ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к химерному цитокиновому рецептору (CCR) и клетке, которая экспрессирует такой химерный цитокиновый рецептор, и, дополнительно, химерному антигенному рецептору на клеточной поверхности.
УРОВЕНЬ ТЕХНИКИ
Химерные антигенные рецепторы (CARs)
Описан ряд иммунотерапевтических агентов для применения при лечении рака, включая терапевтические моноклональные антитела (mAbs), привлекающие T-клетки би-специфические активаторы и химерные антигенные рецепторы (CARs).
Химерные антигенные рецепторы представляют собой белки, которые прививают специфичность моноклонального антитела (mAb) к эффекторной функции Т-клетки. Их обычной формой является форма белка трансмембранного домена 1 типа с антиген-распознающим аминоконцом, спейсером, трансмембранным доменом, все они соединены с составным эндодоменом, который передает Т-клеточные сигналы выживаемости и активации.
Наиболее распространенной формой этих молекул являются слияния одноцепочечных вариабельных фрагментов (scFv), полученных из моноклональных антител, которые распознают целевой антиген, связанные с помощью спейсера и трансмембранного домена с эндодоменом сигнализации. Такие молекулы приводят к активации Т-клетки в ответ на распознавание scFv ее мишени. Когда Т-клетки экспрессируют такой CAR, они распознают и уничтожают клетки-мишени, которые экспрессируют целевой антиген. Было разработано несколько CARs против связанных с опухолью антигенов, и подходы адаптивного переноса с использованием таких Т-клеток, экспрессирующих CAR, в настоящее время находятся в клинических испытаниях для лечения различных видов рака.
Подходы к лечению рака предстательной железы, основанные на CAR
Рак предстательной железы является вторым наиболее распространенным раком у мужчин во всем мире и шестой по значимости причиной смерти от рака. В глобальном масштабе ежегодно насчитывается около 1,100,000 новых случаев заболевания и 300,000 смертей, составляющие 4% всех смертей от рака. Согласно оценкам, 1 из каждых 6 мужчин будет диагностирован с такой болезнью в течение своей жизни.
Первоначальное лечение рака предстательной железы может состоять из хирургического вмешательства, облучения или гормональной терапии, или любой комбинации каждого из них. Гормональная терапия заключается в снижении уровня тестостерона, мужского гормона, который подпитывает неконтролируемый рост клеток. Химиотерапия, как правило, предназначена для распространенных злокачественных опухолей.
Когда увеличивается рак предстательной железы, несмотря на снижение уровня тестостерона с помощью гормональной терапии, возможности лечения ограничены. Как правило, противораковая вакцина «sipuleucel-T» (Provenge®) - терапевтическая противораковая вакцина на основе, предназначенная для индуцирования иммунного ответа, направленного дендритных клеток против антигена простатической кислой фосфатазы ((PAP)), радиофармпрепарат (такой как хлорид радия-223), вторичная гормональная терапия (например, абиратерон или энзалутамид) и/или химиотерапия (доцетаксел и кабазитаксел) добавляются к гормональной терапии последовательно. Хотя каждый из этих методов лечения может замедлять рост рака в течение нескольких месяцев и ослаблять симптомы, вызванные этим заболеванием, болезнь, в конечном итоге, становится устойчивой к ним.
Доклинически, два антигена, связанные с раком предстательной железы, были нацелены на терапии, основанные на CAR-Т-клетках: простатоспецифический мембранный антиген (PSMA) и антиген стволовых клеток предстательной железы (PSCA).
Мыши, обработанные PSCA-CAR-сконструированными Т-клетками, показали замедленный рост опухоли (Hillerdal et al (2014) BMC Cancer 14:30; and Abate-Daga et al (2014) 25:1003-1012). Хотя клетки показали высокую цитотоксичность in vitro, рост опухоли in vivo задерживался, но опухоленесущие мыши не были вылечены.
Это может быть связано с тем, что CAR-Т-клетки in vivo стремятся преодолеть микроокружение злокачественной карциномы. В частности, CAR-Т-клетки могут быть не способными к внедрению и распространению в пределах ложа опухоли рака предстательной железы.
Жизнеспособность и активность CAR-Т-клеток может быть усилена путем введения цитокинов или CAR-Т-клеток, продуцирующих цитокины конститутивно. Однако эти подходы имеют ограничения: системное введение цитокинов может быть токсичным; конститутивное продуцирование цитокинов может привести к неконтролируемой пролиферации и трансформации (Nagarkatti et al (1994) PNAS 91:7638-7642; Hassuneh et al (1997) Blood 89:610-620).
Следовательно, существует потребность в альтернативных CAR-Т-клеточных подходах, которые способствуют приживлению и экспансии Т-клеток для противодействия комплексному воздействию вредоносного микроокружения опухоли.
Токсичность на мишень вне опухоли
Это сравнительно редко присутствие одного антигена эффективно для описания рака, что может привести к недостаточной специфичности.
Большинство видов рака нельзя отличить от нормальных тканей на основе одного антигена. Следовательно, возникает значительная токсичность «на мишень вне опухоли», когда нормальные ткани повреждаются терапией. Например, при нацеливании CD20 на лечение В-клеточной лимфомы ритуксимабом исчерпан весь пул нормальных В-клеток, а при нацеливании CD52 для лечения хронического лимфоцитарного лейкоза истощен весь лимфатический пул, при нацеливании CD33 для лечения острого миелоидного лейкоза поврежден весь миелоидный пул и т.д.
Прогнозируемая проблема токсичности «на мишень вне опухоли» подтверждена клиническими испытаниями. Например, подход, нацеленный на ERBB2, вызвал смерть пациента с метастатическим раком толстой кишки в легкие и печень. ERBB2 избыточно экспрессируется при раке толстой кишки у некоторых пациентов, но он также экспрессируется в нескольких здоровых тканях, включая сердце и здоровую сосудистую систему.
Следовательно, существует потребность в улучшенных подходах к терапии рака, при которых такая токсичность «на мишень вне опухоли» уменьшается или устраняется.
ОПИСАНИЕ ФИГУР
Фигура 1: Схематическая диаграмма, суммирующая структуру различных цитокиновых рецепторов, типы клеток, которые продуцируют цитокины, и типы клеток, которые экспрессируют цитокиновые рецепторы.
Фигура 2: Схематическая диаграмма, показывающая предполагаемый химерный цитокиновый рецептор
(a) Цитокин IL2 и IL7-цитокиновые рецепторы сигнализируют через общую гамма-цепь и цитокиновую специфическую альфа/бета-цепь.
(b) Одно осуществление химерного цитокинового рецептора заключается в замене эктодомена цитокиновой альфа/бета- и гамма-цепи различными scFvs (или любым другим подходящим связующим), которые распознают различные эпитопы PSA.
(c) Альтернативный подход заключается в замене эктодоменов альфа/бета и гамма VH/VL PSA-специфического антитела, где VH и VL участвуют в связывании, так что связывание объединяет их.
Фигура 3: Усилитель сигнализации цитокинов на основе агрегации.
Схематическая диаграмма, показывающая химерный цитокиновый рецептор и CAR-комбинирующую систему. Клетка содержит два химерных цитокиновых рецептора, которые связывают различные эпитопы с одним и тем же растворимым лигандом. В отсутствие растворимого лиганда (например, PSA), но в присутствии антигена клеточной поверхности (например, PSMA) сигнализация происходит посредством CAR. В присутствии растворимого лиганда происходит агрегация двух химерных цитокиновых рецепторов, что приводит к усилению сигнала на основе цитокинов.
Фигура 4: Теоретическая карта конструкции для химерного цитокинового рецептора/CAR-комбинирующей системы, показанной на Фиг. 3.
Фигура 5: Схематическая диаграмма, иллюстрирующая пример структуры для химерного трансмембранного белка по настоящему изобретению. Химерный трансмембранный белок включает домен димеризации и эндодомен цитокинового рецептора. Показанный вариант осуществления имеет структуру типа «Fab», поскольку домен димеризации содержит константные области тяжелой и легкой цепи антитела. Константная димеризация между данными доменами объединяет общую γ-цепь рецептора IL2 либо с β-цепью рецептора IL-2, либо с α-цепью рецептора IL-7, что приводит к конститутивной цитокиновой сигнализации.
Фигура 6: IL-2 сигнализация химерным трансмембранным белком.
Два химерных трансмембранных белка, имеющих общую структуру, показанную на Фиг. 5, тестировали на их способность индуцировать сигнализацию IL-2. Один химерный трансмембранный белок содержал эндодомен рецептора IL2, а другой содержал эндодомен рецептора IL-7. Сигнализация IL-2 была протестирована с использованием мышиной клеточной линии CTLL2, рост которой зависит от сигнализации IL-2. В качестве положительного контроля клетки CTLL2 культивировали с 100 ед/мл мышиного IL2. Клетки, экспрессирующие химерный трансмембранный белок, содержащий эндодомен рецептора IL2 (Fab_IL2эндо), поддерживали выживаемость и рост клеток CTLL2, тогда как клетки, экспрессирующие химерный трансмембранный белок, содержащий рецептор IL-7 (Fab_IL7эндо), не делали этого.
Фигура 7: Схематическая диаграмма, показывающая группу PSA химерных цитокиновых рецепторов
Группа химерных цитокиновых рецепторов (CCRs), нацеленная на PSA, была разработана с использованием scFvs, полученных из двух антител, которые связываются с различными эпитопами PSA: 5D5A5 и 5D3D11.
Верхняя левая группа: CCR с эндодоменом IL-2R, имеющим A5 на цепи с β-цепью IL2R и D11 на цепи с общей γ-цепью;
Верхняя правая группа: CCR с эндодоменом IL7R, имеющим A5 на цепи с α-цепью IL7R и D11 на цепи с общей γ-цепью;
Нижняя левая группа: CCR с эндодоменом IL-2R, имеющим D11 на цепи с β-цепью IL2R и A5 на цепи с общей γ-цепью; и
Нижняя правая группа: CCR с эндодоменом IL-7R, имеющим D11 в цепи с α-цепью IL7R и A5 на цепи с общей γ-цепью.
Отрицательный контроль был также создан для каждого CCR, в котором цепь IL2Rγ была заменена жестким линкером.
Фигура 8: Сигнализация IL2 из клеток, экспрессирующих PSA химерные цитокиновые рецепторы, в присутствии PSA - пролиферация CTLL2.
Клетки CTLL2 трансдуцировали с помощью конструкций, экспрессирующих некоторые из PSA рецепторов, показанных на Фиг. 7. Клетки культивировали в отсутствие и в присутствии IL2 (положительный контроль), а также в отсутствие и присутствии 5 нг/мл или 5 мкг/мл PSA. Пролиферацию CTLL2 оценивали через 3 и 7 д.
Анти-PSA химерный цитокиновый рецептор с эндодоменом IL2 поддерживал пролиферацию клеток CTLL2 в отсутствие IL2 и в присутствии PSA, но не рецептор, имеющий эндодомен IL7R, или любые из CCRs, содержащие жесткий линкер в месте общей γ-цепи.
Фигура 9: Сигнализация IL2 из клеток, экспрессирующих PSA химерный цитокиновый рецептор в присутствии PSA - CTLL2 STAT5-фосфорилирование
Клетки CTLL2 либо оставляли нетрансдуцированными (WT); или трансдуцировали вектором, экспрессирующим CCR против PSA (D11-CD8STK-IL2Rg_A5-Шарнир-IL2Rb) или эквивалентной конструкцией, имеющей жесткий линкер вместо общей γ-цепи (D11-CD8STK-RL_A5-Шарнир-IL2Rb). Клетки инкубировали с 500 мкМ перванадата или 500 нг/мл PSA в течение 1 или 4 ч. Затем фосфорилирование Y694 STAT5 исследовали с использованием проточной цитометрии.
СУЩНОСТЬ АСПЕКТОВ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали «химерные цитокиновые рецепторы» (CCR), которые прививают связывающую специфичность связывающей молекулы, отличной от цитокина, эндодомену цитокинового рецептора. Коэкспрессия такого CCR с химерным антигенным рецептором (CAR) помогает CAR-Т-клетке внедряться и распространяться в микроокружении злокачественной опухоли. Требование, которое должно присутствовать к лиганду для CCR, а также к лиганду для CAR, добавляет еще один уровень избирательного подхода и помогает предотвратить токсичность на мишень вне опухоли.
Например, была разработана клетка, которая со-экспрессирует CAR с химерным цитокиновым рецептором, который обнаруживает PSA и передает IL2/15 или IL7 сигнал CAR-Т-клетке. Таким образом, CAR-Т-клетка избирательно стимулируется для пролиферации только в микроокружении рака предстательной железы, и, в отсутствие PSA (т.е. после того, как пациент находится в состоянии ремиссии), цитокиновая стимуляция теряется.
В первом аспекте настоящее изобретение относится к химерному цитокиновому рецептору (CCR), содержащему:
экзодомен, который связывается с лигандом, выбранным из фактора, секретируемого опухолью, хемокина и антигена клеточной поверхности; а также
эндодомен цитокинового рецептора.
В первом варианте осуществления первого аспекта изобретения химерный цитокиновый рецептор включает два полипептида:
(i) первый полипептид, который включает:
(а) первый антигенсвязывающий домен, который связывает первый эпитоп лиганда
(b) первую цепь эндодомена цитокинового рецептора; а также
(ii) второй полипептид, который включает:
(а) второй антигенсвязывающий домен, который связывает второй эпитоп лиганда
(b) вторую цепь эндодомена цитокинового рецептора. На Фиг. 2b показана такая схема.
Каждый из первого и второго антигенсвязывающих доменов может быть, например, одноцепочечными вариабельными фрагментами (scFvs) или однодоменными связующими.
Во втором варианте осуществления первого аспекта изобретения химерный цитокиновый рецептор, который включает два полипептида:
(i) первый полипептид, который включает:
(a) вариабельный домен тяжелой цепи (VH)
(b) первую цепь эндодомена цитокинового рецептора; а также
(ii) второй полипептид, который включает:
(a) вариабельный домен легкой цепи (VL)
(b) вторую цепь эндодомена цитокинового рецептора.
На Фиг. 2с показана такая схема.
Первая и вторая цепи для эндодоменов цитокинового рецептора могут быть различными и могут быть выбраны из эндодомена α-, β- и γ-цепей цитокинового рецептора I типа.
В альтернативном варианте, первая и вторая цепи для эндодоменов цитокинового рецептора могут быть одинаковыми, и могут быть выбраны из эндодомена α-, β- и γ-цепей цитокинового рецептора I типа.
Например, эндодомен цитокинового рецептора может включать:
(i) эндодомен β-цепи рецептора IL-2
(ii) эндодомен α-цепи рецептора IL-7;
(iii) эндодомен α-цепи рецептора IL-15; или
(iv) общий эндодомен γ-цепи рецептора.
Эндодомен цитокинового рецептора может включать (i), (ii) или (iii); и (iv).
Лиганд может представлять собой секретируемый опухолью фактор, например, секретируемый опухолью фактор, выбранный из: простатоспецифического антигена (PSA), карциноэмбрионального антигена (СЕА), фактора роста эндотелия сосудов (VEGF) и СА125.
Лиганд может быть хемокином, например, хемокином, выбранным из хемокина, выбранного из: CXCL12, CCL2, CCL4, CCL5 и CCL22.
Лиганд может быть молекулой клеточной поверхности, такой как трансмембранный белок. Лигандом может быть, например, CD22.
Во втором аспекте настоящее изобретение относится к клетке, которая содержит химерный цитокиновый рецептор, в соответствии с первым аспектом изобретения.
Клетка может содержать первый химерный цитокиновый рецептор и второй химерный цитокиновый рецептор, которые связывают различные эпитопы на одном лиганде.
Клетка может содержать первый химерный цитокиновый рецептор, который включает эндодомен α- или β-цепи цитокинового рецептора I типа, и второй химерный цитокиновый рецептор, который включает в себя эндодомен γ-цепи цитокинового рецептора I типа, так что, когда первый химерный цитокиновый рецептор и второй химерный цитокиновый рецептор связывают лиганд, происходит комбинированная сигнализация через α-/β-цепь и γ-цепь.
Клетка может также содержать химерный антигенный рецептор, например, химерный антигенный рецептор, который связывает ассоциированный с опухолью антиген клеточной поверхности.
Химерный антигенный рецептор может связывать антиген клеточной поверхности, ассоциированный с раком предстательной железы, такой как антиген стволовых клеток предстательной железы (PSCA) или простатоспецифический мембранный антиген (PSMA).
Если CCR распознает поверхностный клеточный антиген, CCR и CAR могут распознавать антигены клеточной поверхности, которые коэкспрессируются на одной и той же целевой (например, опухолевой) клетке. Например, для В-клеточных злокачественных опухолей CAR может распознавать антиген клеточной поверхности, такой как CD19, и CCR может распознавать молекулу, которая со-экспрессируется на поверхности клетки-мишени, такая как CD22, тем самым усиливая приживление.
В третьем аспекте настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей химерный цитокиновый рецептор (CCR), в соответствии с первым аспектом изобретения.
В четвертом аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, которая содержит первую последовательность нуклеиновой кислоты, кодирующую первый CCR, и вторую последовательность нуклеиновой кислоты, кодирующую второй CCR, конструкцию нуклеиновой кислоты, имеющую структуру:
AgB1-спейсер1-TM1-эндо1-коэкспр-AbB2-спейсер2-ТМ2-эндо2
в которой
AgB1 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен первого CCR;
спейсер 1 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер первого CCR;
TM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен первого CCR;
эндо 1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен первого CCR;
коэкспр представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию обоих CCRs;
AgB2 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен второго CCR;
спейсер 2 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер второго CCR;
TM2 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен второго CCR;
эндо 2 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен второго CCR.
Конструкция нуклеиновой кислоты также может кодировать химерный антигенный рецептор (CAR). В данном варианте осуществления изобретения конструкция нуклеиновой кислоты может иметь структуру:
(i) CCRAgB1-CCRспейсер1-CCRTM1-CCRэндо1-коэкспр1-CCRAgB2-CCRспейсер2-CCRTM2-CCRэндо2-коэкспр2-CARAgB-CARспейсер-CARTM-CARэндо;
(ii) CCRAgB1-CCRспейсер1-CCRTM1-CCRэндо1-коэкспр1-CARAgB-CARспейсер-CARTM-CARэндо-коэкспр2-CCRAgB2-CCRспейсер2-CCRTM2-CCRendo2; или
(iii) CARAgB-CARспейсер-CARTM-CARэндо-коэкспр1-CCRAgB1-CCRспейсер1-CCRTM1-CCRэндо1-коэкспр2-CCRAgB2-CCRспейсер2-CCRTM2-CCRэндо2;
в котором
CCRAgB1 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен первого CCR;
CCRспейсер1 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер первого CCR;
CCRTM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен первого CCR;
CCRэндо1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен первого CCR;
CCRAgB2 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен второго CCR;
CCRспейсер2 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер второго CCR;
CCRTM2 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен второго CCR;
CCRэндо2 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен второго CCR;
Коэкспр1 и коэкспр2 представляют собой последовательности нуклеиновой кислоты, обеспечивающие коэкспрессию двух фланкирующих последовательностей;
CARAgB представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен CAR;
CARспейсер представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер CAR;
CARTM представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен CAR; а также
CARэндо представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен CAR.
Любая или все последовательности коэкспр, коэкспр1, коэкспр2 могут кодировать последовательность, содержащую саморасщепляющийся пептид.
Альтернативные кодоны могут быть использованы в областях последовательности, кодирующих одни и те же или подобные аминокислотные последовательности, чтобы избежать гомологической рекомбинации.
В пятом аспекте настоящее изобретение относится к вектору, содержащему конструкцию нуклеиновой кислоты, в соответствии с четвертым аспектом изобретения.
Вектор может быть, например, ретровирусным вектором или лентивирусным вектором, или транспозоном.
В шестом аспекте настоящее изобретение относится к набору, который содержит:
i) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую первый CCR, в соответствии с первым аспектом изобретения; а также
ii) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую второй CCR, в соответствии с вторым аспектом изобретения.
Набор может также содержать вектор, содержащий последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор.
Набор может содержать:
i) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую CCR, в соответствии с первым аспектом изобретения; а также
ii) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор.
В седьмом аспекте настоящее изобретение относится к способу изготовления клетки, в соответствии с вторым аспектом изобретения, который включает стадию введения: последовательности нуклеиновой кислоты, в соответствии с третьим аспектом изобретения; конструкции нуклеиновой кислоты, в соответствии с четвертым аспектом изобретения; вектора, в соответствии с пятым аспектом изобретения; или набора векторов, в соответствии с шестым аспектом изобретения, в клетку.
Клетка может быть из образца, полученного от субъекта.
В восьмом аспекте предлагается фармацевтическая композиция, содержащая множество клеток, в соответствии со вторым аспектом изобретения.
В девятом аспекте предлагается способ лечения и/или профилактики заболевания, который включает стадию введения фармацевтической композиции, в соответствии с восьмым аспектом изобретения, субъекту.
Способ может содержать следующие этапы:
(i) выделение образца, содержащего клетки, от субъекта;
(ii) трансдукцию или трансфекцию клеток с помощью последовательности нуклеиновой кислоты, в соответствии с третьим аспектом изобретения; конструкцию нуклеиновой кислоты, в соответствии с четвертым аспектом изобретения; вектор, в соответствии с пятым аспектом изобретения; или набор векторов в соответствии с шестым аспектом изобретения; а также
(iii) введение клеток из (ii) субъекту.
Образец может быть образцом, содержащим Т-клетки.
Болезнь может быть раком.
Также предлагается фармацевтическая композиция, в соответствии с восьмым аспектом изобретения, для использования при лечении и/или профилактике заболевания.
Также предусматривается применение клетки, в соответствии со вторым аспектом изобретения, при изготовлении лекарственного средства для лечения и/или профилактики заболевания.
Дальнейшие аспекты изобретения обобщены в следующих пронумерованных пунктах:
1. Химерный трансмембранный белок, содержащий:
домен димеризации; а также
эндодомен цитокинового рецептора.
2. Химерный трансмембранный белок, в соответствии с п. 1, где домен димеризации содержит участок димеризации константного домена тяжелой цепи (CH) и константного домена легкой цепи (CL).
3. Химерный трансмембранный белок, в соответствии с любым из предшествующих пунктов, который включает два полипептида:
(i) первый полипептид, который включает:
(а) первый домен димеризации; а также
(b) первую цепь эндодомена цитокинового рецептора; а также
(ii) второй полипептид, который включает:
(а) второй домен димеризации, который димеризуется с первым доменом димеризации; а также
(b) вторую цепь эндодомена цитокинового рецептора.
4. Химерный трансмембранный белок, в соответствии с п. 3, где первый и второй участки димеризации либо димеризуются спонтанно, либо в присутствии химического индуктора димеризации (CID).
5. Химерный трансмембранный белок, в соответствии с п. 2, 3 или 4, который содержит два полипептида:
(i) первый полипептид, который включает:
(a) константный домен тяжелой цепи (CH)
(b) первую цепь эндодомена цитокинового рецептора; а также
(ii) второй полипептид, который включает:
(a) константный домен легкой цепи (CL)
(b) вторую цепь эндодомена цитокинового рецептора.
6. Химерный трансмембранный белок, в соответствии с п. 5, где первая и вторая цепи для эндодоменов цитокинового рецептора различны и выбраны из эндодомена α-, β-, γ-цепей цитокинового рецептора I типа.
7. Химерный трансмембранный белок, в соответствии с п. 5, где первая и вторая цепи для эндодоменов цитокинового рецептора являются одинаковыми и выбранными из эндодомена α-, β-, γ-цепей цитокинового рецептора I типа.
8. Химерный трансмембранный белок, в соответствии с любым из предшествующих пунктов, где эндодомен цитокинового рецептора содержит:
(i) эндодомен β-цепи рецептора IL-2
(ii) эндодомен α-цепи рецептора IL-7; или
(iii) эндодомен α-цепи рецептора IL-15; и/или
(iv) общий эндодомен γ-цепи рецептора.
9. Химерный трансмембранный белок, в соответствии с п. 5, где первый полипептид включает вариабельный домен тяжелой цепи (VH) и константный домен тяжелой цепи (CH); и второй полипептид включает вариабельный домен легкой цепи (VL) и константный домен легкой цепи (CL).
10. Химерный трансмембранный белок, в соответствии с п. 9, который включает экзодомен Fab.
11. Клетка, которая содержит химерный трансмембранный белок, в соответствии с любым предыдущим пунктом.
12. Клетка, в соответствии с п. 11, которая также включает химерный антигенный рецептор.
13. Клетка, в соответствии с п. 12, где химерный антигенный рецептор связывает ассоциированный с опухолью антиген клеточной поверхности.
14. Последовательность нуклеиновой кислоты, кодирующая химерный трансмембранный белок, в соответствии с любым из пп. 1-10.
15. Конструкция нуклеиновой кислоты, которая содержит первую последовательность нуклеиновой кислоты, кодирующую первый полипептид, как определено в п. 3, и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид, как определено в п. 3, конструкция нуклеиновой кислоты, имеющая структуру:
Dim1-TM1-эндо1-коэкспр-Dim2-TM2-эндо2
в которой
Dim1 представляет собой последовательность нуклеиновой кислоты, кодирующую первый домен димеризации;
TM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен первого полипептида;
эндо 1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен первого полипептида;
коэкспр представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию обоих CCRs;
Dim2 представляет собой последовательность нуклеиновой кислоты, кодирующую второй домен димеризации;
TM2 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный эндодомен второго полипептида;
эндо 2 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен второго полипептида.
16. Конструкция нуклеиновой кислоты, в соответствии с п. 15, которая также кодирует химерный антигенный рецептор (CAR).
17. Конструкция нуклеиновой кислоты, в соответствии с п. 15 или 16, где коэкспр кодирует последовательность, содержащую саморасщепляющийся пептид.
18. Конструкция нуклеиновой кислоты, в соответствии с пп. 15-17, где альтернативные кодоны используются в областях последовательности, кодирующих одни и те же или подобные аминокислотные последовательности, чтобы избежать гомологической рекомбинации.
19. Вектор, содержащий конструкцию нуклеиновой кислоты, в соответствии с любым из пп. 15-18.
20. Ретровирусный вектор или лентивирусный вектор, или транспозон, в соответствии с п. 19.
21. Набор, который включает:
i) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую первый полипептид, как определено в п. 3; а также
ii) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую второй полипептид, как определено в п. 3.
22. Набор, в соответствии с п. 21, который также включает вектор, содержащий последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор.
23. Набор, который включает:
i) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую химерный трансмембранный белок, в соответствии с любым из пп. 1-10; а также
ii) вектор, содержащий последовательность нуклеиновой кислоты, кодирующую рецептор химерного антигена.
24. Способ изготовления клетки, в соответствии с любым из пп. 11-13, который включает стадию введения: последовательности нуклеиновой кислоты, в соответствии с п. 14; конструкции нуклеиновой кислоты, в соответствии с любым из пп. 15-18; вектора, в соответствии с п. 19 или 20; или набора векторов, в соответствии с любым из пп. 21-23, в клетку.
25. Способ, в соответствии п. 24, где клетка из образца, полученного от субъекта.
26. Фармацевтическая композиция, содержащая множество клеток, в соответствии с любым из пп. 11-13.
27. Способ лечения и/или профилактики заболевания, который включает стадию введения фармацевтической композиции, в соответствии с п. 26, субъекту.
28. Способ по п. 27, который включает следующие этапы:
(i) выделение клеточного образца у субъекта;
(ii) трансдукция или трансфекция клеток с помощью последовательности нуклеиновой кислоты, в соответствии с п. 14; конструкции нуклеиновой кислоты, в соответствии с любым из пп. 15-18; вектора, в соответствии с п. 19 или 20; или набора векторов, в соответствии с любыми пп. 21-23; а также
(iii) введение клеток из (ii) субъекту.
29. Способ, в соответствии с п. 28, где образец представляет собой образец, содержащий Т-клетки.
30. Способ, в соответствии с п. 28 или 29, в котором заболевание представляет собой рак.
31. Фармацевтическая композиция, в соответствии с п. 26, для применения при лечении и/или профилактике заболевания.
32. Применение клетки, в соответствии с любым из пп. 11-13, при изготовлении лекарственного средства для лечения и/или профилактики заболевания.
ПОДРОБНОЕ ОПИСАНИЕ
ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР (CCR)
Химерный цитокиновый рецептор (CCR) представляет собой молекулу, которая включает эндодомен цитокинового рецептора и гетерологичный лиганд-связывающий экзодомен. Гетерологичный экзодомен связывает лиганд, отличный от цитокина, для которого цитокиновый рецептор, из которого был получен эндодомен, является селективным. Таким образом, можно модифицировать специфичность лиганда цитокинового рецептора путем прививания гетерологичной связывающей специфичности.
Химерный цитокиновый рецептор содержит:
(i) лиганд-связывающий экзодомен;
(ii) опциональный спейсер;
(iii) трансмембранный домен; а также
(iv) эндодомен цитокинового рецептора.
ЦИТОКИНОВЫЕ РЕЦЕПТОРЫ И СИГНАЛИЗАЦИЯ
Многие клеточные функции регулируются членами суперсемейства цитокиновых рецепторов. Сигнализация с помощью данных рецепторов зависит от их связи с Янус-киназами (JAKs), которые соединяют связывание лиганда с фосфорилированием тирозина сигнальных белков, вовлеченных в рецепторный комплекс. Среди них - переносчики сигнала и активаторы транскрипции (STATs), семейство факторов транскрипции, которые способствуют разнообразию цитокиновых ответов.
Когда химерный цитокиновый рецептор по изобретению связывает его лиганд, может быть инициирован один или несколько следующих внутриклеточных сигнальных путей:
(i) путь JAK-STAT
(ii) путь MAP-киназы; а также
(iii) путь фосфоинозитида-3-киназы (PI3K).
Система JAK-STAT состоит из трех основных компонентов: (1) рецептора (2) Янус-киназы (JAK) и (3) переносчика сигнала и активатора транскрипции (STAT).
JAKs, которые обладают активностью тирозинкиназы, связываются с цитокиновыми рецепторами клеточной поверхности. Связывание лиганда с рецептором запускает активацию JAKs. С повышенной активностью киназы они фосфорилируют тирозиновые остатки на рецепторе и создают сайты для взаимодействия с белками, которые содержат фосфотирозин-связывающие SH2-домены. STATs, обладающие доменами SH2, способными связывать данные фосфотирозиновые остатки, привлекаются к рецепторам, и сами являются тирозинфосфорилированными JAKs. Эти фосфотирозины затем действуют как сайты связывания для доменов SH2 других STATs, опосредуя их димеризацию. Различные STATs образуют гетеро- или гомодимеры. Активированные STAT-димеры накапливаются в ядре клетки и активируют транскрипцию их целевых генов.
ЭНДОДОМЕН ЦИТОКИНОВОГО РЕЦЕПТОРА
Химерный цитокиновый рецептор по настоящему изобретению содержит эндодомен, который вызывает клеточную сигнализацию «цитокинового типа» (либо отдельно, либо в присутствии другого химерного цитокинового рецептора), когда экзодомен связывает его лиганд.
Эндодомен может представлять собой эндодомен цитокинового рецептора.
Эндодомен может быть получен из цитокинового рецептора I типа. Цитокиновые рецепторы I типа имеют общий аминокислотный мотив (WSXWS) во внеклеточной части, прилегающей к клеточной мембране.
Эндодомен может быть получен из цитокинового рецептора II типа. Цитокиновые рецепторы II типа включают те, которые связывают интерфероны I типа и II типа, и те, которые связывают члены семейства интерлейкина-10 (интерлейкин-10, интерлейкин-20 и интерлейкин-22).
Цитокиновые рецепторы I типа:
(i) рецепторы интерлейкина, такие как рецепторы для IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-12, IL13, IL -15, IL-21, IL-23 и IL-27;
(ii) рецепторы колониестимулирующих факторов, такие как рецепторы для эритропоэтина, GM-CSF и G-CSF; а также
(iii) рецептор гормона/рецептор нейропептида, такой как рецептор гормона и рецептор пролактина.
Члены семейства цитокиновых рецепторов I типа включают различные цепи, некоторые из которых вовлечены в взаимодействие лиганд/цитокин и другие, участвующие в передаче сигнала. Например, рецептор IL-2 содержит α-, β-, γ-цепь.
Общая гамма-цепь рецептора IL-2 (также известная как CD132) разделяется между рецептором IL-2, рецептором IL-4, рецептором IL-7, рецептором IL-9, рецептором IL-13 и рецептором IL-15.
IL-2
IL-2 связывается с рецептором IL-2, который имеет три формы, генерируемые различными комбинациями трех различных белков, часто называемых «цепями»: α, β и γ; данные субъединицы также являются частями рецепторов для других цитокинов. β- и γ-цепи IL-2R являются членами семейства цитокинового рецептора I типа.
Три цепи рецептора экспрессируются отдельно и различно на разных типах клеток и могут собираться в разных комбинациях и порядках для получения рецепторов IL-2 с низкой, средней и высокой аффиностью.
α-цепь связывает IL-2 с низкой аффиностью, комбинация β и γ вместе образуют комплекс, который связывает IL-2 со средней аффиностью, главным образом на Т-клетках памяти и NK-клетках; и все три цепи рецептора образуют комплекс, который связывает IL-2 с высокой аффиностью (Kd~10-11 М) на активированных Т-клетках и регуляторных Т-клетках.
Три цепи рецептора IL-2 охватывают клеточную мембрану и распространяются в клетку, тем самым доставляя биохимические сигналы во внутреннюю среду клетки. Альфа-цепь не участвует в передаче сигналов, но бета-цепь объединена с тирозинфосфатазой JAK1. Аналогично, комплексы гамма-цепи - с другой тирозинкиназой, называемой JAK3. Эти ферменты активируются связыванием IL-2 с внешними доменами IL-2R.
Сигнализация IL-2 способствует дифференцировке Т-клеток в эффекторные Т-клетки и в Т-клетки памяти, когда исходные Т-клетки также стимулируются антигеном. Благодаря роли в развитии иммунологической памяти Т-клеток, которая зависит от экспансии числа и функции антигензависимых клонов Т-клеток, они также играют ключевую роль в долгосрочном иммунитете, опосредованном клетками.
Химерный цитокиновый рецептор по настоящему изобретению может содержать β-цепь рецептора IL-2 и/или γ-цепь рецептора IL-2 (т.е. общую).
Аминокислотные последовательности для эндодоменов β-цепи IL-2 и общей γ-цепи показаны как SEQ ID No. 1 и 2.
SEQ ID No. 1: эндодомен, полученный из общей гамма-цепи человека:
ERTMPRIPTLKNLEDLVTEYHGNFSAWSGVSKGLAESLQPDYSERLCLVSEIPPKGGALGEGPGASPCNQHSPYWAPPCYTLKPET
SEQ ID No. 2: эндодомен, полученный из IL-2Rβ человека:
NCRNTGPWLKKVLKCNTPDPSKFFSQLSSEHGGDVQKWLSSPFPSSSFSPGGLAPEISPLEVLERDKVTQLLLQQDKVPEPASLSSNHSLTSCFTNQGYFFFHLPDALEIEACQVYFTYDPYSEEDPDEGVAGAPTGSSPQPLQPLSGEDDAYCTFPSRDDLLLFSPSLLGGPSPPSTAPGGSGAGEERMPPSLQERVPRDWDPQPLGPPTPGVPDLVDFQPPPELVLREAGEEVPDAGPREGVSFPWSRPPGQGEFRALNARLPLNTDAYLSLQELQGQDPTHLV
Термин «полученный из» означает, что эндодомен химерного цитокинового рецептора по изобретению имеет такую же последовательность, что и последовательность дикого типа эндогенной молекулы, или соответствующий вариант, который сохраняет способность образовывать комплекс с JAK-1 или JAK-3 и активировать один из сигнальных путей, упомянутых выше.
«Вариантная» последовательность, имеющая, по меньшей мере, 80, 85, 90, 95, 98 или 99% идентичности последовательности с последовательностью дикого типа (например, SEQ ID No. 1 или 2), при условии, что вариантная последовательность сохраняет функцию последовательности дикого типа, то есть способность образовывать комплекс с JAK-1 или JAK-3 и активировать, например, сигнальный путь JAK-STAT.
Процент идентичности между двумя полипептидными последовательностями может быть легко определен с помощью таких программ, как BLAST, которая свободно доступна на http://blast.ncbi.nlm.nih.gov.
IL-7
Рецептор интерлейкина-7 состоит из двух цепей: α-цепи рецептора интерлейкина-7 (CD127) и общей γ-цепи рецептора (CD132). Общая γ-цепь рецепторов разделяется различными цитокинами, включая интерлейкин-2, -4, -9 и -15. Рецептор интерлейкина-7 экспрессируется на различных типах клеток, включая наивные и Т-клетки памяти.
Рецептор интерлейкина-7 играет решающую роль в развитии лимфоцитов, особенно в V(D)J-рекомбинации. IL-7R также контролирует доступность области генома, которая содержит ген Т-клеточного гамма-рецептора, посредством STAT5 и ацетилирования гистонов. Исследования нокаута у мышей предполагают, что блокирующий апоптоз является важной функцией данного белка при дифференцировке и активации Т-лимфоцитов.
Химерный цитокиновый рецептор по настоящему изобретению может содержать α-цепь рецептора IL-7 и/или γ-цепь рецептора IL-7 (т.е. общую), или соответствующий вариант.
Аминокислотная последовательность для эндодомена α-цепи IL-7 показана как SEQ ID No. 3.
SEQ ID No. 3 - эндодомен, полученный из IL-7Rα человека:
KKRIKPIVWPSLPDHKKTLEHLCKKPRKNLNVSFNPESFLDCQIHRVDDIQARDEVEGFLQDTFPQQLEESEKQRLGGDVQSPNCPSEDVVITPESFGRDSSLTCLAGNVSACDAPILSSSRSLDCRESGKNGPHVYQDLLLSLGTTNSTLPPPFSLQSGILTLNPVAQGQPILTSLGSNQEEAYVTMSSFYQNQ
IL-15
Интерлейкин 15 (IL-15) представляет собой цитокин со структурным сходством с IL-2. IL-15, как и IL-2, связывается и сигнализирует через комплекс, состоящий из бета-цепи рецептора IL-2/IL-15 (CD122) и общей гамма-цепи (gamma-C, CD132). IL-15 секретируется мононуклеарными фагоцитами (и некоторыми другими клетками) вследствие вирусной инфекции. IL-15 индуцирует клеточную пролиферацию естественных клеток-киллеров.
Рецептор интерлейкина-15 состоит из альфа-субъединицы рецептора интерлейкина-15 и имеет общие бета-и гамма-субъединицы с рецептором IL-2.
СПЕЙСЕР
Химерный цитокиновый рецептор по настоящему изобретению может включать спейсер для соединения антигенсвязывающего домена с трансмембранным доменом и пространственного отделения антигенсвязывающего домена от эндодомена. Гибкий спейсер позволяет антигенсвязывающему домену ориентироваться в разных направлениях для обеспечения связывания антигена.
Когда клетка по настоящему изобретению содержит два или более химерных цитокиновых рецептора, спейсеры могут быть одинаковыми или разными. Когда клетка по настоящему изобретению содержит химерный цитокиновый рецептор (CCR) и химерный антигенный рецептор (CAR), спейсер CCR и CAR могут отличаться, например, иметь разную длину. Спейсер CAR может быть длиннее спейсера или каждого CCR.
Последовательность спейсера может, например, содержать область Fc IgG1, шарнир IgG1 или «ствол» CD8. Линкер может также содержать альтернативную последовательность, которая имеет сходные свойства длины и/или расстояния между доменами как область Fc IgG1, шарнир IgG1 или «ствол» CD8.
Спейсер IgG1 человека может быть изменен для удаления мотивов Fc-связывания.
Примеры аминокислотных последовательностей для данных спейсеров приведены ниже:
SEQ ID No. 4 (шарнир-CH2CH3 IgG1 человека)
AEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKD
SEQ ID No. 5 («ствол» CD8 человека):
TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDI
SEQ ID No. 6 (шарнир IgG1человека):
AEPKSPDKTHTCPPCPKDPK
ТРАНСМЕМБРАННЫЙ ДОМЕН
Трансмембранный домен представляет собой последовательность CCR, которая охватывает мембрану. Он может включать гидрофобную альфа-спираль. Трансмембранный домен может быть получен из CD28, который обеспечивает достаточную стабильность рецептора.
В альтернативном варианте, трансмембранный домен может быть получен из цитокинового рецептора, например, того же цитокина, из которого получен эндодомен.
Трансмембранный домен может быть, например, получен из IL-2R, IL-7R или IL-15R.
SEQ ID No. 7 - трансмембранный, полученных из общей гамма-цепи человека:
VVISVGSMGLIISLLCVYFWL
SEQ ID No. 8 - трансмембранный, полученный из IL-2Rβ человека:
IPWLGHLLVGLSGAFGFIILVYLLI
SEQ ID No. 9 - трансмембранный, полученный из IL-7Rα человека:
PILLTISILSFFSVALLVILACVLW
SEQ ID No. 10 - трансмембранный, полученный из IL-15Rα человека:
AISTSTVLLCGLSAVSLLACYL
ЛИГАНДСВЯЗЫВАЮЩИЙ ЭКЗОДОМЕН
Лигандсвязывающий домен содержит антигенсвязывающий домен. Антигенсвязывающий домен связывает целевой лиганд для CCR, т.е. секретируемый опухолью фактор или хемокин, или антиген клеточной поверхности.
В данной области техники известно множество антигенсвязывающих доменов, в том числе на основе антигенсвязывающего сайта антитела, миметиков антител и Т-клеточных рецепторов. Например, антигенсвязывающий домен может содержать: одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела; связывающий домен из природного рецептора для целевого антигена; пептид с достаточной аффинностью к целевому лиганду; однодоменное связующее, такое как верблюжье; искусственное одно-связующее, такое как дарпин; или одноцепочечное, полученное из Т-клеточного рецептора.
Термин «лиганд» используется синонимично с «антигеном» для обозначения объекта, который специфически распознается и связывается антигенсвязывающим доменом CCR.
Когда лиганд является секретируемым опухолью фактором, антигенсвязывающий домен может содержать основанный на иммуноглобулине антигенсвязывающий сайт, такой как scFv или однодоменное связующее.
Когда лиганд является хемокином, антигенсвязывающий домен может включать хемокиносвязывающую часть природного рецептора для хемокина.
ЛИГАНД
CCR по настоящему изобретению связывает лиганд.
Лиганд может быть растворимым лигандом, таким как секретируемый опухолью фактор или хемокин.
В альтернативном варианте, лиганд может быть лигандом, связанным с мембраной, таким как антиген клеточной поверхности.
Термин «растворимый лиганд» используется для обозначения лиганда или антигена, который не является частью клетки или прикрепленным к ней, но свободно перемещается во внеклеточном пространстве, например, в физиологической жидкости представляющей интерес ткани. Растворимый лиганд может существовать в бесклеточном состоянии в сыворотке, плазме или другой физиологической жидкости индивидуума.
Растворимый лиганд может быть связан с наличием или патологией конкретного заболевания, такого как рак.
Растворимый лиганд может быть частью секретом рака, т.е. группой факторов, секретируемых опухолью, будь то от раковых стволовых клеток, клеток, отличных от стволовых, или окружающей стромы. Растворимый лиганд может быть секретируемым или распространенным опухолевыми клетками (см. следующий раздел).
Растворимый лиганд может быть характерным для заболевания или пораженной ткани. Он может быть найден исключительно или на более высоком уровне у субъекта, имеющего заболевание, по сравнению со здоровым субъектом; или в больной ткани против здоровой ткани. Растворимый лиганд может быть экспрессирован, по меньшей мере, в 2-кратном, 5-кратном, 10-кратном, 100-кратном, 1000-кратном, 10,000-кратном или 100,000-кратном размере выше уровня у субъекта, имеющего заболевание, против здорового субъекта; или в больной ткани против здоровой ткани.
Термины «антиген клеточной поверхности» и «лиганд клеточной поверхности» используется синонимично с «мембранносвязанным антигеном» и «мембранносвязанным лигандом», что означает лиганд, который прикреплен или экспрессируется на поверхности клетки. Лиганд клеточной поверхности может быть, например, трансмембранным белком.
Клетка, на которой обнаружен лиганд клеточной поверхности, может быть клеткой-мишенью, такой как раковая клетка.
Лиганд клеточной поверхности может быть связан с наличием или патологией конкретного заболевания, такого как рак. В качестве альтернативы, лиганд клеточной поверхности может быть характерным для клеточного типа клетки-мишени (например, В-клетки), не обязательно связанного с болезненным состоянием.
Если лиганд клеточной поверхности характерен для заболевания или пораженной ткани, он может быть обнаружен исключительно или на более высоком уровне в соответствующих клетках субъекта, имеющего заболевание, против здорового субъекта; или в пораженной ткани против здоровой ткани. Лиганд клеточной поверхности может быть экспрессирован, по меньшей мере, в 2-кратном, 5-кратном, 10-кратном, 100-кратном, 1000-кратном, 10,000-кратном размере выше уровня в клетках субъекта, имеющего заболевание, против здорового субъекта; или в больной ткани против здоровой ткани.
СЕКРЕТИРУЕМЫЙ ОПУХОЛЬЮ ФАКТОР
Лиганд, распознаваемый CCR, может быть растворимым лигандом, секретируемым или распространенным из опухоли.
Данный «секретируемый опухоль фактор» может представлять собой, например, простатоспецифический антиген (PSA), карциноэмбриональный антиген (СЕА), фактор роста эндотелия сосудов (VEGF) или раковый антиген-125 (CA-125).
Секретируемый опухоль фактор может быть растворимым лигандом, который не является цитокином. Таким образом, CCR по настоящему изобретению прививает специфичность связывания для лиганда, отличного от цитокинового, к эндодомену цитокинового рецептора.
ПРОСТАТОСПЕЦИФИЧЕСКИЙ АНТИГЕН (PSA)
Растворимый лиганд может представлять собой антиген простатоспецифический антиген (PSA).
Простатоспецифический антиген (PSA), также известный как гамма-семинопротеин или калликреин-3 (KLK3), представляет собой гликопротеиновый фермент, кодируемый геном KLK3 у людей. PSA является членом семейства пептидазы, связанного с калликреином, и секретируется эпителиальными клетками предстательной железы.
PSA присутствует в небольших количествах в сыворотке мужчин со здоровой простатой, но повышен у индивидуумов с раком предстательной железы и другими расстройствами предстательной железы.
PSA представляет собой гликопротеин, образованный 237 остатками, и активируемый KLK2. Его физиологическая роль заключается в разжижении компонентов сгустков спермы, ведущих к высвобождению сперматозоидов. При раке PSA может участвовать в процессах неопластического роста и метастаза.
PSA представляет собой химотрипсиноподобную сериновую протеазу с типичной триадой His-Asp-Ser и каталитическим доменом, подобным таковому других пептидаз, связанных с калликреином. Кристаллическая структура PSA была получена i) в комплексе с моноклональным антителом (mAb) 8G8F5 и ii) в сэндвич-комплексе с двумя mAb 5D5A5 и 5D3D11 (Stura et al (J. Mol. Biol. (2011) 414:530-544).
Известны различные моноклональные антитела, включая клоны 2G2-B2, 2D8-E8, IgG1/K, описанные в Bavat et al Avicenna J. Med. Biotechnol. 2015, 7:2-7; and Leinonen (2004) 289:157-67.
CCR по настоящему изобретению может, например, содержать 6 CDRs или VH и/или VL-домен(ы) из PSA-связывающего mAb, такого как 8G8F5, 5D5A5 или 5D3D11.
В тех случаях, когда CCR содержит две антигенсвязывающие специфичности, связывающие различные эпитопы на PSA, можно использовать, например, 5D3D11, и один может быть основан, например, на 5D5A5.
Ниже приведены аминокислотные последовательности для 5D3D11 и 5D5A5 VH и VL. Области, определяющие комплементарности (CDRs), выделены жирным шрифтом.
5D3D11 VH (SEQ ID No. 11)
QVQLQQSGPELVKPGASVKISCKVSGYAISSSWMNWVKQRPGQGLEWIGRIYPGDGDTKYNGKFKDKATLTVDKSSSTAYMQLSSLTSVDSAVYFCARDGYRYYFDYWGQGTSVTVSS
5D3D11 VL (SEQ ID No. 12)
DIVMTQTAPSVFVTPGESVSISCRSSKSLLHSNGNTYLYWFLQRPGQSPQLLIYRMSNLASGVPDRFSGSGSGTDFTLRISRVEAEDVGVYYCMQHLEYPVTFGAGTKVEIK
5D5A5 VH (SEQ ID No. 13)
QVQLQQSGAELAKPGASVKMSCKTSGYSFSSYWMHWVKQRPGQGLEWIGYINPSTGYTENNQKFKDKVTLTADKSSNTAYMQLNSLTSEDSAVYYCARSGRLYFDVWGAGTTVTVSS
5D5A5 VL (SEQ ID No. 14)
DIVLTQSPPSLAVSLGQRATISCRASESIDLYGFTFMHWYQQKPGQPPKILIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDVATYYCQQTHEDPYTFGGGTKLEIK
ScFv на основе 5D5A5 (SEQ ID No. 15)
QVQLQQSGAELAKPGASVKMSCKTSGYSFSSYWMHWVKQRPGQGLEWIGYINPSTGYTENNQKFKDKVTLTADKSSNTAYMQLNSLTSEDSAVYYCARSGRLYFDVWGAGTTVTVSSGGGGSGGGGSGGGGSGGGGSDIVLTQSPPSLAVSLGQRATISCRASESIDLYGFTFMHWYQQKPGQPPKILIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDVATYYCQQTHEDPYTFGGGTKLEIK
ScFv на основе 5D3D11 (SEQ ID No. 16)
QVQLQQSGPELVKPGASVKISCKVSGYAISSSWMNWVKQRPGQGLEWIGRIYPGDGDTKYNGKFKDKATLTVDKSSSTAYMQLSSLTSVDSAVYFCARDGYRYYFDYWGQGTSVTVSSGGGGSGGGGSGGGGSGGGGSDIVMTQTAPSVFVTPGESVSISCRSSKSLLHSNGNTYLYWFLQRPGQSPQLLIYRMSNLASGVPDRFSGSGSGTDFTLRISRVEAEDVGVYYCMQHLEYPVTFGAGTKVEIK
Когда клетка содержит два CCRs, антигенсвязывающий домен первого CCR может содержать 6 CDRs из 5D5A5, и антигенсвязывающий домен второго CCR может содержать 6 CDRs из 5D3D11.
Антигенсвязывающий домен первого CCR может содержать домен(ы) VH и/или VL из 5D5A5 или соответствующий вариант; и антигенсвязывающий домен второго CCR может содержать домен(ы) VH и/или VL из 5D3D11, или соответствующий вариант. Варианты доменов VH и VL могут иметь, по меньшей мере, 80, 90, 95 или 99% идентичности с приведенными выше последовательностями при условии, что они сохраняют активность PSA-связывания.
Клетка, экспрессирующая CCR, которая связывает PSA, может быть полезна при лечении рака предстательной железы.
КАРЦИНОЭМБРИОНАЛЬНОЕ АНТИГЕН (CEA)
Растворимый лиганд может представлять собой СЕА.
Карциноэмбриональный антиген (СЕА) описывает набор высокосвязанных гликопротеинов, участвующих в клеточной адгезии. CEA обычно продуцируется в желудочно-кишечных тканях во время развития плода, но продуцирование прекращается до рождения. Поэтому СЕА обычно присутствует только при очень низких уровнях в крови здоровых взрослых людей. Тем не менее, уровни в сыворотке повышаются при некоторых типах рака, а это означает, что это может использоваться в опухолевого маркера при клинических испытаниях.
CEA представляют собой гликопротеины, заякоренные на клеточной поверхности гликозил фосфатидилинозитолом (GPI), чьи специализированные сиалофукозилированные гликоформы служат в качестве функционального L-селектина карциномы толстой кишки и лигандов Е-селектина, которые могут иметь решающее значение для метастатического распространения клеток карциномы толстой кишки. Иммунологически они характеризуются как члены кластера дифференцировки CD66.
СЕА и родственные гены составляют семейство СЕА, принадлежащее к суперсемейству иммуноглобулинов. У людей карциноэмбриональное семейство антигенов состоит из 29 генов, 18 из которых обычно экспрессируются. Ниже приведен список генов человека, которые кодируют карциноэмбриональные связанные с антигеном клеточные адгезионные белки: CEACAM1, CEACAM3, CEACAM4, CEACAM5, CEACAM6, CEACAM7, CEACAM8, CEACAM16, CEACAM18, CEACAM19, CEACAM20, CEACAM21.
Различные антитела, которые нацелены на СЕА, описаны в патентном документе WO 2011/034660.
Клетка, экспрессирующая CCR против CEA, может быть полезна при лечении, например, колоректального рака.
ФАКТОР РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF)
Растворимый лиганд может представлять собой VEGF.
Фактор роста эндотелия сосудов (VEGF) представляет собой сигнальный белок, продуцируемый клетками, который стимулирует васкулогенез и ангиогенез. Это часть системы, которая восстанавливает подачу кислорода к тканям, когда кровообращение является недостаточным. Концентрация в сыворотке VEGF является высокой при бронхиальной астме и сахарном диабете. Нормальная функция VEGF заключается в создании новых кровеносных сосудов во время эмбрионального развития, новых кровеносных сосудов после травмы, мышц после тренировки, и новых сосудов (коллатеральное кровообращение) для обхода заблокированных сосудов.
Когда VEGF экспрессируется избыточно, это может способствовать заболеванию. Солидные раковые образования не могут расти за пределами ограниченного размера без адекватного кровоснабжения; раки, которые могут экспрессировать VEGF, способны расти и метастазировать.
VEGF является суб-семейством тромбоцитарных факторов роста семейства факторов роста цистинового узла. Они являются важными сигнальными белками, участвующими в васкулогенезе (de novo образование эмбриональной системы кровообращения) и ангиогенезе (рост кровеносных сосудов из ранее существовавшей сосудистой сети).
У млекопитающих семейство VEGF содержит пять членов: VEGF-A, фактор роста плаценты (PGF), VEGF-B, VEGF-C и VEGF-D.
Известны различные антитела к VEGF, такие как бевацизумаб (авастин) и ранибизумаб (люцентис).
РАКОВЫЙ АНТИГЕН 125 (CA-125)
CA-125 связан с раком яичников и является наиболее часто используемым биомаркером для выявления рака яичников. В то время как CA-125 наиболее известен как маркер рака яичников, он также может быть повышен при других раках, включая рак эндометрия, рак фаллопиевой трубы, рак легких, рак молочной железы и рак желудочно-кишечного тракта.
Последовательность CA-125 человека (также известного как муцин-16) доступна в NCBI, Код No. 078966.
Известен ряд CA125-связывающих моноклональных антител, включая OC125 и M11 (Nustad et al 1996, Tumour Biol. 17:196-329). В этом исследовании изучали специфичность 26 моноклональных антител против антигена CA 125. Было обнаружено, что антиген CA 125 содержит только два основных антигенных домена, которые классифицируют антитела как OC125-подобные (группа A) или M11-подобные (группа B).
Химерный цитокиновый рецептор по настоящему изобретению может включать антигенсвязывающий домен от такого антитела. Клетка, содержащая такой CCR, может быть полезна при лечении, например, рака яичников.
Секретируемый опухоль фактор (или, если в связанной с мембраной форме - трансмембранный белок), может быть выбран из следующего открытого списка:
Перестройки гена ALK и избыточная экспрессия, дающая мутированные формы белков ALK
Альфа-фетопротеин (AFP)
Бета-2-микроглобулин (В2М)
Бета-человеческий хорионический гонадотропин (бета-hCG)
Мутации BRAF V600, дающие мутированный белок B-REF
C-kit/CD117
CA15-3/CA27,29
CA19-9
Кальцитонин
CD20
Хромогранин A (CgA)
Фрагмент цитокератина 21-1
Анализ мутации гена EGFR
Рецептор эстрогена (ER)/прогестероновый рецептор (PR)
Фибрин/фибриноген
HE4
Амплификация гена HER2/neu или избыточная экспрессия белка
Иммуноглобулины
Анализ мутации гена KRAS
Лактатдегидрогеназа
Нейронспецифическая энолаза (NSE)
Белок ядерного матрикса 22
Лиганд запрограммированной смерти клетки 1 (PD-L1)
Тиреоглобулин
Урокиназный активатор плазминогена (uPA) и ингибитор активатора плазминогена (PAI-1)
ХЕМОКИН
Хемокины представляют собой хемотаксические цитокины. Клеточная миграция руководствуется хемокиновыми градиентами, заключенными и иммобилизованными во внеклеточном матриксе. Положительно заряженные хемокины, такие как CXCL12, связываются с отрицательно заряженными молекулами ECM. Данные градиенты обеспечивают направление раковых клеток и хоуминг иммунных клеток. Действие на Т-клетки, по-видимому, является ингибирующим для хоуминга цитотоксических Т-клеток, в то время как, как представляется, привлекаются регуляторные Т-клетки.
Хемокины составляют приблизительно 8-10 килодальтон по массе и имеют четыре цистеиновых остатка в консервативных участках, которые являются ключевыми для формирования их трехмерной формы.
Некоторые хемокины считаются провоспалительными и могут быть индуцированы во время иммунного ответа для привлечения клеток иммунной системы в место инфекции, тогда как другие считаются гомеостатическими и участвуют в контроле миграции клеток при нормальных процессах поддержания или развития тканей.
Хемокины были классифицированы на четыре основных подсемейства: CXC, CC, CX3C и XC. Все эти белки проявляют свои биологические эффекты, взаимодействуя с трансмембранными рецепторами, сопряженными с G-белком, называемыми хемокиновыми рецепторами, которые селективно обнаруживаются на поверхности клеток-мишеней.
Основная роль хемокинов заключается в том, чтобы действовать как хемоаттрактант, направляющий миграцию клеток. Клетки, которые притягиваются хемокинами, следуют за сигналом увеличения концентрации хемокина в направлении источника хемокина. Некоторые хемокины контролируют клетки иммунной системы во время процессов иммунного надзора, например, направляют лимфоциты в лимфатические узлы, чтобы они могли скринировать распространение патогенов путем взаимодействия с антигенпрезентирующими клетками, находящимися в данных тканях. Другие хемокины являются воспалительными и высвобождаются из широкого спектра клеток в ответ на бактериальную инфекцию, вирусы и другие агенты. Их секреция часто стимулируется провоспалительными цитокинами, такими как интерлейкин 1. Воспалительные хемокины функционируют в основном как хемоаттрактанты для лейкоцитов, рекрутирующие моноциты, нейтрофилы и другие эффекторные клетки из крови в места инфицирования или повреждения тканей. Некоторые воспалительные хемокины активируют клетки, чтобы инициировать иммунный ответ или способствовать заживлению ран. Они секретируются многими различными типами клеток и служат для направления клеток как врожденной иммунной системы, так и адаптивной иммунной системы.
CC-хемокины
Белки СС-хемокина (или β-хемокина) имеют два граничащих цистеина (аминокислоты) вблизи аминоконца. Для млекопитающих было зарегистрировано, по крайней мере, 27 различных члена данной подгруппы, называемых лигандами CC-хемокина (CCL) от -1 до -28; CCL10 является точно таким же, как и CCL9. Хемокины этого подсемейства обычно содержат четыре цистеина (C4-CC-хемокины), но небольшое количество CC-хемокинов имеет шесть цистеинов (C6-CC-хемокины). C6-CC-хемокины включают CCL1, CCL15, CCL21, CCL23 и CCL28. CC-хемокины индуцируют миграцию моноцитов и других типов клеток, таких как NK-клетки и дендритные клетки.
Примеры CC-хемокинов включают моноцитарный хемоаттрактантный белок-1 (MCP-1 или CCL2), который индуцирует моноциты для выхода из кровотока и входа в окружающую ткань для превращения в тканевые макрофаги.
CCL5 (или RANTES) привлекает клетки, такие как Т-клетки, эозинофилы и базофилы, которые экспрессируют рецептор CCR5.
CXC-хемокины
Два N-концевых цистеина CXC-хемокинов (или α-хемокинов) разделены одной аминокислотой, представленной в данном обозначении как «X». У млекопитающих было описано 17 различных CXC-хемокинов, которые подразделяются на две категории: те, у которых определенная аминокислотная последовательность (или мотив) глутаминовой кислоты-лейцина-аргинина (или сокращенно ELR) непосредственно перед первым цистеином CXC-мотива (ELR-положительные), и те, у которых нет ELR-мотива (ELR-отрицательные). ELR-положительные CXC-хемокины специфически индуцируют миграцию нейтрофилов и взаимодействуют с хемокиновыми рецепторами CXCR1 и CXCR2.
C-хемокины
Третья группа хемокинов известна как C-хемокины (или γ-хемокины) и отличается от всех других хемокинов тем, что имеет только два цистеина; один N-концевой цистеин и один цистеин ниже. Для данной подгруппы были описаны два хемокина, и называются XCL1 (лимфотактин-α) и XCL2 (лимфотактин-β).
CX3C-хемокин
CX3C-хемокины имеют три аминокислоты между двумя цистеинами. Единственный CX3C-хемокин, обнаруженный к настоящему времени, называется фракталкин (или CX3CL1). Он секретируется и привязывается к поверхности клетки, которая его экспрессирует, тем самым служа как в качестве хемоаттрактанта, так и адгезионной молекулы.
Хемокиновые рецепторы являются рецепторами, сопряженными с G-белком, содержащими 7 трансмембранных доменов, которые обнаружены на поверхности лейкоцитов. К настоящему времени было охарактеризовано приблизительно 19 различных хемокиновых рецепторов, которые разделяются на четыре семейства в зависимости от типа хемокина, который они связывают; CXCR, которые связывают CXC-хемокины, CCR, которые связывают CC-хемокины, CX3CR1, которые связывают один CX3C-хемокин (CX3CL1) и XCR1, который связывает два XC-хемокина (XCL1 и XCL2). Они разделяют многие структурные особенности; они являются сходными по размеру (с примерно 350 аминокислотами), имеют короткий кислотный N-концевой конец, семь спиральных трансмембранных доменов с тремя внутриклеточными и тремя внеклеточными гидрофильными петлями и внутриклеточный С-конец, содержащий сериновые и треониновые остатки, важные для рецепторного регулирования. Первые две внеклеточные петли хемокиновых рецепторов имеют консервативный цистеиновый остаток, который позволяет образовывать дисульфидный мостик между данными петлями. G-белки соединяются с С-терминальным концом хемокинового рецептора, чтобы обеспечить внутриклеточную сигнализацию после активации рецептора, тогда как N-концевой домен хемокинового рецептора определяет специфичность связывания лиганда.
CXCL12
CXCL12 является в высокой степени хемотаксическим для лимфоцитов. CXCL12 играет важную роль в ангиогенезе путем привлечения эндотелиальных клеток-предшественников (EPCs) из костного мозга через CXCR4-зависимый механизм. Именно эта функция CXCL12 делает его очень важным фактором в канцерогенезе и неоваскуляризации, связанной с прогрессированием опухоли. CXCL12 также играет роль в метастазировании опухолей, в тех случаях, когда раковые клетки, которые экспрессируют рецептор CXCR4, притягиваются к тканям мишени метастазов, которые секретируют лиганд CXCL12.
Рецептором для CXCL12 является CXCR4. CCR по настоящему изобретению может включать CXCL12-связывающий домен из CXCR4, связанный с эндодоменом, полученным из цитокинового рецептора, такого как рецептор IL-2.
CXCR4-связанная экспрессия IL2 будет поддерживать приживление терапевтической Т-клетки для лечения рака. При множественной миеломе клетка, экспрессирующая такой CCR, может мобилизовать клетки и изменить окружение костного мозга. Такие клетки также используют при лечении солидных раковых образований путем модификации микроокружения солидной опухоли.
Последовательность аминокислот для CXCR4 показана ниже как SEQ ID No. 17
SEQ ID No. 17
1 msiplpllqi ytsdnyteem gsgdydsmke pcfreenanf nkiflptiys iifltgivgn
61 glvilvmgyq kklrsmtdky rlhlsvadll fvitlpfwav davanwyfgn flckavhviy
121 tvnlyssvli lafisldryl aivhatnsqr prkllaekvv yvgvwipall ltipdfifan
181 vseaddryic drfypndlwv vvfqfqhimv glilpgivil scyciiiskl shskghqkrk
241 alkttvilil affacwlpyy igisidsfil leiikqgcef entvhkwisi tealaffhcc
301 lnpilyaflg akfktsaqha ltsvsrgssl kilskgkrgg hssvsteses ssfhss
CXCR7 также связывает CXCL12.
CCL2
Хемокиновый (C-C-мотив) лиганд 2 (CCL2) также упоминается как моноцитарный хемотаксический белок 1 (MCP1) и малый индуцибельный цитокин A2. CCL2 привлекает моноциты, Т-клетки памяти и дендритные клетки в места воспаления, вызванные либо повреждением ткани, либо инфекцией.
CCR2 и CCR4 представляют собой два рецептора клеточной поверхности, которые связывают CCL2.
CCR2 имеет аминокислотную последовательность, показанную как SEQ ID No. 18
SEQ ID No. 18
1 mlstsrsrfi rntnesgeev ttffdydyga pchkfdvkqi gaqllpplys lvfifgfvgn
61 mlvvlilinc kklkcltdiy llnlaisdll flitlplwah saanewvfgn amcklftgly
121 higyfggiff iilltidryl aivhavfalk artvtfgvvt svitwlvavf asvpgiiftk
181 cqkedsvyvc gpyfprgwnn fhtimrnilg lvlpllimvi cysgilktll rcrnekkrhr
241 avrviftimi vyflfwtpyn ivillntfqe ffglsncest sqldqatqvt etlgmthcci
301 npiiyafvge kfrslfhial gcriaplqkp vcggpgvrpg knvkvttqgl ldgrgkgksi
361 grapeaslqd kega
CCR4 имеет аминокислотную последовательность, показанную как SEQ ID No. 19.
SEQ ID No. 19
1 mnptdiadtt ldesiysnyy lyesipkpct kegikafgel flpplyslvf vfgllgnsvv
61 vlvlfkykrl rsmtdvylln laisdllfvf slpfwgyyaa dqwvfglglc kmiswmylvg
121 fysgiffvml msidrylaiv havfslrart ltygvitsla twsvavfasl pgflfstcyt
181 ernhtycktk yslnsttwkv lssleinilg lviplgimlf cysmiirtlq hcknekknka
241 vkmifavvvl flgfwtpyni vlfletlvel evlqdctfer yldyaiqate tlafvhccln
301 piiyfflgek frkyilqlfk tcrglfvlcq ycgllqiysa dtpsssytqs tmdhdlhdal
CCR по настоящему изобретению может содержать сайт связывания CCL2 в CCR2 или CCR4 в лигандсвязывающем домене.
АНТИГЕН КЛЕТОЧНОЙ ПОВЕРХНОСТИ
Лиганд может представлять собой антиген клеточной поверхности, такой как трансмембранный белок.
Антиген клеточной поверхности может быть CD22.
CD22 или кластер дифференцировки-22, является молекулой, принадлежащей к семейству лектинов SIGLEC. Он обнаруживается на поверхности зрелых В-клеток и, в меньшей степени, на некоторых незрелых В-клетках. В общем, CD22 является регуляторной молекулой, которая предотвращает сверхактивацию иммунной системы и развитие аутоиммунных заболеваний.
CD22 представляет собой трансмембранный белок, связывающий сахар, который специфически связывает сиаловую кислоту с доменом иммуноглобулина (Ig), расположенным на его N-конце. Присутствие доменов Ig делает CD22 членом суперсемейства иммуноглобулинов. CD22 функционирует как ингибирующий рецептор для сигнализации B-клеточного рецептора (BCR).
Повышенная экспрессия CD22 наблюдается при неходжкинских и других лимфомах. Известны различные моноклональные антитела, нацеленные на CD22, включая эпратузумаб, инотузумаба озогамицин, m971 и m972.
ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (CAR)
Клетка по настоящему изобретению также может содержать один или несколько химерных антигенных рецепторов. CAR(s) могут быть специфическими для ассоциированного с опухолью антигена.
Типичные CAR представляют собой химерные трансмембранные белки I типа, которые соединяют внеклеточный антигенраспознающий домен (связующее) с внутриклеточным сигнальным доменом (эндодомен). Связующее обычно представляет собой одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела (mAb), но может быть основано на других форматах, которые содержат антителоподобный или основанный на лиганде антигенсвязывающий сайт. Трансмембранный домен закрепляет белок в клеточной мембране и соединяет спейсер с эндодоменом.
Ранние конструкции CAR имели эндодомены, полученные из внутриклеточных частей либо γ-цепи FcεR1, либо CD3ζ. Следовательно, данные рецепторы первого поколения передавали иммунологический сигнал 1, который был достаточным для запуска T-клеточного уничтожения родственных клеток-мишеней, но не смог полностью активировать Т-клетку для пролиферации и выживания. Для преодоления данного ограничения были сконструированы составные эндодомены: слияние внутриклеточной части костимулирующей молекулы Т-клеток с молекулой CD3ζ дает в результате рецепторы второго поколения, которые одновременно могут передавать активирующий и костимулирующий сигнал после распознавания антигена. Наиболее распространенным костимулирующим доменом является CD28. Это обеспечивает наиболее сильный костимулирующий сигнал, а именно иммунологический сигнал 2, который вызывает пролиферацию Т-клеток. Также были описаны некоторые рецепторы, которые включают эндодомены семейства рецепторов TNF, такие как близкородственные OX40 и 41BB, которые передают сигналы выживания. В настоящее время описаны еще более сильные CAR третьего поколения, которые имеют эндодомены, способные передавать сигналы активации, пролиферации и выживания.
Нуклеиновые кислоты, кодирующие CAR, могут быть перенесены в Т-клетки, используя, например, ретровирусные векторы. Таким образом, может быть получено большое количество антигенспецифических Т-клеток для адоптивного переноса клеток. Когда CAR связывает целевой антиген, это приводит к передаче активирующего сигнала в Т-клетку, на которой он экспрессируется. Таким образом, CAR направляет специфичность и цитотоксичность Т-клетки в клетки, экспрессирующие целевой антиген.
Клетка по настоящему изобретению может содержать один или несколько CAR(s).
CAR(s) может включать антигенсвязывающий домен, домен спейсера, трансмембранный домен и эндодомен. Эндодомен может включать или ассоциировать с доменом, который передает сигналы активации Т-клеток.
CAR-АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН
Антигенсвязывающий домен представляет собой часть CAR, которая распознает антиген.
В данной области техники известно множество антигенсвязывающих доменов, в том числе на основе антигенсвязывающего сайта антитела, миметиков антител и Т-клеточных рецепторов. Например, антигенсвязывающий домен может содержать: одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела; природный лиганд целевого антигена; пептид с достаточной аффинностью к мишени; однодоменное связующее, такое как верблюжье; искусственное одно-связующее, такое как дарпин; или одноцепочечное, полученное из Т-клеточного рецептора.
Термин «лиганд» используется синонимично с «антигеном» для обозначения объекта, который специфически распознается и связывается антигенсвязывающим доменом CAR.
АНТИГЕН КЛЕТОЧНОЙ ПОВЕРХНОСТИ
CAR может распознавать антиген клеточной поверхности, то есть объект, такой как трансмембранный белок, который экспрессируется на поверхности целевой клетки, такой как опухолевая клетка.
CAR может специфически связывать ассоциированный с опухолью антиген клеточной поверхности.
Известны различные связанные с опухолью антигены (TAA), некоторые из которых показаны в Таблице 1. Антигенсвязывающий домен, используемый в настоящем изобретении, может быть доменом, который способен связывать TAA, как указано здесь.
Таблица 1
Когда CAR распознает В-клеточную лимфому или антиген лейкемии (такой как CD19, CD20, CD52, CD160 или CD5), CCR может распознавать другой B-клеточный антиген, такой как CD22.
АНТИГЕНЫ, АССОЦИИРОВАННЫЕ С РАКОМ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
CAR может специфически связывать антиген клеточной поверхности, ассоциированный с раком, такой как антиген стволовых клеток предстательной железы (PSCA) или простатоспецифический мембранный антиген (PSMA).
PSCA представляет собой заякоренный гликозилфосфатидилинозитолом гликопротеин клеточной мембраны. Он активируется в большей части случаев рака предстательной железы и также обнаруживается при раке мочевого пузыря и поджелудочной железы.
Известны различные анти-PSCA-антитела, такие как 7F5 (Morgenroth et al (Prostate (2007) 67:1121-1131); 1G8 (Hillerdal et al (2014) BMC Cancer 14:30); and Ha1-4.117 (Abate-Daga et al (2014) 25:1003-1012).
CCR-экспрессирующая клетка по изобретению может также экспрессировать CAR-анти-PSCA, который может содержать антигенсвязывающий домен на основе одного из этих антител.
PSMA представляет собой цинк-металлофермент, который находится в мембранах. PSMA экспрессируется в высокой степени в предстательной железе человека, что в сто раз больше, чем экспрессия в большинстве других тканей. При раке он повышается в экспрессии, и был назван вторым по величине повышающимся геном при раке предстательной железы с увеличением в 8-12 раз, по сравнению с нераковой предстательной железой. В дополнение к экспрессии в предстательной железе человека и раке простаты, PSMA также оказывается высокоэкспрессируемым при опухолевой неоваскуляризации, но не нормальной васкуляризации всех типов солидных опухолей, таких как почки, груди, толстой кишки и т. д.
Известны различные анти-PSMA-антитела, такие как 7E11, J591, J415 и Hybritech PEQ226,5 и PM2J004,5, каждое из которых связывает различный эпитоп PSMA (Chang et al (1999) Cancer Res 15:3192-8).
CCR-экспрессирующая клетка по изобретению может также экспрессировать анти-PSMA CAR, который может содержать антигенсвязывающий домен на основе одного из этих антител.
Например, CCR может содержать scFv на основе J591, имеющий последовательность, показанную как SEQ ID No. 20.
SEQ ID No. 20 (J591 scFv)
EVQLQQSGPELKKPGTSVRISCKTSGYTFTEYTIHWVKQSHGKSLEWIGNINPNNGGTTYNQKFEDKATLTVDKSSSTAYMELRSLTSEDSAVYYCAAGWNFDYWGQGTTLTVSSGGGGSGGGGSGGGGSDIVMTQSHKFMSTSVGDRVSIICKASQDVGTAVDWYQQKPGQSPKLLIYWASTRHTGVPDRFTGSGSGTDFTLTITNVQSEDLADYFCQQYNSYPLTFGAGTMLDLKR
CAR-ТРАНСМЕМБРАННЫЙ ДОМЕН
Трансмембранный домен представляет собой последовательность CAR, которая охватывает мембрану. Он может содержать гидрофобную альфа-спираль. CAR-трансмембранный домен может быть получен из CD28, который обеспечивает достаточную стабильность рецептора.
CAR-СИГНАЛЬНЫЙ ПЕПТИД
CAR и CCR, описанные здесь, могут содержать сигнальный пептид, так что когда он/они экспрессируется в клетке, такой как Т-клетка, образующийся белок направляется в эндоплазматический ретикулум, а затем на поверхность клетки, где он экспрессируется.
Ядро сигнального пептида может содержать длинный участок гидрофобных аминокислот, который имеет тенденцию образовывать одну альфа-спираль. Сигнальный пептид может начинаться с короткого положительно заряженного участка аминокислот, что помогает обеспечить правильную топологию полипептида во время транслокации. В конце сигнального пептида обычно имеется участок аминокислот, который распознается и расщепляется сигнальной пептидазой. Сигнальная пептидаза может расщепляться во время или после завершения транслокации, чтобы генерировать свободный сигнальный пептид и зрелый белок. Затем свободные сигнальные пептиды расщепляются специфическими протеазами.
Сигнальный пептид может находиться на N-конце молекулы.
Сигнальный пептид может содержать последовательность, показанную как SEQ ID No. 21, 22 или 23, или соответствующий вариант, имеющий 5, 4, 3, 2 или 1 аминокислотные мутации (вставки, замены или добавления), при условии, что сигнальный пептид по-прежнему функционирует для того, чтобы вызвать экспрессию клеточной поверхности CAR.
SEQ ID No. 21: MGTSLLCWMALCLLGADHADG
Сигнальный пептид SEQ ID No. 21 является компактным и высокоэффективным, и получаемым из бета-цепи TCR. Предполагается, что после терминального глицина будет произведено около 95% расщепления, что обеспечит эффективное удаление сигнальной пептидазой.
SEQ ID No. 22: MSLPVTALLLPLALLLHAARP
Сигнальный пептид SEQ ID No. 22 получают из IgG1.
SEQ ID No. 23: MAVPTQVLGLLLLWLTDARC
Сигнальный пептид SEQ ID No. 23 получают из CD8a.
CAR-ЭНДОДОМЕН
Эндодомен представляет собой часть типичного CAR, расположенного на внутренней стороне клеточной мембраны.
Эндодомен представляет собой передающую сигналы часть типичного CAR. После распознавания антигена антигенсвязывающим доменом отдельные молекулы CAR-молекулы кластера, нативные CD45 и CD148 исключаются из синапса, и сигнал передается в клетку.
CAR-эндодомен может представлять собой или включать внутриклеточный сигнальный домен. В альтернативном варианте осуществления изобретения эндодомен предлагаемого CAR может быть способен взаимодействовать с внутриклеточной сигнальной молекулой, которая присутствует в цитоплазме, что приводит к сигнализации.
Внутриклеточный сигнальный домен или отдельная внутриклеточная сигнальная молекула может представлять собой или содержать Т-клеточный сигнальный домен.
Наиболее часто используемым сигнальным компонентом домена является компонент эндодомена CD3-дзета, который содержит 3 ITAMs. Он передает сигнал активации в Т-клетку после того, как антиген связан. CD3-дзета может не обеспечивать полностью компетентный сигнал активации, и может потребоваться дополнительная костимулирующая сигнализация. Например, химерные CD28 и OX40 могут использоваться с CD3-дзета для передачи сигнала пролиферации/выживания, или все три могут использоваться вместе.
CAR может включать только эндодомен CD3-дзета, эндодомен CD3-дзета вместе с или CD28, или OX40, или эндодомен CD28 и OX40, и эндодомен CD3-дзета.
CAR-эндодомен может содержать один или несколько из следующих: эндодомен ICOS, эндодомен CD27, эндодомен BTLA, эндодомен CD30, эндодомен GITR и эндодомен HVEM.
Эндодомен может содержать последовательность, показанную как SEQ ID No. 24-32, или соответствующий вариант, имеющий, по меньшей мере, 80% идентичности последовательности.
SEQ ID No. 24 - эндодомен CD3-Z RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID No. 25 - эндодомены CD28 и CD3-дзета
SKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID No. 26 - эндодомены CD28, OX40 and CD3-дзета
SKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKIRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID No. 27 - эндодомен ICOS
CWLTKKKYSSSVHDPNGEYMFMRAVNTAKKSRLTDVTL
SEQ ID No. 28 - эндодомен CD27
QRRKYRSNKGESPVEPAEPCHYSCPREEEGSTIPIQEDYRKPEPACSP
SEQ ID No. 29 - эндодомен BTLA
RRHQGKQNELSDTAGREINLVDAHLKSEQTEASTRQNSQVLLSETGIYDNDPDLCFRMQEGSEVYSNPCLEENKPGIVYASLNHSVIGPNSRLARNVKEAPTEYASICVRS
SEQ ID No. 30 - эндодомен CD30
HRRACRKRIRQKLHLCYPVQTSQPKLELVDSRPRRSSTQLRSGASVTEPVAEERGLMSQPLMETCHSVGAAYLESLPLQDASPAGGPSSPRDLPEPRVSTEHTNNKIEKIYIMKADTVIVGTVKAELPEGRGLAGPAEPELEEELEADHTPHYPEQETEPPLGSCSDVMLSVEEEGKEDPLPTAASGK
SEQ ID No. 31 - эндодомен GITR
QLGLHIWQLRSQCMWPRETQLLLEVPPSTEDARSCQFPEEERGERSAEEKGRLGDLWV
SEQ ID No. 32 - эндодомен HVEM
CVKRRKPRGDVVKVIVSVQRKRQEAEGEATVIEALQAPPDVTTVAVEETIPSFTGRSPNH
Вариантная последовательность может иметь, по меньшей мере, 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с SEQ ID No. 24-32 при условии, что последовательность обеспечивает эффективный внутриклеточный сигнальный домен.
НУКЛЕИНОВАЯ КИСЛОТА
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей CCR по изобретению.
Нуклеиновая кислота может иметь структуру:
AgB-спейсер-TM-эндо
в котором
AgB1 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен CCR;
спейсер 1 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер CCR;
TM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен CCR;
эндо 1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен CCR.
КОНСТРУКЦИЯ НУКЛЕИНОВОЙ КИСЛОТЫ
Настоящее изобретение также относится к конструкции нуклеиновой кислоты, которая включает первую последовательность нуклеиновой кислоты, кодирующую первый CCR, как определено в связи с первым аспектом изобретения; и вторую последовательность нуклеиновой кислоты, кодирующую второй CCR, как определено в связи с первым аспектом изобретения.
Конструкция нуклеиновой кислоты может иметь следующую структуру:
AgB1-спейсер1-TM1-эндо1-коэкспр-AgB2-спейсер2-ТМ2-эндо2
в которой
AgB1 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен первого CCR;
спейсер 1 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер первого CCR;
TM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен первого CCR;
эндо 1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен первого CCR;
коэкспр представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию обоих CCRs
AgB2 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен второго CCR;
спейсер 2 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер второго CCR;
TM2 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен второго CCR;
эндо 2 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен второго CCR.
Когда конструкция нуклеиновой кислоты экспрессируется в клетке, такой как Т-клетка, она кодирует полипептид, который расщепляется на сайте расщепления таким образом, что первый и второй CCRs коэкспрессируются на поверхности клетки.
Первый и второй CCRs могут связывать различные эпитопы с одним и тем же антигеном.
Первый и второй CCRs могут иметь комплементарные эндодомены, например, один, полученный из α- или β-цепи цитокинового рецептора, и один, полученный из γ-цепи того же цитокинового рецептора.
Настоящее изобретение также относится к конструкции нуклеиновой кислоты, кодирующей CCR по изобретению и CAR. Такая конструкция может иметь структуру:
CCRAgB-CCRспейсер-CCRTM-CCRэндо-коэкспр-CARAgB-CARспейсер-CARTM-CARэндо
или
CARAgB-CARспейсер-CARTM-CARэндо-коэкспр-CCRAgB-CCRспейсер-CCRTM-CCRэндо
в которой
CCRAgB представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен CCR;
CCRспейсер представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер CCR;
CCRTM представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен CCR;
CCRэндо представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен CCR;
коэкспр представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию как CCR, так и CAR
CARAgB представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен CAR;
CARспейсер представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер CAR;
CARTM представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен CAR; а также
CARэндо представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен CAR.
Настоящее изобретение также относится к конструкции нуклеиновой кислоты, кодирующей первый и второй CCR по изобретению и CAR. Первый и второй CCR могут связывать отдельные эпитопы с одним и тем же антигеном. Такая конструкция может иметь структуру:
(i) CCRAgB1-CCRспейсер1-CCRTM1-CCRэндо1-коэкспр1-CCRAgB2-CCRспейсер2-CCRTM2-CCRэндо2-коэкспр2-CARAgB-CARспейсер-CARTM-CARэндо;
(ii) CCRAgB1-CCRспейсер1-CCRTM1-CCRэндо1-коэкспр1-CARAgB-CARспейсер-CARTM-CARэндо-коэкспр2-CCRAgB2-CCRспейсер2-CCRTM2-CCRэндо2; или
(iii) CARAgB-CARспейсер-CARTM-CARэндо-коэкспр1-CCRAgB1-CCRспейсер1-CCRTM1-CCRэндо1-коэкспр2-CCRAgB2-CCRспейсер2-CCRTM2-CCRэндо2;
в которой
CCRAgB1 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен первого CCR;
CCRспейсер1 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер первого CCR;
CCRTM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен первого CCR;
CCRэндо1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен первого CCR;
CCRAgB2 представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен второго CCR;
CCRспейсер2 представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер второго CCR;
CCRTM2 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен второго CCR;
CCRэндо2 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен второго CCR;
Коэкспр1 и коэкспр2 являются последовательностями нуклеиновой кислоты, обеспечивающими коэкспрессию двух фланкирующих последовательностей;
CARAgB представляет собой последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен CAR;
CARспейсер представляет собой последовательность нуклеиновой кислоты, кодирующую спейсер CAR;
CARTM представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен CAR; а также
CARэндо представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен CAR.
Используемые в данном документе термины «полинуклеотид», «нуклеотид» и «нуклеиновая кислота» предназначены для того, чтобы быть синонимами друг друга.
Специалисту должно быть понятно, что многочисленные различные полинуклеотиды и нуклеиновые кислоты могут кодировать один и тот же полипептид в результате вырождения генетического кода. Кроме того, следует понимать, что квалифицированные специалисты могут, используя принятые методы, делать нуклеотидные замены, которые не влияют на полипептидную последовательность, кодируемую описанными здесь полинуклеотидами, чтобы отражать использование кодонов какого-либо конкретного организма-хозяина, в котором должны быть экспрессированы полипептиды.
Нуклеиновые кислоты, в соответствии с изобретением могут содержать ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Они также могут быть полинуклеотидами, которые включают в себя синтетические или модифицированные нуклеотиды. В данной области техники известен ряд различных типов модификации олигонуклеотидов. К ним относятся метилфосфонатные и фосфоротиоатные остовы, добавление акридиновых или полилизиновых цепей на 3'- и/или 5'-концах молекулы. Следует понимать, что для целей использования, как описано в данном документе, полинуклеотиды могут быть модифицированы любым способом, доступным в данной области техники. Такие модификации могут быть выполнены с целью повышения активности in vivo или периода полураспада целевых полинуклеотидов.
Термины «вариант», «гомолог» или «производное» по отношению к нуклеотидной последовательности включают любое замещение, изменение, модификацию, замену, делецию или добавление одной (или более) нуклеиновой кислоты из или к последовательности.
В вышеприведенной структуре «коэкспр» представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию как первого, так и второго CARs. Это может быть последовательность, кодирующая сайт расщепления, так что конструкция нуклеиновой кислоты включает два или более CCR, или CCR и CAR, объединенных сайтом(ами) расщепления. Сайт расщепления может быть саморасщепляющимся, так что при получении полипептида он немедленно расщепляется на отдельные пептиды без необходимости какой-либо внешней расщепляющей активности.
Участком расщепления может быть любая последовательность, которая позволяет отделить первый и второй CCRs или CCR и CAR.
Термин «расщепление» используется в данном документе из соображений удобства, но сайт расщепления может привести к тому, что пептиды будут разделяться на отдельные объекты по механизму, отличному от классического расщепления. Например, для саморасщепляющегося пептида 2A вируса ящура (FMDV) (см. ниже) были предложены различные модели для учета «расщепляющей» активности: протеолиз протеиназой клетки-хозяина, аутопротеолиз или трансляционный эффект (Donnelly et al (2001) J. Gen. Virol. 82:1027-1041). Точный механизм такого «расщепления» не важен для целей по настоящего изобретения, если сайт расщепления при размещении между последовательностями нуклеиновых кислот, которые кодируют белки, вызывает экспрессию белков в виде отдельных объектов.
Сайт расщепления может быть сайтом расщепления фурина.
Фурин представляет собой фермент, который относится к семейству субтилизин-подобных пропротеинконвертаз. Членами этого семейства являются пропротеинконвертазы, которые превращают латентные белки-предшественники в их биологически активные продукты. Фурин представляет собой зависимую от кальция сериновую эндопротеазу, которая может эффективно расщеплять белки-предшественники на их парных сайтах процессинга основных аминокислот. Примеры фуриновых субстратов включают пропаратиреоидный гормон, предшественник трансформирующего ростового фактора бета 1, проальбумин, про-бета-секретазу, металлопротеиназу матрикса мембранного типа 1, бета-субъединицу про-фактора роста нервов и фактора фон Виллебранда. Фурин расщепляет белки сразу после основной целевой аминокислотной последовательности (канонически, Arg-X-(Arg/Lys)-Arg'), и обогащается в аппарате Гольджи.
Участком расщепления может быть сайт расщепления вируса гравировки табака (TEV).
Протеаза TEV представляет собой в высокой степени сиквенс-специфичную цистеиновую протеазу, которая представляет собой химотрипсин-подобные протеазы. Она очень специфична для целевого сайта расщепления и поэтому часто используется для контролируемого расщепления слитых белков как in vitro, так и in vivo. Консенсусным сайтом расщепления TEV является ENLYFQ\S (где «'\'» обозначает расщепленную пептидную связь). Клетки млекопитающих, такие как клетки человека, не экспрессируют протеазу TEV. Таким образом, в вариантах осуществления изобретения, в которых настоящая конструкция нуклеиновой кислоты включает сайт расщепления TEV и экспрессируется в клетке млекопитающего - экзогенная протеаза TEV также должна экспрессироваться в клетке млекопитающего.
Сайт расщепления может кодировать саморасщепляющийся пептид.
«Саморасщепляющийся пептид» относится к пептиду, который функционирует так, что, когда продуцируется полипептид, содержащий белки и саморасщепляющийся пептид, его немедленно «расщепляют» или разделяют на отдельные и дискретные первый и второй полипептиды без необходимости любой внешней расщепляющей активности.
Саморасщепляющийся пептид может представлять собой саморасщепляющийся пептид 2А из афто- или кардиовируса. Первичное расщепление 2A/2B афто- и кардиовирусов опосредуется 2A «расщеплением» на собственном C-конце. В афтовирусах, таких как вирусы ящура (FMDV) и вирус лошадиного ринита A, область 2A представляет собой короткий участок около 18 аминокислот, который, вместе с N-концевым остатком белка 2B (консервативный пролиновый остаток) представляет собой автономный элемент, способный опосредовать «расщепление» на собственном С-конце (Donelly et al (2001), как указано выше).
«2А-подобные» последовательности были обнаружены в пикорнавирусах, отличных от афто- или кардиовирусов, вирусов насекомых, подобных пикоравирусам, ротавирусов типа С и повторяющихся последовательностей в пределах трипаносом, и бактериальной последовательности (Donelly et al (2001), как указано выше). Сайт расщепления может содержать одну из этих 2А-подобных последовательностей, такую как:
YHADYYKQRLIHDVEMNPGP (SEQ ID No. 33)
HYAGYFADLLIHDIETNPGP (SEQ ID No. 34)
QCTNYALLKLAGDVESNPGP (SEQ ID No. 35)
ATNFSLLKQAGDVEENPGP (SEQ ID No. 36)
AARQMLLLLSGDVETNPGP (SEQ ID No. 37)
RAEGRGSLLTCGDVEENPGP (SEQ ID No. 38)
TRAEIEDELIRAGIESNPGP (SEQ ID No. 39)
TRAEIEDELIRADIESNPGP (SEQ ID No. 40)
AKFQIDKILISGDVELNPGP (SEQ ID No. 41)
SSIIRTKMLVSGDVEENPGP (SEQ ID No. 42)
CDAQRQKLLLSGDIEQNPGP (SEQ ID No. 43)
YPIDFGGFLVKADSEFNPGP (SEQ ID No. 44)
Сайт расщепления может содержать 2A-подобную последовательность, показанную как SEQ ID No. 38 (RAEGRGSLLTCGDVEENPGP).
Настоящее изобретение также относится к набору, содержащему одну или несколько последовательностей нуклеиновой кислоты, кодирующих первый и второй CCRs, в соответствии с первым аспектом настоящего изобретения, или один или несколько CCR(s), в соответствии с изобретением, и один или несколько CAR(s).
SEQ ID NOS 45 и 46 предоставляют полные аминокислотные последовательности слияния между анти-PSMA CAR и анти-PSA CCR. Заголовки даны для обозначения каждой части последовательности, но на практике различные элементы соединены, предоставляя одну непрерывную последовательность.
Конструкция нуклеиновой кислоты по изобретению может кодировать слитый белок, как показано в SEQ ID No. 45 или 46.
SEQ ID No. 45 - Иллюстративная конструкция с бета-цепью IL-2R
Сигнальная последовательность, полученная из CD8a человека:
MSLPVTALLLPLALLLHAA
scFv aPSMA (J591 H/L)
EVQLQQSGPELKKPGTSVRISCKTSGYTFTEYTIHWVKQSHGKSLEWIGNINPNNGGTTYNQKFEDKATLTVDKSSSTAYMELRSLTSEDSAVYYCAAGWNFDYWGQGTTLTVSSGGGGSGGGGSGGGGSDIVMTQSHKFMSTSVGDRVSIICKASQDVGTAVDWYQQKPGQSPKLLIYWASTRHTGVPDRFTGSGSGTDFTLTITNVQSEDLADYFCQQYNSYPLTFGAGTMLDLKR
Линкер
SDPA
Спейсер IgG1Fc человека (HCH2CH3pvaa):
EPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Трансмембранный, полученный из CD28 человека:
FWVLVVVGGVLACYSLLVTVAFIIFWV
Эндодомен, полученный из TCRz:
RRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
2A пептид из капсидного белка вируса Thosea asigna:
RAEGRGSLLTCGDVEENPGP
Сигнальная последовательность, полученная из мышиной каппа VIII:
METDTLILWVLLLLVPGSTG
scFv aPSA (5D5A5 H/L):
QVQLQQSGAELAKPGASVKMSCKTSGYSFSSYWMHWVKQRPGQGLEWIGYINPSTGYTENNQKFKDKVTLTADKSSNTAYMQLNSLTSEDSAVYYCARSGRLYFDVWGAGTTVTVSSGGGGSGGGGSGGGGSGGGGSDIVLTQSPPSLAVSLGQRATISCRASESIDLYGFTFMHWYQQKPGQPPKILIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDVATYYCQQTHEDPYTFGGGTKLEIK
Линкер:
SDPA
CD8aSTK спейсер человека:
TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDI
Трансмембранный, полученный из общей гамма-цепи человека:
VVISVGSMGLIISLLCVYFWL
Эндодомен, полученный из общей гамма-цепи человека:
ERTMPRIPTLKNLEDLVTEYHGNFSAWSGVSKGLAESLQPDYSERLCLVSEIPPKGGALGEGPGASPCNQHSPYWAPPCYTLKPET
2A пептид из полипротеина вируса лошадиного ринита A:
QCTNYALLKLAGDVESNPGP
Сигнальная последовательность, полученная из мышиной каппа VIII:
METDTLILWVLLLLVPGSTG
scFv aPSA (5D3D11 H/L):
QVQLQQSGPELVKPGASVKISCKVSGYAISSSWMNWVKQRPGQGLEWIGRIYPGDGDTKYNGKFKDKATLTVDKSSSTAYMQLSSLTSVDSAVYFCARDGYRYYFDYWGQGTSVTVSSGGGGSGGGGSGGGGSGGGGSDIVMTQTAPSVFVTPGESVSISCRSSKSLLHSNGNTYLYWFLQRPGQSPQLLIYRMSNLASGVPDRFSGSGSGTDFTLRISRVEAEDVGVYYCMQHLEYPVTFGAGTKVEIK
Линкер:
SDPA
Спейсер CD28STK человека:
KIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKP
Трансмембранный, полученный из IL-2Rβ человека:
IPWLGHLLVGLSGAFGFIILVYLLI
Эндодомен, полученный из IL-2Rβ человека:
NCRNTGPWLKKVLKCNTPDPSKFFSQLSSEHGGDVQKWLSSPFPSSSFSPGGLAPEISPLEVLERDKVTQLLLQQDKVPEPASLSSNHSLTSCFTNQGYFFFHLPDALEIEACQVYFTYDPYSEEDPDEGVAGAPTGSSPQPLQPLSGEDDAYCTFPSRDDLLLFSPSLLGGPSPPSTAPGGSGAGEERMPPSLQERVPRDWDPQPLGPPTPGVPDLVDFQPPPELVLREAGEEVPDAGPREGVSFPWSRPPGQGEFRALNARLPLNTDAYLSLQELQGQDPTHLV
SEQ ID No. 46 - Иллюстративная конструкция с альфа-цепью IL-7R
Сигнальная последовательность, полученная из CD8a человека:
MSLPVTALLLPLALLLHAA
scFv aPSMA (J591 H/L)
EVQLQQSGPELKKPGTSVRISCKTSGYTFTEYTIHWVKQSHGKSLEWIGNINPNNGGTTYNQKFEDKATLTVDKSSSTAYMELRSLTSEDSAVYYCAAGWNFDYWGQGTTLTVSSGGGGSGGGGSGGGGSDIVMTQSHKFMSTSVGDRVSIICKASQDVGTAVDWYQQKPGQSPKLLIYWASTRHTGVPDRFTGSGSGTDFTLTITNVQSEDLADYFCQQYNSYPLTFGAGTMLDLKR
Линкер
SDPA
Спейсер IgG1Fc человека (HCH2CH3pvaa):
EPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Трансмембранный, полученный из CD28 человека:
FWVLVVVGGVLACYSLLVTVAFIIFWV
Эндодомен, полученный из TCRz:
RRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
2A пептид из капсидного белка вируса Thosea asigna:
RAEGRGSLLTCGDVEENPGP
Сигнальная последовательность, полученная из мышиной каппа VIII:
METDTLILWVLLLLVPGSTG
scFv aPSA (5D5A5 H/L):
QVQLQQSGAELAKPGASVKMSCKTSGYSFSSYWMHWVKQRPGQGLEWIGYINPSTGYTENNQKFKDKVTLTADKSSNTAYMQLNSLTSEDSAVYYCARSGRLYFDVWGAGTTVTVSSGGGGSGGGGSGGGGSGGGGSDIVLTQSPPSLAVSLGQRATISCRASESIDLYGFTFMHWYQQKPGQPPKILIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDVATYYCQQTHEDPYTFGGGTKLEIK
Линкер:
SDPA
Спейсер CD8aSTK человека:
TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDI
Трансмембранный, полученный из общей гамма-цепи человека:
VVISVGSMGLIISLLCVYFWL
Эндодомен, полученный из общей гамма-цепи человека:
ERTMPRIPTLKNLEDLVTEYHGNFSAWSGVSKGLAESLQPDYSERLCLVSEIPPKGGALGEGPGASPCNQHSPYWAPPCYTLKPET
2A пептид из полипротеина вируса лошадиного ринита A:
QCTNYALLKLAGDVESNPGP
Сигнальная последовательность, полученная из мышиной каппа VIII:
METDTLILWVLLLLVPGSTG
scFv aPSA (5D3D11 H/L):
QVQLQQSGPELVKPGASVKISCKVSGYAISSSWMNWVKQRPGQGLEWIGRIYPGDGDTKYNGKFKDKATLTVDKSSSTAYMQLSSLTSVDSAVYFCARDGYRYYFDYWGQGTSVTVSSGGGGSGGGGSGGGGSGGGGSDIVMTQTAPSVFVTPGESVSISCRSSKSLLHSNGNTYLYWFLQRPGQSPQLLIYRMSNLASGVPDRFSGSGSGTDFTLRISRVEAEDVGVYYCMQHLEYPVTFGAGTKVEIK
Линкер:
SDPA
Спейсер CD28STK человека:
KIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKP
Трансмембранный, полученный из IL-7Rα человека:
PILLTISILSFFSVALLVILACVLW
Эндодомен, полученный из IL-7Rα человека:
KKRIKPIVWPSLPDHKKTLEHLCKKPRKNLNVSFNPESFLDCQIHRVDDIQARDEVEGFLQDTFPQQLEESEKQRLGGDVQSPNCPSEDVVITPESFGRDSSLTCLAGNVSACDAPILSSSRSLDCRESGKNGPHVYQDLLLSLGTTNSTLPPPFSLQSGILTLNPVAQGQPILTSLGSNQEEAYVTMSSFYQNQ
ВЕКТОР
Настоящее изобретение также относится к вектору или набору векторов, который содержит одну или несколько последовательность(ей) нуклеиновой кислоты, кодирующих один или несколько CCR(s) в соответствии с первым аспектом изобретения и, дополнительно, один или несколько CAR(s). Такой вектор может быть использован для введения последовательности(ей) нуклеиновой кислоты в клетку-хозяина, так что она экспрессирует CCR, в соответствии с первым аспектом изобретения.
Вектор может представлять собой, например, плазмидный или вирусный вектор, такой как ретровирусный вектор или лентивирусный вектор, или вектор на основе транспозона или синтетической мРНК.
Вектор может быть способен трансфицировать или трансдуцировать Т-клетку или NK-клетку.
КЛЕТКА
Настоящее изобретение относится к клетке, которая содержит один или несколько CCR(s) по изобретению и, дополнительно, один из нескольких CAR(s).
Клетка может содержать нуклеиновую кислоту или вектор по настоящему изобретению.
Клетка может быть цитолитической иммунной клеткой, такой как Т-клетка или NK-клетка.
Т-клетки или Т-лимфоциты являются типом лимфоцитов, которые играют центральную роль в клеточно-опосредованном иммунитете. Их можно отличить от других лимфоцитов, таких как В-клетки и естественные клетки-киллеры (NK-клетки), наличием Т-клеточного рецептора (ТКР) на поверхности клетки. Существуют различные типы Т-клеток, как описано ниже.
Вспомогательные Т-хелперы (TH-клетки) содействуют другим лейкоцитам в иммунологических процессах, включая созревание В-клеток в плазмоциты и В-клетки памяти, и активацию цитотоксических Т-клеток и макрофагов. TH-клетки экспрессируют на поверхности CD4. TH-клетки активируются, когда они представлены с пептидными антигенами молекулами MHC класса II на поверхности антигенпредставляющих клеток (APCs). Эти клетки могут дифференцироваться в один из нескольких подтипов, включая TH1, TH2, TH3, TH17, Th9 или TFH, которые секретируют различные цитокины для содействия различным типам иммунных реакций.
Цитолитические Т-клетки (ТС-клетки или CTLs) уничтожают зараженные вирусом клетки и опухолевые клетки и также участвуют в отторжении трансплантата. CTLs экспрессируют CD8 на поверхности. Данные клетки распознают свои мишени путем связывания с антигеном, ассоциированным с МНС класса I, который присутствует на поверхности всех ядросодержащих клеток. С использованием IL-10, аденозина и других молекул, секретируемых регуляторными Т-клетками, клетки CD8+ могут быть инактивированы до неактивного состояния анергии, которые предотвращают аутоиммунные заболевания, такие как экспериментальный аутоиммунный энцефаломиелит.
Т-клетки памяти являются подмножеством антигенспецифических Т-клеток, которые сохраняются в течение длительного времени после того, как устранилась инфекция. Они быстро расширяются до большого числа эффекторных Т-клеток после повторного воздействия на их родственный антиген, тем самым обеспечивая иммунную систему «памятью» против прошлых инфекций. Т-клетки памяти содержат три подтипа: Т-клетки центральной памяти (TCM-клетки) и два типа Т-клеток эффекторной памяти (ТЕМ-клетки и TEMRA-клетки). Клетки памяти могут быть либо CD4+, либо CD8+. Т-клетки памяти обычно экспрессируют белок клеточной поверхности CD45RO.
Регуляторные Т-клетки (Treg-клетки), ранее известные как супрессорные Т-клетки, имеют решающее значение для поддержания иммунологической толерантности. Их основная роль заключается в том, чтобы остановить иммунитет, опосредуемый Т-клеткой, к концу иммунной реакции и подавить автореактивные Т-клетки, которые избежали процесса негативного отбора в тимусе.
Были описаны два основных класса клеток Treg-клеток CD4+: естественные Treg-клетки и адаптивные Treg-клетки.
Естественные Treg-клетки (также известные как Treg-клетки CD4+CD25+FoxP3+) возникают в тимусе и связаны с взаимодействием между развивающимися Т-клетками как с миелоидными (CD11c+), так и с плазмоцитоидными (CD123+) дендритными клетками, которые были активированы TSLP. Естественные Treg-клетки можно отличить от других Т-клеток наличием внутриклеточной молекулы, называемой FoxP3. Мутации гена FOXP3 могут препятствовать развитию регуляторных Т-клеток, вызывая фатальное аутоиммунное заболевание IPEX.
Адаптивные Treg-клетки (также известные как Tr1-клетки или Th3-клетки) могут возникать при нормальном иммунном ответе.
Клеткой может быть натуральная клетка-киллер (или NK-клетка). NK-клетки являются частью врожденной иммунной системы. NK-клетки обеспечивают быструю реакцию на врожденные сигналы от зараженных вирусом клеток независимо от MHC.
NK-клетки (принадлежащие к группе врожденных лимфоидных клеток) определяются как большие гранулярные лимфоциты (LGL) и составляют третий вид клеток, дифференцированных от общего лимфоидного предшественника, продуцирующего B- и T-лимфоциты. Известно, что NK-клетки дифференцируются и созревают в костном мозге, лимфатическом узле, селезенке, миндалинах и тимусе, где они затем выходят в кровоток.
CCR-экспрессирующие клетки по изобретению могут быть любыми типами клеток, упомянутыми выше.
Т- или NK-клетки, в соответствии с первым аспектом изобретения, могут быть созданы ex vivo или из собственной периферической крови пациента (1-я группа), или при трансплантации гемопоэтических стволовых клеток из периферической крови донора (2-я группа), или периферической крови от несвязанного донора (3-я группа).
В качестве альтернативы, Т- или NK-клетки, в соответствии с первым аспектом изобретения, могут быть получены из ex vivo дифференцировки индуцибельных клеток-предшественников или эмбриональных клеток-предшественников в Т- или NK-клетки. В альтернативном варианте, может быть использована иммортализованная линия Т-клеток, которая сохраняет свою литическую функцию и может действовать как терапевтическая.
Во всех этих вариантах осуществления CCR-экспрессирующие клетки генерируются путем введения ДНК или РНК, кодирующей один или каждый CCR(s) одним из множества способов, включая трансдукцию с вирусным вектором, трансфекцию с помощью ДНК или РНК.
Клеткой по изобретению может быть ex vivo T- или NK-клетка от субъекта. Т- или NK-клетки могут быть из образца мононуклеарной клетки периферической крови (РВМС). T- или NK-клетки могут быть активированы и/или размножены перед трансдуцированием нуклеиновой кислотой, кодирующей молекулы, обеспечивающие CCR, в соответствии с первым аспектом изобретения, например, путем обработки анти-CD3 моноклональным антителом.
Т- или NK-клетку по изобретению можно изготовить путем:
(i) выделения образца, содержащего Т- или NK-клетку, у субъекта или других источников, перечисленных выше; а также
(ii) трансдукции или трансфекции Т- или NK-клеток одной или несколькими последовательностью(ями) нуклеиновой кислоты, кодирующими CCR.
Затем Т- или NK-клетки могут быть очищены, например, выбраны на основе экспрессии антигенсвязывающего домена антигенсвязывающего полипептида.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Настоящее изобретение также относится к фармацевтической композиции, содержащей множество клеток, в соответствии с изобретением.
Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый носитель, разбавитель или эксципиент. Фармацевтическая композиция может опционально содержать один или несколько дополнительных фармацевтически активных полипептидов и/или соединений. Такая композиция может, например, быть в форме, подходящей для внутривенной инфузии.
СПОСОБ ЛЕЧЕНИЯ
Настоящее изобретение относится к способу лечения и/или профилактики заболевания, которое включает стадию введения клеток по настоящему изобретению (например, в фармацевтической композиции, как описано выше) субъекту.
Способ лечения заболевания относится к терапевтическому применению клеток по настоящему изобретению. При этом клетки могут вводиться субъекту, уже имеющему заболевание или предпосылку для того, чтобы облегчить, уменьшить или улучшить, по меньшей мере, один симптом, связанный с заболеванием, и/или замедлить, уменьшить или заблокировать прогрессирование заболевания.
Способ предупреждения заболевания относится к профилактическому применению клеток по настоящему изобретению. При этом такие клетки могут вводиться субъекту, который еще не заражен данным заболеванием, и/или который не проявляет каких-либо симптомов заболевания, чтобы уменьшить или предупредить причину заболевания, или уменьшить или предотвратить развитие, по меньшей мере, одного симптома, связанного с болезнью. У субъекта может быть предрасположенность или, как считается, риск развития заболевания.
Способ может включать в себя стадии:
(i) выделение образца, содержащего Т- или NK-клетки;
(ii) трансдуцирование или трансфекцию таких клеток нуклеотидной последовательностью или вектором, предоставляемыми настоящим изобретением;
(iii) введение клеток из (ii) субъекту.
Образец, содержащий Т- или NK-клетки, может быть получен от субъекта или из других источников, например, как описано выше. Т- или NK-клетки могут быть выделены из собственной периферической крови субъекта (1-я группа) или трансплантации гемопоэтических стволовых клеток из периферической крови донора (2-я группа), или периферической крови от несвязанного донора (3-я группа).
Настоящее изобретение относится к клетке, экспрессирующей CCR по настоящему изобретению, для применения при лечении и/или профилактике заболевания.
Изобретение также относится к применению клетки, экспрессирующей CCR по настоящему изобретению, при изготовлении лекарственного средства для лечения и/или профилактики заболевания.
Заболевание, подлежащее лечению и/или предотвращению с помощью способов по настоящему изобретению, может представлять собой раковое заболевание, такое как рак мочевого пузыря, рак молочной железы, рак толстой кишки, рак эндометрия, рак почки (клетки почечного эпителия), лейкемия, рак легких, меланома, неходжкинская лимфома, рак поджелудочной железы, рак предстательной железы и рак щитовидной железы.
Когда лиганд, распознаваемый CCR, представляет собой PSA, рак может быть раком предстательной железы.
Клетки по настоящему изобретению могут быть способны уничтожать целевые клетки, такие как раковые клетки. Целевая клетка может быть охарактеризована наличием секретируемого опухолью лиганда или хемокинового лиганда рядом с целевой клеткой. Целевая клетка может быть охарактеризована наличием растворимого лиганда вместе с экспрессией ассоциированного с опухолью антигена (ТАА) на поверхности целевой клетки.
Клетки и фармацевтические композиции по настоящему изобретению могут быть использованы для лечения и/или профилактики заболеваний, описанных выше.
Клетки и фармацевтические композиции по настоящему изобретению могут быть использованы для любого из способов, описанных выше.
ХИМЕРНЫЙ ТРАНСМЕМБРАННЫЙ БЕЛОК
Настоящее изобретение также относится к химерному трансмембранному белку, включающему домен димеризации; и эндодомен цитокинового рецептора.
Димеризация может происходить спонтанно, и в этом случае химерный трансмембранный белок будет конститутивно активным. В альтернативном варианте, димеризация может происходить только в присутствии химического индуктора димеризации (CID), и в этом случае трансмембранный белок вызывает цитокиновую сигнализацию только в присутствии CID.
Подходящие домены димеризации и CID описаны в патентном документе WO025/50771, содержание которого включено в настоящий документ посредством ссылки.
Например, один домен димеризации может включать домен, связывающий рапамицин FK-связывающего белка 12 (FKBP12), другой может включать FKBP12-рапамицинсвязывающий (FRB) домен mTOR; и CID может быть рапамицином или его производным.
Один домен димеризации может включать Fc506 (такролимус)-связывающий домен FK-связывающего белка 12 (FKBP12), а другой домен димеризации может включать циклоспоринсвязывающий домен цилкофилина A; и CID может быть FK506/циклоспорином или его производным.
Один домен димеризации может включать эстрогенсвязывающий домен (EBD), а другой домен димеризации может включать стрептавидинсвязывающий домен; и CID может представлять собой слитый белок эстрона/биотина или его производное.
Один домен димеризации может содержать глюкокортикоидсвязывающий домен (GBD), а другой домен димеризации может содержать домен, связывающий дигидрофолатредуктазу (DHFR); и CID может представлять собой слитый белок дексаметазона/метотрексата или его производное.
Один домен димеризации может включать домен, связывающий O6-алкилгуанин-ДНК алкилтрансферазу (AGT), а другой домен димеризации может включать домен, связывающий дигидрофолатредуктазу (DHFR); и CID может представлять собой слитый белок производного O6-бензилгуанина/метотрексата или его производное.
Один домен димеризации может включать домен рецептора ретиноевой кислоты, а другой домен димеризации может включать домен рецептора экодизона; и CID может быть RSL1 или его производным.
Если домен димеризации спонтанно гетеродимеризуется, он может быть основан на домене димеризации антитела. В частности, он может содержать участок димеризации константного домена тяжелой цепи (CH) и константного домена легкой цепи (CL). «Участок димеризации» константного домена является частью последовательности, которая образует межцепочечную дисульфидную связь.
Химерный цитокиновый рецептор может содержать Fab-участок антитела как экзодомен, например, как показано схематически на Фиг. 5.
Химерный трансмембранный белок может включать два полипептида:
(i) первый полипептид, который включает:
(a) первый домен димеризации; а также
(b) первую цепь эндодомена цитокинового рецептора; а также
(ii) второй полипептид, который включает:
(а) второй домен димеризации, который димеризуется с первым доменом димеризации; а также
(b) вторую цепь эндодомена цитокинового рецептора.
Вышеуказанные разделы, определяющие эндодомен цитокинового рецептора химерного цитокинового рецептора, также применимы к химерному трансмембранному белку по настоящему изобретению.
Вышеупомянутые разделы, относящиеся к нуклеиновым кислотам, векторам, наборам, клеткам, фармацевтическим композициям и способам, также относятся к химерному трансмембранному белку по настоящему изобретению.
Изобретение будет далее описано посредством Примеров, которые предназначены для оказания содействия специалисту в данной области техники в осуществлении изобретения и никоим образом не предназначены для ограничения объема изобретения.
ПРИМЕРЫ
Пример 1 - Тестирование in vitro
Т-клетки трансдуцируют либо с помощью PSMA-специфического CAR, либо трансдуцируют конструкцией, которая коэкспрессирует PSMA-специфический CAR с PSA-специфическим CCR. Т-клетки совместно культивируют с экспрессирующими PSMA целевыми клетками, которые секретируют или не секретируют PSA. Это совместное культивирование проводят в присутствии или отсутствии экзогенного IL2. Это совместное культивирование проводят при различных соотношениях эффектор-мишень. Это совместное культивирование повторяют последовательно с Т-клетками, стимулированными повторными клетками-мишенями. Определяют пролиферацию Т-клеток и уничтожение целевых клеток. Таким образом, можно измерять содействие CCR пролиферации и выживанию Т-клеток. Кроме того, возможность применения для повторения возможности последовательного действия повторного назначения препарата.
Пример 2 - Тестирование in vivo
Мышам NSG приживляют клеточную линию рака простаты человека, которая экспрессирует PSMA и секретирует PSA, и которая экспрессирует люциферазу светлячка. Т-клетки трансдуцируют либо с помощью PSMA-специфического CAR, либо трансдуцируют конструкцией, которая коэкспрессирует CARMA-специфический CAR с PSA-специфическим CCR. Т-клетки вводят мышам. Опухолевую нагрузку можно измерять серийно с использованием биолюминесцентной визуализации и оценивать Т-клеточный ответ на CAR. Мышей внутри каждой группы могут быть умерщвлены в разные контрольные моменты времени, и опухолевая нагрузка непосредственно измеряется макроскопическими измерениями и иммуногистохимией. Кроме того, приживление/размножение Т-клеток в ложе опухоли или в лимфоидных тканях, таких как лимфатические узлы, селезенка и костный мозг, измеряется проточной цитометрией указанных тканей.
Пример 3 - Создание и тестирование конститутивно активной цитокиновой сигнальной молекулы
Конститутивно активный цитокиновый сигнальный химерный трансмембранный белок получали путем связывания эндодоменов цитокинового рецептора с экзодоменом типа «Fab» (Фиг. 5). Эта структура использует компоненты природной димеризации антител, а именно домен димеризации из константных участков тяжелой и легкой цепей. Химерный трансмембранный белок имеет две цепи; первый полипептид, который включает легкую κ-цепь антитела и общую γ-цепь рецептора IL2 в качестве эндодомена; и второй полипептид, который включает тяжелую цепь CH1 антитела и эндодомен, который включает либо: β-цепь рецептора IL2 (дающую конститутивно активную IL2-сигнальную молекулу); или рецептор IL7 (дающий конститутивно активную IL7-сигнальную молекулу). Конститутивно активные цитокиновые сигнальные химерные трансмембранные белки, протестированные в данном исследовании, включали вариабельные участки тяжелой и легкой цепи scFv. Данные домены не требуются для димеризации. Сигнал не зависит от связывания антигена, и структура в равной степени может быть «без верхушки» (как показано на Фиг. 5) или содержать другой объект, такой как метка с белком.
Последовательности нуклеиновой кислоты, кодирующие эти два полипептида, клонировали внутри рамки, отделенной 2А-пептидной кодирующей последовательностью.
CTLL-2 (ATCC® TIB-214™) представляют собой мышиные цитотоксические Т-лимфоциты, рост которых зависит от IL-2. В отсутствие IL-2 клетки подвергаются апоптозу. Клетки CTLL-2 трансдуцировали вектором, экспрессирующим химерный белок, включающий эндодомен IL2-рецептора (Fab_IL2эндо), или вектор, экспрессирующий химерный белок, включающий эндодомен рецептора IL7 (Fab_IL7эндо), или оставляли нетрансдуцированными (WT). В качестве положительного контроля клетки всех трех типов совместно культивировали с 100 ед/мл мышиного IL2. Пролиферацию клеток оценивали через 3 и 7 д культивирования, и результаты показаны на Фиг. 6.
Нетрансдуцированные клетки CTLL2 вместе с клетками CTLL2, трансдуцированными либо с помощью конструкции (Fab_IL2эндо или Fab_IL7эндо), размножали в присутствии 100 ед/мл мышиного IL2 (Фиг. 6, левая группа). Однако в отсутствии экзогенно добавленного IL2 выживали и пролиферировали только клетки, трансдуцированные конструкцией, имеющей эндодомен IL2R (Fab_IL2эндо). Это показывает, что химерный трансмембранный рецептор предоставляет клеткам CTLL2 необходимый IL2-сигнал.
Пример 4 - Получение и тестирование химерного цитокинового рецептора против PSA
Группа химерный цитокиновый рецепторов, нацеленных на PSA, была разработана с использованием scFvs, полученных из двух антител, которые связываются с различными эпитопами PSA: 5D5A5 и 5D3D11. Кристаллическая структура PSA была получена в сэндвич-комплексе с этими двумя эпитопами (Stura et al (2011), как указано выше).
Схематические рисунки, иллюстрирующие некоторые из группы CCRs, показаны на Фиг. 7.
Группа включала следующие конструкции:
A5-CD8stk-IL2Rg_D11-Шарнир-IL2Rb: CCR с эндодоменом IL-2R, имеющим A5 на цепи с общей γ-цепью и D11 на цепи с β-цепью IL2R;
D11-CD8stk-IL2Rg_A5-Шарнир-IL2Rb: CCR с эндодоменом IL-2R, имеющим D11 на цепи с общей γ-цепью и A5 на цепи с β-цепью IL2R;
D11-CD8stk-RL_A5-Шарнир-IL2Rb: конструкция отрицательного контроля, которая эквивалентна D11-CD8stk-IL2Rg_A5-Шарнир-IL2Rb, но в которой цепь IL2Rγ заменена жестким линкером;
D11-CD8stk-IL2Rg_A5-Шарнир-IL7Ra: CCR с эндодоменом IL-7R, имеющим D11 на цепи с общей γ-цепью и A5 на цепи с α-цепью IL7R; а также
D11-CD8stk-RL_A5-Шарнир-IL7Ra: конструкция отрицательного контроля, которая эквивалентна D11-CD8stk-IL2Rg_A5-Шарнир-IL7Ra, но в которой цепь IL2Rγ заменена жестким линкером;
Клетки CTLL2 трансдуцировали векторами, экспрессирующими данные конструкции. Клетки культивировали в присутствии или в отсутствие IL2 (в присутствии IL2 в качестве положительного контроля), а также в присутствии или в отсутствие 5 нг/мл или 5 мкг/мл PSA. Пролиферацию клеток CTLL2 оценивали через 3 и 7 д, и результаты показаны на Фиг. 8.
Клетки CTLL2, экспрессирующие CCR, с эндодоменом IL7, не поддерживали выживаемость и пролиферацию клеток CTLL2 (Фиг. 8, последние две группы). Присутствие мышиного IL-2 в этих клетках поддерживало рост и пролиферацию клеток CTLL2 на день 3, но на день 7 большинство клеток подверглось апоптозу.
Анти-PSA химерные цитокиновые рецепторы с эндодоменом ILR2 поддерживали выживаемость и пролиферацию клеток CTLL2 в отсутствие IL2 и присутствии PSA как при 5 нг/мл, так и 5 мкг/мл (Фиг. 8, первая группа), с 5 мкг/мл, дающей большую выживаемость и пролиферацию, особенно на день 7.
Оба анти-PSA химерных цитокиновых рецептора с эндодоменом IL2R, т.е. A5-CD8stk-IL2Rg_D11-Шарнир-IL2Rb и D11-CD8stk-IL2Rg_A5-Шарнир-IL2Rb, что указывает на то, что относительное позиционирование двух PSA-связывающих доменов: 5D5A5 и 5D3D11 не имеет значения для функции.
Замена общей γ-цепи жестким линкером отменила способность CCR поддерживать выживаемость и пролиферацию клеток CTLL2 (Фиг. 8, третья группа).
В качестве еще одного регистрируемого показателя для сигнализации IL2 фосфорилирование Y694 STAT5 исследовали с использованием проточной цитометрии.
Клетки CTLL2 были либо нетрансдуцированными (WT); трансдуцированы с помощью конструкций PSA CCR, имеющих эндодомен IL2R (D11-CD8STK-IL2Rg_A5-Шарнир-IL2Rb); или трансдуцированы с эквивалентной конструкцией отрицательного контроля, в которой IL2Rγ-цепь заменена жестким линкером (D11-CD8STK-RL_A5-Шарнир-IL2Rb). Клетки инкубировали в течение ночи в отсутствие экзогенно добавленного IL-2. На следующий день клетки инкубировали или с перванадатом при концентрации 500 мкМ (положительный контроль, который ингибирует фосфатазу и приводит к фосфорилированию STAT5), или с 500 нг/мл PSA в течение 1 или 4 ч. После инкубации клетки фиксировали, пермеабилизировали и анализировали с помощью проточной цитометрии.
Результаты показаны на Фиг. 9. В клетках, экспрессирующих PSA CCR, присутствие PSA со временем приводит к увеличению фосфорилирования STAT5 (Фиг. 9, центральная группа). Такое увеличение фосфорилирования не наблюдалось у нетрансдуцированных клеток CTLL2 или клеток CTLL2, трансдуцированных с помощью эквивалентной конструкции, в которой IL2Rγ-цепь заменена жестким линкером (Фиг. 9, правая группа).
Эти результаты согласуются с данными выживаемости/пролиферации CTLL2, показанными на Фиг. 8, и демонстрируют, что химерный цитокиновый рецептор против растворимого лиганда (здесь - PSA) можно использовать для активации цитокиновой сигнализации в Т-клетках.
Все публикации, упомянутые в вышеуказанной спецификации, включены в настоящий документ посредством ссылки. Различные модификации и варианты описанных способов и системы изобретения будут очевидны для специалистов в данной области техники без отклонения от объема и сущности изобретения. Хотя изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявляемое изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. Более того, различные модификации описанных способов для осуществления изобретения, которые очевидны для специалистов в области молекулярной биологии или связанных областей, предназначены для охвата следующих пунктов формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Autolus Ltd
<120> ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР
<130> P108038PCT
<150> GB1514875.2
<151> 2015-08-20
<160> 55
<170> PatentIn версия 3.5
<210> 1
<211> 86
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен, полученный из общей гамма-цепи рецептора IL-2
<400> 1
Glu Arg Thr Met Pro Arg Ile Pro Thr Leu Lys Asn Leu Glu Asp Leu
1 5 10 15
Val Thr Glu Tyr His Gly Asn Phe Ser Ala Trp Ser Gly Val Ser Lys
20 25 30
Gly Leu Ala Glu Ser Leu Gln Pro Asp Tyr Ser Glu Arg Leu Cys Leu
35 40 45
Val Ser Glu Ile Pro Pro Lys Gly Gly Ala Leu Gly Glu Gly Pro Gly
50 55 60
Ala Ser Pro Cys Asn Gln His Ser Pro Tyr Trp Ala Pro Pro Cys Tyr
65 70 75 80
Thr Leu Lys Pro Glu Thr
85
<210> 2
<211> 286
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен, полученный из общей бета-цепи рецептора IL-2
<400> 2
Asn Cys Arg Asn Thr Gly Pro Trp Leu Lys Lys Val Leu Lys Cys Asn
1 5 10 15
Thr Pro Asp Pro Ser Lys Phe Phe Ser Gln Leu Ser Ser Glu His Gly
20 25 30
Gly Asp Val Gln Lys Trp Leu Ser Ser Pro Phe Pro Ser Ser Ser Phe
35 40 45
Ser Pro Gly Gly Leu Ala Pro Glu Ile Ser Pro Leu Glu Val Leu Glu
50 55 60
Arg Asp Lys Val Thr Gln Leu Leu Leu Gln Gln Asp Lys Val Pro Glu
65 70 75 80
Pro Ala Ser Leu Ser Ser Asn His Ser Leu Thr Ser Cys Phe Thr Asn
85 90 95
Gln Gly Tyr Phe Phe Phe His Leu Pro Asp Ala Leu Glu Ile Glu Ala
100 105 110
Cys Gln Val Tyr Phe Thr Tyr Asp Pro Tyr Ser Glu Glu Asp Pro Asp
115 120 125
Glu Gly Val Ala Gly Ala Pro Thr Gly Ser Ser Pro Gln Pro Leu Gln
130 135 140
Pro Leu Ser Gly Glu Asp Asp Ala Tyr Cys Thr Phe Pro Ser Arg Asp
145 150 155 160
Asp Leu Leu Leu Phe Ser Pro Ser Leu Leu Gly Gly Pro Ser Pro Pro
165 170 175
Ser Thr Ala Pro Gly Gly Ser Gly Ala Gly Glu Glu Arg Met Pro Pro
180 185 190
Ser Leu Gln Glu Arg Val Pro Arg Asp Trp Asp Pro Gln Pro Leu Gly
195 200 205
Pro Pro Thr Pro Gly Val Pro Asp Leu Val Asp Phe Gln Pro Pro Pro
210 215 220
Glu Leu Val Leu Arg Glu Ala Gly Glu Glu Val Pro Asp Ala Gly Pro
225 230 235 240
Arg Glu Gly Val Ser Phe Pro Trp Ser Arg Pro Pro Gly Gln Gly Glu
245 250 255
Phe Arg Ala Leu Asn Ala Arg Leu Pro Leu Asn Thr Asp Ala Tyr Leu
260 265 270
Ser Leu Gln Glu Leu Gln Gly Gln Asp Pro Thr His Leu Val
275 280 285
<210> 3
<211> 195
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен, полученный из альфа-цепи рецептора IL-7
<400> 3
Lys Lys Arg Ile Lys Pro Ile Val Trp Pro Ser Leu Pro Asp His Lys
1 5 10 15
Lys Thr Leu Glu His Leu Cys Lys Lys Pro Arg Lys Asn Leu Asn Val
20 25 30
Ser Phe Asn Pro Glu Ser Phe Leu Asp Cys Gln Ile His Arg Val Asp
35 40 45
Asp Ile Gln Ala Arg Asp Glu Val Glu Gly Phe Leu Gln Asp Thr Phe
50 55 60
Pro Gln Gln Leu Glu Glu Ser Glu Lys Gln Arg Leu Gly Gly Asp Val
65 70 75 80
Gln Ser Pro Asn Cys Pro Ser Glu Asp Val Val Ile Thr Pro Glu Ser
85 90 95
Phe Gly Arg Asp Ser Ser Leu Thr Cys Leu Ala Gly Asn Val Ser Ala
100 105 110
Cys Asp Ala Pro Ile Leu Ser Ser Ser Arg Ser Leu Asp Cys Arg Glu
115 120 125
Ser Gly Lys Asn Gly Pro His Val Tyr Gln Asp Leu Leu Leu Ser Leu
130 135 140
Gly Thr Thr Asn Ser Thr Leu Pro Pro Pro Phe Ser Leu Gln Ser Gly
145 150 155 160
Ile Leu Thr Leu Asn Pro Val Ala Gln Gly Gln Pro Ile Leu Thr Ser
165 170 175
Leu Gly Ser Asn Gln Glu Glu Ala Tyr Val Thr Met Ser Ser Phe Tyr
180 185 190
Gln Asn Gln
195
<210> 4
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> шарнир-CH2CH3 IgG1 человека
<400> 4
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp
225 230
<210> 5
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> «ствол» CD8 человека
<400> 5
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile
35 40 45
<210> 6
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> шарнир IgG1 человека
<400> 6
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Lys Asp Pro Lys
20
<210> 7
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> трансмембранный, полученный из общей гамма-цепи человека
<400> 7
Val Val Ile Ser Val Gly Ser Met Gly Leu Ile Ile Ser Leu Leu Cys
1 5 10 15
Val Tyr Phe Trp Leu
20
<210> 8
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> трансмембранный, полученный из IL-2R-бета человека
<400> 8
Ile Pro Trp Leu Gly His Leu Leu Val Gly Leu Ser Gly Ala Phe Gly
1 5 10 15
Phe Ile Ile Leu Val Tyr Leu Leu Ile
20 25
<210> 9
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> трансмембранный, полученный из IL-7R-альфа человека
<400> 9
Pro Ile Leu Leu Thr Ile Ser Ile Leu Ser Phe Phe Ser Val Ala Leu
1 5 10 15
Leu Val Ile Leu Ala Cys Val Leu Trp
20 25
<210> 10
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> трансмембранный, полученный из IL-15-альфа человека
<400> 10
Ala Ile Ser Thr Ser Thr Val Leu Leu Cys Gly Leu Ser Ala Val Ser
1 5 10 15
Leu Leu Ala Cys Tyr Leu
20
<210> 11
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность 5D3D11 VH
<400> 11
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Ala Ile Ser Ser Ser
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Tyr Pro Gly Asp Gly Asp Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Val Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Asp Gly Tyr Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 12
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность 5D3D11 VL
<400> 12
Asp Ile Val Met Thr Gln Thr Ala Pro Ser Val Phe Val Thr Pro Gly
1 5 10 15
Glu Ser Val Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Tyr Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Arg Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln His
85 90 95
Leu Glu Tyr Pro Val Thr Phe Gly Ala Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 13
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность 5D5A5 VH
<400> 13
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Ser Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Thr Gly Tyr Thr Glu Asn Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Arg Leu Tyr Phe Asp Val Trp Gly Ala Gly Thr Thr
100 105 110
Val Thr Val Ser Ser
115
<210> 14
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность 5D5A5 VL
<400> 14
Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Ile Asp Leu Tyr
20 25 30
Gly Phe Thr Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Ile Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asn
65 70 75 80
Pro Val Glu Ala Asp Asp Val Ala Thr Tyr Tyr Cys Gln Gln Thr His
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 15
<211> 248
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ScFv на основе 5D5A5
<400> 15
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Ser Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Thr Gly Tyr Thr Glu Asn Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Arg Leu Tyr Phe Asp Val Trp Gly Ala Gly Thr Thr
100 105 110
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser
130 135 140
Pro Pro Ser Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys
145 150 155 160
Arg Ala Ser Glu Ser Ile Asp Leu Tyr Gly Phe Thr Phe Met His Trp
165 170 175
Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Ile Leu Ile Tyr Arg Ala
180 185 190
Ser Asn Leu Glu Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser
195 200 205
Arg Thr Asp Phe Thr Leu Thr Ile Asn Pro Val Glu Ala Asp Asp Val
210 215 220
Ala Thr Tyr Tyr Cys Gln Gln Thr His Glu Asp Pro Tyr Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Lys
245
<210> 16
<211> 250
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ScFv на основе 5D3D11
<400> 16
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Ala Ile Ser Ser Ser
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Tyr Pro Gly Asp Gly Asp Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Val Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Asp Gly Tyr Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln
130 135 140
Thr Ala Pro Ser Val Phe Val Thr Pro Gly Glu Ser Val Ser Ile Ser
145 150 155 160
Cys Arg Ser Ser Lys Ser Leu Leu His Ser Asn Gly Asn Thr Tyr Leu
165 170 175
Tyr Trp Phe Leu Gln Arg Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr
180 185 190
Arg Met Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
195 200 205
Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala Glu
210 215 220
Asp Val Gly Val Tyr Tyr Cys Met Gln His Leu Glu Tyr Pro Val Thr
225 230 235 240
Phe Gly Ala Gly Thr Lys Val Glu Ile Lys
245 250
<210> 17
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность CXCR4
<400> 17
Met Ser Ile Pro Leu Pro Leu Leu Gln Ile Tyr Thr Ser Asp Asn Tyr
1 5 10 15
Thr Glu Glu Met Gly Ser Gly Asp Tyr Asp Ser Met Lys Glu Pro Cys
20 25 30
Phe Arg Glu Glu Asn Ala Asn Phe Asn Lys Ile Phe Leu Pro Thr Ile
35 40 45
Tyr Ser Ile Ile Phe Leu Thr Gly Ile Val Gly Asn Gly Leu Val Ile
50 55 60
Leu Val Met Gly Tyr Gln Lys Lys Leu Arg Ser Met Thr Asp Lys Tyr
65 70 75 80
Arg Leu His Leu Ser Val Ala Asp Leu Leu Phe Val Ile Thr Leu Pro
85 90 95
Phe Trp Ala Val Asp Ala Val Ala Asn Trp Tyr Phe Gly Asn Phe Leu
100 105 110
Cys Lys Ala Val His Val Ile Tyr Thr Val Asn Leu Tyr Ser Ser Val
115 120 125
Leu Ile Leu Ala Phe Ile Ser Leu Asp Arg Tyr Leu Ala Ile Val His
130 135 140
Ala Thr Asn Ser Gln Arg Pro Arg Lys Leu Leu Ala Glu Lys Val Val
145 150 155 160
Tyr Val Gly Val Trp Ile Pro Ala Leu Leu Leu Thr Ile Pro Asp Phe
165 170 175
Ile Phe Ala Asn Val Ser Glu Ala Asp Asp Arg Tyr Ile Cys Asp Arg
180 185 190
Phe Tyr Pro Asn Asp Leu Trp Val Val Val Phe Gln Phe Gln His Ile
195 200 205
Met Val Gly Leu Ile Leu Pro Gly Ile Val Ile Leu Ser Cys Tyr Cys
210 215 220
Ile Ile Ile Ser Lys Leu Ser His Ser Lys Gly His Gln Lys Arg Lys
225 230 235 240
Ala Leu Lys Thr Thr Val Ile Leu Ile Leu Ala Phe Phe Ala Cys Trp
245 250 255
Leu Pro Tyr Tyr Ile Gly Ile Ser Ile Asp Ser Phe Ile Leu Leu Glu
260 265 270
Ile Ile Lys Gln Gly Cys Glu Phe Glu Asn Thr Val His Lys Trp Ile
275 280 285
Ser Ile Thr Glu Ala Leu Ala Phe Phe His Cys Cys Leu Asn Pro Ile
290 295 300
Leu Tyr Ala Phe Leu Gly Ala Lys Phe Lys Thr Ser Ala Gln His Ala
305 310 315 320
Leu Thr Ser Val Ser Arg Gly Ser Ser Leu Lys Ile Leu Ser Lys Gly
325 330 335
Lys Arg Gly Gly His Ser Ser Val Ser Thr Glu Ser Glu Ser Ser Ser
340 345 350
Phe His Ser Ser
355
<210> 18
<211> 374
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность рецептора клеточной поверхности CCR2
<400> 18
Met Leu Ser Thr Ser Arg Ser Arg Phe Ile Arg Asn Thr Asn Glu Ser
1 5 10 15
Gly Glu Glu Val Thr Thr Phe Phe Asp Tyr Asp Tyr Gly Ala Pro Cys
20 25 30
His Lys Phe Asp Val Lys Gln Ile Gly Ala Gln Leu Leu Pro Pro Leu
35 40 45
Tyr Ser Leu Val Phe Ile Phe Gly Phe Val Gly Asn Met Leu Val Val
50 55 60
Leu Ile Leu Ile Asn Cys Lys Lys Leu Lys Cys Leu Thr Asp Ile Tyr
65 70 75 80
Leu Leu Asn Leu Ala Ile Ser Asp Leu Leu Phe Leu Ile Thr Leu Pro
85 90 95
Leu Trp Ala His Ser Ala Ala Asn Glu Trp Val Phe Gly Asn Ala Met
100 105 110
Cys Lys Leu Phe Thr Gly Leu Tyr His Ile Gly Tyr Phe Gly Gly Ile
115 120 125
Phe Phe Ile Ile Leu Leu Thr Ile Asp Arg Tyr Leu Ala Ile Val His
130 135 140
Ala Val Phe Ala Leu Lys Ala Arg Thr Val Thr Phe Gly Val Val Thr
145 150 155 160
Ser Val Ile Thr Trp Leu Val Ala Val Phe Ala Ser Val Pro Gly Ile
165 170 175
Ile Phe Thr Lys Cys Gln Lys Glu Asp Ser Val Tyr Val Cys Gly Pro
180 185 190
Tyr Phe Pro Arg Gly Trp Asn Asn Phe His Thr Ile Met Arg Asn Ile
195 200 205
Leu Gly Leu Val Leu Pro Leu Leu Ile Met Val Ile Cys Tyr Ser Gly
210 215 220
Ile Leu Lys Thr Leu Leu Arg Cys Arg Asn Glu Lys Lys Arg His Arg
225 230 235 240
Ala Val Arg Val Ile Phe Thr Ile Met Ile Val Tyr Phe Leu Phe Trp
245 250 255
Thr Pro Tyr Asn Ile Val Ile Leu Leu Asn Thr Phe Gln Glu Phe Phe
260 265 270
Gly Leu Ser Asn Cys Glu Ser Thr Ser Gln Leu Asp Gln Ala Thr Gln
275 280 285
Val Thr Glu Thr Leu Gly Met Thr His Cys Cys Ile Asn Pro Ile Ile
290 295 300
Tyr Ala Phe Val Gly Glu Lys Phe Arg Ser Leu Phe His Ile Ala Leu
305 310 315 320
Gly Cys Arg Ile Ala Pro Leu Gln Lys Pro Val Cys Gly Gly Pro Gly
325 330 335
Val Arg Pro Gly Lys Asn Val Lys Val Thr Thr Gln Gly Leu Leu Asp
340 345 350
Gly Arg Gly Lys Gly Lys Ser Ile Gly Arg Ala Pro Glu Ala Ser Leu
355 360 365
Gln Asp Lys Glu Gly Ala
370
<210> 19
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> последовательность рецептора клеточной поверхности CCR4
<400> 19
Met Asn Pro Thr Asp Ile Ala Asp Thr Thr Leu Asp Glu Ser Ile Tyr
1 5 10 15
Ser Asn Tyr Tyr Leu Tyr Glu Ser Ile Pro Lys Pro Cys Thr Lys Glu
20 25 30
Gly Ile Lys Ala Phe Gly Glu Leu Phe Leu Pro Pro Leu Tyr Ser Leu
35 40 45
Val Phe Val Phe Gly Leu Leu Gly Asn Ser Val Val Val Leu Val Leu
50 55 60
Phe Lys Tyr Lys Arg Leu Arg Ser Met Thr Asp Val Tyr Leu Leu Asn
65 70 75 80
Leu Ala Ile Ser Asp Leu Leu Phe Val Phe Ser Leu Pro Phe Trp Gly
85 90 95
Tyr Tyr Ala Ala Asp Gln Trp Val Phe Gly Leu Gly Leu Cys Lys Met
100 105 110
Ile Ser Trp Met Tyr Leu Val Gly Phe Tyr Ser Gly Ile Phe Phe Val
115 120 125
Met Leu Met Ser Ile Asp Arg Tyr Leu Ala Ile Val His Ala Val Phe
130 135 140
Ser Leu Arg Ala Arg Thr Leu Thr Tyr Gly Val Ile Thr Ser Leu Ala
145 150 155 160
Thr Trp Ser Val Ala Val Phe Ala Ser Leu Pro Gly Phe Leu Phe Ser
165 170 175
Thr Cys Tyr Thr Glu Arg Asn His Thr Tyr Cys Lys Thr Lys Tyr Ser
180 185 190
Leu Asn Ser Thr Thr Trp Lys Val Leu Ser Ser Leu Glu Ile Asn Ile
195 200 205
Leu Gly Leu Val Ile Pro Leu Gly Ile Met Leu Phe Cys Tyr Ser Met
210 215 220
Ile Ile Arg Thr Leu Gln His Cys Lys Asn Glu Lys Lys Asn Lys Ala
225 230 235 240
Val Lys Met Ile Phe Ala Val Val Val Leu Phe Leu Gly Phe Trp Thr
245 250 255
Pro Tyr Asn Ile Val Leu Phe Leu Glu Thr Leu Val Glu Leu Glu Val
260 265 270
Leu Gln Asp Cys Thr Phe Glu Arg Tyr Leu Asp Tyr Ala Ile Gln Ala
275 280 285
Thr Glu Thr Leu Ala Phe Val His Cys Cys Leu Asn Pro Ile Ile Tyr
290 295 300
Phe Phe Leu Gly Glu Lys Phe Arg Lys Tyr Ile Leu Gln Leu Phe Lys
305 310 315 320
Thr Cys Arg Gly Leu Phe Val Leu Cys Gln Tyr Cys Gly Leu Leu Gln
325 330 335
Ile Tyr Ser Ala Asp Thr Pro Ser Ser Ser Tyr Thr Gln Ser Thr Met
340 345 350
Asp His Asp Leu His Asp Ala Leu
355 360
<210> 20
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv на основе J591
<400> 20
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Arg Ile Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Thr Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Glu Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Gly Trp Asn Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
100 105 110
Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Ser Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser
130 135 140
Val Gly Asp Arg Val Ser Ile Ile Cys Lys Ala Ser Gln Asp Val Gly
145 150 155 160
Thr Ala Val Asp Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu
165 170 175
Leu Ile Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe
180 185 190
Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Thr Asn Val
195 200 205
Gln Ser Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Asn Ser Tyr
210 215 220
Pro Leu Thr Phe Gly Ala Gly Thr Met Leu Asp Leu Lys Arg
225 230 235
<210> 21
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид, полученный из бета-цепи TCR
<400> 21
Met Gly Thr Ser Leu Leu Cys Trp Met Ala Leu Cys Leu Leu Gly Ala
1 5 10 15
Asp His Ala Asp Gly
20
<210> 22
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид, полученный из IgG1
<400> 22
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 23
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид, полученный из CD8a
<400> 23
Met Ala Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr
1 5 10 15
Asp Ala Arg Cys
20
<210> 24
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен CD3 Z
<400> 24
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 25
<211> 152
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомены CD28 и CD3-дзета
<400> 25
Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro
1 5 10 15
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
20 25 30
Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala
35 40 45
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
50 55 60
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
65 70 75 80
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
85 90 95
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
100 105 110
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
115 120 125
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
130 135 140
His Met Gln Ala Leu Pro Pro Arg
145 150
<210> 26
<211> 188
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомены CD28, OX40 и CD3-дзета
<400> 26
Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro
1 5 10 15
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
20 25 30
Arg Asp Phe Ala Ala Tyr Arg Ser Arg Asp Gln Arg Leu Pro Pro Asp
35 40 45
Ala His Lys Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu
50 55 60
Glu Gln Ala Asp Ala His Ser Thr Leu Ala Lys Ile Arg Val Lys Phe
65 70 75 80
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
85 90 95
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
100 105 110
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
115 120 125
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
130 135 140
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
145 150 155 160
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
165 170 175
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
180 185
<210> 27
<211> 38
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен ICOS
<400> 27
Cys Trp Leu Thr Lys Lys Lys Tyr Ser Ser Ser Val His Asp Pro Asn
1 5 10 15
Gly Glu Tyr Met Phe Met Arg Ala Val Asn Thr Ala Lys Lys Ser Arg
20 25 30
Leu Thr Asp Val Thr Leu
35
<210> 28
<211> 48
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен CD27
<400> 28
Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser Pro Val Glu Pro
1 5 10 15
Ala Glu Pro Cys His Tyr Ser Cys Pro Arg Glu Glu Glu Gly Ser Thr
20 25 30
Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro Ala Cys Ser Pro
35 40 45
<210> 29
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен BTLA
<400> 29
Arg Arg His Gln Gly Lys Gln Asn Glu Leu Ser Asp Thr Ala Gly Arg
1 5 10 15
Glu Ile Asn Leu Val Asp Ala His Leu Lys Ser Glu Gln Thr Glu Ala
20 25 30
Ser Thr Arg Gln Asn Ser Gln Val Leu Leu Ser Glu Thr Gly Ile Tyr
35 40 45
Asp Asn Asp Pro Asp Leu Cys Phe Arg Met Gln Glu Gly Ser Glu Val
50 55 60
Tyr Ser Asn Pro Cys Leu Glu Glu Asn Lys Pro Gly Ile Val Tyr Ala
65 70 75 80
Ser Leu Asn His Ser Val Ile Gly Pro Asn Ser Arg Leu Ala Arg Asn
85 90 95
Val Lys Glu Ala Pro Thr Glu Tyr Ala Ser Ile Cys Val Arg Ser
100 105 110
<210> 30
<211> 188
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен CD30
<400> 30
His Arg Arg Ala Cys Arg Lys Arg Ile Arg Gln Lys Leu His Leu Cys
1 5 10 15
Tyr Pro Val Gln Thr Ser Gln Pro Lys Leu Glu Leu Val Asp Ser Arg
20 25 30
Pro Arg Arg Ser Ser Thr Gln Leu Arg Ser Gly Ala Ser Val Thr Glu
35 40 45
Pro Val Ala Glu Glu Arg Gly Leu Met Ser Gln Pro Leu Met Glu Thr
50 55 60
Cys His Ser Val Gly Ala Ala Tyr Leu Glu Ser Leu Pro Leu Gln Asp
65 70 75 80
Ala Ser Pro Ala Gly Gly Pro Ser Ser Pro Arg Asp Leu Pro Glu Pro
85 90 95
Arg Val Ser Thr Glu His Thr Asn Asn Lys Ile Glu Lys Ile Tyr Ile
100 105 110
Met Lys Ala Asp Thr Val Ile Val Gly Thr Val Lys Ala Glu Leu Pro
115 120 125
Glu Gly Arg Gly Leu Ala Gly Pro Ala Glu Pro Glu Leu Glu Glu Glu
130 135 140
Leu Glu Ala Asp His Thr Pro His Tyr Pro Glu Gln Glu Thr Glu Pro
145 150 155 160
Pro Leu Gly Ser Cys Ser Asp Val Met Leu Ser Val Glu Glu Glu Gly
165 170 175
Lys Glu Asp Pro Leu Pro Thr Ala Ala Ser Gly Lys
180 185
<210> 31
<211> 58
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен GITR
<400> 31
Gln Leu Gly Leu His Ile Trp Gln Leu Arg Ser Gln Cys Met Trp Pro
1 5 10 15
Arg Glu Thr Gln Leu Leu Leu Glu Val Pro Pro Ser Thr Glu Asp Ala
20 25 30
Arg Ser Cys Gln Phe Pro Glu Glu Glu Arg Gly Glu Arg Ser Ala Glu
35 40 45
Glu Lys Gly Arg Leu Gly Asp Leu Trp Val
50 55
<210> 32
<211> 60
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен HVEM
<400> 32
Cys Val Lys Arg Arg Lys Pro Arg Gly Asp Val Val Lys Val Ile Val
1 5 10 15
Ser Val Gln Arg Lys Arg Gln Glu Ala Glu Gly Glu Ala Thr Val Ile
20 25 30
Glu Ala Leu Gln Ala Pro Pro Asp Val Thr Thr Val Ala Val Glu Glu
35 40 45
Thr Ile Pro Ser Phe Thr Gly Arg Ser Pro Asn His
50 55 60
<210> 33
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 33
Tyr His Ala Asp Tyr Tyr Lys Gln Arg Leu Ile His Asp Val Glu Met
1 5 10 15
Asn Pro Gly Pro
20
<210> 34
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 34
His Tyr Ala Gly Tyr Phe Ala Asp Leu Leu Ile His Asp Ile Glu Thr
1 5 10 15
Asn Pro Gly Pro
20
<210> 35
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 35
Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 36
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 36
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
1 5 10 15
Pro Gly Pro
<210> 37
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 37
Ala Ala Arg Gln Met Leu Leu Leu Leu Ser Gly Asp Val Glu Thr Asn
1 5 10 15
Pro Gly Pro
<210> 38
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 38
Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 39
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 39
Thr Arg Ala Glu Ile Glu Asp Glu Leu Ile Arg Ala Gly Ile Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 40
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 40
Thr Arg Ala Glu Ile Glu Asp Glu Leu Ile Arg Ala Asp Ile Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 41
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 41
Ala Lys Phe Gln Ile Asp Lys Ile Leu Ile Ser Gly Asp Val Glu Leu
1 5 10 15
Asn Pro Gly Pro
20
<210> 42
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 42
Ser Ser Ile Ile Arg Thr Lys Met Leu Val Ser Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 43
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 43
Cys Asp Ala Gln Arg Gln Lys Leu Leu Leu Ser Gly Asp Ile Glu Gln
1 5 10 15
Asn Pro Gly Pro
20
<210> 44
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления, 2A-подобная последовательность
<400> 44
Tyr Pro Ile Asp Phe Gly Gly Phe Leu Val Lys Ala Asp Ser Glu Phe
1 5 10 15
Asn Pro Gly Pro
20
<210> 45
<211> 1722
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> конструкция слияния с бета-цепью IL-2R
<400> 45
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Thr Ser Val Arg Ile Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Glu Tyr Thr Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu
50 55 60
Glu Trp Ile Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Thr Tyr Asn
65 70 75 80
Gln Lys Phe Glu Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Ala Gly Trp Asn Phe Asp Tyr Trp Gly Gln Gly Thr
115 120 125
Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser His Lys Phe Met
145 150 155 160
Ser Thr Ser Val Gly Asp Arg Val Ser Ile Ile Cys Lys Ala Ser Gln
165 170 175
Asp Val Gly Thr Ala Val Asp Trp Tyr Gln Gln Lys Pro Gly Gln Ser
180 185 190
Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro
195 200 205
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
210 215 220
Thr Asn Val Gln Ser Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr
225 230 235 240
Asn Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Met Leu Asp Leu Lys
245 250 255
Arg Ser Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys
260 265 270
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
275 280 285
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val
290 295 300
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
305 310 315 320
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
325 330 335
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
340 345 350
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
355 360 365
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
370 375 380
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
385 390 395 400
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
405 410 415
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
420 425 430
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
435 440 445
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
450 455 460
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
465 470 475 480
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Phe Trp Val Leu
485 490 495
Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val
500 505 510
Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg Ser Ala
515 520 525
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
530 535 540
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
545 550 555 560
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
565 570 575
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
580 585 590
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
595 600 605
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
610 615 620
His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly Ser Leu
625 630 635 640
Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Glu Thr Asp
645 650 655
Thr Leu Ile Leu Trp Val Leu Leu Leu Leu Val Pro Gly Ser Thr Gly
660 665 670
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
675 680 685
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Ser Phe Ser Ser Tyr
690 695 700
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
705 710 715 720
Gly Tyr Ile Asn Pro Ser Thr Gly Tyr Thr Glu Asn Asn Gln Lys Phe
725 730 735
Lys Asp Lys Val Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
740 745 750
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
755 760 765
Ala Arg Ser Gly Arg Leu Tyr Phe Asp Val Trp Gly Ala Gly Thr Thr
770 775 780
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
785 790 795 800
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser
805 810 815
Pro Pro Ser Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys
820 825 830
Arg Ala Ser Glu Ser Ile Asp Leu Tyr Gly Phe Thr Phe Met His Trp
835 840 845
Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Ile Leu Ile Tyr Arg Ala
850 855 860
Ser Asn Leu Glu Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser
865 870 875 880
Arg Thr Asp Phe Thr Leu Thr Ile Asn Pro Val Glu Ala Asp Asp Val
885 890 895
Ala Thr Tyr Tyr Cys Gln Gln Thr His Glu Asp Pro Tyr Thr Phe Gly
900 905 910
Gly Gly Thr Lys Leu Glu Ile Lys Ser Asp Pro Ala Thr Thr Thr Pro
915 920 925
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
930 935 940
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
945 950 955 960
Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Val Val Ile Ser Val Gly
965 970 975
Ser Met Gly Leu Ile Ile Ser Leu Leu Cys Val Tyr Phe Trp Leu Glu
980 985 990
Arg Thr Met Pro Arg Ile Pro Thr Leu Lys Asn Leu Glu Asp Leu Val
995 1000 1005
Thr Glu Tyr His Gly Asn Phe Ser Ala Trp Ser Gly Val Ser Lys
1010 1015 1020
Gly Leu Ala Glu Ser Leu Gln Pro Asp Tyr Ser Glu Arg Leu Cys
1025 1030 1035
Leu Val Ser Glu Ile Pro Pro Lys Gly Gly Ala Leu Gly Glu Gly
1040 1045 1050
Pro Gly Ala Ser Pro Cys Asn Gln His Ser Pro Tyr Trp Ala Pro
1055 1060 1065
Pro Cys Tyr Thr Leu Lys Pro Glu Thr Gln Cys Thr Asn Tyr Ala
1070 1075 1080
Leu Leu Lys Leu Ala Gly Asp Val Glu Ser Asn Pro Gly Pro Met
1085 1090 1095
Glu Thr Asp Thr Leu Ile Leu Trp Val Leu Leu Leu Leu Val Pro
1100 1105 1110
Gly Ser Thr Gly Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu
1115 1120 1125
Val Lys Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Val Ser Gly
1130 1135 1140
Tyr Ala Ile Ser Ser Ser Trp Met Asn Trp Val Lys Gln Arg Pro
1145 1150 1155
Gly Gln Gly Leu Glu Trp Ile Gly Arg Ile Tyr Pro Gly Asp Gly
1160 1165 1170
Asp Thr Lys Tyr Asn Gly Lys Phe Lys Asp Lys Ala Thr Leu Thr
1175 1180 1185
Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu
1190 1195 1200
Thr Ser Val Asp Ser Ala Val Tyr Phe Cys Ala Arg Asp Gly Tyr
1205 1210 1215
Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
1220 1225 1230
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1235 1240 1245
Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Thr Ala
1250 1255 1260
Pro Ser Val Phe Val Thr Pro Gly Glu Ser Val Ser Ile Ser Cys
1265 1270 1275
Arg Ser Ser Lys Ser Leu Leu His Ser Asn Gly Asn Thr Tyr Leu
1280 1285 1290
Tyr Trp Phe Leu Gln Arg Pro Gly Gln Ser Pro Gln Leu Leu Ile
1295 1300 1305
Tyr Arg Met Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser
1310 1315 1320
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val
1325 1330 1335
Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln His Leu Glu
1340 1345 1350
Tyr Pro Val Thr Phe Gly Ala Gly Thr Lys Val Glu Ile Lys Ser
1355 1360 1365
Asp Pro Ala Lys Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp
1370 1375 1380
Asn Glu Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His
1385 1390 1395
Leu Cys Pro Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Ile Pro
1400 1405 1410
Trp Leu Gly His Leu Leu Val Gly Leu Ser Gly Ala Phe Gly Phe
1415 1420 1425
Ile Ile Leu Val Tyr Leu Leu Ile Asn Cys Arg Asn Thr Gly Pro
1430 1435 1440
Trp Leu Lys Lys Val Leu Lys Cys Asn Thr Pro Asp Pro Ser Lys
1445 1450 1455
Phe Phe Ser Gln Leu Ser Ser Glu His Gly Gly Asp Val Gln Lys
1460 1465 1470
Trp Leu Ser Ser Pro Phe Pro Ser Ser Ser Phe Ser Pro Gly Gly
1475 1480 1485
Leu Ala Pro Glu Ile Ser Pro Leu Glu Val Leu Glu Arg Asp Lys
1490 1495 1500
Val Thr Gln Leu Leu Leu Gln Gln Asp Lys Val Pro Glu Pro Ala
1505 1510 1515
Ser Leu Ser Ser Asn His Ser Leu Thr Ser Cys Phe Thr Asn Gln
1520 1525 1530
Gly Tyr Phe Phe Phe His Leu Pro Asp Ala Leu Glu Ile Glu Ala
1535 1540 1545
Cys Gln Val Tyr Phe Thr Tyr Asp Pro Tyr Ser Glu Glu Asp Pro
1550 1555 1560
Asp Glu Gly Val Ala Gly Ala Pro Thr Gly Ser Ser Pro Gln Pro
1565 1570 1575
Leu Gln Pro Leu Ser Gly Glu Asp Asp Ala Tyr Cys Thr Phe Pro
1580 1585 1590
Ser Arg Asp Asp Leu Leu Leu Phe Ser Pro Ser Leu Leu Gly Gly
1595 1600 1605
Pro Ser Pro Pro Ser Thr Ala Pro Gly Gly Ser Gly Ala Gly Glu
1610 1615 1620
Glu Arg Met Pro Pro Ser Leu Gln Glu Arg Val Pro Arg Asp Trp
1625 1630 1635
Asp Pro Gln Pro Leu Gly Pro Pro Thr Pro Gly Val Pro Asp Leu
1640 1645 1650
Val Asp Phe Gln Pro Pro Pro Glu Leu Val Leu Arg Glu Ala Gly
1655 1660 1665
Glu Glu Val Pro Asp Ala Gly Pro Arg Glu Gly Val Ser Phe Pro
1670 1675 1680
Trp Ser Arg Pro Pro Gly Gln Gly Glu Phe Arg Ala Leu Asn Ala
1685 1690 1695
Arg Leu Pro Leu Asn Thr Asp Ala Tyr Leu Ser Leu Gln Glu Leu
1700 1705 1710
Gln Gly Gln Asp Pro Thr His Leu Val
1715 1720
<210> 46
<211> 1631
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> конструкция слияния с альфа-цепью IL-7R
<400> 46
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Thr Ser Val Arg Ile Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Glu Tyr Thr Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu
50 55 60
Glu Trp Ile Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Thr Tyr Asn
65 70 75 80
Gln Lys Phe Glu Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Ala Gly Trp Asn Phe Asp Tyr Trp Gly Gln Gly Thr
115 120 125
Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser His Lys Phe Met
145 150 155 160
Ser Thr Ser Val Gly Asp Arg Val Ser Ile Ile Cys Lys Ala Ser Gln
165 170 175
Asp Val Gly Thr Ala Val Asp Trp Tyr Gln Gln Lys Pro Gly Gln Ser
180 185 190
Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro
195 200 205
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
210 215 220
Thr Asn Val Gln Ser Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr
225 230 235 240
Asn Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Met Leu Asp Leu Lys
245 250 255
Arg Ser Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys
260 265 270
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
275 280 285
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val
290 295 300
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
305 310 315 320
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
325 330 335
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
340 345 350
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
355 360 365
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
370 375 380
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
385 390 395 400
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
405 410 415
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
420 425 430
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
435 440 445
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
450 455 460
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
465 470 475 480
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Phe Trp Val Leu
485 490 495
Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val
500 505 510
Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg Ser Ala
515 520 525
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
530 535 540
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
545 550 555 560
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
565 570 575
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
580 585 590
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
595 600 605
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
610 615 620
His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly Ser Leu
625 630 635 640
Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Glu Thr Asp
645 650 655
Thr Leu Ile Leu Trp Val Leu Leu Leu Leu Val Pro Gly Ser Thr Gly
660 665 670
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
675 680 685
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Ser Phe Ser Ser Tyr
690 695 700
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
705 710 715 720
Gly Tyr Ile Asn Pro Ser Thr Gly Tyr Thr Glu Asn Asn Gln Lys Phe
725 730 735
Lys Asp Lys Val Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
740 745 750
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
755 760 765
Ala Arg Ser Gly Arg Leu Tyr Phe Asp Val Trp Gly Ala Gly Thr Thr
770 775 780
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
785 790 795 800
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser
805 810 815
Pro Pro Ser Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys
820 825 830
Arg Ala Ser Glu Ser Ile Asp Leu Tyr Gly Phe Thr Phe Met His Trp
835 840 845
Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Ile Leu Ile Tyr Arg Ala
850 855 860
Ser Asn Leu Glu Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser
865 870 875 880
Arg Thr Asp Phe Thr Leu Thr Ile Asn Pro Val Glu Ala Asp Asp Val
885 890 895
Ala Thr Tyr Tyr Cys Gln Gln Thr His Glu Asp Pro Tyr Thr Phe Gly
900 905 910
Gly Gly Thr Lys Leu Glu Ile Lys Ser Asp Pro Ala Thr Thr Thr Pro
915 920 925
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
930 935 940
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
945 950 955 960
Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Val Val Ile Ser Val Gly
965 970 975
Ser Met Gly Leu Ile Ile Ser Leu Leu Cys Val Tyr Phe Trp Leu Glu
980 985 990
Arg Thr Met Pro Arg Ile Pro Thr Leu Lys Asn Leu Glu Asp Leu Val
995 1000 1005
Thr Glu Tyr His Gly Asn Phe Ser Ala Trp Ser Gly Val Ser Lys
1010 1015 1020
Gly Leu Ala Glu Ser Leu Gln Pro Asp Tyr Ser Glu Arg Leu Cys
1025 1030 1035
Leu Val Ser Glu Ile Pro Pro Lys Gly Gly Ala Leu Gly Glu Gly
1040 1045 1050
Pro Gly Ala Ser Pro Cys Asn Gln His Ser Pro Tyr Trp Ala Pro
1055 1060 1065
Pro Cys Tyr Thr Leu Lys Pro Glu Thr Gln Cys Thr Asn Tyr Ala
1070 1075 1080
Leu Leu Lys Leu Ala Gly Asp Val Glu Ser Asn Pro Gly Pro Met
1085 1090 1095
Glu Thr Asp Thr Leu Ile Leu Trp Val Leu Leu Leu Leu Val Pro
1100 1105 1110
Gly Ser Thr Gly Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu
1115 1120 1125
Val Lys Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Val Ser Gly
1130 1135 1140
Tyr Ala Ile Ser Ser Ser Trp Met Asn Trp Val Lys Gln Arg Pro
1145 1150 1155
Gly Gln Gly Leu Glu Trp Ile Gly Arg Ile Tyr Pro Gly Asp Gly
1160 1165 1170
Asp Thr Lys Tyr Asn Gly Lys Phe Lys Asp Lys Ala Thr Leu Thr
1175 1180 1185
Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu
1190 1195 1200
Thr Ser Val Asp Ser Ala Val Tyr Phe Cys Ala Arg Asp Gly Tyr
1205 1210 1215
Arg Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
1220 1225 1230
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1235 1240 1245
Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Thr Ala
1250 1255 1260
Pro Ser Val Phe Val Thr Pro Gly Glu Ser Val Ser Ile Ser Cys
1265 1270 1275
Arg Ser Ser Lys Ser Leu Leu His Ser Asn Gly Asn Thr Tyr Leu
1280 1285 1290
Tyr Trp Phe Leu Gln Arg Pro Gly Gln Ser Pro Gln Leu Leu Ile
1295 1300 1305
Tyr Arg Met Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser
1310 1315 1320
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val
1325 1330 1335
Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln His Leu Glu
1340 1345 1350
Tyr Pro Val Thr Phe Gly Ala Gly Thr Lys Val Glu Ile Lys Ser
1355 1360 1365
Asp Pro Ala Lys Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp
1370 1375 1380
Asn Glu Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His
1385 1390 1395
Leu Cys Pro Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Pro Ile
1400 1405 1410
Leu Leu Thr Ile Ser Ile Leu Ser Phe Phe Ser Val Ala Leu Leu
1415 1420 1425
Val Ile Leu Ala Cys Val Leu Trp Lys Lys Arg Ile Lys Pro Ile
1430 1435 1440
Val Trp Pro Ser Leu Pro Asp His Lys Lys Thr Leu Glu His Leu
1445 1450 1455
Cys Lys Lys Pro Arg Lys Asn Leu Asn Val Ser Phe Asn Pro Glu
1460 1465 1470
Ser Phe Leu Asp Cys Gln Ile His Arg Val Asp Asp Ile Gln Ala
1475 1480 1485
Arg Asp Glu Val Glu Gly Phe Leu Gln Asp Thr Phe Pro Gln Gln
1490 1495 1500
Leu Glu Glu Ser Glu Lys Gln Arg Leu Gly Gly Asp Val Gln Ser
1505 1510 1515
Pro Asn Cys Pro Ser Glu Asp Val Val Ile Thr Pro Glu Ser Phe
1520 1525 1530
Gly Arg Asp Ser Ser Leu Thr Cys Leu Ala Gly Asn Val Ser Ala
1535 1540 1545
Cys Asp Ala Pro Ile Leu Ser Ser Ser Arg Ser Leu Asp Cys Arg
1550 1555 1560
Glu Ser Gly Lys Asn Gly Pro His Val Tyr Gln Asp Leu Leu Leu
1565 1570 1575
Ser Leu Gly Thr Thr Asn Ser Thr Leu Pro Pro Pro Phe Ser Leu
1580 1585 1590
Gln Ser Gly Ile Leu Thr Leu Asn Pro Val Ala Gln Gly Gln Pro
1595 1600 1605
Ile Leu Thr Ser Leu Gly Ser Asn Gln Glu Glu Ala Tyr Val Thr
1610 1615 1620
Met Ser Ser Phe Tyr Gln Asn Gln
1625 1630
<210> 47
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальная последовательность, полученная из CD8a человека
<400> 47
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala
<210> 48
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 48
Ser Asp Pro Ala
1
<210> 49
<211> 231
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> спейсер IgG1Fc человека (HCH2CH3pvaa)
<400> 49
Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
20 25 30
Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val
35 40 45
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
50 55 60
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
65 70 75 80
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
85 90 95
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
100 105 110
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
115 120 125
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
130 135 140
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
145 150 155 160
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
165 170 175
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
180 185 190
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
195 200 205
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
210 215 220
Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 50
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> трансмебранный, полученный из CD28 человека
<400> 50
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 51
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> эндодомен, полученный из TCRz
<400> 51
Arg Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln
1 5 10 15
Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu
20 25 30
Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly
35 40 45
Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
50 55 60
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
65 70 75 80
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
85 90 95
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
100 105 110
Arg
<210> 52
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальная последовательность, полученная из мышиной каппа VIII
<400> 52
Met Glu Thr Asp Thr Leu Ile Leu Trp Val Leu Leu Leu Leu Val Pro
1 5 10 15
Gly Ser Thr Gly
20
<210> 53
<211> 40
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> спейсер CD28STK человека
<400> 53
Lys Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser
1 5 10 15
Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro
20 25 30
Leu Phe Pro Gly Pro Ser Lys Pro
35 40
<210> 54
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сайт расщепления фурина
<220>
<221> прочий_признак
<222> (2)..(2)
<223> Xaa может быть любой встречающейся в природе аминокислотой
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (3)..(3)
<223> Xaa может быть Arg или Lys
<400> 54
Arg Xaa Xaa Arg
1
<210> 55
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> консенсусный сайт расщепления вируса гравировки табака (TEV)
<400> 55
Glu Asn Leu Tyr Phe Gln Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КЛЕТКА | 2014 |
|
RU2717984C2 |
| КЛЕТКА | 2014 |
|
RU2732236C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| КЛЕТКА | 2015 |
|
RU2768019C2 |
| СИГНАЛЬНАЯ СИСТЕМА | 2015 |
|
RU2731638C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
| СВЯЗЫВАЮЩИЙ ДОМЕН | 2019 |
|
RU2829934C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2016 |
|
RU2752880C2 |
| ИНДУЦИБЕЛЬНЫЕ ХИМЕРНЫЕ ЦИТОКИНОВЫЕ РЕЦЕПТОРЫ | 2019 |
|
RU2826155C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИММУНОТЕРАПИИ | 2013 |
|
RU2832516C2 |


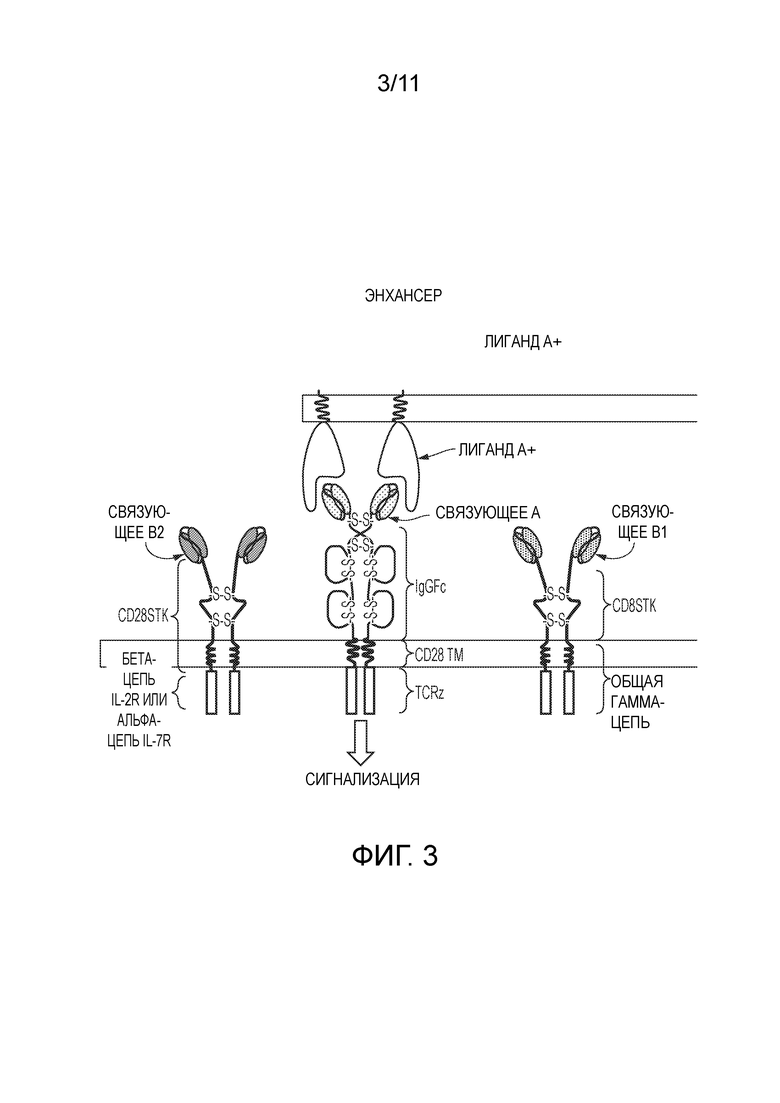
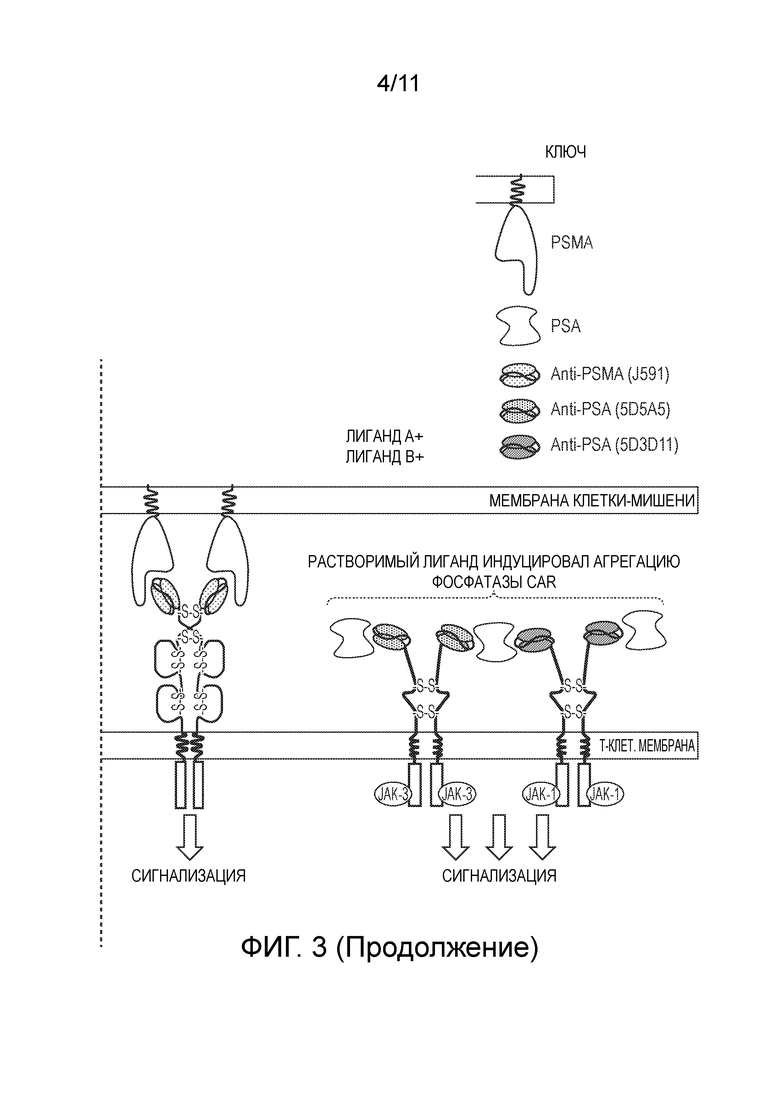
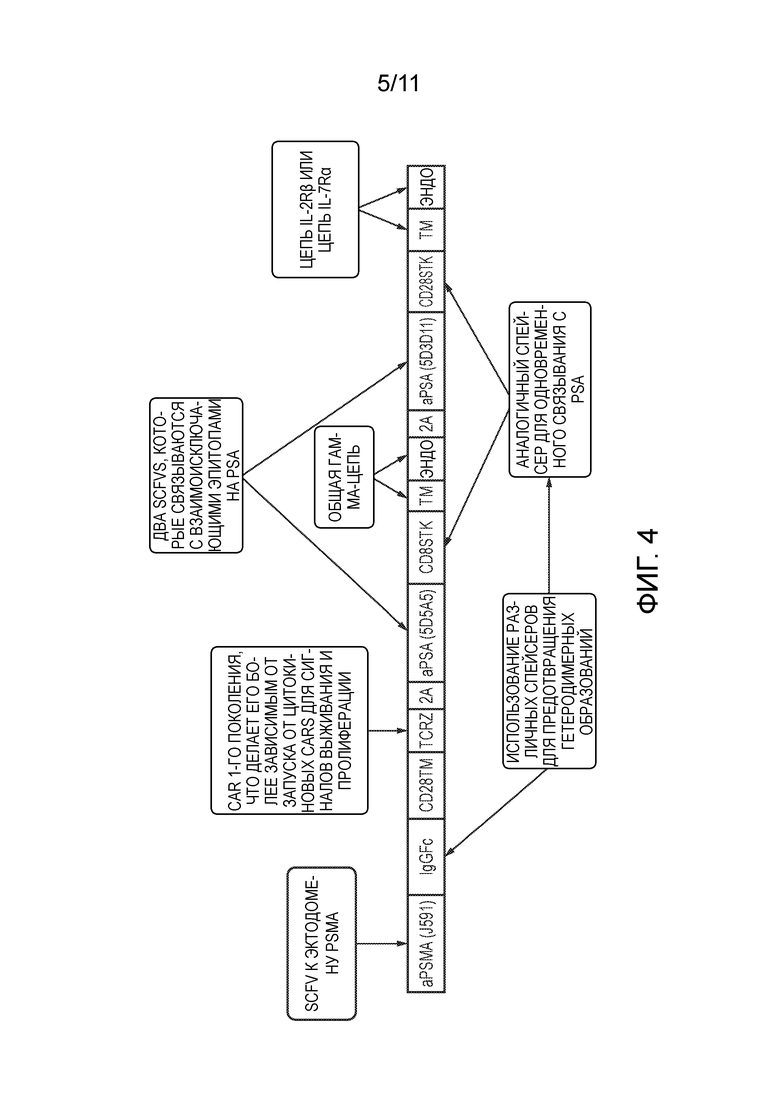
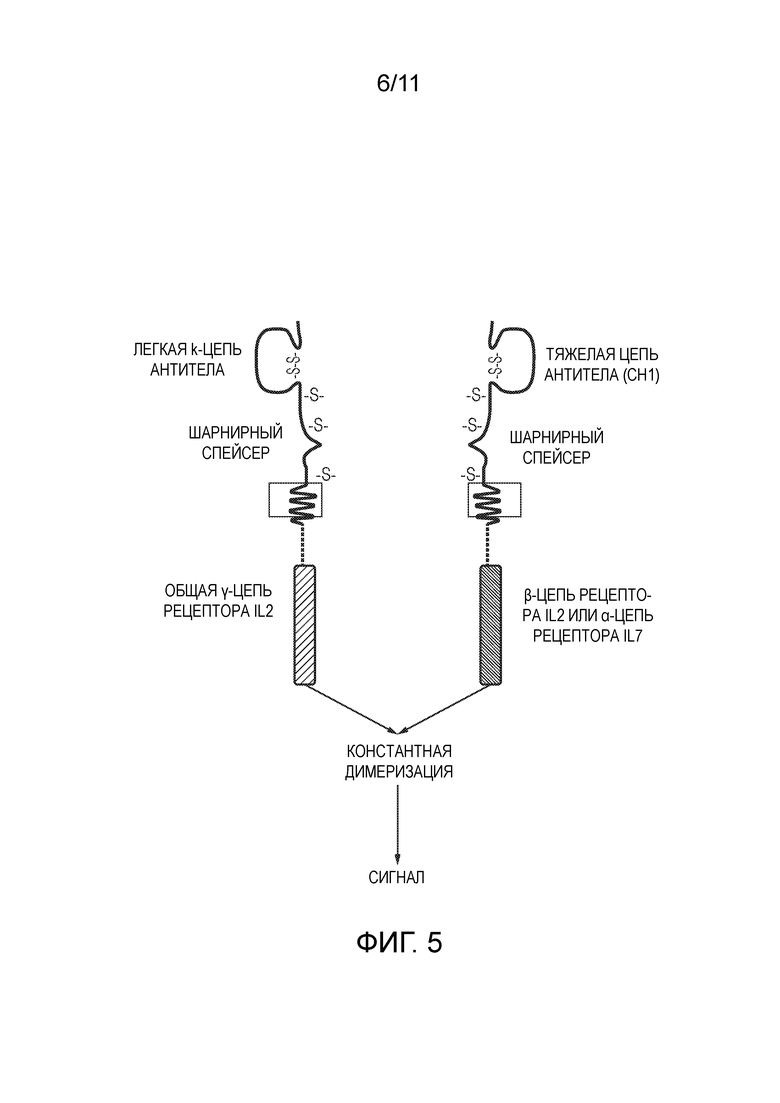





Изобретение относится к области биотехнологии, конкретно к химерным цитокиновым рецепторам (CCR), и может быть использовано в медицине для лечения рака. Предложена конструкция нуклеиновой кислоты для получения клетки, которая экспрессирует химерный трансмембранный белок, причем указанная конструкция нуклеиновой кислоты кодирует химерный трансмембранный белок и содержит первую последовательность нуклеиновой кислоты, кодирующую первый полипептид, содержащий домен димеризации константного домена тяжелой цепи (CH) и первую цепь эндодомена цитокинового рецептора типа I; и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид, содержащий домен димеризации константного домена легкой цепи (CL), который спонтанно гетеродимеризуется с указанным доменом димеризации константного домена тяжелой цепи (CH), и вторую цепь эндодомена цитокинового рецептора типа I. При этом указанный химерный трансмембранный белок обеспечивает конститутивно активный цитокиновый сигнал в указанной клетке. 5 н. и 12 з.п. ф-лы, 9 ил., 1 табл., 4 пр.
1. Конструкция нуклеиновой кислоты для получения клетки, которая экспрессирует химерный трансмембранный белок, причем указанная конструкция нуклеиновой кислоты кодирует химерный трансмембранный белок и содержит:
(i) первую последовательность нуклеиновой кислоты, кодирующую первый полипептид, содержащий:
(а) домен димеризации константного домена тяжелой цепи (CH); и
(b) первую цепь эндодомена цитокинового рецептора типа I; и
(ii) вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид, содержащий:
(а) домен димеризации константного домена легкой цепи (CL), который спонтанно гетеродимеризуется с указанным доменом димеризации константного домена тяжелой цепи (CH); и
(b) вторую цепь эндодомена цитокинового рецептора типа I,
причём указанный химерный трансмембранный белок обеспечивает конститутивно активный цитокиновый сигнал в указанной клетке.
2. Конструкция нуклеиновой кислоты по п. 1, отличающаяся тем, что указанная конструкция нуклеиновой кислоты имеет следующую структуру:
Dim1-TM1-эндо1-коэкспр-Dim2-TM2-эндо2,
в которой
Dim1 представляет собой последовательность нуклеиновой кислоты, кодирующую домен димеризации константного домена тяжелой цепи (CH);
TM1 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен первого полипептида;
эндо 1 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен первого полипептида;
коэкспр представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию обоих полипептидов;
Dim2 представляет собой последовательность нуклеиновой кислоты, кодирующую домен димеризации константного домена легкой цепи (CL);
TM2 представляет собой последовательность нуклеиновой кислоты, кодирующую трансмембранный домен второго полипептида;
эндо 2 представляет собой последовательность нуклеиновой кислоты, кодирующую эндодомен второго полипептида.
3. Конструкция нуклеиновой кислоты по п. 1 или 2, которая дополнительно содержит последовательность нуклеиновой кислоты, которая кодирует химерный антигенный рецептор (CAR).
4. Конструкция нуклеиновой кислоты по любому из пп. 1-3, где коэкспр кодирует последовательность, содержащую саморасщепляющийся пептид.
5. Конструкция нуклеиновой кислоты по любому из пп. 1-4, где альтернативные кодоны используют в областях последовательности, кодирующих одни и те же или подобные аминокислотные последовательности, чтобы избежать гомологической рекомбинации.
6. Конструкция нуклеиновой кислоты по любому из пп. 1-5, отличающаяся тем, что эндодомен цитокинового рецептора в химерном трансмембранном белке содержит:
(i) эндодомен β-цепи рецептора IL-2
(ii) эндодомен α-цепи рецептора IL-7; или
(iii) эндодомен α-цепи рецептора IL-15; и/или
(iv) общий эндодомен γ-цепи рецептора.
7. Конструкция нуклеиновой кислоты по любому из пп. 1-6, отличающаяся тем, что первый полипептид в химерном трансмембранном белке включает вариабельный домен тяжелой цепи (VH) и константный домен тяжелой цепи (CH); и второй полипептид включает вариабельный домен легкой цепи (VL) и константный домен легкой цепи (CL).
8. Конструкция нуклеиновой кислоты по п. 7, отличающаяся тем, что кодируемый химерный трансмембранный белок включает экзодомен Fab.
9. Вектор для получения клетки, которая экспрессирует химерный трансмембранный белок, где указанный вектор содержит конструкцию нуклеиновой кислоты по любому из пп. 1-8, причём указанный химерный трансмембранный белок способен обеспечивать цитокиновый сигнал в указанной клетке.
10. Вектор по п. 9, где указанный вектор представляет собой ретровирусный вектор, или лентивирусный вектор, или транспозон.
11. Цитолитическая иммунная клетка для адоптивного переноса клеток, которая содержит конструкцию нуклеиновой кислоты по любому из пп. 1-8 или вектор по п. 9 или 10, причём указанная клетка экспрессирует химерный трансмембранный белок, который обеспечивает конститутивно активный цитокиновый сигнал в указанной клетке.
12. Клетка в соответствии с п. 11, которая дополнительно содержит химерный антигенный рецептор.
13. Клетка в соответствии с п. 12, где химерный антигенный рецептор связывает ассоциированный с опухолью антиген клеточной поверхности.
14. Фармацевтическая композиция для адоптивного переноса клеток, содержащая эффективное количество клеток в соответствии с любым из пп. 11-13.
15. Способ лечения рака, который включает стадию введения фармацевтической композиции в соответствии с п. 14 или эффективного количества клеток по любому из пп. 11-13 субъекту.
16. Способ по п. 15, который включает следующие этапы:
(i) выделение образца, содержащего клетки, у субъекта;
(ii) трансдукция или трансфекция клеток: конструкцией нуклеиновой кислоты в соответствии с любым из пп. 1-8 или вектором в соответствии с п. 9 или 10; и
(iii) введение клеток из (ii) субъекту.
17. Способ по п. 16, где образец представляет собой образец, содержащий Т-клетки.
| SOGO T | |||
| et al., Selective expansion of genetically modified T cells using an antibody/interleukin-2 receptor chimera, Journal of immunological methods, 2008, v | |||
| Ленточный тормозной башмак | 1922 |
|
SU337A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| et al., Reversal of Tumor Immune Inhibition Using a Chimeric Cytokine Receptor, MOL | |||
| THERAPY, 2014, v | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| et al., Selective | |||

