ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится к штамму для получения L-глутаминовой кислоты в высокой концентрации и к способу получения L-глутаминовой кислоты с его применением.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
L-глутаминовая кислота представляет собой типичную аминокислоту, получаемую посредством ферментации, и имеет своеобразный характерный вкус и поэтому является важной аминокислотой, широко применяемой в области пищевой промышленности, а также в области медицины и других областях кормов для животных. Известно, что способы получения L-глутаминовой кислоты включают способ с применением микроорганизма рода Corynebacterium, Escherichia coli, или микроорганизмов рода Bacillus, Streptomyces, Penicillium, Klebsiella, Erwinia, Pantoea, и так далее (патенты US 3220929 и 6682912).
В настоящее время проводятся исследования по созданию микроорганизмов и технологии процесса ферментации, которые бы позволяли получать L-глутаминовую кислоту с высокой эффективностью. Например, у микроорганизма рода Corynebacterium для улучшения выхода продуцируемой L-глутаминовой кислоты в основном применяется подход, специфический для целевого материала, такой как повышение экспрессии гена, кодирующего фермент, задействованный в биосинтезе аминокислоты, или удаление гена, ненужного для биосинтеза аминокислоты.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ Техническая задача
Авторы данного изобретения предприняли множество попыток получения L-глутаминовой кислоты с высоким выходом и в результате подтвердили, что способность продуцировать L-глутаминовую кислоту повышалась при инактивации белка VKOR (белок семейства витамин К-эпоксид-редуктазы), тем самым завершив данное изобретение.
Техническое решение
Задачей данного изобретения является создание микроорганизма рода Corynebacterium, в котором белок VKOR (белок семейства витамин К-эпоксид-редуктазы) инактивирован.
Другой задачей данного изобретения является разработка способа получения L-глутаминовой кислоты, включающего: культивирование в среде микроорганизма рода Corynebacterium, в котором белок VKOR инактивирован.
Еще одной задачей данного изобретения является разработка композиции для получения L-глутаминовой кислоты, включающей: микроорганизм рода Corynebacterium, в котором белок VKOR инактивирован; среду для его культивирования или их комбинацию.
Еще одной задачей данного изобретения является разработка способа получения микроорганизма рода Corynebacterium, включающего инактивацию белка VKOR.
Еще одной задачей данного изобретения является разработка композиции для получения L-глутаминовой кислоты, включающей: микроорганизм рода Corynebacterium, в котором белок VKOR инактивирован; среду для его культивирования или их комбинацию.
Еще одной задачей данного изобретения является разработка способа получения микроорганизма рода Corynebacterium, включающего инактивацию белка VKOR.
Еще одной задачей данного изобретения является разработка применения L-глутаминовой кислоты, произведенной микроорганизмом рода Corynebacterium, в котором белок VKOR инактивирован.
Полезные эффекты
Продуцирующий L-глутаминовую кислоту микроорганизм рода Corynebacterium, в котором белок VKOR по данному изобретению инактивирован, может продуцировать L-глутаминовую кислоту с высоким выходом и поэтому может эффективно применяться для промышленного производства L-глутаминовой кислоты.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее данное изобретение будет описано более подробно. При этом, каждое описание и воплощение, раскрытое в данном изобретении, также применимо к другим описаниям и воплощениям, соответственно. Таким образом, все комбинации различных элементов, описанных в данном изобретении, входят в объем данного изобретения. Более того, объем данного изобретения не ограничивается конкретным описанием, приведенным ниже. Кроме того, специалистам в данной области будут очевидны или найдены в ходе рутинных экспериментов множество эквивалентов конкретных аспектов изобретения, описанных в данном документе. Более того, предполагается, что данное изобретение будет охватывать такие эквиваленты.
В одном аспекте данного изобретения предложен микроорганизм рода Corynebacterium, в котором белок VKOR (белок семейства витамин K-эпоксид-редуктазы) инактивирован.
В данном описании термин «VKOR (белок семейства витамин K-эпоксид-редуктазы)» относится к ферменту, обладающему активностью, восстанавливающей витамин K-2,3-эпоксид и витамин K в витамин K-гидрохинон (Vitamins & Hormones Volume 78, 2008, p.103-130).
В одном примере данной заявки белок VKOR может происходить из микроорганизма. Микроорганизм, в частности, может происходить от микроорганизма рода Corynebacterium. Более конкретно, он может происходить от Corynebacterium glutamicum, Corynebacterium deserti, Corynebacterium crudilactis, Corynebacterium efficiens, Corynebacterium callunae и так далее, без ограничения.
Аминокислотная последовательность белка VKOR может кодироваться геном VKOR. Например, ген VKOR может представлять собой NCgl0775, происходящий из Corynebacterium glutamicum АТСС13032, или BBD29_04485, происходящий из Corynebacterium glutamicum АТСС13869, без ограничения.
Белок VKOR по данному изобретению может включать или состоять из полипептида, представленного аминокислотной последовательностью, обладающей гомологией 80% или более с аминокислотной последовательностью SEQ ID NO: 1. Кроме того, белок VKOR по данному изобретению может иметь аминокислотную последовательность, обладающую гомологией 80% или более с аминокислотной последовательностью SEQ ID NO:1, или может по существу состоять из аминокислотной последовательности. В частности, белок может состоять из полипептида, представленного аминокислотной последовательностью SEQ ID NO: 1.
Аминокислотная последовательность SEQ ID NO:1 может быть получена из известной базы данных NIH Genbank. В данном изобретении аминокислотная последовательность SEQ ID NO:1 может включать аминокислотную последовательность, гомологичную или идентичную по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,7% или 99,9% аминокислотной последовательности SEQ ID NO: 1. Кроме того, очевидно, что белки, имеющие аминокислотную последовательность с делецией, модификацией, заменой, консервативной заменой или добавлением части последовательности, могут входить в объем данного изобретения, при условии, что их аминокислотная последовательность обладает такой гомологией или идентичностью, и демонстрируют эффективность, соответствующую таковой белка, включающего аминокислотную последовательность SEQ ID NO: 1.
Например, он может включать добавления или делеции последовательности, мутации естественного происхождения, молчащие мутации или консервативные замены на N-конце, С-конце и/или внутри аминокислотной последовательности, которые не изменяют функцию белка по данному изобретению.
В данном описании термин «консервативная замена» относится к замене одной аминокислоты на другую аминокислоту, имеющую схожую структуру и/или химические свойства. Такие аминокислотные замены, как правило, могут возникать на основании сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатка. Как правило, консервативные замены могут оказывать незначительное влияние или не оказывать влияния на активность белка или полипептида.
В данном описании термин «гомология» или «идентичность» относится к степени родства двух заданных аминокислотных последовательностей или нуклеотидных последовательностей и может выражаться в процентах. Термины «гомология» и «идентичность» зачастую могут использоваться взаимозаменяемо друг с другом.
Гомология или идентичность последовательностей консервативных полипептидов или полинуклеотидов может быть установлена при помощи стандартных алгоритмов выравнивания, для которых можно использовать значения штрафов за открытие гэпа, установленные в используемой программе по умолчанию. Обычно предполагается, что по существу гомологичные или идентичные последовательности гибридизуются в условиях умеренной или высокой строгости по всей длине последовательностей или в ее части. Очевидно, что сюда также входит гибридизация с полинуклеотидами, содержащими общие кодоны или вырожденные кодоны в гибридизующихся полинуклеотидах.
Установить, обладают ли две любые полинуклеотидные последовательности гомологией, сходством или идентичностью, можно, например, при помощи известного компьютерного алгоритма, такого как программа «FASTA» с использованием параметров по умолчанию (Pearson et al., (1988) Proc. Natl. Acad. Sci. USA 85:2444). В альтернативном варианте, это можно установить при помощи алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), используемого в программе Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277) (версии 5.0.0 или более поздней) (пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F., et al., J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed, Academic Press, San Diego, 1994 и CARILLO et al (1988) SIAM J Applied Math 48:1073). Например, гомологию, сходство или идентичность можно устанавливать при помощи BLAST или ClustalW Национального Центра Биотехнологической Информации (NCBI).
Гомологию, сходство или идентичность полипептидов или полинуклеотидов можно устанавливать путем сравнения информации о последовательности с применением, например, компьютерной программы GAP, такой как Needleman et al. (1970), J Mol Biol. 48:443, как изложено Smith and Waterman, Adv. Appl. Math (1981) 2:482. Вкратце, программа GAP определяет гомологию, сходство или идентичность как значение, полученное путем деления количества одинаковых выровненных символов (то есть нуклеотидов или аминокислот) на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать (1) одномерную матрицу сравнения (содержащую значение 1 для идентичных и 0 для неидентичных символов) и взвешенную матрицу сравнения, предложенную Gribskov et al. (1986), Nucl. Acids Res. 14:6745, как описано в Атласе белковых последовательностей и структур под ред. Schwartz and Dayhoff, Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или подстановочную матрицу EDNAFULL (EMBOSS версия NCBI NUC4.4)); (2) штраф 3,0 за каждый гэп и дополнительный штраф 0,10 за каждый символ в каждом гэпе (или штраф за открытие гэпа 10 и штраф за продолжение гэпа 0,5); и (3) отсутствие штрафа за концевые гэпы.
В данном изобретении полинуклеотид, кодирующий белок VKOR, может представлять собой ген VKOR. В одном примере ген VKOR может включать ген NCgl0775 или BBD29_04485 и так далее, как описано выше.
В данном описании «полинуклеотид», представляющий собой полимер нуклеотидов, состоящий из мономеров нуклеотидов, соединенных в длинную цепочку ковалентными связями, представляет собой цепь ДНК или РНК, имеющую по меньшей мере определенную длину. Более конкретно, он может относиться к фрагменту полинуклеотида, кодирующему белок.
Полинуклеотид, кодирующий белок VKOR по данному изобретению, может включать в себя нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID NO: 1. В качестве примера данного изобретения полинуклеотид по данному изобретению может иметь или включать последовательность SEQ ID NO: 2. Кроме того, полинуклеотид по данному изобретению может состоять или по существу состоять из последовательности SEQ ID NO: 2. В частности, белок VKOR может кодироваться полинуклеотидом, представленным в нуклеотидной последовательности SEQ ID NO: 2.
Вследствие вырожденности кодонов или с учетом предпочтения кодонов у организма, в котором будет экспрессироваться белок VKOR, полинуклеотид по данному изобретению может претерпевать различные модификации в кодирующей области в таком объеме, который не меняет аминокислотную последовательность белка VKOR. В частности, полинуклеотид по данному изобретению может иметь или включать в себя нуклеотидную последовательность, обладающую гомологией или идентичностью 70% или более, 75% или более, 76% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более или 98% или более с последовательностью SEQ ID NO:2, или может состоять или по существу состоять из нуклеотидной последовательности, обладающей гомологией или идентичностью 70% или более, 75% или более, 76% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более или 98% или более с последовательностью SEQ ID NO:2, без ограничения.
Кроме того, полинуклеотид по данному изобретению может включать в себя зонд, который может быть получен из известной генной последовательности, например, любой последовательности, которая может гибридизоваться в строгих условиях с последовательностью, комплементарной всей или части полинуклеотидной последовательности по данному изобретению, без ограничения. «Строгие условия» относятся к условиям, при которых обеспечивается специфическая гибридизация между полинуклеотидами. Такие условия, в частности, описаны в литературе (J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F. M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, 9.50-9.51, 11.7-11.8). Например, строгие условия могут включать условия, при которых гены, обладающие высокой гомологией или идентичностью 70% или более, 75% или более, 76% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более, гибридизуются друг с другом, а полинуклеотиды, обладающие гомологией или идентичностью меньше указанных, не гибридизуются друг с другом, или условия отмывки при гибридизации по Саузерну, представляющие собой однократную, двукратную или трехкратную отмывку при концентрации соли и температуре, соответствующих 60°С, 1X SSC (стандартный солевой раствор), 0,1% SDS (додецилсульфат натрия), в частности, 60°С, 0,1X SSC, 0,1% SDS, и более конкретно, 68°С, 0,1XSSC, 0,1% SDS.
Для гибридизации необходимо, чтобы две нуклеиновых кислоты содержали комплементарные последовательности, хотя, в зависимости от строгости гибридизации, допускаются несовпадения между основаниями. Термин «комплементарный» используется для описания взаимодействия между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в случае ДНК аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, полинуклеотид по данному изобретению может включать выделенные фрагменты нуклеотидов, комплементарные полной последовательности, а также по существу схожие с ними нуклеиновокислотные последовательности.
В частности, полинуклеотиды, обладающие гомологией или идентичностью с полинуклеотидами по данному изобретению, можно выявить с применением условий гибридизации, включающих стадию гибридизации при значении Tm 55°С в описанных выше условиях. Кроме того, значение Tm может быть 60°С, 63°С или 65°С, без ограничения, и может быть соответствующим образом скорректировано специалистом в области техники в зависимости от его задачи.
Надлежащая строгость для гибридизации полинуклеотидов зависит от длины полинуклеотидов и степени комплементарности, и эти переменные хорошо известны в области техники (например, Sambrook et al.).
В данном описании термин «микроорганизм (или штамм)» включает все микроорганизмы дикого типа или микроорганизмы, генетически модифицированные естественным или искусственным образом, и он может представлять собой микроорганизм, в котором конкретный механизм ослаблен или усилен вследствие вставки чужеродного гена или усиления или инактивации активности эндогенного гена, и может представлять собой микроорганизм, включающий генетическую модификацию для продуцирования желаемого полипептида, белка или продукта.
В данном описании термин «инактивация» активности полипептида представляет собой всеобъемлющее понятие, включающее как сниженную активность, так и отсутствие активности по сравнению с эндогенной активностью. Инактивация может использоваться взаимозаменяемо с такими терминами, как ослабление, дефицит, снижение, уменьшение, сокращение, аттенюация и так далее.
Инактивация может также включать случай, когда активность самого полипептида снижена или устранена по сравнению с активностью полипептида, исходно присущей микроорганизму, вследствие мутации полинуклеотида, кодирующего полипептид; случай, когда общий уровень внутриклеточной активности и/или концентрации (уровень экспрессии) полипептида снижены по сравнению с природным штаммом вследствие ингибирования экспрессии гена полинуклеотида, кодирующего полипептид, или ингибирования трансляции в полипептид и так далее; случай, когда полинуклеотид совсем не экспрессируется, и/или случай, когда активность полипептида не проявляется даже несмотря на экспрессию полинуклеотида. В данном описании термин «эндогенная активность» относится к активности конкретного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм или микроорганизм дикого типа, когда признак изменен вследствие генетической модификации, вызванной факторами естественного или искусственного происхождения, и может использоваться взаимозаменяемо с «активностью до модификации». Выражение «активность полипептида «инактивирована, ослаблена, находится в дефиците, уменьшена, снижена, сокращена или аттенуирована» по сравнению с его эндогенной активностью» означает, что активность полипептида снижена по сравнению с активностью конкретного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм.
Инактивация активности полипептида может осуществляться любым способом, известным в области техники, без ограничения, и может достигаться путем применения различных способов, хорошо известных в области техники (например, Nakashima N et at., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014;15(2):2773-2793, Sambrook et al. Molecular Cloning 2012 и так далее).
В частности, инактивация активности полипептида в данном изобретении может достигаться посредством:
1) делеции части гена или всего гена, кодирующего полипептид;
2) модификации области, регулирующей экспрессию (последовательности, регулирующей экспрессию) таким образом, что экспрессия гена, кодирующего полипептид, уменьшается;
3) модификации аминокислотной последовательности, составляющей полипептид (например, делеции/замены/добавления одной или более аминокислот в аминокислотную последовательность), таким образом, что активность полипептида устраняется или ослабляется;
4) модификации последовательности гена, кодирующего полипептид, таким образом, что активность полипептида устраняется или ослабляется (например, делеции/замены/добавления одного или более нуклеотидов в нуклеотидную последовательность гена полипептида, для кодирования полипептида, модифицированного таким образом, что активность полипептида устраняется или ослабляется);
5) модификации нуклеотидной последовательности, кодирующей инициирующий кодон, последовательность Шайна-Дальгарно или 5'-нетранслируемую область (5'-UTR) транскрипта гена, кодирующего полипептид;
6) внедрения антисмыслового олигонуклеотида (например, антисмысловой РНК), которая комплементарно связывается с транскриптом гена, кодирующего полипептид;
7) добавления последовательности, комплементарной последовательности Шайна-Дальгарно, к переднему концу последовательности Шайна-Дальгарно гена, кодирующего полипептид, для образования вторичной структуры, тем самым ингибирования присоединения рибосомы;
8) инженерии на основе транскрипции в обратном направлении (RTE), представляющей собой добавление промотора, который будет транскрибироваться в обратном направлении, к 3' концу открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид; или
9) комбинации двух или более способов, выбранных из (1) - (8), без конкретного ограничения.
Например,
Способ 1) делеции части или всего гена, кодирующего полипептид, может быть реализован путем делеции всего полинуклеотида, кодирующего эндогенный целевой полипептид в составе хромосомы, или замену полинуклеотида полинуклеотидом или маркерным геном, имеющим делецию части нуклеиновых кислот.
Способ 2) модификации области, регулирующей экспрессию (последовательности, регулирующей экспрессию), может быть реализован путем индукции модификации в области, регулирующей экспрессию (последовательности, регулирующую экспрессию), путем делеции, вставки, неконсервативной замены или консервативной замены или их комбинации; или путем замены последовательности последовательностью, обладающей более слабой активностью. Область, регулирующая экспрессию, может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность для регуляции транскрипции и трансляции, без ограничения.
Способы 3) и 4) модификации аминокислотной последовательности или полинуклеотидной последовательности могут быть реализованы путем индукции модификации последовательности в результате делеции, вставки, неконсервативной или консервативной замены в аминокислотной последовательности полипептида или в полинуклеотидной последовательности, кодирующей полипептид, или их комбинации для ослабления активности полипептида, или путем замены последовательности аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной для проявления более слабой активности, или аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной, чтобы не обладать активностью, без ограничения. Например, можно ингибировать или ослаблять экспрессию гена путем внедрения мутации в полинуклеотидную последовательность для образования стоп-кодона, без ограничения.
Способ 5) модификации нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-нетранслируемую область транскрипта гена, кодирующего полипептид, можно реализовать, например, путем замены нуклеотидной последовательности нуклеотидной последовательностью, кодирующей другой инициирующий кодон, имеющий более низкую степень экспрессии полипептида по сравнению с эндогенным инициирующим кодоном, без ограничения.
Способ 6) внедрения антисмыслового олигонуклеотида (например, антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего полипептид, можно найти в литературе (Weintraub, Н. et al., Antisense-RNA as a molecular tool for genetic analysis, Reviews - Trends in Genetics, Vol. 1(1) 1986).
Способ 7) добавления последовательности, комплементарной последовательности Шайна-Дальгарно, к переднему концу последовательности Шайна-Дальгарно гена, кодирующего полипептид, для образования вторичной структуры, тем самым ингибирования присоединения рибосомы, может быть реализован путем ингибирования трансляции мРНК или уменьшения ее скорости.
8) Инженерия на основе транскрипции в обратном направлении (RTE) при которой к 3' концу открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид, добавляют промотор, который будет транскрибироваться в обратном направлении, может быть реализован путем образования антисмыслового нуклеотида, комплементарного транскрипту гена, кодирующего полипептид, для ослабления активности.
В данном описании термин «усиление» активности полипептида означает, что активность полипептида увеличена по сравнению с его эндогенной активностью.
Усиление может использоваться взаимозаменяемо с такими терминами, как активация, повышение, сверхэкспрессия и увеличение и так далее. В частности, термины активация, усиление, повышение, сверхэкспрессия и увеличение могут включать как случаи, когда проявляется активность, которая исходно отсутствовала, так и усиление активности по сравнению с эндогенной активностью или активностью до модификации. «Эндогенная активность» относится к активности конкретного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм, когда признак изменен вследствие генетической модификации, вызванной факторами естественного или искусственного происхождения, и может использоваться взаимозаменяемо с «активностью до модификации». «Усиление», «повышение», «сверхэкспрессия» или «увеличение» активности полипептида по сравнению с его эндогенной активностью означает, что активность и/или концентрация (уровень экспрессии) усилена по сравнению с таковой конкретного полипептида, которой исходно обладал родительский штамм до трансформации или немодифицированный микроорганизм.
Усиление может достигаться посредством внедрения чужеродного полипептида или посредством усиления активности и/или концентрации (уровня экспрессии) эндогенного полипептида. Усиление активности полипептида можно подтвердить по увеличению уровня активности полипептида, уровня экспрессии или количества продукта, выделенного из полипептида.
Для усиления активности полипептида могут применяться различные способы, хорошо известные в области техники, без ограничения, при условии, что они позволяют усилить активность целевого полипептида по сравнению с таковой у микроорганизма до модификации. В частности, можно применять генетическую инженерию и/или белковую инженерию, хорошо известные специалистам в области молекулярной биологии, без ограничения (например, Sitnicka et al. Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol. 2. 1-16, Sambrook et al. Molecular Cloning 2012 и так далее).
В частности, усиление полипептида в данном изобретении может достигаться посредством:
1) увеличения внутриклеточного числа копий полинуклеотида, кодирующего полипептид;
2) замены на хромосоме последовательности, регулирующей экспрессию гена, кодирующего полипептид, последовательностью, обладающей сильной активностью;
3) модификации нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR транскрипта гена, кодирующего полипептид;
4) модификации аминокислотной последовательности полипептида таким образом, чтобы активность полипептида усиливалась;
5) модификации полинуклеотидной последовательности, кодирующей полипептид, таким образом, чтобы активность полипептида усиливалась (например, модификации полинуклеотидной последовательности гена полипептида, чтобы кодировать полипептид, модифицированный для усиления активности полипептида);
6) внедрения чужеродного полипептида, обладающего активностью полипептида, или чужеродного полинуклеотида, кодирующего его;
7) оптимизации кодонов полинуклеотида, кодирующего полипептид;
8) анализа третичной структуры полипептида и, таким образом, выбора и модификации экспонированного сайта или его химической модификации; или
9) комбинации двух или более, выбранных из приведенных выше (1) - (8), без конкретного ограничения.
Более конкретно,
Способ 1) увеличения числа внутриклеточных копий полинуклеотида, кодирующего полипептид, может быть реализован путем внедрения в клетку-хозяина вектора, функционально связанного с полинуклеотидом, кодирующим полипептид, и способного реплицироваться и функционировать независимо от клетки-хозяина. В качестве альтернативы способ может достигаться путем внедрения одной копии или двух копий полинуклеотидов, кодирующих полипептид, в хромосому клетки-хозяина. Внедрение в хромосому может осуществляться путем внедрения в клетку-хозяина вектора, который способен встраивать полинуклеотид в хромосому клетки-хозяина, без ограничения.
Способ 2) замены области, регулирующей экспрессию (или последовательности, регулирующей экспрессию) гена, кодирующего полипептид, на хромосоме последовательностью, имеющей сильную активность, может быть реализован путем индукции модификации последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации для дополнительного усиления активности области, регулирующей экспрессию, или путем замены последовательности последовательностью, обладающей более высокой активностью. Область, регулирующая экспрессию, может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, и последовательность, регулирующую терминацию транскрипции и трансляции, без ограничения. В одном примере способ может включать замену исходного промотора сильным промотором, без ограничения.
Примеры сильного промотора могут включать промоторы CJ1 - CJ7 (US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13(sm3) (US 10584338 В2), промотор O2 (US 10273491 В2), промотор tkt, промотор уссА и так далее, без ограничения.
Способ 3) модификации нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-нетранслируемую область транскрипта гена, кодирующего полипептид, может быть реализован, например, путем замены нуклеотидной последовательности нуклеотидной последовательностью, кодирующей другой инициирующий кодон, имеющий более высокую степень экспрессии полипептида по сравнению с эндогенным инициирующим кодоном, без ограничения.
Способы 4) и 5) модификации аминокислотной последовательности или полинуклеотидной последовательности могут быть реализованы путем индукции модификации последовательности за счет делеции, вставки, неконсервативной или консервативной замены в аминокислотной последовательности полипептида или в полинуклеотидной последовательности, кодирующей полипептид, или их комбинации для усиления активности полипептида, или путем замены последовательности аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной для проявления более высокой активности, или аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной, чтобы усиливать активность, без ограничения. В частности, замена может осуществляться путем встраивания полинуклеотида в хромосому посредством гомологичной рекомбинации, без ограничения. Используемый при этом вектор может дополнительно включать в себя селективный маркер для подтверждения встраивания в хромосому. Селективный маркер является таким, как описано выше.
Способ 6) внедрения чужеродного полинуклеотида, обладающего активностью полипептида, может быть реализован путем внедрения в клетку-хозяина чужеродного полинуклеотида, кодирующего полипептид, демонстрирующий такую же или схожую активность с полипептидом. Чужеродный полинуклеотид может применяться без ограничения, независимо от его происхождения или последовательности, при условии, что он демонстрирует такую же или схожую активность с полипептидом. Внедрение может осуществлять специалист в области техники, выбрав подходящий способ трансформации, известный в области техники, а экспрессия внедренного полинуклеотида в клетке-хозяине обеспечивает продуцирование полипептида, тем самым увеличивая его активность.
Способ 7) оптимизации кодонов полинуклеотида, кодирующего полипептид, может быть реализован путем оптимизации кодонов эндогенного полинуклеотида для увеличения транскрипции или трансляции в клетке-хозяине, или путем оптимизации его кодонов таким образом, что в клетке-хозяине может достигаться оптимизированная транскрипция и трансляция чужеродного полинуклеотида.
Кроме того, способ 8) анализа третичной структуры полипептида и, таким образом, выбор и модификация экспонированного сайта или его химическая модификация может быть реализован, например, путем сравнения информации о последовательности анализируемого полипептида с базой данных, в которой хранится информация о последовательностях известных белков, для определения белков-кандидатов в матрицы в зависимости от степени сходства последовательностей и, таким образом, подтверждения структуры на основе информации, таким образом, выбора и трансформации или модификации экспонированного сайта, подлежащего модификации или химической модификации.
Такое усиление активности полипептида может означать, что активность или концентрация (уровень экспрессии) полипептида увеличены по сравнению с активностью или концентрацией полипептида, экспрессируемого в микроорганизме дикого типа или микроорганизме до модификации, или что количество продукта, получаемого из полипептида, увеличено, без ограничения.
Модификация всех полинуклеотидов в микроорганизме по данному изобретению или их части может быть реализована (а) посредством гомологичной рекомбинации с применением вектора для встраивания в хромосому микроорганизма или редактирования генома с применением модифицированных нуклеаз (например, CRTSPR-Cas9) и/или (б) может быть индуцирована излучением, таким как ультрафиолетовое излучение и радиоактивное излучение и так далее, и/или химической обработкой, без ограничения. Способ модификации части гена или всего гена может включать способ с применением технологии рекомбинации ДНК. Например, делеция всего гена или его части может быть реализована путем инъекции в микроорганизм нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, гомологичную целевому гену, для индукции гомологичной рекомбинации. Инъецированная нуклеотидная последовательность или вектор может включать доминантный селективный маркер, без ограничения.
Вектор по данному изобретению может включать ДНК-конструкцию, содержащую нуклеотидную последовательность полинуклеотида, кодирующего целевой полипептид, функционально связанную с подходящей областью, регулирующей экспрессию (последовательностью, регулирующей экспрессию), чтобы обеспечивалась экспрессия целевого полипептида в подходящей клетке-хозяине. Последовательность, регулирующая экспрессию, может включать промотор, способный инициировать транскрипцию, любую последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы на мРНК, и последовательность для регуляции терминации транскрипции и трансляции. После трансформации подходящей клетки-хозяина вектор может реплицироваться или функционировать независимо от генома хозяина или может интегрироваться в его геном.
Вектор, используемый в данном изобретении, не ограничивается каким-либо конкретным, и можно применять любой известный в области техники вектор. Примеры обычно используемых векторов могут включать естественные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A и так далее, а в качестве плазмидного вектора использовать векторы на основе pDC, pBR, pUC, pBluescriptII, pGEM, pTZ, pCL и pET и так далее. В частности, можно использовать вектор pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и так далее.
В одном примере полинуклеотид, кодирующий целевой полипептид, может быть встроен в хромосому при помощи вектора для внутриклеточного встраивания в хромосому. Встраивание полинуклеотида в хромосому может быть реализовано любым известным в области техники способом, например, посредством гомологичной рекомбинации, без ограничения. Вектор может дополнительно включать в себя селективный маркер для подтверждения встраивания в хромосому. Селективный маркер используют для отбора клеток, трансформированных вектором, то есть для подтверждения, встроилась ли целевая молекула нуклеиновой кислоты, и могут применяться маркеры, обеспечивающие фенотип, позволяющий осуществлять отбор, такой как устойчивость к лекарственным средствам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия поверхностных белков. В среде, обработанной селективным агентом, могут выживать или демонстрировать иные фенотипы только клетки, экспрессирующие селективный маркер, таким образом можно осуществлять отбор трансформированных клеток.
В данном описании термин «трансформация» относится к внедрению вектора, который включает в себя полинуклеотид, кодирующий целевой полипептид, в клетку-хозяина или микроорганизм, таким образом, что полипептид, кодируемый полинуклеотидом, может экспрессироваться в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения, встроен ли он в хромосому клетки-хозяина и находится в ней или находится за пределами хромосомы, и можно включать оба варианта, при условии, что после трансформации полинуклеотид может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид может включать ДНК и/или РНК, которые кодируют целевой полипептид. Полинуклеотид может быть внедрен в любой форме, при условии, что он может быть внедрен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть внедрен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую в себя все элементы, необходимые для ее автономной экспрессии. Экспрессионная кассета обычно может включать в себя промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции. Экспрессионная кассета может быть в форме самореплицирующегося экспрессирующего вектора. Кроме того, полинуклеотид может быть внедрен в клетку-хозяина как таковой и быть функционально связан с последовательностями, необходимыми для экспрессии в клетке-хозяине, без ограничения.
Кроме того, в данном описании термин «функционально связанный» означает, что последовательность полинуклеотида функционально связана с последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой полипептид по данному изобретению.
Микроорганизм по данному изобретению может представлять собой микроорганизм, в котором белок VKOR, или полинуклеотид, кодирующий белок, инактивирован, или микроорганизм (то есть рекомбинантный микроорганизм), который генетически модифицирован при помощи вектора таким образом, что белок VKOR или полинуклеотид, кодирующий белок, инактивирован, без ограничения. Вектор является таким, как описано выше.
Микроорганизм по данному изобретению может представлять собой микроорганизм, обладающий способностью продуцировать L-глутаминовую кислоту.
Микроорганизм по данному изобретению может представлять собой микроорганизм, обладающий способностью продуцировать L-глутаминовую кислоту естественным образом, или микроорганизм, которому способность продуцировать L-глутаминовую кислоту была придана посредством инактивации белка VKOR или полинуклеотида, кодирующего белок, в родительском штамме, не обладающем способностью продуцировать L-глутаминовую кислоту, без ограничения.
В одном Примере рекомбинантный микроорганизм по данному изобретению представляет собой микроорганизм, трансформированный вектором таким образом, что белок VKOR или полинуклеотид, кодирующий белок, инактивирован, и, таким образом, белок VKOR или полинуклеотид, кодирующий белок, инактивирован, и может включать все микроорганизмы, способные продуцировать L-глутаминовую кислоту благодаря инактивации белка VKOR или полинуклеотида, кодирующего белок.
В контексте данного изобретения рекомбинантный микроорганизм по данному изобретению может представлять собой микроорганизм, в котором способность продуцировать L-глутаминовую кислоту повышена благодаря инактивации белка VKOR или кодирующего его полинуклеотида в естественном микроорганизме дикого типа или микроорганизме для продуцирования L-глутаминовой кислоты посредством включения белка VKOR или полинуклеотида, кодирующего белок, по сравнению с естественным микроорганизмом дикого типа или микроорганизмом для продуцирования L-глутаминовой кислоты благодаря включению белка VKOR или полинуклеотида, кодирующего белок, без ограничения. В одном Примере немодифицированный микроорганизм, в котором белок VKOR не инактивирован, представляющий собой целевой штамм, с которым проводят сравнение, чтобы установить увеличение способности продуцировать L-глутаминовую кислоту, может представлять собой штамм Corynebacterium glutamicum АТСС13869, в котором произведена делеция гена odhA, известный как продуцирующий L-глутаминовую кислоту штамм, или штамм Corynebacterium glutamicum BL2, известный как продуцирующий L-глутаминовую кислоту штамм, подвергнутый NTG-индуцированному мутагенезу (KFCC11074, патент Кореи 10-0292299), без ограничения.
В одном Примере рекомбинантный штамм, обладающий увеличенной продуцирующей способностью, может обладать способностью продуцировать L-глутаминовую кислоту, повышенной приблизительно на 1% или более, в частности приблизительно на 1% или более, приблизительно на 2,5% или более, приблизительно на 5% или более, приблизительно на 6% или более, приблизительно на 7% или более, приблизительно на 8% или более, приблизительно на 9% или более, приблизительно на 10% или более, приблизительно на 14,3% или более, приблизительно на 20% или более, приблизительно на 28,6% или более, приблизительно на 30% или более, приблизительно на 31,9% или более, приблизительно на 33,3% или более, приблизительно на 37,7% или более, приблизительно на 40% или более, приблизительно на 42,9% или более, приблизительно на 46,3% или более, приблизительно на 50% или более или приблизительно на 52,4% или более (верхняя граница не ограничена кокой-либо конкретной, например, приблизительно 200% или менее, приблизительно 150% или менее, приблизительно 100% или менее, приблизительно 50% или менее, приблизительно 40% или менее, приблизительно 30% или менее, приблизительно 20% или менее или приблизительно 15% или менее) по сравнению с таковой у родительского штамма до модификации или у немодифицированного микроорганизма, без ограничения, при условии, что она повышена на положительное значение по сравнению с продуцирующей способностью у родительского штамма до модификации или немодифицированного микроорганизма. В другом примере микроорганизм, обладающий повышенной продуцирующей способностью, может обладать способностью продуцировать L-глутаминовую кислоту, повышенной приблизительно в 1,01 раз или более, приблизительно в 1,02 раз или более, приблизительно в 1,03 раз или более, приблизительно в 1,05 раз или более, приблизительно в 1,06 раз или более, приблизительно в 1,07 раз или более, приблизительно в 1,08 раз или более, приблизительно в 1,09 раз или более, приблизительно в 1,1 раз или более, приблизительно в 1,14 раз или более, приблизительно в 1,28 раз или более, приблизительно в 1,32 раз или более, приблизительно в 1,33 раз или более, приблизительно в 1,37 раз или более, приблизительно в 1,43 раз или более, приблизительно в 1,46 раз или более или приблизительно в 1,52 раз или более (верхняя граница не ограничена какой-либо конкретной, например, приблизительно в 10 раз или менее, приблизительно в 5 раз или менее, приблизительно в 3 раза или менее или приблизительно в 2 раза или менее) по сравнению с таковой у родительского штамма до модификации или немодифицированного микроорганизма, без ограничения.
В данном описании термин «немодифицированный микроорганизм» не исключает штамма, содержащего мутацию, которая может возникнуть в микроорганизме естественным путем, и может относиться к штамму дикого типа или к самому штамму естественного типа или к штамму до изменения его признака вследствие генетической модификации, вызванной естественными или искусственными факторами. Например, немодифицированный микроорганизм может относиться к штамму, в котором белок VKOR, описанный в данном изобретении, или полинуклеотид, кодирующий белок, не инактивирован, или к штамму до его инактивации. «Немодифицированный микроорганизм» может использоваться взаимозаменяемо со «штаммом до модификации», «микроорганизмом до модификации», «немутированным штаммом», «немодифицированным штаммом», «немутированным микроорганизмом» или «референсным микроорганизмом».
В одном воплощении микроорганизм по данному изобретению может представлять собой Corynebacterium glutamicum, Corynebacterium stationis, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiem, Corynebacterium callunae, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium ammoniagenes, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens и, в частности, Corynebacterium glutamicum, без ограничения.
В другом примере рекомбинантный микроорганизм по данному изобретению может представлять собой микроорганизм, у которого активность части белков пути биосинтеза L-глутаминовой кислоты дополнительно усилена, или активность части белков пути распада L-глутаминовой кислоты дополнительно ослаблена, благодаря чему усилена способность продуцировать L-глутаминовую кислоту.
В частности, микроорганизм по данному изобретению может представлять собой микроорганизм, в котором белок OdhA дополнительно инактивирован или осуществлена дополнительная делеция гена odhA. Более конкретно, микроорганизм по данному изобретению может представлять собой Corynebacterium glutamicum, где белок OdhA инактивирован в Corynebacterium glutamicum АТСС13869 или микроорганизм, где осуществлена делеция гена odhA в Corynebacterium glutamicum АТСС13869. Белок OdhA может включать аминокислотную последовательность (SEQ ID NO: 32) последовательности с регистрационным номером NCBI WP_060564343.1, а ген odhA может включать нуклеотидную последовательность NCBI GenBank BBD29_06050, но данное изобретение не ограничивается указанным.
Однако, инактивация белка OdhA или делеция гена odhA является только примером, и данное изобретение не ограничивается указанным. Кроме того, микроорганизм по данному изобретению может представлять собой микроорганизм, у которого усилена активность белков различных известных путей биосинтеза L-глутаминовой кислоты или инактивирована активность белков пути распада.
В другом аспекте данного изобретения предложен способ получения L-глутаминовой кислоты, включающий культивирование в среде микроорганизма рода Corynebacterium, в котором белок VKOR инактивирован.
Способ получения L-глутаминовой кислоты по данному изобретению может включать стадию культивирования в среде микроорганизма, в котором белок VKOR или полинуклеотид, кодирующий белок, инактивирован, или микроорганизм рода Corynebacterium, который генетически модифицирован при помощи вектора таким образом, что белок VKOR или полинуклеотид, кодирующий белок, инактивирован.
В данном описании термин «культивирование» означает, что микроорганизм по данному изобретению выращивают в надлежащим образом контролируемых внешних условиях. Процесс культивирования по данному изобретению можно осуществлять в подходящей культуральной среде и условиях культивирования, известных в области техники. Такой способ культивирования может быть легко адаптирован для применения специалистами в области техники в зависимости от выбранного микроорганизма. В частности, культивирование может представлять собой периодическое культивирование, непрерывное культивирование и/или непрерывное культивирование с подпиткой, без ограничения.
В данном описании термин «среда» относится к смеси веществ, содержащих в качестве основного ингредиента питательные вещества, необходимые для культивирования микроорганизма по данному изобретению, и она обеспечивает питательные вещества и факторы роста наряду с водой, которая незаменима для выживания и роста. В частности, среда и другие условия культивирования, применяемые для культивирования микроорганизма по данному изобретению, может представлять собой любую среду, используемую при стандартном культивировании микроорганизмов, без конкретного ограничения. При этом, микроорганизм по данному изобретению можно культивировать в аэробных условиях в стандартной среде, содержащей надлежащий источник углерода, источник азота, источник фосфора, неорганические соединения, аминокислоты и/или витамины, при этом регулируя температуру, рН и так далее.
В частности, культуральная среда для микроорганизма рода Corynebacterium может быть найдена в литературе ("Manual of Methods for General Bacteriology" by the American Society for Bacteriology (Washington D.C., USA, 1981)).
В данном изобретении источник углерода может включать углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, сукроза, мальтоза и так далее; сахароспирты, такие как маннит, сорбит и так далее, органические кислоты, такие как пировиноградная кислота, молочная кислота, лимонная кислота и так далее, аминокислоты, такие как глутаминовая кислота, метионин, лизин и так далее. Кроме того, источник углерода может включать природные органические питательные вещества, такие как гидролизат крахмала, меласса, сырая меласса, рисовые отруби, маниок, меласса тростниково-сахарного производства, жидкий кукурузный экстракт и так далее. В частности, можно использовать такие углеводы как глюкоза и стерилизованная обработанная меласса (то есть меласса, в которой сахара конвертированы в восстанавливающие) и, кроме того, можно использовать различные другие источники углерода в соответствующем количестве, без ограничения. Такие источники углерода можно использовать по отдельности или в виде комбинации двух или более видов, без ограничения.
Источник азота может включать неорганические источники азота, такие как аммоний, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и так далее; аминокислоты, такие как глутаминовая кислота, метионин, глутамин и так далее, и органические источники азота, такие как пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, мальтозный экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыба или продукты ее переработки, обезжиренный соевый жмых или продукты его переработки и так далее. Такие источники углерода можно использовать по-отдельности или в виде комбинации двух или более видов, без ограничения.
Источники фосфора могут включать дигидрофосфат калия, гидрофосфат калия или соответствующие натрий-содержащие соли и так далее. Примеры неорганического соединения могут включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и так далее. Кроме того, можно включать аминокислоты, витамины и/или соответствующие предшественники и так далее. Такие ингредиенты или предшественники можно добавлять в среду при периодическом культивировании или непрерывном культивировании, но такие источники фосфора не ограничиваются перечисленными.
В процессе культивирования микроорганизма по данному изобретению рН среды можно корректировать путем добавления в среду надлежащим образом такого соединения, как гидроксид аммония, гидроксид калия, аммоний, фосфорная кислота, серная кислота и так далее. Кроме того, для предупреждения образования пены в ходе культивирования можно добавлять пеногаситель, такой как сложный полигликолевый эфир жирной кислоты. Кроме того, для поддержания аэробного состояния среды в среду можно вводить кислород или кислород-содержащий газ, либо для поддержания анаэробного или микроаэробного состояния среды в среду можно вводить азот, водород или диоксид углерода без введения газа, однако газ не ограничивается указанными.
Температура среды при культивировании по данному изобретению может находиться в диапазоне от 20°С до 45°С, и в частности, в диапазоне от 25°С до 40°С, и культивирование можно осуществлять в течение от приблизительно 10 до 160 часов, без ограничения.
L-глутаминовая кислота, продуцируемая при культивировании по данному изобретению, может высвобождаться в среду или оставаться в клетках.
Способ получения L-глутаминовой кислоты по данному изобретению может дополнительно включать стадию получения микроорганизма по данному изобретению, стадию приготовления среды для культивирования микроорганизма или их комбинацию (независимо от порядка, в любом порядке), например, перед стадией культивирования.
Способ получения L-глутаминовой кислоты по данному изобретению может дополнительно включать стадию выделения L-глутаминовой кислоты из культуральной среды (среды, в которой выращивали культуру) или культивируемого микроорганизма. После стадии культивирования может быть дополнительно включена стадия выделения.
На стадии выделения необходимую L-глутаминовую кислоту можно собирать, используя способ культивирования микроорганизма по данному изобретению, например, используя подходящий способ, известный в области техники, подходящий для периодического культивирования, непрерывного культивирования или культивирования с подпиткой. Например, может применяться центрифугирование, фильтрация, обработка средством для осаждения кристаллизованного белка (высаливание), экстракция, разрушение ультразвуком, ультрафильтрация, диализ, различные виды хроматографии, такие как хроматография молекулярных сит (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, аффинная хроматография и так далее, ВЭЖХ (высокоэффективная жидкостная хроматография) или их комбинация, и необходимую L-глутаминовую кислоту можно выделять из среды или микроорганизмов при помощи соответствующих способов, известных в области техники.
Кроме того, способ получения L-глутаминовой кислоты по данному изобретению может дополнительно включать процесс очистки, который можно осуществлять при помощи подходящего способа, известного в области техники. В одном примере, когда способ получения L-глутаминовой кислоты по данному изобретению включает как стадию выделения, так и стадию очистки, стадию выделения и стадию очистки можно осуществлять непрерывно или с перерывами, независимо от порядка, или можно осуществлять одновременно или можно объединять в одну стадию, без ограничения.
В способе по данному изобретению белок VKOR, полинуклеотиды, векторы и микроорганизмы и так далее являются такими, как описано в других аспектах.
В еще одном аспекте данного изобретения предложена композиция для получения L-глутаминовой кислоты, включающая микроорганизм рода Corynebacterium, в котором белок VKOR инактивирован; среду для его культивирования или их комбинацию.
Композиция по данному изобретению может дополнительно включать подходящие эксципиенты, обычно используемые для получения L-глутаминовой кислоты, и такие эксципиенты могут представлять собой, например, консервант, увлажняющее вещество, диспергирующее вещество, суспендирующее вещество, буфер, стабилизатор или изотоническое вещество и так далее, без ограничения.
В еще одном аспекте данного изобретения предложен способ получения микроорганизма рода Corynebacterium, включающий инактивацию белка VKOR.
В еще одном аспекте данного изобретения предложено применение L-глутаминовой кислоты, произведенной микроорганизмом рода Corynebacterium, в котором белок VKOR инактивирован.
Белок VKOR, инактивация, микроорганизм рода Corynebacterium и так далее являются такими, как описано в других аспектах.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее данное изобретение будет описано более подробно на Примерах. Однако, данные Примеры являются лишь предпочтительными Примерами, приведенными в целях иллюстрации и, следовательно, объем данного изобретения не ограничивается указанными Примерами. При этом, технические признаки, которые не описаны в данном документе, могут быть в достаточной степени поняты и легко реализованы специалистами в области техники, к которой относится данное изобретение, или в схожей области техники.
Пример 1. Конструирование библиотеки случайных мутаций с применением транспозона
Пример 1-1. Конструирование штамма Corynebacterium glutamicum, обладающего способностью продуцировать L-глутаминовую кислоту, происходящего из Corynebacterium glutamicum дикого типа
Для получения штамма, обладающего способностью производить L-глутаминовую кислоту, происходящего из Corynebacterium glutamicum дикого типа АТСС13869, получали штамм Corynebacterium glutamicum ATCC13869ΔodhA, в котором осуществлялась делеция гена odhA на основе описания, известного из уровня техники (Appl Environ Microbiol. 2007 Feb;73(4): 1308-19. Epub 2006 Dec 8.).
В частности, для делеции гена odhA получали области, расположенные в направлении против хода и по ходу транскрипции относительно гена odhA, при помощи ПЦР с использованием набора праймеров с SEQ ID NO:3 и SEQ ID NO:4 и SEQ ID NO: 5 и SEQ ID NO:6, взяв за основу хромосомную ДНК Corynebacterium glutamicum дикого типа АТСС13869 как матрицу. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР при следующих условиях ПЦР-амплификации: денатурация при 95°С в течение 5 мин; затем 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 55°С в течение 30 сек и полимеризацию при 72°С в течение 60 сек; и затем полимеризация при 72°С в течение 5 мин.
Амплифицированные области, расположенные против хода и по ходу транскрипции от odhA, и вектор pDCM2 для трансформации хромосомы, расщепленные рестрикционным ферментом SmaI, клонировали, используя метод сборки по Гибсону (D.G. Gibson et al., NATURE METHODS, Vol.6 No. 5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix) с получением рекомбинантного вектора, и полученный вектор обозначали pDCM2-ΔodhA. Клонирование осуществляли путем смешивания реагента для сборки по Гибсону и каждого из генных фрагментов в рассчитанном молярном количестве с последующей инкубацией при 50°С в течение 1 часа.
Полученным таким образом вектором pDCM2-ΔodhA трансформировали штамм Corynebacterium glutamicum АТСС13869 посредством электропорации и затем подвергали вторичному кроссинговеру с получением таким образом штамма, в хромосоме которого осуществлена делеция гена odhA. Делецию гена подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO: 7 и SEQ ID NO:8 и обозначали полученный штамм ATCC13869ΔodhA.
Использованные последовательности праймеров приведены в Таблице 1 ниже.

Пример 1-2. Конструирование библиотеки случайных мутаций с применением транспозона
Для получения штамма, обладающего повышенной способностью продуцировать L-глутаминовую кислоту, конструировали векторную библиотеку следующим образом.
Осуществляли трансформацию плазмидами, полученными с применением набора EZ-Tn5™<R6Kγori/KAN-2>Tnp Transposome™ (Epicentre), взяв за основу Corynebacterium glutamicum ATCC13869ΔodhA в качестве родительского штамма, посредством электропорации (Appl. Microbiol. Biothcenol.(1999) 52:541-545), и осуществляли посев газоном на чашку с комплексной средой, содержащей канамицин (25 мг/л) с получением таким образом приблизительно 5000 колоний.
Чашка с комплексной средой (рН 7.0)
Глюкоза 10 г, пептон 10 г, мясной экстракт 5 г, дрожжевой экстракт 5 г, сердечно-мозговая вытяжка 18,5 г, NaCl 2,5 г, мочевина 2 г, сорбит 91 г, агар 20 г (на литр дистиллированной воды).
Пример 2. Скрининг библиотеки случайных мутаций с применением транспозона
Каждой из приблизительно 5000 колоний, полученных в Примере 1-2, инокулировали 300 мкл следующей селективной среды и культивировали в 96-луночном планшете при 37°С при 1000 об/мин в течение приблизительно 48 часов.
Селективная среда (рН 7,0)
Сахар-сырец 5%, 1 М фосфатный буфер (рН 8,0) 100 мл, HSM (гидролизованный соевый жмых) 0,096%, сульфат аммония 2,25%, дигидрофосфат калия 0,1%, сульфат магния 0,04%, сульфат железа 10 мг/л, тиамин-HCL 0,2 мг/л, биотин 0,3 мг/л (на литр дистиллированной воды)
После завершения культивирования измеряли L-глутаминовую кислоту при помощи YSI (биохимический анализатор YSI 2900). Отбирали 10 колоний как мутантные штаммы, демонстрирующие высокие значения L-глутаминовой кислоты по сравнению с родительским штаммом Corynebacterium glutamicum АТСС13869ΔodhA. Другие колонии демонстрировали значения L-глутаминовой кислоты, схожие или более низкие, чем у Corynebacterium glutamicum АТСС13869ΔodhA, который использовали в качестве родительского штамма.
Отобранные 10 штаммов вновь культивировали таким же образом, как указано выше, и в результате отбирали один наилучший мутантный штамм ATCC13869ΔodhA/mt-8 с усиленной способностью продуцировать L-глутаминовую кислоту по сравнению с родительским штаммом Corynebacterium glutamicum ATCC13869ΔodhA.
Пример 3. Установление причин увеличения способности отобранного мутантного штамма продуцировать L-глутаминовую кислоту
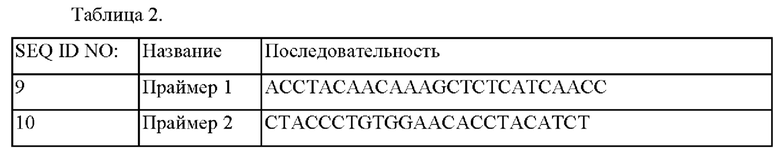
Взяв за основу ATCC13869ΔodhA/mt-8, отобранный в Примере 2, осуществляли анализ гена, делецию которого осуществляли посредством случайного встраивания транспозона, с применением праймера 1 (SEQ ID NO: 9) и праймера 2 (SEQ ID NO: 10) и набора согласно инструкциям производителя. В результате анализа на основе нуклеотидных последовательностей, зарегистрированных в NIH GenBank подтверждали, что ген (BBD29_04485), включающий полинуклеотидную последовательность SEQ ID NO:2, был инактивирован.
Последовательности использованных для этого праймеров приведены в Таблице 2 ниже.
Пример 4. Конструирование рекомбинантного вектора для инактивации белка VKOR
Пример 4-1. Конструирование рекомбинантного вектора для делеции гена BBD29_04485, кодирующего белок VKOR
Как подтверждено в Примере 3, получали рекомбинантный вектор с делецией гена для проверки, усиливалась ли способность продуцировать L-глутаминовую кислоту при делеции гена BBD29_04485 (SEQ ID NO: 2), кодирующего белок VKOR, на хромосоме штамма рода Corynebacterium.
Для этого вначале синтезировали праймеры с SEQ ID NO: 11-14 для получения фрагмента для делеции гена.
В частности, получали фрагменты гена BBD29_04485 при помощи ПЦР с использованием набора праймеров с SEQ ID NO: 11 и SEQ ID NO: 12, и SEQ ID NO: 13 и SEQ ID NO: 14, взяв за основу хромосомную ДНК Corynebacterium glutamicum дикого типа АТСС13869 как матрицу. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР при следующих условиях ПЦР-амплификации: денатурация при 95°С в течение 5 мин; затем 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 55°С в течение 30 сек и полимеризацию при 72°С в течение 60 сек; и затем полимеризация при 72°С в течение 5 мин.
Амплифицированные фрагменты гена и вектор pDCM2 для трансформации хромосомы, расщепленные рестрикционным ферментом SmaI, клонировали, используя метод сборки по Гибсону, с получением рекомбинантного вектора, и полученный вектор обозначали pDCM2-ΔBBD29_04485. Клонирование осуществляли путем смешивания реагента для сборки по Гибсону и каждого из генных фрагментов в рассчитанном молярном количестве с последующей инкубацией при 50°С в течение 1 часа.
Последовательности использованных для этого праймеров показаны в Таблице 3 ниже.

Пример 4-2. Конструирование рекомбинантных векторов для изменения инициирующего кодона BBD29_04485, кодирующего белок VKOR
Для проверки, усиливалась ли способность продуцировать L-глутаминовую кислоту, когда в хромосоме штамма рода Corynebacterium инициирующий кодон гена BBD29_04485 изменяли на каждый из TTG и CTG, вначале получали рекомбинантные векторы для изменения инициирующего кодона.
Для этого вначале синтезировали праймеры с SEQ ID NO: 15-20 для получения фрагмента для изменения инициирующего кодона гена.
В частности, получали фрагменты гена при помощи ПЦР с использованием наборов праймеров с SEQ ID NO:15 и SEQ ID NO:16, SEQ ID NO:17 и SEQ ID NO:18, SEQ ID NO:15 и SEQ ID NO:19, и SEQ ID NO: 18 и SEQ ID NO:20, взяв за основу хромосомную ДНК Corynebacterium glutamicum дикого типа АТСС13869 как матрицу. В качестве полимеразы использовали ДНК-полимер азу Solg™ Pfu-X и проводили ПЦР при следующих условиях ПЦР-амплификации: денатурация при 95°С в течение 5 мин; затем 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 55°С в течение 30 сек и полимеризацию при 72°С в течение 60 сек; и затем полимеризация при 72°С в течение 5 мин.
Амплифицированные фрагменты гена и вектор pDCM2 для трансформации хромосомы, расщепленные рестрикционным ферментом SmaI, клонировали, используя метод сборки по Гибсону с получением рекомбинантных векторов, и полученные векторы обозначали вектор pDCM2-ΔBBD29_04485::BBD29_04485(g1t) и вектор pDCM2-ΔBBD29_04485::BBD29_04485(g1c). Клонирование осуществляли путем смешивания реагента для сборки по Гибсону и каждого из генных фрагментов в рассчитанном молярном количестве с последующей инкубацией при 50°С в течение 1 часа.
Последовательности использованных для этого праймеров приведены в Таблице 4 ниже.

Пример 4-3. Конструирование рекомбинантных векторов для изменения сайта связывания рибосомы (RBS) BBD29_ 04485, кодирующего белок VKOR
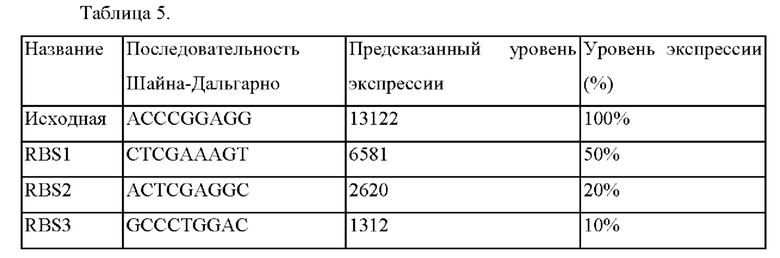
Для ослабления гена BBD29_04485 на хромосоме штамма рода Corynebacterium предсказывали последовательность Шайна-Дальгарно (SD) по нуклеотидным последовательностям, содержащим 35 пар оснований перед геном и 35 пар оснований от N-конца открытой рамки считывания (ORF).
На основе вышеупомянутых нуклеотидных последовательностей при помощи калькулятора RBS (github) всего было предсказано три группы кандидатных последовательностей связывания рибосомы, а именно RBS1, RBS2, и RBS3. В результате было предсказано, что экспрессия трех групп кандидатных последовательностей связывания рибосомы снижалась на 50%, 20% и 10%, соответственно, по сравнению с существующими последовательностями связывания рибосомы.
Предсказанные последовательности связывания рибосомы показаны в Таблице 5.
Для проверки, усиливалась ли способность продуцировать L-глутаминовую кислоту, когда ген был ослаблен за счет изменения последовательности связывания рибосомы BBD29_04485 в хромосоме штамма рода Corynebacterium, получали рекомбинантные векторы для изменения последовательностей связывания рибосомы для каждой из трех предсказанных последовательностей связывания рибосомы.
Для этого вначале синтезировали праймеры с SEQ ID NO:21-28 для получения фрагмента для изменения последовательностей связывания рибосомы в гене.
В частности, получали фрагменты гена при помощи ПЦР с использованием наборов праймеров с SEQ ID NO:21 и SEQ ID NO:22, SEQ ID NO:23 и SEQ ID NO:24, SEQ ID NO:21 и SEQ ID NO:25, SEQ ID NO:24 и SEQ ID NO:26, SEQ ID NO:21 и SEQ ID NO:27, и SEQ ID NO:24 и SEQ ID NO:28, взяв за основу хромосомную ДНК Corynebacterium glutamicum дикого типа АТСС13869 как матрицу. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР при следующих условиях ПЦР-амплификации: денатурация при 95°С в течение 5 мин; затем 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 55°С в течение 30 сек и полимеризацию при 72°С в течение 60 сек; и затем полимеризация при 72°С в течение 5 мин.
Амплифицированные фрагменты гена и вектор pDCM2 для трансформации хромосомы, расщепленные рестрикционным ферментом SmaI, клонировали, используя метод сборки по Гибсону, с получением рекомбинантных векторов, и полученные векторы обозначали вектор pDCM2-ΔRBS(wt)::RBS1, вектор pDCM2-ΔRBS(wt)::RBS2 и вектор pDCM2-ΔRBS(wt)::RBS3. Клонирование осуществляли путем смешивания реагента для сборки по Гибсону и каждого из генных фрагментов в рассчитанном молярном количестве с последующей инкубацией при 50°С в течение 1 часа.
Последовательности использованных для этого праймеров показаны в Таблице 6 ниже.
Пример 5. Конструирование штамма, обладающего способностью продуцировать L-глутаминовую кислоту, происходящего из Corynebacterium glutamicum дикого типа, в котором BBD29_04485, кодирующий белок VKQR, ослаблен
Влияние на способность продуцировать L-глутаминовую кислоту подтверждали путем внедрения вектора pDCM2-ΔBBD29_04485, вектора pDCM2-ΔBBD29_04485::BBD29_04485(g1t), вектора pDCM2-ΔBBD29_04485::BBD29_04485(g1c), вектора pDCM2-ΔRBS(wt)::RBS1, вектора pDCM2-ΔRBS(wt)::RBS2 и вектора pDCM2-ΔRBS(wt)::RBS3, полученных в Примере 4, взяв за основу штамм ATCC13869ΔodhA, полученный в Примере 1.
Вначале вектором pDCM2-ΔBBD29_04485 трансформировали штамм Corynebacterium glutamicum ATCC13869ΔodhA посредством электропорации и подвергали вторичному кроссинговеру, с получением таким образом штамма, в хромосоме которого была делеция гена BBD29_04485. Манипуляцию с геном подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO:29 и SEQ ID NO:30, которые позволяют амплифицировать гомологичные рекомбинантные области выше и ниже по ходу транскрипции соответственно, и полученный штамм обозначали СА02-1624.
Затем вектором pDCM2-ABBD29_04485::BBD29_04485(g1t) и вектором pDCM2-ABBD29_04485::BBD29_04485(g1c) трансформировали штамм Corynebacterium glutamicum АТСС13869ΔodhA посредством электропорации и подвергали вторичному кроссинговеру, получая таким образом штаммы, в которых инициирующий кодон гена BBD29_04485 был изменен с GTG на TTG и CTG, соответственно.
Манипуляцию с геном подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO:31 и SEQ ID NO:30, которые позволяют амплифицировать гомологичные рекомбинантные области выше и ниже по ходу транскрипции соответственно, и полученные штаммы обозначали СА02-1625 и СА02-1630.
Затем вектором pDCM2-ΔRBS(wt)::RBS1, вектором pDCM2-ΔRBS(wt)::RBS2 и вектором pDCM2-ΔRBS(wt)::RBS3 трансформировали штамм Corynebacterium glutamicum ATCC13869ΔodhA посредством электропорации и подвергали вторичному кроссинговеру, получая таким образом штаммы, в хромосоме которых последовательность связывания рибосомы гена BBD29_04485 была изменена на RBS1, RBS2 и RBS3, соответственно.
Манипуляцию с геном подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO:31 и SEQ ID NO:30, которые позволяют амплифицировать гомологичные рекомбинантные области выше и ниже по ходу транскрипции соответственно, и полученные штаммы обозначали СА02-1631, СА02-1632 и СА02-1633.
Последовательности использованных для этого праймеров показаны в Таблице 7 ниже.

Для подтверждения способности продуцировать L-глутаминовую кислоту всего культивировали 6 штаммов, СА02-1624, СА02-1625, СА02-1630, СА02-1631, СА02-1632 и СА02-1633, следующим образом, взяв за основу штамм АТСС13869ΔodhA в качестве контрольного.
Вышеупомянутыми штаммами инокулировали чашку с комплексной средой, представляющей собой среду для затравочной культуры, и культивировали при температуре 30°С в течение 20 часов. Затем вышеупомянутыми штаммами инокулировали 25 мл следующей среды для продуцирования, находящейся в конической колбе с дефлекторами объемом 250 мл, при помощи петли для инокуляции и затем культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 40 часов. После окончания культивирования измеряли продуцирование L-глутаминовой кислоты при помощи высокоэффективной жидкостной хроматографии (ВЭЖХ), и результаты измерений показаны в Таблице 8 ниже.
Среда для затравочной культуры
Глюкоза 1%, мясной экстракт 0,5%, полипептон 1%, хлорид натрия 0,25%, дрожжевой экстракт 0,5%, агар 2%, мочевина 0,2%, рН 7,2.
Среда для продуцирования
Сахар-сырец 6%, карбонат кальция 5%, сульфат аммония 2,25%, дигидрофосфат калия 0,1%, сульфат магния 0,04%, сульфат железа 10 мг/л, тиамин-HCL 0,2 мг/л, биотин 50 мкг/л
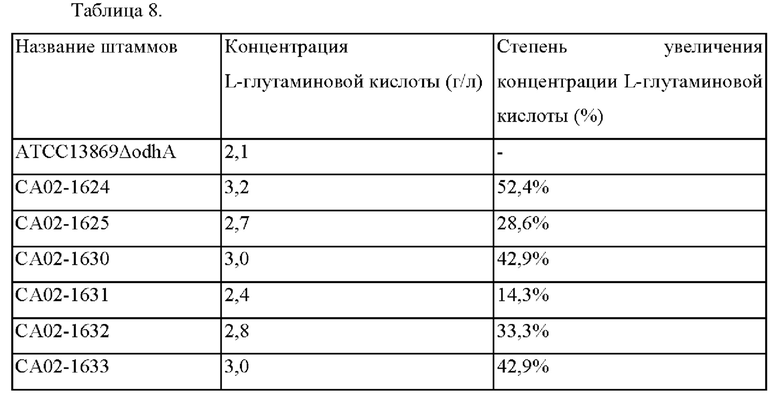
Как показано в Таблице 8, подтверждали, что концентрация L-глутаминовой кислоты повышалась у всех из штамма СА02-1624, имевшего делецию гена BBD29_04485, штаммов СА02-1625 и СА02-1630, у которых инициирующий кодон гена BBD29_04485 был ослаблен на TTG и CTG, и штаммов СА02-1631, СА02-1632 и СА02-1633, у которых последовательность связывания рибосомы гена BBD29_04485 была ослаблена на RBS1, RBS2 и RBS3, по сравнению со штаммом АТСС13869ΔodhA, происходящим от дикого типа.
На основании указанных результатов можно отметить, что способность продуцировать L-глутаминовую кислоту усиливалась при инактивации активности белка VKOR, независимо от конкретного способа.
Из этих штаммов СА02-1624 депонировали 13 января 2021 в Центре культур микроорганизмов Кореи в соответствии с Будапештским Соглашением под регистрационным номером KCCM12928P.
Пример 6. Подтверждение влияния делеции BBD29_04485, кодирующего белок VKOR, на штамме Corynebacterium glutamicum, подвергнутом мутагенезу, индуцированному N-метил-N'-нитро-N-нитрозогуанидином (NTG)
Кроме штаммов, происходящих из Corynebacterium sp.дикого типа, для проверки, оказывает ли ген такое же влияние на штамм, происходящий из Corynebacterium sp., подвергнутый NTG-индуцированному мутагенезу, обладающий повышенной способностью продуцировать L-глутаминовую кислоту, аттенуирующее влияние гена, подтвержденное в Примере 5, подтверждали на штамме Corynebacterium glutamicum BL2, известном как продуцирующий L-глутаминовую кислоту штамм, подвергнутый NTG-индуцированному мутагенезу (KFCC11074, патент Кореи №10-0292299).
В частности, влияние на способность продуцировать L-глутаминовую кислоту подтверждали путем внедрения в штамм KFCC11074 всего 3 векторов, вектора pDCM2-ΔBBD29_04485, вектора pDCM2-ΔBBD29_04485::BBD29_04485(glc) и вектора pDCM2-ΔRBS(wt)::RBS3, продемонстрировавших в Примере 5 увеличение концентрации L-глутаминовой кислоты на 40% или более.
Вначале вектором pDCM2-ΔBBD29 04485 трансформировали штамм KFCC11074 посредством электропорации и подвергали вторичному кроссинговеру, получая таким образом штамм, в хромосоме которого была делеция гена BBD29_04485.
Манипуляцию с геном подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO:29 и SEQ ID NO:30, которые позволяют амплифицировать гомологичные рекомбинантные области выше и ниже по ходу транскрипции соответственно, и полученный штамм обозначали СА02-1634.
Затем вектором pDCM2-ΔBBD29_04485::BBD29_04485(g1c) трансформировали штамм KFCC11074 посредством электропорации и подвергали вторичному кроссинговеру, получая таким образом штамм, у которого инициирующий кодон гена BBD29_04485 был изменен с GTG на CTG.
Манипуляцию с геном подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO:31 и SEQ ID NO:30, которые позволяют амплифицировать гомологичные рекомбинантные области выше и ниже по ходу транскрипции соответственно, и полученный штамм обозначали СА02-1635.
Затем вектором pDCM2-ΔRBS(wt)::RBS3 трансформировали штамм KFCC11074 посредством электропорации и подвергали вторичному кроссинговеру, получая таким образом штамм, в хромосоме которого последовательность связывания рибосомы гена BBD29_04485 была изменена на RBS3.
Манипуляцию с геном подтверждали при помощи секвенирования генома и ПЦР с использованием SEQ ID NO:31 и SEQ ID NO:30, которые позволяют амплифицировать гомологичные рекомбинантные области выше и ниже по ходу транскрипции, соответственно, и полученный штамм обозначали СА02-1636.
Для подтверждения способности продуцировать L-глутаминовую кислоту всего культивировали 3 полученных штамма, СА02-1634, СА02-1635 и СА02-1636, следующим образом, взяв за основу штамм KFCC11074 в качестве контрольного.
Вышеупомянутыми штаммами инокулировали чашку с комплексной средой, представляющей собой среду для затравочной культуры, и культивировали при температуре 30°С в течение 20 часов. Затем вышеупомянутыми штаммами инокулировали 25 мл следующей среды для продуцирования, находящейся в конической колбе с дефлекторами объемом 250 мл, при помощи петли для инокуляции и затем культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 40 часов. После окончания культивирования измеряли продуцирование L-глутаминовой кислоты при помощи высокоэффективной жидкостной хроматографии (ВЭЖХ), и результаты измерений показаны в Таблице 9 ниже. Среда для затравочной культуры
Глюкоза 1%, мясной экстракт 0,5%, полипептон 1%, хлорид натрия 0,25%, дрожжевой экстракт 0,5%, агар 2%, мочевина 0,2%, рН 7,2.
Среда для продуцирования
Сахар-сырец 6%, карбонат кальция 5%, сульфат аммония 2,25%, дигидрофосфат калия 0,1%, сульфат магния 0,04%, сульфат железа 10 мг/л, тиамин-HCL 0,2 мг/л, биотин 500 мкг/л
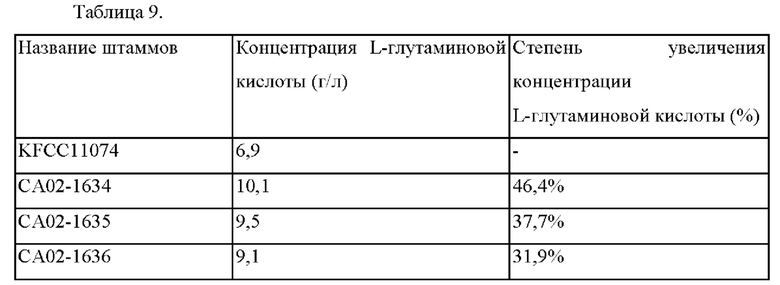
Как показано в Таблице выше, подтверждали, что концентрация L-глутаминовой кислоты у СА02-1634, имевшего делецию гена BBD29_04485, повышалась приблизительно на 46,4% по сравнению со штаммом KFCC11074. Кроме того, подтверждали, что концентрация L-глутаминовой кислоты у СА02-1635, у которого инициирующий кодон гена BBD29_04485 был ослаблен на СТО, повышалась приблизительно на 37,7%. Далее, подтверждали, что концентрация L-глутаминовой кислоты у СА02-1636, у которого последовательность связывания рибосомы гена BBD29_04485 была ослаблена на RBS3, повышалась приблизительно на 31,9%.
Специалистам в области техники, к которой относится данное изобретение, очевидно, что возможны и другие частные воплощения данного изобретения, без отклонения от его сущности или существенных признаков. Описанные воплощения следует считать только иллюстративными во всех аспектах, но не ограничивающими изобретение. Таким образом, объем данного изобретения определяется приведенной ниже формулой изобретения, а не описанием, приведенным выше. Все изменения, охватываемые значениями и диапазоном эквивалентности формулы изобретения, следует считать входящими в объем данного изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> ШТАММ ДЛЯ ПОЛУЧЕНИЯ ВЫСОКОКОНЦЕНТРИРОВАННОЙ L-ГЛУТАМИНОВОЙ КИСЛОТЫ И
СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ С ЕГО ПРИМЕНЕНИЕМ
<130> OPA21393
<150> KR 10-2021-0085738
<151> 2021-06-30
<160> 32
<170> KoPatentIn 3.0
<210> 1
<211> 196
<212> PRT
<213> Unknown
<220>
<223> VKOR
<400> 1
Met Ser Thr Glu Ile His Asn Ala Pro Pro Lys Ala Pro Thr Trp Leu
1 5 10 15
Gly Trp Val Leu Met Ile Gly Gly Ile Ile Gly Leu Ile Leu Ser Val
20 25 30
Ile Ile Met Ala Glu Lys Leu Ala Ile Leu Glu Asp Pro Gly His Ile
35 40 45
Thr Ser Cys Asp Phe Asn Ala Val Leu Ala Cys Gly Asp Val Met Arg
50 55 60
Ser Gly Gln Ala Asn Ala Phe Gly Ile Pro Asn Pro Leu Ile Gly Ile
65 70 75 80
Ala Gly Phe Ala Ala Val Ala Ile Ile Gly Ala Gly Ile Leu Ala Gly
85 90 95
Gly Arg Phe Arg Gly Trp Phe Trp Phe Gly Ala Gln Ala Gly Leu Thr
100 105 110
Phe Ala Met Met Phe Cys His Trp Leu Ala Tyr Gln Ser Met Ser Val
115 120 125
Ile Arg Ala Leu Cys Pro Tyr Cys Met Gly Val Trp Thr Val Ser Ile
130 135 140
Ile Met Phe Val Leu Val Thr Ala Trp Asn Val Lys Thr Phe Ser Gly
145 150 155 160
Ser Asp Ser Thr Phe Val Asn Ala Leu Tyr Lys Tyr Lys Trp Val Ile
165 170 175
Ala Ile Val Trp Leu Leu Leu Ile Ala Ala Ala Ala Val Trp Ser Phe
180 185 190
Arg Tyr Met Phe
195
<210> 2
<211> 591
<212> DNA
<213> Unknown
<220>
<223> VKOR
<400> 2
gtgtctaccg aaatccacaa cgccccaccc aaggccccaa cttggcttgg ctgggtgctc 60
atgatcggcg gaataatcgg cctcattttg tcggtgatca tcatggccga aaaacttgcc 120
atcctcgagg atcccggtca catcaccagc tgcgatttca atgcagtcct agcttgtggc 180
gatgtcatgc gttccggcca agctaacgcg ttcggcatcc cgaatccgct catcggcatc 240
gccggtttcg ccgctgtcgc catcatcggc gccggcatcc tcgcgggcgg caggttccgc 300
ggttggttct ggttcggcgc ccaggccgga ctcacttttg ccatgatgtt ctgccactgg 360
ctcgcctacc aatccatgtc cgtcatccgc gcgctctgcc cttactgcat gggcgtgtgg 420
accgtatcga tcatcatgtt cgtgctggtc actgcatgga atgtgaaaac tttcagcggc 480
tccgacagca cgttcgtcaa cgcactgtac aaatacaagt gggtcatcgc gatcgtctgg 540
ctgctgctca tcgcagccgc agctgtgtgg tcattccgct acatgttcta g 591
<210> 3
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> odhA_up_F
<400> 3
tgaattcgag ctcggtaccc ttgaacggaa ttgggtgg 38
<210> 4
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> odhA_up_R
<400> 4
cccaggtggc atcggtacct tcacccagcg ccacgcag 38
<210> 5
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> odhA_down_F
<400> 5
cgctgggtga aggtaccgat gccacctggg ttggtcaag 39
<210> 6
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> odhA_down_R
<400> 6
gtcgactcta gaggatcccc ggacaaggaa tggagaga 38
<210> 7
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> odhA_del_F
<400> 7
cttaccgttg ttgccctt 18
<210> 8
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> odhA_del_R
<400> 8
ctccttcacc cacatcatt 19
<210> 9
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 1
<400> 9
acctacaaca aagctctcat caacc 25
<210> 10
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 2
<400> 10
ctaccctgtg gaacacctac atct 24
<210> 11
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_F
<400> 11
aattcgagct cggtacccgt gagtccaacc aggtcgaag 39
<210> 12
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_R
<400> 12
ctgaaagcct taaatgctgg cttgatttct tgtgctgtg 39
<210> 13
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_F
<400> 13
acaagaaatc aagccagcat ttaaggcttt caggccg 37
<210> 14
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_R
<400> 14
cgactctaga ggatccccgt atcgacaagg gtgagtgatg 40
<210> 15
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_F_2
<400> 15
ttcgagctcg gtacccgttg atgatcaggg aaggctgt 38
<210> 16
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_R_2
<400> 16
tggatttcgg tagacaaggg gagacctccg ggtgggaa 38
<210> 17
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_F_2
<400> 17
acccggaggt ctccccttgt ctaccgaaat ccacaacg 38
<210> 18
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_R_2
<400> 18
actctagagg atcccctcgg tcaggtgtgt gtaaatag 38
<210> 19
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_R_3
<400> 19
tggatttcgg tagacagggg gagacctccg ggtgggaa 38
<210> 20
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_F_3
<400> 20
acccggaggt ctccccctgt ctaccgaaat ccacaacg 38
<210> 21
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_F_4
<400> 21
ttcgagctcg gtaccccagc atcttcttct tcgccgaa 38
<210> 22
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_R_4
<400> 22
gtagacacgg ggagaacttt cgaggggaaa acgtttgttg 40
<210> 23
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_F_4
<400> 23
aacaaacgtt ttcccctcga aagttctccc cgtgtctacc 40
<210> 24
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_R_4
<400> 24
actctagagg atcccccttt tctgagctgg aggaacaa 38
<210> 25
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_R_5
<400> 25
gtagacacgg ggagagcctc gagtgggaaa acgtttgttg 40
<210> 26
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_F_5
<400> 26
aacaaacgtt ttcccactcg aggctctccc cgtgtctacc 40
<210> 27
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_up_R_6
<400> 27
gtagacacgg ggagagtcca gggcgggaaa acgtttgttg 40
<210> 28
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_down_F_6
<400> 28
aacaaacgtt ttcccgccct ggactctccc cgtgtctacc 40
<210> 29
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_del_F
<400> 29
ataatccacc aaaactgcc 19
<210> 30
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_del_R
<400> 30
aggtgtgtgt aaataggga 19
<210> 31
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> BBD29_04485_seq_F
<400> 31
gtggttgtct tcgatagt 18
<210> 32
<211> 1205
<212> PRT
<213> Unknown
<220>
<223> OdhA
<400> 32
Met Phe Gln Gln Phe Gln Lys Asp Pro Lys Ser Val Asp Lys Glu Trp
1 5 10 15
Arg Glu Leu Phe Glu Ala Gln Gly Gly Pro Asn Ala Thr Pro Ala Thr
20 25 30
Thr Glu Ala Gln Pro Ser Ala Pro Lys Glu Ser Ala Lys Pro Ala Pro
35 40 45
Lys Ala Ala Pro Ala Ala Lys Ala Ala Pro Arg Val Glu Thr Lys Pro
50 55 60
Ala Ala Lys Thr Ala Pro Lys Ala Lys Glu Ser Ser Val Pro Gln Gln
65 70 75 80
Pro Lys Leu Pro Glu Pro Gly Gln Thr Pro Ile Arg Gly Ile Phe Lys
85 90 95
Ser Ile Ala Lys Asn Met Asp Ile Ser Leu Glu Ile Pro Thr Ala Thr
100 105 110
Ser Val Arg Asp Met Pro Ala Arg Leu Met Phe Glu Asn Arg Ala Met
115 120 125
Val Asn Asp Gln Leu Lys Arg Thr Arg Gly Gly Lys Ile Ser Phe Thr
130 135 140
His Ile Ile Gly Tyr Ala Met Val Lys Ala Val Met Ala His Pro Asp
145 150 155 160
Met Asn Asn Ser Tyr Asp Val Ile Asp Gly Lys Pro Thr Leu Ile Val
165 170 175
Pro Glu His Ile Asn Leu Gly Leu Ala Ile Asp Leu Pro Gln Lys Asp
180 185 190
Gly Ser Arg Ala Leu Val Val Ala Ala Ile Lys Glu Thr Glu Lys Met
195 200 205
Asn Phe Ser Glu Phe Leu Ala Ala Tyr Glu Asp Ile Val Thr Arg Ser
210 215 220
Arg Lys Gly Lys Leu Thr Met Asp Asp Tyr Gln Gly Val Thr Val Ser
225 230 235 240
Leu Thr Asn Pro Gly Gly Ile Gly Thr Arg His Ser Val Pro Arg Leu
245 250 255
Thr Lys Gly Gln Gly Thr Ile Ile Gly Val Gly Ser Met Asp Tyr Pro
260 265 270
Ala Glu Phe Gln Gly Ala Ser Glu Asp Arg Leu Ala Glu Leu Gly Val
275 280 285
Gly Lys Leu Val Thr Ile Thr Ser Thr Tyr Asp His Arg Val Ile Gln
290 295 300
Gly Ala Val Ser Gly Glu Phe Leu Arg Thr Met Ser Arg Leu Leu Thr
305 310 315 320
Asp Asp Ser Phe Trp Asp Glu Ile Phe Asp Ala Met Asn Val Pro Tyr
325 330 335
Thr Pro Met Arg Trp Ala Gln Asp Val Pro Asn Thr Gly Val Asp Lys
340 345 350
Asn Thr Arg Val Met Gln Leu Ile Glu Ala Tyr Arg Ser Arg Gly His
355 360 365
Leu Ile Ala Asp Thr Asn Pro Leu Ser Trp Val Gln Pro Gly Met Pro
370 375 380
Val Pro Asp His Arg Asp Leu Asp Ile Glu Thr His Ser Leu Thr Ile
385 390 395 400
Trp Asp Leu Asp Arg Thr Phe Ser Val Gly Gly Phe Gly Gly Lys Glu
405 410 415
Thr Met Thr Leu Arg Glu Val Leu Ser Arg Leu Arg Ala Ala Tyr Thr
420 425 430
Leu Lys Val Gly Ser Glu Tyr Thr His Ile Leu Asp Arg Asp Glu Arg
435 440 445
Thr Trp Leu Gln Asp Arg Leu Glu Ala Gly Met Pro Lys Pro Thr Gln
450 455 460
Ala Glu Gln Lys Tyr Ile Leu Gln Lys Leu Asn Ala Ala Glu Ala Phe
465 470 475 480
Glu Asn Phe Leu Gln Thr Lys Tyr Val Gly Gln Lys Arg Phe Ser Leu
485 490 495
Glu Gly Ala Glu Ala Leu Ile Pro Leu Met Asp Ser Ala Ile Asp Thr
500 505 510
Ala Ala Gly Gln Gly Leu Asp Glu Val Val Ile Gly Met Pro His Arg
515 520 525
Gly Arg Leu Asn Val Leu Phe Asn Ile Val Gly Lys Pro Leu Ala Ser
530 535 540
Ile Phe Asn Glu Phe Glu Gly Gln Met Glu Gln Gly Gln Ile Gly Gly
545 550 555 560
Ser Gly Asp Val Lys Tyr His Leu Gly Ser Glu Gly Gln His Leu Gln
565 570 575
Met Phe Gly Asp Gly Glu Ile Lys Val Ser Leu Thr Ala Asn Pro Ser
580 585 590
His Leu Glu Ala Val Asn Pro Val Met Glu Gly Ile Val Arg Ala Lys
595 600 605
Gln Asp Tyr Leu Asp Lys Gly Val Asp Gly Lys Thr Val Val Pro Leu
610 615 620
Leu Leu His Gly Asp Ala Ala Phe Ala Gly Leu Gly Ile Val Pro Glu
625 630 635 640
Thr Ile Asn Leu Ala Lys Leu Arg Gly Tyr Asp Val Gly Gly Thr Ile
645 650 655
His Ile Val Val Asn Asn Gln Ile Gly Phe Thr Thr Thr Pro Asp Ser
660 665 670
Ser Arg Ser Met His Tyr Ala Thr Asp Tyr Ala Lys Ala Phe Gly Cys
675 680 685
Pro Val Phe His Val Asn Gly Asp Asp Pro Glu Ala Val Val Trp Val
690 695 700
Gly Gln Leu Ala Thr Glu Tyr Arg Arg Arg Phe Gly Lys Asp Val Phe
705 710 715 720
Ile Asp Leu Val Cys Tyr Arg Leu Arg Gly His Asn Glu Ala Asp Asp
725 730 735
Pro Ser Met Thr Gln Pro Lys Met Tyr Glu Leu Ile Thr Gly Arg Glu
740 745 750
Thr Val Arg Ala Gln Tyr Thr Glu Asp Leu Leu Gly Arg Gly Asp Leu
755 760 765
Ser Asn Glu Asp Ala Glu Ala Val Val Arg Asp Phe His Asp Gln Met
770 775 780
Glu Ser Val Phe Asn Glu Val Lys Glu Gly Gly Lys Lys Gln Ala Glu
785 790 795 800
Ala Gln Thr Gly Ile Thr Gly Ser Gln Lys Leu Pro His Gly Leu Glu
805 810 815
Thr Asn Ile Ser Arg Glu Glu Leu Leu Glu Leu Gly Gln Ala Phe Ala
820 825 830
Asn Thr Pro Glu Gly Phe Asn Tyr His Pro Arg Val Ala Pro Val Ala
835 840 845
Lys Lys Arg Val Ser Ser Val Thr Glu Gly Gly Ile Asp Trp Ala Trp
850 855 860
Gly Glu Leu Leu Ala Phe Gly Ser Leu Ala Asn Ser Gly Arg Leu Val
865 870 875 880
Arg Leu Ala Gly Glu Asp Ser Arg Arg Gly Thr Phe Thr Gln Arg His
885 890 895
Ala Val Ala Ile Asp Pro Ala Thr Ala Glu Glu Phe Asn Pro Leu His
900 905 910
Glu Leu Ala Gln Ser Lys Gly Asn Asn Gly Lys Phe Leu Val Tyr Asn
915 920 925
Ser Ala Leu Thr Glu Tyr Ala Gly Met Gly Phe Glu Tyr Gly Tyr Ser
930 935 940
Val Gly Asn Glu Asp Ser Val Val Ala Trp Glu Ala Gln Phe Gly Asp
945 950 955 960
Phe Ala Asn Gly Ala Gln Thr Ile Ile Asp Glu Tyr Val Ser Ser Gly
965 970 975
Glu Ala Lys Trp Gly Gln Thr Ser Lys Leu Ile Leu Leu Leu Pro His
980 985 990
Gly Tyr Glu Gly Gln Gly Pro Asp His Ser Ser Ala Arg Ile Glu Arg
995 1000 1005
Phe Leu Gln Leu Cys Ala Glu Gly Ser Met Thr Val Ala Gln Pro Ser
1010 1015 1020
Thr Pro Ala Asn His Phe His Leu Leu Arg Arg His Ala Leu Ser Asp
1025 1030 1035 1040
Leu Lys Arg Pro Leu Val Ile Phe Thr Pro Lys Ser Met Leu Arg Asn
1045 1050 1055
Lys Ala Ala Ala Ser Ala Pro Glu Asp Phe Thr Glu Val Thr Lys Phe
1060 1065 1070
Gln Ser Val Ile Asp Asp Pro Asn Val Ala Asp Ala Ala Lys Val Lys
1075 1080 1085
Lys Val Met Leu Val Ser Gly Lys Leu Tyr Tyr Glu Leu Ala Lys Arg
1090 1095 1100
Lys Glu Lys Asp Gly Arg Asp Asp Ile Ala Ile Val Arg Ile Glu Met
1105 1110 1115 1120
Leu His Pro Ile Pro Phe Asn Arg Ile Ser Glu Ala Leu Ala Gly Tyr
1125 1130 1135
Pro Asn Ala Glu Glu Val Leu Phe Val Gln Asp Glu Pro Ala Asn Gln
1140 1145 1150
Gly Pro Trp Pro Phe Tyr Gln Glu His Leu Pro Glu Leu Ile Pro Asn
1155 1160 1165
Met Pro Lys Met Arg Arg Val Ser Arg Arg Ala Gln Ser Ser Thr Ala
1170 1175 1180
Thr Gly Val Ala Lys Val His Gln Leu Glu Glu Lys Gln Leu Ile Asp
1185 1190 1195 1200
Glu Ala Phe Glu Ala
1205
<---
Изобретение относится к биотехнологии. Предложен микроорганизм рода Corynebacterium для продуцирования L-глутаминовой кислоты, в котором белок VKOR (белок семейства витамин K-эпоксид-редуктазы) инактивирован, где инактивация белка VKOR означает сниженную активность или отсутствие активности по сравнению с эндогенной активностью. Также предложен способ получения L-глутаминовой кислоты, с использованием указанного микроорганизма. Изобретение обеспечивает продуцирование L-глутаминовой кислоты с высоким выходом. 2 н. и 13 з.п. ф-лы, 9 табл., 6 пр.
1. Микроорганизм рода Corynebacterium для продуцирования L-глутаминовой кислоты, в котором белок VKOR (белок семейства витамин K-эпоксид-редуктазы) инактивирован, где инактивация белка VKOR означает сниженную активность или отсутствие активности по сравнению с эндогенной активностью.
2. Микроорганизм по п. 1, где белок VKOR происходит из рода Corynebacterium.
3. Микроорганизм по п. 1, где белок VKOR состоит из полипептида, представленного аминокислотной последовательностью, обладающей идентичностью 80% или более с аминокислотной последовательностью SEQ ID NO: 1.
4. Микроорганизм по п. 1, где белок VKOR кодируется полинуклеотидом, представленным нуклеотидной последовательностью SEQ ID NO: 2.
5. Микроорганизм по п. 1, где инактивация белка VKOR означает делецию или ослабление полинуклеотида, представленного нуклеотидной последовательностью SEQ ID NO: 2.
6. Микроорганизм по п. 1, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
7. Микроорганизм по п. 1, обладающий повышенной способностью продуцировать L-глутаминовую кислоту по сравнению с родительским штаммом или штаммом дикого типа, в котором белок VKOR не инактивирован.
8. Микроорганизм по п. 1, содержащий белок OdhA, который дополнительно удален.
9. Способ получения L-глутаминовой кислоты, включающий культивирование в среде микроорганизма рода Corynebacterium, в котором белок VKOR инактивирован; и где инактивация белка VKOR означает сниженную активность или отсутствие активности по сравнению с эндогенной активностью.
10. Способ по п. 9, где белок VKOR происходит из рода Corynebacterium.
11. Способ по п. 9, где белок VKOR состоит из полипептида, представленного аминокислотной последовательностью, обладающей идентичностью 80% или более с аминокислотной последовательностью SEQ ID NO: 1.
12. Способ по п. 9, где белок VKOR кодируется полинуклеотидом, представленным нуклеотидной последовательностью SEQ ID NO: 2.
13. Способ по п. 9, где инактивация белка VKOR означает делецию или ослабление полинуклеотида, представленного нуклеотидной последовательностью SEQ ID NO: 2.
14. Способ по п. 9, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
15. Способ по п. 9, где микроорганизм содержит белок OdhA, который дополнительно удален.
| KR 20090069571 A, 01.07.2009 | |||
| REARDON-ROBINSON M | |||
| E | |||
| et al | |||
| Disulfide-bond-forming pathways in Gram-positive bacteria | |||
| Journal of Bacteriology | |||
| Токарный резец | 1924 |
|
SU2016A1 |
| Складная решетчатая мачта | 1919 |
|
SU198A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ГАЗОГЕНЕРАТОР ДЛЯ ДРОВ, ТОРФА И КИЗЯКА | 1923 |
|
SU746A1 |
| Нагревательный прибор | 1929 |
|
SU22856A1 |
| Приспособление для выгрузки шахтных печей | 1930 |
|
SU21799A1 |
| Найдено по адресу: | |||

