Область техники
Настоящее изобретение относится к области биотехнологии, а именно к выделенному альтернативному внутриклеточному сигнальному домену химерного антигенного рецептора (CAR) и включающему данный сигнальный домен химерному антигенному рецептору (CAR). Изобретение также относится к нуклеиновой кислоте, кодирующей альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора, и нуклеиновой кислоте, кодирующей химерный антигенный рецептор с вышеуказанным сигнальным доменом, вектору экспрессии, вектору доставки, а также генетически модифицированной клетке, которая включает вышеуказанный химерный антигенный рецептор, и способу получения данной клетки.
Уровень техники
Химерные антигенные рецепторы (CAR) представляют собой рекомбинантные белковые молекулы, которые нацелены на клеточные поверхностные антигены (Sadelain M, Brentjens R, Rivière I. The promise and potential pitfalls of chimeric antigen receptors. Curr Opin Immunol [Internet]. 2009;21:215–23. Available from: http://linkinghub.elsevier.com/retrieve/pii/S0952791509000211).
CAR в общем случае состоит из нескольких доменов, выполняющих специфические функции. Внеклеточный домен CAR чаще всего представлен одноцепочечным вариабельным фрагментом антитела, специфичным к опухолевому антигену (single-chain variable fragment, scFv) и обеспечивает независимое от молекул главного комплекса гистосовместимости распознавание мишени. Внутриклеточные домены отвечает за передачу сигнала при связывании с антигеном-мишенью и эффективную активацию экспрессирующих CAR иммунных клеток. Внутриклеточная часть рецептора состоит из CD3ζ-цепи (CD3-дзета цепи) и дополнительных ко-стимуляторных доменов, чаще всего производных природных белков CD28 и 4-1ВВ. Установлено, что наличие ко-стимуляторного домена в составе CAR рецептора существенно повышает активность CAR экспрессирующих иммунных клеток по сравнению с CAR, внутриклеточная часть которых состоит только из CD3 ζ-цепи (CD3-дзета цепи) ( Maher J, Brentjens RJ, Gunset G, Rivière I, Sadelain M. Human T-lymphocyte cytotoxicity and proliferation directed by a single chimeric TCRζ /CD28 receptor. Nat Biotechnol [Internet]. 2002;20:70–5. Available from: http://www.nature.com/articles/nbt0102-70; Imai C, Mihara K, Andreansky M, Nicholson IC, Pui C-H, Geiger TL, et al. Chimeric receptors with 4-1BB signaling capacity provoke potent cytotoxicity against acute lymphoblastic leukemia. Leukemia [Internet]. 2004;18:676–84. Available from: http://www.nature.com/articles/2403302; Kowolik CM, Topp MS, Gonzalez S, Pfeiffer T, Olivares S, Gonzalez N, et al. CD28 costimulation provided through a CD19-specific chimeric antigen receptor enhances in vivo persistence and antitumor efficacy of adoptively transferred T cells. Cancer Res. 2006;66:10995–1004).
Оба варианта дизайна CAR рецептора показали выраженную противоопухолевую активность и высокую эффективность в доклинических и клинических исследованиях (Sadelain M, Brentjens R, Rivi?re I. The Basic Principles of Chimeric Antigen Receptor Design. Cancer Discov [Internet]. 2013; 3:388-98. Available from: http://cancerdiscovery.aacrjournals.org/lookup/doi/10.1158/2159-8290.CD-12-0548). Показано, что существуют существенные различия между CAR рецепторами, содержащими ко-стимуляторный домен CD28 и 4-1BB.
Результаты клинических исследований, полученные при лечении лимфопролиферативных заболеваний при помощи CD19-специфичной CAR-T терапии, показали, что 4-1BB - CD3ζ (CD3-дзета цепи) CAR обеспечивает более длительную персистенцию CAR-T лимфцитов in vivo (более 6 месяцев для большинства пациентов) и его использование позволяет получить менее выраженный синдром выброса цитокинов чем в случае применения CD28 - CD3ζ CAR (CD3-дзета цепи) ( Davila ML, Riviere I, Wang X, Bartido S, Park J, Curran K, et al. Efficacy and Toxicity Management of 19-28z CAR T Cell Therapy in B Cell Acute Lymphoblastic Leukemia. Sci Transl Med [Internet]. 2014;6:224ra25-224ra25. Available from: http://stm.sciencemag.org/cgi/doi/10.1126/scitranslmed.3008226; Maude SL. Chimeric Antigen Receptor T Cells for Sustained Remissions in Leukemia. N Engl J Med [Internet]. 2014;371:1507–17. Available from: http://www.nejm.org/doi/abs/10.1056/NEJMoa1407222%5Cnhttp://www.ncbi.nlm.nih.gov/pubmed/25317870; Zhang T, Cao L, Xie J, Shi N, Zhang Z, Luo Z, et al. Efficiency of CD19 chimeric antigen receptor-modified T cells for treatment of B cell malignancies in phase I clinical trials: a meta-analysis. Oncotarget [Internet]. 2015;6:33961–71. Available from: http://www.oncotarget.com/fulltext/5582).
По совокупности данных CD28 - CD3ζ (CD3-дзета цепи) ) CAR рецептор может обеспечить более раннюю, чем в случае использования 4-1BB - CD3ζ (CD3-дзета цепи) CAR, активацию CAR-T лимфоцитов за счет выброса цитокинов и выраженный цитотоксический эффект (van der Stegen SJC, Hamieh M, Sadelain M. The pharmacology of second-generation chimeric antigen receptors. Nat Rev Drug Discov [Internet]. Nature Publishing Group; 2015;14:499–509. Available from: http://www.nature.com/doifinder/10.1038/nrd4597), что, в свою очередь, может обеспечить повышенную терапевтическую эффективность.
Умеренное снижение уровня активации CAR-T клеток и выброса цитокинов, при сохранении скорости передачи активационного сигнала через ко-стимуляторный домен на основе CD28 без потери цитотоксической активности позволят существенно улучшить терапевтические свойства адоптивной иммунотерапии онкологических заболеваний на основе экспрессирующих CD28 - CD3ζ (CD3-дзета цепи) CAR рецептор иммунных клеток.
Взаимодействие природных Т-клеточных рецепторов (TCR) с компонентами главного комплекса гистосовместимости (MHC) приводит к последующему связыванию CD8 корецептора и активации тирозинкиназы LcK. Она фосфорилирует пептидные последовательности - иммунорецепторные тирозин-основные активационные мотивы (immunoreceptor tyrosine-based activation motifs (ITAMs)) в составе CD3ζ (CD3-дзета цепи). Киназа Zap-70 взаимодействует с фосфорилированными по двум сайтам ITAM в составе CD3ζ (CD3-дзета цепи) с помощью Src-homology 2 (SH2) доменов. Связывание Zap-70 с CD3ζ (CD3-дзета цепи) приводит к изменению конформации киназы и ее автофософрилированию по аминокислотным остаткам Y315 и Y319. Lck опосредвоанное фосфорилирование и транс-автофосфорилирование приводит к активации Zap70. После активации Zap70 перестает взаимодействовать с CD3ζ (CD3-дзета цепи) и ассоциирует с плазматической мембраной, где, в свою очередь, фосфорилирует другие субстратные белки, включая LAT20 [Yamasaki S, Takamatsu M, Iwashima M, The kinase, SH3, and SH2 domains of Lck play critical roles in T-cell activation after ZAP-70 membrane localization, Mol Cell Biol. 1996 Dec; 16(12):7151-60; Visco C, Magistrelli G, Bosotti R, Perego R, Rusconi L, Toma S, Zamai M, Acuto O, Isacchi A, Activation of Zap-70 tyrosine kinase due to a structural rearrangement induced by tyrosine phosphorylation and/or ITAM binding, Biochemistry. 2000 Mar 14;39(10):2784-91].
Таким образом, Zap-70 является важнейшим участником сигнального пути CAR, поэтому регуляция активности этой киназы может представлять один из путей регуляции уровня активации CAR-T, который непосредственно влияет на их жизнеспособность, персистенцию и функциональную активность. Многочисленные исследования позволили выявить основные структурные и функциональные элементы системы взаимодействия ZAP70 и ITAM мотивов [Katz ZB, Novotná L, Blount A, Lillemeier BF, A cycle of Zap70 kinase activation and release from the TCR amplifies and disperses antigenic stimuli, Nat Immunol. 2017 Jan;18(1):86-95; Deindl S, Kadlecek TA, Cao X, Kuriyan J, Weiss A, Stability of an autoinhibitory interface in the structure of the tyrosine kinase ZAP-70 impacts T cell receptor response, Proc Natl Acad Sci U S A. 2009 Dec 8;106(49):20699-704; Szabo M, Czompoly T, Kvell K, Talaber G, Bartis D, Nemeth P, Berki T, Boldizsar F, Fine-tuning of proximal TCR signaling by ZAP-70 tyrosine residues in Jurkat cells, Int Immunol. 2012 Feb;24(2):79-87].
Регуляция активности CAR представляется возможной через создание гибридных пептидных субстратов для Zap-70, отличных от природных ITAM мотивов в составе активационного домена CAR на основе CD3ζ (CD3-дзета цепи). Одним из кандидатов являются фрагменты аминокислотной последовательности интегрина бета 3 (ITGB3) (cytoplasmic tail of integrin beta 3; ITGB3) [Liu XY, Timmons S, Lin YZ, Hawiger J., Identification of a functionally important sequence in the cytoplasmic tail of integrin beta 3 by using cell-permeable peptide analogs, Proc Natl Acad Sci U S A. 1996 Oct 15;93(21):11819-24; Durrant TN, van den Bosch MT, Hers I, Integrin αIIbβ3 outside-in signaling, Blood. 2017 Oct 5;130(14):1607-1619].
Авторами изобретения было неожиданно установлено, что гибридные сигнальные домены цитоплазматической части химерного антигенного рецептора (CAR), включающие вместо одного или двух иммунорецепторных тирозин-основных активационных мотивов (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, в составе внутриклеточного сигнального домена химерного антигенного рецептора (CAR) (например, CD3ζ (CD3-дзета)), пептидные фрагменты аминокислотной последовательности интегрина бета 3 (ITGB3) стимулируют CAR зависимую активацию генетически модифицированных иммунных клеток, экспрессирующих соответствующую генетическую конструкцию.
Краткое описание изобретения
В одном из аспектов настоящее изобретение относится к выделенному альтернативному внутриклеточному сигнальному домену химерного антигенного рецептора (CAR), где один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что один иммунорецепторный тирозин-основный активационный мотив (ITAM), выбранный из ITAM1, ITAM2 и ITAM3, заменен на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 заменен на ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM2 заменен на ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM3 заменен на ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM2 и ITAM3 заменены на два ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 и ITAM3 заменены на два ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 и ITAM2 заменены на два ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 имеет аминокислотную последовательность APAYKQGQNQLYNELNLGRREEYDVLDKR (SEQ ID NO:1).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM2 имеет аминокислотную последовательность PRRKNPQEGLYNELQKDKMAEAYSEIGM (SEQ ID NO:2).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM3 имеет аминокислотную последовательность ERRRGKGHDGLYQGLSTATKDTYDALHMQ (SEQ ID NO:3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT SEQ ID NO:4) или аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT SEQ ID NO:4) с любой 1 или 2 консервативными заменами.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO: 4) с любой 1 консервативной заменой.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO: 4) с любыми 2 консервативными заменами.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO: 4).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO: 5) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 5 и имеет отличия по сравнению с SEQ ID NO: 5 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:6) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 6 и имеет отличия по сравнению с SEQ ID NO: 6 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:7) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO:7 и имеет отличия по сравнению с SEQ ID NO: 7 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:8) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 8 и имеет отличия по сравнению с SEQ ID NO: 8 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:9) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 9 и имеет отличия по сравнению с SEQ ID NO: 9 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:10) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 10 и имеет отличия по сравнению с SEQ ID NO: 10 только в неконсервативных аминокислотных остатках.
В одном из аспектов настоящее изобретение относится к выделенному альтернативному внутриклеточному сигнальному домену химерного антигенного рецептора (CAR), который включает один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и один или два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3), при этом общее количество ITAM и ITGB3 равно трем.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и один фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM2, ITAM3 и ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный который включает ITAM1, ITAM3 и ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM1, ITAM2 и ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает один иммунорецепторный тирозин-основный активационный мотив (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM1 и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM2 и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM3 и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 имеет аминокислотную последовательность APAYKQGQNQLYNELNLGRREEYDVLDKR (SEQ ID NO:1).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM2 имеет аминокислотную последовательность PRRKNPQEGLYNELQKDKMAEAYSEIGM (SEQ ID NO:2).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM3 имеет аминокислотную последовательность ERRRGKGHDGLYQGLSTATKDTYDALHMQ (SEQ ID NO:3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO:4) или аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO:4) с любой 1 или 2 консервативными заменами.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO:4) с любой 1 консервативной заменой.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO:4) с любыми 2 консервативными заменами.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность DTANNPLYKEATSTFTNITYRGT (SEQ ID NO:4).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:5) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 5 и имеет отличия по сравнению с SEQ ID NO: 5 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:6) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 6 и имеет отличия по сравнению с SEQ ID NO: 6 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:7) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 7 и имеет отличия по сравнению с SEQ ID NO: 7 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность
RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:8) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 8 и имеет отличия по сравнению с SEQ ID NO: 8 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность
RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:9) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 9 и имеет отличия по сравнению с SEQ ID NO: 9 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:10) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 10 и имеет отличия по сравнению с SEQ ID NO: 10 только в неконсервативных аминокислотных остатках.
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует вышеуказанный альтернативный внутриклеточный сигнальный домен.
В одном из аспектов настоящее изобретение относится к выделенному химерному антигенному рецептору (CAR), содержащему вышеуказанный альтернативный внутриклеточный сигнальный домен.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что содержит:
а) антигенсвязывающий домен, который специфичен к опухолевому антигену;
б) трансмембранный домен;
в) ко-стимуляторный домен;
г) вышеуказанный альтернативный внутриклеточный сигнальный домен.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен, который специфичен к опухолевому антигену, представляет собой Fab или scFv.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен, который специфичен к опухолевому антигену, представляет собой scFv.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен специфичен к опухолевому антигену, который выбран из группы: CD19, CD20, CD123 или BCMA.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен специфичен к опухолевому антигену CD19.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен, который специфичен к опухолевому антигену CD19, имеет аминокислотную последовательность DIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSS (SEQ ID NO: 11).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что трансмембранный домен представляет собой трансмембранный домен белка, выбранного из группы, состоящей из альфа, бета или дзета-цепи T-клеточного рецептора, CD28, CD3-эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что где трансмембранный домен представляет собой CD8.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что трансмембранный домен CD8 имеет аминокислотную последовательность представленную IYIWAPLAGTCGVLLLSLVITLYC (SEQ ID NO: 12).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен представляет собой функциональный сигнальный домен белка, выбранного из группы, состоящей из OX40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278) и 4-1BB (CD137).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен представляет собой CD28 или 4- 1BB.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен представляет собой CD28.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен CD28 имеет аминокислотную последовательность AAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:13).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен представляет собой 4- 1BB.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен 4- 1BB имеет аминокислотную последовательность представленную KRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL (SEQ ID NO: 14).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что дополнительно содержит лидерную последовательность.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) имеет аминокислотную последовательность представленную аминокислотной последовательностью, которую выбирают из группы: SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности, которую выбирают из группы: SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, и имеет отличия по сравнению с аминокислотной последовательностью, которую выбирают из группы: SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, только в неконсервативных аминокислотных остатках.
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITGB3- ITAM2- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:15).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITAM1- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:16).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITAM1- ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:17).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITGB3- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:18).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITAM1- ITGB3- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:19).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITGB3 - ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:20).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITGB3- ITAM2- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:21).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITAM1- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:22).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITAM1- ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:23).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITGB3- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:24).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITAM1- ITGB3- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:25).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITGB3 - ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:26).
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует вышеуказанный выделенный химерный антигенный рецептор (CAR).
В одном из аспектов настоящее изобретение относится к экспрессионному вектору, содержащему вышеуказанную нуклеиновую кислоту.
В одном из аспектов настоящее изобретение относится к вектору доставки, содержащему вышеуказанную нуклеиновую кислоту.
В некоторых вариантах вектор выбирают из группы, состоящей из ДНК, РНК, плазмиды, лентивирусного вектора, аденоассоциированного вирусного вектора, аденовирусного вектора или ретровирусного вектора.
В некоторых вариантах вектор дополнительно содержит промотор.
В некоторых вариантах вектор представляет собой in vitro транскрибируемый вектор.
В некоторых вариантах последовательность нуклеиновой кислоты дополнительно содержит поли(A)-последовательность.
В некоторых вариантах последовательность нуклеиновой кислоты дополнительно содержит 3'UTR.
В одном из аспектов настоящее изобретение относится к способу получения генетически модифицированной клетки, которая содержит вышеуказанный химерный антигенный рецептор (CAR), включающий трансформирование клетки вышеуказанным вектором.
В одном из аспектов настоящее изобретение относится к генетически модифицированной клетке, которая содержит вышеуказанный химерный антигенный рецептор (CAR), которую получают вышеуказанным способом.
В некоторых вариантах клетка представляет собой Т-лимфоцит.
В некоторых вариантах клетка представляет собой CD8+ T-клетку.
В некоторых вариантах клетка представляет собой NK-клетку.
В некоторых вариантах клетку используют в качестве лекарственного средства.
В некоторых вариантах клетку используют в качестве лекарственного средства при опухолевых заболеваниях.
В некоторых вариантах клетку используют в качестве лекарственного средства при заболеваниях, где опухоль представляет собой рак.
Краткое описание чертежей
Фиг. 1. Карта вектора pCDH-19CAR-T-ITGB3-ITGB3-eGFP.
Фиг. 2. Схематическое изображение экспрессионной кассеты 19CAR-T-ITGB3-ITGB3-eGFP.
Фиг. 3. Карта вектора pCDH-19CAR-T-ITGB3-ITGB3-ITGB3-eGFP.
Фиг. 4. Схематическое изображение экспрессионной кассеты 19CAR-T-ITGB3-ITGB3-ITGB3-eGFP.
Фиг. 5. Карта вектора pCDH-19CAR-T-ITAM1-ITGB3-ITGB3-eGFP.
Фиг. 6. Схематическое изображение экспрессионной кассеты 19CAR-T-ITAM1-ITGB3-ITGB3-eGFP.
Фиг. 7. Карта вектора pCDH-19CAR-T-ITAM1-ITAM2-ITGB3-eGFP.
Фиг. 8. Схематическое изображение экспрессионной кассеты 19CAR-T-ITAM1- ITAM2-ITGB3-eGFP.
Фиг. 9. Карта вектора pCDH-CAR-T-GFP.
Фиг. 10. Схематическое изображение экспрессионной кассеты pCDH-CAR-T-GFP.
Фиг. 11. Карта вектора pCDH-19CAR-T-ITAM1-ITGB3-ITAM2-eGFP.
Фиг. 12. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T-ITAM1-ITGB3-ITAM2-eGFP.
Фиг. 13. Карта вектора pCDH-19CAR-T -ITGB3-ITAM2-ITGB3-eGFP.
Фиг. 14. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T -ITGB3-ITAM2-ITGB3-eGFP.
Фиг. 15. Карта вектора pCDH-19CAR-T_CD28-ITGB3-ITGB3-eGFP.
Фиг. 16. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T_CD28-ITGB3-ITGB3-eGFP.
Фиг. 17. Карта вектора pCDH-19CAR-T-CD28_ITGB3-ITGB3-ITGB3-eGFP.
Фиг. 18. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T-CD28_ITGB3-ITGB3-ITGB3-eGFP.
Фиг. 19. Карта вектора pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITGB3-eGFP.
Фиг. 20. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITGB3-eGFP.
Фиг. 21. Карта вектора pCDH-19CAR-T-CD28_ITAM1-ITAM2-ITGB3-eGFP.
Фиг. 22. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T-CD28_ITAM1-ITAM2-ITGB3-eGFP.
Фиг. 23. Карта вектора pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITAM2-eGFP.
Фиг. 24. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITAM2-eGFP.
Фиг. 25. Карта вектора pCDH-19CAR-T-CD28_ITGB3-ITAM2-ITGB3-eGFP.
Фиг. 26. Схематическое изображение экспрессионной кассеты pCDH-19CAR-T-CD28_ ITGB3-ITAM2-ITGB3-eGFP.
Фиг. 27. Уровень экспрессии GFP в жизнеспособных Т-лимфоцитах после трансдукции лентивирусными препаратами, 72 часа, %.
Фиг. 28. Оценка цитотоксической активности CAR-T-лимфоцитов экспрессирующих контрольный CAR-ITAMx3 и экспериментальный CAR-ITAMx2-ITGB3x1 при со-культивировании с клетками-мишенями.
A. - Динамика изменения процентного соотношения Т-лимфоцитов / CAR-T-лимфоцитов и клеток Raji при со-культивировании 0 часов и 72 часа;
В. - Концентрация IFN-γ в культуральной среде при со-культивировании Т-лимфоцитов / CAR-Т-лимфоцитов и клеток-мишеней, 72 часа.;
С. - Концентрация IL2 в культуральной среде при со-культивировании Т-лимфоцитов / CAR-Т-лимфоцитов и клеток-мишеней, 72 часа.
Фиг. 29. Сравнение функциональной активности генетических конструкций контрольного CAR-ITAMx3 и экспериментального CAR-ITAMx2-ITGB3x1 по способности активировать NFAT сигнальный каскад в репортерной клеточной линии JurkatΔTCRab NFAT-GFP, 72 часа.
Описание изобретения
Определения и общие методы
Если иное не определено в настоящем документе, научные и технические термины, используемые в связи с настоящим изобретением, будут иметь значения, которые обычно понятны специалистам в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментативные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
Под «млекопитающим» понимается любое животное, классифицируемое как млекопитающее, в том числе приматы, люди, грызуны, собачьи, кошачьи, крупный рогатый скот, мелкий рогатый скот, лошади, свиньи и т.д.
«Выделенный» означает измененный или удаленный из природного состояния. Например, нуклеиновая кислота или пептид, в природе присутствующие в животном, не являются «выделенными», но те же нуклеиновая кислота или пептид, частично или полностью отделенные от материалов, сопутствующих им в их природном состоянии, являются «выделенными». Выделенная нуклеиновая кислота или белок могут существовать, по существу, в очищенной форме или могут существовать в неприродном окружении, таком как, например, генетически модифицированной клетке.
Термин «идентичность» или «гомологичность» следует толковать как означающее процентное содержание остатков аминокислот в кандидатной последовательности, которые идентичны остаткам соответствующей последовательности, с которой ее сравнивают, после сравнения последовательностей и введения «брешей», если необходимо достичь максимального процента идентичности для полной последовательности и не учитывая любые консервативные замещения как часть идентичности последовательности. Ни N- или С-концевой удлиняющей, ни инсерционные сегменты не следует толковать как уменьшающие идентичность или гомологичность. Методы и компьютерные программы для сравнения хорошо известны. Идентичность последовательности можно определить, используя программное обеспечение для анализа последовательности (например, Sequence Analysis Software Package, Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Ave., Madison, WI 53705). Данное программное обеспечение подходит для подобных последовательностей путем определения степени гомологичности для разнообразных замещений, делеций (элиминирований) и других модификаций.
Как применяют в настоящем описании, термин «антиген» или «Ag» определяют как молекулу, провоцирующую иммунный ответ. Этот иммунный ответ может включать продукцию антител или активацию специфичных иммунологически компетентных Т-клеток, или и то, и другое. Специалистам в этой области будет понятно, что любая макромолекула, включая практически все белки или пептиды, может служить в качестве антигена. Кроме того, антигены можно получать из рекомбинантной или геномной ДНК. Специалистам в этой области будет понятно, что любая ДНК, содержащая нуклеотидные последовательности или неполную нуклеотидную последовательность, кодирующую белок, вызывающий иммунный ответ, таким образом, кодирует «антиген» в том виде, в котором этот термин используют в настоящем описании. Кроме того, специалисту в этой области будет понятно, что антиген необязательно кодирует исключительно полноразмерная нуклеотидная последовательность гена. Вполне очевидно, что настоящее изобретение, в качестве неограничивающих примеров, относится к применению неполных нуклеотидных последовательностей нескольких генов, и что эти нуклеотидные последовательности располагаются в различных комбинациях для стимуляции желаемого иммунного ответа. Кроме того, специалистам в этой области будет понятно, что антиген совершенно необязательно кодирует «ген». Вполне очевидно, что антиген можно синтезировать или его можно получать из биологического образца. Такой биологический образец может включать, в качестве неограничивающих примеров, образец ткани, образец опухоли, клетку или биологическую жидкость.
«Заболевание» является состоянием здоровья животного, где животное не может поддерживать гомеостаз, и где, если заболевание не облегчают, то здоровье животного продолжает ухудшаться.
Наоборот, «нарушение» у животного является состоянием здоровья, при котором животное способно поддерживать гомеостаз, но состояние здоровья животного является менее благоприятным, чем оно бы было в отсутствие нарушения. Если его не лечить, нарушение не обязательно приводит к ухудшению состояния здоровья животного.
Как применяют в настоящем описании, термин «злокачественное новообразование» определяют как заболевание, отличающееся быстрым и неконтролируемым ростом аномальных клеток. Злокачественные клетки могут распространяться местно или через кровоток и лимфатическую систему в другие части организма. Примеры различных злокачественных новообразований включают, в качестве неограничивающих примеров, рак молочной железы, рак предстательной железы, рак яичников, рак шейки матки, рак кожи, рак поджелудочной железы, колоректальный рак, рак почки, рак печени, злокачественные новообразования головного мозга, лимфому, лейкоз, рак легких, миелому, множестввенную миелому и т.п.
Злокачественные новообразования, которые можно лечить, включают опухоли, не васкуляризованные или, по существу, не васкуляризованные, а также васкуляризованные опухоли.
Злокачественные новообразования могут включать несолидные опухоли (такие как гематологические опухоли, например, лейкозы и лимфомы) или могут включать солидные опухоли. Типы злокачественных новообразований, подлежащих лечению с помощью CAR по изобретению, включают, в качестве неограничивающих примеров, карциному, бластому и саркому, и конкретные лейкозы или лимфоидные злокачественные новообразования, доброкачественные и злокачественные опухоли и злокачественные новообразования, например, саркомы, карциномы и меланомы. Также включены опухоли/злокачественные новообразования взрослых и опухоли/злокачественные новообразования детей.
Гематологическими злокачественными новообразованиями являются злокачественные новообразования крови или костного мозга. Примеры гематологических (или гематогенных) злокачественных новообразований включают лейкозы, включая острые лейкозы (такие как острый лимфоцитарный лейкоз, острый миелоцитарный лейкоз, острый миелогенный лейкоз и миелобластный, промиелоцитарный, миеломоноцитарный, моноцитарный и эритролейкоз), хронические лейкозы (такие как хронический миелоцитарный (гранулоцитарный) лейкоз, хронический миелогенный лейкоз и хронический лимфоцитарный лейкоз), истинную полицитемию, лимфому, болезнь Ходжкина, неходжкинскую лимфому (медленно растущую и высокозлокачественную формы), множественную миелому, макроглобулинемия Вальденстрема, болезнь тяжелых цепей, миелодиспластический синдром, волосатоклеточный лейкоз и миелодисплазию.
Солидные опухоли являются аномальными опухолями ткани, как правило, не содержащими кисты или области жидкости. Солидные опухоли могут являться доброкачественными или злокачественными. Различные типы солидных опухолей называют по типу образующих их клеток (например, саркомы, карциномы и лимфомы). Примеры солидных опухолей, таких как саркомы и карциномы, включают фибросаркому, миксосаркому, липосаркому, хондросаркому, остеосаркому и другие саркомы, синовиому, мезотелиому, опухоль Юинга, лейомиосаркому, рабдомиосаркому, карциному толстого кишечника, лимфолейкоз, рак поджелудочной железы, рак молочной железы, рак легких, рак яичников, рак предстательной железы, печеночноклеточную карциному, плоскоклеточную карциному, базальноклеточную карциному, аденокарциному, карциному потовых желез, медуллярный рак щитовидной железы, папиллярный рак щитовидной железы, феохромоцитому, карциному сальных желез, папиллярную карциному, папиллярные аденокарциномы, медуллярную карциному, бронхогенную карциному, почечноклеточную карциному, гепатому, карциному желчных протоков, хориокарциному, опухоль Вильмса, рак шейки матки, опухоль яичка, семиному, карциному мочевого пузыря, меланому и опухоли ЦНС (такие как глиома (такая как глиома ствола головного мозга и смешанная глиома), глиобластома (также известная как мультиформная глиобластома) астроцитома, лимфома ЦНС, герминома, медуллобластома, шваннома, краниофарингиома, эпендимома, пинеалома, гемангиобластома, невринома слухового нерва, олигодендроглиома, менингиома, нейробластома, ретинобластома и метастазы в головном мозге).
Как применяют в настоящем описании, «эндогенный» относится к любому материалу из организма, клетки, ткани или системы или материалу, продуцируемому в организме, клетке, ткани или системе.
Как применяют в настоящем описании, термин «экзогенный» относится к любому материалу, вводимому в организм, клетку, ткань или систему, или продуцируемому вне организма, клетки, ткани или системы.
Как применяют в настоящем описании, термин «экспрессия» определяют как транскрипцию и/или трансляцию конкретной нуклеотидной последовательности, запускаемую ее промотором.
Термин «гиперэкспрессированный» опухолевый антиген или «гиперэкспрессия» опухолевого антигена предназначен для обозначения аномального уровня экспрессии опухолевого антигена в клетке из очага заболевания, такого как солидная опухоль в конкретной ткани или органе пациента, относительно уровня экспрессии в нормальной клетке из этой ткани или органа. Пациентов с солидными опухолями или гематологическими злокачественными новообразованиями, отличающимися гиперэкспрессией опухолевого антигена, можно определять стандартными анализами, известными в этой области.
Термины «пациент», «индивидуум» и т.п. используют в настоящем описании взаимозаменяемо, и они относятся к любому животному или его клеткам in vitro или in situ, поддающимся воздействию способами, представленными в настоящем описании. В конкретных неограничивающих вариантах осуществления пациент или индивидуум является человеком.
Как применяют в настоящем описании, термин «полинуклеотид» определяют как цепь нуклеотидов. Кроме того, нуклеиновые кислоты являются полимерами нуклеотидов. Таким образом, как применяют в настоящем описании, термины нуклеиновые кислоты и полинуклеотиды являются взаимозаменяемыми. Специалист в этой области имеет общие знания о том, что нуклеиновые кислоты являются полинуклеотидами, которые можно гидролизовать до мономерных «нуклеотидов». Мономерные нуклеотиды можно гидролизовать в нуклеозиды. Как применяют в настоящем описании, полинуклеотиды включают, в качестве неограничивающих примеров, все последовательности нуклеиновой кислоты, получаемые любыми способами, доступными в этой области, включая, в качестве неограничивающих примеров, рекомбинантные способы, т.е. клонирование последовательностей нуклеиновой кислоты из рекомбинантной библиотеки или генома клетки, использование обычной технологии клонирования и ПЦР и т.п., и способами синтеза.
Как применяют в настоящем описании, термины «пептид», «полипептид» и «белок» используют взаимозаменяемо, и они относятся к соединению, состоящему из аминокислотных остатков, ковалентно связанных пептидными связями. Белок или пептид должен содержать по меньшей мере две аминокислоты, и не существует ограничений по максимальному количеству аминокислот, которые может содержать последовательность белка или пептида. Полипептиды включают любой пептид или белок, содержащий две или более аминокислот, соединенных друг с другом пептидными связями. Как применяют в настоящем описании, термин относится и к коротким цепям, также общепринято обозначаемым в этой области, например, как пептиды, олигопептиды и олигомеры, и к более длинным цепям, как правило, обозначаемым в этой области как белки, множество типов которых существует. «Полипептиды» включают, помимо прочего, например, биологически активные фрагменты, по существу, гомологичные полипептиды, олигопептиды, гомодимеры, гетеродимеры, варианты полипептидов, модифицированные полипептиды, производные, аналоги, слитные белки. Полипептиды включают природные пептиды, рекомбинантные пептиды, синтетические пептиды или их комбинацию.
Подробное описание изобретения
Выделенный альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR)
Внутриклеточный сигнальный домен CAR или, иначе, цитоплазматический домен по изобретению отвечает за активацию по меньшей мере одной нормальной эффекторной функции клетки иммунной системы, в которую помещают CAR. Термин «эффекторная функция» относится к специализированной функции клетки. Эффекторная функция Т-клетки, например, может являться цитолитической активностью или хелперной активностью, включая секрецию цитокинов. Таким образом, термин «внутриклеточный сигнальный домен» относится к части белка, передающей сигнал эффекторной функции и заставляющей клетку осуществлять специализированную функцию. Хотя, как правило, можно использовать целый внутриклеточный сигнальный домен, во многих случаях использование целой цепи не является необходимым. В случае если используют укороченную часть внутриклеточного сигнального домена, такую укороченную часть можно использовать вместо интактной цепи при условии, что она передает сигнал эффекторной функции. Таким образом, термин «внутриклеточный сигнальный домен» предназначен для включения любой укороченной части внутриклеточного сигнального домена, достаточной для передачи сигнала эффекторной функции.
Предпочтительные примеры внутриклеточных сигнальных доменов для применения в CAR по изобретению включают цитоплазматические последовательности Т-клеточного рецептора (TCR) и ко-рецепторов, действующих совместно для инициации передачи сигнала после связывания антигенного рецептора, а также любое производное или вариант этих последовательностей и любую синтетическую последовательность, обладающую той же функциональной способностью.
Известно, что сигналов, генерируемых только через TCR, недостаточно для полной активации Т-клетки, и что также необходим вторичный или костимуляторный сигнал. Таким образом, можно сказать, что активацию Т-клетки опосредуют два различных класса цитоплазматических последовательностей, опосредующих передачу сигнала: инициирующие антигензависимую первичную активацию через TCR (первичные цитоплазматические последовательности, опосредующие передачу сигнала) и действующие антиген-независимым образом для обеспечения вторичного или костимуляторного сигнала (вторичные цитоплазматические последовательности, опосредующие передачу сигнала).
Первичные цитоплазматические последовательности, опосредующие передачу сигнала, регулируют первичную активацию комплекса TCR стимулирующим или ингибиторным образом. Первичные цитоплазматические последовательности, опосредующие передачу сигнала, действующие стимулирующим образом, могут содержать сигнальные мотивы, известные как иммунорецепторные тирозиновые активационные мотивы (иммунорецепторный тирозин-основный активационный мотив) или ITAM.
Примеры ITAM, включающие первичные цитоплазматические последовательности, опосредующие передачу сигнала, включают полученные из TCR-дзета, FcR-гамма, FcR-бета, CD3-гамма, CD3-дельта, CD3-эпсилон, CD8, CD22, CD79a, CD79b и CD66d. Особенно предпочтительно, если цитоплазматическая молекула, опосредующая передачу сигнала, в CAR содержит цитоплазматическую последовательность, опосредующую передачу сигнала, полученную из CD3-дзета.
Цитоплазматический домен CAR можно конструировать содержащим сигнальный домен CD3-дзета сам по себе или в комбинации с любыми другими желаемыми цитоплазматическими доменами, применимыми в условиях CAR.
Сигнальный домен CD3-дзета имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR.
Вышеуказанный сигнальный домен содержит 3 ITAM (иммунорецепторные тирозиновые активационные мотивы), а именно ITAM1, ITAM2 и ITAM3.
ITAM1 имеет аминокислотную последовательность, представленную APAYKQGQNQLYNELNLGRREEYDVLDKR (SEQ ID NO:1).
ITAM2 имеет аминокислотную последовательность, представленную PRRKNPQEGLYNELQKDKMAEAYSEIGM (SEQ ID NO:2).
ITAM3 имеет аминокислотную последовательность, представленную ERRRGKGHDGLYQGLSTATKDTYDALHMQ (SEQ ID NO:3).
В одном из аспектов настоящее изобретение относится к выделенному альтернативному внутриклеточному сигнальному домену химерного антигенного рецептора (CAR), где один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что один иммунорецепторный тирозин-основный активационный мотив (ITAM), выбранный из ITAM1, ITAM2 и ITAM3, заменен на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 заменен на ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM2 заменен на ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM3 заменен на ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM2 и ITAM3 заменены на два ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 и ITAM3 заменены на два ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITAM1 и ITAM2 заменены на два ITGB3.
В одном из аспектов настоящее изобретение относится к выделенному альтернативному внутриклеточному сигнальному домену химерного антигенного рецептора (CAR), который включает один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и один или два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3), при этом общее количество ITAM и ITGB3 равно трем.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и один фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM2, ITAM3 и ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный который включает ITAM1, ITAM3 и ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM1, ITAM2 и ITGB3.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает один иммунорецепторный тирозин-основный активационный мотив (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM1 и и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM2 и и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что включает ITAM3 и и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность представленную DTANNPLYKEATSTFTNITYRGT SEQ ID NO:4) или аминокислотную последовательность представленную DTANNPLYKEATSTFTNITYRGT SEQ ID NO:4) с любой 1 или 2 консервативными заменами.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность представленную DTANNPLYKEATSTFTNITYRGT (SEQ ID NO: 4) с любой 1 консервативной заменой.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность представленную DTANNPLYKEATSTFTNITYRGT (SEQ ID NO: 4) с любыми 2 консервативными заменами.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) характеризуется тем, что ITGB3 имеет аминокислотную последовательность представленную DTANNPLYKEATSTFTNITYRGT (SEQ ID NO: 4).
Под аминокислотными консервативными заменами подразумевается замена одной аминокислоты на другую сходную по структуре. Такая замена не приводит к изменению свойств ITGB3.
Консервативные замены показаны в таблице А под заголовком «предпочтительные замены». Если такие замены приводят к изменению биологической активности, то могут быть введены дополнительные существенные изменения, названные «примерами заменам» в таблице А.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO: 5) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 5 и имеет отличия по сравнению с SEQ ID NO: 5 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:6) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 6 и имеет отличия по сравнению с SEQ ID NO: 6 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:7) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 7 и имеет отличия по сравнению с SEQ ID NO: 7 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:8) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 8 и имеет отличия по сравнению с SEQ ID NO: 8 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPR (SEQ ID NO:9) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 9 и имеет отличия по сравнению с SEQ ID NO: 9 только в неконсервативных аминокислотных остатках.
В некоторых вариантах альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) имеет аминокислотную последовательность RVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:10) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 10 и имеет отличия по сравнению с SEQ ID NO: 10 только в неконсервативных аминокислотных остатках.
Химерный антигенный рецептор (CAR)
В одном из аспектов настоящее изобретение относится к выделенному химерному антигенному рецептору (CAR), содержащему вышеуказанный альтернативный внутриклеточный сигнальный домен.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что содержит:
а) антигенсвязывающий домен, который специфичен к опухолевому антигену;
б) трансмембранный домен;
в) ко-стимуляторный домен;
г) вышеуказанный альтернативный внутриклеточный сигнальный домен.
Настоящее изобретение относится к химерному антигенному рецептору (CAR), содержащему внеклеточный и внутриклеточный домен. Внеклеточный домен содержит мишенеспецифичный связывающий элемент, иначе обозначаемый как антигенсвязывающий домен. Внутриклеточный домен или, иначе, цитоплазматический домен содержит костимуляторную сигнальную область и часть дзета-цепи. Костимуляторная сигнальная область относится к части CAR, содержащей внутриклеточный домен костимуляторной молекулы. Костимуляторные молекулы являются молекулами клеточной поверхности, иными, чем антигенные рецепторы или их лиганды, необходимые для эффективного ответа лимфоцитов на антиген.
Между внеклеточным доменом и трансмембранным доменом CAR или между цитоплазматическим доменом и трансмембранным доменом CAR можно встраивать спейсерный домен. Как применяют в настоящем описании, термин «спейсерный домен», как правило, означает любой олиго- или полипептид, функционирующий, связывая трансмембранный домен с внеклеточным доменом или цитоплазматическим доменом в полипептидной цепи. Спейсерный домен может содержать до 300 аминокислот, предпочтительно от 10 до 100 аминокислот и наиболее предпочтительно от 25 до 50 аминокислот.
В одном из вариантов осуществления CAR по изобретению содержит внеклеточный домен, содержащий антигенсвязывающий домен; трансмембранный домен и цитоплазматический домен.
В одном из вариантов осуществления используют трансмембранный домен, который в природе ассоциирован с одним из доменов в CAR.
В другом варианте осуществления трансмембранный домен можно выбирать или модифицировать посредством замены аминокислоты для избежания связывания таких доменов с трансмембранными доменами одинаковых или различных белков на поверхности мембраны для минимизации взаимодействий с другими компонентами рецепторного комплекса.
Антигенсвязывающий домен
Термин «антигенсвязывающий домен» антитела или «антигенсвязывающая часть» антитела или «антигенсвязывающий фрагмент» (или просто «часть антитела» или «фрагмент антитела»), как использовано в данном описании, относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может выполняться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, включенных в термин «антигенсвязывающая часть» антитела включают:
(i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1;
(ii) F(ab’)2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области;
(iii) Fd- фрагмент, состоящий из доменов VH и CH1;
(iv) Fv-фрагмент, состоящий из доменов VL и VH в едином плече антитела,
(v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH/VHH; и
(vi) выделенная определяющая комплементарность область (CDR).
Кроме того, две области Fv-фрагмента, VL и VH, кодируются разными генами, они могут быть соединены при помощи рекомбинантных способов с использованием синтетического линкера, который дает возможность получать их в виде единой белковой цепи, в которой области VL и VH спариваются с образованием одновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие одноцепочечные молекулы также включены в термин «антигенсвязывающая часть» антитела. Такие фрагменты антител получают с использованием общепринятых способов, известных специалистам в данной области, и эти фрагменты подвергают скринингу таким же образом, как и интактные антитела.
Термин «вариабельный» относится к тому факту, что определенные сегменты вариабельных доменов широко отличаются в последовательности среди антител. Домен V опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность неравномерно распределяется на участке вариабельных доменов из 110 аминокислот. Напротив, V области состоят из инвариантных фрагментов, называемых каркасными областями (FR) из 15-30 аминокислот, разделенных более короткими участками чрезвычайной вариабельности, называемых «гипервариабельными областями» или CDR. Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR, в основном принимающих конфигурацию бета-листов, связанных тремя гипервариабельными областями, которые образуют петли, связывающие, и в некоторых случаях являющиеся частью бета-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются вместе в тесной близости с помощью FR и с гипервариабельными областями другой цепи вносят вклад в образование антигенсвязывающего сайта антител.
Термин «гипервариабельная область» по данному описанию относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Обычно гипервариабельная область содержит аминокислотные остатки из «области, определяющей комплементарность» или «CDR», и/или такие остатки из «гипервариабельной петли».
В некоторых случаях может также быть предпочтительным изменение одного или более остатков аминокислот CDR-участков с целью повышения аффинности связывания с целевым эпитопом. Это известно, как «созревание аффиности» и в некоторых случаях может выполняться в связи с гуманизацией, например, в ситуациях, когда гуманизация антитела приводит к снижению специфичности или аффинности связывания, и не представляется возможным в достаточной степени улучшить специфичность или аффинность связывания с помощью только обратных мутаций. Различные методы созревания аффинности известны в данной области техники, например, способ in vitro сканирующего насыщающего мутагенеза, описанный Burks et al., Proc Natl Acad Sci USA, 94:412-417 (1997), и способ пошагового in vitro созревания аффинности, предложенный Wu et al., Proc Natl Acad Sci USA 95:6037 6042 (1998).
Антигенсвязывающий домен антитела по данному изобретению, «которое связывает» целевой антиген, представляет собой антигенсвязывающий домен антитела, которое связывает антиген с достаточной аффинностью так, что антигенсвязывающий домен антитела можно применять в качестве диагностического и/или терапевтического агента при таргетировании на белок или клетку, или ткань, экспрессирующую антиген, и в незначительной степени перекрестно реагирует с другими белками. По данным аналитических методов: сортинга флуоресцентно-активированных клеток (FACS), радиоиммунопреципитации (RIA) или ИФА (ELISA), степень связывания антигенсвязывающего домена антитела с белком, не являющимся «мишенью» (с «нецелевым белком»), составляет менее 10% от связывания антигенсвязывающего домена антитела с конкретным белком-мишенью. По отношению к связыванию антигенсвязывающего домена антитела с молекулой-мишенью термин «специфическое связывание» или выражения «специфически связывается с» или «специфический к» конкретному полипептиду или эпитопу на конкретном полипептиде-мишени означает связывание, которое заметно (измеримо) отличается от неспецифического взаимодействия.
В одном из вариантов осуществления CAR по изобретению содержит мишенеспецифичный связывающий элемент, иначе обозначаемый как антигенсвязывающий домен. Выбор домена зависит от типа и количества лигандов, определяющих поверхность клетки-мишени. Например, антигенсвязывающий домен можно выбирать для распознавания лиганда, действующего как поверхностный клеточный маркер на клетках-мишенях, ассоциированных с конкретным состоянием заболевания. Таким образом, примеры маркеров клеточной поверхности, которые могут действовать как лиганды для домена антигенсвязывающего домена в CAR по изобретению, включают маркеры, ассоциированные с злокачественными клетками.
В одном из вариантов осуществления CAR по изобретению можно конструировать для воздействия на интересующий опухолевый антиген посредством конструирования желаемого антигенсвязывающего домена, специфически связывающегося с антигеном на опухолевой клетке. В контексте настоящего изобретения «опухолевый антиген», или «антиген гиперпролиферативного нарушения» или «антиген, ассоциированный с гиперпролиферативным нарушением» относится к антигенам, типичным для конкретных гиперпролиферативных нарушений, таких как злокачественное новообразование. Антигены, представленные в настоящем описании, включают исключительно в качестве примера. Список не предназначен для ограничения, и дополнительные примеры будут очевидны специалистам в этой области.
Опухолевые антигены являются белками, продуцируемыми опухолевыми клетками, вызывающими иммунный ответ, конкретно, опосредуемые Т-клетками иммунные ответы. Выбор антигенсвязывающего домена по изобретению будет зависеть от конкретного типа злокачественного новообразования, подлежащего лечению. Опухолевые антигены хорошо известны в этой области и включают, например, глиома-ассоциированный антиген, карциноэмбриональный антиген (СЕА), р-хорионический гонадотропин человека, альфа-фетопротеин (AFP), лектин-реактивный AFP, тиреоглобулин, RAGE-1, MN-CA IX, теломеразная обратная транскриптаза человека, RU1, RU2 (AS), кишечная карбоксилэстераза, mut hsp70-2, M-CSF, простаза, простатический специфический антиген (PSA), PAP, NY-ESO-1, LAGE-1a, p53, простеин, PSMA, Her2/neu, сурвивин и теломераза, антиген карциномы простаты-1 (РСТА-1), MAGE, ELF2M, эластаза нейтрофилов, эфрин В2, CD22, инсулиноподобный фактор роста (IGF)-I, IGF-II, рецептор IGF-I и мезотелин.
В одном из вариантов осуществления опухолевый антиген содержит один или несколько эпитопов опухолевых антигенов, ассоциированных со злокачественной опухолью. Злокачественные опухоли экспрессирует ряд белков, которые могут служить в качестве антигенов-мишеней для иммунного ответа. Эти молекулы включают, в качестве неограничивающих примеров, тканеспецифические антигены, такие как MART-1, тирозиназа и GP 100 при меланоме и простатическая кислая фосфатаза (РАР) и простатический специфический антиген (PSA) при раке предстательной железы. Другие молекулы-мишени принадлежат к группе участвующих в трансформации молекул, таких как онкоген HER-2/Neu/ErbB-2. Еще одной группой антигенов-мишеней являются онкофетальные антигены, такие как карциноэмбриональный антиген (СЕА). При В-клеточной лимфоме иммуноглобулин опухолеспецифичного идиотипа образует, в сущности, опухолеспецифичный иммуноглобулиновый антиген, уникальный для отдельной опухоли. Антигены В-клеточной дифференцировки, такие как CD19, CD20 и CD37 являются другими кандидатами для антигенов-мишеней при В-клеточной лимфоме. Некоторые из этих антигенов (СЕА, HER-2, CD19, CD20, идиотип) с ограниченным успехом используют в качестве мишеней для пассивной иммунотерапии с использованием моноклональных антител.
Тип опухолевого антигена, описываемого в изобретении, также может являться опухолеспецифичным антигеном (TSA) или ассоциированным с опухолью антигеном (ТАА). TSA является уникальным для опухолевых клеток и не присутствует на других клетках в организме. ТАА-антиген не является уникальным для опухолевых клеток и также экспрессируется на нормальных клетках в условиях, препятствующих индуцированию состояния иммунологической толерантности к антигену. Экспрессия антигена в опухоли может происходить в условиях, позволяющих иммунной системе отвечать на антиген. ТАА могут являться антигенами, экспрессируемыми на нормальных клетках при развитии плода, когда иммунная система является незрелой и не способна отвечать, или они могут являться антигенами, присутствующими в норме на крайне низких уровнях на нормальных клетках, но экспрессируемыми на гораздо более высоких уровнях на опухолевых клетках.
Неограничивающие примеры TSA- или ТАА-антигенов включают следующие: антигены дифференцировки, такие как MART-1/MelanA (MART-1), gp100 (Pmel 17), тирозиназа, TRP-1, TRP-2 и опухолеспецифичные мультилинейные антигены, такие как MAGE-1, MAGE-3, BAGE, GAGE-1, GAGE-2, р15; гиперэкспрессируемые эмбриональные антигены, такие как СЕА; гиперэкспрессируемые онкогены и мутантные гены опухолевых супрессоров, такие как р53, Ras, HER-2/neu; уникальные опухолевые антигены, образующиеся в результате хромосомных транслокаций, такие как BCR-ABL, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR; и вирусные антигены, такие как антигены EBVA вируса Эпштейн-Барр и антигены Е6 и Е7 папилломавируса человека (HPV). Другие большие антигены на основе белков включают TSP-180, MAGE-4, MAGE-5, MAGE-6, RAGE, NY-ESO, p185erbB2, p180erbB-3, c-met, nm-23H1, PSA, TAG-72, CA 19-9, CA 72-4, CAM 17.1, NuMa, K-ras, бета-катенин, CDK4, Mum-1, p15, p16, 43-9F, 5T4, 791Tgp72, альфа-фетопротеин, бета-HCG, BCA225, BTAA, CA 125, CA 15-3/CA 27.29/BCAA, CA 195, CA 242, CA-50, CAM43, CD68/P1, CO-029, FGF-5, G250, Ga733/EpCAM, HTgp-175, M344, MA-50, MG7-Ag, MOV18, NB/70K, NY-CO-1, RCAS1, SDCCAG16, ТА-90/Мас-2-связывающий белок/циклофилин С-ассоциированный белок, TAAL6, TAG72, TLP и TPS.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен специфичен к опухолевому антигену, который выбран из группы: CD19, CD20, CD123 или BCMA.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен специфичен к опухолевому антигену CD19.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен, который специфичен к опухолевому антигену, представляет собой Fab или scFv.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен, который специфичен к опухолевому антигену, представляет собой scFv.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что антигенсвязывающий домен, который специфичен к опухолевому антигену CD19, имеет аминокислотную последовательность представленную DIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSS (SEQ ID NO: 11) или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 11 и имеет отличия по сравнению с SEQ ID NO: 11 только в неконсервативных аминокислотных остатках.
Антигенсвязывающий домен с аминокислотной последовательностью SEQ ID NO: 11 состоит из:
а) вариабельного домена легкой цепи с аминокислотной последовательностью DIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEIT;
б) линкера с аминокислотной последовательностью GSTSGSGKPGSGEGSTKG;
в) вариабельного домена тяжелой цепи с аминокислотной последовательностью EVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSS.
Ко-стимуляторный домен
Ко-стимуляторный домен или костимуляторная сигнальная область относится к части CAR, содержащей внутриклеточный домен костимуляторной молекулы. Костимуляторная молекула является молекулой клеточной поверхности, иной, чем антигенный рецептор или его лиганды, необходимые для эффективного ответа лимфоцитов на антиген. Примеры таких молекул включают CD27, CD28, 4-1ВВ (CD137), ОХ40, CD30, ICAM-1, CDS, CD40, PD-1, ICOS (CD278), функционально-ассоциированный антиген лимфоцитов-1 (LFA-1, CD11a/CD18), CD2, CD7, LIGHT, NKG2C, В7-Н3 и лиганд, специфически связывающийся с CD83, и т.п.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен представляет собой функциональный сигнальный домен белка, выбранного из группы, состоящей из OX40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278) и 4-1BB (CD137).
Цитоплазматические последовательности, опосредующие передачу сигнала, в цитоплазматической сигнальной части CAR, можно соединять друг с другом в случайном или специальном порядке. Необязательно, соединение может образовывать короткий олиго- или полипептидный линкер, предпочтительно от 2 до 10 аминокислот в длину. В частности, глицин-сериновый дуплет является подходящим линкером.
В одном из вариантов осуществления цитоплазматический домен CAR конструируют содержащим вышеуказанный альтернативный внутриклеточный сигнальный домен и ко-стимуляторный сигнальный домен CD28.
В другом варианте осуществления цитоплазматический домен CAR конструируют содержащим вышеуказанный альтернативный внутриклеточный сигнальный домен и ко-стимуляторный сигнальный домен 4-1ВВ.
В еще одном варианте осуществления цитоплазматический домен конструируют содержащим вышеуказанный альтернативный внутриклеточный сигнальный домен и ко-стимуляторный домен CD28 и 4-1ВВ.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен CD28 имеет аминокислотную последовательность представленную AAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:13).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что ко-стимуляторный домен 4- 1BB имеет аминокислотную последовательность представленную KRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL (SEQ ID NO: 14).
Трансмембранный домен
Что касается трансмембранного домена, CAR можно конструировать содержащим трансмембранный домен, слитный с внеклеточным доменом CAR. В одном из вариантов осуществления используют трансмембранный домен, в природе ассоциированный с одним из доменов в CAR. В некоторых случаях трансмембранный домен можно выбирать или модифицировать заменой аминокислоты для избежания связывания таких доменов с трансмембранными доменами тех же или других белков на поверхности мембраны для минимизации взаимодействий с другими компонентами рецепторного комплекса.
Трансмембранный домен можно получать из природного или синтетического источника. Если источник является природным, домен можно получать из любого мембраносвязанного или трансмембранного белка. Трансмембранные области для конкретного использования в этом изобретении можно получать из (т.е. получать содержащими, по меньшей мере, трансмембранные области) альфа-, бета- или дзета-цепи Т-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, CD154. Альтернативно, трансмембранный домен может являться синтетическим, в этом случае он будет содержать, преимущественно, гидрофобные остатки, такие как лейцин и валин. Предпочтительно на каждом конце синтетического трансмембранного домена будут обнаруживать триплет фенилаланина, триптофана и валина.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что трансмембранный домен представляет собой трансмембранный домен белка, выбранного из группы, состоящей из альфа, бета или дзета-цепи T-клеточного рецептора, CD28, CD3-эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что где трансмембранный домен представляет собой CD8.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что трансмембранный домен CD8 имеет аминокислотную последовательность IYIWAPLAGTCGVLLLSLVITLYC (SEQ ID NO: 12).
Необязательно, короткий олиго- или полипептидный линкер, предпочтительно от 2 до 10 аминокислот в длину может образовывать связь между трансмембранным доменом и цитоплазматическим сигнальным доменом CAR. В частности глицин-сериновый дуплет представляет собой подходящий линкер.
В некоторых случаях трансмембранный домен CAR по изобретению содержит шарнирный домен CD8. В одном из вариантов осуществления шарнирный домен CD8 имеет аминокислотную последовательность TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACD (SEQ ID NO: 35).
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) характеризуется тем, что дополнительно содержит лидерную последовательность.
В некоторых вариантах выделенный химерный антигенный рецептор (CAR) имеет аминокислотную последовательность представленную аминокислотной последовательностью, которую выбирают из группы: SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности, которую выбирают из группы: SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, и имеет отличия по сравнению с аминокислотной последовательностью, которую выбирают из группы: SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, только в неконсервативных аминокислотных остатках.
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITGB3- ITAM2- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:15).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITAM1- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:16).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITAM1- ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:17).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITGB3- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:18).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITAM1- ITGB3- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:19).
Химерный антигенный рецептор 19CAR-T- CD28 - altcd3дзета (ITGB3 - ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCAAAIEVMYPPPYLDNEKSNGTIIHVKGKHLCPSPLFPGPSKPFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:20).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITGB3- ITAM2- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:21).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITAM1- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:22).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITAM1- ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:23).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITGB3- ITGB3- ITAM3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRGSAAA (SEQ ID NO:24).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITAM1- ITGB3- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKDTANNPLYKEATSTFTNITYRGTKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:25).
Химерный антигенный рецептор 19CAR-T- 41bb - altcd3дзета (ITGB3 - ITAM2- ITGB3) имеет аминокислотную последовательность MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADDTANNPLYKEATSTFTNITYRGTRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGDTANNPLYKEATSTFTNITYRGTALPPRGSAAA (SEQ ID NO:26).
Молекулы нуклеиновых кислот
Термины «нуклеиновая кислота», «нуклеиновая последовательность» или «нуклеиновокислотная последовательность», «полинуклеотид», «олигонуклеотид», «полинуклеотидная последовательность» и «нуклеотидная последовательность», которые используются равнозначно в данном описании, обозначают четкую последовательность нуклеотидов, модифицированных или не модифицированных, определяющую фрагмент или участок нуклеиновой кислоты, содержащую или не содержащую неприродные нуклеотиды и являющуюся либо двухцепочечной ДНК или РНК, либо одноцепочечной ДНК или РНК, либо продуктами транскрипции указанных ДНК.
Здесь также следует упомянуть, что данное изобретение не относится к нуклеотидным последовательностям в их природной хромосомной среде, т.е. в природном состоянии. Последовательности данного изобретения были выделены и/или очищены, т.е. были взяты прямо или косвенно, например, путем копирования, при этом их среда была по меньшей мере частично модифицирована. Таким образом, также здесь следует подразумевать изолированные нуклеиновые кислоты, полученные путем генетической рекомбинации, например, с помощью принимающих клеток (клеток-хозяев), или полученные путем химического синтеза.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена от по меньшей мере одной молекулы нуклеиновой кислоты-примеси, с которой она обычно связана в естественном источнике нуклеиновой кислоты белка (полипептида). Выделенная молекула нуклеиновой кислоты отличается от той формы или набора, в которых она находится в естественных условиях. Таким образом, выделенная молекула нуклеиновой кислоты отличается от молекулы нуклеиновой кислоты, существующей в клетках в естественных условиях. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, находящуюся в клетках, в которых в норме происходит экспрессия белка (полипептида), например, в случае, если молекула нуклеиновой кислоты имеет локализацию в хромосоме, отличную от ее локализации в клетках в естественных условиях.
Термин нуклеотидная последовательность охватывает его комплемент, если не указано иное. Таким образом, нуклеиновую кислоту, имеющую определенную последовательность следует понимать, как охватывающие ее комплементарную цепь с ее комплементарной последовательностью.
В контексте настоящего изобретения используют следующие аббревиатуры для повсеместно встречающихся оснований нуклеиновых кислот. «А» относится к аденину, «С» относится к цитозину, «G» относится к гуанину, «Т» относится к тимину (5-метилурацил), не встречается в РНК , занимает место урацила в ДНК, и «U» относится к урацилу (занимает место тимина в РНК).
Если не указано иначе, «нуклеотидная последовательность, кодирующая аминокислотную последовательность» включает все нуклеотидные последовательности, являющиеся вырожденными версиями друг друга и кодирующие одну и ту же аминокислотную последовательность. Фраза «нуклеотидная последовательность, кодирующая белок или РНК, также может включать интроны в случае, если нуклеотидная последовательность, кодирующая белок, в некоторых версиях может содержать интроны.
«Кодирующий» относится к неотъемлемому свойству конкретных последовательностей нуклеотидов в полинуклеотиде, таком как ген, кДНК или мРНК, служить матрицами для синтеза других полимеров и макромолекул в биологических процессах, имеющих определенную последовательность нуклеотидов (т.е. рРНК, тРНК и мРНК) или определенную последовательность аминокислот и, в результате, биологические свойства. Таким образом, ген кодирует белок, если в результате транскрипции и трансляции мРНК, соответствующей этому гену, в клетке или другой биологической системе образуется белок. И кодирующая цепь, нуклеотидная последовательность которой является идентичной последовательности мРНК и которую, как правило, представляют в списках последовательностей, и некодирующую цепь, используемую в качестве матрицы для транскрипции гена или кДНК, можно обозначать как кодирующую белок или другой продукт этого гена или кДНК.
Последовательности нуклеиновой кислоты, кодирующие желаемые молекулы, можно получать с использованием известных в этой области рекомбинантных способов, таких как, например, с помощью скрининга библиотек из клеток, экспрессирующих ген, выделяя ген из вектора, о котором известно, что он его включает, или выделяя его напрямую из содержащих его клеток и тканей с использованием стандартных способов. Альтернативно, интересующий ген чаще можно получать синтетически, чем клонировать.
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует вышеуказанный альтернативный внутриклеточный сигнальный домен.
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует вышеуказанный выделенный химерный антигенный рецептор (CAR), который включает вышеуказанный альтернативный внутриклеточный сигнальный домен.
Вектор
Термин «вектор» при использовании в настоящем документе означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она соединена. В некоторых вариантах осуществления изобретения вектор представляет собой плазмиду, т.е. кольцевую двухцепочечную молекулу ДНК, в которую могут быть встроены с помощью реакции ферментативного лигирования дополнительные сегменты ДНК. В некоторых вариантах осуществления изобретения вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть встроены с помощью реакции ферментативного лигирования в вирусный геном. В некоторых вариантах осуществления изобретения векторы способны к автономной репликации в генетически модифицированной клетке, в которую они введены (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации и эписомные векторы млекопитающих). В других вариантах осуществления изобретения векторы (например, неэписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина, и таким образом реплицируются вместе с геном хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально соединены. Такие векторы упоминаются в данном документе как «рекомбинантные экспрессирующие векторы» (или просто «экспрессирующие векторы»).
Термин «последовательность контроля экспрессии», используемый в данном описании, означает полинуклеотидные последовательности, которые необходимы для воздействия на экспрессию и процессинг кодирующих последовательностей, к которым они встроены с помощью реакции ферментативного лигирования. Контролирующие экспрессию последовательности включают соответствующие последовательности инициации транскрипции, терминации, промотора и энхансера; эффективные сигналы процессинга РНК, такие как сплайсинг и сигналы полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, которые повышают эффективность трансляции (т.е. консенсусная последовательность Козака); последовательности, которые повышают стабильность белка; и, при желании, последовательности, которые усиливают секрецию белка. Характер таких контролирующих последовательностей различается в зависимости от организма-хозяина. В эукариотических клетках присутствуют промоторы, сигналы полиаденилирования и энхансеры. Термин «контролирующие последовательности» включает, как минимум, все компоненты, наличие которых имеет важное значение для экспрессии и процессинга, и может также включать дополнительные компоненты, чье присутствие является полезным, например, лидирующие последовательности и последовательности слившихся клеток.
Нуклеиновая кислота «функционально связана», если она находится в функциональной связи с другой нуклеотидной последовательностью. Например, ДНК предпоследовательности или секреторной лидерной последовательности функционально связывают с ДНК полипептида, если он экспрессируется в виде предпротеина, который принимает участие в секреции полипептида; промотор или энхансер функционально связывают с кодирующей последовательностью, если он оказывает воздействие на транскрипцию последовательности; сайт связывания рибосомы функционально связывают с кодирующей последовательностью, если он расположен так, что может облегчать трансляцию. Как правило, «функционально связан» обозначает, что связанные последовательности ДНК являются смежными, а в случае секреторной лидерной последовательности являются смежными и находятся в рамке считывания. Однако энхансеры не обязательно должны быть смежными.
Под «вектором доставки» или «векторами доставки» понимается любой вектор доставки, который можно использовать в настоящем изобретении для того, чтобы ввести в контакт с клеткой (т.е. «контактирование») или доставить внутрь клеток или внутриклеточных компартментов (т.е. «введение») средства/химические вещества и молекулы (белки или нуклеиновые кислоты), нужные в настоящем изобретении. Он включает, но без ограничения, липосомные векторы доставки, вирусные векторы доставки, лекарственные векторы доставки, химические носители, полимерные носители, липоплексы, полиплексы, дендримеры, микропузырьки (ультразвуковые контрастные средства), наночастицы, эмульсии или другие соответствующие векторы переноса. Данные векторы доставки делают возможной доставку молекул, химических веществ, макромолекул (генов, белков) или других векторов, таких как плазмиды, пептиды. В этих случаях векторы доставки представляют собой молекулярные носители. Под «вектором доставки» или «векторами доставки» также понимаются способы доставки для осуществления трансфекции.
«Экспрессирующий вектор» относится к вектору, содержащему рекомбинантный полинуклеотид, содержащий последовательности, контролирующие экспрессию, функционально связанные с подлежащей экспрессии нуклеотидной последовательностью. Экспрессирующий вектор содержит соответствующие цис-активные элементы для экспрессии; другие элементы для экспрессии можно восполнять элементами генетически модифицированной клетки или в системе экспрессии in vitro. Экспрессирующие векторы включают все векторы, известные в этой области, такие как космиды, плазмиды (например, безоболочечные или содержащиеся в липосомах) и вирусы (например, лентивирусы, ретровирусы, аденовирусы и адено-ассоциированные вирусы), включающие рекомбинантный полинуклеотид.
Вкратце, экспрессии природных или синтетических нуклеиновых кислот, кодирующих CAR, как правило, достигают функциональным соединением нуклеиновой кислоты, кодирующей полипептид CAR или его части, с промотором, и встраиванием конструкции в экспрессирующий вектор. Векторы могут подходить для репликации и интеграции в эукариотические клетки. Типичные клонирующие векторы содержат терминаторы транскрипции и трансляции, инициирующие последовательности и промоторы, применимые для регуляции экспрессии желаемой последовательности нуклеиновой кислоты.
Как применяют в настоящем описании, термин «промотор» определяют как последовательность ДНК, распознаваемую синтетическим аппаратом клетки или встраиваемым синтетическим аппаратом, необходимым для инициации конкретной транскрипции полинуклеотидной последовательности.
Как применяют в настоящем описании, термин «промотор/регуляторная последовательность» означает последовательность нуклеиновой кислоты, необходимую для экспрессии продукта гена, функционально связанного с промотором/регуляторной последовательностью. В некоторых случаях эта последовательность может являться коровой последовательностью промотора, а в других случаях эта последовательность также может включать энхансерную последовательность и другие регуляторные элементы, необходимые для экспрессии продукта гена. Промотор/регуляторная последовательность, например, может являться последовательностью, экспрессирующей продукт гена тканеспецифическим образом.
«Конститутивный» промотор является нуклеотидной последовательностью, которая, если функционально связана с полинуклеотидом, кодирующим или определяющим продукт гена, вызывает продукцию продукта гена в клетке в большинстве или всех физиологических условиях в клетке.
«Индуцибельный» промотор является нуклеотидной последовательностью, которая, если функционально связана с полинуклеотидом, кодирующим или определяющим продукт гена, вызывает продукцию продукта гена в клетке, по существу, только когда индуцирующий фактор, соответствующий промотору, присутствует в клетке.
«Тканеспецифический» промотор является нуклеотидной последовательностью, которая, если функционально связана с полинуклеотидом, кодирующим или определяющим продукт гена, вызывает продукцию продукта гена в клетке, по существу, только когда клетка представляет собой клетку ткани типа, соответствующего промотору.
Одним из примеров подходящего промотора является промоторная последовательность предраннего цитомегаловируса (CMV). Эта промоторная последовательность является сильной конститутивной промоторной последовательностью, способной запускать высокие уровни экспрессии любой полинуклеотидной последовательности, функционально с ней связанной. Другим примером подходящего промотора является промотор фактора элонгации-1а (EF-1a). Однако также можно использовать другие конститутивные промоторные последовательности, включая, в качестве неограничивающих примеров, ранний промотор вируса обезьян 40 (SV40), вирус опухоли молочной железы мыши (MMTV), промотор в области длинных концевых повторов (LTR) вируса иммунодефицита человека (ВИЧ), промотор MoMuLV, промотор вируса лейкоза птиц, предранний промотор вируса Эпштейна-Барр, промотор вируса саркомы Рауса, а также промоторы генов человека, в качестве неограничивающих примеров, такие как, промотор гена актина, промотор гена миозина, промотор гена гемоглобина и промотор гена креатинкиназы. Кроме того, изобретение не должно ограничиваться использованием конститутивных промоторов. Индуцибельные промоторы также включены как часть изобретения. Использование индуцибельного промотора представляет молекулярный «переключатель», способный запускать экспрессию полинуклеотидной последовательности, с которой он функционально связан, если такая экспрессия является желательной, или прекращать экспрессию, если экспрессия нежелательна. Примеры индуцибельных промоторов включают, в качестве неограничивающих примеров, промотор металлотионеинов, промоторы генов глюкокортикоидов, промотор гена прогестерона и промотор гена тетрациклина.
Дополнительные промоторные элементы, например, энхансеры, регулируют частоту инициации транскрипции. Как правило, они локализуются в области на 30-110 п.н. выше стартового участка, хотя недавно показано, что ряд промоторов содержит функциональные элементы ниже стартового участка. Часто спейсинг между промоторными элементами является гибким таким образом, что предотвращают функционирование промотора, когда элементы инвертированы или перемещены относительно друг друга. В промоторе тимидинкиназы (tk) спейсинг между промоторными элементами можно повышать до 50 п.н. до начала снижения активности. Считают, что в зависимости от промотора отдельные элементы могут функционировать совместно или независимо, активируя транскрипцию.
Настоящее изобретение относится к конструкции ДНК, содержащей последовательности CAR, где последовательность содержит последовательность нуклеиновой кислоты антигенсвязывающего домена, функционально связанного с последовательностью нуклеиновой кислоты внутриклеточного домена. Пример внутриклеточного домена, который можно использовать в CAR по изобретению, включает, в качестве неограничивающих примеров, вышеуказанный альтернативный внутриклеточный сигнальный домен, ко-стимуляторный домен CD28 и/или 4-1ВВ и т.п., в некоторых случаях CAR может содержать любую комбинацию вышеуказанный альтернативный внутриклеточный сигнальный домен, ко-стимуляторный домен CD28, ко-стимуляторный домен 4-1ВВ и т.п.
В одном из вариантов осуществления CAR по изобретению содержит scFv против CD19, шарнирный и трансмембранный домен CD8 человека и ко-стимуляторный домен 4-1ВВ и вышеуказанный альтернативный внутриклеточный сигнальный домен.
В одном из вариантов осуществления CAR по изобретению содержит scFv против CD19, шарнирный и трансмембранный домен CD8 человека и ко-стимуляторный домен CD28 и вышеуказанный альтернативный внутриклеточный сигнальный домен.
Экспрессирующие конструкции по настоящему изобретению также можно использовать для иммунизации нуклеиновыми кислотами и генной терапии с использованием стандартных способов доставки генов. В этой области известны способы доставки генов. См., например, патенты США № 5399346, 5580859, 5589466, включенные в настоящее описании в качестве ссылки в полном объеме. В другом варианте осуществления изобретение относится к вектору для генной терапии.
Нуклеиновую кислоту можно клонировать в некоторые типы векторов. Например, нуклеиновую кислоту можно клонировать в вектор, включая, в качестве неограничивающих примеров, плазмиду, фагмиду, фаговое производное, вирус животных и космиду. Конкретные интересующие векторы включают экспрессирующие векторы, реплицирующиеся векторы, векторы для получения зондов и векторы для секвенирования.
Кроме того, экспрессирующий вектор можно доставлять в клетку в форме вирусного вектора. Технология вирусных векторов хорошо известна в этой области, и ее описывают, например, в Sambrook et al. (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York) и других руководствах по вирусологии и молекулярной биологии. Вирусы, применимые в качестве векторов, включают, в качестве неограничивающих примеров, ретровирусы, аденовирусы, адено-ассоциированные вирусы, вирус герпеса и лентивирусы. В основном, подходящий вектор содержит участок начала репликации, функциональный по меньшей мере в одном организме, промоторную последовательность, подходящие участки эндонуклеаз рестрикции и один или несколько селективных маркеров (например, WO 01/96584; WO 01/29058 и патент США № 6326193).
Для передачи генов в клетки млекопитающих разрабатывают ряд систем на основе вирусов. Например, ретровирусы представляют собой подходящую платформу для систем доставки генов. Выбранный ген можно встраивать в вектор и упаковывать в ретровирусные частицы с использованием известных в этой области способов. Затем рекомбинантный вирус можно выделять и доставлять в клетки индивидуума in vivo или ex vivo. В этой области известен ряд ретровирусных систем. В некоторых вариантах осуществления используют аденовирусные векторы. В этой области известен ряд аденовирусных векторов. В одном из вариантов осуществления используют лентивирусные векторы.
Настоящее изобретение также относится к векторам, в которые встраивают ДНК по настоящему изобретению. Векторы на основе ретровирусов, таких как лентивирус, являются подходящими инструментами для достижения длительной передачи генов, т.к. они делают возможной длительную стабильную интеграцию трансгена и его размножение в дочерних клетках. Лентивирусные векторы обладают дополнительным преимуществом по сравнению с векторами, полученными из онкоретровирусов, таких как вирусы лейкемии мышей, состоящим в том, что ими можно трансдуцировать непролиферирующие клетки, такие как гепатоциты. Они также обладают дополнительным преимуществом - низкой иммуногенностью.
Как применяют в настоящем описании, «лентивирус» относится к роду семейства Retroviridae. Лентивирусы уникальны среди ретровирусов, т.к. способны инфицировать неделящиеся клетки; они могут доставлять значительное количество генетической информации в виде ДНК в клетку-хозяина, таким образом, они представляют собой один из наиболее эффективных способов реализации вектора для доставки гена. ВИЧ, SIV и FIV являются примерами лентивирусов. Векторы, полученные из лентивирусов, представляют средства достижения значительных уровней передачи генов in vivo.
В целях оценки экспрессии полипептида CAR или его частей экспрессирующий вектор для встраивания в клетку также может содержать ген селективного маркера, или репортерный ген или оба для облегчения определения и селекции экспрессирующих клеток из популяции клеток, которые необходимо трансфицировать или инфицировать с помощью вирусных векторов. В других аспектах селективный маркер может нести отдельный фрагмент ДНК и его можно использовать в способе котрансфекции. И селективные маркеры, и репортерные гены можно фланкировать соответствующими регуляторными последовательностями, делая возможной экспрессию в клетках-хозяевах. Применимые селективные маркеры включают, например, гены устойчивости к антибиотикам, такие как neo и т.п.
Для определения потенциально трансфицированных клеток и для оценки функциональности регуляторных последовательностей используют репортерные гены. В основном, репортерный ген является геном, не присутствующим или экспрессируемым в организме или ткани и кодирующим полипептид, экспрессия которых проявляется в некотором легко определяемом свойстве, например, ферментативной активности. Экспрессию репортерного гена анализируют в подходящий момент времени после встраивания ДНК в клетки-реципиенты. Подходящие репортерные гены могут включать гены, кодирующие люциферазу, бета-галактозидазу, хлорамфеникол ацетилтрансферазу, секретируемую щелочную фосфатазу, или ген зеленого флуоресцентного белка (GFP) (например, Ui-Tei et al., 2000 FEBS Letters 479: 79-82). Подходящие системы экспрессии хорошо известны и их можно получать с использованием известных способов или из коммерческого источника. В основном, конструкцию с минимальной 5'-фланкирующей областью, демонстрирующую наибольший уровень экспрессия репортерного гена, определяют как промотор. Такие промоторные области можно соединять с репортерным геном и использовать для оценки средств на способность модулировать запускаемую промотором транскрипцию.
Настоящее изобретение относится к векторам, содержащим молекулы нуклеиновых кислот, которые кодируют вышеуказанный химерный антигенный рецептор (CAR) по изобретению.
В одном из аспектов настоящее изобретение относится к экспрессионному вектору, содержащему вышеуказанную нуклеиновую кислоту, которая кодирует вышеуказанный химерный антигенный рецептор (CAR) по изобретению.
В одном из аспектов настоящее изобретение относится к вектору доставки, содержащему вышеуказанную нуклеиновую кислоту, которая кодирует вышеуказанный химерный антигенный рецептор (CAR) по изобретению.
В некоторых вариантах вектор выбирают из группы, состоящей из ДНК, РНК, плазмиды, лентивирусного вектора, адено-ассоциированного вирусного вектора, аденовирусного вектора или ретровирусного вектора.
В некоторых вариантах вектор дополнительно содержит промотор.
В некоторых вариантах вектор представляет собой in vitro транскрибируемый вектор.
В некоторых вариантах последовательность нуклеиновой кислоты дополнительно содержит поли(A)-последовательность.
В некоторых вариантах последовательность нуклеиновой кислоты дополнительно содержит 3'UTR.
Клетки-хозяева и способы их получения
Термин «генетически модифицированная клетка» (или «рекомбинантная клетка-хозяин») при использовании в данном документе означает клетку, в которую введен рекомбинантный экспрессионный вектор. Настоящее изобретение относится к «генетически модифицированным клеткам», которые могут включать, например, вектор в соответствии с настоящим изобретением, описанным выше. Следует понимать, что «генетически модифицированная клетка» и «рекомбинантная клетка-хозяин» означают не только конкретную заявленную клетку, но также и потомство такой клетки. Поскольку модификации могут проходить в последующих поколениях вследствие мутации или воздействий окружающей среды, такое потомство не может, на самом деле, быть идентичным родительской клетке, но такие клетки по-прежнему включены в объем термина «генетически модифицированная клетка» при использовании в настоящем документе.
В одном из аспектов настоящее изобретение относится к способу получения «генетически модифицированной клетки», которая содержит вышеуказанный химерный антигенный рецептор (CAR), включающий трансформирование клетки вышеуказанным вектором.
В этой области известны способы встраивания и экспрессии генов в клетке. Что касается экспрессирующего вектора, вектор легко можно встраивать в клетку-хозяина, например, клетку млекопитающего, бактерии, дрожжей или насекомых, любым способом, известным в этой области. Например, экспрессирующий вектор можно трансфицировать в клетку-хозяина физическими, химическими или биологическими способами.
Физические способы введения полинуклеотида в клетку-хозяина включают осаждение фосфатом кальция, липофекцию, бомбардировку частицами, микроинъекции, электропорацию и т.п. Способы получения клеток, содержащих векторы и/или экзогенные нуклеиновые кислоты, хорошо известны в этой области. См., например, Sambrook et al. (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York). Предпочтительным способом введения полинуклеотида в клетку-хозяина является трансфекция с помощью фосфата кальция.
Биологические способы введения интересующего полинуклеотида в клетку-хозяина включают использование ДНК- и РНК-векторов. Вирусные векторы и, особенно, ретровирусные векторы стали наиболее широко используемым способом встраивания генов в клетки млекопитающих, например, клетки человека. Другие вирусные векторы можно получать на основе лентивируса, поксвирусов, вируса простого герпеса I, аденовирусов и аденоассоциированных вирусов и т.п. См., например, патенты США № 5350674 и 5585362.
Химические способы введения полинуклеотида в клетку-хозяина включают системы коллоидных дисперсий, такие как комплексы макромолекул, нанокапсулы, микросферы, бусы и системы на основе липидов, включая эмульсии «масло-в-воде», мицеллы, смешанные мицеллы и липосомы. Примером коллоидной системы для применения в качестве средства доставки in vitro и in vivo является липосома (например, искусственное мембранное средство).
В случае если используют невирусные системы доставки, примером средства доставки является липосома. Использование липидных составов предназначено для введения нуклеиновых кислот в клетку-хозяина (in vitro, ex vivo или in vivo). В другом аспекте нуклеиновую кислоту можно соединять с липидом. Нуклеиновую кислоту, соединенную с липидом, можно инкапсулировать в водной внутренней части липосомы, можно распределять внутри липидного бислоя липосомы, прикреплять к липосоме с помощью линкерной молекулы, соединенной и с липосомой, и с олигонуклеотидом, заключать в липосому, получать ее комплекс с липосомой, диспергировать в растворе, содержащем липид, смешивать с липидом, комбинировать с липидом, она может содержаться в виде суспензии в липиде, содержаться или образовывать комплекс с мицеллой, или ее можно иначе соединять с липидом. Композиции, ассоциированные с липидами, липидами/ДНК или липидами/экспрессирующим вектором, не ограничены любой конкретной структурой в растворе. Например, они могут присутствовать в бислойной структуре, в виде мицеллы или со «сложенной» структурой. Они также могут просто распределяться в растворе, возможно, образуя агрегаты, не являющиеся однородными по размеру или форме. Липиды являются жирными веществами, которые могут представлять собой природные или синтетические липиды. Например, липиды включают жировые капли, в природе присутствующие в цитоплазме, а также класс соединений, содержащих длинноцепочечные алифатические углеводороды и их производные, такие как жирные кислоты, спирты, амины, аминоспирты и альдегиды.
Пригодные для использования липиды можно получать из коммерческих источников. Например, димиристилфосфатидилхолин («DMPC») можно приобретать в Sigma, St. Louis, МО; дицетил фосфат («DCP») можно приобретать в K & K Laboratories (Plainview, NY); холестерин («Choi») можно приобретать в Calbiochem-Behring; димиристил фосфатидилглицерин («DMPG») и другие липиды можно приобретать в Avanti Polar Lipids, Inc. (Birmingham, AL). Стоковые растворы липидов в хлороформе или хлороформ/метаноле можно хранить приблизительно при -20°С. В качестве растворителя используют исключительно хлороформ, т.к. он испаряется легче, чем метанол. «Липосома» представляет собой общий термин, включающий множество одно- и многослойных липидных средств, получения посредством образования замкнутых липидных бислоев или агрегатов. Липосомы можно охарактеризовать как имеющие везикулярную структуру с фосфолипидной бислойной мембраной и внутренней водной средой. Многослойные липосомы имеют многочисленные липидные слои, разделенные водной средой. Они образуются спонтанно, когда фосфолипиды суспендируют в избытке водного раствора. Липидные компоненты подвергаются самоперестройке до образования замкнутых структур и удерживают воду и растворенные вещества между липидными бислоями (Ghosh et al., 191 Glycobiology 5; 505-10). Однако также включают композиции, имеющие в растворе структуры, иные, чем нормальная везикулярная структура. Например, липиды могут иметь мицеллярную структуру или просто существовать как неоднородные агрегаты молекул липидов. Также включают комплексы липофектамин-нуклеиновая кислота.
Независимо от способа, используемого для введения экзогенных нуклеиновых кислот в клетку-хозяина или иного воздействия на клетку ингибитора по настоящему изобретению, для подтверждения наличия рекомбинантной последовательности ДНК в клетке-хозяине можно осуществлять множество анализов. Такие анализы включают, например, «молекулярно-биологические» анализы, хорошо известные специалистам в этой области, такие как Саузерн- и «нозерн»-блоттинг, RT-ПЦР и ПЦР; «биохимические» анализы, такие как определение наличия или отсутствия конкретного пептида, например, иммунологическими способами (ELISA и вестерн-блоттингом), или посредством анализов, представленных в настоящем описании, для определения средств, включенных в объем изобретения.
В одном из аспектов настоящее изобретение относится к генетически модифицированной клетке, которая содержит вышеуказанный химерный антигенный рецептор (CAR), которую получают вышеуказанным способом.
Воплощение согласно изобретению также предусматривает генетически модифицированную клетку, содержащую любой из рекомбинантных экспрессионных векторов, описанных в данном документе. Используемый в данном документе термин «генетически модифицированная клетка» относится к любому типу клеток, которые могут содержать рекомбинантный экспрессионный вектор согласно изобретению. Генетически модифицированная клетка может быть эукариотической, например, клеткой растения, животного, гриба или водоросли, либо она может быть прокариотической клеткой, например, клеткой бактерии или простейшего. Генетически модифицированная клетка может быть культивированной клеткой или первичной клеткой, т.е. выделенной непосредственно из организма, например, человека. Генетически модифицированная клетка может быть прикрепленной клеткой или суспензионной клеткой, т.е. клеткой, которая растет в суспензии. Подходящие клетки-хозяева известны в данной области и включают, например, клетки Е. coli DH5α, клетки яичников китайского хомячка, клетки обезьяны VERO, клетки COS, клетки HEK293 и т.п. Для амплификации или репликации рекомбинантного экспрессионного вектора клетка-хозяин может быть прокариотической клеткой, например, клеткой DH5α. Для получения рекомбинантного CAR генетически модифицированная клетка может быть клеткой млекопитающего. Генетически модифицированная клетка может быть клеткой человека. Притом что генетически модифицированная клетка может быть клеткой любого типа, может происходить из любого типа ткани и может находиться на любой стадии развития, генетически модифицированная клетка может быть лимфоцитом периферической крови (PBL) или мононуклеарной клеткой периферической крови (РВМС). Генетически модифицированная клетка может быть Т-клеткой. Генетически модифицированная клетка может быть NK-клеткой.
Для целей данного изобретения Т-клетка может быть любой Т-клеткой, такой как культивированная Т-клетка, например, первичная Т-клетка или Т-клетка из культивируемой Т-клеточной линии, например, Jurkat, SupT1 и т.д., или Т-клетка, полученная от млекопитающего. Если Т-клетка получена от млекопитающего, то она может быть получена из различных источников, включая, но не ограничиваясь ими, кровь, костный мозг, лимфатические узлы, вилочковую железу или другие ткани или жидкости. Т-клетки также могут быть обогащенными или очищенными. Т-клетка могут быть человеческой Т-клеткой. Т-клетка может быть Т-клеткой, выделенной из организма человека. Т-клетка может быть Т-клеткой любого типа и может находиться на любой стадии развития, включая, но не ограничиваясь ими, CD4+/CD8+-двойные положительные Т-клетки, CD4+Т-хелперы, например, клетки Th1 и Th2, CD8+Т-клетки (например, цитотоксические Т-клетки), инфильтрирующие опухоль клетки, Т-клетки памяти, наивные Т-клетки и т.п. Т-клетка может быть CD8+Т-клеткой или CD4+Т-клеткой.
Т-клетки можно получать из ряда источников, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфоузла, пуповинную кровь, ткань тимуса, ткань из места инфекции, асцитическую жидкость, плевральный выпот, ткань селезенки и опухоли. В определенных вариантах осуществления настоящего изобретения можно использовать любое количество Т-клеточных линий, доступных в этой области. В определенных вариантах осуществления настоящего изобретения Т-клетки можно получать из единицы крови, забранной у индивидуума, с использованием любого количества способов, известных специалистам в этой области, таких как разделение с использованием фиколла. В одном из предпочтительных вариантов осуществления клетки из циркулирующей крови индивидуума получают аферезом. Как правило, продукт афереза содержит лимфоциты, включая Т-клетки, моноциты, гранулоциты, В-клетки, другие ядерные лейкоциты, эритроциты и тромбоциты. В одном из вариантов осуществления клетки, собранные аферезом, можно промывать для удаления фракции плазмы и для помещения клеток в соответствующий буфер или среду для последующих этапов обработки. В одном из вариантов осуществления изобретения клетки промывают фосфатно-солевым буфером (PBS). В альтернативном варианте осуществления промывочный раствор не содержит кальций и может не содержать магний или может не содержать многие, если не все, бивалентные катионы. Кроме того, неожиданно, исходные этапы активации в отсутствие кальция приводят к усиленной активации. Как будет очевидно специалистам в этой области, этап промывания можно осуществлять известными в этой области способами, такими как использование полуавтоматической «проточной» центрифуги (например, клеточного процессора Cobe 2991, Baxter CytoMate или Haemonetics Cell Saver 5) по инструкциям производителя. После промывания клетки можно ресуспендировать во множестве биосовместимых буферов, таких как, например, несодержащий Са2+, несодержащий Mg2+ PBS, плазмалита А или другой физиологический раствор с буфером или без него. Альтернативно, можно удалять нежелательные компоненты образца после афереза и напрямую ресуспендировать клетки в средах для культивирования.
В некоторых вариантах генетически модифицированная клетка представляет собой Т-лимфоцит.
В некоторых вариантах генетически модифицированная клетка представляет собой CD8+ T-клетку.
В некоторых вариантах генетически модифицированная клетка представляет собой NK-клетку.
В некоторых вариантах генетически модифицированную клетку используют в качестве лекарственного средства.
В некоторых вариантах генетически модифицированную клетку используют в качестве лекарственного средства при опухолевых заболеваниях.
В некоторых вариантах генетически модифицированную клетку используют в качестве лекарственного средства при заболеваниях, где опухоль представляет собой рак.
Примеры
Для наилучшего понимания изобретения приводятся следующие примеры. Эти примеры приведены только в иллюстративных целях и не должны толковаться как ограничивающие сферу применения изобретения в любой форме.
Все публикации, патенты и патентные заявки, указанные в этой спецификации включены в данный документ путем отсылки. Хотя вышеупомянутое изобретение было довольно подробно описано путем иллюстрации и примера в целях исключения двусмысленного толкования, специалистам в данной области на основе идей, раскрытых в данном изобретении, будет вполне понятно, что могут быть внесены определенные изменения и модификации без отклонения от сущности и объема прилагаемых вариантов осуществления изобретения.
Материалы и общие методы
Методы рекомбинантной ДНК
Для манипуляций с ДНК использовали стандартные методы, описанные у Sambrook J. и др., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии использовали согласно инструкциям производителей.
Синтез генов
Требуемые сегменты генов получали из олигонуклеотидов, созданных путем химического синтеза. Генные сегменты длиной от 300 до 4000 т.п.н., которые фланкированы уникальными сайтами рестрикции, собирали путем отжига и лигирования олигонуклеотидов, включая ПЦР-амплификацию и последующее клонирование через указанные сайты рестрикции. Последовательности ДНК субклонированных генных фрагментов подтверждали путем секвенирования ДНК.
Слияние генов, содержащих описанные цепи антитела, как указано ниже, осуществляли с помощью ПЦР и/или синтеза и сборки генов с использованием известных методов и процедур рекомбинации путем соединения соответствующих сегментов нуклеиновых кислот, например, с использованием уникальных сайтов рестрикции в соответствующих векторах. Субклонированные нуклеотидные последовательности подтверждали секвенированием ДНК. Для кратковременных трансфекций создавали большие количества плазмид посредством получения плазмид из трансформированных культур E. coli.
Определение последовательностей ДНК
Последовательности ДНК определяли путем секвенирования по Сенгеру.
Анализ последовательностей ДНК и белков и обработка данных о последовательностях
Применяли пакет программ Infomax's Vector NTI Advance suite, версия 8.0 и SnapGene Viewer для создания, картирования, анализа, аннотирования и иллюстрации последовательностей.
Протокол клонирования для получения вектора с ко-стимуляторным доменом 4-1BB
Фрагмент из трех повторов ITGB3 собирали из синтетических олигонуклеотидов (10 штук по 59 п.о. каждый), в результате двух раундов амплификации получали ампликон размером 308 п.о. Далее амплифицировали неизменяемую часть scFv (Leader-SVL-GS-linker-SVH-4-1BB CD3), к которой подшивали методом сплайсинг-ПЦР (SOE, Splicing by overlap extension) заданное количество фрагментов ITGB3 (1, 2 или 3, в зависимости от конструкционных особенностей конкретного вектора). Полученный продукт клонировали в заранее линеаризованный вектор pCDH-eGFP по сайтам XbaI/NotI. Валидировали полученные конструкции секвенированием всей последовательности кассеты.
Протокол клонирования для получения вектора с ко-стимуляторным доменом CD28
Фрагмент из трех повторов ITGB3 собирали из синтетических олигонуклеотидов (10 штук по 59 п.о. каждый), в результате двух раундов амплификации получали ампликон размером 308 п.о. Далее амплифицировали неизменяемую часть scFv (Leader-SVL-GS-linker-SVH-CD28-CD3), к которой подшивали методом сплайсинг-ПЦР (SOE, Splicing by overlap extension) заданное количество фрагментов ITGB3 (1, 2 или 3, в зависимости от конструкционных особенностей конкретного вектора). Полученный продукт клонировали в заранее линеаризованный вектор pCDH-eGFP по сайтам XbaI/NotI. Валидировали полученные конструкции секвенированием всей последовательности кассеты.
Пример 1
Получение последовательностей синтетических генов ITGB3.
Для получения блока из трех повторяющихся фрагментов ITGB3 использовали синтетические олигонуклеотиды по 60 п.о. каждый, образующие полностью перекрывающуюся последовательность участка гена, входящего в состав химерной структуры CD8 4-1BB altCD3 дзета или CD8 CD28 altCD3 дзета. Сборку гена производили методом двухраундовой ПЦР.
Пример 2
Последовательности кодон-оптимизированных фрагментов ITGB3.
Для совокупного расположения фрагментов ITGB3 в составе кассеты 19CAR-T и предотвращения возможной рекомбинации в гене были использованы 3 различных варианта нуклеотидных последовательностей, кодирующих ITGB3. Варианты таких последовательностей приведены ниже.
Вариант 1
gatactgccaacaacccactttacaaagaagctacatccaccttcaccaatataacctacagaggtact
D T A N N P L Y K E A T S T F T N I T Y R G T
Вариант 2
gacaccgctaacaatcctctgtataaagaggcaaccagcacttttacaaacattacttataggggaacc
D T A N N P L Y K E A T S T F T N I T Y R G T
Вариант 3
gatacagcaaataaccccttgtacaaggaagccacttctacattcactaatatcacctatcgcggcaca
D T A N N P L Y K E A T S T F T N I T Y R G T
Пример 3
Клонирование различных вариантов последовательности кассеты 19CAR-T-ITGB3 и 19CAR-T-CD28-ITGB3.
Для клонирования кассеты 19CAR-T-ITGB3-ITGB3-ITGB3 (фиг. 4) получали неизменяемый участок гена путем амплификации в ПЦР, используя в качестве матрицы вектор pCDH-CAR-T-EGFP, с участием специфических олигонуклеотидных праймеров, ограничивающих неизменяемую часть кассеты 19CAR-T. Методом сплайсинг-ПЦР (SOE, Splicing by overlap extension) производили сшивку участков гена 19CAR-T и блока из трех ITGB3 фрагментов.
Для клонирования кассеты 19CAR-T-ITGB3-ITGB3 (фиг. 2) аналогично получали участок гена путем амплификации в ПЦР, используя в качестве матрицы валидированную pCDH-19CAR-T-ITGB3-ITGB3-ITGB3, ограниченный специфическими праймерами и включающий в себя повтор ITGB3-ITGB3.
Для клонирования кассеты 19CAR-T-ITAM1-ITGB3-ITGB3 (фиг. 6) получали неизменяемый участок гена путем амплификации в ПЦР используя в качестве матрицы вектор pCDH-CAR-T-EGFP, с участием специфических олигонуклеотидных праймеров, ограничивающих неизменяемую часть 19CAR-T-ITAM1. Методом сплайсинг-ПЦР производили сшивку участков гена с блоком ITGB3-ITGB3.
Для клонирования кассеты 19CAR-T-ITAM1-ITAM2-ITGB3 (фиг. 8) получали неизменяемый участок гена путем амплификации в ПЦР используя в качестве матрицы вектор pCDH-CAR-T-EGFP, с участием специфических олигонуклеотидных праймеров, ограничивающих неизменяемую часть 19CAR-T-ITAM1-ITAM2. Методом сплайсинг-ПЦР производили сшивку участков гена с блоком ITGB3.
Для клонирования кассеты 19CAR-T-ITGB3-ITAM2-ITGB3 (фиг. 14) получали неизменяемый участок гена путем амплификации в ПЦР используя в качестве матрицы вектор pCDH-CAR-T-EGFP, с участием специфических олигонуклеотидных праймеров, ограничивающих неизменяемую часть с 19CAR-T. Методом сплайсинг-ПЦР производили сшивку участков гена с блоками из двух ITGB3 и ITAM2.
Для клонирования кассеты 19CAR-T-ITAM1-ITGB3-ITAM2 (фиг. 12) получали неизменяемый участок гена путем амплификации в ПЦР используя в качестве матрицы вектор pCDH-CAR-T-EGFP, с участием специфических олигонуклеотидных праймеров, ограничивающих неизменяемую часть 19CAR-T-ITAM1. Методом сплайсинг-ПЦР производили сшивку участков гена с блоком ITGB3 и ITAM2.
Аналогичным образом были получены кассеты с ко-стимуляторным доменом CD28, а именно:
экспрессионная кассета pCDH-19CAR-T_CD28-ITGB3-ITGB3-eGFP (Фиг. 16);
экспрессионная кассета pCDH-19CAR-T-CD28_ITGB3-ITGB3-ITGB3-eGFP (Фиг. 18);
экспрессионная кассета pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITGB3-eGFP (Фиг. 20);
экспрессионная кассета pCDH-19CAR-T-CD28_ITAM1-ITAM2-ITGB3-eGFP (Фиг. 22);
экспрессионная кассета pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITAM2-eGFP (Фиг. 24);
экспрессионная кассета pCDH-19CAR-T-CD28_ITGB3-ITAM2-ITGB3-eGFP (Фиг. 26).
Полученные кассеты интегрировали в состав плазмидного вектора pCDH-eGFP по специфическим сайтам рестрикции, куда в той же рамке считывания помещены элементы T2A и открытая рамка считывания зелёного флуоресцентного белка (eGFP) в качестве маркера экспрессии.
В результате были получены вектора, где CAR включает ко-стимуляторный домен 4-1BB и альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора с различным набором ITGB3 и ITAM, а именно:
вектор pCDH-19CAR-T-ITGB3-ITGB3-eGFP (Фиг. 1);
вектор pCDH-19CAR-T-ITGB3-ITGB3-ITGB3-eGFP (Фиг. 3);
вектор pCDH-19CAR-T-ITAM1-ITGB3-ITGB3-eGFP (Фиг. 5);
вектор pCDH-19CAR-T-ITAM1-ITAM2-ITGB3-eGFP (Фиг. 7);
вектор pCDH-19CAR-T-ITAM1-ITGB3-ITAM2-eGFP (Фиг. 11;.
вектор pCDH-19CAR-T -ITGB3-ITAM2-ITGB3-eGFP (Фиг. 13);
А также были получены вектора, где CAR включает ко-стимуляторный домен CD28 и альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора с различным набором ITGB3 и ITAM, а именно:
вектор pCDH-19CAR-T_CD28-ITGB3-ITGB3-eGFP (Фиг. 15);
вектор pCDH-19CAR-T-CD28_ITGB3-ITGB3-ITGB3-eGFP (Фиг. 17);
вектор pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITGB3-eGFP (Фиг. 19);
вектор pCDH-19CAR-T-CD28_ITAM1-ITAM2-ITGB3-eGFP (Фиг. 21);
вектор pCDH-19CAR-T-CD28_ITAM1-ITGB3-ITAM2-eGFP (Фиг. 23);
вектор pCDH-19CAR-T-CD28_ ITGB3-ITAM2-ITGB3-eGFP (Фиг. 25).
Пример 4
Выделение и активация Т-лимфоцитов.
У условно здорового добровольного донора было забрано 27 мл венозной периферической крови в стандартные пробирки Vacutainer, содержащие стерильный раствор гепарина натрия. Для получения фракции мононуклеаров в стерильную 15 мл центрифужную пробирку (Corning, 430791) переносили 3 мл раствора фиколла (градиент плотностью 1,077 г/л) (ПанЭко, Р052п) на который наслаивалась венозная кровь. Пробирки закручивались крышками и центрифугировали 18 минут при 1500 об/мин при температуре 24°С. Слой мононуклеарных клеток был собран с границы раздела фаз. Для получения популяции Т-лимфоцитов (CD3+) была произведена иммуномагнитная сепарация с негативной селекцией с использованием набора EasySep Human T Cell Enrichment Kit (Stemcell, 19051) согласно инструкции производителя. Был проведен один раунд иммуномагнитной сепарации.
Полученные Т-лимфоциты были сконцентрированы в 1 мл среды для культивирования (RPMI-1640 c L-глутамином (Биолот, 1.3.4.1) + 10% cыворотки крови эмбриональной телячьей (Gibco, 26140079) + 100U IL2 (Ронколейкин, ООО «НПК»БИОТЕХ»)). Концентрация Т-лимфоцитов была подсчитана при помощи счетчика клеток и анализатора жизнеспособности клеток Countess® II FL (Thermofisher, AMQAF1000). 12×106 Т-лимфоцитов были перенесены в культуральный флакон Т75 для суспензионных клеточных культур (Eppendorf, 0030711025) с 15 мл среды для культивирования. Для активации Т-лимфоцитов были добавлены 300 мкл Dynabeads Human T-Activator CD3/CD28 for T Cell Expansion and Activation (Thermofisher, 11132D). Культуральный флакон был помещен на 72 часа в СО2 инкубатор (37°С, 5% СО2).
Все работы выполнялись в стерильных условиях в ламинарных шкафах второго класса биобезопасности.
Пример 5
Трансдукция активированных Т-лимфоцитов лентивирусным препаратом.
Для трансдукции Т-лимфоцитов человека были использованы два лентивирусных препарата: первый содержащий гены anti-CD19-CAR-ITAMх3 и GFP (контроль) и второй препарат (экспериментальный), который выбирают из группы векторов, полученных в примере 3 (Фиг. 1, Фиг. 3, Фиг. 5, Фиг. 7, Фиг. 11, Фиг. 13, Фиг. 15, Фиг. 17, Фиг. 19, Фиг. 21, Фиг. 23, Фиг. 25). В обоих препаратах методом ИФА была определена концентрация лентивирусных частиц (LVP/мл). Трансдукция была поставлена в формате 15 мл биореактора (TPP, 87017). В качестве активатора трансдукции был применен протамина сульфат (ООО Эллара) в концентрации 50 мкг/мл. 5×105 активированных Т-лимфоцитов, лентивирусный препарат (в количестве 2×105 LVP/клетку), протамина сульфат и 10 мл культуральной среды вносили в биореактор, биореактор оставляли на 5 часов в шейкере Multitron (110 RPM; 37°C; 5% CO2). Через 5 часов биореактор доставали и центрифугировали 5 минут при 1500 об/мин, 24°С. Надосадок удаляли, осадок ресуспендировали в 2 мл культуральной среды, переносили в культуральный 6-луночный планшет для суспензионных культур (Eppendorf, 0030720016) и помещали в CO2 инкубатор (37°С, 5% СО2). Анализ трансдукции проводили через 72 часа. Эффективность трансдукции Т-лимфоцитов оценивалась по уровню сигнала репортерного белка GFP, оценка эффективности проводилась методом проточной цитометрии. Для анализа жизнеспособности клеток применялся краситель пропидий йодид.
Цитотоксическая активность полученных CAR-T-лимфоцитов оценивалась в присутствии клеток-мишеней, экспрессирующих рецептор CD19, при прямом со-культивировании. В лунку 48-луночного планшета для суспензионных культур (Eppendorf, 0030723015) в 1 мл культуральной среды (RPMI-1640 c L-глутамином (Биолот, 1.3.4.1) + 10% cыворотки крови эмбриональной телячьей (Gibco, 26140079)) переносились по 10000 CAR-T лимфоцитов и 5000 клеток-мишеней (Raji). Время инкубации составило 72 часа в СО2 инкубаторе (37°С, 5% СО2). Анализ проводили на проточном цитометре, оценивалось соотношение CD3 положительных и CD3 негативных клеток.
Для анализа количества цитокинов в культуральной среде был использован набор для определения цитокинов BD Cytometric Bead Array (CBA) Human Th1/Th2 Cytokine Kit II (BD, 551809). Анализ проводился согласно инструкции производителя.
Наличие CAR на поверхности клетки при взаимодействии с антигеном вызывает активацию NFAT сигнального каскада. Использование репортерной клеточной линии JurkatΔTCRab NFAT-GFP позволило провести скрининг генетических конструкций anti-CD19-CAR-ITAMх3 (контроль) и препарата экспериментальной генетической конструкции, которую выбирают из группы векторов на Фиг. 1, Фиг. 3, Фиг. 5, Фиг. 7, Фиг. 11, Фиг. 13, Фиг. 15, Фиг. 17, Фиг. 19, Фиг. 21, Фиг. 23, Фиг. 25). Трансфекция репортерной клеточной линии осуществлялась методом электропорации (электропоратор: Neon, Thermo Scientific, США). В качестве клеток-мишеней, экспрессирующих рецептор CD19, использовали клетки Raji, инкубация составляла 6 часов в СО2 инкубаторе (37°С, 5% СО2). Анализ проводили на проточном цитометре, оценивалась интенсивность свечения репортного белка GFP (фиг. 27).
Пример 6
Результаты.
С помощью трансдукции рекомбинантными лентивирусными векторами были получены CAR-T, экспрессирующие контрольный CAR-ITAMx3, и экспериментальные CAR-T, которые экспрессировали один из вариантов CAR (в соответствии с выбранным вектором из группы векторов, указанных на Фиг. 1, Фиг. 3, Фиг. 5, Фиг. 7, Фиг. 11, Фиг. 13, Фиг. 15, Фиг. 17, Фиг. 19, Фиг. 21, Фиг. 23, Фиг. 25), а именно:
CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITGB3x2);
CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITGB3x3);
CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITAMx2-ITGB3x1);
CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITAMx1-ITGB3x2);
CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITGB3x2);
CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITGB3x3);
CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITAMx2-ITGB3x1);
CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITAMx1-ITGB3x2).
Уровень экспрессии GFP в экспериментальных клетках (17,6%) был сопоставим с уровнем GFP в контроле (15,63%).
При прямом со-культивировании экспериментальных CAR-T, где CAR включает сигнальный домен с только ITGB3x2 или только ITGB3x3 («CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITGB3x2)»; «CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITGB3x3)»; «CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITGB3x2)»; «CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITGB3x3)») с CD19+ клетками-мишенями линии Raji (2:1) наблюдалась низкая цитотоксическая активность по сравнению с контрольными CAR-ITAMx3.
Поэтому вышеуказанные CAR-T, где CAR включает сигнальный домен с только ITGB3x2 или только ITGB3x3 были отбракованы и не являются предметом настоящего изобретения.
При прямом со-культивировании различных экспериментальных CAR-ITAMx2-ITGB3x1 («CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITAMx1-ITGB3x2)»; «CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITAMx2-ITGB3x1)») с CD19+ клетками-мишенями линии Raji (2:1) наблюдалась сопоставимая цитотоксическая активность с контрольными CAR-ITAMx3. Культивирование обоих вариантов CAR клеток сопровождалось секрецией характерных для CAR-T лимфоцитов провоспалительных цитокинов IL-2 и IFN-γ (фиг. 28 (A, B, C)).
Аналогично вышеуказанному, при прямом со-культивировании различных экспериментальных CAR-ITAMx1-ITGB3x2 («CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITAMx1-ITGB3x2)»; «CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITAMx1-ITGB3x2)») с CD19+ клетками-мишенями линии Raji (2:1) с CD19+ клетками-мишенями линии Raji (2:1) наблюдалась сопоставимая цитотоксическая активность с контрольными CAR-ITAMx3.
Добавление клеток-мишеней (Raji CD19+) к репортерной клеточной линии JurkatΔTCRab NFAT-GFP, экспрессирующей контрольный CAR-ITAMx3 и различные экспериментальные CAR-ITAMx1-ITGB3x2 («CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITAMx1-ITGB3x2)»; «CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITAMx1-ITGB3x2)»), вызывает CAR опосредованную активацию NFAT сигнального каскада. Таким образом, наличие синтетического активационного домена в составе CAR рецептора не приводит к существенному падению CAR зависимой активации клеток, экспрессирующих соответствующую генетическую конструкцию (фиг. 29).
Аналогично вышеуказанному, добавление клеток-мишеней (Raji CD19+) к репортерной клеточной линии JurkatΔTCRab NFAT-GFP, экспрессирующей контрольный CAR-ITAMx3 и различные экспериментальные CAR-ITAMx1-ITGB3x2 («CAR-(ко-стимуляторным доменом 4-1BB) - (сигнальный домен с ITAMx1-ITGB3x2)»; «CAR-(ко-стимуляторным доменом CD28) - (сигнальный домен с ITAMx1-ITGB3x2)»), вызывает CAR опосредованную активацию NFAT сигнального каскада. Таким образом, наличие синтетического активационного домена в составе CAR рецептора не приводит к существенному падению CAR зависимой активации клеток, экспрессирующих соответствующую генетическую конструкцию.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ООО "АНАБИОН"
<120> Выделенный альтернативный внутриклеточный сигнальный домен химерного
антигенного рецептора и включающий его химерный антигенный рецептор
<160> 35
<170> BiSSAP 1.3.6
<210> 1
<211> 29
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITAM1
<400> 1
Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
1 5 10 15
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
20 25
<210> 2
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITAM2
<400> 2
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
1 5 10 15
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
20 25
<210> 3
<211> 29
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITAM3
<400> 3
Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser
1 5 10 15
Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
20 25
<210> 4
<211> 23
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITGB3
<400> 4
Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr
1 5 10 15
Asn Ile Thr Tyr Arg Gly Thr
20
<210> 5
<211> 106
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITAM1- ITAM2- ITGB3
<400> 5
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Asp Thr
65 70 75 80
Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile
85 90 95
Thr Tyr Arg Gly Thr Ala Leu Pro Pro Arg
100 105
<210> 6
<211> 106
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITGB3- ITAM2- ITAM3
<400> 6
Arg Val Lys Phe Ser Arg Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu
1 5 10 15
Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr
20 25 30
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
35 40 45
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
50 55 60
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
65 70 75 80
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
85 90 95
Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105
<210> 7
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITAM1- ITGB3- ITAM3
<400> 7
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr
50 55 60
Asn Ile Thr Tyr Arg Gly Thr Lys Gly Glu Arg Arg Arg Gly Lys Gly
65 70 75 80
His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr
85 90 95
Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105
<210> 8
<211> 101
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITAM1- ITGB3- ITGB3
<400> 8
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr
50 55 60
Asn Ile Thr Tyr Arg Gly Thr Lys Gly Asp Thr Ala Asn Asn Pro Leu
65 70 75 80
Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr
85 90 95
Ala Leu Pro Pro Arg
100
<210> 9
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITGB3- ITAM2- ITGB3
<400> 9
Arg Val Lys Phe Ser Arg Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu
1 5 10 15
Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr
20 25 30
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
35 40 45
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
50 55 60
Tyr Ser Glu Ile Gly Met Lys Gly Asp Thr Ala Asn Asn Pro Leu Tyr
65 70 75 80
Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Ala
85 90 95
Leu Pro Pro Arg
100
<210> 10
<211> 101
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ITGB3-ITGB3- ITAM3
<400> 10
Arg Val Lys Phe Ser Arg Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu
1 5 10 15
Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr
20 25 30
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Asp Thr Ala Asn Asn Pro
35 40 45
Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly
50 55 60
Thr Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln
65 70 75 80
Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
85 90 95
Ala Leu Pro Pro Arg
100
<210> 11
<211> 245
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность антигенсвязывающего домена,
который специфичен к опухолевому антигену CD19
<400> 11
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr His Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr Gly Ser Thr Ser Gly
100 105 110
Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Lys
115 120 125
Leu Gln Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln Ser Leu Ser
130 135 140
Val Thr Cys Thr Val Ser Gly Val Ser Leu Pro Asp Tyr Gly Val Ser
145 150 155 160
Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu Glu Trp Leu Gly Val Ile
165 170 175
Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys Ser Arg Leu
180 185 190
Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln Val Phe Leu Lys Met Asn
195 200 205
Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Ala Lys His Tyr
210 215 220
Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser
225 230 235 240
Val Thr Val Ser Ser
245
<210> 12
<211> 24
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность трансмембранного домена CD8
<400> 12
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr Leu Tyr Cys
20
<210> 13
<211> 222
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ко-стимуляторнного домена CD28
<400> 13
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
1 5 10 15
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
20 25 30
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
35 40 45
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
50 55 60
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
65 70 75 80
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
85 90 95
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
100 105 110
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
115 120 125
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
130 135 140
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
145 150 155 160
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
165 170 175
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
180 185 190
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
195 200 205
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
210 215 220
<210> 14
<211> 42
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность ко-стимуляторнного домена 4- 1BB
<400> 14
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 15
<211> 669
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- CD28 - cd3dzeta (ITGB3-
ITAM2- ITAM3)
<400> 15
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
340 345 350
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
355 360 365
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
370 375 380
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
385 390 395 400
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
405 410 415
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
420 425 430
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
435 440 445
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
450 455 460
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
465 470 475 480
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
485 490 495
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
500 505 510
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
515 520 525
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
530 535 540
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Val
545 550 555 560
Lys Phe Ser Arg Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu Tyr Lys
565 570 575
Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Arg Gly
580 585 590
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
595 600 605
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
610 615 620
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
625 630 635 640
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
645 650 655
His Met Gln Ala Leu Pro Pro Arg Gly Ser Ala Ala Ala
660 665
<210> 16
<211> 670
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- CD28 - cd3dzeta (ITAM1-
ITGB3- ITAM3)
<400> 16
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
340 345 350
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
355 360 365
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
370 375 380
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
385 390 395 400
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
405 410 415
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
420 425 430
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
435 440 445
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
450 455 460
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
465 470 475 480
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
485 490 495
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
500 505 510
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
515 520 525
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
530 535 540
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Val
545 550 555 560
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn
565 570 575
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
580 585 590
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Asp Thr
595 600 605
Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile
610 615 620
Thr Tyr Arg Gly Thr Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
625 630 635 640
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
645 650 655
Leu His Met Gln Ala Leu Pro Pro Arg Gly Ser Ala Ala Ala
660 665 670
<210> 17
<211> 669
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- CD28 - cd3dzeta (ITAM1-
ITAM2- ITGB3)
<400> 17
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
340 345 350
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
355 360 365
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
370 375 380
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
385 390 395 400
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
405 410 415
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
420 425 430
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
435 440 445
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
450 455 460
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
465 470 475 480
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
485 490 495
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
500 505 510
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
515 520 525
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
530 535 540
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Val
545 550 555 560
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn
565 570 575
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
580 585 590
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
595 600 605
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
610 615 620
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Asp Thr Ala Asn
625 630 635 640
Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr
645 650 655
Arg Gly Thr Ala Leu Pro Pro Arg Gly Ser Ala Ala Ala
660 665
<210> 18
<211> 664
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- CD28 - cd3dzeta (ITGB3-
ITGB3- ITAM3)
<400> 18
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
340 345 350
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
355 360 365
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
370 375 380
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
385 390 395 400
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
405 410 415
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
420 425 430
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
435 440 445
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
450 455 460
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
465 470 475 480
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
485 490 495
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
500 505 510
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
515 520 525
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
530 535 540
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Val
545 550 555 560
Lys Phe Ser Arg Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu Tyr Lys
565 570 575
Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Arg Gly
580 585 590
Arg Asp Pro Glu Met Gly Gly Lys Asp Thr Ala Asn Asn Pro Leu Tyr
595 600 605
Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Lys
610 615 620
Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu
625 630 635 640
Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu
645 650 655
Pro Pro Arg Gly Ser Ala Ala Ala
660
<210> 19
<211> 664
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- CD28 - cd3dzeta (ITAM1-
ITGB3- ITGB3)
<400> 19
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
340 345 350
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
355 360 365
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
370 375 380
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
385 390 395 400
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
405 410 415
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
420 425 430
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
435 440 445
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
450 455 460
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
465 470 475 480
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
485 490 495
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
500 505 510
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
515 520 525
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
530 535 540
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Val
545 550 555 560
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn
565 570 575
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
580 585 590
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Asp Thr
595 600 605
Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile
610 615 620
Thr Tyr Arg Gly Thr Lys Gly Asp Thr Ala Asn Asn Pro Leu Tyr Lys
625 630 635 640
Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Ala Leu
645 650 655
Pro Pro Arg Gly Ser Ala Ala Ala
660
<210> 20
<211> 663
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- CD28 - cd3dzeta (ITGB3
- ITAM2- ITGB3)
<400> 20
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Ala Ala Ala Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu
340 345 350
Lys Ser Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro
355 360 365
Ser Pro Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val
370 375 380
Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe
385 390 395 400
Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp
405 410 415
Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr
420 425 430
Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val
435 440 445
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
450 455 460
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
465 470 475 480
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
485 490 495
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
500 505 510
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
515 520 525
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
530 535 540
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Val
545 550 555 560
Lys Phe Ser Arg Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu Tyr Lys
565 570 575
Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Arg Gly
580 585 590
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
595 600 605
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
610 615 620
Glu Ile Gly Met Lys Gly Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu
625 630 635 640
Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Ala Leu Pro
645 650 655
Pro Arg Gly Ser Ala Ala Ala
660
<210> 21
<211> 489
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- 41bb - cd3dzeta (ITGB3-
ITAM2- ITAM3)
<400> 21
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser
385 390 395 400
Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Arg Gly Arg Asp Pro Glu
405 410 415
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
420 425 430
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
435 440 445
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
450 455 460
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
465 470 475 480
Leu Pro Pro Arg Gly Ser Ala Ala Ala
485
<210> 22
<211> 490
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- 41bb - cd3dzeta (ITAM1-
ITGB3- ITAM3)
<400> 22
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Asp Thr Ala Asn Asn Pro
420 425 430
Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly
435 440 445
Thr Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln
450 455 460
Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
465 470 475 480
Ala Leu Pro Pro Arg Gly Ser Ala Ala Ala
485 490
<210> 23
<211> 489
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- 41bb - cd3dzeta (ITAM1-
ITAM2- ITGB3)
<400> 23
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Asp Thr Ala Asn Asn Pro Leu Tyr
450 455 460
Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Ala
465 470 475 480
Leu Pro Pro Arg Gly Ser Ala Ala Ala
485
<210> 24
<211> 484
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T - 41bb - cd3dzeta
(ITGB3- ITGB3- ITAM3)
<400> 24
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser
385 390 395 400
Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Arg Gly Arg Asp Pro Glu
405 410 415
Met Gly Gly Lys Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr
420 425 430
Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Lys Gly Glu Arg Arg
435 440 445
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr
450 455 460
Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Gly
465 470 475 480
Ser Ala Ala Ala
<210> 25
<211> 484
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- 41bb - cd3dzeta (ITAM1-
ITGB3- ITGB3)
<400> 25
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Asp Thr Ala Asn Asn Pro
420 425 430
Leu Tyr Lys Glu Ala Thr Ser Thr Phe Thr Asn Ile Thr Tyr Arg Gly
435 440 445
Thr Lys Gly Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser
450 455 460
Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Ala Leu Pro Pro Arg Gly
465 470 475 480
Ser Ala Ala Ala
<210> 26
<211> 483
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность 19CAR-T- 41bb - cd3dzeta (ITGB3
- ITAM2- ITGB3)
<400> 26
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser
20 25 30
Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr
85 90 95
Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln
100 105 110
Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
115 120 125
Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
130 135 140
Thr Lys Gly Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
145 150 155 160
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
165 170 175
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
180 185 190
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
195 200 205
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
210 215 220
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
225 230 235 240
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser
385 390 395 400
Thr Phe Thr Asn Ile Thr Tyr Arg Gly Thr Arg Gly Arg Asp Pro Glu
405 410 415
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
420 425 430
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
435 440 445
Lys Gly Asp Thr Ala Asn Asn Pro Leu Tyr Lys Glu Ala Thr Ser Thr
450 455 460
Phe Thr Asn Ile Thr Tyr Arg Gly Thr Ala Leu Pro Pro Arg Gly Ser
465 470 475 480
Ala Ala Ala
<210> 27
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность CD3 дзета
<400> 27
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 28
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность СD8 hinge
<400> 28
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 29
<211> 22
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность лидерного пептида
<400> 29
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro
20
<210> 30
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность вариабельного домена легкой цепи
(VL) антигенсвязывающего домена, который специфичен к опухолевому
антигену CD19
<400> 30
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr His Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
100 105
<210> 31
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность вариабельного домена тяжелой
цепи (VH) антигенсвязывающего домена, который специфичен к
опухолевому антигену CD19
<400> 31
Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu Pro Asp Tyr
20 25 30
Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 32
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность линкера
<400> 32
Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Thr
1 5 10 15
Lys Gly
<210> 33
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность T2A
<400> 33
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
1 5 10 15
Gly Pro
<210> 34
<211> 239
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность eGFP
<400> 34
Met Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu
1 5 10 15
Val Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly
20 25 30
Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile
35 40 45
Cys Thr Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu Val Thr Thr
50 55 60
Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys
65 70 75 80
Gln His Asp Phe Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu
85 90 95
Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu
100 105 110
Val Lys Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly
115 120 125
Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr
130 135 140
Asn Tyr Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys Asn
145 150 155 160
Gly Ile Lys Val Asn Phe Lys Ile Arg His Asn Ile Glu Asp Gly Ser
165 170 175
Val Gln Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly
180 185 190
Pro Val Leu Leu Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu
195 200 205
Ser Lys Asp Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe
210 215 220
Val Thr Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys
225 230 235
<210> 35
<211> 45
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность шарнирного домена CD8
<400> 35
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2018 |
|
RU2770002C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2016 |
|
RU2752880C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНОВ | 2015 |
|
RU2745705C2 |
| CD19-СПЕЦИФИЧЕСКИЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2014 |
|
RU2727447C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ M971 | 2018 |
|
RU2770411C1 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ К BCMA | 2015 |
|
RU2747457C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА FLT3 | 2018 |
|
RU2820859C2 |
| Т-КЛЕТКИ С КОСТИМУЛИРУЮЩИМ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ, НАЦЕЛЕННЫЕ НА IL13Rα2 | 2015 |
|
RU2749922C2 |
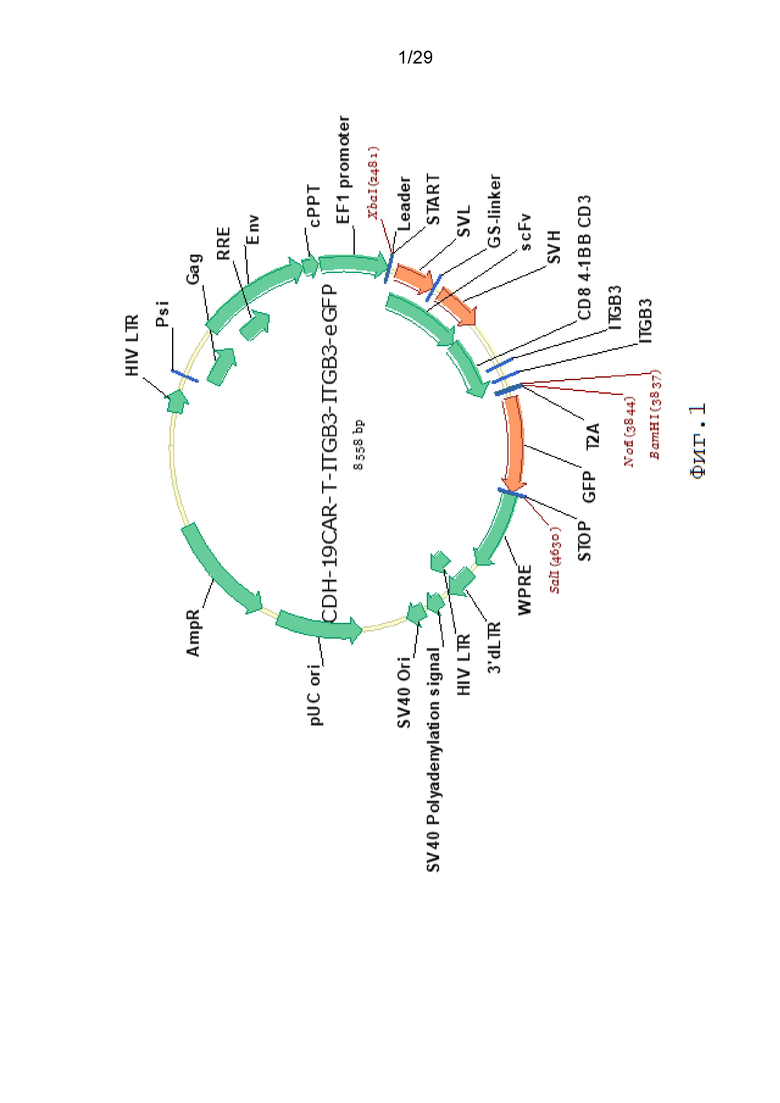
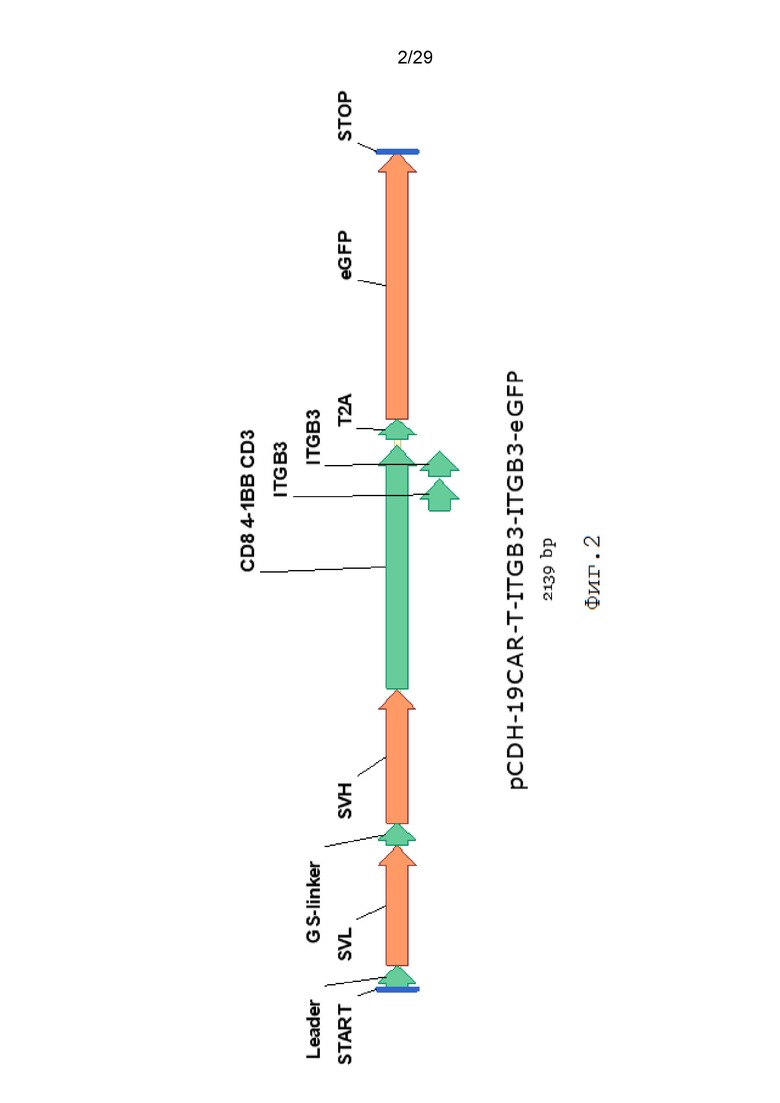
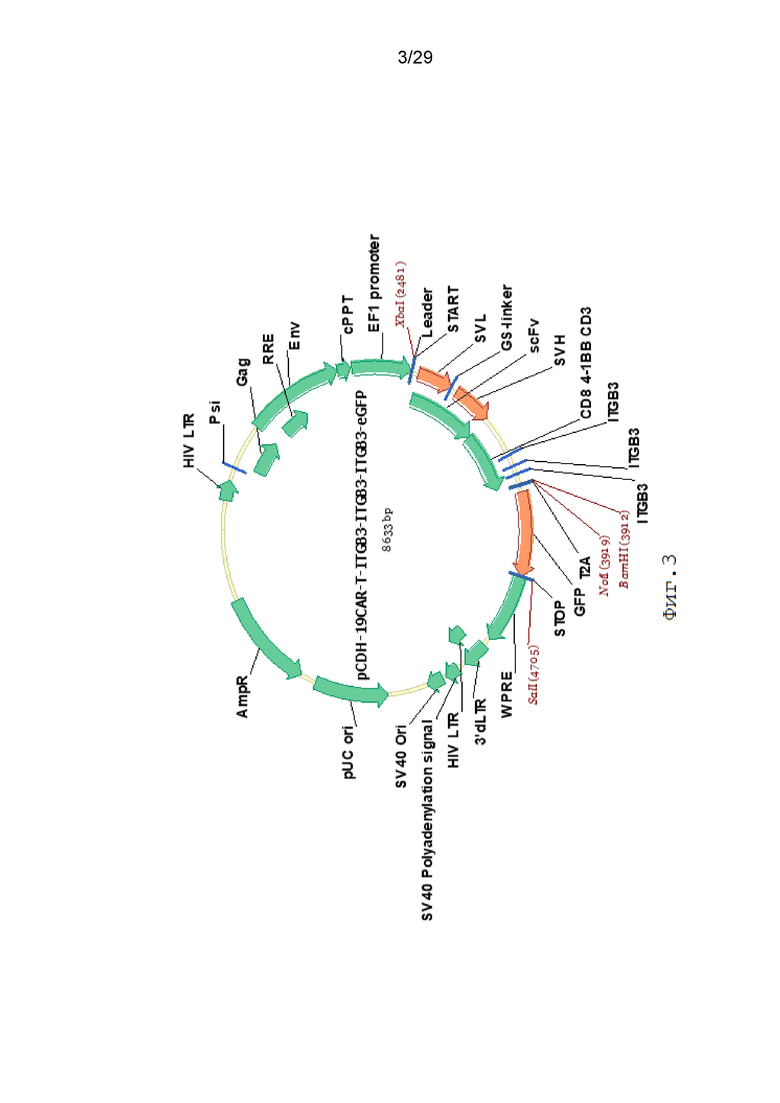
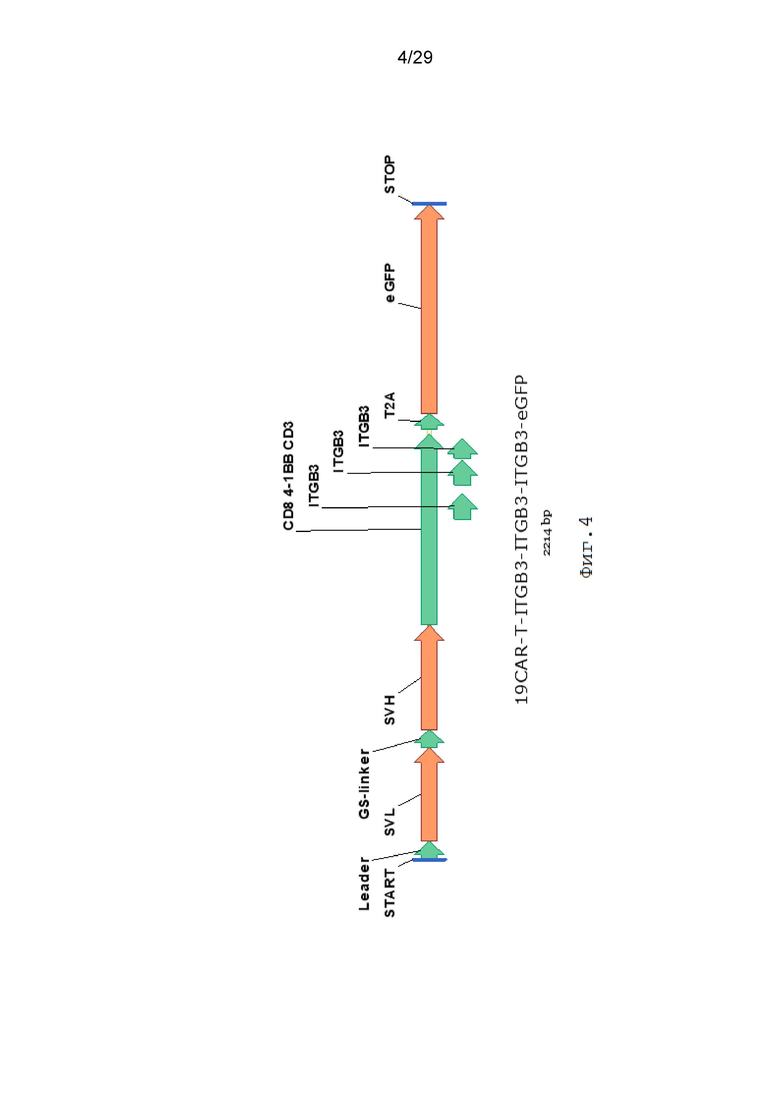
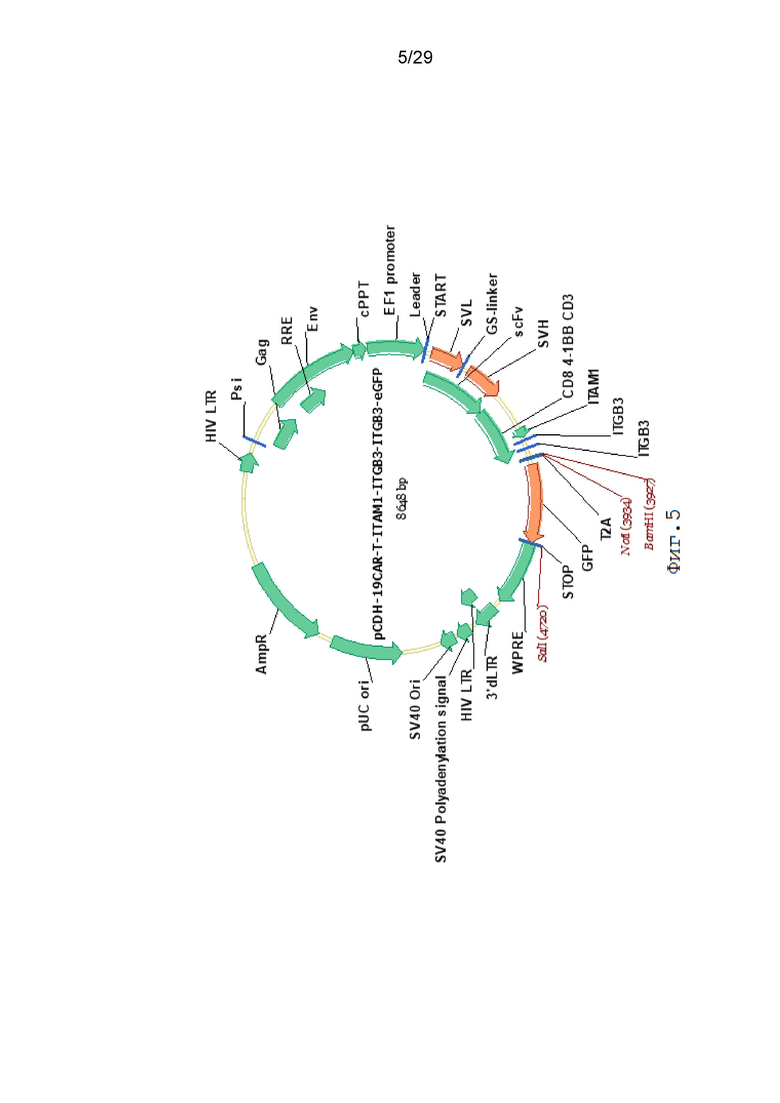
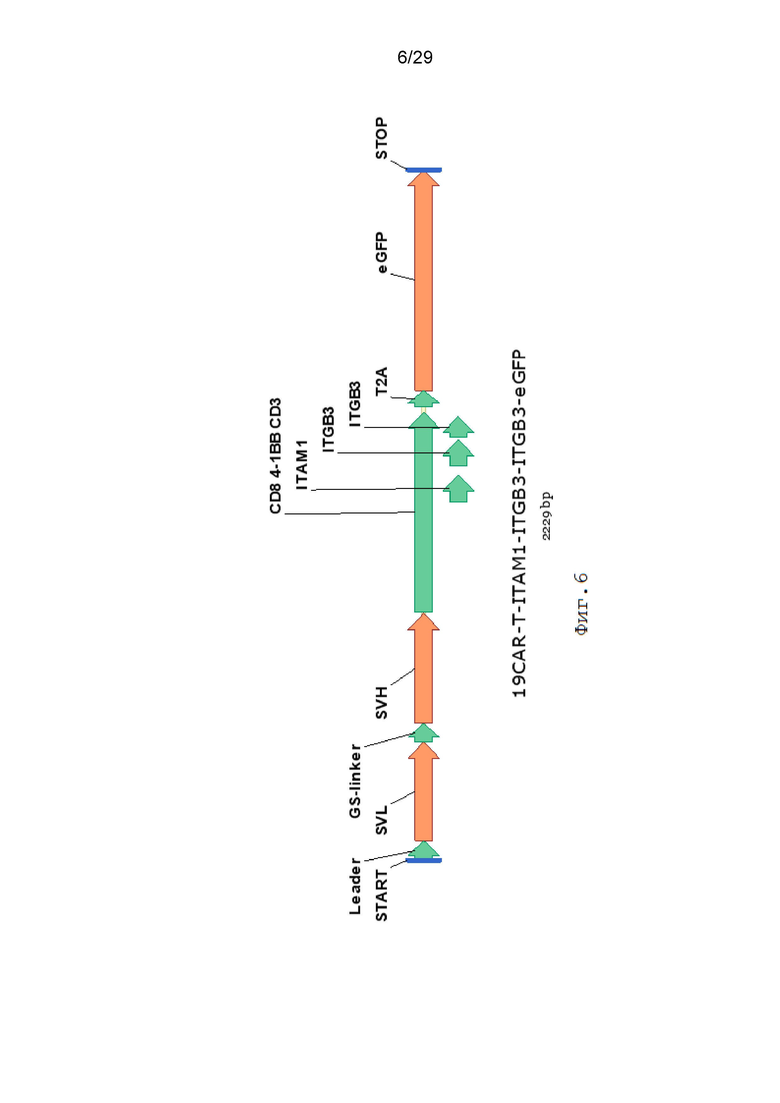
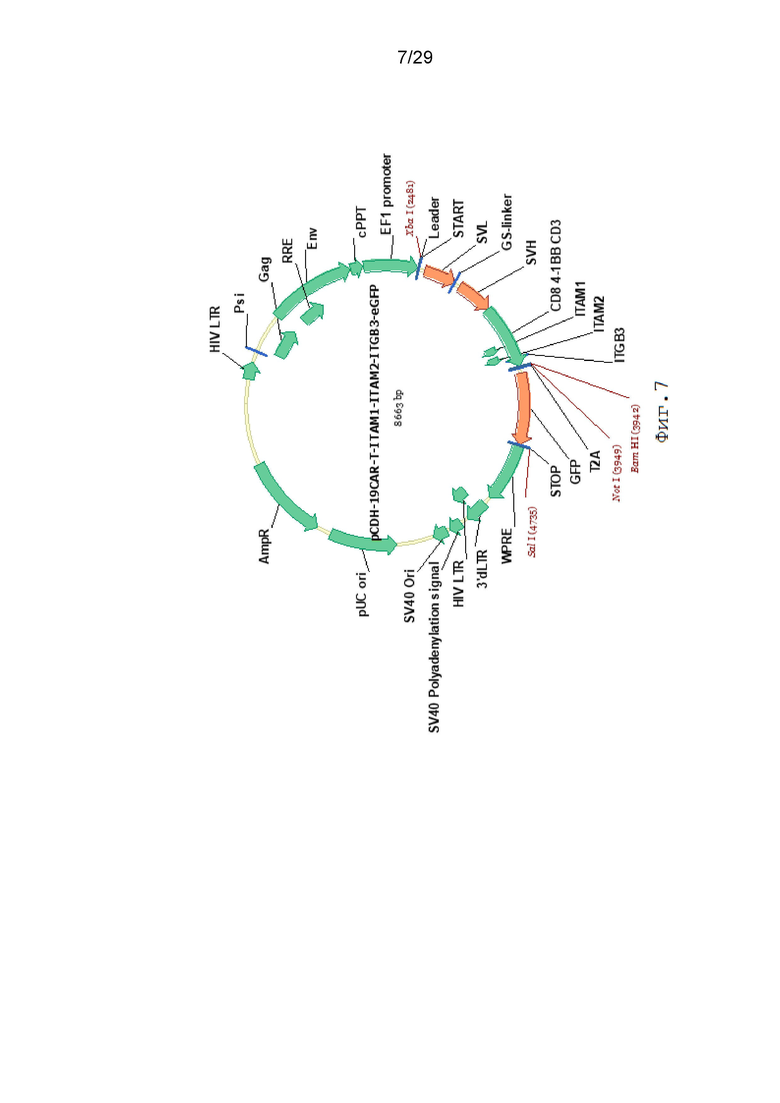
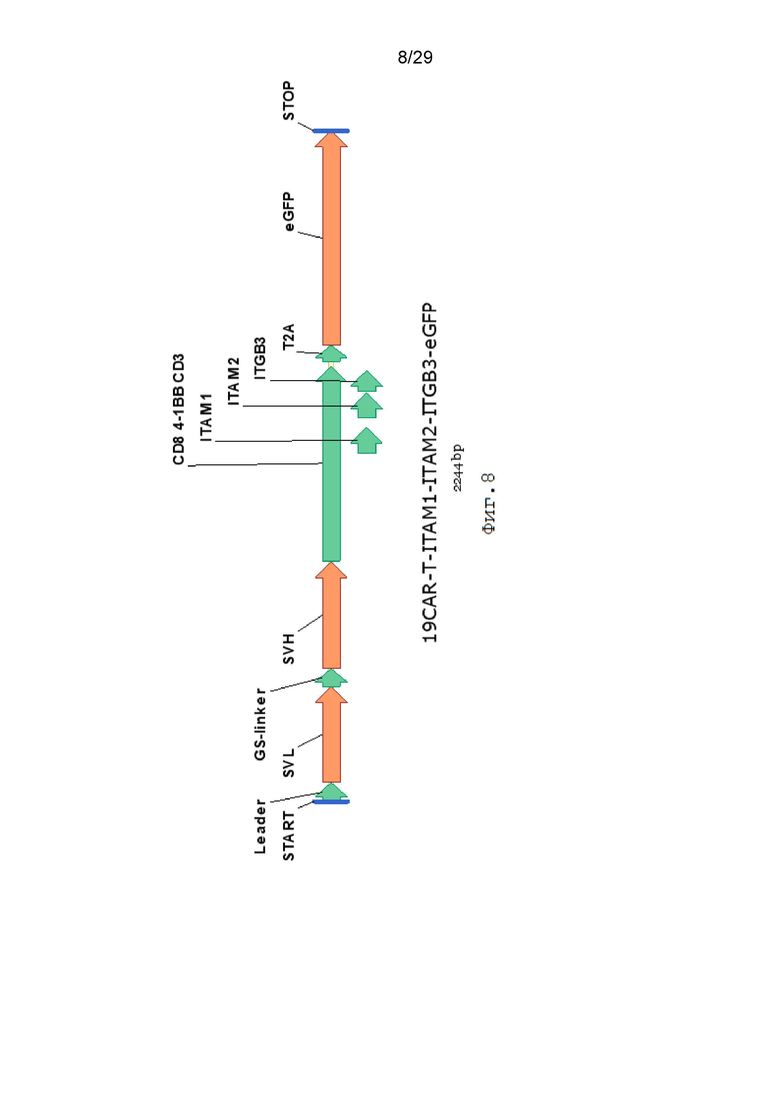
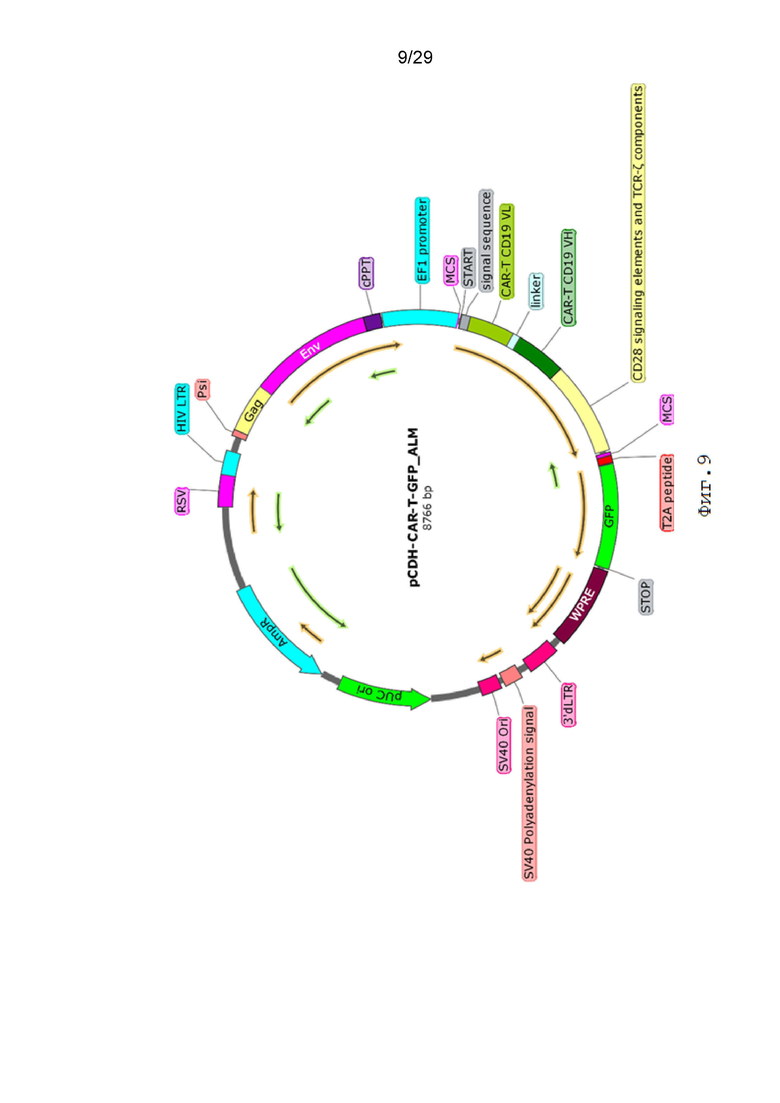
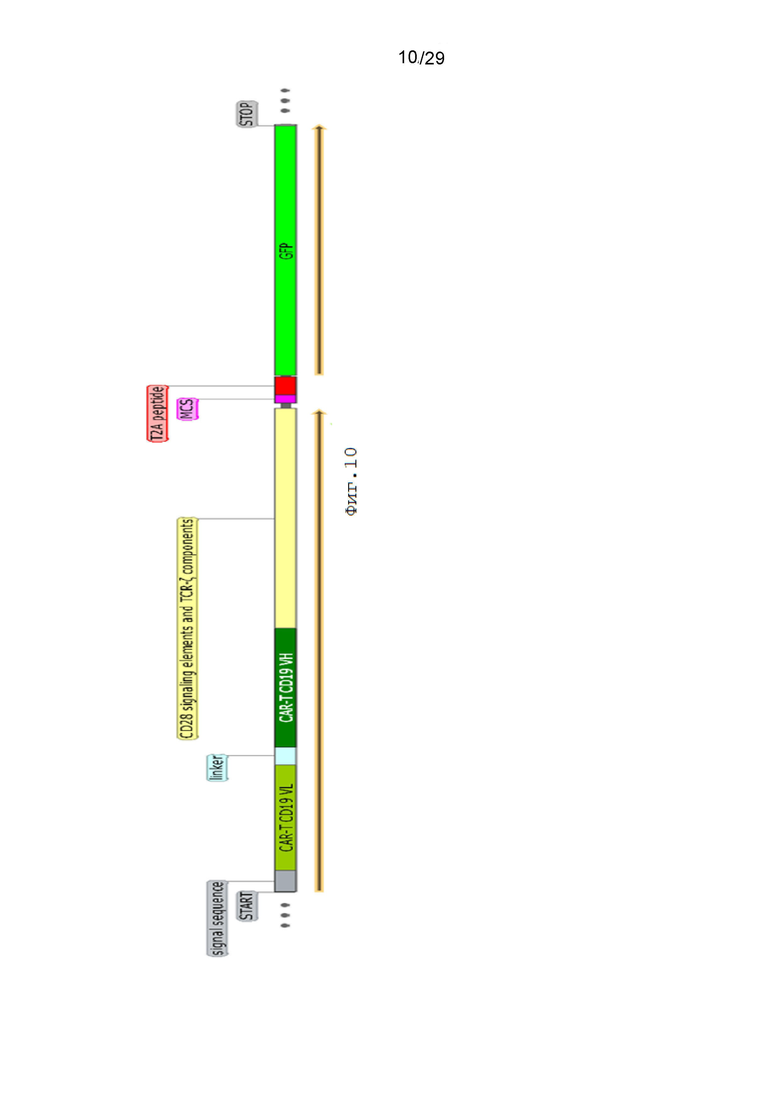
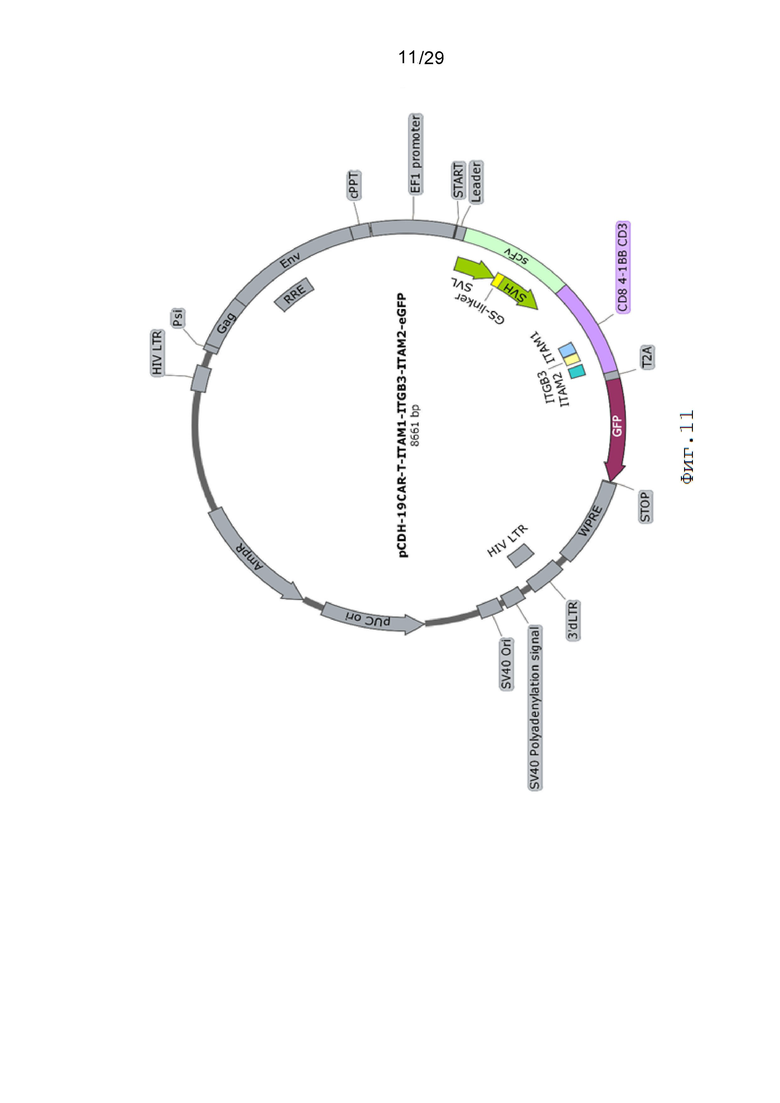
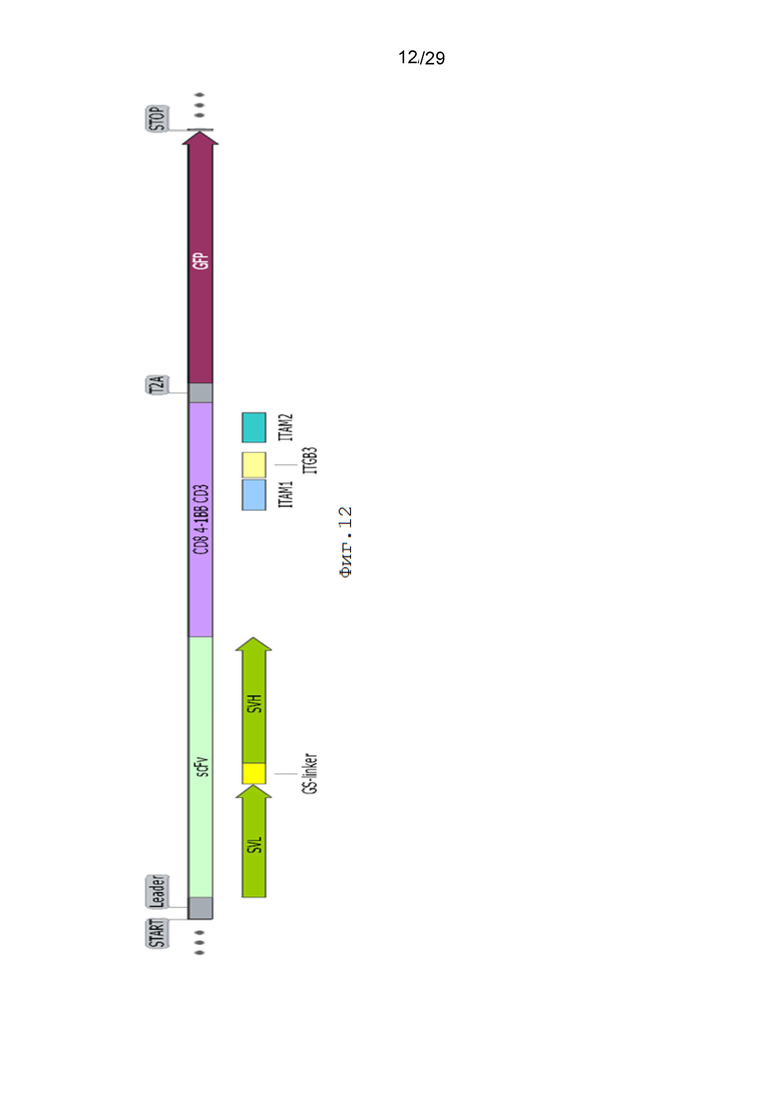
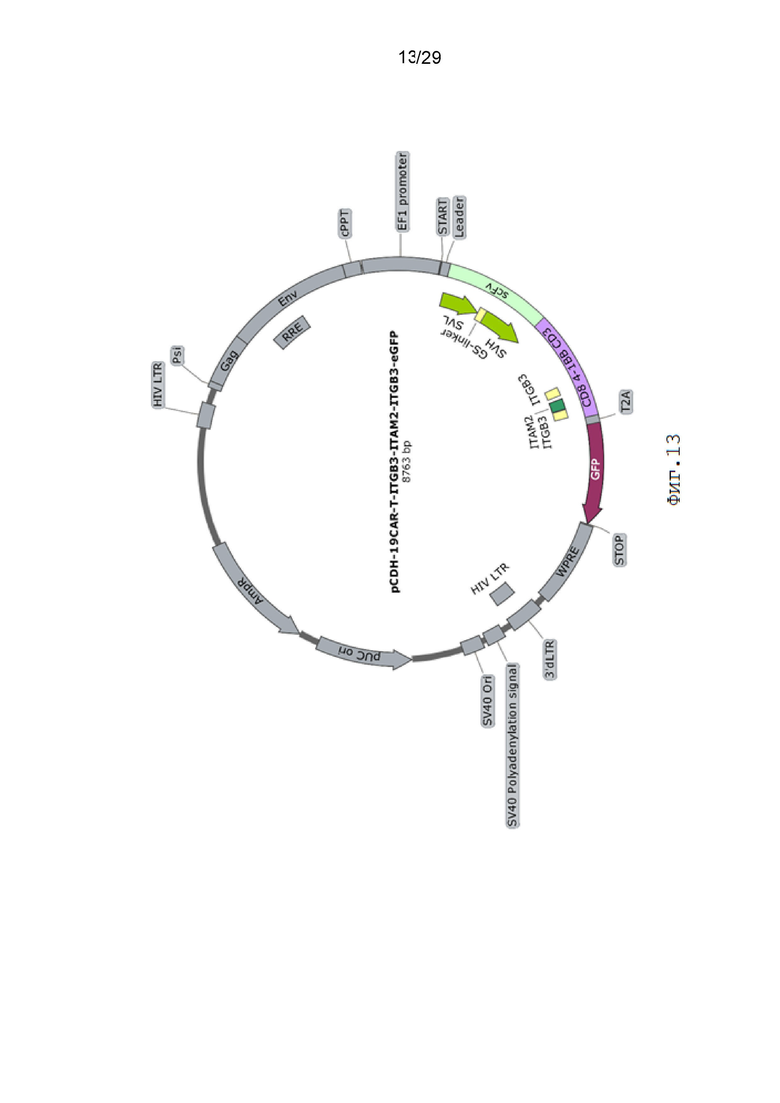
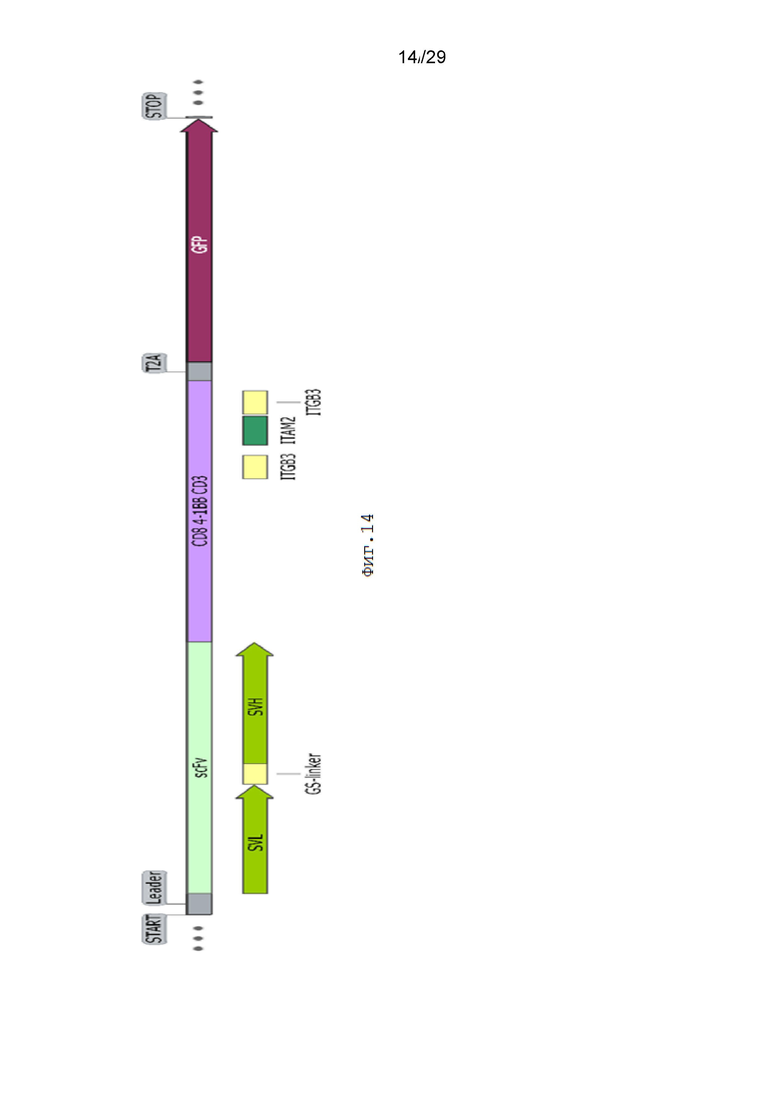
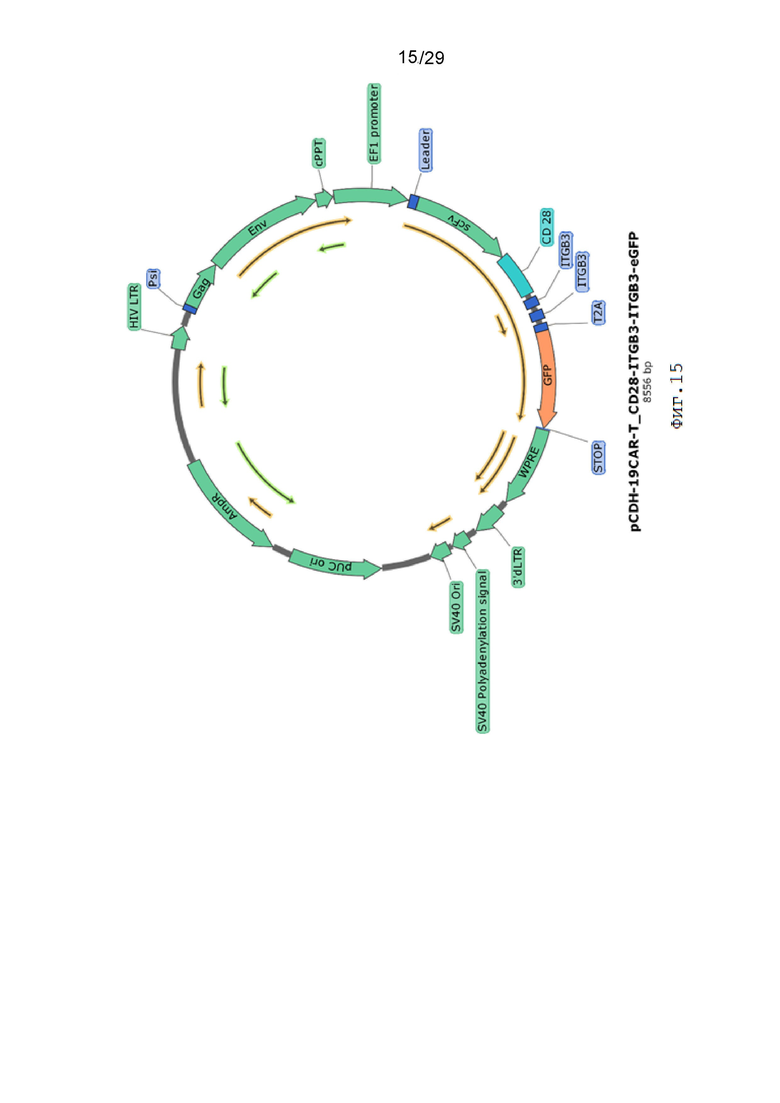
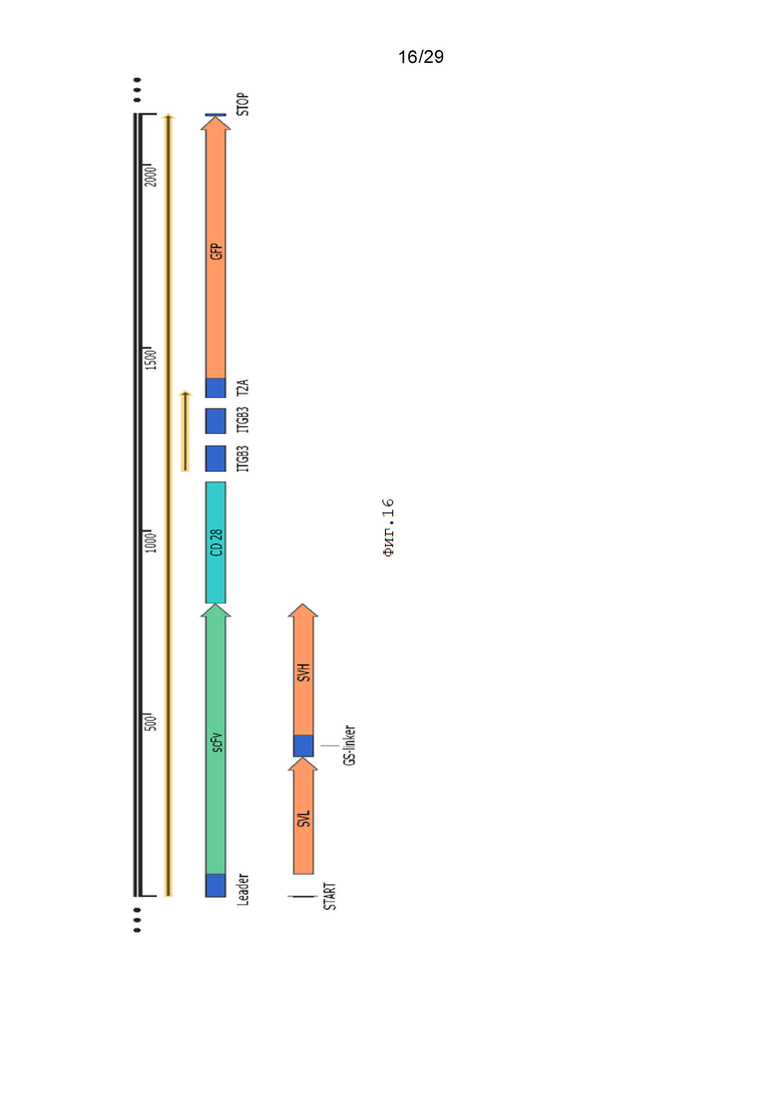
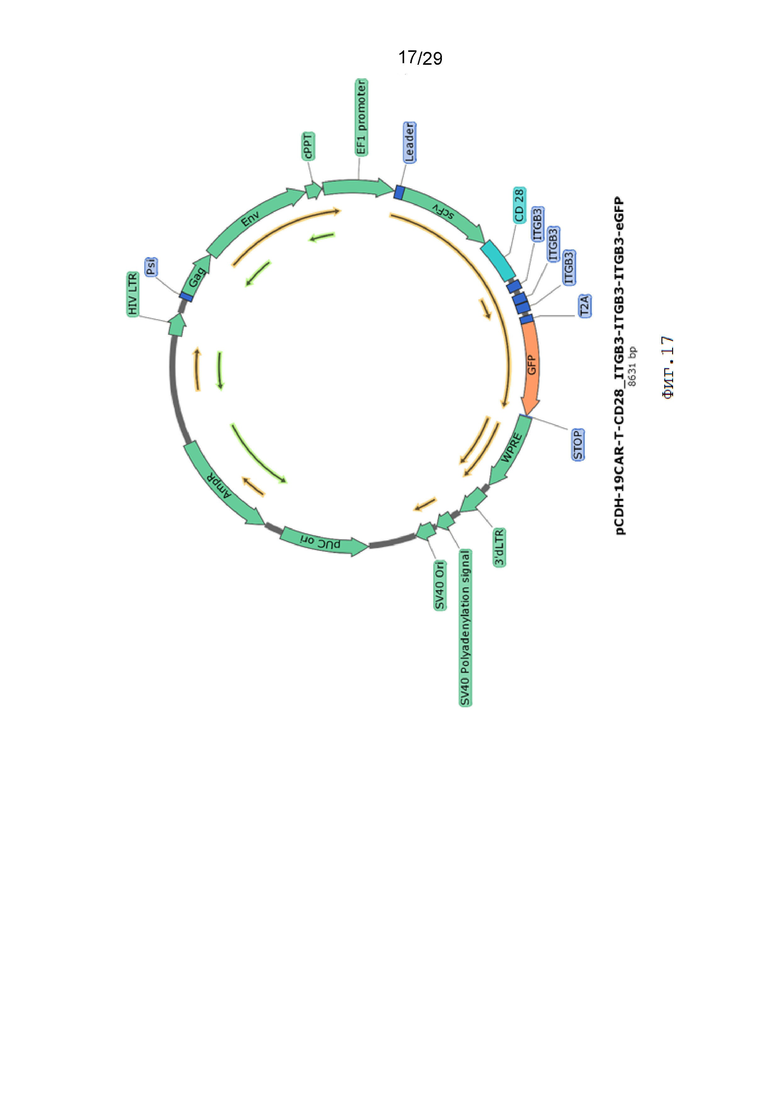
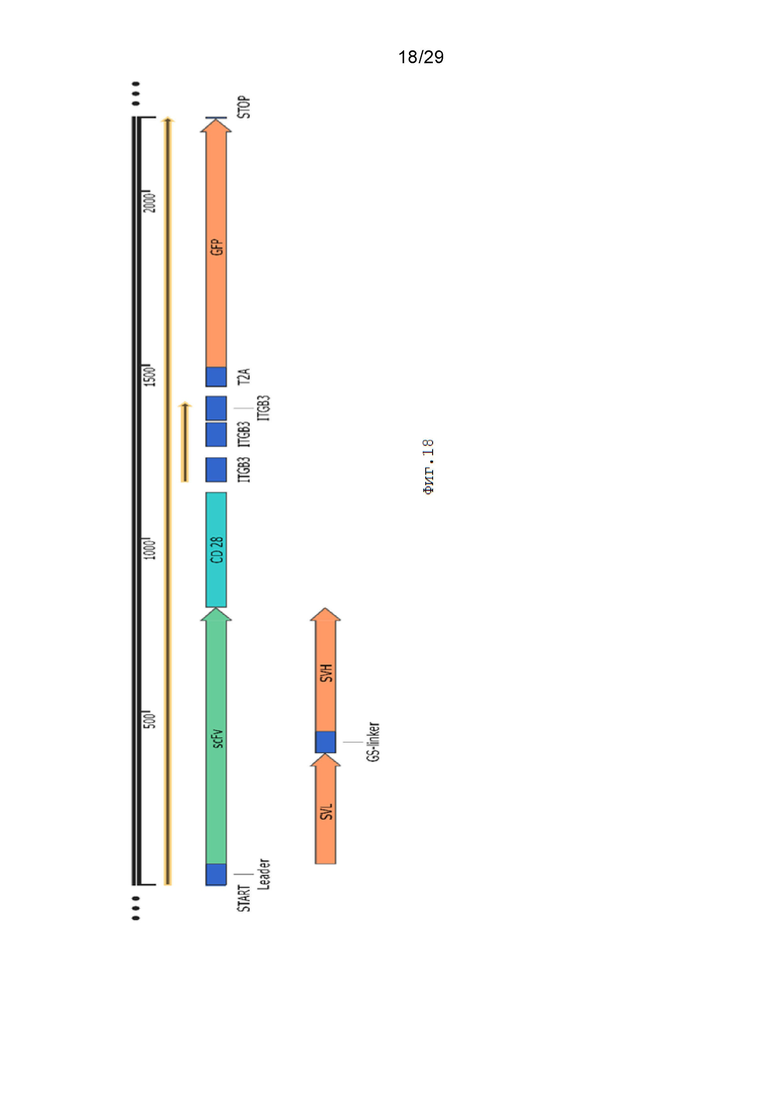
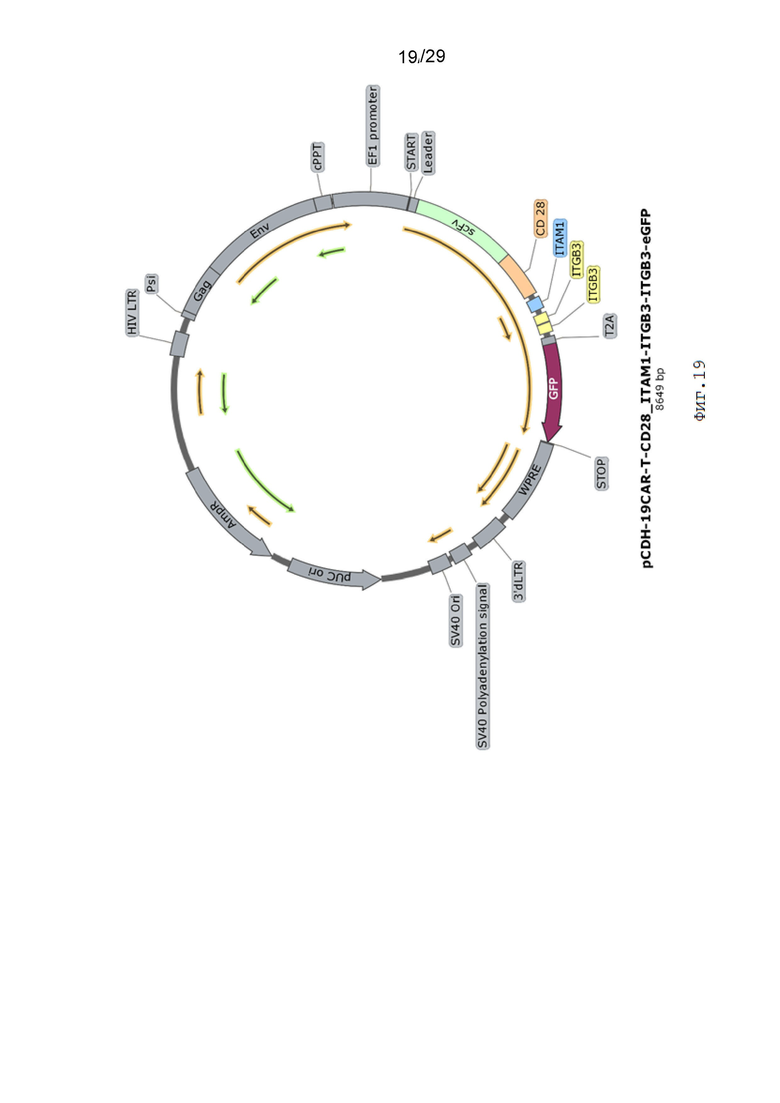
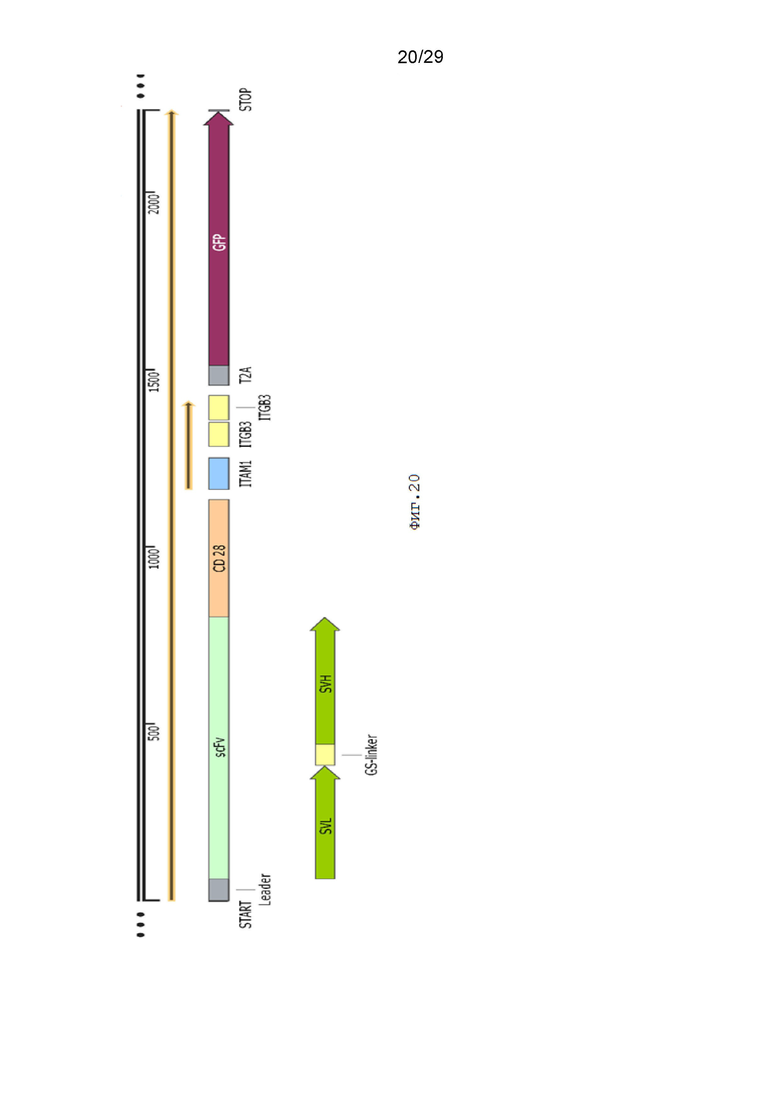
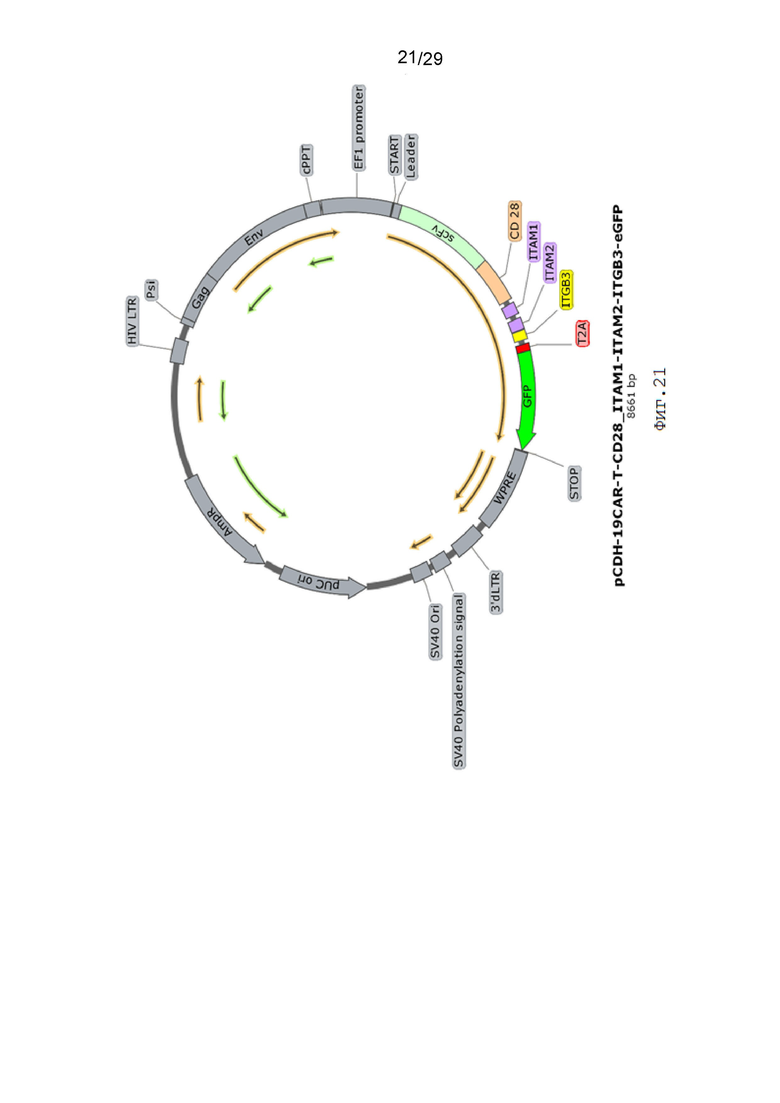
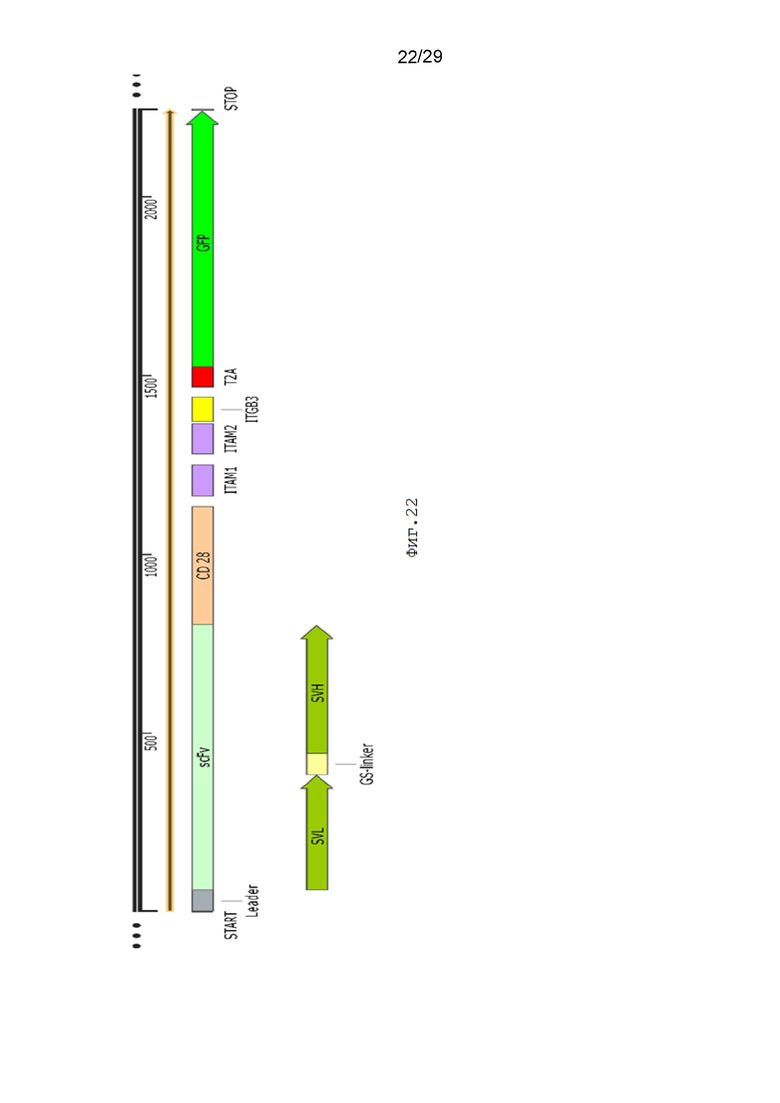
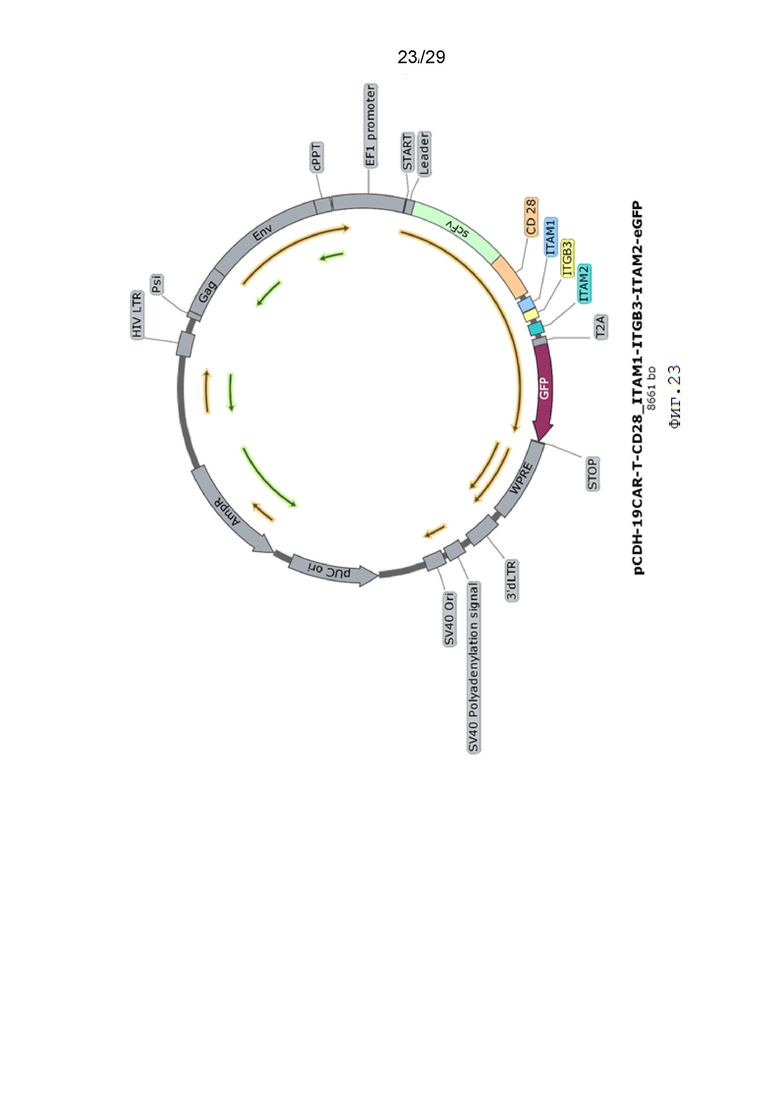

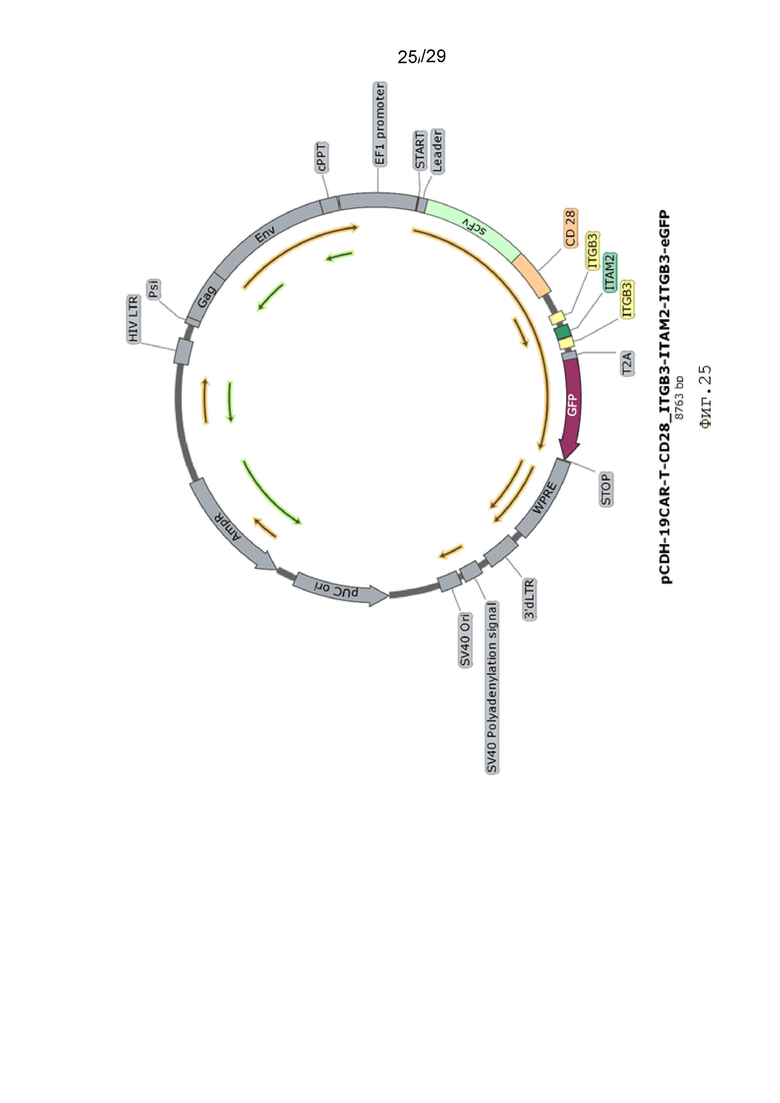
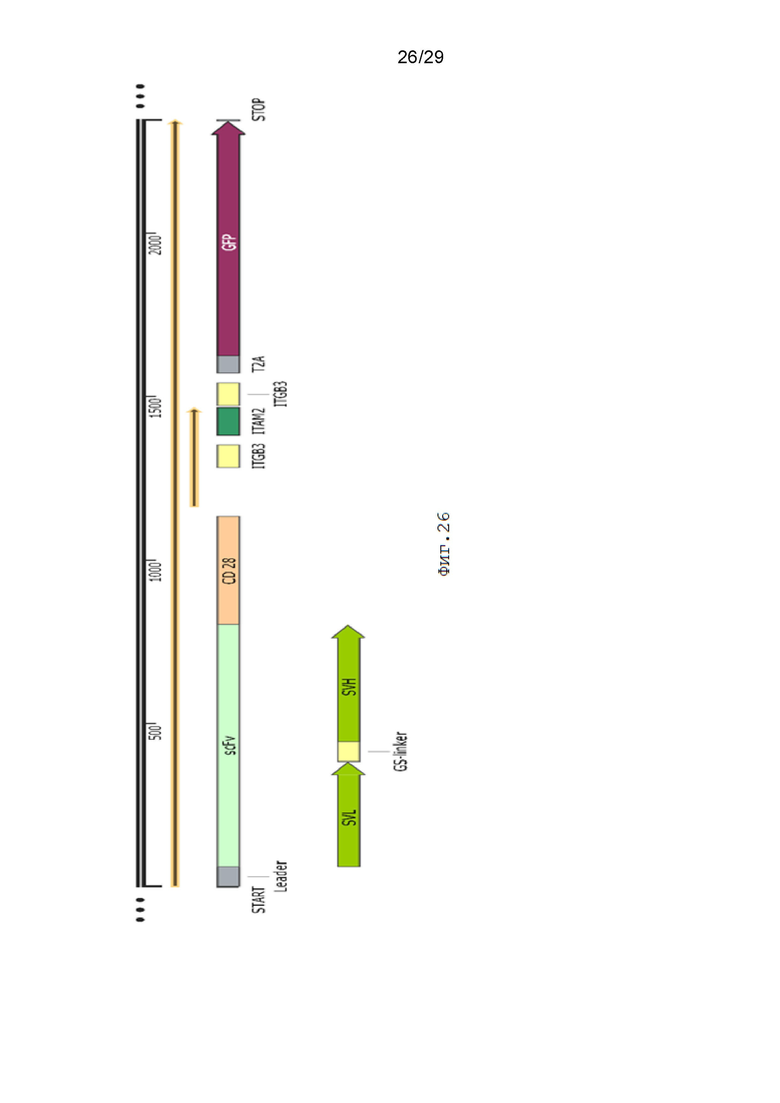
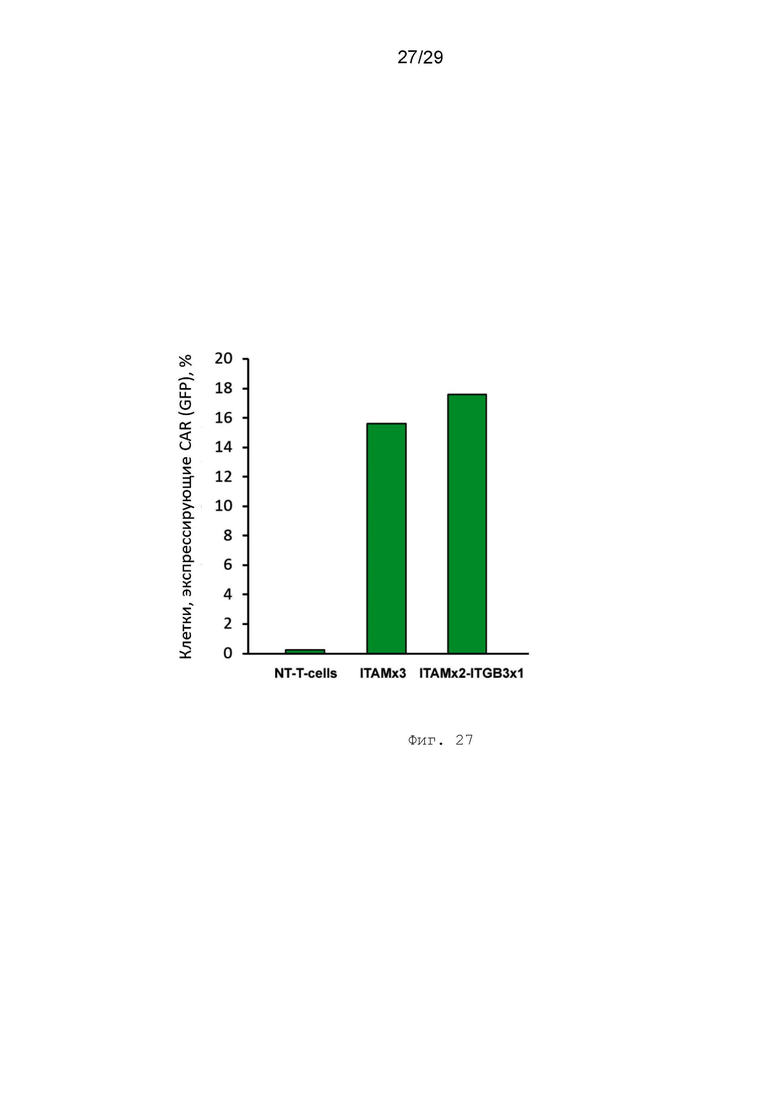
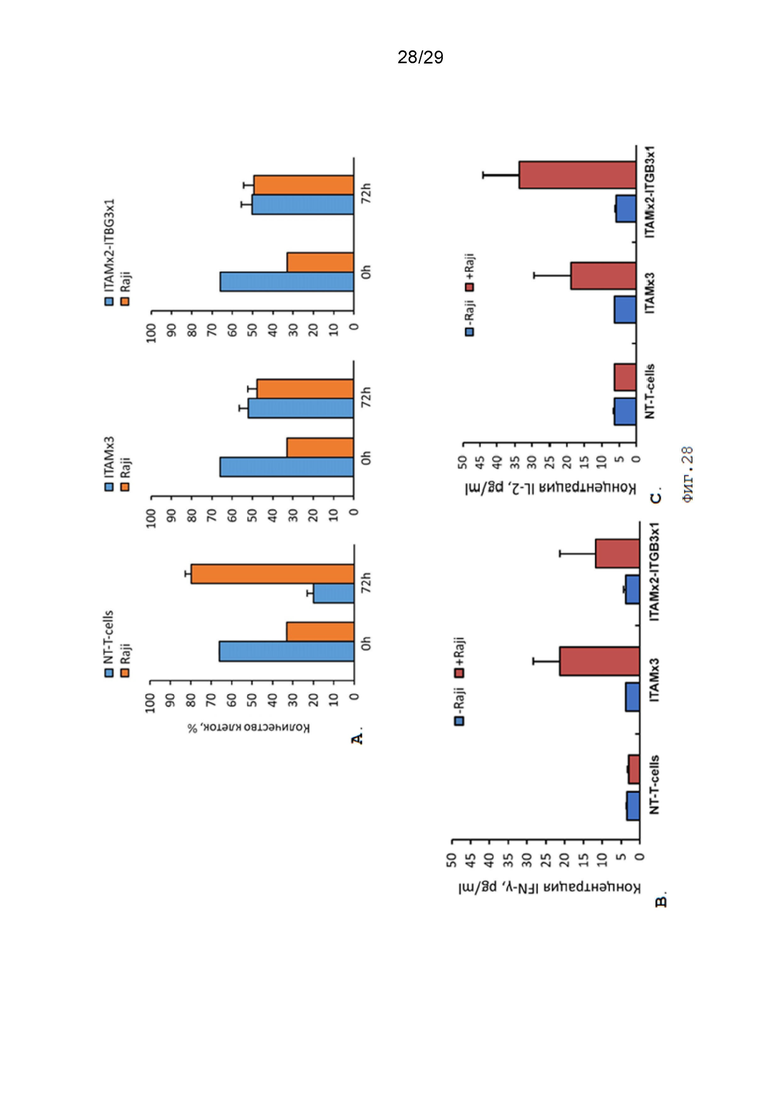
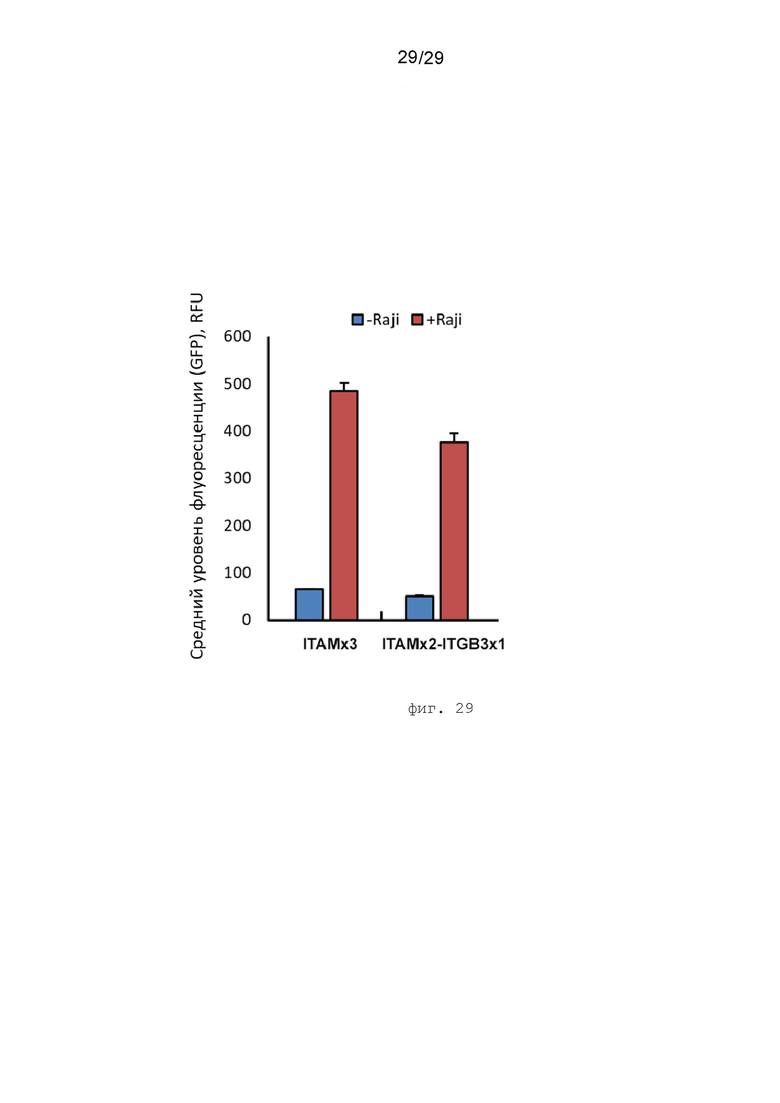
Изобретение относится к области биотехнологии, конкретно к получению химерных антигенных рецепторов (CAR) с модифицированным сигнальным доменом CD3-дзета, и может быть использовано в медицине. Полученный CAR, где в сигнальном домене CD3-дзета один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3), может быть использован для создания модифицированных нацеленных на опухолевый антиген Т-клеток, используемых в эффективной иммунотерапии рака. 8 н. и 56 з. п. ф-лы, 29 ил., 1 табл., 6 пр.
1. Выделенный альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR), где в сигнальном домене CD3-дзета один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3), где фрагмент ITGB3 имеет аминокислотную последовательность, представленную SEQ ID NO: 4.
2. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 1, где один иммунорецепторный тирозин-основный активационный мотив (ITAM), выбранный из IT AMI, ITAM2 и ITAM3, заменен на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
3. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 2, где ITAM1 заменен на ITGB3.
4. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 2, где ITAM2 заменен на ITGB3.
5. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 2, где ITAM3 заменен на ITGB3.
6. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 1, где два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, заменены на фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
7. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 6, где ITAM2 и ITAM3 заменены на два ITGB3.
8. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 6, где IT AMI и ITAM3 заменены на два ITGB3.
9. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 6, где IT AMI и ITAM2 заменены на два ITGB3.
10. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 1, где ITAM1 имеет аминокислотную последовательность, представленную SEQ ID NO: 1.
11. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 1, где ITAM2 имеет аминокислотную последовательность, представленную SEQ ID NO: 2.
12. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 1, где ITAM3 имеет аминокислотную последовательность, представленную SEQ ID NO: 3.
13. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 2, который имеет аминокислотную последовательность, представленную SEQ ID NO: 5.
14. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 2, который имеет аминокислотную последовательность, представленную SEQ ID NO: 6.
15. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 2, который имеет аминокислотную последовательность, представленную SEQ ID NO: 7.
16. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 6, который имеет аминокислотную последовательность, представленную SEQ ID NO: 8.
17. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 6, который имеет аминокислотную последовательность, представленную SEQ ID NO: 9.
18. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 6, который имеет аминокислотную последовательность, представленную SEQ ID NO: 10.
19. Выделенный альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR), который включает один или два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и один или два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3), при этом общее количество ITAM и ITGB3 равно трем, и где фрагмент ITGB3 имеет аминокислотную последовательность, представленную SEQ ID NO: 4,
где ITAM1 имеет аминокислотную последовательность, представленную SEQ ID NO: 1,
где ITAM2 имеет аминокислотную последовательность, представленную SEQ ID NO: 2,
где ITAM3 имеет аминокислотную последовательность, представленную SEQ ID NO: 3.
20. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 19, который включает два иммунорецепторных тирозин-основных активационных мотива (ITAM), выбранных из ITAM1, ITAM2 и ITAM3, и один фрагмент аминокислотной последовательности интегрина бета 3 (ITGB3).
21. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 20, который включает ITAM2, ITAM3 и ITGB3.
22. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 20, который включает ITAM1, ITAM3 и ITGB3.
23. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 20, который включает ITAM1, ITAM2 и ITGB3.
24. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 19, который включает один иммунорецепторный тирозин-основный активационный мотив (ITAM), выбранный из ITAM1, ITAM2 и ITAM3, и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
25. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 24, который включает ITAM1 и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
26. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 24, который включает ITAM2 и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
27. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 24, который включает ITAM3 и два фрагмента аминокислотной последовательности интегрина бета 3 (ITGB3).
28. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 20, который имеет аминокислотную последовательность, представленную SEQ ID NO: 5.
29. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 20, который имеет аминокислотную последовательность, представленную SEQ ID NO: 6.
30. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 20, который имеет аминокислотную последовательность, представленную SEQ ID NO: 7.
31. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 24, который имеет аминокислотную последовательность, представленную SEQ ID NO: 8.
32. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 24, который имеет аминокислотную последовательность, представленную SEQ ID NO: 9.
33. Альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора (CAR) по п. 24, который имеет аминокислотную последовательность, представленную SEQ ID NO: 10.
34. Выделенная нуклеиновая кислота, которая кодирует альтернативный внутриклеточный сигнальный домен по пп. 1-33.
35. Выделенный химерный антигенный рецептор (CAR), который специфичен к опухолевому антигену, содержащий альтернативный внутриклеточный сигнальный домен по пп. 1-33.
36. Выделенный химерный антигенный рецептор (CAR) по 35, содержащий:
а) антигенсвязывающий домен, который специфичен к опухолевому антигену;
б) трансмембранный домен;
в) ко-стимуляторный домен;
г) альтернативный внутриклеточный сигнальный домен по пп. 1-33.
37. Выделенный химерный антигенный рецептор (CAR) по п. 36, где антигенсвязывающий домен, который специфичен к опухолевому антигену, представляет собой Fab или scFv.
38. Выделенный химерный антигенный рецептор (CAR) по п. 37, где антигенсвязывающий домен, который специфичен к опухолевому антигену, представляет собой scFv.
39. Выделенный химерный антигенный рецептор (CAR) по п. 36, где антигенсвязывающий домен специфичен к опухолевому антигену, который выбран из группы: CD 19, CD20, CD 123 или ВСМА.
40. Выделенный химерный антигенный рецептор (CAR) по п. 39, где антигенсвязывающий домен специфичен к опухолевому антигену CD 19.
41. Выделенный химерный антигенный рецептор (CAR) по п. 40, где антигенсвязывающий домен, который специфичен к опухолевому антигену CD 19, имеет аминокислотную последовательность, представленную SEQ ID NO: 11.
42. Выделенный химерный антигенный рецептор (CAR) по п. 36, где трансмембранный домен представляет собой трансмембранный домен белка, выбранного из группы, состоящей из альфа-, бета- или дзета-цепи Т-клеточного рецептора, CD28, СО3-эпсилон, CD45, CD4, CD5, CD8, CD9, CD 16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154.
43. Выделенный химерный антигенный рецептор (CAR) по п. 42, где трансмембранный домен представляет собой CD8.
44. Выделенный химерный антигенный рецептор (CAR) по п. 43, где трансмембранный домен CD8 имеет аминокислотную последовательность, представленную SEQ ID NO: 12.
45. Выделенный химерный антигенный рецептор (CAR) по п. 36, где ко-стимуляторный домен представляет собой функциональный сигнальный домен белка, выбранного из группы, состоящей из ОХ40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278) и 4-1 ВВ (CD137).
46. Выделенный химерный антигенный рецептор (CAR) по п. 45, где ко-стимуляторный домен представляет собой CD28 или 4-1 ВВ.
47. Выделенный химерный антигенный рецептор (CAR) по п. 46, где ко-стимуляторный домен представляет собой CD28.
48. Выделенный химерный антигенный рецептор (CAR) по п. 47, где ко-стимуляторный домен CD28 имеет аминокислотную последовательность, представленную SEQ ID NO: 13.
49. Выделенный химерный антигенный рецептор (CAR) по п. 45, где ко-стимуляторный домен представляет собой 4-1 ВВ.
50. Выделенный химерный антигенный рецептор (CAR) по п. 49, где ко-стимуляторный домен 4-1 ВВ имеет аминокислотную последовательность, представленную SEQ ID NO: 14.
51. Выделенный химерный антигенный рецептор (CAR) по п. 35, дополнительно содержащий лидерную последовательность.
52. Выделенный химерный антигенный рецептор (CAR) по п. 35, который имеет аминокислотную последовательность, представленную аминокислотной последовательностью, которую выбирают из группы: SEQ ID NO: 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, или имеет аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности, которую выбирают из группы: SEQ ID NO: 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, и имеет отличия по сравнению с аминокислотной последовательностью, которую выбирают из группы: SEQ ID NO: 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26, только в неконсервативных аминокислотных остатках.
53. Выделенная нуклеиновая кислота, которая кодирует выделенный химерный антигенный рецептор (CAR) по пп. 35-52.
54. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 53.
55. Вектор по п. 54, выбранный из группы, состоящей из ДНК, РНК, плазмиды, лентивирусного вектора, аденоассоциированного вирусного вектора, аденовирусного вектора или ретровирусного вектора.
56. Вектор по п. 54, дополнительно содержащий промотор.
57. Вектор по п. 54, представляющий собой in vitro транскрибируемый вектор.
58. Вектор по п. 54, в котором последовательность нуклеиновой кислоты дополнительно содержит поли(А)-последовательность.
59. Вектор по п. 54, в котором последовательность нуклеиновой кислоты дополнительно содержит 3'UTR.
60. Способ получения рекомбинантной клетки-хозяина млекопитающего, экспрессирующей химерный антигенный рецептор (CAR) по пп. 35-52, включающий трансформирование клетки вектором по пп. 54-59.
61. Рекомбинантная клетка-хозяин млекопитающего, которая экспрессирует химерный антигенный рецептор (CAR) по пп. 35-52, которую получают способом по п. 60.
62. Клетка по п. 61, где клетка представляет собой Т-лимфоцит.
63. Клетка по п. 61, где клетка представляет собой CD8+ Т-клетку.
64. Клетка по п. 61, где клетка представляет собой NK-клетку.
| WO 2015187528, 10.12.2015 | |||
| VAN KUPPEVELT T | |||
| H | |||
| et al., An alternative cytoplasmic domain of the integrin beta 3 subunit, Proceedings of the National Academy of Sciences, 1989, V | |||
| Пюпитр для работы на пишущих машинах | 1922 |
|
SU86A1 |
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| KUMAR C | |||
| S | |||
| et al., Cloning and Characterization of a Novel Integrin β3Subunit, Journal of Biological Chemistry, 1997, V | |||
| Паровоз с приспособлением для автоматического регулирования подвода и распределения топлива в его топке | 1919 |
|
SU272A1 |
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |

