения этой проблемы авторы настоящего изобретения разработали «супер-CAR» на основе суперспирального спейсера, который рекрутирует несколько цепей TCR-дзета для каждого взаимодействия с антигеном.
Внутриклеточная часть полипептида, составляющая суперспиральную структуру CAR, содержит множество гетеродимеризующихся доменов, каждый из которых способен взаимодействовать с одним или несколькими внутриклеточными сигнальными компонентами, которые содержат один или несколько внутриклеточных сигнальных доменов.
В конструкциях, показанных на фиг. 15, внутриклеточная димеризация представляет собой димеризацию между стыковочные доменом (DDD1) и якорным доменом (AD1). Каждый полипептид, составляющий CAR с суперспиральным спейсером, состоит из четырех отдельных AD1-доменов. Поэтому, суперспиральный CAR, содержащий 5 полипептидов, будет содержать 20 AD1-доменов.
Суперспиральный супер-CAR был протестирован в сочетании с различными сигнальными компонентами, имеющими 0 или 2 копии сигнального домена TCR-дзета. Поскольку DDD1 связывает AD1 со стехиометрией 2:1, эти сигнальные домены дают 0 и 80 копий домена TCR-дзета, соответственно, для каждого связывающегося с мишенью компонента 5-полипептидного суперспирального CAR.
В качестве контроля использовали классический гомодимерный анти-CDl9-CAR (фиг.15: анти-CD19-IgGFc-Z) с таким же антигенсвязывающим доменом.
Мышиную Т-клеточную линию BW5 трансдуцировали каждым CAR и активировали клетками SupT1, экспрессирующими соответствующий антиген (CD19), при различных концентрациях: низкой, средней и высокой. Эти клетки SupT1 были сконструированы для экспрессии CD19 на различном уровне путем использования субоптимальных сигнальных пептидов и/или введения мотивов удерживания в цитоплазме, полученных из Tyrp-1 (встраивается близко к мембране) или аденовирусного гликопротеина Е3-19k (встраивается на С-конец). Высвобождение IL-2 измеряли после введения антигена.
Результаты показаны на фиг. 16. Было обнаружено, что суперспиральный супер-CAR, содержащий 80 копий TCR-дзета на 5-членный суперспиральный CAR, давал намного больший ответ на антиген, чем эквивалентный классический CAR, содержащий две копии TCR-дзета на молекулу.
Все публикации, указанные в приведенном выше описании, включены путем ссылки. Различные модификации и вариации описанных способов и системы по изобретению будут очевидны специалистам в данной области техники, не выходя за пределы объема и сущности настоящего изобретения. Несмотря на то что изобретение описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. В самом деле, предполагается, что различные модификации описанных способов осуществления изобретения, которые очевидны специалистам в данной области молекулярной биологии, клеточной биологии или в смежных областях, входят в объем нижеследующей формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCL Business PLC
<120> ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР
<130> P106296PCT
<150> GB 1504840.8
<151> 2015-03-23
<160> 48
<170> PatentIn версии 3.5
<210> 1
<211> 45
<212> Белок
<213> Искусственная последовательность
<220>
<223> Суперспиральный домен
<400> 1
Asp Leu Gly Pro Gln Met Leu Arg Glu Leu Gln Glu Thr Asn Ala Ala
1 5 10 15
Leu Gln Asp Val Arg Glu Leu Leu Arg Gln Gln Val Arg Glu Ile Thr
20 25 30
Phe Leu Lys Asn Thr Val Met Glu Cys Asp Ala Cys Gly
35 40 45
<210> 2
<211> 21
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид получен из цепи TCR-бета
<400> 2
Met Gly Thr Ser Leu Leu Cys Trp Met Ala Leu Cys Leu Leu Gly Ala
1 5 10 15
Asp His Ala Asp Gly
20
<210> 3
<211> 21
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид получен из IgG1
<400> 3
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 4
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид получен из CD8a
<400> 4
Met Ala Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr
1 5 10 15
Asp Ala Arg Cys
20
<210> 5
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен CD3 Z
<400> 5
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 6
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомены CD28 и CD3-дзета
<400> 6
Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro
1 5 10 15
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
20 25 30
Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala
35 40 45
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
50 55 60
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
65 70 75 80
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
85 90 95
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
100 105 110
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
115 120 125
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
130 135 140
His Met Gln Ala Leu Pro Pro Arg
145 150
<210> 7
<211> 188
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомены CD28, OX40 и CD3-дзета
<400> 7
Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro
1 5 10 15
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
20 25 30
Arg Asp Phe Ala Ala Tyr Arg Ser Arg Asp Gln Arg Leu Pro Pro Asp
35 40 45
Ala His Lys Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu
50 55 60
Glu Gln Ala Asp Ala His Ser Thr Leu Ala Lys Ile Arg Val Lys Phe
65 70 75 80
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
85 90 95
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
100 105 110
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
115 120 125
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
130 135 140
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
145 150 155 160
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
165 170 175
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
180 185
<210> 8
<211> 38
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен ICOS
<400> 8
Cys Trp Leu Thr Lys Lys Lys Tyr Ser Ser Ser Val His Asp Pro Asn
1 5 10 15
Gly Glu Tyr Met Phe Met Arg Ala Val Asn Thr Ala Lys Lys Ser Arg
20 25 30
Leu Thr Asp Val Thr Leu
35
<210> 9
<211> 48
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен CD27
<400> 9
Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser Pro Val Glu Pro
1 5 10 15
Ala Glu Pro Cys His Tyr Ser Cys Pro Arg Glu Glu Glu Gly Ser Thr
20 25 30
Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro Ala Cys Ser Pro
35 40 45
<210> 10
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен BTLA
<400> 10
Arg Arg His Gln Gly Lys Gln Asn Glu Leu Ser Asp Thr Ala Gly Arg
1 5 10 15
Glu Ile Asn Leu Val Asp Ala His Leu Lys Ser Glu Gln Thr Glu Ala
20 25 30
Ser Thr Arg Gln Asn Ser Gln Val Leu Leu Ser Glu Thr Gly Ile Tyr
35 40 45
Asp Asn Asp Pro Asp Leu Cys Phe Arg Met Gln Glu Gly Ser Glu Val
50 55 60
Tyr Ser Asn Pro Cys Leu Glu Glu Asn Lys Pro Gly Ile Val Tyr Ala
65 70 75 80
Ser Leu Asn His Ser Val Ile Gly Pro Asn Ser Arg Leu Ala Arg Asn
85 90 95
Val Lys Glu Ala Pro Thr Glu Tyr Ala Ser Ile Cys Val Arg Ser
100 105 110
<210> 11
<211> 188
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен CD30
<400> 11
His Arg Arg Ala Cys Arg Lys Arg Ile Arg Gln Lys Leu His Leu Cys
1 5 10 15
Tyr Pro Val Gln Thr Ser Gln Pro Lys Leu Glu Leu Val Asp Ser Arg
20 25 30
Pro Arg Arg Ser Ser Thr Gln Leu Arg Ser Gly Ala Ser Val Thr Glu
35 40 45
Pro Val Ala Glu Glu Arg Gly Leu Met Ser Gln Pro Leu Met Glu Thr
50 55 60
Cys His Ser Val Gly Ala Ala Tyr Leu Glu Ser Leu Pro Leu Gln Asp
65 70 75 80
Ala Ser Pro Ala Gly Gly Pro Ser Ser Pro Arg Asp Leu Pro Glu Pro
85 90 95
Arg Val Ser Thr Glu His Thr Asn Asn Lys Ile Glu Lys Ile Tyr Ile
100 105 110
Met Lys Ala Asp Thr Val Ile Val Gly Thr Val Lys Ala Glu Leu Pro
115 120 125
Glu Gly Arg Gly Leu Ala Gly Pro Ala Glu Pro Glu Leu Glu Glu Glu
130 135 140
Leu Glu Ala Asp His Thr Pro His Tyr Pro Glu Gln Glu Thr Glu Pro
145 150 155 160
Pro Leu Gly Ser Cys Ser Asp Val Met Leu Ser Val Glu Glu Glu Gly
165 170 175
Lys Glu Asp Pro Leu Pro Thr Ala Ala Ser Gly Lys
180 185
<210> 12
<211> 58
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен GITR
<400> 12
Gln Leu Gly Leu His Ile Trp Gln Leu Arg Ser Gln Cys Met Trp Pro
1 5 10 15
Arg Glu Thr Gln Leu Leu Leu Glu Val Pro Pro Ser Thr Glu Asp Ala
20 25 30
Arg Ser Cys Gln Phe Pro Glu Glu Glu Arg Gly Glu Arg Ser Ala Glu
35 40 45
Glu Lys Gly Arg Leu Gly Asp Leu Trp Val
50 55
<210> 13
<211> 60
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен HVEM
<400> 13
Cys Val Lys Arg Arg Lys Pro Arg Gly Asp Val Val Lys Val Ile Val
1 5 10 15
Ser Val Gln Arg Lys Arg Gln Glu Ala Glu Gly Glu Ala Thr Val Ile
20 25 30
Glu Ala Leu Gln Ala Pro Pro Asp Val Thr Thr Val Ala Val Glu Glu
35 40 45
Thr Ile Pro Ser Phe Thr Gly Arg Ser Pro Asn His
50 55 60
<210> 14
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 14
Tyr His Ala Asp Tyr Tyr Lys Gln Arg Leu Ile His Asp Val Glu Met
1 5 10 15
Asn Pro Gly Pro
20
<210> 15
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 15
His Tyr Ala Gly Tyr Phe Ala Asp Leu Leu Ile His Asp Ile Glu Thr
1 5 10 15
Asn Pro Gly Pro
20
<210> 16
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 16
Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 17
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 17
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
1 5 10 15
Pro Gly Pro
<210> 18
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 18
Ala Ala Arg Gln Met Leu Leu Leu Leu Ser Gly Asp Val Glu Thr Asn
1 5 10 15
Pro Gly Pro
<210> 19
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 19
Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 20
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 20
Thr Arg Ala Glu Ile Glu Asp Glu Leu Ile Arg Ala Gly Ile Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 21
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 21
Thr Arg Ala Glu Ile Glu Asp Glu Leu Ile Arg Ala Asp Ile Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 22
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 22
Ala Lys Phe Gln Ile Asp Lys Ile Leu Ile Ser Gly Asp Val Glu Leu
1 5 10 15
Asn Pro Gly Pro
20
<210> 23
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 23
Ser Ser Ile Ile Arg Thr Lys Met Leu Val Ser Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 24
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 24
Cys Asp Ala Gln Arg Gln Lys Leu Leu Leu Ser Gly Asp Ile Glu Gln
1 5 10 15
Asn Pro Gly Pro
20
<210> 25
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> 2A-подобная последовательность
<400> 25
Tyr Pro Ile Asp Phe Gly Gly Phe Leu Val Lys Ala Asp Ser Glu Phe
1 5 10 15
Asn Pro Gly Pro
20
<210> 26
<211> 647
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность ORF анти-CD33 COMP CAR
<400> 26
Met Gly Thr Ser Leu Leu Cys Trp Met Ala Leu Cys Leu Leu Gly Ala
1 5 10 15
Asp His Ala Asp Ala Cys Pro Tyr Ser Asn Pro Ser Leu Cys Ser Gly
20 25 30
Gly Gly Gly Ser Glu Leu Pro Thr Gln Gly Thr Phe Ser Asn Val Ser
35 40 45
Thr Asn Val Ser Pro Ala Lys Pro Thr Thr Thr Ala Cys Pro Tyr Ser
50 55 60
Asn Pro Ser Leu Cys Ser Gly Gly Gly Gly Ser Pro Ala Pro Arg Pro
65 70 75 80
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
85 90 95
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
100 105 110
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
115 120 125
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg
130 135 140
Asn Arg Arg Arg Val Cys Lys Cys Pro Arg Pro Val Val Arg Ala Glu
145 150 155 160
Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly
165 170 175
Pro Met Ala Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu
180 185 190
Thr Asp Ala Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
195 200 205
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu
210 215 220
Asp Ile Tyr Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala
225 230 235 240
Pro Lys Leu Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro
245 250 255
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile
260 265 270
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr
275 280 285
Lys Asn Tyr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
290 295 300
Arg Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
305 310 315 320
Ser Gly Gly Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly
325 330 335
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
340 345 350
Ser Gly Phe Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala
355 360 365
Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly
370 375 380
Ser Thr Tyr Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
385 390 395 400
Asp Asn Ala Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
405 410 415
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly
420 425 430
Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
435 440 445
Met Asp Pro Ala Gly Ser Asp Leu Gly Pro Gln Met Leu Arg Glu Leu
450 455 460
Gln Glu Thr Asn Ala Ala Leu Gln Asp Val Arg Glu Leu Leu Arg Gln
465 470 475 480
Gln Val Arg Glu Ile Thr Phe Leu Lys Asn Thr Val Met Glu Cys Asp
485 490 495
Ala Cys Gly Ser Gly Lys Lys Asp Pro Lys Phe Trp Val Leu Val Val
500 505 510
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
515 520 525
Ile Ile Phe Trp Val Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp
530 535 540
Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
545 550 555 560
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
565 570 575
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
580 585 590
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
595 600 605
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
610 615 620
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
625 630 635 640
Met Gln Ala Leu Pro Pro Arg
645
<210> 27
<211> 638
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность ORF анти-ROR-1 COMP CAR
<400> 27
Met Gly Thr Ser Leu Leu Cys Trp Met Ala Leu Cys Leu Leu Gly Ala
1 5 10 15
Asp His Ala Asp Ala Cys Pro Tyr Ser Asn Pro Ser Leu Cys Ser Gly
20 25 30
Gly Gly Gly Ser Glu Leu Pro Thr Gln Gly Thr Phe Ser Asn Val Ser
35 40 45
Thr Asn Val Ser Pro Ala Lys Pro Thr Thr Thr Ala Cys Pro Tyr Ser
50 55 60
Asn Pro Ser Leu Cys Ser Gly Gly Gly Gly Ser Pro Ala Pro Arg Pro
65 70 75 80
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
85 90 95
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
100 105 110
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
115 120 125
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg
130 135 140
Asn Arg Arg Arg Val Cys Lys Cys Pro Arg Pro Val Val Arg Ala Glu
145 150 155 160
Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly
165 170 175
Pro Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val
180 185 190
Pro Gly Ser Thr Gly Gln Ser Val Lys Glu Ser Glu Gly Asp Leu Val
195 200 205
Thr Pro Ala Gly Asn Leu Thr Leu Thr Cys Thr Ala Ser Gly Ser Asp
210 215 220
Ile Asn Asp Tyr Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly
225 230 235 240
Leu Glu Trp Ile Gly Phe Ile Asn Ser Gly Gly Ser Thr Trp Tyr Ala
245 250 255
Ser Trp Val Lys Gly Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val
260 265 270
Asp Leu Lys Met Thr Ser Leu Thr Thr Asp Asp Thr Ala Thr Tyr Phe
275 280 285
Cys Ala Arg Gly Tyr Ser Thr Tyr Tyr Gly Asp Phe Asn Ile Trp Gly
290 295 300
Pro Gly Thr Leu Val Thr Ile Ser Ser Gly Gly Gly Gly Ser Gly Gly
305 310 315 320
Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Val Met Thr Gln Thr Pro
325 330 335
Ser Ser Thr Ser Gly Ala Val Gly Gly Thr Val Thr Ile Asn Cys Gln
340 345 350
Ala Ser Gln Ser Ile Asp Ser Asn Leu Ala Trp Phe Gln Gln Lys Pro
355 360 365
Gly Gln Pro Pro Thr Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser
370 375 380
Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Glu Tyr Thr
385 390 395 400
Leu Thr Ile Ser Gly Val Gln Arg Glu Asp Ala Ala Thr Tyr Tyr Cys
405 410 415
Leu Gly Gly Val Gly Asn Val Ser Tyr Arg Thr Ser Phe Gly Gly Gly
420 425 430
Thr Glu Val Val Val Lys Arg Ser Asp Pro Ala Gly Ser Asp Leu Gly
435 440 445
Pro Gln Met Leu Arg Glu Leu Gln Glu Thr Asn Ala Ala Leu Gln Asp
450 455 460
Val Arg Glu Leu Leu Arg Gln Gln Val Arg Glu Ile Thr Phe Leu Lys
465 470 475 480
Asn Thr Val Met Glu Cys Asp Ala Cys Gly Ser Gly Lys Lys Asp Pro
485 490 495
Lys Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
500 505 510
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Arg Val
515 520 525
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
530 535 540
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
545 550 555 560
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
565 570 575
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
580 585 590
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
595 600 605
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
610 615 620
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
625 630 635
<210> 28
<211> 1944
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК-последовательность ORF анти-CD33 COMP CAR
<400> 28
atgggcacca gcctgctgtg ctggatggcc ctgtgcctgc tgggcgccga ccacgccgat 60
gcctgcccct acagcaaccc cagcctgtgc agcggaggcg gcggcagcga gctgcccacc 120
cagggcacct tctccaacgt gtccaccaac gtgagcccag ccaagcccac caccaccgcc 180
tgtccttatt ccaatccttc cctgtgtagc ggagggggag gcagcccagc ccccagacct 240
cccaccccag cccccaccat cgccagccag cctctgagcc tgagacccga ggcctgccgc 300
ccagccgccg gcggcgccgt gcacaccaga ggcctggatt tcgcctgcga tatctacatc 360
tgggccccac tggccggcac ctgtggcgtg ctgctgctga gcctggtgat caccctgtac 420
tgcaaccacc gcaaccgcag gcgcgtgtgc aagtgcccca ggcccgtggt gagagccgag 480
ggcagaggca gcctgctgac ctgcggcgac gtggaggaga acccaggccc catggccgtg 540
cccactcagg tcctggggtt gttgctactg tggcttacag atgccagatg tgacatccag 600
atgacacagt ctccatcttc cctgtctgca tctgtcggag atcgcgtcac catcacctgt 660
cgagcaagtg aggacattta ttttaattta gtgtggtatc agcagaaacc aggaaaggcc 720
cctaagctcc tgatctatga tacaaatcgc ttggcagatg gggtcccatc acggttcagt 780
ggctctggat ctggcacaca gtatactcta accataagta gcctgcaacc cgaagatttc 840
gcaacctatt attgtcaaca ctataagaat tatccgctca cgttcggtca ggggaccaag 900
ctggaaatca aaagatctgg tggcggaggg tcaggaggcg gaggcagcgg aggcggtggc 960
tcgggaggcg gaggctcgag atctgaggtg cagttggtgg agtctggggg cggcttggtg 1020
cagcctggag ggtccctgag gctctcctgt gcagcctcag gattcactct cagtaattat 1080
ggcatgcact ggatcaggca ggctccaggg aagggtctgg agtgggtctc gtctattagt 1140
cttaatggtg gtagcactta ctatcgagac tccgtgaagg gccgattcac tatctccagg 1200
gacaatgcaa aaagcaccct ctaccttcaa atgaatagtc tgagggccga ggacacggcc 1260
gtctattact gtgcagcaca ggacgcttat acgggaggtt actttgatta ctggggccaa 1320
ggaacgctgg tcacagtctc gtctatggat cccgccggga gcgacctggg ccctcagatg 1380
ctgcgggagc tgcaggagac aaatgccgcc ctgcaggacg tgcgcgagct gctgagacag 1440
caggtgcggg agattacatt cctgaagaac accgtgatgg agtgcgatgc ctgcggatct 1500
gggaagaagg accccaagtt ctgggtcctg gtggtggtgg gaggcgtgct ggcctgttac 1560
tctctcctgg tgaccgtggc cttcatcatc ttttgggtgc gctcccgggt gaagttttct 1620
cgctctgccg atgccccagc ctatcagcag ggccagaatc agctgtacaa tgaactgaac 1680
ctgggcaggc gggaggagta cgacgtgctg gataagcgga gaggcagaga ccccgagatg 1740
ggcggcaaac cacggcgcaa aaatccccag gagggactct ataacgagct gcagaaggac 1800
aaaatggccg aggcctattc cgagatcggc atgaagggag agagaagacg cggaaagggc 1860
cacgacggcc tgtatcaggg attgtccacc gctacaaaag atacatatga tgccctgcac 1920
atgcaggccc tgccacccag atga 1944
<210> 29
<211> 1914
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК-последовательность ORF анти-ROR-1 COMP CAR
<400> 29
atgggcacca gcctgctgtg ctggatggcc ctgtgcctgc tgggcgccga ccacgccgat 60
gcctgcccct acagcaaccc cagcctgtgc agcggaggcg gcggcagcga gctgcccacc 120
cagggcacct tctccaacgt gtccaccaac gtgagcccag ccaagcccac caccaccgcc 180
tgtccttatt ccaatccttc cctgtgtagc ggagggggag gcagcccagc ccccagacct 240
cccaccccag cccccaccat cgccagccag cctctgagcc tgagacccga ggcctgccgc 300
ccagccgccg gcggcgccgt gcacaccaga ggcctggatt tcgcctgcga tatctacatc 360
tgggccccac tggccggcac ctgtggcgtg ctgctgctga gcctggtgat caccctgtac 420
tgcaaccacc gcaaccgcag gcgcgtgtgc aagtgcccca ggcccgtggt gagagccgag 480
ggcagaggca gcctgctgac ctgcggcgac gtggaggaga acccaggccc catggagacc 540
gacaccctgc tgctgtgggt gctgctgctg tgggtgcccg gcagcaccgg ccagagcgtg 600
aaggagagcg agggcgacct ggtgacccca gccggcaacc tgaccctgac ctgcaccgcc 660
agcggcagcg acatcaacga ctaccccatc agctgggtgc ggcaggcccc aggcaagggc 720
ctggagtgga tcggcttcat caacagcggc ggcagcacct ggtacgccag ctgggtgaag 780
ggccggttca ccatcagccg gaccagcacc accgtggacc tgaagatgac cagcctgacc 840
accgacgaca ccgccaccta cttctgcgcc agaggctaca gcacctacta cggcgacttc 900
aacatctggg gacccggcac cctggtgacc atcagcagcg gaggcggagg gtctgggggc 960
ggcggtagcg gcggaggagg gagcgagctg gtgatgaccc agaccccaag cagcaccagc 1020
ggcgccgtgg gcggcaccgt gaccatcaac tgccaggcca gccagagcat cgacagcaac 1080
ctggcctggt tccagcagaa gcctggccag ccacccaccc tgctgatcta ccgggccagc 1140
aacctggcca gcggcgtgcc cagccggttc agcggcagcc ggagcggcac cgagtacacc 1200
ctgaccatca gcggcgtgca gcgggaggac gccgccacct actactgcct gggcggagtg 1260
ggcaacgtga gctaccggac cagcttcggc ggaggcaccg aggtggtggt gaagcggtcg 1320
gatcccgccg ggagcgacct gggccctcag atgctgcggg agctgcagga gacaaatgcc 1380
gccctgcagg acgtgcgcga gctgctgaga cagcaggtgc gggagattac attcctgaag 1440
aacaccgtga tggagtgcga tgcctgcgga tctgggaaga aggaccccaa gttctgggtc 1500
ctggtggtgg tgggaggcgt gctggcctgt tactctctcc tggtgaccgt ggccttcatc 1560
atcttttggg tgcgctcccg ggtgaagttt tctcgctctg ccgatgcccc agcctatcag 1620
cagggccaga atcagctgta caatgaactg aacctgggca ggcgggagga gtacgacgtg 1680
ctggataagc ggagaggcag agaccccgag atgggcggca aaccacggcg caaaaatccc 1740
caggagggac tctataacga gctgcagaag gacaaaatgg ccgaggccta ttccgagatc 1800
ggcatgaagg gagagagaag acgcggaaag ggccacgacg gcctgtatca gggattgtcc 1860
accgctacaa aagatacata tgatgccctg cacatgcagg ccctgccacc caga 1914
<210> 30
<211> 57
<212> Белок
<213> Искусственная последовательность
<220>
<223> Кинезиновый моторный белок: параллельный гомодимер
<400> 30
Met His Ala Ala Leu Ser Thr Glu Val Val His Leu Arg Gln Arg Thr
1 5 10 15
Glu Glu Leu Leu Arg Cys Asn Glu Gln Gln Ala Ala Glu Leu Glu Thr
20 25 30
Cys Lys Glu Gln Leu Phe Gln Ser Asn Met Glu Arg Lys Glu Leu His
35 40 45
Asn Thr Val Met Asp Leu Arg Gly Asn
50 55
<210> 31
<211> 50
<212> Белок
<213> Искусственная последовательность
<220>
<223> Дельта-антиген гепатита D: параллельный гомодимер
<400> 31
Gly Arg Glu Asp Ile Leu Glu Gln Trp Val Ser Gly Arg Lys Lys Leu
1 5 10 15
Glu Glu Leu Glu Arg Asp Leu Arg Lys Leu Lys Lys Lys Ile Lys Lys
20 25 30
Leu Glu Glu Asp Asn Pro Trp Leu Gly Asn Ile Lys Gly Ile Ile Gly
35 40 45
Lys Tyr
50
<210> 32
<211> 61
<212> Белок
<213> Искусственная последовательность
<220>
<223> Коровый белок бокса С/D sRNP архей: анти-параллельный гетеродимер
<400> 32
Arg Tyr Val Val Ala Leu Val Lys Ala Leu Glu Glu Ile Asp Glu Ser
1 5 10 15
Ile Asn Met Leu Asn Glu Lys Leu Glu Asp Ile Arg Ala Val Lys Glu
20 25 30
Ser Glu Ile Thr Glu Lys Phe Glu Lys Lys Ile Arg Glu Leu Arg Glu
35 40 45
Leu Arg Arg Asp Val Glu Arg Glu Ile Glu Glu Val Met
50 55 60
<210> 33
<211> 31
<212> Белок
<213> Искусственная последовательность
<220>
<223> Маннозосвязывающий белок А: параллельный гомотример
<400> 33
Ala Ile Glu Val Lys Leu Ala Asn Met Glu Ala Glu Ile Asn Thr Leu
1 5 10 15
Lys Ser Lys Leu Glu Leu Thr Asn Lys Leu His Ala Phe Ser Met
20 25 30
<210> 34
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Суперспиральный серин-богатый белок 1: параллельный гомотример
<400> 34
Glu Trp Glu Ala Leu Glu Lys Lys Leu Ala Ala Leu Glu Ser Lys Leu
1 5 10 15
Gln Ala Leu Glu Lys Lys Leu Glu Ala Leu Glu His Gly
20 25
<210> 35
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> Полипептид-высвобождающий фактор 2: антипараллельный гетеротример, цепь А
<400> 35
Ile Asn Pro Val Asn Asn Arg Ile Gln Asp Leu Thr Glu Arg Ser Asp
1 5 10 15
Val Leu Arg Gly Tyr Leu Asp Tyr
20
<210> 36
<211> 51
<212> Белок
<213> Искусственная последовательность
<220>
<223> Полипептид-высвобождающий фактор 2: антипараллельный гетеротример, цепь B
<400> 36
Val Val Asp Thr Leu Asp Gln Met Lys Gln Gly Leu Glu Asp Val Ser
1 5 10 15
Gly Leu Leu Glu Leu Ala Val Glu Ala Asp Asp Glu Glu Thr Phe Asn
20 25 30
Glu Ala Val Ala Glu Leu Asp Ala Leu Glu Glu Lys Leu Ala Gln Leu
35 40 45
Glu Phe Arg
50
<210> 37
<211> 51
<212> Белок
<213> Искусственная последовательность
<220>
<223> SNAP-25 и SNARE: параллельный гетеротетрамер, цепь А
<400> 37
Ile Glu Thr Arg His Ser Glu Ile Ile Lys Leu Glu Asn Ser Ile Arg
1 5 10 15
Glu Leu His Asp Met Phe Met Asp Met Ala Met Leu Val Glu Ser Gln
20 25 30
Gly Glu Met Ile Asp Arg Ile Glu Tyr Asn Val Glu His Ala Val Asp
35 40 45
Tyr Val Glu
50
<210> 38
<211> 67
<212> Белок
<213> Искусственная последовательность
<220>
<223> SNAP-25 и SNARE: параллельный гетеротетрамер, цепь B
<400> 38
Ala Leu Ser Glu Ile Glu Thr Arg His Ser Glu Ile Ile Lys Leu Glu
1 5 10 15
Asn Ser Ile Arg Glu Leu His Asp Met Phe Met Asp Met Ala Met Leu
20 25 30
Val Glu Ser Gln Gly Glu Met Ile Asp Arg Ile Glu Tyr Asn Val Glu
35 40 45
His Ala Val Asp Tyr Val Glu Arg Ala Val Ser Asp Thr Lys Lys Ala
50 55 60
Val Lys Tyr
65
<210> 39
<211> 69
<212> Белок
<213> Искусственная последовательность
<220>
<223> SNAP-25 и SNARE: параллельный гетеротетрамер, цепь C
<400> 39
Glu Leu Glu Glu Met Gln Arg Arg Ala Asp Gln Leu Ala Asp Glu Ser
1 5 10 15
Leu Glu Ser Thr Arg Arg Met Leu Gln Leu Val Glu Glu Ser Lys Asp
20 25 30
Ala Gly Ile Arg Thr Leu Val Met Leu Asp Glu Gln Gly Glu Gln Leu
35 40 45
Glu Arg Ile Glu Glu Gly Met Asp Gln Ile Asn Lys Asp Met Lys Glu
50 55 60
Ala Glu Lys Asn Leu
65
<210> 40
<211> 51
<212> Белок
<213> Искусственная последовательность
<220>
<223> SNAP-25 и SNARE: параллельный гетеротетрамер, цепь D
<400> 40
Ile Glu Thr Arg His Ser Glu Ile Ile Lys Leu Glu Asn Ser Ile Arg
1 5 10 15
Glu Leu His Asp Met Phe Met Asp Met Ala Met Leu Val Glu Ser Gln
20 25 30
Gly Glu Met Ile Asp Arg Ile Glu Tyr Asn Val Glu His Ala Val Asp
35 40 45
Tyr Val Glu
50
<210> 41
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Lac-репрессор: параллельный гомотетрамер
<400> 41
Ser Pro Arg Ala Leu Ala Asp Ser Leu Met Gln Leu Ala Arg Gln Val
1 5 10 15
Ser Arg Leu Glu
20
<210> 42
<211> 136
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аполипопротеин Е: антипараллельный гетеротетрамер
<400> 42
Ser Gly Gln Arg Trp Glu Leu Ala Leu Gly Arg Phe Trp Asp Tyr Leu
1 5 10 15
Arg Trp Val Gln Thr Leu Ser Glu Gln Val Gln Glu Glu Leu Leu Ser
20 25 30
Ser Gln Val Thr Gln Glu Leu Arg Ala Leu Met Asp Glu Thr Met Lys
35 40 45
Glu Leu Lys Ala Tyr Lys Ser Glu Leu Glu Glu Gln Leu Thr Ala Arg
50 55 60
Leu Ser Lys Glu Leu Gln Ala Ala Gln Ala Arg Leu Gly Ala Asp Met
65 70 75 80
Glu Asp Val Cys Gly Arg Leu Val Gln Tyr Arg Gly Glu Val Gln Ala
85 90 95
Met Leu Gly Gln Ser Thr Glu Glu Leu Arg Val Arg Leu Ala Ser His
100 105 110
Leu Arg Lys Leu Arg Lys Arg Leu Leu Arg Asp Ala Asp Asp Leu Gln
115 120 125
Lys Arg Leu Ala Val Tyr Gln Ala
130 135
<210> 43
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гетеродимеризующийся домен
<400> 43
Gln Leu Glu Lys Glu Leu Gln Ala Leu Glu Lys Glu Asn Ala Gln Leu
1 5 10 15
Glu Trp Glu Leu Gln Ala Leu Glu Lys Glu Leu Ala Gln
20 25
<210> 44
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гетеродимеризующийся домен
<400> 44
Gln Leu Glu Lys Lys Leu Gln Ala Leu Lys Lys Lys Asn Ala Gln Leu
1 5 10 15
Lys Trp Lys Leu Gln Ala Leu Lys Lys Lys Leu Ala Gln
20 25
<210> 45
<211> 44
<212> Белок
<213> Искусственная последовательность
<220>
<223> Димеризующийся и стыковочный домен (DDD1)
<400> 45
Ser His Ile Gln Ile Pro Pro Gly Leu Thr Glu Leu Leu Gln Gly Tyr
1 5 10 15
Thr Val Glu Val Leu Arg Gln Gln Pro Pro Asp Leu Val Glu Phe Ala
20 25 30
Val Glu Tyr Phe Thr Arg Leu Arg Glu Ala Arg Ala
35 40
<210> 46
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Якорный домен (AD1)
<400> 46
Val Gln Ile Glu Tyr Leu Ala Lys Gln Ile Val Asp Asn Ala Ile Gln
1 5 10 15
Gln Ala
<210> 47
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Укороченный COMP
<400> 47
Gln Gln Val Arg Glu Ile Thr Phe Leu Lys Asn Thr Val Met Glu Cys
1 5 10 15
Asp Ala Cys Gly
20
<210> 48
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Консенсусный сайт расщепления протеазы из вируса гравировки табака (TEV)
<400> 48
Glu Asn Leu Tyr Phe Gln Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР | 2016 |
|
RU2826122C1 |
| СИГНАЛЬНАЯ СИСТЕМА | 2015 |
|
RU2731638C2 |
| КЛЕТКА | 2014 |
|
RU2717984C2 |
| КЛЕТКА | 2014 |
|
RU2732236C2 |
| КЛЕТКА | 2015 |
|
RU2768019C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
| ХИМЕРНЫЙ БЕЛОК | 2016 |
|
RU2746755C2 |
| СВЯЗЫВАЮЩИЙ ДОМЕН | 2019 |
|
RU2829934C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| Т-КЛЕТКИ С КОСТИМУЛИРУЮЩИМ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ, НАЦЕЛЕННЫЕ НА IL13Rα2 | 2015 |
|
RU2749922C2 |


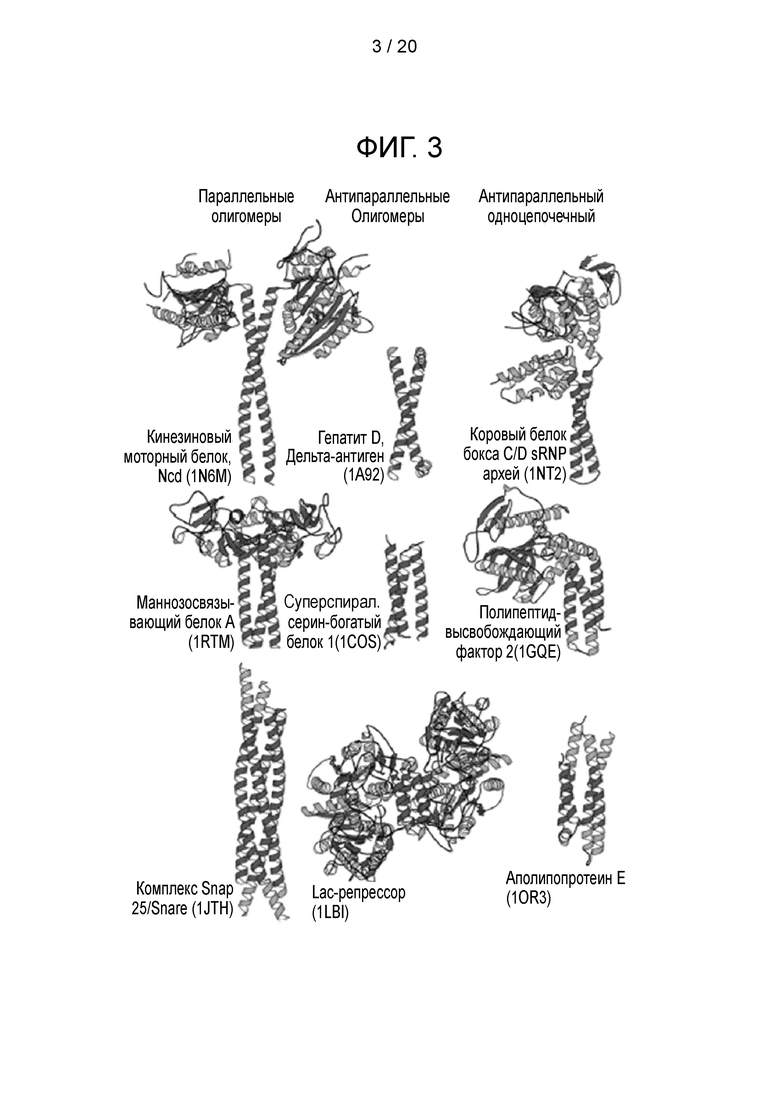

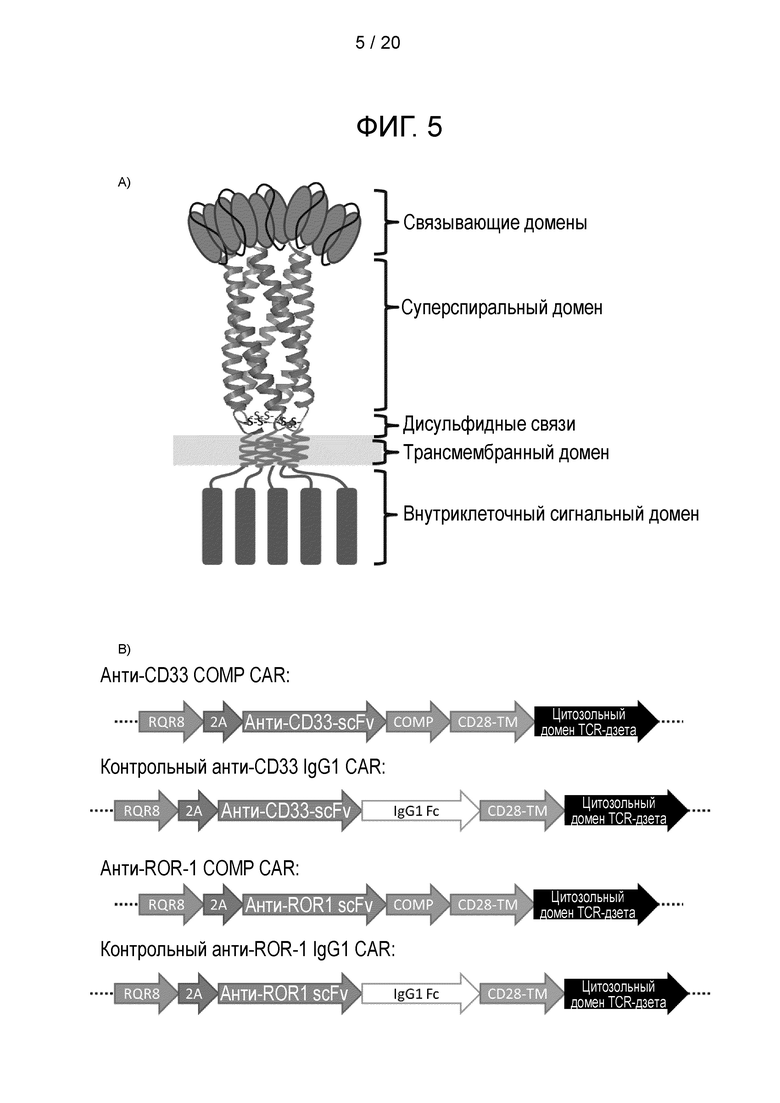


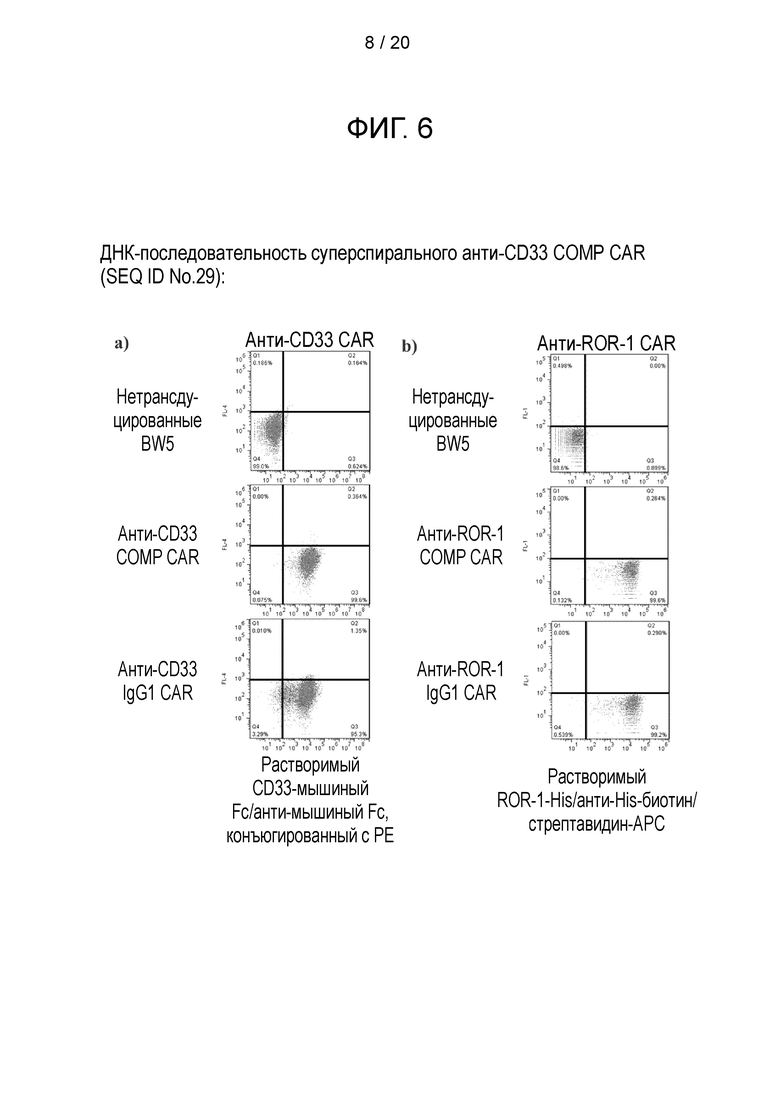
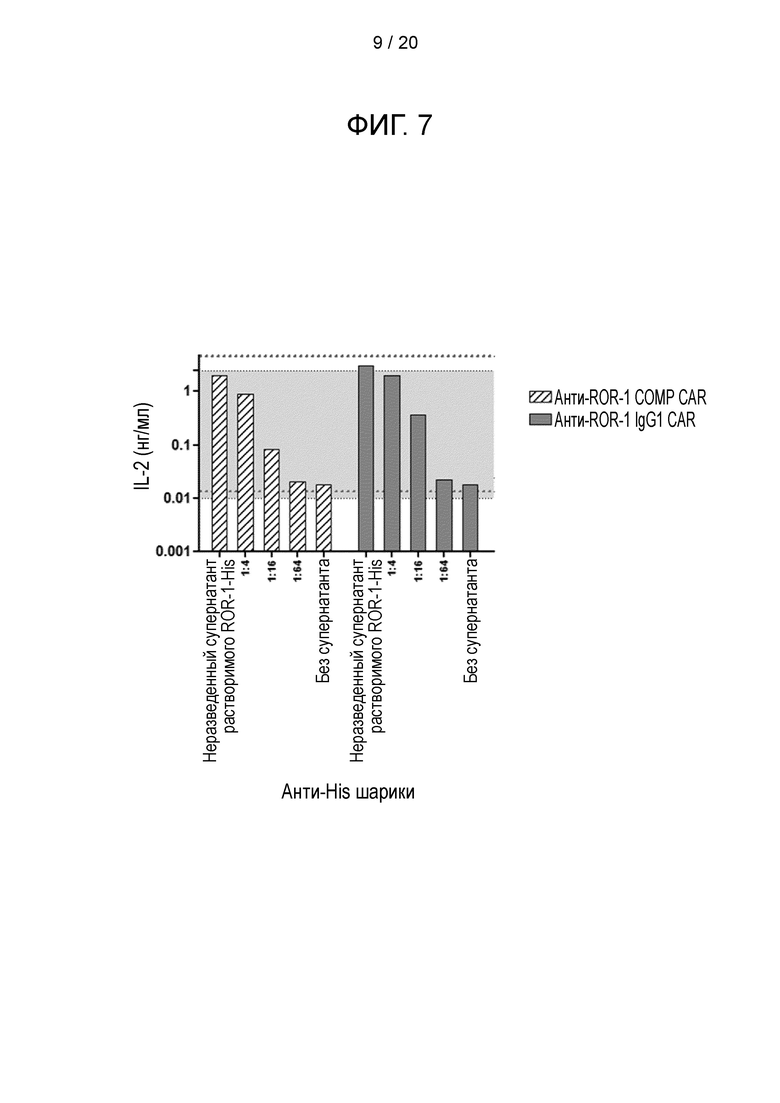
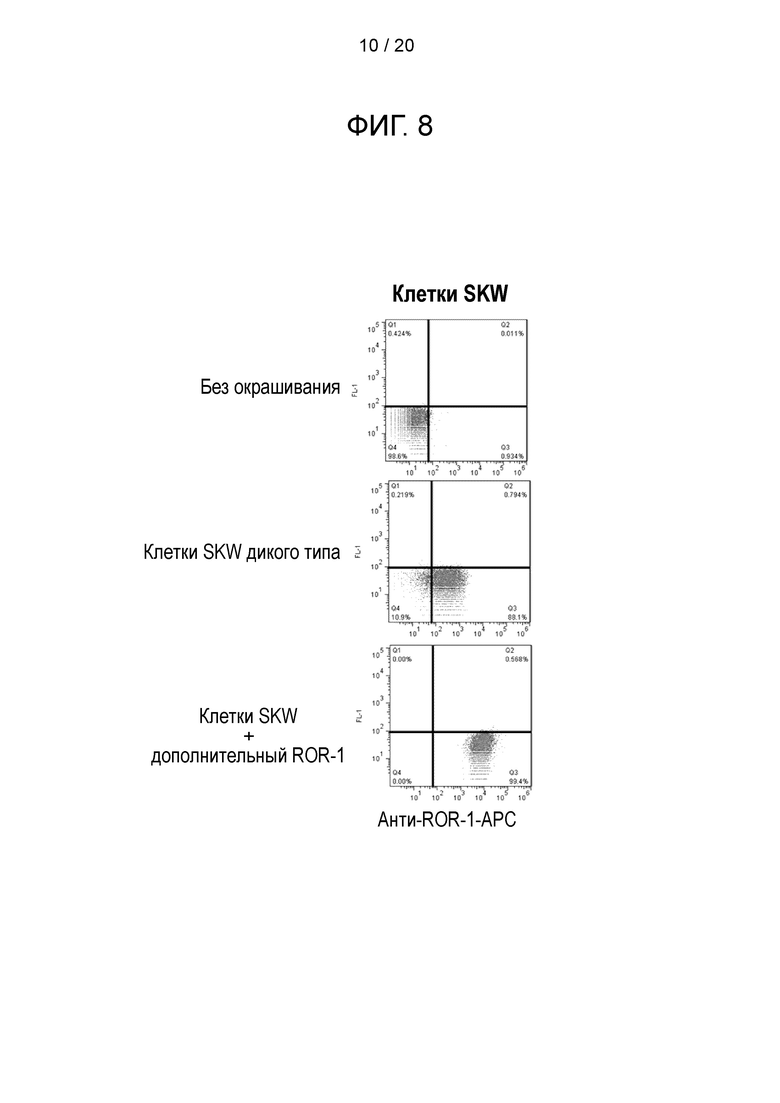
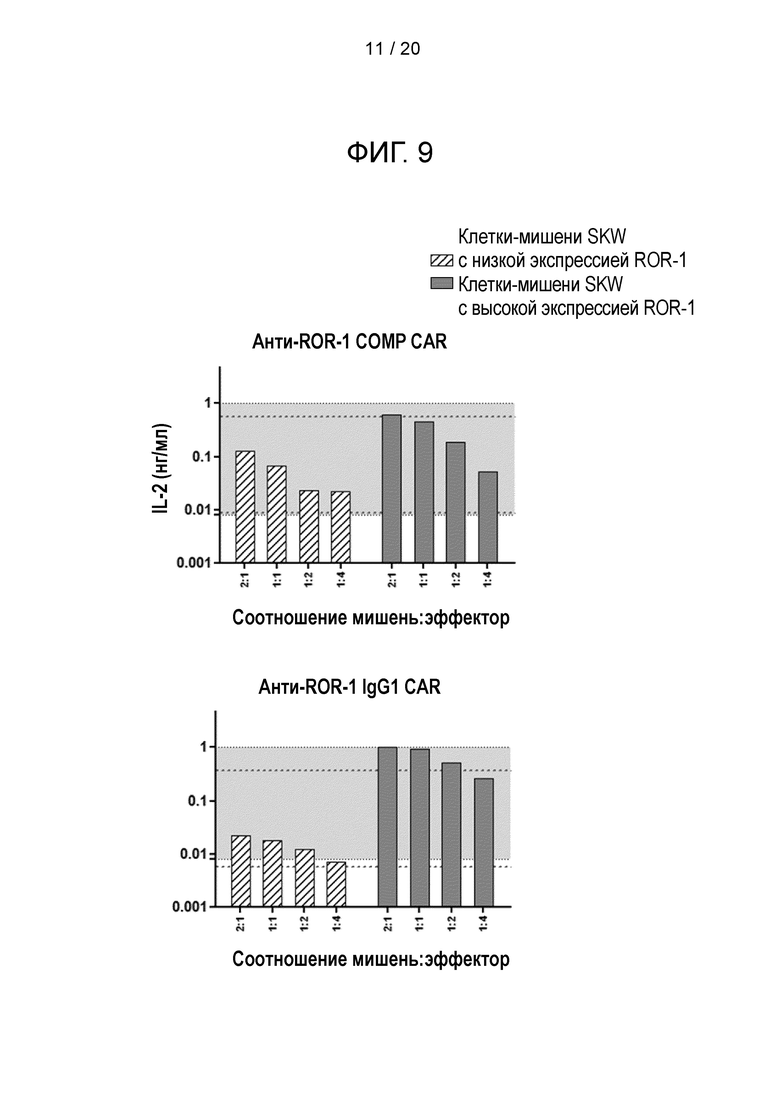
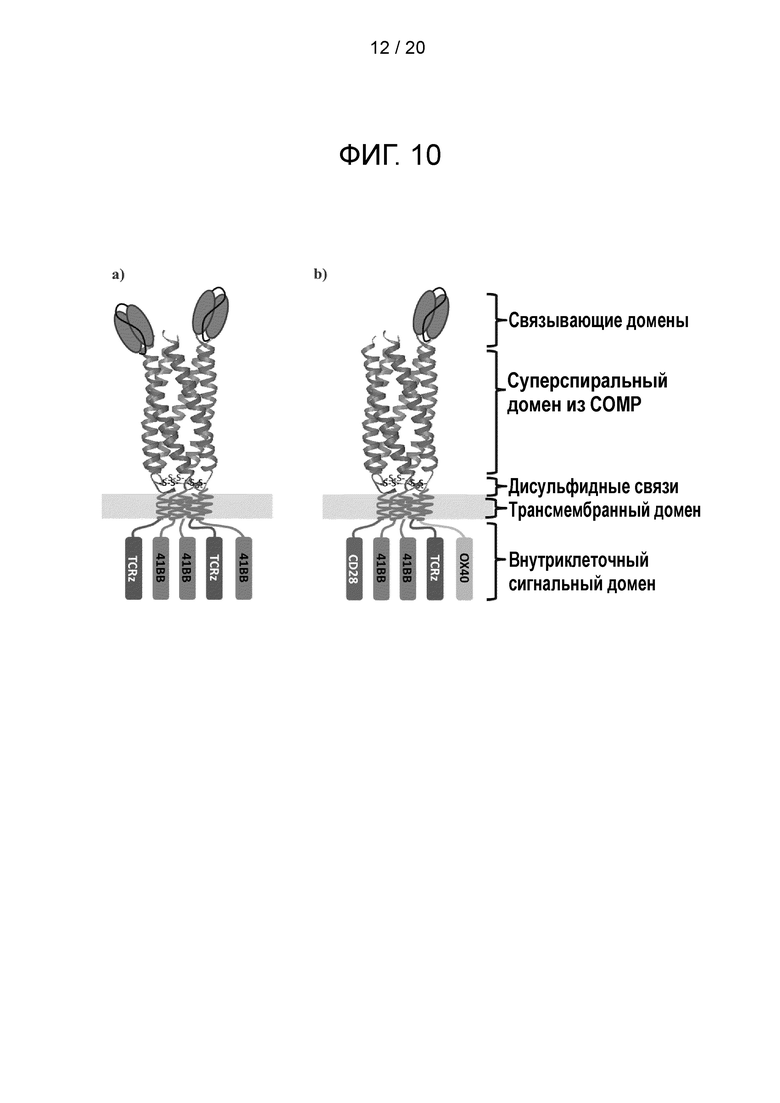
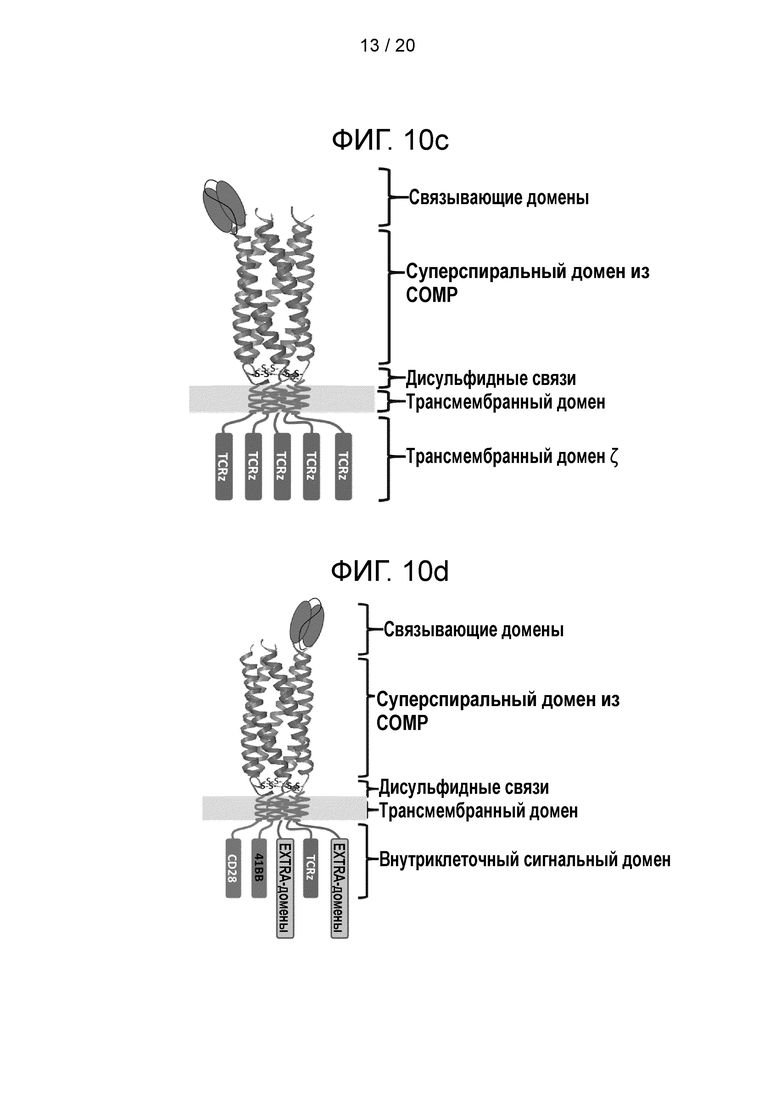
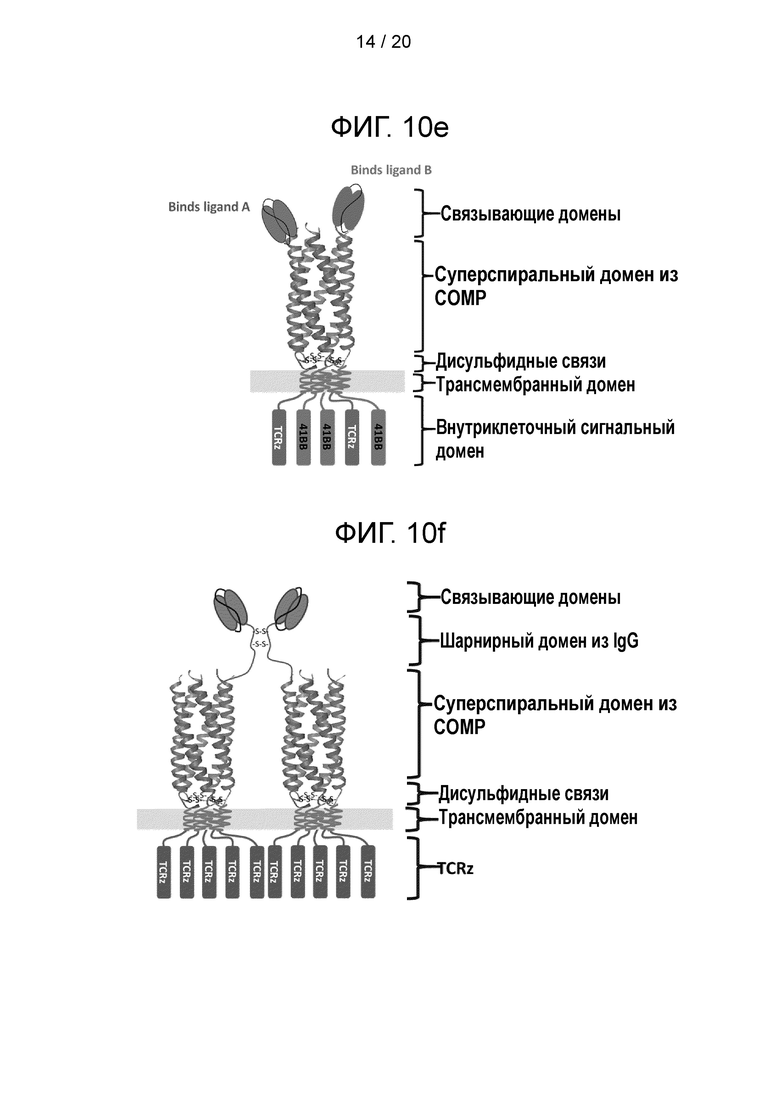
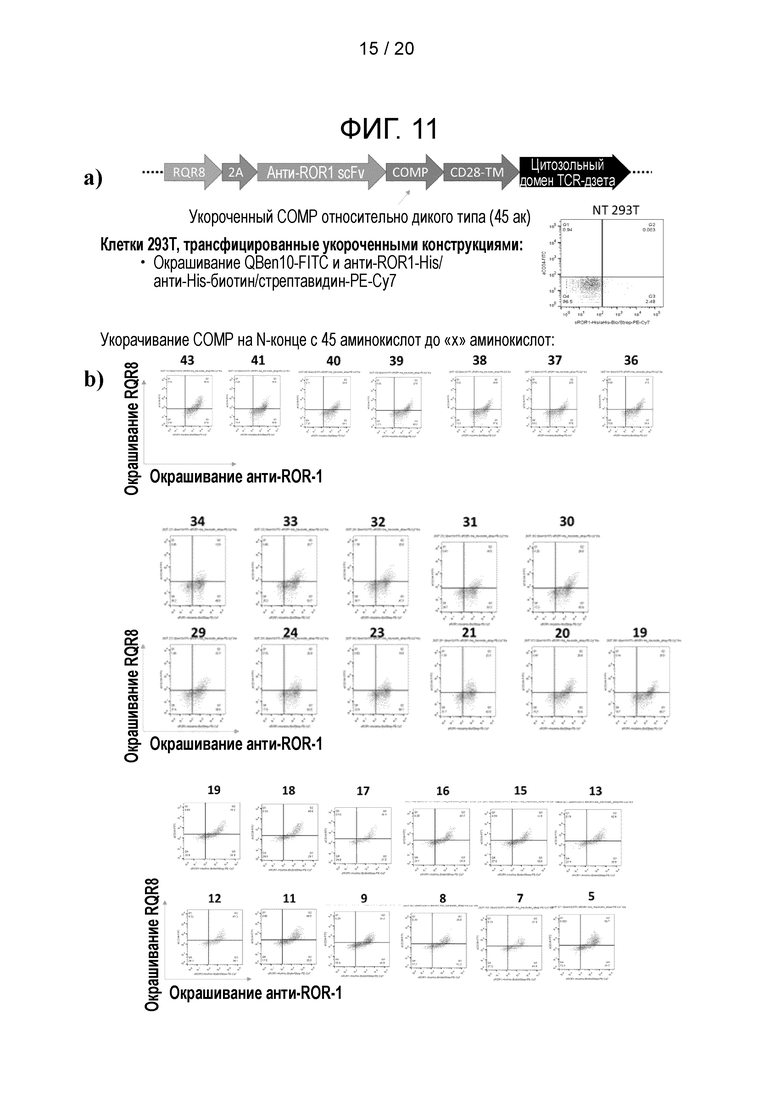
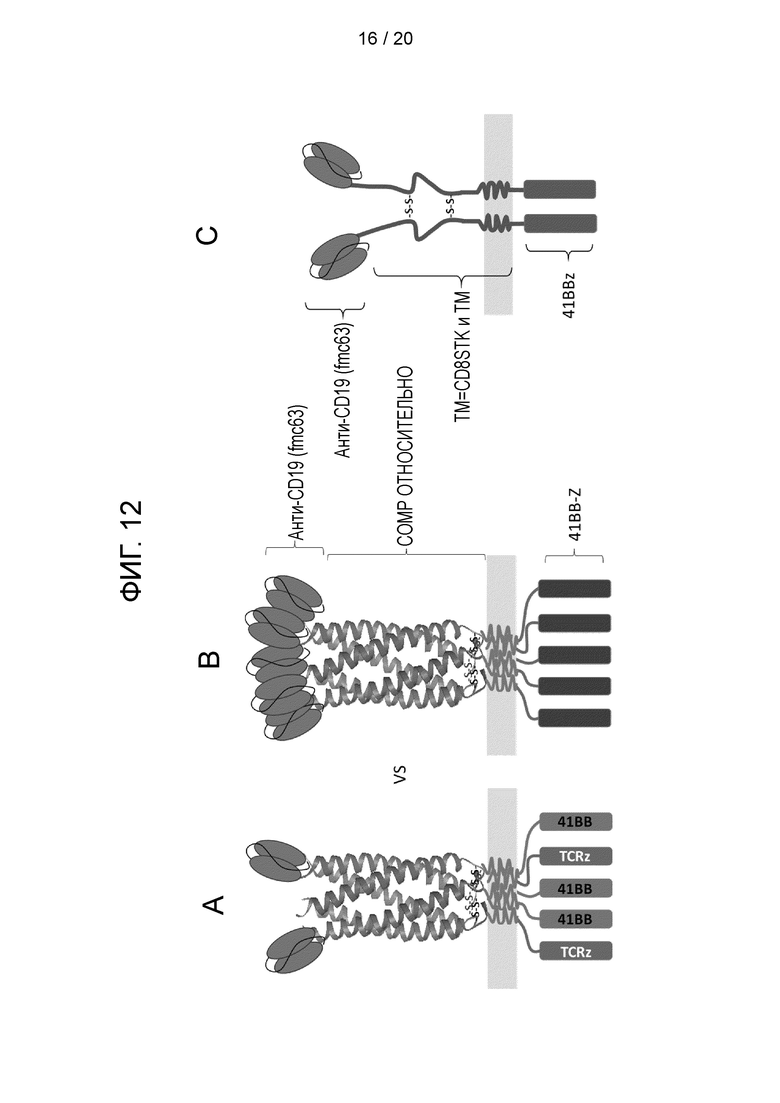
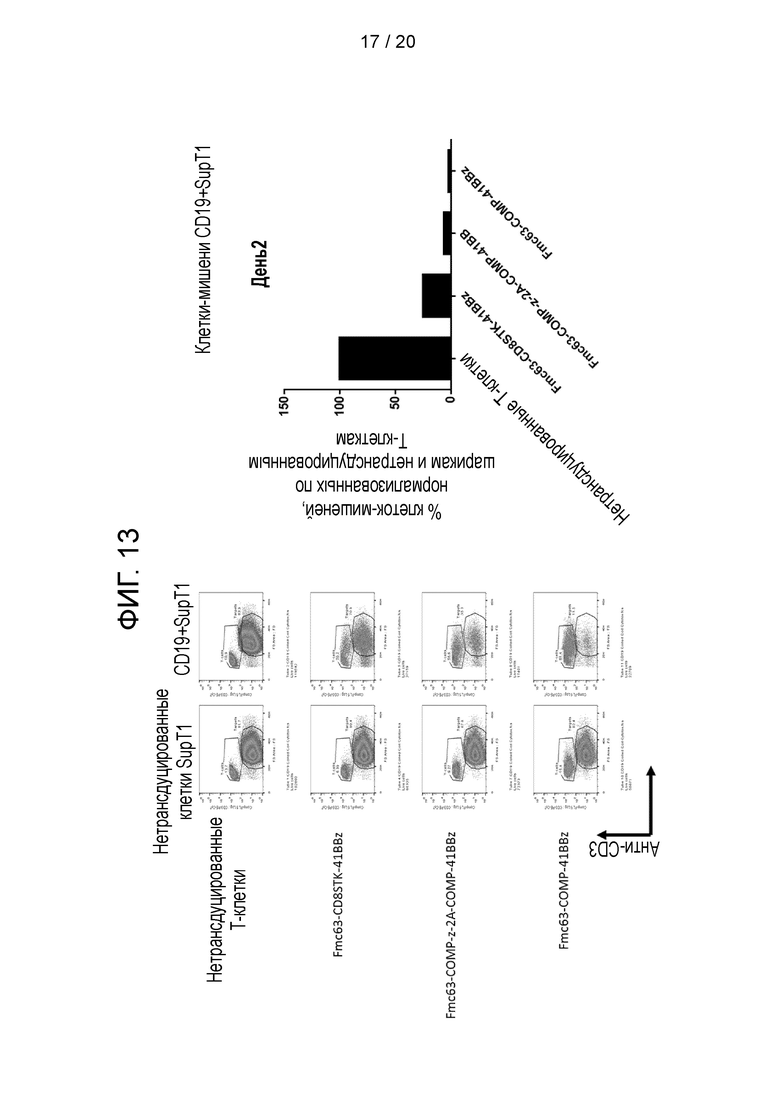
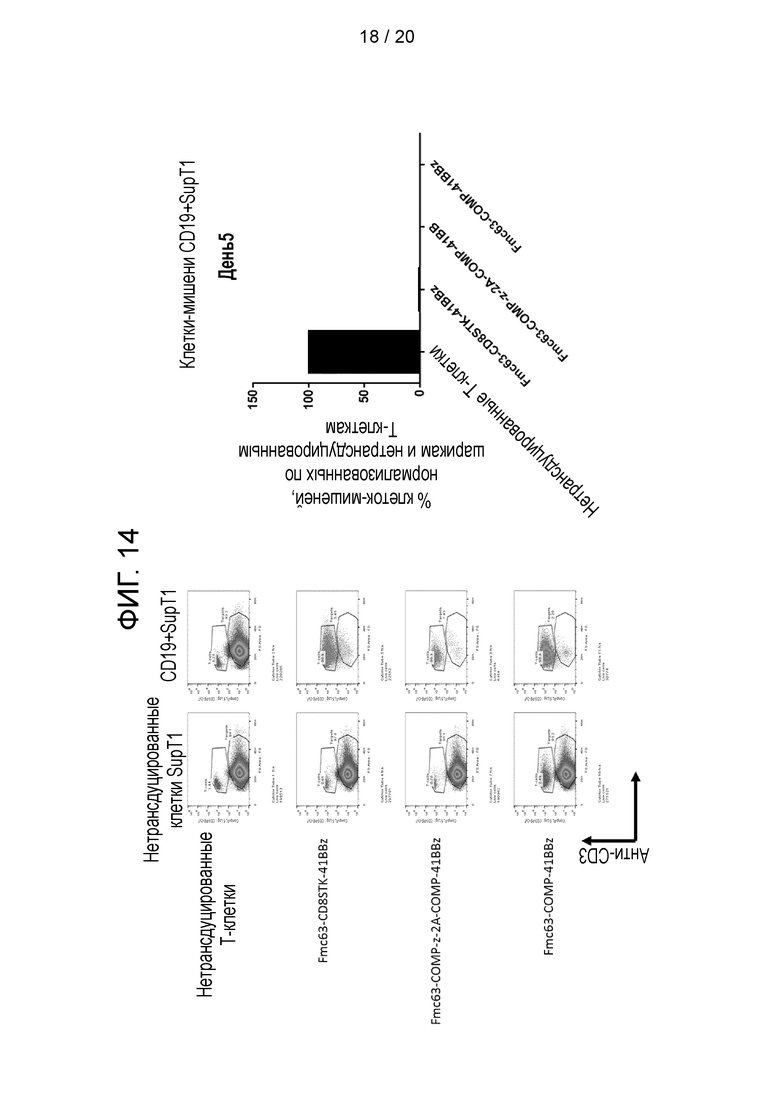
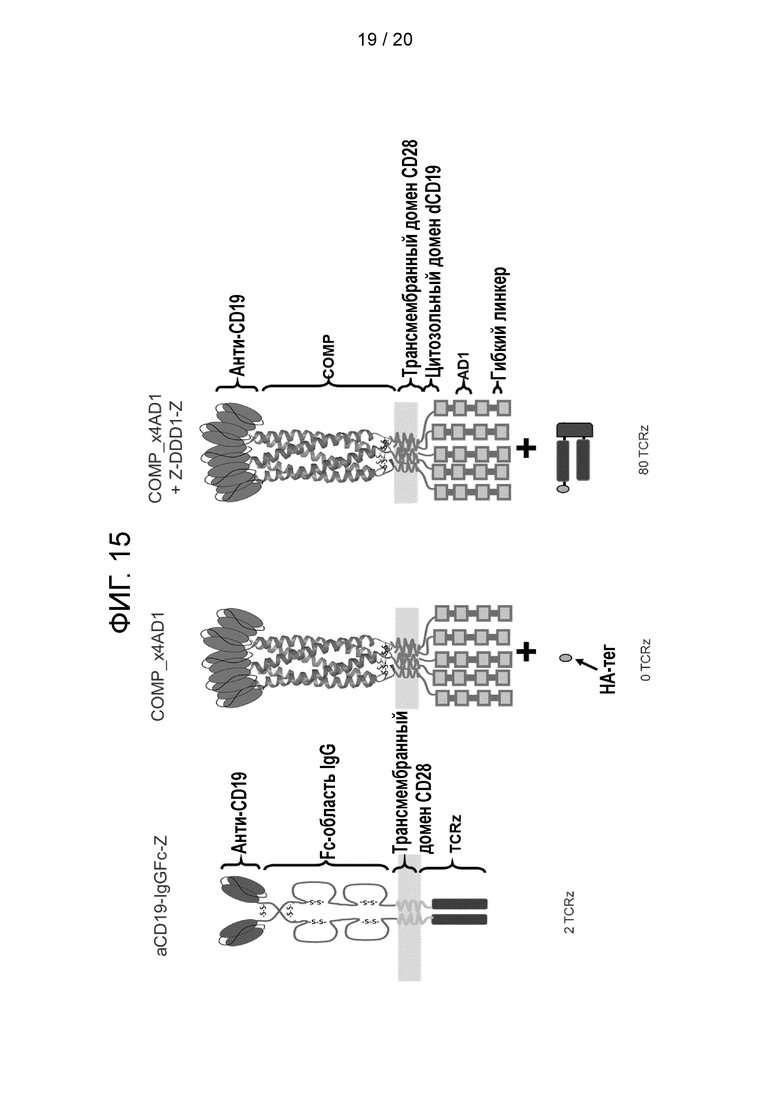
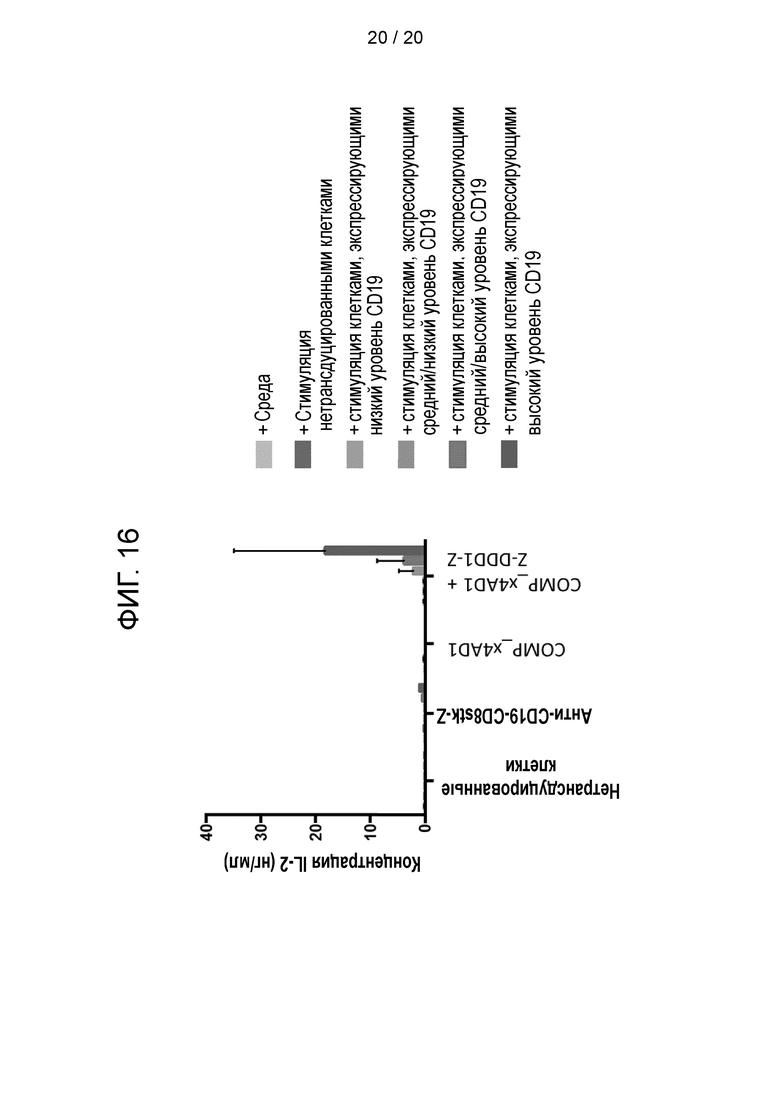
Изобретение относится к области биотехнологии, конкретно к получению формирующего химерный антигенный рецептор (CAR) полипептида, содержащего антигенсвязывающий домен, суперспиральный спейсерный домен, трансмембранный домен, и эндодомен, и может быть использовано в медицине. Мультимерный CAR, образованный путем объединения множества формирующих CAR полипептидов посредством ассоциации их суперспиральных спейсерных доменов, может быть использован для эффективной иммунотерапии рака. 11 н. и 26 з.п. ф-лы, 16 ил., 5 табл., 7 пр.
1. Формирующий химерный антигенный рецептор (CAR) полипептид для получения мультимерного CAR, содержащий в следующем порядке:
(i) антигенсвязывающий домен;
(ii) суперспиральный спейсерный домен;
(iii) трансмембранный домен; и
(iv) эндодомен.
2. Формирующий CAR полипептид по п.1, отличающийся тем, что указанный антигенсвязывающий домен связывается с ROR-1, TYRP-1 или ВСМА.
3. Формирующий CAR полипептид по любому из пп.1-2, отличающийся тем, что указанный суперспиральный спейсерный домен происходит из олигомерного матриксного белка хряща (COMP), маннозосвязывающего белка А, суперспирального серин-богатого белка 1, полипептид-высвобождающего фактора 2, SNAP-25, SNARE, Lac-репрессора или аполипопротеина Е.
4. Формирующий CAR полипептид по п.3, отличающийся тем, что указанный суперспиральный спейсерный домен содержит одну из последовательностей SEQ ID NO: 1 или 30-42.
5. Формирующий CAR полипептид по любому из пп.1-2, 4, отличающийся тем, что указанный эндодомен содержит по меньшей мере один из следующего: эндодомен CD3-дзета, эндодомен CD28, эндодомен 41BB и эндодомен OX40.
6. Формирующий CAR полипептид по любому из пп.1-2, 4, отличающийся тем, что указанный эндодомен содержит последовательность SEQ ID NO: 7.
7. Мультимерный химерный антигенный рецептор (CAR), который содержит от трех до семи формирующих CAR полипептидов по любому из пп.1-6 для запуска опосредованной CAR передачи сигналов в присутствии антигена-мишени на клетке-мишени.
8. Мультимерный CAR по п.7, отличающийся тем, что указанные формирующие CAR полипептиды содержат различные эндодомены.
9. Мультимерный CAR по п.7, содержащий по меньшей мере два формирующих CAR полипептида, имеющих различные антигенсвязывающие домены.
10. Мультимерный химерный антигенный рецептор (CAR), содержащий по меньшей мере один формирующий CAR полипептид по п.1 или 2 и по меньшей мере один вспомогательный полипептид, при этом каждый вспомогательный полипептид содержит в следующем порядке:
(a) суперспиральный спейсерный домен;
(b) трансмембранный домен; и
(c) эндодомен,
при этом общее число формирующих CAR полипептидов и вспомогательных полипептидов в составе указанного мультимерного CAR составляет от трех до семи, и при этом указанный мультимерный CAR предназначен для запуска опосредованной CAR передачи сигнала в присутствии антигена-мишени на клетке-мишени.
11. Мультимерный CAR по п.10, отличающийся тем, что указанный суперспиральный спейсерный домен происходит из олигомерного матриксного белка хряща (COMP), маннозосвязывающего белка А, суперспирального серин-богатого белка 1, полипептид-высвобождающего фактора 2, SNAP-25, SNARE, Lac-репрессора или аполипопротеина Е.
12. Мультимерный CAR по п.11, отличающийся тем, что указанный суперспиральный спейсерный домен содержит одну из последовательностей SEQ ID NO: 1 или 30-42.
13. Мультимерный CAR по любому из пп.10-12, отличающийся тем, что указанный эндодомен содержит по меньшей мере один из следующего: эндодомен CD3-дзета, эндодомен CD28, эндодомен 41BB и эндодомен OX40.
14. Мультимерный CAR по любому из пп.10-12, отличающийся тем, что указанный эндодомен содержит последовательность SEQ ID NO: 7.
15. Мультимерный CAR по любому из пп.10-12, отличающийся тем, что указанный формирующий(е) CAR полипептид(ы) и/или вспомогательный(е) полипептид(ы) содержат различные эндодомены.
16. Мультимерный CAR по любому из пп.10-12, содержащий по меньшей мере два формирующих CAR полипептида, имеющих различные антигенсвязывающие домены.
17. Экспрессионная конструкция нуклеиновой кислоты для экспрессии формирующего CAR полипептида, которая содержит последовательность нуклеиновой кислоты, кодирующую формирующий CAR полипептид по любому из пп.1-6.
18. Экспрессионная конструкция нуклеиновой кислоты для экспрессии формирующего CAR полипептида и вспомогательного полипептида, которая содержит первую последовательность нуклеиновой кислоты, кодирующую по меньшей мере один формирующий CAR полипептид согласно любому из пп.1-6; и вторую последовательность нуклеиновой кислоты, кодирующую по меньшей мере один вспомогательный полипептид, при этом каждый вспомогательный полипептид содержит в следующем порядке:
(a) суперспиральный спейсерный домен;
(b) трансмембранный домен; и
(c) эндодомен.
19. Экспрессионная конструкция по п.18, отличающаяся тем, что указанный суперспиральный спейсерный домен происходит из олигомерного матриксного белка хряща (COMP), маннозосвязывающего белка А, суперспирального серин-богатого белка 1, полипептид-высвобождающего фактора 2, SNAP-25, SNARE, Lac-репрессора или аполипопротеина Е.
20. Экспрессионная конструкция по п.19, отличающаяся тем, что указанный суперспиральный спейсерный домен содержит одну из последовательностей SEQ ID NO: 1 или 30-42.
21. Экспрессионная конструкция по любому из пп.18-20, отличающаяся тем, что указанный эндодомен содержит по меньшей мере один из следующего: эндодомен CD3-дзета, эндодомен CD28, эндодомен 41BB и эндодомен OX40.
22. Экспрессионная конструкция по любому из пп.18-20, отличающаяся тем, что указанный эндодомен содержит последовательность SEQ ID NO: 7.
23. Экспрессионная конструкция по любому из пп.18-20, отличающаяся тем, что указанный(е) формирующий(е) CAR полипептид(ы) и/или вспомогательный(е) полипептид(ы) содержат различные эндодомены.
24. Экспрессионная конструкция по любому из пп.18-20, содержащая по меньшей мере два формирующих CAR полипептида, имеющих различные антигенсвязывающие домены.
25. Экспрессионный вектор для получения цитотоксической Т-клетки, которая способна уничтожать клетку-мишень, экспрессирующую антиген-мишень, который содержит конструкцию нуклеиновой кислоты по п.17 или 18.
26. Экспрессионный вектор по п.25, который представляет собой ретровирусный вектор или лентивирусный вектор или транспозон.
27. Цитотоксическая Т-клетка, которая способна уничтожать клетку-мишень, экспрессирующую антиген-мишень, при этом указанная Т-клетка экспрессирует мультимерный CAR по любому из пп.7-16.
28. Цитотоксическая Т-клетка по п.27, отличающаяся тем, что указанный антиген-мишень представляет собой антиген, который присутствует в низкой плотности на клетке-мишени.
29. Фармацевтическая композиция для лечения рака посредством уничтожения клеток-мишеней, которые экспрессируют антиген-мишень, которая содержит Т-клетку по п.27 или 28 в эффективном количестве.
30. Т-клетка по п.27 или 28 для применения при лечении рака.
31. Т-клетка по п.30, где рак представляет собой хронический лимфоцитарный лейкоз (CLL), меланому или миелому.
32. Применение Т-клетки по п.30 в производстве лекарственного средства для лечения рака.
33. Применение по п.32, где рак представляет собой хронический лимфоцитарный лейкоз (CLL), меланому или миелому.
34. Способ лечения рака, который включает стадию введения субъекту Т-клетки по п.30.
35. Способ по п.34, где рак представляет собой хронический лимфоцитарный лейкоз (CLL), меланому или миелому.
36. Способ получения Т-клетки по п.27 или 28, который включает стадию введения в клетку ex vivo экспрессионного вектора по п.25 или 26.
37. Способ по п.36, где клетка происходит из образца, выделенного из субъекта.
| WO 2014127261 A1, 21.08.2014 | |||
| GUEST R | |||
| D | |||
| et al | |||
| The role of extracellular spacer regions in the optimal design of chimeric immune receptors: evaluation of four different scFvs and antigens, Journal of immunotherapy, 2005, V | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| CHANG H | |||
| C | |||
| et al | |||
| A general method for facilitating heterodimeric pairing between two proteins: | |||

