ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к модифицированной гомосериндегидрогеназе. Конкретно, настоящее изобретение относится к модифицированной гомосериндегидрогеназе, имеющей полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотные замены выполняют путем замены аминокислоты в положении 285 на изолейцин; аминокислоты в положении 398 на глутамин; или аминокислот в обоих этих положениях на изолейцин и глутамин соответственно. Кроме того, настоящее изобретение относится к способу получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, с использованием модифицированной гомосериндегидрогеназы, к композиции для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, к способу повышения способности продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, или к применению модифицированной гомосериндегидрогеназы.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Из L-аминокислот, L-треонина, L-изолейцина и L-метионина, обычно используют гомосерин, продуцируемый гомосериндегидрогеназой (называемой ниже «Hom»; EC:1.1.1.3) из аспартат-полуальдегида (называемого ниже «ASA»). Поэтому для получения аминокислот методом ферментации необходимо поддерживать активность ферментов, используемых в пути их биосинтеза, на определенном уровне или выше, и были проведены интенсивные исследования этого вопроса.
В частности, известно, что активность гомосериндегидрогеназы, действующей в точке разветвления путей биосинтеза L-лизина и L-треонина, регулируется L-треонином и L-изолейцином. Недавно появилось несколько сообщений о Hom, десенсибилизированной к ингибированию L-треонином по принципу обратной связи, и о способе получения L-треонина с использованием такой Hom. В 1991 г. Eikmann et al. в Германии сообщили о Hom, десенсибилизированной путем замены глицина, представляющего собой аминокислотный остаток в положении 378 Hom, на глутамат (Eikmanns BJ et al., Appl. Microbial Biotechnol. 34: 617-622, 1991); и в 1991 г. Archer et al. сообщили о том, что десенсибилизация происходит при повреждении C-конца Hom в результате мутации со сдвигом рамки считывания (Archer JA et al., Gene 107: 53-59, 1991).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Авторы настоящего изобретения провели исследование десенсибилизации к ингибированию треонином по принципу обратной связи и, в результате, обнаружили, что новый ген, кодирующий модифицированую Hom, выделен и что способность продуцировать L-аминокислоты улучшена у микроорганизма, трансдуцированного этим новым геном, тем самым завершив настоящее изобретение.
Техническое решение
Задача настоящего изобретения состоит в том, чтобы предложить модифицированную гомосериндегидрогеназу, имеющую полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотные замены выполняют путем замены аминокислоты в положении 285 на другую аминокислоту, аминокислоты в положении 398 на другую аминокислоту или аминокислот в обоих этих положениях на другие аминокислоты.
Другая задача настоящего изобретения состоит в том, чтобы предложить полинуклеотид, кодирующий модифицированную дегидрогеназу.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить микроорганизм рода Corynebacterium, содержащий модифицированную гомосериндегидрогеназу.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить способ получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, включающий культивирование микроорганизма в среде и выделение гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, из микроорганизма или среды.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить композицию для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, содержащую модифицированную гомосериндегидрогеназу или микроорганизм, содержащий модифицированную гомосериндегидрогеназу по настоящему изобретению.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить способ повышения способности продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, включающий экспрессию модифицированной гомосериндегидрогеназы по настоящему изобретению в микроорганизме рода Corynebacterium.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить применение модифицированной гомосериндегидрогеназы для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, по настоящему изобретению.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить применение полинуклеотида для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, по настоящему изобретению.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить применение микроорганизма рода Corynebacterium для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, по настоящему изобретению.
Еще одна задача настоящего изобретения состоит в том, чтобы предложить применение композиции для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, по настоящему изобретению.
Полезные эффекты
Модифицированную гомосериндегидрогеназу по настоящему изобретению можно широко использовать для крупномасштабного получения эффективного гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, благодаря десенсибилизации к ингибированию конечным продуктом по принципу обратной связи по сравнению с природным или диким типом.
Наилучший вариант осуществления изобретения
Далее настоящее изобретение будет описано подробно. Между тем, каждое из пояснений и типичных воплощений, раскрытых здесь, может быть применено к другим пояснениям и типичным воплощениям. То есть, все комбинации различных факторов, раскрытых здесь, входят в объем настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать конкретным описанием изобретения, предложенным ниже.
Для решения указанных выше задач согласно одному аспекту настоящего изобретения предложена модифицированная гомосериндегидрогеназа, имеющая полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотные замены выполняют путем замены аминокислоты в положении 285 или аминокислоты в положении 398 другой аминокислотой или их комбинацией.
Конкретно, согласно настоящему изобретению предложен вариант гомосериндегидрогеназы, имеющий полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотные замены выполняют путем замены аминокислоты в положении 285 на изолейцин, аминокислоты в положении 398 на глутамин или аминокислот в обоих этих положениях на изолейцин и глутамин соответственно. Более конкретно, согласно настоящему изобретению предложена модифицированная гомосериндегидрогеназа, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 285 заменена на изолейцин, аминокислота в положении 398 заменена на глутамин или аминокислоты в обоих этих положениях заменены на изолейцин и глутамин соответственно.
В настоящем изобретении гомосериндегидрогеназа (EC:1.1.1.3) относится к ферменту, катализирующему синтез гомосерина, общего промежуточного продукта в биосинтезе метионина, треонина и изолейцина у растений и микроорганизмов. Гомосериндегидрогеназа может быть включена в настоящее изобретение независимо от ее происхождения при условии, что она обладает указанной выше ферментативной активностью, и в качестве гомосериндегидрогеназы может быть использован фермент, имеющий происхождение от любого организма (растений, микроорганизмов и так далее). Конкретно, гомосериндегидрогеназа может иметь происхождение от микроорганизма рода Corynebacterium, и, более конкретно, может иметь происхождение от Corynebacterium glutamicum. Например, гомосериндегидрогеназа может представлять собой белок, содержащий аминокислотную последовательность SEQ ID NO: 1. «Белок, имеющий аминокислотную последовательность SEQ ID NO: 1» можно использовать взаимозаменяемо с термином «белок, имеющий аминокислотную последовательность SEQ ID NO: 1» или «белок, состоящий из аминокислотной последовательности SEQ ID NO: 1».
В настоящем изобретении для способа получения гомосериндегидрогеназы можно использовать множество методов, хорошо известных в данной области. Примеры таких методов включают методики синтеза генов, включающие оптимизацию кодонов, с получением белков с высокой эффективностью в микроорганизме рода Corynebacterium, который часто используют для экспрессии белков, и способы скрининга источников полезных ферментов с использованием биоинформационных способов на основе метагенома микроорганизмов, но такие способы не ограничены указанными методами.
В настоящем изобретении белок, обладающий активностью гомосериндегидрогеназы, не исключает мутацию, которая может произойти в результате добавления бессмысленной последовательности выше или ниже аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, например аминокислотной последовательности SEQ ID NO: 1, или ее естественной мутации, или молчащей мутации в ней. Кроме того, белок, активность которого идентична или соответствует активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1, соответствует белку, обладающему активностью гомосериндегидрогеназы по настоящему изобретению. В качестве конкретного примера, белок, обладающий активностью гомосериндегидрогеназы по настоящему изобретению, может представлять собой белок, состоящий из аминокислотной последовательности SEQ ID NO: 1, или аминокислотной последовательности по меньшей мере на 80%, 90%, 95% или 97% гомологичной SEQ ID NO: 1.
Кроме того, несмотря на описание «белок или полипептид, содержащий аминокислотную последовательность определенной SEQ ID NO: » в данном описании изобретения, очевидно, что объем настоящего изобретения может также включать любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или добавлением части последовательности при условии, что этот белок имеет аминокислотную последовательность с любой из указанных выше степеней гомологии и демонстрирует эффект, соответствующий указанному выше белку. Например, в настоящем изобретении белок, обладающий активностью гомосериндегидрогеназы, может представлять собой гомосериндегидрогеназу, имеющую происхождение от Corynebacterium glutamicum. Более конкретно, белок, обладающий активностью гомосериндегидрогеназы, может представлять собой аминокислотную последовательность (SEQ ID NO: 1) гомосериндегидрогеназы, имеющей происхождение от Corynebacterium glutamicum ATCC13032, аминокислотную последовательность (SEQ ID NO: 49) гомосериндегидрогеназы, имеющей происхождение от Corynebacterium glutamicum ATCC14067, или аминокислотную последовательность (SEQ ID NO: 50) гомосериндегидрогеназы, имеющей происхождение от Corynebacterium glutamicum ATCC13869. Поскольку гомосериндегидрогеназы, имеющие указанные выше последовательности, гомологичны друг другу на 80%, 90%, 95%, 97% или более, и поскольку эти гомосериндегидрогеназы демонстрируют эффекты, соответствующие эффектам гомосериндегидрогеназы, очевидно, что они включены в белок, обладающий активностью гомосериндегидрогеназы, по настоящему изобретению.
При использовании здесь термин «гомология» относится к проценту идентичности двух полинуклеотидных или полипептидных группировок. Гомология относится к степени совпадения с заданной аминокислотной последовательностью или нуклеотидной последовательностью и может быть выражена в процентах. В настоящем изобретении гомология последовательностей, обладающих активностью, идентичной или сходной с заданной аминокислотной последовательностью или нуклеотидной последовательностью, выражена как «% гомологии». Гомология последовательностей от одной группировки до другой может быть определена при помощи методов, известных в данной области. Например, гомология может быть подтверждена с использованием стандартного программного обеспечения, то есть BLAST 2.0, для расчета таких параметров, как баллы, идентичность и сходство, или путем сравнения последовательностей в экспериментах с Саузерн-гибридизацией, при этом подходящие условия гибридизации могут быть определены методом, известным специалистам в данной области (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
При использовании здесь термины «модификация», «модифицированный» или «вариант» относятся к культуре или индивидууму, демонстрирующему наследуемое или ненаследуемое изменение одного стабилизированного фенотипа. Конкретно, термин «вариант» может быть предназначен для обозначения варианта, активность которого эффективно повышена в результате того, что одна или более аминокислот в аминокислотной последовательности, соответствующей белку, обладающему активностью гомосериндегидрогеназы, модифицированы по сравнению с диким типом, нативным или немодифицированным, или варианта, у которого ингибирование изолейцином, треонином или их производным по принципу обратной связи устранено, или варианта, у которого и повышение активность, и ингибирование по принципу обратной связи устранены.
В настоящем изобретении термин «модифицированная гомосериндегидрогеназа» можно использовать взаимозаменяемо с термином «вариант гомосериндегидрогеназы». С другой стороны, такой вариант может не встречаться в природе.
Конкретно, модифицированная гомосериндегидрогеназа по настоящему изобретению может представлять собой модифицированный белок, имеющий полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотные замены выполняют путем замены аминокислоты в положении 285 изолейцином, аминокислоты в положении 398 глутамином или их комбинацией. Аминокислотная последовательность белка, обладающего активностью гомосериндегидрогеназы, описана выше и может представлять собой, например, аминокислотную последовательность SEQ ID NO: 1. Кроме того, аминокислота в положении 285 может являться такой, что треонин заменен изолейцином, и аминокислота в положении 398 может являться такой, что аргинин заменен глутамином.
Более того, модифицированная гомосериндегидрогеназа по настоящему изобретению может представлять собой модифицированный белок, имеющий полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотную замену выполняют путем замены аминокислоты в положении 378 на триптофан. Конкретно, модифицированная гомосериндегидрогеназа по настоящему изобретению может представлять собой модифицированный белок, имеющий полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы, где аминокислотную замену выполняют путем замены аминокислоты в положении 285 на изолейцин, аминокислоты в положении 398 на глутамин или их комбинации; в этой модифицированной гомосериндегидрогеназе аминокислота в положении 378 может быть дополнительно заменена триптофаном. Более конкретно, аминокислота в положении 378 может быть такой, что глицин заменен триптофаном.
Еще более конкретно, модифицированная гомосериндегидрогеназа по настоящему изобретению представляет собой модифицированный белок, имеющий полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, где аминокислотную замену выполняют путем замены аминокислоты в положении 285 на изолейцин, аминокислоты в положении 398 на глутамин, или их комбинации. Например, модифицированная гомосериндегидрогеназа по настоящему изобретению может представлять собой белок, содержащий аминокислотную последовательность SEQ ID NO: 10, 11, 12 или 13. Кроме того, может иметь место мутация, которая может произойти в результате добавления бессмысленной последовательности выше или ниже этой аминокислотной последовательности, естественная мутация или молчащая мутация в ней также не исключены. Кроме того, белок, активность которого идентична или соответствует активности модифицированной гомосериндегидрогеназы, соответствует белку, обладающему активностью гомосериндегидрогеназы по настоящему изобретению. В качестве конкретного примера, модифицированная гомосериндегидрогеназа по настоящему изобретению может представлять собой белок, состоящий из аминокислотной последовательности SEQ ID NO: 10, 11, 12 или 13, или белок, гомологичный по меньшей мере на 80%, 90%, 95% или 97% указанной выше аминокислотной последовательности. Кроме того, несмотря на описание как «белок или полипептид, имеющий аминокислотную последовательность определенной SEQ ID NO: » в настоящем описании изобретения, очевидно, что объем настоящего изобретения может также включать любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или добавлением части последовательности при условии, что этот белок имеет аминокислотную последовательность с любой из указанных выше степеней гомологии и демонстрирует эффект, соответствующий указанному выше белку.
Кроме того, модифицированная гомосериндегидрогеназа по настоящему изобретению представляет собой модифицированную гомосериндегидрогеназу, имеющую полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности белка, обладающего активностью гомосериндегидрогеназы. Очевидно, что объем настоящего изобретения включает любой белок, имеющий модификацию, где аминокислота в положении 285 или 398 заменена другой аминокислотой, и демонстрирующий эффект, соответствующий гомосериндегидрогеназе.
Кроме того, в отличие от белка дикого типа, или нативного белка, или немодифицированного белка, обладающего активностью гомосериндегидрогеназы, модифицированная гомосериндегидрогеназа по настоящему изобретению может представлять собой гомосериндегидрогеназу, ингибирование которой конечным продуктом, то есть изолейцином, треонином, метионином или гомосерином, их производным или аналогом, устранено или десенсибилизировано. При использовании здесь термин «ингибирование по принципу обратной связи» означает, что конечный продукт метаболизма препятствует реакции на более ранней стадии. Поэтому, когда ингибирование гомосериндегидрогеназы по принципу обратной связи устранено или десенсибилизировано, продуктивность по гомосерину или L-аминокислоте, имеющей происхождение от гомосерина, может быть улучшена по сравнению с отсутствием устранения или десенсибилизации ингибирования по принципу обратной связи.
L-аминокислота, имеющая происхождение от гомосерина, относится к L-аминокислоте, которая может быть биологически синтезирована с использованием L-гомосерина в качестве предшественника и не ограничена, при условии, что она представляет собой вещество, которое может быть биологически синтезировано из L-гомосерина. L-аминокислота, имеющая происхождение от гомосерина, может включать не только L-аминокислоту, имеющую происхождение от гомосерина, но и ее производное. Например, L-аминокислота, имеющая происхождение от гомосерина, может представлять собой, без ограничения, L-треонин, L-изолейцин, O-ацетилгомосерин, O-сукцинил-L-гомосерин, O-фосфо-L-гомосерин, L-метионин и/или L-глицин. Более конкретно, L-аминокислота, имеющая происхождение от гомосерина, может представлять собой, без ограничения ими, L-треонин, L-изолейцин, O-ацетилгомосерин, O-сукцинил-L-гомосерин и/или L-метионин.
Согласно другому аспекту настоящего изобретения предложен полинуклеотид, кодирующий модифицированную гомосериндегидрогеназу.
Гомосериндегидрогеназа и вариант являются такими как описано выше.
При использовании здесь термин «полинуклеотид» представляет собой нуклеотидный полимер, состоящий из нуклеотидных мономеров, ковалентно связанных с образованием цепи, и его примерами являются цепи ДНК или РНК, имеющие предопределенную или большую длину, и, более конкретно, он относится к фрагменту полинуклеотида, кодирующему модифицированную гомосериндегидрогеназу. Полинуклеотид, кодирующий модифицированный белок по настоящему изобретению, может быть включен, без ограничения, при условии, что он имеет полинуклеотидную последовательность, кодирующую модифицированный белок, обладающий активностью гомосериндегидрогеназы по настоящему изобретению.
В настоящем изобретении полинуклеотид, кодирующий аминокислотную последовательность варианта гомосериндегидрогеназы, может, без ограничения ими, иметь происхождение конкретно от микроорганизма рода Corynebacterium и более конкретно от Corynebacterium glutamicum.
Кроме того, в кодирующей области полинуклеотида, кодирующего белок, могут быть выполнены различные модификации без изменения аминокислотной последовательности белка из-за вырожденности кодонов или в соответствии с кодонами, предпочтительными в организме, в котором предстоит проводить экспрессию белка. Конкретно, полинуклеотид может представлять собой полинуклеотид, содержащий полинуклеотидную последовательность, кодирующую белок, или полинуклеотидную последовательность, по меньшей мере на 80%, 90%, 95% или 97% гомологичную указанной выше полинуклеотидной последовательности. Кроме того, очевидно, что объем настоящего изобретения может также включать полинуклеотидную последовательность с делецией, модификацией, заменой или добавлением части последовательности при условии, что она представляет собой полинуклеотидную последовательность, кодирующую белок, имеющий указанные выше степени гомологии и демонстрирующий эффект, по существу идентичный или соответствующий указанному выше белку. Полинуклеотид, кодирующий белок, имеющий активность гомосериндегидрогеназы по настоящему изобретению, может иметь полинуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 1. Например, полинуклеотид может иметь полинуклеотидную последовательность SEQ ID NO: 48, без ограничения ей. Кроме того, полинуклеотид, кодирующий модифицированную гомосериндегидрогеназу по настоящему изобретению, может иметь полинуклеотидную последовательность, кодирующую полипептид, содержащий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, и, конкретно, может иметь полинуклеотидную последовательность, кодирующую SEQ ID NO: 10, 11, 12 или 13. Например, полинуклеотид может иметь полинуклеотидную последовательность SEQ ID NO: 6, 7, 8 или 9, без ограничения ими.
Кроме того, зонд, который может быть получен из известной последовательности гена, например любой последовательности, которая гибридизируется с последовательностью, комплементарной всей или части полинуклеотидной последовательности, в жестких условиях, кодирующей белок, обладающий активностью гомосериндегидрогеназы по настоящему изобретению, может также быть включена без ограничения. Термин «жесткие условия» означает условия, в которых возможна специфичная гибридизация полинуклеотидов. Такие условия подробно описаны в литературе (например, в J. Sambrook et al., ниже). Жесткие условия могут включать, например, условия, в которых высокогомологичные гены, гомологичные на 80% или более, конкретно гомологичные на 90% или более, более конкретно гомологичные на 95% или более, еще более конкретно гомологичные на 97% или более, еще более конкретно гомологичные на 99% или более, гибридизируются друг с другом, а менее гомологичные гены не гибридизируются друг с другом, или обычные условия промывки при Саузерн-гибридизации, то есть однократная промывка, особенно двукратная или трехкратная промывка при концентрации соли и температуре, соответствующих 60°C, 1×SSC (хлорид натрия/цитрат натрия), 0,1% SDS (додецилсульфат натрия), конкретно 60°C, 0,1×SSC, 0,1% SDS, и более конкретно 68°C, 0,1×SSC, 0,1% SDS. Для гибридизации необходимо, чтобы два полинуклеотида содержали комплементарные последовательности, однако, в зависимости от жесткости гибридизации, возможны несоответствия оснований. Термин «комплементарный» использован для описания взаимоотношения нуклеотидных оснований, которые могут гибридизироваться друг с другом. Например, применительно к ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, настоящее изобретение может также включать выделенный нуклеотидный фрагмент, комплементарный полноразмерной последовательности, а также нуклеотидную последовательность, по существу сходную с ним. Конкретно, полинуклеотид, имеющий гомологию, может быть определен с использованием условий гибридизации, включающих стадию гибридизации при значении Tm 55°C в условиях, описанных выше. Кроме того, значение Tm может составлять, без ограничения ими, 60°C, 63°C или 65°C, и специалисты в данной области могут надлежащим образом его контролировать в зависимости от поставленной задачи. Подходящая жесткость гибридизации полинуклеотидов зависит от длины полинуклеотидов и степени их комплементарности, и эти показатели хорошо известны в данной области (см. Sambrook et al., ниже, 9.50-9.51, 11.7-11.8).
Согласно еще одному аспекту настоящего изобретения предложен микроорганизм, содержащий модифицированную гомосериндегидрогеназу. Конкретно, согласно настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, содержащий модифицированную гомосериндегидрогеназу. Кроме того, согласно настоящему изобретению предложен микроорганизм рода Corynebacterium, продуцирующий L-аланин, содержащий модифицированную гомосериндегидрогеназу. Однако изобретение не ограничено указанным микроорганизмом.
Гомосериндегидрогеназа и вариант являются такими, как описано выше.
Конкретно, микроорганизм, содержащий модифицированную гомосериндегидрогеназу по настоящему изобретению, относится к микроорганизму, который по своей природе обладает способностью продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, или к микроорганизму, исходному штамму которого, не обладавшему способностью продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, была придана способность продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина. Конкретно, микроорганизм, содержащий гомосериндегидрогеназу, может представлять собой, без ограничения им, микроорганизм, экспрессирующий модифицированную гомосериндегидрогеназу, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 285 заменена изолейцином, аминокислота в положении 398 заменена глутамином или аминокислоты в обоих этих положениях заменены изолейцином и глутамином соответственно. Микроорганизм может представлять собой клетку или микроорганизм, содержащий полинуклеотид, кодирующий модифицированную гомосериндегидрогеназу или способный экспрессировать модифицированный полипептид при трансформации вектором, содержащим полинуклеотид, кодирующий модифицированную гомосериндегидрогеназу. Для целей настоящего изобретения клетка-хозяин или микроорганизм может представлять собой любой микроорганизм, способный продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, содержащий модифицированный полипептид.
Микроорганизм, содержащий модифицированную гомосериндегидрогеназу по настоящему изобретению, обладает улучшенной способностью продуцировать гомосерин, L-аминокислоту, имеющую происхождение от гомосерина, или L-аланин по сравнению с диким типом или микроорганизмом, содержащим белок, обладающий активностью модифицированной гомосериндегидрогеназы. Таким образом, гомосерин, L-аминокислота, имеющая происхождение от гомосерина, или L-аланин могут быть получены из этого микроорганизма с высоким выходом.
В настоящем изобретении нет конкретных ограничений в отношении типа микроорганизма, содержащего модифицированную гомосериндегидрогеназу, и он может представлять собой Enterobacter sp., Escherichia sp., Erwinia sp., Serratia sp., Pseudomonas sp., Providencia sp., Corynebacterium sp. или Brevibacterium sp. Более конкретно, микроорганизм может представлять собой микроорганизм рода Corynebacterium.
В настоящем изобретении «микроорганизм рода Corynebacterium» может конкретно представлять собой, без ограничения ими, Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens и так далее. Более конкретно, в настоящем изобретении микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum.
В то же время, микроорганизм, содержащий модифицированную гомосериндегидрогеназу, может представлять собой, без ограничения им, микроорганизм, в который введен вектор, содержащий полинуклеотид, кодирующий вариант гомосериндегидрогеназы.
При использовании здесь термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, где целевой белок функционально связан с подходящей контрольной последовательностью, так, что он может быть экспрессирован в подходящем хозяине. Контрольная последовательность включает промотор, способный инициировать транскрипцию, любую операторную последовательность для контроля транскрипции, последовательность, кодирующую подходящий домен связывания рибосом с мРНК, и последовательность, контролирующую терминацию транскрипции и трансляции. После трансформации подходящей клетки-хозяина вектор может быть реплицирован или функционировать независимо от генома хозяина или может быть интегрирован в сам геном хозяина.
Вектор, используемый в настоящем изобретении, конкретно не ограничен при условии, что он способен реплицироваться в клетке-хозяине, и может быть использован любой вектор, известный в данной области. Примеры обычных векторов могут включать естественную или рекомбинантную плазмиду, космиду, вирус и бактериофаг. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, M13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A; и в качестве плазмидного вектора могут быть использованы векторы типа pBR, типа pUC, типа pBluescriptII, типа pGEM, типа pTZ, типа pCL и типа pET. Конкретно, могут быть использованы векторы pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 и pCC1BAC, но вектор не ограничен ими.
Вектор, который может быть использован в настоящем изобретении, конкретно не ограничен, и может быть использован любой известный вектор экспрессии. Кроме того, полинуклеотид, кодирующий целевой белок в хромосоме, может быть введен с использованием вектора для хромосомной вставки. Введение полинуклеотида в хромосому может быть выполнено любым методом, известным в данной области (например посредством гомологичной рекомбинации), без ограничения ими. Может быть дополнительно включен селективный маркер для подтверждения успешного введения гена в хромосому. Селективный маркер предназначен для скрининга клеток, трансформированных вектором, иными словами, для определения, произошло ли введение целевой полинуклеотидной молекулы. Могут быть использованы маркеры, обеспечивающие селектируемые фенотипы, такие как резистентность к лекарственным препаратам, ауксотрофия, резистентность к цитотоксическим агентам или экспрессия поверхностных белков. В среде, обработанной селективным агентом, могут выжить только клетки, экспрессирующие селективный маркер, или клетки демонстрируют отличный от других фенотип, благодаря чему посредством этого метода могут быть выбраны успешно трансформированные клетки.
При использовании здесь термин «трансформация» относится к введению вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, так, что белок, кодируемый полинуклеотидом, экспрессируется в клетке-хозяине. При условии, что трансформированный полинуклеотид может экспрессироваться в клетке- хозяине, он может быть или интегрирован, или помещен в хромосому клетки-хозяина либо может существовать вне хромосомы. Кроме того, полинуклеотид включает ДНК и РНК, кодирующую целевой белок. Полинуклеотид может быть введен в любой форме, при условии, что он может быть введен в клетку-хозяина и экспрессирован в ней. Например, полинуклеотид может быть введен в клетку-хозяина в виде экспрессионной кассеты, представляющей собой генную конструкцию, содержащую все элементы, необходимые для ее автономной экспрессии. Экспрессионная кассета может содержать промотор, функционально связанный с полинуклеотидом, терминатор транскрипции, сайты связывания рибосом или терминатор трансляции. Экспрессионная кассета может быть представлена в форме самореплицируемого вектора экспрессии. Кроме того, полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностями, необходимыми для экспрессии в клетке-хозяине, без ограничения им. Метод трансформации включает любой метод введения полинуклеотида в клетку-хозяина и может быть осуществлен путем выбора подходящей стандартной методики, известной в данной области, в зависимости от клетки-хозяина. Примеры данного метода включают, без ограничения ими, электропорацию, преципитацию с фосфатом кальция (Ca(H2PO4)2, CaHPO4 или Ca3(PO4)2), преципитацию с хлоридом кальция (CaCl2), микроинъекцию, методику с полиэтиленгликолем (ПЭГ), методику с DEAE-декстраном, методику с катионными липосомами, методику с ацетатом лития и DMSO и так далее.
Кроме того, термин «функциональная связь» означает, что полинуклеотидная последовательность функционально связана с промоторной последовательностью, инициирующей и опосредующей транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению. Функциональная связь может быть получена путем применением рекомбинантной генетической методики, известной в данной области, а сайт-специфичное расщепление или связывание ДНК может быть проведено с использованием известной лиазы и лигазы, без ограничения ими.
Микроорганизм, содержащий модифицированную гомосериндегидрогеназу, может представлять собой микроорганизм, трансформированный, чтобы содержать модифицированную гомосериндегидрогеназу в микроорганизме рода Corynebacterium. Например, микроорганизм рода Corynebacterium может включать, без ограничения ими, штамм, резистентный к 2-амино-3-гидроксивалерату (AHV); штамм, продуцирующий L-треонин, благодаря замене лейцина, представляющего собой аминокислоту в положении 377 аспартаткиназы (LysC), на лизин для прекращения ингибирования по принципу обратной связи LysC, которая является первым важным ферментом в пути биосинтеза треонина; штамм, продуцирующий L-изолейцин, благодаря замене аминокислоты в положении 323 гена ilvA, кодирующего L-треониндегидратазу (первый важный фермент в пути биосинтеза изолейцина), в штамме, продуцирующем L-треонин, на аланин (Appl. Enviro. Microbiol., Dec. 1996, p. 4345-4351); штамм, продуцирующий O-ацетилгомосерин, благодаря инактивации O-ацетилгомосерин(тиол)-лиазы, вовлеченной в путь расщепления O-ацетилгомосерина, и цистатионин-гамма-синтазы; или штамм, продуцирующий метионин, благодаря инактивации транскрипционных регуляторных факторов метионина и цистеина.
Согласно еще одному аспекту настоящего изобретения предложен способ получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, включающий культивирование микроорганизма в среде и выделение гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, из микроорганизма или среды.
Как описано выше, микроорганизм может представлять собой микроорганизм рода Corynebacterium, содержащий вариант гомосериндегидрогеназы по настоящему изобретению, и, более конкретно, может представлять собой Corynebacterium glutamicum. Кроме того, микроорганизм рода Corynebacterium или Corynebacterium glutamicum может представлять собой микроорганизм, продуцирующий гомосерин или L-аминокислоту, имеющую происхождение от гомосерина. L-аминокислота, имеющая происхождение от гомосерина, может включать не только L-аминокислоту, имеющую происхождение от гомосерина, но и ее производное. Например, L-аминокислота, имеющая происхождение от гомосерина, может представлять собой, без ограничения ими, L-треонин, L-изолейцин, O-ацетилгомосерин, O-сукцинил-L-гомосерин, O-фосфо-L-гомосерин, L-метионин и/или L-глицин. Более конкретно, L-аминокислота, имеющая происхождение от гомосерина, может представлять собой, без ограничения ими, L-треонин, L-изолейцин, O-ацетилгомосерин, O-сукцинил-L-гомосерин и/или L-метионин. Кроме того, микроорганизм рода Corynebacterium или Corynebacterium glutamicum может представлять собой микроорганизм, продуцирующий L-аланин.
Гомосерин или L-аминокислота, имеющая происхождение от гомосерина, может представлять собой культуральную среду с гомосерином или L-аминокислотой, имеющей происхождение от гомосерина, продуцируемую микроорганизмом, описанным в настоящем изобретении, супернатант культуры, ее обработанный продукт или его очищенную форму. Специалистам в данной области очевидно, что гомосерин или L-аминокислота, имеющая происхождение от гомосерина, включает не только ее нейтральную форму, но также и ее соль.
Способ получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, может быть легко определен специалистами в данной области в оптимизированных условиях культивирования и условиях ферментативной активности, известных в данной области.
В указанном выше способе культивирование микроорганизма может быть выполнено в периодическом процессе, непрерывном процессе, процессе с подпиткой культуры и так далее, известном в данной области, но способ культивирования не ограничен данными способами. В частности, применительно к условиям культивирования, pH культуры может быть скорректирован до подходящего значения pH (например от pH 5 до pH 9, конкретно от pH 6 до pH 8, и наиболее конкретно, с использованием подходящего основного соединения (например гидроксида натрия, гидроксида калия или аммиака) или кислого соединения (например фосфорной кислоты или серной кислоты)), и аэробные условия в культуре можно поддерживать путем введения в культуру кислорода или кислородсодержащей газовой смеси. Температура при культивировании обычно может находиться в диапазоне от 20°C до 45°C и, конкретно, от 25°C до 40°C в течение приблизительно 10-160 часов, но условия культивирования не ограничены этими значениями. Треонин, изолейцин или ацетилгомосерин, продуцируемые посредством описанного выше культивирования, могут секретироваться в культуру или могут оставаться в клетках.
Кроме того, примеры источников углерода для использования в культуральной среде могут включать, без ограничения ими, сахара и углеводы (например глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу); масла и жиры (например соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло); жирные кислоты (например пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту); спирты (например глицерин и этанол); и органические кислоты (например уксусную кислоту). Эти источники углерода могут быть использованы, без ограничения ими, по отдельности или в комбинации. Примеры источников азота для использования в культуральной среде могут включать азотсодержащие органические соединения (например пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) и так далее. Эти источники азота, без ограничения ими, могут быть использованы по отдельности или в комбинации. Примеры источников фосфора для использования в культуральной среде могут включать, без ограничения ими, дигидрофосфат калия, гидрофосфат калия, соответствующие натрийсодержащие соли и так далее. Кроме того, в среду могут также быть включены соли металлов (например сульфат магния или сульфат железа), аминокислоты, витамины и так далее, являющиеся незаменимыми ростостимулирующими веществами.
В настоящем изобретении способ выделения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, продуцируемой на стадии культивирования, может быть осуществлен посредством сбора целевого продукта из культуральной жидкости с использованием подходящего метода, известного в данной области. Например, могут быть применены такие методы, как центрифугирование, фильтрация, анионообменная хроматография, кристаллизация, ВЭЖХ и так далее, и нужное вещество, которым является гомосерин или L-аминокислота, имеющая происхождение от гомосерина, может быть выделено из культуры или культивированного микроорганизма с использованием подходящего метода, известного в данной области. Кроме того, выделение может включать дополнительный процесс очистки и может быть выполнено с использованием подходящего метода, известного в данной области. Для повышения выхода целевого продукта до/после стадии культивирования или стадии выделения может быть включен дополнительный метод.
Согласно еще одному аспекту настоящего изобретения предложена композиция для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, содержащая модифицированную гомосериндегидрогеназу или микроорганизм, содержащий модифицированную гомосериндегидрогеназу, по настоящему изобретению.
Композиция для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, относится к композиции, способной продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, содержащей: модифицированную гомосериндегидрогеназу, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 285 заменена на изолейцин, аминокислота в положении 398 заменена на глутамин, или аминокислоты в обоих этих положениях заменены на изолейцин и глутамин соответственно; полинуклеотид, кодирующий модифицированную гомосериндегидрогеназу; или микроорганизм, содержащий такой полинуклеотид. Например, полинуклеотид может содержать, без ограничения ею, дополнительную структуру, способную управлять полинуклеотидом. Например, полинуклеотид может быть представлен в форме, включенной в вектор, так что функционально связанный ген может экспрессироваться в клетке-хозяине, в которую он введен.
Кроме того, композиция может дополнительно содержать любой подходящий эксципиент, обычно используемый в композициях для получения гомосерина или L-аминокислот, имеющих происхождение от гомосерина. Например, эксципиент может представлять собой, без ограничения ими, консервант, увлажнитель, диспергирующий агент, суспендирующий агент, буфер, стабилизатор, изотонический агент и так далее.
Согласно еще одному аспекту настоящего изобретения предложен способ повышения способности продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, у микроорганизма, включающий замену аминокислоты в положении 285 на изолейцин, аминокислоты в положении 398 на глутамин, или аминокислот в обоих этих положениях на изолейцин и глутамин соответственно, в аминокислотной последовательности SEQ ID NO: 1, обладающей активностью гомосериндегидрогеназы.
Термины «гомосериндегидрогеназа» и «гомосерин или L-аминокислота, имеющая происхождение от гомосерина», описаны выше.
Согласно еще одному аспекту настоящего изобретения предложено применение модифицированной гомосериндегидрогеназы для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
Согласно еще одному аспекту настоящего изобретения предложено применение полинуклеотида, кодирующего модифицированную гомосериндегидрогеназу, для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
Согласно еще одному аспекту настоящего изобретения предложено применение микроорганизма рода Corynebacterium, содержащего модифицированную гомосериндегидрогеназу, для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
Согласно еще одному аспекту настоящего изобретения предложено применение композиции для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
Вариант осуществления изобретения
Ниже настоящее изобретение будет описано подробно со следующими типичными воплощениями. Однако типичные воплощения раскрыты здесь исключительно в иллюстративных целях, и их не следует истолковывать как ограничение объема настоящего изобретения.
Пример 1: Скрининг AHV-резистентных микроорганизмов, полученных посредством искусственной модификации
В данном Примере эксперимент по приданию резистентности к 2-амино-3-гидроксивалерату (называемому ниже «AHV»), являющемуся аналогом L-треонина, проводили с использованием Corynebacterium glutamicum KFCC10881 (корейский патент №0159812) в качестве исходного штамма с целью устранения ингибирования гомосериндегидрогеназы (называемой ниже «Hom», EC:1.1.1.3) L-треонином по принципу обратной связи.
Модификацию создавали методом искусственной модификации с использованием N-метил-N'-нитро-N-нитрозогуанидина (называемого ниже «NTG»). Штамм KFCC10881, культивированный в посевной среде в течение 18 часов, инокулировали в 4 мл посевной среды и затем культивировали до достижения OD660 примерно 1,0. Культуральную среду центрифугировали с выделением клеток и затем клетки промывали два раза 50 мМ трис-малатным буфером (pH 6,5) и суспендировали в конечном объеме 4 мл того же буфера. В суспензию клеток добавляли раствор NTG (2 мг/мл в 0,05 М трис-малатном буфере (pH 6,5)) до конечной концентрации 150 мг/л и затем оставляли ее стоять при комнатной температуре в течение 20 минут. Затем клетки выделяли центрифугированием и промывали два раза тем же буфером для удаления NTG. После последней промывки клетки суспендировали в 4 мл 20%-го раствора глицерина и затем хранили при -70°C до использования. Штаммы, обработанные NTG, высевали на минимальную среду, содержащую 3 г/л AHV, после чего получали 155 AHV-резистентных штаммов KFCC10881, используя описанную выше методику.
Посевная среда (pH 7,0)
Глюкоза 20 г; пептон 10 г; дрожжевой экстракт 5 г; мочевина 1,5 г; KH2PO4 4 г; K2HPO4 8 г; MgSO4⋅7H2O 0,5 г; биотин 100 мкг; тиамин-HCl 1000 мкг; пантотенат кальция 2000 мкг; никотинамид 2000 мкг (на 1 л дистиллированной воды).
Минимальная среда (pH 7,2)
Глюкоза 5 г; KH2PO4 1 г; (NH4)2SO4 5 г; MgSO4⋅7H2O 0,4 г; NaCl 0,5 г; биотин 200 мкг; тиамин-HCl 100 мкг; пантотенат кальция 100 мкг; никотинамид 0,03 г; мочевина 2 г; Na2B4O7⋅10H2O 0,09 мг; (NH4)6Mo7O27⋅4H2O 0,04 мг; ZnSO4⋅7H2O 0,01 мг; CuSO4⋅5H2O, MnCl2⋅4H2O 0,01 мг; FeCl3⋅6H2O 1 мг; CaCl2 0,01 мг (на 1 л дистиллированной воды).
Пример 2: Анализпродуцированию L-треонина AHV-резистентными штаммами KFCC10881
Анализ способности продуцировать L-треонин проводили на 155 AHV-резистентных штаммах, полученных в Примере 1. 155 штаммов, полученных в Примере 1, высевали в колбы с угловыми перегородками (250 мл), содержащие посевную среду (25 мл), и затем культивировали с покачиванием при 30°C и 200 об/мин в течение 20 часов. Посевную культуральную среду (1 мл) вносили в колбу с угловыми перегородками (250 мл), содержащую описанную ниже среду для продуцированию L-треонина (24 мл), и затем культивировали с покачиванием при 30°C и 200 об/мин в течение 48 часов.
Среда для продуцирования L-треонина (pH 7,2)
Глюкоза 30 г; KH2PO4 2 г; мочевина 3 г; (NH4)2SO4 40 г; пептон 2,5 г; CSL (Sigma) 5 г (10 мл); MgSO4⋅7H2O 0,5 г; лейцин 400 мг; CaCO3 20 г (на 1 л дистиллированной воды).
После культивирования количество различных продуцируемых аминокислот измеряли с использованием ВЭЖХ. Концентрации аминокислот в культуральных средах 22 штаммов, продемонстрировавших очень хорошую способность продуцировать L-треонин, из 155 штаммов, включенных в эксперимент, показаны в Таблице 1. 22 штаммам-кандидатам, подтвержденным описанной выше методикой, были присвоены названия от KFCC10881-1 до KFCC10881-22.

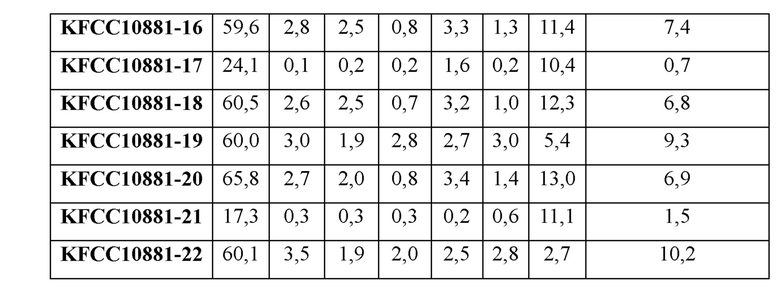
Как показано в Таблице 1, количества L-треонина, L-гомосерина, L-глицина, L-аланина и L-изолейцина, продуцируемых 22 типами штаммов, обладающих резистентностью к AHV, было увеличены по сравнению с контрольной группой, в то время как количество L-лизина было уменьшено.
Начальной точкой разделения путей биосинтеза L-треонина и L-лизина является аспартат-полуальдегид (называемый далее «ASA»). То есть, количество продуцируемого L-лизина уменьшается по мере увеличения количества продуцируемого L-треонина. Соответственно, количества гомосерина (Hse), L-глицина (Gly) и L-изолейцина (Ile), которые могут быть побочными продуктами пути биосинтеза L-треонина, могут увеличиваться по мере увеличения количества продуцируемого L-треонина, и таким образом было также подтверждено общее количество этих продуцируемых аминокислот (Thr + Hse + Gly + Ile).
Таким образом, среди AHV-резистентных штаммов, описанных выше, 4 типа штаммов (KFCC10881-1, KFCC10881-14, KFCC10881-19 и KFCC10881-22) с уменьшенным количеством продуцируемого L-лизина, увеличенным количеством продуцируемого L-треонина и увеличенным общим количеством продуцируемых Thr + Hse + Gly + Ile, были выбраны как наиболее удачные AHV-резистентные штаммы.
Пример 3: Анализ нуклеотидных последовательностей штаммов, обладающих очень хорошей способностью продуцировать треонин, имеющий происхождение от KFCC10881
Чтобы проанализировать нуклеотидные последовательности ферментов биосинтеза L-треонина у штаммов, выбранных в Примере 1 выше, был проведен следующий эксперимент. На основании информации о генах, представленной в Киотской энциклопедии генов и геномов (Kyoto Encyclopedia of Genes and Genomes, KEGG) была получена каждая из нуклеотидной последовательности hom (SEQ ID NO: 1, NCgl1136), кодирующей гомосериндегидрогеназу Corynebacterium glutamicum ATCC13032, и нуклеотидной последовательности thrB (SEQ ID NO: 2, ген № NCgl1137), кодирующей гомосеринкиназу. Известно, что hom и thrB состоят из оперонной структуры (Peoples et al., Mol. Biol. 2(1):63-72, 1988).
Для получения фрагмента ДНК, содержащего оперон hom-thrB выбранных штаммов, проводили ПЦР с использованием геномной ДНК штаммов в качестве матрицы и комбинации праймеров SEQ ID NO: 3 и SEQ ID NO: 4. В качестве полимеразы для ПЦР-реакции использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene). Условия ПЦР были следующими: денатурация при 96°C в течение 30 секунд; отжиг при 52°C в течение 30 секунд; и полимеризация при 72°C в течение 3 минут; в общей сложности были повторены 30 циклов. В результате, удалось амплифицировать фрагмент гена (2778 п.о.; SEQ ID NO: 5), который включает нуклеотидную последовательность (300 п.о.), содержащую промоторный сайт выше кодона инициации SEQ ID NO: 1, и 200 п.о. ниже кодона терминации SEQ ID NO: 2.
Нуклеотидная последовательность была определена с использованием полученного выше праймера и анализатора ABI PRISM 3730XL Analyzer (тип с 96 капиллярами; Applied Biosystems). В нуклеотидной последовательности, соответствующей hom, в опероне hom-thrB KFCC10881-1, цитозин, представляющий собой нуклеотид в положении 854 SEQ ID NO: 1, был мутирован в тимин, и, таким образом, кодон ACT, кодирующий треониновый остаток, был мутирован в кодон ATT, кодирующий изолейциновый остаток (ниже «модификация T285I»; SEQ ID NO: 6). Кроме того, в нуклеотидной последовательности, соответствующей оперону hom-thrB KFCC10881-14, гуанин, представляющий собой нуклеотид в положении 1193 SEQ ID NO: 1, был мутирован на аденин, и, таким образом, кодон CGA, кодирующий аргининовый остаток, был мутирован в кодон CAA, кодирующий глутаминовый остаток (ниже «модификация R398Q»; SEQ ID NO: 7). Кроме того, в нуклеотидной последовательности, соответствующей оперону hom-thrB KFCC10881-19, гуанин, представляющий собой нуклеотид в положении 1132 SEQ ID NO: 1, был мутирован на цитозин, и, таким образом, кодон GGG, кодирующий глициновый остаток, был мутирован на кодон TGG, кодирующий триптофановый остаток (ниже «модификация G378W»; SEQ ID NO: 8). Кроме того, в нуклеотидной последовательности, соответствующей оперону hom-thrB KFCC10881-22, гуанин, представляющий собой нуклеотид в положении 1132 SEQ ID NO: 1, был мутирован на аденин, а гуанин, представляющий собой нуклеотид в положении 1134 SEQ ID NO: 1, был мутирован на цитозин, и, таким образом, кодон GGG, кодирующий глициновый остаток, был мутирован на кодон AGC, кодирующий сериновый остаток (ниже «модификация G378S»; SEQ ID NO: 9). В то же время, в thrB, соответствующем SEQ ID NO: 2, модификаций не было.
По результатам анализов нуклеотидных последовательностей, описанных выше, удалось подтвердить, что ингибирование L-треонином по принципу обратной связи было десенсибилизировано, так как в Hom (SEQ ID NO: 10), экспрессируемой в KFCC10881-1, треонин, представляющий собой аминокислотный остаток в положении 285, был мутирован на изолейцин (модификация T285I); в Hom (SEQ ID NO: 11), экспрессируемой в KFCC10881-14, аргинин, представляющий собой аминокислотный остаток в положении 398, был мутирован на глутамин (модификация R398Q); в Hom (SEQ ID NO: 12), экспрессируемой в KFCC10881-19, глицин, представляющий собой аминокислотный остаток в положении 378, был мутирован на триптофан (модификация G378W); и в Hom (SEQ ID NO: 13), экспрессируемой в KFCC10881-22, глицин, представляющий собой аминокислотный остаток в положении 378, был мутирован на серин (модификация G378S).
Пример 4: Получение новых штаммов, в которые введена гомосериндегидрогеназа
Праймеры SEQ ID NO: 14 и SEQ ID NO: 15 были приготовлены для получения штаммов, где варианты (T285I, R398Q, G378W и G378S), указанные в Примере 2, были введены в штаммы дикого типа.
Для получения штаммов, в которые были введены каждая из модификаций hom T285I, R398Q, G378W и G378S, проводили ПЦР с использованием геномной ДНК, выделенной из каждого из штаммов KFCC10811-1, KFCC10811-14, KFCC10811-19 и KFCC10811-22, в качестве матрицы и с использованием праймеров SEQ ID NO: 14 и SEQ ID NO: 15. В качестве полимеразы для ПЦР использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene). Условия ПЦР были следующими: денатурация при 95°C в течение 30 секунд; отжиг при 55°C в течение 30 секунд; и полимеризация при 72°C в течение 2 минут; и в общей сложности повторяли 28 циклов. В результате был получен фрагмент гена (1668 п.о.), включающий промоторный сайт (примерно 100 п.о.) гена hom (1338 п.о.). Амплифицированный продукт очищали с использованием набора PCR Purification (QUIAGEN) и затем использовали в качестве вводимого фрагмента ДНК для получения вектора. В тоже время, после обработки рестриктазой SmaI, отношение молярной концентрации (М) вектора pDZ, обработанного нагреванием до 65°C в течение 20 минут, к вводимому фрагменту ДНК, амплифицированному посредством описанной выше ПЦР, было установлено как 1:2, после чего проводили клонирование с использованием набора Infusion Cloning Kit (TaKaRa), следуя руководству по его применению. Таким образом, были получены векторы, то есть pDZ-T285I, pDZ-R398Q, pDZ-G378W и pDZ-G378S, для введения модификаций T285I, R398Q, G378W и G378S в хромосому.
Corynebacterium glutamicum ATCC13032 трансформировали каждым из полученных векторов посредством электропорации. После вторичного кроссовера были получены штаммы, в хромосомы которых были введены каждый из модифицированных нуклеотидов. Используя комбинацию указанных ниже праймеров и ПЦР-методики MASA (Mutant Allele Specific Amplification, специфичная амплификация мутантных аллелей) (Takeda et al., Hum. Mutation, 2, 112-117 (1993)) проводили первичную оценку правильности замен, выбирая амплифицированные штаммы в комбинации праймеров, соответствующих каждой из модифицированных последовательностей (CTR-T285I: SEQ ID NO: 16 и SEQ ID NO: 17; CTR-R398Q: SEQ ID NO: 16 и SEQ ID NO: 18; CTR-G378W: SEQ ID NO: 16 и SEQ ID NO: 19; и CTR-G378S: SEQ ID NO: 16 и SEQ ID NO: 20). Кроме того, были проведены анализы последовательностей hom выбранных штаммов для вторичного подтверждения правильности замен с использованием SEQ ID NO: 16 и SEQ ID NO: 21 и путем анализа модифицированных последовательностей таким же образом, как в Примере 2. Штаммы с заменами каждым из модифицированных нуклеотидов были названы CTR-T285I, CTR-R398Q, CTR-G378W и CTR-G378S соответственно.
Пример 5: Измерение активности гомосериндегидрогеназы
В полученных штаммах измеряли активность фермента Hom. Штамм дикого типа ATCC13032 в контрольной группе и CTR-T285I, CTR-R398Q, CTR-G378W и CTR-G378S, полученные в Примере 4, инокулировали в 25 мл посевной среды и затем культивировали до достижения поздней логарифмической фазы роста. Клетки выделяли центрифугированием, промывали два раза 0,1 М калий-фосфатным буфером (pH 7,6) и, наконец, суспендировали в 2 мл того же буфера, содержащего глицерин в концентрации 30%. Суспензию клеток подвергали физическому разрушению обычным методом встряхивания со стеклянными гранулами в течение 10 минут, и затем отбирали супернатант от двух центрифугирований (13000 об/мин, 4°C, 30 минут) и использовали его в качестве неочищенного экстракта для измерения активности фермента Hom. Для измерения активности Hom в реакционный раствор добавляли раствор кофермента (0,1 мл) для измерения ферментативной активности (калий-фосфатный буфер (pH 7,0), 25 мМ NADPH, 5 мМ аспартат-полуальдегида) и затем проводили взаимодействие при 30°C. Активность Hom (ЕД) определяли как количество мкмоль NADPH, потребляемых за минуту, в зависимости от присутствия L-треонина (0 мМ, 10 мМ), и полученные результаты по ферментативной активности показаны в Таблице 2 ниже.
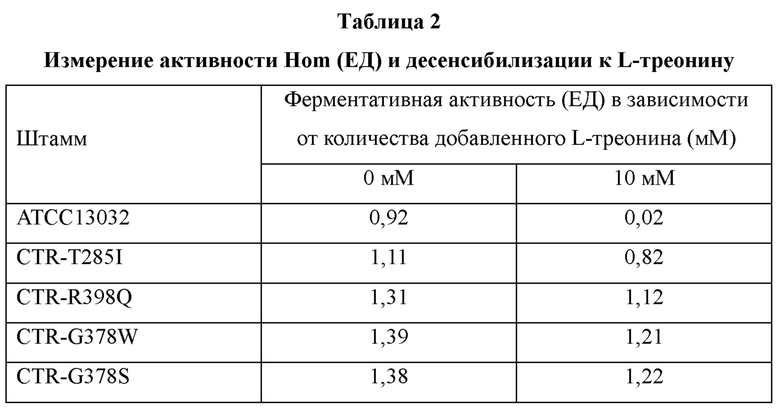
В результате эксперимента было подтверждено, что ингибирование активности
Hom, содержащей каждую из модификаций T285I, R398Q, G378W и G378S, было снижено в условиях содержания 10 мМ L-треонина, в отличие от Hom дикого типа, то есть происходила десенсибилизацию к L-треонину.
При мер 6: Получение и анализ штамма микроорганизма рода Corynebacterium , обладающего продуктивностью по L-треонину
Штаммы, продуцирующие L-треонин, были получены из Corynebacterium glutamicum ATCC13032 дикого типа. Конкретно, для устранения ингибирования аспартаткиназы (LysC), которая является важным, первым действующим ферментом в пути биосинтеза треонина, по принципу обратной связи, лейцин, представляющий собой аминокислоту в положении 377 LysC, был заменен на лизин (SEQ ID NO: 22).
Более конкретно, для получения штаммов, в которые введена модификация LysC (L377K), проводили ПЦР с использованием хромосомы ATCC13032 в качестве матрицы и праймеров SEQ ID NO: 23 и 24 или SEQ ID NO: 25 и 26. В качестве полимеразы для ПЦР использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene). Условия ПЦР были следующими: денатурация при 95°C в течение 30 секунд; отжиг при 55°C в течение 30 секунд; и полимеризация при 72°C в течение 1 минуты; и в общей сложности повторяли 28 циклов. В результате были получены фрагмент ДНК (515 п.о.) в 5'-восходящей области и фрагмент ДНК (538 п.о.) в 3'-нисходящей области с сайтом модификации гена lysC в центре. Проводили ПЦР с двумя амплифицированными фрагментами ДНК в качестве матриц, используя праймеры SEQ ID NO: 23 и SEQ ID NO: 26. После денатурации при 95°C в течение 5 минут проводили ПЦР в течение в общей сложности 28 циклов в следующих условиях: денатурация при 95°C в течение 30 секунд; отжиг при 55°C в течение 30 секунд; и полимеризация при 72°C в течение 2 минут. Затем проводили реакцию полимеризации при 72°C в течение 5 минут. В результате был амплифицирован фрагмент ДНК (1023 п.о.), включающий модификацию гена lysC, который кодирует вариант аспартаткиназы, в котором лейцин в положении 377 заменен на лизин. Амплифицированный продукт очищали с использованием набора PCR Purification kit (QUIAGEN) и использовали в качестве вводимого фрагмента ДНК для получения вектора. В тоже время, после обработки рестриктазой SmaI, отношение молярной концентрации (М) вектора pDZ, обработанного нагреванием при 65°C в течение 20 минут, к вводимому фрагменту ДНК, амплифицированному при помощи описанной выше ПЦР, было установлено как 1:2, после чего проводили клонирование с использованием набора Infusion Cloning Kit (TaKaRa), следуя руководству по его применению. Таким образом, был получен вектор pDZ-L377K для введения модификации L377K в хромосому.
ATCC13032 трансформировали полученным вектором посредством электропорации. После вторичного кроссовера был получен штамм, в котором каждая из нуклеотидных модификаций заменена модифицированными нуклеотиды, и штамм был назван CJP1.
Для ясного подтверждения изменения в продуцировании L-треонина данным штаммом, в ген, кодирующий гомосериндегидрогеназу, вводили каждую из модификаций, указанных в Примере 4. Конкретно, для введения каждой из модификаций T285I, R398Q, G378W и G378S в штамм CTR-L377K, CJP1 трансформировали каждым из векторов pDZ-T285I, pDZ-R398Q, pDZ-G378W и pDZ-G378S, полученных в Примере 4, посредством электропорации и затем получали штаммы, где каждая из нуклеотидных модификаций в хромосоме заменена модифицированными нуклеотидами, посредством вторичного кроссовера таким же образом, как в Примере 4. Штаммы с замещением каждым из модифицированных нуклеотидов были названы CJP1-T285I, CJP1-R398Q, CJP1-G378W и CJP1-G378S.
Штаммы CJP1-T285I и CJP1-R398Q были депонированы в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM), являющемся международным органом по депонированию согласно Будапештскому соглашению, 26 сентября 2017 г. под регистрационными номерами KCCM12119P и KCCM12120P соответственно.
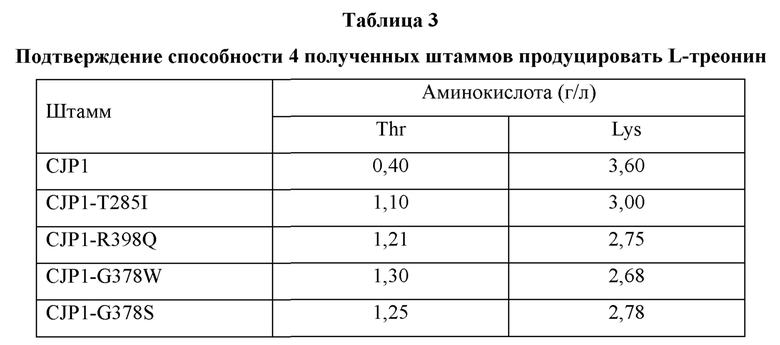
В результате, у штаммов, в которые были введены каждая из модификаций, количество продуцируемого L-лизина уменьшилось, а количество продуцируемого L-треонина увеличилось на 0,7-0,9 г/л по сравнению со штаммом CJP1.
В то же время, для получения штамма, одновременно содержащего модификации T285I и R398Q, штамм CJP1-T285I трансформировали вектором pDZ-R398Q, после чего получали штамм CJP1-T285I,R398Q таким же методом, как описано выше. Кроме того, для получения штамма, одновременно содержащего модификации G378W и R398Q, штамм CJP1-G378W трансформировали вектором pDZ-R398Q, после чего получали штамм CJP1-G378W,R398Q таким же методом, как описано выше. Кроме того, для получения штамма, одновременно содержащего модификации T285I и G378W, штамм CJP1-T285I трансформировали вектором pDZ-G378W, после чего получали штамм CJP1-T285I,G378W таким же методом, как описано выше. Анализ способности продуцировать L-треонин проводили методом, описанным в Примере 2, и его результаты показаны в Таблице 4 ниже.

В результате было подтверждено, что при введении двух типов модификаций по настоящему изобретению способность продуцировать треонин была выше, чем у штамма CJP1-G378W, показавшим в Примерах наибольшую способность продуцировать треонин. Количество треонина, продуцируемого штаммами, в которые были введены две модификации, было на 1,1-1,7 г/л больше, чем у штамма CJP1, являющегося контрольной группой, и таким образом было подтверждено значительное улучшение эффекта десенсибилизации Hom.
Пример 7: Получение и анализ штамма микроорганизма рода Corynebacterium , продуцирующего L-изолейцин
С целью получения штаммов, продуцирующих изолейцин, был получен вектор для усиления экспрессии модифицированного гена ilvA (V323A) (Appl. Enviro. Microbiol., Dec. 1996, p. 4345-4351), кодирующего известную L-треониндегидратазу (первый фермент в пути биосинтеза изолейцина), у штаммов, полученных в Примере 6.
Конкретно, для получения вектора для введения модификации, нацеленного на ген ilvA, были сконструированы пара праймеров (SEQ ID NO: 27 и 28) для амплификации 5'- восходящей области и пара праймеров (SEQ ID NO: 29 и 30) для амплификации 3'- нисходящей области с сайтом модификации в центре. В каждый конец праймеров SEQ ID NO: 27 и 30 были введены сайты рестрикции BamHI (подчеркнуты), и праймеры SEQ ID NO: 28 и 29 были сконструированы таким образом, что модификация с заменой нуклеотидов (подчеркнута) располагается в области, где следует индуцировать кроссовер.
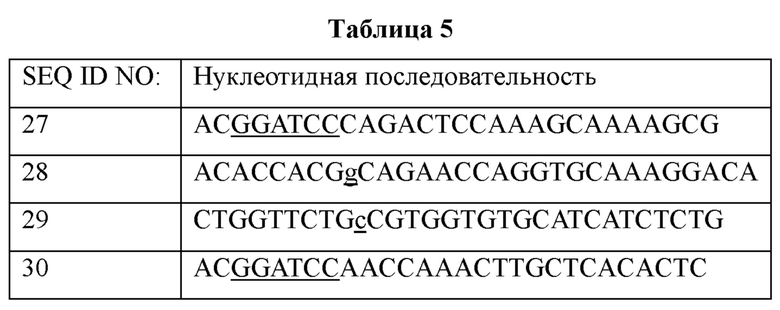
ПЦР проводили с использованием хромосомы дикого типа в качестве матрицы и праймеров SEQ ID NO: 27, 28, 29 и 30. После денатурации при 95°C в течение 5 минут проводили ПЦР в течение в общей сложности 30 циклов в следующих условиях: денатурация при 95°C в течение 30 секунд; отжиг при 55°C в течение 30 секунд; и полимеризация при 72°C в течение 30 секунд. Затем проводили реакцию полимеризации при 72°C в течение 7 минут. В результате были получены фрагмент ДНК (627 п.о.) 5'- восходящей области и фрагмент ДНК (608 п.о.) 3'- нисходящей области с сайтом модификации гена ilvA в центре.
Проводили ПЦР с двумя амплифицированными фрагментами ДНК в качестве матриц, используя праймеры SEQ ID NO: 27 и SEQ ID NO: 30. После денатурации при 95°C в течение 5 минут проводили ПЦР в течение в общей сложности 30 циклов в следующих условиях: денатурация при 95°C в течение 30 секунд; отжиг при 55°C в течение 30 секунд; и полимеризация при 72°C в течение 60 секунд. Затем проводили реакцию полимеризации при 72°C в течение 7 минут. В результате был амплифицирован фрагмент ДНК (1217 п.о.), включающий модификацию гена ilvA, кодирующий вариант IlvA, где валин в положении 323 заменен аланином. Вектор pECCG117 (корейский патент №10-0057684) и фрагмент ДНК (1011 п.о.) обрабатывали рестриктазой BamHI, лигировали с использованием ДНК-лигазы и затем клонировали с получением плазмиды. Полученная таким образом плазмида была названа pECCG117-ilvA(V323A).
Вектор pECCG117-ilvA(V323A) вводили в каждый из штаммов CJP1-T285I,R398Q, CJP1-G378W,R398Q и CJP1-T285I,G378W, полученных в Примере 6, посредством электропорации и наносили мазками на селективную среду, содержащую канамицин (25 мг/л), с получением трансформированных штаммов. Полученные таким образом трансформированные штаммы культивировали тем же методом культивирования в колбах, что и в Примере 2, и анализировали концентрации L-изолейцина в культуральных средах. Результаты анализа показаны в Таблице 6.

В результате было подтверждено, что у штамма, содержащего модификацию hom(G378W), концентрация L-изолейцина, была на 0,2 г/л выше, чем у контрольного штамма. Кроме того, у штамма, включающего модификацию hom, в который были введены одновременно две модификации, способность продуцировать L-изолейцин была дополнительно улучшена на 0,3-0,5 г/л по сравнению с контрольным штаммом. Кроме того, среди полученных штаммов, штамм CJP1-T285I,R398Q/pECCG117-ilvA(V323A), содержащий обе модификации T285I и R398Q, продуцировал L-изолейцин в количестве 1,1 г/л.
Пример 8: Получение и оценка штамма, продуцирующего O -ацетилгомосерин (OAH) с заменой модифицированной Hom
8-1. Получение штамма ATCC13032 с заменой модифицированной Hom
В штамм ATCC13032 вводили модификации двух типов (T285I и R398Q) таким же образом, как в Примере 7, и полученный таким образом штамм был назван ATCC13032::HomFBR.
8-2. Делеция гена metB
В данном примере получали ген metB, кодирующий цистатионин-гамма-синтазу в пути расщепления O-ацетилгомосерина, посредством ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum ATCC13032 в качестве матрицы. Информацию о нуклеотидной последовательности metB (регистрационный номер в NCBI Ncgl2360; SEQ ID NO: 31) получали из базы GenBank Национальных институтов здравоохранения США (National Institutes of Health, NIH GenBank). Затем на основании этой информации синтезировали праймеры (SEQ ID NO: 32 и 33), содержащие N-концевую и линкерную последовательность гена metB, и праймеры (SEQ ID NO: 34 и 35), содержащие C-концевую и линкерную последовательность гена metB. Проводили ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum ATCC13032 в качестве матрицы и олигонуклеотидов с нуклеотидными последовательностями SEQ ID NO: 32 и 33 и SEQ ID NO: 34 и 35 в качестве праймеров. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene). Условия ПЦР были следующими: денатурация при 96°C в течение 30 секунд; отжиг при 53°C в течение 30 секунд; и полимеризация при 72°C в течение 1 минуты; и проводили в общей сложности 30 циклов. В результате были получены амплифицированный ген (500 п.о.), содержащий N-конец и линкер гена metB, и амплифицированный ген (500 п.о.), содержащий C-конец и линкер гена metB.
Проводили ПЦР с использованием двух полученных таким образом амплифицированных генов в качестве матрицы в течение в общей сложности 10 циклов в следующих условиях: денатурация при 96°C в течение 60 секунд; отжиг при 50°C в течение 60 секунд; и полимеризация при 72°C в течение 1 минуты. Затем добавляли нуклеотидные последовательности SEQ ID NO: 32 и 35, после чего проводили в общей сложности 20 циклов. В результате был получен амплифицированный ген -ΔmetB (1000 п.о.), представляющий собой кассету инактивации metB, содержащую N-конец-линкер-C-конец гена metB. Ген metB, полученный посредством ПЦР, обрабатывали рестриктазами XbaI и SalI, включенными на концах, и затем клонировали в вектор pDZ (KR 0924065), с обработанными рестриктазами XbaI и SalI, посредством лигирования. В результате был получен рекомбинантный вектор pDZ-ΔmetB, в который окончательно клонирована кассета инактивации metB.
Полученным таким образом вектором pDZ-ΔmetB трансформировали Corynebacterium glutamicum ATCC13032 и ATCC13032::HomFBR. После вторичного кроссовера были получены Corynebacterium glutamicum ATCC13032 ΔmetB и ATCC13032::HomFBR ΔmetB, в хромосомах которых инактивирован ген metB. В завершение, инактивацию гена metB подтверждали, проводя ПЦР с использованием праймеров SEQ ID NO: 32 и 25, и затем его сравнивали с ATCC13032, в котором ген metB не инактивирован.
8-3. Делеция гена metY
В данном Примере получали ген metY, кодирующий O-ацетилгомосерин(тиол)- лиазу в пути распада O-ацетилгомосерина, посредством ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum ATCC13032 в качестве матрицы. Информацию о нуклеотидной последовательности metY (регистрационный номер в NCBI Ncgl0625; SEQ ID NO: 36) получали из базы GenBank Национальных институтов здравоохранения США (NIH GenBank). Затем на основании этой информации синтезировали праймеры (SEQ ID NO: 37 и 38), содержащие N-концевую и линкерную последовательность гена metY, и праймеры (SEQ ID NO: 39 и 40), содержащие C-концевую и линкерную последовательность гена metY.
Проводили ПЦР с хромосомной ДНК Corynebacterium glutamicum ATCC13032 в качестве матрицы, используя олигонуклеотиды с нуклеотидными последовательностями SEQ ID NO: 39 и 40 в качестве праймеров. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene). Условия ПЦР были следующими: денатурация при 96°C в течение 30 секунд; отжиг при 53°C в течение 30 секунд; и полимеризация при 72°C в течение 1 минуты; повторяли в общей сложности 30 циклов. В результате, были получены амплифицированный ген (500 п.о.), содержащий N-конец и линкер гена metY, и амплифицированный ген (500 п.о.), содержащий C-конец и линкер гена metY. Проводили ПЦР с использованием двух полученных таким образом амплифицированных генов в качестве матрицы в течение в общей сложности 10 циклов в следующих условиях: денатурация при 96°C в течение 60 секунд; отжиг при 50°C в течение 60 секунд; и полимеризация при 72°C в течение 1 минуты. Затем добавляли нуклеотидные последовательности SEQ ID NO: 37 и 40, после чего повторяли в общей сложности 20 циклов. В результате был получен амплифицированный ген ΔmetY (1000 п.о.), представляющий собой кассету инактивации metY, содержащую N-конец- линкер-C-конец гена metY.
Ген metY, полученный посредством ПЦР, обрабатывали рестриктазами XbaI и SalI, включенными на концах, и затем клонировали в вектор pDZ (KR 2008-0025355) с обработкой рестриктаз XbaI и SalI, посредством лигирования. В результате был получен рекомбинантный вектор pDZ-ΔmetY, в который окончательно клонирована кассета инактивации metY.
Полученным таким образом вектором pDZ-ΔmetY трансформировали штаммы Corynebacterium glutamicum ATCC13032, ATCC13032::HomFBR, ATCC13032 ΔmetB и ATCC13032::HomFBR ΔmetB. После вторичного кроссовера были получены Corynebacterium glutamicum ATCC13032 ΔmetY, ATCC13032::HomFBR ΔmetY, ATCC13032 ΔmetB ΔmetY и ATCC13032::HomFBR ΔmetB ΔmetY, в хромосомах которых инактивирован ген metY. В завершение, инактивацию гена metY подтверждали, проводя ПЦР с использованием праймеров SEQ ID NO: 37 и 40 и последующее сравнение с ATCC13032, в котором ген metY не инактивирован.
8-4. Получение и оценка штамма, продуцирующего O -ацетилгомосерин
Проводили сравнение способности продуцировать O-ацетилгомосерин у штаммов ATCC13032, ATCC13032 ΔmetB, ATCC13032 ΔmetY, ATCC13032 ΔmetB ΔmetY, ATCC13032::HomFBR, ATCC13032::HomFBR ΔmetB, ATCC13032::HomFBR ΔmetY и ATCC13032::HomFBR ΔmetB ΔmetY, полученных в Примерах 8-1, 8-2 и 8-3, с делецией генов metB, metY, metBY и модификацией гена hom.
Конкретно, отдельные колонии культивировали на твердой среде LB в течение ночи в инкубаторе при 32°C и по одной петле каждой из отдельных колоний высевали на среду для определения титров O-ацетилгомосерина (25 мл), культивируя затем полученные культуры при 32°C и 250 об/мин в течение 42-64 часов. O-ацетилгомосерин из каждой полученной культуры анализировали посредством ВЭЖХ, и результаты анализа показаны в Таблице 7 ниже.
Среда для продуцирования O -ацетилгомосерина (pH 7,2)
Глюкоза 30 г; KH2PO4 2 г; мочевина 3 г; (NH4)2SO4 40 г; пептон 2,5 г; CSL (Sigma) 5 г (10 мл); MgSO4⋅7H2O 0,5 г; метионин 400 мг; лейцин 400 мг; CaCO3 20 г (на 1 л дистиллированной воды).
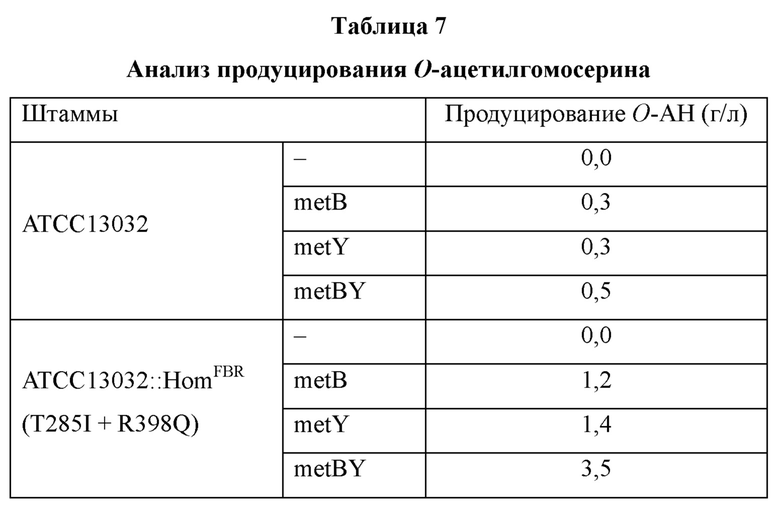
В результате, как показано в Таблице 7 выше, при культивировании контрольного штамма Corynebacterium glutamicum ATCC13032 накопления O-ацетилгомосерина не происходило, в то время как у штаммов ATCC13032 ΔmetB, ATCC13032 ΔmetY и ATCC13032 ΔmetB ΔmetY с инактивированными генами metB, metY и metBY, происходило накопление O-ацетилгомосерина в количестве 0,3 г/л, 0,3 г/л и 0,5 г/л соответственно.
Кроме того, в случае штамма ATCC13032::HomFBR, в котором ген hom заменен на мутантную форму, и штаммов ATCC13032::HomFBR ΔmetB, ATCC13032::HomFBR ΔmetY и ATCC13032::HomFBR ΔmetB ΔmetY, в которых инактивированы гены metB, metY и metBY соответственно, было подтверждено, что O-ацетилгомосерин накапливается в количестве 1,2 г/л, 1,4 г/л и 3,5 г/л для каждого их этих штаммов.
Таким образом, изложенные выше результаты подтвердили возможность значительного повышения количества продуцируемой целевой аминокислоты, предшественником которой является гомосерин, путем использования модифицированной hom по настоящему изобретению.
Пример 9: Получение и оценка штамма, продуцирующего метионин (Met)
При мер 9-1: Получение рекомбинантного вектора для делеции гена mcbR
В данном Примере с целью получения штаммов, продуцирующих метионин, был получен вектор для инактивации гена mcbR (J. Biotechnol. 103:51-65, 2003), кодирующего известные метиониновые и цистеиновые транскрипционные регуляторные белки у штаммов, полученных в Примере 6.
Конкретно, рекомбинантный плазмидный вектор получали, используя описанный ниже метод для нокаута гена mcbR в хромосоме Corynebacterium ATCC13032. Исходя из нуклеотидных последовательностей, представленных в базе GenBank Национальных институтов здравоохранения США (NIH GenBank), были получены ген mcbR и окружающая его последовательность (SEQ ID NO: 41) Corynebacterium glutamicum.
Для делеции гена mcbR проводили ПЦР с хромосомной ДНК Corynebacterium glutamicum ATCC13032 в качестве матрицы, используя праймеры SEQ ID NO: 42 и 43 и SEQ ID NO: 44 и 45. После денатурации при 95°C в течение 5 минут проводили ПЦР в течение в общей сложности 30 циклов в следующих условиях: денатурация при 95°C в течение 30 секунд; отжиг при 53°C в течение 30 секунд; и полимеризация при 72°C в течение 30 секунд. В результате были получены фрагменты ДНК (700 п.о.).
Вектор pDZ (корейский патент №10-0924065), который не может быть клонирован в Corynebacterium glutamicum, и амплифицированные фрагменты гена mcbR обрабатывали рестриктазой SmaI для введения в хромосому. Затем их лигировали с использованием ДНК-лигазы, после чего трансформировали клетки E. coli DH5α, затем наносили мазками на твердую среду LB, содержащую канамицин (25 мг/л).
Проводили отбор трансформированных вектором колоний, в которые посредством ПЦР введены делетированные фрагменты целевых генов, и получали плазмиду, используя метод экстракции плазмид. Полученная таким образом плазмида была названа pDZ-ΔmcbR.
Пример 9-2: Получение и оценка штамма микроорганизма рода Corynebacterium , продуцирующего L-метионин
Каждый из штаммов CJP1-G378W, CJP1-T285I,R398Q, CJP1-G378W,R398Q, CJP1-T285I,G378W и CJP1, полученных в Примере 6 посредством гомологичной рекомбинации хромосом, трансформировали вектором pDZ-ΔmcbR, полученным в Примере 9, с применением электропорации (van der Rest et al., Appl. Microbiol. Biotechnol. 52:541-545, 1999). Затем проводили вторичную рекомбинацию на твердой среде, содержащей X-gal. Штаммы с делецией гена mcbR подтверждали методом ПЦР трансформированных штаммов Corynebacterium glutamicum, в которых была проведена вторичная рекомбинация, с использованием праймеров SEQ ID NO: 46 и 47. Эти рекомбинантные штаммы были названы Corynebacterium glutamicum CJP1-G378W/ΔmcbR, CJP1-T285I,R398Q/ΔmcbR, CJP1-G378W,R398Q/ΔmcbR, CJP1-T285I,G378W/ΔmcbR и CJP1/ΔmcbR соответственно.
Для анализа способности продуцировать L-метионин у полученных штаммов CJP1-G378W/ΔmcbR, CJP1-T285I,R398Q/ΔmcbR, CJP1-G378W,R398Q/ΔmcbR и CJP1-T285I,G378W/ΔmcbR эти штаммы культивировали вместе с их исходным штаммом Corynebacterium glutamicum CJP1/ΔmcbR следующим образом.
Corynebacterium glutamicum CJP1/ΔmcbR и штаммы по изобретению (Corynebacterium glutamicum CJP1-G378W/ΔmcbR, CJP1-T285I,R398Q/ΔmcbR, CJP1-G378W,R398Q/ΔmcbR и CJP1-T285I,G378W/ΔmcbR) инокулировали в колбы с угловыми перегородками (250 мл), содержащие посевную среду, указанную ниже (25 мл), и затем культивировали с покачиванием при 30°C и 200 об/мин в течение 20 часов. После этого посевную культуральную среду (1 мл) вносили в колбу с угловыми перегородками (250 мл), содержащую описанную ниже продукционную среду (24 мл), и затем культивировали с покачиванием при 30°C и 200 об/мин в течение 48 часов. Состав посевной среды и продукционной среды является следующим.
Посевная среда (pH 7,0)
Глюкоза 20 г; пептон 10 г; дрожжевой экстракт 5 г; мочевина 1,5 г; KH2PO4 4 г; K2HPO4 8 г; MgSO4⋅7H2O 0,5 г; биотин 100 мкг; тиамин-HCl 1000 мкг; пантотенат кальция 2000 мкг; никотинамид 2000 мкг (на 1 л дистиллированной воды).
Продукционная среда (pH 8,0)
Глюкоза 50 г; (NH4)2S2O3 12 г; дрожжевой экстракт 5 г; KH2PO4 1 г; MgSO4⋅7H2O 1,2 г; биотин 100 мкг; тиамин-HCl 1000 мкг; пантотенат кальция 2000 мкг; никотинамид 3000 мкг; CaCO3 0 г (на 1 л дистиллированной воды).
После культивирования с использованием способа культивирования, описанного выше, анализировали концентрацию L-метионина в каждой культуральной среде, и результаты анализа показаны в Таблице 8.
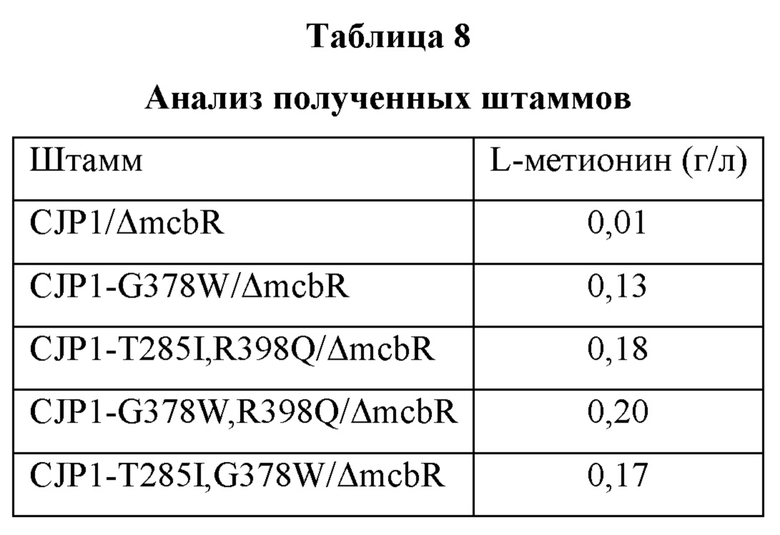
В результате было подтверждено, что у штамма, содержащего модификацию hom G378W, способность продуцировать L-метионин была на 0,12 г/л выше, чем у контрольного штамма. Кроме того, было подтверждено, что у штаммов, содержащих модификацию hom, в которые были введены одновременно две модификации, способность продуцировать L-метионин была на 0,16-0,19 г/л выше, чем у контрольного штамма.
Изложенные выше результаты подтвердили, что количество продуцируемого L-метионина может быть значительного увеличено путем использования модифицированной hom по настоящему изобретению.
Несмотря на то, что настоящее изобретение описано со ссылкой на конкретные наглядные воплощения, специалистам в области, к которой относится настоящее изобретение, очевидно, что оно может быть воплощено в других конкретных формах без отклонения от технической сущности или существенных характеристик настоящего изобретения. Таким образом, описанные выше воплощения следует считать иллюстративными и не ограничивающими во всех аспектах. Кроме того, объем настоящего изобретения определен приложенной формулой изобретения, но не его подробным описанием, и следует понимать, что все модификации или варианты, имеющие происхождение от значений и объема настоящего изобретения и его эквивалентов, включены в объем прилагаемой формулы изобретения.
Регистрационный номер
Наименование органа по депонированию: Корейский центр культур микроорганизмов (Korean Culture Center of Microorganisms) (международный орган по депонированию)
Депозитарный номер: KCCM12119P
Дата депонирования: 26 сентября 2017 г.
Наименование органа по депонированию: Корейский центр культур микроорганизмов (Korean Culture Center of Microorganisms) (международный орган по депонированию)
Депозитарный номер: KCCM12120P
Дата депонирования: 26 сентября 2017 г.
Изобретение относится к биотехнологии, в частности к модифицированной гомосериндегидрогеназе и к способу получения L-аминокислоты, имеющей происхождение от гомосерина, с использованием такой гомосериндегидрогеназы. Изобретение позволяет получать аминокислоты с высокой степенью эффективности. 9 н. и 8 з.п. ф-лы, 8 табл., 9 пр.
1. Модифицированная гомосериндегидрогеназа, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 285 заменена на изолейцин; аминокислота в положении 398 заменена на глутамин; или аминокислоты в обоих этих положениях заменены на изолейцин и глутамин соответственно.
2. Модифицированная гомосериндегидрогеназа по п. 1, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 378 дополнительно заменена на триптофан.
3. Полинуклеотид, кодирующий модифицированную гомосериндегидрогеназу по любому из пп. 1, 2.
4. Микроорганизм рода Corynebacterium для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, содержащий модифицированную гомосериндегидрогеназу, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 285 заменена на изолейцин; аминокислота в положении 398 заменена на глутамин; или аминокислоты в обоих этих положениях заменены на изолейцин и глутамин соответственно.
5. Микроорганизм по п. 4, где микроорганизм рода Corynebacterium представляет собой микроорганизм рода Corynebacterium, продуцирующий гомосерин или L-аминокислоту, имеющую происхождение от гомосерина.
6. Микроорганизм по п. 5, где L-аминокислота, имеющая происхождение от гомосерина, представляет собой по меньшей мере одну, выбранную из группы, состоящей из L-треонина, L-изолейцина, O-ацетилгомосерина и L-метионина.
7. Микроорганизм по п. 4, где микроорганизм рода Corynebacterium продуцирует L-аланин.
8. Микроорганизм по п. 4, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
9. Способ получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, включающий:
культивирование микроорганизма рода Corynebacterium, содержащего модифицированную гомосериндегидрогеназу, где в аминокислотной последовательности SEQ ID NO: 1 аминокислота в положении 285 заменена на изолейцин; аминокислота в положении 398 заменена на глутамин; или аминокислоты в обоих этих положениях заменены на изолейцин и глутамин соответственно, в среде; и
выделение гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, из микроорганизма или среды.
10. Способ по п. 9, где L-аминокислота, имеющая происхождение от гомосерина, представляет собой по меньшей мере одну, выбранную из группы, состоящей из L-треонина, L-изолейцина, O-ацетилгомосерина и L-метионина.
11. Композиция для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина, содержащая модифицированную гомосериндегидрогеназу по п. 1 или микроорганизм по п. 4 и эксципиент.
12. Композиция по п. 11, где L-аминокислота, имеющая происхождение от гомосерина, представляет собой по меньшей мере одну, выбранную из группы, состоящей из L-треонина, L-изолейцина, O-ацетилгомосерина и L-метионина.
13. Способ повышения способности продуцировать гомосерин или L-аминокислоту, имеющую происхождение от гомосерина, у микроорганизма рода Corynebacterium, включающий замену аминокислоты в положении 285 на изолейцин; аминокислоты в положении 398 на глутамин; или аминокислот в обоих этих положениях на изолейцин и глутамин соответственно, в аминокислотной последовательности SEQ ID NO: 1, обладающей активностью гомосериндегидрогеназы.
14. Способ по п. 13, где L-аминокислота, имеющая происхождение от гомосерина, представляет собой по меньшей мере одну, выбранную из группы, состоящей из L-треонина, L-изолейцина, O-ацетилгомосерина и L-метионина.
15. Применение модифицированной гомосериндегидрогеназы по п. 1 для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
16. Применение полинуклеотида по п. 3 для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
17. Применение микроорганизма рода Corynebacterium по п. 4 для получения гомосерина или L-аминокислоты, имеющей происхождение от гомосерина.
| МОДИФИЦИРОВАННЫЙ ПОЛИПЕПТИД, ОБДАДАЮЩИЙ ГОМОСЕРИНАЦЕТИЛТРАНСФЕРАЗНОЙ АКТИВНОСТЬЮ, И ЭКСПРЕССИРУЮЩИЙ ЕГО МИКРООРГАНИЗМ | 2011 |
|
RU2557411C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ МЕТОДОМ ФЕРМЕНТАЦИИ, ШТАММ БАКТЕРИИ ESCHERICHIA COLI - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 1999 |
|
RU2207376C2 |
| DE 204326 T1, 02.07.1987 | |||
| DE 602005021342 D1, 01.07.2010. | |||

