1. Область изобретения
Настоящее изобретение относится к микроорганизму, продуцирующему L-метионин, в который введен белок, кодируемый чужеродным геном metZ, и способу получения L-метионина с использованием этого микроорганизма.
2. Уровень техники
L-метионин, одну из незаменимых аминокислот в организме, применяют в качестве корма, медицинского исходного вещества, такого как синтетическое исходное вещество для медицинских растворов, медицинских расходных материалов и так далее, и пищевой добавки. Метионин представляет собой важную аминокислоту, вовлеченную в реакцию переноса метальной группы в организме, и играющую роль в обеспечении серой.
В химическом синтезе метионина используют главным образом способ получения метионина в форме смеси L- и D-изомеров путем гидролиза 5-(β-метилмеркаптоэтил)гидантоина. Однако такой химический синтез приводит к получению смешанной формы из L- и D-изомеров.
Между тем, L-метионин также можно получать биологическим способом. Более конкретно, один способ получения L-метионина с использованием микроорганизмов заключается в получении метионина путем прямого сульфгидрилирования с использованием О-ацилгомосерина (О-ацетилгомосерина или О-сукцинилгомосерина) и сероводорода в качестве субстратов. Например, известно, что фермент, кодируемый геном metY у Corynebacterium, выполняет функцию прямого сульфгидрилирования. Другой способ получения L-метионина с помощью микроорганизмов заключается в получении метионина путем транссульфурации с использованием О-ацилгомосерина (О-ацетилгомосерина или O-сукцинилгомосерина) и цистеина в качестве субстратов. Например, известно что фермент, кодируемый геном metB у Corynebacterium, выполняет функцию транссульфурации. Однако, существуют недостатки, которые заключаются в том, что фермент, кодируемый metB, продуцирует много побочных продуктов, и ген metY получает ингибирование по типу обратной связи, и следовательно, его трудно применить для получения L-метионина в промышленном масштабе (Kromer JO et al., J Bacteriol 188(2):609-618, 2006; Yeom HJ et al., J Microbiol Biotechnol 14(2):373-378, 2004 и так далее).
Автор настоящего изобретения поставил задачу разработать белок, который может заменить указанный белок, и в результате обнаружил, что микроорганизм, в который введен белок, кодируемый геном metZ, продуцирует L-метионин с высоким выходом, тем самым выполняя настоящее изобретение.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задача настоящего изобретения заключается в предоставлении микроорганизма, продуцирующего L-метионин, в который введен белок, кодируемый чужеродным геном metZ.
Задача настоящего изобретения также заключается в предоставлении способа получения L-метионина, включающего культивирование микроорганизма в среде, содержащей тиосульфат.
Задача настоящего изобретения также заключается в предоставлении композиции для получения L-метионина, содержащей этот микроорганизм и тиосульфат.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 представлено изображение плазмиды pDCM2.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ
Далее настоящее изобретение описано более подробно. Между тем, каждое описание и воплощение, раскрытое в настоящем описании, также можно применить к другим описаниям и воплощениям. То есть, все комбинации различных элементов, раскрытых в настоящем описании, попадают в объем настоящего изобретения. Далее, объем настоящего изобретения не ограничен конкретным описанием, раскрытым здесь ниже.
Далее, специалист в данной области техники понимает или способен определить с помощью только рутинных экспериментов множество воплощений, эквивалентных определенным воплощениям описания, раскрытым здесь. Далее, эти эквивалентные воплощения следует интерпретировать как попадающие в объем настоящего изобретения.
В одном аспекте настоящего изобретения предложен микроорганизм, продуцирующий L-метионин, в который введен белок, кодируемый чужеродным геном metZ.
В другом аспекте настоящего изобретения предложен способ получения L-метионина, включающий культивирование микроорганизма, продуцирующего L-метионин в среде, содержащей тиосульфат.
Как его используют здесь, термин "ген metZ" обозначает ген, кодирующий фермент, вовлеченный в сульфгидрацию с использованием ацилгомосерина в качестве субстрата.
Как его используют здесь, термин "ацилгомосерин" относится к соединению, в котором ацильная группа присоединена к гомосерину, и включает как сукцинилгомосерин, так и ацетилгомосерин. Например, ацилгомосерин может представлять собой О-сукцинилгомосерин или О-ацетилгомосерин, но не ограничиваясь ими.
Как его используют здесь, фермент, кодируемый геном metZ, может представлять собой сукцинилгомосерин-сульфгидрилазу, ацетилгомо серии-сульфгидрилазу или фермент, вовлеченный в сульфгидрацию с использованием О-сукцинилгомосерина в качестве субстрата, но не ограничиваясь ими.
Как его используют здесь, термин "сульфгидрация" можно использовать взаимозаменяемо с термином "сульфгидрилирование", и он относится к реакции, которая предоставляет сульфгидрильную группу (-SH) для определенной молекулы. В отношении задачи настоящего изобретения этот термин может относиться к реакции в процессе синтеза метионина, но не ограничиваясь этим. Фермент, вовлеченный в "сульфгидрацию" также можно называть "сульфгидрилазой", но не ограничиваясь этим.
При традиционном ферментировании метионина фермент, экспрессируемый геном metZ, использовали в следующих реакциях in vitro:
CH3SH + O-ацетил-L-гомосерин => ацетат + метионин
CH3SH + О-сукцинил-L-гомосерин => сукцинат + метионин
Другими словами, в способе получения метионина, включающем первую стадию получения предшественника метионина с использованием микроорганизма, и вторую стадию проведения ферментативной реакции in vitro путем добавления метилмеркаптана и метионин-конвертирующего фермента в ферментируемый раствор, содержащий предшественник метионина, фермент, экспрессируемый геном metZ, использовали в качестве метионин-конвертирующего фермента in vitro (смотрите US 2010-0184164 A1).
Между тем, при ферментировании метионина в микроорганизме рода Corynebacterium используют два пути сульфгидрации (этапы сульфгидрилирования) (Hwang BJ et al., J Bacteriol 184(5): 1277-1286, 2002). Один из них представляет собой превращение (9-ацетилгомосерина (ацетилгомосерин: АН) в цистатионин с помощью фермента, кодируемого геном metB. В этом случае цистеин используют в качестве источника серы. Другими словами, реакция, при которой происходит превращение ацилгомосерина и цистеина в качестве взаимодействующих соединений в цистатионин, носит название "транссульфурация", и фермент, вовлеченный в эту реакцию, носит название "транссульфураза". Другой путь представляет собой превращение О-ацетилгомосерина в гомоцистеин с помощью фермента, кодируемого геном metY. В этом случае в качестве источника серы используют неорганическое соединение серы, такое как сероводород и так далее. В этой реакции, в которой происходит превращение ацилгомосерина и сероводорода в качестве реагирующих веществ в гомоцистеин, не образуется цистатионин в качестве промежуточного соединения в процессе получения гомоцистеина, который является предшественником метионина, в отличие от описанной выше транссульфурации. Эту реакцию называют прямым сульфгидрилированием.
Другими словами, путь сульфгидрилирования может относиться к пути реакции, в которой ацилгомосерин превращается в другое соединение путем реакции с источником серы, и по большому счету может быть разделен на транссульфурацию и прямую сульфгидрацию.
Однако, у штаммов Corynebacterium оба фермента, вовлеченные в сульфгидрацию, имеют недостатки. Например, при использовании ацетилгомосерина и гомоцистеина белок, кодируемый геном metB, продуцирует побочный продукт гомолантионин дополнительно к цистатионину (Kromer JO et al., J Bacteriol 188(2):609-618, 2006). Далее, известно, что метионин ингибирует ген metY по типу обратной связи (Yeom HJ et al., J Microbiol Biotechnol 14(2):373-378, 2004).
Согласно настоящему изобретению чужеродный ген metZ вводят в штамм Corynebacterium для биологического получения метионина путем только одностадийной реакции, и показано, что введение гена metZ полезно применять при ферментировании метионина.
В пути синтеза метионина, в который вовлечен белок, кодируемый геном metZ по настоящему изобретению, образование побочных продуктов может быть уменьшено. Побочный продукт может представлять собой гомолантионин. Уменьшение образования побочных продуктов может относиться к уменьшению образования побочных продуктов по сравнению с образованием побочных продуктов у микроорганизма дикого типа или в пути синтеза, в который вовлечен белок, кодируемый геном metB, но не ограничиваясь этим.
Следовательно, микроорганизм по настоящему изобретению, в который введен чужеродный ген metZ, и способ получения метионина, включающий культивирование этого микроорганизма, могут демонстрировать уменьшение образования побочных продуктов по сравнению с микроорганизмом, продуцирующим метионин, в который не введен чужеродный ген metZ, и способом получения метионина с использованием этого микроорганизма. Белок, кодируемый геном metZ, по настоящему изобретению, может не регулироваться метионином по типу обратной связи.
Белок, кодируемый геном metZ, по настоящему изобретению представляет собой O-ацилгомосеринсульфгидрилазу, которая может использовать сероводород в качестве источника серы, и также является О-ацилгомосеринтранссульфуразой, которая в качестве источника серы может использовать цистеин. Более конкретно, белок может представлять собой O-ацетилгомосеринсульфгидрилазу, О-ацетилгомосеринтранссульфуразу, O-сукцинилгомосеринсульфгидрилазу или О-сукцинилгомосеринтранссульфуразу. Следовательно, белок, кодируемый геном metZ, по настоящему изобретению может представлять собой белок, обладающий активностью O-ацилгомосеринсульфгидрилазы, и в частности, он может представлять собой белок, обладающий одной или более чем одной активностью из О-ацетилгомосеринсульфгидрилазы, O-ацетилгомосеринтранссульфуразы, О-сукцинилгомосеринсульфгидрилазы и O-сукцинилгомосеринтранссульфуразы.
Например, чужеродный ген metZ по настоящему изобретению может представлять собой ген, имеющий происхождение из микроорганизмов, отличных от микроорганизма, продуцирующего L-метионин, в который введен этот ген, или может быть отличным от гена, исходно присутствующего в микроорганизме, продуцирующем L-метионин, в который введен этот ген. В частности, ген может представлять собой ген, называемый metZ, имеющий происхождение из Chromobacterium violaceum, Hyphomonas neptunium или Rhodobacter sphaeroides, но не ограничиваясь ими. Относительно задачи настоящего изобретения ген может включать любой ген без ограничения, при условии, что он может усиливать способность продуцировать L-метионин. Последовательность гена metZ доступна из известной базы данных GenBank от NCBI (the National Center for Biotechnology Information, Нацилнальный центр биотехнологической информации) и в качестве способа получения соответствующей последовательности применимы различные способы, известные в данной области техники.
В настоящем изобретении белок, кодируемый чужеродным metZ, может включать любую одну или более чем одну последовательность, выбранную из группы, состоящей из полипептидных последовательностей SEQ ID NO: 60, 61 и 62; и аминокислотных последовательностей (полипептидных последовательностей), на 90% или более гомологичных или идентичных SEQ ID NO: 60, 61 и 62, но не ограничиваясь ими. Например, белок может включать полипептидную последовательность, на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 97,5%, 97,7%, 97,8%, 98%, 98,5%, 98,7%, 98,8%, 99%, 99,5%, 99,7%, 99,8% или менее чем на 100% гомологичную или идентичную любой полипептидной последовательности из SEQ ID NO: 60, 61 и 62. Например, белок может включать любую полипептидную последовательность из SEQ ID NO: 66-71 и последовательность, выбранную из полипептидных последовательностей, на 90% или более гомологичных SEQ ID NO: 66-71, но не ограничиваясь ими.
Ген metZ по настоящему изобретению может включать любую одну или более чем одну последовательность, выбранную из группы, состоящей из полинуклеотидных последовательностей SEQ ID NO: 63, 64 и 65; и полинуклеотидной последовательности, на 90% или более гомологичной или идентичной SEQ ID NO: 63, 64 и 65, но не ограничиваясь ими. Например, ген может включать любую одну или более чем одну полинуклеотидную последовательность, выбранную из SEQ ID NO: 63, 64 и 65, и полинуклеотидную последовательность на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 97,5%, 97,7%, 97,8%, 98%, 98,5%, 98,7%, 98,8%, 99%, 99,5%, 99,7%, 99,8% или менее чем на 100% гомологичную или идентичную SEQ ID NO: 63, 64 и 65.
Как его используют здесь, термин "полинуклеотид" относится к цепи ДНК, имеющей заданную длину или длину более заданной, представляющий собой длинноцепочечный полимер из нуклеотидов, образованный путем соединения нуклеотидных мономеров ковалентными связями.
В настоящем изобретении при условии, что ген metZ включает полинуклеотид, кодирующий белок, кодируемый одной или более чем одной полинуклеотидной последовательностью, выбранной из SEQ ID NO: 63, 64 и 65, или полинуклеотид, кодирующий белок, имеющий эффективность, соответствующую белку, имеющему одну или более чем одну из аминокислотных последовательностей SEQ ID NO: 60, 61 и 62, очевидно, что любой полинуклеотид, кодирующий аминокислотную последовательность, в которой часть последовательности удалена, модифицирована, заменена или добавлена, также может попадать в объем настоящего изобретения.
Например, ген metZ может представлять собой ген, кодирующий аминокислотную последовательность, имеющую частичную замену, например, 1-20 аминокислот в любой из аминокислотных последовательностей SEQ ID NO: 60, 61 и 62. В другом воплощении ген metZ может представлять собой последовательность, кодирующую аминокислотную последовательность, имеющую вставку последовательностей из 20, 19, 18, 17, 16, 15, 14, 13, 12 или 11 или менее аминокислот перед аминокислотной последовательностью/после этой аминокислотной последовательности. Еще в одном воплощении ген metZ может представлять собой последовательность, кодирующую аминокислотную последовательность, включающую все вышеописанные замены и вставки, но не ограничиваясь ими.
Далее, зонд, который может быть получен из известной нуклеотидной последовательности, например, полинуклеотид, который гибридизуется с последовательностью, комплементарной полной полинуклеотидной последовательности или ее части в строгих условиях, также может быть включен без ограничения.
Другими словами, хотя в настоящем описании он описан как "полинуклеотид, включающий нуклеотидную последовательность с определенным номером последовательности", "полинуклеотид, состоящий из нуклеотидной последовательности с определенным номером последовательности" или "полинуклеотид, имеющий нуклеотидную последовательность с определенным номером последовательности", очевидно, что любой полинуклеотид, кодирующий аминокислотную последовательность, в которой часть последовательности вырезана, модифицирована, заменена, заменена консервативной заменой или добавлена, может быть включен в объем настоящего изобретения при условии, что он обладает такой же активностью как полипептид, кодируемый нуклеотидной последовательностью, состоящей из полинуклеотида с определенным номером последовательности, или соответствующей активностью. Например, это может быть случай, где на N-конце и/или С-конце аминокислотной последовательности добавлена последовательность, которая не изменяет функцию белка, природная мутация, молчащая мутация или консервативная замена.
Как его используют здесь, термин "консервативная замена" относится к замене аминокислоты другой аминокислотой, имеющей похожие структурные и/или химические свойства. Такая аминокислотная замена обычно может происходить на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатка.
Как его используют здесь, термин "гомология" или "идентичность" относится к степени родства между двумя данными нуклеотидными последовательностями, которая может быть выражена в процентах.
Термины "гомология" и "идентичность" часто можно использовать взаимозаменяемо друг с другом.
Гомологию или идентичность последовательностей консервативных полинуклеотидов можно определить с помощью стандартных алгоритмов выравнивания и можно применять штраф за пропуск в последовательности, установленный по умолчанию в используемой программе. По существу, обычно ожидают, что гомологичные или идентичные последовательности гибридизуются по всей длине или по меньшей мере примерно на 50%, 60%, 70%, 80% или 90% от полной длины последовательностей в умеренно строгих или очень строгих условиях. Также в гибридизующихся полинуклеотидах рассматривают полинуклеотиды, содержащие вырожденные кодоны вместо обычных кодонов.
Обладают ли две любые полинуклеотидные последовательности гомологией, сходством или идентичностью, можно определить с помощью известного компьютерного алгоритма, такого как программа "FASTA" согласно Pearson et al. (1988) Proc. Natl. Acad. Sci. USA 85:2444 с использованием параметров по умолчанию. Альтернативно, это можно определить с помощью алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), который выполняют с помощью программы "Needleman" пакета "EMBOSS" (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al.., 2000, Trends Genet. 16:276-277) (версия 5.0.0 или более поздняя) (пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S.F., et al., J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994 и CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, гомологию, сходство или идентичность можно определить с помощью "BLAST" или "ClustalW" от Национального центра биотехнологической информации (США).
Гомологию, сходство или идентичность полинуклеотидов можно определить путем сравнения информации о последовательностях с использованием, например, компьютерной программы "GAP", например как описано в Needleman et al. (1970), J Mol Biol. 48:443, как раскрыто в Smith and Waterman, Adv. Appl. Math (1981) 2:482. Коротко, программа GAP определяет гомологию, сходство или идентичность как величину, полученную путем деления числа одинаково выровненных символов (то есть нуклеотидов или аминокислот) на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) бинарную матрицу сравнения (содержащую значение 1 для идентичных символов и 0 для не идентичных) и взвешенную матрицу сравнения от Gribskov et al. (1986) Nucl. Acids Res. 14:6745, как раскрыто в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979) (альтернативно, матрицу замен EDNAFULL (версия NCBI NUC4.4 от EMBOSS); (2) штраф 3,0 за каждый пропуск и дополнительно штраф 0,10 за каждый символ в каждом пропуске (или штраф за открытие пропуска 10 и штраф за продление пропуска 0,5); и (3) отсутствие штрафа за окончание пропуска.
Далее, обладают ли две полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью друг с другом, можно определить путем сравнения последовательностей в эксперименте с гибридизацией по Саузерну в строгих условиях как определено, и определенные подходящие условия гибридизации известны в данной области техники и могут быть определены способом, хорошо известным специалисту в данной области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
Далее, в полинуклеотид по настоящему изобретению могут быть внесены различные модификации в кодирующей области при условии, что они не изменяют полипептидную последовательность вследствие вырожденности генетического кода или с учетом кодонов, предпочтительных для организма, в котором полинуклеотид должен экспрессироваться. Далее, зонд, который может быть получен из известной последовательности гена, например, любой полинуклеотидной последовательности, которая может гибридизоваться с последовательностью, комплементарной полной нуклеотидной последовательности или ее части в строгих условиях, и который может повышать продуктивность по L-метионину, в то же время не являясь последовательностью, в природе присутствующей в микроорганизме, в который вводят эту последовательность, может быть включен без ограничения. Термин "строгие условия" относится к условиям, обеспечивающим специфическую гибридизацию между полинуклеотидами. Такие условия специально описаны в различных литературных источниках (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989) и хорошо известны в данной области техники. Например, строгие условия могут включать условия, в которых гены, обладающие высокой степенью гомологии или идентичности, составляющей 40% или выше, в частности, 70% или выше, 80% или выше, 85% или выше, 90% или выше, более конкретно 95% или выше, более конкретно 97% или выше, и особенно конкретно 99% или выше, гибридизуются друг с другом, и гены, обладающие более низкой степенью гомологии или идентичности, чем перечисленные выше значения гомологии или идентичности, не гибридизуются друг с другом, или обычные условия промывки для гибридизации по Саузерну, то есть, однократная промывка, в частности двукратная или трехкратная промывка при концентрации соли и температуре, соответствующей 60°С, 1×SSC, 0,1% SDS, в частности 60°С, 0,1×SSC, 0,1% SDS и более конкретно 68°С, 0,1×SSC, 0,1% SDS.
Для гибридизации необходимо, чтобы два полинуклеотида имели комплементарные последовательности, хотя несоответствия между основаниями возможны в зависимости от строгости условий гибридизации. Термин "комплементарный" используют для описания взаимосвязи между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в отношении ДНК, аденозин комплементарен тимину, и цитозин комплементарен гуанину. Следовательно, полинуклеотид по настоящему изобретению может включать выделенные полинуклеотидные фрагменты, комплементарные полной последовательности, а также полинуклеотидные последовательности по существу сходные с ними.
В частности, полинуклеотиды, обладающие гомологией или идентичностью, можно выявить с использованием условий гибридизации, включающих этап гибридизации при значении Tm 55°С в вышеописанных условиях. Далее, значение Tm может составлять 60°С, 63°С или 65°С, но не ограничиваясь ими, и может быть подходящим образом отрегулировано специалистом в данной области техники в зависимости от его задач.
Подходящая степень строгости условий для гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов, и эти переменные хорошо известны в данной области техники (смотрите Sambrook et al., выше, 9.50 9.51, 11.7-11.8).
Как его используют здесь, термин "введение белка" обозначает, что микроорганизм проявляет активность определенного белка, которой он изначально не обладал, или обозначает, что микроорганизм проявляет повышенную активность по сравнению с его собственной активностью или активностью этого белка до модификации. Например, это может означать введение определенного белка, введение в хромосому микроорганизма полинуклеотид а, кодирующего этот определенный белок, или введение в микроорганизм вектора, содержащего полинуклеотид, кодирующий этот определенный белок, что таким образом дает возможность проявления его активности. В настоящем изобретении введение белка также может быть выражено как повышение активности белка у микроорганизма, не обладающего определенной активностью белка.
Введение белка может быть выполнено путем введения чужеродного полинуклеотид а, кодирующего белок, проявляющий активность, идентичную/сходную с активностью вышеописанного белка, или путем введения кодон-оптимизированного варианта его полинуклеотида в клетку-хозяина. Любую чужеродную полинуклеотидную последовательность можно использовать без ограничения по ее происхождению или последовательности при условии, что она демонстрирует активность, идентичную/сходную с активностью вышеописанного белка. Далее, чужеродный полинуклеотид можно вводить в клетку-хозяина после оптимизации его кодонов таким образом, чтобы обеспечить оптимизированную транскрипцию и трансляцию в клетке-хозяине. Это введение можно проводить путем известного способа трансформации, подходящим образом выбранного специалистом в данной области техники, и белок может быть получен путем экспрессии введенного полинуклеотида в клетке-хозяине, и в результате его активность может быть увеличена.
Повышение активности введенного белка может быть выполнено путем:
1) увеличения числа внутриклеточных копий гена или полинуклеотида, кодирующего белок,
2) замены области регуляции экспрессии гена в хромосоме, кодирующей белок, на последовательность, обладающую высокой активностью,
3) модификации нуклеотидной последовательности стартового кодона или 5'-UTR (нетранслируемой)-области белка,
4) модификации нуклеотидной последовательности на хромосоме для повышения активности белка, или
5) комбинации этих способов, но не ограничиваясь этим.
Как его используют здесь, термин "вектор" относится к ДНК-конструкции, содержащей целевую полинуклеотидную последовательность, функционально связанную с подходящей регуляторной последовательностью, так что целевой ген может быть введен в подходящего хозяина. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любую последовательность оператора для контроля транскрипции, последовательность, кодирующую подходящий домен связывания с рибосомой мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После трансформации подходящей клетки-хозяина вектор может реплицироваться или функционировать независимо от генома хозяина и сам может интегрироваться в геном хозяина. Например, целевой полинуклеотид в хромосоме может быть заменен на модифицированный полинуклеотид с помощью вектора для вставки в хромосому. Вставку полинуклеотида в хромосому можно выполнять с помощью любого способа, известного в данной области техники, например, гомологичной рекомбинацией, но не ограничиваясь этим.
Вектор по настоящему изобретению специально не ограничен, можно использовать любой вектор, известный в данной области техники. Примеры обычно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее, и в качестве плазмидного вектора можно использовать векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и так далее. В частности, можно использовать векторы pDZ, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
Как его используют здесь, термин "трансформация" относится к процессу введения вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, что таким образом дает возможность экспрессии белка, кодируемого этим полинуклеотидом, в клетке-хозяине. При условии, что трансформирующий полинуклеотид может экспрессироваться в клетке-хозяине, не имеет значения, вставлен ли он в хромосому клетки-хозяина и локализован в ней или находится вне хромосомы, и оба случая могут быть включены. Дополнительно, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид может быть введен в любой форме при условии, что он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой генетическую конструкцию, включающую все элементы, необходимые для самостоятельной экспрессии. Обычно экспрессионная кассета может включать промотор, функционально связанный с полинуклеотидом, терминатор транскрипции, домен связывания с рибосомой и терминатор трансляции. Экспрессионная кассета может быть в форме экспрессионного вектора, способного к саморепликации. Дополнительно, полинуклеотид может быть введен в клетку-хозяина как таковой и быть функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине, но не ограничиваясь этим.
Далее, как его используют здесь, термин "функционально связанный" относится к функциональной связи между вышеописанной последовательностью гена и последовательностью промотора, которая инициирует транскрипцию и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению.
Способ трансформации вектором по настоящему изобретению включает любой способ введения нуклеиновой кислоты в клетку, и он может быть выполнен путем выбора подходящего стандартного способа, известного в данной области техники, в зависимости от клетки-хозяина. Например, способ может включать электропорацию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), микроинъекцию, полиэтиленгликолевый (PEG) метод, DEAE (диэтиламиноэтил)-декстрановый метод, катионно-липосомный метод, метод с ацетатом лития-DMSO (диметилсульфоксид) и так далее, но не ограничиваясь ими.
Микроорганизм по настоящему изобретению может включать микроорганизм дикого типа и микроорганизм, генетически модифицированный естественным путем или искусственно. Любой микроорганизм, в который введен или включен чужеродный ген metZ, как описано в настоящем изобретении, может быть включен без ограничения.
Микроорганизм может представлять собой микроорганизм, продуцирующий L-метионин, включающий один или более чем один чужеродный ген metZ по настоящему изобретению; кодируемый им белок и вектор, включающий ген metZ.
Как его используют здесь, термин "микроорганизм, продуцирующий L-метионин" включает все микроорганизмы дикого типа или микроорганизмы, модифицированные естественным путем или искусственно, и он может представлять собой микроорганизм, в котором определенный механизм ослаблен или усилен вследствие вставки чужеродного гена или повышения либо инактивации активности эндогенного гена, и он может представлять собой микроорганизм, включающий генетическую модификацию для продукции желаемого L-метионина.
Микроорганизм, продуцирующий L-метионин, может представлять собой микроорганизм, включающий белок, кодируемый чужеродным геном metZ, по настоящему изобретению для получения повышенной способности продуцировать L-метионин по сравнению с родительским штаммом или немодифицированным микроорганизмом.
Как его используют здесь, термин "штамм до модификации" или "микроорганизм до модификации" может относиться к самому штамму дикого типа или штамму природного типа, не исключая штамм, включающий мутацию, которая может произойти у микроорганизма естественным путем, или к штамму до изменения его признака, который изменен вследствие генетического изменения, вызванного природными или искусственными факторами. Термины "штамм до модификации" или "микроорганизм до модификации" можно использовать взаимозаменяемо с терминами "немодифицированный штамм", "штамм немодифицированного типа", "немодифицированный микроорганизм", "микроорганизм немодифицированного типа" или "референтный микроорганизм". Альтернативно, они могут обозначать микроорганизм, в котором уровни экспрессии генов, вовлеченных в путь биосинтеза L-метионина не регулируются, или микроорганизм, в который изначально отсутствующий ген metZ не введен.
Микроорганизм, продуцирующий L-метионин, по настоящему изобретению может представлять собой микроорганизм, обладающий повышенной способностью продуцировать L-метионин вследствие повышения активности части белков в пути биосинтеза L-метионина или вследствие снижения активности части белков в пути расщепления L-метионина.
В частности, примеры белков или генов, экспрессию которых можно регулировать для усиления пути биосинтеза L-метионина или для ослабления/инактивации пути расщепления L-метионина, являются следующими: белки, репрезентативные гены, кодирующие эти белки, и репрезентативные номера ЕС описаны по порядку. Названия белков начинаются с заглавной буквы, и названия генов обозначены курсивом. Например, путь биосинтеза L-аминокислоты может быть усилен, или путь расщепления L-аминокислоты может быть ослаблен путем повышения активности части одного(одной) или более чем одного(одной) белка или системы, выбранного(ной) из: тиосульфат-сульфотрансферазы, такой как Rdl2p, GlpE, PspE, YgaP, ThiI, YbbB, SseA, YnjE, YceA, YibN, NCgl0671, NCgll369, NCgl2616, NCgl0053, NCgl0054, NCG12678, NCgl2890 и так далее; сульфитредуктазы, су si; транспортной системы тиосульфат/сульфат, cysPUWA (ЕС 3.6.3.25); 3'-фосфоаденозин-5'-фосфосульфатредуктазы, cysH (ЕС 1.8.4.8); сульфитредуктазы, cysJI (ЕС 1.8.1.2); цистеинсинтазы A, cysK (ЕС 2.5.1.47); цистеинсинтазы В, cysM (ЕС 2.5.1.47); серинацетилтрансферазы, cysE (ЕС 2.3.1.30); системы расщепления глицина, gcvTHP-Ipd (ЕС 2.1.2.10, ЕС 1.4.4.2, ЕС 1.8.1.4); липоилсинтазы, lipA (ЕС 2.8.1.8); липоат-протеинлигазы, lipB (ЕС 2.3.1.181); фосфоглицератдегидрогеназы, serA (ЕС 1.1.1.95); 3-фосфосеринфосфатазы, serB (ЕС 3.1.3.3); 3-фосфосерин/фосфогидрокситреонин-амино трансферазы, serC (ЕС 2.6.1.52); серингидроксиметилтрансферазы, glyA (ЕС 2.1.2.1); аспартокиназы I (ЕС 2.7.2.4); гомосериндегидрогеназы I, thrA (ЕС 1.1.1.3); аспартаткиназы, lysC (ЕС 2.7.2.4); гомосериндегидрогеназы, horn (ЕС 1.1.1.3); гомосерин-О-ацетилтрансферазы, metX (ЕС 2.3.1.31); гомосерин-О-сукцинилтрансферазы, metA (ЕС 2.3.1.46); цистатионин-галша-синтазы, metB (ЕС 2.5.1.48); β-C-S-лиазы, aecD (ЕС 4.4.1.8, бета-лиазы); цистатионин-бета-лиазы, metC (ЕС 4.4.1.8); В12-независимой гомоцистеин-S-метилтрансферазы, metE (ЕС 2.1.1.14); метионинсинтазы, metH (ЕС 2.1.1.13); метилентетрагидрофолат-редуктазы, metF (ЕС 1.5.1.20); экспортера L-метионина BrnFE; экспортера валина YgaZH (В2682, В2683), ygaZH (B2682, B2683); экспортера YjeH, b4141; пиридиннуклеотид-трансгидрогеназы PntAB, pntAB (ЕС 1.6.1.2) и фосфоенолпируваткарбоксилазы, Рус (ЕС 4.1.1.31), или путем сверхэкспрессии полинуклеотидов, кодирующих эти белки. Альтернативно, активность одного или более чем одного белка, выбранного из группы, состоящей из глюкозо-6-фосфат-изомеразы, pgi (ЕС 5.3.1.9); гомосеринкиназы, thrB (ЕС 2.7.1.39); S-аденозилметионин-синтазы, metK (ЕС 2.5.1.6); дигидродипиколинат-синтазы, dapA (ЕС 4.2.1.52); фосфоенолпируваткарбоксилазы, рек (ЕС 4.1.1.49); формилтетрагидрофолат-гидролазы, purU (ЕС 3.5.1.10); пируваткиназы I, pykF (ЕС 2.7.1.40); пируваткиназы II, pykA (ЕС 2.7.1.40); цистатионин-у-лиазы, cg3086 (ЕС 4.4.1.1); цистатионин-β-синтазы, cg2344 (ЕС 4.2.1.22); регуляторного белка Cg3031, cg3031; белка-репрессора биосинтеза метионина и цистеина McbR, mcbR; белка-репрессора транскрипции L-метионина (Met), metJ; транспортера L-метионина MetQNI, metQ, metN, metl; N-ацилтрансферазы, yncA; fnrS малой РНК (sRNA) и транспортера L-метионина, metP, может быть инактивирована или снижена, или экспрессия генов, кодирующих эти белки, может быть супрессирована или устранена.
В одном конкретном воплощении микроорганизм, продуцирующий L-метионин, по настоящему изобретению дополнительно к введению metZ может включать одну или более чем одну генетическую модификацию, выбранную из группы, состоящей из ослабления или инактивации активности цистатионин-гамма-синтазы; ослабления или инактивации активности O-ацетилгомосерин-сульфгидрилазы; ослабления или инактивации активности белка-репрессора биосинтеза метионина-цистеина; повышения активности метионинсинтазы и повышения активности сульфитредуктазы. Альтернативно, генетическая модификация может включать одну или более чем одну модификацию, выбранную из группы, состоящей из делеции/игибирования экспрессии гена metB; делеции гена metY; делеции/игибирования экспрессии гена mcbR и усиления экспрессии генов metH и cysI. Например, ген metB, ген metY, ген mcbR, ген metH и ген cysI могут включать полинуклеотидную последовательность по меньшей мере на 70%, 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% гомологичную или идентичную полинуклеотидной последовательности SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 1, SEQ ID NO: 39 и SEQ ID NO: 40, соответственно, но не ограничиваясь ими. Вышеприведенное описание, касающееся гомологии или идентичности, также применимо к генам metB, metY, mcbR, metH и cysI.
Однако эти гены приведены только в качестве примера, и гены не ограничены ими, и микроорганизм может представлять собой микроорганизм, в котором активность различных известных белков пути биосинтеза L-метионина повышена или активность белков пути расщепления L-метионина инактивирована или снижена.
Как его используют здесь, термин "повышение" активности полипептида или белка обозначает увеличение активности полипептида или белка по сравнению с его собственной активностью. Термин "повышение" можно использовать взаимозаменяемо с термином "повышающая регуляция", "сверхэкспрессия", "увеличение" и так далее. Здесь, "увеличение" может включать как проявление изначально отсутствующей активности, так и проявление улучшенной активности, по сравнению с собственной активностью или активностью до модификации. Термин "собственная активность" обозначает активность определенного полипептида или белка, изначально присутствующего у родительского штамма до трансформации или у немодифицированного микроорганизма, когда признак изменен вследствие генетической модификации, вызванной природным или искусственным фактором. Этот термин можно использовать взаимозаменяемо с термином "активность до модификации". Выражение "активность полипептида или белка повышена или увеличена по сравнению с собственной активностью" обозначает, что активность улучшена по сравнению с активностью определенного полипептида или белка, изначально присутствующей у родительского штамма до трансформации или у немодифицированного микроорганизма. "Увеличение активности" может быть достигнуто путем введения чужеродного полипептида или белка или путем повышения активности собственного полипептида или белка, в частности, путем повышения активности собственного полипептида или белка. Повышена ли активность полипептида или белка, можно определить по степени активности соответствующего полипептида или белка, уровню его экспрессии или увеличению количества продукта соответствующего белка.
В данной области техники хорошо известны различные способы, которые можно применять для повышения активности полипептида или белка, и способ не ограничен при условии, что он может повышать активность желаемого полипептида или белка по сравнению с таковой у микроорганизма до модификации. Способ может представлять собой способ использования генетической инженерии и/или белковой инженерии, хорошо известный специалисту в данной области техники, представляющий собой рутинный способ молекулярной биологии (Sitnicka et al. "Functional Analysis of Genes". Advances in Cell Biology. 2010, Vol. 2. 1-16; Sambrook et al. Molecular Cloning 2012 и так далее), но не ограничиваясь этим.
Такой способ повышения активности полипептида или белка с использованием генетической инженерии может быть выполнен, например, с помощью:
1) способа увеличения числа внутриклеточных копий гена или полинуклеотида, кодирующего полипептид или белок;
2) способа замены области регуляции экспрессии гена на хромосоме, кодирующей полипептид или белок, на последовательность, имеющую высокую активность,
3) способа модификации нуклеотидной последовательности стартового кодона или области 5'-UTR полипептида или белка,
4) способа модификации полинуклеотидной последовательности на хромосоме для увеличения активности полипептида или белка,
5) способа введения чужеродного полинуклеотида, проявляющего активность полипептида или белка, или введения варианта полинуклеотида, полученного путем оптимизации кодонов полинуклеотида, или
6) комбинации этих способов, но не ограничиваясь этим.
Способ повышения активности полипептида или белка с использованием белковой инженерии может быть выполнен, например, с помощью способа анализа третичной структуры полипептида или белка, выбора его экспонированного участка и затем изменения или химической модификации этого участка, но не ограничиваясь этим.
1) Способ увеличения числа внутриклеточных копий гена или полинуклеотида, кодирующего полипептид или белок, может быть выполнен любым способом, известным в данной области техники, например, путем введения в клетку-хозяин а вектора, функционально связанного с геном или полинуклеотидом, кодирующим соответствующий полипептид или белок, и обладающего способностью реплицироваться и функционировать независимо от клетки-хозяина. Альтернативно, он может быть выполнен путем введения в клетку-хозяина вектора, функционально связанного с геном и обладающего способностью вставлять ген или полинуклеотид в хромосому клетки-хозяина, но не ограничиваясь этим. Вектор является таким же, как описано выше.
2) Способ замены области регуляции экспрессии гена (или последовательности регуляции экспрессии) на хромосоме, кодирующей полипептид или белок, на последовательность, имеющую высокую активность, может быть выполнен любым способом, известным в данной области техники, например, путем введения мутации в последовательность путем делеции, вставки, неконсервативной или консервативной замены в нуклеотидной последовательности или их комбинации для дополнительного повышения активности области регуляции экспрессии или путем замены последовательности на нуклеотидную последовательность, имеющую более высокую активность. Область регуляции экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания с рибосомой, последовательность, регулирующую терминацию транскрипции и трансляции и так далее, но специально не ограничиваясь этим. Этим способом можно специфически присоединять сильный гетерологичный промотор вместо исходного промотора, но не ограничиваясь этим.
Примеры известных сильных промоторов могут включать промоторы cj1 - cj7 (Патент США No. 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор фага лямбда PR, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13(sm3) (Патент США No. 10584338 В2), промотор O2 (Патент США No. 10273491 В2), промотор tkt, промотор уссА и так далее, но не ограничиваясь ими.
3) Способ модификации нуклеотидной последовательности стартового кодона или области 5'-UTR полипептида или белка может быть выполнен любым способом, известным в данной области техники, например, путем замены собственного стартового кодона полипептида или белка на другой стартовый кодон, обеспечивающий более высокий уровень экспрессии полипептида или белка, чем собственный стартовый кодон, но не ограничиваясь этим.
4) Способ модификации полинуклеотидной последовательности на хромосоме для увеличения активности полипептида или белка может быть выполнен любым способом, известным в данной области техники, например, путем индуцирования мутации в последовательности регуляции экспрессии путем делеции, вставки, неконсервативной или консервативной замены нуклеотидной последовательности или их комбинации для дополнительного повышения активности полинуклеотидной последовательности или путем замены последовательности на полинуклеотидную последовательность, которая улучшена так, чтобы иметь более высокую активность. Замена может представлять собой специфическую вставку гена в хромосому путем гомологичной рекомбинации, но не ограничиваясь этим.
Вектор, который можно использовать здесь, может дополнительно включать селективный маркер для идентификации вставки в хромосому. Селективный маркер является таким же как описано выше.
5) Способ введения чужеродного полинуклеотида, проявляющего активность полипептида или белка, может быть выполнен любым способом, известным в данной области техники, например, путем введения в клетку-хозяина чужеродного полинуклеотида, кодирующего полипептид или белок, проявляющий активность, идентичную/похожую на таковую полипептида или белка, или кодон-оптимизированного варианта его полинуклеотида. Любой чужеродный полинуклеотид можно использовать без ограничения по его происхождению или последовательности при условии, что он проявляет активность, идентичную/похожую на таковую полипептида или белка. Далее чужеродный полинуклеотид можно вводить в клетку-хозяина после оптимизации кодонов, так чтобы происходила его оптимизированная транскрипция и трансляция в клетке-хозяине. Введение может быть выполнено известным способом трансформации, подходящим образом выбранным специалистом в данной области техники, и полинуклеотид, введенный в клетку-хозяина, экспрессируется с получением полипептида или белка, и в результате его активность может быть повышена.
Наконец, (6) может быть выполнена комбинация способов путем применения любого одного или более чем одного способа из (1)-(5).
Такое повышение активности полипептида или белка может представлять собой увеличение активности или концентрации соответствующего полипептида или белка относительно активности или концентрации полипептида или белка, экспрессируемого в штамме дикого типа или штамме микроорганизма до модификации, или увеличение количества продукта, полученного из соответствующего полипептида или белка, но не ограничиваясь этим.
Как его используют здесь, термин "инактивация" или "ослабление" активности полипептида или белка обозначает понятие, включающее все случаи, когда активность уменьшена, или активность отсутствует, по сравнению с собственной активностью. Термины "инактивация" или "ослабление" можно использовать взаимозаменяемо с терминами "понижающая регуляция", "уменьшение", "снижение" и так далее. Понятия "инактивация" или "ослабление" также могут включать случай, когда активность самого белка снижена или устранена вследствие мутации в гене, кодирующем белок, и так далее, по сравнению с активностью белка, изначально присутствующей у микроорганизма, случай, когда общий уровень внутриклеточной активности белка ниже, чем таковой у нативного штамма вследствие ингибирования экспрессии или ингибирования трансляции гена, кодирующего этот белок, случай, когда ген совсем не экспрессируется, и случай, когда белок не проявляет активность, несмотря на то, что экспрессируется. Термин "собственная активность" обозначает активность определенного полипептида или белка, изначально присущую родительскому штамму до трансформации или немодифицированному микроорганизму, когда признак изменен вследствие генетической модификации, вызванной природным или искусственным фактором. Этот термин можно использовать взаимозаменяемо с термином "активность до модификации". Выражение "активность полипептида или белка снижена по сравнению с собственной активностью" обозначает, что активность снижена по сравнению с активностью определенного полипептида или белка, изначально присущей родительскому штамму до трансформации или немодифицированному организму.
Инактивация или ослабление активности белка может быть достигнута(то) различными способами, хорошо известными в данной области техники (Nakashima N et at, "Bacterial cellular engineering by genome editing and gene silencing". Int J Mol Sci. 2014; 15(2):2773-2793; Sambrook et al. Molecular Cloning 2012 и так далее), но не ограничиваясь ими.
Примеры способа могут включать:
1) способ делетирования всего гена, кодирующего белок, или его части;
2) способ модификации области регуляции экспрессии (или последовательности регуляции экспрессии), с тем чтобы уменьшить экспрессию гена, кодирующего белок;
3) способ модификации последовательности гена, кодирующей белок, с тем чтобы устранить или ослабить активность белка;
4) способ введения антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующего белок;
5) способ добавления последовательности, комплементарной последовательности Шайна-Дальгарно, выше по ходу транскрипции от последовательности Шайна-Дальгарно гена, кодирующего белок, для образования вторичной структуры, что таким образом ингибирует присоединение рибосомы; и
6) способ с использованием технологии обратной транскрипции (RTE) с добавлением промотора на 3'-конце открытой рамки считывания (ORF) полинуклеотидной последовательности гена, кодирующего белок, с тем чтобы обеспечить обратную транскрипцию этого гена; и способ может быть выполнен путем их комбинирования, но специально не ограничиваясь этим.
В частности, способ делетирования всего гена, кодирующего белок, или его части может быть выполнен путем замены полинуклеотида, кодирующего желаемый эндогенный белок в хромосоме, на полинуклеотид, имеющий частичную делецию нуклеотидной последовательности или маркерный ген, с помощью вектора для вставки в хромосому микроорганизма. Этот способ делетирования всего полинуклеотида или его части может быть выполнен, например, с помощью способа делетирования полинуклеотида путем гомологичной рекомбинации, но не ограничиваясь этим.
Далее, способ делетирования всего гена или его части может быть выполнен путем индуцирования мутации с использованием излучения, такого как УФ (ультрафиолет), или химического соединения с последующим отбором штамма с делецией целевого гена среди полученных мутантов. Способ делетирования гена включает способ с использованием технологии рекомбинантной ДНК. Технологию рекомбинантной ДНК можно применять, например, путем введения нуклеотидной последовательности или вектора, включающего нуклеотидную последовательность, обладающую гомологией с целевым геном, в микроорганизм для индукции гомологичной рекомбинации. Дополнительно, нуклеотидная последовательность или вектор, подлежащая(щий) введению, может включать доминантный селективный маркер, но не ограничиваясь этим.
Дополнительно, способ модификации последовательности регуляции экспрессии может быть выполнен путем применения различных способов, хорошо известных в данной области техники. Примеры способа могут быть выполнены путем индукции мутации в области регуляции экспрессии (или последовательности регуляции экспрессии) путем делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного снижения активности области регуляции экспрессии (или последовательности регуляции экспрессии) или путем замены последовательности на полинуклеотидную последовательность, имеющую более низкую активность. Область регуляции экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания с рибосомой, последовательность, регулирующую терминацию транскрипции и трансляции и так далее, но не ограничиваясь этим.
Дополнительно, способ модификации последовательности гена может быть выполнен путем индукции мутации в последовательности путем делеции, вставки, неконсервативной или консервативной замены в последовательности гена или их комбинации для дополнительного снижения активности полипептида или путем замены последовательности на последовательность гена, улучшенную так, чтобы снизить активность, или на последовательность гена, улучшенную так, чтобы устранить активность, но не ограничиваясь этим.
Например, экспрессия гена может быть супрессирована или ослаблена путем введения мутации в последовательность гена для образования стоп-кодона.
Однако вышеописанный способ приведен только для примера, и способы повышения или инактивации активности белка и способы управления генами известны в данной области техники и, следовательно, микроорганизм, продуцирующий L-метионин, может быть получен путем применения различных известных способов.
Микроорганизм по настоящему изобретению может представлять собой микроорганизм рода Corynebacterium.
В настоящем изобретении термин "микроорганизм рода Corynebacterium" может включать любые микроорганизмы рода Corynebacterium, в частности, Corynebacterium glutamicum, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium stationis, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium ammoniagenes, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens и более конкретно Corynebacterium glutamicum.
Среда и другие условия культивирования, используемые для культивирования микроорганизма по настоящему изобретению, могут представлять собой любую среду, обычно используемую для культивирования микроорганизмов рода Corynebacterium, без какого-либо специального ограничения. В частности, микроорганизм по настоящему изобретению можно культивировать в аэробных или анаэробных условиях в обычной среде, содержащей подходящий источник углерода, источник азота, источник фосфора, неорганическое соединение, аминокислоту и/или витамин и так далее, при регулировании температуры, рН и так далее.
В настоящем изобретении источник углерода может включать углеводы, такие как глюкоза, фруктоза, сахароза, мальтоза и так далее; сахарные спирты, такие как маннит, сорбит и так далее; органические кислоты, такие как пировиноградная кислота, молочная кислота, лимонная кислота и так далее; аминокислоты, такие как глутаминовая кислота, метионин, лизин и так далее. Дополнительно можно использовать природные органические питательные вещества, такие как гидролизат крахмала, меласса, сырая меласса, рисовые отруби, маниока, меласса сахарного тростника, жидкий кукурузный экстракт и так далее, и в частности, можно использовать такие углеводы как глюкоза, стерилизованная предварительно обработанная меласса (то есть меласса, превращенная в редуцирующий сахар). Дополнительно подходящее количество различных других источников углерода можно использовать без ограничения. Эти источники углерода можно использовать по отдельности или в комбинации из двух или более чем двух из них, но не ограничиваясь этим.
Источник азота может включать неорганические источника азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и так далее; аминокислоты, такие как глутаминовая кислота, метионин, глутамин и так далее; и органические источники азота, такие как пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыба или продукты ее переработки, обезжиренный соевый жмых или продукты его переработки и так далее. Эти источники азота можно использовать по отдельности или в комбинации из двух или более чем двух из них, но не ограничиваясь этим.
Источник фосфора может включать дигидрофосфат калия, гидрофосфат калия или соответствующие натрийс о держащие соли и так далее. Неорганическое соединение может включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и так далее.
Источник серы может включать алкансульфонат, такой как метансульфонат и этансульфонат, органические и неорганические серосодержащие соединения, такие как сульфат, сульфит, сероводород, такой как H2S, сульфид, производные сульфида, смесь органического и неорганического серосодержащего соединения и тиосульфата, такого как тиогликолят, тиоцианат и/или тиомочевина, либо источник серы может не включать другие вещества, кроме тиосульфата, но не ограничиваясь этим.
Способ получения L-метионина по настоящему изобретению может включать культивирование микроорганизма в среде, содержащей тиосульфат. В частности, микроорганизм по настоящему изобретению может представлять собой микроорганизм, содержащий чужеродный metZ и использующий тиосульфат в качестве источника серы. Тиосульфат можно использовать в качестве источника серы для микроорганизма, но не ограничиваясь этим.
В качестве неорганического соединения можно использовать хлорид натрия, хлорид кальция, хлорид железа, карбонат кальция и так далее. Дополнительно среда может включать витамины и/или подходящие предшественники и так далее. Среду или предшественник можно добавлять в культуральную среду периодическим или непрерывным способом, но не ограничиваясь этим.
В настоящем изобретении рН культуральной среды можно регулировать во время культивирования микроорганизма путем добавления в культуральную среду подходящим образом такого соединения как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота, серная кислота и так далее. Далее во время культивирования может быть добавлен пеногаситель, такой как полигликоле вый сложный эфир жирной кислоты, для предупреждения образования пены. Дополнительно, кислород или кислородсодержащий газ можно вводить в культуральную среду для поддержания аэробного состояния культуральной среды; или можно не вводить газ, или можно вводить азот, водород или углекислый газ в культуральную среду для поддержания анаэробного или микро аэробно го состояния, но не ограничиваясь этим.
Температура культуральной среды может составлять 25°С-40°С, и более конкретно, 28°С-37°С, но не ограничиваясь этим. Культивирование можно продолжать до получения желаемого количества полезных веществ и, в частности, в течение от 1 часа до 100 часов, но не ограничиваясь этим.
Способ получения метионина по настоящему раскрытию может включать выделение L-метионина из микроорганизма или среды.
Целевые серосодержащие аминокислоты или производные серосодержащих аминокислот можно выделять из среды подходящим способом, известным в данной области техники, в соответствии со способом культивирования микроорганизма по настоящему изобретению, например, периодическим, непрерывным способом культивирования или периодическим способом культивирования с подпиткой. Например, такие способы как центрифугирование, фильтрование, обработка осадителем, вызывающим кристаллизацию белка (способ высаливания), экстракция, обработка ультразвуком, ультрафильтрация, диализ, различные виды хроматографии, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, аффинная хроматография и так далее, и HPLC (высокоэффективная жидкостная хроматография) можно использовать по отдельности или в комбинации, но способы не ограничены этим.
Способ может включать дополнительный процесс очистки. В процессе очистки можно использовать подходящий способ очистки, известный в данной области техники.
Еще в одном аспекте настоящего раскрытия предложена композиция для получения L-метионина, включающая микроорганизм и тиосульфат.
Композиция по настоящему раскрытию может дополнительно включать любые подходящие эксципиенты, обычно используемые в композициях для получения L-метионина, и эти эксципиенты могут включать, например, консерванты, увлажняющие вещества, диспергирующие вещества, суспендирующие вещества, буферные вещества, стабилизирующие вещества, изотонические вещества и так далее, но не ограничиваясь ими.
Еще в одном аспекте настоящего раскрытия предложен способ получения микроорганизма, продуцирующего L-метионин, включающий стадию введения в микроорганизм белка, кодируемого чужеродным геном metZ.
Еще в одном аспекте настоящего раскрытия предложено применение микроорганизма, в который введен белок, кодируемый чужеродным геном metZ, для продуцирования L-метионина.
Микроорганизм, чужеродный ген metZ, и кодируемый им белок, и введение этого белка являются такими как описано выше.
Здесь и далее настоящее изобретение описано более подробно со ссылкой на примеры воплощений. Однако эти примеры воплощений предназначены исключительно для иллюстративных целей и не предназначены никаким образом ограничивать объем настоящего изобретения.
Референсный Пример 1: Получение плазмиды
Плазмида (pDCM2, Фиг. 1, SEQ ID NO: 81) была сконструирована для вставки и замены гена в хромосоме Corynebacterium, и плазмиду синтезировали с использованием сервиса синтеза генов от Bionics Со. Плазмида была сконструирована так, чтобы включать сайт рестрикции, который легко использовать для клонирования, со ссылкой на статью, в которой рассматривается общеизвестная система sacB (Gene, 145 (1994) 69-73). Синтезированная таким образом плазмида pDCM2 имела следующие характеристики:
1) способность к саморепликации в Е. coli, но отсутствие способности к саморепликации в Corynebacterium вследствие наличия ориджина репликации, который работает только в Е. coli;
2) наличие в плазмиде pDCM2 гена устойчивости к канамицину в качестве селективного маркера;
3) наличие в плазмиде pDCM2 гена левансахаразы (sacB) в качестве вторичного положительного селективного маркера; и
4) плазмида pDCM2 не оставляет никакой генетической информации, имеющей происхождение из нее, в конечном полученном штамме.
Пример 1: Получение рекомбинантного вектора для делеции гена mcbR
В этом примере для получения штамма, продуцирующего метионин, для получения вектора для инактивации mcbR, кодирующего ранее раскрытый белок-репрессор биосинтеза метионина-цистеина (J. Biotechnol. 103:51-65, 2003), использовали штамм АТСС13032 дикого типа.
Более подробно, для делетирования гена mcbR из хромосомы Corynebacterium АТСС13032 был получен рекомбинантный плазмидный вектор следующим способом. На основе нуклеотидных последовательностей, опубликованных в GenBank от Национального института здравоохранения США (NIH), был получен ген mcbR и окружающая его последовательность (SEQ ID NO: 1) Corynebacterium glutamicum.
Для получения делетированного гена mcbR проводили ПЦР на матрице хромосомной ДНК Corynebacterium glutamicum АТСС13032 с использованием праймеров SEQ ID NO: 2 и 3 и SEQ ID NO: 4 и 5 (Таблица 1).
ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, с последующей полимеризацией при 72°С в течение 7 минут. В результате были получены фрагменты ДНК 700 п.н., соответственно.
Вектор pDCM2, не реплицируемый в Corynebacterium glutamicum, и амплифицированные фрагменты гена mcbR обрабатывали рестриктазой smal для введения в хромосому. После реакции клонирования изотермической сборкой продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB (лизогенная среда), содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставкой делетированных фрагментов целевых генов, отбирали путем ПЦР, и методом выделения плазмид была получена плазмида, затем получившая название pDCM2-AmcbR.
Пример 2: Получение и культивирование штамма с делецией гена mcbR
Штамм АТСС13032 трансформировали путем гомологичной рекомбинации на хромосоме с помощью вектора pDC-AmcbR, полученного в Примере 1, путем электропорации (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). После этого проводили вторичную рекомбинацию на твердой среде, содержащей сахарозу. Трансформированный штамм Corynebacterium glutamicum, в котором была завершена вторичная рекомбинация, подвергали ПЦР с использованием SEQ ID NO: 6 и 7 (Таблица 2) для идентификации штамма с делецией гена mcbR. Этот рекомбинантный штамм получил название Corynebacterium glutamicum СМ02-0618.
Этот штамм СМ02-0618 был депонирован в Корейском центре культур микроорганизмов, международном депозитарии, 4 января 2019 года, в соответствии с положениями Будапештского договора и присвоением номера доступа No. KCCM12425P.
Для анализа способности полученного штамма СМ02-0618 продуцировать L-метионин штамм культивировали вместе с его родительским штаммом, штаммом Corynebacterium glutamicum АТСС13032, следующим образом.
Corynebacterium glutamicum АТСС 13032 и Corynebacterium glutamicum СМ02-0618 по настоящему раскрытию инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 25 мл среды, описанной далее, соответственно, и затем культивировали со встряхиванием при 30°С при 200 об./мин в течение 20 часов. После этого 1 мл посевной культуральной среды инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 24 мл продукционной среды, и затем культивировали со встряхиванием при 30°С при 200 об./мин в течение 48 часов. Состав посевной среды и продукционной среды является следующим. В продукционной среде (NH4)2S2O3, представляющий собой тип тиосульфата, использовали в качестве источника серы.
<Посевная среда (рН 7,0)>
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на основе 1 л дистиллированной воды).
<Продукционная среда (рН 8,0)>
50 г глюкозы, 12 г (NH4)2S2O3, 5 г дрожжевого экстракта, 1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3, 1 мкг цианокобаламина (Витамина В12) (на основе 1 л дистиллированной воды).
После культивирования вышеописанным способом анализировали концентрацию L-метионина в культуральной среде, и она представлена в Таблице 3 ниже.

В результате в штамме, из которого был удален только mcbR, наблюдали продукцию L-метионина.
Пример 3-1: Получение векторов для введения трех чужеродных генов metZ
Была сделана попытка преодолеть недостатки существующего способа биосинтеза метионина при одновременном повышении продукции метионина путем введения чужеродного metZ в штамм Corynebacterium. Более подробно, были получены векторы для введения metZ, полученные из Chromobacterium violaceum, Hyphomonas neptunium, Rhodobacter sphaeroides.
Более подробно, рекомбинантные плазмидные векторы для дополнительной вставки каждого из трех типов чужеродных генов metZ в хромосому Corynebacterium АТСС13032 были получены следующими способами.
Сначала для вставки metZ был получен вектор для удаления Ncgl1021 (транспозаза). На основе нуклеотидных последовательностей, опубликованных в GenBank от Национального института здравоохранения США (NIH), были получены Ncgl1021 и окружающая его последовательность (SEQ ID NO: 8) из Corynebacterium glutamicum. Для получения делетированного гена Ncgl1021 проводили ПЦР на матрице хромосомной ДНК Corynebacterium glutamicum АТСС13032 с использованием праймеров SEQ ID NO: 9 и 10 и SEQ ID NO: 11 и 12 (Таблица 4).
ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, с последующей полимеризацией при 72°С в течение 7 минут. В результате были получены соответствующие фрагменты ДНК. Вектор pDCM2, не реплицируемый в Corynebacterium glutamicum, и амплифицированные фрагменты гена Ncgl1021 обрабатывали рестриктазой smal для введения в хромосому. После реакции клонирования изотермической сборкой продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставкой делетированных фрагментов целевых генов, отбирали путем ПЦР, и методом выделения плазмид была получена плазмида, затем получившая название pDCM2-ANcgl1021.
Для получения генов metZ (имеющих происхождение из Chromobacterium violaceum, Hyphomonas neptunium и Rhodobacter sphaeroides) проводили ПЦР на хромосомной ДНК каждого из Chromobacterium violaceum, Hyphomonas neptunium и Rhodobacter sphaeroides в качестве матрицы с использованием праймеров SEQ ID NO: 13 и 14, SEQ ID NO: 15 и 16 и SEQ ID NO: 17 и 18. Для экспрессии трех типов генов metZ, соответственно, использовали промотор Pspl1, и Pspl1 подвергали ПЦР с использованием ранее раскрытой векторной ДНК spll-GFP (KR 10-1783170 В1) в качестве матрицы и праймеров SEQ ID NO: 19 и 20, SEQ ID NO: 19 и 21, SEQ ID NO: 19 и 22 (Таблица 5). ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, с последующей полимеризацией при 72°С в течение 7 минут.

В результате были получены три типа чужеродных генных фрагментов metZ (SEQ ID NO: 63 - 65) и соответствующие промоторные фрагменты spl1 для экспрессии трех типов генов metZ, соответственно. Вектор pDCM2-ΔNcgl1021, не реплицируемый в Corynebacterium glutamicum, обрабатывали рестриктазой scaI. После реакции клонирования изотермической сборкой амплифицированных промоторных фрагментов spll с фрагментами metZ в соответствии с каждым штаммом, каждым продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставкой целевого гена, отбирали путем ПЦР, и методом выделения плазмид были получены в общей сложности три типа плазмид, затем получившие названия pDCM2-ΔNcgl1021-PsplCvimetZ (Chromobacterium violaceum metZ), pDCM2-ΔNcgl1021-PsplHnemetZ (Hyphomonas neptunium metZ) и pDCM2-ΔNcgl1021-PsplRspmetZ (Rhodobacter sphaeroides metZ), соответственно.
Пример 3-2: Получение вектора для введения гена metZ
Дополнительно были получены векторы для последовательностей ранее известных шести генов metZ, имеющих происхождение из Rhodobacter sphaeroides. Таким же способом как в Примере 3-1 (смотрите US 2013-0273614 A1 и US 2018-0355389 A1), были получены векторы с генами metZ, каждый из которых кодирует аминокислотные последовательности SEQ ID NO: 66-71, соответственно. Гены metZ получили названия RspmetZ_long, RspmetZ_3, RspmetZ_65, RspmetZ_104, RspmetZ_196 и RspmetZ_3_65_104, соответственно, и праймеры, используемые для введения каждого гена, были следующими.

Сначала, для получения RspmetZlong проводили ПЦР на матрице хромосомной ДНК Rhodobacter sphaeroides с использованием праймеров SEQ ID NO: 19 и 22 и SEQ ID NO: 72 и 18.
В результате были получены генный фрагмент и промоторный фрагмент spl1.
Вектор pDCM2-ΔNcgl1021, не реплицируемый в Corynebacterium glutamicum, обрабатывали рестриктазой scaI. После реакции клонирования изотермической сборкой амплифицированных промоторных фрагментов spl1 с фрагментами metZ в соответствии с каждым штаммом, каждым продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставкой целевого гена, отбирали путем ПЦР, и методом выделения плазмид была получена плазмида, затем получившая название pDCM2-ΔNcgl1021-PsplRspmetZ_long.
Для получения RspmetZ_3, RspmetZ_65, RspmetZ_104, RspmetZ_196 и RspmetZ_3_65_104 проводили ПЦР на матрице вектора pDCM2-ΔNcgl1021-PsplRspmetZ long с использованием SEQ ID NO: 72 и 73, 74 и 18 (RspmetZ_3), SEQ ID NO: 72 и 75, 76 и 18 (RspmetZ_65), SEQ ID NO: 72 и 77, 78 и 18 (RspmetZ_104), SEQ ID NO: 71 и 79, 80 и 18 (RspmetZ 196), SEQ ID NO: 72 и 73, 74 и 75, 76 и 77, 77 и 18 (RspmetZ_3_65 104). ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, с последующей полимеризацией при 72°С в течение 7 минут.
В результате было получено в общей сложности десять фрагментов.

Вектор pDCM2-ΔNcgl1021, не реплицируемый в Corynebacterium glutamicum, обрабатывали рестриктазой scaI. После реакции клонирования изотермической сборкой RspmetZ_3 и фрагмента 1, фрагмента 2, RspmetZ_65 и фрагмента 3, фрагмента 4, RspmetZ_104 и фрагмента 5, фрагмента 6, RspmetZ_196 и фрагмента 7, фрагмента 8, RspmetZ_3_65_104 и фрагмента 1, фрагмента 11, фрагмента 12, фрагмента 6, затем каждым продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставкой целевого гена отбирали путем ПЦР, и методом выделения плазмид были получены в общей сложности шесть типов плазмид, затем получивших названия pDCM2-ΔNcgl1021-PsplRspmetZ_long, pDCM2-ΔNcgl1021-PsplRspmetZ_3, pDCM2-ΔNcgl1021-PsplRspmetZ_65, PDCM2-ΔNcgl1021-PsplRspmetZ_104, pDCM2-ΔNcgl1021-PsplRspmetZ_196, PDCM2-ΔNcgl1021-PsplRspmetZ_3_65_104, соответственно.
Пример 4: Получение и культивирование штамма с введенным чужеродным metZ
Девять типов чужеродных генов metZ были введены в штамм СМ02-0618, представляющий собой штамм, продуцирующий метионин, полученный в Примере 2, соответственно.
Более подробно, штаммом СМ02-0618, представляющим собой штамм, продуцирующий метионин, полученный в Примере 2, трансформировали путем гомологичной рекомбинации на хромосоме векторами pDCM2-ANcgl1021, pDCM2-ΔNcgl1021-PsplCvimetZ, pDCM2-ΔNcgl1021-PsplHnemetZ, pDCM2-ΔNcgl1021-PsplRspmetZ, pDCM2-ΔNcgl1021-PsplRspmetZ_long, pDCM2-ΔNcgl1021-PsplRspmetZ_3, pDCM2-ΔNcgl1021-PsplRspmetZ_65, pDCM2-ΔNcgl1021-PsplRspmetZ_104, pDCM2-ΔNcgl1021-PsplRspmetZ_196 и pDCM2-ΔNcgl1021-PsplRspmetZ_3_65_104, полученными в Примере 3 путем электропорации, соответственно (van der Rest et al., ApplMicrobiol Biotechnol 52:541-545, 1999).
После этого проводили вторичную рекомбинацию на твердой среде, содержащей сахарозу. Трансформированный штамм Corynebacterium glutamicum, в котором была завершена вторичная рекомбинация, подвергали ПЦР с использованием SEQ ID NO: 23 и 24 для выявления штамма с делецией гена Ncgl1021 и штамма с делецией Ncgl1021 и вставкой metZ. Рекомбинантные штаммы, каждый из которых получен путем введения одного из девяти типов векторов в СМ02-0618, получили названия CM02-0618/ANcgl021 и СМ02-0757, СМ02-0758, СМ02-0759-1, СМ02-0759-2, СМ02-0759-3, СМ02-0759-4, СМ02-0759-5 и СМ02-0759, соответственно.
Для изучения способности полученных штаммов продуцировать L-метионин каждый штамм культивировали вместе с их родительским штаммом СМ02-0618 следующим образом.
Каждый из штаммов инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 25 мл описанной ниже среды, и затем культивировали со встряхиванием при 30°С при 200 об./мин в течение 20 часов. После этого 1 мл посевной культуральной среды инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 24 мл продукционной среды, и затем культивировали со встряхиванием при 30°С при 200 об./мин в течение 48 часов. Состав посевной среды и продукционной среды является следующим.
<Посевная среда (рН 7,0)>
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на основе 1 л дистиллированной воды)
<Продукционная среда (рН 8,0)>
50 г глюкозы, 12 г (NH4)2S2O3, 5 г дрожжевого экстракта, 1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3, 1 мкг кобаламина (Витамина В12) (на основе 1 л дистиллированной воды).
Далее, для сравнения с источником серы, используемым существующим метионин-превращающим ферментом, источник серы, используемый в продукционной среде, был заменен с тиосульфата (S2O3) на метилмеркаптан (CH3SH), и штаммы культивировали таким же образом. После завершения культивирования каждого штамма измеряли концентрацию L-метионина в каждой культуральной среде, и результаты представлены в Таблице 8.

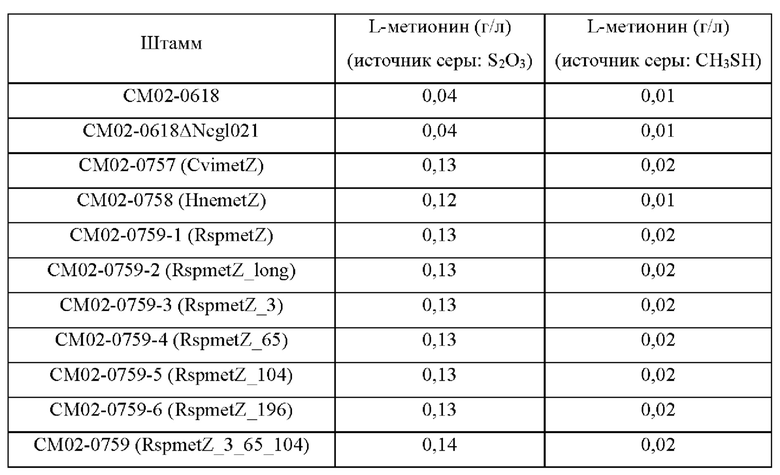
В результате при введении девяти типов генов metZ, соответственно, продуктивность по L-метионину повышалась на 266% или более по сравнению с контрольным штаммом, показывая, что чужеродный metZ по настоящему раскрытию значительно повышает продуктивность по L-метионину путем сульфгидрилирования, и в частности, эффективность является высокой по сравнению с таковой при использовании метилмеркаптана в качестве источника серы в существующем способе. Это можно объяснить так, что в отличие от metB и metY Corynebacterium glutamicum, чужеродный metZ по настоящему раскрытию не претерпевал ингибирования по типу обратной связи, и таким образом выход метионина увеличился.
Штаммы СМ02-0757, СМ02-0758 и СМ02-0759 депонировали в Корейском центре культур микроорганизмов, международном депозитарии, 2 мая 2019 года в соответствии с положениями Будапештского договора и присвоением номеров доступа No. KCCM12506P, KCCM12507P, KCCM12508P, соответственно.
Пример 5: Получение рекомбинантного вектора для делеции генов metB и metY
Для изучения функции (активности) белка, кодируемого metZ, по настоящему раскрытию и для сравнения его активности с активностью metY и metB, metB и metY, присутствующие у Corynebacterium glutamicum, были делетированы, соответственно. Более подробно, для делетирования генов metB и metY, соответственно, следующим способом были получены рекомбинантные плазмидные векторы. На основе нуклеотидных последовательностей, опубликованных в GenBank от Национального института здоравоохранения США (NIH), были получены гены metB и metY и окружающие их последовательности (SEQ ID NO: 25 и 26) из Corynebacterium glutamicum.
Для осуществления делетирования генов metB и metY, соответственно, проводили ПЦР с хромосомной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы с использованием праймеров SEQ ID NO: 27 и 28 и SEQ ID NO: 29 и 30 (metB) и праймеров SEQ ID NO: 31 и 32 и SEQ ID NO: 33 и 34 (metY) (Таблица 9).
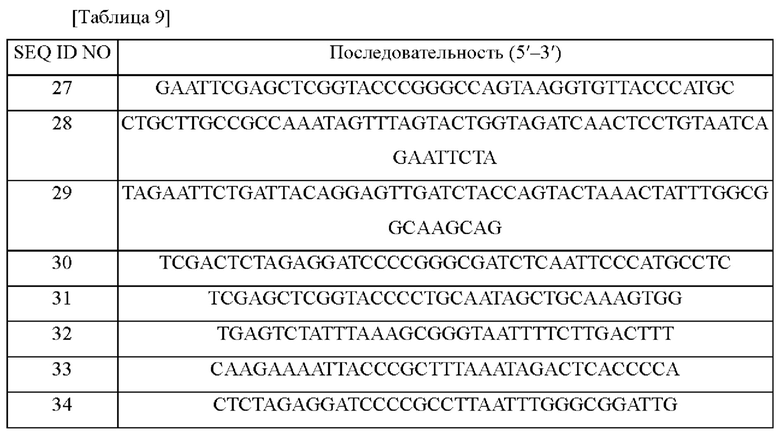
ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, с последующей полимеризацией при 72°С в течение 7 минут. В результате были получены фрагменты ДНК 700 п.н., соответственно.
Вектор pDCM2, не реплицируемый в Corynebacterium glutamicum, и амплифицированные фрагменты генов metB и metY обрабатывали рестриктазой smaI для введения в хромосому, соответственно. После лигирования с помощью ДНК-лигазы затем продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные каждым вектором со вставками делетированных фрагментов целевого гена, отбирали путем ПЦР, и каждую плазмиду получали методом выделения плазмид, и затем полученные плазмиды получили названия pDCM2-ΔmetB и pDCM2-ΔmetY, соответственно.
Пример 6: Получение штаммов с делецией генов metB или metY из трех типов штаммов с усиленным metZ и культивирование полученных штаммов
Штаммы СМ02-0618, СМ02-0757, СМ02-0758 и СМ02-0759 трансформировали путем гомологичной рекомбинации на хромосоме векторами pDCM2-ΔmetB и pDCM2-ΔmetY, полученными, как описано выше, путем электропорации, соответственно (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Затем проводили вторичную рекомбинацию на твердой среде, содержащей сахарозу. Трансформированные штаммы Corynebacterium glutamicum, в которых вторичная рекомбинация была завершена, исследовали на наличие делеции генов metB и metY с использованием SEQ ID NO: 35 и 36 (metB) и SEQ ID NO: 37 и 38 (metY) (Таблица 10), соответственно.
Эти рекомбинантные штаммы получили названия Corynebacterium glutamicum CM02-0618/ΔmetB CM02-0757/ΔmetB, CM02-0758/ΔmetB, CM02-0759/ΔmetB, CM02-0618/ΔmetY, CM02-0757/ΔmetY, CM02-0758/ΔmetY, CM02-0759/ΔmetY, соответственно.
Для анализа способности полученных штаммов CM02-0618/ΔmetB, СМ02-0757/ ΔmetB, CM02-0758/ΔmetB, CM02-0759/ΔmetB, CM02-0618/ΔmetY, CM02-0757/ΔmetY, CM02-0758/ΔmetY и CM02-0759/ΔmetY продуцировать L-метионин их культивировали следующим способом, соответственно.
СМ02-0618, СМ02-0757, СМ02-0758 и СМ02-0759 в качестве родительских штаммов и CM02-0618/ΔmetB, СМ02-0757/ ΔmetB, CM02-0758/ΔmetB, СМ02-0759/ ΔmetB, CM02-0618/ΔmetY, CM02-0757/ΔmetY, CM02-0758/ΔmetY и CM02-0759/AmetY, полученные как указано выше, инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 25 мл среды, описанной ниже, соответственно, и затем культивировали с встряхиванием при 30°С при 200 об./мин в течение 20 часов. После этого 1 мл каждой культуральной среды инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 24 мл продукционной среды, и затем культивировали с встряхиванием при 30°С при 200 об./мин в течение 48 часов. Состав посевной среды и продукционной среды является следующим.
<Посевная среда (рН 7,0)>
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на основе 1 л дистиллированной воды)
<Продукционная среда (рН 8,0)>
50 г глюкозы, 12 г (NH4)2S2O3, 5 г дрожжевого экстракта, 1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3, 1 мкг кобаламина (витамина В12) (на основе 1 л дистиллированной воды).
Анализировали концентрации L-метионина и побочного продукта гомолантионина в каждой культуральной среде, культивируемой описанным выше способом культивирования,, и они представлены в Таблице 11.
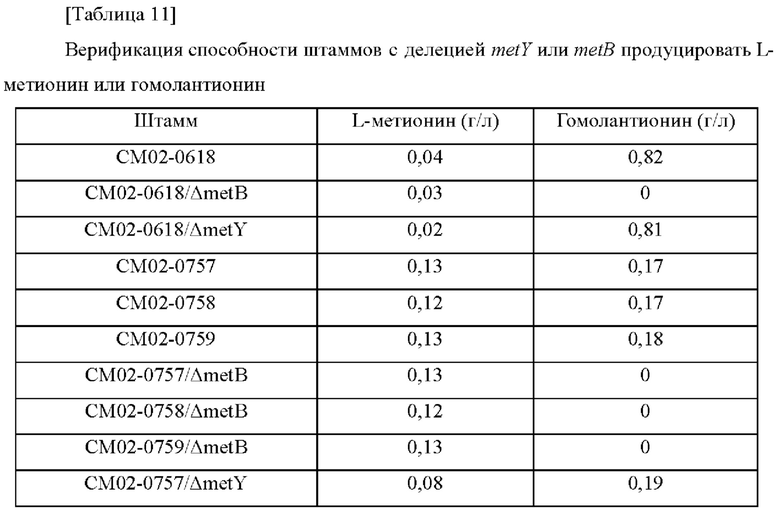

Эти результаты подтверждают, что чужеродный metZ выполняет такую же функцию как metB, то есть продуцирует метионин путем транссульфурации, используя цистеин в качестве источника серы. Другими словами, даже несмотря на делецию metB или metY, продуцирование метионина может поддерживаться с высоким выходом посредством использования metZ, и следовательно, этот ген может быть заменен на чужеродный metZ для компенсации недостатков metB и/или metY у Corynebacterium.
Далее, в штамме, в котором присутствует metB, и в который введен чужеродный metZ, продукция метионина была повышена, а продукция гомолантионина составила только примерно 20% относительно контроля (СМ02-0618), указывая на то, что продукция побочного продукта, гомолантионина, была снижена при усилении metZ. Гомолантионин представляет собой вещество, синтезированное путем потребления О-ацетилгомосерина, и, следовательно, продуцирование гомолантионина снижает продукцию метионина. Чужеродный metZ, который ингибирует продуцирование побочного продукта, компенсирует недостатки metB путем ингибирования продукции побочного продукта и усиления синтеза метионина.
Вышеприведенные результаты впервые выявили, что metZ по настоящему раскрытию может опосредовать транссульфурацию путем использования цистеина, и metZ по настоящему раскрытию можно использовать не только для увеличения продукции метионина, но также для компенсации недостатков metB у штамма рода Corynebacterium.
Пример 7: Получение рекомбинантного вектора для одновременного усиления metH и cysI
В настоящем примере для получения штамма, продуцирующего метионин, в котором mcbR не делетирован, был получен вектор для усиления metH, кодирующего метионинсинтазу (Ncgll450), и cysI, кодирующего сульфитредуктазу (Ncgl2718), одновременно.
Более подробно, для дополнительной вставки генов metH и cysI в хромосому Corynebacterium АТСС 13032 следующим способом был получен рекомбинантный плазмидный вектор. На основе нуклеотидных последовательностей, опубликованных в GenBank от Национального института здравоохранения США (NIH), был получен ген metH и окружающая его последовательность (SEQ ID NO: 39) и ген cysI и окружающая его последовательность (SEQ ID NO: 40) из Corynebacterium glutamicum.
Для получения генов metH и cysI проводили ПЦР с хромосомной ДНК Corynebacterium glutamicum АТСС 13032 в качестве матрицы с использованием праймеров SEQ ID NO: 41 и 42 и SEQ ID NO: 43 и 44. Промотор Pcj7 использовали для усиления экспрессии гена metH и промотор Pspl1 использовали для усиления экспрессии гена cysI. Для получения каждого промотора проводили ПЦР: для Pcj7 на хромосомной ДНК Corynebacterium ammoniagenes АТСС6872 в качестве матрицы с использованием SEQ ID NO: 45 и 46 и для Pspl1 с использованием ДНК ранее раскрытого вектора spl1-GFP (KR 10-1783170 В1) в качестве матрицы с использованием SEQ ID NO: 47 и 48. Последовательности используемых праймеров представлены в Таблице 12 ниже.

ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 4 минут, с последующей полимеризацией при 72°С в течение 7 минут. В результате были получены фрагменты ДНК metH и cysI, промотора Pcj7 и промотора Pspl1, соответственно.
Вектор pDCM2-ΔNcgl1021, не реплицируемый в Corynebacterium glutamicum, обрабатывали рестриктазой scaI, и четыре амплифицированных фрагмента ДНК обрабатывали scaI для введения в хромосому. После реакции IST (иодат-сульфит-тиосульфат) продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором, в который были вставлены делетированные фрагменты целевого гена, отбирали путем ПЦР, и методом выделения плазмид получали плазмиду, которая затем получила название pDCM2-ANcgl1021-Pcj7metH-Pspl1cysI.
Пример 8: Получение штамма для одновременного усиления metH и cysI и продуцирование L-метионина с использованием этого штамма
Штамм АТСС 13032 трансформировали путем гомологичной рекомбинации на хромосоме векторами pDCM2-ANcgl1021 и pDCM2-ANcgl1021-Pcj7metH-Pspl1cysI, полученными как описано выше путем электропорации (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999), соответственно. После этого проводили вторичную рекомбинацию на твердой среде, содержащей сахарозу. Трансформированные штаммы Corynebacterium glutamicum, в которых была завершена вторичная рекомбинация, исследовали на наличие делеции гена Ncgl1021 и вставки гена Pcj7-metH-Pspl1cysI с использованием SEQ ID NO: 23 и 24, соответственно. Рекомбинантные штаммы получили названия 1303 2/ΔNcgl1021 и СМ02-0753, соответственно.
Для исследования способности полученных штаммов 13032/ΔNcgl1021 и СМ02-0753 продуцировать L-метионин их культивировали вместе с родительским штаммом Corynebacterium glutamicum АТСС 13032 следующим способом, соответственно.
Corynebacterium glutamicum АТСС13032 и штаммы 13032/ΔNcgl1021 и СМ02-0753, полученные как описано выше, инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 25 мл описанной ниже среды, соответственно, и затем культивировали с встряхиванием при 30°С при 200 об./мин в течение 20 часов. После этого 1 мл каждой посевной среды инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 24 мл продукционной среды, и затем культивировали с встряхиванием при 30°С при 200 об./мин в течение 48 часов. Состав посевной среды и продукционной среды является следующим.
<Посевная среда (рН 7,0)>
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина-HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на основе 1 л дистиллированной воды)
<Продукционная среда (рН 8,0)>
50 г глюкозы, 12 г (NH4)2S2O3, 5 г дрожжевого экстракта, 1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3, 1 мкг кобаламина (витамина В12) (на основе 1 л дистиллированной воды).
Анализировали концентрации L-метионина в каждой культуральной среде, культивируемой вышеописанным способом культивирования, и они представлены в Таблице 13.

В результате было подтверждено, что способность продуцировать L-метионин штамма, сверхэкспрессирующего metH и cysI, в котором все еще присутствует mcbR, была повышена на 0,03 г/л по сравнению с контрольным штаммом.
Пример 9-1: Получение вектора для усиления генов metZ в сайтах, отличных от сайта Ncgl1021
Для вставки трех типов чужеродных генов metZ в сайты, отличные от существующего сайта на хромосоме Corynebacterium АТСС13032, рекомбинантные плазмидные векторы были получены следующим способом.
Сначала для вставки metZ был получен вектор для делеции Ncgl2748 (транспозаза). На основе нуклеотидных последовательностей, опубликованных в GenBank от Национального института здравоохранения США (NIH), были получены Ncgl2748 и окружающая его последовательность (SEQ ID NO: 49) из Corynebacterium glutamicum. Для осуществления делетирования гена Ncgl2748 проводили ПЦР на хромосомной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы с использованием праймеров SEQ ID NO: 50 и 51 и SEQ ID NO: 52 и 53 (Таблица 14). ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, с последующей полимеризацией при 72°С в течение 7 минут. В результате были получены соответствующие фрагменты ДНК.
Вектор pDCM2, не реплицируемый в Corynebacterium glutamicum, и амплифицированные генные фрагменты Ncgl2748 обрабатывали рестриктазой smaI для введения в хромосому. После реакции 1ST продуктом затем трансформировали Е. coli DH5a с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставками делетированных целевых генных фрагментов, отбирали путем ПЦР, и методом выделения плазмид получали плазмиду, которая затем получила название PDCM2-ANcgl2748.
Для получения трех типов генов metZ (Chromobacterium violaceum, Hyphomonas neptunium, Rhodobacter sphaeroides) проводили ПЦР с полученными векторами pDCM2-ΔNcgl1021-PsplCvimetZ (metZ Chromobacterium violaceum), pDCM2-ΔNcgl1021-PsplHnemetZ (metZ Hyphomonas neptunium) и pDCM2-ΔNcgl1021-PsplRspmetZ (metZ Rhodobacter sphaeroides) в качестве матрицы с использованием SEQ ID NO: 54 и 55, 54 и 56, и 54 и 57 (Таблица 15). ПЦР проводили в следующих условиях: денатурация при 95°С в течение 5 минут, в общей сложности 30 циклов, состоящих из денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд, с последующей полимеризацией при 72°С в течение 7 минут. В результате были получены три фрагмента ДНК.

Вектор pDCM2-ANcgl2748, не реплицируемый в Corynebacterium glutamicum, обрабатывали рестриктазой scaI. Затем, после реакции 1ST с амплифицированными фрагментами в соответствии с каждым штаммом, продуктом трансформировали Е. coli DH5α с последующим нанесением полученных клеток на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором со вставками целевого гена, отбирали путем ПЦР, и были получены в общей сложности три плазмиды методом выделения плазмид, которые затем получили названия pDCM2-ANcgl2748-PsplCvimetZ (Chromobacterium violaceum metZ), pDCM2-ANcgl2748-PsplHnemetZ (Hyphomonas neptunium metZ), pDCM2-ANcgl2748-PsplRspmetZ (Rhodobacter sphaeroides metZ), соответственно.
Пример 9-2: Получение вектора для усиления генов metZ в сайтах, отличных от сайта Ncgl1021
Дополнительно для исследования того, повышается ли также продукция метионина у штамма, в который введен ген metZ, имеющий 99% или более гомологии последовательности, были дополнительно получены векторы для пяти генов metZ из Примеров 3-2.
Векторы, в каждый из которых введен один из шести генов metZ, были получены таким же способом как в Примере 9-1.
Всего были получены шесть векторов, под названиями pDCM2-ANcgl2748-PsplRspmetZlong, ΔNcgl2748-PsplRspmetZ_3, pDCM2-ANcgl2748-PsplRspmetZ_65, pDCM2-ΔNcgl2748-PsplRspmetZ 104, pDCM2-ANcgl2748-PsplRspmetZ_196 и pDCM2-ΔNcgl2748-PsplRspmetZ_3_65_104, соответственно, следующим образом. Для получения этих векторов ANcgl102l-PsplRspmetZ_long pDCM2-ΔNcgl1021-PsplRspmetZ_3, pDCM2-ANcgl 1021-PsplRspmetZ_65, pDCM2-ΔNcgl1021-PsplRspmetZ_104, pDCM2-ANcgl1021-PsplRspmetZ_196 и pDCM2-ΔNcgl1021-PsplRspmetZ_3_65_104, полученные в Примере 4, использовали в качестве ДНК-матрицы, соответственно, и обычно использовали праймеры SEQ ID NO: 54 и 57. Другие методики были такими же как в Примере 9-1.
Пример 10: Разработка штамма, усиленного чужеродным metZ, на основе имеющего mcbR штамма, продуцирующего L-метионин, и получение L-метионина с использованием этого штамма
Штамм СМ02-0753 трансформировали путем гомологичной рекомбинации на хромосоме векторами pDCM2-ANcgl2748, pDCM2-ANcgl2748-PsplCvimetZ, pDCM2-ΔNcgl2748-PsplHnemetZ, pDCM2-ΔNcgl2748-PsplRspmetZ, pDCM2-ΔNcgl2748-PsplRspmetZ long, pDCM2-ΔNcgl2748-PsplRspmetZ_3, pDCM2-ΔNcgl2748-PsplRspmetZ_65, pDCM2-ΔNcgl2748-PsplRspmetZ_104, PDCM2-ΔNcgl2748-PsplRspmetZ_196 и pDCM2-ΔNcgl2748-PsplRspmetZ_3_65_104, полученными как описано выше путем электропорации, соответственно (van der Rest et al., Appl Microbiol Biotechnol 52:541 545, 1999). После этого проводили вторичную рекомбинацию на твердой среде, содержащей сахарозу. Трансформированные штаммы Corynebacterium glutamicum, в которых была завершена вторичная рекомбинация, исследовали на вставку каждого чужеродного гена metZ в сайте Ncgl2748 с использованием SEQ ID NO: 58 и 59 (Таблица 16), соответственно.
Эти рекомбинантные штаммы получили названия Corynebacterium glutamicum 13032/ANcgl2748, СМ02-0765, СМ02-0766, СМ02-0767-1, СМ02-0767-2, СМ02-0767-3, СМ02-0767-4, СМ02-0767-5, СМ02-0767-6, СМ02-0767, соответственно.
Для исследования способности полученных штаммов продуцировать L-метионин их культивировали вместе с родительским штаммом Corynebacterium glutamicum СМ02-0753 следующим способом, соответственно.
Каждый штамм инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 25 мл описанной ниже среды, соответственно, и затем культивировали со встряхиванием при 30°С и 200 об./мин в течение 20 часов. После этого 1 мл каждой посевной среды инокулировали в колбу с угловыми перегородками на 250 мл, содержащую 24 мл продукционной среды, и затем культивировали со встряхиванием при 30°С при 200 об./мин в течение 48 часов. Состав посевной среды и продукционной среды является следующим.
<Посевная среда (рН 7,0)>
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на основе 1 л дистиллированной воды)
<Продукционная среда (рН 8,0)>
50 г глюкозы, 12 г (NH4)2S2O3, 5 г дрожжевого экстракта, 1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамин-HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3, 1 мкг кобаламина (витамина В12) (на основе 1 л дистиллированной воды).
Анализировали концентрации L-метионина в каждой культуральной среде, культивированной вышеописанным способом культивирования, и они представлены в Таблице 17.

В результате было подтверждено, что при введении чужеродных генов metZ в штамм, продуцирующий метионин, имеющий mcbR, соответственно, выход метионина также увеличивался.
СМ02-0765, СМ02-0766 и СМ02-0767 были депонированы в Корейском центре культур микроорганизмов, международном депозитарии, 2 мая 2019 года, соответствии с положениями Будапештского договора и присвоением номеров доступа No. КССМ12509Р, КССМ12510Р и КССМ12511Р, соответственно.
На основе вышеприведенного описания специалист в данной области техники понимает, что настоящее изобретение может быть выполнено в различных конкретных формах без изменения его технической идеи или существенных признаков. В этом отношении следует понимать, что вышеприведенное описание не является ограничивающим, а является иллюстративным во всех аспектах. Объем настоящего изобретения определяет прилагаемая формула изобретения, а не предшествующее ей описание и, следовательно, подразумевается, что формула изобретения охватывает все изменения и модификации, попадающие в объем формулы изобретения, или их эквиваленты.
Технический эффект изобретения
Микроорганизм, в который введена активность белка, кодируемого metZ, по настоящему раскрытию демонстрирует высокий выход, потому что он производит меньше побочных продуктов по сравнению с metY, и микроорганизм не получает регуляцию по типу братной связи в отличие от metB, получающему ингибирование метионином по типу обратной связи, и следовательно продуцирует L-метионин с высоким выходом, что делает полезным его применение в промышленном производстве L-метионина.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ L-МЕТИОНИН, В КОТОРЫЙ ВВЕДЕН БЕЛОК,
КОДИРУЕМЫЙ ЧУЖЕРОДНЫМ ГЕНОМ metZ, И СПОСОБ ПОЛУЧЕНИЯ L-МЕТИОНИНА
С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА
<130> OPA20069
<150> KR 10-2019-0134797
<151> 2019-10-28
<160> 81
<170> KoPatentIn 3.0
<210> 1
<211> 2642
<212> DNA
<213> Corynebacterium glutamicum
<400> 1
ctcccgcgca ctgctgcaat ccgcaccgtg cccaatgatg gtggttcgcc cacctgagaa 60
gattaagaag tagtttcttt taagtttcga tgccccggtt tcctgatttt gtgcagggag 120
gccggggcat tggtgtttgc gggttagttc gggccattcg aaagggagaa accaagggca 180
gccagacaga cgtgccaaga atctggattt ccgccaggtt ttggcacgcc cgtctggttt 240
aggcaatgag ataccgaaca cacgtgccaa aagttcggct ttttcgccga tcttgtcacg 300
cctgcctggt ttgtcttgta aagagtgatt tcatggccga gactcctaaa agtttgacct 360
cacaggattg cttctaaggg cctctccaat ctccactgag gtacttaatc cttccgggga 420
attcgggcgc ttaaatcgag aaattaggcc atcacctttt aataacaata caatgaataa 480
ttggaatagg tcgacacctt tggagcggag ccggttaaaa ttggcagcat tcaccgaaag 540
aaaaggagaa ccacatgctt gccctaggtt ggattacatg gatcattatt ggtggtctag 600
ctggttggat tgcctccaag attaaaggca ctgatgctca gcaaggaatt ttgctgaaca 660
tagtcgtcgg tattatcggt ggtttgttag gcggctggct gcttggaatc ttcggagtgg 720
atgttgccgg tggcggcttg atcttcagct tcatcacatg tctgattggt gctgtcattt 780
tgctgacgat cgtgcagttc ttcactcgga agaagtaatc tgctttaaat ccgtagggcc 840
tgttgatatt tcgatatcaa caggcctttt ggtcattttg gggtggaaaa agcgctagac 900
ttgcctgtgg attaaaacta tacgaaccgg tttgtctata ttggtgttag acagttcgtc 960
gtatcttgaa acagaccaac ccgaaaggac gtggccgaac gtggctgcta gcgcttcagg 1020
caagagtaaa acaagtgccg gggcaaaccg tcgtcgcaat cgaccaagcc cccgacagcg 1080
tctcctcgat agcgcaacca accttttcac cacagaaggt attcgcgtca tcggtattga 1140
tcgtatcctc cgtgaagctg acgtggcgaa ggcgagcctc tattcccttt tcggatcgaa 1200
ggacgccttg gttattgcat acctggagaa cctcgatcag ctgtggcgtg aagcgtggcg 1260
tgagcgcacc gtcggtatga aggatccgga agataaaatc atcgcgttct ttgatcagtg 1320
cattgaggaa gaaccagaaa aagatttccg cggctcgcac tttcagaatg cggctagtga 1380
gtaccctcgc cccgaaactg atagcgaaaa gggcattgtt gcagcagtgt tagagcaccg 1440
cgagtggtgt cataagactc tgactgattt gctcactgag aagaacggct acccaggcac 1500
cacccaggcg aatcagctgt tggtgttcct tgatggtgga cttgctggat ctcgattggt 1560
ccacaacatc agtcctcttg agacggctcg cgatttggct cggcagttgt tgtcggctcc 1620
acctgcggac tactcaattt agtttcttca ttttccgaag gggtatcttc gttgggggag 1680
gcgtcgataa gccccttctt tttagcttta acctcagcgc gacgctgctt taagcgctgc 1740
atggcggcgc ggttcatttc acgttgcgtt tcgcgcctct tgttcgcgat ttctttgcgg 1800
gcctgttttg cttcgttgat ttcggcagta cgggttttgg tgagttccac gtttgttgcg 1860
tgaagcgttg aggcgttcca tggggtgaga atcatcaggg cgcggttttt gcgtcgtgtc 1920
cacaggaaga tgcgcttttc tttttgtttt gcgcggtaga tgtcgcgctg ctctaggtgg 1980
tgcactttga aatcgtcggt aagtgggtat ttgcgttcca aaatgaccat catgatgatt 2040
gtttggagga gcgtccacag gttgttgctg acccaataga gtgcgattgc tgtggggaat 2100
ggtcctgtga ggccaaggga cagtgggaag atcggcgcga ggatcgacat cacgatcatg 2160
aacttcagca tgccgttaga gaatccggat gcgtaatcgt tggtttggaa gctgcggtac 2220
atggacatcg ccatgttgat tgcggtgagg attgcggctg tgatgaacag tggcaaaacg 2280
aaactaagaa cttccgcctg cgtggtgctc aaatatttta gctgctcagt gggcatcgaa 2340
acataagcgg gcagaggcac attgctcacg cgaccagcga ggaaagattc cacttcctca 2400
ggagttagga agccgatcga ctggaagacg ggattttcca aaccaccttc agggcgagcc 2460
atgcggagaa gtgcccagta aagaccaagg acaatcggta tctggatcag cccaggcaca 2520
caacctgcca gcgggttaat gccgtattcc ttattcaaat cattctggcg cttctgcaac 2580
tcccgaatgg acgcttcatc gtactttccc ttgtattctt cccggagcgc agcgcggtga 2640
gg 2642
<210> 2
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 2
tcgagctcgg tacccctgcc tggtttgtct tgta 34
<210> 3
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 3
cggaaaatga agaaagttcg gccacgtcct ttcgg 35
<210> 4
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 4
aggacgtggc cgaactttct tcattttccg aaggg 35
<210> 5
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 5
ctctagagga tccccgtttc gatgcccact gagca 35
<210> 6
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 6
aatctggatt tccgccaggt 20
<210> 7
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 7
cttcctaact cctgaggaag 20
<210> 8
<211> 3311
<212> DNA
<213> Corynebacterium glutamicum
<400> 8
ctcattccag cgtcacgacg ttccgaaggt actggttacc tggcattggg cactaccgtt 60
tctgcagcac ttggaccagc cctagcactt tttgtcctag gaacatttga ttacgacatg 120
ctgtttatcg tggtcttggc aacctcggtc atctctttga tcgccgtcgt gttcatgtac 180
tttaagacca gcgaccctga gccttctggg gaaccagcca agttcagctt caaatctatt 240
atgaacccaa agatcatccc catcggcatc tttatcttgc ttatttgctt tgcttactct 300
ggcgtcattg cctacatcaa cgcatttgct gaagaacgcg atctgattac gggtgctgga 360
ttgttcttca ttgcctacgc agtatcaatg tttgtgatgc gcagcttcct tggcaaactg 420
caggaccgtc gcggagacaa cgtcgttatt tactttggat tgttcttctt cgttatttcc 480
ttgacgattt tgtcctttgc cacttccaac tggcacgttg tgttgtccgg agtcattgca 540
ggtctgggat acggcacttt gatgccagca gtgcagtcca tcgctgttgg tgtagtagac 600
aaaaccgaat tcggtacggc cttctccact ttgttcctgt ttgtggactt aggttttggc 660
tttggaccta ttatcctggg agcagtttct gcggcaattg gtttcggacc tatgtatgca 720
gcactggcag gtgtgggtgt gattgccgga atcttctacc tgttcacaca cgctcgcacc 780
gatcgagcta agaatggctt tgttaaacac ccagagcctg tcgctttagt tagctagttc 840
tttcagcttt ccctcccgat cagcgtaaac cggcccttcc ggttttgggg tacatcacag 900
aacctgggct agcggtgtag acccgaaaat aaacgagcct tttgtcaggg ttaaggttta 960
ggtatctaag ctaaccaaac accaacaaaa ggctctaccc atgaagtcta ccggcaacat 1020
catcgctgac accatctgcc gcactgcgga actaggactc accatcaccg gcgcttccga 1080
tgcaggtgat tacaccctga tcgaagcaga cgcactcgac tacacctcca cctgcccaga 1140
atgctcccaa cctggggtgt ttcgtcatca cacccaccgg atgctcattg atttacccat 1200
cgtcgggttt cccaccaaac tgtttatccg tctacctcgc taccgctgca ccaaccccac 1260
atgtaagcaa aagtatttcc aagcagaact aagctgcgct gaccacggta aaaaggtcac 1320
ccaccgggtc acccgctgga ttttacaacg ccttgctatt gaccggatga gtgttcacgc 1380
aaccgcgaaa gcacttgggc tagggtggga tttaacctgc caactagccc tcgatatgtg 1440
ccgtgagctg gtctataacg atcctcacca tcttgatgga gtgtatgtca ttggggtgga 1500
tgagcataag tggtcacata atagggctaa gcatggtgat gggtttgtca ccgtgattgt 1560
cgatatgacc gggcatcggt atgactcacg gtgtcctgcc cggttattag atgtcgtccc 1620
aggtcgtagt gctgatgctt tacggtcctg gcttggctcc cgcggtgaac agttccgcaa 1680
tcagatacgg atcgtgtcca tggatggatt ccaaggctac gccacagcaa gtaaagaact 1740
cattccttct gctcgtcgcg tgatggatcc attccatgtt gtgcggcttg ctggtgacaa 1800
gctcaccgcc tgccggcaac gcctccagcg ggagaaatac cagcgtcgtg gtttaagcca 1860
ggatccgttg tataaaaacc ggaagacctt gttgaccacg cacaagtggt tgagtcctcg 1920
tcagcaagaa agcttggagc agttgtgggc gtatgacaaa gactacgggg cgttaaagct 1980
tgcgtggctt gcgtatcagg cgattattga ttgttatcag atgggtaata agcgtgaagc 2040
gaagaagaaa atgcggacca ttattgatca gcttcgggtg ttgaaggggc cgaataagga 2100
actcgcgcag ttgggtcgta gtttgtttaa acgacttggt gatgtgttgg cgtatttcga 2160
tgttggtgtc tccaacggtc cggtcgaagc gatcaacgga cggttggagc atttgcgtgg 2220
gattgctcta ggtttccgta atttgaacca ctacattctg cggtgcctta tccattcagg 2280
gcagttggtc cataagatca atgcactcta aaacaggaag agcccgtaaa cctctgacta 2340
gcgtcaccct ctgattaagg cgaccgcgga tttaagagca gaggctgcca cgagcgcatc 2400
ttcacggctg tgtgttgtac taaaagtaca gcgcacagcc gttcgtgctt gatcctcctc 2460
aagccccaac gccagcaaca catgggatac ctctccggaa ccacaggcag aaccagggga 2520
gcacacaatg ccttggcgtt ccaattccag aagaacagtt tcagatccta tgctgtcgaa 2580
gagaaaagat gcgtgtccat caatgcgcat cctaggatgt ccagtcaggt gtgctcccgg 2640
gatagtgaga acttcctcga tgaattcgcc aagatctgga taggattccg ccctggccaa 2700
ttccaaggca gtggcaaagg cgatagcccc cgcaacgttt tccgtgccac tacgccgccc 2760
tttttcctgg ccgccgccat ggattaccgg ctccagggga agctttgacc ataacactcc 2820
aatcccttta ggcgcaccga atttatgacc cgacaaactt aacgcgtcaa ctcccaagtc 2880
aaaggttaaa tgtgcagctt gcactgcatc ggtgtgaaaa ggcgtactgc ttaccgccgc 2940
caactcagct atcggctgaa tggttcccac ctcattgttg gcataaccaa tgctgatcaa 3000
tgtggtgtcc ggcctgactg ctttgcggag accctccggg gagatcagcc cagtgtgatc 3060
gggggatagg taggtgatct cgaaatcatg aaacctttca agataagcag cagtttctag 3120
gacactgtca tgctcgatcg gggtggtgat gaggtgccgg ccacgaggat tagctaagca 3180
cgctcctttg atagcgaggt tgttggcttc tgatccaccc gacgtaaacg tcacctgtgt 3240
ggggcgtcct ccgataatgc gggccacccg agttcgagca tcctccagcc ccgcagaggc 3300
gagtcttccc a 3311
<210> 9
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 9
acccggggat cctctagaat gtttgtgatg cgcag 35
<210> 10
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 10
gtcagagagt acttacgctg atcgggaggg aaagc 35
<210> 11
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 11
atcagcgtaa gtactctctg actagcgtca ccctc 35
<210> 12
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 12
ctgcaggtcg actctagaaa agggattgga gtgtt 35
<210> 13
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 13
atcaaaacag atatcatggc atccgacgcg ccgca 35
<210> 14
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 14
cgctagtcag agagtttagt caaggccccg caaca 35
<210> 15
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 15
atcaaaacag atatcatggc ggatgcaccc ggcgg 35
<210> 16
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 16
cgctagtcag agagttcaca agctgttaag cgaag 35
<210> 17
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 17
atcaaaacag atatcatgac gaaggactgg aagac 35
<210> 18
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 18
cgctagtcag agagttcaga tcaccgcgag cgcct 35
<210> 19
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 19
ccgatcagcg taagtggcgc ttcatgtcaa caatc 35
<210> 20
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 20
cgcgtcggat gccatgatat ctgttttgat ctcct 35
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 21
gggtgcatcc gccatgatat ctgttttgat ctcct 35
<210> 22
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 22
ccagtccttc gtcatgatat ctgttttgat ctcct 35
<210> 23
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 23
aatggtccag gagctcat 18
<210> 24
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 24
gatcacctac ctatcccc 18
<210> 25
<211> 3161
<212> DNA
<213> Corynebacterium glutamicum
<400> 25
cggtggacga tgagggaaaa cgtttcctcg ccgcccacag aatcataaaa attttctgag 60
gttgtcatgg gtaccagtct aagccctggc cttacgccag taaggtgtta cccatgcgcg 120
aactagcact caacatggcc ggcgtcaccg tgcggcgcgg cgagaaattg cttctcgacg 180
atatctccct ctcaattccg caagggtcgc actgggccgt acttggtcca aatggcgccg 240
gtaaaaccac catgctgaag atcgcagcca ccttgctgta cccatcggaa ggcaccgtgg 300
acatcctggg gcatcgcttt ggtcgggtgg atactcgtga gctgcggaaa acaatcggcc 360
tggtggaccc gaagcaaaga tttaccaacc tgccggccca cgaaattgtg ctgtcggggt 420
taaccgcctc caacgggttg ttgccacggt ggtcggcttc ggcttcggag ttggagcggt 480
gcgctttgat gttggagttg gtgggcatga cagcgcgtgc cgatcgttac tgggccgata 540
tgagccaggg cgaaaaagcc cgcaccctga ttgctcgtgc gctgattatc tcaccgaccc 600
tactgctgct tgatgaaccc accaccggcc ttgacctgcc cggacgtgaa actttgctca 660
gtgtgattga tggtttgcga gccgctcttc ctggtctgac gacagtgatg atcacccacc 720
acgtcgaaga gatcgccgcc tccacgacag atatcctcat gatcaaggac gcccgcatac 780
tggcttcggg gactgtttca gaagtgatga ctcctgaaaa tttgggcgcg ctgtatgaca 840
tgtcggtgtc gttggaaact gtgcgcagcc ggtggttcgc gttcgatgct ctgcattaaa 900
aggggctagt tttacacaaa agtggacagc ttggtctatc attgccagaa gaccggtcct 960
tttagggcca tagaattctg attacaggag ttgatctacc ttgtcttttg acccaaacac 1020
ccagggtttc tccactgcat cgattcacgc tgggtatgag ccagacgact actacggttc 1080
gattaacacc ccaatctatg cctccaccac cttcgcgcag aacgctccaa acgaactgcg 1140
caaaggctac gagtacaccc gtgtgggcaa ccccaccatc gtggcattag agcagaccgt 1200
cgcagcactc gaaggcgcaa agtatggccg cgcattctcc tccggcatgg ctgcaaccga 1260
catcctgttc cgcatcatcc tcaagccggg cgatcacatc gtcctcggca acgatgctta 1320
cggcggaacc taccgcctga tcgacaccgt attcaccgca tggggcgtcg aatacaccgt 1380
tgttgatacc tccgtcgtgg aagaggtcaa ggcagcgatc aaggacaaca ccaagctgat 1440
ctgggtggaa accccaacca acccagcact tggcatcacc gacatcgaag cagtagcaaa 1500
gctcaccgaa ggcaccaacg ccaagctggt tgttgacaac accttcgcat ccccatacct 1560
gcagcagcca ctaaaactcg gcgcacacgc agtcctgcac tccaccacca agtacatcgg 1620
aggacactcc gacgttgttg gcggccttgt ggttaccaac gaccaggaaa tggacgaaga 1680
actgctgttc atgcagggcg gcatcggacc gatcccatca gttttcgatg catacctgac 1740
cgcccgtggc ctcaagaccc ttgcagtgcg catggatcgc cactgcgaca acgcagaaaa 1800
gatcgcggaa ttcctggact cccgcccaga ggtctccacc gtgctctacc caggtctgaa 1860
gaaccaccca ggccacgaag tcgcagcgaa gcagatgaag cgcttcggcg gcatgatctc 1920
cgtccgtttc gcaggcggcg aagaagcagc taagaagttc tgtacctcca ccaaactgat 1980
ctgtctggcc gagtccctcg gtggcgtgga atccctcctg gagcacccag caaccatgac 2040
ccaccagtca gctgccggct ctcagctcga ggttccccgc gacctcgtgc gcatctccat 2100
tggtattgaa gacattgaag acctgctcgc agatgtcgag caggccctca ataaccttta 2160
gaaactattt ggcggcaagc agcttttcaa tataagcaat gcgagcctcc accatgtagc 2220
cgaagagttc gtcagaagtt gagacggact cttcgactgc tttacgggtc agtggcgctt 2280
ccacatctgg gttctcatca agccatggct taggaaccgg agcaaacaca tccggctttt 2340
cgccctctgg acgattgtca aaggtgtagt cagaagtcag ggtgaagctg aagacatcag 2400
ccatcatcat ctcccggatg atggttgctt ccttgaggga cagcccgata tcagtcaaga 2460
acttcaagga ctcctccgca cccgcgattc gcagtgggga agtgccctga gtagagatct 2520
gttcatccag cgcgaccaga agaacacgtg gagtctcacg gaattggtcg cgcaatgagc 2580
tccacagcgt atgaatagat tgccgccaat tgtccggatc aagatcgggc accttgatat 2640
catcgatgat gcgcacccag acgcgatcaa tgatttcttg acgatttagc acatggttat 2700
acagtgcgcg aggggtgaca cccatgtctc gggcgaggcg gttcatggtc acggcagcga 2760
atccttcgcg gccggcaatg tttaaagtgc gctccactat ggattcgacg gaaaggatac 2820
gttgggtggg gcgtccagga cgacgtcccg tggaagtggc cgccagagtt gacgctacgg 2880
ctggtttcat agtttcgcta ggcatgttat atgacgttac gcctttttct acaagacaac 2940
cagcgttttc agcgagatac tggacatatc aactaaaatc cctgaataaa acatctaaca 3000
tgggttttat acagaaaatt catacgaaag gttgatcatg aagaagaaga ttgcggtcgt 3060
taccggagcg accggaggca tgggaattga gatcgtcaaa gacctctccc gcgaccacat 3120
tgtctacgcc ttgggccgaa atccagagca tctggcagct c 3161
<210> 26
<211> 3314
<212> DNA
<213> Corynebacterium glutamicum
<400> 26
gataccggca gctccaccga ccgtgcccat ttcatcacga accatctggc caagtgagcg 60
tccacgccta cgagtagaca cccacagcac taggtagtcc tgcactgcac cggcgaaaat 120
cacaccgagg ataatccaca aggtgcctgg caggtagccc atctgcgcgg ccatgacagg 180
tccaaccaat ggaccggcac ctgcaatagc tgcaaagtgg tggccaaaaa gcacacgacg 240
atccgttggg acatagtcct tgccgtcatt aacgtattcc gccggggttg ctcgctgatc 300
tttcggctta acaactttgt attcaatcag tcgggcatag aaagaaaacg caatgatata 360
ggaaccaact gccgccaaaa ccagccacac agagttgatt gtttcgccac gggagaaagc 420
gattgctccc caacccaccg ccgcgataac cccaaagaca aggagaccaa cgcgggcggt 480
cggtgacatt ttaggggact tcttcacgcc tactggaagg tcagtagcgt tgctgtacac 540
caaatcatcg tcattgatgt tgtcagtctg ttttatggtc acgatcttta ctgttttctc 600
ttcgggtcgt ttcaaagcca ctatgcgtag aaacagcggg cagaaactgt gtgcagaaat 660
gcatgcagaa aaaggaaagt tcggccagat gggtgtttct gtatgccgat gatcggatct 720
ttgacagctg ggtatgcgac aaatcaccga gagttgttaa ttcttaacaa tggaaaagta 780
acattgagag atgatttata ccatcctgca ccatttagag tggggctagt cataccccca 840
taaccctagc tgtacgcaat cgatttcaaa tcagttggaa aaagtcaaga aaattacccg 900
agaataaatt tataccacac agtctattgc aatagaccaa gctgttcagt agggtgcatg 960
ggagaagaat ttcctaataa aaactcttaa ggacctccaa atgccaaagt acgacaattc 1020
caatgctgac cagtggggct ttgaaacccg ctccattcac gcaggccagt cagtagacgc 1080
acagaccagc gcacgaaacc ttccgatcta ccaatccacc gctttcgtgt tcgactccgc 1140
tgagcacgcc aagcagcgtt tcgcacttga ggatctaggc cctgtttact cccgcctcac 1200
caacccaacc gttgaggctt tggaaaaccg catcgcttcc ctcgaaggtg gcgtccacgc 1260
tgtagcgttc tcctccggac aggccgcaac caccaacgcc attttgaacc tggcaggagc 1320
gggcgaccac atcgtcacct ccccacgcct ctacggtggc accgagactc tattccttat 1380
cactcttaac cgcctgggta tcgatgtttc cttcgtggaa aaccccgacg accctgagtc 1440
ctggcaggca gccgttcagc caaacaccaa agcattcttc ggcgagactt tcgccaaccc 1500
acaggcagac gtcctggata ttcctgcggt ggctgaagtt gcgcaccgca acagcgttcc 1560
actgatcatc gacaacacca tcgctaccgc agcgctcgtg cgcccgctcg agctcggcgc 1620
agacgttgtc gtcgcttccc tcaccaagtt ctacaccggc aacggctccg gactgggcgg 1680
cgtgcttatc gacggcggaa agttcgattg gactgtcgaa aaggatggaa agccagtatt 1740
cccctacttc gtcactccag atgctgctta ccacggattg aagtacgcag accttggtgc 1800
accagccttc ggcctcaagg ttcgcgttgg ccttctacgc gacaccggct ccaccctctc 1860
cgcattcaac gcatgggctg cagtccaggg catcgacacc ctttccctgc gcctggagcg 1920
ccacaacgaa aacgccatca aggttgcaga attcctcaac aaccacgaga aggtggaaaa 1980
ggttaacttc gcaggcctga aggattcccc ttggtacgca accaaggaaa agcttggcct 2040
gaagtacacc ggctccgttc tcaccttcga gatcaagggc ggcaaggatg aggcttgggc 2100
atttatcgac gccctgaagc tacactccaa ccttgcaaac atcggcgatg ttcgctccct 2160
cgttgttcac ccagcaacca ccacccattc acagtccgac gaagctggcc tggcacgcgc 2220
gggcgttacc cagtccaccg tccgcctgtc cgttggcatc gagaccattg atgatatcat 2280
cgctgacctc gaaggcggct ttgctgcaat ctagctttaa atagactcac cccagtgctt 2340
aaagcgctgg gtttttcttt ttcagactcg tgagaatgca aactagacta gacagagctg 2400
tccatataca ctggacgaag ttttagtctt gtccacccag aacaggcggt tattttcatg 2460
cccaccctcg cgccttcagg tcaacttgaa atccaagcga tcggtgatgt ctccaccgaa 2520
gccggagcaa tcattacaaa cgctgaaatc gcctatcacc gctggggtga ataccgcgta 2580
gataaagaag gacgcagcaa tgtcgttctc atcgaacacg ccctcactgg agattccaac 2640
gcagccgatt ggtgggctga cttgctcggt cccggcaaag ccatcaacac tgatatttac 2700
tgcgtgatct gtaccaacgt catcggtggt tgcaacggtt ccaccggacc tggctccatg 2760
catccagatg gaaatttctg gggtaatcgc ttccccgcca cgtccattcg tgatcaggta 2820
aacgccgaaa aacaattcct cgacgcactc ggcatcacca cggtcgccgc agtacttggt 2880
ggttccatgg gtggtgcccg caccctagag tgggccgcaa tgtacccaga aactgttggc 2940
gcagctgctg ttcttgcagt ttctgcacgc gccagcgcct ggcaaatcgg cattcaatcc 3000
gcccaaatta aggcgattga aaacgaccac cactggcacg aaggcaacta ctacgaatcc 3060
ggctgcaacc cagccaccgg actcggcgcc gcccgacgca tcgcccacct cacctaccgt 3120
ggcgaactag aaatcgacga acgcttcggc accaaagccc aaaagaacga aaacccactc 3180
ggtccctacc gcaagcccga ccagcgcttc gccgtggaat cctacttgga ctaccaagca 3240
gacaagctag tacagcgttt cgacgccggc tcctacgtct tgctcaccga cgccctcaac 3300
cgccacgaca ttgg 3314
<210> 27
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 27
gaattcgagc tcggtacccg ggccagtaag gtgttaccca tgc 43
<210> 28
<211> 57
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 28
ctgcttgccg ccaaatagtt tagtactggt agatcaactc ctgtaatcag aattcta 57
<210> 29
<211> 57
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 29
tagaattctg attacaggag ttgatctacc agtactaaac tatttggcgg caagcag 57
<210> 30
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 30
tcgactctag aggatccccg ggcgatctca attcccatgc ctc 43
<210> 31
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 31
tcgagctcgg tacccctgca atagctgcaa agtgg 35
<210> 32
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 32
tgagtctatt taaagcgggt aattttcttg acttt 35
<210> 33
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 33
caagaaaatt acccgcttta aatagactca cccca 35
<210> 34
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 34
ctctagagga tccccgcctt aatttgggcg gattg 35
<210> 35
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 35
ttcctggtct gacgacagtg 20
<210> 36
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 36
gatgtcttca gcttcaccct g 21
<210> 37
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 37
ccgaggataa tccacaaggt 20
<210> 38
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 38
cgaagcgttc gtcgatttct 20
<210> 39
<211> 5666
<212> DNA
<213> Corynebacterium glutamicum
<400> 39
tgtcatgctt ccggaggtgc gcagggctcg agactccgga aagctatttg ccactccgat 60
gtttgggtca ctcgacgaga tacgtgctga tcacctaatt tggtgcacag ggtttcggcc 120
ggcgattagg ccagttcgtc aacttctcaa acacggacaa ccaaaggttc ctggtcttta 180
tttagtaggc tacggagatt ggacgggacc tgggtctgcg actatcacag gggtcgggct 240
ttatgccaag cgagcagcca aagagattgc cgcgtcagtc ggcaaagtcg ttaaatagtt 300
tgaaggctaa gaacttaatg ttaaagcgaa aattgttttg acacctcaac taatgcagcg 360
atgcgttctt tccagaatgc tttcatgaca gggatgctgt cttgatcagg caggcgtctg 420
tgctggatgc cgaagctgga tttattgtcg cctttggagg tgaagttgac gctcactcga 480
gaatcatcgg ccaaccattt ggcattgaat gttctaggtt cggaggcgga ggttttctca 540
attagtgcgg gatcgagcca ctgcgcccgc aggtcatcgt ctccgaagag cttccacact 600
ttttcgaccg gcaggttaag ggttttggag gcattggccg cgaacccatc gctggtcatc 660
ccgggtttgc gcatgccacg ttcgtattca taaccaatcg cgatgccttg agcccaccag 720
ccactgacat caaagttgtc cacgatgtgc tttgcgatgt gggtgtgagt ccaagaggtg 780
gcttttacgt cgtcaagcaa ttttagccac tcttcccacg gctttccggt gccgttgagg 840
atagcttcag gggacatgcc tggtgttgag ccttgcggag tggagtcagt catgcgaccg 900
agactagtgg cgctttgcct gtgttgctta ggcggcgttg aaaatgaact acgaatgaaa 960
agttcgggaa ttgtctaatc cgtactaagc tgtctacaca atgtctactt cagttacttc 1020
accagcccac aacaacgcac attcctccga atttttggat gcgttggcaa accatgtgtt 1080
gatcggcgac ggcgccatgg gcacccagct ccaaggcttt gacctggacg tggaaaagga 1140
tttccttgat ctggaggggt gtaatgagat tctcaacgac acccgccctg atgtgttgag 1200
gcagattcac cgcgcctact ttgaggcggg agctgacttg gttgagacca atacttttgg 1260
ttgcaacctg ccgaacttgg cggattatga catcgctgat cgttgccgtg agcttgccta 1320
caagggcact gcagtggcta gggaagtggc tgatgagatg gggccgggcc gaaacggcat 1380
gcggcgtttc gtggttggtt ccctgggacc tggaacgaag cttccatcgc tgggccatgc 1440
accgtatgca gatttgcgtg ggcactacaa ggaagcagcg cttggcatca tcgacggtgg 1500
tggcgatgcc tttttgattg agactgctca ggacttgctt caggtcaagg ctgcggttca 1560
cggcgttcaa gatgccatgg ctgaacttga tacattcttg cccattattt gccacgtcac 1620
cgtagagacc accggcacca tgctcatggg ttctgagatc ggtgccgcgt tgacagcgct 1680
gcagccactg ggtatcgaca tgattggtct gaactgcgcc accggcccag atgagatgag 1740
cgagcacctg cgttacctgt ccaagcacgc cgatattcct gtgtcggtga tgcctaacgc 1800
aggtcttcct gtcctgggta aaaacggtgc agaataccca cttgaggctg aggatttggc 1860
gcaggcgctg gctggattcg tctccgaata tggcctgtcc atggtgggtg gttgttgtgg 1920
caccacacct gagcacatcc gtgcggtccg cgatgcggtg gttggtgttc cagagcagga 1980
aacctccaca ctgaccaaga tccctgcagg ccctgttgag caggcctccc gcgaggtgga 2040
gaaagaggac tccgtcgcgt cgctgtacac ctcggtgcca ttgtcccagg aaaccggcat 2100
ttccatgatc ggtgagcgca ccaactccaa cggttccaag gcattccgtg aggcaatgct 2160
gtctggcgat tgggaaaagt gtgtggatat tgccaagcag caaacccgcg atggtgcaca 2220
catgctggat ctttgtgtgg attacgtggg acgagacggc accgccgata tggcgacctt 2280
ggcagcactt cttgctacca gctccacttt gccaatcatg attgactcca ccgagccaga 2340
ggttattcgc acaggccttg agcacttggg tggacgaagc atcgttaact ccgtcaactt 2400
tgaagacggc gatggccctg agtcccgcta ccagcgcatc atgaaactgg taaagcagca 2460
cggtgcggcc gtggttgcgc tgaccattga tgaggaaggc caggcacgta ccgctgagca 2520
caaggtgcgc attgctaaac gactgattga cgatatcacc ggcagctacg gcctggatat 2580
caaagacatc gttgtggact gcctgacctt cccgatctct actggccagg aagaaaccag 2640
gcgagatggc attgaaacca tcgaagccat ccgcgagctg aagaagctct acccagaaat 2700
ccacaccacc ctgggtctgt ccaatatttc cttcggcctg aaccctgctg cacgccaggt 2760
tcttaactct gtgttcctca atgagtgcat tgaggctggt ctggactctg cgattgcgca 2820
cagctccaag attttgccga tgaaccgcat tgatgatcgc cagcgcgaag tggcgttgga 2880
tatggtctat gatcgccgca ccgaggatta cgatccgctg caggaattca tgcagctgtt 2940
tgagggcgtt tctgctgccg atgccaagga tgctcgcgct gaacagctgg ccgctatgcc 3000
tttgtttgag cgtttggcac agcgcatcat cgacggcgat aagaatggcc ttgaggatga 3060
tctggaagca ggcatgaagg agaagtctcc tattgcgatc atcaacgagg accttctcaa 3120
cggcatgaag accgtgggtg agctgtttgg ttccggacag atgcagctgc cattcgtgct 3180
gcaatcggca gaaaccatga aaactgcggt ggcctatttg gaaccgttca tggaagagga 3240
agcagaagct accggatctg cgcaggcaga gggcaagggc aaaatcgtcg tggccaccgt 3300
caagggtgac gtgcacgata tcggcaagaa cttggtggac atcattttgt ccaacaacgg 3360
ttacgacgtg gtgaacttgg gcatcaagca gccactgtcc gccatgttgg aagcagcgga 3420
agaacacaaa gcagacgtca tcggcatgtc gggacttctt gtgaagtcca ccgtggtgat 3480
gaaggaaaac cttgaggaga tgaacaacgc cggcgcatcc aattacccag tcattttggg 3540
tggcgctgcg ctgacgcgta cctacgtgga aaacgatctc aacgaggtgt acaccggtga 3600
ggtgtactac gcccgtgatg ctttcgaggg cctgcgcctg atggatgagg tgatggcaga 3660
aaagcgtggt gaaggacttg atcccaactc accagaagct attgagcagg cgaagaagaa 3720
ggcggaacgt aaggctcgta atgagcgttc ccgcaagatt gccgcggagc gtaaagctaa 3780
tgcggctccc gtgattgttc cggagcgttc tgatgtctcc accgatactc caaccgcggc 3840
accaccgttc tggggaaccc gcattgtcaa gggtctgccc ttggcggagt tcttgggcaa 3900
ccttgatgag cgcgccttgt tcatggggca gtggggtctg aaatccaccc gcggcaacga 3960
gggtccaagc tatgaggatt tggtggaaac tgaaggccga ccacgcctgc gctactggct 4020
ggatcgcctg aagtctgagg gcattttgga ccacgtggcc ttggtgtatg gctacttccc 4080
agcggtcgcg gaaggcgatg acgtggtgat cttggaatcc ccggatccac acgcagccga 4140
acgcatgcgc tttagcttcc cacgccagca gcgcggcagg ttcttgtgca tcgcggattt 4200
cattcgccca cgcgagcaag ctgtcaagga cggccaagtg gacgtcatgc cattccagct 4260
ggtcaccatg ggtaatccta ttgctgattt cgccaacgag ttgttcgcag ccaatgaata 4320
ccgcgagtac ttggaagttc acggcatcgg cgtgcagctc accgaagcat tggccgagta 4380
ctggcactcc cgagtgcgca gcgaactcaa gctgaacgac ggtggatctg tcgctgattt 4440
tgatccagaa gacaagacca agttcttcga cctggattac cgcggcgccc gcttctcctt 4500
tggttacggt tcttgccctg atctggaaga ccgcgcaaag ctggtggaat tgctcgagcc 4560
aggccgtatc ggcgtggagt tgtccgagga actccagctg cacccagagc agtccacaga 4620
cgcgtttgtg ctctaccacc cagaggcaaa gtactttaac gtctaacacc tttgagaggg 4680
aaaactttcc cgcacattgc agatcgtgcc actttaacta aggttgacgg catgattaag 4740
gcgattttct gggacatgga cggcacgatg gtggactctg agccacagtg gggcattgct 4800
acctacgagc tcagcgaagc catgggccgc cgcctcaccc cggagctccg ggaactcacc 4860
gtcggctcga gcctgccgcg caccatgcgc ttatgcgcag agcacgcagg cattacattg 4920
agcgacgcgg actacgagcg ctaccgggct ggcatgttcg cccgggtcca tgagcttttc 4980
gacgaatccc tcgtcccaaa tccaggcgtc accgaactcc tgacagagtt gaaggccctc 5040
gagatcccca tgttggtcac caccaacaca gagcgcgatc tcgcgacccg ttcagtcgca 5100
gccgtgggaa atgagttctt catcggttct atcgctggtg atgaagtccc aacagcaaag 5160
ccagcccccg acatgtacct cgaagcagca cgacgtgtgg gctttgaccc atcagagtgc 5220
ctcgtgttcg aagattccta caacggcatg ctgggcgctg ttactgcagg ttgccgcgtc 5280
attggtctgc acccagaaga agtccaagcg ccagaaggtg tagtgccttt gcgttccctc 5340
cacggtaaaa actctttcga aggtgtcacc gctgagatgg tcactgcctg gtaccaccag 5400
atcgagccgg caggtgtcgc aaaataaaac caggtggggg agtgaaatta ttcgactaat 5460
atcctccccc aaacacacat tgataactgt tgtgtggaag aatgtaccga gtgaagacat 5520
ttgactcgct gtacgaagaa cttcttaacc gtgctcagac ccgccctgaa gggtctggaa 5580
ccgtggccgc cttggataaa ggcatccatc atctaggtaa gaaggtcatc gaagaagccg 5640
gagaggtctg gattgcagcc gagtat 5666
<210> 40
<211> 3613
<212> DNA
<213> Corynebacterium glutamicum
<400> 40
tcctgtgggg tgaacttgac ctgtgctggg ccacgacgtc cgaaaacgtg cacttcagtg 60
gccttgtttt ctttgaggga gtcgtagacg ttgtcggaaa tttcggtgac tttgagctcg 120
tcgcctgtct tagccaggat gcgggctacg tcgaggccga cgttaccaac gccgataaca 180
gcgacggact gtgcagacag atcccaggag cgctcgaagc gtgggttgcc gtcgtagaag 240
ccaacgaact cgccggcacc gaaggagcct tctgcttcaa ttccggggat gttgaggtcg 300
cggtctgcaa ctgcgccggt ggagaacacg actgcatcgt agtagtcgcg gagttcttcg 360
acggtgatgt ctttgccgat ttcaatgtta ccgagcaggc gcaggcgtgg cttgtccaac 420
acgttgtgca gggacttaac gatgcccttg atgcgtgggt ggtctggagc aacgccgtaa 480
cggatgagtc cgaacggtgc aggcatttgc tcgaaaaggt caacgaacac ttcgcgctct 540
tcattgcgga tgaggaggtc ggatgcgtaa atgccagcag ggccagctcc gatgacggct 600
acgcgcaggg gagttgtcat gtgtttgaag ttgcctttcg tgagcccttt tatggaaaca 660
agggtgtgaa aatcaagtag ttaaaggtgt ttcaagtcca ggctgtttaa cactcctaga 720
ccgcttggtc tgtaaacgta gcagcgaaat gcgacaatgc gaagactttt gcttaattaa 780
attcaaactc catgaaaaaa ctagacagat cggtctatta tattcacggt gaacctaacc 840
taatatcccc aggttaattc atttaaacgg gcattaggtg actccattgc tttcagtctc 900
atgaatctaa tggttggtct agacagagcg gtacgtctaa gtttgcggat agatcaaacc 960
gagtgacatg tacttcacta gctctttaag gattaactcc ccatgacaac aaccaccgga 1020
agtgcccggc cagcacgtgc cgccaggaag cctaagcccg aaggccaatg gaaaatcgac 1080
ggcaccgagc cgcttaacca tgccgaggaa attaagcaag aagaacccgc ttttgctgtc 1140
aagcagcggg tcattgatat ttactccaag cagggttttt cttccattgc accggatgac 1200
attgccccac gctttaagtg gttgggcatt tacacccagc gtaagcagga tctgggcggt 1260
gaactgaccg gtcagcttcc tgatgatgag ctgcaggatg agtacttcat gatgcgtgtg 1320
cgttttgatg gcggactggc ttcccctgag cgcctgcgtg ccgtgggtga aatttctagg 1380
gattatgctc gttccaccgc ggacttcacc gaccgccaga acattcagct gcactggatt 1440
cgtattgaag atgtgcctgc gatctgggag aagctagaaa ccgtcggact gtccaccatg 1500
cttggttgcg gtgacgttcc acgtgttatc ttgggctccc cagtttctgg cgtagctgct 1560
gaagagctga tcgatgccac cccggctatc gatgcgattc gtgagcgcta cctagacaag 1620
gaagagttcc acaaccttcc tcgtaagttt aagactgcta tcactggcaa ccagcgccag 1680
gatgttaccc acgaaatcca ggacgtttcc ttcgttcctt cgattcaccc agaattcggc 1740
ccaggatttg agtgctttgt gggcggtggc ctgtccacca acccaatgct tgctcagcca 1800
cttggttctt ggattccact tgatgaggtt ccagaagtgt gggctggcgt cgccggaatt 1860
ttccgcgact acggcttccg acgcctgcgt aaccgtgctc gcctcaagtt cttggtggca 1920
cagtggggta ttgagaagtt ccgtgaagtt cttgagaccg aatacctcga gcgcaagctg 1980
atcgatggcc cagttgttac caccaaccct ggctaccgtg accacattgg cattcaccca 2040
caaaaggacg gcaagttcta cctcggtgtg aagccaaccg ttggacacac caccggtgag 2100
cagctcattg ccattgctga tgttgcagaa aagcacggca tcaccaggat tcgtaccacg 2160
gcggaaaagg aactgctctt cctcgatatt gagagaaaga accttactac cgttgcacgc 2220
gacctggatg aaatcggact gtactcttca ccttccgagt tccgccgcgg catcatttcc 2280
tgcaccggct tggagttctg caagcttgcg cacgcaacca ccaagtcacg agcaattgag 2340
cttgtcgacg aactggaaga gcgcctcggc gatttggatg ttcccatcaa gattgcactg 2400
aacggttgcc ctaactcttg tgcacgcacc caggtttccg acatcggatt caagggacag 2460
accgtcactg atgctgacgg caaccgcgtt gaaggtttcc aggttcacct gggcggttcc 2520
atgaacttgg atccaaactt cggacgcaag ctcaagggcc acaaggttat tgccgatgaa 2580
gtgggagagt acgtcactcg cgttgttacc cacttcaagg aacagcgcca cgaggacgag 2640
cacttccgcg attgggtcca gcgggccgct gaggaagatt tggtgtgagt cttcggagga 2700
aacccaatcc caaccgcaac caccctctgt actgcccata ctgcgcggga gaagttcttt 2760
tccccgatga gcaaacagaa ttcgcgtggt tgtgtgcgga ttgcaccaga gtttttgaag 2820
tgaaatatca cggccaggac gatccagtgc acaggccagc accagcaaag tccacatcgc 2880
aagcattaaa agaatctctc gaaagacaca aaagaggtga gtcgcaacaa tgagctttca 2940
actagttaac gccctgaaaa atactggttc ggtaaaagat cccgagatct cacccgaagg 3000
acctcgcacg accacaccgt tgtcaccaga ggtagcaaaa cataacgagg aactcgtcga 3060
aaagcatgct gctgcgttgt atgacgccag cgcgcaagag atcctggaat ggacagccga 3120
gcacgcgccg ggcgctattg cagtgacctt gagcatggaa aacaccgtgc tggcggagct 3180
ggctgcgcgg cacctgccgg aagctgattt cctctttttg gacaccggtt accacttcaa 3240
ggagaccctt gaagttgccc gtcaggtaga tgagcgctat tcccagaagc ttgtcaccgc 3300
gctgccgatc ctcaagcgca cggagcagga ttccatttat ggtctcaacc tgtaccgcag 3360
caacccagcg gcgtgctgcc gaatgcgcaa agttgaaccg ctggcggcgt cgttaagccc 3420
atacgctggc tggatcaccg gcctgcgccg cgctgatggc ccaacccgtg ctcaagcccc 3480
tgcgctgagc ttggatgcca ccggcaggct caagatttct ccaattatca cctggtcatt 3540
ggaggaaacc aacgagttca ttgcggacaa caacctcatc gatcacccac ttacccatca 3600
gggttatcca tca 3613
<210> 41
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 41
caacgaaagg aaacaatgtc tacttcagtt acttc 35
<210> 42
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 42
tagtcagaga gtgatttaga cgttaaagta ctttg 35
<210> 43
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 43
atcaaaacag atatcatgac aacaaccacc ggaag 35
<210> 44
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 44
cgctagtcag agagttcaca ccaaatcttc ctcag 35
<210> 45
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 45
ccgatcagcg taagtagaaa catcccagcg ctact 35
<210> 46
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 46
aactgaagta gacattgttt cctttcgttg ggtac 35
<210> 47
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 47
tactttaacg tctaaggtac cggcgcttca tgtca 35
<210> 48
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 48
ggtggttgtt gtcatgatat ctgttttgat ctcct 35
<210> 49
<211> 3261
<212> DNA
<213> Corynebacterium glutamicum
<400> 49
cctcactggc gaacacggcc gactcctgga acagcgcaac atggcatgga cgaaactcaa 60
cgaaatccca ggtgtcagct gtgtgaaacc aatgggagct ctatacgcgt tccccaagct 120
cgaccccaac gtgtacgaaa tccacgacga cacccaactc atgctggatc ttctccgtgc 180
cgagaaaatc ctcatggttc agggcactgg cttcaactgg ccacatcacg atcacttccg 240
agtggtcacc ctgccatggg catcccagtt ggaaaacgca attgagcgcc tgggtaactt 300
cctgtccact tacaagcagt agtagttgtt aggattcacc acgaatctca ggatttttga 360
gattcgtggt gaatttttgc gttttccagt caggctcctg caactttcgg accgatttca 420
gaggggcgga gctggtttgt ggtggatcct tgaaatggaa cctcgcagga agctttcagg 480
aagaccaagt tgggcctagg ggtggcggga ttgcaaaaat ccgtccccgg ttcgccatga 540
aatgctgatt ttgatcgaat ctttgcgcta actgtagggc gggttcaggg ggtgaatgca 600
ccacgagcaa cccgaagggt gcgaagtggg cattcgtaga acaatcccag aggaaagccg 660
tacggctttc ctcgacatga tcaatcaagg tatgtcaggt cttgctgcgt ctacagcggt 720
cggggtcagt gaattcaccg ggcgaaagtg ggcgaaggcc gccggggtga aactgacccg 780
cggcccgcga ggtggcaatg cttttgacac cgccgagaaa cttgagattg cagccagcat 840
gctagagaaa ggatgcctac cccgagaaat cggcgagtat gtcggcatga ctcgggccaa 900
tatatcccta tggcgcaaac aaggcccaga caagcttcgc caacgcgcag ccaccttgcg 960
caccggcaag cgagcagctg aattcatcca cgccccggtg atgggccctt attatgggcc 1020
acgcacactc catcaagtgt tgcgtgagga ctacacaaca ctgtttgacg agttatctgc 1080
gttggggttg ccagcacagg tgtgtggggc cttacttcat cttgctccac caccatcatt 1140
acgcttttct tatatgtcgt gtgtagtgcc gttatttgct gatgaaatca aagtcgtagg 1200
acaaggcaca cgattatcgt tagaagagaa aatgatgatc caacgtttcc atgacaccgg 1260
ggtcagtgca gcagaaatcg gtcgacgcct gggtcggtgt cggcaaacaa tttccaggga 1320
acttcgacgt ggtcaagatg atgatggacg ttatcgtgca cgcgactcct atgaaggtgc 1380
gatcaggaaa ctagcgcgtc cgaaaacacc gaaacttgat gccaatcgta ggcttcgggc 1440
tgtggtggtc gaggcgttga ataataaatt atctccggag cagatttctg gtcttttagc 1500
caccgagcat gctaacgata gctctatgca gattagtcat gaaactattt accaggcgtt 1560
atatgttcaa ggtaaagggg cgttgcgtga tgaattgaag gtggagaaat ttcttcgtac 1620
cggtcggaag ggacgtaaac cgcagtcgaa gttgccatcg agaggtaagc cgtgggtgga 1680
gggtgcgttg attagtcaac gcccagcaga agttgctgat cgtgctgtgc ctgggcactg 1740
ggagggcgat ttagtaattg gtggtgaaaa ccaagcgaca gcgttggtga cgttggtgga 1800
gcgcacgagc cggttgacgt tgattaagcg gttgggggtt aatcatgagg cgtcgactgt 1860
gacggatgcg ttggtggaga tgatgggtga tttgccgcag gcgttgcgtc ggagtttgac 1920
gtgggatcag ggtgtggaga tggcagagca tgcgcggttt agcgtggtga ccaagtgtcc 1980
ggtgtttttc tgtgatcctc attcgccgtg gcagcgtggg tcgaatgaga atacgaatgg 2040
attggtcagg gattttttcc cgaagggcac taattttgct aaagtaagtg acgaagaagt 2100
tcagcgggca caggatctgc tgaattaccg gccgcggaaa atgcatggtt ttaaaagcgc 2160
gacgcaggta tatgaaaaaa tcgtagttgg tgcatccacc gattgaattc gcctaggaga 2220
ttgtacgaaa attcgttcgg ctttcggatt tcctggcgat ctgagacgag aagttgaaca 2280
gctaacctgc agaaaccttg caagaatcac aacagcccca atggcctcaa aagtcacgcc 2340
ctcagaatcg ctgccaggcg tctaaatccc ctaaaacggg acaataggtc actgggcgat 2400
cccaagccct taaaacgtga tccttaaata cccactgtcc tctattctgg gttaggcttc 2460
actgggtaaa agtgcctgcc tatgcctgaa acttgagcat ggcaacagca aggagacacc 2520
gtgggaaaac atgcagctga aacatcggaa ccgaagaaaa attcaccgtg gcgcattggt 2580
ttgttgacgt ttttgatttc ttcagttgtc gtgacgctgg tgggcatggt gatgctgtgg 2640
ccggattctg atgatgtggt gttggcggat aacttttcgc agacgtttgc gggaaatcat 2700
gagcaggtgg atggaacgat cacgctcgtt gataattctg cgtgtaattc gccagacacc 2760
ggccgagttt ttgcggaaag ccccacgatt tctgcggagc cggcaacgtt ggagtgcgtg 2820
cgtgcactcg tagacatcac atcgggtgcc aatgaggggc agaaaactca gctgatcact 2880
tacgcgcaac ctggtgatcc ggagttttcc gagggcgaca agatccgcat ggtggaaaca 2940
ccggatacaa atggcgagat catctacacc tttgctgatt accagcgcgg accggcgttg 3000
atcatttggg gtgtggttct cattgtggcg atgggagctt tcgcggcgtg gcgaggtgtg 3060
cgtgcgctgg ttggtttggt cgtcaccttg ggaattgttg gtattttctt gctgccagga 3120
ttggccagcg ggcacgatgc gatgtggttg gcgctggtgt gtggcgcggc gatcttgttg 3180
attgtggtgc cgatggttca cggaatcaac tggaaatcgg cagctgcgtt ggcgggcacg 3240
ctggtggcat tgttgttgtc g 3261
<210> 50
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 50
gtacccgggg atcctctaga cctgggtaac ttcctgtcca 40
<210> 51
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 51
caggttagca gtacttctca agtttctcgg cggtg 35
<210> 52
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 52
aacttgagaa gtactgctaa cctgcagaaa ccttg 35
<210> 53
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 53
gcctgcaggt cgactctaga ctccgcagaa atcgtggggc 40
<210> 54
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 54
cgccgagaaa cttgagaagt ggcgcttcat gtcaa 35
<210> 55
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 55
ctgcaggtta gcagtttagt caaggccccg caaca 35
<210> 56
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 56
ctgcaggtta gcagttcaca agctgttaag cgaag 35
<210> 57
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 57
ctgcaggtta gcagttcaga tcaccgcgag cgcct 35
<210> 58
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 58
ttctccgtgc cgagaaaatc 20
<210> 59
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 59
gtagatgatc tcgccatttg 20
<210> 60
<211> 394
<212> PRT
<213> Chromobacterium violaceum
<400> 60
Met Ala Ser Asp Ala Pro His Leu Pro Leu His Pro Glu Thr Leu Ala
1 5 10 15
Ile Arg Ala Gly Leu Glu Thr Ser Gln Phe Asn Glu His Ser Gln Gly
20 25 30
Leu Phe Leu Thr Ser Ser Phe Thr Tyr Glu Ser Ala Ala Gln Ala Ala
35 40 45
Ala Met Phe Leu Gly Glu Ile Asp Gly Tyr Thr Tyr Ser Arg Phe Thr
50 55 60
Asn Pro Thr Val Ala Ala Phe Gln His Arg Leu Ala Gln Met Glu Gly
65 70 75 80
Gly Glu Arg Ala Ile Ala Thr Ala Thr Gly Met Ala Ala Ile Gln Ala
85 90 95
Ile Met Met Thr Leu Leu Gln Ala Gly Asp His Ile Val Ser Ser Gln
100 105 110
Ser Leu Phe Gly Ser Thr Thr Asn Leu Phe Ala Asn Gln Leu Ala Lys
115 120 125
Phe Ala Val Ala Thr Asp Phe Val Asp Ala Arg Asp Leu Ser Ala Trp
130 135 140
Arg Glu Ala Leu Arg Pro Asn Thr Lys Leu Leu Phe Leu Glu Thr Pro
145 150 155 160
Ser Asn Pro Leu Thr Glu Val Ala Asp Ile Ala Ala Ile Ala Asp Ile
165 170 175
Ala His Ala His Gly Ala Leu Leu Val Val Asp Asn Ser Phe Cys Ser
180 185 190
Pro Ala Leu Gln Gln Pro Leu Lys Leu Gly Ala Asp Leu Val Met His
195 200 205
Ser Ala Thr Lys Phe Ile Asp Gly His Gly Arg Val Met Gly Gly Ala
210 215 220
Val Val Gly Ser Asp Lys Leu Val Glu Gln Val Tyr Leu His Val Arg
225 230 235 240
Ala Ala Gly Pro Ser Leu Ala Pro Phe Asn Ala Trp Thr Leu Leu Ser
245 250 255
Gly Leu Glu Thr Leu His Leu Arg Met Glu Lys His Ser Ala Asn Ala
260 265 270
Leu Glu Leu Ala Arg Trp Leu Glu Ala Gln Pro Asn Val Glu Arg Val
275 280 285
Tyr Tyr Pro Gly Leu Glu Ser His Pro Gln His Glu Leu Ala Leu Arg
290 295 300
Gln Gln Lys Ser Gly Gly Ala Val Val Ser Phe Val Val Lys Gly Gly
305 310 315 320
Arg Lys Ala Ala Trp Lys Val Val Asp Ala Val Arg Val Ile Ser Arg
325 330 335
Thr Ala Asn Leu Gly Asp Val Lys Thr Thr Leu Thr His Pro Ala Ser
340 345 350
Thr Thr His Ala Arg Val Thr Gln Glu Ala Arg Glu Arg Ala Gly Ile
355 360 365
Val Glu Gly Leu Leu Arg Val Ser Val Gly Leu Glu Asn Val Arg Asp
370 375 380
Leu Gln Gln Asp Leu Leu Arg Gly Leu Asp
385 390
<210> 61
<211> 399
<212> PRT
<213> Unknown
<220>
<223> Hyphomonas neptunium
<400> 61
Met Ala Asp Ala Pro Gly Gly Asp Lys Lys Gly Trp Lys Pro Ala Thr
1 5 10 15
Gln Ala Val Arg Gly Gly Leu Met Arg Ser Gln His Gly Glu Ile Ser
20 25 30
Glu Ala Leu Tyr Leu Thr Ser Gly Tyr Ala Tyr Asp Ser Ala Glu Gln
35 40 45
Ala Met Arg Arg Met Ala Gly Glu Glu Glu Gly Phe Val Tyr Ser Arg
50 55 60
Tyr Gly Ser Pro Thr Asn Glu Met Leu Gln Gln Arg Leu Ala Leu Ile
65 70 75 80
Glu Gly Ala Glu Ala Cys Arg Val Thr Gly Ser Gly Met Gly Ala Ile
85 90 95
Ser Ser Ala Ile Leu Ala Pro Leu Lys Ala Gly Asp Arg Val Val Ala
100 105 110
Ala Thr Ala Leu Phe Gly Ser Cys Arg Trp Ile Ile Ala Asn Gln Met
115 120 125
Pro Lys Phe Gly Ile Glu Ala Val Phe Val Asp Gly Ala Asp Leu Asp
130 135 140
Ala Trp Lys Arg Glu Ile Asp Lys Gly Cys Gln Leu Val Leu Ile Glu
145 150 155 160
Ser Pro Ala Asn Pro Leu Leu Asp Gly Val Asp Ile Glu Ala Val Ala
165 170 175
Arg Leu Ala Lys Ala Ala Gly Ala Leu Leu Val Val Asp Asn Val Phe
180 185 190
Ala Thr Pro Val Leu Gln Arg Pro Leu Glu Met Gly Ala Asp Val Ile
195 200 205
Ala Tyr Ser Ala Thr Lys His Met Asp Gly Gln Gly Arg Val Leu Leu
210 215 220
Gly Ala Ile Leu Thr Asp Ala Lys Arg Met Ser Asp Val Tyr Asp Pro
225 230 235 240
Trp Leu Arg His Met Gly Pro Ala Ala Ser Pro Phe Asn Ala Trp Val
245 250 255
Val Leu Lys Gly Leu Glu Thr Met Gln Leu Arg Val Glu Ala Gln Ser
260 265 270
Arg Thr Ala Ala Arg Leu Ala Asp Val Leu Ala Asp His Pro Ala Val
275 280 285
Asn Ala Val Arg Tyr Pro His Arg Lys Asp His Pro His Tyr Glu Val
290 295 300
His Lys Arg Gln Met Lys Ser Gly Gly Thr Leu Leu Ala Leu Ser Leu
305 310 315 320
Lys Gly Gly Gln Asp Ala Ala Phe Arg Phe Leu Asn Gly Leu Gln Leu
325 330 335
Val Asp Ile Cys Asn Asn Leu Gly Asp Thr Lys Ser Leu Ala Cys His
340 345 350
Pro Ser Thr Thr Thr His Arg Ala Leu Ser Asp Glu Asp Gln Ala Ala
355 360 365
Met Gly Leu Asp Arg Ser Trp Val Arg Leu Ser Val Gly Leu Glu Asp
370 375 380
Ala Asp Asp Leu Glu Ala Asp Leu Leu Ala Ser Leu Asn Ser Leu
385 390 395
<210> 62
<211> 393
<212> PRT
<213> Rhodobacter sphaeroides
<400> 62
Met Thr Lys Asp Trp Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser
1 5 10 15
Arg Arg Ser Gln Tyr Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln
20 25 30
Gly Phe Val Tyr Asp Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu
35 40 45
Thr Gly Ala Asp Glu Phe Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr
50 55 60
Arg Met Phe Glu Glu Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala
65 70 75 80
Phe Ala Thr Ala Ser Gly Met Ala Ala Ile His Gly Val Leu Thr Ser
85 90 95
Ile Val Arg Ala Gly Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly
100 105 110
Ser Cys Ile Tyr Ile Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu
115 120 125
Val Thr Phe Val Asp Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val
130 135 140
Arg Pro Gly Thr Lys Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr
145 150 155 160
Leu Glu Val Ala Asp Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val
165 170 175
Gly Ala Leu Val Ile Val Asp Asn Val Phe Ala Thr Pro Val Phe Ser
180 185 190
Thr Ala Val Arg Gln Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys
195 200 205
His Ile Asp Gly Gln Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser
210 215 220
Gln Ala Phe Ile Arg Lys Val Leu Glu Pro Phe Met Lys His Thr Gly
225 230 235 240
Gly Ser Met Ser Pro Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala
245 250 255
Thr Leu Asp Leu Arg Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile
260 265 270
Ala Arg Ala Leu Glu Gly His Pro Gln Leu Gly Arg Val Ile His Pro
275 280 285
Ala Leu Glu Ser His Pro Gln His Glu Met Ala Lys Ala Gln Met Glu
290 295 300
Arg Pro Gly Thr Met Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala
305 310 315 320
Ala Phe Arg Phe Leu Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn
325 330 335
Leu Gly Asp Ala Arg Ser Ile Ala Thr His Pro Ala Thr Thr Thr His
340 345 350
Gln Arg Leu Ser Asp Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly
355 360 365
Leu Val Arg Leu Ser Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala
370 375 380
Asp Leu Lys Gln Ala Leu Ala Val Ile
385 390
<210> 63
<211> 1185
<212> DNA
<213> Chromobacterium violaceum
<400> 63
atggcatccg acgcgccgca tcttccgctg caccctgaaa ccctggccat ccgggccggg 60
ttggaaacca gccagttcaa cgagcacagc cagggcctgt tcctgacgtc cagcttcacc 120
tacgaatcgg ccgcgcaggc ggcggcgatg ttcctgggcg agatcgacgg ctacacctat 180
tcccgcttca ccaatccgac cgtcgccgcg ttccagcata ggctggcgca gatggagggc 240
ggggagcgcg ccatcgccac cgccaccggc atggcggcga tccaggccat catgatgact 300
ttgctgcagg ctggcgacca catcgtgtcg tcgcaaagcc tgttcggctc caccaccaat 360
ctgttcgcca accagttggc caagttcgcc gtggccaccg acttcgtcga cgcgcgcgac 420
ctgtccgcct ggcgggaggc gctgcggccg aacaccaagc tgctgttcct ggagacgccg 480
tccaatccct tgaccgaagt ggccgacatc gcggccatcg ccgacatcgc ccacgcgcat 540
ggcgcgctgc tggtggtgga caacagcttc tgttcgccgg ccttgcagca gccgttgaaa 600
ctgggcgccg atctggtcat gcattccgcc accaagttca tcgacggcca tggccgggtg 660
atgggcgggg cggtggtcgg cagcgacaag ctggtcgagc aggtctattt gcacgtgcgc 720
gccgccggtc cctcgctggc gccgttcaat gcctggacgc tgctgtccgg tttggagacg 780
ctgcacctgc ggatggagaa gcacagcgcc aacgcgctgg agctggcgcg ctggctggag 840
gcgcagccca atgtggagcg cgtctattac ccgggcctgg agagccaccc ccagcacgag 900
ctggcgctgc gccagcagaa gagcggcgga gcggtggtgt ccttcgtggt caagggcggc 960
cgcaaggccg cgtggaaagt ggtggacgcg gtcagggtga tctcgcgcac cgccaatctg 1020
ggcgatgtga aaaccaccct cactcatccg gccagcacca cccacgcccg cgtgacgcag 1080
gaggcgcgcg agcgcgccgg catcgtcgag gggctgttgc gcgtcagcgt cggcctggaa 1140
aatgtacggg accttcaaca agatctgttg cggggccttg actaa 1185
<210> 64
<211> 1200
<212> DNA
<213> Unknown
<220>
<223> Hyphomonas neptunium
<400> 64
atggcggatg cacccggcgg cgacaagaag ggctggaagc ctgcgaccca ggcggtacgc 60
ggcggcctga tgcggtccca gcatggggag atttccgagg cgctgtatct gacctccggc 120
tacgcttacg actcggccga gcaggcgatg cgccggatgg cgggcgagga agaaggcttc 180
gtctattccc gctatggcag cccgaccaat gagatgctgc aacagcgcct cgcgctgatt 240
gaaggcgccg aagcgtgccg ggtgacgggc tctggcatgg gcgcgatttc gtcggccatc 300
ctggcgccgc ttaaagcggg cgaccgggtg gtggcggcga ccgcgctgtt tggctcgtgc 360
cgctggatca ttgccaacca gatgccgaag tttggcatcg aggcagtgtt cgtggacggg 420
gccgatcttg atgcttggaa gcgcgagatc gacaagggct gccagctggt gctgatcgaa 480
agcccggcca atccgttgct cgacggcgtg gacatcgaag cggtcgccag gctcgccaag 540
gcggcgggcg cgctgctggt ggtggacaat gtgtttgcca cgccggtgct tcagcggccg 600
ctggaaatgg gcgccgatgt gatcgcctat tcggccacca aacatatgga cgggcagggc 660
cgcgttctgc tgggcgcgat cctgacggac gccaagcgga tgagtgatgt gtatgatccg 720
tggctgcgcc atatggggcc ggcggcctcg ccgtttaacg cctgggtagt gctgaagggc 780
cttgagacga tgcagctgcg cgtggaagcg cagagccgca cggcggcgcg gctggcggat 840
gttctggccg atcatccggc ggtcaatgcc gtgcgctatc cccaccgcaa ggatcacccg 900
cattatgagg tgcacaagcg ccagatgaaa tcgggcggca cgctgctcgc gctgtcgctc 960
aagggcgggc aggacgcggc gttccgcttc ctcaacgggc tgcagctggt cgacatctgc 1020
aacaaccttg gcgatacgaa atcgctggcc tgtcatccct ccaccacgac gcaccgcgcg 1080
ttgagtgatg aggatcaggc ggcgatgggg cttgaccgca gctgggtccg gctctctgtt 1140
ggtcttgaag acgcagatga tctggaagct gatcttctcg cttcgcttaa cagcttgtga 1200
1200
<210> 65
<211> 1182
<212> DNA
<213> Rhodobacter sphaeroides
<400> 65
atgacgaagg actggaagac aaggacgcaa ctcgtccacg ggggcagccg ccggagccag 60
tatggcgaaa tggccgaggc gatcttcctg acccagggct tcgtctacga ctcggccgaa 120
caggccgaag cgcgcttcat cgagaccggc gccgacgaat tcatctatgc ccgctacggc 180
aaccccacga cgcgcatgtt cgaagagcgc atcgcggccg tcgagggcac cgaggatgcg 240
ttcgccaccg cctcgggcat ggccgcgatc cacggcgtgc tcacctcgat cgtgcgggcg 300
ggcgatcatc tggtggcggc gcgcgctctg ttcggctcct gcatctacat cctcgaggag 360
gtgctgggcc gattcggcgt cgaggtgacc ttcgtcgacg gcaccgatct cgatcagtgg 420
cgcgcggcgg tgcggcccgg cacgaaggcc gtgttcttcg agtcggtctc gaatccgacg 480
ctcgaggtgg ccgatatcgg cgccatcgcc gagatcgccc atgccgtggg cgcgctcgtc 540
atcgtggaca atgtcttcgc gacgcccgtc ttctcgacgg cggtgcggca gggcgcggat 600
gtggtgatct attcggccac caagcacatc gacgggcaag ggcgcgcgct cggcggcgtg 660
gtctgcgcct cgcaggcctt catccgcaag gtgctcgaac ccttcatgaa gcacaccggc 720
ggctcgatga gccccttcaa cgcctggctc atgctgaacg ggatggcgac gctcgacctg 780
cgctgccgcg cgatggccga cacggccgag aagatcgccc gcgcgctcga gggccatccg 840
cagctcggcc gcgtgatcca tcccgcgctg gaaagccacc cgcagcacga gatggccaag 900
gcgcagatgg agcgtcccgg cacgatgatc gcgctcgacc tcgccggggg caaggaggcg 960
gccttccgct tcctcgacgc cctgaggatc gtgaagatct ccaacaatct gggcgatgcc 1020
cgctcgatcg cgacccaccc ggcaacgacc acccaccagc gtctttccga cgcgcagaag 1080
gcccatctcg gcatcacgcc cgggctcgtg cggctgtcgg tggggctcga ggatgcggac 1140
gacctgatcg ccgatctgaa acaggcgctc gcggtgatct ga 1182
<210> 66
<211> 404
<212> PRT
<213> Artificial Sequence
<220>
<223> RspmetZ_long
<400> 66
Met Gly Ile Ala Phe Arg Glu Gly Arg Thr Gly Met Thr Lys Asp Trp
1 5 10 15
Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser Arg Arg Ser Gln Tyr
20 25 30
Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln Gly Phe Val Tyr Asp
35 40 45
Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu Thr Gly Ala Asp Glu
50 55 60
Phe Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr Arg Met Phe Glu Glu
65 70 75 80
Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala Phe Ala Thr Ala Ser
85 90 95
Gly Met Ala Ala Ile His Gly Val Leu Thr Ser Ile Val Arg Ala Gly
100 105 110
Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly Ser Cys Ile Tyr Ile
115 120 125
Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu Val Thr Phe Val Asp
130 135 140
Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val Arg Pro Gly Thr Lys
145 150 155 160
Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr Leu Glu Val Ala Asp
165 170 175
Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val Gly Ala Leu Val Ile
180 185 190
Val Asp Asn Val Phe Ala Thr Pro Val Phe Ser Thr Ala Val Arg Gln
195 200 205
Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys His Ile Asp Gly Gln
210 215 220
Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser Gln Ala Phe Ile Arg
225 230 235 240
Lys Val Leu Glu Pro Phe Met Lys His Thr Gly Gly Ser Met Ser Pro
245 250 255
Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala Thr Leu Asp Leu Arg
260 265 270
Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile Ala Arg Ala Leu Glu
275 280 285
Gly His Pro Gln Leu Gly Arg Val Ile His Pro Ala Leu Glu Ser His
290 295 300
Pro Gln His Glu Met Ala Lys Ala Gln Met Glu Arg Pro Gly Thr Met
305 310 315 320
Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala Ala Phe Arg Phe Leu
325 330 335
Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn Leu Gly Asp Ala Arg
340 345 350
Ser Ile Ala Thr His Pro Ala Thr Thr Thr His Gln Arg Leu Ser Asp
355 360 365
Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly Leu Val Arg Leu Ser
370 375 380
Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala Asp Leu Lys Gln Ala
385 390 395 400
Leu Ala Val Ile
<210> 67
<211> 404
<212> PRT
<213> Artificial Sequence
<220>
<223> RspmetZ_3
<400> 67
Met Gly Asn Ala Phe Arg Glu Gly Arg Thr Gly Met Thr Lys Asp Trp
1 5 10 15
Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser Arg Arg Ser Gln Tyr
20 25 30
Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln Gly Phe Val Tyr Asp
35 40 45
Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu Thr Gly Ala Asp Glu
50 55 60
Phe Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr Arg Met Phe Glu Glu
65 70 75 80
Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala Phe Ala Thr Ala Ser
85 90 95
Gly Met Ala Ala Ile His Gly Val Leu Thr Ser Ile Val Arg Ala Gly
100 105 110
Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly Ser Cys Ile Tyr Ile
115 120 125
Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu Val Thr Phe Val Asp
130 135 140
Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val Arg Pro Gly Thr Lys
145 150 155 160
Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr Leu Glu Val Ala Asp
165 170 175
Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val Gly Ala Leu Val Ile
180 185 190
Val Asp Asn Val Phe Ala Thr Pro Val Phe Ser Thr Ala Val Arg Gln
195 200 205
Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys His Ile Asp Gly Gln
210 215 220
Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser Gln Ala Phe Ile Arg
225 230 235 240
Lys Val Leu Glu Pro Phe Met Lys His Thr Gly Gly Ser Met Ser Pro
245 250 255
Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala Thr Leu Asp Leu Arg
260 265 270
Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile Ala Arg Ala Leu Glu
275 280 285
Gly His Pro Gln Leu Gly Arg Val Ile His Pro Ala Leu Glu Ser His
290 295 300
Pro Gln His Glu Met Ala Lys Ala Gln Met Glu Arg Pro Gly Thr Met
305 310 315 320
Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala Ala Phe Arg Phe Leu
325 330 335
Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn Leu Gly Asp Ala Arg
340 345 350
Ser Ile Ala Thr His Pro Ala Thr Thr Thr His Gln Arg Leu Ser Asp
355 360 365
Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly Leu Val Arg Leu Ser
370 375 380
Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala Asp Leu Lys Gln Ala
385 390 395 400
Leu Ala Val Ile
<210> 68
<211> 404
<212> PRT
<213> Artificial Sequence
<220>
<223> RspmetZ_65
<400> 68
Met Gly Ile Ala Phe Arg Glu Gly Arg Thr Gly Met Thr Lys Asp Trp
1 5 10 15
Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser Arg Arg Ser Gln Tyr
20 25 30
Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln Gly Phe Val Tyr Asp
35 40 45
Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu Thr Gly Ala Asp Glu
50 55 60
Tyr Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr Arg Met Phe Glu Glu
65 70 75 80
Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala Phe Ala Thr Ala Ser
85 90 95
Gly Met Ala Ala Ile His Gly Val Leu Thr Ser Ile Val Arg Ala Gly
100 105 110
Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly Ser Cys Ile Tyr Ile
115 120 125
Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu Val Thr Phe Val Asp
130 135 140
Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val Arg Pro Gly Thr Lys
145 150 155 160
Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr Leu Glu Val Ala Asp
165 170 175
Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val Gly Ala Leu Val Ile
180 185 190
Val Asp Asn Val Phe Ala Thr Pro Val Phe Ser Thr Ala Val Arg Gln
195 200 205
Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys His Ile Asp Gly Gln
210 215 220
Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser Gln Ala Phe Ile Arg
225 230 235 240
Lys Val Leu Glu Pro Phe Met Lys His Thr Gly Gly Ser Met Ser Pro
245 250 255
Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala Thr Leu Asp Leu Arg
260 265 270
Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile Ala Arg Ala Leu Glu
275 280 285
Gly His Pro Gln Leu Gly Arg Val Ile His Pro Ala Leu Glu Ser His
290 295 300
Pro Gln His Glu Met Ala Lys Ala Gln Met Glu Arg Pro Gly Thr Met
305 310 315 320
Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala Ala Phe Arg Phe Leu
325 330 335
Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn Leu Gly Asp Ala Arg
340 345 350
Ser Ile Ala Thr His Pro Ala Thr Thr Thr His Gln Arg Leu Ser Asp
355 360 365
Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly Leu Val Arg Leu Ser
370 375 380
Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala Asp Leu Lys Gln Ala
385 390 395 400
Leu Ala Val Ile
<210> 69
<211> 404
<212> PRT
<213> Artificial Sequence
<220>
<223> RspmetZ_104
<400> 69
Met Gly Ile Ala Phe Arg Glu Gly Arg Thr Gly Met Thr Lys Asp Trp
1 5 10 15
Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser Arg Arg Ser Gln Tyr
20 25 30
Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln Gly Phe Val Tyr Asp
35 40 45
Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu Thr Gly Ala Asp Glu
50 55 60
Phe Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr Arg Met Phe Glu Glu
65 70 75 80
Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala Phe Ala Thr Ala Ser
85 90 95
Gly Met Ala Ala Ile His Gly Ala Leu Thr Ser Ile Val Arg Ala Gly
100 105 110
Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly Ser Cys Ile Tyr Ile
115 120 125
Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu Val Thr Phe Val Asp
130 135 140
Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val Arg Pro Gly Thr Lys
145 150 155 160
Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr Leu Glu Val Ala Asp
165 170 175
Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val Gly Ala Leu Val Ile
180 185 190
Val Asp Asn Val Phe Ala Thr Pro Val Phe Ser Thr Ala Val Arg Gln
195 200 205
Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys His Ile Asp Gly Gln
210 215 220
Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser Gln Ala Phe Ile Arg
225 230 235 240
Lys Val Leu Glu Pro Phe Met Lys His Thr Gly Gly Ser Met Ser Pro
245 250 255
Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala Thr Leu Asp Leu Arg
260 265 270
Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile Ala Arg Ala Leu Glu
275 280 285
Gly His Pro Gln Leu Gly Arg Val Ile His Pro Ala Leu Glu Ser His
290 295 300
Pro Gln His Glu Met Ala Lys Ala Gln Met Glu Arg Pro Gly Thr Met
305 310 315 320
Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala Ala Phe Arg Phe Leu
325 330 335
Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn Leu Gly Asp Ala Arg
340 345 350
Ser Ile Ala Thr His Pro Ala Thr Thr Thr His Gln Arg Leu Ser Asp
355 360 365
Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly Leu Val Arg Leu Ser
370 375 380
Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala Asp Leu Lys Gln Ala
385 390 395 400
Leu Ala Val Ile
<210> 70
<211> 404
<212> PRT
<213> Artificial Sequence
<220>
<223> RspmetZ_196
<400> 70
Met Gly Ile Ala Phe Arg Glu Gly Arg Thr Gly Met Thr Lys Asp Trp
1 5 10 15
Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser Arg Arg Ser Gln Tyr
20 25 30
Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln Gly Phe Val Tyr Asp
35 40 45
Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu Thr Gly Ala Asp Glu
50 55 60
Phe Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr Arg Met Phe Glu Glu
65 70 75 80
Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala Phe Ala Thr Ala Ser
85 90 95
Gly Met Ala Ala Ile His Gly Val Leu Thr Ser Ile Val Arg Ala Gly
100 105 110
Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly Ser Cys Ile Tyr Ile
115 120 125
Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu Val Thr Phe Val Asp
130 135 140
Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val Arg Pro Gly Thr Lys
145 150 155 160
Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr Leu Glu Val Ala Asp
165 170 175
Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val Gly Ala Leu Val Ile
180 185 190
Val Asp Asn Ile Phe Ala Thr Pro Val Phe Ser Thr Ala Val Arg Gln
195 200 205
Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys His Ile Asp Gly Gln
210 215 220
Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser Gln Ala Phe Ile Arg
225 230 235 240
Lys Val Leu Glu Pro Phe Met Lys His Thr Gly Gly Ser Met Ser Pro
245 250 255
Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala Thr Leu Asp Leu Arg
260 265 270
Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile Ala Arg Ala Leu Glu
275 280 285
Gly His Pro Gln Leu Gly Arg Val Ile His Pro Ala Leu Glu Ser His
290 295 300
Pro Gln His Glu Met Ala Lys Ala Gln Met Glu Arg Pro Gly Thr Met
305 310 315 320
Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala Ala Phe Arg Phe Leu
325 330 335
Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn Leu Gly Asp Ala Arg
340 345 350
Ser Ile Ala Thr His Pro Ala Thr Thr Thr His Gln Arg Leu Ser Asp
355 360 365
Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly Leu Val Arg Leu Ser
370 375 380
Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala Asp Leu Lys Gln Ala
385 390 395 400
Leu Ala Val Ile
<210> 71
<211> 404
<212> PRT
<213> Artificial Sequence
<220>
<223> RspmetZ_3_65_104
<400> 71
Met Gly Asn Ala Phe Arg Glu Gly Arg Thr Gly Met Thr Lys Asp Trp
1 5 10 15
Lys Thr Arg Thr Gln Leu Val His Gly Gly Ser Arg Arg Ser Gln Tyr
20 25 30
Gly Glu Met Ala Glu Ala Ile Phe Leu Thr Gln Gly Phe Val Tyr Asp
35 40 45
Ser Ala Glu Gln Ala Glu Ala Arg Phe Ile Glu Thr Gly Ala Asp Glu
50 55 60
Tyr Ile Tyr Ala Arg Tyr Gly Asn Pro Thr Thr Arg Met Phe Glu Glu
65 70 75 80
Arg Ile Ala Ala Val Glu Gly Thr Glu Asp Ala Phe Ala Thr Ala Ser
85 90 95
Gly Met Ala Ala Ile His Gly Ala Leu Thr Ser Ile Val Arg Ala Gly
100 105 110
Asp His Leu Val Ala Ala Arg Ala Leu Phe Gly Ser Cys Ile Tyr Ile
115 120 125
Leu Glu Glu Val Leu Gly Arg Phe Gly Val Glu Val Thr Phe Val Asp
130 135 140
Gly Thr Asp Leu Asp Gln Trp Arg Ala Ala Val Arg Pro Gly Thr Lys
145 150 155 160
Ala Val Phe Phe Glu Ser Val Ser Asn Pro Thr Leu Glu Val Ala Asp
165 170 175
Ile Gly Ala Ile Ala Glu Ile Ala His Ala Val Gly Ala Leu Val Ile
180 185 190
Val Asp Asn Val Phe Ala Thr Pro Val Phe Ser Thr Ala Val Arg Gln
195 200 205
Gly Ala Asp Val Val Ile Tyr Ser Ala Thr Lys His Ile Asp Gly Gln
210 215 220
Gly Arg Ala Leu Gly Gly Val Val Cys Ala Ser Gln Ala Phe Ile Arg
225 230 235 240
Lys Val Leu Glu Pro Phe Met Lys His Thr Gly Gly Ser Met Ser Pro
245 250 255
Phe Asn Ala Trp Leu Met Leu Asn Gly Met Ala Thr Leu Asp Leu Arg
260 265 270
Cys Arg Ala Met Ala Asp Thr Ala Glu Lys Ile Ala Arg Ala Leu Glu
275 280 285
Gly His Pro Gln Leu Gly Arg Val Ile His Pro Ala Leu Glu Ser His
290 295 300
Pro Gln His Glu Met Ala Lys Ala Gln Met Glu Arg Pro Gly Thr Met
305 310 315 320
Ile Ala Leu Asp Leu Ala Gly Gly Lys Glu Ala Ala Phe Arg Phe Leu
325 330 335
Asp Ala Leu Arg Ile Val Lys Ile Ser Asn Asn Leu Gly Asp Ala Arg
340 345 350
Ser Ile Ala Thr His Pro Ala Thr Thr Thr His Gln Arg Leu Ser Asp
355 360 365
Ala Gln Lys Ala His Leu Gly Ile Thr Pro Gly Leu Val Arg Leu Ser
370 375 380
Val Gly Leu Glu Asp Ala Asp Asp Leu Ile Ala Asp Leu Lys Gln Ala
385 390 395 400
Leu Ala Val Ile
<210> 72
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 72
atcaaaacag atatcatggg tatcgcgttt cgtga 35
<210> 73
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 73
ccttcacgaa acgcgttacc catgatatct g 31
<210> 74
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 74
cagatatcat gggtaacgcg tttcgtgaag g 31
<210> 75
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 75
tagcgggcat agatgtattc gtcggcgccg g 31
<210> 76
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 76
ccggcgccga cgaatacatc tatgcccgct a 31
<210> 77
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 77
acgatcgagg tgagcgcgcc gtggatcgcg g 31
<210> 78
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 78
ccgcgatcca cggcgcgctc acctcgatcg t 31
<210> 79
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 79
cgggcgtcgc gaagatattg tccacgatga c 31
<210> 80
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 80
gtcatcgtgg acaatatctt cgcgacgccc g 31
<210> 81
<211> 5803
<212> DNA
<213> Artificial Sequence
<220>
<223> pDCM2
<400> 81
gttcgcttgc tgtccataaa accgcccagt ctagctatcg ccatgtaagc ccactgcaag 60
ctacctgctt tctctttgcg cttgcgtttt cccttgtcca gatagcccag tagctgacat 120
tcatccgggg tcagcaccgt ttctgcggac tggctttcta cgtgttccgc ttcctttagc 180
agcccttgcg ccctgagtgc ttgcggcagc gtgaagctag cttttatcgc cattcgccat 240
tcaggctgcg caactgttgg gaagggcgat cggtgcgggc ctcttcgcta ttacgccagc 300
tggcgaaagg gggatgtgct gcaaggcgat taagttgggt aacgccaggg ttttcccagt 360
cacgacgttg taaaacgacg gccagtgaat tcgagctcgg tacccgggga tcctctagag 420
tcgacctgca ggcatgcaag cttggcgtaa tcatggtcat agctgtttcc tgtgtgaaat 480
tgttatccgc tcacaattcc acacaacata cgagccggaa gcataaagtg taaagcctgg 540
ggtgcctaat gagtgagcta actcacatta attgcgttgc gctcactgcc cgctttccag 600
tcgggaaacc tgtcgtgcca gctgcattaa tgaatcggcc aacgcgcggg gagaggcggt 660
ttgcgtattg ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc ggtcgttcgg 720
ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac agaatcaggg 780
gataacgcag gaaagaacat gtgagcaaaa ggccagcaaa aggccaggaa ccgtaaaaag 840
gccgcgttgc tggcgttttt ccataggctc cgcccccctg acgagcatca caaaaatcga 900
cgctcaagtc agaggtggcg aaacccgaca ggactataaa gataccaggc gtttccccct 960
ggaagctccc tcgtgcgctc tcctgttccg accctgccgc ttaccggata cctgtccgcc 1020
tttctccctt cgggaagcgt ggcgctttct caatgctcac gctgtaggta tctcagttcg 1080
gtgtaggtcg ttcgctccaa gctgggctgt gtgcacgaac cccccgttca gcccgaccgc 1140
tgcgccttat ccggtaacta tcgtcttgag tccaacccgg taagacacga cttatcgcca 1200
ctggcagcag ccactggtaa caggattagc agagcgaggt atgtaggcgg tgctacagag 1260
ttcttgaagt ggtggcctaa ctacggctac actagaagga cagtatttgg tatctgcgct 1320
ctgctgaagc cagttacctt cggaaaaaga gttggtagct cttgatccgg caaacaaacc 1380
accgctggta gcggtggttt ttttgtttgc aagcagcaga ttacgcgcag aaaaaaagga 1440
tctcaagaag atcctttgat cttttctacg gggtctgacg ctcagtggaa cgaaaactca 1500
cgttaaggga ttttggtcat gagattatca aaaaggatct tcacctagat ccttttgggg 1560
tgggcgaaga actccagcat gagatccccg cgctggagga tcatccagcc ctgatagaaa 1620
cagaagccac tggagcacct caaaaacacc atcatacact aaatcagtaa gttggcagca 1680
tcacccgacg cactttgcgc cgaataaata cctgtgacgg aagatcactt cgcagaataa 1740
ataaatcctg gtgtccctgt tgataccggg aagccctggg ccaacttttg gcgaaaatga 1800
gacgttgatc ggcacgtaag aggttccaac tttcaccata atgaaataag atcactaccg 1860
ggcgtatttt ttgagttatc gagattttca ggagctgata gaaacagaag ccactggagc 1920
acctcaaaaa caccatcata cactaaatca gtaagttggc agcatcaccc gacgcacttt 1980
gcgccgaata aatacctgtg acggaagatc acttcgcaga ataaataaat cctggtgtcc 2040
ctgttgatac cgggaagccc tgggccaact tttggcgaaa atgagacgtt gatcggcacg 2100
taagaggttc caactttcac cataatgaaa taagatcact accgggcgta ttttttgagt 2160
tatcgagatt ttcaggagct ctttggcatc gtctctcgcc tgtcccctca gttcagtaat 2220
ttcctgcatt tgcctgtttc cagtcggtag atattccaca aaacagcagg gaagcagcgc 2280
ttttccgctg cataaccctg cttcggggtc attatagcga ttttttcggt atatccatcc 2340
tttttcgcac gatatacagg attttgccaa agggttcgtg tagactttcc ttggtgtatc 2400
caacggcgtc agccgggcag gataggtgaa gtaggcccac ccgcgagcgg gtgttccttc 2460
ttcactgtcc cttattcgca cctggcggtg ctcaacggga atcctgctct gcgaggctgg 2520
ccggctaccg ccggcgtaac agatgagggc aagcggatgg ctgatgaaac caagccaacc 2580
aggaagggca gcccacctat caaggtgtac tgccttccag acgaacgaag agcgattgag 2640
gaaaaggcgg cggcggccgg catgagcctg tcggcctacc tgctggccgt cggccagggc 2700
tacaaaatca cgggcgtcgt ggactatgag cacgtccgcg agggcgtccc ggaaaacgat 2760
tccgaagccc aacctttcat agaaggcggc ggtggaatcg aaatctcgtg atggcaggtt 2820
gggcgtcgct tggtcggtca tttcgaaaaa ggttaggaat acggttagcc atttgcctgc 2880
ttttatatag ttcantatgg gattcacctt tatgttgata agaaataaaa gaaaatgcca 2940
ataggatatc ggcattttct tttgcgtttt tatttgttaa ctgttaattg tccttgttca 3000
aggatgctgt ctttgacaac agatgttttc ttgcctttga tgttcagcag gaagctcggc 3060
gcaaacgttg attgtttgtc tgcgtagaat cctctgtttg tcatatagct tgtaatcacg 3120
acattgtttc ctttcgcttg aggtacagcg aagtgtgagt aagtaaaggt tacatcgtta 3180
ggcggatcaa gatccatttt taacacaagg ccagttttgt tcagcggctt gtatgggcca 3240
gttaaagaat tagaaacata accaagcatg taaatatcgt tagacgtaat gccgtcaatc 3300
gtcatttttg atccgcggga gtcagtgaac aggtaccatt tgccgttcat tttaaagacg 3360
ttcgcgcgtt caatttcatc tgttactgtg ttagatgcaa tcagcggttt catcactttt 3420
ttcagtgtgt aatcatcgtt tagctcaatc ataccgagag cgccgtttgc taactcagcc 3480
gtgcgttttt tatcgctttg cagaagtttt tgactttctt gacggaagaa tgatgtgctt 3540
ttgccatagt atgctttgtt aaataaagat tcttcgcctt ggtagccatc ttcagttcca 3600
gtgtttgctt caaatactaa gtatttgtgg cctttatctt ctacgtagtg aggatctctc 3660
agcgtatggt tgtcgcctga gctgtagttg ccttcatcga tgaactgctg tacattttga 3720
tacgtttttc cgtcaccgtc aaagattgat ttataatcct ctacaccgtt gatgttcaaa 3780
gagctgtctg atgctgatac gttaacttgt gcagttgtca gtgtttgttt gccgtaatgt 3840
ttaccggaga aatcagtgta gaataaacgg atttttccgt cagatgtaaa tgtggctgaa 3900
cctgaccatt cttgtgtttg gtcttttagg atagaatcat ttgcatcgaa tttgtcgctg 3960
tctttaaaga cgcggccagc gtttttccag ctgtcaatag aagtttcgcc gactttttga 4020
tagaacatgt aaatcgatgt gtcatccgca tttttaggat ctccggctaa tgcaaagacg 4080
atgtggtagc cgtgatagtt tgcgacagtg ccgtcagcgt tttgtaatgg ccagctgtcc 4140
caaacgtcca ggccttttgc agaagagata tttttaattg tggacgaatc aaattcagaa 4200
acttgatatt tttcattttt ttgctgttca gggatttgca gcatatcatg gcgtgtaata 4260
tgggaaatgc cgtatgtttc cttatatggc ttttggttcg tttctttcgc aaacgcttga 4320
gttgcgcctc ctgccagcag tgcggtagta aaggttaata ctgttgcttg ttttgcaaac 4380
tttttgatgt tcatcgttca tgtctccttt tttatgtact gtgttagcgg tctgcttctt 4440
ccagccctcc tgtttgaaga tggcaagtta gttacgcaca ataaaaaaag acctaaaata 4500
tgtaaggggt gacgccaaag tatacacttt gccctttaca cattttaggt cttgcctgct 4560
ttatcagtaa caaacccgcg cgatttactt ttcgacctca ttctattaga ctctcgtttg 4620
gattgcaact ggtctatttt cctcttttgt ttgatagaaa atcataaaag gatttgcaga 4680
ctacgggcct aaagaactaa aaaatctatc tgtttctttt cattctctgt attttttata 4740
gtttctgttg catgggcata aagttgcctt tttaatcaca attcagaaaa tatcataata 4800
tctcatttca ctaaataata gtgaacggca ggtatatgtg atgggttaaa aaggatcacc 4860
ccagagtccc gctcagaaga actcgtcaag aaggcgatag aaggcgatgc gctgcgaatc 4920
gggagcggcg ataccgtaaa gcacgaggaa gcggtcagcc cattcgccgc caagctcttc 4980
agcaatatca cgggtagcca acgctatgtc ctgatagcgg tccgccacac ccagccggcc 5040
acagtcgatg aatccagaaa agcggccatt ttccaccatg atattcggca agcaggcatc 5100
gccatgggtc acgacgagat cctcgccgtc gggcatccgc gccttgagcc tggcgaacag 5160
ttcggctggc gcgagcccct gatgctcttc gtccagatca tcctgatcga caagaccggc 5220
ttccatccga gtacgtgctc gctcgatgcg atgtttcgct tggtggtcga atgggcaggt 5280
agccggatca agcgtatgca gccgccgcat tgcatcagcc atgatggata ctttctcggc 5340
aggagcaagg tgagatgaca ggagatcctg ccccggcact tcgcccaata gcagccagtc 5400
ccttcccgct tcagtgacaa cgtcgagaca gctgcgcaag gaacgcccgt cgtggccagc 5460
cacgatagcc gcgctgcctc gtcttggagt tcattcaggg caccggacag gtcggtcttg 5520
acaaaaagaa ccgggcgccc ctgcgctgac agccggaaca cggcggcatc agagcagccg 5580
attgtctgtt gtgcccagtc atagccgaat agcctctcca cccaagcggc cggagaacct 5640
gcgtgcaatc catcttgttc aatcatgcga aacgatcctc atcctgtctc ttgatcagat 5700
cttgatcccc tgcgccatca gatccttggc ggcaagaaag ccatccagtt tactttgcag 5760
ggcttcccaa ccttaccaga gggcgcccca gctggcaatt ccg 5803
<---
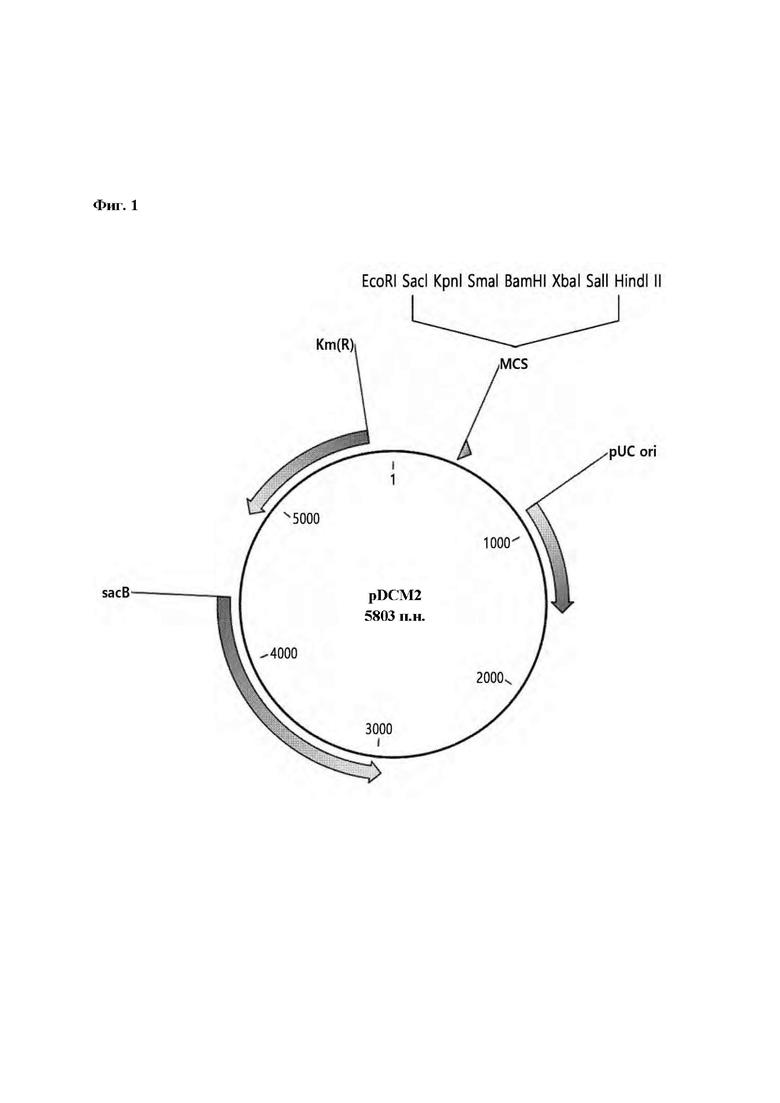
Изобретение относится к биотехнологии. Предложен способ получения L-метионина, включающий культивирование микроорганизма, в который введен белок, кодируемый чужеродным геном metZ, в среде, содержащей тиосульфат. Также предложена композиция для получения L-метионина, содержащая микроорганизм, продуцирующий L-метионин, в который введен белок, кодируемый чужеродным геном metZ, и тиосульфат. Изобретение обеспечивает повышение выхода метионина. 2 н. и 7 з.п. ф-лы, 1 ил., 17 табл., 10 пр.
1. Способ получения L-метионина, включающий культивирование микроорганизма, в который введен белок, кодируемый чужеродным геном metZ, в среде, содержащей тиосульфат.
2. Способ получения L-метионина по п. 1, где белок обладает активностью O-ацилгомосеринтранссульфуразы.
3. Способ получения L-метионина по п. 1, где белок имеет происхождение из Chromobacterium violaceum, Hyphomonas neptunium или Rhodobacter sphaeroides.
4. Способ получения L-метионина по п. 1, где белок включает любой один или более чем один белок, выбранный из группы, состоящей из полипептидных последовательностей SEQ ID NO: 60, 61 и 62 и полипептидных последовательностей, гомологичных или идентичных SEQ ID NO: 60, 61 и 62 на 90% или более.
5. Способ получения L-метионина по п. 1, где микроорганизм включает одну или более чем одну генетическую модификацию, выбранную из группы, состоящей из ослабления или инактивации активности цистатионингаммасинтазы, ослабления или инактивации активности O-ацетилгомосеринсульфгидрилазы, ослабления или инактивации активности белка-репрессора биосинтеза метионина-цистеина, повышения активности метионинсинтазы и повышения активности сульфитредуктазы.
6. Способ получения L-метионина по п. 1, где микроорганизм представляет собой микроорганизм рода Corynebacterium.
7. Способ получения L-метионина по п. 6, где микроорганизм представляет собой Corynebacterium glutamicum.
8. Способ получения L-метионина по п. 1, включающий выделение L-метионина из микроорганизма или среды.
9. Композиция для получения L-метионина, содержащая микроорганизм, продуцирующий L-метионин, в который введен белок, кодируемый чужеродным геном metZ, и тиосульфат.
| US 9080155 B2, 14.04.2015 | |||
| US 9029105 B2, 12.05.2015 | |||
| US 20180223319 A1, 09.08.2018 | |||
| US 2009298137 A1, 03.12.2009 | |||
| TATE R | |||
| et al | |||
| The Rhizobium etli metZ gene is essential for methionine biosynthesis and nodulation of Phaseolus vulgaris | |||
| Mol Plant Microbe Interact | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| РЕКОМБИНАНТНЫЕ МИКРООРГАНИЗМЫ, ПРОДУЦИРУЮЩИЕ МЕТИОНИН | 2006 |
|
RU2447146C2 |

