ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к соединениям, предназначенным для использования в генной терапии заболеваний глаз. Более конкретно, изобретение относится к вирусным векторам, в частности, аденоассоциированным вирусным (AAV) векторам, предназначенным для использования в лечении или предотвращении пигментного ретинита (RP), которые обеспечивают доставку изоформы ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) в глаза.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Пигментный ретинит (RP) представляет собой фенотипически связанную группу наследственных дистрофий сетчатки, приводящих к постепенному ослаблению зрения. От RP страдает примерно 1 из 3000-4000 людей.
Ранние симптомы RP включают ухудшение ночного и периферического зрения. По мере прогрессирования заболевания также могут быть затронуты четкость зрения, острота зрения и цветовое восприятие. Возраст, в котором впервые появляются симптомы RP, варьируется, однако, как правило, находится в диапазоне от 10 и 30 лет, и скорость ухудшения зрения различается у разных людей.
RP обычно бывает вызван прогрессирующей дегенерацией палочковидных фоторецепторных клеток. Однако пигментный эпителий сетчатки (RPE) и колбочковидные фоторецепторные клетки также могут дегенерировать при прогрессировании заболевания.
RP может быть вызван, например, мутациями в одном из множества разных генов, связанных со здоровьем и функцией глаз. Из всех разновидностей RP, связанных с дефектом единственного гена, X-связанное заболевание, развивающееся в результате дефектов в гене регулятора ГТФазы пигментного ретинита (RPGR), является наиболее распространенным.
X-связанный пигментный ретинит (XLRP) считают самой тяжелой формой пигментного ретинита. У субъектов, страдающих XLRP, развивается ограничение периферического зрительного поля и ночная слепота в течение первых двух десятилетий жизни. Кроме того, это состояние характеризуется ощущениями вспышек света в глазах, непроизвольным маятниковым движением глаз, нарушением цветового зрения и уменьшением остроты центрального зрения. В результате, пациенты, как правило, становятся инвалидами по зрению еще до окончания средней школы.
Ген RPGR в высокой степени подвержен мутациям, что увеличивает вероятность появления вызывающих заболевание мутаций in vivo. Однако такая подверженность мутациям также создает проблемы при получении векторов, кодирующих RPGR, для использования в генной терапии. Действительно, предыдущие стратегии для разработки заместительной генной терапии для XLRP потерпели неудачу в результате таких мутаций: лишь совсем недавно многообещающая исследовательская программа была отложена в результате обнаружения мутации в области ORF15 трансгенной кассеты. В другой программе альтернативно сплайсированные варианты RPGR были обнаружены методом вестерн-блоттинга (Wu, Z. et al. (2015) Hum. Mol. Genet. 24: 3956-3970).
В настоящее время не существует одобренных терапевтических методов для предотвращения развития RP или для улучшения зрения после начала заболевания. Соответственно, сохраняется насущная потребность в способах лечения RP, в частности, способах предотвращения угасания зрительной функции или улучшения зрения заболевших людей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения неожиданно обнаружили, что последовательность гена изоформы ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) может быть изменена для повышения стабильности последовательности, например, за счет уменьшения частоты, или предотвращения возникновения, мутаций при репликации (то есть, повышения точности репликации). Авторы настоящего изобретения также неожиданно обнаружили, что риск того, что модификация повлияет на функцию гена, может быть уменьшен. Кроме того, авторы настоящего изобретения продемонстрировали, что модифицированные гены успешно лечат пигментный ретинит в животных моделях.
Соответственно, в одном аспекте изобретение относится к полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15), при этом кодирующая RPGRORF15 нуклеотидная последовательность была кодон-оптимизирована для повышения точности репликации последовательности.
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована для уменьшения частоты образования альтернативно сплайсированных вариантов.
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована, чтобы свести к минимуму или предотвратить создание новых сайтов CpG в сравнении с последовательностью дикого типа. В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит менее 220, 210, 200, 190, 170, 165, 160, 150, 140, 130, 120, 110 или 100 сайтов CpG.
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована путем замены некоторых или всех кодонов GGN кодонами GGC (там, где кодоны GGN не были исходно кодонами GGC).
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована, чтобы свести к минимуму или предотвратить введение новых тиминовых нуклеотидов в богатые пуринами (то есть, богатые GA) области кодирующей RPGRORF15 нуклеотидной последовательности, например, область экзона ORF 15 (которая соответствует нуклеотидам 1754-3459 в RPGRORF15, например, нуклеотидам 1754-3459 в SEQ ID NO: 2).
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована для уменьшения числа сайтов GT (то есть, потенциальных донорских сайтов сплайсинга) в сравнении с последовательностью дикого типа. В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит менее 120, 115, 110, 105, 100, 90, 80, 70, 60, 50, 40, 30, 20 или 10 сайтов GT. В предпочтительном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит менее 105 сайтов GT.
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована, чтобы избежать создания аномальных сигналов полиA (например, AATAAA).
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована для уменьшения числа пуриновых нуклеотидов в сравнении с последовательностью дикого типа. Последовательность дикого типа может представлять собой, например, последовательность SEQ ID NO: 2.
Предпочтительно, число пуриновых нуклеотидов уменьшено путем замены пуриновых нуклеотидов пиримидиновыми нуклеотидами.
В одном варианте осуществления число пуриновых нуклеотидов уменьшено в богатых пуринами (то есть, богатых GA) областях кодирующей RPGRORF15 нуклеотидной последовательности, например, области экзона ORF 15 (которая соответствует нуклеотидам 1754-3459 в RPGRORF15, например, нуклеотидам 1754-3459 в SEQ ID NO: 2).
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2505, 2520, 2547, 2574, 2622, 2634, 2661, 2673, 2706, 2712, 2763, 2775, 2796, 2811, 2817, 2832, 2838, 2871, 2886, 2892, 2898, 2910, 2922, 2931, 2937, 2958, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 в SEQ ID NO: 2. Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению не содержит пуриновые нуклеотиды по меньшей мере в 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или во всех из этих нуклеотидных положений. Предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность с по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 1. Более предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 1.
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению получена из SEQ ID NO: 2 путем замены пуриновых нуклеотидов в положениях 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2505, 2520, 2547, 2574, 2622, 2634, 2661, 2673, 2706, 2712, 2763, 2775, 2796, 2811, 2817, 2832, 2838, 2871, 2886, 2892, 2898, 2910, 2922, 2931, 2937, 2958, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 пиримидиновыми нуклеотидами (например, тимином или цитозином). Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. В другом варианте осуществления пуриновые нуклеотиды были заменены по меньшей мере в 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или во всех из этих нуклеотидных положений. Предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность с по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 1. Более предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 1.
В соответствии с изобретением, число пуриновых нуклеотидов может быть уменьшено на по меньшей мере 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5% или 5% от числа пуриновых нуклеотидов в последовательности дикого типа (например, SEQ ID NO: 2).
В соответствии с изобретением, число пуриновых нуклеотидов может быть уменьшено на 0,5-10%, 0,5-7,5%, 0,5-5%, 0,5-4,5%, 0,5-4%, 0,5-3,5%, 0,5-3%, 1-5%, 1-4,5%, 1-4%, 1-3,5% или 1-3% от числа пуриновых нуклеотидов в последовательности дикого типа (например, SEQ ID NO: 2).
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована для уменьшения числа адениновых нуклеотидов в сравнении с последовательностью дикого типа. Последовательность дикого типа может представлять собой, например, последовательность SEQ ID NO: 2. В другом варианте осуществления число адениновых нуклеотидов было уменьшено на по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190 или 200. В одном варианте осуществления аденины в положениях, соответствующих нуклеотидам 58, 114, 123, 129, 181, 213, 219, 226, 237, 285, 306, 309, 315, 324, 330, 339, 400, 444, 456, 478, 480, 594, 606, 618, 697, 744, 807, 852, 877, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1194, 1197, 1221, 1249, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1413, 1446, 1452, 1464, 1474, 1482, 1519, 1542, 1584, 1608, 1653, 1674, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1854, 1881, 1893, 1902, 1909, 1929, 1960, 1987, 1992, 1998, 2020, 2047, 2049, 2062, 2067, 2076, 2167, 2169, 2190, 2208, 2229, 2259, 2298, 2323, 2361, 2373, 2382, 2403, 2410, 2412, 2448, 2457, 2472, 2476, 2622, 2661, 2673, 2712, 2775, 2811, 2832, 2886, 2910, 2931, 2958, 3033, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3306, 3321, 3369, 3405 и/или 3438 в SEQ ID NO: 2, заменены пиримидиновыми нуклеотидами (например, тимином или цитозином). Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению не содержит пуриновый нуклеотид по меньшей мере в одном положении, предпочтительно по меньшей мере в 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170 или 180 положениях, предпочтительно во всех положениях, соответствующих положениям, где пуриновый нуклеотид в SEQ ID NO: 2 (wtRPGR) выравнивается с пиримидиновым нуклеотидом в SEQ ID NO: 3 (coRPGR), например, при выравнивании последовательностей на Фигуре 2 (wtRPGR, «исходная»; coRPGR, «оптимизированная»).
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 33, 58, 59, 114, 123, 129, 181, 182, 213, 219, 226, 227, 237, 267, 285, 306, 309, 315, 324, 330, 339, 400, 401, 444, 456, 478, 480, 510, 594, 606, 618, 639, 697, 726, 744, 777, 807, 852, 877, 879, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1184, 1194, 1197, 1221, 1249, 1251, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1373, 1413, 1446, 1452, 1464, 1474, 1475, 1482, 1519, 1520, 1542, 1584, 1590, 1599, 1608, 1653, 1668, 1674, 1689, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2505, 2520, 2547, 2574, 2622, 2634, 2661, 2673, 2706, 2712, 2763, 2775, 2796, 2811, 2817, 2832, 2838, 2871, 2886, 2892, 2898, 2910, 2922, 2931, 2937, 2958, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 в SEQ ID NO: 2. Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению не содержит пуриновые нуклеотиды по меньшей мере в 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180 или во всех из этих нуклеотидных положений. Предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность с по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 1. Более предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 1.
Следует отметить, что в данной заявке нуклеотидные положения идентифицированы как положения, «соответствующие» конкретным положениям в SEQ ID NO: 2. Это не следует интерпретировать как указание на то, что последовательности по настоящему изобретению должны включать последовательности, приведенные в SEQ ID NO: 2. В настоящем документе нуклеотиды кодирующей RPGRORF15 нуклеотидной последовательности пронумерованы в соответствии с соглашением по нумерации, на основании чего 5′-аденин в SEQ ID NO: 2 считают нуклеотидом 1. Обозначение отдельных нуклеотидов и положений мутаций, описанных в настоящем документе, соответствует данному соглашению по нумерации. Квалифицированный специалист с легкостью сможет определять аналогичные положения в гомологичных последовательностях путем выравнивания последовательностей с SEQ ID NO: 2. Пример такого выравнивания приведен на Фигуре 2.
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению получена из SEQ ID NO: 2 путем замены пуриновых нуклеотидов в положениях 33, 58, 59, 114, 123, 129, 181, 182, 213, 219, 226, 227, 237, 267, 285, 306, 309, 315, 324, 330, 339, 400, 401, 444, 456, 478, 480, 510, 594, 606, 618, 639, 697, 726, 744, 777, 807, 852, 877, 879, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1184, 1194, 1197, 1221, 1249, 1251, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1373, 1413, 1446, 1452, 1464, 1474, 1475, 1482, 1519, 1520, 1542, 1584, 1590, 1599, 1608, 1653, 1668, 1674, 1689, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2505, 2520, 2547, 2574, 2622, 2634, 2661, 2673, 2706, 2712, 2763, 2775, 2796, 2811, 2817, 2832, 2838, 2871, 2886, 2892, 2898, 2910, 2922, 2931, 2937, 2958, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 пиримидиновыми нуклеотидами (например, тимином или цитозином). Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. В другом варианте осуществления пуриновые нуклеотиды были заменены по меньшей мере в 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180 или во всех из этих нуклеотидных положений. Предпочтительно, пуриновые нуклеотиды были заменены во всех из этих нуклеотидных положений. Предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность с по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 1. Более предпочтительно, нуклеотидная последовательность кодирует аминокислотную последовательность SEQ ID NO: 1.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность является такой, что нуклеотид, соответствующий положению 33 в SEQ ID NO: 2, представляет собой T, нуклеотид, соответствующий положению 58, представляет собой T, нуклеотид, соответствующий положению 59, представляет собой C, нуклеотид, соответствующий положению 114, представляет собой C, нуклеотид, соответствующий положению 123, представляет собой T, нуклеотид, соответствующий положению 129, представляет собой C, нуклеотид, соответствующий положению 181, представляет собой T, нуклеотид, соответствующий положению 182, представляет собой C, нуклеотид, соответствующий положению 213, представляет собой C, нуклеотид, соответствующий положению 219, представляет собой T, нуклеотид, соответствующий положению 226, представляет собой T, нуклеотид, соответствующий положению 227, представляет собой C, нуклеотид, соответствующий положению 237, представляет собой C, нуклеотид, соответствующий положению 267, представляет собой C, нуклеотид, соответствующий положению 285, представляет собой C, нуклеотид, соответствующий положению 306, представляет собой C, нуклеотид, соответствующий положению 309, представляет собой C, нуклеотид, соответствующий положению 315, представляет собой C, нуклеотид, соответствующий положению 324, представляет собой C, нуклеотид, соответствующий положению 330, представляет собой C, нуклеотид, соответствующий положению 339, представляет собой C, нуклеотид, соответствующий положению 400, представляет собой T, нуклеотид, соответствующий положению 401, представляет собой C, нуклеотид, соответствующий положению 444, представляет собой C, нуклеотид, соответствующий положению 456, представляет собой T, нуклеотид, соответствующий положению 478, представляет собой C, нуклеотид, соответствующий положению 480, представляет собой C, нуклеотид, соответствующий положению 510, представляет собой C, нуклеотид, соответствующий положению 594, представляет собой C, нуклеотид, соответствующий положению 606, представляет собой C, нуклеотид, соответствующий положению 618, представляет собой C, нуклеотид, соответствующий положению 639, представляет собой C, нуклеотид, соответствующий положению 697, представляет собой C, нуклеотид, соответствующий положению 726, представляет собой T, нуклеотид, соответствующий положению 744, представляет собой C, нуклеотид, соответствующий положению 777, представляет собой T, нуклеотид, соответствующий положению 807, представляет собой C, нуклеотид, соответствующий положению 852, представляет собой C, нуклеотид, соответствующий положению 877, представляет собой C, нуклеотид, соответствующий положению 879, представляет собой C, нуклеотид, соответствующий положению 888, представляет собой T, нуклеотид, соответствующий положению 891, представляет собой C, нуклеотид, соответствующий положению 921, представляет собой C, нуклеотид, соответствующий положению 930, представляет собой C, нуклеотид, соответствующий положению 960, представляет собой C, нуклеотид, соответствующий положению 1042, представляет собой C, нуклеотид, соответствующий положению 1050, представляет собой C, нуклеотид, соответствующий положению 1116, представляет собой T, нуклеотид, соответствующий положению 1140, представляет собой T, нуклеотид, соответствующий положению 1183, представляет собой T, нуклеотид, соответствующий положению 1184, представляет собой C, нуклеотид, соответствующий положению 1194, представляет собой T, нуклеотид, соответствующий положению 1197, представляет собой C, нуклеотид, соответствующий положению 1221, представляет собой T, нуклеотид, соответствующий положению 1249, представляет собой C, нуклеотид, соответствующий положению 1251, представляет собой C, нуклеотид, соответствующий положению 1257, представляет собой T, нуклеотид, соответствующий положению 1273, представляет собой C, нуклеотид, соответствующий положению 1276, представляет собой C, нуклеотид, соответствующий положению 1281, представляет собой C, нуклеотид, соответствующий положению 1290, представляет собой T, нуклеотид, соответствующий положению 1293, представляет собой C, нуклеотид, соответствующий положению 1357, представляет собой C, нуклеотид, соответствующий положению 1372, представляет собой T, нуклеотид, соответствующий положению 1373, представляет собой C, нуклеотид, соответствующий положению 1413, представляет собой C, нуклеотид, соответствующий положению 1446, представляет собой C, нуклеотид, соответствующий положению 1452, представляет собой C, нуклеотид, соответствующий положению 1464, представляет собой T, нуклеотид, соответствующий положению 1474, представляет собой T, нуклеотид, соответствующий положению 1475, представляет собой C, нуклеотид, соответствующий положению 1482, представляет собой C, нуклеотид, соответствующий положению 1519, представляет собой T, нуклеотид, соответствующий положению 1520, представляет собой C, нуклеотид, соответствующий положению 1542, представляет собой T, нуклеотид, соответствующий положению 1584, представляет собой T, нуклеотид, соответствующий положению 1590, представляет собой C, нуклеотид, соответствующий положению 1599, представляет собой T, нуклеотид, соответствующий положению 1608, представляет собой C, нуклеотид, соответствующий положению 1653, представляет собой C, нуклеотид, соответствующий положению 1668, представляет собой C, нуклеотид, соответствующий положению 1674, представляет собой T, нуклеотид, соответствующий положению 1689, представляет собой C, нуклеотид, соответствующий положению 1692, представляет собой T, нуклеотид, соответствующий положению 1734, представляет собой T, нуклеотид, соответствующий положению 1761, представляет собой C, нуклеотид, соответствующий положению 1776, представляет собой C, нуклеотид, соответствующий положению 1788, представляет собой C, нуклеотид, соответствующий положению 1803, представляет собой T, нуклеотид, соответствующий положению 1824, представляет собой C, нуклеотид, соответствующий положению 1845, представляет собой C, нуклеотид, соответствующий положению 1854, представляет собой C, нуклеотид, соответствующий положению 1881, представляет собой T, нуклеотид, соответствующий положению 1893, представляет собой T, нуклеотид, соответствующий положению 1902, представляет собой C, нуклеотид, соответствующий положению 1909, представляет собой T, нуклеотид, соответствующий положению 1910, представляет собой C, нуклеотид, соответствующий положению 1929, представляет собой T, нуклеотид, соответствующий положению 1932, представляет собой C, нуклеотид, соответствующий положению 1960, представляет собой C, нуклеотид, соответствующий положению 1987, представляет собой T, нуклеотид, соответствующий положению 1988, представляет собой C, нуклеотид, соответствующий положению 1992, представляет собой C, нуклеотид, соответствующий положению 1998, представляет собой T, нуклеотид, соответствующий положению 2020, представляет собой C, нуклеотид, соответствующий положению 2031, представляет собой C, нуклеотид, соответствующий положению 2047, представляет собой C, нуклеотид, соответствующий положению 2049, представляет собой C, нуклеотид, соответствующий положению 2062, представляет собой C, нуклеотид, соответствующий положению 2067, представляет собой T, нуклеотид, соответствующий положению 2076, представляет собой C, нуклеотид, соответствующий положению 2121, представляет собой C, нуклеотид, соответствующий положению 2167, представляет собой C, нуклеотид, соответствующий положению 2169, представляет собой C, нуклеотид, соответствующий положению 2190, представляет собой C, нуклеотид, соответствующий положению 2193, представляет собой C, нуклеотид, соответствующий положению 2208, представляет собой C, нуклеотид, соответствующий положению 2229, представляет собой C, нуклеотид, соответствующий положению 2259, представляет собой C, нуклеотид, соответствующий положению 2283, представляет собой C, нуклеотид, соответствующий положению 2298, представляет собой C, нуклеотид, соответствующий положению 2323, представляет собой C, нуклеотид, соответствующий положению 2343, представляет собой C, нуклеотид, соответствующий положению 2346, представляет собой C, нуклеотид, соответствующий положению 2361, представляет собой C, нуклеотид, соответствующий положению 2373, представляет собой C, нуклеотид, соответствующий положению 2382, представляет собой C, нуклеотид, соответствующий положению 2388, представляет собой C, нуклеотид, соответствующий положению 2403, представляет собой C, нуклеотид, соответствующий положению 2409, представляет собой C, нуклеотид, соответствующий положению 2410, представляет собой C, нуклеотид, соответствующий положению 2412, представляет собой C, нуклеотид, соответствующий положению 2448, представляет собой C, нуклеотид, соответствующий положению 2457, представляет собой C, нуклеотид, соответствующий положению 2472, представляет собой C, нуклеотид, соответствующий положению 2476, представляет собой C, нуклеотид, соответствующий положению 2478, представляет собой C, нуклеотид, соответствующий положению 2505, представляет собой C, нуклеотид, соответствующий положению 2520, представляет собой C, нуклеотид, соответствующий положению 2547, представляет собой C, нуклеотид, соответствующий положению 2574, представляет собой C, нуклеотид, соответствующий положению 2622, представляет собой C, нуклеотид, соответствующий положению 2634, представляет собой C, нуклеотид, соответствующий положению 2661, представляет собой C, нуклеотид, соответствующий положению 2673, представляет собой C, нуклеотид, соответствующий положению 2706, представляет собой C, нуклеотид, соответствующий положению 2712, представляет собой C, нуклеотид, соответствующий положению 2763, представляет собой C, нуклеотид, соответствующий положению 2775, представляет собой C, нуклеотид, соответствующий положению 2796, представляет собой C, нуклеотид, соответствующий положению 2811, представляет собой C, нуклеотид, соответствующий положению 2817, представляет собой C, нуклеотид, соответствующий положению 2832, представляет собой C, нуклеотид, соответствующий положению 2838, представляет собой C, нуклеотид, соответствующий положению 2871, представляет собой C, нуклеотид, соответствующий положению 2886, представляет собой C, нуклеотид, соответствующий положению 2892, представляет собой C, нуклеотид, соответствующий положению 2898, представляет собой C, нуклеотид, соответствующий положению 2910, представляет собой C, нуклеотид, соответствующий положению 2922, представляет собой C, нуклеотид, соответствующий положению 2931, представляет собой C, нуклеотид, соответствующий положению 2937, представляет собой C, нуклеотид, соответствующий положению 2958, представляет собой C, нуклеотид, соответствующий положению 2970, представляет собой C, нуклеотид, соответствующий положению 2997, представляет собой C, нуклеотид, соответствующий положению 3033, представляет собой C, нуклеотид, соответствующий положению 3039, представляет собой C, нуклеотид, соответствующий положению 3069, представляет собой C, нуклеотид, соответствующий положению 3075, представляет собой C, нуклеотид, соответствующий положению 3117, представляет собой C, нуклеотид, соответствующий положению 3129, представляет собой C, нуклеотид, соответствующий положению 3156, представляет собой C, нуклеотид, соответствующий положению 3166, представляет собой C, нуклеотид, соответствующий положению 3235, представляет собой C, нуклеотид, соответствующий положению 3246, представляет собой C, нуклеотид, соответствующий положению 3273, представляет собой C, нуклеотид, соответствующий положению 3285, представляет собой C, нуклеотид, соответствующий положению 3306, представляет собой C, нуклеотид, соответствующий положению 3321, представляет собой C, нуклеотид, соответствующий положению 3369, представляет собой T, нуклеотид, соответствующий положению 3399, представляет собой C, нуклеотид, соответствующий положению 3405, представляет собой T и/или нуклеотид, соответствующий положению 3438, представляет собой C.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 1689 и/или 3438 в SEQ ID NO: 2. Нуклеотид в положении 1689 может быть заменен пиримидином, предпочтительно, C. Аденин в положении 3438 может быть заменен пиримидином, предпочтительно, C.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность получена из SEQ ID NO: 2 путем замены аденинового нуклеотида в положении 3405 пиримидиновым нуклеотидом, без введения динуклеотида CpG.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность содержит адениновый нуклеотид в положении, соответствующем положению 3405 в SEQ ID NO: 2.
В одном варианте осуществления кодируемый RPGRORF15 белок представляет собой человеческий RPGRORF15.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность содержит один или более нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность содержит один или более нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 48 в SEQ ID NO: 2, A в положении, соответствующем положению 57 в SEQ ID NO: 2, T в положении, соответствующем положению 60 в SEQ ID NO: 2, T в положении, соответствующем положению 69 в SEQ ID NO: 2, A в положении, соответствующем положению 72 в SEQ ID NO: 2, C в положении, соответствующем положению 171 в SEQ ID NO: 2, C в положении, соответствующем положению 189 в SEQ ID NO: 2, T в положении, соответствующем положению 441 в SEQ ID NO: 2, T в положении, соответствующем положению 537 в SEQ ID NO: 2, C в положении, соответствующем положению 546 в SEQ ID NO: 2, T в положении, соответствующем положению 786 в SEQ ID NO: 2, T в положении, соответствующем положению 792 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, A в положении, соответствующем положению 990 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, A в положении, соответствующем положению 3355 в SEQ ID NO: 2, G в положении, соответствующем положению 3357 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3403 в SEQ ID NO: 2, C в положении, соответствующем положению 3404 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит последовательность, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 1;
(b) нуклеотидной последовательности, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 3; и
(c) нуклеотидной последовательности SEQ ID NO: 3,
предпочтительно, при этом белок, кодируемый нуклеотидной последовательностью, по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность, которая имеет по меньшей мере 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 1, предпочтительно, при этом аминокислотная последовательность по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
Кроме того, настоящее изобретение относится к полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) или ее функциональный вариант, имеющий по меньшей мере 80% идентичности с SEQ ID NO: 1, при этом нуклеотидная последовательность содержит один или более, предпочтительно все, из нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2. Полинуклеотид может содержать 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17 из указанных нуклеотидов.
Изобретение также относится к полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) или ее функциональный вариант, имеющий по меньшей мере 80% идентичности с SEQ ID NO: 1, при этом нуклеотидную последовательность выбирают из группы, состоящей из:
(a) нуклеотидной последовательности, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 3, при этом указанная последовательность содержит один или более, предпочтительно все, из нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 48 в SEQ ID NO: 2, A в положении, соответствующем положению 57 в SEQ ID NO: 2, T в положении, соответствующем положению 60 в SEQ ID NO: 2, T в положении, соответствующем положению 69 в SEQ ID NO: 2, A в положении, соответствующем положению 72 в SEQ ID NO: 2, C в положении, соответствующем положению 171 в SEQ ID NO: 2, C в положении, соответствующем положению 189 в SEQ ID NO: 2, T в положении, соответствующем положению 441 в SEQ ID NO: 2, T в положении, соответствующем положению 537 в SEQ ID NO: 2, C в положении, соответствующем положению 546 в SEQ ID NO: 2, T в положении, соответствующем положению 786 в SEQ ID NO: 2, T в положении, соответствующем положению 792 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, A в положении, соответствующем положению 990 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, A в положении, соответствующем положению 3355 в SEQ ID NO: 2, G в положении, соответствующем положению 3357 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3403 в SEQ ID NO: 2, C в положении, соответствующем положению 3404 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2; или (b) нуклеотидной последовательности SEQ ID NO: 3. Полинуклеотид может содержать 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33 или 34 из указанных нуклеотидов.
Предпочтительно, функциональный вариант по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит нуклеотидную последовательность, которая имеет по меньшей мере 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 3, предпочтительно, при этом белок, кодируемый нуклеотидной последовательностью, по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
Предпочтительно, белок, кодируемый кодирующей RPGRORF15 нуклеотидной последовательностью по изобретению, обеспечивает такое же, или улучшенное, функционирование клеток сетчатки и/или зрительной функции, что и белок, представленный SEQ ID NO: 1.
В предпочтительном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению кодирует белок с SEQ ID NO: 1.
Полинуклеотид по изобретению может иметь по существу такую же, или повышенную, точность репликации в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
Полинуклеотид по изобретению может иметь повышенную стабильность и/или быть менее подвержен мутациям, возникающим в процессе циклов репликации полинуклеотида, в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
Полинуклеотид по изобретению может обеспечивать более высокие скорости трансляции и/или экспрессии белка в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
Полинуклеотид по изобретению может приводить к уменьшению или предотвращению образования альтернативно сплайсированных вариантов и/или укороченных белков в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
В предпочтительном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению имеет по существу такую же, или повышенную, точность репликации, что и SEQ ID NO: 3. Точность репликации может быть измерена любым из множества методов, известных специалистам в данной области, например, методами, описанными в настоящем документе.
В предпочтительном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит нуклеотидную последовательность SEQ ID NO: 3.
Кодирующая RPGRORF15 нуклеотидная последовательность может содержать менее 165 динуклеотидов CpG и/или содержит не более двух CpG-островков. CpG-островки можно легко идентифицировать обычными методами, например, с использованием EMBOSS Cpgplot (http://www.ebi.ac.uk/Tools/seqstats/emboss_cpgplot/help/).
Полинуклеотид по изобретению может кодировать RPGRorf15, который укорочен в сравнении с последовательностью дикого типа. При этом, кодон-оптимизацию, описанную в настоящем документе, применяют к соответствующим фрагментам укороченной последовательности. Предпочтительно, укороченный RPGRorf15 обладает способностью сохранять функцию RPGR.
Кодирующая RPGRorf15 последовательность по изобретению может кодировать укороченный RPGRorf15, описанный в WO 2016/001693.
В одном варианте осуществления кодирующая RPGRorf15 последовательность по изобретению укорочена за счет удаления некоторых или всех из нуклеотидов, соответствующих положениям 2485-2940 в SEQ ID NO: 2. В соответствии с данным вариантом осуществления, кодирующая RPGRORF15 нуклеотидная последовательность предпочтительно не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 33, 58, 59, 114, 123, 129, 181, 182, 213, 219, 226, 227, 237, 267, 285, 306, 309, 315, 324, 330, 339, 400, 401, 444, 456, 478, 480, 510, 594, 606, 618, 639, 697, 726, 744, 777, 807, 852, 877, 879, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1184, 1194, 1197, 1221, 1249, 1251, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1373, 1413, 1446, 1452, 1464, 1474, 1475, 1482, 1519, 1520, 1542, 1584, 1590, 1599, 1608, 1653, 1668, 1674, 1689, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2958, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 в SEQ ID NO: 2. Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. Предпочтительная кодирующая RPGRORF15 нуклеотидная последовательность в соответствии с данным вариантом осуществления содержит или состоит из последовательности, приведенной в SEQ ID NO: 9.
Полинуклеотид по изобретению может содержать:
(a) нуклеотидную последовательность, приведенную в SEQ ID NO: 9;
(b) нуклеотидную последовательность, содержащую последовательность SEQ ID NO: 3, но с делецией, соответствующей (i) последовательности SEQ ID NO: 10, (ii) последовательности SEQ ID NO: 10 и вплоть до 75 дополнительных нуклеотидов, фланкирующих SEQ ID NO: 10 с одной или с обеих сторон SEQ ID NO: 10 в последовательности SEQ ID NO: 3, или (iii) 390 или более последовательных нуклеотидов из SEQ ID NO: 10; или
(c) нуклеотидную последовательность по пункту (a) или (b), но с укорочением с одного или с обоих из его 5' и 3'-концов длиной до 150 нуклеотидов с каждого конца.
Предпочтительно, делеция составляет по меньшей мере 400, 420 или 450 последовательных нуклеотидов из SEQ ID NO: 10.
ATGAGAGAGCCAGAGGAGCTGATGCCAGACAGTGGAGCAGTGTTTACATTCGGAAAATCTAAGTTCGCTGAAAATAACCCAGGAAAGTTCTGGTTTAAAAACGACGTGCCCGTCCACCTGTCTTGTGGCGATGAGCATAGTGCCGTGGTCACTGGGAACAATAAGCTGTACATGTTCGGGTCCAACAACTGGGGACAGCTGGGGCTGGGATCCAAATCTGCTATCTCTAAGCCAACCTGCGTGAAGGCACTGAAACCCGAGAAGGTCAAACTGGCCGCTTGTGGCAGAAACCACACTCTGGTGAGCACCGAGGGCGGGAATGTCTATGCCACCGGAGGCAACAATGAGGGACAGCTGGGACTGGGGGACACTGAGGAAAGGAATACCTTTCACGTGATCTCCTTCTTTACATCTGAGCATAAGATCAAGCAGCTGAGCGCTGGCTCCAACACATCTGCAGCCCTGACTGAGGACGGGCGCCTGTTCATGTGGGGAGATAATTCAGAGGGCCAGATTGGGCTGAAAAACGTGAGCAATGTGTGCGTCCCTCAGCAGGTGACCATCGGAAAGCCAGTCAGTTGGATTTCATGTGGCTACTATCATAGCGCCTTCGTGACCACAGATGGCGAGCTGTACGTCTTTGGGGAGCCCGAAAACGGAAAACTGGGCCTGCCTAACCAGCTGCTGGGCAATCACCGGACACCCCAGCTGGTGTCCGAGATCCCTGAAAAAGTGATCCAGGTCGCCTGCGGGGGAGAGCATACAGTGGTCCTGACTGAGAATGCTGTGTATACCTTCGGACTGGGCCAGTTTGGCCAGCTGGGGCTGGGAACCTTCCTGTTTGAGACATCCGAACCAAAAGTGATCGAGAACATTCGCGACCAGACTATCAGCTACATTTCCTGCGGAGAGAATCACACCGCACTGATCACAGACATTGGCCTGATGTATACCTTTGGCGATGGACGACACGGGAAGCTGGGACTGGGACTGGAGAACTTCACTAATCATTTTATCCCCACCCTGTGTTCTAACTTCCTGCGGTTCATCGTGAAACTGGTCGCTTGCGGCGGGTGTCACATGGTGGTCTTCGCTGCACCTCATAGGGGCGTGGCTAAGGAGATCGAATTTGACGAGATTAACGATACATGCCTGAGCGTGGCAACTTTCCTGCCATACAGCTCCCTGACTTCTGGCAATGTGCTGCAGAGAACCCTGAGTGCAAGGATGCGGAGAAGGGAGAGGGAACGCTCTCCTGACAGTTTCTCAATGCGACGAACCCTGCCACCTATCGAGGGAACACTGGGACTGAGTGCCTGCTTCCTGCCTAACTCAGTGTTTCCACGATGTAGCGAGCGGAATCTGCAGGAGTCTGTCCTGAGTGAGCAGGATCTGATGCAGCCAGAGGAACCCGACTACCTGCTGGATGAGATGACCAAGGAGGCCGAAATCGACAACTCTAGTACAGTGGAGTCCCTGGGCGAGACTACCGATATCCTGAATATGACACACATTATGTCACTGAACAGCAATGAGAAGAGTCTGAAACTGTCACCAGTGCAGAAGCAGAAGAAACAGCAGACTATTGGCGAGCTGACTCAGGACACCGCCCTGACAGAGAACGACGATAGCGATGAGTATGAGGAAATGTCCGAGATGAAGGAAGGCAAAGCTTGTAAGCAGCATGTCAGTCAGGGGATCTTCATGACACAGCCAGCCACAACTATTGAGGCTTTTTCAGACGAGGAAGTGGAGATCCCCGAGGAAAAAGAGGGCGCAGAAGATTCCAAGGGGAATGGAATTGAGGAACAGGAGGTGGAAGCCAACGAGGAAAATGTGAAAGTCCACGGAGGCAGGAAGGAGAAAACAGAAATCCTGTCTGACGATCTGACTGACAAGGCCGAGGTGTCCGAAGGCAAGGCAAAATCTGTCGGAGAGGCAGAAGACGGACCAGAGGGACGAGGGGATGGAACCTGCGAGGAAGGCTCAAGCGGGGCTGAGCATTGGCAGGACGAGGAACGAGAGAAGGGCGAAAAGGATAAAGGCCGCGGGGAGATGGAACGACCTGGAGAGGGCGAAAAAGAGCTGGCAGAGAAGGAGGAATGGAAGAAAAGGGACGGCGAGGAACAGGAGCAGAAAGAAAGGGAGCAGGGCCACCAGAAGGAGCGCAACCAGGAGATGGAAGAGGGCGGCGAGGAAGAGCATGGCGAGGGAGAAGAGGAAGAGGGCGATAGAGAAGAGGAAGAGGAAAAAGAAGGCGAAGGGAAGGAGGAAGGAGAGGGCGAGGAAGTGGAAGGCGAGAGGGAAAAGGAGGAAGGAGAACGGAAGAAAGAGGAAAGAGCCGGCAAAGAGGAAAAGGGCGAGGAAGAGGGCGATCAGGGCGAAGGCGAGGAGGAAGAGACCGAGGGCCGCGGGGAAGAGAAAGAGGAGGGAGGAGAGGTGGAGGGCGGAGAGGTCGAAGAGGGAAAGGGCGAGCGCGAAGAGGGGGAGGAAGAGGAAGGCGAAGGAGAAGGCGAGGAAGAAGAGGGAGAGGAGGAAGGCGAGGAGGAAGGAGAGGGGGAGGAGGAGGGAGAAGGCGAGGGCGAAGAAGAAGAAGAGGGAGAAGTGGAGGGCGAAGTCGAGGGGGAGGAGGGAGAAGGGGAAGGGGAGGAAGAAGAGGGCGAAGAAGAAGGCGAGGAAAGAGAAAAAGAGGGAGAAGGCGAGGAAAACCGGAGAAATAGGGAAGAGGAGGAAGAGGAAGAGGGAAAGTACCAGGAGACAGGCGAAGAGGAAAACGAGCGGCAGGATGGCGAGGAATATAAGAAAGTGAGCAAGATCAAAGGATCCGTCAAGTACGGCAAGCACAAAACCTATCAGAAGAAAAGCGTGACCAACACACAGGGGAATGGAAAAGAGCAGAGGAGTAAGATGCCTGTGCAGTCAAAACGGCTGCTGAAGAATGGCCCATCTGGAAGTAAAAAATTCTGGAACAATGTGCTGCCCCACTATCTGGAACTGAAATAA (SEQ ID NO: 9)
GAAGAGGAAGAGGGCGAGGGCGAGGAAGAAGAGGGCGAGGGGGAAGAAGAGGAGGGAGAGGGCGAAGAGGAAGAGGGGGAGGGAAAGGGCGAAGAGGAAGGAGAGGAAGGGGAGGGAGAGGAAGAGGGGGAGGAGGGCGAGGGGGAAGGCGAGGAGGAAGAAGGAGAGGGGGAAGGCGAAGAGGAAGGCGAGGGGGAAGGAGAGGAGGAAGAAGGGGAAGGCGAAGGCGAAGAGGAGGGAGAAGGAGAGGGGGAGGAAGAGGAAGGAGAAGGGAAGGGCGAGGAGGAAGGCGAAGAGGGAGAGGGGGAAGGCGAGGAAGAGGAAGGCGAGGGCGAAGGAGAGGACGGCGAGGGCGAGGGAGAAGAGGAGGAAGGGGAATGGGAAGGCGAAGAAGAGGAAGGCGAAGGCGAAGGCGAAGAAGAGGGCGAAGGGGAGGGCGAGGAGGGCGAAGGCGAA (SEQ ID NO: 10)
Кодирующая RPGRorf15 последовательность по изобретению может кодировать укороченный RPGRorf15, описанный в WO 2016/014353.
В одном варианте осуществления кодирующая RPGRorf15 последовательность по изобретению укорочена за счет удаления некоторых или всех из нуклеотидов, соответствующих положениям 2086-3027 в SEQ ID NO: 2.
В соответствии с данным вариантом осуществления, кодирующая RPGRORF15 нуклеотидная последовательность предпочтительно не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 33, 58, 59, 114, 123, 129, 181, 182, 213, 219, 226, 227, 237, 267, 285, 306, 309, 315, 324, 330, 339, 400, 401, 444, 456, 478, 480, 510, 594, 606, 618, 639, 697, 726, 744, 777, 807, 852, 877, 879, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1184, 1194, 1197, 1221, 1249, 1251, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1373, 1413, 1446, 1452, 1464, 1474, 1475, 1482, 1519, 1520, 1542, 1584, 1590, 1599, 1608, 1653, 1668, 1674, 1689, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 в SEQ ID NO: 2. Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. Предпочтительная кодирующая RPGRORF15 нуклеотидная последовательность в соответствии с данным вариантом осуществления содержит или состоит из последовательности, приведенной в SEQ ID NO: 11.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность содержит SEQ ID NO: 3, но с делецией, соответствующей некоторой части, или всей, последовательности SEQ ID NO: 12.
ATGAGAGAGCCAGAGGAGCTGATGCCAGACAGTGGAGCAGTGTTTACATTCGGAAAATCTAAGTTCGCTGAAAATAACCCAGGAAAGTTCTGGTTTAAAAACGACGTGCCCGTCCACCTGTCTTGTGGCGATGAGCATAGTGCCGTGGTCACTGGGAACAATAAGCTGTACATGTTCGGGTCCAACAACTGGGGACAGCTGGGGCTGGGATCCAAATCTGCTATCTCTAAGCCAACCTGCGTGAAGGCACTGAAACCCGAGAAGGTCAAACTGGCCGCTTGTGGCAGAAACCACACTCTGGTGAGCACCGAGGGCGGGAATGTCTATGCCACCGGAGGCAACAATGAGGGACAGCTGGGACTGGGGGACACTGAGGAAAGGAATACCTTTCACGTGATCTCCTTCTTTACATCTGAGCATAAGATCAAGCAGCTGAGCGCTGGCTCCAACACATCTGCAGCCCTGACTGAGGACGGGCGCCTGTTCATGTGGGGAGATAATTCAGAGGGCCAGATTGGGCTGAAAAACGTGAGCAATGTGTGCGTCCCTCAGCAGGTGACCATCGGAAAGCCAGTCAGTTGGATTTCATGTGGCTACTATCATAGCGCCTTCGTGACCACAGATGGCGAGCTGTACGTCTTTGGGGAGCCCGAAAACGGAAAACTGGGCCTGCCTAACCAGCTGCTGGGCAATCACCGGACACCCCAGCTGGTGTCCGAGATCCCTGAAAAAGTGATCCAGGTCGCCTGCGGGGGAGAGCATACAGTGGTCCTGACTGAGAATGCTGTGTATACCTTCGGACTGGGCCAGTTTGGCCAGCTGGGGCTGGGAACCTTCCTGTTTGAGACATCCGAACCAAAAGTGATCGAGAACATTCGCGACCAGACTATCAGCTACATTTCCTGCGGAGAGAATCACACCGCACTGATCACAGACATTGGCCTGATGTATACCTTTGGCGATGGACGACACGGGAAGCTGGGACTGGGACTGGAGAACTTCACTAATCATTTTATCCCCACCCTGTGTTCTAACTTCCTGCGGTTCATCGTGAAACTGGTCGCTTGCGGCGGGTGTCACATGGTGGTCTTCGCTGCACCTCATAGGGGCGTGGCTAAGGAGATCGAATTTGACGAGATTAACGATACATGCCTGAGCGTGGCAACTTTCCTGCCATACAGCTCCCTGACTTCTGGCAATGTGCTGCAGAGAACCCTGAGTGCAAGGATGCGGAGAAGGGAGAGGGAACGCTCTCCTGACAGTTTCTCAATGCGACGAACCCTGCCACCTATCGAGGGAACACTGGGACTGAGTGCCTGCTTCCTGCCTAACTCAGTGTTTCCACGATGTAGCGAGCGGAATCTGCAGGAGTCTGTCCTGAGTGAGCAGGATCTGATGCAGCCAGAGGAACCCGACTACCTGCTGGATGAGATGACCAAGGAGGCCGAAATCGACAACTCTAGTACAGTGGAGTCCCTGGGCGAGACTACCGATATCCTGAATATGACACACATTATGTCACTGAACAGCAATGAGAAGAGTCTGAAACTGTCACCAGTGCAGAAGCAGAAGAAACAGCAGACTATTGGCGAGCTGACTCAGGACACCGCCCTGACAGAGAACGACGATAGCGATGAGTATGAGGAAATGTCCGAGATGAAGGAAGGCAAAGCTTGTAAGCAGCATGTCAGTCAGGGGATCTTCATGACACAGCCAGCCACAACTATTGAGGCTTTTTCAGACGAGGAAGTGGAGATCCCCGAGGAAAAAGAGGGCGCAGAAGATTCCAAGGGGAATGGAATTGAGGAACAGGAGGTGGAAGCCAACGAGGAAAATGTGAAAGTCCACGGAGGCAGGAAGGAGAAAACAGAAATCCTGTCTGACGATCTGACTGACAAGGCCGAGGTGTCCGAAGGCAAGGCAAAATCTGTCGGAGAGGCAGAAGACGGACCAGAGGGACGAGGGGATGGAACCTGCGAGGAAGGCTCAAGCGGGGCTGAGCATTGGCAGGACGAGGAACGAGAGAAGGGCGAAAAGGATAAAGGCCGCGGGGAGATGGAACGACCTGGAGAGGGCGAAAAAGAGGAAGGCGAGGGCGAAGAAGAAGAAGAGGGAGAAGTGGAGGGCGAAGTCGAGGGGGAGGAGGGAGAAGGGGAAGGGGAGGAAGAAGAGGGCGAAGAAGAAGGCGAGGAAAGAGAAAAAGAGGGAGAAGGCGAGGAAAACCGGAGAAATAGGGAAGAGGAGGAAGAGGAAGAGGGAAAGTACCAGGAGACAGGCGAAGAGGAAAACGAGCGGCAGGATGGCGAGGAATATAAGAAAGTGAGCAAGATCAAAGGATCCGTCAAGTACGGCAAGCACAAAACCTATCAGAAGAAAAGCGTGACCAACACACAGGGGAATGGAAAAGAGCAGAGGAGTAAGATGCCTGTGCAGTCAAAACGGCTGCTGAAGAATGGCCCATCTGGAAGTAAAAAATTCTGGAACAATGTGCTGCCCCACTATCTGGAACTGAAATAA (SEQ ID NO: 11)
CTGGCAGAGAAGGAGGAATGGAAGAAAAGGGACGGCGAGGAACAGGAGCAGAAAGAAAGGGAGCAGGGCCACCAGAAGGAGCGCAACCAGGAGATGGAAGAGGGCGGCGAGGAAGAGCATGGCGAGGGAGAAGAGGAAGAGGGCGATAGAGAAGAGGAAGAGGAAAAAGAAGGCGAAGGGAAGGAGGAAGGAGAGGGCGAGGAAGTGGAAGGCGAGAGGGAAAAGGAGGAAGGAGAACGGAAGAAAGAGGAAAGAGCCGGCAAAGAGGAAAAGGGCGAGGAAGAGGGCGATCAGGGCGAAGGCGAGGAGGAAGAGACCGAGGGCCGCGGGGAAGAGAAAGAGGAGGGAGGAGAGGTGGAGGGCGGAGAGGTCGAAGAGGGAAAGGGCGAGCGCGAAGAGGAAGAGGAAGAGGGCGAGGGCGAGGAAGAAGAGGGCGAGGGGGAAGAAGAGGAGGGAGAGGGCGAAGAGGAAGAGGGGGAGGGAAAGGGCGAAGAGGAAGGAGAGGAAGGGGAGGGAGAGGAAGAGGGGGAGGAGGGCGAGGGGGAAGGCGAGGAGGAAGAAGGAGAGGGGGAAGGCGAAGAGGAAGGCGAGGGGGAAGGAGAGGAGGAAGAAGGGGAAGGCGAAGGCGAAGAGGAGGGAGAAGGAGAGGGGGAGGAAGAGGAAGGAGAAGGGAAGGGCGAGGAGGAAGGCGAAGAGGGAGAGGGGGAAGGCGAGGAAGAGGAAGGCGAGGGCGAAGGAGAGGACGGCGAGGGCGAGGGAGAAGAGGAGGAAGGGGAATGGGAAGGCGAAGAAGAGGAAGGCGAAGGCGAAGGCGAAGAAGAGGGCGAAGGGGAGGGCGAGGAGGGCGAAGGCGAAGGGGAGGAAGAGGAAGGCGAAGGAGAAGGCGAGGAAGAAGAGGGAGAGGAGGAAGGCGAGGAGGAAGGAGAGGGGGAGGAGGAGGGA (SEQ ID NO: 12)
В другом варианте осуществления кодирующая RPGRorf15 последовательность по изобретению укорочена за счет удаления некоторых или всех из нуклеотидов, соответствующих положениям 2584-2961 в SEQ ID NO: 2.
В соответствии с данным вариантом осуществления, кодирующая RPGRORF15 нуклеотидная последовательность предпочтительно не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 33, 58, 59, 114, 123, 129, 181, 182, 213, 219, 226, 227, 237, 267, 285, 306, 309, 315, 324, 330, 339, 400, 401, 444, 456, 478, 480, 510, 594, 606, 618, 639, 697, 726, 744, 777, 807, 852, 877, 879, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1184, 1194, 1197, 1221, 1249, 1251, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1373, 1413, 1446, 1452, 1464, 1474, 1475, 1482, 1519, 1520, 1542, 1584, 1590, 1599, 1608, 1653, 1668, 1674, 1689, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2505, 2520, 2547, 2574, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 в SEQ ID NO: 2. Предпочтительно, нуклеотиды в этих положениях соответствуют тем, которые находятся в SEQ ID NO: 3. Предпочтительная кодирующая RPGRORF15 нуклеотидная последовательность в соответствии с данным вариантом осуществления содержит или состоит из последовательности, приведенной в SEQ ID NO: 13.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность содержит SEQ ID NO: 3, но с делецией, соответствующей части, или всей, последовательности SEQ ID NO: 14.
ATGAGAGAGCCAGAGGAGCTGATGCCAGACAGTGGAGCAGTGTTTACATTCGGAAAATCTAAGTTCGCTGAAAATAACCCAGGAAAGTTCTGGTTTAAAAACGACGTGCCCGTCCACCTGTCTTGTGGCGATGAGCATAGTGCCGTGGTCACTGGGAACAATAAGCTGTACATGTTCGGGTCCAACAACTGGGGACAGCTGGGGCTGGGATCCAAATCTGCTATCTCTAAGCCAACCTGCGTGAAGGCACTGAAACCCGAGAAGGTCAAACTGGCCGCTTGTGGCAGAAACCACACTCTGGTGAGCACCGAGGGCGGGAATGTCTATGCCACCGGAGGCAACAATGAGGGACAGCTGGGACTGGGGGACACTGAGGAAAGGAATACCTTTCACGTGATCTCCTTCTTTACATCTGAGCATAAGATCAAGCAGCTGAGCGCTGGCTCCAACACATCTGCAGCCCTGACTGAGGACGGGCGCCTGTTCATGTGGGGAGATAATTCAGAGGGCCAGATTGGGCTGAAAAACGTGAGCAATGTGTGCGTCCCTCAGCAGGTGACCATCGGAAAGCCAGTCAGTTGGATTTCATGTGGCTACTATCATAGCGCCTTCGTGACCACAGATGGCGAGCTGTACGTCTTTGGGGAGCCCGAAAACGGAAAACTGGGCCTGCCTAACCAGCTGCTGGGCAATCACCGGACACCCCAGCTGGTGTCCGAGATCCCTGAAAAAGTGATCCAGGTCGCCTGCGGGGGAGAGCATACAGTGGTCCTGACTGAGAATGCTGTGTATACCTTCGGACTGGGCCAGTTTGGCCAGCTGGGGCTGGGAACCTTCCTGTTTGAGACATCCGAACCAAAAGTGATCGAGAACATTCGCGACCAGACTATCAGCTACATTTCCTGCGGAGAGAATCACACCGCACTGATCACAGACATTGGCCTGATGTATACCTTTGGCGATGGACGACACGGGAAGCTGGGACTGGGACTGGAGAACTTCACTAATCATTTTATCCCCACCCTGTGTTCTAACTTCCTGCGGTTCATCGTGAAACTGGTCGCTTGCGGCGGGTGTCACATGGTGGTCTTCGCTGCACCTCATAGGGGCGTGGCTAAGGAGATCGAATTTGACGAGATTAACGATACATGCCTGAGCGTGGCAACTTTCCTGCCATACAGCTCCCTGACTTCTGGCAATGTGCTGCAGAGAACCCTGAGTGCAAGGATGCGGAGAAGGGAGAGGGAACGCTCTCCTGACAGTTTCTCAATGCGACGAACCCTGCCACCTATCGAGGGAACACTGGGACTGAGTGCCTGCTTCCTGCCTAACTCAGTGTTTCCACGATGTAGCGAGCGGAATCTGCAGGAGTCTGTCCTGAGTGAGCAGGATCTGATGCAGCCAGAGGAACCCGACTACCTGCTGGATGAGATGACCAAGGAGGCCGAAATCGACAACTCTAGTACAGTGGAGTCCCTGGGCGAGACTACCGATATCCTGAATATGACACACATTATGTCACTGAACAGCAATGAGAAGAGTCTGAAACTGTCACCAGTGCAGAAGCAGAAGAAACAGCAGACTATTGGCGAGCTGACTCAGGACACCGCCCTGACAGAGAACGACGATAGCGATGAGTATGAGGAAATGTCCGAGATGAAGGAAGGCAAAGCTTGTAAGCAGCATGTCAGTCAGGGGATCTTCATGACACAGCCAGCCACAACTATTGAGGCTTTTTCAGACGAGGAAGTGGAGATCCCCGAGGAAAAAGAGGGCGCAGAAGATTCCAAGGGGAATGGAATTGAGGAACAGGAGGTGGAAGCCAACGAGGAAAATGTGAAAGTCCACGGAGGCAGGAAGGAGAAAACAGAAATCCTGTCTGACGATCTGACTGACAAGGCCGAGGTGTCCGAAGGCAAGGCAAAATCTGTCGGAGAGGCAGAAGACGGACCAGAGGGACGAGGGGATGGAACCTGCGAGGAAGGCTCAAGCGGGGCTGAGCATTGGCAGGACGAGGAACGAGAGAAGGGCGAAAAGGATAAAGGCCGCGGGGAGATGGAACGACCTGGAGAGGGCGAAAAAGAGCTGGCAGAGAAGGAGGAATGGAAGAAAAGGGACGGCGAGGAACAGGAGCAGAAAGAAAGGGAGCAGGGCCACCAGAAGGAGCGCAACCAGGAGATGGAAGAGGGCGGCGAGGAAGAGCATGGCGAGGGAGAAGAGGAAGAGGGCGATAGAGAAGAGGAAGAGGAAAAAGAAGGCGAAGGGAAGGAGGAAGGAGAGGGCGAGGAAGTGGAAGGCGAGAGGGAAAAGGAGGAAGGAGAACGGAAGAAAGAGGAAAGAGCCGGCAAAGAGGAAAAGGGCGAGGAAGAGGGCGATCAGGGCGAAGGCGAGGAGGAAGAGACCGAGGGCCGCGGGGAAGAGAAAGAGGAGGGAGGAGAGGTGGAGGGCGGAGAGGTCGAAGAGGGAAAGGGCGAGCGCGAAGAGGAAGAGGAAGAGGGCGAGGGCGAGGAAGAAGAGGGCGAGGGGGAAGAAGAGGAGGGAGAGGGCGAAGAGGAAGAGGGGGAGGGAAAGGGCGAAGAGGAAGGAGAAGGCGAGGAAGAAGAGGGAGAGGAGGAAGGCGAGGAGGAAGGAGAGGGGGAGGAGGAGGGAGAAGGCGAGGGCGAAGAAGAAGAAGAGGGAGAAGTGGAGGGCGAAGTCGAGGGGGAGGAGGGAGAAGGGGAAGGGGAGGAAGAAGAGGGCGAAGAAGAAGGCGAGGAAAGAGAAAAAGAGGGAGAAGGCGAGGAAAACCGGAGAAATAGGGAAGAGGAGGAAGAGGAAGAGGGAAAGTACCAGGAGACAGGCGAAGAGGAAAACGAGCGGCAGGATGGCGAGGAATATAAGAAAGTGAGCAAGATCAAAGGATCCGTCAAGTACGGCAAGCACAAAACCTATCAGAAGAAAAGCGTGACCAACACACAGGGGAATGGAAAAGAGCAGAGGAGTAAGATGCCTGTGCAGTCAAAACGGCTGCTGAAGAATGGCCCATCTGGAAGTAAAAAATTCTGGAACAATGTGCTGCCCCACTATCTGGAACTGAAATAA (SEQ ID NO: 13)
GGAGAGGAAGGGGAGGGAGAGGAAGAGGGGGAGGAGGGCGAGGGGGAAGGCGAGGAGGAAGAAGGAGAGGGGGAAGGCGAAGAGGAAGGCGAGGGGGAAGGAGAGGAGGAAGAAGGGGAAGGCGAAGGCGAAGAGGAGGGAGAAGGAGAGGGGGAGGAAGAGGAAGGAGAAGGGAAGGGCGAGGAGGAAGGCGAAGAGGGAGAGGGGGAAGGCGAGGAAGAGGAAGGCGAGGGCGAAGGAGAGGACGGCGAGGGCGAGGGAGAAGAGGAGGAAGGGGAATGGGAAGGCGAAGAAGAGGAAGGCGAAGGCGAAGGCGAAGAAGAGGGCGAAGGGGAGGGCGAGGAGGGCGAAGGCGAAGGGGAGGAAGAGGAAGGCGAA (SEQ ID NO: 14)
Кодирующая RPGRORF15 нуклеотидная последовательность по изобретению может быть функционально связана с полинуклеотидом, содержащим элемент промотора, способный управлять экспрессией RPGRORF15 или его функционального варианта в палочковидных и колбочковидных фоторецепторных клетках человека.
Кодирующая RPGRORF15 нуклеотидная последовательность по изобретению может быть функционально связана с промотором родопсин-киназы (GRK1), предпочтительно промотором GRK1 человека.
Предпочтительно, помимо промотора, никакие дополнительные энхансерные элементы не используют для контроля экспрессии RPGRORF15. Таким образом, в одном варианте осуществления нуклеотидная последовательность по изобретению не содержит элемент энхансера. В одном варианте осуществления нуклеотидная последовательность не содержит посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE).
В другом аспекте изобретение относится к вирусному вектору, содержащему полинуклеотид по изобретению.
В одном варианте осуществления вирусный вектор представляет собой аденоассоциированный вирусный (AAV), ретровирусный, лентивирусный или аденовирусный вектор.
В предпочтительном варианте осуществления вирусный вектор имеет форму вирусной частицы.
В предпочтительном варианте осуществления вирусный вектор представляет собой AAV вектор.
AAV вектор может относиться к любому серотипу (например, содержать геном и/или капсидный белок любого серотипа AAV) при условии, что вектор способен инфицировать или трансдуцировать клетки глаза.
В одном варианте осуществления AAV вектор содержит геном AAV серотипа 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11. В другом варианте осуществления AAV вектор содержит геном AAV серотипа 2, 4, 5 или 8. Предпочтительно, AAV вектор содержит геном AAV серотипа 2.
В одном варианте осуществления AAV векторная частица содержит капсидный белок AAV серотипа 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11. В другом варианте осуществления AAV векторная частица содержит капсидный белок AAV серотипа 2, 4, 5 или 8. Предпочтительно, AAV векторная частица содержит капсидный белок AAV серотипа 8. Капсидный белок AAV серотипа 8 может, например, представлять собой AAV8/Y733F мутантный капсидный белок.
В одном варианте осуществления AAV векторная частица содержит геном AAV2 и капсидные белки AAV2 (AAV2/2); геном AAV2 и капсидные белки AAV5 (AAV2/5) или геном AAV2 и капсидные белки AAV8 (AAV2/8). Предпочтительно, AAV векторная частица содержит геном AAV2 и капсидные белки AAV8 (AAV2/8).
AAV векторная частица по изобретению может быть химерным, перетасованным или капсид-модифицированным производным одного или более природных AAV. В частности, AAV векторная частица может содержать последовательности капсидных белков из разных серотипов, клад, клонов или изолятов AAV в одном и том же векторе (то есть, представлять собой псевдотипированный вектор). Таким образом, в одном варианте осуществления AAV вектор имеет форму псевдотипированной AAV векторной частицы.
В другом аспекте изобретение относится к системе продуцирования вирусного вектора, содержащей набор полинуклеотидов, кодирующих компоненты, необходимые для продуцирования вирусного вектора, при этом геном вирусного вектора содержит полинуклеотид по изобретению.
В одном варианте осуществления вирусный вектор представляет собой аденоассоциированный вирусный (AAV), ретровирусный, лентивирусный или аденовирусный вектор. Предпочтительно, вирусный вектор представляет собой AAV вектор.
В другом аспекте изобретение относится к конструкции ДНК для применения в системе продуцирования вирусного вектора по изобретению, содержащей полинуклеотид по изобретению.
В другом аспекте изобретение относится к клетке, продуцирующей вирусный вектор, которая содержит полинуклеотид, систему продуцирования вирусного вектора или конструкцию ДНК по изобретению. Клетка, продуцирующая вирусный вектор, может, например, представлять собой клетку HEK293, HEK293T, Sf9, C12 или HeLa. Предпочтительно, клетка, продуцирующая вирусный вектор, представляет собой клетку HEK293 или HEK293T.
В другом аспекте изобретение относится к способу продуцирования вирусного вектора, включающему введение полинуклеотида по изобретению в клетку и культивирование клетки в условиях, подходящих для продуцирования вирусного вектора.
В другом аспекте изобретение относится к вирусному вектору, получаемому с применением продуцирующей вирусный вектор клетки по изобретению или способа по изобретению.
В другом аспекте изобретение относится к клетке, трансфицированной полинуклеотидом по изобретению или трансдуцированной вирусным вектором по изобретению.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей полинуклеотид, вирусный вектор или клетку по изобретению в сочетании с фармацевтически приемлемым носителем, разбавителем или эксципиентом.
В другом аспекте изобретение относится к полинуклеотиду или вирусному вектору по изобретению для применения с целью лечения или предотвращения пигментного ретинита. Изобретение также относится к полинуклеотиду или вирусному вектору по изобретению для применения с целью уменьшения гибели фоторецепторных клеток у субъекта, страдающего от, или имеющего риск развития, пигментного ретинита.
В одном варианте осуществления пигментный ретинит представляет собой X-связанный пигментный ретинит.
В одном варианте осуществления полинуклеотид или вирусный вектор вводят в глаз субъекта субретинальной, прямой ретинальной или интравитреальной инъекцией. Предпочтительно, полинуклеотид или вирусный вектор вводят в глаз субъекта субретинальной инъекцией. Субретинальную инъекцию можно выполнять методом двухэтапной субретинальной инъекции, описанным в настоящем документе.
Предпочтительно, субретинальная инъекция включает этапы:
(a) введения раствора субъекту субретинальной инъекцией в количестве, эффективном для по меньшей мере частичного отслоения сетчатки, с образованием субретинального пузыря, при этом раствор не содержит полинуклеотид или вирусный вектор; и
(b) введения лекарственной композиции субретинальной инъекцией в пузырь, образованный на этапе (a), при этом лекарственная композиция содержит полинуклеотид или вирусный вектор.
В одном варианте осуществления AAV вектор вводят субъекту одной дозой.
AAV вектор может, например, находиться в суспензии в концентрации примерно 1-2×109, 1-2×1010, 1-2×1011, 1-2×1012 или 1-2×1013 геномных частиц (гч) в мл. Таким образом, дозу AAV вектора примерно 2×1010 гч можно, например, вводить путем инъекции примерно 10 мкл суспензии AAV вектора в концентрации примерно 2×1012 гч в мл. Квалифицированный специалист с легкостью сможет корректировать дозу, объем и концентрацию AAV вектора по мере необходимости.
Объем вводимого AAV вектора может составлять, например, примерно 1-500 мкл, например, примерно 10-500, 50-500, 100-500, 200-500, 300-500, 400-500, 50-250, 100-250, 200-250, 50-150, 1-100 или 1-10 мкл. Объем может составлять, например, примерно 1, 2, 5, 10, 50, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мкл. Предпочтительно, инъецируемый объем композиции AAV вектора составляет примерно 100 мкл.
В одном варианте осуществления AAV вектор вводят в дозе по меньшей мере 2×107, 2×108, 2×109, 2×1010, 2×1011 или 2×1012 гч в каждый глаз. В другом варианте осуществления AAV вектор вводят в дозе примерно 1-2×107, 1-2×108, 1-2×109, 1-2×1010, 1-2×1011 или 1-2×1012 гч в каждый глаз. Предпочтительно, AAV вектор вводят в дозе примерно 2×1011 гч в каждый глаз, предпочтительно, субретинальной инъекцией.
В одном варианте осуществления дегенерация фоторецепторных клеток вследствие пигментного ретинита по существу предотвращается на всем протяжении жизни субъекта. Фоторецепторные клетки могут включать колбочковидные клетки и/или палочковидные клетки, предпочтительно колбочковидные клетки и палочковидные клетки. В другом варианте осуществления менее примерно 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% от количества фоторецепторных клеток (например, колбочковидных клеток и/или палочковидных клеток), находящихся в получающем лечение глазу в момент введения полинуклеотида или вирусного (например, AAV) вектора, впоследствии дегенерируют в результате пигментного ретинита на протяжении жизни субъекта. Предпочтительно, выжившие клетки остаются функциональными.
В одном варианте осуществления зрительная функция по существу восстанавливается или сохраняется в получающем лечение глазу. Зрительная функция (например, определяемая в тесте для оценки зрительной функции, описанном в настоящем документе) может, например, восстанавливаться в пораженном глазу до примерно того же уровня, который имел место до начала пигментного ретинита. Альтернативно, зрительная функция может, например, сохраняться примерно на том же уровне у здорового субъекта, имеющего риск развития пигментного ретинита, или у субъекта, уже страдающего от пигментного ретинита (например, по существу никакого ухудшения или дальнейшего ухудшения зрительной функции не происходит вследствие пигментного ретинита после введения полинуклеотида или вирусного (например, AAV) вектора по изобретению).
Без применения лечения большинство или все палочковидные клетки могут дегенерировать (например, погибать) с течением времени вследствие пигментного ретинита. Колбочковидные клетки также могут дегенерировать при прогрессировании заболевания.
В другом аспекте изобретение относится к способу лечения или предотвращения пигментного ретинита, включающему введение полинуклеотида или вирусного вектора по изобретению субъекту, который нуждается в этом. Изобретение также относится к способу уменьшения гибели фоторецепторных клеток у субъекта, страдающего от, или имеющего риск развития, пигментного ретинита, включающему введение полинуклеотида или вирусного вектора по изобретению субъекту.
Режим (например, способ и дозировка) и эффект введения, а также субъект, получающий лечение, могут быть такими, как описано в настоящем документе.
Изобретение также относится к применению полинуклеотида или вирусного вектора по изобретению для уменьшения или предотвращения образования альтернативно сплайсированных вариантов и/или укороченных белков RPGRORF15 в сравнении с вектором или полинуклеотидом, содержащим ген RPGRORF15 дикого типа.
Изобретение также относится к применению полинуклеотида или вирусного вектора по изобретению для повышения стабильности и/или точности репликации нуклеотидной последовательности, содержащей ген RPGRORF15, в сравнении с нуклеотидной последовательностью, содержащей ген RPGRORF15 дикого типа.
Изобретение также относится к применению полинуклеотида или вирусного вектора по изобретению для повышения скоростей трансляции и/или экспрессии белка RPGRORF15 в сравнении с вектором или полинуклеотидом, содержащим ген RPGRORF15 дикого типа.
Изобретение также относится к применению полинуклеотида или вирусного вектора по изобретению для увеличения продуцирования кодирующей RPGRORF15 нуклеотидной последовательности в сравнении с нуклеотидной последовательностью, содержащей ген RPGRORF15 дикого типа.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1
Сравнение геномных последовательностей RPGR человека, мыши и собаки.
Фигура 2
Сравнение кодон-оптимизированных (верх) и дикого типа (низ) последовательностей RPGRORF15. Различия в первичных последовательностях выделены красным цветом и подчеркнуты.
Фигура 3
Сравнение эффективности клонирования и точности последовательности между дикого типа (A, B) и кодон-оптимизированными (C, D) последовательностями RPGR в стандартном клонирующем векторе.
Сравнение выхода плазмид из минипрепаратов (E) и мегапрепаратов (G), а также сравнение чистоты образцов (F) между одинаковыми последовательностями.
Фигура 4
Сравнение точности последовательности между дикого типа (A) и кодон-оптимизированной (B) последовательностями RPGR в стандартном клонирующем векторе.
Фигура 5
Жидкостная хроматография с тандемной масс-спектрометрией (ЖХ/МС-МС) белкового продукта кодон-оптимизированной последовательности RPGR позволила идентифицировать примерно 80% аминокислот (аминокислоты, которые не удалось подтвердить, выделены красным цветом).
Фигура 6
Вестерн-блоты, подтверждающие идентичные молекулярные массы (220 кДа) пептидов, полученных с дикого типа и кодон-оптимизированной последовательностей RPGR (wtRPGR и coRPGR, соответственно) (верхняя панель).
График различий в интенсивности сигналов вестерн-блоттинга показывает примерно в 4 раза более высокое продуцирование белка RPGR с последовательности coRPGR (нижняя панель).
Фигура 7
Обзор эффекта воздействия при использовании AAV.coRPGR в мышиной модели с естественной мутацией в RPGR (C57BL/6Rd9/Boc).
Вестерн-блоттинг суммарных лизатов сетчатки через 6 месяцев после инъекции показывает наличие белкового продукта RPGR массой примерно 200 кДа в получавшем инъекцию глазу (TE), но не в глазу, не получавшем инъекцию (UE), у мышей C57BL/6Rd9/Boc (A). L, белки-маркеры молекулярной массы; +, положительный контроль (белковый лизат клеток HEK293T, трансфицированных экспрессионной плазмидой coRPGR); -, отрицательный контроль (белковый лизат не трансфицированных клеток HEK293T).
Иммуногистохимический анализ нефиксированных срезов замороженных тканей через 6 месяцев после инъекции (B). Верхняя панель демонстрирует отсутствие окрашивания на RPGR (зеленый) в не получавшем инъекцию глазу. В получавшем инъекцию глазу (низ) видно, что окрашенный RPGR совместно локализован с цилиарным белком Rpgrip (красный). Масштабная полоска=20 мкм.
ЭРГ при однократных вспышках света разной интенсивности после темновой адаптации (ТА) глаз через 3 (C) и 6 (D) месяцев после инъекции. Кривые красного цвета соответствуют средним значениям для получавшего инъекцию глаза (TE), кривые черного цвета отражают результаты для не получавшего инъекцию глаза (UE). Планки погрешностей показывают 95% доверительный интервал.
Фигура 8
Идентичность и структурная стабильность плазмиды, плазмида pAAV.RK.coRPGR получена из RCB. Размеры фрагментов после расщепления эндонуклеазами рестрикции плазмидной ДНК, полученной из RCB, для pAAV.RK.coRPGR. Ожидаемая картина при расщеплении ферментом XmnI: 11+11+161+211+2681+4006 п.н.; SmaI: 11+11+161+211+2681+4006 п.н. (фрагменты размером 11 п.н. проходят через агарозный гель и их невозможно визуализировать).
Набор маркеров, отличающихся 1 т.п.н. (PlasmidFactory, каталожный № MSM-865-50), 300 нг
Дорожка 1 ДНК RCB1729-151023, нерасщепленная, 250 нг
Дорожка 2 ДНК RCB1729-151023, нерасщепленная, 250 нг
Дорожка 3 ДНК RCB1729-151023, расщепленная XmnI, 250 нг
Дорожка 4 ДНК RCB1729-151023, расщепленная SmaI, 250 нг.
Фигура 9
Идентичность и структурная стабильность плазмиды для 100 мг препарата плазмиды pAAV.RK.coRPGR. Размеры фрагментов после расщепления эндонуклеазами рестрикции для плазмиды pAAV.RK.RPGR. Ожидаемая картина при расщеплении ферментом XmnI: 11+11+161+211+2681+4006 п.н. (фрагменты размером 11 п.н. проходят через агарозный гель и их невозможно визуализировать).
Набор маркеров, отличающихся 1 т.п.н. (PlasmidFactory, каталожный № MSM-865-50), 300 нг
Дорожка 1 pAAV.RK.coRPGR, 250 нг
Дорожка 2 pAAV.RK.coRPGR, 250 нг
Набор маркеров, отличающихся 1 т.п.н. (PlasmidFactory, каталожный № MSM-865-50), 300 нг
Дорожка 3 pAAV.RK.coRPGR, расщепленная XmaI, 250 нг
Дорожка 4 pAAV.RK.co.RPGR, расщепленная XmaI, 250 нг
Набор маркеров, отличающихся 1 т.п.н. (PlasmidFactory, каталожный № MSM-865-50), 300 нг.
Фигура 10
Концентрация плазмидной ДНК, определенная на основании A260, для 100 мг препарата плазмиды pAAV.RK.coRPGR. Рассчитанная усредненная концентрация: 1,0 мг мл-1.
Фигура 11
Чистота ДНК. Метод: УФ-сканирование при длине волны от 220 нм до 320 нм. Определение чистоты плазмиды методом УФ-сканирования при длине волны от 220 нм до 320 нм для 100 мг препарата плазмиды pAAV.RK.coRPGR.
Фигура 12
Вестерн-блот анализ экспрессии RPGRORF15 в клетках HEK293. (A) Экспрессию RPGRORF15 (черная стрелка) определяли в клетках, трансфицированных либо CAG.coRPGRORF15 (ко), либо CAG.wtRPGRORF15 (дт), в сравнении с не трансфицированными образцами (Н/Т) в трех независимых экспериментах. (B) Коробчатая диаграмма показывает уровни экспрессии RPGRORF15, измеренные методом денситометрии и выраженные в условных единицах (УЕ), после нормирования на эндогенный контроль (β-актин).
Фигура 13
Вестерн-блот анализ экспрессии RPGRORF15 в клетках HEK293T. (A) Экспрессию RPGRORF15 (черная стрелка) определяли в клетках, трансфицированных либо CAG.coRPGRORF15 (ко), либо CAG.wtRPGRORF15 (дт), в сравнении с не трансфицированными образцами (Н/Т). Укороченный белок (белая стрелка) обнаруживали в клетках, трансфицированных последовательностью дт. (B) Гистограмма показывает уровень экспрессии RPGRORF15, измеренный методом денситометрии, после нормирования на эндогенный контроль (β-актин) в каждом образце.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Различные предпочтительные признаки и варианты осуществления изобретения далее описаны с помощью неограничивающих примеров.
При осуществлении на практике изобретения будут использованы, если нет иных указаний, общепринятые методы химии, биохимии, молекулярной биологии, микробиологии и иммунологии, которые находятся в пределах компетенции специалиста в данной области. Такие методы описаны в литературе. Смотри, например, Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2-е издание, Cold Spring Harbor Laboratory Press; Ausubel, F.M. et al. (1995, и периодические дополнения) Current Protocols in Molecular Biology, Ch. 9, 13 и 16, John Wiley & Sons; Roe, B., Crabtree, J. and Kahn, A. (1996) DNA Isolation and Sequencing: Essential Techniques, John Wiley & Sons; Polak, J.M. and McGee, J.O'D. (1990) In Situ Hybridization: Principles and Practice, Oxford University Press; Gait, M.J. (1984) Oligonucleotide Synthesis: A Practical Approach, IRL Press; и Lilley, D.M. and Dahlberg, J.E. (1992) Methods in Enzymology: DNA Structures Part A: Synthesis and Physical Analysis of DNA, Academic Press. Каждый из этих литературных источников общего характера включен в настоящий документ посредством ссылки.
В одном аспекте изобретение относится к полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15), при этом кодирующая RPGRORF15 нуклеотидная последовательность была кодон-оптимизирована для повышения точности репликации последовательности.
Регулятор ГТФазы пигментного ретинита (RPGR)
Регулятор ГТФазы пигментного ретинита (RPGR), вероятно, действует как рилизинг-фактор гуанин-нуклеотидов и имеет важное значение для нормального зрения.
Результаты исследований свидетельствуют о том, что RPGR играет определенную роль в образовании ресничек, возможно, за счет участия в организации микротрубочек и регуляции транспорта. Реснички представляют собой пальцеобразные выросты на поверхности клетки, которые могут быть вовлечены в целый ряд видов биологической активности, включая сигнализацию и движение клеток. Реснички также необходимы для целого ряда сенсорных восприятий, включая слух, обоняние и зрение, и они являются ключевыми органеллами фоторецепторных клеток.
Несколько изоформ белка RPGR экспрессируются с гена RPGR. Изоформа, содержащая экзон ORF15, имеет особое значение для настоящего изобретения, и ее иногда называют ORF15. В настоящем документе данную изоформу называют RPGRORF15. RPGRORF15 экспрессируется преимущественно в сетчатке, в частности, в фоторецепторных клетках, в то время как другие изоформы экспрессируются в других зонах и, вероятно, также участвуют в образовании и/или функции ресничек.
Примером аминокислотной последовательности человеческого белка RPGRORF15 дикого типа является:
MREPEELMPDSGAVFTFGKSKFAENNPGKFWFKNDVPVHLSCGDEHSAVVTGNNKLYMFGSNNWGQLGLG
SKSAISKPTCVKALKPEKVKLAACGRNHTLVSTEGGNVYATGGNNEGQLGLGDTEERNTFHVISFFTSEH
KIKQLSAGSNTSAALTEDGRLFMWGDNSEGQIGLKNVSNVCVPQQVTIGKPVSWISCGYYHSAFVTTDGE
LYVFGEPENGKLGLPNQLLGNHRTPQLVSEIPEKVIQVACGGEHTVVLTENAVYTFGLGQFGQLGLGTFL
FETSEPKVIENIRDQTISYISCGENHTALITDIGLMYTFGDGRHGKLGLGLENFTNHFIPTLCSNFLRFI
VKLVACGGCHMVVFAAPHRGVAKEIEFDEINDTCLSVATFLPYSSLTSGNVLQRTLSARMRRRERERSPD
SFSMRRTLPPIEGTLGLSACFLPNSVFPRCSERNLQESVLSEQDLMQPEEPDYLLDEMTKEAEIDNSSTV
ESLGETTDILNMTHIMSLNSNEKSLKLSPVQKQKKQQTIGELTQDTALTENDDSDEYEEMSEMKEGKACK
QHVSQGIFMTQPATTIEAFSDEEVEIPEEKEGAEDSKGNGIEEQEVEANEENVKVHGGRKEKTEILSDDL
TDKAEVSEGKAKSVGEAEDGPEGRGDGTCEEGSSGAEHWQDEEREKGEKDKGRGEMERPGEGEKELAEKE
EWKKRDGEEQEQKEREQGHQKERNQEMEEGGEEEHGEGEEEEGDREEEEEKEGEGKEEGEGEEVEGEREK
EEGERKKEERAGKEEKGEEEGDQGEGEEEETEGRGEEKEEGGEVEGGEVEEGKGEREEEEEEGEGEEEEG
EGEEEEGEGEEEEGEGKGEEEGEEGEGEEEGEEGEGEGEEEEGEGEGEEEGEGEGEEEEGEGEGEEEGEG
EGEEEEGEGKGEEEGEEGEGEGEEEEGEGEGEDGEGEGEEEEGEWEGEEEEGEGEGEEEGEGEGEEGEGE
GEEEEGEGEGEEEEGEEEGEEEGEGEEEGEGEGEEEEEGEVEGEVEGEEGEGEGEEEEGEEEGEEREKEG
EGEENRRNREEEEEEEGKYQETGEEENERQDGEEYKKVSKIKGSVKYGKHKTYQKKSVTNTQGNGKEQRS
KMPVQSKRLLKNGPSGSKKFWNNVLPHYLELK (SEQ ID NO: 1).
В одном варианте осуществления нуклеотидной последовательностью, кодирующей человеческий белок RPGRORF15 дикого типа, является:
ATGAGGGAGCCGGAAGAGCTGATGCCCGATTCGGGTGCTGTGTTTACATTTGGGAAAAGTAAATTTGCTG
AAAATAATCCCGGTAAATTCTGGTTTAAAAATGATGTCCCTGTACATCTTTCATGTGGAGATGAACATTC
TGCTGTTGTTACCGGAAATAATAAACTTTACATGTTTGGCAGTAACAACTGGGGTCAGTTAGGATTAGGA
TCAAAGTCAGCCATCAGCAAGCCAACATGTGTCAAAGCTCTAAAACCTGAAAAAGTGAAATTAGCTGCCT
GTGGAAGGAACCACACCCTGGTGTCAACAGAAGGAGGCAATGTATATGCAACTGGTGGAAATAATGAAGG
ACAGTTGGGGCTTGGTGACACCGAAGAAAGAAACACTTTTCATGTAATTAGCTTTTTTACATCCGAGCAT
AAGATTAAGCAGCTGTCTGCTGGATCTAATACTTCAGCTGCCCTAACTGAGGATGGAAGACTTTTTATGT
GGGGTGACAATTCCGAAGGGCAAATTGGTTTAAAAAATGTAAGTAATGTCTGTGTCCCTCAGCAAGTGAC
CATTGGGAAACCTGTCTCCTGGATCTCTTGTGGATATTACCATTCAGCTTTTGTAACAACAGATGGTGAG
CTATATGTGTTTGGAGAACCTGAGAATGGGAAGTTAGGTCTTCCCAATCAGCTCCTGGGCAATCACAGAA
CACCCCAGCTGGTGTCTGAAATTCCGGAGAAGGTGATCCAAGTAGCCTGTGGTGGAGAGCATACTGTGGT
TCTCACGGAGAATGCTGTGTATACCTTTGGGCTGGGACAATTTGGTCAGCTGGGTCTTGGCACTTTTCTT
TTTGAAACTTCAGAACCCAAAGTCATTGAGAATATTAGGGATCAAACAATAAGTTATATTTCTTGTGGAG
AAAATCACACAGCTTTGATAACAGATATCGGCCTTATGTATACTTTTGGAGATGGTCGCCACGGAAAATT
AGGACTTGGACTGGAGAATTTTACCAATCACTTCATTCCTACTTTGTGCTCTAATTTTTTGAGGTTTATA
GTTAAATTGGTTGCTTGTGGTGGATGTCACATGGTAGTTTTTGCTGCTCCTCATCGTGGTGTGGCAAAAG
AAATTGAATTCGATGAAATAAATGATACTTGCTTATCTGTGGCGACTTTTCTGCCGTATAGCAGTTTAAC
CTCAGGAAATGTACTGCAGAGGACTCTATCAGCACGTATGCGGCGAAGAGAGAGGGAGAGGTCTCCAGAT
TCTTTTTCAATGAGGAGAACACTACCTCCAATAGAAGGGACTCTTGGCCTTTCTGCTTGTTTTCTCCCCA
ATTCAGTCTTTCCACGATGTTCTGAGAGAAACCTCCAAGAGAGTGTCTTATCTGAACAGGACCTCATGCA
GCCAGAGGAACCAGATTATTTGCTAGATGAAATGACCAAAGAAGCAGAGATAGATAATTCTTCAACTGTA
GAAAGCCTTGGAGAAACTACTGATATCTTAAACATGACACACATCATGAGCCTGAATTCCAATGAAAAGT
CATTAAAATTATCACCAGTTCAGAAACAAAAGAAACAACAAACAATTGGGGAACTGACGCAGGATACAGC
TCTTACTGAAAACGATGATAGTGATGAATATGAAGAAATGTCAGAAATGAAAGAAGGGAAAGCATGTAAA
CAACATGTGTCACAAGGGATTTTCATGACGCAGCCAGCTACGACTATCGAAGCATTTTCAGATGAGGAAG
TAGAGATCCCAGAGGAGAAGGAAGGAGCAGAGGATTCAAAAGGAAATGGAATAGAGGAGCAAGAGGTAGA
AGCAAATGAGGAAAATGTGAAGGTGCATGGAGGAAGAAAGGAGAAAACAGAGATCCTATCAGATGACCTT
ACAGACAAAGCAGAGGTGAGTGAAGGCAAGGCAAAATCAGTGGGAGAAGCAGAGGATGGGCCTGAAGGTA
GAGGGGATGGAACCTGTGAGGAAGGTAGTTCAGGAGCAGAACACTGGCAAGATGAGGAGAGGGAGAAGGG
GGAGAAAGACAAGGGTAGAGGAGAAATGGAGAGGCCAGGAGAGGGAGAGAAGGAACTAGCAGAGAAGGAA
GAATGGAAGAAGAGGGATGGGGAAGAGCAGGAGCAAAAGGAGAGGGAGCAGGGCCATCAGAAGGAAAGAA
ACCAAGAGATGGAGGAGGGAGGGGAGGAGGAGCATGGAGAAGGAGAAGAAGAGGAGGGAGACAGAGAAGA
GGAAGAAGAGAAGGAGGGAGAAGGGAAAGAGGAAGGAGAAGGGGAAGAAGTGGAGGGAGAACGTGAAAAG
GAGGAAGGAGAGAGGAAAAAGGAGGAAAGAGCGGGGAAGGAGGAGAAAGGAGAGGAAGAAGGAGACCAAG
GAGAGGGGGAAGAGGAGGAAACAGAGGGGAGAGGGGAGGAAAAAGAGGAGGGAGGGGAAGTAGAGGGAGG
GGAAGTAGAGGAGGGGAAAGGAGAGAGGGAAGAGGAAGAGGAGGAGGGTGAGGGGGAAGAGGAGGAAGGG
GAGGGGGAAGAGGAGGAAGGGGAGGGGGAAGAGGAGGAAGGAGAAGGGAAAGGGGAGGAAGAAGGGGAAG
AAGGAGAAGGGGAGGAAGAAGGGGAGGAAGGAGAAGGGGAGGGGGAAGAGGAGGAAGGAGAAGGGGAGGG
AGAAGAGGAAGGAGAAGGGGAGGGAGAAGAGGAGGAAGGAGAAGGGGAGGGAGAAGAGGAAGGAGAAGGG
GAGGGAGAAGAGGAGGAAGGAGAAGGGAAAGGGGAGGAGGAAGGAGAGGAAGGAGAAGGGGAGGGGGAAG
AGGAGGAAGGAGAAGGGGAAGGGGAGGATGGAGAAGGGGAGGGGGAAGAGGAGGAAGGAGAATGGGAGGG
GGAAGAGGAGGAAGGAGAAGGGGAGGGGGAAGAGGAAGGAGAAGGGGAAGGGGAGGAAGGAGAAGGGGAG
GGGGAAGAGGAGGAAGGAGAAGGGGAGGGGGAAGAGGAGGAAGGGGAAGAAGAAGGGGAGGAAGAAGGAG
AGGGAGAGGAAGAAGGGGAGGGAGAAGGGGAGGAAGAAGAGGAAGGGGAAGTGGAAGGGGAGGTGGAAGG
GGAGGAAGGAGAGGGGGAAGGAGAGGAAGAGGAAGGAGAGGAGGAAGGAGAAGAAAGGGAAAAGGAGGGG
GAAGGAGAAGAAAACAGGAGGAACAGAGAAGAGGAGGAGGAAGAAGAGGGGAAGTATCAGGAGACAGGCG
AAGAAGAGAATGAAAGGCAGGATGGAGAGGAGTACAAAAAAGTGAGCAAAATAAAAGGATCTGTGAAATA
TGGCAAACATAAAACATATCAAAAAAAGTCAGTTACTAACACACAGGGAAATGGGAAAGAGCAGAGGTCC
AAAATGCCAGTCCAGTCAAAACGACTTTTAAAAAACGGGCCATCAGGTTCCAAAAAGTTCTGGAATAATG
TATTACCACATTACTTGGAATTGAAGTAA (SEQ ID NO: 2).
Более 300 мутаций в гене RPGR имеют отношение к X-связанному пигментному ретиниту (XLRP). Кроме того, мутации RPGR имеют место примерно в 70% случаев XLRP. Большинство ассоциированных с XLRP мутаций RPGR имеют место в экзоне ORF15 (который соответствует нуклеотидам 1754-3459 в RPGRORF15, например, нуклеотидам 1754-3459 в SEQ ID NO: 2) и, как правило, приводят к образованию укороченного, нефункционального белка. Мутация в RPGR, по всей вероятности, приводит к нарушению нормальной функции ресничек фоторецепторных клеток, однако неясно, каким образом это приводит к постепенной утрате фоторецепторов и, как следствие, проблемам со зрением, характерным для этого заболевания.
RPGR, как правило, связан с развитием RP из-за высоко мутагенной области в богатой пуринами области экзона 15 гена RPGR. Ожидается, что аналогичные проблемы возникнут при клонировании в AAV векторе, если для этой цели использовать нуклеотидную последовательность RPGR дикого типа. Для решения этой проблемы авторы изобретения использовали кодон-оптимизацию для добавления пиримидиновых нуклеотидов с целью прерывания повторяющихся последовательностей GA в экзоне 15 гена RPGR и предотвращения образования других потенциальных сайтов сплайсинга в дт кДНК, которые могут быть причиной большой доли описанных укороченных вариантов RPGR (Wu et al., Human Molecular Genetics 2015: 24(14); 3956-70). Последовательность coRPGR по настоящему изобретению была разработана для устранения последовательностей CpG, скрытых сайтов сплайсинга и аномальных сигналов полиA.
Кодон-оптимизация
Кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована относительно последовательности гена дикого типа, например, нуклеотидной последовательности SEQ ID NO: 2.
Вследствие этого, кодон-оптимизированный ген RPGR по настоящему изобретению, полученный с использованием стратегии оптимизации, раскрытой в настоящем документе, является более стабильным, чем последовательность кДНК дикого типа, и при этом устраняются проблемы, связанные с дт RPGR, которые могут приводить к образованию альтернативно сплайсированных вариантов и укороченных белков в случае, если последовательность дт RPGR будет вновь введена в аппарат транскрипции методами генной терапии (Wu et al., Human Molecular Genetics 2015: 24(14); 3956-70).
Кодон-оптимизация основана на том преимуществе, которое дает вырожденность генетического кода, что позволяет изменять нуклеотидную последовательность, при этом сохраняя ту же самую аминокислотную последовательность закодированного белка.
Как правило, кодон-оптимизацию выполняют с целью увеличения или уменьшения экспрессии закодированного белка. Это достигается за счет адаптации использования кодонов в нуклеотидной последовательности к характерному использованию кодонов в клетке конкретного типа, таким образом, используется преимущество предпочтительного использования кодонов в клетке и, соответственно, преимущество относительно большей представленности конкретных тРНК в клетке данного типа. За счет изменения кодонов в нуклеотидной последовательности таким образом, что они становятся адаптированы к относительному избытку соответствующих тРНК, можно добиваться увеличения экспрессии. И наоборот, можно добиваться уменьшения экспрессии за счет выбора кодонов, для которых, как известно, соответствующие тРНК являются более редкими в клетке конкретного типа.
Однако кодон-оптимизация по изобретению не предназначена конкретно для оказания влияния на уровни экспрессии в клетке. Вместо этого, в изобретении использовано преимущество, которое дает вырожденность генетического кода, для модификации предрасположенной к мутациям нуклеотидной последовательности RPGRORF15 дикого типа (например, SEQ ID NO: 2) с целью создания последовательности, отличающейся повышенной точностью репликации (то есть, менее предрасположенной к мутациям, возникающим во время циклов репликации полинуклеотида, например, в процессе клонирования или в процессе естественного клеточного цикла) в сравнении с последовательностью дикого типа (например, SEQ ID NO: 2).
Кроме того, авторы настоящего изобретения показали, что: (i) в случае кодон-оптимизированной последовательности может быть предотвращено образование альтернативно сплайсированных вариантов, которые наблюдались в конструкциях не кодон-оптимизированной кДНК RPGR человека; и (ii) уровни экспрессии белка с кодон-оптимизированной последовательности превышают таковые в случае последовательности дикого типа, что делает конструкцию более эффективной для общей трансляции (без привязки к конкретной теории, это может быть следствием сокращения числа повторяющихся кодонов GAN, которые используют клеточный пул тРНК для глутамата и глицина).
Область ORF15 дикого типа содержит намного меньше нуклеотидов T и C, чем можно предсказать на основании генома. Например, последовательность из 750 пар нуклеотидов между положениями 2410 и 3160 совсем не содержит нуклеотидов C в последовательности дикого типа. Это приводит к появлению большого количества повторяющихся последовательностей, которые могут ошибочно рекомбинировать в процессе клонирования и продуцирования вектора. Поскольку большинство кодонов начинаются с G в положении 1 в этой области, в стратегии кодон-оптимизации по настоящему изобретению было проведено добавление нуклеотидов C в положение 3 предшествующего кодона с учетом ограничения числа динуклеотидов CpG. Причина этого заключается в том, что слишком большое количество этих динуклеотидов может приводить к идентификации трансгенной ДНК как неприродной, результатом чего может стать подверженность основанному на метилировании сайленсингу нуклеотида C. Например, в последовательности из 750 пар нуклеотидов между положениями 2410 и 3160 в SEQ ID NO: 3 было добавлено в общей сложности 45 нуклеотидов C в процессе кодон-оптимизации, что привело к появлению 45 динуклеотидов CpG (6,00%). Это выгодно отличается от предсказанной частоты встречаемости данных динуклеотидов в последовательности дикого типа (6,25%).
Дополнительная причина ограничения количества динуклеотидов CpG при кодон-оптимизации последовательности RPGRORF15 заключается в том, что эукариотические клетки имеют эволюционно сохраняющиеся механизмы врожденного иммунитета, которые служат им (или окружающим клеткам) в качестве защитных механизмов, например, от вирусных инфекций (Willett et al., Frontiers in immunology 2013;4:261). Неметилированные CpG-островки могут быть идентифицированы клетками-хозяевами как вирусная ДНК, в результате будут запущены пути (сигнализации Toll-/Nod-/RIG I-подобного рецептора и так далее), потенциально приводящие к программируемой клеточной гибели (Krieg et al. The Journal of laboratory and clinical medicine 1996;128:128-133).
Кроме того, неметилированные динуклеотиды CpG могут стимулировать иммунный ответ, приводящий к воспалению.
Помимо этого, кодон-оптимизация в отношении C была применена, где это возможно, к четырехкратно вырожденному кодону «GGN», кодирующему глицин. Это было выполнено, поскольку любое последующее метилирование нуклеотида C динуклеотида CpG в трансгене и последующее дезаминирование в тимин (T), даже если и произойдет, не приведет к изменению белковой последовательности RPGR, поскольку как GGC, так и GGT, кодируют глицин.
Вставка нуклеотидов T в богатую GA область RPGR также была ограничена в нуклеотидной последовательности по изобретению, во избежание создания аномальных сигналов полиA (например, AATAAA) и возможных донорских сайтов сплайсинга (GT) в процессе кодон-оптимизации по настоящему изобретению. Следует избегать создания донорских сайтов сплайсинга, поскольку данная область содержит множество акцепторных последовательностей сплайсинга (AG) с повторяющимися пиримидиновыми нуклеотидами G и потенциальными A-нуклеотидными точками ветвления в 5′-направлении. Схема кодон-оптимизации была тщательно смоделирована in silico с целью определения оптимальной модификации для уменьшения числа GA-повторов, а также для уменьшения риска аномального сплайсинга и создания преждевременных сигналов полиA. Вследствие этого, кодон-оптимизированный ген по настоящему изобретению, созданный с использованием стратегии оптимизации, раскрытой в настоящем документе, является более стабильным, чем последовательность кДНК дикого типа, в случае которой могут возникать альтернативно сплайсированные варианты и укороченные белки при ее повторном введении в аппарат транскрипции методами генной терапии (Wu et al., Human Molecular Genetics 2015: 24(14); 3956-70). Таким образом, кодон-оптимизированный ген по настоящему изобретению позволяет избегать вызывающих заболевание проблем, связанных с дт RPGR.
Принципы, которые были разработаны и подтверждены авторами настоящего изобретения, можно адаптировать и применять для создания вариантов SEQ ID NO: 3, имеющих аналогичные полезные характеристики. Соответственно, изобретение не следует рассматривать, как ограниченное конкретной последовательностью SEQ ID NO: 3.
Точность репликации можно определять любым из целого ряда методов, известных специалисту в данной области.
Например, кодирующую RPGRORF15 нуклеотидную последовательность можно амплифицировать методом ПЦР и лигировать в стандартный клонирующий вектор. Продукт лигирования затем можно вводить трансформацией в линию клеток (например, стандартного штамма E. coli) и ряд полученных колоний трансформантов можно анализировать для определения содержащейся в них нуклеотидной последовательности гена RPGRORF15. Результаты секвенирования можно сравнивать между разными кодирующими RPGRORF15 нуклеотидными последовательностями (например, в том числе, эталонной ожидаемой последовательностью) для определения доли протестированных колоний, которые содержат мутантные и не мутантные последовательности.
В одном варианте осуществления менее 9, например, менее 8, 7, 6, 5, 4, 3 или 2, мутаций присутствуют в кодирующей RPGRORF15 нуклеотидной последовательности, когда полинуклеотид, содержащий последовательность (например, полинуклеотид, выделенный из колонии клеток, такой как колония E. coli, например, путем выделения плазмиды, содержащей последовательность, из колонии) анализируют путем секвенирования полинуклеотида, например, секвенирования методом Сэнгера. Предпочтительно, при секвенировании кодирующей RPGRORF15 нуклеотидной последовательности обнаруживают 0 мутаций.
В другом варианте осуществления менее 25%, например, менее 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1%, протестированных клонов (то есть, полинуклеотидов, содержащих кодирующую RPGRORF15 нуклеотидную последовательность по изобретению, выделенных из колонии клеток, например, плазмид, выделенных из колонии E. coli) содержат кодирующую RPGRORF15 нуклеотидную последовательность, имеющую по меньшей мере одну мутацию, например, при анализе 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 100, 200, 300, 400 или 500 клонов. Предпочтительно, 0% протестированных клонов содержат кодирующую RPGRORF15 нуклеотидную последовательность, имеющую по меньшей мере одну мутацию.
В одном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению была кодон-оптимизирована для уменьшения числа пуриновых нуклеотидов в сравнении с последовательностью дикого типа (например, SEQ ID NO: 2).
Аденин и гуанин являются двумя пуриновыми нуклеотидами, присутствующими в кодирующей RPGRORF15 нуклеотидной последовательности дикого типа.
В одном варианте осуществления число пуриновых нуклеотидов уменьшено в богатых пуринами (то есть, богатых GA) областях кодирующей RPGRORF15 нуклеотидной последовательности, например, области экзона ORF 15.
В одном варианте осуществления число пуриновых нуклеотидов уменьшено на по меньшей мере 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5% или 5% от числа пуриновых нуклеотидов в последовательности дикого типа (например, SEQ ID NO: 2).
В другом варианте осуществления число пуриновых нуклеотидов уменьшено на 0,5-10%, 0,5-7,5%, 0,5-5%, 0,5-4,5%, 0,5-4%, 0,5-3,5%, 0,5-3%, 1-5%, 1-4,5%, 1-4%, 1-3,5% или 1-3% от числа пуриновых нуклеотидов в последовательности дикого типа (например, SEQ ID NO: 2).
Примером кодон-оптимизированной последовательности RPGRORF15 по изобретению является:
ATGAGAGAGCCAGAGGAGCTGATGCCAGACAGTGGAGCAGTGTTTACATTCGGAAAATCTAAGTTCGCTG
AAAATAACCCAGGAAAGTTCTGGTTTAAAAACGACGTGCCCGTCCACCTGTCTTGTGGCGATGAGCATAG
TGCCGTGGTCACTGGGAACAATAAGCTGTACATGTTCGGGTCCAACAACTGGGGACAGCTGGGGCTGGGA
TCCAAATCTGCTATCTCTAAGCCAACCTGCGTGAAGGCACTGAAACCCGAGAAGGTCAAACTGGCCGCTT
GTGGCAGAAACCACACTCTGGTGAGCACCGAGGGCGGGAATGTCTATGCCACCGGAGGCAACAATGAGGG
ACAGCTGGGACTGGGGGACACTGAGGAAAGGAATACCTTTCACGTGATCTCCTTCTTTACATCTGAGCAT
AAGATCAAGCAGCTGAGCGCTGGCTCCAACACATCTGCAGCCCTGACTGAGGACGGGCGCCTGTTCATGT
GGGGAGATAATTCAGAGGGCCAGATTGGGCTGAAAAACGTGAGCAATGTGTGCGTCCCTCAGCAGGTGAC
CATCGGAAAGCCAGTCAGTTGGATTTCATGTGGCTACTATCATAGCGCCTTCGTGACCACAGATGGCGAG
CTGTACGTCTTTGGGGAGCCCGAAAACGGAAAACTGGGCCTGCCTAACCAGCTGCTGGGCAATCACCGGA
CACCCCAGCTGGTGTCCGAGATCCCTGAAAAAGTGATCCAGGTCGCCTGCGGGGGAGAGCATACAGTGGT
CCTGACTGAGAATGCTGTGTATACCTTCGGACTGGGCCAGTTTGGCCAGCTGGGGCTGGGAACCTTCCTG
TTTGAGACATCCGAACCAAAAGTGATCGAGAACATTCGCGACCAGACTATCAGCTACATTTCCTGCGGAG
AGAATCACACCGCACTGATCACAGACATTGGCCTGATGTATACCTTTGGCGATGGACGACACGGGAAGCT
GGGACTGGGACTGGAGAACTTCACTAATCATTTTATCCCCACCCTGTGTTCTAACTTCCTGCGGTTCATC
GTGAAACTGGTCGCTTGCGGCGGGTGTCACATGGTGGTCTTCGCTGCACCTCATAGGGGCGTGGCTAAGG
AGATCGAATTTGACGAGATTAACGATACATGCCTGAGCGTGGCAACTTTCCTGCCATACAGCTCCCTGAC
TTCTGGCAATGTGCTGCAGAGAACCCTGAGTGCAAGGATGCGGAGAAGGGAGAGGGAACGCTCTCCTGAC
AGTTTCTCAATGCGACGAACCCTGCCACCTATCGAGGGAACACTGGGACTGAGTGCCTGCTTCCTGCCTA
ACTCAGTGTTTCCACGATGTAGCGAGCGGAATCTGCAGGAGTCTGTCCTGAGTGAGCAGGATCTGATGCA
GCCAGAGGAACCCGACTACCTGCTGGATGAGATGACCAAGGAGGCCGAAATCGACAACTCTAGTACAGTG
GAGTCCCTGGGCGAGACTACCGATATCCTGAATATGACACACATTATGTCACTGAACAGCAATGAGAAGA
GTCTGAAACTGTCACCAGTGCAGAAGCAGAAGAAACAGCAGACTATTGGCGAGCTGACTCAGGACACCGC
CCTGACAGAGAACGACGATAGCGATGAGTATGAGGAAATGTCCGAGATGAAGGAAGGCAAAGCTTGTAAG
CAGCATGTCAGTCAGGGGATCTTCATGACACAGCCAGCCACAACTATTGAGGCTTTTTCAGACGAGGAAG
TGGAGATCCCCGAGGAAAAAGAGGGCGCAGAAGATTCCAAGGGGAATGGAATTGAGGAACAGGAGGTGGA
AGCCAACGAGGAAAATGTGAAAGTCCACGGAGGCAGGAAGGAGAAAACAGAAATCCTGTCTGACGATCTG
ACTGACAAGGCCGAGGTGTCCGAAGGCAAGGCAAAATCTGTCGGAGAGGCAGAAGACGGACCAGAGGGAC
GAGGGGATGGAACCTGCGAGGAAGGCTCAAGCGGGGCTGAGCATTGGCAGGACGAGGAACGAGAGAAGGG
CGAAAAGGATAAAGGCCGCGGGGAGATGGAACGACCTGGAGAGGGCGAAAAAGAGCTGGCAGAGAAGGAG
GAATGGAAGAAAAGGGACGGCGAGGAACAGGAGCAGAAAGAAAGGGAGCAGGGCCACCAGAAGGAGCGCA
ACCAGGAGATGGAAGAGGGCGGCGAGGAAGAGCATGGCGAGGGAGAAGAGGAAGAGGGCGATAGAGAAGA
GGAAGAGGAAAAAGAAGGCGAAGGGAAGGAGGAAGGAGAGGGCGAGGAAGTGGAAGGCGAGAGGGAAAAG
GAGGAAGGAGAACGGAAGAAAGAGGAAAGAGCCGGCAAAGAGGAAAAGGGCGAGGAAGAGGGCGATCAGG
GCGAAGGCGAGGAGGAAGAGACCGAGGGCCGCGGGGAAGAGAAAGAGGAGGGAGGAGAGGTGGAGGGCGG
AGAGGTCGAAGAGGGAAAGGGCGAGCGCGAAGAGGAAGAGGAAGAGGGCGAGGGCGAGGAAGAAGAGGGC
GAGGGGGAAGAAGAGGAGGGAGAGGGCGAAGAGGAAGAGGGGGAGGGAAAGGGCGAAGAGGAAGGAGAGG
AAGGGGAGGGAGAGGAAGAGGGGGAGGAGGGCGAGGGGGAAGGCGAGGAGGAAGAAGGAGAGGGGGAAGG
CGAAGAGGAAGGCGAGGGGGAAGGAGAGGAGGAAGAAGGGGAAGGCGAAGGCGAAGAGGAGGGAGAAGGA
GAGGGGGAGGAAGAGGAAGGAGAAGGGAAGGGCGAGGAGGAAGGCGAAGAGGGAGAGGGGGAAGGCGAGG
AAGAGGAAGGCGAGGGCGAAGGAGAGGACGGCGAGGGCGAGGGAGAAGAGGAGGAAGGGGAATGGGAAGG
CGAAGAAGAGGAAGGCGAAGGCGAAGGCGAAGAAGAGGGCGAAGGGGAGGGCGAGGAGGGCGAAGGCGAA
GGGGAGGAAGAGGAAGGCGAAGGAGAAGGCGAGGAAGAAGAGGGAGAGGAGGAAGGCGAGGAGGAAGGAG
AGGGGGAGGAGGAGGGAGAAGGCGAGGGCGAAGAAGAAGAAGAGGGAGAAGTGGAGGGCGAAGTCGAGGG
GGAGGAGGGAGAAGGGGAAGGGGAGGAAGAAGAGGGCGAAGAAGAAGGCGAGGAAAGAGAAAAAGAGGGA
GAAGGCGAGGAAAACCGGAGAAATAGGGAAGAGGAGGAAGAGGAAGAGGGAAAGTACCAGGAGACAGGCG
AAGAGGAAAACGAGCGGCAGGATGGCGAGGAATATAAGAAAGTGAGCAAGATCAAAGGATCCGTCAAGTA
CGGCAAGCACAAAACCTATCAGAAGAAAAGCGTGACCAACACACAGGGGAATGGAAAAGAGCAGAGGAGT
AAGATGCCTGTGCAGTCAAAACGGCTGCTGAAGAATGGCCCATCTGGAAGTAAAAAATTCTGGAACAATG
TGCTGCCCCACTATCTGGAACTGAAATAA (SEQ ID NO: 3)
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит последовательность, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 1;
(b) нуклеотидной последовательности, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 3; и
(c) нуклеотидной последовательности SEQ ID NO: 3,
предпочтительно, при этом белок, кодируемый нуклеотидной последовательностью, по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность, которая имеет по меньшей мере 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 1, предпочтительно, при этом аминокислотная последовательность по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
В другом варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит нуклеотидную последовательность, которая имеет по меньшей мере 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, или 99,9% идентичности с SEQ ID NO: 3, предпочтительно, при этом белок, кодируемый нуклеотидной последовательностью, по существу сохраняет естественную функцию белка, представленного SEQ ID NO: 1.
В предпочтительном варианте осуществления кодирующая RPGRORF15 нуклеотидная последовательность по изобретению содержит нуклеотидную последовательность SEQ ID NO: 3.
Предпочтительно, кодирующая RPGRORF15 нуклеотидная последовательность по изобретению кодирует белок, который способствует аналогичному или более эффективному предотвращению:
(a) клинических проявлений изменений пигмента сетчатки, которые связаны с RP;
(b) гибели фоторецепторных клеток (например, колбочковидных клеток, предпочтительно, колбочковидных и палочковидных клеток); и/или
(c) ослабления зрительной функции
у субъекта, страдающего от, или имеющего риск развития, пигментного ретинита, в сравнении с белком SEQ ID NO: 1.
В настоящем документе символ нуклеотида «N» указывает на то, что любой нуклеотид может находиться в данном положении (например, G, A, T или C), в соответствии с условными обозначениями, принятыми IUPAC-IUB.
Пигментный ретинит
Пигментный ретинит (RP) представляет собой фенотипически связанную группу наследственных дистрофий сетчатки, которые обычно бывают вызваны прогрессирующей дегенерацией палочковидных фоторецепторных клеток. Пигментный эпителий сетчатки (RPE) и колбочковидные фоторецепторные клетки также могут дегенерировать при прогрессировании заболевания.
X-связанный пигментный ретинит (XLRP), форма заболевания, имеющая сцепленный с X-хромосомой характер наследования (то есть, гены, связанные с заболеванием, расположены на X-хромосоме), считается наиболее тяжелой формой пигментного ретинита.
Клинические проявления RP включают изменения в пигменте сетчатки, которые могут сопровождаться ослаблением артериол и атрофией зрительного нерва. Изменения в сетчатке могут происходить в результате диспергирования и агрегации пигмента сетчатки. Вследствие этого может возникать спектр внешних проявлений от зернистости или крапчатости до четко различимых фокальных агрегатов, напоминающих костные иглы. Могут появляться пигментные скопления черного или темно-коричневого цвета звездчатой формы. Кроме того, может иметь место пигментация, ограниченная одним квадрантом сетчатки, аномалии, которые выглядят расходящимися от диска, и изменения, ассоциированные с тяжелой васкулопатией.
Методы лечения и предотвращения RP, описанные в настоящем документе, могут приводить к уменьшению или предотвращению проявления фенотипа RP, описанного выше. Они могут обеспечивать защиту фоторецепторных клеток, таких как колбочковидные клетки, от дегенерации. Предпочтительно, такое лечение защищает как колбочковидные, так и палочковидные клетки, от дегенерации.
Число палочек и колбочек может оценивать квалифицированный специалист в клинике такими методами, как сканирование с использованием адаптивной оптики, аутофлуоресценция и оптическая когерентная томография (ОКТ).
Предпочтительно, лечение RP позволяет сохранять или улучшать зрительную функцию.
Визуализацию внешнего вида сетчатки и оценку зрительной функции может с легкостью выполнять специалист в данной области. Например, тестирование зрительной функции, которое может проводить квалифицированный специалист, включает проверку наилучшей корригированной остроты зрения, проверку поля зрения, микропериметрию, проверку цветового зрения, темновую адаптометрию, электроретинографию и определение частоты слияния мельканий для колбочек. Используемое в настоящем документе выражение «сохранение или улучшение зрительной функции» следует понимать, как сохранение по существу такого же уровня, или повышение уровня, зрения при оценке в одном или более из таких тестов зрительной функция, когда сравнивают зрение в подвергнутом лечению глазу до и после применения способов по изобретению.
Структура глаза
Лекарственные средства, описанные в настоящем документе, могут быть доставлены в глаз млекопитающего, предпочтительно глаз человека, для лечения или предотвращения пигментного ретинита (RP).
Специалист по лечению заболеваний глаз имеет подробное и глубокое понимание структуры глаза. Однако далее приведено описание структур глаза, имеющих непосредственное отношение к изобретению.
Сетчатка
Сетчатка представляет собой многослойную мембрану, которая выстилает внутреннюю заднюю камеру глаза и воспринимает зрительные образы окружающего мира, которые передаются в мозг через зрительный нерв. В порядке расположения от внутренней к наружной части глаза сетчатка содержит слои нейросенсорной части сетчатки и пигментного эпителия сетчатки, с хориоидом, находящимся за пигментным эпителием сетчатки.
Нейросенсорная часть сетчатки и фоторецепторные клетки
В нейросенсорной части сетчатки находятся фоторецепторные клетки, которые непосредственно воспринимают свет. Она состоит из следующих слоев: внутренняя пограничная мембрана (ILM); слой нервных волокон; слой ганглиозных клеток; внутренний сетчатый слой; внутренний ядерный слой; внешний сетчатый слой; внешний ядерный слой (ядра фоторецепторов); внешняя пограничная мембрана (ELM) и фоторецепторы (внутренний и внешний сегменты) палочки и колбочки.
Квалифицированный специалист хорошо понимает устройство фоторецепторных клеток. Вкратце, фоторецепторные клетки представляют собой специализированные нейроны, расположенные в сетчатке, которые превращают свет в биологические сигналы. Фоторецепторные клетки включают палочковидные и колбочковидные клетки, которые по-разному распределены в сетчатке.
Палочковидные клетки распределены в основном во внешних зонах сетчатки. Они отличаются высокой чувствительностью и отвечают за зрение при низком уровне освещения. В нормальной сетчатке человека находится в среднем примерно 125 миллионов палочковидных клеток.
Колбочковидные клетки встречаются по всей сетчатке, но особенно сильно сконцентрированы в ямке, углублении в нейросенсорной части сетчатки, которая отвечает за четкое центральное зрение. Колбочковидные клетки менее чувствительны, чем палочковидные клетки. В нормальной сетчатке человека находится в среднем примерно 6-7 миллионов колбочковидных клеток.
Пигментный эпителий сетчатки
Пигментный эпителий сетчатки (RPE) представляет собой пигментированный слой клеток, находящийся непосредственно с внешней стороны нейросенсорной части сетчатки. RPE выполняет ряд функций, включая транспорт питательных веществ и других веществ к фоторецепторным клеткам, а также поглощение рассеянного света для улучшения зрительного восприятия.
Хориоид
Хориоид представляет собой сосудистый слой, расположенный между RPE и наружной склерой глаза. Сосудистая система хориоида позволяет доставлять кислород и питательные вещества к сетчатке.
Векторы
Вектор представляет собой инструмент, обеспечивающий или облегчающий перенос какого-либо компонента из одной среды в другую. В соответствии с изобретением, и в качестве примера, некоторые векторы, используемые в методах рекомбинантных нуклеиновых кислот, позволяют переносить компоненты, такие как сегмент нуклеиновой кислоты (например, сегмент гетерологичной ДНК, такой как сегмент гетерологичной кДНК), в клетку-мишень. Вектор может служить для целей сохранения гетерологичной нуклеиновой кислоты (например, ДНК или РНК) в клетке, облегчения репликации вектора, содержащего сегмент нуклеиновой кислоты, или облегчения экспрессии белка, кодируемого сегментом нуклеиновой кислоты.
Векторы могут быть не вирусными и вирусными. Примеры векторов, используемых в методах рекомбинантных нуклеиновых кислот, включают, но не ограничиваются ими, плазмиды, хромосомы, искусственные хромосомы и вирусы. Вектор также может представлять собой, например, «голую» нуклеиновую кислоту (например, ДНК или РНК). В самой простой форме вектор может сам представлять собой интересующий нуклеотид.
В одном аспекте изобретение относится к вектору, содержащему полинуклеотид по изобретению.
Используемые по изобретению векторы могут представлять собой, например, плазмидные или вирусные векторы и могут включать промотор для экспрессии полинуклеотида и, необязательно, регулятор промотора.
Вирусные векторы
В предпочтительном варианте осуществления вектор по изобретению представляет собой вирусный вектор. Предпочтительно, вирусный вектор имеет форму вирусной векторной частицы.
Вирусный вектор может представлять собой, например, аденоассоциированный вирусный (AAV), ретровирусный, лентивирусный или аденовирусный вектор.
Специалист в данной области способен выбирать подходящий вирус для использования в качестве вектора по изобретению, например, на основании размера и вида трансгена, который предстоит доставлять, и типа клетки-мишени. Кроме того, методы получения и модификации вирусных векторов и вирусных векторных частиц, например, полученных из AAV, ретровирусов, лентивирусов или аденовирусов, хорошо известны в данной области и могут быть легко адаптированы квалифицированным специалистом для нужной цели.
Аденоассоциированные вирусные (AAV) векторы
В предпочтительном варианте осуществления вектор по изобретению представляет собой аденоассоциированный вирусный (AAV) вектор. Предпочтительно, AAV вектор имеет форму AAV частицы.
AAV вектор может содержать геном AAV или его производное.
Геном AAV представляет собой полинуклеотидную последовательность, которая кодирует функции, необходимые для продуцирования AAV частицы. Эти функции включают те, которые задействованы в цикле репликации и упаковки AAV в клетке-хозяине, включая капсидирование генома AAV в AAV частицу. Природные AAV являются дефектными по репликации и нуждаются в предоставлении хелперных функций извне для завершения цикла репликации и упаковки. Соответственно, геном AAV вектора по изобретению, как правило, является репликативно-дефектным.
Геном AAV может находиться в одноцепочечной форме, либо смысловой, либо антисмысловой, или, альтернативно, в двухцепочечной форме. Использование двухцепочечной формы позволяет обходить этап репликации ДНК в клетке-мишени и, следовательно, ускорять экспрессию трансгена.
Геном AAV может быть из любого природного серотипа, изолята или клады AAV. Таким образом, геном AAV может представлять собой полный геном природного AAV. Как известно специалисту в данной области, существующие в природе AAV могут быть классифицированы в соответствии с разными биологическими системами.
Обычно AAV определяют по их серотипу. Серотип означает вариантный подвид AAV, который вследствие его профиля экспрессии поверхностных антигенов капсида имеет определенную реакционную способность, которая может быть использована, чтобы отличать его от других вариантных подвидов. Как правило, вирус, имеющий конкретный AAV серотип, не имеет эффективную перекрестную реактивность с нейтрализующими антителами, специфичными для другого AAV серотипа.
AAV серотипы включают AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10 и AAV11, а также рекомбинантные серотипы, такие как Rec2 и Rec3, недавно идентифицированные в мозгу приматов. Любые из этих AAV серотипов можно использовать по изобретению. Таким образом, в одном варианте осуществления изобретения AAV векторная частица представляет собой AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, Rec2 или Rec3 AAV векторную частицу.
Обзоры AAV серотипов можно найти в публикациях Choi et al. (2005) Curr. Gene Ther. 5: 299-310 и Wu et al. (2006) Molecular Therapy 14: 316-27. Последовательности геномов AAV или элементов геномов AAV, включая последовательности ITR, гены rep или cap, для использованию по изобретению можно найти под следующими регистрационными номерами для полных последовательностей генома AAV: аденоассоциированный вирус 1 NC_002077, AF063497; аденоассоциированный вирус 2 NC_001401; аденоассоциированный вирус 3 NC_001729; аденоассоциированный вирус 3B NC_001863; аденоассоциированный вирус 4 NC_001829; аденоассоциированный вирус 5 Y18065, AF085716; аденоассоциированный вирус 6 NC_001862; птичий AAV ATCC VR-865 AY186198, AY629583, NC_004828; птичий AAV штамма DA-1 NC_006263, AY629583; бычий AAV NC_005889, AY388617.
AAV также можно определять по кладам или клонам. Это относится к филогенетическим отношениям природных AAV и, как правило, к филогенетической группе AAV, представителей которой можно проследить до общего предка, группа также включает всех их потомков. Кроме того, AAV можно определять по конкретному изоляту, то есть, генетическому изоляту конкретного AAV, имеющемуся в природе. Термин «генетический изолят» означает популяцию AAV, которая подверглась ограниченному генетическому смешиванию с другими природными AAV, за счет чего создалась четко отличающаяся на генетическом уровне популяция.
Квалифицированный специалист может выбирать соответствующий серотип, клад, клон или изолят AAV для использования по изобретению на основании общих знаний. Например, показано, что капсид AAV5 эффективно трансдуцирует колбочковидные фоторецепторные клетки приматов, о чем свидетельствует успешная коррекция наследственного дефекта цветового зрения (Mancuso et al. (2009) Nature 461: 784-7).
AAV серотип определяет тканевую специфичность инфицирования (или тропизм) AAV вируса. Соответственно, предпочтительными AAV серотипами для использования в AAV, вводимых пациентам в соответствии с изобретением, являются те, которые имеют естественный тропизм в отношении, или высокую эффективность инфицирования, клеток-мишеней в глазу. В одном варианте осуществления AAV серотипы для использования по изобретению являются такими, которые инфицируют клетки нейросенсорной части сетчатки, пигментного эпителия сетчатки и/или хориоида.
Как правило, геном AAV природного серотипа, изолята или клады AAV содержит по меньшей мере одну последовательность инвертированного концевого повтора (ITR). Последовательность ITR действует в цис-положении, обеспечивая функциональную точку начала репликации, и делает возможным интегрирование вектора в геном и вырезание вектора из генома клетки. В предпочтительных вариантах осуществления одна или более последовательностей ITR фланкируют нуклеотидную последовательность, кодирующую RPGRORF15. Геном AAV, как правило, также содержит гены для упаковки, такие как гены rep и/или cap, которые обеспечивают выполнение функций упаковки для AAV частицы. Ген rep кодирует один или более из белков Rep78, Rep68, Rep52 и Rep40 или их вариантов. Ген cap кодирует один или более капсидных белков, таких как VP1, VP2 и VP3 или их варианты. Эти белки составляют капсид AAV частицы. Варианты капсида обсуждаются ниже.
Промотор будет функционально связан с каждым из генов белков-упаковщиков. Конкретные примеры таких промоторов включают промоторы p5, p19 и p40 (Laughlin et al. (1979) Proc. Natl. Acad. Sci. USA 76: 5567-5571). Например, промоторы p5 и p19, как правило, используют для экспрессии гена rep, в то время как промотор p40, как правило, используют для экспрессии гена cap.
Как описано выше, геном AAV, используемый в векторе по изобретению, может представлять собой полный геном природного AAV. Например, вектор, содержащий полный геном AAV, может быть использован для получения AAV вектора или векторной частицы in vitro. Однако, хотя такой вектор, в принципе, можно вводить пациентам, это редко осуществляют на практике. Предпочтительно, геном AAV дериватизируют для введения пациентам. Такая дериватизация является стандартной в данной области, и изобретение охватывает применение любого известного производного генома AAV, а также производных, которые могут быть получены при использовании методов, известных в данной области. Дериватизация генома AAV и капсида AAV описана в Coura and Nardi (2007) Virology Journal 4: 99, и в публикациях Choi et al. и Wu et al., ссылки на которые приведены выше.
Производные генома AAV включают любые укороченные или модифицированные формы генома AAV, которые допускают экспрессию трансгена с вектора по изобретению in vivo. Как правило, можно значительно укорачивать геном AAV, чтобы он содержал минимальную вирусную последовательность, но при этом сохранял описанную выше функцию. Это предпочтительно из соображений безопасности для уменьшения риска рекомбинации вектора с вирусом дикого типа, а также чтобы избежать инициирования клеточного иммунного ответа из-за присутствия белков вирусных генов в клетке-мишени.
Как правило, производное будет включать по меньшей мере одну последовательность инвертированного концевого повтора (ITR), предпочтительно, более одного ITR, например, два ITR или более. Один или более из ITR могут происходить из геномов AAV разных серотипов, или могут представлять собой химерные или мутантные ITR. Предпочтительным мутантным ITR является такой, который имеет делецию trs (сайт концевого разрешения). Эта делеция допускает постоянную репликацию генома, с образованием одноцепочечного генома, содержащего как кодирующую, так и комплементарную последовательности, то есть, самокомплементарного генома AAV. Это позволяет обходить этап репликации ДНК в клетке-мишени и также способствует ускоренной экспрессии трансгена.
Один или более ITR, предпочтительно, будут фланкировать нуклеотидную последовательность, кодирующую RPGRORF15, с каждого конца. Включение одного или более ITR является предпочтительным для содействия образованию конкатемеров вектора по изобретению в ядре клетки-хозяина, например, после превращения одноцепочечной векторной ДНК в двухцепочечную ДНК под действием ДНК-полимераз клетки-хозяина. Образование таких эписомных конкатемеров защищает векторную конструкцию на протяжении жизни клетки-хозяина, тем самым способствуя продолжительной экспрессии трансгена in vivo.
В предпочтительных вариантах осуществления элементы ITR будут единственными последовательностями в производном, сохранившимися из природного генома AAV. Таким образом, производное, предпочтительно, не будет содержать гены rep и/или cap из природного генома, а также любые другие последовательности из природного генома. Это предпочтительно из соображений, приведенных выше, а также для уменьшения вероятности интеграции вектора в геном клетки-хозяина. Кроме того, уменьшение размера генома AAV способствует увеличению гибкости при включении других элементов последовательности (таких, как регуляторные элементы) в вектор в дополнение к трансгену.
Таким образом, следующие части могут быть удалены в производном по изобретению: одна последовательность инвертированного концевого повтора (ITR), гены, участвующие в репликации (rep) и гены, кодирующие белки капсида (cap). Однако в некоторых вариантах осуществления производные могут дополнительно включать один или более генов rep и/или cap, или другие вирусные последовательности генома AAV. Природный AAV интегрируется с высокой частотой в конкретный сайт на человеческой хромосоме 19 и отличается очень низкой частотой случайной интеграции, так что сохранение у вектора способности к интегрированию может допускаться при терапевтическом применении.
Если производное содержит капсидные белки, то есть, VP1, VP2 и/или VP3, производное может представлять собой химерное, перетасованное или капсид-модифицированное производное одного или более природных AAV. В частности, изобретение включает использование последовательностей капсидных белков из AAV разных серотипов, клад, клонов или изолятов в одном векторе (то есть, псевдотипированном векторе).
Химерные, перетасованные или капсид-модифицированные производные, как правило, будут выбраны для обеспечения одной или более желательных функциональных характеристик вирусного вектора. Так, эти производные могут отличаться повышенной эффективностью доставки генов, сниженной иммуногенностью (гуморальной или клеточной), измененным диапазоном тропизма и/или улучшенным нацеливанием на клетки конкретного типа в сравнении с AAV вектором, содержащим природный геном AAV, например, AAV2. Повышенная эффективность доставки генов может быть достигнута за счет улучшения связывания с рецептором или корецептором на клеточной поверхности, более эффективной интернализации, более эффективного перемещения внутри клетки и в ядро, улучшенной декапсидации вирусной частицы и более эффективного превращения одноцепочечного генома в двухцепочечную форму. Повышенная эффективность также может относиться к изменению диапазона тропизма или нацеливанию на конкретную клеточную популяцию для того, чтобы доза вектора не разбавлялась за счет введения в ткани, не предназначенные для этого.
Химерные капсидные белки включают те, которые получены путем рекомбинации между двумя или более кодирующими капсид последовательностями природных серотипов AAV. Это можно осуществлять, например, методом «спасения» генетического маркера, в котором неинфекционные последовательности капсида одного серотипа совместно трансфицируют с последовательностями капсида другого серотипа и используют направленную селекцию для отбора последовательностей капсида, имеющих желаемые свойства. Последовательности капсида разных серотипов могут быть изменены путем гомологичной рекомбинации внутри клетки, с получением новых химерных капсидных белков.
Химерные капсидные белки также включают те, которые получены путем генетической модификации последовательностей капсидных белков с переносом конкретных доменов, поверхностных петель или конкретных аминокислотных остатков капсидных белков между двумя или более капсидными белками, например, между двумя или более капсидными белками разных серотипов.
Перетасованные или химерные капсидные белки также могут быть получены путем перетасовки ДНК за счет ПЦР с пониженной точностью. Гибридные гены капсида AAV могут быть получены путем случайной фрагментации последовательностей родственных генов AAV, например, тех, которые кодируют капсидные белки, нескольких разных серотипов, с последующей повторной сборкой фрагментов в самоинициируемой полимеразной реакции, что также может вызывать кроссинговеры в областях гомологии последовательностей. Библиотеку гибридных генов AAV, созданных таким способом за счет перетасовки генов капсидных белков нескольких серотипов, можно подвергать скринингу для идентификации вирусных клонов, обладающих желательными функциональными характеристиками. Аналогично, можно использовать ПЦР с пониженной точностью для случайных мутаций генов капсидных белков AAV с целью создания разнообразной библиотеки вариантов, которые затем можно отбирать на наличие желательных качеств.
Последовательности генов капсидных белков также можно генетически модифицировать для внесения конкретных делеций, замен или вставок в сравнении с последовательностью дикого типа. В частности, гены капсидных белков можно модифицировать путем вставки последовательности постороннего белка или пептида в открытую рамку считывания кодирующей последовательности капсида, либо на N- и/или C-конце кодирующей последовательности капсида.
Посторонний белок или пептид, предпочтительно, может быть тем, который действует как лиганд для клетки конкретного типа, тем самым обеспечивая улучшенное связывание с клеткой-мишенью или повышая специфичность нацеливания вектора на конкретную популяцию клеток. Пример может включать использование пептида RGD для блокирования поглощения в пигментный эпителий сетчатки и за счет этого повышения трансдукции окружающих тканей сетчатки (Cronin et al. (2008) ARVO Abstract: D1048). Посторонний белок также может быть белком, облегчающим очистку вирусной частицы в процессе производства, то есть, эпитопным или аффинным маркером. Сайт вставки, как правило, можно выбирать таким образом, чтобы не препятствовать выполнению других функций вирусной частицы, например, интернализации, перемещения вирусной частицы. Квалифицированный специалист может определять подходящие сайты для вставки на основании общих знаний. Конкретные сайты описаны в публикации Choi et al., ссылка на которую приведена выше.
Изобретение также охватывает создание последовательностей генома AAV в ином порядке и конфигурации, чем у природного генома AAV. Изобретение также охватывает замену одной или более последовательностей, или генов, AAV последовательностями из другого вируса или химерными генами, состоящими из последовательностей более чем одного вируса. Такие химерные гены могут состоять из последовательностей для двух или более родственных вирусных белков из разных видов вирусов.
Вектор по изобретению может принимать форму нуклеотидной последовательности, содержащей геном AAV, или его производное, и последовательность, кодирующую трансген RPGRORF15 или его вариант.
AAV частицы по изобретению включают транскапсидированные формы, в которых геном AAV, или его производное, содержащий ITR одного серотипа, упакован в капсид другого серотипа. AAV частицы по изобретению также включают мозаичные формы, в которых вирусный капсид состоит из смеси не модифицированных капсидных белков двух или более разных серотипов. AAV частица также включает химически модифицированные формы, несущие лиганды, адсорбированные на поверхности капсида. Например, такие лиганды могут включать антитела для нацеливания на конкретный рецептор клеточной поверхности.
Таким образом, например, AAV частицы по изобретению включают частицы с геномом AAV2 и капсидными белками AAV2 (AAV2/2), частицы с геномом AAV2 и капсидными белками AAV5 (AAV2/5), а также частицы с геномом AAV2 и капсидными белками AAV8 (AAV2/8).
Ретровирусные и лентивирусные векторы
В другом варианте осуществления изобретения вирусный вектор представляет собой ретровирусный вектор.
Ретровирусы и лентивирусы были адаптированы для использования в качестве генно-терапевтических векторов для широкого диапазона задач.
Ретровирусы могут быть, в целом, разделены на две категории, «простые» и «сложные». Ретровирусы также можно подразделять на семь групп. Пять из этих групп представляют собой ретровирусы с онкогенным потенциалом. Остальные две группы представляют собой лентивирусы и спумавирусы. Обзор этих ретровирусов приведен в Coffin, J.M. et al. (1997) Retroviruses, pp. 758-763, Cold Spring Harbor Laboratory Press, Eds: J.M. Coffin, S.M. Hughes, H.E. Varmus.
Ретровирусный вектор, используемый по изобретению, может быть получен, или может быть получаем, из любого подходящего ретровируса. Идентифицировано большое количество разных ретровирусов, и квалифицированный специалист способен выбирать соответствующий ретровирус для конкретной цели. Примеры включают: вирус лейкоза мышей (MLV), вирус T-клеточного лейкоза человека (HTLV), вирус опухоли молочной железы мышей (MMTV), вирус саркомы Рауса (RSV), вирус саркомы Фуджинами (FuSV), вирус лейкоза мышей Молони (MoMLV), вирус остеосаркомы мышей FBR (FBR MSV), вирус саркомы мышей Молони (Mo-MSV), вирус лейкоза мышей Абельсона (A-MLV), вирус миелоцитоматоза птиц 29 (MC29) и вирус эритробластоза птиц (AEV). Подробный список ретровирусов можно найти в Coffin, J.M. et al. (1997) Retroviruses, pp. 758-763, Cold Spring Harbor Laboratory Press, Eds: J.M. Coffin, S.M. Hughes, H.E. Varmus.
В другом варианте осуществления изобретения вирусный вектор представляет собой лентивирусный вектор.
Лентивирусные векторы составляют часть большой группы ретровирусных векторов. Подробный список лентивирусов можно найти в Coffin, J.M. et al. (1997) Retroviruses, pp. 758-763, Cold Spring Harbor Laboratory Press, Eds: J.M. Coffin, S.M. Hughes, H.E. Varmus. Вкратце, лентивирусы можно разделить на группы вирусов приматов и не приматов. Примеры лентивирусов приматов включают, но не ограничиваются ими: вирус иммунодефицита человека (HIV), возбудитель синдрома аутоиммунодефицита человека (СПИД); и вирус иммунодефицита обезьян (SIV). Группа лентивирусов не приматов включает прототип «медленного вируса» Висна-Маеди (VMV), а также родственный вирус артрита-энцефалита коз (CAEV), вирус инфекционной анемии лошадей (EIAV) и описанный совсем недавно вирус иммунодефицита кошек (FIV) и вирус иммунодефицита крупного рогатого скота (BIV).
Аденовирусные векторы
В другом варианте осуществления изобретения вирусный вектор представляет собой аденовирусный вектор.
Аденовирус представляет собой двухцепочечный линейный ДНК вирус, который не проходит промежуточную стадию РНК. Существует более 50 разных серотипов аденовирусов человека, которые разделяют на 6 подгрупп на основании гомологии генетических последовательностей. Природной мишенью аденовируса является эпителий дыхательных путей и желудочно-кишечного тракта, как правило, инфицирование вызывает лишь слабые симптомы. Серотипы 2 и 5 (с 95% гомологии последовательностей) наиболее часто используют в аденовирусных векторных системах, и они обычно ассоциированы с инфекциями верхних дыхательных путей у молодых людей.
Аденовирусы были использованы в качестве векторов для генной терапии и для экспрессии гетерологичных генов. Большой (36 т.п.н.) геном может принимать постороннюю вставку ДНК размером до 8 т.п.н. и способен эффективно реплицироваться в комплементирующих линиях клеток, с продуцированием высоких титров вплоть до 1012. Таким образом, аденовирус представляет собой одну из наилучших систем для изучения экспрессии генов в первичных не реплицирующихся клетках.
Для экспрессии вирусных или чужеродных генов с генома аденовируса нет необходимости в реплицирующейся клетке. Аденовирусные векторы проникают в клетки за счет опосредованного рецепторами эндоцитоза. Находясь внутри клетки, аденовирусные векторы редко интегрируются в хромосомы хозяина. Вместо этого, они функционируют эписомно (независимо от генома хозяина) в виде линейного генома в ядре клетки-хозяина. Таким образом, использование рекомбинантных аденовирусов устраняет проблемы, связанные со случайной интеграцией в геном хозяина.
Промоторы и регуляторные последовательности
Вектор по изобретению также может включать элементы, делающие возможной экспрессию трансгена RPGRORF15 in vitro или in vivo. Они могут быть названы последовательностями контроля экспрессии. Таким образом, вектор, как правило, содержит последовательности контроля экспрессии (например, включая последовательность промотора), функционально связанные с нуклеотидной последовательностью, кодирующей трансген.
Можно использовать любой подходящий промотор, который может быть с легкостью выбран специалистом в данной области. Последовательность промотора может быть конститутивно активной (то есть, функциональной в любых условиях клетки-хозяина) или, альтернативно, может быть активной лишь в конкретных условиях клетки-хозяина, таким образом обеспечивая направленную экспрессию трансгена в клетках конкретного типа (например, тканеспецифичный промотор). Промотор может демонстрировать индуцируемую экспрессию в ответ на присутствие другого фактора, например, фактора, присутствующего в клетке-хозяине. В любом случае, если вектор вводят для терапии, предпочтительно, чтобы промотор был функциональным в условиях, имеющих место в клетке-хозяине.
В некоторых вариантах осуществления предпочтительно, если промотор обеспечивает специфичную для клеток сетчатки экспрессию, чтобы экспрессия трансгена имела место только в популяции клеток сетчатки. Таким образом, экспрессия с промотора может быть специфичной для клеток сетчатки, например, ограничиваться только клетками нейросенсорной части сетчатки и пигментного эпителия сетчатки.
Предпочтительные промоторы включают промотор бета-актина кур (CBA), необязательно, в сочетании с энхансерным элементом цитомегаловируса (CMV). Примером промотора для использования по изобретению является гибридный промотор CBA/CAG, например, промотор, используемый в экспрессионной кассете rAVE (GeneDetect.com). Другой пример промотора для использования по изобретению имеет последовательность:
ATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTG
GAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTG
ACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTG
GCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTCGAGGTGAGCCCCACGTTCTGCTTCACTCTC
CCCATCTCCCCCCCCTCCCCACCCCCAATTTTGTATTTATTTATTTTTTAATTATTTTGTGCAGCGATGG
GGGCGGGGGGGGGGGGGGGGCGCGCGCCAGGCGGGGCGGGGCGGGGCGAGGGGCGGGGCGGGGCGAGGCG
GAGAGGTGCGGCGGCAGCCAATCAGAGCGGCGCGCTCCGAAAGTTTCCTTTTATGGCGAGGCGGCGGCGG
CGGCGGCCCTATAAAAAGCGAAGCGCGCGGCGGGCGGGAGTCGCTGCGCGCTGCCTTCGCCCCGTGCCCC
GCTCCGCCGCCGCCTCGCGCCGCCCGCCCCGGCTCTGACTGACCGCGTTACTCCCACAGGTGAGCGGGCG
GGACGGCCCTTCTCCTCCGGGCTGTAATTAGCGCTTGGTTTAATGACGGCTTGTTTCTTTTCTGTGGCTG
CGTGAAAGCCTTGAGGGGCTCCGGGAGGGCCCTTTGTGCGGGGGGAGCGGCTCGGGGCTGTCCGCGGGGG
GACGGCTGCCTTCGGGGGGGACGGGGCAGGGCGGGGTTCGGCTTCTGGCGTGTGACCGGCGGCTCTAGAG
CCTCTGCTAACCATGTTCATGCCTTCTTCTTTTTCCTACAGCTCCTGGGCAACGTGCTGGTTATTGTGCT
GTCTCATCATTTTGGCAAAGAATT (SEQ ID NO: 4)
Примеры промоторов на основе человеческих последовательностей, которые будут индуцировать специфичную для сетчатки экспрессию гена, включают промоторы родопсин-киназы для палочек и колбочек (Allocca et al. (2007) J. Virol. 81: 11372-80), PR2.1 только для колбочек (Mancuso et al. (2009) Nature 461: 784-7) и/или RPE65 для пигментного эпителия сетчатки (Bainbridge et al. (2008) N. Engl. J. Med. 358: 2231-9). В некоторых вариантах осуществления кодирующий RPGRORF15 полинуклеотид функционально связан с промотором (предпочтительно человеческой) родопсин-киназы (GRK1), который может содержать нуклеотидную последовательность SEQ ID NO: 7 или ее функциональный вариант, имеющий по меньшей мере 90 или 95% идентичности.
GGGCCCCAGAAGCCTGGTGGTTGTTTGTCCTTCTCAGGGGAAAAGTGAGGCGGCCCCTTGGAGGAAGGGGCCGGGCAGAATGATCTAATCGGATTCCAAGCAGCTCAGGGGATTGTCTTTTTCTAGCACCTTCTTGCCACTCCTAAGCGTCCTCCGTGACCCCGGCTGGGATTTAGCCTGGTGCTGTGTCAGCCCCGGG (SEQ ID NO: 7)
В других вариантах осуществления элементом промотора, с которым кодирующий RPGRORF15 полинуклеотид функционально связан, является промотор (предпочтительно человеческого) интерфоторецепторного ретинол-связывающего белка (IRBP), который может содержать нуклеотидную последовательность SEQ ID NO: 8 или ее функциональный вариант, имеющий по меньшей мере 90 или 95% идентичности.
agcacagtgtctggcatgtagcaggaactaaaataatggcagtgattaatgttatgatatgcagacacaacacagcaagataagatgcaatgtaccttctgggtcaaaccaccctggccactcctccccgatacccagggttgatgtgcttgaattagacaggattaaaggcttactggagctggaagccttgccccaactcaggagtttagccccagaccttctgtccaccagc (SEQ ID NO: 8)
Предпочтительно, помимо промотора, никакие дополнительные элементы не используют для контроля экспрессии RPGRORF15.
Однако вектор по изобретению также может содержать одну или более дополнительных регуляторных последовательностей, которые могут действовать до или после транскрипции. Регуляторная последовательность может быть частью природного локуса трансгена или может быть гетерологичной регуляторной последовательностью. Вектор по изобретению может содержать части 5'-UTR или 3'-UTR из природного транскрипта трансгена.
Регуляторные последовательности представляют собой любые последовательности, которые способствуют экспрессии трансгена, то есть, действуют, увеличивая экспрессию транскрипта, улучшая экспорт мРНК из ядра или повышая ее стабильность. Такие регуляторные последовательности включают, например, энхансерные элементы, посттранскрипционные регуляторные элементы и сайты полиаденилирования. Предпочтительным сайтом полиаденилирования является сигнал поли-A бычьего гормона роста, который может быть таким, как приведено ниже:
TCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCC
TTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGA
GTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAG
CAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTTCTGAGGCGGAAAGAACCAGCTGGGG (SEQ ID NO: 5)
В контексте вектора по изобретению такие регуляторные последовательности могут действовать в цис-положении. Однако изобретение также охватывает использование действующих в транс-положении регуляторных последовательностей, расположенных на дополнительных генетических конструкциях.
Предпочтительным посттранскрипционным регуляторным элементом для использования в векторе по изобретению является посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE) или его вариант. Пример последовательности WPRE приведен ниже:
ATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCT
ATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCC
TTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGT
GCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGAC
TTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGG
GCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAATCATCGTCCTTTCCTTGGCTGCTCG
CCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGA
CCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGT
CGGATCTCCCTTTGGGCCGCCTCCCCGC (SEQ ID NO: 6)
Изобретение охватывает использование любой вариантной последовательности WPRE, которая приводит к увеличению экспрессии трансгена в сравнении с вектором без WPRE. Предпочтительно, вариантные последовательности имеют по меньшей мере 70% идентичности с SEQ ID NO: 6 на всем протяжении последовательности, более предпочтительно, 75%, 80%, 85%, 90%, и более предпочтительно, по меньшей мере 95%, 96% 97%, 98% или 99% идентичности SEQ ID NO: 6 на всем протяжении последовательности.
Другой регуляторной последовательностью, которую можно использовать в векторе по изобретению, является последовательность, обладающая способностью к связыванию с ядерным матриксом (SAR). Дополнительные регуляторные последовательности может с легкостью выбирать специалист в данной области.
Способ введения
В одном варианте осуществления изобретения вирусный (например, AAV) вектор вводят в глаз субъекта субретинальной, прямой ретинальной или интравитреальной инъекцией.
Специалист в данной будет обладать соответствующими знаниями и умением для выполнения субретинальных, прямых ретинальных или интравитреальных инъекций субъекту.
Предпочтительно, вирусный (например, AAV) вектор вводят субретинальной инъекцией.
Субретинальная инъекция
Субретинальные инъекции представляют собой инъекции в субретинальное пространство, то есть, под нейросенсорную часть сетчатки. При субретинальной инъекции инъецируемый материал направляется в область, и создает пространство, между слоями фоторецепторных клеток и пигментного эпителия сетчатки (RPE).
Когда инъекцию выполняют через небольшое рассечение сетчатки, может возникать отслоение сетчатки. Отслоившийся приподнятый слой сетчатки, создаваемый инъецированным материалом, называют «пузырем».
Отверстие, создаваемое субретинальной инъекцией, должно быть достаточно небольшим, чтобы инъецируемый раствор в значительной степени не выбрасывался повторно в полость стекловидного тела после введения. Такой обратный заброс был бы особенно проблематичным при инъекции лекарственного средства, поскольку эффекты лекарственного средства были бы направлены не на целевую зону. Предпочтительно, инъекция создает самозатягивающуюся точку ввода в нейросенсорной части сетчатки, то есть, после удаления инъекционной иглы отверстие, создаваемое иглой, затягивается, так что очень малое, или по существу никакое, количество инъецированного материала не вытекает через отверстие.
Для облегчения этого процесса коммерчески доступны специальные иглы для субретинальных инъекций (например, тефлоновая игла для субретинальных инъекций DORC 41G, Dutch Ophthalmic Research Center International BV, Zuidland, The Netherlands). Эти иглы разработаны для выполнения субретинальных инъекций.
Если в процессе инъекции сетчатка не повреждается, и при условии использования достаточно мелкой иглы, по существу весь инъецированный материал остается локализованным между отслоенной нейросенсорной частью сетчатки и RPE в зоне локализованного отслоения сетчатки (то есть, не выбрасывается обратно в полость стекловидного тела). Действительно, типичная персистенция пузыря на протяжении короткого времени указывает на то, что обычно очень немного инъецированного материала попадает в стекловидное тело. Пузырь может исчезать в течение длительного времени по мере абсорбции инъецированного материала.
Перед операцией можно проводить визуализацию структур глаза, в частности, сетчатки, например, с использованием оптической когерентной томографии.
Двухэтапная субретинальная инъекция
Вектор по изобретению может быть доставлен с повышенной точностью и безопасностью при использовании двухэтапного способа, в котором создается локализованное отслоение сетчатки за счет субретинальной инъекции первого раствора. Первый раствор не содержит вектор. Затем используют вторую субретинальную инъекцию для доставки лекарственного средства, содержащего вектор, в субретинальную жидкость пузыря, созданного в результате первой субретинальной инъекции. Поскольку инъекционную доставку лекарственного средства не используют для отслоения сетчатки, на втором этапе можно вводить конкретный объем раствора.
В одном варианте осуществления изобретения вирусный (например, AAV) вектор доставляют путем:
(a) введения раствора субъекту субретинальной инъекцией в количестве, эффективном для по меньшей мере частичного отслоения сетчатки, с образованием субретинального пузыря, при этом раствор не содержит вектор; и
(b) введения лекарственной композиции субретинальной инъекцией в пузырь, образованный на этапе (a), при этом лекарственная композиция содержит вектор.
Объем раствора, инъецируемого на этапе (a) для по меньшей мере частичного отслоения сетчатки, может составлять, например, примерно 10-1000 мкл, например, примерно 50-1000, 100-1000, 250-1000, 500-1000, 10-500, 50-500, 100-500, 250-500 мкл. Объем может составлять, например, примерно 10, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 мкл.
Объем лекарственной композиции, инъецируемой на этапе (b), может составлять, например, примерно 10-500 мкл, например, примерно 50-500, 100-500, 200-500, 300-500, 400-500, 50-250, 100-250, 200-250 или 50-150 мкл. Объем может составлять, например, примерно 10, 50, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мкл. Предпочтительно, объем лекарственной композиции, инъецируемой на этапе (b), составляет 100 мкл. Большие объемы могут увеличивать риск растяжения сетчатки, в то время как меньшие объемы трудно рассмотреть.
Раствор, не содержащий лекарственное средство (то есть, «первый раствор» этапа (a)), может быть сформулирован аналогично раствору, который содержит лекарственное средство, как описано ниже. Предпочтительный раствор, не содержащий лекарственное средство, представляет собой сбалансированный солевой раствор (BSS) или аналогичный буферный раствор, совпадающий по значению pH и осмоляльности с субретинальным пространством.
Визуализация сетчатки в процессе операции
При определенных обстоятельствах, например, на последних стадиях дегенерации сетчатки, рассмотреть сетчатку бывает сложно, поскольку она является тонкой, прозрачной и ее сложно увидеть на фоне нарушенного и сильно пигментированного эпителия, на котором она располагается. Использование синего витального красителя (например, Brilliant Peel®, Geuder; MembraneBlue-Dual®, Dorc) может облегчать нахождение отверстия в сетчатке, сделанного во время процедуры отслоения сетчатки (то есть, на этапе (a) в способе двухэтапной субретинальной инъекции по изобретению), чтобы лекарственное средство можно было ввести через то же самое отверстие без риска обратного выброса в полость стекловидного тела.
Использование синего витального красителя также позволяет выявлять любые области сетчатки, где имеет место утолщение внутренней пограничной мембраны или эпиретинальной мембраны, поскольку инъекция через любую из этих структур затрудняла бы чистый доступ в субретинальное пространство. Кроме того, сокращение любой из этих структур в промежуточный послеоперационный период могло бы приводить к растяжению входного отверстия в сетчатке, что могло бы привести к обратному выбросу лекарственного средства в полость стекловидного тела.
Фармацевтические композиции и инъекционные растворы
Лекарственные средства, например, векторы, по изобретению можно формулировать в фармацевтические композиции. Эти композиции могут содержать, помимо лекарственного средства, фармацевтически приемлемый носитель, разбавитель, эксципиент, буфер, стабилизатор или другие материалы, хорошо известные в данной области. Такие материалы должны быть нетоксичными и не должны уменьшать эффективность активного ингредиента. Точную природу носителя или другого материала может определять квалифицированный специалист в зависимости от пути введения, например, субретинальной, прямой ретинальной или интравитреальной инъекции.
Фармацевтическая композиция, как правило, находится в жидкой форме. Жидкие фармацевтические композиции обычно содержат жидкий носитель, такой как вода, вазелиновое масло, животные или растительные масла, минеральное масло или синтетическое масло. Можно включать физиологический солевой раствор, растворы хлорида магния, декстрозы или других сахаридов, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль. В некоторых случаях можно использовать сурфактант, такой как 0,001% раствор плюрониловой кислоты (PF68).
Для инъекции в поврежденную зону активный ингредиент может быть в форме водного раствора, который является апирогенным и имеет соответствующее значение pH, изотоничность и стабильность. Квалифицированный специалист способен готовить соответствующие растворы с использованием, например, изотоничных сред, таких как хлорид натрия для инъекций, раствор Рингера для инъекций или раствор Рингера лактат для инъекций. По мере необходимости можно включать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки.
Для отсроченного высвобождения лекарственное средство можно включать в фармацевтическую композицию, которая сформулирована для замедленного высвобождения, например, микрокапсулы, образованные из биосовместимых полимеров, или системы липосомного носителя, методами, известными в данной области.
Способ лечения
Следует понимать, что в настоящем документе все ссылки на лечение включают радикальное, паллиативное и профилактическое лечение; хотя в контексте изобретения ссылка на предотвращение обычно ассоциируется с профилактическим лечением. Лечение также может включать остановку прогрессирования степени тяжести заболевания.
Лечение млекопитающих, в частности, людей, является предпочтительным. Однако в объем изобретения входит лечение как людей, так и животных.
Варианты, производные, аналоги, гомологи и фрагменты
Помимо конкретных белков и нуклеотидов, упомянутых в настоящем документе, изобретение также охватывает использование их вариантов, производных, аналогов, гомологов и фрагментов.
В контексте изобретения вариант какой-либо конкретной последовательности представляет собой последовательность, в которой конкретная последовательность остатков (либо аминокислотных, либо нуклеотидных остатков) была изменена таким образом, что указанный полипептид или полинуклеотид по существу сохраняет свою функцию. Вариантная последовательность может быть получена путем добавления, делеции, замещения, модификации, замены и/или варьирования по меньшей мере одного остатка, присутствующего в природном белке.
В настоящем документе термин «производное», используемый применительно к белкам или полипептидам по изобретению, включает последовательность с любым замещением, вариацией, модификацией, заменой, делецией и/или добавлением одного (или более) аминокислотных остатков в (или из) последовательности, при условии, что полученный белок или полипептид по существу сохраняет по меньшей мере одну из своих эндогенных функций.
Используемый в настоящем документе термин «аналог» применительно к полипептидам или полинуклеотидам включает любой миметик, то есть, химическое соединение, которое обладает по меньшей мере одной из эндогенных функций полипептидов или полинуклеотидов, которые он имитирует.
Как правило, аминокислотные замены могут состоять, например, из 1, 2 или 3-10 или 20 замен, при условии, что модифицированная последовательность по существу сохраняет необходимую активность или способность. Для аминокислотных замен могут быть использованы неприродные аналоги.
Белки, используемые по изобретению, также могут иметь делеции, вставки или замены аминокислотных остатков, которые приводят к «молчащему» изменению, с получением в результате функционально эквивалентного белка. Намеренные аминокислотные замены могут быть произведены на основании сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков, при условии сохранения эндогенной функции. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин; и аминокислоты с незаряженными полярными головными группами, имеющие сходные показатели гидрофильности, включают аспарагин, глутамин, серин, треонин и тирозин.
Консервативные замены могут быть произведены, например, в соответствии с приведенной ниже таблицей. Аминокислоты в одном и том же блоке во второй колонке и, предпочтительно, в одной и той же строке в третьей колонке могут быть заменены друг на друга:
Используемый в настоящем документе термин «гомолог» означает фрагмент, имеющий определенную гомологию с аминокислотной последовательностью дикого типа и нуклеотидной последовательностью дикого типа. Термин «гомология» может быть приравнен к термину «идентичность».
Гомологичная последовательность может включать аминокислотную последовательность, которая может быть по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% идентичной, предпочтительно по меньшей мере на 95% или 97%, или 99% идентичной конкретной последовательности. Как правило, гомологи будут содержать точно такие активные центры и так далее, что и конкретная аминокислотная последовательность. Хотя гомологию также можно считать сходством (то есть, аминокислотные остатки, имеющие сходные химические свойства/функции), в контексте изобретения предпочтительно выражать гомологию в терминах идентичности последовательностей.
Гомологичная последовательность может включать нуклеотидную последовательность, которая может быть по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85% или 90% идентичной, предпочтительно по меньшей мере на 95% или 97%, или 99% идентичной конкретной последовательности. Хотя гомологию также можно считать сходством, в контексте изобретения предпочтительно выражать гомологию в терминах идентичности последовательностей.
Предпочтительно, ссылка на последовательность, которая имеет некоторый процент идентичности с любой из SEQ ID NOs, приведенных в настоящем документе, относится к последовательности, которая имеет указанный процент идентичности по всей длине указанной последовательности SEQ ID NO.
Сравнения для определения гомологии можно проводить визуально или, чаще, при помощи широкодоступных программ сравнения последовательностей. Такие коммерчески доступные компьютерные программы могут рассчитывать процент гомологии или идентичности между двумя или более последовательностями.
Процент гомологии можно рассчитывать для непрерывных последовательностей, то есть, одну последовательность выравнивают с другой последовательностью и каждую аминокислоту в одной последовательности непосредственно сравнивают с соответствующей аминокислотой в другой последовательности, по одному остатку за раз. Это называют выравниванием «без пробелов». Как правило, такие выравнивания без пробелов выполняют только на протяжении относительно небольшого числа остатков.
Хотя этот метод является очень простым и логичным, в нем не учитывается, что, например, в остальном идентичной паре последовательностей, одна вставка или делеция в нуклеотидной последовательности может препятствовать выравниванию следующих далее кодонов, что, таким образом, потенциально приводит к сильному уменьшению процента гомологии при выполнении глобального выравнивания. Вследствие этого, большинство методов сравнения последовательностей разработаны для создания оптимальных выравниваний, при которых учитываются возможные вставки и делеции без ненужного уменьшения показателя общей гомологии. Это достигается за счет вставки «пробелов» при выравнивании последовательностей с целью максимального увеличения локальной гомологии.
Однако в этих более сложных методах присуждаются штрафы «за внесение пробела» для каждого пробела, возникающего при выравнивании, следовательно, при одинаковом количестве идентичных аминокислот выравниваемая последовательность с наименьшим возможным количеством пробелов, что отражает большую степень родства между двумя сравниваемыми последовательностями, будет иметь более высокий балл, чем последовательность с большим количеством пробелов. Как правило, используют «аффинные штрафы за пробел», которые включают относительно высокий штраф за внесение пробела и меньший штраф за каждый последующий остаток в пробеле. Эта система наиболее часто используется для расчета баллов за пробелы. Высокие штрафы за пробелы, безусловно, будут приводить к оптимизированным выравниваниям с меньшим количеством пробелов. Большинство программ выравнивания последовательностей позволяют изменять штрафы за пробелы. Однако предпочтительно использовать параметры по умолчанию в таких программах сравнения последовательностей. Например, при использовании пакета программ GCG Wisconsin Bestfit штраф за пробел по умолчанию для аминокислотных последовательностей составляет: -12 за внесение пробела и -4 за каждое удлинение.
Таким образом, для расчета максимального процента гомологии, во-первых, необходимо произвести оптимальное выравнивание, учитывая штрафы за пробелы. Подходящей компьютерной программой для выполнения такого выравнивания является пакет программ GCG Wisconsin Bestfit (Университет штата Висконсин, США; Devereux et al. (1984) Nucleic Acids Res. 12: 387). Примеры других программ, которые могут выполнять сравнение последовательностей, включают, но не ограничиваются ими, пакет программ BLAST (смотри Ausubel et al. (1999), там же - Ch. 18), FASTA (Atschul et al. (1990) J. Mol. Biol. 403-410) и набор инструментов сравнения GENEWORKS. Как BLAST, так и FASTA, доступны для офлайн и онлайн поиска (смотри Ausubel et al. (1999), там же, страницы 7-58 до 7-60). Однако для некоторых приложений предпочтительно использовать программу GCG Bestfit. Для сравнения белковых и нуклеотидных последовательностей также доступен и другой инструмент, называемый BLAST 2 Sequences (смотри FEMS Microbiol. Lett. (1999) 174: 247-50; FEMS Microbiol. Lett. (1999) 177: 187-8).
Хотя окончательный процент гомологии может быть измерен с точки зрения идентичности, сам по себе процесс выравнивания, как правило, не основан на сравнении пар по принципу «все или ничего». Вместо этого, как правило, используют матрицу нормированных весов замен, которая присваивает баллы каждому парному сравнению на основании химического сходства или эволюционного расстояния. Примером такой часто используемой матрицы является матрица BLOSUM62 - матрица по умолчанию для набора программ BLAST. В программах GCG Wisconsin, как правило, используют либо общепринятые параметры по умолчанию, либо пользовательскую таблицу сравнения символов, при наличии (смотри руководство пользователя для подробной информации). Для некоторых приложений предпочтительно использовать общепринятые параметры по умолчанию для пакета программ GCG или, в случае других программ, матрицу по умолчанию, такую как BLOSUM62.
После выполнения программой оптимального выравнивания можно рассчитывать процент гомологии, предпочтительно, процент идентичности последовательностей. Программа, как правило, выполняет это в виде части процесса сравнения последовательностей и выдает числовой результат.
«Фрагменты» полноразмерного RPGRORF15 также представляют собой варианты, и термин, как правило, относится к выбранной области полипептида или полинуклеотида, которая представляет интерес либо с точки зрения функции, либо, например, для использования в анализе. Таким образом, «фрагмент» означает аминокислотную или нуклеотидную последовательность, которая является частью полноразмерного полипептида или полинуклеотида.
Такие варианты можно получать с использованием стандартных методов рекомбинантной ДНК, таких как сайт-направленный мутагенез. Если предстоит делать вставки, можно создавать синтетическую ДНК, кодирующую вставку вместе с 5' и 3'-фланкирующими областями, соответствующими природной последовательности с каждой стороны от сайта вставки. Фланкирующие области будут содержать удобные сайты рестрикции, соответствующие сайтам в природной последовательности, так что последовательность может быть разрезана соответствующим ферментом(ами) и синтетическая ДНК лигирована в место разреза. Затем ДНК экспрессируется в соответствии с изобретением, приводя к образованию закодированного белка. Эти методы являются лишь примерами множества стандартных методов, известных в данной области, для манипуляций с последовательностями ДНК, и другие известные методы также могут быть использованы.
Все публикации, упомянутые в приведенной выше спецификации, включены в настоящий документ посредством ссылки. Различные модификации и вариации описанных продуктов, систем, вариантов применения, процессов и способов по изобретению, не выходящие за пределы объема и сущности изобретения, будут очевидны для специалистов в данной области. Хотя изобретение было описано на примере конкретных предпочтительных вариантов осуществления, следует понимать, что заявленное изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. Действительно, различные модификации описанных вариантов осуществления изобретения, которые очевидны для специалистов в области биохимии и биотехнологии или родственных областях, должны входить в объем прилагаемой формулы изобретения.
ПРИМЕРЫ
ПРИМЕР 1
Пример 1A - Модельные системы на мышах для X-связанного пигментного ретинита
Некоторые виды животных, например, мыши и собаки, имеют гены, гомологичные гену RPGR человека (сравните выделенные желтым цветом последовательности экзонов на Фигуре 1). Вследствие этого, животные таких видов потенциально могут служить моделями для X-связанного пигментного ретинита, вызываемого мутациями в человеческом RPGR.
Были получены две мышиные модели для тестирования безопасности и эффективности терапевтического вектора в данном исследовании. В обеих моделях имели место изменения в гомологичном гене Rpgr:
1. Линия Rpgr -/-: эта линия была создана путем направленного разрушения частей экзона 4 до части экзона 6 за счет последовательности, содержащей гены, кодирующие β-галактозидазу и маркер устойчивости к неомицину (Hong D.H. et al. (2000) Proc. Natl. Acad. Sci. USA 97: 3649-3654).
2. Линия Rd9: для этой линии характерна природная инсерционная мутация со вставкой 32 п.н., которая приводит к сдвигу рамки считывания (Thompson D.A. et al. (2012) PLoS One 7: e35865).
В обеих мышиных моделях, Rpgr -/- и Rd9, отсутствует экспрессия белка Rpgr, но лишь с очень небольшим фенотипическим проявлением (Thompson D.A. et al. (2012) PLoS One 7: e35865).
Пример 1B - Кодон-оптимизация RPGR
Кодон-оптимизация RPGR
Синтетическую последовательность RPGRORF15 получали с использованием кодон-оптимизации (coRPGR, «оптимизированная; SEQ ID NO: 3) для стабилизации высоко мутагенной богатой пуринами области (Фигура 2).
Характеризация кодон-оптимизированного гена RPGR
Сравнение эффективности клонирования и точности последовательностей проводили между последовательностью дикого типа и кодон-оптимизированной последовательностью в стандартном клонирующем векторе (Фигура 3A-D).
При использовании рестрикционного анализа было обнаружено лишь 3 клона из 24, успешно демонстрирующие наличие последовательности дикого типа (wtRPGR; Фигура 3B, успешные клоны отмечены звездочками). Напротив, 18 из 24 клонов успешно демонстрировали наличие правильной кодон-оптимизированной последовательности (coRPGR; Фигура 3D).
Было обнаружено, что выход плазмиды из минипрепаратов был выше при использовании последовательности coRPGR, чем последовательности wtRPGR (Фигура 3E), даже при сохранении аналогичного уровня чистоты образца (Фигура 3F). В подтверждение этого результата, общий выход плазмиды из мегапрепаратов (Фигура 3G) также был выше при использовании последовательности coRPGR.
Анализ частоты мутаций RPGR в процессе клонирования
Было установлено, что секвенирование методом Сэнгера полученных клонов было легче выполнимо и отличалось лучшим общим отношением сигнала к шуму при использовании последовательности coRPGR, поскольку, как показано, неоднозначное определение нуклеотидов было менее вероятным (Фигура 4).
Из-за множественных G-отрезков и богатого пуринами характера wtRPGR пришлось выполнить 34 отдельные реакции секвенирования для покрытия примерно 4 т.п.н. последовательности ДНК. В области ORF15 было обнаружено 6 потенциальных мутаций (2 делеции и 4 вставки), а также 74 неоднозначных определений нуклеотидов с потенциалом для дополнительных точечных мутаций.
Напротив, последовательность coRPGR было легче секвенировать и чистое подтверждение (по меньшей мере 2×) ее последовательности было достигнуто с использованием лишь 17 реакций.
Количественный анализ точности последовательности
Все четыре независимо полученных мегапрепарата плазмид wtRPGRORF15 и coRPGRORF15 были секвенированы в сертифицированной по ISO 9001, GLP-аккредитованной лаборатории (Source BioScience, UK), в качестве коммерческого подрядчика, методом Сэнгера, что является золотым стандартом для валидации последовательностей ДНК (Sanger F. et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463-5467).
Сравнение данных секвенирования с опубликованными данными для эталонной последовательности (NCBI Gene ID: 6103) позволило выявить в среднем 20 мутаций (преимущественно однонуклеотидные делеции, вставки и точечные мутации) в секвенированной последовательности wtRPGRORF15, но не в coRPGRORF15.
Результаты этого анализа отчетливо демонстрируют гораздо большую стабильность последовательности у кодон-оптимизированной последовательности RPGR.
Анализ белка - продукта трансгена coRPGR
Была проведена жидкостная хроматография с тандемной масс-спектрометрией (ЖХ/МС-МС) для подтверждения белковой последовательности продукта трансгена coRPGR (Фигура 5).
Примерно 80% аминокислот RPGR были непосредственно идентифицированы. Лишь для части области, в высокой степени богатой глутаминовой кислотой и повторениями, около C-конца белка не удалось провести пептидный анализ (Фигура 5, последовательность выделена красным цветом). Причиной этого было отсутствие специфических фрагментов для создания олигомеров для спектрометрии. Однако идеальное совпадение C-концевой последовательности свидетельствует о том, что был получен полноразмерный белок без сдвига рамки считывания.
Кроме того, вестерн-блот анализ продемонстрировал идентичные молекулярные массы полученных из wtRPGR и coRPGR пептидов из трансфицированных клеток (Фигура 6). Это подтверждает вывод об отсутствии различий (например, вставок или делеций), которые были внесены при использовании подхода кодон-оптимизации.
Вестерн-блот анализ также четко показал, что кодон-оптимизация позволяет достигать более высокой скорости трансляции и продуцирования белка (Фигура 6).
Вывод
В целом, кодон-оптимизированная последовательность RPGRORF15 (coRPGR) продемонстрировала превосходную точность последовательности и повышенные уровни экспрессии в сравнении с последовательностью дикого типа и, таким образом, была признана потенциально способной повысить терапевтический потенциал заместительной генной терапии для XLRP.
Пример 1C - AAV-опосредованная in vivo доставка кодон-оптимизированной последовательности RPGR
Последовательность coRPGR была упакована в вектор AAV2/8, который был использован для введения RPGR в фоторецепторные клетки мышей, у которых отсутствовала экспрессия RPGR (линии мышей Rd9 и Rpgr -/-). Более подробно, трансгенная кассета содержала промотор родопсин-киназы и консенсусную последовательность Kozak выше полинуклеотидной последовательности coRPGR, а также последовательность полиA из бычьего гормона роста ниже кодирующей последовательности.
coRPGR приводит к экспрессии белка RPGR у мышей Rd9 и Rpgr -/-
Вестерн-блот анализ показал, что введение мышам Rd9 и Rpgr -/- вектора AAV.coRPGR приводит к экспрессии белка RPGR (Фигура 7A). Кроме того, иммуногистохимическое окрашивание на RPGR было проведено на получавших и не получавших инъекцию глазах у мышей Rd9 и Rpgr -/- и продемонстрировало правильную локализацию белкового продукта (Фигура 7B).
Генное усиление за счет coRPGR приводит к терапевтическому эффекту у мышей Rd9 и Rpgr -/-
Модельных мышей линий Rd9 и Rpgr -/- подвергали генной терапии только для одного глаза и использовали метод электроретинографии (ЭРГ) для объективной оценки функции в получавших инъекцию глазах в сравнении с не получавшими инъекцию глазами.
Изучение амплитуд a- и b-волн при однократной вспышке света разной интенсивности после темновой адаптации (Фигура 7C,D) показало статистически значимые величины амплитуд (p<0,05) в получавших инъекцию группах в сравнении с не получавшими инъекцию группами в двух временных точках после воздействия.
В заключение, приведенные выше данные, которые описаны более подробно в примерах 4 и 5, показывают, что кодон-оптимизированная последовательность человеческого RPGR продуцирует белок RPGR дикого типа с предсказанным размером и аминокислотной последовательностью. Кроме того, кодон-оптимизация приводит к более высокой точности последовательности и повышенным уровням экспрессии. При использовании в методах заместительной генной терапии в моделях с наследуемым отсутствием экспрессии белка Rpgr последовательность coRPGR обеспечивает экспрессию RPGR. Более того, RPGR локализуется в физиологической зоне действия в соединительных ресничках и способен улучшать структуру (длину внутреннего и внешнего сегмента) и функцию сетчатки, как показывают исследования методом ЭРГ.
ПРИМЕР 2 - Производство плазмиды pAAV.RK.coRPGR в промышленном масштабе
Пример 2a
Получение бактериального клеточного банка для плазмиды, кодирующей кодон-оптимизированную последовательность RPGR
Бактериальный штамм K-12 Escherichia coli, XL10 Gold, с генотипом TetrD(mcrA)183 D(mcrCB-hsdSMR-mrr)173 endA1 supE44 thi-1 recA1 gyrA96 relA1 lac Hte [F´ proAB lacIqZDM15 Tn10 (Tetr) Amy Camr] трансформировали плазмидой pAAV.RK.coRPGR и высевали на чашки с агаром с добавлением ампициллина для отбора одиночных колоний, содержащих плазмидную ДНК с кодон-оптимизированной последовательностью RPGR. Одиночную колонию выбирали, инокулировали в культуральную среду и наращивали в маломасштабной суспензионной культуре. Когда клетки находились в середине логарифмической фазы роста, их собирали и ресуспендировали в криоконсервационной среде, содержащей глицерин, после чего примерно пятьдесят 1,5-мл аликвот разливали в 1,8-мл криопробирки и замораживали при -80°C, получая бактериальный клеточный банк, RCB pAAV.RK.coRPGR E.coli XL10 Gold.
Клеточный банк размораживали, плазмидную ДНК получали путем экстрагирования из минипрепаратов (Birnboim H.C. and Doly J. 1979; A rapid alkaline extraction procedure for screening recombinant plasmid DNA in 1979 Nov 24;7(6):1513-23) и тестировали на идентичность и структурную стабильность плазмиды путем расщепления эндонуклеазами рестрикции Xmn1 и Sma1, с последующим анализом методом электрофореза в 0,8% агарозном геле с окрашиванием бромидом этидия в концентрации 0,5 мг/мл (Фигура 8). Фигура 8 четко демонстрирует ожидаемую картину рестрикционных фрагментов из стабильной и структурно интактной плазмиды pAAV.RK.coRPGR, что свидетельствует о стабильном сохранении и репродуцировании плазмидной ДНК в процессе наращивания и производства клеточного банка в культуре.
Клеточный банк также оценивали на выход плазмиды после культивирования во встряхиваемых колбах с бульоном и получали 598 мкг плазмидной ДНК на грамм сырой клеточной массы. Был достигнут высокий выход надлежащей плазмиды.
Также не было обнаружено свидетельств потери плазмиды при тестировании сегрегационной стабильности плазмиды путем высевания методом реплик колоний на содержащие антибиотик и не содержащие антибиотик чашки с агаром. Результаты показали 100% сохранение плазмиды.
Плазмидную ДНК из клеточного банка секвенировали методом Сэнгера (Sanger F. et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463-5467), анализ полученной последовательности показал, что кодон-оптимизированная последовательность RPGR сохранялась со 100% точностью при сравнении с теоретической контрольной последовательностью кодон-оптимизированной последовательности RPGR. Опять-таки отсутствовало какое-либо свидетельство нестабильности.
RCB pAAV.RK.coRPGR E.coli XL10 Gold также тестировали на бактериальную чистоту (было продемонстрировано отсутствие бактериального загрязнения) и на отсутствие литических и лизогенных бактериофагов (не обнаружено). Видовая принадлежность RCB pAAV.RK.coRPGR E.coli XL10 Gold также была подтверждена биохимическим методом идентификации в тесте API-20E (BioMerieux).
Вывод
Кодон-оптимизация гена RPGR привела к повышению стабильности гена RPGR, что позволило создать пригодный для промышленного использования бактериальный клеточный банк, при этом была показана 100% точность последовательности относительно эталонной кодон-оптимизированной последовательности, 100% сегрегационная стабильность плазмиды и хороший выход плазмиды.
Пример 2b
Получение высококачественной плазмиды, кодирующей кодон-оптимизированную последовательность RPGR, в промышленном масштабе
Высококачественную плазмидную ДНК производили и очищали (Schmeer et al. (2014) Pharmaceutical Grade Large-Scale Plasmid DNA Manufacturing Process: 219-242) из бактериального клеточного банка RCB pAAV.RK.coRPGR в E. coli XL10 Gold, полученного в примере 2a, как описано вкратце. Одну пробирку с бактериальным клеточным банком размораживали, наращивали и культивировали в промышленном масштабе, затем бактериальную клеточную массу собирали центрифугированием. Плазмидную ДНК экстрагировали из бактериальной биомассы путем щелочного лизиса бактериальных клеток, полученную растворимую плазмидную ДНК отделяли от нерастворимого белка и находящейся в комплексе геномной ДНК методом центрифугирования и фильтрования. Плазмидную ДНК дополнительно очищали хроматографией в несколько этапов. Полностью очищенную плазмидную ДНК окончательно переводили в буфер формулирования путем осаждения и проточной фильтрации вдоль потока, а также мембранной фильтрации, с получением 100 мг высокоочищенной плазмидной ДНК с концентрацией 1,0 мг/мл в 10 мМ Tris+1 мМ ЭДТА, pH 8,0 (Фигура 10), которая обладала достаточной степенью чистоты (Фигура 11) для использования в дальнейших производственных процессах получения rAAV векторов.
Очищенную плазмиду тестировали на идентичность и структурную стабильность плазмиды путем расщепления эндонуклеазой рестрикции Xmn1 и последующего анализа методом электрофореза в 0,8% агарозном геле с окрашиванием бромидом этидия в концентрации 0,5 мг/мл (Фиг. 10). Фигура 11 четко демонстрирует ожидаемую картину рестрикционных фрагментов из стабильной и структурно интактной плазмиды pAAV.RK.coRPGR, что свидетельствует о стабильном сохранении и репродуцировании плазмидной ДНК в процессе наращивания клеточной культуры и очистки плазмиды.
Очищенную плазмиду секвенировали методом Сэнгера (Sanger F. et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463-5467), анализ полученной последовательности показал, что кодон-оптимизированная последовательность RPGR сохранялась со 100% точностью при сравнении с теоретической эталонной последовательностью кодон-оптимизированной последовательности RPGR.
Вывод
Кодон-оптимизация гена RPGR привела к повышению стабильности гена RPGR, что позволило получать высокочистую плазмидную ДНК в достаточном количестве (100 мг) и качестве для использования в дальнейших производственных процессах получения rAAV вектора.
Пример 2c
Получение rAAV2/8, кодирующего кодон-оптимизированную последовательность RPGR, в промышленном масштабе
Высококачественную плазмидную ДНК, полученную в примере 2b, использовали вместе с хелперной плазмидой pDP8.ape (PlasmidFactory, Bielefeld, Germany и Grimm D, Kay MA, Kleinschmidt JA. Helper virus-free, optically controllable, and two-plasmid-based production of adeno-associated virus vectors of serotypes 1 to 6. Mol Ther 2003;7:839-850) для производства rAAV8/2 вектора для использования in vivo методом крупномасштабной временной трансфекции с последующей очисткой (Lock et al. 2010; Human Gene Therapy: 1259-1271). Вкратце, клетки HEK293 выращивали в виде прикрепленной к субстрату культуры в модифицированной по способу Дульбекко среде Игла (DMEM) с добавлением 10% эмбриональной бычьей сыворотки (ЭБС) в условиях 37°C и 5% CO2 до получения достаточного количества клеток для засевания достаточного количества многослойных сосудов для культивирования клеток. Неочищенный AAV получали путем временной трансфекции с использованием фосфата кальция прикрепленных клеток HEK293, растущих в многослойных сосудах для культивирования клеток, плазмидой pAAV.RK.coRPGR и хелперной плазмидой pDP8.ape для получения rAAV частиц, секретируемых в ростовую среду. rAAV частицы собирали путем удаления среды из сосудов для культивирования клеток и фильтровали для удаления клеточного детрита. Сначала rAAV концентрировали методом проточной фильтрации вдоль потока (TFF) и ультрафильтрации, и частично очищали методом диафильтрации с использованием того же оборудования для TFF. Затем rAAV очищали ультрацентрифугированием в прерывистом градиенте иодиксанола и ионообменной хроматографией на колонке. Очищенный rAAV затем переводили в конечный буфер формулирования в концентрации 3,55 x1012 гч/мл дополнительной ультрафильтрацией и диафильтрацией на базе TFF. Второй препарат rAAV с более низкой концентрацией 1,00 x1012 гч/мл был получен путем разбавления более концентрированной лекарственной формы буфером формулирования. Обе лекарственные формы разливали по пробиркам 50-мкл аликвотами и хранили при ≤60°C.
Вывод
Кодон-оптимизация гена RPGR привела к повышению стабильности гена RPGR, что позволило получить очищенный rAAV вектор в достаточном количестве и качестве для использования в исследованиях с in vivo дозированием.
Пример 2d
Целью данных исследований была разработка метода in vivo доставки генно-терапевтического вектора AAV8-RPGR, вводимого путем субретинальной инъекции, предпочтительным клиническим путем введения (ROA). Было проведено соответствующее требованиям GLP исследование с однократным введением дозы AAV8-RPGR путем субретинальной инъекции мышам C57B/6J, с последующим анализом через 4 недели и 26 недель.
Линия мышей C57B/6J с пигментацией глаз была выбрана в качестве подходящего вида животных для данных исследований с in vivo доставкой препарата по следующим причинам. Во-первых, животные этой линии имеют пигментированные глаза, что позволяет очень точно имитировать процедуру введения, используемую в клинических условиях. Во-вторых, животные трансгенного варианта этой линии, мышиная модель с нокаутом RPGR, демонстрируют биологический ответ на изучаемый продукт AAV8-RPGR, аналогичный тому, который ожидается у человека. В-третьих, человеческий и мышиный белки RPGR имеют высокую гомологию аминокислотных последовательностей, и успешное нацеливание AAV8-RPGR и трансдукция ткани сетчатки мышей ранее было продемонстрировано в in vivo фармакологических исследованиях.
Животные получали одну субретинальную инъекцию 1 мкл растворителя (BSS Plus, Alcon Pharma) или AAV8-RPGR (в двух разных дозах гч/глаз) в оба глаза. Была проведена подробная оценка любых токсических эффектов, включая оценку массы тела, клинических признаков токсичности, в том числе потребления пищи, клинической патологии и гистопатологии, а также подробное и регулярное офтальмологическое обследование глазного яблока, внешних структур глаза, переднего сегмента глаза, в основном роговицы и хрусталика, а также внутренних структур, включая глазное дно. Кроме того, были проведены регулярные тонометрические оценки и электроретинографические оценки. Метод электроретинографии (ЭРГ) позволяет регистрировать электрические потенциалы, возникающие в сетчатке после световой стимуляции при разной интенсивности, длинах волн и продолжительности воздействия света. Электроретинограмма представляет совокупную активность миллионов клеток сетчатки на всем протяжении от пигментного эпителия до внутреннего ядерного слоя. Этот метод используют для оценки функции сетчатки и обнаружения дегенерации сетчатки на ранних стадиях.
Генная терапия с использованием AAV8-RPGR хорошо переносилась мышами и не было отмечено никаких серьезных побочных реакций на инъекции. Любые отклонения были временными и совпадали с процедурами дозирования и анестезирования, они также имели место в группах введения растворителя.
Для получения информации о распределении доставляемого вектора AAV8-RPGR животных в группах введения делили на подгруппы, в которых животных умерщвляли в дни 8, 29 и 183 после инъекции. Кроме того, прижизненные образцы крови собирали в дни 2, 15 и 92 в каждой из подгрупп для определения уровней вектора в специфичном для AAV8-RPGR анализе методом количественной ПЦР (кПЦР). Животные получали одну субретинальную инъекцию 1 мкл растворителя (BSS Plus, Alcon Pharma) или AAV8-RPGR в оба глаза.
Образцы для кПЦР из нескольких тканей/жидкостей организма (кровь, костный мозг, слезная жидкость, головной мозг, глаз, сердце, внутриглазная жидкость и жидкость стекловидного тела, почка, печень, легкое, лимфатический узел, зрительный нерв, сетчатка, слюна, яичко, селезенка, моча), собранные у животных при аутопсии в дни 8, 29 и 183, будут проанализированы на количественное содержание векторной ДНК. Количественное определение векторной ДНК в этих тканях/жидкостях будет проведено методом кПЦР по окончании исследования. Данные результаты подтвердят успешную доставку гена методом генной терапии и позволят картировать его распределение в тканях мышей.
ПРИМЕР 3 - In vitro анализ белкового продукта трансгена coRPGRORF15
В данном примере анализировали эффект кодон-оптимизации на повышение уровней экспрессии RPGR и улучшение ситуации с синтезом альтернативно сплайсированных вариантов. Для этой цели использовали клетки 293 эмбриональной почки человека (HEK293) и клетки HEK293, экспрессирующие T-антиген вируса обезьян 40 (SV40) (HEK293T). Эти клетки трансфицировали конструкциями плазмид CAG.coRPGRORF15 и CAG.wtRPGRORF15 и обрабатывали для обнаружения трансгена, как описано ниже.
Материалы и методы
Клетки HEK293 и HEK293T высевали при плотности 4 и 2,5×105 клеток/мл, соответственно, в 6-луночные планшеты и культивировали в среде DMEM, содержащей 10% инактивированной нагреванием эмбриональной бычьей сыворотки (ЭБС) и 1% смеси пенициллин/стрептомицин, в условиях 37°C и 5% CO2 до достижения ими конфлюэнтности более 70%. Один микрограмм плазмидной ДНК (CAG.coRPGRORF15 и CAG.wtRPGRORF15) доставляли в клетки с использованием реагента для трансфекции Mirus TransIT®-LT1 (Geneflow Ltd., Lichfield, UK) в не содержащей сыворотку/антибиотик среде. Трансфицированные клетки инкубировали при 37°C в течение 48 часов.
Анализ экспрессии трансгена проводили на уровне белка. Через сорок восемь часов после трансфекции клетки промывали в фосфатно-солевом буфере (PBS), с последующим лизисом клеток и солюбилизацией белка в буфере для радио-иммунопреципитационного анализа (RIPA) (Sigma-Aldrich Company Ltd., Dorset, UK) с минитаблеткой не содержащего ЭДТА коктейля ингибиторов протеаз cOmplete™ (Roche Products Ltd., Welwyn Garden City, UK). Клетки в осадке разрушали обработкой ультразвуком, и клеточные фрагменты удаляли центрифугированием при 13000 g и 4°C в течение 10 минут. Общее содержание белка в супернатанте количественно определяли с использованием набора для анализа белка с бицинхониновой кислотой (BCA) Pierce™ (Thermo Scientific) в соответствии с инструкциями производителя.
Экспрессию белка оценивали вестерн-блот анализом. Тридцать мкг суммарного белка денатурировали в 2 x буфере Лэммли (Sigma-Aldrich) в течение 5 минут при 95°C и разделяли в 7,5% полиакриламидном геле с додецилсульфатом натрия (Criterion™ TGX™ Precast Gels, Bio-Rad Laboratories Ltd., Hemel Hempstead, UK) электрофорезом при 100 В в течение 2 часов. Образцы белка, разделенные методом SDS-ПААГ, переносили на мембраны из поливинилиден дифторида (PVDF) (Trans-Blot Turbo™ Midi PVDF, Bio-Rad) с использованием системы Trans-Blot Turbo™ Transfer Starter System (Bio-Rad). Мембраны блокировали 3% раствором бычьего сывороточного альбумина (BSA) в PBS, содержащим 0,1% Tween 20, в течение 45 минут и инкубировали с первичным антителом при комнатной температуре в течение 1 часа. RPGR в качестве целевого белка и β-актин в качестве контроля нагрузки идентифицировали с использованием следующих первичных антител: кроличьих поликлональных антител против RPGR (1:500, Sigma-Aldrich) и мышиного моноклонального антитела против β-актина (1:30000, Ambion Inc., Thermo Scientific, Nortumberland, UK). Полосы выявляли при помощи конъюгированных с пероксидазой хрена вторичных антител, используя обнаруживающий реагент ECL. Уровни белка RPGR количественно определяли денситометрией с использованием Image Studio Lite (версия 5.2) и нормировали на β-актин.
Результаты
Вестерн-блот анализ суммарных белковых лизатов показал наличие преобладающей полосы с массой примерно 220 кДа, соответствующей белку RPGRORF15 (Фигуры 12A и 13A), в клетках HEK293 и HEK293T, трансфицированных плазмидами CAG.coRPGRORF15 и CAG.wtRPGRORF15. Количественное определение интенсивности полосы RPGRORF15, нормированной на интенсивность полосы β-актина, показало, что экспрессия RPGR выше в клетках HEK293, трансфицированных кодон-оптимизированной плазмидой, 2,79 [1,1-17,0] условных единиц (УЕ) (среднее [Q1 - Q3]), чем в клетках, трансфицированных последовательностью дикого типа, 0,36 [0,08-4,88] УЕ. Аналогично, клетки HEK293T, трансфицированные кодон-оптимизированной последовательностью, демонстрировали повышенные уровни экспрессии RPGR, 4,23 ± 0,11 УЕ (среднее ± стандартное отклонение) в сравнении с клетками, трансфицированными последовательностью дикого типа, 3,01 ± 0,07 УЕ (среднее ± стандартное отклонение) (Фигуры 12B и 13B).
Помимо экспрессированной целой последовательности RPGRORF15 в случае обоих плазмидных конструкций, дополнительная четкая полоса с молекулярной массой 80 кДа была обнаружена в клетках HEK293T, трансфицированных последовательностью RPGRORF15 дикого типа. Этот результат показал, что кодон-оптимизация повышает стабильность последовательности, тем самым уменьшая образование альтернативно сплайсированных форм.
Вывод
Данные результаты подтверждают, что увеличение индекса адаптации кодонов (CAI) кодирующей последовательности RPGRORF15 за счет введения синонимичных основных кодонов приводит к более высоким уровням экспрессии трансгена, при этом уменьшается синтез укороченных белков за счет повышения стабильности и точности последовательности.
ПРИМЕР 4
Генная терапия с использованием RPGR направлена на восстановление экспрессии RPGR в клетках-мишенях, которые имеют вызывающие заболевание мутации в RPGR, приводящие к полной потере RPGR (нулевые мутации) или образованию нефункционального белка. Один из путей достижения этого заключается во введении правильной копии нуклеотидной последовательности, кодирующей RPGR, которая затем транслируется в белок RPGR собственным аппаратом трансляции клеток-мишеней. Такую нуклеотидную последовательность можно вводить путем трансдукции при помощи рекомбинантного AAV.
В настоящем документе авторы изобретения представили данные из пилотного исследования, разработанного для изучения эффективности трансдукции рекомбинантных AAV векторов AAV2/8.RK.coRPGR и AAV2/8.RK.wtRPGR после субретинальной инъекции мышам. Для того, чтобы модель генной терапии с использованием RPGR больше напоминала клинические условия, авторы изобретения использовали мышей Rpgr-/y, у которых отсутствует экспрессия Rpgr.
Rpgr является мышиным гомологом RPGR, гена, затронутого в большинстве случаев X-связанного пигментного ретинита, и, таким образом, трансгенные мыши Rpgr-/y имитируют ситуацию с нулевыми мутациями у пациентов-людей на генетическом уровне. Кодон-оптимизированный RPGR отличается более высокими уровнями экспрессии, чем RPGR дикого типа, и, что важно, обеспечивает большую точность последовательности при образовании идентичного белкового продукта RPGR. Рекомбинантные AAV векторы AAV2/8.RK.coRPGR и AAV2/8.RK.wtRPGR способны трансдуцировать подобные фоторецепторам клетки in vitro.
Целью данного исследования было изучение того, будет ли RPGR экспрессироваться in vivo после субретинальной инъекции AAV2/8.RK.coRPGR или AAV2/8.RK.wtRPGR, а также будет ли RPGR локализован в соединительных ресничках в фоторецепторных клетках, наследуемым образом лишенных экспрессии Rpgr.
Материалы и методы
Растворы рекомбинантных AAV, содержащие AAV2/8.RK.coRPGR и AAV2/8.RK.wtRPGR, получали и оценивали на качество и титр. Для контроля на эффект хирургического вмешательства и трансдукцию фоторецепторных клеток вектором AAV2/8 использовали третью конструкцию с геном GFP в качестве гена репортера под контролем промотора CAG (AAV2/8.CAG.GFP). Проводили количественную оценку этих растворов векторов методом кПЦР для расчета числа векторных геномов/мл. Аликвоты, хранящиеся при -80°C, размораживали на льду непосредственно перед использованием и разбавляли в сбалансированном солевом растворе (BSS®) для витреоретинальной хирургии (Alcon Laboratories, Camberley, UK) с 0,001% плюроника® PF68 (BASF, Ludwigshafen, Germany) для субретинальной доставки 1×1010 вг в объеме 2 мкл.
Для данного пилотного исследования использовали мышей Rpgr-/y, поскольку у них отсутствует экспрессия Rpgr, но сохранены соединительные реснички для потенциальной локализации продукта трансгена RPGR. Мышей анестезировали для субретинальной инъекции 2 мкл раствора AAV под верхнюю половину сетчатки.
Через три недели после хирургической операции получавших инъекцию мышей Rpgr-/y вновь анестезировали для in vivo визуализации сетчатки методом конфокальной сканирующей лазерной офтальмоскопии (кСЛО) для изучения картины аутофлуоресценции у животных, которые получали AAV2/8.RK.coRPGR, AAV2/8.RK.wtRPGR, и флуоресценции GFP в качестве показателя эффективности трансдукции у животных, получавших AAV2/8.CAG.eGFP.
Сразу после визуализации мышей умерщвляли и быстро энуклеировали. Целые глаза быстро обрабатывали для иммуногистохимических исследований без фиксации. Вкратце, 16-мкм срезы нефиксированных образцов сетчатки окрашивали красителем Hoechst 33342 и поликлональными антителами, направленными против аминокислот 379-509 в RPGR (Sigma, HPA001593). Антитела осла против иммуноглобулинов кролика, конъюгированные с AlexaFluor 488 (Invitrogen), использовали в качестве вторичного антитела для указанного обнаружения RPGR. Глазные срезы рассматривали под большим увеличением (x63 с масляной иммерсией) на конфокальном микроскопе (Zeiss LSM710) для изучения экспрессии и локализации RPGR в фоторецепторных клетках мышей Rpgr-/y, получавших инъекцию. Не получавшие инъекцию мыши служили в качестве отрицательного контроля для проверки специфичности анализа.
Исход хирургической операции
Все животные получили предназначенную для них дозу раствора AAV в субретинальное пространство и хорошо перенесли анестезию. Это стало возможным благодаря хирургической процедуре, при которой сначала уменьшают внутриглазное давление (IOP) за счет парацентеза передней камеры глаза перед созданием отслоения половины сетчатки при помощи суспензии вектора. Этот метод позволяет осуществлять доставку объема до 2 мкл без развития отека роговицы и/или ограничения внутриглазной циркуляции жидкости из-за высокого IOP. В то же время, пониженное IOP уменьшает риск обратного выброса субретинальной жидкости через инъекционный канал (то есть, в хориоидальную циркуляцию или глазную орбиту).
Оценка состояния глазного дна у получавших инъекцию мышей через 24 ч после операции показала полное повторное прикрепление сетчатки во всех случаях. Визуализация сетчатки продемонстрировала безопасную доставку и экспрессию гена репортера. Через три недели визуализация методом кСЛО показала хорошую оптическую среду с четко просматриваемым глазным дном при визуализации в инфракрасном свете. Визуализация аутофлуоресценции показала гиперфлуоресцентные точки в получавших и не получавших инъекцию глазах у мышей Rpgr-/y, которым вводили AAV2/8.RK.coRPGR или AAV2/8.RK.wtRPGR. У мышей, которым вводили вектор AAV2/8.CAG.GFP, наблюдали сильную и повсеместную флуоресценцию за счет GFP. Это указывало на надежную экспрессию трансгена и вероятность того, что другие рекомбинантные AAV векторы имели достаточно времени для экспрессии трансгена.
Проводили конфокальную сканирующую лазерную офтальмоскопию у (не)получавших инъекцию мышей Rpgr-/y. Выполняли регистрацию изображений в инфракрасном свете с фокальной плоскостью на внутренней части сетчатки. Были получены изображения в режиме аутофлуоресценции. Как не получавшие инъекцию, так и получавшие инъекцию AAV.RK.wtRPGR или AAV.RK.coRPGR, мыши демонстрировали прерывистую картину гиперфлуоресценции. При тех же параметрах чувствительности в глазу, в который вводили AAV.CAG.GFP, наблюдали распространенную и сильную флуоресценцию GFP за пределами зоны отслоения верхней половины сетчатки, что указывало на успешную трансдукцию клеток за пределами участка субретинальной доставки вектора.
Иммуногистохимическое исследование показало экспрессию RPGR
Специфический сигнал наблюдали в срезах глаз у получавших инъекцию мышей, но не у мышей, которые не получали инъекцию. Этот сигнал возникал вследствие связывания антитела с эпитопом RPGR и коррелировал с экспрессией RPGR в результате трансдукции AAV вектора, несущего RPGR в качестве трансгена. Наиболее мощный сигнал был виден в срезах из глаз, в которые вводили AAV2/8.RK.coRPGR, AAV, несущий кодон-оптимизированную последовательность coRPGR.
Результаты иммуногистохимического исследования показали наличие RPGR у получавших инъекцию мышей Rpgr-/y. В не получавшем инъекцию глазу у животных Rpgr-/y отсутствовало окрашивание на RPGR. Однократное введение AAV.RK.wtRPGR приводило к появлению детектируемого сигнала, большая часть которого была локализована в области между внутренним и внешним сегментами. У мышей Rpgr-/y, получавших кодон-оптимизированный вектор, наблюдалась наибольшая экспрессия RPGR с типичной картиной окрашивания маркера соединительных ресничек в виде запятых.
Обсуждение
Хирургический метод, использованный в данном исследовании, позволял безопасно вводить до 2 мкл в субретинальное пространство мышам. Отслоившаяся половина сетчатки самопроизвольно вновь прикреплялась в пределах 24 ч у всех животных и какие-либо осложнения в газах отсутствовали. В то же время этот метод предотвращал (временно) образование отека роговицы и/или прекращение внутриглазной циркуляции жидкости, что можно наблюдать после субретинальных инъекций без предварительного парацентеза. Оптическая среда оставалась прозрачной в течение следующих 3 недель и отсутствовали признаки внутриглазной патологии, такие как клеточные инфильтраты, передние/задние синехии радужки или образование катаракты. Визуализация сетчатки показала высокие уровни экспрессии трансгена GFP в контрольной группе животных, получавших AAV2/8.CAG.GFP. Интересно, что репортерный белок GFP был виден по всей сетчатке, даже при том, что зона непосредственного введения была ограничена верхней половиной сетчатки. Это указывало на по меньшей мере некоторую степень трансдукции клеток за пределами зоны отслоения. Экспрессия GFP находилась под управлением неспецифического промотора CAG, который обеспечивает повсеместную экспрессию трансгена, не ограниченную фоторецепторными клетками. В этом заключается отличие от векторов, несущих последовательность coRPGR по настоящему изобретению, в которых промотор родопсин-киназы обеспечивает специфическую для фоторецепторных клеток экспрессию coRPGR.
Глаза, в которые вводили AAV2/8.RK.coRPGR или AAV2/8.RK.wtRPGR, имели нормальную сосудистую сеть сетчатки и слой нервных волокон, о чем свидетельствовали изображения в инфракрасном свете, сфокусированные на внутренней части сетчатки. Напротив, у мышей, получавших вектор AAV2/8.CAG.eGFP, наблюдали сильную и повсеместную флуоресценцию, вызываемую GFP. В нефиксированных срезах была продемонстрирована экспрессия и локализация RPGR в соединительных ресничках у мышей Rpgr-/y.
Это является первым свидетельством успешной доставки вектора с кодон-оптимизированной последовательностью RPGR при помощи AAV в качестве генной терапии в мышиной модели RP. В отличие от предыдущих исследований, в которых мутации при создании вектора, используемого для экспрессии трансгена, вызывали образование альтернативных белковых продуктов, данная работа продемонстрировала экспрессию и правильную локализацию RPGR с кодон-оптимизированной последовательностью, что привело к образованию белкового продукта RPGR дикого типа.
ПРИМЕР 5
Безопасность и эффективность генной терапии с использованием coRPGR
Разработка метода генной терапии для XLRP оставалась сложной задачей по ряду причин. Одной из причин является богатая пуринами повторяющаяся последовательность RPGRORF15, что осложняет клонирование без возникновения спонтанных мутаций. Подтверждение целостности последовательности при секвенировании методом Сэнгера также является проблематичным, поскольку частые полигуаниновые отрезки заставляют ДНК-полимеразы задерживаться или останавливаться.
Второй проблемой является умеренно выраженный фенотип в мышиных моделях заболевания, таких как мыши Rpgr/y и C57BL/6JRd9/Boc. При относительно небольших структурных и функциональных различиях между этими моделями заболевания и контролями дикого типа сложно добиться уровня статистической значимости в группе лечения.
Для решения первой проблемы авторы изобретения использовали принцип кодон-оптимизации для изменения первичной нуклеотидной последовательности кодирующей RPGRORF15 последовательности. Поскольку были использованы только синонимичные кодоны, полученная аминокислотная последовательность оставалась неизменной, в то время как кодон-оптимизированная конструкция RPGR демонстрировала превосходную стабильность последовательности и эффективность трансляции в сравнении с конструкцией RPGR дикого типа. Это преимущество было очевидно in vitro при использовании трансфекции плазмидами и экспериментов по трансдукции AAV конструкциями. Те же AAV конструкции затем были использованы в пилотном исследовании для демонстрации их способности трансдуцировать фоторецепторные клетки (пример 4). Полученный белок RPGR был локализован в соединительных ресничках, его физиологическом компартменте в фоторецепторных клетках.
Для решения второй проблемы с небольшими различиями между группами животных, имеющих и не имеющих разрушительные мутации Rpgr, авторы изобретения выполнили расчеты размера выборки и мощности исследования на основе соответствующих объективных и количественных показателей результатов.
Цель данной работы заключалась в проверке эффективности AAV.RK.coRPGR в качестве генно-терапевтического средства для XLRP3 в двух релевантных животных моделях (мыши Rpgr-/y и C57BL/6JRd9/Boc) и в изучении любых потенциальных токсических эффектов у животных дикого типа (C57BL/6J). Дизайн исследования был выбран для получения надежных статистических данных, которые потенциально могут служить основой для получения одобрения регулирующих органов на проведение фазы I клинических испытаний.
Материалы и методы
Расчеты размера выборки и мощности исследования
Расчеты размера выборки и мощности исследования выполняли с использованием алгоритма на основе JavaScript, предоставленного Rollin Brant с кафедры биостатистики отдела эпидемиологии и биостатистики Университета Калифорнии, Сан-Франциско. В качестве первичного критерия эффективности использовали амплитуду a-волны [мкВ] после однократного стимулирования адаптированного к темноте глаза вспышкой света при 0,01 кд.с/м2, что отражает физиологию подавляющего большинства фоторецепторных клеток в сетчатке мыши, целевой популяции клеток в данном исследовании. Эти электроретинографические (ЭРГ) ответы были измерены, и средние значения и стандартные отклонения для n=4 на группу (мышей C57BL/6J, C57BL/6JRd9/Boc и Rpgr-/y) были рассчитаны в Excel. Перед расчетом размера выборки рассчитывали разницу между средней амплитуды a-волны у мышей C57BL/6J (целевое нормальное значение) и C57BL/6JRd9/Boc (исходное значение до лечения), а также эту разницу у мышей C57BL/6J и Rpgr-/y, и ½ от этого значения прибавляли к текущему среднему значению у соответствующих животных моделей заболевания, предполагая 50%-е улучшение вследствие лечения. Это позволило получить значения для μ0 (среднее в группе без лечения) и μ1 (среднее в группе с лечением). Значение для σ (стандартное отклонение для совокупности выборки) было принято равным стандартному отклонению амплитуды a-волны в соответствующей модели заболевания. Предполагалось одностороннее тестирование, частота ошибок I рода была принята за α=0,05 и желаемую мощность исследования считали равной 90% для расчета размера выборки.
Затем использовали сопоставимый размер выборки для расчета мощности исследования для обнаружения токсических эффектов у мышей C57BL/6J. Для этого, потерю 10% от средней амплитуды a-волны в начале исследования считали токсическим эффектом и вводили стандартное отклонение исходного значения как σ.
Дизайн исследования
ЭРГ ответы были выбраны в качестве первичного критерия эффективности, поскольку они являются объективным и количественным биомаркером функции сетчатки, который связан с процессом заболевания, и подходящим показателем для определения потенциального терапевтического и/или токсического эффекта предмета исследования, AAV.RK.coRPGR. Из-за относительно высокой вариабельности между отдельными членами группы была выбрана парадигма внутрииндивидуального тестирования: один глаз будет получать AAV.RK.coRPGR (настоящий лекарственный препарат), при этом контралатеральный глаз будет служить в качестве контроля. Для того, чтобы отслеживать естественный процесс заболевания и иметь в качестве контроля инъекцию неактивного вещества (AAV.control) в таком дизайне исследования, проводили два параллельных исследования: одно (обязательное) открытое исследование с односторонним лечением рандомизированных глаз при помощи AAV.RK.coRPGR использовали для сравнения эффекта лечения с естественным процессом заболевания.
Второе исследование имело дизайн слепого исследования со случайным выбором глаз, получавших AAV.RK.coRPGR или AAV.control. Последнее исследование использовали для контролирования эффекта хирургической операции и воздействия AAV. Все 129 животных были подвергнуты воздействию после отлучения от матерей в постнатальный день P22 ± 2 и тестированию в трех последующих временных точках: постнатальный месяц 2 (PM2), PM4 и PM6 до момента умерщвления. ЭРГ регистрировали во всех трех временных точках и, кроме того, проводили кСЛО в последней временной точке PM6.
Дизайн исследования для определения эффективности и токсичности
Мышам Rpgr-/y, C57BL/6JRd9/Boc и C57BL/6J выполняли либо двусторонние инъекции в слепом режиме для тестирования AAV.RK.coRPGR против AAV.control (верх), либо одностороннюю инъекцию только AAV.RK.coRPGR. Все глаза случайным образом распределяли для получения инъекции настоящего или фиктивного препарата, либо для получения и не получения инъекции. Хирургическую операцию выполняли в постнатальный день 22 (P22), с последующей электроретинографией (ЭРГ) в постнатальный месяц 2 (PM2), PM4 и PM6. В PM6 проводили сканирующую лазерную офтальмоскопию (СЛО) перед умерщвлением и обработкой глаз для гистологического или вестерн-блот анализа.
Мыши C57BL/6J дикого типа
В общей сложности n=47 C57BL/6J мышей были использованы в двух исследованиях для изучения потенциальных токсических эффектов субретинальной доставки AAV2/8.RK.coRPGR. 24 животным были проведены односторонние инъекции в глаз в открытом исследовании, при этом глаз выбирали случайным образом. Инъекция представляла собой одну субретинальную инъекцию 1,5×109 вг AAV2/8.RK.coRPGR, разведенных в BSS® с 0,001% PF-68. Остальные 23 животных получали двусторонние инъекции в слепом и рандомизированном режиме. Инъекция в группе получения настоящего препарата была такой, как описано выше, при этом контрольная группа получала 1,5×109 вг AAV2/8.control, разведенных в той же среде (BSS® с 0,001% PF-68). Оба вектора описаны подробно в примере 4.
Субретинальные инъекции выполняли животным после отлучении от матери в возрасте 3 недель. Хирург использовал систему оценки в баллах (0-10) для определения качества субретинальной инъекции, при этом показатель 0/10 соответствовал отсутствию доставки вектора и 10/10 соответствовал полной доставке вектора без какого-либо осложнения, такого как внешнее (например, подконъюнктивальное) или внутреннее (например, внутриретинальное/витреальное) кровотечение в процессе или после субретинальной инъекции. Показатель 9/10 присваивали, например, в случае полной доставки вектора, но с незначительным подконъюнктивальным кровотечением. Неудачными считали инъекции с показателем ≤7, и они были исключены из исследования.
За хирургическим вмешательством следовало продолжительное наблюдение внутрииндивидуальных различий кривых ЭРГ в постнатальный месяц два (PM2) и PM4. В PM6 проводили дополнительную визуализацию сетчатки методом СЛО сразу после записи кривых ЭРГ. Мышей умерщвляли в последней временной точке (PM6), и глаза извлекали для обнаружения трансгена в вестерн-блот, или иммуногистохимическом, анализе.
Мыши C57BL/6JRd9/Boc
В общей сложности n=36 мышей C57BL/6JRd9/Boc были использованы в обоих исследованиях. 19 животным были проведены односторонние инъекции в глаз в открытом исследовании, в то время как 17 животных получали в слепом и рандомизированном режиме двусторонние инъекции настоящего или контрольного препарата, как описано выше для мышей C57BL/6J. Это исследование было проведено для оценки эффективности основанной на AAV заместительной генной терапии для гена RPGR. Последующее наблюдение было таким, как описано выше.
Мыши Rpgr-/y
Мышей Rpgr-/y использовали для оценки эффективности основанной на AAV заместительной генной терапии для гена RPGR во второй животной модели XLRP. Для этого в общей сложности n=46 животным были проведены инъекции в двух отдельных исследованиях, описанных выше. 25 животным были проведены односторонние инъекции в глаз в открытом исследовании, в то время как 21 животное получало инъекции в оба глаза (настоящий препарат и контроль), как описано выше. Обследование в период последующего наблюдения проводили таким же образом, как описано выше для мышей C57BL/6J.
Результаты
Расчеты размера выборки и мощности исследования
Рассчитанный размер выборки был основан на данных ЭРГ, полученных для четырех глаз (от двух животных) для каждой группы (C57BL/6J, C57BL/6JRd9/Boc и Rpgr-/y mice). Средняя амплитуда a-волны из n=9 испытаний была зарегистрирована для каждого глаза.
После расчетов значений μ0, μ1 и сигма на основании приведенных выше данных, рекомендуемые размеры выборки находились в диапазоне от 16 до 21. У мышей C57BL/6JRd9/Boc наблюдали наименьшую амплитуду (65 ± 11 мкВ; среднее ± стандартное отклонение) в сравнении с контролем дикого типа (81 ± 7 мкВ) и, вследствие этого, необходимый размер выборки, по оценкам, составлял лишь n=16. Рекомендуемый размер выборки для Rpgr-/y составлял n=21 из-за более высокой амплитуды в начале исследования (728 мкВ). Расчеты мощности исследования показали, что потеря 10% амплитуды от исходного значения у мышей дикого типа была бы обнаружена с мощностью 100% при использовании размера группы n=20.
Мыши C57BL/6J дикого типа
Шесть глаз были исключены из исследования вследствие неудачного хирургического вмешательства и один глаз был исключен вследствие ранее существующих изменений переднего сегмента глаза. В результате этого общий размер группы составил n=19 для исследования с односторонним введением и n=22 для исследования с двусторонним введением. Средние значения [и диапазон] документированного качества хирургической операции были очень сходными во всех группах: 9,0 [8-10] для открытого исследования, 9,5 [8-10] для группы введения настоящего препарата и 9,3 [8-10] для контрольной группы. Все мыши быстро восстанавливались после операции.
Ни одна из зарегистрированных серий сигналов при разной интенсивности света в условиях адаптации как к темноте, так и к свету, не показала существенных различий между получавшей и не получавшей инъекцию группами, или между получавшей настоящий препарат и контрольной группами в любой временной точке. Результаты визуализации сетчатки методом сканирующей лазерной офтальмоскопии (СЛО) также подтвердили, что токсический эффект на структуры сетчатки отсутствовал вплоть до 6 месяцев после субретинального введения AAV.RK.coRPGR.
Кривые ЭРГ у мышей C57BL/6J после двусторонней субретинальной инъекции AAV.RK.coRPGR или AAV.control были получены в PM2, в PM4 и в PM6, последней протестированной временной точке. Факториальный анализ ANOVA для повторных измерений подтвердил нулевую гипотезу (отсутствие разницы) во всех анализах.
Вестерн-блот анализ суммарных лизатов сетчатки подтвердил экспрессию трансгена RPGR в получавшем инъекцию глазу, но не в контрольном глазу. Полоса имела предсказанную молекулярную массу, и какие-либо лишние полосы отсутствовали. Иммуногистохимический анализ нефиксированных срезов замороженных тканей также продемонстрировал экспрессию трансгена RPGR и его совместную локализацию с естественным Rpgrip в получавшем инъекцию глазу, но не оставленном без инъекции контрольном глазу.
Окрашивание гематоксилином и эозином (H&E) срезов сетчатки из получавших (TE) и не получавших инъекцию (UE) глаз продемонстрировало нормальную анатомию сетчатки в обеих группах. Ни в одной группе не были обнаружены какие-либо признаки воспаления или дегенерации в каком-либо из срезов.
Был проведен вестерн-блот анализ суммарных лизатов сетчатки из получавших инъекцию глаз в сравнении с не получавшими инъекцию глазами у мышей C57BL/6J, C57BL/6JRd9/Boc и Rpgr-/y. Экспрессия RpgrORF15 имела место только в получавших инъекцию глазах, о чем свидетельствовала полоса на уровне, соответствующем 200 кДа. Наиболее сильно выраженная полоса имела место у мышей C57BL/6JRd9/Boc, следующей по интенсивности была полоса у C57BL/6J, при этом образец от Rpgr-/y демонстрировал наиболее слабую экспрессию. GAPDH использовали в качестве контроля нагрузки (красный цвет). Нативные клетки HEK293T использовали в качестве отрицательного контроля (ок) и клетки HEK293T, трансфицированные экспрессионной плазмидой coRPGR, использовали в качестве положительного контроля.
Иммуногистохимическое исследование мышей C57BL/6J. Никакой экспрессии RPGR не было обнаружено в контрольном получавшем инъекцию глазу. Введение AAV.coRPGR приводило к экспрессии RPGR и совместной локализации человеческого RPGR с Rpgrip.
Окрашивание гематоксилином и эозином глазных срезов от мышей C57BL/6J показало отсутствие признаков воспаления или дегенерации во всех срезах. Были видны структуры в нормальном состоянии.
Мыши C57BL/6JRd9/Boc
Одну мышь из открытого исследования с односторонней инъекцией пришлось исключить из-за ранее существующего инфильтрата роговицы с сопутствующим микрофтальмом в не получавшем инъекцию контрольном глазу. Микрофтальм мог привести к ложно низким показателям ЭРГ в не получавшем инъекцию глазу. В результате этого общее количество животных составило n=18 в исследовании с односторонней инъекцией и n=17 животных в исследовании с двусторонними инъекциями. Ни одно из получавших инъекции животных не было исключено из-за осложнений во время операции, и все животные получили инъекции категории «хорошо» или «отлично» со средним баллом (диапазон составлял 8-10 во всех группах) 9,7 в открытом исследовании, 9,1 в группе получения настоящего препарата и 9,0 в контрольной группе.
Во время длительного последующего наблюдения два животных из исследования с односторонней инъекцией не оправились от анестезии в PM4. В общей сложности 102 набора данных двусторонней ЭРГ в трех временных точках были успешно получены и сохранены для последующего анализа. В открытом исследовании с односторонней инъекцией факториальный анализ ANOVA для повторных измерений подтвердил нулевую гипотезу (отсутствие разницы) в первой временной точке (PM2), но продемонстрировал тенденцию в сторону больших амплитуд для получавших инъекцию глаз при более высоких интенсивностях света в серии экспериментов как с адаптацией к темноте, так и к свету. В PM4 получавшие инъекцию глаза реагировали с гораздо более высокими амплитудами в серии с разными интенсивностями света после темновой адаптации как в случае a-волны (p=0,001), так и b-волны (p=0,002). Ответы адаптированных к свету глаз существенно не отличались. Эта картина сохранялась до последней временной точки (PM6).
В слепом исследовании с двусторонними инъекциями нулевая гипотеза была подтверждена во всех временных точках. Однако в получавших настоящий препарат глазах опять-таки всегда наблюдалась тенденция в сторону более высоких амплитуд, особенно при высоких интенсивностях стимула.
Результаты визуализации сетчатки методом сканирующей лазерной офтальмоскопии (СЛО) также подтверждали отсутствие токсического эффекта на структуру сетчатки вплоть до 6 месяцев после субретинального введения AAV.RK.coRPGR. Однако аутофлуоресцентная визуализация выявила изменение в проявлении заболевания у получавших инъекцию мышей C57BL/6JRd9/Boc: авторы изобретения ранее показали, что отсутствие экспрессии Rpgr у мышей связано с присутствием гиперфлуоресцентных точек во всей сетчатке, видимых при анализе аутофлуоресценции, хотя мыши дикого типа имели лишь минимальный гомогенный сигнал аутофлуоресценции. Интересно, что визуализация в получавших настоящий препарат глазах в PM6 показала уменьшение количества этих гиперфлуоресцентных точек в верхней половине сетчатки, куда исходно был введен вектор. Поля, соответствующие половинам сетчатки, значительно отличались в получавших инъекцию глазах: 13 ± 14 (среднее ± стандартное отклонение) в получавших инъекцию верхних половинах сетчатки против 59 ± 40 в ипсилатеральных нижних половинах сетчатки (p=0,005; n=6, парный критерий Стьюдента). Сравнение верхней половины сетчатки получавших инъекцию глаз с контралатеральной не получавшей инъекцию сетчаткой приводило к аналогичному результату (p=0,037; n=6, парный критерий Стьюдента).
Вестерн-блот анализ суммарных лизатов сетчатки подтвердил экспрессию трансгена RPGR в получавшем инъекцию глазу, но не в контрольном глазу. Полоса соответствовала предсказанной молекулярной массе, и какие-либо лишние полосы отсутствовали. Иммуногистохимический анализ нефиксированных срезов замороженных тканей также продемонстрировал экспрессию трансгена RPGR и его совместную локализацию с естественным Rpgrip в получавшем инъекцию глазу, но не в контрольном глазу.
Мыши Rpgr-/y
Одну мышь из открытого исследования с односторонней инъекцией пришлось исключить из-за ранее существующего микрофтальма, который был замечен только после хирургического вмешательства, в не получавшем инъекцию контралатеральном глазу и привел бы к ложно низким показателям ЭРГ в не получавшем инъекцию глазу. Кроме того, одно животное из исследования с двусторонними инъекциями пришлось исключить из-за осложнений во время хирургического вмешательства (частичная интравитреальная инъекция с интравитреальным кровотечением). В результате этого общее количество животных составило n=24 в исследовании с односторонней инъекцией и n=20 животных в исследовании с двусторонними инъекциями. Рейтинг успеха хирургического вмешательства был высоким во всех группах: средние баллы [диапазон] составляли 9,3 [8-10] в открытом исследовании, 9,5 [9-10] в группе получения настоящего препарата и 9,4 [8-10] в контрольной группе.
Иммуногистохимическое исследование мышей C57BL/6JRd9/Boc. Верхние панели демонстрируют отсутствие экспрессии RPGR в получавшем контрольную инъекцию глазу. Введение AAV.coRPGR приводило к экспрессии RPGR и его совместной локализации с Rpgrip. Масштабная полоска соответствует 20 мкм. Во время длительного последующего наблюдения одно животное в группе двусторонних инъекций в PM2 и два животных в группе односторонней инъекции в PM4 не оправились от анестезии. В общей сложности 128 наборов данных двусторонней ЭРГ в трех временных точках были успешно получены и сохранены для последующего анализа.
В открытом исследовании с односторонней инъекцией отсутствовало видимое различие между глазами в самой ранней временной точке (PM2), но сильный эффект воздействия был виден в ЭРГ ответах адаптированных к темноте глаз в PM4 (p<0,001) и PM6 (p<0,001), как в случае амплитуд a-, так и b-волн. Фотопические b-волновые ответы были значительно больше в PM6 (p=0,004). В слепом исследовании с двусторонними инъекциями были продемонстрированы значительно повышенные амплитуды b-волновых ответов после темновой адаптации в PM6, и амплитуда b-волн после световой адаптации была значительно увеличена в PM4 (но не в PM6).
В глазах у мышей Rpgr-/y, получавших AAV.RK.coRPGR, наблюдалось уменьшение количества гиперфлуоресцентных точек в верхней половине сетчатки, что также имело место в получавших инъекцию глазах у мышей C57BL/6JRd9/Boc. Напротив, в не получавших инъекцию или получавших фиктивный препарат глазах у мышей Rpgr-/y повсеместно наблюдались гиперфлуоресцентные точки, ассоциированные с нулевой мутацией Rpgr.
Вестерн-блот анализ суммарных лизатов сетчатки подтвердил экспрессию трансгена RPGR в получавшем инъекцию глазу, но не в контрольном глазу. Полоса соответствовала предсказанной молекулярной массе, и какие-либо лишние полосы отсутствовали. Иммуногистохимический анализ нефиксированных срезов замороженных тканей также продемонстрировал экспрессию трансгена RPGR и его совместную локализацию с естественным Rpgrip в получавшем инъекцию глазу, но не в контрольном глазу. ЭРГ кривые у мышей Rpgr-/y после односторонней субретинальной инъекции AAV.RK.coRPGR.
ЭРГ кривые у мышей Rpgr-/y после двусторонней субретинальной инъекции AAV.RK.coRPGR или AAV.control. Введение AAV.RK.coRPGR приводило к значительному улучшению амплитуд ЭРГ после темновой адаптации. Эффект воздействия на амплитуды b-волны после световой адаптации не сохранялся в PM6.
С использованием визуализации методом сканирующей лазерной офтальмоскопии в режиме инфракрасного света фокальная плоскость была установлена на внутренней части сетчатки или внешней части сетчатки. Четкое отличие картин аутофлуоресценции между получавшими лечение и получавшими фиктивный препарат или контрольными глазами: в получавшем лечение глазу отмечено меньше гиперфлуоресцентных точек в верхней половине сетчатки - зоне введения AAV.RK.coRPGR.
Иммуногистохимическое исследование мышей Rpgr-/y. Никакой экспрессии RPGR не было обнаружено в контрольном получавшем инъекцию глазу. Введение AAV.coRPGR приводило к экспрессии RPGR и совместной локализации человеческого RPGR с Rpgrip. Масштабная полоска соответствует 20 мкм.
Обсуждение
Заместительная терапия для гена RPGR вызывала интерес со времени определения RPGR, как генетической причины XLRP3. Основной причиной того, что она все еще остается целью, не доведенной до стадии клинических испытаний, является тот факт, что RPGR представляет собой сложный ген с высокой предрасположенностью к мутационным изменениям. Это вызвало серьезную задержку в получении набора данных в поддержку заявок на клинические испытания. И даже в случае одобрения регулирующими органами испытаний на безопасность, производство подходящего для клинического применения AAV для генной терапии гена RPGR будет представлять серьезную проблему.
Приведенные данные показывают отсутствие токсических эффектов у животных дикого типа, C57BL/6J, и показывают эффективность векторной конструкции нового типа для генной терапии гена RPGR в двух релевантных животных моделях (мыши Rpgr-/y и C57BL/6JRd9/Boc). Этот вектор на основе AAV содержит кодон-оптимизированную кодирующую последовательность RPGRORF15, которая делает конструкцию более генетически стабильной, при этом приводя к образованию идентичного белкового продукта, RPGRORF15. Результаты тщательных in vitro анализов и пилотного исследования на мышах Rpgr-/y (пример 4), полученные в соответствии с настоящим изобретением, показывают, что одно субретинальное введение AAV.RK.coRPGR является безопасным и способно приводить к остановке или замедлению прогрессирования дегенерации сетчатки из-за мутаций в Rpgr.
Безопасность
Для изучения потенциальных токсических эффектов AAV.RK.coRPGR авторы изобретения тестировали одну и ту же дозу (1,5×109 вг в 1,5 мкл) на сорока одной мыши C57BL/6J. Девятнадцать мышей получали инъекцию только в один глаз, в то время как 22 мыши получали двустороннюю инъекцию либо AAV.RK.coRPGR, либо AAV.control, в слепом режиме. Не было отмечено никаких существенных различий между показателями функции сетчатки, полученными методом ЭРГ, в любой временной точке любого исследования в группе получения настоящего препарата и группе получения контрольного препарата или не получавшей инъекцию группе. Кроме того, визуализация сетчатки in vivo свидетельствовала о том, что отсутствовало влияние инъекции AAV.RK.coRPGR на структуру сетчатки. Окрашивание H&E показало отсутствие токсических эффектов инъекции, а иммуногистохимический и вестерн-блот анализы продемонстрировали экспрессию и локализацию RPGRORF15 у получавших инъекцию животных, но не у животных, не получавших инъекцию или получавших контрольную инъекцию. Суммируя приведенные результаты, одна инъекция AAV.RK.coRPGR мышам дикого типа не приводила к каким-либо токсическим эффектам и характеризовалась хорошим профилем безопасности. Вследствие этого, авторы изобретения сделали вывод о том, что введение AAV.RK.coRPGR было безопасным.
Эффективность
Терапевтический эффект AAV.RK.coRPGR был продемонстрирован в двух хорошо охарактеризованных мышиных моделях XLRP3: трансгенных мышах Rpgr-/y и мышах C57BL/6JRd9/Boc, имеющих природную мутацию в гене Rpgr. Обе модели характеризуются отсутствием экспрессии RpgrORF15 в сетчатке и, вследствие этого, были выбраны в качестве релевантных животных моделей XLRP3. Однако использование данных животных моделей имеет некоторые недостатки. Наиболее важно, что заболевание имеет неожиданно слабое фенотипическое проявление, что приводит к необходимости использования в исследовании относительно больших групп животных для достижения необходимой статистической мощи исследования. Вследствие этого, авторы изобретения проводили исследования в общей сложности на 80 животных и предоставили надежное доказательство эффективности, о чем свидетельствует значительное улучшение электрофизиологических показателей у мышей Rpgr-/y и C57BL/6JRd9/Boc.
Эффект инъекции не был явным в первой временной точке исследования, наиболее вероятно, вследствие медленного прогрессирования заболевания в обеих животных моделях. Однако введение AAV.RK.coRPGR было связано со значительным улучшением показателей ЭРГ в обеих животных моделях в PM4 и PM6. Это улучшение показателей было более очевидным при использовании вспышек света разной интенсивности после темновой адаптации, что отражает суммарный потенциал палочковидных фоторецепторных клеток в диапазоне света меньшей интенсивности (однократные вспышки с интенсивностью до 0,01 кд.с/м2). Считается, что вспышки с большей интенсивностью стимулируют смешанный колбочко-палочковый ответ. Наибольшая разница между получавшими лечение и получавшими фиктивный препарат/не получавшими инъекцию глазами была заметна при интенсивностях света примерно 1 кд.с/м2, это указывало на то, что как палочковидные, так и колбочковидные фоторецепторные клетки могли стать более эффективными в результате трансдукции AAV.RK.coRPGR.
Подводя итоги, введение AAV.RK.coRPGR было безопасным и эффективным. Успешная трансдукция AAV.RK.coRPGR фоторецепторных клеток у мышей дикого типа не приводила к токсическим эффектам, ассоциированным с экспрессией RPGRORF15. Более того, лечение в животных моделях XLRP3 приводило к статистически значимому улучшению показателей ЭРГ в получавших инъекцию глазах, но не в глазах, не получавших инъекцию. Эти данные показали, что кодон-оптимизированную кодирующую RPGRORF15 последовательность по настоящему изобретению можно использовать для безопасного лечения глаз.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Oxford University Innovation Limited
<120> ЛЕЧЕНИЕ ПИГМЕНТНОГО РЕТИНИТА
<130> P107084PCT
<150> GB 1516066.6
<151> 2015-09-10
<160> 14
<170> PatentIn version 3.5
<210> 1
<211> 1152
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Arg Glu Pro Glu Glu Leu Met Pro Asp Ser Gly Ala Val Phe Thr
1 5 10 15
Phe Gly Lys Ser Lys Phe Ala Glu Asn Asn Pro Gly Lys Phe Trp Phe
20 25 30
Lys Asn Asp Val Pro Val His Leu Ser Cys Gly Asp Glu His Ser Ala
35 40 45
Val Val Thr Gly Asn Asn Lys Leu Tyr Met Phe Gly Ser Asn Asn Trp
50 55 60
Gly Gln Leu Gly Leu Gly Ser Lys Ser Ala Ile Ser Lys Pro Thr Cys
65 70 75 80
Val Lys Ala Leu Lys Pro Glu Lys Val Lys Leu Ala Ala Cys Gly Arg
85 90 95
Asn His Thr Leu Val Ser Thr Glu Gly Gly Asn Val Tyr Ala Thr Gly
100 105 110
Gly Asn Asn Glu Gly Gln Leu Gly Leu Gly Asp Thr Glu Glu Arg Asn
115 120 125
Thr Phe His Val Ile Ser Phe Phe Thr Ser Glu His Lys Ile Lys Gln
130 135 140
Leu Ser Ala Gly Ser Asn Thr Ser Ala Ala Leu Thr Glu Asp Gly Arg
145 150 155 160
Leu Phe Met Trp Gly Asp Asn Ser Glu Gly Gln Ile Gly Leu Lys Asn
165 170 175
Val Ser Asn Val Cys Val Pro Gln Gln Val Thr Ile Gly Lys Pro Val
180 185 190
Ser Trp Ile Ser Cys Gly Tyr Tyr His Ser Ala Phe Val Thr Thr Asp
195 200 205
Gly Glu Leu Tyr Val Phe Gly Glu Pro Glu Asn Gly Lys Leu Gly Leu
210 215 220
Pro Asn Gln Leu Leu Gly Asn His Arg Thr Pro Gln Leu Val Ser Glu
225 230 235 240
Ile Pro Glu Lys Val Ile Gln Val Ala Cys Gly Gly Glu His Thr Val
245 250 255
Val Leu Thr Glu Asn Ala Val Tyr Thr Phe Gly Leu Gly Gln Phe Gly
260 265 270
Gln Leu Gly Leu Gly Thr Phe Leu Phe Glu Thr Ser Glu Pro Lys Val
275 280 285
Ile Glu Asn Ile Arg Asp Gln Thr Ile Ser Tyr Ile Ser Cys Gly Glu
290 295 300
Asn His Thr Ala Leu Ile Thr Asp Ile Gly Leu Met Tyr Thr Phe Gly
305 310 315 320
Asp Gly Arg His Gly Lys Leu Gly Leu Gly Leu Glu Asn Phe Thr Asn
325 330 335
His Phe Ile Pro Thr Leu Cys Ser Asn Phe Leu Arg Phe Ile Val Lys
340 345 350
Leu Val Ala Cys Gly Gly Cys His Met Val Val Phe Ala Ala Pro His
355 360 365
Arg Gly Val Ala Lys Glu Ile Glu Phe Asp Glu Ile Asn Asp Thr Cys
370 375 380
Leu Ser Val Ala Thr Phe Leu Pro Tyr Ser Ser Leu Thr Ser Gly Asn
385 390 395 400
Val Leu Gln Arg Thr Leu Ser Ala Arg Met Arg Arg Arg Glu Arg Glu
405 410 415
Arg Ser Pro Asp Ser Phe Ser Met Arg Arg Thr Leu Pro Pro Ile Glu
420 425 430
Gly Thr Leu Gly Leu Ser Ala Cys Phe Leu Pro Asn Ser Val Phe Pro
435 440 445
Arg Cys Ser Glu Arg Asn Leu Gln Glu Ser Val Leu Ser Glu Gln Asp
450 455 460
Leu Met Gln Pro Glu Glu Pro Asp Tyr Leu Leu Asp Glu Met Thr Lys
465 470 475 480
Glu Ala Glu Ile Asp Asn Ser Ser Thr Val Glu Ser Leu Gly Glu Thr
485 490 495
Thr Asp Ile Leu Asn Met Thr His Ile Met Ser Leu Asn Ser Asn Glu
500 505 510
Lys Ser Leu Lys Leu Ser Pro Val Gln Lys Gln Lys Lys Gln Gln Thr
515 520 525
Ile Gly Glu Leu Thr Gln Asp Thr Ala Leu Thr Glu Asn Asp Asp Ser
530 535 540
Asp Glu Tyr Glu Glu Met Ser Glu Met Lys Glu Gly Lys Ala Cys Lys
545 550 555 560
Gln His Val Ser Gln Gly Ile Phe Met Thr Gln Pro Ala Thr Thr Ile
565 570 575
Glu Ala Phe Ser Asp Glu Glu Val Glu Ile Pro Glu Glu Lys Glu Gly
580 585 590
Ala Glu Asp Ser Lys Gly Asn Gly Ile Glu Glu Gln Glu Val Glu Ala
595 600 605
Asn Glu Glu Asn Val Lys Val His Gly Gly Arg Lys Glu Lys Thr Glu
610 615 620
Ile Leu Ser Asp Asp Leu Thr Asp Lys Ala Glu Val Ser Glu Gly Lys
625 630 635 640
Ala Lys Ser Val Gly Glu Ala Glu Asp Gly Pro Glu Gly Arg Gly Asp
645 650 655
Gly Thr Cys Glu Glu Gly Ser Ser Gly Ala Glu His Trp Gln Asp Glu
660 665 670
Glu Arg Glu Lys Gly Glu Lys Asp Lys Gly Arg Gly Glu Met Glu Arg
675 680 685
Pro Gly Glu Gly Glu Lys Glu Leu Ala Glu Lys Glu Glu Trp Lys Lys
690 695 700
Arg Asp Gly Glu Glu Gln Glu Gln Lys Glu Arg Glu Gln Gly His Gln
705 710 715 720
Lys Glu Arg Asn Gln Glu Met Glu Glu Gly Gly Glu Glu Glu His Gly
725 730 735
Glu Gly Glu Glu Glu Glu Gly Asp Arg Glu Glu Glu Glu Glu Lys Glu
740 745 750
Gly Glu Gly Lys Glu Glu Gly Glu Gly Glu Glu Val Glu Gly Glu Arg
755 760 765
Glu Lys Glu Glu Gly Glu Arg Lys Lys Glu Glu Arg Ala Gly Lys Glu
770 775 780
Glu Lys Gly Glu Glu Glu Gly Asp Gln Gly Glu Gly Glu Glu Glu Glu
785 790 795 800
Thr Glu Gly Arg Gly Glu Glu Lys Glu Glu Gly Gly Glu Val Glu Gly
805 810 815
Gly Glu Val Glu Glu Gly Lys Gly Glu Arg Glu Glu Glu Glu Glu Glu
820 825 830
Gly Glu Gly Glu Glu Glu Glu Gly Glu Gly Glu Glu Glu Glu Gly Glu
835 840 845
Gly Glu Glu Glu Glu Gly Glu Gly Lys Gly Glu Glu Glu Gly Glu Glu
850 855 860
Gly Glu Gly Glu Glu Glu Gly Glu Glu Gly Glu Gly Glu Gly Glu Glu
865 870 875 880
Glu Glu Gly Glu Gly Glu Gly Glu Glu Glu Gly Glu Gly Glu Gly Glu
885 890 895
Glu Glu Glu Gly Glu Gly Glu Gly Glu Glu Glu Gly Glu Gly Glu Gly
900 905 910
Glu Glu Glu Glu Gly Glu Gly Lys Gly Glu Glu Glu Gly Glu Glu Gly
915 920 925
Glu Gly Glu Gly Glu Glu Glu Glu Gly Glu Gly Glu Gly Glu Asp Gly
930 935 940
Glu Gly Glu Gly Glu Glu Glu Glu Gly Glu Trp Glu Gly Glu Glu Glu
945 950 955 960
Glu Gly Glu Gly Glu Gly Glu Glu Glu Gly Glu Gly Glu Gly Glu Glu
965 970 975
Gly Glu Gly Glu Gly Glu Glu Glu Glu Gly Glu Gly Glu Gly Glu Glu
980 985 990
Glu Glu Gly Glu Glu Glu Gly Glu Glu Glu Gly Glu Gly Glu Glu Glu
995 1000 1005
Gly Glu Gly Glu Gly Glu Glu Glu Glu Glu Gly Glu Val Glu Gly
1010 1015 1020
Glu Val Glu Gly Glu Glu Gly Glu Gly Glu Gly Glu Glu Glu Glu
1025 1030 1035
Gly Glu Glu Glu Gly Glu Glu Arg Glu Lys Glu Gly Glu Gly Glu
1040 1045 1050
Glu Asn Arg Arg Asn Arg Glu Glu Glu Glu Glu Glu Glu Gly Lys
1055 1060 1065
Tyr Gln Glu Thr Gly Glu Glu Glu Asn Glu Arg Gln Asp Gly Glu
1070 1075 1080
Glu Tyr Lys Lys Val Ser Lys Ile Lys Gly Ser Val Lys Tyr Gly
1085 1090 1095
Lys His Lys Thr Tyr Gln Lys Lys Ser Val Thr Asn Thr Gln Gly
1100 1105 1110
Asn Gly Lys Glu Gln Arg Ser Lys Met Pro Val Gln Ser Lys Arg
1115 1120 1125
Leu Leu Lys Asn Gly Pro Ser Gly Ser Lys Lys Phe Trp Asn Asn
1130 1135 1140
Val Leu Pro His Tyr Leu Glu Leu Lys
1145 1150
<210> 2
<211> 3459
<212> ДНК
<213> Homo sapiens
<400> 2
atgagggagc cggaagagct gatgcccgat tcgggtgctg tgtttacatt tgggaaaagt 60
aaatttgctg aaaataatcc cggtaaattc tggtttaaaa atgatgtccc tgtacatctt 120
tcatgtggag atgaacattc tgctgttgtt accggaaata ataaacttta catgtttggc 180
agtaacaact ggggtcagtt aggattagga tcaaagtcag ccatcagcaa gccaacatgt 240
gtcaaagctc taaaacctga aaaagtgaaa ttagctgcct gtggaaggaa ccacaccctg 300
gtgtcaacag aaggaggcaa tgtatatgca actggtggaa ataatgaagg acagttgggg 360
cttggtgaca ccgaagaaag aaacactttt catgtaatta gcttttttac atccgagcat 420
aagattaagc agctgtctgc tggatctaat acttcagctg ccctaactga ggatggaaga 480
ctttttatgt ggggtgacaa ttccgaaggg caaattggtt taaaaaatgt aagtaatgtc 540
tgtgtccctc agcaagtgac cattgggaaa cctgtctcct ggatctcttg tggatattac 600
cattcagctt ttgtaacaac agatggtgag ctatatgtgt ttggagaacc tgagaatggg 660
aagttaggtc ttcccaatca gctcctgggc aatcacagaa caccccagct ggtgtctgaa 720
attccggaga aggtgatcca agtagcctgt ggtggagagc atactgtggt tctcacggag 780
aatgctgtgt atacctttgg gctgggacaa tttggtcagc tgggtcttgg cacttttctt 840
tttgaaactt cagaacccaa agtcattgag aatattaggg atcaaacaat aagttatatt 900
tcttgtggag aaaatcacac agctttgata acagatatcg gccttatgta tacttttgga 960
gatggtcgcc acggaaaatt aggacttgga ctggagaatt ttaccaatca cttcattcct 1020
actttgtgct ctaatttttt gaggtttata gttaaattgg ttgcttgtgg tggatgtcac 1080
atggtagttt ttgctgctcc tcatcgtggt gtggcaaaag aaattgaatt cgatgaaata 1140
aatgatactt gcttatctgt ggcgactttt ctgccgtata gcagtttaac ctcaggaaat 1200
gtactgcaga ggactctatc agcacgtatg cggcgaagag agagggagag gtctccagat 1260
tctttttcaa tgaggagaac actacctcca atagaaggga ctcttggcct ttctgcttgt 1320
tttctcccca attcagtctt tccacgatgt tctgagagaa acctccaaga gagtgtctta 1380
tctgaacagg acctcatgca gccagaggaa ccagattatt tgctagatga aatgaccaaa 1440
gaagcagaga tagataattc ttcaactgta gaaagccttg gagaaactac tgatatctta 1500
aacatgacac acatcatgag cctgaattcc aatgaaaagt cattaaaatt atcaccagtt 1560
cagaaacaaa agaaacaaca aacaattggg gaactgacgc aggatacagc tcttactgaa 1620
aacgatgata gtgatgaata tgaagaaatg tcagaaatga aagaagggaa agcatgtaaa 1680
caacatgtgt cacaagggat tttcatgacg cagccagcta cgactatcga agcattttca 1740
gatgaggaag tagagatccc agaggagaag gaaggagcag aggattcaaa aggaaatgga 1800
atagaggagc aagaggtaga agcaaatgag gaaaatgtga aggtgcatgg aggaagaaag 1860
gagaaaacag agatcctatc agatgacctt acagacaaag cagaggtgag tgaaggcaag 1920
gcaaaatcag tgggagaagc agaggatggg cctgaaggta gaggggatgg aacctgtgag 1980
gaaggtagtt caggagcaga acactggcaa gatgaggaga gggagaaggg ggagaaagac 2040
aagggtagag gagaaatgga gaggccagga gagggagaga aggaactagc agagaaggaa 2100
gaatggaaga agagggatgg ggaagagcag gagcaaaagg agagggagca gggccatcag 2160
aaggaaagaa accaagagat ggaggaggga ggggaggagg agcatggaga aggagaagaa 2220
gaggagggag acagagaaga ggaagaagag aaggagggag aagggaaaga ggaaggagaa 2280
ggggaagaag tggagggaga acgtgaaaag gaggaaggag agaggaaaaa ggaggaaaga 2340
gcggggaagg aggagaaagg agaggaagaa ggagaccaag gagaggggga agaggaggaa 2400
acagagggga gaggggagga aaaagaggag ggaggggaag tagagggagg ggaagtagag 2460
gaggggaaag gagagaggga agaggaagag gaggagggtg agggggaaga ggaggaaggg 2520
gagggggaag aggaggaagg ggagggggaa gaggaggaag gagaagggaa aggggaggaa 2580
gaaggggaag aaggagaagg ggaggaagaa ggggaggaag gagaagggga gggggaagag 2640
gaggaaggag aaggggaggg agaagaggaa ggagaagggg agggagaaga ggaggaagga 2700
gaaggggagg gagaagagga aggagaaggg gagggagaag aggaggaagg agaagggaaa 2760
ggggaggagg aaggagagga aggagaaggg gagggggaag aggaggaagg agaaggggaa 2820
ggggaggatg gagaagggga gggggaagag gaggaaggag aatgggaggg ggaagaggag 2880
gaaggagaag gggaggggga agaggaagga gaaggggaag gggaggaagg agaaggggag 2940
ggggaagagg aggaaggaga aggggagggg gaagaggagg aaggggaaga agaaggggag 3000
gaagaaggag agggagagga agaaggggag ggagaagggg aggaagaaga ggaaggggaa 3060
gtggaagggg aggtggaagg ggaggaagga gagggggaag gagaggaaga ggaaggagag 3120
gaggaaggag aagaaaggga aaaggagggg gaaggagaag aaaacaggag gaacagagaa 3180
gaggaggagg aagaagaggg gaagtatcag gagacaggcg aagaagagaa tgaaaggcag 3240
gatggagagg agtacaaaaa agtgagcaaa ataaaaggat ctgtgaaata tggcaaacat 3300
aaaacatatc aaaaaaagtc agttactaac acacagggaa atgggaaaga gcagaggtcc 3360
aaaatgccag tccagtcaaa acgactttta aaaaacgggc catcaggttc caaaaagttc 3420
tggaataatg tattaccaca ttacttggaa ttgaagtaa 3459
<210> 3
<211> 3459
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность RPGRORF15
<400> 3
atgagagagc cagaggagct gatgccagac agtggagcag tgtttacatt cggaaaatct 60
aagttcgctg aaaataaccc aggaaagttc tggtttaaaa acgacgtgcc cgtccacctg 120
tcttgtggcg atgagcatag tgccgtggtc actgggaaca ataagctgta catgttcggg 180
tccaacaact ggggacagct ggggctggga tccaaatctg ctatctctaa gccaacctgc 240
gtgaaggcac tgaaacccga gaaggtcaaa ctggccgctt gtggcagaaa ccacactctg 300
gtgagcaccg agggcgggaa tgtctatgcc accggaggca acaatgaggg acagctggga 360
ctgggggaca ctgaggaaag gaataccttt cacgtgatct ccttctttac atctgagcat 420
aagatcaagc agctgagcgc tggctccaac acatctgcag ccctgactga ggacgggcgc 480
ctgttcatgt ggggagataa ttcagagggc cagattgggc tgaaaaacgt gagcaatgtg 540
tgcgtccctc agcaggtgac catcggaaag ccagtcagtt ggatttcatg tggctactat 600
catagcgcct tcgtgaccac agatggcgag ctgtacgtct ttggggagcc cgaaaacgga 660
aaactgggcc tgcctaacca gctgctgggc aatcaccgga caccccagct ggtgtccgag 720
atccctgaaa aagtgatcca ggtcgcctgc gggggagagc atacagtggt cctgactgag 780
aatgctgtgt ataccttcgg actgggccag tttggccagc tggggctggg aaccttcctg 840
tttgagacat ccgaaccaaa agtgatcgag aacattcgcg accagactat cagctacatt 900
tcctgcggag agaatcacac cgcactgatc acagacattg gcctgatgta tacctttggc 960
gatggacgac acgggaagct gggactggga ctggagaact tcactaatca ttttatcccc 1020
accctgtgtt ctaacttcct gcggttcatc gtgaaactgg tcgcttgcgg cgggtgtcac 1080
atggtggtct tcgctgcacc tcataggggc gtggctaagg agatcgaatt tgacgagatt 1140
aacgatacat gcctgagcgt ggcaactttc ctgccataca gctccctgac ttctggcaat 1200
gtgctgcaga gaaccctgag tgcaaggatg cggagaaggg agagggaacg ctctcctgac 1260
agtttctcaa tgcgacgaac cctgccacct atcgagggaa cactgggact gagtgcctgc 1320
ttcctgccta actcagtgtt tccacgatgt agcgagcgga atctgcagga gtctgtcctg 1380
agtgagcagg atctgatgca gccagaggaa cccgactacc tgctggatga gatgaccaag 1440
gaggccgaaa tcgacaactc tagtacagtg gagtccctgg gcgagactac cgatatcctg 1500
aatatgacac acattatgtc actgaacagc aatgagaaga gtctgaaact gtcaccagtg 1560
cagaagcaga agaaacagca gactattggc gagctgactc aggacaccgc cctgacagag 1620
aacgacgata gcgatgagta tgaggaaatg tccgagatga aggaaggcaa agcttgtaag 1680
cagcatgtca gtcaggggat cttcatgaca cagccagcca caactattga ggctttttca 1740
gacgaggaag tggagatccc cgaggaaaaa gagggcgcag aagattccaa ggggaatgga 1800
attgaggaac aggaggtgga agccaacgag gaaaatgtga aagtccacgg aggcaggaag 1860
gagaaaacag aaatcctgtc tgacgatctg actgacaagg ccgaggtgtc cgaaggcaag 1920
gcaaaatctg tcggagaggc agaagacgga ccagagggac gaggggatgg aacctgcgag 1980
gaaggctcaa gcggggctga gcattggcag gacgaggaac gagagaaggg cgaaaaggat 2040
aaaggccgcg gggagatgga acgacctgga gagggcgaaa aagagctggc agagaaggag 2100
gaatggaaga aaagggacgg cgaggaacag gagcagaaag aaagggagca gggccaccag 2160
aaggagcgca accaggagat ggaagagggc ggcgaggaag agcatggcga gggagaagag 2220
gaagagggcg atagagaaga ggaagaggaa aaagaaggcg aagggaagga ggaaggagag 2280
ggcgaggaag tggaaggcga gagggaaaag gaggaaggag aacggaagaa agaggaaaga 2340
gccggcaaag aggaaaaggg cgaggaagag ggcgatcagg gcgaaggcga ggaggaagag 2400
accgagggcc gcggggaaga gaaagaggag ggaggagagg tggagggcgg agaggtcgaa 2460
gagggaaagg gcgagcgcga agaggaagag gaagagggcg agggcgagga agaagagggc 2520
gagggggaag aagaggaggg agagggcgaa gaggaagagg gggagggaaa gggcgaagag 2580
gaaggagagg aaggggaggg agaggaagag ggggaggagg gcgaggggga aggcgaggag 2640
gaagaaggag agggggaagg cgaagaggaa ggcgaggggg aaggagagga ggaagaaggg 2700
gaaggcgaag gcgaagagga gggagaagga gagggggagg aagaggaagg agaagggaag 2760
ggcgaggagg aaggcgaaga gggagagggg gaaggcgagg aagaggaagg cgagggcgaa 2820
ggagaggacg gcgagggcga gggagaagag gaggaagggg aatgggaagg cgaagaagag 2880
gaaggcgaag gcgaaggcga agaagagggc gaaggggagg gcgaggaggg cgaaggcgaa 2940
ggggaggaag aggaaggcga aggagaaggc gaggaagaag agggagagga ggaaggcgag 3000
gaggaaggag agggggagga ggagggagaa ggcgagggcg aagaagaaga agagggagaa 3060
gtggagggcg aagtcgaggg ggaggaggga gaaggggaag gggaggaaga agagggcgaa 3120
gaagaaggcg aggaaagaga aaaagaggga gaaggcgagg aaaaccggag aaatagggaa 3180
gaggaggaag aggaagaggg aaagtaccag gagacaggcg aagaggaaaa cgagcggcag 3240
gatggcgagg aatataagaa agtgagcaag atcaaaggat ccgtcaagta cggcaagcac 3300
aaaacctatc agaagaaaag cgtgaccaac acacagggga atggaaaaga gcagaggagt 3360
aagatgcctg tgcagtcaaa acggctgctg aagaatggcc catctggaag taaaaaattc 3420
tggaacaatg tgctgcccca ctatctggaa ctgaaataa 3459
<210> 4
<211> 934
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность промотора
<400> 4
attgacgtca ataatgacgt atgttcccat agtaacgcca atagggactt tccattgacg 60
tcaatgggtg gagtatttac ggtaaactgc ccacttggca gtacatcaag tgtatcatat 120
gccaagtacg ccccctattg acgtcaatga cggtaaatgg cccgcctggc attatgccca 180
gtacatgacc ttatgggact ttcctacttg gcagtacatc tacgtattag tcatcgctat 240
taccatggtc gaggtgagcc ccacgttctg cttcactctc cccatctccc ccccctcccc 300
acccccaatt ttgtatttat ttatttttta attattttgt gcagcgatgg gggcgggggg 360
gggggggggg cgcgcgccag gcggggcggg gcggggcgag gggcggggcg gggcgaggcg 420
gagaggtgcg gcggcagcca atcagagcgg cgcgctccga aagtttcctt ttatggcgag 480
gcggcggcgg cggcggccct ataaaaagcg aagcgcgcgg cgggcgggag tcgctgcgcg 540
ctgccttcgc cccgtgcccc gctccgccgc cgcctcgcgc cgcccgcccc ggctctgact 600
gaccgcgtta ctcccacagg tgagcgggcg ggacggccct tctcctccgg gctgtaatta 660
gcgcttggtt taatgacggc ttgtttcttt tctgtggctg cgtgaaagcc ttgaggggct 720
ccgggagggc cctttgtgcg gggggagcgg ctcggggctg tccgcggggg gacggctgcc 780
ttcggggggg acggggcagg gcggggttcg gcttctggcg tgtgaccggc ggctctagag 840
cctctgctaa ccatgttcat gccttcttct ttttcctaca gctcctgggc aacgtgctgg 900
ttattgtgct gtctcatcat tttggcaaag aatt 934
<210> 5
<211> 270
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность сигнала поли-A бычьего гормона роста
<400> 5
tcgctgatca gcctcgactg tgccttctag ttgccagcca tctgttgttt gcccctcccc 60
cgtgccttcc ttgaccctgg aaggtgccac tcccactgtc ctttcctaat aaaatgagga 120
aattgcatcg cattgtctga gtaggtgtca ttctattctg gggggtgggg tggggcagga 180
cagcaagggg gaggattggg aagacaatag caggcatgct ggggatgcgg tgggctctat 240
ggcttctgag gcggaaagaa ccagctgggg 270
<210> 6
<211> 588
<212> ДНК
<213> Искусственная последовательность
<220>
<223> иллюстративная последовательность посттранскрипционного регуляторного
элемента вируса гепатита сурков (WPRE)
<400> 6
atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac tatgttgctc 60
cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt gcttcccgta 120
tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat gaggagttgt 180
ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca acccccactg 240
gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc cccctcccta 300
ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg gctcggctgt 360
tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct tggctgctcg 420
cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct tcggccctca 480
atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt ccgcgtcttc 540
gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgc 588
<210> 7
<211> 199
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность промотора родопсин-киназы (GRK1)
<400> 7
gggccccaga agcctggtgg ttgtttgtcc ttctcagggg aaaagtgagg cggccccttg 60
gaggaagggg ccgggcagaa tgatctaatc ggattccaag cagctcaggg gattgtcttt 120
ttctagcacc ttcttgccac tcctaagcgt cctccgtgac cccggctggg atttagcctg 180
gtgctgtgtc agccccggg 199
<210> 8
<211> 235
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность промотора интерфоторецепторного ретинол-связывающего
белка (IRBP)
<400> 8
agcacagtgt ctggcatgta gcaggaacta aaataatggc agtgattaat gttatgatat 60
gcagacacaa cacagcaaga taagatgcaa tgtaccttct gggtcaaacc accctggcca 120
ctcctccccg atacccaggg ttgatgtgct tgaattagac aggattaaag gcttactgga 180
gctggaagcc ttgccccaac tcaggagttt agccccagac cttctgtcca ccagc 235
<210> 9
<211> 3003
<212> ДНК
<213> Искусственная последовательность
<220>
<223> предпочтительная кодирующая RPGRORF15 нуклеотидная последовательность
<400> 9
atgagagagc cagaggagct gatgccagac agtggagcag tgtttacatt cggaaaatct 60
aagttcgctg aaaataaccc aggaaagttc tggtttaaaa acgacgtgcc cgtccacctg 120
tcttgtggcg atgagcatag tgccgtggtc actgggaaca ataagctgta catgttcggg 180
tccaacaact ggggacagct ggggctggga tccaaatctg ctatctctaa gccaacctgc 240
gtgaaggcac tgaaacccga gaaggtcaaa ctggccgctt gtggcagaaa ccacactctg 300
gtgagcaccg agggcgggaa tgtctatgcc accggaggca acaatgaggg acagctggga 360
ctgggggaca ctgaggaaag gaataccttt cacgtgatct ccttctttac atctgagcat 420
aagatcaagc agctgagcgc tggctccaac acatctgcag ccctgactga ggacgggcgc 480
ctgttcatgt ggggagataa ttcagagggc cagattgggc tgaaaaacgt gagcaatgtg 540
tgcgtccctc agcaggtgac catcggaaag ccagtcagtt ggatttcatg tggctactat 600
catagcgcct tcgtgaccac agatggcgag ctgtacgtct ttggggagcc cgaaaacgga 660
aaactgggcc tgcctaacca gctgctgggc aatcaccgga caccccagct ggtgtccgag 720
atccctgaaa aagtgatcca ggtcgcctgc gggggagagc atacagtggt cctgactgag 780
aatgctgtgt ataccttcgg actgggccag tttggccagc tggggctggg aaccttcctg 840
tttgagacat ccgaaccaaa agtgatcgag aacattcgcg accagactat cagctacatt 900
tcctgcggag agaatcacac cgcactgatc acagacattg gcctgatgta tacctttggc 960
gatggacgac acgggaagct gggactggga ctggagaact tcactaatca ttttatcccc 1020
accctgtgtt ctaacttcct gcggttcatc gtgaaactgg tcgcttgcgg cgggtgtcac 1080
atggtggtct tcgctgcacc tcataggggc gtggctaagg agatcgaatt tgacgagatt 1140
aacgatacat gcctgagcgt ggcaactttc ctgccataca gctccctgac ttctggcaat 1200
gtgctgcaga gaaccctgag tgcaaggatg cggagaaggg agagggaacg ctctcctgac 1260
agtttctcaa tgcgacgaac cctgccacct atcgagggaa cactgggact gagtgcctgc 1320
ttcctgccta actcagtgtt tccacgatgt agcgagcgga atctgcagga gtctgtcctg 1380
agtgagcagg atctgatgca gccagaggaa cccgactacc tgctggatga gatgaccaag 1440
gaggccgaaa tcgacaactc tagtacagtg gagtccctgg gcgagactac cgatatcctg 1500
aatatgacac acattatgtc actgaacagc aatgagaaga gtctgaaact gtcaccagtg 1560
cagaagcaga agaaacagca gactattggc gagctgactc aggacaccgc cctgacagag 1620
aacgacgata gcgatgagta tgaggaaatg tccgagatga aggaaggcaa agcttgtaag 1680
cagcatgtca gtcaggggat cttcatgaca cagccagcca caactattga ggctttttca 1740
gacgaggaag tggagatccc cgaggaaaaa gagggcgcag aagattccaa ggggaatgga 1800
attgaggaac aggaggtgga agccaacgag gaaaatgtga aagtccacgg aggcaggaag 1860
gagaaaacag aaatcctgtc tgacgatctg actgacaagg ccgaggtgtc cgaaggcaag 1920
gcaaaatctg tcggagaggc agaagacgga ccagagggac gaggggatgg aacctgcgag 1980
gaaggctcaa gcggggctga gcattggcag gacgaggaac gagagaaggg cgaaaaggat 2040
aaaggccgcg gggagatgga acgacctgga gagggcgaaa aagagctggc agagaaggag 2100
gaatggaaga aaagggacgg cgaggaacag gagcagaaag aaagggagca gggccaccag 2160
aaggagcgca accaggagat ggaagagggc ggcgaggaag agcatggcga gggagaagag 2220
gaagagggcg atagagaaga ggaagaggaa aaagaaggcg aagggaagga ggaaggagag 2280
ggcgaggaag tggaaggcga gagggaaaag gaggaaggag aacggaagaa agaggaaaga 2340
gccggcaaag aggaaaaggg cgaggaagag ggcgatcagg gcgaaggcga ggaggaagag 2400
accgagggcc gcggggaaga gaaagaggag ggaggagagg tggagggcgg agaggtcgaa 2460
gagggaaagg gcgagcgcga agagggggag gaagaggaag gcgaaggaga aggcgaggaa 2520
gaagagggag aggaggaagg cgaggaggaa ggagaggggg aggaggaggg agaaggcgag 2580
ggcgaagaag aagaagaggg agaagtggag ggcgaagtcg agggggagga gggagaaggg 2640
gaaggggagg aagaagaggg cgaagaagaa ggcgaggaaa gagaaaaaga gggagaaggc 2700
gaggaaaacc ggagaaatag ggaagaggag gaagaggaag agggaaagta ccaggagaca 2760
ggcgaagagg aaaacgagcg gcaggatggc gaggaatata agaaagtgag caagatcaaa 2820
ggatccgtca agtacggcaa gcacaaaacc tatcagaaga aaagcgtgac caacacacag 2880
gggaatggaa aagagcagag gagtaagatg cctgtgcagt caaaacggct gctgaagaat 2940
ggcccatctg gaagtaaaaa attctggaac aatgtgctgc cccactatct ggaactgaaa 3000
taa 3003
<210> 10
<211> 456
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность делеции
<400> 10
gaagaggaag agggcgaggg cgaggaagaa gagggcgagg gggaagaaga ggagggagag 60
ggcgaagagg aagaggggga gggaaagggc gaagaggaag gagaggaagg ggagggagag 120
gaagaggggg aggagggcga gggggaaggc gaggaggaag aaggagaggg ggaaggcgaa 180
gaggaaggcg agggggaagg agaggaggaa gaaggggaag gcgaaggcga agaggaggga 240
gaaggagagg gggaggaaga ggaaggagaa gggaagggcg aggaggaagg cgaagaggga 300
gagggggaag gcgaggaaga ggaaggcgag ggcgaaggag aggacggcga gggcgaggga 360
gaagaggagg aaggggaatg ggaaggcgaa gaagaggaag gcgaaggcga aggcgaagaa 420
gagggcgaag gggagggcga ggagggcgaa ggcgaa 456
<210> 11
<211> 2517
<212> ДНК
<213> Искусственная последовательность
<220>
<223> предпочтительная кодирующая RPGRORF15 нуклеотидная последовательность
<400> 11
atgagagagc cagaggagct gatgccagac agtggagcag tgtttacatt cggaaaatct 60
aagttcgctg aaaataaccc aggaaagttc tggtttaaaa acgacgtgcc cgtccacctg 120
tcttgtggcg atgagcatag tgccgtggtc actgggaaca ataagctgta catgttcggg 180
tccaacaact ggggacagct ggggctggga tccaaatctg ctatctctaa gccaacctgc 240
gtgaaggcac tgaaacccga gaaggtcaaa ctggccgctt gtggcagaaa ccacactctg 300
gtgagcaccg agggcgggaa tgtctatgcc accggaggca acaatgaggg acagctggga 360
ctgggggaca ctgaggaaag gaataccttt cacgtgatct ccttctttac atctgagcat 420
aagatcaagc agctgagcgc tggctccaac acatctgcag ccctgactga ggacgggcgc 480
ctgttcatgt ggggagataa ttcagagggc cagattgggc tgaaaaacgt gagcaatgtg 540
tgcgtccctc agcaggtgac catcggaaag ccagtcagtt ggatttcatg tggctactat 600
catagcgcct tcgtgaccac agatggcgag ctgtacgtct ttggggagcc cgaaaacgga 660
aaactgggcc tgcctaacca gctgctgggc aatcaccgga caccccagct ggtgtccgag 720
atccctgaaa aagtgatcca ggtcgcctgc gggggagagc atacagtggt cctgactgag 780
aatgctgtgt ataccttcgg actgggccag tttggccagc tggggctggg aaccttcctg 840
tttgagacat ccgaaccaaa agtgatcgag aacattcgcg accagactat cagctacatt 900
tcctgcggag agaatcacac cgcactgatc acagacattg gcctgatgta tacctttggc 960
gatggacgac acgggaagct gggactggga ctggagaact tcactaatca ttttatcccc 1020
accctgtgtt ctaacttcct gcggttcatc gtgaaactgg tcgcttgcgg cgggtgtcac 1080
atggtggtct tcgctgcacc tcataggggc gtggctaagg agatcgaatt tgacgagatt 1140
aacgatacat gcctgagcgt ggcaactttc ctgccataca gctccctgac ttctggcaat 1200
gtgctgcaga gaaccctgag tgcaaggatg cggagaaggg agagggaacg ctctcctgac 1260
agtttctcaa tgcgacgaac cctgccacct atcgagggaa cactgggact gagtgcctgc 1320
ttcctgccta actcagtgtt tccacgatgt agcgagcgga atctgcagga gtctgtcctg 1380
agtgagcagg atctgatgca gccagaggaa cccgactacc tgctggatga gatgaccaag 1440
gaggccgaaa tcgacaactc tagtacagtg gagtccctgg gcgagactac cgatatcctg 1500
aatatgacac acattatgtc actgaacagc aatgagaaga gtctgaaact gtcaccagtg 1560
cagaagcaga agaaacagca gactattggc gagctgactc aggacaccgc cctgacagag 1620
aacgacgata gcgatgagta tgaggaaatg tccgagatga aggaaggcaa agcttgtaag 1680
cagcatgtca gtcaggggat cttcatgaca cagccagcca caactattga ggctttttca 1740
gacgaggaag tggagatccc cgaggaaaaa gagggcgcag aagattccaa ggggaatgga 1800
attgaggaac aggaggtgga agccaacgag gaaaatgtga aagtccacgg aggcaggaag 1860
gagaaaacag aaatcctgtc tgacgatctg actgacaagg ccgaggtgtc cgaaggcaag 1920
gcaaaatctg tcggagaggc agaagacgga ccagagggac gaggggatgg aacctgcgag 1980
gaaggctcaa gcggggctga gcattggcag gacgaggaac gagagaaggg cgaaaaggat 2040
aaaggccgcg gggagatgga acgacctgga gagggcgaaa aagaggaagg cgagggcgaa 2100
gaagaagaag agggagaagt ggagggcgaa gtcgaggggg aggagggaga aggggaaggg 2160
gaggaagaag agggcgaaga agaaggcgag gaaagagaaa aagagggaga aggcgaggaa 2220
aaccggagaa atagggaaga ggaggaagag gaagagggaa agtaccagga gacaggcgaa 2280
gaggaaaacg agcggcagga tggcgaggaa tataagaaag tgagcaagat caaaggatcc 2340
gtcaagtacg gcaagcacaa aacctatcag aagaaaagcg tgaccaacac acaggggaat 2400
ggaaaagagc agaggagtaa gatgcctgtg cagtcaaaac ggctgctgaa gaatggccca 2460
tctggaagta aaaaattctg gaacaatgtg ctgccccact atctggaact gaaataa 2517
<210> 12
<211> 942
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность делеции
<400> 12
ctggcagaga aggaggaatg gaagaaaagg gacggcgagg aacaggagca gaaagaaagg 60
gagcagggcc accagaagga gcgcaaccag gagatggaag agggcggcga ggaagagcat 120
ggcgagggag aagaggaaga gggcgataga gaagaggaag aggaaaaaga aggcgaaggg 180
aaggaggaag gagagggcga ggaagtggaa ggcgagaggg aaaaggagga aggagaacgg 240
aagaaagagg aaagagccgg caaagaggaa aagggcgagg aagagggcga tcagggcgaa 300
ggcgaggagg aagagaccga gggccgcggg gaagagaaag aggagggagg agaggtggag 360
ggcggagagg tcgaagaggg aaagggcgag cgcgaagagg aagaggaaga gggcgagggc 420
gaggaagaag agggcgaggg ggaagaagag gagggagagg gcgaagagga agagggggag 480
ggaaagggcg aagaggaagg agaggaaggg gagggagagg aagaggggga ggagggcgag 540
ggggaaggcg aggaggaaga aggagagggg gaaggcgaag aggaaggcga gggggaagga 600
gaggaggaag aaggggaagg cgaaggcgaa gaggagggag aaggagaggg ggaggaagag 660
gaaggagaag ggaagggcga ggaggaaggc gaagagggag agggggaagg cgaggaagag 720
gaaggcgagg gcgaaggaga ggacggcgag ggcgagggag aagaggagga aggggaatgg 780
gaaggcgaag aagaggaagg cgaaggcgaa ggcgaagaag agggcgaagg ggagggcgag 840
gagggcgaag gcgaagggga ggaagaggaa ggcgaaggag aaggcgagga agaagaggga 900
gaggaggaag gcgaggagga aggagagggg gaggaggagg ga 942
<210> 13
<211> 3081
<212> ДНК
<213> Искусственная последовательность
<220>
<223> предпочтительная кодирующая RPGRORF15 нуклеотидная последовательность
<400> 13
atgagagagc cagaggagct gatgccagac agtggagcag tgtttacatt cggaaaatct 60
aagttcgctg aaaataaccc aggaaagttc tggtttaaaa acgacgtgcc cgtccacctg 120
tcttgtggcg atgagcatag tgccgtggtc actgggaaca ataagctgta catgttcggg 180
tccaacaact ggggacagct ggggctggga tccaaatctg ctatctctaa gccaacctgc 240
gtgaaggcac tgaaacccga gaaggtcaaa ctggccgctt gtggcagaaa ccacactctg 300
gtgagcaccg agggcgggaa tgtctatgcc accggaggca acaatgaggg acagctggga 360
ctgggggaca ctgaggaaag gaataccttt cacgtgatct ccttctttac atctgagcat 420
aagatcaagc agctgagcgc tggctccaac acatctgcag ccctgactga ggacgggcgc 480
ctgttcatgt ggggagataa ttcagagggc cagattgggc tgaaaaacgt gagcaatgtg 540
tgcgtccctc agcaggtgac catcggaaag ccagtcagtt ggatttcatg tggctactat 600
catagcgcct tcgtgaccac agatggcgag ctgtacgtct ttggggagcc cgaaaacgga 660
aaactgggcc tgcctaacca gctgctgggc aatcaccgga caccccagct ggtgtccgag 720
atccctgaaa aagtgatcca ggtcgcctgc gggggagagc atacagtggt cctgactgag 780
aatgctgtgt ataccttcgg actgggccag tttggccagc tggggctggg aaccttcctg 840
tttgagacat ccgaaccaaa agtgatcgag aacattcgcg accagactat cagctacatt 900
tcctgcggag agaatcacac cgcactgatc acagacattg gcctgatgta tacctttggc 960
gatggacgac acgggaagct gggactggga ctggagaact tcactaatca ttttatcccc 1020
accctgtgtt ctaacttcct gcggttcatc gtgaaactgg tcgcttgcgg cgggtgtcac 1080
atggtggtct tcgctgcacc tcataggggc gtggctaagg agatcgaatt tgacgagatt 1140
aacgatacat gcctgagcgt ggcaactttc ctgccataca gctccctgac ttctggcaat 1200
gtgctgcaga gaaccctgag tgcaaggatg cggagaaggg agagggaacg ctctcctgac 1260
agtttctcaa tgcgacgaac cctgccacct atcgagggaa cactgggact gagtgcctgc 1320
ttcctgccta actcagtgtt tccacgatgt agcgagcgga atctgcagga gtctgtcctg 1380
agtgagcagg atctgatgca gccagaggaa cccgactacc tgctggatga gatgaccaag 1440
gaggccgaaa tcgacaactc tagtacagtg gagtccctgg gcgagactac cgatatcctg 1500
aatatgacac acattatgtc actgaacagc aatgagaaga gtctgaaact gtcaccagtg 1560
cagaagcaga agaaacagca gactattggc gagctgactc aggacaccgc cctgacagag 1620
aacgacgata gcgatgagta tgaggaaatg tccgagatga aggaaggcaa agcttgtaag 1680
cagcatgtca gtcaggggat cttcatgaca cagccagcca caactattga ggctttttca 1740
gacgaggaag tggagatccc cgaggaaaaa gagggcgcag aagattccaa ggggaatgga 1800
attgaggaac aggaggtgga agccaacgag gaaaatgtga aagtccacgg aggcaggaag 1860
gagaaaacag aaatcctgtc tgacgatctg actgacaagg ccgaggtgtc cgaaggcaag 1920
gcaaaatctg tcggagaggc agaagacgga ccagagggac gaggggatgg aacctgcgag 1980
gaaggctcaa gcggggctga gcattggcag gacgaggaac gagagaaggg cgaaaaggat 2040
aaaggccgcg gggagatgga acgacctgga gagggcgaaa aagagctggc agagaaggag 2100
gaatggaaga aaagggacgg cgaggaacag gagcagaaag aaagggagca gggccaccag 2160
aaggagcgca accaggagat ggaagagggc ggcgaggaag agcatggcga gggagaagag 2220
gaagagggcg atagagaaga ggaagaggaa aaagaaggcg aagggaagga ggaaggagag 2280
ggcgaggaag tggaaggcga gagggaaaag gaggaaggag aacggaagaa agaggaaaga 2340
gccggcaaag aggaaaaggg cgaggaagag ggcgatcagg gcgaaggcga ggaggaagag 2400
accgagggcc gcggggaaga gaaagaggag ggaggagagg tggagggcgg agaggtcgaa 2460
gagggaaagg gcgagcgcga agaggaagag gaagagggcg agggcgagga agaagagggc 2520
gagggggaag aagaggaggg agagggcgaa gaggaagagg gggagggaaa gggcgaagag 2580
gaaggagaag gcgaggaaga agagggagag gaggaaggcg aggaggaagg agagggggag 2640
gaggagggag aaggcgaggg cgaagaagaa gaagagggag aagtggaggg cgaagtcgag 2700
ggggaggagg gagaagggga aggggaggaa gaagagggcg aagaagaagg cgaggaaaga 2760
gaaaaagagg gagaaggcga ggaaaaccgg agaaataggg aagaggagga agaggaagag 2820
ggaaagtacc aggagacagg cgaagaggaa aacgagcggc aggatggcga ggaatataag 2880
aaagtgagca agatcaaagg atccgtcaag tacggcaagc acaaaaccta tcagaagaaa 2940
agcgtgacca acacacaggg gaatggaaaa gagcagagga gtaagatgcc tgtgcagtca 3000
aaacggctgc tgaagaatgg cccatctgga agtaaaaaat tctggaacaa tgtgctgccc 3060
cactatctgg aactgaaata a 3081
<210> 14
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность делеции
<400> 14
ggagaggaag gggagggaga ggaagagggg gaggagggcg agggggaagg cgaggaggaa 60
gaaggagagg gggaaggcga agaggaaggc gagggggaag gagaggagga agaaggggaa 120
ggcgaaggcg aagaggaggg agaaggagag ggggaggaag aggaaggaga agggaagggc 180
gaggaggaag gcgaagaggg agagggggaa ggcgaggaag aggaaggcga gggcgaagga 240
gaggacggcg agggcgaggg agaagaggag gaaggggaat gggaaggcga agaagaggaa 300
ggcgaaggcg aaggcgaaga agagggcgaa ggggagggcg aggagggcga aggcgaaggg 360
gaggaagagg aaggcgaa 378
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕННОЕ РЕДАКТИРОВАНИЕ ГЛУБОКИХ ИНТРОННЫХ МУТАЦИЙ | 2016 |
|
RU2759335C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ НАРУШЕНИЙ И ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С RDH12 | 2017 |
|
RU2764920C2 |
| ГЕННАЯ ТЕРАПИЯ | 2016 |
|
RU2740038C2 |
| Кодон-оптимизированная последовательность нуклеотидов, кодирующая hAIPL1, и её содержащий экспрессионный вектор | 2021 |
|
RU2785621C1 |
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ МИОЦИЛИНОВОЙ (MYOC) ГЛАУКОМЫ | 2015 |
|
RU2718047C2 |
| ВИРИОНЫ АДЕНОАССОЦИИРОВАННОГО ВИРУСА С ВАРИАНТНЫМ КАПСИДОМ И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2017 |
|
RU2742724C1 |
| КОДОН-ОПТИМИЗИРОВАННЫЙ ФАКТОР КОМПЛЕМЕНТА I | 2019 |
|
RU2824048C2 |
| СПОСОБЫ УСИЛЕНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ РЕКОМБИНАНТНОГО АДЕНОАССОЦИИРОВАННОГО ВУРУСА, ПРОДУЦИРУЕМОГО БАКУЛОВИРУСНОЙ СИСТЕМОЙ | 2017 |
|
RU2762948C2 |
| ВИРИОНЫ АДЕНОАССОЦИИРОВАННОГО ВИРУСА С ВАРИАНТНЫМ КАПСИДОМ И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2012 |
|
RU2611202C2 |
| ФАКТОР КОМПЛЕМЕНТА I И КОФАКТОР ФАКТОРА КОМПЛЕМЕНТА I, КОДИРУЮЩИЕ ИХ ВЕКТОРЫ И ПРИМЕНЕНИЕ В ТЕРАПЕВТИЧЕСКИХ ЦЕЛЯХ | 2019 |
|
RU2809247C2 |
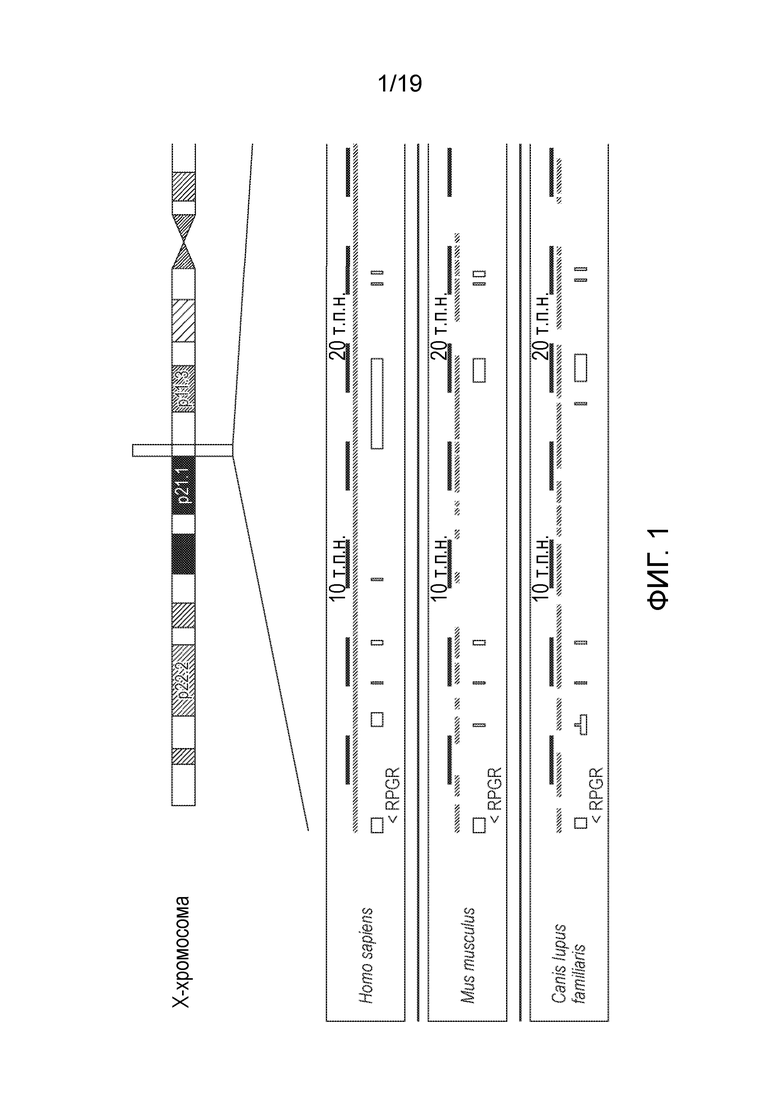
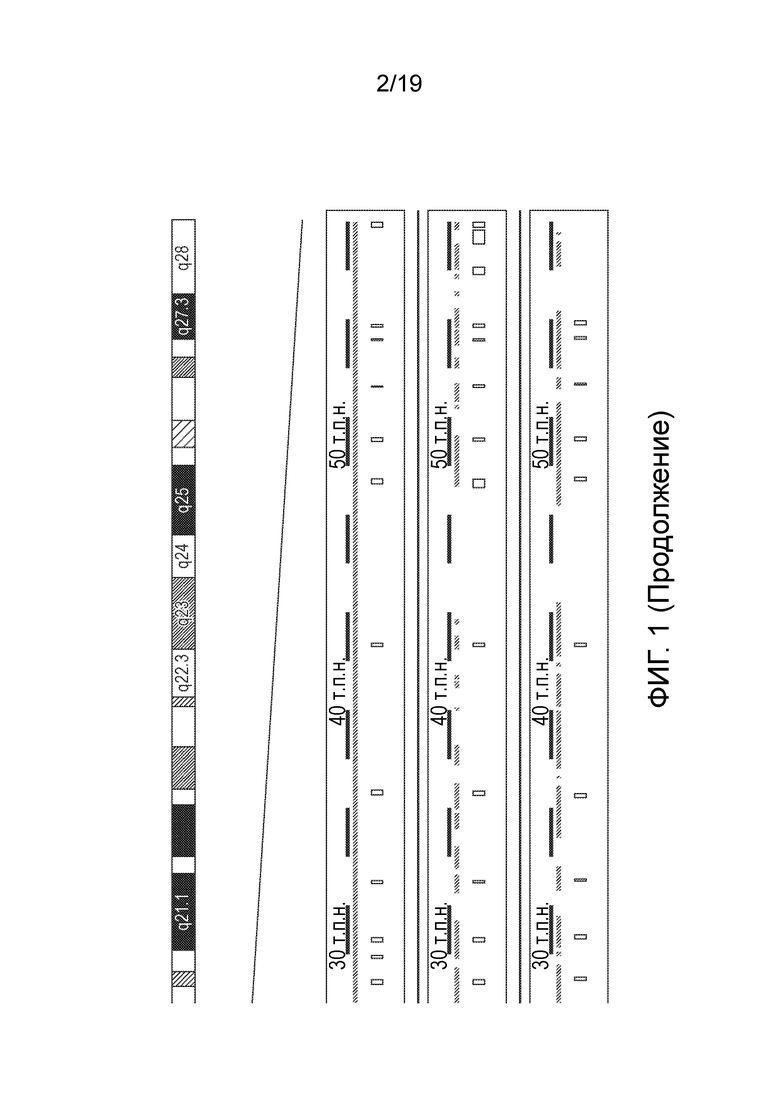




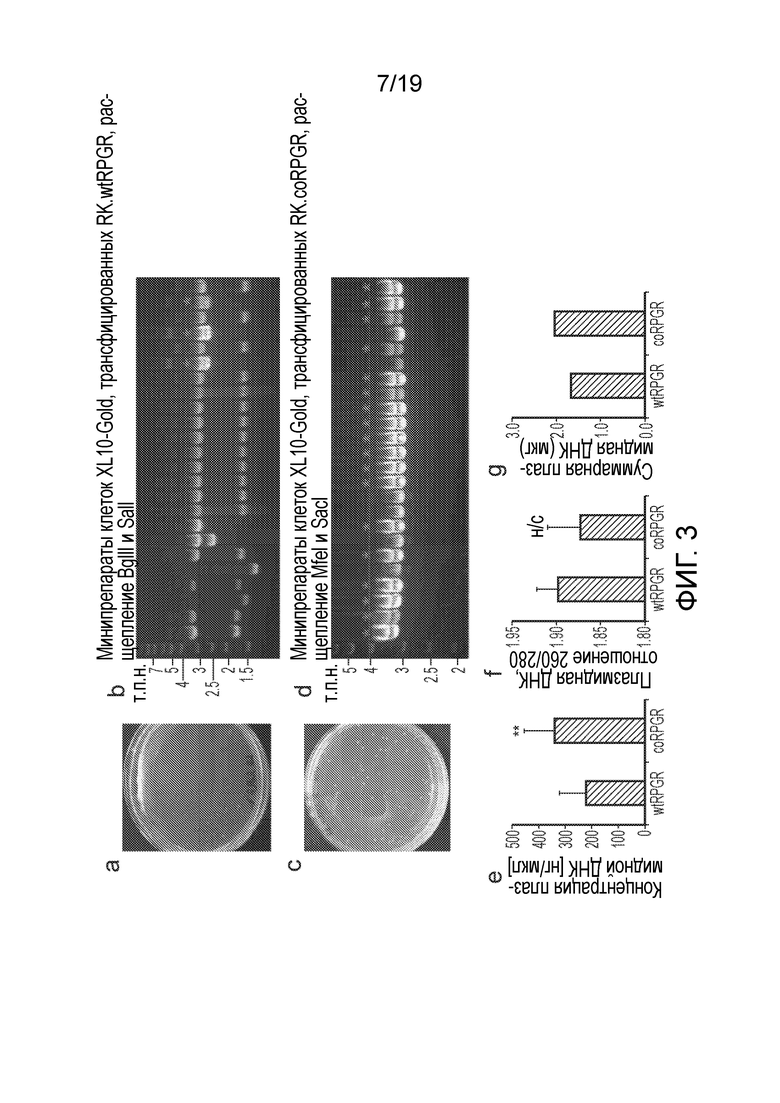
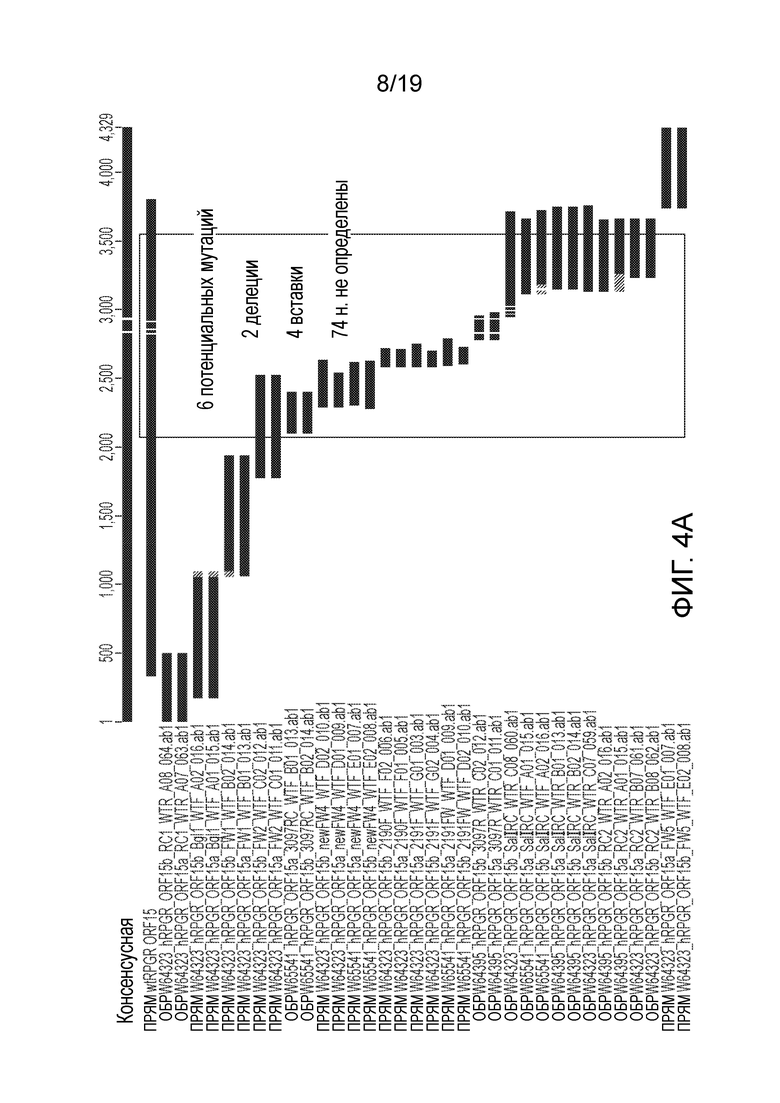

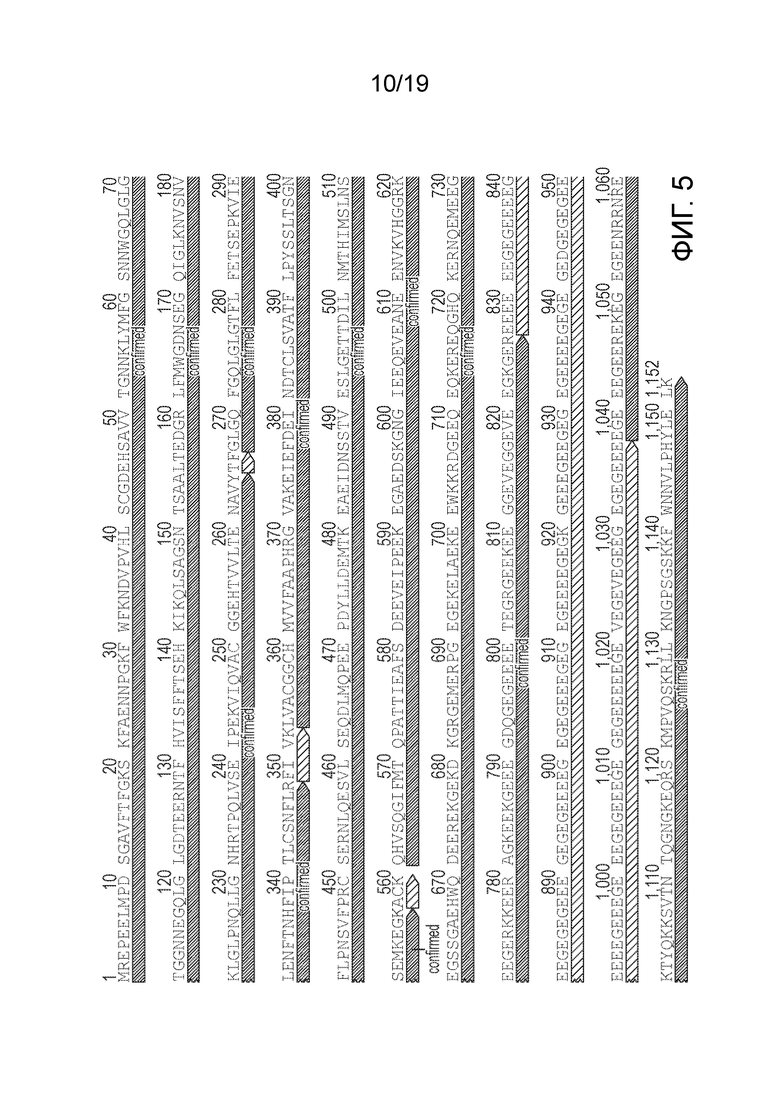
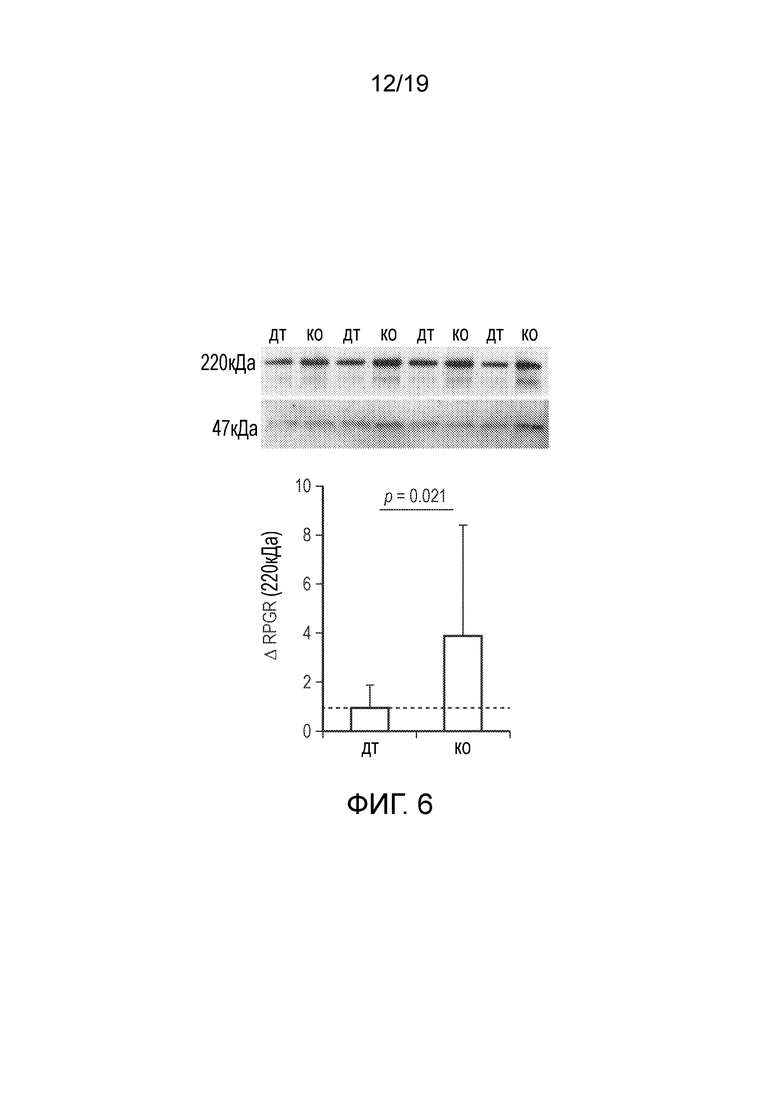
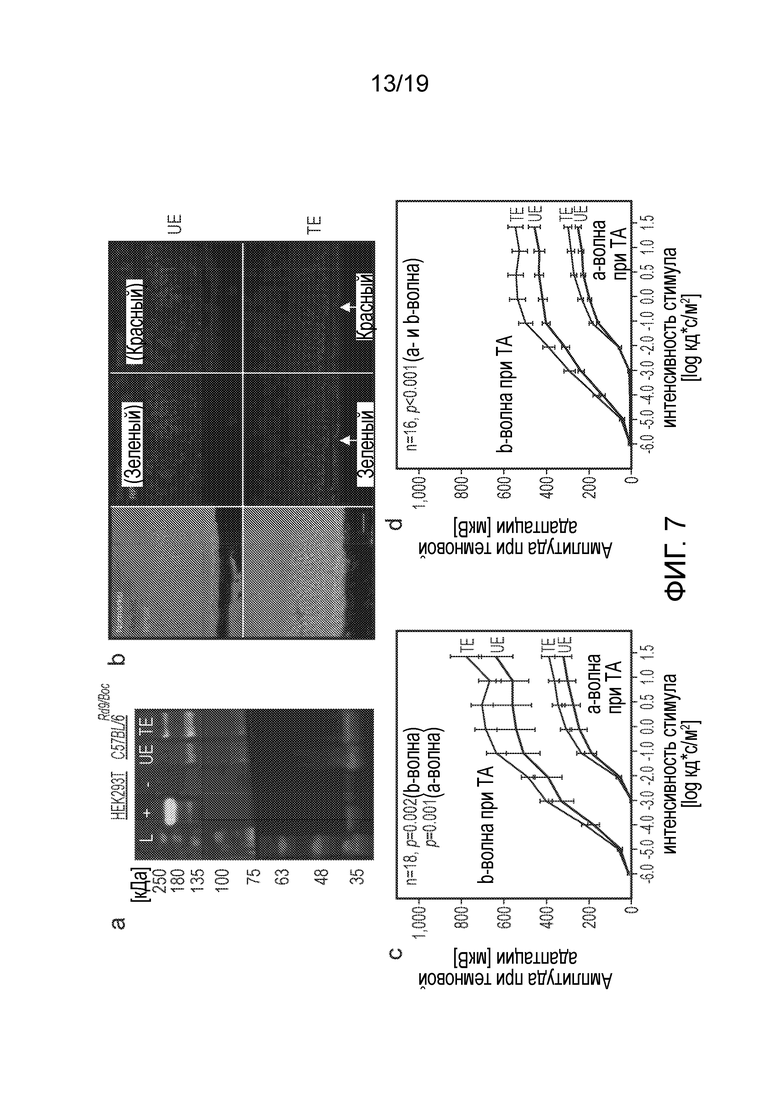
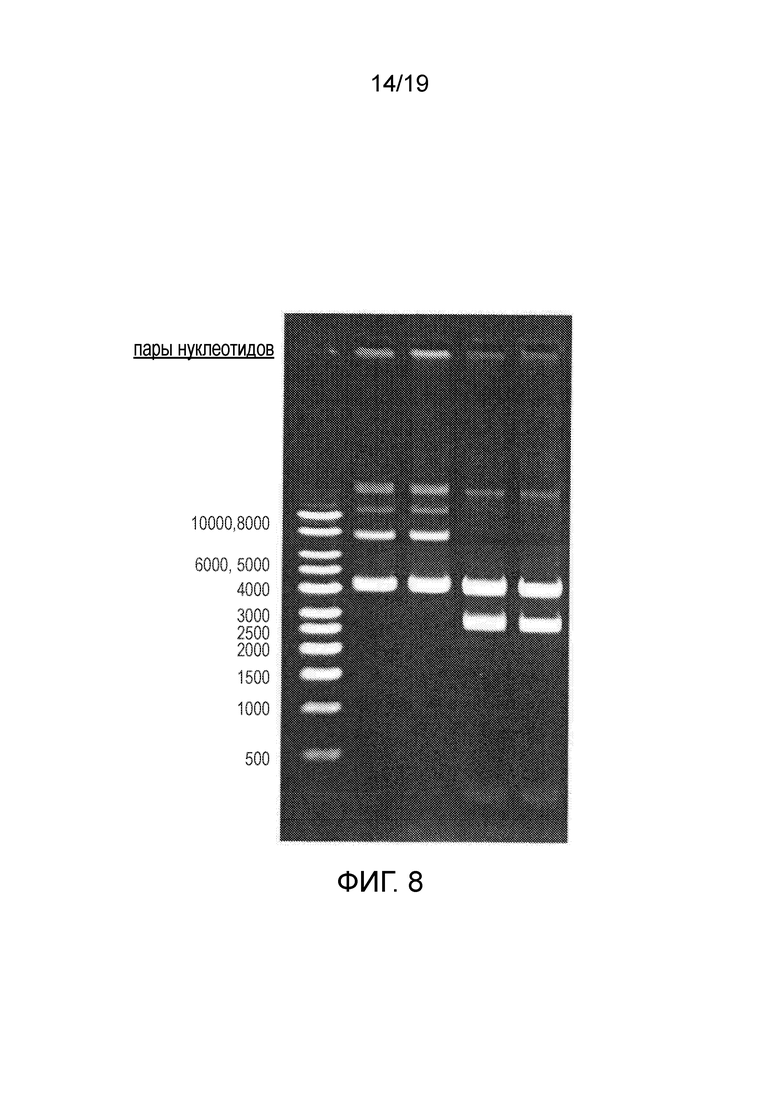
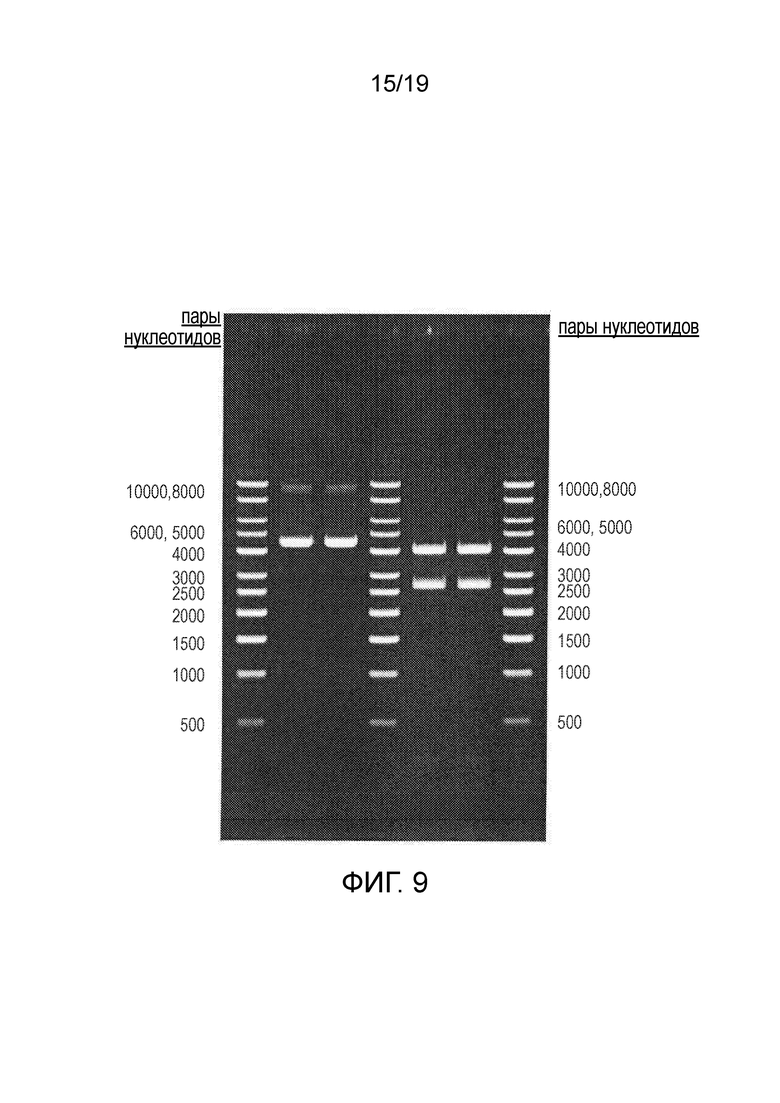
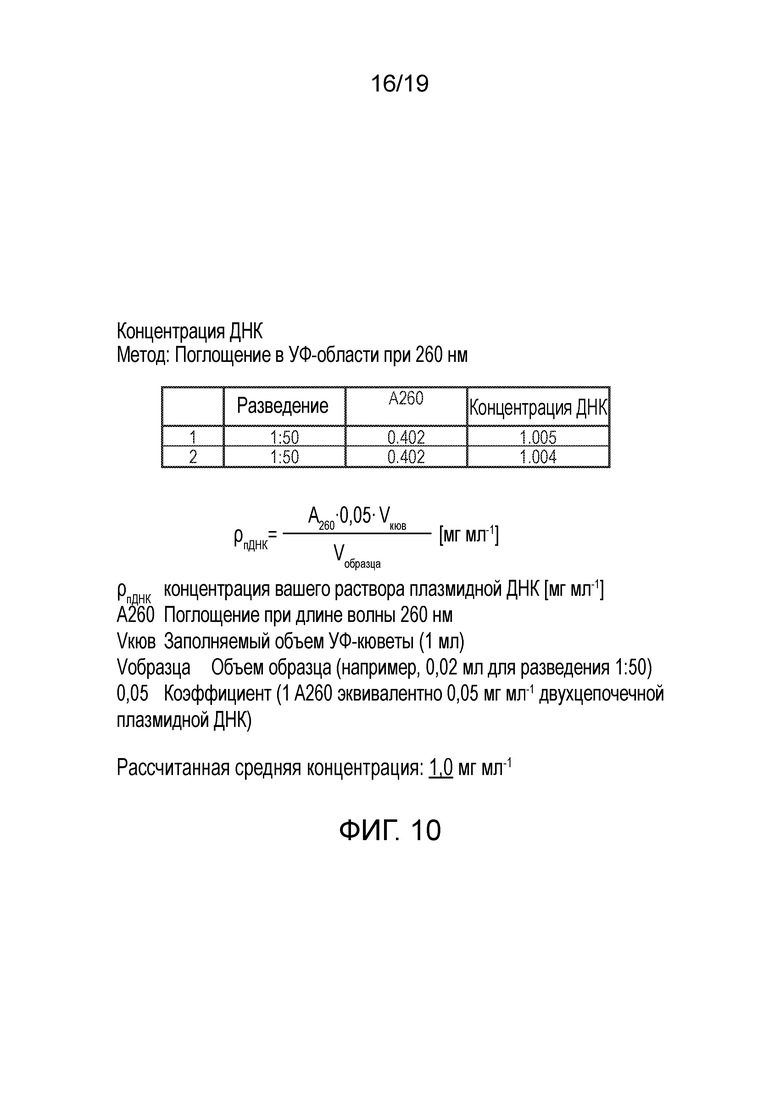
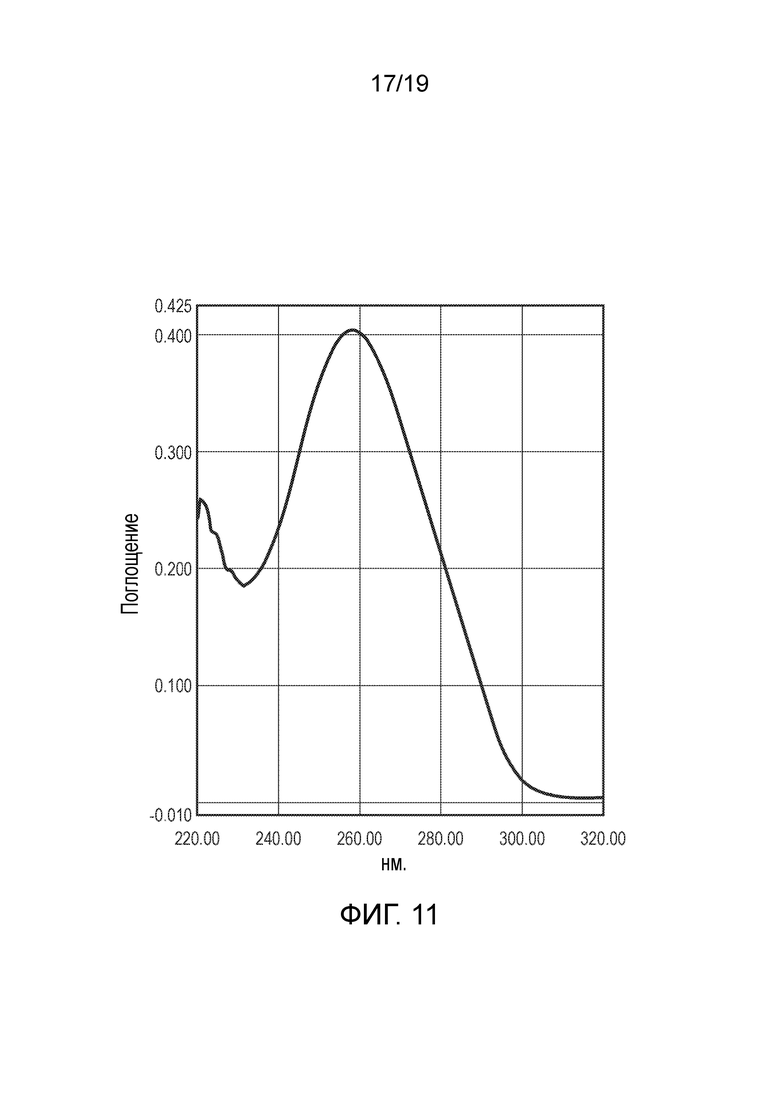
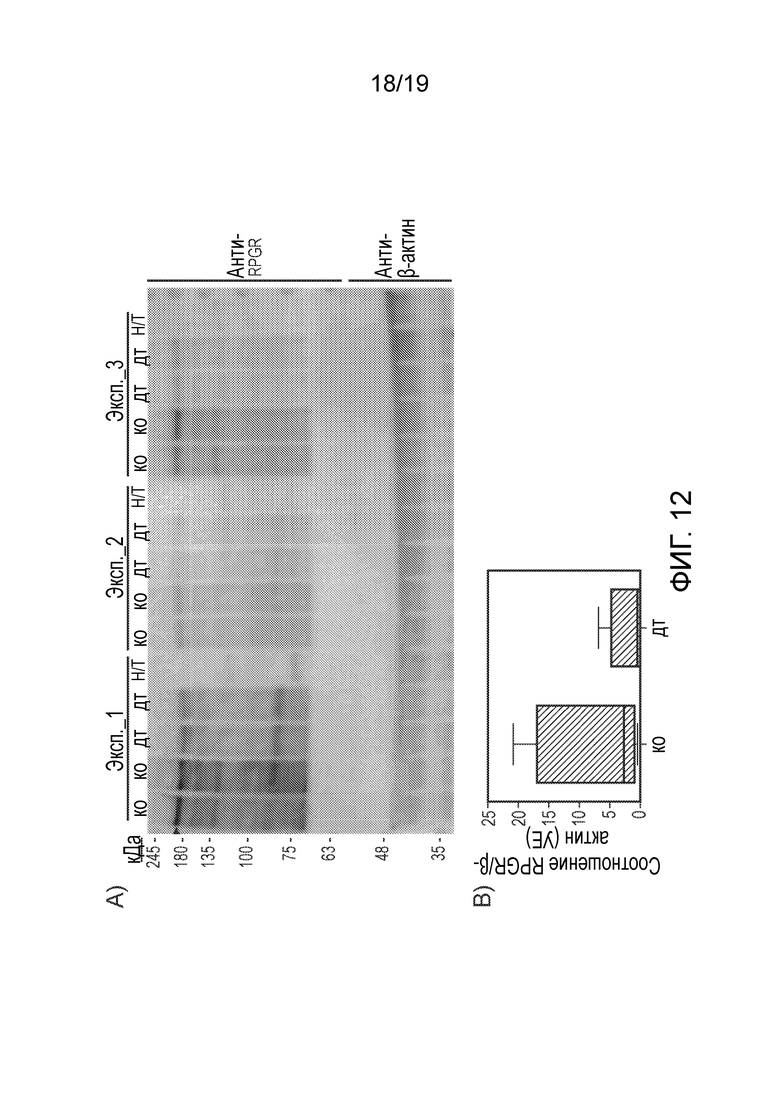
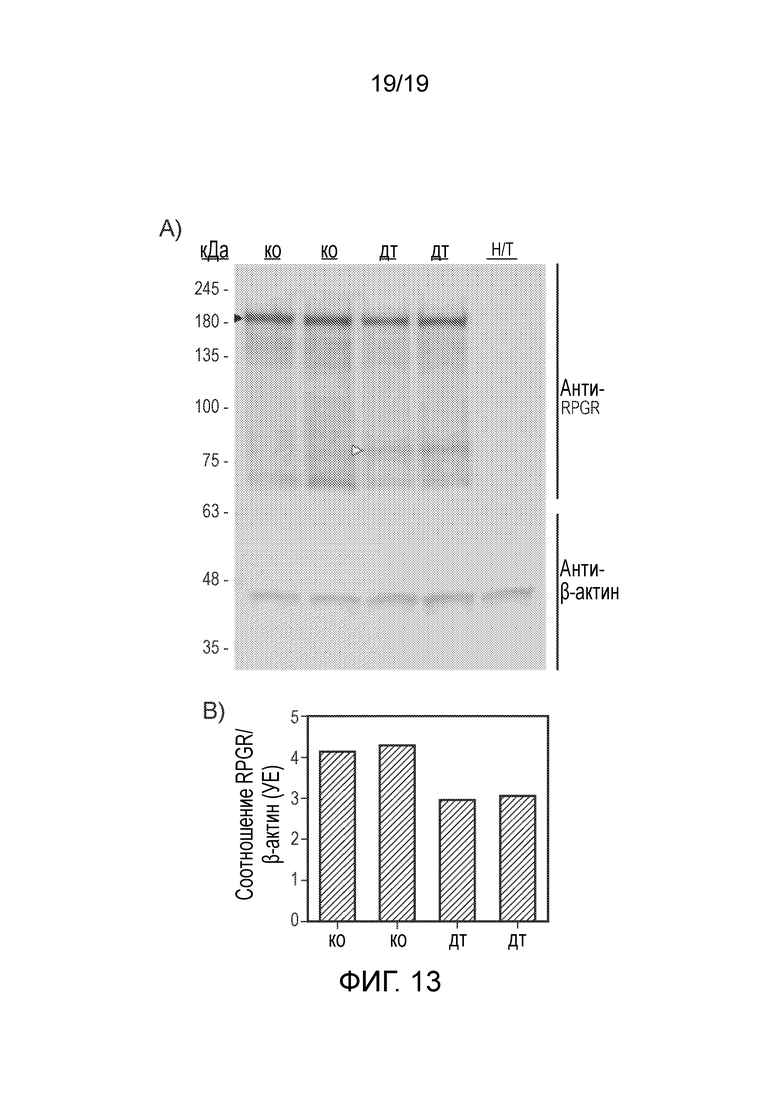
Изобретение относится к биотехнологии. Описан полинуклеотид, кодирующий изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15), содержащий нуклеотидную последовательность, которая является кодон-оптимизированной для повышения точности репликации последовательности, где нуклеотидная последовательность содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 1. Изобретение расширяет арсенал средств для лечения или предотвращения пигментного ретинита. 16 н. и 32 з.п. ф-лы, 13 ил., 5 пр.
1. Полинуклеотид, кодирующий изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15), содержащий нуклеотидную последовательность, которая является кодон-оптимизированной для повышения точности репликации последовательности, где нуклеотидная последовательность содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 1.
2. Полинуклеотид по п. 1, отличающийся тем, что нуклеотидная последовательность была кодон-оптимизирована для уменьшения числа пуриновых нуклеотидов в сравнении с последовательностью дикого типа.
3. Полинуклеотид по п. 1 или 2, отличающийся тем, что закодированный белок RPGRORF15 представляет собой человеческий RPGRORF15.
4. Полинуклеотид по любому из пп. 1-3, отличающийся тем, что нуклеотидная последовательность не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 33, 58, 59, 114, 123, 129, 181, 182, 213, 219, 226, 227, 237, 267, 285, 306, 309, 315, 324, 330, 339, 400, 401, 444, 456, 478, 480, 510, 594, 606, 618, 639, 697, 726, 744, 777, 807, 852, 877, 879, 888, 891, 921, 930, 960, 1042, 1050, 1116, 1140, 1183, 1184, 1194, 1197, 1221, 1249, 1251, 1257, 1273, 1276, 1281, 1290, 1293, 1357, 1372, 1373, 1413, 1446, 1452, 1464, 1474, 1475, 1482, 1519, 1520, 1542, 1584, 1590, 1599, 1608, 1653, 1668, 1674, 1689, 1692, 1734, 1761, 1776, 1788, 1803, 1824, 1845, 1854, 1881, 1893, 1902, 1909, 1910, 1929, 1932, 1960, 1987, 1988, 1992, 1998, 2020, 2031, 2047, 2049, 2062, 2067, 2076, 2121, 2167, 2169, 2190, 2193, 2208, 2229, 2259, 2283, 2298, 2323, 2343, 2346, 2361, 2373, 2382, 2388, 2403, 2409, 2410, 2412, 2448, 2457, 2472, 2476, 2478, 2505, 2520, 2547, 2574, 2622, 2634, 2661, 2673, 2706, 2712, 2763, 2775, 2796, 2811, 2817, 2832, 2838, 2871, 2886, 2892, 2898, 2910, 2922, 2931, 2937, 2958, 2970, 2997, 3033, 3039, 3069, 3075, 3117, 3129, 3156, 3166, 3235, 3246, 3273, 3285, 3306, 3321, 3369, 3399, 3405 и/или 3438 в SEQ ID NO: 2.
5. Полинуклеотид по любому из пп. 1-4, отличающийся тем, что нуклеотидная последовательность не содержит пуриновые нуклеотиды в положениях, соответствующих нуклеотидам 1689 и/или 3438 в SEQ ID NO: 2.
6. Полинуклеотид по любому из пп. 1-5, отличающийся тем, что нуклеотидная последовательность содержит нуклеотид T в положении, соответствующем положению 3405 в SEQ ID NO: 2.
7. Полинуклеотид по любому из предшествующих пунктов, отличающийся тем, что нуклеотидная последовательность не содержит пуриновые нуклеотиды по меньшей мере в 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180 или во всех из нуклеотидных положений, перечисленных в п. 4.
8. Полинуклеотид по любому из предшествующих пунктов, отличающийся тем, что нуклеотидная последовательность не содержит пуриновые нуклеотиды по меньшей мере во всех из нуклеотидных положений, перечисленных в п. 4.
9. Полинуклеотид по любому из пп. 1-8, отличающийся тем, что нуклеотидная последовательность содержит последовательность, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 3; или
(b) нуклеотидной последовательности SEQ ID NO: 3.
10. Полинуклеотид по п. 9(a), при этом указанная аминокислотная последовательность имеет по меньшей мере 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичности с SEQ ID NO: 1.
11. Полинуклеотид по любому из предшествующих пунктов, при этом нуклеотидная последовательность содержит один или более нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2.
12. Полинуклеотид по любому из предшествующих пунктов, при этом нуклеотидная последовательность содержит один или более нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 48 в SEQ ID NO: 2, A в положении, соответствующем положению 57 в SEQ ID NO: 2, T в положении, соответствующем положению 60 в SEQ ID NO: 2, T в положении, соответствующем положению 69 в SEQ ID NO: 2, A в положении, соответствующем положению 72 в SEQ ID NO: 2, C в положении, соответствующем положению 171 в SEQ ID NO: 2, C в положении, соответствующем положению 189 в SEQ ID NO: 2, T в положении, соответствующем положению 441 в SEQ ID NO: 2, T в положении, соответствующем положению 537 в SEQ ID NO: 2, C в положении, соответствующем положению 546 в SEQ ID NO: 2, T в положении, соответствующем положению 786 в SEQ ID NO: 2, T в положении, соответствующем положению 792 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, A в положении, соответствующем положению 990 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, A в положении, соответствующем положению 3355 в SEQ ID NO: 2, G в положении, соответствующем положению 3357 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3403 в SEQ ID NO: 2, C в положении, соответствующем положению 3404 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2.
13. Полинуклеотид, кодирующий изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) или ее функциональный вариант, имеющий по меньшей мере 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичности с SEQ ID NO: 1, содержащий нуклеотидную последовательность, которая содержит один или более нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2.
14. Полинуклеотид, кодирующий изоформу ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) или ее функциональный вариант, имеющий по меньшей мере 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичности с SEQ ID NO: 1, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из:
(a) нуклеотидной последовательности, которая имеет по меньшей мере 80% идентичности с SEQ ID NO: 3, при этом указанная последовательность содержит один или более нуклеотидов, выбранных из: C в положении, соответствующем положению 30 в SEQ ID NO: 2, T в положении, соответствующем положению 33 в SEQ ID NO: 2, A в положении, соответствующем положению 48 в SEQ ID NO: 2, A в положении, соответствующем положению 57 в SEQ ID NO: 2, T в положении, соответствующем положению 60 в SEQ ID NO: 2, T в положении, соответствующем положению 69 в SEQ ID NO: 2, A в положении, соответствующем положению 72 в SEQ ID NO: 2, C в положении, соответствующем положению 171 в SEQ ID NO: 2, C в положении, соответствующем положению 189 в SEQ ID NO: 2, T в положении, соответствующем положению 441 в SEQ ID NO: 2, T в положении, соответствующем положению 537 в SEQ ID NO: 2, C в положении, соответствующем положению 546 в SEQ ID NO: 2, T в положении, соответствующем положению 786 в SEQ ID NO: 2, T в положении, соответствующем положению 792 в SEQ ID NO: 2, A в положении, соответствующем положению 966 в SEQ ID NO: 2, A в положении, соответствующем положению 969 в SEQ ID NO: 2, A в положении, соответствующем положению 990 в SEQ ID NO: 2, T в положении, соответствующем положению 1011 в SEQ ID NO: 2, T в положении, соответствующем положению 1014 в SEQ ID NO: 2, T в положении, соответствующем положению 1029 в SEQ ID NO: 2, A в положении, соответствующем положению 1299 в SEQ ID NO: 2, C в положении, соответствующем положению 1689 в SEQ ID NO: 2, A в положении, соответствующем положению 3355 в SEQ ID NO: 2, G в положении, соответствующем положению 3357 в SEQ ID NO: 2, G в положении, соответствующем положению 3363 в SEQ ID NO: 2, T в положении, соответствующем положению 3403 в SEQ ID NO: 2, C в положении, соответствующем положению 3404 в SEQ ID NO: 2, T в положении, соответствующем положению 3405 в SEQ ID NO: 2, A в положении, соответствующем положению 3408 в SEQ ID NO: 2, A в положении, соответствующем положению 3409 в SEQ ID NO: 2, G в положении, соответствующем положению 3410 в SEQ ID NO: 2, G в положении, соответствующем положению 3432 в SEQ ID NO: 2, C в положении, соответствующем положению 3438 в SEQ ID NO: 2, и A в положении, соответствующем положению 3456 в SEQ ID NO: 2; или/и
(b) нуклеотидной последовательности SEQ ID NO: 3.
15. Полинуклеотид по любому из предшествующих пунктов, отличающийся тем, что нуклеотидная последовательность функционально связана с полинуклеотидом, содержащим элемент промотора, который способен управлять экспрессией RPGRORF15 или его функционального варианта в палочковидных и колбочковидных фоторецепторных клетках человека.
16. Полинуклеотид по любому из предшествующих пунктов, отличающийся тем, что нуклеотидная последовательность функционально связана с промотором родопсин-киназы (GRK1), предпочтительно промотором GRK1 человека.
17. Полинуклеотид по п. 16, отличающийся тем, что указанный промотор GRK1 содержит нуклеотидную последовательность SEQ ID NO: 7 или функциональный вариант, имеющий по меньшей мере 90 или 95% идентичности с ней.
18. Полинуклеотид по любому из предшествующих пунктов, отличающийся тем, что нуклеотидная последовательность содержит менее 165 динуклеотидов CpG и/или содержит не более двух CpG-островков.
19. Полинуклеотид по любому из предшествующих пунктов, имеющий по существу такую же, или повышенную, точность репликации в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
20. Полинуклеотид по любому из предшествующих пунктов, имеющий повышенную стабильность и/или менее подверженный мутациям, возникающим в процессе циклов репликации полинуклеотида, в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
21. Полинуклеотид по любому из предшествующих пунктов, обеспечивающий более высокие скорости трансляции и/или экспрессии белка в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
22. Полинуклеотид по любому из предшествующих пунктов, способствующий уменьшению или предотвращению образования альтернативно сплайсированных вариантов и/или укороченных белков в сравнении с эквивалентным полинуклеотидом, содержащим человеческий RPGRORF15 дикого типа с SEQ ID NO: 2.
23. Вирусный вектор для доставки изоформы ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) в глаз, содержащий полинуклеотид по любому из предшествующих пунктов.
24. Вирусный вектор по п. 23, представляющий собой аденоассоциированный вирусный (AAV), ретровирусный, лентивирусный или аденовирусный вектор.
25. Вирусный вектор по п. 23 или 24, представляющий собой AAV вектор.
26. Вирусный вектор по любому из пп. 23-25, имеющий форму вирусной частицы.
27. Вирусный вектор по п. 26, представляющий собой транскапсидированный AAV вектор.
28. Вирусный вектор по п. 26, содержащий геном AAV2 и капсидные белки AAV5 (AAV2/5) или AAV8 (AAV2/8).
29. AAV вектор по п. 25 или 26, представляющий собой AAV вектор, содержащий геном AAV2 и/или капсидный белок AAV8.
30. Система продуцирования вирусного вектора, содержащая набор полинуклеотидов, кодирующих компоненты, необходимые для продуцирования вирусного вектора, при этом система продуцирования вирусного вектора содержит полинуклеотид по любому из пп. 1-22.
31. Конструкция ДНК для применения в системе продуцирования вирусного вектора по п. 30, содержащая полинуклеотид по любому из пп. 1-22.
32. Клетка, продуцирующая вирусный вектор, которая содержит полинуклеотид по любому из пп. 1-22, систему продуцирования вирусного вектора по п. 30 или конструкцию ДНК по п. 31.
33. Способ продуцирования вирусного вектора, включающий введение полинуклеотида по любому из пп. 1-22 в клетку и культивирование клетки в условиях, подходящих для продуцирования вирусного вектора.
34. Вирусный вектор для доставки изоформы ORF15 регулятора ГТФазы пигментного ретинита (RPGRORF15) в глаз, получаемый с применением продуцирующей вирусный вектор клетки по п. 32 или способа по п. 33.
35. Клетка, трансфицированная полинуклеотидом по любому из пп. 1-22 или трансдуцированная вирусным вектором по любому из пп. 23-29 или 34.
36. Способ по п. 33 или клетка по п. 35, при этом указанная клетка представляет собой клетку HEK293 или HEK293T.
37. Фармацевтическая композиция для лечения или предотвращения пигментного ретинита, содержащая полинуклеотид по любому из пп. 1-22, вирусный вектор по любому из пп. 23-29 или 34 или клетку по п. 35 в сочетании с фармацевтически приемлемым носителем, разбавителем или эксципиентом.
38. Применение полинуклеотида по любому из пп. 1-22 или вирусного вектора по любому из пп. 23-29 или 34 для лечения или предотвращения пигментного ретинита.
39. Применение полинуклеотида по любому из пп. 1-22 или вирусного вектора по любому из пп. 23-29 или 34 для уменьшения гибели фоторецепторных клеток у субъекта, страдающего от, или имеющего риск развития, пигментного ретинита.
40. Применение по п. 38 или 39, при этом пигментный ретинит представляет собой X-связанный пигментный ретинит.
41. Применение по любому из пп. 38-40, при этом вирусный вектор вводят в глаз субъекта субретинальной, прямой ретинальной или интравитреальной инъекцией.
42. Применение по любому из пп. 38-41, при этом субретинальная инъекция включает этапы:
(a) введения раствора субъекту субретинальной инъекцией в количестве, эффективном для по меньшей мере частичного отслоения сетчатки, с образованием субретинального пузыря, при этом раствор не содержит полинуклеотид или вирусный вектор; и
(b) введения лекарственной композиции субретинальной инъекцией в пузырь, образованный на этапе (a), при этом лекарственная композиция содержит полинуклеотид или вирусный вектор.
43. Применение по любому из пп. 38-42, при этом вирусный вектор вводят субъекту одной дозой.
44. Применение по любому из пп. 38-43, при этом дегенерация фоторецепторных клеток вследствие пигментного ретинита по существу предотвращается на всем протяжении жизни субъекта.
45. Применение по любому из пп. 38-44, при этом зрительная функция по существу восстанавливается или сохраняется в получающем лечение глазу.
46. Способ лечения или предотвращения пигментного ретинита, включающий введение полинуклеотида по любому из пп. 1-22 или вирусного вектора по любому из пп. 23-29 или 34 субъекту, который нуждается в этом.
47. Способ уменьшения гибели фоторецепторных клеток у субъекта, страдающего от, или имеющего риск развития, пигментного ретинита, включающий введение полинуклеотида по любому из пп. 1-22 или вирусного вектора по любому из пп. 23-29 или 34 субъекту.
48. Применение полинуклеотида по любому из пп. 1-22 или вирусного вектора по любому из пп. 23-29 или 34 для:
(a) уменьшения или предотвращения образования альтернативно сплайсированных вариантов и/или укороченных белков RPGRORF15 в сравнении с вектором или полинуклеотидом, содержащим ген RPGRORF15 дикого типа, и/или
(b) повышения стабильности и/или точности репликации нуклеотидной последовательности, содержащей ген RPGRORF15, в сравнении с нуклеотидной последовательностью, содержащей ген RPGRORF15 дикого типа, и/или
(c) повышения скоростей трансляции и/или экспрессии белка RPGRORF15 в сравнении с вектором или полинуклеотидом, содержащим ген RPGRORF15 дикого типа, и/или
(d) увеличения продуцирования кодирующей RPGRORF15 нуклеотидной последовательности в сравнении с нуклеотидной последовательностью, содержащей ген RPGRORF15 дикого типа.
| МЕСТНЫЙ ГЛАЗНОЙ ПЕПТИДНЫЙ СОСТАВ | 2010 |
|
RU2561585C2 |
| N-СУЛЬФАТИРОВАННЫЕ ОЛИГОСАХАРИДЫ, АКТИВИРУЮЩИЕ РЕЦЕПТОРЫ FGF, ИХ ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ В ТЕРАПИИ | 2010 |
|
RU2559629C2 |
| Способ получения продуктов конденсации сульфоароматических жирных кислот с альдегидами | 1924 |
|
SU8801A1 |
| US 20050048536 А1, 03.03.2005. | |||

