ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет на основании Международной заявки PCT/CN2014/081797, поданной 8 июля 2014 года, которая включена в настоящее описание в качестве ссылки во всей своей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0002] Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, способным связываться с каннабиноидным рецептором 1 (CB1), и к способам применения таких антител и антигенсвязывающих фрагментов.
ОПИСАНИЕ ТЕКСТОВОГО ФАЙЛА, ПРЕДСТАВЛЕННОГО В ЭЛЕКТРОННОМ ВИДЕ
[0003] Содержимое текстового файла, представленного в электронном виде вместе с данным документом, включено в настоящее описание в качестве ссылки во всей полноте: копия перечня последовательностей в машиночитаемом формате (имя файла: 15-343-W03-Seq_List_ST25.txt, дата регистрации: 27 марта 2015 года, размер файла: 793 кБ).
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[0004] Каннабиноидный рецептор 1 (CB1) является членом суперсемейства G-белок-сопряженных рецепторов (GPCR). Рецептор CB1 экспрессируется в центральной нервной системе (ЦНС), легких, печени, жировой ткани и почках, и участвует в развитии многих заболеваний человека, включающих в себя, в числе прочих, ожирение, диабет, фиброз, заболевания печени, сердечно-сосудистые заболевания, рак, боль, спастичность при MS и глаукому. Более конкретно, показано, что рецептор CB1 проявляет вредную активность, например, при ожирении, сахарном диабете, фиброзе, заболеваниях печени, сердечно-сосудистых заболеваниях и раке; и полезную активность при боли, MS-спастичности, глаукоме и др.
[0005] В данной области существует потребность в новых антагонистах и агонистах рецепторов CB1 для применения в терапевтических целях, а также в селективных связывающих средствах для применения в целях диагностики/визуализации. В частности, желательным может быть соединение, специфичное к рецептору CB1, но не способное к проникновению в ЦНС, что может уменьшить потенциальные ЦНС-опосредованные побочные эффекты модуляции рецептора CB1, такие как психиатрические нежелательные явления, связанные с обратным агонистом CB1, римонабантом.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0006] В одном аспекте настоящее изобретение предлагает антитела и их антигенсвязывающие фрагменты, способные связываться с каннабиноидным рецептором 1 (также называемым здесь "рецептор CB1" или "CB1"). В некоторых вариантах осуществления рецептор CB1 представляет собой человеческий рецептор CB1. В некоторых вариантах осуществления антитело или его фрагмент распознает один или несколько внеклеточных эпитопов рецептора CB1. В некоторых вариантах осуществления антитела и их фрагменты, способные связываться с рецептором CB1, представляют собой функциональные антитела или их антигенсвязывающие фрагменты. В некоторых вариантах осуществления антитела или их фрагменты, связывающиеся с рецептором CB1, ингибируют или повышают сигнальную активность рецептора CB1. В некоторых вариантах осуществления настоящего изобретения антитела или их фрагменты, связывающиеся с рецептором CB1, представляют собой антагонистические антитела, которые ингибируют сигнальную активность рецептора CB1. В некоторых вариантах осуществления антитела или их фрагменты, связывающиеся с рецептором CB1, представляют собой агонистические антитела, которые повышают сигнальную активность рецептора CB1. В некоторых вариантах осуществления антитела или их фрагменты, связывающиеся с рецептором CB1, являются модуляторами сигнальной активности рецептора CB1, или аллостерическими модуляторами сигнальной активности рецептора CB1. В некоторых вариантах осуществления антитела или их фрагменты, связывающиеся с рецептором CB1, представляют собой селективные связывающие средства, которые не обладают агонистической или антагонистической активностью. В некоторых вариантах осуществления настоящего изобретения антитела или их фрагменты, связывающиеся с рецептором CB1, представляют собой селективные связывающие средства, не обладающие агонистической или антагонистической активностью, которые можно использовать в целях диагностики и/или визуализации.
[0007] В некоторых вариантах осуществления выделенные антитела или их антигенсвязывающие фрагменты являются, по меньшей мере, столь же эффективными, как и низкомолекулярные модуляторы рецептора CB1, такие как, например, AM6545, AM251 или римонабант. В некоторых вариантах осуществления антитела или их фрагменты обладают ингибирующей или активирующей активностью в отношении рецептора СВ1, которая, по меньшей мере в 2 раза, по меньшей мере, в 3 раза, по меньшей мере, в 4 раза, по меньшей мере, в 5 раз, по меньшей мере в 10 раз, или, по меньшей мере, в 20 раз превышает активность низкомолекулярных AM6545, AM251 или римонабанта. В некоторых вариантах осуществления настоящего изобретения выделенные антитела или их антигенсвязывающие фрагменты ингибируют передачу сигнала, опосредованную агонистом рецептора CB1. В некоторых вариантах осуществления настоящего изобретения ингибирование передачи сигнала, опосредованной агонистом рецептора CB1, измеряют путем определения уровня внутриклеточного цАМФ и/или нижестоящего фосфорилирования ERK.
[0008] В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты имеют преимущество, заключающееся в снижении или отсутствии способности проникать в мозг. В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты обладают более низкой способностью проникать в мозг, чем низкомолекулярные агонисты или антагонисты рецептора СВ1 (такие как AM6545, AM251 или римонабант). В некоторых вариантах осуществления антитела и их фрагменты, связывающиеся с рецептором CB1, оказывают терапевтическое действие, сопровождающееся пониженными побочными эффектами, связанными с центральной нервной системой, по сравнению с низкомолекулярными агонистами или антагонистами рецептора СВ1. Связанные с ЦНС побочные эффекты низкомолекулярного антагониста рецептора CB1 римонабанта включают в себя беспокойство, депрессию, возбуждение, расстройство пищевого поведения, раздражительность, агрессивность и бессонницу (Moreira, 2009, Rev Bras Psiquiatr., 31(2):145-53).
[0009] В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты получают из гибридомных клеточных линий. В других вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты получают из фаговых библиотек.
[0010] В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты обладают сродством к нативному человеческому рецептору СВ1, которое находится, по меньшей мере, в нМ диапазоне. Например, в некоторых вариантах осуществления сродство к рецептору СВ1 составляет примерно 1 мкМ или менее, или примерно 750 нМ или менее, или примерно 500 нМ или менее, или примерно 250 нМ или менее, или примерно 100 нМ или менее, или примерно 75 нМ или менее, или примерно 50 нМ или менее, или примерно 25 нМ или менее, или примерно 10 нМ или менее, или примерно 1 нМ или менее. В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты обладают сродством к человеческому рецептору СВ1, которое составляет примерно от 0,01 нМ до 500 нМ, примерно от 0,02 нМ до 250 нМ, примерно от 0,02 до 200 нМ, примерно от 0,05 до 100 нМ, примерно от 0,05 до 50 нМ.
[0011] Выделенные антитела и их антигенсвязывающие фрагменты настоящего изобретения можно получить из любых видов, включающих в себя, без ограничения, мышей, крыс, кроликов, хомяков, морских свинок, приматов, лам или людей. В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты представляют собой мышиные антитела. В других вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты представляют собой химерные антитела. В следующих вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты представляют собой гуманизированные антитела. В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты представляют собой полностью человеческие антитела.
[0012] В одном варианте осуществления выделенные антитела и их антигенсвязывающие фрагменты представляют собой гуманизированные или химерные антитела P1C4, описанные в настоящем документе. В одном варианте осуществления гуманизированные антитела P1C4 выбраны из группы, состоящей из P1C4-H0, P1C4-H2 и P1C4-H4, описанных в настоящем документе. В одном варианте осуществления выделенные антитела и их антигенсвязывающие фрагменты содержат модификации Fc, которые приводят к снижению, нарушению или устранению эффекторной функции антитела. В другом варианте осуществления выделенные антитела и их антигенсвязывающие фрагменты выбраны из группы, включающей в себя гибрид P1C4-Н0-IgG2-4, P1C4-Н0-IgG2A330S/P331S, P1C4-Н0-IgG4S228P, гибрид P1C4-Н2-IgG2-4, P1C4-H2-IgG2A330S/P331S, P1C4-H2-IgG4S228P, гибрид P1C4-H4-IgG2-4, P1C4-H4-IgG2A330S/P331S, P1C4-H4-IgG4S228P.
[0013] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, 9, 17, 25, 33, 41, 49 и 57. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 10, 18, 26, 34, 42, 50 и 58. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 3, 11, 19, 27, 35, 43, 51 и 59. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит константный участок тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 12, 20, 28, 36, 44, 52 и 60. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5, 13, 21, 29, 37, 45, 53 и 61. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, 14, 22, 30, 38, 46, 54 и 62. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит константный участок легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 15, 23, 31, 39, 47, 55 и 63. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит константный участок легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 8, 16, 24, 32, 40, 48, 56 и 64.
[0014] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 1; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 3; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 5; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 7.
[0015] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 2; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 4; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 6; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 8.
[0016] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 9; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 11; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 13; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 15.
[0017] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 10; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 12; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 14; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 16.
[0018] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 17; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 19; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 21; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 23.
[0019] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 18; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 20; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 22; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 24.
[0020] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 25; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 27; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 29; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 31.
[0021] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 26; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 28; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 30; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 32.
[0022] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 33; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 35; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 37; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 39.
[0023] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 34; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 36; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 38; и константный участок легкой цепи, имеющий аминокислотную последовательность ID NO: 40.
[0024] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 41; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 43; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 45; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 47.
[0025] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 42; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 44; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 46; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 48.
[0026] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 49; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 51; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 53; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 55.
[0027] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 50; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 52; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 54; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 56.
[0028] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 57; константный участок тяжелой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 59; вариабельный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 61; и константный участок легкой цепи, кодируемый нуклеотидной последовательностью SEQ ID NO: 63.
[0029] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 58; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 60; вариабельный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 62; и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 64.
[0030] В одном варианте осуществления настоящее изобретение предлагает выделенное антитело или его фрагмент, нуклеотидная последовательность или аминокислотная последовательность которого, по меньшей мере, на 65%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-351.
[0031] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержат гипервариабельные участки (CDR) тяжелой цепи, независимо выбранные из CDR, присутствующих в вариабельных участках тяжелой цепи SEQ ID NO: 2, 10, 18 и 26. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR легкой цепи, независимо выбранные из CDR, присутствующих в вариабельных участках легкой цепи SEQ ID NO: 6, 14, 22 и 30.
[0032] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент представляет собой гуманизированное антитело, содержащее гипервариабельные участки (CDR) тяжелой цепи, независимо выбранные из CDR, присутствующих в вариабельных участках тяжелой цепи SEQ ID NO: 2, 10, 18 и 26. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент представляет собой гуманизированное антитело, содержащее гипервариабельные участки (CDR) легкой цепи, независимо выбранные из CDR, присутствующих в вариабельных участках легкой цепи SEQ ID NO: 6, 14, 22 и 30.
[0033] В одном варианте осуществления вариабельный участок тяжелой цепи содержит аминокислотную последовательность, по существу состоит из, или состоит из аминокислотной последовательности, выбранной из группы, включающей в себя SEQ ID NO: 339-341. В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи, аминокислотная последовательность которого, по меньшей мере, на 65%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 339-341. В следующем варианте осуществления вариабельный участок тяжелой цепи содержит аминокислотную последовательность, по существу состоит из, или состоит из аминокислотной последовательности, выбранной из группы, включающей в себя SEQ ID NO: 339-341.
[0034] В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность тяжелой цепи, которая, по меньшей мере, на 65%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 343-351. В следующем варианте осуществления тяжелая цепь содержит аминокислотную последовательность, по существу состоит из, или состоит из аминокислотной последовательности, выбранной из группы, включающей в себя SEQ ID NO: 343-351.
[0035] В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность вариабельного участка легкой цепи, которая, по меньшей мере, на 65%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% идентична аминокислотной последовательности SEQ ID NO: 337. В следующем варианте осуществления вариабельный участок тяжелой цепи содержит аминокислотную последовательность, по существу состоит из, или состоит из аминокислотной последовательности SEQ ID NO: 337. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность легкой цепи, которая, по меньшей мере, на 65%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% идентична аминокислотной последовательности SEQ ID NO: 338. В следующем варианте осуществления легкая цепь содержит аминокислотную последовательность, по существу состоит из, или состоит из аминокислотной последовательности SEQ ID NO: 338.
[0036] В одном варианте осуществления настоящее изобретение предлагает гуманизированное выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с СВ1. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок легкой цепи с последовательностью SEQ ID NO: 337, и вариабельный участок тяжелой цепи с последовательностью SEQ ID NO: 339. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок легкой цепи SEQ ID NO: 337 и вариабельный участок тяжелой цепи SEQ ID NO: 340. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельный участок легкой цепи SEQ ID NO: 337 и вариабельный участок тяжелой цепи SEQ ID NO: 341. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит полноразмерную легкую цепь с последовательностью SEQ ID NO: 338 и полноразмерную тяжелую цепь с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 343-351.
[0037] В одном варианте осуществления выделенное антитело или его фрагмент содержит последовательность CDR1 тяжелой цепи, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% гомологичную аминокислотной последовательности SEQ ID NO: 352 (YYWMN). В другом варианте осуществления выделенное антитело или его фрагмент содержит последовательность CDR2 тяжелой цепи, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% гомологичную аминокислотной последовательности SEQ ID NO: 353 (QIYPGDGETKY). В другом варианте осуществления выделенное антитело или его фрагмент содержит последовательность CDR3 тяжелой цепи, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% гомологичную аминокислотной последовательности SEQ ID NO: 354 (SHGNYLPY). В другом варианте осуществления выделенное антитело или его фрагмент содержит последовательность CDR1 легкой цепи, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% гомологичную аминокислотной последовательности SEQ ID NO: 355 (SSYLH). В другом варианте осуществления выделенное антитело или его фрагмент содержит последовательность CDR2 легкой цепи, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% гомологичную аминокислотной последовательности SEQ ID NO: 356 (STSNLAS). В другом варианте осуществления выделенное антитело или его фрагмент содержит последовательность CDR3 легкой цепи, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% гомологичную аминокислотной последовательности SEQ ID NO: 357 (HQYHRSPPTF).
[0038] В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR1, CDR2 и CDR3 тяжелой цепи, имеющие аминокислотные последовательности SEQ ID NO: 352, 353 и 354, соответственно. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR1, CDR2 и CDR3 легкой цепи, имеющие аминокислотные последовательности SEQ ID NO: 355, 356 и 357, соответственно. В следующем варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи, CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, имеющие аминокислотные последовательности SEQ ID NO: 352, 353, 354, 355, 356 и 357, соответственно. В другом варианте осуществления выделенное антитело или его фрагмент является химерным или гуманизированным.
[0039] Специалисту в данной области должно быть известно, что CDR тяжелых и легких цепей описанных здесь антител, можно выбирать независимо друг от друга, или комбинировать с получением антитела или его связывающего фрагмента, содержащего любые CDR1, CDR2 и CDR3 легкой цепи; и любые CDR1, CDR2 и CDR3 тяжелой цепи описанных здесь антител. Специалисту в данной области также следует понимать, что вариабельные участки тяжелых и легких цепей описанных здесь антител можно выбирать независимо друг от друга, или комбинировать с получением антитела или его связывающего фрагмента, содержащего любые тяжелые и легкие цепи описанных здесь антител.
[0040] В одном варианте осуществления описанное здесь антитело, или его антигенсвязывающий фрагмент, представляет собой химерное антитело, или его фрагмент, содержащие CDR тяжелых и легких цепей, выбранные из описанных здесь CDR, или консервативные варианты описанных здесь CDR. В другом варианте осуществления описанное здесь антитело, или его антигенсвязывающий фрагмент, представляет собой гуманизированное антитело, или его фрагмент, содержащие CDR тяжелых и легких цепей, выбранные из описанных здесь CDR, или консервативные варианты описанных здесь CDR. В одном варианте осуществления изобретения описанное здесь антитело, или его антигенсвязывающий фрагмент, содержит легкую цепь и/или тяжелую цепь, имеющую описанную здесь последовательность, или их консервативный вариант. В одном варианте осуществления консервативные варианты, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% гомологичны описанной здесь стандартной последовательности. В одном варианте осуществления консервативные варианты содержат 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислотных замен, инсерций или делеций.
[0041] В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с СВ1 при пониженной эффекторной функции. В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с CB1 и содержит одну или несколько модификаций Fc-участка. В следующем варианте осуществления антитело или его антигенсвязывающий фрагмент связывается с CB1 и имеет аминокислотную последовательность, содержащую одну или несколько мутаций в Fc-участке. В следующем варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит мутацию в положении 228, и/или 330, и/или 331. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит мутацию в положении 228 Fc-участка, где Fc-участок относится к изотипу IgG4. В следующем варианте осуществления мутация представляет собой S228P. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит мутацию в положении 330, и/или в положении 331. В следующем варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит мутацию в положении 330 и/или 331, где Fc-участок относится к изотипу IgG2. В следующем варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит следующие мутации в Fc-участке: A330S и P331S. В другом варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит участок Fc, который представляет собой гибридный Fc-участок. Например, в одном варианте осуществления Fc-участок представляет собой гибридный Fc-участок IgG2/IgG4, где CH1 и шарнирные участки получены из IgG2, а участки СН2 и СН3 получены из IgG4.
[0042] Таким образом, в одном варианте осуществления антитело или его антигенсвязывающий фрагмент представляет собой химерное или гуманизированное антитело, или фрагмент, содержащий CDR тяжелых и легких цепей, выбранные из описанных здесь CDR или консервативных вариантов описанных здесь CDR, где выделенное антитело или его фрагмент содержит Fc-участок, несущий модификации, которые изменяют эффекторные функции антитела. Например, в одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR легкой и тяжелой цепи, имеющие последовательности SEQ ID NO: 352-357, или их консервативные варианты, а также содержит гибридный Fc-участок IgG2-IgG4, Fc-участок IgG2, содержащий аминокислотные мутации в положениях 330 и 331 (например, A330S и P331S), или Fc-участок IgG4, содержащий аминокислотную мутацию в положении 228 (например, S228P).
[0043] В одном варианте осуществления настоящее изобретение предлагает выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с CB1, где значение Kd, характеризующее сродство антитела или фрагмента к рецептору СВ1, составляет примерно 70 нМ или менее, примерно 60 нМ или менее, примерно 50 нМ или менее, примерно 40 нМ или менее, примерно 30 нМ или менее, примерно 25 нМ или менее, примерно 20 нМ или менее, примерно 15 нМ или менее, примерно 10 нМ или менее, примерно 8 нМ или менее, примерно 6 нМ или менее, примерно 5 нМ или менее, примерно 4 нМ или менее, примерно 3 нм или менее, примерно 2 нМ или менее, или примерно 1 нМ или менее. В одном варианте осуществления настоящее изобретение предлагает выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с CB1, где значение Kd, характеризующее сродство антитела или фрагмента к рецептору СВ1, находится в диапазоне примерно от 1 нМ до 100 нМ, примерно от 2 нМ до 75 нМ, примерно от 3 нМ до 50 нМ, примерно от 4 нм до 10 нМ, или значение Kd, характеризующее сродство к рецептору СВ2, составляет примерно 50 нМ, или примерно 40 нМ, или примерно 30 нМ, или примерно 20 нМ, или примерно 10 нМ, или примерно 5 нМ, или примерно 4 нМ, или примерно 3 нМ, или примерно 2 нМ, или примерно 1 нМ.
[0044] В одном варианте осуществления настоящее изобретение предлагает выделенное антитело или его антигенсвязывающий фрагмент, которые являются, по меньшей мере, в 2 раза, по меньшей мере, в 3 раза, по меньшей мере, в 4 раза, по меньшей мере, в 5 раз, по меньшей мере, в 6 раз, по меньшей мере, в 7 раз, по меньшей мере, в 8 раз, по меньшей мере, в 9 раз, по меньшей мере, в 10 раз, по меньшей мере, в 11 раз, по меньшей мере, в 12 раз, по меньшей мере, в 13 раз, по меньшей мере, в 14 раз, или, по меньшей мере, в 15 раз более активными, чем низкомолекулярный римонабант, где активность антитела, или фрагмента, или римонабанта измеряют путем ингибирования передачи сигнала, опосредованной антагонистом рецептора СВ1, в анализе цАМФ. В следующем варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент является гуманизированным.
[0045] В одном варианте осуществления настоящее изобретение предлагает выделенное гуманизированное антитело или его антигенсвязывающий фрагмент, способные связываться с CB1, где антитело или его фрагмент обладает более высоким сродством связывания и/или более высокой активностью, чем соответствующее негуманизированное или химерное антитело, причем гуманизированное антитело, или его фрагмент, и соответствующее негуманизированное или химерное антитело содержат одинаковые CDR тяжелой и легкой цепи. Например, в одном варианте осуществления настоящее изобретение предлагает гуманизированное антитело, или его фрагмент, которые содержат CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, имеющие последовательности SEQ ID NO: 352, 353, 354, 355, 356 и 357, соответственно, где гуманизированное антитело обладает более высоким сродством к рецептору CB1 и/или более высокой активностью в отношении ингибирования агониста рецептора CB1. В одном из вариантов осуществления активность описанных здесь гуманизированных антител и фрагментов, по меньшей мере на 50% выше, по меньшей мере, на 100% выше, по меньшей мере, в 2 раза выше, по меньшей мере, в 3 раза выше, по меньшей мере, в 4 раза выше, по меньшей мере, в 5 раз выше, или, по меньшей мере, в 10 раз выше, чем активность соответствующего негуманизированного или химерного антитела. В другом варианте осуществления активность измеряют по ингибированию CB1 - продукции цАМФ.
[0046] Активность описанных здесь антител против рецептора CB1 можно измерить с помощью любого способа, известного в данной области техники. Например, в одном варианте осуществления активность описанных здесь антител и фрагментов измеряют по уровню внутриклеточного цАМФ или фосфорилирования ERK. Например, активность можно измерить путем определения уровня ингибирования продукции цАМФ с помощью функционального анализа цАМФ (Cisbio), или уровня ингибирования WIN55,212-индуцированного фосфорилирования ERK методом вестерн-блоттинга.
[0047] В некоторых вариантах осуществления настоящее изобретение предлагает антитело, или его антигенсвязывающий фрагмент, способные конкурировать с описанным здесь антителом, или его антигенсвязывающим фрагментом, за связывание с рецептором CB1. В некоторых других вариантах осуществления настоящее изобретение предлагает антитело, или его антигенсвязывающий фрагмент, способные специфически связываться, по существу, с тем же эпитопом на рецепторе CB1, что и описанные здесь антитела или антигенсвязывающие фрагменты. Такие антитела можно идентифицировать с помощью рутинных анализов конкурентного связывания. В некоторых вариантах осуществления конкурентную способность измеряют методами ELISA, проточной цитометрии или поверхностного плазмонного резонанса (SPR).
[0048] В некоторых вариантах осуществления антитела и их фрагменты конъюгируют с одним или несколькими средствами, выбранными из группы, включающей в себя другое терапевтическое средство, цитотоксическое средства, молекулу иммуноадгезии и визуализирующее средство. В некоторых вариантах осуществления визуализирующее средство выбрано из группы, состоящей из радиоактивной метки, фермента, флуоресцентной метки, люминесцентной метки, биолюминесцентной метки, магнитной метки и биотина.
[0049] В одном аспекте изобретение предлагает способы модуляции сигнальной активности рецептора СВ1, включающие в себя приведение клетки, экспрессирующей рецептор CB1, в контакт с описанным здесь антителом или его фрагментом. В некоторых вариантах осуществления предлагаемые способы приводят к ингибированию сигнальной активности рецептора СВ1. В некоторых вариантах осуществления предлагаемые способы приводят к увеличению сигнальной активности рецептора СВ1. В некоторых вариантах осуществления модуляция сигнальной активности рецептора СВ1 является косвенной, например, как под действием аллостерического модулятора. В некоторых вариантах осуществления модуляция сигнальной активности рецептора СВ1 приводит к смещению в сторону передачи сигналов, опосредованной Galpha i/o, от передачи сигналов, опосредованной бета-аррестином.
[0050] В одном аспекте изобретение предлагает способы лечения заболевания или расстройства, отвечающего на антагонистическую или агонистическую активность в отношении рецептора СВ1, у индивидуума, нуждающегося в этом. В некоторых вариантах осуществления способы включают в себя введение индивидууму описанного здесь антитела против рецептора CB1, или его антигенсвязывающего фрагмента. В одном из вариантов осуществления индивидуум представляет собой млекопитающее. В следующем варианте осуществления индивидуум представляет собой человека. В некоторых вариантах осуществления заболевание или расстройство представляет собой ожирение, диабет, дислипидемию, метаболические заболевания, фиброз, неалкогольный стеатогепатит (NASH), заболевание печени, первичный билиарный цирроз, заболевание почек, фиброз почек, хроническое заболевание почек, остеопороз, атеросклероз, сердечно-сосудистое заболевание, рак, воспалительное заболевание, боль, спастичность мышечной системы и глазные болезни, включающие в себя глаукому. В некоторых вариантах осуществления заболевание или расстройство представляет собой, например, ожирение, диабет, фиброз, заболевание печени, сердечно-сосудистое заболевание или рак, а предлагаемый способ приводит к ингибированию активности рецептора СВ1. В некоторых вариантах осуществления заболевание или расстройство представляет собой, например, боль или глаукому, а предлагаемый способ приводит к активации рецептора СВ1, или увеличению активности рецептора СВ1.
[0051] В одном аспекте изобретение предлагает способ детекции рецептора СВ1 в клетке, ткани или организме индивидуума, включающий в себя приведение клетки в контакт с описанным здесь антителом, способным связываться с рецептором СВ1, или его антигенсвязывающим фрагментом. В одном из вариантов осуществления клетка присутствует в организме индивидуума. В другом варианте осуществления клетка присутствует в организме человека. В другом варианте осуществления уровень экспрессии рецептора СВ1 на клетках коррелирует со стадией заболевания. Таким образом, в одном аспекте настоящее изобретение предлагает способы применения антител и их фрагментов, способных специфически связываться с рецептором СВ1, в качестве средств диагностики и/или прогнозирования заболеваний человека. В одном варианте осуществления настоящее изобретение предлагает способы визуализации рецептора СВ1, включающие в себя применение описанных здесь антител против рецептора СВ1 и их фрагментов. В одном варианте осуществления способ детекции рецептора CB1 проводят с использованием описанного здесь антитела против рецептора СВ1, или его фрагмента, способных селективно связываться с рецептором CB1. В следующем варианте осуществления антитело, способное селективно связываться с рецептором CB1, или его фрагмент, не обладает агонистической или антагонистической активностью. В другом варианте осуществления антитело, селективно связывающееся с рецептором CB1, или его фрагмент, не способны к интернализации. В одном варианте осуществления настоящее изобретение предлагает способы диагностики и визуализации, включающие в себя применение антитела против рецептора CB1, или его фрагмента, конъюгированного с визуализирующим средством, таким как, например, радиоактивная метка, фермент, флуоресцентная метка, люминесцентная метка, биолюминесцентная метка, магнитная метка или биотин.
[0052] В одном варианте осуществления настоящее изобретение предлагает клетку-хозяина, экспрессирующую выделенное антитело, или его фрагмент, способные специфически связываться с рецептором CB1. В другом варианте осуществления изобретение предлагает способ получения антитела или его фрагмента, способных специфически связываться с рецептором CB1, где указанный способ включает в себя иммунизацию млекопитающих очищенным рецептором CB1 или его антигенным фрагментом, CB1/липидными комплексами, рецептор CB1-экспрессирующими iCAPS, и/или ДНК, кодирующей рецептор CB1. В другом варианте осуществления в качестве млекопитающих для иммунизации используют мышей. В другом варианте осуществления, перед сбором клеток иммунизированных млекопитающих, животных иммунизируют один, два, три, четыре, пять или более раз очищенным CB1, или его антигенным фрагментом, CB1/липидным комплексом, рецептор CB1-экспрессирующими iCAPS, и/или ДНК, кодирующей рецептор CB1. В следующем варианте осуществления антитело или его фрагмент, способные специфически связываться с рецептором CB1, получают из гибридомной клеточной линии, содержащей клетки, полученные из иммунизированных млекопитающих. В другом варианте осуществления антитело или его фрагмент, способные специфически связываться с рецептором CB1, получают из библиотеки фагового дисплея. В следующем варианте осуществления библиотеку фагового дисплея получают из клеток, выделенных из иммунизированных млекопитающих. В следующем варианте осуществления библиотеку фагового дисплея получают из последовательностей наивных человеческих иммуноглобулинов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0053] На фигуре 1А приведен ряд гистограмм, демонстрирующих связывание P2A12 Bril mAb (левый столбец), PA2LR3-P2D3 (средний столбец) или PA2R3-P1A7 (правый столбец) при 300 нМ (верхний ряд) или 30 нМ (нижний ряд) с клеточной линией Trex-CHO нативный человеческий CB1 (экспрессирующей нативный рецептор CB1, темно-серые линии), клеточной линией Trex-CHO CB1 T210A/партнер по гибридизации (экспрессирующей CB1 на повышенном уровне; средне-серые линии), или исходной клеточной линией Trex-CHO (не экспрессирующей рецептор CB1; светло-серые линии). На фигуре 1B приведен ряд гистограмм, демонстрирующих связывание PA2R3-P1F1 (левый столбец), PA2LR3-P2E5 (средний столбец) или PA2LR3-P3B10 (правый столбец) при 300 нМ (верхний ряд) или 30 нМ (нижний ряд) с клеточной линией Trex-CHO нативный CB1 (экспрессирующей нативный человеческий рецептор CB1, темно-серые линии), клеточной линией Trex-CHO CB1 T210A/партнер по гибридизации (экспрессирующей CB1 на повышенном уровне; средне-серые линии), или исходной клеточной линией Trex-CHO (не экспрессирующей рецептор CB1; светло-серые линии). На фигуре 1C приведен ряд гистограмм, демонстрирующих связывание PA2LR3-P3B8 (левый столбец) или PA2LR3-P1H4 (правый столбец) при 300 нМ (верхний ряд) или 30 нМ (нижний ряд) с клеточной линией Trex-CHO нативный человеческий CB1 (экспрессирующей нативный рецептор CB1, темно-серые линии), клеточной линией Trex-CHO CB1 T210A/партнер по гибридизации (экспрессирующей CB1 на повышенном уровне; средне-серые линии), или исходной клеточной линией Trex-CHO (не экспрессирующей рецептор CB1; светло-серые линии). На фигуре 1D приведен ряд гистограмм, демонстрирующих связывание PA2LR3-P4B1 (левый столбец), PA2LR3-P4B5 (средний столбец) или PA2LR3-P4C6 (правый столбец) при 300 нм (верхний ряд) или 30нм (нижний ряд) с клеточной линией Trex-CHO нативный человеческий CB1 (экспрессирующей нативный рецептор CB1, темно-серые линии), клеточной линией Trex-CHO CB1 T210A/партнер по гибридизации (экспрессирующей CB1 на повышенном уровне; средне-серые линии), или исходной клеточной линией Trex-CHO (не экспрессирующей рецептор CB1; светло-серые линии). На фигуре 1E приведен ряд гистограмм, демонстрирующих связывание PA2LR3-P4G10 (левый столбец) или PA2LR3-P6D7 (правый столбец) при 300 нМ (верхний ряд) или 30 нМ (нижний ряд) с клеточной линией Trex-CHO нативный человеческий CB1 (экспрессирующей нативный рецептор CB1, темно-серые линии), клеточной линией Trex-CHO CB1 T210A/партнер по гибридизации (экспрессирующей CB1 на повышенном уровне; средне-серые линии), или исходной клеточной линией Trex-CHO (не экспрессирующей рецептор CB1; светло-серые линии). На фигуре 1F приведен ряд гистограмм, демонстрирующих связывание PA2LR3-P1G6 (левый столбец), PA2LR3-P1H4 (средний столбец) или PA2LR3-P2B8 (правый столбец) с исходной клеточной линией Trex-CHO (не экспрессирующей рецептор CB1; верхний ряд), клеточной линией Trex-CHO CB1 T210A/партнер по гибридизации (экспрессирующей CB1 на повышенном уровне; средний ряд), или клеточной линией Trex-CHO A156 (экспрессирующей нативный человеческий рецептор CB1; нижний ряд).
[0054] На фигуре 2 показана способность двух антител против рецептора CB1, PA13R3-P1C4 и 36E12B6C2, селективно связываться с клетками, экспрессирующими CB1. На фигуре 2А (демонстрирующей связывание PA13R3-P1C4) и на фигуре 2В (демонстрирующей связывание 36E12B6C2) показано, что оба антитела связываются с A156 (экспрессирующими нативный человеческий рецептор CB1) и, в еще большей степени, с А56 (экспрессирующими рецептор CB1 на повышенном уровне и модифицированными путем мутации T210A и замены ICL3 на партнер по гибридизации), но не связываются с клетками СНО, не экспрессирующими рецептор CB1, клеточной линией, экспрессирующей СВ2, или клеточной линией, экспрессирующей 5HT2b. Экспрессию CB2 (фигура 2C) и 5HT2b (фигура 2D) подтверждают в клеточных линиях, экспрессирующих CB2 и 5HT2b, соответственно.
[0055] На фигуре 3 приведены результаты конкурентного анализа, позволяющие определить, действительно ли 36E12B6C2 и P1C4 связываются с подобными эпитопами. Клетки Trex CHO A156, экспрессирующие нативный человеческий CB1, инкубируют с конкурирующими IgG (IgG или Fab PA13R3-P1C4 и 36E12B6C2) и затем с разными концентрациями окрашивающих IgG (300 нМ или 75 нМ P1C4, фигура 3А; 80 нМ или 25 нМ 36E12B6C2, фигура 3B).
[0056] На фигуре 4 показаны результаты функционального антагонистического анализа цАМФ. Антитела 36E12B2H8 (фигура 4A) и PA13R3-P1C4 (фигура 4В) проявляют антагонистическую активность, равную (36E12B2H8) или превышающую (PA13R3-P1C4) активность используемых в качестве положительного контроля низкомолекулярных ингибиторов CB1 AM251 (фигура 4C), SR141716A (римонабант) (фигура 4D) и AM6545 (фигура 4Е). P2A12 и изотип гибридомных IgG используют в качестве отрицательного контроля (фигуры 4F и 4G).
[0057] На фигуре 5A приведены результаты вестерн-блоттинга, демонстрирующие уровень фосфорилированной ERK (pERK) и общей ERK в клетках Trex-СНО, экспрессирующих нативный человеческий рецептор CB1, после экспрессии рецептора CB1 и обработки контрольным IgG, используемым в качестве положительного контроля низкомолекулярным AM6545, или полученным из фага mAb PA13R3-P1C4 или PA13R3-P1E4, с последующей обработкой 100 нм агониста рецептора CB1, WIN55,212. Вестерн-блоттинг проводят через 10 минут после активации WIN55,212 (левые панели), или через 15 минут после активации WIN55,212 (правые панели). На фигуре 5В приведены результаты вестерн-блоттинга, демонстрирующие уровень pERK и общей ERK в клетках Trex-СНО, экспрессирующих нативный человеческий рецептор CB1, после экспрессии рецептора CB1 и обработки контрольным IgG1, контрольным IgG2, WIN55,212, AM6545 или полученным из гибридом mAb 36E12B2E5, 36E12B6C2 или 36E12B2F2, с последующей активацией под действием WIN55,212.
[0058] На фигуре 6А показаны результаты функционального анализа цАМФ, проводимого в отсутствии CP55940 для оценки потенциальной агонистической активности PA2LR3-P2D3, PA2LR3-P4B1, PA2LR3-P6B12 и PA2LR3-P6G6, по сравнению с контрольными соединениями, изображенными на фигуре 6В: CP55940 (положительный контроль), P2A12 (отрицательный контроль) или PA2R3-P1A7 (отрицательный контроль). На фигуре 6С показаны результаты функционального анализа цАМФ, проводимого в присутствии CP55940 для оценки потенциальной аллостерической модуляторной активности PA2LR3-P2D3, PA2LR3-P4B1, PA2LR3-P6B12 и PA2LR3-P6G6, по сравнению с контрольными соединениями, изображенными на фигуре 6D: CP55940 один (положительный контроль), P2A12 (отрицательный контроль) или PA2R3-P1A7 (отрицательный контроль).
[0059] На фигуре 7 показаны результаты анализа цАМФ, проводимого для оценки обратной агонистической или нейтральный антагонистической активности PA13R3-P1C4 и 36E12B6C2. AM6545 и SR141716A используют в качестве положительного контроля для нейтрального антагониста и обратного агониста, соответственно.
[0060] На фигуре 8А показаны результаты анализа iCAPS методом ELISA с целью определения связывания 36E12B6C2 Fab (верхняя левая панель), или IgG (верхняя правая панель), или P1F7 Fab (нижняя левая панель), или Fab (нижняя правая панель), с rBril-0918, пустыми iCAPS, iCAPS, не экспрессирующими рецептор CB1 (h13h iCAPS), или iCAPS, экспрессирующими человеческий рецептор CB1 (A138 iCAPS и A139 iCAPS). На фигуре 8В показаны результаты анализа iCAPS методом ELISA с целью определения связывания PA13R3-P1C4 Fab (верхняя левая панель), или IgG (верхняя правая панель), или P1F7 Fab (нижняя левая панель), или Fab (нижняя правая панель) с rBril-0918, пустыми iCAPS, iCAPS, не экспрессирующими рецептор CB1 (h13h iCAPS), или iCAPS, экспрессирующими человеческий рецептор CB1 (A138 iCAPS и A139 iCAPS).
[0061] На фигурах 9A и 9B показана интернализация рецептора CB1 после обработки разными средствами. На фигуре 9A показано, что антитела против CB1 не блокируют WIN55,212-индуцированную интернализацию рецептора. На гистограммах, приведенных на фигуре 9А в верхнем ряду, показана поверхностная экспрессия CB1 после обработки WIN55,212, или контролем, или после предварительной обработки CB1-специфическим нейтральным антагонистом AM6545 с последующей обработкой WIN55,212. На гистограммах, приведенных на фигуре 9А в среднем и нижнем рядах, показана поверхностная экспрессия CB1 после предварительной обработки антителами против CB1 PA2LR3-P3A8, PA2LR3-P3F8, PA2LR3-P5B11, PA2LR3-P5E7, PA2LR3-P6B12, PA2LR3-P6G7, PA3R3-P4D5, PA2LR3-P4B1, PA2LR3-P4B5, PA2LR3-P4C6 и PA2LR3-P4G10, или отрицательным контролем P2A12 с последующей обработкой WIN55,212. На фигуре 9B показано, что антитела против CB1 сами по себе не индуцируют интернализацию рецептора CB1. На гистограммах, приведенных на фигуре 9B в верхнем ряду, показана поверхностная экспрессия CB1 после обработки WIN55,212, или контролем, или после предварительной обработки CB1-специфическим нейтральным антагонистом AM6545 с последующей обработкой WIN55,212. На гистограммах, приведенных на фигуре 9B в среднем и нижнем рядах, показана поверхностная экспрессия CB1 после обработки антителами против CB1 PA2LR3-P3A8, PA2LR3-P3F8, PA2LR3-P5B11, PA2LR3-P5E7, PA2LR3-P6B12, PA2LR3-P6G7, PA3R3- P4D5, PA2LR3- P4B1, PA2LR3-P4B5, PA2LR3-P4C6 и PA2LR3-P4G10, или отрицательным контролем P2A12.
[0062] На фигуре 10 показаны результаты измерения цАМФ в функциональном сравнительном анализе антагонистической активности гуманизированных и химерных антител P1C4. Гуманизированное антитело P1C4-H0 обладает антагонистической активностью, подобной активности химерного антитела P1C4. Гуманизированные антитела P1C4-H4 и PIC4-Н2 обладают более высокой антагонистической активностью, чем химерное антитело P1C4, или используемый в качестве положительного контроля низкомолекулярный ингибитор рецептора CB1 римонабант.
[0063] На фигуре 11 показаны результаты измерения сродства связывания, перекрестной реакционноспособности и специфичности гуманизированных антител P1C4 методом проточной цитометрии. Сродство гуманизированных антител P1C4, P1C4-H2 и P1C4-H4, как к клеткам, осуществляющим нативную экспрессию человеческого CB1 (фигура 11А, верхняя панель), так и к клеткам, осуществляющим экспрессию CB1 на повышенном уровне (фигура 11А, нижняя панель), превосходит сродство химерного антитела P1C4. Ни одно из химерных или гуманизированных антител не связывается с мышиными клетками, экспрессирующими мышиный CB1, исходными клетками TRex-CHO или клетками TRex-CHO, экспрессирующими человеческий CB2 (фигура 11B).
[0064] На фигуре 12 показано сродство немеченого антитела P4B5 по сравнению со сродством Vivotag 680 XL-меченного антитела P4B5 к CB1, экспрессированному на клеточной поверхности.
[0065] На фигуре 13A показаны результаты детекции меченого антитела P4B5 в сердце, легких, печени, почках, желудке, кишечнике и мочевом пузыре в моменты времени 0 ч, 1 ч, 5 ч, 24 ч, 48 ч, 72 ч, 96 ч и 144 ч (13A.1); и детекции меченого антитела P4B5 в головном мозге в моменты времени 0 ч, 1 ч, 5 ч, 24 ч, 48 ч, 72 ч, 96 ч и 144 ч (13A.2). На фигуре 13В показаны результаты детекции меченого антитела в головном мозге, включающей в себя детекцию в ткани и в крови (левая панель), по сравнению с результатами детекции меченого антитела в мозге с вычитанием сигнала, поступающего от крови (т.е. только ткань мозга, правая панель).
[0066] На фигуре 14 показаны профили SEC и результаты анализа SDS-PAGE, полученные для гуманизированных вариантов PA13R3-P1C4, экспрессированных в клетках 293 и CHO-K1. На фигуре 14A показаны профиль SEC (вверху) и результаты анализа методом SDS-PAGE (внизу) для одной из партий 293 FreeStyle. На фигуре 14B показаны профиль SEC (вверху) и результаты анализа методом SDS-PAGE (внизу) для одной из партий CHO-K1.
[0067] На фигуре 15 показаны результаты функциональных анализов гуманизированных вариантов PA13R3-P1C4 по сравнению с исходным химерным PA13R3-P1C4 и mAb P2A12, используемым в качестве отрицательного контроля, не специфичным к GPCR антителом изотипа IgG1, где указанные анализы проводят путем измерения уровня цАМФ.
[0068] На фигуре 16 показано сравнение активности гуманизированных вариантов P1C4-h2-IgG2 и P1C4-h2-IgG4 с активностью римонабанта, AM6545 и используемого в качестве отрицательного контроля антитела P2A12-IgG1 в отношении клеток TRex-CHO CB1, стимулированных 1,5 мкМ форсколином (фигура 16A), а также клеток TRex-CHO CB1, стимулированных 5 мкМ форсколином (фигура 16B).
[0069] На фигуре 17 показано влияние возрастающих концентраций P1C4-h2-IgG4 на CP55,940 (фигура 17A) и WIN55,212 (фигура 17C). Также показаны кривые Шильда для каждого вида обработки (фигуры 17B и 17D).
[0070] На фигурах 18А и 18В показаны результаты анализов активации ERK методом вестерн-блоттинга, в которых измеряют способность гуманизированных вариантов PA13R3-P1C4 блокировать WIN55,212-опосредованную активацию ERK.
[0071] На фигуре 19А показаны результаты исследования интернализации рецептора CB1 методом проточной цитометрии в разных индуцирующих условиях в отсутствии ингибитора (панель А), или в присутствии римонабанта (панель B) или гуманизированных вариантов PA13R3-P1C4 (панели С-G). На фигуре 19B показаны результаты такого же анализа интернализации рецептора CB1, в котором исследуют влияние P1C4-h2, клонированного в разных человеческих каркасных участках Fc IgG2 и IgG4.
[0072] На фигурах 20 A-D показаны результаты анализа методом проточной цитометрии, в котором измеряют связывание гуманизированных вариантов антител PA13R3-P1C4 с клетками TRex-СНО, стабильно трансфицированными тетрациклин-индуцируемым человеческим CB1. На фигурах 20 Е-М показаны результаты анализа селективности связывания и перекрестной реакционноспособности гуманизированных вариантов PA13R3-P1C4 в отношении человеческого CB1 по сравнению с человеческим CB2 и мышиным CB1.
[0073] На фигуре 21 показаны результаты анализа методом проточной цитометрии, в котором измеряют связывание антител (показаны в верхней части) с клетками, экспрессирующими разные конструкции CB1 (показаны слева).
[0074] На фигуре 22 показаны результаты анализа антитело-опосредованной цитотоксичности и комплемент-зависимой цитотоксичности гуманизированных вариантов P1C4, P1C4-h2-IgG2 и P1C4-H2-IgG4, в отношении клеток Дауди. На фигурах 22А-С показаны результаты анализа антитело-опосредованной цитотоксичности вариантов P14C. На фигуре 22D показаны результаты анализа комплемент-зависимой цитотоксичности вариантов P14C.
[0075] На фигуре 23 показаны результаты анализа методом вестерн-блоттинга, в котором определяют распознавание денатурированного белка CB1 указанными антителами P1C4, используемыми в качестве первичных антител, или контрольными антителами, с использованием вторичных антител против человеческих (фигура 23A) или мышиных (фигура 23B) иммуноглобулинов. В качестве отрицательного контроля используют очищенный человеческий IgG со вторичным антителом против человеческих иммуноглобулинов (фигура 23А, линия 1) и первичное мышиное антитело со вторичным антителом против кроличьих иммуноглобулинов (фигура 23B, линия 11).
[0076] На фигуре 24 показаны результаты анализа Fab-фрагментов химерных и гуманизированных антител P1C4, инкубированных с клетками, экспрессирующими рецептор CB1, на способность к связыванию методом проточной цитометрии (фигура 24A) и ингибированию продукции цАМФ (фигура 24B).
[0077] На фигуре 25 показано положительное CB1-специфическое окрашивание макрофагов, гепатоцитов и печеночных миофибробластов в образцах, соответствующих ранней стадии NASH (левая панель), фиброзу, обусловленному NASH (средняя панель), и поздней стадии фиброза (правая панель).
[0078] На фигуре 26 демонстрируется отсутствие окрашивания клеток, полученных из нормального образца (средняя панель), или из образца, соответствующего фиброзу, обусловленному NASH (правая панель), изотипическими нерелевантными антителами.
[0079] На фигуре 27 показано отсутствие CB1-специфического окрашивания в нормальных тканях.
[0080] На фигуре 28 показаны результаты измерения методом ОТ-ПЦТ экспрессии коллагена А1(I) в первичных звездчатых клетках печени, обработанных PBS, нефункциональным контрольным антителом и антителами P1C4-h2.
[0081] На фигуре 29 показаны результаты измерения методом ОТ-ПЦТ уровней экспрессии TGFβ в первичных звездчатых клетках печени, обработанных указанными антителами в разных концентрациях и контрольными соединениями.
[0082] На фигуре 30 показаны результаты измерения методом ОТ-ПЦТ уровней экспрессии TIMP1 в первичных звездчатых клетках печени, обработанных указанными антителами в разных концентрациях и контрольными соединениями.
[0083] На фигуре 31 показаны результаты измерения методом ОТ-ПЦТ уровней экспрессии α-SMA в первичных звездчатых клетках печени, обработанных указанными антителами в разных концентрациях и контрольными соединениями.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0084] В одном аспекте настоящее изобретение предлагает антигенсвязывающие белки, такие как антитела и их антигенсвязывающие фрагменты, способные селективно связываться с человеческим каннабиноидным рецептором 1 (СВ1). Антитела и их фрагменты представляют собой функциональные антитела, обладающие агонистической или антагонистической активностью в отношении рецептора СВ1, или способные селективно распознавать CB1, но не обладающие агонистической или антагонистической активностью.
[0085] В данном описании термин "антитело" относится к связывающим белкам, содержащим, по меньшей мере, один антигенсвязывающий домен, и включает в себя моноклональные антитела, их фрагменты и/или варианты, в том числе рекомбинантные полипептиды, гибридные белки и иммуноконъюгаты. Таким образом, термины "антитело", "фрагмент антитела" и "вариант антитела" используются здесь как взаимозаменяемые. Примеры фрагментов антител настоящего изобретения включают в себя, без ограничения, фрагмент Fab, состоящий из доменов VL, VH, CL и CHI; фрагмент Fc, состоящий из доменов VH и CHI; фрагмент Fv, состоящий из VL и VH; фрагмент dAb, состоящий из домена VH; выделенные CDR; двухвалентный фрагмент F(аb')2, содержащий два связанных фрагмента Fab; и одноцепочечные молекулы Fv (scFv). Описанные здесь антитела, способные связываться с рецептором CB1, можно получить из любых видов, включающих в себя, без ограничения, мышей, крыс, кроликов, приматов, лам и людей. Антитела, связывающие рецептор СВ1, могут представлять собой химерные, гуманизированные или полностью человеческие антитела.
[0086] В данном описании термин "полученный", используемый для обозначения молекулы или полипептида, родственных исходному антителу или другому связывающему белку, относится к молекуле, или полипептиду, которые способны специфически связываться с тем же эпитопом, что и исходное антитело, или другой связывающий белок.
[0087] Если конкретно не указано иначе, применение термина в единственном числе также подразумевает его использование во множественном числе. Если специально не указано иначе, единственное число означает "по меньшей мере один". Если конкретно не указано иначе, слово "или" используется в значении "и/или". Значение фразы "по меньшей мере один" эквивалентно значению фразы "один или несколько". Далее, термин "включающий в себя", а также другие формы, такие как "включает в себя" и "включенный", используется как неограничивающий. Кроме того, если конкретно не указано иначе, такие термины, как "элемент" или "компонент" охватывают как элементы или компоненты, содержащие одну единицу, так и элементы или компоненты, содержащие несколько единиц.
[0088] Описанные здесь антитела и их антигенсвязывающие фрагменты являются специфичными к каннабиноидному рецептору 1 (CB1). Под термином "специфичный к" подразумевают, что антитела и их фрагменты связывают рецептор CB1 с большим сродством (т.е. с более низким значением Kd), чем любую другую мишень. Таким образом, антитела и их фрагменты, которые являются селективными в отношении рецептора CB1, связывают рецептор CB1 с большим сродством (т.е. с более низким значением Kd), чем любой другой каннабиноидный рецептор, или любой другой GPCR, или любую другую мишень. Значение Kd, характеризующее сродство антител и их фрагментов или вариантов к рецептору CB1, может находиться в диапазоне примерно от 0,01 нМ до 500 нМ, примерно от 0,02 нМ до 250 нМ, примерно от 0,02 до 200 нМ, примерно от 0,05 до 100 нМ, примерно от 0,05 до 50 нМ. Значение Kd, характеризующее сродство антител и их фрагментов или вариантов к рецептору CB1, может составлять примерно 500 нМ, примерно 250 нм, примерно 200 нм, примерно 150 нм, примерно 100 нм, примерно 75 нМ, примерно 50 нМ, примерно 25 нМ, примерно 10 нМ, примерно 5 нМ, примерно 1 нМ, примерно 500 пМ, примерно 250 пМ, примерно 100 пМ, примерно 50 пМ или примерно 10 пМ. Значение Kd, характеризующее сродство антител и их фрагментов или вариантов к рецептору CB1, может составлять примерно 100 нМ или менее, примерно 75 нМ или менее, примерно 50 нМ или менее, примерно 10 нМ или менее, примерно 1 нМ или менее, примерно 500 пМ или менее, или примерно 100 пМ или менее.
[0089] В данном описании термин "агонист" относится к соединению, которое усиливает сигнальную активность другого соединения или рецепторного участка.
[0090] В данном описании термин "антагонист" относится к соединению, которое ингибирует, уменьшает или предотвращает сигнальную активность другого соединения в участке рецептора и, в более общем смысле, данный термин относится к соединению, которое уменьшает или предотвращает активацию и/или сигнальную активность рецептора.
[0091] "Аллостерический модулятор" представляет собой соединение, которое косвенно модулирует агонистические эффекты другого соединения. Например, аллостерический модулятор может косвенно модулировать агонистический эффект агониста рецептора путем индукции конформационного изменения в структуре белка. Аллостерические модуляторы могут представлять собой положительные (усиливают агонистический эффект соединения-агониста) или отрицательные (уменьшают эффект соединения-агониста) модуляторы.
[0092] В данном описании термин "лечение" относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. Индивидуум, нуждающийся в лечении, представляет собой индивидуума, который уже имеет заболевание или расстройство, или индивидуума, у которого может развиться заболевание или расстройство, и который нуждается в предотвращении, задержке или ослаблении заболевания или расстройства. Описанные здесь способы "лечения" включают в себя введение индивидууму описанного здесь антитела или его антигенсвязывающего фрагмента, например, индивидууму, страдающему от CB1-ассоциированного заболевания или расстройства (такого как фиброзное заболевание), или предрасположенного к такому заболеванию или расстройству, с целью предотвращения, вылечивания, задержки, уменьшения тяжести, или ослабления одного или нескольких симптомов заболевания или расстройства, или рецидивирующего заболевания или расстройства, или с целью увеличения срока жизни индивидуума по сравнению со сроком жизни, ожидаемым в отсутствии такого лечения. В данном описании термин "индивидуум" относится к млекопитающему, включающему в себя грызунов, кошачьих, собачьих и приматов. В соответствии с настоящим изобретением индивидуум предпочтительно представляет собой человека.
[0093] Термин "терапевтически эффективное количество" в данном описании относится к количеству соединения или композиции, которое требуется для достижения терапевтического и/или профилактического эффекта у индивидуума. Терапевтически эффективное количество варьирует в зависимости от индивидуума и болезненного состояния, подлежащего лечению, массы и возраста индивидуума, тяжести болезненного состояния, способа введения и т.п., и может быть легко определено рядовым специалистом в данной области. Вводимая доза описанного здесь антитела или его антигенсвязывающего фрагмента может варьировать, например, примерно от 1 нг до 10000 мг, примерно от 1 мкг до 5000 мг, примерно от 1 мг до 1000 мг, примерно от 10 мг до 100 мг. Режим дозирования можно регулировать, чтобы достичь оптимальный терапевтический ответ. Эффективное количество также представляет собой количество, которое обеспечивает сведение к минимуму любых токсических или вредных эффектов (то есть побочных эффектов) антитела или его антигенсвязывающего фрагмента, или преобладание положительных эффектов над токсическими или вредными эффектами.
I. Модифицированные антитела против CB1
[0094] В некоторых вариантах осуществления описанные здесь антитела против рецептора CB1 могут содержать одну или несколько модификаций. Описанные здесь модифицированные формы антител против рецептора CB1 можно получить с помощью любых методов, известных в данной области.
[0095] В некоторых вариантах осуществления антитела против рецептора CB1 и их фрагменты представляют собой конъюгаты, дополнительно содержащие средство, выбранное из группы, включающей в себя другое терапевтическое средство, цитотоксическое средство, молекулы иммуноадгезии и визуализирующее средство. В некоторых вариантах осуществления визуализирующее средство выбрано из группы, состоящей из радиоактивной метки, фермента, флуоресцентной метки, люминесцентной метки, биолюминесцентной метки, магнитной метки и биотина. В некоторых вариантах осуществления визуализирующее средство представляет собой радиоактивную метку, выбранную из группы, состоящей из: 3H, 14C, 35S, 64Cu, 89Zr, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho и 153Sm. В некоторых вариантах осуществления терапевтическое средство или цитотоксическое средство выбрано из группы, включающей в себя иммунодепрессант, иммуностимулирующее средство, антиметаболит, алкилирующее средство, антибиотик, фактор роста, цитокин, антиангиогенное средство, антимитотическое средство, антрациклин, токсин и апоптотическое средство.
[0096] В одном варианте осуществления описанное здесь выделенное антитело против рецептора CB1 или его антигенсвязывающий фрагмент конъюгируют с антагонистом CB1. Неограничивающие примеры известных антагонистов CB1 включают в себя римонабант, таранабант, VD60, Isis-414930 Antisense CB1, JD5037, AM6545 и ™38837. В одном варианте осуществления описанное здесь выделенное антитело против рецептора CB1, или его антигенсвязывающий фрагмент, конъюгируют с римонабантом. В одном варианте осуществления выделенное антитело, или его антигенсвязывающий фрагмент, конъюгированные с цитотоксическим средством, представляют собой агонисты рецептора CB1. В другом варианте осуществления выделенное антитело, или его антигенсвязывающий фрагмент, конъюгированные с цитотоксическим средством, представляют собой нейтральные средства, связывающие рецептор CB1, которые обеспечивают интернализацию рецептора.
[0097] В другом аспекте изобретения описанное здесь выделенное антитело против рецептора CB1 или его антигенсвязывающий фрагмент конъюгируют с химиотерапевтическим средством. "Химиотерапевтическое средство" представляет собой химическое соединение, используемое для лечения рака. Примеры химиотерапевтических средств включают в себя алкилирующие средства, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, такие как алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (главным образом, буллатацин и буллатацинон); камптотецин (в том числе синтетический аналог топотекан); бриостатин; каллистатин; CC-1065 (в том числе его синтетические аналоги адозелезин, карзелезин и бизелезин); криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (в том числе его синтетические аналоги KW-2189 и CB1-™1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид мехлорэтаминоксида, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, такие как энедииновые антибиотики (например, каличемицин, в особенности каличемицин гамма 11 и каличемицин омега 11 (см., например, Agnew, Chem Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, в том числе динемицин А; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарциностатиновый хромофор и родственные хромопротеиновые энедииновые антибиотические хромофоры), аклациномизины, актиномицин, отрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицйины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, ADRIAMYCIN® доксорубицин (в том числе морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идаруцибин, марцелломицин, митомицины, такие как митомицин C, микофеноловая кислота, ногаламицин, оливомицины, пепломицины, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолона пропионат, эпитиостанол, мепитиостан, тестолактон; средства, угнетающие функции надпочечников, такие как аминоглютетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; алдофосфамида гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; эллиптиния ацетат; эпотилон; этоглюцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтансиноиды, такие как майтансин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, Oreg.); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2"-трихлортриэтиламин; трихотецены (в особенности, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-C"); циклофосфамид; тиотепа; таксоиды, например, паклитаксел TAXOL® (Bristol-Myers Squibb Oncology, Princeton, N.J.), не содержащий кремофор ABRAXANE™, композиция паклитаксела на основе наночастиц рекомбинантного альбумина (American Pharmaceutical Partners, Schaumberg, Illinois), и доксетаксел TAXOTERE® (Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин GEMZAR®; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платину; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин NAVELBINE®; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселоду; ибандронат; СРТ-11; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин, а также фармацевтически приемлемые соли, кислоты или производные любого из указанных выше соединений. Данное определение также включает в себя ингибиторы протеасом, такие как бортезомиб (велкейд), ингибиторы BCL-2, антагонисты IAP (например, имитаторы Smac/xIAP и ингибиторы cIAP, такие как некоторые пептиды, пиридиновые соединения, такие как (S)-N-{6-бензо[1,3]диоксол-5-ил-1-[5-(4-фтор-бензоил)-пиридин-3-ил-метил]-2-оксо-1,2-дигидропиридин-3-ил}-2-метиламинопропионамид, антисмысловая последовательность xIAP), ингибиторы HDAC (HDACI) и ингибиторы киназ (сорафениб). В одном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент, конъюгированные с цитотоксическим средством, являются агонистами рецептора СВ1.
[0098] В некоторых вариантах осуществления связывающий белок конъюгируют непосредственно со средством. В других вариантах осуществления связывающий белок конъюгируют со средством через линкер. Подходящие линкеры включают в себя, без ограничения, описанные здесь аминокислоты и полипептидные линкеры. Линкеры могут быть расщепляемыми или нерасщепляемыми.
[0100] В некоторых вариантах осуществления антитела и их фрагменты представляют собой биспецифические или бифункциональные антитела. Термин "биспецифические антитела" относится к соединениям, которые содержат антигенсвязывающие участки двух антител в одной молекуле. Таким образом, биспецифическое антитело способно связываться одновременно с двумя разными антигенами. Биспецифическое антитело, как правило, представляет собой искусственное гибридное антитело, содержащее две разные пары тяжелая цепь/легкая цепь и два разных связывающих участка или эпитопа. Биспецифические антитела могут представлять собой моноклональные, предпочтительно человеческие или гуманизированные, антитела, которые обладают специфичностью, по меньшей мере, к двум разным антигенам.
[0100] В одном варианте осуществления биспецифическое антитело и/или его фрагмент является специфичным в отношении CB1 и второго антигена-мишени. В некоторых вариантах осуществления биспецифическое антитело и/или его фрагмент является специфичным в отношении CB1 и TGF-β, 2-AG, PDGF-β, IL-6, анандамида (АЕА) или LOXL-2.
[0101] Описанные здесь антитела и их фрагменты могут связываться с одним или несколькими антигенами-мишенями, выбранными из группы, включающей в себя карбоангидразу IX, альфа-фетопротеин, α-актинин-4, A3, антиген, специфичный для антитела А33, ART-4, B7, Ba 733, BAGE, BrE3-антиген, CA125, CAMEL, CAP-1, CASP-8/m, CCCL19, CCCL21, CD1, CD1a, CD2, CD3, CD4, CD5, CD8, CD11A, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD29, CD30, CD32b, CD33, CD37, CD38, CD40, CD40L, CD45, CD46, CD52, CD54, CD55, CD59, CD64, CD66a-e, CD67, CD70, CD74, CD79a, CD80, CD83, CD95, CD126, CD132, CD133, CD138, CD147, CD154, CDC27, CDK-4/m, CDKN2A, CXCR4, специфический антиген-р рака толстой кишки (CSAp), CEA (CEACAM5), CEACAM1, CEACAM6, с-Met, DAM, EGFR, EGFRvIII, EGP-1, EGP-2, ELF2-М, Ер-САМ, Flt-1, Flt-3, рецептор фолиевой кислоты, антиген G250, GAGE, gp100, GROB, HLA-DR, HM1.24, хорионический гонадотропин человека (ХГЧ) и его субъединицы, HER2/neu, HMGB-1, индуцируемый гипоксией фактор (HIF-1), HSP70-2M, HST-2, Ia, IGF-1R, IFN-γ, IFN-α, IFN-β, IL-2, IL-4R, IL-6R, IL-13R, IL-15R, IL-17R, IL-18R, IL-6, IL-8, IL-12, IL-15, IL-17, IL-18, IL-23, IL-25, инсулиноподобный фактор роста-1 (IGF-1), KC4-антиген, KS-1-антиген, KS1-4, Le-Y, LDR/FUT, фактор ингибирования миграции макрофагов (MIF), MAGE, MAGE-3, MART-1, MART-2, NY-ESO-1, TRAG-3, mCRP, MCP-1, MIP-1A, MIP-1B, MIF, MUC1, MUC2, MUC3, MUC4, MUC5, MUM-1/2, MUM-3, NCA66, NCA95, NCA90, антиген, специфичный для антитела РАМ-4, плацентарный фактор роста, p53, PLAGL2, простатическая кислая фосфатаза, PSA, PRAME, PSMA, PIGF, IGF, IGF-1R, IL-6, RS5, RANTES, T101, SAGE, S100, сурвивин, сурвивин-2B, ТАС, TAG-72, тенасцин, рецепторы TRAIL, TNF-α, антиген Tn, антигены Томсена-Фриденрейха, некротические опухолевые антигены, VEGFR, фибронектин ED-B, WT-1, 17-1A-антиген, факторы комплемента С3, C3a, C3b, C5a, С5, маркер ангиогенеза, bcl-2, bcl-6, Kras, cMET, онкогенный маркер и продукт онкогена (см., например, Sensi et al., Clin Cancer Res 2006, 12:5023-32; Parmiani et al., J Immunol 2007, 178:1975-79; Novellino et al. Cancer Immunol Immunother 2005, 54:187-207).
[0102] Способы получения биспецифических антител хорошо известны. Получение биспецифических антител рекомбинантными методами обычно проводят путем совместной экспрессии двух пар тяжелая цепь/легкая цепь иммуноглобулинов, где две тяжелые цепи обладают разной специфичностью (Milstein et al., Nature 305:537 (1983)). Вследствие случайного выбора тяжелых и легких цепей иммуноглобулинов гибридомы (квадромы) потенциально продуцируют смесь десяти разных молекул антител, из которых только одна имеет правильную биспецифическую структуру. Выделение правильной молекулы обычно проводят методом аффинной хроматографии. Аналогичные способы описаны в WO 93/08829 и в Traunecker et al., EMBO J 10:3655 (1991). Другие способы получения биспецифических антител описаны, например, в Kufer et al., Trends Biotech 22:238-244, 2004.
[0103] Вариабельные домены антитела, обладающие желательной специфичностью связывания, можно гибридизовать с последовательностями константных доменов иммуноглобулина. Предпочтительно проводят гибридизацию с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть шарнирного участка и участки CH2 и CH3. Он может содержать первый константный участок тяжелой цепи (CH1), содержащий участок, необходимый для связывания легкой цепи, присутствующей, по меньшей мере, в одном из гибридов. ДНК, кодирующие гибриды тяжелой цепи иммуноглобулина и, при необходимости, легкую цепь иммуноглобулина, вставляют в разные векторы экспрессии, которые используют для совместной трансформации подходящего организма-хозяина. Более подробное описание способов получения биспецифических антител можно найти, например, в Suresh et al., Meth Enzym 121:210 (1986). Разработан ряд рекомбинантных методов, позволяющих осуществлять эффективную продукцию биспецифических антител, как в виде фрагментов антител (Carter et al. (1995), J. Hematotherapy 4: 463-470; Pluckthun et al. (1997) Immunotechology 3: 83-105; Todorovska et al. (2001) J. Immunol. Methods 248: 47-66), так и в виде полноразмерных IgG (Carter (2001) J. Immunol. Methods 248: 7-15).
[0105] Если не указано иначе, для осуществления настоящего изобретения используют традиционные методы молекулярной биологии, клеточной биологии, биохимии и иммунологии, хорошо известные в данной области и описанные, например, в Methods in Molecular Biology, Humana Press; Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989), Current Protocols in Immunology (J. E. Coligan et al., eds., 1991); Immunobiology (C. A. Janeway and P. Travers, 1997); Антитела (P. Finch, 1997); Антитела: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Моноклональные антитела: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Phage display: a laboratory manual (C. Barbas III et al, Cold Spring Harbor Laboratory Press, 2001); и Using антитела: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999).
[0105] В одном аспекте настоящее изобретение предлагает способы получения описанных здесь антител, а также их фрагментов или вариантов, включающие в себя иммунизацию мышей иммуногеном, обеспечивающим иммунный ответ на рецептор CB1, таким как, например, ДНК рецептора CB1, белок рецептора CB1, или липидный комплекс CB1. В некоторых вариантах осуществления мышей иммунизируют 1, 2, 3, 4, 5 или более раз. В некоторых вариантах осуществления мышей иммунизируют ДНК рецептора CB1 и ответ на рецептор CB1 усиливают путем дополнительной иммунизации ДНК, кодирующей рецептор CB1, и/или очищенным белком рецептора CB1, мембранами, содержащими белок рецептора CB1, или iCAPS рецептора CB1. В некоторых вариантах осуществления клетки иммунизированных мышей используют для получения гибридом и фаговых библиотек.
[0106] В некоторых вариантах осуществления продукцию антител против рецептора CB1 осуществляют путем извлечения В-клеток из иммунизированных мышей и использования их для получения гибридомных клеток, продуцирующих антитела против рецептора CB1. Способы получения гибридом хорошо известны в данной области и включают в себя гибридизацию антитело-продуцирующих клеток с клетками миеломы или другими иммортализованными клетками с получением иммортализованной антитело-продуцирующей гибридомной клеточной линии (см., например, Kohler and Milstein, 1975, Nature, 256:495). Моноклональные антитела, продуцируемые клетками гибридомы, можно выделить из супернатанта с помощью обычных методов очистки иммуноглобулинов, таких как преципитация, хроматография, ультрафильтрация, центрифугирование, гель-электрофорез и/или любой другой способ, известный в данной области. Супернатанты или отдельные моноклональные антитела можно протестировать на связывание с рецептором CB1 путем оценки связывания с мембранами, содержащими рецептор CB1, по сравнению с наивными мембранами. Например, супернатанты или выделенные моноклональные антитела можно протестировать на связывание с рецептором CB1 с помощью метода ELISA.
[0107] В другом аспекте изобретение предлагает способы получения описанных здесь антител, а также их фрагментов или вариантов, включающие в себя применение фаговой библиотеки. Рекомбинантные способы получения антител с использованием технологии фагового дисплея известны в данной области (см., например, Winter et al., Annu. Rev. Immunol. 12:433-455 (1994), McCafferty et al., Nature 348: 552-553 (1990), and Clackson et al. Nature 352:624 (1991)). В некоторых вариантах осуществления из селезенки иммунизированных мышей выделяют множество антител против рецептора CB1 и получают случайную комбинаторную библиотеку вариабельных генов, соответствующих данным антителам. В некоторых вариантах осуществления используют не библиотеку фагового дисплея, полученную из клеток иммунизированных мышей, а библиотеку генов вариабельных доменов тяжелой и легкой цепи, полученных из первичных лимфоцитов крови человека.
[0108] В некоторых вариантах осуществления библиотеку фагов подвергают, по меньшей мере, 3 циклам пэннинга с выявлением фагов, связывающихся с рецептором СВ1, после чего связующие компоненты фагов подвергают скринингу на специфическое связывание с рецептором CB1 методом ELISA. Затем выбирают конкретные связующие компоненты и превращают их в полноразмерные антитела.
[0109] В некоторых вариантах осуществления описанные здесь антитела и их фрагменты представляют собой химерные или гуманизированные антитела. Способы получения химерных и гуманизированных антител хорошо известны в данной области и описаны, например, в Lo, Benny, K. C., editor, in Антитело Engineering: Methods and Protocols, volume 248, Humana Press, New Jersey, 2004.
[0110] "Химерное антитело" представляет собой антитело, содержащее, по меньшей мере, часть вариабельного участка тяжелой цепи и, по меньшей мере, часть вариабельного участка легкой цепи, полученные от одного вида; и, по меньшей мере, часть константного участка, полученную от другого вида. Например, в одном из вариантов осуществления химерное антитело может содержать мышиные вариабельные участки и человеческий константный участок. "Гуманизированное антитело" представляет собой антитело, которое содержит гипервариабельные участки (CDR) нечеловеческого антитела; и каркасные участки, а также константные участки человеческого антитела.
[0111] В данном описании термин "CDR" или "гипервариабельный участок" относится к несмежным антигенсвязывающим участкам, присутствующим в вариабельных участках полипептидов как тяжелой, так и легкой цепей. Эти конкретные участки описаны Kabat et al., J. Biol. Chem. 252, 6609-6616 (1977), Kabat et al., Последовательности of protein of immunological interest. (1991), Chothia et al., J. Mol. Biol. 196:901-917 (1987), и MacCallum et al., J. Mol. Biol. 262:732-745 (1996), где определения включают в себя аминокислотные остатки, которые при сравнении друг с другом образуют перекрывающиеся участки, или подгруппы. Определение Kabat основано на вариабельности последовательности. Уникальная нумерация IMGT V-участков всех IG и TR всех видов опирается на высокую консервативность структуры вариабельного участка (Lefranc, Mp et al., Dev comp. Immunol. 27:55-77, 2003). Нумерация IMGT, установленная после выравнивания более 5000 последовательностей, учитывает и объединяет определения каркасных участков и CDR. Определение Clothia основано на расположении структурных петлеобразных участков. Контактное определение (MacCallum et al.) основано на анализе кристаллических структур комплексов и взаимодействий антитело-антиген. Аминокислотные остатки, которые составляют CDR в соответствии с каждым из вышеуказанных определений, приведены для сравнения. В одном варианте осуществления настоящего изобретения термин "CDR" относится к CDR, определенному по системе Kabat. В другом варианте осуществления настоящего изобретения CDR представляет собой CDR, определенный по системе IMGT.
[0112] CDR, как правило, играют важную роль в распознавании эпитопа и связывании антитела. Тем не менее можно осуществить изменения остатков, составляющих CDR, которые не препятствуют способности антитела распознавать и связывать свой эпитоп. Например, можно осуществить изменения, которые не влияют на распознавание эпитопа, но увеличивают сродство антитела к эпитопу. Было проведено несколько исследований с целью анализа влияния введения одной или нескольких аминокислотных замен в разных положениях последовательности антитела с использованием информации о первичной последовательности антитела, его свойствах, таких как способность к связыванию, и уровне экспрессии (Yang et al., 1995, J Mol Biol 254:392-403; Rader et al., 1998, Proc Natl Acad Sci USA 95:8910-8915; и Vaughan et al., 1998, Nature Biotechnology 16, 535-539).
[0113] Таким образом, эквиваленты представляющего интерес антитела можно получить путем изменения последовательностей генов тяжелых и легких цепей по участкам CDR1, CDR2 или CDR3, или по каркасным участкам, с использованием таких методов, как олигонуклеотид-опосредованный сайт-направленный мутагенез, кассетный мутагенез, ПЦР с внесением ошибок, перетасовка ДНК или применение штаммов E. Coli, содержащих гены-мутаторы (Vaughan et al., 1998, Nat Biotech 16:535-539; and Adey et al., 1996, Chap. 16, pp. 277-291, in Phage Display of Peptides and Proteins, eds. Kay et al., Academic Press). Путем изменения нуклеотидной последовательности первичного антитела можно получать антитела с повышенным сродством (Gram et al., 1992, Proc Natl Acad Sci USA 89:3576-3580; Boder et al., 2000, Proc Natl Acad Sci USA 97:10701-10705; Davies & Riechmann, 1996, Immunotech 2:169-179; Thompson et al., 1996, J Mol Biol 256:77-88; Short et al., 2002, J Biol Chem 277:16365-16370; and Furukawa et al., 2001, J Biol Chem 276:27622-27628).
[0114] Например, описанные здесь антитела против CB1 могут содержать CDR, полученные из одного или нескольких мышиных антител, и человеческие каркасные и константные участки. Таким образом, в одном варианте осуществления описанное здесь гуманизированное антитело связывается с тем же эпитопом на CB1, что и мышиное антителао, из которого получены CDR. В данном описании приведены примеры гуманизированных антител. Другие гуманизированные антитела против CB1, содержащие описанные здесь CDR тяжелых и легких цепей, или их варианты, которые можно получить с использованием любой человеческой каркасной последовательности, также также входят в объем настоящего изобретения. В одном варианте осуществления каркасные последовательности, подходящие для применения в настоящем изобретении, включают в себя каркасные последовательности, структурно подобные описанным здесь каркасным последовательностям. В некоторых вариантах осуществления человеческие каркасные участки выбирают на основе гомологии генов исходного антитела и зародышевых человеческих VH и VK. В некоторых вариантах осуществления выбранные каркасные участки обладают наивысшей степенью гомологии по отношению к генам VH и VK исходного антитела, а также их структуру можно рассчитать с помощью компьютерного моделирования или других способов, так, чтобы сохранить структуру CDR, которые, согласно расчетам, присутствуют в исходном антителе.
[0115] Для улучшения свойств описанных здесь антител можно осуществить дополнительные модификации каркасных участков. Такие дополнительные модификации каркасных участков могут включать в себя химические модификации; точечные мутации, вводимые для уменьшения иммуногенности или удаления эпитопов Т-клеток; или обратные мутации с введением остатка, присутствующего в исходной зародышевой последовательности. В одном варианте осуществления настоящего изобретения гуманизированные антитела и их фрагменты содержат человеческие каркасные участки и привитые на них описанные здесь CDR, без дополнительных модификаций вариабельного участка. Гуманизированные антитела, которые не содержат обратных мутаций в человеческом каркасном участке, в данном описании обозначают H0 (например, P1C4-Н0). В другом варианте осуществления настоящего изобретения гуманизированные антитела и их фрагменты содержат человеческие каркасные участки и привитые на них описанные здесь CDR, где аминокислоту в положении 27 и/или 28 каркасного участка тяжелой цепи 1 подвергают обратной мутации. В следующем варианте осуществления аминокислоту в положении 27 подвергают обратной мутации из глицина (G) на Tyr (Y); а аминокислоту в положении 28 подвергают обратной мутации из Thr (T) на глутаминовую кислоту (Е). Гуманизированные антитела, содержащие такие мутации в положениях 27 и 28, в настоящем документе обозначают "Н2" или "H2 (YE)" (например, P1C4-H2 или P1C4-H2 (YE)). В другом варианте осуществления настоящего изобретения гуманизированные антитела и их фрагменты содержат человеческие каркасные участки и привитые на них описанные здесь CDR, где аминокислоту в положении 27 и/или 28 каркасного участка 1 тяжелой цепи и аминокислоту в положении 60 и/или 61 каркасного участка 3 тяжелой цепи подвергают обратной мутации. В следующем варианте осуществления аминокислоту в положении 27 подвергают обратной мутации из глицина (G) на Tyr (Y); аминокислотув положении 28 подвергают обратной мутации из Thr (T) на глутаминовую кислоту (Е); аминокислоту в положении 60 подвергают обратной мутации из Ala (A) на Asn (N); и аминокислоту в положении 61 подвергают обратной мутации из Gln (Q) на глицин (G). Гуманизированные антитела, содержащие такие мутации в положениях 27, 28, 60 и 61, в настоящем документе обозначают "H4" или "H4 (YENG)" (например, P1C4-H4 или P1C4-H4 (YENG)). В одном варианте осуществления настоящего изобретения антитела и антигенсвязывающие фрагменты содержат модификации в каркасном участке, такие как обратные мутации в легкой цепи. Например, в одном варианте осуществления антитела содержат мутацию в положении 45 и/или 47 каркасного участка 2 легкой цепи. В следующем варианте осуществления аминокислоту в положении 45 подвергают мутации из Arg (R) на Lys (K), а аминокислоту в положении 47 подвергают мутации из лейцина (L) на Trp (W). Настоящее изобретение также охватывает гуманизированные антитела, которые способны связываться с CB1 и содержат модификации в каркасном участке, соответствующие описанным здесь примерам модификаций в применении к любой подходящей каркасной последовательности, а также другие каркасные модификации, способные улучшать свойства антител. Описанные здесь антитела против CB1 и их фрагменты могут относится к изотипу IgG1, IgG2, IgG3 или IgG4, или любому их сочетанию. Термин "изотип" относится к классу антител, кодируемых генами константного участка тяжелой цепи. Кроме того, константный участок тяжелой цепи может быть получен из любого вида, включающего в себя, без ограничения, мышей, крыс, кроликов, хомяков, морских свинок, приматов, лам или людей. Например, в одном варианте осуществления антитела против CB1 и их фрагменты настоящего изобретения включают в себя константный участок Fc человеческого IgG1. В другом варианте осуществления антитела против CB1 и их фрагменты содержат константный участок Fc человеческого IgG2, человеческого IgG4, или гибрида IgG2-IgG4.
II. Эффекторные функции и модификации Fc
[0116] В некоторых вариантах осуществления настоящее изобретение предлагает антитела CB1, содержащие вариантные участки Fc. Fc-участок антитела представляет собой часть антитела, которая связывается с рецепторами Fcγ (FcγR) и молекулами комплемента C1q. Fc-участок играет важную роль в осуществлении эффекторных функций антитела. Термин "эффекторные функции", используемый в данном описании в применении к Fc антитела, относится к таким функциям антитела, как, например, связывание C1q; комплемент-зависимая цитотоксичность (CDC); связывание рецептора Fc; антитело-зависимая клеточно-опосредованная цитотоксичность (ADCC); фагоцитоз; опсонизация; трансцитоз; и понижающая регуляция рецепторов клеточной поверхности (например, В-клеточного рецептора). Такие эффекторные функции, для осуществления которых обычно требуется сочетание Fc-участка со связывающим доменом (например, с вариабельным доменом антитела), можно оценить с помощью разных методов, традиционно используемых в данной области для анализа указанных эффекторных функций антитела. Вариантные Fc-участки представляют собой Fc-участки, которые содержат изменения, оказывающие влияние на эффекторные функции. В некоторых вариантах осуществления описанные здесь антитела против CB1 содержат модификации Fc-участка, которые уменьшают, ухудшают или устраняют одну или несколько эффекторных функций. Например, в одном варианте осуществления описанные здесь антитела и их фрагменты обладают способностью связывать CB1 при пониженной, ухудшенной или отсутствующей способности к связыванию C1q и/или CDC и/или ADCC. Модификации Fc могут включать в себя вставки, делеции или замены аминокислот, а также химические модификации. Например, модификации Fc-участка можно осуществлять с целью увеличения или уменьшения связывания комплемента; с целью увеличения или уменьшения антитело-зависимой клеточно-опосредованной цитотоксичности; или с целью изменения характера гликозилирования. В данной области известны разные модификации Fc, которые описаны, например, в Labrijin et al., Nature Biotech 27(8):767-71 (2009); Idusogie, et al. J Immunol 2000; Greenwood et al Eur J Immunol 23:1098-104 (1993); Mueller et al. Mol Immunol 1997; 34:441-52; и Rother et al Nature Biotechnol 2007; 25:1256-64. Чтобы изменить эффекторные функции описанных здесь примеров антител против СВ1, можно использовать любые из известных в данной области модификаций Fc. Кроме того, разработаны разные терапевтические антитела, содержащие модификации Fc-участка, влияющие на эффекторные функции. Такие терапевтические антитела известны в данной области и включают в себя, например, алемтузумаб, бенрализумаб, бевацизумаб, бимекизумаб, кантузумаб, кодритузумаб, далотузумаб, эфализумаб, элотузумаб, энаватузумаб, энокизумаб, этролизумаб, фарлетузумаб, фиклатузумаб, имгатузумаб, итолизумаб, лифастузумаб, лигелизумаб, лоделцизумаб, лорвотузумаб, могамулизумаб, мотавизумаб, обинутузумаб, окаратузумаб, омализумаб, парсатузумаб, патексизумаб, перакизумаб, пертузумаб, пидилизумаб, квилизумаб, ронтализумаб, софитузумаб, соланезумаб, сувизумаб, теплизумаб, тилдракизумаб, тоцилизумаб, трастузумаб, трастузумаба эмтансин, трегализумаб, ведолизумаб, ворсетузумаб, ворсетузумаба мафодотин, иттрий (90Y)-кливатузумаба тетраксетан, анрукинзумаб, дацетузумаб, даклизумаб, этарацизумаб, милатузумаб, озанезумаб, пинатузумаба ведотин, полатузумаба ведотин, тигатузумаб, велтузумаб, абитузумаб, бокоцизумаб, демцизумаб, гевокизумаб, понезумаб, ралпанцизумаб, ромосозумаб, танезумаб, блозозумаб, концизумаб, кренезумаб, ибализумаб, иксекизумаб, лебрикизумаб, олокизумаб, пембролизумаб, симтузумаб, улокаплумаб, вателизумаб и самализумаб (см., например, SEQ ID NO: 358-432). Чтобы изменить эффекторные функции, период полужизни или другие свойства описанных здесь антител против рецептора СВ1, можно использовать любые известные в данной области модификации Fc.
[0117] В одном варианте осуществления антитело против CB1 выполняет эффекторную функцию на пониженном уровне. В следующем варианте осуществления антитело против СВ1 содержит Fc-участок IgG4, несущий мутацию в положении 228. В следующем варианте осуществления в положении 228 аминокислоту серин (S) заменяют на пролин (P) (т.е. S228P). В другом варианте осуществления антитело против CB1 выполняет эффекторную функцию на пониженном уровне и содержит Fc-участок IgG2, несущий мутацию в положении 330 и/или 331. В следующем варианте осуществления в положении 330 аминокислоту аланин (А) заменяют на серин (S), и/или в положении 331 аминокислоту пролин (Р) заменяют на серин (S). В следующем варианте осуществления антитело против CB1 содержит домен Fc IgG2, несущий обе мутации A330S и P331S. В другом варианте осуществления антитело против CB1 содержит гибридный Fc-участок IgG2/IgG4. Например, в одном варианте осуществления антитело CB1 содержит участок CH1 и шарнирный участок, полученные из IgG2, и участки СН2 и СН3, полученные из IgG4.
[0118] Конформационная антигенпрезентирующая система (iCAPS). iCAPS позволяет осуществлять конформационно правильную презентацию очищенных, выделенных, функциональных GPCR. Очищенные GPCR являются стабильными в липидных бислоях, окруженных белковым поясом.
[0119] В одном варианте осуществления настоящее изобретение предлагает выделенную нуклеиновую кислоту, кодирующую одно из описанных здесь антител, или его антигенсвязывающий фрагмент или вариант. В некоторых вариантах осуществления изобретение предлагает вектор, содержащий выделенную нуклеиновую кислоту. В некоторых вариантах осуществления изобретение предлагает клетку-хозяина, трансформированную указанным вектором. В некоторых вариантах осуществления клетка-хозяин представляет собой прокариотическую клетку. В других вариантах осуществления клетка-хозяин представляет собой клетку Escherichia coli. В некоторых вариантах осуществления клетка-хозяин представляет собой эукариотическую клетку. В других вариантах осуществления эукариотическая клетка выбрана из группы, состоящей из клетки протиста, клетки животного, клетки растения и клетки грибка. В некоторых вариантах осуществления клетка-хозяин представляет собой клетку млекопитающего, включающую в себя, без ограничения, 293, COS, NSO и CHO; или клету грибка, такого как Saccharomyces cerevisiae; или клетку насекомого, такую как Sf9. В одном варианте осуществления изобретение предлагает способы получения описанных здесь антител, а также их фрагментов или вариантов, включающие в себя культивирование одной из указанных в данном документе клеток-хозяев в культуральной среде в условиях, обеспечивающих продукцию связывающего белка.
[0120] Примеры рецептор CB1-связывающих антител настоящего изобретения приведены ниже в таблице 1. Другие типичные рецептор CB1-связывающие антитела настоящего изобретения, определяемые по SEQ ID NO, приведены в перечне последовательностей, показанном в таблице 2. Последовательности типичных рецептор CB1-связывающих антител настоящего изобретения приведены в таблице 3.
Таблица 1. Нуклеотидные и аминокислотные последовательности вариабельных участков тяжелой цепи и вариабельных участков легкой цепи типичных рецептор CB1-связывающих антител
тельности
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
тельность константного участка LC
ная последова
тельность константного участка LC
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
ная последова
тельность константного участка LC
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
тельность константного участка LC
ная последова
тельность константного участка LC
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
ная последова
тельность константного участка LC
ная последова
тельность константного участка LC
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
тельность константного участка LC
ная последова
тельность константного участка LC
(химер
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
тельность константного участка LC
ная последова
тельность константного участка LC
(химер
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
тельность константного участка LC
ная последова
тельность константного участка LC
(химер
ная)
тельность вариабель
ного участка тяжелой цепи (HC)
ная последова
тельность вариабель
ного участка HC
тельность константного участка HC
ная последова
тельность константного участка HC
тельность вариабель
ного участка легкой цепи (LC)
ная последова
тельность вариабель
ного участка LC
тельность константного участка LC
ная последова
тельность константного участка LC
Таблица 2. SEQ ID NO других рецептор CB1-связывающих антител
Таблица 3. Последовательности типичных гуманизированных антител
[0121] В некоторых вариантах осуществления предлагаемые в данном описании антитела против рецептора CB1 содержат описанные здесь последовательности или их консервативные варианты. В данном описании термин "консервативные варианты" включает в себя консервативные замены, вставки или делеции аминокислот. Специалистам в данной области известно, что консервативная аминокислотная замена представляет собой замену одной аминокислоты на другую аминокислоту, обладающую подобными структурными или химическими свойствами, например, на аминокислоту, содержащую подобную боковую цепь; и что консервативные замены, вставки или делеции аминокислот приводят к получению последовательности, которая сохраняет билогическую активность исходной последовательности. Примеры консервативных замен описаны в данной области, например, в Watson et al., Molecular Biology of the Gene, The Bengamin/Cummings Publication Company, 4th Ed. (1987).
III. Способы лечения CB1-ассоциированных расстройств
[0122] Описанные здесь антитела и их антигенсвязывающие фрагменты можно вводить человеку в терапевтических целях. В некоторых вариантах осуществления способы лечения включают в себя введение индивидууму описанных здесь антител и их связывающих фрагментов или вариантов.
[0123] В некоторых вариантах осуществления изобретение предлагает способы лечения заболеваний, направленные предпочтительно на периферические рецепторы CB1. "Периферические рецепторы CB1" в соответствии с данным описанием представляют собой рецепторы CB1, локализованные не в мозге, или не в центральной нервной системе (например, рецепторы CB1, находящиеся в пеферической системе). Тогда как термин "общие рецепторы CB1" относится к рецепторам CB1, находящимся в любом участке организма, в том числе, в мозге или в ЦНС.
[0124] В одном варианте осуществления выделенные антитела и их антигенсвязывающие фрагменты можно использовать для лечения разных заболеваний или расстройств, таких как, например, ожирение, диабет, дислипидемия, фиброз, неалкогольный стеатогепатит (NASH), заболевания печени, первичный билиарный цирроз, сердечно-сосудистые заболевания, рак, боль, спастичность при рассеянном склерозе (MS), глаукома, воспалительные заболевания, нефропатии, остеопороз, метаболические расстройства, психиатрические расстройства, неврологические расстройства, нейродегенеративные расстройства, репродуктивные расстройства, заболевания почек, фиброз почек, хронические заболевания почек, атеросклероз, рак, заболевания кожи и др.
[0125] Показано, что сигнальный путь рецептора CB1 обуславливает вредную активность, например, при ожирении, диабете, фиброзе, болезнях печени, сердечно-сосудистых заболеваниях и раке (Kunos et al., 2009, Trends Pharmacol Sci 30:1-7). В одном аспекте описанные здесь антитела против CB1, или их фрагменты, используют для противодействия активности CB1. Соответственно, в другом аспекте изобретение предлагает способы лечения CB1-ассоциированных заболеваний или расстройств путем введения индивидууму, нуждающемуся в этом, фармацевтической композиции, содержащей одно или несколько из описанных здесь антител против CB1, или их антигенсвязывающих фрагментов. В некоторых вариантах осуществления описанные здесь антагонистические антитела против рецептора CB1 и их фрагменты обеспечивают благоприятный эффект при использовании с целью лечения или профилактики ожирения, диабета, фиброза, болезней печени, сердечно-сосудистых заболеваний, зависимостей, таких как никотиновая зависимость, или рака.
[0126] Неалкогольный стеатогепатит (NASH), также известный как неалкогольная жировая болезнь печени (NAFLD), характеризуется нарастанием жировой дегенерации печени, не связанной с избыточным потреблением алкоголя. NASH представляет собой болезнь печени, характеризующуюся воспалением печени и одновременным накоплением жира. NASH, который часто присутствует у людей, страдающих от диабета и ожирения, связан с метаболическим синдромом. NASH представляет собой прогрессирующую форму относительно доброкачественной неалкогольной жировой болезни печени, которая может медленно ухудшаться, вызывая нарастание фиброза в печени, который, в свою очередь, может привести к развитию цирроза (обзор см. в Smith, et al., 2011, Crit Rev Clin Lab Sci., 48(3):97-113). В настоящее время не существует конкретных способов лечения NASH.
[0127] В одном аспекте описанные здесь антитела против CB1 или их фрагменты используют для лечения, профилактики, диагностики или исследования фиброза. Некоторые исследования на мышиных моделях подтверждают участие рецептора CB1 в развитии фиброза, включающего в себя фиброз печени. (См., например, Wei et al., 2014, Exp. Biol. Med. 239(2):183-192; Tam et al., 2010, J. Clin. Invest. 120(8):2953-66; Wan et al., 2014, Cell Metabolism, 19(6):900-1; Takano et al., 2014, Synapse, 68:89-97). Периферические CB1 участвуют в некоторых механизмах, вносящих вклад в развитие NASH и фиброза печени, в том числе стеатоза (жировое перерождение печени), воспаления и повреждения печени (обзор: Mallat et al., 2013, J Hepatology, 59(4):891-896). Показано, что CB1 подвергается повышащей регуляции в активированных звездчатых клетках печени человека (HSC), которые опосредуют фиброз путем трансформации в миофибробласты. (Teixeira-Clerc et al., 2006, Nature Med., 12(6):671-76). CB1 также участвует в диабетической нефропатии. Lin et al., 2014 J. Mol. Med. 92(7):779-92).
[0128] Исследования на мышах с гепатоцит-специфическим и общим нокаутом CB1 подразумевают важную роль CB1 в периферических клетках (гепатоцитах), связанных с некоторыми метаболическими заболеваниями и расстройствами. У мышиной модели индуцированного диетой ожирения как общий нокаут CB1 (CB1-/-), так и гепатоцит-специфический нокаут CB1 (LCB1-/-) приводят к уменьшению стеатоза (жирового перерождения печени) и усилению функции печени, демонстрируя участие CB1, присутствующего в периферических клетках (гепатоцитах), в развитии болезненных патологий, связанных с нелкогольным стеатогепатитом (NASH), диабетом и метаболическим синдромом. (Osei-Hyiaman et al., 2008, J. Clin. Invest., 118(9):3160-3169; Liu et al., 2012, Gastroenterology, 142:1218-1228). Селективный нокдаун CB1 с использованием миРНК (CB1R-GeRPs), обеспечивающей макрофаг-специфический нокдаун CB1, предотвращает прогрессирующую гипергликемию и снижение уровней инсулина и C-пептида в плазме диабетических тучных крыс Цукера (ZDF), которые представляют собой распространенную модель устойчивости к инсулину T2D, гипергликемии и недостаточности бета-клеток (Jourdan et al., 2013, Nature Med. 19(9):1132-1140). У мышиной модели индуцированного алкоголем стеатоза печени как общий нокаут CB1 (CB1-/-), так и гепатоцит-специфический нокаут CB1 (LCB1-/-) приводят к уменьшению стеатоза и усилению функции печени, демонстрируя участие CB1, присутствующего в периферических клетках (гепатоцитах), в развитии болезненной патологии, связанной со стеатозом. (Jeong, et al., 2008, Cell Metabolism, 7:227-235). Показано, что в линиях эпидидимальных белых жировых клеток, полученных от CB1-нокаутных мышей, накопление липидов ниже, чем в контрольных клетках дикого типа (Wagner et al., 2011, Nutrition and Diabetes, 1:e16).
[0129] Исследования, проводимые на разных мышиных моделях заболевания, демонстрируют, что низкомолекулярные антагонисты, действие которых ограничено периферическими рецепторами CB1, могут эффективно подавлять развитие фиброза печени. (См., например, Wei et al., 2014, Exp. Biol. Med. 239(2):183-192; Tam et al., 2010, J. Clin. Invest. 120(8):2953-66; Wan et al., 2014, Cell Metabolism, 19(6):900-1; Takano et al., 2014, Synapse, 68:89-97). Неограничивающие примеры известных антагонистов CB1 включают в себя римонабант, таранабант, VD60, антисмыловую последовательность CB1 Isis-414930, JD5037, AM6545 и ™38837. Показано, что такие антагонисты CB1, как римонабант, ингибируют пролиферацию клеток и уменьшают экспрессию профибротического гена в первичных звездчатых клетках печени человека (HSC), которые опосредуют фиброз путем трансформации в миофибробласты (Patsenker et al., 2011, Mol Med.,17(11-12):1285-1294). На мышиной модели CCl4-индуцированного фиброза печени показано, что антагонист CB1 VD60 (3,4,22-3-деметоксикарбонил-3-гидроксилметил-4-дезацетилвиндолина 3,4-тионокарбонат) ингибирует экспрессию профибротического гена (альфа-коллагена) в активированных звездчатых клетках печени (линия HSC LX-2) и пролиферацию данных клеток, тогда как селективный агонист CB1 ACEA (N-(2-хлорэтил)-5Z,8Z,11Z,14Z-эйкозатетраенамид) предотвращает данный эффект (Wei Y. et al., 2014, Exp. Biol. Med. 239(2):183-192). Показано, что антагонист CB1 JD5037 отменяет эндоканнабиноид-индуцированное ингибирование сигнального пути инсулина. (Cinar et al., 2014, Hepatology, 59(1)143-153). Блокада CB1 под действием римонабанта приводит к обратному развитию индуцированного воспалением нарушения поглощения глюкозы адипоцитами, выделенными из крыс, получавших рацион с высоким содержанием жиров (Miranville et al, 2010, Obesity 18: 2247-2254).
[0130] Исследования на людях также демонстрируют связь периферических рецепторов CB1 с появлением и развитием заболевания. Например, повышающая регуляция CB1 в печени пациентов с NASH и HCV коррелирует с тяжестью стеатоза и фиброза печени. (Auguet et al., 2014, BioMed Res. Intl. Vol. 2014, Article ID 502542). Кроме того, хронический агонизм CB1 (в результате употребления каннабиса) коррелирует с повышенной тяжестью стеатоза и фиброза печени у пациентов с HCV. (Van der Poorten et al., 2010, PlosOne 5, e12841). Кроме того, показано, что блокада CB1 у пациентов, страдающих от ожирения, уменьшает стеатоз печени. (Despres et al., 2009, Arterioscler Thromb Vasc Biol. 29:416-423).
[0131] Известно, что антагонисты CB1 оказывают разное влияние в зависимости от местонахождения рецептора. Например, можно сделать вывод, что эффекты антагонистов CB1 являются тканеспецифичными, поскольку наблюдающиеся у пациентов благоприятные кардиометаболические эффекты римонабанта не зависят от потери массы. (Pi-Sunyeret al, 2006, J Am Coll Cardio. 147: 362A). Кроме того, известно, что римонабант улучшает гликемический контроль у пациентов с диабетом типа 2 (см., например, Hollander et al., 2010, Diabetes Care. 33(3):605-7), однако данный эффект сопровождается значительными психиатрическими побочными эффектами, обусловленными рецепторами CB1, расположенными в ЦНС. (Kunos et al, 2009, Trends Pharmacol Sci 30:1-7; Moreira et al., 2009, Rev Bras Psiquiatr. 31(2):145-53; Pacher et al, 2013, FEBS J. 280(9): 1918-1943). Результаты исследования Rimonabant in Obesity-Lipids (RIO-Lipids) свидетельствуют о том, что антагонист рецептора CB1 римонабант улучшает профиль некоторых метаболических факторов риска (включающих в себя уровень адипонектина) у пациентов с избыточной массой. (См., например, Després et al., 2005, N Engl J Med, 353:2121-2134).
[0132] Показано, что сигнальный путь рецептора CB1 обуславливает благоприятную активность при боли, спастичности MS, глаукоме и других состояниях. (Pacher et al., 2013, FEBS J. 280(9):1918-1943). В некоторых вариантах осуществления описанные здесь агонистические антитела против рецептора CB1 и их фрагменты оказывают благоприятный эффект при применении для лечения или профилактики боли, спастичности MS и глаукомы. Показано, что агонисты CB1 активируют синтез жирных кислот в печени, глюконеогенез и другие метаболические пути. (См., например, Osei-Hyiaman et al, 2005, J. Clin. Invest., 115(5):1298-1305; Chanda et al., 2012, JBC, 287(45):38041-38049).
[0133] Спастичность при рассеянном склерозе (MS) включает в себя ощущение скованности и широкий диапазон непроизвольных мышечных спазмов (длительные мышечные сокращения или внезапные движения). Спастичность представляет собой один из распространенных симптомов MS, тяжесть которого может варьировать от небольшой скованности до болезненных, неконтролируемых судорог конечностей. В отсутствии лечения спастичность может приводить к серьезным осложнениям, включающим в себя контрактуры (сросшиеся или фиксированные суставы) и пролежни. Существующие в настоящее время средства для лечения спастичности MS включают в себя, в числе прочих, баклофен, тизанидин, диазепам, дантролен, фенол. Показано, что рецепторы CB1 участвуют в регуляции спастичности у мышиной модели MS. (Pryce et al., 2007, Br J Pharmacol. 150(4): 519-525).
[0134] Активация рецепторов CB1 вызывает обезболивающие эффекты при некоторых экспериментальных моделях боли, включающих в себя висцеральную боль, возникающую в желудочнокишечном тракте. Показано, что агонисты CB1, такие как WIN55,212-2 и SAB-378, ингибируют связанные с болью ответы на повторяющееся воздействие болевых раздражителей (Brusberg et al., 2009, J. Neuroscience, 29(5):1554-1564; Talwar et al., 2011, CNS Neurol Disord Drug Targets. 10(5):536-44).
[0135] Специалист в данной области может путем рутинного экспериментирования определить эффективное нетоксичное количество антитела (или другого терапевтического средства), необходимое для лечения CB1-ассоциированного заболевания или растройства. Например, терапевтически активное количество полипептида может варьировать в зависимости от таких факторов, как стадия заболевания (например, стадия I или стадия IV), возраст, пол, медицинские осложнения (например, иммуносупрессивные состояния или заболевания) и масса индивидуума, а также способность антитела вызывать желательный ответ у индивидуума. Дозировочный режим можно корректировать, чтобы достичь оптимального терапевтического ответа. Например, ежедневно вводимую дозу можно разделить на несколько приемов, или пропорционально уменьшить, в зависимости от терапевтической ситуации. Однако в большинстве случаев эффективная доза находится в диапазоне примерно от 0,05 до 100 миллиграмм на килограмм массы тела в день, в одном варианте осуществления примерно от 0,5 до 10 миллиграмм на килограмм массы тела в день.
[0136] В некоторых вариантах осуществления описанные здесь антитела против CB1 и их фрагменты используют в способах сочетанной терапии, где человеческие антитела вводят индивидууму в сочетании с другим терапевтическим средством, например, с одним или несколькими другими антителами, которые связываются с другими мишенями (такими как антитела, которые связываются с другими цитокинами или с молекулами клеточной поверхности). Поскольку наличие альтернативных сигнальных путей может приводить к отсутствию ответа на одно антитело, изобретение предлагает разные стратегии сочетанной терапии, включающие в себя применение антител, связывающихся с разными эпитопами или с разными антигенами на одной клетке-мишени. Проведены доклинические испытания таких сочетаний, как антитело против CD20 и антитело против CD22 (Stein et al., Clin Cancer Res 2004, 10:2868-2878), антитело против CD20 и антитело против HLA-DR (Tobin et al., Leuk Lymphoma 2007, 48:944-956), антитело против CD20 и антитело против TRAIL-R1 (Maddipatla et al., Clin Cancer Res 2007, 13:4556-4564), антитело против IGF-1R и антитело против EGFR (Goetsche et al., Int J Cancer 2005, 113:316-328), антитело против IGF-1R и антитело против VEGF (Shang et al., Mol Cancer Ther 2008, 7:2599-2608), или трастузумаб и пертузумаб, направленные на разные участки человеческого EGFR2 (Nahta et al., Cancer Res 2004, 64:2343-2346), демонстрирующие повышенную или синергическую противоопухолевую активность in vitro и in vivo. Преимуществом таких сочетанных способов лечения является возможность введения более низких доз терапевтических средств, позволяющая избежать токсические эффекты или осложнения, которые могут возникать при использовании разных монотерапий.
[0137] Описанные здесь антитела и фрагменты можно вводить в сочетании с любым желательным терапевтическим средством. В некоторых вариантах осуществления в сочетании с описанными здесь антителами и фрагментами можно вводить, например, антитело против LOXL2, антитело против TGFβ, нинтеданиб, ингибитор тирозинкиназы, агонист PPAR, агонист фарнезоидного X-рецептора (FXR), агонист рецептора глюкагоноподобного пептида 1, или ингибитор каспазы.
IV. Фармацевтические композиции
[0138] В другом аспекте изобретение предлагает фармацевтические композиции, содержащие антитело против CB1, или его фрагмент.
[0139] Способы получения и введения индивидууму описанных здесь антител, или их фрагментов, хорошо известны специалистам в данной области, или могут быть легко определены специалистами в данной области. Способ введения описанных здесь антител, или их фрагментов, может представлять собой пероральное введение, парентеральное введение, введение путем ингаляции или местное введение. Термин парентеральное введение в соответствии с данным описанием включает в себя внутривенное, внутриартериальное, внутрибрюшинное, внутримышечное, подкожное, ректальное или вагинальное введение. В некоторых вариантах осуществления используют внутривенный, внутриартериальный, подкожный и внутримышечный способы парентерального введения. В то время, как однозначно подразумевается, что все указанные способы введения входят в объем настоящего изобретения, форма для введения может представлять собой раствор для инъекции, например, раствор для внутривенной или внутриартериальной инъекции, или раствор для капельного вливания. Как правило, подходящая для инъекции фармацевтическая композиция содержит буфер (такой как ацетатный, фосфатный или цитратный буфер), поверхностно-активное вещество (такое как полисорбат), необязательно стабилизирующее средство (такое как человеческий альбумин), и др. Однако в других способах, подходящих для применения в настоящем изобретении, полипептиды можно доставлять непосредственно к участку аберрантной клеточной популяции, повышая уровень воздействия терапевтического средства на целевую ткань.
[0140] Препараты для парентерального введения включают в себя стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и подходящие для введения органические сложные эфиры, такие как этилолеат. Водные носители включают в себя воду, спиртовые/водные растворы, эмульсии или суспензии, в том числе солевые и забеференные среды. Фармацевтически приемлемые носители, подходящие для применения в настоящем изобретении, включают в себя, без ограничения, 0,01-0,1M (например, 0,05M) фосфатный буфер, или 0,8% раствор хлорида натрия. Другие среды, обычно используемые для парентерального введения, включают в себя растворы фосфата натрия, растворы декстрозы в растворе Рингера, растворы декстрозы и хлорида натрия, Рингер-лактат или нелетучие масла. Среды для внутривенного введения включают в себя жидкие и питательные компенсаторные добавки, компенсаторы электролитов, например, на основе раствора Рингера-декстрозы, и т.п. В состав композиций также могут входить консерванты и другие добавки, такие как, например, противомикробные средства, антиоксиданты, хелатирующие средства, инертные газы и т.п. Более конкретно, фармацевтические композиции, подходящие для инъекций, включают в себя стерильные водные растворы (в случае водорастворимых средств) или дисперсии, а также стерильные порошки для приготовления стерильных растворов или дисперсий для инъекций непосредственно перед введением. В таких случаях композиция должна быть стерильной и обладать достаточной текучестью, чтобы легко проходить через шприц. Она должна быть стабильной в условиях производства и хранения и, в одном варианте осуществления, она должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибки. Носитель может представлять собой растворитель или среду для дисперсии, и может содержать, например, воду, этанол, полиол (такой как глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.), и их подходящие смеси. Подходящую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, частиц нужного размера в случае дисперсии и поверхностно-активных веществ. Действие микроорганизмов можно предотвратить путем применения разных противобактериальных и противогрибковых средств, таких как парабены, хлорбутанол, фенол, аскорбиновая кислота, тимеросал и т.п. В некоторых вариантах осуществления в состав композиции вводят изотонические средства, такие как сахара, полиспирты, например, маннит, сорбит, или хлорид натрия. Пролонгированную абсорбцию инъекционных композиций можно достичь путем включения в состав композиции средства, замедляющего абсорбцию, такого как моностеарат алюмия и желатин.
[0141] В любом случае стерильные растворы для инъекций можно получить путем введения активного соединения (например, антитела, отдельно или в сочетании с другими активными средствами) в нужном количестве в подходящий растворитель, при необходимости вместе с одним или несколькими из перечисленных здесь ингредиентов, с последующей стерилизацией фильтрованием. Дисперсии обычно получают путем введения активного соединения в стерильную среду, которая содержит диспергирующую основу и другие необходимые ингредиенты из перечисленных выше. Стерильные порошки для приготовления стерильных инъекционных растворов можно получить путем вакуумной сушки и лиофилизации, причем полученный порошок может содержать активный ингредиент и дополнительный желательный ингредиент, присутствующий в полученном ранее стерилизованном фильтрацией растворе. Препараты для инъекций обрабатывают и помещают в контейнеры, такие как ампулы, мешки, бутылки, шприцы или флаконы, и герметично закрывают в асептических условиях с помощью известных в данной области методов. Кроме того, препараты можно упаковывать и продавать в виде наборов, таких как описанные в одновременно рассматриваемых патентах США № 09/259337 и № 09/259338, каждый из которых включен в настоящее описание в качестве ссылки. Такие изделия промышленного производства в одном варианте осуществления содержат этикетки или вкладыши в упаковку, указывающие, что предлагаемые композиции можно использовать для лечения индивидуума, страдающего от аутоиммунного или неопластического расстройства, или предрасположенного к такому расстройству.
[0142] Дозы описанных здесь стабилизированных антител, или их фрагментов, эффективные для лечения вышеуказанных состояний, варьируют в зависимости от многих разных факторов, включающих в себя способ введения, участок-мишень, физиологическое состояние пациента, независимо от того, является ли пациент человеком или животным, применение других лекарственных средств, а также от того, с какой целью, профилактической или терапевтической, проводится лечение. Как правило, пациент представляет собой человека, однако с помощью указанных способов также можно лечить и отличных от человека млекопитающих, в том числе, трансгенных млекопитающих. Используемые для лечения дозы можно титровать с помощью рутинных методов, известных специалистам в данной области, с целью оптимизации безопасности и эффективности.
[0143] Используемая для пассивной иммунизации доза описанного здесь антитела может варьировать, например, примерно от 0,0001 до 100 мг/кг, более предпочтительно от 0,01 до 5 мг/кг (например, она может составлять 0,02 мг/кг, 0,25 мг/кг, 0,5 мг/кг, 0,75 мг/кг, 1 мг/кг, 2 мг/кг и т.д.) массы тела хозяина. Например, доза может составлять 1 мг/кг массы тела или 10 мг/кг массы тела, или она может находиться в диапазоне 1-10 мг/кг, или в конкретных вариантах осуществления она может составлять, по меньшей мере, 1 мг/кг. Дозы, соответствующие промежуточным значениям указанных здесь диапазонов, также входят в объем настоящего описания.
[0144] Такие дозы можно вводить индивидуумам ежедневно, через день, еженедельно или по любой другой схеме, установленной на основании анализа опытных данных. Один из примеров схемы лечения включает в себя введение нескольких доз в течение длительного периода, например, в течение, по меньшей мере, шести месяцев. Другие примеры схемы лечения включают в себя введение один раз каждые две недели, или один раз в месяц, или один раз каждые 3-6 месяцев. Примеры режимов дозирования включают в себя введение 1-10 мг/кг или 15 мг/кг каждый день, 30 мг/кг через день или 60 мг/кг еженедельно. В некоторых способах одновременно вводят два или более моноклональных антител, обладающих разной специфичностью связывания, причем доза каждого вводимого антитела может находиться в одном из указанных диапазонов.
[0145] Описанные здесь антитела или их фрагменты можно вводить многократно. Интервалы между введением отдельных доз могут составлять, например, один день, неделю, месяц или год. Интервалы также могут быть неодинаковыми и могут определяться результатами измерения уровня полипептида или молекулы-мишени в крови пациента. В некоторых способах дозу корректируют, чтобы достичь определенной концентрации антитела или токсина в плазме, такой как 1-1000 мкг/мл или 25-300 мкг/мл. Альтернативно антитела или их фрагменты можно вводить в составе композиции, обеспечивающей замедленное высвобождение, при этом введение можно осуществлять с меньшей частотой. Доза и частота введения варьируют в зависимости от периода полужизни антитела в организме пациента. Как правило, наиболее продолжительным периодом полужизни обладают гуманизированные антитела, после них следуют химерные антитела и нечеловеческие антитела. В одном варианте осуществления описанные здесь антитела или их фрагменты можно вводить в неконъюгированном виде. В другом варианте осуществления описанные здесь антитела можно вводить несколько раз в конъюгированном виде. В следующем варианте осуществления описанные здесь антитела, или их фрагменты можно вводить в неконъюгированном виде, затем в конъюгированном виде, или наоборот.
[0146] Доза и частота введения могут варьировать в зависимости от того, является ли лечение профилактическим или терапевтическим. При профилактическом применении композиции, содержащие антитела настоящего изобретения или их смеси, вводят пациенту, еще не находящемуся в болезненном состоянии, чтобы повысить устойчивость пациента. Используемое для этого количество определяют как "профилактически эффективная доза". При таком применении точное количество, опять же, зависит от состояния здоровья пациента и его иммунной системы в целом, однако обычно оно варьирует от 0,1 до 25 мг на дозу, предпочтительно от 0,5 до 2,5 мг на дозу. Относительно низкие дозы вводят с относительно маленькими интервалами в течение длительного периода времени. Некоторые пациенты продолжают получать лечение в течение всей оставшейся жизни.
[0147] При терапевтическом применении иногда требуется введение относительно высоких доз (например, содержащих примерно от 1 до 400 мг антитела/кг, причем в случае радиоиммуноконъюгатов дозы обычно составляют от 5 до 25 мг, а в случае молекул, конъюгированных с цитотоксином-лекарственным средством, используют более высокие дозы) с относительно короткими интервалами до уменьшения или прекращения развития заболевания, или, в конкретных вариантах осуществления, до частичного или полного улучшения симптомов заболевания у пациента. Затем лекарственное средство можно вводить пациенту в профилактическом режиме.
[0148] В одном варианте осуществления для лечения индивидуума можно использовать молекулу нуклеиновой кислоты (например, в составе вектора), кодирующую описанный здесь полипептид. Дозы нуклеиновых кислот, кодирующих полипептиды, варьируют примерно от 10 нг до 1 г, от 100 нг до 100 мг, от 1 мкг до 10 мг, или они находятся в диапазоне 30-300 мкг на пациента. В случае инфекционных вирусных векторов можно использовать дозы, соответствующие содержанию 10-100 или более вирионов в одной дозе.
[0149] Терапевтические средства можно доставлять путем парентерального, местного, внутривенного, перорального, подкожного, внутриартериального, внутричерепного, внутрибрюшинного, интраназального или внутримышечного введения с профилактическими или терапевтическими целями. Для введения описанного здесь антитела можно использовать внутримышечную инъекцию или внутривенную инфузию. В некоторых способах терапевтические антитела или их фрагменты вводят непосредственно в черепную коробку. В некоторых способах антитела или их фрагменты вводят в составе композиций или устройств, обеспечивающих замедленное высвобождение, таких как устройство Medipad™.
[0150] Описанные здесь средства необязательно вводят в сочетании с другими средствами, которые могут обеспечить эффективное лечение расстройства или состояния, требующего такого лечения (например, профилактического или терапевтического). В качестве других средств можно использовать средства, известные в данной области и традиционно используемые для лечения конкретного расстройства.
[0151] Хотя в клинических экспериментах чаще всего используют 131I и 90Y, в данной области известны и другие радиоактивные метки, которые можно использовать в подобных целях. Другие радиоизотопы также используют для визуализации. Например, другие радиоизотопы, подходящие для применения в настоящем изобретении, включают в себя, без ограничения, 123I, 125I, 32P, 57Co, 64Cu, 67Cu, 77Br, 81Rb, 81Kr, 87Sr, 113In, 127Cs, 129Cs, 132I, 197Hg, 203Pb, 206Bi, 177Lu, 186Re, 212Pb, 212Bi, 47Sc, 105Rh, 109Pd, 153Sm, 188Re, 199Au, 225Ac, 211A, 213Bi. В настоящем изобретении можно использовать альфа-, гамма- и бета-излучатели. Кроме того, с учетом настоящего описания специалист в данной области может легко определить, без излишнего экспериментирования, какие радионуклиды можно использовать в выбранной схеме лечения. В данной связи другие радионуклиды, уже используемые для клинической диагностики, включают в себя 125I, 123I, 99Tc, 43K, 52Fe, 67Ga, 68Ga, а также 111In. Для потенциального применения в направленной иммунотерапии антитела можно метить разными радионуклидами (Peirersz et al. Immunol. Cell Biol. 65: 111, 1987). Такие радионуклиды включают в себя 188Re и 186Re, а также, в меньшей степени, 199Au и 67Cu. В патенте США № 5460785, который включен в настоящееописание в качестве ссылки, можно найти дополнительную информацию, касающуюся таких радиоизотопов.
[0152] Как указано выше, описанные здесь антитела, или их фрагменты, можно вводить в фармацевтически эффективном количестве для лечения заболеваний млекопитающих in vivo. В данной связи следует понимать, что описанные антитела, или их фрагменты вводят в состав композиций, облегчающих введение и обеспечивающих стабильность активного средства. В некоторых вариантах осуществления фармацевтические композиции настоящего изобретения содержат фармацевтически приемлемые, нетоксичные, стерильные носители, такие как физиологический раствор, нетоксичные буферы, консерванты и т.п. В целях настоящего изобретения фармацевтически эффективное количество описанного здесь антитела, конъюгированного или не конъюгированного с терапевтическим средством, представляет собой количество, достаточное для обеспечения эффективного связывания с мишенью, а также для достижения улучшения, например, уменьшения симптомов заболевания или расстройства, или для детекции вещества или клетки. В случае опухолевых клеток полипептид в некоторых вариантах осуществления может взаимодействовать с выбранными иммунореактивными антигенами, присутствующими на неопластических или иммунореактивных клетках, и обеспечивать повышенную гибель данных клеток. Разумеется, описанные здесь фармацевтические композиции можно вводить в виде одной или нескольких доз, с достижением фармацевтически эффективного количества полипептида.
[0153] В соответствии с объемом настоящего изобретения описанные здесь антитела можно вводить человеку или другому животному с помощью указанных выше способов лечения в количестве, достаточном для достижения терапевтического или профилактического эффекта. Описанные здесь полипептиды можно вводить человеку или другому животному в виде традиционной лекарственной формы, полученной путем объединения описанного здесь антитела с традиционным фармацевтически приемлемым носителем или разбавителем с помощью известных методов. Специалистам в данной области известно, что тип и свойства фармацевтически приемлемого носителя или разбавителя зависят от количества активного ингредиента, с которым его объединяют, способа введения и других хорошо известных факторов. Специалистам в данной области также следует понимать, что смесь, содержащая один или несколько полипептидов настоящего изобретения, может оказаться особенно эффективной.
[0154] Изобретение также предлагает способ лечения состояния, вызванного повышенной экспрессией CB1, или повышенной чувствительностью к CB1, включающий в себя пероральное, парентеральное с использованием раствора для инъекции, ингаляционное, или местное введение пациенту или другому индивидууму фармацевтически эффективного количества антитела против CB1.
[0155] Кроме того, изобретение предлагает способ применения фармацевтически эффективного количества антитела против CB1 в производстве лекарственного средства для лечения состояния, вызванного повышенной экспрессией CB1, или повышенной чувствительностью к CB1, включающий в себя пероральное, парентеральное с использованием раствора для инъекции, ингаляционное, или местное введение пациенту или другому индивидууму.
[0156] В некоторых вариантах осуществления описанные здесь выделенные антитела и их антигенсвязывающие фрагменты обладают преимуществом, заключающимся в минимальном проникновении в мозг. В некоторых вариантах осуществления выделенные антитела и их фрагменты обладают высокой селективностью в отношении рецептора CB1 и не проникают через гематоэнцефалитический барьер, или обладают пониженной способностью проникать через гематоэнцефалитический барьер по сравнению с низкомолекулярными соединениями, специфичными к рецептору CB1, что приводит к минимизации побочных эффектов, связанных с ЦНС. В других вариантах осуществления выделенные антитела и их фрагменты после внутривенной инъекции не проникают через гематоэнцефалитический барьер, или обладают пониженной способностью проникать через гематоэнцефалитический барьер по сравнению с низкомолекулярными соединениями, специфичными к рецептору CB1, такими как римонабант.
[0157] В одном варианте осуществления описанные здесь антитела и их связывающие фрагменты или варианты можно вводить индивидууму с помощью по меньшей мере одного способа, выбранного из парентерального, подкожного, внутримышечного, внутривенного, внутрисуставного, внутрибронхиального, внутрибрюшного, внутрикапсульного, внутрихрящевого, внутриполостного, внутриклеточного, внутримозжечкового, интрацеребровентрикулярного, внутритолстокишечного, внутрицервикального, внутрижелудочкового, внутрипеченочного, внутримиокардиального, внутрикостного, внутритазового, внутриперикардиального, внутрибрюшинного, внутриплеврального, внутрипростатного, внутрилегочного, внутриректального, внутрипочечного, интраретинального, интраспинального, внутрисиновиального, внутригрудного, интратимпанального, внутриматочного, интравезикального, интравитреального, болюсного, подконъюнктивального, вагинального, ректального, буккального, подъязычного, интраназального и трансдермального введения.
[0158] Настоящее изобртеение предлагает выделенные антитела и их антигенсвязывающие фрагменты, нуклеиновые кислоты, кодирующие такие антитела или фрагменты, а также композиции, содержащие указанные выделенные антитела, фрагменты и нуклеиновые кислоты. Кроме того, настоящее изобретение предлагает фармацевтические композиции, содержащие выделенные антитела, или их фрагменты, или нуклеиновые кислоты, кодирующие такие антитела или фрагменты, а также один или несколько из фармацевтически приемлемых носителей. Фармацевтически приемлемые носители включают в себя, например, вспомогательные вещества, разбавители, капсулирующие вещества, наполнители, буферы и другие средства.
описание конкертных аспектов и вариантов осуществления
[0159] Разные описанные здесь аспекты и варианты их осуществления можно объединять друг с другом. Кроме того, любой из описанных выше аспектов и вариантов их осуществления можно объединить с любым из описанных ниже конкретных аспектов и вариантов их осуществления.
[0160] Ниже приведены некоторые конкретные аспекты и варианты осуществления, которые служат для дополнительной иллюстрации настоящего изобретения:
[0161] 1. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или фрагмент обладает сродством к рецептору CB1, характеризующимся Kd примерно 1 мкМ или менее.
[0162] 2. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент обладает сродством к рецептору CB1, характеризующимся Kd примерно 100 нМ или менее.
[0163] 3. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент обладает сродством к рецептору CB1, характеризующимся Kd примерно 10 нМ или менее.
[0164] 4. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент обладает сродством к рецептору CB1, характеризующимся Kd примерно 1 нМ или менее.
[0165] 5. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент связывается с внеклеточным эпитопом на рецепторе CB1.
[0166] 6. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент связывается с человеческим рецептором CB1.
[0167] 7. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент ингибирует сигнальную активность рецептора CB1.
[0168] 8. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 7, где антитело или фрагмент обладает ингибирующей активностью в отношении сигнального пути рецептора CB1, которая по величине, по меньшей мере, эквивалентна активности низкомолекулярного римонабанта, где величину активности измеряют по ингибированию передачи сигнала, опосредованной агонистом рецептора CB1, с помощью анализа цАМФ.
[0169] 9. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 7, где антитело или фрагмент обладает ингибирующей активностью в отношении сигнального пути рецептора CB1, которая, по меньшей мере, в 3 раза превышает активность низкомолекулярного римонабанта, где величину активности измеряют по ингибированию передачи сигнала, опосредованной агонистом рецептора CB1, с помощью анализа цАМФ.
[0170] 10. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 7, где антитело или фрагмент обладает ингибирующей активностью в отношении сигнального пути рецептора CB1, которая по величине, по меньшей мере, эквивалентна активности низкомолекулярного римонабанта, где величину активности измеряют по ингибированию фосфорилирования ERK, опосредованного агонистом рецептора CB1.
[0171] 11. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 7, где антитело или фрагмент обладает ингибирующей активностью в отношении сигнального пути рецептора CB1, которая, по меньшей мере, в 3 раза превышает активность низкомолекулярного римонабанта, где величину активности измеряют по ингибированию фосфорилирования ERK, опосредованного агонистом рецептора CB1.
[0172] 12. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент активирует рецептор CB1, или повышает активность рецептора CB1.
[0173] 13. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент представляет собой аллостерический модулятор рецептора CB1.
[0174] 14. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент представляет собой обратный агонист рецептора CB1.
[0175] 15. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент является мышиным.
[0176] 16. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент является химерным.
[0177] 17. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент является гуманизированным.
[0178] 18. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент селективно связывается с CB1.
[0179] 19. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело конъюгировано со средством, таким как другое терапевтическое средство, цитотоксическое средство, молекула иммуноадгезии или визуализирующее средство.
[0180] 20. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 19, где средство представляет собой другое терапевтическое средство, цитотоксическое средство, молекулу иммуноадгезии или визуализирующее средство.
[0181] 21. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 20, где терапевтическое средство представляет собой римонабант.
[0182] 22. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 20, где визуализирующее средство выбрано из группы, состоящей из радиоактивной метки, фермента, флуоресцентной метки, люминесцентной метки, биолюминесцентной метки, магнитной метки и биотина.
[0183] 23. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 18, где антитело не обладает агонистической или антагонистической активностью.
[0184] 24. Антитело или его антигенсвязывающий фрагмент, способные конкурировать за связывание с рецептором CB1 с антителом или антигенсвязывающим фрагментом по варианту осуществления 1.
[0185] 25. Антитело или его антигенсвязывающий фрагмент, способные специфически связываться по существу с тем же эпитопом на рецепторе CB1, что и антитело или антигенсвязывающий фрагмент по варианту осуществления 1.
[0186] 26. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело, или его антигенсвязывающий фрагмент, содержит: вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 10, 18 и 26; и вариабельный участок легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, 14, 22 и 30.
[0187] 27. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или фрагмент содержит константный участок тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 12, 20 и 28, и, константный участок легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 8, 16, 24 и 32.
[0188] 28. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или его фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 2; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 4; вариабельной участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 6, и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 8.
[0189] 29. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или его фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 10; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 12; вариабельной участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 14, и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 16.
[0190] 30. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или его фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 18; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 20; вариабельной участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 22, и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 24.
[0191] 31. Выделенное антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где антитело или его фрагмент содержит вариабельный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 26; константный участок тяжелой цепи, имеющий аминокислотную последовательность SEQ ID NO: 28; вариабельной участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 30, и константный участок легкой цепи, имеющий аминокислотную последовательность SEQ ID NO: 32.
[0192] 32. Способ противодействия CB1, включающий в себя приведение клетки, экспрессирующей рецептор CB1, в контакт с антителом или связывающим фрагментом по варианту осуществления 1.
[0193] 33. Способ усиления активности CB1, включающий в себя приведение клетки, экспрессирующей рецептор CB1, в контакт с антителом или связывающим фрагментом по варианту осуществления 1.
[0194] 34. Способ лечения заболевания или расстройства, отвечающего на антагонистическую или агонистическую активность в отношении рецептора CB1, у индивидуума, нуждающегося в этом, где указанный способ включает в себя введение индивидууму антитела или его антигенсвязывающего фрагмента по варианту осуществления 1.
[0195] 35. Способ по варианту осуществления 34, где индивидуум представляет собой человека.
[0196] 36. Способ детекции CB1, включающий в себя приведение клетки в контакт с антителом или антигенсвязывающим фрагментом по варианту осуществления 1.
[0197] 37. Способ по варианту осуществления 34, где заболевание или расстройство выбрано из группы, состоящей из ожирения, диабета, дислипидемии, метаболических заболеваний, фиброза, неалкогольного стеатогепатита (NASH), заболевания печени, первичного билиарного цирроза печени, заболевания почек, фиброза почек, хронического заболевания почек, остеопороза, атеросклероза, сердечно-сосудистого заболевания, рака и воспалительного заболевания.
[0198] 38. Способ по варианту осуществления 34, где заболевание или расстройство выбрано из группы, состоящей из боли, спастичности при рассеянном склерозе и глаукомы.
[0199] 39. Способ по варианту осуществления 37, где заболевание или расстройство представляет собой фиброз.
[0200] 40. Способ по варианту осуществления 37, где антитело или антигенсвязывающий фрагмент противодействует CB1.
[0201] 41. Способ по варианту осуществления 38, где антитело или антигенсвязывающий фрагмент способствует проявлению активности CB1.
[0202] 42. Способ диагностики заболевания или расстройства, связанного с CB1, включающий в себя приведение клетки в контакт с антителом или антигенсвязывающим фрагментом по варианту осуществления 1.
[0203] 43. Способ определения прогноза для индивидуума, у которого диагностировано заболевание или расстройство, связанное с CB1, включающий в себя измерение экспрессии CB1 путем приведения клетки в контакт с антителом или его фрагментом по варианту осуществления 1.
[0204] 44. Способ по варианту осуществления 42-43, где заболевание или расстройство выбрано из группы, состоящей из ожирения, диабета, дислипидемии, метаболических заболеваний, фиброза, NASH, заболевания печени, первичного билиарного цирроза, заболевания почек, фиброза почек, хронического заболевания почек, остеопороза, атеросклероза, сердечно-сосудистого заболевания, рака, воспалительного заболевания, боли, спастичности при MS и глазных заболеваний, включающих в себя глаукому.
[0205] 45. Способ по варианту осуществления 44, где заболевание или расстройство представляет собой фиброз.
[0206] 46. Способ по варианту осуществления 36, где выделенное антитело, или его антигенсвязывающий фрагмент, конъюгированы с визуализирующим средством.
[0207] 47. Способ по варианту осуществления 46, где визуализирующее средство выбрано из группы, состоящей из радиоактивной метки, фермента, флуоресцентной метки, люминесцентной метки, биолюминесцентной метки, магнитной метки и биотина.
[0208] 48. Способ по варианту осуществления 42-43, где клетка присутствует в организме индивидуума.
[0209] 49. Способ по варианту осуществления 48, где индивидуум представляет собой человека.
[0210] 50. Клетка-хозяин, экспрессирующая выделенное антитело или фрагмент по варианту осуществления 1.
[0211] 51. Способ получения антитела или его фрагмента, способного специфически связываться с рецептором CB1, где указанный способ включает в себя иммунизацию млекопитающих очищенным рецептором CB1 или его антигенным фрагментом, CB1/липидными комплексами, и/или ДНК, кодирующей рецептор CB1.
[0212] 52. Способ по варианту осуществления 51, где антитело или его фрагмент получают из гибридомной клеточной линии, полученной с использованием клеток иммунизированных млекопитающих.
[0213] 53. Способ по варианту осуществления 51, где антитело или его фрагмент получают из фаговой библиотеки.
[0214] 54. Способ получения антитела или его фрагмента, способного специфически связываться с CB1, где указанный способ включает в себя получение фаговой библиотеки, содержащей вариабельные участки тяжелых и легких цепей из человеческих первичных лимфоцитов крови, и пэннинг фаговой библиотеки по связыванию с рецептором CB1.
[0215] 55. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит CDR1, CDR2 и CDR3 тяжелой цепи, имеющие последовательности SEQ ID NO: 352, 353 и 354, соответственно.
[0216] 56. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит CDR1, CDR2 и CDR3 легкой цепи, имеющие последовательности SEQ ID NO: 355, 356 и 357, соответственно.
[0217] 57. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит CDR1, CDR2 и CDR3 тяжелой цепи, имеющие последовательности, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% гомологичные аминокислотным последовательностям SEQ ID NO: 352, 353 и 354, соответственно.
[0218] 58. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит CDR1, CDR2 и CDR3 легкой цепи, имеющие последовательности, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% гомологичные аминокислотным последовательностям SEQ ID NO: 355, 356 и 357, соответственно.
[0219] 59. Антитело или его антигенсвязывающий фрагмент, способные специфически связываться с тем же эпитопом, что и антитело или антигенсвязывающий фрагмент по любому из перечисленных или раскрытых здесь вариантов осуществления.
[0220] 60. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит: CDR1 тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 443-463; CDR2 тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 464-577, и CDR3 тяжелой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 578-625.
[0221] 61. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит: CDR1 легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 626-661; CDR2 легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 662-742, и CDR3 легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 742-824.
[0222] 62. Выделенное антитело или его фрагмент по варианту осуществления 1, где антитело или его антигенсвязывающий фрагмент представляет собой гуманизированное антитело.
[0223] 63. Раскрытое в данном документе выделенное антитело, или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент содержит участок Fc человеческого IgG1.
[0224] 64. Выделенное антитело, или его антигенсвязывающий фрагмент по любому из предшествующих вариантов осуществления, где антитело или его антигенсвязывающий фрагмент содержит модифицированный участок Fc.
[0225] 65. Выделенное антитело, или его антигенсвязывающий фрагмент по варианту осуществления 64, где антитело или его антигенсвязывающий фрагмент содержит участок Fc, выбранный из группы, состоящей из Fc-участка гибрида IgG2/IgG4, Fc-участка IgG2, содержащего мутации A330S и P331S, и Fc-участка IgG4, содержащего мутацию S228P.
[0226] 66. Выделенное антитело или его антигенсвязывающий фрагмент, способные связываться с каннабиоидным рецептором 1 (CB1), где антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность вариабельного участка тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 339-341, и, необязательно, вариабельный участок легкой цепи с последовательностью SEQ ID NO: 337.
[0227] 67. Выделенное антитело, или его антигенсвязывающий фрагмент по варианту осуществления 64, где антитело или его антигенсвязывающий фрагмент содержит константный участок тяжелой цепи с последовательностью SEQ ID NO: 342.
[0228] 68. Выделенное антитело или его антигенсвязывающий фрагмент по варианту осуществления 66, где антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 343-351, и аминокислотную последовательность легкой цепи SEQ ID NO: 338.
[0229] 69. Выделенное антитело или его антигенсвязывающий фрагмент, содержащие вариабельный участок тяжелой цепи с последовательностью SEQ ID NO: 341 и константный участок тяжелой цепи с последовательностью SEQ ID NO: 433, SEQ ID NO: 434 или SEQ ID NO: 435.
[0230] 70. Выделенное антитело или его антигенсвязывающий фрагмент, содержащие вариабельный участок тяжелой цепи с последовательностью SEQ ID NO: 340 и константный участок тяжелой цепи с последовательностью SEQ ID NO: 433, SEQ ID NO: 434 или SEQ ID NO: 435.
[0231] 71. Выделенное антитело или его фрагмент, нуклеотидная последовательность или аминокислотная последовательность которого, по меньшей мере, на 65%, по меньшей мере, на 70%, по меньшей мере, на 75%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO 1-351 и SEQ ID NO 436-824.
[0232] 72. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент, способные связываться с CB1, где антитело или фрагмент обладает сродством к рецептору CB1, характеризующимся Kd примерно 100 нМ или менее.
[0233] 73. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 72, где антитело или фрагмент обладает сродством к рецептору CB1, характеризующимся Kd примерно 5 нМ или менее.
[0234] 74. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент, способные связываться с CB1, где антитело или фрагмент обладает ингибирующей активностью в отношении рецептора CB1, которая, по меньшей мере, в 10 раз превышает активность низкомолекулярного римонабанта, где величину активности измеряют по ингибированию передачи сигнала, опосредованной агонистом рецептора CB1, с помощью анализа цАМФ.
[0235] 75. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 74, где антитело или фрагмент обладает ингибирующей активностью в отношении рецептора CB1, которая, по меньшей мере, в 5 раз превышает активность низкомолекулярного римонабанта, где величину активности измеряют по ингибированию передачи сигнала, опосредованной агонистом рецептора CB1, с помощью анализа цАМФ.
[0236] 76. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 74, где антитело или фрагмент обладает ингибирующей активностью в отношении рецептора CB1, которая по величине, по меньшей мере, эквивалентна активности низкомолекулярного римонабанта, где величину активности измеряют по ингибированию передачи сигнала, опосредованной агонистом рецептора CB1, с помощью анализа цАМФ.
[0237] 77. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент, способные связываться с CB1, где антитело или его фрагмент обладает более высокой активностью, чем соответствующее негуманизированное или химерное антитело, причем гуманизированное антитело, или его фрагмент, и соответствующее негуманизированное или химерное антитело содержат одинаковые CDR тяжелой и легкой цепи, где активность измеряют по ингибированию передачи сигнала, опосредованной агонистом рецептора CB1, с помощью анализа цАМФ.
[0238] 78. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 77, где гуманизированное антитело или его фрагмент обладает ингибирующей активностью в отношении рецептора CB1, которая, по меньшей мере, в 2 раза превышает активность соответствующего негуманизированного или химерного антитела или фрагмента.
[0239] 79. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 78, где гуманизированное антитело или его фрагмент обладает ингибирующей активностью в отношении рецептора CB1, которая, по меньшей мере, в 3 раза превышает активность соответствующего негуманизированного или химерного антитела или фрагмента.
[0240] 80. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 77, где гуманизированное антитело или его фрагмент обладает ингибирующей активностью в отношении рецептора CB1, которая, по меньшей мере, в 5 раз превышает активность соответствующего негуманизированного или химерного антитела или фрагмента.
[0241] 81. Раскрытое в данном описании выделенное гуманизированное антитело или его антигенсвязывающий фрагмент, где способность антитела или фрагмента проникать в мозг снижена или отсутствует.
[0242] 82. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 81, где способность антитела или фрагмента проникать в мозг снижена по сравнению с низкомолекулярным агонистом или антагонистом рецептора CB1.
[0243] 83. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 81, где антитело или фрагмент обладает пониженными побочными эффектами, связанными с центральной нервной системой (ЦНС), по сравнению с низкомолекулярным агонистом или антагонистом рецептора CB1.
[0244] 84. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по варианту осуществления 83, где низкомолекулярный агонист или антагонист рецептора CB1 представляет собой AM6545, AM251, таранабант или римонабант.
[0245] 85. Способ лечения заболевания или расстройства, отвечающего на антагонистическую или агонистическую активность в отношении рецептора CB1, у индивидуума, нуждающегося в этом, где указанный способ включает в себя введение индивидууму антитела или его антигенсвязывающего фрагмента по любому из вариантов осуществления 55-84.
[0246] 86. Способ по варианту осуществления 85, где индивидуум представляет собой человека.
[0247] 87. Способ по варианту осуществления 85, где заболевание или расстройство выбрано из группы, состоящей из ожирения, диабета, дислипидемии, метаболических заболеваний, фиброза, NASH, заболевания печени, первичного билиарного цирроза, заболевания почек, фиброза почек, хронического заболевания почек, остеопороза, атеросклероза, сердечно-сосудистого заболевания, рака, воспалительного заболевания, боли, спастичности при MS и глазных заболеваний, включающих в себя глаукому.
[0248] 88. Способ по варианту осуществления 85, где способность антитела или антигенсвязывающего фрагмента проникать в мозг снижена или отсутствует.
[0249] Хотя выше изобретение описано с приведением некоторых подробностей в качестве иилюстраций и примеров для облегчения понимания, для рядового специалиста в данной области должно быть очевидно в свете разъяснений, приведенных в настоящем изобретении, что можно осуществить некоторые изменения и модификации, не отступая от сущности и объема прилагаемой формулы изобретения. Нижеследующие примеры приведены только для иллюстрации, но не для ограничения. Специалистам в данной области хорошо известны разные некритические параметры, которые можно изменить или модифицировать с получением практически таких же результатов.
ПРИМЕРЫ
Пример 1. Иммунизация мышей с целью получения антител против рецептора CB1
[0250] Последовательность кДНК и праймеры для клонирования человеческого рецептора CB1 получают на основе стандартной последовательности NM_001160258 из электронного архива по биологии и медицине NCBI. Рецептор CB1 получают путем временной экспрессии, осуществляемой в клетках 293 с использованием липофектамина, или путем получения стабильных клеточных линий. Для получения стабильной клеточной линии конструкцию нативного полноразмерного человеческого рецептора CB1 pcDNA4/TO трансфицируют в тетрациклин-индуцируемую систему Trex-CHO и клетки Trex-293. Клетки культивируют в присутствии антибиотиков зеоцина и бластицидина. Клоны, экспрессирующие CB1 после индукции тетрациклином, идентифицируют путем FACS-окрашивания антителом против рецептора CB1, R&D (клон 368302). Получают мембраны, которые используют для иммунизации либо непосредственно, либо после солюбилизации мембранных белков в детергенте с последующей очисткой. Очищенный рецептор CB1 стабилизируют в липидном бислое. CB1/липидный комплекс обозначают как iCAPS рецептора CB1.
[0251] Мышей Balb/c иммунизируют в течение 2 циклов ДНК, кодирующей рецептор CB1, с последующими иммунизациями мембранами, содержащими рецептор CB1, или iCAPS, содержащими рецептор CB1. Образцы крови берут до и после иммунизации, сыворотку тестируют методом ELISA на связывание с мембранами, содержащими рецептор CB1, по сравнению с наивными мембранами. Когда мышиная сыворотка дает положительный сигнал на мембранах, содержащих рецептор CB1, по сравнению с наивными мембранами, мышей умерщвляют, селезенки удаляют и используют для получения гибридом и фаговых библиотек.
Пример 2. Извлечение лимфоцитов, выделение B-клеток, гибридизация и выбор гибридом, связывающихся с рецептором CB1
[0252] Из иммунизированных мышей удаляют селезенки и получают суспензии отдельных клеток. Чтобы получить суспензии отдельных клеток, селезенки переносят на сито, помещенное сверху конической центрифужной пробирки объемом 50 мл, и с помощью поршня от шприца на 3 мл выдавливают клетки из селезенки. Красные кровяные клетки лизируют охлажденным на льду буфером ACK в течение 10 мин, добавляют 5 мл DMEM и проводят центрифугирование. Данную стадию повторяют один раз, после чего клетки ресуспендируют в среде DMEM с получением концентрации 2×107/мл.
[0253] Извлечение и получение клеток миеломы: клетки SP2/0 высевают при плотности примерно 5×104 клеток/мл и пересаживают каждые 2 дня. Перед гибридизацией исходные клетки миеломы собирают путем центрифугирования в конической центрифужной пробирке объемом 50 мл при комнатной температуре (RT) при 500 g в течение 10 минут. Клетки промывают 3 раза путем добавления 30 мл среды, не содержащей сыворотку, и повторяют центрифугирование. Супернатант удаляют пипеткой, клеточный осадок ресуспендируют в 25 мл среды и доводят до концентрации жизнеспособных клеток 2×107/мл.
[0254] Слияние клеток: Клетки миеломы и клетки селезенки смешивают в соотношении 1:5. Смесь клеток центрифугируют при 1000 об/мин в течение 5 мин, после чего супернатант отбрасывают, получая клеточный осадок. Смесь клеток дважды промывают рабочим буфером для гибридизации и центрифугируют с получением клеточного осадка. Клеточный осадок осторожно ресуспендируют в буфере для гибридизации до конечной концентрации клеток 8×107/мл. Смеси клеток помещают в ячейку с электродами и проводят электрослияние. После слияния клетки оставляют в покое в течение 5 мин, промывают 1× средой DMEM, к гибридизованным клеткам добавляют нагретую среду HAT до конечной концентрации 0,5×106 B-клеток/мл. Затем в каждую лунку 96-луночного планшета добавляют 100 мкл клеточной суспензии (5×104 B-клеток). Средняя эффективность слияния составляет 1 гибридома/1×104 B-клеток, следовательно, в соответствии с данной методикой, в каждой лунке содержится 5 гибридом.
[0255] Культивирование гибридомных клеток. Гибридомы культивируют в инкубаторе для культивирования клеток при 5% CO2, 37°C. Рост гибридом проверяют ежедневно. Обычно колонии можно увидеть через 5 дней. Среду заменяют на свежую DMEM на 7 день перед положительным скринингом.
[0256] Положительный скрининг: через 7-9 дней после слияния клеток, когда колонии увеличиваются, 100 мкл супернатанта из каждой гибридома-содержащей лунки переносят в отдельную лунку нового 96-луночного планшета и анализируют методом ELISA с использованием белка CB1.
Пример 3. Получение и скрининг фаговых библиотек
[0257] Экстракция общей РНК: Ткань селезенки собирают в RNAlater. Маленький кусочек замороженной ткани (~30 мг) гомогенизируют в ступке, охлажденной в жидком N2, и перемалывают до состояния мелкого порошка. Добавляют реагент TRIzol®, энергично встряхивая вручную в течение 30 секунд. 1 мл раствора переносят в микроцентрифужную пробирку объемом 1,5 мл при комнатной температуре 2-3 мин и оставляют стоять при комнатной температуре в течение нескольких минут. К смеси добавляют 0,2 мл хлороформа на 1 мл раствора и энергично встряхивают вручную в течение 30 секунд. Смесь инкубируют при комнатной температуре в течение 5 мин и затем центрифугируют при 12000 g в течение 20 мин при 4°C. Водную фазу удаляют и переносят в новую пробирку. В пробирку добавляют равный объем изопропилового спирта и перемешивают в течение 30 секунд. После инкубации при комнатной температуре в течение 5 мин смесь центрифугируют при 12000 g в течение 15 мин при 4°C. Осадок промывают путем добавления 500 мкл 75% этанола и центрифугируют при 12000 g в течение 15 мин при 4°C. Полученный осадок общей РНК сушат на воздухе и растворяют в 50 мкл не содержащей РНКазы воде на 1 мкг предполагаемой РНК. OD измеряют при 260 нм и 280 нм в образце РНК, разведенном в соотношении 1:10.
[0258] Получение кДНК: Синтез первой цепи кДНК проводят с помощью коммерческого набора (Invitrogen, № по каталогу: 18080-051), коротко говоря, 20 мкг общей РНК смешивают с 5 мкМ oligo(dT)20 и 1 мМ dNTP в DEPC-обработанной воде объемом 40 мкл, инкубируют при 65°C в течение 5 мин и затем добавляют 80 мкл буфера при КТ, содержащего 5 мМ MgCl2, 10 мкМ DTT, 16 единиц RNaseOUT и 80 единиц обратной транскриптазы Superscript III. Полученную смесь инкубируют при 50°C в течение 50 мин, инактивируют нагреванием, после чего добавляют 4 мкл РНКазы H, чтобы удалит остаток РНК. кДНК используют для последующего конструирования библиотеки.
[0259] Библиотеку химерных Fab конструируют следующим образом: Вариабельные участки тяжелой цепи или легкой цепи амплифицируют с использованием праймеров, специфичных для тяжелой цепи или легкой цепи, представляющих несколько зародышевых семейств, описанных в Barbas et al, и полученной выше мышиной кДНК в качестве матрицы. Константные участки человеческой тяжелой цепи и легкой цепи, Ch1 и CL1, соответственно, амплифицируют из существующего клона pCOM3xTT. Вариабельный участок и константный участок тяжелой цепи соединяют, используя ПЦР с перекрывающимися праймерами. Полученные тяжелую цепь и легкую цепь соединяют, снова используя ПЦР с перекрывающимися праймерами, и получают ДНК-фрагмент химерного Fab, который клонируют в модифицированном векторе pCOM3x в виде фрагмента SfiI, вставленного путем лигирования. Библиотеку лигированной ДНК очищают и используют для трансформации высокоэффективных компетентных клеток SS320. Общее число полученных уникальных трансформантов составляет, по меньшей мере, 5×107.
[0260] Проводят пэннинг путем двойного вычитания фаговой библиотеки, полученной из иммунизированных мышей, в иммунопробирках Maxisorp (Thermo Scientific), покрытых пустыми iCAPS, с последующим двойным вычитанием на Dynabeads MyOne Streptavidin T1 (Life Technologies), покрытых биотинилированным белком Bril. Все стадии вычитания длятся 30 минут. В это время iCAPS, содержащие биотинилированный рецептор CB1, наносят на Dynabeads MyOne Strepavidin T1 (Life Technologies). Затем полученный после вычитания фаговый пул и CB1-содержащие iCAPS на гранулах смешивают, наряду с небиотинилированным белком Bril и пустыми iCAPS для конкуренции, и инкубируют в течение одного часа при комнатной температуре при вращении. Гранулы отделяют от связывающей смеси с помощью магнита и промывают несколько раз, чтобы удалить несвязанные фаги (5 раз в цикле 1, 10 в каждом из циклов 2 и 3). Связанные фаги элюируют дважды глициновым буфером, pH 2,2, каждый раз по 10 минут. Элюаты объединяют, нейтрализуют Tris-HCl, pH 8,0, и используют для инфицирования клеток TG1 с получением фага, который подвергают следующему циклу пэннинга. После 3 циклов пэннинга отбирают отдельные колонии и подвергают их скринингу методом ELISA для моноклональных фагов.
[0261] Для скрининга средств, связывающих фаги, отдельные колонии TG1, инфицированных фагами, отобранными в результате пэннинга, помещают в 96-луночные планшеты, содержащие среду 2YT/карбенициллин/глюкоза, и встряхивают в течение ночи при 30°C. На следующий день небольшой объем насыщенной культуры переносят на свежую среду 2YT/карбенициллин/глюкоза и встряхивают при 37°C до достижения значения OD600 0,6-0,7. Затем культуру инфицируют херперным фагом KO7. Инфицированные клетки TG1 центрифугируют, ресуспендируют в среде 2YT/карбенициллин/канамицин и встряхивают при 30°C в течение ночи. В это время 96-луночные планшеты Maxisorp (Thermo Scientific) покрывают стрептавидином (Wako) при 4°C в течение ночи. На третий день биотинилированные антигены улавливают на планшетах, покрытых стрептавидином, которые затем блокируют 3% обезжиренным молоком в PBST. Клетки TG1 снова центрифугируют. Фаг-содержащие супернатанты блокируют 3% обезжиренным молоком, затем загружают на планшеты ELISA и инкубируют в течение одного часа при комнатной температуре. После трех циклов промывания PBST в планшеты добавляют HRP-меченое мышиное антитело против M13 (GE Healthcare), разведенное 1:2000 3% обезжиренным молоком в PBST и инкубируют еще час при комнатной температуре. Планшеты промывают три раза PBST и затем проявляют ™B (Biopanda). Реакцию останавливают путем добавления в планшеты HCl. Поглощение при 450 нм прочитывают на ридере для микропланшет Emax precision (Molecular Devices). Клоны, специфически связывающиеся с iCAPS, отбирают и подвергают дальнейшей характеризации и секвенированию.
[0262] Выделяют колонии E. coli, несущие плазмиды, которые продуцируют Fab, презентированные на фаге. Плазмидную ДНК экстрагируют и секвенируют, получая информацию по ДНК, кодирующей Fab. Конструируют специфические праймеры, чтобы амплифицировать V-участок тяжелой цепи, продукт ПЦР клонируют в pTT5-HCV3, модифицированной версии вектора экспрессии pTT5, ранее обработанной ферментами рестрикции ApaI и SacI, перед константным участком человеческой тяжелой цепи, путем непрерывного клонирования. Кроме того, конструируют специфические праймеры, чтобы амплифицировать поноразмерный участок легкой цепи фрагмента Fab. Фрагмент, полученный методом ПЦР, клонируют в pTT5-IL2-LCC, модифицированном векторе pTT5, обработанном EcoRI и NotI, путем непрерывного клонирования. Последовательности 2 полученных плазмид подтверждают секвенированием.
Пример 4. Секвенирование гибридом
[0263] Клетки гибридомы (1×107) собирают и экстрагируют общую РНК с помощью реагента Tri по способу, описанному выше для ткани селезенки. кДНК получают с помощью набора SuperScript III в соответствии с инструкцией производителя, как описано выше. Проводят ПЦР с использованием полученной кДНК в качестве матрицы и праймеров VhRevU и VhForU, получая продукт ПЦР размером 300 п.о., который очищают с помощью набора для очистки продуктов ПЦР и секвенируют, используя такие же праймеры. Также проводят реакцию ПЦР, используя праймеры VkRev7 и VkFor, специфичные для V-участка легкой цепи (только для вариабельного участка), или праймеры KappaFor (специфичные для полноразмерной легкой цепи каппа). Очищенный продукт ПЦР подвергают реакциям секвенирования, получая последовательность ДНК.
Пример 5. Экспрессия и анализ IgG
[0264] Экспрессия IgG: Две плазмиды, полученные на основе pTT5, одна из которых содержит ДНК тяжелой цепи, а другая - ДНК легкой цепи, совместно трансфицируют в клетки HEK293F, чтобы обеспечить экспрессию IgG. За 24 часа до трансфекции клетки 293F разбавляют до плотности 8×105 клеток/мл. В день трансфекции плотность клеток поддерживают на уровне 1,1-1,3×106 клеток/мл. Для трансфекции используют один мкг плазмидной дНК на 1 мл суспензионной культуры клеток. 80 мкг ДНК разводят в 4 мл свежей среды 293F freestyle. 240 мкг реагента для трансфекции полиэтиленимина (PEI) разводят в конечном объеме 4 мл среды 293F freestyle. После инкубации в течение 3 минут 4 мл ДНК тщательно смешивают с 4 мл PEI. 8 мл смеси ДНК и PEI инкубируют в течение 15 минут при комнатной температуре и медленно добавляют к 80 мл суспензионной культуры клеток 293F. Клетки инкубируют на платформе с орбитальным вращением со скоростью 130 об/мин при 37°C и 5% CO2 в течение 4 дней, после чего собирают.
[0265] Очистка IgG: Слой, содержащий белок A, объемом 0,4 мл помещают в колонку объемом 1 мл и промывают 10 мл dH2O и 10 мл PBS, pH 8,0. Суспензии трансфицированных клеток 293F центрифугируют при 4000 об/мин в течение 45 минут при 4°C. Осадки отбрасывают, супернатант доводят до pH 8,0 на льду и загружают на приготовленную колонку с белком A. После окончания загрузки супернатанта колонку промывают 5 мл PBS, pH 8,0, и элюируют 4 мл 0,1 M буфера цитрат Na-HCl, pH 3,5. Элюированные образцы, содержащие IgG, нейтрализуют 200 мкл 1,5M буфера Tris-HCl, pH 8,8, и концентрируют с использованием концентратора 4 мл, 30 кДа. 4,5 мл PBS помещают в концентратор и центрифугируют. В конечном счете, после замены среды IgG хранят в PBS. IgG детектируют по OD280 и определяют их чистоту методами SDS-PAGE и SEC.
[0266] Значения концентраций, объемов и выходов PA13R3-P1C4, полученные в четырех разных экспериментах, показаны в таблице 4. Значения концентраций, объемов и количеств разных клонов показаны ниже в таблице 5.
Таблица 4. Экспрессия PA13R3-P1C4
Таблица 5. Экспрессия клонов
Пример 6. Анализ связывания IgG с CB1-содержащими iCAPS методом ELISA
[0267] Очищенные IgG тестируют на связывание с очищенным рецептором CB1 (CB1-содержащая конформационная антиген-презентирующая система (iCAPS)) методом ELISA. Небиотинилированные антигены (например, пустые iCAPS) наносят непосредственно на планшеты Maxisorp. Первичные антитела, представляющие собой серийные разведения очищенных IgG 1:3, инкубируют на планшете в течение 1 часа. После 3 циклов промывания PBST, в качестве вторичных антител добавляют HRP-меченные козлиные антитела против мышиных IgG (Abmart) или HRP-меченные козлиные антитела против человеческих IgG (Sigma), в зависимости от вида IgG, и инкубируют еще час. Планшеты промывают три раза PBST и затем проявляют ™B (Biopanda). Реакцию останавливают путем добавления HCl. Поглощение при 450 нм прочитывают на ридере для микропланшетов Emax precision (Molecular Devices). Результаты анализа связывания приведены в таблице 6.
Таблица 6. Связывание с CB1-содержащими iCAPS
Пример 7. Анализ связывания IgG методом FACS
[0268] Исходные клетки TRex CHO, TRex CHO A56, экспрессирующие CB1 (CB1 T210A/партнер по гибридизации) на повышенном уровне, и TRex CHO A156, экспрессирующие нативный человеческий CB1, собирают из колб. 100 мкл клеточной суспензии, содержащей 1×106 клеток/мл, инкубируют с первичными антителами IgG. В качестве вторичных антител используют PE-конъюгированные антитела против человеческих IgG и против мышиных IgG, разведенные 1:200. FITC-меченные антитела против человеческого Fab разводят 1:32. Клетки дважды промывают 200 мкл буфера FACS, переносят в пробирки BD Falcon объемом 5 мл и анализируют методом проточной цитометрии. Связывание очищенных IgG изначально тестируют при концентрациях 30 нМ и 300 нМ. Идентифицируют ряд связывающих соединений, показанных на фигурах 1A-1F.
[0269] Проводят дополнительный анализ исходных клеток TRex CHO, клеток TRex CHO A56, экспрессирующих CB1 на повышенном уровне, и клеток TRex CHO A156, экспрессирующих нативный человеческий CB1, в котором используют 5HT2B, мышиный CB1 и человеческий CB2, чтобы определить специфичность связывания IgG. 100 мкл клеточной суспензии, содержащей 1×106 клеток/мл, инкубируют с первичными антителами IgG в 3-кратных серийных разведениях, начиная от 1 мкМ до 0,5 нМ в течение 30 минут на льду. Клетки дважды промывают 200 мкл буфера FACS, затем инкубируют со вторичным антителом в течение 30 минут на льду. После двух промываний 200 мкл буфера FACS клетки переносят в пробирки BD Falcon объемом 5 мл и анализируют методом FACS.
[0270] Кроличье моноклональное антитело против CB1 Santa Cruz и FITC-конъюгированное вторичное антитело против кроличьих IgG используют для детекции экспрессии мышиного CB1. Мышиные моноклональные антитела против CB2 R&D и человеческий IgG P2C2 используют для подтверждения экспрессии CB2 и 5HT2B соответственно. В качестве вторичных антител используют антитела против мышиных IgG и антитела против человеческих IgG, конъюгированные с PE.
[0271] Чтобы выбрать связывающие соединения, строят полные кривые связывания для рецептора CB1 на основе тестирования в диапазоне концентраций. Готовят трехкратные серийные разведения от 1 мкМ до 0,1 мкМ. Селективность определяют путем измерения связывания с клетками, экспрессирующими 5HT2B или CB2, по сравнению с клетками A156 (экспрессирующими нативный человеческий рецептор CB1) или A56 (экспрессирующими рецептор CB1 на повышенном уровне и содержащими модификацию T210A и замену ICL3 на партнера по гибридизации), используя метод проточной цитометрии. Как показано на фигуре 2C и фигуре 2D, экспрессию CB2 и 5HT2B подтвержают с использованием мышиных моноклональных антител против CB2 и P2C2 против человеческих IgG, соответственно. Для детекции CB2 и 5HT2B используют PE-конъюгированные антитела против мышиных IgG (дя детекции антител против CB2) и PE-конъюгированные антитела против человеческих IgG, соответственно. Антитела PA13R3-P1C4 и 36E12B6C2 селективно связываются с CB1, как показано на фигуре 2A и фигуре 2B. Кроме того, в таблице 7 приведены концентрации и константы диссоциации (Kd) для каждой из нескольких партий PA13R3-P1C4 и 36E12B6C2.
[0272] Результаты исследования демонстрируют, что IgG и Fab PA13R3-P1C4 и 36E12B6C2 связываются как с A56, так и с A156, но не связываются с исходной линией TRex CHO и не обладают перекрестной активностью в отношении 5ht2b, человеческого CB2 или мышиного CB1.
Таблица 7. Результаты анализа разных партий антител методом проточной цитометрии
Пример 8. Конкурентный анализ
[0273] Чтобы определить, связываются ли 36E12B6C2 и P1C4 с подобными эпитопами, используют клетки TRex CHO A156, экспрессирующие нативный человеческий CB1. Для окрашивания используют концентрации P1C4 и 36E12B6C2, соответствующие EC80 и EC50. Для анализа конкуренции используют избыток IgG, Fab PA13R3-P1C4 и 36E12B6C2. 100 мкл суспензии клеток A156 с плотностью 1×106 клеток/мл инкубируют с конкурирующими IgG в течение 30 минут на льду, после чего к смеси добавляют окрашивающие IgG и инкубируют еще 30 минут на льду. Дважды промывают 200 мкл буфера FACS и затем клетки инкубируют со вторичным антителом в течение 30 минут на льду. PE-конъюгированные антитела против человеческих IgG и против мышиных IgG разводят 1:200. Клетки дважды промывают 200 мкл буфера FACS, переносят в пробирки BD Falcon объемом 5 мл и анализируют методом FACS.
[0274] Результаты иследования демонстрируют, что Fab и IgG PA13R3-P1C4 конкурируют с 36E12B6C2 за связывание с CB1, позволяя предположить, что PA13R3-P1C4 и 36E12B6C2 связываются с перекрывающимися эпитопами (фигуры 3A и B). Соединение 36E12B6C2, используемое в концентрациях от 100 нМ до 500 нМ, также способно конкурировать с PA13R3-P1C4 за связывание с CB1.
Пример 9. Функциональный анализ цАМФ
[0275] Чтобы измерить антагонистическую активность антител, проводят функциональный анализ цАМФ. Функциональный анализ цАМФ (Cisbio) проводят на белом 384-луночном низкообъемном планшете (Greiner). В каждую лунку планшета высевают 8000 клеток TRex CHO, стабильно экспрессирующих CB1, после чего проводят инкубацию с антагонистом в разных концентрациях (варьирующих от 10 мкМ до 0 мкМ) при комнатной температуре в течение 10 мин. Для стимуляции смеси к клеткам добавляют 5 мкМ форсколин (Sigma Aldrich) и 9 мкМ каннабиноид CP55940 (Sigma Aldrich), после чего инкубируют в течение 30 мин при комнатной температуре, чтобы активировать CB1. После инкубации в течение 30 мин добавляют 5 мкл цАМФ-d2 (разведение 1:39, конъюгат и буфер для лизиса получают от Cisbio) и 5 мкл анти-цАМФ-криптата (разведение 1:9, конъюгат и буфер для лизиса получают от Cisbio) для стимуляции клеток и инкубируют в течение часа. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism.
[0276] Как показано на фигуре 4, два антитела, 36E12B2H8 (полученное с использованием гибридом) и PA13R3-P1C4 (полученное с использованием фагов) обладают антагонистической активностью, равной (36E12B2H8) или превышающей (PA13R3-PIC4) активность низкомолекулярных соединений, используемых в качестве положительных контролей (таких как обратные агонисты рецептора CB1 SR141716A (римонабант) и AM251 и нейтральный антагонист AM6545), со значениями IC50 350±28 нМ и 90±13 нМ, соответственно.
Пример 10: Анализ активации ERK
[0277] Чтобы подтвердить антагонистическую активность mAb, определяют активацию ERK как часть сигнального пути рецептора CB1. За два дня до эксперимента клетки Trex-CHO, экспрессирующие рецептор CB1, высевают в 6-луночные планшеты при плотности 500000 клеток/лунку. Через 24 часа добавляют 1 мкг/мл тетрациклина, чтобы индуцировать экспрессию рецептора CB1. В течение, по меньшей мере, двух часов перед началом эксперимента клетки держат на бессывороточной среде. К культуральной среде добавляют IgG в концентрации 300 нМ и через 30 минут клетки стимулируют агонистом рецептора CB1 WIN55,212 (100 нМ) в течение 10 и 15 минут. Собирают клеточные лизаты и определяют уровень активации ERK методом вестерн-блоттинга. Антитела против ERK и против фосфо-ERK получают от Cell Signaling Inc.
[0278] Обработка агонистом рецептора CB1 WIN55,212 индуцирует активацию ERK, что подтверждает увеличение сигнала фосфорилированной ERK. Общую ERK используют в качестве контроля загрузки в методе вестерн-блоттинга, чтобы подтвердить равную загрузку образцов. Как показано на фигуре 5A, полученное с использованием фагов антитело PA13R3-P1C4 (300 нМ), но не контрольный IgG (неспецифическое связывающее соединение) или PA13R3-P1E4, блокирует индуцированное WIN55,212 (100 нМ) фосфорилирование ERK. Как показано на фигуре 5B, полученные с использованием гибридом антитела 36E12B2E5, 36E12B6C2 и 36E12B6F2, но не контрольный IgG, блокируют WIN55,212-индуцированное фосфорилирование ERK. AM6545 (нейтральный антагонист) используют в качестве положительного контроля, как показано на фигурах 5A и 5B.
Пример 11. Функциональные анализы цАМФ
[0279] Функциональный анализ агонистической активности путем измерения цАМФ проводят на белом 384-луночном низкообъемном планшете (Greiner). В каждую лунку планшета высевают 8000 клеток TRex CHO, стабильно экспрессирующих CB1, после чего проводят инкубацию с агонистом в разных концентрациях (варьирующих от 1,5 мкМ до 0 мкМ) при комнатной температуре в течение 10 мин. Для тестирования агонистической активности (фигуры 6A и 6B) к смеси, стимулирующей клетки, добавляют 5 мкМ форсколин (Sigma Aldrich) и инкубируют в течение 30 мин при комнатной температуре. Чтобы определить активность положительного аллостерического модулятора (фигуры 6C и 6D), к смеси, стимулирующей клетки, добавляют 5 мкМ форсколин (Sigma Aldrich) и 1 мкМ CP55940, после чего смесь инкубируют в течение 30 мин при комнатной температуре.
[0280] После инкубации в течение 30 мин добавляют 5 мкл цАМФ-d2 (разведение 1:39, конъюгат и буфер для лизиса получают от Cisbio) и 5 мкл анти-цАМФ-криптата (разведение 1:9, конъюгат и буфер для лизиса получают от Cisbio) для стимуляции клеток и инкубируют в течение часа. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism.
[0281] Результаты скрининга на агонистическую активность путем измерения цАМФ позволяют идентифицировать четыре потенциальных агонистических IgG, включающих в себя PA2LR3-P2D3, PA2LR3-P4B1, PA2LR3-P6G7 и PA2LR3-P6B12. В частности, PA2LR3-P2D3, PA2LR3-P4B1 и PA2LR3-P6G7 характеризуются EC50 >300 нМ, как показано на фигурах 6A и 6B. Кроме того, PA2LR3-P6B12 является потенциальным аллостерическим модулятором с EC50 примерно 1000 нМ в присутствии CP55940, как показано на фигуре 6C. Положительные и отрицательные контроли показаны на фигурах 6B и 6D.
[0282] Анализ цАМФ также проводят для дополнительной характеризации PA13R3-P1C4 и 36E12B6C2. Функциональный анализ с измерением цАМФ (Cisbio) проводят на белом 384-луночном низкообъемном планшете (Greiner). В каждую лунку планшета высевают 8000 клеток TRex CHO, стабильно экспрессирующих CB1, после чего проводят инкубацию с антагонистами, включающими в себя AM6545, SR141716A, PA12R3-P1C4 и 36E12B6C2, в концентрациях, варьирующих от 3 мкМ до 0 мкМ в течение 10 минут при комнатной температуре. После инкубации в течение 10 минут к смеси, стимулирующей клетки, добавляют 5 мкМ форсколин (Sigma Aldrich) и инкубируют в течение 30 мин при комнатной температуре. Чтобы количественно определить продукцию цАМФ, добавляют 5 мкл цАМФ-d2 и 5 мкл анти-цАМФ-криптата для стимуляции клеток и инкубируют в течение часа. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism. Результаты исследования показаны на фигуре 7. AM6545 и SR141716A были ранее охарактеризованы как нейтральный антагонист и обратный агонист, соответственно. Результаты функциональных анализов с измерением цАМФ свидетельствуют о том, что характер ингибирования PA12R3-P1C4 и 36E12B6C2 подобен характеру ингибирования SR141716A, позволяя предположить, что PA12R3-P1C4 и 36E12B6C2 являются обратными агонистами.
Пример 12. Анализ связывания iCAPS методом ELISA
[0283] Анализ связывания методом ELISA проводят, чтобы оценить связывание IgG или Fab антитела против рецептора CB1 с iCAPS, экспрессирующими CB1 (A138, содержащие замену нативной последовательности ICL3 на белковую последовательность 1, и A139, содержащие замену нативной последовательности ICL3 на BRIL), iCAPS, экспрессирующими 5HT2B (h13h), пустыми iCAPS, или rBril-0918. IgG и Fab включают в себя IgG 36E12B6C2, Fab 36E12B6C2, IgG PA13R3-P1C4 и Fab PA13R3-P1C4, которые тестируют по сравнению с используемым в качестве отрицательного контроля BRIL-связывающим IgG P1F7 и Fab P1F7. В случае IgG антитела, вторичное антитело, используемое для детекции связывающих соединений, представляет собой конъюгат антитело против мышиного IgG-HRP; в случае Fab вторичное антитело, используемое для детекции связывающих соединений, представляет собой конъюгат антитело против человеческого IgG-HRP. Результаты исследования приведены на фигурах 8A и 8B, а также в приведенной ниже таблице 8. В случае соединений, связывающих и iCAPS A138, и iCAPS A139, Fab 36E12B6C2 характеризуется более высокими значениями EC50, чем IgG 36E12B6C2. С другой стороны, IgG и Fab PA13R3-P1C4 имеют примерно одинаковые значения EC50 для связывания с A139 и A138. Ни одно из антител или Fab против CB1 не связывается с rBril-0918, пустыми iCAPs, или iCAPS, экспрессирующими 5HT2B. Контрольное mAb P1F7 распознает BRIL и, следовательно, может связываться с A139, содержащим гибрид BRIL в ICL3, но не с A138, не содержащим BRIL.
Таблица 8. Значения EC50, определенные методом ELISA для IgG и Fab 36E12B6C2 и PA13R3-P1C4
Пример 13. Исследование интернализации рецептора CB1
[0284] Влияние антитела против CB1 на WIN55,212 (CB1-специфичный агонист)-индуцированную интернализацию CB1 анализируют методом проточной цитометрии. Клетки TRex-CHO, стабильно экспрессирующие CB1, высевают в 6-луночный планшет с плотностью 5×105 клеток/лунку. В культуральную среду добавляют тетрациклин (1 мкг/мл) и инкубируют в течение 24 часов, чтобы индуцировать экспрессию CB1. В день эксперимента клетки держат в бессывороточной среде в течение 2 часов. Затем клетки предварительно инкубируют с антителом против CB1 (300 нМ), AM6545 (нейтральный антагонист CB1) и отрицательным контролем (соединение, связывающее BRIL) в течение получаса. Затем в культуральную среду добавляют агонист CB1 (1 мкМ WIN55,212) и инкубируют 1 час, чтобы индуцировать интернализацию рецептора. Экспрессированный на поверхности CB1 окрашивают мышиным моноклональным антителом против N-концевого участка CB1, полученным от R&D, и методом проточной цитометрии определяют среднюю интенсивность флуоресценции (MFI). Результаты исследования приведены на фигуре 9A. После обработки агонистом CB1 WIN55,212 значение MFI уменьшается по сравнению с контролем, что позволяет предположить наличие интернализации CB1 (фигура 9A, верхний ряд гистограмм). Предварительная обработка CB1-специфичным нейтральным антагонистом AM6545 блокирует WIN55,212-индуцированную интернализацию рецептора CB1 (фигура 9A, верхний ряд гистограмм). Предварительная обработка антителами против CB1 (300 нМ) не влияет на WIN55,212-индуцированную интернализацию рецептора (фигура 9A, средний и нижний ряды гистограмм).
[0285] Также исследуют влияние антитела против CB1 на интернализацию рецептора. После культивирования в бессывороточной среде в течение 2 часов к культуральной среде добавляют 300 nM антитела против CB1, отрицательный контроль P2A12 (соединение, связывающее BRIL), агонист CB1 WIN55,212 и инкубируют в течение 1 часа. Клетки собирают и окрашивают мышиным моноклональным антителом против N-концевого участка CB1 (R&D). Результаты исследования приведены на фигуре 9B. Опять же, WIN55212 индуцирует интернализацию рецептора CB1, которая блокируется предварительной обработкой нейтральным антагонистом CB1 AM6545 (фигура 9B, верхний ряд гистограмм). Антитела против CB1 не влияют на поверхностную экспрессию CB1, позволяя предположить, что антитела против CB1 не индуцируют интернализацию рецептра (фигура 9B, средний и нижний ряды гистограмм).
Пример 14. Специфическая активность гуманизированных антител против CB1
[0286] Получают гуманизированные антитела P1C4 и тестируют их на активность, специфичность и сродство. Для получения гуманизированных антител P1C4 выбирают человеческие каркасные участки на основе гомологии P1C4 и генов человеческих зародышевых VH и VK. Каркасные участки, обладающие наивысшей степенью гомологии в отношении участков VH и VK P1C4, подвергают отбору на основе компьютерного моделирования по способности поддерживать структуру CDR, предположительно присутствующую в P1C4.
Получают следующие гуманизированные антитела: (1) P1C4-H0-IgG; (2) P1C4-H2(YE)- IgG (содержащее мутации G27Y и T28E в вариабельном участке тяжелой цепи); и (3) P1C4-H4(YENG)- IgG (содержащее мутации G27Y, T28E, A60N и Q61G в вариабельном участке тяжелой цепи)
[0287] Активность химерного антитела PA13R3-P1C4 и гуманизированного антитела PA13R3-P1C4 определяют путем анализа цАМФ. Функциональный анализ с измерением цАМФ (Cisbio) проводят на белых 384-луночных низкообъемных планшетах (Greiner). В каждую лунку планшета высевают клетки TRex-CHO, стабильно экспрессирующие нативный человеческий CB1, с плотностью 8000 клеток/лунку и затем проводят инкубацию с римонабантом (SR141716A), химерным PA12R3-P1C4, PA12R3-P1C4 H0 (без обратной мутации), PA12R3-P1C4 H2 (YE), PA12R3-P1C4 H4 (YENG) и P2A12 (отрицательный контроль) в концентрациях, варьирующих от 1 мкМ до 0 мкМ, в течение 10 минут при комнатной температуре. Через 10 минут к стимулирующей смеси добавляют 5 мкМ форсколин (Sigma Aldrich) и инкубируют в течение 30 мин при комнатной температуре. Чтобы количественно определить продукцию цАМФ, добавляют 5 мкл цАМФ-d2 и 5 мкл анти-цАМФ-криптата и инкубируют в течение часа. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism.
[0288] Результаты исследования приведены на фигуре 10 и ниже в таблице 9. Функциональный анализ с измерением цАМФ показывает, что гуманизированные P1C4-H2 и P1C4-H4 имеют IC50 21 нМ и 17 нМ, соответственно. Таким образом, среди тестируемых антител гуманизированные антитела H1C4-H2 и P1C4-H4 обладают даже большей активностью, измеряемой по ингибированию цАМФ, чем соответствующее химерное антитело.
Таблица 9. IC50 химерных и гуманизированных антител против CB1
химерное
Число мутаций
(YE)
(YENG)
[0289] Сродство связывания, перекрестную реакционноспособность и специфичность гуманизированных антител P1C4 определяют методом проточной цитометрии с использованием исходных клеток TRex CHO, клеток TRex CHO A56, экспрессирующих CB1 на повышенном уровне (T210A/партнер по гибридизации), клеток TRex CHO A156, экспрессирующих нативный человеческий CB1, исходных клеток Trex (не экспрессирующих CB1), мышиных клеток, экспрессирующих CB1, и клеток, стабильно экспрессирующих человеческий CB2. 100 мкл суспензии, содержащей 1×106 клеток/мл, инкубируют с химерным PA13R3-P1C4, гуманизированными P1C4-H0 (не содержит мутации), P1C4-H2 (YE), P1C4-H4 (YENG) или P2A12 (контрольный) IgG в 3-кратных серийных разведениях от 300 нМ до 0,5 нМ в течение 30 минут на льду. Затем клетки дважды промывают 200 мкл буфера FACS и инкубируют с PE-конъюгированным вторичным антителом против человеческих IgG (Southern Biotech) в течение 30 минут на льду. Клетки дважды промывают 200 мкл буфера FACS, переносят в пробирки BD Falcon объемом 5 мл и анализируют методом проточной цитометрии (BD FACScalibur).
Таблица 10. Сродство связывания и перекрестная реакционноспособность гуманизированных антител P1C4
CB1 T210A/партнер по гибридизации
(контроль)
[0290] Результаты исследования показаны на фигуре 11 и в приведенной выше таблице 10. Гуманизированные антитела против CB1 связываются с клетками A56 и с клетками A156. Оба гуманизированные антитела P1C4-H2 и PIC4-H4 связываются с клетками TRex CHO A56, экспрессирующими CB1 на повышенном уровне, а также с клетками A156, экспрессирующими нативный человеческий CB1, с более высоким сродством, чем химерное антитело P1C4 (фигура 11A). Кроме того, ни одно из тестируемых антител не обладает перекрестной реакционноспособностью в отношении мышиного CB1 или человеческого CB2 (фигура 11B).
Пример 15. Пониженные эффекторные функции антител против CB1
[0291] Конструируют и тестируют антагонистические антитела против CB1, обладающие пониженными эффекторными функциями. Получают антагонистические антитела против CB1, содержащие одну или несколько из перечисленных ниже модификаций Fc: (1) константный участок IgG4, содержащий замену серина на пролин в положении 228 (S228P); (2) константный участок IgG2, содержащий замену аланина на серин в положении 330 (A330S) и замену пролина на серин в положении 331 (P331S); и (3) гибридный константный участок IgG2/IgG4.
[0292] Последовательности полученных гуманизированных антител против CB1, содержащих 0, 2 или 4 обратные мутации в каркасных участках, описаны в SEQ ID NO: 343-351. Антитела тестируют на связывание с рецепторами Fcγ и компонентом комплемента C1q методом ELISA. Полученные антитела против CB1 также тестируют на способность активировать первичные человеческие иммунные клетки in vitro. А именно, антитела против CB1, содержащие одну или несколько модификаций Fc, тестируют на способность активировать иммунные клетки, например, путем определения их способности образовывать поперечные связи или индуцировать экспрессию маркеров активации. Результаты исследования демонстрируют, что антагонистические антитела против CB1 обладают более низкой способностью связывать FcγR и/и C1q, и/или индуцировать активацию иммунных клеток, чем соответствующее антагонистическое антитело против CB1, не содержащее модификации константного участка.
Пример 16. Исследование биораспределения
[0293] В данном исследовании определяют биораспределение антитела против CB1 P4B5 у мышей in vivo. Антитело P4B5 метят Vivotag 680 XL (Perkin Elmer) и вводят бесшерстным мышам (n=4 на группу) в.в. в дозе 5 мг/кг или 25 мг/кг. Визуализацию всего организма с измерением флуоресценции в разных тканях проводят методом флуоресцентной томографии (FMT) в следующие моменты времени: 0 часов (0 ч), 1 ч, 5 ч, 24 ч, 48 ч, 72 ч, 96 ч и 144 ч. Как показано на фигуре 12, меченное P4B5 связывается с CB1 с таким же сродством, что и немеченное P4B5. EC50 немеченного P4B5 составляет 60,5 нМ, а EC50 антитела P4B5, меченного Vivotag 680 XL, составляет 57,8 нМ.
[0294] Результаты исследования приведены на фигурах 13A и 13B. Меченное антитело детектируют на протяжении определенного периода времени, как показано на фигурах 13A.1 и A.2, на которых приведены результаты, полученные от типичной мыши, получающей повышенную дозу антитела (25 мг/кг). Однако после вычитания фонового сигнала крови антитело против CB1 не детектируют в мозге (фигура 13B), делая вывод, что антитело не протикает через гематоэнцефалический барьер после в.в. инъекции.
Пример 17. Экспрессия и анализ IgG гуманизированных вариантов PA13R3-P1C4
[0295] Среди разных подклассов человеческих IgG Fc-участки подклассов IgG2 и IgG4 отличаются низким связыванием с эффекторными молекулами, такими как активирующие FcγR или компонент комплемента 1q (C1q), которое обуславлявает пониженную эффекторную активность. Чтобы минимизировать активацию иммунных эффекторных функций, гуманизированные антитела основных серий P1C4-H2 и P1C4-H4 клонируют в 3 вариантах человеческих каркасных участков, Fc IgG2, IgG4 и гибрида IgG2 и IgG4, с целью дополнительной характеризации.
[0296] Последовательности вариабельного участка тяжелой цепи гуманизированного P1C4-H2 (SEQ ID NO: 340), вариабельного участка тяжелой цепи гуманизированного P1C4-H4 (SEQ ID NO: 341) и легкой цепи гуманизированного P1C4 (SEQ ID NO: 338) приведены ниже в таблице 11. Жирным шрифтом выделены остатки, введенные в результате обратных мутаций, а подчеркиванием выделены участки CDR. Для получения вариантов Fc разных семейств IgG используют три последовательности константных участков тяжелой цепи. Последовательности константного участка тяжелой цепи IgG2 (SEQ ID NO: 433), константного участка тяжелой цепи IgG4 (SEQ ID NO: 434), гибридного константного участка тяжелой цепи IgG2/4 (SEQ ID NO: 435) показаны ниже в таблице 11.
[0297] Варианты антител экспрессируют в клетках 293FreeStyle, CHO-S и CHO-K1 с последующей очисткой в разных партиях и в разные моменты времени.
[0298] Для проведения экспрессии в 293 FreeStyle отдельные плазмиды pTT5, кодирующие последовательности тяжелой и легкой цепей, совместно трансфицируют в клетки FreeStyle HEK293F, чтобы обеспечить экспрессию полноразмерного антитела IgG. Клетки культивируют при 37°C в атмосфере 5% CO2 в среде для экспрессии FreeStyle 293. За двадцать четыре часа до трансфекции клетки разбавляют до плотности 8×105 клеток/мл. Чтобы получить раствор для трансфекции, 80 мкг ДНК (40 мкг ДНК, кодирующей легкую цепь,+40 мкг ДНК, кодирующей тяжелую цепь) и 240 мкг полиэтиленимина (PEI) разводят в 8 мл среды Freestyle 293F, тщательно перемешивают, фильтруют через фильтровальные насадки для шприца с размером пор 0,2 мкм в коническую пробирку объемом 50 мл и инкубируют в течение 15 минут при 22°C. Восемь мл раствора для трансфекции медленно добавляют к 80 мл клеточной культуры 293F (разбавленной средой для экспрессии FreeStyle 293 до плотности 1,1-1,3×106 клеток/мл) и инкубируют при 37°C в 5% CO2 при вращении со скоростью 130 об/мин в течение 4 дней. Супернатанты трансфицированных клеточных культур собирают путем центрифугирования при 4000 об/мин в течение 45 минут при 4°C, доводят до pH 8,0 0,1 M NaOH и держат на льду до очистки белка.
[0299] Для проведения экспрессии в CHO-S отдельные плазмиды pTT5, кодирующие последовательности тяжелой и легкой цепей, совместно трансфицируют в клетки CHO-S, чтобы обеспечить экспрессию полноразмерного антитела IgG. Клетки культивируют в среде CD-CHO при 37°C и 5% CO2. За двадцать четыре часа до трансфекции клетки разбавляют до плотности 0,6-0,7×106 клеток/мл в среде CD-CHO. В день трансфекции клетки разбавляют до плотности 1,1-1,3×106 клеток/мл средой CD-CHO во встряхиваемых колбах объемом 250 мл. К каждой встряхиваемой колбе объемом 250 мл добавляют 80 мл клеток. Восемьдесят мкг плазмидной ДНК (40 мкг ДНК, кодирующей легкую цепь,+40 мкг ДНК, кодирующей тяжелую цепь) разводят в конечном объеме 4 мл среды CD-CHO и фильтруют через фильтровальные насадки для шприца с размером пор 0,2 мкм в коническую пробирку объемом 50 мл. В отдельной конической пробирке объемом 50 мл 80 мкл реагента FreeStyle Max разводят в конечном объеме 4 мл среды CD-CHO. 2 полученные смеси инкубируют при 22°C в течение 3 минут, после чего их объединяют, смешивают и инкубируют еще 15 минут при 22°C. 8 мл смеси ДНК/реагент для трансфекции медленно добавляют к 80 мл клеточной культуры в колбе объемом 250 мл. В результате достигается конечная плотность ~1,0×106 клеток/мл. Затем культуральные колбы инкубируют на платформе с орбитальным вращением со скоростью 133 об/мин при 37°C и 5% CO2 в течение 6 дней. Культуральные супернатанты собирают путем центрифугирования (Allegra X-15R, Beckman) при 4000 g в течение 40 минут при 4°C, доводят до pH 8,0 0,1 M NaOH и держат на льду до очистки белка.
[0300] Очистку IgG, полученных с использованием 293 FreeStyle и CHO-S, проводят следующим образом: супернатанты загружают на колонки с белком A (объем слоя 0,4 мл), предварительно уравновешенные PBS pH 7,4, и оставляют стекать под действием силы тяжести. Колонки промывают 5 мл PBS pH 7,4, после чего белок элюируют 4 мл буфера 0,1 M Na цитрат-HCl pH 3,5. Элюированное вещество нейтрализуют 200 мкл буфера 1,5 M Tris-HCl pH 8,8, концентрируют и заменяют буфер на PBS, pH 7.4, с помощью концентратора Amicon, 30 кДа, 4 мл (Millipore), в соответствии с инструкциями производителя, с достижением конечного объема примерно 0,5-1 мл. Концентрацию белка определяют по поглощению при 280 нм, а чистоту определяют методами SDS-PAGE и SEC.
[0301] Экспрессию белка в CHO-K1 и последующую очистку проводят с помощью методов, запатентованных контрактной исследовательской организацией (CRO). Коротко говоря, для трансфекции используют клетки CHO-K1. IgG очищают с помощью гранул MabSelect™ SuRe™, стадии промывания проводят с использованием PBS в модификации Дульбекко (Lonza BE17-512Q). IgG элюируют 0,1 M глицином, pH 3,5.
[0302] Во всех тестах, анализах SDS-PAGE и SEC, проводимых с целью QC (контроля качества) белка, используют 3 мкг антитела. Критерии удовлетворяющие требованиям QC, включают в себя чистоту >90% по результатам SDS PAGE и мономерный пик >90% в методе SEC. Аликвоты очищенного белка IgG, прошедшего тесты QC, помещают в пробирки с завинчивающимися крышками в количестве 100 мкл/пробирку при концентрации ~5 мг/мл. Аликвоты быстро замораживают в жидком азоте и хранят при -80°C.
Выход белка в клетках 293FreeStyle варьирует от низкого до 53 мг/л. Выход в клетках CHO-S является низким и составляет примерно 1 мг/л. Выход в клетках CHO-K1 варьирует от 198 мг/л до 350 мг/л. Результаты SDS-PAGE свидетельствуют о наличии интактного белка, элюирующегося в невосстанавливающих условииях как белок с массой примерно 150 кДа, и 2 полос, соответствующих тяжелой цепи и легкой цепи, при отсутствии видимой деградации или агрегации. В качестве примеров профили SEC и результаты SDS-PAGE для одной из партий 293 FreeStyle приведены на фигуре 14A, и для одной из партий CHO-K1 - на фигуре 14B. Результаты очистки белков, экспрессированных с использованием партий 293FreeStyle и CHO-S, приведены в таблице 12.
Таблица 12. Результаты очистки белка mAb PA13R3-P1C4
Пример 18. Функциональный анализ гуманизированных вариантов PA13R3-P1C4 путем измерения цАМФs
[0303] Коммерчески доступный набор (Cisbio) для проведения иммуноанализа в конкурентном формате, содержащий криптат-меченое антитело против цАМФ и d2-меченый цАМФ, используют для характеристики гуманизированных вариантов антител PA13R3-P1C4 путем измерения изменений уровней внутриклеточного цАМФ в клетках TRex-CHO, стабильно экспрессирующих CB1. Число клеток, концентрацию форсколина и концентрацию CP55,940 оптимизируют в соответствии с инструкциями производителя. Антагонистический функциональный анализ цАМФ проводят в белых 384-луночных низкообъемных планшетах (Greiner). Клетки TRex-CHO, экспрессирующие человеческий CB1, высевают с плотностью восемь тысяч клеток/лунку в бессывороточной среде Hamʹs F12 и инкубируют с mAb или контрольным соединением в разных концентрациях при 22°C в течение 10 минут. Затем к клеткам добавляют пяти мкМ форсколин (при концентрации EC20, Sigma Aldrich) и 9 нМ CP55,940 (при EC80, Sigma Aldrich) и инкубируют в течение 30 минут при 22°C для повышениям активности аденилатциклазы и активации сигнального пути CB1, соответственно. Затем к клеткам добавляют пять мкл цАМФ-d2 (разведение 1:39, конъюгат и буфер для лизиса получают от Cisbio) и 5 мкл анти-цАМФ-криптата (разведение 1:9, конъюгат и буфер для лизиса получают от Cisbio) и инкубируют в течение 1 часа при 22°C. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism.
[0304] В данном анализе активность гуманизированных антител PA13R3-P1C4 сравнивают с активностью исходного химерного PA13R3-P1C4, римонабанта, низкомолекулярного обратного агониста CB1, и mAb P2A12, неспецифичного к GPCR антитела изотипа IgG1, используемого в качестве отрицательного контроля. mAb PA13R3-P1C4 и его гуманизированные варианты дозо-зависимо ингибируют CP55,940-индуцированное уменьшение уровня внутриклеточного цАМФ, тогда как используемое в качестве отрицательного контроля mAb P2A12 не оказывает никаких эффектов (фигура 15). Средние значения IC50±SD гуманизированных вариантов PA13R3-P1C4 приведены в таблице 13.
Таблица 13. Значения IC50 гуманизированных вариантов P1C4, определенные путем антагонистического анализа цАМФ
*p <0,05, **p <0,005; и в сравнении с римонабантом #p <0,05, ##p <0,02.
[0305] Гуманизированные варианты P1C4-h2 и h4 являются более активными (в 1,6-3,3 раза), чем химерное mAb PA13R3-P1C4 по результатам антагонистического анализа цАМФ (p<0,005). Гуманизированные варианты P1C4-h2 и P1C4-h4 обладают сравнимой активностью, тогда как среди разных изотипов гуманизированных вариантов PA13R3-P1C4 более высокая активность чаще наблюдается у вариантов IgG1 и IgG4, чем у вариантов IgG2 и IgG2/4.
[0306] Авторы изобретения дополнительно характеризуют механизм антагонизма гуманизированного варианта антитела PA13R3-P1C4, в частности, могут ли такие антитела вести себя как обратные агонисты или нейтральный антагонисты CB1. Римонабант (SR141716A), известный обратный агонист CB1, и AM6545, известный нейтральный антагонист CB1, используют в качестве стандартных соединений.
[0307] Чтобы определить, действительно ли гуманизированный вариант антитела PA13R3-P1C4 действует как обратный агонист или нейтральный антагонист, проводят анализ цАМФ в отсутствии экзогенного агониста. Форсколин, неспецифический активатор аденилилциклазы, добавляют в аналитическую среду, чтобы повысить базовый уровень цАМФ до пределов детекции. Агонистический функциональный анализ цАМФ (Cisbio) проводят в белых 384-луночных низкообъемных планшетах (Greiner). Клетки TRex-CHO, экспрессирующие человеческий CB1, высевают с плотностью восемь тысяч клеток/лунку в бессывороточной среде Hamʹs F12 и инкубируют с mAb или контрольным соединением в разных концентрациях при 22°C в течение 10 минут. Затем к клеткам добавляют пяти мкМ форсколин (Sigma Aldrich) и инкубируют в течение 30 минут при 22°C. Затем к клеткам добавляют 5 мкл цАМФ-d2 (разведение 1:39, конъюгат и буфер для лизиса получают от Cisbio) и 5 мкл анти-цАМФ-криптата (разведение 1:9, конъюгат и буфер для лизиса получают от Cisbio) и инкубируют в течение 1 часа. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism.
[0308] Авторы настоящего изобретения сравнивают активность гуманизированных вариантов P1C4-h2-IgG2 и P1C4-h2-IgG4 с активностью римонабанта, AM6545 и антитела P2A12-IgG1, используемого в качестве отрицательного контроля, в клетках TRex-CHO CB1, стимулированных 1,5 мкМ форсколином (фигура 16A), а также в клетках TRex-CHO CB1, стимулированных 5 мкМ форсколином (фигура 16B). Результаты демонстрируют, что P1C4-h2-IgG1, P1C4-h2-IgG2, P1C4-h2-IgG4 и римонабант дозо-зависимо повышают уровень цАМФ, что указывает на механизм обратного агонизма. И наоборот, AM6545, нейтральный антагонист CB1, и антитело P2A12-IgG1, используемое в качестве отрицательного контроля, не влияют на уровень цАМФ.
[0309] Проводя анализ кривых Шильда, чтобы определить равновесную константу диссоциации (KB), которая является показателем сродства антагониста к рецептору независимо от природы и концентрации используеого агониста. Кривые доза-ответ для агонистов CB1 CP55,940 и WIN55,212 в анализе с использованием антагониста цАМФ HTRF получают в присутствии разных концентраций P1C4-h2-IgG4.
[0310] На фигурах 17A-D показано влияние увеличивающихся концентраций P1C4-h2-IgG4 на CP55,940- и WIN55,212-индуцированную активность CB1, измеряемое путем анализа цАМФ (соответственно). Отношение доз (R) рассчитывают на основании EC50 CP55,940 или WIN55,212, концентрации, до которой нужно повысить количество агонистов CB1 (CP55,940 или WIN55,212), чтобы получить такой же ответ в присутствии P1C4-h2-IgG4, как и в его отсутствии. В приведенных ниже таблицах 14 и 15 показаны значения наклона кривых Шильда и равновесные константы диссоциации (KB), измеренные в 4 разных экспериментах.
Таблица 14. Значения наклона кривых Шильда и равновесные константы диссоциации для CP55,940
Таблица 15. Значения наклона кривых Шильда и равновесные константы диссоциации для WIN55,212
Пример 19. Анализ активации ERK под действием гуманизированных вариантов PA13R3-P1C4
[0311] В примере 10 и на фигуре 5A показано, что исходное антитело PA13R3-P1C4 блокирует WIN55,212-индуцированную активацию ERK. Чтобы подтвердить, что гуманизированные варианты PA13R3-P1C4 также блокируют WIN55,212-индуцированную активацию ERK, авторы изобретения тестируют способность указанных антител ингибировать WIN55,212-индуцированное фосфорилирование ERK.
[0312] За два дня до эксперимента клетки Trex-CHO, экспрессирующие рецептор CB1, высевают в 6-луночные планшеты при плотности 500000 клеток/лунку. Через 24 часа добавляют 1 мкг/мл тетрациклина, чтобы индуцировать экспрессию рецептора CB1. В течение, по меньшей мере, двух часов перед началом эксперимента клетки держат на бессывороточной среде. К культуральной среде добавляют очищенные IgG в концентрации 300 нМ и через 30 минут клетки стимулируют агонистом рецептора CB1 WIN55,212 (100 нМ) в течение 10 и 15 минут. Собирают клеточные лизаты и определяют уровень активации ERK методом вестерн-блоттинга. Антитела против ERK и против фосфо-ERK получают от Cell Signaling Inc.
[0313] Как показано на фигуре 18A, обработка агонистом CB1 WIN55,212 повышает уровень фосфорилированной ERK, позволяя предположить, что сигналы рецептора CB1 опосредуются путем ERK. Предварительная обработка CB1-специфическим антагонистом римонабантом ингибирует WIN55,212-индуцированную активацию ERK. Подобно римонабанту, предварительная обработка антителами против CB1 P1C4-h2-IgG1 и P1C4-h4-IgG1 ингибирует WIN55,212-индуцированную активацию ERK (фигура 18A). P1C4-h0-IgG1 не ингибирует WIN55,212-индуцированную активацию ERK. Это согласуется с результатами антагонистического анализа цАМФ, которые демонстрируют, что P1C4-h0-IgG1 является менее активным, чем другие гуманизированные варианты P1C4, и позволяет предположить, что указанное антитело не обладает способностью блокировать WIN55,212-индуцированную активацию ERK.
[0314] Также определяют влияние P1C4-h2, содержащих каркасные участки IgG2 и IgG4, на WIN55,212-активируемый путь ERK. Подобно химерному PA13R3-P1C4 и гуманизированному P1C4-h2-IgG1, предварительная обработка антителами против CB1 P1C4-h2-IgG2 и P1C4-h2-IgG4 блокирует WIN55,212-индуцированное фосфорилирование ERK (фигура 18B). Не специфичное к GPCR mAb P2A12 не блокирует WIN55,212-индуцированную активацию ERK. Полученные результаты свидетельствуют о том, что указанные Fc каркасные участки не влияют на антагонистические свойства PA13R3-P1C4.
Пример 20. Исследование влияния гуманизированных вариантов PA13R3-P1C4 на интернализацию рецептора CB1
[0315] Влияние PA13R3-P1C4 и гуманизированных вариантов антител на интернализацию рецептора CB1 анализируют методом проточной цитометрии в присутствии или отсутствии агонистических или антагонистических соединений.
[0316] В день эксперимента клетки держат в бессывороточной среде в течение 2 часов. Затем клетки предварительно инкубируют с антителом против CB1 (300 нМ), AM6545 (нейтральный антагонист CB1) и отрицательным контролем (соединение, связывающее BRIL) в течение получаса. Затем в культуральную среду добавляют агонист CB1 (1 мкМ WIN55,212) и инкубируют 1 час, чтобы индуцировать интернализацию рецептора. Экспрессированный на поверхности CB1 окрашивают мышиным моноклональным антителом против N-концевого участка CB1, полученным от R&D, и методом проточной цитометрии (Guava) определяют среднюю интенсивность флуоресценции (MFI).
[0317] Тетрациклин индуцирует экспрессию CB1 в клетках TRex-293, что подтверждает увеличение поверхностного окрашивания мышиным моноклональным антителом, специфичным к N-концевому участку CB1 (фигура 19A, панель A, линия, обозначенная точками). Обработка тетрациклином в присутствии агониста CB1 WIN55,212 уменьшает поверхностное окрашивание, свидетельствуя о снижении количества CB1 на клеточной поверхности в результате интернализации (фигура 19A, панель A, пунктирная линия). Предварительная обработка CB1-специфичным антагонистом римонабантом (фигура 19A, панель B, сплошная черная линия) ингибирует агонист-индуцированное уменьшение окрашивания CB1 на клеточной поверхности. Подобно римонабанту, предварительная обработка антителами против CB1 (PA13R3-P1C4 (фигура 19A, панель D, сплошная черная линия), P1C4-h2-IgG1 (фигура 19A, панель F, сплошная черная линия) и P1C4-h4-IgG1 (фигура 19A, панель G, сплошная черная линия)) ингибирует WIN55,212-индуцированную интернализацию рецептора CB1. P1C4-h0-IgG1 (фиг. 19A, панель E, сплошная черная линия) и используемое в качестве отрицательного контроля антитело P2A12-IgG1 (фигура 19A, панель C, сплошная черная линия) не ингибируют WIN55,212-индуцированную интернализацию CB1. Согласно результатам антагонистического анализа цАМФ и анализа активаци ERK, P1C4-h0-IgG1 является менее активным, чем другие гуманизированные варианты P1C4, причем его неспособность блокировать WIN55,212-индуцированную интернализацию рецептора может быть обусловлена высокой скоростью диссоциации P1C4-h0-IgG1.
[0318] Среди разных подклассов человеческих IgG Fc-участки подклассов IgG2 и IgG4 отличаются низким связыванием с эффекторными молекулами, такими как активирующие FcγR или компонент комплемента 1q (C1q), которое обуславлявает пониженную эффекторную активность. Соответственно, P1C4-h2 клонируют в человеческих каркасных участках Fc IgG2 и IgG4, поскольку при терапевтическом применении активация иммунных эффекторных функций является нежелательной (как описано в примере 17). Затем гуманизированные варианты P1C4-h2-IgG1, P1C4-h4-IgG1, P1C4-h2-IgG2 и P1C4-h2-IgG4 анализируют на способность блокировать интернализацию рецептора CB1. Как указано выше, экспрессию CB1 в клетках TRex-293, индуцированную тетрациклином, подтверждают путем увеличения окрашивания поверхности мышиным моноклональным антителом, специфичным к N-концевому участку CB1 (фигура 19B, панель A, линия, отмеченная точками). Обработка тетрациклином в присутствии агониста CB1 WIN55,212 уменьшает поверхностное окрашивание, свидетельствуя о снижении количества CB1 на клеточной поверхности в результате интернализации (фигура 19B, панель A, пунктирная линия). Предварительная обработка CB1-специфичным антагонистом римонабантом (фигура 19B, панель B, сплошная черная линия) ингибирует агонист-индуцированное уменьшение окрашивания CB1 на клеточной поверхности. Подобно римонабанту, предварительная обработка антителами против CB1 (PA13R3-P1C4 (фигура 19B, панель D, сплошная черная линия), P1C4-h2-IgG1 (фигура 19B, панель F, сплошная черная линия) и P1C4-h4-IgG1 (фигура 19B, панель G, сплошная черная линия)) ингибирует WIN55,212-индуцированную интернализацию рецептора CB1. P1C4-h0-IgG1 (фиг. 19B, панель E, сплошная черная линия) и используемое в качестве отрицательного контроля антитело P2A12-IgG1 (фигура 19B, панель C, сплошная черная линия) не ингибируют WIN55,212-индуцированную интернализацию CB1.
Пример 21. Анализ связывания гуманизированных вариантов антител PA13R3-P1C4 методом проточной цитометрии
[0319] Сродство гуманизированных вариантов PA13R3-P1C4 определяют методом проточной цитометрии с использованием клеток TRex CHO, стабильно трансфицированных CB1. Собирают исходные клетки TRex CHO и клетки TRex-CHO, стабильно трансфицированные конструкциями, обеспечивающими экспрессию тетрациклин-индуцируемого человеческого CB1, человеческого CB2 или мышиного CB1. Чтобы определить связывание тестируемых антител, сто микролитров суспензии клеток с плотностью 1×106 клеток/мл инкубируют с тестируемыми антителами в диапазоне концентраций от 1 мкМ до 1,3 нМ в течение 30 минут на льду. Затем клетки центрифугируют при 4°C и 1600 об/мин в течение 3 минут; супернатант удаляют отсасыванием, после чего клетки промывают 200 мкл буфера FACS. Процедуру промывания повторяют дважды. После последнего промывания клетки ресуспендируют в буфере FACS, содержащем вторичные PE-конъюгированные антитела против Fc человеческих IgG (в разведении 1:200), и инкубируют при 4°C в течение 30 минут. Затем клетки дважды промывают 200 мкл буфера FACS и анализируют методом проточной цитометрии (Guava). Анализ результатов и измерение сродства (KD) проводят с помощью программного обеспечения GraphPad Prism. Средние значения константы диссоциации (KD) гуманизированных вариантов PA13R3-P1C4 определяют, используя результаты, по меньшей мере, 4 разных экспериментов, проводимых с использованием, по меньшей мере, 2 разных партий белка (фигуры 20A-D и таблица 16).
Таблица 16. Средние значения константы диссоциации (KD) гуманизированных вариантов PA13R3-P1C4
*p <0,05, **p <0,01, ***p <0,001.
[0320] У гуманизированных вариантов, за исключением h2-IgG2, h2-IgG2/4 и h4-IgG2/4, наблюдается статистически значимое увеличение сродства к человеческому CB1 по сравнению с исходным химерным антителом PA13R3-P1C4. Средние значения константы диссоциации для каждого соединения определяют с использованием, по меньшей мере, двух разных препаратов белка (за исключением P1C4-h2-IgG1, который существует в виде только одного препарата). Сравнивают константы диссоциации, полученные для разных партий одного соединения (таблица 16). Варианты P1C4-h2 и P1C4-h4 обладают сходными значениями сродства. Однако существует слабая тенденция, которая не является статистически значимой, заключающаяся в том, что варианты IgG1 обладают более высоким сродством, чем варианты IgG2, IgG4 и IgG2/4.
[0321] P1C4-h0-IgG1 имеет самую низкое измеренное значение константы диссоциации (KD 24 нМ). Однако максимальный сигнал FACS (средняя интенсивность флуоресценции) у P1C4-h0-IgG1 меньше, чем у других вариантов P1C4. Это может свидетельствовать о более высокой скорости диссоциации P1C4-h0-IgG1.
[0322] Селективность связывания и перекрестную реакционноспособность PA13R3-P1C4 и его гуманизированных вариантов также характеризуют методом проточной цитометрии с использованием клеток TRex CHO, стабильно трансфицированных представляющими интерес GPCR. А именно, определяют селективность связывания с CB1 по сравнению с CB2. Также определяют перекрестную реакционноспособность в отношении мышиного CB1. Чтобы детектировать экспрессию мышиного CB1, 100 мкл суспензии клеток с плотностью 1×106 клеток/мл инкубируют с кроличьим поликлональным антителом против CB1 (Santa Cruz) в концентрации 1 мкг/100 мкл на льду в течение 30 мин. Мышиное моноклональное антитело против CB2, полученное от R&D Systems, используют для подтверждения экспрессии CB2. 100 мкл суспензии клеток с плотностью 1×106 клеток/мл инкубируют с антителом против CB2 в концентрации 0,5 мкг/100 мкл на льду в течение 30 минут. После инкубации клетки центрифугируют при 4°C и 1600 об/мин в течение 3 минут; супернатант удаляют отсасыванием, после чего клетки промывают 200 мкл буфера FACS. Процедуру промывания повторяют дважды. После последнего промывания клетки ресуспендируют в буфере FACS, содержащем FITC-конъюгированные вторичные антитела против кроличьих IgG (в разведении 1:200) для детекции мышиного CB1, и PE-конъюгированные вторичные антитела против мышиных IgG (в разведении 1:200) для детекции CB2, и инкубируют при 4°C в течение 30 минут. После инкубации в течение 30 минут со вторичными антителами клетки центрифугируют при 4°C и 1600 об/мин в течение 3 минут; супернатант отсасывают, чтобы удалить избыток вторичных антител. Клетки дважды промывают 200 мкл буфера FACS и анализируют методом проточной цитометрии (Guava). Строят кривые связывания очищенных IgG с учетом всех концентраций от 1 мкМ до 1,3 нМ. Анализ результатов и измерение сродства (KD) проводят с помощью программного обеспечения GraphPad Prism.
[0323] Значения селективности связывания и перекрестной реакционноспособности гуманизированных вариантов PA13R3-P1C4 в отношении человеческого CB1 по сравнению с человеческим CB2 и мышиным CB1 показаны на фигурах 20E-M. Подобно PA13R3-P1C4, гуманизированные варианты селективно связываются с человеческим CB1 по сравнению с человеческим CB2. Связывание с мышиным CB1 практически не наблюдается при концентрациях до 1 мкМ, свидетельствуя об отсутствии перекрестной реакционноспособности в отношении данных соединений, несмотря на высокую степень идентичности аминокислотных последовательностей внеклеточных доменов человеческого и мышиного CB1 (идентичность 97%).
Пример 22. Влияние перестановки ELC2 на связывание FACS
[0324] Чтобы протестировать влияние мутаций ECL2 на способность P1C4 связывать CB1, экспрессированный на клеточной поверхности, авторы изобретения получают конструкцию, обеспечивающую экспрессию CB1 в клетках путем направленного мутагенеза. Коротко говоря, проводят ПЦР, используя 2 олигонуклеотида, Apollo_ECL2_h2m_F (CTGCAATCTGTTTGCTCAGACATTTTCCCACTCATTGATGAAACCTACCT) (SEQ ID NO: 826) и Apollo_ECL2_h2m_R (GGAAAATGTCTGAGCAAACAGATTGCAGTTTCTTGCAGTTCCAGCCCAGG) (SEQ ID NO: 827), в качестве праймеров и pcDNA4TO-человеческий CB1 в качестве матрицы. Посредством данной реакции в ECL2 вводят мутации E→K и H→L, получая последовательность ECL2 человеческого CB1, идентичную последовательности ECL2 мышиного CB1. 50 мкл реакционной смеси для ПЦР содержат 10 мкл буфера 5×PCR, 2 мкл dNTP (каждый в концентрации 10 мМ), 0,25 мкл каждого из обратного и прямого праймеров (100 мМ исходный раствор), 50 нг ДНК-матрицы, 1 мкл DMSO и 1 мкл полимеразы Phusion (NEB). Циклы ПЦР, которые включают в себя 95°C в течение 30 секунд, 55°C в течение 1 минуты, 72°C в течение 7 минут, повторяют 16 раз. После окончания реакции к продукту ПЦР добавляют 1 мкл DpnI (20 ед/мкл). Продукт ПЦР инкубируют при 37°C в течение 1 часа и затем 1 мкл используют для трансформации Dh5α E. coli. Полученные трансформанты культивируют в чашках и отдельные колонии подвергают секвенированию, чтобы подтвердить наличие мутации. Полученную конструкцию называют pcDNA4TO-CB1 с перестановкой человеческий/мышиный ECL2-IRES-GFP.
[0325] Чтобы подтвердить, что участки E→K и H→L в ECL2 играют важную роль в связывании PA13R3-P1C4 с CB1, методом проточной цитометрии проводят анализ связывания P1C4-h4-IgG1 с клетками TRex-CHO, временно трансфицированными конструкциями, обеспечивающими экспрессию человеческого CB1, мышиного CB1 или CB1 с перестановкой человеческий/мышиный ECL2.
[0326] Клетки TRex-CHO выращивают в культуральной среде Hamʹs F12, содержащей 10% фетальной бычьей сыворотки, 1% пенициллина и стрептомицина и 10 мкг/мл бластицидина. За день до трансфекции клетки пересаживают и высевают в 6-луночный планшет в количестве 0,5×106 клеток на лунку, содержащую 2 мл культуральной среды. В день трансфекции в клетки вводят pcDNA4TO-человеческий CB1, pcDNA4TO-мышиный CB1-IRES-GFP, pcDNA4TO-CB1 с перестановкой человеческий/мышиный ECL2-IRES-GFP или pcDNA4TO-GFP в качестве отрицательного контроля, используя липофектамин 2000 в соответствии с инструкциями Life Technologiesʹ. На следующий день после трансфекции клетки обрабатывают тетрациклином, 1 мкг/мл, в течение 24 часов, чтобы индуцировать экспрессию CB1. В день эксперимента среду отсасывают; клетки промывают один раз DPBS и инкубируют с буфером для диссоциации клеток в течение 5 минут. Клетки собирают и центрифугируют при 1600 об/мин в течение 3 минут. Супернатанты удаляют отсасыванием, после чего клетки промывают DPBS. Число клеток определяют с помощью цитометра Bio-Rad T10. Клетки центрифугируют при 1600 об/мин в течение 3 минут; супернатант отсасывают, чтобы удалить DPBS, после чего клетки ресуспендируют в буфере FACS (3% FBS в DPBS, 0,09% азида натрия) до плотности 1×106 клеток/мл. Чтобы определить связывание mAb PA13R3-P1C4 с клетками, экспрессирующими человеческий CB1, мышиный CB1, CB1 с перестановкой человеческий/мышиный ECL2, а также с контрольными клетками, 100 мкл суспензии, содержащей 1×106 клеток/мл, инкубируют с P1C4-h4-IgG1, mAb против CB1, R&D, или P2A12 в течение 30 минут на льду. Вторичные PE-конъюгированные mAb против человеческих IgG или PE-конъюгированные mAb против мышиных IgG разводят 1:200 буфером FACS. Клетки, окрашенные только вторичным антителом, используют в качестве отрицательного контроля. После инкубации со вторичным антителом клетки дважды промывают 200 мкл буфера FACS и анализируют методом проточной цитометрии (Guava).
[0327] Как показано на фигуре 21, P1C4-h4-IgG1 связывается с человеческим CB1, но не с мышиным CB1 и не с CB1, содержащим перестановку человеческий/мышиный ECL2. Единственным различием между человеческим CB1 и CB1, содержащим перестановку человеческий/мышиный ECL2, является замена остатков E→K и H→L в ECL2. Результаты измерения связывания методом проточной цитометрии свидетельствуют о том, что ECL2 играет ключевую роль в связывании P1C4. Моноклональное антитело против CB1, R&D, используют в качестве положительного контроля, демонстрирующего экспрессию CB1. P2A12 и pcDNA4TO-GFP используют в качестве контроля окрашивания и пустого контрольного вектора, соответственно.
Пример 23. Анализ эффекторных функций ADCC и CDC
[0328] Чтобы подтвердить предполагаемое отсутствие эффекторных функций у вариантов P1C4-h2-IgG2 и P1C4-h2-IgG4 гуманизированного P1C4, проводят анализы антителозависимой клеточноопосредованной цитотоксичности (ADCC) и комплементзависимой цитотоксичности (CDC) с использованием клеток Дауди.
[0329] Чтобы провести анализ ADCC, концентрацию клеток-мишеней Дауди доводят до 12,5×104 клеток/мл средой ADCC. В каждую лунку круглодонного 96-луночного планшета добавляют 80 мкл клеточной суспензии (1×104 жизнеспособных клеток). Двадцать мкл серийных разведений антитела в среде ADCC распределяют в лунки с тройными повторами. Конечные концентрации ритуксимаба и Anti-hel-hIgG1 составляют: 0,128 нг/мл, 0,64 нг/мл, 3,2 нг/мл, 16 нг/мл, 80 нг/мл, 0,4 мкг/мл, 2 мкг/мл; конечные концентрации Anti-hel-hIgG2, Anti-hel-hIgG4, P1C4-h2-IgG2 и P1C4-h2-IgG4 составляют: 3,2 нг/мл, 16 нг/мл, 80 нг/мл, 0,4 мкг/мл, 2 мкг/мл, 10 мкг/мл, 50 мкг/мл. Планшшет инкубируют при 22-25°C в течение 30 минут. Концентрацию PBMC доводят средой ADCC так, чтобы после добавления 100 мкл клеток PBMC к клеткам-мишеням, отношение эффекторные клетки:клетки-мишени составляло 25:1 и 50:1, соответственно. Планшет центрифугируют при 250 g в течение 4 минут и затем инкубируют при 37°C. За 45 минут до сбора супернатанта 20 мкл лизирующего раствора из набора CytoTox 96 (Promega) добавляют в контрольные лунки максимального лизиса. После инкубации в течение 5 часов планшет центрифугируют при 250 g в течение 4 минут. 50 мкл супернатанта из каждой лунки переносят в чистый плоскодонный 96-луночный аналитический планшет. В каждую лунку добавляют 50 мкл субстрата из набора CytoTox 96 (Promega) и перемешивают в течение 30 секунд. Планшет инкубируют при комнатной температуре в течение 30 минут, после чего в каждую лунку добавляют 50 мкл останавливающего раствора и перемешивают в течение 30 секунд. Поглощение считывают при 490 нм. Анализ полученных данных: % лизиса рассчитывают с помощью GraphPad Prism 5.0 с получением IC50 по следующей формуле:
% лизиса=[(экспериментальное высвобождение)- (мишени+PBMC)]/[(максимальный лизис)-среднее(только мишени)]×100
[0330] Ритуксимаб индуцирует эффект ADCC в клетках Дауди со значением IC50 1,19 нг/мл при соотношении E/T 50:1, 0,92 нг/мл при соотношении E/T 25:1. Как показано на фигурах 22A-C варианты Fc антител P1C4 не обладают активностью ADCC в клетках Дауди при соотношении E/T 50:1 и 25:1.
[0331] Чтобы провести анализ CDC, клетки-мишени Дауди собирают и доводят их концентрацию до 80×104 клеток/мл средой для культивирования клеток. Суспензию клеток вносят в плоскодонный 96-луночный белый планшет в объеме 25 мкл/лунку. Затем во все лунки с двойными повторами добавляют серийные разведения P1C4, ритуксимаба, Anti-HEL-hIgG1 и Anti-HEL-hIgG2 в среде ADCC, в объеме 12,5 мкл/лунку. Конечные концентрации ритуксимаба и Anti-hel-hIgG1 составляют: 0,128 нг/мл, 0,64 нг/мл, 3,2 нг/мл, 16 нг/мл, 80 нг/мл, 0,4 мкг/мл, 2 мкг/мл; конечные концентрации Anti-hel-hIgG2, Anti-hel-hIgG4, P1C4-H2-IgG2 и P1C4-H2-IgG4 составляют: 3,2 нг/мл, 16 нг/мл, 80 нг/мл, 0,4 мкг/мл, 2 мкг/мл, 10 мкг/мл, 50 мкг/мл. Планшеты инкубируют в вытяжном шкафу в течение 15 минут. В планшеты, содержащие клетки, добавляют человеческий комплемент, разбавленный средой ADCC, в объеме 12,5 мкл/лунку с достижением конечной концентрации 10%. В лунки максимального лизиса добавляют 5 мкл лизирующего раствора. Конечный объем во всех лунках доводят до 50 мкл средой ADCC. Планшеты инкубируют при 37°C в течение 2 часов и затем к клеткам добавляют 50 мкл/лунку раствора CTG. Планшеты встряхивают на шейкере для микропланшет в течение 2 минут со скоростью 200 и затем инкубируют при комнатной температуре в течение 10 минут. Люминесцентный сигнал считывают с помощью Envision. Анализ полученных данных: % цитотоксичности и IC50 рассчитывают с помощью GraphPad Prism 5.0 по следующей формуле:
% цитотоксичности=[(экспериментальное значение)- среднее(клетки+комплемент)]/[среднее(максимальный лизис)-среднее(только клетки)]×100
[0332] CDC-активность антител тестируют на клетках Дауди, лизис клеток Дауди под действием PBMC оценивают с помощью анализа клеточного титра Glo, конечная концентрация комплемента составляет 10%. Ритуксимаб индуцирует CDC-активность в отношении клеток Дауди с IC50 399 нг/мл. Как показано на фигуре 22D, варианты Fc антитела P1C4 не проявляют CDC-активности в отношении клеток Дауди.
Пример 24. Распознавание денатурированного белка CB1 антителами P1C4
[0333] В данном исследовании методом вестерн-блоттинга определяют, способны ли PA13R3-P1C4 и его гуманизированные варианты распознавать эпитоп (эпитопы) на денатурированном белке CB1 (His-меченый, укороченный по N/C-концам CB1). Очищенный человеческий белок CB1 (750 нг на полосу) смешивают с SDS-содержащим восстанавливающим буфером, содержащим бета-меркаптоэтанол (Double Helix). Денатурированный рекомбинантный белок CB1 загружают на 12% восстанавливающие гели SDS-PAGE, после чего белок разделяют при 120 В в течение одного часа и переносят на мембраны PVDF (предварительно замоченные в метаноле) под действием электрического тока при 300 мА в течение 70 минут. Мембраны блокируют 5% NFDM/PBS-T в течение одного часа при 22°C и затем проводят иммуноблоттинг с использованием тестируемых антител (2 мкг/мл) в 5% NFDM/PBS-T в течение ночи at 4°C. В качестве положительных контролей используют коммерческое мышиное антитело против His и мышиное антитело против CB1 (R&D Systems), а в качестве отрицательного контроля используют кроличье антитело против CB1 (Cayman), котрое распознает C-концевой участок, удаленный из рекомбинантного белка CB1, используемого в данном исследовании.
[0334] После инкубации в течение ночи мембраны при 22°C промывают три раза PBS, содержащим 0,5% твин-20 (PBS-T). К соответствующим мембранам добавляют вторичные AP-конъюгированные антитела против человеческих IgG (1:5000), или против мышиных IgG, или против кроличьих IgG, в 5% NFDM/PBS-T и инкубируют в течение 1 часа при 22°C. Мембраны промывают PBS-T три раза по 5 минут. После последнего промывания развивают окрашивание путем инкубации с раствором субстрата NBT/BCIP при 22°C и затем реакцию останавливают путем промывания мембран проточной водой.
[0335] Результаты вестерн-блоттинга демонстрируют, что используемые в качестве положительного контроля антитела позволяют детектировать денатурированный белок CB1 с правильной кажущейся молекулярной массой 63 кДа (фигура 23B, полосы 9 и 10). Полосы, соответствующие используемому в качестве отрицательного контроля антителу, специфичному к C-концу (фигура 23B, полоса 11), PA13R3-P1C4 (фигура 23A, полоса 2) или гуманизированным вариантам антител (фигура 23A, полосы 3-6), не детектируются. Испльзуемые в данном эксперименте вторичные антитела против человеческого Fc позволяют детектировать очищенный человеческий IgG (фигура 23A, полоса 1). Результаты свидетельствуют о том, что PA13R3-P1C4 и гуманизированные варианты антител не способны распознавать денатурированный и развернутый белок CB1. Наряду с результатами, полученными методом проточной цитометрии, результаты данного эксперимента подтвержают, что PA13R3-P1C4 и гуманизированные варианты антител распознают конформационные, но не линейные эпитопы.
Пример 25. Связывание химерных и гуманизированных Fab P1C4, определяемое путем измерения цАМФ и методом FACS
[0336] Чтобы определить сродство Fab PA13R3-P1C4, получают полные кривые связывания с рецептором CB1 путем тестирования диапазона концентраций с использованием клеток TRex-CHO, стабильно трансфицированных конструкцией, обеспечивающей тетрациклин-индуцируемую экспрессию CB1, методом проточной цитометрии. Получают трехкратные серийные разведения от 3 мкМ до 0,1 мкМ. Для детекции Fab PA13R3-P1C4 используют FITC-конъюгированные антитела против человеческих IgG. Fab PA13R3-P1C4 связываются с клетками TRex-CHO CB1 в дозо-зависимой манере (фигура 24A и таблица 17).
[0337] Функциональный анализ с измерением цАМФ проводят, чтобы определить антагонистическую активность Fab P1C4. Функциональный анализ цАМФ (Cisbio) проводят на белом 384-луночном низкообъемном планшете (Greiner). В каждую лунку планшета высевают 8000 клеток TRex CHO, стабильно экспрессирующих CB1, после чего проводят инкубацию с Fab P1C4 в разных концентрациях при комнатной температуре в течение 10 мин. Для стимуляции смеси к клеткам добавляют 5 мкМ форсколин (Sigma Aldrich) и 9 мкМ каннабиноид CP55940 (Sigma Aldrich), после чего инкубируют в течение 30 мин при комнатной температуре, чтобы активировать CB1. После инкубации в течение 30 мин добавляют 5 мкл цАМФ-d2 (разведение 1:39, конъюгат и буфер для лизиса получают от Cisbio) и 5 мкл анти-цАМФ-криптата (разведение 1:9, конъюгат и буфер для лизиса получают от Cisbio) для стимуляции клеток и инкубируют в течение часа. Сигнал FRET детектируют с помощью планшет-ридера Envision для нескольких меток (Perkin Elmer) при длине волны возбуждения анти-цАМФ-криптата 620 нм и длине волны испускания 665 нм. Анализ результатов проводят с помощью GraphPad Prism. Результаты приведены на фигуре 24B и в таблице 17. Показанные средние значения IC50 и константы диссоциации (Kd) ± SD получают с использованием результатов, по меньшей мере, 2 разных экспериментов.
Таблица 17. Значения IC50 и константы диссоциации Fab PA13R3-P1C4
Пример 26. Определение биофизических характеристик гуманизированных вариантов P1C4
[0338] Авторы характеризуют стабильность и растворимость гуманизированных вариантов Fc PA13R3-P1C4 путем проведения тестов с измерением стабильности при низких и высоких pH и с определением максимальной растворимости. Авторы также характеризуют стабильность указанных молекул в ускоренных условиях, в человеческой сыворотке и в сыворотке отличных от человека приматов, после нескольких циклов замораживания/оттаивания, а также в условиях сдвига pH.
[0339] Чтобы охарактеризовать pH-стабильность гуманизированных вариантов P1C4, 200 мкл каждого антитела (~5 мг/мл) и PBS в качестве контроля вводят в диализную кассету Pur-A-Lyzer Maxi 12000 (Sigma, Cat# PURX12015-1KT). Белки диализуют против 2 л буфера с pH 3 (0,1 M уксусная кислота, доведенная до pH 3 NaOH) или буфера с pH 9 (0,2 M глицин, доведенный до pH 9 NaOH), соответственно, при 4°C в течение ночи. Образцы белков извлекают из диализных кассет, помещают в предварительно взвешенные пустые пробирки Эппендорфа и проверяют на наличие видимого осадка. Измеряют массу образца. Чтобы подтвердить успешное протекание диализа, берут 3 мкл диализованного образца и проверяют с помощью индикаторной бумаги для определения pH. Аликвоты образцов объемом пятьдесят мкл используют для анализа SEC. Оставшуюся часть держат в инкубаторе при 40°C в течение 48 часов. Затем образцы снова проверяют на наличие видимого осадка. После инкубации в течение 48 часов шесть мкл образца вводят на колонку TSK G3000SWXL SEC и рассчитывают концентрацию белка по площади пиков SEC (после инкубации при 40°C). Степень извлечения белка определяют по следующей формуле: количество до диализа (об.×конц.)/количество после диализа (об.×конц.) ×100%.
[0340] После диализа против буферов с pH 3 и pH 9 и последующей инкубации в течение 48 часов при 40°C осадок в растворах 4 IgG не наблюдается. Степень извлечения всех четырех IgG составляет примерно 71-83%. Низкая степень извлечения может быть связана с тем, что часть образца остается в диализной кассете. Рассчитанные по профилям SEC количества мономеров IgG составляют более 99% для всех 4 диализованных IgG. После диализа против буфера pH 3 и последующей инкубации в течение 48 часов при 40°C IgG дают более широкие пики SEC, позволяющие предположить более высокую гетерогенность IgG после длительной инкубации в буфере с pH 3. Способность антител влиять на уровень цАМФ и связываться с клетками, экспрессирующими CB1, измеряют с помощью способов, описанных в примерах 11 и 7, результаты приведены в таблицах 18 и 19, соответственно. Результаты демонстрируют, что после инкубации при pH 3 активность P1C4-h2-IgG4, P1C4-h4-IgG2 и P1C4-h4-IgG4 в отношении цАМФ уменьшается, как и активность P1C4-h4-IgG2 после инкубации при pH 9.
Таблица 18. pH-стабильность гуманизированных вариантов P1C4: растворимость
Таблица 19. pH-стабильность гуманизированных вариантов P1C4: активность в отношении цАМФ
[0341] Растворимость гуманизированных вариантов P1C4 характеризуют путем концентрирования 400 мкл IgG (~5 мг/мл) фильтрованием на центрифуге (Amicon Ultra-0,5 мл 30K) при 14000 g и 4°C до ~100 мкл. В центрифужные фильтры добавляют еще 200 мкл IgG и концентрируют при 14000 g и 4°C до ~100 мкл. Затем в центрифужные фильтры добавляют еще порцию IgG объемом 200 мкл и концентрируют при 14000 g и 4°C до ~100 мкл (всего 800 мкл концентрируют до 100 мкл). Концентрированный белок проверяют на наличие видимого осадка путем перемешивания пипетированием. Затем концнетрирование продолжают при 14000 g и 4°C до достижения объема менее 50 мкл. Центрифужные фильтры переворачивают и помещают в заранее взвешенные пустые пробирки. Центрифужные фильтры центрифугируют при 1000 g в течение 5 минут при 4°C и собирают концентрированный образец. Пробирки взвешивают, чтобы определить объем образцов. Если наблюдается образование осадка, пробирки центрифугируют при 14000 g в течение 10 минут при 4°C. Супернатанты удаляют, помещают в новые пробирки Эппендорфа объемом 1,5 мл и инкубируют при комнатной температуре в течение 24 часов. Затем осадок осаждают центрифугированием при 14000 g в течение 10 минут при 4°C и супернатанты переносят в новые пробирки Эппендорфа объемом 1,5 мл. Чтобы провести анализ методом SEC, 6 мкл супернатанта концентрированного образца вводят на колонку TSK G3000SWXL SEC и концентрацию белка рассчитывают по площади пика SEC. Степень извлечения белка рассчитывают по следующей формуле: количество до конц. (об.×конц.)/количество после конц. (об.×конц.) ×100%.
[0342] Четыре варианта Fc концентрируют до достижения концентрации белка более 85 мг/мл. Степень извлечения белка составляет более 99%. Наблюдается небольшой видимый осадок P1C4-h2-IgG4 при 93,9 мг/мл до и после инкубации в течение 24 часов при комнатной температуре. В образцах 3 других IgG при концентрации выше 85 мг/мл после инкубации в течение 24 часов при комнатной температуре видимый осадок отсутствует. Содержание мономеров IgG, рассчитанное по профилю SEC, составляет более 96% для всех 4 концентрированных IgG по сравнению с исходным уровнем ~99%. Результаты приведены в таблице 20.
Таблица 20. Растворимость гуманизированных вариантов P1C4
[0343] Кроме того, определяют стабильность гуманизированных вариантов P1C4 в ускоренных условиях. В пробирку Эппендорфа объемом 1,5 мл помещают 100 мкл образца белка (~5 мг/мл). Пробирку закрывают парафильмом. Используют две пробирки, одну держат при 4°C, а другую - при 40°C в течение 33 дней. Берут аликвоту объемом 20 мкл, чтобы проверить образец на наличие видимого осдка и провести анализ SEC. Анализ SEC проводят путем введения 6 мкл образца на колонку TSK G3000SWXL SEC с последующим определением концентрации белка по площади пика SEC. Степень извлечения белка рассчитывают по следующей формуле: количество до инкуб. (об.×конц.)/количество после инкуб. (об.×конц.) ×100%.
[0344] После инкубации в течение 33 дней при 4°C и 40°C осадок не наблюдается. Содержание мономеров IgG, рассчитанное по профилю SEC, составляет более 98% для всех 4 IgG после инкубации как при 4°C, так и при 40°C. Степень извлечения, рассчитанная по профилю SEC, составляет более 96% для всех 4 IgG после инкубации как при 4°C, так и при 40°C (таблица 21). После инкубации в течение 33 дней при 4°C и 40°C изменение активности вариантов Fc P1C4 не наблюдается, за исключением P1C4-h4-IgG2, у которого IC50 находится вне стандартного диапазона, свидетельствуя о небольшом снижении активности (таблица 22).
Таблица 21. Стабильность в ускоренных условиях: растворимость
Таблица 22. Стабильность в ускоренных условиях: активность
[0345] Определяют стабильность гуманизированных вариантов P1C4 в сыворотке. Человеческую сыворотку получают от Sigma. Сыворотку отличных от человека приматов собирают в Crown Bioscience. В пробирке Эппендорфа смешивают 950 мкл сыворотки и 50 мкл IgG до конечной концентрации 250 мкг/мл (~1,67 мкМ) и инкубируют при 37°C. Образцы объемом 200 мкл берут через 0, 24, 48 и 72 часа и методом проточной цитометрии анализируют связывание с клетками, экспрессирующими CB1. Готовят 3-кратные серийные разведения образцов тестируемых IgG с исходной концентрацией 500 nM и используют их для определения KD методом проточной цитометрии. Результаты, приведенные в таблицах 23 и 24, свидетельствуют об отсутствии изменений сродства после инкубации в течение 24 часов с человеческой сывороткой или сывороткой NHP при 37°C. Значительные изменения KD отсутствуют в образцах, за исключением P1C4-h2-IgG2, у которого наблюдается уменьшение сродства к клеткам, экспрессирующим CB1, после инкубации в течение 48 часов с человеческой сывороткой или сывороткой NHP. Кроме того, у P1C4-h4-IgG2 также наблюдается уменьшение сродства к клеткам, экспрессирующим CB1, после инкубации в течение 48 часов.
Таблица 23. Стабильность гуманизированных вариантов P1C4 в человеческой сыворотке, 24 часа
Таблица 24. Стабильность гуманизированных вариантов P1C4 в сыворотке NHP
[0346] Стабильность гуманизированных вариантов P1C4 после замораживания/оттаивания характеризуют следующим образом. Аликвоту объемом 100 мкл замороженного исходного раствора каждого из гуманизированных вариантов Fc P1C4 отаивают на водяной бане при 22°C, затем быстро замораживают в жидком азоте. Замороженный образец держат при -80°C в течение, по меньшей мере, 20 минут, после чего его снова отаивают на водяной бане при 22°C. Образцы подвергают 10 указанным циклам замораживания/оттаивания. Наличие осадка проверяют визуально. Аликвоты образца объемом 20 мкл берут для анализа методом SEC после 1, 5 и 10 циклов замораживания/оттаивания. Для анализа методом SEC 6 мкл инкубированного образца вводят на колонку TSK G3000SWXL SEC. Концентрацию белка определяют по площади пика SEC. Степень извлечения белка рассчитывают по следующей формуле: количество до З/О (об.×конц.)/количество после З/О (об.×конц.) ×100%.
[0347] Результаты демонстрируют, что после 10 циклов замораживания/оттаивания содержание мономеров IgG выше 97% в образцах всех 4 тестируемых вариантов IgG. Степень извлечения белка составляет более 96% для всех IgG. В образце, содержащем P1C4-H2-IG4, наблюдается небольшое помутнение после 1-ого цикла замораживания/оттаивания, однако заметный осадок или значительное уменьшение концентрации белка не наблюдается. Результаты приведены в таблицах 25-28. Образцы белков, извлеченных после 5 циклов замораживания/оттаивания, также тестируют на активность с помощью антагонистического анализа цАМФ, получая значения IC50, которые демонстрируют отсутствие существенных изменений в активности и находятся в нормальном диапазоне (таблица 29).
Таблица 25. Стабильность P1C4-H2-IgG2 после замораживания/оттаивания
Таблица 26. Стабильность P1C4-H2-IgG4 после замораживания/оттаивания
Таблица 27. Стабильность P1C4-H4-IgG2 после замораживания/оттаивания
Таблица 28. Стабильность P1C4-H4-IgG4 после замораживания/оттаивания
Таблица 29. Стабильность гуманизированных вариантов после замораживания/оттаивания: активность в отношении цАМФ после 5 циклов
[0348] Чтобы определить стабильность 4 вариантов Fc P1C4 при сдвиге pH, 0,2 мл смолы MabSelect помещают в колонку объемом 10 мл и уравновешивают 10 мл DPBS, pH 7,4. На колонку загружают 150 мкл образца IgG (~5 мг/мл). Колонку промывают 1,6 мл DPBS. Затем IgG элюируют 1,6 мл цитрата натрия (pH ~3,5). Измеряют концнетрацию белка. Затем белок концентрируют до ~3 мг/мл и используют для функционального анализа. Видимый осадок отсутствует. Полученные образцы центрифугируют при 14000 g в течение 10 минут при 4°C. Супернатант переносят в новую пробирку Эппендорфа и измеряют концентрацию белка. Полученный образец проверяют на агрегацию методом SEC и подвергают функциональному тестированию с помощью антагонистического анализа с измерением цАМФ. Профили SEC свидетельствуют о том, что содержание мономеров IgG составляет более 95% после сдвига pH до 3,5. Результаты антагонистического анализа цАМФ, приведенные в таблице 30, демонстрируют, что P1C4-h2-IgG4, P1C4-h4-IgG2 и P1C4-h4-IgG4 являются стабильными при pH 3,5, а измеренные значения IC50 попадают в допустимый диаппазон. P1C4-h2-IgG2 ведет себя при pH 3,5 по-другому и имеет значение IC50 ниже допустимого диапазона. Данный результат согласуется с результатами тестирования pH-стабильности.
Таблица 30. Стабильность гуманизированных вариантов P1C4 при сдвиге pH
Пример 27. CDR-мутагенез PA13R3-P1C4
[0349] Чтобы исследовать роль каждого аминокислотного положения в каждом CDR, проводят сайт-направленный насыщающий мутагенез по всем положениям CDR Fab химерного P1C4. Синтезируют мутагенные праймеры, содержащие NNS или специфический кодон, растворяют их в буфере трис-EDTA и разбавляют до 10 мкМ рабочим раствором. Двадцать пять мкл реакционных смесей ПЦР помещают в 96-луночные планшеты, используя высокоточную ДНК-полимеразу. Полученные продукты ПЦР обрабатывают путем добавления 0,8 мкл DpnI (20 ед/мкл) в каждую лунку при 37°C в течение 5 часов. DpnI-обработанный продукт ПЦР (2 мкл) используют для трансформации 30 мкл Dh5α-компетентных клеток E. coli. ДНК выделяют из трансформантов с использованием miniprep и секвенируют, чтобы идентифицировать желательные мутации. Плазмидную ДНК из желательных клонов используют для трансформации BL21 (CodonPlus)-компетентных клеток E. coli. Отдельные колонии используют для экспрессии белка Fab.
[0350] Чтобы осуществить экспрессию Fab, выбранные колонии помещают в 96-луночный планшет, содержащий 100 мкл среды SB и культивируют в течение ночи. На следующий день 10 мкл из каждой лунки используют для инокуляции 500 мкл среды ZYM, содержащей 50 мкг/мл канамицина, в 96-луночном планшете с глубокими лунками. Планшет закрывают воздухопроницаемым покрытием для планшетов и встряхивают в течение 36 часов при 25°C на шейкере при 450 об\мин.
[0351] Чтобы получить образцы для ELISA, 96-луночный планшет с глубокими лунками центрифугируют при 3000 об/мин в настольной центрифуге Beckman в течение 20 минут (~2050 rcf) при 4°C. 100 мкл супернатантов переносят из планшета для экспрессии в планшет для разведения, содержащий 200 мкл PBS, pH 7,4, и тщательно перемешивают.
[0352] Экспрессию Fab измеряют методом ELISA. 96-луночные планшеты ELISA с половинным объемом лунок (Corning, 3690) непосредственно покрывают 2 мкг/мл антитела против his (Sigma H1029 2 мг/мл) в концентрации 50 мкл/лунку в течение ночи при 4°C. Затем планшет промывают 6 раз 150 мкл PBST. Каждую лунку блокируют 175 мкл/лунку 3% молоко/PBS, pH 7,4, в течение 1 часа при 22°C. Затем планшет промывают 6 раз PBST, в каждую лунку добавляют 25 мкл/лунку Fab-содержащей культуральной среды, разведенной в 3 раза PBS, pH 7,4, и инкубируют в течение 1 часа при 22°C. Планшет снова промывают 6 раз PBST и затем добавляют 50 мкл/лунку HRP-меченных антител против Fab человеческих IgG (0293 Sigma), разведенных 1:10000 3% молоком, и инкубируют при 22°C в течение 1 часа. Для развития окраски планшет промывают 6 раз PBST и добавляют 50 мкл/лунку субстрата ™B. Реакцию останавливают путем добавления 50 мкл/лунку 1N HCl и прочитывают OD450 на ридере BioTek.
[0353] Для измерения связывания iCAPS методом ELISA 96-луночные планшеты ELISA с половинным объемом лунок покрывают 25 мкл/лунку раствора стрептавидина в PBS, 2 мкг/мл, pH 7,4, в течение ночи при 4°C. Затем планшет промывают 6 раз 150 мкл PBST, переворачивая его в промежутке между промываниями. В планшет добавляют 25 мкл/лунку iCAPS CB1, 5 мкг/мл, и инкубируют в течение 1 часа. Промывают 6 раз PBST, после чего каждую лунку блокируют 175 мкл/лунку 3% молоко/PBS, pH 7,4, в течение 1 часа при 22°C. Затем планшет промывают 6 раз PBST и добавляют в каждую лунку 25 мкл/лунку Fab-содержащей культуральной среды, разбавленной в 3 раза PBS, pH 7,4, и инкубируют в течение 1 часа при 22°C. Планшет снова промывают 6 раз PBST и затем добавляют 50 мкл/лунку антител против человеческих Fab (0293 Sigma), разведенных 1:10000 3% молоком, и инкубируют при 22°C в течение 1 часа. Для развития окраски планшет промывают 6 раз PBST и добавляют 50 мкл/лунку субстрата ™B. Реакцию останавливают путем добавления 50 мкл/лунку 1N HCl и прочитывают OD450 на ридере BioTek.
[0354] Каждый клон анализируют с тройными повторами. Чтобы рассчитать относительное связывание с iCAPS, нормализованное по экспрессии Fab, сигнал ELISA, соответствующий связыванию iCAPS, делят на величину экспрессии Fab, определенную методом ELISA, и сравнивают с результатами, полученными от исходного клона. Допустимыми считают изменения, содержащиеся в клонах, которые сохраняют, по меньшей мере, 50% способности исходных клонов специфически связываться с iCAPS. Указанные допустимые изменения приведены в таблице 31 и таблице 32. Последовательности CDR, содержащие указанные мутации, показаны в таблице 34.
Таблица 31. Допустимые мутации в CDR тяжелой цепи PA13R3-P1C4
Таблица 32. Допустимые мутации в CDR легкой цепи PA13R3-P1C4
Пример 28. Иммуноокрашивание CB1 в образцах ткани печени антителом P1C4-h2-IgG4, конъюгированным с HRP.
[0355] P1C4-h2-IgG4 антитело метят с помощью набора для конъюгации Lightning-Link HRP (Innova Bioscience, 701-0010) в соответствии с инструкциями производителя. Обработанные парафильмом срезы образцов печени человека обрабатывают растворителем Clearene (Leica Biosystems) в течение 5 минут и затем этанолом в уменьшающихся концентрациях, до конечной концентрации 50%. Затем срезы помещают в метанол/пероксид водорода на 15 минут, после чего их быстро промывают PBS. После этого срезы обрабатывают раствором лимонной кислоты в физрастворе (Vector, H-3300), нагревая для извлечения антигена, и помещают в предварительно нагретый PBS (350 мл) и трипсин (2,45 мл) на водяной бане при 37°C в течение 20 минут. После промывания PBS срезы блокируют казеином (Vector, SP-5020) в течение 1 часа при комнатной температуре. Затем добавляют разведенное 1:100 HRP-конъюгированное антитело P1C4-h2-IgG4 или изотипическое контрольное антитело и инкубируют в течение ночи при 4°C. После инкубации срезы промывают PBS и обрабатывают 3 каплями Vector ABC Tertiary (Vector, PK-7100) при 22°C в течение 45 минут. Срезы 3 раза промывают PBS и добавляют смесь DAB (Vector, SK-4100), инкубируют в течение 5-10 минут и снова быстро промывают PBS. Затем срезы подвергают контрастному окрашиванию гематоксилином Майера (TCS Biosciences, HS315) в течение 1 минуты с последующей обработкой водой, возрастающими концентрациями этанола (от 50% до 100%), 2 циклами растворителя Clearene, после чего их заключают в Pertex (Leica Biosystems).
[0356] Результаты демонстрируют наличие положительного CB1-специфичного окрашивания макрофагов, гепатоцитов и миофибробластов печени в образцах, соответствующих раннему NASH (фигура 25, левая панель), фиброзу NASH (фигура 25, средняя панель) и позднему фиброзу (фигура 25, правая панель). При использовании изотипических контрольных иррелевантных антител (фигура 26) или образцов нормальной ткани (фигура 27) окрашивание не наблюдается.
Пример 29. Измерение влияния антитела против CB1 на генетические маркеры фиброза в первичных звездчатых клетках печени человека
[0357] Первичные звездчатые клетки печени (HSCs) выделяют из ткани печени, полученной от 3 здоровых доноров. После 2-3 пассажей в DMEM+10% FBS клетки активируют путем адгезии на пластике, помещают в среду, содержащую 0,5% сыворотки, и держат в ней в течение ночи. Затем клетки обрабатывают в течение 6 или 24 часов римонабантом (антагонист CB1), P1C4-h2-IgG4 и нефункциональным контрольным антителом в разных концентрациях. Ингибирование профиля экспрессии профибротического гена, включающего в себя экспрессию α-SMA, проколлагена A1(I), TIMP1 и TGFβ, измеряют методом ОТ-ПЦР, результаты приводят в виде графиков.
[0358] Результаты демонстрируют значительное уменьшение экспрессии проколлагена A1(I) после обработки HSC антителом P1C4-h2, но не нефункциональным контрольным антителом и не PBS (фигура 28). Также наблюдается значительное уменьшение экспрессии TGFβ (фигура 29) и TIMP1 (фигура 30) по сравнению с PBS и нефункциональным контрольным антителом. Кроме того, наблюдается значительное уменьшение экспрессии α-SMA в клетках, обработанных P1C4-IgG1 или IgG4 по сравнению с клетками, обработанными PBS или нефункциональным связывающим соединением (фигура 31).
Пример 30: Количественное определение антитела против CB1 в цереброспинальной жидкости (CSF) яванских макак
[0359] Яванским макакам (2 самца и 2 самки/группу) вводят P1C4-h2-IgG4 в соответствии со схемой, приведенной в таблице 33, и в указанные моменты времени собирают CSF. Количество P1C4-h2-IgG4 определяют методом ELISA, используя 96-луночные планшеты, покрытые антителом против ID в концентрации 1 мкг/мл, с последующим добавлением CSF и детекцией с использованием HRP-конъюгированного антитела против IgG (Abcam) и окрашивающего реагента.
[0360] Как показано в таблице 33, антитело детектируют только на очень низком уровне, или совсем не детектируют. В группе, получающей самую высокую дозу, в CSF детектируют менее 0,1% от введенного количества, что свидетельствует об очень низком проникновении антитела в ЦНС.
Таблица 33. Количественное определение антитела против CB1 в CSF яванских макак
BLQ=ниже предела количественного определения
Пример 31. Измерение влияния антитела против CB1 на метаболические и сердечнососудистые факторы у яванских макак
[0361] Программа RIO демонстрирует кардиометаболические эффекты лечения римонабантом с использованием нескольких факторов. См., например, Pi-Sunyeret al., 2006, J Am Coll Cardio, l47:362A. Также определяют влияние описанных здесь антител против CB1, как таковых, на аналогичные кардиометаболические факторы.
[0362] Страдающим от ожирения яванским макакам и макакам-резус еженедельно п.к. вводят антитело против CB1 P1C4-h2-IgG4 в дозе 3 мг/кг или 0,3 мг/кг. В качестве отрицательного контроля ряду приматов вводят либо (1) один фармацевтический носитель; либо (2) контрольное антитело, которое заведомо не связывает CB1. Наблюдают влияние на потребление пищи, массу тела, чувствительность к инсулину, уровни триглицеридов и другие факторы сердечнососудистого риска у приматов.
[0363] У приматов, получающих антитело против CB1 P1C4-h2-IgG4 в дозе 3 мг/кг или 0,3 мг/кг, наблюдается снижение уровней триглицеридов и других факторов сердечнососудистого риска. У приматов, получающих антитело против CB1 P1C4-h2-IgG4 в дозе 3 мг/кг или 0,3 мг/кг, также наблюдается улучшение чувствительности к инсулину. У приматов, которым вводят либо контрольное антитело, либо только носитель, отсутствуют улучшения каких-либо из указанных факторов.
Специалисты в данной области могут осуществлять многочисленные модификации и вариации настоящего изобретения, описанного с использованием приведенных выше иллюстративных примеров. Следовательно, к настоящему изобретению можно применять лишь ограничения, накладываемые прилагающейся формулой изобретения. Все цитирующиеся здесь ссылки включены в настоящее описание в качестве ссылки во всей их полноте и во всех целях.
Таблица 34. Последовательности допустимых мутаций в CDR PA13R3-P1C4
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> RuiYi, Inc.
KRETZ-ROMMEL, Anke
SHI, Lei
FERRINI, Roger
YANG, Teddy
Xu, Fei
CAMPION, Brian
<120> АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ЧЕЛОВЕЧЕСКИМ КАННАБИНОИДНЫМ РЕЦЕПТОРОМ 1 (CB1)
<130> 15-343-WO3
<150> PCT/CN2014/074199
<151> 2014-03-27
<150> PCT/CN2014/081797
<151> 2014-07-08
<160> 827
<170> PatentIn version 3.5
<210> 1
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3-P1C4
<400> 1
gaggtccagc tgcagcagtc tggggctgag ctggtgaggc ctggggtctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccaa cacagcctat 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 2
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3-P1C4
<400> 2
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Val
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 3
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3-P1C4
<400> 3
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 4
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3-P1C4
<400> 4
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 5
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3-P1C4
<400> 5
gatattgttc tcacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 6
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3-P1C4
<400> 6
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 7
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3-P1C4
<400> 7
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 8
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3-P1C4
<400> 8
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 9
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи 36E12B6C2
<400> 9
caggttcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttcagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcacctca ccagcctaac gtctgaggac tctgcggtct atttctgtgc aagatcgggg 300
ggtaacccct ttgctttctg gggccaaggg actctggtca ctgtctctgc a 351
<210> 10
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи 36E12B6C2
<400> 10
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Gln Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met His Leu Thr Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser Gly Gly Asn Pro Phe Ala Phe Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 11
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи 36E12B6C2
<400> 11
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 12
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи 36E12B6C2
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 13
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи 36E12B6C2
<400> 13
gatatccaga tgacacagac ttcatcctcc ctgtctgcct ctctgggaga cagagtcacc 60
ttcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca 120
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcacatca 180
aggttccgtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa 240
gaagacgttg ccacttactt ttgccaacag ggtcatacgc ttccgtggtc gttcggtgga 300
ggcaccaagc tggaaatcaa a 321
<210> 14
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи 36E12B6C2
<400> 14
Asp Ile Gln Met Thr Gln Thr Ser Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Phe Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Thr Ser Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Val Ala Thr Tyr Phe Cys Gln Gln Gly His Thr Leu Pro Trp
85 90 95
Ser Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 15
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи 36E12B6C2
<400> 15
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttag 324
<210> 16
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи 36E12B6C2
<400> 16
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 17
<211> 351
<212> ДНК
<213> Mus sp.
<400> 17
caggttcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttcagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcacctca ccagcctaac gtctgaggac tctgcggtct atttctgtgc aagatcgggg 300
ggtaacccct ttgctttctg gggccaaggg actctggtca ctgtctctgc a 351
<210> 18
<211> 117
<212> Белок
<213> Mus sp.
<400> 18
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Gln Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met His Leu Thr Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser Gly Gly Asn Pro Phe Ala Phe Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 19
<211> 993
<212> ДНК
<213> Mus sp.
<400> 19
gctaaaacaa cagccccatc ggtctatcca ctggcccctg tgtgtggaga tacaactggc 60
tcctcggtga ctctaggatg cctggtcaag ggttatttcc ctgagccagt gaccttgacc 120
tggaactctg gatccctgtc cagtggtgtg cacaccttcc cagctgtcct gcagtctgac 180
ctctacaccc tcagcagctc agtgactgta acctcgagca cctggcccag ccagtccatc 240
acctgcaatg tggcccaccc ggcaagcagc accaaggtgg acaagaaaat tgagcccaga 300
gggcccacaa tcaagccctg tcctccatgc aaatgcccag cacctaacct cttgggtgga 360
ccatccgtct tcatcttccc tccaaagatc aaggatgtac tcatgatctc cctgagcccc 420
atagtcacat gtgtggtggt ggatgtgagc gaggatgacc cagatgtcca gatcagctgg 480
tttgtgaaca acgtggaagt acacacagct cagacacaaa cccatagaga ggattacaac 540
agtactctcc gggtggtcag tgccctcccc atccagcacc aggactggat gagtggcaag 600
gagttcaaat gcaaggtcaa caacaaagac ctcccagcgc ccatcgagag aaccatctca 660
aaacccaaag ggtcagtaag agctccacag gtatatgtct tgcctccacc agaagaagag 720
atgactaaga aacaggtcac tctgacctgc atggtcacag acttcatgcc tgaagacatt 780
tacgtggagt ggaccaacaa cgggaaaaca gagctaaact acaagaacac tgaaccagtc 840
ctggactctg atggttctta cttcatgtac agcaagctga gagtggaaaa gaagaactgg 900
gtggaaagaa atagctactc ctgttcagtg gtccacgagg gtctgcacaa tcaccacacg 960
actaagagct tctcccggac tccgggtaaa tga 993
<210> 20
<211> 330
<212> Белок
<213> Mus sp.
<400> 20
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 21
<211> 321
<212> ДНК
<213> Mus sp.
<400> 21
gatatccaga tgacacagac ttcatcctcc ctgtctgcct ctctgggaga cagagtcacc 60
ttcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca 120
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcacatca 180
aggttccgtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa 240
gaagacgttg ccacttactt ttgccaacag ggtcatacgc ttccgtggtc gttcggtgga 300
ggcaccaagc tggaaatcaa a 321
<210> 22
<211> 107
<212> Белок
<213> Mus sp.
<400> 22
Asp Ile Gln Met Thr Gln Thr Ser Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Phe Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Thr Ser Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Val Ala Thr Tyr Phe Cys Gln Gln Gly His Thr Leu Pro Trp
85 90 95
Ser Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 23
<211> 324
<212> ДНК
<213> Mus sp.
<400> 23
cgggcagatg ctgcaccaac tgtatccatc ttcccaccat ccagtgagca gttaacatct 60
ggaggtgcct cagtcgtgtg cttcttgaac aacttctacc ccaaagacat caatgtcaag 120
tggaagattg atggcagtga acgacaaaat ggcgtcctga acagttggac tgatcaggac 180
agcaaagaca gcacctacag catgagcagc accctcacgt tgaccaagga cgagtatgaa 240
cgacataaca gctatacctg tgaggccact cacaagacat caacttcacc cattgtcaag 300
agcttcaaca ggaatgagtg ttag 324
<210> 24
<211> 107
<212> Белок
<213> Mus sp.
<400> 24
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 25
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3- P4B5
<400> 25
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 26
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3- P4B5
<400> 26
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 27
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3- P4B5
<400> 27
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 28
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3- P4B5
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 29
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3- P4B5
<400> 29
gacattgttc tcaaccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccgct cacgttcggt 300
gctgggacca aactggaaat aaaa 324
<210> 30
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3- P4B5
<400> 30
Asp Ile Val Leu Asn Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 31
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3- P4B5
<400> 31
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 32
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3- P4B5
<400> 32
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 33
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P2D3
<400> 33
gaggttcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtagctatt ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 34
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P2D3
<400> 34
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Ser Tyr Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 35
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P2D3
<400> 35
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 36
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P2D3
<400> 36
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 37
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P2D3
<400> 37
gatattgagc tggcccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atatcctgca gagccagtga aagtgttgat agttatggca atagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatc ttgcatccaa cctagaatct 180
ggggtccctg ccaggttcag cggcagtggg tctagggcag acttcaccct caccattgat 240
cctgtggagg ctgatgatgc tgcaacctat tactgtctac aatatgctag ttctcctcct 300
acgttcggtg ctgggaccaa actggaaata aaa 333
<210> 38
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P2D3
<400> 38
Asp Ile Glu Leu Ala Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Ala Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Tyr Ala
85 90 95
Ser Ser Pro Pro Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 39
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P2D3
<400> 39
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 40
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P2D3
<400> 40
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 41
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P4B1
<400> 41
gaggtccagc ttcagcaatc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggctt tgcattcagt aactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaacttc 180
aatggaaagt tcaagggtag agccatactg actgcagaca tatcctccaa cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 42
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P4B1
<400> 42
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Phe Ala Phe Ser Asn Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Phe Asn Gly Lys Phe
50 55 60
Lys Gly Arg Ala Ile Leu Thr Ala Asp Ile Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 43
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P4B1
<400> 43
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 44
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P4B1
<400> 44
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 45
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P4B1
<400> 45
caaattgtgt tgacacagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccgct cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 46
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P4B1
<400> 46
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 47
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P4B1
<400> 47
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 48
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P4B1
<400> 48
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 49
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P6B12
<400> 49
gaggtccagc ttcagcaatc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 50
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P6B12
<400> 50
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 51
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P6B12
<400> 51
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 52
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P6B12
<400> 52
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 53
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P6B12
<400> 53
caaattgtac tcacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttcccccct cgcgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 54
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P6B12
<400> 54
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Leu Ala Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 55
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P6B12
<400> 55
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 56
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P6B12
<400> 56
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 57
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P6G7
<400> 57
gaggttcagc ttcagcaatc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggctt tgcattcagt aactactgga tgaactgggt gaagcagagg 120
cccggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaacttc 180
aatggaaagt tcaagggtag agccatactg actgcagaca tatcctccaa cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 58
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P6G7
<400> 58
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Phe Ala Phe Ser Asn Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Phe Asn Gly Lys Phe
50 55 60
Lys Gly Arg Ala Ile Leu Thr Ala Asp Ile Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 59
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P6G7
<400> 59
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 60
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P6G7
<400> 60
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 61
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P6G7
<400> 61
gatattgtgc taactcagtc tccagcaatc atgtccgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacctcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagcatcatc gttccccacc cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 62
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P6G7
<400> 62
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln His His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 63
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P6G7
<400> 63
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 64
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P6G7
<400> 64
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 65
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P1G6
<400> 65
gaggtgcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatgggaagt tcaagggtaa agccacactg actgtagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 66
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P1G6
<400> 66
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 67
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P1G6
<400> 67
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 68
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P1G6
<400> 68
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 69
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P1G6
<400> 69
gacattgttc tctcccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag ccgaatggag 240
gctgaagatg ctgccactta ttactgccag cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 70
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P1G6
<400> 70
Asp Ile Val Leu Ser Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 71
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P1G6
<400> 71
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 72
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P1G6
<400> 72
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 73
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P1H4
<400> 73
gaggttcaac tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgtattcggt aactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatgggaagt tcaaaggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctgac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 74
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P1H4
<400> 74
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Val Phe Gly Asn Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 75
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P1H4
<400> 75
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 76
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P1H4
<400> 76
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 77
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P1H4
<400> 77
caaattgttc tcactcagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgtcaa cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 78
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P1H4
<400> 78
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 79
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P1H4
<400> 79
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 80
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P1H4
<400> 80
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 81
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P2B8
<400> 81
gaggtccagc ttcagcaatc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 82
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P2B8
<400> 82
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 83
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P2B8
<400> 83
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 84
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P2B8
<400> 84
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 85
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P2B8
<400> 85
gatattgtaa tgacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtataagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttcccccct cgcgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 86
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P2B8
<400> 86
Asp Ile Val Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Ile Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Leu Ala Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 87
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P2B8
<400> 87
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 88
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P2B8
<400> 88
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 89
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P2E5
<400> 89
gaggttcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattccgt gactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtgc tactaactac 180
attggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctgac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccacggg actctggtca ctgtctctgc a 351
<210> 90
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P2E5
<400> 90
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Arg Asp Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Ala Thr Asn Tyr Ile Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly His Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 91
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P2E5
<400> 91
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 92
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P2E5
<400> 92
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 93
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P2E5
<400> 93
caaattgttc tcactcagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttattctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgtcaa cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 94
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P2E5
<400> 94
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 95
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P2E5
<400> 95
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 96
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P2E5
<400> 96
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 97
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3A8
<400> 97
gaggtccagc ttcagcaatc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagg tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 98
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3A8
<400> 98
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Val
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 99
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3A8
<400> 99
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 100
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3A8
<400> 100
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 101
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3A8
<400> 101
gacattgtga tgacccaatc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccag cagtggagta gtaacccgct cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 102
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3A8
<400> 102
Asp Ile Val Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 103
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3A8
<400> 103
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 104
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3A8
<400> 104
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 105
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3B10
<400> 105
gaggtgcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 106
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3B10
<400> 106
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 107
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3B10
<400> 107
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 108
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3B10
<400> 108
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 109
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3B10
<400> 109
gacattgtga taactcagtc tccagcaatc gtgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttattctc tcacaatcag cagcatggag 240
gctgaagatg ccgccactta ttactgtcaa cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggaaat caaa 324
<210> 110
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3B10
<400> 110
Asp Ile Val Ile Thr Gln Ser Pro Ala Ile Val Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 111
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3B10
<400> 111
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 112
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3B10
<400> 112
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 113
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3B8
<400> 113
gaggtccagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggctt tgcattcagt aactactgga tgaactgggt gaagcagagg 120
cccggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaacttc 180
aatggaaagt tcaagggtag agccatactg actgcagaca tatcctccaa cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 114
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3B8
<400> 114
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Phe Ala Phe Ser Asn Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Phe Asn Gly Lys Phe
50 55 60
Lys Gly Arg Ala Ile Leu Thr Ala Asp Ile Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 115
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3B8
<400> 115
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 116
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3B8
<400> 116
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 117
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3B8
<400> 117
gatattgtga tgacacagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat ggcacatcca acctggcttc tggagtccct 180
gttcgcttca gtggcagtgg atctgggacc tcttattctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgtcaa cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggagct gaaac 325
<210> 118
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3B8
<400> 118
Asp Ile Val Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Gly Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 119
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3B8
<400> 119
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 120
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3B8
<400> 120
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 121
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3F8
<400> 121
gaggttcagc ttcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt aactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaatt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 122
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P3F8
<400> 122
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Asn Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 123
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3F8
<400> 123
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 124
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P3F8
<400> 124
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 125
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3F8
<400> 125
gatattgttc tcacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccag cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggagtt gaaa 324
<210> 126
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P3F8
<400> 126
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 127
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3F8
<400> 127
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 128
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P3F8
<400> 128
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 129
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P4C6
<400> 129
gaggtgcagc tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 130
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P4C6
<400> 130
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 131
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P4C6
<400> 131
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 132
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P4C6
<400> 132
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 133
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P4C6
<400> 133
gacattgtga taactcagtc tccagcaatc gtgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttattctc tcacaatcag cagcatggag 240
gctgaagatg ccgccactta ttactgtcaa cagtggagta gttacccact cacgttcggt 300
gctgggacca agctggaaat caaa 324
<210> 134
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P4C6
<400> 134
Asp Ile Val Ile Thr Gln Ser Pro Ala Ile Val Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 135
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P4C6
<400> 135
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 136
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P4C6
<400> 136
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 137
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P4G10
<400> 137
gaggttcagc tgcagcagtt tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag tacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 138
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P4G10
<400> 138
Glu Val Gln Leu Gln Gln Phe Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 139
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P4G10
<400> 139
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 140
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P4G10
<400> 140
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 141
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P4G10
<400> 141
gacattgttc tcacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccag cagtggagta gtaacccgct cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 142
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P4G10
<400> 142
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro
85 90 95
Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 143
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P4G10
<400> 143
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 144
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P4G10
<400> 144
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 145
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P5E7
<400> 145
gaggtccagc tccagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt aactactgga tgaactgggt gaagcagagg 120
cccggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaacttc 180
aatggaaagt tcaagggtag agccatactg actgcagaca tatcctccaa cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 146
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P5E7
<400> 146
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Asn Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Phe Asn Gly Lys Phe
50 55 60
Lys Gly Arg Ala Ile Leu Thr Ala Asp Ile Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 147
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P5E7
<400> 147
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 148
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P5E7
<400> 148
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 149
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P5E7
<400> 149
gacattgtgc tcactcagtc tccaaccacc atggctgcat ctcccgggga gaagatcact 60
atcacctgca gtgccagctc aagtataagt tccaattact tgcattggta tcagcagaag 120
ccaggattct cccctaaact cttgatttat aggacatcca ctctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccaca cacgttcggc 300
tcggggacca aactggaaat caaa 324
<210> 150
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P5E7
<400> 150
Asp Ile Val Leu Thr Gln Ser Pro Thr Thr Met Ala Ala Ser Pro Gly
1 5 10 15
Glu Lys Ile Thr Ile Thr Cys Ser Ala Ser Ser Ser Ile Ser Ser Asn
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Phe Ser Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Thr Ser Thr Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
His Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 151
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P5E7
<400> 151
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 152
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P5E7
<400> 152
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 153
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P6D7
<400> 153
gaggtgaagc tggtggagtc tgggggaggt ttagtgcagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt agctatacca tgtcttgggt tcgccagact 120
ccagagaaga ggctggagtg ggtcgcatac attagtaatg gtggtggtag cacctactat 180
ccagacactg taaagggccg attcaccatc tccagagaca atgccaagaa caccctgtac 240
ctgcaaatga gcagtctgaa gtctgaggac acggccatgt attactgtgc aagacatcga 300
gatggtaact acgactactg gggccaaggc accacggtca ccgtctcctc a 351
<210> 154
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P6D7
<400> 154
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Asn Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg His Arg Asp Gly Asn Tyr Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Val Thr Val Ser Ser
115
<210> 155
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P6D7
<400> 155
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 156
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P6D7
<400> 156
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 157
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P6D7
<400> 157
gatattgtga taactcagga tgaactctcc aaccctgtca cttctggaga atcagtttcc 60
atctcctgca ggtctagtaa gagtctccta tataaggatg ggaagacata cttgaattgg 120
tttctgcaga gaccaggaca atctcctcag ctcctgatct atttgatgtc cacccgtgca 180
tcaggagtct cagaccggtt tagtggcagt gggtcaggaa cagatttcac cctggaaatc 240
agtaaagtga aggctgagga tgtgggtgtg tattactgtc aacaacttgt agagtatccg 300
ctcacgttcg gtgctgggac aaagttggaa ataaaa 336
<210> 158
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P6D7
<400> 158
Asp Ile Val Ile Thr Gln Asp Glu Leu Ser Asn Pro Val Thr Ser Gly
1 5 10 15
Glu Ser Val Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu Tyr Lys
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Met Ser Thr Arg Ala Ser Gly Val Ser
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Glu Ile
65 70 75 80
Ser Lys Val Lys Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Leu
85 90 95
Val Glu Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 159
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P6D7
<400> 159
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 160
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P6D7
<400> 160
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 161
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P1A7
<400> 161
ggggttcaac tgcagcagtc tggggctgag ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggctt tgcattcagt tactactgga tgaactgggt gaagcagagg 120
cccggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaacttc 180
aatggaaagt tcaagggtag agccatactg actgcagaca tatcctccaa cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccgaggg actctggtca ctgtctctgc a 351
<210> 162
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2LR3-P1A7
<400> 162
Gly Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Phe Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Phe Asn Gly Lys Phe
50 55 60
Lys Gly Arg Ala Ile Leu Thr Ala Asp Ile Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 163
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P1A7
<400> 163
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 164
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2LR3-P1A7
<400> 164
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 165
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P1A7
<400> 165
gatattgtgc taactcagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aaatgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcggt 300
gctgggacca agctggagct gaaa 324
<210> 166
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2LR3-P1A7
<400> 166
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Asn Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 167
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P1A7
<400> 167
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 168
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2LR3-P1A7
<400> 168
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 169
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2R3-P1F1
<400> 169
gaggtccagc tgcagcagtc tggagctgag ctgatgaagc ctggggcctc agtgaagata 60
tcctgcaagg ctactggcta cacattcagt agctactgga tagagtgggt aaagcagagg 120
cctggacatg gccttgagtg gattggagag attttacctg gaagtggtag tactaactac 180
aatgagaagt tcaagggcaa ggccacattc actgcagata catcctccaa cacagcctat 240
atgcaactca gcagcctgac atctgaggac tctgccgtct attactgtgc aagaggggca 300
tggttactat cctactggta cttcgatgtc tggggcgcgg ggacctcagt caccgtctcc 360
tca 363
<210> 170
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA2R3-P1F1
<400> 170
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Met Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Thr Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Trp Ile Glu Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Leu Pro Gly Ser Gly Ser Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Phe Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Trp Leu Leu Ser Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 171
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2R3-P1F1
<400> 171
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 172
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA2R3-P1F1
<400> 172
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 173
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2R3-P1F1
<400> 173
gacattcaga tgatgcagtc tacatcctcc ctgtctgcct ctctgggaga cagagtcacc 60
atcagttgca gtgcaagtca gggcattagc aattatttaa actggtatca gcagaaacca 120
gatggaactg ttaaactcct gatctattac acatcaagtt tacactcagg agtcccatca 180
aggttcagtg gcagtggatc aggcacacag ttttctctga agatcaacag cctgcagcct 240
gaagattttg ggagttatta ctgtcaacat cattatggta ctccgtacac gttcggaggg 300
gggaccaagc tggagctgaa a 321
<210> 174
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA2R3-P1F1
<400> 174
Asp Ile Gln Met Met Gln Ser Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Ser Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Ser Leu Lys Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Ser Tyr Tyr Cys Gln His His Tyr Gly Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 175
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2R3-P1F1
<400> 175
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 176
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA2R3-P1F1
<400> 176
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 177
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-A7
<400> 177
gaggtccagc tccagcaatc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 178
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-A7
<400> 178
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 179
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-A7
<400> 179
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 180
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-A7
<400> 180
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 181
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-A7
<400> 181
gatattgttc tcaaccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
ataacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccct 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagtatggag 240
gctgaagatg ctgccactta ttactgccag cagtatcata cttacccacc cacgttcggc 300
tcggggacaa agttggaaat aaaa 324
<210> 182
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-A7
<400> 182
Asp Ile Val Leu Asn Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr His Thr Tyr Pro
85 90 95
Pro Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 183
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-A7
<400> 183
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ccgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgcccc ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 184
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-A7
<400> 184
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Pro Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 185
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-C3
<400> 185
gaggtccagc tccagcaatc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagttca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaaccacc ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 186
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-C3
<400> 186
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Phe Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn His Leu Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 187
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-C3
<400> 187
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 188
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-C3
<400> 188
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 189
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-C3
<400> 189
gatattgttc tctcccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagc tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctgggtttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gcggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca aactggaaat caaa 324
<210> 190
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-C3
<400> 190
Asp Ile Val Leu Ser Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Val Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 191
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-C3
<400> 191
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 192
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-C3
<400> 192
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 193
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-D10
<400> 193
gaggtccagc tccagcaatc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccaa cacagcctat 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtac aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 194
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-D10
<400> 194
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Thr Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 195
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-D10
<400> 195
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 196
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-D10
<400> 196
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 197
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-D10
<400> 197
gacattgttc tctcccagtc tccagcaatc atgtctgcat ctccagggga acgggtcacc 60
atgacctgca ccgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 198
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-D10
<400> 198
Asp Ile Val Leu Ser Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 199
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-D10
<400> 199
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 200
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-D10
<400> 200
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 201
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-D11
<400> 201
gaggttcagc tgcagcagtc tggacctgag ctgatgaagc ctggggcttc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttcctg gggccaaggg actctggtca ctgtctctgc a 351
<210> 202
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-D11
<400> 202
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Met Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Ser Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 203
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-D11
<400> 203
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 204
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-D11
<400> 204
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 205
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-D11
<400> 205
gacattgttc tctcccagtc tccagcaatc atgtccgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 206
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-D11
<400> 206
Asp Ile Val Leu Ser Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 207
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-D11
<400> 207
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 208
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-D11
<400> 208
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 209
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-F6
<400> 209
gaggtccagc tgcagcagtc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactaca ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 210
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P3-F6
<400> 210
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Ile Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 211
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-F6
<400> 211
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 212
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P3-F6
<400> 212
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 213
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-F6
<400> 213
gatattgttc tctcccagtc tccagcaatc atgtctgcat ctctagggga gcgtgtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 214
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P3-F6
<400> 214
Asp Ile Val Leu Ser Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 215
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-F6
<400> 215
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 216
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P3-F6
<400> 216
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 217
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-C4
<400> 217
gaggttcaac tgcagcagtc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcaat tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 218
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-C4
<400> 218
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Asn Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 219
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-C4
<400> 219
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 220
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-C4
<400> 220
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 221
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-C4
<400> 221
gacattgagc tcacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atatcctgca gagccagtga aagtgttgat agttatggca atagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatc ttgcatccaa cctagaatct 180
ggggtccctg ccaggttcag tggcagtggg tctaggacag acttcaccct caccattgat 240
cctgtggagg ctgatgatgc tgcaacctat tactgtcagc aaaataatga ggatcctccg 300
acgttcggtg gagggaccaa gctggaaata aaa 333
<210> 222
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-C4
<400> 222
Asp Ile Glu Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 223
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-C4
<400> 223
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 224
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-C4
<400> 224
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 225
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-F8
<400> 225
gaggtccagc ttcagcagtc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 226
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-F8
<400> 226
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 227
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-F8
<400> 227
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 228
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-F8
<400> 228
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 229
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-F8
<400> 229
gacattgtgc tcacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atatcctgca gagccagtga aagtgttgat agttatggca atagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatc ttgcatccaa cctagaacct 180
ggggtccctg ccaggttcag tggcagtggg tctaggacag acttcaccct caccattgat 240
cctgtggagg ctgatgatgc tgcaacctat tactgtcagc aaaataatga ggatcctccg 300
acgttcggtg gagggaccaa gctggagctg aaa 333
<210> 230
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-F8
<400> 230
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Pro Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 231
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-F8
<400> 231
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 232
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-F8
<400> 232
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 233
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-G11
<400> 233
gaggtgcagc tccagcagtc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 234
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-G11
<400> 234
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 235
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-G11
<400> 235
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 236
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-G11
<400> 236
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 237
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-G11
<400> 237
gatattgtgt taacacagtc tcctgcttcc ttagctgtgt ctctggggca gagggccacc 60
atctcatgca gggccagcca aagtgtcagt acatctcgct atagttatat gcactggtac 120
caacagaaac caggacagcc acccaaactc ctcatcaagt atgcatccaa cctagaatct 180
ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccac 240
cctgtggagg aggaggatat tgcaacatat tactgtcagc acagttggga gattcccacg 300
ttcggctcgg ggaccaagct ggagctgaaa 330
<210> 238
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-G11
<400> 238
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Ser Thr Ser
20 25 30
Arg Tyr Ser Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Lys Tyr Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ile Ala Thr Tyr Tyr Cys Gln His Ser Trp
85 90 95
Glu Ile Pro Thr Phe Gly Ser Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 239
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-G11
<400> 239
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 240
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-G11
<400> 240
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 241
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-H10
<400> 241
gaggttcaac tgcagcagtc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaacttc 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaaccacc tagcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 242
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA13R3P4-H10
<400> 242
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Phe Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn His Leu Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 243
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-H10
<400> 243
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 244
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA13R3P4-H10
<400> 244
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 245
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-H10
<400> 245
gatattgttc tcaaccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 246
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA13R3P4-H10
<400> 246
Asp Ile Val Leu Asn Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 247
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-H10
<400> 247
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg tgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 248
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA13R3P4-H10
<400> 248
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 249
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA15R3P3-A6
<400> 249
gaggttcaac tgcagcagtc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 250
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA15R3P3-A6
<400> 250
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 251
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA15R3P3-A6
<400> 251
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 252
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA15R3P3-A6
<400> 252
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 253
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA15R3P3-A6
<400> 253
gatattgtaa tgacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 254
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA15R3P3-A6
<400> 254
Asp Ile Val Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 255
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA15R3P3-A6
<400> 255
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 256
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA15R3P3-A6
<400> 256
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 257
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA15R3P3-A7
<400> 257
gaggttcagc tgcagcagtc tggacctgag ctggtgaagc ctgggtcttc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaaccacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 258
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA15R3P3-A7
<400> 258
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn His Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 259
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA15R3P3-A7
<400> 259
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 260
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA15R3P3-A7
<400> 260
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 261
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA15R3P3-A7
<400> 261
gacattgttc tcacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
ctgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacttcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agttggaaat aaaa 324
<210> 262
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA15R3P3-A7
<400> 262
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Leu Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 263
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA15R3P3-A7
<400> 263
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 264
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA15R3P3-A7
<400> 264
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 265
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA15R3P3-C9
<400> 265
gaggtccagc tgcagcaatc tggggctgag ctgctgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgaattcagt tactactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga aactaagtac 180
aatggaaagt tcaagggtaa agccacactg actgcagaca aatcctccaa cacagcctat 240
atgcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcccat 300
ggtaactacc ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 266
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA15R3P3-C9
<400> 266
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Leu Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 267
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA15R3P3-C9
<400> 267
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 268
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA15R3P3-C9
<400> 268
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 269
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA15R3P3-C9
<400> 269
ggcattctga tgacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtataagt tccaattact tgcattggta tcagcagaag 120
ccaggattct cccctaaact cttgatttat aggacatcca atctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtttcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 270
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA15R3P3-C9
<400> 270
Gly Ile Leu Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Ile Ser Ser Asn
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Phe Ser Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Phe His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 271
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA15R3P3-C9
<400> 271
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 272
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA15R3P3-C9
<400> 272
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 273
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P2G6
<400> 273
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 274
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P2G6
<400> 274
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 275
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P2G6
<400> 275
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 276
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P2G6
<400> 276
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 277
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P2G6
<400> 277
gatattgagc tgacccaatc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta tttctgccag cagtggagta gtttcccgct cacgttcggt 300
ggagggacca agctggagct gaaa 324
<210> 278
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P2G6
<400> 278
Asp Ile Glu Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Phe Cys Gln Gln Trp Ser Ser Phe Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 279
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P2G6
<400> 279
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 280
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P2G6
<400> 280
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 281
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1A6
<400> 281
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 282
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1A6
<400> 282
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 283
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1A6
<400> 283
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 284
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1A6
<400> 284
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 285
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1A6
<400> 285
gatattgttc tcaaccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagcatcatc gttccccacc cacgttcgga 300
ggggggacaa agttggaaat aaaa 324
<210> 286
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1A6
<400> 286
Asp Ile Val Leu Asn Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln His His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 287
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1A6
<400> 287
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 288
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1A6
<400> 288
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 289
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1B5
<400> 289
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 290
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1B5
<400> 290
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 291
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1B5
<400> 291
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 292
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1B5
<400> 292
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 293
<211> 334
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1B5
<400> 293
gatattgtgc taacacagtc tcctgcttcc ttagctgtat ctctggggca gagggccacc 60
atctcatgca gggccagcaa aagtgtcagt acatctggct atagttatat gcactggttc 120
cagcagaaac caggacagcc acccaaactc ctcatcaagt atgcatccaa cctagaatct 180
ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat 240
cctgtggagg aggaggatgc tgcaacctat tactgtcagc acggttggga gcttccattc 300
acgttcggct cggggaccaa gctggagctg aaac 334
<210> 294
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1B5
<400> 294
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Lys Ser Val Ser Thr Ser
20 25 30
Gly Tyr Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Lys Tyr Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys Gln His Gly Trp
85 90 95
Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 295
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1B5
<400> 295
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 296
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1B5
<400> 296
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 297
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1E5
<400> 297
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 298
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1E5
<400> 298
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 299
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1E5
<400> 299
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 300
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1E5
<400> 300
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 301
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1E5
<400> 301
gacattgtga tgacccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
ctgacctgca ctgccagttc aagtgttagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacttcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttccccacc cacgttcgga 300
ggggggacca agctggagct gaaa 324
<210> 302
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1E5
<400> 302
Asp Ile Val Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Leu Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 303
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1E5
<400> 303
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 304
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1E5
<400> 304
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 305
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1H5
<400> 305
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 306
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA16R3-P1H5
<400> 306
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 307
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1H5
<400> 307
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 308
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA16R3-P1H5
<400> 308
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 309
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1H5
<400> 309
gatattgtga tgacccagtc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atatcctgca gagccagtga aagtgttgat agttatggca atagttttat gcactggtac 120
cagcagaaag caggacagcc acccaaactc ctcatctatc ttgcatccaa cctagaatct 180
ggggtccctg ccaggttcag tggcagtggg tctaggacag acttcaccct caccattgat 240
cctgtggagg ctgatgatgc tgcaacctat tactgtcaac aacattatag tactcctccg 300
acgttcggtg gagggacaaa gttggaaata aaa 333
<210> 310
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA16R3-P1H5
<400> 310
Asp Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Ala Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln His Tyr
85 90 95
Ser Thr Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 311
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1H5
<400> 311
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 312
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA16R3-P1H5
<400> 312
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 313
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA18R3-P1D8
<400> 313
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 314
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA18R3-P1D8
<400> 314
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 315
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA18R3-P1D8
<400> 315
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 316
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA18R3-P1D8
<400> 316
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 317
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA18R3-P1D8
<400> 317
gatattgtga taactcagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tacactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccattta ttactgccag cagtggaatt atcctcctat cacgttcggc 300
tcggggacca agctggaaat caaa 324
<210> 318
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA18R3-P1D8
<400> 318
Asp Ile Val Ile Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Ile Tyr Tyr Cys Gln Gln Trp Asn Tyr Pro Pro
85 90 95
Ile Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 319
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA18R3-P1D8
<400> 319
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 320
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA18R3-P1D8
<400> 320
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 321
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA18R3-P1E5
<400> 321
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 322
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA18R3-P1E5
<400> 322
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 323
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA18R3-P1E5
<400> 323
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 324
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA18R3-P1E5
<400> 324
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 325
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA18R3-P1E5
<400> 325
gacattgtaa tgacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
ataacctgca gtgccagctc aagtataagt tccaattact tgcattggta tcagcagaag 120
ccaggattct cccctaaact cttgatttat aggacatcca atctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaattgg caccatggag 240
gctgaagatg ttgccactta ctactgccag cagggtagta gtataccatt cacgttcggc 300
tcggggacca agctggaaat aaaa 324
<210> 326
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA18R3-P1E5
<400> 326
Asp Ile Val Met Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Ile Ser Ser Asn
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Phe Ser Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Gly Thr Met Glu
65 70 75 80
Ala Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Ser Ile Pro
85 90 95
Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 327
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA18R3-P1E5
<400> 327
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 328
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA18R3-P1E5
<400> 328
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 329
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA18R3-P1H5
<400> 329
gaggttcagc tgcagcagtc tggggctgaa ctggtgaggc ctgggtcctc agtgaagatt 60
tcctgcaagg cttctggcta tgcattcagt tattactgga tgaactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggacag atttatcctg gagatggtga tactaactac 180
agtggaaggt tcaagggtaa agccacactg actgcagaca aatcctccag cacagcctac 240
attcagctca gcagcctaac atctgaggac tctgcggtct atttctgtgc aagatcgcac 300
ggtaactatt ttccttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 330
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи PA18R3-P1H5
<400> 330
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Ile Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Phe Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 331
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA18R3-P1H5
<400> 331
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtca 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 332
<211> 204
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи PA18R3-P1H5
<400> 332
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200
<210> 333
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA18R3-P1H5
<400> 333
gacattgttc tcaaccagtc tccagcaatc atgtctgcat ctctagggga acgggtcacc 60
atgacctgca ctgccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
ccaggatcct cccccaaact ctggatttat agcacatcca acctggcttc tggagtccca 180
gctcgcttca gtggcagtgg gtctgggacc tcttactctc tcacaatcag cagcatggag 240
gctgaagatg ctgccactta ttactgccac cagtatcatc gttcccacac gttcggaggg 300
gggaccaaac tggaaatcaa a 321
<210> 334
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи PA18R3-P1H5
<400> 334
Asp Ile Val Leu Asn Gln Ser Pro Ala Ile Met Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Thr Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Tyr His Arg Ser His
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 335
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA18R3-P1H5
<400> 335
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagcttgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 336
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок легкой цепи PA18R3-P1H5
<400> 336
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Leu
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 337
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок легкой цепи гуманизированного P1C4
<400> 337
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 338
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> Полноразмерная легкая цепь гуманизированного P1C4
<400> 338
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 339
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи гуманизированного P1C4-H0
<400> 339
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 340
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи гуманизированного P1C4-H2
<400> 340
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 341
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариабельный участок тяжелой цепи гуманизированного P1C4-H4
<400> 341
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 342
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи гуманизированного P1C4
<400> 342
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 343
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гибрид IgG2-4 гуманизированного P1C4 H0
<400> 343
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 344
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG2A330S/P331S гуманизированного P1C4 H0
<400> 344
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
290 295 300
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 345
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG4S228P гуманизированного P1C4 H0
<400> 345
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 346
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гибрид IgG2-4 гуманизированного P1C4 H2
<400> 346
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 347
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG2A330S/P331S гуманизированного P1C4 H2
<400> 347
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
290 295 300
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 348
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG4S228P гуманизированного P1C4 H2
<400> 348
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 349
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гибрид IgG2-4 гуманизированного P1C4 H4
<400> 349
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 350
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG2A330S/P331S гуманизированного P1C4 H4
<400> 350
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
290 295 300
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 351
<211> 444
<212> Белок
<213> IgG4S228P гуманизированного P1C4 H4
<400> 351
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 352
<211> 5
<212> Белок
<213> Mus sp.
<400> 352
Tyr Tyr Trp Met Asn
1 5
<210> 353
<211> 11
<212> Белок
<213> Mus sp.
<400> 353
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 354
<211> 8
<212> Белок
<213> Mus sp.
<400> 354
Ser His Gly Asn Tyr Leu Pro Tyr
1 5
<210> 355
<211> 5
<212> Белок
<213> Mus sp.
<400> 355
Ser Ser Tyr Leu His
1 5
<210> 356
<211> 7
<212> Белок
<213> Mus sp.
<400> 356
Ser Thr Ser Asn Leu Ala Ser
1 5
<210> 357
<211> 10
<212> Белок
<213> Mus sp.
<400> 357
His Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 358
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> алемтузумаб моноклональное антитело
<400> 358
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Phe Ile Arg Asp Lys Ala Lys Gly Tyr Thr Thr Glu Tyr Asn Pro
50 55 60
Ser Val Lys Gly Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln
65 70 75 80
Phe Ser Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Gly His Thr Ala Ala Pro Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 359
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> бенрализумаб моноклональное антитело
<400> 359
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Val Ile His Trp Val Arg Gln Arg Pro Gly Gln Gly Leu Ala Trp Met
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Gly Thr Lys Tyr Asn Glu Arg Phe
50 55 60
Lys Gly Lys Val Thr Ile Thr Ser Asp Arg Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Leu Cys
85 90 95
Gly Arg Glu Gly Ile Arg Tyr Tyr Gly Leu Leu Gly Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 360
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> бевастизумаб моноклональное антитело
<400> 360
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Ala Asp Phe
50 55 60
Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Pro His Tyr Tyr Gly Ser Ser His Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 361
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> бимекизумаб моноклональное антитело
<400> 361
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Asn Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Tyr Glu Gly Arg Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Pro Gln Tyr Tyr Glu Gly Ser Ile Tyr Arg Leu Trp Phe
100 105 110
Ala His Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 362
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> кантузумаб равтансин моноклональное антитело
<400> 362
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Asp Tyr Thr Phe Thr Tyr Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asp Thr Thr Thr Gly Glu Pro Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ile Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Lys Ser Leu Lys Ser Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Pro Tyr Asn Trp Tyr Phe Asp Val Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 363
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> кодритузумаб моноклональное антитело
<400> 363
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 364
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> далотузумаб моноклональное антитело
<400> 364
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Gly Gly
20 25 30
Tyr Leu Trp Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Asp Gly Thr Asn Asn Tyr Lys Pro Ser Leu
50 55 60
Lys Asp Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Val Phe Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 365
<211> 391
<212> Белок
<213> Искусственная последовательность
<220>
<223> эфализумаб моноклональное антитело
<400> 365
Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys
1 5 10 15
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
20 25 30
Val Tyr Tyr Cys Ala Arg Gly Ile Tyr Phe Tyr Gly Thr Thr Tyr Phe
35 40 45
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
50 55 60
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
65 70 75 80
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
85 90 95
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
100 105 110
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
115 120 125
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
130 135 140
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
145 150 155 160
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
165 170 175
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
180 185 190
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
195 200 205
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
210 215 220
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
225 230 235 240
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
245 250 255
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
260 265 270
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
275 280 285
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
290 295 300
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
305 310 315 320
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
325 330 335
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
340 345 350
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
355 360 365
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
370 375 380
Leu Ser Leu Ser Pro Gly Lys
385 390
<210> 366
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> элотузумаб моноклональное антитело
<400> 366
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Asp Gly Asn Tyr Trp Tyr Phe Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 367
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> энаватузумаб моноклональное антитело
<400> 367
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Glu Ile Arg Leu Lys Ser Asp Asn Tyr Ala Thr His Tyr Ala Glu
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Gly Tyr Tyr Ala Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 368
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> энокизумаб моноклональное антитело
<400> 368
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Tyr Tyr
20 25 30
Trp Ile Glu Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Glu Ile Leu Pro Gly Ser Gly Thr Thr Asn Pro Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Asp Tyr Tyr Gly Ser Asp Tyr Val Lys Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 369
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> этролизумаб моноклональное антитело
<400> 369
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Phe Ile Thr Asn Asn
20 25 30
Tyr Trp Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Ser Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Phe Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Gly Ser Ser Gly Tyr Phe Asp Phe Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 370
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> фарлетузумаб моноклональное антитело
<400> 370
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Met Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Arg His Gly Asp Asp Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 371
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> фиклатузумаб моноклональное антитело
<400> 371
Gln Val Gln Leu Val Gln Pro Gly Ala Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Asn Gly His Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Val Gly Ser Ile Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 372
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> имгатузумаб моноклональное антитело
<400> 372
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Lys Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Phe Asn Pro Asn Ser Gly Tyr Ser Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ser Pro Gly Gly Tyr Tyr Val Met Asp Ala Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 373
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> итолизумаб моноклональное антитело
<400> 373
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Lys Phe Ser Arg Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Tyr Ile Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Asp Leu Asp Tyr Phe Asp Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 374
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> лифастузумаб ведотин моноклональное антитело
<400> 374
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Asp Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Gly Arg Val Ala Phe His Thr Tyr Tyr Pro Asp Ser Met
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Arg Gly Phe Asp Val Gly His Phe Asp Phe Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 375
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> лигелизумаб моноклональное антитело
<400> 375
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Met Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Trp Tyr
20 25 30
Trp Leu Glu Trp Val Arg Gln Ala Pro Gly His Gly Leu Glu Trp Met
35 40 45
Gly Glu Ile Asp Pro Gly Thr Phe Thr Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ala Arg Val Thr Phe Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Ser His Phe Ser Gly Ser Asn Tyr Asp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 376
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> лоделцизумаб моноклональное антитело
<400> 376
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Thr Met
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Glu His Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Tyr Tyr Tyr Tyr Asn Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 377
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> лорвотузумаб мертансин моноклональное антитело
<400> 377
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Phe Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Arg Lys Gly Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 378
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> могамулизумаб моноклональное антитело
<400> 378
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Ala Ser Thr Tyr Ser Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Gly Arg His Ser Asp Gly Asn Phe Ala Phe Gly Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 379
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> мотавизумаб моноклональное антитело
<400> 379
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ala
20 25 30
Gly Met Ser Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Asp Ile Trp Trp Asp Asp Lys Lys His Tyr Asn Pro Ser
50 55 60
Leu Lys Asp Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Lys Val Thr Asn Met Asp Pro Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Asp Met Ile Phe Asn Phe Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 380
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> обнитузумаб моноклональное антитело
<400> 380
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 381
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> окаратузумаб моноклональное антитело
<400> 381
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Arg Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Tyr Pro Leu Thr Gly Asp Thr Ser Tyr Asn Gln Lys Ser
50 55 60
Lys Leu Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Ser Thr Tyr Val Gly Gly Asp Trp Gln Phe Asp Val Trp Gly
100 105 110
Lys Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Ile Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Gln Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly
450
<210> 382
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> омализумаб моноклональное антитело
<400> 382
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Ser Ile Thr Tyr Asp Gly Ser Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 383
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> парсатузумаб моноклональное антитело
<400> 383
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ile Asp Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Leu Asp Asn Ser Gly Thr His Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Val Tyr His Asp Tyr Asp Asp Tyr Ala Met Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 384
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> патеклизумаб моноклональное антитело
<400> 384
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Val Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Tyr Asn Asn Pro Tyr Asn Ala Gly Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ser Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Pro Thr Met Leu Pro Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 385
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> перакизумаб моноклональное антитело
<400> 385
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Thr Met Leu Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Lys Ser Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Asp Tyr Gly Ser Ser Tyr Gly Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 386
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> пертузумаб моноклональное антитело
<400> 386
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His
225
<210> 387
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> пидилизумаб моноклональное антитело
<400> 387
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Thr Asp Ser Gly Glu Ser Thr Tyr Ala Glu Glu Phe
50 55 60
Lys Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Val Asn Thr Ala Tyr
65 70 75 80
Leu Gln Ile Thr Ser Leu Thr Ala Glu Asp Thr Gly Met Tyr Phe Cys
85 90 95
Val Arg Val Gly Tyr Asp Ala Leu Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 388
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> квилизумаб моноклональное антитело
<400> 388
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Ile Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Ser Asp Leu Ala Tyr Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asn Trp Asp Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 389
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> ронтализумаб моноклональное антитело
<400> 389
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Ile Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Asn Pro Asp Tyr Asp Ile Thr Asn Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Leu Asp Lys Ser Lys Arg Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Trp Ile Ser Asp Phe Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 390
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> софитузумаб ведотин моноклональное антитело
<400> 390
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Ile Thr Asn Asp
20 25 30
Tyr Ala Trp Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Tyr Ile Ser Tyr Ser Gly Tyr Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Thr Ser Gly Leu Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 391
<211> 442
<212> Белок
<213> Искусственная последовательность
<220>
<223> соланезумаб моноклональное антитело
<400> 391
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ser Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Gln Ile Asn Ser Val Gly Asn Ser Thr Tyr Tyr Pro Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Gly Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
340 345 350
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 392
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> сувизумаб моноклональное антитело
<400> 392
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Ser
20 25 30
Trp Ile Gly Trp Phe Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Gly Gly Gly Tyr Thr Asn Tyr Asn Glu Ile Phe
50 55 60
Lys Gly Lys Ala Thr Met Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Gly Ile Pro Gly Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 393
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> теплизумаб моноклональное антитело
<400> 393
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Val
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Ala Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 394
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> тилдракизумаб моноклональное антитело
<400> 394
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Thr Tyr
20 25 30
Trp Met Thr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Phe Pro Ala Ser Gly Ser Ala Asp Tyr Asn Glu Lys Phe
50 55 60
Glu Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Gly Gly Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 395
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> тоцилизумаб моноклональное антитело
<400> 395
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 396
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> трастузумаб моноклональное антитело
<400> 396
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 397
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> трастузумаб эмтансин моноклональное антитело
<400> 397
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 398
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> трегализумаб моноклональное антитело
<400> 398
Glu Glu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Asp Cys
20 25 30
Arg Met Tyr Trp Leu Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Ser Val Lys Ser Glu Asn Tyr Gly Ala Asn Tyr Ala Glu
50 55 60
Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ser Ala Ser Tyr Tyr Arg Tyr Asp Val Gly Ala Trp Phe Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 399
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> ведолизумаб моноклональное антитело
<400> 399
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Gly Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp Pro Ser Glu Ser Asn Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Ile Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Asp Gly Trp Asp Tyr Ala Ile Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Ala Gly
225 230 235 240
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 400
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> ворсетузумаб моноклональное антитело
<400> 400
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Ala Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Asp Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 401
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> ворсетузумаб мафодотин моноклональное антитело
<400> 401
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Ala Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Asp Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 402
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> иттрий 90 Y кливатузумаб тетраксетан моноклональное антитело
<400> 402
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Phe Pro Ser Tyr
20 25 30
Val Leu His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Gly Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Arg Asp Thr Ser Ile Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Phe Gly Gly Ser Tyr Gly Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 403
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> анрукинзумаб моноклональное антитело
<400> 403
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ile Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Asn Thr Tyr Tyr Pro Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Leu Asp Gly Tyr Tyr Phe Gly Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Leu Gly Ala Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 404
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> дацетузумаб моноклональное антитело
<400> 404
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 405
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> даклизумаб моноклональное антитело
<400> 405
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Arg Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Thr Gly Tyr Thr Glu Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Ile Thr Ala Asp Glu Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Gly Val Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 406
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> этарацизумаб моноклональное антитело
<400> 406
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Val Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Leu Asp Thr Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Leu His Gly Ser Phe Ala Ser Trp Gly Gln Gly Thr Thr
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 407
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> милатузумаб моноклональное антитело
<400> 407
Gln Val Gln Leu Gln Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Val Asn Trp Ile Lys Gln Ala Pro Gly Gln Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Thr Gly Glu Pro Thr Phe Asp Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ser Arg Ser Arg Gly Lys Asn Glu Ala Trp Phe Ala Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 408
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> озанезумаб моноклональное антитело
<400> 408
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Lys Ala Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Glu Leu Met Gln Gly Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Ala Gly Ala Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 409
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> пинатузумаб ведотин моноклональное антитело
<400> 409
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Glu Phe Ser Arg Ser
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Ser Gly Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Ser Ser Trp Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 410
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> полатузумаб ведотин моноклональное антитело
<400> 410
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Trp Ile Glu Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Leu Pro Gly Gly Gly Asp Thr Asn Tyr Asn Glu Ile Phe
50 55 60
Lys Gly Arg Ala Thr Phe Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Arg Val Pro Ile Arg Leu Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 411
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> тигатузумаб моноклональное антитело
<400> 411
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Val Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Asp Ser Met Ile Thr Thr Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 412
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> велтузумаб моноклональное антитело
<400> 412
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Phe Tyr Tyr Cys
85 90 95
Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 413
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> абитузумаб моноклональное антитело
<400> 413
Gln Val Gln Leu Gln Gln Ser Gly Gly Glu Leu Ala Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Arg Ser Gly Tyr Thr Glu Tyr Asn Glu Ile Phe
50 55 60
Arg Asp Lys Ala Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Phe Leu Gly Arg Gly Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Pro Lys Ser Ser Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Ala Gln Ser Thr Phe Arg Val Val Ser
290 295 300
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 414
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> бокоцизумаб моноклональное антитело
<400> 414
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Glu Ile Ser Pro Phe Gly Gly Arg Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Pro Leu Tyr Ala Ser Asp Leu Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 415
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> демцизумаб моноклональное антитело
<400> 415
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ala Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Ser Ser Tyr Asn Gly Ala Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Asp Tyr Asp Val Gly Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 416
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> гевокизумаб моноклональное антитело
<400> 416
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Gly Asp Glu Ser Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Ser Leu Lys Ile Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe
85 90 95
Cys Ala Arg Asn Arg Tyr Asp Pro Pro Trp Phe Val Asp Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Thr
180 185 190
Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val
210 215 220
Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Met Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val
290 295 300
Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 417
<211> 442
<212> Белок
<213> Искусственная последовательность
<220>
<223> понезумаб моноклональное антитело
<400> 417
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Tyr Thr Glu Ala Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Thr Gly Asn Thr Lys Tyr Ala Pro Arg Leu
50 55 60
Gln Asp Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Leu Tyr Ser Leu Pro Val Tyr Trp Gly Gln Gly Thr Thr Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 418
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> ралпанцизумаб моноклональное антитело
<400> 418
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Glu Ile His Pro Ser Gly Gly Arg Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Ser Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Pro Leu Tyr Ala Ser Asp Leu Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 419
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> ромозозумаб моноклональное антитело
<400> 419
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Glu Ile Asn Pro Asn Ser Gly Gly Ala Gly Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Asp Asp Ile Tyr Asp Asp Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys
210 215 220
Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val
290 295 300
Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 420
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> танезумаб моноклональное антитело
<400> 420
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ile Gly Tyr
20 25 30
Asp Leu Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Gly Asp Gly Thr Thr Asp Tyr Asn Ser Ala Val Lys
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Gly Tyr Trp Tyr Ala Thr Ser Tyr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys
210 215 220
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
290 295 300
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 421
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> блосозумаб моноклональное антитело
<400> 421
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Val Ser Gly Phe Pro Ile Lys Asp Thr
20 25 30
Phe Gln His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Trp Ser Asp Pro Glu Ile Gly Asp Thr Glu Tyr Ala Ser Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Glu Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Gly Asp Thr Thr Tyr Lys Phe Asp Phe Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 422
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> концизумаб моноклональное антитело
<400> 422
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 423
<211> 438
<212> Белок
<213> Искусственная последовательность
<220>
<223> кренезумаб моноклональное антитело
<400> 423
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Ser Ile Asn Ser Asn Gly Gly Ser Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Gly Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
115 120 125
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
180 185 190
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
210 215 220
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
245 250 255
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
260 265 270
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
275 280 285
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
305 310 315 320
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
325 330 335
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
340 345 350
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
355 360 365
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
370 375 380
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
385 390 395 400
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
405 410 415
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
420 425 430
Leu Ser Leu Ser Leu Gly
435
<210> 424
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> ибализумаб моноклональное антитело
<400> 424
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Val Ile His Trp Val Arg Gln Lys Pro Gly Gln Gly Leu Asp Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Gly Thr Asp Tyr Asp Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ser Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Lys Asp Asn Tyr Ala Thr Gly Ala Trp Phe Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys
<210> 425
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> иксекизумаб моноклональное антитело
<400> 425
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asp Tyr
20 25 30
His Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asn Pro Met Tyr Gly Thr Thr Asp Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Asp Tyr Phe Thr Gly Thr Gly Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 426
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> лебрикизумаб моноклональное антитело
<400> 426
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ala Tyr
20 25 30
Ser Val Asn Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Ala Met Ile Trp Gly Asp Gly Lys Ile Val Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Gly Asp Gly Tyr Tyr Pro Tyr Ala Met Asp Asn Trp Gly Gln Gly Ser
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 427
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> олокизумаб моноклональное антитело
<400> 427
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Asn Asp Tyr
20 25 30
Phe Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Met Arg Asn Lys Asn Tyr Gln Tyr Gly Thr Tyr Tyr Ala Glu
50 55 60
Ser Leu Glu Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Ser Tyr Tyr Gly Phe Thr Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 428
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> пембролизумаб моноклональное антитело
<400> 428
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Phe Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 429
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> симтузумаб моноклональное антитело
<400> 429
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Tyr Tyr
20 25 30
Leu Ile Glu Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Asn Pro Gly Ser Gly Gly Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Asn Trp Met Asn Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 430
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> улокаплумаб моноклональное антитело
<400> 430
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ala Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Arg Ser Arg Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Gly Gln Pro Pro Tyr Tyr Tyr Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser
130 135 140
Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys
195 200 205
Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Leu Gly
450
<210> 431
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> вателизумаб моноклональное антитело
<400> 431
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Ile His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ala Arg Gly Phe Thr Asn Tyr Asn Ser Ala Leu Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Asn Asp Gly Val Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 432
<211> 442
<212> Белок
<213> Искусственная последовательность
<220>
<223> самализумаб моноклональное антитело
<400> 432
Gln Val Gln Leu Gln Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asp Tyr
20 25 30
Ile Ile Leu Trp Val Arg Gln Asn Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly His Ile Asp Pro Tyr Tyr Gly Ser Ser Asn Tyr Asn Leu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Gln Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Ser Lys Arg Asp Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 433
<211> 326
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи IgG2
<400> 433
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 434
<211> 327
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи IgG4
<400> 434
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 435
<211> 327
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константный участок тяжелой цепи гибрида IgG2/4
<400> 435
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 436
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-H2-IgG2
<400> 436
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
290 295 300
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 437
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-H2-IgG4
<400> 437
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 438
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-H2-IgG2/4
<400> 438
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 439
<211> 443
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-H4-IgG2
<400> 439
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
290 295 300
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 440
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-H4-IgG4
<400> 440
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 441
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-H4-IgG2/4
<400> 441
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Ser Tyr Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr Asn Gly Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser His Gly Asn Tyr Leu Pro Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn
180 185 190
Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 442
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> P1C4-Lc гуманизированный
<400> 442
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Tyr His Arg Ser Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 443
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 443
His Tyr Trp Met Asn
1 5
<210> 444
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 444
Ile Tyr Trp Met Asn
1 5
<210> 445
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 445
Trp Tyr Trp Met Asn
1 5
<210> 446
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 446
Tyr Phe Trp Met Asn
1 5
<210> 447
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 447
Tyr Lys Trp Met Asn
1 5
<210> 448
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 448
Tyr Asn Trp Met Asn
1 5
<210> 449
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 449
Tyr Tyr Ala Met Asn
1 5
<210> 450
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 450
Tyr Tyr Phe Met Asn
1 5
<210> 451
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 451
Tyr Tyr Trp Ala Asn
1 5
<210> 452
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 452
Tyr Tyr Trp Glu Asn
1 5
<210> 453
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 453
Tyr Tyr Trp Phe Asn
1 5
<210> 454
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 454
Tyr Tyr Trp Leu Asn
1 5
<210> 455
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 455
Tyr Tyr Trp Asn Asn
1 5
<210> 456
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 456
Tyr Tyr Trp Gln Asn
1 5
<210> 457
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 457
Tyr Tyr Trp Thr Asn
1 5
<210> 458
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 458
Tyr Tyr Trp Val Asn
1 5
<210> 459
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 459
Tyr Tyr Trp Met Ile
1 5
<210> 460
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 460
Tyr Tyr Trp Met Lys
1 5
<210> 461
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 461
Tyr Tyr Trp Met Leu
1 5
<210> 462
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 462
Tyr Tyr Trp Met Ser
1 5
<210> 463
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 тяжелой цепи
<400> 463
Tyr Tyr Trp Met Trp
1 5
<210> 464
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 464
Asp Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 465
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 465
Glu Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 466
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 466
Gln Ala Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 467
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 467
Gln Asp Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 468
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 468
Gln Glu Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 469
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 469
Gln Phe Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 470
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 470
Gln Gly Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 471
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 471
Gln His Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 472
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 472
Gln Lys Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 473
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 473
Gln Leu Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 474
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 474
Gln Asn Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 475
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 475
Gln Gln Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 476
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 476
Gln Arg Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 477
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 477
Gln Ser Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 478
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 478
Gln Thr Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 479
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 479
Gln Trp Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 480
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 480
Gln Tyr Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 481
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 481
Gln Ile Phe Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 482
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 482
Gln Ile Tyr Ala Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 483
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 483
Gln Ile Tyr Phe Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 484
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 484
Gln Ile Tyr His Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 485
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 485
Gln Ile Tyr Lys Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 486
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 486
Gln Ile Tyr Pro Ala Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 487
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 487
Gln Ile Tyr Pro Cys Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 488
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 488
Gln Ile Tyr Pro Asp Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 489
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 489
Gln Ile Tyr Pro Glu Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 490
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 490
Gln Ile Tyr Pro Phe Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 491
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 491
Gln Ile Tyr Pro His Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 492
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 492
Gln Ile Tyr Pro Ile Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 493
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 493
Gln Ile Tyr Pro Lys Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 494
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 494
Gln Ile Tyr Pro Leu Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 495
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 495
Gln Ile Tyr Pro Met Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 496
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 496
Gln Ile Tyr Pro Gln Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 497
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 497
Gln Ile Tyr Pro Arg Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 498
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 498
Gln Ile Tyr Pro Ser Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 499
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 499
Gln Ile Tyr Pro Thr Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 500
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 500
Gln Ile Tyr Pro Val Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 501
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 501
Gln Ile Tyr Pro Trp Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 502
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 502
Gln Ile Tyr Pro Tyr Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 503
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 503
Gln Ile Tyr Pro Gly Ile Gly Glu Thr Lys Tyr
1 5 10
<210> 504
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 504
Gln Ile Tyr Pro Gly Leu Gly Glu Thr Lys Tyr
1 5 10
<210> 505
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 505
Gln Ile Tyr Pro Gly Met Gly Glu Thr Lys Tyr
1 5 10
<210> 506
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 506
Gln Ile Tyr Pro Gly Asn Gly Glu Thr Lys Tyr
1 5 10
<210> 507
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 507
Gln Ile Tyr Pro Gly Pro Gly Glu Thr Lys Tyr
1 5 10
<210> 508
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 508
Gln Ile Tyr Pro Gly Gln Gly Glu Thr Lys Tyr
1 5 10
<210> 509
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 509
Gln Ile Tyr Pro Gly Val Gly Glu Thr Lys Tyr
1 5 10
<210> 510
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 510
Gln Ile Tyr Pro Gly Trp Gly Glu Thr Lys Tyr
1 5 10
<210> 511
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 511
Gln Ile Tyr Pro Gly Tyr Gly Glu Thr Lys Tyr
1 5 10
<210> 512
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 512
Gln Ile Tyr Pro Gly Asp Ala Glu Thr Lys Tyr
1 5 10
<210> 513
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 513
Gln Ile Tyr Pro Gly Asp Asp Glu Thr Lys Tyr
1 5 10
<210> 514
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 514
Gln Ile Tyr Pro Gly Asp Glu Glu Thr Lys Tyr
1 5 10
<210> 515
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 515
Gln Ile Tyr Pro Gly Asp Phe Glu Thr Lys Tyr
1 5 10
<210> 516
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 516
Gln Ile Tyr Pro Gly Asp His Glu Thr Lys Tyr
1 5 10
<210> 517
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 517
Gln Ile Tyr Pro Gly Asp Ile Glu Thr Lys Tyr
1 5 10
<210> 518
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 518
Gln Ile Tyr Pro Gly Asp Lys Glu Thr Lys Tyr
1 5 10
<210> 519
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 519
Gln Ile Tyr Pro Gly Asp Leu Glu Thr Lys Tyr
1 5 10
<210> 520
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 520
Gln Ile Tyr Pro Gly Asp Met Glu Thr Lys Tyr
1 5 10
<210> 521
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 521
Gln Ile Tyr Pro Gly Asp Asn Glu Thr Lys Tyr
1 5 10
<210> 522
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 522
Gln Ile Tyr Pro Gly Asp Pro Glu Thr Lys Tyr
1 5 10
<210> 523
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 523
Gln Ile Tyr Pro Gly Asp Gln Glu Thr Lys Tyr
1 5 10
<210> 524
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 524
Gln Ile Tyr Pro Gly Asp Arg Glu Thr Lys Tyr
1 5 10
<210> 525
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 525
Gln Ile Tyr Pro Gly Asp Val Glu Thr Lys Tyr
1 5 10
<210> 526
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 526
Gln Ile Tyr Pro Gly Asp Thr Glu Thr Lys Tyr
1 5 10
<210> 527
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 527
Gln Ile Tyr Pro Gly Asp Val Glu Thr Lys Tyr
1 5 10
<210> 528
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 528
Gln Ile Tyr Pro Gly Asp Tyr Glu Thr Lys Tyr
1 5 10
<210> 529
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 529
Gln Ile Tyr Pro Gly Asp Gly Ala Thr Lys Tyr
1 5 10
<210> 530
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 530
Gln Ile Tyr Pro Gly Asp Gly Asp Thr Lys Tyr
1 5 10
<210> 531
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 531
Gln Ile Tyr Pro Gly Asp Gly Met Thr Lys Tyr
1 5 10
<210> 532
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 532
Gln Ile Tyr Pro Gly Asp Gly Gln Thr Lys Tyr
1 5 10
<210> 533
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 533
Gln Ile Tyr Pro Gly Asp Gly Val Thr Lys Tyr
1 5 10
<210> 534
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 534
Gln Ile Tyr Pro Gly Asp Gly Tyr Thr Lys Tyr
1 5 10
<210> 535
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 535
Gln Ile Tyr Pro Gly Asp Gly Glu Ala Lys Tyr
1 5 10
<210> 536
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 536
Gln Ile Tyr Pro Gly Asp Gly Glu Asp Lys Tyr
1 5 10
<210> 537
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 537
Gln Ile Tyr Pro Gly Asp Gly Glu Glu Lys Tyr
1 5 10
<210> 538
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 538
Gln Ile Tyr Pro Gly Asp Gly Glu Phe Lys Tyr
1 5 10
<210> 539
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 539
Gln Ile Tyr Pro Gly Asp Gly Glu Gly Lys Tyr
1 5 10
<210> 540
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 540
Gln Ile Tyr Pro Gly Asp Gly Glu His Lys Tyr
1 5 10
<210> 541
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 541
Gln Ile Tyr Pro Gly Asp Gly Glu Ile Lys Tyr
1 5 10
<210> 542
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 542
Gln Ile Tyr Pro Gly Asp Gly Glu Lys Lys Tyr
1 5 10
<210> 543
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 543
Gln Ile Tyr Pro Gly Asp Gly Glu Gln Lys Tyr
1 5 10
<210> 544
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 544
Gln Ile Tyr Pro Gly Asp Gly Glu Arg Lys Tyr
1 5 10
<210> 545
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 545
Gln Ile Tyr Pro Gly Asp Gly Glu Ser Lys Tyr
1 5 10
<210> 546
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 546
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Tyr
1 5 10
<210> 547
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 547
Gln Ile Tyr Pro Gly Asp Gly Glu Val Lys Tyr
1 5 10
<210> 548
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 548
Gln Ile Tyr Pro Gly Asp Gly Glu Tyr Lys Tyr
1 5 10
<210> 549
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 549
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Asp Tyr
1 5 10
<210> 550
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 550
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Glu Tyr
1 5 10
<210> 551
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 551
Gln Ile Tyr Pro Gly Asp Gly Glu Thr His Tyr
1 5 10
<210> 552
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 552
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Ile Tyr
1 5 10
<210> 553
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 553
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Leu Tyr
1 5 10
<210> 554
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 554
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Met Tyr
1 5 10
<210> 555
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 555
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Asn Tyr
1 5 10
<210> 556
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 556
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Pro Tyr
1 5 10
<210> 557
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 557
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Gln Tyr
1 5 10
<210> 558
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 558
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Arg Tyr
1 5 10
<210> 559
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 559
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Ser Tyr
1 5 10
<210> 560
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 560
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Thr Tyr
1 5 10
<210> 561
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 561
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Val Tyr
1 5 10
<210> 562
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 562
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Trp Tyr
1 5 10
<210> 563
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 563
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Tyr Tyr
1 5 10
<210> 564
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 564
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Cys
1 5 10
<210> 565
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 565
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Asp
1 5 10
<210> 566
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 566
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Glu
1 5 10
<210> 567
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 567
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Phe
1 5 10
<210> 568
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 568
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Gly
1 5 10
<210> 569
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 569
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys His
1 5 10
<210> 570
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 570
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Ile
1 5 10
<210> 571
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 571
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Leu
1 5 10
<210> 572
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 572
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Asn
1 5 10
<210> 573
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 573
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Pro
1 5 10
<210> 574
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 574
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Gln
1 5 10
<210> 575
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 575
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Arg
1 5 10
<210> 576
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 576
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Thr
1 5 10
<210> 577
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 тяжелой цепи
<400> 577
Gln Ile Tyr Pro Gly Asp Gly Glu Thr Lys Trp
1 5 10
<210> 578
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> NHGNYLPY
<400> 578
Asn His Gly Asn Tyr Leu Pro Tyr
1 5
<210> 579
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> THGNYLPY
<400> 579
Thr His Gly Asn Tyr Leu Pro Tyr
1 5
<210> 580
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 580
Tyr His Gly Asn Tyr Leu Pro Tyr
1 5
<210> 581
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 581
Ser His Gly Asn Ala Leu Pro Tyr
1 5
<210> 582
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 582
Ser His Gly Asn Cys Leu Pro Tyr
1 5
<210> 583
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 583
Ser His Gly Asn Phe Leu Pro Tyr
1 5
<210> 584
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 584
Ser His Gly Asn His Leu Pro Tyr
1 5
<210> 585
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 585
Ser His Gly Asn Asn Leu Pro Tyr
1 5
<210> 586
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 586
Ser His Gly Asn Ser Leu Pro Tyr
1 5
<210> 587
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 587
Ser His Gly Asn Tyr Asp Pro Tyr
1 5
<210> 588
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 588
Ser His Gly Asn Tyr Glu Pro Tyr
1 5
<210> 589
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 589
Ser His Gly Asn Tyr Phe Pro Tyr
1 5
<210> 590
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 590
Ser His Gly Asn Tyr Gly Pro Tyr
1 5
<210> 591
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 591
Ser His Gly Asn Tyr His Pro Tyr
1 5
<210> 592
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 592
Ser His Gly Asn Tyr Ile Pro Tyr
1 5
<210> 593
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 593
Ser His Gly Asn Tyr Lys Pro Tyr
1 5
<210> 594
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 594
Ser His Gly Asn Tyr Met Pro Tyr
1 5
<210> 595
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 595
Ser His Gly Asn Tyr Asn Pro Tyr
1 5
<210> 596
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 596
Ser His Gly Asn Tyr Gln Pro Tyr
1 5
<210> 597
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 597
Ser His Gly Asn Tyr Ser Pro Tyr
1 5
<210> 598
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 598
Ser His Gly Asn Tyr Trp Pro Tyr
1 5
<210> 599
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 599
Ser His Gly Asn Tyr Tyr Pro Tyr
1 5
<210> 600
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 600
Ser His Gly Asn Tyr Leu Ala Tyr
1 5
<210> 601
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 601
Ser His Gly Asn Tyr Leu Glu Tyr
1 5
<210> 602
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 602
Ser His Gly Asn Tyr Leu Phe Tyr
1 5
<210> 603
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 603
Ser His Gly Asn Tyr Leu Gly Tyr
1 5
<210> 604
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 604
Ser His Gly Asn Tyr Leu His Tyr
1 5
<210> 605
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 605
Ser His Gly Asn Tyr Leu Lys Tyr
1 5
<210> 606
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 606
Ser His Gly Asn Tyr Leu Leu Tyr
1 5
<210> 607
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 607
Ser His Gly Asn Tyr Leu Gln Tyr
1 5
<210> 608
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 608
Ser His Gly Asn Tyr Leu Arg Tyr
1 5
<210> 609
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 609
Ser His Gly Asn Tyr Leu Ser Tyr
1 5
<210> 610
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 610
Ser His Gly Asn Tyr Leu Val Tyr
1 5
<210> 611
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 611
Ser His Gly Asn Tyr Leu Trp Tyr
1 5
<210> 612
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 612
Ser His Gly Asn Tyr Leu Tyr Tyr
1 5
<210> 613
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 613
Ser His Gly Asn Tyr Leu Pro Ala
1 5
<210> 614
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 614
Ser His Gly Asn Tyr Leu Pro Asp
1 5
<210> 615
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 615
Ser His Gly Asn Tyr Leu Pro Glu
1 5
<210> 616
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 616
Ser His Gly Asn Tyr Leu Pro Phe
1 5
<210> 617
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 617
Ser His Gly Asn Tyr Leu Pro Gly
1 5
<210> 618
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 618
Ser His Gly Asn Tyr Leu Pro His
1 5
<210> 619
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 619
Ser His Gly Asn Tyr Leu Pro Ile
1 5
<210> 620
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 620
Ser His Gly Asn Tyr Leu Pro Lys
1 5
<210> 621
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 621
Ser His Gly Asn Tyr Leu Pro Leu
1 5
<210> 622
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 622
Ser His Gly Asn Tyr Leu Pro Met
1 5
<210> 623
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 623
Ser His Gly Asn Tyr Leu Pro Arg
1 5
<210> 624
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 624
Ser His Gly Asn Tyr Leu Pro Ser
1 5
<210> 625
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 тяжелой цепи
<400> 625
Ser His Gly Asn Tyr Leu Pro Val
1 5
<210> 626
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 626
Ala Ser Tyr Leu His
1 5
<210> 627
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 627
Asp Ser Tyr Leu His
1 5
<210> 628
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 628
Phe Ser Tyr Leu His
1 5
<210> 629
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 629
Leu Ser Tyr Leu His
1 5
<210> 630
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 630
Met Ser Tyr Leu His
1 5
<210> 631
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 631
Arg Ser Tyr Leu His
1 5
<210> 632
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 632
Thr Ser Tyr Leu His
1 5
<210> 633
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 633
Val Ser Tyr Leu His
1 5
<210> 634
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 634
Trp Ser Tyr Leu His
1 5
<210> 635
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 635
Tyr Ser Tyr Leu His
1 5
<210> 636
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 636
Ser Ala Tyr Leu His
1 5
<210> 637
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 637
Ser Asp Tyr Leu His
1 5
<210> 638
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 638
Ser Glu Tyr Leu His
1 5
<210> 639
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 639
Ser Phe Tyr Leu His
1 5
<210> 640
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 640
Ser Gly Tyr Leu His
1 5
<210> 641
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 641
Ser His Tyr Leu His
1 5
<210> 642
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 642
Ser Lys Tyr Leu His
1 5
<210> 643
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 643
Ser Met Tyr Leu His
1 5
<210> 644
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 644
Ser Asn Tyr Leu His
1 5
<210> 645
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 645
Ser Pro Tyr Leu His
1 5
<210> 646
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 646
Ser Gln Tyr Leu His
1 5
<210> 647
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 647
Ser Val Tyr Leu His
1 5
<210> 648
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 648
Ser Trp Tyr Leu His
1 5
<210> 649
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 649
Ser Ser Phe Leu His
1 5
<210> 650
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 650
Ser Ser Tyr Phe His
1 5
<210> 651
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 651
Ser Ser Tyr His His
1 5
<210> 652
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 652
Ser Ser Tyr Ile His
1 5
<210> 653
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 653
Ser Ser Tyr Lys His
1 5
<210> 654
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 654
Ser Ser Tyr Asn His
1 5
<210> 655
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 655
Ser Ser Tyr Pro His
1 5
<210> 656
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 656
Ser Ser Tyr Gln His
1 5
<210> 657
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 657
Ser Ser Tyr Arg His
1 5
<210> 658
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 658
Ser Ser Tyr Ser His
1 5
<210> 659
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 659
Ser Ser Tyr Tyr His
1 5
<210> 660
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 660
Ser Ser Tyr Trp His
1 5
<210> 661
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR1 легкой цепи
<400> 661
Ser Ser Tyr Tyr His
1 5
<210> 662
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 662
Ala Thr Ser Asn Leu Ala Ser
1 5
<210> 663
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 663
His Thr Ser Asn Leu Ala Ser
1 5
<210> 664
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 664
Asn Thr Ser Asn Leu Ala Ser
1 5
<210> 665
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 665
Arg Thr Ser Asn Leu Ala Ser
1 5
<210> 666
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 666
Ser Ala Ser Asn Leu Ala Ser
1 5
<210> 667
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 667
Ser Asp Ser Asn Leu Ala Ser
1 5
<210> 668
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 668
Ser Phe Ser Asn Leu Ala Ser
1 5
<210> 669
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 669
Ser Gly Ser Asn Leu Ala Ser
1 5
<210> 670
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 670
Ser His Ser Asn Leu Ala Ser
1 5
<210> 671
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 671
Ser Leu Ser Asn Leu Ala Ser
1 5
<210> 672
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 672
Ser Met Ser Asn Leu Ala Ser
1 5
<210> 673
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 673
Ser Ser Ser Asn Leu Ala Ser
1 5
<210> 674
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 674
Ser Val Ser Asn Leu Ala Ser
1 5
<210> 675
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 675
Ser Trp Ser Asn Leu Ala Ser
1 5
<210> 676
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 676
Ser Tyr Ser Asn Leu Ala Ser
1 5
<210> 677
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 677
Ser Thr Asp Asn Leu Ala Ser
1 5
<210> 678
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 678
Ser Thr Phe Asn Leu Ala Ser
1 5
<210> 679
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 679
Ser Thr His Asn Leu Ala Ser
1 5
<210> 680
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 680
Ser Thr Ile Asn Leu Ala Ser
1 5
<210> 681
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 681
Ser Thr Lys Asn Leu Ala Ser
1 5
<210> 682
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 682
Ser Thr Leu Asn Leu Ala Ser
1 5
<210> 683
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 683
Ser Thr Asn Asn Leu Ala Ser
1 5
<210> 684
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 684
Ser Thr Gln Asn Leu Ala Ser
1 5
<210> 685
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 685
Ser Thr Tyr Asn Leu Ala Ser
1 5
<210> 686
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 686
Ser Thr Ser Asp Leu Ala Ser
1 5
<210> 687
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 687
Ser Thr Ser Glu Leu Ala Ser
1 5
<210> 688
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 688
Ser Thr Ser Phe Leu Ala Ser
1 5
<210> 689
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 689
Ser Thr Ser Gly Leu Ala Ser
1 5
<210> 690
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 690
Ser Thr Ser His Leu Ala Ser
1 5
<210> 691
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 691
Ser Thr Ser Ile Leu Ala Ser
1 5
<210> 692
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 692
Ser Thr Ser Lys Leu Ala Ser
1 5
<210> 693
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 693
Ser Thr Ser Gln Leu Ala Ser
1 5
<210> 694
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 694
Ser Thr Ser Arg Leu Ala Ser
1 5
<210> 695
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 695
Ser Thr Ser Ser Leu Ala Ser
1 5
<210> 696
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 696
Ser Thr Ser Val Leu Ala Ser
1 5
<210> 697
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 697
Ser Thr Ser Asn Asp Ala Ser
1 5
<210> 698
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 698
Ser Thr Ser Asn Glu Ala Ser
1 5
<210> 699
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 699
Ser Thr Ser Asn Phe Ala Ser
1 5
<210> 700
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 700
Ser Thr Ser Asn Gly Ala Ser
1 5
<210> 701
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 701
Ser Thr Ser Asn His Ala Ser
1 5
<210> 702
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 702
Ser Thr Ser Asn Ile Ala Ser
1 5
<210> 703
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 703
Ser Thr Ser Asn Lys Ala Ser
1 5
<210> 704
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 704
Ser Thr Ser Asn Met Ala Ser
1 5
<210> 705
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 705
Ser Thr Ser Asn Asn Ala Ser
1 5
<210> 706
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 706
Ser Thr Ser Asn Pro Ala Ser
1 5
<210> 707
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 707
Ser Thr Ser Asn Gln Ala Ser
1 5
<210> 708
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 708
Ser Thr Ser Asn Val Ala Ser
1 5
<210> 709
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 709
Ser Thr Ser Asn Trp Ala Ser
1 5
<210> 710
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 710
Ser Thr Ser Asn Tyr Ala Ser
1 5
<210> 711
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 711
Ser Thr Ser Asn Leu Asp Ser
1 5
<210> 712
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 712
Ser Thr Ser Asn Leu Phe Ser
1 5
<210> 713
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 713
Ser Thr Ser Asn Leu Gly Ser
1 5
<210> 714
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 714
Ser Thr Ser Asn Leu Ile Ser
1 5
<210> 715
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 715
Ser Thr Ser Asn Leu Lys Ser
1 5
<210> 716
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 716
Ser Thr Ser Asn Leu Gln Ser
1 5
<210> 717
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 717
Ser Thr Ser Asn Leu Arg Ser
1 5
<210> 718
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 718
Ser Thr Ser Asn Leu Ser Ser
1 5
<210> 719
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 719
Ser Thr Ser Asn Leu Val Ser
1 5
<210> 720
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 720
Ser Thr Ser Asn Leu Trp Ser
1 5
<210> 721
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 721
Ser Thr Ser Asn Leu Ala Ala
1 5
<210> 722
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 722
Ser Thr Ser Asn Leu Ala Asp
1 5
<210> 723
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 723
Ser Thr Ser Asn Leu Ala Phe
1 5
<210> 724
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 724
Ser Thr Ser Asn Leu Ala Gly
1 5
<210> 725
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 725
Ser Thr Ser Asn Leu Ala His
1 5
<210> 726
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 726
Ser Thr Ser Asn Leu Ala Lys
1 5
<210> 727
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 727
Ser Thr Ser Asn Leu Ala Leu
1 5
<210> 728
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 728
Ser Thr Ser Asn Leu Ala Arg
1 5
<210> 729
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 729
Ser Thr Ser Asn Leu Ala Thr
1 5
<210> 730
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 730
Ser Thr Ser Asn Leu Ala Val
1 5
<210> 731
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 731
Ser Thr Ser Asn Leu Ala Trp
1 5
<210> 732
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность CDR2 легкой цепи
<400> 732
Ser Thr Ser Asn Leu Ala Ser Gly
1 5
<210> 733
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 733
Ser Thr Ser Asn Leu Ala Ser Ala
1 5
<210> 734
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 734
Ser Thr Ser Asn Leu Ala Ser Phe
1 5
<210> 735
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 735
Ser Thr Ser Asn Leu Ala Ser Ile
1 5
<210> 736
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 736
Ser Thr Ser Asn Leu Ala Ser Asn
1 5
<210> 737
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 737
Ser Thr Ser Asn Leu Ala Ser Pro
1 5
<210> 738
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 738
Ser Thr Ser Asn Leu Ala Ser Arg
1 5
<210> 739
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 739
Ser Thr Ser Asn Leu Ala Ser Ser
1 5
<210> 740
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 740
Ser Thr Ser Asn Leu Ala Ser Thr
1 5
<210> 741
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 741
Ser Thr Ser Asn Leu Ala Ser Val
1 5
<210> 742
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR2 легкой цепи
<400> 742
Ser Thr Ser Asn Leu Ala Ser Tyr
1 5
<210> 743
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 743
Ala Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 744
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 744
Cys Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 745
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 745
Asp Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 746
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 746
Glu Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 747
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 747
Phe Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 748
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 748
Ile Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 749
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 749
Lys Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 750
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 750
Leu Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 751
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 751
Asn Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 752
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 752
Arg Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 753
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 753
Ser Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 754
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 754
Thr Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 755
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 755
Val Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 756
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 756
Trp Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 757
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 757
Tyr Gln Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 758
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 758
His Ala Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 759
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 759
His Cys Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 760
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 760
His Glu Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 761
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 761
His Gly Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 762
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 762
His Asn Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 763
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 763
His Ser Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 764
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 764
His Thr Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 765
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 765
His Val Tyr His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 766
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 766
His Gln Ala His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 767
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 767
His Gln Phe His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 768
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 768
His Gln Gly His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 769
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 769
His Gln His His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 770
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 770
His Gln Gln His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 771
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 771
His Gln Trp His Arg Ser Pro Pro Thr Phe
1 5 10
<210> 772
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 772
His Gln Tyr Ala Arg Ser Pro Pro Thr Phe
1 5 10
<210> 773
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 773
His Gln Tyr Glu Arg Ser Pro Pro Thr Phe
1 5 10
<210> 774
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 774
His Gln Tyr Gly Arg Ser Pro Pro Thr Phe
1 5 10
<210> 775
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 775
His Gln Tyr Lys Arg Ser Pro Pro Thr Phe
1 5 10
<210> 776
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 776
His Gln Tyr Leu Arg Ser Pro Pro Thr Phe
1 5 10
<210> 777
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 777
His Gln Tyr Asn Arg Ser Pro Pro Thr Phe
1 5 10
<210> 778
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 778
His Gln Tyr Gln Arg Ser Pro Pro Thr Phe
1 5 10
<210> 779
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 779
His Gln Tyr Ser Arg Ser Pro Pro Thr Phe
1 5 10
<210> 780
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 780
His Gln Tyr Thr Arg Ser Pro Pro Thr Phe
1 5 10
<210> 781
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 781
His Gln Tyr Val Arg Ser Pro Pro Thr Phe
1 5 10
<210> 782
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 782
His Gln Tyr Trp Arg Ser Pro Pro Thr Phe
1 5 10
<210> 783
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 783
His Gln Tyr His Ala Ser Pro Pro Thr Phe
1 5 10
<210> 784
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 784
His Gln Tyr His Cys Ser Pro Pro Thr Phe
1 5 10
<210> 785
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 785
His Gln Tyr His Asp Ser Pro Pro Thr Phe
1 5 10
<210> 786
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 786
His Gln Tyr His Glu Ser Pro Pro Thr Phe
1 5 10
<210> 787
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 787
His Gln Tyr His Phe Ser Pro Pro Thr Phe
1 5 10
<210> 788
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 788
His Gln Tyr His Ile Ser Pro Pro Thr Phe
1 5 10
<210> 789
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 789
His Gln Tyr His Leu Ser Pro Pro Thr Phe
1 5 10
<210> 790
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 790
His Gln Tyr His Met Ser Pro Pro Thr Phe
1 5 10
<210> 791
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 791
His Gln Tyr His Asn Ser Pro Pro Thr Phe
1 5 10
<210> 792
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 792
His Gln Tyr His Gln Ser Pro Pro Thr Phe
1 5 10
<210> 793
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 793
His Gln Tyr His Val Ser Pro Pro Thr Phe
1 5 10
<210> 794
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 794
His Gln Tyr His Trp Ser Pro Pro Thr Phe
1 5 10
<210> 795
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 795
His Gln Tyr His Tyr Ser Pro Pro Thr Phe
1 5 10
<210> 796
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 796
His Gln Tyr His Arg Ala Pro Pro Thr Phe
1 5 10
<210> 797
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 797
His Gln Tyr His Arg Cys Pro Pro Thr Phe
1 5 10
<210> 798
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 798
His Gln Tyr His Arg Glu Pro Pro Thr Phe
1 5 10
<210> 799
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 799
His Gln Tyr His Arg Phe Pro Pro Thr Phe
1 5 10
<210> 800
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 800
His Gln Tyr His Arg Gly Pro Pro Thr Phe
1 5 10
<210> 801
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 801
His Gln Tyr His Arg Ile Pro Pro Thr Phe
1 5 10
<210> 802
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 802
His Gln Tyr His Arg Met Pro Pro Thr Phe
1 5 10
<210> 803
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 803
His Gln Tyr His Arg Pro Pro Pro Thr Phe
1 5 10
<210> 804
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 804
His Gln Tyr His Arg Arg Pro Pro Thr Phe
1 5 10
<210> 805
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 805
His Gln Tyr His Arg Thr Pro Pro Thr Phe
1 5 10
<210> 806
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 806
His Gln Tyr His Arg Val Pro Pro Thr Phe
1 5 10
<210> 807
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 807
His Gln Tyr His Arg Trp Pro Pro Thr Phe
1 5 10
<210> 808
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 808
His Gln Tyr His Arg Tyr Pro Pro Thr Phe
1 5 10
<210> 809
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 809
His Gln Tyr His Arg Ser Lys Pro Thr Phe
1 5 10
<210> 810
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 810
His Gln Tyr His Arg Ser Trp Pro Thr Phe
1 5 10
<210> 811
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 811
His Gln Tyr His Arg Ser Tyr Pro Thr Phe
1 5 10
<210> 812
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 812
His Gln Tyr His Arg Ser Pro Asp Thr Phe
1 5 10
<210> 813
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 813
His Gln Tyr His Arg Ser Pro His Thr Phe
1 5 10
<210> 814
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 814
His Gln Tyr His Arg Ser Pro Pro Asp Phe
1 5 10
<210> 815
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 815
His Gln Tyr His Arg Ser Pro Pro Phe Phe
1 5 10
<210> 816
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 816
His Gln Tyr His Arg Ser Pro Pro Gly Phe
1 5 10
<210> 817
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 817
His Gln Tyr His Arg Ser Pro Pro Ile Phe
1 5 10
<210> 818
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 818
His Gln Tyr His Arg Ser Pro Pro Leu Phe
1 5 10
<210> 819
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 819
His Gln Tyr His Arg Ser Pro Pro Met Phe
1 5 10
<210> 820
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 820
His Gln Tyr His Arg Ser Pro Pro Asn Phe
1 5 10
<210> 821
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 821
His Gln Tyr His Arg Ser Pro Pro Gln Phe
1 5 10
<210> 822
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 822
His Gln Tyr His Arg Ser Pro Pro Arg Phe
1 5 10
<210> 823
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 823
His Gln Tyr His Arg Ser Pro Pro Ser Phe
1 5 10
<210> 824
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 824
His Gln Tyr His Arg Ser Pro Pro Val Phe
1 5 10
<210> 825
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутантная последовательность CDR3 легкой цепи
<400> 825
His Gln Tyr His Arg Ser Pro Pro Tyr Phe
1 5 10
<210> 826
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Apollo_ECL2_h2m_F
<400> 826
ctgcaatctg tttgctcaga cattttccca ctcattgatg aaacctacct 50
<210> 827
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Apollo_ECL2_h2m_R
<400> 827
ggaaaatgtc tgagcaaaca gattgcagtt tcttgcagtt ccagcccagg 50
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЫДЕЛЕННОЕ АНТИТЕЛО (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, ЭКСПРЕССИОННАЯ КАССЕТА (ВАРИАНТЫ), ПЛАЗМИДА (ВАРИАНТЫ), КЛЕТКА-ХОЗЯИН, ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, НАБОР, СПОСОБ ЛЕЧЕНИЯ БОЛЬНОГО, ПОДВЕРЖЕННОГО РИСКУ ИЛИ СТРАДАЮЩЕГО ОТ ИНФЕКЦИИ E.COLI, И СПОСОБ ДИАГНОСТИКИ ОПРЕДЕЛЕНИЯ ИНФЕКЦИЙ E.COLI | 2014 |
|
RU2724530C2 |
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| ЛИГАНДСВЯЗЫВАЮЩАЯ МОЛЕКУЛА С РЕГУЛИРУЕМОЙ ЛИГАНДСВЯЗЫВАЮЩЕЙ АКТИВНОСТЬЮ | 2017 |
|
RU2803067C2 |
| АНТИ-LILRB1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2801535C1 |
| СКОНСТРУИРОВАННЫЕ PH-ЗАВИСИМЫЕ АНТИТЕЛА К CD3, А ТАКЖЕ СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2832079C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732042C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СОДЕРЖАЩАЯ ДВА АНТИГЕНСВЯЗЫВАЮЩИХ ДОМЕНА, КОТОРЫЕ СВЯЗАНЫ ДРУГ С ДРУГОМ | 2019 |
|
RU2838581C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2827926C2 |
| АНТИТЕЛА К ТАУ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2787779C2 |
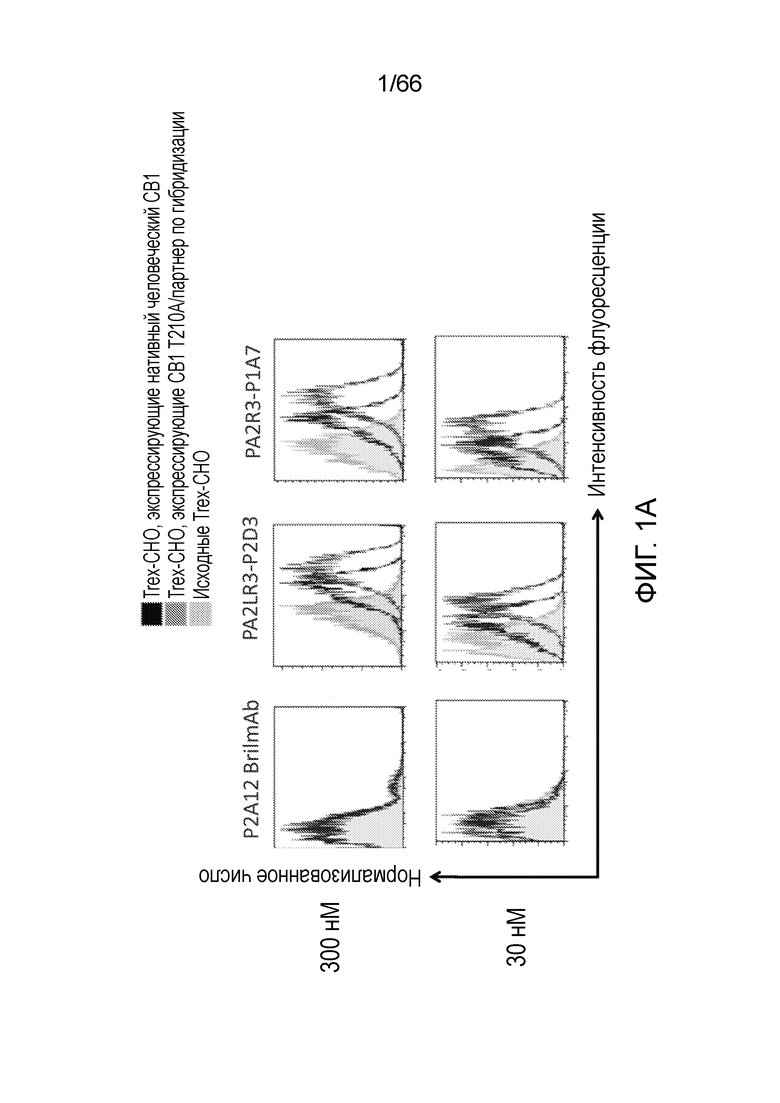
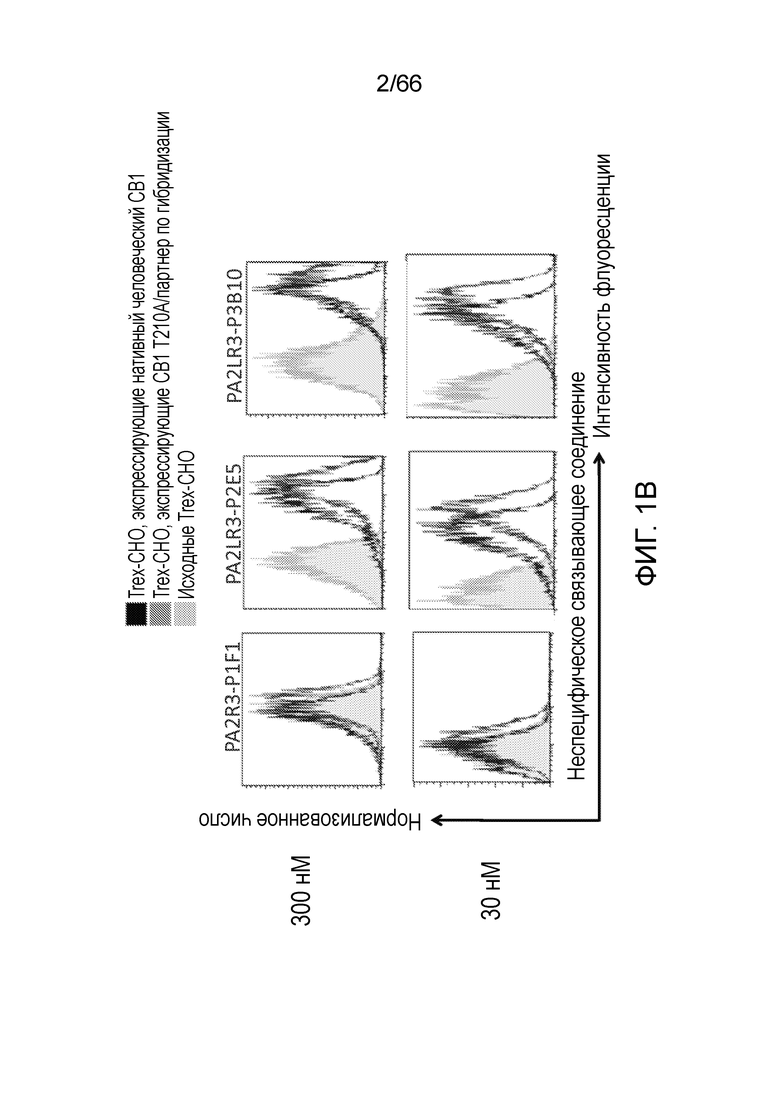
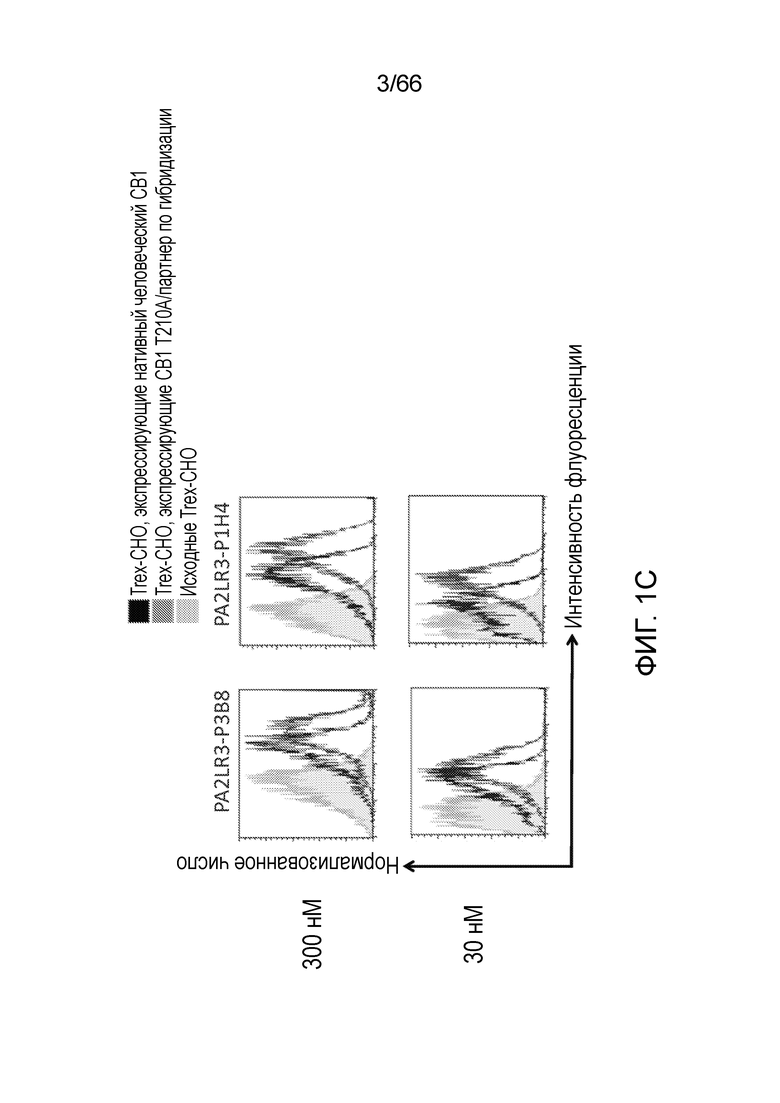
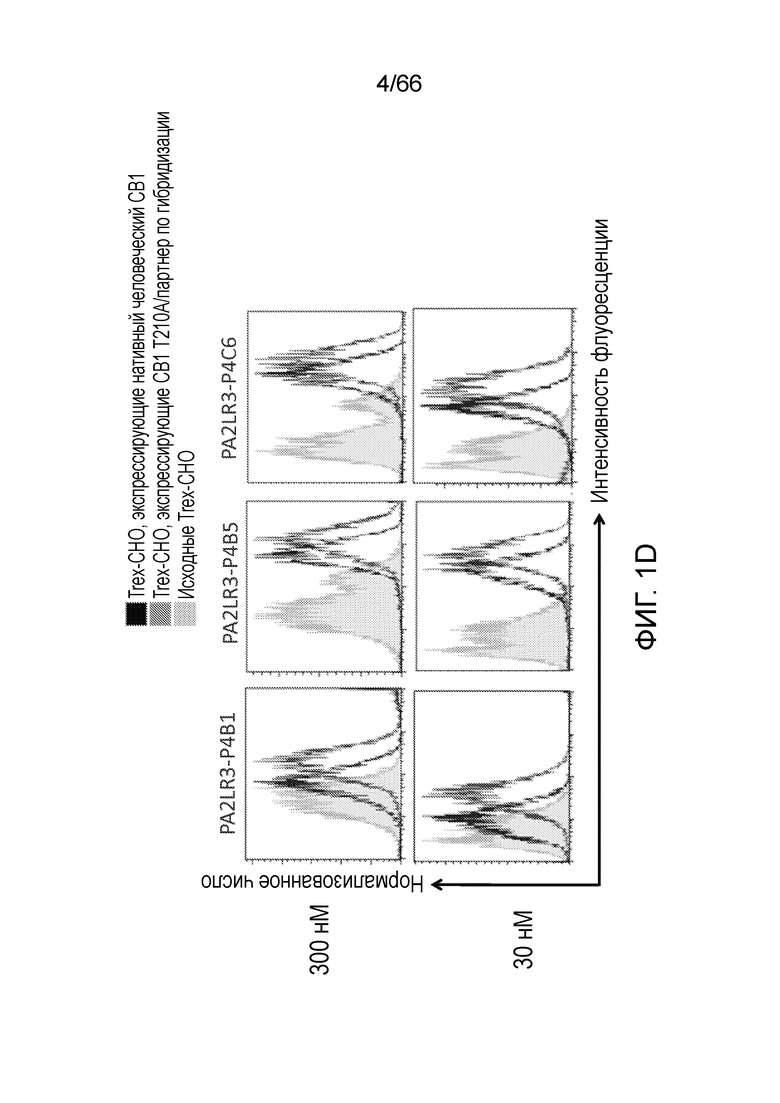
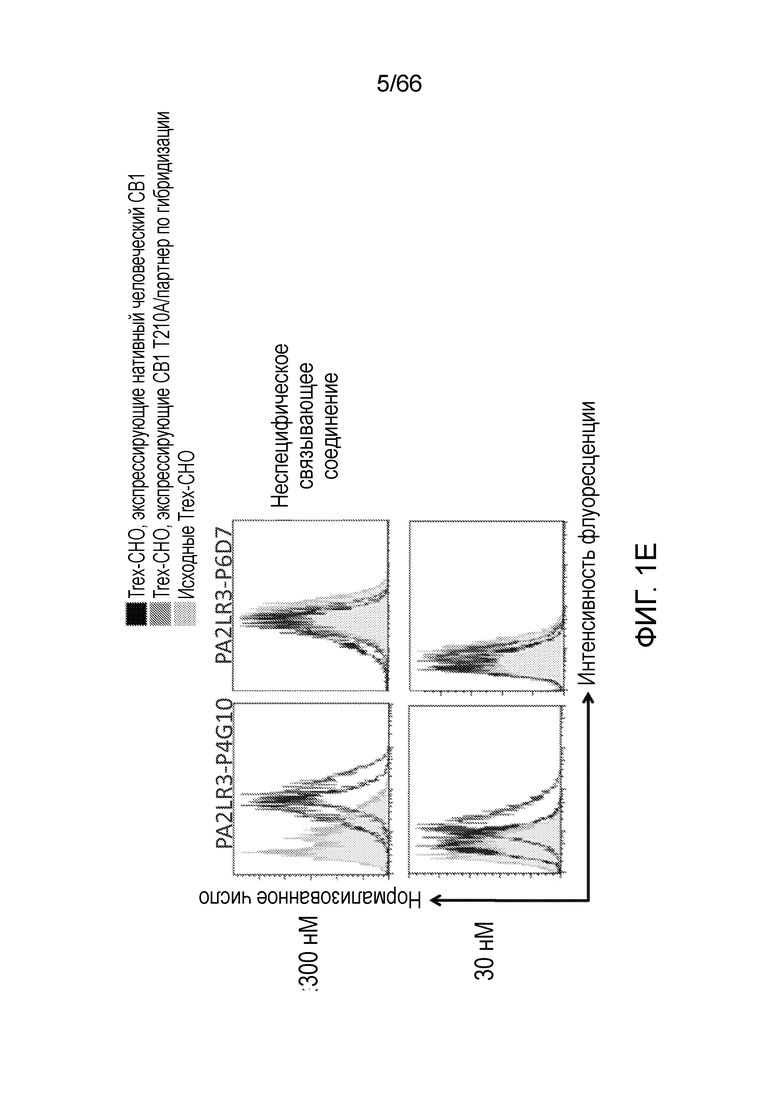
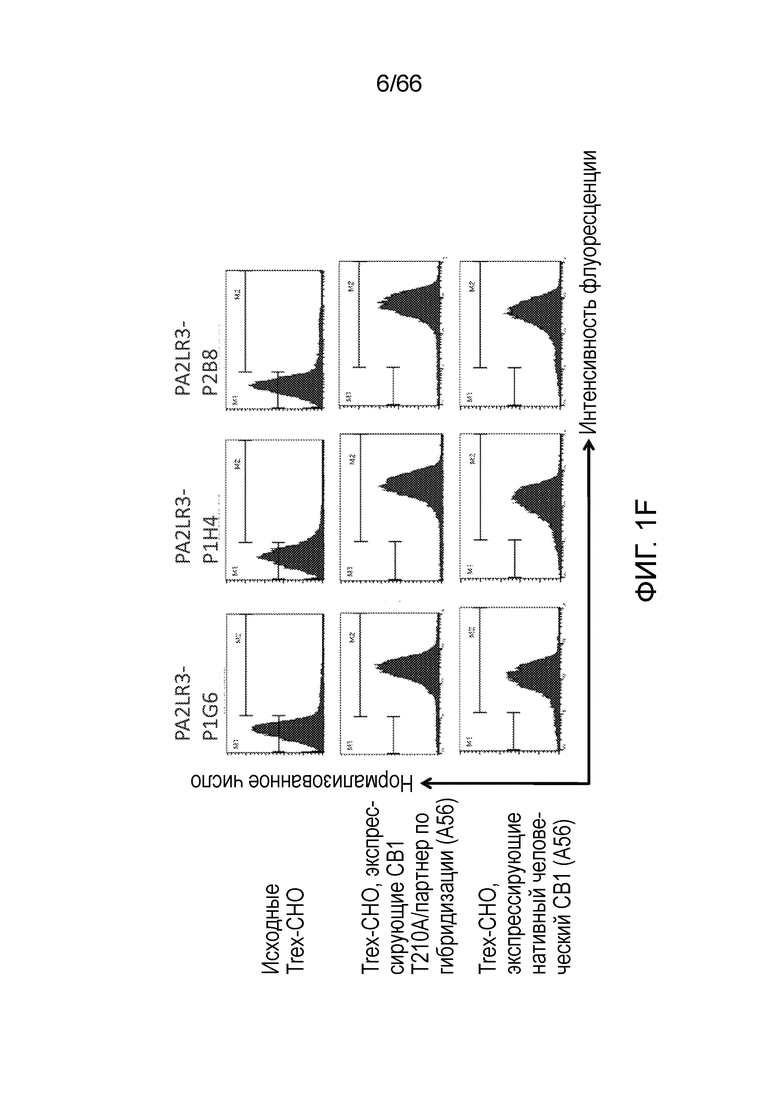
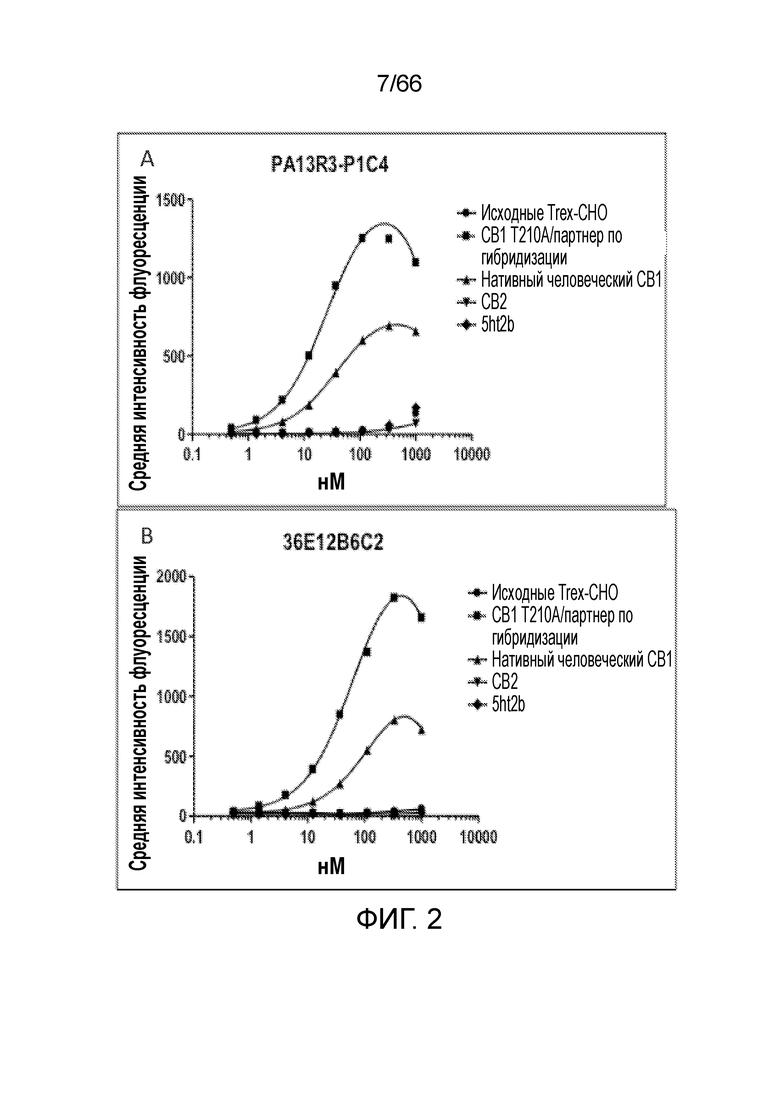
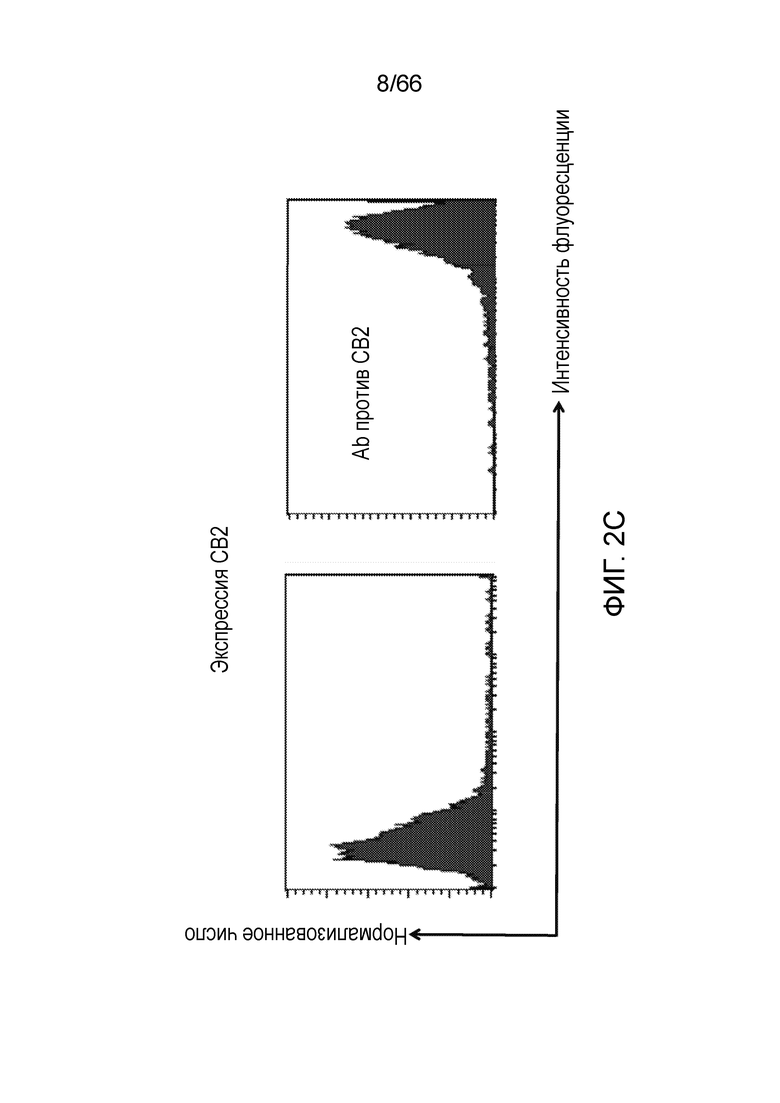
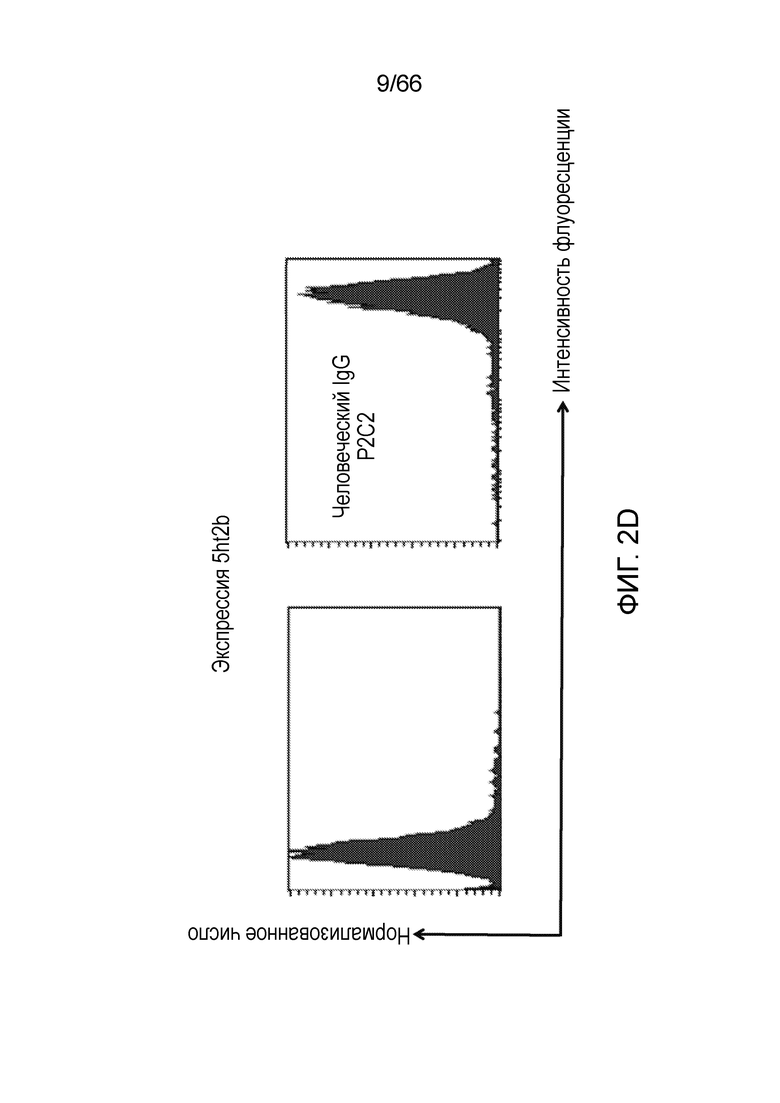
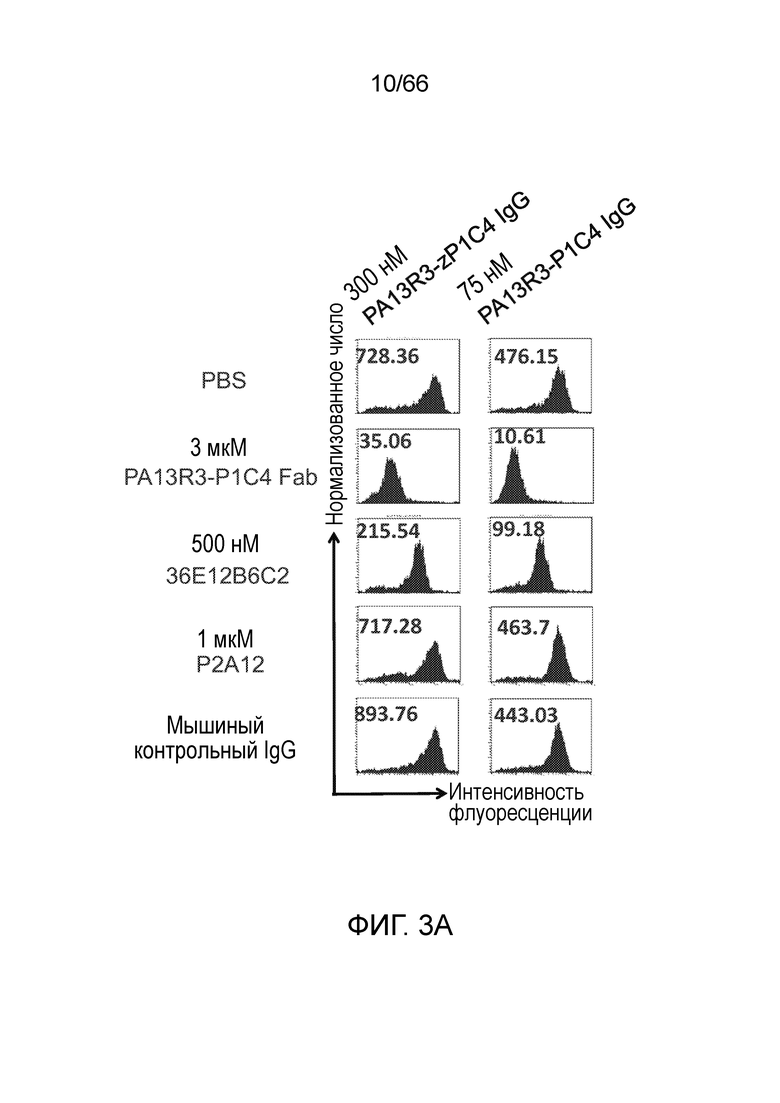
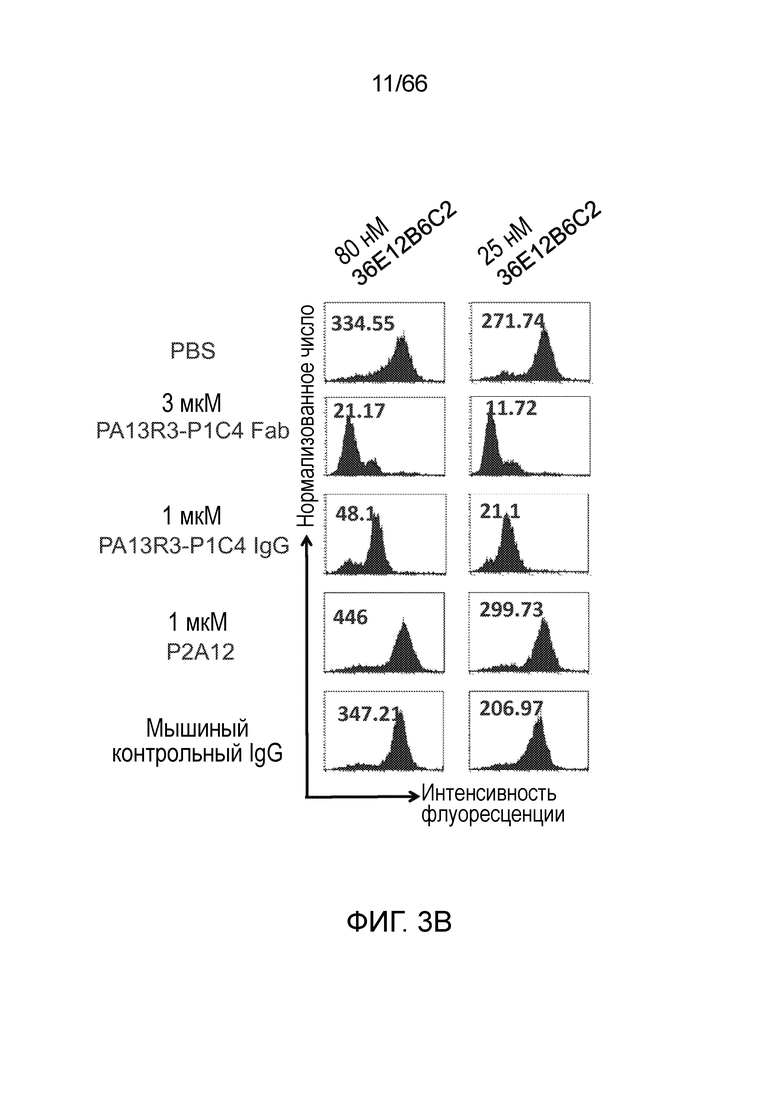
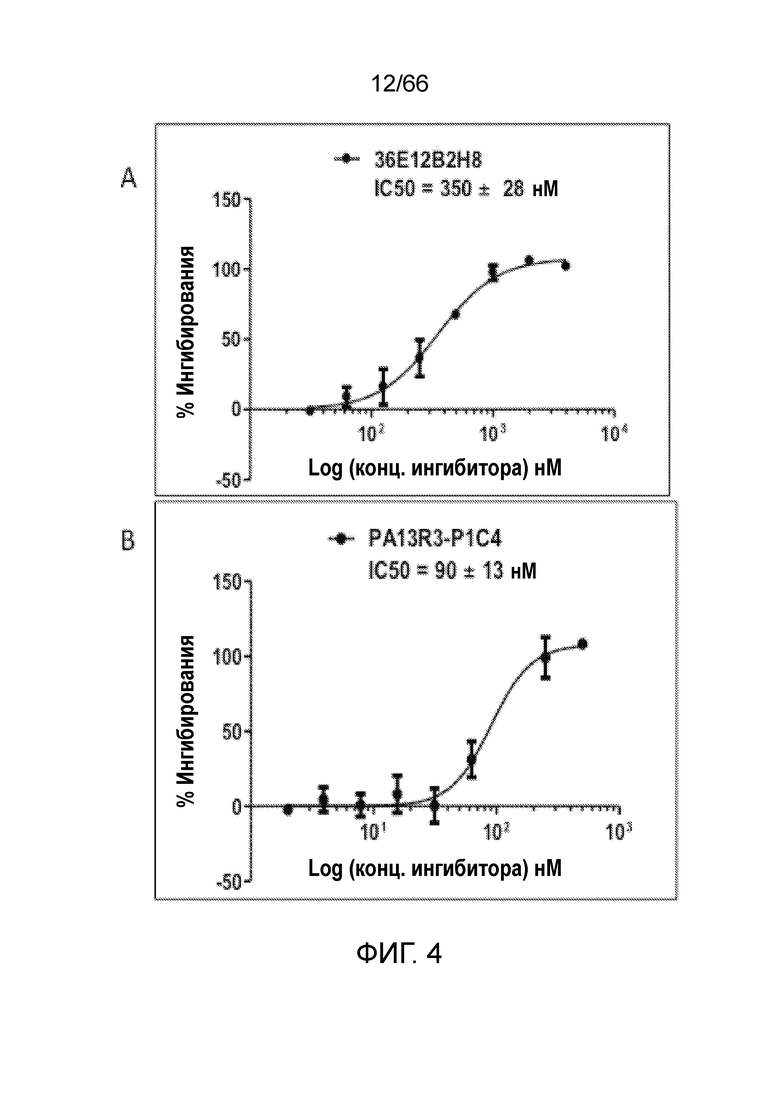
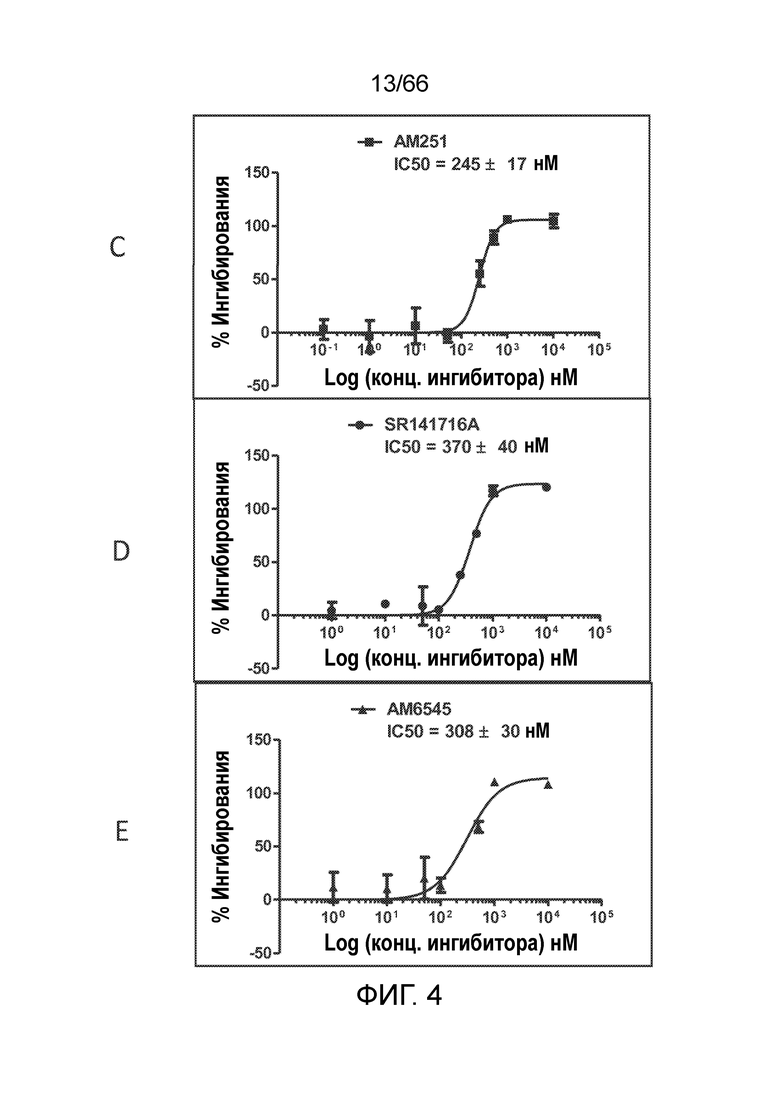
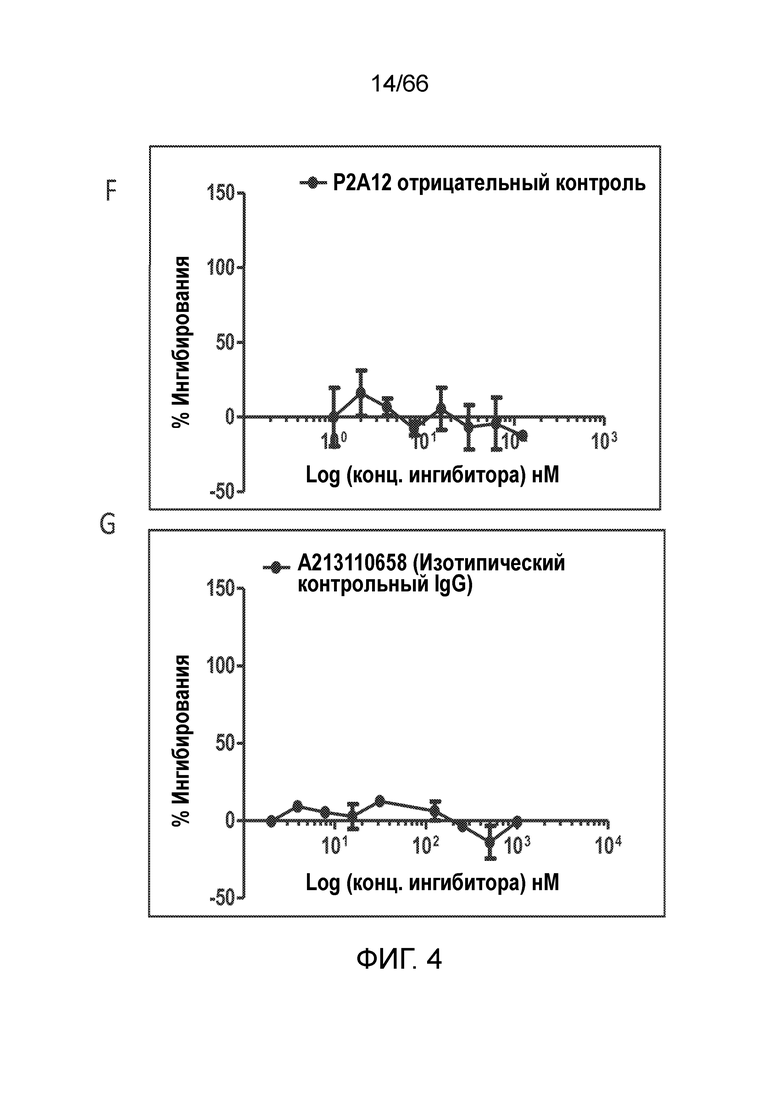
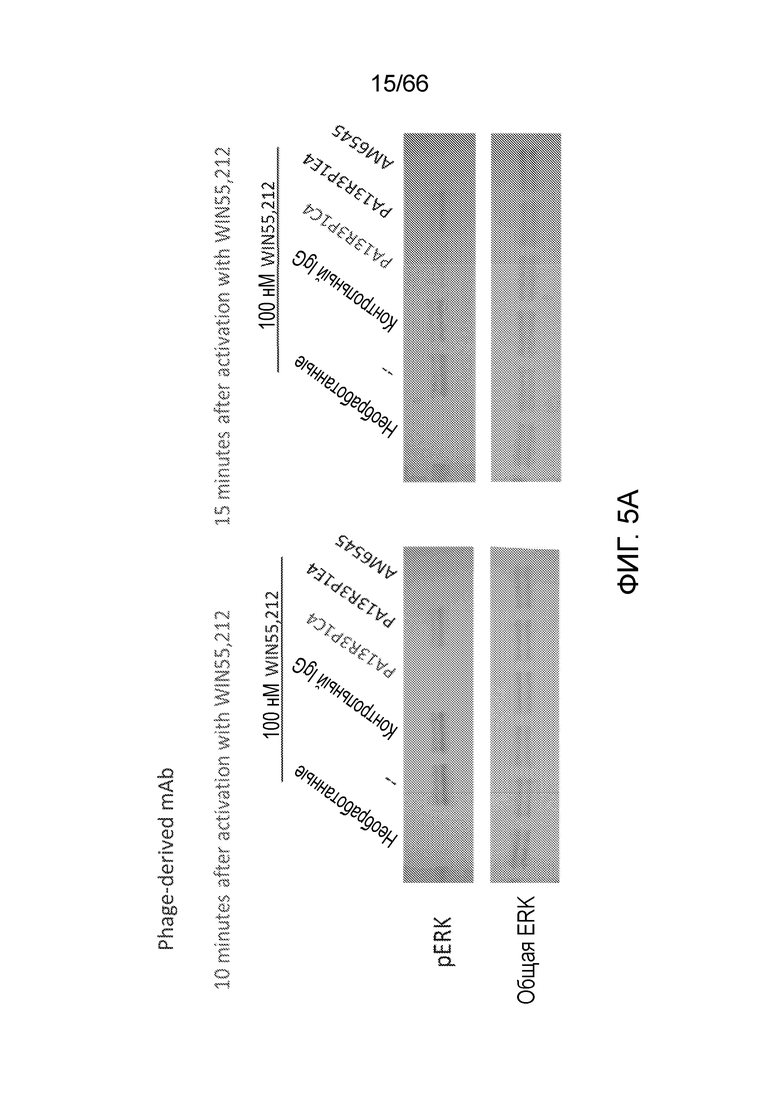
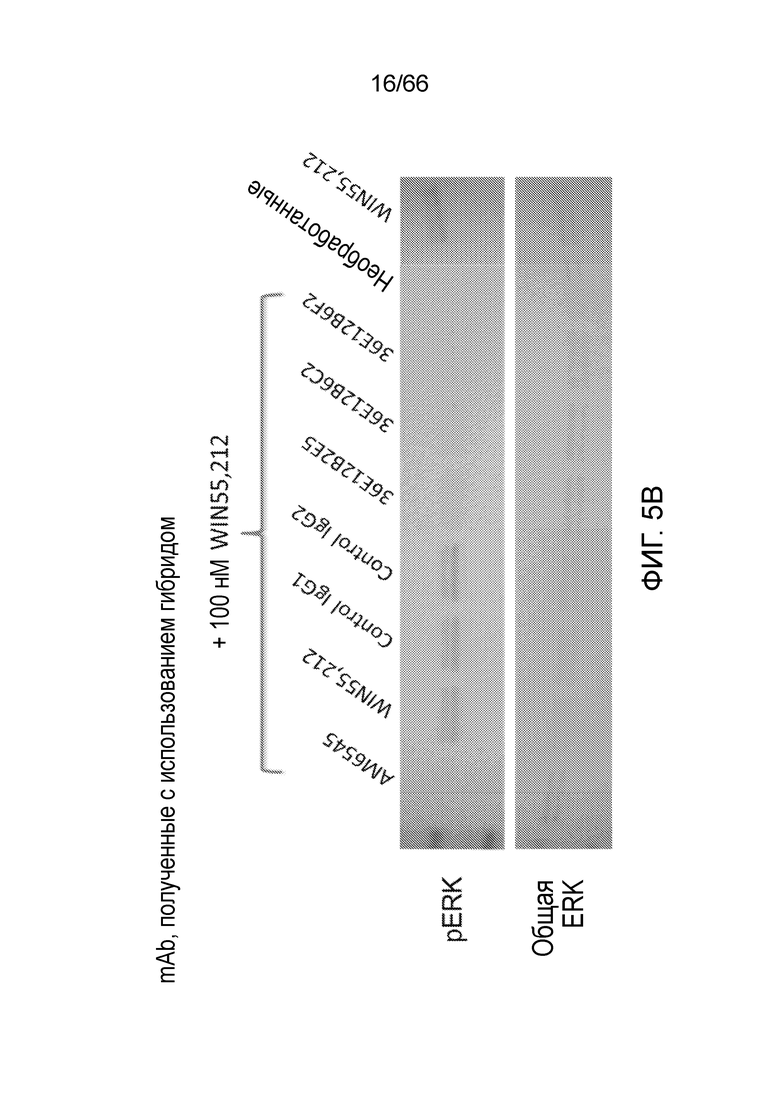
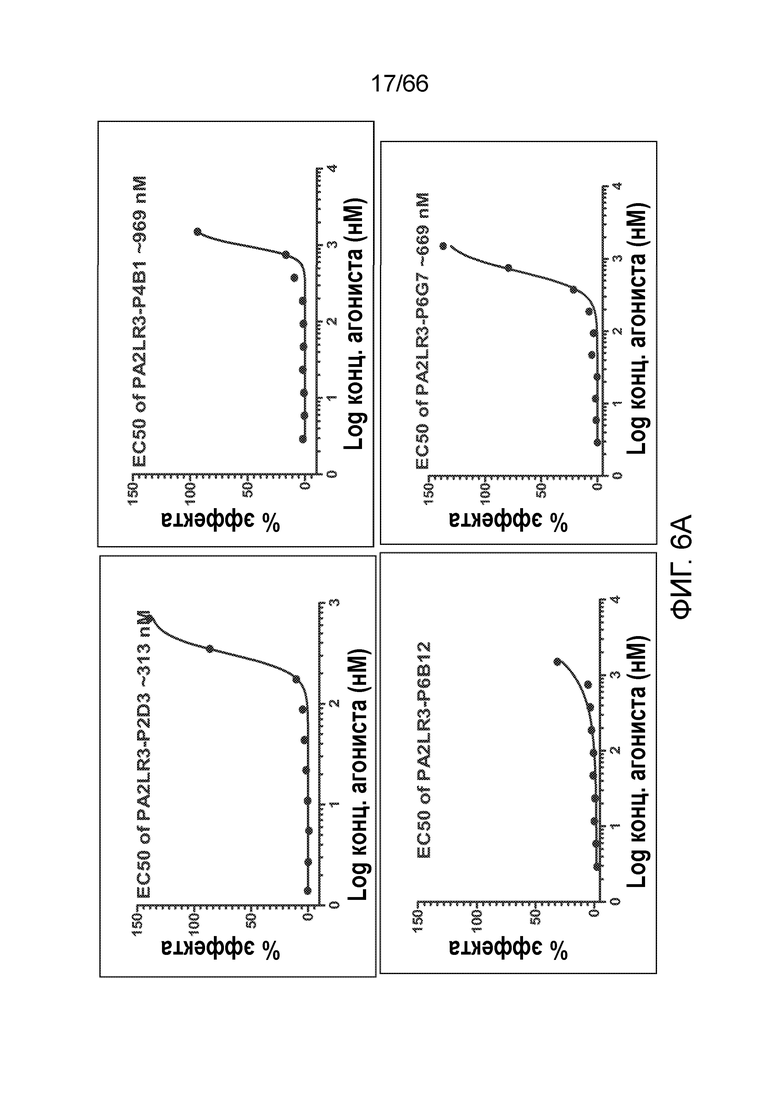

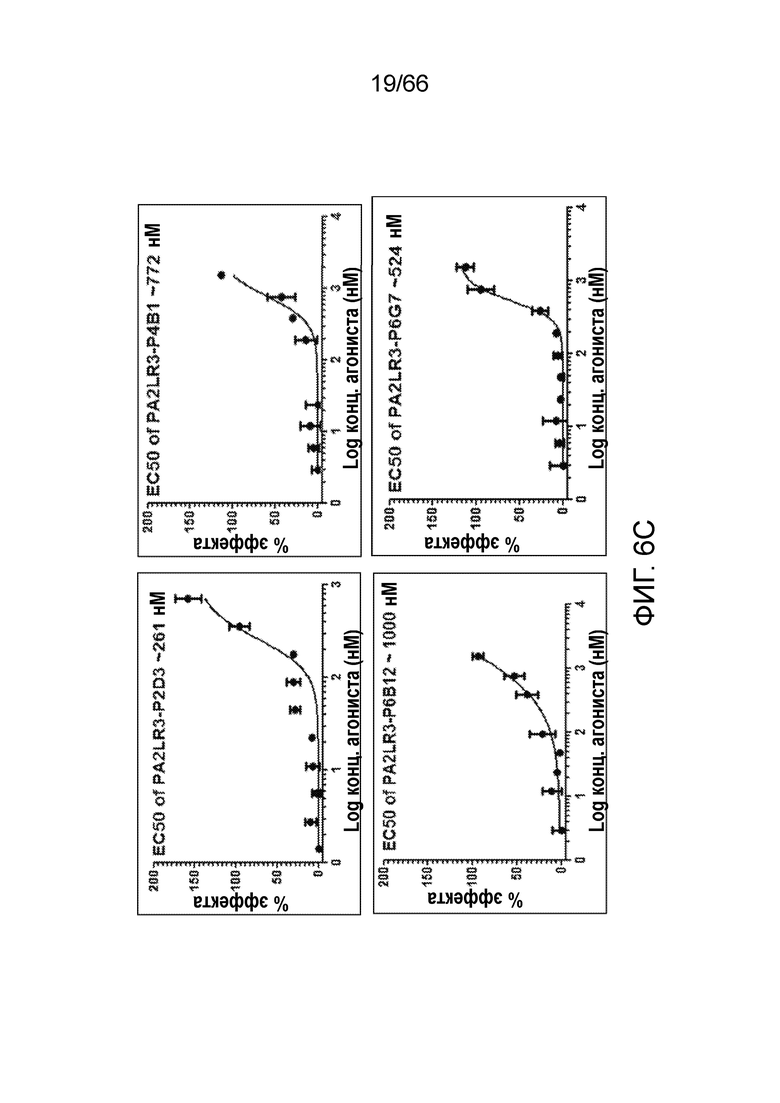

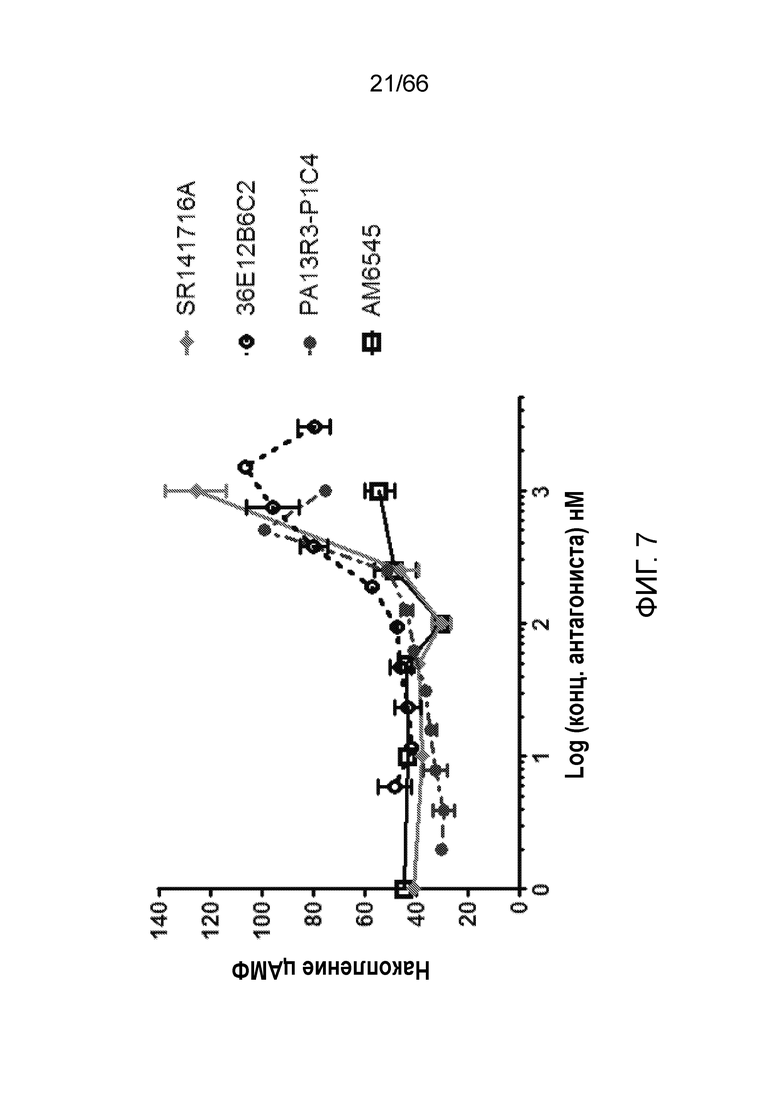
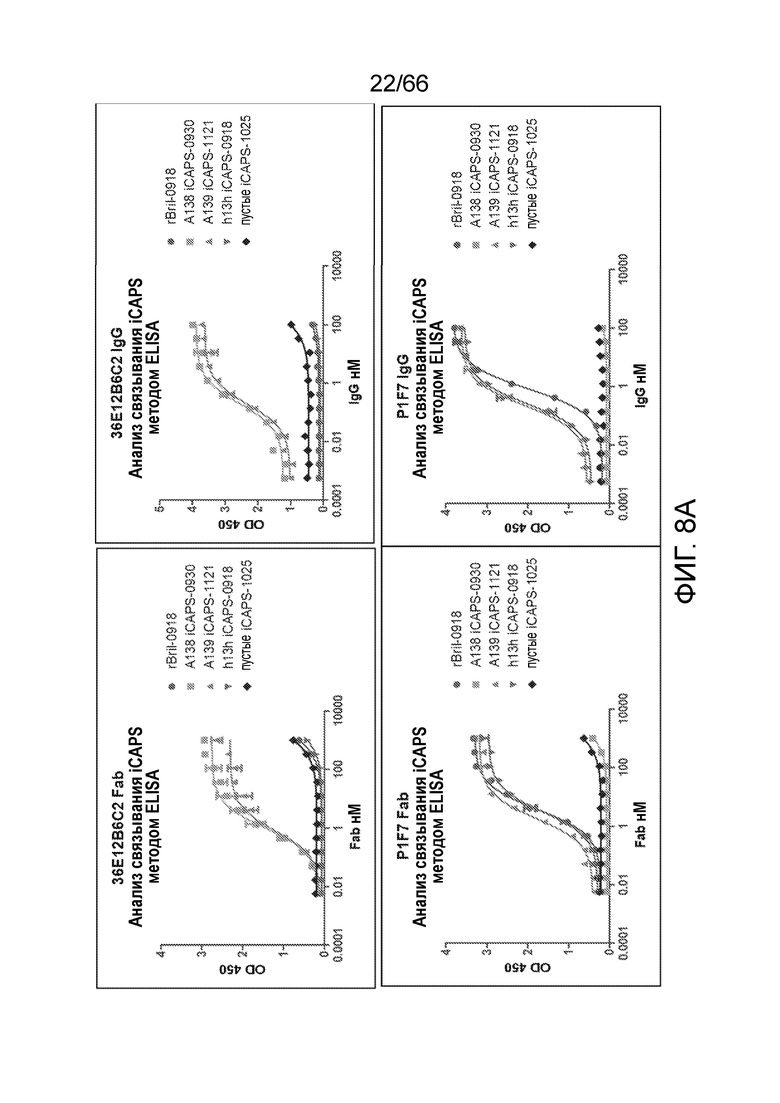
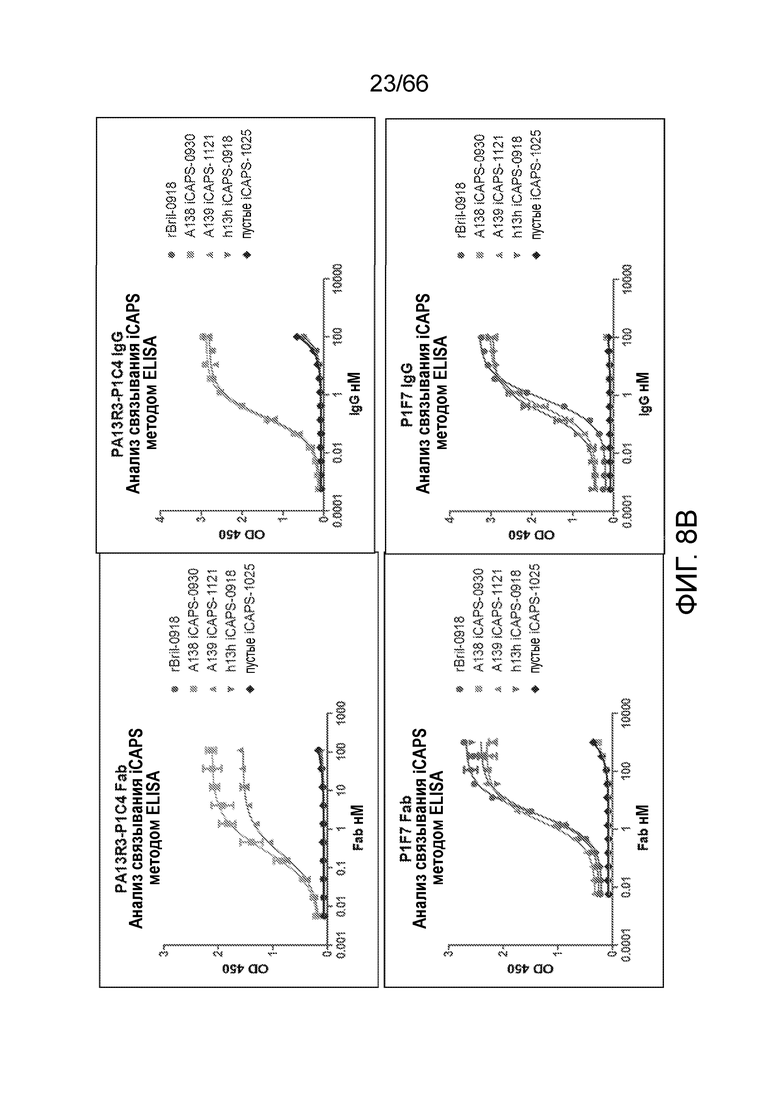


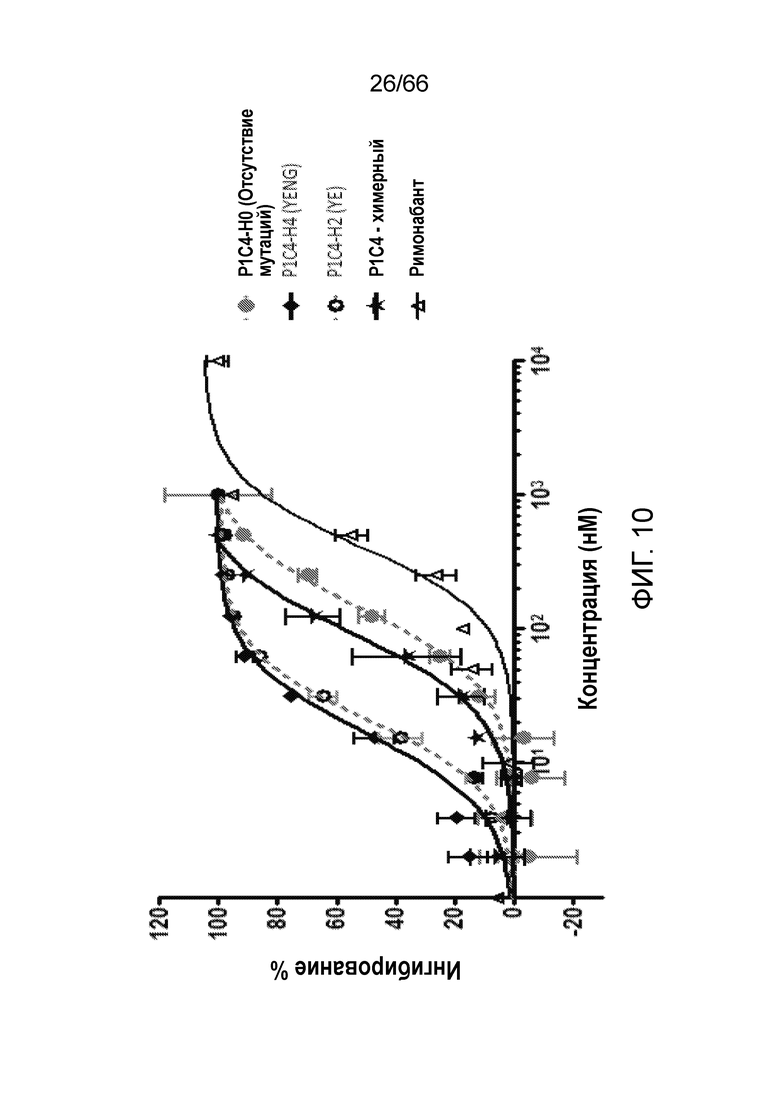
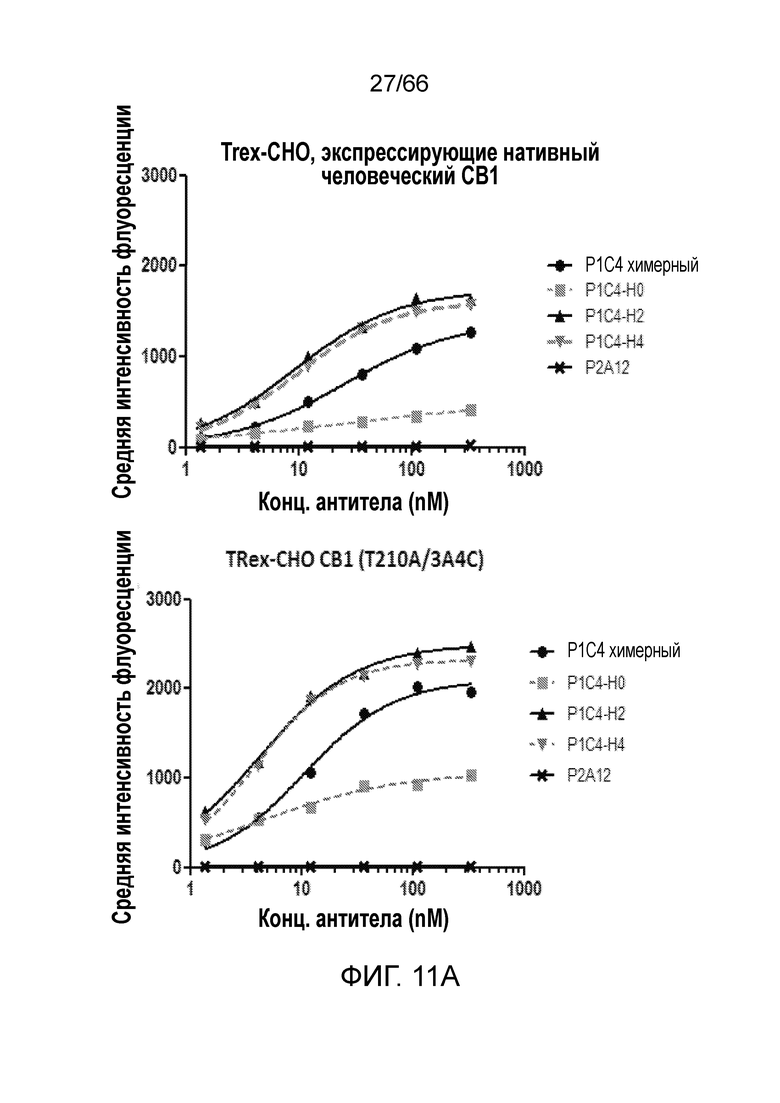
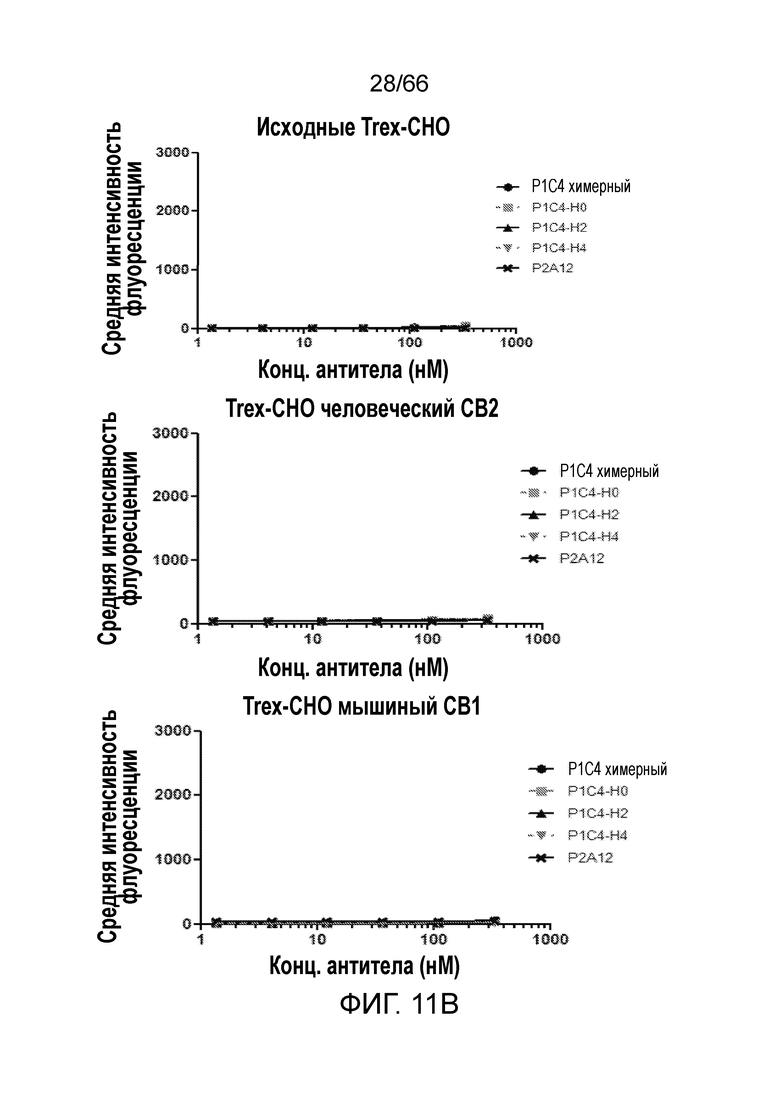
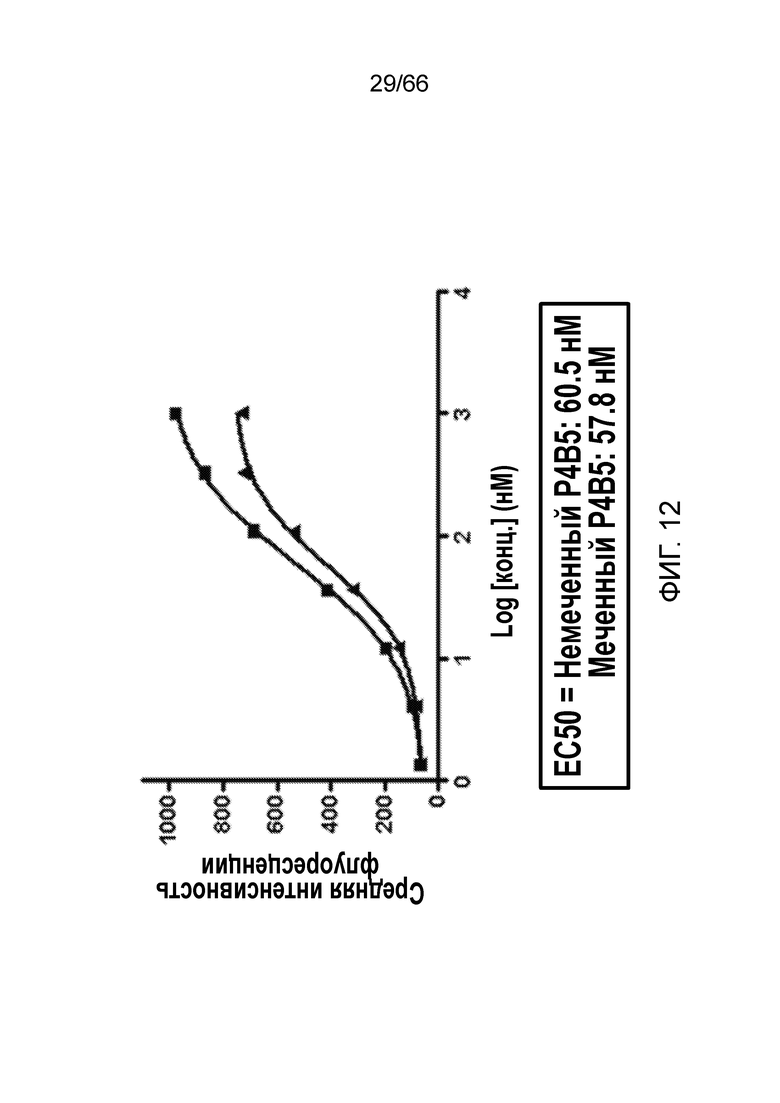

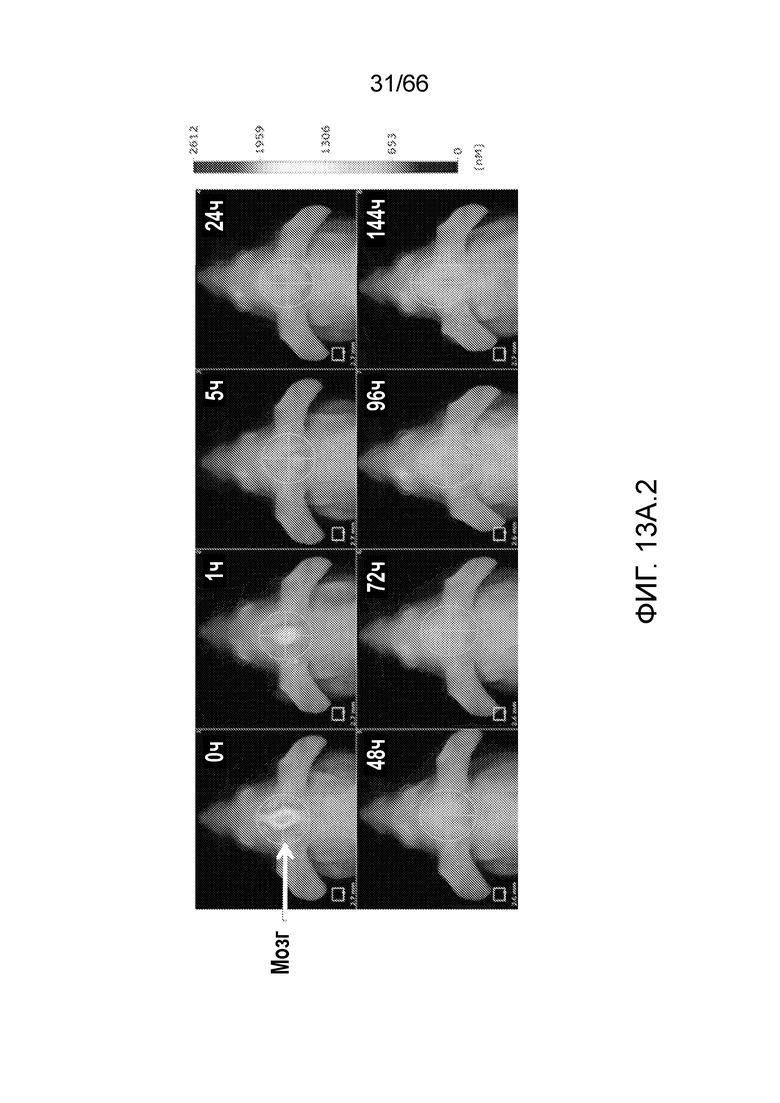
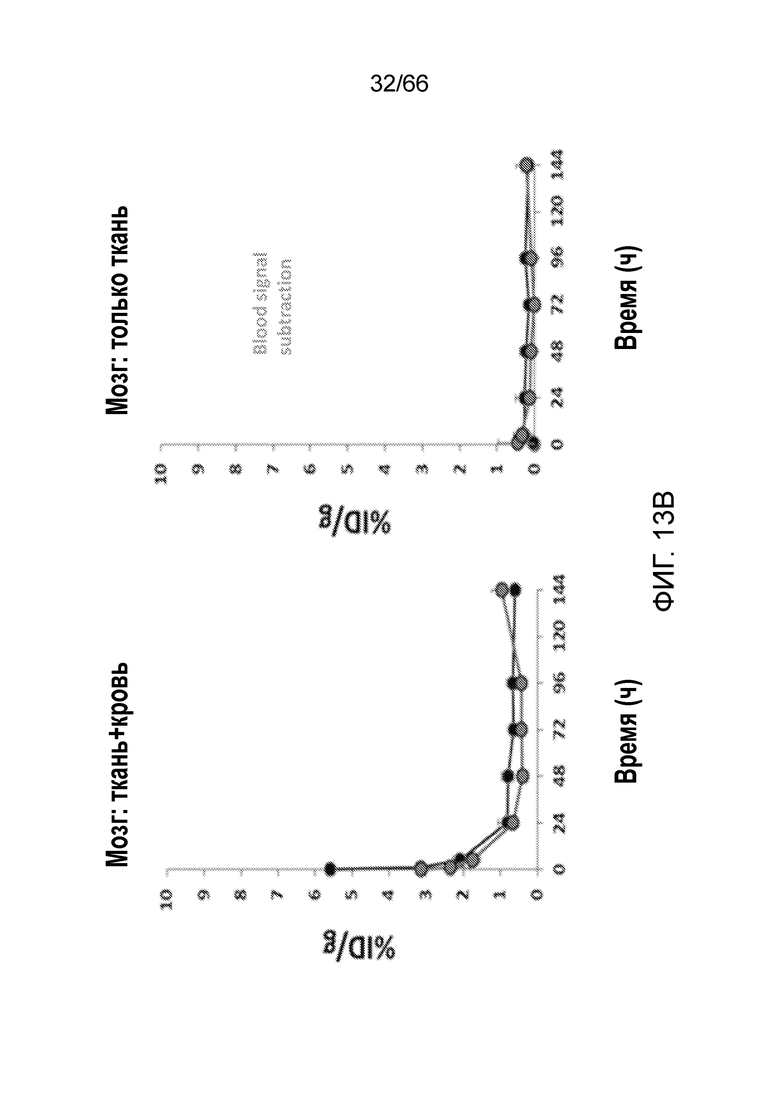
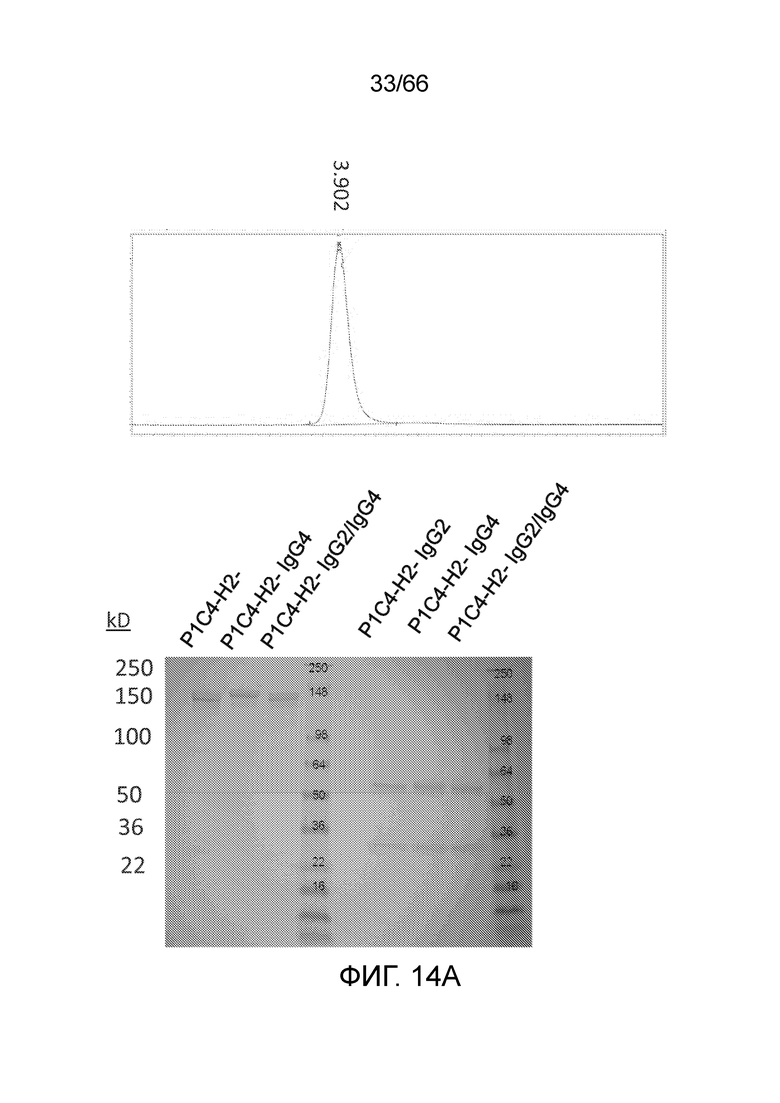
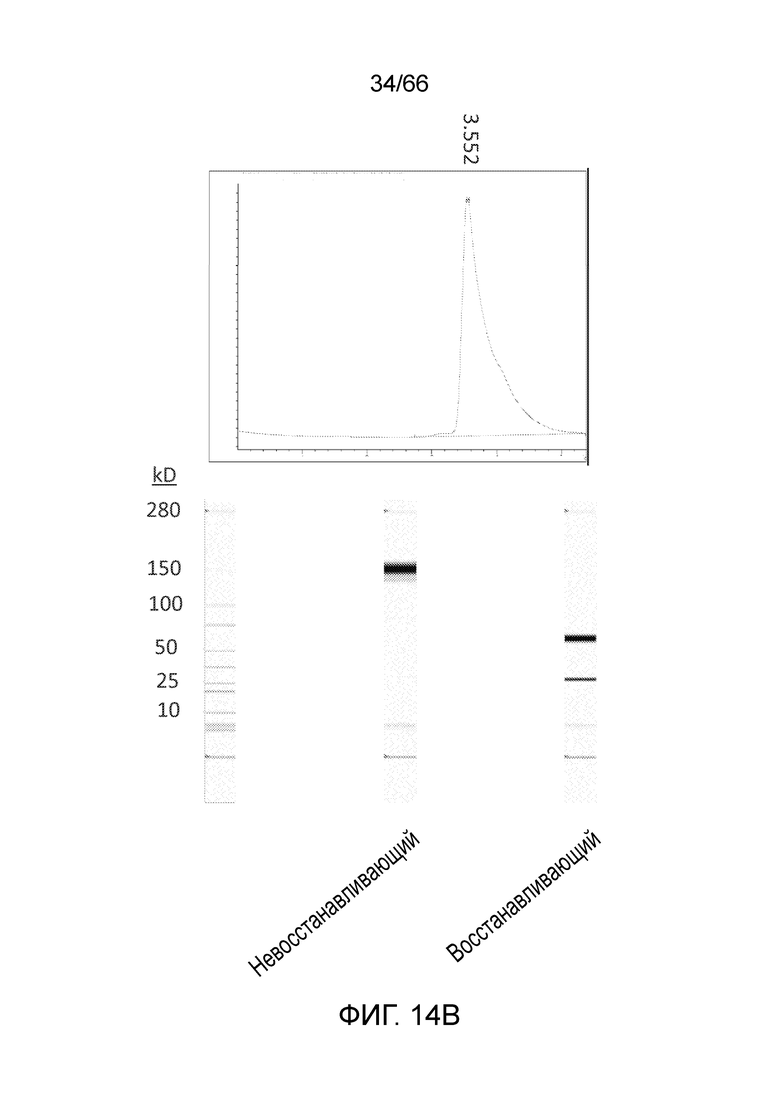
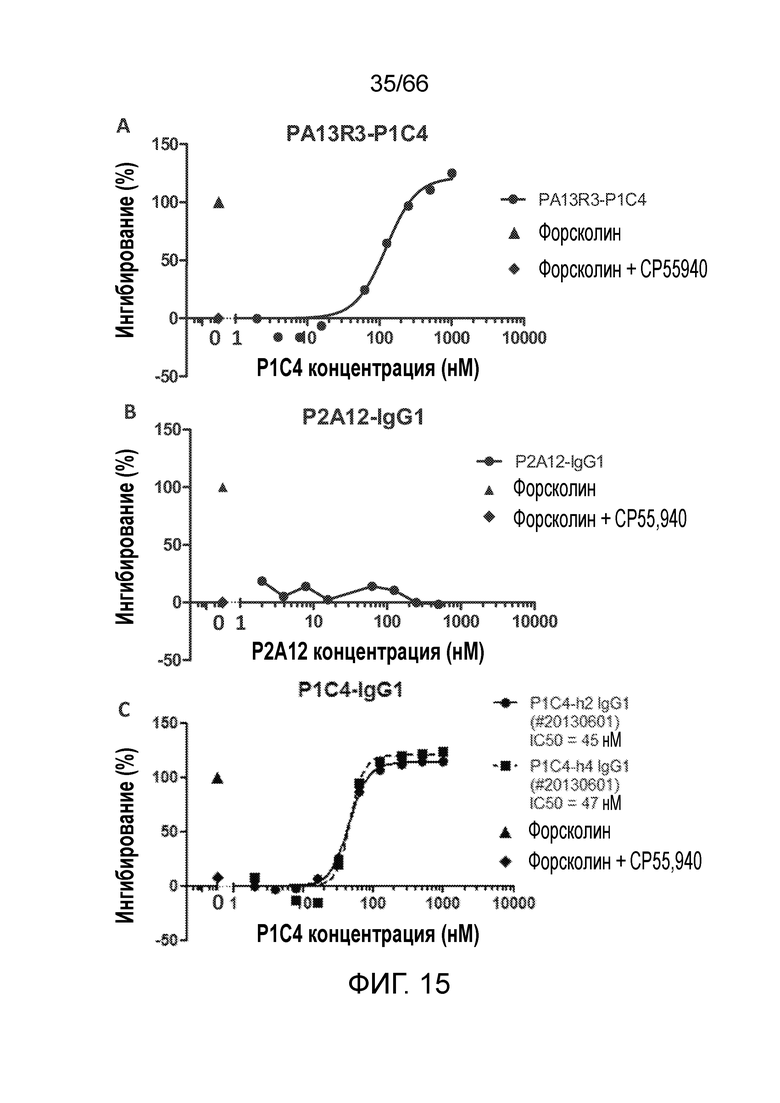

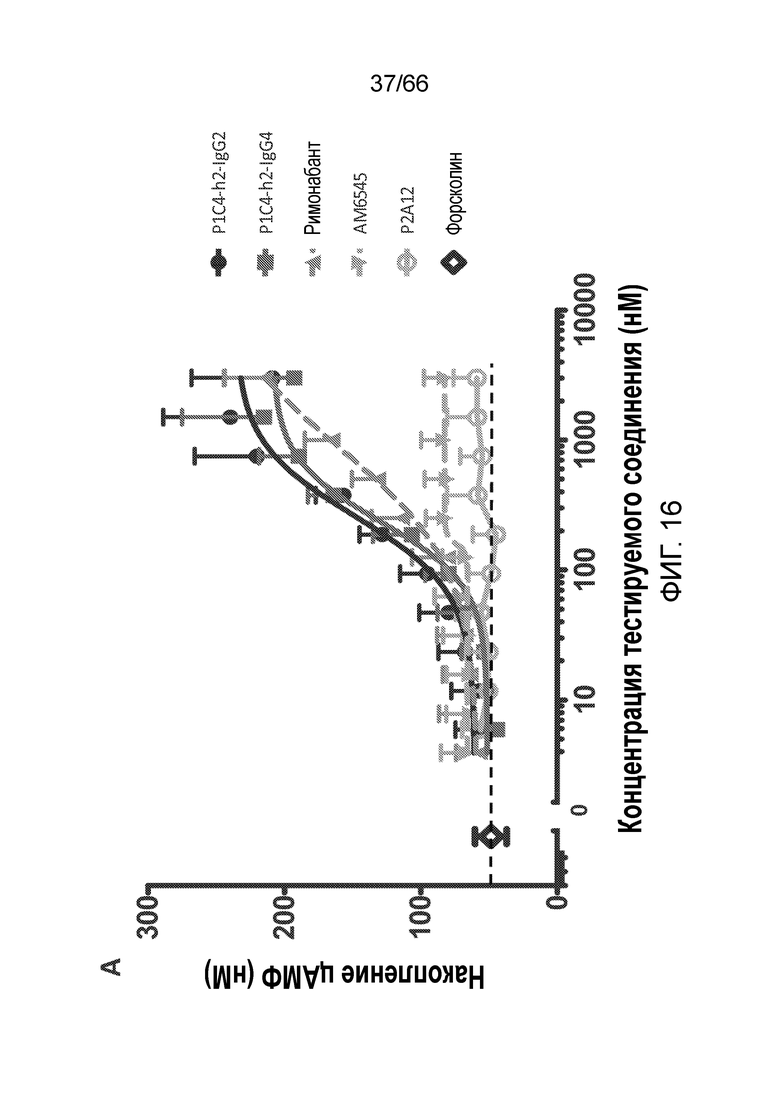
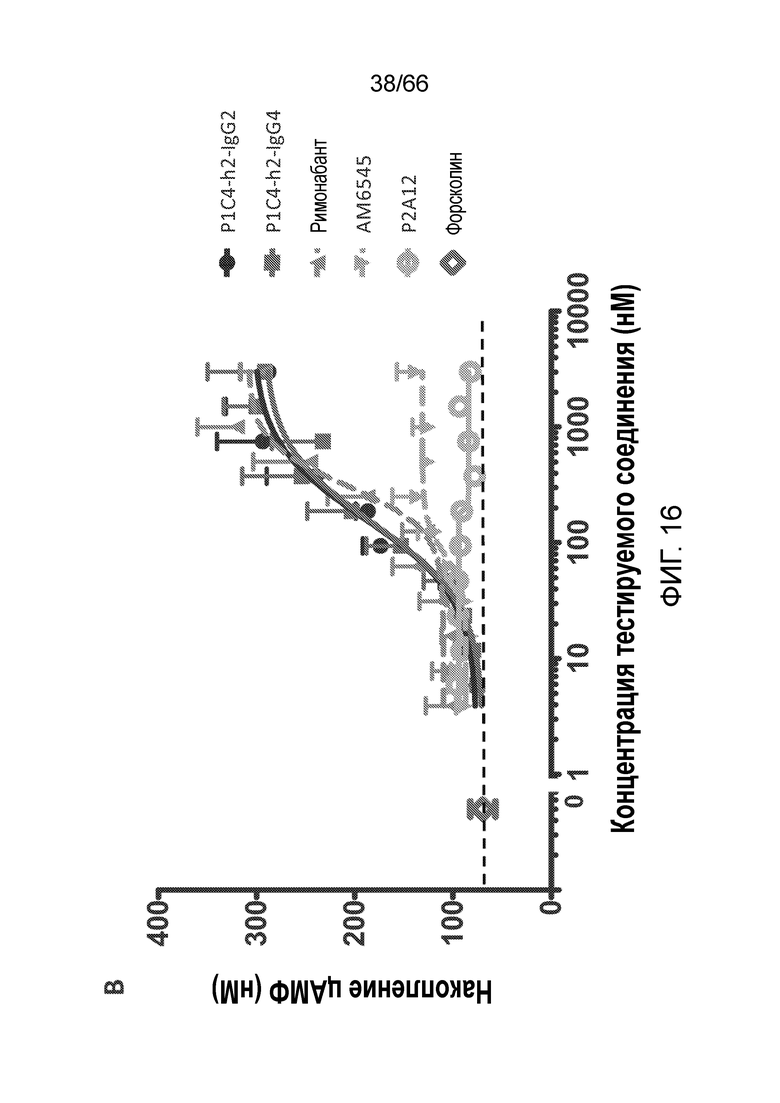
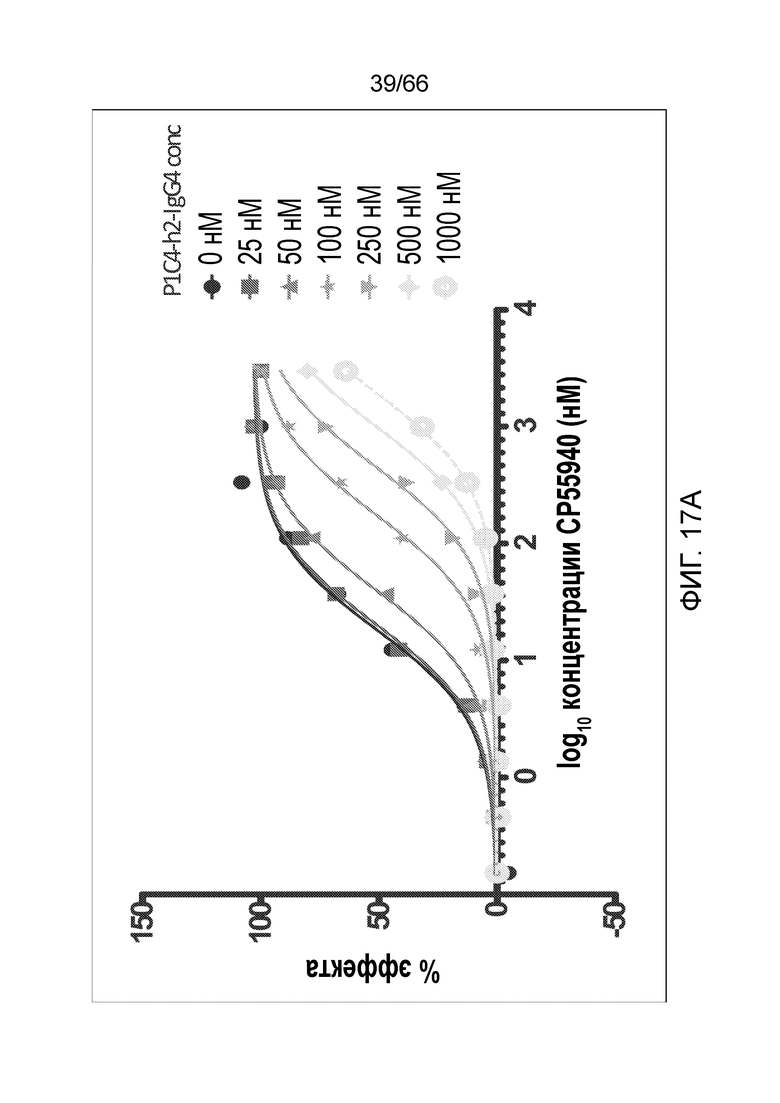
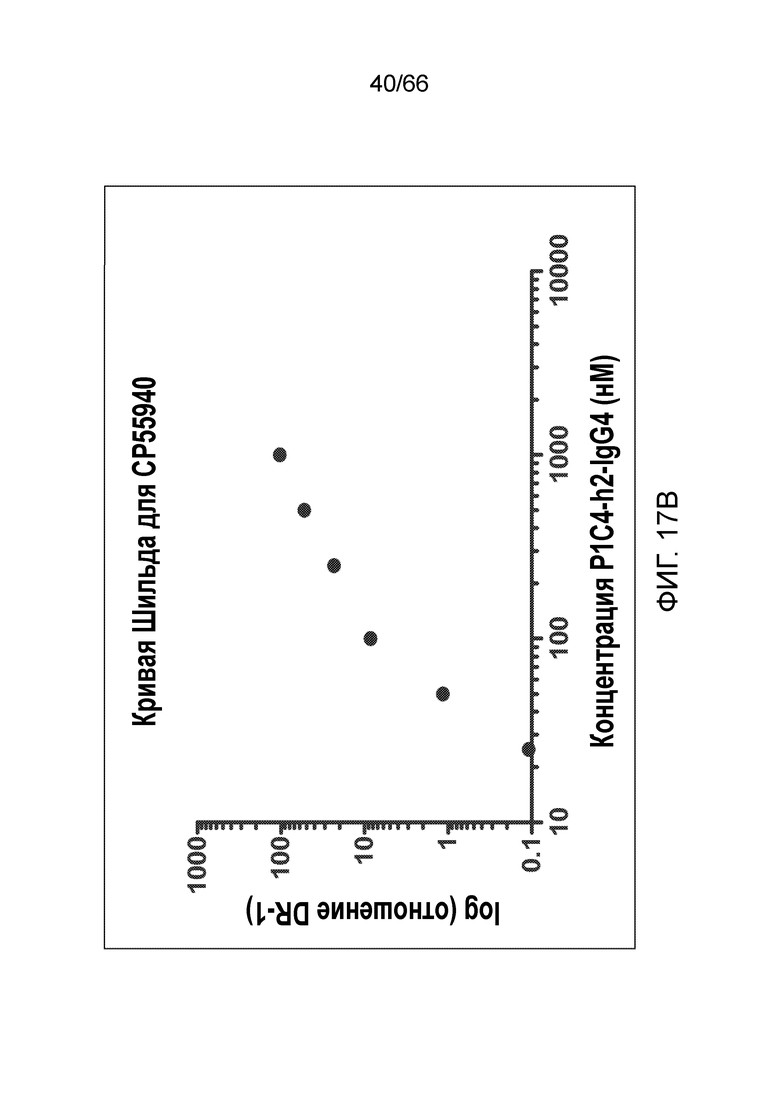
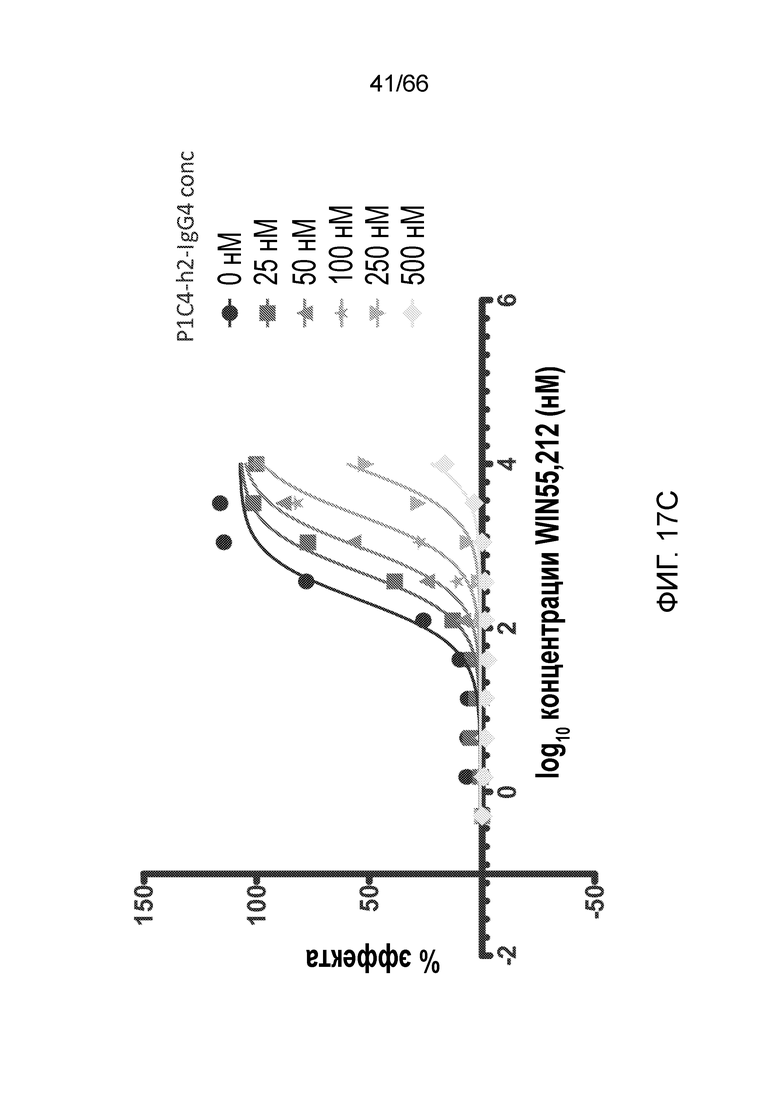
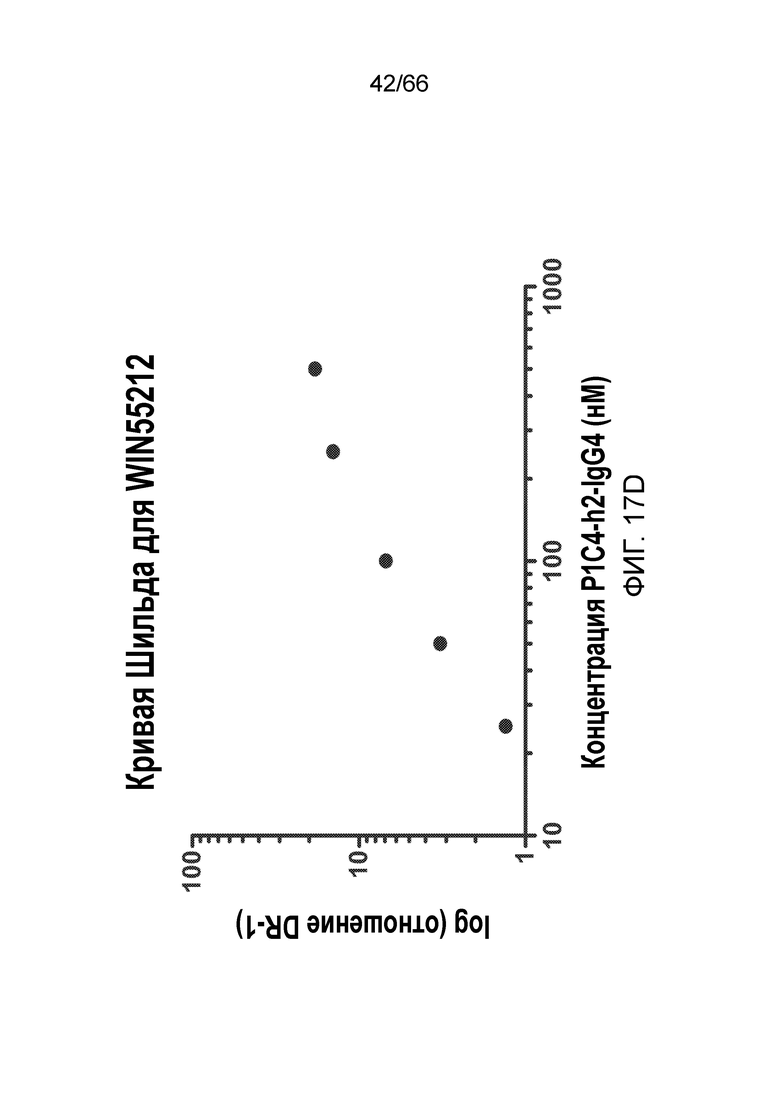
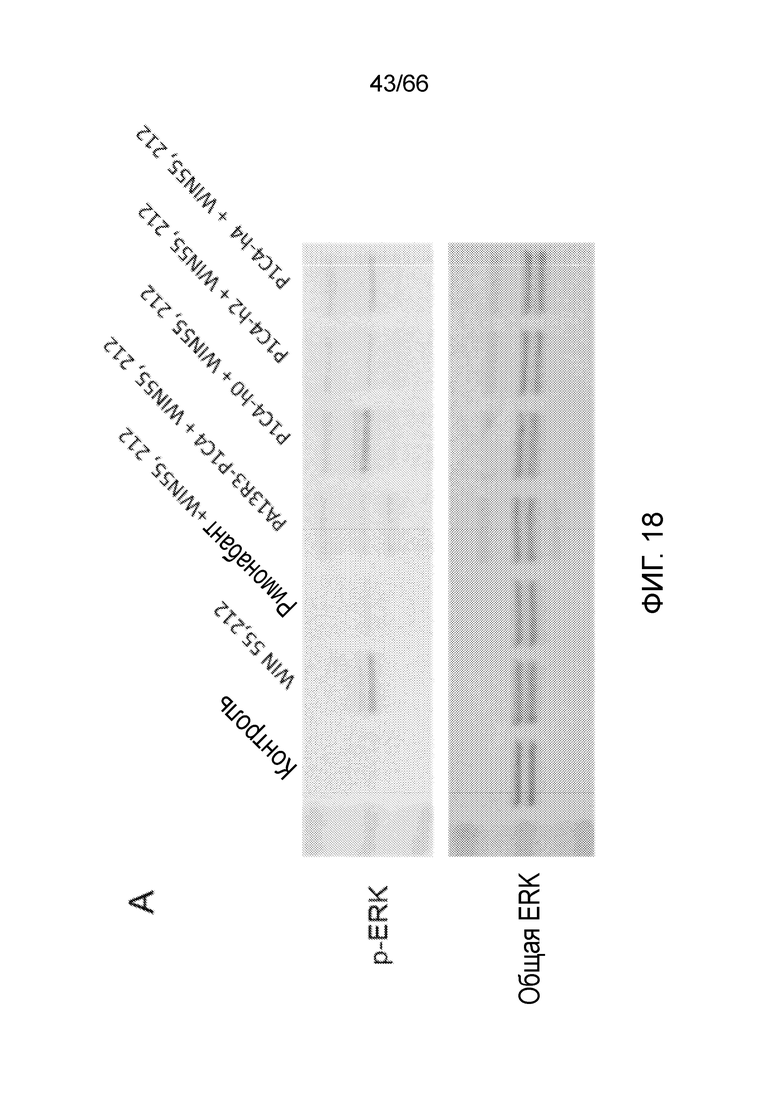
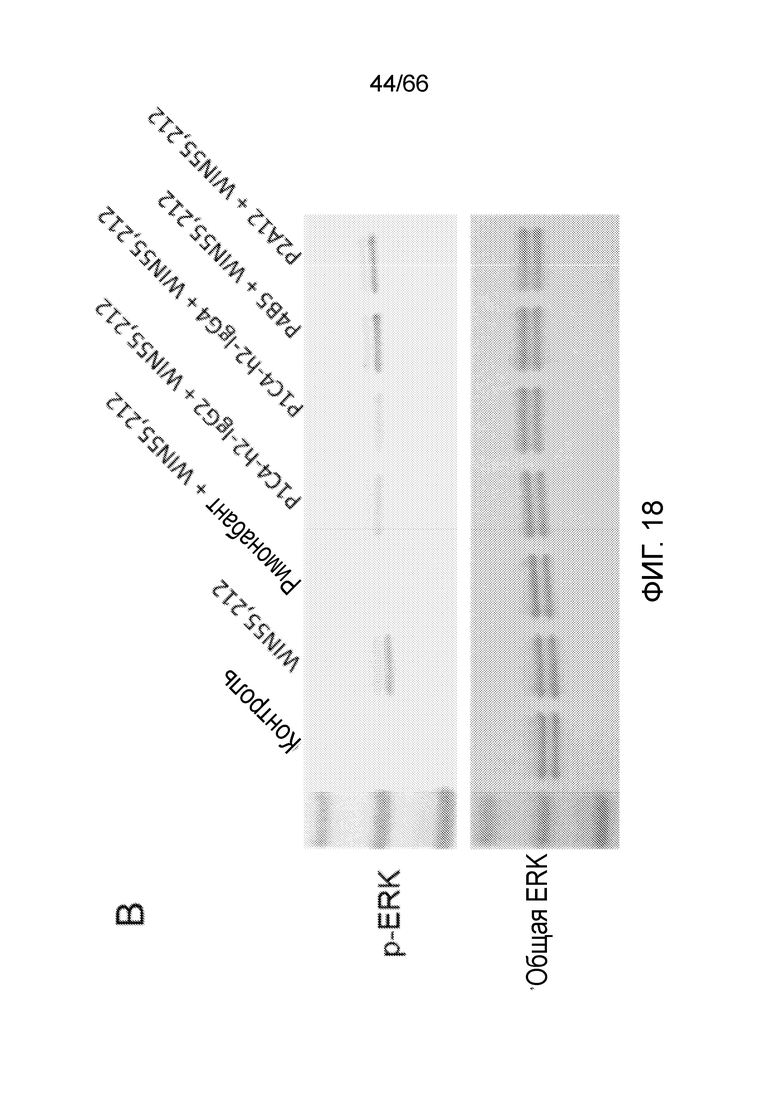
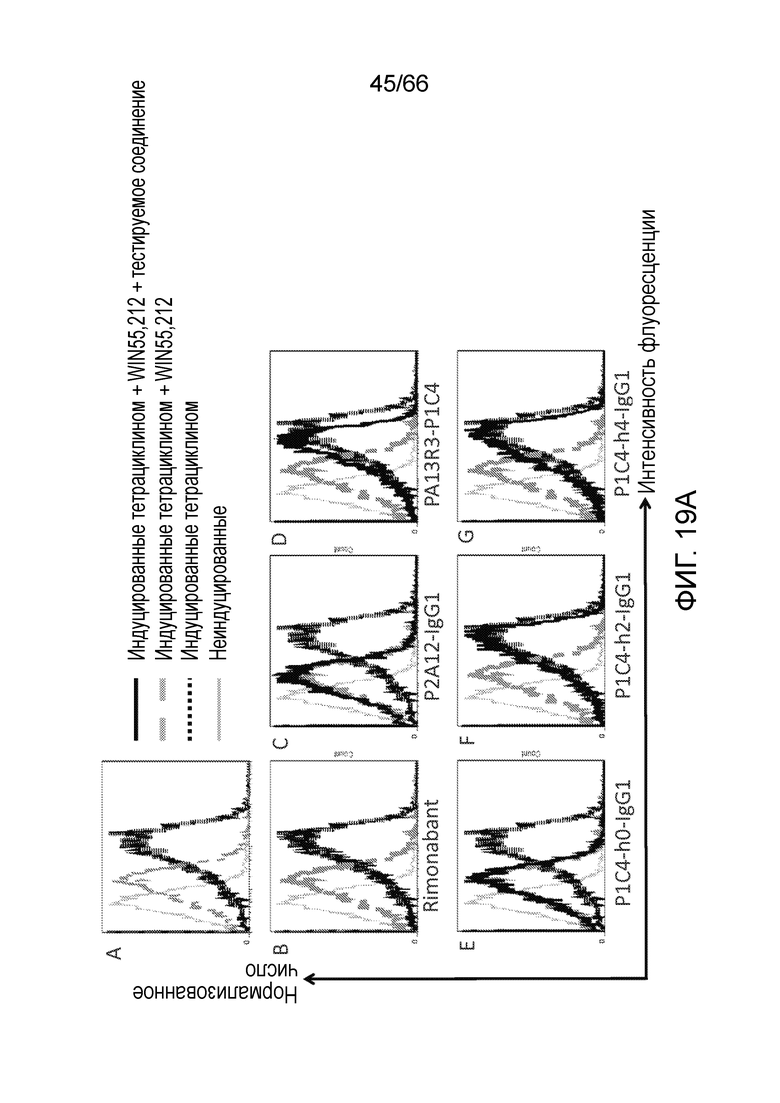

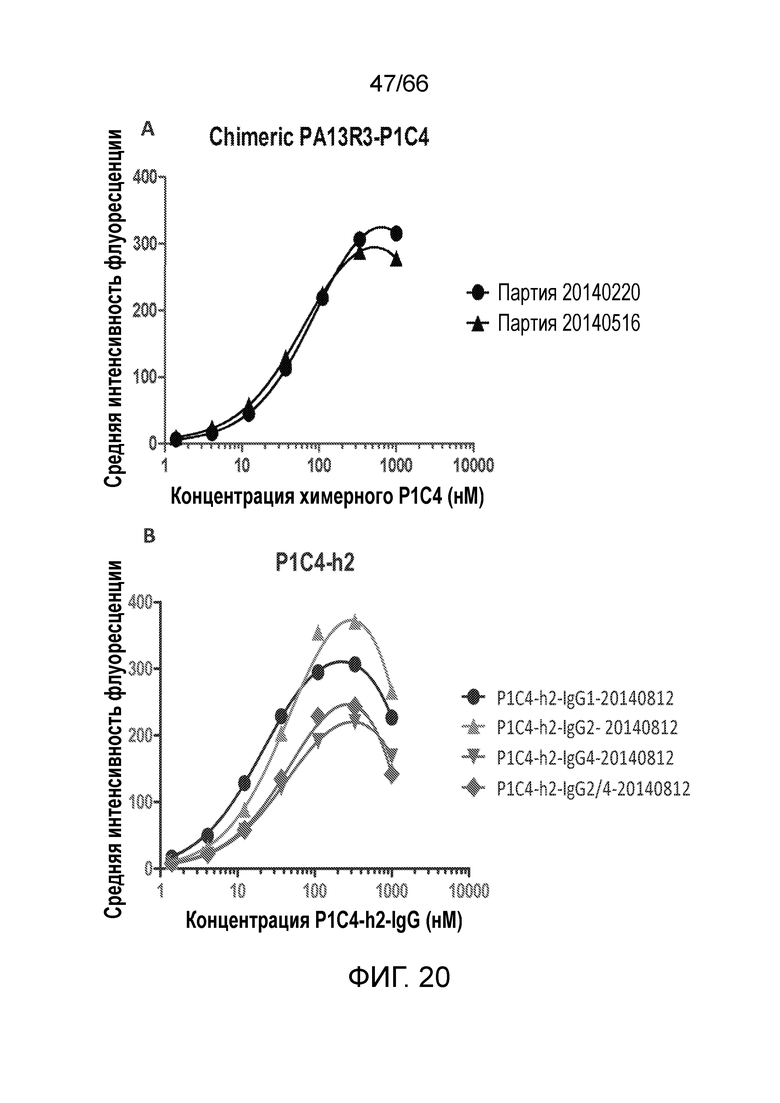
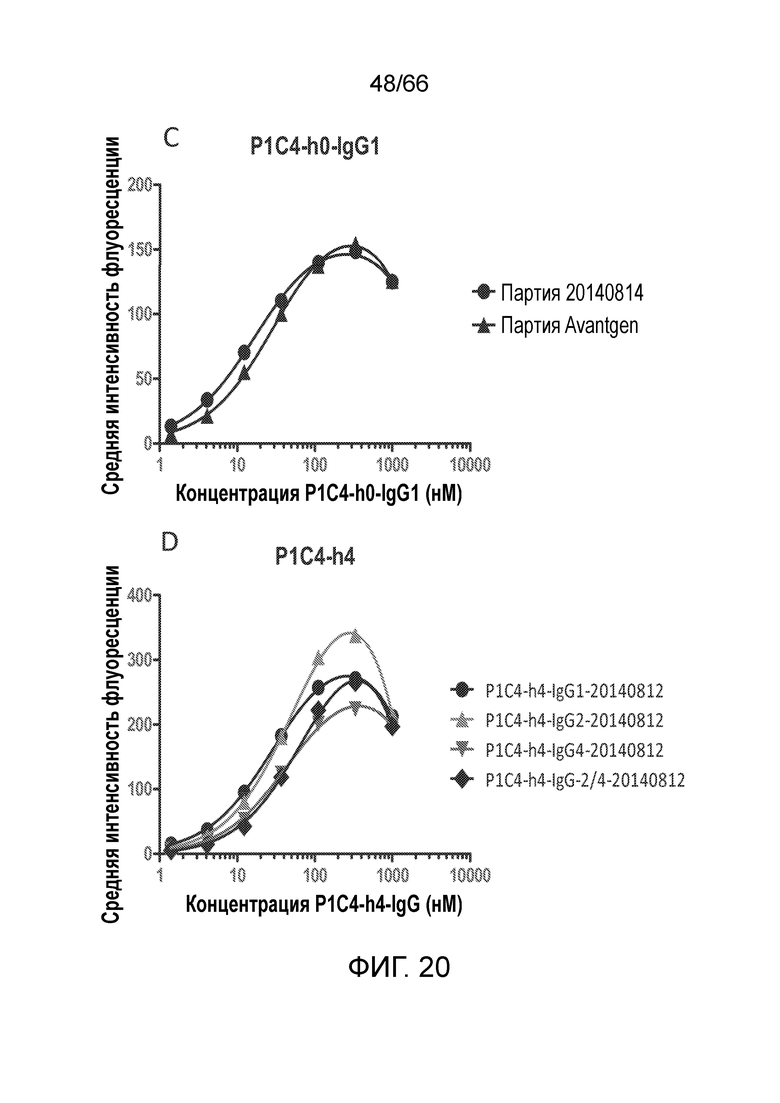
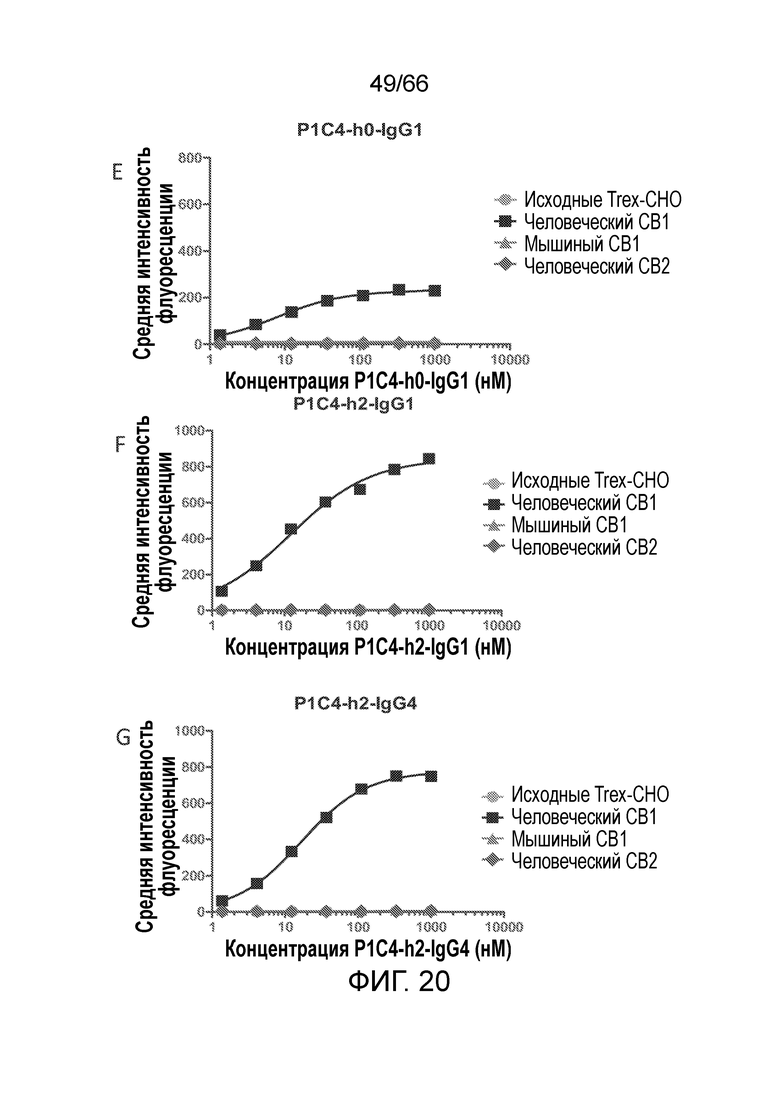
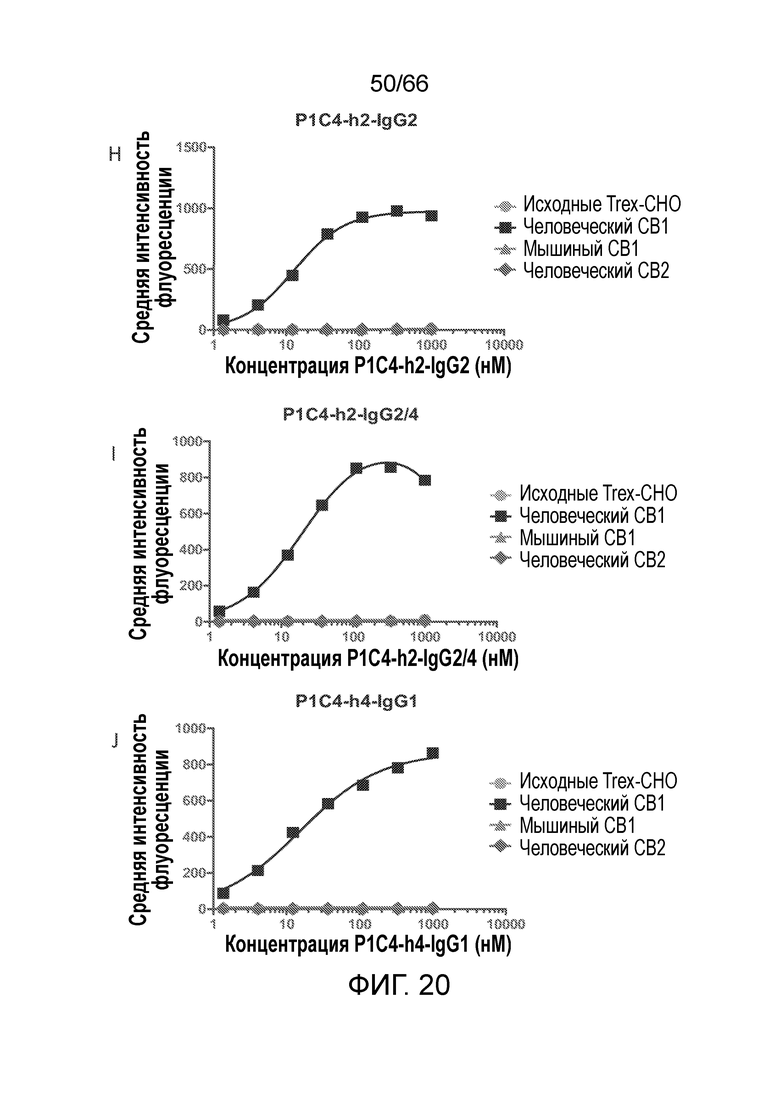
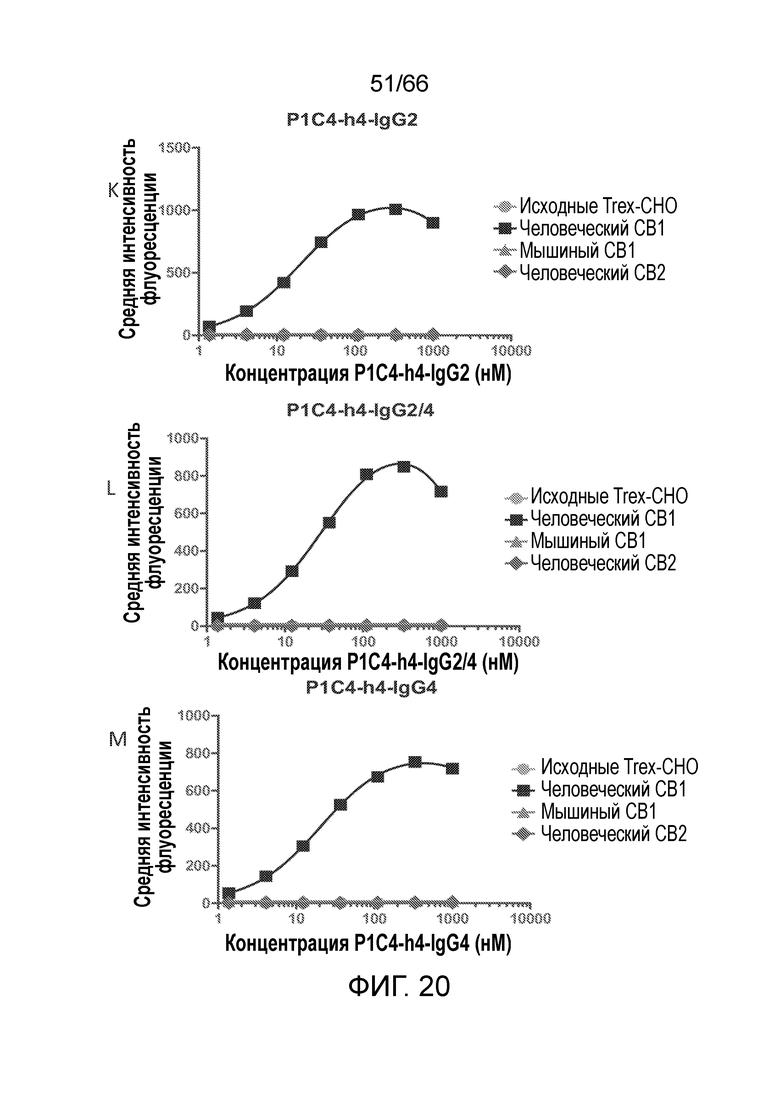
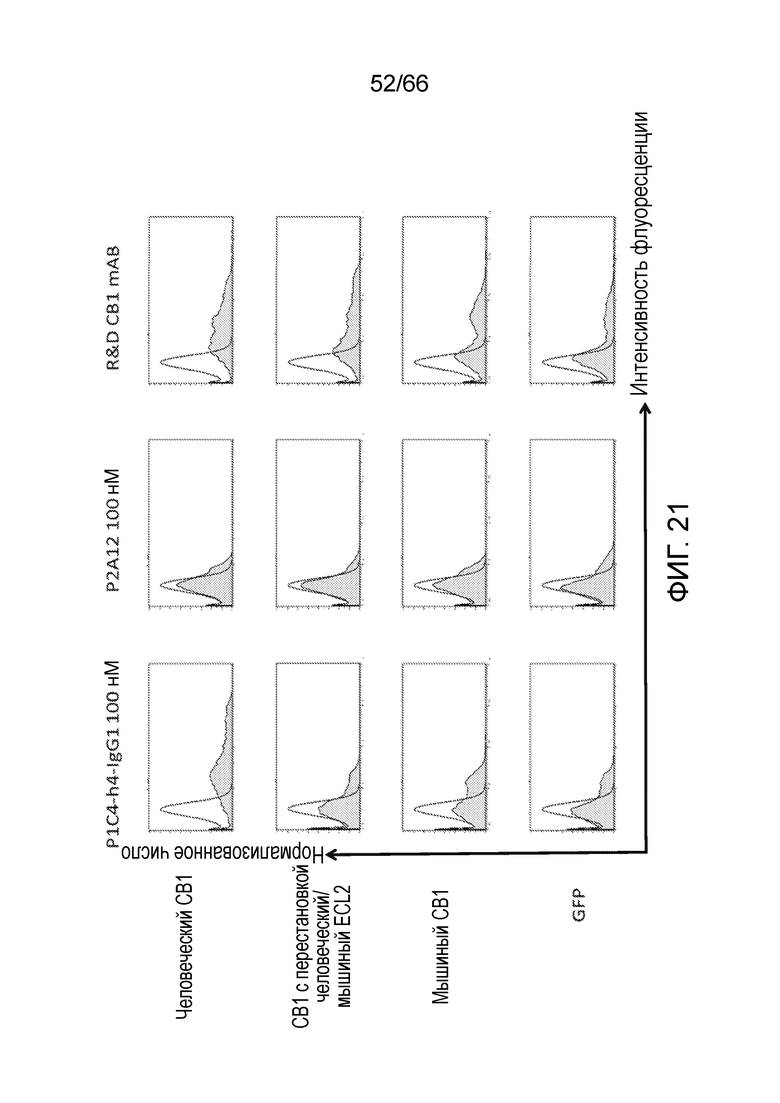

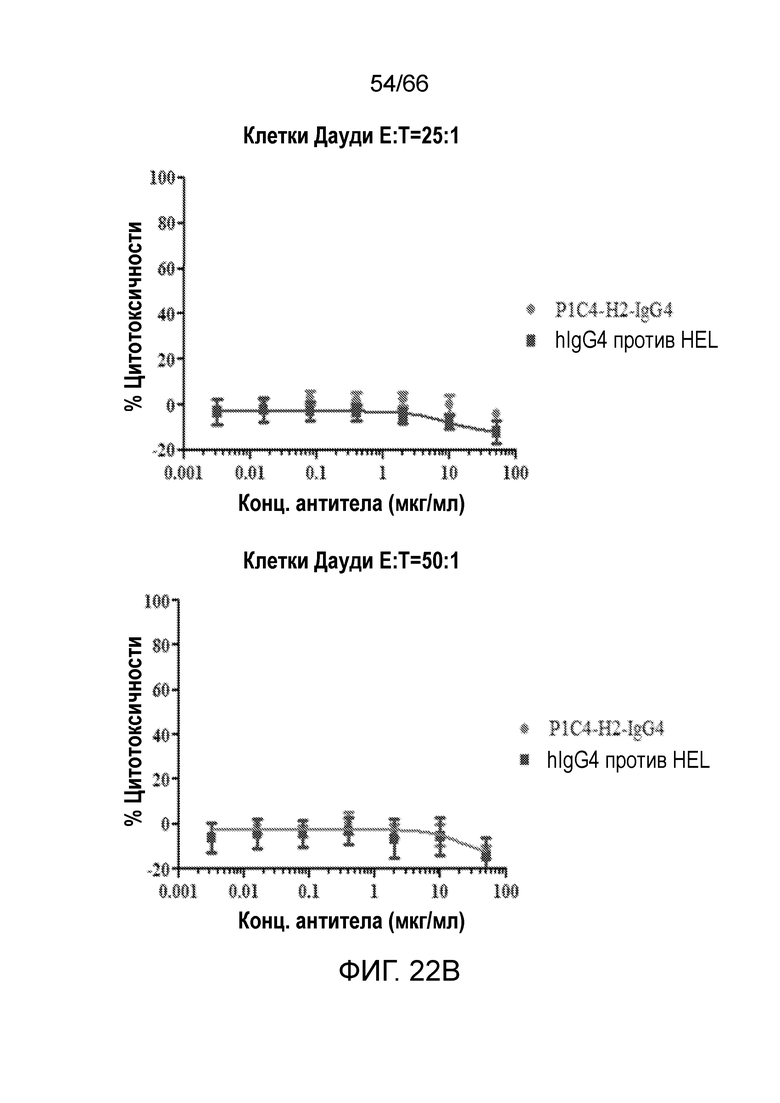
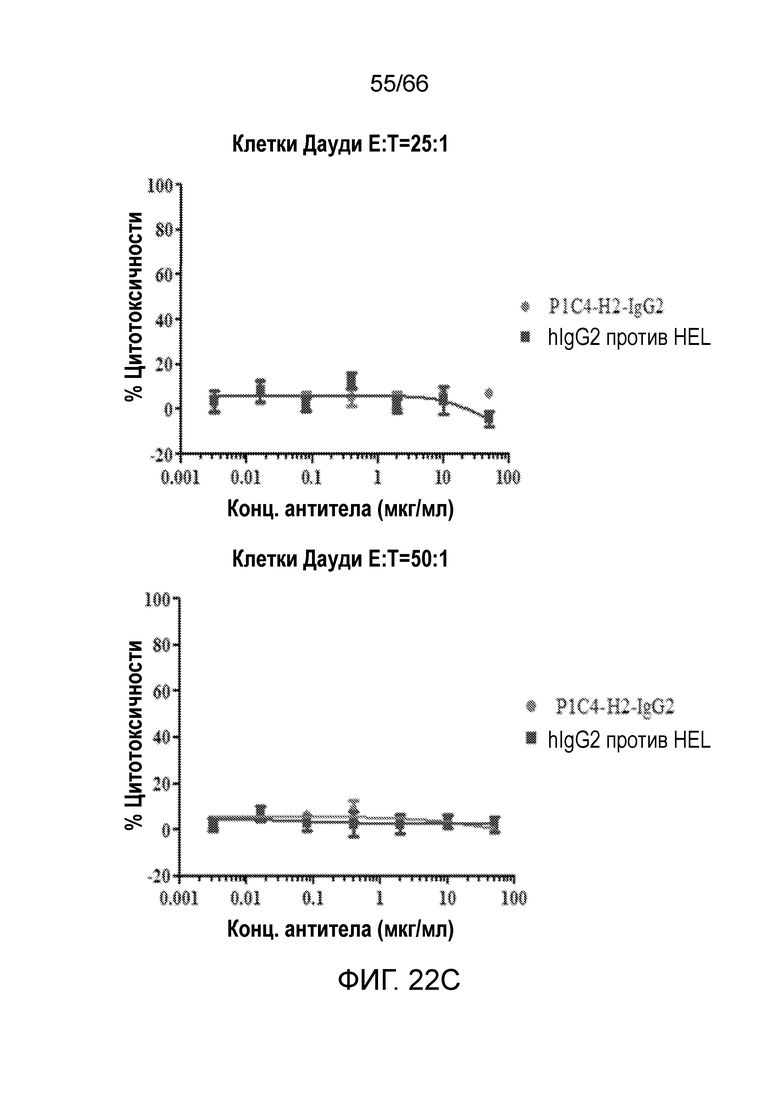
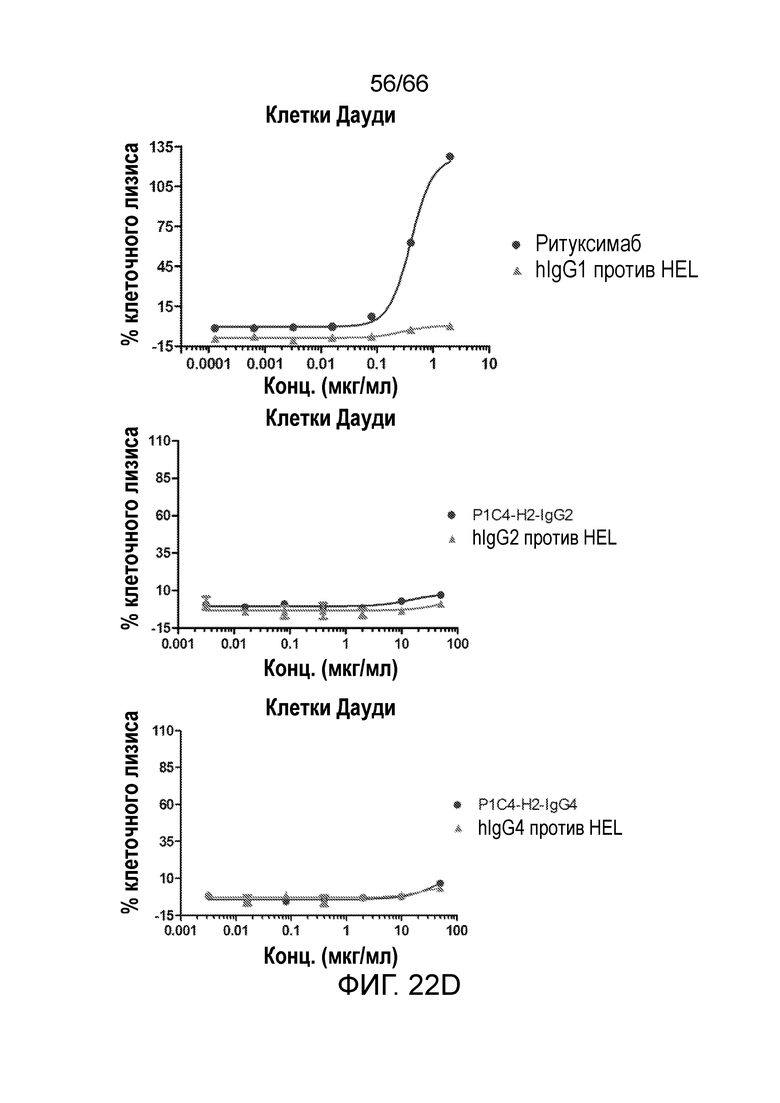
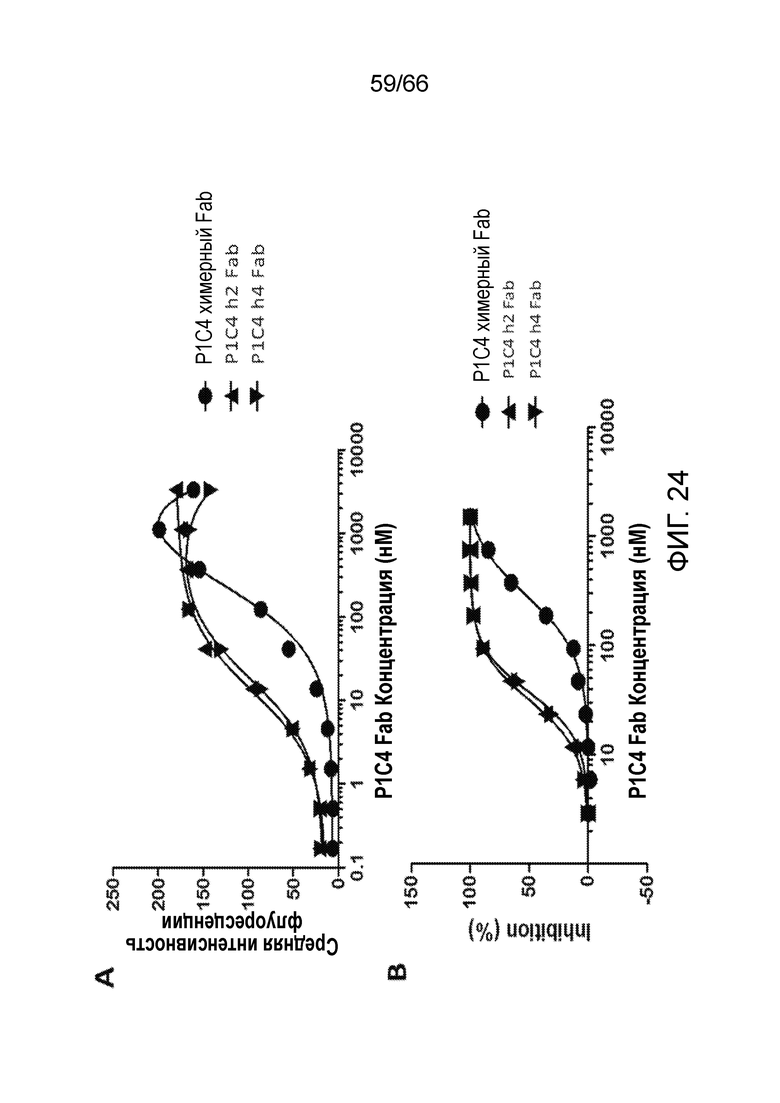
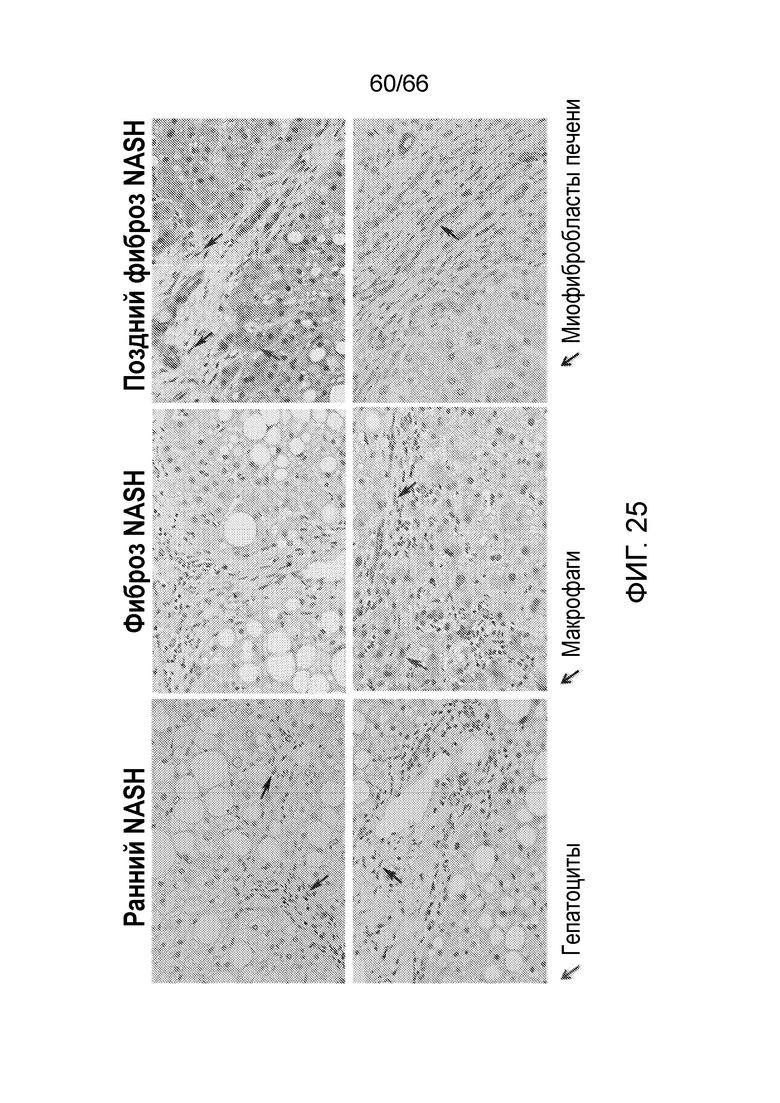

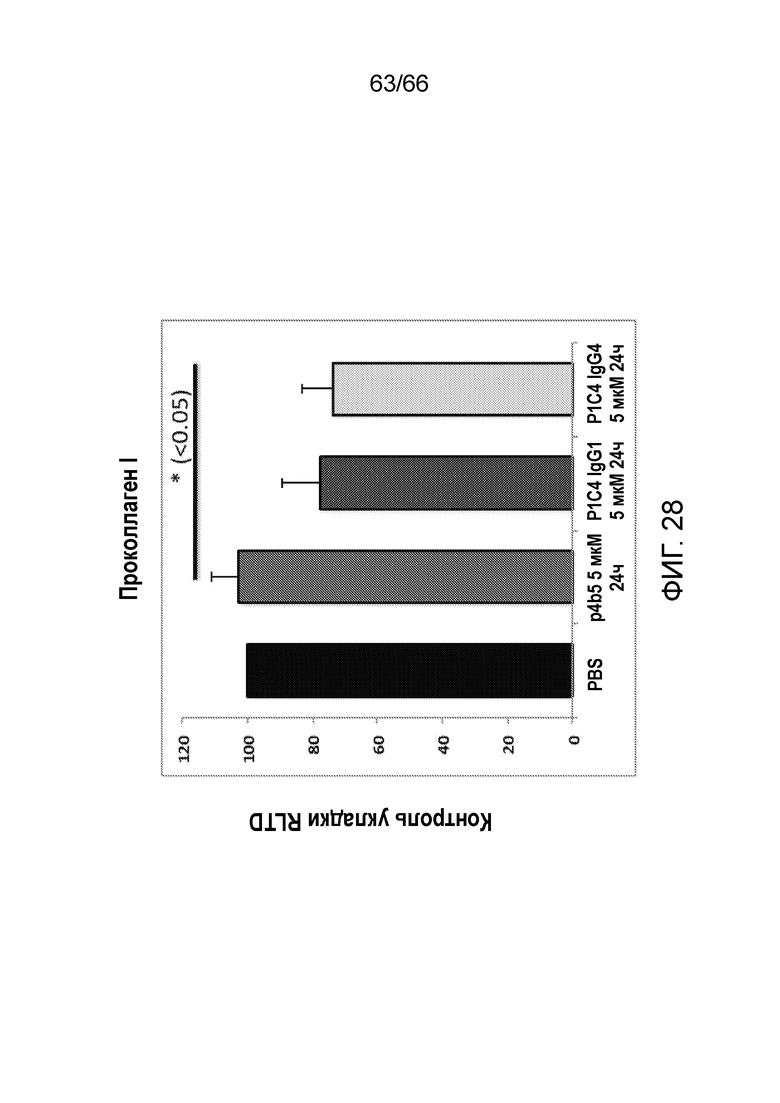
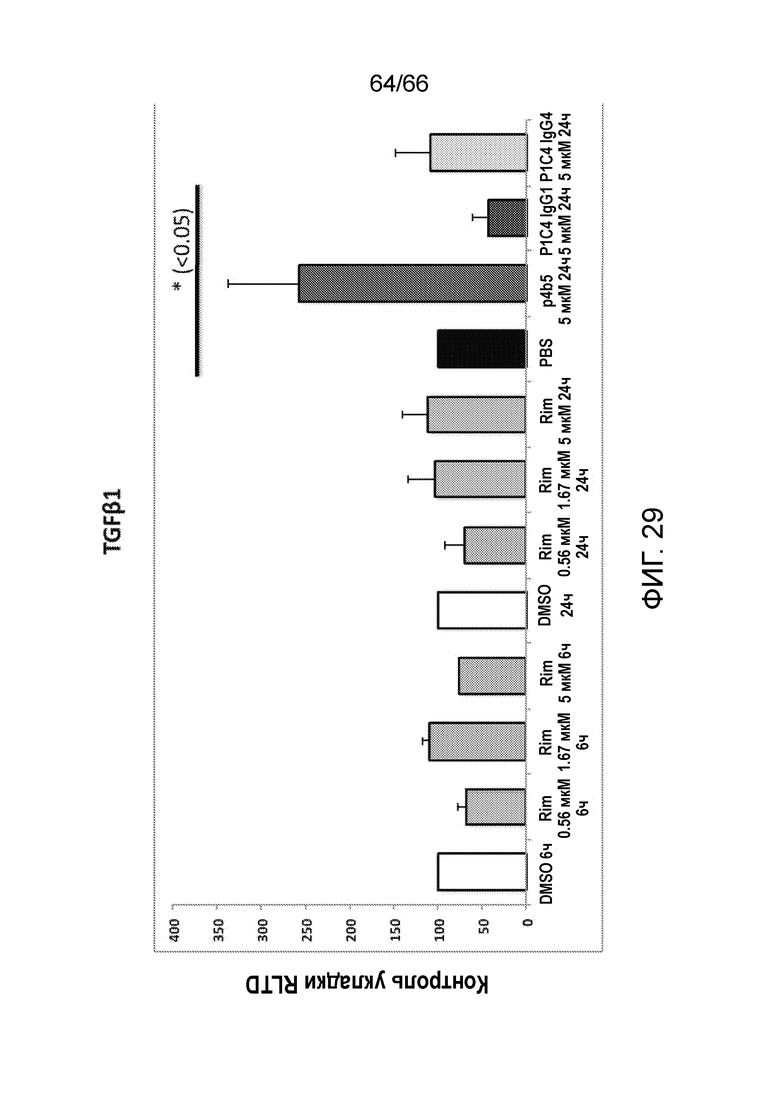

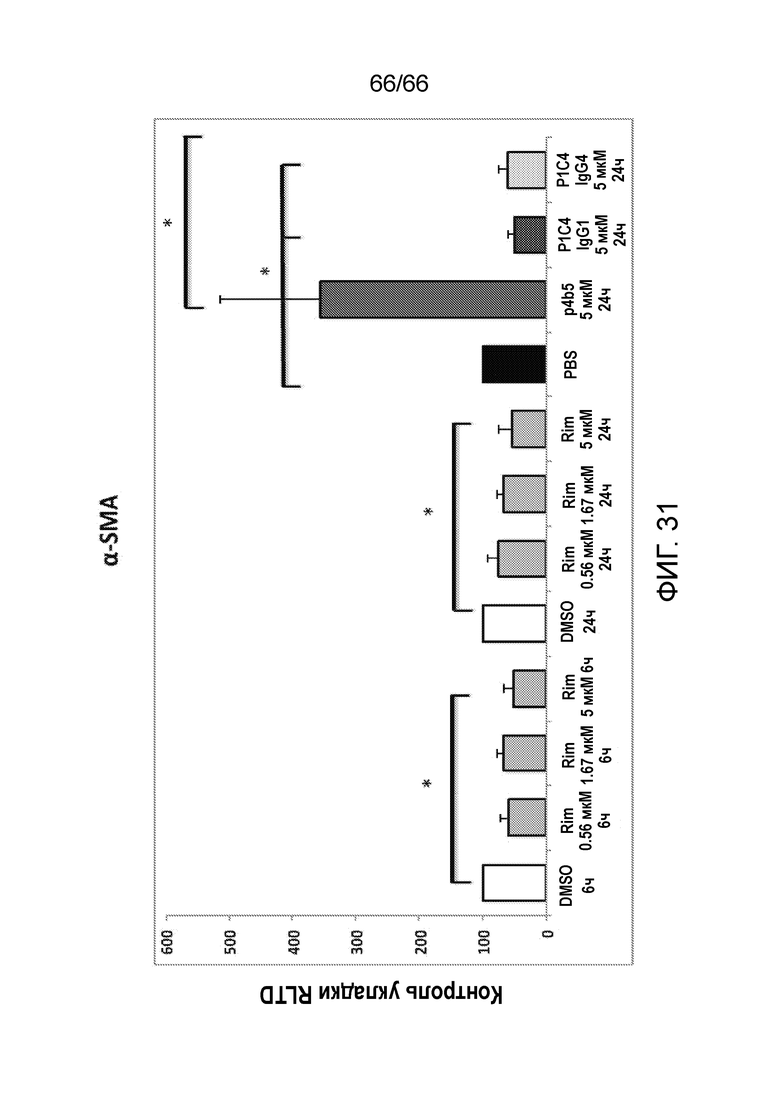
Изобретение относится к области биохимии, в частности к антителу или его антигенсвязывающему фрагменту, обладающему обратным антагонизмом, которое специфично связывается с конформационным эпитопом 1 каннабиноидного рецептора 1 (CB1) человека. Также раскрыты конъюгат, содержащий указанное антитело, клетка-хозяин, экспрессирующая указанное антитело. Также раскрыты способы придания рецептору СВ1 свойств антагониста или агониста, лечения, диагностики. Изобретение позволяет эффективно лечить заболевания, ассоциированные с каннабиноидным рецептором 1 (CB1). 13 н. и 21 з.п. ф-лы, 31 ил., 34 табл., 31 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, обладающие обратным антагонизмом, которое специфично связывается с конформационным эпитопом 1 каннабиноидного рецептора 1 (CB1) человека, полученное с использованием рецептора CB1 человека, имеющего домен ECL2 с последовательностью GWNCEKLQSDIFPHIDE.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, где антитело или фрагмент имеет аффинность связывания Kd в отношении рецептора CB1 человека, составляющую примерно 1 мкМ или меньше.
3. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, где антитело или фрагмент ингибирует сигнальную активность рецептора CB1.
4. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, где антитело или фрагмент активирует или усиливает активность рецептора CB1.
5. Конъюгат для нацеливания на рецептор CB1 человека, содержащий выделенное антитело или его антигенсвязывающий фрагмент по п.1, где антитело или его антигенсвязывающий фрагмент конъюгировано с агентом, например, дополнительным терапевтическим средством, цитотоксическим средством, молекулой иммуноадгезии или агентом для визуализации.
6. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:2, 10, 18 и 26; и вариабельную область легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 14, 22 и 30.
7. Выделенное антитело или его антигенсвязывающий фрагмент по п.6, где антитело или его антигенсвязывающий фрагмент содержит константную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:4, 12, 20 и 28, и константную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO:8, 16, 24 и 32.
8. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:2; и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:6.
9. Выделенное антитело или его антигенсвязывающий фрагмент по п.8, где антитело или его антигенсвязывающий фрагмент содержит константную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:4; и константную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8.
10. Способ придания рецептору CB1 свойств антагониста, включающий приведение клетки, экспрессирующей рецептор CB1, в контакт с антителом или его антигенсвязывающим фрагментом по любому из пп.1-4 и 6-9.
11. Способ придания рецептору CB1 свойств агониста, включающий приведение клетки в контакт с антителом или связывающим фрагментом по любому из пп.1-4 и 6-9.
12. Способ лечения заболевания или расстройства, отвечающего на антагонизм или агонизм рецептора CB1, у больного, который включает введение больному антитела или его антигенсвязывающего фрагмента по любому из пп.1-4 и 6-9.
13. Способ по п.12, где заболевание или расстройство выбрано из группы, состоящей из ожирения, диабета, дислипидемии, метаболических заболеваний, фиброза, неалкогольного стеатогепатита (NASH), болезни печени, первичного билиарного цирроза, болезни почек, фиброза почек, хронической болезни почек, остеопороза, атеросклероза, сердечно-сосудистого заболевания, злокачественного новообразования и воспалительного заболевания.
14. Способ по п.12, где заболевание или расстройство выбрано из группы, состоящей из боли, спастичности при рассеянном склерозе и глаукомы.
15. Способ диагностики заболевания или расстройства, связанного с CB1 человека, включающий приведение клетки в контакт с антителом или его антигенсвязывающим фрагментом по пп.1-4 и 6-9.
16. Способ детекции рецептора CB1, включающий приведение клетки в контакт с антителом или его антигенсвязывающим фрагментом по пп.1-4 и 6-9.
17. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с каннабиноидным рецептором 1 (CB1), которое содержит СDR1, CDR2 и CDR3 тяжелой цепи с SEQ ID NO:352, 353 и 354 соответственно, где каждая CDR1, CDR2 и CDR3 тяжелой цепи может содержать одну аминокислотную замену, и CDR1, CDR2 и CDR3 легкой цепи с SEQ ID NO:355, 356 и 357 соответственно, где каждая CDR1, CDR2 и CDR3 легкой цепи может содержать одну аминокислотную замену.
18. Выделенное антитело или его антигенсвязывающий фрагмент по п.17, которое содержит:
CDR1 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:443-463;
CDR2 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:464-577, и
CDR3 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:578-625.
19. Выделенное антитело или антигенсвязывающий фрагмент по п.17, которое содержит:
CDR1 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:626-661;
CDR2 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:662-742, и
CDR3 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:742-824.
20. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп.1-4, 6-9 и 17-19, которое представляет собой гуманизированное антитело.
21. Выделенное антитело или его антигенсвязывающий фрагмент по п.20, которое содержит Fc-фрагмент IgG1 человека.
22. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп.1-4, 6-9 и 17-19, которое содержит модифицированный Fc-фрагмент.
23. Выделенное антитело или его антигенсвязывающий фрагмент, способное специфически связывается с каннабиноидным рецептором 1 (CB1), которое содержит аминокислотную последовательность вариабельного участка тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO:339-341, и вариабельный участок легкой цепи с последовательностью SEQ ID NO:337.
24. Выделенное антитело или его антигенсвязывающий фрагмент по п.23, содержащее вариабельный участок тяжелой цепи, содержащий SEQ ID NO:341, и константный участок тяжелой цепи, содержащий SEQ ID NO:433, SEQ ID NO:434 или SEQ ID NO:435.
25. Выделенное антитело или его антигенсвязывающий фрагмент по п.23, которое специфически связывается с каннабиноидным рецептором 1 (СВ1), содержащее вариабельный участок тяжелой цепи, содержащий SEQ ID NO:340, и константный участок тяжелой цепи, содержащий SEQ ID NO:433, SEQ ID NO:434 или SEQ ID NO:435.
26. Клетка-хозяин, содержащая нуклеиновую кислоту, кодирующую выделенное антитело или его антигенсвязывающий фрагмент по пп.17-25, экспрессирующая выделенное антитело или его антигенсвязывающий фрагмент.
27. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент по п.1, аффинность связывания Кd которого в отношении рецептора CB1 составляет примерно 100 нМ или менее.
28. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент по п.27, которое обладает ингибирующей активностью в отношении рецептора CB1, которая по меньшей мере в 2 раза превышает активность соответствующего негуманизированного или химерного антитела или фрагмента.
29. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по любому из пп.27, 28, где способность антитела или антигенсвязывающего фрагмента проникать в мозг снижена или отсутствует.
30. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по п.29, у которого понижены побочные эффекты на центральную нервную систему (ЦНС) по сравнению с низкомолекулярным агонистом или антагонистом рецептора CB1.
31. Выделенное гуманизированное антитело или антигенсвязывающий фрагмент по п.30, где низкомолекулярный агонист или антагонист рецептора CB1 представляет собой AM6545, AM251, таранабант или римонабант.
32. Способ лечения заболевания или расстройства, отвечающего на антагонизм или агонизм рецептора CB1, у больного, включающий введение больному антитела или его антигенсвязывающего фрагмента по любому из пп.18-31.
33. Способ по п.32, где заболевание или расстройство выбрано из группы, состоящей из ожирения, диабета, дислипидемии, метаболических заболеваний, фиброза, NASH, болезни печени, первичного билиарного цирроза, болезни почек, фиброза почек, хронического заболевания почек, остеопороза, атеросклероза, сердечно-сосудистого заболевания, злокачественного новообразования, воспалительного заболевания, боли, спастичности при рассеянном склерозе и глазных болезней, включая глаукому.
34. Способ по п.32, где способность антитела или его антигенсвязывающего фрагмента проникать в мозг снижена или отсутствует.
| LIM J.C | |||
| and et.al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| MOROZOV Y | |||
| M | |||
| and et.al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

