ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к химерному белку, пригодному для адоптивной клеточной терапии (ACT). Химерный белок может действовать в виде гена самоубийства, обеспечивая удаление клеток, экспрессирующих химерный белок. Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей такой химерный белок, клетке, содержащей такую нуклеиновую кислоту и их терапевтическим применениям.
ПРЕДШЕСТВУЮЩИЙ ИЗОБРЕТЕНИЮ УРОВЕНЬ ТЕХНИКИ
Адоптивная клеточная терапия
Адоптивная иммунотерапия представляет собой общепризнанный и развивающийся терапевтический подход. В случае трансплантации аллогенных гемопоэтических стволовых клеток (HSCT) для лечения рецидивов злокачественных гематологических новообразований часто назначают инфузии донорских лимфоцитов (DLI). Инфильтрирующие опухоли лимфоциты (TIL) эффективны при лечении метастатической меланомы. Генетическая инженерия Т-клеток существенно увеличила область и эффективность Т-клеточной терапии: перенос Т-клеточных рецепторов обеспечивает направленное взаимодействие с внутриклеточными опухолевыми антигенами, в то время как химерные антигенные рецепторы (CAR) обеспечивают направленное взаимодействие с поверхностными антигенами злокачественных опухолей или линиеспецифическими антигенами. Клинический ответ наблюдали при обоих подходах, и в настоящее время проводят многочисленные дополнительные исследования.
При адоптивной иммунотерапии могут происходить неблагоприятные побочные явления. Основным и опасным осложнением DLI является реакция "трансплантат против хозяина" (РТПХ). Введение сконструированных Т-клеток также приводило к токсичности. Например, сообщалось о направленной внеопухолевой токсичности в исследованиях переноса нативных Т-клеточных рецепторов к антигенам меланомы; Т-клетки перенаправленные к антигену почечноклеточной карциномы карбоангидразе IX (CAIX) проявляли неожиданную гепатотоксичность. Сообщалось о синдромах активации иммунной системы после терапии CAR к CD19. Наконец, индуцированный вектором инсерционный мутагенез приводит к теоретическому риску лимфопролиферативных нарушений. Частота и тяжесть этих токсических проявлений являются непредсказуемыми. Кроме того, в отличие от терапевтических белковых молекул или низкомолекулярных соединений, неблагоприятные побочные явления которых, как правило, ослабевают при полувыведении терапевтических соединений, Т-клетки приживаются и реплицируются, потенциально приводя к растущей и быстро развивающейся токсичности. Гены самоубийства
Ген самоубийства представляет собой генетически кодируемый механизм, который обеспечивает селективное уничтожение адоптивно перенесенных клеток, таких как Т-клетки, в условиях неприемлемой токсичности. В клинических исследованиях тестировали два гена самоубийства: ген тимидинкиназы вируса простого герпеса (HSV-TK) и индуцибельной каспазы 9 (iCasp9).
Ген происходящей из вируса простого герпеса I тимидинкиназы (HSV-TK) использовали в качестве переключателя на программу самоубийства in vivo при инфузиях донорных Т-клеток для лечения рецидивирующих злокачественных новообразований и лимфопролиферации вследствие вируса Эпштейна-Барр (EBV) после трансплантации гемопоэтических стволовых клеток. Однако уничтожение Т-клеток, вызывающих реакцию "трансплантат против хозяина", являлось неполным, а применение для активации HSV-TK ганцикловира (или аналогов) в качестве пролекарственного средства исключает введение ганцикловира в качестве противовирусного лекарственного средства при цитомегаловирусных инфекциях. Кроме того, направляемый HSV-TK иммунный ответ приводил к устранению трансдуцированных HSV-TK клеток даже у пациентов с иммуносупрессией вследствие вируса иммунодефицита человека и трансплантации костного мозга, что ставит под угрозу продолжительное существование и, таким образом, эффективность инфузируемых Т-клеток.
В исходной молекуле iCasp9 использовали механизм активации каспазы 9. Все, что необходимо для активации каспазы 9, - это преодоление энергетического барьера гомодимеризации каспазы 9. Гомодимер претерпевает конформационные изменения, и протеолитический домен одного из пары димеров, становится активным. Физиологически это происходит посредством связывания домена CARD каспазы 9 с APAF-1. В iCasp9 домен APAF-1 замещен модифицированным FKBP12, который подвергнут мутированию для селективного связывания химического индуктора димеризации (CID). Присутствие CID приводит к гомодимеризации и активации. iCasp9 основана на модифицированной каспазе 9 человека, слитой со связывающим FK506 белком человека (FKBP) (Straathof et al. (2005) Blood 105:4247-4254). Это дает возможность обуславливаемой димеризации в присутствии низкомолекулярного соединения CID, известного как АР 1903. АР1903 представляет собой экспериментальное лекарственное средство и считается биологически инертным, так как он не взаимодействует с FKBP12 дикого типа. Однако клиническая практика с использованием этого средства ограничена очень малым количеством пациентов (Di Stasi, A. et al. (2011) N. Engl. J. Med. 365, 1673-1683 и Iuliucci, J. D. et al. (2001) J. Clin. Pharmacol. 41, 870-879). Также АР1903 является относительно большой и полярной молекулой, и маловероятно, что она пересекает гематоэнцефалический барьер.
В альтернативном подходе исполнительные каспазы можно активировать низкомолекулярными соединениями с использованием комплексной стратегии, включающей введение в каспазу 3, или 6, или 7 участков протеолиза вируса гравировки табака (TeV) и коэкспрессию с разделенной протеазой TEV, воссоединяющейся в присутствии рапамицина (Morgan et al (2014) Methods Enzymol. 544:179-213). Это является неудовлетворительной стратегией для клинически пригодного переключателя на программу самоубийства по ряду причин: во-первых, необходимы три раздельных белка, являющиеся очень сложными: модифицированная каспаза и два компонента разделенной протеазы TEV, соответственно; во-вторых, компоненты TeV являются ксеногенными и, вероятно, иммуногенными; наконец, эта стратегия активирует только чувствительные к протеазе молекулы каспаз, которые находятся ниже по каскаду и менее чувствительны, чем находящиеся выше каспазы.
Описан ген самоубийства на основе активации FAS посредством CID (Amara et al (1999) Hum. Gene Ther. 10, 2651-2655). Это также зависит от этого CID для активации, и так как это не активирует апоптоз напрямую, возможна устойчивость (вследствие устойчивости FAS).
Привлекательной альтернативой могла бы являться система гомодимеризации на основе стандартного фармацевтического средства, заменяющего необходимость в экспериментальном CID. Однако доступных гомодимеризующихся низкомолекулярных фармацевтических средств нет.
Предложены другие гены самоубийства, например, полноразмерный CD20 при экспрессии на Т-клетках может делать Т-клетки чувствительными к лизису терапевтическим антителом к CD20 ритуксимабом (Introna, М. et al. (2000) Hum. Gene Ther. 11, 611-620). По этой теме распознавания антителами описаны дополнительные гены самоубийства, например: RQR8 делает Т-клетки чувствительными к CD20, но является более компактным, чем полноразмерная молекула CD20 (Philip, В. et al. (2014) Blood doi:10.1182/blood-2014-01-545020); укороченная версия EGFR (huEGFRt) делает клетки чувствительными к лизису mAb к EGFR (Wang, X. et al. (2011) Blood 118, 1255-1263); и эпитопная метка myc, экспрессируемая на клеточной поверхности, делает клетки чувствительными к лизису антителом к myc (Kieback et al. (2008) Proc. Natl. Acad. Sci. U. S. A. 105, 623-628). Основным ограничением этих подходов с применением антител является их зависимость от биодоступности терапевтического антитела в высокой локальной концентрации для действия. Например, известно, что литические антитела не очень эффективны против заболевания с массированным поражением, а ограничением основанных на антителах генах самоубийства является то, что находящиеся в организме клетки, где не будет достигаться высокие концентрации антител, могут быть устойчивыми. Кроме того, в определенных ситуациях, например, при тяжелом синдроме активации макрофагов или цитокиновом шторме, индуцированном Т-клетками с CAR; дополнительная активация иммунной системы, индуцируемая моноклинальным антителом, может являться неблагоприятной для активации суицидального гена при клинической ситуации, которую пытаются лечить.
Таким образом, существует необходимость в альтернативном гене самоубийства, который не ассоциирован с указанными выше недостатками.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 - Рисунки, демонстрирующие различные подходы к RapCasp9. (а) Двойная конструкция, где две молекулы экспрессированы раздельно. Каждая молекула содержит каталитический домен Casp9, слитый с FKBP12 или FRB, соответственно. (b) Единая конструкция, где FKBP12 и FRB слиты непосредственно друг с другом, а затем слиты с каталитическим доменом Casp9 гибким линкером. В этой ориентации самогетеродимеризация не должна быть возможной, (с) Единая конструкция, где каталитический домен каспазы 9 фланкирован FRB и FKBP12. Здесь самогетеродимеризация происходить может, поэтому этот хорошего функционирования этого варианта не ожидают, (d) Двойная конструкция, где каталитический домен каспазы 9 слит с FKBP12, и коэкспрессируют отдельный малый белок, представляющий собой слияние двух копий FRB.
Фигура 2 - Демонстрация того, что гетеродимеризующим средством можно активировать каспазу 9. Т-клетки подвергали трансдукции только eGFP (фигура 2а) или совместной трансдукции с FKBP12-dCasp9 (с коэкспрессией eGFP) и FRB-dCasp9 (с коэкспрессией eBFP2) (фигура 2b). Т-клетки намеренно трансдуцировали только частично так, чтобы нетрансдуцированные Т-клетки могли служить в качестве внутренних контролей. Затем Т-клетки подвергали действию снижающихся концентраций рапамицина. Через 48 часов клетки окрашивали аннексином-V и 7AAD и анализировали посредством проточной цитометрии, анализируя долю жизнеспособных клеток, экспрессирующих флуоресцентные белки. Устранение Т-клеток, экспрессирующих eGFP и eBFP2 проходило очень эффективно даже в присутствии наименьшей концентрации рапамицина.
Фигура 3 - Функционирование вариантов RapCasp9. Т-клетки подвергали трансдукции (а) только eGFP; (b) подвергали двойной трансдукции FKBP12-Casp9 и FRB-Casp9 с коэкспрессией eGFP и eBFP2, соответственно; (с) подвергали трансдукции FRB-FKBP12-Casp9 и (d) подвергали трансдукции FRB-Casp9-FKBP12 и (е) FBP12-Casp9-2A-FRB-FRBw. Трансдукции подвергали только часть клеток, клетки с непрошедшей трансдукцией служили в качестве внутреннего отрицательного контроля. Т-клетки в течение 48 часов подвергали действию 2,5 нМ рапамицина. Затем Т-клетки окрашивали аннексином-V и 7AAD и анализировали посредством проточной цитометрии. Представлены eGFP по сравнению с eBFP2 на жизнеспособных клетках, как определено посредством окрашивания аннексином-V и 7AAD.
Фигура 4 - Рапамицин и рапалоги. А) Рапамицин; В) C-20-метилаллилрапамицин (MaRap); С) С16(S)-бутилсульфонамидорапамицин (C16-BS-Rap); D) C16-(S)-3-метилиндолрапамицин (C16-iRap) и Е) С16-(S)-7-метилиндолрапамицин (АР21976/С16-AiRap).
Фигура 5 - Обзор конструкций, тестируемых в примере 3.
Фигура 6 - Краткое описание стратегии гейтирования для примера 3.
Фигуры 7, 8 и 9 - Исследование, демонстрирующее уничтожение клеток Jurkat, трансфицированных конструкциями, представленными на фигуре 5, после инкубации с различными концентрациями рапамицина.
Фигура 10 - График для обобщения данных FACS, представленных на фигурах 7, 8 и 9.
Фигура 11 - График сравнения уничтожения клеток Jurkat в присутствии рапамицина и темсиролимуса.
СУЩНОСТЬ АСПЕКТОВ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали новый ген самоубийства, кодирующий продукт, который димеризуется в присутствии химического индуктора димеризации (CID), такого как рапамицин или аналог рапамицина.
Рапамицин и аналоги рапамицина индуцируют гетеродимеризацию, образуя область взаимодействия между доменом FRB mTOR и FKBP12. Эта ассоциация приводит к тому, что FKBP12 блокирует доступ к активному центру mTOR, ингибируя его функцию. Хотя mTOR является очень большим белком, известен и можно использовать определенный небольшой участок mTOR, необходимый для взаимодействия с рапамицином.
Авторы настоящего изобретения продемонстрировали, что для индукции гомодимеризации каспазы можно использовать гетеродимеризацию, опосредованную рапамицином. В частности, они неожиданно показали, что можно получить мультидоменную молекулу, которая содержит (i) домен FRB mTOR; (ii) FKBP12 и (iii) каспазу, и использовать гетеродимеризацию между доменом FRB одной копии молекулы и домен FKBP12 другой копии молекулы, обеспечивая гомодимеризацию доменов каспазы.
Таким образом, в первом варианте осуществления первого аспекта изобретения настоящее изобретение относится к химерному белку формулы: Ht1-Ht2-Casp где Casp представляет собой домен каспазы;
Ht1 представляет собой первый домен гетеродимеризации; и
Ht2 представляет собой второй домен гетеродимеризации,
и где в присутствии химического индуктора димеризации (CID) пара идентичных химерных белков взаимодействует так, что Ht1 одного химерного белка гетер одимеризуется с Ht2 другого химерного белка, обуславливая гомодимеризацию двух доменов каспазы.
Конфигурация является такой, что Ht1 не гетеродимеризуется в какой-либо значительной степени с Ht2 того же химерного белка.
Домен каспазы может содержать инициаторную каспазу, выбранную из следующей группы: каспаза-8, каспаза-9 и каспаза-10, или исполнительную каспазу, выбранную из каспазы-3 и каспазы-7.
В мультидоменном белке по этому первому варианту осуществления первого аспекта изобретения один домен гетеродимеризации может содержать связывающий FK506 белок (FKBP), а другой домен гетеродимеризации может содержать домен FRB mTOR.
Для этой комбинации доменов гетеродимеризации подходящим CID является рапамицин или аналог рапамицина.
Во втором варианте осуществления первого аспекта изобретения предоставлены химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий связывающий FK506 белок (FKBP12), и химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий домен FRB mTOR.
В третьем варианте осуществления этого аспекта изобретения предоставлены два белка:
Ht1-Casp и Ht2-Ht2
где Ht1-Casp представляет собой химерный белок, содержащий домен каспазы (Casp) и первый домен гетеродимеризации (Ht1); a Ht2-Ht2 представляет собой обеспечивающий взаимодействие белок, содержащий два или более вторых доменов гетеродимеризации (Ht2); и
где в присутствии химического индуктора димеризации (CID) пара химерных белков Ht1-Casp9 взаимодействует так, что Ht1 каждого химерного белка гетер одимеризуется с доменом Ht2 обеспечивающего взаимодействие белка, обуславливая гомодимеризацию двух доменов каспазы.
В четвертом варианте осуществления этого аспекта изобретения предоставлен химерный белок формулы:
Ht1-Casp-Ht2
где
Casp представляет собой домен каспазы;
Ht1 представляет собой первый домен гетеродимеризации; и
Ht2 представляет собой второй домен гетеродимеризации
и где в присутствии химического индуктора димеризации (CID) пара идентичных химерных белков взаимодействует так, что Ht1 одного химерного белка гетер одимеризуется с Ht2 другого химерного белка, обуславливая гомодимеризацию двух доменов каспазы.
В этом четвертом варианте осуществления первого аспекта изобретения, когда один домен гетеродимеризации содержит связывающий FK506 белок (FKBP), а другой домен гетеродимеризации содержит домен FRB mTOR, и CID представляет собой рапамицин или его производное, тогда для обеспечения гомодимеризации двух доменов каспазы можно использовать концентрации менее 5 нм, например, 1-3 нм или приблизительно 1 нм.
Химерный белок может содержать домен каспазы, слитый с FKBP12, а обеспечивающий взаимодействие белок может представлять собой слияние двух или более доменов FRB. Эти два или более домена FRB служат в качестве области взаимодействия, сближая два домена FKBP12-Casp.
Во втором аспекте настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей химерный белок по первому аспекту изобретения.
Нуклеиновая кислота может находиться в форме конструкции нуклеиновой кислоты, содержащей множество последовательностей нуклеиновой кислоты. Например, конструкция может содержать одну или несколько последовательностей нуклеиновых кислот по второму аспекту изобретения и последовательность нуклеиновой кислоты, кодирующую Т-клеточный рецептор (TCR) или химерный антигенный рецептор (CAR). Конструкция нуклеиновой кислоты может содержать:
i) первую последовательность нуклеиновой кислоты, кодирующую химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий связывающий FK506 белок (FKBP);
ii) вторую последовательность нуклеиновой кислоты, кодирующую химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий домен FRB mTOR.
Также предоставлена конструкция нуклеиновой кислоты структуры:
Ht1-Casp-coexpr-Ht2-Ht2 где:
Casp представляет собой последовательность нуклеиновой кислоты, кодирующую домен каспазы;
Ht1 представляет собой последовательность нуклеиновой кислоты, кодирующую первый домен гетеродимеризации;
Ht2 представляет собой последовательность нуклеиновой кислоты, кодирующую втор домен гетеродимеризации; и
coexpr представляет собой последовательность нуклеиновой кислоты, обеспечивающую коэкспрессию Ht1-Casp и Ht2-Ht2,
где экспрессия конструкции нуклеиновой кислоты приводит к получению химерного белка Ht1-Casp и обеспечивающего взаимодействие белка Ht2-Ht2, и где в присутствии химического индуктора димеризации (CID) пара химерных белков Ht1-Casp взаимодействует так, что Ht1 каждого химерного белка гетеродимеризуется с доменом Ht2 обеспечивающего взаимодействие белка, обуславливая гомодимеризацию двух доменов каспазы.
Ht1 может содержать связывающий FK506 белок (FKBP), а Ht2 может содержать домен FRB mTOR.
Конструкция нуклеиновой кислоты также может содержать последовательность нуклеиновой кислоты, кодирующую Т-клеточный рецептор (TCR) или химерный антигенный рецептор (CAR).
В третьем аспекте настоящее изобретение относится к вектору, содержащему последовательность нуклеиновой кислоты или конструкцию нуклеиновой кислоты по второму аспекту изобретения.
Вектор, который также может содержать представляющий интерес нуклеотид, такой как нуклеотидная последовательность, кодирующая химерный антигенный рецептор или Т-клеточный рецептор, так, что когда вектор используют для трансдукции клетки-мишени, клетка-мишень коэкспрессирует химерный белок по первому аспекту изобретения и химерный антигенный рецептор или Т-клеточный рецептор.
В четвертом аспекте настоящее изобретение относится к клетке, которая экспрессирует химерный белок по первому аспекту изобретения.
Клетка может содержать:
i) первый химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий связывающий FK506 белок (FKBP); и
ii) второй химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий домен FRB mTOR.
Также предоставлена клетка, экспрессирующая два белка: Ht1-Casp и Ht2-Ht2
где Ht1-Casp представляет собой химерный белок, содержащий домен каспазы (Casp) и первый домен гетеродимеризации (Ht1); a Ht2-Ht2 представляет собой обеспечивающий взаимодействие белок, содержащий два вторых домена гетеродимеризации (Ht2); и
где в присутствии химического индуктора димеризации (CID) пара химерных белков Ht1-Casp9 взаимодействует так, что Ht1 каждого химерного белка гетеродимеризуется с доменом Ht2 обеспечивающего взаимодействие белка, обуславливая гомодимеризацию двух доменов каспазы.
Клетка может содержать последовательность нуклеиновой кислоты или конструкцию по второму аспекту изобретения.
Клетка может представлять собой, например, гемопоэтическую стволовую клетку, лимфоцит или Т-клетку.
Также предоставлен способ получения клетки по четвертому аспекту изобретения, включающий этап трансдукции или трансфекции клетки вектором по третьему аспекту изобретения.
Также предоставлен способ удаления клеток по четвертому аспекту изобретения, включающий этап воздействия на клетки химическим индуктором димеризации (CID).
CID может представлять собой рапамицин или аналог рапамицина.
Также предоставлен способ профилактики или лечения заболевание у индивидуума, включающий этап введения индивидууму клеток по четвертому аспекту изобретения.
Способ может включать следующие этапы:
(i) трансдукции или трансфекции образца клеток, выделенных у индивидуума, вектором по второму аспекту изобретения, и
(ii) введение трансдуцированных/трансфицированных клеток пациенту. Способ может предназначаться для лечения рака.
Также предоставлен способ профилактики и/или лечения у индивидуума патологического иммунного ответа, вызванного введением индивидууму клеток по четвертому аспекту изобретения, включающий этап введения индивидууму рапамицина или аналога рапамицина.
Патологический иммунный ответ можно выбирать из следующей группы: реакция "трансплантат против хозяина"; направленная внеопухолевая токсичность; синдром активации иммунной системы и лимфопролиферативные нарушения.
Способ лечения или профилактики заболевания у индивидуума может включать следующие этапы:
(i) введение индивидууму клеток по четвертому аспекту изобретения;
(ii) наблюдение индивидуума на развитие патологического иммунного ответа; и
(iii) введение индивидууму рапамицина или аналога рапамицина, если индивидуум демонстрирует признаки развития или наличие развитого патологического иммунного ответа.
Также предоставлены клетки по четвертому аспекту изобретения для применения в трансплантации гемопоэтических стволовых клеток, инфузии лимфоцитов или адоптивном переносе клеток.
Также предоставлен рапамицин или аналог рапамицина для применения в профилактике или лечении патологического иммунного ответа, вызванного введением индивидууму клеток по четвертому аспекту изобретения.
Таким образом, настоящее изобретение относится к гену самоубийства, который обеспечивает селективное уничтожение адоптивно инфундированных клеток в случае неприемлемой токсичности, и который активируется рапамицином и/или его аналогами.
Рапамицин представляет собой стандартное фармацевтическое средство с хорошо известными свойствами, превосходной биодоступностью и объемом распределения и являющееся широкодоступным. Также рапамицин фактически не ухудшает подвергаемое лечению состояние, так как он представляет собой иммуносупрессор, он с большой вероятностью оказывает положительное воздействие на нежелательную токсичность, также как его функция гена самоубийства.
ПОДРОБНОЕ ОПИСАНИЕ
ХИМЕРНЫЙ БЕЛОК
Настоящее изобретение относится к химерному белку, который действует в качестве гена самоубийства. Клетки, экспрессирующие химерный белок, можно удалять in vivo или in vitro посредством введения химического индуктора димеризации (CID), такого как рапамицин или аналог рапамицина.
Химерный белок может иметь формулу:
Ht1-Ht2-Casp
в которой
Casp представляет собой домен каспазы;
Ht1 представляет собой первый домен гетеродимеризации; и
Ht2 представляет собой второй домен гетеродимеризации.
Химерный белок может иметь формулу:
Ht1-Ht2-L-Casp
в которой Casp, Ht1 и Ht2 являются такими, как определено выше, a L представляет собой необязательный линкер.
Конфигурация должна быть такой, чтобы Ht1 не гетеродимеризовался в значительной степени с Ht2 той же молекулы химерного белка, но когда два химерных белка сближаются в присутствии химического индуктора димеризации (CID) Ht1 одного химерного белка гетеродимеризовался с Ht2 другого химерного белка, обуславливая гомодимеризацию двух доменов каспазы.
Конфигурация является такой, что Ht1 не гетеродимеризуется в значительной степени с Ht2 того же химерного белка. Например, в клетке, экспрессирующей химерный белок по этому варианту осуществления первого аспекта изобретения, присутствие CID должно обуславливать большую долю димеризации между двумя химерными белками, чем гетеродимеризацию в пределах одного и того же химерного белка. Количество химерных белков, которые гетеродимеризуется в пределах одной и той же молекулы в клетке или группе клеток или в растворе, может составлять менее 50%, 40%, 30%, 20%, 10%, 5% или 1% от количества химерных белков, которые в присутствии CID димеризуются с другой молекулой химерного белка.
Химерный белок может содержать последовательность, представленную как SEQ ID NO: 1.
SEQ ID NO: l (FRB-FKBP12-L3-dCasp9)
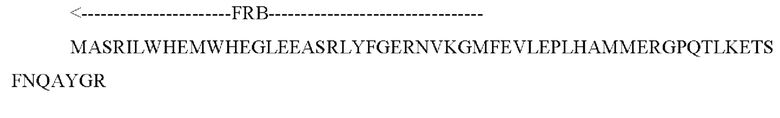

В указанной выше последовательности "FKBP12" относится к последовательности FKBP12; "dCasp9" относится к каталитическому домену Casp9; "L1" представляет собой линкер с одним повтором; "FMD-2A" представляет собой пептид ERAV, подобный пептиду 2А ящура; "FRB" представляет собой домен FRB mTOR; "L3" представляет собой линкер с двумя повторами; и "FRBw" представляет собой FRB с вырожденными кодонами.
Во втором варианте осуществления изобретение относится к "двухмолекулярной" системе гена самоубийства, в которой CID представляет собой рапамицин или аналог рапамицина.
Таким образом, настоящее изобретение также относится к i) химерному белку, содержащему домен каспазы и домен гетеродимеризации, содержащий связывающий FK506 белок (FKBP 12); и ii) химерному белку, содержащему домен каспазы и домен гетеродимеризации, содержащий домен FRB mTOR.
Когда клетка, такая как Т-клетка, экспрессирует оба этих химерных белка, присутствие рапамицина или аналога рапамицина вызывает гетеродимеризацию содержащего FKBP домена или i) с содержащим FRB доменом или ii), таким образом, обуславливая гомодимеризацию доменов каспазы из i) и ii).
В этом варианте осуществления изобретения химерный белок может содержать последовательность, представленную как SEQ ID NO: 2 или 3.


В третьем варианте осуществления изобретение относится к альтернативному "двухмолекулярному" подходу с меньшей областью узнавания, чем во втором варианте осуществления. Здесь, Ht1 слит с каспазой, и коэкспрессируют вторую молекулу, содержащую Ht2-Ht2. В присутствии CID Ht2-Ht2 сближает две молекулы Ht1-Casp.На практике, это можно реализовать посредством коэкспрессии FKBP12-Casp9 с FRB-FRB и активации рапамицином. В целях удобства эти компоненты можно коэкспрессировать с пептидом, подобным пептиду 2А ящура. Вторая кодирующая Ht2 (например, FRB) последовательность может представлять собой последовательность с вырожденными кодонами для предотвращения рекомбинации.


В указанной выше последовательности: "FKBP 12" относится к FKBP12; "dCasp9" представляет собой каталитический домен Casp9; "L1" представляет собой линкер с одним повтором; "FMD-2A" представляет собой пептид ERAV, подобный пептиду ящура 2А; "FRB" представляет собой домен FRB mTOR; "L2" представляет собой линкер с двумя повторами; и "FRBw" представляет собой FRB с вырожденными кодонами.
КАСПАЗА
Каспазы или цистеин-аспарагиновые протеазы или цистеин-зависимые направляемые аспартатом протеазы представляют собой семейство цистеиновых протеаз, которые играют существенные роли в апоптозе.
У людей идентифицировано двенадцать каспаз. Существует два типа апоптотических каспаз: инициаторные каспазы и исполнительные каспазы. Инициаторные каспазы, такие как каспаза-2, каспаза-8, каспаза-9 и каспаза-10, расщепляют неактивные про-формы исполнительных каспаз, таким образом, активируя их. Затем исполнительные каспазы, такие как каспаза-3, каспаза-6 и каспаза-7, расщепляют другие белковые субстраты в клетке, запуская процессы апоптоза.
Домен каспазы химерного белка по первому аспекту настоящего изобретения может содержать инициаторную каспазу, выбранную из каспазы-2; каспазы-8, каспазы-9 и каспазы-10; или исполнительную каспазу, выбранную из каспазы-3, каспазы-6 и каспазы-7.
В частности, домен каспазы химерного белка по первому аспекту настоящего изобретения может содержать каспазу-9. Каспаза 9 представляет собой ключевую инициаторную каспазу так как ее активация представляет собой очень чувствительный пусковой фактор индукции апоптоза. Кроме того, гомодимеризация является всем, что необходимо для активации, в отличие от гомодимеризации и протеолитического расщепления.
Полноразмерная каспаза-9 имеет последовательность, представленную как SEQ ID NO: 5.

Каспаза-9 может быть укороченной, например, с удалением домена активации и рекрутирования каспазы. Укороченная каспаза-9 представлена как SEQ ID NO: 6


Химерный белок по первому аспекту изобретения может содержать SEQ ID NO: 5 или SEQ ID NO: 6 или их фрагмент или вариант, который сохраняет способность к гомодимеризации и, таким образом, инициации апоптоза.
Вариант последовательности каспаза-9 может обладать последовательностью, по меньшей мере на 80%, 85%, 90%, 95%, 98% или 99% идентичной с SEQ ID NO: 5 или 6.
Процент идентичности двух полипептидных последовательностей можно легко определять такими программами как BLAST, которые свободно доступны на http://blast.ncbi.nlm.nih.gov.
Протеаза каспаза 9 in vivo является центральным участником многокомпонентного каскада, известного как апоптосома, который контролирует удаление клеток при эмбриогенезе и физиологических ответы, которые инициируют гибель клеток, а также летальные повреждения клеток, такие как ионизирующая радиация или химиотерапевтические лекарственные средства. Функцией каспазы 9 является образование активных форм каспаз 3 и 7 посредством ограниченного протеолиза, и, таким образом, в передаче сигнала апоптоза в исполнительную фазу. Однако каспаза 9 необычна в ряду своих ближайших родственников тем, что в каталитическую форму латентный зимоген преобразует не протеолиз большой и малой субъединиц. Фактически, этим является гомодимеризация, необходимая для активации.
ДОМЕНЫ ГЕТЕРОДИМЕРИЗАЦИИ
Макролиды рапамицин и FK506 действуют, индуцируя гетеродимеризацию клеточных белков. Каждое лекарственное средство с высокой аффинностью связывается с белком FKBP12, образуя комплекс лекарственное средство-белок, которые затем связывается и инактивирует mTOR/FRAP и кальциневрин, соответственно. Связывающий FKBP-рапамицин (FRB) домен mTOR определен и применяется как выделенная молекула белка из 98 аминокислот, которую можно сливать с представляющим интерес белком. Затем рапамицин может индуцировать сближение слияний с FRB с FKBP12 или белками, слитыми с FKBP12.
В контексте настоящего изобретения один из доменов гетеродимеризации (Ht1 или Ht2) может представлять собой или содержать FRB или его вариант, а другой домен гетеродимеризации (Ht2 или Ht1) может представлять собой или содержать FKBP12 или его вариант.
Рапамицин обладает несколькими свойствами идеального димеризующего средства: он обладает высокой аффинностью (KD<1 нМ) к FRB при связывании с FKBP12, и он высокоспецифичен к домену FRB mTOR. Рапамицин представляет собой эффективный терапевтический иммуносупрессор с приемлемыми фармакокинетическим и фармакодинамическим профилями у млекопитающих. В соответствии с клинической ситуацией также можно использовать фармакологические аналоги рапамицина с другими фармакокинетическими и -динамическими свойствами, такие как эверолимус, темсиролимус и дефоролимус (Ben jamin et al, Nature Reviews, Drug Discovery, 2011).
Для предотвращения связывания и инактивации рапамицином эндогенного mTOR, поверхность рапамицина, которая контактирует с FRB можно модифицировать. Компенсирующая мутация домена FRB с формированием вогнутости, которая адаптирует "выпуклый" рапамицин восстанавливает димеризующие взаимодействия только с мутантным FRB, а не с эндогенным белком mTOR.
В Bayle et al. (Chem Bio; 2006; 13; 99-107) описаны различные аналоги рапамицина или "рапалоги" и соответствующие им модифицированные связывающие FRB домены. Например, в Bayle et al. (2006) описаны рапалоги: C-20-метилаллилрапамицин (MaRap), С16(S)-бутилсульфонамидорапамицин (C16-BS-Rap) и С16-(S)-7-метилиндолрапамицин (AP21976/C16-AiRap), как представлено на фигуре 3, в комбинации с соответствующими комплементарными связывающими домены для каждого. Другие рапамицины/рапалоги включают сиролимус и такролимус.
Домены гетеродимеризации химерного белка могут представлять собой или содержать одну из последовательностей, представленных как SEQ ID NO:7-SEQ ID NO: 11 или их вариант.

SEQ ID No 10 - участок FRB mTOR с заменой T на H по остатку 2098 и с W на F по остатку остаток 2101 полноразмерного mTOR, который связывает рапамицин со сниженной по сравнению с диким типом аффинностью
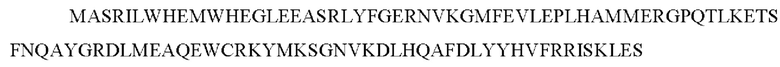
SEQ ID No 11 - участок FRB mTOR с заменой к K на Р по остатку 2095 полноразмерного mTOR, который связывает рапамицин со сниженной аффинностью
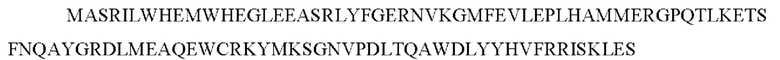
Варианты последовательностей могут обладать последовательностями по меньшей мере на 80%, 85%, 90%, 95%, 98% или 99% идентичными с SEQ ID NO:7-11, при условии, что последовательности обеспечивают эффективную систему димеризации. Т.е., при условии, что последовательности способствуют достаточный уровень колокализации двух химерных белков, чтобы обеспечить гомодимеризацию двух доменов каспазы.
Домен FRB "дикого типа", представленный как SEQ ID NO: 8, содержит аминокислоты 2025-2114 mTOR человека. С использованием система нумерации аминокислот mTOR человека, последовательность химерного белка FRB по изобретению может содержать замену аминокислоты по одной или нескольким из следующих положений: 2095, 2098, 2101.
Вариант FRB, используемый в химерном белке по изобретению может содержать одну из следующих аминокислоты в положениях 2095, 2098 и 2101:
2095: K, Р, Тили А
2098: Т, L, Н или F
2101: W или F
В Bayle et al (как выше) описаны следующие варианты FRB, указанные в соответствии с аминокислотами в положениях 2095, 2098 и 2101 (см. таблицу 1): KTW, PLF, KLW, PLW, TLW, ALW, PTF, ATF, TTF, KLF, PLF, TLF, ALF, KTF, KHF, KFF, KLF. Эти варианты способны к связыванию с рапамицином и рапалогами в различной степени, как представлено в таблице 1 и на фигуре 5А Bayle et al. Химерный белок по изобретению может содержать один из этих вариантов FRB.
ЛИНКЕР
В пространственно разделенные домен каспазы и домен(ы) гетеродимеризации можно включать линкер.
В первом варианте осуществления первого аспекта настоящего изобретения химерный белок содержит два домена гетеродимеризации, которые расположены в конфигурации так, что они не могут гетеродимеризоваться друг с другом в одной молекуле в присутствии CID, но Ht1 одной молекулы может гетеродимеризоваться с Ht2 другой химерной молекулы с такими же доменами гетеродимеризации (фигура 1В). В конструкции, где Ht1 и Ht2 фланкируют домен каспазы (Ht1-Casp-Ht2), активация уступала конструкциям, где Ht1 и Ht2 были связаны вместе, указывая на важность предотвращения непродуктивного связывания Ht1 и Ht2 одной молекулы при одном CID.
В этом варианте осуществления линкер (L1) должен обеспечивать достаточную гибкость, чтобы могла проходить гомодимеризация каталитических доменов, но не настолько большую гибкость, чтобы не преодолеть энергетический барьер гомодимеризации (фигура 1). Например, длина линкера может составлять менее 15, менее 10 или 5-15 или 5-10 аминокислот.
Во втором варианте осуществления первого аспекта настоящего изобретения химерный белок содержит один домен гетеродимеризации, который в присутствии CID способен к гетеродимеризации с комплементарным доменом гетеродимеризации второго химерного белка.
В альтернативной конфигурации в одной молекуле можно размещать два домена гетеродимеризации с длинным линкером (L2), обеспечивая конструкцию формулы: Ht1-Casp1-L2-Ht2-Casp2
Домены НТ и Casp на любой стороне линкера могут находиться в любом порядке.
В этом варианте осуществления линкер L2 может обеспечивать достаточную гибкость так, что первый домен гетеродимеризации может гетеродимеризоваться со вторым доменом гетеродимеризации; и так, что домен каспазы в части молекулы, соответствующей "первому химерному белку" может гомодимеризоваться с доменом каспазы в части молекулы, соответствующей "второму химерному белку".
В третьем варианте осуществления первого аспекта изобретения Casp слит с одним доменом гетеродимеризации, а вторая молекула представляет собой слияние двух или более копий другого домена гетеродимеризации. Две молекулы можно коэкспрессировать. В этом случае, вторая молекула действует в качестве области взаимодействия, сближая два или более доменов Casp в присутствии CID. В этом случае две или более копии доменов гетеродимеризации должны быть слиты таким образом, чтобы обеспечить достаточное для их активации сближение доменов Casp9.
Обеспечивающий взаимодействие белок может являться мультимерным, содержа более двух доменов Ht2. Например, в одном обеспечивающим взаимодействие белке можно комбинировать множество доменов Ht2 с использованием мультимеризующего линкера, такого как суперспиральный домен.
В этом варианте осуществления обеспечивающий взаимодействие белок может иметь формулу Ht2-L2-Ht2 или Ht2-L2, в которой L2 представляет собой суперспиральный домен.
Суперспираль представляет собой структурный мотив, в котором от двух до семи альфа-спиралей скручены вместе, подобно нитям каната. Структура суперспиральных доменов хорошо известна в данной области. Например, как описано в Lupas & Gruber (Advances in Protein Chemistry; 2007; 70; 37-38).
Как правило, суперспирали, содержат повторяющийся шаблон, hxxhcxc, гидрофобных (h) и заряженных (с) аминокислотных остатков, обозначаемый как семичленный повтор. Как правило, положения в семичленном повторе, обозначают abcdefg, где a и d представляют собой гидрофобные положения, часто являясь занятыми изолейцином, лейцином или валином. Сворачивание последовательности с этим повторяющимся шаблоном в альфа-спиральную вторичную структуру заставляет гидрофобные остатки выстраиваться в виде "полосы", которая медленно обвивается вокруг спирали в направлении влево, формируя амфипатическую структуру. Наиболее предпочтительным способом расположения двух таких спиралей в цитоплазме является закручивание гидрофобных цепей друг с другом, зажатыми между гидрофильными аминокислотами. Таким образом, происходит углубление гидрофобных поверхностей, которое обеспечивает термодинамическую движущую силу для олигомеризации. Укладка в суперспиральном внутреннем слое является исключительно плотной, почти с полным вандерваальсовским контактом между боковыми цепями остатков a и d.
Примеры белков, которые содержат суперспиральный домен, в качестве неограничивающих примеров включают моторный белок кинезин, дельта-антиген гепатита D, коровый белок sRNP C/D-бокса архебактерий, олигомерный матриксный белок хряща (СОМР), связывающий маннозу белок А, суперспиральный богатый серином белок 1, полипептидный фактор высвобождения 2, SNAP-25, SNARE, репрессор Lac или аполипопротеин Е.
ХИМИЧЕСКИЙ ИНДУКТОР ДИМЕРИЗАЦИИ (CID)
Химический индуктор димеризации (CID) может представлять собой любую молекулу, которая индуцирует гетеродимеризацию Ht1 и Ht2 на раздельных химерных молекулах с теми же доменами Ht1 и Ht2.
CID может представлять собой рапамицин или аналоги рапамицина ("рапалоги") с улучшенными или отличающимися фармакодинамическими или фармакокинетическими свойствами по сравнению с рапамицином, но с тем же общим механизмом действия. CID может представлять собой измененный рапамицин со сконструированной специфичностью к комплементарным FKBP12 или FRB - например, как представлено на фигуре 4. В Bayle et al. (2006, как выше) описаны различные рапалоги, функционализированные по С16 и/или С20.
Примеры таких рапалогов первой категории включают сиролимус, эверолимус, темсиролимус и дефоролимус. Примеры рапалогов второй категории включают С-20-метилаллилрапамицин (MaRap); С16(S)-бутилсульфонамидорапамицин (C16-BS-Rap); С16-(S)-3-метилиндолрапамицин (C16-iRap); и С16-(S)-7-метилиндолрапамицин (AP21976/C16-AiRap).
Домены гомодимеризации каспазы в присутствии CID могут обеспечивать активацию каспазы, приводящую к активности, которая в 2, 5, 10, 50, 100, 1000 или 10000 раз превосходит активность каспазы в отсутствие CID.
Рапамицин представляет собой сильнодействующее иммуносупрессорное средство. Аналоги рапамицина (рапалоги) находятся в ежедневном клиническом применении. Современные рапалоги обладают превосходной биодоступностью и объемами распределения. Хотя они являются сильнодействующими иммуносупрессорными средствами, короткий период дозирования (для активации гена самоубийства) должен обуславливать минимальное побочное действие. Кроме того, в отличие от введения mAb, фармакологическое действие рапамицина и аналогов может быть выгодным в клинических ситуациях, где необходима активация генов самоубийства, таки как внеопухолевая токсичность или синдромы гиперактивации иммунной системы.
ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ
Второй аспект изобретения относится к последовательности нуклеиновой кислоты, кодирующей химерный белок по изобретению.
Как используют в настоящем документе, термины "полинуклеотид", "нуклеотид" и "нуклеиновая кислота" являются синонимами друг с другом.
Специалисту понятно, что в результате вырожденности генетического кода один и тот же полипептид могут кодировать множество различных полинуклеотидов и нуклеиновых кислот. Кроме того, следует понимать, что для соответствия частоте использования кодонов любого конкретного организма-хозяина, в котором необходимо экспрессировать полипептиды, специалисты общепринятыми способами могут проводит замены нуклеотидов, которые не влияют на полипептидную последовательность, кодируемую полинуклеотидами, описываемыми в настоящем документе.
Нуклеиновые кислоты по второму аспекту изобретения могут содержать ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Также они могут представлять собой полинуклеотиды, которые содержат синтетические или модифицированные нуклеотиды. В данной области известен ряд модификаций олигонуклеотидов различного типа. Они включают метилфосфонатные и тиофосфатные каркасы, добавление акридиновых или полилизиновых цепей на 3'- и/или 5'-концах молекулы. Следует понимать, что для целей использования, описанных в настоящем документе, полинуклеотиды можно модифицировать любым способом, доступным в данной области. Такие модификации можно проводить для увеличения активности или продолжительность жизни представляющих интерес полинуклеотидов in vivo.
Термины "вариант", "гомолог" или "производное" в отношении нуклеотидной последовательности включают любую вариацию, модификацию, замену, делецию или добавление одной (или более) нуклеиновой кислоты из последовательности или в ней.
В первом варианте осуществления этого аспекта изобретения предоставлена нуклеиновая кислота, кодирующая химерный белок формулы:
Ht1-Ht2-L-Casp
где
Ht1 представляет собой первый домен гетеродимеризации; и Ht2 представляет собой второй домен гетеродимеризации. L представляет собой необязательный линкер; Casp представляет собой домен каспазы;
Последовательность нуклеиновой кислоты может кодировать последовательность химерного белка, представленную как SEQ ID NO: l, или ее вариант.
Например, нуклеотидная последовательность может содержать последовательность, представленную как SEQ ID NO: 12


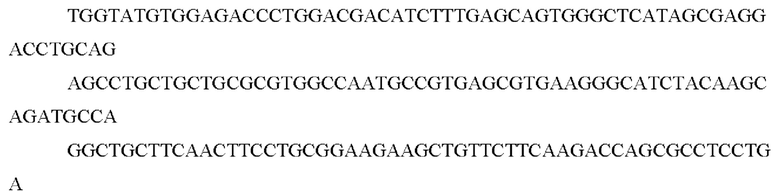
Во втором варианте осуществления этого аспекта изобретения предоставлена последовательность нуклеиновой кислоты, кодирующая химерный белок формулы: Ht1-L-Casp где Ht1 представляет собой домен гетеродимеризации. L представляет собой необязательный линкер; и Casp представляет собой домен каспазы;
Последовательность нуклеиновой кислоты может кодировать последовательность химерного белка, представленную как SEQ ID NO: 2 или 3 или ее вариант.
Например, нуклеотидная последовательность может содержать последовательность, представленную как SEQ ID NO: 13 или 14



В этом втором варианте осуществления последовательности нуклеиновой кислоты можно предоставлять в форме конструкций, кодирующих оба химерных белка. Конструкция может кодировать полипротеин формулы: Ht1-L2-Casp-coexpr-Ht2-L2-Casp где
Ht1 представляет собой первый домен гетеродимеризации;
L1 и L2 представляют собой необязательные линкеры, которые могут являться одинаковыми или различными;
Соехрг представляет собой последовательность, обеспечивающую коэкспрессию двух белков: Ht1-L1-Casp и Ht2-L2-Casp;
Ht2 представляет собой второй домен гетеродимеризации; и
Casp представляет собой домен каспазы.
Когда присутствуют последовательности нуклеиновых кислот, кодирующие одни и те же или сходные последовательности, такие как два домена каспазы, одна из последовательностей может содержать вырожденные кодоны во избежание гомологичной рекомбинации.
В третьем варианте осуществления предоставлена последовательность нуклеиновой кислоты, кодирующая последовательность следующей формулы: Ht1-Casp-coexpr-Ht2-Ht2 где
Casp представляет собой домен каспазы;
Ht1 представляет собой первый домен гетеродимеризации;
Coexpr представляет собой последовательность, обеспечивающую коэкспрессию белков Ht1-Casp и Ht2-Ht2, такую как участок расщепления; и
Ht2 представляет собой второй домен гетеродимеризации, который гетеродимеризуется с Ht1 в присутствии химического индуктора димеризации (CID).
В последовательности, кодирующей второй белок, Ht2-Ht2, одна из последовательностей, кодирующих Ht2 может содержать вырожденные кодоны во избежание гомологичной рекомбинации.
Конструкция нуклеиновой кислоты по третьему варианту осуществления может содержать последовательность, представленную как SEQ ID NO: 15.



Последовательности нуклеиновой кислоты с высокой степенью сходства, такие как последовательности каспаз или последовательности FRB могут содержать вырожденные кодоны во избежание рекомбинации.
КОНСТРУКЦИЯ НУКЛЕИНОВОЙ КИСЛОТЫ
Изобретение также относится к конструкции нуклеиновой кислоты, содержащей: i) первую последовательность нуклеиновой кислоты, кодирующую химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий связывающий FK506 белок (FKBP); и
ii) вторую последовательность нуклеиновой кислоты, кодирующую химерный белок, содержащий домен каспазы и домен гетеродимеризации, содержащий домен FRB mTOR.
Изобретение также относится к конструкции нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, кодирующую один или несколько химерных белков и дополнительную необходимую последовательность нуклеиновой кислоты (NOI). Например, NOI может кодировать Т-клеточный рецептор (TCR) или химерный антигенный рецептор (CAR).
Последовательности нуклеиновой кислоты можно связывать последовательностью, обеспечивающей коэкспрессию двух или более последовательностей нуклеиновой кислоты. Например, конструкция может содержать последовательность внутреннего промотора, внутреннего участка связывания рибосомы (IRES) или последовательность, кодирующую участок расщепления. Участок расщепления может являться саморасщепляемым так, что при продукции полипептида он сразу же расщепляется на отдельные белки без необходимости в какой либо внешнем расщепляющем действии.
Известны различные саморасщепляющиеся участки, включая саморасщепляющийся пептид вируса ящура (FMDV) 2а с последовательностью, представленной как SEQ ID NO: 16 или 17:
SEQ ID NO: 16
RAEGRGSLLTCGDVEENPGP или
SEQ ID No 17
QCTNYALLKLAGDV SNPGP.
Последовательность для коэкспрессии может представлять собой внутренний участок связывания рибосомы (IRES). Последовательность для коэкспрессии может представлять собой внутренний промотор.
Т-КЛЕТОЧНЫЙ РЕЦЕПТОР (TCR)
Т-клеточный рецептор или TCR представляет собой молекулу, находящуюся на поверхности Т-клеток, которая отвечает за распознавание антигенов, связанных с молекулами главного комплекса гистосовместимости (МНС). Связывание TCR и антигена происходит с относительно низкой аффинностью и является вырожденным: многие TCR распознают один и тот же антиген и один и тот TCR распознает множество антигенов.
TCR состоит из два различных белковых цепей, т.е. он представляет собой гетеродимер. У 95% Т-клеток он состоит из альфа- (α) и бета- (β) цепей, тогда как у 5% Т-клеток он состоит из гамма- и дельта- (γ/δ) цепей. Это отношение изменяется в течение онтогенеза и при болезнях.
Когда TCR связывается с антигенным пептидом и МНС (пептид/МНС), Т-лимфоцит активируется путем ряда биохимических событий, опосредуемых ассоциированными ферментами, корецепторами, специализированными адаптерными молекулами и активированными или высвобожденными факторами транскрипции.
Конструкция нуклеиновой кислоты или вектор по настоящему изобретению может содержать последовательность нуклеиновой кислоты, кодирующую α-цепь TCR, β-цепь TCR, γ-цепь TCR или δ-цепь TCR. Например, она может содержать последовательность нуклеиновой кислоты, кодирующую α-цепь TCR, и последовательность нуклеиновой кислоты, кодирующую β-цепь TCR; или последовательность нуклеиновой кислоты, кодирующую γ-цепь TCR, или последовательность нуклеиновой кислоты, кодирующую δ-цепь TCR. Два последовательности нуклеиновой кислоты может связывать последовательность, обеспечивающая коэкспрессию двух цепей TCR, такую как последовательность внутреннего промотора, IRES или участка расщепления, такого как саморасщепляющийся участок.
ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (CAR)
Необходимая последовательность нуклеиновой кислоты (NOI) может кодировать химерный антигенный рецептор (CAR).
Классические CAR представляют собой химерные трансмембранные белки I типа, которые связывают внеклеточный распознающий антиген домен (связывающее средство) с внутриклеточным сигнальным доменом (эндодомен). Как правило, связывающее средство представляет собой одноцепочечный фрагмент вариабельной области (scFv), получаемый из моноклонального антитела (mAb), но оно может основываться на других форматах, содержащих антигенсвязывающий участок, таких как лиганд. Для изоляции связывающего средство от мембраны и для обеспечения подходящей ориентации необходимым может являться разделительный домен. Общий используемый разделительный домен представляет собой Fc IgG1. В зависимости от антигена достаточными могут являться более компактные разделители, например, стебель из CD8α и даже только шарнир IgG1. Трансмембранный домен заякоревает белок в клеточной мембране и связывает разделитель с эндодоменом, который может содержать или ассоциироваться с внутриклеточным сигнальным доменом.
Ранние конструкции CAR содержали внутриклеточные сигнальные домены, получаемые из внутриклеточных частей γ-цепи FcεR1 или CD3ξ. Таким образом, эти рецепторы первого поколения передавали иммунологический сигнал 1, которого было достаточно для инициации уничтожения Т-клеткой распознаваемых ею клеток-мишеней, но недостаточно для полной активации Т-клетки для пролиферации и выживания. Для преодоления этого ограничения сконструированы составные сигнальные домены: слияние внутриклеточной части костимулирующей Т-клетку молекулы с внутриклеточной частью CD3ξ приводит к рецепторам второго поколения, которые после распознавания антигена могут передавать активирующий и костимулирующий сигнал одновременно. Наиболее широко используемый костимулирующий домен представляет собой домен CD28. Он обеспечивает наиболее сильный костимулирующий сигнал, а именно иммунологический сигнал 2, который инициирует пролиферацию Т-клеток. Также описаны определенные рецепторы, которые содержат эндодомены представителей семейства рецептора TNF, таких как близкородственные ОХ40 и 41ВВ, которые передают сигналы выживания. В настоящее время описаны даже более сильнодействующие CAR третьего поколения, которые содержат внутриклеточные сигнальные домены, способные к передаче сигналов активации, пролиферации и выживания.
Кодирующие CAR нуклеиновые кислоты можно переносить в Т-клетки, например, с использованием ретровирусных векторов. Таким образом, можно получать большое количество антиген-специфичных Т-клеток для адоптивного клеточного переноса. Когда CAR связывает антиген-мишень, это приводит к передаче активирующего сигнала Т-клетке, на которой он экспрессирован. Таким образом, CAR обуславливает специфичность и цитотоксичность Т-клетки в направлении клетки, экспрессирующей антиген-мишень.
ВЕКТОР
В третьем аспекте настоящее изобретение относится к вектору, содержащему последовательность нуклеиновой кислоты или конструкцию нуклеиновой кислоты по изобретению.
Настоящее изобретение также относится к вектору или набору векторов, содержащих одну или несколько последовательностей нуклеиновых кислот или конструкций нуклеиновых кислот по изобретению и необязательно одну или несколько дополнительных необходимых последовательностей нуклеиновых кислот (NOI). Такой вектор можно использовать для введения последовательностей нуклеиновых кислот или конструкций нуклеиновых кислот в клетки-хозяева так, чтобы они экспрессировали один или несколько химерных белков по первому аспекту изобретения и необязательно один или несколько других необходимых белков (POI). Набор также может содержать CID.
Например, вектор может представлять собой плазмиду или вирусный вектор, такой как ретровирусный вектор или лентивирусный вектор, или вектор на основе транспозона или синтетическую иРНК.
Вектор может быть способным к трансфекции или трансдукции Т-клетки.
Например, NOI может кодировать химерный антигенный рецептор или Т-клеточный рецептор так, что когда вектор используют для трансдукции клетки-мишени, клетка-мишень коэкспрессирует химерный белок и химерный антигенный рецептор или Т-клеточный рецептор.
КЛЕТКА
Настоящее изобретение также относится к клетке, содержащей химерный белок по первому аспекту изобретения.
Клетка может экспрессировать химерный белок с двумя доменами гетеродимеризации по первому варианту осуществления первого аспекта настоящего изобретения.
Клетка может экспрессировать два химерных белка; где один содержит домен каспазы и домен гетеродимеризации, содержащий связывающий FK506 белок (FKBP); и один содержит домен каспазы и домен гетеродимеризации, содержащий домен FRB mTOR, по второму варианту осуществления первого аспекта изобретения.
Также предоставлена клетка, экспрессирующая два белка:
Ht1-Casp и Ht2-Ht2
где Ht1-Casp представляет собой химерный белок, содержащий домен каспазы (Casp) и первый домен гетеродимеризации (Ht1); и Ht2-Ht2 представляет собой обеспечивающий взаимодействие белок, содержащий два вторых домена гетеродимеризации (Ht2) так, что в присутствии химического индуктора димеризации (CID) пара химерных белков Ht1-Casp9 взаимодействует так, что Ht1 каждого химерного белка гетеродимеризуется с доменом Ht2 обеспечивающего взаимодействие белка, обуславливая гомодимеризацию двух доменов каспазы (см. фигуру 1d).
Например, клетка может представлять собой клетку иммунной системы, такую как Т-клетка или естественную киллерную (NK) клетку.
Клетка может представлять собой стволовую клетку, такую как гемопоэтическая стволовая клетка.
Т-клетки или Т-лимфоциты являются типом лимфоцитов, который играет центральную роль в клеточном иммунитете. Их можно отличать от других лимфоцитов, таких как В-клетки и естественные киллерные клетки (NK клетки), по присутствию на клеточной поверхности Т-клеточного рецептора (TCR). Существуют различные типы Т-клеток, как изложено ниже.
Т-хелперные клетки (ТН клетки) помогают другим лейкоцитам в иммунологических процессах, включая созревание В-клеток в плазматические клетки и В-клетки памяти и активацию цитотоксических Т-клеток и макрофагов. ТН клетки экспрессируют на своей поверхности CD4. ТН клетки активируются при презентации им пептидных антигенов молекулами МНС класса II на поверхности антигенпрезентирующих клеток (АРС). Эти клетки могут дифференцироваться в один из несколько подтипов, включая TH1, TH2, TH3, TH17, TH9 или TFH, которые секретируют различные цитокины, содействуя различным типам иммунного ответа.
Цитолитические Т-клетки (ТС клетки или CTL) разрушают инфицированные вирусами клетки и опухолевые клетки, а также вовлечены в отторжение трансплантата. CTL экспрессируют на своей поверхности CD8. Эти клетки распознают свои мишени, связывая антиген, ассоциированный с МНС класса I, который презентирован на поверхности всех ядросодержащих клеток. Под действием IL-10, аденозина и других молекул, секретируемых регуляторными Т-клетками, CD8+ клетки можно инактивировать до анергического состояния, которое предотвращает аутоиммунные заболевания, такие как экспериментальный аутоиммунный энцефаломиелит.
Т-клетки памяти представляют собой субпопуляцию антиген-специфичных Т-клеток, которые длительно персистируют после устранения инфекции. Они быстро размножаются до больших количеств эффекторных Т-клеток после повторной презентации распознаваемого ими антигена, таким образом, обеспечивая иммунную систему "памятью" против прошедших инфекций. Т-клетки памяти включают три подтипа: центральные Т-клетки памяти (ТСМ клетки) и два типа эффекторных Т-клеток памяти (ТЕМ клетки и TEMRA клетки). Клетки память могут представлять собой CD4+ клетки или CD8+ клетки. Как правило, Т-клетки памяти экспрессируют белок клеточной поверхности CD45RO.
Регуляторные Т-клетки (клетки Treg), ранее известные как супрессорные Т-клетки, являются ключевыми для поддержания иммунологической толерантности. Их основной ролью является подавление Т-клеточного иммунитета в направлении окончания иммунного ответа и супрессия аутореактивных Т-клеток, которые избежали процесса негативного отбора в тимусе.
Описано два основных класса CD4+ клеток Treg - естественные клетки Treg и адаптивные клетки Treg.
Естественные клетки Treg (также известные как CD4+CD25+FoxP3+ клетки Treg) образуются в тимусе и связаны со взаимодействиями между развивающимися Т-клетками с миелоидными (CD11c+) и плазмацитоидными (CD123+) дендритными клетками, которые активированы TSLP. Естественные клетки Treg можно отличить от других Т-клеток по наличию внутриклеточной молекулы, называемой FoxP3. Мутации гена FOXP3 могут предотвращать развитие регуляторных Т-клеток, вызывая летальное аутоиммунное заболевание IPEX.
Адаптивные клетки Treg (также известные как клетки Tr1 или клетки Th3) могут образовываться при нормальном иммунном ответе.
Естественные киллерные клетки (или NK клетки) представляют собой тип цитолитических клеток, которые формируют часть врожденной иммунной системы. NK клетки обеспечивают быстрый ответ на врожденные сигналы от инфицированных вирусами клеток независимым от МНС способом.
NK клетки (принадлежащие к группе врожденных лимфоидных клеток) определяют как большие гранулярные лимфоциты (LGL), и они составляют третий класс клеток, дифференцирующихся из общего лимфоидного предшественника, порождающего В- и Т-лимфоциты. Известно, что NK клетки are дифференцируются и созревают в костном мозге, лимфоузлах, селезенке, миндалинах и тимусе, откуда они потом попадают в кровоток.
Стволовые клетки представляют собой недифференцированные клетки, которые могут дифференцироваться в специализированные клетки. У млекопитающих существуют два основных типа стволовых клеток: эмбриональные стволов клетки, которые отделены от внутренней клеточной массы бластоцисты, и взрослые стволовые клетки, которые находятся в различных тканях. В организмах взрослых стволовые клетки и клетки-предшественники действуют в качестве системы репарации организма, пополняя ткани взрослого. В развивающихся эмбрионах стволовые клетки могут дифференцироваться во все специализированные клетки эктодермы, энтодермы и мезодермы (см. индуцированные плюрипотентные стволовые клетки), но также сохраняют нормальные кругооборот регенерирующих органов, таких как кровь, кожа или ткани кишечника.
У людей существуют три известных доступных источника аутологичных взрослых стволовых клеток:
1. Костный мозг, который требует извлечения посредством сбора, т.е. сверления кости.
2. Жировая ткань, которая требует извлечения посредством липосакции.
3. Кровь, которая требует извлечения посредством афереза, где кровь берут у донора и пропускают через устройство, которое извлекает стволовые клетки и возвращает другие части other крови донору.
Взрослые стволов клетки часто используют в медицинских способах лечения, например, при трансплантации костного мозга. В настоящее время стволовые клетки можно выращивать искусственно и трансформировать (дифференцировать) в специализированные типы клеток с характеристиками, соответствующими клетками различных тканей, таких как мышцы или нервы. Для получения специализированных типов клеток для клеточной терапии также можно использовать линии эмбриональных клеток и аутологичные эмбриональные стволовые клетки, получаемые посредством переноса ядер или дедифференцировки соматических клеток.
Гемопоэтические стволовые клетки (HSC) представляют собой клетки крови, которые служат источником всех остальных клеток крови и происходят из мезодермы. Они находятся в красном костном мозге, который находится в сердцевине большинства костей.
Они служат источником миелоидных (моноциты и макрофаги, нейтрофилы, базофилы, эозинофилы, эритроциты, мегакариоциты/тромбоциты, дендритные клетки) и лимфоидных линий дифференцировки (Т-клетки, В-клетки, NK-клетки). Гемопоэтическая ткань содержит клетки со способностью к долговременной и кратковременной регенерации и коммитированные плюрипотентные, олигопотентные и унипотентные предшественники.
HSC представляют собой гетерогенную популяцию. Существует три класса стволовых клеток, отличающихся соотношением лимфоидного и миелоидного потомства (L/M) в крови. Смещенные в направлении миелоидной линии (My-bi) HSC обладают низким отношением L/M (от 0 до 3), тогда как смещенные в направлении лимфоидной линии (Ly-bi) HSC демонстрируют высокое отношение (>10). Третья категория состоит из сбалансированных (Bala) HSC, отношение L/M которых составляет от 3 до 10. Только смещенные в направлении миелоидной линии и сбалансированные HSC обладают свойствами длительного самоподдержания.
Экспрессирующие химерные белки клетки по изобретению могут являться клетками любого типа из указанных выше.
Т- или NK клетки, экспрессирующие один или несколько химерных белков по первому аспекту изобретения, можно получать ex vivo из собственной периферической крови пациента (1 партия), или в условиях трансплантата гемопоэтических стволовых клеток из периферической крови донора (2 партия), или периферической крови несвязанного донора (3 часть).
Альтернативно, Т- или NK клетки, экспрессирующие один или несколько химерных белков по первому аспекту изобретения, можно получать на основе дифференцировки ex vivo индуцибельных клеток-предшественников или эмбриональных клеток-предшественников в Т-клетки. Альтернативно можно использовать линию иммортализованных Т-клеток, которые сохраняют свою литическую функцию и могут действовать в качестве терапевтического средства.
Во всех этих варианты осуществления экспрессирующие химерный белок(ки) клетки получают посредством введения ДНК или РНК, кодирующих один или каждый из химерных белков и необязательно NOI, например, посредством трансдукция вирусным вектором или трансфекции ДНК или РНК.
Клетка по изобретению может представлять собой Т- или NK клетку индивидуума ex vivo. Т- или NK клетка можно получать из образца мононуклеарных клеток периферической крови (РВМС). Т- или NK клетки, до трансдукции в них нуклеиновой кислоты, кодирующей один или несколько химерных белков по первому аспекту изобретения, можно активировать и/или размножать, например, обрабатывая моноклональным антителом к CD3.
Т- или NK клетки по изобретению можно получать посредством:
(i) выделения содержащего Т- или NK клетки образца у индивидуума или из других источников, перечисленных выше; и
(ii) трансдукции или трансфекции Т- или NK клеток одной или несколькими последовательностями нуклеиновых кислот по второму аспекту изобретения.
Настоящее изобретение также относится к набору, содержащему Т- или NK клетки, содержащие один или несколько химерных белков по первому аспекту изобретения, и CID.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Настоящее изобретение также относится к фармацевтической композиции, содержащей множество клеток по четвертому аспекту изобретения. Дополнительно фармацевтическая композиция может содержать фармацевтически приемлемые носитель, разбавитель или эксципиент. Фармацевтическая композиция необязательно может содержать один или несколько дополнительных фармацевтически активных полипептидов и/или соединений. Такой состав может находиться, например, в форме, подходящей для внутривенной инфузии.
СПОСОБЫ
Изобретение также относится к способу получения клеток по четвертому аспекту изобретения, включающему этап трансдукции или трансфекции клеток вектором по третьему аспекту изобретения.
Вектор может представлять собой, например, ретровирусный или лентивирусный вектор.
Изобретение также относится к способу удаления клеток по четвертому аспекту изобретения, включающему этап воздействия на клетки CID, таким как рапамицин или аналог рапамицина. Клетки можно подвергать воздействию CID in vivo или in vitro. Удаление клеток можно вызывать апоптозом, индуцированным активацией каспазы, после индуцированной CID гомодимеризации доменов каспазы.
CID можно вводить в форме фармацевтической композиции. Дополнительно фармацевтическая композиция может содержать фармацевтически приемлемые носитель, разбавитель или эксципиент. Необязательно фармацевтическая композиция может содержать один или несколько дополнительных фармацевтически активных полипептидов и/или соединений. Такой состав может находиться, например, в форме, подходящей для внутривенной инфузии.
Изобретение также относится к способу профилактики и/или лечения патологического иммунного ответа у индивидуума, вызванного введением индивидууму клеток по четвертому аспекту изобретения, где способ включает этап введения индивидууму CID, такого как рапамицин или аналог рапамицина.
Патологический иммунный ответ можно выбирать из следующей группы: реакция "трансплантат против хозяина"; направленная внеопухолевая токсичность; синдром активации иммунной системы и лимфопролиферативные нарушения.
Изобретение также относится к способу лечения или профилактики заболевания у индивидуума, где способ включает этап введения индивидууму клеток по четвертому аспекту изобретения. Клетки могут находиться в форме фармацевтической композиции, как определено выше.
Способ может включать следующие этапы:
(i) трансдукции или трансфекции образца клеток, выделенных у индивидуума, вектором по третьему аспекту изобретения, и
(ii) введение трансдуцированных/трансфицированных клеток пациенту.
Способ лечения заболевания относится к терапевтическому применению клеток по настоящему изобретению. По настоящему документу клетки можно вводить индивидууму с существующим заболеванием или состоянием для снижения, ослабления или улучшения состояния по меньшей мере одного симптома, ассоциированного с заболеванием, и/или для замедления, снижения или блокировки прогрессирования заболевания.
Способ профилактики заболевание относится к профилактическому применению клеток иммунной системы по настоящему изобретению. По настоящему документу такие клетки можно вводить индивидууму, который еще не страдает заболеванием и/или который не демонстрирует никаких симптомов заболевания, для профилактики или ослабления причины заболевания или для снижения или предотвращения развития по меньшей мере одного симптома, ассоциированного с заболеванием. Индивидуум может обладать предрасположенностью к заболеванию или, его можно рассматривать, как подверженного риску развития заболевания.
Способы лечения заболевания, предоставляемые по настоящему изобретению, могут включать мониторинг прогрессирования заболевания и мониторинг любой токсической активности и регулирование дозы CID, вводимой индивидууму с обеспечением приемлемых уровней прогрессирования заболевания и токсической активности.
Мониторинг прогрессирования заболевания означает оценку симптомов, ассоциированных с заболеванием с течением времени с определением их ослабления/снижения или увеличения/ухудшения.
Токсическая активность относится к неблагоприятным воздействиям, вызываемым клетками по изобретению после их введения индивидууму. Токсическая активность может включать, например, иммунологическую токсичность, желчную токсичность и респираторный дистресс-синдром.
В частности изобретение относится к способу лечения заболевание у индивидуума, который включает следующие этапы:
(i) введение индивидууму клеток по четвертому аспекту изобретения;
(ii) мониторинг развития у индивидуума патологического иммунного ответа; и
(iii) введение индивидууму рапамицина или аналога рапамицина, если индивидуум демонстрирует признаки развития или наличия развитого патологического иммунного ответа.
Настоящее изобретение относится к клеткам по настоящему изобретению для применения в лечении и/или профилактики заболевания.
Например, клетки могут предназначаться для применения в трансплантации гемопоэтических стволовых клеток, инфузии лимфоцитов или адоптивном переносе клеток.
Изобретение также относится к применению клеток по настоящему изобретению в производстве лекарственного средства для лечения и/или профилактики заболевания.
Настоящее изобретение также относится к средству CID, способному к индукции димеризации химерного белка по первому аспекту изобретения для применения в лечении и/или профилактики токсической активности.
Настоящее изобретение также относится к средству CID для применения в активации в клетке пары доменов каспазы химерных белков по первому аспекту изобретения.
Заболевание для лечения и/или профилактики клетками и способами по настоящему изобретению может представлять собой инфекцию, такую как вирусная инфекция.
Способы по изобретению также могут предназначаться для контроля патогенетического иммунного ответа, например, при аутоиммунных заболеваниях, аллергиях и отторжения по типу "трансплантат против хозяина".
Когда клетки по изобретению экспрессируют TCR или CAR, их можно применять для лечения онкологического заболевания, такого как рак мочевого пузыря, рак молочной железы, рак толстого кишечника, рак эндометрия, рак почки (почечная клетка), лейкоз, рак легких, меланома, неходжкинская лимфома, рак поджелудочной железы, рак предстательной железы и рак щитовидной железы.
Экспрессирующие TCR/CAR клетки по настоящему изобретению могут обладать способностью к уничтожению клеток-мишеней, таких как раковые клетки.
Изобретение также относится к рапамицину или аналогу рапамицина для применения в профилактике или лечении патологического иммунного ответа, вызываемого введением индивидууму клеток по четвертому аспекту изобретения.
Клетки по настоящему изобретению можно использовать в любой клеточной терапии, при которой пациенту вводят модифицированные или немодифицированные клетки. Пример клеточной терапии представляет собой адоптивный перенос Т-клеток после трансплантации CD34+ стволовых клеток. Введение Т-клеток после переноса стволовых клеток помогает ускорить восстановление иммунной системы у пациента-реципиента. Когда совпадающий родственный или неродственный донор недоступен, или заболевание является слишком агрессивным для широкого поиска доноров, эффективным может являться использование гаплоидентично го по семейству HLA донора. Такие доноры могут представлять собой родителей, сибсов или родственников второй линии. Такие инфузии могут усиливать восстановление иммунитета и, таким образом, снижать вирусные инфекции и устранять рецидивирующие лейкозные клетки. Однако совместное существования в трансплантате стволовых клеток донора аллореактивных Т-клеток может вызывать реакцию "трансплантат против хозяина" (РТПХ), при которой клетки донора реагируют против реципиента, что может прогрессирующе поражать кожу, кишечник, печень и другие органы реципиента.
Другие примеры клеточной терапии включают использование нативных клеток или клеток, генетически сконструированных для экспрессии гетерологичного гена. Такое лечение используют при многих нарушениях, включая нарушения крови, но это лечение может оказывать негативное побочное действие. В другом способе для лечения нарушения можно использовать незрелые клетки-предшественники, которые могут дифференцироваться во множество типов зрелых клеток, таких как, например, мезенхимальные стромальные клетки, заменяя функцию поврежденных клеток. По настоящему изобретению предоставлен быстрый и эффективный механизм устранения возможного негативного действия клеток донора, используемых в клеточной терапии.
Настоящее изобретение относится к способу уменьшения эффекта реакции "трансплантат против хозяина" у пациента-человека после трансплантации Т-клеток донора, включающему трансфекцию или трансдукцию Т-клеток донора-человека в культуре клеток донора вектором по настоящему изобретению; введение трансдуцированных или трансфицированных Т-клеток донора пациенту; затем детекцию наличия или отсутствия у пациента реакции "трансплантат против хозяина"; и введение химического индуктора димеризации (CID) тому пациенту, у которого детектирована реакция "трансплантат против хозяина". Т-клетки могут представлять собой клетки, не подвергавшиеся аллодеплеции.
Настоящее изобретение относится к способу трансплантации стволовых клеток, включающему введение пациенту-человеку трансплантата гаплоидентичных стволовых клеток и введение пациенту гаплоидентичных Т-клеток донора, где Т-клетки в культуре гаплоидентичных клеток донора трансфицируют или трансдуцируют вектором по изобретению.
Клетки в культуре клеток донора могут представлять собой не подвергавшиеся аллодеплеции Т-клетки донора-человека.
Настоящее изобретение также относится к способу трансплантации стволовых клеток, включающему введение пациенту-человеку трансплантата гаплоидентичных стволовых клеток и введение пациенту не подвергавшихся аллодеплеции гаплоидентичных Т-клеток донора, где Т-клетки в культуре гаплоидентичных клеток донора трансфицируют или трансдуцируют вектором по изобретению.
Трансплантат гаплоидентичных стволовых клеток может представлять собой трансплантат гаплоидентичных CD34+ стволовых клеток. Т-клетки донора-человека могут являться гаплоидентичными Т-клеткам пациента. Пациент может страдать заболеванием или нарушением, которое можно облегчать трансплантацией стволовых клеток. Пациент может страдать онкологическим заболеванием, таким как солидная опухоль, или рак крови или костного мозга. Пациент может страдать заболеванием крови или костного мозга. Пациент может страдать серповидноклеточной анемией или метахроматической лейкодистрофией.
Культуру клеток донора можно получать из образца костного мозга или из периферической крови. Культуру клеток донора можно получать из мононуклеарных клеток периферической крови донора. В определенных вариантах осуществления Т-клетки донора из культуры клеток донора перед трансфекцией или трансдукцией подвергают аллодеплеции. Трансдуцированные или трансфицированные Т-клетки перед введением пациенту можно культивировать в присутствии IL-2.
Далее изобретение дополнительно описано посредством примеров, которые предназначены для помощи специалисту в данной области в осуществлении изобретения и никоим образом не предназначены для ограничения объема изобретения.
ПРИМЕРЫ
Пример 1 - Получение Т-клеток, экспрессирующих химерные белки Т-клетки подвергали трансдукции различными конструкциями. Для получения двухмолекулярной rapCasp9 (фигура 1а) Т-клетки подвергали трансдукции двумя векторами: одним, кодирующим FKBP12-Casp9 с коэкспрессией зеленого флуоресцентного белка eGFP посредством внутренней последовательности связывания рибосомы, и другим, кодирующим FRB-Casp9 с коэкспрессией синего флуоресцентного белка eBFP2. Для получения одномолекулярной rapCasp9 (фигура 1b) Т-клетки подвергали трансдукции только одним вектором, кодирующим соответствующую rapCasp9 с коэкспрессией eGFP. Конструкция, обеспечивающая FKBP12-Casp9 и FRB-FRBw, кодировала трицистронную кассету, посредством которой коэкспрессировали FKBP12-Casp9 и FRB-FRBw с использованием подобного FMD-2A пептид, и с помощью IRES коэкспрессировали eGFP. Т-клетки намеренно трансдуцировали только частично с тем, чтобы в культуре клеток часть клеток оставалась нетрансдуцированной так, чтобы служить в качестве внутреннего отрицательного контроля. В качестве дополнительного контроля, для исключения неспецифического действия рапамицина на трансдуцированные клетки, Т-клетки подвергали трансдукции вектором, кодирующим только eGFP.
Пример 2 - Тестирование удаления экспрессирующих химерный белок клеток рапамицином
Т-клетки подвергали действию различных концентраций рапамицина и инкубировали в течение 48 часов. После этого Т-клетки окрашивали аннексином-V и 7AAD и анализировали посредством проточной цитометрии. Посредством гейтирования по жизнеспособным клеткам и распознавания популяции клеток, экспрессирующих флуоресцентные белки, можно легко определять жизнеспособность трансдуцированных и нетрансдуцированных популяций. Подход с двойной FRB-Casp9 и FKBP12-Casp9 обуславливал эффективное удалению только двойных позитивных клеток, как и ожидалось. Конструкция FKBP12-FRB-Casp9 обуславливала эффективное удаление одиночно-позитивных клеток. Конструкция FKBP12-Casp9-FRB обуславливала минимальное удаление. FKBP12-Casp9/FRB-FRBw обуславливала эффективное удаление одиночно-позитивных клеток. Контроль не приводил к специфическому удалению (фигуры 2 и 3).
Пример 3 - Тестирование расширенного набора конструкций
Авторы получали конструкции, представленные на фигуре 5, и трансдуцировали в клетки Jurkat. Трансдуцированные клетки смешивали с нетрансдуцированными (NT) клетками с получением популяции с положительными и отрицательными по наличию конструкции клетками. Добавляли рапамицин с концентрациями 0, 1, 10, 100 и 1000 нМ и клетки инкубировали в течение 24 часов. После сбора клетки окрашивали PI и аннексином V и анализировали FACS. Результаты представлены на фигурах 6 до 9 и обобщены на фигуре 10.
Конструкция с конфигурацией, как определено по первому варианту осуществления первого аспекта изобретения, а именно МР20244, в этом анализе функционировала очень хорошо, обеспечивая очень эффективное уничтожение трансфицированных клеток при всех концентрациях рапамицина, указанных выше, включая 1 нМ.
Пара конструкций с конфигурацией, как определено по второму варианту осуществления первого аспекта изобретения, а именно МР20206 и МР20207, также функционировала очень хорошо, обеспечивая очень эффективное уничтожение транс фицированных клеток при всех концентрациях рапамицина, указанных выше, включая 1 нМ.
Конструкция с конфигурацией, как определено по третьему варианту осуществления первого аспекта изобретения, а именно МР20265, также функционировала хорошо, обеспечивая определенное уничтожение при 1 нМ рапамицина и эффективное уничтожение при концентрациях рапамицина 10 нМ и выше.
Конструкции с конфигурацией, как определено по четвертому варианту осуществления первого аспекта изобретения, а именно МР20263, МР20264 и МР21067, функционировали хорошо при 1 нМ рапамицина, но при более высоких концентрациях рапамицина уничтожение было менее эффективным.
Пример 4 - Тестирование конструкций с темсиролимусом
В эксперименте, эквивалентном эксперименту, описанному в пример 3, клетки, экспрессирующие конструкции, представленные на фигуре 5, обрабатывали рапамицином и темсиролимусом, аналогом рапамицина.
Как и в эксперименте, описанном в примере 3, трансдуцированные клетки Jurkat смешивали с нетрансдуцированными (NT), получая популяцию, содержащую клетки, экспрессирующие конструкции, и нетрансдуцированные клетки.
Клетки с концентрацией 2×105 клеток на лунку оставляли необработанными или обрабатывали рапамицином или темсиролимусом в следующих концентрациях: 0,01, 0,1, 1,10 нМ (рапамицина или темсиролимуса)
Клетки инкубировали в течение 24 часов и затем окрашивали аннексином V и PI и анализировали FACS. Результаты приведены на фигуре 11.
В присутствии темсиролимуса, при использовании различных конструкций, приведенных на фигуре 5, наблюдали картину уничтожения клеток Jurkat, эквивалентную той, которую наблюдали ранее в присутствии рапамицина.
В частности, конструкция МР20244, с конфигурацией, как определено по первому варианту осуществления первого аспекта изобретения; и пара конструкций МР20206 и МР20207, с конфигурацией, как определено по второму варианту осуществления первого аспекта изобретения, функционировали хорошо. Обе обуславливали эффективное уничтожение трансфицированных клеток при всех концентрациях темсиролимуса, приведенных выше, включая 1 нМ.
Все публикации, указанные в приведенном выше описании включены в настоящий документ в качестве ссылки. Специалистам в данной области очевидны различные модификации и вариации описываемых способов и системы по изобретению без отклонений от объема и сущности изобретения. Хотя изобретение описано в отношении конкретных предпочтительных вариантов осуществления, следует понимать, что заявляемое изобретение не следует ненадлежащим образом ограничивать такими конкретными вариантами осуществления. Фактически, для включения в объем приводимой ниже формулы изобретения предназначены различные модификации описываемых способов осуществления изобретения, которые очевидны специалистам в молекулярной биологии или родственных областей.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCL Business PLC
Syncona Partners LLP
<120> Химерный белок
<130> P106629PCT
<150> GB 1503133.9
<151> 2015-02-24
<160> 17
<170> PatentIn version 3.5
<210> 1
<211> 517
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный белок FRB-FKBP12-L3-dCasp9
<400> 1
Met Ala Ser Arg Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu
1 5 10 15
Glu Ala Ser Arg Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe
20 25 30
Glu Val Leu Glu Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr
35 40 45
Leu Lys Glu Thr Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu
50 55 60
Ala Gln Glu Trp Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp
65 70 75 80
Leu Leu Gln Ala Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser
85 90 95
Lys Leu Glu Tyr Ser Gly Gly Gly Ser Leu Glu Gly Val Gln Val Glu
100 105 110
Thr Ile Ser Pro Gly Asp Gly Arg Thr Phe Pro Lys Arg Gly Gln Thr
115 120 125
Cys Val Val His Tyr Thr Gly Met Leu Glu Asp Gly Lys Lys Phe Asp
130 135 140
Ser Ser Arg Asp Arg Asn Lys Pro Phe Lys Phe Met Leu Gly Lys Gln
145 150 155 160
Glu Val Ile Arg Gly Trp Glu Glu Gly Val Ala Gln Met Ser Val Gly
165 170 175
Gln Arg Ala Lys Leu Thr Ile Ser Pro Asp Tyr Ala Tyr Gly Ala Thr
180 185 190
Gly His Pro Gly Ile Ile Pro Pro His Ala Thr Leu Val Phe Asp Val
195 200 205
Glu Leu Leu Lys Leu Glu Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Gly Gly Gly Gly Ser Gly Val Asp Gly Phe Gly Asp Val Gly Ala
225 230 235 240
Leu Glu Ser Leu Arg Gly Asn Ala Asp Leu Ala Tyr Ile Leu Ser Met
245 250 255
Glu Pro Cys Gly His Cys Leu Ile Ile Asn Asn Val Asn Phe Cys Arg
260 265 270
Glu Ser Gly Leu Arg Thr Arg Thr Gly Ser Asn Ile Asp Cys Glu Lys
275 280 285
Leu Arg Arg Arg Phe Ser Ser Leu His Phe Met Val Glu Val Lys Gly
290 295 300
Asp Leu Thr Ala Lys Lys Met Val Leu Ala Leu Leu Glu Leu Ala Gln
305 310 315 320
Gln Asp His Gly Ala Leu Asp Cys Cys Val Val Val Ile Leu Ser His
325 330 335
Gly Cys Gln Ala Ser His Leu Gln Phe Pro Gly Ala Val Tyr Gly Thr
340 345 350
Asp Gly Cys Pro Val Ser Val Glu Lys Ile Val Asn Ile Phe Asn Gly
355 360 365
Thr Ser Cys Pro Ser Leu Gly Gly Lys Pro Lys Leu Phe Phe Ile Gln
370 375 380
Ala Cys Gly Gly Glu Gln Lys Asp His Gly Phe Glu Val Ala Ser Thr
385 390 395 400
Ser Pro Glu Asp Glu Ser Pro Gly Ser Asn Pro Glu Pro Asp Ala Thr
405 410 415
Pro Phe Gln Glu Gly Leu Arg Thr Phe Asp Gln Leu Asp Ala Ile Ser
420 425 430
Ser Leu Pro Thr Pro Ser Asp Ile Phe Val Ser Tyr Ser Thr Phe Pro
435 440 445
Gly Phe Val Ser Trp Arg Asp Pro Lys Ser Gly Ser Trp Tyr Val Glu
450 455 460
Thr Leu Asp Asp Ile Phe Glu Gln Trp Ala His Ser Glu Asp Leu Gln
465 470 475 480
Ser Leu Leu Leu Arg Val Ala Asn Ala Val Ser Val Lys Gly Ile Tyr
485 490 495
Lys Gln Met Pro Gly Cys Phe Asn Phe Leu Arg Lys Lys Leu Phe Phe
500 505 510
Lys Thr Ser Ala Ser
515
<210> 2
<211> 402
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный белок FKBP12-dCasp9
<400> 2
Met Leu Glu Gly Val Gln Val Glu Thr Ile Ser Pro Gly Asp Gly Arg
1 5 10 15
Thr Phe Pro Lys Arg Gly Gln Thr Cys Val Val His Tyr Thr Gly Met
20 25 30
Leu Glu Asp Gly Lys Lys Phe Asp Ser Ser Arg Asp Arg Asn Lys Pro
35 40 45
Phe Lys Phe Met Leu Gly Lys Gln Glu Val Ile Arg Gly Trp Glu Glu
50 55 60
Gly Val Ala Gln Met Ser Val Gly Gln Arg Ala Lys Leu Thr Ile Ser
65 70 75 80
Pro Asp Tyr Ala Tyr Gly Ala Thr Gly His Pro Gly Ile Ile Pro Pro
85 90 95
His Ala Thr Leu Val Phe Asp Val Glu Leu Leu Lys Leu Glu Ser Gly
100 105 110
Gly Gly Ser Gly Val Asp Gly Phe Gly Asp Val Gly Ala Leu Glu Ser
115 120 125
Leu Arg Gly Asn Ala Asp Leu Ala Tyr Ile Leu Ser Met Glu Pro Cys
130 135 140
Gly His Cys Leu Ile Ile Asn Asn Val Asn Phe Cys Arg Glu Ser Gly
145 150 155 160
Leu Arg Thr Arg Thr Gly Ser Asn Ile Asp Cys Glu Lys Leu Arg Arg
165 170 175
Arg Phe Ser Ser Leu His Phe Met Val Glu Val Lys Gly Asp Leu Thr
180 185 190
Ala Lys Lys Met Val Leu Ala Leu Leu Glu Leu Ala Gln Gln Asp His
195 200 205
Gly Ala Leu Asp Cys Cys Val Val Val Ile Leu Ser His Gly Cys Gln
210 215 220
Ala Ser His Leu Gln Phe Pro Gly Ala Val Tyr Gly Thr Asp Gly Cys
225 230 235 240
Pro Val Ser Val Glu Lys Ile Val Asn Ile Phe Asn Gly Thr Ser Cys
245 250 255
Pro Ser Leu Gly Gly Lys Pro Lys Leu Phe Phe Ile Gln Ala Cys Gly
260 265 270
Gly Glu Gln Lys Asp His Gly Phe Glu Val Ala Ser Thr Ser Pro Glu
275 280 285
Asp Glu Ser Pro Gly Ser Asn Pro Glu Pro Asp Ala Thr Pro Phe Gln
290 295 300
Glu Gly Leu Arg Thr Phe Asp Gln Leu Asp Ala Ile Ser Ser Leu Pro
305 310 315 320
Thr Pro Ser Asp Ile Phe Val Ser Tyr Ser Thr Phe Pro Gly Phe Val
325 330 335
Ser Trp Arg Asp Pro Lys Ser Gly Ser Trp Tyr Val Glu Thr Leu Asp
340 345 350
Asp Ile Phe Glu Gln Trp Ala His Ser Glu Asp Leu Gln Ser Leu Leu
355 360 365
Leu Arg Val Ala Asn Ala Val Ser Val Lys Gly Ile Tyr Lys Gln Met
370 375 380
Pro Gly Cys Phe Asn Phe Leu Arg Lys Lys Leu Phe Phe Lys Thr Ser
385 390 395 400
Ala Ser
<210> 3
<211> 392
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный белок FRB-dCasp9
<400> 3
Met Ala Ser Arg Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu
1 5 10 15
Glu Ala Ser Arg Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe
20 25 30
Glu Val Leu Glu Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr
35 40 45
Leu Lys Glu Thr Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu
50 55 60
Ala Gln Glu Trp Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp
65 70 75 80
Leu Leu Gln Ala Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser
85 90 95
Lys Leu Glu Tyr Ser Gly Gly Gly Ser Gly Val Asp Gly Phe Gly Asp
100 105 110
Val Gly Ala Leu Glu Ser Leu Arg Gly Asn Ala Asp Leu Ala Tyr Ile
115 120 125
Leu Ser Met Glu Pro Cys Gly His Cys Leu Ile Ile Asn Asn Val Asn
130 135 140
Phe Cys Arg Glu Ser Gly Leu Arg Thr Arg Thr Gly Ser Asn Ile Asp
145 150 155 160
Cys Glu Lys Leu Arg Arg Arg Phe Ser Ser Leu His Phe Met Val Glu
165 170 175
Val Lys Gly Asp Leu Thr Ala Lys Lys Met Val Leu Ala Leu Leu Glu
180 185 190
Leu Ala Gln Gln Asp His Gly Ala Leu Asp Cys Cys Val Val Val Ile
195 200 205
Leu Ser His Gly Cys Gln Ala Ser His Leu Gln Phe Pro Gly Ala Val
210 215 220
Tyr Gly Thr Asp Gly Cys Pro Val Ser Val Glu Lys Ile Val Asn Ile
225 230 235 240
Phe Asn Gly Thr Ser Cys Pro Ser Leu Gly Gly Lys Pro Lys Leu Phe
245 250 255
Phe Ile Gln Ala Cys Gly Gly Glu Gln Lys Asp His Gly Phe Glu Val
260 265 270
Ala Ser Thr Ser Pro Glu Asp Glu Ser Pro Gly Ser Asn Pro Glu Pro
275 280 285
Asp Ala Thr Pro Phe Gln Glu Gly Leu Arg Thr Phe Asp Gln Leu Asp
290 295 300
Ala Ile Ser Ser Leu Pro Thr Pro Ser Asp Ile Phe Val Ser Tyr Ser
305 310 315 320
Thr Phe Pro Gly Phe Val Ser Trp Arg Asp Pro Lys Ser Gly Ser Trp
325 330 335
Tyr Val Glu Thr Leu Asp Asp Ile Phe Glu Gln Trp Ala His Ser Glu
340 345 350
Asp Leu Gln Ser Leu Leu Leu Arg Val Ala Asn Ala Val Ser Val Lys
355 360 365
Gly Ile Tyr Lys Gln Met Pro Gly Cys Phe Asn Phe Leu Arg Lys Lys
370 375 380
Leu Phe Phe Lys Thr Ser Ala Ser
385 390
<210> 4
<211> 651
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный белок FKBP12-dCasp9-2A-FRB-FRBw
<400> 4
Met Leu Glu Gly Val Gln Val Glu Thr Ile Ser Pro Gly Asp Gly Arg
1 5 10 15
Thr Phe Pro Lys Arg Gly Gln Thr Cys Val Val His Tyr Thr Gly Met
20 25 30
Leu Glu Asp Gly Lys Lys Phe Asp Ser Ser Arg Asp Arg Asn Lys Pro
35 40 45
Phe Lys Phe Met Leu Gly Lys Gln Glu Val Ile Arg Gly Trp Glu Glu
50 55 60
Gly Val Ala Gln Met Ser Val Gly Gln Arg Ala Lys Leu Thr Ile Ser
65 70 75 80
Pro Asp Tyr Ala Tyr Gly Ala Thr Gly His Pro Gly Ile Ile Pro Pro
85 90 95
His Ala Thr Leu Val Phe Asp Val Glu Leu Leu Lys Leu Glu Ser Gly
100 105 110
Gly Gly Ser Gly Val Asp Gly Phe Gly Asp Val Gly Ala Leu Glu Ser
115 120 125
Leu Arg Gly Asn Ala Asp Leu Ala Tyr Ile Leu Ser Met Glu Pro Cys
130 135 140
Gly His Cys Leu Ile Ile Asn Asn Val Asn Phe Cys Arg Glu Ser Gly
145 150 155 160
Leu Arg Thr Arg Thr Gly Ser Asn Ile Asp Cys Glu Lys Leu Arg Arg
165 170 175
Arg Phe Ser Ser Leu His Phe Met Val Glu Val Lys Gly Asp Leu Thr
180 185 190
Ala Lys Lys Met Val Leu Ala Leu Leu Glu Leu Ala Gln Gln Asp His
195 200 205
Gly Ala Leu Asp Cys Cys Val Val Val Ile Leu Ser His Gly Cys Gln
210 215 220
Ala Ser His Leu Gln Phe Pro Gly Ala Val Tyr Gly Thr Asp Gly Cys
225 230 235 240
Pro Val Ser Val Glu Lys Ile Val Asn Ile Phe Asn Gly Thr Ser Cys
245 250 255
Pro Ser Leu Gly Gly Lys Pro Lys Leu Phe Phe Ile Gln Ala Cys Gly
260 265 270
Gly Glu Gln Lys Asp His Gly Phe Glu Val Ala Ser Thr Ser Pro Glu
275 280 285
Asp Glu Ser Pro Gly Ser Asn Pro Glu Pro Asp Ala Thr Pro Phe Gln
290 295 300
Glu Gly Leu Arg Thr Phe Asp Gln Leu Asp Ala Ile Ser Ser Leu Pro
305 310 315 320
Thr Pro Ser Asp Ile Phe Val Ser Tyr Ser Thr Phe Pro Gly Phe Val
325 330 335
Ser Trp Arg Asp Pro Lys Ser Gly Ser Trp Tyr Val Glu Thr Leu Asp
340 345 350
Asp Ile Phe Glu Gln Trp Ala His Ser Glu Asp Leu Gln Ser Leu Leu
355 360 365
Leu Arg Val Ala Asn Ala Val Ser Val Lys Gly Ile Tyr Lys Gln Met
370 375 380
Pro Gly Cys Phe Asn Phe Leu Arg Lys Lys Leu Phe Phe Lys Thr Ser
385 390 395 400
Ala Ser Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val
405 410 415
Glu Ser Asn Pro Gly Pro Gly Val Gln Val Glu Thr Ile Ser Pro Gly
420 425 430
Asp Gly Arg Thr Phe Pro Lys Arg Gly Gln Thr Cys Val Val His Tyr
435 440 445
Thr Gly Met Leu Glu Asp Gly Lys Lys Val Asp Ser Ser Arg Asp Arg
450 455 460
Asn Lys Pro Phe Lys Phe Met Leu Gly Lys Gln Glu Val Ile Arg Gly
465 470 475 480
Trp Glu Glu Gly Val Ala Gln Met Ser Val Gly Gln Arg Ala Lys Leu
485 490 495
Thr Ile Ser Pro Asp Tyr Ala Tyr Gly Ala Thr Gly His Pro Gly Ile
500 505 510
Ile Pro Pro His Ala Thr Leu Val Phe Asp Val Glu Leu Leu Lys Leu
515 520 525
Glu Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Met Leu Glu Gly
530 535 540
Val Gln Val Glu Thr Ile Ser Pro Gly Asp Gly Arg Thr Phe Pro Lys
545 550 555 560
Arg Gly Gln Thr Cys Val Val His Tyr Thr Gly Met Leu Glu Asp Gly
565 570 575
Lys Lys Phe Asp Ser Ser Arg Asp Arg Asn Lys Pro Phe Lys Phe Met
580 585 590
Leu Gly Lys Gln Glu Val Ile Arg Gly Trp Glu Glu Gly Val Ala Gln
595 600 605
Met Ser Val Gly Gln Arg Ala Lys Leu Thr Ile Ser Pro Asp Tyr Ala
610 615 620
Tyr Gly Ala Thr Gly His Pro Gly Ile Ile Pro Pro His Ala Thr Leu
625 630 635 640
Val Phe Asp Val Glu Leu Leu Lys Leu Glu Ser
645 650
<210> 5
<211> 416
<212> Белок
<213> Homo sapiens
<400> 5
Met Asp Glu Ala Asp Arg Arg Leu Leu Arg Arg Cys Arg Leu Arg Leu
1 5 10 15
Val Glu Glu Leu Gln Val Asp Gln Leu Trp Asp Ala Leu Leu Ser Ser
20 25 30
Glu Leu Phe Arg Pro His Met Ile Glu Asp Ile Gln Arg Ala Gly Ser
35 40 45
Gly Ser Arg Arg Asp Gln Ala Arg Gln Leu Ile Ile Asp Leu Glu Thr
50 55 60
Arg Gly Ser Gln Ala Leu Pro Leu Phe Ile Ser Cys Leu Glu Asp Thr
65 70 75 80
Gly Gln Asp Met Leu Ala Ser Phe Leu Arg Thr Asn Arg Gln Ala Ala
85 90 95
Lys Leu Ser Lys Pro Thr Leu Glu Asn Leu Thr Pro Val Val Leu Arg
100 105 110
Pro Glu Ile Arg Lys Pro Glu Val Leu Arg Pro Glu Thr Pro Arg Pro
115 120 125
Val Asp Ile Gly Ser Gly Gly Phe Gly Asp Val Gly Ala Leu Glu Ser
130 135 140
Leu Arg Gly Asn Ala Asp Leu Ala Tyr Ile Leu Ser Met Glu Pro Cys
145 150 155 160
Gly His Cys Leu Ile Ile Asn Asn Val Asn Phe Cys Arg Glu Ser Gly
165 170 175
Leu Arg Thr Arg Thr Gly Ser Asn Ile Asp Cys Glu Lys Leu Arg Arg
180 185 190
Arg Phe Ser Ser Pro His Phe Met Val Glu Val Lys Gly Asp Leu Thr
195 200 205
Ala Lys Lys Met Val Leu Ala Leu Leu Glu Leu Ala Gln Gln Asp His
210 215 220
Gly Ala Leu Asp Cys Cys Val Val Val Ile Leu Ser His Gly Cys Gln
225 230 235 240
Ala Ser His Leu Gln Phe Pro Gly Ala Val Tyr Gly Thr Asp Gly Cys
245 250 255
Pro Val Ser Val Glu Lys Ile Val Asn Ile Phe Asn Gly Thr Ser Cys
260 265 270
Pro Ser Leu Gly Gly Lys Pro Lys Leu Phe Phe Ile Gln Ala Cys Gly
275 280 285
Gly Glu Gln Lys Asp His Gly Phe Glu Val Ala Ser Thr Ser Pro Glu
290 295 300
Asp Glu Ser Pro Gly Ser Asn Pro Glu Pro Asp Ala Thr Pro Phe Gln
305 310 315 320
Glu Gly Leu Arg Thr Phe Asp Gln Leu Asp Ala Ile Ser Ser Leu Pro
325 330 335
Thr Pro Ser Asp Ile Phe Val Ser Tyr Ser Thr Phe Pro Gly Phe Val
340 345 350
Ser Trp Arg Asp Pro Lys Ser Gly Ser Trp Tyr Val Glu Thr Leu Asp
355 360 365
Asp Ile Phe Glu Gln Trp Ala His Ser Glu Asp Leu Gln Ser Leu Leu
370 375 380
Leu Arg Val Ala Asn Ala Val Ser Val Lys Gly Ile Tyr Lys Gln Met
385 390 395 400
Pro Gly Cys Phe Asn Phe Leu Arg Lys Lys Leu Phe Phe Lys Thr Ser
405 410 415
<210> 6
<211> 282
<212> Белок
<213> Homo sapiens
<400> 6
Gly Phe Gly Asp Val Gly Ala Leu Glu Ser Leu Arg Gly Asn Ala Asp
1 5 10 15
Leu Ala Tyr Ile Leu Ser Met Glu Pro Cys Gly His Cys Leu Ile Ile
20 25 30
Asn Asn Val Asn Phe Cys Arg Glu Ser Gly Leu Arg Thr Arg Thr Gly
35 40 45
Ser Asn Ile Asp Cys Glu Lys Leu Arg Arg Arg Phe Ser Ser Leu His
50 55 60
Phe Met Val Glu Val Lys Gly Asp Leu Thr Ala Lys Lys Met Val Leu
65 70 75 80
Ala Leu Leu Glu Leu Ala Gln Gln Asp His Gly Ala Leu Asp Cys Cys
85 90 95
Val Val Val Ile Leu Ser His Gly Cys Gln Ala Ser His Leu Gln Phe
100 105 110
Pro Gly Ala Val Tyr Gly Thr Asp Gly Cys Pro Val Ser Val Glu Lys
115 120 125
Ile Val Asn Ile Phe Asn Gly Thr Ser Cys Pro Ser Leu Gly Gly Lys
130 135 140
Pro Lys Leu Phe Phe Ile Gln Ala Cys Gly Gly Glu Gln Lys Asp His
145 150 155 160
Gly Phe Glu Val Ala Ser Thr Ser Pro Glu Asp Glu Ser Pro Gly Ser
165 170 175
Asn Pro Glu Pro Asp Ala Thr Pro Phe Gln Glu Gly Leu Arg Thr Phe
180 185 190
Asp Gln Leu Asp Ala Ile Ser Ser Leu Pro Thr Pro Ser Asp Ile Phe
195 200 205
Val Ser Tyr Ser Thr Phe Pro Gly Phe Val Ser Trp Arg Asp Pro Lys
210 215 220
Ser Gly Ser Trp Tyr Val Glu Thr Leu Asp Asp Ile Phe Glu Gln Trp
225 230 235 240
Ala His Ser Glu Asp Leu Gln Ser Leu Leu Leu Arg Val Ala Asn Ala
245 250 255
Val Ser Val Lys Gly Ile Tyr Lys Gln Met Pro Gly Cys Phe Asn Phe
260 265 270
Leu Arg Lys Lys Leu Phe Phe Lys Thr Ser
275 280
<210> 7
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен FKBP12
<400> 7
Met Gly Val Gln Val Glu Thr Ile Ser Pro Gly Asp Gly Arg Thr Phe
1 5 10 15
Pro Lys Arg Gly Gln Thr Cys Val Val His Tyr Thr Gly Met Leu Glu
20 25 30
Asp Gly Lys Lys Phe Asp Ser Ser Arg Asp Arg Asn Lys Pro Phe Lys
35 40 45
Phe Met Leu Gly Lys Gln Glu Val Ile Arg Gly Trp Glu Glu Gly Val
50 55 60
Ala Gln Met Ser Val Gly Gln Arg Ala Lys Leu Thr Ile Ser Pro Asp
65 70 75 80
Tyr Ala Tyr Gly Ala Thr Gly His Pro Gly Ile Ile Pro Pro His Ala
85 90 95
Thr Leu Val Phe Asp Val Glu Leu Leu Lys Leu Glu
100 105
<210> 8
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> участок FRB mTOR дикого типа
<400> 8
Met Ala Ser Arg Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu
1 5 10 15
Glu Ala Ser Arg Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe
20 25 30
Glu Val Leu Glu Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr
35 40 45
Leu Lys Glu Thr Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu
50 55 60
Ala Gln Glu Trp Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp
65 70 75 80
Leu Thr Gln Ala Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser
85 90 95
Lys Leu Glu Ser
100
<210> 9
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> FRB с заменой T на L в положении 2098, которое
обеспечивает связывание с AP21967
<400> 9
Met Ala Ser Arg Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu
1 5 10 15
Glu Ala Ser Arg Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe
20 25 30
Glu Val Leu Glu Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr
35 40 45
Leu Lys Glu Thr Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu
50 55 60
Ala Gln Glu Trp Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp
65 70 75 80
Leu Leu Gln Ala Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser
85 90 95
Lys Leu Glu Ser
100
<210> 10
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> участок FRB mTOR с заменой T на H в положении 2098 и
с W на F в положении 2101 полноразмерного mTOR
<400> 10
Met Ala Ser Arg Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu
1 5 10 15
Glu Ala Ser Arg Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe
20 25 30
Glu Val Leu Glu Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr
35 40 45
Leu Lys Glu Thr Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu
50 55 60
Ala Gln Glu Trp Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp
65 70 75 80
Leu His Gln Ala Phe Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser
85 90 95
Lys Leu Glu Ser
100
<210> 11
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> участок FRB segment mTOR с заменой K на P в положении 2095
полноразмерного mTOR
<400> 11
Met Ala Ser Arg Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu
1 5 10 15
Glu Ala Ser Arg Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe
20 25 30
Glu Val Leu Glu Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr
35 40 45
Leu Lys Glu Thr Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu
50 55 60
Ala Gln Glu Trp Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Pro Asp
65 70 75 80
Leu Thr Gln Ala Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser
85 90 95
Lys Leu Glu Ser
100
<210> 12
<211> 1554
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FRB-FKBP12-L3-Casp9
<400> 12
atggcttcta gaatcctctg gcatgagatg tggcatgaag gcctggaaga ggcatctcgt 60
ttgtactttg gggaaaggaa cgtgaaaggc atgtttgagg tgctggagcc cttgcatgct 120
atgatggaac ggggccccca gactctgaag gaaacatcct ttaatcaggc ctatggtcga 180
gatttaatgg aggcccaaga gtggtgcagg aagtacatga aatcagggaa tgtcaaggac 240
ctcctccaag cctgggacct ctattatcat gtgttccgac gaatctcaaa gctcgagtat 300
agcggcggcg gcagcctgga gggcgtgcag gtggagacca tcagcccagg cgacggcaga 360
accttcccca agagaggcca gacctgcgtg gtgcactata ccggcatgct ggaggacggc 420
aagaagttcg acagcagccg cgaccgcaat aagcccttca agttcatgct gggcaagcag 480
gaggtgatca gaggctggga ggagggcgtg gcccagatga gcgtgggcca gagagccaag 540
ctgaccatca gccccgacta cgcctatggc gccaccggcc accccggcat catcccaccc 600
cacgccaccc tggtgtttga tgtggagctg ctgaagctgg agtccggcgg aggcgggtct 660
ggaggaggcg gcagcggcgg cggcgggtca ggcgtggatg gcttcggcga cgtgggagcc 720
ctggagagcc tgagaggcaa cgccgatctg gcctacatcc tgagcatgga gccctgtggc 780
cactgcctga tcatcaacaa cgtgaacttc tgccgggaga gcggcctgcg gacccggacc 840
ggcagcaaca tcgactgcga gaagctgagg aggcgcttct cctccctgca ctttatggtg 900
gaggtgaaag gcgatctgac tgccaagaaa atggtgctgg ccctgctgga gctggcccag 960
caggaccacg gagccctgga ttgctgtgtg gtggtgatcc tgtcccacgg ctgccaggcc 1020
agccacctgc agttccccgg agccgtgtac ggcaccgacg gctgtcccgt gtccgtggag 1080
aagatcgtga acatcttcaa cggcacctcc tgcccctccc tgggcggcaa gcccaagctg 1140
ttctttatcc aggcctgtgg cggcgagcag aaggaccacg gctttgaggt ggccagcacc 1200
tcccccgagg acgagagccc aggcagcaac cccgagcccg acgccacccc cttccaggag 1260
ggcctgcgca ccttcgacca gctggacgcc atcagcagcc tgcccacccc cagcgacatc 1320
ttcgtgagct acagcacctt tcccggcttc gtgagctggc gcgatcccaa gtccggctct 1380
tggtatgtgg agaccctgga cgacatcttt gagcagtggg ctcatagcga ggacctgcag 1440
agcctgctgc tgcgcgtggc caatgccgtg agcgtgaagg gcatctacaa gcagatgcca 1500
ggctgcttca acttcctgcg gaagaagctg ttcttcaaga ccagcgcctc ctga 1554
<210> 13
<211> 1209
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FKBP12-dCasp9
<400> 13
atgctggagg gcgtgcaggt ggagaccatc agcccaggcg acggcagaac cttccccaag 60
agaggccaga cctgcgtggt gcactatacc ggcatgctgg aggacggcaa gaagttcgac 120
agcagccgcg accgcaataa gcccttcaag ttcatgctgg gcaagcagga ggtgatcaga 180
ggctgggagg agggcgtggc ccagatgagc gtgggccaga gagccaagct gaccatcagc 240
cccgactacg cctatggcgc caccggccac cccggcatca tcccacccca cgccaccctg 300
gtgtttgatg tggagctgct gaagctggag tccggaggcg gctccggcgt ggatggcttc 360
ggcgacgtgg gagccctgga gagcctgaga ggcaacgccg atctggccta catcctgagc 420
atggagccct gtggccactg cctgatcatc aacaacgtga acttctgccg ggagagcggc 480
ctgcggaccc ggaccggcag caacatcgac tgcgagaagc tgaggaggcg cttctcctcc 540
ctgcacttta tggtggaggt gaaaggcgat ctgactgcca agaaaatggt gctggccctg 600
ctggagctgg cccagcagga ccacggagcc ctggattgct gtgtggtggt gatcctgtcc 660
cacggctgcc aggccagcca cctgcagttc cccggagccg tgtacggcac cgacggctgt 720
cccgtgtccg tggagaagat cgtgaacatc ttcaacggca cctcctgccc ctccctgggc 780
ggcaagccca agctgttctt tatccaggcc tgtggcggcg agcagaagga ccacggcttt 840
gaggtggcca gcacctcccc cgaggacgag agcccaggca gcaaccccga gcccgacgcc 900
acccccttcc aggagggcct gcgcaccttc gaccagctgg acgccatcag cagcctgccc 960
acccccagcg acatcttcgt gagctacagc acctttcccg gcttcgtgag ctggcgcgat 1020
cccaagtccg gctcttggta tgtggagacc ctggacgaca tctttgagca gtgggctcat 1080
agcgaggacc tgcagagcct gctgctgcgc gtggccaatg ccgtgagcgt gaagggcatc 1140
tacaagcaga tgccaggctg cttcaacttc ctgcggaaga agctgttctt caagaccagc 1200
gcctcctga 1209
<210> 14
<211> 1179
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FRB-dCasp9
<400> 14
atggcttcta gaatcctctg gcatgagatg tggcatgaag gcctggaaga ggcatctcgt 60
ttgtactttg gggaaaggaa cgtgaaaggc atgtttgagg tgctggagcc cttgcatgct 120
atgatggaac ggggccccca gactctgaag gaaacatcct ttaatcaggc ctatggtcga 180
gatttaatgg aggcccaaga gtggtgcagg aagtacatga aatcagggaa tgtcaaggac 240
ctcctccaag cctgggacct ctattatcat gtgttccgac gaatctcaaa gctcgagtat 300
agcggcggcg gcagcggcgt ggatggcttc ggcgacgtgg gagccctgga gagcctgaga 360
ggcaacgccg atctggccta catcctgagc atggagccct gtggccactg cctgatcatc 420
aacaacgtga acttctgccg ggagagcggc ctgcggaccc ggaccggcag caacatcgac 480
tgcgagaagc tgaggaggcg cttctcctcc ctgcacttta tggtggaggt gaaaggcgat 540
ctgactgcca agaaaatggt gctggccctg ctggagctgg cccagcagga ccacggagcc 600
ctggattgct gtgtggtggt gatcctgtcc cacggctgcc aggccagcca cctgcagttc 660
cccggagccg tgtacggcac cgacggctgt cccgtgtccg tggagaagat cgtgaacatc 720
ttcaacggca cctcctgccc ctccctgggc ggcaagccca agctgttctt tatccaggcc 780
tgtggcggcg agcagaagga ccacggcttt gaggtggcca gcacctcccc cgaggacgag 840
agcccaggca gcaaccccga gcccgacgcc acccccttcc aggagggcct gcgcaccttc 900
gaccagctgg acgccatcag cagcctgccc acccccagcg acatcttcgt gagctacagc 960
acctttcccg gcttcgtgag ctggcgcgat cccaagtccg gctcttggta tgtggagacc 1020
ctggacgaca tctttgagca gtgggctcat agcgaggacc tgcagagcct gctgctgcgc 1080
gtggccaatg ccgtgagcgt gaagggcatc tacaagcaga tgccaggctg cttcaacttc 1140
ctgcggaaga agctgttctt caagaccagc gcctcctga 1179
<210> 15
<211> 1956
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность FKBP12-Casp9-2A-FRB-FRBw
<400> 15
atgctggagg gcgtgcaggt ggagaccatc agcccaggcg acggcagaac cttccccaag 60
agaggccaga cctgcgtggt gcactatacc ggcatgctgg aggacggcaa gaagttcgac 120
agcagccgcg accgcaataa gcccttcaag ttcatgctgg gcaagcagga ggtgatcaga 180
ggctgggagg agggcgtggc ccagatgagc gtgggccaga gagccaagct gaccatcagc 240
cccgactacg cctatggcgc caccggccac cccggcatca tcccacccca cgccaccctg 300
gtgtttgatg tggagctgct gaagctggag tccggaggcg gctccggcgt ggatggcttc 360
ggcgacgtgg gagccctgga gagcctgaga ggcaacgccg atctggccta catcctgagc 420
atggagccct gtggccactg cctgatcatc aacaacgtga acttctgccg ggagagcggc 480
ctgcggaccc ggaccggcag caacatcgac tgcgagaagc tgaggaggcg cttctcctcc 540
ctgcacttta tggtggaggt gaaaggcgat ctgactgcca agaaaatggt gctggccctg 600
ctggagctgg cccagcagga ccacggagcc ctggattgct gtgtggtggt gatcctgtcc 660
cacggctgcc aggccagcca cctgcagttc cccggagccg tgtacggcac cgacggctgt 720
cccgtgtccg tggagaagat cgtgaacatc ttcaacggca cctcctgccc ctccctgggc 780
ggcaagccca agctgttctt tatccaggcc tgtggcggcg agcagaagga ccacggcttt 840
gaggtggcca gcacctcccc cgaggacgag agcccaggca gcaaccccga gcccgacgcc 900
acccccttcc aggagggcct gcgcaccttc gaccagctgg acgccatcag cagcctgccc 960
acccccagcg acatcttcgt gagctacagc acctttcccg gcttcgtgag ctggcgcgat 1020
cccaagtccg gctcttggta tgtggagacc ctggacgaca tctttgagca gtgggctcat 1080
agcgaggacc tgcagagcct gctgctgcgc gtggccaatg ccgtgagcgt gaagggcatc 1140
tacaagcaga tgccaggctg cttcaacttc ctgcggaaga agctgttctt caagaccagc 1200
gcctcccagt gcaccaatta tgctttgctt aagctggcag gcgatgtgga atcaaacccg 1260
ggtcctgggg tacaggtgga gaccatctct cctggcgacg ggagaacatt tcctaaaagg 1320
ggccaaacat gcgtggttca ctataccggt atgctggagg atggcaaaaa agtagactcc 1380
agccgggata gaaacaaacc ctttaagttc atgctgggta agcaggaagt tatacggggc 1440
tgggaagagg gagtagctca gatgtctgtg ggccagaggg ccaagctgac catctcaccg 1500
gactacgcct acggcgctac cggccaccct ggcattatac caccccatgc aactctcgtc 1560
ttcgatgttg agttgctcaa actggaatca ggcggaggcg ggtctggagg aggcggcagc 1620
atgctggagg gcgtgcaggt ggagaccatc agcccaggcg acggcagaac cttccccaag 1680
agaggccaga cctgcgtggt gcactatacc ggcatgctgg aggacggcaa gaagttcgac 1740
agcagccgcg accgcaataa gcccttcaag ttcatgctgg gcaagcagga ggtgatcaga 1800
ggctgggagg agggcgtggc ccagatgagc gtgggccaga gagccaagct gaccatcagc 1860
cccgactacg cctatggcgc caccggccac cccggcatca tcccacccca cgccaccctg 1920
gtgtttgatg tggagctgct gaagctggag tcctga 1956
<210> 16
<211> 20
<212> Белок
<213> вирус ящура
<400> 16
Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 17
<211> 20
<212> Белок
<213> вирус ящура
<400> 17
Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР | 2016 |
|
RU2826122C1 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| СИГНАЛЬНАЯ СИСТЕМА | 2015 |
|
RU2731638C2 |
| ВАРИАНТЫ, КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ ЭНДОНУКЛЕАЗЫ CBLB | 2018 |
|
RU2779097C2 |
| НОВЫЕ АНТИ-IL13 АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2020 |
|
RU2752833C1 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| ИНДУЦИБЕЛЬНЫЕ ХИМЕРНЫЕ ЦИТОКИНОВЫЕ РЕЦЕПТОРЫ | 2019 |
|
RU2826155C2 |
| АКТИВИРУЕМЫЕ ПОЛИПЕПТИДЫ ИНТЕРЛЕЙКИНА 12 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2826183C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
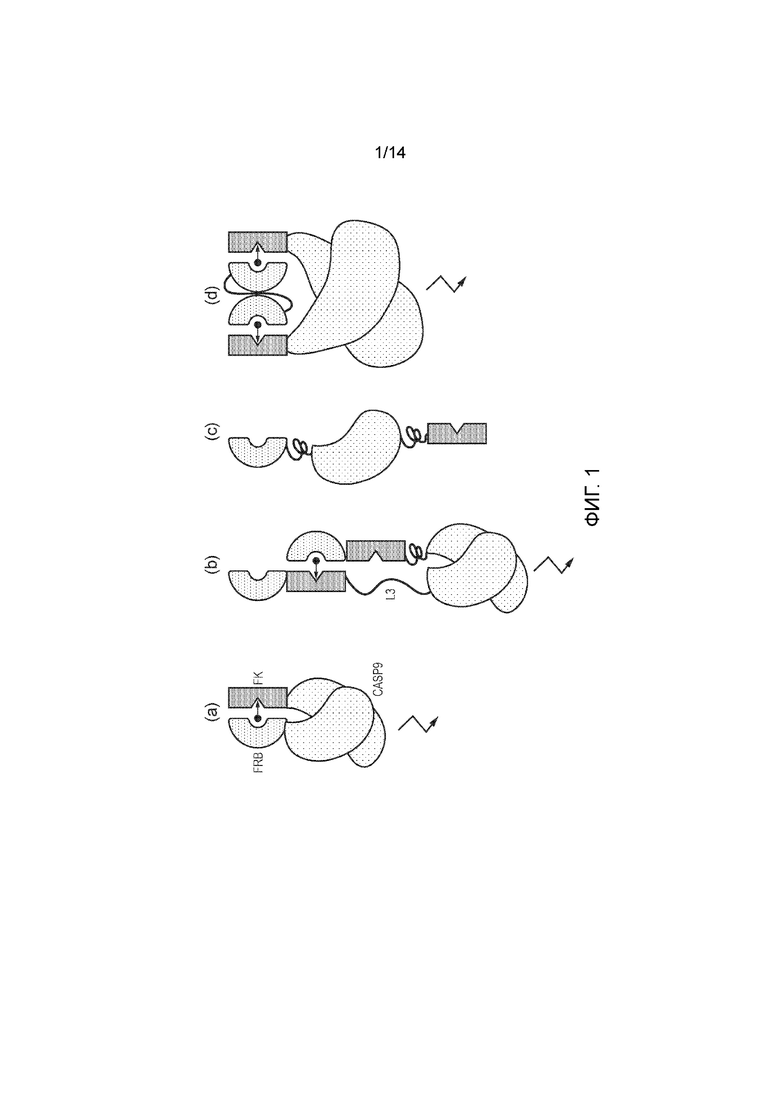
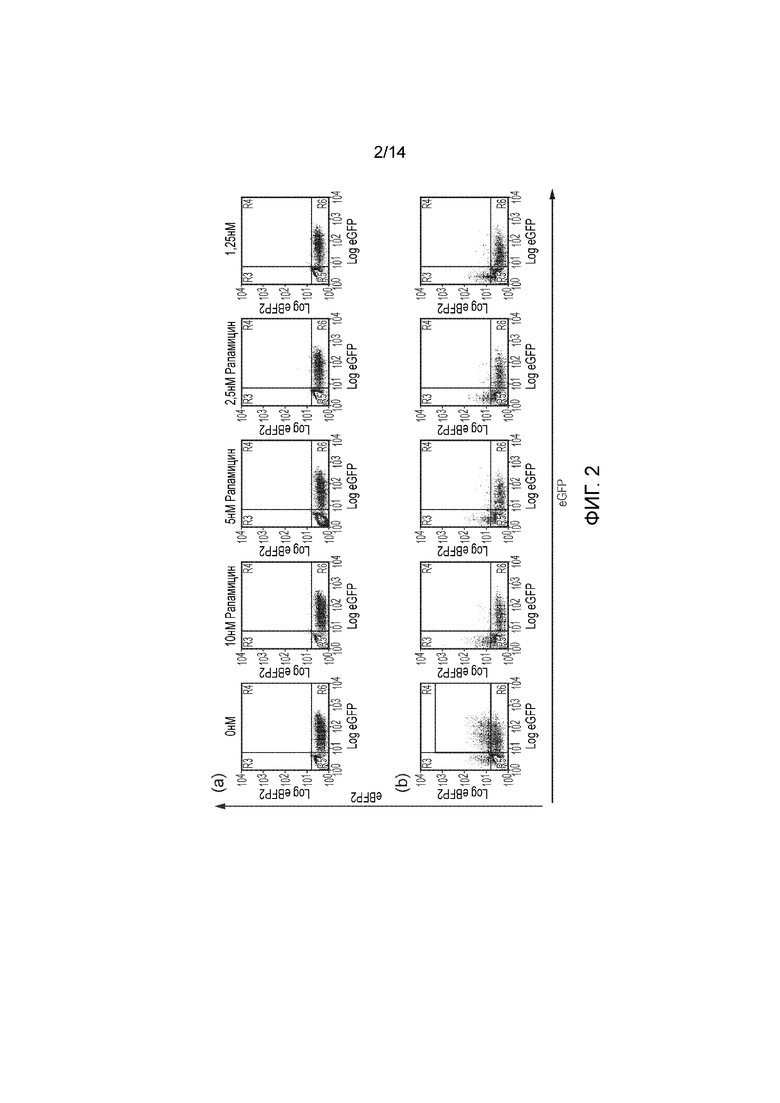
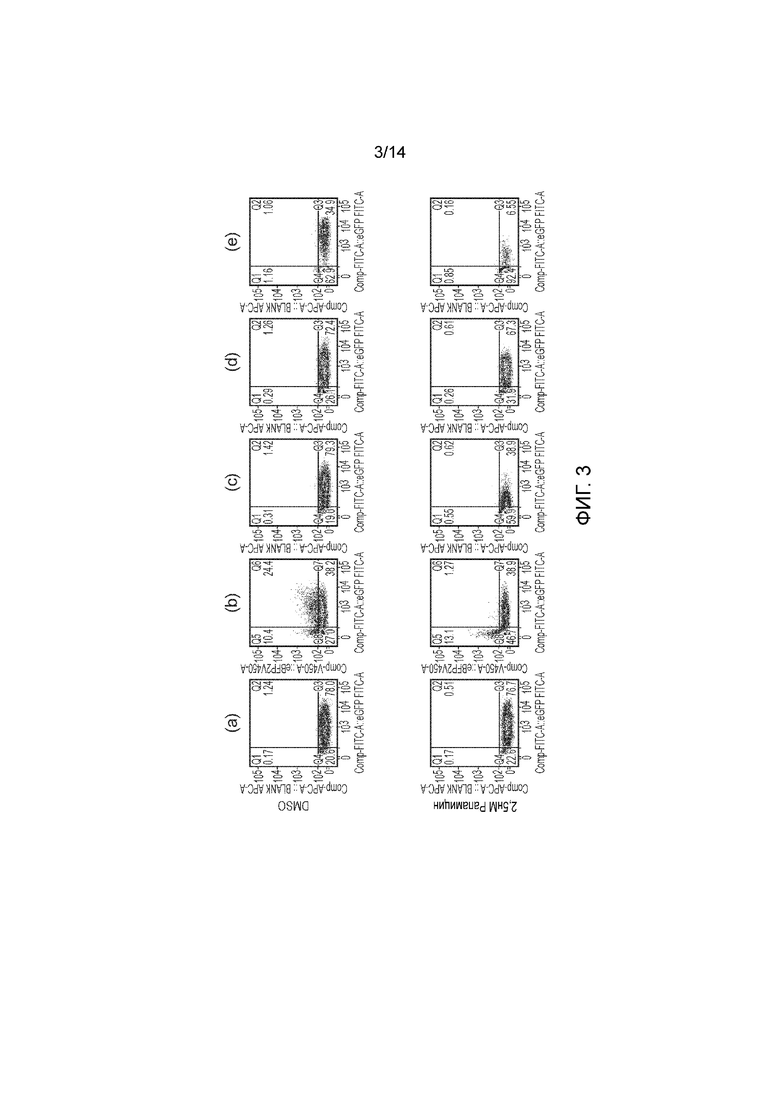
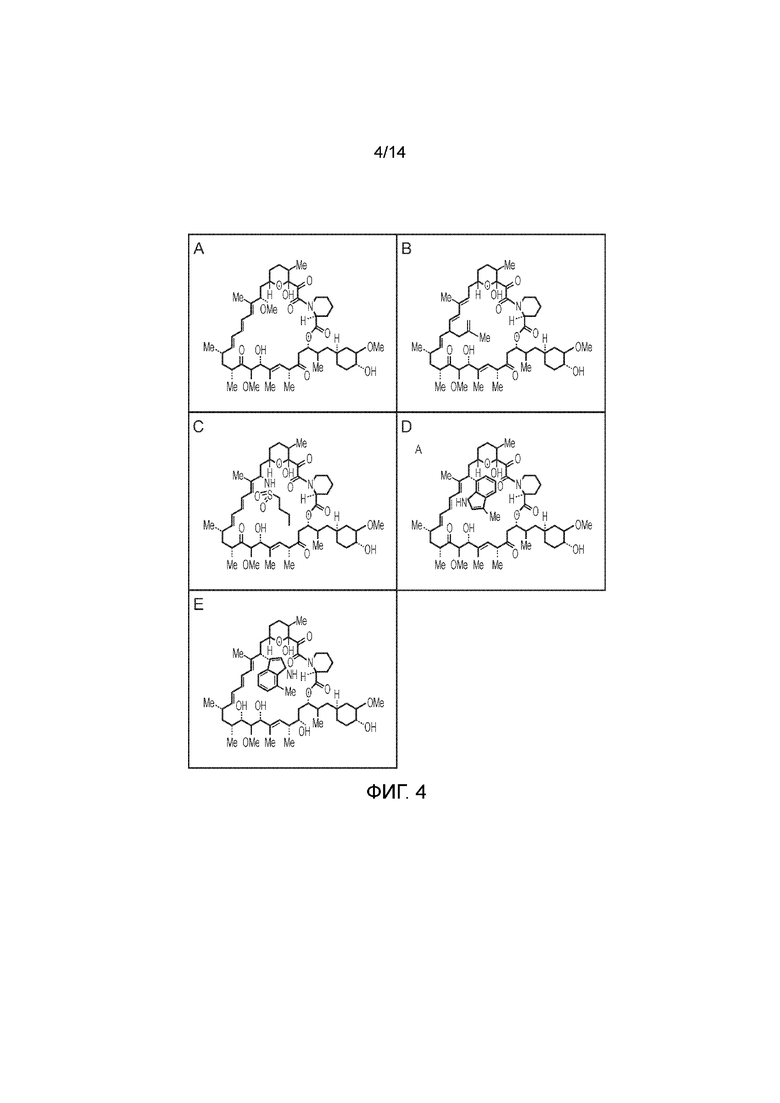
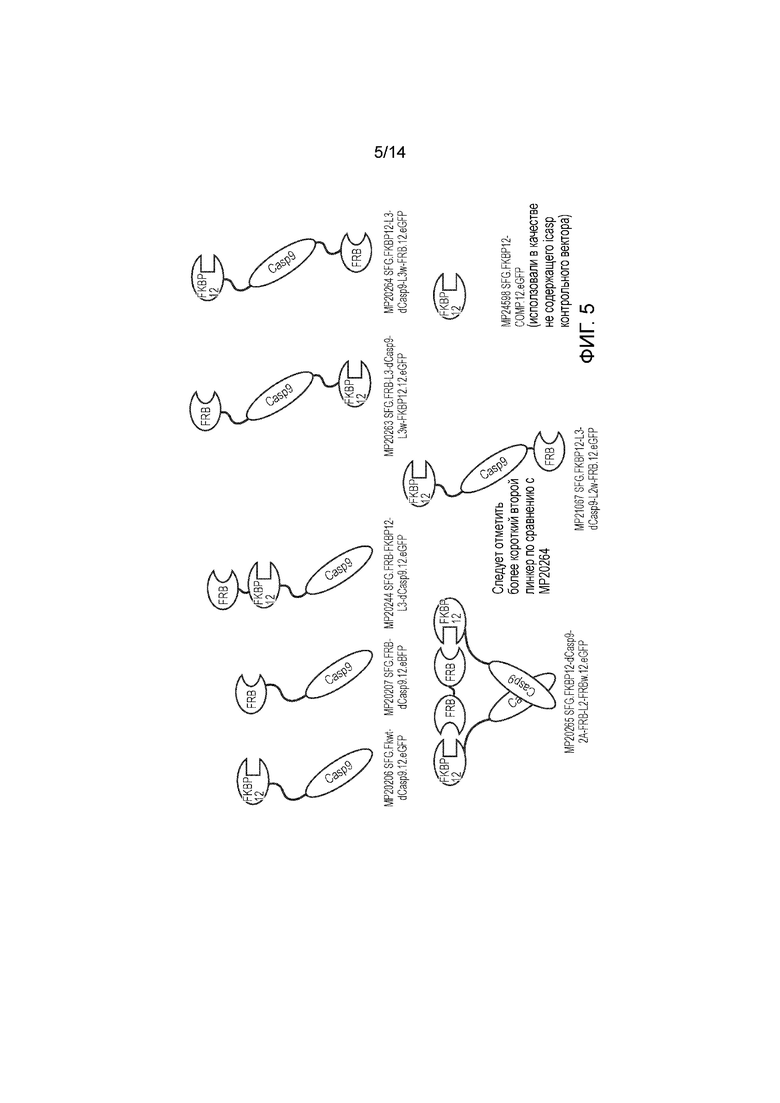

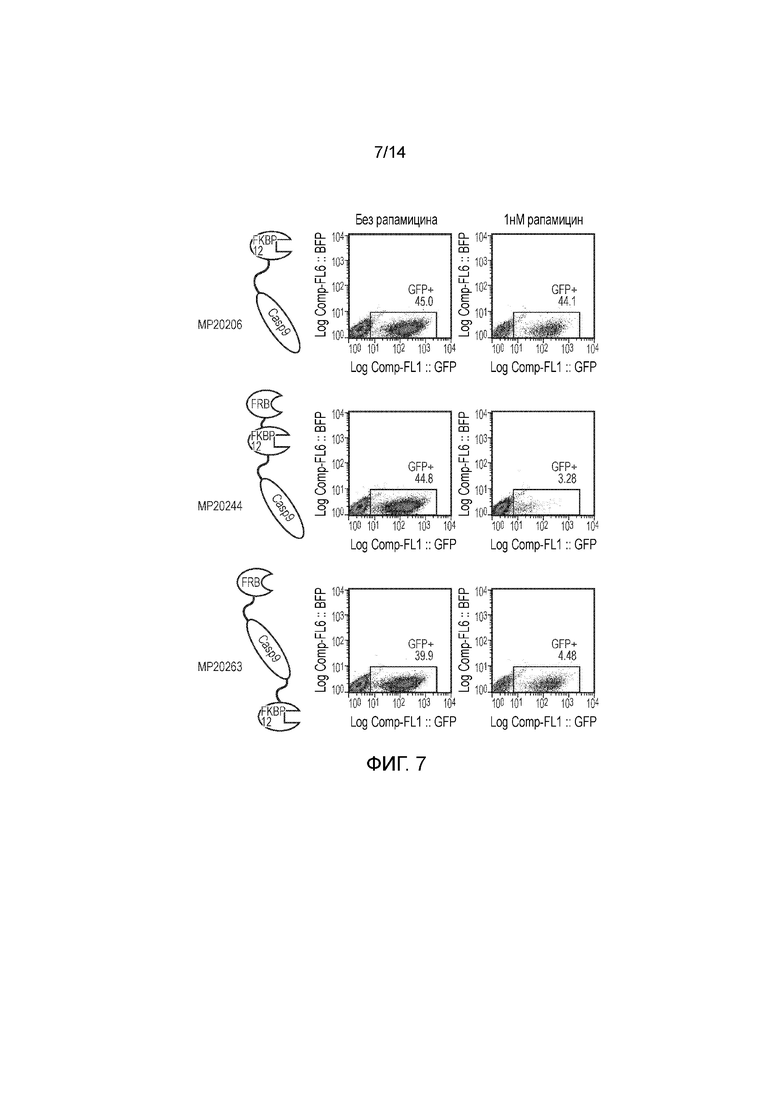
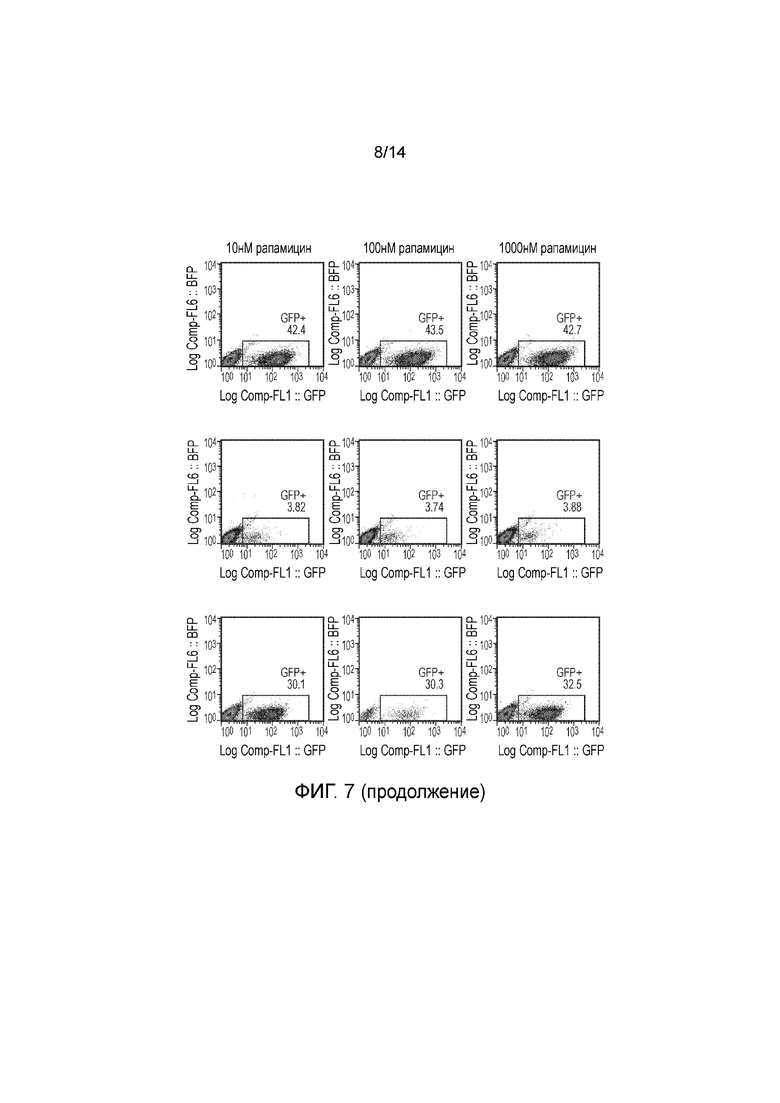
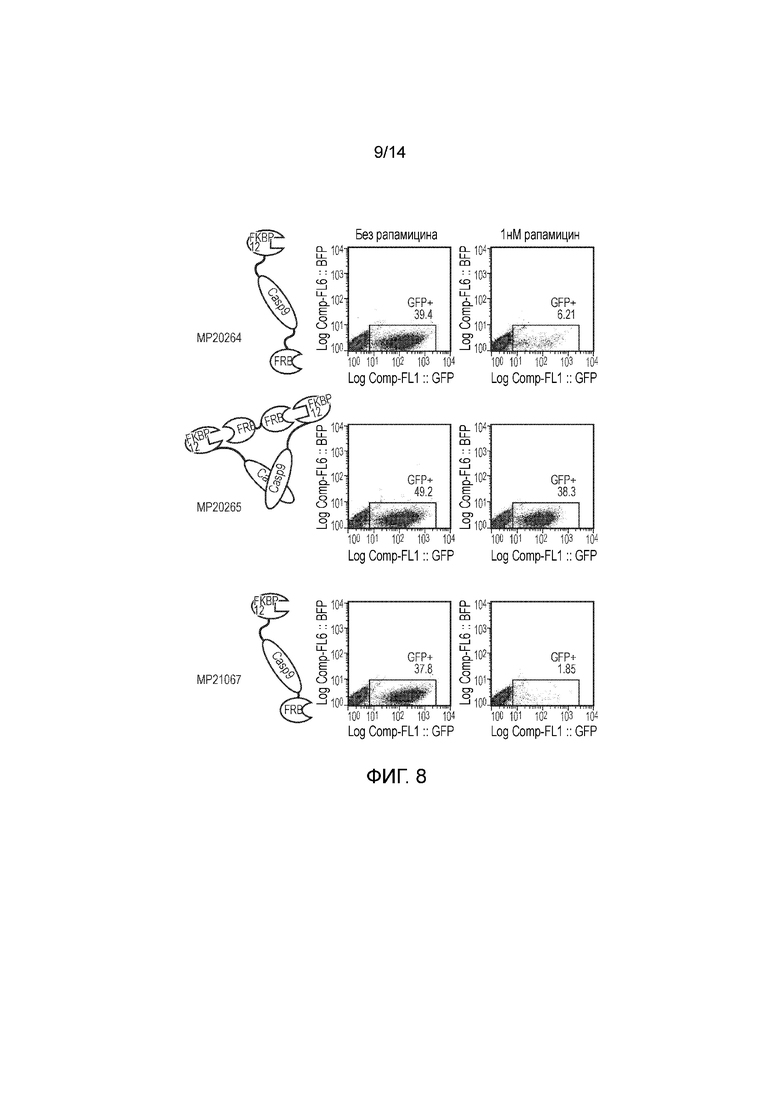
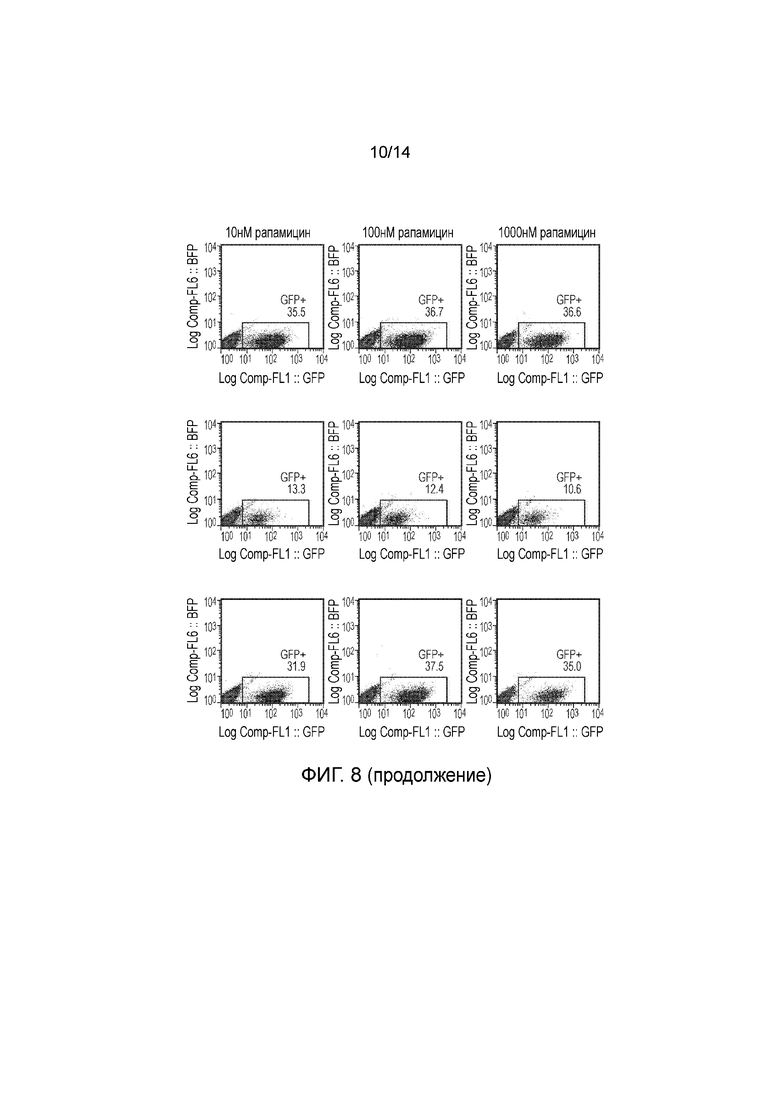
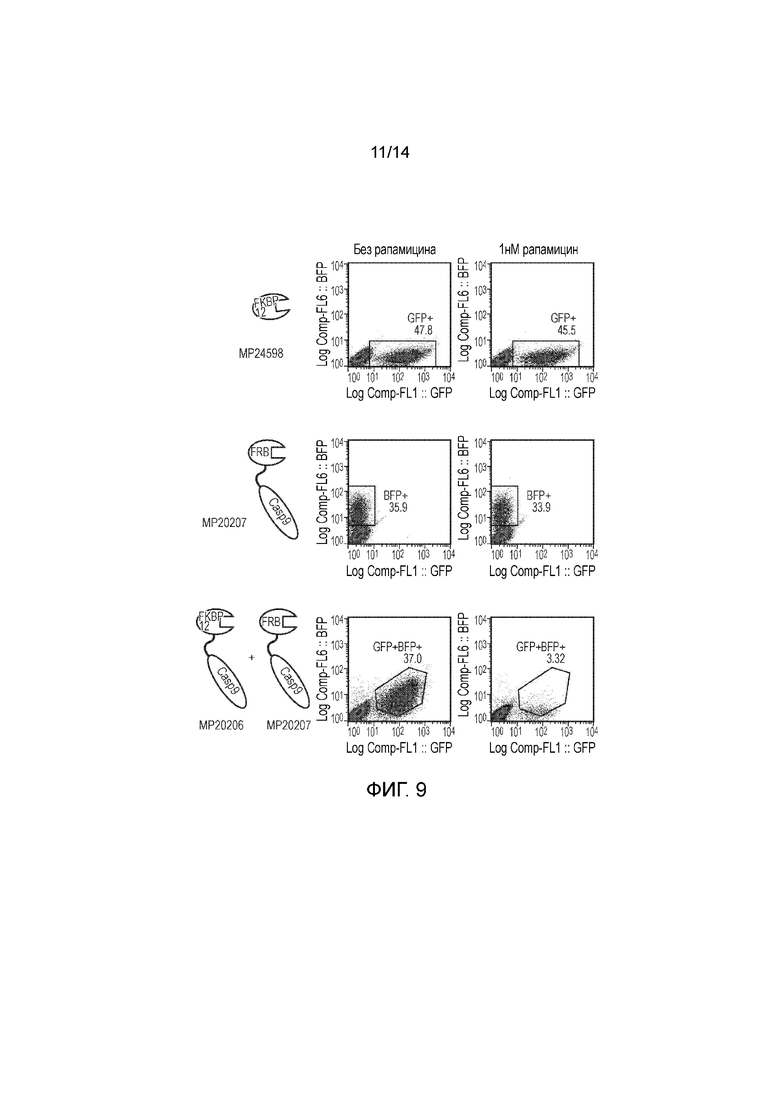
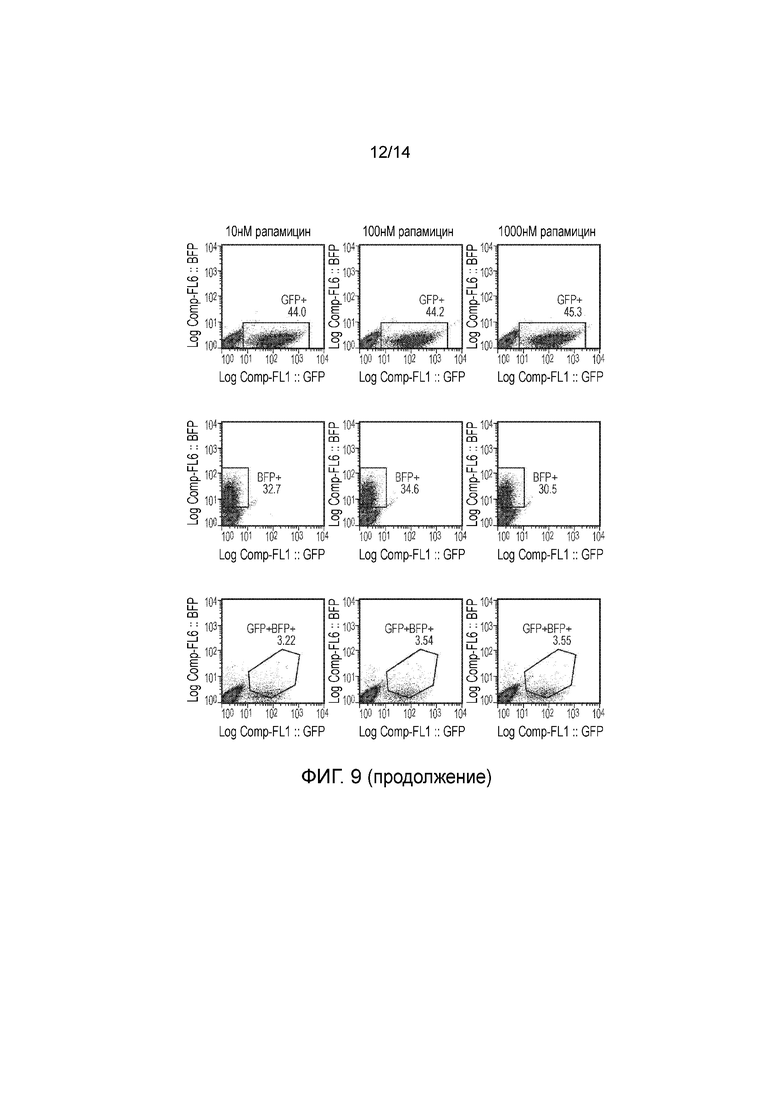
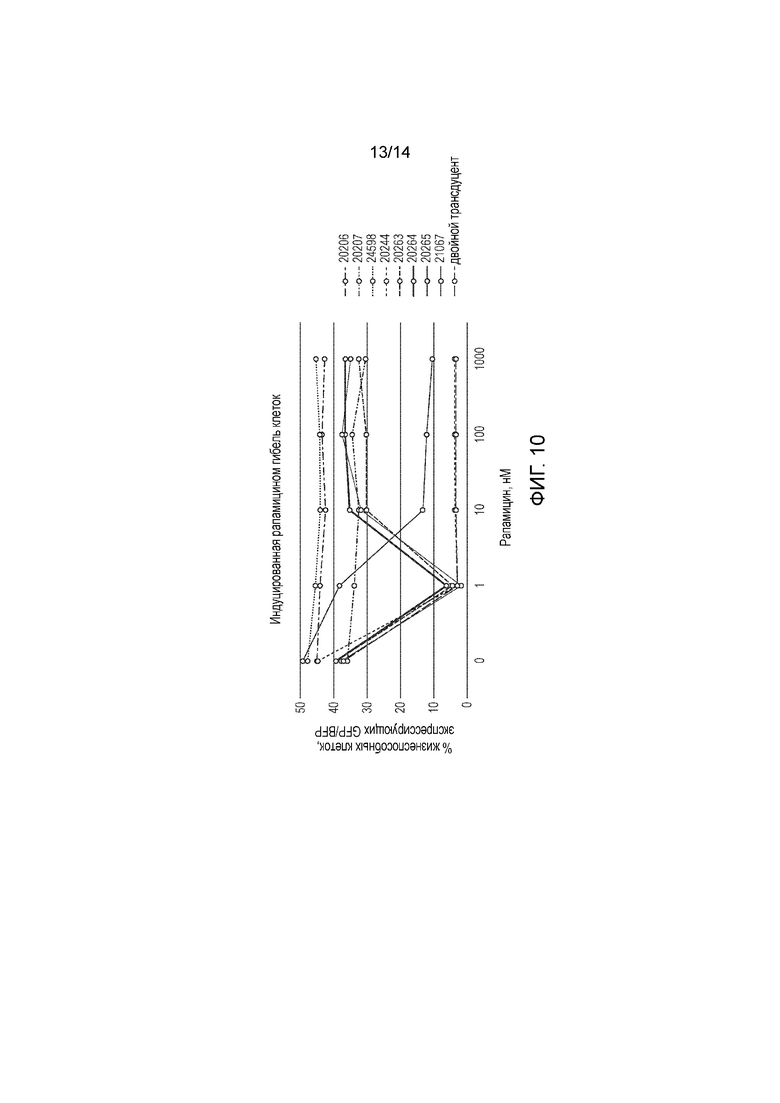
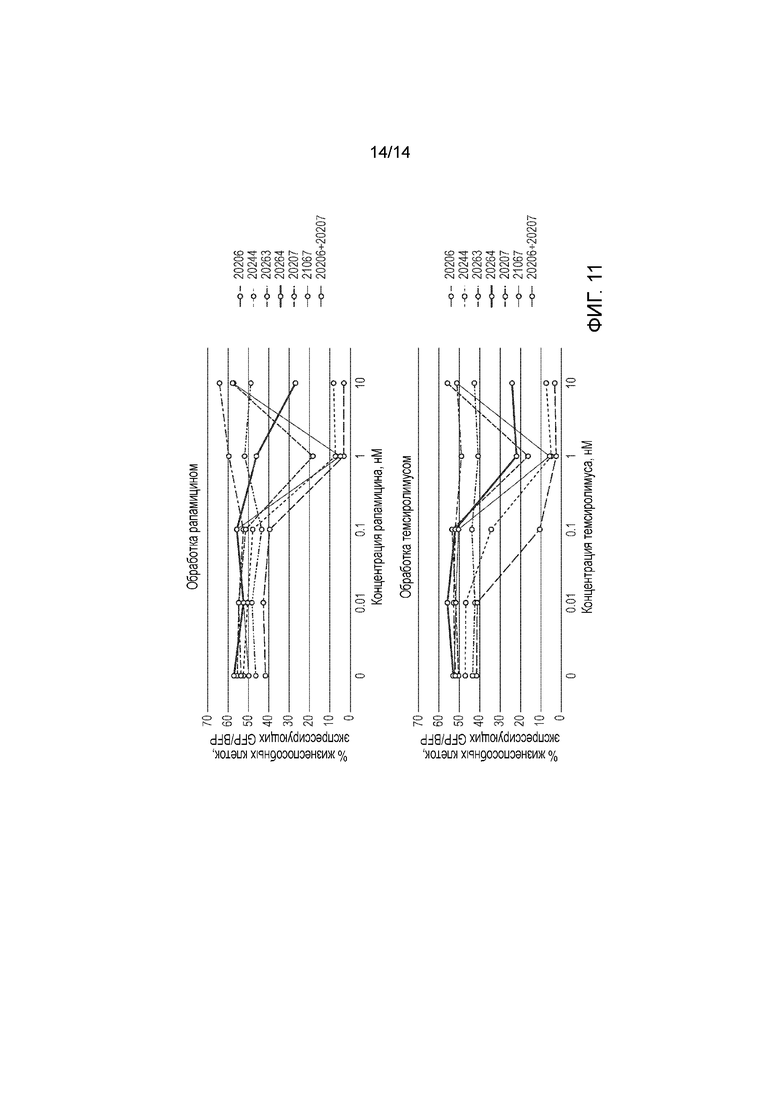
Изобретение относится к области биотехнологии, конкретно к химерным белкам для индукции апоптоза клетки, и может быть использовано в клеточной терапии. Предложен химерный белок, который характеризуется определенной формулой и содержит первый домен гетеродимеризации, второй домен гетеродимеризации и домен каспазы, причем в присутствии химического индуктора димеризации (CID) пара идентичных химерных белков взаимодействует так, что первый домен гетеродимеризации одного химерного белка гетеродимеризуется со вторым доменом гетеродимеризации другого химерного белка, обуславливая гомодимеризацию и активацию двух доменов каспазы. Причём один домен гетеродимеризации из указанных первого и второго доменов гетеродимеризации содержит связывающий FK506 белок (FKBP), а другой домен гетеродимеризации из указанных первого и второго доменов гетеродимеризации содержит домен FRB mTOR и при этом CID представляет собой рапамицин или аналог рапамицина. Изобретение обеспечивает эффективную индукцию апоптоза в клетке, включающей указанный химерный белок, при воздействии рапамицином или его аналогом. 14 н. и 12 з.п. ф-лы, 4 пр., 11 ил.
1. Химерный белок для индукции апоптоза клетки, который содержит первый домен гетеродимеризации, второй домен гетеродимеризации и домен каспазы, причем в присутствии химического индуктора димеризации (CID) пара идентичных химерных белков взаимодействует так, что первый домен гетеродимеризации одного химерного белка гетеродимеризуется со вторым доменом гетеродимеризации другого химерного белка, обуславливая гомодимеризацию и активацию двух доменов каспазы, и при этом указанный химерный белок имеет формулу, выбранную из:
Ht1-Ht2-L1-Casp;
Ht1-Casp1-L2-Ht2-Casp2;
Ht1-Casp1-L2-Casp2-Ht2;
Casp1-Ht1-L2-Ht2-Casp2;
или
Casp1-Ht1-L2 -Casp2-Ht2,
где
Casp представляет собой домен каспазы;
Casp1 и Casp2 представляют собой домены каспазы, независимо выбранной из инициаторных каспаз: каспазы-2; каспазы-8, каспазы-9 и каспазы-10; или исполнительных каспаз: каспазы-3, каспазы-6 и каспазы-7;
L1 представляет собой гибкий пептидный линкер или связь;
L2 представляет собой длинный гибкий линкер;
Ht1 представляет собой первый домен гетеродимеризации; и
Ht2 представляет собой второй домен гетеродимеризации, при этом
L1 обеспечивает достаточную гибкость, чтобы указанные каталитические домены могли гомодимеризоваться; и
L2 придает достаточную гибкость таким образом, что в присутствии химического индуктора димеризации (CID) указанный первый домен гетеродимеризации может гетеродимеризоваться с указанным вторым доменом гетеродимеризации таким образом, что домены Casp1 и Casp2 могут гомодимеризоваться;
причём один домен гетеродимеризации из указанных первого и второго доменов гетеродимеризации содержит связывающий FK506 белок (FKBP), а другой домен гетеродимеризации из указанных первого и второго доменов гетеродимеризации содержит домен FRB mTOR и при этом CID представляет собой рапамицин или аналог рапамицина.
2. Химерный белок по п. 1, где Ht1 не гетеродимеризуется с Ht2 того же химерного белка.
3. Химерный белок по п. 1 или 2, где домен каспазы содержит инициаторную каспазу, выбранную из следующей группы: каспаза-8, каспаза-9 и каспаза-10.
4. Химерный белок по п. 1 или 2, где домен каспазы содержит исполнительную каспазу, выбранную из каспазы-3 и каспазы-7.
5. Химерный белок по любому из пп. 1-4, где Ht1 содержит FRB и Ht2 содержит FKBP.
6. Молекула нуклеиновой кислоты, кодирующая химерный белок по любому из пп. 1-5.
7. Конструкция нуклеиновой кислоты, содержащая молекулу нуклеиновой кислоты по п. 6 для экспрессии химерного белка согласно любому из пп. 1-5 в клетке.
8. Конструкция нуклеиновой кислоты по п.7, отличающаяся тем, что указанная конструкция дополнительно содержит представляющий интерес полинуклеотид, где указанный представляющий интерес полинуклеотид кодирует химерный антигенный рецептор или T-клеточный рецептор, так что химерный белок по любому из пп.1-5 и химерный антигенный рецептор или T-клеточный рецептор совместно экспрессируются в клетке.
9. Конструкция нуклеиновой кислоты по п. 8, отличающаяся тем, что последовательности молекулы нуклеиновой кислоты по п. 6 и представляющего интерес полинуклеотида соединены последовательностью, обеспечивающей возможность их коэкспрессии.
10. Конструкция нуклеиновой кислоты по п. 8, отличающаяся тем, что указанная последовательность, обеспечивающая возможность коэкспрессии, представляет собой последовательность внутреннего промотора, внутреннего участка связывания рибосомы (IRES) или последовательность, кодирующую участок расщепления.
11. Вектор для трансдукции молекулы нуклеиновой кислоты, которая кодирует химерный белок согласно любому из пп. 1-5, в клетку, содержащий конструкцию нуклеиновой кислоты по любому из пп. 7-10.
12. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п. 6.
13. Вектор по п. 12, где указанный вектор дополнительно содержит представляющий интерес полинуклеотид, причём указанный представляющий интерес полинуклеотид кодирует химерный антигенный рецептор или T-клеточный рецептор так, что когда вектор используют для трансдукции клетки-мишени, клетка-мишень коэкспрессирует химерный белок по любому из пп. 1-5 и химерный антигенный рецептор или T-клеточный рецептор.
14. Клетка млекопитающего для применения в клеточной терапии, экспрессирующая химерный белок по любому из пп. 1-5, при этом указанная клетка содержит конструкцию нуклеиновой кислоты по любому из пп. 7-10 или вектор по любому из пп. 11-13.
15. Клетка по п. 12, которая представляет собой гемопоэтическую стволовую клетку, лимфоцит или T-клетку.
16. Способ получения клетки по любому из пп. 14-15, включающий этап трансдукции или трансфекции клетки млекопитающего конструкцией нуклеиновой кислоты по любому из пп. 7-10 или вектором по любому из пп. 11-13.
17. Способ индукции апоптоза в клетке по любому из пп. 14, 15, включающий этап воздействия на указанную клетку химическим индуктором димеризации (CID), где CID представляет собой рапамицин или аналог рапамицина.
18. Способ профилактики или лечения заболевания у индивидуума, включающий этап введения указанному индивидууму клетки по любому из пп. 14 или 15.
19. Способ по п.18, включающий следующие этапы:
(i) трансдукции или трансфекции образца клеток, выделенных у индивидуума, вектором по любому из пп. 11-13, и
(ii) введения трансдуцированных/трансфицированных клеток указанному пациенту.
20. Способ по п. 19 для лечения рака.
21. Способ профилактики или лечения у индивидуума патологического иммунного ответа, вызванного введением указанному индивидууму клетки по любому из пп. 14, 15, включающий этап введения индивидууму рапамицина или аналога рапамицина.
22. Способ по п. 21, где патологический иммунный ответ выбран из следующей группы: реакция "трансплантат против хозяина"; направленная внеопухолевая токсичность; синдром активации иммунной системы и лимфопролиферативные нарушения.
23. Способ лечения заболевания у индивидуума по п. 18, который включает следующие этапы:
(i) введение указанному индивидууму клетки по любому из пп. 14 или 15;
(ii) мониторинг развития у указанного индивидуума патологического иммунного ответа; и
(iii) введение указанному индивидууму рапамицина или аналога рапамицина, если указанный индивидуум демонстрирует признаки развития или наличие развитого патологического иммунного ответа.
24. Применение клетки по любому из пп. 14 или 15 в трансплантации гемопоэтических стволовых клеток, инфузии лимфоцитов или адоптивном переносе клеток.
25. Применение рапамицина или аналога рапамицина в профилактике или лечении патологического иммунного ответа, вызванного введением индивидууму клетки по любому из пп. 14 или 15.
26. Вектор для трансдукции молекулы нуклеиновой кислоты, которая кодирует химерный белок по любому из пп. 1-5 в клетку, при этом указанный вектор содержит молекулу нуклеиновой кислоты по п. 6.
| WO 9641865 A1, 27.12.1996 | |||
| US 2004040047 A1, 26.02.2004 | |||
| ADRIAN FEGAN et al., Chemically Controlled Protein Assembly: Techniques and Applications, CHEMICAL REVIEWS, 2010, v | |||
| Прибор, автоматически записывающий пройденный путь | 1920 |
|
SU110A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| МНОГОШОМПОЛЬНАЯ ПАПИРОСО-НАБИВНАЯ МАШИНА | 1925 |
|
SU3315A1 |
| WO 2014197638 A1, 11.12.2014 | |||
| DI STASI A | |||
| et al., Inducible Apoptosis as a Safety Switch for Adoptive Cell Therapy, NEW ENGLAND JOURNAL OF | |||

