Область техники
Настоящее изобретение относится к новому промотору и способу получения целевого вещества с его использованием.
Предшествующий уровень техники
Относящиеся к коринебактериям микроорганизмы являются промышленными микроорганизмами, наиболее широко и традиционно использующимися для производства аминокислот и родственных нуклеиновым кислотам веществ. Относящиеся к коринебактериям микроорганизмы представляют собой грамположительные бактерии, которые главным образом используются для производства химических веществ, находящих различные применения в области кормов для животных, лекарственных средств, медикаментов, пищевых продуктов и тому подобного, в том числе аминокислот и нуклеиновых кислот различных видов, и которым для роста требуется биотин. Для этих бактерий характерно изгибание под прямым углом в процессе деления клеток («щелкающее деление»), и одним из их преимуществ является то, что они характеризуются низкой активностью в отношении разложения продуцируемых метаболитов.
Среди продуктов, продуцируемых относящимися к коринебактериям микроорганизмами, L-аминокислоты являются основными структурными единицами белка и используются в качестве важного материала для производства фармацевтического сырья, пищевых добавок, кормов для животных, биологически активных добавок, пестицидов, дезинфицирующих средств и тому подобного. Ввиду этого, получение аминокислот в промышленных масштабах стало экономически важным промышленным процессом.
С целью разработки эффективного способа получения аминокислот были проведены различные исследования; например, были предприняты усилия в отношении разработки микроорганизмов или технологий способа ферментации для получения аминокислот с высокой эффективностью. В частности, были разработаны способы получения, характерные для целевых веществ, такие как усиление экспрессии генов, кодирующих ферменты, вовлеченные в биосинтез аминокислот, или делетирование генов, не являющихся необходимыми для биосинтеза аминокислот в штаммах рода Cory nebacterium (US 8030036 B2 и так далее). В дополнение к этим способам также были разработаны способ удаления генов, которые не вовлечены в продуцирование аминокислот, и способ удаления генов, функции которых, связанные с продуцированием аминокислот, конкретно не известны. Однако, по-прежнему сохраняется растущая потребность в изучении способов эффективного получения аминокислот с высоким выходом.
Чтобы разработать характеризующиеся высоким титром штаммы таких относящихся к коринебактериям микроорганизмов с использованием генно-инженерного конструирования или конструирования метаболических путей, необходимо в этих микроорганизмах выполнить избирательную регуляцию экспрессии генов, вовлеченных в различные метаболические пути. Для осуществления такой регуляции важно регулировать активность промотора, представляющего собой регуляторный ген, на котором происходит инициация транскрипции посредством связывания РНК-полимеразы с молекулой ДНК.
Описание
Техническая проблема
Авторы настоящего изобретения приложили усилия по разработке промотора, проявляющего сильную индуцирующую экспрессию активность, и в результате авторы настоящего изобретения модифицировали промотор гена ilvC в хромосоме Corynebacterium посредством замены нуклеотидов и установили, что модифицированный промотор может усиливать экспрессию функционально связанного с ним гена, тем самым завершая настоящее изобретение.
Техническое решение
Согласно настоящему изобретению предложен полинуклеотид, обладающий промоторной активностью, в котором по меньшей мере один нуклеотид заменен на другой нуклеотид в нуклеотидной последовательности с SEQ ID NO: 1.
Согласно настоящему изобретению предложен промотор, включающий этот полинуклеотид.
Согласно настоящему изобретению предложен вектор, включающий этот промотор и ген, кодирующий целевой белок.
Согласно настоящему изобретению предложен микроорганизм рода Corynebacterium, включающий этот полинуклеотид.
Согласно настоящему изобретению предложен способ получения целевого вещества, включающий культивирование микроорганизма рода Corynebacterium в среде.
Согласно настоящему изобретению предложен способ усиления экспрессии целевого гена, включающий функциональное связывание промотора с данным целевым геном.
Согласно настоящему изобретению предложено применение в качестве промотора полинуклеотида, в котором по меньшей мере один нуклеотид заменен на другой нуклеотид в нуклеотидной последовательности с SEQ ID NO: 1.
Полезные эффекты
Полинуклеотиды, обладающие активностью новых промоторов по настоящему изобретению, можно использовать для усиления экспрессии соединенного с ними целевого гена, и поэтому они могут быть с успехом применены для получения целевых веществ.
Краткое описание графических материалов
На ФИГ. 1-4 показаны промоторные области гена ilvC валин-продуцирующих штаммов.
На ФИГ. 5 и 6 показаны промоторные области гена ilvC изолейцин-продуцирующих штаммов.
На ФИГ. 7 и 8 показаны промоторные области гена ilvC лейцин-продуцирующих штаммов.
Подробное описание изобретения
Далее настоящее изобретение будет описано подробно. Каждое описание и воплощение какого-либо аспекта, описанного в данной заявке, могут быть применены для описания и воплощения другого аспекта применительно к перекрывающемуся содержимому. Помимо этого, все комбинации различных элементов, раскрытые в подробном описании настоящего изобретения, находятся в пределах объема настоящего изобретения. Кроме того, объем настоящего изобретения не должен ограничиваться конкретным приведенным ниже описанием.
Кроме того, специалисты в данной области техники распознают или смогут установить, используя не более чем стандартные эксперименты, многие эквиваленты конкретных воплощений, описанных в данной заявке. Подразумевается, что такие эквиваленты охватываются настоящим изобретением.
Согласно одному из аспектов настоящего изобретения предложен полинуклеотид, в котором по меньшей мере один нуклеотид заменен на другой нуклеотид в нуклеотидной последовательности с SEQ ID NO: 1 и который обладает промоторной активностью.
Использованный в данном описании термин «полинуклеотид» относится к нуклеотидному полимеру, состоящему из нуклеотидных мономеров, соединенных ковалентной связью в длинную цепь, такую как нить ДНК, имеющую предварительно заданную или большую длину.
Использованный в данном описании термин «полинуклеотид, обладающий промоторной активностью», относится к области ДНК, находящейся вблизи сайта, который вовлечен в транскрипцию целевого гена, в том числе сайта, с которым связывается РНК-полимераза, энхансер или тому подобное, для осуществления экспрессии целевого гена, расположенного от него вниз по течению. Для задач настоящего изобретения этот полинуклеотид можно использовать в качестве усиленного промотора для общего применения. Данный полинуклеотид может быть сконструирован с возможностью регуляции экспрессии целевого гена, функционально с ним связанного, и регуляции продуцирования и/или активности белка, кодируемого этим целевым геном, и может быть сконструирован с возможностью усиления продуцирования и/или активности целевого продукта (биологически активного вещества, например, по меньшей мере одного вещества, выбранного из группы, состоящей из аминокислот, нуклеиновых кислот, витаминов, белков, жирных кислот и органических кислот), в образовании которого участвует данный белок, в клетках, по сравнению с традиционными промоторами или клеточными эндогенными промоторами, но этим не ограничиваясь.
В одном из воплощений полинуклеотид, обладающий промоторной активностью по настоящему изобретению, может быть использован в качестве промотора, способного усиливать экспрессию изомероредуктазы ацетогидроксикислот. Такой полинуклеотид может представлять собой полинуклеотид, вовлеченный в усиление процесса продуцирования или в увеличение количества образованных аминокислот, включая, конкретно, аминокислоты с разветвленной цепью, более конкретно, лейцин, валин и изолейцин, но этим не ограничиваясь, и полинуклеотидные последовательности, обладающие промоторной активностью, включены без ограничения.
В настоящем изобретении SEQ ID NO: 1 представляет собой последовательность, обладающую промоторной активностью, и нуклеотидная последовательность SEQ ID NO: 1 может быть идентифицирована с использованием известной базы данных Genbank Национального центра биотехнологической информации (NCBI) и может происходить из Corynebacterium sp., но этим не ограничивается. Любая последовательность, которая обладает такой же активностью, что и данная нуклеотидная последовательность, может быть включена без ограничения. При этом SEQ ID NO: 1 может представлять собой промотор изомероредуктазы ацетогидроксикислот. Однако данная последовательность этим не ограничивается.
Использованный в данном описании термин «изомероредуктаза ацетогидроксикислот» относится к ферменту, вовлеченному в биосинтез L-аминокислоты с разветвленной цепью. Что касается пути биосинтеза L-аминокислот с разветвленной цепью, то сначала синтаза ацетогидроксикислот катализирует декарбоксилирование пировиноградной кислоты и реакцию конденсации пировиноградной кислоты с другой молекулой пировиноградной кислоты с образованием ацетомолочной кислоты, предшественника валина, или декарбоксилирование пировиноградной кислоты и реакцию конденсации пировиноградной кислоты с 2-кетобутиратом с образованием ацетогидроксибутирата, предшественника изолейцина. Изомероредуктаза ацетогидроксикислот переводит реакцию на следующую стадию, используя образующиеся в результате этого ацетомолочную кислоту или ацетогидроксибутират в качестве субстрата, с образованием тем самым L-валина, L-лейцина и L-изолейцина. Конкретно, изомеризация, катализируемая изомероредуктазой ацетогидроксикислот, происходит в реакции с ацетомолочной кислотой или ацетогидроксибутиратом, полученной(ым) в реакции с участием синтазы ацетогидроксикислот, а затем, в результате реакции восстановления, из каждого субстрата образуется (2R)-2,3-дигидрокси-3-изовалерат или (2R,3R)-2,3-дигидрокси-3-метилвалерат. (2R)-2,3-Дигидрокси-3-изовалерат подвергается реакциям, катализируемым дегидратазой дигидроксикислот и трансаминазой В, с образованием L-валина и после этого подвергается реакциям, катализируемым дегидратазой дигидроксикислот, 2-изопропилмалатсинтазой, изопропилмалатизомеразой, 3-изопропилмалатдегидрогеназой и трансаминазой В, с образованием L-лейцина. (2R,3R)-2,3-Дигидрокси-3-метилвалерат подвергается реакциям, катализируемым дегидратазой дигидроксикислот и трансаминазой В, с образованием L-изолейцина. Таким образом, изомероредуктаза ацетогидроксикислот является важным ферментом в пути биосинтеза L-аминокислот с разветвленной цепью.
Полинуклеотид, обладающий промоторной активностью по настоящему изобретению, означает полинуклеотид, в котором по меньшей мере один нуклеотид заменен на другой нуклеотид в нуклеотидной последовательности SEQ ID NO: 1 и/или нуклеотидной последовательности, имеющей по меньшей мере 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологии или идентичности с SEQ ID NO: 1. Гомологичные или идентичные нуклеотидные последовательности могут представлять собой последовательности, имеющие гомологию или идентичность в указанном выше диапазоне, за исключением последовательности, имеющей 100% идентичности, или могут представлять собой последовательности, имеющие менее 100% идентичности.
В частности, полинуклеотид, обладающий промоторной активностью, может включать полинуклеотид, обладающий промоторной активностью, в котором по меньшей мере один нуклеотид заменен на другой нуклеотид в нуклеотидной последовательности SEQ ID NO: 1, или может состоять из полинуклеотид а, обладающего промоторной активностью, в котором по меньшей мере один нуклеотид заменен на другой нуклеотид в нуклеотидной последовательности SEQ ID NO: 1.
Полинуклеотидом, обладающим промоторной активностью, может быть полинуклеотид, представленный общей формулой X-Y-Z, где 1) X представляет собой CNGN; 2) Y представляет собой CTAATTN; и 3) Z представляет собой С ATGTGTGTGGTAN A AN; и 4) N выбран из аденина (А), тимина (Т), гуанина (G) или цитозина (С). В полинуклеотид ной последовательности этой общей формулы Z может состоять из SEQ ID NO: 2, Y может состоять из SEQ ID NO: 3, или X может состоять из SEQ ID NO: 4.
В частности, в этой общей формуле X может быть представлен в виде CN1GN2, Y может быть представлен в виде CTAATTN3, и Z может быть представлен в виде CATGTGTGTGGTATAAT, где каждый из N1, N2 и N3 представляет собой любое, выбранное из аденина (А), тимина (Т), гуанина (G) или цитозина (С).
Более конкретно, в этой общей формуле 1) N1 может представлять собой цитозин (С) или гуанин (G), 2) N2 может представлять собой аденин (А) или тимин (Т), 3) N3 может представлять собой аденин (А) или гуанин (G), либо 4) может иметь место комбинация замен (1)-(3), без ограничения этим.
В одном из воплощений в полинуклеотид ной последовательности этой общей формулы, в Z, 14-ый нуклеотид N и 17-ый нуклеотид могут представлять собой тимин (Т) в нуклеотидной последовательности, представленной в SEQ ID NO: 2, при этом в X, 2-ой нуклеотид N может представлять собой цитозин (С) или гуанин (G), а 4-ый нуклеотид N может представлять собой аденин (А) или тимин (Т) в нуклеотидной последовательности, представленной в SEQ ID NO: 4, или X может представлять собой любую из последовательностей с SEQ ID NO: 8-11; и в Y, 7-ой нуклеотид N может представлять собой аденин (А) или гуанин (G) в нуклеотидной последовательности, представленной в SEQ ID NO: 3. В частности, в Z, 14-ый нуклеотид N и 17-ый нуклеотид могут представлять собой тимин (Т) в нуклеотидной последовательности, представленной в SEQ ID NO: 2; X может представлять собой любую из последовательностей с SEQ ID NO: 8-11; и Y может представлять собой последовательность SEQ ID NO: 6 или 7.
В еще одном воплощении полинуклеотид может иметь любую полинуклеотидную последовательность, выбранную из SEQ ID NO: 13-20.
Хотя в настоящем изобретении полинуклеотид описан с использованием выражения «полинуклеотид, имеющий нуклеотидную последовательность, представленную под конкретным порядковым номером» или «полинуклеотид, включающий нуклеотидную последовательность, представленную под конкретным порядковым номером», очевидно, что полинуклеотид, имеющий полинуклеотидную последовательность, содержащую делецию, модификацию, замену или добавление в какой-либо своей части, также может быть использован в настоящем изобретении, при условии, что данный полинуклеотид имеет активность, идентичную или соответствующую таковой у полинуклеотида, состоящего из нуклеотидной последовательности соответствующего порядкового номера. Например, очевидно, что такие выражения не исключают добавления несмысловых последовательностей вверх по течению или вниз по течению от нуклеотидной последовательности с соответствующим порядковым номером, природной мутации или молчащей мутации, при условии, что полинуклеотид имеет активность, идентичную или эквивалентную таковой у данного полинуклеотид а, и нуклеотидные последовательности, имеющие такие добавление или мутацию в последовательности, также находятся в пределах объема настоящего изобретения.
Понятия гомологии или идентичности относятся к степени сходства между двумя указанными нуклеотидными последовательностями и могут быть выражены в процентном отношении.
Термины «гомология» и «идентичность» часто могут быть использованы взаимозаменяемо.
Гомологию или идентичность последовательностей для консервативных полинуклеотидов можно определить при сочетанном применении стандартного алгоритма выравнивания и штрафов за разрыв по умолчанию, установленных используемой программой. По существу, гомологичные или идентичные последовательности, как правило, могут гибридизоваться друг с другом на протяжении всех их последовательностей или на протяжении по меньшей мере примерно 50%, 60%, 70%, 80% или 90% всей длины этих последовательностей в условиях умеренной или высокой жесткости. Среди полинуклеотидов, подлежащих гибридизации, также учитывается полинуклеотид, содержащий вырожденный кодон вместо какого-либо кодона.
Тот факт, имеются ли между какими-либо двумя полинуклеотидными последовательностями гомология, сходство или идентичность, можно определить с применением известного компьютерного алгоритма, такого как программа в формате «FASTA», использующая параметры по умолчанию как в работе Pearson et al. (1988) Proc. Natl. Acad. Sci. USA, 85: 2444. Альтернативно, это можно определить, используя алгоритм Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol., 48: 443-453), который реализован в программе Нидлмана из Европейского пакета открытого программного обеспечения по молекулярной биологии (EMBOSS) (Rice et al., 2000, Trends Genet., 16: 276-277) (версия 5.0.0 или более поздние версии) (в том числе пакет программ GCG (Devereux J., et al., Nucleic Acids Research, 12: 387 (1984)), BLASTP (основное средство поиска путем локального выравнивания (BLAST) для белков, BLASTN (BLAST для нуклеотидов), FASТА (Atschul S.F. et al., J. MOLEC. BIOL., 215: 403 (1990); Guide to Huge Computers, Martin J. Bischop (ed)., Academic Press, San Diego, 1994, и CARILLO et al., (1988) SIAM J. Applied Math., 48: 1073). Например, гомологию, сходство или идентичность можно определить, используя BLAST из базы данных Национального центра биотехнологической информации или ClustalW (кластерный анализ W множественных выравниваний).
Гомологию, сходство или идентичность полинуклеотидов можно определить путем сравнения информации о последовательностях, используя компьютерную программу GAP, например, Needleman et al. (1970), J. Mol. Biol, 48: 443), которая описана в Smith and Waterman, Adv. Appl. Math., (1981) 2: 482. Кратко, с использованием программы GAP гомологию, сходство или идентичность определяют в виде числа, полученного в результате деления числа аналогично выравненных символов (т.е. нуклеотидов или аминокислот) на общее число данных символов в более короткой из этих двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) бинарную матрицу сравнения (содержащую значение 1 в случае идентичности и значение 0 в случае отсутствия идентичности) и матрицу взвешенного сравнения согласно Gribskov et al. (1986) Nucl. Acids Res., 14: 6745 или как описано в работе Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979) (или матрицу замен EDNAFULL (EMBOSS версия NCBI NUC4.4)); (2) штраф 3,0 за каждый разрыв и дополнительный штраф 0,10 за каждый символ в каждом разрыве (или штраф за внесение разрыва 10 и штраф за удлинение разрыва 0,5); и (3) отсутствие штрафа за внесение концевых разрывов. Таким образом, термин «гомология» или «идентичность», использованный в данном описании, указывает на соответствие между последовательностями.
Помимо этого, любая полинуклеотидная последовательность, которая может в жестких условиях гибридизоваться с зондом, который может быть получен на основании последовательности известного гена, например, последовательности, комплементарной части или всей описанной выше полинуклеотидной последовательности, и которая обладает такой же активностью, может быть включена без ограничения. Термин «жесткие условия» относится к условиям, в которых возможно осуществление специфической гибридизации между полинуклеотидами. Такие условия, в частности, описаны в литературе (например, J. Sambrook и др., выше). Например, данные условия могут включать условия, при которых гены, имеющие высокую степень гомологии или идентичности, как например, гены, имеющие по меньшей мере 40%, конкретно по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85% или по меньшей мере 90%, более конкретно по меньшей мере 95%, еще конкретнее по меньшей мере 97% и даже еще более конкретно по меньшей мере 99% гомологии или идентичности, гибридизуются друг с другом, но гены, имеющие более низкую степень гомологии или идентичности, чем в указанных выше диапазонах, не гибридизуются друг с другом; либо условия могут включать типичные условия промывки для гибридизации по Саузерну, то есть когда промывку проводят один раз, в частности, два раза или три раза, при концентрации соли и температуре, соответствующих 60°С, 1xSSC (раствор хлорида и цитрата натрия) и 0,1% додецилсульфата натрия (SDS), конкретно, 60°С, 0,1xSSC и 0,1% SDS и более конкретно 68°С, 0,1xSSC и 0,1% SDS.
Для гибридизации необходимо, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя в зависимости от жесткости условий гибридизации могут быть возможны ошибочные спаривания между основаниями. Термин «комплементарные» используется для описания взаимосвязи между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, что касается ДНК, то аденин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, настоящее изобретение может включать не только по существу одинаковые нуклеиновокислотные последовательности, но также выделенные фрагменты нуклеиновых кислот, комплементарные полной последовательности.
В частности, полинуклеотиды, имеющие гомологию или идентичность, можно обнаружить при значении температуры плавления (Тпл) 55°С с использованием условий гибридизации, включающих стадию гибридизации и применение описанных выше условий. Кроме того, значение Тпл может составлять 60°С, 63°С или 65°С, но этим не ограничивается, и может соответствующим образом регулироваться специалистом в данной области техники сообразно задаче.
Соответствующая жесткость гибридизации полинуклеотидов зависит от длины полинуклеотидов и степени их комплементарности, и такие показатели хорошо известны в данной области техники (см. Sambrook и др., выше, 9.50-9.51, 11.7-11.8).
Согласно другому аспекту настоящего изобретения предложен промотор, включающий полинуклеотид по настоящему изобретению.
Использованный в данном описании термин «промотор» относится к нетранслируемой нуклеотидной последовательности, которая локализована «вверх по течению» относительно кодирующей области, содержит сайт связывания РНК-полимеразы и обладает активностью в отношении инициации транскрипции целевого гена с образованием матричной РНК (мРНК), то есть, относится к участку ДНК, с которым связывается РНК-полимераза и тем самым инициирует транскрипцию данного гена. Промотор может быть расположен в 5'-области сайта инициации транскрипции, приводящей к образованию мРНК.
Промотор по настоящему изобретению может обладать промоторной активностью, которая усилена по сравнению с традиционными промоторами. То есть, промотор может усиливать экспрессию целевого гена, а также экспрессию и активность белка, кодируемого данным целевым геном.
Для задач настоящего изобретения в отношении усиления экспрессии целевой ген может быть соответствующим образом изменен в зависимости от продукта, который должен быть получен, то есть, «целевого продукта», и промотор может быть использован как универсальный промотор для усиления экспрессии целевого гена.
Термин «целевой ген», для задач настоящего изобретения, относится к гену, экспрессия которого регулируется последовательностью промотора по настоящему изобретению. Белок, кодируемый целевым геном, может быть назван «целевым белком», а ген, кодирующий «целевой белок», может быть назван «целевым геном». Например, целевым геном для промотора может быть ген, кодирующий изомероредуктазу ацетогидроксикислот, то есть он может представлять собой ген ilvC, но этим не ограничивается.
Полинуклеотид, кодирующий целевой белок, может иметь различные модификации в своей кодирующей области в тех пределах, при которых полинуклеотидная последовательность не изменяется ввиду вырожденности кодонов или с учетом кодонов, предпочтительных для микроорганизма, в котором данный полинуклеотид должен быть экспрессирован. Описание данной полинуклеотид ной последовательности приведено выше.
Согласно еще одному аспекту настоящего изобретения предложен вектор, включающий промотор по настоящему изобретению.
Согласно еще одному аспекту настоящего изобретения предложен вектор, включающий промотор по настоящему изобретению и ген, кодирующий целевой белок.
В частности, такой вектор может представлять собой вектор, при этом целевым белком является изомероредуктаза ацетогидроксикислот, но этим не ограничивается.
Использованный в данном описании термин «вектор» относится к конструкции ДНК, содержащей нуклеотидную последовательность, представляющую собой целевой полинуклеотид, которая функционально связана с соответствующей контролирующей экспрессию областью или контролирующей экспрессию последовательностью, чтобы осуществить экспрессию целевого полинуклеотида в соответствующем хозяине. Контролирующая экспрессию последовательность может включать в себя промотор, способный инициировать транскрипцию, любую последовательность оператора для контроля такой транскрипции, последовательность для кодирования соответствующего сайта связывания рибосомы с мРНК и последовательности для контроля терминации транскрипции и трансляции и, в частности, контролирующая экспрессию последовательность может включать в себя промотор по настоящему изобретению. Вектор после трансформации соответствующего хозяина может реплицироваться или функционировать независимо от генома хозяина либо может быть интегрирован в сам геном.
Например, замену целевого полинуклеотида в хромосоме можно осуществлять с использованием вектора для вставки в хромосому клетки. Вставка полинуклеотида в хромосому может быть осуществлена с использованием любого метода, известного в данной области техники, например, гомологической рекомбинации, но этим не ограничивается. Вектор может дополнительно включать селективный маркер для идентификации вставки в хромосому. Селективный маркер используют для отбора трансформированных вектором клеток, то есть для подтверждения того, что молекула целевого нуклеотида была успешно встроена, и могут быть использованы маркеры для придания селектируемых фенотипов, таких как лекарственная устойчивость, ауксотрофия, устойчивость к цитотоксическим лекарственным средствам, и маркеры экспрессии поверхностных белков. В условиях, когда обрабатывают агентами для осуществления селекции, только клетки, способные экспрессировать селективные маркеры, могут выживать или проявлять другие фенотипические признаки, на основании чего может быть осуществлен отбор трансформированных клеток.
Особых ограничений в выборе вектора, используемого в настоящей заявке, нет, и может быть использован любой известный в данной области техники вектор. Примеры обычно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фаговых векторов или космидных векторов могут быть использованы pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и им подобные, а в качестве плазмидных векторов могут быть использованы векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL и pET. В частности, могут быть использованы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC или им подобные векторы.
Согласно еще одному аспекту настоящего изобретения предложен микроорганизм Corynebacterium sp., содержащий полинуклеотид, обладающий промоторной активностью по настоящему изобретению.
Согласно еще одному аспекту настоящего изобретения предложен микроорганизм Corynebacterium sp., содержащий полинуклеотид по настоящему изобретению и ген, кодирующий целевой белок.
Использованный в данном описании термин «микроорганизм» охватывает все микроорганизмы дикого типа либо природно или искусственно генетически модифицированные микроорганизмы и относится к микроорганизму, в котором конкретный механизм ослаблен или усилен благодаря вставке экзогенного гена либо усилению или ослаблению активности эндогенного гена. Микроорганизм по настоящему изобретению может включать микроорганизм, в который введен полинуклеотид, обладающий промоторной активностью по настоящему изобретению, или который включает данный полинуклеотид, без ограничения.
В частности, такой микроорганизм представляет собой микроорганизм, полученный путем трансформации вектором, содержащим полинуклеотид, обладающий промоторной активностью по настоящему изобретению, и ген, кодирующий целевой белок, либо микроорганизм, включающий полинуклеотид, обладающий промоторной активностью, и ген, кодирующий целевой белок, либо включающий содержащий их вектор. В частности, такой микроорганизм может представлять собой микроорганизм, включающий полинуклеотид, обладающий промоторной активностью, и ген, кодирующий целевой белок, и таким образом, микроорганизм обладает способностью продуцировать целевой белок или целевой продукт, в продуцирование которого вовлечен целевой белок, но этим не ограничивается. Данный микроорганизм может представлять собой микроорганизм, обладающий природной способностью продуцировать целевой белок или целевой продукт, либо микроорганизм, полученный путем придания родительскому штамму, не обладавшему способностью продуцировать целевой белок или целевой продукт, способности продуцировать целевой белок или целевой продукт, но этим не ограничивается.
Использованный в данном описании термин «микроорганизм, продуцирующий целевой белок или целевой продукт»» охватывает все микроорганизмы дикого типа либо природно или искусственно генетически модифицированные микроорганизмы и относится к микроорганизму, в котором конкретный механизм ослаблен или усилен благодаря вставке экзогенного гена либо усилению или ослаблению активности эндогенного гена, при этом микроорганизм может иметь генетическую мутацию для продуцирования целевого белка или продукта. Соответствующий микроорганизм может представлять собой: микроорганизм, генетически модифицированный в отношении любого из перечисленного, а именно, целевого белка, кодирующего этот белок полинуклеотида и вектора, включающего данный полинуклеотид; микроорганизм, модифицированный с целью экспрессии белка или кодирующего этот белок полинуклеотида; рекомбинантный микроорганизм, экспрессирующий целевой белок или кодирующий этот белок полинуклеотид; или рекомбинантный микроорганизм, характеризующийся активностью целевого белка, но этим не ограничивается.
Использованный в данном описании термин «трансформация» указывает на то, что полинуклеотид или вектор, содержащий полинуклеотид по настоящему изобретению, и полинуклеотид, кодирующий целевой белок, вводят в клетку-хозяина или в микроорганизм, что дает возможность целевому белку экспрессироваться в клетке-хозяине. Может быть включена любая клетка-хозяин при условии, что целевой белок может экспрессироваться в клетке-хозяине независимо от того, встроен ли полинуклеотид или вектор, введенный в клетку-хозяина или в микроорганизм, и локализован ли он в хромосоме клетки-хозяина или расположен вне хромосомы. Полинуклеотид может быть введен в любой форме при условии, что данный полинуклеотид может быть введен и экспрессирован в этой клетке-хозяине. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, содержащую все факторы, необходимые для самостоятельной экспрессии. Обычно экспрессионная кассета может включать в себя промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции, а промотор может представлять собой полинуклеотид, обладающий промоторной активностью по настоящему изобретению. Экспрессионная кассета может представлять собой экспрессирующий вектор, обладающий способностью к самостоятельной репликации. Помимо этого, полинуклеотид, кодирующий целевой белок, может быть функционально связан с полинуклеотидом по настоящему изобретению и в таком виде введен в клетку-хозяина, но этим не ограничивается.
Использованный в данном описании термин «функционально связанный» означает функциональную связь между последовательностью гена и последовательностью промотора, с использованием которой инициируется и опосредуется транскрипция полинуклеотида, кодирующего целевой белок. Промоторная последовательность может представлять собой промотор, предложенный в настоящем изобретении.
Способ трансформации вектором по настоящему изобретению включает любой метод введения нуклеиновой кислоты в клетку, и в зависимости от клетки-хозяина может быть выбран и осуществлен любой подходящий стандартный метод, известный в данной области техники. Примерами данного метода могут служить электропорация, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), микроинъекция, метод с применением полиэтиленгликоля (ПЭГ), метод с применением DEAE-декстрана (диэтиламиноэтил-декстрана), метод с применением катионных липосом, метод с применением ацетата лития-DMSO (диметилсульфоксид) и тому подобные, но этим не ограничивается.
Для задач настоящего изобретения микроорганизм представляет собой микроорганизм, продуцирующий целевой белок или целевой продукт, при этом в результате введения полинуклеотида по настоящему изобретению данный микроорганизм обладает повышенной способностью продуцировать целевой белок или целевой продукт.
В одном из воплощений микроорганизм, включающий полинуклеотид по настоящему изобретению, может представлять собой микроорганизм, который обуславливает наличие повышенной активности у целевого белка в результате замены по меньшей мере одного нуклеотида на другой нуклеотид в полинуклеотид ной последовательности SEQ ID NO: 1, но этим не ограничивается.
В частности, данный микроорганизм представляет собой микроорганизм, включающий полинуклеотид, который обладает промоторной активностью в результате замены по меньшей мере одного нуклеотида на другой нуклеотид в полинуклеотидной последовательности SEQ ID NO: 1, при этом полинуклеотид, обладающий промоторной активностью, может быть представлен общей формулой Х-Y-Z, в которой X представляет собой CNGN; Y представляет собой CTAATTN; и Z представляет собой CATGTGTGTGGTANAAN; и N выбран из аденина (А), тимина (Т), гуанина (G) или цитозина (С). Данный полинуклеотид описан выше.
В одном из воплощений микроорганизм по настоящему изобретению может характеризоваться повышенной активностью целевого белка в результате трансформации вектором, включающим полинуклеотид по настоящему изобретению и ген, кодирующий целевой белок.
В настоящем изобретении микроорганизм, продуцирующий целевой белок или целевой продукт, либо микроорганизм, обладающий способностью продуцировать целевой белок или целевой продукт, может представлять собой микроорганизм, в котором некоторые гены, вовлеченные в путь биосинтеза целевого белка или продукта, усилены или ослаблены, либо некоторые гены, вовлеченные в путь разложения целевого белка или целевого продукта, усилены или ослаблены.
Например, когда целевой белок представляет собой белок, вовлеченный в продуцирование аминокислоты с разветвленной цепью, тогда таким микроорганизмом может быть микроорганизм, обладающий природной способностью продуцировать аминокислоту с разветвленной цепью, либо микроорганизм, полученный путем придания родительскому штамму, не обладавшему способностью продуцировать аминокислоту с разветвленной цепью, способности продуцировать аминокислоту с разветвленной цепью, но этим не ограничивается.
В одном из воплощений, когда целевой белок представляет собой изомероредуктазу ацетогидроксикислот, данный микроорганизм может представлять собой клетку или микроорганизм, в котором полинуклеотид по настоящему изобретению функционально связан с геном, кодирующим изомероредуктазу ацетогидроксикислот для повышения активности изомероредуктазы ацетогидроксикислот, и в таких случаях клетка-хозяин или микроорганизм может представлять собой микроорганизм, способный продуцировать аминокислоту с разветвленной цепью с использованием целевого белка.
В данном описании выражение «микроорганизм, способный продуцировать аминокислоту с разветвленной цепью», может быть использовано взаимозаменяемо с выражениями «микроорганизм, продуцирующий аминокислоту с разветвленной цепью» и «микроорганизм, обладающий способностью продуцировать аминокислоту с разветвленной цепью».
Использованный в данном описании термин «аминокислота с разветвленной цепью» относится к аминокислоте с разветвленной алкильной группой в ее боковой цепи, которая включает валин, лейцин и изолейцин. В частности, в настоящем изобретении аминокислота с разветвленной цепью может представлять собой L-аминокислоту с разветвленной цепью, и данная L-аминокислота с разветвленной цепью может представлять собой L-валин, L-изолейцин или L-лейцин, но этим не ограничивается.
Использованный в данном описании термин «микроорганизм, продуцирующий аминокислоту с разветвленной цепью», охватывает все микроорганизмы дикого типа либо природно или искусственно генетически модифицированные микроорганизмы и относится к микроорганизму, в котором конкретный механизм ослаблен или усилен благодаря вставке экзогенного гена либо усилению или ослаблению активности эндогенного гена, при этом микроорганизм может иметь генетическую мутацию или повышенную активность в отношении продуцирования целевой аминокислоты с разветвленной цепью. Для задач настоящего изобретения микроорганизм, продуцирующий аминокислоту с разветвленной цепью, может представлять собой микроорганизм, который обладает усиленной способностью продуцировать целевую аминокислоту с разветвленной цепью в результате введения полинуклеотида, обладающего промоторной активностью по настоящему изобретению, и в частности, данный микроорганизм может представлять собой микроорганизм рода Corynebacterium. В частности, микроорганизм, продуцирующий аминокислоту с разветвленной цепью, или микроорганизм, обладающий способностью продуцировать аминокислоту с разветвленной цепью, может представлять собой микроорганизм, в котором некоторые из генов, вовлеченных в путь биосинтеза аминокислоты с разветвленной цепью, усилены или ослаблены, либо некоторые из генов, вовлеченных в путь разложения аминокислоты с разветвленной цепью, усилены или ослаблены. Например, микроорганизм, продуцирующий аминокислоту с разветвленной цепью, может обладать повышенной экспрессией гена ilvC, кодирующего изомероредуктазу ацетогидроксикислот, обусловленной введением полинуклеотида, обладающего промоторной активностью, предложенного в настоящем изобретении, но этим не ограничивается.
Использованный в данном описании термин «микроорганизм рода Corynebacterium, продуцирующий аминокислоту с разветвленной цепью», может относиться к микроорганизму рода Corynebacterium, обладающему способностью продуцировать аминокислоту с разветвленной цепью, природному или полученному в результате модификации. Конкретно, микроорганизм рода Corynebacterium, продуцирующий аминокислоту с разветвленной цепью по настоящему изобретению, может представлять собой микроорганизм рода Corynebacterium, который содержит ген ilvC, кодирующий изомероредуктазу ацетогидроксикислот, и обладает повышенной способностью продуцировать аминокислоту с разветвленной цепью в результате усиления промоторной активности гена ilvC. Более конкретно, микроорганизм рода Corynebacterium, продуцирующий аминокислоту с разветвленной цепью по настоящему изобретению, может представлять собой микроорганизм рода Corynebacterium, который содержит полинуклеотид, обладающий промоторной активностью по настоящему изобретению, или обладает усиленной способностью продуцировать аминокислоту с разветвленной цепью в результате трансформации вектором, содержащим данный полинуклеотид и ген, кодирующий целевой белок.
«Микроорганизм рода Corynebacterium, обладающий способностью продуцировать аминокислоту с разветвленной цепью», относится к микроорганизму, обладающему усиленной способностью продуцировать аминокислоту с разветвленной цепью по сравнению с родительским штаммом до трансформации или немодифицированным микроорганизмом. «Немодифицированный микроорганизм» относится к штамму дикого типа как таковому, к микроорганизму, не содержащему ген, кодирующий изомероредуктазу ацетогидроксикислот, или к микроорганизму, не содержащему полинуклеотидную последовательность по настоящему изобретению или не трансформированному вектором, содержащим полинуклеотид по настоящему изобретению и ген, кодирующий целевой белок.
«Родительский штамм» может представлять собой микроорганизм рода Corynebacterium, продуцирующий аминокислоту с разветвленной цепью. В частности, родительский штамм может представлять собой микроорганизм, продуцирующий аминокислоту с разветвленной цепью с природной или искусственной генетической модификацией. Например, родительский штамм может представлять собой штамм, обладающий улучшенной способностью продуцировать L-валин, обусловленной введением модификации (ilvN(A42V); Biotechnology and Bioprocess Engineering, June 2014, Volume 19, Issue 3, pp. 456-467) в микроорганизм рода Corynebacterium, или штамм, обладающий улучшенной способностью продуцировать L-изолейцин, обусловленной введением варианта lysC(L377K) и варианта hom(G378E) (Appl. Microbiol. Biotechnol., 45, 612-620 (1996)) в микроорганизм рода Corynebacterium, и введением модификации ilvA(V383A) (World J. Microbiol. Biotechnol. (2015), 31: 1369-1377) в ген, кодирующий L-треониндегидратазу. Помимо этого родительский штамм может представлять собой штамм, обладающий улучшенной способностью продуцировать L-лейцин, обусловленной введением модификации (leuA(R558H, G561D); US 2020-0032305 A1) в микроорганизм рода Corynebacterium, но этим не ограничивается.
В настоящем изобретении «микроорганизм рода Corynebacterium» может включать все микроорганизмы рода Corynebacterium. Конкретно, их примерами могут быть Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens, Corynebacterium stationis, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium callunae, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris, Corynebacterium flavescens и тому подобное, но этим не ограничиваясь.
Согласно еще одному аспекту настоящего изобретения предложен способ получения целевого вещества, включающий культивирование микроорганизма рода Corynebacterium в среде.
Конкретно, целевое вещество может представлять собой аминокислоту и, более конкретно, аминокислоту с разветвленной цепью, но этим не ограничивается.
В данном способе культивирование микроорганизма может быть выполнено с использованием известных методов стационарного культивирования, непрерывного культивирования, периодического культивирования с подпиткой или тому подобного, но этим не ограничивается. Условия культивирования особенно не ограничиваются, но подведение рН до соответствующего значения (например, рН 5-9, конкретно, рН 6-8 и наиболее конкретно рН 6,8) может быть достигнуто с использованием основного соединения (например, гидроксида натрия, гидроксида калия или аммиака) или кислотного соединения (например, фосфорной кислоты или серной кислоты), а аэробные условия можно поддерживать, добавляя в культуру кислород или кислород-содержащую газовую смесь. Температуру культивирования можно поддерживать при 20°С - 45°С и, конкретно, при 25°С - 40°С, и культивирование можно проводить в течение примерно 10-160 часов, но данные условия этим не ограничиваются. Полученная посредством культивирования аминокислота может высвобождаться в среду или может оставаться в клетках без высвобождения.
В применяемой для культивирования среде в качестве источника углерода, по отдельности или в комбинации, могут быть использованы сахара и углеводы (например, глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, крахмал и целлюлоза), масла и жиры (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновая кислота, стеариновая кислота и линолевая кислота), спирты (например, глицерин и этанол), органические кислоты (например, уксусная кислота) и тому подобное, но источник углерода этим не ограничивается. В качестве источника азота можно использовать, по отдельности или в комбинации, азотсодержащее органическое соединение (например, пептон, дрожжевой экстракт, мясной экстракт, экстракт солода, кукурузный экстракт, соевую муку и мочевину) или неорганическое соединение (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) и тому подобное, но источник азота этим не ограничивается. В качестве источника фосфора можно использовать, по отдельности или в комбинации, дигидрофосфат калия, гидрофосфат дикалия, соответствующую им натрий-содержащую соль и тому подобное, но источник фосфора этим не ограничивается. Помимо этого, среда может содержать необходимые способствующие росту вещества, такие как соли других металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
Способ получения целевого вещества по настоящему изобретению может дополнительно включать извлечение целевого вещества из среды.
Для извлечения целевого вещества, образованного на стадии культивирования, данное целевое вещество может быть собрано из среды с использованием подходящего метода, известного в данной области техники, соответствующего способу культивирования. Например, можно использовать центрифугирование, фильтрацию, анионообменную хроматографию, кристаллизацию, высокоэффективную жидкостную хроматографию (HPLC) и тому подобное, и целевое вещество может быть извлечено из среды или микроорганизма с применением соответствующего метода, известного в данной области техники.
Кроме того, стадия извлечения может включать процесс очистки, который может быть осуществлен с использованием соответствующего метода, известного в данной области техники. Например, когда целевое вещество представляет собой аминокислоту, то извлеченная аминокислота может быть в очищенной форме или может представлять собой содержащий аминокислоту бульон после ферментации микроорганизма (Introduction to Biotechnology and Genetic Engineering, A. J. Nair., 2008).
Согласно другому аспекту настоящего изобретения предложен способ усиления экспрессии целевого гена, включающий функциональное связывание промотора, содержащего полинуклеотид по настоящему изобретению, с целевым геном.
Полинуклеотид, целевой ген, промотор и тому подобное описаны выше.
Согласно другому аспекту настоящего изобретения предложено применение, в качестве промотора, полинуклеотида, содержащего замену по меньшей мере одного нуклеотида на другой нуклеотид в нуклеотидной последовательности SEQ ID NO: 1.
Данный полинуклеотид описан выше.
Способ осуществления изобретения
Далее настоящее изобретение будет описано подробно со ссылкой на типичные воплощения. Однако, эти типичные воплощения приведены для конкретной иллюстрации настоящего изобретения, и объем настоящего изобретения ими не ограничивается.
Пример 1. Отбор мутантного штамма с повышенной способностью продуцировать валин посредством внесения случайных мутаций
Пример 1-1. Случайный мутагенез с использованием УФ-облучения
Для отбора мутантных штаммов с повышенной способностью продуцировать валин штамм Corynebacterium glutamicum из Корейского центра культур микроорганизмов под номером KCCM11201P (US 8465962 В2), который представляет собой валин-продуцирующий штамм, высевали на питательную среду, содержащую агар, и культивировали при 30°С в течение 36 часов. Полученные таким образом несколько сотен колоний подвергали ультрафиолетовому (УФ) облучению при комнатной температуре для осуществления случайного мутагенеза в геноме данного штамма.
Питательная среда (рН 7,2)
Глюкоза - 10 г, мясной сок - 5 г, полипептон - 10 г, хлорид натрия - 2,5 г, дрожжевой экстракт - 5 г, агар - 20 г, и мочевина - 2 г (из расчета на 1 л дистиллированной воды).
Пример 1-2. Тестирование титра после ферментации на подвергнутых мутагенезу штаммах и отбор штаммов
Для отбора мутантных штаммов с повышенной способностью продуцировать L-валин по сравнению со штаммом Corynebacterium glutamicum KCCM11201P, использованным в качестве родительского штамма, на подвергнутых мутагенезу штаммах проводили тестирование титра после ферментации. Каждую колонию пересевали на питательную среду и затем каждым штаммом инокулировали содержимое конической колбы с перегородками емкостью 250 мл, содержащей 25 мл среды для продуцирования, и культивировали со встряхиванием при 30°С и 200 об./мин в течение 72 часов. После этого, используя HPLC, определяли концентрацию L-валина, и установленные концентрации L-валина занесены в Таблицу 1.
Питательная среда (рН 7,2)
Глюкоза - 10 г, мясной сок - 5 г, полипептон - 10 г, хлорид натрия - 2,5 г, дрожжевой экстракт - 5 г, агар - 20 г, и мочевина - 2 г (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
Глюкоза - 100 г, сульфат аммония - 40 г, соевый белок - 2,5 г, кукурузный экстракт высушенный - 5 г, мочевина - 3 г, фосфат калия двухосновный - 1 г, сульфата магния гептагидрат - 0,5 г, биотин - 100 мкг, тиамин-HCl - 1 мг, пантотенат кальция - 2 мг, никотинамид - 3 мг, карбонат кальция - 30 г (из расчета на 1 л дистиллированной воды).
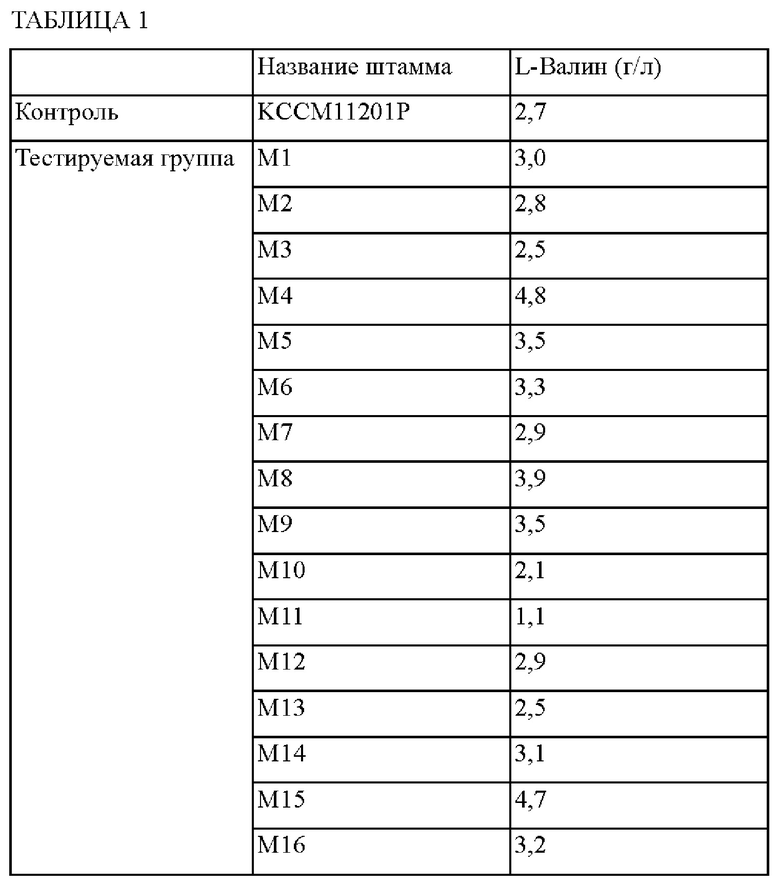
С учетом данных Таблицы 1 отбирали штаммы М4 и M15, для которых продуцируемые количества валина возрастали на 178% и 174%, соответственно, по сравнению со штаммом KCCM11201P в качестве контроля.
Пример 2. Исследование модификации посредством секвенирования генов
Проводили секвенирование основных генов пути биосинтеза валина в штаммах М4 и M15 с усиленной способностью к продуцированию валина и их сравнение с подобными генами штамма KCCM11201P и штаммов Corynebacterium glutamicum дикого типа из Американской коллекции типовых культур с номерами АТСС14067, АТСС13032 и АТСС13869. В результате было установлено, что штаммы М4 и М15 содержали одинаковые мутации в определенных положениях промоторной области гена ilvC, кодирующего изомероредуктазу ацетогидроксикислот (AHAIR) (ФИГ. 1). Конкретно, в М4 и M15 14-ый нуклеотид G и 17-ый нуклеотид С были заменены на Т в последовательности промоторной области, включающей последовательность, приведенную в SEQ ID NO: 5. Последовательность, приведенная в SEQ ID NO: 5, представляет собой последовательность, которая обычно содержится в промоторной области гена ilvC штаммов Corynebacterium glutamicum дикого типа (АТСС 14067, АТСС13032 и АТСС13869). В приведенных далее примерах исследовали, влияют ли данные мутации на количество аминокислот, продуцируемое микроорганизмами рода Corynebacterium.
Пример 3. Получение штаммов с внесенными мутациями и исследование способности продуцировать валин
Пример 3-1. Получение штаммов с внесением мутаций в Corynebacterium glutamicum КССМ11201Р и оценка способности продуцировать валин
Пример 3-3-1. Получение штаммов
Чтобы выполнить замены 14-ого и 17-ого нуклеотидов на Т в полинуклеотидной последовательности, приведенной в SEQ ID NO: 5, конструировали вектор, содержащий целевые мутации, для внесения этих мутаций в валин-продуцирующий штамм Corynebacterium glutamicum KCCM11201P.
Конкретно, геномную ДНК из АТСС14067, штамма Corynebacterium glutamicum дикого типа, выделяли, используя мини-набор для выделения общей ДНК G-spin (Intron, №по каталогу 17045) в соответствии с приложенным к набору протоколом. Полимеразную цепную реакцию (ПНР) проводили, используя геномную ДНК в качестве матрицы. Чтобы сконструировать вектор для внесения мутаций в промоторную область гена ilvC, получали фрагменты ДНК (А и В), используя пару праймеров - праймер 1 (SEQ ID NO: 21) и праймер 2 (SEQ ID NO: 22), и пару праймеров - праймер 3 (SEQ ID NO: 23) и праймер 4 (SEQ ID NO: 24), соответственно.
Проводили ПЦР с перекрывающимися праймерами, используя эти два фрагмента в качестве матрицы вместе с праймером 1 (SEQ ID NO: 21) и праймером 4 (SEQ ID NO: 24) и получая ПЦР-продукт размером приблизительно 1,4 тысячи пар оснований (т.п.о.) (далее именуемый как «фрагмент с внесенными мутациями»). Использованные праймеры показаны в Таблице 2.


Полученные фрагменты с внесенными мутациями обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MA) и затем лигировали с применением вектора pDZ (WO 2008-033001 A1), обработанного этим же ферментом рестрикции, и лигазы Т4 (New England Biolabs, Beverly, MA). Полученным геном трансформировали клетки Е coli DH5α, затем проводили их отбор на среде Лурия-Бертани (LB), содержащей канамицин, и с использованием набора для очистки плазмидной ДНК DNA-spin (Intron) получали ДНК, чтобы сконструировать таким образом рекомбинантную плазмиду pDZ-ilvC(Pm3)-14067. Выполняли эту же методику, но используя, вместо геномной ДНК штамма АТСС14067, ДНК штаммов АТСС13869 и АТСС13032 Corynebacterium glutamicum дикого типа, и в результате этого были сконструированы рекомбинантные плазмиды, обозначенные как pDZ-ilvC(Pm3)-13869 и pDZ-ilvC(Pm3)-13032, соответственно.
Одной из этих трех сконструированных, как описано выше, рекомбинантных плазмид, плазмидой pDZ-ilvC(Pm3)-14067, трансформировали L-валин-продуцирующий штамм Corynebacterium glutamicum KССМ11201Р посредством гомологичной рекомбинации с участием хромосомы (van der Rest et al., Appl. Microbiol. Biotechnol, 52: 541-545, 1999). Штамм, в хромосому которого встроился данный вектор посредством рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин в концентрации 25 мг/л. После этого подвергнутый трансформации штамм Corynebacterium glutamicum, в котором произошла рекомбинация с участием двух нитей ДНК (secondary recombination), подвергали ПЦР с использованием праймера 1 и праймера 4 с целью конструирования штамма KCCM11201P-ilvC(Pm3) с мутациями, внесенными в промотор гена ilvC в хромосоме (ФИГ. 2). Рекомбинантный штамм обозначали как Corynebacterium glutamicum СА08-1063, был депонирован на международном уровне в международном депозитарии Корейского центра культур микроорганизмов (KCCM) 21 августа 2019 г. в соответствии с положениями Будапештского договора и ему был присвоен номер доступа KCCM12574P.
Пример 3-1-2. Оценка способности продуцировать валин
Чтобы сравнить способность продуцировать валин среди валин-продуцирующих штаммов Corynebacterium glutamicum KCCM11201P и KCCM11201P-ilvC(Pm3), проводили оценку титра после ферментации. Каждый штамм пересевали на питательную среду и затем инокулировали содержимое конической колбы с перегородками емкостью 250 мл, содержащей 25 мл среды для продуцирования, и культивировали со встряхиванием при 30°С и 200 об./мин в течение 72 часов. После этого, используя HPLC, определяли концентрацию L-валина, и установленные концентрации L-валина занесены в Таблицу 3.
Питательная среда (рН 7,2)
Глюкоза - 10 г, мясной сок - 5 г, полипептон - 10 г, хлорид натрия - 2,5 г, дрожжевой экстракт - 5 г, агар - 20 г, и мочевина - 2 г (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
Глюкоза - 100 г, сульфат аммония - 40 г, соевый белок - 2,5 г, кукурузный экстракт высушенный - 5 г, мочевина - 3 г, фосфат калия двухосновный - 1 г, сульфата магния гептагидрат - 0,5 г, биотин - 100 мкг, тиамин-HCl - 1 мг, пантотенат кальция - 2 мг, никотинамид - 3 мг, карбонат кальция - 30 г (из расчета на 1 л дистиллированной воды).

Как видно из приведенных выше результатов, способность штамма KCCM11201P-ilvC(Pm3) продуцировать L-валин увеличилась на 11% по сравнению с таковой в случае контроля. Следовательно, способность продуцировать L-валин может быть улучшена посредством внесения мутации в промотор гена ilvC.
Пример 3-2. Получение штаммов с внесением мутации в Corynebacterium glutamicum CJ7V и оценка способности продуцировать валин
Пример 3-2-1. Получение валин-продуцирующего штамма CJ7V
Для выяснения того, будет ли наблюдаться такой же эффект, который описан выше, для других продуцирующих L-валин штаммов Corynebacterium glutamicum, в штамм Corynebacterium glutamicum дикого типа АТСС14067 вносили мутацию одного вида (ilvN(A42V); Biotechnology and Bioprocess Engineering, June 2014, Volume 19, Issue 3, pp. 456-467) с целью получения штаммов, обладающих улучшенной способностью продуцировать L-валин.
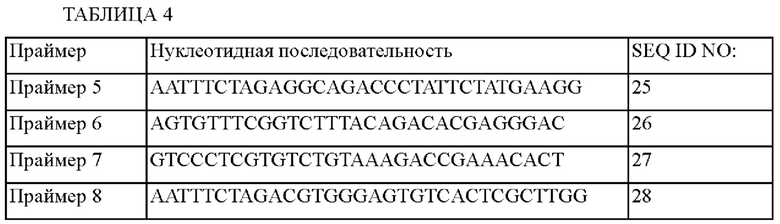
Конкретно, геномную ДНК из АТСС14067, штамма Corynebacterium glutamicum дикого типа, выделяли, используя мини-набор для выделения общей ДНК G-spin (Intron, № по каталогу 17045) в соответствии с приложенным к набору протоколом. ПЦР проводили, используя геномную ДНК в качестве матрицы. Чтобы сконструировать вектор для внесения мутации A42V в ген ilvC, получали фрагменты (А и В), используя пару праймеров - праймер 5 (SEQ ID NO: 25) и праймер 6 (SEQ ID NO: 26), и пару праймеров - праймер 7 (SEQ ID NO: 27) и праймер 8 (SEQ ID NO: 28), соответственно. ПЦР проводили в условиях, приведенных ниже: денатурация при 94°С в течение 5 минут, 25 циклов с денатурацией при 94°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Использованные праймеры показаны в Таблице 4.
В результате для обоих фрагментов А и В могут быть получены полинуклеотиды размером 537 пар оснований (п.о.). Проводили ПЦР с перекрывающимися праймерами, используя эти два фрагмента в качестве матрицы вместе с праймером 5 (SEQ ID NO: 25) и праймером 8 (SEQ ID NO: 28) и получая ПЦР-продукт размером приблизительно 1044 п.о. (далее именуемый как «фрагмент с внесенными мутациями»).
Полученные фрагменты с внесенными мутациями обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MA) и затем лигировали с применением вектора pDZ, обработанного этим же ферментом рестрикции, и лигазы Т4 (New England Biolabs, Beverly, MA). Полученным геном трансформировали клетки Е coli DH5α, затем проводили их отбор на среде LB, содержащей канамицин, и с использованием набора для очистки плазмидной ДНК DNA-spin (Intron) получали ДНК. Вектор, предназначенный для внесения A42V в ген ilvN, обозначали как pDZ-ilvN(A42V).
После этого рекомбинантной плазмидой pDZ-ilvN(A42V), сконструированной так, как описано выше, трансформировали штамм Corynebacterium glutamicum дикого типа АТСС14067 посредством гомологичной рекомбинации с участием хромосомы (van der Rest et al., Appl. Microbiol. Biotechnol, 52: 541-545, 1999). Штамм, в хромосому которого встроился данный вектор посредством рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин в концентрации 25 мг/л. После этого трансформированный штамм Corynebacterium glutamicum, в котором произошла рекомбинация с участием двух нитей ДНК, подвергали ПЦР с использованием праймера 5 и праймера 8 с целью осуществления амплификации генного фрагмента и затем проводили идентификацию штамма с внесенной мутацией путем секвенирования гена. Рекомбинантный штамм обозначали как Corynebacterium glutamicum CJ7V.
Пример 3-2-2. Оценка способности продуцировать валин
Плазмидой pDZ-ilvC(Pm3)-14067 трансформировали штамм Corynebacterium glutamicum CJ7V, обладающий способностью продуцировать L-валин, полученный в примере 3-2-1, тем же способом, что и в примере 3-1, с получением в результате этого штамма с мутацией в промоторе гена ilvC, который обозначали как CJ7V-ilvC(Pm3) (ФИГ. 3). Для сравнения способности к продуцированию L-валина среди полученных штаммов эти штаммы культивировали, концентрацию L-валина определяли тем же методом, что и в примере 3-1, и установленные концентрации L-валина занесены в Таблицу 5.
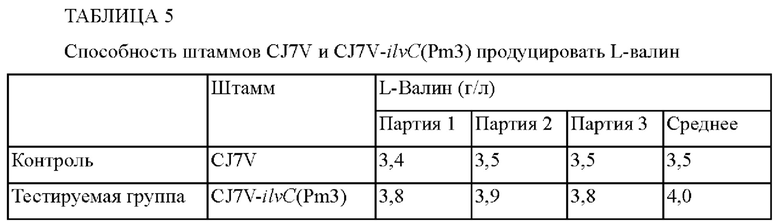
Как видно из приведенных выше результатов, способность штамма CJ7V-ilvC(Pm3) продуцировать L-валин увеличилась на 14% по сравнению с таковой в случае контроля. Следовательно, вновь было установлено, что способность продуцировать L-валин может быть улучшена посредством внесения мутации в промотор гена ilvC.
Пример 3-3. Получение штаммов с внесением мутации в Corynebacterium glutamicum CJ8V и оценка способности продуцировать L-валин
Пример 3-3-1. Получение валин-продуцирующего штамма CJ8V
Для выяснения того, будет ли наблюдаться такой же эффект, который описан выше, для других продуцирующих L-валин штаммов Corynebacterium glutamicum, в штамм Corynebacterium glutamicum дикого типа АТСС13869 вносили мутацию одного вида (ilvN(A42V)) с целью получения штаммов, обладающих улучшенной способностью продуцировать L-валин, тем же способом, что и в примере 3-2, и рекомбинантный штамм обозначали как Corynebacterium glutamicum CJ8V.
Пример 3-3-2. Оценка способности продуцировать валин
Получали штаммы CJ8V с внесенной мутацией в промотор гена ilvC штамма Corynebacterium glutamicum, обладающие способностью продуцировать L-валин, как в примере 3-3-1. Каждым из рекомбинантных векторов pDZ-ilvC(Pm3)-14067 и pDZ-ilvC(Pm3)-13869, полученных в примере 3-1-1, трансформировали штамм CJ8V (van der Rest et al., Appl. Microbiol. Biotechnol, 52: 541-545, 1999). Штамм, в хромосому которого встроился данный вектор посредством рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин в концентрации 25 мг/л. После этого трансформированный штамм Corynebacterium glutamicum, в котором произошла рекомбинация с участием двух нитей ДНК, подвергали ПЦР с использованием праймера 1 и праймера 4 с целью конструирования штаммов CJ8V-ilvC(Pm3) и CJ8V-ilvC(Pm3)-2 с мутацией, которая была внесена в промотор гена ilvC в хромосоме (ФИГ. 4). Среди этих рекомбинантных штаммов штамм CJ8V-ilvC(Pm3)-2 обозначали как Corynebacterium glutamicum СА08-2034, депонировали на международном уровне в международном депозитарии Корейского центра культур микроорганизмов (KCCM) 21 августа 2019 г. в соответствии с положениями Будапештского договора, и ему был присвоен номер доступа KCCM12575P.
Для сравнения способности полученных штаммов продуцировать L-валин эти штаммы культивировали и концентрацию L-валина определяли тем же методом, что и в примере 3-1, и установленные концентрации L-валина занесены в Таблицу 6.

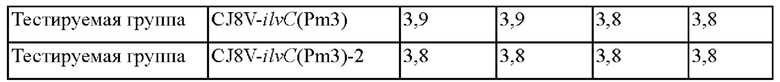
Как видно из приведенных выше результатов, способность каждого из штаммов CJ8V-ilvC(Pm3) и CJ8V-ilvC(Pm3)-2 продуцировать L-валин увеличилась на 8,6% по сравнению с таковой в случае контроля. Следовательно, вновь было установлено, что способность продуцировать L-валин может быть улучшена посредством внесения мутации в промотор гена ilvC.
Пример 4. Получение изолейщш-продуцирующих штаммов и оценка продуцирующей способности
Пример 4-1. Получение штаммов с мутацией в промоторе гена ilvC, внесенной в продуцирующий L-изолейцин штамм Corynebacterium glutamicum KCCM11248P
Получали штаммы на основе L-изолейцин-продуцирующего штамма Corynebacterium glutamicum KCCM11248P (патент Кореи №10-1335789), в которые посредством гомологичной рекомбинации с участием хромосомы тем же способом, что и в примере 3, были введены рекомбинантные плазмиды pDZ-ilvC(Pm3)-14067 и pDZ-ilvC(Pm3)-13869, сконструированные в примере 3-1, и эти штаммы обозначали как KCCM11248P::ilvC(Pm3) и KCCM11248P::ilvC(Pm3)-2, соответственно (ФИГ. 5). Полученные штаммы культивировали способом, описанным ниже, и затем сравнивали по способности продуцировать изолейцин.
Каждым штаммом инокулировали содержимое конической колбы с перегородками емкостью 250 мл, содержащей 25 мл среды для посева, и культивировали со встряхиванием при 200 об./мин в течение 20 часов при 30°С. Затем, используя 1 мл посевной культуры, инокулировали содержимое конической колбы с перегородками емкостью 250 мл, содержащей 24 мл среды для продуцирования, и культивировали со встряхиванием при 200 об./мин в течение 48 часов при 30°С. Составы среды для посева и среды для продуцирования приведены ниже.
Среда для посева (рН 7,0)
Глюкоза - 20 г, пептон - 10 г, дрожжевой экстракт - 5 г, мочевина - 1,5 г, KH2PO4 - 4 г, K2HPO4 - 8 г, MgSO4 7H2O - 0,5 г, биотин - 100 мкг, тиамин-HCl - 1000 мкг, пантотенат кальция - 2000 мкг, никотинамид - 2000 мкг (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
Глюкоза - 50 г, (NH4)2SO4 - 12,5 г, соевый белок - 2,5 г, кукурузный экстракт высушенный - 5 г мочевина - 3 г, KH2PO4 - 1 г, MgSO4 7H2O - 0,5 г, биотин - 100 мкг, тиамина гидрохлорид - 1000 мкг, пантотенат кальция - 2000 мкц никотинамид - 3000 мкц СаСО3 - 30 г (из расчета на 1 л дистиллированной воды).
По окончании культивирования определяли способность продуцировать L-изолейцин. Концентрации L-изолейцина в культуральных средах для каждого тестируемого штамма показаны ниже в Таблице 7.
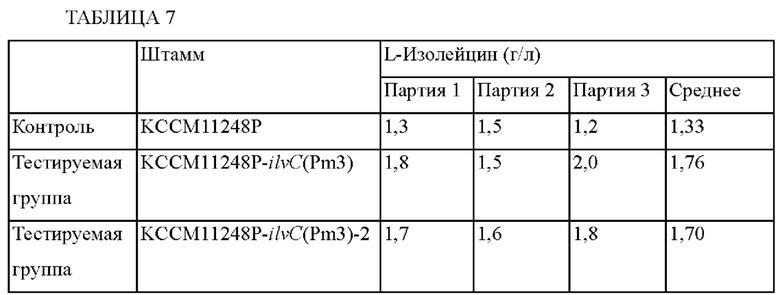
Как показано в приведенной выше Таблице 7, концентрации L-изолейцина, продуцируемого штаммами KCCM11248P::ilvC(Pm3) и KCCM11248::ilvC(Pm3)-2, в которые была внесена мутация, усиливающая промотор гена ilvC, увеличились примерно на 32,3% и 27,8%, соответственно, по сравнению с таковыми для L-изолейцин-продуцирующего штамма KCCM11248P. Таким образом, было установлено, что способность продуцировать L-изолейцин улучшилась в результате внесения мутации в промотор гена ilvC. Приведенные выше результаты показывают, что внесение мутации в промотор гена ilvC L-изолейцин-продуцирующих штаммов рода Corynebacterium является эффективным в отношении продуцирования L-изолейцина.
Пример 4-2. Получение L-изолейцин-продуцирующего штамма с мутацией в промоторе гена ilvC, внесенной в штамм Corynebacterium glutamicum дикого типа АТСС13032, и оценка способности продуцировать L-изолейцин
Чтобы исследовать влияние внесения мутации в промотор гена ilvC на способность продуцировать L-изолейцин, получали штаммы путем введения варианта lysC(L377K) (KR10-2019-0003019 А) и варианта hom(G378E) (Appl. Microbiol. Biotechnol., 45, 612-620 (1996)) в штамм Corynebacterium glutamicum АТСС 13032 (далее, штамм дикого типа (WT)), а мутацию ilvA(V383A) (World J. Microbiol. Biotechnol. (2015), 31: 1369-1377) вносили в известный ген, кодирующий L-треониндегидратазу, и сравнивали способность продуцировать L-изолейцин. Использованные праймеры показаны в Таблице 8.
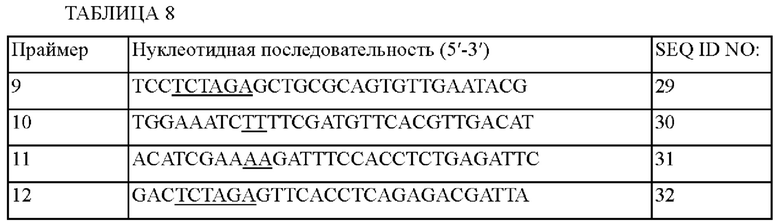
Пример 4-2-1. Внесение мутации L377K
Проводили ПЦР, используя хромосому WT в качестве матрицы вместе с парой праймеров 9 и 10 или праймеров 11 и 12. ПЦР проводили в условиях, приведенных ниже: денатурация при 95°С в течение 5 минут, 30 циклов с денатурацией при 95°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 30 секунд, а затем полимеризация при 72°С в течение 7 минут. В результате получали фрагмент ДНК размером 509 п.о., соответствующий области, расположенной в 5'-направлении вверх по течению по отношению к мутации в гене lysC, и фрагмент ДНК размером 520 п.о., соответствующий области, расположенной в 3'-направлении вниз по течению по отношению к этой мутации, соответственно.
Проводили ПЦР, используя эти два амплифицированных фрагмента ДНК в качестве матрицы вместе с парой праймеров 9 и 12. ПЦР проводили в условиях, приведенных ниже: денатурация при 95°С в течение 5 минут, 30 циклов с денатурацией при 95°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. В результате был амплифицирован фрагмент ДНК размером 1011 п.о., содержащий мутацию в гене lysC, кодирующем вариант аспартаткиназы, в котором лейцин в положении 377 был заменен на лизин.
Вектор pDZ, который не может реплицироваться в Corynebacterium glutamicum, и фрагмент ДНК размером 1011 п.о. обрабатывали ферментом рестрикции XbaI и лигировали с использованием ДНК-лигазы, а затем клонировали, получая плазмиду, которую обозначали как pDZ-lysC(L377K).
Вектор pDZ-lysC(L377K), полученный так, как описано выше, вводили в штамм WT, применяя метод электропорации (Appl. Microbiol. Biotechnol. (1999), 52: 541-545), и затем на селективной среде, содержащей канамицин в концентрации 25 мг/л, отбирали трансформированные штаммы. Получали штамм, в котором мутация с заменой нуклеотида была внесена в ген lysC путем встраивания этого фрагмента ДНК в хромосому с применением процесса рекомбинации с участием двух нитей ДНК (кроссинговера).
Пример 4-2-2. Внесение мутации G378E
Чтобы сконструировать вектор для внесения мутации hom(G378E), проводили ПЦР, используя геномную ДНК WT в качестве матрицы вместе с парой праймеров 13 и 14 и парой праймеров 15 и 16. ПЦР проводили в условиях, приведенных ниже: денатурация при 95°С в течение 5 минут, 30 циклов с денатурацией при 95°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 30 секунд, а затем полимеризация при 72°С в течение 7 минут. В результате получали фрагмент ДНК размером 220 п.о., соответствующий области, расположенной в 5'-направлении вверх по течению по отношению к мутации в гене hom, и фрагмент ДНК размером 220 п.о., соответствующий области, расположенной в 3'-направлении вниз по течению по отношению к этой мутации, соответственно. Проводили ПЦР, используя эти два продукта ДНК в качестве матрицы вместе с парой праймеров 13 и 16. ПЦР проводили в условиях, приведенных ниже: денатурация при 95°С в течение 5 минут, 30 циклов с денатурацией при 95°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. В результате был амплифицирован фрагмент ДНК размером 440 п.о., содержащий мутацию в гене horn. Использованные праймеры показаны в Таблице 9.

Использованный ранее вектор pDZ и фрагмент ДНК размером 440 п.о. обрабатывали ферментом рестрикции XbaI, лигировали с использованием ДНК-лигазы и затем клонировали, получая плазмиду, которую обозначали как pDZ-hom(G378E).
Полученный вектор pDZ-hom(G378E) вводили в штамм WT::lysC(L377K), приготовленный как в примере 4-2-1, применяя метод электропорации, а затем на селективной среде, содержащей канамицин в концентрации 25 мг/л, отбирали трансформированные штаммы. Получали штамм WT::lysC(L377K)-hom(G378E), в котором мутация с заменой нуклеотида была внесена в ген horn путем встраивания этого фрагмента ДНК в хромосому с применением процесса рекомбинации с участием двух нитей ДНК (кроссинговера).
Пример 4-2-3. Внесение мутации в промотор гена ilvC
С применением тех же способов, которые описаны в приведенных выше примерах, получали штаммы, в которых рекомбинантные плазмиды pDZ-ilvC(Pm3)-14067 и pDZ-ilvC(Pm3)-13032, полученные как в примере 3-1, были введены в штамм WT::lysC(L377K)-hom(G378E), полученный как в примере 4-2-2 посредством гомологичной рекомбинации с участием хромосомы, и эти штаммы обозначали как WT::lysC(L377K)-hom(G378E)-ilvC(Pm3) и WT::lysC(L377K)-hom(G378E)-ilvC(Pm3)-3, соответственно.
Пример 4-2-4. Внесение мутации в ген ilvA
Чтобы сконструировать вектор для внесения в ген ilvA известной ранее мутации liv4(V383A) (World J. Microbiol. Biotechnol, (2015) 31: 1369-1377), были разработаны одна пара праймеров (праймеры 17 и 18) для амплификации области, расположенной в 5'-направлении вверх по течению по отношению к положению мутации, и одна пара праймеров (праймеры 19 и 20) для амплификации области, расположенной в 3'-направлении вниз по течению по отношению к этой мутации, соответственно. Сайт для фермента BamHI (подчеркнут) был встроен в один конец каждого из праймеров 17 и 20, а мутация с заменой нуклеотида (подчеркнута) была расположена в сайте, предназначенном для проведения кроссинговера, в праймерах 18 и 19. Использованные праймеры показаны в Таблице 10.

Проводили ПЦР, используя хромосому WT в качестве матрицы вместе с парой праймеров 17 и 19 или парой праймеров 19 и 20. ПЦР проводили в условиях, приведенных ниже: денатурация при 95°С в течение 5 минут, 30 циклов с денатурацией при 95°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 30 секунд, а затем полимеризация при 72°С в течение 7 минут. В результате получали фрагмент ДНК размером 627 п.о., соответствующий области, расположенной в 5'-направлении вверх по течению по отношению к мутации в гене ilvA, и фрагмент ДНК размером 608 п.о., соответствующий области, расположенной в 3'-направлении вниз по течению по отношению к этой мутации, соответственно. Проводили ПЦР, используя эти два амплифицированных фрагмента ДНК в качестве матрицы вместе с парой праймеров 17 и 20. ПЦР проводили в условиях, приведенных ниже: денатурация при 95°С в течение 5 минут, 30 циклов с денатурацией при 95°С по 30 секунд, отжигом при 55°С в течение 30 секунд и полимеризацией при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. В результате был амплифицирован фрагмент ДНК размером 1217 п.о., содержащий мутацию в гене ilvA, кодирующем вариант IlvA, в котором валин в положении 383 был заменен на аланин.
Вектор pECCG117 (патент Кореи №10-0057684) и фрагмент ДНК размером 1011 п.о. обрабатывали ферментом рестрикции BamHI, лигировали с использованием ДНК лигазы и затем клонировали, получая плазмиду, которую обозначали как pECCG117-ilvA(V383A).
Получали штаммы путем введения вектора pECCG117-ilvA(V383A) в штаммы ATCC13032::hom(G378E)-lysC(L377K)-ilvC(Pm3) и АТСС13032::hom(G378E)-lysC(L377K)-ilvC(Pm3)-3, соответственно, и эти штаммы обозначали как ATCC13032::hom(G378E)-lysC(L377K)-ilvC(Pm3)/pECCG117-ilvA(V383A) и ATCC13032::hom(G378E)-lysC(L377K)-ilvC(Pm3)-3/pECCG117-ilvA(V383A), соответственно (ФИГ. 6). Помимо этого, также получали штамм путем внесения в штамм ATCC13032::-hom(G378E)-lysC(L377K) только мутации ilvA(V383A) в качестве контроля. Пример 4-2-5. Оценка способности продуцировать изо лейцин Штаммы культивировали тем же способом, который приведен в примере 4-1, и определяли концентрацию L-изолейцина в культуре.
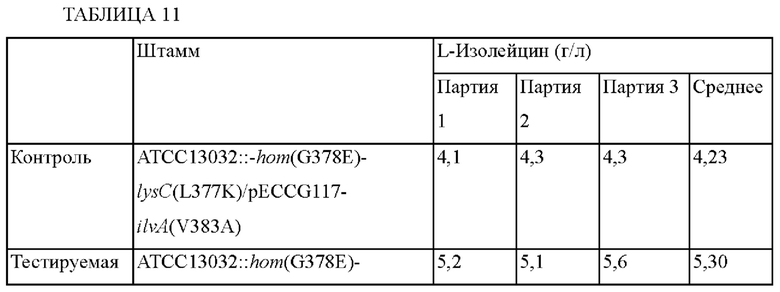

Как показано в приведенной выше Таблице 11, концентрации L-изолейцина в случае штаммов ATCC13032::hom(G378E)-lysC(L377K)-ilvC(Pm3)/pECCG117-ilvA(V383A) и ATCC13032::hom(G378E)-lysC(L377K)-ilvC(Pm3)-3/pECCG117-ilvA(V383A), каждый из которых содержит мутацию в гене ilvC, увеличились примерно на 25% и 24%, соответственно, по сравнению с таковой для штамма дикого типа ATCC13032::-hom(G378E)-lysC(L377K)/pECCG117-ilvA(V383A). Указанные выше результаты показывают, что внесение мутации в промотор гена ilvC L-изолейцин-продуцирующих штаммов рода Corynebacterium является эффективным в плане продуцирования L-изолейцина.
Пример 5. Получение лейцин-продуцирующих штаммов и оценка продуцирующей способности
Пример 5-1. Получение штамма с мутацией в промоторе гена ilvC, внесенной в L-изолейцин-продуцирующие штаммы Corynebacterium glutamicum KCCM11661P и KCCM11662P, и оценка способности продуцировать лейцин
Рекомбинантной плазмидой pDZ-ilvC(Pm3)-14067, сконструированной как в примере 3-1, трансформировали штаммы Corynebacterium glutamicum KCCM11661P (US 10351859 B2) и KCCM11662P (US 10351859 В2), которые являются L-лейцин-продуцирующими штаммами, посредством гомологичной рекомбинации с участием хромосомы (van der Rest et al.,Appl. Microbiol. Biotechnol., 52: 541-545, 1999). Штамм, в хромосому которого встроился данный вектор посредством рекомбинации гомологичных последовательностей, отбирали на среде, содержащей канамицин в концентрации 25 мг/л. После этого трансформированный штамм Corynebacterium glutamicum, в котором произошла рекомбинация с участием двух нитей ДНК, подвергали ПЦР с использованием праймера 1 и праймера 4 с целью конструирования штаммов с мутацией, которая была внесена в промотор гена ilvC в хромосоме. Рекомбинантные штаммы обозначали как Corynebacterium glutamicum KCCM11661P-ilvC(Pm3) и KCCM11662P-ilvC(Pm3), соответственно (ФИГ. 7). Чтобы сравнить способность продуцировать лейцин среди лейцин-продуцирующих штаммов Corynebacterium glutamicum KCCM11661P-ilvC(Pm3) и KCCM11662P-ilvC(Pm3), проводили оценку титра после ферментации. Каждый штамм пересевали на питательную среду и затем каждым штаммом инокулировали содержимое конической колбы с перегородками емкостью 250 мл, содержащей 25 мл среды для продуцирования, и культивировали со встряхиванием при 30°С и 200 об./мин в течение 72 часов. После этого, используя HPLC, определяли концентрацию L-лейцина, и установленные концентрации L-лейцина занесены в приведенную ниже Таблицу 12.
Питательная среда (рН 7,2)
Глюкоза - 10 г, мясной сок - 5 г, полипептон - 10 г, хлорид натрия - 2,5 г, дрожжевой экстракт - 5 г, агар - 20 г, и мочевина - 2 г (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
Глюкоза - 50 г, сульфат аммония - 20 г, кукурузный экстракт высушенный - 20 г, фосфат калия двухосновный - 1 г, сульфата магния гептагидрат - 0,5 г, биотин - 100 мкг, тиамин-HCl - 1 мг, карбонат кальция - 15 г (из расчета на 1 л дистиллированной воды).
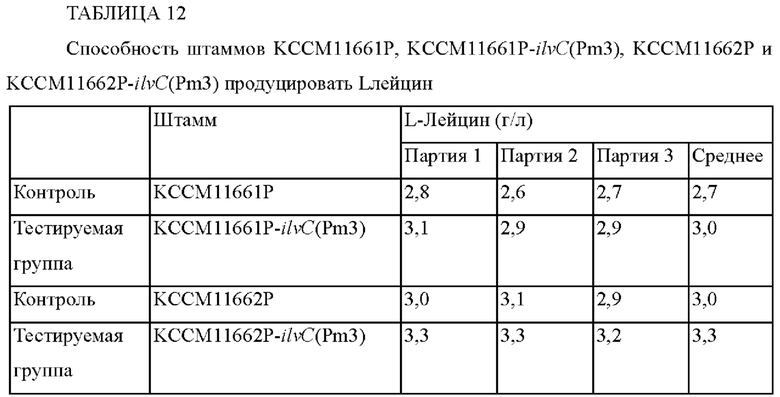
Как видно из приведенных выше результатов, способность штаммов KCCM11661P-ilvC(Pm3) и KCCM11662P-ilvC(Pm3) продуцировать L-лейцин увеличилась на 11% и 10% по сравнению с таковой в случае контроля. Таким образом, было установлено, что способность продуцировать L-лейцин может быть улучшена посредством внесения мутации в промотор гена ilvC.
Пример 5-2. Получение штамма с мутацией, внесенной в лейцин-продуцирующий штамм Corynebacterium glutamicum CJL8001, и оценка способности продуцировать L-лейцин
Для выяснения того, будет ли наблюдаться такой же эффект, который описан выше, для других продуцирующих L-лейцин штаммов Corynebacterium glutamicum, в штамм Corynebacterium glutamicum дикого типа АТСС 13032 вносили мутацию одного вида (leuA(R558H, G561D); US 2020-0032305 A1) с целью получения штаммов, обладающих улучшенной способностью продуцировать L-лейцин.
Конкретно, рекомбинантной плазмидой pDZ-leuA(R558H, G561D), сконструированной в упомянутом выше патенте, трансформировали штамм Corynebacterium glutamicum дикого типа АТСС130332 посредством гомологичной рекомбинации с участием хромосомы (van der Rest et al., Appl. Microbiol. Biotechnol, 52: 541-545, 1999). После этого, с целью идентификации штамма с внесенной в ген мутацией осуществляли секвенирование трансформированного штамма Corynebacterium glutamicum, в котором произошла рекомбинация с участием двух нитей ДНК. Рекомбинантный штамм обозначали как Corynebacterium glutamicum CJL8001.
Наконец, векторами pDZ-ilvC(Pm3)-14067 и pDZ-ilvC(Pm3)-13032 трансформировали штамм Corynebacterium glutamicum CJL8001, обладающий способностью продуцировать L-лейцин, тем же способом, что и в примере 5-1, с получением в результате этого штаммов CJL8001-ilvC(Pm3) и CJL8001-ilvC(Pm3)-3 с мутациями, которые были внесены в ген ilvC (ФИГ. 8), при этом штамм CJL8001-ilvC(Pm3)-3 обозначали как Corynebacterium glutamicum СА13-8101, был депонирован на международном уровне в международном депозитарии Корейского центра культур микроорганизмов (KCCM) 21 августа 2019 г. в соответствии с положениями Будапештского договора и ему был присвоен номер доступа KCCM12576P.
Для сравнения способности полученных штаммов продуцировать L-лейцин эти штаммы культивировали и концентрацию L-лейцина определяли тем же методом, что и в примере 5-1, и установленные концентрации L-лейцина занесены в Таблицу 13.
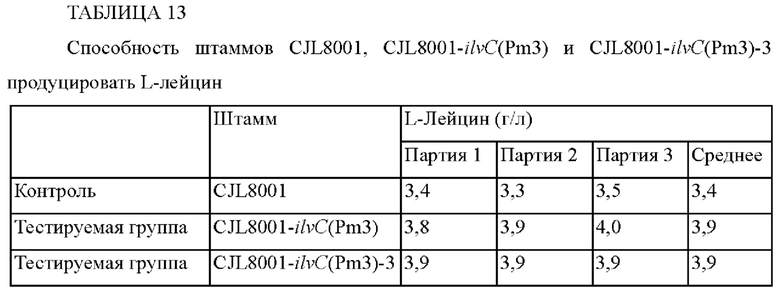
Как видно из приведенных выше результатов, способность штаммов CJL8001-ilvC(Pm3) и CJL8001-ilvC(Pm3)-3 продуцировать L-лейцин увеличилась на 15% по сравнению с таковой в случае контроля. Таким образом, вновь было установлено, что способность продуцировать L-лейцин может быть улучшена посредством внесения мутации в промотор гена ilvC микроорганизма рода Corynebacterium glutamicum.
Из приведенного выше описания специалист в данной области техники, к которой относится настоящее изобретение, сможет понять, что настоящее изобретение может быть реализовано в других конкретных формах без отклонения от его технической сущности или существенных признаков. Таким образом, описанные выше воплощения следует рассматривать как иллюстративные и не ограничивающие объем настоящего изобретения. Объем настоящего изобретения следует понимать в том смысле, что все изменения или модификации, вытекающие из определений и объемов формулы изобретения, и их эквиваленты включены в объем данного изобретения.
--->
<110> CJ CheilJedang Corporation
<120> НОВЫЙ ПРОМОТОР И СПОСОБ ПОЛУЧЕНИЯ ЖЕЛАЕМОГО ВЕЩЕСТВА С ЕГО
ИСПОЛЬЗОВАНИЕМ
<130> OPA20073-PCT
<150> KR 10-2019-0108263
<151> 2019-09-02
<160> 40
<170> KoPatentIn 3.0
<210> 1
<211> 28
<212> DNA
<213> Unknown
<220>
<223> Corynebacterium glutamicum
<400> 1
ccgtctaatt gcatgtgtgt ggtagaac 28
<210> 2
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_общая формула Z
<400> 2
catgtgtgtg gtanaan 17
<210> 3
<211> 7
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_общая формула Y
<400> 3
ctaattn 7
<210> 4
<211> 4
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_общая формула X
<400> 4
cngn 4
<210> 5
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула Z
<400> 5
catgtgtgtg gtagaac 17
<210> 6
<211> 7
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула Y
<400> 6
ctaattg 7
<210> 7
<211> 7
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула Y
<400> 7
ctaatta 7
<210> 8
<211> 4
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула X
<400> 8
ccgt 4
<210> 9
<211> 4
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула X
<400> 9
cgga 4
<210> 10
<211> 4
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула X
<400> 10
cggt 4
<210> 11
<211> 4
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула X
<400> 11
ccga 4
<210> 12
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор_формула Z
<400> 12
catgtgtgtg gtataat 17
<210> 13
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 13
ccgtctaatt gcatgtgtgt ggtataat 28
<210> 14
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 14
ccgtctaatt acatgtgtgt ggtataat 28
<210> 15
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 15
cggactaatt gcatgtgtgt ggtataat 28
<210> 16
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 16
cggactaatt acatgtgtgt ggtataat 28
<210> 17
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> promoter
<400> 17
ccgactaatt gcatgtgtgt ggtataat 28
<210> 18
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 18
ccgactaatt acatgtgtgt ggtataat 28
<210> 19
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 19
cggtctaatt gcatgtgtgt ggtataat 28
<210> 20
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> промотор
<400> 20
cggtctaatt acatgtgtgt ggtataat 28
<210> 21
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 21
ctattctaga gtgatgaatc tgcagcagaa gatc 34
<210> 22
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 22
gacaactaca ttattattat accacacaca tgca 34
<210> 23
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 23
tgcatgtgtg tggtataata ataatgtagt tgtc 34
<210> 24
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 24
ctattctaga gaagaggtcg gtgacggtct cagc 34
<210> 25
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 25
aatttctaga ggcagaccct attctatgaa gg 32
<210> 26
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 26
agtgtttcgg tctttacaga cacgagggac 30
<210> 27
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 27
gtccctcgtg tctgtaaaga ccgaaacact 30
<210> 28
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 28
aatttctaga cgtgggagtg tcactcgctt gg 32
<210> 29
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 29
tcctctagag ctgcgcagtg ttgaatacg 29
<210> 30
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 30
tggaaatctt ttcgatgttc acgttgacat 30
<210> 31
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 31
acatcgaaaa gatttccacc tctgagattc 30
<210> 32
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 32
gactctagag ttcacctcag agacgatta 29
<210> 33
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 33
tcctctagac tggtcgcctg atgttctac 29
<210> 34
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 34
gccaaaacct ccacgcgatc 20
<210> 35
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 35
atcgcgtgga ggttttggct 20
<210> 36
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 36
gactctagat tagtcccttt cgaggcgga 29
<210> 37
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 37
acggatccca gactccaaag caaaagcg 28
<210> 38
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 38
gcgcttgagg tactctgcca gcgtgatgtc 30
<210> 39
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 39
gacatcacgc tggcagagta cctcaagcgc 30
<210> 40
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 40
acggatccaa ccaaacttgc tcacactc 28
<---


Изобретение относится к биотехнологии. Предложен полинуклеотид, обладающий промоторной активностью. Также предложены промотор, содержащий указанный полинуклеотид, экспрессирующий вектор, содержащий указанный промотор, микроорганизм рода Corynebacterium для получения целевого вещества, содержащий указанный полинуклеотид, способ получения целевого вещества с использованием микроорганизма. Также предложен способ усиления экспрессии целевого гена, включающий функциональное связывание промотора, содержащего указанный полинуклеотид с целевым геном, и экспрессию целевого гена в клетке-хозяине. Изобретение обеспечивает усиление экспрессии целевых генов при получении целевых веществ. 6 н. и 6 з.п. ф-лы, 8 ил., 13 табл., 5 пр.
1. Полинуклеотид, обладающий промоторной активностью, содержащий нуклеотидную последовательность, представленную приведенной ниже общей формулой 1:
общая формула 1
X-Y-Z,
где X представляет собой CN1GN2,
Y представляет собой CTAATTN3 и
Z представляет собой CATGTGTGTGGTATAAT,
где каждый N1 в X представляет собой цитозин (C) или гуанин (G), N2 в X представляет собой аденин (A) или тимин (T) и N3 в Y представляет собой аденин (A) или гуанин (G).
2. Полинуклеотид по п. 1, где X представляет собой любую из последовательностей SEQ ID NO: 8-11 и Y представляет собой SEQ ID NO: 6 или 7.
3. Полинуклеотид по п. 1, где полинуклеотид имеет любую полинуклеотидную последовательность, выбранную из SEQ ID NO: 13-20.
4. Промотор, содержащий полинуклеотид по любому из пп. 1-3.
5. Экспрессирующий вектор, содержащий промотор по п. 7 и ген, кодирующий целевой белок, где промотор функционально связан с геном, кодирующим целевой белок.
6. Вектор по п. 5, где целевой белок представляет собой изомерoредуктазу ацетогидроксикислот.
7. Микроорганизм рода Corynebacterium для получения целевого вещества, содержащий полинуклеотид по любому из пп. 1-3.
8. Микроорганизм по п. 7, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
9. Способ получения целевого вещества, включающий:
культивирование микроорганизма рода Corynebacterium по п. 7 в среде; и
извлечение целевого вещества из этой среды.
10. Способ по п. 9, где целевое вещество представляет собой аминокислоту.
11. Способ по п. 10, где аминокислота представляет собой L-аминокислоту с разветвленной цепью.
12. Способ усиления экспрессии целевого гена, включающий функциональное связывание промотора, содержащего полинуклеотид по любому из пп. 1-3, с целевым геном и экспрессию целевого гена в клетке-хозяине.
| Прибор для определения экспозиции при печатании на фотографических бумагах | 1927 |
|
SU9232A1 |
| Corynebacterium glutamicum acetohydroxy acid synthase (ilvB) and (ilvN) genes, and acetohydroxy acid isomeroreductase (ilvC) gene, complete cds Найдено онлайн: https://www.ncbi.nlm.nih.gov/nuccore/L09232.1/ Дата обращения 19.10.2022 | |||
| WO 2014195154 A1, 11.12.2014 | |||
| EP 1860193 A1, 28.11.2007 | |||
| САМОИНДУЦИРУЕМАЯ ЭКСПРЕССИОННАЯ СИСТЕМА И ЕЕ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ПОЛЕЗНЫХ МЕТАБОЛИТОВ С ПОМОЩЬЮ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2013 |
|
RU2549708C2 |

