СПОСОБЫ ЛЕЧЕНИЯ РАССТРОЙСТВ, СВЯЗАННЫХ С FGF21
По данной заявке испрашивается приоритет по предварительной заявке США № 62/200445, поданной 3 августа 2015 года, которая включена в настоящее описание посредством ссылки в полном объеме.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
В настоящей заявке содержится список последовательностей, который представлен в электронном виде в формате ASCII, и он полностью включен в настоящее описание посредством ссылки. Указанная копия ASCII, созданная 28 июля 2016 года, называется PAT056954_SL.txt и имеет размер 124487 байтов.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к миметическим антителам против фактора роста фибробластов 21 (FGF21). Также описаны способы лечения расстройств, связанных с FGF21, таких как ожирение, сахарный диабет 1 типа и 2 типа, панкреатит, дислипидемия, неалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперсулинемия, непереносимость глюкозы, гипергликемия, метаболический синдром и другие нарушения обмена веществ, а также способы снижения смертности и заболеваемости критически больных пациентов.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Семейство фактора роста фибробластов (FGF) характеризуется 22 генетически отличными, гомологичными лигандами, которые сгруппированы в семь подсемейств. Согласно опубликованной литературе, семейство FGF в настоящее время состоит по меньшей мере из двадцати трех членов, от FGF-1 до FGF-23 (Reuss et al. (2003) Cell Tissue Res. 313: 139-157).
Фактор роста фибробластов 21 (FGF21) был выделен из эмбрионов мыши и наиболее близок к FGF19 и FGF23. Это подсемейство FGF регулирует разнообразные физиологические процессы, необычные для классических FGF, а именно: гомеостаз энергии и желчных кислот, метаболизм глюкозы и липидов, а также гомеостаз фосфата и витамина D. Более того, в отличие от классических FGF, это подсемейство действует эндокринным путем (Moore, D. D. (2007) Science 316, 1436-8). Как было описано, FGF21 предпочтительно экспрессируется в печени (Nishimura et al. (2000) Biochimica et Biophysica Acta, 1492: 203-206; патентная публикация WO01/36640; и патентная публикация WO01/18172), и рассматривается для лечения ишемического сосудистого заболевания, заживления ран и заболеваний, связанных с потерей функции клеток легких, бронхов или альвеол, и многих других расстройств.
FGF21 был идентифицирован как мощный метаболический регулятор. Системное введение FGF21 грызунам и макакам-резусам с ожирением, вызванным питанием, или генетическим ожирением и диабетом оказывает сильное антигипергликемическое и триглицеридное действие и вызывает снижение массы тела (Coskun, T, et al. (2008) Endocrinology 149:6018-6027; Kharitonenkov, A. et al. (2005) Journal of Clinical Investigation 115:1627-1635; Kharitonenkov, A., et al. (2007) Endocrinology 148:774-781; Xu, J, et al. (2009) Diabetes 58:250-259). FGF21 представляет собой полипептид, состоящий из 209 аминокислот, который содержит лидерную последовательность из 28 аминокислот. Аминокислотная последовательность FGF21 человека приблизительно на 79% идентична FGF21 мыши и приблизительно на 80% идентична FGF21 крысы.
У млекопитающих FGF опосредуют свое действие посредством набора из четырех рецепторов FGF, FGFR1-4, которые, в свою очередь, экспрессированы во множественных вариантах сплайсинга. Каждый рецептор FGF содержит внутриклеточный тирозинкиназный домен, который активируется при связывании лиганда, что ведет к следующим стадиям сигнальных путей, включая MAPK (Erk1/2), RAF1, AKT1 и STAT (Kharitonenkov, A. et al. (2008) BioDrugs 22:37-44). В некоторых сообщениях было высказано предположение, что варианты сплайсинга с «c»-репортером FGFR1-3 проявляют специфическую аффинность в отношении β-клото и могут действовать в качестве эндогенных рецепторов FGF21 (Kurosu et al., 2007 J. Biol. Chem. 282:26687-26695); Ogawa et al., 2007 Proc. Natl. Acad. Sci. USA 104: 7432-7437; Kharitonenkov et al., 2008 J. Cell Physiol. 215, 1-7). В клетках 3T3-L1 и белой жировой ткани FGFR1 является, безусловно, самым распространенным рецептором, и поэтому наиболее вероятно, что основными функциональными рецепторами FGF21 в этой ткани являются комплексы β-клото-FGFR1c.
Хотя FGF21 активирует рецепторы FGF и молекулы в последующих стадиях сигнального пути, включая FRS2a и ERK, прямое взаимодействие FGFR и FGF21 не было обнаружено. Кроме того, различные нежировые клетки не реагируют на FGF21, хотя они экспрессируют множество изоформ FGFR. Все эти данные свидетельствуют о том, что кофактор должен опосредовать сигнальный путь FGF21 через FGFR. В исследованиях был обнаружен бета-клото (β-клото), который имеет высокий уровень экспрессии в печени, адипоцитах и поджелудочной железе в качестве детерминанта клеточного ответа на FGF21 (Kurosu, H. et al. (2007) J Biol Chem 282, 26687-95). Комплекс β-клото-FGFR, но не отдельно FGFR, связывается с FGF21 in vitro (Kharitonenkov, A., et al. (2008) J Cell Physiol 215, 1-7). FGF21 связывается с β-клото в комплексе с FGFR1c, 2c или 3c; но не связывается с β-клото в комплексе с FGFR4 (Owen et al., 2015, Trends in Endocrinology 26: 22-29). Аналогичный механизм был идентифицирован в системе FGF23-клото-FGFR (Urakawa, I. et al. (2006) Nature 444, 770-4).
Биоактивность FGF21 была впервые идентифицирована в анализе поглощения глюкозы адипоцитами мышей 3T3-L1 (Kharitonenkov, A. et al. (2005) J Clin Invest 115, 1627-35). Впоследствии было показано, что FGF21 индуцирует инсулиннезависимое поглощение глюкозы и экспрессию GLUT1. Было также показано, что FGF21 улучшает гипергликемию в ряде моделей диабета на грызунах. Кроме того, было обнаружено, что трансгенные мыши со сверхэкспрессией FGF21, устойчивы к нарушениям метаболизма, вызванными питанием, включая снижение массы тела и жировой массы и повышение чувствительности к инсулину (Badman, MK et al. (2007) Cell Metab 5, 426-37). Введение FGF21 страдающим диабетом приматам, не относящимся к человеку (NHP), приводило к снижению натощак уровня глюкозы, уровням триглицеридов, инсулина и глюкагона и приводило к значительному улучшению профилей липопротеинов, включая увеличение холестерина ЛПВП на 80% (Kharitonenkov, A. et al. (2007) Endocrinology 148, 774-81). Важно отметить, что гипогликемия не отмечалась в любой момент этого исследования NHP. Другие исследования определили FGF21 в качестве важного эндокринного гормона, который помогает контролировать адаптацию к состоянию голодания. Это обеспечивает ранее отсутствующую связь, ниже PPARα, с помощью которого печень взаимодействует с остальной частью организма при регулировании биологии энергетического гомеостаза. Объединенные данные наблюдений о том, что FGF21 регулирует жировой метаболизм (липолиз), процессы в печени (окисление жирных кислот и кетогенез) и мозге (отсутствие активности) определяют его в качестве основного эндокринного регулятора ответа на голодание (Kharitonenkov, A. & Shanafelt, A. B. (2008) BioDrugs 22, 37-44).
Проблема непосредственного использования FGF21 в качестве биотерапевтического средства заключается в том, что его период полувыведения очень короткий. У мышей период полувыведения FGF21 человека составляет от 0,5 до 1 часа, а у яванских макак период полувыведения составляет от 2 до 3 часов. Кроме того, если в фармацевтических составах или препаратах используется нативный тип FGF21, то на его стабильность оказывают негативное воздействие консерванты, например, м-крезол.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к миметическим антителам FGF21, то есть моноклональным антителам, которые связываются с бета-клото (β-клото) и активируют рецепторный комплекс фактора роста фибробластов человека 21 (в дальнейшем иногда называемый «FGF21») и FGF21-опосредованный сигнальный путь (например, FGF21-рецептор-зависимый сигнальный путь), к его антигенсвязывающим фрагментам, а также к фармацевтическим композициям и способам лечения, которые их включают.
Антигенсвязывающие фрагменты (миметика FGF21, β-клото-связывающие антитела) по изобретению могут представлять собой молекулы с FGF21-подобной активностью и селективностью, но с добавлением терапевтически желаемых характеристик, таких как стабильность белка, низкая иммуногенность, легкость получения и желаемый период полувыведения in vivo.
Моноклональные миметические антитела FGF21 по изобретению, их антигенсвязывающие фрагменты и фармацевтические композиции, содержащие их, могут быть использованы при лечении расстройств, связанных с FGF21, таких как ожирение, сахарный диабет 2 типа, сахарный диабет 1 типа, панкреатит, дислипидемия, неалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперсулинемия, непереносимость глюкозы, гипергликемия, метаболический синдром, гипертензия, сердечно-сосудистое заболевание, атеросклероз, периферическая артериальная болезнь, инсульт, сердечная недостаточность, ишемическая болезнь сердца, заболевание почек, диабетические осложнения, нейропатия, гастропарез и другие нарушения обмена веществ, и для снижения смертности и заболеваемости критически больных пациентов.
Выделенные миметические антитела FGF21 или антигенсвязывающие фрагменты, описанные в настоящем документе, связывают β-клото со значением равновесной константой диссоциации (KD) менее чем или равной 100 пМ. Например, выделенные антитела или антигенсвязывающие фрагменты, описанные в настоящем документе, могут связываться с человеческим β-клото со значением KD менее чем или равным 100 пМ, менее чем или равным 50 пМ, менее чем или равным 45 пМ, менее чем или равным 40 пМ, менее чем или равным 35 пМ, менее чем или равным 25 пМ, или менее чем или равным 15 пМ. Более конкретно, выделенные антитела или антигенсвязывающие фрагменты, описанные в настоящем документе, также могут связываться с человеческим β-клото со значением KD менее чем или равным 10 пМ, определенные с помощью титриметрического анализа равновесного раствора (SET); а также могут активировать рецепторный комплекс FGFR1c_β-клото яванских макак со значением EC50 менее чем или равным 50 нМ, определенные с помощью анализы pERK клеток.
Настоящее изобретение относится к выделенному антителу или его антигенсвязывающим фрагментам, которые связываются с β-клото человека и яванского макака. Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые связывают β-клото и активируют рецепторный комплекс FGF21 и FGF21-опосредованный сигнальный путь (например, FGF21-рецептор-зависимый сигнальный путь). В конкретных аспектах выделенное антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, не активируют рецепторные комплексы FGFR2c_β-клото, FGFR3c_β-клото или FGFR4_β-клото человека.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые связывают β-клото и далее конкурируют за связывание с антителом, как показано в таблице 1. Настоящее изобретение, кроме того, относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые связывают тот же эпитоп, что и антитело, как показано в таблице 1.
Как описано в настоящем документе, «конкуренция» между антителами и/или их антигенсвязывающими фрагментами указывает на то, что как антитела (так и их связывающие фрагменты) связываются с одним и тем же эпитопом β-клото (например, как определено анализом конкурентного связывания, любым из способов, хорошо известных специалистам в данной области). Антитело или его антигенсвязывающий фрагмент также «конкурируют» с антителом β-клото или антигенсвязывающим фрагментом по изобретению (например, NOV001 или NOV002), если указанное конкурирующее антитело или его антигенсвязывающий фрагмент связывают один и тот же эпитоп β-клото или перекрывающий эпитоп β-клото, что и антитело или антигенсвязывающий фрагмент по изобретению. Как используется в настоящем документе, конкурирующее антитело или его антигенсвязывающий фрагмент также могут включать такие, которые (i) пространственно блокируют антитело или антигенсвязывающий фрагмент по изобретению от связывания с его мишенью (например, если указанное конкурирующее антитело связываются с соседним, неперекрывающимся β-клото и/или эпитопом β-клото и физически препятствует связыванию антитела или антигенсвязывающего фрагмента по изобретению с его мишенью); и/или (ii) связываются с отличным, неперекрывающимся эпитопом β-клото и вызывают конформационное превращение в белок β-клото, так что указанный белок больше не может связываться с антителом или антигенсвязывающим фрагментом β-клото по изобретению таким образом, чтобы это не сопровождалось указанным конформационным изменением.
Аффинность связывания выделенных антител и антигенсвязывающих фрагментов, описанных в настоящем документе, может быть определена с помощью титрования равновесного раствора (SET). Методы SET известны в данной области техники и более подробно описаны далее. Альтернативно, аффинность связывания выделенных антител или фрагментов, описанных в настоящем документе, может быть определена с помощью анализа Biacore. Методы кинетических анализов Biacore известны в данной области техники и более подробно описаны далее.
Выделенные миметические антитела FGF21 или их антигенсвязывающие фрагменты могут быть использованы для увеличения активации рецепторного комплекса FGF21, и, таким образом, каскада реакций сигнального пути FGF21.
Выделенные миметические антитела FGF21 или их антигенсвязывающие фрагменты, как описано в настоящем документе, могут быть моноклональными антителами, человеческими или гуманизированными антителами, химерными антителами, одноцепочечными антителами, фрагментами Fab, фрагментами Fv, фрагментами F(ab')2 или фрагментами scFv и/или изотипами IgG.
Выделенные миметические антитела FGF21 или их антигенсвязывающие фрагменты, как описано в настоящем документе, также могут включать каркасный участок, в котором аминокислота заменена на каркасный участок антитела из соответствующих последовательностей VH или VL зародышевой линии человека.
Другой аспект изобретения включает выделенное антитело или его антигенсвязывающие фрагменты, имеющие полные последовательности тяжелых и легких цепей Fabs, описанные в таблице 1. Более конкретно, выделенное антитело или его антигенсвязывающие фрагменты могут иметь последовательности тяжелых и легких цепей Fab NOV001, NOV002, NOV003, NOV004.
Еще один аспект изобретения включает выделенное антитело или его антигенсвязывающие фрагменты, имеющие последовательности вариабельной области тяжелой и легкой цепей Fabs, описанные в таблице 1. Более конкретно, выделенное антитело или его антигенсвязывающий фрагмент могут иметь последовательность вариабельной области тяжелой и легкой цепи Fab NOV001, NOV002, NOV003, NOV004.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые включают CDR1 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 3, 23, 43 и 63; CDR2 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 4, 24, 44 и 64; и CDR3 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 5, 25, 45 и 65, где выделенное антитело или его антигенсвязывающие фрагменты связываются с человеческим β-клото. В другом аспекте такое выделенное антитело или его антигенсвязывающие фрагменты, кроме того, включают CDR1 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73; CDR2 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74; и CDR3 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые включают CDR1 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73; CDR2 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74; и CDR3 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75, где выделенное антитело или его антигенсвязывающие фрагменты связываются с человеческим β-клото.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые связывают β-клото, имеющий HCDR1, HCDR2 и HCDR3 и LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat, где HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 3, 4 и 5, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 13, 14 и 15; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 23, 24 и 25, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 33, 34 и 35; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 43, 44 и 45, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 53, 54 и 55; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 63, 64 и 65, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 73, 74 и 75.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые связывают β-клото, имеющий HCDR1, HCDR2 и HCDR3 и LCDR1, LCDR2 и LCDR3, определенные с помощью Chothia, где HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 6, 7 и 8, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 16, 17 и 18; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 26, 27 и 28, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 36, 37 и 38; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 46, 47 и 48, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 56, 57 и 58; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NO: 66, 67 и 68, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NO: 76, 77 и 78.
В одном аспекте изобретения выделенное антитело или его антигенсвязывающие фрагменты включают последовательность вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49 и 69. Выделенное антитело или антигенсвязывающий фрагмент, кроме того, могут включать последовательность вариабельной области легкой цепи, где вариабельная область тяжелой цепи и вариабельная область легкой цепи объединены с образованием антигенсвязывающего сайта для β-клото. В частности, последовательность вариабельной области легкой цепи может быть выбрана из SEQ ID NO: 19, 39, 59 и 79, где указанные выделенное антитело или его антигенсвязывающие фрагменты связывают бета-клото.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые включают последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79, где указанные выделенное антитело или его антигенсвязывающие фрагменты связываются с β-клото человека. Выделенное антитело или антигенсвязывающий фрагмент, кроме того, могут содержать последовательность вариабельной области тяжелой цепи, где вариабельная область легкой цепи и вариабельная область тяжелой цепи объединены с образованием антигенсвязывающего сайта β-клото.
В частности, выделенное антитело или его антигенсвязывающие фрагменты которое связывает β-клото, могут иметь вариабельные области тяжелой и легкой цепей, содержащие последовательности SEQ ID NO: 9 и 19; 29 и 39; 49 и 59; или 69 и 79, соответственно.
Настоящее изобретение относится далее к выделенному антителу или его антигенсвязывающие фрагментам, которые включают вариабельную область тяжелой цепи, имеющую последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 9, 29, 49 и 69, где указанное антитело связывается с β-клото. В одном аспекте, выделенное антитело или его антигенсвязывающие фрагменты также включают вариабельную область легкой цепи, имеющую последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79. В следующем аспекте изобретения выделенное антитело или антигенсвязывающий фрагмент имеют HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat и как описано в таблице 1.
Настоящее изобретение относится также к выделенному антителу или его антигенсвязывающим фрагментам, имеющим вариабельную область легкой цепи, имеющую последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79, где указанное антитело связывает β-клото.
В другом аспекте изобретения выделенное антитело или его антигенсвязывающие фрагменты, которые связываются с β-клото, могут иметь тяжелую цепь, содержащую последовательность SEQ ID NO: 11, 31, 51 или 71. Выделенное антитело также может включать легкую цепь, которая может быть объединена с тяжелой цепью с образованием антигенсвязывющего сайта к β-клото человека. В частности, легкая цепь может иметь последовательность, содержащую SEQ ID NO: 21, 41, 61 или 81. В частности, выделенное антитело или его антигенсвязывающие фрагменты, которые связывают β-клото, могут иметь тяжелую цепь и легкую цепь, содержащие последовательности SEQ ID NO: 11 и 21; 31 и 41; 51 и 61; или 71 и 81, соответственно.
Настоящее изобретение далее относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые включают тяжелую цепь, имеющую последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 9, 29, 49 или 69, где указанное антитело связывается с β-клото. В одном аспекте выделенное антитело или его антигенсвязывающие фрагменты также включают легкую цепь, имеющую последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 21, 41, 61 или 81.
Настоящее изобретение далее относится также к выделенному антителу или его антигенсвязывающим фрагментам, которые включают легкую цепь, имеющую последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 21, 41, 61 или 81, где указанное антитело связывает β-клото.
Настоящее изобретение относится также к композициям, содержащим выделенное антитело или его антигенсвязывающие фрагменты, описанные в настоящем документе, в том числе к композициям антител в комбинации с фармацевтически приемлемым носителем. А именно, настоящее изобретение, более того, включает фармацевтические композиции, содержащие антитело или его антигенсвязывающие фрагменты из таблицы 1, такие как, например, антитело NOV001, NOV002, NOV003, NOV004. Настоящее изобретение относится также к фармацевтическим композициям, содержащим комбинацию двух или более выделенных антител или их антигенсвязывающих фрагментов из таблицы 1.
Настоящее изобретение относится также к выделенной последовательности нуклеиновой кислоты, кодирующей вариабельную тяжелую цепь, имеющую последовательность, выбранную из SEQ ID NO: 9, 29, 49 и 69. В частности, нуклеиновая кислота имеет последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 10, 30, 50 и 70. В следующем аспекте изобретения последовательность представляет собой SEQ ID NO: 10, 30, 50 и 70.
Настоящее изобретение относится также к выделенной последовательности нуклеиновой кислоты, кодирующей вариабельную легкую цепь, имеющую последовательность, выбранную из SEQ ID NO: 20, 40, 60 и 80. В частности, нуклеиновая кислота имеет последовательность, которая по меньшей мере, на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 20, 40, 60 и 80. В следующем аспекте изобретения последовательность представляет собой SEQ ID NO: 20, 40, 60 и 80.
Настоящее изобретение относится также к выделенной нуклеиновой кислоте, содержащей последовательность, кодирующую полипептид, который включает вариабельную область легкой цепи, имеющей последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 20, 40, 60 и 80.
Настоящее изобретение относится также к вектору, который включает одну или несколько молекул нуклеиновых кислот, описанных в настоящем документе.
Настоящее изобретение относится также к выделенной клетке-хозяину, которая включают последовательность рекомбинантной ДНК, кодирующую тяжелую цепь антитела, описанного выше, и вторую последовательность рекомбинантной ДНК, кодирующую легкую цепь антитела, описанного выше, где указанные последовательности ДНК функционально связаны с промотором и способны экспрессироваться в клетке-хозяине. Предполагается, что антитело может быть моноклональным антителом человека. Также предполагается, что клетка-хозяин представляет собой клетку млекопитающего, отличную от человека.
Изобретение также относится к активации рецептора фактора роста фибробластов 21 (FGF21) и, таким образом, опосредованного FGF21 сигнального пути (например, FGF21-рецептор-зависимый сигнальный путь), где способ включает стадию контактирования клетки с эффективным количеством композиции, содержащей выделенное антитело или антигенсвязывающие фрагменты, описанные в настоящем документе.
Предполагается, что клетка является клеткой человека. Кроме того, предполагается, что клетка находится в субъекте. В одном варианте осуществления предполагается, что клетка является адипоцитом. В других вариантах осуществления клетка может быть одной или несколькими из следующих: гепатоциты, клетки поджелудочной железы, эндотелиальные клетки, клетки мышц или почек. Кроме того, предполагается, что субъект является человеком.
Изобретение также относится к способу лечения, улучшения или предотвращения связанного с FGF21 расстройства у субъекта, где способ включает стадию введения субъекту эффективного количества композиции, содержащей антитело или его антигенсвязывающие фрагменты, описанные выше в настоящем документе. В одном аспекте связанное с FGF21 расстройство представляет собой ожирение. В одном аспекте связанное с FGF21 расстройство является диабетом 2 типа. Предполагается, что субъектом является человек.
Любое из вышеперечисленных выделенных антител или их антигенсвязывающих фрагментов может быть моноклональным антителом или его антигенсвязывающими фрагментами.
Неограничивающие варианты осуществления изобретения описаны в следующих аспектах:
1. Выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом внутри внеклеточного домена β-клото.
2. Выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где эпитоп содержит одну или несколько SEQ ID NO, показанных в таблице 2.
3. Выделенное миметическое антитело FGF21 или его антигенсвязывающий фрагмент, которые связываются с β-клото, где указанные антитело или фрагмент увеличивают активность β-клото и FGFR1c.
4. Выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с белком β-клото человека с KD менее чем или равной 10 пМ, определенные с помощью титриметрического анализа равновесного раствора (SET).
5. Выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит один или несколько аминокислотных остатков 246-265, 536-550, 834-857 и 959-986 последовательности β-клото (SEQ ID NO: 262).
6. Выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с одним или несколькими эпитопами β-клото, где указанные эпитопы содержат один или несколько аминокислотных остатков 646-670, 696-700 и 646-689 последовательности β-клото (SEQ ID NO: 262).
7. Выделенное антитело или его антигенсвязывающий фрагмент, которые активируют рецепторный комплекс FGFR1c_β-клото яванского макака с EC50 менее чем или равной 50 нМ, определенные с помощью анализов pERK клеток.
8. Выделенное антитело или антигенсвязывающий фрагмент аспекта 1, где указанные антитело или фрагмент содержат, по меньшей мере, один участок, определяющий комплементарность, который по меньшей мере на 95% идентичен по меньшей мере одной из CDR, приведенных в таблице 1.
9. Выделенное антитело или антигенсвязывающий фрагмент аспекта 1, где указанные антитело или фрагмент содержат, по меньшей мере, один участок, определяющий комплементарность, который по меньшей мере на 98% идентичен по меньшей мере одной из CDR, приведенных в таблице 1.
10. Выделенное антитело или антигенсвязывающий фрагмент аспекта 1, где указанные антитело или фрагмент содержат, по меньшей мере, один участок, определяющий комплементарность, который по меньшей мере на 99% идентичен по меньшей мере одной из CDR, приведенных в таблице 1.
11. Выделенное антитело или антигенсвязывающий фрагмент по любому из предшествующих аспектов, где указанные антитело или фрагмент содержат CDR1 тяжелой цепи, CDR2 тяжелой цепи и CDR3 тяжелой цепи из таблицы 1, и/или CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи из таблицы 1.
12. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где указанные антитело или фрагмент содержат CDR1, CDR2 и CDR3 из таблицы 1, и где вариант имеет по меньшей мере от одного до четырех аминокислотных изменений в одном из CDR1, CDR2 или CDR3.
13. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR3 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 5, 25, 45 и 65.
14. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат VH, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49, 69 или аминокислотной последовательности, которая на 90% им идентична; и VL, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79 или аминокислотной последовательности, которая на 90% им идентична.
15. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат VH, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49, и 29 или аминокислотной последовательности, которая на 95% им идентична; и VL, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79 или аминокислотной последовательности, которая на 95% им идентична.
16. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат VH, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49, и 69 или аминокислотной последовательности, которая на 97% им идентична; и VL, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79 или аминокислотной последовательности, которая на 97% им идентична.
17. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат последовательность вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49 и 69.
18. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79.
19. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат последовательность вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49 и 69; и последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79.
20. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент выбраны из группы, состоящей из антитела или фрагмента, содержащих последовательность вариабельной области тяжелой цепи с SEQ ID NO: 9 и последовательность вариабельной области легкой цепи с SEQ ID NO: 19, антитела или фрагмента, содержащих последовательность вариабельной области тяжелой цепи с SEQ ID NO: 29 и последовательность вариабельной области легкой цепи с SEQ ID NO: 39; антитела или фрагмента, содержащих последовательность вариабельной области тяжелой цепи с SEQ ID NO: 49 и последовательность вариабельной области легкой цепи с SEQ ID NO: 59; и антитела или фрагмента, содержащих последовательность вариабельной области тяжелой цепи с SEQ ID NO: 69 и последовательность вариабельной области легкой цепи с SEQ ID NO: 79.
21. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 3, 23, 43 и 63; CDR2 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 4, 24, 44 и 64; CDR3 тяжелой цепи, выбранную из группы, состоящей из 5, 25, 45 и 65; CDR1 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73; CDR2 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74; и CDR3 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75.
22. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 6, 26, 46 и 66; CDR2 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 7, 27, 47 и 67; CDR3 тяжелой цепи, выбранную из группы, состоящей из 8, 28, 48 и 68; CDR1 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 16, 36, 56 и 76; CDR2 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 17, 37, 57 и 77; и CDR3 легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 18, 38, 58 и 78.
23. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 3; CDR2 тяжелой цепи с SEQ ID NO: 4; CDR3 тяжелой цепи с SEQ ID NO: 5; CDR1 легкой цепи с SEQ ID NO: 13; CDR2 легкой цепи с SEQ ID NO: 14; и CDR3 легкой цепи с SEQ ID NO: 15.
24. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 23; CDR2 тяжелой цепи с SEQ ID NO: 24; CDR3 тяжелой цепи с SEQ ID NO: 25; CDR1 легкой цепи с SEQ ID NO: 33; CDR2 легкой цепи с SEQ ID NO: 34; и CDR3 легкой цепи с SEQ ID NO: 35.
25. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 43; CDR2 тяжелой цепи с SEQ ID NO: 44; CDR3 тяжелой цепи с SEQ ID NO: 45; CDR1 легкой цепи с SEQ ID NO: 53; CDR2 легкой цепи с SEQ ID NO: 54; и CDR3 легкой цепи с SEQ ID NO: 55.
26. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 63; CDR2 тяжелой цепи с SEQ ID NO: 64; CDR3 тяжелой цепи с SEQ ID NO: 65; CDR1 легкой цепи с SEQ ID NO: 73; CDR2 легкой цепи с SEQ ID NO: 74; и CDR3 легкой цепи с SEQ ID NO: 75.
27. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 6; CDR2 тяжелой цепи с SEQ ID NO: 7; CDR3 тяжелой цепи с SEQ ID NO: 8; CDR1 легкой цепи с SEQ ID NO: 16; CDR2 легкой цепи с SEQ ID NO: 17; и CDR3 легкой цепи с SEQ ID NO: 18.
28. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 26; CDR2 тяжелой цепи с SEQ ID NO: 27; CDR3 тяжелой цепи с SEQ ID NO: 28; CDR1 легкой цепи с SEQ ID NO: 36; CDR2 легкой цепи с SEQ ID NO: 37; и CDR3 легкой цепи с SEQ ID NO: 38.
29. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 46; CDR2 тяжелой цепи с SEQ ID NO: 47; CDR3 тяжелой цепи с SEQ ID NO: 48; CDR1 легкой цепи с SEQ ID NO: 56; CDR2 легкой цепи с SEQ ID NO: 57; и CDR3 легкой цепи с SEQ ID NO: 58.
30. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент содержат CDR1 тяжелой цепи с SEQ ID NO: 66; CDR2 тяжелой цепи с SEQ ID NO: 67; CDR3 тяжелой цепи с SEQ ID NO: 68; CDR1 легкой цепи с SEQ ID NO: 76; CDR2 легкой цепи с SEQ ID NO: 77; и CDR3 легкой цепи с SEQ ID NO: 78.
31. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или фрагмент связываются с тем же эпитопом, что и выделенное антитело или фрагмент согласно любому из аспектов 12-30.
32. Выделенное антитело или его антигенсвязывающий фрагмент, где антитело или фрагмент конкурируют за связывание с β-клото с выделенным антителом или фрагментом согласно любому из аспектов 12-30.
33. Выделенное антитело или антигенсвязывающий фрагмент по любому аспекту 1-7, где антитело или фрагмент выбраны из группы, состоящей из NOV001, NOV002, NOV003 и NOV004.
34. Фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент согласно любому из вышеупомянутых аспектов и фармацевтически приемлемый носитель.
35. Способ лечения метаболического нарушения, включающий введение субъекту, страдающему метаболическим нарушением, эффективного количества фармацевтической композиции, содержащий антитело или антигенсвязывающий фрагмент согласно любому из аспектов 1-30.
36. Способ аспекта 35, где субъект страдает одним или несколькими из следующих заболеваний: ожирение, сахарный диабет 1 типа и 2 типа, панкреатит, дислипидемия, неалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперсулинемия, непереносимость глюкозы, гипергликемия и метаболический синдром.
37. Способ аспекта 35, где субъект страдает одним или несколькими из следующих заболеваний: ожирение, диабет и дислипидемия.
38. Способ лечения сердечно-сосудистого нарушения, включающий введение субъекту, страдающему сердечно-сосудистым нарушением, эффективного количества фармацевтической композиции, содержащий антитело или фрагмент согласно любому предшествующему аспекту.
39. Способ аспекта 38, где субъект страдает одним или несколькими из следующих заболеваний: атеросклероз, периферическая артериальная болезнь, инсульт, сердечная недостаточность и ишемическая болезнь сердца.
40. Антитело или его антигенсвязывающий фрагмент согласно любому из аспектов 1-30 для применения в качестве лекарственного средства.
41. Нуклеиновая кислота, кодирующая одно или несколько антител согласно любому предшествующему аспекту.
42. Нуклеиновая кислота, содержащая последовательность, которая по меньшей мере на 90% идентична последовательностям, представленным в таблице 1.
43. Нуклеиновая кислота, содержащая последовательность, которая по меньшей мере на 95% идентична последовательностям, представленным в таблице 1.
44. Вектор, содержащий нуклеиновую кислоту согласно аспекту 41.
45. Клетка-хозяин, содержащий вектор аспекта 44.
46. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент согласно любому из аспектов 1-30 для использования при лечении метаболических нарушений.
ОПРЕДЕЛЕНИЯ
Если не указано иначе, все технические и научные термины, используемые в настоящем документе, имеют то же значение, которое обычно понимается специалистами в данной области техники, к которой относится настоящее изобретение.
Как используется в настоящем документе, термин «FGF21» относится к члену семейства белков факторов роста фибробластов (FGF). Аминокислотная последовательность FGF21 (номер доступа в базе данных GenBank: NP_061986.1) представлена в виде SEQ ID NO: 1, соответствующая полинуклеотидная последовательность которой представлена в виде SEQ ID NO: 2 (номер контрольной последовательности в базе данных NCBI: NM_019113.2).
Как используется в настоящем документе, термин «рецептор FGF21» относится к рецептору для FGF21 (Kharitonenkov, A, et al. (2008) Journal of Cellular Physiology 215:1-7; Kurosu, H, et al. (2007) JBC 282:26687-26695; Ogawa, Y, et al. (2007) PNAS 104:7432-7437).
Термин «полипептид FGF21» относится к природному полипептиду, экспрессируемому у людей. Для целей настоящего описания термин «полипептид FGF21» может быть использован взаимозаменяемо по отношению к любому полноразмерному полипептиду FGF21, например, SEQ ID NO: 1, который состоит из 209 аминокислотных остатков, и который кодируется нуклеотидной последовательностью SEQ ID NO: 2; любой зрелой форме полипептида, который состоит из 181 аминокислотных остатков, и в котором 28 аминокислотных остатков на аминотерминальном конце полноразмерного полипептида FGF21 (то есть которые составляют сигнальный пептид) были удалены, и их вариантам.
Термин «антитело», как он используется в настоящем документе означает полное антитело и любой антигенсвязывающий фрагмент (то есть «антигенсвязывающую часть») или одну его цепь. Полное антитело представляет собой гликопротеин, содержащий, по меньшей мере, две тяжелые (H) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначаемой в настоящем документе как VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов: CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой в настоящем документе как VL) и константной области легкой цепи. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой в настоящем документе как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. Области VH и VL можно далее подразделить на области гипервариабельности, которые называются участками, определяющими комплементарность (CDR), которые чередуются с более консервативными областями, называемыми каркасными участками (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Термин «антигенсвязывающая часть» или «антигенсвязывающий фрагмент» антитела, как он используется в настоящем документе, относится к одному или нескольким фрагментам интактного антитела, которые сохраняют способность специфически связываться с данным антигеном (например, β-клото). Антигенсвязывающие функции антитела могут быть осуществлены фрагментами интактного антитела. Примеры связывающих фрагментов, охватываемых терминами антигенсвязывающая часть или антигенсвязывающий фрагмент антитела, включают фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; фрагмент F(ab)2, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; Fd-фрагмент, состоящий из доменов VH и CH1; Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; фрагмент одного домена (dAb) (Ward et al., 1989 Nature 341: 544-546), который состоит из домена VH или домена VL; и изолированный участок, определяющий комплементарность (CDR).
Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их можно объединить с использованием рекомбинантных методов с помощью синтетического пептидного линкера, который позволяет сделать их в виде единой белковой цепочки, в которой пара областей VL и VH образует одновалентные молекулы (известные как одноцепочечные Fv (scFv); смотри, например, Bird et al., 1988 Science 242:423-426; и Huston et al., 1988 Proc. Natl. Acad. Sci. 85:5879-5883). Такие одноцепочечные антитела включают одно или несколько антигенсвязывающей частей или фрагментов антитела. Фрагменты этих антител получают традиционными методами, известными специалистам в соответствующей области; их проверка на применимость осуществляется так же, как и в случае интактных антител.
Антигенсвязывающие фрагменты также могут быть включены в однодоменные антитела, макситела, минитела, интратела, диатела, триатела, тетратела, v-NAR и bis-scFv (смотри, например, Hollinger and Hudson, 2005, Nature Biotechnology, 23, 9, 1126-1136). Антигенсвязывающие части антител могут быть привиты в каркасы на основе полипептидов, таких как фибронектин типа III (Fn3) (смотри, патент США № 6703199, в котором описаны мономеры полипептидной цепи фибронектина).
Антигенсвязывающие фрагменты могут быть встроены в одноцепочечные молекулы, содержащие пару тандемных сегментов Fv (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антигенсвязывающих областей (Zapata et al. (1995) Protein Eng. 8(10):1057-1062; и патент США № 5641870).
Как используется в настоящем документе, термин «аффинность» относится к силе взаимодействия между антителом и антигеном на отдельных сайтах антигена. Внутри каждого сайта антигена вариабельная область «плеча» антитела взаимодействует через слабые нековалентные силы с антигеном на многочисленных участках; чем больше взаимодействий, тем сильнее аффинность. Как используется в настоящем документе, термин «высокая аффинность» для антитела или его антигенсвязывающих фрагментов (например, Fab-фрагмента) обычно относится к антителу или антигенсвязывающему фрагменту, имеющему KD 10-9 M или менее.
Термин «аминокислота» относится к природным и синтетическим аминокислотам, а также к аминокислотным аналогам и аминокислотным миметикам, которые функционируют аналогично природным аминокислотам. Природными аминокислотами являются такие, которые кодируются генетическим кодом, а также аминокислоты, которые затем модифицируются, например, гидроксипролин, γ-карбоксиглутамат и O-фосфосерин. Аминокислотные аналоги относятся к соединениям, которые имеют ту же основную химическую структуру, что и природная аминокислота, то есть содержат альфа-углерод, связанный с водородом, карбоксильную группу, аминогруппу и группу R, например, гомосерин, норлейцин, метионинсульфоксид, метионин-метил-сульфоний. Такие аналоги имеют модифицированные R-группы (например, норлейцин) или модифицированные пептидные скелеты, но сохраняют ту же основную химическую структуру, что и природная аминокислота. Аминокислотные миметики относятся к химическим соединениям, которые имеют структуру, которая отличается от общей химической структуры аминокислоты, но они функционирует аналогично природной аминокислоте.
Термин «специфичность связывания», как он используется в настоящем документе, относится к способности отдельного антигенсвязывающего центра антитела взаимодействовать только с одной антигенной детерминантой.
Фраза «специфически (или селективно) связывается» с антителом (например, антитело, связывающееся с β-клото) относится к реакции связывания, при помощи которой определяют присутствие родственного антигена (например, β-клото человека или β-клото яванского макака) в гетерогенной популяции белков и других биологических компонентов. Фразы «антитело, распознающее антиген» и «антитело, специфичное к антигену», взаимозаменяемы в настоящем документе с термином «антитело, которое специфически связывается с антигеном».
Термин «FGF21-опосредованный» или аналогичный термин относится к тому факту, что рецептор FGF21 и/или антитела по изобретению опосредуют клеточный ответ и сигнальный путь FGF21 при связывании с β-клото, тем самым вызывая различные физиологические эффекты, в том числе, но этим не ограничиваясь, уменьшение одного или нескольких из следующих факторов: триглицериды в плазме, плазменный инсулин, глюкоза в плазме, потребление пищевых продуктов и вес тела.
«Расстройство, связанное с FGF21», «состояние, связанное с FGF21», «заболевание или состояние, связанное с FGF21» или аналогичные термины, используемые в настоящем документе, относятся к любому числу состояний или заболеваний, при которых для предотвращения, диагностики и/или лечения требуется активация сигнального пути FGF21 (например, путем активации сигнального пути рецептора FGF21). Они могут включать в себя состояния, заболевания или расстройства, характеризующиеся аберрантным сигнальным путем FGF21 (например, аберрантной активацией FGF21-опосредованного сигнального пути и/или сигнального пути FGF21-рецептора). Эти состояния включают, но этим не ограничиваются, метаболические, эндокринные и сердечно-сосудистые нарушения, такие как ожирение, сахарный диабет 1 типа и 2 типа, панкреатит, дислипидемия, неалкогольная жировая болезнь печени (NAFLD), неалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперсулинемия, непереносимость глюкозы, гипергликемия, метаболический синдром, острый инфаркт миокарда, гипертензия, сердечно-сосудистое заболевание, атеросклероз, периферическая артериальная болезнь, инсульт, сердечная недостаточность, ишемическая болезнь сердца, заболевание почек, диабетические осложнения, нейропатия, гастропарез, заболевания, связанные с тяжелыми инактивирующими мутациями в рецепторе инсулина, и другие метаболические нарушения, и для снижения смертности и заболеваемости критически больных пациентов.
«Сахарный диабет 2 типа» представляет собой состояние, характеризующееся избыточным выработкой глюкозы, несмотря на доступность инсулина, и уровни циркулирующей глюкозы остаются чрезмерно высоким в результате недостаточного клиренса глюкозы.
«Сахарный диабет 1 типа» представляет собой состояние, характеризующееся высоким уровнем глюкозы в крови, вызванным полным отсутствием инсулина. Это происходит, когда иммунная система организма атакует инсулин-продуцирующие бета-клетки в поджелудочной железе и разрушает их. Поджелудочная железа тогда мало или вообще не вырабатывает инсулин.
«Панкреатит» представляет собой воспаление поджелудочной железы.
«Дислипидемия» является расстройством метаболизма липопротеинов, включая избыточный синтез или дефицит липопротеинов. Дислипидемии могут проявляться в повышении общего уровня холестерина, концентрации липопротеинов низкой плотности (ЛПНП) и триглицеридов, а также в снижении концентрации холестерина липопротеинов высокой плотности (ЛПВП) в крови.
«Неалкогольный стеатогепатит (NASH)» представляет собой заболевание печени, не связанное с потреблением алкоголя, характеризующееся жировым изменением гепатоцитов, сопровождающееся внутридольковым воспалением и фиброзом.
«Непереносимость глюкозы» или «неудовлетворительная толерантность к глюкозе» (IGT) является преддиабетическим состоянием дисгликемии, которое связано с повышенным риском сердечно-сосудистой патологии. Предварительное диабетическое состояние препятствует тому, чтобы у субъекта эффективно перемещалась глюкоза в клетки и использовалась в качестве эффективного источника топлива, что приводит к повышенным уровням глюкозы в крови и некоторой степени резистентности к инсулину.
«Гипергликемия» определяется как избыток сахара (глюкозы) в крови.
«Гипогликемия», также называемая низким уровнем сахара в крови, возникает, когда уровень глюкозы в крови падает слишком низко, чтобы обеспечить достаточную энергию для деятельности вашего организма.
«Гиперинсулинемия» определяется как более высокий, чем обычно, уровень инсулина в крови.
«Резистентность к инсулину» определяется как состояние, при котором нормальное количество инсулина вызывает субнормальный биологический ответ.
«Ожирение», с точки зрения человеческого субъекта, можно определить как вес тела более чем на 20 процентов выше идеального веса тела для данной группы (R. H. Williams, Textbook of Endocrinology, 1974, p. 904-916). Его также можно определить как индекс массы тела (ИМТ, определяемый как вес человека в килограммах, деленный на квадрат его высоты в метрах (кг/м2)) больше или равный 30.
«Метаболический синдром» можно определить как группу по меньшей мере трех следующих признаков: абдоминальный жир - у большинства мужчин - 40-дюймовая талия или больше; высокий уровень сахара в крови - по меньшей мере 110 миллиграммов на децилитр (мг/дл) после голодания; высокие триглицериды - по меньшей мере 150 мг/дл в крови; низкий уровень ЛПВП - менее 40 мг/дл; и артериальное давление 130/85 мм рт.ст. или выше.
«Гипертензия» или высокое кровяное давление представляет собой временное или стабильное повышение системного артериального давления до уровня, который может вызвать сердечно-сосудистые повреждения или другие неблагоприятные последствия. Гипертензия была условно определена как систолическое артериальное давление выше 140 мм рт. ст. или диастолическое артериальное давление выше 90 мм рт.ст.
«Сердечно-сосудистые заболевания» представляют собой болезни, связанные с сердцем или кровеносными сосудами.
«Периферическая артериальная болезнь» возникает, когда в артериях, которые переносят кровь к голове, органам и конечностям, накапливаются бляшки. Со временем бляшка может затвердеть и сузить артерии, которые ограничивают поток богатой кислородом крови к органам и другим частям тела.
«Атеросклероз» представляет собой сосудистое заболевание, характеризующееся нерегулярно распределенными липидными отложениями в интиме крупных и средних артерий, вызывая сужение артериальных просветов и в конечном итоге приводя к фиброзу и кальцификации. Повреждения обычно являются очаговыми и прогрессируют медленно и с перерывами. Ограничение кровотока объясняет большинство клинических проявлений, которые различаются в зависимости от распределения и тяжести поражений.
«Инсульт» представляет собой острое клиническое событие, связанное с нарушением мозгового кровообращения, которое длится более 24 часов. Инсульт включает необратимое повреждение головного мозга, тип и тяжесть симптомов которого зависят от местоположения и объема ткани головного мозга, циркуляция которой была нарушена.
«Сердечная недостаточность», также называемая застойной сердечной недостаточностью, является состоянием, при котором сердце не может больше накачивать достаточное количество крови для остальной части тела.
«Ишемическая болезнь сердца», также называемая болезнью коронарной артерии, представляет собой сужение мелких кровеносных сосудов, которые снабжают сердце кровью и кислородом.
«Болезнь почек» или нефропатия представляет собой любое заболевание почек. Диабетическая нефропатия является основной причиной заболеваемости и смертности у людей с сахарным диабетом 1 или 2 типа.
«Диабетические осложнения» представляют собой проблемы, вызванные высоким уровнем глюкозы в крови, проявляемые другими функциями организма, такими как почки, нервы (невропатии), стопы (язвы стопы и плохое кровообращение) и глаза (например, ретинопатии). Диабет также увеличивает риск сердечных заболеваний и заболеваний костей и суставов. Другие долгосрочные осложнения диабета включают проблемы с кожей, проблемы с пищеварением, сексуальную дисфункцию и проблемы с зубами и деснами.
«Нейропатии» представляют собой заболевания, связанные с черепными нервами или периферической или вегетативной нервной системой.
«Гастропарез» представляет собой слабость перистальтики желудка, что приводит к задержке опорожнения кишечника.
Критически больные пациенты, охватываемые настоящим изобретением, обычно испытывают нестабильное гиперметаболическое состояние. Это нестабильное метаболическое состояние обусловлено изменениями метаболизма субстрата, что может привести к относительным недостаткам в некоторых питательных веществах. Как правило, наблюдается повышенное окисление жира и мышц.
Кроме того, критически больные пациенты, предпочтительно, являются пациентами, которые испытывают системный синдром воспалительного ответа или респираторный дистресс. Снижение заболеваемости означает снижение вероятности того, что у критически больного пациента возникнут дополнительные болезни, состояния или симптомы или уменьшится тяжесть дополнительных заболеваний, состояний или симптомов. Например, снижение заболеваемости может быть связано с уменьшением частоты бактериемии или сепсиса или осложнений, связанных с множественной недостаточностью органов.
Термин «консервативно модифицированный вариант» относится как к аминокислотам, так и к последовательностям нуклеиновых кислот. Что касается конкретных последовательностей нуклеиновых кислот, то консервативно модифицированные варианты относятся к таким нуклеиновым кислотам, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или, если нуклеиновая кислота не кодирует аминокислотную последовательность, то к по существу идентичным последовательностям. Из-за вырожденности генетического кода большое количество функционально идентичных нуклеиновых кислот кодируют каждый конкретный белок. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, если аланин указан с помощью кодона, кодон может быть изменен на любой описанный соответствующий кодон без изменения кодируемого полипептида. Такие варианты нуклеиновой кислоты являются «молчащими вариантами», которые являются одним из видов консервативно модифицированных вариантов. В настоящем документе каждая последовательность нуклеиновой кислоты, которая кодирует полипептид, также описывает все возможные молчащие варианты нуклеиновой кислоты. Специалисту в данной области понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который в обычной ситуации является единственным кодоном для метионина, и TGG, который в обычной ситуации является единственным кодоном для триптофана) может быть модифицирован с получением функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, который кодирует полипептид, подразумевается в каждой описанной последовательности.
Для полипептидных последовательностей «консервативно модифицированные варианты» включают отдельные замены, делеции или добавления к полипептидной последовательности, которые приводят к замене аминокислоты на химически подобную аминокислоту. Таблицы консервативных замен, которые дают функционально сходные аминокислоты, хорошо известны в данной области. Такие консервативно модифицированные варианты дополняют и не исключают полиморфные варианты, межвидовые гомологи и аллели по изобретению. Следующие восемь групп содержат аминокислоты, которые являются консервативными заменами друг друга: 1) аланин (А), глицин (G); 2) аспарагиновая кислота (D), глутаминовая кислота (E); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (M), валин (V); 6) фенилаланин (F), тирозин (Y), триптофан (W); 7) серин (S), треонин (Т) и 8) цистеин (C), метионин (M) (смотри, например, Creighton, Proteins (1984)). В некоторых вариантах осуществления термин «консервативные модификации в последовательности» используется для обозначения аминокислотных модификаций, которые не оказывают существенного влияния или не изменяют характеристики связывания антитела, содержащего эту аминокислотную последовательность.
Термин «эпитоп» означает белковую детерминанту, способную специфически связываться с антителом. Эпитопы обычно состоят из химически активных групп молекул, расположенных на поверхности, таких как аминокислоты или боковые цепи сахаров, и обычно имеют определенные характеристики трехмерной структуры, а также определенные характеристики заряда. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первым, в отличие от последнего, теряется в присутствии денатурирующих растворителей.
Термин «антитело человека», как используется в настоящем документе, предполагает включение антител с вариабельными областями, в которых как каркасный участок, так и области CDR получены из последовательностей человеческого происхождения. Кроме того, если антитело содержит константную область, то константная область также получена из таких последовательностей человека, например, последовательности зародышевой линии человека или мутированные варианты последовательностей зародышевой линии человека. Антитела человека по изобретению могут включать аминокислотные остатки, не кодируемые последовательностями человека (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro или с помощью соматической мутации in vivo).
Термин «моноклональное антитело человека» относится к антителам, проявляющим единственную специфичность связывания, которые имеют вариабельные области, в которых как каркасные участки, так и области CDR получены из последовательностей человека. В одном из вариантов осуществления моноклональные антитела человека получают с помощью гибридомы, которая включает В-клетку, полученную у трансгенного животного, не являющегося человеком, например, у трансгенной мыши, с геномом, содержащим трансген тяжелой цепи и трансген легкой цепи человека, слитым с иммортализованной клеткой.
«Гуманизированное» антитело представляет собой антитело, которое сохраняет реакционную способность антитела, не являющегося человеческим, но которое является менее иммуногенным для людей. Такой эффект может быть достигнут, например, за счет сохранения областей CDR, не являющихся человеческими, и заменой оставшихся частей антитела на их человеческие аналоги (то есть константная область, а также каркасные части вариабельной области). Смотри, например, Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855, 1984; Morrison and Oi, Adv. Immunol., 44: 65-92, 1988; Verhoeyen et al., Science, 239: 1534-1536, 1988; Padlan, Molec. Immun., 28:489-498, 1991; и Padlan, Molec. Immun., 31:169-217, 1994. Другие примеры биоинженерных технологий включают, но не ограничиваются ими, технологию Xoma, описанную в патенте США 5766886.
Термины «идентичная» или процентная «идентичность» в контексте двух или более нуклеиновых кислот или полипептидных последовательностей относятся к двум или более последовательностям или последовательностям, которые являются одинаковыми. Две последовательности «по существу идентичны», если две последовательности имеют определенный процент одинаковых аминокислотных остатков или нуклеотидов (то есть 60% идентичность, возможно, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичность в определенной области или, если не указано, по всей последовательности) при сравнении и выравнивании для максимального совпадения в окне сравнения или обозначенной области, определенных с помощью одного из следующих алгоритмов сравнения последовательностей, либо путем выравнивания вручную и при визуальной проверке. Необязательно, идентичность существует в области, длина которой равна по меньшей мере приблизительно 50 нуклеотидов (или 10 аминокислот) или, более предпочтительно, в области, длина которой равна от 100 до 500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот).
Для сравнения последовательностей обычно одна последовательность служит эталонной последовательностью, с которой сравнивают исследуемые последовательности. При использовании алгоритма сравнения последовательностей исследуемая и эталонная последовательности вводятся в компьютер, обозначаются координаты последовательности, при необходимости, и указываются параметры программы с алгоритмом для работы с последовательностями. Можно использовать параметры программы по умолчанию или указать альтернативные параметры. По алгоритму сравнения последовательностей затем рассчитывается процент идентичности исследуемых последовательностей в сравнении с эталонной последовательностью на основании параметров программы.
«Окно сравнения», как используется в настоящем документе, включает указание на сегмент любого из множества непрерывных положений, выбранных из группы, состоящей из от 20 до 600, обычно от около 50 до около 200, более обычно от около 100 до около 150, в которых последовательность можно сравнивать с эталонной последовательностью с таким же количеством непрерывных положений после оптимального выравнивания двух последовательностей. Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Оптимальное выравнивание последовательностей для сравнения может быть проведено, например, с помощью алгоритма локальной гомологии Smith and Waterman (1970) Adv. Appl. Math. 2:482c, с помощью алгоритма гомологичного выравнивания Needleman and Wunsch, (1970) J. Mol. Biol. 48:443, 1970, с помощью способа поиска сходства Pearson and Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444, 1988, с помощью этих алгоритмов, реализованных в компьютерных программах (GAP, BESTFIT, FASTA и TFASTA в программном пакете Wisconsin Genetics Software Package от компании Genetics Computer Group, 575 Science Dr., Madison, WI), или с помощью выравнивания вручную и визуальной проверки (смотри, например, Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (Ringbou ed., 2003)).
Двумя примерами алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, описанные в Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977; и Altschul et al., J. Mol. Biol. 215:403-410, 1990, соответственно. Программное обеспечение для осуществления анализов BLAST является общедоступным на сайте Национального центра по биотехнологической информации. Этот алгоритм включает, во-первых, идентификацию пар последовательностей с наибольшей балльной оценкой (HSP) путем определения коротких слов длиной W в искомой последовательности, которые либо совпадают, либо удовлетворяют некоторому положительному порогу показателя Т при сравнении со словом той же длины в базе данных последовательностей. Т называют порогом показателя сходства слов (Altschul et al., выше). Эти начальные совпадения соседних слов выполняют роль затравки на начальном поиске более длинных HSP, их содержащих. Совпадения слов распространяют в оба направления по всей длине каждой последовательности по мере увеличения суммирующейся оценки выравнивания. Сумму баллов рассчитывают с помощью параметра М (балл за совпадение остатков, всегда >0) и параметра N (штраф за несовпадение остатков, всегда <0) для нуклеотидных последовательностей. Для аминокислотных последовательностей матрица замен используется для расчета суммарного балла. Продление слова в каждом направлении прекращается, если: суммарный балл выравнивания опускается ниже величины Х от его полученного максимального значения; суммарный балл достигает нуля или ниже из-за накопления одного или нескольких остатков с отрицательным баллом; либо же при достижении конца последовательности. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) по умолчанию используют длину слова (W), равную 11, ожидание (E), равное 10, M=5, N=-4 и сравнение обеих цепей. Для аминокислотных последовательностей в программе BLASTP по умолчанию используют длину слова, равную 3 и ожидание (E), равное 10, и матрицу замен BLOSUM62 (смотри, Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89: 10915, 1989) (B), равное 50, ожидание (E), равное 10, M=5, N=-4 и сравнение обеих цепей.
Алгоритм BLAST также выполняет статистический анализ сходности двух последовательностей (смотри, например, Karlin and Altschul, Proc. Natl. Acad. Sci. USA, 90: 5873-5787, 1993). Одно измерение сходности, проводимое алгоритмом BLAST, заключается в определении наименьшей суммарной вероятности P(N)), которая указывает на вероятность, при которой совпадение двух нуклеотидных или аминокислотных последовательностей будет происходить случайно. Например, нуклеиновая кислота считается сходной эталонной последовательности, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой меньше приблизительно 0,2, более предпочтительно, меньше приблизительно 0,01 и, наиболее предпочтительно, меньше приблизительно 0,001.
Процент идентичности двух аминокислотных последовательностей также может быть определен с использованием алгоритма Е. Мейерса и В. Миллера (Comput. Appl. Biosci., 4: 11-17, 1988), который включен в программу ALIGN (версия 2.0), используя таблицу веса остатков PAM120, штраф за длину пробела, равный 12, и штраф за пробел, равный 4. Кроме того, процент идентичности двух аминокислотных последовательностей может быть определен с использованием Нидлмана и Вунша (J. Mol, Biol. 48: 444-453, 1970), который включен в программу GAP в программный пакет GCG (доступен во всемирной паутине на сайте gcg.com), используя либо матрицу Blossom 62, либо матрицу PAM250, а также вес пробела 16, 14, 12, 10, 8, 6 или 4 и размер длины 1, 2, 3, 4, 5 или 6.
Помимо процента идентичности последовательностей, как указано выше, еще одно указание на то, что две последовательности нуклеиновой кислоты или полипептиды по существу идентичны, состоит в том, что полипептид, кодируемый первой нуклеиновой кислотой, перекрестно реагирует при иммунных реакциях с антителами, продуцируемыми против полипептида, кодируемого второй нуклеиновой кислотой, как описано далее. Таким образом, полипептид обычно по существу идентичен второму полипептиду, например, если два пептида отличаются только консервативными заменами. Еще одно указание на то, что две последовательности нуклеиновых кислот по существу идентичны, состоит в том, что две молекулы или их комплементарные варианты гибридизуются друг с другом в жестких условиях, как описано далее. Еще одно указание на то, что две последовательности нуклеиновых кислот по существу идентичны, состоит в том, что для амплификации последовательности можно использовать одни и те же праймеры.
Термин «выделенное антитело» относится к антителу, которое по существу не содержит других антител с другими антигенными специфичностями (например, выделенное антитело, которое специфически связывается с β-клото, по существу не содержит антител, которые специфически связываются с антигенами, отличными от β-клото). Однако выделенное антитело, которое специфически связывается с β-клото, может иметь перекрестную реактивность в отношении других антигенов. Кроме того, выделенное антитело может быть по существу свободным от другого клеточного вещества и/или химических веществ.
Термин «изотип» относится к классу антител (например, IgM, IgE, IgG, таких как IgG1 или IgG4), которые соответствуют генам константной области тяжелой цепи. Изотип также включает модифицированные варианты одного из этих классов, в которых были сделаны модификации с изменением функции Fc, например, для усиления или уменьшения эффекторных функций или связывания с рецепторами Fc.
Термин «kассоц» или «ka», как он используется в настоящем документе, относится к обозначению скорости ассоциации конкретной реакции антитело-антиген, причем термин «kдис» или «kd», как используется в настоящем документе, относится к скорости диссоциации конкретной реакции антитело-антиген. Термин «KD», как он используется в настоящем документе, относится к константе диссоциации, которую получают из соотношения kd к ka (то есть kd/ka) и выражают в виде молярной концентрации (М). Значения KD для антител могут быть определены способами, хорошо известными в данной области. Способы определения KD антитела включают измерение поверхностного плазмонного резонанса с помощью биосенсорной системы, такой как система Biacore®, или измерение аффинности в растворе путем титрования раствора (SET).
Термины «моноклональное антитело» или «композиция моноклональных антител», как используется в настоящем документе, относятся к препарату молекул антител одного молекулярного состава. Композиция моноклональных антител демонстрирует одну специфичность связывания и аффинность к определенному эпитопу.
Термин «нуклеиновая кислота» используется в настоящем документе взаимозаменяемо с термином «полинуклеотид» и относится к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам в одно- или двухцепочечной форме. Термин включает нуклеиновые кислоты, содержащие известные нуклеотидные аналоги или модифицированные остатки каркаса или связи, которые являются синтетическими, природными и неприродными, которые обладают связывающими свойствами, аналогичными эталонной нуклеиновой кислоте, и которые метаболизируются способом, сходным с эталонными нуклеотидами. Примеры таких аналогов включают, но ими не ограничиваются, фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептид-нуклеиновые кислоты (PNA).
Если не указано иное, конкретная последовательность нуклеиновой кислоты также подразумевает их консервативно модифицированные варианты (например, замены из-за вырожденности кодонов) и комплементарные последовательности, а также конкретно указанную последовательность. В частности, как подробно описано далее, замены из-за вырожденности кодонов могут быть получены путем создания последовательностей, в которых третье положение одного или нескольких (или всех) выбранных кодонов заменено смешанным основанием и/или дезоксиинозиновыми остатками (Batzer et al., Nucleic Acid Res. 19:5081, 1991; Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); и Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)).
Термин «функционально связанный» относится к функциональному взаимодействию двух или более полинуклеотидных сегментов (например, ДНК). Обычно термин относится к функциональному взаимодействию последовательности, регулирующей транскрипцию, с транскрибируемой последовательностью. Например, промоторная или энхансерная последовательность функционально связана с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой экспрессирующей системе. Как правило, промоторные последовательности, регулирующие транскрипцию, которые функционально связаны с транскрибируемой последовательностью, физически непрерывны с транскрибируемой последовательностью, то есть они являются цис-действующими. Однако некоторые последовательности, регулирующие транскрипцию, такие как энхансеры, не обязательно должны быть физически связаны или находиться в непосредственной близости от кодирующих последовательностей, транскрипцию которых они усиливают.
Как используется в настоящем документе, термин «оптимизированный» означает, что нуклеотидная последовательность была изменена в отношении кодирования аминокислотной последовательности, используя кодоны, которые являются предпочтительными для продуцирующей клетки или организма, главным образом, для эукариотической клетки, например, клетка Pichia, клетка яичника китайского хомячка (CHO) или клетка человека. Оптимизированную нуклеотидную последовательность конструируют таким образом, чтобы полностью или максимально возможно сохранить аминокислотную последовательность, изначально кодируемую исходной нуклеотидной последовательностью, которая также называется «родительской» последовательностью. В настоящем документы оптимизированные последовательности сконструированы так, что имеют кодоны, которые являются предпочтительными для клеток млекопитающих. Однако в настоящем документе также предусмотрена оптимизированная экспрессия этих последовательностей в других эукариотических клетках или прокариотических клетках. Аминокислотные последовательности, кодируемые оптимизированными нуклеотидными последовательностями, также называются оптимизированными.
Термины «полипептид» и «белок» используются в настоящем документе взаимозаменяемо и обозначают полимер из аминокислотных остатков. Эти термины применимы к аминокислотным полимерам, в которых один или несколько аминокислотных остатков является синтетическим химический миметиком соответствующей природной аминокислоты, а также к природным полимерам из аминокислот и к неприродному полимеру из аминокислот. Если не указано иное, конкретная полипептидная последовательность также предусматривает консервативно модифицированные варианты.
Термин «рекомбинантное антитело человека», как используется в настоящем документе, включает все антитела человека, которые получены, экспрессированы, созданы или выделены рекомбинантными способами, например, антитела, выделенные у животного (например, мышь), которое является трансгенным или трансхромосомным по генам иммуноглобулина человека, или из полученной из них гибридомы, антитела, выделенные из клетки-хозяина, трансформированной с тем, чтобы экспрессировать антитело человека, например, из трансфектомы, антитела, выделенные из рекомбинантной комбинаторной библиотеки антител человека, и антитела, полученные, экспрессированные, созданные или выделенные любым другим способом, которые включают сплайсинг всей или части последовательности гена иммуноглобулина человека, с другими последовательностями ДНК. Такие рекомбинантные антитела человека имеют вариабельные области, в которых каркасный участок и области CDR получены из последовательностей иммуноглобулина зародышевой линии человека. Однако в некоторых вариантах осуществления такие рекомбинантные антитела человека могут быть подвергнуты in vitro мутагенезу (или, если используется животное, трансгенное по последовательностям Ig человека, то in vivo соматическому мутагенезу), и, таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые при том, что они выделены и относятся к последовательностям VH и VL зародышевой линии человека, могут не существовать в природе в репертуаре зародышевой линии человеческого антитела in vivo.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») относится к клетке, в которую введен рекомбинантный вектор экспрессии. Следует понимать, что такие термины относятся не только к конкретной заданной клетке, но и к потомству такой клетки. Поскольку в последующих поколениях могут произойти некоторые изменения из-за мутаций или воздействия окружающей среды, то такое потомство может фактически быть не идентичным родительской клетке, но сохраняется в объеме термина «клетка-хозяин», как используется в настоящем документе.
Термин «субъект» включает человека или животных, отличных от человека. Животные, отличные от человека, включают всех позвоночных животных (например, млекопитающих и не млекопитающих), таких как приматы, отличные от человека (например, яванский макак), овцы, собаки, коровы, куры, амфибии и рептилии. Кроме указанных случаев, термины «пациент» или «субъект» используются в настоящем документе взаимозаменяемо. Как используется в настоящем документ, термин «яванский» относятся к яванскому макаку (Macaca fascicularis).
Как используется в настоящем документе, термин «лечить» или «лечение» какого-либо заболевания или расстройства (например, расстройства, связанного с FGF21) относится в одном варианте осуществления к облегчению заболевания или расстройства (то есть, к замедлению или прекращению или к уменьшению развития заболевания, или по крайней мере одного из его клинических симптомов). В еще одном варианте осуществления «лечить» или «лечение» относятся к облегчению или улучшению по меньшей мере одного физического параметра, включая параметры, которые не могут быть ощутимыми самим пациентом. В еще одно варианте осуществления «лечить» или «лечение» относятся к изменению заболевания или расстройства либо физически (например, стабилизация ощутимого симптома), либо физиологически (например, стабилизация физического параметра), либо и физически и физиологически. В еще одном варианте осуществления изобретения, «лечить» или «лечение» относятся к профилактике или задержке начала или развития, или к прогрессированию заболевания или расстройства.
«Профилактика», поскольку она относится к показаниям, описанным в настоящем документе, включая, например, расстройство, связанное с FGF21, означает любое действие, которое предотвращает или замедляет ухудшение, например, связанных с FGF21 параметров заболевания, как описано ниже, у пациента с риском такого ухудшения.
Термин «вектор» предназначен для обозначения полинуклеотидной молекулы, способной переносить другой полинуклеотид, с которым она связана. Одним из типов векторов является «плазмида», которая относится к циклической двухцепочечной петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, такой как аденоассоциированный вирусный вектор (AAV или AAV2), в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, куда они введены (например, бактериальные вектор с бактериальной точкой начала репликации и эписомальные векторы млекопитающих). Другие векторы (например, неэпизомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяин и, таким образом, реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы упоминаются в настоящем документе как «рекомбинантные векторы экспрессии» (или просто «векторы экспрессии»).
Обычно векторы экспрессии, используемые в методах рекомбинантных ДНК, часто находятся в виде плазмид. В настоящем описании «плазмида» и «вектор» могут использоваться взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако в изобретении подразумеваются и другие формы векторов экспрессии, такие как вирусные векторы (например, ретровирусы, дефектные по репликации, аденовирусы и аденоассоциированные вирусы), которые выполняют аналогичные функции.
«Модуляция активности FGF21», как используется в настоящем документе, относится к увеличению или уменьшению активности FGF21, что может быть результатом, например, взаимодействия средства с полинуклеотидом или полипептидом FGF21, активацией сигнального пути FGF21 и/или активацией сигнального пути, опосредованного FGF21 (например, сигнальный путь, зависящий от рецептора FGF21), и тому подобное. Например, модуляция биологической активности относится к увеличению или уменьшению биологической активности. Активность FGF21 может оценена способами, включающими, но ими не ограничивающимися, анализ уровня глюкозы в крови, инсулина, триглицеридов или холестерина у субъекта; оценкой уровней полипептидов бета-клото и/или рецепторов FGF (например, FGFR-1c); или оценкой активации сигнального пути, опосредуемого FGF21 (например, сигнального пути, зависящего от рецептора FGF21).
Сравнение активности FGF21 также может быть осуществлено, например, путем измерения уровней биомаркера FGF21 в следующих стадиях сигнального пути и измерения усиления сигнального пути FGF21. Активность также можно оценить путем измерения: сигнального пути клетки; киназной активности; поглощения глюкозы адипоцитами; колебаний уровней инсулина, триглицеридов или холестерина в крови; изменений уровней липидов или триглицеридов в печени; взаимодействия между FGF21 и/или бета-клото и рецептором FGF21; или фосфорилирования рецептора FGF21. В некоторых вариантах осуществления фосфорилирование рецептора FGF21 может быть фосфорилированием тирозина. В некоторых вариантах осуществления модуляция активности FGF21 может вызывать модуляцию фенотипа, связанного с FGF21.
«Биомаркер в последующих стадиях сигнального пути FGF21», как используется в настоящем документе, представляет собой ген или продукт гена, или измеримые показатели гена или продукта гена. В некоторых вариантах осуществления ген или активность, являющаяся маркером последующих стадий сигнального пути FGF21, показывают изменения в уровне экспрессии, или в сосудистой ткани. В некоторых вариантах осуществления активность маркера последующих стадий сигнального пути изменяется в присутствии модулятора FGF21. В некоторых вариантах осуществления маркеры в последующих стадиях сигнального пути демонстрируют изменение в уровнях экспрессии, если на FGF21 воздействует модулятор FGF21 по настоящему изобретению. Маркеры в последующих стадиях сигнального пути FGF21 включают, но ими не ограничиваются, поглощение глюкозы или 2-дезокси-глюкозы, pERK и уровни других фосфорилированных или ацетилированных белков или NAD.
Как используется в настоящем документе термин «активировать» относится к увеличению, активации или стимуляции активности или количества. Например, в контексте настоящего изобретения модуляторы FGF21 могут увеличивать активность бета-клото и/или рецептора FGF21. В одном варианте осуществления FGFR-1c может быть активирован в ответ на модулятор FGF21. Активирование также может относиться к активности, связанной с FGF21, такой как, например, способность понижать уровни глюкозы, инсулина, триглицерида или холестерина в крови; понижать уровни липидов или триглицеридов в печени; уменьшать вес тела; улучшать толерантность к глюкозе, расход энергии или чувствительность к инсулину; или вызывать фосфорилирование рецептора FGF21; или повышать уровень маркера в последующих стадиях сигнального пути FGF21. Рецептор FGF21 может представлять собой β-клото и FGFR-1c. Активирование может составлять по меньшей мере 25%, по меньшей мере 50%, по меньшей мере 75%, по меньшей мере 100%, по меньшей мере 150%, по меньшей мере 200%, по меньшей мере 250%, по меньшей мере 400% или по меньшей мере 500% по сравнению с контролем.
Как используется в настоящем документе, термин «модулятор» относится к композиции, которая модулирует один или несколько физиологических или биохимических событий, связанных с расстройством, связанным с FGF21, таким как сахарный диабет типа 1 или типа 2, или метаболическое состояние, такое как ожирение. Указанные события включают, но ими не ограничиваются, способность снижать уровни глюкозы, инсулина, триглицерида или холестерина в крови; понижать уровни липидов или триглицеридов в печени; уменьшать вес тела; и улучшать толерантность к глюкозе, расход энергии или чувствительность к инсулину.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: Анализ связывания FGF21-миметических антител NOV001, NOV002 и NOV004 с β-клото человека титрованием раствора.
Фигура 2: Активация pERK клеток FGFR1c_β-клото_HEK293 A) человека и B) японского макака с помощью NOV002 и NOV004.
Фигура 3: Профиль NOV002 и NOV004 активации pERK клеток A) FGFR2c_β-клото, B) FGFR3c_β-клото и C) FGFR4_β-клото HEK293 человека. FGF21 использовали в качестве положительного контроля для активации FGFR2c_β-клото или FGFR3c_β-клото. FGF19 использовали в качестве положительного контроля активации FGFR4_β-клото.
Фигура 4: Профиль NOV002 и NOV004 активации FGF23 с использованием клеток HEK293, трансфицированных α-клото, Egr1-люциферазой и люмиферазой Renilla. FGF23 использовался в качестве положительного контроля.
Фигура 5: Профиль NOV002 и NOV004 перекрестной реактивности мыши с использованием обратного захвата 2-DOG адипоцитами 3T3-L1. FGF21 использовали в качестве положительного контроля.
Фигура 6: Профили «концентрация-время» NOV002 и NOV004 после внутривенной инъекции крысам.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение основано, в частности, на обнаружении молекул антител, которые специфически связываются с β-клото и приводят к активации рецепторов FGF, например FGFR1c, и активации FGF21-опосредованного сигнального пути (например, сигнального пути, зависимого от FGF21-рецептора). Изобретение относится как к полной форме антител IgG, так и к их антигенсвязывающим фрагментам, таким как Fab-фрагменты (например, антитела NOV001, NOV002, NOV003 и NOV004).
Соответственно, настоящее изобретение относится к антителам, которые специфически связываются с β-клото (например, β-клото человека и яванского макака), фармацевтическими композициями, способами получения и способами применения таких антител и композиций.
Белки FGF21
Настоящее изобретение относится к миметическим mAb FGF21 (то есть моноклональным антителам, которые связываются с бета-клото (β-клото)), которые могут индуцировать FGF21-опосредованный сигнальный путь (например, опосредованный FGF21-рецептором сигнальный путь), как определено в настоящем документе. In vivo зрелая форма FGF21 является активной формой молекулы. Последовательность дикого типа FGF21 имеет номер последовательности NCBI NP_061986.1 и описание может быть найдено в таких выданных патентах, как, например, US 6716626B1, выдан компании Chiron Corporation (SEQ ID NO: 1).


Соответствующая последовательность мРНК, кодирующая полноразмерный полипептид FGF21 (номер контрольной последовательности NCBI NM_019113.2), показана ниже (SEQ ID NO: 2):
1 ctgtcagctg aggatccagc cgaaagagga gccaggcact caggccacct gagtctactc
61 acctggacaa ctggaatctg gcaccaattc taaaccactc agcttctccg agctcacacc
121 ccggagatca cctgaggacc cgagccattg atggactcgg acgagaccgg gttcgagcac
181 tcaggactgt gggtttctgt gctggctggt cttctgctgg gagcctgcca ggcacacccc
241 atccctgact ccagtcctct cctgcaattc gggggccaag tccggcagcg gtacctctac
301 acagatgatg cccagcagac agaagcccac ctggagatca gggaggatgg gacggtgggg
361 ggcgctgctg accagagccc cgaaagtctc ctgcagctga aagccttgaa gccgggagtt
421 attcaaatct tgggagtcaa gacatccagg ttcctgtgcc agcggccaga tggggccctg
481 tatggatcgc tccactttga ccctgaggcc tgcagcttcc gggagctgct tcttgaggac
541 ggatacaatg tttaccagtc cgaagcccac ggcctcccgc tgcacctgcc agggaacaag
601 tccccacacc gggaccctgc accccgagga ccagctcgct tcctgccact accaggcctg
661 ccccccgcac tcccggagcc acccggaatc ctggcccccc agccccccga tgtgggctcc
721 tcggaccctc tgagcatggt gggaccttcc cagggccgaa gccccagcta cgcttcctga
781 agccagaggc tgtttactat gacatctcct ctttatttat taggttattt atcttattta
841 tttttttatt tttcttactt gagataataa agagttccag aggagaaaaa aaaaaaaaaa
901 aaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa
У зрелой последовательности FGF21 отсутствует лидерная последовательность, и она может также включать другие модификации полипептида, такие как протеолитический процессинг аминоконца (с лидерной последовательностью или без нее) и/или карбоксильного конца, отщепление меньшего полипептида от более крупного предшественника, N-связанное и/или O-связанное гликозилирование и другие посттрансляционные модификации, понятные специалистам в данной области. Характерный пример зрелой последовательности FGF21 имеет следующую последовательность (SEQ ID NO: 83, в которой представлены аминокислотные положения 29-209 последовательности полноразмерного белка FGF21 (номер контрольной последовательности NCBI NP_061986.1):
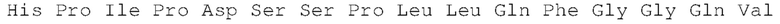

Соответствующая кДНК-последовательность, кодирующая зрелый полипептид FGF21 (SEQ ID NO: 83), показана ниже (SEQ ID NO: 84):
1 caccccatcc ctgactccag tcctctcctg caattcgggg gccaagtccg gcagcggtac
61 ctctacacag atgatgccca gcagacagaa gcccacctgg agatcaggga ggatgggacg
121 gtggggggcg ctgctgacca gagccccgaa agtctcctgc agctgaaagc cttgaagccg
181 ggagttattc aaatcttggg agtcaagaca tccaggttcc tgtgccagcg gccagatggg
240 gccctgtatg gatcgctcca ctttgaccct gaggcctgca gcttccggga gctgcttctt
301 gaggacggat acaatgttta ccagtccgaa gcccacggcc tcccgctgca cctgccaggg
360 aacaagtccc cacaccggga ccctgcaccc cgaggaccag ctcgcttcct gccactacca
421 ggcctgcccc ccgcactccc ggagccaccc ggaatcctgg ccccccagcc ccccgatgtg
481 ggctcctcgg accctctgag catggtggga ccttcccagg gccgaagccc cagctacgct
541 tcctga
Миметические антитела FGF21 и антигенсвязывающие фрагменты
Настоящее изобретение относится к антителам, которые специфически связываются с β-клото. В некоторых вариантах осуществлений настоящее изобретение относится к антителам, которые специфически связываются с β-клото человека и яванского макака. Антитела по изобретению включают, но этим не ограничиваются моноклональные антитела человека и Fabs, выделенные, как описано в примерах.
Последовательность β-клото дикого типа имеет номер последовательности NCBI NP_783864.1, и описание ее может быть найдено в такой литературе, как Xu, et al. (2007) J Biol Chem. 282 (40): 29069-72, и Lin, et al. (2007) J Biol Chem. 282 (37): 27277-84. Полноразмерная кДНК, кодирующая β-клото человека, имеет номер доступа GenBank NM_175737). Последовательность белка выглядит следующим образом (SEQ ID NO: 262).
1 mkpgcaagsp gnewiffstd eittryrntm sngglqrsvi lsalillrav tgfsgdgrai
61 wsknpnftpv nesqlflydt fpknffwgig tgalqvegsw kkdgkgpsiw dhfihthlkn
121 vsstngssds yiflekdlsa ldfigvsfyq fsiswprlfp dgivtvanak glqyystlld
181 alvlrniepi vtlyhwdlpl alqekyggwk ndtiidifnd yatycfqmfg drvkywitih
241 npylvawhgy gtgmhapgek gnlaavytvg hnlikahskv whnynthfrp hqkgwlsitl
301 gshwiepnrs entmdifkcq qsmvsvlgwf anpihgdgdy pegmrkklfs vlpifseaek
361 hemrgtadff afsfgpnnfk plntmakmgq nvslnlreal nwikleynnp riliaengwf
421 tdsrvktedt taiymmknfl sqvlqairld eirvfgytaw slldgfewqd aytirrglfy
481 vdfnskqker kpkssahyyk qiirengfsl kestpdvqgq fpcdfswgvt esvlkpesva
541 sspqfsdphl yvwnatgnrl lhrvegvrlk trpaqctdfv nikkqlemla rmkvthyrfa
601 ldwasvlptg nlsavnrqal ryyrcvvseg lklgisamvt lyypthahlg lpepllhadg
661 wlnpstaeaf qayaglcfqe lgdlvklwit inepnrlsdi ynrsgndtyg aahnllvaha
721 lawrlydrqf rpsqrgavsl slhadwaepa npyadshwra aerflqfeia wfaeplfktg
781 dypaamreyi askhrrglss salprlteae rrllkgtvdf calnhfttrf vmheqlagsr
841 ydsdrdiqfl qditrlsspt rlavipwgvr kllrwvrrny gdmdiyitas giddqaledd
901 rlrkyylgky lqevlkayli dkvrikgyya fklaeekskp rfgfftsdfk akssiqfynk
961 vissrgfpfe nsssrcsqtq entectvclf lvqkkplifl gccffstlvl llsiaifqrq
1021 krrkfwkakn lqhiplkkgk rvvs
Настоящее изобретение относится к антителам, которые специфически связывают белок β-клото (например, β-клото человека и яванского макака), где антитела содержат домен VH, имеющий аминокислотную последовательность SEQ ID NO: 9, 29, 49 или 69. Настоящее изобретение также относится к антителам, которые специфически связываются с белком β-клото, где антитела содержат CDR VH, имеющие аминокислотную последовательность любой из CDR VH, перечисленных в таблице 1, ниже. В частности, настоящее изобретение относится к антителам, которые специфически связываются с белком β-клото (например, β-клото человека и яванского макака), где антитела содержат (или, альтернативно, состоят из) одну, две, три или более CDR VH, имеющие аминокислотную последовательность любой из CDR VH, перечисленных в таблице 1, ниже.
Настоящее изобретение относится к антителам, которые специфически связываются с белком β-клото, где указанные антитела содержат домен VL, имеющий аминокислотную последовательность SEQ ID NO: 19, 39, 59 или 79. Настоящее изобретение также относится к антителам, которые специфически связываются с белком β-клото (например, β-клото человека и яванского макака), где указанные антитела, содержащие CDR VL, имеют аминокислотную последовательность любой из CDR VL, перечисленных в таблице 1, ниже. В частности, настоящее изобретение относится к антителам, которые специфически связываются с белком β-клото (например, β-клото человека и яванского макака), где указанные антитела содержат (или, альтернативно, состоят из) одну, две, три или более CDR VL, имеющих аминокислотную последовательность любой из CDR VL, перечисленных в таблице 1, ниже.
Другие антитела по изобретению включают аминокислоты, которые были мутированы, но по меньшей мере на 60, 70, 80, 85, 90 или 95 процентов идентичны в областях CDR тем областям CDR, которые изображены в последовательностях, описанных в таблице 1. В некоторых вариантах осуществления они включает мутантные аминокислотные последовательности, в которых не более 1, 2, 3, 4 или 5 аминокислот были мутированы в областях CDR по сравнению с областями CDR, изображенными в последовательности, описанной в таблице 1.
Настоящее изобретение также относится к последовательности нуклеиновых кислот, которые кодируют VH, VL, полноразмерную тяжелую цепь и полноразмерную легкую цепь антител, которые специфически связываются с белком β-клото (например, β-клото человека и яванского макака). Такие последовательности нуклеиновых кислот могут быть оптимизированы для экспрессии в клетках млекопитающих (например, в таблице 1 показаны оптимизированные последовательности нуклеиновых кислот для тяжелой цепи и легкой цепи антител по изобретению).
Таблица 1. Примеры миметических антител FGF21 и Fabs
(IgG1 LALA версия NOV003)
(IgG1 LALA версия NOV004)
(IgG1 DAPA версия NOV001)
(IgG1 DAPA версия NOV002)
Другие антитела по изобретению включают такие, где аминокислоты или нуклеиновые кислоты, кодирующие аминокислоты, были мутированы, но по меньшей мере на 60, 65, 70, 75, 80, 85, 90 или 95 процентов идентичны последовательностям, описанным в таблице 1. Некоторые варианты осуществления включают мутантные аминокислотные последовательности, где не более чем 1, 2, 3, 4 или 5 аминокислот были мутированы в вариабельных областях по сравнению с вариабельными областями, изображенными в последовательности, описанной в таблице 1, при сохранении по существу той же антигенсвязывающей активности.
Поскольку каждое из указанных антител может связываться с β-клото, последовательности VH, VL, полноразмерной легкой цепи и полноразмерной тяжелой цепи (аминокислотные последовательности и нуклеотидные последовательности, кодирующие аминокислотные последовательности) могут быть «смешаны и сопоставлены» для создания других связывающих β-клото антител по изобретению. Такие «смешанные и сопоставленные» связывающие β-клото антитела могут быть исследованы с использованием анализов связывания, известных в данной области (например, ELISA и других анализов, описанных в разделе примеров). Когда эти цепи смешивают и сопоставляют, последовательность VH конкретного VH/VL-спаривания должна быть заменена структурно подобной последовательностью VH. Точно так же последовательность полноразмерной тяжелой цепи конкретного спаривания «полноразмерная тяжелая цепь/полноразмерная легкая цепь» должна быть заменена структурно подобной последовательностью полноразмерной тяжелой цепи. Аналогично, последовательность VL конкретного VH/VL-спаривания должна быть заменена структурно подобной последовательностью VL. Точно так же последовательность полноразмерной легкой цепи конкретного спаривания «полноразмерная тяжелая цепь/полноразмерная легкая цепь» должна быть заменена структурно подобной последовательностью полноразмерной легкой цепи.
Соответственно, в одном аспекте настоящее изобретение относится к выделенному антителу или его антигенсвязывающей области, имеющим: вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49 и 69, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 и 79, где антитело специфически связывается с β-клото (например, β-клото человека и яванского макака).
Более конкретно, в некоторых аспектах настоящее изобретение относится к выделенному антителу или его антигенсвязывающей области, имеющим вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, содержащие аминокислотные последовательности, выбранные из SEQ ID NO: 9 и 19; 29 и 39; 49 и 59; или 69 и 79, соответственно.
В другом аспекте настоящее изобретение относится к (i) выделенному антителу, имеющему: полноразмерную тяжелую цепь, содержащую аминокислотную последовательность, которая была оптимизирована для экспрессии в клетке млекопитающего, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49 или 69, и полноразмерную легкую цепь, содержащую аминокислотную последовательность, которая была оптимизирована для экспрессии в клетке млекопитающего, выбранную из группы, состоящей из SEQ ID NO: 21, 41, 61 или 81; или (ii) функциональный белок, содержащий его антигенсвязывающую часть. Более конкретно, в некоторых аспектах настоящее изобретение относится к выделенному антителу или его антигенсвязывающей области, имеющим тяжелую цепь и легкую цепь, содержащей аминокислотные последовательности, выбранные из SEQ ID NO: 9 и 19; 29 и 39; 49 и 59; или 69 и 79, соответственно.
Термины «область, определяющая комплементарность», и «CDR», как они используются в настоящем документе, относятся к последовательности аминокислот в вариабельных областях антитела, которые придают антигенную специфичность и аффинность связывания. Обычно в каждой вариабельной области тяжелой цепи (HCDR1, HCDR2, HCDR3) имеются три CDR и в каждой вариабельной области легкой цепи (LCDR1, LCDR2, LCDR3) имеются три CDR.
Точные границы аминокислотной последовательности данной CDR могут быть легко определены с использованием какой-либо из числа хорошо известных систем, в том числе описанных в работах Kabat et al. (1991), «Sequences of Proteins of Immunological Interest», 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (система нумерации «Kabat»), Al-Lazikani et al., (1997) JMB 273, 927-948 (система нумерации «Chothia»), и системы нумерации ImMunoGenTics (IMGT) (Lefranc, M.-P., The Immunologist, 7, 132-136 (1999); Lefranc, M.-P. et al., Dev. Comp. Immunol., 27, 55-77 (2003) (система нумерации «IMGT»). Например, для классических форм, согласно Kabat, аминокислотные остатки CDR в вариабельном домене тяжелой цепи (VH) пронумерованы 31-35 (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3); и аминокислотные остатки CDR в вариабельном домене легкой цепи (VL) пронумерованы 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3). Согласно Chothia, аминокислоты CDR в VH пронумерованы 26-32 (HCDR1), 52-56 (HCDR2) и 95-102 (HCDR3); и аминокислотные остатки в VL пронумерованы 26-32 (LCDR1), 50-52 (LCDR2) и 91-96 (LCDR3). Объединяя определения CDR как по Kabat, так и по Chothia, CDR состоят из аминокислотных остатков 26-35 (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3) в VH человека, и из аминокислотных остатков 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в VL человека. Согласно IMGT, аминокислотные остатки CDR в VH пронумерованы предположительно 26-35 (CDR1), 51-57 (CDR2) и 93-102 (CDR3), и аминокислотные остатки CDR в VL пронумерованы предположительно 27-32 (CDR1), 50-52 (CDR2) и 89-97 (CDR3) (нумерация согласно «Kabat»). Согласно IMGT области CDR антитела могут быть определены с использованием программы IMGT/DomainGap Align.
Например, согласно Kabat, аминокислотные остатки CDR антитела NOV001 в вариабельном домене тяжелой цепи (VH) пронумерованы 31-35 (HCDR1), 50-65 (HCDR2) и 99-108 (HCDR3); и аминокислотные остатки CDR в вариабельном домене легкой цепи (VL) пронумерованы 24-39 (LCDR1), 55-61 (LCDR2) и 94-102 (LCDR3). Согласно Chothia аминокислоты CDR в VH пронумерованы 26-32 (HCDR1), 52-57 (HCDR2) и 99-108 (HCDR3); и аминокислотные остатки в VL пронумерованы 26-39 (LCDR1), 55-57 (LCDR2) и 96-101 (LCDR3). Объединяя определения e CDR определения как по Kabat, так и по Chothia, CDR состоят из аминокислотных остатков 26-35 (HCDR1), 50-65 (HCDR2) и 99-108 (HCDR3) в VH человека и аминокислотных остатков 24-39 (LCDR1), 55-61 (LCDR2) и 94-102 (LCDR3) в VL человека.
В другом аспекте настоящее изобретение относится к β-клото связывающим антителам, которые содержат CDR1, CDR2 и CDR3 тяжелых цепей и легких цепей, как описано в таблице 1, или их комбинации. Аминокислотные последовательности CDR1 VH антител показаны в SEQ ID NO: 3, 23, 43 и 63. Аминокислотные последовательности CDR2 VH антител показаны в SEQ ID NO: 4, 24, 44 и 64. Аминокислотные последовательности CDR3 VH антител показаны в SEQ ID NO: 5, 25, 45 и 65. Аминокислотные последовательности CDR1 VL антител показаны в SEQ ID NO: 13, 33, 53 и 73. Аминокислотные последовательности CDR2 VL антител показаны в SEQ ID NO: 14, 34, 54 и 74. Аминокислотные последовательности CDR3 антител VL показаны в SEQ ID NO: 15, 35, 55 и 75. Эти области CDR обозначаются с использованием системы Kabat.
Альтернативно, определенные с помощью системы Chothia (Al-Lazikani et al., (1997) JMB 273,927-948), аминокислотные последовательности CDR1 VH антител показаны в SEQ ID NO: 6, 26, 46 и 66. Аминокислотные последовательности CDR2 VH антител показаны в SEQ ID NO: 7, 27, 47 и 67. Аминокислотные последовательности CDR3 VH антител показаны в SEQ ID NO: 8, 28, 48 и 68. Аминокислотные последовательности CDR1 VL антител показаны в SEQ ID NO: 16, 36, 56 и 76. Аминокислотные последовательности CDR2 VL антител показаны в SEQ ID NO: 17, 37, 57 и 77. Аминокислотные последовательности CDR3 VL антител показаны в SEQ ID NO: 18, 38, 58 и 78.
Альтернативно, определенные с помощью комбинированной системы, аминокислотные последовательности CDR1 VH антител показаны в SEQ ID NO: 263 и 268. Аминокислотные последовательности CDR2 VH антител показаны в SEQ ID NO: 4, 24, 44 и 64. Аминокислотные последовательности CDR VH3 антител показаны в SEQ ID NO: 5, 25 и 45. Аминокислотные последовательности CDR1 VL антител показаны в SEQ ID NO: 13, 33, 53 и 73. Аминокислотные последовательности CDR2 VL антител показаны в SEQ ID NO: 14, 34, 54 и 74. Аминокислотные последовательности CDR3 VL антител показаны в SEQ ID NO: 15, 35, 55 и 75.
Альтернативно, как определено с использованием системы IMGT, аминокислотные последовательности CDR1 VH антител показаны в SEQ ID NO: 264 и 269. Аминокислотные последовательности CDR2 VH антител показаны в SEQ ID NO: 265 и 270. Аминокислотные последовательности CDR VH3 антител показаны в SEQ ID NO: 266 и 271. Аминокислотные последовательности CDR1 VL антител показаны в SEQ ID NO: 267 и 272. Аминокислотные последовательности CDR2 VL антител показаны в SEQ ID NO: 17, 273 и 57. Аминокислотные последовательности CDR3 VL антител показаны в SEQ ID NO: 15, 35, 55 и 75.
Учитывая, что каждое из указанных антител может связываться с β-клото, и что антигенсвязывающая специфичность обеспечивается в основном областями CDR1, 2 и 3, последовательности CDR1, 2 и 3 VH и последовательности CDR1, 2 и 3 VL могут быть «смешаны и сопоставлены» (то есть CDR из разных антител могут быть смешаны и сопоставлены, хотя каждое антитело предпочтительно содержит CDR1, 2 и 3 VH и CDR1, 2 и 3 VL для создания других связывающих β-клото веществ по изобретению. Такие «смешанные и сопоставленные» связывающие β-клото антитела могут быть исследованы с использованием анализов связывания, известных в данной области и описанных в примерах (например, анализы связывания ELISA, SET, Biacore®). Когда последовательности CDR VH смешивают и сопоставляют, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VH должна быть заменена структурно подобной(ыми) последовательностью(ями) CDR. Подобным образом, когда последовательности CDR VL смешивают и сопоставляют, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VL должна быть заменена структурно подобной(ыми) последовательностью(ями) CDR. Специалисту в данной области техники будет очевидно, что новые последовательности VH и VL могут быть созданы путем замещения одной или нескольких последовательностей CDR областей VH и/или VL на структурно подобные последовательности из последовательностей CDR, показанных в настоящем документе для моноклональных антител по настоящему изобретению. В дополнение к вышесказанному, в одном варианте осуществления антигенсвязывающие фрагменты антител, описанные в настоящем документе, могут содержать CDR1, 2, и 3 VH или CDR 1, 2, и 3 VL, где фрагмент связывается с β-клото в виде единственного вариабельного домена.
В некоторых вариантах осуществления изобретения антитела или их антигенсвязывающие фрагменты могут иметь последовательности тяжелых и легких цепей Fabs, описанные в таблице 1. Более конкретно, антитело или его антигенсвязывающие фрагменты могут иметь последовательность тяжелых и легких Fab NOV001, NOV002, NOV003, NOV004.
В некоторых вариантах осуществления изобретения антитело или антигенсвязывающий фрагмент, которые специфически связывают β-клото, содержат CDR1 вариабельной области тяжелой цепи, CDR2 и CDR3 Fab NOV001, NOV002, NOV003 или NOV004, и CDR1 вариабельной области легкой цепи, CDR2 и CDR3 Fab NOV001, NOV002, NOV003 или NOV004, например, представленные в таблице 1.
В других вариантах осуществления изобретения антитело или антигенсвязывающий фрагмент, которые специфически связывают β-клото, содержат CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные с помощью Kabat и описанные в таблице 1. В еще других вариантах осуществления изобретения антитело или антигенсвязывающий фрагмент, которые специфически связывают β-клото, содержат CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные с помощью Chothia и описанные в таблице 1. В еще других вариантах осуществления изобретения антитело или антигенсвязывающий фрагмент, которые специфически связывают β-клото, содержат CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные с помощью объединенных Kabat и Chothia и описанные в таблице 1. В еще других вариантах осуществления по изобретению антитело или антигенсвязывающий фрагмент, которые специфически связывают β-клото, содержат CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные с помощью IMGT и описанные в таблице 1.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 3; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 4; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 5; CDR1 вариабельной области легкой цепи с SEQ ID NO: 13; CDR2 вариабельной области легкой цепи с SEQ ID NO: 14; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 15.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 23; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 24; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 25; CDR1 вариабельной области легкой цепи с SEQ ID NO: 33; CDR2 вариабельной области легкой цепи с SEQ ID NO: 34; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 35.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 43; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 44; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 45; CDR1 вариабельной области легкой цепи с SEQ ID NO: 53; CDR2 вариабельной области легкой цепи с SEQ ID NO: 54; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 55.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 63; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 64; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 65; CDR1 вариабельной области легкой цепи с SEQ ID NO: 73; CDR2 вариабельной области легкой цепи с SEQ ID NO: 74; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 75.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 6; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 7; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 8; CDR1 вариабельной области легкой цепи с SEQ ID NO: 16; CDR2 вариабельной области легкой цепи с SEQ ID NO: 17; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 18.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 26; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 27; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 28; CDR1 вариабельной области легкой цепи с SEQ ID NO: 36; CDR2 вариабельной области легкой цепи с SEQ ID NO: 37; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 38.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 46; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 47; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 48; CDR1 вариабельной области легкой цепи с SEQ ID NO: 56; CDR2 вариабельной области легкой цепи с SEQ ID NO: 57; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 58.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 66; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 67; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 68; CDR1 вариабельной области легкой цепи с SEQ ID NO: 76; CDR2 вариабельной области легкой цепи с SEQ ID NO: 77; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 78.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 263; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 4; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 5; CDR1 вариабельной области легкой цепи с SEQ ID NO: 13; CDR2 вариабельной области легкой цепи с SEQ ID NO: 14; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 15.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 268; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 24; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 25; CDR1 вариабельной области легкой цепи с SEQ ID NO: 33; CDR2 вариабельной области легкой цепи с SEQ ID NO: 34; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 35.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 263; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 44; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 45; CDR1 вариабельной области легкой цепи с SEQ ID NO: 53; CDR2 вариабельной области легкой цепи с SEQ ID NO: 54; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 55.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 268; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 64; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 25; CDR1 вариабельной области легкой цепи с SEQ ID NO: 73; CDR2 вариабельной области легкой цепи с SEQ ID NO: 74; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 75.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 264; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 265; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 266; CDR1 вариабельной области легкой цепи с SEQ ID NO: 267; CDR2 вариабельной области легкой цепи с SEQ ID NO: 17; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 15.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 269; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 270; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 271; CDR1 вариабельной области легкой цепи с SEQ ID NO: 272; CDR2 вариабельной области легкой цепи с SEQ ID NO: 273; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 35.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 264; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 265; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 266; CDR1 вариабельной области легкой цепи с SEQ ID NO: 267; CDR2 вариабельной области легкой цепи с SEQ ID NO: 57; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 55.
В конкретном варианте осуществления настоящее изобретение включает антитело, которое специфически связывается с β-клото, содержащее CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 269; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 270; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 271; CDR1 вариабельной области легкой цепи с SEQ ID NO: 272; CDR2 вариабельной области легкой цепи с SEQ ID NO: 273; и CDR3 вариабельной области легкой цепи с SEQ ID NO: 75.
В некоторых вариантах осуществления настоящее изобретение включает антитела или антигенсвязывающие фрагменты, которые специфически связываются с β-клото, описанные в таблице 1. В предпочтительном варианте осуществления антитело или антигенсвязывающий фрагмент, которые связывают β-клото и активируют рецепторный комплекс FGF21, представляют собой Fab NOV001, NOV002, NOV003, NOV004.
Как используется в настоящем документе, антитело человека содержит вариабельные области тяжелой или легкой цепи или полноразмерные тяжелые или легкие цепи, которые являются «продуктом» или «получены из» конкретной последовательности зародышевой линии, если вариабельные области или полноразмерные цепи антитела получены из системы, в которой используются гены иммуноглобулина зародышевой линии человека. Такие системы включают в себя иммунизацию трансгенной мыши, несущей гены иммуноглобулина человека, с представляющим интерес антигеном или скрининг библиотеки генов иммуноглобулина человека, отображаемой на фаге с представляющим интерес антигеном. Антитело человека, которое является «продуктом» или «производным» последовательности иммуноглобулина зародышевой линии человека, можно идентифицировать как таковое, сравнивая аминокислотную последовательность антитела человека с аминокислотными последовательностями иммуноглобулинов зародышевой линии человека и выбирая последовательность иммуноглобулина зародышевой линии человека, которая является наиболее близкой последовательностью (то есть с наибольшим % идентичности) к последовательности антитела человека.
Антитело человека, которое является «продуктом» или «производным» определенной последовательности иммуноглобулина зародышевой линии человека, может обнаруживать аминокислотные отличия по сравнению с последовательностью зародышевой линии, например, из-за природных соматических мутаций или преднамеренного введения ориентированных на сайт мутаций. Однако в каркасных участках VH или VL выбранное антитело человека в аминокислотной последовательности обычно по меньшей мере на 90% идентично аминокислотной последовательности, кодируемой геном иммуноглобулина человека, и содержит аминокислотные остатки, которые идентифицируют антитело человека как человеческое по сравнению с аминокислотными последовательностями иммуноглобулина зародышевой линии других видов (например, последовательности зародышевой линии мышей). В некоторых случаях антитело человека в аминокислотной последовательности может быть по меньшей мере на 60%, 70%, 80%, 90% или по меньшей мере на 95% или даже по меньшей мере на 96%, 97%, 98% или 99% идентично аминокислотной последовательности, кодируемой геном иммуноглобулина зародышевой линии.
Обычно рекомбинантное антитело человека будет обнаруживать не более 10 аминокислотных отличий от аминокислотной последовательности, кодируемой геном иммуноглобулина зародышевой линии человека в каркасных участках VH или VL. В некоторых случаях антитело человека может обнаруживать не более 5 или даже не более 4, 3, 2 или 1 аминокислотных отличий от аминокислотной последовательности, кодируемой геном иммуноглобулина зародышевой линии. Примеры генов иммуноглобулина зародышевой линии человека включают, но ими не ограничиваются, фрагменты вариабельного домена в зародышевой линии, описанные далее, а также DP47 и DPK9.
Гомологичные антитела
В еще одном варианте осуществления настоящее изобретение относится к антителу, или его антигенсвязывающему фрагменту, содержащим аминокислотные последовательности, которые являются гомологичными по отношению к последовательностям, описанным в таблице 1, и антитело связывается с белком β-клото (например, β-клото человека и яванского макака) и сохраняет желаемые функциональные свойства тех антител, которые описаны в таблице 1.
Например, настоящее изобретение относится к выделенному антителу или его функциональному антигенсвязывающему фрагменту, содержащим вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 9, 29, 49 или 69; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 19, 39, 59 или 79; и антитело специфически связывается с β-клото (например, β-клото человека и яванского макака). В некоторых аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat, например, SEQ ID NO: 3, 4, 5, 13, 14 и 15, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Chothia, например, SEQ ID NO: 6, 7, 8, 16, 17 и 18, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Combined, например, SEQ ID NO: 263, 4, 5, 13, 14 и 15, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью IMGT, например, SEQ ID NO: 264, 265, 266, 267, 17 и 15, соответственно.
Например, настоящее изобретение относится к выделенному антителу или его функциональному антигенсвязывающему фрагменту, содержащим вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 9; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 19; и антитело специфически связывается с β-клото (например, β-клото человека и яванского макака). В некоторых аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat, например, SEQ ID NO: 3, 4, 5, 13, 14 и 15, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Chothia, например, SEQ ID NO: 6, 7, 8, 16, 17 и 18, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Combined, например, SEQ ID NO: 263, 4, 5, 13, 14 и 15, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью IMGT, например, SEQ ID NO: 264, 265, 266, 267, 17 и 15, соответственно.
Например, настоящее изобретение относится к выделенному антителу или его функциональному антигенсвязывающему фрагменту, содержащим вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 29; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 39; и антитело специфически связывается с β-клото (например, β-клото человека и яванского макака). В некоторых аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat, например, SEQ ID NO: 23, 24, 25, 33, 34 и 35, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Chothia, например, SEQ ID NO: 26, 27, 28, 36, 37 и 38, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Combined, например, SEQ ID NO: 268, 24, 25, 33, 34 и 35, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью IMGT, например, SEQ ID NO: 269, 270, 271, 272, 273 и 35, соответственно.
Например, настоящее изобретение относится к выделенному антителу или его функциональному антигенсвязывающему фрагменту, содержащим вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 49; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 59; и антитело специфически связывается с β-клото (например, β-клото человека и яванского макака). В некоторых аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat, например, SEQ ID NO: 43, 44, 45, 53, 54 и 55, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Chothia, например, SEQ ID NO: 46, 47, 48, 56, 57 и 58, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Combined, например, SEQ ID NO: 263, 44, 45, 53, 54 и 55, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью IMGT, например, SEQ ID NO: 264, 265, 266, 267, 57 и 55, соответственно.
Например, настоящее изобретение относится к выделенному антителу или его функциональному антигенсвязывающему фрагменту, содержащим вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 69; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, 95%, 96%, 97%, 98% или по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 79; и антитело специфически связывается с β-клото (например, β-клото человека и яванского макака). В некоторых аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Kabat, например, SEQ ID NO: 63, 64, 65, 73, 74 и 75, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Chothia, например, SEQ ID NO: 66, 67, 68, 76, 77 и 78, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью Combined, например, SEQ ID NO: 268, 64, 25, 73, 74 и 75, соответственно. В некоторых других аспектах изобретения последовательности тяжелых и легких цепей дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные с помощью IMGT, например, SEQ ID NO: 269, 270, 271, 272, 273 и 75, соответственно.
В других вариантах осуществления аминокислотные последовательности VH и/или VL могут быть на 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, представленным в таблице 1. В других вариантах осуществления аминокислотные последовательности VH и/или VL могут быть идентичны, за исключением аминокислотной замены в не более чем 1, 2, 3, 4 или 5 аминокислотных положениях. Антитело, имеющее области VH и VL, имеющие высокую (то есть 80% или больше) идентичность к тем областям VH и VL, которые описаны в таблице 1, могут быть получены путем мутагенеза (например, сайт-направленный или ПЦР-опосредованный мутагенез) молекул нуклеиновых кислот, кодирующих SEQ ID NO: 10, 30, 50 или 70 и SEQ ID NO: 20, 40, 60 или 80, соответственно, с последующим исследованием кодированного измененного антитела на сохранение функций с использованием функциональных анализов, описанных в настоящем документе.
В других вариантах осуществления аминокислотные последовательности полноразмерной тяжелой цепи и/или полноразмерной легкой цепи могут быть на 50% 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, представленным в таблице 1. Антитело, имеющее полноразмерную тяжелую цепь и полноразмерная легкую цепь, имеющие высокую (то есть 80% или больше) идентичность к полноразмерным тяжелым цепям какой-либо SEQ ID NO: 9, 29, 49 или 69, и полноразмерным легким цепям какой-либо SEQ ID NO: 19, 39, 59 или 79, могут быть получены путем мутагенеза (например, сайт-направленный или ПЦР-опосредованный мутагенез) молекул нуклеиновых кислот, кодирующих такие полипептиды, с последующим исследованием кодированного измененного антитела на сохранение функций с использованием функциональных анализов, описанных в настоящем документе. Антитело, имеющее полноразмерную тяжелую цепь и полноразмерную легкую цепь, имеющее высокую (то есть 80% или больше) идентичность к полноразмерным тяжелым цепям какой-либо SEQ ID NO: 11, 31, 51 или 71, и полноразмерным легким цепям какой-либо SEQ ID NO: 21, 41, 61 или 81, могут быть получены путем мутагенеза (например, сайт-направленный или ПЦР-опосредованный мутагенез) молекул нуклеиновых кислот, кодирующих такие полипептиды, с последующим исследованием кодированного измененного антитела на сохранение функций с использованием функциональных анализов, описанных в настоящем документе.
В других вариантах осуществления нуклеотидные последовательности полноразмерной тяжелой цепи и/или полноразмерной легкой цепи могут быть на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, представленным в таблице 1.
В других вариантах осуществления нуклеотидные последовательности вариабельных областей тяжелой цепи и/или вариабельных областей легкой цепи могут быть на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, представленным в таблице 1.
В некоторых вариантах осуществлений тяжелая цепь и/или легкая цепь CDR могут быть на 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, представленным в таблице 1.
Как используется в настоящем документе, процент идентичности между двумя последовательностями является функцией количества идентичных позиций, разделяемых последовательностями (то есть, % идентичности равен число одинаковых позиций/общее число позиций×100), принимая во внимание количество пробелов и размер каждого пробела, которые необходимо ввести для оптимального выравнивания двух последовательностей. Сравнение последовательностей и определение процентной идентичности между двумя последовательностями может быть выполнено с использованием математического алгоритма, как описано в приведенных далее неограничивающих примерах.
Дополнительно или альтернативно белковые последовательности по настоящему изобретению могут, кроме того, использоваться в качестве «запрашиваемой последовательности» для выполнения поиска по открытым базам данных, например, для идентификации связанных последовательностей. Например, такие поиски могут выполняться с использованием программы BLAST (версия 2.0) Altschul, et al., 1990 J.Mol. Biol. 215:403-10.
Антитела с консервативными модификациями
В некоторых вариантах осуществления антитело по изобретению имеет вариабельный домен тяжелой цепи, содержащий последовательности CDR1, CDR2 и CDR3, и вариабельный домен легкой цепи, содержащий последовательности CDR1, CDR2 и CDR3, где одна или несколько этих последовательностей CDR имеют специфические аминокислотные последовательности на основе антител, описанных в настоящем документе, или их консервативные модификации, и где антитела сохраняют желаемые функциональные свойства связывающих β-клото антител по изобретению.
Соответственно, настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, состоящим из вариабельной области тяжелой цепи, содержащей последовательности CDR1, CDR2 и CDR3, и вариабельной области легкой цепи, содержащей последовательности CDR1, CDR2 и CDR3, где аминокислотные последовательности вариабельной области тяжелой цепи CDR1 выбраны из группы, состоящей из SEQ ID NO: 3, 23, 43 и 63 и их консервативных модификаций; аминокислотные последовательности вариабельной области тяжелой цепи CDR2 выбраны из группы, состоящей из SEQ ID NO: 4, 24, 44, и 64 и их консервативных модификаций; аминокислотные последовательности вариабельной области тяжелой цепи CDR3 выбраны из группы, состоящей из SEQ ID NO: 5, 25, 45 и 65 и их консервативных модификаций; аминокислотные последовательности вариабельной области легкой цепи CDR1 выбраны из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73 и их консервативных модификаций; аминокислотные последовательности вариабельной области легкой цепи CDR2 выбраны из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74 и их консервативных модификаций; аминокислотные последовательности вариабельной области легкой цепи CDR3 выбраны из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75 и их консервативных модификаций; и антитело или его антигенсвязывающие фрагменты специфически связываются с β-клото.
Соответственно, настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, состоящим из вариабельной области тяжелой цепи, содержащей последовательности CDR1, CDR2 и CDR3, и вариабельной области легкой цепи, содержащий последовательности CDR1, CDR2 и CDR3, где аминокислотные последовательности вариабельной области тяжелой цепи CDR1 выбраны из группы, состоящей из SEQ ID NO: 263 и 268 и их консервативных модификаций; аминокислотные последовательности вариабельной области тяжелой цепи CDR2 выбраны из группы, состоящей из SEQ ID NO: 4, 24, 44 и 64 и их консервативных модификаций; аминокислотные последовательности вариабельной области тяжелой цепи CDR3 выбраны из группы, состоящей из SEQ ID NO: 5, 25 и 45 и их консервативных модификаций; аминокислотные последовательности вариабельной области легкой цепи CDR1 выбраны из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73 и их консервативных модификаций; аминокислотные последовательности вариабельной области легкой цепи CDR2 выбраны из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74 и их консервативных модификаций; аминокислотные последовательности вариабельной области легкой цепи CDR3 выбраны из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75 и их консервативных модификаций; и антитело или его антигенсвязывающие фрагменты специфически связываются с β-клото.
В других вариантах осуществления антитело по изобретению оптимизировано для экспрессии в клетке млекопитающего, имеет последовательность полноразмерной тяжелой цепи и последовательность полноразмерной легкой цепи, где одна или несколько из указанных последовательности имеют специфические аминокислотные последовательности, основанных на антителах, описанных в настоящем документе, или их консервативных модификациях, и где антитела сохраняют желаемые функциональные свойства связывающих β-клото антител по изобретению. Соответственно, настоящее изобретение относится к выделенному антителу, оптимизированному для экспрессии в клетке млекопитающего, состоящему из полноразмерной тяжелой цепи и полноразмерной легкой цепи, где полноразмерная тяжелая цепь имеет аминокислотные последовательности, выбранные из группы SEQ ID NO: 11, 31, 51 или 71 и их консервативных модификаций; и полноразмерная легкая цепь имеет аминокислотные последовательности, выбранные из группы SEQ ID NO: 21, 41, 61, или 81 и их консервативных модификаций; и антитело специфически связывается с β-клото (например, β-клото человека и яванского макака).
Антитела, которые связываются с одним и тем же эпитопом
Настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающие β-клото антитела, описанные в таблице 1 (например, NOV001, NOV002, NOV003 или NOV004). В конкретном аспекте такие антитела и антигенсвязывающие фрагменты способны увеличивать активность β-клото и FGFR1c. Следовательно, могут быть идентифицированы дополнительные антитела по их способности конкурировать (например, конкурентно ингибировать связывание статистически значимым образом) с другими антителами по изобретению в анализах связывания с β-клото (например, описанных в примерах). Способность исследуемого антитела ингибировать связывание антител по настоящему изобретению с β-клото указывает на то, что исследуемое антитело может конкурировать с этим антителом за связывание с β-клото; такое антитело может, в соответствии с неограничивающей теорией, связываться с тем же или соответствующим (например, структурно подобным или пространственно проксимальным) эпитопом на белке β-клото в качестве антитела, с которым он конкурирует. В конкретном варианте осуществления антитело, которое связывается с одним и тем же эпитопом на β-клото в качестве антител по настоящему изобретению, представляет собой моноклональное антитело человека. Такие моноклональные антитела человека могут быть получены и выделены, как описано в настоящем документе. Как используется в настоящем документе, антитело «конкурирует» за связывание, когда конкурирующее антитело ингибирует связывание β-клото антителом или антигенсвязывающим фрагментом по изобретению более чем на 50% (например, на 80%, на 85%, на 90%, на 95%, на 98% или на 99%) в присутствии эквимолярной концентрации конкурирующего антитела. В конкретном варианте осуществления антитело, которое связывается с одним и тем же эпитопом на β-клото в качестве антител по настоящему изобретению, представляет собой моноклональное антитело человека. Такие моноклональные антитела человека могут быть получены и выделены, как описано в настоящем документе.
В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело NOV001. В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело NOV002. В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело NOV003. В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело NOV004.
В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело, которое содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 9, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 19.
В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело, которое содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 29, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 39.
В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело, которое содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 49, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 59.
В конкретном аспекте настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и связывающее β-клото антитело, которое содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 69, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 79.
В конкретных аспектах в настоящем документе предложены выделенные антитела или их антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где эпитоп содержит одну или несколько SEQ ID NO, показанных в таблице 2. В конкретном аспекте такие антитела и антигенсвязывающие фрагменты способны увеличить активность β-клото и FGFR1c.
В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит один или несколько аминокислотных остатков 246-265 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит один или несколько аминокислотных остатков 536-550 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит один или несколько аминокислотных остатков 834-857 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит один или несколько аминокислотных остатков 959-986 последовательности β-клото (SEQ ID NO: 262).
В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит один или несколько аминокислотных остатков 246-265, 536-550, 834-857 и 959-986 последовательности β-клото (SEQ ID NO: 262). В конкретных аспектах в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит два или более аминокислотных остатков 246-265, 536-550, 834-857 и 959-986 последовательности β-клото (SEQ ID NO: 262). В конкретных аспектах в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит три или более аминокислотных остатков 246-265, 536-550, 834-857 и 959-986 последовательности β-клото (SEQ ID NO: 262). В конкретных аспектах в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где указанный эпитоп содержит аминокислотные остатки 246-265, 536-550, 834-857 и 959-986 последовательности β-клото (SEQ ID NO: 262).
В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с одним или несколькими эпитопами β-клото, где указанные эпитопы содержат один или несколько аминокислотных остатков 646-670 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с одним или несколькими эпитопами β-клото, где указанные эпитопы содержат один или несколько аминокислотных остатков 696-700 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с одним или несколькими эпитопами β-клото, где указанные эпитопы содержат один или несколько аминокислотных остатков 646-689 последовательности β-клото (SEQ ID NO: 262).
В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с одним или несколькими эпитопами β-клото, где указанные эпитопы содержат один, или два, или три, или четыре, или пять, или более аминокислотных остатков 646-670, 696-700 и 646-689 последовательности β-клото (SEQ ID NO: 262). В некотором аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с двумя или более эпитопами β-клото, где указанные эпитопы содержат один или несколько аминокислотных остатков 646-670, 696-700 и 646-689 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с тремя или более эпитопами β-клото, где указанные эпитопы содержат один или несколько аминокислотных остатков 646-670, 696-700 и 646-689 последовательности β-клото (SEQ ID NO: 262). В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые связываются с тремя или более эпитопами β-клото, где указанные эпитопы содержат аминокислотные остатки 646-670, 696-700 и 646-689 последовательности β-клото (SEQ ID NO: 262).
В конкретных аспектах в настоящем документе предложены выделенные антитела или их антигенсвязывающий фрагмент, которые связываются с эпитопом β-клото, где эпитоп содержит одну или несколько SEQ ID NO, показанных в таблице 2, где указанные антитела и антигенсвязывающие фрагменты способны увеличить активность β-клото и FGFR1c, и где указанные антитела и антигенсвязывающие фрагменты способны защищать путем водородно-дейтериевого обмена (HDx) один или несколько пептидов β-клото, что характеризуются изменением включения дейтерия в диапазоне от -0,5 до -2,1, например как указано в таблице 2.
В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые защищают, как определено водородно-дейтериевым обменом (HDx), один, два, три, четыре, пять или более следующих пептидов β-клото (SEQ ID NO: 262): 245-266, 246-265, 343-349, 344-349, 421-429, 488-498, 509-524, 536-550, 568-576, 646-669, 646-670, 696-700, 773-804, 834-857 и 959-986 aa.
В конкретном аспекте в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые увеличивают активность β-клото и FGFR1c, где антитело или его антигенсвязывающий фрагмент защищают, как определено водородно-дейтериевым обменом (HDx), один, два, три, четыре, пять или более пептидов из следующих, указанных в таблице 2: SEQ ID NO: 109, 110, 111, 112, 113, 125, 126, 127, 128, 129, 141, 142, 143, 156, 157, 158, 159, 160, 161, 163, 164, 165, 167, 168, 169, 170, 171, 172, 184, 185, 186, 187, 188, 195, 196, 197, 198, 204, 212, 213, 214, 215, 216, 217, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 256, 257, 258, 259, 260 и 261.
В некоторых аспектах, в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые увеличивают активность β-клото и FGFR1c, где антитело или его антигенсвязывающий фрагмент защищают, как определено водородно-дейтериевым обменом (HDx), шесть, семь, восемь, девять, десять или более пептидов из следующих, указанных в таблице 2: SEQ ID NO: 109, 110, 111, 112, 113, 125, 126, 127, 128, 129, 141, 142, 143, 156, 157, 158, 159, 160, 161, 163, 164, 165, 167, 168, 169, 170, 171, 172, 184, 185, 186, 187, 188, 195, 196, 197, 198, 204, 212, 213, 214, 215, 216, 217, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 256, 257, 258, 259, 260 и 261.
В некоторых аспектах, в настоящем документе предложено выделенное антитело или его антигенсвязывающий фрагмент, которые увеличивают активность β-клото и FGFR1c, где антитело или его антигенсвязывающий фрагмент не содержат остатков 701 (Tyr) или 703 (Arg) β-клото человека (SEQ ID NO: 262).
Сконструированные и модифицированные антитела
Антитело по изобретению, кроме того, может быть получено с использованием антитела, имеющего одну или несколько последовательностей VH и/или VL, показанных в настоящем документе в качестве исходного материала для конструирования модифицированного антитела, причем модифицированное антитело может обладать измененными свойствами по сравнению с исходным антителом. Антитело может быть сконструировано путем модификации одного или нескольких остатков в одной или обеих вариабельных областях (то есть VH и /или VL), например, в пределах одной или нескольких областей CDR и/или в пределах одного или нескольких каркасных участков. Дополнительно или альтернативно, антитело может быть сконструировано путем модификации остатков пределах константной(ых) области(ей), например, c изменением эффекторной(ых) функции(ий) антитела.
Одним из видов конструирования вариабельных областей, который может быть осуществлен, является прививка CDR. Антитела взаимодействуют с мишеневыми антигенами преимущественно через аминокислотные остатки, которые расположены в шести областях, определяющих комплементарность, тяжелых и легких цепей (CDR). По этой причине аминокислотные последовательности в CDR отличаются сильнее среди разных антител, чем последовательности вне CDR. Поскольку последовательности CDR ответственны за большинство взаимодействий антитело-антиген, рекомбинантные антитела, которые копируют свойства специфических природных антител, могут быть экспрессированы путем конструирования векторов экспрессии, которые включают последовательности CDR из конкретного природного антитела, привитого на последовательности каркасных участков из другого антитела с отличными свойствами (смотри, например, Riechmann, L. et al., 1998 Nature 332:323-327; Jones, P. et al., 1986 Nature 321:522-525; Queen, C. et al., 1989 Proc. Natl. Acad., USA 86: 10029-10033; патент США № 5225539, Winter, и патенты США №№ 5530101, 5585089, 5693762 и 6180370, Queen et al.).
Соответственно, другой вариант осуществления изобретения относится к выделенному антителу или его антигенсвязывающему фрагменту, содержащим вариабельную область тяжелой цепи, содержащей последовательности CDR1, имеющие аминокислотную последовательность, выбранную из последовательностей HCDR1, представленных в таблице 1; последовательности CDR2, имеющие аминокислотную последовательность, выбранную из последовательностей HCDR2, представленных в таблице 1; последовательности CDR3, имеющие аминокислотную последовательность, выбранную из последовательностей HCDR3, представленных в таблице 1; и вариабельную область легкой цепи, содержащей последовательности CDR1, имеющие аминокислотную последовательность, выбранную из последовательностей LCDR1, представленных в таблице 1; последовательности CDR2, имеющие аминокислотную последовательность, выбранную из последовательностей LCDR2, представленных в таблице 1; и последовательности CDR3, состоящие из аминокислотной последовательности, выбранной из последовательностей LCDR3, представленных в таблице 1. Таким образом, такие антитела содержат последовательности CDR VH и VL моноклональных антител, но могут содержать различные каркасные последовательности из этих антител.
Соответственно, другой вариант осуществления изобретения относится к выделенному антителу или его антигенсвязывающему фрагменту, содержащим вариабельную область тяжелой цепи, содержащей последовательности CDR1, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, 23, 43 и 63; последовательности CDR2, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 24, 44 и 64; последовательности CDR3, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 25, 45 и 65, соответственно; и вариабельную область легкой цепи, содержащей последовательности CDR1, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73; последовательности CDR2, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74; и последовательности CDR3, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75, соответственно. Таким образом, такие антитела содержат последовательности CDR VH и VL моноклональных антител, но могут содержать различные каркасные последовательности из этих антител.
Такие каркасные последовательности могут быть получены из публичных баз данных ДНК или опубликованных ссылок, которые включают последовательности генов антител в клетках зародышевой линии. Например, последовательности ДНК зародышевой линии для генов вариабельной области тяжелой и легкой цепей человека могут быть найдены в базе данных последовательности зародышевой линии человека «VBase» (доступной во всемирной паутине по адресу mrc-cpe.cam.ac.uk/vbase), а также в работах Kabat, E. A., et al., 1991 Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242; Tomlinson, I. M., et al., 1992 J. Mol. Biol. 227:776-798; и Cox, J. P. L. et al., 1994 Eur. J Immunol. 24:827-836; содержание каждой из которых явно включено в настоящий документ посредством ссылки.
Примером последовательностей каркасных участков для использования в антителах по изобретению являются последовательности, которые структурно подобны последовательностям каркасных участков, используемых выбранными антителами по изобретению, например, консенсусным последовательностям и/или последовательностям каркасных участков, используемых моноклональными антителами по изобретению. Последовательности CDR1, 2 и 3 VH и последовательности CDL1, 2 и 3 VL могут быть привиты на каркасные участки, последовательность которых идентична последовательности гена иммуноглобулина зародышевой линии, из которой получают каркасную последовательность, или последовательности CDR могут быть привиты на каркасные участки, которые содержат одну или несколько мутаций по сравнению с последовательностями зародышевой линии. Например, было обнаружено, что в некоторых случаях мутирование остатков на каркасных участках дает преимущества поддержания или усиления антигенсвязывающей способности антитела (смотри, например, патенты США 5530101, 5585089, 5693762 и 6180370, Queen et al). Каркасные участки, которые могут быть использованы в качестве скаффолдов, на которых можно построить антитела и антигенсвязывающие фрагменты, описанные в настоящем документе, включают, но не ограничиваются ими, VH1A, VH1B, VH3, Vk1, Vl2 и Vk2. Дополнительные каркасные участки известны в данной области и могут быть найдены, например, в базе данных vBase во всемирной паутине на сайте vbase.mrc-cpe.cam.ac.uk/index.php?& MMN_position=1:1.
Соответственно, вариант осуществления изобретения относится к выделенным связывающим β-клото антителам или их антигенсвязывающим фрагментам, содержащим вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 29, 49 или 69, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей, и, кроме того, содержащую вариабельную область легкой цепи, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 19, 39, 59 или 79, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей.
Соответственно, вариант осуществления изобретения относится к выделенным связывающим β-клото антителам или их антигенсвязывающим фрагментам, содержащим вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 9, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей, и, кроме того, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 19, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей.
В конкретном аспекте вариант осуществления изобретения относится к выделенным связывающим β-клото антителам или их антигенсвязывающим фрагментам, содержащим вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 29, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей, и, кроме того, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 39, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей.
В конкретном аспекте вариант осуществления изобретения относится к выделенным связывающим β-клото антителам или их антигенсвязывающим фрагментам, содержащим вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 49, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей, и, кроме того, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 59, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей.
В конкретном аспекте вариант осуществления изобретения относится к выделенным связывающим β-клото антителам или их антигенсвязывающим фрагментам, содержащим вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 69, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей, и, кроме того, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 79, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок в каркасном участке таких последовательностей.
Другой тип модификации вариабельной области состоит в мутировании аминокислотных остатков в областях VH и/или CDR1 VL, CDR2 и/или CDR3, тем самым улучшая одно или несколько связывающих свойств (например, аффинность) интересующего антитела, известное как «созревание аффинности». Сайт-направленный мутагенез или ПЦР-опосредованный мутагенез могут быть выполнены на введение мутации(ий), и влияние на связывание антитела или другое представляющее интерес функциональное свойство может быть оценено в анализах in vitro или in vivo, описанных в настоящем документе и приведенных в примерах. Могут быть введены консервативные модификации (как обсуждалось выше). Мутации могут представлять собой аминокислотные замены, добавления или делеции. Кроме того, обычно изменяют не более одного, двух, трех, четырех или пяти остатков в области CDR.
Известно, что некоторые мотивы аминокислотных последовательностей подвергаются посттрансляционной модификации (ПТМ), такой как гликозилирование (то есть NxS/T, x любая аминокислота, кроме P), окисление свободных цистеинов, дезамидирование (например, NG) или изомеризация (например, DG). Если они присутствуют в регионах CDR, то эти мотивы идеально удаляются с помощью сайт-направленного мутагенеза с повышением гомогенности продукта.
Процесс созревания аффинности подробно описан в данной области. Среди большого числа систем дисплея фаговый дисплей (Smith GP (1985) Science 228: 1315-1317) и дисплей на эукариотических клетках, таких как дрожжи (Boder ET и Wittrup KD (1997) Nature Biotechnology 15: 553-557), по-видимому, являются наиболее часто используемыми системами для выбора реакции антитело-антиген. Преимущества этих систем дисплея состоят в том, что они подходят для широкого спектра антигенов, и что жесткость выбора может быть легко отрегулирована. В фаговом дисплее могут быть представлены фрагменты scFv или Fab, а в дрожжевых клетках полноразмерный IgG. Эти обычно используемые методы позволяют выбирать желаемые варианты антител из более крупных библиотек с разбросами более 10E7. Библиотеки с меньшим разнообразием, например 10E3, могут быть тестированы с помощью микроэкспрессии и ELISA.
Нецелевые или случайные библиотеки вариантов антител могут быть получены, например, с помощью ПЦР с внесением ошибок (Cadwell RC and Joyce GF (1994) Mutagenic PCR. Methods Appl. 3: S136-S140), и они обеспечивают очень простой, но иногда ограниченный подход. Другой стратегией является CDR направленная диверсификация кандидата антител. Одно или несколько положений в одном или более CDR могут быть специфическими целями с использованием, например, вырожденных олигонуклеотидов (Thompson J et al. (1996) J.Mol.Biol. 256: 77-88), тринуклеотидного мутагенеза (TRIM) (Kayushin AL et al. (1996) Nucleic Acids Res. 24: 3748-3755) или любого другого подхода, известного в данной области.
Соответственно, в другом варианте осуществления настоящее изобретение относится к выделенным связывающим β-клото антителам или их антигенсвязывающие фрагментам, состоящим из вариабельной области тяжелой цепи, имеющей область CDR1 VH, имеющую аминокислотную последовательность, выбранную из группы, имеющей SEQ ID NO: 3, 23, 43 и 63, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 3, 23, 43 и 63; область CDR2 VH, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 24, 44 и 64, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 4, 24, 44 и 64; область CDR VH3, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 25, 45 и 65, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 5, 25, 45 и 65; область CDR1 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 13, 33, 53 и 73; область CDR2 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 14, 34, 54 и 74; и область CDR3 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 15, 35, 55 и 75.
Соответственно, в другом варианте осуществления настоящее изобретение относится в выделенным связывающим β-клото антителам или их антигенсвязывающие фрагментам, состоящим из вариабельной области тяжелой цепи, имеющей область CDR1 VH, имеющую аминокислотную последовательность, выбранную из группы, имеющей SEQ ID NO: 6, 26, 46 и 66, или аминокислотную последовательность, имеющей одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 6, 26, 46 и 66; область CDR2 VH, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, 27, 47 и 67, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 7, 27 47, и 67; область CDR VH3, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 8, 28, 48 и 68, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 8, 28, 48 и 68; область CDR1 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящую из SEQ ID NO: 13, 33, 53 и 73, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 13, 33, 53 и 73; область CDR2 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 14, 34, 54 и 74; и область CDR3 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или вставок по сравнению с SEQ ID NO: 15, 35, 55 и 75.
Трансплантация антигенсвязывающих доменов в альтернативные каркасные участки или скаффолды
Широкое разнообразие каркасных участков антител/иммуноглобулина или скаффолдов может быть использовано при условии, что полученный полипептид содержит по меньшей мере одну связывающую область, которая специфически связывается с β-клото. Такие каркасные участки или скаффолды включают 5 основных идиотипов иммуноглобулинов человека или их фрагментов и включают иммуноглобулины других видов животных, предпочтительно, имеющих гуманизированные части. Особый интерес в этом отношении представляют антитела только с тяжелой цепью, такие были обнаружены у верблюдов. Специалисты в данной области продолжают открывать и разрабатывать новые каркасные участки, скаффолды и фрагменты.
В одном аспекте изобретение относится к получению антител не на основе иммуноглобулинов с использованием неиммуноглобулиновых скаффолдов, на которые могут быть трансплантированы CDR по изобретению. Могут быть использованы известные или еще не найденные неиммуноглобулиновые каркасные участки и скаффорды, при условии, что они содержат связывающую область, специфическую к целевому белку β-клото. Известные неиммуноглобулиновые каркасные участки или скаффорды включают, но ими не ограничиваются, фибронектин (Compound Therapeutics, Inc., Waltham, MA), анкирин (Molecular Partners AG, Zurich, Switzerland), доменные антитела (Domantis, Ltd., Cambridge, MA, и Ablynx nv, Zwijnaarde, Belgium), липокалин (Pieris Proteolab AG, Freising, Germany), небольшие модульные иммунофармацевтические препараты (Trubion Pharmaceuticals Inc., Сиэтл, WA), макситела (Avidia, Inc., Mountain View, CA), белок A (Affibody AG, Швеция) и аффилин (гамма-кристаллин или убиквитин) (Scil Proteins GmbH, Halle, Germany).
Фибронектиновые скаффолды основаны на домене фибронектина типа III (например, десятый модуль фибронектина типа III (домен 10 Fn3)). Домен фибронектина типа III имеет 7 или 8 бета-цепей, распределенных между двумя бета-листами, которые сами упаковываются друг против друга с образованием ядра белка и дополнительно содержат петли (аналогичные CDR), которые соединяют бета-цепи друг с другом и доступны воздействию растворителя. Существует по крайней мере три таких петли на каждом краю сэндвича с бета-листами, где край является границей белка перпендикулярно направлению бета-цепей (смотри, US 6818418). Эти скаффолды на основе фибронектина не являются иммуноглобулином, хотя общая укладка очень близка укладе самого маленького функционального фрагмента антитела, вариабельной области тяжелой цепи, которая содержит всю единицу распознавания антигена в IgG верблюда и ламы. Из-за такой структуры неиммуноглобулиновое антитело имитирует антигенсвязывающие свойства, которые сходны по своей природе и аффинности свойствам антител. Эти скаффолды могут использоваться в стратегии рандомизации петель и перестановки in vitro, которые аналогичны процессу созревания аффинности антител in vivo. Эти молекулы на основе фибронектина могут использоваться в качестве скаффолдов, где области петель молекулы могут быть заменены CDR по изобретению, используя стандартные методики клонирования.
Анкириновая методика основана на использовании белков с модулями повторов, выделенными из анкирина, в качестве скаффолдов, несущих вариабельные области, которые могут использоваться для связывания с различными мишенями. Анкириновый модуль повтора представляет собой полипептид длиной в 33 аминокислоты, состоящий из двух антипараллельных α-спиралей и β-витка. Связывание вариабельных областей обычно оптимизируют, используя рибосомный дисплей.
Авимеры получают из природного белка, содержащего A-домен, такого как LRP-1. Эти домены используются по своей природе для белок-белкового взаимодействия, и у человека более 250 белков основаны на A-доменах. Авимеры состоят из ряда различных «A-доменных» мономеров (2-10), связанных посредством аминокислотных линкеров. Можно создавать авимеры, которые могут связываться с мишеневым антигеном, используя методику, описанную, например, в публикациях патентных заявок США №№ 20040175756, 20050053973, 20050048512 и 20060008844.
Аффинные лиганды аффитела представляют собой низкомолекулярные простые белки, состоящие из трехспирального пучка на основе скаффолда одного из IgG-связывающих доменов белка A. Белок A представляет собой поверхностный белок бактерии Staphylococcus aureus. Этот домен скаффолда состоит из 58 аминокислот, 13 из которых рандомизированы с получением библиотек аффител с большим числом вариантов лиганда (смотри, например, US 5831012). Молекулы аффител копируют антитела, они имеют молекулярную массу 6 кДа в сравнении с молекулярной массой антител, которая составляет 150 кДа. Несмотря на свой небольшой размер, сайт связывания молекул аффител аналогичен сайту связывания антитела.
Антикалины представляют собой продукты, разработанные компанией Pieris ProteoLab AG. Их получают из липокалинов, широко распространенной группы низкомолекулярных и устойчивых белков, которые обычно участвуют в физиологическом транспорте или хранении химически чувствительных или нерастворимых соединений. Некоторые природные липокалины встречаются в тканях или жидкостях организма человека. Архитектура белка напоминает иммуноглобулины с гипервариабельными петлями в верхней части жесткого каркасного участка. Однако в отличие от антител или их рекомбинантных фрагментов липокалины состоят из одной полипептидной цепи с 160-180 аминокислотными остатками, которые лишь немного больше одного домена иммуноглобулина. Набор из четырех петель, которые образуют связывающий карман, демонстрирует выраженную структурную пластичность и допускает множество боковых цепей. Таким образом, сайт связывания может быть изменен в запатентованном процессе и распознавать заданные молекулы-мишени различной формы с высокой аффинностью и специфичностью. Один белок из семейства липокалинов, билин-связывающий белок (BBP) Pieris Brassicae, был использован для разработки антикалинов путем мутагенизации набора из четырех петель. Примером патентной заявки, описывающей антикалины, является публикация РСТ WO 199916873.
Молекулы аффилина представляют собой низкомолекулярные неиммуноглобулиновые белки, которые разработаны таким образом, что специфически связываются с белками и низкомолекулярными молекулам. Новые молекулы аффилина могут быть очень быстро выбраны из двух библиотек, каждая из которых основана на отличающемся скаффолд-белке человека. Молекулы аффилина не имеют никакой структурной гомологии с белками иммуноглобулина. В настоящее время используются два аффилиновых скаффолда, один из которых представляет собой гамма кристаллин, структурный белок хрусталика глаза человека, а другой представляет собой белки семейства «убиквитина». Оба скаффолда человека имеют низкий молекулярный вес, стабильны при высокой температуре и по существу устойчивы к изменениям рН и к денатурирующим агентам. Такая высокая стабильность обусловлена, главным образом, благодаря широкой структуре бета-листов белков. Примеры белков, полученных из гамма кристаллина, описаны в WO200104144, а примеры белков, подобных «убиквитину», описаны в WO2004106368.
Миметики эпитопов белков (PEM) представляют собой среднего размера циклические пептидоподобные молекулы (MW 1-2кДа) подобные вторичной структуре белков бета-шпильке, главной вторичной структуре, участвующей в «белок-белковых» взаимодействиях.
Настоящее изобретение относится к полностью человеческим антителам, которые специфически связываются с белком β-клото. По сравнению с химерными или гуманизированными антителами связывающие человеческие β-клото антитела по изобретению обладают дополнительной сниженной антигенностью при введении индивидам.
Верблюжьи антитела
Белки антител, полученные из членов семейства верблюдов и дромадеров (Camelus bactrianus и Calelus dromaderius), включая новые члены, например, виды лам (Lama paccos, Lama glama и Lama vicugna), были охарактеризованы по размеру, структурной сложности и антигенности в отношении людей. Было обнаружено, что некоторые антитела IgG из этого семейства млекопитающих по своей природе лишены легких цепей и, следовательно, структурно отличаются от типичной четырехцепочечной четвертичной структуры, имеющей две тяжелые и две легкие цепи, антител других животных. Смотри PCT/EP93/02214 (WO 94/04678, опубликована 3 марта 1994).
Область верблюжьего антитела, которая является низкомолекулярным единичным вариабельным доменом, идентифицированным как VHH, может быть получена с помощью генной инженерии с получением низкомолекулярного белка, обладающего высокой аффинностью в отношении мишени, что дает низкомолекулярный белок, полученный из антитела, известный как «верблюжье нанотело». Смотри патент США № 5759808, выдан 2 июня 1998; смотри также Stijlemans, B. et al., 2004 J Biol Chem 279: 1256-1261; Dumoulin, M. et al., 2003 Nature 424: 783-788; Pleschberger, M. et al. 2003 Bioconjugate Chem 14: 440-448; Cortez-Retamozo, V. et al. 2002 Int J Cancer 89: 456-62; и Lauwereys, M. et al. 1998 EMBO J 17: 3512-3520. Полученные методами генной инженерии библиотеки верблюжьих антител и фрагментов антител являются коммерчески доступными, например, от компании Ablynx, Ghent, Belgium. Как и в случае с другими антителами нечеловеческого происхождения, аминокислотная последовательность верблюжьего антитела может быть рекомбинантно изменена с получением последовательности, которая наиболее близка к последовательности человека, то есть нанотела могут быть «гуманизированы». Таким образом, природная низкая антигенность верблюжьих антител в отношении людей может быть дополнительно снижена.
Верблюжьи нанотела имеют молекулярную массу, равную примерно одной десятой молекулы IgG человека, и белок имеет физический диаметр всего в несколько нанометров. Одним из следствий низкомолекулярного размера является способность верблюжьих нанотел связываться с антигенными сайтами, которые функционально невидимы для более крупных белков антител, то есть верблюжьи нанотела могут быть использованы в качестве реагентов, обнаруживающих антигены, которые в противном случае являются скрытыми, используя классические иммунологические методики, и, насколько это возможно, в качестве терапевтических средств. Таким образом, еще одним следствием низкомолекулярного размера является способность верблюжьих нанотел ингибировать в результате связывания со специфическим сайтом в бороздке или узкой щели мишеневого белка и, следовательно, они могут выполнять свои функции ближе к функциям классического низкомолекулярного лекарственного средства, чем классического антитела.
Низкий молекулярный вес и компактный размер также приводят к тому, что верблюжьи нанотела являются чрезвычайно термостабильными, устойчивыми к экстремальным значениям рН и к действию протеолитических ферментов, а также являются слабо антигенными. Еще одним следствием этого является то, что верблюжьи нанотела легко перемещаются из системы кровообращения в ткани и даже пересекают гематоэнцефалический барьер и могут лечить расстройства, поражающие нервную ткань. Нанотела могут дополнительно облегчить транспортировку лекарственных средств через гематоэнцефалический барьер. Смотри патентную заявку США 20040161738, опубликованную 19 августа 2004 года. Эти особенности в сочетании с низкой антигенностью в отношении людей указывают на большой терапевтический потенциал. Кроме того, эти молекулы могут быть полностью экспрессированы в прокариотических клетках, таких как E. coli, и они экспрессируются в виде слитых белков с бактериофагом и являются функциональными.
Соответственно, отличительной особенностью настоящего изобретения является верблюжье антитело или нанотело, обладающие высокой аффинностью к β-клото. В некоторых вариантах осуществления настоящего изобретения верблюжье антитело или нанотело продуцируется естественным образом у верблюда, то есть продуцируется верблюдом после иммунизации β-клото или его пептидным фрагментом, используя методики, описанные в настоящем документе для других антител. Альтернативно, верблюжье нанотело, связывающееся с β-клото, сконструировано, то есть получено путем отбора, например, из библиотеки фагового дисплея, соответственным образом мутированного белка нанотела верблюда, используя методики пэннинга с β-клото в качестве мишени, как описано в приведенных в настоящем документе примерах. Сконструированные нанотела могут быть, кроме того, получены с помощью методов генной инженерии с тем, чтобы их период полураспада в организме реципиента составлял от 45 минут до двух недель. В конкретном варианте осуществления верблюжье антитело или нанотело получают путем прививки последовательностей CDR тяжелой или легкой цепи антител человека по изобретению на нанотело или последовательности каркасного участка однодоменных антител, как описано, например, в PCT/EP93/02214.
Биспецифические молекулы и мультивалентные антитела
В еще одном аспекте настоящее изобретение относится к биспецифическим или мультиспецифическим молекулам, содержащим антитело, связывающее β-клото, или его фрагмент по изобретению. Антитело по изобретению или его антигенсвязывающие области могут быть изменены или связаны с другой функциональной молекулой, например, другим пептидом или белком (например, другое антитело или лиганд рецептора) с образованием биспецифической молекулы, которая связывается, по меньшей мере, с двумя различными сайтами связывания или молекулами-мишенями. Антитело по изобретению фактически может быть изменено или связано с более чем одной другой функциональной молекулой с получением мультиспецифических молекул, которые связываются с более чем двумя различными сайтами связывания и/или молекулами-мишенями; такие мультиспецифические молекулы также предусмотрены термином «биспецифическая молекула», как используется в настоящем документе. Для создания биспецифической молекулы по изобретению, антитело по изобретению может быть функционально связано (например, путем химической связи, генетического слияния, не ковалентной ассоциации или иным образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, фрагмент антитела, пептид или связывающий миметик, с получением биспецифической молекулы.
Соответственно, настоящее изобретение включает биспецифические молекулы, содержащие по меньшей мере одну первую специфичность связывания в отношении β-клото и вторую специфичность связывания в отношении второго мишеневого эпитопа. Например, второй мишеневый эпитоп является еще одним эпитопом β-клото, отличным от первого мишеневого эпитопа.
Кроме того, по изобретению, когда биспецифическая молекула является мультиспецифической, молекула может дополнительно включать третью специфичность связывания, кроме первого и второго мишеневого эпитопа.
В одном варианте осуществления биспецифические молекулы по изобретению включают в качестве специфичности связывания по меньшей мере одно антитело или его фрагмент антитела, включая, например, Fab, Fab', F (ab')2, Fv или одноцепочечный Fv. Антитело также может представлять собой димер легкой цепи или тяжелой цепи, или любой его минимальный фрагмент, такой как Fv, или одноцепочечную конструкцию, как описано Ladner et al., патент США № 4946778.
Диатела представляют собой бивалентные биспецифические молекулы, в которых домены VH и VL экспрессированы на одной полипептидной цепи, соединенной линкером, который является слишком коротким, чтобы обеспечить спаривание двух доменов в одну цепь. Домены VH и VL спариваются с комплементарными доменами другой цепи, тем самым образуя два антигенсвязывающих сайта (смотри, например, Holliger et al., 1993 Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak et al., 1994 Structure 2:1121-1123). Диатела могут быть получены путем экспрессии двух полипептидных цепей либо со структурой VHA-VLB, либо VHB-VLA (конфигурация VH-VL), либо VLA-VHB и VLB-VHA (конфигурация VL-VH) в пределах одной и той же клетки. Большинство из них могут быть экспрессированы в растворимой форме в бактериях. Одноцепочечные дитела (scDb) получают путем соединения двух образующих диатело полипептидных цепей с линкером, содержащим приблизительно 15 аминокислотных остатков (смотри, Holliger and Winter, 1997 Cancer Immunol. Immunother., 45(3-4):128-30; Wu et al., 1996 Immunotechnology, 2(1):21-36). scDb может быть экспрессировано в бактериях в растворимой активной мономерной форме (смотри, Holliger and Winter, 1997 Cancer Immunol. Immunother., 45(34): 128-30; Wu et al., 1996 Immunotechnology, 2(1):21-36; Pluckthun and Pack, 1997 Immunotechnology, 3(2): 83-105; Ridgway et al., 1996 Protein Eng., 9(7):617-21). Диатело может быть слито с Fc с образованием «ди-диатела» (смотри, Lu et al., 2004 J. Biol. Chem., 279 (4): 2856-65).
Другими антителами, которые могут быть использованы в биспецифических молекулах по изобретению, являются мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы могут быть получены путем конъюгирования составляющих специфичность связывания, используя способы, известные в данной области. Например, каждую специфичность связывания биспецифической молекулы можно получить независимо от другой, а затем конъюгировать их друг с другом. Если специфичность связывания относится к белкам или пептидам, то для ковалентного конъюгирования можно использовать ряд связывающих или перекрестно связывающих агентов. Примеры перекрестно-связывающих агентов включают белок А, карбодиимид, N-сукцинимидил-S-ацетил-тиоацетат (SATA), 5,5'-дитиобис(2-нитробензойную кислоту) (DTNB), о-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (смотри, например, Karpovsky et al., 1984 J. Exp. Med. 160:1686; Liu, MA et al., 1985 Proc. Natl. Acad. Sci. USA 82:8648). Другие методы включают способы, описанные в работах Paulus, 1985 Behring Ins. Mitt. No. 78,118-132; Brennan et al., 1985 Science 229:81-83), и Glennie et al., 1987 J. Immunol. 139: 2367-2375). Конъюгирующими агентами являются SATA и сульфо-SMCC, оба доступны от компании Pierce Chemical Co. (Rockford, IL).
Если специфичность связывания относится к антителам, то они могут быть конъюгированы посредством сульфгидрильного связывания С-концевых шарнирных областей двух тяжелых цепей. В конкретном варианте осуществления шарнирная область модифицирована так, что перед конъюгированием содержит нечетное количество сульфгидрильных остатков, например, один.
Альтернативно, обе специфичности связывания могут быть закодированы в одном и том же векторе и экспрессированы и собраны в одной и той же клетке-хозяине. Этот способ особенно эффективен, если биспецифическая молекула представляет собой mAb×mAb, mAb×Fab, Fab×F(ab')2 или лиганд×слитый белок Fab. Биспецифическая молекула по изобретению может представлять собой одноцепочечную молекулу, содержащую одноцепочечное антитело и эпитоп связывания, или одноцепочечную биспецифическую молекулу, содержащую два связывающих эпитопа. Биспецифические молекулы могут содержать по меньшей мере две одноцепочечные молекулы. Способы получения биспецифических молекул описаны, например, в патенте США № 5260203, патенте США № 5132405, патенте США № 5091513, патенте США № 5476786, патенте США № 5013653, патенте США № 5258498 и патенте США № 5482858.
Связывание биспецифических молекул с их специфическими мишенями может быть подтверждено, например, иммуноферментным анализом с ферментным связыванием (ELISA), радиоиммуноанализом (REA), анализом FACS, биоанализом (например, ингибирование роста) или Вестерн-блот анализом. Каждый из этих анализов обычно определяет наличие комплексов белок-антитело, представляющих интерес, используя меченый реагент (например, антитело), специфический в отношении представляющего интерес комплекса.
В еще одном аспекте настоящее изобретение относится к мультивалентным соединениям, содержащим по меньшей мере две идентичные или различные антигенсвязывающие части антител по изобретению, связывающихся с β-клото. Антигенсвязывающие части могут быть соединены вместе посредством слияния белка или ковалентной или не ковалентной связи. Альтернативно, способы соединения описаны для биспецифических молекул. Тетравалентные соединения могут быть получены, например, с помощью перекрестного связывания антител по изобретению с антителом, которое связывается с константными областями антител по изобретению, например, с Fc или шарнирной областью.
Тримеризующий домен описан, например, в патенте компании BOREAN, EP 1012280B1. Модули пентамеризации описаны, например, в PCT/EP97/05897.
Антитела с увеличенным периодом полувыведения
Настоящее изобретение относится к антителам, которые специфически связываются с белком бета-клото, имеющим увеличенный период полувыведения in vivo.
На период полувыведения белка in vivo могут влиять множество факторов. Например, фильтрация почками, метаболизм в печени, деградация протеолитическими ферментами (протеазами) и иммунные ответы (например, нейтрализация белка антителами и поглощение макрофагами и дендритными клетками). Для увеличения периода полувыведения антител по настоящему изобретению можно использовать множество стратегий. Например, путем химической связывания с полиэтиленгликолем (ПЭГ), reCODE ПЭГ, скаффолдом антитела, полисиаловой кислотой (PSA), гидроксиэтилкрахмалом (HES), связывающими альбумин лигандами и углеводными защитными группами; путем генетического слияния с белками, связывающимися с белками сыворотки, такими как альбумин, IgG, FcRn и трансферрин; путем сочетания (генетически или химически) с другими связывающими фрагментами, которые связываются с белками сыворотки, такими как наночастицы, Fab, DARPin, авимеры, аффитела и антикалины; путем генетического слияния с rPEG, альбумина, домена альбумина, альбуминсвязывающих белков и Fc; или путем включения в наноносители, препараты с замедленным высвобождением или медицинские устройства.
Для увеличения циркуляции антител in vivo в сыворотке, инертные полимерные молекулы, такие как высокомолекулярный ПЭГ, могут быть присоединены к антителам или к их фрагменту с мультифункциональным линкером или без него либо через сайт-специфическую конъюгацию ПЭГ с N- или С-концом антител, либо через эпсилон-аминогруппы, присутствующие на остатках лизина. Для антитела антитело или его фрагмент обычно подвергают взаимодействию с полиэтиленгликолем (ПЭГ), таким как реакционноспособный сложный эфир или альдегидное производное ПЭГ, в условиях, когда одна или несколько групп ПЭГ присоединяются к антителу или фрагменту антитела. Пэгилирование может быть осуществлено реакцией ацилирования или реакцией алкилирования с реакционноспособной молекулой ПЭГ (или аналогичным реакционноспособным водорастворимым полимером). Используемый в настоящем документе термин «полиэтиленгликоль» подразумевает любую форму ПЭГ, которая была использована для образования производных других белков, таких как моно (C1-C10)алкокси- или арилокси-полиэтиленгликоль или полиэтиленгликоль-малеимид. В некоторых вариантах осуществления антитело, подлежащее пэгилированию, представляет собой агликозилированное антитело. Можно использовать линейную или разветвленную полимеризационную дериватизацию, которая приводит к минимальной потере биологической активности. Степень конъюгации можно тщательно контролировать с помощью SDS-PAGE и масс-спектрометрии для обеспечения правильной конъюгации молекул ПЭГ с антителами. Непрореагировавший ПЭГ можно отделить от конъюгатов антитело-ПЭГ с помощью гель-фильтрации или путем ионообменной хроматографии. ПЭГ-производные антител могут быть исследованы на активность связывания, а также на эффективность in vivo с использованием методов, хорошо известных специалистам в данной области, например, с помощью описанных в настоящем документе иммуноанализов. Способы пэгилирования белков известны в данной области и могут быть применены к антителам по изобретению. Смотри, например, ЕР 0154316, Nishimura et al., и ЕР 0401384, Ishikawa et al.
Другие технологии модифицирования пэгилированием включают биоинженерную технологию с независимым восстановлением химической активности (ReCODE PEG), которая встраивает химически активные боковые цепи в белки при биосинтезе посредством восстанавливающей системы, которая включает в себя tRNA синтетазу и tRNA. Эта технология позволяет включать более 30 новых аминокислот в белки при биосинтезе в клетках E. coli, дрожжей и млекопитающих. tRNA встраивает неприродную аминокислоту в любом месте, где расположен ампер-кодон, превращая амбер из стоп-кодона в кодон, который сигнализирует о включении химически активной специфической аминокислоты.
Рекомбинантную технологию пэгилирования (rPEG) также можно использовать для удлинения времени полувыведения из сыворотки крови. Эта технология включает в себя генетическое слияние неструктурированного белкового хвоста длиной 300-600 аминокислот с существующим фармацевтическим белком. Поскольку кажущаяся молекулярная масса такой неструктурированной белковой цепи примерно в 15 раз больше, чем ее фактическая молекулярная масса, время полувыведения из сыворотке крови белка значительно увеличивается. В отличие от традиционного ПЭГилирования, при котором требуется химическая конъюгация и повторная очистка, процесс производства значительно упрощается, и продукт является гомогенным.
Полисиалирование представляет собой еще одну технологию, в которой используется природный полимер полисиаловая кислота (PSA) для продления активной жизни и улучшения стабильности терапевтических пептидов и белков. PSA представляет собой полимер сиаловой кислоты (сахар). При использовании для доставки белка и терапевтического пептидного лекарственного средства полисиаловая кислота обеспечивает защитное микроокружение при конъюгации. Это повышает активную жизнь терапевтического белка в кровообращении и препятствует его распознаванию иммунной системой. Полимер PSA находится в организме человека в естественном состоянии. Его используют некоторые бактерии, которые в процессе эволюции в течение миллионов лет покрыли им свои стенки. Эти природно полисиалилированные бактерии затем стали способными в силу своей молекулярной мимикрии разрушать защитную систему организма. PSA, природная технология абсолютной невидимости, может быть легко продуцирована такими бактериями в больших количествах и с заданными физическими характеристиками. Бактериальная PSA полностью неиммуногенна, даже при связывании с белками, поскольку она химически идентична PSA организма человека.
Еще одна технология включает использование производных гидроксиэтилкрахмала («HES»), связанных с антителами. HES является модифицированным природным полимером, полученным из крахмала восковой кукурузы, и он может метаболизироваться под действием ферментов организма. Растворы HES обычно вводят при замещении недостаточного объема крови и для улучшения реологических свойств крови. HESилирование антитела позволяет увеличить период полувыведения, повышая стабильность молекулы, а также уменьшая почечный клиренс, что приводит к увеличению биологической активности. Изменяя различные параметры, такие как молекулярная масса HES, можно получить индивидуальный широкий спектр конъюгатов антител HES.
Антитела с повышенным периодом полувыведения in vivo также могут быть созданы введением одной или нескольких модификаций аминокислот (то есть заменами, вставками или делецией) в константном домене IgG или его связывающем фрагменте FcRn (предпочтительно, Fc или шарнирный фрагмент домена Fc). Смотри, например, международную публикацию № WO 98/23289, международную публикацию № WO 97/34631 и патент США № 6277375.
Кроме того, антитела могут быть конъюгированы с альбумином (например, альбумином сыворотки человека, HSA), чтобы повысить стабильность антитела или фрагмента антитела in vivo или повысить период полураспада in vivo. Эти способы хорошо известны в данной области, смотри, например, международные публикации №№ WO 93/15199, WO 93/15200 и WO 01/77137, а также европейский патент № EP 413622. Кроме того, в контексте биспецифического антитела, как описано выше, специфичность антитела может быть сконструирована таким образом, что один связывающий домен антитела связывается с FGF21, а второй связывающий домен антитела связывается с сывороточным альбумином, предпочтительно, с HSA.
Стратегии повышения периода полувыведения особенно эффективны в нанотелах, связывающих агентов на основе фибронектина, и в других антителах или белках, для которых желательно повышение in vivo периода полураспада.
Конъюгаты антител
Настоящее изобретение относится к антителам или их фрагментам, которые специфически связываются с белком β-клото, рекомбинантно конденсированным или химически конъюгированным (включая как ковалентные, так и нековалентные конъюгации) с гетерологичным белком или полипептидом (или его фрагментом, предпочтительно, с полипептидом с по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислоты) для генерирования слитых белков. В частности, настоящее изобретение относится к слитым белкам, содержащим антигенсвязывающий фрагмент антитела, описанного в настоящем документе, (например, фрагмент Fab, фрагмент Fd, фрагмент Fv, фрагмент F(ab)2, домен VH, CDR VH, домена VL или a VL CDR) и к гетерологичному белку, полипептиду или пептид. Способы слияния или конъюгирования белков, полипептидов или пептидов с антителом или фрагментом антитело известны в данной области. Смотри, например, патенты США №№ 5336603, 5622929, 5359046, 5349053, 5447851 и 5112946; Европейские патенты №№ EP 307434 и EP 367166; международные публикации №№ WO 96/04388 и WO 91/06570; Ashkenazi et al., 1991, Proc. Natl. Acad. Sci. USA 88: 10535-10539; Zheng et al., 1995, J. Immunol. 154:5590-5600; и Vil et al., 1992, Proc. Natl. Acad. Sci. USA 89:11337- 11341.
Дополнительные слитые белки могут быть получены с помощью методов перетасовки генов, перестановки мотивов, перетасовки экзонов и/или перетасовки кодонов (в совокупности называемых «ДНК шаффлингом»). ДНК шаффлинг может быть использован для изменения активности антител по изобретению или их фрагментов (например, антител или их фрагментов с более высокой аффинностью и более низкой скоростью диссоциации). Смотри, в основном, патенты США №№ 5605793, 5811238, 5830721, 5834252 и 5837458; Patten et al., 1997, Curr. Opinion Biotechnol. 8:724-33; Harayama, 1998, Trends Biotechnol. 16(2):76-82; Hansson, et al., 1999, J. Mol. Biol. 287:265-76; и Lorenzo и Blasco, 1998, Biotechniques 24(2):308-313 (каждый из этих патентов и публикаций полностью включен в настоящее описание посредством ссылки). Антитела или их фрагменты, или кодированные антитела или их фрагменты, могут быть изменены путем случайного мутагенеза методом ПЦР сниженной точности, случайной нуклеотидной вставкой или другими способами перед рекомбинацией. Полинуклеотид, кодирующий антитело или его фрагмент, который специфически связывается с белком β-клото, может быть рекомбинирован с одним или несколькими компонентами, мотивами, частями, частями, доменами, фрагментами и т. д. одной или более гетерологичных молекул.
Более того, для облегчения очистки антитела или их фрагменты могут быть слиты с маркерными последовательностями, такими как пептид. В предпочтительных вариантах осуществления аминокислотная последовательность маркера представляет собой гексагистидиновый пептид (SEQ ID NO: 274), такой как таг, представленный в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311), помимо прочих, многие из которых являются коммерчески доступными. Как описано в работе Gentz et al., 1989, Proc. Natl. Acad. Sci. USA 86: 821-824, например, гексагистидин (SEQ ID NO: 274) обеспечивает удобную очистку слитого белка. Другие пептидные таги, используемые для очистки, включают, но не ограничиваются ими, метку гемагглютинина («HA»), которая соответствует эпитопу, полученному из белка гемагглютинина гриппа (Wilson et al., 1984, Cell 37:767), и маркер «flag».
В других вариантах осуществления антитела по настоящему изобретению или их фрагменты конъюгированы с диагностическим или детектируемым агентом. Такие антитела могут быть использованы для мониторинга или прогнозирования начала, развития, прогрессирования и/или тяжести заболевания или расстройства в рамках процедуры клинических испытаний, таких как определение эффективности конкретной терапии. Такой диагноз и обнаружение могут быть осуществлены путем связывания антитела с детектируемыми веществами, включая, но не ограничиваясь ими, различные ферменты, такие как, но не ограничиваясь ими, пероксидаза хрена, щелочная фосфатаза, бета-галактозидаза или ацетилхолинэстераза; простетические группы, такие как, но не ограничиваясь ими, стрептавинилбвитин и авидин/биотин; флуоресцентные вещества, такие как, но не ограничиваясь ими, умбеллиферон, флуоресцеин, изотиотионат флуоресцеина, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; люминесцентные материалы, такие как, но не ограничиваясь ими, люминол; биолюминесцентные материалы, такие как, но не ограничиваясь ими, люцифераза, люциферин и аэкорин; радиоактивные вещества, такие как, но не ограничиваясь ими, йод (131I, 125I, 123I и 121I), углерод (14C), сера (35S), тритий (3H), индий (115In, 113In, 112In и 111In), технеций (99Tc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 188Re, 142 Pr, 105Rh, 97Ru, 68Ge, 57Co, 65Zn, 85Sr, 32P, 153Gd, 169Yb, 51Cr, 54Mn, 75Se, 113Sn и 117Tin; и позитрон-эмиттирующие металлы, используемые в различных видах позитронно-эмиссионной томографии, и нерадиоактивные ионы парамагнитных металлов.
Настоящее изобретение, кроме того, охватывает применение антител или их фрагментов, конъюгированных с терапевтическим фрагментом. Антитело или его фрагмент могут быть конъюгированы с терапевтическим фрагментом, таким как цитотоксин, например, цитостатический или цитоцидный агент, терапевтический агент или ион радиоактивного металла, например, альфа-излучатели. Цитотоксин или цитотоксический агент включает любой агент, который оказывает негативные воздействия на клетки.
Кроме того, антитело или его фрагмент могут быть конъюгированы с терапевтическим фрагментом или лекарственным фрагментом, который модифицирует данный биологический ответ. Следует понимать, что терапевтические фрагменты или лекарственные фрагменты не должны быть ограничены классическими химическими терапевтическими средствами. Например, лекарственный фрагмент может представлять собой белок, пептид или полипептид, обладающий желательной биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин А, псевдомонас экзотоксин, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, фактор роста тромбоцитов, активатор тканевого плазминогена, апоптотический агент, антиангиогенный агент или, модификатор биологического ответа, такой как, например, лимфокин.
Более того, антитело может быть конъюгировано с терапевтическими фрагментами, такими как ион радиоактивного металла, такие как альфа-излучатели, такие как 213Bi или макроциклические хелаторы, используемые для конъюгации радиометаллических ионов, включая, но этим не ограничиваясь, 131In, 131LU, 131Y, 131Ho, 131Sm, с полипептидами. В некоторых вариантах осуществления макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N,N',Nʺ,Nʺ'-тетрауксусную кислоту (DOTA), которая может быть присоединена к антителу посредством линкерной молекулой. Такие линкерные молекулы широко известны в данной области и описаны в работах Denardo et al., 1998, Clin Cancer Res. 4(10):2483-90; Peterson et al., 1999, Bioconjug. Chem. 10(4):553-7; и Zimmerman et al., 1999, Nucl. Med. Biol. 26(8):943-50, все включены в качестве ссылки во всей их полноте.
Методики конъюгирования терапевтических фрагментов с антителами хорошо известны, смотри, например, Arnon et al., «Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy», in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds. ), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., «Antibodies For Drug Delivery», in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review», Monoclonal Antibodies 84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); «Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy», Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., 1982, Immunol. Rev. 62: 119-58.
Антитела также могут быть присоединены к твердым подложкам, которые особенно эффективны для иммунологических анализов или очистки мишеневого антигена. Такие твердые подложки включают, но не ограничиваются ими, стекло, целлюлозу, полиакриламид, найлон, полистирол, поливинилхлорид или полипропилен.
Способы получения антител по изобретению
Нуклеиновые кислоты, кодирующие антитела
Настоящее изобретение относится к по существу очищенным молекулам нуклеиновых кислот, которые кодируют полипептиды, содержащие сегменты или домены цепей связывающих β-клото антител, описанных выше. Некоторые из нуклеиновых кислот по изобретению содержат нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, показанную в SEQ ID NO: 10, 30, 50 или 70, и/или нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, показанную в SEQ ID NO: 20, 40, 60 или 80. В конкретном варианте осуществления молекулы нуклеиновой кислоты являются такими, которые указаны в таблице 1. Некоторые другие молекулы нуклеиновой кислоты по изобретению содержат нуклеотидные последовательности, которые по существу идентичны (например, по меньшей мере на 65, 80%, 95% или 99%) нуклеотидным последовательностям, указанным в таблице 1. При экспрессии из соответствующих векторов экспрессии полипептиды, кодируемые этими полинуклеотидами, могут проявлять антигенсвязывающую способность FGF21.
Также в настоящем изобретении предложены полинуклеотиды, которые кодируют, по меньшей мере, одну область CDR и обычно все три области CDR из тяжелой или легкой цепи связывающего β-клото антитела, описанного выше. Некоторые другие полинуклеотиды кодируют всю или по существу всю последовательность вариабельной области тяжелой цепи и/или легкой цепи связывающего β-клото антитела, описанного выше. Из-за вырождения кода различные последовательности нуклеиновых кислот кодируют каждую из аминокислотных последовательностей иммуноглобулина.
Молекулы нуклеиновых кислот по изобретению могут кодировать как вариабельный участок, так и константную область антитела. Некоторые из последовательностей нуклеиновых кислот по изобретению включают нуклеотиды, кодирующие последовательность тяжелой цепи, которая по существу идентична (например, по меньшей мере на 80%, 90% или 99%) последовательности тяжелой цепи, представленной в SEQ ID NO: 11, 31, 51 или 71. Некоторые другие последовательности нуклеиновых кислот, содержащие нуклеотид, кодирующий последовательность легкой цепи, которая по существу идентична (например, по меньшей мере на 80%, 90% или 99%) последовательности легкой цепи, представлены в SEQ ID NO: 21, 41, 61 или 81.
Полинуклеотидные последовательности могут быть получены путем синтеза твердофазной ДНК de novo или путем ПЦР-мутагенеза существующей последовательности (например, последовательностей, описанных в примерах ниже), кодирующих связывающее β-клото антитело или его связывающий фрагмент. Прямой химический синтез нуклеиновых кислот может быть осуществлен способами, известными в данной области, такими как фосфотриэфирный метод Narang et al., 1979, Meth. Enzymol. 68:90; фосфодиэфирный метод Brown et al., Meth. Enzymol. 68:109, 1979; диэтилфосфорамидитный метод Beaucage et al., Tetra. Lett., 22:1859, 1981; и метод твердой подложки, патент США № 4458066. Введение мутаций в полинуклеотидную последовательность посредством ПЦР может быть выполнено, как описано, например, в методах ПЦР: Principles and Applications for DNA Amplification, H.A. Erlich (Ed.), Freeman Press, NY, NY, 1992; протоколы PCR: A Guide to Methods and Applications, Innis et al. (Ed.), Academic Press, San Diego, CA, 1990; Mattila et al., Nucleic Acids Res. 19:967, 1991; и Eckert et al., PCR Methods and Applications 1:17, 1991.
Также в изобретении предложены векторы экспрессии и клетки-хозяева для продуцирования описанных выше связывающих β-клото антител. Для экспрессии полинуклеотидов, кодирующих цепи связывающего β-клото антитела или связывающих фрагментов, могут быть использованы различные экспрессирующие векторы. Для получения антител в клетке-хозяине млекопитающих могут быть использованы как вирусные, так и невирусные экспрессирующие векторы. Невирусные векторы и системы включают плазмиды, эписомальные векторы, обычно с экспрессионной кассетой для экспрессии белка или РНК, и искусственные хромосомы человека (смотри, например, Harrington et al., Nat Genet 15:345, 1997). Например, невирусные векторы, используемые для экспрессии связывающих FGF21 полинуклеотидов и полипептидов в клетках млекопитающих (например, человека), включают pThioHis A, B & C, pcDNA3.1/His, pEBVHis A, B & C (Invitrogen, San Diego, CA), векторы MPSV и множество других векторов, известных в данной области для экспрессии других белков. Используемые вирусные векторы включают векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, векторов на основе SV40, папилломавирусов, вируса Эпштейна Барра HBP, векторы на основе вируса осповакцины и вируса леса Семлики (SFV). См., Brent et al., выше; Smith, Annu. Rev. Microbiol. 49:807, 1995; и Rosenfeld et al., Cell 68:143, 1992.
Выбор вектора экспрессии зависит от предполагаемых клеток-хозяев, в которых должен быть экспрессирован вектор. Обычно векторы экспрессии содержат промотор и другие регуляторные последовательности (например, энхансеры), которые функционально связаны с полинуклеотидами, кодирующими цепь или фрагмент связывающего β-клото антитела. В некоторых вариантах осуществления используется индуцибельный промотор для предотвращения экспрессии вставленных последовательностей, за исключением условий, вызывающих индуцирование. Индуцируемые промоторы включают, например, арабинозу, lacZ, промотор металлотионеина или промотор теплового шока. Культуры трансформированных организмов могут быть увеличены в неиндуцирующих условиях без смещения популяции в сторону кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. В дополнение к промоторам также могут потребоваться или быть желательными другие регуляторные элементы для эффективной экспрессии цепи связывающего β-клото антитела или фрагмента. Эти элементы обычно включают кодон инициации ATG и соседний сайт связывания рибосом или другие последовательности. Кроме того, эффективность экспрессии может быть повышена за счет включения энхансеров, подходящих для используемой клеточной системы (смотри, например, Scharf et al., Results Probl. Cell Differ. 20:125, 1994; и Bittner et al., Meth. Enzymol., 153: 516, 1987). Например, энхансер SV40 или энхансер CMV можно использовать для усиления экспрессии в клетках-хозяевах млекопитающих.
Векторы экспрессии также могут обеспечивать положение сигнальной последовательности секреции для образования слитого белка с полипептидами, кодируемыми встроенными последовательностями антител, связывающих FGF21. Чаще всего встроенные последовательности связывающего β-клото антитела соединяют с сигнальными последовательностями перед включением в вектор. Векторы, используемые для получения последовательностей, кодирующих вариабельные домены легких и тяжелых цепей связывающих β-клото антител иногда также кодируют константные области или их части. Такие векторы обеспечивают экспрессию вариабельных областей в виде слитых белков с константными областями, что приводит к продукции интактных антител или их фрагментов. Обычно такие константные области являются человеческими.
Клетки-хозяева, содержащие и экспрессирующие цепи связывающего β-клото антитела, могут быть либо прокариотическими, либо эукариотическими. E. coli представляет собой один из прокариотических хозяев, которые могут использоваться для клонирования и экспрессии полинуклеотидов по настоящему изобретению. Другие микробные клетки-хозяева, подходящие для использования, включают бациллы, такие как Bacillus subtilis, и другие энтеробактерии, такие как Salmonella, Serratia и различные виды Pseudomonas. В этих прокариотических хозяевах также можно создавать векторы экспрессии, которые обычно содержат последовательности контроля экспрессии, совместимые с клеткой-хозяином (например, точка начала репликации). Кроме того, может присутствовать любое количество различных известных промоторов, таких как лактозная промоторная система, триптофановая (trp) промоторная система, бета-лактамазная промоторная система или промоторная система фага лямбда. Промоторы обычно контролируют экспрессию, необязательно, с помощью последовательности оператора и имеют последовательности сайтов связывания с рибосомами и тому подобное для инициирования и завершения транскрипции и трансляции. Другие микробы, такие как дрожжи, также могут быть использованы для экспрессии полипептидов, связывающих FGF21, по изобретению. Также можно использовать клетки насекомых в сочетании с векторами бакуловируса.
В некоторых предпочтительных вариантах осуществления для экспрессии и получения связывающих β-клото полипептидов по настоящему изобретению используют клетки млекопитающих-хозяева. Например, они могут представлять собой либо гибридомную клеточную линию, экспрессирующей эндогенные гены иммуноглобулина (например, клон гибридомы миеломы 1D6.C9, как описано в примерах), либо линию клеток млекопитающих, несущих экзогенный экспрессирующий вектор (например, клетки миеломы SP2/0, приведенные далее). Они включают любую нормальную смертную или нормальную или патологическую иммортализованную клетку животного или человека. Например, был разработан ряд подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, включая клеточные линии СНО, различные клеточные линии Cos, клетки HeLa, клеточные линии миеломы, трансформированные B-клетки и гибридомы. Использование клеточной культуры ткани млекопитающих для экспрессии полипептидов обсуждалось в основном, например, в обзоре Winnacker, FROM GENES TO CLONES, VCH Publishers, N.Y., N.Y., 1987. Экспрессирующие векторы для клеток млекопитающих-хозяев могут включать последовательности контроля экспрессии, такие как точка начала репликации, промотор и энхансер (смотри, например, Queen, et al., Immunol. Rev. 89: 49-68, 1986), и необходимые сайты процессинга информации, такие как сайты связывания рибосом, сплайс-сайты РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Эти векторы экспрессии обычно содержат промоторы, полученные из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфическими в отношении конкретного типа клеток, специфическими для стадии и/или модулируемыми или регулируемыми. Используемые промоторы включают, но не ограничиваются ими, промотор металлотионеина, конститутивный главный поздний промотор аденовируса, индуцируемый дексаметазоном промотор MMTV, промотор SV40, промотор MRP polIII, конститутивный промотор MPSV, индуцируемый тетрациклином промотор CMV (такой как человеческий средне-ранний промотор CMV), конститутивный промотор CMV и комбинации промотор-энхансер, известные в области техники.
Способы введения векторов экспрессии, содержащие представляющие интерес полинуклеотидные последовательности, различаются в зависимости от типа клеточного хозяина. Например, трансфекция хлорида кальция обычно используется для прокариотических клеток, тогда как обработка фосфатом кальция или электропорация может быть использована для других клеточных хозяев (смотри, главным образом Sambrook, et al., выше). Другие методы включают, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические методы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, голую ДНК, искусственные вирионы, слияние структурного белка вируса герпеса VP22 (Elliot and O'Hare, Cell 88: 223, 1997), усиленное агентом поглощение ДНК и трансдукцию ex vivo. Для длительного высокопродуктивного получения рекомбинантных белков часто требуется стабильная экспрессия. Например, клеточные линии, которые стабильно экспрессируют цепи связывающего β-клото антитела или связывающие фрагменты, могут быть получены с использованием векторов экспрессии по изобретению, которые содержат вирусные точки начала репликации или эндогенные элементы экспрессии и ген селектируемого маркера. После введения вектора клетки оставляют расти в течение 1-2 дней в обогащенной среде перед тем, как среду меняют на селективную. Целью селектируемого маркера является придать устойчивость к отбору, и его присутствие обеспечивает рост клеток, которые успешно экспрессируют введенные последовательности в селективных средах. Можно обеспечить пролиферацию устойчивых стабильно трансфицированных клеток с использованием методов тканевых культур, подходящих к типу клеток.
Получение моноклональных антител
Моноклональные антитела (mAb) могут быть получены различными способами, включая обычный метод получения моноклональных антител, например, стандартный метод гибридизации соматических клеток, Kohler and Milstein, 1975 Nature 256: 495. Могут быть использованы многие способы получения моноклонального антитела, например, вирусной или онкогенной трансформацией В-лимфоцитов.
Семейства животных для получения гибридом включают семейства мышей, крыс и кроликов. Получение гибридомы у мышей представляет собой хорошо установившийся метод. Протоколы иммунизации и способы выделения иммунизированных спленоцитов для слияния известны в данной области. Сливающиеся клетки (например, клетки мышиной миеломы) и методы слияния также известны.
Химерные или гуманизированные антитела по настоящему изобретению могут быть получены на основе последовательности мышиного моноклонального антитела, полученного, как описано выше. ДНК, кодирующую тяжелые и легкие цепи иммуноглобулинов, может быть получена из представляющей интерес мышиной гибридомы и сконструирована так, что содержит немышиный (например, человеческий) иммуноглобулин, используя стандартные способы молекулярной биологии. Например, для создания химерного антитела вариабельные области мыши могут быть связаны с константными областями человека с использованием способов, известных в данной области (смотри, например, патент США № 4816567, Cabilly et al.). Для создания гуманизированного антитела мышиные области CDR могут быть вставлены в рамку считывания человека с использованием способов, известных в данной области. См., например, патент США № 5225539, Winter, и патенты США № 5530101; 5585089; 5693762 и 6180370, Queen et al.
В конкретном варианте осуществления антитела по изобретению представляют собой моноклональные антитела человека. Такие моноклональные антитела человека, направленные против β-клото, могут быть образованы с использованием трансгенных или трансхромосомных мышей, несущих скорее части иммунной системы человека, чем мышиной системы. Эти трансгенные и трансхромосомные мыши включают мышей, которые называются в настоящем документе мышами HuMAb и мышами KM, соответственно, и в совокупности упоминаются здесь как «мыши с Ig человека».
Мышь HuMAb® (Medarex, Inc.) содержит минилокусы гена иммуноглобулина человека, которые кодируют неупорядоченные последовательности тяжелой цепи (μ и γ) и легкой цепи ĸ иммуноглобулинов человека, вместе с нацеленными мутациями, которые инактивируют эндогенные локусы цепей μ и ĸ (смотри, например, Lonberg, et al., 1994 Nature 368(6474): 856-859). Таким образом, мыши демонстрируют пониженную экспрессию IgM мыши или ĸ, и в ответ на иммунизацию введенные трансгены тяжелой и легкой цепей человека подвергаются переключению класса и соматической мутации с генерированием моноклонального IgGĸ с высокой аффинностью (Lonberg, N. et al., 1994, выше; рассмотрено N., 1994 Handbook of Experimental Pharmacology 113:49-101; Lonberg, N. and Huszar, D., 1995 Intern. Rev. Immunol.13: 65-93, и Harding, F. and Lonberg, N., 1995 Ann. N. Y. Acad. Sci. 764: 536-546). Получение и использование мышей HuMAb и геномные модификации, которые несут такие мыши, дополнительно описаны Taylor, L. et al., 1992 Nucleic Acids Research 20:6287-6295; Chen, J. et at., 1993 International Immunology 5: 647-656; Tuaillon et al., 1993 Proc. Natl. Acad. Sci. USA 94:3720-3724; Choi et al., 1993 Nature Genetics 4:117-123; Chen, J. et al., 1993 EMBO J. 12: 821-830; Tuaillon et al., 1994 J. Immunol. 152:2912-2920; Taylor, L. et al., 1994 International Immunology 579-591; и Fishwild, D. et al., 1996 Nature Biotechnology 14: 845-851, содержание которых полностью включено в качестве ссылки во всей их полноте. См., кроме того, патенты США №№ 5545806; 5569825; 5625126; 5633425; 5789650; 5877397; 5661016; 5814318; 5874299 и 5770429; все Lonberg и Kay; патент США № 5545807, Surani et al.; публикации PCT №№ WO 92103918, WO 93/12227, WO 94/25585, WO 97113852, WO 98/24884 и WO 99/45962, все Lonberg и Kay; и публикация PCT № WO 01/14424, Korman et al.
В другом варианте осуществления антитела человека по изобретению могут быть индуцированы с использованием мыши, которая несет последовательности человеческого иммуноглобулина на трансгены и трансхомосомы, например, мыши, которая несет трансген тяжелой цепи человека и трансхромосому легкой цепи человека. Такие мыши, называемые в настоящем документе «мышами KM», подробно описаны в публикации PCT WO 02/43478, Ishida et al.
Кроме того, в данной области доступны альтернативные трансгенные системы животных, экспрессирующие гены иммуноглобулина человека, и они могут быть использованы для индукции связывающих β-клото антител по изобретению. Например, может быть использована альтернативная трансгенная система, называемая ксеномышью (Abgenix, Inc.). Такие мыши описаны, например, в патентах США №№ 5939598; 6075181; 6114598; 6150584 и 6162963, Kucherlapati et al.
Кроме того, альтернативные трансхромосомные системы животных, экспрессирующие гены иммуноглобулина человека, доступны в данной области и могут быть использованы для индукции связывающих β-клото антител по изобретению. Например, могут использоваться мыши, несущие как трансхромосому тяжелой цепи человека, так и транхромосому легкой цепи человека, называемые «мышами TC»; такие мыши описаны Tomizuka et al., 2000 Proc. Natl. Acad. Sci. USA 97: 722-727. Кроме того, в данной области были описаны коровы, несущие трансхромосомы тяжелой и легкой цепей человека (Kuroiwa et al., 2002 Nature Biotechnology 20:889-894), и они могут быть использованы для индукции антител, связывающих FGF21, по изобретению.
Моноклональные антитела человека по изобретению также могут быть получены с использованием способов фагового дисплея для скрининга библиотек генов иммуноглобулинов человека. Такие способы фагового дисплея для выделения антител человека являются установленными в данной области техники или описаны в примерах ниже. См., например: патенты США №№ 5223409, 5403484 и 5571698, Ladner et al.; патенты США №№ 5427908 и 5580717, Dower et al.; патенты США №№ 5969108 и 6172197, McCafferty et al.; и патенты США №№ 5885793, 6521404, 6544731, 6555313, 6582915 и 6593081, Griffiths et al.
Моноклональные антитела человека по изобретению также могут быть получены с использованием мышей SCID, в которых иммунные клетки человека были реконструированы таким образом, что после иммунизации может быть вызвана гуморальная (антительная) реакция человека. Такие мыши описаны, например, в патентах США №№ 5476996 и 5698767, Wilson et al.
Конструирование каркаса или Fc
Сконструированные антитела по настоящему изобретению включают в себя антитела, в которых модификации были произведены в каркасных остатках в VH и/или VL, например, для улучшения свойств данного антитела. Обычно такие модификации каркасных участков производят для уменьшения иммуногенности антитела. Например, одним подходом является «обратное мутирование» одного или нескольких каркасных остатков в соответствующую последовательность зародышевой линии. Более конкретно, антитело, которое подверглось соматической мутации, может содержать каркасные остатки, которые отличаются от последовательности зародышевой линии, из которой получено антитело. Такие остатки могут быть идентифицированы путем сравнения последовательностей каркасных участков антитела с последовательностями зародышевой линии, из которых получено антитело. Для возврата последовательностей каркасного участка в их конфигурацию зародышевой линии, соматические мутации могут быть «обратно мутированы» в последовательность зародышевой линии, например, путем сайт-направленного мутагенеза. Такие «обратно мутированные» антитела также должны охватываться изобретением.
Другой тип модификации каркаса включает мутацию одного или нескольких остатков в каркасном участке или даже в пределах одной или нескольких областей CDR для удаления эпитопов T-клеток для уменьшения таким образом возможной иммуногенности антитела. Этот подход называют также как «деиммунизация», и он более подробно описан в публикации патента США № 20030153043, Carr et al.
В качестве дополнения или альтернативы модификациям, произведенным в каркасных участках или областях CDR, антитела по настоящему изобретению могут быть сконструированы таким образом, что они включают в себя модификации в Fc-области, обычно для изменения одного или нескольких функциональных свойств антитела, таких как период полувыведения в сыворотке, фиксация комплемента, связывание рецептора Fc и/или антиген-независимая клеточно опосредованная цитотоксичность. Кроме того, антитело по изобретению может быть химически модифицировано (например, к антителу могут быть присоединены один или несколько химических фрагментов) или может быть модифицировано с изменением его гликозилирования, опять же для изменения одного или нескольких функциональных свойств данного антитела. Каждый из этих вариантов осуществления более подробно описан далее. Нумерация остатков в Fc-области является схемой нумерации EU по Кабату.
В одном варианте осуществления шарнирная область CH1 модифицирована таким образом, что количество цистеиновых остатков в шарнирной области изменено, например, увеличено или уменьшено. Этот подход более подробно описан в патенте США № 5677425, Bodmer et al. Количество цистеиновых остатков в шарнирной области CH1 изменяют, например, для облегчения сборки легких и тяжелых цепей или для увеличения или уменьшения стабильности антитела.
В другом варианте осуществления шарнирную область Fc антитела мутируют для уменьшения биологического периода полувыведения антитела. Более конкретно, одну или несколько аминокислотных мутаций вводят в область контакта доменов CH2-CH3 Fc-шарнирного фрагмента, таким образом, что что у антитела нарушается связывание со стафилококковым белком A (SpA) по сравнению со связыванием нативного Fc-шарнирного домена с SpA. Этот подход более подробно описан в патенте США № 6165745, Ward et al.
В другом варианте осуществления антитело модифицируют для увеличения его биологического периода полураспада. Возможны различные подходы. Например, могут быть введены одна или несколько из следующих мутаций: T252L, T254S, T256F, описанные в патент США № 6277375, Ward. Альтернативно, чтобы увеличить биологический период полураспада, антитело может быть изменено в области CH1 или CL таким образом, что оно содержит эпитоп связывания рецептора реутилизации, образованного из двух петель СН2-домена Fc-области IgG, как описано в патентах США №№ 5869046 и 6121022, Presta et al.
Еще в одном варианте осуществления Fc-область изменяют путем замены по меньшей мере одного аминокислотного остатка на другой аминокислотный остаток для изменения эффекторных функций антитела. Например, одна или несколько аминокислот могут быть заменены другим аминокислотным остатком таким образом, что антитело будет иметь измененную аффинность в отношении эффекторного лиганда, но сохранит антигенсвязывающую способность исходного антитела. Эффекторным лигандом, в отношении которого аффинность изменяется, может быть, например, рецептор Fc или C1-компонент комплемента. Этот подход более подробно описан в патентах США №№ 5624821 и 5648260, оба Winter et al.
В другом варианте осуществления одна или несколько аминокислот, выбранных из аминокислотных остатков, могут быть заменены другим аминокислотным остатком таким образом, что антитело имеет измененное связывание С1q и/или уменьшенную или устраненную комплементзависимую цитотоксичность (CDC). Этот подход более подробно описан в патенте США № 6194551, Idusogie et al.
В другом варианте осуществления один или несколько аминокислотных остатков изменяют, чтобы таким образом изменить способность антитела фиксировать комплемент. Этот подход описан более подробно в публикации PCT WO 94/29351, Bodmer et al.
В еще одном варианте осуществления область Fc модифицируют для увеличения способности антитела опосредовать зависимую от антитела клеточную цитотоксичность (ADCC) и/или для увеличения аффинности антитела в отношении рецептора Fcγ путем модификации одной или нескольких аминокислот. Этот подход описан более подробно в публикации PCT WO 00/42072, Presta. Кроме того, сайты связывания на IgG1 человека для FcγRl, FcγRII, FcγRIII и FcRn были картированы, и были описаны варианты с улучшенным связыванием (смотри, Shields, R.L. et al., 2001 J. Biol. Chen. 276: 6591-6604).
В еще одном варианте осуществления модифицируют гликозилирование антитела. Например, может быть получено агликозированное антитело (то есть антитело без гликозилирования). Гликозилирование может быть изменено, например, для увеличения аффинности антитела к «антигену». Такие углеводные модификации могут быть выполнены, например, путем изменения одного или нескольких сайтов гликозилирования в последовательности антитела. Например, могут быть произведены одна или несколько аминокислотных замен, которые приводят к элиминации одного или нескольких сайтов гликозилирования каркаса вариабельной области, чтобы таким образом элиминировать гликозилирование в этом сайте. Такое агликозилирование может увеличивать аффинность антитела к антигену. Такой подход более подробно описан в патентах США №№ 5714350 и 6350861, Co et al.
Дополнительно или альтернативно может быть получено антитело, которое имеет измененный тип гликозилирования, такое как гипофукозилированное антитело, имеющее уменьшенные количества остатков фукозила, или антитело, имеющее увеличенные бисекционные структуры GlcNac. Было продемонстрировано, что такие измененные картины гликозилирования повышают ADCC-способность антител. Такие углеводные модификации могут быть осуществлены, например, путем экспрессии антитела в клетке-хозяине с измененным аппаратом гликозилирования. Клетки с измененным аппаратом гликозилирования были описаны в данной области и могут быть использованы в качестве клеток-хозяев, в которых экспрессируются рекомбинантные антитела по изобретению, в результате чего получают антитело с измененным гликозилированием. Например, в ЕР 1176195, Hang et al., описана клеточная линия с функционально разрушенным геном FUT8, который кодирует фукозилтрансферазу таким образом, что антитела, экспрессируемые в такой клеточной линии, проявляют гипофукозилирование. В публикации РСТ WO 03/035835, Presta, описан вариант клеточной линии CHO, клетки Lecl3, с уменьшенной способностью присоединять фукозу к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых в этой клетке-хозяине (смотри, также, Shields, R.L. et al., 2002 J. Biol. Chem. 277: 26733-26740). В публикации РСТ WO 99/54342, Umana et al., описаны клеточные линии, сконструированные для экспрессии гликопротеин-модифицирующих гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминилтрансфераза III (GnTIII)) таким образом, что антитела, экспрессируемые в сконструированных клеточных линиях, демонстрируют повышенные бисекционные структуры GlcNac, что приводит к увеличению активности ADCC в антителах (смотри, также Umana et al., 1999 Nat. Biotech. 17:176-180).
Способы конструирования измененных антител
Как обсуждалось выше, связывающие β-клото антитела, имеющие последовательности VH и VL или полноразмерные последовательности тяжелой и легкой цепей, показанные в настоящем документе, могут быть использованы для создания новых связывающих β-клото антител путем модификации полноразмерных последовательностей тяжелых цепей и/или легких цепей, последовательностей VH и или VL или константной(ых) области(ей), присоединенной(ых) к ним. Таким образом, в другом аспекте изобретения структурные особенности связывающего β-клото антитела по изобретению используются для создания структурно родственных связывающих β-клото антител, которые сохраняют по меньшей мере одно функциональное свойство антител по изобретению, такое как связывание с человеческим β-клото, а также активирование одного или нескольких функциональных свойств комплекса FGF21-рецептор (например, активируя передачу сигналов FGF21-рецепторов).
Например, одна или несколько областей CDR антител по настоящему изобретению или их мутации могут быть комбинированы рекомбинантно с известными каркасными участками и/или другими CDR с созданием дополнительных рекомбинантно сконструированных связывающих β-клото антител по изобретению, как обсуждалось выше. Другие типы модификаций включают модификации, которые описаны в предыдущем разделе. Исходным материалом для способа конструирования является одна или несколько последовательностей VH и/или VL, представленные в настоящем документе, или одна или несколько областей CDR. Чтобы создать сконструированное антитело, нет необходимости действительно получать (то есть экспрессировать в виде белка) антитело, имеющее одну или несколько последовательностей VH и/или VL, представленных в настоящем документе, или одну или несколько их областей CDR. Предпочтительно, используют информацию, содержащуюся в последовательности(ях), в качестве исходного материала для создания последовательности(ей) «второй генерации», полученной(ых) из начальной(ых) последовательности(ей), и затем последовательность(ти) «второй генерации» получают и экспрессируют в виде белка.
Таким образом, в другом варианте осуществления настоящее изобретение относится к способу получения связывающего β-клото антитела, состоящего из последовательности вариабельной области тяжелой цепи антитела, имеющей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO: 3, 23, 43 и 63, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO: 4, 24, 44 и 64, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO: 5, 25, 45 и 65; и последовательности вариабельной области легкой цепи антитела, имеющей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO: 13, 33, 53 и 73, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO: 14, 34, 54 и 74, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO: 15, 35, 55 и 75; изменяя по меньшей мере один аминокислотный остаток в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела для создания по меньшей мере одной измененной последовательности антитела; и экспрессируя измененную последовательность антитела в виде белка.
Соответственно, в другом варианте осуществления настоящее изобретение относится к способу получения связывающего β-клото антитела, состоящего из последовательности вариабельной области тяжелой цепи антитела, имеющей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO: 6, 26, 46 и 66, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO: 7, 27, 47 и 67, и/или последовательности CDR3, выбранную из группы, состоящей из SEQ ID NO: 8, 28, 48 и 68; и последовательности вариабельной области легкой цепи антитела, имеющей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO: 16, 36, 56 и 76, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO: 17, 37, 57 и 77, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO: 18, 38, 58 и 78; изменяя по меньшей мере один аминокислотный остаток в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела для создания по меньшей мере одной измененной последовательности антитела; и экспрессируя измененную последовательность антитела в виде белка.
Соответственно, в другом варианте осуществления настоящее изобретение относится к способу получения связывающего β-клото антитела, оптимизированного для экспрессии в клетке млекопитающего, состоящего из последовательности тяжелой цепи полноразмерного антитела, имеющей последовательность, выбранную из группы SEQ ID NO: 11, 31, 51 или 71; и последовательности легкой цепи полноразмерного антитела, имеющей последовательность, выбранную из группы 21, 41, 61 или 81; изменяя по меньшей мере один аминокислотный остаток в последовательности тяжелой цепи полноразмерного антитела и/или последовательности легкой цепи полноразмерного антитела для создания по меньшей мере одной измененной последовательности антитела; и экспрессируя измененную последовательность антитела в виде белка. В одном варианте осуществления изменение тяжелой или легкой цепи находится в каркасном участке тяжелой или легкой цепи.
Измененная последовательность антител также может быть получена путем скрининга библиотек антител, имеющих фиксированные последовательности CDR3 или минимальные существенные связывающие детерминанты, как описано в US2005/0255552, и разнообразие последовательностей CDR1 и CDR2. Скрининг может проводиться в соответствии с любым способом скрининга, подходящим для скрининга антител из библиотек антител, таким как способ фагового дисплея.
Для получения и экспрессии измененной последовательности антитела могут быть использованы стандартные методы молекулярной биологии. Антитело, кодируемое измененной(ыми) последовательностью(ами) антитела, представляет собой антитело, которое сохраняет один, несколько или все функциональные свойства связывающих β-клото антител, описанных в настоящем документе, причем функциональные свойства включают, но не ограничиваются ими, специфическое связывание с β-клото человека, яванского макака, крысы и/или мыши; и антитело активирует передачу сигналов, опосредованную FGF21, например, передачу сигналов, зависящую от FGF21-рецептора, в анализе клеток FGFR1c_ β-клото_HEK293 pERK.
В некоторых вариантах способов конструирования антител по настоящему изобретению мутации могут вводиться случайным образом или селективно на протяжении всей или части кодирующей последовательности связывающего β-клото антитела, и полученные модифицированные связывающие β-клото антитела могут быть подвергнуты скринингу на связывающую активность и/или другие функциональные свойства, как описано в настоящем документе. Мутационные способы были описаны в данной области техники. Например, в публикации PCT WO 02/092780, Short, описаны способы создания и скрининга мутаций антител с использованием насыщенного мутагенеза, сборки синтетическим лигированием или их комбинации. Альтернативно, в публикации PCT WO 03/074679, Lazar et al., описаны способы с использованием компьютерных программ при скрининге для оптимизации физико-химических свойств антител.
В некоторых вариантах осуществления изобретения антитела были сконструированы с удалением сайтов дезамидирования. Известно, что дезамидирование вызывает структурные и функциональные изменения в пептиде или белке. Дезамидирование может привести к снижению биологической активности, а также к изменениям фармакокинетики и антигенности белкового препарата. (Anal Chem. 2005 Mar 1;77(5):1432-9).
В некоторых вариантах осуществления изобретения антитела были сконструированы для увеличения pI и улучшения их лекарственно-подобных свойств. pI белка является ключевым фактором, определяющим общие биофизические свойства молекулы. Известно, что антитела, которые имеют низкие pI, менее растворимы, менее стабильны и склонны к агрегации. Кроме того, очистка антител с низким pI является сложной и может быть проблематичной, особенно при масштабировании для клинического применения. Увеличение pI антител против β-клото, или Fab, по изобретению улучшало их растворимость, позволяя вводить антибиотики при более высоких концентрациях (>100 мг/мл). Состав антител с высокими концентрациями (например, >100 мг/мл) имеет преимущество в том, что можно вводить более высокие дозы антител в глаза пациентов посредством интравитреальных инъекций, что, в свою очередь, позволит снизить частоту дозирования, что является значительным преимуществом при лечении хронических заболеваний, включая сердечно-сосудистые заболевания. Более высокие pI могут также увеличивать опосредованную FcRn рециркуляцию антитела варианта IgG, что позволяет лекарственному средству оставаться в организме в течение более длительного времени, требуя меньше инъекций. Наконец, из-за более высоких значений pI значительно улучшается общая стабильность антител, что приводит к увеличению срока хранения и биоактивности in vivo. Предпочтительно, pI больше или равно 8,2.
Функциональные свойства измененных антител могут быть оценены с использованием стандартных анализов, доступных в данной области и/или описанных в настоящем документе, таких как приведенные в примерах (например, ELISA).
Профилактическое и терапевтическое применение
Антитела, которые связывают β-клото, описанные в настоящем документе, в терапевтически полезной концентрации могут быть использованы для лечения заболевания или расстройства, ассоциированного с патологической сигнализацией FGF21 (например, патологической активация передачи сигналов, опосредованной FGF21 и/или передачи сигналов рецепторами FGF21), путем введения субъекту, нуждающемуся в этом, эффективного количества антител или антигенсвязывающих фрагментов по изобретению. Настоящее изобретение относится к способу лечения метаболических нарушений, связанных с FGF21, путем введения субъекту, нуждающемуся в этом, эффективного количества антител по изобретению. Настоящее изобретение относится к способу лечения связанных с FGF21 сердечно-сосудистых заболеваний путем введения субъекту, нуждающемуся в этом, эффективного количества антител по изобретению.
Антитела по изобретению могут быть использованы, помимо прочего, для превентативного лечения, предотвращения и улучшения состояний или расстройств, связанных с FGF21, включая, но этим не ограничиваясь, метаболические, эндокринные и сердечно-сосудистые расстройства, таких как ожирение, сахарный диабет типа 1 и типа 2, панкреатит, дислипидемию, неалкогольную жировую болезнь печени (NAFLD), неалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперсулинемию, непереносимость глюкозы, гипергликемию, метаболический синдром, острый инфаркт миокарда, гипертензию, сердечно-сосудистое заболевание, атеросклероз, периферическую артериальную болезнь, инсульт, сердечную недостаточность, ишемическую болезнь сердца, заболевание почек, диабетические осложнения, нейропатию, гастропарез, заболевания, связанные с тяжелыми инактивирующими мутациями в рецепторе инсулина, и другие метаболические нарушения, и для снижения смертности и заболеваемости критически больных пациентов.
Антитела по изобретению могут также использоваться в комбинации с другими агентами для профилактики, лечения или облегчения расстройств, связанных с FGF21. Например, можно использовать статиновую терапию в комбинации с миметическими антителами FGF21 и антигенсвязывающими фрагментами по изобретению для лечения пациентов с сердечно-сосудистыми или метаболическими нарушениями.
В конкретных аспектах в настоящем документе предложен способ снижения веса тела (например, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, по меньшей мере на 10%, по меньшей мере на 12%, по меньшей мере на 15% или по меньшей мере на 20%), включающий введение субъекту, нуждающемуся в этом, эффективного количества фармацевтической композиции, содержащей описанные в настоящем документе антитело или антигенсвязывающий фрагмент, связывающие β-клото и способные увеличивать активность β-клото и FGFR1c.
В конкретных аспектах в настоящем документе предложен способ снижения аппетита или потребления пищи (например, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, при по меньшей мере на 10%, по меньшей мере на 12%, по меньшей мере на 15% или по меньшей мере на 20%), включающий введение субъекту, нуждающемуся в этом, эффективного количества фармацевтической композиции, содержащей описанные в настоящем документе антитело или антигенсвязывающий фрагмент, связывающие β-клото и способные увеличивать активность β-клото и FGFR1c.
В конкретных аспектах приведенные в настоящем документе аспекты представляют собой способ снижения (например, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, по меньшей мере на 10% по меньшей мере на 12%, по меньшей мере на 15% или по меньшей мере на 20%) концентрации триглицеридов в плазме (ТГ) или концентрации общего холестерина в плазме (ТС) у пациента, включая введение нуждающемуся в этом субъекту эффективного количества фармацевтической композиции, содержащей описанные в настоящем документе антитело или антигенсвязывающий фрагмент, связывающие β-клото и способные увеличивать активность β-клото и FGFR1c.
В конкретных аспектах субъект страдает от метаболического расстройства, такого как ожирение, сахарный диабет 1-го типа и типа 2, панкреатит, дислипидемия, безалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперинсулинемия, непереносимость глюкозы, гипергликемия и метаболический синдром. В конкретных аспектах субъект страдает сердечно-сосудистым расстройством.
Фармацевтические композиции
Изобретение относится к фармацевтическим композициям, содержащим связывающие β-клото антитела (интактные или связывающие фрагменты) в составе вместе с фармацевтически приемлемым носителем. Композиции могут дополнительно содержать один или несколько других терапевтических агентов, которые пригодны для лечения или профилактики, например, сердечно-сосудистых заболеваний. Фармацевтически приемлемые носители усиливают или стабилизируют композицию, либо могут быть использованы для облегчения изготовления композиции. Фармацевтически приемлемые носители включают растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и абсорбирующие агенты и тому подобное, являющиеся физиологически совместимыми.
Фармацевтическая композиция по настоящему изобретению может быть введена различными способами, известными в данной области. Путь и/или способ введения варьируется в зависимости от желаемых результатов. Предпочтительно, чтобы введение было интравитреальным, внутривенным, внутримышечным, внутрибрюшинным или подкожным, или вводимым проксимально к сайту мишени. Фармацевтически приемлемый носитель должен быть подходящим для интравитреального, внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от способа введения активное вещество, то есть антитело, биспецифическое и мультиспецифическое действующее вещество, может быть покрыто материалом для защиты соединения от действия кислот и других естественных условий, которые могут инактивировать соединение.
Композиция должна быть стерильной и жидкой. Соответствующую текучесть можно поддерживать, например, с помощью покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и используя поверхностно-активные вещества. Во многих случаях предпочтительно включать в композиции изотонические агенты, например сахара, полиспирты, такие как маннит или сорбит, и хлорид натрия. Продолжительное поглощение инъекционных композиций может быть обеспечено путем включения в состав агента, который задерживает абсорбцию, например моностеарат алюминия или желатин.
Фармацевтические композиции по изобретению могут быть получены в соответствии со способами, хорошо известными и обычно применяемыми в данной области техники. Смотри, например, Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20th ed., 2000; и Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978. Фармацевтические композиции предпочтительно изготавливают в условиях надлежащей производственной практики GMP. Обычно в фармацевтических композициях по изобретению используют терапевтически эффективную дозу или эффективную дозу связывающего β-клото антитела. Связывающие β-клото антитела вводят в состав фармацевтически приемлемых лекарственных форм обычными способами, известными специалистам в данной области. Схемы дозирования регулируют для обеспечения оптимального желаемого ответа (например, терапевтического ответа). Например, может быть введен один болюс, несколько разделенных доз могут быть введены в течение длительного периода временем, или доза может быть пропорционально уменьшена или увеличена, как показано необходимостью терапевтической ситуации. Особенно удобно составлять парентеральные композиции в единичной дозированной форме для удобства введения и единообразия дозировки. Используемая в настоящем документе форма дозировки относится к физически дискретным единицам, подходящим в качестве унифицированных доз для пациентов, подлежащих лечению; каждая единица содержит предопределенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, в сочетании с необходимым фармацевтическим носителем.
Фактические уровни дозировки активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать, чтобы получить количество активного ингредиента, которое эффективно для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения, не являясь токсичным для пациента. Выбранный уровень дозировки зависит от множества фармакокинетических факторов, включая активность конкретных соединений по настоящему изобретению или их сложных эфиров, солей или амидов, пути введения, время введения, скорость экскреции конкретного применяемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или вещества, используемые в комбинации с конкретными применяемыми соединениями, возраст, пол, вес, состояние, общее состояние здоровья и предысторию болезни пациента, подлежащего лечению, и подобные факторы.
Врач или ветеринар могут начать с доз антител по изобретению, используемых в фармацевтической композиции, на уровнях ниже, чем требуется для достижения желаемого терапевтического эффекта, и постепенно увеличивать дозировку до достижения желаемого эффекта. Обычно эффективные дозы композиций по настоящему изобретению для лечения сердечно-сосудистых нарушений, описанных в настоящем документе, варьируются в зависимости от множества различных факторов, включая способы введения, сайт-мишень, физиологическое состояние пациента, независимо от того, является ли пациент человеком или животных, других назначаемых препаратов, и того, является ли лечение профилактическим или терапевтическим. Лечебные дозы должны быть оттитрованы для оптимизации безопасности и эффективности. В конкретном варианте осуществления для системного введения с антителом доза составляет от примерно 0,0001 до 100 мг/кг или от 0,01 до 15 мг/кг массы тела хозяина. В конкретном варианте осуществления для внутривизорного введения с антителом доза может варьироваться от 0,1 мг/глаз до 5 мг/глаз. Например, 0,1 мг/мл, 0,2 мг/мл, 0,3 мг/мл, 0,4 мг/мл, 0,5 мг/мл, 0,6 мг/мл, 0,7 мг/мл, 0,8 мг/мл, 0,9 мг/мл, 1,0 мг/мл, 1,1 мг/мл, 1,2 мг/мл, 1,3 мг/мл, 1,4 мг/мл, 1,5 мг/мл, 1,6 мг/мл, 1,7 мг/мл, 1,8 мг/мл, 1,9 мг/мл, 2,0 мг/мл, 2,1 мг/мл, 2,2 мг/мл, 2,3 мг/мл, 2,4 мг/мл, 2,5 мг/мл, 2,6 мг/мл, 2,7 мг/мл, 2,8 мг/мл, 2,9 мг/мл, 3,0 мг/мл, 3,1 мг/мл, 3,2 мг/мл, 3,3 мг/мл, 3,4 мг/мл, 3,5 мг/мл, 3,6 мг/мл, 3,7 мг/мл, 3,8 мг/мл, 3,9 мг/мл, 4,0 мг/мл, 4,1 мг/мл, 4,2 мг/мл, 4,3 мг/мл, 4,4 мг/мл, 4,5 мг/мл, 4,6 мг/мл, 4,7 мг/мл, 4,8 мг/мл, 4,9 мг/мл, или 5,0 мг/мл. Примерный режим лечения предусматривает системное введение один раз каждые две недели или один раз в месяц или один раз каждые 3-6 месяцев или по мере необходимости (PRN).
Антитело обычно вводят несколько раз. Интервалы между разовыми дозами могут быть недельными, ежемесячными или ежегодными. Интервалы могут также быть нерегулярными, как показано путем измерения уровня связывающего β-клото антитела в крови у пациента. Кроме того, альтернативные интервалы дозировки могут определяться врачом, и может быть эффективным ежемесячное введение или по мере необходимости. В некоторых способах системного введения дозу корректируют для достижения концентрации антител в плазме 1-1000 мкг/мл и в некоторых способах 25-500 мкг/мл. Альтернативно, антитело можно вводить в виде препарата с замедленным высвобождением, и в этом случае требуется менее частое введение. Дозировка и частота варьируются в зависимости от периода полувыведения антитела у пациента. Обычно гуманизированные антитела показывают более длительный период полувыведения, чем химерные антитела и нечеловеческие антитела. Дозировка и частота введения могут варьироваться в зависимости от того, является ли лечение профилактическим или терапевтическим. При профилактическом применении относительно низкая дозировка вводится относительно нечасто в течение длительного периода времени. Некоторые пациенты продолжают получать лечение всю оставшуюся жизнь. При терапевтическом применении иногда требуется относительно высокая дозировка с относительно короткими интервалами, пока прогрессирование заболевания не будет уменьшено или прекращено, и, предпочтительно, до тех пор, пока у пациента не проявится частичное или полное улучшение симптомов заболевания. После этого пациенту можно назначать профилактический режим.
ПРИМЕРЫ
Следующие примеры приведены для дополнительной иллюстрации настоящего изобретения, но не для ограничения его объема. Другие варианты изобретения будут очевидны для обычного специалиста в данной области и охватываются прилагаемой формулой изобретения.
Пример 1: Получение клеток FGFR1c_β-клото_300.19 человека для использования в качестве антигена
Клетки 300.19, которые стабильно экспрессировали FGFR1c человека (1-386 aa) и β-клото, получали для использования в качестве полного клеточного антигена. Полноразмерную кДНК, кодирующая β-клото человека (номер доступа GenBank NM_175737), клонировали в сайты EcoRI и EcoRV pEF1/Myc-His B (Invitrogen номер в катологе #V92120). кДНК, кодирующую аминокислоты 1-386 FGFR1c человека (номер доступа GenBank NM_023106), клонировали в сайты BamHI и NotI pEF6/Myc-His B (Invitrogen, номер в каталоге V96220). В обе конструкции включали последовательность Козака (CACC) сразу перед старт-кодоном, а стоп-кодон добавляли перед тагом Myc-His в векторе. Клетки мышей pre-B 300-19 совместно трансфицировали с плазмидами β-клото и FGFR1c путем электропорации, используя устройство Amaxa Nucleofector и набор Nucleofector (Lonza, номер в каталоге #VCA-1003). В течение 3 недель отбирали стабильные клоны, используя 1 мг/мл генетицина (Invitrogen, номер в каталоге #10131) и 8 мкг/мл бластицидина (Invitrogen, номер в каталоге #46-1120).
Пример 2: Получение клеток FGFR_β-клото HEK293 для использования в клеточных анализах
Для проверки специфичности связывания, функциональной активности или ортологичной перекрестной реактивности антител к β-клото получали клетки HEK293, стабильно экспрессирующие FGFR1c_β-клото человека, FGFR2c_β-клото человека, FGFR3c_β-клото человека, FGFR4_β-клото человека или FGFR1c_β-клото яванского макака, используя стандартные методы трансфекции Lipofectamine 2000 и методы селекции клонов клеток.
Использовали следующие плазмиды экспрессии млекопитающих, кодирующие полноразмерные кДНК β-клото человека (NM_175737), FGFR1c человека (NM_023106), FGFR2c человека (NP_001138387), FGFR3c человека (NP_000133) или FGFR4 человека (NP_998812): для β-клото яванского макака полноразмерная последовательность была ПЦР амплифицирована из кДНК жировой ткани яванского макака (BioChain, номер в каталоге #C1534003-Cy) с праймерами на основе последовательностей β-клото человека и резус-обезьян, и клонирована. кДНК FGFR1c яванского макака клонировали из кДНК жировой ткани яванского макака (BioChain, номер в каталоге #C1534003-Cy), используя праймеры на основе последовательности FGFR1 человека (#NM_023106) и, как было показано, она на 100% идентична по аминокислотному уровню FGFR1c человека. Таким образом, конструкцию кДНК FGFR1c человека, описанную выше, использовали для получения клеток HEK293, которые стабильно экспрессировали β-клото яванского макака и FGFR1c человека (#NM_023106), так как аминокислотные последовательности FGFR1c человека и яванского макака идентичны.
Пример 3: Измерение активации рецептора FGFR_β-клото с использованием клеточного анализа pERK
Для культивирования клеток и для измерения уровней фосфо-ERK 1/2 (pERK) использовали стандартные методики. Коротко, клетки HEK293, стабильно экспрессирующие FGFR1c_β-клото человека, FGFR2c_β-клото человека, FGFR3c_β-клото человека, FGFR4_β-клото человека или FGFR1c_β-клото яванского макака поддерживали в среде DMEM (Invitrogen, 11995) содержащей 10% FBS (Hyclone, SH30071), бластицидин (Invitrogen, A1113902) и генетицин (Invitrogen, 10131035) при 37°C в 5% CO2. Клетки высевали в 384-луночные планшеты, покрытые поли-D-лизином, (BD Biosciences, 354663) и инкубировали в течение ночи при 37°C в 5% CO2, после чего переводили на сывороточное голодание.
Супернатанты гибридомы или антитела против β-клото разводили в среде Freestyle 293, и в планшет добавляли различные концентрации антител. После инкубации клетки промывали, затем лизировали буфером для лизиса. Клеточные лизаты переносили в 384-луночный аналитический планшет (PerkinElmer, Cat. #6008280) и для измерения уровней фосфо-ERK 1/2 использовали набор AlphaScreen SureFireTM pERK 1/2 (Perkin Elmer, TGRES10K). Планшеты считывали на многоканальном ридере EnVision 2104 (Perkin Elmer), используя стандартные настройки AlphaScreen. Данные «доза-ответ» графически отображали как кратность активности pERK по сравнению с основной концентрацией против концентрации белка для определения значений EC50, используя уравнение Y=нижнее+(верхнее-нижнее)/(1+10^((LogEC50-X)×HillSlope)) и программное обеспечение GraphPad Prism 5.
Пример 4. Получение моноклональных антител
Антитела против β-клото человека получали в мышах Balb/c (штамм Jackson Laboratory: BALB/cJ) или Bcl2 22 wehi (штамм Jackson Laboratory: C.Cg-Tg(BCL2)22Wehi/J) путем иммунизации цельными клетками, по существу как описано в работе Dreyer et al. (2010) (Dreyer AM et al. (2010) BMC Biotechnology 10:87).
Коротко, 1×107 клеток FGFR1c_β-клото_300.19 человека вводили мышам Balb/c шесть раз с интервалами от 10 до 30 дней. Первые инъекции цельных клеток проводили вместе с полным адъювантом Фрейнда (Sigma-Aldrich F5881). Клетки и адъювант не смешивали, а вводили подкожно отдельно в двух близких, но разных местах. После этого проводили внутрибрюшинные инъекции тех же клеток либо с системой адъювантов Sigma (Sigma-Aldrich S6322), либо без адъюванта.
Используя мышей Bcl2 22 wehi, 1×107 клеток FGFR1c_β-клото_300.19 человека вводили этим животным четыре раза с интервалом в семь дней. Первые инъекции проводили вместе с полным адъювантом Фрейнда (Sigma-Aldrich F5881). Клетки и адъювант вводили отдельно двумя наборами в два близких, но различных места, подкожно. Последующие инъекции клеток проводили подкожно без адъюванта.
Иммунные реакции у иммунизированных мышей измеряли в анализе клеток, активированных флуоресценцией (FACS). Сыворотку иммунизированных мышей, разведенную в 1000 или 10000 раз, использовали для окрашивания клеток FGFR1c_β-клото-HEK человека и FGFR1c_β-клото-300.19 человека, а затем вторым аллоцикоцианиновым (APC) антителом против IgG мыши (Jackson ImmunoResearch Cat #115-136-071). Флуоресценцию считывали на проточном цитометре Becton Dickinson LSRII или Foressa. Для слияния методом электрошока выбирали четырех мыши с самым высоким титром.
Пример 5: Скрининг, субклонирование и селекция гибридомы
2×108 спленоцитов и 5×107 слитых клеток-партнеров F0 (ATCC, CRL-1646) промывали в среде Cytofusion (LCM-C, Cyto Pulse Sciences) и проводили слияние, используя генератор Hybrimune Waveform (Cyto Pulse Sciences, модель CEEF-50B) в соответствии с описанием производителя с пиковым импульсом в 600 вольт. Клетки высевали в 384 луночные планшеты с рассчитанной плотностью 3000 клеток F0 на лунку и культивировали в среде для отбора HAT (Sigma-Aldrich, номер в каталоге H0262).
Первый скрининг осуществляли с использованием высокопроизводительной платформы FACS (Anderson, Paul. Automated Hybridoma Screening, Expansion, Archiving and Antibody Purification. In: 3rd Annual 2014 SLAS Conference. Jan. 18-22, 2014, San Diego, CA). Коротко, супернатанты гибридомы инкубировали с клеточными линиями, стабильно экспрессирующими FGFR1c_β-клото человека и не экспрессирующими его, и связывание антитела определяли с помощью второго антитела против IgG мыши APC (Jackson ImmunoReseach Cat#115-136-071).
Антитела из каждого супернатанта гибридомы исследовали на одновременное связывание с четырьмя линиями клеток со штрих-кодом: родительские клетки 300.19, клетки FGFR1c_β-клото_300.19 человека, родительские клетки HEK 293 и клетки FGFR1c-β-клото-HEK 293 человека. При первичном скрининге было выбрано 348 совпадений. Первичные совпадения распределяли в 96-луночные планшеты, и связывание снова подтверждали на клетках FGFR1c_β-клото-HEK 293 человека с помощью FACS, с выходом 122 подтвержденных совпадений. Строили профиль клеточной активации рецепторного комплекса FGFR1c_β-клото человека, используя анализ фосфо-ERK 1/2, описанный в примере 2, для супернатантов, содержащих HAT (гипоксантин-аминоптерин-тимидин), 115 совпадений, связывание которых повторно подтверждалось с помощью FACS.
Гибридомы с наивысшей клеточной активностью фосфо-ERK 1/2 в супернатантах увеличивали в объеме, и IgG очищали от их супернатантов. Строили профиль клеточной активности рецепторного комплекса FGFR1c_β-клото человека, используя анализ фосфо-ERK 1/2, описанный в примере 2, для очищенных IgG из 74 гибридом. Строили профиль ортологичной перекрестной реактивности для рецепторного комплекса FGFR1c_β-клото яванского макака и селективности для рецепторных комплексов FGFR2c_β-клото человека и FGFR3c_β-клото человека, используя анализ фосфо-ERK 1/2, описанный в примере 2, для IgG из гибридомы с лучшей эффективностью активации фосфор-ERK 1/2 рецепторного комплекса FGFR1c_β-клото человека. На основе результатов этих профилей отбирали клоны гибридом, 99G09 и 127F19 для построения дополнительного профиля.
Для оценки сигнального пути 99G09 и 127F19 в клетках, экспрессирующих α-клото, клетки HEK293 трансфицировали α-клото, Egr1-люциферазой и люмиферазой Renilla. Коротко, клетки HEK293 культивировали в DMEM, 10% FBS и высевали в количестве 30000 клеток/лунка и трансфицировали клото, Egr-1-luc и TK-Rennila, используя Lipofectamine 2000. На следующий день FGF23, FGF21, 99G09 и 127F19 разводили до указанной концентрации в DMEM с добавлением 0,1% FBS, и добавляли к трансфицированным клеткам в течение ночи. Активность люциферазы определяли с помощью набора для люциферазного анализа Dual-Glo (Promega, E2920) в соответствии с инструкцией производителя. Как и ожидалось, FGF23, для сигнального пути которого требуется экспрессия α-клото, показал сильную экспрессию люциферазы. Однако ни в FGF21, ни в 99G09, ни в 127F19 не было значительной экспрессии люциферазы, что предполагает, что α-клото не действует как корецептор для FGF21 или антител-миметиков против FGF21.
Пример 6: Гуманизация и созревание аффинности моноклональных антител
Гуманизация
Процесс гуманизации хорошо описан в данной области (Jones PT et al. (1986) Nature 321: 522-525; Queen C et al. (1989) PNAS USA 86: 10029-10033; Riechmann L et al. (1988) Nature 33:323-327; Verhoeyen M et al. (1988) Science 239: 1534-1536). Термин гуманизация описывает перенос антигенсвязывающего сайта антитела, отличного от человека, например, мышиного антитела, на акцепторный каркас человека, например, последовательности зародышевой линии человека (Retter I et al. (2005). Nucleic Acids Res. 33:D671-D674.).
Основной целью гуманизации антитела является минимизация риска развития иммуногенного ответа на антитело у людей (Rebello PR et al. (1999) Transplantation 68: 1417-1420). Антигенсвязывающий сайт содержит определяющие комплементарность области (CDR) (Chothia C and Lesk AM (1987) Journal of Molecular Biology 196: 901-917; Kabat E et al. (1991) Anon. 5th Edition ed; NIH Publication No. 91: 3242) и положения вне CDR, то есть в каркасном участке вариабельных доменов (VL и VH), которые напрямую или косвенно влияют на связывание. Остатки в каркасном участке, которые могут непосредственно влиять на связывание, могут, например, находится в так называемой «внешней» петлевой области, расположенной между CDR2 и CDR3. Остатки, которые косвенно влияют на связывание, находятся, например, в так называемых областях Верньера (Foote J, Winter G. (1992) Journal of Molecular Biology 224: 4 87-499). Считается, что они обеспечивают конформацию CDR. Эти положения вне CDR учитываются при выборе подходящего акцепторного каркасного участка для минимизации числа отклонений конечного гуманизированного антитела от акцепторной последовательности, полученной из зародышевой линии человека, в каркасных участках.
Оптимизация последовательности при созревании аффинности
Известно, что некоторые мотивы аминокислотных последовательностей подвергаются посттрансляционной модификации (ПТМ), такой как гликозилирование (то есть NxS/T, x любая кислота, но не P), окисление свободных цистеинов, дезамидирование (например, NG) или изомеризация (например, DG). Если они присутствуют в регионах CDR, эти мотивы идеально удаляются с помощью сайт-направленного мутагенеза, чтобы повысить однородность продукта.
Процесс созревания аффинности подробно описан в данной области. Среди большого числа систем дисплея фаговый дисплей (Smith GP, 1985, Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 228: 1315-1317) и дисплей на эукариотических клетках, таких как дрожжи (Boder ET and Wittrup KD, 1997, Yeast surface display for screening combinatorial polypeptide libraries. Nature Biotechnology 15: 553-557), по-видимому, являются наиболее часто используемыми системами для выбора реакции антитело-антиген. Преимущества этих систем дисплея состоят в том, что они подходят для широкого спектра антигенов, и что жесткость выбора может быть легко отрегулирована. В фаговом отображении могут быть отображены фрагменты scFv или Fab, а в дрожжевых клетках - полноразмерный IgG. Эти обычно применяемые методы позволяют выбирать желаемые варианты антител из более крупных библиотек с разбросами более 10E7. Библиотеки с меньшим разнообразием, например, 10E3, могут быть исследованы с помощью микроэкспрессии и ELISA. Нецелевые или случайные библиотеки вариантов антител могут быть получены, например, с помощью ПЦР с внесением ошибок (Cadwell RC and Joyce GF, 1994, Mutagenic PCR. PCR Methods Appl. 3: S136-S140), и они обеспечивают очень простой, но иногда ограниченный подход. Другой стратегией является CDR направленная диверсификация кандидата антител. Одно или несколько положений в одном или более CDR могут быть специфическими целями с использованием, например, вырожденных олигонуклеотидов (Thompson J et al., 1996, Affinity maturation of a high-affinity human monoclonal antibody against the third hypervariable loop of human immunodeficiency virus: use of phage display to improve affinity and broaden strain reactivity. J.Mol.Biol. 256: 77-88), тринуклеотидного мутагенеза (TRIM) (Kayushin AL et al., 1996, A convenient approach to the synthesis of trinucleotide phosphoramidites--synthons for the generation of oligonucleotide/peptide libraries. Nucleic Acids Res. 24: 3748-3755) или любого другого подхода, известного в данной области.
Получение плазмид экспрессии
Последовательности ДНК, кодирующие гуманизированные домены VL и VH, были заказаны у компании GeneArt (Life Technologies Inc. Regensburg, Germany), включая оптимизацию кодонов для человека. Последовательности, кодирующие домены VL и VH, субклонировали путем вырезания и встраивания из векторов, полученных от компании GeneArt, в векторы экспрессии, подходящие для секреции в клетках млекопитающих. Тяжелую и легкую цепи клонировали в отдельные векторы экспрессии для обеспечения котрансфекции. Элементы вектора экспрессии включали промотор (энхансерный промотор цитомегаловируса (CMV)), сигнальную последовательность для облегчения секреции, сигнал полиаденилирования и терминатор транскрипции (ген бычьего гормона роста (BGH)), элемент, обеспечивающий эписомальную репликацию и репликацию в прокариотах (например, точка начала репликации SV40 и ColE1 или другие известные в данной области) и элементы, обеспечивающие селекцию (ген устойчивости к ампициллину и маркер зеоцин).
Экспрессия и очистка кандидатных гуманизированных антител
Эмбриональные клетки почки человека, конститутивно экспрессирующие большой Т-антиген SV40 (HEK293-T ATCC11268), являются одной из предпочтительных линий клеток-хозяев для временной экспрессии гуманизированных и/или оптимизированных белков IgG. Трансфекцию проводят с помощью PEI (полиэтиленимин, линейный MW 25000, Polysciences, США номер в каталоге 23966) в качестве трансфекционного реагента. Основной раствор PEI готовили путем тщательного растворения 1 г PEI в 900 мл воды со степенью очистки для клеточной культуры при комнатной температуре (RT).
Очистку проводили в две стадии, в результате были получены гуманизированные mAb со зрелой аффиностью из гибридом мыши. NOV001 представляет собой гуманизированное mAb со зрелой аффиностью, полученное из гибридомы мыши 99G09. NOV002 представляет собой гуманизированное mAb со зрелой аффиностью, полученное из гибридомы мыши 127F19. Изотипы IgG1 L234A/L235A (LALA) или IgG1κ D265A/P329A (DAPA) были выбраны в качестве профилактических средств для снижения способности антитела стимулировать антителозависимую клеточную цитотоксичность (ADCC) и комплементарную цитотоксичность (CDC) (Hezareh M et al. (2001) Journal of Virology 75: 12161-12168). NOV001 является изотипом IgG1 (LALA), а NOV003 является изотипом IgG1 (DAPA) того же mAb. NOV002 является изотипом IgG1 (LALA), а NOV004 является изотипом IgG1 (DAPA) того же mAb.
Пример 7: Характеристика моноклональных антител in vitro
Анализ SET, как описано в примере 5, использовали для оценки KD mAb против β-клото человека. Значение KD NOV001, NOV002 и NOV004 составляет примерно 42 пМ, 8 пМ и 9 пМ, соответственно (фигура 1). Анализ pERK, как описано в примере 3, использовали для создания профиля активности mAb в отношении рецептора FGFR_β-клото. NOV002 активировал рецепторный комплекс FGFR1c_β-клото человека и яванского макака с EC50, равным примерно 5 нМ и 37 нМ, соответственно (фигура 2). NOV004 активировал рецепторный комплекс FGFR1c_β-клото человека и яванского макака с EC50, равным примерно 6 нМ и 40 нМ, соответственно (фигура 2). NOV002 и NOV004 не активировали рецепторные комплексы FGFR2c_β-клото, FGFR3c_β-клото или FGFR4_β-клото (фигура 3). Строили профили MAb в отношении активности FGF23, как описано в примере 5. NOV002 и NOV004 не проявляли активность в отношении FGF23 (фигура 4). Строили профиль MAb в отношении перекрестной реактивности к рецепторному комплексу FGFR_β-клото мыши, как описано в примере 5. MAb NOV002 и NOV004 не вызывали захват глюкозы адипоцитами 3T3L1 мыши (фигура 5).
Пример 8: Картирование эпитопа по обмену водород-дейтерий внеклеточного домена β-клота человека с NOV001 и NOV002
Обмен водород-дейтерий (HDx) в сочетании с масс-спектрометрией (MS) (Woods VL et al. (2001) High Resolution, High-Throughput Amide Deuterium Exchange-Mass Spectrometry (DXMS) Determination of Protein Binding Site Structure and Dynamics: Utility in Pharmaceutical Design. J. Cell. Biochem. Supp.; 84(37): 89-98) использовали для картирования предполагаемых сайтов связывания NOV001 и NOV002 с внеклеточным доменом β-клото человека (ECD) (52-997aa). В остове HDx водороды амида белков заменяли на дейтерий. Этот процесс чувствителен к структуре/динамическим свойствам белка и доступности растворителей и, следовательно, способен сообщать о местах, в которых происходит снижение захвата дейтерия при связывании с лигандом. Цель этих экспериментов состояла в идентификации возможных эпитопов и понимании динамики ECD β-клото человека при связывании с NOV001 или NOV002. Изменения в захвате дейтерия чувствительны как к прямому связыванию, так и к аллостерическим событиям.
Эксперименты HDx-MS проводили способами, аналогичными описанным в литературе (Chalmers MJ et al. (2006) Probing protein ligand interactions by automated hydrogen/deuterium exchange mass spectrometry. Anal. Chem.; 78(4): 1005-1014). Эксперименты проводили на платформе Waters HDx-MS, которая включает автосамплер LEAP, nanoACQUITY UPLC и масс-спектрометр Synapt G2. Дейтериевым буфером, используемым для мечения белкового каркаса β-клото человека ECD (52-997aa) был D-PBS с pH 7,2; общий процент дейтерия в растворе составлял 95%. Белок β-клото человека (52-997aa) заказывали у компании R&D System (5889-KB-050). Проводили диализ белка в буфере PBS pH 7,2 в течение ночи перед анализом HDx-MS. Для экспериментов с мечением дейтерием β-клото человека ECD (52-997aa) в отсутствие антител, 300 пмоль β-клото человека ECD (52-997aa) объемом 13 мкл разводили в 100 мкл дейтериевого буфера (95% дейтерия) в охлажденной пробирке и инкубировали в течение 15 минут на ротаторе при 4°C. Затем реакцию маркировки гасили 100 мкл охлажденного буфера при 2°C в течение пяти минут с последующим введением в систему LC-MS для автоматического ферментативного переваривания пепсином и пептидного анализа.
Для экспериментов с мечением дейтерием β-клото человека ECD (52-997aa) в присутствии NOV001 или NOV002 350 пмоль NOV001 или NOV002 сначала иммобилизировали на гранулы Thermo Protein G Plus и перекрестно связывали с DSS (дисукцинимидил субератом). Для проведения экспериментов по маркировке гранулы антител, содержащие антитело 350 пмоль, инкубировали с ECD (52-997aa) человека с размером частиц 300 пмоль в течение 20 минут при 4°C. Через 20 минут гранулы промывали 200 мкл PBS-буфера. Затем добавляли 200 мкл охлажденного дейтериевого буфера (84,1% дейтерия) и комплекс инкубировали в течение 15 минут при 4°C. Через 15 минут дейтериевый буфер центрифугировали, и реакцию маркировки гасили 200 мкл охлажденного на льду буфера в течение 4,5 минут. После центрифугирования образца в течение 30 секунд в центрифуге потушенный раствор вводили в систему LC-MS для автоматического ферментативного переваривания пепсином и пептидного анализа.
Все эксперименты по обмену дейтерия гасили с использованием 0,5М TCEP (трис(2-карбоксиэтил)фосфин) и 3М мочевины (pH=2,6). После гашения обмениваемый антиген подвергали он-лайн ферментативному перевариванию пепсином, используя колонку с иммобилизованным пепсином Poroszyme (2,1×30 мм) при 12°C с последующим захватом на колонке для захвата Waters Vanguard HSS T3. Пептиды элюировали из улавливающей колонны и разделяли на колонке Waters BEH C18 1×100 мм (поддерживали при 1°C) со скоростью потока 40 мкл/мин с использованием бинарного 8,4-минутного градиента от 2 до 35% В (подвижная фаза А составляла 99,9% воды и 0,1% муравьиной кислоты, подвижная фаза В составляла 99,9% ацетонитрила и 0,1% муравьиной кислоты).
Для β-клото-ECD человека (52-997aa) 82% последовательности контролировали экспериментами по обмену с дейтерием, как описано в настоящем документе. Для дифференциальных экспериментов между NOV001 или NOV002 связанными и несвязанными β-клото ECD, информативным является исследовать изменение включения дейтерия между двумя состояниями. В таблице 2 отрицательное значение указывает на то, что комплекс NOV001- или NOV002-β-клото ECD подвергается меньшему поглощению дейтерия по сравнению с β-клото ECD. Снижение захвата дейтерия может быть связано с защитой антигена от обменного дейтерия с помощью антитела или путем стабилизации водородной связи. Напротив, положительное значение указывает на то, что комплекс подвергается более активному захвату дейтерия по сравнению с β-клото ECD. Увеличение поглощения дейтерия может быть связано с дестабилизацией водородных сетей (то есть локализованным разворачиванием белка). При изучении дифференциального изменения обмена дейтерия между двумя различными состояниями, такими как апо-β-клото-ECD и комплекс NOV001- или NOV002-β-клото ECD, подход обычно используется для определения того, являются ли изменения значительными. Как правило, если разница больше 0,5 Да, то разница считается значительной (Houde D, et al. (2010) J. Pharma. Sci.; 100(6): 2071-2086).
В таблице 2 приведено изменение включения дейтерия для комплекса NOV001- или NOV002-β-клото ECD по сравнению с β-клото ECD. Используя значительную отсечку со значением -0,5 Да (Houde D, et al. (2010) The Utility of Hydrogen/Deuterium Exchange Mass Spectrometry in Biopharmaceutical Comparability Studies. J. Pharma. Sci.; 100(6): 2071-2086). Следующие пептиды значительно защищены в комплексе NOV001-β-клото ECD: 245-266, 344-349, 421-429, 488-498, 509-524, 536-550, 568-576, 646-670, 696- 700, 773-804, 834-857 и 959-986aa. Следующие пептиды значительно защищены в комплексе NOV002-β-клото ECD: 246-265, 343-349, 421-429, 488-498, 509-524, 536-550, 568-576, 646-669, 773- 804, 834-857 и 959-986aa.
Сравнивая значения защиты перекрывающихся пептидов, можно дополнительно улучшить разрешение данных обмена с дейтерием. Например, пептид 329-342aa защищен только -0,51 Да в комплексе NOV001-β-клото ECD, тогда как более крупный пептид 330-348aa защищен -1,48 Да. Следовательно, можно сделать вывод, что существенно защищенная область более крупного пептида должна быть областью 343-347aa. Осуществление этого анализа по данным показывает, что области 246-265, 536-550, 834-857 и 959-986aa наиболее сильно защищены при связывании NOV002 или NOV001 с β-клото ECD (например, к указанным областям внутри SEQ ID NO: 262).
Кроме того, большое число областей, кроме наиболее сильно защищенных областей, указанных ранее, защищенных NOV001, защищены в NOV002. Эти области защиты распределены по длине многих областей в пространстве линейной последовательности, хотя в области С-конца ECD имеется больше областей защиты. Напротив, существует несколько областей, которые различаются между двумя этими антителами. В NOV001 две области имеют специальную защиту: 646-670aa и 697-700aa, а для NOV002 одна область, 646-689aa, защищена специфическим образом. В целом, проведение различия между эпитопами NOV002 и NOV001, используя только HDx-MS на β-клото ECD, было сложным. Данные HDx показывают, что эпитопы весьма схожи.
В таблице 2 показано влияние связывания NOV001 и NOV002 на включение дейтерия β-клото человека (53-997aa). Для каждого пептида, обнаруженного методом масс-спектрометрии, показано изменение включения дейтерия (в дальтонах) для комплекса NOV001-β-клото по сравнению с β-клото ECD и комплексу NOV002-β-клото по сравнению с β-клото ECD.
Таблица 2: Охват пептидом HDx-MS внеклеточного домена β-клото человека (52-997aa)
Пример 9: Фармакокинетические профили моноклональных антител у крыс
Животные и условия содержания
Уход за животными и кормление соответствовало Руководству по уходу и использованию лабораторных животных (Институт лабораторных животных, Национальный исследовательский совет). Все процедуры регулировались стандартами, установленными Министерством здравоохранения и социальных служб США, и выполнялись в соответствии с протоколом CV9054, утвержденным Комитетом по уходу и использованию животных в Восточном Ганновере «Новартис». Самцов крыс Спрага-Доули (n=3/группа) размещали в клетках с твердым дном на стойке, оборудованной для автоматической подачи воды ad libitum, содержали при 12-часовом цикле свет/темнота (с 6:00 до 18:00), и обеспечивали свободный доступ к стандартному корму для грызунов (Harlan-Teklad, Frederick, MD, cat # 8604). Виварий содержали при температуре в диапазоне 68 и 76°F при влажности от 30 до 70%.
Получение NOV002 или NOV004 и дозирование
Исходные растворы NOV002 (12,00 мг/мл) и NOV004 (16,0 мг/мл) в 20мМ гистидиновом буфере (рН 6,0) приобретали в замороженном состоянии, хранили при -80°C и оттаивали при низких температурах перед использованием. При утреннем введении дозы оба антитела разводили до 2 мг/мл в 20 мМ гистидиновом буфере и соответствующие объемы вносили в шприцы для введения дозы (5 мл/кг) и выдерживали при комнатной температуре перед введением. Животных помещали в фиксирующий бокс и вводили NOV002 или NOV004 путем внутривенной (IV) инъекции в хвостовую вену (10 мг/кг).
Сбор образцов крови
Образцы крови собирали на -3 день (фоновый уровень), 0 день (через 1 и 6 ч после приема) и в 1, 2, 4, 7, 14, 21 и 28 дни после введения дозы. Все временные точки рассчитывали от конца введения дозы в 0 день. В каждой временной точке собирали приблизительно 0,2 мл (200 мкл) крови в пробирки для сбора/разделения BD Microtainer с EDTA (Becton, Dickinson, Company, Franklin Lakes, NJ, номер в каталоге 365973). Для остановки кровотечения применяли давление с марлей. Образцы центрифугировали в течение 10 мин при 20 817×g, а затем ~100 мкл плазмы переносили в ряд пробирок объемом 0,2 мл Thermo-strip tube (Thermo-Scientific, Pittsburg, PA, номер в каталоге AB-0451) и замораживали при 80°C. После каждого сбора крыс возвращали в их домашнюю клетку.
Измерение общих концентраций NOV002 или NOV004 в плазме
IgG человека (т.е. NOV002 или NOV004) в плазме крысы определяли количественно с использованием индивидуального сэндвич-иммуноанализа с моноклональным антителом мыши против IgG человека (R10Z8E9) в качестве иммобилизованного антитела и антитела козла против IgG человека с меткой HRP в качестве детектирующего антитела. Иммобилизованное антитело (2 мкг/мл в PBS, 30 мкл/лунка) наносили на 384-луночные микротитровальные планшеты белого цвета (Greiner Bio-One; Monroe, NC; номер в каталоге 781074). Планшеты инкубировали в течение ночи при комнатной температуре (RT) без встряхивания. После аспирации раствора для покрытия без промывания в каждую лунку добавляли 90 мкл 1×молочный разбавитель/блокирующий раствор (KPL, Gaithersburg, MD, cat no. 50-82-01) и планшеты инкубировали в течение 2 часов при комнатной температуре. В конце инкубирования раствор отсасывали, и планшеты хранили в пакетах из фольги с осушителем при -80°C.
В день анализа 16 стандартных концентраций NOV002 или NOV004 в диапазоне от 0,15-600 нг/мл получали путем серийного разведения в казеиновом буфере, включая буферный отрицательный контроль. Все образцы исследования разбавляли 1:50 вручную в казеиновом буфере и затем последовательно разводили в 5 раз с использованием Freedom EVO (Tecan, Mannedorf, Switzerland) в общей сложности в пять разведений. Планшеты инкубировали в течение 2 ч при комнатной температуре, встряхивая со скоростью 300 об/мин, а затем промывали 3 раза фосфатным буфером для промывки (90 мкл/лунка). В каждый планшет помещали антитело козла против IgG человека, меченное HRP (400 нг/мл в казеиновом буфере, 30 мкл/лунка), и планшеты инкубировали в течение 1 часа при комнатной температуре при встряхивании со скоростью 300 об/мин. Планшеты промывали 3 раза фосфатным буфером для промывки (90 мкл/лунку), а затем добавляли хемилюминесцентный субстрат KPL LumiGLO (30 мкл/лунка, номер в каталоге 54-61-00). Хемилюминесценцию сразу считывали на ридере для планшетов SpectraMax M5 (Molecular Devices) на всех длинах волн с временем интегрирования 50 мсек. Концентрации Fc человека (нг/мл) в образцах плазмы интерполировали из стандартной кривой NOV002 или NOV004, умножали на коэффициенты разбавления и преобразовывали в концентрации в нМ.
Животные показали среднее значение Cmax 1207 и 1115 нМ через 1 ч после IV введения NOV002 или NOV004, соответственно. NOV002 и NOV004 показали эквивалентные профили PK у крыс Спрага-Доули (фигура 6).
Пример 10: Исследование яванских макак с ожирением
Изучали эффекты NOV002 на потребление пищи, вес тела и биомаркеры плазмы у яванских макак с ожирением.
Протоколы:
Пяти самцам яванских макак делали две подкожные инъекции (s.c.) дозами по 1 мг/кг NOV002, вводимыми с интервалом в одну неделю (0 и 7 дни исследования), и потребление пищи, вес тела и биомаркеры плазмы оценивали в течение более 100 дней после введения дозы. Для каждой дозы животных обездвиживали в их домашней клетке, собирали образцы крови, а затем каждому животному вводили подкожно дозу в 1 мг/кг NOV002. Измерения потребления пищи начинали за 1 неделю до введения первой дозы и продолжали в ходе исследования. Пищу при исследовании взвешивали перед кормлением и каждый день делили на две равные части. На следующее утро оставшуюся пищу собирали и взвешивали. Количество гранул (по 1 г каждый), уроненных в поддон, подсчитывали и добавляли к весу оставшейся пищи. Ежедневное потребление пищи рассчитывали как вес пищи, получаемой за вычетом собранной пищи. Потребление фруктов и овощей не измерялось. Массу тела до приема пищи измеряли в повторе три раза в неделю (перед сбором крови или введением дозы), используя динамический объект в масштабе.
Измерение концентрации NOV002 в плазме
Fc IgG человека в плазме яванского макака определяли количественно с использованием иммуноанализа сэндвич типа на основе ELISA. В качестве иммобилизованного антитела использовали антитела мыши против IgG1 человека, моноклональное антитело мыши против IgG человека. 384-луночные планшеты Greiner белого цвета покрывали 2 мкг/мл IgG1 мыши против IgG человека (30 мкл/лунка) и инкубировали в течение ночи при комнатной температуре (RT). Покрывающее антитело отсасывали и добавляли 1×молочный блокатор (KPL #50-82-01) при 90 мкл/лунку в течение 2 ч при комнатной температуре. Блокирующий раствор отсасывали, и планшеты хранили при -80°C в мешках для планшетов с осушителем до анализа. В день анализа планшеты и реагенты доводили до RT. Готовили стандарты, разбавляя очищенный IgG от 4000 до 16 пМ в стандартном казеиновом разбавителе для образцов и включая буферный контроль. Образцы разбавляли в повторе 1:50, 1:250, 1:1250 и 1:6250 в том же разбавителе, что и стандарты, а затем стандарты, разбавленные образцы и контрольные образцы добавляли на планшет в течение 2 часов при комнатной температуре (рабочий объем для всех стадий 30 мкл/лунка). Затем планшеты промывали 3 раза промывочным буфером на основе фосфата. К планшету добавляли гамма антитело против Fc человека, меченное пероксидазой хрена (HRP), в течение 1 часа при комнатной температуре, и затем планшеты 3 раза промывали промывочным буфером на основе фосфата. К планшету добавляли хемилюминесцентный субстрат, и планшет сразу же считывали на люминесцентном ридере для планшетов.
Стандарты NOV002 анализировали в трех повторах на планшет. Образцы разведенной плазмы анализировали в повторе. Неизвестные образцы интерполировали из стандартной кривой IgG. С использованием программного обеспечения SoftMax Pro Software v5.4.1 выполняли построение кривой по точкам, обратное исчисление, % нормализации и интерполяцию концентраций образцов. Сигнал, генерируемый стандартами IgG, наносили на график и подгоняли, используя логистическую кривую с подгоном и 4-мя параметрами. Концентрации Fc (pM) в образцах плазмы интерполировали из стандартной кривой NOV002 и умножали на коэффициенты разбавления. Нижний предел количественной оценки анализа (LLOQ) составил 31,250 пМ, а верхний предел количественной оценки (ULOQ) составил 4000 пМ. LLOQ и ULOQ определяли как нижнюю и верхнюю стандартную концентрацию со 100% -ной нормализацией ±20% и CV <20%, а затем умножали на коэффициенты разбавления.
Обнаружение антител против лекарственного средства
Образцы плазмы разбавляли 1:5 в буфере LowCross (Boca Scientific; Boca Raton, FL; номер в каталоге 100500). Получали реакционную смесь, содержащую 0,6 мкг/мл NOV002, меченного биотином, и 0,6 мкг/мл NOV002, меченного дигоксигенином, в буфере LowCross. Разведенную плазму (80 мкл) объединяли с 160 мкл реакционной смеси в 96-луночных планшетах с U-образным дном (BD Falcon, Billerica, MA, номер в каталоге 351177). Края планшетов запечатывали парафином, и планшеты инкубировали на платформе-шейкере при 37°C в течение 2 часов (150 об/мин, с защитой от света). Аликвоту каждой смеси (100 мкл) затем переносили в дублированные лунки 96-луночных планшетов, покрытых стрептавидином (Roche; номер в каталоге 11734776001), которые сначала промывали 3 раза буфером для промывки, состоящим из 1×PBS, содержащим 0,05% (об./об.) Tween-20 (300 мкл на лунку). Планшеты герметизировали и затем инкубировали при комнатной температуре на платформе-шейкере в течение 1 часа (300 об/мин, с защитой от света). Планшеты промывали 3 раза буфером для промывки (300 мкл на лунку), а затем к каждой лунке добавляли 100 мкл фрагмента антидигоксигенин-пероксидазы-POD Fab (Roche, номер в каталоге 11633716001), разбавленного 1:2500 в буфере LowCross. Планшеты герметизировали, инкубировали при комнатной температуре на платформе-шейкере в течение 45 минут (300 об/мин, с защитой от света), и затем промывали 3 раза буфером для промывки (300 мкл на лунку). Однокомпонентный субстрат TMB One Component HRP Microwell Substrate (Bethyl Laboratories, Montgomery TX; номер в каталоге E102; 100 мкл/лунку) добавляли в каждую лунку, и в течение 9-10 мин появлялся синий цвет, защищали от света. Цветную реакцию останавливали, добавляя 100 мкл 0,18н H2SO4 в каждую лунку, планшеты слегка встряхивали и желтый цвет измеряли при OD450.
Измерение концентрации глюкозы в плазме
Концентрацию глюкозы в плазме измеряли с использованием анализа Autokit Glucose (Wako Chemicals; Richmond, VA; номер в каталоге 439-90901). Стандартную кривую получали разведением калибратора до стандартов 500, 200, 100, 50, 20 и 0 мг/дл. Аналитический реагент (300 мкл), предварительно нагретый до 37°C, добавляли к 2 мкл плазмы, стандартам и контрольным образцам на прозрачном 96-луночной плоскодонном планшете (Thermo Scientific; номер в каталоге 269620). Планшет перемешивали на планшетном шейкере в течение 30 сек и затем инкубировали при 37°C в течение 5 мин. Через 20 сек перемешивания планшет считывали при 505/600 нм, используя устройство Molecular Devices SPECTRAmax PLUS 384 (Sunnyvale, CA). Концентрации образцов глюкозы рассчитывали по сравнению со стандартной кривой.
Измерение концентрации инсулина в плазме
Концентрации инсулина в плазме определяли с использованием набора Millipore Human Insulin Specific RIA Kit (Billerica, MA; номер в каталоге HI-14K) в соответствии с инструкциями производителя. Соответствующие количества буфера для анализа, стандартов или образца разбавленной плазмы смешивали с 125I-инсулином и антителу к инсулину в 5 мл, 75×12 мм пробирках PP SARSTEDT (номер в каталоге 55526). Пробирки встряхивали, закрывали и инкубировали в течение 20 ч при комнатной температуре. После инкубации добавляли 1 мл с температурой 4°C осаждающего реагента и пробирки встряхивали и инкубировали в течение 30 мин при 4°C. Все пробирки центрифугировали в течение 30 мин (3000 об/мин при 4°C), супернатанты декантировали и гранулы подсчитывали на автоматическом гамма-счетчике PerkinElmer WIZARD2 (модель № 2470, PerkinElmer, Waltham, MA). Концентрации инсулина рассчитывали по сравнению со стандартной кривой, полученной с использованием известных количеств инсулина.
Измерение концентрации триглицеридов в плазме
Концентрации триглицеридов (ТГ) в плазме измеряли с использованием набора Triglyceride (GPO) Liquid Reagent (Pointe Scientific, Canton, MI, номер в каталоге T7532-500). Предварительно подогретый аналитический реагент (300 мкл, 37°C) добавляли к 5 мкл плазмы на прозрачном 96-луночной планшете с плоским дном (Thermo Fisher Scientific; Tewksbury, MA; номер в каталоге 269620). Планшет перемешивали на шейкере для планшетов в течение 30 сек и затем инкубировали при 37°C в течение 5 мин. Через 20 сек перемешивания измеряли поглощение при 500 нм на ридере для планшетов SPECTRAmax PLUS. Концентрации TG рассчитывали по сравнению с калибровочной кривой, полученной с использованием известных величин стандарта TG (Pointe Scientific; номер в каталоге T7531-STD).
Измерение концентрации холестерина в плазме:
Общий уровень холестерина (ТС) в плазме определяли количественно с использованием набора Cholesterol (Liquid) Reagent (Pointe Scientific, номер в каталоге C7510-500). Предварительно подогретый аналитический реагент (200 мкл, 37°C) добавляли к 10 мкл плазмы в прозрачном 96-луночном планшете с плоским дном (Thermo Fisher Scientific; номер в каталоге 269620). Планшет перемешивали на планшетном шейкере в течение 30 сек и затем инкубировали при 37°C в течение 5 мин. Через 20 сек перемешивания измеряли поглощение при 500 нм на ридере для планшетов SPECTRAmax PLUS. Концентрации холестерина рассчитывали по сравнению с калибровочной кривой, полученной с использованием известных количеств стандарта холестерина (Stanbio Laboratory; Boerne, TX; номер в каталоге 1012-030).
Измерение концентрации холестерина липопротеинов высокой плотности в плазме
Для определения концентраций холестерина липопротеина высокой плотности (ЛПВП) 50 мкл образца плазмы объединяли с 50 мкл реагента, осаждающего холестерин (Wako Chemicals; Richmond, VA; номер в каталоге 431-52501), в 0,5 мл микроцентрифужной пробирке и быстро перемешивали на Vortex. Пробирку оставляли при комнатной температуре в течение 10 мин и затем центрифугировали при 2000×g в течение 10 мин при 4°C. После центрифугирования удаляли приблизительно половину супернатанта (содержащего часть холестерина ЛПВП исходного образца плазмы) и использовали 10 мкл для анализа холестерина, описанного выше.
Измерение концентрации β-гидроксибутирата в плазме
β-Гидроксибутират (β-HB) в плазме измеряли, используя набор β-Hydroxybutyrate LiquiColor Test (Stanbio Laboratory; номер в каталоге 2440-058). Аналитический реагент R1 (215 мкл, предварительно нагретый до 37°C) добавляли к 20 мкл образца контроля качества или плазмы в прозрачный 96-луночный планшет с плоским дном (Thermo Fisher Scientific, № 269620). Планшет перемешивали на шейкере для планшетов в течение 30 сек и затем инкубировали при 37°C в течение 5 мин. Предварительно считываемое поглощение измеряли при 505 нм на ридере для планшетов SPECTRAmax PLUS. В каждую лунку добавляли реагент R2 (35 мкл, предварительно нагретый до 37°C), и планшет снова перемешивали на шейкере для планшетов в течение 30 сек и инкубировали при 37°C в течение 5 мин. После 20-секундного перемешивания конечное поглощение измеряли при 505 нм, из которого вычитали предварительно считываемое значение. Концентрации β-HB рассчитывали по сравнению с калибровочной кривой, полученной с использованием известных количеств калибратора β-HB (Wako Diagnostics; Richmond, VA; номер в каталоге 412-73791).
Статистический анализ
Статистический анализ проводился с использованием GraphPad Prism (версия 6.05, GraphPad Software, La Jolla, CA). Данные о потреблении пищи для каждого животного были нормализованы в процентах от базовой линии (рассчитаны как среднее в дни от -6 до 0), и затем были рассчитаны средние групповые значения±стандартные ошибки среднего (SEM); каждый день сравнивали с днем 0 с помощью непараметрического теста Фридмана с последующими множественными сравнениями с помощью критерия Данна. Вес тела представлен как среднее групповое значение±стандартные ошибки среднего (SEM), рассчитанный как процент от базовой линии (рассчитывается как среднее в дни -7, -5, -3 и 0). Вес необработанного тела и данные биомаркера плазмы также анализировали с помощью непараметрического теста Фридмана с последующими множественными сравнениями с помощью критерия Данна. P <0,05 считалось значительным.
Результаты
Перед введением дозы NOV002 животных подвергали скринингу на ранее присутствовавшие антитела против лекарственных средств (ADA) и было обнаружено, что все обезьяны были негативны по ADA. Чтобы определить, может ли NOV002 снизить потребление пищи и вес тела обезьян с ожирением (средний исходный вес тела=12,4±0,9 кг, который был рассчитан как средний в дни от -6 до 0), каждое животное получило две подкожные дозы 1 мг/кг NOV002 с интервалом в неделю в дни исследований 0 и 7. NOV002 значительно снизил потребление пищи по сравнению с исходным уровнем, при этом среднее пиковое снижение до ~44% от базовой линии произошло на 32-й день после введения дозы. Потребление пищи было значительно ниже в дни 18, 22, 24, 25, 26, 29 и 32 после дозирования в сравнении с днем 0 по тесту Фридмана с пост-тестом Данна). У всех пяти обезьян отмечалось четкое снижение потребления пищи, но величина и продолжительность снижения потребления пищи варьировались по всей группе.
Вес тела также измерялся на протяжении всего исследования, и наблюдалось пиковое среднее изменение массы тела -8,9% на 67-й день после введения дозы. Вес тела был значительно ниже в дни с 37 по 70 после дозирования против дня 0 по тесту Фридмана с пост-тестом Данна. Подобно эффектам приема пищи, вариабельность и продолжительность изменений веса тела наблюдались после лечения 1 мг/кг NOV002, которые соответствовали ответам на потребление пищи отдельными животными: обезьяны с наибольшим уменьшением потребления пищи потеряли больше всего веса тела во время исследования.
В дополнение к оценке влияния NOV002 на потребление пищи и вес тела, было показано влияние на биомаркеры плазмы липидного и углеводного обмена. NOV002 уменьшал как концентрацию ТГ, так и ТС в плазме, со значительной разницей на 32-й день после дозирования в сравнении с исходными уровнями (значение в дни -7, -3 и 0, тест Фридмана с пост-тестом Данна).
В таблице 3 представлены все изменения биомаркера плазмы на 32-й день по сравнению с исходным уровнем. По сравнению с исходным уровнем NOV002 в течение какого-либо времени во время исследования NOV002 существенно не изменял уровень холестерина липопротеина высокой плотности плазмы (HDL), β-гидроксибутирата (β-HB), глюкозы или инсулина, но NOV002 значительно увеличивал уровни адипонектина в плазме в день 35 после введения дозы.
Таблица 3 Улучшенные с помощью NOV002 уровни биомаркера в плазме
TC (мг/дл)
HDL (мг/дл)
β-HB (мкM)
Глюкоза (мг/дл)
Инсулин (мкЕд/мл)
Адипонектин (кмг/мл)3
126±11
53±9
53±5
80±3
271±57
2,9±0,6
96±6*
53±5
115±31
76±7
124±12
6,0±1,8*
-22±5
6±12
137±77
-10±9
-40±17
94±22
Значения представляют собой среднее групповое значение±SEM.
1Исходные значения отражают среднее значение в дни -7, -3 и 0.
2Процентное изменение было рассчитано для каждого индивидуума, а затем усреднено для среднего значения группы±SEM.
3Уровни адипонектина измеряли только в подмножестве образцов, поэтому данные взяты на дни исследования 0 и 35.
*P <0,05 по сравнению с исходным уровнем по непараметрическому тесту Фридмана с пост-тестом Данна.
Образование ADA также исследовали в различные моменты времени (например, 1, 2, 3, 4, 7 день (до введения дозы и 6 ч после приема), два раза в неделю от 11 до 81 дня, а затем один раз в неделю после этого) на протяжении всего исследования. Ни одна из обезьян не показала ADA к NOV002 при предварительном скрининге, но у трех животных во время исследования развился ADA к NOV002. PK профили NOV002, по-видимому, не отличались между животными с положительными ADA и отрицательными ADA.
NOV002 эффективно снижало потребление пищи и массу тела у яванских макак с ожирением. У животных, получавших две подкожные дозы 1 мг/кг NOV002, наблюдалось следующее: снижение веса тела на 8,9% на день 67 после дозирования, хотя снижение потребления пищи и массы тела варьировалось между отдельными животными. Потеря веса тела сопровождалась уменьшением концентраций ТГ и ТС в плазме, демонстрируя положительные эффекты NOV002 на профили липидов плазмы. Эти данные показывают, что NOV002 эффективен при снижении потребления пищи, веса тела и концентраций TG и ТС в плазме; что NOV002, а также его варианты, такие как NOV004, другие антитела против β-клото, содержащие CDR NOV002 или NOV004, и другие антитела против β-клото, которые связываются с тем же эпитопом, что и NOV002 (например, антитела, связывающиеся с одним или более защищенными пептидами, указанными в таблице 2) или которые конкурируют с NOV002, будут являться новой эффективной терапией для лечения метаболических нарушений, таких как ожирение.
Включение путем ссылки
Все цитируемые в настоящем документе ссылки, включая патенты, заявки на патент, научные статьи, учебники и тому подобное, а также ссылки, приведенные в них, в той мере, в которой они еще не охвачены, включены в настоящий документ в качестве ссылки во всей их полноте.
Эквиваленты
Вышеупомянутое письменное описание считается достаточным, чтобы дать возможность специалисту в данной области техники осуществить на практике изобретение. Вышеприведенное описание и примеры подробно описывают некоторые предпочтительные варианты осуществления изобретения и описывают наилучший режим, предложенный изобретателями. Однако будет понятно, что независимо от того, насколько подробно изложено вышеизложенное в тексте, изобретение может быть осуществлено на практике многими способами, и изобретение должно толковаться в соответствии с прилагаемой формулой изобретения и любыми ее эквивалентами.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
<120> СПОСОБЫ ЛЕЧЕНИЯ РАССТРОЙСТВ, СВЯЗАННЫХ С FGF21
<130> PAT056954
<140>
<141>
<150> 62/200,445
<151> 2015-08-03
<160> 274
<170> PatentIn версия 3.5
<210> 1
<211> 209
<212> Белок
<213> Homo sapiens
<400> 1
Met Asp Ser Asp Glu Thr Gly Phe Glu His Ser Gly Leu Trp Val Ser
1 5 10 15
Val Leu Ala Gly Leu Leu Leu Gly Ala Cys Gln Ala His Pro Ile Pro
20 25 30
Asp Ser Ser Pro Leu Leu Gln Phe Gly Gly Gln Val Arg Gln Arg Tyr
35 40 45
Leu Tyr Thr Asp Asp Ala Gln Gln Thr Glu Ala His Leu Glu Ile Arg
50 55 60
Glu Asp Gly Thr Val Gly Gly Ala Ala Asp Gln Ser Pro Glu Ser Leu
65 70 75 80
Leu Gln Leu Lys Ala Leu Lys Pro Gly Val Ile Gln Ile Leu Gly Val
85 90 95
Lys Thr Ser Arg Phe Leu Cys Gln Arg Pro Asp Gly Ala Leu Tyr Gly
100 105 110
Ser Leu His Phe Asp Pro Glu Ala Cys Ser Phe Arg Glu Leu Leu Leu
115 120 125
Glu Asp Gly Tyr Asn Val Tyr Gln Ser Glu Ala His Gly Leu Pro Leu
130 135 140
His Leu Pro Gly Asn Lys Ser Pro His Arg Asp Pro Ala Pro Arg Gly
145 150 155 160
Pro Ala Arg Phe Leu Pro Leu Pro Gly Leu Pro Pro Ala Leu Pro Glu
165 170 175
Pro Pro Gly Ile Leu Ala Pro Gln Pro Pro Asp Val Gly Ser Ser Asp
180 185 190
Pro Leu Ser Met Val Gly Pro Ser Gln Gly Arg Ser Pro Ser Tyr Ala
195 200 205
Ser
<210> 2
<211> 938
<212> ДНК
<213> Homo sapiens
<400> 2
ctgtcagctg aggatccagc cgaaagagga gccaggcact caggccacct gagtctactc 60
acctggacaa ctggaatctg gcaccaattc taaaccactc agcttctccg agctcacacc 120
ccggagatca cctgaggacc cgagccattg atggactcgg acgagaccgg gttcgagcac 180
tcaggactgt gggtttctgt gctggctggt cttctgctgg gagcctgcca ggcacacccc 240
atccctgact ccagtcctct cctgcaattc gggggccaag tccggcagcg gtacctctac 300
acagatgatg cccagcagac agaagcccac ctggagatca gggaggatgg gacggtgggg 360
ggcgctgctg accagagccc cgaaagtctc ctgcagctga aagccttgaa gccgggagtt 420
attcaaatct tgggagtcaa gacatccagg ttcctgtgcc agcggccaga tggggccctg 480
tatggatcgc tccactttga ccctgaggcc tgcagcttcc gggagctgct tcttgaggac 540
ggatacaatg tttaccagtc cgaagcccac ggcctcccgc tgcacctgcc agggaacaag 600
tccccacacc gggaccctgc accccgagga ccagctcgct tcctgccact accaggcctg 660
ccccccgcac tcccggagcc acccggaatc ctggcccccc agccccccga tgtgggctcc 720
tcggaccctc tgagcatggt gggaccttcc cagggccgaa gccccagcta cgcttcctga 780
agccagaggc tgtttactat gacatctcct ctttatttat taggttattt atcttattta 840
tttttttatt tttcttactt gagataataa agagttccag aggagaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaa 938
<210> 3
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 3
Asp Tyr Tyr Ile Asn
1 5
<210> 4
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 4
Arg Ile His Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe Gln
1 5 10 15
Gly
<210> 5
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 5
Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp
1 5 10
<210> 6
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 6
Gly Tyr Thr Phe Thr Asp Tyr
1 5
<210> 7
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 7
His Pro Gly Ser Gly Asn
1 5
<210> 8
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 8
Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp
1 5 10
<210> 9
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 9
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile His Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 10
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 10
caggtgcagc tggtgcagtc tggcgccgaa gtgaagaaac ccggcagcag cgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc gactactaca tcaactgggt gcgccaggcc 120
cctggacagg gcctggaatg gatgggcaga atccaccccg gctccggcaa cacctactac 180
aacgagaagt tccagggcag agtgaccctg accgccgaca agagcaccag caccgcctac 240
atggaactga gcagcctgcg gagcgaggac accgccgtgt actactgtgc catcctgctg 300
ctgcggagct acggcatgga tgattggggc cagggcacca ccgtgaccgt cagctca 357
<210> 11
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile His Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 12
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 12
caggtgcagc tggtgcagtc tggcgccgaa gtgaagaaac ccggcagcag cgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc gactactaca tcaactgggt gcgccaggcc 120
cctggacagg gcctggaatg gatgggcaga atccaccccg gctccggcaa cacctactac 180
aacgagaagt tccagggcag agtgaccctg accgccgaca agagcaccag caccgcctac 240
atggaactga gcagcctgcg gagcgaggac accgccgtgt actactgtgc catcctgctg 300
ctgcggagct acggcatgga tgattggggc cagggcacca ccgtgaccgt cagctcagct 360
agcaccaagg gccccagcgt gttccccctg gcccccagca gcaagagcac cagcggcggc 420
acagccgccc tgggctgcct ggtgaaggac tacttccccg agcccgtgac cgtgtcctgg 480
aacagcggag ccctgacctc cggcgtgcac accttccccg ccgtgctgca gagcagcggc 540
ctgtacagcc tgtccagcgt ggtgacagtg cccagcagca gcctgggcac ccagacctac 600
atctgcaacg tgaaccacaa gcccagcaac accaaggtgg acaagagagt ggagcccaag 660
agctgcgaca agacccacac ctgccccccc tgcccagccc cagaggcagc gggcggaccc 720
tccgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagcag gacccccgag 780
gtgacctgcg tggtggtgga cgtgagccac gaggacccag aggtgaagtt caactggtac 840
gtggacggcg tggaggtgca caacgccaag accaagccca gagaggagca gtacaacagc 900
acctacaggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaggaa 960
tacaagtgca aggtctccaa caaggccctg ccagccccca tcgaaaagac catcagcaag 1020
gccaagggcc agccacggga gccccaggtg tacaccctgc ccccctcccg ggaggagatg 1080
accaagaacc aggtgtccct gacctgtctg gtgaagggct tctaccccag cgacatcgcc 1140
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc cccagtgctg 1200
gacagcgacg gcagcttctt cctgtacagc aagctgaccg tggacaagtc caggtggcag 1260
cagggcaacg tgttcagctg cagcgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagagcctga gcctgtcccc cggcaag 1347
<210> 13
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 13
Lys Ser Ser Gln Ser Ile Val His Ser Ser Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 14
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 14
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 15
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 15
Phe Gln Gly Ser His Ile Pro Tyr Thr
1 5
<210> 16
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 16
Ser Gln Ser Ile Val His Ser Ser Gly Asn Thr Tyr
1 5 10
<210> 17
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 17
Lys Val Ser
1
<210> 18
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 18
Gly Ser His Ile Pro Tyr
1 5
<210> 19
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 19
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val His Ser
20 25 30
Ser Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Ile Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 20
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 20
gacgtggtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atctcctgca agagcagcca gagcatcgtg cacagcagcg gcaacaccta cctggaatgg 120
tatctgcaga agcccggcca gagcccccag ctgctgatct acaaggtgtc caaccggttc 180
agcggcgtgc ccgacagatt ttctggcagc ggctccggca ccgacttcac cctgaagatc 240
tcccgggtgg aagccgagga cgtgggcgtg tactactgtt ttcaaggctc ccacatcccc 300
tacaccttcg gccagggcac caagctggaa atcaag 336
<210> 21
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 21
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val His Ser
20 25 30
Ser Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Ile Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 22
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 22
gacgtggtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atctcctgca agagcagcca gagcatcgtg cacagcagcg gcaacaccta cctggaatgg 120
tatctgcaga agcccggcca gagcccccag ctgctgatct acaaggtgtc caaccggttc 180
agcggcgtgc ccgacagatt ttctggcagc ggctccggca ccgacttcac cctgaagatc 240
tcccgggtgg aagccgagga cgtgggcgtg tactactgtt ttcaaggctc ccacatcccc 300
tacaccttcg gccagggcac caagctggaa atcaagcgta cggtggccgc tcccagcgtg 360
ttcatcttcc cccccagcga cgagcagctg aagagcggca ccgccagcgt ggtgtgcctg 420
ctgaacaact tctacccccg ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgaccctgag caaggccgac tacgagaagc ataaggtgta cgcctgcgag 600
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgc 657
<210> 23
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 23
Ser Gly Tyr Thr Trp His
1 5
<210> 24
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 24
Tyr Ile His Tyr Ser Val Tyr Thr Asn Tyr Asn Pro Ser Val Lys Gly
1 5 10 15
<210> 25
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 25
Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val
1 5 10
<210> 26
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 26
Gly Tyr Ser Ile Thr Ser Gly Tyr
1 5
<210> 27
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 27
His Tyr Ser Val Tyr
1 5
<210> 28
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 28
Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val
1 5 10
<210> 29
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Thr Trp His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Leu Ser Tyr Ile His Tyr Ser Val Tyr Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ala Lys Asn Ser Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 30
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 30
gaggtgcagc tggtggaatc tggcggcgga ctcgtgaagc ctggcggctc tctgagactg 60
agctgtgccg tgtccggcta cagcatcacc agcggctaca cctggcattg ggtgcgccag 120
gcccctggca aaggactgga atggctgtcc tacatccact acagcgtgta caccaactac 180
aaccccagcg tgaagggccg gttcaccatc agcagagaca ccgccaagaa cagcttctac 240
ctgcaaatga acagcctgcg ggccgaggac accgccgtgt actactgtgc cagacggacc 300
accagcctgg aacggtactt cgacgtgtgg ggccagggca cactcgtgac cgtcagctca 360
<210> 31
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 31
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Thr Trp His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Leu Ser Tyr Ile His Tyr Ser Val Tyr Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ala Lys Asn Ser Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 32
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 32
gaggtgcagc tggtggaatc tggcggcgga ctcgtgaagc ctggcggctc tctgagactg 60
agctgtgccg tgtccggcta cagcatcacc agcggctaca cctggcattg ggtgcgccag 120
gcccctggca aaggactgga atggctgtcc tacatccact acagcgtgta caccaactac 180
aaccccagcg tgaagggccg gttcaccatc agcagagaca ccgccaagaa cagcttctac 240
ctgcaaatga acagcctgcg ggccgaggac accgccgtgt actactgtgc cagacggacc 300
accagcctgg aacggtactt cgacgtgtgg ggccagggca cactcgtgac cgtcagctca 360
gctagcacca agggccccag cgtgttcccc ctggccccca gcagcaagag caccagcggc 420
ggcacagccg ccctgggctg cctggtgaag gactacttcc ccgagcccgt gaccgtgtcc 480
tggaacagcg gagccctgac ctccggcgtg cacaccttcc ccgccgtgct gcagagcagc 540
ggcctgtaca gcctgtccag cgtggtgaca gtgcccagca gcagcctggg cacccagacc 600
tacatctgca acgtgaacca caagcccagc aacaccaagg tggacaagag agtggagccc 660
aagagctgcg acaagaccca cacctgcccc ccctgcccag ccccagaggc agcgggcgga 720
ccctccgtgt tcctgttccc ccccaagccc aaggacaccc tgatgatcag caggaccccc 780
gaggtgacct gcgtggtggt ggacgtgagc cacgaggacc cagaggtgaa gttcaactgg 840
tacgtggacg gcgtggaggt gcacaacgcc aagaccaagc ccagagagga gcagtacaac 900
agcacctaca gggtggtgtc cgtgctgacc gtgctgcacc aggactggct gaacggcaag 960
gaatacaagt gcaaggtctc caacaaggcc ctgccagccc ccatcgaaaa gaccatcagc 1020
aaggccaagg gccagccacg ggagccccag gtgtacaccc tgcccccctc ccgggaggag 1080
atgaccaaga accaggtgtc cctgacctgt ctggtgaagg gcttctaccc cagcgacatc 1140
gccgtggagt gggagagcaa cggccagccc gagaacaact acaagaccac ccccccagtg 1200
ctggacagcg acggcagctt cttcctgtac agcaagctga ccgtggacaa gtccaggtgg 1260
cagcagggca acgtgttcag ctgcagcgtg atgcacgagg ccctgcacaa ccactacacc 1320
cagaagagcc tgagcctgtc ccccggcaag 1350
<210> 33
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 33
Gln Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 34
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 34
Tyr Thr Ser Arg Leu Gln Ser
1 5
<210> 35
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 35
Gln Gln Gly Asn Thr Leu Pro Tyr Thr
1 5
<210> 36
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 36
Ser Gln Asp Ile Ser Asn Tyr
1 5
<210> 37
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 37
Tyr Thr Ser
1
<210> 38
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 38
Gly Asn Thr Leu Pro Tyr
1 5
<210> 39
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 39
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Ala Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 40
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 40
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgtc aggccagcca ggacatcagc aactacctga actggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactac accagcagac tgcagagcgg cgtgcccagc 180
agatttaccg gctctggaag cggagccgac tacaccttca ccatcagctc cctgcagccc 240
gaggatatcg ctacctactt ctgtcagcaa ggcaacaccc tgccttacac cttcggccag 300
ggcaccaagc tggaaatcaa g 321
<210> 41
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 41
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Ala Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 42
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 42
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgtc aggccagcca ggacatcagc aactacctga actggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactac accagcagac tgcagagcgg cgtgcccagc 180
agatttaccg gctctggaag cggagccgac tacaccttca ccatcagctc cctgcagccc 240
gaggatatcg ctacctactt ctgtcagcaa ggcaacaccc tgccttacac cttcggccag 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag cggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 43
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 43
Asp Tyr Tyr Ile Asn
1 5
<210> 44
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 44
Arg Ile His Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe Gln
1 5 10 15
Gly
<210> 45
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 45
Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp
1 5 10
<210> 46
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 46
Gly Tyr Thr Phe Thr Asp Tyr
1 5
<210> 47
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 47
His Pro Gly Ser Gly Asn
1 5
<210> 48
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 48
Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp
1 5 10
<210> 49
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 49
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile His Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 50
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 50
caagtccaac tcgtccagtc cggagccgaa gtgaaaaagc cgggctcatc agtgaaggtg 60
tcctgcaagg cgtcgggcta caccttcacc gactactaca tcaactgggt gcgccaggcc 120
ccgggacagg gtctggaatg gatggggagg attcaccccg gatcgggaaa cacctactac 180
aacgagaagt tccagggcag agtgaccctg actgccgaca agtccacgtc cactgcctac 240
atggaactgt cgtccctgcg gtccgaggat accgccgtgt actattgtgc gatcctgctg 300
ttgcggagct acgggatgga tgactgggga cagggtacca ctgtgactgt gtccagc 357
<210> 51
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 51
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile His Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 52
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 52
caagtccaac tcgtccagtc cggagccgaa gtgaaaaagc cgggctcatc agtgaaggtg 60
tcctgcaagg cgtcgggcta caccttcacc gactactaca tcaactgggt gcgccaggcc 120
ccgggacagg gtctggaatg gatggggagg attcaccccg gatcgggaaa cacctactac 180
aacgagaagt tccagggcag agtgaccctg actgccgaca agtccacgtc cactgcctac 240
atggaactgt cgtccctgcg gtccgaggat accgccgtgt actattgtgc gatcctgctg 300
ttgcggagct acgggatgga tgactgggga cagggtacca ctgtgactgt gtccagcgct 360
agcaccaagg gcccctccgt gttccctctg gccccttcca gcaagtctac ctccggcggc 420
acagctgctc tgggctgcct ggtcaaggac tacttccctg agcctgtgac agtgtcctgg 480
aactctggcg ccctgacctc tggcgtgcac accttccctg ccgtgctgca gtcctccggc 540
ctgtactccc tgtcctccgt ggtcacagtg ccttcaagca gcctgggcac ccagacctat 600
atctgcaacg tgaaccacaa gccttccaac accaaggtgg acaagcgggt ggagcctaag 660
tcctgcgaca agacccacac ctgtcctccc tgccctgctc ctgaactgct gggcggccct 720
tctgtgttcc tgttccctcc aaagcccaag gacaccctga tgatctcccg gacccctgaa 780
gtgacctgcg tggtggtggc cgtgtcccac gaggatcctg aagtgaagtt caattggtac 840
gtggacggcg tggaggtgca caacgccaag accaagcctc gggaggaaca gtacaactcc 900
acctaccggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaagag 960
tacaagtgca aagtctccaa caaggccctg gccgccccta tcgaaaagac aatctccaag 1020
gccaagggcc agcctaggga accccaggtg tacaccctgc cacccagccg ggaggaaatg 1080
accaagaacc aggtgtccct gacctgtctg gtcaagggct tctacccttc cgatatcgcc 1140
gtggagtggg agtctaacgg ccagcctgag aacaactaca agaccacccc tcctgtgctg 1200
gactccgacg gctccttctt cctgtactcc aaactgaccg tggacaagtc ccggtggcag 1260
cagggcaacg tgttctcctg ctccgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagtccctgt ccctgtctcc cggcaag 1347
<210> 53
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 53
Lys Ser Ser Gln Ser Ile Val His Ser Ser Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 54
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 54
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 55
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 55
Phe Gln Gly Ser His Ile Pro Tyr Thr
1 5
<210> 56
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 56
Ser Gln Ser Ile Val His Ser Ser Gly Asn Thr Tyr
1 5 10
<210> 57
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 57
Lys Val Ser
1
<210> 58
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 58
Gly Ser His Ile Pro Tyr
1 5
<210> 59
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 59
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val His Ser
20 25 30
Ser Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Ile Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 60
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 60
gatgtcgtga tgacccagac tccgctgtcc ctgtccgtga cccctggaca gcccgcgtct 60
atctcgtgca agagctccca gtccattgtg cattcaagcg ggaacaccta tctggagtgg 120
tacctccaga agcctggcca gagcccacag ctgctgatct acaaagtgtc gaacagattc 180
tccggtgtcc cggaccggtt ctccggctcg ggaagcggca ctgactttac actgaagatc 240
tcacgggtgg aagccgagga cgtgggagtg tactactgtt tccaagggtc ccacattccc 300
tacaccttcg gccaaggaac taagctggaa atcaag 336
<210> 61
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 61
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val His Ser
20 25 30
Ser Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Ile Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 62
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 62
gatgtcgtga tgacccagac tccgctgtcc ctgtccgtga cccctggaca gcccgcgtct 60
atctcgtgca agagctccca gtccattgtg cattcaagcg ggaacaccta tctggagtgg 120
tacctccaga agcctggcca gagcccacag ctgctgatct acaaagtgtc gaacagattc 180
tccggtgtcc cggaccggtt ctccggctcg ggaagcggca ctgactttac actgaagatc 240
tcacgggtgg aagccgagga cgtgggagtg tactactgtt tccaagggtc ccacattccc 300
tacaccttcg gccaaggaac taagctggaa atcaagcgta cggtggccgc tcccagcgtg 360
ttcatcttcc cccccagcga cgagcagctg aagagcggca ccgccagcgt ggtgtgcctg 420
ctgaacaact tctacccccg ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgaccctgag caaggccgac tacgagaagc ataaggtgta cgcctgcgag 600
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgc 657
<210> 63
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 63
Ser Gly Tyr Thr Trp His
1 5
<210> 64
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 64
Tyr Ile His Tyr Ser Val Tyr Thr Asn Tyr Asn Pro Ser Val Lys Gly
1 5 10 15
<210> 65
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 65
Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val
1 5 10
<210> 66
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 66
Gly Tyr Ser Ile Thr Ser Gly Tyr
1 5
<210> 67
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 67
His Tyr Ser Val Tyr
1 5
<210> 68
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 68
Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val
1 5 10
<210> 69
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Thr Trp His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Leu Ser Tyr Ile His Tyr Ser Val Tyr Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ala Lys Asn Ser Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 70
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 70
gaagtccaac tcgtcgaatc cggcggcgga ctggtcaagc cgggaggatc gctgagactg 60
tcgtgcgcag tgtcagggta cagcatcacc tccggttaca cctggcactg ggtcagacag 120
gcgccgggaa aaggcctgga atggctgtcc tacattcatt actccgtgta cactaactac 180
aacccctcag tgaaggggcg gttcaccatc tcccgggaca ctgccaagaa tagcttctat 240
ctgcaaatga actccctgcg ggccgaggat accgccgtgt actactgcgc gaggcgcacc 300
acgtccctgg agcgctactt tgacgtgtgg ggccagggta ccctcgtgac tgtgtcctcg 360
<210> 71
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 71
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Thr Trp His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Leu Ser Tyr Ile His Tyr Ser Val Tyr Thr Asn Tyr Asn Pro Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ala Lys Asn Ser Phe Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 72
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 72
gaagtccaac tcgtcgaatc cggcggcgga ctggtcaagc cgggaggatc gctgagactg 60
tcgtgcgcag tgtcagggta cagcatcacc tccggttaca cctggcactg ggtcagacag 120
gcgccgggaa aaggcctgga atggctgtcc tacattcatt actccgtgta cactaactac 180
aacccctcag tgaaggggcg gttcaccatc tcccgggaca ctgccaagaa tagcttctat 240
ctgcaaatga actccctgcg ggccgaggat accgccgtgt actactgcgc gaggcgcacc 300
acgtccctgg agcgctactt tgacgtgtgg ggccagggta ccctcgtgac tgtgtcctcg 360
gctagcacca agggcccctc cgtgttccct ctggcccctt ccagcaagtc tacctccggc 420
ggcacagctg ctctgggctg cctggtcaag gactacttcc ctgagcctgt gacagtgtcc 480
tggaactctg gcgccctgac ctctggcgtg cacaccttcc ctgccgtgct gcagtcctcc 540
ggcctgtact ccctgtcctc cgtggtcaca gtgccttcaa gcagcctggg cacccagacc 600
tatatctgca acgtgaacca caagccttcc aacaccaagg tggacaagcg ggtggagcct 660
aagtcctgcg acaagaccca cacctgtcct ccctgccctg ctcctgaact gctgggcggc 720
ccttctgtgt tcctgttccc tccaaagccc aaggacaccc tgatgatctc ccggacccct 780
gaagtgacct gcgtggtggt ggccgtgtcc cacgaggatc ctgaagtgaa gttcaattgg 840
tacgtggacg gcgtggaggt gcacaacgcc aagaccaagc ctcgggagga acagtacaac 900
tccacctacc gggtggtgtc cgtgctgacc gtgctgcacc aggactggct gaacggcaaa 960
gagtacaagt gcaaagtctc caacaaggcc ctggccgccc ctatcgaaaa gacaatctcc 1020
aaggccaagg gccagcctag ggaaccccag gtgtacaccc tgccacccag ccgggaggaa 1080
atgaccaaga accaggtgtc cctgacctgt ctggtcaagg gcttctaccc ttccgatatc 1140
gccgtggagt gggagtctaa cggccagcct gagaacaact acaagaccac ccctcctgtg 1200
ctggactccg acggctcctt cttcctgtac tccaaactga ccgtggacaa gtcccggtgg 1260
cagcagggca acgtgttctc ctgctccgtg atgcacgagg ccctgcacaa ccactacacc 1320
cagaagtccc tgtccctgtc tcccggcaag 1350
<210> 73
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 73
Gln Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 74
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 74
Tyr Thr Ser Arg Leu Gln Ser
1 5
<210> 75
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 75
Gln Gln Gly Asn Thr Leu Pro Tyr Thr
1 5
<210> 76
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 76
Ser Gln Asp Ile Ser Asn Tyr
1 5
<210> 77
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 77
Tyr Thr Ser
1
<210> 78
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 78
Gly Asn Thr Leu Pro Tyr
1 5
<210> 79
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 79
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Ala Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 80
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 80
gatattcaga tgactcagag cccctcctcg ctctccgcct ccgtggggga tcgcgtgaca 60
atcacctgtc aagcgtccca ggacatctca aactacctga actggtatca gcagaagcca 120
gggaaggccc cgaagctgct gatctactac acttcgcggc tgcagtccgg cgtgccgtca 180
cggttcactg gctcgggctc cggagcagac tacaccttca ccattagcag cctgcagccc 240
gaggacatcg ctacctactt ttgccaacaa ggaaacaccc tgccttacac cttcggacag 300
ggtactaagc tggaaatcaa a 321
<210> 81
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Ala Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 82
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 82
gatattcaga tgactcagag cccctcctcg ctctccgcct ccgtggggga tcgcgtgaca 60
atcacctgtc aagcgtccca ggacatctca aactacctga actggtatca gcagaagcca 120
gggaaggccc cgaagctgct gatctactac acttcgcggc tgcagtccgg cgtgccgtca 180
cggttcactg gctcgggctc cggagcagac tacaccttca ccattagcag cctgcagccc 240
gaggacatcg ctacctactt ttgccaacaa ggaaacaccc tgccttacac cttcggacag 300
ggtactaagc tggaaatcaa acgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag cggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 83
<211> 181
<212> Белок
<213> Homo sapiens
<400> 83
His Pro Ile Pro Asp Ser Ser Pro Leu Leu Gln Phe Gly Gly Gln Val
1 5 10 15
Arg Gln Arg Tyr Leu Tyr Thr Asp Asp Ala Gln Gln Thr Glu Ala His
20 25 30
Leu Glu Ile Arg Glu Asp Gly Thr Val Gly Gly Ala Ala Asp Gln Ser
35 40 45
Pro Glu Ser Leu Leu Gln Leu Lys Ala Leu Lys Pro Gly Val Ile Gln
50 55 60
Ile Leu Gly Val Lys Thr Ser Arg Phe Leu Cys Gln Arg Pro Asp Gly
65 70 75 80
Ala Leu Tyr Gly Ser Leu His Phe Asp Pro Glu Ala Cys Ser Phe Arg
85 90 95
Glu Leu Leu Leu Glu Asp Gly Tyr Asn Val Tyr Gln Ser Glu Ala His
100 105 110
Gly Leu Pro Leu His Leu Pro Gly Asn Lys Ser Pro His Arg Asp Pro
115 120 125
Ala Pro Arg Gly Pro Ala Arg Phe Leu Pro Leu Pro Gly Leu Pro Pro
130 135 140
Ala Leu Pro Glu Pro Pro Gly Ile Leu Ala Pro Gln Pro Pro Asp Val
145 150 155 160
Gly Ser Ser Asp Pro Leu Ser Met Val Gly Pro Ser Gln Gly Arg Ser
165 170 175
Pro Ser Tyr Ala Ser
180
<210> 84
<211> 546
<212> ДНК
<213> Homo sapiens
<400> 84
caccccatcc ctgactccag tcctctcctg caattcgggg gccaagtccg gcagcggtac 60
ctctacacag atgatgccca gcagacagaa gcccacctgg agatcaggga ggatgggacg 120
gtggggggcg ctgctgacca gagccccgaa agtctcctgc agctgaaagc cttgaagccg 180
ggagttattc aaatcttggg agtcaagaca tccaggttcc tgtgccagcg gccagatggg 240
gccctgtatg gatcgctcca ctttgaccct gaggcctgca gcttccggga gctgcttctt 300
gaggacggat acaatgttta ccagtccgaa gcccacggcc tcccgctgca cctgccaggg 360
aacaagtccc cacaccggga ccctgcaccc cgaggaccag ctcgcttcct gccactacca 420
ggcctgcccc ccgcactccc ggagccaccc ggaatcctgg ccccccagcc ccccgatgtg 480
ggctcctcgg accctctgag catggtggga ccttcccagg gccgaagccc cagctacgct 540
tcctga 546
<210> 85
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 85
Phe Ser Gly Asp Gly Arg Ala Ile Trp
1 5
<210> 86
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 86
Phe Leu Tyr Asp Thr Phe Pro Lys Asn Phe Phe
1 5 10
<210> 87
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 87
Phe Phe Trp Gly Ile Gly Thr
1 5
<210> 88
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 88
Phe Phe Trp Gly Ile Gly Thr Gly Ala Leu
1 5 10
<210> 89
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 89
Gly Ile Gly Thr Gly Ala Leu Gln
1 5
<210> 90
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 90
Gln Val Glu Gly Ser Trp Lys Lys Asp Gly Lys Gly Pro Ser Ile Trp
1 5 10 15
Asp His Phe
<210> 91
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 91
Leu Glu Lys Asp Leu Ser Ala
1 5
<210> 92
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 92
Leu Glu Lys Asp Leu Ser Ala Leu
1 5
<210> 93
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 93
Ser Ala Leu Asp Phe Ile Gly Val Ser Phe Tyr Gln Phe Ser Ile Ser
1 5 10 15
Trp Pro Arg Leu Phe Pro Asp Gly Ile Val Thr Val Ala Asn
20 25 30
<210> 94
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 94
Ser Ile Ser Trp Pro Arg Leu
1 5
<210> 95
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 95
Ser Ile Ser Trp Pro Arg Leu Phe Pro Asp Gly Ile Val Thr
1 5 10
<210> 96
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 96
Phe Pro Asp Gly Ile Val Thr
1 5
<210> 97
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 97
Val Ala Asn Ala Lys Gly Leu Gln
1 5
<210> 98
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 98
Val Ala Asn Ala Lys Gly Leu Gln Tyr
1 5
<210> 99
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 99
Leu Val Leu Arg Asn Ile Glu Pro Ile Val Thr
1 5 10
<210> 100
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 100
Val Leu Arg Asn Ile Glu Pro Ile Val Thr
1 5 10
<210> 101
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 101
Arg Asn Ile Glu Pro Ile Val Thr
1 5
<210> 102
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 102
Arg Asn Ile Glu Pro Ile Val Thr Leu
1 5
<210> 103
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 103
Leu Tyr His Trp Asp Leu Pro Leu Ala Leu
1 5 10
<210> 104
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 104
Ile Phe Asn Asp Tyr Ala Thr
1 5
<210> 105
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 105
Phe Asn Asp Tyr Ala Thr Tyr Cys
1 5
<210> 106
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 106
Cys Phe Gln Met Phe Gly Asp Arg Val Lys Tyr
1 5 10
<210> 107
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 107
Phe Gln Met Phe Gly Asp Arg Val Lys
1 5
<210> 108
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 108
Phe Gln Met Phe Gly Asp Arg Val Lys Tyr
1 5 10
<210> 109
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 109
Val Ala Trp His Gly Tyr Gly Thr Gly Met His Ala Pro Gly Glu Lys
1 5 10 15
Gly Asn Leu
<210> 110
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 110
Ala Trp His Gly Tyr Gly Thr Gly Met His Ala Pro Gly Glu Lys Gly
1 5 10 15
Asn Leu
<210> 111
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 111
Trp His Gly Tyr Gly Thr Gly Met His Ala Pro Gly Glu Lys Gly Asn
1 5 10 15
Leu
<210> 112
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 112
Trp His Gly Tyr Gly Thr Gly Met His Ala Pro Gly Glu Lys Gly Asn
1 5 10 15
Leu Ala Ala
<210> 113
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 113
His Gly Tyr Gly Thr Gly Met His Ala Pro Gly Glu Lys Gly Asn Leu
1 5 10 15
<210> 114
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 114
Tyr Thr Val Gly His Asn Leu Ile Lys Ala
1 5 10
<210> 115
<211> 29
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 115
Tyr Thr Val Gly His Asn Leu Ile Lys Ala His Ser Lys Val Trp His
1 5 10 15
Asn Tyr Asn Thr His Phe Arg Pro His Gln Lys Gly Trp
20 25
<210> 116
<211> 30
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 116
Tyr Thr Val Gly His Asn Leu Ile Lys Ala His Ser Lys Val Trp His
1 5 10 15
Asn Tyr Asn Thr His Phe Arg Pro His Gln Lys Gly Trp Leu
20 25 30
<210> 117
<211> 28
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 117
Val Gly His Asn Leu Ile Lys Ala His Ser Lys Val Trp His Asn Tyr
1 5 10 15
Asn Thr His Phe Arg Pro His Gln Lys Gly Trp Leu
20 25
<210> 118
<211> 27
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 118
Gly His Asn Leu Ile Lys Ala His Ser Lys Val Trp His Asn Tyr Asn
1 5 10 15
Thr His Phe Arg Pro His Gln Lys Gly Trp Leu
20 25
<210> 119
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 119
Leu Ile Lys Ala His Ser Lys Val Trp His Asn Tyr Asn Thr His Phe
1 5 10 15
Arg Pro His Gln Lys Gly Trp Leu
20
<210> 120
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 120
Ser Ile Thr Leu Gly Ser His
1 5
<210> 121
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 121
Ser Ile Thr Leu Gly Ser His Trp Ile Glu Pro Asn Arg Ser Glu Asn
1 5 10 15
Thr Met Asp
<210> 122
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 122
Ile Phe Lys Cys Gln Gln Ser Met Val
1 5
<210> 123
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 123
Phe Lys Cys Gln Gln Ser Met
1 5
<210> 124
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 124
Lys Cys Gln Gln Ser Met Val
1 5
<210> 125
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 125
Phe Ala Asn Pro Ile His Gly Asp Gly Asp Tyr Pro Glu Gly
1 5 10
<210> 126
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 126
Phe Ala Asn Pro Ile His Gly Asp Gly Asp Tyr Pro Glu Gly Met Arg
1 5 10 15
Lys Lys Leu
<210> 127
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 127
Phe Ala Asn Pro Ile His Gly Asp Gly Asp Tyr Pro Glu Gly Met Arg
1 5 10 15
Lys Lys Leu Phe
20
<210> 128
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 128
Leu Pro Ile Phe Ser Glu Ala Glu Lys His Glu Met Arg Gly Thr
1 5 10 15
<210> 129
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 129
Pro Ile Phe Ser Glu Ala Glu Lys His Glu Met Arg Gly Thr Ala Asp
1 5 10 15
<210> 130
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 130
Pro Ile Phe Ser Glu Ala Glu Lys His Glu Met Arg Gly Thr Ala Asp
1 5 10 15
Phe
<210> 131
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 131
Ser Glu Ala Glu Lys His Glu Met Arg Gly Thr Ala Asp Phe
1 5 10
<210> 132
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 132
Ala Glu Lys His Glu Met Arg Gly Thr Ala Asp Phe
1 5 10
<210> 133
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 133
Glu Lys His Glu Met Arg Gly Thr Ala Asp Phe
1 5 10
<210> 134
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 134
Phe Ala Phe Ser Phe Gly Pro Asn Asn Phe
1 5 10
<210> 135
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 135
Phe Ala Phe Ser Phe Gly Pro Asn Asn Phe Lys Pro Leu Asn Thr
1 5 10 15
<210> 136
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 136
Phe Gly Pro Asn Asn Phe Lys Pro Leu Asn Thr
1 5 10
<210> 137
<211> 12
<212> Белок
<213> Искусственная последовательност0ь
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 137
Phe Gly Pro Asn Asn Phe Lys Pro Leu Asn Thr Met
1 5 10
<210> 138
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 138
Asn Leu Arg Glu Ala Leu Asn
1 5
<210> 139
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 139
Ile Lys Leu Glu Tyr Asn Asn Pro Arg Ile Leu
1 5 10
<210> 140
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 140
Glu Tyr Asn Asn Pro Arg Ile Leu
1 5
<210> 141
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 141
Phe Thr Asp Ser Arg Val Lys Thr Glu Asp Thr Thr Ala
1 5 10
<210> 142
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 142
Asp Ser Arg Val Lys Thr Glu Asp Thr Thr Ala
1 5 10
<210> 143
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 143
Asp Thr Thr Ala Ile Tyr Met Met Lys Asn Phe
1 5 10
<210> 144
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 144
Tyr Met Met Lys Asn Phe Leu
1 5
<210> 145
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 145
Met Met Lys Asn Phe Leu Ser Gln Val Leu Gln Ala
1 5 10
<210> 146
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 146
Met Lys Asn Phe Leu Ser Gln Val Leu Gln Ala
1 5 10
<210> 147
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 147
Gln Ala Ile Arg Leu Asp Glu
1 5
<210> 148
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 148
Asp Glu Ile Arg Val Phe Gly Tyr Thr Ala
1 5 10
<210> 149
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 149
Ile Arg Val Phe Gly Tyr Thr Ala
1 5
<210> 150
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 150
Ile Arg Val Phe Gly Tyr Thr Ala Trp Ser Leu
1 5 10
<210> 151
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 151
Tyr Thr Ala Trp Ser Leu Leu
1 5
<210> 152
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 152
Asp Gly Phe Glu Trp Gln Asp Ala
1 5
<210> 153
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 153
Phe Glu Trp Gln Asp Ala Tyr Thr
1 5
<210> 154
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 154
Tyr Thr Ile Arg Arg Gly Leu Phe
1 5
<210> 155
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 155
Thr Ile Arg Arg Gly Leu Phe
1 5
<210> 156
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 156
Asn Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala His Tyr
1 5 10 15
<210> 157
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 157
Asn Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala His Tyr Tyr
1 5 10 15
Lys Gln Ile Ile Arg Glu
20
<210> 158
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 158
Asn Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala His Tyr Tyr
1 5 10 15
Lys Gln Ile Ile Arg Glu Asn Gly
20
<210> 159
<211> 25
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 159
Asn Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala His Tyr Tyr
1 5 10 15
Lys Gln Ile Ile Arg Glu Asn Gly Phe
20 25
<210> 160
<211> 26
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 160
Asn Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala His Tyr Tyr
1 5 10 15
Lys Gln Ile Ile Arg Glu Asn Gly Phe Ser
20 25
<210> 161
<211> 21
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 161
Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala His Tyr Tyr Lys
1 5 10 15
Gln Ile Ile Arg Glu
20
<210> 162
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 162
Tyr Lys Gln Ile Ile Arg Glu Asn Gly
1 5
<210> 163
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 163
Phe Ser Leu Lys Glu Ser Thr Pro Asp Val Gln Gly Gln Phe Pro Cys
1 5 10 15
Asp
<210> 164
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 164
Ser Leu Lys Glu Ser Thr Pro Asp Val Gln Gly Gln Phe Pro Cys Asp
1 5 10 15
<210> 165
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 165
Leu Lys Glu Ser Thr Pro Asp Val Gln Gly Gln Phe Pro Cys Asp
1 5 10 15
<210> 166
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 166
Phe Ser Trp Gly Val Thr Glu
1 5
<210> 167
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 167
Ser Val Leu Lys Pro Glu Ser Val Ala Ser Ser Pro Gln Phe Ser Asp
1 5 10 15
Pro His Leu
<210> 168
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 168
Lys Pro Glu Ser Val Ala Ser Ser Pro Gln Phe Ser Asp Pro His Leu
1 5 10 15
<210> 169
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 169
Val Arg Leu Lys Thr Arg Pro Ala Gln Cys
1 5 10
<210> 170
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 170
Phe Val Asn Ile Lys Lys Gln Leu Glu Met
1 5 10
<210> 171
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 171
Val Asn Ile Lys Lys Gln Leu Glu Met
1 5
<210> 172
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 172
Val Asn Ile Lys Lys Gln Leu Glu Met Leu
1 5 10
<210> 173
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 173
Asn Ile Lys Lys Gln Leu Glu Met
1 5
<210> 174
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 174
Leu Ala Arg Met Lys Val Thr His Tyr Arg
1 5 10
<210> 175
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 175
Leu Ala Arg Met Lys Val Thr His Tyr Arg Phe
1 5 10
<210> 176
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 176
Ala Arg Met Lys Val Thr His Tyr Arg Phe
1 5 10
<210> 177
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 177
Ala Leu Asp Trp Ala Ser Val Leu
1 5
<210> 178
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 178
Tyr Arg Cys Val Val Ser Glu Gly
1 5
<210> 179
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 179
Val Val Ser Glu Gly Leu Lys Leu Gly Ile Ser Ala
1 5 10
<210> 180
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 180
Gly Leu Lys Leu Gly Ile Ser Ala Met
1 5
<210> 181
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 181
Leu Lys Leu Gly Ile Ser Ala
1 5
<210> 182
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 182
Leu Lys Leu Gly Ile Ser Ala Met
1 5
<210> 183
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 183
Ile Ser Ala Met Val Thr Leu Tyr Tyr Pro Thr
1 5 10
<210> 184
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 184
Leu Tyr Tyr Pro Thr His Ala His Leu Gly Leu Pro Glu Pro Leu Leu
1 5 10 15
<210> 185
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 185
Tyr Tyr Pro Thr His Ala His Leu Gly Leu Pro Glu Pro Leu Leu
1 5 10 15
<210> 186
<211> 28
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 186
Tyr Tyr Pro Thr His Ala His Leu Gly Leu Pro Glu Pro Leu Leu His
1 5 10 15
Ala Asp Gly Trp Leu Asn Pro Ser Thr Ala Glu Ala
20 25
<210> 187
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 187
His Ala Asp Gly Trp Leu Asn Pro Ser Thr Ala Glu Ala
1 5 10
<210> 188
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 188
His Ala Asp Gly Trp Leu Asn Pro Ser Thr Ala Glu Ala Phe
1 5 10
<210> 189
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 189
Ala Phe Gln Ala Tyr Ala Gly Leu
1 5
<210> 190
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 190
Phe Gln Ala Tyr Ala Gly Leu Cys
1 5
<210> 191
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 191
Gln Ala Tyr Ala Gly Leu Cys
1 5
<210> 192
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 192
Gln Ala Tyr Ala Gly Leu Cys Phe
1 5
<210> 193
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 193
Cys Phe Gln Glu Leu Gly Asp
1 5
<210> 194
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 194
Cys Phe Gln Glu Leu Gly Asp Leu
1 5
<210> 195
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 195
Trp Ile Thr Ile Asn Glu Pro Asn Arg Leu
1 5 10
<210> 196
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 196
Trp Ile Thr Ile Asn Glu Pro Asn Arg Leu Ser Asp
1 5 10
<210> 197
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 197
Trp Ile Thr Ile Asn Glu Pro Asn Arg Leu Ser Asp Ile
1 5 10
<210> 198
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 198
Ile Thr Ile Asn Glu Pro Asn Arg
1 5
<210> 199
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 199
Val Ala His Ala Leu Ala Trp Arg Leu
1 5
<210> 200
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 200
Ala His Ala Leu Ala Trp Arg Leu
1 5
<210> 201
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 201
His Ala Leu Ala Trp Arg Leu
1 5
<210> 202
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 202
Tyr Asp Arg Gln Phe Arg Pro Ser Gln Arg Gly Ala Val Ser
1 5 10
<210> 203
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 203
Tyr Asp Arg Gln Phe Arg Pro Ser Gln Arg Gly Ala Val Ser Leu
1 5 10 15
<210> 204
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 204
Asp Arg Gln Phe Arg Pro Ser Gln Arg Gly Ala Val Ser
1 5 10
<210> 205
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 205
Ser Leu His Ala Asp Trp Ala Glu Pro Ala Asn Pro Tyr Ala Asp Ser
1 5 10 15
His Trp Arg Ala Ala Glu Arg Phe
20
<210> 206
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 206
His Ala Asp Trp Ala Glu Pro Ala Asn Pro Tyr Ala Asp Ser His Trp
1 5 10 15
<210> 207
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 207
His Ala Asp Trp Ala Glu Pro Ala Asn Pro Tyr Ala Asp Ser His Trp
1 5 10 15
Arg Ala
<210> 208
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 208
His Ala Asp Trp Ala Glu Pro Ala Asn Pro Tyr Ala Asp Ser His Trp
1 5 10 15
Arg Ala Ala Glu Arg Phe
20
<210> 209
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 209
Trp Ala Glu Pro Ala Asn Pro Tyr Ala Asp Ser His Trp Arg Ala Ala
1 5 10 15
Glu Arg Phe
<210> 210
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 210
Pro Ala Asn Pro Tyr Ala Asp Ser His Trp Arg Ala Ala Glu Arg Phe
1 5 10 15
<210> 211
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 211
Pro Tyr Ala Asp Ser His Trp Arg Ala Ala Glu Arg Phe
1 5 10
<210> 212
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 212
Phe Ala Glu Pro Leu Phe Lys Thr Gly Asp Tyr Pro Ala Ala
1 5 10
<210> 213
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 213
Phe Ala Glu Pro Leu Phe Lys Thr Gly Asp Tyr Pro Ala Ala Met
1 5 10 15
<210> 214
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 214
Lys Thr Gly Asp Tyr Pro Ala Ala Met
1 5
<210> 215
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 215
Arg Glu Tyr Ile Ala Ser Lys His Arg Arg Gly Leu Ser Ser Ser Ala
1 5 10 15
Leu
<210> 216
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 216
Arg Glu Tyr Ile Ala Ser Lys His Arg Arg Gly Leu Ser Ser Ser Ala
1 5 10 15
Leu Pro Arg Leu
20
<210> 217
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 217
Tyr Ile Ala Ser Lys His Arg Arg Gly Leu Ser Ser Ser Ala Leu
1 5 10 15
<210> 218
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 218
Pro Arg Leu Thr Glu Ala Glu
1 5
<210> 219
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 219
Pro Arg Leu Thr Glu Ala Glu Arg Arg Leu Leu Lys Gly Thr Val Asp
1 5 10 15
Phe
<210> 220
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 220
Ala Glu Arg Arg Leu Leu Lys Gly Thr Val Asp Phe
1 5 10
<210> 221
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 221
Glu Arg Arg Leu Leu Lys Gly Thr Val Asp Phe
1 5 10
<210> 222
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 222
Arg Arg Leu Leu Lys Gly Thr Val Asp Phe
1 5 10
<210> 223
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 223
Cys Ala Leu Asn His Phe Thr Thr Arg Phe
1 5 10
<210> 224
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 224
Val Met His Glu Gln Leu Ala Gly Ser Arg Tyr Asp Ser Asp Arg Asp
1 5 10 15
<210> 225
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 225
Val Met His Glu Gln Leu Ala Gly Ser Arg Tyr Asp Ser Asp Arg Asp
1 5 10 15
Ile Gln Phe
<210> 226
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 226
His Glu Gln Leu Ala Gly Ser Arg Tyr Asp Ser Asp Arg Asp Ile Gln
1 5 10 15
Phe
<210> 227
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 227
Ala Gly Ser Arg Tyr Asp Ser Asp Arg Asp
1 5 10
<210> 228
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 228
Ala Gly Ser Arg Tyr Asp Ser Asp Arg Asp Ile Gln Phe
1 5 10
<210> 229
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 229
Gly Ser Arg Tyr Asp Ser Asp Arg Asp Ile Gln Phe
1 5 10
<210> 230
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 230
Leu Gln Asp Ile Thr Arg Leu Ser Ser Pro Thr Arg
1 5 10
<210> 231
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 231
Leu Gln Asp Ile Thr Arg Leu Ser Ser Pro Thr Arg Leu
1 5 10
<210> 232
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 232
Ile Thr Arg Leu Ser Ser Pro Thr Arg Leu
1 5 10
<210> 233
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 233
Thr Arg Leu Ser Ser Pro Thr Arg
1 5
<210> 234
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 234
Ser Pro Thr Arg Leu Ala Val
1 5
<210> 235
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 235
Ser Pro Thr Arg Leu Ala Val Ile Pro Trp Gly Val Arg Lys Leu
1 5 10 15
<210> 236
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 236
Ala Val Ile Pro Trp Gly Val Arg Lys Leu
1 5 10
<210> 237
<211> 21
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 237
Ala Val Ile Pro Trp Gly Val Arg Lys Leu Leu Arg Trp Val Arg Arg
1 5 10 15
Asn Tyr Gly Asp Met
20
<210> 238
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 238
Ala Val Ile Pro Trp Gly Val Arg Lys Leu Leu Arg Trp Val Arg Arg
1 5 10 15
Asn Tyr Gly Asp Met Asp
20
<210> 239
<211> 23
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 239
Ala Val Ile Pro Trp Gly Val Arg Lys Leu Leu Arg Trp Val Arg Arg
1 5 10 15
Asn Tyr Gly Asp Met Asp Ile
20
<210> 240
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 240
Ile Pro Trp Gly Val Arg Lys
1 5
<210> 241
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 241
Ile Pro Trp Gly Val Arg Lys Leu Leu Arg Trp Val Arg Arg Asn Tyr
1 5 10 15
Gly Asp Met Asp
20
<210> 242
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 242
Leu Arg Trp Val Arg Arg Asn Tyr Gly Asp Met Asp Ile
1 5 10
<210> 243
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 243
Ile Tyr Ile Thr Ala Ser Gly Ile Asp Asp Gln Ala Leu Glu Asp
1 5 10 15
<210> 244
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 244
Tyr Ile Thr Ala Ser Gly Ile Asp Asp Gln Ala Leu
1 5 10
<210> 245
<211> 25
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 245
Tyr Ile Thr Ala Ser Gly Ile Asp Asp Gln Ala Leu Glu Asp Asp Arg
1 5 10 15
Leu Arg Lys Tyr Tyr Leu Gly Lys Tyr
20 25
<210> 246
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 246
Glu Asp Asp Arg Leu Arg Lys Tyr Tyr Leu Gly Lys Tyr
1 5 10
<210> 247
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 247
Asp Arg Leu Arg Lys Tyr Tyr Leu Gly Lys Tyr
1 5 10
<210> 248
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 248
Asp Arg Leu Arg Lys Tyr Tyr Leu Gly Lys Tyr Leu Gln Glu
1 5 10
<210> 249
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 249
Ile Asp Lys Val Arg Ile Lys Gly
1 5
<210> 250
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 250
Ile Asp Lys Val Arg Ile Lys Gly Tyr
1 5
<210> 251
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 251
Ile Asp Lys Val Arg Ile Lys Gly Tyr Tyr Ala
1 5 10
<210> 252
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 252
Ile Asp Lys Val Arg Ile Lys Gly Tyr Tyr Ala Phe
1 5 10
<210> 253
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 253
Asp Lys Val Arg Ile Lys Gly Tyr Tyr Ala
1 5 10
<210> 254
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 254
Lys Val Arg Ile Lys Gly Tyr Tyr Ala
1 5
<210> 255
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 255
Lys Val Arg Ile Lys Gly Tyr Tyr Ala Phe
1 5 10
<210> 256
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 256
Phe Lys Leu Ala Glu Glu Lys Ser Lys Pro Arg Phe Gly Phe
1 5 10
<210> 257
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 257
Phe Lys Leu Ala Glu Glu Lys Ser Lys Pro Arg Phe Gly Phe Phe
1 5 10 15
<210> 258
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 258
Lys Leu Ala Glu Glu Lys Ser Lys Pro Arg Phe Gly Phe Phe
1 5 10
<210> 259
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 259
Phe Lys Ala Lys Ser Ser Ile Gln Phe
1 5
<210> 260
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 260
Tyr Asn Lys Val Ile Ser Ser Arg Gly Phe Pro Phe Glu Asn Ser Ser
1 5 10 15
Ser Arg Cys Ser Gln Thr Gln Glu
20
<210> 261
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 261
Ser Ser Arg Cys Ser Gln Thr Gln Glu Asn Thr Glu Cys Thr
1 5 10
<210> 262
<211> 1044
<212> Белок
<213> Homo sapiens
<400> 262
Met Lys Pro Gly Cys Ala Ala Gly Ser Pro Gly Asn Glu Trp Ile Phe
1 5 10 15
Phe Ser Thr Asp Glu Ile Thr Thr Arg Tyr Arg Asn Thr Met Ser Asn
20 25 30
Gly Gly Leu Gln Arg Ser Val Ile Leu Ser Ala Leu Ile Leu Leu Arg
35 40 45
Ala Val Thr Gly Phe Ser Gly Asp Gly Arg Ala Ile Trp Ser Lys Asn
50 55 60
Pro Asn Phe Thr Pro Val Asn Glu Ser Gln Leu Phe Leu Tyr Asp Thr
65 70 75 80
Phe Pro Lys Asn Phe Phe Trp Gly Ile Gly Thr Gly Ala Leu Gln Val
85 90 95
Glu Gly Ser Trp Lys Lys Asp Gly Lys Gly Pro Ser Ile Trp Asp His
100 105 110
Phe Ile His Thr His Leu Lys Asn Val Ser Ser Thr Asn Gly Ser Ser
115 120 125
Asp Ser Tyr Ile Phe Leu Glu Lys Asp Leu Ser Ala Leu Asp Phe Ile
130 135 140
Gly Val Ser Phe Tyr Gln Phe Ser Ile Ser Trp Pro Arg Leu Phe Pro
145 150 155 160
Asp Gly Ile Val Thr Val Ala Asn Ala Lys Gly Leu Gln Tyr Tyr Ser
165 170 175
Thr Leu Leu Asp Ala Leu Val Leu Arg Asn Ile Glu Pro Ile Val Thr
180 185 190
Leu Tyr His Trp Asp Leu Pro Leu Ala Leu Gln Glu Lys Tyr Gly Gly
195 200 205
Trp Lys Asn Asp Thr Ile Ile Asp Ile Phe Asn Asp Tyr Ala Thr Tyr
210 215 220
Cys Phe Gln Met Phe Gly Asp Arg Val Lys Tyr Trp Ile Thr Ile His
225 230 235 240
Asn Pro Tyr Leu Val Ala Trp His Gly Tyr Gly Thr Gly Met His Ala
245 250 255
Pro Gly Glu Lys Gly Asn Leu Ala Ala Val Tyr Thr Val Gly His Asn
260 265 270
Leu Ile Lys Ala His Ser Lys Val Trp His Asn Tyr Asn Thr His Phe
275 280 285
Arg Pro His Gln Lys Gly Trp Leu Ser Ile Thr Leu Gly Ser His Trp
290 295 300
Ile Glu Pro Asn Arg Ser Glu Asn Thr Met Asp Ile Phe Lys Cys Gln
305 310 315 320
Gln Ser Met Val Ser Val Leu Gly Trp Phe Ala Asn Pro Ile His Gly
325 330 335
Asp Gly Asp Tyr Pro Glu Gly Met Arg Lys Lys Leu Phe Ser Val Leu
340 345 350
Pro Ile Phe Ser Glu Ala Glu Lys His Glu Met Arg Gly Thr Ala Asp
355 360 365
Phe Phe Ala Phe Ser Phe Gly Pro Asn Asn Phe Lys Pro Leu Asn Thr
370 375 380
Met Ala Lys Met Gly Gln Asn Val Ser Leu Asn Leu Arg Glu Ala Leu
385 390 395 400
Asn Trp Ile Lys Leu Glu Tyr Asn Asn Pro Arg Ile Leu Ile Ala Glu
405 410 415
Asn Gly Trp Phe Thr Asp Ser Arg Val Lys Thr Glu Asp Thr Thr Ala
420 425 430
Ile Tyr Met Met Lys Asn Phe Leu Ser Gln Val Leu Gln Ala Ile Arg
435 440 445
Leu Asp Glu Ile Arg Val Phe Gly Tyr Thr Ala Trp Ser Leu Leu Asp
450 455 460
Gly Phe Glu Trp Gln Asp Ala Tyr Thr Ile Arg Arg Gly Leu Phe Tyr
465 470 475 480
Val Asp Phe Asn Ser Lys Gln Lys Glu Arg Lys Pro Lys Ser Ser Ala
485 490 495
His Tyr Tyr Lys Gln Ile Ile Arg Glu Asn Gly Phe Ser Leu Lys Glu
500 505 510
Ser Thr Pro Asp Val Gln Gly Gln Phe Pro Cys Asp Phe Ser Trp Gly
515 520 525
Val Thr Glu Ser Val Leu Lys Pro Glu Ser Val Ala Ser Ser Pro Gln
530 535 540
Phe Ser Asp Pro His Leu Tyr Val Trp Asn Ala Thr Gly Asn Arg Leu
545 550 555 560
Leu His Arg Val Glu Gly Val Arg Leu Lys Thr Arg Pro Ala Gln Cys
565 570 575
Thr Asp Phe Val Asn Ile Lys Lys Gln Leu Glu Met Leu Ala Arg Met
580 585 590
Lys Val Thr His Tyr Arg Phe Ala Leu Asp Trp Ala Ser Val Leu Pro
595 600 605
Thr Gly Asn Leu Ser Ala Val Asn Arg Gln Ala Leu Arg Tyr Tyr Arg
610 615 620
Cys Val Val Ser Glu Gly Leu Lys Leu Gly Ile Ser Ala Met Val Thr
625 630 635 640
Leu Tyr Tyr Pro Thr His Ala His Leu Gly Leu Pro Glu Pro Leu Leu
645 650 655
His Ala Asp Gly Trp Leu Asn Pro Ser Thr Ala Glu Ala Phe Gln Ala
660 665 670
Tyr Ala Gly Leu Cys Phe Gln Glu Leu Gly Asp Leu Val Lys Leu Trp
675 680 685
Ile Thr Ile Asn Glu Pro Asn Arg Leu Ser Asp Ile Tyr Asn Arg Ser
690 695 700
Gly Asn Asp Thr Tyr Gly Ala Ala His Asn Leu Leu Val Ala His Ala
705 710 715 720
Leu Ala Trp Arg Leu Tyr Asp Arg Gln Phe Arg Pro Ser Gln Arg Gly
725 730 735
Ala Val Ser Leu Ser Leu His Ala Asp Trp Ala Glu Pro Ala Asn Pro
740 745 750
Tyr Ala Asp Ser His Trp Arg Ala Ala Glu Arg Phe Leu Gln Phe Glu
755 760 765
Ile Ala Trp Phe Ala Glu Pro Leu Phe Lys Thr Gly Asp Tyr Pro Ala
770 775 780
Ala Met Arg Glu Tyr Ile Ala Ser Lys His Arg Arg Gly Leu Ser Ser
785 790 795 800
Ser Ala Leu Pro Arg Leu Thr Glu Ala Glu Arg Arg Leu Leu Lys Gly
805 810 815
Thr Val Asp Phe Cys Ala Leu Asn His Phe Thr Thr Arg Phe Val Met
820 825 830
His Glu Gln Leu Ala Gly Ser Arg Tyr Asp Ser Asp Arg Asp Ile Gln
835 840 845
Phe Leu Gln Asp Ile Thr Arg Leu Ser Ser Pro Thr Arg Leu Ala Val
850 855 860
Ile Pro Trp Gly Val Arg Lys Leu Leu Arg Trp Val Arg Arg Asn Tyr
865 870 875 880
Gly Asp Met Asp Ile Tyr Ile Thr Ala Ser Gly Ile Asp Asp Gln Ala
885 890 895
Leu Glu Asp Asp Arg Leu Arg Lys Tyr Tyr Leu Gly Lys Tyr Leu Gln
900 905 910
Glu Val Leu Lys Ala Tyr Leu Ile Asp Lys Val Arg Ile Lys Gly Tyr
915 920 925
Tyr Ala Phe Lys Leu Ala Glu Glu Lys Ser Lys Pro Arg Phe Gly Phe
930 935 940
Phe Thr Ser Asp Phe Lys Ala Lys Ser Ser Ile Gln Phe Tyr Asn Lys
945 950 955 960
Val Ile Ser Ser Arg Gly Phe Pro Phe Glu Asn Ser Ser Ser Arg Cys
965 970 975
Ser Gln Thr Gln Glu Asn Thr Glu Cys Thr Val Cys Leu Phe Leu Val
980 985 990
Gln Lys Lys Pro Leu Ile Phe Leu Gly Cys Cys Phe Phe Ser Thr Leu
995 1000 1005
Val Leu Leu Leu Ser Ile Ala Ile Phe Gln Arg Gln Lys Arg Arg
1010 1015 1020
Lys Phe Trp Lys Ala Lys Asn Leu Gln His Ile Pro Leu Lys Lys
1025 1030 1035
Gly Lys Arg Val Val Ser
1040
<210> 263
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 263
Gly Tyr Thr Phe Thr Asp Tyr Tyr Ile Asn
1 5 10
<210> 264
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 264
Gly Tyr Thr Phe Thr Asp Tyr Tyr
1 5
<210> 265
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 265
Ile His Pro Gly Ser Gly Asn Thr
1 5
<210> 266
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 266
Ala Ile Leu Leu Leu Arg Ser Tyr Gly Met Asp Asp
1 5 10
<210> 267
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 267
Gln Ser Ile Val His Ser Ser Gly Asn Thr Tyr
1 5 10
<210> 268
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 268
Gly Tyr Ser Ile Thr Ser Gly Tyr Thr Trp His
1 5 10
<210> 269
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 269
Gly Tyr Ser Ile Thr Ser Gly Tyr Thr
1 5
<210> 270
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 270
Ile His Tyr Ser Val Tyr Thr
1 5
<210> 271
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 271
Ala Arg Arg Thr Thr Ser Leu Glu Arg Tyr Phe Asp Val
1 5 10
<210> 272
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 272
Gln Asp Ile Ser Asn Tyr
1 5
<210> 273
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 273
Tyr Thr Ser
1
<210> 274
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
6xHis tag
<400> 274
His His His His His His
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА-МИМЕТИКИ FGF21 И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2774368C2 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ FC-РЕЦЕПТОР-ПОДОБНОГО БЕЛКА 5, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2774158C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| АНТИ-CLDN АНТИТЕЛО, ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБ ЕГО ОБНАРУЖЕНИЯ | 2020 |
|
RU2801315C2 |

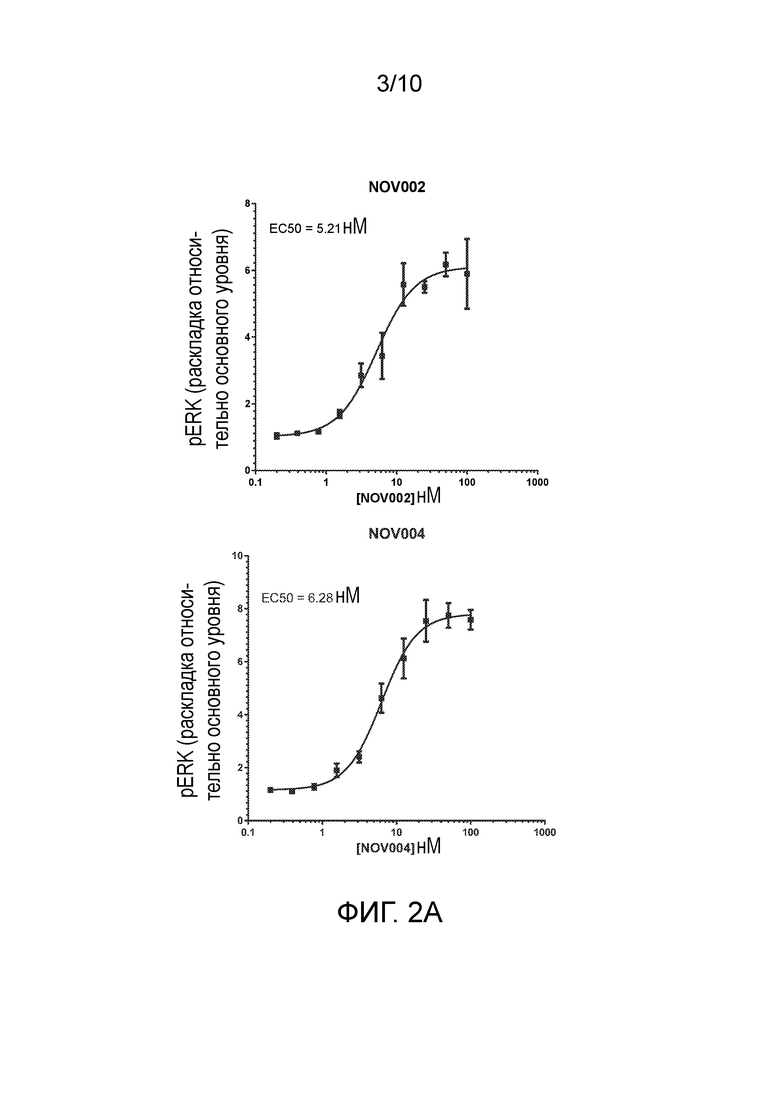







Изобретение относится к области биохимии, в частности к антителу или к его антигенсвязывающему фрагменту, которое специфически связывается с последовательностью β-клото. Также раскрыты нуклеиновая кислота, кодирующая указанная антитело; вектор, содержащий указанную нуклеиновую кислоту; клетка-хозяин, содержащий указанный вектор; фармацевтическая композиция, содержащая указанное антитело. Раскрыто применение указанного антитела в качестве лекарственного средства для лечения FGF21-связанного заболевания. Изобретение позволяет эффективно лечить заболевания, ассоциированные с FGF21. 5 н. и 9 з.п. ф-лы, 11 ил., 3 табл., 10 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, которое(ый) специфически связывается с последовательностью β-клото (SEQ ID NO: 262), и указанное антитело или его антигенсвязывающий фрагмент содержит одно из следующих:
(i) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 3, 4 и 5, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 13, 14 и 15;
(ii) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 6, 7 и 8, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 16, 17 и 18;
(iii) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 263, 4 и 5, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 13, 14 и 15;
(iv) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 264, 265 и 266, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 267, 17 и 15;
(v) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 23, 24 и 25, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи CDR содержат SEQ ID NO: 33, 34 и 35;
(vi) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 26, 27 и 28, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 36, 37 и 38;
(vii) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 268, 24 и 25, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 33, 34 и 35; и
(viii) вариабельную область тяжелой цепи, содержащую три CDR тяжелой цепи, где три CDR тяжелой цепи содержат SEQ ID NO: 269, 270 и 271, и вариабельную область легкой цепи, содержащую три CDR легкой цепи, где три CDR легкой цепи содержат SEQ ID NO: 272, 273 и 35.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, которое(ый) связывается с белком β-клото человека с KD менее чем или равной 10 пМ, определенные с помощью титриметрического анализа равновесного раствора (SET).
3. Выделенное антитело или его антигенсвязывающий фрагмент по п.1 или 2, которое(ый) активирует рецепторный комплекс FGFR1c_бета-клото яванского макака с EC50 менее чем или равной 50 нМ, определенные с помощью анализов pERK клеток.
4. Выделенное антитело или антигенсвязывающий фрагмент по любому из пп.1-3, где антитело или фрагмент содержит VH, содержащий одну или несколько из SEQ ID NO: 9, 29, 49, 69 или аминокислотную последовательность, которая на 90% ей идентична; и VL, содержащую одну или несколько SEQ ID NO: 19, 39, 59 и 79 или аминокислотную последовательность, которая на 90% ей идентична.
5. Выделенное антитело или антигенсвязывающий фрагмент по п.4, где антитело или фрагмент содержит VH, содержащую одну или несколько из SEQ ID NO: 9, 29, 49 и 29 или аминокислотную последовательность, которая на 95% или 97% ей идентична; и VL, содержащую одну или несколько из SEQ ID NO: 19, 39, 59 и 79 или аминокислотную последовательность, которая на 95% или 97% ей идентична.
6. Выделенное антитело или антигенсвязывающий фрагмент по любому из пп.1-5, где антитело или фрагмент представляет собой:
- антитело или фрагмент, содержащее(ий) последовательность вариабельной области тяжелой цепи SEQ ID NO: 9 и последовательность вариабельной области легкой цепи SEQ ID NO: 19, и
- антитело или фрагмент, содержащее(ий) последовательность вариабельной области тяжелой цепи SEQ ID NO: 29 и последовательность вариабельной области легкой цепи SEQ ID NO: 39.
7. Выделенное антитело или антигенсвязывающий фрагмент по любому из пп.1-6, где антитело или фрагмент содержит:
(i) CDR1 тяжелой цепи c SEQ ID NO: 3; CDR2 тяжелой цепи c SEQ ID NO: 4; CDR3 тяжелой цепи c SEQ ID NO: 5; CDR1 легкой цепи c SEQ ID NO: 13; CDR2 легкой цепи c SEQ ID NO: 14; и CDR3 легкой цепи c SEQ ID NO: 15;
(ii) CDR1 тяжелой цепи c SEQ ID NO: 23; CDR2 тяжелой цепи c SEQ ID NO: 24; CDR3 тяжелой цепи c SEQ ID NO: 25; CDR1 легкой цепи c SEQ ID NO: 33; CDR2 легкой цепи c SEQ ID NO: 34; и CDR3 легкой цепи c SEQ ID NO: 35;
(iii) CDR1 тяжелой цепи c SEQ ID NO: 43; CDR2 тяжелой цепи c SEQ ID NO: 44; CDR3 тяжелой цепи c SEQ ID NO: 45; CDR1 легкой цепи c SEQ ID NO: 53; CDR2 легкой цепи c SEQ ID NO: 54; и CDR3 легкой цепи c SEQ ID NO: 55;
(iv) CDR1 тяжелой цепи c SEQ ID NO: 63; CDR2 тяжелой цепи c SEQ ID NO: 64; CDR3 тяжелой цепи c SEQ ID NO: 65; CDR1 легкой цепи c SEQ ID NO: 73; CDR2 легкой цепи c SEQ ID NO: 74; и CDR3 легкой цепи c SEQ ID NO: 75;
(v) CDR1 тяжелой цепи c SEQ ID NO: 6; CDR2 тяжелой цепи c SEQ ID NO: 7; CDR3 тяжелой цепи c SEQ ID NO: 8; CDR1 легкой цепи c SEQ ID NO: 16; CDR2 легкой цепи c SEQ ID NO: 17; и CDR3 легкой цепи c SEQ ID NO: 18;
(vi) CDR1 тяжелой цепи c SEQ ID NO: 26; CDR2 тяжелой цепи c SEQ ID NO: 27; CDR3 тяжелой цепи c SEQ ID NO: 28; CDR1 легкой цепи c SEQ ID NO: 36; CDR2 легкой цепи c SEQ ID NO: 37; и CDR3 легкой цепи c SEQ ID NO: 38;
(vii) CDR1 тяжелой цепи c SEQ ID NO: 46; CDR2 тяжелой цепи c SEQ ID NO: 47; CDR3 тяжелой цепи c SEQ ID NO: 48; CDR1 легкой цепи c SEQ ID NO: 56; CDR2 легкой цепи c SEQ ID NO: 57; и CDR3 легкой цепи c SEQ ID NO: 58; или
(viii) CDR1 тяжелой цепи c SEQ ID NO: 66; CDR2 тяжелой цепи c SEQ ID NO: 67; CDR3 тяжелой цепи c SEQ ID NO: 68; CDR1 легкой цепи c SEQ ID NO: 76; CDR2 легкой цепи c SEQ ID NO: 77; и CDR3 легкой цепи c SEQ ID NO: 78.
8. Фармацевтическая композиция для лечения FGF21-связанного заболевания, содержащая фармацевтически приемлемое количество антитела или его антигенсвязывающего фрагмента по любому из пп.1-7 и фармацевтически приемлемый носитель.
9. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-7 в качестве лекарственного средства для лечения FGF21-связанного заболевания.
10. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-7 или фармацевтическая композиция по п.8 для применения в способе лечения матаболического расстройства у индивида, нуждающегося в таком лечении.
11. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-7 или фармацевтическая композиция по п.8 для применения в способе лечения одного или нескольких следующих состояний у индивида, нуждающегося в таком лечении: ожирение, сахарный диабет типа 1 и типа 2, панкреатит, дислипидемия, неалкогольный стеатогепатит (NASH), резистентность к инсулину, гиперинсулинемия, толерантность к глюкозе, гипергликемия и метаболический синдром.
12. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-7 или фармацевтическая композиция по п.8 для применения в способе лечения одного или нескольких следующих состояний у индивида, нуждающегося в таком лечении: атеросклероз, болезни периферических артерий, инсульт, сердечная недостаточность и коронарная болезнь сердца.
13. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому пп.1-7, где нуклеиновая кислота содержит последовательность, которая по меньшей мере на 90% идентична одной из нуклеиновых последовательностей, указанных в Таблице 1.
14. Вектор для экспрессии антитела или его антигенсвязывающего фрагмента по любому из пп.1-7, содержащий нуклеиновую кислоту по п.13.
| Предохранительное приспособление при фрезерных деревообделочных станках | 1930 |
|
SU21425A1 |
| КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ РАССЕЯННОГО СКЛЕРОЗА | 2008 |
|
RU2492234C2 |
| WO 2013184958 A1, 12.12.2013 | |||
| US 9006400 B2, 14.04.2015. | |||

