Настоящее изобретение относится к способу получения L-метионина, в котором микроорганизм культивируют в присутствии L-гомосерина и метилмеркаптана, его соли или диметилдисульфида, в результате чего L-метионин накапливается в культуральной среде.
Аминокислоту метионин в настоящее время получают в промышленных масштабах по всему миру в больших количествах, и она имеет важное коммерческое значение. Метионин используют во многих областях, а именно в фармацевтических продуктах, спортивно-оздоровительных продуктах, а особенно в качестве кормовой добавки во многих кормовых продуктах для различных сельскохозяйственных животных, для которых может применяться как рацемическая, так и энантиомерно чистая форма метионина.
В промышленном масштабе метионин получают химическим путем посредством реакции Бушера-Берга, которая является вариантом Синтеза Штрекера. В данном случае реагируют исходные вещества метилмеркаптопропиональдегид (полученный из акролеина и метилмеркаптана), цианид водорода, аммиак и диоксид углерода с получением 5-(2-метилмеркаптоэтил)гидантоина (гидантоин метионина), при этом данное вещество впоследствии гидролизуется щелочью с получением метионината щелочного металла и метионина, а затем метионин высвобождается путем нейтрализации кислотой (EP 0780370 А2). Для получения метионина могут также применяться различные другие способы, например, реакция амидокарбонилирования, гидролиз белков или ферментация вырабатывающих метионин микроорганизмов. В химическом синтезе метионин получают в виде рацемической смеси D- и L-метионина, при этом L-метионин или его предшественники с L-конфигурацией, например, L-гомосерин, можно получать при ферментации подходящих микроорганизмов.
L-гомосерин, потенциальный предшественник L-метионина (H.J. Teas et al., J. Biol. Chem. 1948, 172: 651-658), можно получать как химическим путем (M.D. Armstrong, J. Am. Chem. Soc., Vol. 70, 1756-1759, 1948), так и посредством ферментации с помощью микроорганизмов (см., например, US 3598701, US 6303348 B1, EP 0994190 A2, EP 1149911 A2, WO 2004/067757 A1).
У Hateley et al. раскрыт способ, в котором L-метионин получают химическим путем, начиная с L-гомосерина (WO 2007/085514 A2).
Lievense удалось доказать, что штаммы микроорганизмов, лишенные активности метилазы гомоцистеина, выработка L-гомосерина которыми была активирована и которые были трансформированы плазмидой, кодирующей L-гомосеринацетилтрансферазу и O-ацетил-L-гомосеринсульфгидрилазу (O-ацетилгомосерин-(тиол)-лиаза), вырабатывали L-метионин, превышающий их собственную потребность, в присутствии метилмеркаптана в отличие от исходного штамма (E. coli, C. glutamicum или B. flavum) (WO 93/17112 A1).
Bolten et al. (J. Microbiol. Biotechnol. (2010), 20(8), 1196-1203) смогли продемонстрировать, что (дикий тип) C. glutamicum способен расти в присутствии метилмеркаптана и его димерной формы, диметилдисульфида, в качестве единственного источника серы вместо сульфата, наиболее распространенного источника серы для культивирования микроорганизмов, и исследовали основные пути и ферменты. Они продемонстрировали, что MetY (O-ацетил-L-гомосеринсульфогидрилаза) отвечает на замену ацетильной группы O-ацетил-L-гомосерина на меркаптогруппу метилмеркаптана или диметилдисульфида непосредственно с получением L-метионина. С целью увеличения выработки L-метионина авторы данного изобретения предложили амплификацию не только MetY, но также и других ферментов при биосинтезе L-метионина.
Zelder et al. в заявке (WO 2007/011939 A2) продемонстрировали, что L-метионин можно получать из микроорганизмов, таких как E. coli и C. glutamicum, путем культивирования микроорганизмов, характеризующихся дерегулирующим действием O-ацетил-L-гомосеринсульфгидрилазы или O-сукцинил-L-гомосеринсульфгидрилазы и/или L-гомосеринацетилтрансферазы или L-гомосеринсукцинилтрансферазы в присутствии сульфидного соединения с метиловым покрытием, такого как диметилдисульфид или диметилтрисульфид.
Kim et al. предложили двухстадийный биотехнологический способ получения L-метионина (WO 2008/013432 A1). На первом этапе в настоящем документе вначале получают предшественник L-метионина, O-сукцинил-L-гомосерина или O-ацетил-L-гомосерина с помощью рекомбинантных микроорганизмов, которые накапливаются в культуральной жидкости. На последующей второй стадии предшественник L-метионина вводят в реакцию с метилмеркаптаном в присутствии белка, характеризующегося O-сукцинил-L-гомосеринсульфгидрилазной активностью или O-ацетил-L-гомосеринсульфгидрилазной активностью, или в присутствии микроорганизма, вырабатывающего данный белок или перерабатывающего клетку данного микроорганизма, с получением L-метионина и соответствующей карбоновой кислоты, т. е. ацетата или сукцината.
Однако в данной ферментативной реакции эквимолярные количества ацетата или сукцината образованы в дополнение к L-метионину. При выборе O-ацетил-L-гомосерина в качестве предшественника L-метионина, например, это приводит к высоким концентрациям ацетата в ходе реакции, в частности, в промышленном масштабе. При низком внешнем значении pH недиссоциированные молекулы ацетата способны проникать в клетку через мембрану и депротонироваться в ней, что приводит к падению внутреннего значения pH цитоплазмы и приводит к нарушению гомеостаза клеточного pH (I.R. Booth, Microbiological Reviews 49, No. 4 (1985), 359-378). Кроме того, ацетат не могут полностью удалить из продукта, содержащего L-метионин, приемлемыми усилиями. Соответственно, Hong et al. (WO 2012/091479 A2) предлагают множество способов удаления и получения относительно большого количества ацетата, образующегося на второй стадии способа получения L-метионина из продукта, содержащего L-метионин.
Целью настоящего изобретения является обеспечение способа получения L-метионина в микроорганизме, в котором ацетат, образующийся в результате превращения O-ацетил-L-гомосерина в L-метионин, в значительной степени повторно используется тем же микроорганизмом.
Данной цели достигают с помощью способа получения L-метионина, где микроорганизм, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью и O-ацетил-L-гомосеринсульфгидрилазной активностью, культивируют в культуральной среде, содержащей L-гомосерин и источник серы, при этом источник серы выбран из группы, состоящей из метилмеркаптана (MC), соли метилмеркаптана и диметилдисульфида (DMDS), в результате чего L-метионин накапливается в культуральной среде.
Как правило, ферментативные активности в микроорганизмах осуществлялись путем экспрессии соответствующего гена, кодирующего соответствующий фермент. Так называемые промоторы располагаются выше гена. Промотор представляет собой последовательность ДНК, состоящую из приблизительно 40-50 пар оснований, и которые составляют сайт связывания для холофермента РНК-полимеразы и отправную точку транскрипции (M. Pátek et al., Microbial Biotechnology, 6 (2013), 103-117), в результате чего можно влиять на активность экспрессии контролируемого полинуклеотида или гена. Выражение “функциональная связь” понимают в значении последовательное расположение промотора с геном, который приводит к транскрипции гена.
Микроорганизм может также быть рекомбинантным и характеризоваться повышенной L-гомосерин-O-ацетилтрансферазной активностью и повышенной O-ацетил-L-гомосеринсульфгидрилазной активностью.
Повышенные ферментативные активности у микроорганизмов могут осуществляться, например, за счет мутации соответствующего эндогенного гена. Ферментативные активности также могут усиливаться при повышении экспрессии соответствующего гена, например, при увеличении числа копий гена и/или при усилении регуляторных факторов генов. Усиление таких регуляторных факторов, которые положительно влияют на экспрессию генов, может, например, достигаться с помощью модифицирования промоторной последовательности, расположенной выше структурного гена, для увеличения эффективности промотора или путем полной замены указанного промотора на более эффективный промотор.
В способе по настоящему изобретению как L-гомосерин-O-ацетилтрансферазная активность, так и O-ацетил-L-гомосеринсульфгидрилазная активность предпочтительно усиливаются при повышенной экспрессии гена, который кодирует белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью, или белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью. Повышенной экспрессии генов предпочтительно достигают путем увеличения числа копий гена, кодирующего белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью, или белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью, и/или с помощью функциональной связи в каждом случае гена, кодирующего белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью, или белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью по отношению к сильному промотору.
Подходящие сильные промоторы или способы получения таких промоторов для повышения экспрессии известны из литературы (например, S. Lisser and H. Margalit, Nucleic Acid Research, 1993, Vol. 21, No. 7, 1507-1516; M. Pátek and J. Nesvera in H. Yukawa and M Inui (eds.), Corynebacterium glutamicum, Microbiology Monographs 23, Springer Verlag Berlin Heidelberg 2013, 51-88; B. J. Eikmanns et al., Gene, 102 (1991) 93-98). Например, нативные промоторы могут быть оптимизированы путем изменения промоторной последовательности в направлении известных консенсусных последовательностей с повышением экспрессии генов, функционально связанных с данными промоторами (M. Patek et al., Microbiology (1996), 142, 1297-1309; M. Patek et al., Microbial Biotechnology 6 (2013), 103-117). С увеличением экспрессии гена, кодирующего белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью (metX), или гена, кодирующего белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью (metY), например, подходящим является промотор tacI (PtacI) (H.A. deBoer et al., Proc. Natl. Acad. Sci. USA, Vol. 80, 21-25, January 1983, Biochemistry). Последовательность PtacI показана под номером 5 последовательности (SEQ ID No. 5).
Конститутивные промоторы также являются подходящими для сверхэкспрессии, при которой ген, кодирующий ферментативную активность, экспрессируется непрерывно под контролем промотора, такого как, например, зависимый от глюкозы промотор deo. Химически индуцированные промоторы также являются подходящими, а именно tac, lac или trp. Наиболее широко распространенной системой для индукции промоторов является lac-оперон E. coli. В данном случае в качестве индуктора применяют лактозу или изопропил-ß-D-тиогалактопиранозид (IPTG). В качестве индукторов общепризнанными также являются системы, применяющие арабинозу (например, система pBAD) или рамнозу (например, E. coli KRX). Система для физической индукции представляет собой, например, индуцируемую температурой промоторную систему холодового шока на основе промотора E. coli cspA из Takara или Lambda PL, а также осмотически индуцируемые промоторы, например, osmB (например, WO 95/25785 A1).
В способе по настоящему изобретению рекомбинантный микроорганизм выбран из группы, состоящей из Enterobacteriaceae и Corynebacteriaceae, например штамма Escherichia coli (E. coli), например непатогенного штамма E. coli K-12 MG1655 (DSM 18039), или штамма Corynebacterium glutamicum (C. glutamicum), например ATCC13032, или штамма Corynebacterium humireducens (C. humireducens), например DSM 45392.
В способе по настоящему изобретению L-гомосерин-O-ацетилтрансферазная активность обеспечивается, например, ферментом MetX, который получают из Corynebacterium glutamicum или из C. humireducens. У Kim et al. (EP 2657345 A1; EP 2657250 A2) или Ochrombel et al. (WO 2015/165746 A1) раскрыты примеры подходящих ферментов, характеризующихся L-гомосерин-O-ацетилтрансферазной активностью. Фермент MetX, применяемый в описанных ниже экспериментальных примерах, характеризуется аминокислотной последовательностью в соответствии с последовательностью номер 2 (SEQ ID No. 2). Соответствующая нуклеотидная последовательность для гена metX показана под последовательностью номер 1 (SEQ ID No. 1). Последовательность получают из C. glutamicum (ATTC13032) NC_003450.
O-ацетил-L-гомосеринсульфгидрилазная активность, подходящая для способа по настоящему изобретению, представляет собой, например, фермент MetY, который получают из Corynebacterium glutamicum или C. humireducens. У Möckel et al. (WO 02/18613 A1), Kröger et al. (WO 2007/024933 A2) или Kim et al. (EP 2 657 345 A1) раскрыты примеры ферментов, характеризующихся O-ацетил-L-гомосеринсульфгидрилазной активностью по настоящему изобретению. Фермент MetY, применяемый в описанных ниже экспериментальных примерах, характеризуется аминокислотной последовательностью в соответствии с последовательностью номер 4 (SEQ ID No. 4). Соответствующая нуклеотидная последовательность для гена metY показана под последовательностью номер 3 (SEQ ID No. 3). Последовательность получают из C. glutamicum (ATTC13032) NC_003450.
L-Гомосерин транспортируется в микроорганизмы с помощью импортеров аминокислот с разветвленной цепью, например системы LIV у Escherichia coli (B.A. Templeton and M. A. Savageau, JOURNAL OF BACTERIOLOGY, Vol. 117, No. 3, Mar. 1974, p. 1002-1009). У Corynebacterium glutamicum (C. glutamicum) также существует гомологичная транспортная система, кодируемая с помощью BrnQ of cgl2310 (A. Tauch et al., Arch Microbiol 169 (1998): 303-312).
В клетке L-гомосерин активируется по его гидроксильной группе с помощью переноса ацетильной группы ацетилкофермента A (ацетил-CoA) с получением O-ацетил-L-гомосерина с помощью (гетерологичной) гомосерин-O-ацетилтрансферазы (MetX). Затем O-ацетил-L-гомосерин превращают в L-метионин и ацетат в присутствии источника восстановленной серы, а именно метилмеркаптана (MC) и пиридоксаль-5'-фосфата (PLP) с помощью (гетерологичной) сульфгидрилазы (MetY). При этом O-ацетил-L-гомосерин является одним из природных промежуточных продуктов биосинтеза метионина у corynebacteria, при этом биосинтез метионина у enterobacteria протекает с помощью промежуточного продукта O-сукцинил-L-гомосерина (см., например, Figge R (2007) Methionine biosynthesis in Escherichia coli and Corynebacterium glutamicum. В Wendisch VF (ed) Amino acid biosynthesis - pathways, regulation and metabolic engineering. Microbiology Monographs, vol 5. Springer, Berlin, pp 163-193). Следовательно, L-гомосерин-O-ацетилтрансферазная и O-ацетил-L-гомосеринсульфгидрилазная активности сначала должны гетерологично вводится в enterobacteria, а именно в E. coli, при этом данные ферментативные активности по своей природе уже присутствуют в corynebacteria, а именно в C. glutamicum. Каждый из соответствующих гомологичных или гетерологичных генов, кодирующих соответствующие ферменты, может усиливаться с помощью описанных вначале мер (таких как увеличение числа копий обоих генов и/или применения сильных промоторов).
У enterobacteria, а именно у E. coli, усиленная L-гомосерин-O-ацетилтрансферазная и O-ацетил-L-гомосеринсульфгидрилазная активности могут предоставляться путем трансформации с помощью подходящих векторов, содержащих последовательности генов metX (например SEQ ID No. 1) и metY (например SEQ ID No. 3), которые в любом случае находятся выше сильного промотора (например PtacI). Примером такой конструкции является последовательность в соответствии с последовательностью номер 6 (SEQ ID No. 6).
Ацетат, высвобожденный путем превращения O-ацетил-L-гомосерина в L-метионин в присутствии метилмеркаптана (MC) и пиридоксаль-5'-фосфата (PLP), а также (гетерологичной) сульфгидрилазы (MetY), затем используется снова для синтеза ацетил-CoA с потреблением ATP в цитоплазме E. coli (аналогично Bacillus subtilis) с помощью индуцируемой ацетатом ацетил-CoA-синтетазы (Acs), которая активируется, в частности, в стационарной фазе или в анаэробных условиях с помощью регулятора CsrA (S. Kumari et al., JOURNAL OF BACTERIOLOGY, Vol. 177, No. 10, May 1995, p. 2878-2886).
В отличие от E. coli избыток ацетата у C. glutamicum превращается исключительно в ATP-зависимой реакции в ацетилфосфат с помощью ацетаткиназы (AK), которая в итоге вступает в реакцию с помощью фосфотрансацетилазы (PTA) в присутствии CoA с получением ацетил-CoA. Соответствующие гены ack и pta организованы в C. glutamicum в оперон, регулируемый ацетатом на транскрипционном уровне (R. Gerstmeir et al. Journal of Biotechnology 104 (103) 99-122).
L-Метионин выводится из клетки E. coli с помощью экспортера YjeH (Q. Liu et al., Appl Environ Microbiol 81 (2015) p. 7753-7766). Кроме того, ген ygaZH у E. coli кодирует экспортер метионина (WO 2015/028675 A1). L-Метионин выводится из клетки C. glutamicum в культуральную среду с помощью частиц экспортера BrnFE (C. Trötschel et al., JOURNAL OF BACTERIOLOGY, June 2005, p. 3786-3794).
На фигуре 1 показана карта плазмиды pMW218.
На фигуре 2 показана карта плазмиды pMW218_Ptac-metX_Ptac-metY.
На фигуре 3 показано каталитическое превращение L-гомосерина и ацетил-CoA с помощью клеточных гомогенатов MG1655/pMW218 или MG1655/pMW218_Ptac-metX_Ptac-metY.
На фигуре 4 показано каталитическое превращение метилмеркаптида натрия в присутствии O-ацетилгомосерина и пиридоксаль-5'-фосфата (PLP) с помощью фермента O-ацетилгомосеринсульфгидрилазы (MetY). Сравнение клеточных гомогенатов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY показано с помощью используемых соответствующих общих концентраций белка.
Примеры
1) Получение энтеробактерии, гетерологично экспрессирующей гены L-гомосерина-O-ацетилтрансферазы и сульфгидрилазы видов коринебактерий
На основе геномной последовательности Corynebacterium glutamicum (ATCC13032) NC_003450 синтезировали последовательности генов metX (SEQ ID No. 1) и metY (SEQ ID No. 3), которые кодируют L-гомосерин-O-ацетилтрансферазу с аминокислотной последовательностью в соответствии с SEQ ID No. 2 и O-ацетил-L-гомосеринсульфгидрилазу с аминокислотной последовательностью в соответствии с SEQ ID No. 4, соответственно, вместе с находящимся выше промотором PtacI (SEQ ID No. 5) (H.A. deBoer et al., Proc. Natl. Acad. Sci. USA, Vol. 80, 21-25, January 1983, Biochemistry) от Life Technologies Invitrogen GeneArt (Германия) (SEQ ID No. 6).
В этой последовательности SEQ ID No. 6 промотор PtacI находится в пределах пары оснований 407-447, последовательность гена metX находится в пределах 502-1638, промотор PtacI в пределах 1645-1685, и последовательность гена metY в пределах 1742-3055.
Затем клонирование данной синтетической последовательности выполняли с помощью сайтов рестрикции BssHII и BglI в последовательности вектора pMW218 (номер доступа: AB005477) (Nippon Gene, Тояма, Япония) (фиг. 1). Плазмида pMW218_Ptac-metX_Ptac-metY образуется из нее (фиг. 2). Для анализа плазмиды pMW218_Ptac-metX_Ptac-metY дополнительно выполняли секвенирование ДНК с помощью Eurofins MWG Operon. Полученные последовательности ДНК проверяли на точность с применением программного обеспечения Clone Manager, чтобы подтвердить последовательность SEQ ID No. 6.
Плазмиды pMW218 и pMW218_Ptac-metX_Ptac-metY трансформировали в каждом случае в штамме Escherichia coli K-12 MG1655 (DSM № 18039). Затем трансформанты культивировали в чашках с агаровой средой LB с 50 мкг/мл канамицина, так что смогли получить штаммы MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY. В каждом случае была выбрана колония, которую инокулировали в каждом случае в 10 мл среды LB с 50 мкг/мл канамицина и культивировали при 37°C, 200 об./мин. в течение 6 часов. Затем в 10 мл питательной среды A [25 г/л сульфата аммония; 1 г/л магния сернокислого семиводного; 2 г/л дигидрофосфата калия; 0,03 г/л гептагидрата железа; 0,02 г/л моногидрата сульфата марганца; 20 г/л моногидрата глюкозы; 30 г/л карбоната кальция; 0,05 г/л канамицина; 0,025 г/л пиридоксаль-5'-фосфата (PLP); 0,0024 г/л изопропил-β-D-тиогалактопиранозида (IPTG)] инокулировали 200 мкл культуры для роста клеток и инкубировали при 37°C, 200 об./мин. В течение 16 ч. Данные клеточные культуры разбавляли 10 мл свежей среды A в 100-мл колбе при OD 2 и дополнительно культивировали при идентичных условиях до достижения OD приблизительно 5 (приблизительно 3-4 ч.). Затем эти клетки, которые находились в экспоненциальной фазе роста и характеризовались гомосерин-O-ацетилтрансферазной (MetX) и сульфгидрилазной (MetY) активностью, можно применять для биотрансформации. Под биотрансформацией понимают превращение вещества, при котором используют целые живые клетки, фиксированные клетки или изолированные свободные или связанные с носителем ферменты или комбинацию вышеуказанного.
2) Определение ферментативной активности L-гомосерин-O-ацетилтрансферазы и ацетил-L-гомосеринсульфогидрилазы
В 10 мл среды LB с 50 мкг/мл канамицина в каждом случае инокулировали одну колонию штаммов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY и культивировали при 37°C, 200 об./мин. в течение 6 часов. Затем в 10 мл среды A (см. пример 1) инокулировали 200 мкл культуры для роста клеток и инкубировали при 37°C, 200 об./мин. в течение 16 ч. Затем каждую клеточную культуру собирали (8 мл приведенной к OD=1), надосадочную жидкость удаляли с помощью центрифугирования (20 мин., 4000 об./мин., 4°C) и дважды промывали осадок клеток с помощью 800 мкл 0,1 M калий-фосфатного буфера (pH 7,5) и отбирали в 1 мл буфера. Выполняли механическое разрушение клеток на устройстве FastPrep FP120 (QBiogene, Гейдельберг), где клетки трижды встряхивали в течение 2 с при 6,5 м/с в сосудах для расщепления с 300 мг стеклянных гранул (∅ 0,2-0,3 мм). Затем центрифугировали неочищенный экстракт при 12000 об./мин., 4°C, 20 мин. для удаления нерасщепленных клеток и клеточного дебриса. Общее количество белка определяли с использованием количественного анализа для белка Bio-Rad (Bio-Rad, США). Затем использовали клеточный гомогенат для ферментативного определения цитоплазматической активности L-гомосерин-O-ацетилтрансферазы и ацетил-L-гомосеринсульфгидрилазы.
2a) Определение цитоплазматической активности MetX (L-гомосерин-O-ацетилтрансферазы)
Реакцией, которую катализирует фермент L-гомосерин-O-ацетилтрансфераза (MetX) [EC2.3.1.31], является превращение L-гомосерина и ацетил-CoA в O-ацетил-L-гомосерин и CoA. С помощью частиц раствора DTNB (5,5'-дитиобис-2-нитробензойной кислоты, "Ellmans reagent", Sigma Aldrich, Германия) можно регистрировать ход реакции путем измерения поглощения при 412 нм, поскольку DTNB обеспечивает формирование желтого вещества с группой SH в CoA (S. Yamagata Journal of Bacteriology 169, No. 8 (1987) 3458-3463). Фотометрический ферментативный анализ MetX проводили при 37°C, в котором предварительно осуществляли калибровку с применением концентраций CoA в диапазоне 0-200 мкM. Каждое получение проводили в 0,2 мл реакционной смеси со 100 мМ калий-фосфатного буфера (pH 7,5), 0,65 мМ DNTB [100 мкл 1,3 мМ исходного DTNB], 0,13 мМ ацетил-CoA [30 мкл 0,886 мМ исходного ацетил-CoA, Sigma Aldrich, Германия], 10 мМ L-гомосерина [20 мкл 100 мМ исходного L-гомосерина, Sigma Aldrich, Германия] и указывали концентрацию белка 0,012 мг/мл или 0,024 мг/мл соответствующего клеточного гомогената.
Поскольку ацетил-CoA применяется в клетке для различных типов биосинтеза, в цитоплазме присутствуют различные ферменты, которые катализируют расщепление ацетил-CoA до CoA, так что необходимо учитывать разницу между клеточными гомогенатами с MetX и без него.
В результате ферментативного анализа было обнаружено, что увеличение поглощения DNTB клеточным гомогенатом в MG1655/pMW218_Ptac-metX_Ptac-metY постоянно превышало таковое в MG1655/pMW218 с течением времени (фиг. 3). Это подтверждает, что MetX в данном документе является каталитически активным в качестве дополнительного фермента. Сравнение углов наклона исходных линейных участков на зарегистрированных кривых демонстрирует активности в клеточном гомогенате MG1655/pMW218 около 580 мкмоль/мин. на г общего белка и в MG1655/pMW218_Ptac-metX_Ptac-metY около 730 мкмоль/мин. на г общего белка. Таким образом, различие заключается в удельной активности L-гомосерин-О-ацетилтрансферазы (MetX) при около 150 единицах на 1 г общего белка (1 единица = 1 мкмоль превращения субстрата/мин.).
2b) Определение цитоплазматической активности MetY (O-ацетил-L-гомосеринсульфогидрилазы)
Реакцией, которую катализирует фермент O-ацетил-L-гомосеринсульфгидрилаза (MetY) [EC 2.5.1.49], является превращение O-ацетил-L-гомосерина с метантиолом (MC) в присутствии пиридоксаль-5'-фосфата (PLP) с получением L-метионина и ацетата. Как описано в примере 2a, ход данной реакции можно определять с помощью измерения поглощения DTNB при 412 нм, поскольку DTNB реагирует с группой SH непрореагировавшего метилмеркаптана с получением желтого вещества. С этой целью получали два штамма MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY в виде клеточных гомогенатов, как описано выше, и измеряли уменьшение или превращение субстрата метилмеркаптида натрия в последующем ферментативном анализе.
Каждое получение проводили при 37°C в 1 мл реакционной смеси с помощью 100 мМ калий-фосфатного буфера (pH 7,5), 2 мМ метилмеркаптида натрия (NaMC) [10 мкл 200 мМ исходного NaMC], 3 мМ OAH HCl [30 мкл 100 мМ исходного OAH HCl] и 0,01 мМ PLP [10 мкл 1 мМ исходного PLP] с соответствующим клеточным гомогенатом при общей концентрации белка, составляющей 0,012 г/л; 0,024 г/л или 0,048 г/л. После ограниченной по времени ферментативной реакции проводили фотометрическое измерение содержания NaMC с помощью DTNB, где предварительно осуществляли калибровку с применением концентраций MC в диапазоне 0-200 мкM. С этой целью добавляли 180 мкл раствора DTNB (4 мг/мл) в каждые 20 мкл ферментативной реакционной смеси и впоследствии измеряли при 412 нм.
Присутствие клеточного гомогената штамма MG1655/pMW218_Ptac-metX_Ptac-metY приводило к снижению подвергнутого катализу NaMC значительно быстрее, чем в присутствии клеточного гомогената MG1655/pMW218, вследствие ферментативной активности MetY, в зависимости от концентрации общего белка (фигура 4). Снижение количества NaMC, происходящее подобным образом, но более слабо, в препаратах с клеточным гомогенатом MG1655/pMW218 не зависит от количества применяемого белка. Идентичное снижение также может наблюдаться в препарате без клеточного гомогената и происходить вследствие химически зависимого газовыделения метилмеркаптана, происходящего из растворов. На основании разницы в углах наклона в линейном диапазоне можно рассчитать удельную активность сульфгидрилазы MetY, составляющую около 1500 единиц на г общего белка (1 единица = 1 мкмоль превращения субстрата/мин.), которая находится на том же уровне, что и у фермента MetX.
3) Определение клеточной биотрансформации L-гомосерина и метилмеркаптида натрия с получением L-метионина
Культивировали штаммы MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY, как описано в примере 1, а затем в экспоненциальной фазе каждый препарат доводили до OD600 при около 7.
Затем осуществляли биотрансформацию в 100-мл встряхиваемых флаконах при 37°C, 200 об./мин. в течение периода времени, составляющего 0, 2, 4 и 24 ч. Каждое получение проводили в 10 мл среды A с помощью 6,5 г/л L-гомосерина [500 мкл исходного гомосерина в концентрации 100 г/л] (Sigma Aldrich, Германия), 3 г/л NaMC [500 мкл исходного 6% NaMC] и 12 г/л KH2PO4 [600 мкл исходного KH2PO4 в концентрации 200 г/л].
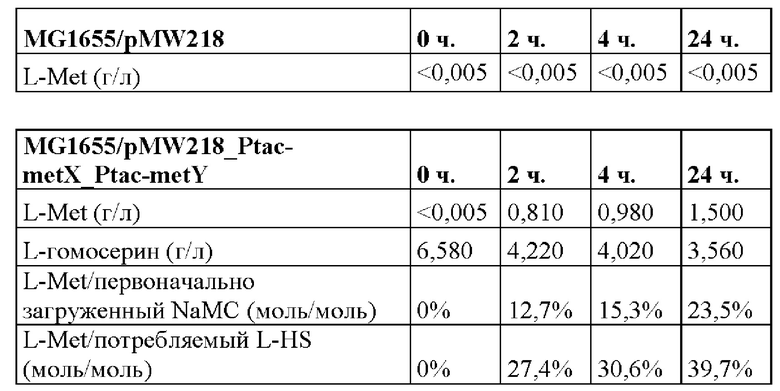
Проводили превращение L-гомосерина с помощью метилмеркаптана с получением L-метионина с применением штамма MG1655/pMW218_Ptac-metX_Ptac-metY, при этом L-метионин не синтезировали с применением штамма MG1655/pMW218 (таблица 1). Различные выходы, основанные на количествах первоначально загруженного NaMC и количествах потребляемого L-гомосерина, основаны на равной стехиометрии обоих субстратов, присутствующих вначале, а также после естественного испарения метилмеркаптана.
Таблица 1. Сравнение биотрансформации 6,5 г/л синтетического L-гомосерина и 3 г/л метилмеркаптида натрия с применением штаммов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY. Показан титр L-метионина во времени, и соответствующие выходы, основанные на количестве NaMC, пульсирующего вначале, и количестве потребляемого L-гомосерина (L-HS).
Кроме того, осуществляли биотрансформацию с применением штамма MG1655/pMW218_Ptac-metX_Ptac-metY, в котором применяли дейтерированный NaMC (D3CSNa) вместо исходного NaMC от Sigma Aldrich. Данное получали путем введения CD3SD (Sigma-Aldrich, 98 атом.% D) в эквимолярное количество водного раствора гидроксида натрия. (В качестве альтернативы его можно получить в соответствии с J. Voss et al., Phosphorous, Sulfur and Silicon and the Related Elements, 2012, 187, 382 из тиомочевины и CD3I.) Анализ раствора через 24 ч. выполнения реакции с помощью LC-MS демонстрирует соотношение метионина к метионину-d-3, составляющее 1:200. Таким образом, может быть обнаружено, что метионин, образующийся в результате биотрансформации, образуется исключительно путем включения подаваемого извне метилмеркаптана.
4) Превращение L-гомосерина, полученного посредством ферментации, в L-метионин путем биотрансформации
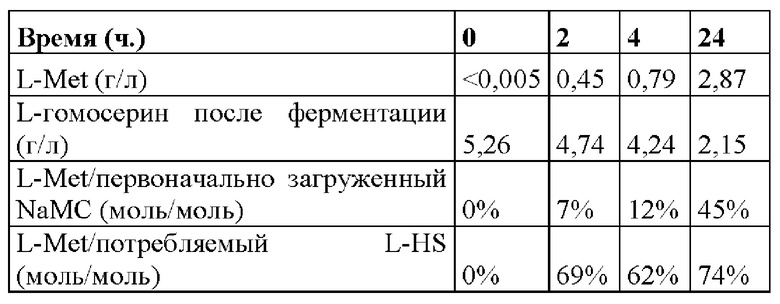
На основе биотрансформации синтетического L-гомосерина в L-метионин, проведенной в примере 3а, также была исследована биотрансформация L-гомосерина, полученного посредством ферментации. Концентрация L-гомосерина в бульоне, полученного посредством ферментации, составляла10 г/л. Культивировали штамм MG1655/pMW218_Ptac-metX_Ptac-metY, как в примере 1, и проводили биотрансформацию в экспоненциальной фазе при OD 5 в присутствии 5 г/л L-гомосерина, полученного посредством ферментации и как описано в примере 3a, в течение 2, 4 и 24 ч. Как показано в таблице 2, через два часа после биотрансформации превращалось около 7%, через четыре часа около 12% и через 24 часа около 45% субстрата L-гомосерина или NaMC в L-метионин, что отражалось в максимальном титре, составляющем около 2,9 г/л L-метионина.
Таблица 2. L-метионин и связанные с ним выходы, полученные путем биотрансформации 5 г/л L-гомосерина, полученного посредством ферментации, и 3 г/л метилмеркаптида натрия штаммом MG1655/pMW218_Ptac-metX_Ptac-metY.
5) Клеточная рециркуляция ацетата во время биотрансформации
Для исследования количества ацетата, образованного при биотрансформации L-гомосерина и метилмеркаптана, записывали препараты с применением штаммов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY в присутствии 5 г/л L-гомосерина, и 3 г/л метилмеркаптида натрия, и 12 г/л KH2PO4 [600 мкл исходного KH2PO4 в концентрации 200 г/л] в течение четырех часов в соответствии с содержанием в них ацетата.
Получали штаммы MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY, как описано в примере 1, чтобы использовать экспоненциальную культуру с исходным значением OD, составляющим около 3, для соответствующих 10 мл препаратов в 100-мл флаконах, как описано в примере 3b.
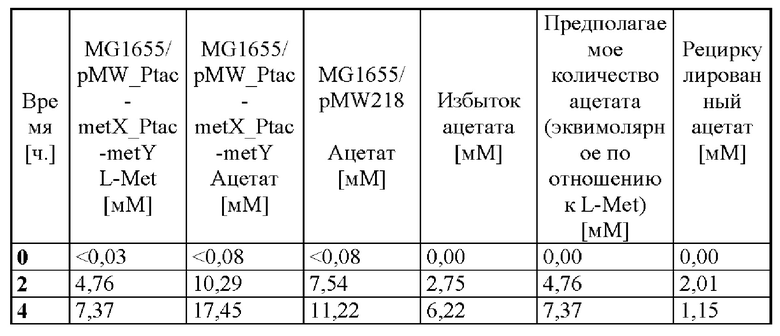
В таблице 3 записаны концентрации ацетата, которые образованы во время биотрансформации 5 г/л L-гомосерина и 3 г/л метилмеркаптида натрия с помощью штаммов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY. L-метионин образуется только в препарате с помощью штамма, гетерологично экспрессирующего гены metX и metY, тогда как в препарате с контрольным штаммом MG1655/pMW218 не обнаружено L-метионина.
В течение первых четырех часов около 11 мМ ацетата образовывалось благодаря экспериментальным параметрам в контрольном препарате, тогда как при биотрансформации образовывалось около 17 мМ ацетата и 7 мМ L-метионина. Избыток ацетата, измеренный в биотрансформированном препарате, который приводит к разности, составляет, таким образом, 6 мМ. Благодаря дополнительному синтезу метионина с эквимолярным продуцированием ацетата и метионина, который не описан для неклеточных систем (WO 2008/013432 A1), это значение будет 7 мМ. Таким образом, можно определять клеточную рециркуляцию ацетата, образованного при синтезе L-метионина при биотрансформации. Таким образом, дополнительный ацетат, полученный при биотрансформации, очевидно, частично рециркулируется с помощью ацетил-CoA-синтетазы (Acs) до ацетил-CoA.
Таблица 3. Образование и рециркуляция ацетата в биотрансформированных препаратах с помощью штаммов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY в присутствии 5 г/л L-гомосерина и 3 г/л метилмеркаптида натрия
6) Добавление L-гомосерина извне
Получали штаммы MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY, как описано в примере 1, чтобы использовать экспоненциальную культуру с исходным значением OD, составляющим около 3, для соответствующих 10 мл препаратов в 100-мл флаконах, как описано в примере 3b. К соответствующим препаратам, к которым сначала ничего не добавляли, после момента времени 0 ч. добавляли 5 г/л L-гомосерина, добавляли 3 г/л метилмеркаптида натрия с 12 г/л KH2PO4 [600 мкл исходного 200 г/л KH2PO4] и добавляли 5 г/л L-гомосерина с 3 г/л метилмеркаптида натрия и 12 г/л KH2PO4 [600 мкл исходного 200 г/л KH2PO4].
В препаратах определяли титры L-метионина и L-гомосерина в моменты времени 0, 2, 4 и 6 ч.
Таблица 4. Титры L-метионина и L-гомосерина после добавления извне 5 г/л L-гомосерина (L-HS) к биотрансформированным препаратам с применением штаммов MG1655/pMW218 и MG1655/pMW218_Ptac-metX_Ptac-metY в присутствии и в отсутствии 3 г/л метилмеркаптида натрия (Na-MC).

7) Биотрансформация L-гомосерина и диметилдисульфида (DMDS) или L-гомосерина и метилмеркаптида натрия (NaMC) с получением L-метионина
Культивировали штамм MG1655/pMW218_Ptac-metX_Ptac-metY, как описано в примере 1, а затем в экспоненциальной фазе каждый препарат доводили до OD600 около 10.
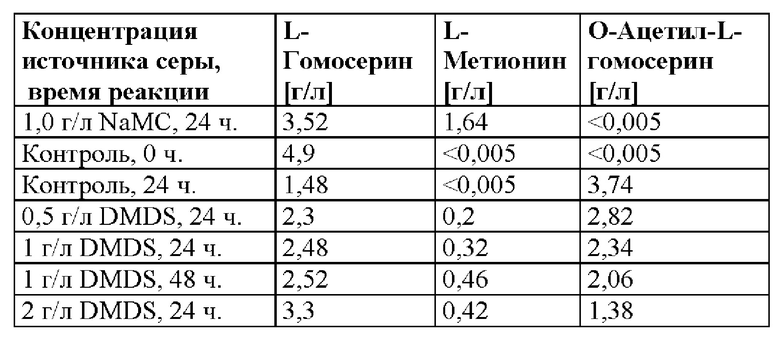
Затем осуществляли биотрансформацию в 100 мл встряхиваемых флаконах при 37°C, 200 об./мин. в течение периода времени, составляющего 0, 24 и 48 ч. Каждое получение проводили в 10 мл среды A (см. пример 1) с помощью 5,0 г/л L-гомосерина и 12 г/л KH2PO4 [600 мкл 200 г/л исходного KH2PO4] и с некоторым количеством источника серы (т. е. NaMC или DMDS), как представлено в таблице 5. Контроль не содержал какого-либо источника серы (т. е. без NaMC и без DMDS).
Таблица 5. Сравнение результатов биотрансформации с помощью NaMC, контроля (без источника серы) и DMDS при различных концентрациях и периодах времени реакции.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Эвоник Дегусса Гмбх
<120> WHCB
<130> .
<160> 6
<170> PatentIn, версия 3.5
<210> 1
<211> 1137
<212> ДНК
<213> Corynebacterium glutamicum
<220>
<221> CDS
<222> (1)..(1137)
<223> metX
<400> 1
atg ccc acc ctc gcg cct tca ggt caa ctt gaa atc caa gcg atc ggt 48
Met Pro Thr Leu Ala Pro Ser Gly Gln Leu Glu Ile Gln Ala Ile Gly
1 5 10 15
gat gtc tcc acc gaa gcc gga gca atc att aca aac gct gaa atc gcc 96
Asp Val Ser Thr Glu Ala Gly Ala Ile Ile Thr Asn Ala Glu Ile Ala
20 25 30
tat cac cgc tgg ggt gaa tac cgc gta gat aaa gaa gga cgc agc aat 144
Tyr His Arg Trp Gly Glu Tyr Arg Val Asp Lys Glu Gly Arg Ser Asn
35 40 45
gtc gtt ctc atc gaa cac gcc ctc act gga gat tcc aac gca gcc gat 192
Val Val Leu Ile Glu His Ala Leu Thr Gly Asp Ser Asn Ala Ala Asp
50 55 60
tgg tgg gct gac ttg ctc ggt ccc ggc aaa gcc atc aac act gat att 240
Trp Trp Ala Asp Leu Leu Gly Pro Gly Lys Ala Ile Asn Thr Asp Ile
65 70 75 80
tac tgc gtg atc tgt acc aac gtc atc ggt ggt tgc aac ggt tcc acc 288
Tyr Cys Val Ile Cys Thr Asn Val Ile Gly Gly Cys Asn Gly Ser Thr
85 90 95
gga cct ggc tcc atg cat cca gat gga aat ttc tgg ggt aat cgc ttc 336
Gly Pro Gly Ser Met His Pro Asp Gly Asn Phe Trp Gly Asn Arg Phe
100 105 110
ccc gcc acg tcc att cgt gat cag gta aac gcc gaa aaa caa ttc ctc 384
Pro Ala Thr Ser Ile Arg Asp Gln Val Asn Ala Glu Lys Gln Phe Leu
115 120 125
gac gca ctc ggc atc acc acg gtc gcc gca gta ctt ggt ggt tcc atg 432
Asp Ala Leu Gly Ile Thr Thr Val Ala Ala Val Leu Gly Gly Ser Met
130 135 140
ggt ggt gcc cgc acc cta gag tgg gcc gca atg tac cca gaa act gtt 480
Gly Gly Ala Arg Thr Leu Glu Trp Ala Ala Met Tyr Pro Glu Thr Val
145 150 155 160
ggc gca gct gct gtt ctt gca gtt tct gca cgc gcc agc gcc tgg caa 528
Gly Ala Ala Ala Val Leu Ala Val Ser Ala Arg Ala Ser Ala Trp Gln
165 170 175
atc ggc att caa tcc gcc caa att aag gcg att gaa aac gac cac cac 576
Ile Gly Ile Gln Ser Ala Gln Ile Lys Ala Ile Glu Asn Asp His His
180 185 190
tgg cac gaa ggc aac tac tac gaa tcc ggc tgc aac cca gcc acc gga 624
Trp His Glu Gly Asn Tyr Tyr Glu Ser Gly Cys Asn Pro Ala Thr Gly
195 200 205
ctc ggc gcc gcc cga cgc atc gcc cac ctc acc tac cgt ggc gaa cta 672
Leu Gly Ala Ala Arg Arg Ile Ala His Leu Thr Tyr Arg Gly Glu Leu
210 215 220
gaa atc gac gaa cgc ttc ggc acc aaa gcc caa aag aac gaa aac cca 720
Glu Ile Asp Glu Arg Phe Gly Thr Lys Ala Gln Lys Asn Glu Asn Pro
225 230 235 240
ctc ggt ccc tac cgc aag ccc gac cag cgc ttc gcc gtg gaa tcc tac 768
Leu Gly Pro Tyr Arg Lys Pro Asp Gln Arg Phe Ala Val Glu Ser Tyr
245 250 255
ttg gac tac caa gca gac aag cta gta cag cgt ttc gac gcc ggc tcc 816
Leu Asp Tyr Gln Ala Asp Lys Leu Val Gln Arg Phe Asp Ala Gly Ser
260 265 270
tac gtc ttg ctc acc gac gcc ctc aac cgc cac gac att ggt cgc gac 864
Tyr Val Leu Leu Thr Asp Ala Leu Asn Arg His Asp Ile Gly Arg Asp
275 280 285
cgc gga ggc ctc aac aag gca ctc gaa tcc atc aaa gtt cca gtc ctt 912
Arg Gly Gly Leu Asn Lys Ala Leu Glu Ser Ile Lys Val Pro Val Leu
290 295 300
gtc gca ggc gta gat acc gat att ttg tac ccc tac cac cag caa gaa 960
Val Ala Gly Val Asp Thr Asp Ile Leu Tyr Pro Tyr His Gln Gln Glu
305 310 315 320
cac ctc tcc aga aac ctg gga aat cta ctg gca atg gca aaa atc gta 1008
His Leu Ser Arg Asn Leu Gly Asn Leu Leu Ala Met Ala Lys Ile Val
325 330 335
tcc cct gtc ggc cac gat gct ttc ctc acc gaa agc cgc caa atg gat 1056
Ser Pro Val Gly His Asp Ala Phe Leu Thr Glu Ser Arg Gln Met Asp
340 345 350
cgc atc gtg agg aac ttc ttc agc ctc atc tcc cca gac gaa gac aac 1104
Arg Ile Val Arg Asn Phe Phe Ser Leu Ile Ser Pro Asp Glu Asp Asn
355 360 365
cct tcg acc tac atc gag ttc tac atc taa tag 1137
Pro Ser Thr Tyr Ile Glu Phe Tyr Ile
370 375
<210> 2
<211> 377
<212> БЕЛОК
<213> Corynebacterium glutamicum
<400> 2
Met Pro Thr Leu Ala Pro Ser Gly Gln Leu Glu Ile Gln Ala Ile Gly
1 5 10 15
Asp Val Ser Thr Glu Ala Gly Ala Ile Ile Thr Asn Ala Glu Ile Ala
20 25 30
Tyr His Arg Trp Gly Glu Tyr Arg Val Asp Lys Glu Gly Arg Ser Asn
35 40 45
Val Val Leu Ile Glu His Ala Leu Thr Gly Asp Ser Asn Ala Ala Asp
50 55 60
Trp Trp Ala Asp Leu Leu Gly Pro Gly Lys Ala Ile Asn Thr Asp Ile
65 70 75 80
Tyr Cys Val Ile Cys Thr Asn Val Ile Gly Gly Cys Asn Gly Ser Thr
85 90 95
Gly Pro Gly Ser Met His Pro Asp Gly Asn Phe Trp Gly Asn Arg Phe
100 105 110
Pro Ala Thr Ser Ile Arg Asp Gln Val Asn Ala Glu Lys Gln Phe Leu
115 120 125
Asp Ala Leu Gly Ile Thr Thr Val Ala Ala Val Leu Gly Gly Ser Met
130 135 140
Gly Gly Ala Arg Thr Leu Glu Trp Ala Ala Met Tyr Pro Glu Thr Val
145 150 155 160
Gly Ala Ala Ala Val Leu Ala Val Ser Ala Arg Ala Ser Ala Trp Gln
165 170 175
Ile Gly Ile Gln Ser Ala Gln Ile Lys Ala Ile Glu Asn Asp His His
180 185 190
Trp His Glu Gly Asn Tyr Tyr Glu Ser Gly Cys Asn Pro Ala Thr Gly
195 200 205
Leu Gly Ala Ala Arg Arg Ile Ala His Leu Thr Tyr Arg Gly Glu Leu
210 215 220
Glu Ile Asp Glu Arg Phe Gly Thr Lys Ala Gln Lys Asn Glu Asn Pro
225 230 235 240
Leu Gly Pro Tyr Arg Lys Pro Asp Gln Arg Phe Ala Val Glu Ser Tyr
245 250 255
Leu Asp Tyr Gln Ala Asp Lys Leu Val Gln Arg Phe Asp Ala Gly Ser
260 265 270
Tyr Val Leu Leu Thr Asp Ala Leu Asn Arg His Asp Ile Gly Arg Asp
275 280 285
Arg Gly Gly Leu Asn Lys Ala Leu Glu Ser Ile Lys Val Pro Val Leu
290 295 300
Val Ala Gly Val Asp Thr Asp Ile Leu Tyr Pro Tyr His Gln Gln Glu
305 310 315 320
His Leu Ser Arg Asn Leu Gly Asn Leu Leu Ala Met Ala Lys Ile Val
325 330 335
Ser Pro Val Gly His Asp Ala Phe Leu Thr Glu Ser Arg Gln Met Asp
340 345 350
Arg Ile Val Arg Asn Phe Phe Ser Leu Ile Ser Pro Asp Glu Asp Asn
355 360 365
Pro Ser Thr Tyr Ile Glu Phe Tyr Ile
370 375
<210> 3
<211> 1314
<212> ДНК
<213> Corynebacterium glutamicum
<220>
<221> CDS
<222> (1)..(1314)
<223> metY
<400> 3
atg cca aag tac gac aat tcc aat gct gac cag tgg ggc ttt gaa acc 48
Met Pro Lys Tyr Asp Asn Ser Asn Ala Asp Gln Trp Gly Phe Glu Thr
1 5 10 15
cgc tcc att cac gca ggc cag tca gta gac gca cag acc agc gca cga 96
Arg Ser Ile His Ala Gly Gln Ser Val Asp Ala Gln Thr Ser Ala Arg
20 25 30
aac ctt ccg atc tac caa tcc acc gct ttc gtg ttc gac tcc gct gag 144
Asn Leu Pro Ile Tyr Gln Ser Thr Ala Phe Val Phe Asp Ser Ala Glu
35 40 45
cac gcc aag cag cgt ttc gca ctt gag gat cta ggc cct gtt tac tcc 192
His Ala Lys Gln Arg Phe Ala Leu Glu Asp Leu Gly Pro Val Tyr Ser
50 55 60
cgc ctc acc aac cca acc gtt gag gct ttg gaa aac cgc atc gct tcc 240
Arg Leu Thr Asn Pro Thr Val Glu Ala Leu Glu Asn Arg Ile Ala Ser
65 70 75 80
ctc gaa ggt ggc gtc cac gct gta gcg ttc tcc tcc gga cag gcc gca 288
Leu Glu Gly Gly Val His Ala Val Ala Phe Ser Ser Gly Gln Ala Ala
85 90 95
acc acc aac gcc att ttg aac ctg gca gga gcg ggc gac cac atc gtc 336
Thr Thr Asn Ala Ile Leu Asn Leu Ala Gly Ala Gly Asp His Ile Val
100 105 110
acc tcc cca cgc ctc tac ggt ggc acc gag act cta ttc ctt atc act 384
Thr Ser Pro Arg Leu Tyr Gly Gly Thr Glu Thr Leu Phe Leu Ile Thr
115 120 125
ctt aac cgc ctg ggt atc gat gtt tcc ttc gtg gaa aac ccc gac gac 432
Leu Asn Arg Leu Gly Ile Asp Val Ser Phe Val Glu Asn Pro Asp Asp
130 135 140
cct gag tcc tgg cag gca gcc gtt cag cca aac acc aaa gca ttc ttc 480
Pro Glu Ser Trp Gln Ala Ala Val Gln Pro Asn Thr Lys Ala Phe Phe
145 150 155 160
ggc gag act ttc gcc aac cca cag gca gac gtc ctg gat att cct gcg 528
Gly Glu Thr Phe Ala Asn Pro Gln Ala Asp Val Leu Asp Ile Pro Ala
165 170 175
gtg gct gaa gtt gcg cac cgc aac agc gtt cca ctg atc atc gac aac 576
Val Ala Glu Val Ala His Arg Asn Ser Val Pro Leu Ile Ile Asp Asn
180 185 190
acc atc gct acc gca gcg ctc gtg cgc ccg ctc gag ctc ggc gca gac 624
Thr Ile Ala Thr Ala Ala Leu Val Arg Pro Leu Glu Leu Gly Ala Asp
195 200 205
gtt gtc gtc gct tcc ctc acc aag ttc tac acc ggc aac ggc tcc gga 672
Val Val Val Ala Ser Leu Thr Lys Phe Tyr Thr Gly Asn Gly Ser Gly
210 215 220
ctg ggc ggc gtg ctt atc gac ggc gga aag ttc gat tgg act gtc gaa 720
Leu Gly Gly Val Leu Ile Asp Gly Gly Lys Phe Asp Trp Thr Val Glu
225 230 235 240
aag gat gga aag cca gta ttc ccc tac ttc gtc act cca gat gct gct 768
Lys Asp Gly Lys Pro Val Phe Pro Tyr Phe Val Thr Pro Asp Ala Ala
245 250 255
tac cac gga ttg aag tac gca gac ctt ggt gca cca gcc ttc ggc ctc 816
Tyr His Gly Leu Lys Tyr Ala Asp Leu Gly Ala Pro Ala Phe Gly Leu
260 265 270
aag gtt cgc gtt ggc ctt cta cgc gac acc ggc tcc acc ctc tcc gca 864
Lys Val Arg Val Gly Leu Leu Arg Asp Thr Gly Ser Thr Leu Ser Ala
275 280 285
ttc aac gca tgg gct gca gtc cag ggc atc gac acc ctt tcc ctg cgc 912
Phe Asn Ala Trp Ala Ala Val Gln Gly Ile Asp Thr Leu Ser Leu Arg
290 295 300
ctg gag cgc cac aac gaa aac gcc atc aag gtt gca gaa ttc ctc aac 960
Leu Glu Arg His Asn Glu Asn Ala Ile Lys Val Ala Glu Phe Leu Asn
305 310 315 320
aac cac gag aag gtg gaa aag gtt aac ttc gca ggc ctg aag gat tcc 1008
Asn His Glu Lys Val Glu Lys Val Asn Phe Ala Gly Leu Lys Asp Ser
325 330 335
cct tgg tac gca acc aag gaa aag ctt ggc ctg aag tac acc ggc tcc 1056
Pro Trp Tyr Ala Thr Lys Glu Lys Leu Gly Leu Lys Tyr Thr Gly Ser
340 345 350
gtt ctc acc ttc gag atc aag ggc ggc aag gat gag gct tgg gca ttt 1104
Val Leu Thr Phe Glu Ile Lys Gly Gly Lys Asp Glu Ala Trp Ala Phe
355 360 365
atc gac gcc ctg aag cta cac tcc aac ctt gca aac atc ggc gat gtt 1152
Ile Asp Ala Leu Lys Leu His Ser Asn Leu Ala Asn Ile Gly Asp Val
370 375 380
cgc tcc ctc gtt gtt cac cca gca acc acc acc cat tca cag tcc gac 1200
Arg Ser Leu Val Val His Pro Ala Thr Thr Thr His Ser Gln Ser Asp
385 390 395 400
gaa gct ggc ctg gca cgc gcg ggc gtt acc cag tcc acc gtc cgc ctg 1248
Glu Ala Gly Leu Ala Arg Ala Gly Val Thr Gln Ser Thr Val Arg Leu
405 410 415
tcc gtt ggc atc gag acc att gat gat atc atc gct gac ctc gaa ggc 1296
Ser Val Gly Ile Glu Thr Ile Asp Asp Ile Ile Ala Asp Leu Glu Gly
420 425 430
ggc ttt gct gca atc tag 1314
Gly Phe Ala Ala Ile
435
<210> 4
<211> 437
<212> БЕЛОК
<213> Corynebacterium glutamicum
<400> 4
Met Pro Lys Tyr Asp Asn Ser Asn Ala Asp Gln Trp Gly Phe Glu Thr
1 5 10 15
Arg Ser Ile His Ala Gly Gln Ser Val Asp Ala Gln Thr Ser Ala Arg
20 25 30
Asn Leu Pro Ile Tyr Gln Ser Thr Ala Phe Val Phe Asp Ser Ala Glu
35 40 45
His Ala Lys Gln Arg Phe Ala Leu Glu Asp Leu Gly Pro Val Tyr Ser
50 55 60
Arg Leu Thr Asn Pro Thr Val Glu Ala Leu Glu Asn Arg Ile Ala Ser
65 70 75 80
Leu Glu Gly Gly Val His Ala Val Ala Phe Ser Ser Gly Gln Ala Ala
85 90 95
Thr Thr Asn Ala Ile Leu Asn Leu Ala Gly Ala Gly Asp His Ile Val
100 105 110
Thr Ser Pro Arg Leu Tyr Gly Gly Thr Glu Thr Leu Phe Leu Ile Thr
115 120 125
Leu Asn Arg Leu Gly Ile Asp Val Ser Phe Val Glu Asn Pro Asp Asp
130 135 140
Pro Glu Ser Trp Gln Ala Ala Val Gln Pro Asn Thr Lys Ala Phe Phe
145 150 155 160
Gly Glu Thr Phe Ala Asn Pro Gln Ala Asp Val Leu Asp Ile Pro Ala
165 170 175
Val Ala Glu Val Ala His Arg Asn Ser Val Pro Leu Ile Ile Asp Asn
180 185 190
Thr Ile Ala Thr Ala Ala Leu Val Arg Pro Leu Glu Leu Gly Ala Asp
195 200 205
Val Val Val Ala Ser Leu Thr Lys Phe Tyr Thr Gly Asn Gly Ser Gly
210 215 220
Leu Gly Gly Val Leu Ile Asp Gly Gly Lys Phe Asp Trp Thr Val Glu
225 230 235 240
Lys Asp Gly Lys Pro Val Phe Pro Tyr Phe Val Thr Pro Asp Ala Ala
245 250 255
Tyr His Gly Leu Lys Tyr Ala Asp Leu Gly Ala Pro Ala Phe Gly Leu
260 265 270
Lys Val Arg Val Gly Leu Leu Arg Asp Thr Gly Ser Thr Leu Ser Ala
275 280 285
Phe Asn Ala Trp Ala Ala Val Gln Gly Ile Asp Thr Leu Ser Leu Arg
290 295 300
Leu Glu Arg His Asn Glu Asn Ala Ile Lys Val Ala Glu Phe Leu Asn
305 310 315 320
Asn His Glu Lys Val Glu Lys Val Asn Phe Ala Gly Leu Lys Asp Ser
325 330 335
Pro Trp Tyr Ala Thr Lys Glu Lys Leu Gly Leu Lys Tyr Thr Gly Ser
340 345 350
Val Leu Thr Phe Glu Ile Lys Gly Gly Lys Asp Glu Ala Trp Ala Phe
355 360 365
Ile Asp Ala Leu Lys Leu His Ser Asn Leu Ala Asn Ile Gly Asp Val
370 375 380
Arg Ser Leu Val Val His Pro Ala Thr Thr Thr His Ser Gln Ser Asp
385 390 395 400
Glu Ala Gly Leu Ala Arg Ala Gly Val Thr Gln Ser Thr Val Arg Leu
405 410 415
Ser Val Gly Ile Glu Thr Ile Asp Asp Ile Ile Ala Asp Leu Glu Gly
420 425 430
Gly Phe Ala Ala Ile
435
<210> 5
<211> 41
<212> ДНК
<213> Гибридный промотор
<220>
<221> промотор
<222> (1)..(41)
<223> PtacI
<400> 5
gagctgttga caattaatca tcggctcgta taatgtgtgg a 41
<210> 6
<211> 3255
<212> ДНК
<213> синтезированная последовательность
<400> 6
cgcgcatgcc cgacggcgag gatctcgtcg tgacccatgg cgatgcctgc ttgccgaata 60
tcatggtgga aaatggccgc ttttctggat tcatcgactg tggccggctg ggtgtggcgg 120
accgctatca ggacatagcg ttggctaccc gtgatattgc tgaagagctt ggcggcgaat 180
gggctgaccg cttcctcgtg ctttacggta tcgccgctcc cgattcgcag cgcatcgcct 240
tctatcgcct tcttgacgag ttcttctgag cgggactctg gggttcggca cacagcccag 300
cttggagcga acgacctaca ccgaactgag atacctacag cgtgagctat gagaaagcgc 360
ccaatacgca aaccgcctct ccccgcgcgt tggccgattc attatggagc tgttgacaat 420
taatcatcgg ctcgtataat gtgtggaatt gtgagcggat aacaatttca cacaggaaac 480
agaattcaaa ggaggacaac catgcccacc ctcgcgcctt caggtcaact tgaaatccaa 540
gcgatcggtg atgtctccac cgaagccgga gcaatcatta caaacgctga aatcgcctat 600
caccgctggg gtgaataccg cgtagataaa gaaggacgca gcaatgtcgt tctcatcgaa 660
cacgccctca ctggagattc caacgcagcc gattggtggg ctgacttgct cggtcccggc 720
aaagccatca acactgatat ttactgcgtg atctgtacca acgtcatcgg tggttgcaac 780
ggttccaccg gacctggctc catgcatcca gatggaaatt tctggggtaa tcgcttcccc 840
gccacgtcca ttcgtgatca ggtaaacgcc gaaaaacaat tcctcgacgc actcggcatc 900
accacggtcg ccgcagtact tggtggttcc atgggtggtg cccgcaccct agagtgggcc 960
gcaatgtacc cagaaactgt tggcgcagct gctgttcttg cagtttctgc acgcgccagc 1020
gcctggcaaa tcggcattca atccgcccaa attaaggcga ttgaaaacga ccaccactgg 1080
cacgaaggca actactacga atccggctgc aacccagcca ccggactcgg cgccgcccga 1140
cgcatcgccc acctcaccta ccgtggcgaa ctagaaatcg acgaacgctt cggcaccaaa 1200
gcccaaaaga acgaaaaccc actcggtccc taccgcaagc ccgaccagcg cttcgccgtg 1260
gaatcctact tggactacca agcagacaag ctagtacagc gtttcgacgc cggctcctac 1320
gtcttgctca ccgacgccct caaccgccac gacattggtc gcgaccgcgg aggcctcaac 1380
aaggcactcg aatccatcaa agttccagtc cttgtcgcag gcgtagatac cgatattttg 1440
tacccctacc accagcaaga acacctctcc agaaacctgg gaaatctact ggcaatggca 1500
aaaatcgtat cccctgtcgg ccacgatgct ttcctcaccg aaagccgcca aatggatcgc 1560
atcgtgagga acttcttcag cctcatctcc ccagacgaag acaacccttc gacctacatc 1620
gagttctaca tctaatagac gcgtgagctg ttgacaatta atcatcggct cgtataatgt 1680
gtggaattgt gagcggataa caatttcacg cgtttaatta acacgagtac tggaaaacta 1740
aatgccaaag tacgacaatt ccaatgctga ccagtggggc tttgaaaccc gctccattca 1800
cgcaggccag tcagtagacg cacagaccag cgcacgaaac cttccgatct accaatccac 1860
cgctttcgtg ttcgactccg ctgagcacgc caagcagcgt ttcgcacttg aggatctagg 1920
ccctgtttac tcccgcctca ccaacccaac cgttgaggct ttggaaaacc gcatcgcttc 1980
cctcgaaggt ggcgtccacg ctgtagcgtt ctcctccgga caggccgcaa ccaccaacgc 2040
cattttgaac ctggcaggag cgggcgacca catcgtcacc tccccacgcc tctacggtgg 2100
caccgagact ctattcctta tcactcttaa ccgcctgggt atcgatgttt ccttcgtgga 2160
aaaccccgac gaccctgagt cctggcaggc agccgttcag ccaaacacca aagcattctt 2220
cggcgagact ttcgccaacc cacaggcaga cgtcctggat attcctgcgg tggctgaagt 2280
tgcgcaccgc aacagcgttc cactgatcat cgacaacacc atcgctaccg cagcgctcgt 2340
gcgcccgctc gagctcggcg cagacgttgt cgtcgcttcc ctcaccaagt tctacaccgg 2400
caacggctcc ggactgggcg gcgtgcttat cgacggcgga aagttcgatt ggactgtcga 2460
aaaggatgga aagccagtat tcccctactt cgtcactcca gatgctgctt accacggatt 2520
gaagtacgca gaccttggtg caccagcctt cggcctcaag gttcgcgttg gccttctacg 2580
cgacaccggc tccaccctct ccgcattcaa cgcatgggct gcagtccagg gcatcgacac 2640
cctttccctg cgcctggagc gccacaacga aaacgccatc aaggttgcag aattcctcaa 2700
caaccacgag aaggtggaaa aggttaactt cgcaggcctg aaggattccc cttggtacgc 2760
aaccaaggaa aagcttggcc tgaagtacac cggctccgtt ctcaccttcg agatcaaggg 2820
cggcaaggat gaggcttggg catttatcga cgccctgaag ctacactcca accttgcaaa 2880
catcggcgat gttcgctccc tcgttgttca cccagcaacc accacccatt cacagtccga 2940
cgaagctggc ctggcacgcg cgggcgttac ccagtccacc gtccgcctgt ccgttggcat 3000
cgagaccatt gatgatatca tcgctgacct cgaaggcggc tttgctgcaa tctagggccg 3060
gccgtttaaa ccctgcaggt ccgggacctg caggcatgca agcttggcac tggccgtcgt 3120
tttacaacgt cgtgactggg aaaaccctgg cgttacccaa cttaatcgcc ttgcagcaca 3180
tccccctttc gccagctggc gtaatagcga agaggcccgc accgatcgcc cttcccaaca 3240
gttgcgcagc ctgaa 3255
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ХИМИЧЕСКИХ ПРОДУКТОВ ТОНКОГО СИНТЕЗА С ПОМОЩЬЮ CORYNEBACTERIUM, СЕКРЕТИРУЮЩЕЙ МОДИФИЦИРОВАННЫЕ α-1,6-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2763317C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ПОМОЩЬЮ КОРИНЕБАКТЕРИЙ С ПРИМЕНЕНИЕМ СИСТЕМЫ РАСЩЕПЛЕНИЯ ГЛИЦИНА | 2015 |
|
RU2713298C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И ВЕКТОРЫ ДЛЯ ГЕНОТЕРАПИИ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА И ДРУГИХ СОСТОЯНИЙ | 2015 |
|
RU2745567C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| Антитела против белка р17 ВИЧ-1 субтипа А | 2019 |
|
RU2727673C1 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| СПОСОБ УПРАВЛЕНИЯ РИТМОМ СЕРДЦА И СОКРАЩЕНИЕМ ОТДЕЛЬНЫХ КАРДИОМИОЦИТОВ ПРИ ПОМОЩИ ТЕРМОГЕНЕТИКИ | 2022 |
|
RU2802995C1 |
Изобретение относится к способу получения L-метионина. Способ предусматривает культивирование микроорганизма, который представляет собой Escherichia coli, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью и O-ацетил-L-гомосеринсульфгидрилазной активностью, в культуральной среде, содержащей L-гомосерин и источник серы, выбранный из группы, состоящей из метилмеркаптана и соли метилмеркаптана. В результате чего L-метионин накапливается в культуральной среде. Изобретение обеспечивает получение L-метионина в микроорганизме, в котором ацетат, образующийся в результате превращения O-ацетил-L-гомосерина в L-метионин, в значительной степени повторно используется тем же микроорганизмом. 5 з.п. ф-лы, 4 ил., 5 табл.
1. Способ получения L-метионина, где микроорганизм, который представляет собой Escherichia coli, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью и O-ацетил-L-гомосеринсульфгидрилазной активностью, культивируют в культуральной среде, содержащей L-гомосерин и источник серы, выбранный из группы, состоящей из метилмеркаптана и соли метилмеркаптана, в результате чего L-метионин накапливается в культуральной среде.
2. Способ по п. 1, где микроорганизм является рекомбинантным, и при этом L-гомосерин-O-ацетилтрансферазная активность и O-ацетил-L-гомосеринсульфгидрилазная активность являются усиленными при повышенной экспрессии гена, который кодирует белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью, и белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью.
3. Способ по п. 2, где повышенную экспрессию гена обеспечивают путем увеличения числа копий гена, кодирующего белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью, и белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью.
4. Способ по п. 2 или 3, где повышенную экспрессию гена обеспечивают с помощью функциональной связи гена, кодирующего белок, характеризующийся L-гомосерин-O-ацетилтрансферазной активностью, и белок, характеризующийся O-ацетил-L-гомосеринсульфгидрилазной активностью, с сильным промотором.
5. Способ по любому из пп. 1-4, где L-гомосерин-O-ацетилтрансферазная активность обеспечивается ферментом MetX, происходящим из Corynebacterium glutamicum.
6. Способ по любому из пп. 1-5, где O-ацетил-L-гомосеринсульфгидрилазная активность обеспечивается ферментом MetY, происходящим из Corynebacterium glutamicum.
| WO 2007011939 A2, 25.01.2007 | |||
| US 3598701 A, 10.08.1971 | |||
| РЕКОМБИНАНТНЫЕ МИКРООРГАНИЗМЫ, ПРОДУЦИРУЮЩИЕ МЕТИОНИН | 2006 |
|
RU2447146C2 |
| KROMER J.O | |||
| et al | |||
| Metabolic pathway analysis for rational design of L-methionine production by Escherichia coli and Corynebacterium glutamicum | |||
| Metab Eng | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |

