ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к сигнальной системе химерного антигенного рецептора.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Традиционно, антигенспецифические T-клетки создавали путем избирательного размножения T-клеток периферической крови, естественным образом специфичных для целевого антигена. Однако сложно, и довольно часто невозможно, отбирать и размножать большие количества T-клеток, специфичных для большинства раковых антигенов. Генная терапия с использованием интегрирующих векторов позволяет найти решение этой проблемы, поскольку трансгенная экспрессия химерного антигенного рецептора (CAR) позволяет получать большое количество T-клеток, специфичных для любого поверхностного антигена, за счет ex vivo трансдукции вирусным вектором большой популяции T-клеток периферической крови.
Химерные антигенные рецепторы представляют собой белки, которые придают специфичность моноклонального антитела (мАт) эффекторной функции T-клетки. Их обычной формой является белок с трансмембранным доменом типа I, имеющий узнающий антиген амино- конец, спейсер, трансмембранный домен, все связанные с комбинированным эндодоменом, который передает сигналы выживания и активации T-клеток (смотри фигуру 1A).
Наиболее распространенными формами этих молекул являются слитые белки из одноцепочечных вариабельных фрагментов (scFv), полученных из моноклональных антител, узнающих целевой антиген, которые слиты через спейсер и трансмембранный домен с сигнальным эндодоменом. Такие молекулы обеспечивают активацию T-клетки в ответ на узнавания фрагментом scFv его мишени. Когда T-клетки экспрессируют такой CAR, они узнают и уничтожают клетки-мишени, экспрессирующие целевой антиген. Несколько CAR были разработаны против опухоль-ассоциированных антигенов, и подходы с адоптивным переносом таких CAR-экспрессирующих T-клеток в настоящее время проходят клинические испытания для лечения различных форм рака.
В процессе исследований CAR был отмечен целый ряд проявлений токсичности, и существует вероятность дополнительных потенциальных форм токсичности. Такие формы токсичности включают иммунологическую токсичность, вызываемую длительной интенсивной активацией CAR T-клеток, что приводит к развитию синдрома активации макрофагов (MAS) и токсичности «неопухолевых мишеней» то есть, узнаванию целевого антигена на нормальных тканях.
Считается, что причиной MAS является персистентная вызываемая антигеном активация и пролиферация T-клеток, которые, в свою очередь, секретируют большое количество воспалительных цитокинов, что приводит к гиперактивации макрофагов и, в цикле положительной прямой связи, к иммунной активации. Характерной особенностью является большой всплеск концентрации IL-6 в сыворотке, и синдром может приводить к тяжелому системному заболеванию, требующему госпитализации в отделении интенсивной терапии.
О токсичности неопухолевых мишеней сообщалось в случае других CAR, например, у группы пациентов, получающих лечение CAR против антигена CAIX почечно-клеточного рака, развивалась внезапная и ограничивающая лечение токсичность желчных кислот. Сообщалось о двух случаях смерти, связанных с исследованиями CAR: один пациент умер от респираторного дистресс-синдрома, случившегося сразу же после инфузии большой дозы T-клеток с анти-ERBB2 CAR 3-го поколения; другой пациент погиб в другом исследовании в результате возможного цитокинового шторма после лечения CLL с анти-CD19 CAR второго поколения.
Такие формы токсичности очень трудно предвидеть, даже после тщательно проведенных исследованиях на животных или работы с приматами, отличными от человека. Важно, что, в отличие от малых молекул и биологических препаратов, CAR T-клетки не имеют периода полувыведения и невозможно прекратить введение и ожидать, пока средство распадется/будет выведено из организма. CAR T-клетки являются автономными и могут приживаться и пролиферировать. Таким образом, токсичность может быть внезапной и быстро прогрессирующей.
Суицидные гены представляют собой генетически экспрессируемые элементы, продукты которых при определенных условиях могут разрушать экспрессирующие их клетки. Примеры включают тимидинкиназу вируса простого герпеса, которая делает клетки чувствительными к ганцикловиру; индуцируемую каспазу 9, которая делает клетки чувствительными к низкомолекулярному гомодимеризатору, а также CD20 и RQR8, которые делают клетки чувствительными к ритуксимабу.
Данная технология придает определенную безопасность CAR T- клеточной терапии, однако существуют ограничения. Во-первых, это двоякий подход, при котором все CAR T-клетки погибают при добавлении суицидного средства. Кроме того, лекарственные терапевтические средства часто имеют терапевтическое окно. В случае суицидного гена эффективность препарата невозможно корректировать таким образом, чтобы обеспечивать эффективность в сочетании с переносимой токсичностью. Во-вторых, неясно, поможет ли суицидный ген при некоторых формах иммунотоксичности, описанных выше: например, ко времени возникновения синдрома активации макрофагов может уже отсутствовать необходимость в постоянном присутствии CAR T-клеток и суицидный ген более не будет полезен. Более острые синдромы секреции цитокинов, по всей вероятности, развиваются слишком быстро, чтобы суицидный ген мог действовать. Таким образом, существует потребность в альтернативных способах контроля CAR T-клеток, не связанных с недостатками и проблемами, упомянутыми выше.
ОПИСАНИЕ ФИГУР
Фигура 1 - a) Схематическая диаграмма, изображающая классический CAR. (b) - (d): Разные поколения и пермутации эндодоменов CAR: (b) исходные конструкции передают только сигналы ITAM через эндодомен FcR1-γ или CD3ζ, в то время как разработанные позже конструкции передают дополнительно (c) один или (d) два костимулирующих сигнала в одном и том же комбинированном эндодомене.
Фигура 2 - Структуры TetR и TiP. (a) Последовательность TiP, присоединенная к амино-концу произвольного белка; (b) Полученная методом кристаллографии структура TiP, взаимодействующего с TetR (из PDB 2NS8 и статьи Luckner et al. (J. Mol. Biol. 368, 780-790 (2007)). Можно видеть, что TiP расположен глубоко в гомодимере TetR, связываясь со многими остатками, с которыми связывается тетрациклин.
Фигура 3 - (a) Проходящий через мембрану рецепторный компонент содержит внеклеточный антигенсвязывающий домен, трансмембранный домен и внутриклеточный линкер с TetR. Отдельная молекула, сигнальный компонент, представляет собой внутриклеточный белок, полученный путем слияния TiP с одним или несколькими T- клеточными сигнальными доменами. В отсутствие тетрациклина или аналогов тетрациклина рецепторный и сигнальный компоненты взаимодействуют и в присутствии узнаваемого антигена система передает сигнал. (b) В присутствии тетрациклина или аналогов тетрациклина TiP вытесняется с TetR и рецептор не может передавать сигналы даже в присутствии узнаваемого антигена.
Фигура 4 - Внутриклеточный домен линкера, полученный из CD4.
Фигура 5 - Тестовый конструкт с eGFP для демонстрации функционирования системы. (a) бицистронный конструкт, экспрессируемый в виде единого транскрипта, который саморасщепляется в сайте 2A, образуя: TiP, слитый с eGFP; и CAR с TetR в качестве его эндодомена. (b) Флуоресцентная микрофотография клеток SupT1, экспрессирующих данный конструкт в отсутствие тетрациклина. Флуоресценцию eGFP можно отчетливо видеть на клеточной мембране; (c) Флуоресцентная микрофотография тех же клеток, но в присутствии тетрациклина. Видно, что в этом случае eGFP находится в цитоплазме, из чего следует, что тетрациклин вытеснил TiP.
Фигура 6 - Исходный конструкт TetCAR и контроль (a) бицистронный конструкт, экспрессируемый в виде единого транскрипта, который саморасщепляется в сайте 2A, образуя: сигнальный компонент, который содержит TiP, слитый через гибкий линкер с эндодоменом CD3-дзета; и рецепторный компонент, который содержит узнающий CD33 фрагмент scFv, спейсер, полученный из Fc- домена IgG1, полученный из CD4 трансмембранный и внутриклеточный домен; и TetR. (b) также был создан контрольный конструкт, который был идентичен, за исключением того, что TiP отсутствовал в сигнальном компоненте. (c) показана аннотированная аминокислотная последовательность основного TetCAR.
Фигура 7 - Действие исходного конструкта TetR в сравнении с контролем. (a) TetCAR был экспрессирован в T-клетках BW5. Эти T- клетки стимулировали клетками SupT1 дикого типа или клетками SupT1, генетически модифицированными для экспрессии CD33, в отсутствие тетрациклина или в присутствии тетрациклина в возрастающих концентрациях. T-клетки, стимулированные клетками SupT1 дикого типа, не активируются как в присутствии, так и отсутствие тетрациклина; T-клетки, стимулированные клетками SupT1, экспрессирующими CD33, активируются в отсутствие тетрациклина, однако активация быстро ингибируется в присутствии тетрациклина, при этом активация полностью ингибируется в присутствии 100 нМ тетрациклина. (b) Контрольный TetCAR без домена TiP был трансдуцирован в клетки BW5. Опять-таки, эти T-клетки были стимулированы клетками SupT1 дикого типа или клетками SupT1, генетически модифицированными для экспрессии CD33, в отсутствие тетрациклина или в присутствии тетрациклина в возрастающих концентрациях. Отсутствие элемента TiP в сигнальном компоненте приводило к отсутствию сигнализации в любых условиях.
Фигура 8 - TetCAR с двойным доменом TetR. TetR экспрессирован в виде одной цепи с двумя соединенными вместе доменами TetR. При использовании доменов TetR с разной аффинностью к тетрациклину (и, следовательно, TiP) кинетика опосредованного тетрациклином вытеснения TiP может модулировать уровни сигнализации.
Фигура 9 - Сигнальная система TetCAR, в которой использовано несколько сигнальных компонентов, содержащих одиночные эндодомены. Один CAR экспрессируется со многими разными сигнальными компонентами, все из которых содержат TiP на их амино-конце, но разные индивидуальные сигнальные домены, в отличие от комбинированного сигнального домена. Они случайным образом взаимодействуют с рецепторным компонентом. Отсутствие стерического взаимодействия между разными сигнальными доменами и их вторыми мессенджерами улучшает их функцию.
Фигура 10 - Сигнальная система TetCAR, в которой использовано несколько сигнальных компонентов, содержащих одиночные эндодомены и разные домены TiP. Каждый сигнальный компонент содержит индивидуальный сигнальный домен. Каждый сигнальный компонент также содержит TiP, однако все TiP имеют разную аффинность к домену TetR. Таким образом, можно варьировать стехиометрию взаимодействий между CAR и сигнальными доменами. В приведенном примере сигнальная система сконструирована таким образом, что OX40 > CD3-дзета > CD28.
Фигура 11 - Сигнальная система TetCAR, в которой использовано несколько рецепторных компонентов и несколько сигнальных компонентов, при этом каждый сигнальный компонент содержит одиночный эндодомен.
Фигура 12 - Сигнализация TetCAR в первичных клетках (a) Различные протестированные конструкты: (i) классический CAR; (ii) TetCAR; (iii) контрольный TetCAR с делетированным доменом TiP. (b) нетрансдуцированные клетки и клетки SupT1.CD19, окрашенные на CD19; (c) нетрансдуцированные T-клетки и T-клетки, трансдуцированные разными конструктами CAR, окрашенные анти-Fc антителами.
Фигура 13 - Секреция интерферона-гамма из нетрансдуцированных T-клеток и T-клеток, трансдуцированных разными конструктами CAR ((i) Классический CAR первого поколения, (ii) TetCAR и (iii) контрольный TetCAR), стимулированных клетками SupT1, клетками SupT1.CD19 при разных концентрациях тетрациклина.
Фигура 14 - Уничтожение клеток-мишеней. Использовали анализ с высвобождением хрома для демонстрации уничтожения клеток-мишеней (SupT1.CD19) в отсутствие тетрациклина. Пояснение: (i) - обычный CAR; (ii) -TetCAR; (iii) - контрольный TetCAR (без TiP на эндодомене).
КРАТКОЕ ИЗЛОЖЕНИЕ АСПЕКТОВ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения установили, что можно разделять антиген-узнающие и сигнальные компоненты CAR для создания системы, в которой сигнализацию можно быстро ингибировать/прекращать, несмотря на продолжающееся связывание антигена с антиген-узнающим компонентом системы CAR. Такое ингибирование сигнализации происходит в присутствии средства, например, малой молекулы, ингибирующего совместную локализацию и взаимодействие, которые без него имели бы место между внеклеточным антигенсвязывающим компонентом (называемым в настоящем документе рецепторным компонентом) и компонентом внутриклеточного сигналинга в составе CAR.
Таким образом, в первом аспекте настоящее изобретение относится к системе химерного антигенного рецептора (CAR), включающей:
(i) рецепторный компонент, содержащий антигенсвязывающий домен, трансмембранный домен и первый связывающий домен; и
(ii) компонент внутриклеточного сигналинга, содержащий сигнальный домен и второй связывающий домен, который специфически связывает первый связывающий домен рецепторного компонента;
при этом связывание первого и второго связывающих доменов нарушается в присутствии средства таким образом, что в отсутствие средства рецепторный компонент и сигнальный компонент гетеродимеризуются и связывание антигенсвязывающего домена с антигеном приводит к сигнализации через сигнальный домен, в то время как в присутствии средства рецепторный компонент и сигнальный компонент не гетеродимеризуются и связывание антигенсвязывающего домена с антигеном не приводит к сигнализации через сигнальный домен.
Рецепторный компонент может содержать линкер между трансмембранным доменом и первым связывающим доменом.
Линкер может содержать или состоять из последовательности, приведенной в SEQ ID NO: 3.
Первый связывающий домен может представлять собой репрессорный белок Tet (TetR) или его вариант, и второй связывающий домен может представлять собой TetR-индуцирующий пептид (TiP, описанный в статье Klotzsche et al; The Journal of biological chemistry; 2005; 280(26); 24591-9) (TiP); или наоборот. В этом случае средство может представлять собой тетрациклин, доксицилин или миноциклин, либо их аналоги.
Рецепторный компонент может содержать два первых связывающих домена, которые представляют собой домены TetR. Два домена TetR могут быть разделены линкером. Домены TetR могут иметь разную аффинность для средства.
Система CAR по первому аспекту изобретения может включать множественные рецепторные компоненты, узнающие разные антигены.
Первые связывающие домены множественных рецепторных компонентов могут отличаться по связыванию со вторым связывающим доменом сигнального компонента таким образом, что все антигены вызывают сигналы разной силы.
Первые связывающие домены множественных рецепторных компонентов могут отличаться по связыванию со средством таким образом, что все антигены вызывают сигналы разной силы в присутствии средства.
Сигнальный домен сигнального компонента может представлять собой один эндодомен, выбранный из эндодомена CD3-дзета, эндодомена CD28, эндодомена 41BB и эндодомена OX40.
Сигнальный домен сигнального компонента может представлять собой по меньшей мере один из эндодомена CD3-дзета, эндодомена CD28, эндодомена 41BB и эндодомена OX40.
Система CAR по первому аспекту изобретения может включать несколько сигнальных компонентов, каждый из которых содержит сигнальный домен и второй связывающий домен, при этом все вторые связывающие домены узнают одинаковые первые связывающие домены рецепторного компонента, однако сигнальные домены представляют собой разные эндодомены.
Несколько сигнальных компонентов могут содержать несколько вторых связывающих доменов, все из которых независимо узнают первый связывающий домен рецепторного компонента с разной аффинностью.
Во втором аспекте настоящее изобретение относится к рецепторному компоненту, подходящему для использования в системе CAR по первому аспекту изобретения, содержащему антигенсвязывающий домен, трансмембранный домен и первый связывающий домен.
В третьем аспекте настоящее изобретение относится к сигнальному компоненту, подходящему для использования в системе CAR по первому аспекту изобретения, содержащему сигнальный домен и второй связывающий домен.
В четвертом аспекте настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей рецепторный компонент по второму аспекту изобретения.
В пятом аспекте настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей сигнальный компонент по третьему аспекту изобретения.
В шестом аспекте настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей систему CAR по первому аспекту изобретения, при этом рецепторный компонент и сигнальный компонент совместно экспрессируются в виде саморасщепляемого пептида, который расщепляется между рецепторным компонентом и сигнальным компонентом после трансляции.
В седьмом аспекте настоящее изобретение относится к вектору, содержащему последовательность нуклеиновой кислоты по четвертому- шестому аспектам изобретения.
В восьмом аспекте настоящее изобретение относится к ретровирусному вектору или лентивирусному вектору, или транспозону, содержащему вектор по седьмому аспекту изобретения.
В девятом аспекте настоящее изобретение относится к T-клетке или NK-клетке, экспрессирующей рецепторный компонент по второму аспекту изобретения и сигнальный компонент по третьему аспекту изобретения.
T-клетка или NK-клетка может содержать нуклеиновую кислоту по четвертому-шестому аспектам изобретения или вектор по седьмому или восьмому аспекту изобретения.
В десятом аспекте настоящее изобретение относится к фармацевтической композиции, содержащей множество T-клеток или NK- клеток по девятому аспекту изобретения.
В одиннадцатом аспекте настоящее изобретение относится к фармацевтической композиции по десятому аспекту изобретения для использования в лечении и/или предотвращения заболевания.
В двенадцатом аспекте настоящее изобретение относится к способу лечения и/или предотвращения заболевания, включающему этап введения субъекту фармацевтической композиции по десятому аспекту изобретения.
Способ по двенадцатому аспекту изобретения может включать следующие этапы:
(i) выделение образца, содержащего T-клетки или NK-клетки;
(ii) трансдукция или трансфекция T- или NK-клеток последовательностью нуклеиновой кислоты по любому из четвертого- шестого аспектов изобретения или вектором по седьмому или восьмому аспекту изобретения; и
(iii) введение T-клеток или NK-клеток по пункту (ii) субъекту.
Способ может включать введение T-клеток/NK-клеток субъекту, при этом эти T-клетки/NK-клетки ранее были получены от субъекта и трансдуцированы/трансфицированы последовательностью нуклеиновой кислоты по любому из четвертого-шестого аспектов изобретения или вектором по седьмому или восьмому аспекту изобретения.
Способ по двенадцатому аспекту изобретения может включать мониторинг токсической активности у субъекта и включает этап введения субъекту средства, используемого в системе CAR по первому аспекту изобретения, для уменьшения неблагоприятных токсических эффектов.
Способ может включать мониторинг прогрессирования заболевания и/или мониторинг токсической активности у субъекта и включает этап введения субъекту средства, используемого в системе CAR по первому аспекту изобретения, для обеспечения приемлемых уровней прогрессирования заболевания и/или токсической активности.
При использовании фармацевтической композиции по десятому аспекту изобретения или способа по двенадцатому аспекту изобретения заболевание может представлять собой рак.
В тринадцатом аспекте настоящее изобретение относится к использованию фармацевтической композиции по десятому аспекту изобретения в производстве лекарственного средства для лечения и/или предотвращения заболевания.
В четырнадцатом аспекте настоящее изобретение относится к набору, включающему нуклеиновую кислоту по четвертому-шестому аспектам изобретения или вектор по седьмому или восьмому аспекту изобретения.
В пятнадцатом аспекте настоящее изобретение относится к способу получения T- или NK-клетки по девятому аспекту изобретения, включающему этап введения последовательности нуклеиновой кислоты по четвертому-шестому аспектам изобретения или вектора по седьмому или восьмому аспекту изобретения в T- или NK клетку.
T- или NK-клетка может быть клеткой из образца, полученного от субъекта.
В шестнадцатом аспекте настоящее изобретение относится к способу ингибирования системы CAR по первому аспекту изобретения у субъекта, в организме которого находится T- или NK-клетка по девятому аспекту изобретения, включающему этап введения средства субъекту.
Таким образом, по настоящему изобретению предложена система CAR, в которой сигнализация может быть ингибирована в присутствии средства, например, малой молекулы, которое предотвращает совместную локализацию рецепторного компонента и сигнального компонента. Это позволяет обратимо прекращать сигнализацию CAR и, следовательно, активность CAR-клеток, контролируемым образом, чтобы избежать потенциальных токсических эффектов, связанных с неослабевающей сигнализацией CAR. Кроме того, настоящая система также позволяет фармакологически контролировать активность CAR клеток и регулировать приемлемый баланс между достижением желаемого терапевтического эффекта и избеганием нежелательной токсичности.
ПОДРОБНОЕ ОПИСАНИЕ
ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (CAR)
Классические CAR, которые схематически изображены на фигуре 1, представляют собой химерные трансмембранные белки типа I, в которых внеклеточный антиген-узнающий (связывающий) домен соединен с доменом внутриклеточного сигналинга (эндодоменом). Связывающий домен, как правило, представляет собой одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела (мАт), но он может быть основан и на других форматах, содержащих антитело- подобный сайт связывания антигена. Спейсерный домен может быть необходим для отделения связывающего домена от мембраны и создания возможности подходящей ориентации. Обычным используемым спейсерным доменом является Fc из IgG1. Могут быть достаточными и более компактные спейсеры, например, стебель из CD8α и даже только шарнирная область IgG1, в зависимости от антигена. Трансмембранный домен служит якорем для белка в клеточной мембране и соединяет спейсер с эндодоменом.
Ранние конструкции CAR имели эндодомены, полученные из внутриклеточных частей либо γ-цепи FcεR1, либо CD3ζ. Следовательно, эти рецепторы первого поколения передавали иммунологический сигнал 1, достаточный для инициации уничтожения T-клетками узнаваемых клеток-мишеней, но не способный полностью активировать T-клетки для пролиферации и выживания. Чтобы преодолеть это ограничение, были сконструированы комбинированные эндодомены: в результате слияния внутриклеточной части T-клеточной костимулирующей молекулы с таковой от CD3ζ были получены рецепторы второго поколения, способные передавать одновременно активирующий и костимулирующий сигнал после узнавания антигена. Наиболее часто используемым костимулирующим доменом является домен CD28. Он обеспечивает наиболее сильный костимулирующий сигнал - а именно, иммунологический сигнал 2, который инициирует пролиферацию T- клеток. Также были описаны некоторые рецепторы, содержащие эндодомены из семейства рецепторов TNF, например, из близкородственных OX40 и 41BB, которые передают сигналы выживания. В настоящее время описаны еще более эффективные CAR третьего поколения, которые имеют эндодомены, способные передавать сигналы активации, пролиферации и выживания.
Кодирующие CAR нуклеиновые кислоты могут быть перенесены в T- клетки с использованием, например, ретровирусных векторов. Таким способом можно создавать большое число антигенспецифических T- клеток для адоптивного переноса клеток. Когда CAR связывает целевой антиген, это приводит к передаче активирующего сигнала T- клетке, на которой он экспрессирован. Таким образом, CAR управляет специфичностью и цитотоксичностью T-клетки в отношении клеток, экспрессирующих целевой антиген.
В первом аспекте настоящее изобретение относится к системе CAR, в которой антиген-узнающий/антигенсвязывающий домен и трансмембранный домен находятся на первой молекуле (называемой в настоящем документе «рецепторный компонент»), которая расположена на клеточной мембране. Домен внутриклеточного сигналинга находится на второй, внутриклеточной, молекуле (называемой в настоящем документе «компонент сигналинга»).
Важно отметить, что рецепторный компонент содержит первый связывающий домен и сигнальный компонент содержит второй связывающий домен, который специфически связывается с первым связывающим доменом рецепторного компонента. Таким образом, связывание первого связывающего домена со вторым связывающим доменом вызывает гетеродимеризацию и совместную локализацию рецепторного компонента и сигнального компонента. Когда антиген связывается с антигенсвязывающим доменом рецепторного компонента, через сигнальный компонент осуществляется сигнализация.
Первый или второй связывающий домен также способен связывать дополнительное средство, помимо соответствующего связывающего домена. Дополнительное средство может представлять собой, например, малую молекулу. Связывание между средством и первым или вторым связывающим доменом характеризуется более высокой аффинностью, чем связывание между первым связывающим доменом и вторым связывающим доменом. Таким образом, в случае присутствия данного средства оно предпочтительно связывается с первым или вторым связывающим доменом и ингибирует/нарушает гетеродимеризацию между рецепторным компонентом и сигнальным компонентом. Когда антиген связывается с антигенсвязывающим доменом рецепторного компонента в присутствии дополнительного средства, сигнализация через сигнальный компонент отсутствует.
В частности, в присутствии средства рецепторный компонент и сигнальный компонент случайным образом рассредоточены и связывание антигена антигенсвязывающим доменом рецепторного компонента не приводит к сигнализации через сигнальный компонент.
Используемый в настоящем документе термин «совместная локализация» или «гетеродимеризация» рецепторного и сигнального компонентов аналогичен связыванию/рекрутингу сигнального компонента и рецепторного компонента путем связывания первого связывающего домена рецепторного компонента и второго связывающего домена сигнального компонента.
Можно сказать, что связывание антигена рецепторным компонентом в присутствии средства приводит к «непродуктивной» сигнализации через сигнальный компонент. Такая сигнализация не приводит к активации клеток, например, активации T-клеток. Можно сказать, что связывание антигена рецепторным компонентом в отсутствие средства приводит к «продуктивной» сигнализации через сигнальный компонент. Такая сигнализация приводит к активации T- клеток, запуская, например, уничтожение клетки-мишени и T- клеточную активацию.
Связывание антигена рецепторным компонентом в отсутствие средства может приводить к сигнализации через сигнальный компонент, которая в 2, 5, 10, 50, 100, 1000 или 10000 раз выше, чем сигнализация, которая имеет место, когда антиген связывается рецепторным компонентом в присутствии средства.
Сигнализацию через сигнальный компонент можно определять различными способами, известными в данной области. Такие способы включают анализ сигнальной трансдукции, например, анализ уровня определенных протеинтирозинкиназ (PTK), разрушения фосфатидилинозитол-4,5-бифосфата (PIP2), активации протеинкиназы C (PKC) и повышения внутриклеточной концентрации ионов кальция. Также можно использовать функциональные показатели, такие как клональная экспансия T-клеток, повышающая регуляция маркеров активации на клеточной поверхности, дифференциация в эффекторные клетки и индукция цитотоксичности или секреции цитокинов. В качестве иллюстрации, в настоящих примерах авторы изобретения определяли уровни интерлейкина-2 (IL-2), продуцируемого T- клетками, экспрессирующими рецепторный компонент и сигнальный компонент системы CAR в соответствии с настоящим изобретением, при связывании антигена с рецепторным компонентом в присутствии средства в различных концентрациях.
ПЕРВЫЙ СВЯЗЫВАЮЩИЙ ДОМЕН, ВТОРОЙ СВЯЗЫВАЮЩИЙ ДОМЕН И СРЕДСТВО
Первый связывающий домен, второй связывающий домен и средство настоящей системы CAR могут представлять собой любое сочетание молекул/пептидов/доменов, которые делают возможной избирательную совместную локализацию и димеризацию рецепторного компонента и сигнального компонента в отсутствие средства.
Как таковые, первый связывающий домен и второй связывающий домен способны к специфическому связыванию.
Сигнальная система по настоящему изобретению не ограничена конфигурацией определенной системы димеризации. Рецепторный компонент может содержать либо первый связывающий домен, либо второй связывающий домен конкретной системы димеризации при условии, что сигнальный компонент содержит соответствующий, комплементарный связывающий домен, который позволяет рецепторному компоненту и сигнальному компоненту совместно локализоваться в отсутствие средства.
Первый связывающий домен и второй связывающий домен могут представлять собой пептидный домен и пептид-связывающий домен; или наоборот. Пептидный домен и пептид-связывающий домен могут представлять собой любое сочетание пептидов/доменов, которые способны к специфическому связыванию.
Средство представляет собой молекулу, например, малую молекулу, которая способна специфически связываться с первым связывающим доменом или вторым связывающим доменом с более высокой аффинностью, чем аффинность связывания между первым связывающим доменом и вторым связывающим доменом.
Например, система связывания может быть основана на системе пептид:пептид-связывающий домен. Первый или второй связывающий домен может представлять собой пептид-связывающий домен, и другой связывающий домен может представлять собой пептидомиметик, который связывает пептид-связывающий домен с более низкой аффинностью, чем пептид. Использование пептида в качестве средства нарушает связывание пептидомиметика с пептид-связывающим доменом в результате конкурентного связывания. Пептидомиметик может иметь аминокислотную последовательность, сходную с последовательностью пептида «дикого типа», но с одним или более изменениями аминокислот для снижения аффинности связывания с пептид-связывающим доменом.
Например, средство может связывать первый связывающий домен или второй связывающий домен с аффинностью, по меньшей мере в 10, 20, 50, 100, 1000 или 10000 раз превышающей аффинность связывания между первым связывающим доменом и вторым связывающим доменом.
Средство может представлять собой любую фармацевтически приемлемую молекулу, которая предпочтительно связывает первый связывающий домен или второй связывающий домен с более высокой аффинностью, чем аффинность связывания между первым связывающим доменом и вторым связывающим доменом.
Средство может быть доставлено в цитоплазму клетки-мишени и быть доступным для внутриклеточного связывания.
Средство может быть способно к пересечению гематоэнцефалического барьера.
Системы малых молекул для контроля совместной локализации пептидов известны в данной области, например, система из репрессора Tet (TetR), взаимодействующего с TetR белка (TiP) и тетрациклина (Klotzsche et al.; J. Biol. Chem. 280, 24591-24599 (2005); Luckner et al.; J. Mol. Biol. 368, 780-790 (2007)).
Система репрессора Tet (TetR)
Оперон Tet является хорошо известным биологическим опероном, который был адаптирован для использования в клетках млекопитающих. TetR связывает тетрациклин в виде гомодимера и претерпевает конформационное изменение, которое затем модулирует связывание ДНК молекулами TetR. В статье Klotzsche et al. (выше) описан полученный методом фагового дисплея пептид, который активирует TetR. Этот белок (взаимодействующий с TetR белок/TiP) имеет сайт связывания в TetR, который перекрывается, но не является идентичным сайту связывания тетрациклина (Luckner et al.; выше). Таким образом TiP и тетрациклин конкурируют за связывание TetR.
В настоящей системе CAR первый связывающий домен рецепторного компонента может представлять собой TetR или TiP, при условии, что второй связывающий домен сигнального компонента представляет собой соответствующий, комплементарный партнер по связыванию. Например, если первый связывающий домен рецепторного компонента представляет собой TetR, второй связывающий домен сигнального компонента представляет собой TiP. Если первый связывающий домен рецепторного компонента представляет собой TiP, второй связывающий домен сигнального компонента представляет собой TetR.
Например, первый связывающий домен или второй связывающий домен может содержать последовательность, приведенную в SEQ ID NO: 1 или SEQ ID NO: 2: SEQ ID NO: 1 – TetR
MSRLDKSKVINSALELLNEVGIEGLTTRKLAQKLGVEQPTLYWHVKNKRALLDALAIEMLDRHHTHFCPLEGESWQDFLRNNAKSFRCALLSHRDGAKVHLGTRPTEKQYETLENQLAFLCQQ GFSLENALYALSAVGH
SEQ ID NO: 2 - TiP
MWTWNAYAFAAPSGGGS
Для того, чтобы функционировать, TetR должен образовывать гомодимер. Таким образом, если первый связывающий домен на рецепторном компоненте представляет собой TetR, рецепторный компонент может содержать линкер между трансмембранным доменом и первым связывающим доменом (TetR). Линкер позволяет TetR образовывать гомодимер с TetR из соседнего рецепторного компонента и ориентироваться в правильном направлении.
Линкер может иметь последовательность, приведенную в SEQ ID NO: 3.
SEQ ID NO: 3 - модифицированный эндодомен CD4
ALIVLGGVAGLLLFIGLGIFFCVRCRHRRRQAERMAQIKRVVSEKKTAQAPHRFQKTCSPI
Альтернативно, линкер может содержать другую линкерную последовательность, которая имеет сходную длину и/или способность разделять домены в пространстве, как у последовательности, приведенной в SEQ ID NO: 3.
Линкер может иметь по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с SEQ ID NO: 3 при условии, что он выполняет функцию обеспечения способности TetR образовывать гомодимер с TetR из соседнего рецепторного компонента и ориентироваться в правильном направлении.
Одним потенциальным недостатком системы TetR/TiP является то, что TetR является ксеногенным и иммуногенным. Вследствие этого, последовательность TetR может представлять собой вариант, который является менее иммуногенным, но сохраняет способность специфически связывать TiP.
Если первый и второй связывающие домены представляют собой TetR или TiP, либо их варианты, средство может представлять собой тетрациклин, доксицилин, миноциклин или их аналоги.
«Аналог» означает вариант тетрациклина, доксицилина или миноциклина, который сохраняет способность специфически связываться с TetR.
Другие сочетания связывающих доменов и средств, которые можно использовать в настоящей системе CAR, известны в данной области. Например, в системе CAR можно использовать систему связывания на основе стрептавидина/биотина.
Стрептавидин-связывающий эпитоп
Первый или второй связывающий домен может представлять собой один или более стрептавидин-связывающих эпитопов. Другой связывающий домен может представлять собой миметик биотина.
Стрептавидин представляет собой 52,8-кДа белок из бактерии Streptomyces avidinii. Гомотетрамеры стрептавидина имеют очень высокую аффинность для биотина (витамин B7 или витамин H), с константой диссоциации (Kd) ~ 10-15 M. Миметик биотина имеет более низкую аффинность для стрептавидина, чем биотин дикого типа, так что сам биотин можно использовать в качестве средства, нарушающего или предотвращающего гетеродимеризацию между доменом стрептавидина и доменом миметика биотина. Миметик биотина может связывать стрептавидин, например, с Kd от 1 нМ до 100 мкМ.
Домен «миметика биотина» может, например, содержать короткую пептидную последовательность (например, 6-20, 6-18, 8-18 или 8-15 аминокислот), которая специфически связывается со стрептавидином.
Миметик биотина может содержать последовательность, приведенную в таблице 1.
Таблица 1. Пептидомиметики биотина
Короткий наномаркер
DVEAWLGAR (SEQ ID NO: 5)
17 нМ
WSHPQFEK (SEQ ID NO: 7)
AECHPQGPPCIEGRK (SEQ ID NO: 10)
Миметик биотина можно выбирать из следующей группы: стрепмаркер II, фланкированный cc-стрепмаркер и cc-стрепмаркер.
Домен стрептавидина может представлять собой стрептавидин, имеющий последовательность, приведенную в SEQ ID No. 11, либо ее фрагмент или вариант, который сохраняет способность связывать биотин.
Полноразмерный стрептавидин содержит 159 аминокислот. N- и C- концы полноразмерного белка из 159 остатков подвергаются процессингу, с образованием более короткого «корового» стрептавидина, обычно состоящего из остатков 13-139; удаление N- и C-концов необходимо для высокой аффинности связывания биотина.
Последовательность «корового» стрептавидина (остатки 13-139) приведена в SEQ ID No. 11
SEQ ID No. 11
EAGITGTWYNQLGSTFIVTAGADGALTGTYESAVGNAESRYVLTGRYDSAPATDGSGTALGWTVAWKNNYRNAHSATTWSGQYVGGAEARINTQWLLTSGTTEANAWKSTLVGHDTFTKVKPSAAS
В природе стрептавидин существует в виде гомотетрамера. Вторичная структура мономера стрептавидина состоит из восьми антипараллельных бета-тяжей, которые сворачиваются, образуя третичную структуру антипараллельного бета-цилиндра. Сайт связывания биотина расположен на одном конце каждого β-цилиндра. Четыре идентичных мономера стрептавидина (то есть, четыре идентичных β-цилиндра) соединяются, с образованием тетрамерной четвертичной структуры стрептавидина. Сайт связывания биотина в каждом цилиндре образован остатками из внутренней части цилиндра, вместе с консервативным остатком Trp120 из соседней субъединицы. Таким образом, каждая субъединица вносит вклад в сайт связывания на соседней субъединице и, следовательно, тетрамер также можно рассматривать как димер из функциональных димеров.
Домен стрептавидина системы CAR по настоящему изобретению может состоять по существу из мономера, димера или тетрамера стрептавидина.
Последовательность мономера, димера или тетрамера стрептавидина может содержать всю или часть последовательности, приведенной в SEQ ID No. 11, или ее вариант, сохраняющий способность связывать биотин.
Вариант последовательности стрептавидина может иметь по меньшей мере 70, 80, 90, 95 или 99% идентичности с SEQ ID No. 11 или ее функциональной частью. Вариант стрептавидина может содержать одну или более из следующих аминокислот, которые принимают участие в связывании биотина: остатки Asn23, Tyr43, Ser27, Ser45, Asn49, Ser88, Thr90 и Asp128. Вариант стрептавидина может, например, содержать все 8 из этих остатков. Если вариант стрептавидина присутствует в связывающем домене в виде димера или тетрамера, он может также содержать остаток Trp120, который участвует в связывании биотина со стороны соседней субъединицы.
Низкомолекулярные средства, которые нарушают белок-белковые взаимодействия, уже давно разрабатываются для фармацевтических целей (информация в обзоре Vassilev et al; Small-Molecule Inhibitors of Protein-Protein Interactions ISBN: 978-3-642-17082- 9). В описанной системе CAR можно использовать такую малую молекулу. Белки или пептиды, взаимодействие которых нарушается (или соответствующие фрагменты этих белков), можно использовать в качестве первого и/или второго связывающих доменов, и малую молекулу можно использовать в качестве средства, которое ингибирует активацию CAR. Такую систему можно изменять путем изменения малой молекулы и белков таким образом, что система функционирует, как описано, но малая молекула лишена нежелательной фармакологической активности (например, таким образом, как это описано в статье Rivera et al. (Nature Med; 1996; 2; 1028-1032).
Список белков/пептидов, взаимодействие которых может быть нарушено с помощью такого средства, как малая молекула, приведен в таблице 2. Эти рассматриваемые белок-белковые взаимодействия (PPI) можно использовать в системе CAR по настоящему изобретению. Дополнительную информацию по данным PPI можно найти в статье White et al., 2008 (Expert Rev. Mol. Med. 10:e8).
Таблица 2
Вторые связывающие домены, которые конкурентно связываются с тем же первым связывающим доменом, что и средства, описанные выше, и, таким образом, могут быть использованы для совместной локализации рецепторного компонента и сигнального компонента настоящей сигнальной системы в отсутствие средства, можно идентифицировать с использованием методик и способов, которые хорошо известны в данной области. Например, такие вторые связывающие домены можно идентифицировать путем дисплея библиотеки однодоменного VHH.
Первый связывающий домен и/или второй связывающий домен настоящей сигнальной системы могут представлять собой вариант(ы), которые способны специфически связываться с соответствующим связывающим доменом и, таким образом, облегчать совместную локализацию рецепторного компонента и сигнального компонента.
Вариантные последовательности могут иметь по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с последовательностью дикого типа при условии, что эти последовательности обеспечивают эффективную систему димеризации. То есть, при условии, что последовательности способствуют достаточной совместной локализации рецепторного и сигнального компонентов в отсутствие средства для продуктивной сигнализации при связывании антигенсвязывающего домена с антигеном.
Настоящее изобретение также относится к способу ингибирования системы CAR по первому аспекту изобретения, включающему этап введения средства. Как описано выше, введение средства приводит к нарушению совместной локализации рецепторного компонента и сигнального компонента, так что сигнализация через сигнальный компонент ингибируется даже при связывании антигена с антигенсвязывающим доменом.
Первый и второй связывающие домены могут способствовать сигнализации через систему CAR, пропорциональной концентрации присутствующего средства. Таким образом, хотя средство связывает первый связывающий домен или второй связывающий домен с более высокой аффинностью, чем аффинность связывания между первым и вторым связывающими доменами, совместная локализация рецепторного и сигнального компонентов может быть не полностью нарушена в присутствии низких концентраций средства. Например, средство в низких концентрациях может приводить к снижению общего уровня сигнализации в ответ на антиген без того, чтобы полностью ингибировать ее. Конкретные концентрации средства будут отличаться в зависимости от необходимого уровня сигнализации, а также конкретных связывающих доменов и средств. Уровни сигнализации и корреляцию с концентрацией средства можно определять методами, известными в данной области, как описано выше.
РЕЦЕПТОРНЫЙ КОМПОНЕНТ
Настоящее изобретение относится к рецепторному компоненту, содержащему антигенсвязывающий домен, необязательный домен спейсера, трансмембранный домен и первый связывающий домен. При экспрессии в клетке рецепторный компонент располагается на клеточной мембране. В данном случае, антигенсвязывающий домен молекулы располагается на внеклеточной стороне мембраны и первый связывающий домен располагается на внутриклеточной стороне мембраны.
Таким образом, рецепторный компонент обеспечивает антигенсвязывающую функцию системы CAR по настоящему изобретению.
АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН
Антигенсвязывающий домен представляет собой часть классического CAR, которая узнает антиген. В сигнальной системе по настоящему изобретению антигенсвязывающий домен расположен в рецепторном компоненте.
Многочисленные антигенсвязывающие домены известны в данной области, включая те, которые основаны на антигенсвязывающем сайте антител, миметиков антител и T-клеточных рецепторов. Например, антигенсвязывающий домен может представлять собой: одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального 23 антитела; естественный лиганд целевого антигена; пептид с достаточной аффинностью для целевого антигена; однодоменный связывающий фрагмент, такой как антитело верблюдовых; искусственный одиночный связывающий фрагмент, такой как дарпин, или одноцепочечный фрагмент, полученный из T-клеточного рецептора.
Известны различные опухоль-ассоциированные антигены (TAA), приведенные в следующей далее таблице 1. Антигенсвязывающий домен, используемый по настоящему изобретению, может представлять собой домен, способный связывать TAA, как указано в таблице.
Таблица 1
ТРАНСМЕМБРАННЫЙ ДОМЕН
Трансмембранный домен представляет собой последовательность классического CAR, которая пересекает мембрану. В сигнальной системе по настоящему изобретению трансмембранный домен расположен в рецепторном компоненте. Он может содержать гидрофобную альфа- спираль. Трансмембранный домен может быть получен из CD28, который придает хорошую стабильность рецептору.
СИГНАЛЬНЫЙ ПЕПТИД Рецепторный компонент системы CAR по настоящему изобретению может содержать сигнальный пептид, так что, когда рецепторный компонент экспрессируется в клетке, такой как T-клетка, формирующийся белок направляется к эндоплазматическому ретикулуму, а затем на клеточную поверхность, где он экспрессируется.
Коровая часть сигнального пептида может содержать длинную последовательность из гидрофобных аминокислот, которая имеет тенденцию к образованию одиночной альфа-спирали. Сигнальный пептид может начинаться с короткой положительно заряженной последовательности аминокислот, которая помогает обеспечивать правильную топологию полипептида в процессе транслокации. На конце сигнального пептида, как правило, имеется последовательность аминокислот, которая узнается и расщепляется сигнальной пептидазой. Расщепление сигнальной пептидазой может происходить либо в процессе, либо после завершения транслокации, с образованием свободного сигнального пептида и зрелого белка. Затем свободные сигнальные пептиды расщепляются специфическими протеазами.
Сигнальный пептид может находиться на амино-конце молекулы.
Сигнальный пептид может содержать последовательность, приведенную в SEQ ID NO: 12, 13 или 14, либо ее вариант, имеющий 5, 4, 3, 2 или 1 аминокислотные мутации (вставки, замены или добавления), при условии, что сигнальный пептид по-прежнему функционирует, обеспечивая экспрессию CAR на клеточной поверхности.
SEQ ID NO: 12: MGTSLLCWMALCLLGADHADG
Сигнальный пептид SEQ ID NO: 12 является компактным и высокоэффективным. По прогнозам, процент расщепления будет составлять примерно 95% после концевого остатка глицина, приводя к эффективному удалению с помощью сигнальной пептидазы.
SEQ ID NO: 13: MSLPVTALLLPLALLLHAARP
Сигнальный пептид SEQ ID NO: 13 получен из IgG1.
SEQ ID NO: 14: MAVPTQVLGLLLLWLTDARC
Сигнальный пептид SEQ ID NO: 14 получен из CD8.
ДОМЕН СПЕЙСЕРА
Система CAR, описанная в настоящем документе, может содержать последовательность спейсера для связывания антигенсвязывающего домена с трансмембранным доменом в рецепторном компоненте. Гибкий спейсер позволяет антигенсвязывающему домену ориентироваться в разных направлениях для облегчения связывания.
Последовательность спейсера может, например, представлять собой Fc-область IgG1, шарнир IgG1 или стебель CD8 человека или стебель CD8 мыши. Альтернативно, спейсер может представлять собой другую линкерную последовательность, имеющую сходную длину и/или способность разделять домены в пространстве, как у Fc-области IgG1, шарнира IgG1 или стебля CD8. Спейсер из IgG1 человека может быть изменен для удаления Fc-связывающих фрагментов.
Примеры аминокислотных последовательностей для этих спейсеров приведены ниже:
SEQ ID NO: 15 (шарнир-CH2CH3 IgG1 человека)
AEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKD
SEQ ID NO: 16 (стебель CD8 человека):
TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDI
SEQ ID NO: 17 (шарнир IgG1 человека): AEPKSPDKTHTCPPCPKDPK
SEQ ID NO: 18 (эктодомен CD2)
KEITNALETWGALGQDINLDIPSFQMSDDIDDIKWEKTSDKKKIAQFRKEKETFKEKDTYKLFKNGTLKIKHLKTDDQDIYKVSIYDTKGKNVLEKIFDLKIQERVSKPKISWTCINTTLTCEVMNGTDPELNLYQDGKHLKLSQRVITHKWTTSLSAKFKCTAGNKVSKESSVEPVSCPEKGLD
SEQ ID NO: 19 (эктодомен CD34)
SLDNNGTATPELPTQGTFSNVSTNVSYQETTTPSTLGSTSLHPVSQHGNEATTNITETTVKFTSTSVITSVYGNTNSSVQSQTSVISTVFTTPANVSTPETTLKPSLSPGNVSDLSTTSTSLATSPTKPYTSSSPILSDIKAEIKCSGIREVKLTQGICLEQNKTSSCAEFKKDRGEGLARVLCGEEQADADAGAQVCSLLLAQSEVRPQCLLLVLANRTEISSKLQLMKKHQSDLKKLGILDFTEQDVASHQSYSQKT
РЕЦЕПТОРНЫЙ КОМПОНЕНТ, СОДЕРЖАЩИЙ НЕСКОЛЬКО ПЕРВЫХ СВЯЗЫВАЮЩИХ ДОМЕНОВ
Рецепторный компонент может содержать несколько первых связывающих доменов и, таким образом, быть способным к рекрутингу более одного сигнального компонента.
Несколько первых связывающих доменов могут присутствовать в одном внутриклеточном домене рецепторного компонента. Рецепторный компонент может содержать соответствующее количество трансмембранных доменов, так что каждый первый связывающий домен располагается на внутриклеточной стороне клеточной мембраны. Например, рецепторный компонент может содержать 3, 5, 7, 9, 11 или более трансмембранных доменов. В этом случае один рецепторный компонент может рекрутировать множество сигнальных компонентов, усиливая сигнализацию в ответ на антиген.
Все из первых связывающих доменов могут представлять собой варианты, имеющие разную аффинность для второго связывающего домена сигнального компонента.
МНОЖЕСТВЕННЫЕ РЕЦЕПТОРНЫЕ КОМПОНЕНТЫ
В другом варианте осуществления изобретения система CAR может включать два или более рецепторных компонентов, узнающих разные антигены, но содержащих одинаковые внутриклеточные первые связывающие домены. Такая система CAR была бы способна узнавать множество антигенов (фигура 11). Это может быть полезным, например, для преодоления избегания опухолью иммунного надзора. В дополнительном связанном аспекте изобретения первые связывающие домены рецепторных компонентов отличаются по остаткам, которые определяют их аффинность для второго связывающего домена сигнального компонента. В этом случае систему CAR можно регулировать таким образом, что сигнализация в ответ на один антиген будет сильнее или слабее, чем в ответ на другой (фигура 11). Это может быть полезным, например, при нацеливании на два опухолевых антигена одновременно, когда один из них экспрессируется с более высокой плотностью, чем другой. Ответ на такой антиген можно регулировать в сторону понижения, чтобы избежать токсичности, связанной с избыточной стимуляцией.
Способы, подходящие для изменения аминокислотных остатков первого или второго связывающего домена таким образом, чтобы изменялась аффинность связывания между двумя доменами, известны в данной области и включают замену, добавление и удаление аминокислот с использованием как направленного, так и случайного мутагенеза. Способы определения аффинности связывания между первым связывающим доменом и вторым связывающим доменом также хорошо известны в данной области и включают предсказание белок-белковых взаимодействий методами биоинформатики, аффинный электрофорез, поверхностный плазмонный резонанс, интерферометрию биослоя, двойную поляризационную интерферометрию, статическое рассеяние света и динамическое рассеяние света.
СИГНАЛЬНЫЙ КОМПОНЕНТ
Настоящее изобретение также относится к сигнальному компоненту, содержащему сигнальный домен и второй связывающий домен. Сигнальный компонент представляет собой растворимую молекулу и, следовательно, располагается в цитоплазме при экспрессии в клетке, например, T-клетке.
Никакой сигнализации не происходит через сигнальный домен сигнального компонента, если он не локализуется совместно с рецепторным компонентом, предложенным по настоящему изобретению. Такая совместная локализация имеет место только в отсутствие средства, описанного выше.
ДОМЕН ВНУТРИКЛЕТОЧНОГО СИГНАЛИНГА
Домен внутриклеточного сигналинга представляет собой передающую сигнал часть классического CAR. В сигнальной системе по настоящему изобретению домен внутриклеточного сигналинга (сигнальный домен) расположен в компоненте сигналинга. В отсутствие средства связанный с мембраной рецепторный компонент и компонент внутриклеточного сигналинга сближаются. После узнавания антигена рецепторы группируются, природные молекулы CD45 и CD148 исключаются из синапса и сигнал передается клетке.
Как таковой, сигнальный домен сигнального компонента аналогичен эндодомену классической молекулы CAR.
Наиболее часто используемым сигнальным доменом сигнального компонента является эндодомен CD3-дзета, который содержит 3 ITAM. Он передает сигнал активации T-клетке после связывания антигена. CD3-дзета может не обеспечивать генерирование достаточного сигнала активации и может потребоваться дополнительная костимулирующая сигнализация. Например, химерные CD28 и OX40 можно использовать вместе с CD3-дзета для передачи сигнала пролиферации/выживания, или можно использовать все три вместе (проиллюстрировано на фигуре 1B).
Сигнальный компонент, описанный в настоящем документе, содержит сигнальный домен, он может содержать только эндодомен CD3-дзета, эндодомен CD3-дзета с эндодоменом или CD28, или OX40, либо эндодомены CD28 и OX40, и эндодомен CD3-дзета (фигура 3A).
Сигнальный компонент системы CAR по настоящему изобретению может содержать последовательность, приведенную в SEQ ID NO: 20, 21 или 22, либо ее вариант, имеющий по меньшей мере 80% идентичности последовательности.
SEQ ID NO: 20 - эндодомен CD3-дзета
RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID NO: 21 - эндодомены CD28 и CD3-дзета
SKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRG KGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID NO: 22 - эндодомены CD28, OX40 и CD3-дзета
SKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKIRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR.
Вариантная последовательность может иметь по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с SEQ ID NO: 20, 21 или 22 при условии, что эта последовательность образует эффективный внутриклеточный сигнальный домен.
МНОЖЕСТВЕННЫЕ СИГНАЛЬНЫЕ КОМПОНЕНТЫ
Сигнальная система по первому аспекту настоящего изобретения может включать несколько сигнальных компонентов, каждый из которых содержит сигнальный домен и второй связывающий домен, при этом все вторые связывающие домены связываются одинаковыми первыми связывающими доменами рецепторного компонента, но сигнальные домены представляют собой разные эндодомены (фигура 9). В этом случае несколько разных эндодоменов могут быть активированы одновременно. Это является преимуществом по сравнению с комбинированным сигнальным доменом, поскольку каждый сигнальный домен остается свободным от других сигнальных доменов.
Если каждый сигнальный компонент содержит второй связывающий домен, который отличается по остаткам, изменяющим его аффинность для первого связывающего домена рецепторного компонента, сигнальные компоненты, содержащие разные сигнальные домены, связываются с первым связывающим доменом с разной кинетикой (фигура 10). Это обеспечивает лучший контроль над сигнализацией в ответ на связывание антигена рецепторным компонентом, поскольку разные сигнальные компоненты рекрутируются рецепторным компонентом с разной кинетикой/динамикой. Это является преимуществом, поскольку в отличие от фиксированного равного соотношения сигнала, передаваемого комбинированным эндодоменом, оптимальный сигнал T- клеточной активации может требовать разных иммунологических сигналов в разных пропорциях.
НУКЛЕИНОВАЯ КИСЛОТА
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей рецепторный компонент по второму аспекту, и нуклеиновой кислоте, кодирующей сигнальный компонент по третьему аспекту. Используемые в настоящем документе термины «полинуклеотид», «нуклеотид» и «нуклеиновая кислота» являются синонимами. Специалист в данной области понимает, что многочисленные различные полинуклеотиды и нуклеиновые кислоты могут кодировать один и тот же полипептид в результате вырожденности генетического кода. Кроме того, следует понимать, что квалифицированный специалист может, используя обычные методы, производить нуклеотидные замены, не влияющие на полипептидную последовательность, кодируемую полинуклеотидами, описанными в настоящем документе, которые отражают использование кодонов в конкретном организме-хозяине, в котором предстоит экспрессировать полипептиды.
Нуклеиновые кислоты по изобретению могут представлять собой ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Они также могут представлять собой полинуклеотиды, которые содержат в своем составе синтетические или модифицированные нуклеотиды. Множество разных видов модификаций олигонуклеотидов известно в данной области. Сюда относятся метилфосфонатные и фосфоротиоатные каркасы, а также добавление акридиновых или полилизиновых цепей на 3' и/или 5'-концах молекулы. Следует понимать, что для целей применения, описанного в настоящем документе, полинуклеотиды могут быть модифицированы любым способом, известным в данной области. Такие модификации можно осуществлять для повышения активности или продолжительности существования интересующих полинуклеотидов in vivo.
Термины «вариант», «гомолог» или «производное» применительно к нуклеотидной последовательности включают любое замещение, вариацию, модификацию, замену, делецию или добавление одного (или более) нуклеотидов в последовательности.
Нуклеиновая кислота по изобретению может представлять собой нуклеиновую кислоту, которая кодирует как рецепторный компонент, так и сигнальный компонент.
Продуктом нуклеиновой кислоты может являться полипептид, содержащий рецепторный компонент и сигнальный компонент, соединенные сайтом расщепления. Сайт расщепления может быть саморасщепляющимся, так что, когда полипептид продуцируется, он немедленно расщепляется на рецепторный компонент и сигнальный компонент без необходимости во внешней расщепляющей активности.
Известны различные саморасщепляющиеся сайты, включая саморасщепляющийся пептид 2a вируса ящура (FMDV), который имеет последовательность, приведенную ниже:
SEQ ID NO: 23
RAEGRGSLLTCGDVEENPGP.
или
SEQ ID NO: 24
QCTNYALLKLAGDVESNPGP.
Продуктом нуклеиновой кислоты может являться полипептид, содержащий последовательность, приведенную в SEQ ID NO: 25.
SEQ ID NO: 25
MWTWNAYAFAAPSGGGSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRДНКKSTLYLQMNSLRAЭДТАVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHN HYTQKSLSLSPGKKDPM……SGGGGSMSRLDKSKVINSALELLNEVGIEGLTTRKLAQKLGVEQPTLYWHVKNKRALLDALAIEMLDRHHTHFCPLEGESWQDFLRNNAKSFRCALLSHRDGAKVHLGTRPTEKQYETLENQLAFLCQQGFSLENALYALSAVGHFTLGCVLEDQEHQVAKEERETPTTDSM PPLLRQAIELFDHQGAEPAFLFGLELIICGLEKQLKCESGS
где … указывает на положение, в которое может быть включена последовательность антигенсвязывающего домена. Можно включать любой антигенсвязывающий домен, например, scFV, описанный в настоящем документе.
Совместно экспрессируемой последовательностью может быть последовательность внутренней посадки рибосомы (IRES). Совместно экспрессируемой последовательностью может быть внутренний промотор.
Настоящее изобретение также относится к набору, включающему нуклеиновую кислоту, кодирующую рецепторный компонент по второму аспекту, и/или нуклеиновую кислоту, кодирующую сигнальный компонент по третьему аспекту.
ВЕКТОР
Настоящее изобретение также относится к вектору, или набору векторов, содержащему одну или более последовательностей нуклеиновой кислоты, кодирующих рецепторный компонент по второму аспекту и/или сигнальный компонент по третьему аспекту изобретения. Такой вектор можно использовать для введения последовательности(ей) нуклеиновой кислоты в клетку-хозяина таким образом, чтобы она экспрессировала рецепторный компонент и сигнальный компонент системы CAR по первому аспекту изобретения.
Вектор может представлять собой, например, плазмиду или вирусный вектор, такой как ретровирусный вектор или лентивирусный вектор, или вектор на основе транспозона, или синтетическую мРНК.
Вектор может быть способен трансфицировать или трансдуцировать T-клетку или NK-клетку.
ЦИТОЛИТИЧЕСКАЯ ИММУННАЯ КЛЕТКА
Настоящее изобретение также относится к иммунной клетке, содержащей систему CAR по первому аспекту изобретения.
Цитолитическая иммунная клетка может содержать нуклеиновую кислоту или вектор по настоящему изобретению.
Цитолитическая иммунная клетка может содержать рецепторный компонент и сигнальный компонент по настоящему изобретению.
Цитолитическая иммунная клетка может содержать по меньшей мере один сигнальный компонент по настоящему изобретению. Например, цитолитическая иммунная клетка может содержать один, два, три, четыре, пять, вплоть до множества сигнальных компонентов по настоящему изобретению. Цитолитическая иммунная клетка может содержать по меньшей мере один рецепторный компонент по настоящему изобретению. Например, цитолитическая иммунная клетка может содержать один, два, три, четыре, пять, вплоть до множества рецепторных компонентов по настоящему изобретению.
Цитолитические иммунные клетки могут представлять собой T- клетки или T-лимфоциты, которые являются лимфоцитами, играющими центральную роль в опосредованном клетками иммунитете. Их можно отличать от других лимфоцитов, таких как B-клетки и клетки - естественные киллеры (NK-клетки), по присутствию T-клеточного рецептора (TCR) на клеточной поверхности. Существуют различные типы T-клеток, как описано ниже.
Клетки-помощники T-хелперы (TH-клетки) помогают другим белым кровяным клеткам в иммунологических процессах, включая созревание B-клеток в плазматические клетки и B-клетки памяти, а также активацию цитотоксических T-клеток и макрофагов. TH-клетки экспрессируют CD4 на своей поверхности. TH-клетки становятся активированными после представления им пептидных антигенов молекулами MHC класса II на поверхности антигенпредставляющих клеток (APC). Эти клетки могут дифференцироваться в один из нескольких подтипов, включая TH1, TH2, TH3, TH17, Th9 или TFH, которые секретируют разные цитокины, способствующие проявлениям иммунных ответов разных типов.
Цитолитические T-клетки (TC-клетки или CTL) разрушают инфицированные вирусами клетки и клетки опухолей, а также играют роль в отторжении трансплантата. CTL экспрессируют CD8 на своей поверхности. Эти клетки узнают свои мишени, связываясь с антигеном, ассоциированным с молекулами MHC класса I, которые присутствуют на поверхности всех ядросодержащих клеток. Посредством IL-10, аденозина и другие молекул, секретируемых регуляторными T-клетками, CD8+ клетки могут быть инактивированы и переведены в состояние анергии, что предотвращает аутоиммунные заболевания, такие как экспериментальный аутоиммунный энцефаломиелит.
T-клетки памяти представляют собой подгруппу антигенспецифических T-клеток, которые сохраняются долгое время после того, как инфекция была побеждена. Они быстро размножаются до больших количеств эффекторных T-клеток при повторной стимуляции узнаваемым ими антигеном, таким образом, обеспечивая иммунную систему «памятью» относительно прошлых инфекций. T-клетки памяти включают три подтипа: центральные T-клетки памяти (TCM-клетки) и два типа эффекторных T-клеток памяти (TEM-клетки и TEMRA-клетки). Клетки памяти могут представлять собой либо CD4+, либо CD8+ клетки. T-клетки памяти, как правило, экспрессируют на клеточной поверхности белок CD45RO.
Регуляторные T-клетки (Treg-клетки), ранее известные как супрессорные T-клетки, имеют важное значение для поддержания иммунологической толерантности. Их основная роль заключается в выключении опосредованного T-клетками иммунитета к концу иммунного ответа и в подавлении аутореактивных T-клеток, которые избежали процесса негативной селекции в тимусе.
Описано два основных класса CD4+ Treg-клеток - естественные Treg-клетки и адаптивные Treg-клетки.
Естественные Treg-клетки (также известные как CD4+CD25+FoxP3+ Treg-клетки) образуются в тимусе, и была установлена их связь с взаимодействием между развивающимися T-клетками и миелоидными (CD11c+), а также плазмацитоидными (CD123+) дендритными клетками, которые были активированы TSLP. Естественные Treg-клетки можно отличать от других T-клеток по присутствию внутриклеточной молекулы, называемой FoxP3. Мутации гена FOXP3 могут предотвращать развитие регуляторных T-клеток, вызывая смертельное аутоиммунное заболевание IPEX.
Адаптивные Treg-клетки (также известные как Tr1-клетки или Th3-клетки) могут возникать в процессе обычного иммунного ответа.
Клетки - естественные киллеры (или NK-клетки) являются типом цитолитических клеток, которые составляют часть врожденной иммунной системы. NK-клетки обеспечивают быстрые ответы на характерные сигналы от инфицированных вирусом клеток независимым от MHC образом.
NK-клетки (принадлежащие к группе врожденных лимфоидных клеток) представляют собой крупные зернистые лимфоциты (LGL) и составляют третий вид клеток, дифференцирующихся из общего лимфоидного предшественника, из которого происходят B- и T- лимфоциты. Известно, что NK-клетки дифференцируются и созревают в костном мозге, лимфатических узлах, селезенке, миндалинах и тимусе, откуда они затем поступают в систему циркуляции.
CAR клетки по изобретению могут относиться к любому из типов клеток, указанных выше.
T- или NK-клетки, экспрессирующие молекулы системы CAR по первому аспекту изобретения, могут быть созданы ex vivo либо из собственных клеток периферической крови пациента (1-я сторона), либо из трансплантата гемопоэтических стволовых клеток из периферической крови донора (2-я сторона), либо из периферической крови от несвязанного донора (3-я сторона).
Альтернативно, T- или NK-клетки, экспрессирующие молекулы системы CAR по первому аспекту изобретения, могут быть получены путем ex vivo дифференциации индуцируемых клеток-предшественников или эмбриональных клеток-предшественников в T-клетки. Альтернативно, можно использовать линию иммортализованных T- клеток, которые сохраняют свою литическую функцию и могут действовать в качестве терапевтического средства.
Во всех этих вариантах осуществления CAR клетки получают путем введения ДНК или РНК, кодирующей рецепторный компонент и сигнальный компонент, одним из множества методов, включая трансдукцию вирусным вектором, а также трансфекцию ДНК или РНК.
CAR клетка по изобретению может представлять собой ex vivo T- или NK-клетку, полученную от субъекта. T или NK-клетка может быть клеткой из образца мононуклеарных клеток периферической крови (МКПК). T или NK-клетки можно активировать и/или наращивать перед проведением трансдукции нуклеиновой кислотой, кодирующей молекулы, составляющие систему CAR по первому аспекту изобретения, например, путем обработки анти-CD3 моноклональными антителами.
T или NK-клетка по изобретению может быть получена путем:
(i) получения содержащего T- или NK-клетки образца от субъекта или из других источников, перечисленных выше; и
(ii) трансдукции или трансфекции T- или NK-клеток одной или более последовательностями нуклеиновой кислоты, кодирующими рецепторный компонент и/или сигнальный компонент системы CAR по второму и третьему аспектам изобретения.
Затем T или NK-клетки можно очищать, например, отбирать на основании экспрессии антигенсвязывающего домена антигенсвязывающего полипептида.
Настоящее изобретение также относится к набору, включающему T или NK-клетку, содержащую систему CAR по первому аспекту изобретения.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Настоящее изобретение также относится к фармацевтической композиции, содержащей множество цитолитических иммунных клеток, экспрессирующих компоненты системы CAR по первому аспекту изобретения. Кроме того, фармацевтическая композиция может содержать фармацевтически приемлемый носитель, разбавитель или эксципиент. Фармацевтическая композиция может, необязательно, содержать один или более дополнительных фармацевтически активных полипептидов и/или соединений. Такой препарат может, например, быть в форме, подходящей для внутривенной инфузии.
СПОСОБ ЛЕЧЕНИЯ
Настоящее изобретение относится к способу лечения и/или предотвращения заболевания, включающему этап введения субъекту цитолитических иммунных клеток по настоящему изобретению (например, в фармацевтической композиции, описанной выше).
Способ лечения заболевания относится к терапевтическому применению цитолитических иммунных клеток по настоящему изобретению. В соответствии с настоящим документом, клетки можно вводить субъекту, имеющему заболевание или состояние, с целью уменьшения, ослабления или облегчения по меньшей мере одного симптома, ассоциированного с заболеванием, и/или с целью замедления, уменьшения или блокирования прогрессирования заболевания.
Способ предотвращения заболевания относится к профилактическому применению цитолитических иммунных клеток по настоящему изобретению. В соответствии с настоящим документом, такие клетки можно вводить субъекту, у которого еще не начало развиваться заболевание и/или у которого еще не проявляются какие- либо симптомы заболевания, с целью предотвращения или устранения причины заболевания, либо уменьшения или предотвращения развития по меньшей мере одного симптома, ассоциированного с заболеванием. Субъект может иметь предрасположенность или, предположительно, иметь риск развития заболевания.
Способ может включать этапы:
(i) выделения содержащего T- или NK-клетки образца;
(ii) проведения трансдукции или трансфекции таких клеток последовательностью нуклеиновой кислоты или вектором, предложенными по настоящему изобретению;
(iii) введения клеток по пункту (ii) субъекту.
Содержащий T- или NK-клетки образец можно получать от субъекта или из других источников, например, описанных выше. T- или NK-клетки можно выделять из собственной периферической крови субъекта (1-я сторона), либо из трансплантата гемопоэтических стволовых клеток из периферической крови донора (2-я сторона), либо из периферической крови от несвязанного донора (3-я сторона).
Способы, предложенные по настоящему изобретению для лечения заболевания, могут включать мониторинг прогрессирования заболевания и любой токсической активности и введение средства, подходящего для использования в системе CAR по первому аспекту изобретения, с целью ингибирования сигнализации CAR и, таким образом, уменьшения или ослабления каких-либо неблагоприятных токсических эффектов.
Способы, предложенные по настоящему изобретению для лечения заболевания, могут включать мониторинг прогрессирования заболевания и мониторинг любой токсической активности, и корректировку дозы средства, вводимого субъекту, с целью обеспечения приемлемых уровней прогрессирования заболевания и токсической активности.
Мониторинг прогрессирования заболевания означает оценку симптомов, ассоциированных с заболеванием, с течением времени для определения того, имеет ли место их уменьшение/ослабление или усиление/ухудшение.
Токсические активности означают неблагоприятные эффекты, вызываемые CAR клетками по изобретению после их введения субъекту. Токсические активности могут включать, например, иммунологическую токсичность, токсичность желчных кислот и респираторный дистресс-синдром.
Уровень сигнализации через сигнальную систему по первому аспекту изобретения и, следовательно, уровень активации CAR клеток, экспрессирующих сигнальную систему, можно корректировать путем изменения количества присутствующего средства или количества времени, в течение которого средство присутствует. В настоящем способе уровень активации CAR клеток можно увеличивать путем уменьшения дозы средства, вводимого субъекту, или уменьшения частоты такого введения. И наоборот, уровень активации CAR клеток можно уменьшать путем увеличения дозы средства или частоты его введения субъекту.
Более высокие уровни активации CAR клеток, вероятно, будут связаны с уменьшением прогрессирования заболевания, но возрастанием токсических активностей, в то время как более низкие уровни активации CAR клеток, вероятно, будут связаны с возрастанием прогрессирования заболевания, но уменьшением токсических активностей.
Настоящее изобретение также относится к способу лечения и/или предотвращения заболевания у субъекта, в организме которого присутствуют клетки по изобретению, включающему этап введения субъекту средства, подходящего для использования в системе CAR по первому аспекту изобретения. По существу, данный способ включает введение соответствующего средства субъекту, в организме которого уже присутствуют CAR клетки по настоящему изобретению.
По существу, дозу средства, вводимого субъекту, или частоту введения можно изменять для обеспечения приемлемых уровней как прогрессирования заболевания, так и токсической активности. Конкретный уровень прогрессирования заболевания и токсической активности, считающийся «приемлемым», будет варьироваться в зависимости от конкретных обстоятельств и должен быть оценен, исходя из них. Настоящее изобретение относится к способу изменения уровня активации CAR клеток с целью достижения такого приемлемого уровня.
Средство можно вводить в форме фармацевтической композиции. Фармацевтическая композиция, кроме того, может содержать фармацевтически приемлемый носитель, разбавитель или эксципиент. Фармацевтическая композиция может, необязательно, содержать один или более дополнительных фармацевтически активных полипептидов и/или соединений. Такой препарат может, например, быть в форме, подходящей для внутривенной инфузии.
Настоящее изобретение относится к CAR клетке по настоящему изобретению для использования в лечении и/или предотвращении заболевания.
Изобретение также относится к использованию CAR клетки по настоящему изобретению в производстве лекарственного средства для лечения и/или предотвращения заболевания.
Настоящее изобретение также относится к средству, подходящему для ингибирования системы CAR по первому аспекту изобретения, которое предназначено для использования в лечении и/или предотвращении заболевания.
Настоящее изобретение также относится к средству, используемому для ингибирования системы CAR по первому аспекту изобретения в CAR клетке.
Изобретение также относится к использованию средства, подходящего для ингибирования системы CAR по первому аспекту изобретения, в производстве лекарственного средства для лечения и/или предотвращения заболевания.
Заболевание, которое предстоит лечить и/или предотвращать способами по настоящему изобретению, может представлять собой инфекцию, например, вирусную инфекцию.
Способы по изобретению также могут быть предназначены для контроля патогенных иммунных ответов, например, при аутоиммунных заболеваниях, аллергии и реакции отторжения «трансплантат против хозяина».
Способы могут быть предназначены для лечения онкологического заболевания, такого как рак мочевого пузыря, рак молочной железы, рак толстого кишечника, рак эндометрия, рак почки (почечно- клеточный рак), лейкоз, рак легкого, меланома, неходжкинская лимфома, рак поджелудочной железы, рак предстательной железы и рак щитовидной железы.
CAR клетки по настоящему изобретению могут быть способны к уничтожению клеток-мишеней, таких как раковые клетки. Клетки- мишени могут быть узнаны по экспрессии TAA, например, экспрессии TAA, приведенных выше в таблице 1.
CAR клетки и фармацевтические композиции по настоящему изобретению могут быть использованы в лечении и/или предотвращении заболеваний, описанных выше.
CAR клетки и фармацевтические композиции по настоящему изобретению могут быть использованы в любом из способов, описанных выше.
Далее изобретение будет дополнительно описано с помощью примеров, которые предназначены служить для облегчения специалисту в данной области осуществления на практике изобретения и не должны каким-либо образом ограничивать объем данного изобретения.
ПРИМЕРЫ
Пример 1 - Функциональность сигнальной системы TetCAR
Бицистронный конструкт был экспрессирован в виде единого транскрипта, который саморасщепляется в сайте 2A, с образованием TiP, слитого с eGFP, и CAR с TetR в качестве его эндодомена (фигура 5a).
Результаты флуоресцентной микроскопии клеток SupT1, экспрессирующих данный конструкт, в отсутствие тетрациклина показали, что флуоресценция eGFP может быть отчетливо видна на клеточной мембране (фигура 5b); в то время как в присутствии тетрациклина eGFP находился в цитоплазме (фигура 5c). Эти данные свидетельствуют о том, что тетрациклин вытеснил TiP из TetR CAR.
Пример 2 - Сигнализация через систему TetCAR
Бицистронный конструкт был экспрессирован в T-клетках BW5 в виде единого транскрипта, который саморасщепляется в сайте 2A, с образованием сигнального компонента, содержащего TiP, слитый через гибкий линкер с эндодоменом CD3-дзета; и рецепторного компонента, содержащего узнающий CD33 фрагмент scFv, спейсер, полученный из Fc-домена IgG1, полученный из CD4 трансмембранный и внутриклеточный домен; и TetR (фигура 6a). Также был экспрессирован контроль, который был идентичен, за исключением того, что TiP отсутствовал в сигнальном компоненте (фигура 6b).
T-клетки BW5 стимулировали клетками SupT1 дикого типа или клетками SupT1, сконструированными для экспрессии CD33, в отсутствие тетрациклина или в присутствии тетрациклина в возрастающих концентрациях. T-клетки, стимулированные клетками SupT1 дикого типа, не активировались ни в присутствии, ни в отсутствие тетрациклина; T-клетки, стимулированные клетками SupT1, экспрессирующими CD33, активировались в отсутствие тетрациклина, однако активация быстро ингибировалась в присутствии тетрациклина, и активация была полностью ингибирована в присутствии 100 нМ тетрациклина (фигура 7a).
Контрольный TetCAR, у которого отсутствовал домен TiP, также трансдуцировали в клетки BW5. Опять-таки, эти T-клетки стимулировали клетками SupT1 дикого типа или клетками SupT1, сконструированными для экспрессии CD33, в отсутствие тетрациклина или в присутствии тетрациклина в возрастающих концентрациях. Отсутствие элемента TiP в сигнальном компоненте приводило к отсутствию сигнализации в любых условиях (фигура 7b).
Пример 3 - Сигнализация системы TetCAR в первичных T-клетках
Клетки SupT1 (которые являются CD19-отрицательными), были сконструированы CD19-положительными, в результате чего были получены положительные и отрицательные по целевому антигену линии клеток, которые имели максимально возможное сходство. Первичные человеческие T-клетки от 3 доноров были трансдуцированы тремя конструктами CAR: (i) «классическим» анти-CD19 CAR 1-го поколения; (ii) анти-CD19 TetCAR 1-го поколения; (iii) контрольным анти-CD19 TetCAR, у которого в эндодомене отсутствовал TiP. Нетрансдуцированные T-клетки и T-клетки, трансдуцированные разными конструктами CAR, стимулировали в соотношении 1:1 либо клетками SupT1, либо клетками SupT1.CD19 в присутствии тетрациклина в разных концентрациях. Супернатант отбирали через 48 часов после стимуляции. Также получали образцы супернатантов с фоновым (только T-клетки) и максимальным (T-клетки, стимулированные ФМА/иономицином) уровнями. Интерферон-гамма определяли в супернатантах методом ELISA (фигура 13). «Классические» CAR T- клетки были активированы клетками SupT1.CD19 независимо от тетрациклина. TetCAR T-клетки были активированы клетками SupT1.CD19, однако активация ингибировалась тетрациклином. Контрольные TetCAR и н/т T-клетки не реагировали на клетки SupT1.CD19.
Пример 4 - Уничтожение клеток-мишеней
После исследований по секреции интерферона-гамма, описанных в примере 3, уничтожение клеток-мишеней было продемонстрировано с использованием метода с высвобождением хрома. Клетки SupT1 и SupT1.CD19 нагружали 51Cr и инкубировали с контрольными и TetCAR T-клетками в течение 4 часов в присутствии или в отсутствие тетрациклина. Лизис клеток-мишеней определяли путем подсчета 51Cr в супернатанте. Результаты представлены на фигуре 14. Показано, что TetCAR T-клетки лизировали клетки-мишени SupT1.CD19 только в отсутствие тетрациклина.
Все публикации, упомянутые в приведенной выше спецификации, включены в настоящий документ посредством ссылки. Для специалистов в данной области будут очевидны различные модификации и вариации описанных способов и системы по изобретению без отклонения от объема и сущности изобретения. Хотя изобретение описано в связи с определенными предпочтительными вариантами осуществления, следует понимать, что заявленное изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. Действительно, различные модификации описанных способов осуществления изобретения, которые очевидны для специалистов в области молекулярной биологии, клеточной иммунологии или смежных областях, должны входить в объем прилагаемой формулы изобретения.
--->
СПИСКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCL Business PLC
<120> Сигнальная система
<130> P105610PCT
<150> GB1415347.2
<151> 2014-08-29
<160> 29
<170> PatentIn version 3.5
<210> 1
<211> 139
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> связывающий домен TetR
<400> 1
Met Ser Arg Leu Asp Lys Ser Lys Val Ile Asn Ser Ala Leu Glu Leu
1 5 10 15
Leu Asn Glu Val Gly Ile Glu Gly Leu Thr Thr Arg Lys Leu Ala Gln
20 25 30
Lys Leu Gly Val Glu Gln Pro Thr Leu Tyr Trp His Val Lys Asn Lys
35 40 45
Arg Ala Leu Leu Asp Ala Leu Ala Ile Glu Met Leu Asp Arg His His
50 55 60
Thr His Phe Cys Pro Leu Glu Gly Glu Ser Trp Gln Asp Phe Leu Arg
65 70 75 80
Asn Asn Ala Lys Ser Phe Arg Cys Ala Leu Leu Ser His Arg Asp Gly
85 90 95
Ala Lys Val His Leu Gly Thr Arg Pro Thr Glu Lys Gln Tyr Glu Thr
100 105 110
Leu Glu Asn Gln Leu Ala Phe Leu Cys Gln Gln Gly Phe Ser Leu Glu
115 120 125
Asn Ala Leu Tyr Ala Leu Ser Ala Val Gly His
130 135
<210> 2
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> связывающий домен TiP
<400> 2
Met Trp Thr Trp Asn Ala Tyr Ala Phe Ala Ala Pro Ser Gly Gly Gly
1 5 10 15
Ser
<210> 3
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Модифицированный линкер эндодомена CD4
<400> 3
Ala Leu Ile Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile Gly
1 5 10 15
Leu Gly Ile Phe Phe Cys Val Arg Cys Arg His Arg Arg Arg Gln Ala
20 25 30
Glu Arg Met Ala Gln Ile Lys Arg Val Val Ser Glu Lys Lys Thr Ala
35 40 45
Gln Ala Pro His Arg Phe Gln Lys Thr Cys Ser Pro Ile
50 55 60
<210> 4
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, длинный наномаркер
<400> 4
Asp Val Glu Ala Trp Leu Asp Glu Arg Val Pro Leu Val Glu Thr
1 5 10 15
<210> 5
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, короткий наномаркер
<400> 5
Asp Val Glu Ala Trp Leu Gly Ala Arg
1 5
<210> 6
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, стрепмаркер
<400> 6
Trp Arg His Pro Gln Phe Gly Gly
1 5
<210> 7
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, стрепмаркер II
<400> 7
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 8
<211> 38
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, SBP-маркер
<400> 8
Met Asp Glu Lys Thr Thr Gly Trp Arg Gly Gly His Val Val Glu Gly
1 5 10 15
Leu Ala Gly Glu Leu Glu Gln Leu Arg Ala Arg Leu Glu His His Pro
20 25 30
Gln Gly Gln Arg Glu Pro
35
<210> 9
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, cc-стрепмаркер
<400> 9
Cys His Pro Gln Gly Pro Pro Cys
1 5
<210> 10
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Имитирующий биотин пептид, фланкированный cc-стрепмаркер
<400> 10
Ala Glu Cys His Pro Gln Gly Pro Pro Cys Ile Glu Gly Arg Lys
1 5 10 15
<210> 11
<211> 126
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Коровая последовательность стрептавидина
<400> 11
Glu Ala Gly Ile Thr Gly Thr Trp Tyr Asn Gln Leu Gly Ser Thr Phe
1 5 10 15
Ile Val Thr Ala Gly Ala Asp Gly Ala Leu Thr Gly Thr Tyr Glu Ser
20 25 30
Ala Val Gly Asn Ala Glu Ser Arg Tyr Val Leu Thr Gly Arg Tyr Asp
35 40 45
Ser Ala Pro Ala Thr Asp Gly Ser Gly Thr Ala Leu Gly Trp Thr Val
50 55 60
Ala Trp Lys Asn Asn Tyr Arg Asn Ala His Ser Ala Thr Thr Trp Ser
65 70 75 80
Gly Gln Tyr Val Gly Gly Ala Glu Ala Arg Ile Asn Thr Gln Trp Leu
85 90 95
Leu Thr Ser Gly Thr Thr Glu Ala Asn Ala Trp Lys Ser Thr Leu Val
100 105 110
Gly His Asp Thr Phe Thr Lys Val Lys Pro Ser Ala Ala Ser
115 120 125
<210> 12
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид
<400> 12
Met Gly Thr Ser Leu Leu Cys Trp Met Ala Leu Cys Leu Leu Gly Ala
1 5 10 15
Asp His Ala Asp Gly
20
<210> 13
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид
<400> 13
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 14
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид
<400> 14
Met Ala Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr
1 5 10 15
Asp Ala Arg Cys
20
<210> 15
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность спейсера, шарнир-CH2CH3 IgG1 человека
<400> 15
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp
225 230
<210> 16
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность спейсера, стебель CD8 человека
<400> 16
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile
35 40 45
<210> 17
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность спейсера, шарнир IgG1 человека
<400> 17
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Lys Asp Pro Lys
20
<210> 18
<211> 185
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность спейсера, эктодомен CD2
<400> 18
Lys Glu Ile Thr Asn Ala Leu Glu Thr Trp Gly Ala Leu Gly Gln Asp
1 5 10 15
Ile Asn Leu Asp Ile Pro Ser Phe Gln Met Ser Asp Asp Ile Asp Asp
20 25 30
Ile Lys Trp Glu Lys Thr Ser Asp Lys Lys Lys Ile Ala Gln Phe Arg
35 40 45
Lys Glu Lys Glu Thr Phe Lys Glu Lys Asp Thr Tyr Lys Leu Phe Lys
50 55 60
Asn Gly Thr Leu Lys Ile Lys His Leu Lys Thr Asp Asp Gln Asp Ile
65 70 75 80
Tyr Lys Val Ser Ile Tyr Asp Thr Lys Gly Lys Asn Val Leu Glu Lys
85 90 95
Ile Phe Asp Leu Lys Ile Gln Glu Arg Val Ser Lys Pro Lys Ile Ser
100 105 110
Trp Thr Cys Ile Asn Thr Thr Leu Thr Cys Glu Val Met Asn Gly Thr
115 120 125
Asp Pro Glu Leu Asn Leu Tyr Gln Asp Gly Lys His Leu Lys Leu Ser
130 135 140
Gln Arg Val Ile Thr His Lys Trp Thr Thr Ser Leu Ser Ala Lys Phe
145 150 155 160
Lys Cys Thr Ala Gly Asn Lys Val Ser Lys Glu Ser Ser Val Glu Pro
165 170 175
Val Ser Cys Pro Glu Lys Gly Leu Asp
180 185
<210> 19
<211> 259
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность спейсера, эктодомен CD34
<400> 19
Ser Leu Asp Asn Asn Gly Thr Ala Thr Pro Glu Leu Pro Thr Gln Gly
1 5 10 15
Thr Phe Ser Asn Val Ser Thr Asn Val Ser Tyr Gln Glu Thr Thr Thr
20 25 30
Pro Ser Thr Leu Gly Ser Thr Ser Leu His Pro Val Ser Gln His Gly
35 40 45
Asn Glu Ala Thr Thr Asn Ile Thr Glu Thr Thr Val Lys Phe Thr Ser
50 55 60
Thr Ser Val Ile Thr Ser Val Tyr Gly Asn Thr Asn Ser Ser Val Gln
65 70 75 80
Ser Gln Thr Ser Val Ile Ser Thr Val Phe Thr Thr Pro Ala Asn Val
85 90 95
Ser Thr Pro Glu Thr Thr Leu Lys Pro Ser Leu Ser Pro Gly Asn Val
100 105 110
Ser Asp Leu Ser Thr Thr Ser Thr Ser Leu Ala Thr Ser Pro Thr Lys
115 120 125
Pro Tyr Thr Ser Ser Ser Pro Ile Leu Ser Asp Ile Lys Ala Glu Ile
130 135 140
Lys Cys Ser Gly Ile Arg Glu Val Lys Leu Thr Gln Gly Ile Cys Leu
145 150 155 160
Glu Gln Asn Lys Thr Ser Ser Cys Ala Glu Phe Lys Lys Asp Arg Gly
165 170 175
Glu Gly Leu Ala Arg Val Leu Cys Gly Glu Glu Gln Ala Asp Ala Asp
180 185 190
Ala Gly Ala Gln Val Cys Ser Leu Leu Leu Ala Gln Ser Glu Val Arg
195 200 205
Pro Gln Cys Leu Leu Leu Val Leu Ala Asn Arg Thr Glu Ile Ser Ser
210 215 220
Lys Leu Gln Leu Met Lys Lys His Gln Ser Asp Leu Lys Lys Leu Gly
225 230 235 240
Ile Leu Asp Phe Thr Glu Gln Asp Val Ala Ser His Gln Ser Tyr Ser
245 250 255
Gln Lys Thr
<210> 20
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный компонент, эндодомен CD3 Z
<400> 20
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 21
<211> 152
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный компонент, эндодомены CD28 и CD3-дзета
<400> 21
Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro
1 5 10 15
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
20 25 30
Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala
35 40 45
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
50 55 60
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
65 70 75 80
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
85 90 95
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
100 105 110
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
115 120 125
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
130 135 140
His Met Gln Ala Leu Pro Pro Arg
145 150
<210> 22
<211> 188
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный компонент, эндодомены CD28, OX40 и CD3-дзета
<400> 22
Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro
1 5 10 15
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
20 25 30
Arg Asp Phe Ala Ala Tyr Arg Ser Arg Asp Gln Arg Leu Pro Pro Asp
35 40 45
Ala His Lys Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu
50 55 60
Glu Gln Ala Asp Ala His Ser Thr Leu Ala Lys Ile Arg Val Lys Phe
65 70 75 80
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
85 90 95
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
100 105 110
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
115 120 125
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
130 135 140
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
145 150 155 160
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
165 170 175
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
180 185
<210> 23
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> саморасщепляющийся сайт, саморасщепляющийся пептид 2a
<400> 23
Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 24
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> саморасщепляющийся сайт, саморасщепляющийся пептид 2a
<400> 24
Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 25
<211> 652
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность химерного антигенного рецептора (CAR)
<400> 25
Met Trp Thr Trp Asn Ala Tyr Ala Phe Ala Ala Pro Ser Gly Gly Gly
1 5 10 15
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
20 25 30
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
35 40 45
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
50 55 60
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
65 70 75 80
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
85 90 95
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
100 105 110
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
115 120 125
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
130 135 140
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
145 150 155 160
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
165 170 175
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
180 185 190
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
195 200 205
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
210 215 220
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
225 230 235 240
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
245 250 255
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
260 265 270
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
275 280 285
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
290 295 300
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
305 310 315 320
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
325 330 335
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
340 345 350
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
355 360 365
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
370 375 380
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
385 390 395 400
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
405 410 415
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
420 425 430
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
435 440 445
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
450 455 460
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
465 470 475 480
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
485 490 495
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
500 505 510
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
515 520 525
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
530 535 540
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
545 550 555 560
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
565 570 575
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
580 585 590
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
595 600 605
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
610 615 620
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
625 630 635 640
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp Pro Met
645 650
<210> 26
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность химерного антигенного рецептора (CAR)
<400> 26
Ser Gly Gly Gly Gly Ser Met Ser Arg Leu Asp Lys Ser Lys Val Ile
1 5 10 15
Asn Ser Ala Leu Glu Leu Leu Asn Glu Val Gly Ile Glu Gly Leu Thr
20 25 30
Thr Arg Lys Leu Ala Gln Lys Leu Gly Val Glu Gln Pro Thr Leu Tyr
35 40 45
Trp His Val Lys Asn Lys Arg Ala Leu Leu Asp Ala Leu Ala Ile Glu
50 55 60
Met Leu Asp Arg His His Thr His Phe Cys Pro Leu Glu Gly Glu Ser
65 70 75 80
Trp Gln Asp Phe Leu Arg Asn Asn Ala Lys Ser Phe Arg Cys Ala Leu
85 90 95
Leu Ser His Arg Asp Gly Ala Lys Val His Leu Gly Thr Arg Pro Thr
100 105 110
Glu Lys Gln Tyr Glu Thr Leu Glu Asn Gln Leu Ala Phe Leu Cys Gln
115 120 125
Gln Gly Phe Ser Leu Glu Asn Ala Leu Tyr Ala Leu Ser Ala Val Gly
130 135 140
His Phe Thr Leu Gly Cys Val Leu Glu Asp Gln Glu His Gln Val Ala
145 150 155 160
Lys Glu Glu Arg Glu Thr Pro Thr Thr Asp Ser Met Pro Pro Leu Leu
165 170 175
Arg Gln Ala Ile Glu Leu Phe Asp His Gln Gly Ala Glu Pro Ala Phe
180 185 190
Leu Phe Gly Leu Glu Leu Ile Ile Cys Gly Leu Glu Lys Gln Leu Lys
195 200 205
Cys Glu Ser Gly Ser
210
<210> 27
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность TiP
<400> 27
Trp Thr Trp Asn Ala Tyr Ala Phe Ala Ala Pro Ser Gly Gly Gly Ser
1 5 10 15
<210> 28
<211> 87
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Leu Ser Asp Ser Gly Gln Val Leu Leu Glu Ser Asn Ile Lys Val Leu
1 5 10 15
Pro Thr Trp Ser Thr Pro Val Gln Pro Met Ala Leu Ile Val Leu Gly
20 25 30
Gly Val Ala Gly Leu Leu Leu Phe Ile Gly Leu Gly Ile Phe Phe Cys
35 40 45
Val Arg Cys Arg His Arg Arg Arg Gln Ala Glu Arg Met Ser Gln Ile
50 55 60
Lys Arg Leu Leu Ser Glu Lys Lys Thr Cys Gln Cys Pro His Arg Phe
65 70 75 80
Gln Lys Thr Cys Ser Pro Ile
85
<210> 29
<211> 926
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность основного TetCAR
<400> 29
Met Trp Thr Trp Asn Ala Tyr Ala Phe Ala Ala Pro Ser Gly Gly Gly
1 5 10 15
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
20 25 30
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
35 40 45
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
50 55 60
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
65 70 75 80
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
85 90 95
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
100 105 110
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
115 120 125
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
130 135 140
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
145 150 155 160
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
165 170 175
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
180 185 190
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
195 200 205
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
210 215 220
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
225 230 235 240
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
245 250 255
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
260 265 270
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
275 280 285
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
290 295 300
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
305 310 315 320
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
325 330 335
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
340 345 350
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
355 360 365
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
370 375 380
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
385 390 395 400
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
405 410 415
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
420 425 430
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
435 440 445
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
450 455 460
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
465 470 475 480
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
485 490 495
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
500 505 510
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
515 520 525
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
530 535 540
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
545 550 555 560
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
565 570 575
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
580 585 590
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
595 600 605
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
610 615 620
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
625 630 635 640
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp Pro Met Ala Leu Ile Val
645 650 655
Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile Gly Leu Gly Ile Phe
660 665 670
Phe Cys Val Arg Cys Arg His Arg Arg Arg Gln Ala Glu Arg Met Ala
675 680 685
Gln Ile Lys Arg Val Val Ser Glu Lys Lys Thr Ala Gln Ala Pro His
690 695 700
Arg Phe Gln Lys Thr Cys Ser Pro Ile Ser Gly Gly Gly Gly Ser Met
705 710 715 720
Ser Arg Leu Asp Lys Ser Lys Val Ile Asn Ser Ala Leu Glu Leu Leu
725 730 735
Asn Glu Val Gly Ile Glu Gly Leu Thr Thr Arg Lys Leu Ala Gln Lys
740 745 750
Leu Gly Val Glu Gln Pro Thr Leu Tyr Trp His Val Lys Asn Lys Arg
755 760 765
Ala Leu Leu Asp Ala Leu Ala Ile Glu Met Leu Asp Arg His His Thr
770 775 780
His Phe Cys Pro Leu Glu Gly Glu Ser Trp Gln Asp Phe Leu Arg Asn
785 790 795 800
Asn Ala Lys Ser Phe Arg Cys Ala Leu Leu Ser His Arg Asp Gly Ala
805 810 815
Lys Val His Leu Gly Thr Arg Pro Thr Glu Lys Gln Tyr Glu Thr Leu
820 825 830
Glu Asn Gln Leu Ala Phe Leu Cys Gln Gln Gly Phe Ser Leu Glu Asn
835 840 845
Ala Leu Tyr Ala Leu Ser Ala Val Gly His Phe Thr Leu Gly Cys Val
850 855 860
Leu Glu Asp Gln Glu His Gln Val Ala Lys Glu Glu Arg Glu Thr Pro
865 870 875 880
Thr Thr Asp Ser Met Pro Pro Leu Leu Arg Gln Ala Ile Glu Leu Phe
885 890 895
Asp His Gln Gly Ala Glu Pro Ala Phe Leu Phe Gly Leu Glu Leu Ile
900 905 910
Ile Cys Gly Leu Glu Lys Gln Leu Lys Cys Glu Ser Gly Ser
915 920 925
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КЛЕТКА | 2014 |
|
RU2717984C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| КЛЕТКА | 2014 |
|
RU2732236C2 |
| Выделенный альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора и включающий его химерный антигенный рецептор | 2019 |
|
RU2742000C2 |
| ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР | 2016 |
|
RU2826122C1 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2016 |
|
RU2752880C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА FLT3 | 2018 |
|
RU2820859C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
| ВАРИАНТЫ РЕЦЕПТОРА ИНТЕРЛЕЙКИНА-2 (IL2R) И ИНТЕРЛЕЙКИНА-2 (IL2) ДЛЯ СПЕЦИФИЧЕСКОЙ АКТИВАЦИИ ИММУННЫХ ЭФФЕКТОРНЫХ КЛЕТОК | 2020 |
|
RU2832154C2 |
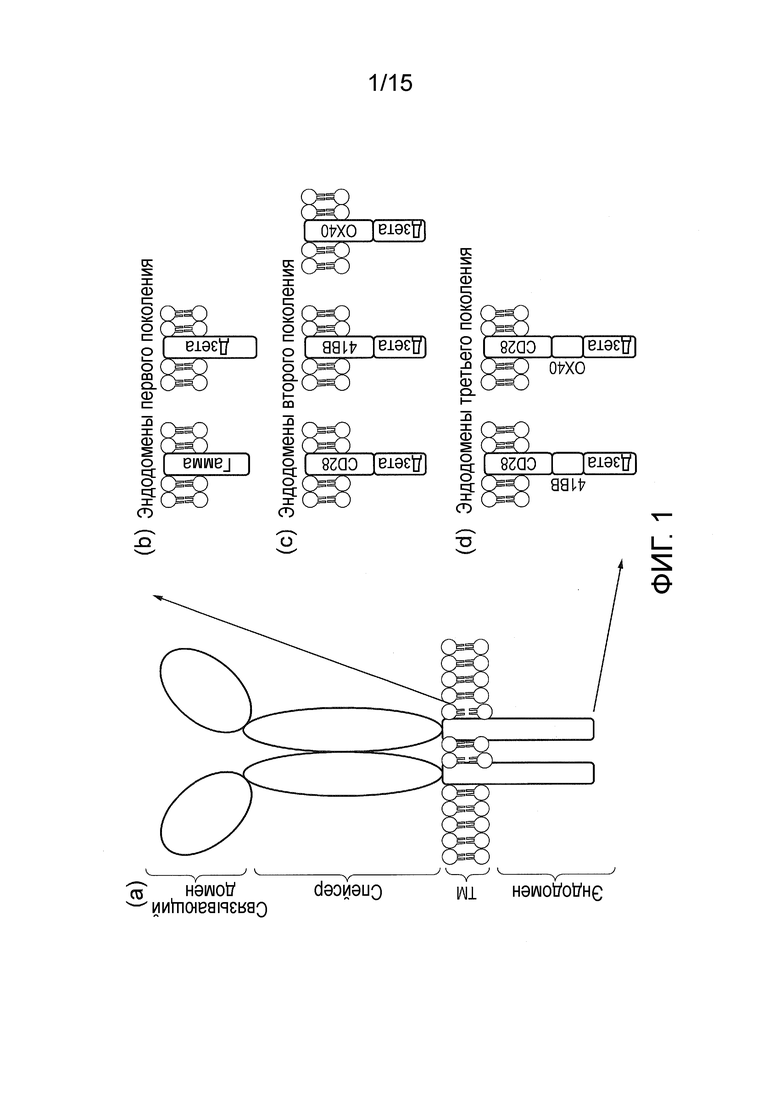
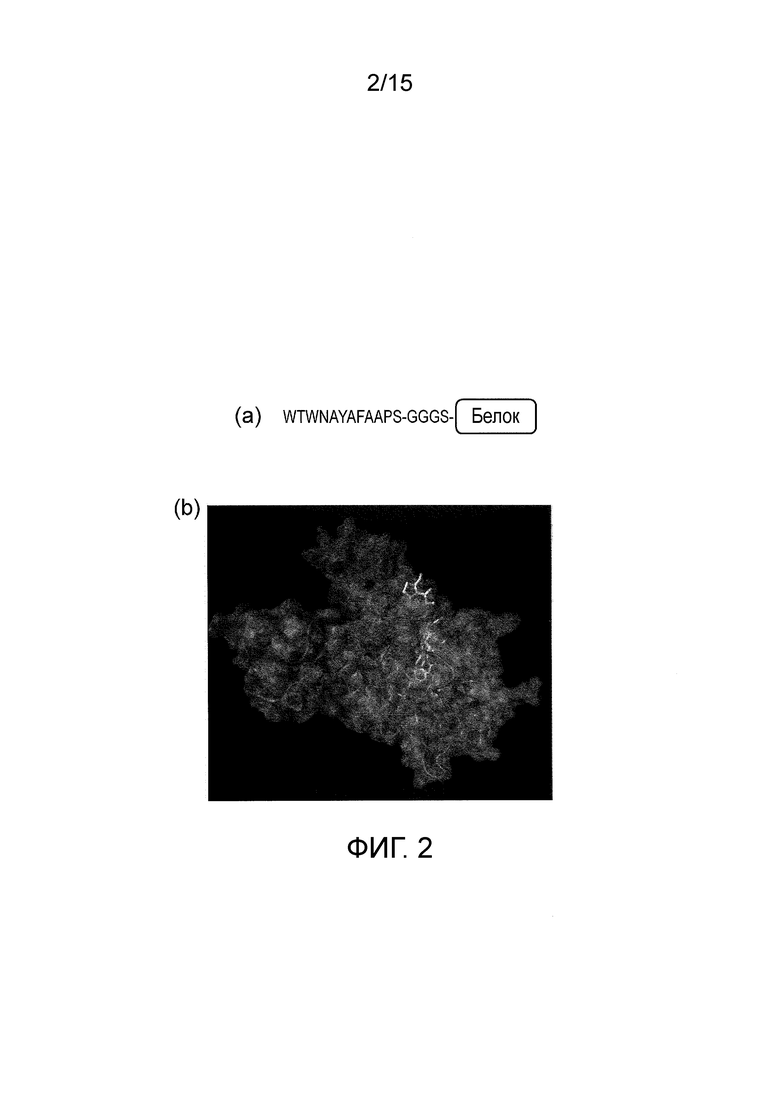
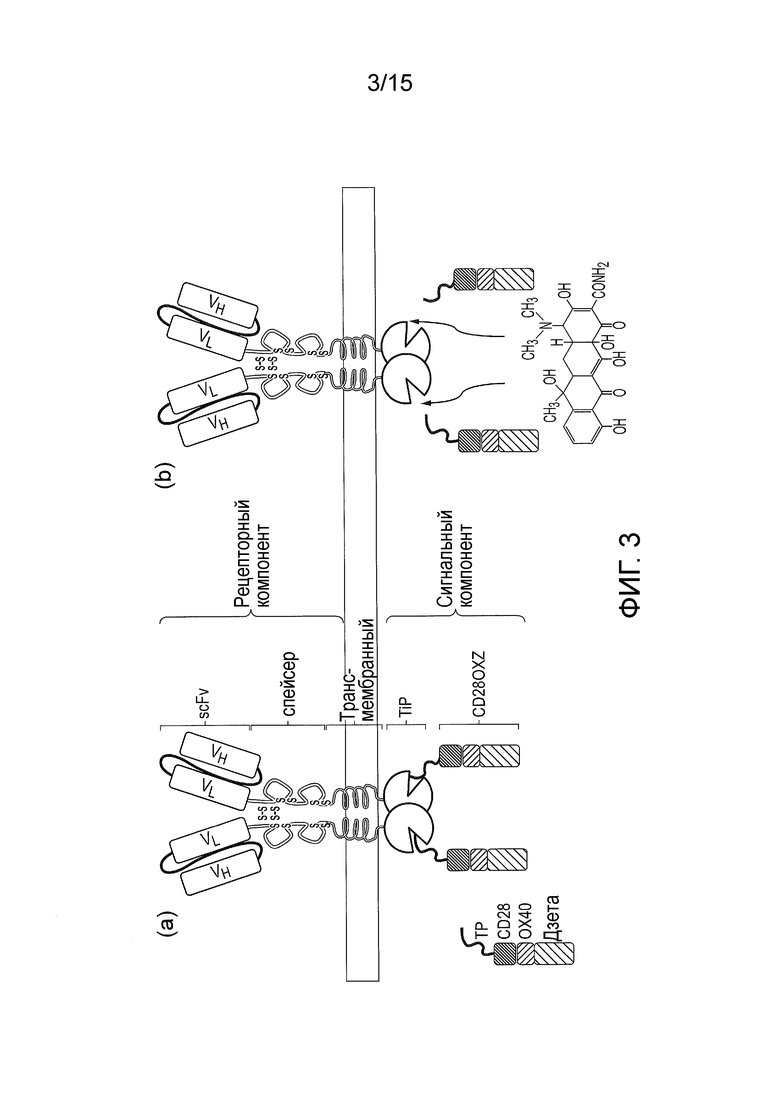
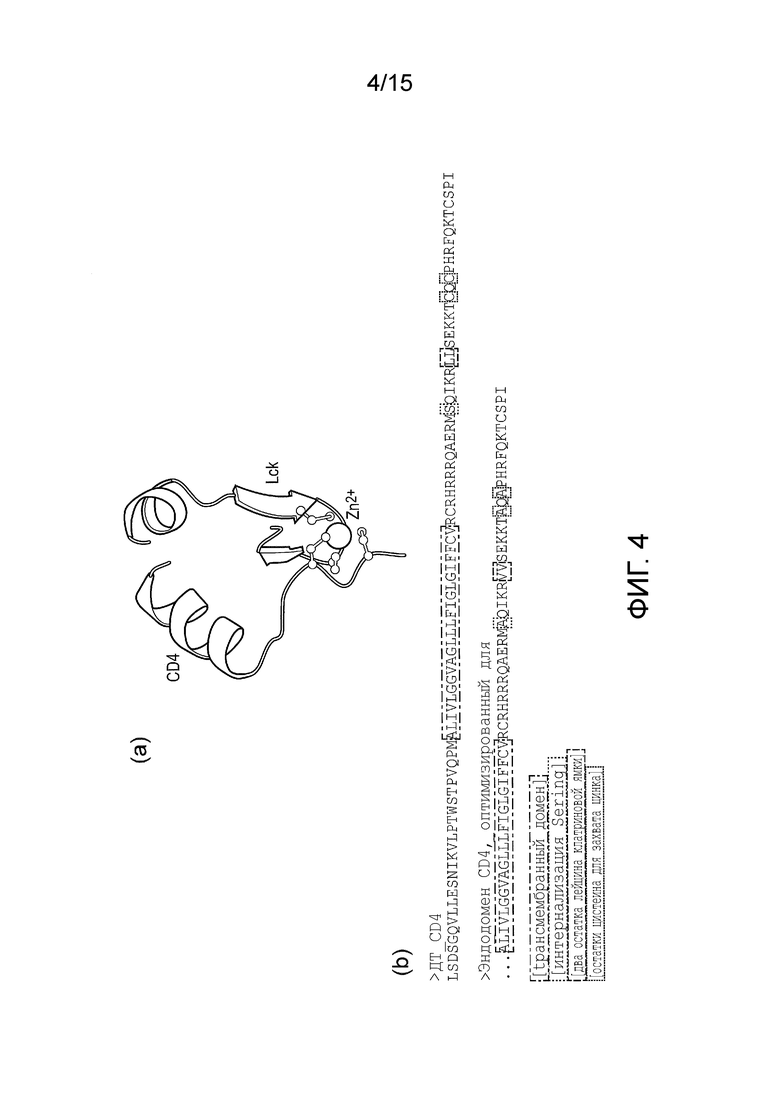
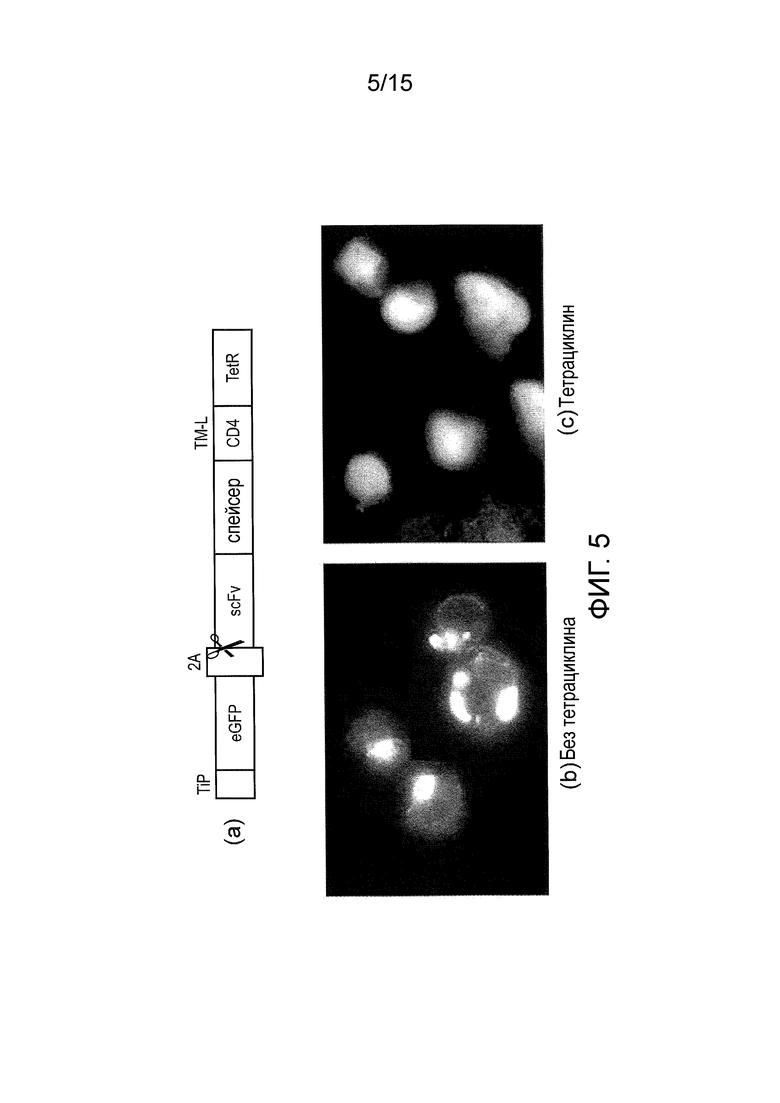
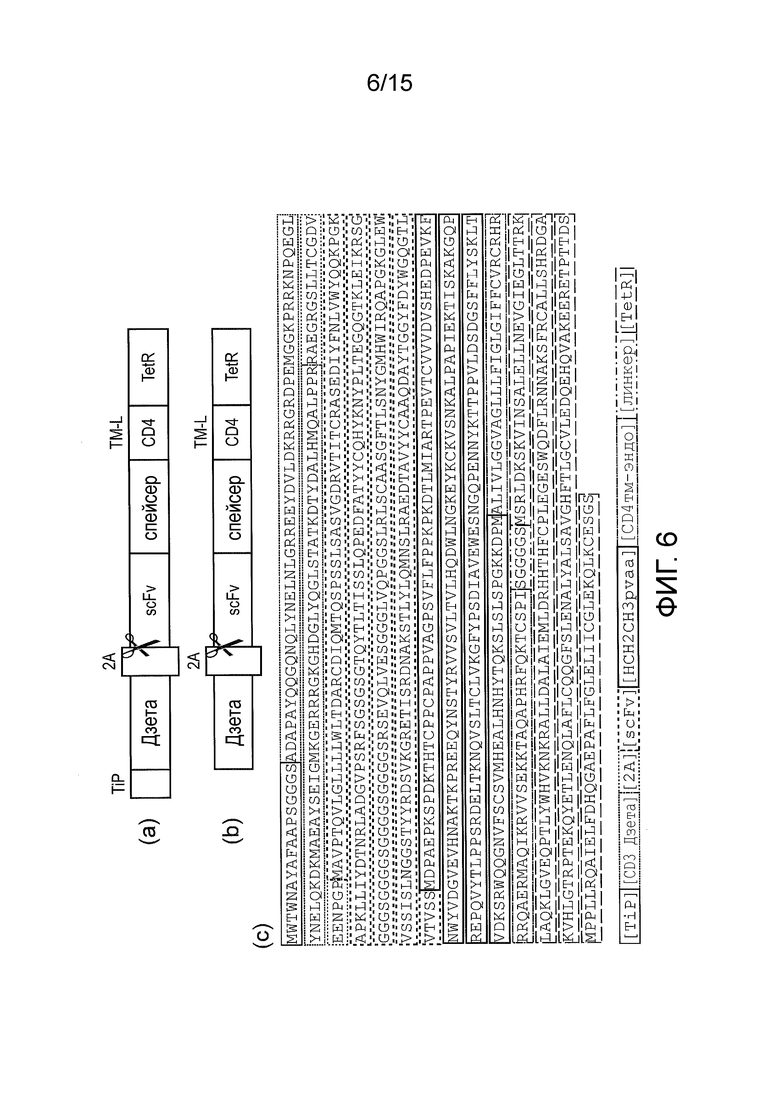
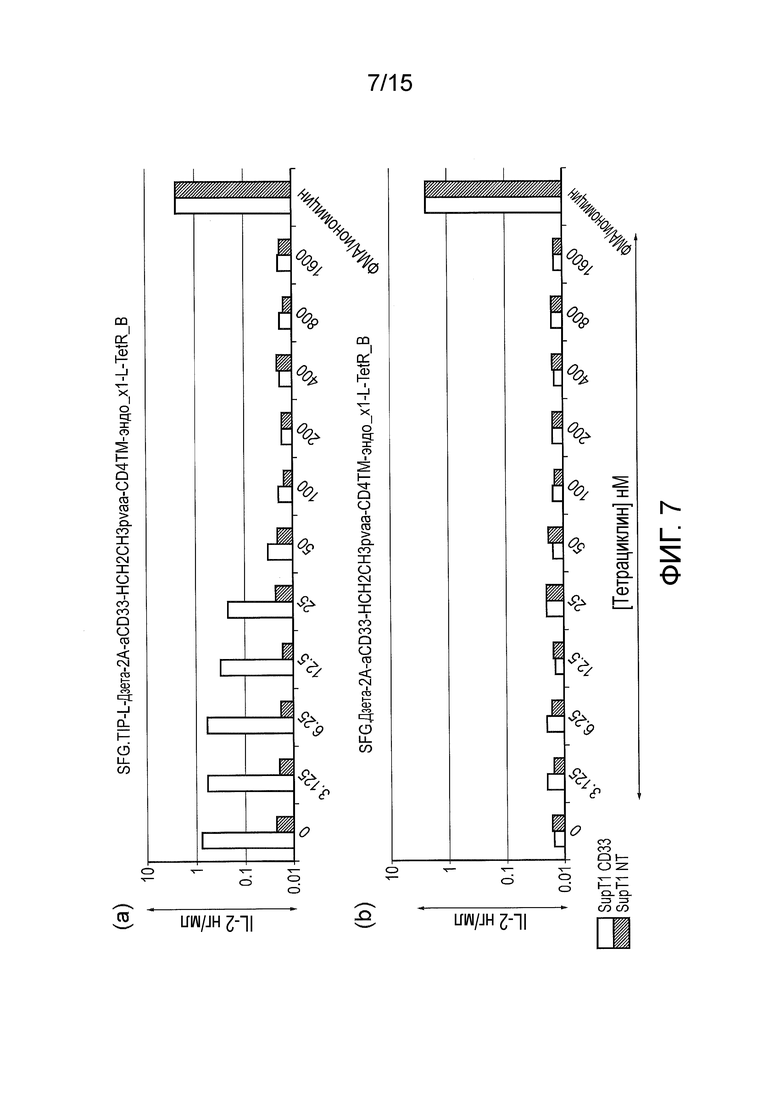
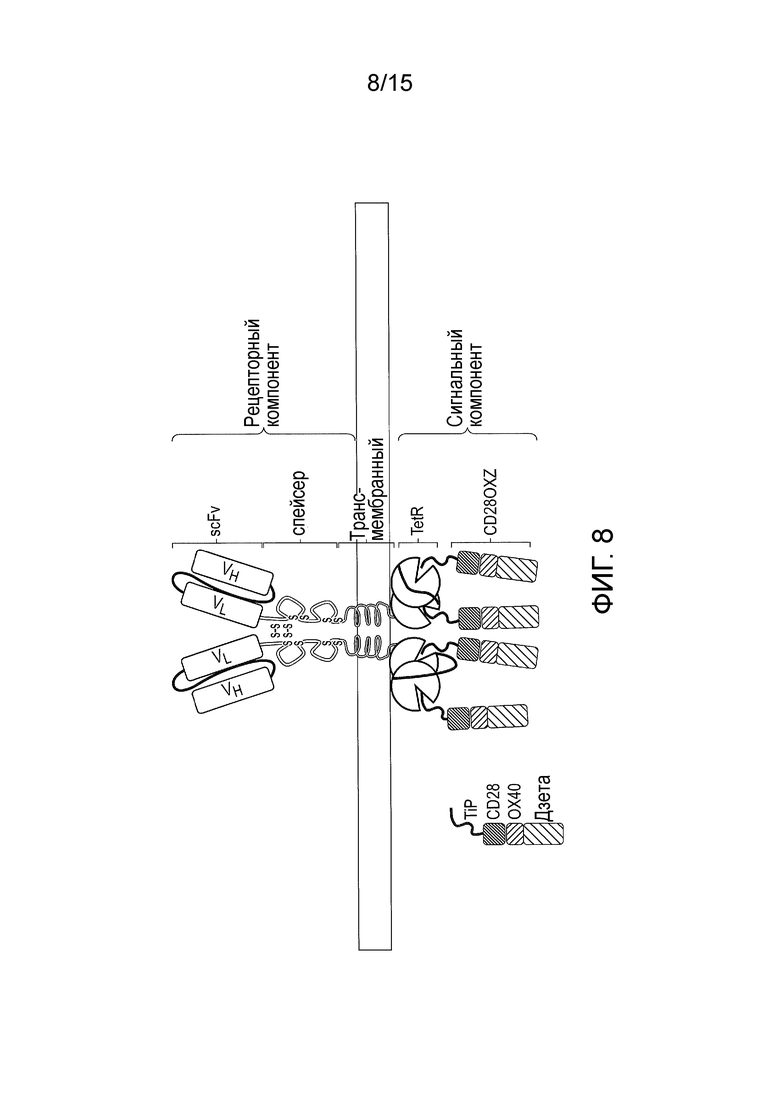
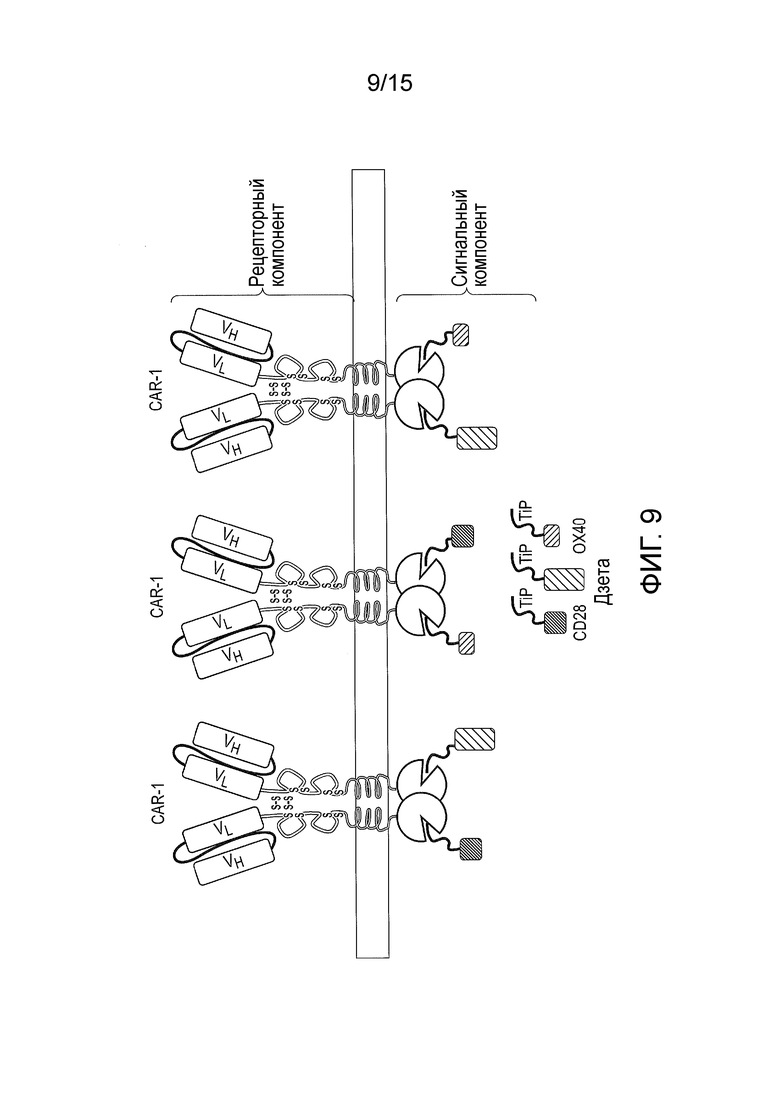
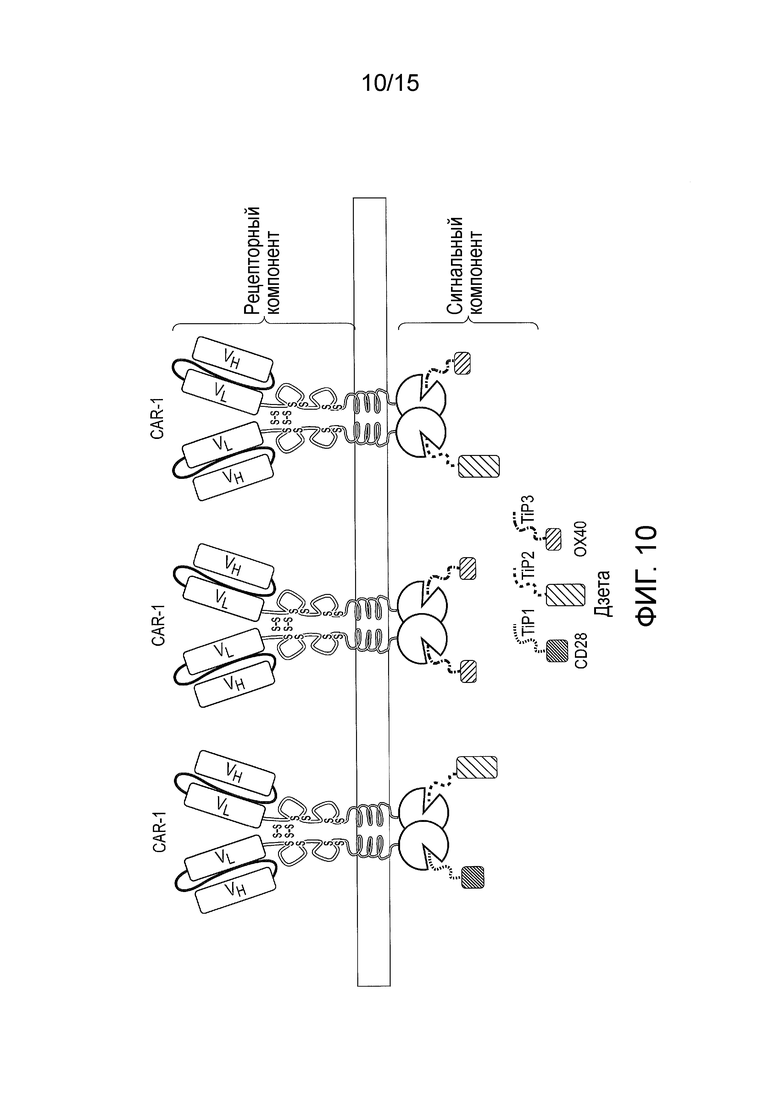
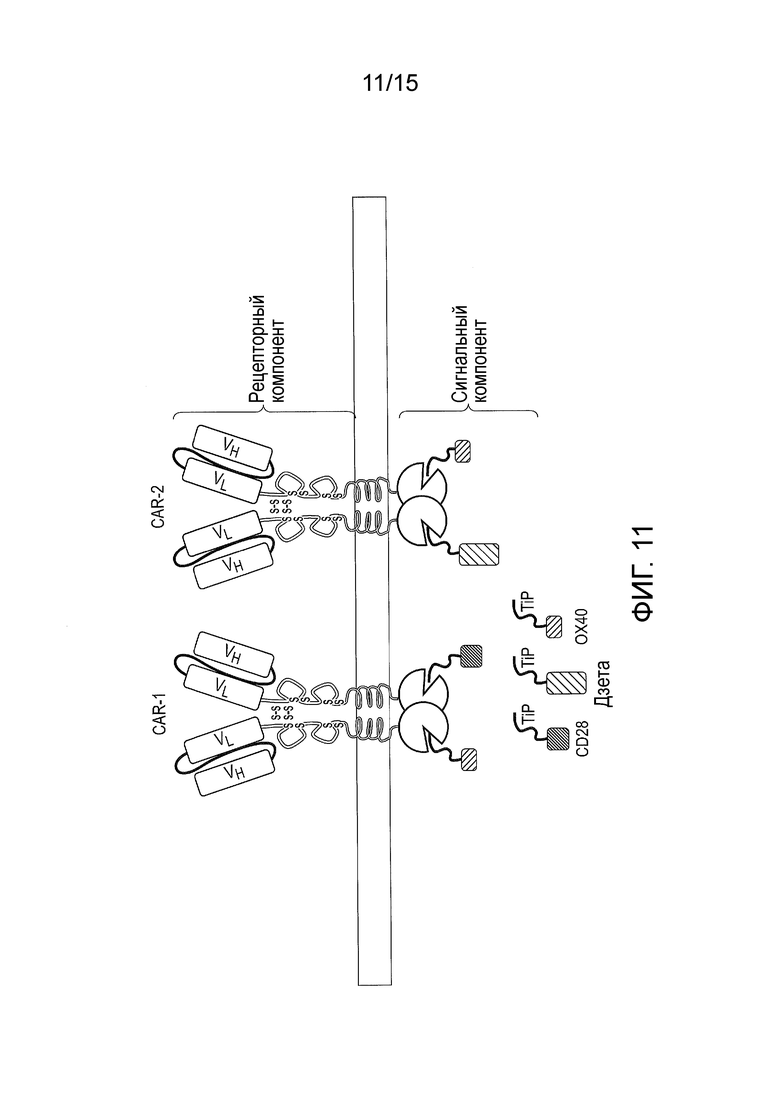
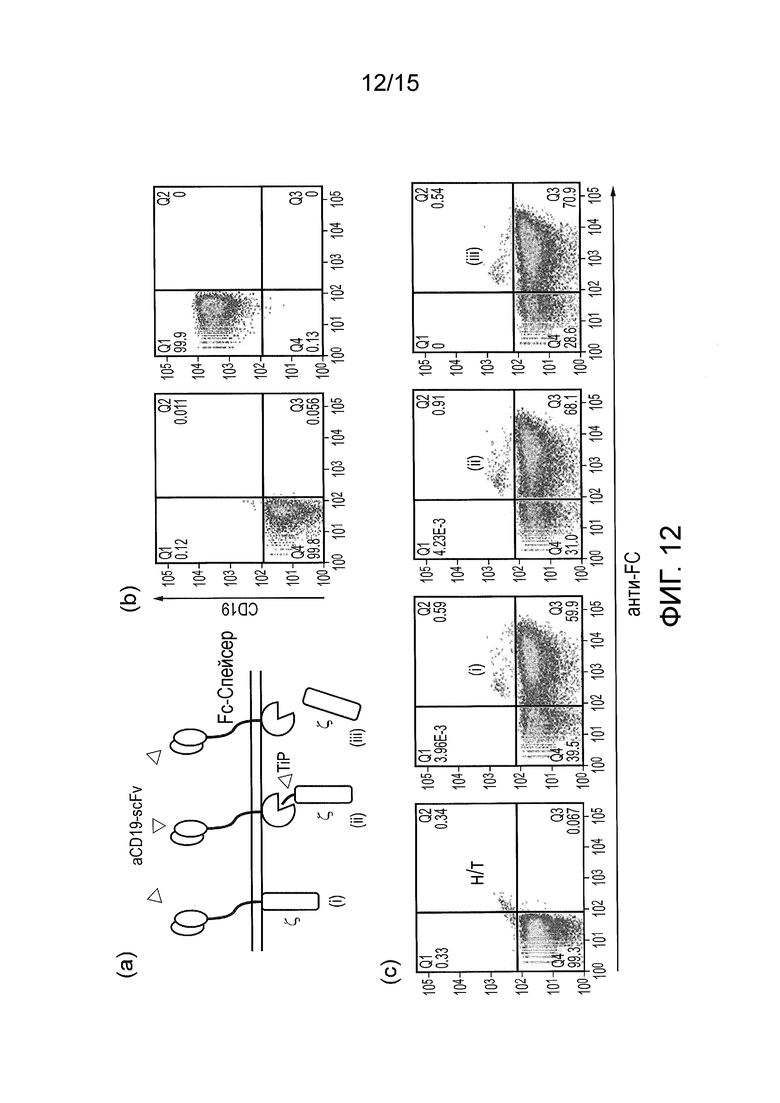
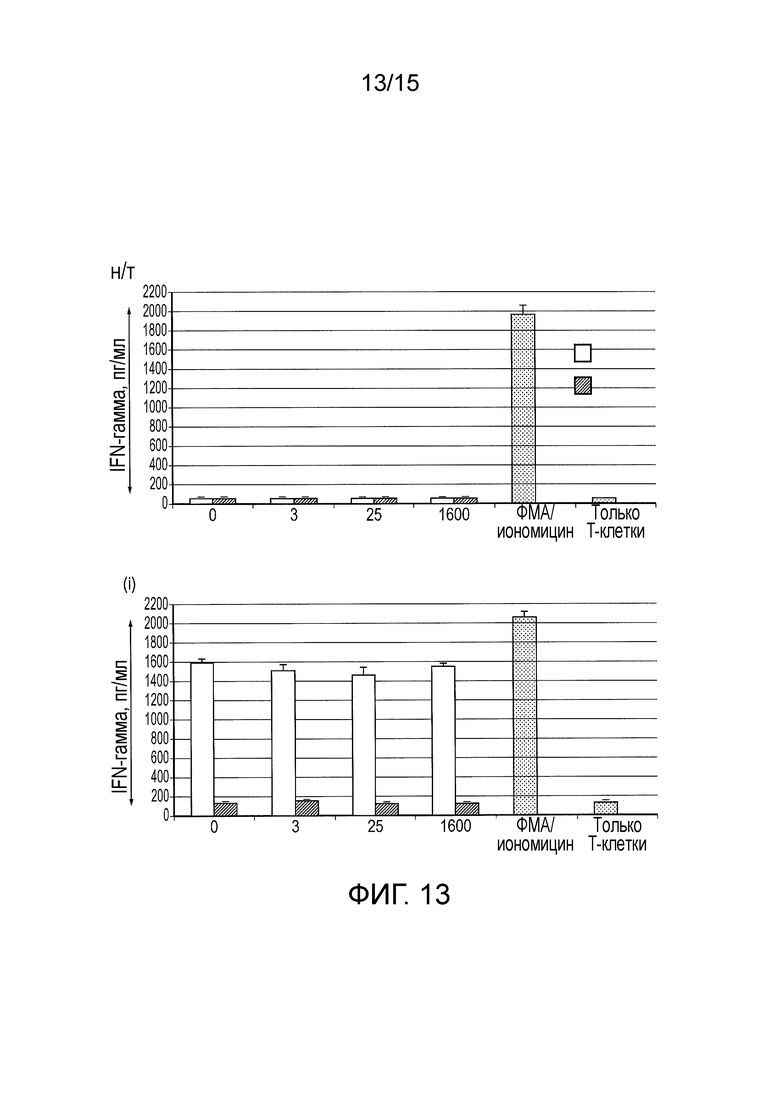
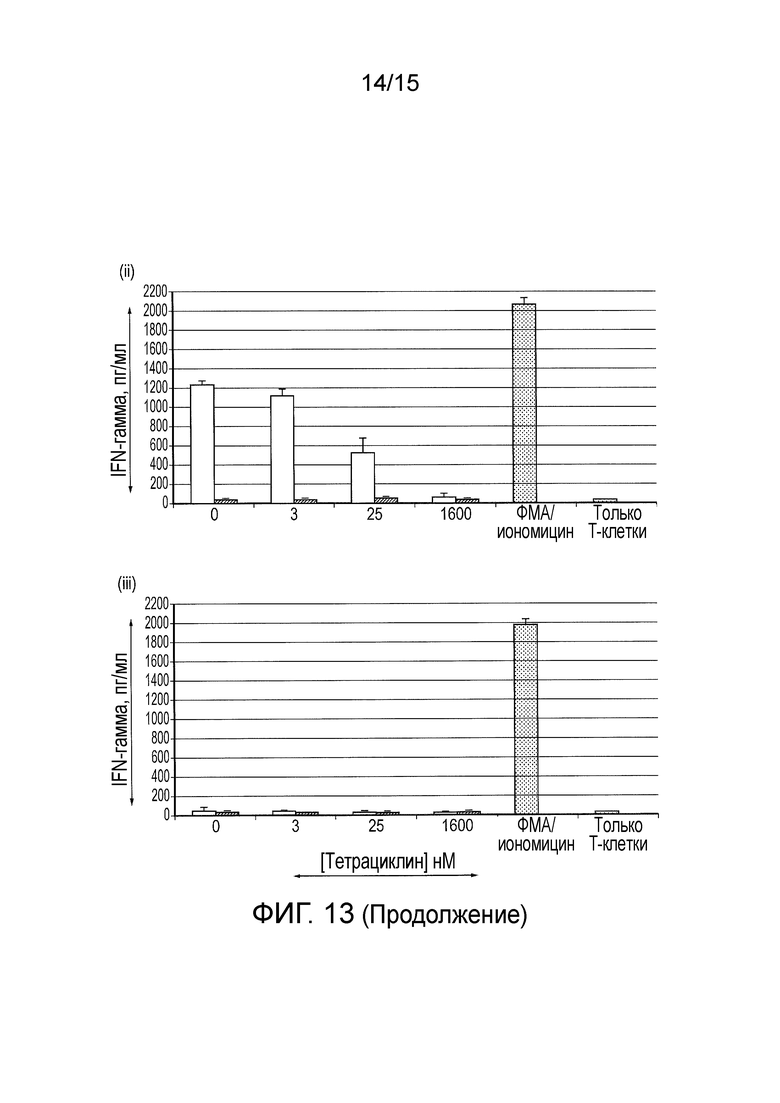
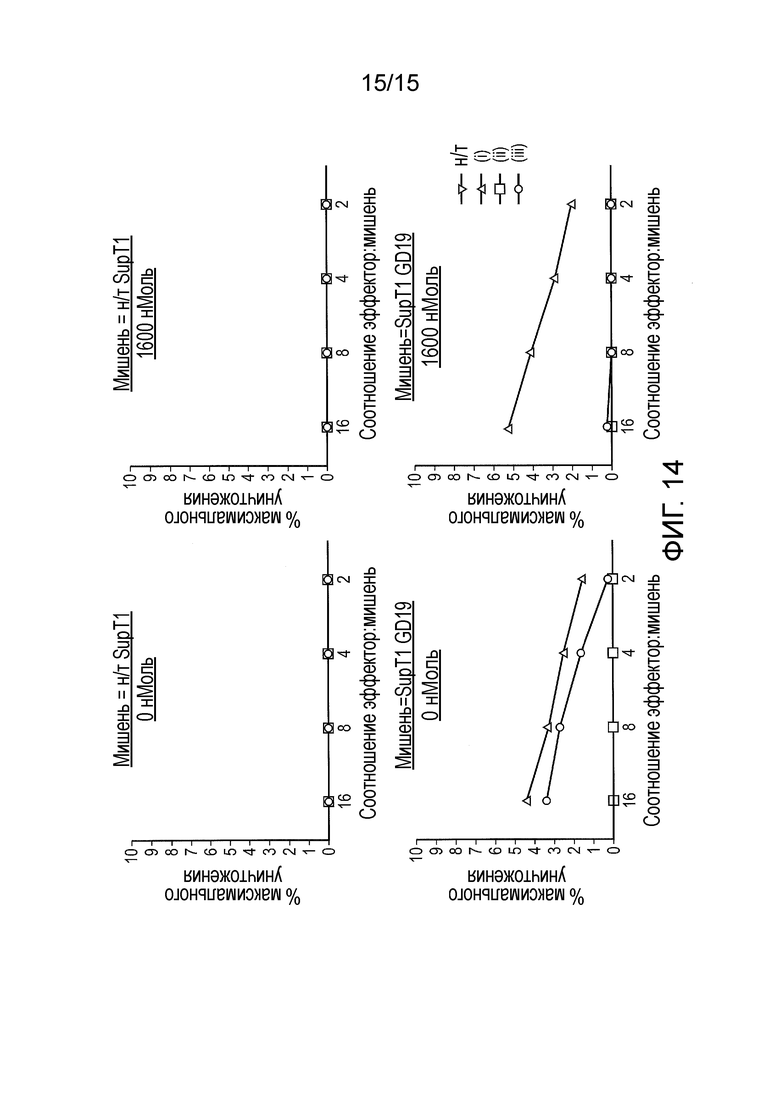
Настоящая группа изобретений относится к иммунологии. Предложена система химерного антигенного рецептора (CAR), которая включает рецепторный компонент, содержащий антигенсвязывающий домен, трансмембранный домен и первый связывающий домен; а также включает компонент внутриклеточного сигналинга, содержащий эндодомен CD3-дзета и второй связывающий домен, который специфически связывает первый связывающий домен, причем связывание первого и второго связывающих доменов может быть нарушено в присутствии определенного средства. Также представлена кодирующая CAR нуклеиновая кислота, экспрессионный вектор, Т- и NK-клетки, их применение, способ получения T- или NK-клетки и способ ингибирования сигнальной системы CAR. Данная группа изобретений может найти применение в производстве лекарственного средства для лечения и/или предотвращения раковых заболеваний. 8 н. и 20 з.п. ф-лы, 3 табл., 4 пр., 14 ил.
1. Система химерного антигенного рецептора (CAR), которая, при экспрессии клеткой, сообщает этой клетке способность уничтожать раковую клетку-мишень, которая экспрессирует антиген-мишень, где указанная система CAR включает:
(i) рецепторный компонент, содержащий антигенсвязывающий домен, который связывается с антигеном-мишенью, трансмембранный домен и первый связывающий домен; и
(ii) компонент внутриклеточного сигналинга, содержащий:
(a) сигнальный домен, содержащий эндодомен CD3-дзета, и (b) второй связывающий домен, который специфически связывает первый связывающий домен рецепторного компонента;
где первый связывающий домен или второй связывающий домен также способны связываться со средством, и где связывание первого или второго связывающего домена с указанным средством имеет большую аффинность, чем связывание первого связывающего домена со вторым связывающим доменом, так что связывание первого и второго связывающих доменов нарушается в присутствии средства таким образом, что в отсутствие средства рецепторный компонент и компонент внутриклеточного сигналинга гетеродимеризуются и связывание антигенсвязывающего домена с антигеном-мишенью приводит к сигнализации через сигнальный домен, в то время как в присутствии средства рецепторный компонент и компонент внутриклеточного сигналинга не гетеродимеризуются и связывание антигенсвязывающего домена с антигеном-мишенью не приводит к сигнализации через сигнальный домен.
2. Система CAR по п. 1, отличающаяся тем, что рецепторный компонент содержит линкер между трансмембранным доменом и первым связывающим доменом.
3. Система CAR по п. 2, отличающаяся тем, что линкер содержит или состоит из последовательности, приведенной в SEQ ID NO: 3.
4. Система CAR по любому из предшествующих пунктов, отличающаяся тем, что первый связывающий домен представляет собой репрессорный белок Tet (TetR) или его вариант, и второй связывающий домен представляет собой индуцирующий транскрипцию пептид (TiP) или его вариант; или отличающаяся тем, что первый связывающий домен представляет собой TiP или его вариант и второй связывающий домен представляет собой TetR или его вариант; и средство представляет собой тетрациклин, доксицилин или миноциклин, либо их аналоги.
5. Система CAR по п. 4, отличающаяся тем, что рецепторный компонент содержит два первых связывающих домена, которые представляют собой домены TetR.
6. Система CAR по п. 5, отличающаяся тем, что два домена TetR разделены линкером.
7. Система CAR по п. 5 или 6, отличающаяся тем, что домены TetR имеют разную аффинность для средства.
8. Система CAR по любому из предшествующих пунктов, отличающаяся тем, что в ней имеются множественные рецепторные компоненты, все из которых узнают разные антигены.
9. Система CAR по п. 8, отличающаяся тем, что связывающие домены множественных рецепторных компонентов отличаются по связыванию со связывающим доменом компонента внутриклеточного сигналинга таким образом, что каждый антиген вызывает разную силу сигнализации.
10. Система CAR по п. 8 или 9, отличающаяся тем, что связывающие домены множественных рецепторных компонентов отличаются по связыванию со средством таким образом, что каждый антиген вызывает разную силу сигнализации в присутствии средства.
11. Система CAR по любому из пп. 1-9, отличающаяся тем, что сигнальный домен компонента внутриклеточного сигналинга содержит эндодомен второго поколения, содержащий эндодомен CD3-дзета вместе с эндодоменом CD28, эндодоменом 4-1BB или эндодоменом OX40.
12. Система CAR по любому из предшествующих пунктов, отличающаяся тем, что сигнальный домен компонента внутриклеточного сигналинга содержит эндодомен третьего поколения, содержащий эндодомен CD3-дзета вместе с эндодоменом CD28 и эндодоменом 4-1BB; или эндодоменом CD28 и эндодоменом OX40.
13. Система CAR по любому из предшествующих пунктов, включающая несколько компонентов внутриклеточного сигналинга, каждый из которых содержит сигнальный домен и связывающий домен, где все связывающие домены узнают одинаковые связывающие домены рецепторного компонента, однако сигнальные домены представляют собой разные эндодомены.
14. Система CAR по п. 13, отличающаяся тем, что несколько компонентов внутриклеточного сигналинга содержат несколько связывающих доменов, каждый из которых независимо узнает связывающий домен рецепторного компонента с разной аффинностью.
15. Нуклеиновая кислота, кодирующая сигнальную систему CAR по любому из пп. 1-14, где рецепторный компонент и компонент внутриклеточного сигналинга совместно экспрессируются в виде саморасщепляющегося пептида, который расщепляется между рецепторным компонентом и компонентом внутриклеточного сигналинга после трансляции.
16. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 15.
17. Вектор по п. 16, который представляет собой ретровирусный вектор или лентивирусный вектор или транспозон.
18. T-клетка или NK-клетка, трансфицированные или трансдуцированные вектором по п. 16 или 17, экспрессирующие сигнальную систему CAR по любому из пп. 1-14.
19. T-клетка или NK-клетка по п. 18, содержащая нуклеиновую кислоту по п. 15 или вектор по п. 16 или 17.
20. T-клетка или NK-клетка по п. 18 для применения в лечении и/или предотвращении инфекции, патогенного иммунного ответа или ракового заболевания.
21. Способ лечения ракового заболевания, включающий этап введения субъекту множества T-клеток или NK-клеток по п. 18 или 19.
22. Способ по п. 21, включающий следующие этапы: (i) выделение образца, содержащего T-клетки или NK-клетки; (ii) трансдукцию или трансфекцию T- или NK-клеток нуклеиновой кислотой по п. 15 или вектором по п. 16 или 17; и (iii) введение T-клеток или NK-клеток по пункту (ii) субъекту.
23. Способ по п. 21 или 22, включающий мониторинг токсической активности у субъекта и включающий этап введения субъекту средства, используемого в сигнальной системе CAR по любому из пп. 1-14, для уменьшения неблагоприятных токсических эффектов.
24. Способ по п. 22 или 23, включающий мониторинг прогрессирования заболевания и/или мониторинг токсической активности у субъекта и включающий этап введения субъекту средства, используемого в сигнальной системе CAR по любому из пп. 1-14, для обеспечения приемлемых уровней прогрессирования заболевания и/или токсической активности.
25. Применение T-клетки или NK-клетки по п. 18 или 19 в производстве лекарственного средства для лечения ракового заболевания.
26. Способ получения T- или NK-клетки по п. 18 или 19, включающий этап введения нуклеиновой кислоты по п. 15 или вектора по п. 16 или 17 в T- или NK-клетку.
27. Способ по п. 26, отличающийся тем, что T- или NK-клетка представляет собой клетку из образца, полученного от субъекта.
28. Способ ингибирования сигнальной системы CAR по любому из пп. 1-14 у субъекта, в организме которого присутствует T- или NK-клетка по п. 18 или 19, включающий этап введения указанного средства субъекту.
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| KLOTZSCHE M | |||
| et al | |||
| "Efficient and exclusive induction of Tet repressor by the oligopeptide Tip results from co-variation of their interaction site." Nucleic acids research, 2007, 35(12): 3945-3952 | |||
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| FEDOROV V.D | |||
| et al., "PD-1-and CTLA-4-based inhibitory chimeric antigen receptors | |||

