Область техники
Настоящее изобретение относится к новому промотору и способу получения целевых веществ с его использованием. Более конкретно, настоящее изобретение относится к новому полинуклеотиду, обладающему промоторной активностью, вектору и клетке-хозяину, включающим его, и к способу получения целевых веществ с использованием микроорганизма.
Предшествующий уровень техники
Процесс получения целевых веществ (например аминокислот) из микроорганизмов изучался различными способами как экологически чистый и безопасный метод производства. Среди них постоянно проводились исследования по получению большого количества целевых веществ в микроорганизмах рода Corynebacterium. Микроорганизмы рода Corynebacterium, в частности Corynebacterium glutamicum, представляют собой грамположительные микроорганизмы, которые широко используют для получения L-аминокислот и других полезных веществ. Для получения L-аминокислот и других полезных веществ проводятся различные исследования по разработке микроорганизмов с высокоэффективным продуцированием и технологиями процессов ферментации.
L-Лизин, являющийся репрезентативным веществом, продуцируемым микроорганизмами рода Corynebacterium, используют в кормах для животных, фармацевтических препаратах для человека и косметической промышленности, и его получают путем ферментации с использованием штаммов Corynebacterium. Микроорганизм, в котором усилены гены, связанные с биосинтезом L-лизина, и способ получения L-лизина с их использованием и т.д. известны в данной области техники (KR 10-0924065 В1).
Кроме того, L-треонин является незаменимой аминокислотой и широко используется в кормах и пищевых добавках, а также в инфузиях для медицины и в качестве сырья для фармацевтических препаратов. Поскольку L-треонина мало в растительном белке, его эффективно используют в качестве добавки к кормам для животных. L-треонин в основном производят путем ферментации с использованием Е. coli или микроорганизмов рода Corynebacterium, получаемых с использованием искусственной мутации или генетической рекомбинации. В данной области техники обычно известен способ с использованием генного рекомбинантного штамма для получения L-треонина путем введения треонинового оперона, происходящего из Е. coli, в Brevibacterium flavum, являющийся штаммом, продуцирующим треонин, и т.д. (TURBA Е., et al., Agric. Biol. Chem. 53:2269-2271, 1989).
O-Ацетилгомосерин представляет собой вещество, используемое в качестве предшественника продукции метионина, и является промежуточным продуктом в пути биосинтеза метионина (WO 2008/013432). О-Ацетил-L-гомосерин синтезируется из L-гомосерина и ацетил-КоА в качестве субстратов с помощью гомосерин-(9-ацетилтрансферазы.
L-Изолейцин имеет общий основной путь биосинтеза с другими аминокислотами с разветвленной цепью, L-валином и L-лейцином. Что касается пути биосинтеза L-изолейцина, в качестве предшественника используется 2-кетобутират, продуцируемый из L-треонина, аминокислоты, получаемой из пирувата и аспарагиновой кислоты, которые образуются в результате гликолиза. Из этих двух предшественников 2-ацето-2-гидроксиацетат синтезируется под действием фермента синтазы ацетогидроксикислот (AHAS), а затем под действием изомероредуктазы ацетогидроксикислот продуцируется 2,3-дигидрокси-3-метилвалерат. Впоследствии под действием дегидратазы дигидроксикислот образуется 2-кето-3-метилвалерат, и в конечном итоге в результате аминотрансферазной реакции образуется L-изолейцин. Кроме того, синтаза ацетогидроксикислот катализирует декарбоксилирование пирувата и реакцию конденсации с другими молекулами пирувата с образованием ацетолактата, предшественника валина и лейцина.
L-Триптофан, одна из незаменимых аминокислот, широко используется в кормовых добавках или в качестве сырья для фармацевтических препаратов, таких как инфузии, и в качестве вещества для здорового питания. В настоящее время L-триптофан в основном производят путем прямой ферментации с использованием микроорганизмов.
L-Тирозин представляет собой аминокислоту и используется в качестве важного материала для фармацевтического сырья, пищевых добавок, кормов для животных, пищевых добавок и т.д. Для производства L-тирозина и других полезных веществ проводят различные исследования по разработке микроорганизмов с высокоэффективным продуцированием и технологиями процессов ферментации. Процесс продуцирования L-тирозина микроорганизмами начинается с 3-дезокси-D-аробино-гептулозонат-7-фосфата (DAHP), образующегося в результате реакции полимеризации фосфоенолпирувата (PEP), являющегося промежуточным продуктом гликолиза, с эритрозо-4-фосфатом (Е4Р), который является промежуточным продуктом пентозофосфатного пути. Затем DAHP подвергается биосинтезу из хоризмата в префенат по обычному пути биосинтеза ароматических соединений и в конечном итоге превращается в L-тирозин посредством пути биосинтеза L-тирозина. Во время этого процесса хоризмат может быть переведен в L-триптофан, а префенат может быть переведен в L-тирозин или L-фенилаланин. Таким образом, когда общий путь биосинтеза ароматических соединений усиливается для увеличения количества вырабатываемого L-тирозина, можно ожидать, что продуцирование L-триптофана и L-фенилаланина также будет увеличиваться одновременно. То есть, чтобы получить L-тирозин, фенилаланин и триптофан продуцируются вместе как побочные продукты, и соответственно необходимо проводить различные исследования, такие как рекомбинация генов, очистка и т.д. Между тем известно, что продукция L-триптофана регулируется репрессорами и аттенюаторами в зависимости от концентрации L-триптофана, продуцируемого микроорганизмами (патент Кореи No. 10-0792095).
L-Фенилаланин является исходным веществом для метилового эфира L-аспартил-L-фенилаланина, который является подсластителем.
При разработке универсального промотора, который не ограничивается конкретными полезными веществами, ожидается, что его можно будет использовать в производстве различных полезных веществ.
При таких обстоятельствах авторы настоящего изобретения разработали полинуклеотид, обладающий новой промоторной активностью, способный продуцировать целевые вещества с высокой продуктивностью в микроорганизмах рода Corynebacterium, и подтвердили, что продуктивность по целевым веществам можно увеличить путем введения полинуклеотида в микроорганизмы, тем самым создав настоящее изобретение.
Описание изобретения
Техническая задача
Авторы настоящего изобретения предприняли значительные усилия для получения нового полинуклеотида, обладающего промоторной активностью, и в результате подтвердили, что промотор целевого гена может быть усилен заменой нуклеотида, и этот усиленный промотор может регулировать экспрессию генов, функционально связанных с промотором, тем самым создав настоящее изобретение. Техническое решение
Одна из задач настоящего изобретения заключается в том, чтобы предложить полинуклеотид, обладающий промоторной активностью, в котором нуклеотиды по положениям 218, 219, 220, 221, 222, 225 и 227 в полинуклеотидной последовательности SEQ ID NO: 1 заменены на другие нуклеотиды.
Другая задача настоящего изобретения заключается в том, чтобы предложить кассету генной экспрессии, включающую: полинуклеотид и ген, кодирующий целевой белок, который функционально связан с полинуклеотидом.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить клетку-хозяин, включающую: полинуклеотид и ген, кодирующий целевой белок, который функционально связан с полинуклеотидом.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить способ получения целевого вещества, включающий: культивирование клетки-хозяина в среде и извлечение целевого вещества из среды.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить применение полинуклеотида, в котором нуклеотиды по положениям 218, 219, 220, 221, 222, 225 и 227 в полинуклеотидной последовательности SEQ ID NO: 1 заменены на другие нуклеотиды, в качестве промотора.
Полезные эффекты
Новый промотор по настоящему изобретению может быть введен в микроорганизм для получения целевых веществ с увеличением продуцирования этих целевых веществ микроорганизмом. В виду повышения выхода продукции в промышленном аспекте можно ожидать такие эффекты, как снижение производственных затрат наряду с удобством производства.
Подробное описание предпочтительных воплощений
Ниже настоящее изобретение будет описано подробно. При этом каждое описание и воплощение, раскрытое здесь, может быть применено к другим описаниям и воплощениям, соответственно. То есть, все комбинации различных элементов, раскрытых здесь, включены в объем настоящего изобретения. Кроме того, объем настоящего изобретения не ограничен подробным описанием, представленным ниже.
Дополнительно, специалисты в данной области техники будут понимать или способны определить, используя только рутинные эксперименты, множество воплощений, эквивалентных конкретным воплощениям настоящего изобретения. Кроме того, подразумевается, что эти эквиваленты включены в объем настоящего изобретения.
В одном аспекте настоящего изобретения предложен полинуклеотид, обладающий промоторной активностью.
В частности, полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может представлять собой полинуклеотид, обладающий промоторной активностью, включающий по меньшей мере одну нуклеотидную замену в полинуклеотидной последовательности SEQ ID NO: 1.
Используемый здесь термин «полинуклеотид» относится к нити ДНК, имеющей определенную минимальную длину как нуклеотидный полимер, в котором нуклеотидные мономеры связаны друг с другом в виде длинной цепи ковалентными связями.
Используемый здесь термин «промотор» относится к нетранслируемой нуклеотидной последовательности, расположенной выше кодирующей области, которая включает сайт связывания полимеразы и обладает активностью инициации транскрипции целевого гена-промотора в мРНК, т.е. области ДНК, которая приводит к инициации транскрипции определенного гена при связывании с ним полимеразы. Промотор может быть расположен в 5'-области сайта инициации транскрипции мРНК.
Используемый здесь термин «полинуклеотид, обладающий промоторной активностью» относится к области ДНК, расположенной в непосредственной близости к области, которая вовлечена в транскрипцию целевого гена, включающей сайт связывания РНК-полимеразы или энхансер, для экспрессии гена, который должен быть функционально связан против хода транскрипции от него, т.е. целевого гена. Для задач настоящего изобретения полинуклеотид может быть использован в качестве промотора для общего применения. Кроме того, полинуклеотид может регулировать (например увеличивать или уменьшать) в клетке, по сравнению с существующим промотором или внутриклеточным промотором, экспрессию целевого гена, функционально связанного с ним, и продукцию и/или активность белка, кодируемого целевым геном, и может регулировать (например увеличивать или уменьшать) продукцию и/или активность целевых продуктов (в качестве биологически активного вещества, например, по меньшей мере одного выбранного из группы, состоящей из аминокислот, нуклеиновых кислот, витаминов, белков, жирных кислот и органических кислот и т.д.), участвующих в продуцировании белков, но не ограничиваясь этим.
Например, полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может быть использован в качестве промотора, способного усиливать экспрессию полипептида, обладающего транскетолазной (tkt) активностью. Кроме того, полинуклеотид может представлять собой полинуклеотид, вовлеченный в увеличение продуцирования или продуцируемого количества лизина, треонина, О-ацетилгомосерина, изолейцина, триптофана, тирозина или фенилаланина.
Полинуклеотид по настоящему изобретению может включать любые полинуклеотидные последовательности, обладающие промоторной активностью, без ограничения.
В одном примере полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может включать полинуклеотид, обладающий промоторной активностью, в котором по меньшей мере один нуклеотид в нуклеотидной последовательности SEQ ID NO: 1 заменен на другой нуклеотид. В частности, в настоящем изобретении полинуклеотид, обладающий промоторной активностью, может представлять собой полинуклеотид, обладающий промоторной активностью, включающий одну или более, две или более, три или более, четыре или более, пять или более, шесть или более или семь или более нуклеотидных замен в полинуклеотидной последовательности SEQ ID NO: 1. В настоящем изобретении полинуклеотид, обладающий промоторной активностью, может быть использован взаимозаменяемо с «модифицированным промотором», и здесь могут быть использованы все вышеописанные термины.
В настоящем изобретении полинуклеотидная последовательность SEQ ID NO: 1 может представлять собой, например, полинуклеотид, обладающий промоторной активностью транскетолазы. Кроме того, полинуклеотид, в котором определенные нуклеотиды заменены в полинуклеотидной последовательности SEQ ID NO: 1, также может представлять собой полинуклеотид, обладающий промоторной активностью транскетолазы, при условии что он обладает промоторной активностью. Полинуклеотидная последовательность SEQ ID NO: 1 может представлять собой репрезентативный полинуклеотид для указания положения мутации, и другие полинуклеотидные последовательности, обладающие промоторной активностью, которые соответствуют ей, также включены в последовательность, которая может вводить мутацию. Например, любые полинуклеотидные последовательности, способные служить промотором для транскетолазы (tkt) или полипептида, обладающего соответствующей активностью, могут быть включены в ряд последовательностей, которые могут вводить мутацию по настоящему изобретению, без ограничения.
Нуклеотидная последовательность SEQ ID NO: 1 может быть подтверждена из NCBI GenBank, известной базы данных, а последовательность, соответствующая SEQ ID NO: 1, как последовательность, которая может служить промотором для транскетолазы, может иметь происхождение из Corynebacterium sp., и, в частности, это может быть последовательность Corynebacterium glutamicum, но последовательность, имеющая активность, равную или превышающую активность полинуклеотида, может быть включена в промотор по настоящему изобретению, без ограничений.
Полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может представлять собой полинуклеотид, в котором промоторная активность усилена заменой нуклеотидов в определенных положениях полинуклеотидной последовательности, обладающей существующей промоторной активностью.
В одном воплощении полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может включать полинуклеотид, обладающий промоторной активностью, в котором по меньшей мере один нуклеотид в нуклеотидной последовательности SEQ ID NO: 1 заменен на другой нуклеотид. В частности, он может состоять из полинуклеотида, обладающего промоторной активностью, в котором по меньшей мере один нуклеотид в нуклеотидной последовательности SEQ ID NO: 1 заменен на другой нуклеотид. В настоящем изобретении полинуклеотид, обладающий промоторной активностью, может быть использован взаимозаменяемо с «модифицированным промотором».
В одном воплощении модифицированный промотор может представлять собой полинуклеотид, обладающий промоторной активностью, включающий замены по меньшей мере одного нуклеотида, выбранного из группы, состоящей из нуклеотидов по положениям 218, 219, 220, 221, 222, 225 и 227, на другие нуклеотиды. В частности, модифицированный промотор может представлять собой промотор, в котором любое одно или более, два или более, три или более, четыре или более, пять или более, шесть или более или семь из вышеуказанных положений или их соответствующие положения заменены на другие нуклеотиды.
Используемый здесь термин «другой нуклеотид» не ограничен, при условии что он отличается от нуклеотида до замены. Например, когда описано, что «нуклеотид по положению 218 в SEQ ID NO: 1 заменен на другой нуклеотид», это означает, что он заменен аденином (А), тимином (Т) или гуанином (G), кроме цитозина (С). Кроме того, если не указано иное, когда описано, что конкретный нуклеотид «замещен», это означает, что нуклеотид замещен другим нуклеотидом, который отличается от нуклеотида до замены.
Между тем, специалисты в данной области техники могут идентифицировать нуклеотид в положении, соответствующем нуклеотидам по положениям 218, 219, 220, 221, 222, 225 и 227 SEQ ID NO: 1 настоящей заявки в любой полинуклеотидной последовательности через выравнивание последовательностей, известное в данной области техники. Кроме того, если в настоящем документе не указано иное, когда описано, что «нуклеотид по определенному положению в конкретной SEQ ID NO», очевидно, что подразумевается включение «нуклеотидов в соответствующем положении» в любые полинуклеотидные последовательности. Следовательно, полинуклеотидные последовательности, в которых любой один или более нуклеотидов выбраны из группы, состоящей из нуклеотидов по положениям, соответствующих 218, 219, 220, 221, 222, 225 и 227 SEQ ID NO: 1 в любой полинуклеотидной последовательности, обладающей промоторной активностью, заменены на другие нуклеотиды, также входят в объем настоящего изобретения.
В одном примере, когда один или более, два или более, три или более, четыре или более, пять или более, шесть или более или семь нуклеотидов по положениям, соответствующим 218, 219, 220, 221, 222, 225 и 227 в полинуклеотидной последовательности SEQ ID NO: 1, заменены на другие нуклеотиды, могут быть получены промоторы с более высокой активностью, чем незамещенные (немодифицированные) промоторные последовательности.
В частности, полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может представлять собой полинуклеотид, в котором нуклеотиды по положениям 218, 219, 220, 221, 222, 225 и 227 в полинуклеотидной последовательности SEQ ID NO: 1 заменены на другие нуклеотиды, но не ограничивается этим.
В конкретном примере полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может представлять собой полинуклеотид, в котором цитозин (С), представляющий собой нуклеотид по положению 218, заменен на тимин (Т), цитозин (С), представляющий собой нуклеотид по положению 219, заменен на гуанин (Г), аденин (А), представляющий собой нуклеотид по положению 220, заменен на тимин (Т), аденин (А), представляющий собой нуклеотид по положению 221, заменен на гуанин (Г), тимин (Т), представляющий собой нуклеотид по положению 222, заменен на гуанин (G), аденин (А), представляющий собой нуклеотид по положению 225, заменен на тимин (Т), и цитозин (С), представляющий собой нуклеотид по положению 227, заменен на аденин (А) в полинуклеотидной последовательности SEQ ID NO: 1.
В более конкретном примере это может быть полинуклеотид, состоящий из нуклеотидной последовательности SEQ ID NO: 2. В частности, полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может представлять собой полинуклеотид, включающий или состоящий (по существу) из полинуклеотидной последовательности SEQ ID NO: 2.
Кроме того, различные модификации полинуклеотидной последовательности могут быть включены в диапазоне, который существенно не снижает активность промотора, не ограничиваясь вышеописанными воплощениями. Например, нуклеотидная последовательность по настоящему изобретению может быть модифицирована с помощью общеизвестного мутагенеза, например, прямой эволюции, сайт-направленного мутагенеза и т.д.
В частности, термин «модификация» относится к фенотипическому изменению, которое является генетически или негенетически стабильным, и может взаимозаменяемо обозначаться термином «модификация» или «мутация» в настоящем документе.
Таким образом, полинуклеотид, обладающий промоторной активностью в настоящем изобретении, может представлять собой полинуклеотидную последовательность SEQ ID NO: 1 или 2 или полинуклеотидную последовательность, имеющую по меньшей мере 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологии или идентичности с SEQ ID NO: 1 или 2. Нуклеотидная последовательность, имеющая гомологию или идентичность, может представлять собой последовательность в вышеуказанном диапазоне, исключая последовательность, имеющую идентичность 100%, или может представлять собой последовательность, имеющую идентичность менее 100%.
В то же время, хотя в настоящем описании используют термины «полинуклеотид, имеющий нуклеотидную последовательность определенной SEQ ID NO» и «полинуклеотид, включающий нуклеотидную последовательность определенной SEQ ID NO», очевидно, что в настоящем изобретении также можно использовать любой полинуклеотид, в котором часть нуклеотидной последовательности делетирована, модифицирована, заменена или добавлена, при условии что этот полинуклеотид обладает активностью, идентичной или соответствующей активности полинуклеотида, состоящего из нуклеотидной последовательности соответствующей SEQ ID NO.
Например, очевидно, что любой полинуклеотид, включающий добавление бессмысленной последовательности внутри или на конце полинуклеотидной последовательности заданной SEQ ID NO или делецию части полинуклеотидной последовательности заданной SEQ ID NO внутри или на ее конце, также может входить в объем настоящего изобретения, при условии что этот полинуклеотид обладает активностью, идентичной или соответствующей активности полинуклеотида по настоящему изобретению.
Как используют здесь, термин «гомология» или «идентичность» относится к степени родства между двумя данными нуклеотидными последовательностями и может быть выражен в процентах.
Термины «гомология» и «идентичность» часто могут быть использованы взаимозаменяемо.
Гомология или идентичность последовательности консервативных полинуклеотидов может быть определена с помощью стандартного алгоритма выравнивания и установленных программой штрафов на введение гэпа по умолчанию. По существу гомологичные или идентичные последовательности могут гибридизоваться друг с другом на по меньшей мере примерно 50%, 60%, 70%, 80% или 90% от всей последовательности или по всей длине последовательности в условиях умеренной или высокой жесткости. Также можно рассматривать полинуклеотиды, включающие вместо кодонов вырожденные кодоны в гибридизуемых полинуклеотидах.
Гомология, подобие или идентичность между двумя данными полинуклеотидными последовательностями могут быть определены с использованием известного компьютерного алгоритма, такого как программа FASTA, (Pearson et al. (1988, Proc. Natl. Acad. Sci. USA 85:2444), используя параметры по умолчанию. Альтернативно, их можно определить, используя алгоритм Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), выполняемый программой Нидлмана в пакете программ от The European Molecular Biology Open Software Suite (EMBOSS) (Rice et al, 2000, Trends Genet. 16:276-277) (версия 5.0.0 или более поздняя) (пакет программ GCG (Devereux, J. et al, Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F. et al, J Molec Biol 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994; and Carillo et al. (1988) SIAM J Applied Math 48:1073). Например, гомологию, подобие или идентичность можно определить с помощью программ BLAST или ClustalW от Национального центра биотехнологической информации (NCBI).
Гомологию, подобие или идентичность полинуклеотидов можно определять путем сравнения информации о последовательности с помощью компьютерной программы GAP, такой как в Needleman et al. (1970J, J Mol Biol 48: 443, как описано в работе Smith and Waterman, Adv. Appl. Math (1981) 2:482. Вкратце, программа GAP определяет гомологию, подобие или идентичность как результат деления количества выровненных символов (т.е. нуклеотидов или аминокислот), являющихся подобными, на общее количество символов в более короткой из двух последовательностей. Параметры программы GAP по умолчанию могут включать: (1) однокомпонентную матрицу сравнений (содержащую значение 1 для идентичности и 0 для отсутствия идентичности) и взвешенную матрицу сравнений или матрицу замен Gribskov et al. (1986) Nucl. Acids Res. 14:6745, раскрытую в Schwartz and DayhofF, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или EDNAFULL (версия EMBOSS NCBI NUC4.4)); (2) штраф 3,0 за каждый гэп и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф на открытие гэпа 10, штраф на удлинение гэпа 0,5); и (3) отсутствие штрафа для концевых гэпов. Таким образом, используемый здесь термин «гомология» или «идентичность» отражает родство между последовательностями.
Кроме того, зонд, который может быть получен из любых известных последовательностей генов, например, любая полинуклеотидная последовательность, которая может гибридизоваться с последовательностью, полностью или частично комплементарной данной полинуклеотидной последовательности, в жестких условиях и обладающая такой же активностью, может быть включена без ограничения. Термин «жесткие условия» относится к условиям, в которых обеспечивается специфическая гибридизация между полинуклеотидами. Такие условия подробно раскрыты в известных документах (например, см. J. Sambrooke? al, Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F. M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York). Например, жесткие условия могут включать осуществление гибридизации генов, имеющих высокую гомологию или идентичность, например гомологию или идентичность 40% или конкретно 70% или более, 80% или более, 85% или более, 90% или более, более конкретно 95% или более, еще более конкретно 97% или более, и наиболее конкретно 99% или более, без осуществления гибридизации генов, имеющих гомологию или идентичность более низкую, чем указанные выше гомологии или идентичности, или выполнение отмывки один раз, конкретно два или три раза, в обычных условиях отмывки для Саузерн-гибридизации с концентрацией соли и температурой 60°С, 1×SSC и 0,1% SDS, конкретно 60°С, 0,1×SSC и 0,1% SDS, и более конкретно 68°С, 0,1×SSC и 0,1% SDS.
Для гибридизации требуется, чтобы два нуклеотида имели комплементарные последовательности, хотя возможно ошибочное спаривание оснований в зависимости от степени жесткости гибридизации. Термин «комплементарный» используют для описания взаимоотношения между нуклеотидными основаниями, способными гибридизоваться друг с другом. Например, что касается ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, настоящее изобретение может включать выделенные нуклеотидные фрагменты, комплементарные целой последовательности, а также нуклеиновокислотные последовательности, по существу подобные ей.
В частности, полинуклеотиды, имеющие гомологию или идентичность, могут быть обнаружены с использованием описанных выше условий гибридизации, включающих стадию гибридизации при значении Tm 55°С. Также, значение Tm может составлять 60°С, 63°С или 65°С, без ограничения ими, и может подходящим образом регулироваться специалистом в данной области техники в соответствии с задачами.
Степень жесткости условий для гибридизации полинуклеотидов зависит от длины полинуклеотидов и степени комплементарности, и эти переменные хорошо известны в данной области техники (Sambrook et al., см. выше, 9.50-9.51, 11.7-11.8).
Полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может быть выделен или получен с использованием стандартных методов молекулярной биологии. Например, он может быть получен с использованием стандартных способов синтеза с использованием автоматического синтезатора ДНК, но не ограничивается этим.
Полинуклеотид, обладающий промоторной активностью, по настоящему изобретению может быть использован в качестве промтора.
Промотор может быть расположен в 5' области сайта инициации транскрипции в мРНК.
Промотор может иметь повышенную или пониженную промоторную активность по сравнению с обычным промотором. То есть, промотор может увеличивать или уменьшать экспрессию и/или активность белка, кодируемого целевым геном, а также экспрессию целевого гена в клетке-хозяине. Для задач настоящего изобретения целевой ген для усиления или ослабления экспрессии может быть изменен в зависимости от продуцируемого продукта, и промотор может быть использован в качестве универсального промотора для усиления или ослабления целевого гена.
В другом аспекте настоящего изобретения предложена кассета генной экспрессии, включающая полинуклеотид и целевой ген.
Полинуклеотид по настоящему изобретению является таким, как описано выше.
Используемый здесь термин «кассета генной экспрессии» относится к единой кассете, которая включает промотор и целевой ген и, таким образом, может экспрессировать целевой ген, функционально связанный с промотором ниже по ходу транскрипции. Такая кассета генной экспрессии может включать различные факторы, которые могут способствовать эффективной экспрессии целевого гена внутри или снаружи кассеты. Кассета генной экспрессии может обычно включать сигнал терминации транскрипции, домен связывания рибосомы и сигнал терминации трансляции, в дополнение к промотору, функционально связанному с целевым геном, но не ограничиваясь этим.
Используемый здесь термин «целевой ген» относится к гену, экспрессия которого должна контролироваться промоторной последовательностью по настоящему изобретению для задач настоящего изобретения. Белок, кодируемый целевым геном, может быть определен как «целевой белок», а ген, кодирующий «целевой белок», может быть определен как «целевой ген».
Кроме того, полинуклеотид, кодирующий целевой белок, может подвергаться различным модификациям в кодирующей области в объеме, который не изменяет последовательность полипептида, из-за вырожденности кодонов или с учетом кодонов, предпочтительных в организме, в котором полинуклеотид по настоящему изобретению подлежит экспрессии. Полинуклеотидная последовательность является такой, как описано выше.
В одном воплощении целевой белок может представлять собой полипептид, обладающий транскетолазной (tkt) активностью. То есть, целевым геном промотора может быть ген, кодирующий полипептид, обладающий транскетолазной (tkt) активностью.
Используемый здесь термин «транскетолаза» представляет собой фермент, который влияет на пентозофосфатный путь и продуцирует В-седогептулозо-7-фосфат и D-глицеральдегид-3-фосфат из В-ксилулозо-5-фосфата и D-рибозо-3-фосфата, и эффект повышения продуктивности полезных веществ, таких как лизин, треонин, О-ацетилгосерин, изолейцин, триптофан, тирозин и фенилаланин, может быть получен путем регулирования ее активности.
Примеры генов, кодирующих транскетолазу, могут включать ген tkt (NCgl1512) Corynebacterium glutamicum АТСС 13032 и т.д., но не ограничиваются этим. Специалисты в данной области техники могут легко получить информацию о гене, кодирующем транскетолазу, из GenBank, известной базы данных и т.д.
Кроме того, аминокислотная последовательность, составляющая транскетолазу, может быть получена из известной базы данных GenBank NCBI. Например, она может иметь происхождение из Corynebacterium glutamicum.
Кроме того, «полипептид, обладающий транскетолазной активностью» по настоящему изобретению включает не только немодифицированную или природную форму транскетолазы дикого типа, но также мутант, обладающий такой же активностью или повышенной активностью.
Используемый здесь термин «модифицированный полипептид» имеет то же значение, что и «вариант», и относится к белку, имеющему одну или более аминокислот, отличающихся от приведенной последовательности консервативными заменами и/или модификациями, так что функции и свойства белка сохраняются.
Варианты отличаются от последовательностей, идентифицированных заменой, делецией или добавлением нескольких аминокислот. Такие варианты обычно могут быть идентифицированы путем модификации одной или более из вышеуказанных аминокислотных последовательностей белка и оценки свойств модифицированного белка. То есть, способность вариантов может быть усилена по сравнению с нативным белком. Кроме того, некоторые варианты могут включать варианты, в которых удалены одна или более частей, таких как N-концевая лидерная последовательность или транс мембранный домен.
Термин «вариант» может быть использован взаимозаменяемо с такими терминами, как модификация, модифицированный белок, модифицированный полипептид, мутант, мутеин, дивергент, вариант и т.д. без ограничений, если эти термины используют для обозначения вариации. Для задач настоящего изобретения вариант может представлять собой вариант, в котором активность модифицированного белка повышена по сравнению с активностью нативного белка дикого типа или немодифицированного белка, но не ограничивается этим.
Используемый здесь термин «консервативная замена» относится к замене аминокислоты на другую аминокислоту, имеющую сходные структурные и/или химические свойства. Вариант может иметь, например, по меньшей мере одну консервативную замену, сохраняя при этом по меньшей мере одну биологическую активность. Такая аминокислотная замена обычно может происходить на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатка.
Кроме того, вариант может также включать делецию или добавление аминокислот, которые оказывают минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью HaN-конце, участвующей в котрансляционном или посттрансляционном переносе белков. Кроме того, полипептид также может быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
Ген, кодирующий полипептид, обладающий транскетолазной активностью, по настоящему изобретению может быть обозначен как «ген tkt».
Ген может иметь происхождение из микроорганизма рода Corynebacterium и, в частности, Corynebacterium glutamicum.
В настоящем изобретении полинуклеотид, кодирующий полипептид, имеющий «ген tkt», т.е. транскетолазную активность, может быть подвергнут различным модификациям в кодирующей области в объеме, который не изменяет последовательность полипептида, вследствие вырожденности кодонов или с учетом кодонов, предпочтительных в организме, в котором должен экспрессироваться данный полипептид.
Полипептид, обладающий транскетолазной активностью, по настоящему изобретению может включать вариантные последовательности и может, в частности, включать варианты модифицированного белка, так чтобы повышенная активность транскетолазы могла быть экспрессирована.
В еще одном аспекте настоящего изобретения предложен рекомбинантный вектор, включающий полинуклеотид, или кассету генной экспрессии, включающую ген, кодирующий целевой белок, функционально связанный с полинуклеотидом.
Полинуклеотид, целевой белок и кассета генной экспрессии по настоящему изобретению являются такими, как описано выше.
В одном воплощении целевой белок может представлять собой полипептид, обладающий транскетолазной активностью.
Используемый здесь термин «вектор» представляет собой молекулу искусственной ДНК, которая содержит генетический материал, обеспечивающий экспрессию целевого гена в подходящей клетке-хозяине, и, в частности, относится к конструкции ДНК, которая включает нуклеотидную последовательность гена, кодирующего целевой белок, функционально связанного с ним.
Используемый здесь термин «функционально связанный» означает, что полинуклеотид, обладающий промоторной активностью, по настоящему изобретению и последовательность гена функционально связаны, так что транскрипция целевого гена может быть инициирована и опосредована. Функциональная связь может быть получена с использованием генетической рекомбинантной технологии, хорошо известной в данной области техники, а сайт-специфическое расщепление ДНК и связывание могут быть получены с использованием ферментов расщепления и связывания и т.д., известных в данной области техники, но не ограничиваясь ими.
Для задач настоящего изобретения регуляторная последовательность может включать полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
При этом регуляторная последовательность может включать промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции такой транскрипции, последовательность, кодирующую соответствующий рибосомо-связывающий домен мРНК, и последовательность для регуляции транскрипции и трансляции. После трансформации в подходящую клетку-хозяина вектор может реплицироваться или функционировать независимо от генома хозяина или может интегрироваться в его геном.
Вектор, используемый в настоящем изобретении, может не иметь конкретных ограничений, при условии что вектор экспрессируется в клетке-хозяине, и клетка-хозяин может быть трансформирована с использованием любого вектора, известного в данной области техники. Примеры обычно используемого вектора могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги.
Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.д.; а в качестве плазмидного вектора могут быть использованы те, которые основаны на pDZ, pBR, pUC, pBluescriptll, pGEM, pTZ, pCL, pET, и т.д., но не ограничены ими. В частности, векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д. могут быть использованы, но вектор не ограничен ими. Встраивание полинуклеотида в хромосому может быть осуществлено способом, хорошо известным в данной области техники, например, гомологичной рекомбинацией.
Поскольку вектор по настоящему изобретению может быть вставлен в хромосому посредством гомологичной рекомбинации, может быть дополнительно включен маркер селекции для подтверждения вставки в хромосому. Маркер селекции используют для отбора трансформированной клетки, т.е. для подтверждения вставки полинуклеотида, и могут быть использованы маркеры, способные обеспечивать селектируемые фенотипы, такие как устойчивость к лекарственным средствам, потребность в питательных веществах, устойчивость к цитотоксическим агентам или экспрессия поверхностных белков. В условиях, когда обрабатывают селективными агентами, только клетки, способные экспрессировать маркеры селекции, могут выживать или экспрессировать другие фенотипические признаки, и, таким образом, трансформированные клетки могут быть отобраны.
Используемый в настоящем документе термин «трансформация» относится к процессу введения вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, тем самым обеспечивая экспрессию белка, кодируемого полинуклеотидом, в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения, встроен ли он в хромосому клетки-хозяина и находится в ней или находится вне хромосомы, при условии что он может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид может включать ДНК и РНК, кодирующие целевой белок, и может быть введен в любой форме, при условии что он может быть введен в клетку-хозяин и экспрессироваться в ней. Например, полинуклеотид может быть введен в виде кассеты экспрессии, которая представляет собой генную конструкцию, включающую все основные элементы, необходимые для самоэкспрессии, или в форме вектора, включающего кассету экспрессии.
Способ трансформации может включать любой способ, с помощью которого можно ввести в клетку ген, кодирующий целевой белок, и трансформацию можно осуществить, выбрав подходящую стандартную методику, известную в данной области техники, в соответствии с клеткой-хозяином. Например, способ может включать электропорацию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), микроинъекцию, метод с полиэтиленгликолем (ПЭГ), метод с ДЭАЭ-декстраном, метод катионных липосом и метод ацетата лития-ДМСО и т.д., но не ограничивается этим.
Еще один аспект настоящего изобретения относится к клетке-хозяину, включающей полинуклеотид и ген, кодирующий целевой белок, функционально связанный с полинуклеотидом.
В частности, клетка-хозяин может представлять собой микроорганизм рода Corynebacterium и, более конкретно, Corynebacterium glutamicum, но не ограничивается этим.
Полинуклеотид и целевой белок являются такими, как описано выше.
Используемый здесь термин «микроорганизм» представляет собой понятие, включающее как микроорганизм дикого типа, так и естественно или искусственно генетически модифицированный микроорганизм, и он может представлять собой микроорганизм, имеющий конкретный ослабленный или усиленный механизм из-за вставки чужеродного гена или усиление или ослабление активности эндогенного гена. В частности, это может быть микроорганизм, включающий полинуклеотид, обладающий промоторной активностью, и целевой белок.
Целевой белок может представлять собой полипептид, обладающий транскетолазной (tkt) активностью. Полинуклеотид, обладающий промоторной активностью, целевой белок, полипептид, обладающий транскетолазной (tkt) активностью, и вектор являются такими, как описано выше.
Микроорганизм может представлять собой микроорганизм рода Corynebacterium и, более конкретно, Corynebacterium glutamicum, но не ограничивается ими.
Микроорганизм может быть микроорганизмом, экспрессирующим транскетолазу, микроорганизмом, экспрессирующим полипептид, обладающий транскетолазной активностью, или микроорганизмом, в который введен полипептид, обладающий транскетолазной активностью, но не ограничиваясь этим.
В настоящем изобретении микроорганизм может включать полинуклеотид, обладающий промоторной активностью, по настоящему изобретению и может конкретно включать полинуклеотид и/или ген, кодирующий целевой белок, который функционально связан с полинуклеотидом. Альтернативно, микроорганизм может включать вектор, содержащий полинуклеотид или ген, кодирующий последовательность, регулирующую экспрессию гена, и целевой белок, но не ограничивается этим. Кроме того, полинуклеотид, ген, кодирующий целевой белок, и вектор могут быть введены в микроорганизм путем трансформации, но не ограничиваются этим. Кроме того, при условии что ген может экспрессироваться, не имеет значения, расположен ли ген, кодирующий полинуклеотид и целевой белок, на хромосоме или вне хромосомы.
Используемый здесь термин «быть экспрессированным/экспрессирующийся» в отношении белка относится к состоянию, при котором целевой белок, например транскетолаза или ее вариант, вводится в микроорганизм, или целевой белок модифицируется для экспрессии в микроорганизме. Когда целевой белок представляет собой белок, присутствующий в микроорганизме, этот термин относится к состоянию, в котором активность белка повышена по сравнению с активностью его эндогенного белка или активностью до его модификации.
В частности, термин «введение белка» означает, что микроорганизм проявляет активность определенного белка, которой микроорганизм изначально не обладал, или микроорганизм проявляет повышенную активность по сравнению с его эндогенной активностью или активностью белка до модификации. Например, это может означать, что в хромосому микроорганизма встраивают полинуклеотид, кодирующий тот или иной белок; или вектор, содержащий полинуклеотид, кодирующий конкретный белок, вводят в микроорганизм и тем самым обеспечивают возможность проявления активности конкретного белка.
Кроме того, термин «повышение активности» означает, что активность конкретного белка, которым обладает микроорганизм, повышается по сравнению с его эндогенной активностью или активностью до его модификации. Термин «эндогенная активность» относится к активности определенного белка, изначально присущего родительскому штамму до модификации, в случае, когда характерная особенность микроорганизма изменена вследствие генетической мутации, вызванной естественным или искусственным фактором.
Для задач настоящего изобретения повышение активности может быть достигнуто за счет использования полинуклеотидной последовательности, обладающей промоторной активностью, по настоящему изобретению в качестве регуляторной последовательности экспрессии целевого белка. Поскольку целевой белок может быть природным или модифицированным белком, как описано выше, регуляторная последовательность экспрессии может представлять собой регуляторную последовательность экспрессии гена, кодирующего вариант белка, или регуляторную последовательность экспрессии гена, кодирующего природный белок на хромосоме.
Кроме того, другие способы повышения активности могут быть использованы в комбинации. Например, в дополнение к использованию полинуклеотида, обладающего промоторной активностью, по настоящему изобретению в качестве регуляторной последовательности экспрессии целевого белка, любой один или более способов, выбранных из группы, состоящей из способа увеличения внутриклеточного числа копий гена, кодирующего целевой белок; способа замены гена, кодирующего природный белок на хромосоме, геном, кодирующим вариант белка; способ дополнительного введения мутации в ген, кодирующий белок, так что активность варианта белка повышается; и способ введения варианта белка в микроорганизм, но способ этим не ограничивается.
Активность целевого белка может быть повышена за счет использования полинуклеотида, обладающего промоторной активностью, по настоящему изобретению в качестве регуляторной последовательности экспрессии целевого белка в микроорганизме.
Например, активность или концентрация соответствующего белка может быть увеличена по меньшей мере на 1%, 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500% и самое большее 1000% или 2000% в зависимости от активности или концентрации белка в штамме микроорганизма дикого типа или немодифицированном, но диапазон увеличения не ограничивается этим.
Используемый здесь термин «немодифицированный микроорганизм» не исключает штамм, содержащий мутацию, которая может встречаться в природе в микроорганизме, и может быть штаммом дикого типа самим по себе, или может включать микроорганизм, не содержащий полинуклеотид, обладающий промоторной активностью, по настоящему изобретению, или микроорганизм, который не трансформирован вектором, содержащим полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
Используемый здесь термин «микроорганизм для получения целевого вещества» включает все микроорганизмы, в которых генетическая модификация происходит естественным или искусственным путем, и это может быть микроорганизм, в котором конкретный механизм ослаблен или усилен за счет вставки чужеродного гена или усиления или инактивации активности эндогенного гена, и может быть микроорганизмом, в котором происходит генетическая модификация для получения желаемого продукта, или активность усилена. Для задач настоящего изобретения микроорганизм для продуцирования целевого вещества может относиться к микроорганизму, способному продуцировать целевое вещество в избытке, по сравнению с микроорганизмом дикого типа или немодифицированным микроорганизмом, включающему полинуклеотид, обладающий промоторной активностью, по настоящему изобретению.
Термин «микроорганизм для продуцирования целевого вещества» может быть использован взаимозаменяемо с терминами «микроорганизм, продуцирующий целевое вещество», «микроорганизм, способный продуцировать целевое вещество», «штамм, продуцирующий целевое вещество», «штамм, способный продуцировать целевое вещество» и т.д.
Целевое вещество может представлять собой аминокислоту, в частности лизин, треонин, O-ацетилгомосерин, изолейцин, триптофан, тирозин или фенилаланин. В более конкретном примере лизин может представлять собой L-лизин, треонин может представлять собой L-треонин, О-ацетилгомосерин может представлять собой О-ацетил-L-гомосерин, изолейцин может представлять собой L-изолейцин, триптофан может представлять собой L-триптофан, тирозин может представлять собой L-тирозин, и фенилаланин может представлять собой L-фенилаланин, но целевое вещество не ограничивается этим.
Для задач настоящего изобретения микроорганизм для продуцирования целевого вещества может представлять собой микроорганизм, у которого повышена способность продуцировать целевое вещество, в частности лизин, треонин, О-ацетилгосерин, изолейцин, триптофан, тирозин или фенилаланин.
При этом микроорганизм для получения целевого вещества может представлять собой микроорганизм дикого типа или рекомбинантный микроорганизм. Рекомбинантный микроорганизм является таким, как описано выше. Микроорганизм может дополнительно включать мутации, такие как усиление путей биосинтеза для повышения способности продуцировать целевые вещества, высвобождение ингибирования по принципу обратной связи или инактивация генов для ослабления путей разложения или путей биосинтеза и т.д., и такие мутации могут, например, быть искусственно вызваны УФ-облучением, но не исключают естественных мутаций.
В частности, микроорганизм для продуцирования целевого вещества может быть мутирован таким образом, чтобы можно было продуцировать целевое вещество. Например, это могут быть те микроорганизмы, в которых микроорганизм, не обладающий способностью продуцировать целевое вещество, мутирован, чтобы иметь способность продуцировать целевое вещество, или это может быть микроорганизм с повышенной способностью к продуцированию. В качестве примера это может быть микроорганизм, в который вводят белок, участвующий в пути биосинтеза, или его вариант, чтобы целевое вещество могло продуцироваться в микроорганизме дикого типа (КР 10-2011994, КР 10-1947959).
Микроорганизм по настоящему изобретению может обладать способностью продуцировать целевое вещество по меньшей мере на 1%, 5%, 10%, 13%, 15%, 16%, 17%, 18%, 19%, 20% или 25% или более по сравнению с микроорганизмом, не включающим полинуклеотид, обладающий промоторной активностью.
В еще одном аспекте настоящего изобретения предложен способ получения целевого вещества, включающий культивирование клетки-хозяина в среде и извлечение целевого вещества из среды.
Клетка-хозяин и целевое вещество являются такими, как описано выше.
Используемый здесь термин «культивирование» означает, что микроорганизмы выращивают в надлежащим образом искусственно контролируемых условиях окружающей среды. В настоящем изобретении способ получения целевого вещества с использованием микроорганизма, включающего полинуклеотид, может быть осуществлен способом, хорошо известным в данной области техники. В частности, культивирование можно проводить непрерывно в периодическом процессе, периодическом процессе с подпиткой или периодическом процессе с повторной подпиткой, но не ограничиваясь этим. Среда, используемая для культивирования, должна соответствовать требованиям конкретного штамма в соответствующем методе. Питательные среды для штаммов, принадлежащих к роду Corynebacterium, уже известны (например, Manual of Methods for General Bacteriology. American Society for Bacteriology. Washington D.C., USA, 1981).
Примеры источников углерода могут включать сахара и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал, целлюлоза и т.д.; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло, кокосовое масло и т.д.; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота, линолевая кислота и т.д.; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Эти источники углерода могут быть использованы по отдельности или в комбинации, но не ограничиваются этим.
Примеры источников азота могут включать пептон, дрожжевой экстракт, мясную подливку, солодовый экстракт, кукурузный экстракт (CSL), соевую муку и мочевину; или неорганические источники азота, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота также могут быть использованы по отдельности или в комбинации, но не ограничиваются этим.
Примеры источников фосфора могут включать дигидрофосфат калия, дигидрофосфат калия или соответствующие натрийсодержащие соли. Кроме того, среда может содержать соли металлов, такие как сульфат магния или сульфат железа, которые необходимы для роста. Наконец, можно использовать необходимые для роста вещества, такие как аминокислоты и витамины. Кроме того, в среде также могут содержаться соответствующие предшественники. Вышеупомянутые вещества могут быть добавлены к культуре периодическим или непрерывным способом соответствующим методом.
Значение рН культуры можно регулировать соответствующим образом с помощью щелочного соединения, такого как гидроксид натрия, гидроксид калия и аммиак, или кислотного соединения, такого как фосфорная кислота и серная кислота. Кроме того, для предотвращения пенообразования может быть добавлен пеногаситель, такой как полигликоле вый эфир жирной кислоты. Для поддержания аэробного состояния культуры в культуру можно вводить кислород или кислородсодержащий газ (например воздух).
Температура культуры (среды) обычно может составлять от 20°С до 45°С, предпочтительно от 25°С до 40°С, и культивирование может продолжаться до тех пор, пока не будет получено желаемое количество целевого вещества, и может быть, в частности, осуществлено в течение от 10 до 160 часов.
Выделение целевого вещества из культуры (среды) может быть проведено путем разделения и извлечения обычным способом, известным в данной области техники. В качестве метода разделения могут быть использованы такие методы, как центрифугирование, фильтрация, хроматография и кристаллизация. Например, надосадочную жидкость, полученную путем удаления биомассы посредством центрифугирования культуры при низкой скорости, можно отделить с помощью ионообменной хроматографии, но не ограничиваясь этим.
Стадия выделения может дополнительно включать процесс очистки.
Еще один аспект настоящего изобретения включает применение полинуклеотида, в котором нуклеотиды по положениям 218, 219, 220, 221, 222, 225 и 227 в полинуклеотидной последовательности SEQ ID NO: 1 заменены на другие нуклеотиды, в качестве промотора.
Полинуклеотид является таким, как описано выше.
Способ осуществления изобретения
Настоящее изобретение будет подробно описано посредством примеров. Однако специалистам в данной области техники должно быть понятно, что эти примеры даны только в иллюстративных целях и не предназначены для ограничения ими объема изобретения.
Пример 1. Подтверждение активности индукции экспрессии целевого гена нового промотора
Пример 1-1. Конструирование библиотеки вариантов промотора tkt с использованием случайного мутагенеза
Сначала, согласно данным GenBank Национального института здравоохранения США (NIH GenBank) была получена нуклеотидная последовательность (SEQ ID NO: 1), содержащая промоторную область гена tkt (регистрационный номер NCBI NCgl1512) Corynebacterium glutamicum дикого типа АТСС 13032. Промотор (Pntkt) гена tkt, состоящий из нуклеотидной последовательности SEQ ID NO: 1, использовали в качестве матрицы с использованием набора для случайного мутагенеза Diversify® PCR (takara), и праймеры SEQ ID NO: 3 и 4 использовали для индуцирования случайного мутагенеза в соответствии с руководством производителя с получением таким образом ПЦР-продукта варианта промотора tkt (Pmtkt), имеющего другую последовательность. Дополнительно проводили ПЦР на ORF (открытая рамка считывания) гена GFP на основе вектора pGFPuv (clontech, США) в качестве матрицы с использованием праймеров SEQ ID NO: 5 и 6. ДНК-полимеразу Solg™ Pfu-X использовали в качестве полимеразы, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут, с получением фрагмента гена, содержащего ORF GFP.
ПЦР-продукт варианта промотора tkt (Pmtkt), амплифицированный продукт, и GFP смешивали с pCES208 (J. Microbiol. Biotechnol. 18:639-647, 2008), челночным вектором Е. coli-Corynebacterium, расщепленным с помощью фермента рестрикции BamHI/Sall, и клонировали с использованием метода сборки Гибсона (DG Gibson et al, NATURE METHODS, Vol.6 No. 5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix) для получения рекомбинантных плазмид, и каждый вектор получил название от pCES208_Pmltkt gfp до pCES208 Pm50tkt gfp.Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
В качестве контроля для подтверждения активности библиотеки вариантов промотора tkt использовали рекомбинантный вектор, в котором промотор (SEQ ID NO: 1) гена tkt дикого типа и GFP связаны. Промоторный ген (Pntkt) гена tkt дикого типа был получен на основе Corynebacterium glutamicum АТСС 13032 дикого типа в качестве матрицы с использованием праймеров SEQ ID NO: 3 и 4. Дополнительно была проведена ПЦР на ORF (открытая рамка считывания) гена GFP на основе вектора pGFPuv в качестве матрицы с использованием праймеров SEQ ID NO: 5 и 6. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд, полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут с получением фрагмента гена, содержащего ORF GFP.
ПЦР-продукт варианта промотора tkt дикого типа (Pntkt), амплифицированный продукт, и GFP смешивали с pCES208, расщепленной ферментом рестрикции BamHI/Sall, и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pCES208_Pntkt_gfp. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Пример 1-2. Конструирование трансформированных штаммов
Вектор pCES208 и библиотеку рекомбинантных векторов, включающую от pCES208_Pmltkt_gfp до pCES208_Pm50tkt_gfp и pCES208_Pntkt_gfp, полученные в Примере 1-1, трансформировали в Corynebacterium glutamicum АТСС13032 посредством электропорации (Appl. Microbiol. Biothcenol. (1999)), и затем трансформированные штаммы были получены в селекционной среде, содержащей 25 мг/л канамицина, и полученные штаммы назвали АТСС 1303 2/pCES208, от ATCC13032/pCES208_Pmltkt_gfp до ATCC13032/pCES208_Pm50tkt_gfp и ATCC13032/pCES208_Pntkt_gfp, соответственно.
Пример 1-3. Отбор варианта промотора tkt
Для подтверждения активности варианта промотора tkt трансформированные штаммы, полученные в Примере 1-2, культивировали следующим образом и измеряли активность GFP.
В частности, каждый из трансформированных штаммов Corynebacterium glutamicum инокулировали в колбу, содержащую 25 мл питательной среды (глюкоза (20 г), сульфат аммония (5 г), дрожжевой экстракт (5 г), мочевина (1,5 г), KH2PO4 (4 г), K2HPO4 (8 г), MgSO4⋅7H2O (0,5 г), биотин (150 мкг), тиамин-HCl (1,5 мг), пантотенат кальция (3 мг) и никотинамид (3 мг) (в расчете на 1 л дистиллированной воды), рН 7,2) и культивировали в инкубаторе при 30°С в течение 20 часов при встряхивании. Бактериальные клетки извлекали центрифугированием (5000 об/мин, 15 минут), дважды промывали буфером 50 мМ Трис-HCl (рН 8,0) и суспендировали в том же буфере. К суспензии добавляли стеклянные шарики в количестве 1,25 г/1,5 мл и разрушали бактериальные клетки с помощью мешалки для шариков в течение 6 минут. Затем полученный продукт подвергали центрифугированию (15000 об/мин, 20 минут) для извлечения из него надосадочной жидкости и количественно определяли концентрации белков по методу Бредфорда. Для равного количества экстрактов бактериальных клеток проводили облучение возбуждающим светом с длиной волны 488 нм в соответствии с методом, предложенным Laure Gory et al. (FEMSMicrobiology Letters 194, 127-133, 2001), и испускаемый свет при 511 нм измеряли с использованием спектрофотометра LS-50B (Perkin-Elmer), и таким образом измеряли уровень экспрессии гена GFP. В результате, по сравнению с уровнем экспрессии гена GFP контрольного штамма ATCC13032/pCES208_Pntkt_gfp был отобран один штамм с наиболее высоким уровнем экспрессии гена GFP (Таблица 1).
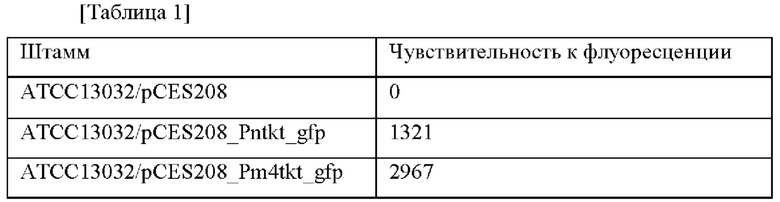
Как показано в Таблице 1, было подтверждено, что промотор Pm4tkt проявлял промоторную активность в Corynebacterium glutamicum и проявлял более высокую чувствительность к флуоресценции, чем промотор tkt дикого типа.
Для подтверждения мутации, введенной в отобранный выше промотор Pm4tkt, проводили ПЦР с использованием праймеров SEQ ID NO: 7 и 8, а затем проводили анализ последовательности. Промотор Pm4tkt сравнивали с SEQ ID NO: 1, которая представляет собой последовательность промотора tkt дикого типа, для подтверждения последовательности варианта промотора tkt, и эта последовательность показана в Таблице 2 ниже.

Пример 2. Конструирование вектора для введения варианта промотора Pm4tkt
Пример 2-1. Конструирование вектора для введения варианта промотора Pm4tkt в Corynebacterium glutamicum АТСС13032
Для конструирования вектора для введения варианта промотора Pm4tkt в Corynebacterium glutamicum АТСС 13032 были получены область выше промотора tkt и нижележащая область, включающая часть ORF tkt, на основе хромосомы Corynebacterium glutamicum АТСС 13032 дикого типа в качестве матрицы с использованием пар праймеров SEQ ID NO: 9 и 10 и SEQ ID NO: 13 и 14. Кроме того, ПЦР-продукты, соответствующие варианту промотора, были получены на основе вектора pCES208_Pm4tkt_gfp в качестве матрицы с использованием SEQ ID NO: 11 и 12. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут с получением таким образом ПЦР-продуктов. Три амплифицированных продукта смешивали с вектором pDCM2 (публикация Корейской патентной заявки №10-2020-0136813), предварительно расщепленным ферментом рестрикции SmaI, и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-Pm4tkt_tkt. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Пример 2-2. Конструирование вектора для введения Pm4tkt_tkt в Corynebacterium glutamicum АТСС13869
Был получен вектор для введения одной копии гена tkt, имеющего вариант промотора Pm4tkt (SEQ ID NO: 2), в Corynebacterium glutamicum АТСС 13869.
Был получен вектор для вставки целевого гена с одновременным удалением положения aroP (гамм а-амино масляная пермеаза, номер доступа GenBank WP_060564335.1) Corynebacterium glutamicum АТСС 13869 в качестве положения, в котором на хромосоме происходит гомологичная рекомбинация.
В частности, фрагменты гена в областях выше и ниже aroP, где происходит гомологичная рекомбинация на хромосомах, были получены с помощью ПЦР на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869 с использованием пар праймеров SEQ ID NO: 15 и 16 и SEQ ID NO: 17 и 18. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут.
Области выше и ниже амплифицированного aroP и вектор pDCM2 для хромосомной трансформации, расщепленный ферментом рестрикции SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-ΔaroP. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
ПЦР проводили с использованием праймеров SEQ ID NO: 19 и 20 на основе pCES208 Pm4tktgfp в качестве матрицы и посредством ПЦР получали фрагмент гена, имеющий вариант промотора Pm4tkt. Дополнительно ORF гена tkt получали с использованием праймеров SEQ ID NO: 21 и 22 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут.
Амплифицированный Pm4tkt, ген tkt и вектор pDCM2-ΔaroP для хромосомной трансформации, расщепленные ферментом рестрикции Seal, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-ΔaroP::Pm4tkt_tkt. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Дополнительно, для сравнения с промотором Pm4tkt получали вектор для добавления одной копии гена tkt в виде собственного промотора (Pntkt) гена tkt. В частности, проводили ПЦР с использованием праймеров SEQ ID NO: 19 и 22 на основе хромосомной ДНК Corynebacterium glutamicum АТСС 13869 в качестве матрицы с получением гена tkt, имеющего собственный промотор гена tkt. Полученный таким образом ген и полученный выше вектор для хромосомной трансформации pDCM2-ΔaroP, расщепленный ферментом рестрикции Seal, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-ΔaroP::Pntkt_tkt. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Пример 2-3. Конструирование вектора для введения варианта промотора Pm4tkt в Corynebacterium glutamicum АТСС13869
Для замены нативного промотора гена tkt, ранее введенного в положение aroP, на вариант промотора Pm4tkt проводили ПЦР на основе вектора pDCM2-ΔaroP::Pm4tkt_tkt, полученного в Примере 2-2, в качестве матрицы с использованием SEQ ID NO: 23 и 24, и был получен фрагмент гена, включающий последовательность Pm4tkt. Полученный таким образом фрагмент гена смешивали с вектором pDCM2, расщепленным ферментом рестрикции SmaI, и клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-ΔPn::Pm4tkt_tkt. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Пример 3. Оценка способности продуцирования целевых продуктов
3-1. Оценка лизин-продуцирующей способности
3-1-1. Конструирование штамма, продуцирующего L-лизин, в который введен вариант промотора tkt
Штамм, трансформированный вариантом промотора tkt, получали с использованием вектора pDCM2-Pm4tkt_tkt, полученного в Примере 2-1. С этой целью вектор трансформировали в Corynebacterium glutamicum CJ3P (US 9556463 В2), который представляет собой штамм, продуцирующий L-лизин, для введения последовательности варианта промотора tkt в хромосому. Штамм CJ3P представляет собой штамм Corynebacterium glutamicum, обладающий способностью продуцировать L-лизин благодаря введению трех типов мутаций (pyc(P458S), hom(V59A) и lysC(T3111)) в штамм дикого типа на основе ранее известных методик.
В частности, вектор, полученный в Примере 2-1, трансформировали в штамм CJ3P посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера штамм, в который был введен вариант промотора tkt с помощью фрагмента ДНК, вставленного в хромосому, был отобран путем проведения ПЦР с использованием праймеров SEQ ID NO: 7 и 8 и секвенирования, и отобранный таким образом штамм был назван Corynebacterium glutamicum CJ3P::Pm4tkt_tkt(CM03-1661) и был депонирован в Корейском центре культур микроорганизмов в соответствии с Будапештским договором 5 апреля 2021 г. под номером доступа KCCM12971P.
3-1-2. Оценка способности штамма с введенным вариантом промотора tkt продуцировать L-лизин
Чтобы оценить способность штамма Corynebacterium glutamicum CJ3P, используемого в качестве исходного штамма, и штамма Corynebacterium glutamicum CJ3P::Pm4tkt_tkt, полученного в Примере 3-1-1, продуцировать L-лизин, эти штаммы культивировали следующим образом и анализировали.
Сначала каждый штамм высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при 30°С в течение 20 часов при 200 об/мин при встряхивании. Затем 1 мл посевного культурального раствора высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 24 мл продукционной среды, и культивировали при 32°С в течение 48 часов при 200 об/мин при встряхивании. Составы питательной среды, среды для высевания и продукционной среды показаны ниже.
<Среда для высевания (рН 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4⋅7H2O 0,5 г, биотин 100 мкг, тиамин-HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (в расчете на 1 л дистиллированной воды)
<Продукционная среда (рН 7,0)>
Глюкоза 45 г, соевый белок 10 г, патока 10 г, (NH4)2SO4 15 г, KH2PO4 0,55 г, MgSO4⋅7H2O 0,6 г, FeSO4⋅7H2O 9 мг, MnSO4⋅5H2O 9 мг, биотин 0,9 мг, тиамин-HCl 4,5 мг, СаСО3 30 г, пантотенат кальция 4,5 мг, никотинамид 30 мг, ZnSO4 0,45 мг, CuSO4 0,45 мг (в расчете на 1 л дистиллированной воды).
После завершения культивирования продукцию L-лизина измеряли с помощью ВЭЖХ. Концентрация L-лизина и степень увеличения концентрации в культуральной среде для штаммов Corynebacterium glutamicum CJ3P и CJ3P::Pm4tkt_tkt показаны в Таблице 3 ниже.
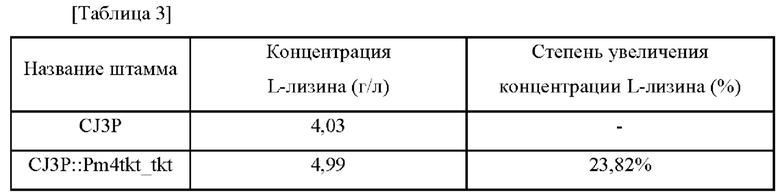
Как показано в Таблице 3, было подтверждено, что концентрация L-лизина в штамме CJ3P::Pm4tkt_tkt, в который был введен вариант промотора tkt, увеличилась на 23,82% по сравнению с исходным штаммом CJ3P.
Пример 3-2. Оценка способности продуцировать треонин 3-2-1. Конструирование штамма, продуцирующего L-треонин Для конструирования штамма, трансформированного вариантом промотора tkt с использованием вектора pDCM2-Pm4tkt_tkt, полученного в Примере 2-1, сначала получали штамм, продуцирующий L-треонин, в который был введен мутант lysC (L377K) (патент Кореи №10-2011994) и мутант horn (R398Q) (патент Кореи №10-1947959), на основе штамма Corynebacterium glutamicum АТСС 13032.
В частности, для получения штамма, продуцирующего L-треонин, сначала получали вектор для введения lysC (L377K). Для получения вектора проводили ПЦР с использованием пар праймеров SEQ ID NO: 25 и 26 и SEQ ID NO: 27 и 28 на основе хромосомы штамма Corynebacterium glutamicum АТСС 13032 дикого типа в качестве матрицы. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут. Амплифицированный продукт и вектор pDCM2 для хромосомной трансформации, расщепленный ферментом рестрикции SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-lysC(L377K). Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный таким образом вектор pDCM2-lysC(L377K) вводили в штамм Corynebacterium glutamicum АТСС 13032 посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера посредством проведения ПЦР с использованием праймеров SEQ ID NO: 29 и 30 и секвенирования был отобран штамм, в котором нуклеотидная мутация была введена в ген lysC с помощью встроенного в хромосому фрагмента ДНК, и отобранный штамм назвали ATCC13032::lysC(L377K).
Кроме того, для получения вектора для введения hom(R398Q) проводили ПЦР с использованием пар праймеров SEQ ID NO: 31 и 32 и SEQ ID NO: 33 и 34 на основе хромосомы штамма Corynebacterium glutamicum АТСС 13032 в качестве матрицы. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут. Амплифицированный продукт и вектор pDCM2 для хромосомной трансформации, расщепленный ферментом рестрикции SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-hom(R398Q). Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный таким образом вектор pDCM2-hom(R398Q) вводили в штамм Corynebacterium glutamicum ATCC13032::lysC(L377K) посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера посредством проведения ПЦР с использованием праймеров SEQ ID NO: 35 и 36 и секвенирования был отобран штамм, в котором нуклеотидная мутация была введена в ген horn с помощью встроенного в хромосому фрагмента ДНК, и отобранный штамм получил название Corynebacterium glutamicum ATCC13032::lysC(L377K)-hom(R398Q).
3-2-2. Конструирование L-треонин-продуцирующего штамма с введенным вариантом промотора tkt
Вектор pDCM2-Pm4tkt_tkt, полученный в Примере 2-1, вводили в штамм Corynebacterium glutamicum ATCC13032::lysC(L377K)-hom(R398Q) посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера штамм, в который вариант промотора tkt был введен с помощью фрагмента ДНК, вставленного в хромосому, был отобран путем проведения ПЦР с использованием праймеров SEQ ID NO: 7 и 8 и секвенирования, и отобранный штамм назвали Corynebacterium glutamicum АТСС13032.::lysC(L377K)-hom(R398Q)-Pm4tkt_tkt.
3-2-3. Оценка способности штамма с введенным вариантом промотора tkt продуцировать L-треонин
Для оценки способности продуцировать L-треонин у Corynebacterium glutamicum ATCC13032::lysC(L377K)-hom(R398Q), используемого в качестве исходного штамма, и ATCC13032::lysC(L377K)-hom(R398Q)-Pm4tkt_tkt, полученного в Примере 3-2-2, эти штаммы культивировали следующим образом и анализировали.
Сначала каждый штамм высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при 30°С в течение 20 часов при 200 об/мин при встряхивании. Затем 1 мл посевного культурального раствора высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 24 мл продукционной среды, и культивировали при 32°С в течение 48 часов при 200 об/мин при встряхивании. Составы питательной среды, среды для высевания и продукционной среды показаны ниже.
<Среда для высевания (рН 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 8 г, MgSO4-7H2O 0,5 г, биотин 100 мкг, тиамин-HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (в расчете на 1 л дистиллированной воды)
<Продукционная среда (рН 7,0)>
Глюкоза 45 г, соевый белок 10 г, патока 10 г, (NH4)2SO4 15 г, KH2PO4 0,55 г, MgSO4⋅7H2O 0,6 г, FeSO4⋅7H2O 9 мг, MnSO4⋅5H2O 9 мг, биотин 0,9 мг, тиамин-HCl 4,5 мг, СаСО3 30 г, пантотенат кальция 4,5 мг, никотинамид 30 мг, ZnSO4 0,45 мг, CuSO4 0,45 мг (в расчете на 1 л дистиллированной воды).
После завершения культивирования продукцию L-треонина измеряли с помощью ВЭЖХ. Концентрация L-треонина и степень увеличения концентрации в культуральной среде для штаммов Corynebacterium glutamicum ATCC13032::lysC(L377K)-hom(R398Q) и ATCC13032::lysC(L377K)-hom(R398Q)-Pm4tkt_tkt показаны в Таблице 4 ниже.


Как показано в Таблице 4, концентрация L-треонина увеличилась на 41,30% в штамме ATCC13032::lysC(L377K)-hom(R398Q)-Pm4tkt_tkt, в который был введен вариант промотора tkt, по сравнению с исходным штаммом ATCC13032::lysC(L377K)-hom(R398Q).
Пример 3-3. Оценка способности продуцировать 0-ацетилгомосерин 3-3-1. Конструирование штамма с введенным вариантом промотора tkt, продуцирующего 0-ацетилгомосерин
Вектор pDCM2-Pm4tkt_tkt, полученный в Примере 2-1, вводили в штамм Corynebacterium glutamicum АТСС 13032 дикого типа посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера штамм, в который вариант промотора tkt был введен с помощью фрагмента ДНК, вставленного в хромосому, был отобран путем проведения ПЦР с использованием праймеров SEQ ID NO: 7 и 8 и секвенирования, и отобранный штамм назвали Corynebacterium glutamicum ATCC13032::Pm4tkt_tkt.
3-3-2. Оценка способности штамма с введенным вариантом промотора tkt продуцировать 0-ацетилгомосерин
Для оценки способности продуцировать О-ацетилгомосерин у Corynebacterium glutamicum АТСС 13032, используемого в качестве исходного штамма, и ATCC13032::Pm4tkt_tkt, полученного в Примере 3-3-1, эти штаммы культивировали следующим образом и анализировали.
Вышеуказанные штаммы инокулировали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при встряхивании при 200 об/мин при 33°С в течение 20 часов.
<Продукционная среда (рН 7,2)>
Глюкоза 30 г, KH2PO4 2 г, мочевина 3 г, (NH4)2SO4 40 г, пептон 2,5 г, CSL (Corn steep liquor, Sigma) 5 г (10 мл), MgSO4 7H2O 0,5 г, СаСО3 20 г (в расчете на 1 л дистиллированной воды).
После завершения культивирования способность продуцировать О-ацетилгомосерина измеряли с помощью ВЭЖХ. Концентрация O-ацетилгомосерина и степень увеличения концентрации в культуральной среде для штаммов Corynebacterium glutamicum Corynebacterium glutamicum ATCC13032 и ATCC13032::Pm4tkt_tkt показаны в Таблице 5 ниже.
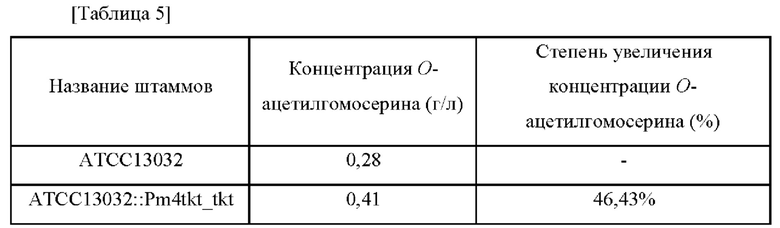
Как показано в Таблице 5, концентрация (3-ацетилгомосерина увеличилась на 46,43% в штамме ATCC13032::Pm4tkt_tkt, в который был введен вариант промотора tkt, по сравнению с исходным штаммом АТСС 13032 дикого типа.
Пример 3-4. Оценка способности продуцировать изолейцин
3-4-1. Конструирование штамма с введенным вариантом промотора tkt, продуцирующего L-изолейцин
Для получения штамма, трансформированного вариантом промотора tkt с использованием вектора pDCM2-Pm4tkt_tkt, полученного в Примере 2-1, сначала этот вектор трансформировали в штамм Corynebacterium glutamicum CJP1 (патент Кореи №10-1996769), и последовательность варианта промотора tkt была введена в его хромосому. После этого вектор, содержащий ген ilvA (V323A) (Appl. Enviro. Microbiol., Dec. 1996, pp.4345-4351), в котором валин, являющийся 323-ей аминокислотой гена ilvA, кодирующего ранее известную L-треониндегидратазу, мутирован в аланин, дополнительно вводили для получения таким образом штамма, продуцирующего L-изолейцин (патент Кореи №10-1996769).
В частности, вектор pDCM2-Pm4tkt_tkt, полученный в Примере 2-1, вводили в штамм CJP1 посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера штамм, в котором вариант промотора tkt был введен с помощью фрагмента ДНК, вставленного в хромосому, был отобран путем проведения ПЦР с использованием праймеров SEQ ID NO: 7 и 8 и секвенирования, и отобранный штамм назвали Corynebacterium glutamicum CJP1::Pm4tkt_tkt.
Каждый вектор pECCG117-ilvA(V323A) вводили в полученный выше штамм CJP1::Pm4tkt_tkt и штамм CJP1, а затем трансформированные штаммы получали в селекционной среде, содержащей 25 мг/л канамицина. Отобранные штаммы назвали Corynebacterium glutamicum CJP1/pECCG117-ilvA(V323A) и
CJP1::Pm4tkt_tkt/pECCG117-ilvA(V323A), соответственно.
3-4-2. Оценка способности штамма с введенным вариантом промотора tkt продуцировать L-изолейцин
Для оценки способности Corynebacterium glutamicum CJP1/pECCG117-ilvA(V323A), используемого в качестве исходного штамма, и штамма CJP1::Pm4tkt_tkt/pECCG117-ilvA(V323A), продуцировать L-изолейцин эти штаммы культивировали следующим образом и анализировали.
Сначала каждый штамм высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при 30°С в течение 20 часов при 200 об/мин при встряхивании. Затем 1 мл посевного культурального раствора высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 24 мл продукционной среды, и культивировали при 32°С в течение 48 часов при 200 об/мин при встряхивании. Составы питательной среды, среды для высевания и продукционной среды показаны ниже.
<Среда для высевания (рН 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, К2НРО4 8 г, MgSO4⋅7H2O 0,5 г, биотин 100 мкг, тиамин-НС1 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (в расчете на 1 л дистиллированной воды).
<Продукционная среда (рН 7,0)>
Глюкоза 45 г, соевый белок 10 г, патока 10 г, (NH4)2SO4 15 г, KH2PO4 0,55 г, MgSO4⋅7H2O 0,6 г, FeSO4⋅7H2O 9 мг, MnSO4⋅5H2O 9 мг, биотин 0,9 мг, тиамин-HCl 4,5 мг, СаСО3 30 г, пантотенат кальция 4,5 мг, никотинамид 30 мг, ZnSO4 0,45 мг, CuSO4 0,45 мг (в расчете на 1 л дистиллированной воды).
После завершения культивирования продукцию L-изолейцина измеряли с помощью ВЭЖХ. Концентрация L-изолейцина и степень увеличения концентрации в культуральной среде для штаммов Corynebacterium glutamicum CJP1/pECCG117-ilvA(V323A) и CJP1::Pm4tkt_tkt/pECCG117-ilvA(V323A) показаны в Таблице 6 ниже.
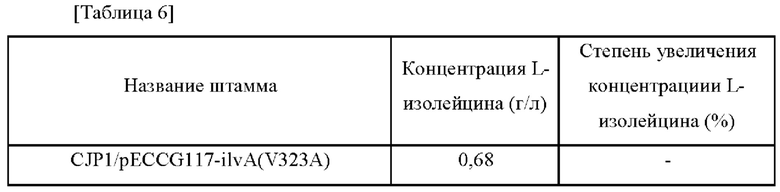

Как показано в Таблице 6, было подтверждено, что концентрация L-изолейцина увеличилась на 57,35% в штамме CJP1::Pm4tkt_tkt/pECCG117-ilvA(V323A), в который был введен вариант промотора tkt, по сравнению со штаммом CJP1/pECCG117-ilvA(V323A).
Пример 3-5. Оценка способности продуцировать триптофан 3-5-1. Конструирование штамма, продуцирующего L-триптофан
Чтобы подтвердить эффект варианта промотора Pm4tkt, сначала был получен штамм, продуцирующий L-триптофан, на основе штамма Corynebacterium glutamicum АТСС13869 дикого типа.
В частности, экспрессию оперона гена биосинтеза L-триптофана усиливали за счет усиления промотора. Кроме того, чтобы снять ингибирование белка TrpE по принципу обратной связи, серии, 38-ая аминокислота trpE, была заменена на аргинин (JOURNAL OF BACTERIOLOGY, Nov. 1987, pp.5330-5332).
Для генетической манипуляции сначала были получены области выше и ниже промотора trpE, где на хромосоме происходит гомологичная рекомбинация. В частности, фрагмент гена в области выше промотора trpE был получен с использованием праймеров SEQ ID NO: 37 и 38, а фрагмент гена в области ниже по ходу транскрипции был получен с использованием праймеров SEQ ID NO: 41 и 42 на основании хромосомной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы с помощью ПЦР. Кроме того, проводили ПЦР с использованием праймеров SEQ ID NO: 39 и 40 на основе синтезированного промотора SPL7 (патент Кореи №10-1783170) и последовательности trpE(S38R) (SEQ ID NO: 43) в качестве матриц. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 58°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут.
Три амплифицированных продукта и вектор pDCM2 для хромосомной трансформации, расщепленный ферментом рестрикции SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-PSPL7 trpE(S38R). Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный выше вектор pDCM2-PSPL7_trpE(S38R) вводили в штамм Corynebacterium glutamicum АТСС13869 дикого типа посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера был получен штамм, в котором промотор trpE был заменен на более сильный промотор SPL7 на хромосоме, а серии, являющийся 38-ой аминокислотой ТгрЕ, заменен на аргинин. Соответствующая генетическая манипуляция была подтверждена с помощью секвенирования генома и ПЦР с использованием SEQ ID NO: 44 и 45, способных амплифицировать области выше и ниже областей гомологичной рекомбинации, в которые был вставлен ген, и полученный продукт назвали Corynebacterium glutamicum ATCC13869::PSPL7 trpE(С38Р).
3-5-2. Конструирование штамма с введенной одной копией гена tkt, продуцирующего L-триптофан
Продуцирование триптофана происходит посредством метаболического пути ароматических аминокислот, и этот метаболический путь начинается с реакции конденсации между фосфоенолпируватом и эритрозо-4-фосфатом. Соответственно, достаточный запас этих двух предшественников необходим для усиления продукции триптофана, а сверхэкспрессия гена tkt необходима для достаточного запаса эритрозо-4-фосфата, который, как известно, является относительно дефицитным. Таким образом, штамм, продуцирующий L-триптофан, был получен с использованием pDCM2-ΔaroP, pDCM2-ΔaroP::Pm4tkt_tkt и pDCM2-ΔaroP::Pntkt_tkt, полученных в Примере 2-2.
Вышеупомянутые три вектора вводили в штамм Corynebacterium glutamicum ATCC13869::PSPL7 trpE(S38R), полученный в Примере 3-5-1, и затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера были получены штаммы, в которых aroP делетирован или ген Pntkt_tkt и ген Pm4tkt_tkt вставлены в положение aroP на хромосоме. Соответствующая генетическая манипуляция была подтверждена секвенированием генома и ПЦР с использованием SEQ ID NO: 46 и 47, а полученные таким образом штаммы назвали ATCC13869::PSPL7 trpE(S38R)ΔaroP, АТСС13869::PSPL7_trpE(S38R)ΔaroP::Pntkt_tkt и ATCC13869::PSPL7_trpE(S38R)ΔaroP::Pm4tkt_tkt, соответственно.
3-5-3. Оценка способности штамма с введенной одной копией гена tkt продуцировать L-триптофан
Для оценки способности продуцировать L-триптофан у Corynebacterium glutamicum ATCC13869::PSPL7_trpE(S38R), используемого в качестве исходного штамма, и ATCC13869::PSPL7 trpE(S38R)ΔaroP, АТСС13869::PSPL7JrpE(S38R)ΔaroP::Pntkt_tkt и ATCC13869::PSPL7_trpE(S38R)ΔaroP::Pm4tkt_tkt, полученных в Примере 3-5-2, эти штаммы культивировали следующим образом и анализировали.
Сначала каждый штамм высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при 30°С в течение 20 часов при 200 об/мин при встряхивании. Затем 1 мл посевного культурального раствора высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 24 мл продукционной среды, и культивировали при 32°С в течение 48 часов при 200 об/мин при встряхивании. Составы питательной среды, среды для высевания и продукционной среды показаны ниже.
<Среда для высевания (рН 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4⋅7H2O 0,5 г, биотин 100 мкг, тиамин-HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (в расчете на 1 л дистиллированной воды)
<Продукционная среда (рН 7,0)>
Глюкоза 30 г, (NH4)2SO4 15 г, MgSO4⋅7H2O 1,2 г, KH2PO4 1 г, дрожжевой экстракт 5 г, биотин 900 мкг, тиамин-HCl 4500 мкг, пантотенат кальция 4500 мкг, СаСО3 30 г (в расчете на 1 л дистиллированной воды).
После завершения культивирования продукцию L-триптофана измеряли с помощью ВЭЖХ. Концентрация L-триптофана и степень увеличения концентрации в культуральной среде для штаммов Corynebacterium glutamicum ATCC13869::PSPL7 trpE(S38R), ATCC13869::PSPL7 JrpE(S38R)ΔaroP,
ATCC13869::PSPL7_trpE(S38R)ΔaroP::Pntkt_tkt и ATCC13869::PSPL7_trpE(S38R)ΔaroP::Pm4tkt_tkt показаны в Таблице 7 ниже.
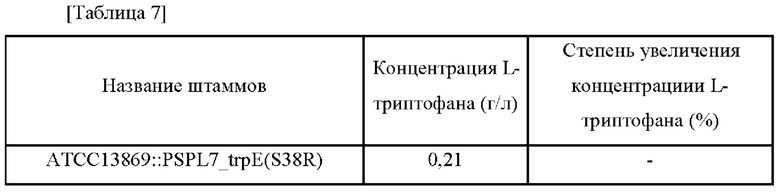
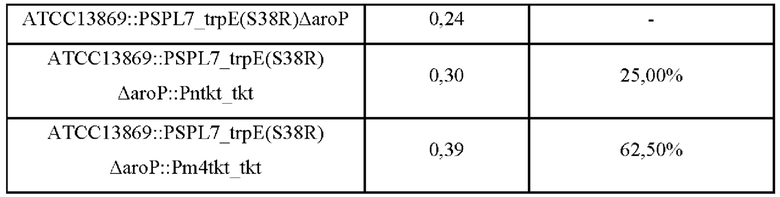
Как показано в Таблице 7, было подтверждено, что концентрация L-триптофана увеличилась на 62,50% в штамме ATCC13869::PSPL71E(S38R)ΔaroP::Pm4tkt_tkt, в который была введена одна копия гена Pm4tkt_tkt, по сравнению со штаммом ATCC13869::PSPL7_trpE(S38R)ΔaroP в качестве контроля.
Пример 3-6. Оценка способности продуцировать тирозин
3-6-1. Конструирование штамма, продуцирующего L-тирозин
Чтобы подтвердить эффект варианта промотора Pm4tkt, трансформацию проводили с использованием штамма, продуцирующего L-тирозин, СМ06-0010 (патент Кореи №10-2134418, депонирован в Корейском центре культур микроорганизмов под номером доступа KCCM12487P) в качестве исходного штамма.
В частности, вектор pDCM2-ΔPn::Pm4tkttkt, полученный в Примере 2-3, вводили в штамм СМ06-0010 посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера был получен штамм, в котором нативный промотор tkt (Pntkt) был заменен на вариант промотора tkt с помощью фрагмента ДНК, вставленного в хромосому. Соответствующая генетическая манипуляция была подтверждена с помощью секвенирования генома и ПЦР с использованием SEQ ID NO: 48 и 49, и полученный таким образом штамм назвали CM06-0010ΔPn::Pm4tkt_tkt.
3-6-2. Оценка способности штамма с введенным вариантом промотора tkt продуцировать L-тирозин
Для оценки способности Corynebacterium glutamicum СМ06-0010, используемого в качестве исходного штамма, и CM06-0010ΔPn::Pm4tkt_tkt, полученного в Примере 3-6-1, продуцировать L-тирозин, эти штаммы культивировали следующим образом и анализировали.
Сначала каждый штамм высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при 30°С в течение 20 часов при 200 об/мин при встряхивании. Затем 1 мл посевного культурального раствора высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 24 мл продукционной среды, и культивировали при 32°С в течение 48 часов при 200 об/мин при встряхивании. Составы питательной среды, среды для высевания и продукционной среды показаны ниже.
<Среда для высевания (рН 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4⋅7H2O 0,5 г, биотин 100 мкг, тиамин-НС1 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (в расчете на 1 л дистиллированной воды).
<Продукционная среда (рН 7,0)>
Глюкоза 30 г, (NH4)2SO4 15 г, MgSO4⋅7H2O 1,2 г, KH2PO4 1 г, дрожжевой экстракт 5 г, биотин 900 мкг, тиамин-HCl 4500 мкг, пантотенат кальция 4500 мкг, СаСО3 30 г (в расчете на 1 л дистиллированной воды).
После завершения культивирования продукцию L-тирозина измеряли с помощью ВЭЖХ. Концентрация L-тирозина и степень увеличения концентрации в культуральной среде для штаммов СМ06-0010 и CM06-0010ΔPn::Pm4tkt_tkt показаны в Таблице 8 ниже.
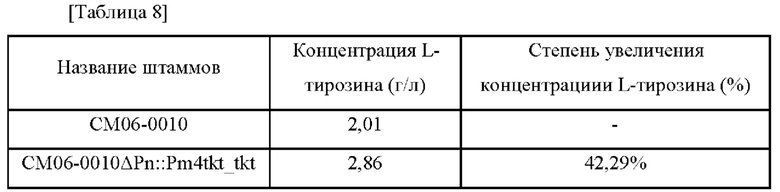
Как показано в Таблице 8, было подтверждено, что концентрация L-тирозина увеличилась на 42,29% в штамме CM06-0010ΔPn::Pm4tkt_tkt, в котором нативный промотор tkt был заменен на вариант Pm4tkt, по сравнению со штаммом СМ06-0010 в качестве контроля.
Пример 3-7. Оценка способности продуцировать фенилаланин 3-7-1. Конструирование штамма, продуцирующего L-фенилаланин Чтобы подтвердить эффект варианта промотора Pm4tkt, сначала получали штамм, продуцирующий L-фенилаланин, на основе штамма Corynebacterium glutamicum АТСС13869 дикого типа (см. патент Кореи №10-2134418).
В частности, ген tyrA, который получает регуляцию по принципу обратной связи с помощью L-тирозина, был заменен на вариант tyrA, который не подвергается регуляции по принципу обратной связи, имеющий происхождение из Е. coli, включающий сильный промотор gap А. Известно, что в белке tyrA, имеющем происхождение Е. coli, обратная связь высвобождается, когда метионин в положении 53 мутируют в изолейцин, а аланин в положении 354 мутируют в валин, и эту форму белка (SEQ ID NO: 50) использовали (Appl Microbiol Biotechnol 75, 103-110 (2007)).
Для генетической манипуляции сначала получали области выше и ниже гена tyrA, в которые ген tyrA должен был быть вставлен путем замены. В частности, фрагмент гена в расположенной выше области гена tyrA был получен с использованием праймеров SEQ ID NO: 51 и 52, а фрагмент гена в расположенной ниже области гена tyrA был получен с использованием праймеров SEQ ID NO: 53 и 54 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы посредством ПЦР. Кроме того, для получения имеющего происхождение из Е. coli варианта гена tyrA, включающего промотор gap А, был получен фрагмент промотора gapA с использованием праймеров SEQ ID NO: 55 и 56 на основе хромосомной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы посредством ПЦР, и фрагмент имеющего происхождение из Е. coli варианта гена tyrA был получен с использованием праймеров SEQ ID NO: 57 и 58 на основе синтетической ДНК варианта tyrA, имеющего происхождение из Е. coli (SEQ ID NO: 50), в качестве матрицы посредством ПЦР. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 60°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут.
Амплифицированные области выше и ниже tyrA, промотор gapA, фрагмент варианта гена tyrA, имеющего происхождение из Е. coli, и вектор pDCM2 для хромосомной трансформации, расщепленный рестрикционным ферментом SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, и полученный продукт назвали pDCM2-ΔtyrA::PgapA-tyrAm. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный таким образом вектор pDCM2-ΔtyrA::PgapA-tyrAm трансформировали в Corynebacterium glutamicum АТСС 13869 дикого типа посредством электропорации, а затем подвергали вторичному кроссоверу для получения штамма, в котором вариант гена tyrA, имеющий происхождение из Е. coli, включающий промотор gap А, был вставлен с одновременной делецией renntyrA. Соответствующая генетическая манипуляция была подтверждена с помощью геномного секвенирования и ПЦР с использованием праймеров SEQ ID NO: 59 и 60, и полученный штамм назвали ATCC13869ΔtyrA::PgapA-tyrAm.
Ген aroG, вовлеченный в первую стадию общего пути биосинтеза ароматических соединений, был усилен путем добавления сильного промотора к варианту aroG с обратной связью, имеющему происхождение из Е. coli. Известно, что в белке aroG, имеющем происхождение из Е. coli, обратная связь высвобождается, когда пролин в положении 150 заменен лейцином, и была использована эта форма белка (SEQ ID NO: 61) (Appl. Environ. Microbiol. 63, 761-762 (1997)).
Для генетической манипуляции были получены области выше и ниже, в которые должен был быть вставлен ген aroG. В частности, был получен вектор с использованием положения BBD29 14470, которое является одной из транспозаз Corynebacterium glutamicum, где на хромосоме происходит гомологичная рекомбинация.
В частности, фрагмент гена в области выше гена BBD29 14470 был получен с использованием праймеров SEQ ID NO: 62 и 63, а фрагмент гена в области ниже гена BBD29 14470 был получен с использованием праймеров SEQ ID NO: 64 и 65 на основе хромосомной ДНК Corynebacterium glutamicum АТСС 13869 в качестве матрицы с помощью ПЦР. Кроме того, для получения имеющего происхождение из Е. coli варианта гена aroG, включающего промотор gapA, был получен фрагмент промотора gapA с использованием праймеров SEQ ID NO: 66 и 67 на основе хромосомной ДНК Corynebacterium glutamicum АТСС 13869 в качестве матрицы посредством ПЦР, и фрагмент гена aroG, имеющего происхождение из£". coli, был получен с использованием праймеров SEQ ID NO: 68 и 69 на основе синтетической ДНК варианта aroG с обратной связью, имеющего происхождение из Е. coli (SEQ ID NO: 61), в качестве матрицы посредством ПЦР. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X, и ПЦР проводили в условиях ПЦР-амплификации: денатурация при 95°С в течение 5 минут, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 60°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 5 минут.
Амплифицированные области выше и ниже гена BBD29 14470, промотор gap А, фрагмент варианта гена aroG, имеющего происхождение из Е. coli, и вектор pDCM2 для хромосомной трансформации, расщепленные ферментом рестрикции SmaI, клонировали с использованием метода сборки Гибсона для получения рекомбинантной плазмиды, а полученный продукт назвали pDCM2-ABBD29_14470::PgapA-aroGm. Клонирование проводили путем смешивания реагента сборки Гибсона и каждого из фрагментов гена в расчетном количестве молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный таким образом вектор pDCM2-ABBD29 14470::PgapA-aroGm трансформировали в штамм АТСС 13869ΔtyrA:: Pgap A-tyr Am посредством электропорации, а затем подвергали вторичному кроссоверу для получения штамма, в который был вставлен имеющий происхождение из Е. coli вариант гена aroG с высвобождением обратной связи, включающий промотор gapA. Соответствующая генетическая манипуляция была подтверждена секвенированием генома и ПЦР с использованием праймеров SEQ ID NO: 70 и 71, и полученный штамм назвали АТСС 13869Δtyr A::Pgap A-tyr AmABBD29 14470:: Pgap A-aroGm.
3-7-2. Конструирование штамма со вставкой одной копии гена tkt, продуцирующего L-фенилаланин
Штамм, продуцирующий L-фенилаланин, получали с использованием pDCM2-ΔaroP, pDCM2-ΔaroP::Pm4tkt_tkt и pDCM2-ΔaroP::Pntkt_tkt, полученных в Примере 2-2. Три вектора вводили в штамм ATCC13869ΔtyrA::PgapA-tyrAmABBD29_14470::PgapA-aroGm, полученный в Примере 3-7-1, посредством электропорации, а затем трансформированный штамм получали в селекционной среде, содержащей 25 мг/л канамицина. Путем вторичного кроссовера были получены штаммы, в которых aroP делетирован или ген Pntkttkt и ген Pm4tkt_tkt вставлены в положение aroP на хромосоме. Соответствующая генетическая манипуляция была подтверждена с помощью секвенирования генома и ПЦР с использованием SEQ ID NO: 46 и 47, а полученные таким образом штаммы назвали АТСС13869ΔtyrA::PgapA-tyrAmABBD29 14470::PgapA-aroGmΔaroP, АТСС 13869ΔtyrA::PgapA-tyrAmA4BBD29 14Pgap A-tyr AmA4BBD29 14Pgap A-tyr AmA4BBD29 14470::PgapA-aroGmΔaroP aroGmΔaroP::Pntkt_tkt и ATCC13869ΔtyrA::PgapA-tyrAmABBD29_14470::PgapA-aroGmΔaroP::Pm4tkt_tkt, соответственно.
3-7-3. Оценка способности штаммов со вставкой одной копии гена tkt продуцировать L-фенилаланин
Для оценки способности Corynebacterium glutamicum продуцировать L-фенилаланин штамм ATCC13869ΔtyrA::PgapA-tyrAm ABBD29_14470::PgapA-aroGm, используемый в качестве исходного штамма, и штаммы АТСС 13869ΔtyrA::PgapA-tyr Am ABBD29 14470:: 6Pgap A3 P 8, ATCCaroGam tyrAm ABBD29 14470::PgapA-aroGmΔaroP::Pntkt_tkt и АТСС13869ΔtyrA:: Pgap A-tyr Am ABBD29_14470::PgapA-aroGmΔaroP::Pm4tkt_tkt, полученные в Примере 3-7-2, культивировали и анализировали следующим образом.
Сначала каждый штамм высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 25 мл среды для высевания, и культивировали при 30°С в течение 20 часов при 200 об/мин при встряхивании. Затем 1 мл посевного культурального раствора высевали в 250-миллилитровую колбу с угловой перегородкой, содержащую 24 мл продукционной среды, и культивировали при 32°С в течение 48 часов при 200 об/мин при встряхивании. Составы питательной среды, среды для высевания и продукционной среды показаны ниже.
<Среда для высевания (рН 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, Л2НРО4 8 г, MgSO4⋅7H2O 0,5 г, биотин 100 мкг, тиамин-HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (в расчете на 1 л дистиллированной воды).
<Продукционная среда (рН 7,0)>
Глюкоза 30 г, (NH4)2SO4 15 г, MgSO4⋅7H2O 1,2 г, KH2PO4 1 г, дрожжевой экстракт 5 г, биотин 900 мкг, тиамин-HCl 4500 мкг, пантотенат кальция 4500 мкг, СаСО3 30 г (в расчете на 1 л дистиллированной воды).
После завершения культивирования продукцию L-фенил ал анина измеряли с помощью ВЭЖХ. Концентрация L-фенилаланина и степень увеличения концентрации в культуральной среде для штаммов Corynebacterium glutamicum ATCC13869ΔtyrA::PgapA-tyrAmABBD29_14470::PgapA-aroGm, ATCC13869ΔtyrA::PgapA-tyrAmABBD29 14470:: Pgap A-aroGmΔaroP, ATCC13869ΔtyrA::PgapA-tyrAmABBD29 14470::PgapA-aroGmΔaroP::Pntkt_tkt и ATCC13869ΔtyrA::PgapA-tyrAmABBD29 14470::PgapA-aroGmΔaroP::Pm4tkt_tkt показаны в Таблице 9 ниже.
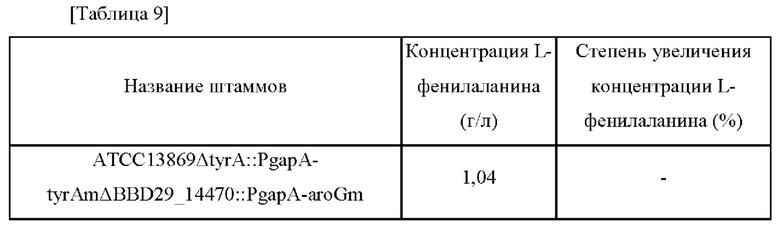
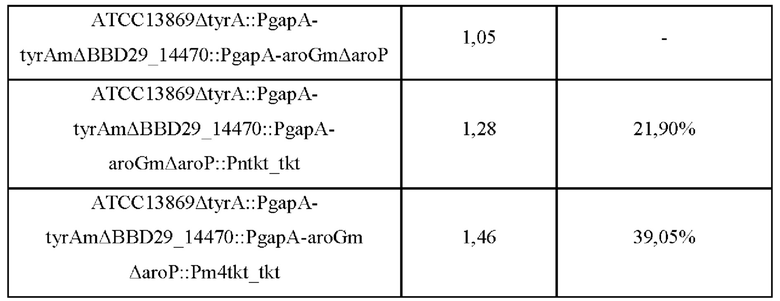
Как показано в Таблице 9, было подтверждено, что концентрация L-фенилаланина увеличилась на 39,05% в штамме АТСС13869ΔtyrA:: Pgap А-tyrAmABBD29_14470::PgapA-aroGmΔaroP::Pm4tkt_tkt, в который была введена одна копия гена Pm4tkt_tkt, по сравнению со штаммом АТСС 13869ΔtyrA::PgapA-tyrAmABBD29 14470::PgapA-aroGmΔaroP в качестве контроля.
Благодаря приведенным выше результатам рекомбинантный микроорганизм, включающий полинуклеотид, обладающий промоторной активностью, по настоящему изобретению, увеличивал продукцию L-лизина, L-треонина, О-ацетилгосерина, L-изолейцина и ароматических аминокислот, таких как L-триптофан, L-тирозин и L-фенилаланин, которые используются в промышленности.
Из вышеизложенного специалист в данной области техники, к которой относится настоящее изобретение, сможет понять, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технических концепций или существенных характеристик настоящего изобретения. В этом отношении репрезентативные варианты осуществления, раскрытые здесь, предназначены только для иллюстративных целей и не должны рассматриваться как ограничивающие объем настоящего изобретения. Таким образом, объем настоящего изобретения определяется прилагаемой формулой изобретения, а не изложенным выше описанием. Все изменения, подпадающие под значение и диапазон эквивалентности пунктов формулы изобретения, должны охватываться объемом настоящего изобретения.
[No. депонирования]
Депозитарный Институт: Корейский центр культур микроорганизмов (Международный Орган Депонирования)
No. доступа: KCCM12971P
Дата депонирования: 20210405
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> НОВЫЙ ПРОМОТОР И ЕГО ПРИМЕНЕНИЕ
<130> OPA21347
<150> KR 10-2021-0064855
<151> 2021-05-20
<160> 71
<170> KoPatentIn 3.0
<210> 1
<211> 300
<212> DNA
<213> Unknown
<220>
<223> промотор
<400> 1
gcttgccgaa catttttctt ttcctttcgg tttttcgaga attttcacct acaaaagccc 60
acgtcacagc tcccagactt aagattgatc acacctttga cacatttgaa ccacagttgg 120
ttataaaatg ggttcaacat cactatggtt agaggtgttg acgggtcaga ttaagcaaag 180
actactttcg gggtagatca cctttgccaa atttgaacca attaacctaa gtcgtagatc 240
tgatcatcgg atctaacgaa aacgaaccaa aactttggtc ccggtttaac ccaggaagga 300
300
<210> 2
<211> 300
<212> DNA
<213> Unknown
<220>
<223> вариант промотора
<400> 2
gcttgccgaa catttttctt ttcctttcgg tttttcgaga attttcacct acaaaagccc 60
acgtcacagc tcccagactt aagattgatc acacctttga cacatttgaa ccacagttgg 120
ttataaaatg ggttcaacat cactatggtt agaggtgttg acgggtcaga ttaagcaaag 180
actactttcg gggtagatca cctttgccaa atttgaatgt ggtatcataa gtcgtagatc 240
tgatcatcgg atctaacgaa aacgaaccaa aactttggtc ccggtttaac ccaggaagga 300
300
<210> 3
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 3
ctctagaact agtggatccg cttgccgaac atttttc 37
<210> 4
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 4
gttcttctcc tttactcatt ccttcctggg ttaaacc 37
<210> 5
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 5
ggtttaaccc aggaaggaat gagtaaagga gaagaac 37
<210> 6
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 6
cccccctcga ggtcgactta tttgtagagc tcatccatg 39
<210> 7
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 7
gcttgccgaa catttttc 18
<210> 8
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 8
tccttcctgg gttaaacc 18
<210> 9
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 9
tgaattcgag ctcggtaccc ccgcacccat ccagcc 36
<210> 10
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 10
gaaaaatgtt cggcaagcaa tacggtgcaa cgtcaa 36
<210> 11
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 11
ttgacgttgc accgtattgc ttgccgaaca tttttc 36
<210> 12
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 12
cagcgtcaag gtggtcaatc cttcctgggt taaacc 36
<210> 13
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 13
ggtttaaccc aggaaggatt gaccaccttg acgctg 36
<210> 14
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 14
gtcgactcta gaggatcccc ggtcgcttgg tgtccttc 38
<210> 15
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 15
cggtacccgg ggatccctgg accacgatcg cc 32
<210> 16
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 16
taatttctat cggcctaagt actgtatcaa ccgtaaaccc 40
<210> 17
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 17
gggtttacgg ttgatacagt acttaggccg atagaaatta 40
<210> 18
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 18
atgcctgcag gtcgacgagg ccgtttgagc cc 32
<210> 19
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 19
ttacggttga tacagtactg cttgccgaac atttttc 37
<210> 20
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 20
cagcgtcaag gtggtcaatc cttcctgggt taaacc 36
<210> 21
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 21
ggtttaaccc aggaaggatt gaccaccttg acgctg 36
<210> 22
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 22
tctatcggcc taagtacttt aaccgttaat ggagtcc 37
<210> 23
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 23
tgaattcgag ctcggtaccc ggagctgctg tccaacgtgg 40
<210> 24
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 24
gtcgactcta gaggatcccc tgggaacttg tcgacgctat 40
<210> 25
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 25
tgaattcgag ctcggtaccc tccaagattt tggtgctgcg 40
<210> 26
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 26
atctcagagg tggaaatctt ttcgatgttc acgttgacat 40
<210> 27
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 27
atgtcaacgt gaacatcgaa aagatttcca cctctgagat 40
<210> 28
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 28
gtcgactcta gaggatcccc gttcacctca gagacgatta 40
<210> 29
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 29
tcctaatgca cagaagctgg 20
<210> 30
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 30
gtggtgcagt tagggttcgc 20
<210> 31
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 31
tgaattcgag ctcggtaccc tttccacacc cgtgttaccg 40
<210> 32
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 32
atcatcgcgc tcttcctgtt ggattgtacg cagggagatt 40
<210> 33
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 33
aatctccctg cgtacaatcc aacaggaaga gcgcgatgat 40
<210> 34
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 34
gtcgactcta gaggatcccc aaccattagc tgcagcaaca 40
<210> 35
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 35
atccaactgc agacgtcgaa 20
<210> 36
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 36
atctgggtgg ccttcaaagg 20
<210> 37
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 37
tgaattcgag ctcggtaccc ggcgttcttg cagtcc 36
<210> 38
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 38
ttgttgacat gaagcgccgg ggcacctacc gaggaa 36
<210> 39
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 39
ttcctcggta ggtgccccgg cgcttcatgt caacaa 36
<210> 40
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 40
gcgcaccgaa ctcttcaaca ccgcgaggga agaaat 36
<210> 41
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 41
atttcttccc tcgcggtgtt gaagagttcg gtgcgc 36
<210> 42
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 42
gtcgactcta gaggatcccc gggtacagct gcagctc 37
<210> 43
<211> 450
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность промотора trpE(S38R)
<400> 43
ggcgcttcat gtcaacaatc tttaacgttt tcaagttcac aagtcgtgtt caaatggtga 60
caagattgga cactgtgctg aattggcacc aagccctcat aaatgataga tctaaatcga 120
atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc atccgctaaa 180
gccccaggaa ccctgtgcag aaagaacaaa taatcgtgaa ttttggcagc aacagcaatt 240
cctgctacaa ttgaaaacgt gcaaaagcat agattattgg aggagatcaa aacaatgagc 300
acgaatcccc atgttttctc cctagatgtc cgctatcacg aggatgcttc tgcgttgttt 360
gcccacttgg gtggcacaac cgcagatgat gcagccctgt tggaacgcgc tgatatcacc 420
accaagaatg gtatttcttc cctcgcggtg 450
<210> 44
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 44
ggcagagcat catacc 16
<210> 45
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 45
ggcatcagtg cgcatatc 18
<210> 46
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 46
ctggaccacg atcgcc 16
<210> 47
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 47
gaggccgttt gagccc 16
<210> 48
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 48
ggagctgctg tccaacgtgg 20
<210> 49
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 49
tgggaacttg tcgacgctat 20
<210> 50
<211> 1122
<212> DNA
<213> Unknown
<220>
<223> ген tyrA
<400> 50
atggttgctg aattgaccgc attacgcgat caaattgatg aagtcgataa agcgctgctg 60
aatttattag cgaagcgtct ggaactggtt gctgaagtgg gcgaggtgaa aagccgcttt 120
ggactgccta tttatgttcc ggagcgcgag gcatctattt tggcctcgcg tcgtgcagag 180
gcggaagctc tgggtgtacc gccagatctg attgaggatg ttttgcgtcg ggtgatgcgt 240
gaatcttact ccagtgaaaa cgacaaagga tttaaaacac tttgtccgtc actgcgtccg 300
gtggttatcg tcggcggtgg cggtcagatg ggacgcctgt tcgagaagat gctgaccctc 360
tcgggttatc aggtgcggat tctggagcaa catgactggg atcgagcggc tgatattgtt 420
gccgatgccg gaatggtgat tgttagtgtg ccaatccacg ttactgagca agttattggc 480
aaattaccgc ctttaccgaa agattgtatt ctggtcgatc tggcatcagt gaaaaatggg 540
ccattacagg ccatgctggt ggcgcatgat ggtccggtgc tggggctaca cccgatgttc 600
ggtccggaca gcggtagcct ggcaaagcaa gttgtggtct ggtgtgatgg acgtaaaccg 660
gaagcatacc aatggtttct ggagcaaatt caggtctggg gcgctcggct gcatcgtatt 720
agcgccgtcg agcacgatca gaatatggcg tttattcagg cactgcgcca ctttgctact 780
tttgcttacg ggctgcacct ggcagaagaa aatgttcagc ttgagcaact tctggcgctc 840
tcttcgccga tttaccgcct tgagctggcg atggtcgggc gactgtttgc tcaggatccg 900
cagctttatg ccgacatcat tatgtcgtca gagcgtaatc tggcgttaat caaacgttac 960
tataagcgtt tcggcgaggc gattgagttg ctggagcagg gcgataagca ggcgtttatt 1020
gacagtttcc gcaaggtgga gcactggttc ggcgattacg tgcagcgttt tcagagtgaa 1080
agccgcgtgt tattgcgtca ggcgaatgac aatcgccagt aa 1122
<210> 51
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 51
ttcgagctcg gtaccctatc aaaaccgagt tcttcc 36
<210> 52
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 52
gtcgttttta ggcctcctga caagtgtggc acatac 36
<210> 53
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 53
tgacaatcgc cagtaatttt atcggctgat gattct 36
<210> 54
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 54
actctagagg atccccaacg cgattgcatt cggctc 36
<210> 55
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 55
gtgccacact tgtcaggagg cctaaaaacg accgag 36
<210> 56
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 56
tcaattcagc aaccatgttg tgtctcctct aaagat 36
<210> 57
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 57
ttagaggaga cacaacatgg ttgctgaatt gaccgc 36
<210> 58
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 58
tcatcagccg ataaaattac tggcgattgt cattcg 36
<210> 59
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 59
cgcagcgagc gaaccg 16
<210> 60
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 60
gttcgaatcc ctccgggc 18
<210> 61
<211> 1053
<212> DNA
<213> Unknown
<220>
<223> ген aroG
<400> 61
atgaattatc agaacgacga tttacgcatc aaagaaatca aagagttact tcctcctgtc 60
gcattgctgg aaaaattccc cgctactgaa aatgccgcga atacggttgc ccatgcccga 120
aaagcgatcc ataagatcct gaaaggtaat gatgatcgcc tgttggttgt gattggccca 180
tgctcaattc atgatcctgt cgcggcaaaa gagtatgcca ctcgcttgct ggcgctgcgt 240
gaagagctga aagatgagct ggaaatcgta atgcgcgtct attttgaaaa gccgcgtacc 300
acggtgggct ggaaagggct gattaacgat ccgcatatgg ataatagctt ccagatcaac 360
gacggtctgc gtatagcccg taaattgctg cttgatatta acgacagcgg tctgccagcg 420
gcaggtgagt ttctcgatat gatcacccca caatatctcg ctgacctgat gagctggggc 480
gcaattggcg cacgtaccac cgaatcgcag gtgcaccgcg aactggcatc agggctttct 540
tgtccggtcg gcttcaaaaa tggcaccgac ggtacgatta aagtggctat cgatgccatt 600
aatgccgccg gtgcgccgca ctgcttcctg tccgtaacga aatgggggca ttcggcgatt 660
gtgaatacca gcggtaacgg cgattgccat atcattctgc gcggcggtaa agagcctaac 720
tacagcgcga agcacgttgc tgaagtgaaa gaagggctga acaaagcagg cctgccagca 780
caggtgatga tcgatttcag ccatgctaac tcgtccaaac aattcaaaaa gcagatggat 840
gtttgtgctg acgtttgcca gcagattgcc ggtggcgaaa aggccattat tggcgtgatg 900
gtggaaagcc atctggtgga aggcaatcag agcctcgaga gcggggagcc gctggcctac 960
ggtaagagca tcaccgatgc ctgcatcggc tgggaagata ccgatgctct gttacgtcaa 1020
ctggcgaatg cagtaaaagc gcgtcgcggg taa 1053
<210> 62
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 62
ttcgagctcg gtaccccccg gcggtatcga ggtagt 36
<210> 63
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 63
ctcggtcgtt tttaggcctc ttaatcaccc gcggggaccc 40
<210> 64
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 64
taaaagcgcg tcgcgggtaa aaacttgtcc cgagggtgag 40
<210> 65
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 65
actctagagg atcccctatc agtcacttcc ctgaga 36
<210> 66
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 66
gggtccccgc gggtgattaa gaggcctaaa aacgaccgag 40
<210> 67
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 67
gttctgataa ttcatgttgt gtctcctcta aagat 35
<210> 68
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 68
ctttagagga gacacaacat gaattatcag aacgacga 38
<210> 69
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 69
ctcaccctcg ggacaagttt ttacccgcga cgcgctttta 40
<210> 70
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 70
ggtaccggca ggtacc 16
<210> 71
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 71
caagggggtc caatcc 16
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Новый экспортер L-триптофана и способ продуцирования L-триптофана с его использованием | 2019 |
|
RU2761871C1 |
| НОВЫЙ ПРОМОТОР И СПОСОБ ПОЛУЧЕНИЯ ЖЕЛАЕМОГО ВЕЩЕСТВА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2020 |
|
RU2794946C1 |
| Новый вариант L-тирозин-экспортирующего белка и способ получения L-тирозина с его использованием | 2021 |
|
RU2808831C1 |
| НОВЫЙ ВАРИАНТ БЕЛКА-ЭКСПОРТЕРА L-ТРИПТОФАНА И СПОСОБ ПОЛУЧЕНИЯ L-ТРИПТОФАНА С ЕГО ПРИМЕНЕНИЕМ | 2020 |
|
RU2763603C1 |
| Новый промотор и его применение | 2021 |
|
RU2787780C1 |
| Новый промотор и его применение | 2021 |
|
RU2787592C1 |
| Новый промотор и его применение | 2022 |
|
RU2831201C2 |
| ВАРИАНТ ПРЕФЕНАТДЕГИДРАТАЗЫ И СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТ С РАЗВЕТВЛЕННОЙ ЦЕПЬЮ С ЕГО ИСПОЛЬЗОВАНИЕМ | 2022 |
|
RU2827315C1 |
| НОВЫЙ ВАРИАНТ ГАММА-АМИНОБУТИРАТПЕРМЕАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ИЗОЛЕЙЦИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2826168C1 |
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ЕГО ИСПОЛЬЗОВАНИЕМ | 2020 |
|
RU2795161C1 |
Настоящее изобретение относится к биотехнологии и микробиологии, в частности к новому промотору и способу получения целевых веществ с его использованием. Более конкретно, настоящее изобретение относится к новому полинуклеотиду, обладающему промоторной активностью, кассете генной экспрессии и клетке-хозяину, включающим его, и способу получения целевых веществ с использованием микроорганизма. Изобретение позволяет продуцировать целевые вещества с высокой продуктивностью в микроорганизмах рода Corynebacterium. 4 н. и 8 з.п. ф-лы, 9 табл., 3 пр.
1. Полинуклеотид, обладающий промоторной активностью, где указанный полинуклеотид состоит из нуклеотидной последовательности SEQ ID NO: 1, в которой нуклеотид по положению 218, цитозин (C), заменен на тимин (T), нуклеотид по положению 219, цитозин (C), заменен на гуанин (G), нуклеотид по положению 220, аденин (A), заменен на тимин (T), нуклеотид по положению 221, аденин (A), заменен на гуанин (G), нуклеотид по положению 222, тимин (T), заменен на гуанин (G), нуклеотид по положению 225, аденин (A), заменен на тимин (T) и нуклеотид по положению 227, цитозин (C), заменен на аденин (A).
2. Полинуклеотид по п. 1, где указанный полинуклеотид состоит из нуклеотидной последовательности SEQ ID NO: 2.
3. Полинуклеотид по п. 1 или 2, где указанный полинуклеотид функционально связан с геном, кодирующим целевой белок.
4. Кассета генной экспрессии, для экспрессии целевого гена, содержащая полинуклеотид по п. 1 или 2 и ген, кодирующий целевой белок, который функционально связан с указанным полинуклеотидом.
5. Кассета генной экспрессии по п. 4, где целевой белок представляет собой транскетолазу.
6. Клетка-хозяин, для продуцирования целевого вещества, содержащая полинуклеотид по п. 1 или 2 и ген, кодирующий целевой белок, который функционально связан с указанным полинуклеотидом.
7. Клетка-хозяин по п. 6, где целевой белок представляет собой транскетолазу.
8. Клетка-хозяин по п. 6, где указанная клетка-хозяин представляет собой микроорганизм рода Corynebacterium.
9. Клетка-хозяин по п. 8, где указанный микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
10. Способ получения целевого вещества, включающий культивирование клетки-хозяина по п. 6 в среде и выделение целевого вещества из среды.
11. Способ по п. 10, где указанное целевое вещество представляет собой аминокислоту.
12. Способ по п. 10, где указанное целевое вещество представляет собой по меньшей мере одно выбранное из группы, состоящей из лизина, треонина, O-ацетилгомосерина, изолейцина, триптофана, тирозина и фенилаланина.
| PATEK M | |||
| et al., Corynebacterium glutamicum promoters: a practical approach, Microb Biotechnol, 2013, vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Клапанный регулятор для паровозов | 1919 |
|
SU103A1 |
| SHANG X | |||
| et al., Native promoters of Corynebacterium glutamicum and its application in L-lysine production, Biotechnol Lett, 2018, vol | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Передвижная комнатная печь | 1922 |
|
SU383A1 |
| RU 2018117175 A, 26.11.2019. | |||

