ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА СВЯЗАННЫЕ ЗАЯВКИ
[0001] В данной заявке заявляется приоритет США по предварительной заявке номер62/093671, поданной 18 декабря 2014 года, содержание которой включено в настоящий документ посредством ссылки в полном объеме и для всех целей.
ЗАЯВЛЕНИЕ ОТНОСИТЕЛЬНО ПРАВ НА ИЗОБРЕТЕНИЕ, ПОЛУЧЕННОЕ В РЕЗУЛЬТАТЕ ИССЛЕДОВАНИЙ, ФИНАНСИРУЕМЫХ ГОСУДАРСТВОМ
[0002] Настоящее изобретение сделано при поддержке правительства в виде грантов №1 RO1 CA133592-01, присужденным Национальными Институтами Здравоохранения, и грантом №ECCS-1231910, присужденный Национальным Научным Фондом. Правительство обладает определенными правами на изобретение.
СПРАВОЧНАЯ ИНФОРМАЦИЯ ПО КАТЕГОРИЯМ «СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ», ТАБЛИЦА, ИЛИ КОМПЬЮТЕРНАЯ ПРОГРАММА, ПРЕДСТАВЛЕНЫ В ПРИЛОЖЕНИИ В ВИДЕ ФАЙЛА ASCII
[0003]Перечень последовательностей записан в файле 48538-526001WO_ST25. TXT, созданный 13 декабря 2015 года, 2828 байт, машинный формат IBM-PC, операционная система MS-Windows, настоящим включена сюда посредством ссылки в полном объеме и для всех целей.
УРОВЕНЬ ТЕХНИКИ
[0004] В секвенировании ДНК, использование синтетических (не природных) молекул является основной стратегией для дифференциации четырех нуклеотидных оснований (A, C, T и G), которые входят в состав ДНК. Эта стратегия была успешно применена к проверенному временем методу секвенсирования Сангера, который использовался в первоначальных попытках определить геном человека.
[0005] Технологии для секвенирования ДНК существуют, но наблюдается коммерческий спрос на новые методы, которые могут повысить скорость, снизить частоту ошибок и снизить сложность, затраты и требования к реагентам. Существует значительный интерес к технологиям, которые могут секвенировать ДНК с использованием электронных схем, поскольку твердотельная электроника может предложить множество преимуществ по скорости, стоимости и сложности.
[0006] В последние годы появились электронные архитектуры, которые управляются путем пропускания ДНК через нанопоры и контроля ионного тока через одну и ту же пору, или путем пропускания ДНК через нанопоры, но с трансдукцией транзита с использованием смежного электрического туннельного перехода. Платформы основаны на прохождении ДНК через нанопоры, поэтому им обеим присущи характерные трудности работы с нанопорами, такие как нестабильность, хрупкость и требования к точной обработке жидкости. Кроме того, прохождение ДНК через нанопоры имеет ограниченное соотношение сигнал-шум, так что на практике любая информация о последовательности должна быть независимо подтверждена с использованием методов флуоресценции. Кроме того, высокая частота ошибок и «скольжение» через нанопоры ограничивают применение, например, для последовательности повторяющихся последовательностей коротких тандемных повторов, необходимых для приложений по идентификации человека.
[0007] Биосенсор представляет собой аналитическое устройство, которое включает элемент биологического распознавания при прямом пространственном контакте с элементом трансдукции. Такая интеграция обеспечивает быстрое и удобное преобразование биологических событий в обнаруживаемые сигналы. Среди различных электрической биосерсорных архитектур, устройства на полевых транзисторах (FET), привлекли большое внимание, поскольку они представляют собой тип биосенсора, который может непосредственно переводить взаимодействия между молекулами-мишенями (например, биологическими молекулами) и поверхностью транзистора, в читаемые электрические сигналы. В стандартном полевом транзисторе ток протекает вдоль проводящего пути (канала), который соединен с двумя электродами, (источник и сток). Проводимость канала между источником и стоком включается и выключается третьим (запорным) электродом, который емкостно связан через тонкий диэлектрический слой. Полевые транзисторы обнаруживают таргетные химические вещества и измеряют химические концентрации для широкого спектра коммерческих применений, включая, например, контроль промышленных процессов, обнаружение утечек, мониторинг стоков, и медицинскую диагностику.
[0008] Например, раскрытое в заявке на патент США № 13/626760 представляет собой электронное устройство, которое достаточно чувствительно для обнаружения на уровне одной молекулы. . Аспекты изобретения выполняются с использованием электропроводящего канала, который имеет единственную сенсибилизирующую молекулу, к нему прикрепленную. Соответственно, устройства, раскрытые в нем, контролируют динамику реакции одной молекулы и могут быть использованы в важных биохимических анализах с одной молекулой, таких как детекторы в реакции секвенирования одной молекулы.
[0009] Таким образом, существует потребность в технике для методов секвенирования ДНК следующего поколения, которые являются более эффективными и более информативными, чем существующие методы. Здесь представлены решения этих и других проблем в данной области техники.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0010] В данном документе представлены, в частности, контуры со смесью природных и неприродных нуклеотидных оснований для определения генетической последовательности образца ДНК. Описаны специальные методы и сокращение практики использования контура для определения генетического кода цепи ДНК.
[0011] Контур позволяет секвенировать ДНК и, кроме того, секвенировать РНК и углеводы. Изобретение предлагает способ недорогого высокоскоростного высокоточного секвенирования ДНК, который может успешно конкурировать с более традиционными методами секвенирования.
[0012] Способы и композиции, представленные в данном документе, могут сопровождать активность одиночных молекул во время ферментативной обработки. Синтетические субстраты, нуклеотиды и флуорофоры могут быть использованы для создания уникальных и различимых сигналов от нити ДНК
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0013]ФИГ. 1A-1C. Электрический мониторинг активности KF с химически модифицированными дезоксиробонуклеотидами dNTPs. ФИГ. 1A:Одиночный наноконтур KF и химически модифицированные dNTP, проверенные на их включение KF. (а) Принципиальная схема однополюсного полевого транзистора на основе однослойных углеродных нанотрубок (SWCNT-FET), нековалентно биоконъюгированного с одной молекулой ДНК-полимеразы I (KF) через одиночный цистеин, введенный в субдомен «пальцы». Пирен-малеимидный линкер (желтый), прикрепленный к SWCNT-FET через π-π укладка и ковалентное присоединение к одиночному цистеину для иммобилизации KF. SWCNT-FET выращивали на SiO2, соединенный с металлическими электродами источника и стока, и пассивированный полимером (ПММА, обозначен красным цветом). ФИГ. 1B:Атомно-силовая микроскопия показывает диаметр 1-2 нм полевого транзистора SWNT с одним креплением KF (7 нм, стрелка). ФИГ. 1C:Химические структуры аналоговых dNTP, описанные в этом документе. Выделены химические модификации нативных dNTP.
[0014] ФИГ. 2A-2F. Изменения в токе во время включения природного и измененного dNTP. ФИГ. 2A:При измерении тока в присутствии poly(dC)42 матрицы и комлемертарного ей природный dGTP, ΔI(t) колебания происходят во время включения каждого снования. Высокие и низкие токи соответствуют открытой и закрытой конформации фермента, соответственно. ФИГ. 2B-2F:Увеличение временного масштаба данных, соответствующих ФИГ. 2A (временное окно от 1,5 до 2,5 сек) иллюстрирует уменьшение событий переключения, соответствующих включению основания dGTP (ФИГ. 2B), α-тио-dGTP (Фиг. 2C), 6-хлор-2APTP (Фиг. 2D), и 2-тио-dCTP (Фиг. 2E-2F). Направо от каждой из ФИГ. 2B-2F, увеличение изображает одиночную ΔI(t) флуктуацию для каждого указанного основания, подчеркивающее разрешение до одного основания, со штрихом, соответствующим интервалу времени 1 мсек.
[0015] ФИГ. 3A-3B. Прямое сравнение распределений вероятностей open, τopen и closed, τclosed, состояний длительности во время включения указанных dNTP из наборов данных >50 с. На Y-осях отложены значения % log вероятности. Как для τclosed (ФИГ. 3A), так и τopen (ФИГ. 3B), матрицу представляет гомополимерный poly(dC)42. На ФИГ. 3A-3B, одно-экспоненциальные подгонки для каждого нуклеотида показаны сплошными линиями.
[0016]ФИГ. 4A-4B. Электронные сигналы, генерируемые при обработке гpoly(dA)42. ФИГ. 4A:Когда KF обрабатывает poly(dA)42 в присутствии природного нуклеотидного дезокситимидинтрифосфата (dTTP), каждое включение пары оснований вызывает отрицательный всплеск тока ΔI<0. ФИГ. 4B:Когда dTTP заменяется неприродным нуклеотидом 2-тио-2'-дезокситимидин-5'-трифосфатом (2-тио-dTTP), включения оснований вызывают положительные импульсы тока ΔI>0.
[0017] ФИГ. 5A-5C. Электронные сигналы, генерируемые при обработке гетерогенных субстратов. ФИГ. 5A:KF обрабатывает гетерогенные субстраты в присутствии всех четырех природных нуклеотидов (dNTP), причем каждое включение пары оснований вызывает отрицательное колебание тока ΔI<0. Индивидуальные колебания могут быть перечислены, как показано, но в целом они не различают один тип основания от другого. ФИГ. 5B:ФИГ. 5B демонстрирует симуляцию одного и того же набора данных с заменой dTTP на 2-тио-dTTP. С тиолированным дезокситимидином, положительные колебания указывают (№ 2, 6, 7) места, где были включены T-нуклеотиды. ФИГ. 5C показывает, что, когда KF обрабатывает гетерогенные субстраты в присутствии природных нуклеотидов (dNTP), смешанных с некоторыми аналогами, полученный шаблон содержит положительные и отрицательные колебания тока, которые могут быть использованы для идентификации выбранного основания. В этом примере показаны данные, полученные с использованием трех нативных нуклеотидов (dATP, dTTP, dCTP), смешанных с 6-Cl-2APTP в качестве аналога для включения G-оснований. Эта информация используется в способе секвенирования олигонуклеотида с помощью нанотрубок.
[0018] ФИГ. 6. На рисунке изображен репрезентативный 15%-ный SDS-PAGE гель KF после суперэкспрессии и очистки. KF очищали до >95% гомогенности и он мигрировал согласно его ожидаемой массе около 68 кДа.
[0019] ФИГ. 7. На рисунке изображен анализ активности на основе флуоресценции, изображающий KF (L790C) (черные круги) и активность KF дикого типа (серые круги) в стационарных условиях. в реакции удлинения праймера, которая происходит в присутствии dATP, dTTP, dCTP и dGTP. Исходные данные были вычтены из фона, который измеряли как активность в отсутствие dNTP.
[0020]На фиг. 8A-8B. На рисунках показан групповой анализ, показывающий включение непрородных аналогов dNTP с матрицами, описанными в этом документе. Продукты полимеризации с аналогами dNTP и матрицей включения A/T (ФИГ. 8A) или матрицей включения G/C (Фиг. 8B) подвергали электрофорезу на 5%-м агарозном геле с высоким разрешением. Реакции отрицательного контроля только с 3 dNTP, опуская dTTP (1), dATP (2), dCTP (8) и dGTP (9), не содержащие dsDNA. Реакции положительного контроля со всеми четырьмя dNTP показали преобразование в dsDNA как с матрицей включения A/T (3), так и с матрицей включения G/C (10). Реакции с аналогами dNTP (4-7 и 11-14) без соответствующего им нативного основания dNTP и содержащие оставшиеся 3 природные dNTP. Напротив матрицы включения A/T α-тио-dTTP (4) и 2-тио-dTTP (5), введенного напротив основания матрицы A, и α-тио-dATP (6) и 6-Cl-2APTP (7) встроенного напротив основания матрицы Т. Напротив матрицы включения G/C, α-тио-dCTP (11) и 2-тио-dCTP (12), встроенного напротив основания матрицы G, и α-тио-dGTP (13) и 6 -Cl-2APTP (14), встроенного напротив основания матрицы C. После визуализации, цвета изображений были инвертированы, а затем изменены на черно-белые.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0021] В данном документе, в частности, предложен способ обнаружения изменения в конформации полимеразы нуклеиновой кислоты; cпособ секвенирования полимеразой нуклеиновой кислоты, в которой обнаруживается изменение конформации полимеразы нуклеиновой кислоты. В вариантах реализации изобретения способы включают обнаружение конформационного изменения полимеразы нуклеиновой кислоты с аналогами нуклеиновой кислоты.
Определения
[0022] Следующие определения включены для понимания настоящего предмета изобретения и для построения прилагаемых патентных заявок. Аббревиатуры, применяемые в данном документе, имеют свое обычное значение в пределах химической и биологической областях техники.
[0023] Если не указано иное значение, все технические и научные термины, используемые в данном документе, имеют то же значение, как их обычно понимает специалист в данной области техники. См., например, Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J. (Wiley & Sons, New York, 1994, and quarterly updates); Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Springs Laboratory, Cold Springs Harbor, N. Y 1989. Любые способы, устройства и материалы, аналогичные или эквивалентные описанным здесь, могут быть использованы в подобной практике. Следующие определения предоставляются для облегчения понимания некоторых терминов, часто используемых в данном документе, и не предназначены для ограничения объема настоящего раскрытия.
[0024] Термин «нуклеиновая кислота» относится к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам в виде одно-, двух- или многоцепочечных форм, или их дополнений. Термин «полинуклеотид» относится к линейной последовательности нуклеотидов. Термин «нуклеотид» обычно относится к одному звену полинуклеотида, т. е. к мономеру. Нуклеотидами могут быть рибонуклеотиды, дезоксирибонуклеотиды или их модифицированные варианты. Примеры рассматриваемых здесь полинуклеотидов включают одно- и двухцепочечную ДНК, одно- и двухцепочечную РНК (включая siRNA) и гибридные молекулы, имеющие смеси одно- и двухцепочечной ДНК и РНК. Нуклеиновые кислоты могут быть линейными или разветвленными. Например, нуклеиновые кислоты могут быть линейной цепью нуклеотидов или нуклеиновые кислоты могут быть разветвленными, например, такие, когда нуклеиновые кислоты содержат одно или несколько плеч или ветвей нуклеотидов. По выбору, разветвленные нуклеиновые кислоты повторно разветвляются с образованием высших упорядоченных структур, таких как дендримеры и тому подобное.
[0025] Нуклеиновые кислоты, включая нуклеиновые кислоты с фосфотиоатным скелетом, могут включать одну или несколько реакционноспособных остатков. Используемый здесь термин реакционноспособный остаток включает любую группу, способную взаимодействовать с другой молекулой, например нуклеиновой кислотой или полипептидом, посредством ковалентных, нековалентных или других взаимодействий. В качестве примера, нуклеиновая кислота может включать реакционноспособный остаток аминокислоты, который реагирует с амиокислотой на белке или полипептиде посредством ковалентного, нековалентного или другого взаимодействия.
[0026] Термины также включают в себя нуклеиновые кислоты, содержащие известные нуклеотидные аналоги или модифицированные остатки или связи со скелетом, которые являются синтетическими, встречающимися в природе, и не встречающимися в природе, которые имеют сходные связывающие свойства, как эталонная нуклеиновая кислота, и которые метаболизируются способом, подобно как талонные нуклеотиды. Примеры таких аналогов включают, без ограничения, производные фосфодиэфира, например, фосфорамидат, фосфорамидат, фосфоротиоат (также известный как фосфотиоат), фосфородитиоат, фосфонокарбоновые кислоты, фосфонокарбоксилаты, фосфоноуксусная кислота, фосфоноформальная кислота, метилфосфонат, борфосфонат или O-метилфосфорамидитные связи (см. Eckstein, Oligonucleotides and Analogues: A Practical Approach, Oxford University Press); и цепи и связи пептидов и нуклеиноых кислот. A Practical Approach, Oxford University Press); пептидные остовы нуклеиновых кислот и связи. Другие аналоговые нуклеиновые кислоты включают группы с положительными скелетами; неионными скелетами, модифицированные сахара и нерибозные скелеты (например, фосфородиамидатные морфолиновые олигонуклеотиды или блокированные нуклеиновые кислоты (LNA)), в том числе в патентах США№№5,235,033 и 5,034,506 и в главах 6 and 7, ASC Symposium Series 580, Carbohydrate Modifications in Antisense Research, Sanghui & Cook, eds. Нуклеиновые кислоты, содержащие один или более карбоциклических сахаров, также включены в одном определении нуклеиновых кислот. Модификации рибозофосфатного остова могут быть осуществлены по целому ряду причин, например, для увеличения стабильности и периода полужизни таких молекул в физиологических средах или в качестве зондов на биочипе. Могут быть приготовлены смеси встречающихся в природе нуклеиновых кислот и аналогов; в альтернативном варианте, могут быть приготовлены смеси различных аналогов нуклеиновых кислот и смеси встречающихся в природе нуклеиновых кислот и аналогов. В вариантах реализации изобретения межнуклеотидные связи в ДНК представляют собой фосфодиэфир, производные фосфодиэфира или их комбинацию.
[0027] Слова «комплементарная» или «комплементарность» относятся к способности нуклеиновой кислоты в полинуклеотиде образовывать пару оснований с другой нуклеиновой кислотой в другом полинуклеотиде. Например, последовательность AGT является комплементарной последовательности TCA. Комплементарность может быть частичной, в которой только некоторые из нуклеиновых кислот соответствуют по парному спариванию, или полной, где все нуклеиновые кислоты спариваются в соответствии с базовым спариванием.
[0028] Термин «гибридизация» и тому подобное относятся в обычном и привычном смысле к образованию двухцепочечной (т. е. дуплексной) нуклеиновой кислоты, включая, например, гибрид ДНК/ДНК, гибрид ДНК/РНК и гибрид РНК/РНК. Понятно, что образование дуплексной нуклеиновой кислоты может происходить через спаривание оснований путем взаимодействий Уотсона-Крика. Фраза «селективно (или специфично) гибридизуется с» относится к связыванию, дуплексированию или гибридизации нуклеиновой кислоты с конкретной нуклеотидной последовательностью с более высокой аффинностью, например, в более жестких условиях, чем с другими нуклеотидными последовательностями (например, с тотальной клеточной или библиотечной ДНК или РНК).
[0029] Как используется в данном документе, конформационное изменение полимеразы нуклеиновой кислоты обнаруживатеся с использованием однополюсного полевого транзистора на основе нанотрубок углерода (SWCNT-FET). Например, наноконтур фрагмента Кленова (KF) включает в себя SWCNT-FET, нековалентно биоконъюгированный с одной молекулой ДНК-полимеразы I (KF) через один цистеин, введенный в субдомен «пальцев». Изменение конформации измеряется сигналами AI(t), создаваемыми наноконтуром KF. Устройство создает непрерывные последовательности отрицательных ΔI(t) колебаний, каждая из которых указывает на образование одной пары оснований, а с инвертированной амплитудой отражает другую конформацию KF (ФИГ. 2C, ФИГ. 2F).
[0030] Как используется в данном документе, в вариантах реализации изобретения, первый нуклеотид или первый нуклеотидный аналог может быть таким же, как второй нуклеотид или второй нуклеотидный аналог, соответственно. первый нуклеотид или первый нуклеотидный аналог могут быть отличными от второго нуклеотида или второго нуклеотидного аналога, соответственно.
[0031] Фраза «условия строгой гибридизации» относится к условиям, при которых нуклеиновая кислота будет гибридизоваться с последовательностью-мишенью, как правило, в сложной смеси нуклеиновых кислот, но ни с какими другими последовательностями. Жесткие условия зависят от последовательностей и будут различаться в разных обстоятельствах. Специфическая гибридизация более длинных последовательностей происходит при более высоких температурах. Расширенное руководство по гибридизации нуклеиновых кислот можно найти в Tijssen, Techniques In Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes, ''Overview of principles of hybridization and the strategy of nucleic acid assays'' (1993 год). Как правило, жесткие условия выбираются таким образом, чтобы быть ниже, чем температура плавления (Tm) на около 5-10°C для определенной последовательности при определенной ионной силе и рН. Tm представляет температуру (при определенных ионной силе, рН и концентрации нуклеиновой кислоты), при которой 50% зондов, комплементарных последовательности-мишени, гибридизируются с последовательностью-мишенью в состоянии равновесия (если последовательности-мишени присутствуют в избытке, при Tm наблюдается занятость 50% зондов в состоянии равновесия). Строгие условия гибридизации также могут быть достигнуты с добавлением дестабилизирующих агентов, таких как формамид. Для селективной или специфической гибридизации положительный сигнал должен быть по меньшей мере в два раза выше фона, предпочтительно в 10 раз выше фоновой гибридизации. Типовые жесткие условия гибридизации включают следующее:50% формамида, 5x SSC и 1% SDS, инкубацию при 42°С или 5x SSC, 1% SDS, инкубацию при 65°С, с промывкой в 0,2x SSC и 0,1% SDS при 65°С. Типичные «умеренно строгие условия гибридизации» включают гибридизацию в буфере с 40% формамида, 1 М NaCl, 1% SDS при 37°C, и промывку в 1X SSC при 45°C. Положительная гибридизация должна быть, по крайней мере, в два раза выше фона. Специалисты со средним уровнем квалификации легко поймут, что альтернативные условия гибридизации и промывки могут быть использованы для обеспечения условий одинаковой жесткости. Дополнительные рекомендации по определению параметров гибридизации приведены в многочисленных ссылках, например, и Current Protocols in Molecular Biology, ed. Xu et al.,John Wiley & Sons.
[0032] В вариантах реализации изобретения, устройство обнаружения одной молекулы 10 может иметь форму транзистора, а именно полевой транзистор (FET) с присоединенными биомолекулами, служащими «затвором» в электрической цепи. В этом варианте реализации изобретения, одна сенсибилизирующая молекула выступает как одномолекулярный затвор для устройства. В этом варианте реализации изобретения, одна сенсибилизирующая молекула выступает как одномолекулярный затвор для устройства. Вариант реализации транзистора может включать в себя два или три терминальных транзистора. Канал проводимости также может быть образован из металлов, оксидов металлов,полупроводников или нанометровых проводников, таких как нанопроводники, графены или однослойные углеродные нанотрубки (SWNT). В одном варианте реализации изобретения, канал проводимости представляет собой одиночный SWNT.
Способы
[0033] Здесь представлен способ обнаружения изменения конформации полимеразы нуклеиновой кислоты. Способ включает контактирование полимеразы нуклеиновой кислоты, нековалентно прикрепленной к однослойной углеродной нанотрубке (SWNT) с нуклеотидом или нуклеотидным аналогом (например, с первым нуклеотидом или нуклеотидным аналогом) и последовательностью матричной нуклеиновой кислоты (например, олигонуклеотидом смысловой цепи или полинуклеотидом) тем самым образуя конформационно измененную полимеразу нуклеиновой кислоты, связанную с нуклеотидом или нуклеотидным аналогом и последовательностью матричной нуклеиновой кислоты. Конформационно измененную полимеразу нуклеиновой кислоты обнаруживают путем измерения изменения электропроводности SWNT между полимеразой нуклеиновой кислоты и конформационно измененной полимеразой нуклеиновой кислоты. Термин «контактирование» и тому подобное относится в обычном и привычном смысле к приведению двух или более объектов в достаточно тесный контакт, таким образом чтобы между объектами могло происходить взаимодействие, например связывание, химическая реакция и тому подобное. Термин «изменение электропроводности» и тому подобное относятся в обычном и привычном смысле к изменению электропроводности, которое может быть измерено способами, известными в данной области техники и раскрытыми здесь. Термин «конформационно измененная полимераза нуклеиновой кислоты» и тому подобное относятся, в обычном и привычном смысле, к изменению вторичной, третичной и/или четвертичной структуры или нуклеиновой кислоты, как известно в данной области.
[0034] Как раскрыто в данном документе, изменение проводимости может быть результатом изменения положения сенсибилизирующей молекулы (например, аминокислоты), которая является частью полимеразы нуклеиновой кислоты, относительно полимеразы нуклеиновой кислоты и конформационно измененной полимеразы нуклеиновой кислоты. Колебания тока могут включать простые увеличения и уменьшения прямоугольной формы. Альтернативно, колебания могут включать любую форму волны, включая формы, которые могут быть треугольными, синусоидальными, или иметь любое количество компонентов Фурье. Амплитуды, длительности и формы этих волн кодируют активность таргетного компонента, и поэтому могут быть проанализированы с использованием компьютера, с тем чтобы выявить кинетику связывания и другие механические и электронные степени свободы. Статистический анализ этих параметров дает представление о кинетической изменчивости, переходах и промежуточных химических состояниях процессов связывания и освобождения мишеней. Степени свободы в сигнале тока различают несколько похожих молекул-мишеней, которые все связываются с одним и тем же сайтом, например между молекулой-мишенью и молекулой-ингибитором сайта связывания. Эти степени свободы также могут различать слабые взаимодействия, такие как распознавание молекул, которые происходят до реального связывания.
[0035] Способ обнаружения изменения конформации полимеразы нуклеиновой кислоты может быть использован как часть способа секвенирования нуклеиновой кислоты (например, ДНК и/или РНК. Таким образом, в некоторых вариантах реализации изобретения, способ дополнительно включает, после обнаружения конформационно измененной полимеразы нуклеиновой кислоты, связанной с первым нуклеотидом или нуклеотидным аналогом, детектирование второго изменения конформации указанной полимеразы нуклеиновой кислоты, позволяя упомянутой конформационно измененной полимеразе нуклеиновой кислоты высвобождать первый нуклеотид или нуклеотидный аналог, тем самым изменяя форму полимеразы нуклеиновой кислоты. Далее, способ включает контактирование полимеразы нуклеиновой кислоты, нековалентно прикрепленной к однослойной углеродной нанотрубке (SWNT), со вторым нуклеотидом или нуклеотидным аналогом, в результате чего образуется конформационно измененная полимераза нуклеиновой кислоты, связанная со вторым нуклеотидом или нуклеотидным аналогом. Конформационно измененную полимеразу нуклеиновой кислоты обнаруживают путем измерения изменения электропроводности SWNT между полимеразой нуклеиновой кислоты и конформационно измененной полимеразой нуклеиновой кислоты.
[0036] В вариантах реализации изобретения, первый и/или второй нуклеотид или нуклеотидный аналог продуцируют уникальный сигнал проводимости, который детектируется. Уникальный сигнал проводимости используется для идентификации упомянутого первого и/или второго нуклеотида или нуклеотидного аналога, тем самым идентифицируя последовательность матричной нуклеиновой кислоты. Термины «сигнал проводимости», «первый сигнал проводимости», «уникальный сигнал проводимости» и тому подобное относятся в обычном и привычном смысле к проводимости объектов, измеренного известными способами, включая способы, описанные в этом документе.
[0037] Полезными в способах, представленных в данном документе, являются контуры углеродных нанотрубок, которые могут работать быстрее, с низкой стоимостью и с гораздо более низкой частотой ошибок, чем более традиционные технологии секвенирования. Представленные здесь композиции и способы обеспечивают значительное улучшение по сравнению с электронными архитектурами на основе нанопор. Во-первых, контур углеродных нанотрубок генерирует электронный сигнал с превосходными шумовыми характеристиками, который не требует независимого подтверждения. Во-вторых, контур нанотрубок выдерживает широкий диапазон условий среды и грубое обращение, таким образом что спецификации по манипулированию жидкостями и общая сложность системы могут быть значительно ослаблены по сравнению с нанопоровыми архитектурами. В-третьих, контур нанотрубок концептуально прост и легко адаптируется для работы в самых различных режимах. В-четвертых, в подходе может применяться фермент высокой специфичности для обеспечения различения пар оснований; оцененные коэффициенты ошибок могут быть такими же низкими, как теоретический максимум для фермента 18×10-6. Такие низкая частота ошибок будет примерно в 10 000 раз лучше по сравнению с имеющимися в настоящее время коммерческими инструментами. Таким образом, в этом документе представлены способы и композиции, которые значительно снижают стоимость, сложность, частоту ошибок и дополнительную нагрузку по общирному повторному секвенированию. Общее описание схем нанотрубок приведено в Приложении А и в заявке на патент США №13/626,760.
[0038] Изобретение в общем и целом предлагает электронное устройство, которое достаточно чувствительно для обнаружения на уровне одной молекулы. Аспекты изобретения выполняются с использованием электропроводящего канала, который имеет единственную сенсибилизирующую молекулу, к нему прикрепленную. Соответственно, устройства, раскрытые в изобретении, контролируют динамику реакции одной молекулы и могут быть использованы в важных биохимических анализах с одной молекулой, таких как детекторы в реакции секвенирования одной молекулы.
[0039] Любой тип канала проводимости, который обычно находится в полевых транзисторах, может быть использован с этим изобретением. Примерные каналы проводимости формируются из металлов, оксидов металлов, полупроводников или проводников нанометрового размера, таких как нанопроводники, графены или однослойные углеродные нанотрубки (SWNT). В вариантах реализации изобретения канал проводимости представляет собой одиночный SWNT.
[0040] В качестве класса материалов SWNT представляют собой полупроводники с электронными запрещенными зонами, которые могут изменяться от одного электронвольта до эффективного нуля. Это изменение приводит к классификации углеродных SWNT как металлических или полуметаллических, а других - как полупроводников. С помощью соединительных электродов, электростатических затворов и других схем управления, полупроводниковые SWNT могут быть сконфигурированы как сенсорные полевые транзисторы, как радиочастотные усилители или как низкотемпературные одноэлектронные транзисторы. Устройство и способ не исключают таких дополнений, поскольку в вариантах реализации изобретения устройство состоит только из двухконтактного проводника SWNT. SWNT являются каналами проводимости, в которых устройства для обнаружения одной молекулы могут быть изготовлены из проводников SWNT любого типа, с или без электродов затвора, а также на стеклянных, пластиковых или кремниевых подложках. Описанное здесь одномолекульное сенсорное устройство может быть одним компонентом внутри полевого транзистора или любым количеством более сложных электронных или оптоэлектронных устройств и схем.
[0041] Одним из аспектов раскрытия является надежное достижение только одной активной сенсибилизирующей молекулы в каждом устройстве. В общем, сенсибилизирующие молекулы будут наноситься на SWNT со средним интервалом, который определяется концентрацией и периодом инкубации, используемыми при приготовлении. Когда это среднее расстояние было эмпирически определено для определенного набора условий, проводник SWNTНа практике, эта длина обычно составляет от 1 до 100 нм, когда чувствительные молекулы непосредственно присоединяются к проводнику SWNT, при этом этот диапазон трудно контролировать с помощью оптической литографии.
[0042] В вариантах реализации, молекулы-линкеры служат в качестве промежуточного посредника, который улучшает контроль над средним разделением сенсибилизирующих молекул. Любой способ, известный в данной области, может быть использован для присоединения одиночной сенсибилизирующей молекулы к проводнику. В вариантах реализации изобретения, молекула-линкер используется для присоединения одной сенсибилизирующей молекулы. В вариантах реализации изобретения, молекула-линкер включает по меньшей мере первую и вторую функциональную группу. Как правило, первая функциональная группа взаимодействует с каналом проводимости (например, однослойная углеродная нанотрубка), а вторая функциональная группа взаимодействует с сенсибилизирующей молекулой. Примеры первых функциональных групп включают пирен, бензол, циклогексан и 2,3-dichloro-5,6-dicyano-1,4-benzoquinone. Примером второй функциональной группы является малеимид. В некоторых вариантах реализации изобретения, в которых канал проводимости представляет собой SWNT, молекула-линкер взаимодействует с боковой стенкой SWNT через pi-pi-укладку.
[0043] Используя линкеры, длина между сенсибилизирующими молекулами может быть резко увеличена до 1 микрометра или более. Сенсибилизирующими молекулами, расположенными на расстоянии 1 микрометр друг от друга, становится возможным использовать стандартные методы литографического маскирования для определения облаток, заполненных проводниками, длиной примерно в 1 микрометр. В качестве альтернативы, учитывая желаемый шаг устройства, заданный дизайном маски, концентрацию сенсибилизирующих молекул и продолжительность инкубации можно варьировать для достижения того же результата, одной молекулы на устройство. Устройства с одной молекулой могут быть получены по меньшей мере в 8 из 10 попыток изготовления, причем все они не нарушают sp2 -характер проводника SWNT.
[0044] Любые сенсибилизирующие молекулы, известные в данной области, могут быть использованы с устройствами в изобретении, а выбор сенсибилизирующей молекулы будет зависеть от молекулы, подлежащей обнаружению, или реакции, подлежащей мониторингу. Типичные сенсибилизирующие молекулы включают фермент, белок, нуклеиновую кислоту, рибозим, аптамер и полисахарид. В некоторых вариантах реализации изобретения, фермент представляет собой лизоцим, протеинкиназу А или ДНК-полимеразу I.
[0045] В других аспектах, в каждом устройстве может потребоваться более одной сенсибилизирующей молекулы для достижения динамического зондирования одной молекулы. Например, при желаемой рабочей температуре или рН, определенный тип сенсибилизирующей молекулы может иметь только 25%-ную вероятность быть химически активной. молекулы (например, четыре) к каждому проводнику, чтобы создать устройство, в котором они могут быть активными. молекулы (например, четыре) к каждому проводнику, чтобы создать устройство, в котором они могут быть активными. Эта, более высокая, плотность прикреплений, легко достигается с использованием схемы, описанной выше, либо путем увеличения длины устройств до соответствующего кратного среднего расстояния разделения молекул, либо путем уменьшения того же разделения путем изменения условий присоединения.
[0046] В вариантах реализации изобретения, одномолекульное сенсорное устройство содержит несколько параллельно соединенных проводников (например, проводники SWNT). Одна активная сенсибилизирующая молекула присоединена к одному из проводников, и способствует динамическому электронному сигналу, который выделяется из параллельной, но статической проводимости немодифицированных проводников. Этот вариант реализации изобретения обеспечивает дополнительную гибкость в разработке синтеза проводника или его размещения, и в успешном изготовлении чувствительных устройств с одной молекулой, с использованием сенсибилизирующих молекул, которые имеют очень низкие вероятности прикрепления.
[0047] В вариантах реализации изобретения, несколько детектирующих устройств с одной молекулой изготавливаются параллельно с использованием сенсибилизирующей молекулы того же типа, с одной сенсибилизирующей молекулой, прикрепленной к одному устройству. В другом варианте реализации изобретения, несколько проводников изготавливаются, а затем подвергаются воздействию различных сенсибилизирующих молекул, для того чтобы получить множество одномолекульных детектирующих устройств, которые сенсибилизированы к различным мишеням. В другом варианте реализации изобретения, устройство для определения одной молекулы реагирует на несколько мишеней посредством сенсибилизирующей молекулы с рядом специфических особенностей.
[0048] В вариантах реализации изобретения, устройство для определения одной молекулы включает в себя первый электрод, и второй электрод. Одностенная углеродная нанотрубка соединена, соответственно, с первым электродом и вторым электродом. Устройство содержит по меньшей мере одну молекулу-линкер, имеющую первую и вторую функциональные группы, по меньшей мере одну молекулу линкера, имеющую первую функциональную группу, нековалентно функционализированную боковой стенкой однослойной углеродной нанотрубки. Одна сенсибилизирующая молекула, имеющая по меньшей мере одну функциональную группу, в значении, что по меньшей мере одна функциональная группа единичной сенсибилизирующей молекулы, является функционализированной второй функциональной группой по меньшей мере одной молекулы-линкера.
[0049] В вариантах реализации изобретения, способ изготовления устройства для обнаружения одной молекулы включает в себя формирование по меньшей мере одной однослойной углеродной нанотрубки на подложке, которая соединена с первым электродом и вторым электродом; нековалентная функционализация боковой стенки однослойного углеродного нанотрубочного устройства, по меньшей мере, одной функциональной группой, по меньшей мере, одной молекулы-линкера, содержащей множество функциональных групп; и функционализация, по меньшей мере, одной из функциональных групп, по меньшей мере одной молекулы линкера с одной или несколькими функциональными группами одной сенсибилизирующей молекулы.
[0050] В вариантах реализации изобретения раскрыт способ использования устройства для обнаружения одной молекулы, имеющего однослойную углеродную нанотрубку (SWNT). SWNT расположен на подложке и соединен с первым электродом и вторым электродом, причем чувствительное устройство имеет единственную сенсибилизирующую молекулу, прикрепленную к SWNT, с использованием молекулы-линкера, нековалентно функционализированной с помощью SWNT. Напряжение подается через SWNT. Сенсибилизирующая молекула подвергается воздействию химической среды. Контролируются колебания тока, протекающего через SWNT.
[0051] В вариантах реализации изобретения раскрыты способы секвенирования нуклеиновой кислоты с использованием устройства для определения одной молекулы. Чувствительное устройство содержит проводящий канал. Проводящий канал может включать однослойную углеродную нанотрубку (SWNT) на подложке, соединенную с первым электродом и вторым электродом. Чувствительное устройство имеет один сенсибилизирующий фермент, закрепленный на канале с использованием молекулы-линкера, нековалентно функционализированной с каналом (например, SWNT). Способ включает воздействие устройства на по меньшей мере один тип нуклеотида; приложение потенциала напряжения по каналу; мониторинг колебаний тока, протекающего через SWNT; и идентификации нуклеотидов, включенных в матрицу нуклеиновой кислоты ферментом, основанным, по меньшей мере, частично на контролируемых флуктуациях тока. Фермент может быть полимеразой или обратной транскриптазой. Нуклеотид может быть нуклеотидным аналогом. В некоторых вариантах реализации изобретения устройство подвергается воздействию более одного типа нуклеотидов за один раз.
[0052] Чувствительное устройство также может быть использовано для определения кинетики обработки белка или фермента. Еще одно применение чувствительного устройства заключается в определении эффектов генетической мутации. Устройства, использующие сенсибилизирующие молекулы или мишени с генетическими мутациями, можно сравнивать с результатами, полученными на аналогичных устройствах с сенсибилизирующими молекулами или мишенями, которые не имеют мутации. В еще одном приложении чувствительные устройства могут использоваться для измерения эффектов лекарств или других небольших молекул на белке, чтобы сделать его активным или неактивным.
[0053] Способ изготовления устройств по изобретению может включать в себя протокол биохимической конъюгации с последующим контролируемым промыванием. Такой способ приводит к тому, что устройства по изобретению имеют одну сенсибилизирующую молекулу и не имеют неспецифического связывания мешающих молекул. В некоторых вариантах реализации изобретения, сенсибилизирующая молекула непосредственно присоединена к проводнику посредством нековалентного взаимодействия. нековалентного присоединения, а другая для универсальной биоконъюгации с сенсибилизирующей молекулой. Одна из схем использования промежуточного линкера обеспечивает химически универсальную платформу для создания устройств по изобретению из широкого класса чувствительных молекул.
[0054] В вариантах реализации изобретения, способ изготовления устройства для обнаружения одной молекулы включает в себя формирование по меньшей мере одной однослойной углеродной нанотрубки на подложке, которая соединена с первым электродом и вторым электродом; нековалентная функционализация боковой стенки однослойного углеродного нанотрубочного устройства, по меньшей мере, одной функциональной группой, по меньшей мере, одной молекулы-линкера, содержащей множество функциональных групп; и функционализация, по меньшей мере, одной из функциональных групп, по меньшей мере одной молекулы линкера с одной или несколькими функциональными группами одной сенсибилизирующей молекулы.
[0055] В вариантах реализации изобретения, устройство обнаружения одной молекулы может иметь форму транзистора, а именно полевого транзистора (FET) с присоединенными биомолекулами, служащими «затвором» в электрической цепи. В этом варианте реализации изобретения, одна сенсибилизирующая молекула выступает как одномолекулярный затвор для устройства. Вариант реализации транзистора может включать в себя два или три терминальных транзистора. Примерные каналы проводимости формируются из металлов, оксидов металлов, полупроводников или проводников нанометрового размера, таких как нанопроводники, графены или однослойные углеродные нанотрубки (SWNT). В одном варианте реализации изобретения, канал проводимости представляет собой одиночный SWNT.
[0056] Как правило, длина SWNT может варьироваться от примерно 0,1 до примерно 10 мкм. Конкретная длина SWNT выбирается таким образом, что статистически большинство изготовленных устройств 10 имеют только одну сенсибилизирующую молекулу, связанную с SWNT. Еще более предпочтительно, длина SWNT, которая подвергается воздействию внешней химической среды, выбирается таким образом, что более 75% изготовленных устройств включают только одну сенсибилизирующую молекулу, связанную с SWNT. В некоторых случаях, это расстояние представляет собой расстояние между первым электродом и вторым электродом.
[0057] Первый электрод и второй электрод могут быть дополнительно покрыты крышкой. Крышка может включать в себя окно, выемку, слот или другой открытый сегмент, который обеспечивает доступ из внешней среды к SWNT. В этом отношении, SWNT может подвергаться воздействию химической среды. Например, открытое окно может быть определено в крышке во время производственного процесса. Защитное покрытие гарантирует, что большая часть поверхности, включая первый и второй электроды, защищена от окружающей среды. Кроме того, в предпочтительном варианте реализации изобретения, длина окна приспособлена для достижения правильной длины устройства. окна можно варьировать для достижения желаемой активной области на SWNT. Например, первый и второй электроды могут быть соединены с SWNT и разделены расстоянием в 2 мкм. Однако окно можно сделать меньшим, чем межэлектродное расстояние. Открытое окно в защитном покрытии подвергает SWNT и прикрепленную сенсибилизирующую молекулу воздействию химической среды. Защитным покрытием может быть любая электрически изолирующая пленка, состоящая из одного или нескольких слоев. Пленочные материалы включают полимеры, оксид алюминия, оксид гафния, диоксид кремния или нитрид кремния. Окно определяется в защитном покрытии с использованием литографических методов. Литографические методы хорошо известны в данной области техники и включают использование любой приемлемой комбинации оптического воздействия, методов электронного луча и положительных или отрицательных сопротивлений.
[0058] В вариантах реализации изобретения, изготовление устройства включает в себя устройства для нанесения покрытия в защитном покрытии из положительного электронного пучка, такого как полиметилметакрилат (PMMA); нанесение литографических рисунков электронным лучом; а затем разработка нанесенных участков для экспонирования активного канала SWNT длиной от 0,5 до 1,0 мкм. В другом варианте реализации изобретения, изготовление устройства включает устройства для нанесения покрытия в защитном покрытии из оксида алюминия; устройства для нанесения покрытий дополнительно на пленку оптического фоторезиста; подвергая желаемые окна воздействию света; разработка нанесенных участков для подвергания узких окон из оксида алюминия; травление оксида алюминия для дальнейшего подвергания лежащих в основе каналов SWNT длиной от 0,5 до 1,0 мкм. Комбинации двух или более слоев материалов в защитном покрытии обеспечивают покрытия, имеющие различные химические свойства.
[0059] Устройство подключено к электронной схеме. Электронная схема используется как для подачи напряжения (например, 50-100 мВ) между первым электродом и вторым электродом, так и для измерения тока по SWNT в зависимости от времени. Электронная схема может быть соединена с компьютером 24, имеющим в нем один или более процессоров, который используется для управления подачей напряжения и тока через устройство, а также для сбора, хранения и анализа данных, генерируемых устройством. Во время работы устройства между первым электродом и вторым электродом подается напряжение (например, постоянное напряжение или комбинация переменного и постоянного напряжения). Ток, проходящий через SWNT, измеряется с использованием электронной схемы, которая может включать в себя измеритель тока с одним или несколькими усилителями.
[0060] Первый электрод, второй электрод и SWNT могут быть расположены на подложке. Подложка может включать любое количество материалов подложки, таких как стекло, пластик или кремний. Один альтернативный вариант реализации изобретения включает в себя изготовление устройства на оптическиВ отличие от полевых транзисторов, и в соответствие с большей частью предшествующего уровня техники, связанной с измерением, устройство не требует электрода затвора или проводящей опорной подложки. Следовательно, устройство может быть изготовлено на широком диапазоне поверхностей, включая прозрачные. Кварц является предпочтительным для процесса изготовления CVD, описанного выше, потому что он совместим с высокими температурами. Стеклянные пластины также можно использовать, если SWNT синтезируются и наносятся на подложку другими способами, такими как нанесение покрытия из раствора, или если устройства изготовлены на пластинах, а затем переносятся на стекло для подложки. В любом случае, использование кварца, стекла, сапфира или другой прозрачной подложки позволяет осуществлять оптический мониторинг устройства. Мониторинг сигнала флуоресценции от прикрепленных молекул хорошо известен в данной области техники, и его лучше всего проводить через прозрачную подложку. Устройство 10, сформированное на кварцевой подложке, обеспечивает независимый мониторинг молекулярной динамики с использованием описанных здесь электрических методов и оптических методов, включая флуоресценцию одной молекулы и smFRET.
[0061] В вариантах реализации изобретения, электрические и оптические сигналы от одной и той же молекулы получаются либо в разное время, либо одновременно. Устройство, чувствительное к единичной молекуле, расположенное на прозрачной подложке (например, кварцевое), дает уникальную возможность, не известную в предшествующем уровне техники, заключающееся в дополнении smFRET независимой одномолекульной технологией. В этом варианте реализации изобретения, SWNT освещается через прозрачную подложку с использованием источника освещения. Люминесцентный свет, который испускается, может быть собран с использованием объектива, который использует масло или воду для контакта с прозрачной подложкой. Флуоресцентный свет можно направлять на счетчик фотонов, используя, например, расщепитель луча.
[0062] Такой двухрежимный мониторинг может калибровать измерения, выполненные одним подходом, такие как электронный мониторинг с измерениями флуоресценции, выполненными на групповом уровне. Одновременный опрос одной молекулы двумя независимыми средствами дает возможность изучить две разные части одной и той же молекулы, например, сравнить часть, которая движется, часть, которая принимает перенос заряда, часть, которая содержит каталитический сайт, или часть, которая поглощает или испускает фотоны. Синхронный мониторинг двух таких частей может определять относительное время и причинность двух событий, таких как движение активного сайта, скоррелированное с конформационными изменениями регулирующего сайта. Кроме того, прозрачная подложка позволяет активировать индуцированную светом активацию функциональности каталитического сайта, или управляемый светом перенос заряда для изучения полученного конформационного изменения. SWNT может, в одном варианте реализации изобретения, быть интегрирован в проточную ячейку или тому подобное, таким образом что жидкость может протекать через SWNT для измерений. Альтернативно, жидкости могут быть выборочно нанесены поверх устройства.
[0063] Устройство может содержать одну линкерную молекулу, содержащую одну или несколько функциональных групп, нековалентно прикрепленных к внешней боковой стенке SWNT. Примеры первых функциональных групп включают пирен, бензол, циклогексан и 2,3-дихлор-5,6-дициано-1,4-бензохинон. Функциональные группы, которые нековалентно прикрепляются к внешней боковой стенке SWNT, хорошо известны в данной области техники, и конкретная конструкция для этой функциональной группы может содержать любую конструкцию, подходящую для использования в настоящем изобретении. Кроме того линкерная молекула(ы) содержат одну или более функциональных групп, функционализированных с другой функциональной группой, присоединенной к сенсибилизирующей молекуле таким образом, чтобы сохранить функциональность сенсибилизирующей молекулы. Пары функциональных групп могут включать азид и алкин, эфир NHS и амин, тиол и алкин, а также тиол и малеимид. Функциональные группы, которые функционально взаимодействуют с другими функциональными группами, хорошо известны в данной области техники, и конкретная конструкция для этой функциональной группы может содержать любую конструкцию, подходящую для использования в настоящем изобретении.
[0064] Устройство может содержать одну сенсибилизирующую молекулу, содержащую одну или несколько функциональных групп, функционализированных одной или несколькими функциональными группами одной из линкерных молекул, таким образом, чтобы сохранить функциональность сенсибилизирующей молекулы. Сенсибилизирующие молекулы настоящего изобретения включают любую молекулу. Предпочтительные сенсибилизирующие молекулы включают молекулы, которые являются химически специфичными в их взаимодействиях с другими молекулами. Более предпочтительно сенсибилизирующие молекулы могут включать полимеры, белки, ДНК, РНК, рибозим и/или аптамер, полисахарид или другую биомолекулу. Сенсибилизирующие молекулы 30 хорошо известны в данной области техники, и могут содержать любую сенсибилизирующую молекулу, подходящую для использования в настоящем изобретении.
[0065] В вариантах реализации изобретения, молекула-линкер может содержать первую функциональную группу, которая присоединяется нековалентно к стенке SWNT, и второй функциональной группы, которая предназначена для присоединения к сенсибилизирующей молекуле. Использование молекулы-линкера позволяет избежать трудностей разработки эффективного прямого присоединения между сенсибилизирующей молекулой и SWNT. В этом варианте реализации изобретения молекула-линкер и сенсибилизирующая молекула являются фактически одним объектом. На практике, достижение и контроль требуемой поверхностной плотности часто требует, чтобы линкерная/ые молекула/ы и сенсибилизирующая молекула были изготовлены в виде двух отдельных растворов, при этом конечная связь между ними выполнялась на непосредственно на SWNT. Сенсибилизирующая молекула может содержать первую функциональную группу и вторую функциональную группу, которая может включать селективную таргетную функциональную группу. Первая функциональная группа сенсибилизирующей молекулы связывается со второй функциональной группой молекулы-линкера. Связыванием может быть любое химическое взаимодействие, известное в данной области техники, например, ковалентное или нековалентное связывание. В вариантах реализации изобретения, связывание осуществляется с помощью ковалентной связи. Вторая функциональная группа предназначена для связывания с молекулой-мишенью или несколькими молекулами-мишенями посредством любого связывающего взаимодействия. Сенсибилизирующая молекула также включает компонент, модулирующий проводимость, который идеально расположен рядом с местом крепления SWNT. Компонент, модулирующий проводимость, не должен находиться в непосредственной близости ко второй функциональной группе, но обе они должны взаимодействовать через механические, аллостерические или электронные средства, таким образом, что взаимодействия сенсибилизирующей молекулы с химической мишенью индуцируют динамические изменения компонента, модулирующего проводимость той же самой сенсибилизирующей молекулы, которая влияет на электронные изменения в SWNT.
[0066] В вариантах реализации изобретения, функциональная группа пирена, нековалентно может присоединяться к поверхности SWNT посредством укладки pi-pi. Единичная сенсибилизирующая молекула может быть связана с SWNT. Типичные электрические характеристики завершенного устройства могут быть измерены водным электролитом, находящимся в прямом контакте с боковой стенкой полупроводниковой SWNT.
[0067] В вариантах реализации изобретения, все три компонента объединяются в единую сенсибилизирующую молекулу. Например, одна аминокислота белка может быть эффективным сайтом для связывания с SWNT, другая аминокислота может иметь поверхностный заряд, который может модулировать проводимость SWNT, а третья аминокислота может служить местом распознавания или связывания для связывающего белок партнера, молекулу-мишень, подлежащую обнаружению. Альтернативно, ковалентный или нековалентный комплекс может быть сконструирован и синтезирован для объединения всех трех компонентов в виде единого сенсибилизирующего агента.
[0068] В вариантах реализации изобретения, различные функциональные компоненты сенсибилизирующей молекулы разделены между двумя или более молекулами, все из которых ковалентно или нековалентно собраны на проводнике SWNT. В этом альтернативном варианте реализации изобретения, компонент модуляции проводимости может быть молекулой, которая присоединена к одной функциональной группе молекулы-линкера, а селективный таргетный химический компонент может быть второй молекулой, которая присоединена к другой функциональной группе того же линкера. Альтернативно, селективный таргетный химический компонент может иметь функциональную группу, которая непосредственно связывается с молекулой, которая содержит компонент, модулирующий проводимость. Это связывание может происходить через ковалентную связь или посредством нековалентного распознавания или стыковки, общих для многих биомолекул. В каждом случае, между компонентами будет достигнута некоторая форма механической, стерической или электрической связи, так что динамика таргетного химического компонента приведет к изменениям компонента, модулирующего проводимость всего комплекса сенсибилизации.
[0069] В вариантах реализации изобретения одномолекульное чувствительное устройство для обнаружения одной молекулы может включать в себя проводник, имеющий одну или более SWNT; одну или более молекул-линкеров, содержащих две или более функциональных групп, из которых одна или более нековалентно связаны с поверхностью SWNT, и одну чувствительную молекулу, которая содержит по меньшей мере одну функциональную группу, которая функционализирована к по меньшей мере одной функциональной группе молекулы-линкера.
[0070] В вариантах реализации изобретения, одномолекульное чувствительное устройство содержит линкерную молекулу, содержащую карбоксилатную группу, и сенсибилизирующую молекулу, содержащую амин. Карбоксилатная функциональная группа молекулы-линкера может быть активирована в качестве реакционноспособного сложного эфира и амидирована с использованием методов, которые хорошо известны в данной области техники. Затем, реакционноспособный сложный эфир может быть ковалентно связан с аминогруппой сенсибилизирующей молекулы, с образованием стабильной амидной связи способом, который хорошо известен в данной области техники.
[0071] В вариантах реализации изобретения, одномолекульное чувствительное устройство содержит молекулу линкера, которая является малеимидом пирена, и сенсибилизирующую молекулу, содержащую реакционноспособную тиольную группу. Малеимидная функциональная группа молекулы-линкера может быть ковалентно связана с тиольной группой сенсибилизирующей молекулы, с образованием стабильной тиоэфирной связи, способом, который хорошо известен в данной области техники.
[0072] В вариантах реализации изобретения, нековалентное одномолекульное чувствительное устройство содержит молекулу линкера, которая представляет собой пириновый малеимид, а сенсибилизирующая молекула представляет собой белок. Другие варианты реализации изобретения включают те, в которых белок является ферментом. В вариантах реализации изобретения, фермент представляет собой ДНК-полимеразу или обратную транскриптазу. Аналогичные выходы одномолекульных чувствительных устройств, использующих каждый из этих ферментов, были достигнуты путем адаптации рН раствора, продолжительности выдержки и условий полоскания, используемых при прикреплении фермента.
[0073] В вариантах реализации изобретения, сенсибилизирующая молекула представляет собой нуклеиновую кислоту (например, ДНК, РНК), рибозим, аптамер, полисахарид или другую биомолекулу. Более предпочтительно, сенсибилизирующие молекулы могут включать полимеры, белки, ДНК, РНК, рибозим и/или аптамер, полисахарид или другую биомолекулу. Любая сенсибилизирующая молекула, которая подвергается изменению конформационной динамики при связывании или воздействии на субстрат или лиганд, подходит для использования в настоящем изобретении. В вариантах реализации изобретения, молекула линкера содержит линкерную молекулу, содержащую, по меньшей мере, одну функциональную группу, которая известна в данной области техники как нековалентно функционализированная на поверхности SWNT, и по меньшей мере одна функциональная группа является функциональной группой, которая известна в данной области техники для образования связей с другой функциональной группой.
[0074] В одном варианте реализации изобретения предполагается применение ДНК- или РНК-полимеразы или обратной транскриптазы в качестве единственной сенситизирующей молекулы, нековалентно присоединенной к SWNT для обеспечения неоптического секвенирования молекул ДНК, кДНК или РНК. Известно, что ферменты, которые катализируют зависящее от матрицы введение dNTP, подвергаются хорошо охарактеризованным конформационным изменениям, которые могут быть использованы для мониторинга специфического включения нуклеотидов, природных или аналоговых dNTP или NTP, в соответствии с описанными здесь способами и устройствами и, таким образом, обеспечивают секвенирование последовательности матричной молекулы. Этот способ безметочного секвенирования имеет преимущества по сравнению с применяемыми в настоящее время методами неоптического секвенирования, поскольку он позволяет распознавать событие специфического включения нуклеотидов из гомогенной смеси из четырех естественных или аналоговых dNTP или NTP, хотя настоящее изобретение является совместимым с практикой протекания отдельных dNTP или аналоговых dNTP или NTP в последовательном и циклическом режиме для целей определения последовательности. Использование обратной транскриптазы в качестве нековалентно связанной сенсибилизирующей молекулы 30 позволяет прямое секвенирование молекул РНК без необходимости проведения промежуточной стадии конверсии в кДНК.
[0075] Поскольку точность правильного включения нуклеотидов имеет одинаковую важность в секвенировании ДНК, РНК или кДНК, альтернативным методом для усиления обнаружения специфического включения правильного dNTP или NTP будет использование аналоговых dNTP или NTP, которые усугубляют конформационную динамику правильного включения нуклеотидов, что обеспечивает точную последовательность. Немеченые аналоговые dNTP или NTP, которые могут быть использованы для усиления кинетической или динамической дискриминации правильного включения нуклеотидов, хорошо известны специалистам в данной области техники, и включают, но не ограничиваются ими, модификации пуринового и пиримидинового оснований (т. е. на позициях C-4 и C-7), дезоксирибозные или рибозные части нуклеотидов, альфа, бета и гамма-фосфаты dNTP или NTP, включая использование тетра или пента-фосфатов, с дополнительными модификациями фосфата, или без них.
[0076] Другие способы повышения точности секвенирования, которые совместимы с настоящим изобретением, которые известны специалисту в данной области, могут быть использованы, включая, но не ограничиваясь этим, однократное считывание одной и той же матричной молекулы. Другие возможности включают использование дважды читаемого формата, в котором пирофосфоролиз используется для считывания одной и той же матричной молекулы во второй раз.
[0077] В вариантах реализации изобретения предлагается способ обнаружения динамики и кинетики устройства обнаружения одной молекулы. Любой способ измерения изменений электропроводности SWNT может использоваться для контроля чувствительного устройства с одной молекулой. В вариантах реализации изобретения, на SWNT подается разность смещения 100 мВ, а ток, протекающий через проводник, измеряется как функция времени с использованием схемы. Химическое связывание распознавание в таргетном компоненте сенсибилизирующей молекулы приводит к изменениям компонента, модулирующего проводимость сенсибилизирующей молекулы, вызывая увеличение и уменьшение измеренного тока. Многократные события связывания и освобождения, которые при усреднении включают химическую кинетику таргетного компонента, создают множественные колебания тока, которые могут быть синхронизированы, подсчитаны, распознаны, анализированы или сохранены с использованием технологий обработки сигналов, которые известны в данной области техники. Колебания тока могут состоять из простых увеличений и уменьшений прямоугольной формы. Альтернативно, колебания могут содержать любую форму волны, включая формы, которые могут быть треугольными, синусоидальными, или иметь любое количество компонентов Фурье. Амплитуды, длительности и формы этих волн кодируют активность таргетного компонента, и поэтому могут быть проанализированы с использованием компьютера 24, с тем чтобы выявить кинетику связывания и другие механические и электронные степени свободы. Статистический анализ этих параметров дает представление о кинетической изменчивости, переходах и промежуточных химических состояниях процессов связывания и освобождения мишеней. Степени свободы в сигнале тока различают несколько похожих молекул-мишеней, которые все связываются с одним и тем же сайтом, например между молекулой-мишенью и молекулой-ингибитором сайта связывания. Эти степени свободытакже могут различать слабые взаимодействия, такие как распознавания молекул, которые происходят до реального связывания.
[0078] В вариантах реализации изобретения, представлена способность различать и контролировать либо ковалентное, либо нековалентное связывание молекул ингибитора. Ингибиторы белковой функции являются коммерчески важными как фармацевтические агенты, включая для антивирусной, противораковой и антибактериальной терапии. Тестирование эффективных ингибиторов является трудоемким и дорогостоящим процессом. Устройство обеспечивает непосредственный мониторинг функции белка с разрешением до одной молекулы, одновременно исследуя белок с любым количеством различных ингибиторов-кандидатов. Используя автоматизированные системы доставки жидкости, хорошо известные в данной области техники, такие как проточная ячейка, растворы ингибиторов-кандидатов могут быть доставлены в устройство один за другим, для идентификации ингибиторов с желаемыми кинетическими свойствами. Альтернативно, ингибиторы-кандидаты могут быть в смесях, либо сразу после синтеза, либо целенаправленно классифицированы по химической структуре или функции или любому другому признаку, чтобы быстро анализировать целые партии молекул-кандидатов.
[0079] Таким образом, будет видно, что способы настоящего раскрытия способны обнаруживать динамику и кинетику одной сенсибилизирующей молекулы. Когда сенсибилизирующая молекула является ферментом, кинетика и динамика включают скорости ферментативного оборота или скорости конформационных движений. Техническое преимущество данного изобретения заключается в том, что динамика и кинетика одной сенсибилизирующей молекулы может быть определена, преодолевая проблемы групповых измерений, возникающие при наличии множественных сенсибилизирующих молекул на SWNT. Кроме того, способы настоящего раскрытия преодолевают проблемы, связанные с предшествующими способами изготовления одномолекулярных устройств, которые создают дефектный сайт на SWNT, который затем функционализирует одну сенсибилизирующую молекулу.
[0080] Варианты реализации изобретения включают в себя способ создания устройства для обнаружения одной молекулы. Способ включает в себя формирование по меньшей мере одной однослойной углеродной нанотрубки на подложке 26, имеющей первый и второй ее концы присоединенные, соответственно, с первым электродом и вторым электродом. Затем, стенка однослойной углеродной нанотрубки устройства нековалентно функционализируется, по меньшей мере, одной функциональной группой, по меньшей мере, одной молекулы линкера, содержащей множество функциональных групп. Одна сенсибилизирующая молекула функционализирована, по меньшей мере, одной функциональной группой по меньшей мере одной молекулы-линкера (например, функциональная группа, которая не является ковалентно функционализированной с помощью SWNT).
[0081] В вариантах реализации изобретения, SWCNT-FET изготавливают и функционализируют с использованием одноцистеинового варианта KF, дефицитного по домену экзонуклеазе (D355A/E357A/L790C/C907S). Очистка KF до >95% обеспечивается его гомогенностью (ФИГ. 6). Анализ на основе флуоресценции подтверждает активность фермента в объеме перед прикреплением (ФИГ. 7). Прикрепление KF к SWCNT-FET осуществляется путем замачивания устройств в растворе N-(1-пиренил)малеимида (1 мМ в этаноле, 30 мин) с последующей инкубацией с KF (300 нМ KF в стандартном буфере активности KF 20 мМ Трис, 50 мМ NaCl, 10 мМ MgCl2, 100 мкM TCEP, pH 8,0). Атомно-силовая микроскопия после сбора данных подтверждает присоединение одной молекулы KF к каждому устройству (ФИГ. 1B). Такие устройства называются просто наноконтурами KF.
[0082] В вариантах реализации изобретения, гомополимерные матрицы poly(dA)42, poly(dT)42, poly(dG)42 или poly(dC)42, смешанные с комплементарными аналогами dNTP, используются для обнаружения конформационного изменения полимеразы, например, ДНК-полимеразы. В вариантах реализации изобретения, каждая матрица слита с праймирующим участком M13, и смешивается с прямым праймером M13 в стехиометрическом соотношении 1: 1; для гибридизации смесь нагревают в термоциклере до 95°С в течение 5-10 мин с последующим охлаждением до 65°С, затем дополнительно охлаждают с градиентом 5°С на каждые пять минут до достижения комнатной температуры. В вариантах реализации изобретения, наноконтуры KF погружают в буфер активности с отожженным матрица-праймер дуплексом, при концентрациях 100 нМ. Природные или аналоговые dNTP добавляются в буфер в избытке, обеспечивая Vmax условия для KF-катализа. Чтобы компенсировать медленное включение аналогов dNTP, в экспериментах применяли более высокую концентрацию аналогов (ФИГ. 1C, например, 100 мкМ), чем нативных dNTP (например, 10 мкМ).
[0083] В вариантах реализации изобретения, измерения состоят из контроля тока источник-сток, I(t), через SWCNT-FET, в то время как присоединенная молекула KF взаимодействовала с окружающей средой. В вариантах реализации изобретения, стоковый электрод смещен на 100 мВ, а электролит, который служит в качестве электрода затвора, удерживается на уровне или около 0 В. Инкубация устройства с любым дуплексом матрица-праймер и его комплементарными dNTP преобразует колебания, ΔI(t), в то время как эти колебания могут отсутствовать в измерениях с некомплементарными dNTP или в контрольных измерениях, не содержащих матрица-праймер или прикрепление KF. В вариантах реализации изобретения, I(t) колебания усиливаются, оцифровываются на частоте 100 кГц, и сохраняются как непрерывные, 600 сек. Между измерениями, наноконтуры KF можно дважды промыть буфером активности, инкубировать в буфере в течение 5 мин, затем дважды промыть буфером перед введением другого нуклеотида и дуплекса матрица-праймер. Каждая молекула KF может контролироваться несколькими аналогами, их соответствующими нативными dNTP, и буфером без нуклеотидов для сбора непосредственно сопоставимых наборов данных, подтверждения типичных активностей KF, и получения колебаний ΔI(t).
[0084] На ФИГ. 2A и 2B показаны примерные сигналы ΔI(t), создаваемые наноконтуром KF, обрабатывающим матрицу poly(dC)42, в присутствии dGTP. В вариантах реализации изобретения, устройство создает непрерывные последовательности отрицательных колебаний ΔI(t), показанные с тремя различными увеличениями. Каждое колебание ΔI(t) указывает на образование одной пары оснований, и кинетические параметры, полученные из наборов данных ΔI(t), согласуются с известным одномолекулярным анализом движений KF и скоростями регистрации сборки KF. В вариантах реализации изобретения, формирование пары G•C или C•G может быть идентичным друг другу; образование пары оснований A•T/T•А также может обеспечить очень сходную кинетику полимеризации, динамику и значения ΔI(t), по сравнению друг с другом. Измерения с природными dNTP могут давать базовые значения для сравнения с аналогами dNTP.
[0085] В вариантах реализации изобретения, коммерчески доступные аналоги dNTP включаются в ДНК через полимеризацию KF как в групповых, так и в одномолекулярных анализах (фиг. 8A-8B). В вариантах реализации изобретения, в измерениях с α-тио-dNTP или dNTPαS, наборы данных ΔI(t), полученных с нуклеотидными аналогами, могут быть похожими на нативные dNTP, но с различными скоростями регистрации (ФИГ. 2C). В вариантах реализации изобретения, при измерении с помощью наноконтуров KF, включение 6-chloro-2-aminopurine-drTP, или 6-Cl-2-APTP, противоположное обеим матрицам, как poly(dC)42, так и poly(dT)42, вызывают сигналы ΔI(t) с инвертированной амплитудой, отражающие другую конформацию KF (ФИГ. 2D). Этот аналог включается медленнее; например, против матрицы poly(dC)42, 6-CL-2APTP вызывал колебания ΔI(t) на 80% от скорости dGTP. Регистрация ΔI(t) саналогами 2-тио-dNTP вызывали смешанное поведение, при котором активность KF вызывала отрицательные колебания ΔI(t) в течение одной минуты, положительные колебания ΔI(t) в течение еще одной минутыи, реже, смеси обоих поведений вдоль одной цепи матрицы (ФИГ. 2E-2F).
[0086] Для нативных dNTPs, константы времени для экспериментального базового тока, τopen также может упоминаться как τhi. Временные константы, представляющие собой событие включения природного dNTP, могут возникать при более низком токе, и называются τ1o. Положительные, отрицательные или смешения как положительных, так и отрицательных колебаний ΔI(t) раскрываются здесь, а временные постоянные для любого направления колебаний называются τclosed. Распределения τopenи τclosed получены из каждой записи данных полимеризации.
[0087] На ФИГ. 3A-3B показаны примерные распределения для включения субстратов dGTP в матрицы poly(dC)42. Распределения от событий τclosed включения нативного и аналогового dGTP были почти неотличимы, за исключением редких событий в хвостах, для которых у нас самая бедная статистика (ФИГ. 3A). Чтобы провести сравнение между натурным и аналоговым dNTP, мы сосредоточились на средней постоянной времени <τ> первичной пуассоновской составляющей этих распределений. Все из средних значений <τclosed>близко совпадали с приблизительно 0,3±0,1 мсек. При сравнении, распределения и средние значения <τopen> явно отличались. Например, KF находился 63,6±2,8 мсек в своей открытой конформации при обработке α-тио-dGTP, что на 56% больше, чем 40,8±0,6 мс, наблюдаемое для нативного dGTP (ФИГ. 3B).
[0088] Кинетические параметры <τclosed>, <τopen> и средняя скорость включения k были проанализированы на четырех гомополимерных матрицах с нативными и аналоговыми dNTP (Таблица 1). Как и в случае, описанном выше, каждая комбинация показала идентичные распределения τclosed со значениями <τclosed> в диапазоне 0,3 ± 0,1 мсек. В то время, как аналогичный эффект наблюдался ранее для всех четырех нативных dNTPs,33 неожиданным было увеличение этого результата для аналогов dNTP, имеющих разные размеры нуклеотидов, электронные свойства, водородную связь или замещение в α-фосфодиэфире.
[0089]С другой стороны, τopen, является более чувствительным к идентичности dNTP. Средняя длительность <τopen> варьировалась от 23 мс с нативным dCTP до 145 мс с α-тио-dATP. Среди четырех нативных dNTP, <τopen> было больше для включения dTTP или dATP, чем для включения dGTP или dCTP. Эта иерархия сохранялась при больших длительностях <τopen>, измеренных для всех четырех α-тио-dNTP. Замена α-тио увеличила <τopen> на 50% в случае dGTP и dCTP, тогда как увеличение было более 100% для dTTP и dATP.
[0090]Средняя скорость KF обработки для включения dNTP рассчитывалась как k=(<τopen>+<τclosed>)-1. В большей степени τopen определяет k, поскольку по меньшей мере в 60 раз больше, чем τclosed- быстрее всего KF включает 2- тио-dCTP быстрее, чем за 30 с-1. Увеличение τopen, описанное выше для α-тио-dNTP, уменьшало k до 15 с-1для α- тио-dCTP и α-тио-dGTP и 7 с-1для α-тио-dATP и α-тио-dTTP. Уровни включения 6-Cl-2APTP наиболее благоприятно сравнивались с самыми медленными темпами, наблюдаемыми для включения нативного dGTP. И наоборот, включение 2-тио-dTTP и 2-тио-dTСP оказалось немного быстрее, чем включение их природных аналогов.
[0091] Аналогичные результаты были воспроизведены с использованием десятков различных молекул KF. Каждый KF был прикреплен к другому SWCNT-FET и измерен независимо. Для сравнения, не гомополимерная матрица, измеренная с аналогами dNTP, привела к аналогичной кинетике (данные не показаны). Как упоминалось ранее, в наших экспериментах применяли 100 мкМ аналогов dNTP для обеспечения условий устойчивого состояния; для сравнения, 10 мкМ α-тио-dATP с матрицей poly(dT)42 не влияли на полимеризацию ДНК. Из-за статического разброса, некоторые молекулы KF обрабатывались быстрее или медленнее, чем среднее значение группы, но без какого-либо существенного изменения относительного сравнения аналоговых и природных dNTP.
[0092] Одномолекулярные эксперименты, проведенные в этом исследовании, иллюстрируют и проливают новый свет на хорошо известную пластичность ДНК-полимераз, таких как KF. Этот класс ферментов может совмещаться даже с сильно измененными входящими dNTP. Однако, мы непосредственно наблюдаем конформационные движения, требуемые ферментом для поддержания точности при взаимодействии с некоторыми измененными dNTP. В отношении отражения пределов для таких применений, известно, что ДНК-полимеразы проявляют сильную чувствительность к незначительным изменениям размера и формы dNTP. Наш анализ пользуется преимуществом от сравнения данных одиночных молекул с природными и аналоговыми dNTP во время многочисленных процессуальных событий включения. Этот анализ начинается с кинетики двух наблюдаемых конформаций ферментов при катализе, которые были отражены в τopen и τclosed.
[0093] События, происходящие во время τopen включают граничивающую скорость стадию распознавания dNTP, которая чувствительна к модификациям как в нуклеотидном основании, так и в скелете. Успешное распознавание и связывание соответствующих нуклеотидов вызывает активацию и закрытие KF. Предыдущие эксперименты на основе FRET с соответствующей ДНК-полимеразой Т7 идентифицировали «полностью открытое» конформационное состояние, вызванное распознаванием несовпадения. Однако, при использовании сайта привязки L790C, SWCNT-FET не регистрирует как движения KF, так и никаких сигналов в присутствии несовпадающих dNTP. Отсутствие промежуточных состояний, или связанных с несовпадениями движений, предполагает, что наш сайт привязки является нечувствительным к этой начальной точке контроля точности. Таким образом, колебание ΔI(t) является результатом каталитически зафиксированной конформации, и не ограничивается просто глобальным движением открытия и закрытия фермента.
[0094] Аналоги dNTP были выбраны по их способности быть включенными в ДНК-матрицы ДНК-полимеразами, и изменениями в размерах, структурах и реакционной способности. Мы исследовали либо замещение в α-фосфате, либо в нуклеотидной основе. Первый тип аналога, например, α-тио-dNTP, замещал немостовой, α-фосфорильный атом кислорода, серой, для введения нового стереоцентра и изменения реактивности на этом критическом участке. Вторая категория аналогов dNTP, замещение, например замещение галогена или серы, на нуклеотидной основе, изменяет размер и электронную структуру пары оснований; некоторые аналоги также изменяют водородную связь, доступную для спаривания основания. Например, 6-Cl-2-APTP (ФИГ. 1C) имеет два профиля водородной связи, что позволяет включать его как в основаниях T, так и в C. По сравнению с dATP, 6-Cl-2-APTP замещает 6-аминогруппу хлором, но вводит 2-аминную функциональную группу, эта конфигурация в конечном итоге обеспечивает такое же количество водородных связей по Уотсону-Крику, комплементарных к T, как и dATP. При использовании в качестве аналога dGTP 6-Cl-2-APTP имеет различную таутомеризацию, которая изменяет N-1 от донора водородной связи до состояния акцептора. В этом случае замена кислорода хлором значительно снижает силу водородного связывания. 41Как и 6-Cl-2APTP, серозамещенные аналоги 2-тио-dTTP и 2-тио-dCTP также образуют более крупные пары оснований благодаря увеличенной длине связи тиокарбонила
[0095] В вариантах реализации изобретения, аналог dNTP или NTP включает химическую модификацию в трифосфатном остатке. В вариантах реализации изобретения, химическая модификация в трифосфатном остатке представляет собой замещение O в α-положении на S. В вариантах реализации изобретения трифосфатный фрагмент любого аналога dNTP или NTP имеет структуру Формулы (I), 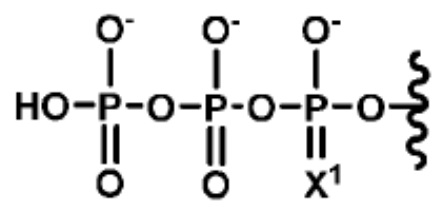 (I), где X-1 является S или O. В вариантах реализации изобретения, аналогом dNTP является α-тио-dATP, α-тио-dGTP, α-тио-dCTP, или α-тио-dTTP. В вариантах реализации изобретения, аналогом NTP является α-тио-ATP, α-тио-GTP, α-тио-CTP, или α-тио-TTP. В вариантах реализации изобретения, аналог dNTP или NTP включает замещение в а-положении трифосфатного остатка, как указано в Формуле (I), и одну или несколько замещений на нуклеотидном основании, как раскрыто здесь и известно в данной области техники.
(I), где X-1 является S или O. В вариантах реализации изобретения, аналогом dNTP является α-тио-dATP, α-тио-dGTP, α-тио-dCTP, или α-тио-dTTP. В вариантах реализации изобретения, аналогом NTP является α-тио-ATP, α-тио-GTP, α-тио-CTP, или α-тио-TTP. В вариантах реализации изобретения, аналог dNTP или NTP включает замещение в а-положении трифосфатного остатка, как указано в Формуле (I), и одну или несколько замещений на нуклеотидном основании, как раскрыто здесь и известно в данной области техники.
[0096] В вариантах реализации изобретения, аналог dNTP или NTP замещен в нуклеотидном основании. В вариантах реализации изобретения, пурин замещен, как показано в Формуле (IIIa), 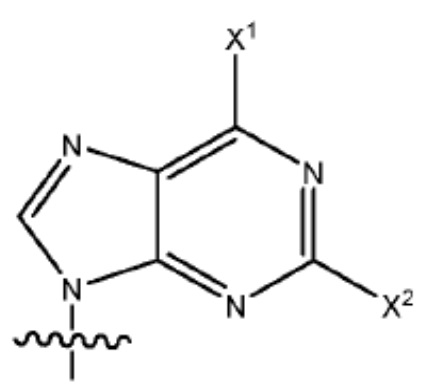 (IIa), где X1представляет собой водород, галоген или -NH2, и X2представляет собой водород или -NH2. В вариантах реализации изобретения, X1является - NH2, а X2 является водородом, обеспечивая dATP или АТФ. В вариантах реализации изобретения, X1 не является -NH 2, а X2является - NH2, обеспечивая 6-замещенный 2-АРТР или его замещенный аналог. В вариантах реализации изобретения, аналогом является 6-Cl-2-APTP, также известный как 6-Cl-dGTP.
(IIa), где X1представляет собой водород, галоген или -NH2, и X2представляет собой водород или -NH2. В вариантах реализации изобретения, X1является - NH2, а X2 является водородом, обеспечивая dATP или АТФ. В вариантах реализации изобретения, X1 не является -NH 2, а X2является - NH2, обеспечивая 6-замещенный 2-АРТР или его замещенный аналог. В вариантах реализации изобретения, аналогом является 6-Cl-2-APTP, также известный как 6-Cl-dGTP.
[0097] В вариантах реализации изобретения, аналог dNTP или NTP включает нуклеотидное основание, которое представляет собой 8-оксогуанин, 2,6-диамино-4-оксо-5-формамидопиримидин, N6-метиладенозин, O6- метилгуанозин, N2-метилгуанозин, 2,6-диаминопурин, индолил, 5-метилиндолил, 5-алкилиндолил (например, 5-этилиндолил), 5-этилениндолил, 5-нитроиндолил, 4-нитроиндолил, 5-фенилиндолил, 5-галогениндолил (например, 5-F-индолил), 5-аминоиндолил или 6-нитроиндолил.
[0098] В вариантах реализации изобретения, dNTP или NTP включает нуклеотидное основание со структурой формулы (IIb) 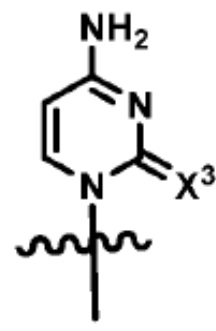 (IIb), где X3является O или S. В вариантах реализации изобретения, X3 представляет собой O, обеспечивая цитозиновое нуклеотидное основание. В вариантах реализации изобретения, X3 представляет собой S, обеспечивая 2-thiо нуклеотидное основание. В вариантах реализации изобретения, нуклеотидное основание представляет собой 2-тио-dCTP или 2-тио-CTP.
(IIb), где X3является O или S. В вариантах реализации изобретения, X3 представляет собой O, обеспечивая цитозиновое нуклеотидное основание. В вариантах реализации изобретения, X3 представляет собой S, обеспечивая 2-thiо нуклеотидное основание. В вариантах реализации изобретения, нуклеотидное основание представляет собой 2-тио-dCTP или 2-тио-CTP.
[0099]В вариантах реализации изобретения, dNTP или NTP включает нуклеотидную группу со структурой Формулы (IIc), 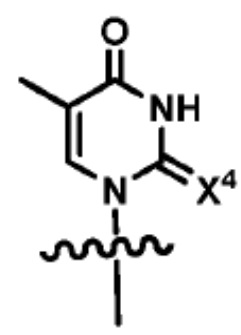 (IIc), где X4 является O или S. В вариантах реализации изобретения, X4 представляет собой O, обеспечивая тиминогенное нуклеотидное основание. В вариантах реализации изобретения, X4 представляет собой S, обеспечивая 2-тио нуклеотидное основание. В вариантах реализации изобретения, нуклеотидное основание представляет собой 2-тио-dTTP или 2-тио-ТТР.
(IIc), где X4 является O или S. В вариантах реализации изобретения, X4 представляет собой O, обеспечивая тиминогенное нуклеотидное основание. В вариантах реализации изобретения, X4 представляет собой S, обеспечивая 2-тио нуклеотидное основание. В вариантах реализации изобретения, нуклеотидное основание представляет собой 2-тио-dTTP или 2-тио-ТТР.
[0100] В вариантах реализации изобретения, dNTP или NTP включает нуклеотидную группу со структурой формулы (IId), 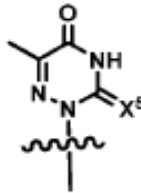 (IId), где X5 является O или S. В вариантах реализации изобретения, X5 является O, обеспечивая 6-азатиминовое нуклеотидное основание. В вариантах реализации изобретения, X5 представляет собой S, обеспечивая 6-аза-2-тио нуклеотидное основание. В вариантах реализации изобретения, нуклеотидное основание представляет собой 6-аза-2-тио-dTTP или 6-аза-2-тио-ТТР.
(IId), где X5 является O или S. В вариантах реализации изобретения, X5 является O, обеспечивая 6-азатиминовое нуклеотидное основание. В вариантах реализации изобретения, X5 представляет собой S, обеспечивая 6-аза-2-тио нуклеотидное основание. В вариантах реализации изобретения, нуклеотидное основание представляет собой 6-аза-2-тио-dTTP или 6-аза-2-тио-ТТР.
[0101] В вариантах реализации изобретения, аналогом является альфа-тио dATP, dTTP, dCTP и dGTP, 2-тио dATP, dTTP, dCTP и dGTP, 2-амино-6-Cl-пурин-2'-дезоксирибозид-трифосфат (называемый 6- Cl dGTP или 6-Cl-2APTP), 4-тио-dTTP, 2-аза-dTTP, 5-фтор-dTTP или гамма-ANS dTTP.
[0102] В вариантах реализации изобретения, различия, наблюдаемые в τopen, в значительной степени отражают механизмы узнавания и связывания неприродных dNTP. Длинные хвосты в распределениях для α-тио-dGTP и 6- Cl-2APTP, по сравнению с нативным dGTP, возможно, отвечали за увеличение <τopen> (ФИГ. 3B). Хвосты могут быть подогнаны ко второй экспоненте с временными константами 200 мсек, примерно в пять раз длиннее чем <τopen> для нативного dGTP. Подобные длинные хвосты можно наблюдать со всеми аналогами dNTP, описанными здесь, что иллюстрирует проблемы, с которыми сталкивается фермент при включении неприродных субстратов. Шаги, отличные от распознавания, потенциально имеют место во время τopen, который представлен в данном документе; образование ковалентной связи является одним из возможных примеров, которые произошли бы слишком быстро, даже с замедленной реактивностью α-тио-dNTP, чтобы их можно было бы определить как такие, которые ограничивают скорость реакции. Более высокие темпы включения, наблюдаемые с 2-тио аналогами, могут быть результатом более стабильного образования пар оснований, эффективно сокращая значения <τopen>. Больший размер атома серы в 2-тио-dCTP на границе водородной связи с основанием матрицы G, по-видимому, не влияет на способность 2-тио-dCTP к эффективному спариванию. В вариантах реализации изобретения, способ обнаружения конформации полимеразы нуклеиновой кислоты обнаруживает повышение эффективности полимеризации с 2-тио-dTTP и 4-тио-dTTP, по сравнению с включением dTTP.
[0103] Аналог 6-Cl-2APTP, с гораздо более слабой водородной связью и, следовательно, неполным спариванием оснований по сравнению с dGTP, иллюстрирует проблемы распознавания спаривания оснований при полимеризации ДНК, катализируемой KF. Более длинные значения <τopen> для включения 6-Cl-2APTP по сравнению с dGTP против матрицы poly(dC)42, иллюстрирует готовность ДНК-полимераз принимать неприродные dNTP, частично за счет удлинения времени, выделенного для распознавания. Значения <τopen>, и, следовательно, скорость включения, наблюдаемая при полимеризации 6-Cl-2APTP против матрицы poly(dC), попала между значениями, измеренными для включения нативных dGTP и dATP против комплементарных гомополимерных матриц. Таким образом, несмотря на измененную таутомеризацию и последующую потерю по меньшей мере одной водородной связи, связывающей пары, по сравнению с dGTP, 6-Cl-2APTP все еще может быть включенным быстрее, чем нативный dATP. В вариантах реализации изобретения, способ обнаружения конформации полимеразы нуклеиновой кислоты обнаруживает, что водородная связь, которая участвует в спаривании в малой канавке, остается неизменной, когда 6-Cl-2APTP считается аналогом dGTP, и может регулировать относительно более высокие скорости, наблюдаемые для dGTP, dCTP, и 6-Cl-2APTP против матрицы poly(dC).
[0104] В вариантах реализации изобретения, см., например, ФИГ. 5A, KF обрабатывает гетерогенные субстраты в присутствии всех четырех природных нуклеотидов (dNTP), причем каждое включение пары оснований вызывает отрицательное колебание тока ΔI<0. Индивидуальные колебания могут быть перечислены, как показано на ФИГ. 5A, но в целом они не различают один тип основания от другого.
[0105]В вариантах реализации изобретения, см., например, ФИГ. 5B, используется 2-тио-dTTP-аналог. С тиолированным дезокситимидином, положительные колебания указывают (№ 2, 6, 7) места, где были включены T-нуклеотиды. Это наблюдение можно распространить на секвенирование РНК путем замены KF на РНК-полимеразу. Процесс, изображенный на ФИГ. 5B идентифицирует все включения T-нуклеотидов в конкретном ДНК-субстрате. Этот процесс можно расширить довсех четырех оснований путем измерения субстрата с каждым из четырех различных тиолированных нуклеотидов.
[0106] В вариантах реализации изобретения, см., например, ФИГ. 5C, когда KF обрабатывает гетерогенные субстраты в присутствии природных нуклеотидов (dNTP), смешанных с некоторыми аналогами, полученная диаграмма содержит положительные и отрицательные колебания тока, которые могут быть использованы для идентификации выбранного основания. В вариантах реализации изобретения, используются три нативных нуклеотида (dATP, dTTP, dCTP), смешанные с 6-Cl-2APTP в качестве аналога G-соединений. Текущие колебания в этом варианте реализации изобретения нумеруются для перечисления 15 событий включения основания. Большинство событий (№ 1, 4, 7, 9, 10, 13, 14, 15) могут состоять из одного отрицательного всплеска тока, который идентифицирует включение одного нативного нуклеотида. В вариантах осуществления пять событий (например, В вариантах реализации изобретения, пять событий (например, № 2, 3, 6, 8, 11; выделены стрелками на ФИГ. 5C) представляют собой положительные импульсы тока, которые идентифицируют включения нуклеотидов 6-Cl-2APTP. В вариантах реализации изобретения, когда 6-Cl-2APTP используется в качестве аналога нативного нуклеотида dGTP, события (например, №2, 3, 6, 8, 11; выделены стрелками на ФИГ. 5С) идентифицируют нуклеотиды G в последовательности ДНК.
[0107] В вариантах реализации изобретения, два из событий (№ 5, 12), показанных на ФИГ. 5C, содержат пару близко расположенных токовых штырей. Эти пары могут указывать на включение одного нативного и одного 6-Cl-2APTP-нуклеотида в быстрой последовательности. Альтернативно, пара пиков может быть уникальным сигналом от 6-Cl-dAPTP, действующий как псевдоаналог dATP.
[0108] В вариантах реализации изобретения, способ обнаружения изменения конформации в полимеразе используется для определения последовательности олигонуклеотида. Полученная информация (например, ФИГ. 5A-5C) сообщает последовательность олигонуклеотидов, так как пики указывают, какой нуклеотид или аналог внедряется, когда полимераза перемещается вдоль ее субстрата.
[0109] В дополнение к распознаванию и связыванию, продолжительные значения <τopen> при включении α-тио-dNTP могут быть результатом пониженной стабильности вновь синтезированной ДНК. Kатализируемая KF обработка гомополимерных матриц может привести к искаженной дцДНК. Кроме того, α-тио-dNTPs особенно подвержены образованию менее стабильных бинарных комплексов с неблагоприятными связями со скелетом ДНК, которые постепенно замедляют каталитическую скорость KF. Более выраженные эффекты на этом этапе, по сравнению с экспериментами с соответствующими нативными dNTP, наблюдались во время α-тио-dATP/α-тио-dTTP по сравнению с включением α-тио-dGTP/α-тио-dCTP. В вариантах реализации изобретения, способ обнаружения конформации полимеразы нуклеиновой кислоты указывает на неустойчивость ДНК, зависящую от последовательности, которая подчеркивает недостаток использования гомополимерных матриц. Альтернативно, это различие может свидетельствовать о том, что α-тио-замещение дополнительно мешает механизму, который вызывает удлинение <τopen> для нативных A•T/T•A пар оснований. Некоторые вариации в τopen, связанные с включением α-тио-dNTP, могут быть результатом слабо ингибирующего Rp стереоизомера (Ki ≈ 30 мкМ), который присутствует в соотношении приблизительно 1:1 со стереоизомером Sp в коммерческом синтезе этого аналога. Это торможение примерно на порядок слабее, чем Km для нативного dNTP,10 и, следовательно, можно ожидать, что он повлияет на значения <τopen> лишь в малой степени.
[0110] Во время τclosed, KF подвергается выраженному конформационному изменению, соответствующему образованию одной фосфодиэфирной связи между входящим нуклеотидом и возникающей dsDNA. В экспериментах с ограниченным субстратом, число колебаний ΔI(t) соответствовало количеству нависающих пар оснований матрицы; таким образом, конформационное изменение в течение τclosed должно происходить для каждого успешного, процессивного включения нуклеотидов. Ранее, короткая и равная длительность <τclosed> для нативных dNTP поддерживала модели, в которых τclosed сама происходила из стадии формирования ковалентной связи. 33 В вариантах реализации изобретения в способе выявления конформации полимеразы нуклеиновых кислот используется три наблюдения с аналогами dNTP. Во-первых, направление колебаний ΔI(t) меняются на противоположные для некоторых аналогов dNTP. Во-вторых, включение аналогов 2-тио-dNTP вызывает смеси как положительных, так и отрицательных колебаний ΔI(t). В-третьих, как показано в Таблице 1, инвариантность в <τclosed> распространяется на все проверенные аналоги, несмотря на замещения на электрофильном α-фосфате или вероятные альтернативные конформации, необходимые для принятия замен в нуклеотидном основании.
[0111] В этой электронной технике, SWCNT-FET, лежащий в основе, является чрезвычайно чувствительным к электростатическому стробированию заряженных поверхностных остатков белка в пределах 1 нм от места прикрепления. Различные варианты того же фермента могут проявлять либо положительные, либо отрицательные колебания ΔI(t), в зависимости от заряда остатков, смежных SWCNT, и направлений их движения. В вариантах реализации изобретения, во время способа обнаружения конформации полимеразы нуклеиновой кислоты, KF и его заряженные остатки, электростатически стробирующие SWCNT-FET, могут оставаться инвариантными. Переменные колебания ΔI(t) могут указывать на то, что остатки, смежные с сайтом присоединения KF, принимают различные движения в ответ на определенные аналоги dNTP в каталитически-компетентном цикле. Такие движения могут быть переданы от активного сайта KF через аллостерию, но они могут не быть движениями образования ковалентных связей. В вариантах реализации изобретения, ковалентная стадия может не проходить по тому же механизму и с той же длительностью <τclosed>, но с двумя противоположными движениями. Вместо этого, соответствующие движения остатков, ответственные за τclosed могут быть независимыми как от начального молекулярного распознавания, так и от химического этапа катализа KF.
[0112] В вариантах реализации изобретения, в способе обнаружения конформации полимеразы нуклеиновой кислоты, KF присоединяется к SWCNT-FET посредством боковой цепи L79°C белка в субдомене «пальцы», связывая электростатические стробирующие движения соответствующих заряженных остатков с каталитически зафиксированными движениями в течение τclosed. Каждое событие τclosed может быть результатом самой O-спирали активного сайта, или конкретного скручивания остатка O-спирали в двух возможных направлениях в течение наблюдаемой стадии успешного включения нуклеотидов. Это предлагаемое скручивание подразумевается путем рассмотрения движений остатков активного сайта на известных стадиях включения нуклеотидов, и их влияния на теоретическую близость заряженных остатков к SWCNT-FET. Например, эксперименты smFRET с KF показывают промежуточную конформацию O-спирали активного узла между открытым и закрытым состояниями; потенциально аналогичная конформация «ajar» наблюдается в кристаллических структурах гомолога KF Bst Pol I. O-спираль С-конца Bst Pol I перегибает путь до замыкания, таким образом, что большой сдвиг эквивалента KF Y766 сопровождается незначительным вращением эквивалента KF F762. Вращение эквивалента KF F762 продолжается до закрытия фермента.
[0113] Путем сравнения кристаллических структур KF и Bst Pol I, идентифицированы смежные с SWCNT-FET заряженные остатки, которые могут перемещаться в ответ на вращение Y766 и F762 в активном центре KF. В вариантах реализации изобретения, в способе обнаружения конформации полимеразы нуклеиновой кислоты, источником колебаний Δ I(t) являются дополнительные движения Y766 и/или F762, после закрытия фермента и включения основания, которые продолжают распространяться на заряженные остатки вблизи SWCNT-FET. Дополнительное конформационное изменение KF, когда зарождающаяся пара оснований переходит на после-вставочный сайт KF, наблюдалось с помощью smFRET после успешного включения нуклеотидов и, возможно, является движением, измеряемым τ closed- существенные взаимодействия, переданные ароматическими остатками активного сайта, могут включать укладку π-π с вновь образованной базовой парой. Такое движение будет оценивать электронную конфигурацию пары оснований и контролировать точность этапа формирования связи, не требуя гидрофильных взаимодействий, которые заменяются заменами аналога dNTP.
[0114] ДНК-полимеразы, в том числе описанные, очень пригодны для мутагенеза, чтобы адаптировать их свойства, в том числе для приложений по секвенированию ДНК. В вариантах реализации изобретения, способ настоящего раскрытия включает мутации, введенные в ДНК-полимеразу для биоконъюгации фермента с углеродной нанотрубкой. В вариантах реализации изобретения, такие мутации изменяют свойства ДНК-полимеразы для более эффективного введения неприродных оснований, и изменения процессивности фермента. В вариантах реализации изобретения, такой мутагенез используется для улучшения электрического считывания и свойств фермента. Например, в вариантах реализации изобретения, мутации вводятся в активный сайт фермента, чтобы принимать специфичные аналоги dNTP с формами или функциональными дополнениями, комплементарные мутациям, сделанным на активном сайте фермента. В вариантах реализации изобретения, ДНК-полимераза мутируется для усиления электрического ответа, получаемого от каждого основания, включенного во время полимеризации ДНК; например, замещение заряженных остатков фермента вблизи углеродной нанотрубки обеспечивало ответы, способные к рационализации, во время движений ферментов.
[0115] Наноконтур KF показывает более значительные колебания Δ I(t) для набора A•T/T•А, чем для набора пар оснований G•С/С•G. Структурные результаты предположили, что основания матриц A и T наиболее глубоко сокрыты в активном сайте ДНК-полимеразы, и поэтому поворот O-спирали может быть максимизирован. E710 и Y766 KF, и другие остатки глутамата и тирозина гомологичного активного сайта, были вовлечены в механизм стабилизации A•T/T•А пар оснований по сравнению с G•С/С•G. В вариантах реализации изобретения, взаимодействие водородной связи между KF E710 и KF Y766 перед включением нуклеотидов может влиять на размер и форму активного сайта, и может играть важную роль на этапе τclosed распознавания аналогового dNTP.
[0116] В вариантах реализации изобретения, аналогичные результаты при включении 2-тио-dTTP и 2-тио-dCTP иллюстрируют преимущественное распознавание KF электронной структуры пары оснований. Хотя замещение серы влияет только на акцептор водородной связи Уотсона-Крика в аналоге 2-тио-dCTP, оба аналога 2-тио-dNTP приводят к смесям положительных и отрицательных колебаний Δ I(t) и, следовательно, одновременно вызывают подобные движения KF во время включения. Замещение серы для аналогов 2-тио-dNTP незначительно по сравнению с более драматическими электронными вариациями, введенными в 6-Cl-2APTP, но фермент реагирует аналогичным, хотя и неэксклюзивным, образом. Наблюдаемые смеси как отрицательных, так и положительныхколебаний ΔI(t) предлагает, что KF обращается как к нативным, так и к альтернативным движениям, соответственно, во время включения 2-тиозамещенных dNTP. Очевидный эффект памяти блокируетфермент в одном движении, или в другом, в течении десятков секунд, что подразумевает дополнительное конформационное изменение, которое энергетически бистабильно в частном случае 2-тио-dNTP.
[0117] В вариантах реализации изобретения, способ обнаружения конформации полимеразы нуклеиновой кислоты включает в себя перемещение зарождающейся ДНК в неактивный домен экзонуклеазы (екзо),как возможный источник положительных колебаний Δ I(t). При плавлении неустойчивого конца праймера из-за несовершенного спаривания оснований, ДНК перемещается в и из неактивного экзодомена, и KF подвергается различным конформационным изменениям. Однако, такие переходы происходят на удалении от места присоединения, и положительные колебания ΔI(t), наблюдаемые здесь, не меняют длительности<τclosed>- соответственно, переход к экзо домену кажется несогласующимся с наблюдением положительных колебаний ΔI(t). Подобно конформационным шагам, которые, как известно, происходят во время распознавания неспаренных dNTP, переход к экзодомену должен происходить в течение τopen. Колебания ΔI(t) настоящего изобретения могут произойти во время начавшегося каталитического цикла, и могут представлять собой адаптируемое движение KF, согласующееся с поворотной O-спиралью, анализирующей электронную целостность вновь сформированной пары оснований ДНК.
[0118] В вариантах реализации изобретения, способ обнаружения конформации полимеразы нуклеиновой кислоты, аналоги dNTP вызывают ограничения включения нуклеотидов ДНК-полимеразами, включая стереохимию на электрофильном фосфате,способность связывания водорода входящего основания, и механизмы проверки точности. Поскольку большинство аналогов dNTP увеличивают среднее значение <τopen> и широту его кинетических распределений, определяющий скорость этап распознавания dNTP очень чувствителен к даже незначительным изменениям структуры подложки. Однако, резкие замены в реактивноспособном сайте образования связи не способны влиять на длительность <τclosed>. Направление колебаний ΔI(t), с другой стороны, переключается на положительные, или на смесь как отрицательных, так и положительных сигналов с аналогами dNTP с модифицированным основанием. Поскольку эти аналоги dNTP имеют функциональные возможности в центре формирования связей, идентичные природным субстратам, драматические изменения в направлении AI(t) могут быть результатом проверки точности KF перед открытием для обработки следующего субстрата. Такие события можно легко отличить от нативных событий включения dNTP, и обеспечить непосредственное наблюдение за ферментом, принимающим неприродные dNTP, путем изменения направления его динамической проверки ошибок.
ПРИМЕРЫ
[0119] Пример 1. Включение аналогов дезоксинуклеозид-трифосфатов с помощью Одномолекулярного Наноконтура с ДНК-полимеразой I (Фрагментом Кленова).
[0120] Введение. Чтобы обеспечить выживание всех известных форм жизни, ДНК-полимеразы должны правильно распознавать дезоксинуклеозидтрифосфатные (dNTP) субстраты и успешно катализировать их включение в новые цепи ДНК. Требуемая точность всех ДНК-полимераз частично зависит от Уотсон-Криковской комплементарности входящих пар оснований dNTP, которые гибридизуются с одноцепочечной матрицей ДНК. [1,2] Однако, неприродные dNTP, неспособные к образованию водородных связей с нативными комплементарными основаниями, также были успешно включены ДНК-полимеразами. Такие аналоги могут образовывать устойчивые пары оснований с формой, подобной каноническим А•T и G•C. [3-5] Исследования с аналогами dNTP выявили требования к стереохимическим, геометрическим, электронным эффектам и гидрофобным взаимодействиям во время включения нуклеотидов. [6-9]
[0121] Результаты экспериментов по включению аналоговых dNTP проясняют критерии выбора нуклеотидов ДНК-полимеразами, включая требования к дальнейшему расширению цепи и эффективному катализу. Например, каталитические скорости полимеризации с аналогами фосфоротиоата определяют стереохимическое предпочтение нуклеофильной атаке праймирующим 3'-ОН на электрофильный а-фосфат dNTP. [6] Небольшие модификации нуклеотидных оснований, включая тио- и гало- замещение в положениях водородной связи, приводят к измененной скорости включения, и демонстрируют потребность в жесткой стерической посадке в активном сайте ДНК-полимеразы. [8,10]]. Уотсон-Криковская геометрия, необходимая для включения неприродных оснований, индуцируется несколькими взаимодействиями между активным центром полимеразы и новой парой оснований. [5]
[0122] Детальная оценка полимеризации неприродных dNTP за пределами одной пары оснований может обеспечить точную кинетическую информацию об этой ненативной полимеразной активности. Кроме того, превращение ассоциированных небольших конформационных изменений при распознавании базы в надежное измерение может выявить новые аспекты ролей для стерических и электронных взаимодействий в полимеризации ДНК. Как сообщается в данном документе, такая информация может быть выяснена одномолекулярными методами.
[0123] Обычные исследования объемных или групповых популяций ферментов не могут наблюдать промежуточные стадии и переходные состояния в механизме реакции. Однако, эксперименты с отдельными молекулами позволяют наблюдать такие состояния, которые иначе усреднялись бы в групповой популяции. [11-13] Эксперименты по полимеризации ДНК, использующие одномолекулярный перенос энергии резонанса Форстера (smFRET), выявили конформационную гибкость и понимание механизма точности фрагмента ДНК-полимеразы I Кленова, названного далее KF. [14-16] Несмотря на свою способность выхватывать новую информацию о динамике ферментов, smFRET требует флуоресцентно меченого белка и/или субстрата. Фотообесцвечивание и поток фотонов между флуорофорами ограничивают как длительность, так и временное разрешение, соответственно, экспериментов с smFRET.
[0124] Недавно мы описали новый подход к одномолекульной энзимологии и применили его к трем ферментам. В этом методе, отдельный белок биоконъюгирован с полевым транзистором на основе одностенной углеродной нанотрубки (SWCNT-FET, ФИГ. 1 A). Подход выявил новые взгляды на количество ступеней, кинетические параметры и процессивность лизоцима T4, фермента, изучаемого более 100 лет. [17,18]. Очень динамичные скорости полинуклеотид-киназы A (PKA) выявили роль фермента в качестве высоко регулируемого молекулярного переключателя. [19] При исследовании KF, конъюгированного с SWCNT-FET, существенные различия между A•T/T•A и G•С/С•G продемонстрировали, что замкнутая конформация фермента зависит от идентичности входящего dNTP. [20] Чувствительность к этой разнице была неожиданной, так как пары оснований Уотсона-Крика имели одинаковые размеры, и это указывало, что метод SWCNT-FET также может реагировать на уникальную кинетику и конформации, связанные с dNTP-аналогами.
[0125] В этом документе, метод SWCNT-FET использовался для распознавания различий между нативными dNTP и аналогами dNTP во время включения KF. Используя тио- и гало-замещенные аналоги dNTP, контролировали полимеризацию ДНК, а затем статистически анализировали, чтобы выявить различия в кинетике включения для некоторых, но не всех, аналогов dNTP. Кроме того, время, в течение которого каждый аналог проводил в открытой и закрытой конформациях фермента, выявляет вариации времени, необходимого для этапов, ограничивающих скорость реакции. Результаты представляют портрет фермента, преодолевающего проблемы молекулярного распознавания при катализе.
[0126] Экспериментальная часть. Профили SWCNT были изготовлены [17] и функционализированы одноцистеиновым вариантом дефицитного по экзонуклеазе KF. Очистка KF до >95% обеспечивала его однородность. Анализ на основе флуоресценции подтвердил активность партии фермента перед прикреплением. [20,21] Атомно-силовая микроскопия подтвердила прикрепление отдельных молекул KF (ФИГ. 1B) после сбора данных с каждого устройства.
[0127] Для каждого измерения, dNTP были выбраны для комплементирования гомополимерных матриц poly(dA)42, poly(dT)42, poly(dG)42, and poly(dC)42. Каждую матрицу сливали с праймирующим сайтом M13 и смешивали с прямым праймером M13 в стехиометрическом соотношении 1:1; для гибридизации смесь нагревали до 95°С в течение 5-10 минут с последующим охлаждением до комнатной температуры. SWCNT FET погружали в стандартный буфер активности ДНК Pol I (20 мМ Трис, 50 мМ NaCl, 10 мМ MgCl2, 100 мкМ TCEP, pH 8,0) с гибридом матрица-праймер в концентрации 100 нМ. Нативные или аналоговые dNTP были добавлены в буфер в избытке, обеспечивая условия Vmaxдля KF. Чтобы компенсировать медленное включение аналогов dNTP, в экспериментах применяли более высокую концентрацию аналогов (ФИГ. 1C, 100 мкМ,Trilink Biotechnologies), чем нативных dNTP (10 мкМ,Fisher).
[0128] Измерения состояли в контроле тока источник-сток I(t) в SWCNT FET, в то время как присоединенная молекула KF взаимодействовала со своей окружающей средой. FET смещен на 100 мВ, а электролит, который служил электродом затвора, удерживался при 0 В. Инкубация устройства с любым нуклеотидом и его комплементарным матрица-праймером трансдуцировала колебания Δ I(t), которые были измерены с помощью текущего предусилителя (Keithley 428), оцифрованы при 100 кГц, и сохранены для последующего анализа. Между измерениями, наноконтуры KF дважды промывали буфером для анализа, инкубировали в буфере в течение 5 минут, затем снова дважды промывали буфером перед введением другого нуклеотида. Каждая молекула KF наблюдалась с многочисленными аналогами, соответствующими природными dNTP, и безнуклеотидным буфером, чтобы собирать прямо сопоставимые наборы данных, подтверждать типичные активности KF, и воспроизводить типы ΔI(t), которые ранее сообщались. [20]
[0129] Результаты. Инкубация наноконтура с нативным dNTP и ее комплементарным матрица-праймером вызывала отрицательные изменения тока, ΔI(t) (ФИГ. 2A). Как сообщалось ранее, быстрые колебания тока происходили только в присутствии KF, dNTP и матрица-праймера. Каждое отклонение ΔI(t) коррелирует с закрытием фермента KF. Таким образом, кинетические параметры, полученные из измерений таких отклонений тока с нативными dNTP, согласуются с предыдущим анализом движений ферментов (например, наФИГ. 2A). [20] Эти измерения предоставили базовые значения для сравнения с аналогами dNTP (Таблица 1).
Таблица 1:Кинетика включения нативного и аналогового dNTP KF. a
(SEQ ID NO:9)
α-тио-дАТФ
145,9 ± 8,4
0,38 ± 0,21
6,8 ± 0,4
(SEQ ID NO:10)
α-тио-дТТФ
2-тио-дТТФ
152,1 ± 6,6
61,1 ± 3,2b
0,29 ± 0,13
0,23 ± 0,14b
6,6 ± 0,3
16,3 ± 0,9b
(SEQ ID NO:11)
α-тио-дТТФ
2-тио-дТТФ
68,8 ± 4,6
32,3 ± 1,1b
0,33 ± 0,19
0,41 ± 0,15b
14,5 ± 1,0
30,6 ± 1,2b
(SEQ ID NO:12)
α-тио-дТТФ
2-тио-дТТФ
63,6 ± 2,8
50,5 ± 1,4
0,21 ± 0,15
0,20 ± 0,12
15,7 ± 0,7
19,7 ± 0,6
а Средние значения ± стандартное отклонение.
b Аналогичные значения наблюдались как для событий переключения вверх, так и для событий переключения вниз.
[0130] Аналоги dNTP изучали либо замещение в α-фосфате, либо изменение в нуклеотидном основании. В первой категории, замена атома фосфорильного кислорода на серу вводит новый стереоцентр в dNTP для создания α-тио-dNTP, также известного как dNTPαS. Коммерчески синтезированные, без контроля над стереохимией, диастереомерные отношения вокруг этого α- фосфора были, вероятно, 1:1. Во второй категории аналогов dNTP, замещение галогена или серы в нуклеотидном основании может привести к образованию большего основания с измененной таутомеризацией и, следовательно, к различной водородной связи. Например, аналоговый 6-хлор-dGTP также известный как 6-Cl-2-APTP) имеет таутомеризацию, отличную от dGTP, что изменяет N-1 от донора водородной связи до акцептора. Кроме того, замена кислорода хлоридом снижает прочность водородных связей. Другие примеры модифицированных нуклеотидов, 2-тио-dTTP и 2-тио-dCTP, заменяют кислород серой для увеличения размера этих оснований. Как ожидалось, что изменения кинетики, приводящие к включению этих аналогов, приводят к отличающимся электронным сигналам, вероятно из-за снижения распознавания фермента подходящего dNTP.
[0131] Аналоги фосфоротиоата (α-тио-dNTP), 6-Cl-dGTP, 2- тио-dTTP и 2-тио-dCTP инкубировали с комплементарными матрица-праймерами. Как наблюдалось с нативными dNTP, отрицательные колебания тока возникали при включении α-тио-dNTP (ФИГ. 2B). Наоборот, 6-Cl-dGTP вызвали положительные колебания тока (ФИГ. 2C). Аналоги, содержащие 2-тио замены, вызвали смесь как отрицательных, так и положительных колебаний тока. Оба α- тио-dGTP и 6-Cl-dGTP-аналоги продуцируют отклонения ΔI(t) в два-три раза реже, чем dGTP.
[0132] Распределения для τopen и τclosed были получены из более чем 50 с данных полимеризации с dGTP и двумя его аналогами (ФИГ. 3A-3B). Данные соответствуют простым распределениям Пуассона с одиночными постоянными времени <τ>. Подгонки распределений для событий τclosed с нативным и аналоговым dGTP лишь слегка отклоняются друг от друга, особенно у хвостов. Большинство событий перекрываются с высокой вероятностью (-50%). Средняя продолжительность замкнутых комплексов в присутствии dGTP, α-тио-dGTP или 6-Cl-dGTP находились в тесном согласии (0,2-0,4 мсек). Для нативного dGTP, единая экспоненциальная подгонка к данным охватывала >90% событий τopen. Однако, аналогичная подгонка с α-тио-dGTP или 6-CldGTP могли охватывать только ≈75% от событий τopen. В открытой конформации KF, только от 20 до 30% аналоговых событий перекрываются с точной постоянной времени, необходимой для нативного dGTP. Кинетика включения с аналогами dGTP значительно отличается от нативной, и их средняя постоянная времени может быть легко различима.
[0133] Этот анализ кинетических параметров τclosed, τopen, а также скорость включения (k) был распространен на другие нативные и аналоговые подложки (Таблица 1). Как описано для α-тио-dGTP и 6- Cl-dGTP, образование фосфодиэфирной связи при закрытии KF, определенное количественно путем τclose, длится от 0,2 до 0,4 мсек для всех рассмотренных dNTP, включая нативные и аналоги. Средняя длительность открытой конформации KF, τopen, для всех α-тиоdNTPs была в 2-3 раза дольше, чем для их нативных dNTP. Например, KF проводит в открытой конформации, τopen, примерно в 2,5 раза дольше, при обработке α-тио-dGTP (63 ± 3 мсек), чем с нативным dGTP (24 ± 1 мсек). Как сообщалось для нативных dNTP,[20] α-тио-dCTP и α-тио-dGTP включались быстрее в зарождающуюся цепь, чем α-тио-dATP и α-тио-dTTP. Значение τopen -доминирующего образования пар оснований A•T/T•А в 2-3 раза больше значения τopen для G•С/С•G.
[0134] Значения τopen и τclosed показывают время, необходимое для полных циклов включения dNTP. Средняя скорость обработки KF рассчитывалась как k=1/(< τopen>+<τclosed>). Учитывая гораздо большее количество времени, проведенного в открытой конформации (>98%), длительность τopen во многом предопределяет скорость ферментативного катализа. Средние скорости обработки KF уменьшались в 2-3 раза для аналоговых α-тио-dNTP и 6-Cl-dGTP, так как значение τopen в 2-3 раза больше. Например, время, затрачиваемое на открытую конформацию KF при обработке 6-Cl-dGTP (<τopen>=50 ± 1 мсек), примерно в два раза больше, чем при обработке dGTP (<τopen>=24 ± 1 мсек), что приводит к скорости 20 S-1, по сравнению с 41 S-1, соответственно. Скорости для 2-тио-dTTP (<k>= 16 S-1,) и 2- тио-dCTP (<k>=36 S-1,) были примерно такими же, как и для их нативных аналогов.
[0135] Обсуждение. Образование фосфодиэфирной связи является нейтральным к аналоговым замещениям, даже при модифицированном α-фосфате. Сера в α-тио-dNTP заменяет не-мостиковый атом кислорода в ключевой электрофильной функциональной группеНесмотря на ослабленную электрофильность фосфата и, как следствие, пониженную реакционную способность тиофосфодиэфира в сравнении с фосфодиэфиром, этап обмена связями по-прежнему является быстрым, а не ограничивающим скорость реакции. [22,23]
[0136] Примерно двадцать пять процентов событий τopen α-тио-dGTP и 6- Cl-dGTP отклоняются от единственной экспоненциальной подгонки к данным, что согласуется с более значительными посторонними событиями, возникающими в связи с тем, что фермент испытывает трудности с закрытием вокруг неприродного dNTP. Более медленные постоянные времени для τopen подтверждают более длительное время, необходимое для ограничивающие скорость процессы распознавания входящего dNTP, присущие модифицированной базе. Однако, существенное перекрытие между распределением для нативного и аналогового dGTP предотвращает мгновенное назначение конформации KF для каждого события.
[0137] Предыдущее исследование определило Spдиастереомер α-тио-dNTP как исключительно предпочтительный субстрат для включения ДНК Pol I. 6Наблюдаемые скорости обработки α-тио-dNTP могут быть обусловлены ингибированием псевдо-субстрата ДНК-полимеразы Rp диастереомером. RP диастереомер α-тио-dTTP слабо связывается с активным центром KF и ингибирует его катализ с Ki ≈ 30 мкМ. Эта константа торможения примерно на порядок слабее, чем Кm для нативного dNTP. 6 Несмотря на то, что нестереохимически контролируемое α-тио-замещение эффективно удаляет половину доступного субстрата, большой избыток α-тио-dGTP обеспечивал то, что такие эффекты были маловероятны как причина резкого удлинения времени, проведенного в открытой конформации фермента. Кроме того, диастереомерные смеси α-тио-dNTP могут также влиять на стабильность ДНК скелета синтезированной нити. [24,25] Однако образование фосфодиэфирной цепи, которое происходит во время замкнутой конформации фермента, которое определяется количественно τclosed, остается неизменной. Поэтому наиболее вероятным виновником замедления кинетики α-тио-dNTP является ингибирование Rpстереоизомером время этапа распознавания основания. Таким образом, KF находит dNTP с правильной стереохимической конфигурацией, отвергая неправильные субстраты, конкурирующие за связывание и ингибирование фермента.
[0138] Предыдущие исследования показали эффективный катализ ДНК-полимеразой α для включения 6- Cl-dGTP в спаривании соединении с матрицей poly(dC). [26]] Хотя 6-Cl-dGTP может быть введена напротив основания Т ДНК-полимеразой Т4 [27, 28], эта способность до настоящего времени не исследовалась в функциональной KF-функциональной наноконтуре. Эксперименты с 6-Cl-dGTP, описанные здесь, иллюстрируют готовность ДНК-полимераз принимать неприродные dNTP с неправильным спариванием Уотсона-Крика. Чтобы успешно включать эти ненативные основания, ДНК-полимераза должна применять альтернативы традиционным критериям распознавания водородной связи для выбора нуклеотидов. Кинетические результаты согласуются с адаптацией ограничивающей скорость стадии, связанной с водородной связью при распознавании dNTP.
[0139] Было выдвинуто предположение, что конформационные изменения после дискриминации больших оснований 6- Cl-dGTP, 2-тио-dTTP и 2-тио-dCTP могут вызывать различия в прохождении заряда на чувствительном SWCNT-FET. К нашему восторгу, были обнаружены уникальные сигналы, когда инкубировали наноконтур KF с 6-Cl-dGTP и poly(dC)42. Положительный отклонения ΔI(t), или «колебания», вероятно, представляют собой другой способ закрытия фермента (ФИГ. 2D). Эти сигналы являются противоположными от результата включения нативного dGTP. 20Как показано в экспериментах с мутантами заряда T4-лизоцима,29 либо отрицательно заряженные функциональные группы аминокислот должны двигаться ближе к нанотрубке, либо положительно заряженные функциональные группы должны двигаться дальше во время включения 6-Cl-dGTP. Для 2-тиозамещенных dNTPs, наблюдалась сложная смесь событий восходящего и нисходящего переключения. Случайное распределение восходящего и нисходящего переключения, связанное с закрытием KF,предполагает, что ферменты применяют более одной конформации для поддержания эффективного катализа.
[0140] Несмотря на большое совпадение в кинетике включения нативного и аналогового dGTP, KF явно обращается к различным конформациям при катализе с 2-тио dCTP, 2-тио dTTP и 6- Cl-dGTP. Эти уникальные сигналы восходящего переключения могут быть вызваны конформационными изменениями, регулируемыми J-спиралью KF во время разделения на домен экзонуклеазы и из него, поскольку KF распознает включение менее совершенного комплементарного основания. [14, 30-32] Однако такое челночное движение должно происходить с очень высокими скоростями, так как не существует разницы между временами закрытия фермента, τclosed, наблюдаемого для 6-Cl-dGTP, 2-тио-dTTP и 2-тио-CTP, по сравнению с нативными dNTP. Потенциал этих различных сигналов, позволяющих дискриминацию входящих dNTP, заслуживает дальнейшего изучения для возможных применений в секвенировании ДНК.
[0141] Вывод. Таким образом, аналоги dNTP бросают вызов ограничениям включения нуклеотидов ДНК-полимеразами, включая стереохимию на электрофильном фосфате, способность связывания водорода входящего основания, и степень закрытия фермента. Поскольку большинство исследованных аналогов уменьшают τopen и, следовательно, скорость фермента, стадия распознавания dNTP, определяющая скорость реакции, представляется как очень чувствительная к незначительным изменениям структуры субстрата. Напротив, даже существенные замены в реакционноспособном сайте формирования связей не влияют на время закрытия KF. Описанные здесь результаты предполагают две возможные стратегии, направленные на конформационное назначение KF на каждом этапе включения аналога dNTP с использованием наноконтура KF. Во-первых, модификации могут ориентироваться на конформацию открытого KF при распознавании и активации dNTP. Однако, на сегодняшний день модифицированные dNTP, включая 6-Cl-dGTP и α-тио-dNTP, могут только замедлять среднюю скорость фермента. Другая стратегия, которая заслуживает дополнительного изучения, влияет на замкнутую конформацию KF. Модифицированные основания, такие как 6-Cl-dGTP, 2-тиоdCTP и 2- тио-dTTP, могут изменять конформацию фермента во время включения, существенно влияя на электронный сигнал, наблюдаемый как увеличение тока, проходящего через SWCNT. Такое восходящее переключение можно легко отличить от включения нативных dNTP и обеспечивает прямое наблюдение за ферментом, который легко приспосабливается к неприродным dNTP. В заключение, конформационно-чувствительные электронные измерения даже хорошо изученного фермента могут выявить новые и неожиданные аспекты движений и динамики фермента.
[0142] Ссылки (Пример 1). [1] Echols, H. ;Goodman, M. F. Annu. Rev. Biochem. 1991, 60, 477; [2] Kunkel, T. A. J. Biol. Chem. 2004, 279, 16895; [3] Goodman, M. F. Proc. Natl. Acad. Sci. 1997, 94, 10493; [4] Kool, E. T. Annu. Rev. Biochem. 2002, 71, 191; [5] Betz, K. ;Malyshev, D. A. ;Lavergne, T. ;Welte, W. ;Diederichs, K. ;Dwyer, T. J. ;Ordoukhanian, P. ;Romesberg, F. E. ;Marx, A; Nat. Chem. Biol. 2012, 8, 612; [6] Burgers, P. M. ;Eckstein, F. J. Biol. Chem. 1979, 254, 6889; [7] Chiaramonte, M. ;Moore, C. L. ;Kincaid, K. ;Kuchta, R. D; Biochemistry 2003, 42, 10472; [8] Kim, T. W. ;Delaney, J. C. ;Essigmann, J. M. ;Kool, E. T. Proc; Natl. Acad. Sci. U. S. A. 2005, 102, 15803; [9] Kincaid, K. ;Beckman, J. ;Zivkovic, A. ;Halcomb, R. L. ;Engels, J. W. ;Kuchta, R. D. Nucleic Acids Res. 2005, 33, 2620; [10] Sintim, H. O. ;Kool, E. T. J. Am. Chem. Soc. 2006, 128, 396; [11] Deniz, A. A. ;Mukhopadhyay, S. ;Lemke, E. A. J. R. Soc; Interface 2008, 5, 15; [12] Lu, H. P. Chem. Soc. Rev. 2014, 43, 1118; [13] Min, W. ;English, B. P. ;Luo, G. ;Cherayil, B. J. ;Kou, S. C. ;Xie, X. S. Acc. Chem. Res. 2005, 38, 923; [14] Christian, T. D. Romano, L. J. ;Rueda, D. Proc. Natl. Acad. Sci; U. S. A. 2009, 106, 21109; [15] Santoso, Y. ;Joyce, C. M. ;Potapova, O. ;Le Reste, L. ;Hohlbein, J. ;Torella, J. P. ;Grindley, N. D. F. Kapanidis, A. N. Proc; Natl. Acad. Sci. U. S. A. 2010, 107, 715; [16] Berezhna, S. Y. ;Gill, J. P. ;Lamichhane, R. ;Millar, D. P. J. Am; Chem. Soc. 2012, 134, 11261; [17] Choi, Y. ;Moody, I. S. ;Sims, P. C. ;Hunt, S. R. ;Corso, B. L. ;Perez, I. ;Weiss, G. A. ;Collins, P. G. Science 2012, 335, 319; [18] Choi, Y. ;Moody, I. S. ;Sims, P. C. ;Hunt, S. R. ;Corso, B. L. ;Seitz, D. E. ;Blaszczak, L. C. ;Blaszcazk, L. C. ;Collins, P. G. ;Weiss, G. A. J. Am. Chem. Soc. 2012, 134, 2032; [19] Sims, P. C. ;Moody, I. S. ;Choi, Y. ;Dong, C. ;Iftikhar, M. ;Corso, B. L. ;Gul, O. T. ;Collins, P. G. ;Weiss, G. A. 2013; [20] Olsen, T. J. ;Choi, Y. ;Sims, P. C. ;Gul, O. T. ;Corso, B. L. ;Dong, C. ;Brown, W. A. ;Collins, P. G. ;Weiss, G. A. J. Am; Chem. Soc. 2013, 135, 7855; [21] Frey, M. W. ;Sowers, L. C. ;Millar, D. P. ;Benkovic, S. J; Biochemistry 1995, 34, 9185; [22] Knowles, J. R. Annu. Rev. Biochem. 1980, 49, 877; [23] Bryant, F. R. ;Johnson, K. A. ;Benkovic, S. J. Biochemistry 1983, 22, 3537; [24] Eckstein, F. ;Jovin, T. M. Biochemistry 1983, 22, 4546; [25] Mizrahi, V. ;Henrie, R. N. Marlier, J. F. Johnson, K. A. ;Benkovic, S. J. Biochemistry 1985, 24, 4010; [26] Patro, J. N. Urban, M. ;Kuchta, R. D. Biochemistry 2009, 48, 180; [27] Devadoss, B. ;Lee, I. ;Berdis, A. J. Biochemistry 2007, 46, 13752; [28] Zhang, X. ;Motea, E. ;Lee, I. ;Berdis, A. J. Biochemistry 2010, 49, 3009; [29] Choi, Y. ;Olsen, T. J. ;Sims, P. C. ;Moody, I. S. ;Corso, B. L. ;Dang, M. N. Weiss, G. A. ;Collins, P. G. Nano Lett. 2013, 13, 625; [30] Mizrahi, V. ;Benkovic, P. ;Benkovic, S. J. Proc. Natl. Acad. Sci; U. S. A. 1986, 83, 5769; [31] Joyce, C. J. Biol. Chem. 1989, 264, 10858; [32] Tuske, S. ;Singh, K; Kaushik, N. ;Modak, M. J. J. Biol. Chem; 2000, 275, 23759;
[0143] Пример 2. Включение аналогов дезоксинуклеозид-трифосфатов с помощью Одномолекулярного Наноконтура-2 с ДНК-полимеразой I (Фрагментом Кленова).
[0144] Описание. Отдельные копии фрагмента Кленова (KF) ДНК-полимеразы I были прикреплены к устройствам с одностенной углеродной нанотрубкой, и электрически измерены в присутствии различных химических кофакторов. Все аспекты изготовления соответствовали протоколу, описанному Olsen et. al.
[0145] Результаты. ФИГ. 4A-4B показано, что когда KF обрабатывает поли poly(dA)42 в присутствии природного нуклеотидного дезокситимидинтрифосфата (dTTP, включение каждой пары оснований вызывает отрицательный всплеск тока ΔI<0. Когда dTTP заменяется неприродным нуклеотидом 2-тио-2'-дезокситимидин-5'-трифосфатом (2-тио-dTTP), включения оснований вызывают положительные импульсы тока ΔI>0.
[0146] На ФИГ. 5А показано, что когда KF обрабатывает гетерогенные субстраты в присутствии всех четырех природных нуклеотидов (dNTP), каждое включение пары оснований вызывает отрицательный всплеск тока ΔI<0. Индивидуальные колебания могут быть перечислены, как показано, но в целом они не различают один тип основания от другого.
[0147] Как показано на ФИГ. 5B, моделирование того же набора данных с заменой dTTP на 2 тио-dTTP. С тиолированным дезокситимидином, положительные колебания теперь указывают (№ 2, 6, 7) места, где были включены T-нуклеотиды. Это наблюдение можно распространить на секвенирование РНК путем замены KF на РНК-полимеразу. Процесс, изображенный на ФИГ. 5B идентифицирует все включения T-нуклеотидов в конкретном ДНК-субстрате. Процесс можно распространить на все четыре основания путем измерения субстрата с каждым из четырех различных тиолированных нуклеотидов.
[0148] На ФИГ. 5C показывает, что, когда KF обрабатывает гетерогенные субстраты в присутствии природных нуклеотидов (dNTP), смешанных с некоторыми аналогами, полученный шаблон содержит положительные и отрицательные колебания тока, которые могут быть использованы для идентификации выбранного основания. В этом примере показаны данные, полученные с использованием трех нативных нуклеотидов (dATP, dTTP, dCTP), смешанных с 6-Cl-2APTP в качестве аналога для включения G-оснований. Текущие колебания в этом варианте реализации изобретения пронумерованы для перечисления 15 событий включения основания. Большинство событий (№ 1, 4, 7, 9, 10, 13, 14, 15) состоят из одного отрицательного всплеска тока, который идентифицирует включение одного нативного нуклеотида. Пять событий (№ 2, 3, 6, 8, 11) выделяются стрелками, поскольку они представляют собой положительные импульсы тока, которые идентифицируют включения нуклеотидов 6-Cl-2APTP. Так как 6-Cl-2APTP используется здесь как аналог нативного dGTP-нуклеотида, эти пять событий идентифицируют G-нуклеотиды в последовательности ДНК.
[0149] Два из событий (№ 5, 12), показанных на ФИГ. 5C, содержат пару близко расположенных импульсов тока. Эти пары могут указывать на включение одного нативного и одного 6-Cl-2APTP-нуклеотида в быстрой последовательности. Альтернативно, пара импульсов может быть уникальным сигналом от 6-Cl-dAPTP, действующим как псевдоаналог dATP.
[0150] Ссылки (Пример 2 и Уровень техники). [1] T. J. Olsen, Y. Choi, P. C. Sims, 0. T. Gul, B. L. Corso, C. Dong,... G. A. Weiss, Electronic Measurements of Single- Molecule Processing by DNA polymerase I (Klenow fragment), I Am. Chem. Soc. 135, 7855 (2013); [2] Y. Choi, 1. S. Moody, P. C. Sims, S. R. Hunt, B. L. Corso, G. A. Weiss, and P. G. Collins, Single-Molecule Lysozyme Dynamics Monitored by an Electronic Circuit, Science 335, 319 (2012) ; [3], Y. Choi, 1. 5. Moody, P. C. Sims, S. R. Hunt, B. L. Corso, D. E. Seitz,.. . G. A. Weiss, Single Molecule Dynamics of Lysozyme Processing Distinguishes Linear and Cross- linked Peptidoglycan Substrates,. 1. Am. Chem. Soc. 134, 2032 (2012); [4], Y. Choi, T. J. Olsen, P. C. Sims, 1. S. Moody, B. L. Corso, M. N. Dang,. P. G. Collins, Dissecting Single-Molecule Signal Transduction in Carbon Nanotube Circuits with Protein Engineering, Nano Lett. 13, 625 (2013) ; [5], P. C. Sims, I. S. Moody, Y. Choi, C. Dong, M. Iftikhar, B. L. Corso,... G. A. Weiss, Electronic Measurements of Single- Molecule Processing by DNA polymerase I (Klenow fragment), I Am. Chem. Soc. 135, 7861 (2013; [6], L. T. C. Franca, E. Wuts and T. B. L. Kist, A review of DNA sequencing techniques, Quarterly Reviews of Biophysics 35, 169 (2002); [7], 5. E. Jacutin, Unnatural Nucleotides for DNA Sequencing. (Texas A & M University, College Station, TX, 1997). [8], T. D. Harris, P. R. Buzby, H. Babcock, E. Beer, J. Bowers, I. Braslavsky,. Z. Xie, Single-Molecule DNA Sequencing of a Viral Genome Science 320 106 (2008); [9], D. Stoddart, A. J. Heron, E. Mikhailova, G. Maglia, and H. Bayley, Single- nucleotide discrimination in immobilized DNA oligonucleotides with a biological nanopore, Proc. Nati. Acad. Sci. U. S. A. 106, 7702 (2009); [10], S. Sorgenfrei, C. -y. Chiu, R. L. Gonzalez, Y. -J. Yu, P. Kim, C. Nuckolls, and K. L. Shepard, Label-free single-molecule detection of DNA-hybridization kinetics with a carbon nanotube field-effect transistor, Nat. Nanotechnol. 6, 126 (2011); [11], T. C. Glenn, Field guide to next- generation DNA sequencers, Molecular Ecology Resources 11, 759 (2011).
[0151] Пример 3:Экспрессия и очистка KF
[0152] Реагенты, приобретенныенакоммерческойоснове, включаютвсебяантибиотики (Fisher Scientific), Ni-IMAC-смолу (Bio-Rad Laboratories), клеточныелинии (Stratagene), дезоксинуклеозидтрифосфаты (Fisher Scientific), дезоксинуклеозидтрифосфатныеаналоги (Trilink Biotechnologies), ферменты (New England Biolabs или Fermentas), олигонуклеотиды (Фишер), агарозувысокогоразрешения (The Nest Group) и 96-луночныефлуоресцентныепланшеты (Nunc). Всеостальныехимикатыбылиприобретенынакоммерческойосновеот Acros Organics, EMD, Fisher Scientific или Sigma Aldrich. Всереагентыиспользовалиськакполученные.
[0153] Плазмиду pET28c, содержащую ген, кодирующий KF (D355 A/E357A/C907S/L790C),1,2 называемый далее как KF, использовали для трансформации CaCl2-компетентных клеток E. coli BL21 (DE3) путем теплового шока. После ночного роста на твердой среде, одну колонию использовали для инокуляции 25 мл среды LB с добавлением 40 (мкг/мл канамицина для роста в жидкой среде в течение ночи при 37 °С при встряхивании. LB (1 л), дополненную 40 мкг/мл канамицина, инокулировали 10 мл ночной культуры и инкубировали при встряхивании при 37 °С в течение нескольких часов. Как только клетки достигали поздней логарифмической фазы (OD600=0,9), экспрессию KF индуцировали добавлением 1 мМ IPTG. Через 3-4 ч экспрессии белка при 37 °С со встряхиванием клетки собирали центрифугированием (6000 об/мин, 20 мин, 4 °С) и ресуспендировали в буфере для лизиса (20 мМ Трис, 50 мМ NaCl, 10 мМ BME, рН 8,0). Клетки лизировали ультразвуком, а клеточный дебрис собирали центрифугированием (15 000 об/мин, 45 мин, 4 °С). После фильтрации через фильтр с порами 0,45 мкм, супернатант лизата оставляли связываться со смолой Ni-IMAC в течение ночи при 4 °С. KF элюировали в лизисном буфере с 250 мМ имидазола, концентрировали, и затем обрабатывали протеазой TEV в течение двух дней при 4 °С. Смесь центрифугировали, и затем фильтровали через фильтр 0,45 мкм перед эксклюзионной хроматографией в TBS (20 мМ Трис, 50 мМ NaCl, 100 мкМ TCEP, pH 7,9) на Bio-Rad Biologic DuoFlow FPLC. Чистоту KF оценивали с помощью SDS-PAGE (ФИГ. 6).
Групповая активность KF и включение dNTP аналогов.
[0154] Олигонуклеотиды, используемые для тестирования активности
[0155] В Таблице 2 перечислены олигонуклеотиды, используемые для тестирования активности KF, включения аналога dNTP, и для измерений с помощью наноконтура. После получения очищенные HPLC олигонуклеотиды солюбилизировали в воде до 100 мкМ. Выделенные жирным шрифтом области указывают на сайт праймирования M13. Выделенные курсивом регионы указывают сайты рестрикции. [2AmPur] указывает на 2-аминопурин.
Таблица 2. Олигонуклеотиды, используемые для измерения активности и электронных измерений.
Групповой анализ активности KF
[0156] Чтобы подтвердить активность варианта KF(L790C) по сравнению с KF дикого типа, ранее описанный анализ был адаптирован следующим образом. 1,3Рандомизированная ДНК-матрица, содержащая как 2-аминопурин (матрица ActAssay в Таблице S1), так и праймирующий участок M13 (подчеркнутый), отжигали праймером M13F путем нагревания смеси до 65 °С и медленного охлаждения до комнатной температуры в течение 1 часа. Сопоставимое снижение флуоресценции наблюдалось для KF (L790C) и KF дикого типа (как 1 мкМ) при инкубации с смесью праймера-матрицы (25 мкМ) и dNTP (250 мкМ). Исходные данные флуоресценции корректировались путем вычитания фона, который измерялся в отсутствие dNTP. Длины волн возбуждения и излучения, использованные в этом эксперименте, составляли 305 и 365 нм, соответственно.
Групповой анализ для включения аналогового dNTP
[0157] Чтобы подтвердить включение аналогов dNTP, рандомизированные матрицы ДНК (Таблица S1) были полимеризованы KF после гибридизации с праймером M13F. Реакции положительного контроля содержали KF (1 мкМ), dNTP или аналоги dNTP (100 мкМ), и матрицу-праймера A/T или G/C (5 мкМ) в 10 мМ Трис, 50 мМ NaCl, 10 мМ MgCl,1 мМ DTT, pH 7,9. Реакции для тестирования включения аналога dNTP содержали 100 мкМ аналога вместо его нативного dNTP, а реакции отрицательного контроля опускали либо аналог, либо его нативный dNTP. Реакции выдерживали при 25 °С в течении 2 ч в термоциклере перед электрофорезом на агарозном геле с высоким разрешением 5% (ФИГ8B).
Варианты реализации изобретения настоящего раскрытия.
[0158] Способ обнаружения изменения конформации полимеразы нуклеиновой кислоты, включающий:
(i) контактирование полимеразы нуклеиновой кислоты, нековалентно прикрепленной к однослойной углеродной нанотрубке (SWNT), с первым нуклеотидом или первым нуклеотидным аналогом, и последовательностью матрицы нуклеиновой кислоты, в результате чего образуется конформационно измененная полимераза нуклеиновой кислоты, связанная с первым нуклеотидом или первым нуклеотидным аналогом, и последовательностью матричной нуклеиновой кислоты;
(ii) обнаружение конформационно измененной полимеразы нуклеиновой кислоты путем измерения первого изменения электропроводности в SWNT между полимеразой нуклеиновой кислоты и конформационно измененной полимеразой нуклеиновой кислоты.
[0159] Способ, в котором указанная полимераза нуклеиновой кислоты контактирует с первым нуклеотидным аналогом.
[0160] Способ дополнительно содержит (iii) идентификацию указанного первого нуклеотидного или первого нуклеотидного аналога на основе первого сигнала, полученного при первом изменении электропроводности.
[0161] Способ, дополнительно включающий(iv) разрешение указанной конформационно измененной полимеразе нуклеиновой кислоты высвобождать указанный первый нуклеотидный или первый нуклеотидный аналог, тем самым реформируя указанную полимеразу нуклеиновой кислоты.
[0162] Способ, дополнительно включающий
(v) контактирование полимеразы нуклеиновой кислоты, нековалентно прикрепленной к однослойной углеродной нанотрубке (SWNT), со вторым нуклеотидом или вторым нуклеотидным аналогом, и указанной последовательностью матрицы нуклеиновой кислоты, в результате чего образуется конформационно измененная полимераза нуклеиновой кислоты, связанная со вторым нуклеотидом или вторым нуклеотидным аналогом, и последовательностью матричной нуклеиновой кислоты; и
(vi) обнаружение конформационно измененной полимеразы нуклеиновой кислоты путем измерения изменения электропроводности в SWNT между полимеразой нуклеиновой кислоты и конформационно измененной полимеразой нуклеиновой кислоты, связанной со вторым нуклеотидом или вторым нуклеотидным аналогом и последовательностью матричной нуклеиновой кислоты.
[0163] Способ формулы изобретения 5, в котором указанная полимераза нуклеиновой кислоты контактирует со вторым нуклеотидным аналогом.
[0164] Способ дополнительно содержит (vii) идентификацию указанного второго нуклеотидного или второго нуклеотидного аналога на основе второго сигнала, полученного вторым изменением электропроводности; и идентификацию последовательности в матричной нуклеиновой кислоте.
[0165] Способ, в котором указанный первый нуклеотидный аналог или указанный второй нуклеотидный аналог гибридизуется с указанной последовательностью матричной нуклеиновой кислоты с не Уотсон-Криковским спариванием оснований.
[0166] Способ, в котором указанный первый нуклеотидный аналог или указанный второй нуклеотидный аналог представляет собой 2-тио dCTP, 2-тио dTTP, или 6-Cl-dGTP, 6-аза-dTTP, α-тио-dATP, или α -тио-dTTP.
[0167] Способ, в котором указанный первый нуклеотидный аналог или указанный второй нуклеотидный аналог модифицирован в трифосфатном фрагменте.
[0168] Способ, в котором указанный трифосфатный фрагмент содержит α-тиозамещение.
[0169] Способ, в котором указанный первый нуклеотидный аналог или указанный второй нуклеотидный аналог является α- тио-dATP, α-тио-dGTP, α-тио-dCTP или α-тио-dTTP.
[0170] Способ, в котором указанный первый нуклеотидный аналог или указанный второй нуклеотидный аналог дополнительно включает замещение в нуклеотидном основании
[0171] Способ, в котором указанный первый нуклеотидный аналог или указанный второй нуклеотидный аналог является α- тио-2-тио-dTTP, α-тио-2-тио-dCTP, α-тио-6-Cl-20APTP, или 6-Cl-2APTP.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Попечители Калифорнийского
Collins, Philip G.
Weiss, Gregory A.
Choi, Yongki
Olsen, Tivoli
<120> ОБНАРУЖЕНИЕ ИЗМЕНЕНИЙ КОНФОРМАЦИИ ПОЛИМЕРАЗЫ НУКЛЕИНОВЫХ КИСЛОТ С
ПОМОЩЬЮ НАНОТРУБКИ
<130> 48538-526001WO
<150> US 62/093671
<151> 2014-12-18
<160> 12
<170> PatentIn, версия 3.5
<210> 1
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 1
tgtaaaacga cggccagt 18
<210> 2
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<220>
<221> модифицированное основание
<222> (19)..(19)
<223> 2-Аминопурин
<400> 2
tcgagctatc tctaaagcag ctaactatcg agctatcgcg aaactggccg tcgttttaca 60
<210> 3
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 3
cctaacgcag atagacgttg tttagagccc gggtcggcca tactggccgt cgttttaca 59
<210> 4
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 4
cctaacgcag atagacgttg tttagagatt taaattcggc cactggccgt cgttttaca 59
<210> 5
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 5
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaactggccg tcgttttaca 60
<210> 6
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 6
tttttttttt tttttttttt tttttttttt tttttttttt ttactggccg tcgttttaca 60
<210> 7
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 7
gggggggggg gggggggggg gggggggggg gggggggggg ggactggccg tcgttttaca 60
<210> 8
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 8
cccccccccc cccccccccc cccccccccc cccccccccc ccactggccg tcgttttaca 60
<210> 9
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 9
tttttttttt tttttttttt tttttttttt tttttttttt tt 42
<210> 10
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 10
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aa 42
<210> 11
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 11
gggggggggg gggggggggg gggggggggg gggggggggg gg 42
<210> 12
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 12
cccccccccc cccccccccc cccccccccc cccccccccc cc 42
<---
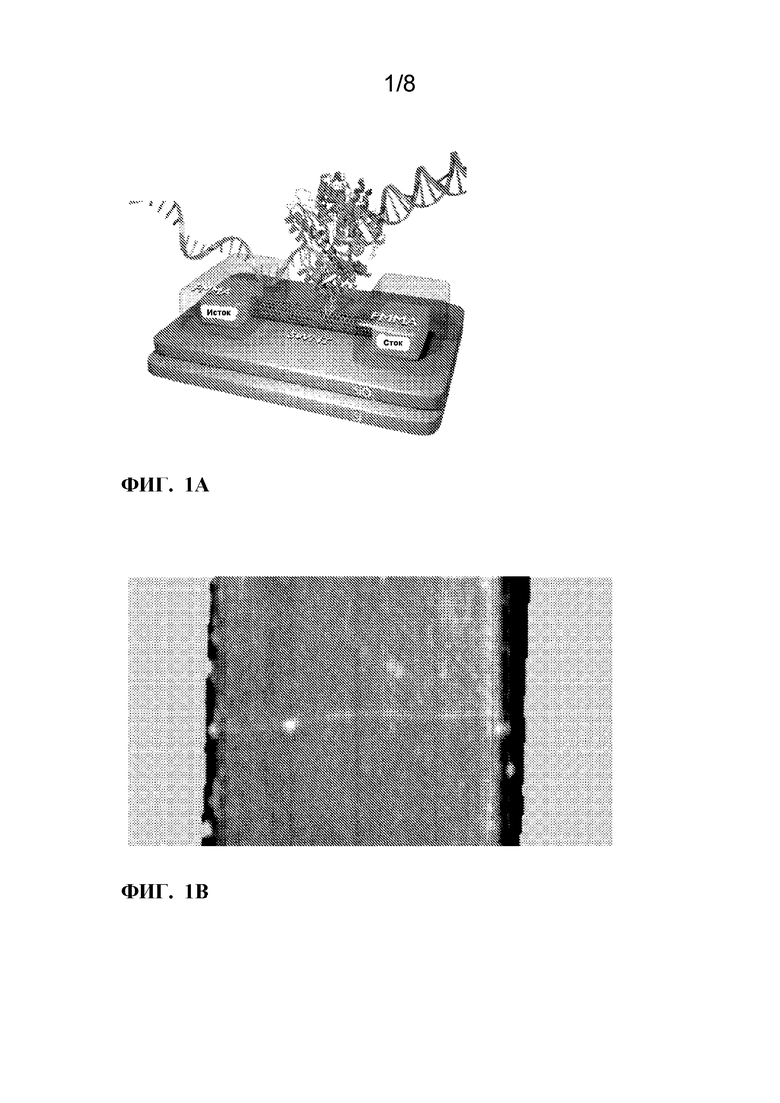
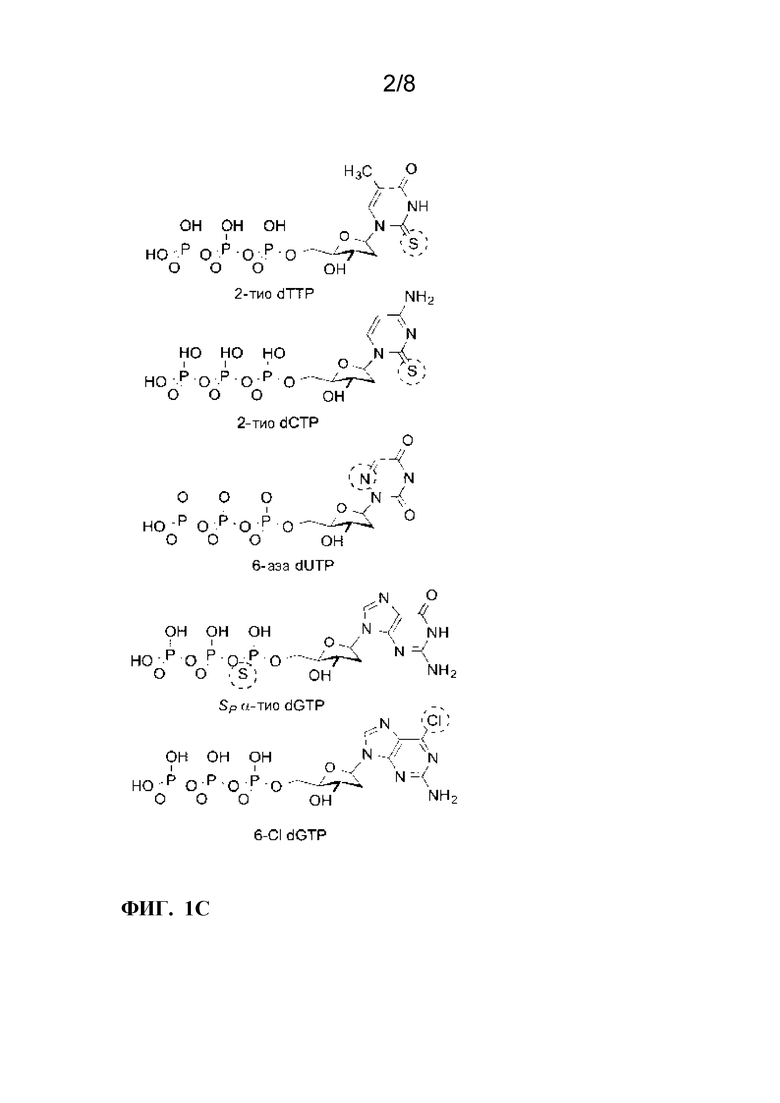
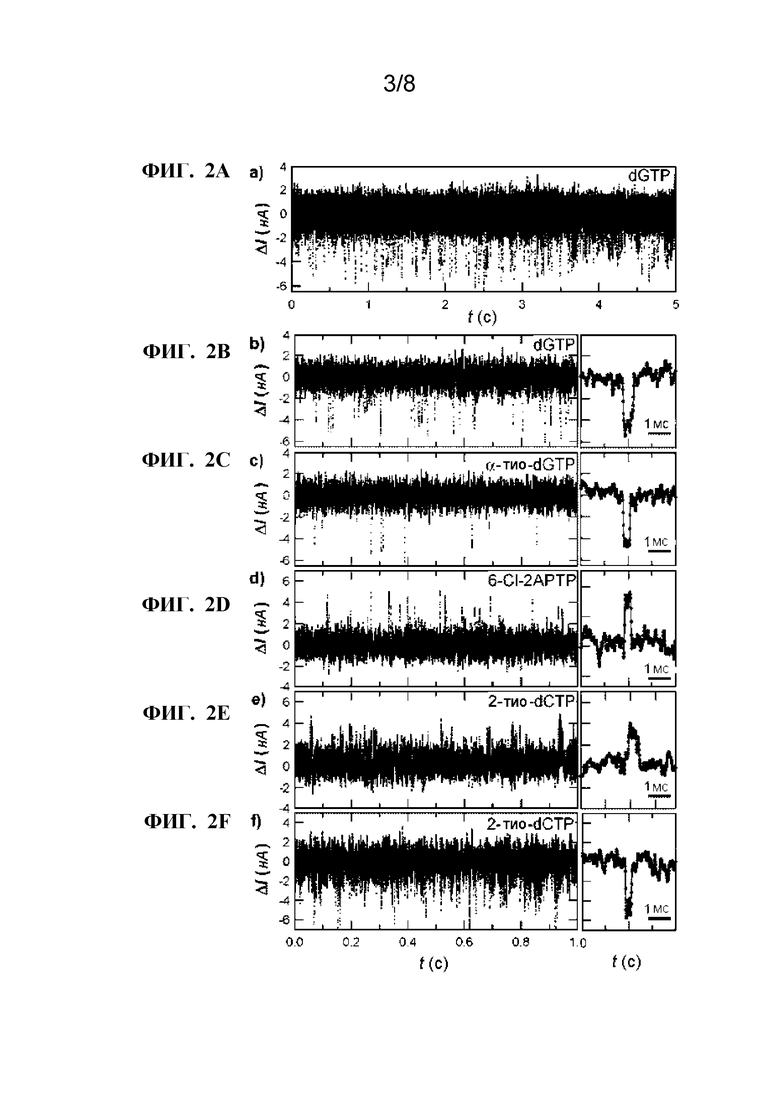
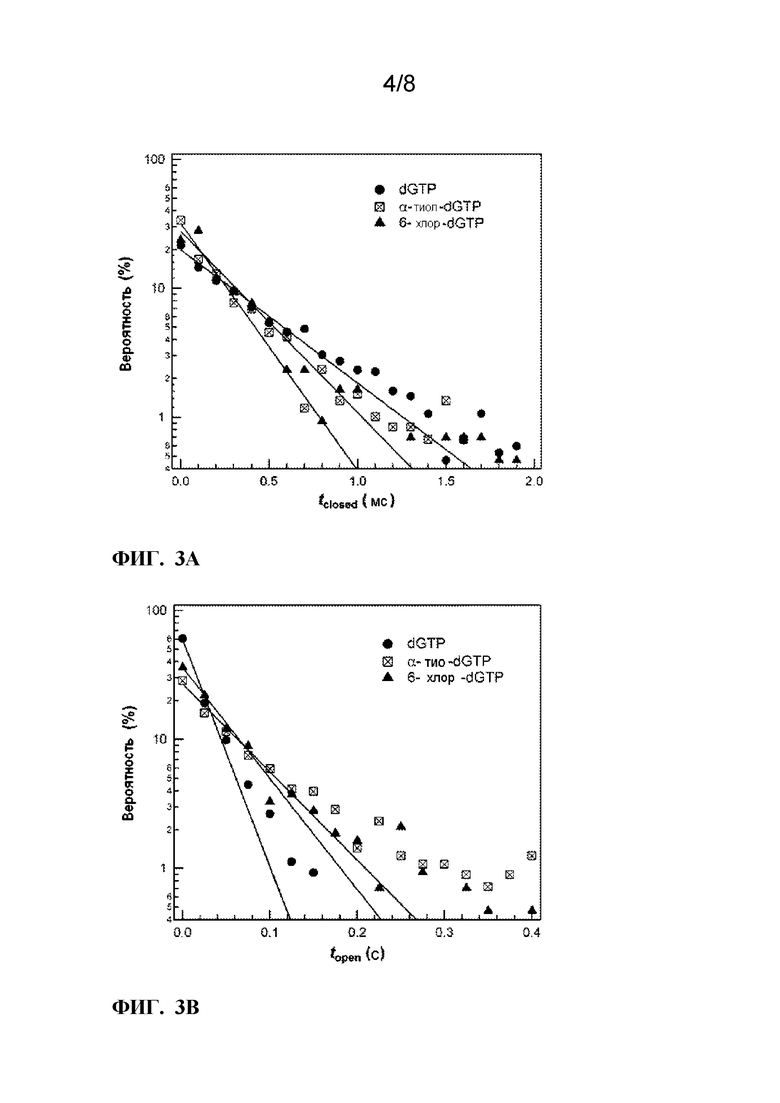
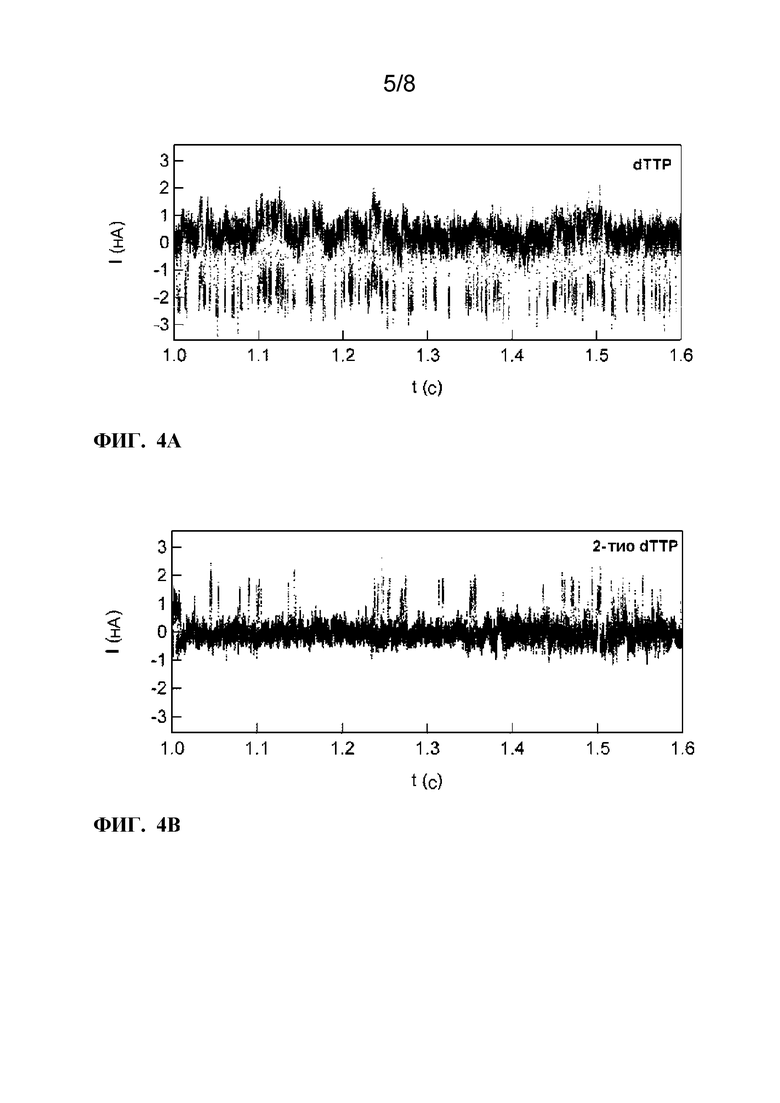
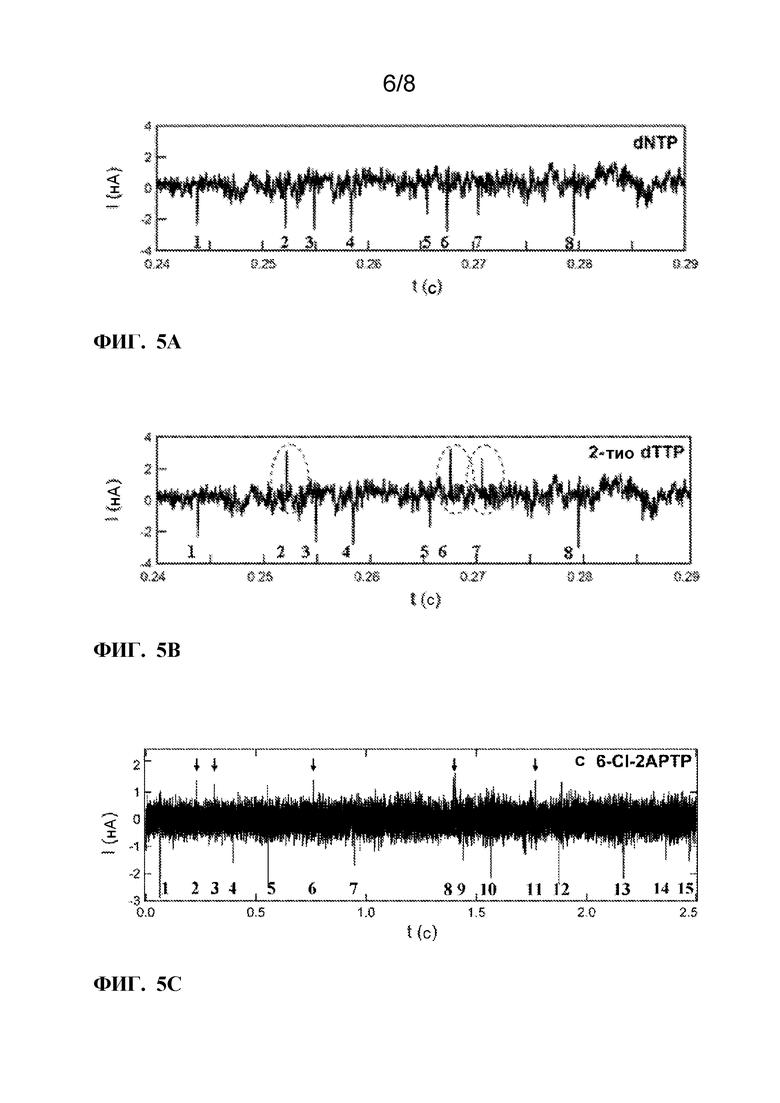

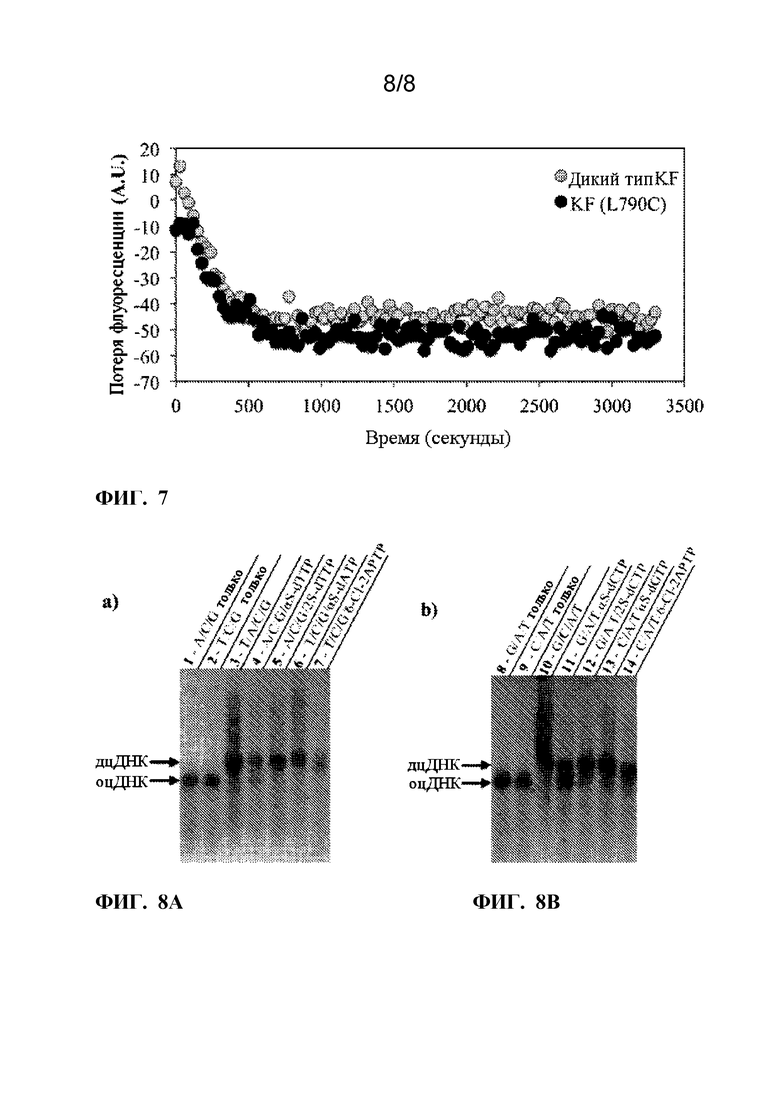
Изобретение относится к области биотехнологии. Предложен способ обнаружения конформационно измененной полимеразы нуклеиновой кислоты. Способ включает приведение прикрепленной к электропроводящему каналу полимеразы нуклеиновой кислоты в контакт с последовательностью матрицы нуклеиновой кислоты и смесью немеченых модифицированных нуклеотидов, обнаружение конформационно измененной полимеразы нуклеиновой кислоты путем измерения изменения электрического сигнала в электропроводящем канале. Причем каждый из немеченых модифицированных нуклеотидов содержит фосфатный остаток и включающее пурин или пиримидин нуклеотидное основание, где по меньшей мере один из немеченых модифицированных нуклеотидов содержит замещение на серу в его фосфатном остатке или где по меньшей мере один из указанных немеченых модифицированных нуклеотидов содержит замещение в положении 6 его пурина или в положении 2 или 6 его пиримидина. Изобретение обеспечивает эффективное и информативное секвенирование ДНК. 19 з.п. ф-лы, 20 ил., 2 табл., 3 пр.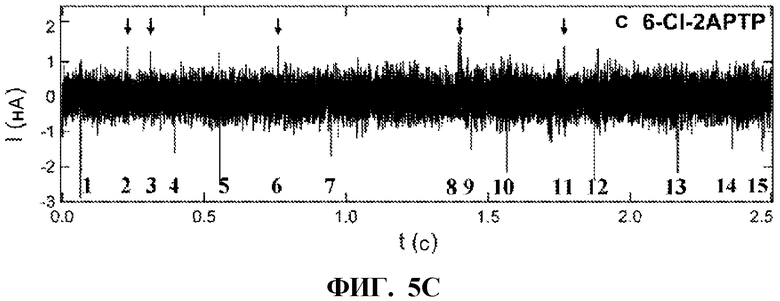
1. Способ обнаружения конформационно измененной полимеразы нуклеиновой кислоты, включающий:
приведение полимеразы нуклеиновой кислоты, прикрепленной к электропроводящему каналу, в контакт с последовательностью матрицы нуклеиновой кислоты и смесью немеченых модифицированных нуклеотидов, где каждый из указанных немеченых модифицированных нуклеотидов содержит фосфатный остаток и нуклеотидное основание, включающее пурин или пиримидин, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов содержит замещение на серу в его фосфатном остатке; или где по меньшей мере один из указанных немеченых модифицированных нуклеотидов содержит замещение в положении 6 его пурина или в положении 2 или 6 его пиримидина, в результате чего образуется конформационно измененная полимераза нуклеиновой кислоты, связанная с одним из нуклеотидов указанной смеси и последовательностью матричной нуклеиновой кислоты; и
обнаружение конформационно измененной полимеразы нуклеиновой кислоты путем измерения изменения электрического сигнала в указанном электропроводящем канале, вызванного конформационным переходом между полимеразой нуклеиновой кислоты и конформационно измененной полимеразой нуклеиновой кислоты.
2. Способ по п. 1, где фосфатный остаток представляет собой трифосфатный остаток, и где атом серы находится в α-положении указанного трифосфатного остатка.
3. Способ по п. 2, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов выбран из группы, состоящей из α-тио-dАТР, α-тио-dGТР, α-тио-dCTP и α-тио-dТТР.
4. Способ по п. 1, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов содержит замещение в его пурине или пиримидине; и указанное замещение выбрано из группы, состоящей из атома галогена, атома азота и атома серы.
5. Способ по п. 4, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на атом галогена в положении 6 пурина, и где указанный немеченый модифицированный нуклеотид представляет собой 6-Cl-dGTP.
6. Способ по п. 4, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на серу в положении 2 пиримидина или где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на азот в положении 6 пиримидина, и где указанный немеченый модифицированный нуклеотид выбран из группы, состоящей из 2-тио-dATP, 2-тио-dGTP, 2-тио-dCTP, 2-тио-CTP, 2-тио-dTTP, 2-тио-TTP, 6-аза-2-тио-dTTP и 6-аза-2-тио-TTP.
7. Способ по п. 4, где указанный по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на азот в положении 6 пиримидина, и где указанный немеченый модифицированный нуклеотид представляет собой 6-аза-dUTP.
8. Способ по п. 1, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на серу в немостовом, α-фосфорильном атоме кислорода; и указанное замещение в положении 2 или 6 пиримидина выбрано из группы, состоящей из атома галогена и второго атома серы.
9. Способ по п. 8, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов выбран из группы, состоящей из α-тио-2-тио-dTTP, α-тио-2-тио-dCTP, α-тио-6-Cl-2-APTP.
10. Способ по п. 1, где указанный электропроводящий канал содержит металл, оксид металла или полупроводник.
11. Способ по п. 1, где указанный электропроводящий канал представляет собой проводник нанометрового размера.
12. Способ по п. 11, где указанный проводник нанометрового размера выбран из группы, состоящей из нанопровода и нанотрубы.
13. Способ по п. 1, где указанный электропроводящий канал представляет собой полупроводниковый нанопровод.
14. Способ по п. 1, дополнительно включающий идентифицирование нуклеотида на основании указанного изменения электрического сигнала.
15. Способ по п. 14, где указанное изменение электрического сигнала повышается или понижается относительно второго изменения электрического сигнала, получаемого, когда нативный нуклеотид приводят в контакт с указанной полимеразой нуклеиновой кислоты.
16. Способ по п. 1, где указанное изменение электрического сигнала представляет собой изменение электропроводности.
17. Способ по п. 1, где указанная полимераза нуклеиновой кислоты ковалентно прикреплена к указанному к электропроводящему каналу.
18. Способ по п. 1, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на серу в немостовом, α-фосфорильном атоме кислорода его фосфатноого остатка; и указанный фосфатный остаток содержит множественные фосфатные группы.
19. Способ по п. 18, где указанный фосфатный остаток выбран из группы, состоящей из три-фосфата, тетра-фосфата и пента-фосфата.
20. Способ по п. 1, где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на серу в немостовом, α-фосфорильном атоме кислорода его фосфатноого остатка; и
где по меньшей мере один из указанных немеченых модифицированных нуклеотидов имеет замещение на галоген, азот или серу в положении 6 его пурина или положении 2 или 6 его пиримидина.
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| СУХОДОЛОВ Н.Г | |||
| и др., Новые материалы, полученные методом Ленгмюра-Блоджетт, и их применение в нанотехнологии и приборостроении (ч.1 гибридные материалы) // Научное приборостроение, т.23, n.1., 2013, стр.86-105. | |||

