ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] Эта заявка испрашивает приоритет международной патентной заявки № PCT/CN2018/078575, поданной 9 марта 2018 г., раскрытие которой полностью включено в настоящий документ ссылкой.
Область техники, к которой относится изобретение
[002] Настоящее изобретение относится к способам осахаривания и/или гидролиза крахмалосодержащих материалов с использованием по меньшей мере одной глюкоамилазы. Глюкоамилазы настоящего изобретения также можно использовать в качестве кормовой добавки для животных, улучшающей усвояемость крахмала. Кроме того, изобретение также относится к способам производства продуктов ферментации, а также продуктам ферментации, полученным этим способом.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[003] Глюкоамилаза (1,4-альфа-D-глюканглюкогидролаза, EC 3.2.1.3) - это фермент, который катализирует высвобождение D-глюкозы из невосстанавливающих концов крахмала или родственных молекул олиго- и полисахаридов. Глюкоамилазы вырабатываются несколькими мицелиальными грибами и дрожжами.
[004] Одним неограничивающим основным применением глюкоамилазы является осахаривание частично переработанного крахмала/декстрина до глюкозы, которая является важным субстратом для многочисленных процессов ферментации. Затем глюкозу можно прямо или косвенно превратить в продукт ферментации с использованием ферментирующего организма. Примеры коммерческих продуктов ферментации включают спирты (например, этанол, метанол, бутанол, 1,3-пропандиол); органические кислоты (например, лимонная кислота, уксусная кислота, итаконовая кислота, молочная кислота, глюконовая кислота, глюконат, молочная кислота, янтарная кислота, 2,5-дикето-D-глюконовая кислота); кетоны (например, ацетон); аминокислоты (например, глутаминовая кислота); газы (например, H2 и CO2) и более сложные соединения.
[005] Глюкоамилазу для коммерческих целей традиционно производят с использованием мицелиальных (нитчатых) грибов, хотя есть сообщения о разнообразных группах микроорганизмов, продуцирующих глюкоамилазу, поскольку они вырабатывают большие количества фермента внеклеточно.
[006] Осахаривание или ферментацию можно проводить при низком pH, поскольку обычно ферментирующие организмы быстро понижают pH, а некоторые лактобациллы могут еще и дальше снижать pH до примерно 3,5. Кроме того, низкие значения pH минимизируют риск загрязнения примесями. Однако коммерчески используемые грибковые глюкоамилазы имеют определенные ограничения, такие как низкая чувствительность к pH или низкая производительность/активность при низких значениях pH (таких, как например, pH ниже 6).
[007] Глюкоамилазы, которые активны при низком pH в присутствии пепсина, могут быть полезны в качестве кормовой добавки для жвачных животных. Ферменты для применения в качестве кормовых добавок для жвачных животных главным образом представляют собой фибролитические ферменты, такие как целлюлазы, бета-глюканазы и гемицеллюлазы (таблица 1 в Beauchemin et al., 2004. A rationale for the development of feed enzyme products for ruminants. Can. J. Anim. Sci. 84: 23-36). Существует ограниченное число сообщений о гидролазах крахмала в применении к жвачным животным.
[008] Усвояемость крахмала в кормах значительно варьируется и зависит от ряда факторов, в том числе физической структуры как крахмала, так и кормовой основы. В публикации заявки на патент США 2005/0037053, опубликованной 17 февраля 2005 г., раскрыто применение фермента, характеризующегося амилолитической активностью и способного к расщеплению устойчивого крахмала в корме, содержащем крахмал для животных с однокамерным желудком, таких как домашняя птица и свиньи. В WO 2008/06881, опубликованной 17 января 2008 г., раскрыто применение бактериальных амилаз в корме для жвачных животных подсемейства бычьих для улучшения удоев молока, кажущейся перевариваемости скармливаемого рациона, перевариваемости сухого вещества кормового продукта, прироста веса и/или коэффициента кормоотдачи. В WO 2015/128366, опубликованной 3 сентября 2015 г., раскрыто применение бактериальных амилаз в комбинации с одной или несколькими протеазами в корме для жвачных животных подсемейства бычьих для улучшения перевариваемости маиса и/или маисового силоса, в частности для улучшения удоев молока, прироста веса и/или коэффициента кормоотдачи. Соответственно, по-прежнему существует потребность в повышении усвояемости крахмала у животных.
[009] Таким образом, существует потребность в поиске новых глюкоамилаз с улучшенной толерантностью к низким значениям pH во время осахаривания и/или ферментации или для увеличения усвояемости крахмала у животных, откармливаемых крахмалосодержащими кормами.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0010] Настоящее изобретение относится к способам осахаривания крахмалосодержащих материалов с использованием по меньшей мере одной глюкоамилазы. Глюкоамилазы настоящего изобретения также можно использовать в качестве кормовой добавки для животных, улучшающей усвояемость крахмала. Аспекты и варианты осуществления способов описаны в следующих независимо пронумерованных абзацах.
1. В одном аспекте предложен способ осахаривания крахмального субстрата, включающий приведение в контакт крахмального субстрата с глюкоамилазой, выбранной из группы, состоящей из:
полипептида, содержащего аминокислотную последовательность с SEQ ID NO: 7, или 8, или 9;
полипептида, который по меньшей мере на 83% идентичен аминокислотной последовательности SEQ ID NO: 7 или 8;
полипептида, который по меньшей мере на 83% идентичен каталитическому домену с последовательностью SEQ ID NO: 7 или 8;
полипептида, содержащего аминокислотную последовательность каталитического домена с SEQ ID NO: 9; или
зрелого полипептида, полученного обработкой полипептида с SEQ ID NO: 1, 2 или 3 сигнальной пептидазой или посттрансляционной модификацией во время секреции из экспрессирующего хозяина;
при этом осахаривание проводят при pH от 2,0 до 6,0.
2. В некоторых вариантах осуществления способа согласно пункту 1 осахаривание крахмального субстрата приводит к получению сиропа с высоким содержанием глюкозы.
3. В некоторых вариантах осуществления способа согласно пункту 1 или 2 содержание глюкозы в сиропе с высоким содержание глюкозы, выбранном из списка, составляет не менее 95,5% глюкозы.
4. В некоторых вариантах осуществления способ согласно любому из пунктов 1-3 дополнительно включает ферментацию сиропа с высоким содержанием глюкозы до конечного продукта.
5. В некоторых вариантах осуществления способа согласно пункту 4 осахаривание и ферментацию проводят как процесс одновременного осахаривания и ферментации (SSF).
6. В некоторых вариантах осуществления способа согласно пункту 4 или 5 конечный продукт представляет собой спирт, например этанол.
7. В некоторых вариантах осуществления способа согласно пункту 4 или 5 конечный продукт представляет собой биохимикат, выбранный из группы, состоящей из аминокислоты, органической кислоты, лимонной кислоты, молочной кислоты, янтарной кислоты, глутамата натрия, глюконовой кислоты, глюконата натрия, глюконата кальция, глюконата калия, глюконо-дельта-лактона, эриторбата натрия, жирной кислоты омега-3, бутанола, лизина, итаконовой кислоты, 1,3-пропандиола, биодизеля и изопрена.
8. В некоторых вариантах осуществления способа согласно любому из пунктов 1-7, где содержание сухого вещества (DS) в крахмальном субстрате составляет от примерно 5% до 99%, от 15% до 50% или 40-99%.
9. В некоторых вариантах осуществления способа согласно любому из пунктов 1-8 крахмальный субстрат выбран из пшеницы, ячменя, кукурузы, ржи, риса, сорго, отрубей, маниоки, дурры, проса, картофеля, батата, тапиоки и любой их комбинации.
10. В некоторых вариантах осуществления способа согласно любому из пунктов 1-9 крахмальный субстрат содержит разжиженный крахмал, желатинизированный крахмал или гранулированный крахмал.
11. В некоторых вариантах осуществления способ согласно любому из пунктов 1-10 дополнительно включает внесение в крахмальный субстрат гексокиназы, ксиланазы, глюкозоизомеразы, ксилозоизомеразы, фосфатазы, фитазы, пуллуланазы, бета-амилазы, альфа-амилазы, глюкоамилазы, протеазы, целлюлазы, гемицеллюлазы, липазы, кутиназы, треалазы, изоамилазы, окисиредуктазы, эстеразы, трансферазы, пектиназы, гидролазы, альфа-глюкозидазы, бета-глюкозидазы или их комбинации.
12. В другом аспекте предложен способ осахаривания и ферментации крахмального субстрата для получения конечного продукта, включающий приведение в контакт крахмального субстрата с глюкоамилазой, выбранной из группы, состоящей из:
полипептида, содержащего аминокислотную последовательность с SEQ ID NO: 7, или 8, или 9;
полипептида, который по меньшей мере на 83% идентичен аминокислотной последовательности SEQ ID NO: 7 или 8;
полипептида, который по меньшей мере на 83% идентичен каталитическому домену с последовательностью SEQ ID NO: 7 или 8;
полипептида, содержащего аминокислотную последовательность каталитического домена с SEQ ID NO: 9; или
зрелого полипептида, полученного обработкой полипептида с SEQ ID NO: 1, 2 или 3 сигнальной пептидазой или посттрансляционной модификацией во время секреции из экспрессирующего хозяина;
при этом осахаривание и ферментацию проводят при pH от 2,0 до 6,0, предпочтительно - от pH 2,0 до 5,0, предпочтительно - от pH 2,0 до 4,0, более предпочтительно - от pH 2,0 до pH 3,0.
13. В некоторых вариантах осуществления способа согласно пункту 12 осахаривание и ферментацию проводят как процесс одновременного осахаривания и ферментации (SSF).
14. В некоторых вариантах осуществления способа согласно пункту 12 или 13 конечный продукт представляет собой спирт, например этанол.
15. В некоторых вариантах осуществления способа согласно пункту 12, где осахаренный и ферментированный крахмальный субстрат характеризуется пониженным уровнем DP3+ и повышенным уровнем DP1 по сравнению с результатом от контакта того же крахмального субстрата с AnGA.
16. В некоторых вариантах осуществления способа согласно пункту 12 или 13 конечный продукт представляет собой биохимикат, выбранный из группы, состоящей из аминокислоты, органической кислоты, лимонной кислоты, молочной кислоты, янтарной кислоты, глутамата натрия, глюконовой кислоты, глюконата натрия, глюконата кальция, глюконата калия, глюконо-дельта-лактона, эриторбата натрия, жирной кислоты омега-3, бутанола, лизина, итаконовой кислоты, 1,3-пропандиола, биодизеля и изопрена.
17. В другом аспекте предложен способ увеличения усвояемости крахмала у животного, который включает внесение по меньшей мере одной глюкоамилазы, выбранной из группы, состоящей из:
полипептида, содержащего аминокислотную последовательность с SEQ ID NO: 7, или 8, или 9;
полипептида, который по меньшей мере на 83% идентичен аминокислотной последовательности SEQ ID NO: 7 или 8;
полипептида, который по меньшей мере на 83% идентичен каталитическому домену с последовательностью SEQ ID NO: 7 или 8;
полипептида, содержащего аминокислотную последовательность каталитического домена с SEQ ID NO: 9; или
зрелого полипептида, полученного обработкой полипептида с SEQ ID NO: 1, 2 или 3 сигнальной пептидазой или посттрансляционной модификацией во время секреции из экспрессирующего хозяина;
в качестве кормовой добавки к корму для животных, где указанная глюкоамилаза (а) проявляет по крайней мере 20% активности или по меньшей мере на 20% более высокую остаточную активность при pH меньше или равном 3 в присутствии пепсина или рубцового сока (например, рубцового сока, содержащего пепсин) по сравнению с активностью ферментов при pH 6 отдельно или в присутствии рубцового сока (например, рубцового сока, содержащего пепсин), и (b) фермент действует вместе с амилазой поджелудочной железы, увеличивая выход глюкозы.
18. В некоторых вариантах осуществления способа согласно пункту 17, где, если животное является жвачным, указанный фермент активен в по меньшей мере двух из трех пищеварительных камер жвачного животного, включая рубец, сычуг и тонкий кишечник.
19. В некоторых вариантах осуществления способа согласно пункту 16 или 17 указанная по меньшей мере одна глюкоамилаза способна гидролизовать сырой крахмал.
20. В другом аспекте предложен полинуклеотид, содержащий нуклеотидную последовательность, которая по меньшей мере на 80% идентична нуклеотидной последовательности SEQ ID NO: 4, 5 или 6.
21. В другом аспекте предложен вектор, содержащий полинуклеотидную последовательность согласно пункту 20, функционально связанную с одной или несколькими управляющими последовательностями, которые регулируют продуцирование кодируемого полипептида в экспрессирующем хозяине, и где указанная регуляторная последовательность гетерологична кодирующей нуклеотидной последовательности, или указанные регуляторная последовательность и кодирующая последовательность не расположены вместе, как это бывает в природе.
22. В другом аспекте предложена рекомбинантная клетка-хозяин, содержащая полинуклеотиды согласно пункту 20.
23. В некоторых вариантах осуществления рекомбинантная клетка-хозяин согласно пункту 22 представляет собой клетку Trichoderma, Aspergillus, Myceliopthora или Saccharomyces.
24. В некоторых вариантах осуществления рекомбинантная клетка-хозяин согласно пункту 22 представляет собой клетку E.coli, Bacillus, Streptomyces или Pseudomonas.
25. В некоторых вариантах осуществления рекомбинантная клетка-хозяин согласно пункту 22 представляет собой этанолопродуцирующие микроорганизмы.
26. В некоторых вариантах осуществления рекомбинантная клетка-хозяин согласно любому из пунктов 22-25 дополнительно экспрессирует и секретирует один или несколько дополнительных ферментов, выбранных из группы, состоящей из протеазы, гемицеллюлазы, целлюлазы, пероксидазы, липолитического фермента, металлолиполитического фермента, ксиланазы, липазы, фосфолипазы, эстеразы, пергидролазы, кутиназы, пектиназы, пектатлиазы, манназы, кератиназы, редуктазы, оксидазы, фенолоксидазы, липоксигеназы, лигниназы, пуллуназы, фитазы, танназы, пентосаназы, маланазы, бета-глюканазы, арабиносидазы, гиалуронидазы, хондроитиназы, лакказы, трансферазы или любой их комбинации.
27. В другом аспекте предложена композиция кормовой добавки или премикс, содержащая по меньшей мере одну глюкоамилазу, выбранную из группы, состоящей из:
полипептида, содержащего аминокислотную последовательность с SEQ ID NO: 7, 8 или 9;
полипептида, который по меньшей мере на 83% идентичен аминокислотной последовательности SEQ ID NO: 7, 8 или 9;
полипептида, который по меньшей мере на 83% идентичен каталитическому домену с последовательностью SEQ ID NO: 7, 8 или 9;
полипептида, который по меньшей мере на 83% идентичен линкеру и каталитическому домену с последовательностью SEQ ID NO: 7, 8 или 9; или
зрелого полипептида, полученного обработкой полипептида с SEQ ID NO: 1, 2 или 3 сигнальной пептидазой или посттрансляционной модификацией во время секреции из экспрессирующего хозяина; и/или
необязательно по меньшей мере один минерал и/или по меньшей мере один витамин.
28. В некоторых вариантах осуществления композиция кормовой добавки или премикса согласно пункту 27 дополнительно включает один или несколько ферментов, выбранных из группы, состоящей из протеазы, амилазы, ксиланазы и фитазы.
29. В некоторых вариантах осуществления композиция кормовой добавки или премикса согласно пункту 27 или пункту 28 дополнительно включает один или несколько пробиотиков, выбранных из группы, состоящей из Bacillus, молочнокислых бактерий и дрожжей.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ И ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0011] На фигуре 1 показано CLUSTALW выравнивание множественных последовательностей TeGA, FraGA1, WcoGA1 и других грибковых глюкоамилаз.
[0012] Следующие последовательности соответствуют §§ 1.821-1.825 37 C.F.R. ("Требования для патентных заявок, содержащих раскрытия нуклеотидных последовательностей и/или аминокислотных последовательностей - Правила для последовательностей") и согласуются со Стандартом ST.25 (2009) Всемирной организации интеллектуальной собственности (WIPO) и требованиями для перечня последовательностей Европейской патентной конвенции (EPC) и Правилами 5.2 и 49.5 (a-bis) Договора о патентной кооперации (РСТ) и разделом 208 и приложением С Административной инструкции. Символы и формат, применяемые для данных относительно нуклеотидных и аминокислотных последовательностей, соответствуют правилам, изложенным в § 1.822 раздела 37 C.F.R.
[0013] SEQ ID NO: 1 - последовательность белка-предшественника FraGA1.
[0014] SEQ ID NO: 2 - последовательность белка-предшественника WcoGA1.
[0015] SEQ ID NO: 3 - последовательность белка-предшественника TeGA.
[0016] SEQ ID NO: 4 - нуклеотидная последовательность гена FraGA1.
[0017] SEQ ID NO: 5 - нуклеотидная последовательность гена WcoGA1.
[0018] SEQ ID NO: 6 - нуклеотидная последовательность гена TeGA.
[0019] SEQ ID NO: 7 - смоделированная последовательность зрелого белка FraGA1.
[0020] SEQ ID NO: 8 - смоделированная последовательность зрелого белка WcoGA1.
[0021] SEQ ID NO: 9 - последовательность зрелого белка TeGA.
[0022] SEQ ID NO: 10 - глюкоамилаза дикого типа из Aspergillus niger, регистрационный номер NCBI - XP_001390530.1.1.
[0023] SEQ ID NO: 11 - глюкоамилаза дикого типа из Trichoderma reesei, регистрационный номер PDB - 2VN4_A.
[0024] SEQ ID NO: 12 представляет собой KZT67263.1 (регистрационный номер NCBI).
[0025] SEQ ID NO: 13 представляет собой XP_002475369.1 (регистрационный номер NCBI).
[0026] SEQ ID NO: 14 представляет собой SEQ ID NO: 1847, описанную в US20090325240.
[0027] SEQ ID NO: 15 представляет собой KZT09226.1 (регистрационный номер NCBI).
[0028] SEQ ID NO: 16 представляет собой SEQ ID NO: 12, описанную в WO2016196202.
[0029] SEQ ID NO: 17 представляет собой SEQ ID NO: 2, описанную в US20170314003.
[0030] SEQ ID NO: 18 представляет собой GAD95639.1 (регистрационный номер NCBI).
[0031] SEQ ID NO: 19 представляет собой SEQ ID NO: 23, описанную в US20170306309.
[0032] SEQ ID NO:20 представляет собой CAC28076.1 (регистрационный номер NCBI).
ПОДРОБНОЕ ОПИСАНИЕ
[0033] Все цитируемые патенты, заявки на патенты и публикации полностью включены в данный документ ссылкой. В данном раскрытии применяется ряд терминов и сокращений. Если четко не указано иное, то применимы следующие определения.
[0034] Термин «включающий» или «содержащий» означает наличие установленных признаков, целых чисел, стадий или компонентов, изложенных в формуле изобретения, но таким образом, что он не исключает наличия или добавления одного или нескольких признаков, целых чисел, стадий, компонентов или их групп. Термин «включающий» или «содержащий» подразумевают как включающий варианты осуществления, охватываемые терминами «по сути состоящий из» и «состоящий из». Аналогично предполагается, что термин «по сути состоящий из» включает варианты осуществления, охватываемые термином «состоящий из». Используемый в данном документе в отношении числового значения термин «примерно» относится к пределам +/- 0,5 от числового значения, если указанный термин иным образом конкретно не определен в контексте. Например, фраза «значение pH, составляющее примерно 6» относится к значениям pH от 5,5 до 6,5, если конкретно не определено иное значение pH.
[0035] Если не указано иное, все используемые технические и научные термины имеют свое обычное значение в соответствующей области науки. Определение многих терминов, описывающих данное изобретение, можно найти в Singleton, et al., Dictionary of Microbiology and Molecular Biology, 2d Ed., John Wiley and Sons, New York (1994) и Hale & Markham, Harper Collins Dictionary of Biology, Harper Perennial, NY (1991).
[0036] Термин «активность глюкоамилазы (1,4-альфа-D-глюканглюкогидролазы, EC 3.2.1.3) в данном контексте означает активность фермента, которая катализирует высвобождение D-глюкозы из невосстанавливающих концов крахмала или родственных молекул олиго- и полисахаридов. Большинство глюкоамилаз представляют собой многодоменные ферменты, состоящие из каталитического домена, связанного с доменом связывания крахмала с помощью О-гликозилированного линкерного участка различной длины. Были определены и описаны кристаллические структуры нескольких глюкоамилаз (см. J. Lee и M. Paetzel 2011. Acta Cryst. Vol. 67 pages 188-192 "Structure of the catalytic domain of glucoamylase from Aspergillus niger" и J. Sauer et al 2000. Biochem. Et Biophys. Acta Vol. 1542 page 275-293 "Glucoamylase: structure/function relationships, and protein engineering".
[0037] Термины «связывающий крахмал домен (SBD)» или «модуль связывания углеводов (CBM)» применяют взаимозаменяемо в данном документе. SBD можно разделить на девять семейств CBM. Являясь источником энергии, крахмал подвергается расщеплению большим числом различных амилолитических ферментов. Однако, лишь примерно 10% из них способны к связыванию и расщеплению сырого крахмала. Такие ферменты обычно содержат отдельную последовательность-структурный модуль, называемый связывающим крахмал доменом, который опосредует присоединение к гранулам крахмала. SBD относится к аминокислотной последовательности, которая связывается преимущественно с субстратом, представляющим собой крахмал (полисахарид), или мальтосахаридом, альфа-, бета- и гамма-циклодекстрином и т. п. Они обычно представляют собой мотивы из примерно 100 аминокислотных остатков, обнаруженные у примерно 10% микробных амилолитических ферментов.
[0038] Термин «каталитический домен (CD)» относится к структурному участку полипептида, который содержит активный центр гидролиза субстрата.
[0039] Термин «гликозидгидролаза» используется взаимозаменяемо с терминами «гликозидаза» и «гликозилгидролаза». Гликозидгидролазы содействуют гидролизу гликозидных связей в сложных сахарах (полисахаридах, таких как, без ограничения, крахмал). Гликозидгидролазы также можно классифицировать как экзо- или эндо-действующие в зависимости от того, действуют ли они на (обычно невосстанавливающем) конце или в середине, соответственно, олиго/полисахаридной цепи. Гликозидгидролазы также можно классифицировать способами, основанными на последовательности или структуре.
[0040] Термин «корм» используется в отношении продуктов, скармливаемых животным при выращивании сельскохозяйственных животных. Термины «корм» и «корм для животных» используются взаимозаменяемо. В предпочтительном варианте осуществления пищевой продукт или корм подходит для скармливания нежвачным и жвачным животным.
[0041] Используемый в данном документе термин «пробиотик» («DFM») является источником живых (жизнеспособных) встречающихся в природе микроорганизмов. Категории DFM включают Bacillus, молочнокислые бактерии и дрожжи. Bacillus представляют собой уникальные грамположительные палочки, которые образуют споры. Типы молочнокислых бактерий включают Bifidobacterium, Lactobacillus и Streptococcus. Дрожжи не являются бактериями. Эти микроорганизмы относятся к растительной группе грибов.
[0042] Термин «пепсин» в данном контексте относится к ферменту, который расщепляет белки на более мелкие пептиды при низких значениях pH в пределах от 2,0 до 3,5, т.е. он представляет собой протеазу, принадлежащую к семейству аспарагиновых пептидаз A1. Он продуцируется и секретируется в желудке и является одним из основных пищеварительных ферментов в пищеварительных системах людей и животных, где он способствует перевариванию белков в пище или корме.
[0043] Термин «жвачное животное» в данном контексте относится к млекопитающему, которое способно получать питательные вещества из пищи растительного происхождения путем ферментирования ее в приспособленном для этого желудке перед перевариванием, главным образом, посредством воздействия микроорганизмов.
[0044] Термин «пищеварительные камеры жвачного животного» в данном контексте относится к рубцу, сетке, книжке, сычугу и тонкой кишке (McDonald et al., 2011, Animal Nutrition (7th Edition), стр. 156-191). Сычуг представляет собой прямой эквивалент однокамерного желудка.
[0045] Термин «гранулированный крахмал» относится к сырому (не подвергнутому тепловой обработке) крахмалу, например гранулированному крахмалу, не прошедшему желатинизацию.
[0046] Термины «фермент, гидролизирующий гранулированный крахмал (GSH)» и «ферменты, имеющие гидролизующую гранулированный крахмал (GSH) активность» используются взаимозаменяемо и относятся к ферментам, которые способны гидролизовать крахмал в гранулированной форме в характерных для пищеварительного тракта условиях, сравнимых с условиями, обнаруживаемыми в пищеварительном тракте животных, и в частности жвачных животных.
[0047] Термин «выделенный» означает вещество в форме или условиях, которые не встречаются в природе. Неограничивающие примеры выделенных веществ включают: (1) любое не встречающееся в природе вещество, (2) любое вещество, включая без ограничения любую клетку-хозяина, фермент, вариант, нуклеиновую кислоту, белок, пептид или кофактор, которое по меньшей мере частично извлечено из одного или нескольких или всех встречающихся в природе компонентов, с которыми оно связано в природе; (3) любое вещество, модифицированное человеком по сравнению с этим веществом в натуральной форме; или (4) любое вещество, модифицированное путем увеличения содержания вещества по отношению к другим составляющим, с которыми оно встречается в природе. Термины «выделенная молекула нуклеиновой кислоты», «выделенный полинуклеотид» и «выделенный фрагмент нуклеиновой кислоты» используются взаимозаменяемо и относятся к полимеру РНК или ДНК, который является одно- или двухцепочечным, необязательно содержащим синтетические, отличные от природных или измененные нуклеотидные основания.
[0048] Термин «очищенный» применительно к нуклеиновым кислотам или полипептидам, как правило, обозначает нуклеиновую кислоту или полипептид, которые фактически не содержат других компонентов, что определяют с помощью аналитических методик, хорошо известных из уровня техники (например, очищенный полипептид или полинуклеотид образует отдельную полосу в электрофоретическом геле, хроматографическом элюате и/или средах, подвергаемых центрифугированию в градиенте плотности). Например, нуклеиновая кислота или полипептид, которые дают фактически одну полосу в электрофоретическом геле, являются «очищенными».
[0049] Термины «пептиды», «белки» и «полипептиды» используются в данном документе взаимозаменяемо и относятся к полимеру из аминокислот, соединенных вместе пептидными связями. "Белок" или "полипептид" содержит полимерную последовательность аминокислотных остатков. В данном раскрытии используется одно- и 3-буквенный код для аминокислот, который определен согласно IUPAC-IUB объединенным комитетом по биохимической номенклатуре (JCBN). Также понятно, что полипептид может быть закодирован несколькими нуклеотидными последовательностями из-за вырожденности генетического кода.
[0050] Термин «зрелая» форма белка, полипептида или фермента относится к функциональной форме белка, полипептида или фермента без сигнальной пептидной последовательности или пропептидной последовательности.
[0051] Термин форма "предшественника" белка или пептида относится к форме белка с пропоследовательностью, функционально связанной с амино- или карбонильным концом белка. Предшественник может также иметь "сигнальную" последовательность, функционально связанную с аминоконцом пропоследовательности.
[0052] Как отмечено выше, регуляторные последовательности могут быть функционально связаны в смысловой или антисмысловой ориентации с представляющими интерес кодирующей последовательностью/геном.
[0053] «Промотор» или «промоторные последовательности» относятся к последовательностям ДНК, которые определяют, где начинается транскрипция гена под действием РНК-полимеразы. Промоторные последовательности обычно располагаются на 5'-конце сайта инициации транскрипции или непосредственно выше него. Промоторы можно получать целиком из нативной или встречающейся в природе последовательности или можно составлять из различных компонент, полученных из различных промоторов, встречающихся в природе, или же они могут содержать сегменты синтетической ДНК. Специалистам в данной области техники будет понятно, что различные промоторы могут управлять экспрессией гена в различных тканях или типах клеток или на различных стадиях развития или в ответ на различные окружающие или физиологические условия («индуцируемые промоторы»).
[0054] Выражение «3'-некодирующие последовательности» относится к последовательностям ДНК, расположенным ниже кодирующей последовательности, и включает последовательности, кодирующие регуляторные сигналы, способные влиять на процессинг мРНК или экспрессию генов, такие как терминация транскрипции.
[0055] Используемый в данном документе термин «трансформация» относится к переносу или введению молекулы нуклеиновой кислоты в организм-хозяин. Молекулу нуклеиновой кислоты можно вводить в виде линейной или кольцевой формы ДНК. Молекула нуклеиновой кислоты может представлять собой плазмиду, которая реплицируется автономно, или она может интегрироваться в геном продуцирующего хозяина. Продуцирующие хозяева, содержащие трансформированную нуклеиновую кислоту, называются «трансформированными», или «рекомбинантными», или «трансгенными» организмами или «трансформантами».
[0056] Используемый в данном документе термин «рекомбинантный» относится к искусственной комбинации двух в ином случае отделенных друг от друга сегментов последовательностей нуклеиновых кислот, например, путем химического синтеза или манипуляции выделенными сегментами нуклеиновых кислот с помощью методик генной инженерии. Например, ДНК, в которую были вставлены один или несколько сегментов или генов естественным образом либо с помощью лабораторной манипуляции из отличающейся молекулы, из другой части той же молекулы или из искусственной последовательности, что привело к введению новой последовательности в ген и, соответственно, в организм. Термины "рекомбинантный", "трансгенный", "трансформированный", "сконструированный" или "модифицированный для экспрессии экзогенного гена" используются в данном документе взаимозаменяемо.
[0057] Термины "рекомбинантная конструкция", "экспрессирующая конструкция", "рекомбинантная экспрессирующая конструкция" и "кассета экспрессии" используются в данном документе взаимозаменяемо. Рекомбинантная конструкция содержит искусственную комбинацию фрагментов нуклеиновых кислот, например регуляторных и кодирующих последовательностей, которые не все встречаются вместе в природе. Например, конструкция может содержать регуляторные последовательности и кодирующие последовательности, которые происходят из разных источников, или регуляторные последовательности и кодирующие последовательности, которые происходят из одного источника, но расположены в порядке, отличающемся от обнаруживаемого в природе. Такую конструкцию можно использовать саму по себе или можно использовать в сочетании с вектором.
[0058] Термин "процент идентичности" представляет собой взаимоотношение между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, определенное путем сравнения последовательностей. В данной области техники «идентичность» также означает степень родства последовательностей полипептидных или полинуклеотидных последовательностей, в зависимости от конкретного случая, определенного по количеству совпадающих нуклеотидов или аминокислот в нитях таких последовательностей. «Идентичность» и «сходство» могут быть легко вычислены с помощью известных способов, в том числе без ограничения тех, которые описаны в: Computational Molecular Biology (Lesk, A. M., ed.) Oxford University Press, NY (1988); Biocomputing: Informatics and Genome Projects (Smith, D. W., ed.) Academic Press, NY (1993); Computer Analysis of Sequence Data, Part I (Griffin, A. M., и Griffin, H. G., eds.) Humana Press, NJ (1994); Sequence Analysis in Molecular Biology (von Heinje, G., ed.) Academic Press (1987); и Sequence Analysis Primer (Gribskov, M. и Devereux, J., eds.) Stockton Press, NY (1991). Способы определения идентичности и сходства запрограммированы в общедоступных компьютерных программах.
[0059] Используемый в данном документе термин «идентичны на %», или "процент тождественности", или "PID" относится к одинаковости белковой последовательности. Процент тождественности может быть определен с использованием стандартных методик, известных из уровня техники. Применимые алгоритмы включают алгоритмы BLAST (см. Altschul et al., J Mol Biol, 215:403-410, 1990; и Karlin and Altschul, Proc Natl Acad Sci USA, 90:5873-5787, 1993). В программе BLAST используются несколько параметров поиска, большинство из которых выставлены на значения по умолчанию. Алгоритм NCBI BLAST находит наиболее подходящие последовательности с точки зрения биологического сходства, но не рекомендуется для запрашиваемых последовательностей из менее чем 20 остатков (Altschul et al., Nucleic Acids Res, 25:3389-3402, 1997; и Schaffer et al., Nucleic Acids Res, 29:2994-3005, 2001). Иллюстративные параметры BLAST по умолчанию для поиска последовательностей нуклеиновой кислоты включают: пороговую длину смежных фрагментов=11; E‐значение для отсечения=10; матрицу замен=NUC.3.1 (соответствие=1, несоответствие = ‐3); штраф за открытие промежутка=5 и штраф за удлинение промежутка=2. Иллюстративные параметры BLAST по умолчанию для поиска аминокислотных последовательностей включают: размер слова=3; E‐значение для отсечения=10; матрицу замен=BLOSUM62; открытие промежутка=11 и продление промежутка=1. Значение процента (%) тождественности аминокислотной последовательности определяется по количеству совпадающих идентичных остатков, деленному на общее количество остатков «эталонной» последовательности, включая любые промежутки, созданные программой для оптимального/максимального выравнивания. Алгоритмы BLAST обращаются к «эталонной» последовательности как к «запрашиваемой» последовательности.
[0060] Используемые в данном документе термины «гомологичные белки» или «гомологичные ферменты» относятся к белкам, которые имеют отчетливое сходство первичной, вторичной и/или третичной структуры. Гомология белка может относиться к сходству в линейной аминокислотной последовательности при выравнивании белков. Гомологичный поиск белковых последовательностей может быть выполнен с использованием BLASTP и PSI-BLAST из NCBI BLAST с порогом (E-значение для отсечки) при 0,001. (Altschul SF, Madde TL, Shaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI BLAST a new generation of protein database search programs. Nucleic Acids Res 1997 Set 1;25(17):3389-402). Используя эту информацию, можно сгруппировать последовательности белков, а также построить филогенетическое дерево с использованием аминокислотных последовательностей. Выравнивание последовательностей и расчеты процента идентичности также можно выполнять, используя программу Megalign, программу AlignX, программное обеспечение EMBOSS Open Software Suite (EMBL-EBI; Rice et al., Trends in Genetics 16, (6):276-277 (2000)) или аналогичные программы. Множественное выравнивание последовательностей также можно выполнить с помощью метода Clustal (например, CLUSTALW) с параметрами по умолчанию. Подходящие параметры для выравниваний белка в CLUSTALW включают следующие: штраф за введение промежутка=15, штраф за удлинение промежутка=0,2, матрица=Gonnet (например, Gonnet250), учет концевых промежутков (ENDGAP) в последовательности белка = -1, расстояние между промежутками (GAPDIST) в белке=4 и размер участка максимального совпадения (KTUPLE) = 1.
[0061] Различные аминокислотные последовательности полипептидов и полинуклеотидные последовательности раскрыты в данном документе в качестве признаков определенных аспектов. В определенных вариантах осуществления могут применяться варианты этих последовательностей, которые по меньшей мере примерно на 70-85%, 85-90% или 90%-95% идентичны последовательностям, раскрываемым в данном документе. В качестве альтернативы, в определенных вариантах осуществления полипептидная последовательность или полинуклеотидная последовательность варианта может быть по меньшей мере на 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична последовательности, раскрываемой в данном документе. Аминокислотная последовательность или полинуклеотидная последовательность варианта имеет аналогичную раскрываемым последовательностям функцию или обладает по меньшей мере примерно 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% функции раскрываемых последовательностей.
[0062] В одном аспекте зрелый полипептид представляет собой аминокислоты с 17 по 567 в SEQ ID NO: 1, с 18 по 569 в SEQ ID NO: 2 и с 28 по 618 в SEQ ID NO: 3 в соответствии с программой SignalP (Nielsen et al., 1997, Protein Engineering 10: 1-6).
[0063] Термин «нуклеиновая кислота» охватывает ДНК, РНК, гетеродуплексы и синтетические молекулы, способные кодировать полипептид. Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными и могут быть изменены химическими методами. Термины «нуклеиновая кислота» и «полинуклеотид» используются взаимозаменяемо. Поскольку генетический код является вырожденным, то можно использовать более одного кодона для кодирования конкретной аминокислоты; при этом композиции и способы по настоящему изобретению охватывают нуклеотидные последовательности, которые кодируют конкретную аминокислотную последовательность. Если не указано иное, то нуклеотидные последовательности представлены в направлении от 5' к 3'.
[0064] Термин «кодирующая последовательность» означает нуклеотидную последовательность, которая непосредственно определяет аминокислотную последовательность ее белкового продукта. Границы кодирующей последовательности обычно определяются открытой рамкой считывания, которая обычно начинается со стартового кодона ATG или альтернативных стартовых кодонов, таких как GTG и TTG, и заканчивается стоп-кодоном, таким как TAA, TAG и TGA. Кодирующая последовательность может быть ДНК, кДНК, синтетической или рекомбинантной нуклеотидной последовательностью.
[0065] «Синтетическую» или «искусственную» молекулу получают посредством химического или ферментативного синтеза in vitro, но без использования организма.
[0066] «Штамм-хозяин» или «клетка-хозяин» представляет собой организм, в который был введен вектор экспрессии, фаг, вирус или другая конструкция ДНК, в том числе полинуклеотид, кодирующий полипептид, представляющий интерес (например, амилазу). Иллюстративными штаммами-хозяевами являются клетки микроорганизмов (например, бактерий, мицелиальных грибов и дрожжей), способные экспрессировать полипептид, представляющий интерес, и/или ферментировать сахариды. Термин «клетка-хозяин» включает протопласты, полученные из клеток.
[0067] Термин «экспрессия» означает процесс, при котором на основе последовательности нуклеиновой кислоты синтезируется полипептид. Данный процесс включает как транскрипцию, так и трансляцию.
[0068] Термин «конечный продукт» относится к спирту, такому как этанол, или биохимикату, выбранному из группы, состоящей из аминокислоты, органической кислоты, лимонной кислоты, молочной кислоты, янтарной кислоты, глутамата натрия, глюконовой кислоты, глюконата натрия, глюконата кальция, глюконата калия, глюконо-дельта-лактона, эриторбата натрия, жирной кислоты омега-3, бутанола, лизина, итаконовой кислоты, 1,3-пропандиола, биодизеля и изопрена.
[0069] Термин «вектор» означает полинуклеотидную последовательность, сконструированную для введения нуклеиновых кислот в клетки одного или нескольких типов. Векторы включают клонирующие векторы, векторы экспрессии, челночные векторы, плазмиды, фаговые частицы, кассеты и т.п.
[0070] «Вектор экспрессии» означает конструкцию ДНК, содержащую последовательность ДНК, кодирующую представляющий интерес полипептид, кодирующая последовательность которого функционально связана с подходящей регуляторной последовательностью, способной воздействовать на экспрессию ДНК в подходящем хозяине. Такие регуляторные последовательности могут включать промотор для воздействия на транскрипцию, необязательно последовательность оператора для контроля транскрипции, последовательность, кодирующую подходящие сайты связывания рибосом на мРНК, энхансеры и последовательности, которые контролируют терминацию транскрипции и трансляции.
[0071] Предполагается, что термин «управляющие последовательности» в данном контексте включает все компоненты, необходимые для экспрессии полинуклеотида, кодирующего полипептид настоящего изобретения. Каждая управляющая последовательность может быть нативной или чужеродной по отношению к нуклеотидной последовательности, кодирующей полипептид, или нативной или чужеродной по отношению друг к другу. Такие управляющие последовательности включают, но не ограничиваются ими, лидерную последовательность, последовательность полиаденилирования, пропептидную последовательность, промотор, последовательность сигнального пептида и терминатор транскрипции. Как минимум, управляющие последовательности включают в себя промотор и сигналы остановки транскрипции и трансляции. Управляющие последовательности могут быть снабжены линкерами с целью введения специфических сайтов рестрикции, облегчающих лигирование управляющей последовательности с кодирующим участком полинуклеотидной последовательности, кодирующей полипептид.
[0072] Термин «функционально связанный» означает, что указанные компоненты взаимосвязаны (включая без ограничения непосредственный контакт), что позволяет им функционировать надлежащим образом. Например, регуляторная последовательность функционально связана с кодирующей последовательностью таким образом, что экспрессия кодирующей последовательности контролируется регуляторными последовательностями.
[0073] «Сигнальная последовательность» представляет собой последовательность аминокислот, присоединенных к N-концевой части белка, которая облегчает секрецию белка из клетки. Зрелая форма внеклеточного белка не содержит сигнальной последовательности, которая отщепляется в процессе секреции.
[0074] «Биологически активный» относится к последовательности, имеющей указанную биологическую активность, такую как ферментативная активность.
[0075] Термин «специфическая активность» означает количество молей субстрата, которое может быть превращено в продукт с помощью фермента или ферментного препарата за единицу времени в определенных условиях. Специфическую активность обычно выражают в единицах (ед)/мг белка.
[0076] Используемый здесь термин «остаточная активность» относится к соотношению активностей, направленных на субстрат, измеренному с инкубированием и без него при определенных условиях (таких как, без ограничения, измененная температура или переменный pH).
[0077] Термины «дикого типа», «исходный» или «эталонный» в применении у полипептиду означают встречающийся в природе полипептид, который не содержит искусственную замену, вставку или делецию в одном или нескольких положениях аминокислот. Аналогичным образом термины «дикого типа», «исходный» или «эталонный» в применении к полинуклеотиду означают встречающийся в природе полинуклеотид, который не содержит искусственную нуклеозидную замену. Однако следует отметить, что полинуклеотид, кодирующий полипептид дикого типа, исходный или эталонный полипептид, не ограничивается встречающимся в природе полинуклеотидом и включает любой полинуклеотид, кодирующий полипептид дикого типа, исходный или эталонный полипептид.
[0078] Термины «термически устойчивый», «термостабильный» и «термостабильность» в применении к ферменту означают способность фермента сохранять активность после воздействия повышенной температуры. Термостабильность фермента, такого, как фермента амилаза, измеряется временем его полужизни (t1/2), выраженным в минутах, часах или днях, в течение которого фермент теряет половину своей активности при воздействии определенных условий. Время полужизни можно рассчитать путем измерения остаточной амилазной активности, например, после воздействия (например, испытания путем воздействия) высокой температуры. Термины «термически устойчивый» и «термостабильный» означает, что по меньшей мере примерно 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% фермента, который присутствовал/был активен в добавке перед нагреванием до указанной температуры, все еще присутствует/активен после охлаждения до комнатной температуры. Предпочтительно по меньшей мере приблизительно 80% фермента, присутствующего и активного в добавке перед нагреванием до указанной температуры, все еще присутствует и активен после охлаждения до комнатной температуры.
[0079] «Пределы значений pH» в применении к ферменту означает пределы значений рН, при которых фермент проявляет каталитическую активность.
[0080] Термины «рН-стабильный» и «рН-стабильность» в применении к ферменту означает способность фермента сохранять активность в широких пределах значений рН в течение предварительно заданного периода времени (например, 15 минут, 30 минут, 1 часа).
[0081] Фраза «одновременные осахаривание и ферментация (SSF)» означает способ получения биохимических веществ, в котором микроорганизм, такой как этанолопродуцирующий микроорганизм, и по меньшей мере один фермент, такой как амилаза, присутствуют на одной и той же стадии способа. SSF включает одновременный гидролиз крахмальных субстратов (гранулированного, разжиженного или солюбилизированного) до сахаридов, в том числе до глюкозы, и ферментацию сахаридов до спирта или другого биохимического вещества или биоматериала в одной емкости реактора.
[0082] «Суспензия» представляет собой водную смесь, содержащую нерастворимые в воде гранулы крахмала.
[0083] Термин «содержание общего сахара» относится к суммарному содержанию растворимого сахара, присутствующего в крахмальной композиции, включая моносахариды, олигосахариды и полисахариды.
[0084] Термин «сухие твердые вещества» (ds) относится к сухим твердым веществам, растворенным в воде, сухим твердым веществам, диспергированным в воде, или их комбинации. Таким образом, сухие твердые вещества включают гранулированный крахмал и продукты его гидролиза, включая глюкозу.
[0085] «Содержание сухих твердых веществ» относится к процентному содержанию как растворенных, так и диспергированных сухих твердых веществ в массовых долях по отношению к воде, в которой эти сухие твердые вещества диспергированы и/или растворены. Первоначальное содержание сухого твердого вещества в крахмале представляет собой массу гранулированного крахмала с поправкой на содержание влаги по отношению к сумме массы гранулированного крахмала и массы воды. Последующее содержание сухого твердого вещества может быть определено из начального содержания, скорректированного с учетом добавленной или потерянной воды, а также привеса в результате гидролиза крахмала. Последующее содержание растворенного сухого твердого вещества можно измерить по показателю преломления, как указано ниже. 8
[0086] Термин «высокий DS» относится к водной суспензии крахмала с массовой долей сухого твердого вещества более 38%.
[0087] «Сухое вещество крахмала» относится к содержанию сухого крахмала в субстрате, таком как крахмальная суспензия, которое может быть определено вычитанием из массы субстрата любого вклада некрахмальных компонентов, таких как белок, клетчатка и вода. Например, если массовая доля воды в суспензии гранулированного крахмала составляет 20%, а массовая доля белка - 1%, то 100 кг гранулированного крахмала содержат 79 кг сухого крахмала. Сухой крахмал можно использовать для определения необходимого количества единиц фермента.
[0088] «Разжиженный» относится к продукту варки (нагревания) и разжижения (уменьшения вязкости) крахмала или крахмальной суспензии (сусла).
[0089] «Разжижение» или «разжижать» относится к процессу, с помощью которого крахмал (или крахмалосодержащие зерна) превращается/превращаются в менее вязкие декстрины с более короткой цепью.
[0090] «Степень полимеризации (DP)» относится к числу (n) единиц ангидроглюкопиранозы в данном сахариде. Примерами DP1 являются моносахариды, такие как глюкоза и фруктоза. Примерами DP2 являются дисахариды, такие как мальтоза и сахароза. DP4+ (>DP3) обозначает полимеры со степенью полимеризации более 3.
[0091] Термин «приведение в контакт» относится к размещению указанных компонентов (включая, помимо прочего, ферменты, субстраты и ферментирующие организмы) в достаточно непосредственной близости, чтобы получить ожидаемый результат, как например, воздействие фермента на субстрат, или ферментацию субстрата ферментирующим организмом. Специалисты в данной области поймут, что «приведение в контакт» можно обеспечить смешиванием растворов.
[0092] «Этанолопродуцирующий микроорганизм» означает микроорганизм, способный превращать сахар или олигосахарид в этанол.
[0093] Термин «биохимикаты» относится к метаболитам микроорганизма, таким как лимонная кислота, молочная кислота, янтарная кислота, глутамат натрия, глюконовая кислота, глюконат натрия, глюконат кальция, глюконат калия, глюконо-дельта-лактон, эриторбат натрия, омега-3 жирная кислота, бутанол, изобутанол, аминокислота, лизин, итаконовая кислота, другие органические кислоты, 1,3-пропандиол, витамины или изопрен или другой биоматериал.
[0094] Термин «примерно» означает ± 15% от указанного значения.
[0095] Следующие сокращения/аббревиатуры имеют приведенные ниже значения, если не указано иное:
EC Комиссия по ферментам
CAZy углеводно-активный фермент
вес/об. вес/объем
вес/вес вес/вес
об./об. объем/объем
вес. % массовая доля в процентах
°C градусы Цельсия
г грамм(ы)
мкг микрограмм
мг миллиграмм
кг килограмм
мкл микролитр
мл миллилитр
мм миллиметр
мкм микрометр
моль моль
ммоль миллимоль
М молярный
мМ миллимолярный
мкм микромолярный
нм нанометр
U или Ед единицы
ppm или м.д. миллионные доли
ч час
[0096] В первом аспекте настоящее изобретение относится к полипептидам, содержащим аминокислотную последовательность, которая предпочтительно по меньшей мере на 83%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% и даже по меньшей мере на 99% идентична аминокислотной последовательности полипептида SEQ ID NO: 7, 8 или 9 и проявляет глюкоамилазную активность.
[0097] В некоторых вариантах осуществления полипептиды настоящего изобретения представляют собой гомологичные полипептиды, содержащие аминокислотные последовательности, отличающиеся десятью аминокислотами, предпочтительно девятью аминокислотами, предпочтительно восемью аминокислотами, предпочтительно семью аминокислотами, предпочтительно шестью аминокислотами, предпочтительно пятью аминокислотами, более предпочтительно четырьмя аминокислотами, еще более предпочтительно тремя аминокислотами, наиболее предпочтительно двумя аминокислотами и еще более предпочтительно одной аминокислотой от полипептида с SEQ ID NO:7, 8 или 9.
[0098] В некоторых вариантах осуществления полипептиды настоящего изобретения представляют собой варианты полипептида с SEQ ID NO: 7, 8 или 9, или их фрагмент, которые проявляют глюкоамилазную активность.
[0099] В некоторых вариантах осуществления полипептиды настоящего изобретения представляют собой каталитические участки, содержащие аминокислоты 17-470 в SEQ ID NO: 1, 18-472 в SEQ ID NO: 2 или 28-500 в SEQ ID NO: 3, смоделированные с помощью ClustalX https://www.ncbi.nlm.nih.gov/pubmed/17846036.
[00100] В некоторых вариантах осуществления полипептиды настоящего изобретения стабильны при низком pH и сохраняют глюкоамилазную активность при низком pH. Полипептиды настоящего изобретения продемонстрировали стабильность при низких pH при значениях pH в пределах от примерно 2,0 до примерно 5,0 (например, от примерно 2,0 до примерно 4,0, от примерно 2,0 до примерно 3,0, от примерно 2,0 до примерно 2,5 и т.д.). Например, при pH от 2,0 до примерно 3,0 полипептиды настоящего изобретения сохраняют большую часть глюкогенной активности в течение продолжительного периода времени при высокой температуре (например, не ниже 40°C, не ниже 50°C, не ниже 55°C, не ниже 60°C, не ниже 65°C, пне ниже 70°C или более высокой температуре), и, например, в течение не менее 4 часов, не менее 17 часов, не менее 24 часов, не менее 48 часов, не менее 72 часов или даже дольше.
[00101] В некоторых вариантах осуществления полипептиды настоящего изобретения обладают лучшими характеристиками осахаривания по сравнению с AnGA (глюкоамилазой из Aspergillus niger) при pH примерно 3 или даже при pH примерно 2, в интервале температур от примерно 30 до примерно 70°C (например, от примерно 30°C до примерно 60°C, от примерно 40°C до примерно 60°C и т.д.) при времени инкубирования не менее 24 часов, не менее 48 часов, не менее 72 часов или даже дольше.
[00102] В некоторых вариантах осуществления полипептиды настоящего изобретения можно применять в способе одновременного осахаривания и ферментации (SSF) или для ферментации при низком рН по сравнению с поступающими в настоящее время в продажу глюкоамилазными продуктами, при pH примерно 3 или даже при pH примерно 2, в интервале температур от примерно 30°C до примерно 70°C (например, от примерно 30°C до примерно 60°C, от примерно 30°C до примерно 50°C и т.д.) при времени инкубирования не менее 17 часов, не менее 24 часов, не менее 48 часов, не менее 72 часов или даже дольше.
[00103] Во втором аспекте настоящие глюкоамилазы содержат консервативную замену одного или нескольких аминокислотных остатков относительно аминокислотной последовательности SEQ ID NO: 7, 8 или 9. Иллюстративные консервативные аминокислотные замены перечислены в таблице 1. Некоторые консервативные мутации можно получать путем генетических манипуляций, а другие - путем введения синтетических аминокислот в полипептид другими способами.
Таблица 1. Консервативные аминокислотные замены
или 5-фенилпролин
D-Met, Met(O), D-Met(O), Val, D-Val
[00104] В некоторых вариантах осуществления настоящая глюкоамилаза включает делецию, замену, вставку или присоединение одного или нескольких аминокислотных остатков относительно аминокислотной последовательности SEQ ID NO: 7, 8 или 9 или гомологичной им последовательности. В некоторых вариантах осуществления настоящие глюкоамилазы получают из аминокислотной последовательности SEQ ID NO: 7, 8 или 9 путем консервативной замены одного или нескольких аминокислотных остатков. В некоторых вариантах осуществления настоящие глюкоамилазы получены из аминокислотной последовательности SEQ ID NO7, 8 или 9 путем делеции, замены, вставки или присоединения одного или нескольких аминокислотных остатков относительно аминокислотной последовательности SEQ ID NO: 7, 8 или 9. Во всех случаях выражение «один или несколько аминокислотных остатков» относится к 10 или менее, т.е. 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотным остаткам. Число аминокислотных замен, делеций и/или вставок в зрелом полипептиде SEQ ID NO: 7, 8 или 9 может составлять не более 10, предпочтительно не более 9, более предпочтительно не более 8, более предпочтительно не более 7, более предпочтительно не более 6, более предпочтительно не более 5, более предпочтительно не более 4, даже более предпочтительно не более 3, наиболее предпочтительно не более 2 и еще более предпочтительно не более 1.
[00105] Альтернативно, характер изменения аминокислот таков, что меняются физико-химические свойства полипептидов. Например, изменения аминокислот могут улучшить термическую стабильность полипептида, изменить специфичность к субстрату, изменить оптимальный pH и т.п.
[00106] Одиночные или множественные аминокислотные замены, делеции и/или вставки можно выполнять и тестировать, используя известные методы мутагенеза, рекомбинации и/или перестановки с последующей соответствующей процедурой отбора, например, описанной у Reidhaar-Olson и Sauer, 1988, Science 241: 53-57; Bowie и Sauer, 1989, Proc. Natl. Acad. Sci. USA 86: 2152-2156; в WO 95/17413; или в WO 95/22625. Другие методы, которые можно использовать, включают ПЦР с внесением ошибок, фаговый дисплей (например, Lowman et al., 1991, Biochem. 30: 10832-10837; U. S. Patent No. 5,223,409; WO 92/06204) и мутагенез, нацеленный на область (Derbyshire et al., 1986, Gene 46: 145; Ner et al., 1988, DNA 7: 127).
[00107] Методы мутагенеза/перестановки можно комбинировать с высокопроизводительными автоматизированными методами отбора для обнаружения активности клонированных, подвергнутых мутагенезу полипептидов, экспрессируемых клетками-хозяевами (Ness et al., 1999, Nature Biotechnology 17: 893-896). Подвергнутые мутагенезу молекулы ДНК, кодирующие активные полипептиды, можно выделять из клеток-хозяев и быстро секвенировать с использованием стандартных в данной области методов. Эти методы позволяют быстро определять важность отдельных аминокислотных остатков в интересующем полипептиде и их можно применять к полипептидам неизвестной структуры.
[00108] Глюкоамилаза может быть «химерным» или «гибридным» полипептидом в том смысле, что она включает по меньшей мере часть из первой глюкоамилазы и по меньшей мере часть из второй амилазы, глюкоамилазы, бета-амилазы, альфа-глюкозидазы или другого разрушающего крахмал фермента, или даже другие гликозилгидролазы, такие как, без ограничения, целлюлазы, гемицеллюлазы и т.д. (включая такие химерные амилазы, которые недавно были «заново открыты» как амилазы с заменой домена). Глюкоамилазы настоящего изобретения могут дополнительно включать гетерологичную сигнальную последовательность - эпитоп для обеспечения отслеживания или очистки и т.п.
[00109] Настоящие глюкоамилазы могут продуцироваться в клетках-хозяевах, например, путем секреции или внутриклеточной экспрессии. Культивированный клеточный материал (например, цельноклеточный бульон), содержащий глюкоамилазу, может быть получен после секреции глюкоамилазы в клеточную среду. По выбору, глюкоамилазу можно изолировать от клеток-хозяев или даже изолировать из клеточного бульона в зависимости от требуемой чистоты конечной глюкоамилазы. Ген, кодирующий глюкоамилазу, можно клонировать и экспрессировать согласно способам, хорошо известным в данном уровне техники. Пригодные клетки-хозяева включают бактериальные, грибковые (включая дрожжи и мицелиальные грибы) и клетки растений (включая водоросли). Особенно полезные клетки-хозяева включают Aspergillus niger, Aspergillus oryzae Trichoderma reesi или Myceliopthora thermophila. Другие клетки-хозяева включают бактериальные клетки, например, Bacillus subtilis или B. licheniformis, а также Streptomyces.
[00110] Кроме того, хозяин может экспрессировать один или несколько вспомогательных ферментов, белков, пептидов. Это может способствовать разжижению, осахариванию, ферментации, SSF и последующим процессам. Кроме того, клетка-хозяин может вырабатывать этанол и другие биохимикаты или биоматериалы помимо ферментов, используемые для расщепления различного сырья. Такие клетки-хозяева можно применять для процессов ферментации или одновременного осахаривания и ферментации для снижения или устранения необходимости вводить ферменты.
[00111] Конструкция ДНК, содержащая нуклеиновую кислоту, кодирующую полипептид глюкоамилазы, может быть сконструирована таким образом, чтобы ее можно было экспрессировать в клетке-хозяине. Из-за известного вырождения генетического кода, с использованием обычных навыков можно разрабатывать и получать разные полинуклеотиды, которые кодируют идентичные аминокислотные последовательности. Также известно, что в зависимости от требуемых клеток-хозяев, перед проведением экспрессии может потребоваться оптимизация кодонов.
[00112] Полинуклеотид, кодирующий полипептид глюкоамилазы настоящего изобретения, может быть встроен в вектор. Векторы можно переносить в клетку-хозяина, используя хорошо известные методики трансформации, такие как раскрыты ниже.
[00113] Подходящим вектором может быть вектор, который можно трансформировать и реплицировать в клетке-хозяине. Например, вектор, содержащий нуклеиновую кислоту, кодирующую полипептид глюкоамилазы настоящего изобретения, можно трансформировать и реплицировать в бактериальной клетке-хозяине как средстве размножения и амплификации вектора. Вектор также можно подходящим образом трансформировать в хозяина экспрессии, так что кодирующий полинуклеотид будет экспрессироваться как функциональный фермент глюкоамилаза.
[00114] Полинуклеотид, кодирующий полипептид глюкоамилазы настоящего изобретения, может быть функционально связан с промотором, который обеспечивает транскрипцию в клетке-хозяине. Промотором может быть любая последовательность ДНК, которая демонстрирует транскрибирующую активность в выбранной клетке-хозяине и может быть получена из генов, кодирующих белки либо гомологичные, либо гетерологичные клетке-хозяину.
[00115] Кодирующая последовательность может быть функционально связана с сигнальной последовательностью. ДНК, кодирующая сигнальную последовательность, может быть последовательностью ДНК, в естественных условиях связанной с представляющим интерес геном глюкоамилазы, подлежащим экспрессии, или может происходить из другого рода или вида, чем глюкоамилаза. Сигнальная последовательность и последовательность промотора, содержащая ДНК-конструкцию или вектор, могут быть введены в грибковую клетку-хозяина и могут быть получены из того же источника. Например, сигнальная последовательность может быть сигнальной последовательностью cbh1 Trichoderma reesei, которая функционально связана с промотором cbh1.
[00116] Вектор экспрессии также может содержать подходящий терминатор транскрипции, а в эукариотах - последовательности полиаденилирования, функционально связанные с последовательностью ДНК, кодирующей глюкоамилазу. Терминирующие и полиаденилирующие последовательности могут быть, соответственно, получены из тех же источников, что и промоторы.
[00117] Вектор также может содержать селектируемый маркер, например, ген, продукт которого комплементарен дефекту в изолированной клетке-хозяине, такой, как ген dal из B. subtilis или B. licheniformis, или ген, который придает устойчивость к антибиотику, такую, как например, устойчивость к ампициллину, канамицину, хлорамфениколу или тетрациклину. Кроме того, вектор может содержать маркеры отбора Aspergillus, такие, как amdS, argB и niaD - маркер, придающий устойчивость к гигромицину, или селективность может быть осуществлена путем совместной трансформации так, как известно в данном уровне техники. См., например, международную заявку PCT WO 91/17243.
[00118] Внутриклеточная экспрессия может быть предпочтительна в некоторых случаях, например, при использовании определенных бактерий или грибов в качестве клеток-хозяев для получения больших количеств альфа-глюкозидазы для последующего обогащения или очистки. Альтернативно, внеклеточное выделение глюкоамилазы в культуральную среду может быть использовано для изготовления культивируемого клеточного материала, содержащего изолированную глюкоамилазу.
[00119] Методики, используемые для лигирования ДНК-конструкции, кодирующей глюкоамилазу, промотор, терминатор и другие элементы, соответственно, и для введения их в подходящий вектор, содержащий информацию, необходимую для репликации, известны специалисту в данной области См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor, 1989, и 3rd ed., 2001.
[00120] Выделенную клетку, содержащую либо ДНК-конструкцию, либо вектор экспрессии предпочтительно применять в качестве клетки-хозяина в рекомбинантном получении глюкоамилазы. Клетку можно удобно трансформировать ДНК-конструкцией, кодирующей фермент, путем внедрения ДНК-конструкции (в одной или нескольких копиях) в хромосому хозяина. Внедрение в общем случае рассматривается как преимущество, поскольку последовательность ДНК с большей вероятностью будет более стабильно поддерживаться в клетке. Включение ДНК-конструкций в хромосому хозяина можно выполнять традиционными методами, например, путем гомологичной или гетерологичной рекомбинации. Альтернативно клетку можно трансформировать вектором экспрессии в зависимости от других типов клеток-хозяев.
[00121] Примерами подходящих бактериальных организмов-хозяев являются грамположительные виды таких бактерий, как Bacillaceae, включая Bacillus subtilis, Bacillus licheniformis, Bacillus lentus, Bacillus brevis, Geobacillus (ранее - Bacillus) stearothermophilus, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus coagulans, Bacillus lautus, Bacillus megaterium и Bacillus thuringiensis; виды Streptomyces, такие, как Streptomyces murinus; виды молочнокислых бактерий, включая Lactococcus sp., такие, как Lactococcus lactis; Lactobacillus sp., включая Lactobacillus reuteri; Leuconostoc sp.; Pediococcus sp.; и Streptococcus sp. Alternatively, штаммы грамотрицательных видов бактерий, принадлежащих к Enterobacteriaceae, влключая E. coli, или к Pseudomonadaceae, которые можно выбирать в качестве организма-хозяина.
[00122] Подходящий дрожжевой организм-хозяин можно выбирать из относящихся к биотехнологическим видам дрожжей, таких как, но не ограничиваясь этими видами дрожжей, Pichia sp., Hansenula sp или виды Kluyveromyces, Yarrowinia, Schizosaccharomyces или виды Saccharomyces, включая Saccharomyces cerevisiae или виды, принадлежащие к Schizosaccharomyces, такие как, например, виды S. pombe. В качестве организма-хозяина можно использовать штамм метилотрофных видов дрожжей Pichia pastoris. Альтернативно, организм-хозяин может представлять собой вид Hansenula.
[00123] Подходящие организмы-хозяева среди мицелиальных грибов включают виды Aspergillus, например, Aspergillus niger, Aspergillus oryzae, Aspergillus tubigensis, Aspergillus awamori или Aspergillus nidulans. Альтернативно, в качестве организма-хозяина можно использовать штаммы видов Fusarium, например, Fusarium oxysporum или видов Rhizomucor, таких как Rhizomucor miehei. Другие пригодные штаммы включают виды Thermomyces и Mucor. Кроме того, в качестве хозяина можно использовать Trichoderma sp. Глюкоамилаза, экспрессированная грибковой клеткой-хозяином, может быть гликозилирована, т.е. содержать остаток гликозила. Характер гликозилирования может быть таким же, как в глюкоамилазе дикого типа, или отличаться от него. Тип и степень гликозилирования может вносить изменения в ферментативные и/или биохимические свойства.
[00124] Удаление генов из экспрессирующих хозяев имеет преимущества, если дефицит гена можно восполнить с помощью трансформированного вектора экспрессии. Можно использовать известные методы для получения грибковой клетки-хозяина, имеющей один или несколько инактивированных генов. Можно удалять любой ген из Trichoderma sp. или другого клонированного мицелиального грибкового хозяина, например, гены cbh1, cbh2, egl1 и egl2. Делеция гена может быть выполнена путем введения формы гена, который необходимо инактивировать, в плазмиду с помощью методов, известных в данной области техники.
[00125] Введение конструкции ДНК или вектора в клетку-хозяина включает такие методики, как трансформация; электропорация; микроинъекция в ядро; трансдукция; трансфекция, например, опосредованная липофекцией и опосредованная DEAE-декстрином трансфекция; инкубировоние с осаждением ДНК с помощью фосфата кальция; высокоскоростная бомбардировка микрочастицами, покрытыми ДНК; и слияние протопластов. В уровне техники известны другие общие методики трансформации. См., например, Sambrook et al. (2001), выше. Экспрессия гетерологичного белка в Trichoderma описана, например, в патенте США № 6022725. О трансформации штаммов Aspergillus также см. в Cao et al. (2000) Science 9: 991-1001. Генетически стабильные трансформанты можно конструировать при помощи векторных систем, в которых нуклеиновая кислота, кодирующая альфа-глюкозидазу, стабильно интегрируется в хромосому клетки-хозяина. Затем трансформанты отбирают и очищают известными методами.
[00126] Способ получения глюкоамилазы может включать культивирование клетки-хозяина в условиях, способствующих продукции фермента, и выделение фермента из клеток и/или культуральной среды.
[00127] Средой, используемой для культивирования клеток, может быть любая традиционная среда, подходящая для выращивания клетки-хозяина и обеспечения экспрессии полипептида глюкоамилазы. Подходящие среды и компоненты сред доступны от коммерческих поставщиков или могут быть получены в соответствии с опубликованными рецептурами (например, описанными в каталогах Американской коллекции типовых культур).
[00128] Любой из способов ферментации, хорошо известных в данной области техники, можно подходящим образом применять для ферментации трансформированного или производного грибкового штамма, описанного выше. В некоторых вариантах осуществления грибковые клетки выращивают в условиях периодической или непрерывной ферментации.
[00129] Методики разделения и концентрирования известны из уровня техники, и традиционные способы можно применять для получения концентрированного раствора или бульона, содержащего полипептид глюкоамилазы настоящего изобретения.
[00130] После ферментации получают ферментационный бульон, удаляют микробные клетки и различные взвешенные твердые вещества, в том числе остаточные сырьевые материалы для ферментации, с помощью традиционных методик разделения с целью получения раствора глюкоамилазы. Обычно применяют фильтрацию, центрифугирование, микрофильтрацию, фильтрацию на вращающемся вакуумном барабане, ультрафильтрацию, центрифугирование с последующей ультрафильтрацией, экстракцию или хроматографию или подобное.
[00131] Иногда для оптимизации извлечения может быть необходимо концентрирование раствора или бульона, содержащего полипептид глюкоамилазу. Применение неконцентрированных растворов или бульона обычно увеличивает время инкубации для сбора осадка обогащенного или очищенного фермента.
[00132] Настоящее изобретение также относится к композициям, содержащим полипептид настоящего изобретения. В некоторых вариантах осуществления полипептид, содержащий аминокислотную последовательность, которая предпочтительно по меньшей мере на 83%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% и даже по меньшей мере на 99% идентична аминокислотной последовательности полипептида SEQ ID NO: 7, 8 или 9, и обладающий глюкоамилазной активностью, также можно использовать в композиции фермента. Предпочтительно композиции составлены для обеспечения желаемых характеристик, таких как слабое окрашивание, слабый запах и приемлемая стабильность при хранении. полипептиды
[00133] Композиция может содержать полипептид настоящего изобретения в качестве основного ферментного компонента, как например, однокомпонентная композиция. Альтернативно, композиция может включать несколько активных ферментов, таких как аминопептидаза, амилаза, карбогидраза, карбоксипептидаза, каталаза, целлюлаза, хитиназа, кутиназа, циклодекстрингликозилтрансфераза, дезоксирибонуклеаза, эстераза, альфа-галактозидаза, бета-галактозидаза, альфа-глюкозидаза, бета-глюкозидаза, бета-амилаза, изоамилаза, галопероксидаза, инвертаза, лакказа, липаза, лизоцим, маннозидаза, оксидаза, пектинолитический фермент, пептидоглутаминаза, пероксидаза, фитаза, полифенолоксидаза, протеолитический фермент, пуллуланаза, рибонуклеаза, трансглютаминаза, ксиланаза или их комбинация, которые можно добавлять в эффективных количествах, хорошо известных специалисту в данной области.
[00134] Полипептидные композиции можно готовить способами, известными в данной области, и они могут иметь форму жидкой или сухой композиции. Например, композиции, содержащие глюкоамилазы настоящего изобретения, могут представлять собой водные или неводные составы, гранулы, порошки, гели, суспензии, пасты и т.д., которые могут дополнительно содержать любой один или несколько дополнительных ферментов, перечисленных в данном документе, наряду с буферами, солями, консервантами, водой, вспомогательными растворителями, поверхностно-активными веществами и т.п. Такие композиции могут работать в комбинации с эндогенными ферментами или другими ингредиентами, уже присутствующими в суспензии, водяной ванне, стиральной машине, пищевом продукте или напитках и т.д., например, эндогенными растительными (включая водорослевые) ферментами, остаточными ферментами из предшествующей стадии переработки и т.п. Полипептид, который должен быть включен в композицию, можно стабилизировать в соответствии со способами, известными в данной области.
[00135] Композиция может представлять собой клетки, экспрессирующие полипептид, включая клетки, способные продуцировать продукт ферментации. Такие клетки могут быть представлены в виде крема или в сухом виде вместе с подходящими стабилизаторами. Такие клетки могут дополнительно экспрессировать дополнительные полипептиды, такие как упомянуты выше.
[00136] Ниже приведены примеры предпочтительных применений полипептидов или композиций данного изобретения. Дозировка полипептидной композиции данного изобретения и другие условия, в которых применяется композиция, могут быть определены на основе методик, известных в данной области.
[00137] Настоящее изобретение также направлено на использование полипептида или композиции настоящего изобретения в процессе разжижения, осахаривания и/или ферментации. Полипептид или композицию можно использовать в одном процессе, например, в процессе разжижения, процессе осахаривания или процессе ферментации. Полипептид или композицию также можно применять в комбинированных процессах, например, в процессе разжижения и осахаривания, в процессе разжижения и ферментации или в процессе осахаривания и ферментации, предпочтительно при переработке крахмала.
[00138] Разжиженный крахмал можно осахаривать с получением сиропа, богатого сахаридами с более низким DP (например, DP1+DP2), с использованием альфа-амилаз и глюкоамилаз, необязательно в присутствии другого фермента(ов). Точный состав продуктов осахаривания зависит от комбинации используемых ферментов, а также от типа перерабатываемого крахмала. Преимущественно получаемый с помощью предложенных глюкоамилаз сироп может содержать массовую долю DP2 от общего количества олигосахаридов в осахаренном крахмале, которая превышает 30%, например, 45%-65% или 55% - 65%. Массовая процентная доля (DP1+DP2) в осахаренном крахмале может превышать примерно 70% и составлять, например, 75% - 85% или 80% - 85%.
[00139] Хотя разжижение обычно осуществляют в виде непрерывного процесса, осахаривание зачастую осуществляют в виде периодического процесса. Условия осахаривания зависят от природы разжиженного вещества и типа доступных ферментов. В некоторых случаях процесс осахаривания может проводиться при температуре примерно 60-65°C и pH примерно 4,0-4,5, например, pH 4,3. Осахаривание можно проводить, например, при температуре от примерно 40°C, примерно 50°C или примерно 55°C до примерно 60°C или примерно 65°C, что требует охлаждения разжиженного вещества. При необходимости также можно отрегулировать pH. Осахаривание обычно проводят в емкостях с мешалкой, заполнение или опорожнение которых может занять несколько часов. При загрузке баков ферменты, как правило, вносят либо в фиксированном соотношении с высушенными твердыми веществами, либо в виде однократной дозы в начале стадии заполнения. Реакцию осахаривания для получения сиропа, как правило, осуществляют в течение примерно 24-72 часов, например, 24-48 часов. Однако обычной практикой является проводить только предварительное осахаривание в течение 40-90 минут при температуре 30-65°C, обычно примерно 60°C, с последующим полным осахариванием при одновременном осахаривании и ферментации (SSF). В одном варианте осуществления способ изобретения включает предварительное осахаривание крахмалосодержащего материала перед процессом одновременного осахаривания и ферментации (SSF). Предварительное осахаривание можно проводить при высокой температуре (например, 50-85°C, предпочтительно 60-75°C) перед переходом к SSF. Предпочтительно осахаривание оптимально проводить в более высоком интервале температур от примерно 30°C до примерно 75°C, например, при 45-75°C или 50-75°C. Путем проведения процесса осахаривания при более высоких температурах процесс можно проводить за более короткий период времени или, альтернативно, процесс можно проводить с использованием более низкой дозировки фермента. Кроме того, риск микробного загрязнения снижается при проведении процесса разжижения и/или осахаривания при более высокой температуре.
[00140] В предпочтительном аспекте настоящего изобретения разжижение и/или осахаривание включает последовательно или одновременно выполняемые процессы разжижения и осахаривания.
[00141] Растворимый гидролизат крахмала, в частности сироп, обогащенный глюкозой, можно ферментировать путем контакта гидролизата крахмала с ферментирующим организмом, обычно при температуре примерно 32°C, например от 30°C до 35°C. «Ферментирующий организм» относится к любому организму, включая бактериальные и грибковые организмы, подходящему для использования в процессе ферментации и способному продуцировать требуемый продукт ферментации. Особенно подходящие ферментирующие организмы способны ферментировать, т.е. преобразовывать сахара, такие как глюкоза или мальтоза, прямо или косвенно в желаемый продукт ферментации. Примеры ферментирующих организмов включают дрожжи, такие как Saccharomyces cerevisiae, и бактерии, например, Zymomonas mobilis, экспрессирующие алкогольдегидрогеназу и пируватдекарбоксилазу. Этанолопродуцирующий микроорганизм может экспрессировать ксилозоредуктазу и ксилитолдегидрогеназу, которые превращают ксилозу в ксилулозу. Усовершенствованные штаммы этанолопродуцирующих микроорганизмов, например, такие, которые способны выдерживать более высокие температуры, известны в данной области техники, и их можно применять. См. Liu et al. ( 2011) Sheng Wu Gong Cheng Xue Bao 27: 1049-56. Дрожжи, поступающие в продажу, включают, например, Red Star™/Lesaffre Ethanol Red (можно приобрести у Red Star/Lesaffre, США), FALI (можно приобрести у Fleischmann's Yeast, подразделения Burns Philp Food Inc., США), SUPERSTART (можно приобрести у Alltech), GERT STRAND (можно приобрести у Gert Strand AB, Швеция), SYNERXIA® ADY (можно приобрести у DuPont), SYNERXIA® THRIVE (можно приобрести у DuPont), FERMIOL (можно приобрести у DSM Specialties). Температура и рН при ферментации будут зависеть от ферментирующего организма. Микроорганизмы, которые путем ферментации могут производить другие метаболиты, такие, как лимонная кислота и молочная кислота, также известны в уровне техники. См., например, Papagianni (2007) Biotechnol. Adv. 25:244-63; John et al. (2009) Biotechnol. Adv. 27:145-52.
[00142] Процессы осахаривания и ферментации можно осуществлять в виде процесса SSF. Процесс SSF можно проводить с грибковыми клетками, которые непрерывно экспрессируют и секретируют глюкоамилазу в ходе всего процесса SSF. Грибковые клетки, экспрессирующие глюкоамилазу, также могут представлять собой ферментирующий микроорганизм, например, этанолопродуцирующий микроорганизм. Таким образом, получение этанола можно осуществлять с использованием грибковой клетки, которая экспрессирует достаточное количество глюкоамилазы для того, чтобы добавлять меньшее количество экзогенного фермента или вообще не добавлять его. Грибковая клетка-хозяин может быть из соответствующим образом модифицированного грибкового штамма. Также можно использовать грибковые клетки-хозяева, которые экспрессируют и секретируют другие ферменты помимо глюкоамилазы. Такие клетки могут экспрессировать амилазу и/или пуллуланазу, фитазу, альфа-глюкозидазу, изоамилазу, бета-амилазу, целлюлазу, другие гемицеллюлазы, протеазу, бета-глюкозидазу, пектиназу, эстеразу, окислительно-восстановительные ферменты, трансферазу или другие ферменты. Вслед за процессом брожения может следовать извлечение этанола.
[00143] В соответствии с настоящим изобретением ферментация включает, без ограничения, процессы ферментации, используемые для получения спиртов (например, этанола, метанола, бутанола); органических кислот (например, лимонной кислоты, уксусной кислоты, итаконовой кислоты, молочной кислоты, глюконовой кислоты); кетонов (например, ацетона); аминокислот (например, глутаминовой кислоты); газов (например, H2 и CO2); антибиотиков (например, пенициллина и тетрациклина); ферментов; витаминов (например, рибофлавина, B12, бета-каротина); и гормонов. В таком предпочтительном варианте осуществления процесс обычно проводят при температуре от 28°C до 36°C, например, от 29°C до 35°C, например, от 30°C до 34°C, например, примерно 32°C, при pH в пределах от 3 до 7, предпочтительно при pH от 3,5 до 6 или более предпочтительно при pH от 4 до 5.
[00144] В других вариантах осуществления низкие значения pH минимизируют риск внесения примесей, поскольку останавливается рост конкурирующих организмов. Процесс ферментации данного изобретения, например ферментацию органических кислот, можно проводить при низком pH, поскольку обычно ферментирующие организмы быстро понижают pH примерно до 4,0-4,5, а некоторые лактобациллы снижают pH даже до примерно 3,5. Содержание свободной молочной кислоты в растворе (например, не менее примерно 50 г/л, предпочтительно - не менее примерно 80 г/л и более предпочтительно - не менее примерно 100 г/л лактата) приводит к относительно низкому pH. Чем выше процентное содержание лактата в форме свободной кислоты, тем ниже pH раствора. Например, если pH среды равен pKa молочной кислоты (примерно 3,8), то содержание лактата в форме свободной кислоты составляет 50%. При pH 4,2 содержание лактата в виде свободной кислоты составляет примерно 31%, а при pH 4,0 и 3,9 - примерно 41% и 47%, соответственно.
[00145] В настоящем изобретении предложено применение глюкоамилазы(глюкоамилаз) данного изобретения для получения глюкозы и других сахаридов из сырого крахмала или гранулированного крахмала. Обычно глюкоамилазу настоящего изобретения либо саму по себе, либо в присутствии альфа-амилазы, можно использовать для гидролиза сырого крахмала (RSH) или гранулированного крахмала (GSH) для получения желаемых сахаров и продуктов ферментации. Гранулированный крахмал солюбилизируют ферментативным гидролизом ниже температуры желатинизации. Сообщалось, что такие «низкотемпературные» системы (известные также как «без варки» или «холодная варка») способны обрабатывать более высокие концентрации сухих твердых веществ, чем традиционные системы (например, вплоть до 45%).
[00146] Процесс «гидролиза сырого крахмала» (RSH) отличается от обычных процессов обработки крахмала, включая последовательное или одновременное осахаривание и ферментацию гранулированного крахмала при температуре желатинизации крахмального субстрата или ниже, обычно в присутствии по меньшей мере глюкоамилазы и/или амилазы. Крахмал, нагретый в воде, начинает желатинизироваться при температуре от 50°C до 75°C, точная температура желатинизации зависит от конкретного крахмала. Например, температура желатинизации может варьироваться в зависимости от вида растений, от конкретного сорта растений, а также от условий роста. В контексте настоящего изобретения температура желатинизации данного крахмала представляет собой температуру, при которой двойное лучепреломление прекращается в 5% гранул крахмала с использованием метода, описанного у Gorinstein. S. и Lii. C., Starch/Starke, Vol. 44 (12) pp. 461-466 (1992).
[00147] Глюкоамилазу данного изобретения также можно использовать в комбинации с ферментом, который гидролизует только альфа-(1,6)-глюкозидные связи в молекулах, содержащих по меньшей мере четыре глюкозильных остатка. Предпочтительно глюкоамилазу данного изобретения использовать в комбинации с пуллуланазой или изоамилазой. Использование изоамилазы и пуллуланазы для расщепления разветвленной структуры крахмала, молекулярные свойства ферментов и потенциальное использование ферментов вместе с глюкоамилазой описано у G. M. A. van Beynum et al., Starch Conversion Technology, Marcel Dekker, New York, 1985, 101-142.
[00148] Термин «продукт ферментации» означает продукт, полученный с помощью способа, предусматривающего процесс ферментации с применением ферментирующего организма. Продукты ферментации, предполагаемые согласно изобретению, включают спирты (например, арабинит, бутанол, этанол, глицерин, метанол, этиленгликоль, пропиленгликоль, бутандиол, глицерин, сорбит и ксилит); органические кислоты (например, уксусную кислоту, ацетоновую кислоту, адипиновую кислоту, аскорбиновую кислоту, лимонную кислоту, 2,5-дикето-D-глюконовую кислоту, муравьиную кислоту, фумаровую кислоту, глюкаровую кислоту, глюконовую кислоту, глюкуроновую кислоту, глутаровую кислоту, 3-гидроксипропионовую кислоту, итаконовую кислоту, молочную кислоту, яблочную кислоту, малоновую кислоту, щавелевую кислоту, щавелевоуксусную кислоту, пропионовую кислоту, янтарную кислоту и ксилоновую кислоту); кетоны (например, ацетон); аминокислоты (например, аспарагиновую кислоту, глутаминовую кислоту, глицин, лизин, серин и треонин); алкан (например, пентан, гексан, гептан, октан, нонан, декан, ундекан и додекан); циклоалкан (например, циклопентан, циклогексан, циклогептан и циклооктан); алкен (например, пентен, гексен, гептен и октен); газы (например, метан, водород (H2), диоксид углерода (CO2) и монооксид углерода (CO)); антибиотики (например, пенициллин и тетрациклин); ферменты; витамины (например, рибофлавин, B12, бета-каротин); и гормоны.
[00149] В предпочтительном аспекте продукт ферментации представляет собой этанол, например топливный этанол; питьевой этанол, т.е. питьевой нейтральный спирт; или промышленный этанол, или продукты, используемые в производстве потребляемого алкоголя (например, пива и вина), молочной промышленности (например, ферментированные молочные продукты), кожевенной и табачной промышленности. Предпочтительные типы пива включают эли, стауты, портеры, лагеры, биттеры, солодовое пиво, крепкое пиво, слабоалкогольное пиво, низкокалорийное пиво или светлое пиво. Предпочтительные используемые способы ферментации включают процессы спиртовой ферментации, которые хорошо известны в данной области. Предпочтительными способами ферментации являются хорошо известные анаэробные процессы ферментации.
[00150] Способы изготовления пива известны в уровне техники. См., например, Wolfgang Kunze (2004) "Technology Brewing and Malting," Research and Teaching Institute of Brewing, Berlin (VLB), 3rd edition. Вкратце способ включает: (a) приготовление браги, (b) фильтрование браги с получением пивного сусла и (c) ферментацию сусла с получением сброженного напитка, такого, как пиво.
[00151] Композицию для пивоварения, содержащую глюкоамилазу, в сочетании с амилазой и, необязательно, пуллуланазой и/или изоамилазой, можно добавлять к браге на стадии (а) выше, т.е. во время приготовления браги. Альтернативно или в дополнение, композицию для пивоварения можно вносить в брагу на стадии (b) выше, т.е. во время фильтрации браги. Альтернативно или в дополнение, композицию для пивоварения можно вносить в сусло на стадии (с) выше, т.е. во время ферментации сусла.
[00152] В еще одном аспекте, описанные здесь глюкоамилазы можно использовать в качестве кормовой добавки для животных для улучшения усвояемости крахмала. В данном документе описан способ увеличения усвояемости крахмала у животного, который включает внесение по меньшей мере одной глюкоамилазы, выбранной из группы, состоящей из:
полипептида, содержащего аминокислотную последовательность с SEQ ID NO: 7, 8 или 9;
полипептида, который по меньшей мере на 83% идентичен аминокислотной последовательности SEQ ID NO: 7, 8 или 9;
полипептида, который по меньшей мере на 83% идентичен каталитическому домену с последовательностью SEQ ID NO: 7, 8 или 9;
полипептида, который по меньшей мере на 83% идентичен линкеру и каталитическому домену с последовательностью SEQ ID NO: 7, 8 или 9; или
зрелого полипептида, полученного обработкой полипептида с SEQ ID NO: 1, 2 или 3 сигнальной пептидазой или посттрансляционной модификацией во время секреции из экспрессирующего хозяина;
в качестве кормовой добавки к корму для животных, где указанная глюкоамилаза (а) проявляет по крайней мере 20% активности или по меньшей мере на 20% более высокую остаточную активность при pH меньше или равном 3 в присутствии пепсина или рубцового сока (например, пепсинсодержащего рубцового сока) по сравнению с активностью ферментов при pH 6 отдельно или в присутствии рубцового сока (например, рубцового сока, содержащего пепсин), и (b) фермент действует вместе с амилазой поджелудочной железы, увеличивая выход глюкозы.
[00153] В другом аспекте, если животное является жвачным, то фермент активен в по меньшей мере двух из трех пищеварительных камер жвачного животного, включая рубец, сычуг и тонкий кишечник. Рубец является самым крупным отделом с объемом, составляющим 150-200 литров (40-50 галлонов).
[00154] Жвачные животные обладают уникальной способностью превращать грубый корм в белок и энергию посредством своих микробных/ферментативных пищеварительных систем. Соответственно, жвачные животные играют важную роль в экологии Земли и в пищевой цепочке.
[00155] Главное различие между жвачным животным и нежвачным животным заключается в том, что жвачное животное имеет четырехкамерный желудок, состоящий из рубца, сетки, книжки и сычуга. Сычуг представляет собой прямой эквивалент однокамерного желудка (McDonald et al., 2011, Animal Nutrition (7th Edition), стр. 156-191).
[00156] В пищеварительной системе присутствуют миллиарды микроорганизмов. Они помогают корове переваривать и усваивать питательные вещества из корма. Для достижения эффективного усваивания корма и высоких удоев молока бактерии должны находиться в оптимальных условиях. Именно бактерии являются теми, кто осуществляет расщепление корма. Кормление коровы, фактически, сопряжено с кормлением микроорганизмов в ее рубце. Процесс ферментации протекает в рубце и сетке. Рубец коровы подобен большому чану для ферментации. Более 200 различных бактерий и 20 типов простейших помогают корове усваивать волокнистые кормовые продукты и небелковые источники азота. Ферментация - это химический процесс, с помощью которого молекулы, такие как глюкоза, расщепляются анаэробно, например, когда микроорганизмы превращают углеводы в летучие жирные кислоты и газы. Данный процесс позволяет корове преобразовывать целлюлозное волокно в энергию. Среди газов, вырабатываемых в рубце во время ферментации (500-1500 литров в день) (150-400 галлонов), 20-40% составляют метан и диоксид углерода. Выработка ферментационных газов представляет собой значительную потерю энергии. Определенные модификаторы ферментации, такие как ионофоры, улучшают энергетическую эффективность жвачных животных путем уменьшения таких потерь энергии на выработку газов.
[00157] Рубец и сетка по сути представляют собой один отдел, но с различными функциями. Хотя большая часть ферментирующего действия происходит в рубце, сетка служит в качестве промежуточной зоны для прохождения в книжку или отрыгивания.
[00158] Идеальное значение pH в рубце составляет от 6 до 7. Микроорганизмы рубца являются наиболее жизнеспособными в пределах данного диапазона. Если значение pH слишком изменчиво, то это может вызвать гибель некоторых типов микроорганизмов, что снижает усваивание корма. Микроорганизмы, которые расщепляют целлюлозу (сено, силос и т.д.) неспособны расти или ферментировать целлюлозу при значениях pH, ниже 6,0. При снижении значения pH в рубце ниже 6 рубец считают ацидозным. Рубцовый ацидоз может быть острым с быстрым сильным падением значения pH. Более распространенным среди высокопроизводительных стад является субклинический ацидоз, который характеризуется хроническими прерывающимися периодами низкого значения pH в рубце.
[00159] Как отмечалось выше, усвояемость крахмала из кормов значительно изменяется в зависимости от ряда факторов, в том числе физической структуры и крахмала, и кормовой основы.
[00160] Было обнаружено, что усвояемость крахмала в рационе животного можно улучшить путем применения по меньшей мере одной глюкоамилазы в качестве кормовой добавки для жвачных животных, где указанная глюкоамилаза: (а) обладает по меньшей мере 20%-ами активности (например, по меньшей мере примерно на 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 80% более высокой активностью, включая все значения между этими процентами) или по крайней мере на 20% более высокой остаточной активностью (например, по крайней мере на примерно 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 80% более высокой остаточной активностью, включая все значения между этими процентами) при рН меньше или равном 3 в присутствии пепсина или рубцового сока (например, содержащего пепсин рубцового сока) по сравнению с активностью ферментов только при pH 6 или в присутствии или рубцового сока (например, содержащего пепсин рубцового сока) и (b) гидролаза действует вместе с пищеварительными ферментами, присутствующими в пищеварительных камерах животного, для повышения выхода глюкозы и повышения усвояемости крахмала.
[00161] Любую из описанных здесь глюкоамилаз можно использовать отдельно или в комбинации по меньшей мере с одним пробиотиком. Категории DFM включают Bacillus, молочнокислые бактерии и дрожжи. Бациллы представляют собой уникальные грамположительные палочки, которые образуют споры. Эти споры очень стабильны и могут выдерживать такие условия окружающей среды, как тепло, влага и определенный диапазон рН. Эти споры прорастают в активные вегетативные клетки при проглатывании животным и могут применяться в молотых и пеллетированных рационах.
[00162] Термины «корм для животных», «корм», «кормовой продукт» и «фураж» используются взаимозаменяемо и могут включать один или несколько кормовых материалов, выбранных из группы, включающей a) зерновые культуры, такие как мелкозерные (например, пшеницу, ячмень, рожь, овес и их комбинации) и/или крупнозерновые, такие как маис или сорго; b) отходы от переработки зерновых культур, такие как кукурузная глютеновая мука, сушеная барда с растворимыми веществами (DDGS) (в частности сушеная барда с растворимыми веществами на основе кукурузы (cDDGS)), пшеничные отруби, пшеничная крупка, пшеничные мелкие отруби, рисовые отруби, рисовая шелуха, овсяная шелуха, ядро кокосового ореха и цитрусовый жом; c) белок, полученный из источников, таких как соя, подсолнечник, арахис, люпин, виды гороха, конские бобы, хлопчатник, канола, рыбная мука, сухой белок плазмы крови, мясо-костная мука, картофельный белок, сыворотка, копра, кунжут; d) масла и жиры, полученные из растительных и животных источников и/или e) минералы и витамины.
[00163] В случае применения в качестве корма, такого как функциональный корм, или при его получении, композицию фермента или кормовой добавки настоящего изобретения можно применять в сочетании с одним или несколькими из приемлемого в питательном отношении носителя, приемлемого в питательном отношении разбавителя, приемлемого в питательном отношении наполнителя, приемлемого в питательном отношении вспомогательного вещества, активного в питательном отношении ингредиента. Например, можно упомянуть по меньшей мере один компонент, выбранный из группы, состоящей из белка, пептида, сахарозы, лактозы, сорбита, глицерина, пропиленгликоля, хлорида натрия, сульфата натрия, ацетата натрия, цитрата натрия, формиата натрия, сорбата натрия, хлорида калия, сульфата калия, ацетата калия, цитрата калия, формиата калия, ацетата калия, сорбата калия, хлорида магния, сульфата магния, ацетата магния, цитрата магния, формиата магния, сорбата магния, метабисульфита натрия, метилпарабена и пропилпарабена.
[00164] В предпочтительном варианте осуществления к композиции на основе фермента или к композиции кормовой добавки по настоящему изобретению примешивают кормовой компонент с образованием кормового продукта. Используемый в данном документе термин «кормовой компонент» означает весь кормовой продукт или его часть. Часть кормового продукта может означать один составляющий компонент кормового продукта или более одного составляющего компонента кормового продукта, например, 2, или 3, или 4, или более. В одном варианте осуществления термин «кормовой компонент» охватывает премикс или составляющие компоненты премикса. Предпочтительно корм может представлять собой фураж или его премикс, комбикорм или его премикс.
[00165] К композиции кормовой добавки в соответствии с настоящим изобретением может быть примешан комбикорм, компонент комбикорма или премикс комбикорма или фураж, компонент фуража или премикс фуража.
[00166] Любой кормовой продукт, описанный в данном документе, может включать один или несколько кормовых материалов, выбранных из группы, содержащей a) злаки, такие как мелкие зерна (например, пшеницу, ячмень, рожь, овес, тритикале и их комбинации) и/или крупные зерна, такие как маис или сорго; b) отходы от переработки зерновых, такие как кукурузная глютеновая мука, влажная барда (в частности кукурузная влажная барда), сушеная барда (DDG) (в частности кукурузная сушеная барда (cDDG)), сушеная барда с растворимыми веществами (DDGS) (в частности кукурузная сушеная барда с растворимыми веществами (cDDGS)), пшеничные отруби, пшеничная крупка, пшеничные мелкие отруби, рисовые отруби, рисовая шелуха, овсяная шелуха, ядро кокосового ореха и цитрусовый жом; c) белок, полученный из таких источников, как соя, подсолнечник, арахис, люпин, виды гороха, конские бобы, хлопчатник, канола, рыбная мука, сухой белок плазмы, мясо-костная мука, картофельный белок, сыворотка, копра, кунжут; d) масла и жиры, полученные из растительных и животных источников; e) минералы и витамины.
[00167] Термин «комбикорм» означает промышленный корм в форме муки, гранулы, орешков, жмыха или крошки. Комбикорма можно смешивать с использованием различного сырья и добавок. Эти смеси составляют в соответствии с конкретными требованиями для целевого животного.
[00168] Комбикорма могут быть полными кормами, которые обеспечивают все суточно необходимые питательные вещества, концентратами, которые обеспечивают часть рациона (белковую, энергетическую), или добавками, которые обеспечивают только дополнительные питательные микроэлементы, такие как минералы и витамины.
[00169] Основными ингредиентами, применяемыми в комбикорме, являются кормовые зерновые, которые включают кукурузу, пшеницу, каноловый шрот, рапсовый шрот, люпин, соевые бобы, сорго, овес и ячмень.
[00170] Предпочтительно премикс, как обозначается в данном документе, может представлять собой композицию, состоящую из микроингредиентов, таких как витамины, минералы, химические консерванты, антибиотики, продукты ферментации и другие необходимые ингредиенты. Премиксы обычно представляют собой композиции, подходящие для смешивания c промышленными рационами.
[00171] В одном варианте осуществления кормовой продукт содержит или состоит из кукурузы, DDGS (такого как cDDGS), пшеницы, пшеничных отрубей или любой их комбинации.
[00172] В одном варианте осуществления кормовой компонент может представлять собой кукурузу, DDGS (например, cDDGS), пшеницу, пшеничные отруби или их комбинацию. В одном варианте осуществления кормовой продукт содержит кукурузу, DDGS (такую как cDDGS) или их комбинацию, или состоит из них.
[00173] Кормовой продукт, описанный в данном документе, может содержать по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50% или по меньшей мере 60% по весу кукурузной и соевой муки или кукурузы и необезжиренной сои, или пшеничной муки, или кормовой муки из жмыха семян подсолнечника.
[00174] Например, кормовой продукт может содержать от примерно 5 до примерно 40% кукурузного DDGS. Для домашней птицы кормовой продукт может содержать в среднем от примерно 7 до примерно 15% кукурузного DDGS. Для домашних свиней (свиней) кормовой продукт может содержать в среднем 5-40% кукурузного DDGS. Он также может содержать кукурузу в виде цельного зерна, и в этом случае кормовой продукт может содержать от примерно 35% до примерно 80% кукурузы.
[00175] Корм может представлять собой одно или несколько из следующего: комбикорм и премикс, в том числе гранулы, орешки или жмых (для крупного рогатого скота); сельскохозяйственную культуру или остатки сельскохозяйственных культур: кукурузу, соевые бобы, сорго, овес, ячмень, копру, солому, мякину, отходы сахарной свеклы; рыбную муку; мясо-костную муку; мелассы; масличный жмых и прессованный жмых; олигосахариды; консервированные кормовые растения: силос; водоросли; семена и зерна, цельные или подготовленные дроблением, измельчением и т.д.; проросшие зерновые и бобовые; экстракт дрожжей.
[00176] Используемый в данном документе термин «корм» охватывает в некоторых вариантах осуществления корм для домашних животных. Корм для домашних животных представляет собой материал растительного или животного происхождения, предназначенный для потребления домашними животными, такой как корм для собак или корм для кошек. Корм для домашних животных, такой как корм для собак и кошек, может быть либо в сухой форме, такой как гранулированный корм для собак, либо во влажной консервированной форме. Корм для кошек может содержать аминокислоту таурин.
[00177] Используемый здесь термин «приведенный в контакт» относится к косвенному или прямому действию глюкоамилазы, как описано в данном документе (или композиции, содержащей глюкоамилазу), на продукт (например, корм). Примеры способов применения, которые можно использовать, включают без ограничения обработку продукта в материале, содержащем композицию кормовой добавки, непосредственное применение путем смешивания композиции кормовой добавки с продуктом, нанесение распылением композиции кормовой добавки на поверхность продукта или погружение продукта в препарат на основе композиции кормовой добавки. В одном варианте осуществления композицию кормовой добавки настоящего изобретения предпочтительно подмешивают к продукту (например, кормовому продукту). В качестве альтернативы композицию на основе кормовой добавки можно включать в эмульсию или сырьевые ингредиенты кормового продукта. Благодаря этому композиция обеспечивает благоприятные воздействия на продуктивность.
[00178] Также возможно, что по меньшей мере одну глюкоамилазу (или композицию фермента, содержащую по меньшей мере одну глюкоамилазу, как описано здесь), описанную в данном документе, можно гомогенизировать, чтобы получать порошок.
[00179] В альтернативном предпочтительном варианте осуществления композицию фермента, содержащую по меньшей мере одну глюкоамилазу, можно получать в виде гранул, как описано в WO2007/044968 (называемых гранулами TPT) или W01997/016076 и W01992/012645, включенных в настоящий документ ссылкой. «TPT» означает технологию термозащиты.
[00180] В другом аспекте, когда композицию кормовой добавки составляют в виде гранул, эти гранулы содержат гидратированную соль, являющуюся барьером, которая покрывает белковое ядро. Преимуществом такого солевого покрытия является улучшенная термоустойчивость, улучшенная стабильность при хранении и защита от других кормовых добавок, неблагоприятно воздействующих каким-либо образом на фермент. Предпочтительно соль, используемая для солевой оболочки, характеризуется активностью воды выше 0,25 или постоянной влажностью выше 60% при 20°C. В некоторых вариантах осуществления солевая оболочка содержит Na2SO4.
[00181] Способ получения по меньшей мере одной глюкоамилазы, как описано здесь (или композиции фермента, содержащей по меньшей мере одну глюкоамилазу, как описано здесь), также может включать дополнительную стадию гранулирования порошка. Порошок можно смешивать с другими компонентами, известными из уровня техники. Порошок или смесь, содержащую порошок, можно продавливать через матрицу, а полученные нити разрезать на подходящие пеллеты различной длины.
[00182] Необязательно стадия пеллетирования может включать обработку паром или стадию кондиционирования перед формированием пеллет. Смесь, содержащую порошок, можно поместить в кондиционирующее устройство, например смеситель с впрыском пара. Смесь нагревают в кондиционирующем устройстве до определенной температуры, например 60-100°C, типичные температуры будут составлять 70°C, 80°C, 85°C, 90°C или 95°C. Время пребывания может варьироваться от секунд до минут и даже часов. Например, 5 секунд, 10 секунд, 15 секунд, 30 секунд, 1 минута, 2 минуты, 5 минут, 10 минут, 15 минут, 30 минут и 1 час. Будет понятно, что гликозидгидролаза, описанная в данном документе (или композиция на основе ферментов, содержащая гликозидгидролазу, описанную в данном документе), подходит для добавления в любой подходящий кормовой материал.
Необязательно кормовой продукт может также содержать дополнительные минералы, такие как, например, кальций, и/или дополнительные витамины. В некоторых вариантах осуществления кормовой продукт представляет собой смесь кукурузной и соевой муки.
[00183] Кормовой продукт обычно получают в кормодробилках, в которых сырьевые материалы сначала измельчают до подходящего размера частиц, а затем смешивают с соответствующими добавками. Затем кормовой продукт может быть получен в виде кашицы или пеллет; причем в отношении последних, как правило, предусматривается способ, при котором температуру повышают до заданного уровня, а затем корм пропускают через матрицу с получением пеллет конкретного размера. Пеллетам дают остыть. Впоследствии можно добавлять жидкие добавки, такие как жир и фермент. Получение кормового продукта также может включать дополнительную стадию, которая включает экструзию или разбухание перед осуществлением пеллетирования, в частности с помощью подходящих методик, которые могут включать по меньшей мере применение пара.
[00184] Композицию кормовой добавки и/или кормовой продукт, содержащий ее, можно применять в любой подходящей форме. Композицию кормовой добавки можно применять в форме твердых или жидких препаратов или их альтернативных вариантов. Примеры твердых препаратов включают порошки, пасты, болюсы, капсулы, пеллеты, таблетки, присыпки и гранулы, которые могут быть смачиваемыми, высушенными распылительной сушкой или лиофилизированными. Примеры жидких препаратов включают без ограничения водные, органические или водно-органические растворы, суспензии и эмульсии.
[00185] Предпочтительно композиция пищевой или кормовой добавки может содержать по меньшей мере один физиологически приемлемый носитель. Физиологически приемлемый носитель предпочтительно выбран из по меньшей мере одного из мальтодекстрина, известняка (карбоната кальция), циклодекстрина, пшеницы или компонента пшеницы, сахарозы, крахмала, Na2SO4, талька, PVA и их смесей. В дополнительном варианте осуществления пищевая или кормовая добавка может дополнительно содержать хелатор ионов металлов. Хелатор ионов металлов может быть выбран из EDTA или лимонной кислоты.
[00186] В некоторых вариантах осуществления содержание гликозидгидролазы, описанной в данном документе, в композиции пищевой или кормовой добавки составляет не менее 0,0001 г/кг, 0,001 г/кг, не менее 0,01 г/кг, не менее 0,1 г/кг, не менее 1 г/кг, не менее 5 г/кг, не менее 7,5 г/кг, не менее 10,0 г/кг, не менее 15,0 г/кг, не менее 20,0 г/кг, не менее 25,0 г/кг. В некоторых вариантах осуществления содержание пищевой или кормовой добавки таково, что при добавлении к пищевому или кормовому материалу содержание гликозидгидролазы, описанной в данном документе, в этом кормовом материале составляет 1-500 мг/кг, 1-100 мг/кг, 2-50 мг/кг или 2-10 мг/кг. В некоторых вариантах осуществления настоящего изобретения пищевой или кормовой материал содержит по меньшей мере 100, 1000, 2000, 3000, 4000, 5000, 10000, 20000, 30000, 50000, 100000, 500000, 1000000 или 2000000 единиц гликозидгидролазы на килограмм кормового или пищевого материала.
[00187] Составы, содержащие любую из гликозидгидролаз, описанных в данном документе, и композиции, описанные в данном документе, можно получать любым подходящим способом, который обеспечивает внесение активных ферментов в состав. Такие составы могут представлять собой жидкость, сухой порошок или гранулу. Предпочтительно композиция кормовой добавки находится в твердой форме, подходящей для добавления на кормовую гранулу или в нее.
[00188] В одном варианте осуществления корм для животных может быть составлен в виде гранул для кормовых композиций, содержащих: ядро; активное средство и по меньшей мере одну оболочку, при этом активное средство гранулы сохраняет не менее 50% активности, не менее 60% активности, не менее 70% активности, не менее 80% активности после воздействия условий, выбранных из одного или нескольких из следующего: а) способа гранулирования корма, b) способа предварительной обработки корма с паровым нагревом, c) хранения, d) хранения в виде ингредиента в негранулированной смеси и e) хранения в виде ингредиента в исходной кормовой смеси или кормовом премиксе, содержащем по меньшей мере одно соединение, выбранное из микроэлементов, органических кислот, восстанавливающих сахаров, витаминов, хлорида холина и соединений, которые придают кислотность или основность исходной кормовой смеси или кормовому премиксу.
[00189] Что касается гранулы, то по меньшей мере одна оболочка может содержать увлажняющий гидратирующий материал, который составляет не менее 55% вес/вес гранулы и/или по меньшей мере одна оболочка может содержать две оболочки. Эти две оболочки могут представлять собой увлажняющую гидратирующую оболочку и влагоизолирующую оболочку. В некоторых вариантах осуществления увлажняющая гидратирующая оболочка может составлять от 25% до 60% вес/вес гранулы, а влагоизолирующая оболочка может составлять от 2% до 15% вес/вес гранулы. Увлажняющую гидратирующую оболочку можно выбирать из неорганических солей, сахарозы, крахмала и мальтодекстрина, а влагоизолирующую оболочку можно выбирать из полимеров, смол, молочной сыворотки и крахмала.
[00190] Гранулу можно получать с помощью способа пеллетирования корма, и при этом способ предварительной обработки корма можно проводить при температуре от 70°C до 95°C в течение нескольких минут, например при температуре от 85°C до 95°C.
[00191] Гранула может иметь влагоизолирующую оболочку, выбранную из полимеров и смол, а увлажняющий гидратирующий материал может представлять собой неорганическую соль. Увлажняющая гидратирующая оболочка может составлять от 25% до 45% вес/вес гранулы, а влагоизолирующая оболочка может составлять от 2% до 10% вес/вес гранулы.
[00192] В качестве альтернативы композиция может быть в виде жидкого состава, подходящего для потребления, при этом предпочтительно такая потребляемая жидкость содержит одно или несколько из следующего: буфер, соль, сорбит и/или глицерин.
[00193] Кроме того, композиция кормовой добавки может быть составлена путем нанесения, например распыления, фермента(ов) на подложку-носитель, такую как, например, молотая пшеница.
[00194] В одном варианте осуществления композиция кормовой добавки может быть составлена в виде премикса. Лишь в качестве примера премикс может содержать один или несколько кормовых компонентов, таких как один или несколько минералов и/или один или несколько витаминов.
[00195] В некоторых вариантах осуществления по меньшей мере одна глюкоамилаза будет находиться в физиологически приемлемом носителе. Подходящими носителями могут быть крупные медленно метаболизируемые макромолекулы, такие как белки, полипептиды, липосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы. Можно применять фармацевтически приемлемые соли, например соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, или соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты. После составления композиции настоящего изобретения можно непосредственно вводить жвачному животному.
[00196] Кроме того, раскрываемые в настоящем документе содержащие глюкоамилазу композиции можно дополнительно составлять с (например, смешивать) с одним или несколькими дополнительными компонентами-ферментами (например, с дополнительными кормовыми ферментами или ферментами пивоварения или соложения, или ферментами обработки зерна или ферментами разделения глютена и крахмала пшеницы).
[00197] Подходящие дополнительные ферменты для использования в композициях, содержащих глюкоамилазу, описанных в данном документе, могут представлять собой один или несколько ферментов, выбранных из группы, состоящей из эндоглюканаз (E.C. 3.2.1.4); целлиобиогидролаз (E.C. 3.2.1.91), β-глюкозидаз (E.C. 3.2.1.21), целлюлаз (E.C. 3.2.1.74), лихеназ (E.C. 3.1.1.73), липаз (E.C. 3.1.1.3), липид-ацилтрансфераз (обычно классифицируемых как E.C. 2.3.1.x), фосфолипаз (E.C. 3.1.1.4, E.C. 3.1.1.32 или E.C. 3.1.1.5), фитаз (например, 6-фитаза (E.C. 3.1.3.26) или 3-фитаза (E.C. 3.1.3.8), альфа-амилаз (E.С. 3.2.1.1), другие ксиланаз (E.C. 3.2.1.8, E.C. 3.2.1.32, E.C. 3.2.1.37, E.C. 3.1.1.72, E.C. 3.1.1.73), глюкоамилаз (E.С. 3.2.1.3), протеаз (например, субтилизин (E.C. 3.4.21.62), или бацилолизин (E.C. 3.4.24.28), или щелочная сериновая протеаза (E.C. 3.4.21.x), или кератиназа (E.C. 3.4.xx)) и/или маннаназ (например, β-маннаназа (E.C. 3.2.1.78)).
[00198] В одном варианте осуществления (например, в применениях корма) другой компонент фермента может представлять собой один или несколько ферментов, выбранных из группы, состоящей из амилазы (включая α-амилазы (E.C. 3.2.1.1), G4-образующие амилазы (E.C. 3.2.1.60), β-амилазы (E.C. 3.2.1.2) и γ-амилазы (E.C. 3.2.1.3); и/или протеазы (например, субтилизин (E.C. 3.4.21.62), или бацилолизин (E.C. 3.4.24.28), или щелочная сериновая протеаза (E.C. 3.4.21.x), или кератиназа (E.C. 3.4.x.x)).
[00199] В одном варианте осуществления (например, применения в кормах) другой компонент фермента может представлять собой комбинацию амилазы (например, α-амилазы (E.C. 3.2.1.1)) и протеазы (например, субтилизина (E.C. 3.4.21.62)).
[00200] В одном варианте осуществления (например, применения в кормах) другим компонентом фермента может быть β-глюканаза, например эндо-1,3(4)-β-глюканаза (E.C. 3.2.1.6).
[00201] В одном варианте осуществления (например, применения в кормах) другим компонентом фермента может быть маннаназа (например, β-маннаназа (E.C. 3.2.1.78)).
[00202] В одном варианте осуществления (например, применения в кормах) другим компонентом фермента может быть липаза (E.C. 3.1.1.3), липид-ацилтрансфераза (обычно классифицируемая как E.C. 2.3.1.x) или фосфолипаза (E.C. 3.1.1.4, E.C. 3.1.1.32 или E.C. 3.1.1.5), предпочтительно липаза (E.C. 3.1.1.3).
[00203] В одном варианте осуществления (особенно применения в кормах) другим компонентом фермента может быть сериновая протеаза (например, E.C. 3.4.21, субтилизин), трипсиноподобные протеазы S1 или S2 или металлопротеаза (например, E.C. 3.4.24, бациллолизин) или аспартил протеаза (например, E.C. 3.4.23).
ПРИМЕРЫ
[00204] Если в данном документе не определено иное, все технические и научные термины, используемые в данном документе, имеют то же самое значение, которое обычно понимает специалист в данной области, к которой принадлежит настоящее изобретение. Singleton, et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY, 2D ED., John Wiley and Sons, New York (1994) и Hale & Marham, THE HARPER COLLINS DICTIONARY OF BIOLOGY, Harper Perennial, N.Y. (1991) предоставляют для специалиста общий словарь многих терминов, используемых в данном раскрытии.
[00205] Настоящее изобретение дополнительно определено в следующих примерах. Следует понимать, что примеры, хотя и показывают определенные варианты осуществления, приводятся исключительно в целях иллюстрации. Из приведенного выше обсуждения и примеров специалист в данной области техники сможет установить существенные характеристики настоящего изобретения и, не отклоняясь от его идеи и объема, сможет осуществить различные изменения и модификации для его адаптации к различным вариантам применения и условиям.
ПРИМЕР 1
Последовательности глюкоамилаз
[00206] Последовательность нуклеиновой кислоты для гена FraGA1 и аминокислотная последовательность гипотетического белка, кодируемого геном FraGA1, были найдены в базе данных NCBI (регистрационный номер NCBI: NW_012133200.1 (ген) и XP_012178139 (белок)).
[00207] Аминокислотная последовательность белка-предшественника FraGA1 представлена как SEQ ID NO: 1.
MLFLLAALGLACSAAAQSTSVSAYIASESPVAKAGVLANIGTEGSLSSGAYSGVVIASPSTVNPDYLYTWVRDSSLTFQALIDQYVYGEDPTLRSLIDEFITAESILQQTTNPSGTVSTGGLGEPKFNINETAFTGPWGRPQRDGPALRSTAIITYATYLWNSGNTSYVSDSLWPIIELDLNYIATYWNFSTFDLWEEIDSSSFWTTAVQHRALRQGITFANLIGQTSPVSNYETQAGDILCFLQTYWNPTGNYMTANTGGGRSGKDSNTVLASVHTFDPDAGCDSTTFQPCSDKALSNLKVYVDSFRSLYAINDGIASDAAVATGRYPEDVYYGGNPWYLCTFAVAEQLYDALIVWSSQGYLEITDLSLAFFQQFDSDVGTGTYDSGSSTYSTLTSAIRTFADGFVLTNAKYTPTNGSLSEEYTSADGTPISAYDLTWSYASALTVFAAEAGTTYGSWGAAGLTVPSTCTSGVAVTFEVDYDTEYGENVYITGSVNALENWSATNALIMSAADYPTWSITVYLPPSTTIQYKYLTQYNGEVTWEDDPNNEITTPASGSMTQVDSWH
[00208] Последовательность нуклеиновой кислоты для гена WcoGA1 и аминокислотная последовательность гипотетического белка, кодируемого геном WcoGA1, были найдены в базе данных JGI (регистрационный номер JGI: Wolco1150090 (ген) и Wolco1149945 (белок)). N-концевой сигнальный пептид был смоделирован с помощью программного обеспечения SignalP версия 4.0 (Nordahl Petersen et al. (2011) Nature Methods, 8:785-786).
[00209] Аминокислотная последовательность белка-предшественника WcoGA1 представлена как SEQ ID NO: 2. MRLSLASVFALAGGALAQTTSVTSYIASESPIAKAGVLANIGADGSLSSGAYSGIVIASPSTVNPNYLYTWTRDSSLTFMELINQYIYGEDDTLRTLIDEFVSAEATLQQVTNPSGTVSTGGLGEPKFNINETAFTGPWGRPQRDGPALRATAIMAYATYLYENGNTSYVTDTLWPIIELDLGYVAEYWNESTFDLWEEIDSSSFFTTAVQHRALRAGVTFANLIGETSDVSNYQENADDLLCFLQSYWNPTGSYVTANTGGGRSGKDANTLLASIHTFDPDAGCNATTFQPCSDKALSNHKVYVDSFRSLYAINDDISSDAAVATGRYPEDVYYNGNPWYLCTLAAAEQLYDSLIVWKAQGYIEVTSLSLAFFQQFDASVSAGTYDSSSDTYTTLLDAVQTYADGFVLMVAQYTPANGSLSEQYAKADGSPTSAYDLTWSFAAALTAFAARDGKTYGSWGAADLSSTCSGSTDTVAVTFEVQYDTQYGENLYITGSVSQLEDWSADDALIMSSADYPTWSITVDLPPSTLIQYKYLTKYNGDVTWEDDPNNEITTPASGSYTQVDSWH
[00210] Аминокислотная последовательность белка-предшественника TeGA из Talaromyces emersonii представлена как SEQ ID NO: 3.
MASLVAGALCILGLTPAAFARAPVAARATGSLDSFLATETPIALQGVLNNIGPNGADVAGASAGIVVASPSRSDPNYFYSWTRDAALTAKYLVDAFNRGNKDLEQTIQQYISAQAKVQTISNPSGDLSTGGLGEPKFNVNETAFTGPWGRPQRDGPALRATALIAYANYLIDNGEASTADEIIWPIVQNDLSYITQYWNSSTFDLWEEVEGSSFFTTAVQHRALVEGNALATRLNHTCSNCVSQAPQVLCFLQSYWTGSYVLANFGGSGRSGKDVNSILGSIHTFDPAGGCDDSTFQPCSARALANHKVVTDSFRSIYAINSGIAEGSAVAVGRYPEDVYQGGNPWYLATAAAAEQLYDAIYQWKKIGSISITDVSLPFFQDIYPSAAVGTYNSGSTTFNDIISAVQTYGDGYLSIVEKYTPSDGSLTEQFSRTDGTPLSASALTWSYASLLTASARRQSVVPASWGESSASSVLAVCSATSATGPYSTATNTVWPSSGSGSSTTTSSAPCTTPTSVAVTFDEIVSTSYGETIYLAGSIPELGNWSTASAIPLRADAYTNSNPLWYVTVNLPPGTSFEYKFFKNQTDGTIVWEDDPNRSYTVPAYCGQTTAILDDSWQ
ПРИМЕР 2
Экспрессия глюкоамилаз
[00211] Последовательности ДНК, кодирующие полноразмерные FraGA1 и WcoGA1, были синтезированы и вставлены в вектор экспрессии pTTT (описано в опубликованной заявке PCT WO2011/063308). Последовательность ДНК, кодирующую TeGA, синтезировали и вставляли в вектор экспрессии pTrex3gM (описано в опубликованной заявке США 2011/0136197 A1).
[00212] Нуклеотидная последовательность гена FraGA1, используемого для экспрессии, представлена ниже как SEQ ID NO: 4.
ATGCTCTTCCTCCTCGCTGCTCTGGGCCTCGCTTGTAGCGCTGCTGCCCAATCCACTTCCGTCTCCGCTTACATCGCCAGCGAGAGCCCCGTTGCCAAAGCCGGTGTTCTGGCTAACATTGGCACTGAAGGTAGCCTGAGCTCCGGTGCCTACTCCGGCGTTGTCATCGCCTCCCCCAGCACCGTCAACCCTGACTACCTCTATACTTGGGTTCGCGACTCCAGCCTCACTTTCCAAGCCCTGATTGACCAGTACGTTTACGGCGAGGACCCCACCCTCCGAAGCCTGATCGACGAGTTCATTACCGCTGAGTCCATCCTGCAGCAAACTACCAACCCCAGCGGCACCGTTAGCACCGGCGGTCTGGGCGAGCCCAAGTTCAACATCAATGAGACTGCCTTTACCGGTCCCTGGGGCCGACCCCAACGCGACGGTCCTGCCCTCCGCAGCACTGCCATCATTACTTATGCCACCTACCTGTGGAACTCCGGTAACACCTCCTACGTTTCCGATTCCCTCTGGCCCATCATCGAACTCGACCTGAATTACATTGCTACCTACTGGAATTTCTCCACTTTTGATCTGTGGGAAGAGATTGACTCCTCCAGCTTCTGGACCACTGCCGTTCAGCATCGAGCCCTGCGCCAGGGTATCACCTTCGCTAATCTGATTGGCCAGACCAGCCCTGTCAGCAACTATGAGACCCAAGCCGGCGATATCCTCTGTTTCCTCCAAACCTATTGGAATCCTACCGGCAACTACATGACCGCCAATACTGGCGGTGGTCGAAGCGGCAAGGACTCCAACACCGTTCTCGCTTCCGTTCATACCTTCGATCCCGATGCCGGCTGTGATAGCACTACTTTTCAACCTTGCTCCGACAAGGCCCTGAGCAACCTCAAGGTCTACGTCGACTCCTTTCGCAGCCTGTACGCCATCAACGACGGTATTGCCTCCGACGCTGCCGTCGCCACCGGCCGCTATCCTGAGGACGTCTACTACGGCGGTAACCCCTGGTACCTCTGCACCTTTGCTGTCGCCGAACAACTCTACGACGCCCTCATCGTCTGGAGCAGCCAGGGCTATCTCGAAATCACTGACCTCAGCCTGGCCTTCTTCCAGCAGTTTGATTCCGATGTCGGTACTGGCACCTACGACAGCGGCTCCAGCACTTACTCCACTCTCACCTCCGCCATCCGAACTTTTGCTGATGGCTTCGTTCTGACCAACGCCAAATACACCCCTACCAATGGTTCCCTGTCCGAGGAGTACACCAGCGCCGATGGCACTCCTATCTCCGCCTATGACCTGACCTGGAGCTACGCCTCCGCTCTGACCGTCTTTGCCGCCGAGGCCGGCACCACTTACGGCTCCTGGGGTGCTGCTGGCCTGACTGTCCCTAGCACCTGCACTAGCGGCGTCGCTGTTACTTTCGAGGTCGATTACGACACCGAGTATGGCGAAAACGTCTATATCACCGGTTCCGTCAATGCCCTGGAAAATTGGTCCGCCACTAATGCTCTGATTATGTCCGCCGCTGACTATCCCACCTGGTCCATCACCGTTTACCTGCCCCCCTCCACCACCATTCAGTATAAGTATCTCACCCAGTACAACGGCGAAGTCACTTGGGAGGACGACCCTAACAACGAGATTACTACCCCTGCTAGCGGTTCCATGACCCAGGTTGACAGCTGGCACtaa
[00213] Нуклеотидная последовательность гена WcoGA1, используемого для экспрессии, представлена ниже как SEQ ID NO: 5.
ATGCGACTGAGCCTGGCCTCCGTTTTTGCTCTCGCCGGTGGTGCCCTCGCCCAGACCACTAGCGTCACCTCCTACATTGCTAGCGAAAGCCCCATTGCCAAAGCCGGTGTTCTCGCTAACATTGGCGCTGACGGCTCCCTGAGCTCCGGTGCTTATTCCGGCATTGTTATCGCCAGCCCCTCCACCGTTAACCCTAACTATCTCTATACCTGGACTCGCGACTCCAGCCTGACCTTCATGGAACTGATCAACCAGTACATCTACGGCGAGGACGATACTCTGCGAACTCTGATTGATGAGTTCGTTTCCGCTGAAGCCACCCTCCAACAGGTCACTAATCCTAGCGGCACTGTCTCCACTGGTGGCCTCGGCGAGCCCAAGTTCAACATCAACGAGACTGCTTTTACTGGTCCCTGGGGCCGACCCCAACGCGATGGCCCTGCCCTGCGCGCTACTGCTATCATGGCCTATGCCACCTACCTGTATGAAAACGGTAATACTAGCTATGTTACTGACACCCTCTGGCCCATCATTGAACTCGACCTCGGTTACGTCGCCGAATATTGGAACGAAAGCACCTTTGATCTCTGGGAGGAAATCGACAGCAGCTCCTTTTTCACTACCGCTGTTCAGCACCGCGCTCTCCGCGCTGGCGTTACCTTCGCCAATCTCATCGGTGAGACCAGCGACGTCAGCAACTACCAGGAAAATGCCGACGACCTCCTCTGCTTCCTCCAAAGCTACTGGAACCCCACTGGCAGCTACGTCACTGCTAACACTGGCGGTGGTCGAAGCGGCAAGGACGCCAACACTCTCCTGGCTAGCATCCACACCTTCGATCCCGACGCTGGCTGCAACGCCACTACCTTTCAACCCTGTTCCGACAAAGCCCTCAGCAATCACAAGGTCTACGTTGACTCCTTCCGCAGCCTCTACGCCATCAATGACGACATTAGCAGCGATGCCGCTGTCGCTACCGGCCGATACCCTGAGGATGTCTACTACAACGGCAACCCCTGGTACCTCTGTACCCTGGCCGCTGCTGAGCAACTCTACGACTCCCTCATCGTCTGGAAGGCCCAAGGCTACATCGAAGTCACCAGCCTCAGCCTCGCCTTTTTTCAACAGTTCGATGCTTCCGTTAGCGCCGGTACTTATGATTCCAGCTCCGACACCTACACCACCCTGCTCGACGCCGTTCAGACCTATGCTGATGGCTTCGTCCTGATGGTCGCTCAGTACACCCCTGCCAACGGTTCCCTCTCCGAGCAGTACGCCAAGGCCGATGGCAGCCCCACTTCCGCCTACGACCTGACTTGGTCCTTTGCTGCTGCCCTCACCGCCTTCGCTGCCCGCGACGGCAAAACCTATGGTAGCTGGGGTGCCGCCGATCTCTCCAGCACCTGCAGCGGTTCCACCGACACTGTCGCCGTCACTTTCGAGGTCCAGTACGACACCCAATATGGTGAAAATCTGTACATTACCGGCAGCGTCTCCCAGCTCGAGGATTGGAGCGCTGATGATGCTCTCATCATGTCCAGCGCCGACTATCCCACCTGGTCCATCACCGTCGATCTGCCCCCTAGCACCCTGATCCAATACAAATACCTCACCAAGTATAACGGCGATGTCACCTGGGAAGACGATCCCAACAACGAAATTACCACTCCTGCCTCCGGCTCCTATACCCAGGTTGACAGCTGGCACtaa
[00214] Нуклеотидная последовательность гена FraGA1, используемого для экспрессии, представлена ниже как SEQ ID NO: 6.
ATGGCCTCCCTGGTTGCTGGTGCTCTGTGCATCCTCGGCCTGACCCCTGCCGCCTTCGCCCGAGCCCCCGTCGCTGCCCGCGCCACTGGCAGCCTCGACAGCTTCCTCGCCACCGAGACCCCTATCGCCCTCCAGGGCGTTCTGAACAACATCGGTCCCAACGGCGCTGACGTCGCCGGTGCTAGCGCCGGTATCGTCGTTGCCAGCCCTAGCCGATCCGACCCCAACTACTTCTACAGCTGGACCCGCGACGCCGCTCTCACCGCTAAGTACCTGGTCGACGCCTTCAATCGCGGCAACAAAGACCTCGAGCAAACCATCCAGCAGTACATCTCCGCTCAGGCCAAGGTCCAGACCATTTCCAACCCCAGCGGCGATCTGAGCACTGGCGGCCTGGGCGAGCCCAAGTTCAACGTCAATGAGACCGCTTTCACTGGCCCCTGGGGCCGACCTCAACGCGATGGCCCTGCTCTCCGAGCCACCGCCCTCATCGCCTATGCTAACTACCTGATCGACAACGGTGAGGCCAGCACTGCCGACGAGATCATCTGGCCCATCGTCCAAAATGACCTCAGCTACATCACCCAATACTGGAACTCCAGCACCTTTGACCTGTGGGAGGAGGTCGAGGGCTCCAGCTTCTTCACCACTGCTGTTCAGCACCGCGCCCTCGTTGAGGGTAATGCCCTGGCCACCCGACTCAATCACACTTGCTCCAACTGCGTCAGCCAGGCCCCCCAGGTCCTCTGCTTTCTCCAGAGCTACTGGACCGGCAGCTACGTCCTGGCCAACTTTGGTGGCAGCGGCCGAAGCGGCAAGGACGTCAACAGCATCCTGGGTTCCATCCACACCTTCGACCCCGCTGGCGGTTGCGACGACTCCACTTTCCAGCCTTGCAGCGCTCGCGCTCTCGCCAACCACAAGGTCGTCACCGATTCCTTCCGCTCCATCTACGCCATCAATTCCGGCATCGCCGAGGGTAGCGCTGTTGCTGTCGGCCGCTACCCCGAGGACGTCTACCAAGGCGGCAATCCCTGGTATCTCGCTACTGCCGCTGCCGCCGAGCAGCTCTATGACGCTATCTATCAGTGGAAAAAGATCGGTAGCATCAGCATTACCGACGTCAGCCTCCCCTTCTTCCAGGACATCTACCCCTCCGCCGCTGTTGGCACCTACAATTCCGGCTCCACCACCTTCAACGACATCATCAGCGCCGTCCAGACTTATGGCGACGGCTACCTGAGCATTGTCGAGAAGTACACCCCCAGCGATGGCAGCCTCACCGAGCAATTCAGCCGCACCGACGGCACCCCCCTGTCCGCTTCCGCCCTCACCTGGAGCTACGCTTCCCTGCTCACCGCCTCCGCTCGCCGCCAGAGCGTCGTTCCCGCTAGCTGGGGCGAGAGCAGCGCCAGCTCCGTCCTGGCCGTCTGCTCCGCTACTAGCGCCACCGGCCCCTACTCCACTGCCACCAACACCGTTTGGCCTTCCAGCGGCTCCGGCAGCTCCACTACCACCTCCAGCGCCCCTTGCACCACCCCTACCAGCGTCGCCGTCACCTTCGACGAGATCGTCAGCACCAGCTACGGCGAGACCATCTATCTGGCTGGCAGCATCCCCGAGCTGGGCAATTGGTCCACCGCCAGCGCTATTCCTCTGCGCGCTGACGCCTACACTAATAGCAACCCTCTGTGGTATGTCACCGTTAACCTCCCTCCCGGCACTAGCTTTGAGTATAAGTTTTTCAAGAACCAGACCGATGGTACTATTGTCTGGGAGGACGACCCCAACCGATCCTACACCGTCCCCGCCTACTGCGGTCAGACTACCGCTATCCTCGACGATTCCTGGCAGtaa
[00215] Плазмиды, кодирующие ферменты FraGA1, WcoGA1 и TeGA, трансформировали в подходящий штамм Trichoderma reesei с использованием трансформации протопластов (Te'o et al., J. Microbiol. Methods 51:393-99, 2002). Трансформанты были отобраны и ферментированы способами, описанными в WO 2016/138315. Надосадочные жидкости этих культур использовали для подтверждения экспрессии белка с помощью анализа SDS-PAGE и анализа активности фермента.
[00216] FraGA1 очищали с помощью аффинной хроматографии на связанной с бета-циклодекстрином сефарозе 6, а затем - гель-фильтрационной хроматографией. WcoGA1 очищали с помощью аффинной хроматографии на связанной с бета-циклодекстрином сефарозе 6, а затем - хроматографией гидрофобных взаимодействий. TeGA очищали хроматографией гидрофобных взаимодействий, а затем - аффинной хроматографией на связанной с бета-циклодекстрином сефарозе 6. Были выполнены анализ активности глюкоамилазы и SDS-PAGE. Фракции, содержащие целевой белок, объединяли и концентрировали, используя устройство Amicon Ultra-15 с 10 K MWCO. Затем концентрированный раствор наносили на колонку Superdex 75 (GE Healthcare) в буфере А. Целевой белок элюировался из колонки в виде единственного пика, который снова объединяли и концентрировали. Чистота очищенного образца была более 90%, и его хранили в 40%-ном глицерине при -80°C до использования.
[00217] Смоделированная последовательность зрелого белка FraGA1 приведена ниже как SEQ ID NO: 7.
QSTSVSAYIASESPVAKAGVLANIGTEGSLSSGAYSGVVIASPSTVNPDYLYTWVRDSSLTFQALIDQYVYGEDPTLRSLIDEFITAESILQQTTNPSGTVSTGGLGEPKFNINETAFTGPWGRPQRDGPALRSTAIITYATYLWNSGNTSYVSDSLWPIIELDLNYIATYWNFSTFDLWEEIDSSSFWTTAVQHRALRQGITFANLIGQTSPVSNYETQAGDILCFLQTYWNPTGNYMTANTGGGRSGKDSNTVLASVHTFDPDAGCDSTTFQPCSDKALSNLKVYVDSFRSLYAINDGIASDAAVATGRYPEDVYYGGNPWYLCTFAVAEQLYDALIVWSSQGYLEITDLSLAFFQQFDSDVGTGTYDSGSSTYSTLTSAIRTFADGFVLTNAKYTPTNGSLSEEYTSADGTPISAYDLTWSYASALTVFAAEAGTTYGSWGAAGLTVPSTCTSGVAVTFEVDYDTEYGENVYITGSVNALENWSATNALIMSAADYPTWSITVYLPPSTTIQYKYLTQYNGEVTWEDDPNNEITTPASGSMTQVDSWH
[00218] Смоделированная последовательность зрелого белка WcoGA1 приведена ниже как SEQ ID NO: 8.
QTTSVTSYIASESPIAKAGVLANIGADGSLSSGAYSGIVIASPSTVNPNYLYTWTRDSSLTFMELINQYIYGEDDTLRTLIDEFVSAEATLQQVTNPSGTVSTGGLGEPKFNINETAFTGPWGRPQRDGPALRATAIMAYATYLYENGNTSYVTDTLWPIIELDLGYVAEYWNESTFDLWEEIDSSSFFTTAVQHRALRAGVTFANLIGETSDVSNYQENADDLLCFLQSYWNPTGSYVTANTGGGRSGKDANTLLASIHTFDPDAGCNATTFQPCSDKALSNHKVYVDSFRSLYAINDDISSDAAVATGRYPEDVYYNGNPWYLCTLAAAEQLYDSLIVWKAQGYIEVTSLSLAFFQQFDASVSAGTYDSSSDTYTTLLDAVQTYADGFVLMVAQYTPANGSLSEQYAKADGSPTSAYDLTWSFAAALTAFAARDGKTYGSWGAADLSSTCSGSTDTVAVTFEVQYDTQYGENLYITGSVSQLEDWSADDALIMSSADYPTWSITVDLPPSTLIQYKYLTKYNGDVTWEDDPNNEITTPASGSYTQVDSWH
[00219] Последовательность зрелого белка FraGA1 приведена ниже как SEQ ID NO: 9.
ATGSLDSFLATETPIALQGVLNNIGPNGADVAGASAGIVVASPSRSDPNYFYSWTRDAALTAKYLVDAFNRGNKDLEQTIQQYISAQAKVQTISNPSGDLSTGGLGEPKFNVNETAFTGPWGRPQRDGPALRATALIAYANYLIDNGEASTADEIIWPIVQNDLSYITQYWNSSTFDLWEEVEGSSFFTTAVQHRALVEGNALATRLNHTCSNCVSQAPQVLCFLQSYWTGSYVLANFGGSGRSGKDVNSILGSIHTFDPAGGCDDSTFQPCSARALANHKVVTDSFRSIYAINSGIAEGSAVAVGRYPEDVYQGGNPWYLATAAAAEQLYDAIYQWKKIGSISITDVSLPFFQDIYPSAAVGTYNSGSTTFNDIISAVQTYGDGYLSIVEKYTPSDGSLTEQFSRTDGTPLSASALTWSYASLLTASARRQSVVPASWGESSASSVLAVCSATSATGPYSTATNTVWPSSGSGSSTTTSSAPCTTPTSVAVTFDEIVSTSYGETIYLAGSIPELGNWSTASAIPLRADAYTNSNPLWYVTVNLPPGTSFEYKFFKNQTDGTIVWEDDPNRSYTVPAYCGQTTAILDDSWQ
ПРИМЕР 3
Стабильность глюкоамилаз при низком pH и в присутствии пепсина
[00220] Стабильность очищенных образцов глюкоамилаз TeGA, FraGA1 и WcoGA1 анализировали в условиях низкого pH и в присутствии пепсина. Ферменты инкубировали с пепсином (Sigma, Кат. № P7000) в 50 мМ глицин-HCl буфере (pH 2,0) и в качестве субстрата для измерения активности использовали 1% (вес./вес.) растворимый крахмал, приготовленный в 50 мМ буфере MES-NaOH (pH 6,5). Глюкоамилазы и пепсин сначала смешивали в соотношениях (вес./вес.) 1:0 или 1:50, при этом глюкоамилазы вводили в размере 100 м.д.; и полученную смесь затем инкубировали при 40°C в течение 30 мин. Между тем, аликвоты 100 м.д. каждой глюкоамилазы инкубировали в 50 мМ буфере MES-NaOH (pH 6,5) при 40°C в течение 30 минут, которые использовали в качестве необработанного контроля. Чтобы начать измерение активности, 10 мкл разведения фермента (доза фермента, выбранная в линейном диапазоне, или только вода в качестве холостого контроля) вносили в 96-луночный микротитровальный планшет (MTP, Corning 3641), содержащий 90 мкл раствора субстрата (1% крахмала). Затем MTP герметично закрывали и инкубировали в течение 10 мин в iEMS (ThermoFisher) при 40°C и 1150 об/мин. Активность глюкоамилазы определяли как скорость высвобождения глюкозы, которую измеряли методом с использованием сопряженной системы глюкозооксидаза/пероксидаза (GOX/HRP) (Anal. Biochem. 105 (1980), 389-397). Как видно из таблицы 2, при использовании высвобождения глюкозы в качестве меры активности фермента, глюкоамилазы FraGa1, WcoGA1 и TeGA стабильны в буфере с pH 2,0 в отсутствие экзогенного пепсина, в отличие от глюкоамилазы дикого типа Trichoderma reesei (TrGA, файл pdb 2VN4_A, SEQ ID NO: 11), который теряет всякую активность при pH 2,0.
ПРИМЕР 4
Глюкогенная активность глюкоамилаз на мальтодекстрине при низком pH
[00221] Активность глюкоамилазы на мальтодекстриновом субстрате измеряли при pH 3 путем анализа композиций сахаров, доза ферментов составляла 5 м.д. Глюкоамилазы: TeGA, FraGA1, WcoGA1, AnGA (глюкоамилаза дикого типа Aspergillus niger, регистрационный номер XP_001390530.1.1, SEQ ID NO: 10) и TrGA (глюкоамилаза дикого типа Trichoderma reesei, pdb файл 2VN4_A, SEQ ID NO: 11) были протестированы на MALTRIN®M040 (Мальтодекстрин с DE4-7), полученном от Staple Flavor & Fragrance Co. Ltd. Реакции инициировали внесением 10 мкл образца очищенных глюкоамилаз в размере 50 м.д. в 90 мкл раствора субстрата Maltrin (5% вес./об.) в 25 мМ буфере глицин/Na-ацетат/HEPES с pH 3,0. Инкубирование проводили в инкубаторе для ПЦР при 55°C в течение 4 и 22 часов, соответственно. Реакции останавливали внесением 50 мкл 0,5 М NaOH и перемешиванием на вибростенде в течение 2 мин. Планшет центрифугировали для сбора надосадочной жидкости, которую затем разбавляли в 20 раз, используя 5 мМ H2SO4. Эти образцы (10 мкл) анализировали с помощью ВЭЖХ на Agilent серии 1200, оснащенном детектором показателя преломления, колонкой Phenomenex Rezex-RFQ Fast Fruit и защитной колонкой Phenomenex Rezex ROA Organic Acid. Применяли подвижную фазу из 5 мМ H2SO4 при расходе 1,0 мл/мин при 85°C. Были идентифицированы пики, соответствующие DP1, 2, 3 и 3+. Были рассчитаны процентные доли площадей пиков DP1 как часть от суммы DP1, DP2, DP3 и DP3 +, которые приведены в таблице 3. Глюкоамилазы FraGA1, WcoGA1 и TeGA проявляли более высокую глюкогенную активность, чем AnGA и TrGA.
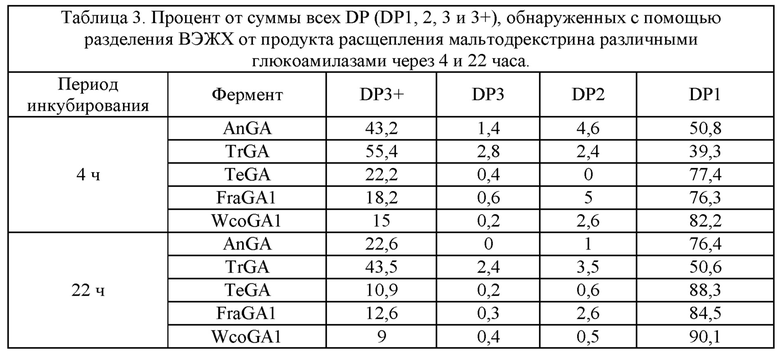
ПРИМЕР 5
Глюкогенная активность глюкоамилаз на разжиженном крахмале, измеренная при низком pH
[00222] Глюкоамилазы FraGA1, WcoGA1, TrGA и AnGA оценивали на разжиженном крахмале для измерения их глюкогенной активности и гидролиза DP3 и DP3+при низком pH. pH разжиженного кукурузного крахмала (32% ds, предварительно обработанного альфа-амилазой) доводили до pH 3,0. Вышеупомянутый субстрат (10 г) и глюкоамилазы (доза 0,25 мг/г сух. в-ва) инкубировали при 32°C и 55°C, соответственно. Реакционные смеси (100 мкл) инкубировали в течение 17, 24, 41, 48, 63 или 72 часов, после чего реакции гасили нагреванием при 100°C в течение 15 минут. После центрифугирования надосадочные жидкости переносили в новые контейнеры и разбавляли в 400 раз, используя 5 мМ H2SO4, для анализа ВЭЖХ с использованием тех же условий, что описаны выше в ПРИМЕРЕ 4. Значения, представленные в таблице 4, представляют собой процентные доли площадей пиков каждого DPn как доли от суммы DP1, DP2, DP3 и DP3 +. Из результатов в таблице 4 видно, что FraGA1 и WcoGA1 проявляли более высокую глюкогенную активность и более быстрый гидролиз DP3+, чем TrGA и AnGA, при дозировании с равным количеством белка (0,25 мг/г сух. в-ва) и инкубировании при 32°C и pH 3. При увеличении температуры инкубирования до 55°C WcoGA1 и FraGA1 очень эффективно гидролизовали DP3+, при этом остаточное содержание DP3+ составляло только 3%, 4% и 5% после 17-часового инкубирования, соответственно, в то время как остаточное содержание в реакциях с TrGA и AnGA составляло 18% и 13%, соответственно.
[00223] Дополнительно оценивали активность FraGA1, WcoGA1, TrGA и AnGA на разжиженном крахмале при pH 2,0. Процедура инкубирования была такой же, как описано выше, за исключением того, что доза ферментов составляла 0,2 мг/г сух. в-ва, а инкубирование проводили при pH 2,0. Образцы отбирали через 4, 21, 29, 45, 53 и 70 ч. При pH 2 FraGA1 и WcoGA1 значительно превосходили TrGA и AnGA по скорости гидролиза DP+ и конечной конверсии DP1, как показано в таблице 5. FraGA1 и WcoGA1 продуцировали ≥90% и ≥72% DP1 после 70 часов инкубирования при 32°C и 55°C, соответственно, тогда как выход от TrGA составлял только 54% и 3%. Эти результаты указывают на то, что FraGA1 и WcoGA1 имеют высокий потенциал для осахаривания и одновременного осахаривания и ферментации (SSF) со значительной активностью при рН 2,0-3,0 и 30-55°С.
ПРИМЕР 6
Оценка осахаривания глюкоамилазами при pH 3,5 и 4,5
[00224] Эффективность осахаривания FraGA1, WcoGA1 и AnGA оценивали при традиционных условиях с pH 4,5 и при более низком pH 3,5. Продуцирование DP1 измеряли путем анализа состава сахара после обработки одинаковой дозой фермента. Инкубирование глюкоамилаз (при дозе 50 мкг/г сух. в-ва) и разжиженного кукурузного крахмала, предварительно обработанного альфа-амилазой (32% сух. в-ва), проводили при 60°C, pH 3,5 и 4,5, а образцы отбирали через 24, 48 и 72 ч. Во всех случаях инкубирование прекращали нагреванием при 100°C в течение 15 мин. После центрифугирования надосадочные жидкости от каждого образца переносили в новые пробирки и разбавляли в 400 раз, используя 5 мМ H2SO4, для анализа ВЭЖХ. Разделение с помощью ВЭЖХ проводили на системе ВЭЖХ Agilent серии 1200 с колонкой Fast Fruit (100 мм × 7,8 мм) при 80°С с изократическим градиентом 5 мМ H2SO4 при расходе 1,0 мл/мин. Продуктовые олигосахариды детектировали с помощью детектора показателя преломления. Глюкогенная активность образцов представлена в таблице 6. Продуцирование DP1 с помощью FraGA1 и WcoGA1 при pH 3,5 после 48-часового инкубирования было сопоставимо с результатами, полученными с использованием AnGA при pH 4,5 после 72-часового инкубирования.
ПРИМЕР 7
Стабильность FraGA1 и WcoGA1 в условиях переваривания жвачными животными
[00225] Стабильность ферментов FraGA1 и WcoGA1 измеряли при pH 2,2, 5,6 и 6,5 в условиях переваривания жвачными животными in vitro, как описано ниже.
Условия анализа при pH 5,6. Реакционная смесь содержала 1 мл рубцового сока, взятого у местных дойных коров (Foulum, Дания), с измеренным pH 5,6, и 20 мкл глюкоамилазы (концентрация белка FraGA1 составляла 5 мг/мл, а концентрация белка WcoGA1 составляла 6,5 мг/мл). Реакционные смеси выдерживали при 40°C в течение 10 ч, встряхивая при 100 об/мин. Реакционная смесь, которую выдерживали при 5°C, служила контролем; в случае холостого опыта ферменты не добавляли. Каждую ферментативную обработку или холостой опыт проводили трижды. В конце периода инкубирования вносили 0,5 мл 10% (мас./мас.) кукурузного крахмала (Sigma, S-4126), и образцы дополнительно инкубировали в течение 140 мин при 40°C, встряхивая при 240 об/мин. Остаточную активность в процентах (%) рассчитывали следующим образом: разность активности после инкубирования с ферментом при 40°C и холостого образца, деленная на активность после инкубирования с ферментом при 5°C минус холостой образец, умноженная на 100.
Условия анализа при pH 6,5. Условия были такими же, как описано выше для pH 5,6, за исключением того, что pH был доведен до pH 6,5 и DTT добавляли до 20 мМ.
Условия анализа при pH 2,2. Рубцовый сок, который был заморожен после взятия, затем оттаивали и центрифугировали перед использованием. Надосадочную жидкость отбрасывали, а осадок суспендировали в очищенной воде в соотношении (осадок к воде) 1: 3. рН суспензии доводили до 2,2. Аликвоты по 0,6 мл вносили в пробирки объемом 2 мл, содержащие 0,4 мл пепсина (2500 Ед. пепсина/мл, Sigma P7000) в 60 мМ глицин-HCl (pH 2,0) и по 20 мкл каждой из глюкоамилаз (концентрация белка FraGA1 составляла 5 мг/мл и концентрация белка WcoGA1 составляла 6,5 мг/мл). Реакционные смеси инкубировали при 40°C в течение 3 ч при встряхивании в режиме 100 об/мин. В конце инкубирования вносили 0,5 мл 15% (вес./вес.) кукурузного крахмала (Sigma, S-4126), суспендированного в 250 мМ MES pH 6,5, и образцы инкубировали в течение 3 ч при 40°C. Остаточная активность в процентах (%) рассчитывалась следующим образом: разность активности после инкубирования с ферментом при 40°C и холостого образца, деленная на активность без предварительного инкубирования (контроль) минус холостой образец, умноженная на 100.
[00226] В конце каждой реакции пробирки для анализа центрифугировали, надосадочные жидкости переносили в планшеты для ПЦР и подвергали термообработке при 95°C в течение 5 мин. Образцы в термообработанных ПЦР пробирках пропускали через 96-луночные фильтровальные планшеты, и 10 мкл аликвоты каждого фильтрата использовали для количественного определения содержания глюкозы с помощью ВЭЖХ, используя колонки для углеводов BioRad (Aminex HPX-87N) и воду в качестве элюента, при 75°C. Содержание глюкозы использовали как меру активности глюкоамилазы на крахмальном субстрате. Как видно из таблицы 7, глюкоамилазы FraGA1 и WcoGA1 сохраняли 60-70% остаточной активности после инкубирования при 40°C, pH 5,6 и pH 6,5 через 10 часов. Также было замечено, что после инкубирования в присутствии свиного пепсина при pH 2,2 и 40°C в течение 3 ч две эти глюкоамилазы сохраняли примерно 90% своей активности.
ПРИМЕР 8
Стабильность и активность FraGA1 и WcoGA1 в условиях переваривания в тонком кишечнике
[00227] Для моделирования условий переваривания в тонком кишечнике одну порцию рубцового сока смешивали с двумя порциями искусственного рубцового сока в соответствии с протоколом, описанным Menke и Steingass (Liu, JX, A. Susenbeth и KH Südekum. 2002. In vitro gas production measurements to evaluate interactions between untreated and chemically treated rice straws, grass hay, and mulberry leaves. Journal of Animal Science 80:517-524). Кукурузную муку добавляли до конечной концентрации 10% (вес./вес.) и доводили до pH 6,5. Один мл этой смеси смешивали с 50 мкл свиного панкреатина (Sigma, P7545, 50 мг/мл маточного раствора панкреатина) и 15 мкл образца глюкоамилазы (0,08 мг FraGA1 или 0,10 мг WcoGA). В холостой образец не добавляли ни панкреатин, ни глюкоамилазу. Реакции проводили при 40°C в течение 3 ч при встряхивании в режиме 240 об/мин. Затем реакционные смеси центрифугировали при 4000 об/мин в течение 10 мин, и 150 мкл надосадочной жидкости переносили в планшет для ПЦР и нагревали при 95°C в течение 5 мин. Аликвоты по 50 мкл переносили на фильтровальный планшет с размером пор 0,45 мкм, добавляли 90 мкл воды, и планшет центрифугировали при 4000 об/мин в течение 15 мин. Фильтрат анализировали на содержание глюкозы (G1), мальтозы (G2) и мальтотриозы (G3) с помощью ВЭЖХ, как описано в Примере 7. Каждую обработку или холостую реакцию проводили трижды, среднее значение приведено в таблице 8. Стд. откл. означает стандартное отклонение.
[00228] Как видно из таблицы 8, свиной панкреатин высвобождает глюкозу, мальтозу и мальтотриозу из кукурузной муки. Оба фермента FraGA1 и WcoGA1 высвобождают глюкозу из сырого крахмала в кукурузной муке. WcoGA1 также способен снижать уровень мальтозы в условиях эксперимента. Комбинация глюкоамилаз с панкреатином образует глюкозу, мальтозу и снижает уровень мальтотриозы. Эти результаты позволяют предположить, что глюкоамилазы могут действовать вместе с альфа-амилазой, присутствующей в панкреатине, даже в присутствии пищеварительных протеаз, присутствующих в свином панкреатине (согласно производителю). Образовавшаяся глюкоза in vivo может абсорбироваться животным непосредственно, в то время как образовавшаяся мальтоза может превращаться в глюкозу с помощью мальтазы-глюкоамилазы, связанной с щеточной каймой, на эпителиальных клетках тонкой кишки. Кукурузная мука из зубовидной кукурузы имела следующий состав: 88,2% сухого вещества, 9,3% сырого протеина, 2,0% кислой очищающей клетчатки, 6,6% нейтральной очищающей клетчатки, обработанной амилазой, 78,5% неволокнистых углеводов и 89,0% общего количества перевариваемых питательных веществ, и измельчали до размера меньше 200 мкм.
ПРИМЕР 9
FraGA1 для улучшения переваривания сухого вещества и газообразования при ферментации кукурузы в рубце
[00229] FraGA1 оценивали по ее действию на усвояемость сухого вещества (DMD), выработку газа, усвояемость крахмала и pH в системе периодической ферментации in vitro с использованием в качестве субстрата зубовидной кукурузы (помол 4 мм). Эффективность FraGA1 оценивали при трех дозах (0, 0,25, 0,5 и 0,75 мг/г субстрата в исходном состоянии) в трех отдельных экспериментах, где каждую комбинацию фермент-доза выполняли в четырех повторностях в каждом эксперименте. Эффективность каждого фермента определяли по типу ферментации в рубце после инкубирования забуференного содержимого рубца с субстратом в установке для ферментации, описанной ранее (Adesogan, AT, Krueger, NK и Kim, SC 2005. Anim. Feed Sci. Technol. 123: 211-223; Krueger, N. A. и A. T. Adesogan. 2008. Anim. Feed Sci. Tech. 145: 84-94; Goering, H. K. и P. J. Van Soest. 1970. Agric. Handbook No. 379. ARS USDA, Вашингтон, округ Колумбия, стр.20). Состав зубчатой кукурузы, используемой в этой системе периодической ферментации in vitro, приведен в Примере 8. Образцы по 0,5 г зубчатой кукурузы, измельченные до 4 мм, взвешивали в 4 одинаковых фильтровальных мешках F57 (ANKOM Technology, Македония, Нью-Йорк). Фермент FraGA1 готовили в 0,1 М цитратно-фосфатном буфере (pH 6,0) и вносили в измельченную зубчатую кукурузу в фильтровальных мешках в количестве 0, 0,25, 0,5 и 0,75 мг ферментного белка на грамм субстрата. Затем фильтровальные мешки герметизировали с помощью Uline Tabletop Poly Bag Sealer (Impulse® типа AIE-200) и сразу же помещали во флаконы для сыворотки объемом 160 мл, содержащие забуференный рубцовый сок (52 мл). Также были включены флаконы с холостой пробой, содержащие только фильтровальный мешок, а также контрольные образцы, содержащие только измельченную зубчатую кукурузу без ферментов помимо забуференного рубцового сока. Затем флаконы укупоривали резиновыми пробками и закрывали алюминиевыми крышками. Их инкубировали в течение 7 ч при 39°C в инкубаторе с принудительной вентиляцией. В конце инкубирования фильтровальные мешки, содержащие остатки зубчатой кукурузы, сушили в сушильном шкафу при 60°C в течение 48 ч и взвешивали для количественного определения DMD. Давление газа внутри флаконов измеряли с помощью пневмодатчика и вычисляли объем газа через 7 ч инкубирования. Следующую формулу применяли для преобразования давления газа в объем газа: объем газа (мл) = (давление газа (фунтов/кв. дюйм) * 4,8843) + 3,1296. Уравнение было написано исходя из условий эксперимента.
[00230] Забуференный рубцовый сок готовили следующим образом: рубцевый сок отбирали у трех лактирующих молочных коров породы Holstein с канюлированием рубца через 2-3 часа после поедания полного смешанного рациона (TMR). Состав ингредиентов TMR, скармливаемых жвачным молочным коровам на основе сухого вещества, состоял из: 38,2% кукурузного силоса, 27,3% измельченной очищенной кукурузы, 14,5% соевой муки с 44% сырого белка, 9,1% мякоти цитрусовых, 4,5% премикса Feedlot, 4,0% сена люцерны в середине цветения, 1,8% энергетической добавки (MS Specialty Nutrition, Данди, Иллинойс) и 0,5% Novasil (BASF, Германия). Отобранный рубцовый сок фильтровали через четыре слоя марли перед смешиванием с предварительно подогретой искусственной слюной (39°C). Состав искусственной слюны включал микроминеральный раствор CaCl2⋅2H2O, MnCl2⋅4H2O, CoCl2⋅6H2O, FeCl2⋅6H2O; макроминеральный раствор Na2HPO4⋅12H2O, KH2PO4, MgSO4⋅7H2O; буферный раствор (NH4)2HCO3 и NaHCO3; триптиказо-пептонный раствор (триптон, Sigma-Aldrich, Сент-Луис, Миссури, США); окислительно-восстановительный индикатор резазурин и восстанавливающий раствор, содержащий цистеин-HCl, 1 М NaOH, Na2S⋅9H2O и дистиллированную воду (Goering, H. K. and P. J. Van Soest. 1970. Agric. Handbook No. 379. ARS USDA, Вашингтон, округ Колумбия, стр.20). Объемное соотношение между рубцовым соком и искусственной слюной составляло 1:2.
[00231] Статистический анализ. Для всех экспериментов в примере 9 собранные данные анализировали с применением процедуры GLIMMIX SAS (версия 9.1; SAS Institute, Кэри, Северная Каролина). Эксперименты разрабатывали в виде полностью рандомизированной блочной схемы, где каждый прогон рассматривался как блок. Дозу использовали в качестве фиксированного эффекта в модели при тестировании ферментов в различных дозах. Переменная отклика включала DMD и газообразование in-vitro. Прогон рассматривали как случайный фактор. Фиксированные параметры модели включали время отбора проб для данных ферментации, измеряемых в разные периоды после инкубирования. Процедуру UNIVARIATE SAS применяли для тестирования остатков на выпадающие значения и нормальность, перед тем как проводили окончательные анализы. Параметры обработок считали статистически значимыми при P < 0,05, тогда как любые тренды определяли при 0,05≤P≤0,10.
[00232] Результаты. По результатам, приведенным в Таблице 9 ниже, видно, что в условиях ферментации в рубце in vitro, описанных здесь, глюкоамилаза FraGA1 увеличивала выработку DMD и газа в зависимости от дозы. Существует тенденция к повышению усвояемости крахмала при дозах 0, 0,25 и 0,5 мг ферментного белка на грамм измельченной зубчатой кукурузы. Вне зависимости от дозы фермента, pH был практически постоянным.
a-c Средние значения в строке с различными верхними индексами отличаются (P < 0,05)
ПРИМЕР 10
Сравнение белковых последовательностей
[00233] Родственные белки были идентифицированы с помощью поиска BLAST (Altschul et al., Nucleic Acids Res., 25:3389-402, 1997) с использованием смоделированных зрелых аминокислотных последовательностей для FraGA1 (SEQ ID NO: 7), WcoGA1 (SEQ ID NO: 8) и TeGA (SEQ ID NO: 9) из баз данных NCBI и Genome Quest Patent с параметрами поиска, установленными на значения по умолчанию, и их подборка приведена в таблицах 10A и 10B (FraGA1), таблицах 11A и 11B (WcoGA1) и таблицах 12A и 12B (TeGA), соответственно. Процентную идентичность (PID) в обоих случаях поиска определяли как число идентичных остатков разделенное на число выровненных остатков при попарном выравнивании. Значение, обозначенное «длина последовательности» в таблицах, соответствует длине (в аминокислотах) для белков, на которые даются ссылки с приведенными номерами доступа, тогда как «выровненная длина» относится к последовательности, используемой для выравнивания и расчета PID.
(несколько последовательностей)
(несколько последовательностей)
[00234] Выравнивание смоделированных зрелых последовательностей FraGA1 (SEQ ID NO: 7); WcoGA1 (SEQ ID NO: 8); TeGA (SEQ ID NO: 9); AnGA (ак 25-640 из XP_001390530.1, SEQ ID NO: 10); TrGA (2VN4_A, SEQ ID NO: 11); KZT67263.1 (ак 19-571 из SEQ ID NO: 12); XP_002475369.1 (ак 18-569 из SEQ ID NO: 13); US20090325240-1847 (ак 17-570 из SEQ ID NO: 14); KZT09226.1 (ак 18-570 из SEQ ID NO: 15); WO2016196202-0012 (SEQ ID NO: 16); US20170314003-0002 (ак 19-575 из SEQ ID NO: 17); GAD95639.1 (ак 38-622 из SEQ ID NO: 18); US20170306309-0023 (SEQ ID NO: 19); и CAC28076.1 (ак 29-618 из SEQ ID NO: 20) выполняли с параметрами по умолчанию с использованием программного обеспечения CLUSTALW (Thompson et al., Nucleic Acids Research, 22:4673-4680, 1994). Множественное выравнивание последовательностей показано на фигуре 1.
Пример 11
Относительная активность глюкоамилаз при pH 1,9, 4,5 и 6,0
[00235] Высвобождение глюкозы из субстрата из кукурузного крахмала измеряли для глюкоамилаз FraGA1 и WcoGA1 в условиях, имитирующих условия, наблюдаемые в пищеварительной системе жвачных животных. Каждая реакционная смесь содержала 0,1 мг очищенного фермента и 10% (вес./об.) кукурузного крахмала (Sigma S-4126) в объеме 1 мл. Для реакций при pH 1,9 в качестве разбавителя использовали 0,1 М глицин-HCl (pH 1,9), и реакционные системы также содержали 1000 единиц свиного пепсина (Sigma P-7000). Для реакций при рН 4,5 в 0,1 М глицине рН доводили до 4,5 с помощью 5 н. NaOH и пепсин не добавляли. Для реакций при pH 6,0 в качестве разбавителя использовали 0,1 М Mes-NaOH (pH 6,0) и пепсин не добавляли.
[00236] Реакции проводили при 40°C, встряхивая в течение 2 ч. Образцы центрифугировали для разделения надосадочной жидкости и непрореагировавших гранул кукурузного крахмала. Надосадочную жидкость нагревали при 99°C в течение 10 минут, фильтровали и 20 мкл фильтрата вводили в ВЭЖХ с использованием колонки Na-Carbohydrate для разделения и количественного определения глюкозы, образованной с использованием стандартного раствора глюкозы с концентрацией 1 мг/мл.
[00237] Результаты реакций приведены в таблице 13. Активность по высвобождению глюкозы выражается в мг высвобождаемой глюкозы на мл реакционной системы. Обе глюкоамилазы показали самую высокую активность при pH 4,5 по сравнению с pH 1,9 и 6,0. Неожиданно было обнаружено, что обе глюкоамилазы проявляли активность при pH 1,9 в присутствии пепсина, и эта активность при pH 1,9 составляла более 80% активности при pH 6,0.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДАНИСКО ЮЭс ИНК.
<120> ГЛЮКОАМИЛАЗЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> NB40933-WO-PCT2
<140> PCT/CN18/078575
<141> 09.03.2018
<160> 9
<170> PatentIn версия 3.5
<210> 1
<211> 567
<212> БЕЛОК
<213> Fibroporia radiculosa
<400> 1
Met Leu Phe Leu Leu Ala Ala Leu Gly Leu Ala Cys Ser Ala Ala Ala
1. 5 10 15
Gln Ser Thr Ser Val Ser Ala Tyr Ile Ala Ser Glu Ser Pro Val Ala
20 25 30
Lys Ala Gly Val Leu Ala Asn Ile Gly Thr Glu Gly Ser Leu Ser Ser
35 40 45
Gly Ala Tyr Ser Gly Val Val Ile Ala Ser Pro Ser Thr Val Asn Pro
50 55 60
Asp Tyr Leu Tyr Thr Trp Val Arg Asp Ser Ser Leu Thr Phe Gln Ala
65 70 75 80
Leu Ile Asp Gln Tyr Val Tyr Gly Glu Asp Pro Thr Leu Arg Ser Leu
85 90 95
Ile Asp Glu Phe Ile Thr Ala Glu Ser Ile Leu Gln Gln Thr Thr Asn
100 105 110
Pro Ser Gly Thr Val Ser Thr Gly Gly Leu Gly Glu Pro Lys Phe Asn
115 120 125
Ile Asn Glu Thr Ala Phe Thr Gly Pro Trp Gly Arg Pro Gln Arg Asp
130 135 140
Gly Pro Ala Leu Arg Ser Thr Ala Ile Ile Thr Tyr Ala Thr Tyr Leu
145 150 155 160
Trp Asn Ser Gly Asn Thr Ser Tyr Val Ser Asp Ser Leu Trp Pro Ile
165 170 175
Ile Glu Leu Asp Leu Asn Tyr Ile Ala Thr Tyr Trp Asn Phe Ser Thr
180 185 190
Phe Asp Leu Trp Glu Glu Ile Asp Ser Ser Ser Phe Trp Thr Thr Ala
195 200 205
Val Gln His Arg Ala Leu Arg Gln Gly Ile Thr Phe Ala Asn Leu Ile
210 215 220
Gly Gln Thr Ser Pro Val Ser Asn Tyr Glu Thr Gln Ala Gly Asp Ile
225 230 235 240
Leu Cys Phe Leu Gln Thr Tyr Trp Asn Pro Thr Gly Asn Tyr Met Thr
245 250 255
Ala Asn Thr Gly Gly Gly Arg Ser Gly Lys Asp Ser Asn Thr Val Leu
260 265 270
Ala Ser Val His Thr Phe Asp Pro Asp Ala Gly Cys Asp Ser Thr Thr
275 280 285
Phe Gln Pro Cys Ser Asp Lys Ala Leu Ser Asn Leu Lys Val Tyr Val
290 295 300
Asp Ser Phe Arg Ser Leu Tyr Ala Ile Asn Asp Gly Ile Ala Ser Asp
305 310 315 320
Ala Ala Val Ala Thr Gly Arg Tyr Pro Glu Asp Val Tyr Tyr Gly Gly
325 330 335
Asn Pro Trp Tyr Leu Cys Thr Phe Ala Val Ala Glu Gln Leu Tyr Asp
340 345 350
Ala Leu Ile Val Trp Ser Ser Gln Gly Tyr Leu Glu Ile Thr Asp Leu
355 360 365
Ser Leu Ala Phe Phe Gln Gln Phe Asp Ser Asp Val Gly Thr Gly Thr
370 375 380
Tyr Asp Ser Gly Ser Ser Thr Tyr Ser Thr Leu Thr Ser Ala Ile Arg
385 390 395 400
Thr Phe Ala Asp Gly Phe Val Leu Thr Asn Ala Lys Tyr Thr Pro Thr
405 410 415
Asn Gly Ser Leu Ser Glu Glu Tyr Thr Ser Ala Asp Gly Thr Pro Ile
420 425 430
Ser Ala Tyr Asp Leu Thr Trp Ser Tyr Ala Ser Ala Leu Thr Val Phe
435 440 445
Ala Ala Glu Ala Gly Thr Thr Tyr Gly Ser Trp Gly Ala Ala Gly Leu
450 455 460
Thr Val Pro Ser Thr Cys Thr Ser Gly Val Ala Val Thr Phe Glu Val
465 470 475 480
Asp Tyr Asp Thr Glu Tyr Gly Glu Asn Val Tyr Ile Thr Gly Ser Val
485 490 495
Asn Ala Leu Glu Asn Trp Ser Ala Thr Asn Ala Leu Ile Met Ser Ala
500 505 510
Ala Asp Tyr Pro Thr Trp Ser Ile Thr Val Tyr Leu Pro Pro Ser Thr
515 520 525
Thr Ile Gln Tyr Lys Tyr Leu Thr Gln Tyr Asn Gly Glu Val Thr Trp
530 535 540
Glu Asp Asp Pro Asn Asn Glu Ile Thr Thr Pro Ala Ser Gly Ser Met
545 550 555 560
Thr Gln Val Asp Ser Trp His
565
<210> 2
<211> 569
<212> БЕЛОК
<213> Wolfiporia cocos
<400> 2
Met Arg Leu Ser Leu Ala Ser Val Phe Ala Leu Ala Gly Gly Ala Leu
1. 5 10 15
Ala Gln Thr Thr Ser Val Thr Ser Tyr Ile Ala Ser Glu Ser Pro Ile
20 25 30
Ala Lys Ala Gly Val Leu Ala Asn Ile Gly Ala Asp Gly Ser Leu Ser
35 40 45
Ser Gly Ala Tyr Ser Gly Ile Val Ile Ala Ser Pro Ser Thr Val Asn
50 55 60
Pro Asn Tyr Leu Tyr Thr Trp Thr Arg Asp Ser Ser Leu Thr Phe Met
65 70 75 80
Glu Leu Ile Asn Gln Tyr Ile Tyr Gly Glu Asp Asp Thr Leu Arg Thr
85 90 95
Leu Ile Asp Glu Phe Val Ser Ala Glu Ala Thr Leu Gln Gln Val Thr
100 105 110
Asn Pro Ser Gly Thr Val Ser Thr Gly Gly Leu Gly Glu Pro Lys Phe
115 120 125
Asn Ile Asn Glu Thr Ala Phe Thr Gly Pro Trp Gly Arg Pro Gln Arg
130 135 140
Asp Gly Pro Ala Leu Arg Ala Thr Ala Ile Met Ala Tyr Ala Thr Tyr
145 150 155 160
Leu Tyr Glu Asn Gly Asn Thr Ser Tyr Val Thr Asp Thr Leu Trp Pro
165 170 175
Ile Ile Glu Leu Asp Leu Gly Tyr Val Ala Glu Tyr Trp Asn Glu Ser
180 185 190
Thr Phe Asp Leu Trp Glu Glu Ile Asp Ser Ser Ser Phe Phe Thr Thr
195 200 205
Ala Val Gln His Arg Ala Leu Arg Ala Gly Val Thr Phe Ala Asn Leu
210 215 220
Ile Gly Glu Thr Ser Asp Val Ser Asn Tyr Gln Glu Asn Ala Asp Asp
225 230 235 240
Leu Leu Cys Phe Leu Gln Ser Tyr Trp Asn Pro Thr Gly Ser Tyr Val
245 250 255
Thr Ala Asn Thr Gly Gly Gly Arg Ser Gly Lys Asp Ala Asn Thr Leu
260 265 270
Leu Ala Ser Ile His Thr Phe Asp Pro Asp Ala Gly Cys Asn Ala Thr
275 280 285
Thr Phe Gln Pro Cys Ser Asp Lys Ala Leu Ser Asn His Lys Val Tyr
290 295 300
Val Asp Ser Phe Arg Ser Leu Tyr Ala Ile Asn Asp Asp Ile Ser Ser
305 310 315 320
Asp Ala Ala Val Ala Thr Gly Arg Tyr Pro Glu Asp Val Tyr Tyr Asn
325 330 335
Gly Asn Pro Trp Tyr Leu Cys Thr Leu Ala Ala Ala Glu Gln Leu Tyr
340 345 350
Asp Ser Leu Ile Val Trp Lys Ala Gln Gly Tyr Ile Glu Val Thr Ser
355 360 365
Leu Ser Leu Ala Phe Phe Gln Gln Phe Asp Ala Ser Val Ser Ala Gly
370 375 380
Thr Tyr Asp Ser Ser Ser Asp Thr Tyr Thr Thr Leu Leu Asp Ala Val
385 390 395 400
Gln Thr Tyr Ala Asp Gly Phe Val Leu Met Val Ala Gln Tyr Thr Pro
405 410 415
Ala Asn Gly Ser Leu Ser Glu Gln Tyr Ala Lys Ala Asp Gly Ser Pro
420 425 430
Thr Ser Ala Tyr Asp Leu Thr Trp Ser Phe Ala Ala Ala Leu Thr Ala
435 440 445
Phe Ala Ala Arg Asp Gly Lys Thr Tyr Gly Ser Trp Gly Ala Ala Asp
450 455 460
Leu Ser Ser Thr Cys Ser Gly Ser Thr Asp Thr Val Ala Val Thr Phe
465 470 475 480
Glu Val Gln Tyr Asp Thr Gln Tyr Gly Glu Asn Leu Tyr Ile Thr Gly
485 490 495
Ser Val Ser Gln Leu Glu Asp Trp Ser Ala Asp Asp Ala Leu Ile Met
500 505 510
Ser Ser Ala Asp Tyr Pro Thr Trp Ser Ile Thr Val Asp Leu Pro Pro
515 520 525
Ser Thr Leu Ile Gln Tyr Lys Tyr Leu Thr Lys Tyr Asn Gly Asp Val
530 535 540
Thr Trp Glu Asp Asp Pro Asn Asn Glu Ile Thr Thr Pro Ala Ser Gly
545 550 555 560
Ser Tyr Thr Gln Val Asp Ser Trp His
565
<210> 3
<211> 618
<212> БЕЛОК
<213> Talaromyces emersonii
<400> 3
Met Ala Ser Leu Val Ala Gly Ala Leu Cys Ile Leu Gly Leu Thr Pro
1. 5 10 15
Ala Ala Phe Ala Arg Ala Pro Val Ala Ala Arg Ala Thr Gly Ser Leu
20 25 30
Asp Ser Phe Leu Ala Thr Glu Thr Pro Ile Ala Leu Gln Gly Val Leu
35 40 45
Asn Asn Ile Gly Pro Asn Gly Ala Asp Val Ala Gly Ala Ser Ala Gly
50 55 60
Ile Val Val Ala Ser Pro Ser Arg Ser Asp Pro Asn Tyr Phe Tyr Ser
65 70 75 80
Trp Thr Arg Asp Ala Ala Leu Thr Ala Lys Tyr Leu Val Asp Ala Phe
85 90 95
Asn Arg Gly Asn Lys Asp Leu Glu Gln Thr Ile Gln Gln Tyr Ile Ser
100 105 110
Ala Gln Ala Lys Val Gln Thr Ile Ser Asn Pro Ser Gly Asp Leu Ser
115 120 125
Thr Gly Gly Leu Gly Glu Pro Lys Phe Asn Val Asn Glu Thr Ala Phe
130 135 140
Thr Gly Pro Trp Gly Arg Pro Gln Arg Asp Gly Pro Ala Leu Arg Ala
145 150 155 160
Thr Ala Leu Ile Ala Tyr Ala Asn Tyr Leu Ile Asp Asn Gly Glu Ala
165 170 175
Ser Thr Ala Asp Glu Ile Ile Trp Pro Ile Val Gln Asn Asp Leu Ser
180 185 190
Tyr Ile Thr Gln Tyr Trp Asn Ser Ser Thr Phe Asp Leu Trp Glu Glu
195 200 205
Val Glu Gly Ser Ser Phe Phe Thr Thr Ala Val Gln His Arg Ala Leu
210 215 220
Val Glu Gly Asn Ala Leu Ala Thr Arg Leu Asn His Thr Cys Ser Asn
225 230 235 240
Cys Val Ser Gln Ala Pro Gln Val Leu Cys Phe Leu Gln Ser Tyr Trp
245 250 255
Thr Gly Ser Tyr Val Leu Ala Asn Phe Gly Gly Ser Gly Arg Ser Gly
260 265 270
Lys Asp Val Asn Ser Ile Leu Gly Ser Ile His Thr Phe Asp Pro Ala
275 280 285
Gly Gly Cys Asp Asp Ser Thr Phe Gln Pro Cys Ser Ala Arg Ala Leu
290 295 300
Ala Asn His Lys Val Val Thr Asp Ser Phe Arg Ser Ile Tyr Ala Ile
305 310 315 320
Asn Ser Gly Ile Ala Glu Gly Ser Ala Val Ala Val Gly Arg Tyr Pro
325 330 335
Glu Asp Val Tyr Gln Gly Gly Asn Pro Trp Tyr Leu Ala Thr Ala Ala
340 345 350
Ala Ala Glu Gln Leu Tyr Asp Ala Ile Tyr Gln Trp Lys Lys Ile Gly
355 360 365
Ser Ile Ser Ile Thr Asp Val Ser Leu Pro Phe Phe Gln Asp Ile Tyr
370 375 380
Pro Ser Ala Ala Val Gly Thr Tyr Asn Ser Gly Ser Thr Thr Phe Asn
385 390 395 400
Asp Ile Ile Ser Ala Val Gln Thr Tyr Gly Asp Gly Tyr Leu Ser Ile
405 410 415
Val Glu Lys Tyr Thr Pro Ser Asp Gly Ser Leu Thr Glu Gln Phe Ser
420 425 430
Arg Thr Asp Gly Thr Pro Leu Ser Ala Ser Ala Leu Thr Trp Ser Tyr
435 440 445
Ala Ser Leu Leu Thr Ala Ser Ala Arg Arg Gln Ser Val Val Pro Ala
450 455 460
Ser Trp Gly Glu Ser Ser Ala Ser Ser Val Leu Ala Val Cys Ser Ala
465 470 475 480
Thr Ser Ala Thr Gly Pro Tyr Ser Thr Ala Thr Asn Thr Val Trp Pro
485 490 495
Ser Ser Gly Ser Gly Ser Ser Thr Thr Thr Ser Ser Ala Pro Cys Thr
500 505 510
Thr Pro Thr Ser Val Ala Val Thr Phe Asp Glu Ile Val Ser Thr Ser
515 520 525
Tyr Gly Glu Thr Ile Tyr Leu Ala Gly Ser Ile Pro Glu Leu Gly Asn
530 535 540
Trp Ser Thr Ala Ser Ala Ile Pro Leu Arg Ala Asp Ala Tyr Thr Asn
545 550 555 560
Ser Asn Pro Leu Trp Tyr Val Thr Val Asn Leu Pro Pro Gly Thr Ser
565 570 575
Phe Glu Tyr Lys Phe Phe Lys Asn Gln Thr Asp Gly Thr Ile Val Trp
580 585 590
Glu Asp Asp Pro Asn Arg Ser Tyr Thr Val Pro Ala Tyr Cys Gly Gln
595 600 605
Thr Thr Ala Ile Leu Asp Asp Ser Trp Gln
610 615
<210> 4
<211> 1704
<212> ДНК
<213> Fibroporia radiculosa
<400> 4
atgctcttcc tcctcgctgc tctgggcctc gcttgtagcg ctgctgccca atccacttcc 60
gtctccgctt acatcgccag cgagagcccc gttgccaaag ccggtgttct ggctaacatt 120
ggcactgaag gtagcctgag ctccggtgcc tactccggcg ttgtcatcgc ctcccccagc 180
accgtcaacc ctgactacct ctatacttgg gttcgcgact ccagcctcac tttccaagcc 240
ctgattgacc agtacgttta cggcgaggac cccaccctcc gaagcctgat cgacgagttc 300
attaccgctg agtccatcct gcagcaaact accaacccca gcggcaccgt tagcaccggc 360
ggtctgggcg agcccaagtt caacatcaat gagactgcct ttaccggtcc ctggggccga 420
ccccaacgcg acggtcctgc cctccgcagc actgccatca ttacttatgc cacctacctg 480
tggaactccg gtaacacctc ctacgtttcc gattccctct ggcccatcat cgaactcgac 540
ctgaattaca ttgctaccta ctggaatttc tccacttttg atctgtggga agagattgac 600
tcctccagct tctggaccac tgccgttcag catcgagccc tgcgccaggg tatcaccttc 660
gctaatctga ttggccagac cagccctgtc agcaactatg agacccaagc cggcgatatc 720
ctctgtttcc tccaaaccta ttggaatcct accggcaact acatgaccgc caatactggc 780
ggtggtcgaa gcggcaagga ctccaacacc gttctcgctt ccgttcatac cttcgatccc 840
gatgccggct gtgatagcac tacttttcaa ccttgctccg acaaggccct gagcaacctc 900
aaggtctacg tcgactcctt tcgcagcctg tacgccatca acgacggtat tgcctccgac 960
gctgccgtcg ccaccggccg ctatcctgag gacgtctact acggcggtaa cccctggtac 1020
ctctgcacct ttgctgtcgc cgaacaactc tacgacgccc tcatcgtctg gagcagccag 1080
ggctatctcg aaatcactga cctcagcctg gccttcttcc agcagtttga ttccgatgtc 1140
ggtactggca cctacgacag cggctccagc acttactcca ctctcacctc cgccatccga 1200
acttttgctg atggcttcgt tctgaccaac gccaaataca cccctaccaa tggttccctg 1260
tccgaggagt acaccagcgc cgatggcact cctatctccg cctatgacct gacctggagc 1320
tacgcctccg ctctgaccgt ctttgccgcc gaggccggca ccacttacgg ctcctggggt 1380
gctgctggcc tgactgtccc tagcacctgc actagcggcg tcgctgttac tttcgaggtc 1440
gattacgaca ccgagtatgg cgaaaacgtc tatatcaccg gttccgtcaa tgccctggaa 1500
aattggtccg ccactaatgc tctgattatg tccgccgctg actatcccac ctggtccatc 1560
accgtttacc tgcccccctc caccaccatt cagtataagt atctcaccca gtacaacggc 1620
gaagtcactt gggaggacga ccctaacaac gagattacta cccctgctag cggttccatg 1680
acccaggttg acagctggca ctaa 1704
<210> 5
<211> 1710
<212> ДНК
<213> Wolfiporia cocos
<400> 5
atgcgactga gcctggcctc cgtttttgct ctcgccggtg gtgccctcgc ccagaccact 60
agcgtcacct cctacattgc tagcgaaagc cccattgcca aagccggtgt tctcgctaac 120
attggcgctg acggctccct gagctccggt gcttattccg gcattgttat cgccagcccc 180
tccaccgtta accctaacta tctctatacc tggactcgcg actccagcct gaccttcatg 240
gaactgatca accagtacat ctacggcgag gacgatactc tgcgaactct gattgatgag 300
ttcgtttccg ctgaagccac cctccaacag gtcactaatc ctagcggcac tgtctccact 360
ggtggcctcg gcgagcccaa gttcaacatc aacgagactg cttttactgg tccctggggc 420
cgaccccaac gcgatggccc tgccctgcgc gctactgcta tcatggccta tgccacctac 480
ctgtatgaaa acggtaatac tagctatgtt actgacaccc tctggcccat cattgaactc 540
gacctcggtt acgtcgccga atattggaac gaaagcacct ttgatctctg ggaggaaatc 600
gacagcagct cctttttcac taccgctgtt cagcaccgcg ctctccgcgc tggcgttacc 660
ttcgccaatc tcatcggtga gaccagcgac gtcagcaact accaggaaaa tgccgacgac 720
ctcctctgct tcctccaaag ctactggaac cccactggca gctacgtcac tgctaacact 780
ggcggtggtc gaagcggcaa ggacgccaac actctcctgg ctagcatcca caccttcgat 840
cccgacgctg gctgcaacgc cactaccttt caaccctgtt ccgacaaagc cctcagcaat 900
cacaaggtct acgttgactc cttccgcagc ctctacgcca tcaatgacga cattagcagc 960
gatgccgctg tcgctaccgg ccgataccct gaggatgtct actacaacgg caacccctgg 1020
tacctctgta ccctggccgc tgctgagcaa ctctacgact ccctcatcgt ctggaaggcc 1080
caaggctaca tcgaagtcac cagcctcagc ctcgcctttt ttcaacagtt cgatgcttcc 1140
gttagcgccg gtacttatga ttccagctcc gacacctaca ccaccctgct cgacgccgtt 1200
cagacctatg ctgatggctt cgtcctgatg gtcgctcagt acacccctgc caacggttcc 1260
ctctccgagc agtacgccaa ggccgatggc agccccactt ccgcctacga cctgacttgg 1320
tcctttgctg ctgccctcac cgccttcgct gcccgcgacg gcaaaaccta tggtagctgg 1380
ggtgccgccg atctctccag cacctgcagc ggttccaccg acactgtcgc cgtcactttc 1440
gaggtccagt acgacaccca atatggtgaa aatctgtaca ttaccggcag cgtctcccag 1500
ctcgaggatt ggagcgctga tgatgctctc atcatgtcca gcgccgacta tcccacctgg 1560
tccatcaccg tcgatctgcc ccctagcacc ctgatccaat acaaatacct caccaagtat 1620
aacggcgatg tcacctggga agacgatccc aacaacgaaa ttaccactcc tgcctccggc 1680
tcctataccc aggttgacag ctggcactaa 1710
<210> 6
<211> 1857
<212> ДНК
<213> Talaromyces emersonii
<400> 6
atggcctccc tggttgctgg tgctctgtgc atcctcggcc tgacccctgc cgccttcgcc 60
cgagcccccg tcgctgcccg cgccactggc agcctcgaca gcttcctcgc caccgagacc 120
cctatcgccc tccagggcgt tctgaacaac atcggtccca acggcgctga cgtcgccggt 180
gctagcgccg gtatcgtcgt tgccagccct agccgatccg accccaacta cttctacagc 240
tggacccgcg acgccgctct caccgctaag tacctggtcg acgccttcaa tcgcggcaac 300
aaagacctcg agcaaaccat ccagcagtac atctccgctc aggccaaggt ccagaccatt 360
tccaacccca gcggcgatct gagcactggc ggcctgggcg agcccaagtt caacgtcaat 420
gagaccgctt tcactggccc ctggggccga cctcaacgcg atggccctgc tctccgagcc 480
accgccctca tcgcctatgc taactacctg atcgacaacg gtgaggccag cactgccgac 540
gagatcatct ggcccatcgt ccaaaatgac ctcagctaca tcacccaata ctggaactcc 600
agcacctttg acctgtggga ggaggtcgag ggctccagct tcttcaccac tgctgttcag 660
caccgcgccc tcgttgaggg taatgccctg gccacccgac tcaatcacac ttgctccaac 720
tgcgtcagcc aggcccccca ggtcctctgc tttctccaga gctactggac cggcagctac 780
gtcctggcca actttggtgg cagcggccga agcggcaagg acgtcaacag catcctgggt 840
tccatccaca ccttcgaccc cgctggcggt tgcgacgact ccactttcca gccttgcagc 900
gctcgcgctc tcgccaacca caaggtcgtc accgattcct tccgctccat ctacgccatc 960
aattccggca tcgccgaggg tagcgctgtt gctgtcggcc gctaccccga ggacgtctac 1020
caaggcggca atccctggta tctcgctact gccgctgccg ccgagcagct ctatgacgct 1080
atctatcagt ggaaaaagat cggtagcatc agcattaccg acgtcagcct ccccttcttc 1140
caggacatct acccctccgc cgctgttggc acctacaatt ccggctccac caccttcaac 1200
gacatcatca gcgccgtcca gacttatggc gacggctacc tgagcattgt cgagaagtac 1260
acccccagcg atggcagcct caccgagcaa ttcagccgca ccgacggcac ccccctgtcc 1320
gcttccgccc tcacctggag ctacgcttcc ctgctcaccg cctccgctcg ccgccagagc 1380
gtcgttcccg ctagctgggg cgagagcagc gccagctccg tcctggccgt ctgctccgct 1440
actagcgcca ccggccccta ctccactgcc accaacaccg tttggccttc cagcggctcc 1500
ggcagctcca ctaccacctc cagcgcccct tgcaccaccc ctaccagcgt cgccgtcacc 1560
ttcgacgaga tcgtcagcac cagctacggc gagaccatct atctggctgg cagcatcccc 1620
gagctgggca attggtccac cgccagcgct attcctctgc gcgctgacgc ctacactaat 1680
agcaaccctc tgtggtatgt caccgttaac ctccctcccg gcactagctt tgagtataag 1740
tttttcaaga accagaccga tggtactatt gtctgggagg acgaccccaa ccgatcctac 1800
accgtccccg cctactgcgg tcagactacc gctatcctcg acgattcctg gcagtaa 1857
<210> 7
<211> 551
<212> БЕЛОК
<213> Fibroporia radiculosa
<400> 7
Gln Ser Thr Ser Val Ser Ala Tyr Ile Ala Ser Glu Ser Pro Val Ala
1. 5 10 15
Lys Ala Gly Val Leu Ala Asn Ile Gly Thr Glu Gly Ser Leu Ser Ser
20 25 30
Gly Ala Tyr Ser Gly Val Val Ile Ala Ser Pro Ser Thr Val Asn Pro
35 40 45
Asp Tyr Leu Tyr Thr Trp Val Arg Asp Ser Ser Leu Thr Phe Gln Ala
50 55 60
Leu Ile Asp Gln Tyr Val Tyr Gly Glu Asp Pro Thr Leu Arg Ser Leu
65 70 75 80
Ile Asp Glu Phe Ile Thr Ala Glu Ser Ile Leu Gln Gln Thr Thr Asn
85 90 95
Pro Ser Gly Thr Val Ser Thr Gly Gly Leu Gly Glu Pro Lys Phe Asn
100 105 110
Ile Asn Glu Thr Ala Phe Thr Gly Pro Trp Gly Arg Pro Gln Arg Asp
115 120 125
Gly Pro Ala Leu Arg Ser Thr Ala Ile Ile Thr Tyr Ala Thr Tyr Leu
130 135 140
Trp Asn Ser Gly Asn Thr Ser Tyr Val Ser Asp Ser Leu Trp Pro Ile
145 150 155 160
Ile Glu Leu Asp Leu Asn Tyr Ile Ala Thr Tyr Trp Asn Phe Ser Thr
165 170 175
Phe Asp Leu Trp Glu Glu Ile Asp Ser Ser Ser Phe Trp Thr Thr Ala
180 185 190
Val Gln His Arg Ala Leu Arg Gln Gly Ile Thr Phe Ala Asn Leu Ile
195 200 205
Gly Gln Thr Ser Pro Val Ser Asn Tyr Glu Thr Gln Ala Gly Asp Ile
210 215 220
Leu Cys Phe Leu Gln Thr Tyr Trp Asn Pro Thr Gly Asn Tyr Met Thr
225 230 235 240
Ala Asn Thr Gly Gly Gly Arg Ser Gly Lys Asp Ser Asn Thr Val Leu
245 250 255
Ala Ser Val His Thr Phe Asp Pro Asp Ala Gly Cys Asp Ser Thr Thr
260 265 270
Phe Gln Pro Cys Ser Asp Lys Ala Leu Ser Asn Leu Lys Val Tyr Val
275 280 285
Asp Ser Phe Arg Ser Leu Tyr Ala Ile Asn Asp Gly Ile Ala Ser Asp
290 295 300
Ala Ala Val Ala Thr Gly Arg Tyr Pro Glu Asp Val Tyr Tyr Gly Gly
305 310 315 320
Asn Pro Trp Tyr Leu Cys Thr Phe Ala Val Ala Glu Gln Leu Tyr Asp
325 330 335
Ala Leu Ile Val Trp Ser Ser Gln Gly Tyr Leu Glu Ile Thr Asp Leu
340 345 350
Ser Leu Ala Phe Phe Gln Gln Phe Asp Ser Asp Val Gly Thr Gly Thr
355 360 365
Tyr Asp Ser Gly Ser Ser Thr Tyr Ser Thr Leu Thr Ser Ala Ile Arg
370 375 380
Thr Phe Ala Asp Gly Phe Val Leu Thr Asn Ala Lys Tyr Thr Pro Thr
385 390 395 400
Asn Gly Ser Leu Ser Glu Glu Tyr Thr Ser Ala Asp Gly Thr Pro Ile
405 410 415
Ser Ala Tyr Asp Leu Thr Trp Ser Tyr Ala Ser Ala Leu Thr Val Phe
420 425 430
Ala Ala Glu Ala Gly Thr Thr Tyr Gly Ser Trp Gly Ala Ala Gly Leu
435 440 445
Thr Val Pro Ser Thr Cys Thr Ser Gly Val Ala Val Thr Phe Glu Val
450 455 460
Asp Tyr Asp Thr Glu Tyr Gly Glu Asn Val Tyr Ile Thr Gly Ser Val
465 470 475 480
Asn Ala Leu Glu Asn Trp Ser Ala Thr Asn Ala Leu Ile Met Ser Ala
485 490 495
Ala Asp Tyr Pro Thr Trp Ser Ile Thr Val Tyr Leu Pro Pro Ser Thr
500 505 510
Thr Ile Gln Tyr Lys Tyr Leu Thr Gln Tyr Asn Gly Glu Val Thr Trp
515 520 525
Glu Asp Asp Pro Asn Asn Glu Ile Thr Thr Pro Ala Ser Gly Ser Met
530 535 540
Thr Gln Val Asp Ser Trp His
545 550
<210> 8
<211> 552
<212> БЕЛОК
<213> Wolfiporia cocos
<400> 8
Gln Thr Thr Ser Val Thr Ser Tyr Ile Ala Ser Glu Ser Pro Ile Ala
1. 5 10 15
Lys Ala Gly Val Leu Ala Asn Ile Gly Ala Asp Gly Ser Leu Ser Ser
20 25 30
Gly Ala Tyr Ser Gly Ile Val Ile Ala Ser Pro Ser Thr Val Asn Pro
35 40 45
Asn Tyr Leu Tyr Thr Trp Thr Arg Asp Ser Ser Leu Thr Phe Met Glu
50 55 60
Leu Ile Asn Gln Tyr Ile Tyr Gly Glu Asp Asp Thr Leu Arg Thr Leu
65 70 75 80
Ile Asp Glu Phe Val Ser Ala Glu Ala Thr Leu Gln Gln Val Thr Asn
85 90 95
Pro Ser Gly Thr Val Ser Thr Gly Gly Leu Gly Glu Pro Lys Phe Asn
100 105 110
Ile Asn Glu Thr Ala Phe Thr Gly Pro Trp Gly Arg Pro Gln Arg Asp
115 120 125
Gly Pro Ala Leu Arg Ala Thr Ala Ile Met Ala Tyr Ala Thr Tyr Leu
130 135 140
Tyr Glu Asn Gly Asn Thr Ser Tyr Val Thr Asp Thr Leu Trp Pro Ile
145 150 155 160
Ile Glu Leu Asp Leu Gly Tyr Val Ala Glu Tyr Trp Asn Glu Ser Thr
165 170 175
Phe Asp Leu Trp Glu Glu Ile Asp Ser Ser Ser Phe Phe Thr Thr Ala
180 185 190
Val Gln His Arg Ala Leu Arg Ala Gly Val Thr Phe Ala Asn Leu Ile
195 200 205
Gly Glu Thr Ser Asp Val Ser Asn Tyr Gln Glu Asn Ala Asp Asp Leu
210 215 220
Leu Cys Phe Leu Gln Ser Tyr Trp Asn Pro Thr Gly Ser Tyr Val Thr
225 230 235 240
Ala Asn Thr Gly Gly Gly Arg Ser Gly Lys Asp Ala Asn Thr Leu Leu
245 250 255
Ala Ser Ile His Thr Phe Asp Pro Asp Ala Gly Cys Asn Ala Thr Thr
260 265 270
Phe Gln Pro Cys Ser Asp Lys Ala Leu Ser Asn His Lys Val Tyr Val
275 280 285
Asp Ser Phe Arg Ser Leu Tyr Ala Ile Asn Asp Asp Ile Ser Ser Asp
290 295 300
Ala Ala Val Ala Thr Gly Arg Tyr Pro Glu Asp Val Tyr Tyr Asn Gly
305 310 315 320
Asn Pro Trp Tyr Leu Cys Thr Leu Ala Ala Ala Glu Gln Leu Tyr Asp
325 330 335
Ser Leu Ile Val Trp Lys Ala Gln Gly Tyr Ile Glu Val Thr Ser Leu
340 345 350
Ser Leu Ala Phe Phe Gln Gln Phe Asp Ala Ser Val Ser Ala Gly Thr
355 360 365
Tyr Asp Ser Ser Ser Asp Thr Tyr Thr Thr Leu Leu Asp Ala Val Gln
370 375 380
Thr Tyr Ala Asp Gly Phe Val Leu Met Val Ala Gln Tyr Thr Pro Ala
385 390 395 400
Asn Gly Ser Leu Ser Glu Gln Tyr Ala Lys Ala Asp Gly Ser Pro Thr
405 410 415
Ser Ala Tyr Asp Leu Thr Trp Ser Phe Ala Ala Ala Leu Thr Ala Phe
420 425 430
Ala Ala Arg Asp Gly Lys Thr Tyr Gly Ser Trp Gly Ala Ala Asp Leu
435 440 445
Ser Ser Thr Cys Ser Gly Ser Thr Asp Thr Val Ala Val Thr Phe Glu
450 455 460
Val Gln Tyr Asp Thr Gln Tyr Gly Glu Asn Leu Tyr Ile Thr Gly Ser
465 470 475 480
Val Ser Gln Leu Glu Asp Trp Ser Ala Asp Asp Ala Leu Ile Met Ser
485 490 495
Ser Ala Asp Tyr Pro Thr Trp Ser Ile Thr Val Asp Leu Pro Pro Ser
500 505 510
Thr Leu Ile Gln Tyr Lys Tyr Leu Thr Lys Tyr Asn Gly Asp Val Thr
515 520 525
Trp Glu Asp Asp Pro Asn Asn Glu Ile Thr Thr Pro Ala Ser Gly Ser
530 535 540
Tyr Thr Gln Val Asp Ser Trp His
545 550
<210> 9
<211> 591
<212> БЕЛОК
<213> Talaromyces emersonii
<400> 9
Ala Thr Gly Ser Leu Asp Ser Phe Leu Ala Thr Glu Thr Pro Ile Ala
1. 5 10 15
Leu Gln Gly Val Leu Asn Asn Ile Gly Pro Asn Gly Ala Asp Val Ala
20 25 30
Gly Ala Ser Ala Gly Ile Val Val Ala Ser Pro Ser Arg Ser Asp Pro
35 40 45
Asn Tyr Phe Tyr Ser Trp Thr Arg Asp Ala Ala Leu Thr Ala Lys Tyr
50 55 60
Leu Val Asp Ala Phe Asn Arg Gly Asn Lys Asp Leu Glu Gln Thr Ile
65 70 75 80
Gln Gln Tyr Ile Ser Ala Gln Ala Lys Val Gln Thr Ile Ser Asn Pro
85 90 95
Ser Gly Asp Leu Ser Thr Gly Gly Leu Gly Glu Pro Lys Phe Asn Val
100 105 110
Asn Glu Thr Ala Phe Thr Gly Pro Trp Gly Arg Pro Gln Arg Asp Gly
115 120 125
Pro Ala Leu Arg Ala Thr Ala Leu Ile Ala Tyr Ala Asn Tyr Leu Ile
130 135 140
Asp Asn Gly Glu Ala Ser Thr Ala Asp Glu Ile Ile Trp Pro Ile Val
145 150 155 160
Gln Asn Asp Leu Ser Tyr Ile Thr Gln Tyr Trp Asn Ser Ser Thr Phe
165 170 175
Asp Leu Trp Glu Glu Val Glu Gly Ser Ser Phe Phe Thr Thr Ala Val
180 185 190
Gln His Arg Ala Leu Val Glu Gly Asn Ala Leu Ala Thr Arg Leu Asn
195 200 205
His Thr Cys Ser Asn Cys Val Ser Gln Ala Pro Gln Val Leu Cys Phe
210 215 220
Leu Gln Ser Tyr Trp Thr Gly Ser Tyr Val Leu Ala Asn Phe Gly Gly
225 230 235 240
Ser Gly Arg Ser Gly Lys Asp Val Asn Ser Ile Leu Gly Ser Ile His
245 250 255
Thr Phe Asp Pro Ala Gly Gly Cys Asp Asp Ser Thr Phe Gln Pro Cys
260 265 270
Ser Ala Arg Ala Leu Ala Asn His Lys Val Val Thr Asp Ser Phe Arg
275 280 285
Ser Ile Tyr Ala Ile Asn Ser Gly Ile Ala Glu Gly Ser Ala Val Ala
290 295 300
Val Gly Arg Tyr Pro Glu Asp Val Tyr Gln Gly Gly Asn Pro Trp Tyr
305 310 315 320
Leu Ala Thr Ala Ala Ala Ala Glu Gln Leu Tyr Asp Ala Ile Tyr Gln
325 330 335
Trp Lys Lys Ile Gly Ser Ile Ser Ile Thr Asp Val Ser Leu Pro Phe
340 345 350
Phe Gln Asp Ile Tyr Pro Ser Ala Ala Val Gly Thr Tyr Asn Ser Gly
355 360 365
Ser Thr Thr Phe Asn Asp Ile Ile Ser Ala Val Gln Thr Tyr Gly Asp
370 375 380
Gly Tyr Leu Ser Ile Val Glu Lys Tyr Thr Pro Ser Asp Gly Ser Leu
385 390 395 400
Thr Glu Gln Phe Ser Arg Thr Asp Gly Thr Pro Leu Ser Ala Ser Ala
405 410 415
Leu Thr Trp Ser Tyr Ala Ser Leu Leu Thr Ala Ser Ala Arg Arg Gln
420 425 430
Ser Val Val Pro Ala Ser Trp Gly Glu Ser Ser Ala Ser Ser Val Leu
435 440 445
Ala Val Cys Ser Ala Thr Ser Ala Thr Gly Pro Tyr Ser Thr Ala Thr
450 455 460
Asn Thr Val Trp Pro Ser Ser Gly Ser Gly Ser Ser Thr Thr Thr Ser
465 470 475 480
Ser Ala Pro Cys Thr Thr Pro Thr Ser Val Ala Val Thr Phe Asp Glu
485 490 495
Ile Val Ser Thr Ser Tyr Gly Glu Thr Ile Tyr Leu Ala Gly Ser Ile
500 505 510
Pro Glu Leu Gly Asn Trp Ser Thr Ala Ser Ala Ile Pro Leu Arg Ala
515 520 525
Asp Ala Tyr Thr Asn Ser Asn Pro Leu Trp Tyr Val Thr Val Asn Leu
530 535 540
Pro Pro Gly Thr Ser Phe Glu Tyr Lys Phe Phe Lys Asn Gln Thr Asp
545 550 555 560
Gly Thr Ile Val Trp Glu Asp Asp Pro Asn Arg Ser Tyr Thr Val Pro
565 570 575
Ala Tyr Cys Gly Gln Thr Thr Ala Ile Leu Asp Asp Ser Trp Gln
580 585 590
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Композиции для получения глюкозных сиропов | 2015 |
|
RU2706297C2 |
| ГЛЮКОАМИЛАЗА TRICHODERMA REESEI И ЕЕ ГОМОЛОГИ | 2005 |
|
RU2394101C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С НЕОНАТАЛЬНЫМ FC-РЕЦЕПТОРОМ (FCRN) ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2830231C2 |
| ВАРИАНТЫ IgG-FC ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2814952C2 |
| СПОСОБ УДАЛЕНИЯ СМОЛИСТЫХ ВЕЩЕСТВ ИЗ РАСТИТЕЛЬНОГО МАСЛА И ЕГО РАФИНИРОВАНИЯ | 2018 |
|
RU2772452C2 |
| АНТАГОНИСТЫ NGF ДЛЯ МЕДИЦИНСКОГО ИСПОЛЬЗОВАНИЯ | 2020 |
|
RU2829812C2 |
| CD47-АНТИГЕНСВЯЗЫВАЮЩАЯ ЕДИНИЦА И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2780859C2 |
| МОЛЕКУЛА РЕЦЕПТОРА IL4/IL13 ДЛЯ ВЕТЕРИНАРНОГО ПРИМЕНЕНИЯ | 2018 |
|
RU2795591C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
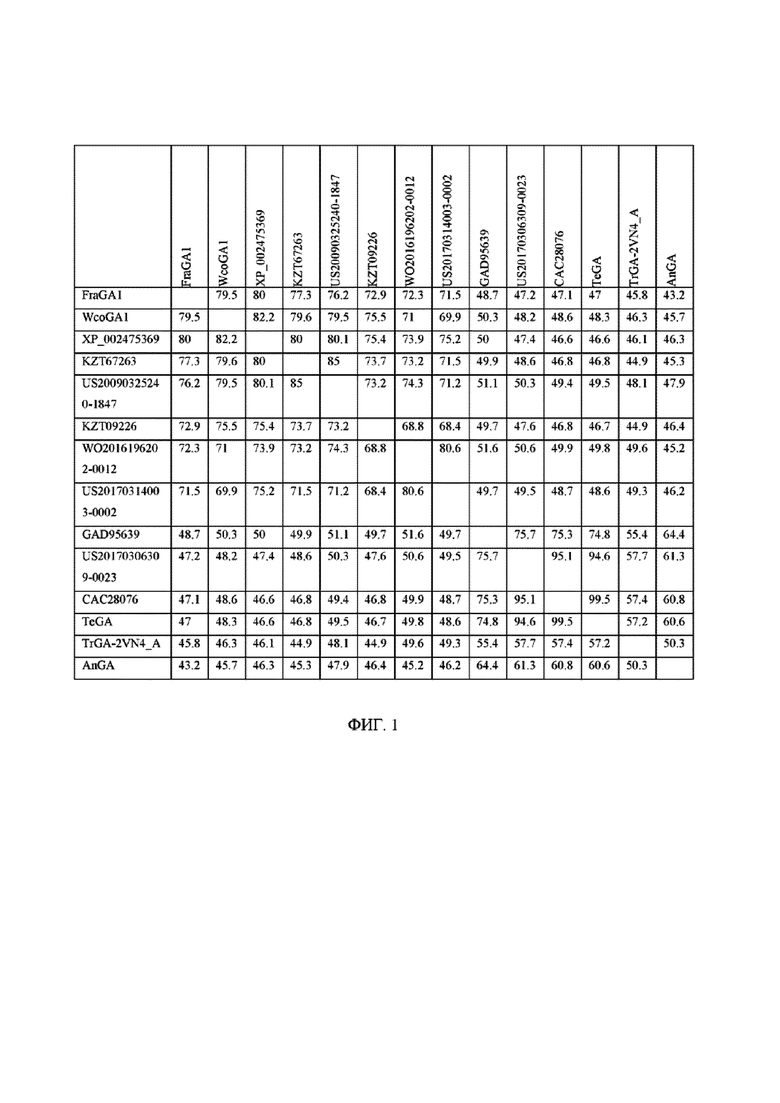
Изобретение относится к биотехнологии. Предложен вектор для трансформации в экспрессирующую клетку-хозяина, содержащий полинуклеотидную последовательность, включающую нуклеотидную последовательность, которая по меньшей мере на 97% идентична нуклеотидной последовательности SEQ ID NO: 4 или 5, при этом нуклеотидная последовательность кодирует функциональный фермент глюкоамилазу. Также предложена рекомбинантная клетка-хозяин для экспрессии представляющего интерес полипептида, содержащая вышеуказанный вектор. Изобретение обеспечивает получение глюкоамилаз с повышенной стабильностью и активностью, улучшенной толерантностью к низким значениям pH. 2 н. и 4 з.п. ф-лы, 1 ил., 13 табл., 11 пр.
1. Вектор для трансформации в экспрессирующую клетку-хозяина, содержащий полинуклеотидную последовательность, включающую нуклеотидную последовательность, которая по меньшей мере на 97% идентична нуклеотидной последовательности SEQ ID NO: 4 или 5, функционально связанную с одной или несколькими управляющими последовательностями, которые регулируют продуцирование кодируемого полипептида в экспрессирующем хозяине, и где указанная регуляторная последовательность гетерологична кодирующей нуклеотидной последовательности, или указанные регуляторная последовательность и кодирующая последовательность не расположены вместе, как это бывает в природе, где нуклеотидная последовательность кодирует функциональный фермент глюкоамилаза.
2. Рекомбинантная клетка-хозяин для экспрессии представляющего интерес полипептида, содержащая вектор по п. 1.
3. Рекомбинантная клетка-хозяин по п. 2, которая представляет собой клетку Trichoderma, Aspergillus, Myceliopthora или Saccharomyces.
4. Рекомбинантная клетка-хозяин по п. 3, которая представляет собой клетку E.coli, Bacillus, Streptomyces или Pseudomonas.
5. Рекомбинантная клетка-хозяин по п. 3, которая представляет собой этанолопродуцирующие микроорганизмы.
6. Рекомбинантная клетка-хозяин согласно любому из пп. 2-5, дополнительно экспрессирующая и секретирующая один или несколько дополнительных ферментов, выбранных из группы, состоящей из протеазы, гемицеллюлазы, целлюлазы, пероксидазы, липолитического фермента, металлолиполитического фермента, ксиланазы, липазы, фосфолипазы, эстеразы, пергидролазы, кутиназы, пектиназы, пектатлиазы, манназы, кератиназы, редуктазы, оксидазы, фенолоксидазы, липоксигеназы, лигниназы, пуллуназы, фитазы, танназы, пентосаназы, маланазы, бета-глюканазы, арабиносидазы, гиалуронидазы, хондроитиназы, лакказы, трансферазы или любой их комбинации.
| US 2011039308 A1, 17.02.2011 | |||
| UniProtKB, J4I862_9APHY, 31.10.2012 | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| UniProtKB, A0A2H3JCA1_WOLCO, 31.01.2018 | |||
| Найдено онлайн: https://www.uniprot.org/uniprotkb/A0A2H3JCA1/entry Дата обращения 09.02.2023 | |||
| WO2016087445 A1, 09.06.2016 | |||
| ГЛЮКОАМИЛАЗА TRICHODERMA REESEI И ЕЕ ГОМОЛОГИ | 2005 |
|
RU2394101C2 |
