ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] Данная заявка заявляет приоритет по заявке на патент США № 62/470605, поданной 13 марта 2017 года, заявке на патент США № 62/470620, поданной 13 марта 2017 года, заявке на патент США № 62/482148, поданной 5 апреля 2017 года, и заявке на патент США № 62/480798, поданной 3 апреля 2017 г., которые включены в данное описание в полном объеме посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[002] Данное изобретение относится к получению гексозных моносахаридов. Более конкретно, данное изобретение относится к способам получения D-гексозы (или гексозы) из сахаридов (например, полисахаридов, олигосахаридов, дисахаридов, сахарозы, D-глюкозы и D-фруктозы), которые включают стадию, на которой фруктозо-6-фосфат превращается в гексозу одной или более ферментативными стадиями.
УРОВЕНЬ ТЕХНИКИ
[003] Гексозы представляют собой моносахариды с шестью атомами углерода. Гексозы могут быть классифицированы по функциональной группе: альдогексозы, имеющие альдегидную группу в положении 1, и кетогексозы, имеющие кетогруппу в положении 2. Альдогексозы (или альдозы) включают аллозу, альтрозу, глюкозу, гулозу, галактозу, идозу, талозу и маннозу. Кетогексозы (или кетозы) включают псикозу (аллулозу), фруктозу, тагатозу и сорбозу. Различные аспекты данных альдогексоз и кетогексоз упоминаются в следующих параграфах.
[004] Например, D-аллоза (далее - аллоза) представляет собой низкокалорийный натуральный подсластитель, который имеет ~80% сладости сахарозы и описана как некалорийный подсластитель и наполнитель. Она представляет собой встречающуюся в природе моносахаридную гексозу, которая присутствует только в небольших количествах в определенных кустарниках и водорослях. Аллоза выгодно отличается несколькими потенциальными медицинскими и сельскохозяйственными преимуществами, включая криопротекторные, антиоксидантные, антигипертензивные, иммунодепрессивные, противовоспалительные, противоопухолевые и противораковые действия. Она также имеет схожую функциональность в пищевых продуктах и напитках с сахарозой. Таким образом, аллоза явно находит применение в пищевой промышленности и производстве напитков. Однако из-за высоких продажных цен аллозы, ее применение в качестве подсластителя ограничено.
[005] В настоящее время аллозу получают преимущественно путем ферментативной изомеризации D-псикозы (WO 2014069537). В целом, метод имеет недостаток из-за более высокой стоимости сырья, дорогостоящего отделения аллозы от D-псикозы и относительно низкого выхода продукта (~23%).
[006] Альтроза является еще одной неестественной альдогексозой и C-3 эпимером маннозы. D-альтрозу ((2S,3R,4R,5R)-2,3,4,5,6-пентагидроксигексаналь) можно применять в качестве субстрата для идентификации, дифференциации и характеристики альдозоизомераз, таких как L-фукозоизомераза, из Caldicellulosiruptor saccharolyticus и D-арабинозоизомераза (d-AI) из Bacillus pallidus (B. pallidus) и Klebsiella pneumoniae. В последнее время сахарные цепи, такие как олигосахариды и полисахариды, которые выполняют функции, пригодные в качестве физиологически активного вещества, привлекли внимание в области высокочистых химических веществ, таких как лекарства и сельскохозяйственные химические вещества. В настоящее время объекты исследований сахарных цепей ограничиваются теми, которые состоят из моносахаридов, присутствующих в природе в больших количествах и легкодоступных для исследователей, таких как D-глюкоза, D-манноза и D-галактоза. Тем не менее, ожидается, что в будущем для изучения синтеза сахарных цепей, выполняющих более полезные функции, потребуются различные моносахариды, помимо тех, которые присутствуют в природе. В этих условиях крайне важно и необходимо разработать метод, который позволяет получать D-альтрозу, которая является редким сахаром, который трудно получить, с высоким выходом при уменьшении количества стадий обработки. Патент США № 5410038.
[007] D-Гулоза пригодна, например, в качестве вспомогательного вещества, хелатирующего агента, фармацевтического промежуточного продукта, чистящего средства для стекла и металлов, пищевой добавки и в качестве добавки для моющих средств. Патент США № 5215591.
[008] D-галактоза (далее галактоза) представляет собой натуральный подсластитель, который имеет ~33% сладости сахарозы и описана как пищевой подсластитель. Она представляет собой встречающуюся в природе моносахаридную гексозу, которая присутствует в молочных продуктах, бобовых, зерновых, орехах, клубнях и овощах. Галактозу применяют в хлебопекарной промышленности для ограничения грубости и кислотности пищевых продуктов. Кроме того, ее применяют в качестве источника энергии для повышения выносливости в индустрии пищевых добавок для спортсменов. В фармацевтической промышленности она является промежуточным звеном для нескольких лекарственных средств, а также ее применяют в качестве модулятора клеточного метаболизма при оптимизации биопродукции терапевтических белков. Кроме того, было показано, что галактоза эффективна в качестве средства борьбы с болезнями растений, вызываемыми определенными патогенами растений, такими как патогены огурца, моркови, картофеля и томатов. Из-за проблем с питанием (например, веганство) и проблем со здоровьем (например, болезнь BSE) источники галактозы, не относящиеся к животным, представляют интерес для промышленности. Таким образом, галактоза, несомненно, имеет множество применений в пище, напитках, физических упражнениях, сельском хозяйстве и фармацевтической промышленности. Однако из-за высоких продажных цен галактозы ее применение было ограничено.
[009] Галактозу получают преимущественно путем гидролиза лактозы (WO 2005039299A3). Этот метод менее желателен из-за более дорогого исходного сырья и дорогостоящего отделения глюкозы от галактозы. В альтернативном варианте, галактоза может быть получена путем гидролиза растительной биомассы (WO 2005001145A1). Этот метод страдает от дорогостоящего отделения галактозы от множества других сахаров, высвобождаемых во время гидролиза биомассы (например, ксилозы, арабинозы, маннозы, глюкозы и рамнозы), и низкого выхода (~4,6% сухой массы обычных источников биомассы составляет галактоза).
[010] Идоза не встречается в природе, но важна ее уроновая кислота, идуроновая кислота. Она является компонентом дерматансульфата и гепарансульфата, которые являются гликозаминогликанами (https://en.wikipedia.org/wiki/Idose - доступно 3/7/18).
[011] Талоза является неестественной альдогексозой, которая растворима в воде и слабо растворяется в метаноле. Она представляет собой C-2 эпимер галактозы и C-4 эпимер маннозы. Талозу можно применять в качестве субстрата для идентификации, дифференциации и характеристики рибозо-5-фосфат-изомеразы из Clostridia.
[012] D-манноза (далее - манноза) - это слабо сладкий природный моносахарид, который содержится во многих фруктах, овощах, растительных материалах и даже в организме человека. Манноза отличается множеством преимуществ для здоровья и фармацевтического применения. Например, маннозу можно применять для лечения синдрома гликопротеина с дефицитом углеводов типа 1b и, чаще, инфекций мочевыводящих путей. Кроме того, манноза является проверенным пребиотиком, не повышает уровень глюкозы в крови и обладает противовоспалительными свойствами. Кроме того, было показано, что она увеличивает выход живого веса у свиней и является широко применяемым вспомогательным увлажняющим средством для ухода за кожей. Таким образом, манноза имеет множество применений в фармацевтике, косметике, производстве напитков, пищевых продуктов, молочной, кондитерской промышленности и животноводстве. Однако из-за высоких продажных цен маннозы ее использование в повседневных продуктах было ограничено.
[013] Маннозу в основном производят путем экстракции из растений. Обычные способы включают кислотный гидролиз, термический гидролиз, ферментативный гидролиз, гидролиз микробной ферментацией и их комбинации. Менее распространенные методы включают химические и биологические преобразования. В целом, данные методы являются проблематичными из-за суровых условий, высоких капитальных затрат, более высоких затрат на сырье, дорогостоящего отделения маннозы от продуктов реакций изомеризации и относительно низких выходов продукта (15-35%).
[014] D-аллулоза (также известная как D-псикоза) (далее псикоза) представляет собой низкокалорийный натуральный подсластитель, который имеет 70% сладости сахарозы, но только 10% калорий. Она представляет собой встречающуюся в природе моносахаридную гексозу, которая присутствует в небольших количествах в пшенице и других растениях. Психоза была одобрена в качестве пищевой добавки Управлением по контролю за продуктами и лекарствами (FDA) в 2012 году, которое объявило ее общепризнанной безопасной (GRAS - generally recognized as safe). Однако из-за высоких продажных цен псикозы ее применение в качестве подсластителя ограничено. Псикоза отличается множеством преимуществ для здоровья: она низкокалорийна (10% сахарозы); имеет очень низкий гликемический индекс 1; она полностью всасывается в тонкой кишке, но не метаболизируется и вместо этого выделяется с мочой и калом; она помогает регулировать уровень сахара в крови путем ингибирования альфа-амилазы, сахаразы и мальтазы; и она имеет такую же функциональность в пищевых продуктах и напитках, что и сахароза. Таким образом, псикоза явно находит применение в пищевой промышленности и производстве напитков.
[015] В настоящее время псикозу получают преимущественно путем ферментативной изомеризации фруктозы (WO 2014049373). В целом, данный способ демонстрирует более высокую стоимость исходного сырья, дорогостоящее отделение пикозы от фруктозы и относительно низкий выход продукта.
[016] Фруктоза представляет собой простой кетоновый моносахарид, встречающийся во многих растениях, где она часто связана с глюкозой с образованием дисахарида, сахарозы. Коммерчески фруктозу получают из сахарного тростника, сахарной свеклы и кукурузы. Основной причиной того, что фруктозу применяют в коммерческих целях в продуктах питания и напитках, помимо ее низкой стоимости, является ее высокая относительная сладость. Она представляет собой самый сладкий из всех встречающихся в природе углеводов. Фруктоза также содержится в производимом подсластителе, кукурузном сиропе с высоким содержанием фруктозы (HFCS - high-fructose corn syrup), который производится путем обработки кукурузного сиропа ферментами, превращающими глюкозу в фруктозу (https://en.wikipedia.org/wiki/Fructose#Physical_and_functional_properties-- доступно 3/7/18).
[017] D-тагатоза (далее тагатоза) представляет собой низкокалорийный натуральный подсластитель, который имеет 92% сладости сахарозы, но только 38% калорий. Она представляет собой встречающуюся в природе моносахаридную гексозу, которая присутствует в небольших количествах в фруктах, какао и молочных продуктах. Тагатоза была одобрена в качестве пищевой добавки Управлением по контролю за продуктами и лекарствами (FDA) в 2003 году, которое объявило ее общепризнанной безопасной (GRAS). Однако из-за высоких продажных цен тагатозы ее применение в качестве подсластителя ограничено. Тагатоза отличается множеством полезных для здоровья свойств: она не вызывает кариеса; она низкокалорийна; она имеет очень низкий гликемический индекс 3; она ослабляет гликемический индекс глюкозы на 20%; она может снизить средний уровень глюкозы в крови; она помогает предотвратить сердечно-сосудистые заболевания, инсульты и другие сосудистые заболевания, повышая уровень холестерина липопротеинов высокой плотности (ЛПВП); и она представляет собой проверенный пребиотик и антиоксидант. Lu и др., Tagatose, a New Antidiabetic and Obesity Control Drug, Diabetes Obes. Metab. 10(2): 109-34 (2008). Таким образом, тагатоза, несомненно, имеет множество применений в фармацевтической, биотехнологической, академической, пищевой промышленности, производстве напитков, диетических добавок и бакалейной промышленности.
[018] Тагатозу получают преимущественно путем гидролиза лактозы с помощью лактазы или кислотного гидролиза с образованием D-глюкозы и D-галактозы (WO 2011150556, CN 103025894, US 5002612, US 6057135 и US 8802843). Затем D-галактозу изомеризуют в D-тагатозу либо химически гидроксидом кальция в щелочных условиях, либо ферментативно изомеразой L-арабинозы в рН нейтральных условиях. Конечный продукт выделяют комбинацией фильтрации и ионообменной хроматографии. Данный процесс проводят в нескольких резервуарах или биореакторах. В целом, данный способ является невыгодным из-за дорогостоящего разделения других сахаров (например, D-глюкозы, D-галактозы и негидролизованной лактозы) и низкого выхода продукта. Разрабатываются несколько методов ферментации микробными клетками, но ни один из них не является практической альтернативой из-за их зависимости от дорогостоящего сырья (например, галактитола и D-псикозы), низкого выхода продукта и дорогостоящего разделения.
[019] Сорбоза ((3R,4S,5R)-1,3,4,5,6-пентагидроксигексан-2-он) представляет собой кетогексозу, обладающую сладостью, эквивалентной сахарозе (столовый сахар), и она представляет собой растительный метаболит, который был обнаружен в природе в винограде в небольших количествах. Было установлено, что D-сорбоза эффективна в качестве средства контроля заболеваний растений, вызванных: Pseudomonas syringae pv. lachrymans и Ralstonia solanacearum. Публикация заявки на патент США № 2016/0037768.
[020] Существует необходимость в разработке экономически эффективных путей синтеза для высокопроизводительного получения гексоз, таких как альдогексозы и альдокетозы, которые обсуждались выше, где, по меньшей мере, одна стадия указанных способов включает энергетически благоприятную химическую реакцию. Кроме того, существует потребность в производственных процессах, где технологические этапы могут проводиться в одном резервуаре или биореакторе и/или где дорогостоящие этапы разделения обходятся или исключаются. Также существует необходимость в способах получения гексоз, которые можно проводить при относительно низкой концентрации фосфата, где фосфат можно рециркулировать, и/или способ не требует использования аденозинтрифосфата (АТФ) в качестве дополнительного источника фосфата. Также существует необходимость в путях получения гексоз, которые не требуют использования дорогостоящего кофермента никотинамид-аденозин динуклеотида (NAD(P)(H)) на любой из стадий реакции.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[021] Описанные в данном документе изобретения в основном относятся к способам получения гексоз из сахаридов ферментативным превращением. Данные изобретения также относятся к гексозам, полученным любым из способов, описанных в данном документе.
[022] Более конкретно, данное изобретение относится к способам получения гексозы, выбранной из аллозы, маннозы, галактозы, фруктозы, альтрозы, талозы, сорбозы, гулозы и идозы, из сахарида, причем данный способ включает: превращение фруктозо-6-фосфата (F6P) в гексозу, катализируемое одним или несколькими ферментами, выбранными из изомеразы, эпимеразы и гексозо-специфической фосфатазы и их смесями.
[023] Способ по данному изобретению для получения аллозы включает превращение F6P в псикозо-6-фосфат (P6P), катализируемое псикозо-6-фосфат-3-эпимеразой (P6PE); превращение P6P в аллозо-6-фосфат (A6P), катализируемое аллозо-6-фосфат-изомеразой (A6PI); и превращение A6P в аллозу, катализируемое аллозо-6-фосфат-фосфатазой (A6PP).
[024] Способ по данному изобретению для получения маннозы включает превращение F6P в маннозо-6-фосфат (M6P), катализируемое маннозо-6-фосфат-изомеразой (M6PI) или фосфоглюкоза/фосфоманнозаизомеразой (PGPMI); и превращение M6P в маннозу, катализируемое маннозо-6-фосфат-фосфатазой (M6PP).
[025] Способ по данному изобретению для получения галактозы включает превращение F6P в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфат-4-эпимеразой (F6PE); превращение T6P в галактозо-6-фосфат (Gal6P), катализируемое галактозо-6-фосфат-изомеразой (Gal6PI); и превращение Gal6P в галактозу, катализируемое галактозо-6-фосфат-фосфатазой (Gal6PP).
[026] Способ по данному изобретению для получения фруктозы включает превращение F6P в фруктозу, катализируемое фруктозо-6-фосфат-фосфатазой (F6PP).
[027] Способ по данному изобретению для получения альтрозы включает превращение F6P в P6P, катализируемое P6PE; превращение P6P в альтрозо-6-фосфат (Alt6P), катализируемое альтрозо-6-фосфат-изомеразой (Alt6PI); и превращение полученного Alt6P в альтрозу, катализируемое альтрозо-6-фосфат-фосфатазой (Alt6PP).
[028] Способ по данному изобретению для получения талозы включает превращение F6P в T6P, катализируемое F6PE; превращение T6P в талозо-6-фосфат (Tal6P), катализируемое талозо-6-фосфат-изомеразой (Tal6PI); и превращение Tal6P в талозу, катализируемое талозо-6-фосфат-фосфатазой (Tal6PP).
[029] Способ по данному изобретению для получения сорбозы включает превращение F6P в T6P, катализируемое F6PE; превращение T6P в сорбозо-6-фосфат (S6P), катализируемое сорбозо-6-фосфат-эпимеразой (S6PE); и превращение S6P в сорбозу, катализируемое сорбозо-6-фосфат-фосфатазой (S6PP).
[030] Способ по данному изобретению для получения гулозы включает превращение F6P в T6P, катализируемое F6PE; превращение S6P в гулозо-6-фосфат (Gul6P), катализируемое гулозо-6-фосфат-изомеразой (Gul6PI); и превращение Gul6P в гулозу, катализируемое гулозо-6-фосфат-фосфатазой (Gul6PP).
[031] Способ по данному изобретению для получения идозы включает превращение F6P в T6P, катализируемое F6PE; превращение T6P в сорбозо-6-фосфат (S6P), катализируемое сорбозо-6-фосфат-эпимеразой (S6PE); превращение S6P в идозо-6-фосфат (I6P), катализируемое идозо-6-фосфат-изомеразой (I6PI); и превращение I6P в идозу, катализируемое идозо-6-фосфат-фосфатазой (I6PP).
[032] Способы получения гексоз в соответствии с данным изобретением могут включать стадию превращения глюкозо-6-фосфата (G6P) в F6P, где указаная стадия катализируется фосфоглюкозоизомеразой (PGI). Данные способы также могут включать стадию превращения глюкозо-1-фосфата (G1P) в G6P, где указанная стадия катализируется фосфоглюкомутазой (PGM). Кроме того, способы в соответствии с данным изобретением могут дополнительно включать стадию превращения сахарида в G1P, где указанная стадия катализируется, по меньшей мере, одним ферментом, и указанный сахарид выбран из группы, состоящей из крахмала или его производного, целлюлозы или ее производного и сахарозы.
[033] Указанный фермент или ферменты, применяемые на стадии превращения сахарида в G1P в способах в соответствии с данным изобретением, могут представлять собой альфа-глюкан-фосфорилазу (αGP), мальтозо-фосфорилазу, сахарозо-фосфорилазу, целлодекстрин-фосфорилазу, целлобиозо-фосфорилазу и/или целлюлозо-фосфорилазу, и их смеси. Когда сахаридом является крахмал или производное крахмала, производное может быть выбрано из группы, состоящей из амилозы, амилопектина, растворимого крахмала, амилодекстрина, мальтодекстрина, мальтозы и глюкозы и их смесей.
[034] Некоторые способы в соответствии с данным изобретением могут дополнительно включать стадию превращения крахмала в производное крахмала, где производное крахмала получают ферментативным гидролизом крахмала или кислотным гидролизом крахмала. Также в способы может быть добавлена 4-глюкантрансфераза (4GT). 4GT можно применять для увеличения выхода гексозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[035] Когда в способах применяют производное крахмала, производное крахмала может быть получено ферментативным гидролизом крахмала, катализируемым изоамилазой, пуллуланазой, альфа-амилазой или их комбинацией.
[036] Способ в соответствии с данным изобретением также может включать стадию превращения фруктозы в F6P, где указанная стадия катализируется по меньшей мере одним ферментом и, необязательно, стадию превращения сахарозы в фруктозу, где указанная стадия катализируется по меньшей мере одним ферментом.
[037] Кроме того, указанные способы получения гексозы в соответствии с данным изобретением могут включать стадию превращения глюкозы в G6P, где указанная стадия катализируется по меньшей мере одним ферментом, и, необязательно, стадию превращения сахарозы в глюкозу, которая катализируется по крайней мере, одним ферментом.
[038] Стадии в каждом из способов синтеза гексозы в соответствии с данным изобретением могут проводиться при температуре в диапазоне от около 40 до около 90°С и при рН в диапазоне от около 5,0 до около 8,0. Их могут проводить в течение от 8 до 48 часов.
[039] Стадии способов в соответствии с данным изобретением могут проводиться в одном биореакторе. Указанные стадии также могут проводиться во множестве биореакторов, расположенных последовательно.
[040] Стадии ферментативного процесса по данному изобретению могут проводиться без АТФ и/или без NAD(Р)(Н). Указанные стадии могут проводиться при концентрации фосфата в диапазоне от около 0,1 до около 150 мМ. Фосфат, применяемый на стадиях фосфорилирования и дефосфорилирования в способах по данному изобретению, может быть переработан. По меньшей мере, одна стадия указанных способов может включать энергетически благоприятную химическую реакцию.
[041] Данное изобретение также относится к аллозе, маннозе, галактозе, фруктозе, альтрозе, талозе, сорбозе, гулозе и идозе, полученным данными способами.
[042] Краткое Описание Графических Материалов
[043] ФИГ. 1 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в аллозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; P6PE, псикозо-6-фосфат-3-эпимераза; A6PI, аллозо-6-фосфат-изомераза; A6PP, аллозо-6-фосфат-фосфатаза.
[044] ФИГ. 2 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в маннозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; PGPMI, бифункциональная фосфоглюкоза/фосфоманнозо-изомераза; M6PI, маннозо-6-фосфат-изомераза; M6PP, маннозо-6-фосфат-фосфатаза.
[045] ФИГ. 3 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в галактозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфат-изомераза; Gal6PI, галактозо-6-фосфат-изомераза; Gal6PP, галактозо-6-фосфат-фосфатаза.
[046] ФИГ. 4 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов во фруктозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; F6PP, фруктозо-6-фосфат-фосфатаза.
[047] ФИГ. 5 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения сахарозы во фруктозу. Используются следующие сокращения: SP, сахарозо-фосфорилаза; PGM, фосфоглюкомутаза; PGI, фосфоглюкоизомераза; F6PP, фруктозо-6-фосфат-фосфатаза.
[048] ФИГ. 6 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в альтрозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; P6PE, псикозо-6-фосфат эпимераза; Alt6PI, альтрозо-6-фосфат-изомераза; Alt6PP, альтрозо-6-фосфат-фосфатаза.
[049] ФИГ. 7 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в талозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфат-эпимераза; Tal6PI, талозо-6-фосфат-изомераза; Tal6PP, талозо-6-фосфат-фосфатаза.
[050] ФИГ. 8 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в сорбозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфат-эпимераза; S6PE, сорбозо-6-фосфат-эпимераза; S6PP, сорбозо-6-фосфат-фосфатаза.
[051] ФИГ. 9 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в гулозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфат-эпимераза; S6PE, сорбозо-6-фосфат-эпимераза; Gul6PI, гулозо-6-фосфат-изомераза; Gul6PP, гулозо-6-фосфат-фосфатаза.
[052] ФИГ. 10 представляет собой схематическую диаграмму, показывающую ферментативный путь превращения крахмала или его производных продуктов в идозу. Используются следующие сокращения: IA, изоамилаза; PA, пуллуланаза; αGP, альфа-глюкан-фосфорилаза или крахмал-фосфорилаза; MP, мальтозо-фосфорилаза; PGM, фосфоглюкомутаза; PPGK, полифосфат-глюкокиназа; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфат-эпимераза; S6PE, сорбозо-6-фосфат-эпимераза; l6PI, идоза 6-фосфат-изомераза; l6PP, идоза 6-фосфат-фосфатаза.
[053] На ФИГ. 11 проиллюстрирована энергия Гиббса реакции между промежуточными продуктами, основанная на энергии Гиббса образования для превращения глюкозо-1-фосфата в другую гексозу.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[054] Описанные в данном документе изобретения предлагают ферментативные пути или способы синтеза гексоз с высоким выходом продукта, в то же время значительно снижая затраты на разделение продуктов и затраты на производство гексозы. В данном документе также описаны гексозы, полученные данными способами.
[055] Способы в соответствии с данным изобретением для получения гексозы из сахарида включают: превращение фруктозо-6-фосфата (F6P) в гексозу, катализируемое одним или несколькими ферментами, где указанная гексоза выбрана из группы, состоящей из аллозы, маннозы, галактозы, фруктозы, альтрозы, талозы, сорбозы, гулозы и идозы; и где указанные ферменты выбраны из группы, состоящей из изомеразы, эпимеразы и гексозо-специфической фосфатазы и их смесей.
[056] Одним из важных преимуществ способов по данному изобретению является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[057] Ионы фосфата, образующиеся на стадии дефосфорилирования, затем могут быть использованы повторно на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность использования повторно фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования гексоз.
[058] Например, концентрации фосфата в реакции в каждом из процессов могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции в каждом из процессов может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ или около 55 мМ.
[059] Низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование фосфатаз высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[060] Кроме того, каждый из описанных в данном документе способов может быть проведен без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Каждый из указанных способов также может быть проведен без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения гексоз включает энергетически выгодную химическую реакцию.
[061] Примеры ферментов, используемых для превращения сахарида в G1P, включают альфа-глюкан-фосфорилазу (αGP, EC 2.4.1.1), мальтозо-фосфорилазу (MP, EC 2.4.1.8), целлодекстрин-фосфорилазу (CDP, EC 2.4.1.49), целлобиозо-фосфорилазу (CBP, EC 2.4.1.20), целлюлозо-фосфорилазу, сахарозо-фосфорилазу (SP, EC 2.4.1.7) и их комбинацию. Выбор фермента или комбинации ферментов зависит от сахарида, используемого в способе.
[062] Сахаридами, используемыми для образования G1P, могут быть полисахариды, олигосахариды и/или дисахариды. Например, сахаридом может быть крахмал, одно или несколько производных крахмала, целлюлоза, одно или несколько производных целлюлозы, сахароза, одно или несколько производных сахарозы или их комбинация.
[063] Крахмал является наиболее широко используемым компонентом накопления энергии в природе и в основном хранится в семенах растений. Натуральный крахмал содержит линейную амилозу и разветвленный амилопектин. Примеры производных крахмала включают амилозу, амилопектин, растворимый крахмал, амилодекстрин, мальтодекстрин, мальтозу, фруктозу и глюкозу. Примеры производных целлюлозы включают редварительно обработанную биомассу, регенерированную аморфную целлюлозу, целлодекстрин, целлобиозу, фруктозу и глюкозу. Производные сахарозы включают фруктозу и глюкозу.
[064] Способы получения F6P из крахмала и его производных, целлюлозы и ее производных, а также сахарозы и ее производных можно найти, например, в публикации международной патентной заявки № WO 2017/059278.
[065] Производные крахмала могут быть получены ферментативным гидролизом крахмала или кислотным гидролизом крахмала. В частности, ферментативный гидролиз крахмала может быть катализирован или усилен изоамилазой (IA, EC. 3.2.1.68), которая гидролизует α-1,6-гликозидные связи; пуллуланаза (PA, EC. 3.2.1.41), которая гидролизует α-1,6-гликозидные связи; 4-α-глюкан-трансфераза (4GT, EC. 2.4.1.25), которая катализирует трансгликозилирование коротких мальтоолигосахаридов с образованием более длинных мальтоолигосахаридов; или альфа-амилаза (ЕС 3.2.1.1), которая расщепляет α-1,4-гликозидные связи.
[066] Кроме того, производные целлюлозы могут быть получены ферментативным гидролизом целлюлозы, катализируемым смесями целлюлаз, кислотами или предварительной обработкой биомассы.
[067] Ферменты, используемые для превращения сахарида в G1P, могут содержать αGP. На данной стадии, когда сахариды включают крахмал, G1P генерируется из крахмала с помощью αGP; когда сахариды содержат растворимый крахмал, амилодекстрин или мальтодекстрин, G1P получают из растворимого крахмала, амилодекстрина или мальтодекстрина с помощью αGP.
[068] Когда сахариды включают мальтозу, а ферменты содержат мальтозо-фосфорилазу, G1P генерируется из мальтозы мальтозо-фосфорилазой. Если сахариды включают сахарозу, а ферменты содержат сахарозо-фосфорилазу, G1P генерируется из сахарозы сахарозо-фосфорилазой.
[069] Когда сахариды включают целлобиозу, а ферменты содержат целлобиозо-фосфорилазу, G1P может быть получен из целлобиозы целлобиозо-фосфорилазой.
[070] Когда сахариды содержат целлодекстрины, а ферменты включают целлодекстрин-фосфорилазу, G1P может быть получен из целлодекстринов целлодекстрин-фосфорилазой.
[071] При превращении сахарида в G1P, когда сахариды включают целлюлозу, а ферменты содержат целлюлозо-фосфорилазу, G1P может быть получен из целлюлозы целлюлозо-фосфорилазой.
[072] В соответствии с данным изобретением, гексозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; и превращение T6P в тагатозу, катализируемое T6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[073] Гексоза может быть получена из сахарозы. Данный способ, например, предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; и превращение T6P в тагатозу, катализируемое T6PP.
[074] Фосфатаза, используемая в способах по данному изобретению, является специфичной для гексозы. Например, аллозо-6-фосфат превращается в аллозу с помощью аллозо-6-фосфат-фосфатазы; маннозо-6-фосфат превращается в маннозу с помощью маннозо-6-фосфат-фосфатазы; галактозо-6-фосфат превращается в галактозу с помощью галактозо-6-фосфат-фосфатазы; фруктозо-6-фосфат превращается во фруктозу с помощью фруктозо-6-фосфат-фосфатазы; альтрозо-6-фосфат превращается в альтрозу с помощью альтрозо-6-фосфат-фосфатазы; талозо-6-фосфат превращается в талозу с помощью талозо-6-фосфат-фосфатазы; сорбозо-6-фосфат превращается в сорбозу с помощью сорбозо-6-фосфат-фосфатазы; гулозо-6-фосфат превращается в гулозу с помощью гулозо-6-фосфат-фосфатазы; и идозо-6-фосфат превращается в идозу с помощью идозо-6-фосфат-фосфатазы. Термин «специфический», как используется в данном документе, означает более высокую специфическую активность для указанной гексозы по сравнению с другими гексозами. Например, аллозо-6-фосфат-фосфатаза обладает более высокой удельной активностью по отношению к аллозо-6-фосфату, чем, например, к сорбозо-6-фосфату или талозо-6-фосфату.
[075] Ионы фосфата, образующиеся на стадии дефосфорилирования гексозы, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода гексозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[076] Способ получения гексозы может включать следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в G6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше. G6P может быть получен из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[077] Данное изобретение предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в гексозу. Могут быть предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в гексозу с использованием бесклеточных ферментных коктейлей.
[078] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в гексозу и повышения растворимости.
[079] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода гексозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода гексозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[080] Кроме того, целлюлоза является наиболее распространенным биоресурсом и основным компонентом клеточных стенок растений. Непищевая лигноцеллюлозная биомасса содержит целлюлозу, гемицеллюлозу и лигнин, а также другие второстепенные компоненты. Чистая целлюлоза, включая Avicel (микрокристаллическая целлюлоза), регенерированная аморфная целлюлоза, бактериальная целлюлоза, фильтровальная бумага и т.д., может быть получена с помощью ряда обработок. Частично гидролизованные целлюлозные субстраты включают водонерастворимые целлодекстрины со степенью полимеризации более 7, водорастворимые целлодекстрины со степенью полимеризации 3-6, целлобиозу, глюкозу и фруктозу.
[081] Целлюлоза и ее производные продукты могут быть преобразованы в гексозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP) соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[082] Несколько ферментов могут быть использованы для гидролиза твердой целлюлозы до водорастворимых целлодекстринов и целлобиозы. Данные ферменты включают эндоглюканазу и целлобиогидролазу, но не включают бета-глюкозидазу (целлобиазу).
[083] Перед гидролизом целлюлозы и образованием G1P целлюлозу и биомассу можно предварительно обработать, чтобы повысить их реакционную способность и снизить степень полимеризации целлюлозных цепей. Способы предварительной обработки целлюлозы и биомассы включают предварительную обработку разбавленной кислотой, фракционирование лигноцеллюлозы растворителем целлюлозы, набухание волокон аммиаком, пропитку водным раствором аммиака, обработку ионной жидкостью и частичный гидролиз с использованием концентрированных кислот, включая соляную кислоту, серную кислоту, фосфорную кислоту и их комбинации.
[084] Полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способам в соответствии с данным изобретением, таким образом увеличивая выход гексозы путем фосфорилирования продукта разложения глюкозы до G6P.
[085] Гексоза может быть получена из глюкозы. Способы получения гексозы могут включать стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK), и превращения G6P в F6P, катализируемого PGI.
[086] Любой пригодный биологически совместимый буферный агент, известный в данной области техники, может быть использован в каждом из способов по данному изобретению, такой как HEPES, PBS, BIS-TRIS, MOPS, DIPSO, Trizma и т.д. Реакционный буфер для способов в соответствии с данным изобретением может иметь рН в диапазоне 5,0-8,0. Более предпочтительно pH реакционного буфера может представлять собой от около 6,0 до около 7,3. Например, pH реакционного буфера может представлять собой 6,0, 6,2, 6,4, 6,6, 6,8, 7,0, 7,2 или 7,3.
[087] Реакционный буфер также может содержать катионы металлов. Примеры ионов металлов включают Mg2+ и Zn2+. Как известно в данной области техники, подходящие соли могут быть использованы для введения желаемого катиона металла.
[088] В каждом из способов по данному изобретению температура реакции, при которой проводят стадии указанных способов, может составлять 37-95°С. Более предпочтительно, указанные стадии могут проводиться при температуре в диапазоне от около 40 до около 90°С. Температура может представлять собой, например, около 40°С, около 45°С, около 50°С, около 55°С, около 60°С, около 65°С, около 70°С, около 75°С, около 80°C, около 85°C или около 90°C. Предпочтительно температура реакции представляет собой около 50°С.
[089] Время реакции каждого из описанных способов может быть отрегулировано по мере необходимости и может варьироваться от около 8 часов до около 48 часов. Например, время реакции может представлять собой около 16 часов, около 18 часов, около 20 часов, около 22 часов, около 24 часов, около 26 часов, около 28 часов, около 30 часов, около 32 часов, около 34 часов, около 36 часов, около 38 часов, около 40 часов, около 42 часов, около 44 часов, около 46 часов или около 48 часов. Более предпочтительно время реакции представляет собой около 24 часов.
[090] Как правило, соотношения единиц фермента, используемых в каждом из описанных способов, составляют от 1:1 до 1:1:1:1:1 (в зависимости от количества катализируемых стадий в способе). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения гексоз. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[091] Каждый из способов в соответствии с данным изобретением может достигать высоких выходов благодаря очень благоприятной константе равновесия для общей реакции. Например, на ФИГ. 11 проиллюстрирована энергия Гиббса реакции между промежуточными продуктами, основанная на энергии Гиббса образования для превращения глюкозо-1-фосфата в гексозу. Энергии Гиббса реакции были получены с использованием http://equilibrator.weizmann.ac.il/. Теоретически, может быть достигнуто до 99% выхода, если исходный материал полностью превращается в промежуточное соединение.
[092] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда гексозу получают из лактозы, глюкозу и другую гексозу(ы) разделяют хроматографией, что приводит к более высоким производственным затратам.
[093] Кроме того, стадия дефосфорилирования гексозы фосфатазой в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, гексозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[094] В некоторых аспектах данного изобретения, для превращения A6P, M6P, F6P или Gal6P в их соответствующие нефосфорилированные формы фосфатазы используют как кофактор двухвалентный металл: предпочтительно магний. В дополнительных аспектах данного изобретения фосфатаза содержит, но не ограничиваясь этим, домен укладки Россмана для катализа; дополнительно содержит, но не ограничиваясь этим, С1 или С2 кэпированный домен для специфичности к субстрату; дополнительно содержит, но не ограничиваясь этим, DxD-сигнатуру в 1-ой β-цепи укладки Россмана для координации магния, где второй Asp представляет собой обычный кислотно-основной катализатор; дополнительно содержит, но не ограничиваясь этим, Thr или Ser на конце 2-ой β-цепи укладки Россмана, что способствует стабильности промежуточных продуктов реакции; дополнительно содержит, но не ограничиваясь этим, Lys на N-конце α-спирали С-терминально относительно 3-ей β-цепи укладки Россмана, что способствует стабильности промежуточных продуктов реакции; и дополнительно содержит, но не ограничиваясь этим, сигнатуру GDxxxD, GDxxxxD, DD или ED в конце 4-ой β-цепи укладки Россмана для координации магния. Данные признаки известны в данной области техники и упоминаются, например, в Burroughs и др., «Evolutionary Genomics of the HAD Superfamily: Understanding the Structural Adaptations and Catalytic Diversity in a Superfamily of Phosphoesterases and Allied Enzymes» J. Mol. Biol. 2006; 361; 1003-1034.
[095] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение гексозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[096] Аллоза
[097] Один вариант реализации данного изобретения представляет собой способ получения аллозы, который включает превращение фруктозо-6-фосфата (F6P) в псикозо-6-фосфат (P6P), катализируемое псикозо-6-фосфат-3-эпимеразой (P6PE), превращение P6P в аллозо-6-фосфат (A6P), катализируемое аллозо-6-фосфат-изомеразой (A6PI), и превращение полученного A6P в аллозу, катализируемое аллозо-6-фосфат-фосфатазой.
[098] Примеры P6PE включают, но не ограничиваются ими, следующие белки, идентифицированные по ID номерам UNIPROT: D9TQJ4, A0A090IXZ8, и P32719. Из них, D9TQJ4 и A0A090IXZ8 получены из термофильных организмов. P32719 получен из мезофильного организма. P32719 на 53% идентичен A0A090IXZ8 и на 55% идентичен D9TQJ4, и каждый белок катализирует эпимеризацию F6P в A6P. Дополнительно, A0A090IXZ8 на 45% идентичен D9TQJ4. И наоборот, другие эпимеразные белки, идентифицированные по ID номерам UNIPROT: A0A101D823, R1AXD6, A0A150LBU8, A0A023CQG9, и H1XWY2, которые имеют степень идентичности с D9TQJ4 45% или менее, не катализируют эпимеризацию F6P в A6P. Примеры P6PE также включают любые гомологи, имеющие по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, или по меньшей мере 99% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[099] Примеры A6PI включают, но не ограничиваются ими, Uniprot ID W4V2C8, с аминокислотной последовательностью, представленной в SEQ ID NO: 1; и Uniprot ID Q67LX4, с аминокислотной последовательностью, представленной в SEQ ID NO: 2. Uniprot ID W4V2C8 и Q67LX4 оба катализируют реакцию A6PI и имеют 56% идентичности аминокислотной последовательности. Таким образом, примеры A6PI также включают любые гомологи, имеющие по меньшей мере 55%, предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0100] A6PI, пригодные для применения в процессе превращения P6P в A6P, содержат укладку Россмана. Мезофильная A6PI, описанная в данной области техники (Mowbray и др., «D-Ribose-5-Phosphate Isomerase B from Escherichia coli is Also a Functional D-Allose-6-phosphate Isomerase, While the Mycobacterium tuberculosis Enzyme is Not» J. Mol. Biol. 2008; 382; 667-679) делит консервативные остатки с термофильной A6PI, описанной в данном изобретении. В некоторых аспектах данного изобретения указанная изомераза содержит, но не ограничиваясь этим, His (мезофильный остаток 10) на С-конце 1-ой β-цепи укладки Россмана для связывания фосфата; дополнительно содержит, но не ограничиваясь этим, Arg (мезофильный остаток 133) на С-конце С-конца α-спирали в отношении 5-ой β-цепи укладки Россмана также для связывания фосфата; дополнительно содержит, но не ограничиваясь этим, His (мезофильный остаток 99) в активном сайте для раскрытия лактона; дополнительно содержит, но не ограничиваясь этим, Cys (мезофильный остаток 66) в активном сайте, действующий в качестве каталитического основания; дополнительно содержит, но не ограничиваясь этим, Thr (мезофильный остаток 68) в активном сайте, действующий в качестве каталитической кислоты; дополнительно содержит, но не ограничиваясь этим, GTG-гидрофобный-G мотив вблизи активного сайта (мезофильные остатки 67-71) для стабилизации высокоэнергетических промежуточных продуктов, и дополнительно содержит, но не ограничиваясь этим, Asn (мезофильный остаток 100) вблизи активного сайта, чтобы также стабилизировать высокоэнергетические промежуточные продукты. A6PI предпочтительно содержит все данные консервативные остатки.
[0101] Примеры A6PP включают, но не ограничиваются ими, следующие белки: Uniprot ID S9SDA3, с аминокислотной последовательностью, представленной в SEQ ID NO: 3; Q9X0Y1, с аминокислотной последовательностью, представленной в SEQ ID NO: 4; I3VT81, с аминокислотной последовательностью, представленной в SEQ ID NO: 5; A0A132NF06, с аминокислотной последовательностью, представленной в SEQ ID NO: 6; и D1C7G9, с аминокислотной последовательностью, представленной в SEQ ID NO: 7. Uniprot ID S9SDA3 и I3VT81 оба катализируют реакцию A6PP и имеют 30% идентичности аминокислотной последовательности. Таким образом, примеры A6PP также включают любые гомологи, имеющие по меньшей мере 30%, предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0102] Предпочтительно, A6PP для превращения A6P в аллозу содержит домен укладки Россмана для катализа, C1 кэпированный домен, сигнатуру DxD в 1-ой β-цепи укладки Россмана, Thr или Ser в конце 2-ой β-цепи укладки Россмана, Lys на N-конце α-спирали С-терминально относительно 3-ей β-цепи укладки Россмана, и сигнатуру ED на конце 4-ой β-цепи укладки Россмана.
[0103] Способ получения аллозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения аллозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения аллозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0104] Таким образом, способ получения аллозы в соответствии с данным изобретением может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с применением одного или более ферментов; (ii) превращение G1P в G6P с применением фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P to F6P с применением фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в P6P с помощью P6PE, (v) превращение P6P в A6P с помощью A6PI, и (vi) превращение A6P в аллозу с помощью A6PP. Пример ферментативного процесса, в котором сахарид представляет собой крахмал, показан на ФИГ. 1.
[0105] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1 (αGP:PGM:PGI:P6PE:A6PI:A6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения аллозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т.д. относительно количества других ферментов.
[0106] Ионы фосфата, образующиеся при дефосфорилировании A6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования аллозы.
[0107] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0108] Низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование A6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0109] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения аллозы включает энергетически выгодную химическую реакцию.
[0110] Аллозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в P6P, катализируемое P6PE; превращение P6P в A6P, катализируемое A6PI, и превращение A6P в аллозу, катализируемое A6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0111] Аллозу также можно получить из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; превращение P6P в A6P, катализируемое A6PI, и превращение A6P в аллозу, катализируемое A6PP.
[0112] Ионы фосфата, образующиеся во время превращения A6P в аллозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода аллозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0113] В определенных вариантах реализации изобретения, способ получения аллозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0114] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0115] Несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в аллозу и повышения растворимости.
[0116] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода аллозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-глюкан-трансферазу (4GT) можно применять для увеличения выхода аллозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0117] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы в аллозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; превращение P6P в A6P, катализируемое A6PI, и превращение A6P в аллозу, катализируемое A6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0118] Несколько ферментов могут быть использованы для гидролиза твердой целлюлозы до водорастворимых целлодекстринов и целлобиозы. Данные ферменты включают эндоглюканазу и целлобиогидролазу, но не включают бета-глюкозидазу (целлобиазу).
[0119] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход аллозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0120] Аллозу также можно получить из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; превращение P6P в A6P, катализируемое A6PI; и превращение A6P в аллозу, катализируемое A6PP.
[0121] В способах по данному изобретению для получения аллозы применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, фруктоза. Когда аллозу получают из псикозы, выходы ниже, чем в данном изобретении, и аллозу необходимо отделять от псикозы с помощью хроматографии, что приводит к более высоким производственным затратам.
[0122] Кроме того, стадия превращения A6P в аллозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, аллозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0123] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение аллозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0124] Конкретным вариантом реализации данного изобретения является аллоза, полученная описанными в данном документе способами получения аллозы.
[0125] Поскольку аллоза обладает аналогичной функциональностью с сахарозой, аллозу, полученную способами по данному изобретению, можно добавлять в любой напиток или пищевой продукт для получения желаемой сладости.
[0126] Аллоза, полученная способами, описанными в данном документе, также может быть применена для усиления действия сильнодействующих подсластителей. В сочетании с одним или более сильнодействующими подсластителями аллоза может оказывать влияние на улучшение органолептических характеристик, таких как ощущение во рту, вкус и послевкусие подслащенного продукта. Использование низкокалорийных подсластителей, таких как сильнодействующие подсластители, в различных пищевых продуктах является обычным явлением в составах пищевых продуктов и напитков. Однако для многих потребителей продукты, которые являются искусственно подслащенными, продаваемые как диетические или легкие версии продуктов, не являются предпочтительными. На протяжении многих лет предпринимались попытки улучшить вкусовые качества данных диетических или легких продуктов путем добавления небольших количеств углеводов. Аллоза, полученная способами по данному изобретению, может не только обеспечивать улучшение качества пищевых продуктов и напитков, особенно в диетических/легких напитках, но также то, что ее использование может быть синергичным с мощными подсластителями, так что оно способно заменить значительные количества сильнодействующих подсластителей, даже когда ее добавляют в концентрациях, которые значительно ниже измеренного порога сладкого вкуса.
[0127] Аллоза, полученная способами, описанными в данном документе, может быть объединена с другими подсластителями, такими как экстракты растения Stevia rebaudiana Bertoni, для приготовления низкокалорийных вариантов пищевых продуктов, таких как мороженое.
[0128] Аллоза, полученная способами, описанными в данном документе, может быть использована в подслащенных готовых к употреблению (ГКУ) сухих завтраках и других продуктах питания, где D-аллоза частично или полностью заменяет сахарозу или другие обычно используемые сахара в качестве глазури.
[0129] Аллоза, полученная способами, описанными в данном документе, может быть использована в качестве подсластителя для пищевых продуктов и напитков в сочетании с сахарными спиртами, такими как эритрит, и питательными подсластителями со значительной калорийностью, такими как фруктоза, сахароза, декстроза, мальтоза, трегалоза, рамноза, кукурузные сиропы и фрукто-олигосахариды.
[0130] Аллоза, полученная способами, описанными в данном документе, также может быть использована как часть композиции, которая усиливает борьбу с болезнями растений.
[0131] Манноза
[0132] Один вариант реализации данного изобретения представляет собой способ для получения маннозы, который включает превращение F6P в маннозо-6-фосфат (M6P), катализируемое маннозо-6-фосфат-изомеразой (M6PI); и превращение M6P в маннозу, катализируемое маннозо-6-фосфат-фосфатазой (M6PP).
[0133] Примеры M6PI включают, но не ограничиваются ими, следующие белки: Uniprot ID A0A1M6TLY7, с аминокислотной последовательностью, представленной в SEQ ID NO: 8; H1XQS6, с аминокислотной последовательностью, представленной в SEQ ID NO: 9; G2Q982, с аминокислотной последовательностью, представленной в SEQ ID NO: 10; и F8F1Z8, с аминокислотной последовательностью, представленной в SEQ ID NO: 11. Uniprot ID G2Q982 и F8F1Z8 оба катализируют реакцию M6PI и имеют 28% идентичности аминокислотной последовательности. Таким образом, примеры M6PI также включают любые гомологи, имеющие по меньшей мере 25%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0134] M6PI, пригодные для применения в процессе превращения F6P в M6P, содержат два домена с ядром из антипараллельных β-цепей, напоминающих укладку купина, и третий домен, состоящий только из α-спиралей. M6PI была структурно охарактеризована в данной области техники (Sagurthi и др. «Structures of mannose-6-phosphate isomerase from Salmonella typhimurium bound to metal atoms and substrate: implications for catalytic mechanism.» Acta Cryst. 2009; D65; 724-732) и делит консервативные остатки с термофильной M6PI, описанной в данном изобретении. В некоторых аспектах данного изобретения указанная изомераза содержит, но не ограничиваясь этим, катион двухвалентного металла, предпочтительно Mg2+ или Zn2+; дополнительно содержит, но не ограничиваясь этим, Glu и два остатка His, предложенные для использования в связывании металлов (PDB 3H1M остатки 134, 99 и 255 соответственно); дополнительно содержит, но не ограничиваясь этим, остатки Asp и Lys, предложенные для кислотно/основного катализа (PDB 3H1M остатки 270 и 132 соответственно); дополнительно содержит, но не ограничиваясь этим, остатки Lys, Pro и Ala, предложенные для связывания фосфата (PDB 3H1M остатки 132, 133 и 267 соответственно). M6PI предпочтительно содержит все данные консервативные остатки.
[0135] Примеры M6PP включают, но не ограничиваются ими, следующие белки: Uniprot ID A0A1A6DSI3, с аминокислотной последовательностью, представленной в SEQ ID NO: 12; A0A1M4UN08, с аминокислотной последовательностью, представленной в SEQ ID NO: 13; и A0A1N6FCW3, с аминокислотной последовательностью, представленной в SEQ ID NO: 14; Uniprot ID A0A1A6DSI3 и A0A1N6FCW3 оба катализируют реакцию M6PP и имеют 35% идентичности аминокислотной последовательности. Таким образом, примеры M6PP также включают любые гомологи, имеющие по меньшей мере 35%, более предпочтительно по меньшей мере 40%, предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0136] Предпочтительно, M6PP для превращения M6P в маннозу содержит домен укладки Россмана для катализа, C1 кэпированный домен, сигнатуру DxD в 1-ой β-цепи укладки Россмана, Thr или Ser на конце 2-ой β-цепи укладки Россмана, Lys на N-конце α-спирали С-терминально относительно 3-ей β-цепи укладки Россмана, и сигнатуру GDxxxD на конце 4-ой β-цепи укладки Россмана.
[0137] Способ получения маннозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения маннозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В других вариантах реализации изобретения, указанный способ получения маннозы включает превращение G6P в F6P в M6P, где данная стадия катализируется бифункциональной фосфоглюкоза/фосфоманнозо-изомеразой (PGPMI). В дополнительных вариантах реализации изобретения, способ получения маннозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0138] Способы по данному изобретению для получения маннозы используют PGPMI, которая превращает G6P или F6P в M6P. Примеры PGPMI включают, но не ограничиваются ими, следующие белки: Uniprot ID D7CPH7, с аминокислотной последовательностью, представленной в SEQ ID NO: 15; A0A085L170, с аминокислотной последовательностью, представленной в SEQ ID NO: 16; и M1E6Z3, с аминокислотной последовательностью, представленной в SEQ ID NO: 17. Uniprot ID A0A085L170 и M1E6Z3 оба катализируют реакцию PGPMI и имеют 28% идентичности аминокислотной последовательности. Таким образом, примеры PGPMI также включают любые гомологи, имеющие по меньшей мере 25%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0139] PGPMI, пригодные для применения в процессе превращения G6P или F6P в M6P, содержат две укладки Россмана. PGPMI была структурно охарактеризована в данной области техники (Swan и др. A «Novel Phosphoglucose Isomerase (PGI)/Phosphomannose Isomerase from the Crenarchaeon Pyrobaculum aerophilum Is a Member of the PGI Superfamily.» J. Biol. Chem. 2004: 279; 39838-39845) и делит консервативные остатки с термофильной PGPMI, описанной в данном изобретении. В некоторых аспектах данного изобретения указанная изомераза содержит, но не ограничиваясь этим, мотив GGS (PDB 1TZB остатки 46-48), где остатки Gly способствуют связыванию субстрата, а остаток Ser связывает фосфат; дополнительно содержит, но не ограничиваясь этим, SYSG-X-T-X-ET-гидрофобный мотив (PDB 1TZB остатки 87-96), который связывает фосфат; дополнительно содержит, но не ограничиваясь этим, остаток Arg (PDB 1TZB остаток 135), который стабилизирует высокоэнергетические промежуточные продукты во время катализа; дополнительно содержит, но не ограничиваясь этим, сигнатуру EN (PDB 1TZB остатки 203-204), где Glu необходим для переноса основного протона в активном сайте; дополнительно содержит, но не ограничиваясь этим, сигнатуру HN (PDB 1TZB остатки 219-220), где His важен для раскрытия/замыкания кольца субстрата во время катализа; и дополнительно содержит, но не ограничиваясь этим, консервативный остаток Lys (PDB 1TZB остаток 298), который важен для раскрытия/замыкания кольца субстрата во время катализа; Функции консервативных остатков проверены в отдельной публикации (Hansen и др. «Bifunctional Phosphoglucose/Phosphomannose Isomerases from the Archaea Aeropyrum pernix and Thermoplasma acidophilum Constitute a Novel Enzyme Family within the Phosphoglucose Isomerase Superfamily.» J. Biol. Chem. 2004; 279; 2262-2272). PGPMI предпочтительно содержит все данные консервативные остатки.
[0140] Следовательно, способ получения маннозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в M6P с помощью маннозо-6-фосфат-изомеразы (M6PI, EC 5.3.1.8), (v) превращение G6P в M6P с помощью бифункциональной фосфоглюкоза/фосфоманнозо-изомеразы (PGPMI, EC 5.3.1.8 и 5.3.1.9) и (vi) превращение M6P в маннозу с помощью M6PP. Пример процесса, в котором сахарид представляет собой крахмал, показан на ФИГ. 2.
[0141] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1 (αGP:PGM:PGI:M6PI:M6PP) или 1:1:1:1 (αGP:PGM:PGPMI:M6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения маннозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т.д. относительно количества других ферментов.
[0142] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0143] Ионы фосфата, образующиеся при дефосфорилировании M6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования маннозы.
[0144] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0145] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование M6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0146] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения маннозы включает энергетически выгодную химическую реакцию.
[0147] Маннозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в M6P, катализируемое M6PI; и превращение M6P в маннозу, катализируемое M6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0148] Маннозу также можно получить из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в M6P, катализируемое M6PI; и превращение M6P в маннозу, катализируемое M6PP. На вышеуказанных стадиях превращение G6P в F6P в M6P может альтернативно катализироваться PGPMI.
[0149] Ионы фосфата, образующиеся во время превращения M6P в маннозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода маннозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0150] В некоторых вариантах реализации изобретения, способ получения маннозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0151] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0152] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в маннозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в маннозу с использованием бесклеточных ферментных коктейлей.
[0153] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в маннозу и повышения растворимости.
[0154] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода маннозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода маннозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0155] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы в маннозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в M6P, катализируемое M6PI; и превращение M6P в маннозу, катализируемое M6PP. В альтернативном варианте, в вышеуказанном пути превращение G6P в F6P в M6P может катализироваться PGPMI. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0156] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход маннозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0157] В других вариантах реализации изобретения манноза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в M6P, катализируемое M6PI; и превращение M6P в маннозу, катализируемое M6PP. В альтернативном варианте, превращение G6P в F6P в M6P может катализироваться PGPMI.
[0158] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, фруктоза. Когда маннозу получают из фруктозы, выходы ниже, чем в данном изобретении, и маннозу необходимо отделять от фруктозы с помощью хроматографии, что приводит к более высоким производственным затратам.
[0159] Кроме того, стадия превращения M6P в маннозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, маннозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0160] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение маннозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0161] Конкретным вариантом реализации данного изобретения является манноза, полученная описанными в данном документе способами получения маннозы.
[0162] Манноза, полученная описанными в данном документе способами, может использоваться, как обсуждалось выше, в различных областях применения: в фармацевтике, косметике, производстве напитков, пищевых продуктов, молочной, кондитерской промышленности и животноводстве.
[0163] Кроме того, манноза, полученная способами, описанными в данном документе, может быть превращена в маннит путем гидрирования. Каталитическое гидрирование маннозы происходит со стехиометрическим выходом и дает маннит. Патент США № 5466795. Маннит широко используется в производстве жевательной резинки, не содержащей сахара, сладостей и фармацевтических вспомогательных веществ. Однако производство высокочистой маннозы чрезвычайно сложно и является дорогостоящим. Соответственно, манноза, полученная вышеупомянутыми способами, может быть преобразована в маннит путем каталитического гидрирования.
[0164] Галактоза
[0165] Один вариант реализации данного изобретения представляет собой способ для получения галактозы, который включает превращение фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфат-4-эпимеразой (F6PE); превращение T6P в галактозо-6-фосфат (Gal6P), катализируемое галактозо-6-фосфат-изомеразой (Gal6PI); и превращение полученного Gal6P в галактозу, катализируемое галактозо-6-фосфат-фосфатазой (Gal6PP).
[0166] Примеры F6PE включают, но не ограничиваются ими, следующие белки: Uniprot ID E8N0N6, E4SEH3, I0I507, H1XRG1, и B5YBD7. Uniprot ID E8N0N6 и I0I507 оба катализируют реакцию F6PE и имеют 27% идентичности аминокислотной последовательности. Таким образом, примеры F6PE также включают любые гомологи, имеющие по меньшей мере 25%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0167] Gal6PI существует как мультимер из двух субъединиц, LacA и LacB. Примеры Gal6PI включают, но не ограничиваются ими, следующую пару субъединиц белка (LacA/LacB): Uniprot ID P23494/P23495, с аминокислотными последовательностями, представленными в SEQ ID NO: 18/SEQ ID NO: 19. Примеры Gal6PI также включают любые гомологи, имеющие по меньшей мере 25%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с вышеупомянутым Uniprot ID для субъединицы LacA и гомологи, имеющие по меньшей мере 25%, по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с вышеупомянутым Uniprot ID для субъединицы LacB.
[0168] Gal6PI, пригодные для применения в процессе превращения T6P в Gal6P, содержат гетеродимер (‘A’ и ‘B’), состоящий из субъединиц с Россман-подобными αβα сэндвич-укладками. Консервативные остатки обсуждаются в данной области техники (Jung и др. «Crystal Structure and Substrate Specificity of D-Galactose-6-Phosphate Isomerase Complexed with Substrates.» PLOS ONE. 2013; 8; e72902). В некоторых аспектах данного изобретения гетеродимер изомеразы содержит, но не ограничиваясь этим, Arg130 и Arg134 в ‘A’ и His9 и Arg39 в ‘B’ для связывания фосфатной группы субстрата; дополнительно содержит, но не ограничиваясь этим, His96 в ‘A’ для раскрытия кольца субстрата; дополнительно содержит, но не ограничиваясь этим, Asn97 в ‘A’ для стабилизации высокоэнергетических промежуточных продуктов; и дополнительно содержит, но ограничиваясь этим, Cys65 и Thr67 в ‘B’ для участия в переносе протона.
[0169] Примеры Gal6PP включают, но не ограничиваются ими, Uniprot ID Q8A2F3, с аминокислотной последовательностью, представленной в SEQ ID NO: 20. Примеры Gal6PP также включают любые гомологи, имеющие по меньшей мере 25%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с вышеупомянутым Uniprot ID.
[0170] Предпочтительно, Gal6PP для превращения Gal6P в галактозу содержит домен укладки Россмана для катализа, C2 кэпированный домен, сигнатуру DxD в 1-ой β-цепи укладки Россмана, Thr или Ser в конце 2-ой β-цепи укладки Россмана, и сигнатуру GDxxxD на конце 4-ой β-цепи укладки Россмана.
[0171] Способ получения галактозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения галактозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения галактозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0172] Следовательно, способ получения галактозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в T6P с помощью F6PE, (v) превращение T6P в Gal6P с помощью Gal6PI (EC 5.3.1.26), и (vi) превращение Gal6P в галактозу с помощью Gal6PP. Пример процесса, в котором сахарид представляет собой крахмал, показан на ФИГ. 3.
[0173] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1 (αGP:PGM:PGI:F6PE:Gal6PI:Gal6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения галактозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0174] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0175] Ионы фосфата, образующиеся при дефосфорилировании Gal6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования галактозы.
[0176] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0177] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование Gal6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0178] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т. е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т. е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения галактозы включает энергетически выгодную химическую реакцию.
[0179] Галактозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; превращение T6P в Gal6P, катализируемое Gal6PI, и превращение Gal6P в галактозу, катализируемое Gal6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0180] Галактоза может быть получена из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в Gal6P, катализируемое Gal6PI, и превращение Gal6P в галактозу, катализируемое Gal6PP.
[0181] Ионы фосфата, образующиеся во время превращения Gal6P в галактозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода галактозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0182] В некоторых вариантах реализации изобретения, способ получения галактозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0183] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0184] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в галактозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в галактозу с использованием бесклеточных ферментных коктейлей.
[0185] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в галактозу и повышения растворимости.
[0186] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода галактозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода галактозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0187] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы в галактозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в Gal6P, катализируемое Gal6PI, и превращение Gal6P в галактозу, катализируемое Gal6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0188] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход галактозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0189] В других вариантах реализации изобретения галактоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в Gal6P, катализируемое Gal6PI; и превращение Gal6P в галактозу, катализируемое Gal6PP.
[0190] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда галактозу получают из биомассы или лактозы, выходы ниже, чем в данном изобретении, и галактозу необходимо отделять от других сахаров с помощью хроматографии, что приводит к более высоким производственным затратам. Кроме того, наш способ осуществляется без участия животных.
[0191] Стадия превращения Gal6P в галактозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, галактозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0192] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение галактозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0193] Конкретным вариантом реализации данного изобретения является галактоза, полученная описанными в данном документе способами получения галактозы.
[0194] Фруктоза
[0195] Один вариант реализации данного изобретения представляет собой способ для получения фруктозы, который включает превращение фруктозо-6-фосфата (F6P) во фруктозу, катализируемое фруктозо-6-фосфат-фосфатазой (F6PP).
[0196] Неограничивающим примером F6PP является Uniprot ID B8CWV3, с аминокислотной последовательностью, представленной в SEQ ID NO: 21. Примеры F6PP также включают любые гомологи, имеющие по меньшей мере 25%, предпочтительно по меньшей мере 30%, более предпочтительно по меньшей мере 35%, более предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 45%, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, или по меньшей мере 94%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с вышеупомянутым Uniprot ID.
[0197] Предпочтительно, F6PP для превращения F6P во фруктозу содержит домен укладки Россмана для катализа, C1 кэпированный домен, сигнатуру DxD в 1-ой β-цепи укладки Россмана, Thr или Ser в конце 2-ой β-цепи укладки Россмана, Lys на N-конце α-спирали С-терминально относительно 3-ей β-цепи укладки Россмана, и сигнатуру ED на конце 4-ой β-цепи укладки Россмана.
[0198] Способ получения фруктозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения фруктозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения фруктозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0199] Следовательно, способ получения фруктозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P во фруктозу с помощью F6PP.
[0200] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1 (αGP:PGM:PGI:F6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения фруктозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0201] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0202] Ионы фосфата, образующиеся при дефосфорилировании F6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования фруктозы.
[0203] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0204] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование F6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0205] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т. е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения фруктозы включает энергетически выгодную химическую реакцию.
[0206] Фруктозу также можно получить из сахарозы через промежуточный продукт F6P. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P во фруктозу, катализируемое F6PP. Пример ферментативного пути представлен на ФИГ. 5
[0207] Ионы фосфата, образующиеся во время превращения F6P во фруктозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода фруктозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0208] В некоторых вариантах реализации изобретения, способ получения фруктозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0209] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0210] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, во фруктозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов во фруктозу с использованием бесклеточных ферментных коктейлей.
[0211] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения во фруктозу и повышения растворимости.
[0212] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода фруктозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода фруктозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0213] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы во фруктозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP) соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P во фруктозу, катализируемое F6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0214] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход фруктозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0215] В других вариантах реализации изобретения фруктоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; и превращение F6P во фруктозу, катализируемое F6PP.
[0216] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда фруктозу получают из биомассы или лактозы, выходы ниже, чем в данном изобретении, и фруктозу необходимо отделять от других сахаров с помощью хроматографии, что приводит к более высоким производственным затратам. Кроме того, наш способ осуществляется без участия животных.
[0217] Стадия превращения F6P во фруктозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, фруктозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0218] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение фруктозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0219] Конкретным вариантом реализации данного изобретения является фруктоза, полученная описанными в данном документе способами получения фруктозы.
[0220] Альтроза
[0221] Один вариант реализации данного изобретения представляет собой способ получения альтрозы, который включает превращение фруктозо-6-фосфата (F6P) в псикозо-6-фосфат (P6P), катализируемое псикозо-6-фосфат-3-эпимеразой (P6PE), превращение P6P в альтрозо-6-фосфат (Alt6P), катализируемое альтрозо-6-фосфат-изомеразой (Alt6PI), и превращение полученного Alt6P в альтрозу, катализируемое альтрозо-6-фосфат-фосфатазой.
[0222] Способ получения альтрозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения альтрозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения альтрозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0223] Следовательно, способ получения альтрозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в P6P с помощью P6PE, (v) превращение P6P в Alt6P с помощью Alt6PI, и (vi) превращение Alt6P в альтрозу с помощью Alt6PP. Пример ферментативного процесса, в котором сахарид представляет собой крахмал, показан на ФИГ. 1.
[0224] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1 (αGP:PGM:PGI:P6PE:Alt6PI:Alt6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения альтрозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0225] Ионы фосфата, образующиеся при дефосфорилировании Alt6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования альтрозы.
[0226] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0227] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование Alt6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0228] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения альтрозы включает энергетически выгодную химическую реакцию.
[0229] Альтрозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в P6P, катализируемое P6PE; превращение P6P в Alt6P, катализируемое Alt6PI, и превращение Alt6P в альтрозу, катализируемое Alt6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0230] Альтрозу также можно получить из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; превращение P6P в Alt6P, катализируемое Alt6PI, и превращение Alt6P в альтрозу, катализируемое Alt6PP.
[0231] Ионы фосфата, образующиеся во время превращения Alt6P в альтрозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода альтрозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0232] В определенных вариантах реализации изобретения, способ получения альтрозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0233] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0234] Несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в альтрозу и повышения растворимости.
[0235] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода альтрозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода альтрозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0236] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы во альтрозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; превращение P6P в Alt6P, катализируемое Alt6PI, и превращение Alt6P в альтрозу, катализируемое Alt6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0237] Несколько ферментов могут быть использованы для гидролиза твердой целлюлозы до водорастворимых целлодекстринов и целлобиозы. Данные ферменты включают эндоглюканазу и целлобиогидролазу, но не включают бета-глюкозидазу (целлобиазу).
[0238] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход альтрозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0239] Альтрозу также можно получить из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; превращение P6P в Alt6P, катализируемое Alt6PI; и превращение Alt6P в альтрозу, катализируемое Alt6PP.
[0240] В способах по данному изобретению для получения альтрозы применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, фруктоза. Когда альтрозу получают из псикозы, выходы ниже, чем в данном изобретении, и альтрозу необходимо отделять от псикозы с помощью хроматографии, что приводит к более высоким производственным затратам.
[0241] Кроме того, стадия превращения Alt6P в альтрозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, альтрозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0242] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение альтрозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0243] Конкретным вариантом реализации данного изобретения является альтроза, полученная описанными в данном документе способами получения альтрозы.
[0244] Талоза
[0245] Один вариант реализации данного изобретения представляет собой способ для получения талозы, который включает превращение фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфат-4-эпимеразой (F6PE); превращение T6P в талозо-6-фосфат (Tal6P), катализируемое талозо-6-фосфат-изомеразой (Tal6PI); и превращение полученного Tal6P в талозу, катализируемое талозо-6-фосфат-фосфатазой (Tal6PP).
[0246] Примеры F6PE включают, но не ограничиваются ими, следующие белки: Uniprot ID E8N0N6, E4SEH3, I0I507, H1XRG1, и B5YBD7. Uniprot ID E8N0N6 и I0I507 оба катализируют реакцию F6PE и имеют 27% идентичности аминокислотной последовательности. Следовательно, примеры F6PE также включают любые гомологи, имеющие по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, или по меньшей мере 99% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0247] Способ получения талозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения талозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения талозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0248] Следовательно, способ получения талозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в T6P с помощью F6PE, (v) превращение T6P в Tal6P с помощью Tal6PI (EC 5.3.1.26), и (vi) превращение Tal6P в талозу с помощью Tal6PP. Пример процесса, в котором сахарид представляет собой крахмал, показан на ФИГ. 3.
[0249] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1 (αGP:PGM:PGI:F6PE:Tal6PI:Tal6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения талозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0250] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0251] Ионы фосфата, образующиеся при дефосфорилировании Tal6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования талозы.
[0252] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0253] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование Tal6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0254] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения талозы включает энергетически выгодную химическую реакцию.
[0255] Талозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; превращение T6P в Tal6P, катализируемое Tal6PI, и превращение Tal6P в талозу, катализируемое Tal6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0256] Талоза может быть получена из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в Tal6P, катализируемое Tal6PI, и превращение Tal6P в талозу, катализируемое Tal6PP.
[0257] Ионы фосфата, образующиеся во время превращения Tal6P в талозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода талозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0258] В некоторых вариантах реализации изобретения, способ получения талозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0259] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0260] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в талозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в талозу с использованием бесклеточных ферментных коктейлей.
[0261] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в талозу и повышения растворимости.
[0262] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода талозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода талозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0263] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы во талозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в Tal6P, катализируемое Tal6PI, и превращение Tal6P в талозу, катализируемое Tal6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0264] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход талозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0265] В других вариантах реализации изобретения талоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в Tal6P, катализируемое Tal6PI; и превращение Tal6P в талозу, катализируемое Tal6PP.
[0266] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда талозу получают из биомассы или лактозы, выходы ниже, чем в данном изобретении, и талозу необходимо отделять от других сахаров с помощью хроматографии, что приводит к более высоким производственным затратам. Кроме того, наш способ осуществляется без участия животных.
[0267] Стадия превращения Tal6P в талозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, талозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0268] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение талозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0269] Конкретным вариантом реализации данного изобретения является талоза, полученная описанными в данном документе способами получения талозы.
[0270] Сорбоза
[0271] Один вариант реализации данного изобретения представляет собой способ для получения сорбозы, который включает превращение фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфат-4-эпимеразой (F6PE); превращение T6P в сорбозо-6-фосфат (S6P), катализируемое сорбозо-6-фосфат-эпимеразой (S6PE); и превращение полученного S6P в сорбозу, катализируемое сорбозо-6-фосфат-фосфатазой (S6PP).
[0272] Примеры F6PE включают, но не ограничиваются ими, следующие белки: Uniprot ID E8N0N6, E4SEH3, I0I507, H1XRG1, и B5YBD7. Uniprot ID E8N0N6 и I0I507 оба катализируют реакцию F6PE и имеют 27% идентичности аминокислотной последовательности. Следовательно, примеры F6PE также включают любые гомологи, имеющие по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, или по меньшей мере 99% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0273] Способ получения сорбозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения сорбозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения сорбозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0274] Следовательно, способ получения сорбозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в T6P с помощью F6PE, (v) превращение T6P в S6P с помощью S6PE (EC 5.3.1.26), и (vi) превращение S6P в сорбозу с помощью S6PP. Пример процесса, в котором сахарид представляет собой крахмал, показан на ФИГ. 3.
[0275] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1 (αGP:PGM:PGI:F6PE:S6PE:S6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения сорбозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т.д. относительно количества других ферментов.
[0276] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0277] Ионы фосфата, образующиеся при дефосфорилировании S6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования сорбозы.
[0278] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0279] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование S6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0280] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения сорбозы включает энергетически выгодную химическую реакцию.
[0281] Сорбозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, и превращение S6P в сорбозу, катализируемое S6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0282] Сорбоза может быть получена из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, и превращение S6P в сорбозу, катализируемое S6PP.
[0283] Ионы фосфата, образующиеся во время превращения S6P в сорбозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода сорбозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0284] В некоторых вариантах реализации изобретения, способ получения сорбозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0285] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0286] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в сорбозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в сорбозу с использованием бесклеточных ферментных коктейлей.
[0287] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в сорбозу и повышения растворимости.
[0288] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода сорбозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода сорбозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0289] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы в сорбозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, и превращение S6P в сорбозу, катализируемое S6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0290] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход сорбозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0291] В других вариантах реализации изобретения сорбоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE; и превращение S6P в сорбозу, катализируемое S6PP.
[0292] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда сорбозу получают из биомассы или лактозы, выходы ниже, чем в данном изобретении, и сорбозу необходимо отделять от других сахаров с помощью хроматографии, что приводит к более высоким производственным затратам. Кроме того, наш способ осуществляется без участия животных.
[0293] Стадия превращения S6P в сорбозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, сорбозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0294] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение сорбозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0295] Конкретным вариантом реализации данного изобретения является сорбоза, полученная описанными в данном документе способами получения сорбозы.
[0296] Гулоза
[0297] Один вариант реализации данного изобретения представляет собой способ для получения гулозы, который включает превращение фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфат-4-эпимеразой (F6PE); превращение T6P в сорбозо-6-фосфат (S6P), катализируемое сорбозо-6-фосфат-эпимеразой (S6PE); превращение полученного S6P в гулозо-6-фосфат (Gul6P), катализируемое гулозо-6-фосфат-изомеразой; и превращение Gul6P в гулозу, катализируемое гулозо-6-фосфат-фосфатазой (Gul6PP).
[0298] Примеры F6PE включают, но не ограничиваются ими, следующие белки: Uniprot ID E8N0N6, E4SEH3, I0I507, H1XRG1, и B5YBD7. Uniprot ID E8N0N6 и I0I507 оба катализируют реакцию F6PE и имеют 27% идентичности аминокислотной последовательности. Следовательно, примеры F6PE также включают любые гомологи, имеющие по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, или по меньшей мере 99% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0299] Способ получения гулозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения гулозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения гулозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0300] Следовательно, способ получения гулозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в T6P с помощью F6PE, (v) превращение T6P в S6P с помощью S6PE (EC 5.3.1.26), (vi) превращение S6P в Gul6P с помощью Gul6PI, и (vii) превращение GulP в гулозу с помощью Gul6PP.
[0301] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1:1 (αGP:PGM:PGI:F6PE:S6PE:Gul6PI:GulPP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения гулозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0302] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0303] Ионы фосфата, образующиеся при дефосфорилировании S6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования гулозы.
[0304] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0305] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование S6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0306] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т. е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т. е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения гулозы включает энергетически выгодную химическую реакцию.
[0307] Гулозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, S6P в Gul6P с помощью Gul6PI, и Gul6P в гулозу с помощью Gul6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0308] Гулоза может быть получена из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, S6P в Gul6P с помощью Gul6PI, и Gul6P в гулозу с помощью Gul6PP.
[0309] Ионы фосфата, образующиеся во время превращения S6P в сорбозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода гулозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0310] В некоторых вариантах реализации изобретения, способ получения гулозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0311] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0312] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в гулозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в гулозу с использованием бесклеточных ферментных коктейлей.
[0313] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в гулозу и повышения растворимости.
[0314] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода гулозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода гулозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0315] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы во гулозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, и превращение S6P в сорбозу, катализируемое S6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0316] В некоторых вариантах реализации изобретения полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход гулозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0317] В других вариантах реализации изобретения гулоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE; S6P в Gul6P с помощью Gul6PI, и Gul6P в гулозу с помощью Gul6PP.
[0318] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда гулозу получают из биомассы или лактозы, выходы ниже, чем в данном изобретении, и гулозу необходимо отделять от других сахаров с помощью хроматографии, что приводит к более высоким производственным затратам. Кроме того, наш способ осуществляется без участия животных.
[0319] Стадия превращения S6P в гулозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, гулозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0320] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение гулозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0321] Конкретным вариантом реализации данного изобретения является гулоза, полученная описанными в данном документе способами получения гулозы.
[0322] Идоза
[0323] Один вариант реализации данного изобретения представляет собой способ для получения идозы, который включает превращение фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфат-4-эпимеразой (F6PE); превращение T6P в сорбозо-6-фосфат (S6P), катализируемое сорбозо-6-фосфат-эпимеразой (S6PE); превращение полученного S6P в идозо-6-фосфат (I6P), катализируемое идозо-6-фосфат-изомеразой; и превращение I6P в идозу, катализируемое идозо-6-фосфат-фосфатазой (I6PP).
[0324] Примеры F6PE включают, но не ограничиваются ими, следующие белки: Uniprot ID E8N0N6, E4SEH3, I0I507, H1XRG1, и B5YBD7. Uniprot ID E8N0N6 и I0I507 оба катализируют реакцию F6PE и имеют 27% идентичности аминокислотной последовательности. Следовательно, примеры F6PE также включают любые гомологи, имеющие по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, или по меньшей мере 99% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0325] Способ получения идозы в соответствии с данным изобретением также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). В других вариантах реализации изобретения, способ получения идозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). В дополнительных вариантах реализации изобретения, способ получения идозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0326] Следовательно, способ получения идозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в T6P с помощью F6PE, (v) превращение T6P в S6P с помощью S6PE (EC 5.3.1.26), (vi) превращение S6P в I6P с помощью I6PI, и (vii) превращение I6P в идозу с помощью I6PP.
[0327] Как правило, соотношения единиц фермента, используемых в описанном способе, составляют 1:1:1:1:1:1:1 (αGP:PGM:PGI:F6PE:S6PE:I6PI:I6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения идозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0328] Одним из важных преимуществ указанных способов является то, что стадии указанного способа могут проводиться в одном биореакторе или реакционном сосуде. В альтернативном варианте, указанные стадии также могут проводиться во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
[0329] Ионы фосфата, образующиеся при дефосфорилировании S6P, затем могут быть переработаны на стадии превращения сахарида в G1P, особенно когда все стадии проводятся в одном биореакторе или реакционном сосуде. Возможность переработки фосфата в описанных способах позволяет использовать нестехиометрические количества фосфата, что поддерживает концентрации фосфата в реакции низкими. Это влияет на общий путь и общую скорость процессов, но не ограничивает активность отдельных ферментов и учитывает общую эффективность процессов образования идозы.
[0330] Например, концентрации фосфата в реакции могут находиться в диапазоне от около 0,1 мМ до около 300 мМ, от около 0,1 мМ до около 150 мМ, от около 1 мМ до около 50 мМ, предпочтительно от около 5 мМ до около 50 мМ, или более предпочтительно от около 10 мМ до около 50 мМ. Например, концентрация фосфата в реакции может быть около 0,1 мМ, около 0,5 мМ, около 1 мМ, около 1,5 мМ, около 2 мМ, около 2,5 мМ, около 5 мМ, около 6 мМ, около 7 мМ, около 8 мМ, около 9 мМ, около 10 мМ, около 15 мМ, около 20 мМ, около 25 мМ, около 30 мМ, около 35 мМ, около 40 мМ, около 45 мМ, около 50 мМ, или около 55 мМ.
[0331] Следовательно, низкая концентрация фосфата приводит к снижению производственных затрат из-за низкого общего содержания фосфата и, следовательно, к снижению стоимости удаления фосфата. Это также предотвращает ингибирование S6PP высокими концентрациями свободного фосфата и снижает вероятность загрязнения фосфатом.
[0332] Кроме того, описанные в данном документе способы могут быть проведены без добавления АТФ в качестве источника фосфата, т.е. без АТФ. Указанные способы также могут быть проведены без добавления NAD(P)(H), т.е. без NAD(P)(H). Другие преимущества также включают тот факт, что по меньшей мере одна стадия описанных способов получения идозы включает энергетически выгодную химическую реакцию.
[0333] Идозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, S6P в I6P с помощью I6PI, и I6P в идозу с помощью I6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0334] Идоза может быть получена из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, S6P в I6P с помощью I6PI, и I6P в идозу с помощью I6PP.
[0335] Ионы фосфата, образующиеся во время превращения S6P в сорбозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода идозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0336] В некоторых вариантах реализации изобретения, способ получения идозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в F6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0337] В других вариантах реализации изобретения, G6P получают из глюкозы и полифосфата натрия с помощью полифосфат-глюкокиназы.
[0338] Данное описание предлагает способы превращения сахаридов, таких как полисахариды и олигосахариды в крахмале, целлюлозе, сахарозе и их производных продуктах, в идозу. В определенных вариантах реализации изобретения, предложены искусственные (не природные) не содержащие АТФ ферментные пути для превращения крахмала, целлюлозы, сахарозы и их производных продуктов в идозу с использованием бесклеточных ферментных коктейлей.
[0339] Как показано выше, несколько ферментов могут быть использованы для гидролиза крахмала для увеличения выхода G1P. Данные ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит много боковых цепей, которые препятствуют действию αGP. Изоамилаза может быть использована для удаления боковых цепей крахмала с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может привести к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляет альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюкан-фосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, поэтому альфа-амилаза используется для разложения крахмала на фрагменты для более быстрого превращения в идозу и повышения растворимости.
[0340] Мальтозо-фосфорилазу (МР) можно использовать для увеличения выхода идозы путем фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу. В альтернативном варианте, 4-гликан-трансферазу (4GT) можно применять для увеличения выхода идозы путем переработки продуктов разложения глюкозы, мальтозы и мальтотриозы в более длинные мальтоолигосахариды; которые могут быть фосфоролитически расщеплены с помощью αGP с получением G1P.
[0341] В определенных вариантах реализации изобретения, целлюлоза и ее производные продукты могут быть преобразованы в идозу с помощью ряда стадий. Данный способ предлагает синтетический путь in vitro, который включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE, и превращение S6P в сорбозу, катализируемое S6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0342] В некоторых вариантах реализации изобретения, полифосфат и полифосфат-глюкокиназа (PPGK) могут быть добавлены к способу, таким образом увеличивая выход идозы путем фосфорилирования продукта разложения глюкозы до G6P.
[0343] В других вариантах реализации изобретения идоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; превращение T6P в S6P, катализируемое S6PE; S6P в I6P с помощью I6PI, и I6P в идозу с помощью I6PP.
[0344] В способах по данному изобретению применяют недорогие исходные материалы и снижают производственные затраты за счет снижения затрат, связанных с разделением сырья и продуктов. Крахмал, целлюлоза, сахароза и некоторые их производные являются менее дорогостоящим сырьем, чем, например, лактоза. Когда идозу получают из биомассы или лактозы, выходы ниже, чем в данном изобретении, и идозу необходимо отделять от других сахаров с помощью хроматографии, что приводит к более высоким производственным затратам. Кроме того, наш способ осуществляется без участия животных.
[0345] Стадия превращения S6P в идозу в соответствии с данным изобретением представляет собой необратимую реакцию фосфатазы, независимо от сырья. Следовательно, идозу получают с очень высоким выходом, при этом эффективно минимизируя последующие затраты на разделение продукта.
[0346] В отличие от способов производства на основе клеток, данное изобретение включает бесклеточное получение идозы, имеет относительно высокие скорости реакции благодаря удалению клеточной мембраны, которая часто замедляет транспорт субстрата/продукта в клетку и из клетки. Также конечный продукт является свободным от богатых питательными веществами ферментационных сред/клеточных метаболитов.
[0347] Конкретным вариантом реализации данного изобретения является идоза, полученная описанными в данном документе способами получения идозы.
[0348] Тагатоза
[0349] Способы получения тагатозы включают превращение F6P в T6P, катализируемое эпимеразой; и превращение T6P в тагатозу, катализируемое фосфатазой.
[0350] Эпимеразы, пригодные для применения в способах превращения F6P в T6P, включают F6PE. Примеры F6PE включают, но не ограничиваются ими, следующие белки: Uniprot ID E8N0N6, E4SEH3, I0I507, H1XRG1, и B5YBD7. Uniprot ID E8N0N6 и I0I507 оба катализируют реакцию F6PE и имеют 27% идентичности аминокислотной последовательности. Следовательно, примеры F6PE также включают любые гомологи, имеющие по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, или по меньшей мере 99% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0351] Фосфатазы, которые превращают T6P в тагатозу (D-тагатозу), T6PP, могут быть использованы в процессе. Примеры T6PP включают, но не ограничиваются ими, следующие белки: Uniprot ID O29805, D2RHV2 и F2KMK2. Uniprot ID O29805 и F2KMK2 оба катализируют реакцию F6PE и имеют 67% идентичности аминокислотной последовательности. Таким образом, примеры T6PP также включают любые гомологи, имеющие по меньшей мере 65%, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и еще наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0352] Способ получения тагатозы также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). Указанный способ получения тагатозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). Более того, способ получения тагатозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0353] Следовательно, способ получения тагатозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в T6P с помощью фруктозо-6-фосфат-эпимеразы (F6PE), и (v) превращение T6P в тагатозу с помощью тагатозо-6-фосфат-фосфатазы (T6PP).
[0354] Как правило, соотношения единиц фермента, используемых в данном способе, составляют 1:1:1:1:1 (αGP:PGM:PGI:F6PE:T6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения тагатозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т.д. относительно количества других ферментов.
[0355] Тагатозу также можно получить из фруктозы. Например, указанный способ включает получение F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в T6P, катализируемое F6PE; и превращение T6P в тагатозу, катализируемое T6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0356] Тагатоза может быть получена из сахарозы. Данный способ предлагает синтетический путь in vitro, который включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; и превращение T6P в тагатозу, катализируемое T6PP.
[0357] Ионы фосфата, образующиеся во время превращения T6P в тагатозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода тагатозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0358] Указанный способ получения тагатозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в G6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0359] Целлюлоза и ее производные продукты могут быть преобразованы в тагатозу с помощью ряда стадий. Данный способ включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; и превращение T6P в тагатозу, катализируемое T6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0360] Тагатоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в T6P, катализируемое F6PE; и превращение T6P в тагатозу, катализируемое T6PP.
[0361] Псикоза
[0362] Способы получения псикозы включают превращение фруктозо-6-фосфата (F6P) в псикозо-6-фосфат (P6P), катализируемое эпимеразой (например, псикозо-6-фосфат-3-эпимеразой, P6PE), и превращение полученного P6P в псикозу, катализируемое фосфатазой (например, псикозо-6-фосфат-фосфатазой, P6PP).
[0363] Примеры P6PE включают, но не ограничиваются ими, следующие белки, идентифицированные по ID номерам UNIPROT: D9TQJ4, A0A090IXZ8, и P32719. Uniprot ID A0A090IXZ8 и D9TQJ4 оба катализируют реакцию P6PE и имеют 45% идентичности аминокислотной последовательности. Таким образом, примеры P6PE также включают любые гомологи, имеющие по меньшей мере 45%, предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0364] Примеры P6PP включают, но не ограничиваются ими, следующие белки: Uniprot ID. A3DC21, Q5LGR4, и Q89ZR1. Uniprot ID A3DC21 и Q89ZR1 оба катализируют реакцию P6PP и имеют 45% идентичности аминокислотной последовательности. Таким образом, примеры P6PP также включают любые гомологи, имеющие по меньшей мере 45%, предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и еще наиболее предпочтительно по меньшей мере 96, 97, 98, 99 или 100% идентичности аминокислотной последовательности с любым из вышеупомянутых Uniprot ID.
[0365] Способ получения псикозы также включает стадию ферментативного превращения глюкозо-6-фосфата (G6P) в F6P, и данная стадия катализируется фосфоглюкоизомеразой (PGI). Указанный способ получения псикозы дополнительно включает стадию превращения глюкозо-1-фосфата (G1P) в G6P, где данная стадия катализируется фосфоглюкомутазой (PGM). Более того, способ получения псикозы также включает стадию превращения сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
[0366] Следовательно, способ получения псикозы по данному изобретению может, например, включать следующие стадии: (i) превращение сахарида в глюкозо-1-фосфат (G1P) с использованием одного или нескольких ферментов; (ii) превращение G1P в G6P с использованием фосфоглюкомутазы (PGM, EC 5.4.2.2); (iii) превращение G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) превращение F6P в P6P с помощью псикозо-6-фосфат-эпимеразы (P6PE), и (v) превращение P6P в псикозу с помощью псикозо-6-фосфат-фосфатазы (P6PP).
[0367] Как правило, соотношения единиц фермента, используемых в данном способе, составляют 1:1:1:1:1 (αGP:PGM:PGI:P6PE:P6PP). Чтобы оптимизировать выход продукта, данные соотношения можно регулировать в любом количестве комбинаций. Например, соотношение 3:1:1:1:1 может быть использовано для максимизации концентрации фосфорилированных промежуточных соединений, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:3 может использоваться для поддержания надежной подачи фосфата для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:3, может быть использовано для дальнейшего увеличения скорости реакции. Следовательно, соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут варьироваться для повышения эффективности получения тагатозы. Например, конкретный фермент может присутствовать в количестве примерно 2х, 3х, 4х, 5х и т. д. относительно количества других ферментов.
[0368] Псикозу также можно получить из фруктозы. Например, указанный способ включает образование F6P из фруктозы и полифосфата, катализируемое полифосфат-фруктокиназой (PPFK); превращение F6P в P6P, катализируемое P6PE; и превращение P6P в псикозу, катализируемое P6PP. Фруктоза может быть получена, например, путем ферментативного превращения сахарозы.
[0369] Псикоза может быть получена из сахарозы. Данный способ включает следующие ферментативные стадии: образование G1P из сахарозы и свободного фосфата, катализируемое сахарозо-фосфорилазой (SP); превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; и превращение P6P в псикозу, катализируемое P6PP.
[0370] Ионы фосфата, образующиеся во время превращения P6P в псикозу, затем могут быть переработаны на стадии превращения сахарозы в G1P. Кроме того, PPFK и полифосфат могут быть использованы для увеличения выхода псикозы путем получения F6P из фруктозы, образующейся при фосфоролитическом расщеплении сахарозы с помощью SP.
[0371] Указанный способ получения псикозы включает следующие стадии: образование глюкозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом, превращение глюкозы в G6P, катализируемое по меньшей мере одним ферментом, образование фруктозы из полисахаридов и олигосахаридов ферментативным гидролизом или кислотным гидролизом и превращение фруктозы в G6P, катализируемое по крайней мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
[0372] Целлюлоза и ее производные продукты могут быть преобразованы в псикозу с помощью ряда стадий. Данный способ включает следующие стадии: образование G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстрин-фосфорилазой (CDP) и целлобиозо-фосфорилазой (CBP), соответственно; превращение G1P в G6P, катализируемое PGM; превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; и превращение P6P в псикозу, катализируемое P6PP. В этом процессе фосфат-ионы могут быть переработаны на стадии превращения целлодекстрина и целлобиозы в G1P.
[0373] Псикоза может быть получена из глюкозы. Указанный способ включает стадии образования G6P из глюкозы и полифосфата, катализируемого полифосфат-глюкокиназой (PPGK); превращение G6P в F6P, катализируемое PGI; превращение F6P в P6P, катализируемое P6PE; и превращение P6P в псикозу, катализируемое P6PP.
ПРИМЕРЫ
[0374] Материалы и методы
[0375] Реактивы
[0376] Все реактивы, включая кукурузный крахмал, растворимый крахмал, мальтодекстрины, глюкозу, фильтровальную бумагу имели квалификацию «химически чистый» или выше и были приобретены в Sigma-Aldrich (Сент-Луис, Миссури, США) или Fisher Scientific (Питтсбург, Пенсильвания, США), если не указано иное. Рестрикционные ферменты, Т4 лигаза и Phusion ДНК-полимераза были приобретены у New England Biolabs (Ипсвич, Массачусетс, США). Олигонуклеотиды были синтезированы либо Integrated ДНК Technologies (Коралвилл, Айова, США) либо Eurofins MWG Operon (Хантсвилл, Алабама, США). Регенерированная аморфная целлюлоза, применяемая в ферментативной очистке, была получена от Avicel PH105 (FMC BioPolymer, Филадельфия, Пенсильвания, США) через ее растворение и регенерацию, как описано в: Ye и др., «Fusion of a family 9 cellulose-binding module improves catalytic potential of Clostridium thermocellum cellodextrin phosphorylase on insoluble cellulose» Appl. Microbiol. Biotechnol. 2011; 92:551-560. Escherichia coli Sig10 (Sigma-Aldrich, Сент-Луис, Миссури, США) применяли в качестве клетки-хозяина для манипулирования ДНК, и E. coli BL21 (DE3) (Sigma-Aldrich, Сент-Луис, Миссури, США) применяли в качестве клетки-хозяина для экспрессии рекомбинантного белка. ZYM-5052 среда, содержащая либо 100 мг л-1 ампициллина либо 50 мг л-1 канамицина, была применена для роста клеток E.coli, и экспрессии рекомбинантного белка. Целлюлаза из Trichoderma reesei (Номер по каталогу: C2730) и пуллуланаза (Номер по каталогу: P1067) были приобретены у Sigma-Aldrich (Сент-Луис, Миссури, США) и получены в Novozymes (Франклинтон, Северная Каролина, США). Мальтозо-фосфорилаза (Номер по каталогу: M8284) была приобретена у Sigma-Aldrich.
[0377] Получение и очистка рекомбинантных ферментов
[0378] Штамм E.coli BL21 (DE3), несущий экспрессирующую белок плазмиду, инкубировали в колбе Эрленмейера на 1 л со 100 мл среды Zym-5052, содержащей либо 100 мг л-1 ампициллина либо 50 мг л-1 канамицина. Клетки выращивали при 37°С с вращательным встряхиванием при 220 оборотах в минуту в течение 16-24 часов. Клетки собирали центрифугированием при 12°С и промывали один раз либо 20 мМ фосфатно-солевым буферным раствором (рН 7,5), содержащим 50 мМ NaCl и 5 мМ MgCl2 (тепловое осаждение и целлюлозосвязывающий модуль), либо 20 мМ фосфатно-солевым буферным раствором (рН 7,5), содержащим 300 мМ NaCl и 5 мМ имидазола (Ni очистка). Клеточную массу ресуспендировали в том же буферном растворе и подвергали лизису с помощью обработки ультразвуком (Fisher Scientific Sonic Dismembrator Model 500; 5 с импульс включен и 10 с выключен, всего 21 мин при амплитуде 50%). После центрифугирования, целевые белки в супернатантах подвергали очистке.
[0379] Три подхода были использованы для очистки различных рекомбинантных белков. His-меченые белки очищали с помощью смолы Ni Sepharose 6 Fast Flow (GE Life Sciences, Марлборо, Массачусетс, США). Слитые белки, содержащие целлюлозосвязывающий модуль (ЦСМ) и интеин для саморасщепления подвергали очистке путем высокоафинной адсорбции на регенерированной аморфной целлюлозе с большой площадью поверхности. Тепловое осаждение при 70-95°С в течение 5-30 мин применяли для очистки гипертермостабильных ферментов. Чистоту рекомбинантных белков исследовали с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПААГ).
[0380] Применяемые ферменты и анализы их активности
[0381] Применяли альфа-глюкан-фосфорилазу (αGP) от Thermotoga maritima (UniProt ID G4FEH8). Активность оценивали в 50 мМ натрий-фосфатном буфере (рН 7,2), содержащем 1 мМ MgCl2 и 30 мМ мальтодекстрина при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ) (Vivaproducts, Inc., Литтлтон, Массачусетс, США). Глюкозо-1-фосфат (G1P) определяли с применением глюкозо-гексокиназа/G6PDH набора для анализа (Sigma Aldrich, № по каталогу GAHK20-1KT) с добавлением 25 Ед/мл фосфоглюкомутазы. Единица (Ед) представляет собой мкмоль/мин.
[0382] Применяли фосфоглюкомутазу (PGM) от Thermococcus kodakaraensis (UniProt ID Q68BJ6). Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 5 мМ G1P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт глюкозо-6-фосфат (G6P) определяли с применением гексокиназа/G6PDH набора для анализа (Sigma Aldrich, № по каталогу GAHK20-1KT).
[0383] Применяли два различных источника фосфоглюкоизомеразы (PGI), от Clostridium thermocellum (UniProt ID A3DBX9) и Thermus thermophilus (UniProt ID Q5SLL6). Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 10 мМ G6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, фруктозо-6-фосфат (F6P), определяли с применением фруктозо-6-фосфат-киназы (F6PK)/пируват-дегидрогеназы (PK)/лактат-дегидрогеназы (LD) сопряженного ферментного анализа, в котором уменьшение оптической плотности при 340 нм указывает на образование F6P. Данная реакционная смесь объемом 200 мкл содержала 50 мМ HEPES (рН 7,2), 5 мМ MgCl2, 10 мМ G6P, 1,5 мМ АТФ, 1,5 мМ фосфоенолпирувата, 200 мкМ NADH, 0,1 Ед PGI, 5 Ед PK и 5 Ед LD.
[0384] Рекомбинантные целлодекстрин-фосфорилаза и целлобиозо-фосфорилаза от С. thermocellum описаны в Ye и др. «Spontaneous high-yield production of hydrogen from cellulosic materials and water catalyzed by enzyme cocktails» ChemSusChem 2009; 2:149-152. Их активности анализировали, как описано.
[0385] Рекомбинантная полифосфат-глюкокиназа от Thermobifida fusca YX описана в Liao и др., «One-step purification and immobilization of thermophilic polyphosphate glucokinase from Thermobifida fusca YX: glucose-6-phosphate generation without ATP» Appl. Microbiol. Biotechnol. 2012; 93:1109-1117. Ее активность анализировали, как описано.
[0386] Рекомбинантная изоамилаза от Sulfolobus tokodaii описана в Cheng и др., «Doubling power output of starch biobattery treated by the most thermostable isoamylase from an archaeon Sulfolobus tokodaii» Scientific Reports 2015; 5:13184. Ее активность анализировали, как описано.
[0387] Рекомбинантная 4-альфа-глюканотрансфераза от Thermococcus litoralis описана в Jeon и др. «4-α-Glucanotransferase from the Hyperthermophilic Archaeon Thermococcus Litoralis» Eur. J. Biochem. 1997; 248:171-178. Ее активность измеряли, как описано.
[0388] Применяли сахарозо-фосфорилазу от Thermoanaerobacterium thermosaccharolyticum (Uniprot ID D9TT09) (Verhaeghe и др. «The quest for a thermostable sucrose phosphorylase reveals sucrose 6'-phosphate phosphorylase as a novel specificity» Appl Microbiol Biotechnol. 2014 Aug;98(16):7027-37). Ее активность измеряли в 50 мМ HEPES буферном растворе (рН 7,5), содержащем 10 мМ сахарозы и 12 мМ органического фосфата. Глюкозо-1-фосфат (G1P) определяли с применением глюкозо-гексокиназа/G6PDH набора для анализа с добавлением 25 Ед/мл фосфоглюкомутазы, как и в случае альфа-глюкан-фосфорилазы.
[0389] Применяли псикозо-6-фосфат-3-эпимеразу (P6PE) от Thermoanaerobacterium thermosaccharolyticum (Uniprot ID D9TQJ4). Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 500 мкМ CoCl2, 1 Ед/мл P6PP, и 10 мМ F6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, псикозо-6-фосфат (P6P), был определен с применением Псикозо-6-фосфат-фосфатазы, обнаруживали высвобождение свободного фосфата. Для обнаружения высвобождения свободного фосфата, 500 мкл раствора, содержащего 0,1 М ацетата цинка и 2 мМ молибдата аммония (рН 5) добавляли к 50 мкл реакционной смеси. Данную смесь перемешивали и затем добавляли 125 мкл 5% аскорбиновой кислоты (рН 5). Полученный раствор перемешивали, затем инкубировали при 30°С в течение 20 мин. Определяли оптическую плотность при 850 нм для обнаружения высвобождения свободного фосфата. Псикозу затем определяли с помощью ВЭЖХ с использованием Agilent Hi-Plex H-колонки (исследуемый и контрольный образцы хроматографируют с 5 мМ H2SO4 при 0,6 мл/мин и 65°С).
[0390] Применяли аллозо-6-фосфат-изомеразу (A6PI) от Clostridium thermocellum (Uniprot ID W4V2C8) с аминокислотной последовательностью, представленной в SEQ ID NO: 1. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 500 мкМ CoCl2, 1 Ед/мл P6PE, 1 Ед/мл A6PP, и 10 мМ F6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, аллозо-6-фосфат (P6P), был определен с применением аллозо-6-фосфат-фосфатазы, обнаруживали высвобождение свободного фосфата, как описано для P6PE. Аллозу определяли с помощью ВЭЖХ таким же образом, как псикозу. Другая A6PI, такая как A6PI от Symbiobacterium thermophilum (Uniprot ID Q67LX4)с аминокислотной последовательностью, представленной в SEQ ID NO: 2, может быть применена.
[0391] Применяли аллозо-6-фосфат-фосфатазу (A6PP) от Rubellimicrobium thermophilum (Uniprot ID S9SDA3) с аминокислотной последовательностью, представленной в SEQ ID NO: 3. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 500 мкМ CoCl2, 1 Ед/мл P6PE, 1 Ед/мл A6PI, и 10 мМ F6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, аллозу, определяли путем обнаружения высвобождения свободного фосфата, как описано для P6PE. Аллозу определяли с помощью ВЭЖХ таким же образом, как псикозу. Другие A6PP, такие как A6PP от Thermotoga maritima (Uniprot ID Q9X0Y1) с аминокислотной последовательностью, представленной в SEQ ID NO: 4, A6PP от Thermoanaerobacterium saccharolyticum (Uniprot ID I3VT81) с аминокислотной последовательностью, представленной в SEQ ID NO: 5, A6PP от Streptomyces thermoautotrophicus (Uniprot ID A0A132NF06) с аминокислотной последовательностью, представленной в SEQ ID NO: 6, и A6PP от Sphaerobacter thermophilus (Uniprot ID D1C7G9) с аминокислотной последовательностью, представленной в SEQ ID NO: 7, могут быть применены.
[0392] Применяли маннозо-6-фосфат-изомеразу (M6PI) от Pseudonocardia thermophila (Uniprot ID A0A1M6TLY7) с аминокислотной последовательностью, представленной в SEQ ID NO: 8. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 1 Ед/мл PGI, 1 Ед/мл M6PP, и 10 мМ F6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, маннозо-6-фосфат (M6P), был определен с применением маннозо-6-фосфат-фосфатазы (M6PP), обнаруживали высвобождение свободного фосфата, как описано для P6PE. Маннозу определяли с помощью ВЭЖХ таким же образом, как псикозу. Другие M6PI такие как M6PI от Caldithrix abyssi (Uniprot ID H1XQS6) с аминокислотной последовательностью, представленной в SEQ ID NO: 9, M6PI от Myceliophthora thermophila (Uniprot ID G2Q982) с аминокислотной последовательностью, представленной в SEQ ID NO: 10, и M6PI от Treponema caldarium (Uniprot ID F8F1Z8) с аминокислотной последовательностью, представленной в SEQ ID NO: 11, могут быть применены.
[0393] Применяли маннозо-6-фосфат-фосфатазу (M6PP) от Tepidimonas fonticaldi (Uniprot ID A0A1A6DSI3) с аминокислотной последовательностью, представленной в SEQ ID NO: 12. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 10 мМ маннозо-6-фосфата при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, маннозу, определяли путем обнаружения высвобождения свободного фосфата, как описано для P6PE. Маннозу определяли с помощью ВЭЖХ таким же образом, как псикозу. Другие M6PP такие как M6PP от Thermomonas hydrothermalis (Uniprot ID A0A1M4UN08) с аминокислотной последовательностью, представленной в SEQ ID NO: 13, и M6PP от Sulfurivirga caldicuralii (Uniprot ID A0A1N6FCW3) с аминокислотной последовательностью, представленной в SEQ ID NO: 14, могут быть применены.
[0394] Была применена бифункциональная фосфоглюкоза/фосфоманнозо-изомераза (PGPMI) от Syntrophothermus lipocalidus (Uniprot ID D7CPH7) с аминокислотной последовательностью, представленной в SEQ ID NO: 15. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 1 Ед/мл M6PP, и 10 мМ G6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, M6P, был определен с применением M6PP, обнаруживали высвобождение свободного фосфата, как описано для P6PE. Маннозу определяли с помощью ВЭЖХ таким же образом, как псикозу. Другие PGPMI такие как PGPMI от Schleiferia thermophila (Uniprot ID A0A085L170) с аминокислотной последовательностью, представленной в SEQ ID NO: 16, и PGPMI от Thermodesulfobium narugense (Uniprot ID M1E6Z3) с аминокислотной последовательностью, представленной в SEQ ID NO: 17, могут быть применены.
[0395] Применяют галактозо-6-фосфат-изомеразу (Gal6PI) от Lactococcus lactis (облигатный димер; Uniprot ID P23494 и P23495 с аминокислотными последовательностями, представленными в SEQ ID NO: 18 и 19, соответственно) (van Rooijen и др. «Molecular Cloning, Characterization, and Nucleotide Sequence of the Tagatose 6-Phosphate Pathway Gene Cluster of the Lactose Operon of Lactococcus Zactis» J. Biol. Chem. 1991;266:7176-7181). Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 1 Ед/мл фруктозо-6-фосфат-4-эпимеразы (F6PE), 1 Ед/мл галактозо-6-фосфат фосфатазы (Gal6PP), и 10 мМ фруктозо-6-фосфата при 37°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, галактоза 6-фосфат (gal6P), был определен с применением Gal6PP, обнаруживали высвобождение свободного фосфата, как описано для P6PE. Галактозу определяли с помощью ВЭЖХ таким же образом, как псикозу.
[0396] Применяли галактозо-6-фосфат-фосфатазу (Gal6PP) от Bacteroides thetaiotaomicron (Uniprot ID Q8A2F3) с аминокислотной последовательностью, представленной в SEQ ID NO: 20. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 10 мМ галактозо-6-фосфата при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, галактозу, определяли путем обнаружения высвобождения свободного фосфата, как описано для P6PE. Галактозу определяли с помощью ВЭЖХ таким же образом, как псикозу.
[0397] Применяли фруктозо-6-фосфат-фосфатазу (F6PP) от Halothermothrix orenii (Uniprot ID B8CWV3) с аминокислотной последовательностью, представленной в SEQ ID NO: 21. Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 10 мМ фруктозо-6-фосфата при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, фруктозу, определяли путем обнаружения высвобождения свободного фосфата, как описано для P6PE. Фруктозу определяли с помощью ВЭЖХ таким же образом, как псикозу.
[0398] Применяли тагатозо-6-фосфат-фосфатазу (T6PP) от Archaeoglobus fugidis (Uniprot ID A0A075WB87). Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 10 мМ T6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Образование тагатозы определяли путем обнаружения высвобождения свободного фосфата, как описано для F6PE.
[0399] Применяли псикозо-6-фосфат фосфатазу (P6PP) от Clostridium thermocellum (UNIPROT ID A3DC21). Активность измеряли в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2, 80 мкМ CoCl2, 1 Ед/мл P6PE, и 10 мМ F6P при 50°С. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, псикозу, определяли путем обнаружения высвобождения свободного фосфата, как описано для P6PE.
[0400] Единицы активности фермента, используемые в каждом Примере ниже, могут быть увеличены или уменьшены, чтобы отрегулировать время реакции по желанию. Например, если кто-либо захочет осуществить Пример 9 в течение 8 ч вместо 24 ч, единицы активности ферментов будут увеличены примерно в 3 раза. И наоборот, если кто-либо захочет осуществить Пример 9 в течение 48 ч вместо 24 ч, единицы активности фермента могут быть снижены примерно в 2 раза. Данные примеры иллюстрируют, как количество единиц активности фермента может быть использовано для увеличения или уменьшения времени реакции при поддержании производительности на постоянном уровне.
[0401] Все продукты
[0402] Пример 1
[0403] Для подтверждения технической осуществимости ферментативного биосинтеза фруктозо-6-фосфата из крахмала, было рекомбинантно экспрессировано три фермента: альфа-глюкан-фосфорилаза из T. maritima (UniProt ID G4FEH8), фосфоглюкомутаза из Thermococcus kodakaraensis (UniProt ID Q68BJ6), и фосфоглюкоизомераза из Clostridium thermocellum (UniProt ID A3DBX9). Рекомбинантные белки были чрезмерно экспрессированы в E. coli BL21 (DE3) и очищены, как описано выше.
[0404] 0,20 мл реакционной смеси, содержащей 10 г/л растворимого крахмала, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,5 мМ ZnCl2, 0,01 Ед αGP, 0,01 Ед PGM, и 0,01 Ед PGI инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, фруктозо-6-фосфат (F6P), определяли с применением фруктозо-6-фосфат-киназы (F6PK)/пируват-дегидрогеназы (PK)/лактат-дегидрогеназы (LD) сопряженного ферментного анализа, в котором уменьшение оптической плотности при 340 нм указывает на образование F6P, как описано выше. Конечная концентрация F6P через 24 ч составляла 3,6 г/л.
[0405] Пример 2
[0406] Такие же тесты, как в Примере 1 (за исключением температуры реакции) были проведены в интервале от 40 до 80°C. Было установлено, что 10 г/л растворимого крахмала образуют 0,9 г/л F6P при 40°C и 3,6 г/л F6P при 80°С после 40 ч реакции. Данные результаты свидетельствуют о том, что повышение температуры реакции для данного набора ферментов позволяет увеличить выход F6P, но слишком высокая температура может ухудшить активность некоторых ферментов.
[0407] Пример 3
[0408] Было обнаружено, что при 80°С соотношение ферментов αGP: PGM: PGI около 1:1:1 приводило к быстрому образованию F6P. Было отмечено, что соотношение ферментов сильно не влияет на конечную концентрацию F6P, если реакция длилась достаточно долго. Однако соотношение ферментов влияет на скорость реакции и общую стоимость ферментов, используемых в системе.
[0409] Пример 4
[0410] 0,20 мл реакционной смеси, содержащей 10 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,5 мМ ZnCl2, 0,01 Ед αGP, 0,01 Ед PGM, и 0,01 Ед PGI инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Продукт, фруктозо-6-фосфат (F6P), определяли с применением фруктозо-6-фосфат-киназы (F6PK)/пируват-дегидрогеназы (PK)/лактат-дегидрогеназы (LD) сопряженного ферментного анализа, в котором уменьшение оптической плотности при 340 нм указывает на образование F6P, как описано выше. Конечная концентрация F6P через 24 ч составляла 3,6 г/л.
[0411] Пример 5
[0412] В эксперименте по получению F6P, для гидролиза целлюлозы при 50°C была применена целлюлаза от Avicel, Sigma. Чтобы удалить бета-глюкозидазу от коммерческой целлюлазы, 10 единиц активности на фильтровальной бумаге/мл целлюлазы смешивали с 10 г/л Avicel на водяной бане со льдом в течение 10 мин. После центрифугирования при 4°С, супернатант, содержащий бета-глюкозидазу, декантировали. Avicel, который был связан с целлюлазой, включающей эндоглюканазу и целлобиогидролазу, ресуспендировали в цитратном буферном растворе (рН 4,8) для гидролиза при 50°С в течение трех дней. Гидролизат целлюлозы смешивали с 5 Ед/мл целлодекстрин-фосфорилазы, 5 Ед/л целлобиозо-фосфорилазы, 5 Ед/мл αGP, 5 Ед/мл PGM, и 5 Ед/мл PGI в 100 мМ HEPES буферном растворе (рН 7,2), содержащем 10 мМ фосфата, 5 мМ MgCl2 и 0,5 мМ ZnCl2. Реакцию проводили при 60°C в течение 72 ч, и были обнаружены высокие концентрации F6P (небольшие количества глюкозы и отсутствие целлобиозы). F6P обнаруживали с помощью сопряженного ферментного анализа, описанного выше. Глюкозу обнаруживали с помощью гексокиназа/G6PDH набора для анализа, как описано выше.
[0413] Пример 6
[0414] Для повышения выхода F6P из Avicel, Avicel был предварительно обработан концентрированной фосфорной кислотой с получением аморфной целлюлозы (РАЦ), как описано в Zhang и др. «A transition from cellulose swelling to cellulose dissolution by o-phosphoric acid: evidence from enzymatic hydrolysis and supramolecular structure» Biomacromolecules 2006; 7:644-648. Чтобы удалить бета-глюкозидазу от коммерческой целлюлазы, 10 единиц активности на фильтровальной бумаге/мл целлюлазы смешивали с 10 г/л РАЦ на водяной бане со льдом в течение 5 мин. После центрифугирования при 4°С, супернатант, содержащий бета-глюкозидазу, декантировали. РАЦ, которая была связана с целлюлазой, включающей эндоглюканазу и целлобиогидролазу, ресуспендировали в цитратном буферном растворе (рН 4,8) для гидролиза при 50°С в течение 12 ч. Гидролизат РАЦ смешивали с 5 Ед/мл целлодекстрин-фосфорилазы, 5 Ед/мл целлобиозо-фосфорилазы, 5 Ед/мл αGP, 5 Ед/мл PGM, и 5 Ед/мл PGI в 100 мМ HEPES буферном растворе (рН 7,2), содержащем 10 мМ фосфата, 5 мМ MgCl2 и 0,5 мМ ZnCl2. Реакцию проводили при 60°C в течение 72 ч. Высокие концентрации F6P и глюкозы были выделены, потому что не были добавлены ферменты для преобразования глюкозы в F6P. F6P обнаруживали с помощью сопряженного ферментного анализа, описанного выше. Глюкозу обнаруживали с помощью гексокиназа/G6PDH набора для анализа, как описано выше.
[0415] Пример 7
[0416] Чтобы дополнительно увеличить выход F6P из РАЦ, были добавлены полифосфат-глюкокиназа и полифосфат. Чтобы удалить бета-глюкозидазу от коммерческой целлюлазы, 10 единиц активности на фильтровальной бумаге/мл целлюлазы смешивали с 10 г/л РАЦ на водяной бане со льдом в течение 5 мин. После центрифугирования при 4°С, супернатант, содержащий бета-глюкозидазу, декантировали. РАЦ, которая была связана с целлюлазой, включающей эндоглюканазу и целлобиогидролазу, ресуспендировали в цитратном буферном растворе (рН 4,8) для гидролиза при 50°С, выдерживали в цитратном буфере (pH 4.8) для гидролиза при 50°С в течение 12 ч. Гидролизат РАЦ смешивали с 5 Ед/мл полифостфат-глюкокиназы, 5 Ед/мл целлодекстрин-фосфорилазы, 5 Ед/мл целлобиозо-фосфорилазы, 5 Ед/мл αGP, 5 Ед/мл PGM, и 5 Ед/мл PGI в 100 мМ HEPES буферном растворе (рН 7,2), содержащем 50 мМ полифосфата, 10 мМ фосфата, 5 мМ MgCl2 и 0,5 мМ ZnCl2. Реакцию проводили при 50°C в течение 72 ч. Высокие концентрации F6P были обнаружены, и теперь присутствовали только небольшие количества глюкозы. F6P обнаруживали с помощью сопряженного ферментного анализа, описанного выше. Глюкозу обнаруживали с помощью гексокиназа/G6PDH набора для анализа, как описано выше.
[0417] Пример 8
[0418] Для того, чтобы определить диапазон концентрации фосфатно-солевого буферного раствора (ФСБ), 0,20 мл реакционной смеси, содержащей 50 г/л мальтодекстрина; 6,25 мМ, 12,5 мМ, 25 мМ, 37,5 мМ, или 50 мМ фосфатно-солевого буферного раствора рН 7,2; 5 мМ MgCl2; 0,1 Ед αGP; 0,1 Ед PGM; и 0,1 Ед PGI инкубировали при 50°С в течение 6 часов. Короткая продолжительность инкубации не обеспечивает достижение завершения, и, следовательно, различия в эффективности может быть четко видно. Образование F6P количественно определяли с применением фруктозо-6-фосфат-киназы (F6PK)/пируват-дегидрогеназы (PK)/лактат-дегидрогеназы (LD) сопряженного ферментного анализа, в котором уменьшение оптической плотности при 340 нм указывает на образование F6P. Соответственно, выход 4,5 г/л, 5,1 г/л, 5,6 г/л, 4,8 г/л или 4,9 г/л F6P был получен для реакций, в которых применяли либо 6,25 мМ, 12,5 мМ, 25 мМ, 37,5 мМ, или 50 мМ фосфатно-солевого буферного раствора рН 7,2 (Таблица 1). Такие результаты указывают на то, что концентрация 25 мМ ФСБ рН 7,2 идеально подходит для данных конкретных условий реакции. Важно отметить, что даже применение 6,25 мМ ФСБ при рН 7,2 приводит к значительному выходу в связи с повторным использованией фосфата. Показано, что описанные способы повторного использования фосфата способствуют поддержанию низкого уровня концентрации фосфата, даже в промышленных масштабах объемной производительности (например, 200-300 г/л мальтодекстрина).
Таблица 1
[0419] Пример 9
[0420] Чтобы определить диапазон рН каскадной реакции, 0,20 мл реакционной смеси, содержащей 50 г/л мальтодекстрина; 50 мМ фосфатно-солевого буферного раствора рН 6,0, 6,2, 6,4, 6,6, 6,8, 7,0, 7,2, или 7,3; 5 мМ MgCl2; 0,02 Ед αGP; 0,02 Ед PGM; и 0,02 Ед PGI инкубировали при 50°С в течение 16 ч. Уменьшение единиц концентраций не обеспечивает достижение завершения реакции, и, следовательно, различия в эффективности может быть четко видно. Образование F6P количественно определяли, как в примере 12. Соответственно, выход 4,0 г/л, 4,1 г/л, 4,2 г/л, 4,1 г/л, 4,4 г/л, 4,1 г/л, 3,8 г/л или 4,0 г/л F6P был получен для реакций, в которых применяли 50 мМ фосфатно-солевого буферного раствора при pH 6,0, 6,2, 6,4, 6,6, 6,8, 7,0, 7,2, или 7,3 (Таблица 2). Такие результаты указывают на то, что рН 6,8 идеально подходит для данных конкретных условий реакции, тем не менее, система работает в широком диапазоне рН.
Таблица 2
[0421] Аллоза
[0422] Пример 10
[0423] Для подтверждения образования аллозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл P6PE, 1 Ед/мл A6PI и 1 Ед/мл A6PP в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 500 мкМ CoCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в аллозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0424] Пример 11
[0425] Для подтверждения образования аллозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 500 мкМ CoCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед A6PI и 0,05 Ед A6PP инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Аллозу определяли с помощью ВЭЖХ, как описано в Примере 10.
[0426] Пример 12
[0427] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 500 мкМ CoCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед A6PI и 0,05 Ед A6PP, которую инкубировали при 50°С в течение 24 ч. Образование аллозы определяли, как в Примере 10.
[0428] Пример 13
[0429] Для дополнительного увеличения выхода аллозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 11.
[0430] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 12), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 500 мкМ CoCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед A6PI, 0,05 Ед A6PP и 0,05 Ед 4GT инкубировали при 50°С в течение 24 ч. Образование аллозы определяли, как в Примере 10.
[0431] Пример 14
[0432] Для дополнительного увеличения выхода аллозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 11.
[0433] Пример 15
[0434] Для дополнительного увеличения выхода аллозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 11.
[0435] Пример 16
[0436] Для получения аллозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 500 мкМ CoCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед P6PE, 0,05 Ед A6PI, и 0,05 Ед A6PP, инкубировали при 50°С в течение 24 ч. Образование аллозы количественно определяли, как в Примере 10.
[0437] Пример 17
[0438] Для получения аллозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 500 мкМ CoCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед A6PI, и 0,05 Ед A6PP, инкубировали при 50°С в течение 24 ч. Образование аллозы количественно определяли, как в Примере 10.
[0439] Пример 18
[0440] Для получения аллозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 500 мкМ CoCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед A6PI, и 0,05 Ед A6PP, инкубировали при 50°С в течение 24 ч. Образование аллозы количественно определяли, как в Примере 10.
[0441] Пример 19
[0442] Для дополнительного увеличения выхода аллозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 18. Образование аллозы количественно определяли, как в Примере 10.
[0443] Манноза
[0444] Пример 20
[0445] Для подтверждения образования маннозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл M6PI/PGPMI и 1 Ед/мл M6PP в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в маннозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0446] Пример 21
[0447] Для подтверждения образования маннозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед M6PI/PGPMI (в случае применения PGPMI, PGI не требуется), и 0,05 Ед M6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Маннозу определяли с помощью ВЭЖХ, как описано в Примере 20.
[0448] Пример 22
[0449] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед M6PI/PGPMI (в случае применения PGPMI, PGI не требуется), и 0,05 Ед M6PP, которую инкубировали при 50°С в течение 24 ч. Образование маннозы определяли, как в Примере 20.
[0450] Пример 23
[0451] Для дополнительного увеличения выхода маннозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 21.
[0452] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 22), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед M6PI/PGPMI (в случае применения PGPMI, PGI не требуется), 0,05 Ед M6PP и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование маннозы определяли, как в Примере 20.
[0453] Пример 24
[0454] Для дополнительного увеличения выхода маннозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 21.
[0455] Пример 25
[0456] Для дополнительного увеличения выхода маннозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 21.
[0457] Пример 26
[0458] Для получения маннозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед M6PI/PGPMI (в случае применения PGPMI, PGI не требуется), и 0,05 Ед M6PP, инкубировали при 50°С в течение 24 ч. Образование маннозы количественно определяли, как в Примере 20.
[0459] Пример 27
[0460] Для получения маннозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед M6PI/PGPMI (в случае применения PGPMI, PGI не требуется), и 0,05 Ед M6PP, инкубировали при 50°С в течение 24 ч. Образование маннозы количественно определяли, как в Примере 20.
[0461] Пример 28
[0462] Для получения маннозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед M6PI/PGPMI (в случае применения PGPMI, PGI не требуется), и 0,05 Ед M6PP, инкубировали при 50°С в течение 24 ч. Образование маннозы количественно определяли, как в Примере 20.
[0463] Пример 29
[0464] Для дополнительного увеличения выхода маннозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 28. Образование маннозы количественно определяли, как в Примере 20.
[0465] Галактоза
[0466] Пример 30
[0467] Для подтверждения образования галактозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл Gal6PI и 1 Ед/мл Gal6PP в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 37°C в течение 3 ч. Превращение F6P в галактозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0468] Пример 31
[0469] Для подтверждения образования галактозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед Gal6PI, и 0,05 Ед Gal6PP, инкубировали при 37°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Галактозу определяли с помощью ВЭЖХ, как описано в Примере 30.
[0470] Пример 32
[0471] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед Gal6PI, и 0,05 Ед Gal6PP, которую инкубировали при 37°С в течение 24 ч. Образование галактозы определяли, как в Примере 30.
[0472] Пример 33
[0473] Для дополнительного увеличения выхода галактозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 31.
[0474] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 12), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед Gal6PI, 0,05 Ед Gal6PP и 0,05 Ед 4GT, инкубировали при 37°С в течение 24 ч. Образование галактозы определяли, как в Примере 30.
[0475] Пример 34
[0476] Для дополнительного увеличения выхода галактозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 31.
[0477] Пример 35
[0478] Для дополнительного увеличения выхода галактозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 31.
[0479] Пример 36
[0480] Для получения галактозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед Gal6PI, и 0,05 Ед Gal6PP, инкубировали при 37°С в течение 24 ч. Образование галактозы количественно определяли, как в Примере 30.
[0481] Пример 37
[0482] Для получения галактозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед Gal6PI, и 0,05 Ед Gal6PP, инкубировали при 37°С в течение 24 ч. Образование галактозы количественно определяли, как в Примере 30.
[0483] Пример 38
[0484] Для получения галактозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед Gal6PI, и 0,05 Ед Gal6PP, инкубировали при 37°С в течение 24 ч. Образование галактозы количественно определяли, как в Примере 30.
[0485] Пример 39
[0486] Для дополнительного увеличения выхода галактозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 38. Образование галактозы количественно определяли, как в Примере 30.
[0487] Пример 40
[0488] Для подтверждения образования галактозы из Gal6P, 10 г/л Gal6P смешивали с 1 Ед/мл Gal6PP в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 1 ч. Превращение Gal6P в галактозу было подтверждено по обнаружению свободного фосфата. Для обнаружения высвобождения свободного фосфата, 500 мкл раствора, содержащего 0,1 М ацетата цинка и 2 мМ молибдата аммония (рН 5) добавляли к 50 мкл реакционной смеси. Данную смесь перемешивали и затем добавляли 125 мкл 5% аскорбиновой кислоты (рН 5). Полученный раствор перемешивали, затем инкубировали при 30°С в течение 20 мин. Определяли оптическую плотность при 850 нм для обнаружения высвобождения свободного фосфата.
[0489] Фруктоза
[0490] Пример 41
[0491] Для подтверждения образования фруктозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл F6PP в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в фруктозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0492] Пример 42
[0493] Для подтверждения образования фруктозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, и 0,05 Ед F6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Фруктозу определяли с помощью ВЭЖХ, как описано в Примере 41.
[0494] Пример 43
[0495] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, и 0,05 Ед F6PP, которую инкубировали при 50°С в течение 24 ч. Образование фруктозы определяли, как в Примере 41.
[0496] Пример 44
[0497] Для дополнительного увеличения выхода фруктозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 42.
[0498] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 12), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PP и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование фруктозы определяли, как в Примере 41.
[0499] Пример 45
[0500] Для дополнительного увеличения выхода фруктозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 42.
[0501] Пример 46
[0502] Для дополнительного увеличения выхода фруктозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 42.
[0503] Пример 47
[0504] Для получения фруктозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, и 0,05 Ед F6PP, инкубировали при 50°С в течение 24 ч. Образование фруктозы количественно определяли, как в Примере 41.
[0505] Пример 48
[0506] Для получения фруктозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 Ед PGM, 0,05 Ед PGI, и 0,05 Ед F6PP, инкубировали при 50°С в течение 24 ч. Образование фруктозы количественно определяли, как в Примере 41.
[0507] Альтроза
[0508] Пример 49
[0509] Для подтверждения образования альтрозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл P6PE, 1 Ед/мл альтрозо-6-фосфат-изомеразы (Alt6PI), и 1 Ед/мл альтрозо-6-фосфат-фосфатазы (Alt6PP) в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в альтрозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0510] Пример 50
[0511] Для подтверждения образования альтрозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед Alt6PI, и 0,05 Ед Alt6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Альтрозу определяли с помощью ВЭЖХ, как описано в Примере 49.
[0512] Пример 51
[0513] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед Alt6PI, и 0,05 Ед Alt6PP, которую инкубировали при 50°С в течение 24 ч. Образование альтрозы определяли, как в Примере 49.
[0514] Пример 52
[0515] Для дополнительного увеличения выхода альтрозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 50.
[0516] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 50), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед Alt6PI, 0,05 Ед Alt6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование альтрозы определяли, как в Примере 49.
[0517] Пример 53
[0518] Для дополнительного увеличения выхода альтрозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 50.
[0519] Пример 54
[0520] Для дополнительного увеличения выхода альтрозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 50.
[0521] Пример 55
[0522] Для получения альтрозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед P6PE, 0,05 Ед Alt6PI, и 0,05 Ед Alt6PP, инкубировали при 50°С в течение 24 ч. Образование альтрозы количественно определяли, как в Примере 49.
[0523] Пример 56
[0524] Для получения альтрозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед Alt6PI, и 0,05 Ед Alt6PP, инкубировали при 50°С в течение 24 ч. Образование альтрозы количественно определяли, как в Примере 49.
[0525] Пример 57
[0526] Для получения альтрозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед Alt6PI, и 0,05 Ед Alt6PP, инкубировали при 50°С в течение 24 ч. Образование альтрозы количественно определяли, как в Примере 49.
[0527] Пример 58
[0528] Для дополнительного увеличения выхода альтрозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 56. Образование альтрозы количественно определяли, как в Примере 49.
[0529] Талоза
[0530] Пример 59
[0531] Для подтверждения образования талозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл F6PE, 1 Ед/мл талозо-6-фосфат-изомеразы (Tal6PI), и 1 Ед/мл талозо-6-фосфат-фосфатазы (Tal6PP) в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в талозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0532] Пример 60
[0533] Для подтверждения образования талозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед Tal6PI, и 0,05 Ед Tal6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Талозу определяли с помощью ВЭЖХ, как описано в Примере 59.
[0534] Пример 61
[0535] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед Tal6PI, и 0,05 Ед Tal6PP, которую инкубировали при 50°С в течение 24 ч. Образование талозы определяли, как в Примере 59.
[0536] Пример 62
[0537] Для дополнительного увеличения выхода талозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 60.
[0538] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 60), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед Tal6PI, 0,05 Ед Tal6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование талозы определяли, как в Примере 59.
[0539] Пример 63
[0540] Для дополнительного увеличения выхода талозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 59.
[0541] Пример 64
[0542] Для дополнительного увеличения выхода талозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 60.
[0543] Пример 65
[0544] Для получения талозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед F6PE, 0,05 Ед Tal6PI, и 0,05 Ед Tal6PP, инкубировали при 50°С в течение 24 ч. Образование талозы количественно определяли, как в Примере 59.
[0545] Пример 66
[0546] Для получения талозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед Tal6PI, и 0,05 Ед Tal6PP, инкубировали при 50°С в течение 24 ч. Образование талозы количественно определяли, как в Примере 59.
[0547] Пример 67
[0548] Для получения талозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед Tal6PI, и 0,05 Ед Tal6PP, инкубировали при 50°С в течение 24 ч. Образование талозы количественно определяли, как в Примере 59.
[0549] Пример 68
[0550] Для дополнительного увеличения выхода талозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 66. Образование талозы количественно определяли, как в Примере 59.
[0551] Сорбоза
[0552] Пример 69
[0553] Для подтверждения образования сорбозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл F6PE, 1 Ед/мл сорбозо-6-фосфат-3-эпимеразы (S6PE), и 1 Ед/мл сорбозо-6-фосфат-фосфатазы (S6PP) в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в сорбозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0554] Пример 70
[0555] Для подтверждения образования сорбозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, и 0,05 Ед S6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Сорбозу определяли с помощью ВЭЖХ, как описано в Примере 68.
[0556] Пример 71
[0557] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, и 0,05 Ед S6PP, которую инкубировали при 50°С в течение 24 ч. Образование сорбозы определяли, как в Примере 69.
[0558] Пример 72
[0559] Для дополнительного увеличения выхода сорбозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 70.
[0560] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 70), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед S6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование сорбозы определяли, как в Примере 69.
[0561] Пример 73
[0562] Для дополнительного увеличения выхода сорбозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 70.
[0563] Пример 74
[0564] Для дополнительного увеличения выхода сорбозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 69.
[0565] Пример 75
[0566] Для получения сорбозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед F6PE, 0,05 Ед S6PE, и 0,05 Ед S6PP, инкубировали при 50°С в течение 24 ч. Образование сорбозы количественно определяли, как в Примере 69.
[0567] Пример 76
[0568] Для получения сорбозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, и 0,05 Ед S6PP, инкубировали при 50°С в течение 24 ч. Образование сорбозы количественно определяли, как в Примере 69.
[0569] Пример 77
[0570] Для получения сорбозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, и 0,05 Ед S6PP, инкубировали при 50°С в течение 24 ч. Образование сорбозы количественно определяли, как в Примере 69.
[0571] Пример 78
[0572] Для дополнительного увеличения выхода сорбозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 76. Образование сорбозы количественно определяли, как в Примере 69.
[0573] Гулоза
[0574] Пример 79
[0575] Для подтверждения образования гулозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл F6PE, 1 Ед/мл S6PE, 1 Ед/мл гулозо-6-фосфат-изомеразы (Gul6PI), и 1 Ед/мл гулозо-6-фосфат-фосфатазы (Gul6PP) в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в гулозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0576] Пример 80
[0577] Для подтверждения образования гулозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед Gul6PI, и 0,05 Ед Gul6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Гулозу определяли с помощью ВЭЖХ, как описано в Примере 79.
[0578] Пример 81
[0579] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед Gul6PI, и 0,05 Ед Gul6PP, которую инкубировали при 50°С в течение 24 ч. Образование гулозы определяли, как в Примере 79.
[0580] Пример 82
[0581] Для дополнительного увеличения выхода гулозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 80.
[0582] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 80), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед Gul6PI, 0,05 Ед Gul6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование гулозы определяли, как в Примере 79.
[0583] Пример 83
[0584] Для дополнительного увеличения выхода гулозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 80.
[0585] Пример 84
[0586] Для дополнительного увеличения выхода гулозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 80.
[0587] Пример 85
[0588] Для получения гулозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед Gul6PI, и 0,05 Ед Gul6PP, инкубировали при 50°С в течение 24 ч. Образование гулозы количественно определяли, как в Примере 79.
[0589] Пример 86
[0590] Для получения гулозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед Gul6PI, и 0,05 Ед Gul6PP, инкубировали при 50°С в течение 24 ч. Образование гулозы количественно определяли, как в Примере 79.
[0591] Пример 87
[0592] Для получения гулозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед Gul6PI, и 0,05 Ед Gul6PP, инкубировали при 50°С в течение 24 ч. Образование гулозы количественно определяли, как в Примере 79.
[0593] Пример 88
[0594] Для дополнительного увеличения выхода гулозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 86. Образование гулозы количественно определяли, как в Примере 79.
[0595] Идоза
[0596] Пример 89
[0597] Для подтверждения образования идозы из F6P, 10 г/л F6P смешивали с 1 Ед/мл F6PE, 1 Ед/мл S6PE, 1 Ед/мл идозо-6-фосфат-изомеразы (I6PI), и 1 Ед/мл идозо-6-фосфат-фосфатазы (I6PP) в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 3 ч. Превращение F6P в идозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый и контрольный образцы хроматографируют в 5 мМ H2SO4 при 0,6 мл/мин и 65°С.
[0598] Пример 90
[0599] Для подтверждения образования идозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед I6PI, и 0,05 Ед I6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Идозу определяли с помощью ВЭЖХ, как описано в Примере 89.
[0600] Пример 91
[0601] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед I6PI, и 0,05 Ед I6PP, которую инкубировали при 50°С в течение 24 ч. Образование идозы определяли, как в Примере 89.
[0602] Пример 92
[0603] Для дополнительного увеличения выхода идозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 90.
[0604] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 90), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед I6PI, 0,05 Ед I6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование идозы определяли, как в Примере 89.
[0605] Пример 93
[0606] Для дополнительного увеличения выхода идозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 90.
[0607] Пример 94
[0608] Для дополнительного увеличения выхода идозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 90.
[0609] Пример 95
[0610] Для получения идозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед I6PI, и 0,05 Ед I6PP, инкубировали при 50°С в течение 24 ч. Образование идозы количественно определяли, как в Примере 89.
[0611] Пример 96
[0612] Для получения идозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед I6PI, и 0,05 Ед I6PP, инкубировали при 50°С в течение 24 ч. Образование идозы количественно определяли, как в Примере 89.
[0613] Пример 97
[0614] Для получения идозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед S6PE, 0,05 Ед I6PI, и 0,05 Ед I6PP, инкубировали при 50°С в течение 24 ч. Образование идозы количественно определяли, как в Примере 89.
[0615] Пример 98
[0616] Для дополнительного увеличения выхода идозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в Примере 96. Образование идозы количественно определяли, как в Примере 89.
[0617] Тагатоза
[0618] Пример 99
[0619] Для подтверждения образования тагатозы из F6P, 2 г/л F6P смешивали с 1 Ед/мл фруктозо-6-фосфат-эпимеразы (F6PE) и 1 Ед/мл тагатозо-6-фосфат-фосфатазы (T6PP) в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2. Реакционную смесь инкубировали при 50°C в течение 16 ч. 100% превращение F6P в тагатозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый образец хроматографировали в 5 мМ H2SO4 при 0,6 мл/мин.
[0620] Пример 100
[0621] Для подтверждения образования тагатозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, и 0,05 Ед T6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Тагатозу обнаруживали и количественно определяли с помощью ВЭЖХ Agilent 1100 series, с применением рефрактометрического детектора и Agilent Hi-Plex H-колонки. Подвижная фаза представляла собой 5 мМ H2SO4, скорость потока составляла 0,6 мл/мин. Полученный выход тагатозы составил 9,2 г/л. Это соответствует 92% от теоретического выхода в связи с пределами деградации мальтодекстрина без участия ферментов, таких как изоамилаза или 4-глюкан-трансфераза. Стандарты различных концентраций тагатозы были использованы для количественного определения нашего выхода.
[0622] Пример 101
[0623] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, и 0,05 Ед T6PP, которую инкубировали при 50°С в течение 24 ч. Образование тагатозы количественно определяли, как в Примере 99. Выход тагатозы был увеличен до 16 г/л с предварительной обработкой мальтодекстрина изоамилазой. Это соответствует 80% от теоретического выхода.
[0624] Пример 102
[0625] Для дополнительного увеличения выхода тагатозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 100.
[0626] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 9), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед F6PE, 0,05 Ед T6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование тагатозы количественно определяли, как в примере 9. Выход тагатозы был увеличен до 17,7 г/л с добавлением 4GT к IA-модифицированному мальтодекстрину. Это соответствует 88,5% от теоретического выхода.
[0627] Пример 103
[0628] Для изучения масштабирования реакции, 20 мл реакционной смеси, содержащей 50 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 99), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 10 Ед αGP, 10 Ед PGM, 10 Ед PGI, 10 Ед F6PE и 10 Ед T6PP, инкубировали при 50°С в течение 24 ч. Образование тагатозы количественно определяли, как в примере 8. Выход тагатозы составил 37,6 г/л в 20 мл смеси содержащей 50 г/л мальтодекстрина. Это соответствует 75% от теоретического выхода. Такие результаты указывают на то, что масштабирование реакции до больших объемов не приводит к существенному снижению выхода.
[0629] Пример 104
[0630] Для дополнительного увеличения выхода тагатозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 100.
[0631] Пример 105
[0632] Для дополнительного увеличения выхода тагатозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 99.
[0633] Пример 106
[0634] Для получения тагатозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед F6PE, и 0,05 Ед T6PP, инкубировали при 50°С в течение 24 ч. Образование тагатозы количественно определяли, как в Примере 100.
[0635] Пример 107
[0636] Для получения тагатозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед F6PE, и 0,05 Ед T6PP, инкубировали при 50°С в течение 24 ч. Образование тагатозы количественно определяли, как в Примере 100.
[0637] Пример 108
[0638] Для получения тагатозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 PGM, 0,05 Ед PGI, 0,05 Ед F6PE, и 0,05 Ед T6PP, инкубировали при 50°С в течение 24 ч. Образование тагатозы количественно определяли, как в Примере 100.
[0639] Пример 109
[0640] Для дополнительного увеличения выхода тагатозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в примере 15. Образование тагатозы количественно определяли, как в Примере 100.
[0641] Псикоза
[0642] Пример 110
[0643] Для подтверждения образования псикозы из F6P, 2 г/л F6P смешивали с 1 Ед/мл P6PE и 1 Ед/мл P6PP в 50 мМ HEPES буферном растворе (рН 7,2), содержащем 5 мМ MgCl2 и 80 мкМ CoCl2. Реакционную смесь инкубировали при 50°C в течение 6 ч. 99% превращение F6P в псикозу было подтверждено с помощью ВЭЖХ (Agilent 1100 series) с применением Agilent Hi-Plex H-колонки и рефрактометрического детектора. Исследуемый образец хроматографировали в 5 мМ H2SO4 при 0,6 мл/мин.
[0644] Пример 111
[0645] Для подтверждения образования псикозы из мальтодекстрина, 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 80 мкМ CoCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, и 0,05 Ед P6PP, инкубировали при 50°С в течение 24 ч. Реакцию останавливали путем фильтрации фермента с помощью Vivaspin 2 концентратора (10000 НОММ). Псикозу обнаруживали и количественно определяли с помощью ВЭЖХ Agilent 1100 series, с применением рефрактометрического детектора и Agilent Hi-Plex H-колонки. Подвижная фаза представляла собой 5 мМ H2SO4, скорость потока составляла 0,6 мл/мин. Стандарты различных концентраций псикозы были использованы для количественного определения нашего выхода.
[0646] Пример 112
[0647] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 5,5), 5 мМ MgCl2, 80 мкМ CoCl2, и 0,1 г/л изоамилазы инкубировали при 80°С в течение 24 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, и 0,05 Ед P6PP, которую инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0648] Пример 113
[0649] Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буферного раствора (рН 4,5), 5 мМ MgCl2, и разбавление Novozymes D6 пуллуланазы 1:200 инкубировали при 50°С в течение 4 ч. Ее использовали для приготовления другой реакционной смеси, содержащей 20 г/л модифицированного пуллуланазой мальтодекстрина, 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 80 мкМ CoCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, и 0,05 Ед P6PP, которую инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0650] Пример 114
[0651] Для дополнительного увеличения выхода псикозы из мальтодекстрина, 0,05 Ед 4-глюкан-трансферазы (4GT) добавляли к реакционной смеси, описанной в Примере 111.
[0652] 0,2 мл реакционной смеси, содержащей 20 г/л модифицированного изоамилазой мальтодекстрина (см. пример 9), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 80 мкМ CoCl2, 0,05 Ед αGP, 0,05 Ед PGM, 0,05 Ед PGI, 0,05 Ед P6PE, 0,05 Ед P6PP, и 0,05 Ед 4GT, инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0653] Пример 115
[0654] Для изучения масштабирования реакции, 20 мл реакционной смеси, содержащей 50 г/л модифицированного изоамилазой мальтодекстрина (см. Пример 10), 50 мМ фосфатно-солевого буферного раствора рН 7,2, 5 мМ MgCl2, 80 мкМ CoCl2, 10 Ед αGP, 10 Ед PGM, 10 Ед PGI, 10 Ед P6PE и 10 Ед P6PP, инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0655] Пример 116
[0656] Для дополнительного увеличения выхода псикозы из мальтодекстрина, 0,05 Ед мальтозо-фосфорилазы добавляли к реакционной смеси, описанной в Примере 110.
[0657] Пример 117
[0658] Для дополнительного увеличения выхода псикозы из мальтодекстрина, 0,05 Ед полифосфат-глюкокиназы и 75 мМ полифосфата добавляли к реакционной смеси, описанной в Примере 111.
[0659] Пример 118
[0660] Для получения псикозы из фруктозы, реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 80 мкМ CoCl2, 0,05 Ед фруктозо-полифосфат-киназы, 0,05 Ед P6PE, и 0,05 Ед P6PP, инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0661] Пример 119
[0662] Для получения псикозы из глюкозы, реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буферного раствора рН 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 80 мкМ CoCl2, 0,05 Ед глюкозо-полифосфат-киназы, 0,05 Ед PGI, 0,05 Ед P6PE, и 0,05 Ед P6PP, инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0663] Пример 120
[0664] Для получения псикозы из сахарозы, реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ фосфатно-солевого буферного раствора рН 7,0, 5 мМ MgCl2, 80 мкМ CoCl2, 0,05 Ед сахарозо-фосфорилазы, 0,05 PGM, 0,05 Ед PGI, 0,05 Ед P6PE, и 0,05 Ед P6PP, инкубировали при 50°С в течение 24 ч. Образование псикозы количественно определяли, как в Примере 111.
[0665] Пример 121
[0666] Для дополнительного увеличения выхода псикозы из сахарозы, 75 мМ полифосфата и 0,05 Ед полифосфат-фруктокиназы добавляли к реакционной смеси, описанной в примере 20. Образование псикозы количественно определяли, как в Примере 111.
[0667] Данное изобретение включает все варианты реализации и вариации, по существу, как описано выше в данном документе и со ссылкой на примеры и фигуры. Хотя различные варианты реализации данного изобретения описаны в данном документе, адаптации и модификации могут быть осуществлены в пределах объема данного изобретения, в соответствии с общеизвестными сведениями специалистов в данной области техники.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Bonumose LLC
<120> ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ГЕКСОЗ
<130> 146.0004-WO00
<150> 62/470605
<151> 2017-03-13
<150> 62/470620
<151> 2017-03-13
<150> 62/482148
<151> 2017-04-05
<150> 62/480798
<151> 2017-04-03
<160> 21
<170> PatentIn версии 3.5
<210> 1
<211> 149
<212> БЕЛОК
<213> Clostridium thermocellum
<400> 1
Met Lys Ile Gly Ile Gly Ser Asp His Gly Gly Tyr Asn Leu Lys Arg
1 5 10 15
Glu Ile Ile Asp Phe Leu Lys Lys Arg Glu Tyr Glu Val Ile Asp Phe
20 25 30
Gly Thr Tyr Gly Thr Asp Ser Val Asp Tyr Pro Asp Phe Gly Leu Lys
35 40 45
Val Ala Glu Ala Val Lys Gly Gly Glu Cys Asp Arg Gly Ile Val Val
50 55 60
Cys Gly Thr Gly Val Gly Ile Ser Ile Ser Ala Asn Lys Val Pro Gly
65 70 75 80
Ile Arg Ala Ala Val Cys Thr Asn Ser Tyr Met Ala Arg Met Ser Arg
85 90 95
Glu His Asn Asp Ala Asn Ile Leu Ala Leu Gly Glu Arg Val Val Gly
100 105 110
Leu Asp Leu Ala Leu Asp Ile Val Asp Thr Trp Leu Lys Ala Glu Phe
115 120 125
Gln Gly Gly Arg His Ser Ala Arg Val Gly Lys Ile Gly Glu Ile Glu
130 135 140
Glu Lys Tyr Ser Lys
145
<210> 2
<211> 154
<212> БЕЛОК
<213> Symbiobacterium thermophilum
<400> 2
Met Arg Ile Ala Ile Gly Asn Asp His Val Gly Thr Glu Met Lys Arg
1 5 10 15
Ala Ile Ala Ala His Leu Glu Ser Leu Gly His Glu Val Val Asn Phe
20 25 30
Gly Thr Asp Ser Thr Glu Arg Thr Asp Tyr Pro Ile Tyr Gly Glu Arg
35 40 45
Val Ala Arg Ala Val Ala Ala Gly Glu Val Asp Cys Gly Ile Leu Ile
50 55 60
Cys Gly Thr Gly Val Gly Ile Ser Leu Ala Ala Asn Lys Val Arg Gly
65 70 75 80
Ile Arg Ala Val Val Cys Ser Glu Pro Tyr Thr Ala Arg Leu Ser Lys
85 90 95
Gln His Asn Asn Thr Asn Ile Leu Ala Phe Gly Ala Arg Val Val Gly
100 105 110
Val Asp Leu Ala Lys Met Ile Val Asp Glu Trp Leu Asn Ala Ser Phe
115 120 125
Glu Gly Gly Arg His Gln Arg Arg Val Asp Met Ile Ala Asp Ile Glu
130 135 140
Arg Arg Glu Glu Cys Gly Pro Glu Gly Cys
145 150
<210> 3
<211> 217
<212> БЕЛОК
<213> Rubellimicrobium thermophilum
<400> 3
Met Thr Ser Arg Tyr Asp Ala Val Val Phe Asp Leu Asp Gly Thr Leu
1 5 10 15
Ile Asp Thr Glu Ser Leu Cys Asn Ala Ala Gly Val Glu Ala Cys Ala
20 25 30
Ala Leu Gly Leu Pro Val Ser Gly Glu Phe Phe Glu Ser Leu Ala Gly
35 40 45
Ile Asp Asp Arg Thr Arg Val Gln Leu Ile Gly Glu His Val Gly Thr
50 55 60
Ala Val Asp Leu Ser Ala Phe Leu Ala Ala Trp Asp Arg Leu Cys Ile
65 70 75 80
Glu Arg Phe Ala Gln Gly Ile Pro Leu Lys Pro Gly Ala Ile Glu Leu
85 90 95
Leu Glu Gln Ile Ala Ala Ala Gly Ile Pro Leu Ala Leu Ala Thr Ser
100 105 110
Ser Arg Arg Gly Pro Ala Glu Asp Lys Leu Arg Met Ala Gly Leu Ala
115 120 125
Arg His Phe Arg Thr Val Val Thr Phe Asp Asp Val Ala Ala Pro Lys
130 135 140
Pro Ala Pro Asp Ala Tyr Leu Leu Ala Val Asp Arg Leu Gly Val Pro
145 150 155 160
Pro Ala Arg Ala Leu Ala Phe Glu Asp Ser Glu Thr Gly Ala Arg Ala
165 170 175
Ala His Ala Ala Gly Leu Thr Val Val Gln Val Pro Asp Leu His Pro
180 185 190
Thr Gln Gly Ala His Ala His His Val Ala Ser Ser Leu Leu Glu Gly
195 200 205
Ala Ala Met Ala Gly Leu Leu Pro Val
210 215
<210> 4
<211> 216
<212> БЕЛОК
<213> Thermotoga maritima
<400> 4
Met Glu Ala Val Ile Phe Asp Met Asp Gly Val Leu Met Asp Thr Glu
1 5 10 15
Pro Leu Tyr Phe Glu Ala Tyr Arg Arg Val Ala Glu Ser Tyr Gly Lys
20 25 30
Pro Tyr Thr Glu Asp Leu His Arg Arg Ile Met Gly Val Pro Glu Arg
35 40 45
Glu Gly Leu Pro Ile Leu Met Glu Ala Leu Glu Ile Lys Asp Ser Leu
50 55 60
Glu Asn Phe Lys Lys Arg Val His Glu Glu Lys Lys Arg Val Phe Ser
65 70 75 80
Glu Leu Leu Lys Glu Asn Pro Gly Val Arg Glu Ala Leu Glu Phe Val
85 90 95
Lys Ser Lys Arg Ile Lys Leu Ala Leu Ala Thr Ser Thr Pro Gln Arg
100 105 110
Glu Ala Leu Glu Arg Leu Arg Arg Leu Asp Leu Glu Lys Tyr Phe Asp
115 120 125
Val Met Val Phe Gly Asp Gln Val Lys Asn Gly Lys Pro Asp Pro Glu
130 135 140
Ile Tyr Leu Leu Val Leu Glu Arg Leu Asn Val Val Pro Glu Lys Val
145 150 155 160
Val Val Phe Glu Asp Ser Lys Ser Gly Val Glu Ala Ala Lys Ser Ala
165 170 175
Gly Ile Glu Arg Ile Tyr Gly Val Val His Ser Leu Asn Asp Gly Lys
180 185 190
Ala Leu Leu Glu Ala Gly Ala Val Ala Leu Val Lys Pro Glu Glu Ile
195 200 205
Leu Asn Val Leu Lys Glu Val Leu
210 215
<210> 5
<211> 218
<212> БЕЛОК
<213> Thermoanaerobacterium saccharolyticum
<400> 5
Met Phe Glu Ala Val Ile Leu Asp Met Asp Gly Val Leu Ile Asp Ser
1 5 10 15
Glu Pro Leu His Ile Gln Leu Glu Glu Glu Ile Phe Lys Glu Ile Gly
20 25 30
Ala Asp Ile Ser Leu Glu Glu His Ile Ser Phe Val Gly Thr Thr Ser
35 40 45
His Tyr Met Trp Glu Tyr Val Lys Asn Lys Cys Asn Val Ser Phe Thr
50 55 60
Val Glu Glu Leu Val Glu Met Asp Arg Lys Arg Tyr Phe Asp Tyr Ile
65 70 75 80
Ser Lys His Asp Gly Ala Val Lys Pro Ile Glu Gly Val Asp Glu Leu
85 90 95
Val Lys Glu Leu Tyr Ser Arg Glu Val Arg Leu Ala Val Ala Ser Ser
100 105 110
Ser Pro Ile Asp Val Ile Glu Leu Val Val Lys Lys Leu His Leu Asn
115 120 125
Asp Tyr Phe Cys Glu Leu Val Ser Gly Asp Phe Val Lys Arg Ser Lys
130 135 140
Pro Tyr Pro Asp Ile Phe Leu Tyr Ala Ala Glu Lys Leu Gly Val Ser
145 150 155 160
Pro Glu Arg Cys Leu Val Val Glu Asp Ser Asn Lys Gly Val Leu Ala
165 170 175
Ala Lys Ser Ala Gly Met Lys Val Ile Gly Phe Ile Asn Pro Asn Ser
180 185 190
Gly Asp Gln Asp Ile Ser Met Ala Asp Met Val Ile Arg Ser Phe Ser
195 200 205
Glu Leu Asn Tyr Glu Lys Leu Gln Asn Ile
210 215
<210> 6
<211> 220
<212> БЕЛОК
<213> Streptomyces thermoautotrophicus
<400> 6
Met Pro Ser Ser Ser Gly Gly Leu Gln Ala Val Phe Phe Asp Met Asp
1 5 10 15
Gly Leu Leu Val Asp Thr Glu Pro Thr Trp His Glu Val Glu Ala Glu
20 25 30
Val Met Ala Glu Tyr Gly Tyr Ala Trp Thr Pro Glu Asp Arg Leu Ala
35 40 45
Cys Leu Gly Gly Pro Met Glu Arg Thr Cys Arg Tyr Met Ile Glu Arg
50 55 60
Cys Gly Ala Asp Ile Thr Val Glu Ala Leu Gly Ala Thr Leu Val Glu
65 70 75 80
Arg Met Ala Leu Arg Val Arg Glu Glu Val Ala Val Gln Pro Gly Ala
85 90 95
Lys Glu Leu Leu Ser Glu Leu Ile Glu Ala Gly Val Pro Arg Ala Leu
100 105 110
Val Ser Ser Ser Phe Arg Val Leu Val Asp Ala Val Leu Asp Ala Val
115 120 125
Gly His Asp Leu Phe Val Val Thr Val Ala Gly Asp Glu Val Ala Arg
130 135 140
Ala Lys Pro His Pro Glu Pro Tyr Leu Thr Ala Ala Ala Arg Leu Gly
145 150 155 160
Val Asp Pro Ala Arg Cys Val Val Leu Glu Asp Ser Pro Pro Gly Val
165 170 175
Ala Ala Ala Glu Ala Ala Gly Cys Leu Val Val Ala Val Pro Ser Val
180 185 190
Ala Pro Leu Glu Pro Ala Pro Arg Arg Leu Val Val Arg Ser Leu Thr
195 200 205
Glu Leu Ser Leu Asp Arg Leu Arg Ala Leu Ile Ala
210 215 220
<210> 7
<211> 219
<212> БЕЛОК
<213> Sphaerobacter thermophilus
<400> 7
Met Ser Gln Gly Val Arg Gly Val Val Phe Asp Leu Asp Gly Leu Leu
1 5 10 15
Val Glu Ser Glu Glu Tyr Trp Glu Gln Ala Arg Arg Glu Phe Val Ser
20 25 30
Arg Tyr Gly Gly Thr Trp Gly Asp Asp Ala Gln Gln Ala Val Met Gly
35 40 45
Ala Asn Thr Arg Gln Trp Ser Arg Tyr Ile Arg Glu Ala Phe Asp Ile
50 55 60
Pro Leu Thr Glu Glu Glu Ile Ala Ala Ala Val Ile Ala Arg Met Gln
65 70 75 80
Glu Leu Tyr His Asp His Leu Pro Leu Leu Pro Gly Ala Ile Pro Ala
85 90 95
Val Arg Ala Leu Ala Asp Arg Tyr Pro Leu Ala Val Ala Ser Ser Ser
100 105 110
Pro Pro Val Leu Ile Arg Phe Val Leu Ala Glu Met Gly Val Ala Glu
115 120 125
Cys Phe Gln Ser Val Thr Ser Ser Asp Glu Val Ala His Gly Lys Pro
130 135 140
Ala Pro Asp Val Tyr His Leu Ala Cys Glu Arg Leu Gly Val Ala Pro
145 150 155 160
Glu Gln Ala Val Ala Phe Glu Asp Ser Thr Ala Gly Ile Ala Ala Ala
165 170 175
Leu Ala Ala Gly Leu Arg Val Ile Ala Val Pro Asn Arg Ser Tyr Pro
180 185 190
Pro Asp Pro Asp Val Leu Arg Arg Ala Asp Leu Thr Leu Pro Ser Leu
195 200 205
Glu Glu Phe Asp Pro Ala Val Leu Glu Gln Trp
210 215
<210> 8
<211> 400
<212> БЕЛОК
<213> Pseudonocardia thermophila
<400> 8
Met Glu Leu Leu Asp Asn Pro Ile Arg Pro Tyr Ala Trp Gly Ser Arg
1 5 10 15
Thr Val Leu Ala Glu Leu Leu Gly His Glu Ser Pro Ser Pro His Pro
20 25 30
Glu Ala Glu Met Trp Leu Gly Ala His Pro Gly Asp Pro Ser Arg Leu
35 40 45
Arg Ser Gly Glu Ser Leu Leu Asp Ala Leu Cys Ala Asp Pro Glu Gly
50 55 60
Leu Leu Gly Ala Asp Arg Ala Arg Lys Trp Asp Gly Lys Leu Pro Phe
65 70 75 80
Leu Leu Lys Val Leu Ala Ala Asp Glu Pro Leu Ser Leu Gln Ala His
85 90 95
Pro Ser Leu Asp Gln Ala Arg Val Gly Phe Glu Arg Glu Glu Arg Ala
100 105 110
Gly Ile Ala Arg Asp Ala Pro Glu Arg Asn Tyr Arg Asp Pro Asn His
115 120 125
Lys Pro Glu Leu Leu Cys Ala Leu Thr Glu Phe His Ala Leu Val Gly
130 135 140
Phe Arg Pro Pro Glu Lys Thr Val Glu Leu Leu Ala Ala Leu Ala Val
145 150 155 160
Pro Glu Leu Asp Ala Tyr Ser Gln Leu Leu Thr Ala Gln Pro Asp Ala
165 170 175
Asn Gly Leu Arg Ala Leu Phe Thr Thr Trp Ile Thr Leu Pro Gln Ser
180 185 190
Val Leu Asp Thr Leu Val Pro Ala Leu Gln Ala Gly Cys Val Arg Leu
195 200 205
Ala Ala Ala Asp Gly Pro Phe Lys Ala Glu Ala Arg Thr Val Leu Glu
210 215 220
Leu Ser Glu Arg Tyr Pro Gly Asp Ala Gly Val Leu Ala Ala Val Leu
225 230 235 240
Leu Asn Arg Val Thr Leu Gln Pro Gly Glu Ala Val Tyr Leu Pro Ala
245 250 255
Gly Asn Leu His Ala Tyr Leu Glu Gly Ala Gly Ile Glu Val Met Ala
260 265 270
Ser Ser Asp Asn Val Leu Arg Gly Gly Leu Thr Pro Lys His Val Asp
275 280 285
Val Pro Glu Leu Leu Arg Val Leu Asp Phe His Ala Ala Val Pro Pro
290 295 300
Val Leu Thr Gly Thr Pro Asp Gly Ala Trp Leu Arg Tyr Asp Thr Pro
305 310 315 320
Phe Glu Glu Phe Leu Leu Arg Arg Leu Glu Gly Asp Pro Ala Ala Gly
325 330 335
Leu Val Ala Val Pro Asp Gly Gly Pro Arg Ile Val Leu Cys Thr Arg
340 345 350
Gly Ala Ala Val Leu Arg Gly Arg Asp Glu Gln Leu Asp Leu His Arg
355 360 365
Gly Ala Ser Ala Trp Leu Gly Ala Asp Asp Thr Gly Leu Thr Val Glu
370 375 380
Ala Val Glu Gln Asn Thr Gln Leu Phe Leu Ala Gly Asp Gly Leu Asp
385 390 395 400
<210> 9
<211> 405
<212> БЕЛОК
<213> Caldithrix abyssi
<400> 9
Met Lys Leu Lys Phe Val Ala Arg Pro Tyr Glu Leu Ile Asn Lys Ile
1 5 10 15
Gln Asn Tyr Ser Trp Gly Thr Arg Asn Glu Gln Ala Phe Ile Pro Arg
20 25 30
Leu Leu Asn Met Ala Val Glu Pro Asp Thr Pro Tyr Ala Glu Leu Trp
35 40 45
Met Gly Thr His Pro Asn Ala Pro Ser Glu Val Val Val Asp Gly Arg
50 55 60
Arg Ile Leu Leu Ser Glu Phe Ile Lys Gln Phe Pro Gln Gln Ile Leu
65 70 75 80
Gly Thr Arg Val Ile Glu Arg Phe Gly Val Gln Leu Pro Phe Leu Phe
85 90 95
Lys Val Leu Ser Ala Ala Glu Ala Leu Ser Ile Gln Ala His Pro Asn
100 105 110
Lys Gln Gln Ala Glu Val Leu His Gln Arg Asp Pro Glu His Tyr Pro
115 120 125
Asp Asp Asn His Lys Pro Glu Ile Ala Ile Ala Leu Asp Glu Leu Thr
130 135 140
Ala Leu Val Gly Phe Arg Ser Leu Lys Glu Met Asp Ala Val Leu Arg
145 150 155 160
Thr Phe Pro Glu Ile Leu Glu Phe Thr Gly Pro Leu Glu Phe Thr Phe
165 170 175
Glu Gly Ala Arg His Glu Glu Gln Glu Asn Arg Gln Lys Phe Arg Gln
180 185 190
Phe Tyr Gln Thr Leu Met Leu Lys Ser Gln Thr His Ala Thr Glu Met
195 200 205
Glu Ala Thr Leu Asn Lys Ile Glu Gln Lys Leu Leu Gln Lys Lys Lys
210 215 220
Arg Thr Glu Arg Asp Glu Trp Phe Leu Lys Leu Lys Lys Lys Tyr Gly
225 230 235 240
Ala Asp Val Gly Leu Phe Ser Ile Tyr Leu Leu Asn Leu Leu His Leu
245 250 255
Lys Lys Gly Gln Gly Val Phe Leu Lys Ala Gly Val Pro His Ala Tyr
260 265 270
Leu Lys Gly Asn Ile Val Glu Cys Met Ala Asn Ser Asp Asn Val Val
275 280 285
Arg Ala Gly Leu Thr Pro Lys Phe Lys Asp Val Lys Thr Leu Ile Glu
290 295 300
Val Leu Thr Tyr Glu Thr Gly Pro Val Glu Ile Tyr Glu Gly Ala Gln
305 310 315 320
Asn Ala Lys Tyr Val Tyr Lys Thr Pro Val Asp Glu Phe Ser Ile Thr
325 330 335
His Val Asn Leu Asp Glu Lys Ser Lys Leu Arg Phe Phe Leu Glu Thr
340 345 350
Val Ser Ile Met Met Val Val Asn Gly Lys Gly Glu Ile Val Phe Asn
355 360 365
Gly Gly Arg Leu Ala Ile Gln Lys Gly Gln Ser Ile Leu Leu Pro Ala
370 375 380
Glu Ile Ala Ser Phe Glu Leu Val Ser Asp Gly Ser Leu Glu Ile Phe
385 390 395 400
Ser Ala Tyr Val Pro
405
<210> 10
<211> 453
<212> БЕЛОК
<213> Myceliophthora thermophila
<400> 10
Met Gln Val Pro Leu Ile Arg Leu Gln Cys Gly Ala Asn Ser Tyr Glu
1 5 10 15
Trp Gly Lys Lys Gly Ser Ser Ser Ala Val Ala Arg Phe Ala Ala Ala
20 25 30
Thr Pro Ser Ser Asp Phe Thr Ile Glu Asp Asp Arg Pro Tyr Ala Glu
35 40 45
Leu Trp Met Gly Thr His Pro Ser Asn Pro Ser Lys Asp Leu Ser Thr
50 55 60
Gly Arg Thr Leu Leu Asp Leu Val Gln Asp Asn Lys Ala Leu Leu Ser
65 70 75 80
Pro Ser Val Ala Ala Arg Tyr Asp Asn Lys Val Pro Phe Leu Phe Lys
85 90 95
Val Leu Ser Ile Asn Lys Ala Leu Ser Ile Gln Ala His Pro Asn Lys
100 105 110
Lys Leu Ala Glu Glu Leu His Arg Lys Asp Pro Lys Asn Tyr Pro Asp
115 120 125
Asp Asn His Lys Pro Glu Met Ala Ile Ala Ile Thr Pro Phe Glu Gly
130 135 140
Leu Cys Gly Phe Arg Pro Leu Gly Glu Ile Ala His Phe Leu Glu Ser
145 150 155 160
Val Pro Pro Leu Arg Gln Leu Val Gly Asp Asp Asn Ala Arg Glu Phe
165 170 175
Ala Gly Ile Val Arg Gln Asn Lys Asp Asn Asp Ser Lys Asp Ala Val
180 185 190
Glu Gln Asn Lys Lys Ala Leu Gln Lys Ile Phe Gly Ala Leu Met Ser
195 200 205
Ser Ser Glu Ala Asp Met Ala Ala Ala Ala Lys Val Leu Val Glu Ser
210 215 220
Ala Ala Thr Ala Gly Ala Asp Phe Ala Ala Gly Gly Val Ala Ala Thr
225 230 235 240
Ser Gly Ser Thr Leu Ala Glu Leu Val Gln Arg Leu His Gly Gln Phe
245 250 255
Gly Ala Asp Tyr Gly Leu Phe Val Leu Phe Phe Leu Asn Phe Val Thr
260 265 270
Leu Gln Pro Gly Glu Ala Leu Phe Leu Arg Ala Asp Asp Ile His Ala
275 280 285
Tyr Val Ser Gly Asp Ile Ile Glu Cys Met Ala Ser Ser Asp Asn Val
290 295 300
Val Arg Ala Gly Phe Thr Pro Lys Phe Lys Asp Val Asp Thr Leu Val
305 310 315 320
Asn Met Leu Thr Tyr Ser Tyr Ala Pro Ile Asp Glu Gln Lys Met Gly
325 330 335
Pro Ser Asp Tyr Pro Tyr Ala Thr Leu Asn Arg Thr Gly Tyr Ser Ser
340 345 350
Gly Ser Thr Ile Ser Phe Tyr Asp Pro Pro Ile Glu Glu Phe Ser Val
355 360 365
Ile Arg Thr Asn Leu Lys Glu Ser Gly Ser Lys Ala Thr Phe Asp Pro
370 375 380
Val Asp Gly Pro Ser Ile Ile Ile Cys Thr Ala Gly Lys Gly Lys Ile
385 390 395 400
Ser Val Gly Pro Thr Ala Gln Glu Val Lys Glu Gly Tyr Val Phe Phe
405 410 415
Val Gly Ala Ser Ala Lys Cys Val Leu Glu Ser Glu Gly Ser Ser Glu
420 425 430
Asp Asp Glu Phe Ile Thr Phe Lys Ala Phe Cys Asp Val Glu Glu His
435 440 445
Arg Gly Ala Ser Leu
450
<210> 11
<211> 410
<212> БЕЛОК
<213> Treponema caldarium
<400> 11
Met Asn Asn Lys Lys Lys Pro Asn Phe Tyr Leu Leu Lys Asn Pro Ile
1 5 10 15
Gln Arg Tyr Ala Trp Gly Ser Lys His Trp Ile Gln Asp Leu Leu Asp
20 25 30
Leu Ser Glu Gln Asp Arg Gln Gly Pro Met Ala Glu Leu Trp Met Gly
35 40 45
Ala His Ser Arg Ser Pro Ser Ile Ala Phe Thr Asp Glu Thr Glu Gln
50 55 60
Pro Leu Asp Lys Leu Ile Gln Glu His Pro Val His Phe Leu Gly Asp
65 70 75 80
Thr Ile Ala His Asp Phe Ser Ser Leu Pro Tyr Leu Phe Lys Ile Leu
85 90 95
Ala Ala Ala Ser Pro Leu Ser Ile Gln Ala His Pro Asp Lys Gln Gln
100 105 110
Ala Glu Gln Gly Phe Ala Arg Glu Ala Lys Ala Gly Ile Pro Leu Ser
115 120 125
Ala Glu Asn Arg Asn Tyr Lys Asp Ser Asn His Lys Pro Glu Ile Ile
130 135 140
Cys Ala Ile Ser Pro Phe Thr Ala Met Cys Gly Phe Arg Thr Gln Ala
145 150 155 160
Glu Ile Ala Glu Leu Leu Ser Leu Leu Asp Val Thr Glu Leu Glu Gln
165 170 175
Ser Leu Val Ala Ile Gln Gln Ile Asp Arg Lys Glu Ala Tyr Arg Asp
180 185 190
Phe Leu Leu Ser Leu Phe Leu Leu Pro Gln Gln Thr Arg Glu Arg Ile
195 200 205
Thr Lys His Ile Gln Ala Lys Leu Pro Lys Leu Glu Gln Lys His Pro
210 215 220
Arg Tyr Ala Lys Glu Trp Glu Leu Ile Asn Leu Phe Cys Thr Leu Tyr
225 230 235 240
Pro Gly Asp Ser Ala Ile Ile Ser Pro Leu Tyr Leu Asn Val Leu Ser
245 250 255
Leu Asn Pro Gly Glu Ala Ile Phe Leu Pro Ala Gly Val Leu His Ala
260 265 270
Tyr Ile His Gly Phe Gly Val Glu Leu Met Ala Asn Ser Asp Asn Val
275 280 285
Leu Arg Gly Gly Leu Thr Pro Lys His Ile Asp Ile Lys Glu Leu Leu
290 295 300
Asn Ile Ile Arg Phe Glu Ser Phe Lys Pro Ala Val Leu Ser Ala Gln
305 310 315 320
Lys Thr Gln Gln Gly Tyr His Ile Tyr Pro Ser Gln Val Arg Glu Phe
325 330 335
Ser Leu Phe His Val Ala Val Thr Met Asp Lys Ala Gln Gln Leu Met
340 345 350
Pro Gly Thr Pro Ile Ile Leu Ile Val Leu Asp Gly Cys Val Ser Ile
355 360 365
Gly Thr Glu Gln Glu Lys Lys Thr Leu Gln Lys Gly Met Ser Val Phe
370 375 380
Leu Pro Ala Glu Arg Glu Gln Leu Ile Leu Glu Gly Ser Ala His Ile
385 390 395 400
Phe Gly Ala Thr Thr Gly Glu Gly Thr Arg
405 410
<210> 12
<211> 223
<212> БЕЛОК
<213> Tepidimonas fonticaldi
<400> 12
Met Thr Arg Trp Arg Gly Ile Arg Ala Val Leu Phe Asp Leu Asp Gly
1 5 10 15
Thr Leu Val Asp Ser Ala Pro Asp Leu Gly His Ala Ala Asp Leu Met
20 25 30
Arg Gln Arg Arg Gly Leu Pro Pro Leu Asp Glu Ala Tyr Tyr Arg Pro
35 40 45
Arg Ala Ser Ser Gly Ala Arg Gly Met Ile Glu Ala Ala Phe Gly Leu
50 55 60
Thr Pro Glu His Pro Glu Phe Glu Ala Tyr Arg Thr Glu Tyr Leu Asp
65 70 75 80
Thr Tyr Gly Gln Val Leu Thr Arg Arg Thr Arg Pro Phe Asp Gly Val
85 90 95
Ala Glu Leu Ile Ala Ala Leu Asp Arg Ala Gln Val Ala Trp Gly Val
100 105 110
Val Thr Asn Lys Val Glu Arg Phe Ala Leu Pro Leu Thr Ala Ala Ile
115 120 125
Pro Leu Phe Ala Thr Ala Ala Thr Val Ile Gly Gly Asp Thr Thr Pro
130 135 140
His Pro Lys Pro His Pro Ala Pro Leu Leu Glu Ala Ala Arg Arg Leu
145 150 155 160
Gln Leu Pro Pro Gln Ala Cys Leu Tyr Val Gly Asp Asp Glu Arg Asp
165 170 175
Ile Val Ala Gly Arg Ala Ala Gly Met Pro Thr Val Ala Ala Arg Tyr
180 185 190
Gly Tyr Leu Gly Val Ala Ala Asp Val Glu Ala Trp Ala Ala Asp Ala
195 200 205
Ile Ile Glu Ser Pro Gln Ala Leu Leu Asn Phe Leu Asp Leu Ala
210 215 220
<210> 13
<211> 221
<212> БЕЛОК
<213> Thermomonas hydrothermalis
<400> 13
Met Ser Ala Arg Arg Phe Pro Pro Leu Val Leu Phe Asp Leu Asp Gly
1 5 10 15
Thr Leu Leu Asp Ser Ala Pro Asp Met Leu Val Thr Val Asn Arg Met
20 25 30
Arg Ala Met Arg Gly Asp Ala Pro Met Ala Leu Asp Ala Leu Arg Pro
35 40 45
His Val Ser Arg Gly Ala Arg Ala Met Ile Ala Ala Ser Phe Pro Ala
50 55 60
Leu Gly Gly Glu Val Pro Ala Glu Met Val Arg Glu Phe Leu Asp Ile
65 70 75 80
Tyr Ala Gln Val Leu Gly Gln His Gly Ala Pro Phe Asp Gly Val Val
85 90 95
Glu Leu Leu Ala Ala Leu Glu Ala Ala Gly Ser Arg Trp Gly Ile Val
100 105 110
Thr Asn Lys Pro Glu Ser Leu Ala Arg Gln Leu Leu Pro Gly Leu Gly
115 120 125
Trp Asp Ala Arg Cys Ala Ile Leu Val Gly Gly Asp Ser Leu Pro Glu
130 135 140
Arg Lys Pro His Pro Leu Pro Leu Leu His Ala Ala Gly Gln Leu Gly
145 150 155 160
Val Ser Cys Gln Asp Cys Ala Tyr Val Gly Asp Asp Arg Arg Asp Ile
165 170 175
Glu Ala Ala Arg Ala Ala Gly Met Arg Ser Val Val Ala Leu Trp Gly
180 185 190
Tyr Arg Leu Pro Asp Glu Asn Pro Gln Asp Trp Gly Gly Asp Ala Leu
195 200 205
Ser Pro Thr Pro Gln Ala Leu Leu Asp Trp Pro Leu Pro
210 215 220
<210> 14
<211> 221
<212> БЕЛОК
<213> Sulfurivirga caldicuralii
<400> 14
Met Arg Glu Phe Asp Cys Val Leu Phe Asp Leu Asp Gly Thr Leu Leu
1 5 10 15
Asp Thr Ser Tyr Asp Phe Ala Trp Ala Leu Asn Thr Leu Gln Lys Gln
20 25 30
Glu Ser Val Pro Leu Thr Pro Tyr Trp Arg Ile Arg Gln Thr Ile Ser
35 40 45
Ser Gly Gly Arg Ala Val Val Lys Leu Gly Phe Pro Asp Ala Asp Asp
50 55 60
Ala Thr Ile Glu Ala Leu Arg Glu Arg Phe Leu Ala Leu Tyr His Glu
65 70 75 80
Asn Ile Ser Val His Thr Asp Leu Phe Pro Gly Leu Glu Lys Val Leu
85 90 95
Thr His Leu Gln Glu Lys Ala Val Pro Trp Gly Ile Val Thr Asn Lys
100 105 110
Pro Ala Trp Leu Thr Asp Lys Leu Leu Gly Glu Leu Asp Leu Pro Ala
115 120 125
Gln Pro Gln Thr Val Val Ser Gly Asp Thr Leu Ala Val Arg Lys Pro
130 135 140
His Pro Glu Pro Met Trp Leu Ala Ala Glu Gln Cys Gly Val Ala Pro
145 150 155 160
Glu Arg Cys Leu Tyr Ile Gly Asp His Pro Arg Asp Ile Glu Ala Pro
165 170 175
Arg Asn Ala Gly Met Gln Ser Ala Ala Ala Leu Tyr Gly Phe Leu Pro
180 185 190
Leu Asp Ala Glu Pro Asp Ser Trp Pro Ala Asp Tyr Arg Tyr His Ala
195 200 205
Pro Ala Asp Ile Leu His His Met Gln Lys Val Phe Pro
210 215 220
<210> 15
<211> 344
<212> БЕЛОК
<213> Syntrophothermus lipocalidus
<400> 15
Met Ala Val Glu Met Gly Pro Glu Met Met Phe Glu Phe Leu Tyr Asn
1 5 10 15
Leu Pro Ala Gln Phe Glu Gly Cys Leu Lys Met Asp Phe Ser Lys Ala
20 25 30
Ser Gly Leu Lys Lys Glu Tyr Ala Asn Ile Val Val Thr Gly Leu Gly
35 40 45
Gly Ser Ala Ile Gly Gly Asp Ile Leu Arg Cys Tyr Cys Gln Ser Arg
50 55 60
Leu Pro Ile Pro Val Val Val Asn Arg Asp Tyr Met Leu Pro Arg Phe
65 70 75 80
Val Gly Pro Asp Ser Leu Val Leu Ala Val Ser Tyr Ser Gly Asn Thr
85 90 95
Glu Glu Thr Leu Ser Ala Tyr Glu Asp Ala Arg Glu Lys Gly Ala Ser
100 105 110
Ile Ile Ala Phe Thr Thr Gly Gly Lys Leu Ala Glu Met Ala Ala Leu
115 120 125
Asp Gly Asn Pro Val Ile Thr Ile Thr Gly Gly Leu Val Pro Arg Ala
130 135 140
Ala Thr Gly Tyr Leu Phe Ala Pro Leu Val Leu Val Leu Glu Arg Leu
145 150 155 160
Gly Leu Val Ser Gly Ala Ser Glu Asp Val Lys Glu Thr Val Thr Val
165 170 175
Leu Thr Gln Leu Arg Glu Glu Ile Glu Pro Gly Arg Glu Glu Asp Ser
180 185 190
Asn Arg Ala Arg Phe Ile Ala Gly Gln Leu Tyr Gln Arg Ile Pro Val
195 200 205
Ile Trp Gly Cys Ser Ser Thr Ser Glu Val Ala Ala Met Arg Trp Lys
210 215 220
Gly Gln Ile Asn Glu Asn Ala Lys Ala Pro Ala Tyr Phe Asn Val Phe
225 230 235 240
Pro Glu Leu Asn His Asn Glu Ile Val Gly Phe Glu Val Pro Glu Asp
245 250 255
Leu Val Lys Lys Leu Ala Val Ile Ile Leu Arg Asp Pro Asp Asp His
260 265 270
Gly Arg Ile Thr Lys Arg Ile Glu Ile Thr Lys Asp Ile Leu Gln Gly
275 280 285
Lys Val Ser Ser Val Ala Glu Val Glu Ala Arg Gly Asn Ser Phe Leu
290 295 300
Ala Lys Thr Tyr Ser Leu Ile Tyr Val Gly Asp Tyr Ala Ser Val Tyr
305 310 315 320
Leu Ala Glu Leu Tyr Gly Ile Asn Pro Thr Pro Val Gln Val Ile Asp
325 330 335
Tyr Leu Lys Ala Arg Met Ala Glu
340
<210> 16
<211> 328
<212> БЕЛОК
<213> Schleiferia thermophila
<400> 16
Met Leu Thr Leu Ile Glu Asn Phe Pro Lys His Leu Val Asp Ala Met
1 5 10 15
Ile Thr Ala Lys Lys Ala Ser Phe Lys Gln Ser Asn Arg Ala Ile Lys
20 25 30
Asn Val Ile Ile Thr Gly Leu Gly Gly Ser Gly Ile Gly Ala Ser Met
35 40 45
Val Gln Asp Leu Leu Ser Pro His Ala Glu Ile Pro Ile Ile Val Asn
50 55 60
Lys Asp Tyr His Leu Pro Ala Phe Ala Asp Glu Asn Thr Leu Val Ile
65 70 75 80
Ala Cys Ser Tyr Ser Gly Glu Thr Glu Glu Thr Leu Ala Ala Leu Ala
85 90 95
Glu Ala Glu Glu His Ser Cys Glu Ile Ala Ile Ile Thr Ser Gly Gly
100 105 110
Thr Leu Leu Gln Met Ala Lys Ser Lys Asn Tyr Asn Tyr Leu Gln Met
115 120 125
Pro Glu Gly Asn Pro Pro Arg Ser Met Ile Gly Tyr Ser Leu Val Tyr
130 135 140
Gln Leu Tyr Met Leu Ala Tyr Tyr Gly Ile Ser Arg Leu Ala Leu Asp
145 150 155 160
Asn Asp Ile Ile Leu Ser Ser Asn Tyr Leu Leu Glu Phe Arg Glu Lys
165 170 175
Ile Gln Ser Gln Ala Arg Tyr Ile Ala Val Arg Leu His Lys Lys Ile
180 185 190
Pro Ala Val Tyr Ala Cys Ser Gly Phe Gly Ser Leu Ala Glu Arg Phe
195 200 205
Arg Gln Gln Leu Asn Glu Asn Ser Lys Met Leu Ala Trp Asn Gly Thr
210 215 220
Val Pro Glu Met Asn His Asn Glu Leu Val Gly Trp Lys Gly Gly Asp
225 230 235 240
Glu His Phe Ala Ala Ile Phe Ile His Thr Pro Phe Asp Asp Asn Arg
245 250 255
Asn Ala Lys Arg Thr Glu Ile Ser Ser Asn Ile Ile Gln Asn Phe Thr
260 265 270
Ser Gly Val Phe His Ile His Ser Glu Gly Glu Thr Pro Leu Arg Ala
275 280 285
Phe Phe Tyr Leu Ile His Ile Thr Asp Trp Ile Ser Tyr Tyr Leu Ser
290 295 300
Glu Leu Asn Gly Val Asp Val Met Asp Ile Ser Ala Ile Asn Gln Leu
305 310 315 320
Lys Gly Glu Leu Ala Asn Phe Asn
325
<210> 17
<211> 349
<212> БЕЛОК
<213> Thermodesulfobium narugense
<400> 17
Met Asp Lys Asn Val Met Asn Ser Tyr Val Ser Asp Ile Ala Tyr His
1 5 10 15
Met Lys Asp Phe Tyr Lys Asp Leu Thr Phe Tyr Lys Asp Gly Lys Ile
20 25 30
Asp Leu Asn Glu Ile Glu Asn Leu Ile Phe Leu Gly Ile Gly Gly Ser
35 40 45
Ala Ile Ser Pro Lys Ile Phe Thr Glu Ile Met Asn Ile Asn Lys Lys
50 55 60
Val Tyr Phe Phe Ser Thr Leu Asn Gly Phe Glu Ala Leu Pro Asp Pro
65 70 75 80
Ser Thr Ser Phe Val Ile Ala Phe Ser Tyr Ser Gly Asn Thr Val Glu
85 90 95
Thr Leu Arg Ser Ile Glu Leu Ile Ala Lys Asp Arg Phe Arg Gly Ile
100 105 110
Gly Ile Ser Ser Gly Gly Lys Ile Val Asp Leu Cys Lys Ser Leu Asn
115 120 125
Trp Gln His Ile Ala Val Pro Lys Gly Arg Ala Pro Arg Ala Ala Met
130 135 140
Pro Phe Thr Leu Ser Ile Leu Phe Lys Leu Ala Leu Ser Lys Gly Trp
145 150 155 160
Thr Glu Tyr Asn Glu Asp Asp Phe Trp Asn Asp Ile Ile Glu Leu Ser
165 170 175
Asn Ser Lys Asn Asn Phe Leu Pro Glu Val Asp Phe Glu Asp Asn Val
180 185 190
Ser Lys Arg Ile Ala Tyr Lys Leu Ala Thr Lys Lys Asn Val Ile Ile
195 200 205
Trp Gly Val Glu Ser Ile Ser Lys Asn Ile Ala Tyr Arg Phe Lys Ser
210 215 220
Gln Leu Glu Glu Asn Ala Lys Gln Leu Ser Tyr Tyr Ser Tyr Leu Pro
225 230 235 240
Glu Ala Ser His Asn Gln Ile Val Pro Ile Ser Leu Val Asp Asn Lys
245 250 255
Glu Glu Tyr Ile Val Leu Ile Phe Arg Ile Pro Gln Leu Glu Ser Val
260 265 270
Leu Val Ser Asn Ile Ile Ser Thr Val Lys Thr Phe Leu Asn Ser Glu
275 280 285
Gly Ile Glu Val Leu Glu Val Phe Gly Ser Gly Lys Asn His Val Leu
290 295 300
Ala Gly Leu Asp Leu Ile Tyr Ser Thr Asp Phe Val Ser Tyr Tyr Leu
305 310 315 320
Ala Leu Leu Lys Gly Ile Glu Pro Glu Pro Ile Glu Pro Ile Ser Arg
325 330 335
Met Lys Val Ile Leu Asn Asp Asn Leu Arg Lys Ala Leu
340 345
<210> 18
<211> 141
<212> БЕЛОК
<213> Lactococcus lactis
<400> 18
Met Ala Ile Val Val Gly Ala Asp Leu Lys Gly Thr Arg Leu Lys Asp
1 5 10 15
Val Val Lys Asn Phe Leu Val Glu Glu Gly Phe Glu Val Ile Asp Val
20 25 30
Thr Lys Asp Gly Gln Asp Phe Val Asp Val Thr Leu Ala Val Ala Ser
35 40 45
Glu Val Asn Lys Asp Glu Gln Asn Leu Gly Ile Val Ile Asp Ala Tyr
50 55 60
Gly Ala Gly Pro Phe Met Val Ala Thr Lys Ile Lys Gly Met Val Ala
65 70 75 80
Ala Glu Val Ser Asp Glu Arg Ser Ala Tyr Met Thr Arg Gly His Asn
85 90 95
Asn Ala Arg Met Ile Thr Val Gly Ala Glu Ile Val Gly Asp Glu Leu
100 105 110
Ala Lys Asn Ile Ala Lys Ala Phe Val Asn Gly Lys Tyr Asp Gly Gly
115 120 125
Arg His Gln Val Arg Val Asp Met Leu Asn Lys Met Cys
130 135 140
<210> 19
<211> 312
<212> БЕЛОК
<213> Lactococcus lactis
<400> 19
Met Arg Ile Ala Ile Gly Cys Asp His Ile Val Thr Asp Val Lys Met
1 5 10 15
Ala Val Ser Glu Phe Leu Lys Ser Lys Gly Tyr Glu Val Leu Asp Phe
20 25 30
Gly Thr Tyr Asp His Val Arg Thr His Tyr Pro Ile Tyr Gly Lys Lys
35 40 45
Val Gly Glu Ala Val Val Ser Gly Gln Ala Asp Leu Gly Val Cys Ile
50 55 60
Cys Gly Thr Gly Val Gly Ile Asn Asn Ala Val Asn Lys Val Pro Gly
65 70 75 80
Val Arg Ser Ala Leu Val Arg Asp Met Thr Ser Ala Leu Tyr Ala Lys
85 90 95
Glu Glu Leu Asn Ala Asn Val Ile Gly Phe Gly Gly Met Ile Thr Gly
100 105 110
Gly Leu Leu Met Asn Asp Ile Ile Glu Ala Phe Ile Glu Ala Glu Tyr
115 120 125
Lys Pro Thr Glu Glu Asn Lys Lys Leu Ile Ala Lys Ile Glu His Val
130 135 140
Glu Thr His Asn Ala His Gln Ala Asp Glu Glu Phe Phe Thr Glu Phe
145 150 155 160
Leu Glu Lys Trp Asp Arg Gly Glu Tyr His Asp Met Ala Ile Val Val
165 170 175
Gly Ala Asp Leu Lys Gly Thr Arg Leu Lys Asp Val Val Lys Asn Phe
180 185 190
Leu Val Glu Glu Gly Phe Glu Val Ile Asp Val Thr Lys Asp Gly Gln
195 200 205
Asp Phe Val Asp Val Thr Leu Ala Val Ala Ser Glu Val Asn Lys Asp
210 215 220
Glu Gln Asn Leu Gly Ile Val Ile Asp Ala Tyr Gly Ala Gly Pro Phe
225 230 235 240
Met Val Ala Thr Lys Ile Lys Gly Met Val Ala Ala Glu Val Ser Asp
245 250 255
Glu Arg Ser Ala Tyr Met Thr Arg Gly His Asn Asn Ala Arg Met Ile
260 265 270
Thr Val Gly Ala Glu Ile Val Gly Asp Glu Leu Ala Lys Asn Ile Ala
275 280 285
Lys Ala Phe Val Asn Gly Lys Tyr Asp Gly Gly Arg His Gln Val Arg
290 295 300
Val Asp Met Leu Asn Lys Met Cys
305 310
<210> 20
<211> 270
<212> БЕЛОК
<213> Bacteroides thetaiotaomicron
<400> 20
Met Lys Tyr Lys Leu Ile Val Leu Asp Leu Asp Gly Thr Leu Thr Asn
1 5 10 15
Ser Lys Lys Glu Ile Ser Ser Arg Asn Arg Glu Thr Leu Ile Arg Ile
20 25 30
Gln Glu Gln Gly Ile Arg Leu Val Leu Ala Ser Gly Arg Pro Thr Tyr
35 40 45
Gly Ile Val Pro Leu Ala Asn Glu Leu Arg Met Asn Glu Phe Gly Gly
50 55 60
Phe Ile Leu Ser Tyr Asn Gly Gly Glu Ile Ile Asn Trp Glu Ser Lys
65 70 75 80
Glu Met Met Tyr Glu Asn Val Leu Pro Asn Glu Val Val Pro Val Leu
85 90 95
Tyr Glu Cys Ala Arg Thr Asn His Leu Ser Ile Leu Thr Tyr Asp Gly
100 105 110
Ala Glu Ile Val Thr Glu Asn Ser Leu Asp Pro Tyr Val Gln Lys Glu
115 120 125
Ala Phe Leu Asn Lys Met Ala Ile Arg Glu Thr Asn Asp Phe Leu Thr
130 135 140
Asp Ile Thr Leu Pro Val Ala Lys Cys Leu Ile Val Gly Asp Ala Gly
145 150 155 160
Lys Leu Ile Pro Val Glu Ser Glu Leu Cys Ile Arg Leu Gln Gly Lys
165 170 175
Ile Asn Val Phe Arg Ser Glu Pro Tyr Phe Leu Glu Leu Val Pro Gln
180 185 190
Gly Ile Asp Lys Ala Leu Ser Leu Ser Val Leu Leu Glu Asn Ile Gly
195 200 205
Met Thr Arg Glu Glu Val Ile Ala Ile Gly Asp Gly Tyr Asn Asp Leu
210 215 220
Ser Met Ile Lys Phe Ala Gly Met Gly Val Ala Met Gly Asn Ala Gln
225 230 235 240
Glu Pro Val Lys Lys Ala Ala Asp Tyr Ile Thr Leu Thr Asn Asp Glu
245 250 255
Asp Gly Val Ala Glu Ala Ile Glu Arg Ile Phe Asn Val Pro
260 265 270
<210> 21
<211> 217
<212> БЕЛОК
<213> Halothermothrix orenii
<400> 21
Met Ile Glu Ala Val Ile Phe Asp Met Asp Gly Val Ile Ile Asn Ser
1 5 10 15
Glu Pro Ile His Tyr Lys Val Asn Gln Ile Ile Tyr Glu Lys Leu Gly
20 25 30
Ile Lys Val Pro Arg Ser Glu Tyr Asn Thr Phe Ile Gly Lys Ser Asn
35 40 45
Thr Asp Ile Trp Ser Phe Leu Lys Arg Lys Tyr Asn Leu Lys Glu Ser
50 55 60
Val Ser Ser Leu Ile Glu Lys Gln Ile Ser Gly Asn Ile Lys Tyr Leu
65 70 75 80
Lys Ser His Glu Val Asn Pro Ile Pro Gly Val Lys Pro Leu Leu Asp
85 90 95
Glu Leu Ser Glu Lys Gln Ile Thr Thr Gly Leu Ala Ser Ser Ser Pro
100 105 110
Glu Ile Tyr Ile Glu Thr Val Leu Glu Glu Leu Gly Leu Lys Ser Tyr
115 120 125
Phe Lys Val Thr Val Ser Gly Glu Thr Val Ala Arg Gly Lys Pro Glu
130 135 140
Pro Asp Ile Phe Glu Lys Ala Ala Arg Ile Leu Gly Val Glu Pro Pro
145 150 155 160
His Cys Val Val Ile Glu Asp Ser Lys Asn Gly Val Asn Ala Ala Lys
165 170 175
Ala Ala Gly Met Ile Cys Ile Gly Tyr Arg Asn Glu Glu Ser Gly Asp
180 185 190
Gln Asp Leu Ser Ala Ala Asp Val Val Val Asp Ser Leu Glu Lys Val
195 200 205
Asn Tyr Gln Phe Ile Lys Asp Leu Ile
210 215
<---
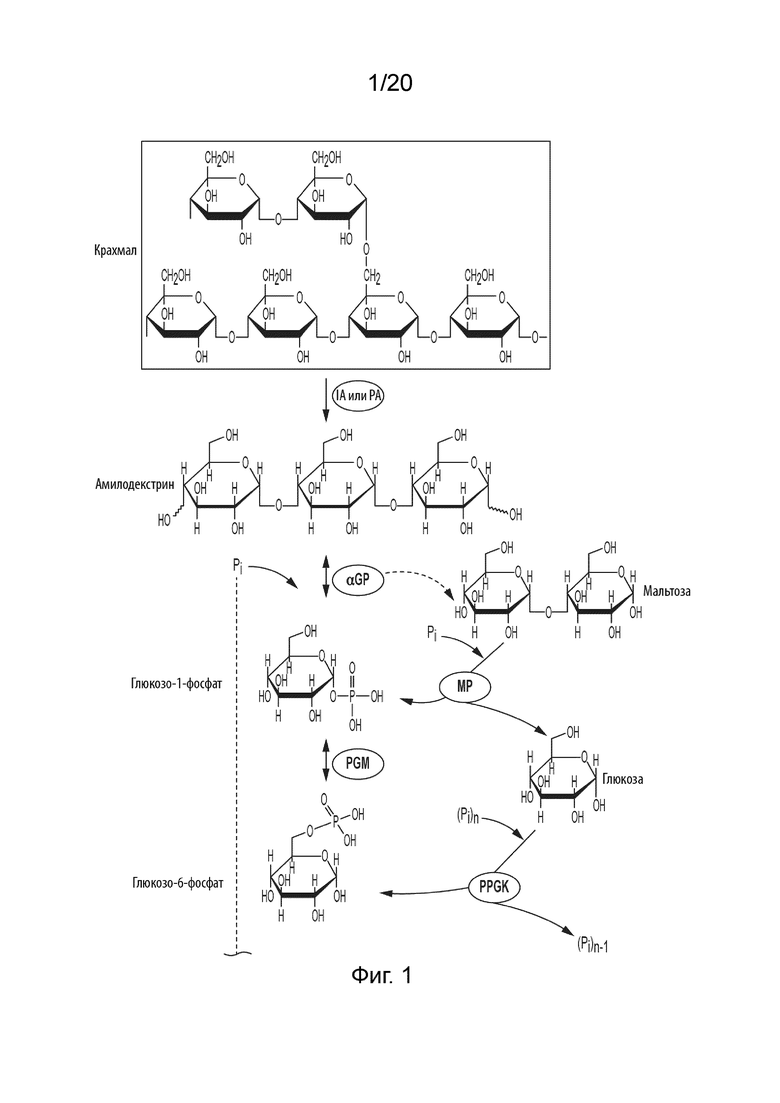
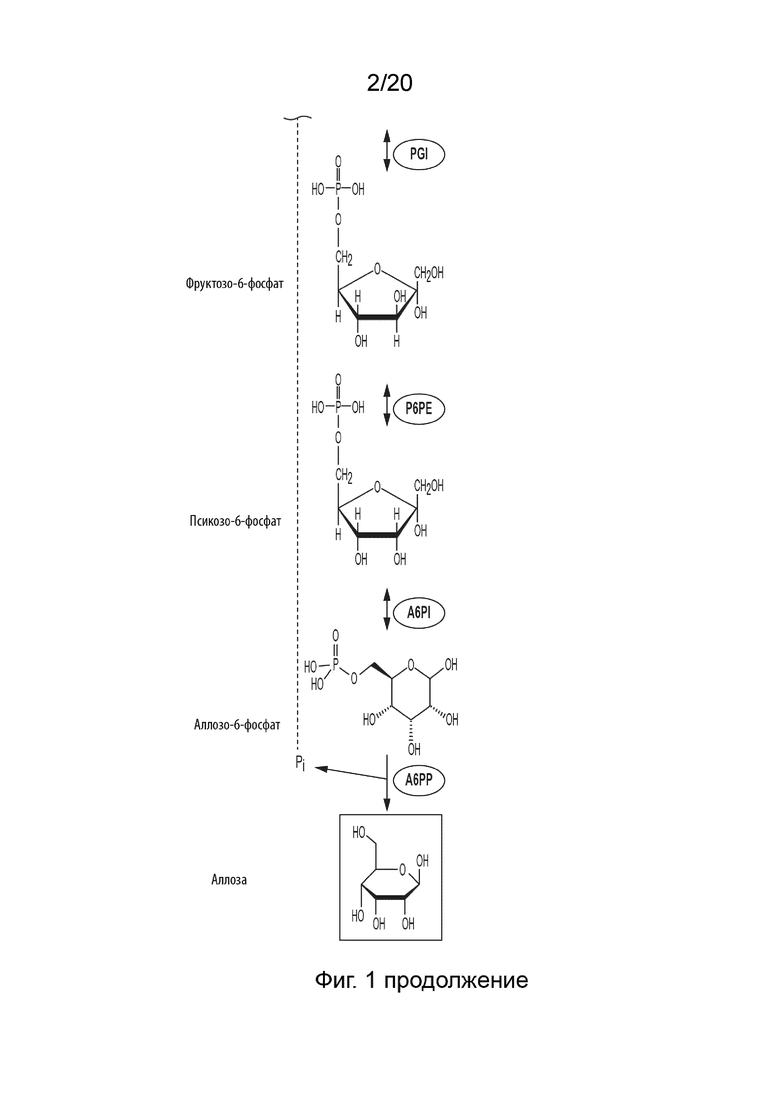
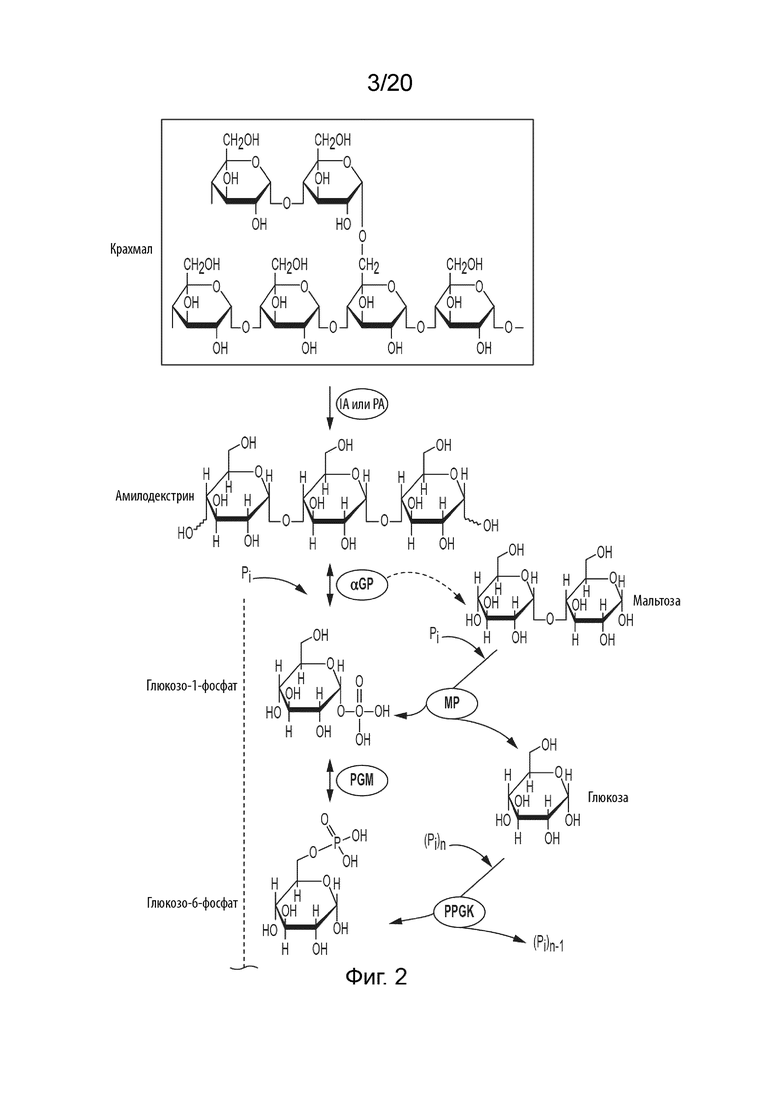
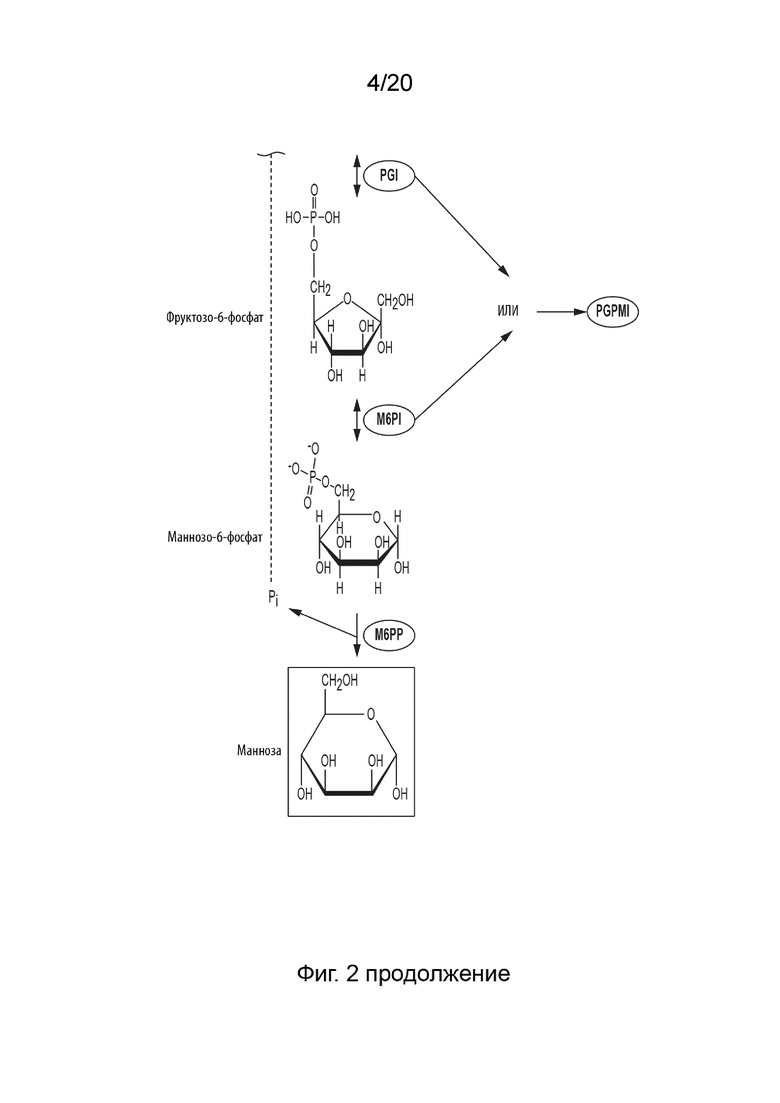
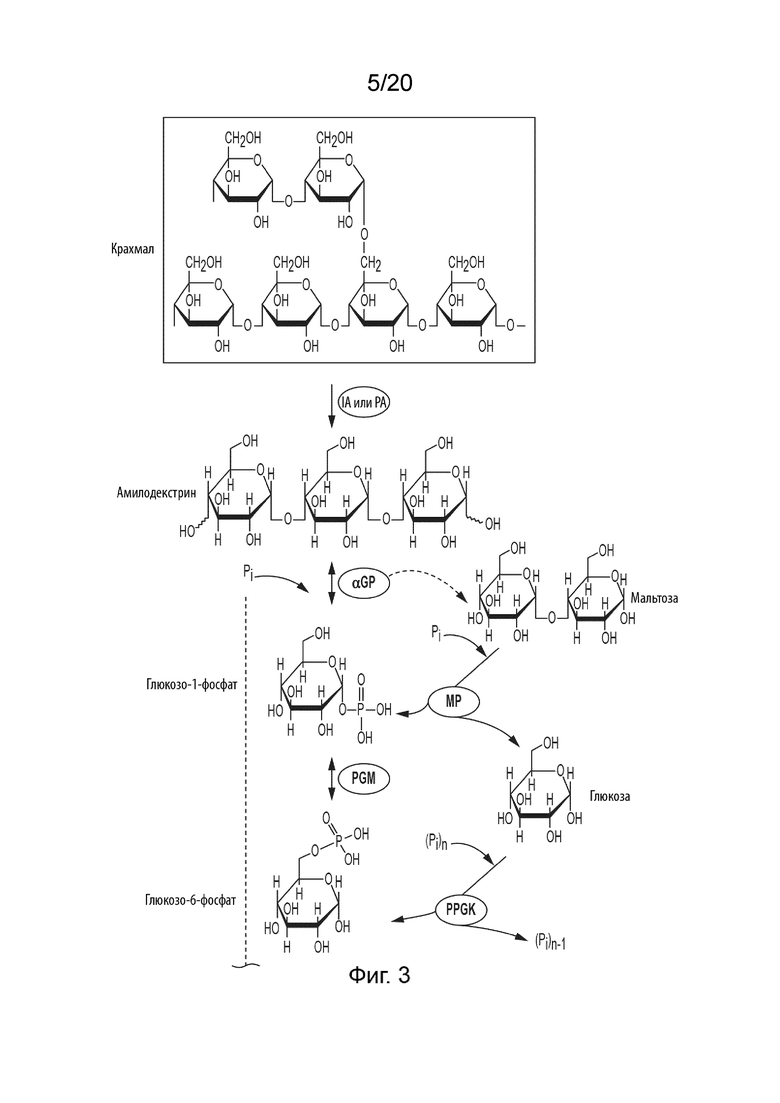
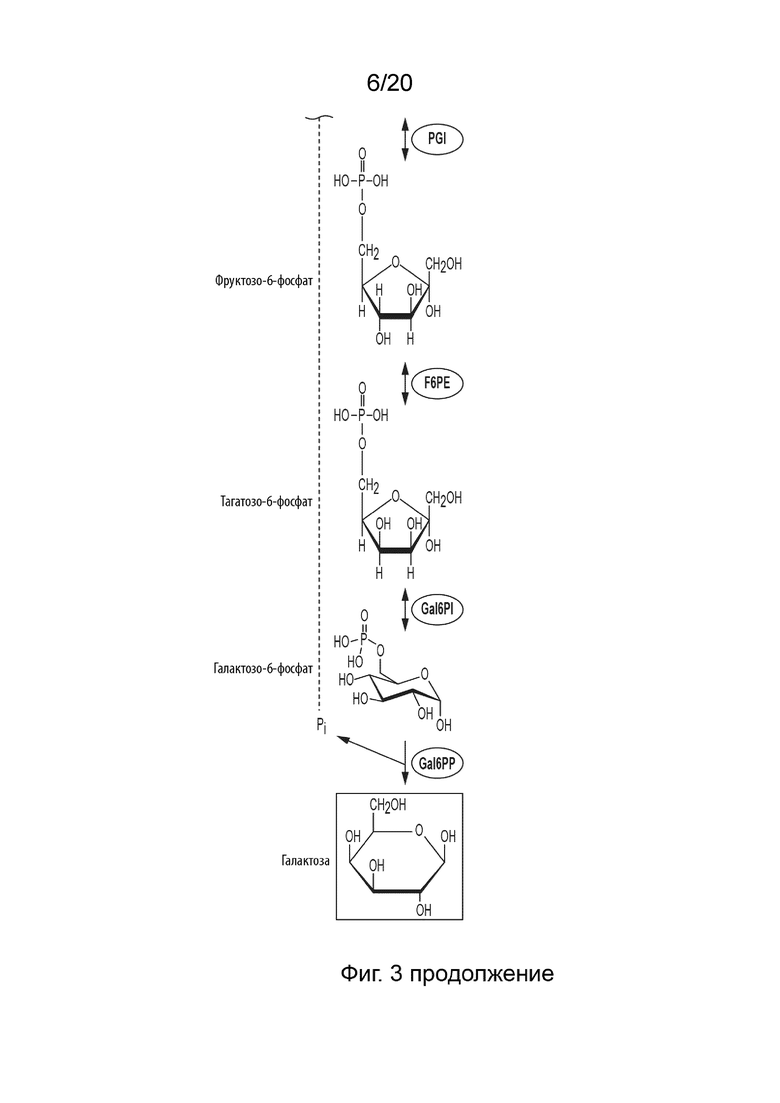
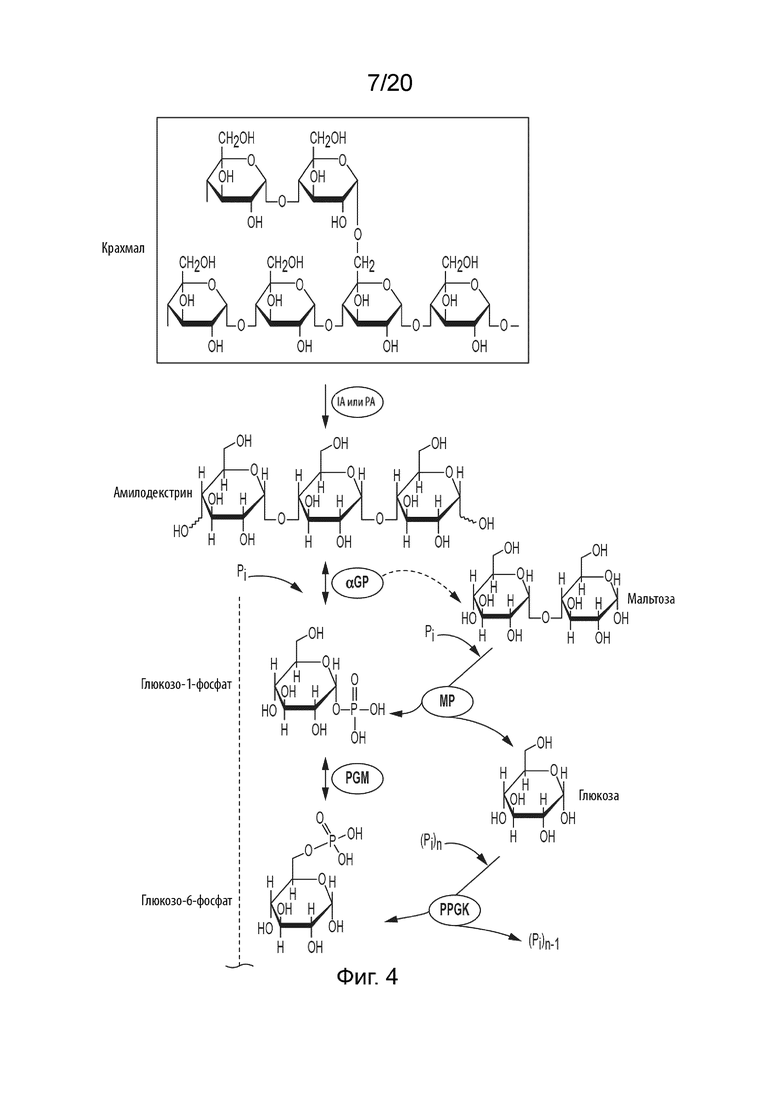
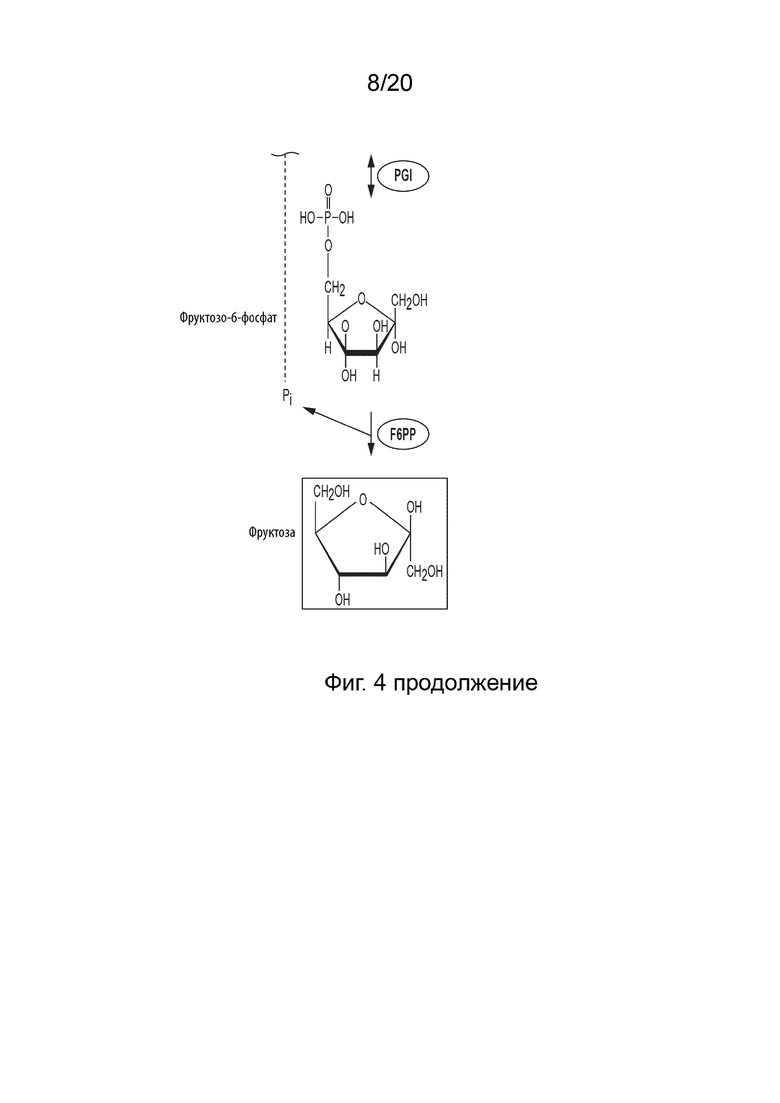
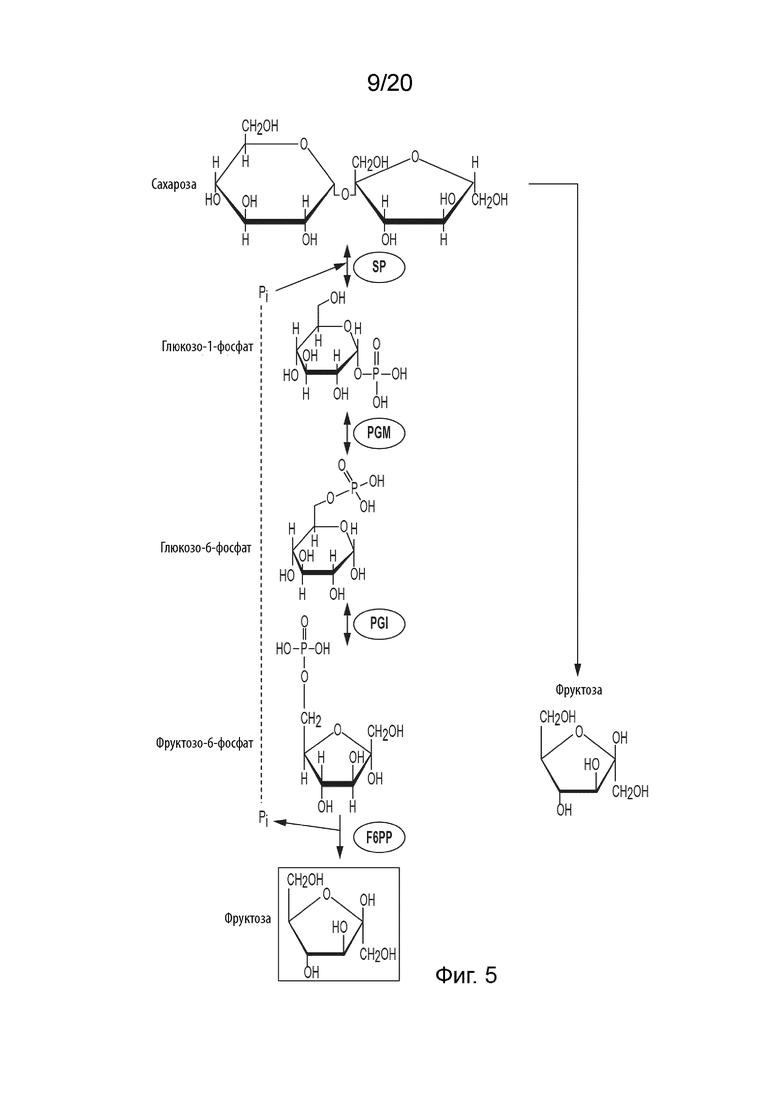
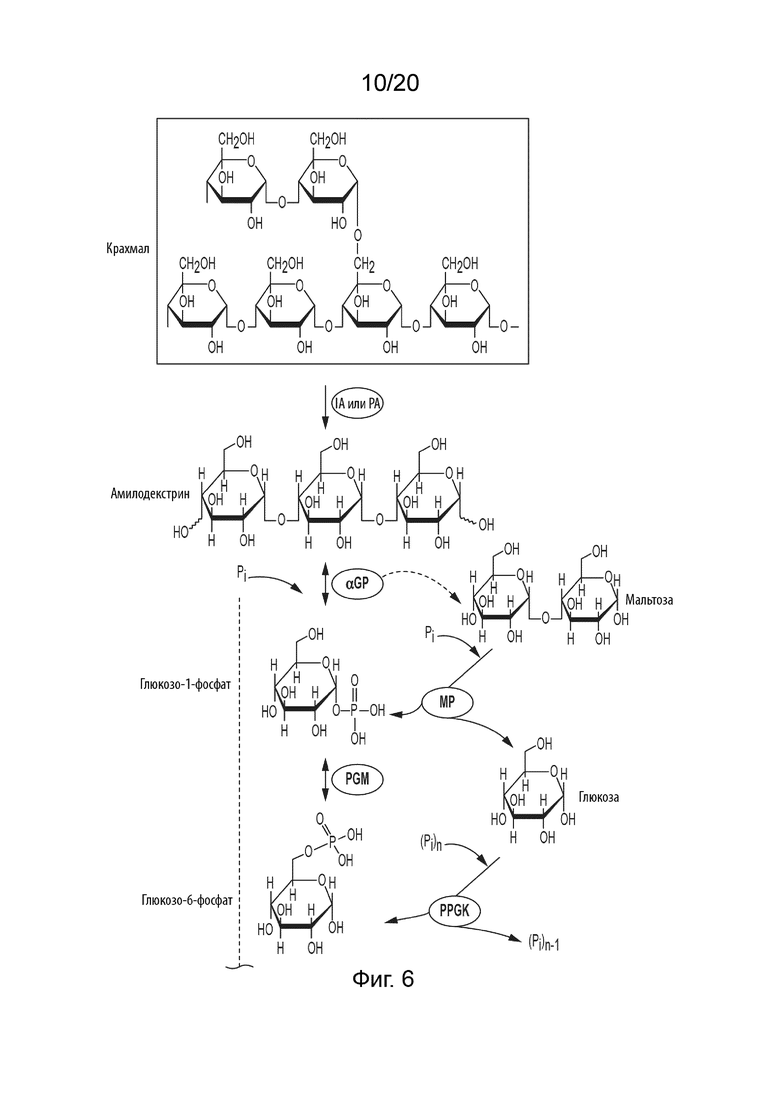
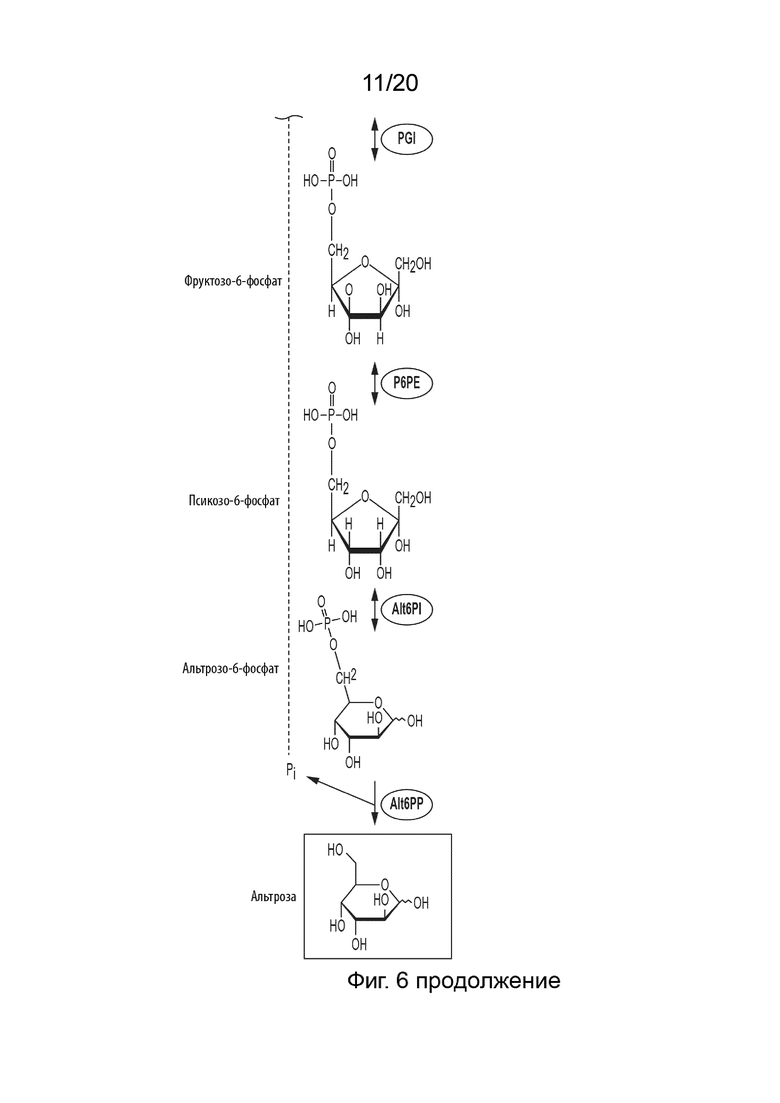
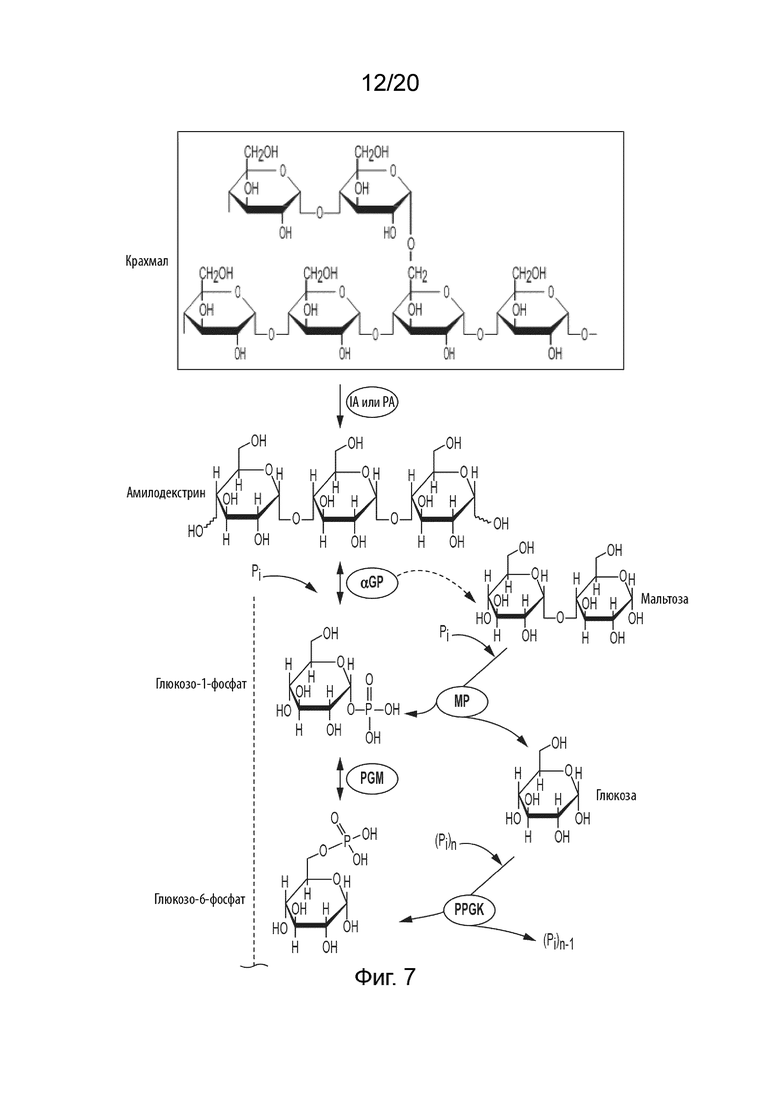
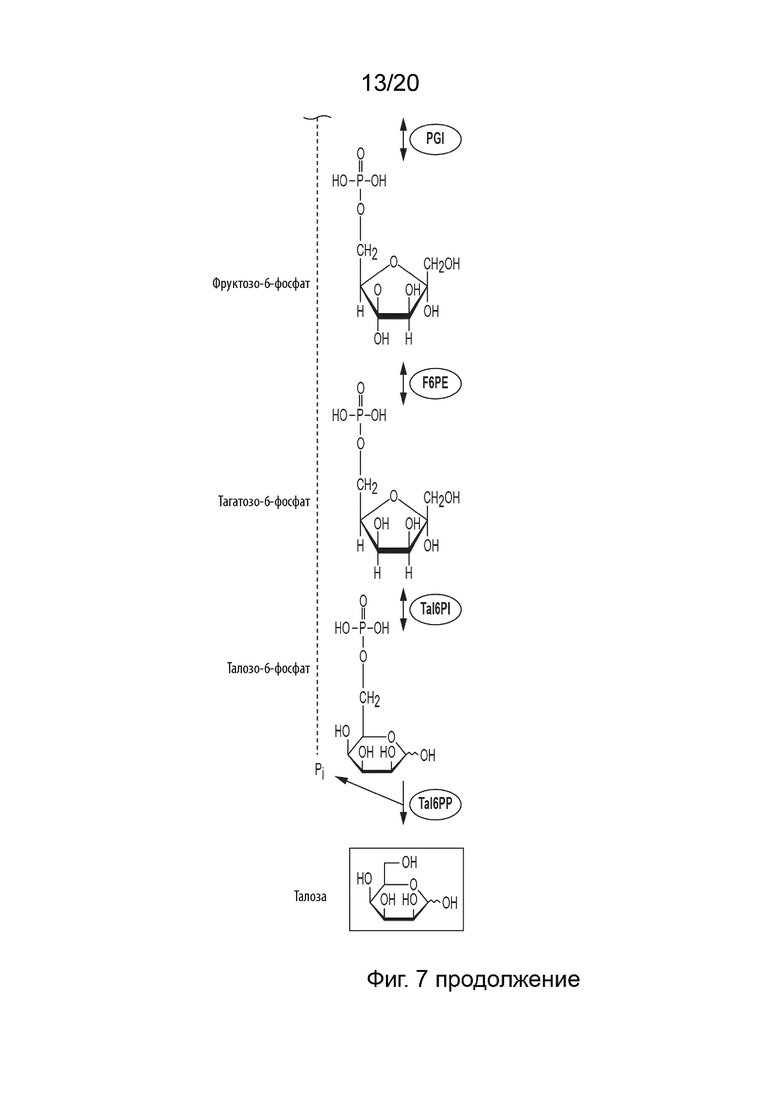
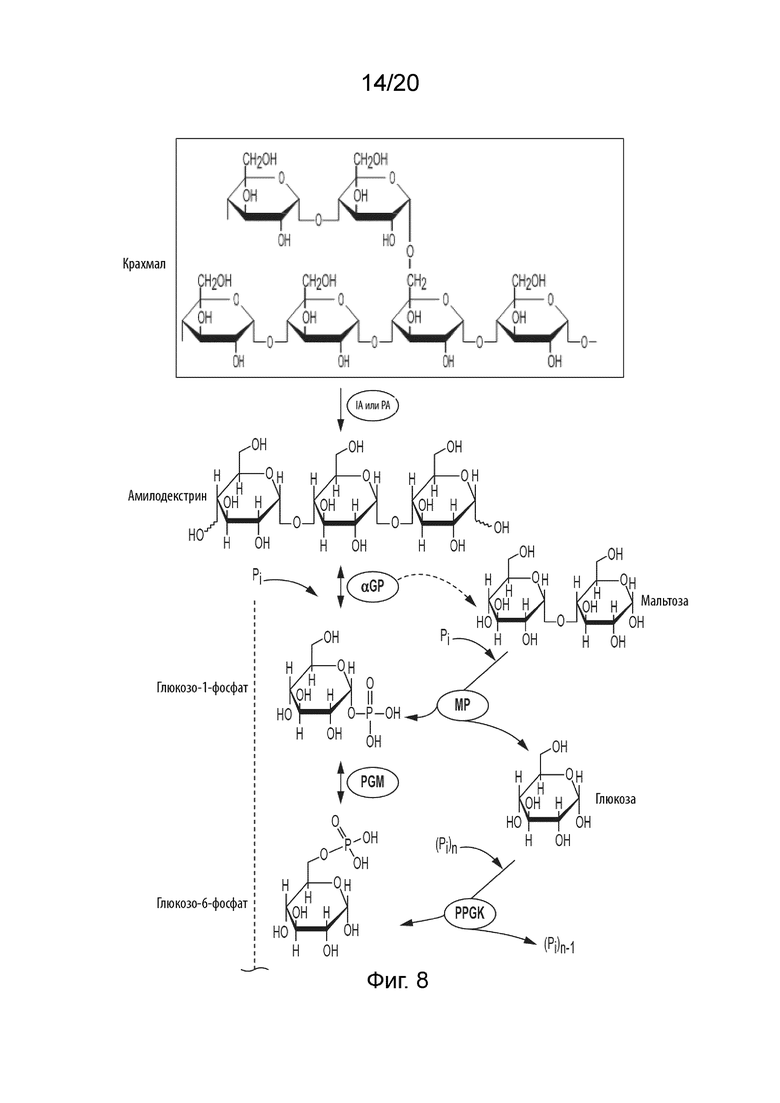
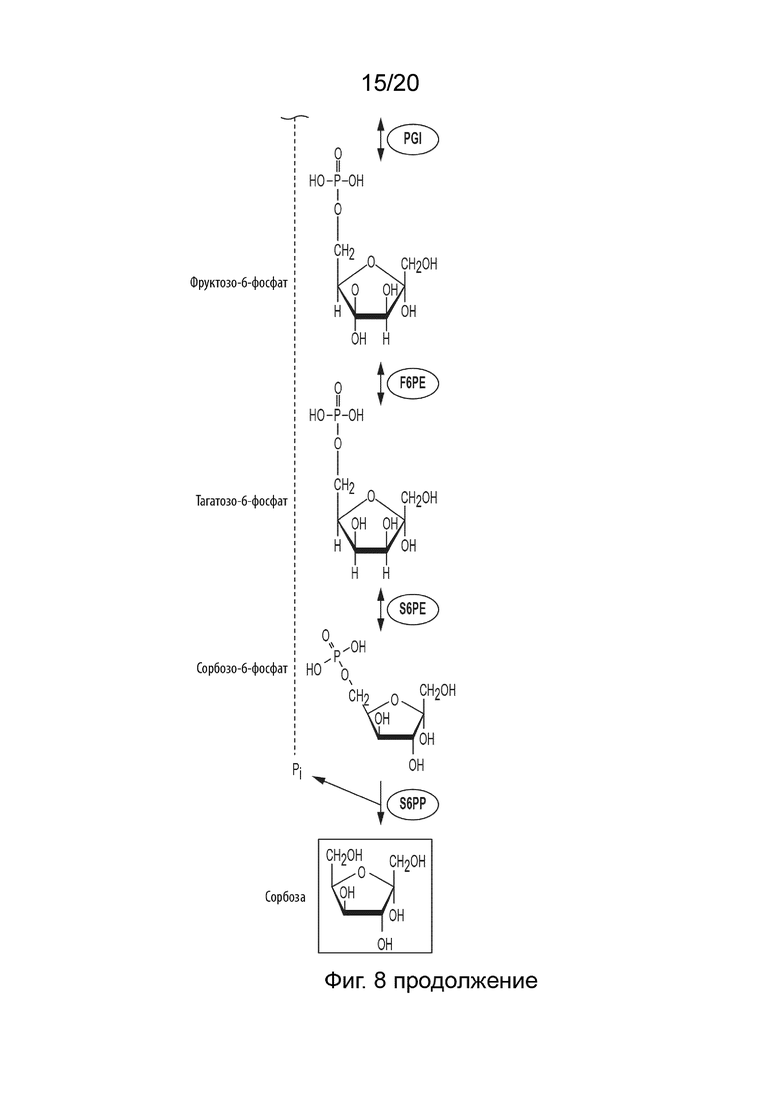
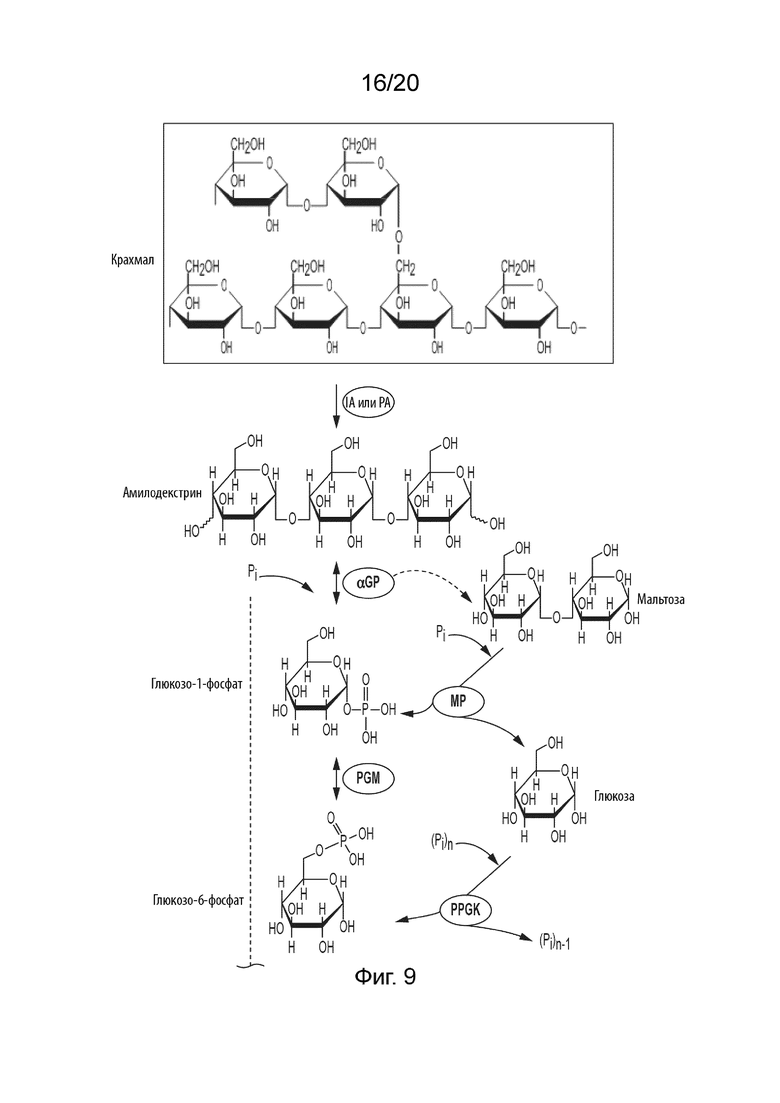
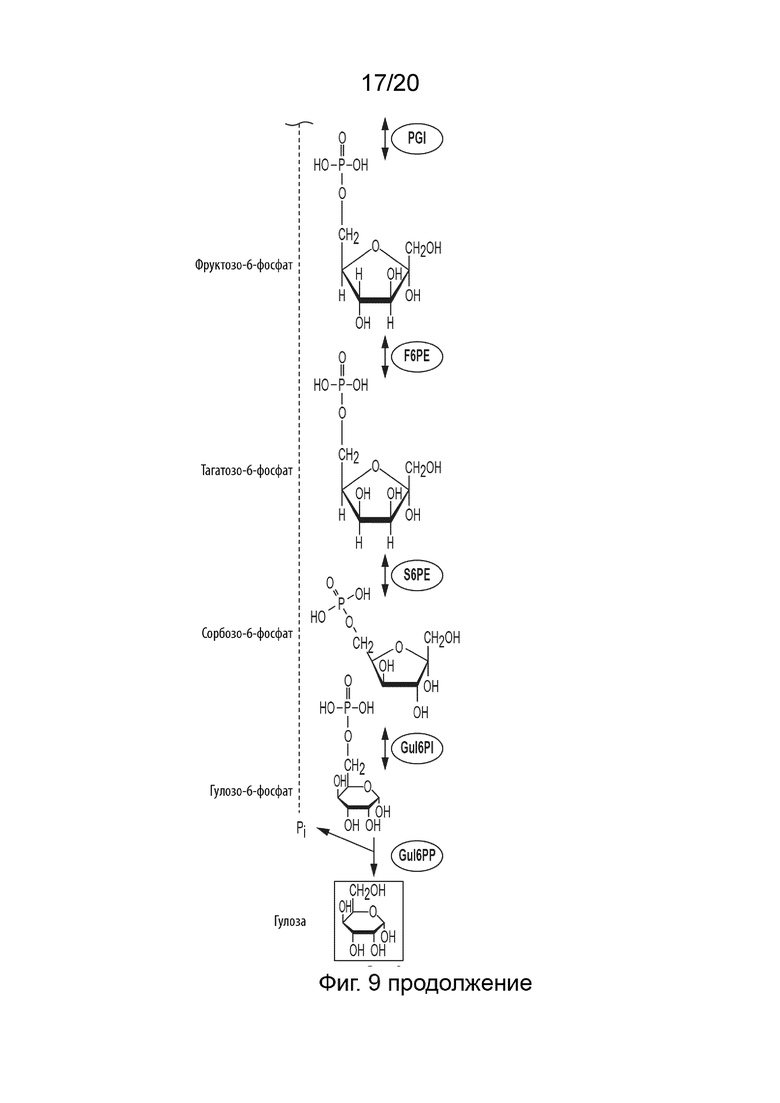
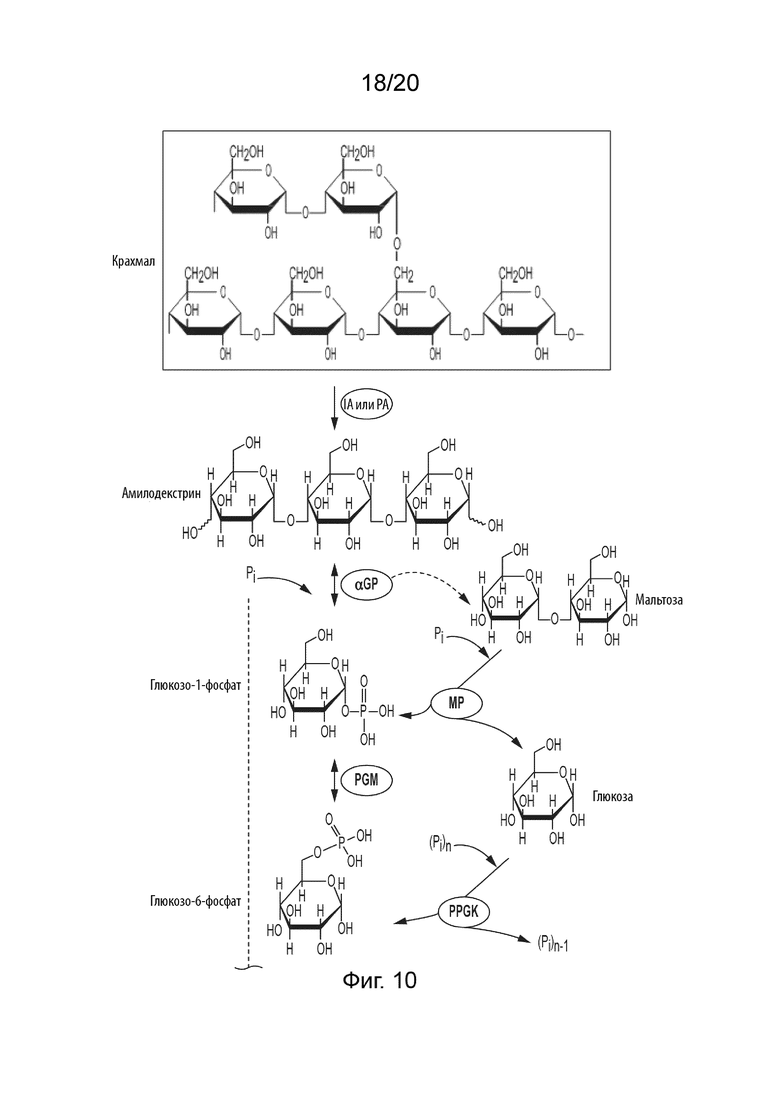
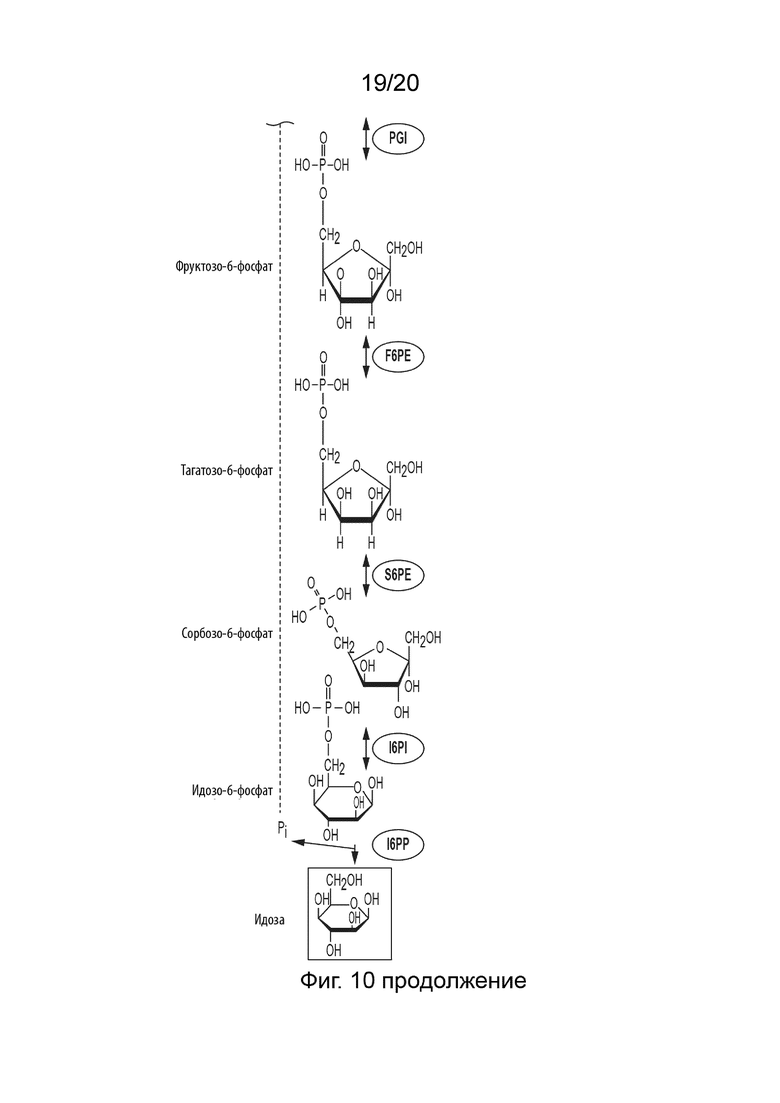
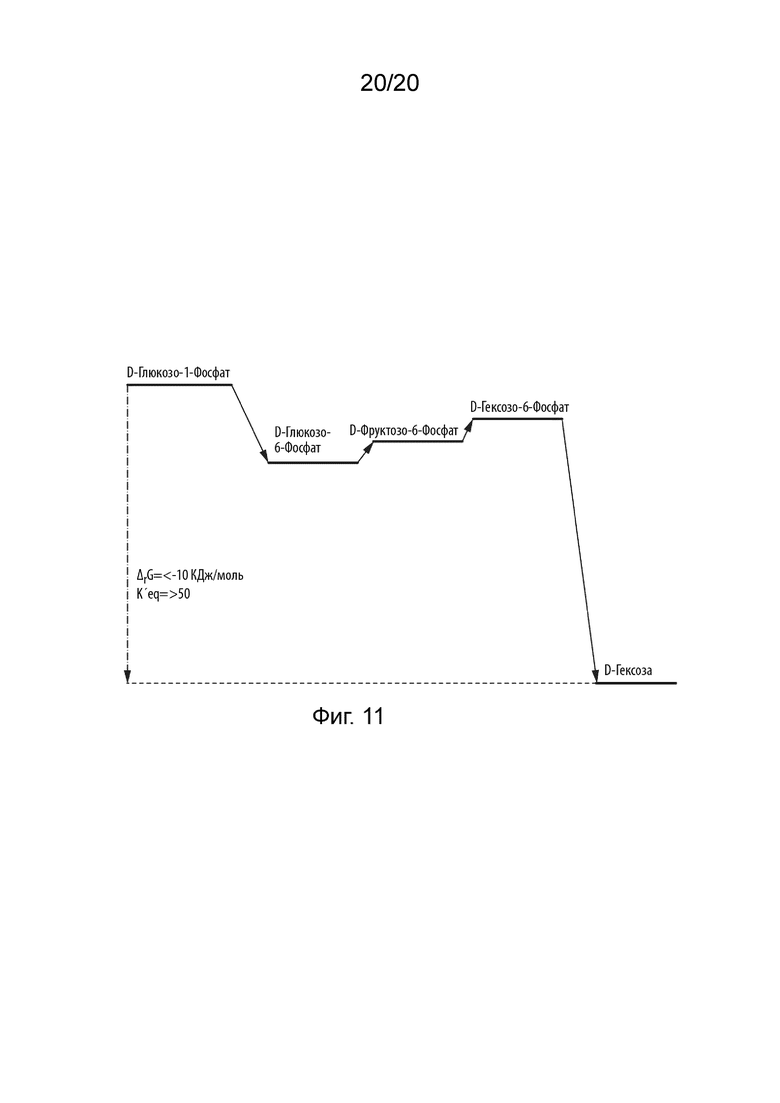
Изобретение относится к биотехнологии и представляет собой способ получения аллозы из сахарида, включающий: превращение фруктозо–6–фосфата (F6P) в псикозо–6–фосфат (P6P), катализируемое псикозо–6–фосфат–3–эпимеразой (P6PE); превращение P6P в аллозо–6–фосфат (A6P), катализируемое аллозо–6–фосфат–изомеразой (A6PI); и превращение A6P в аллозу, катализируемое аллозо–6–фосфат–фосфатазой (A6PP). Изобретение позволяет получать аллозу из сахарида с высокой степенью эффективности. 15 з.п. ф-лы, 2 табл., 121 пр., 11 ил.
1. Способ получения аллозы из сахарида, включающий:
превращение фруктозо–6–фосфата (F6P) в псикозо–6–фосфат (P6P), катализируемое псикозо–6–фосфат–3–эпимеразой (P6PE);
превращение P6P в аллозо–6–фосфат (A6P), катализируемое аллозо–6–фосфат–изомеразой (A6PI); и
превращение A6P в аллозу, катализируемое аллозо–6–фосфат–фосфатазой (A6PP).
2. Способ по п. 1, дополнительно включающий стадию превращения глюкозо–6–фосфата (G6P) в F6P, где указанная стадия катализируется фосфоглюкоизомеразой (PGI).
3. Способ по п. 2, дополнительно включающий стадию превращения глюкозо–1–фосфата (G1P) в G6P, где указанная стадия катализируется фосфоглюкомутазой (PGM).
4. Способ по п. 3, дополнительно включающий стадию превращения сахарида в G1P, причем указанная стадия катализируется, по меньшей мере, одним ферментом, при этоме указанный сахарид выбран из группы, состоящей из крахмала или его производного и сахарозы.
5. Способ по п. 4, отличающийся тем, что по меньшей мере один фермент на стадии превращения сахарида в G1P выбран из группы, состоящей из альфа–глюкан–фосфорилазы (αGP), мальтозо–фосфорилазы, сахарозо–фосфорилазы и их смесей.
6. Способ по п. 4 или 5, отличающийся тем, что сахаридом является крахмал или его производное, выбранное из группы, состоящей из амилозы, амилопектина, растворимого крахмала, амилодекстрина, мальтодекстрина, мальтозы, глюкозы и их смесей.
7. Способ по п. 6, дополнительно включающий стадию превращения крахмала в производное крахмала, где производное крахмала получают ферментативным гидролизом крахмала или кислотным гидролизом крахмала.
8. Способ по п. 6 или 7, отличающийся тем, что в способе добавляют 4–глюкантрансферазу (4GT).
9. Способ по любому из пп. 4-8, отличающийся тем, что производное крахмала получено ферментативным гидролизом крахмала, катализируемым изоамилазой, пуллуланазой, альфа–амилазой или их комбинацией.
10. Способ по п. 1, отличающийся тем, что A6PI содержит аминокислотную последовательность, имеющую по меньшей мере 55% идентичность последовательности с SEQ ID No: 1 или 2, причем указанная A6PI катализирует превращение P6P в A6P.
11. Способ по п. 10, отличающийся тем, что A6PI содержит укладку Россмана для катализа, с His на С–конце 1–й β–цепи укладки Россмана; Arg на С–конце α–спирали С–конца 5–й β–цепи укладки Россмана; His в активном сайте; Cys; Thr в активном сайте; GTG–гидрофобный–G мотив вблизи активного сайта и Asn вблизи активного сайта.
12. Способ по любому из пп. 1 и 10-11, отличающийся тем, что A6PP содержит аминокислотную последовательность, имеющую по меньшей мере 30% идентичность последовательности с любой из SEQ ID No: 3–7, и где указанная A6PP катализирует превращение A6P в аллозу.
13. Способ по п. 12, отличающийся тем, что A6PP содержит домен укладки Россмана для катализа, C1 кэпированный домен, сигнатуру DxD в 1–ой β–цепи укладки Россмана, Thr или Ser на конце 2–ой β–цепи укладки Россмана, Lys на N–конце α–спирали С–терминально относительно 3–ей β–цепи укладки Россмана, и сигнатуру ED на конце 4–ой β–цепи укладки Россмана.
14. Способ по любому из пп. 1-13, отличающийся тем, что стадии указанного способа проводят при температуре в диапазоне от около 40°C до около 70°С, при рН в диапазоне от около 5,0 до около 8,0, и/или в течение от около 8 часов до около 48 часов.
15. Способ по любому из пп. 1-14, отличающийся тем, что стадии указанного способа проводят в одном биореакторе или в множестве биореакторов, расположенных последовательно.
16. Способ по любому из пп. 1-15, отличающийся тем, что стадии указанного способа проводят без АТФ, без NAD(P)(H), при концентрации фосфата от около 0,1 мМ до около 150 мМ, фосфат перерабатывают, и/или, по меньшей мере, одна стадия указанного способа включает энергетически выгодную химическую реакцию.
| US 20170016038 A1, 19.01.2017 | |||
| WO 2017002978 A1, 05.01.2017 | |||
| Способ получения -маннозы | 1978 |
|
SU703104A1 |

