ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к анти-CLDN антителу, настоящее изобретение также относится к фармацевтической композиции, содержащей анти-CLDN антитело, и настоящее изобретение также относится к способу обнаружения того, присутствует ли CLDN в биологическом образце.
УРОВЕНЬ ТЕХНИКИ
Плотный контакт (TJ англ.: tight junction) играет ключевую роль в потоке материала между клетками, поддерживает полярность клеток путем блокирования радиальной диффузии мембранных белков и мембранных липидов. Кроме того, он также участвует в рекрутинге сигнальных молекул, которые регулируют пролиферацию, дифференциацию и движение клеток. Плотный контакт образован клаудином (CLDN), при этом семейство клаудина состоит из более чем 20 белковых молекул, каждая из которых содержит четверной трансмембранный домен и сходные аминокислотные последовательности, но их распределение в тканях является специфичным. Гены CLDN человека распределены парами на разных хромосомах, что подразумевает, что некоторые гены CLDN являются производными от репликации гена.
Молекулярная масса белка CLDN главным образом находится в диапазоне 20-34 кДа, а наибольшее различие заключается в последовательности и размере внутриклеточного С-конца, который содержит мотив связывания домена PDZ, который позволяет белку CLDN непосредственно взаимодействовать с белками в цитоплазме, такими как ZO-1, ZO-2, ZO-3 и MUPP1, имеющими отношение к плотному контакту. Кроме того, эта последовательность содержит сайты пост-транскрипционной модификации, такие как сайты фосфорилирования, которые могут влиять на локализацию и функцию молекул белка. МАРK (Митоген-активированная протеинкиназа) или РKС (протеинкиназа С) способна фосфорилировать CLDN1, а сАМР (циклический AMP) способен индуцировать фосфорилирование CLDN5, и все из них могут промотировать барьерную функцию белка CLDN, в то время как РKА-опосредованное фосфорилирование CLDN16 может усиливать транспорт ионов магния.
CLDN играет ключевую роль в регулировании селективной проницаемости клеточного обходного пути. CLDN2 и CLDN15 участвуют в формировании катионных каналов и катионных пор, в то время как CLDN4/7/10 участвуют в формировании анионных каналов и пор. Белок клаудин экспрессируется на высоком уровне в некоторых клеточных линиях, что влияет на трансэпителиальное электрическое сопротивление и проницаемость. В культивируемых клетках, являющихся производными от эпидермиса, CLDN 1/4/5/7 может увеличивать трансэпителиальное электрическое сопротивление, в то время как CLDN2 и CLDN10 не оказывают такого влияния.
Считают, что мутация гена CLDN имеет отношение к различным заболеваниям. Мутация CLDN1 может приводить к склерозирующему холангиту и ихтиозу, в то время как мутации CLDN16 и CLDN19 считают связанными с гипомагниемией и гиперкальцинурией.
Полагают, что дифференциальная экспрессия белка CLDN связана с различными видами рака. CLDN1 и CLDN7 имеют подавляющую регуляцию при инвазивном раке молочной железы, раке простаты и раке пищевода, тогда как CLDN3/4 обнаруживали в разной степени активированными при раке шейки матки, раке толстой кишки, раке пищевода, раке желудка и других раковых заболеваниях. Sahin et al. обнаружили, что в нормальных тканях изоформа 2 подтипа CLDN18 (CLDN18.2) экспрессировались только в дифференцированных эпидермальных клетках слизистой оболочки желудка, но не в области стволовых клеток желудка, в то время как аномально высокая экспрессия была выявлена при первичном раке желудка и его метастазах. Сообщалось также о высоком уровне экспрессии CLDN18.2 при раке поджелудочной железы, раке пищевода и раке легких. Поскольку CLDN18.2 расположен на поверхности клеточной мембраны, его биологическая функция и характеристики определяют, что он является идеальной терапевтической мишенью. В последние годы также появились моноклональные антитела против этой мишени, а среди них самыми быстроразвивающимися являются IMAB362 (Клаудиксимаб) компании Ganymed. IMAB362 связывает CLDN18.2 на поверхности опухолевых клеток, вызывая антителозависимую цитотоксичность (ADCC) и комплементзависимую цитотоксичность (CDC), тем самым убивая опухолевые клетки. В сочетании с химиотерапией антитела IMAB362 могут также усиливать инфильтрацию Т-лимфоцитов и обеспечивать повышающее регулирование провоспалительных факторов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является предложить анти-CLDN антитело, его фармацевтическую композицию и способ его обнаружения.
В настоящем изобретении приняты следующие технические решения.
Анти-CLDN антитело, которое содержит тяжелую цепь и легкую цепь:
в котором тяжелая цепь антитела содержит одну или несколько областей, определяющих комплементарность (CDR), a CDR тяжелой цепи не более чем на три аминокислоты отличаются от последовательности CDR любой из от SEQ ID No: 1 до SEQ ID No: 7 или от любой от SEQ ID No: 15 до SEQ ID No: 30;
легкая цепь антитела содержит одну или несколько областей, определяющих комплементарность (CDR), a CDR легкой цепи не более чем на три аминокислоты отличаются от последовательности CDR любой из от SEQ ID No: 8 до SEQ ID No: 14 или от любой от SEQ ID No: 31 до SEQ ID No: 46.
Кроме того, анти-CLDN антитело по настоящему изобретению имеет признак, заключающийся в том, что тяжелая цепь антитела выбрана из любой последовательности от SEQ ID No: 1 до SEQ ID No 7.
Кроме того, анти-CLDN антитело по настоящему изобретению имеет признак, заключающийся в том, что легкая цепь антитела выбрана из любой последовательности от SEQ ID No: 8 до SEQ ID No: 14 или от любой от SEQ ID No: 31 до SEQ ID No: 46.
Кроме того, анти-CLDN антитело по настоящему изобретению имеет признак, заключающийся в том, что тяжелая цепь антитела выбрана из любой последовательности от SEQ ID No: 15 до SEQ ID No: 30.
Кроме того, анти-CLDN антитело по настоящему изобретению имеет признак, заключающийся в том, что легкая цепь антитела выбрана из любой последовательности от SEQ ID No: 31 до SEQ ID No: 46.
Кроме того, анти-CLDN антитело по настоящему изобретению имеет признак, заключающийся в том, что тяжелая цепь и легкая цепь антитела образует комбинацию, выбранную из группы, состоящей из:
SEQ ID No: 1 и SEQ ID No: 8, SEQ ID No: 2 и SEQ ID No: 9, SEQ ID No: 3 и SEQ ID No: 10;
SEQ ID No: 4 и SEQ ID No: 11, SEQ ID No: 5 и SEQ ID No: 12, SEQ ID No: 6 и SEQ ID No: 13;
SEQ ID No: 7 и SEQ ID No: 14, SEQ ID No: 15 и SEQ ID No: 31, SEQ ID No: 16 и SEQ ID No: 32;
SEQ ID No: 17 и SEQ ID No: 33, SEQ ID No: 18 и SEQ ID No: 34, SEQ ID No: 19 и SEQ ID No: 35;
SEQ ID No: 20 и SEQ ID No: 36, SEQ ID No: 21 и SEQ ID No: 37, SEQ ID No: 22 и SEQ ID No: 38;
SEQ ID No: 23 и SEQ ID No: 39, SEQ ID No: 24 и SEQ ID No: 40, SEQ ID No: 25 и SEQ ID No: 41;
SEQ ID No: 26 и SEQ ID No: 42, SEQ ID No: 27 и SEQ ID No: 43, SEQ ID No: 28 и SEQ ID No: 44;
SEQ ID No: 29 и SEQ ID No: 45, SEQ ID No: 30 и SEQ ID No: 46.
Настоящее изобретение также предлагает полинуклеотид, кодирующий антитело согласно определению выше в данном документе.
Настоящее изобретение также предлагает фармацевтическую композицию, содержащую любое из упомянутых выше антител.
Настоящее изобретение также предлагает применение любого из вышеупомянутых антител в приготовлении противоопухолевых лекарственных средств.
Настоящее изобретение также предлагает способ обнаружения присутствия или отсутствия CLDN в биологическом образце, который содержит этапы введения антитела, как описано выше, в биологический образец, причем такое антитело имеет обнаруживаемую метку, и этапы обнаружения наличия или отсутствия такой обнаруживаемой метки или обнаружения содержимого такой обнаруживаемой метки.
ПОЛОЖИТЕЛЬНЫЙ ЭФФЕКТ ДАННОГО ИЗОБРЕТЕНИЯ
Анти-CLDN антитело по настоящему изобретению обладает более сильной способностью к связыванию с клетками, чем IMAB362. Более того, антитело по настоящему изобретению демонстрирует лучший эффект ингибирования роста опухоли, чем у IMAB362, по фармакодинамике in vivo у животных.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1а показана способность контрольного антитела QP024025 к связыванию с клетками CHOS
На Фиг. 1b показана способность антитела QP188189, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
На Фиг. 1с показана способность антитела QP190191, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
На Фиг. 1d показана способность антитела QP192193, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
На Фиг. 1е показана способность антитела QP196198, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
На Фиг. 1f показана способность антитела QP199200, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
На Фиг. 1g показана способность антитела QP201202, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
На Фиг. 1h показана способность антитела QP207208, отобранного скринингом из гибридомы, к связыванию с клетками CHOS
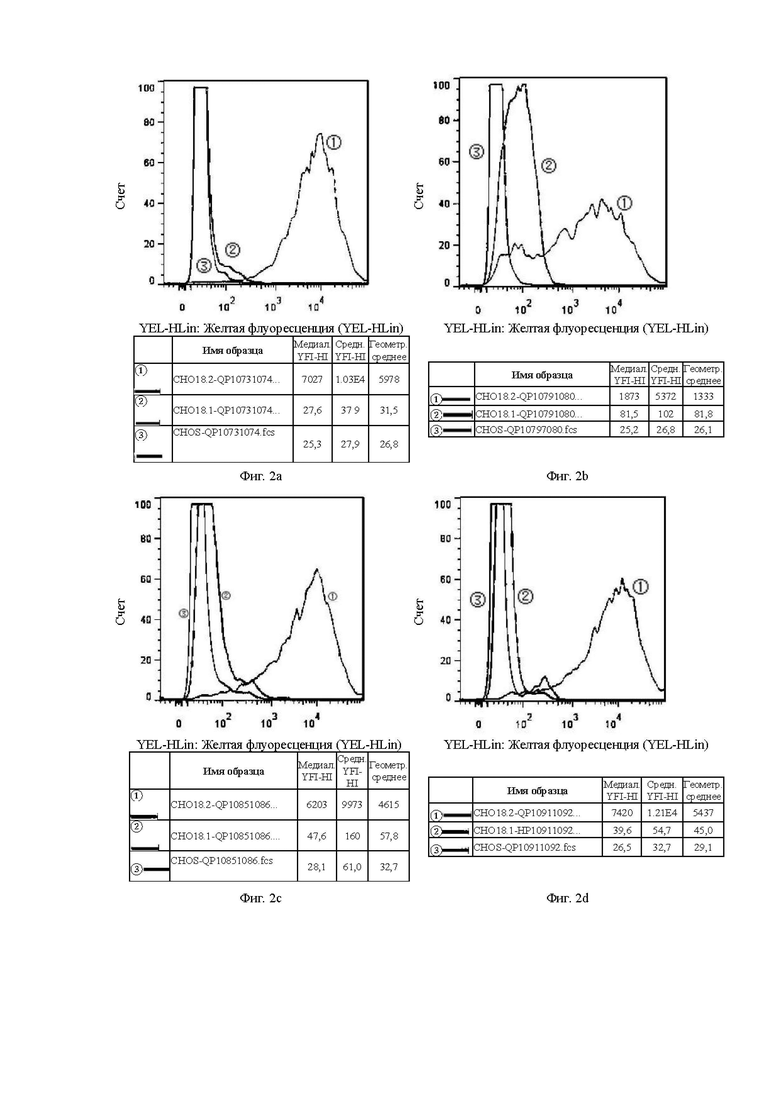
На Фиг. 2а показана способность антитела QP10731074, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 2b показана способность антитела QP10791080, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 2с показана способность антитела QP10851086, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 2d показана способность антитела QP10911092, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
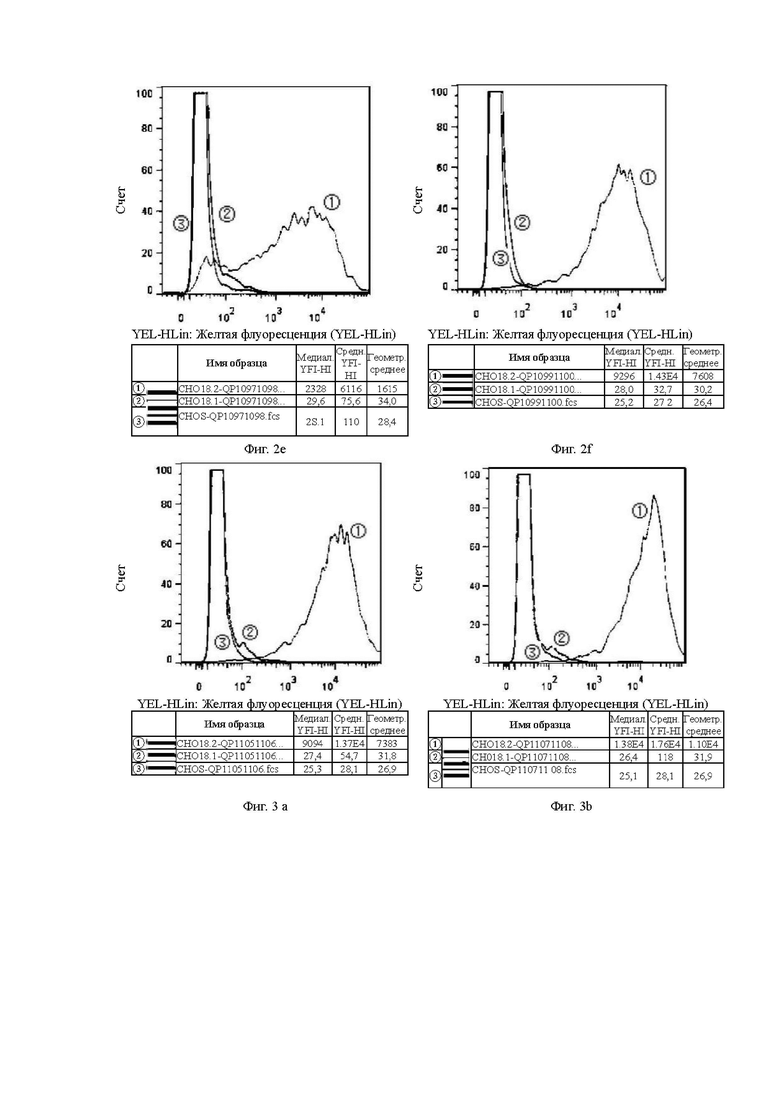
На Фиг. 2е показана способность антитела QP10971098, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 2f показана способность антитела QP10991100, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 3а показана способность антитела QP11051106, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 3b показана способность антитела QP11071108, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
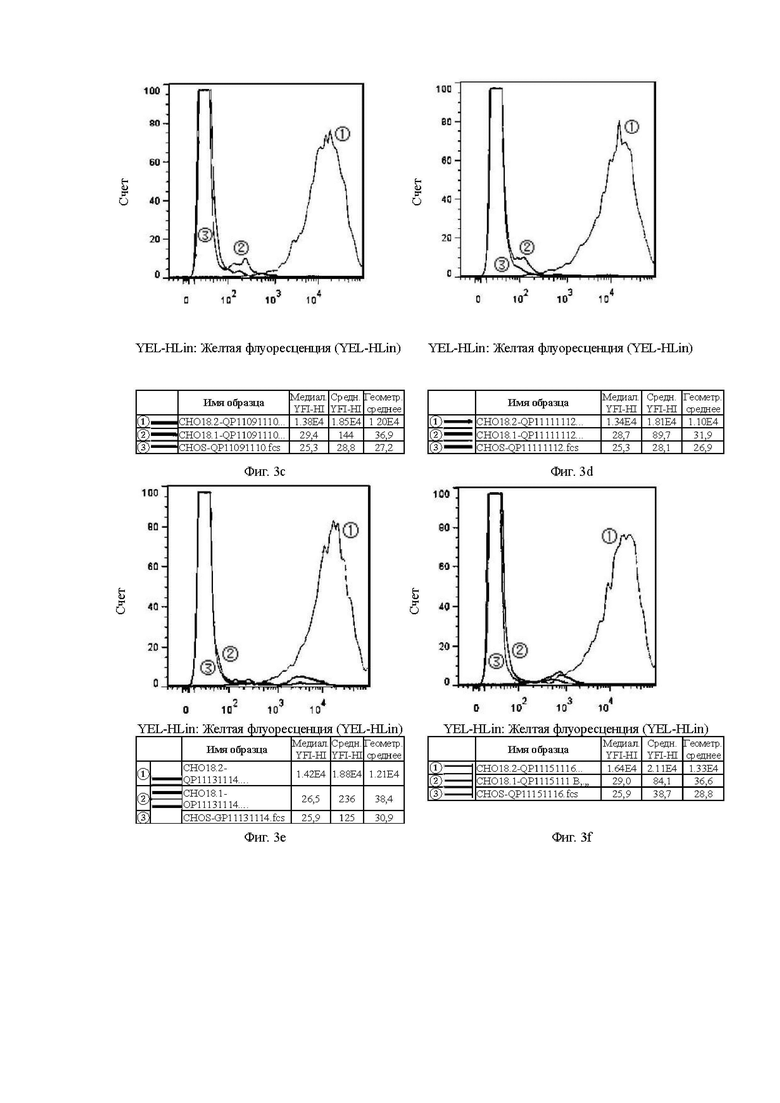
На Фиг. 3с показана способность антитела QP11091110, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 3d показана способность антитела QP11111112, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 3е показана способность антитела QP11131114, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 3f показана способность антитела QP11151116, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
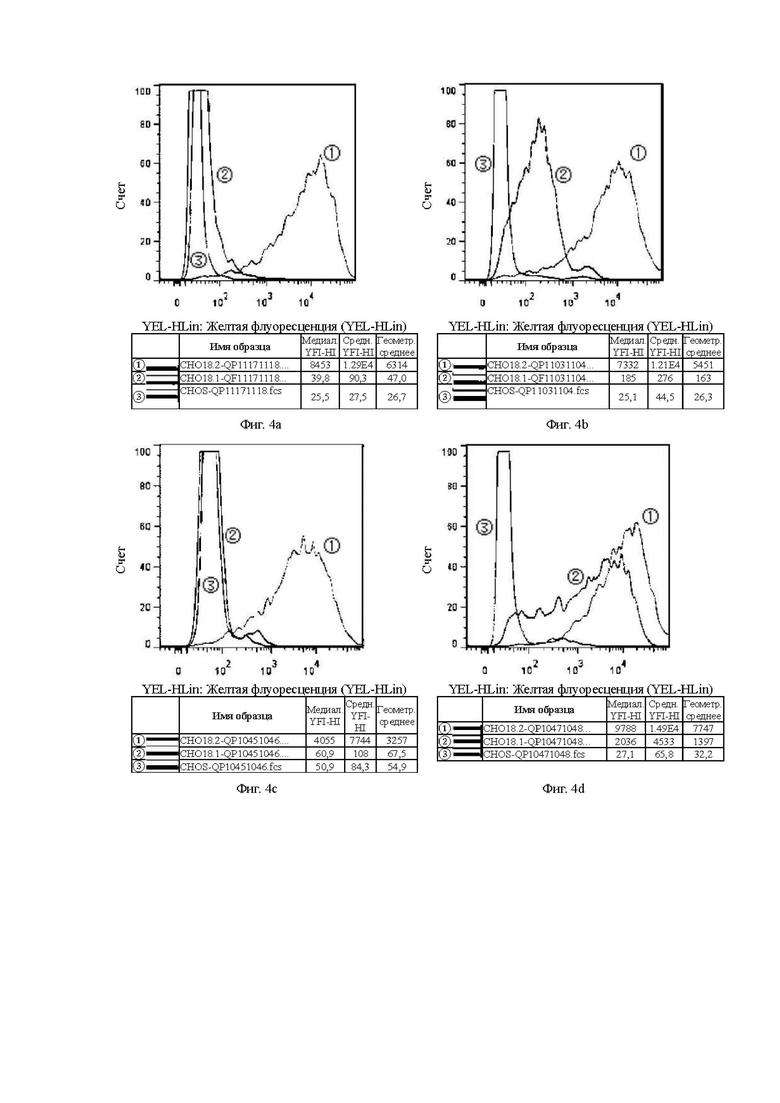
На Фиг. 4а показана способность антитела QP11171118, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 4b показана способность антитела QP11031104, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 4с показана способность антитела QP10451046, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 4d показана способность антитела QP10471048, отобранного скринингом из бактериофага, к связыванию с клетками CHOS.
На Фиг. 5а показана способность контрольного антитела QP024025 к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5b показана способность антитела QP188189, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5с показана способность антитела QP190191, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5d показана способность антитела QP192193, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5е показана способность антитела QP196198, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5f показана способность антитела QP199200, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5g показана способность антитела QP201202, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 5h показана способность антитела QP207208, отобранного скринингом из гибридомы, к связыванию с различными временными клеточными линиями 293Т.
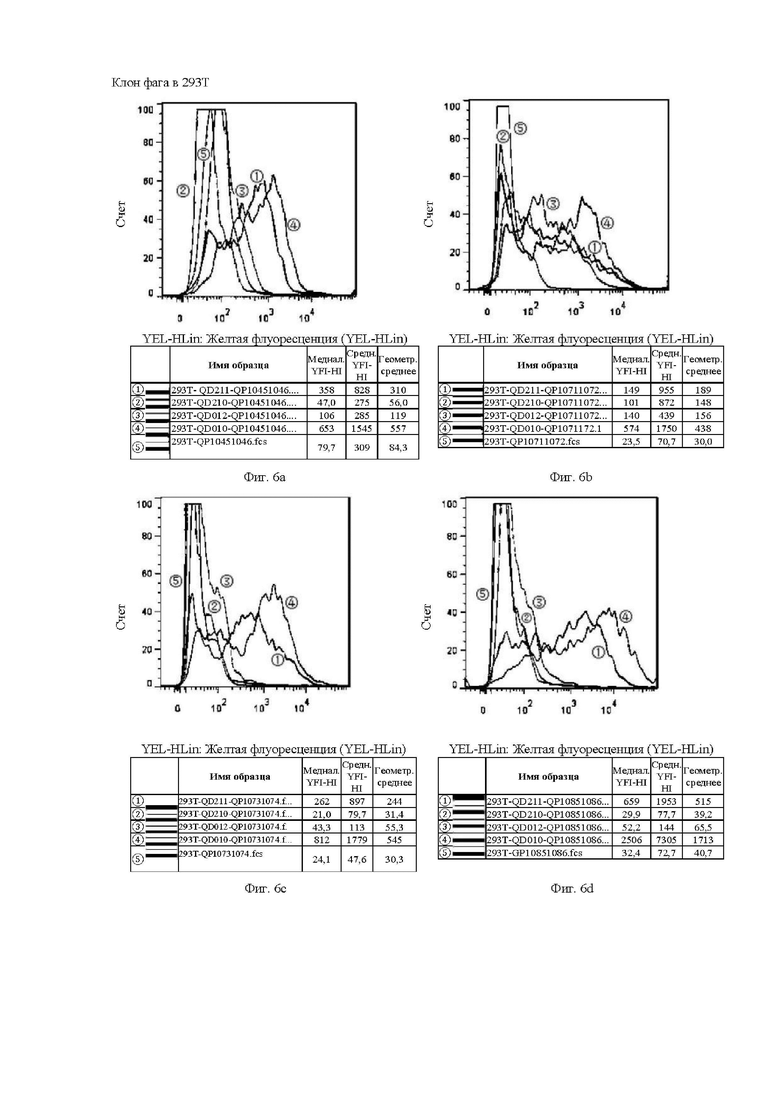
На Фиг. 6а показана способность антитела QP10451046, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 6b показана способность антитела QP10711072, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 6с показана способность антитела QP10731074, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 6d показана способность антитела QP10851086, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
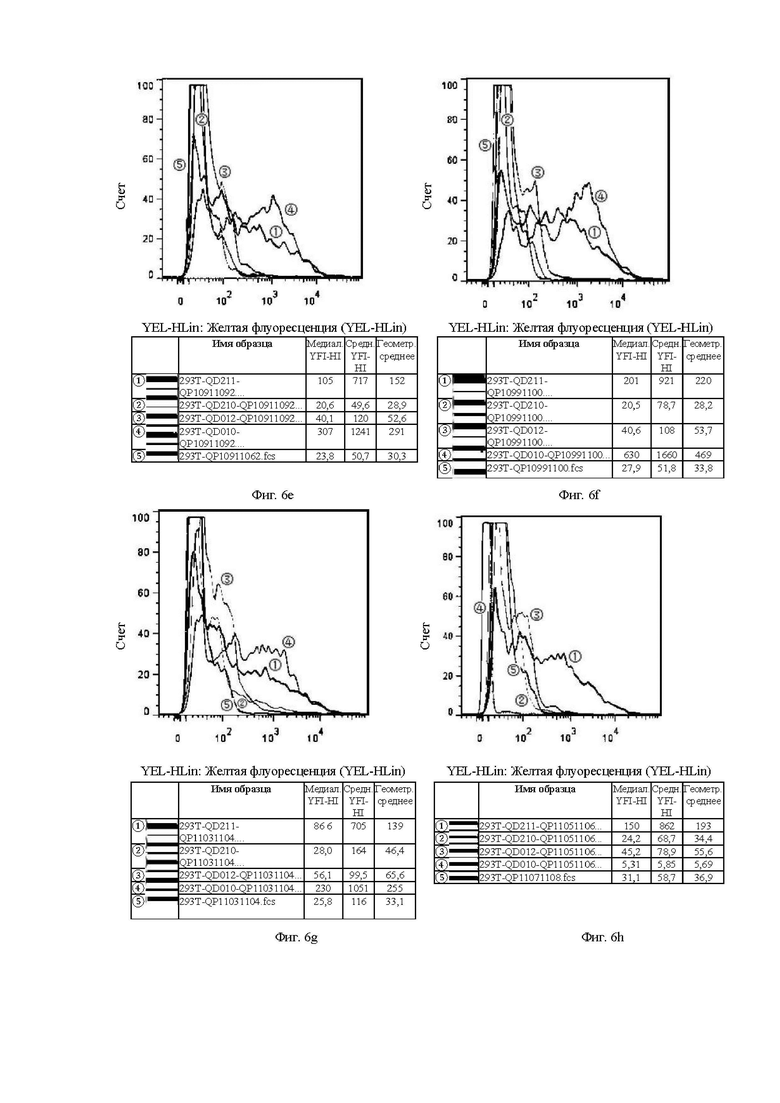
На Фиг. 6е показана способность антитела QP10911092, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 6f показана способность антитела QP10991100, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 6g показана способность антитела QP11031104, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 6h показана способность антитела QP11051106, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
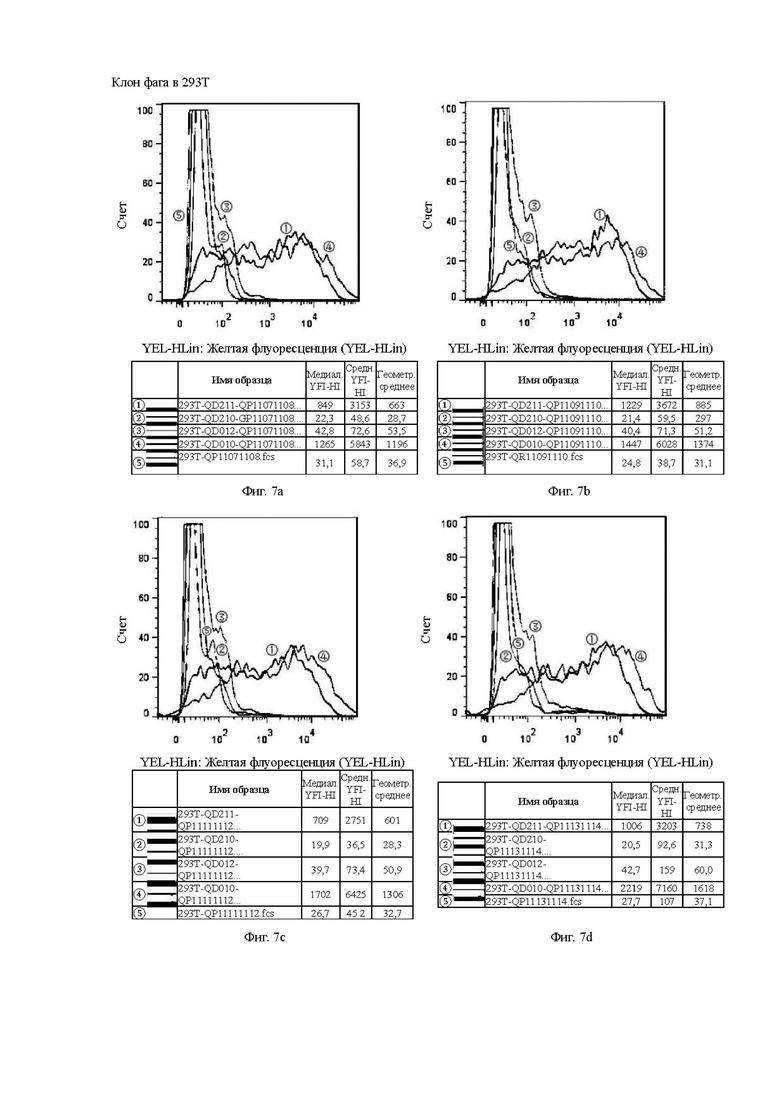
На Фиг. 7а показана способность антитела QP11071108, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 7b показана способность антитела QP11091110, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 7с показана способность антитела QP11111112, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 7d показана способность антитела QP11131114, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
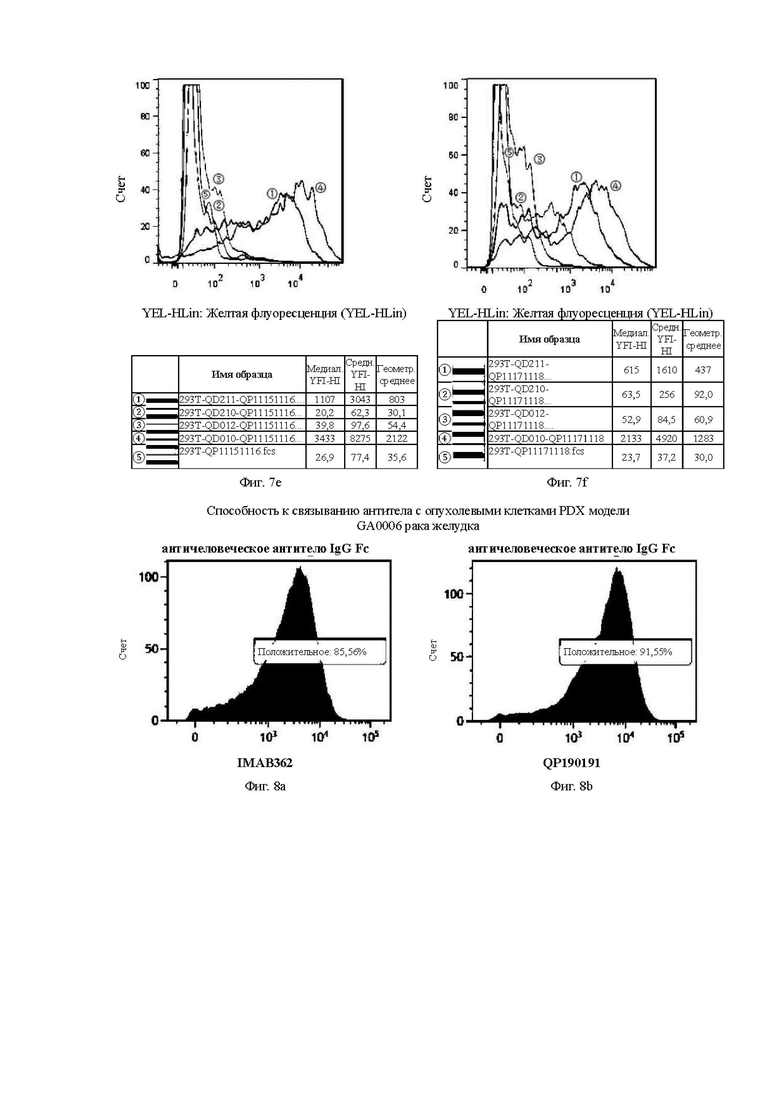
На Фиг. 7е показана способность антитела QP11151116, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 7f показана способность антитела QP11171118, отобранного скринингом из бактериофага, к связыванию с различными временными клеточными линиями 293Т.
На Фиг. 8а показана кривая контрольной группы IMAB362 в эксперименте по способности к связыванию контрольного антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 8b показана кривая QP190191 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
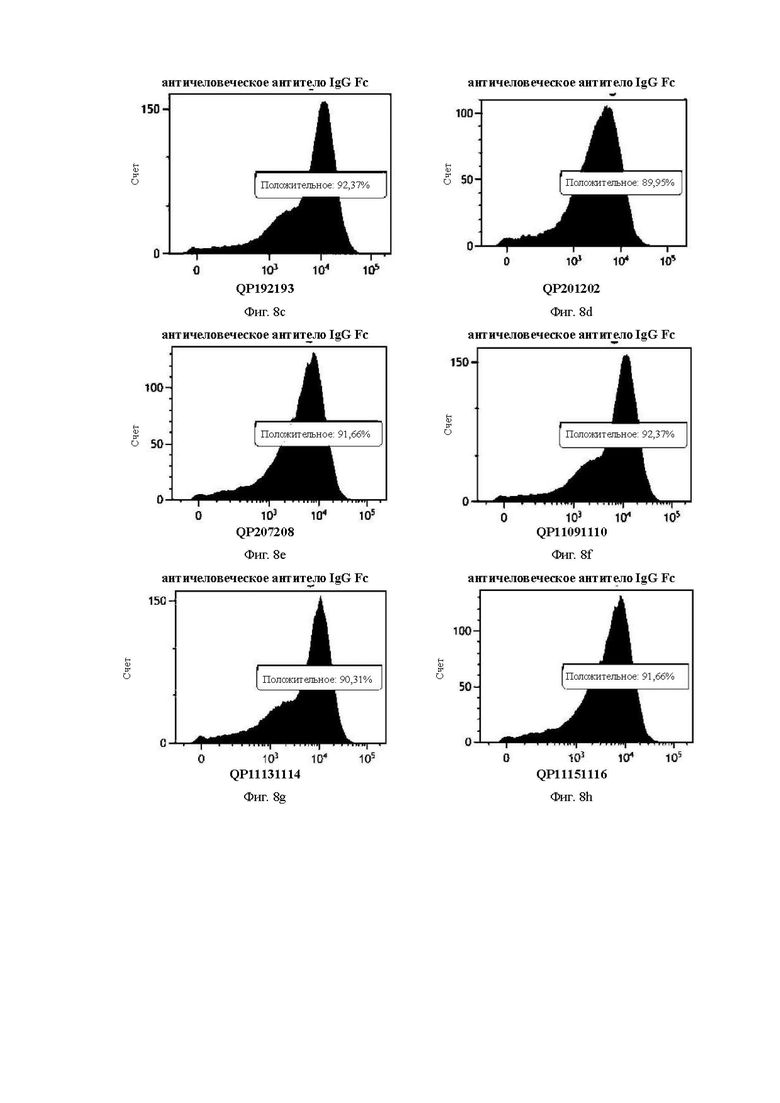
На Фиг. 8с показана кривая QP192193 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 8d показана кривая QP201202 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 8е показана кривая QP207208 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 8f показана кривая QP11091110 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 8g показана кривая QP11131114 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 8h показана кривая QP11151116 в эксперименте по способности к связыванию антитела с опухолевыми клетками рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
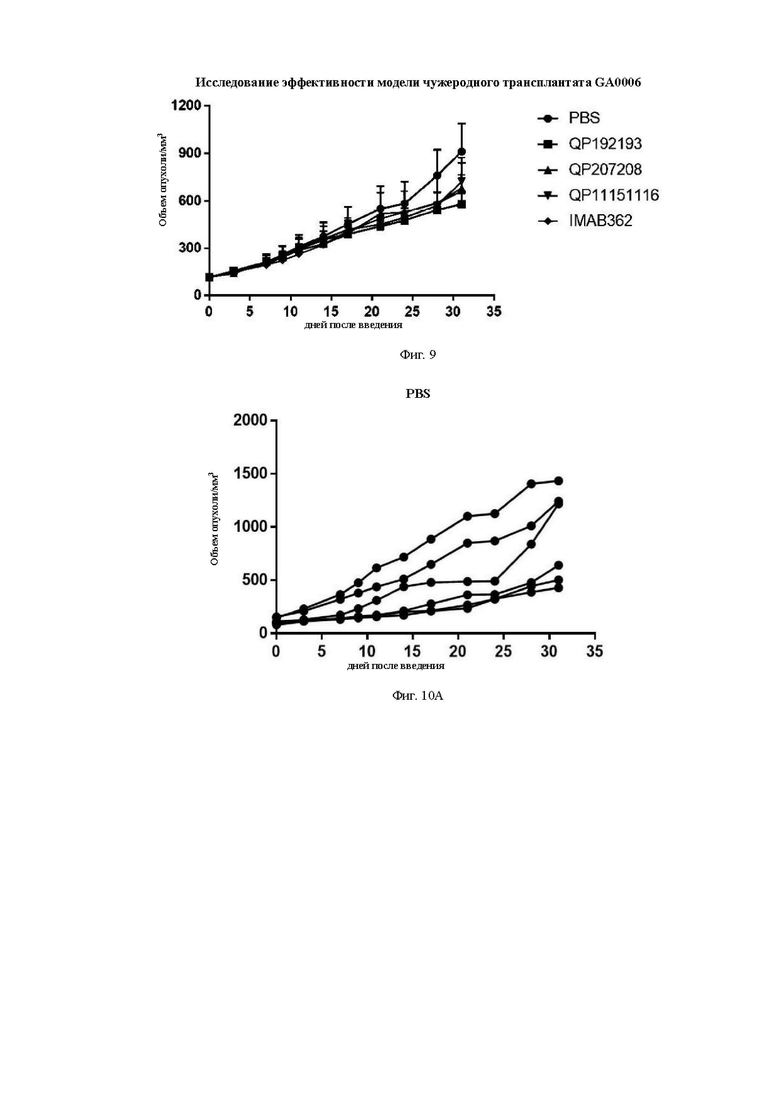
На Фиг. 9 показан фармакодинамический тест антитела против рака желудка PDX (ксенотрансплантата, полученного у пациента) модели GA0006.
На Фиг. 10А показана кривая роста опухоли для группы фосфатно-солевого буфера (PBS) (отрицательный контроль) после группировки.
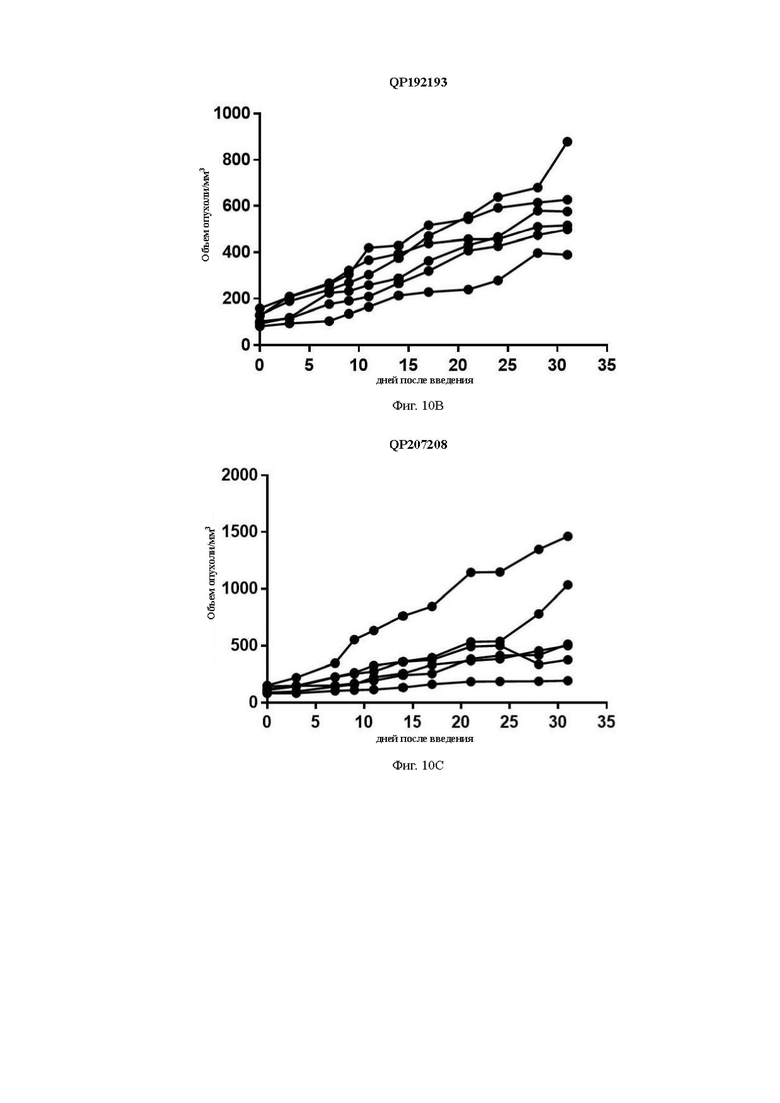
На Фиг. 10В показана кривая роста опухоли для группы QP192193 после группировки.
На Фиг. 10С показана кривая роста опухоли для группы QP207208 после группировки.
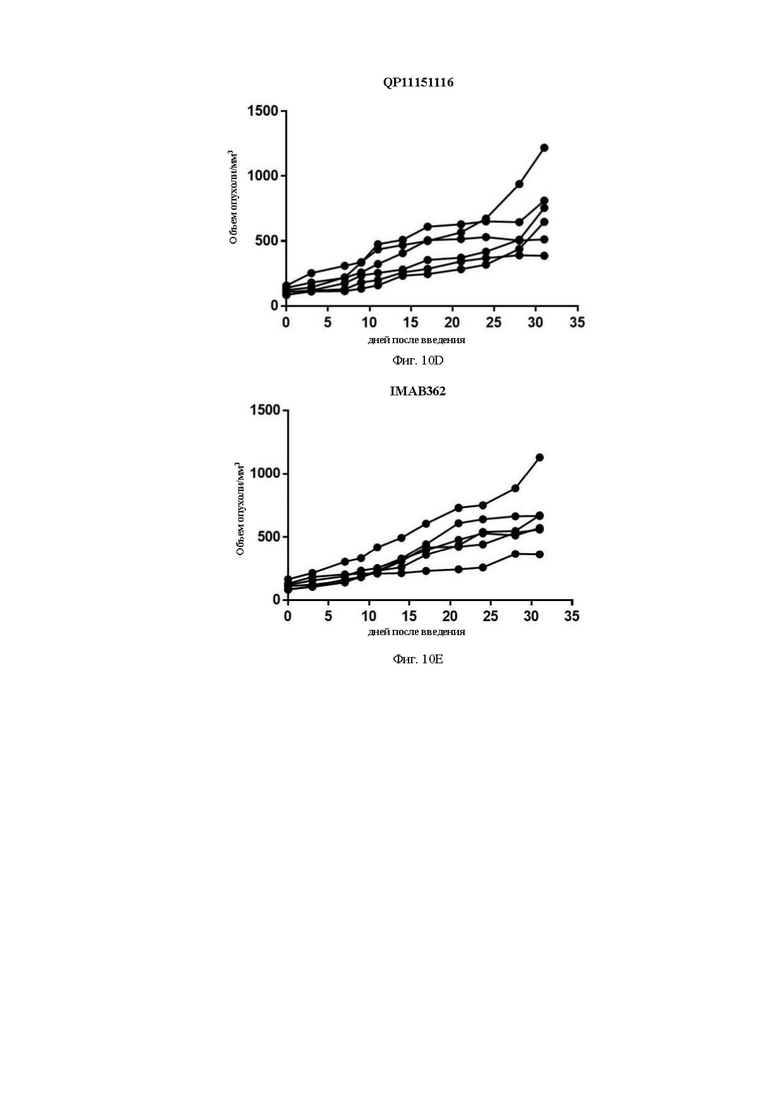
На Фиг. 10D показана кривая роста опухоли для группы QP11151116 после группировки.
На Фиг. 10Е показана кривая роста опухоли для группы контрольного антитела IMAB362 после группировки.
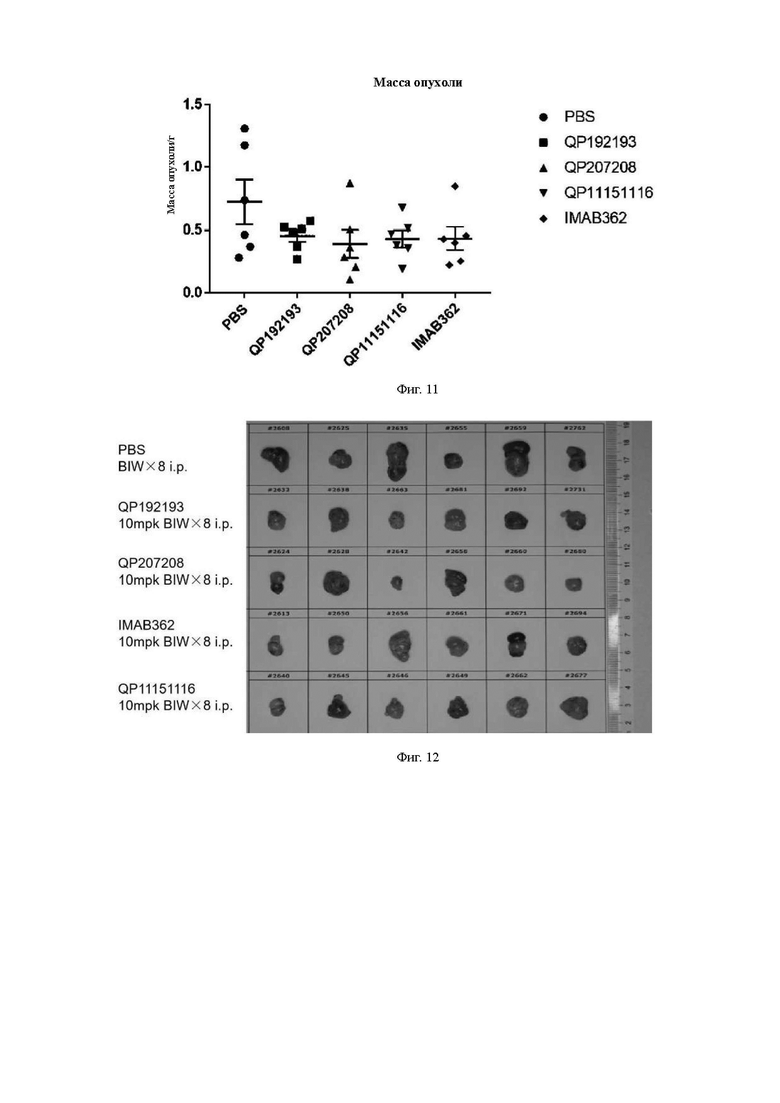
На Фиг. 11 показана масса опухоли на 31-й день в каждой группе мышей с добавлением каждого антитела по настоящему изобретению и контрольного антитела.
На Фиг. 12 показана таблица с реальными снимками объема опухоли каждой экспериментальной группы.
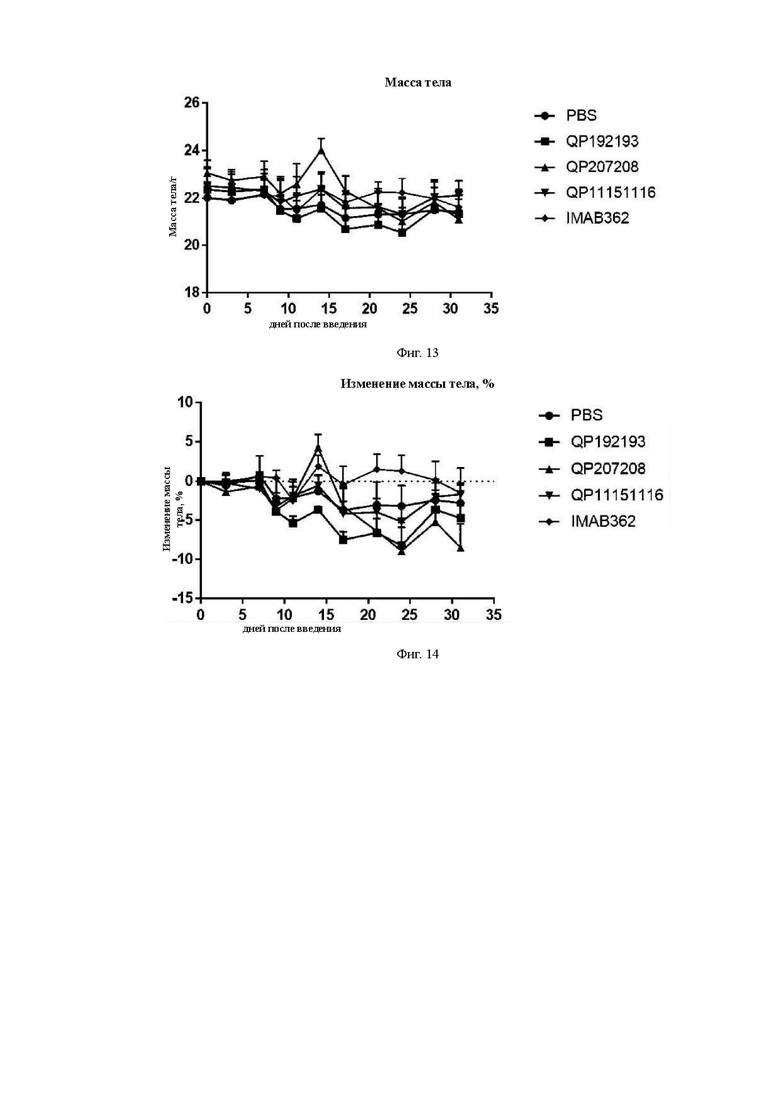
На Фиг. 13 показана кривая массы тела мышей в каждой группе.
На Фиг. 14 показана кривая скорости изменения массы тела мышей в каждой группе.
ПОДРОБНОЕ ОПИСАНИЕ
Конкретные варианты осуществления настоящего изобретения будут разъяснены ниже со ссылкой на прилагаемые графические материалы.
Антитела по настоящему изобретению включают в себя, но не ограничиваются ими, антитела человека. В различных способах скрининга антител, предусмотренных настоящим изобретением, обычно предпочтительным является более удобный отбор антитела путем скрининга гибридомы. Однако антиген этой мишени (клаудин 18.2) получить сложно, поэтому для отбора антитела также используют фаговую библиотеку. Последовательности анти-клаудин 18.2 антител, отобранных скринингом в этом варианте осуществления, являются следующими:
Для антител, отобранных скринингом с помощью гибридомы, области, определяющие комплементарность (CDR) в вариабельной области тяжелой цепи (VH), показаны в подчеркнутых частях последовательностей.
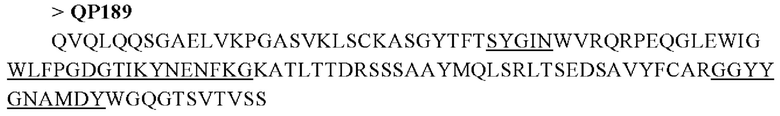

Последовательность легкой цепи:
Области, определяющие комплементарность (CDR) в вариабельном участке (VL) легкой цепи, показаны в подчеркнутых частях последовательностей.


Для антител, отобранных скринингом с помощью фаговой библиотеки, области, определяющие комплементарность (CDR) в вариабельном участке (VH) тяжелой цепи, показаны в подчеркнутых частях последовательностей


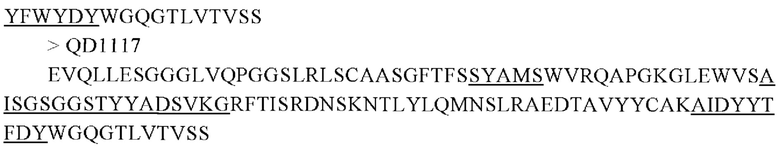
Последовательность легкой цепи:
Области, определяющие комплементарность (CDR) в вариабельном участке (VL) легкой цепи, показаны в подчеркнутых частях последовательностей.


Эксперимент по способности антитела к связыванию:
Эксперимент 1: Обнаружение способности к связыванию антитела с линией опухолевых клеток с помощью методики сортировки способом проточной цитофлуориметрии (FACS)
a) В 96-луночный планшет высевали 2×105 клеток на лунку, центрифугировали при 1000× об/мин в течение 5 минут. Клетки промывали один раз 1× фосфатно-солевым буфером (PBS) с отсасыванием излишка надосадочной жидкости;
b) Для блокирования клеток добавляли 3% раствор бычьего сывороточного альбумина в фосфатно-солевом буфере (BSA-PBS), и клетки блокировали при 4°С в течение 60 минут.
c) Тестируемое антитело разбавляли блокирующим раствором до 5 мкг/мл, добавляли в лунки и инкубировали при 4°С в течение 60 минут.
d) Антитело отсасывали, а клетки в каждой лунке трижды промывали с помощью 220 мкл 1×PBS.
e) В каждую лунку добавляли 50 мкл FC-специфического конъюгированного с фикоэритрином R (РЕ) античеловеческого вторичного антитела (разведение 1: 200), и инкубировали в темноте при температуре 4°С в течение 40 минут.
f) Антитело отсасывали, а клетки в каждой лунке четыре раза промывали с помощью 1×PBS.
g) Выполняли обнаружение в рабочем положении.
Эксперимент 2: Обнаружение способности к связыванию антитела с опухолевыми клетками ткани PDX (ксенотрансплантата) с помощью методики сортировки способом проточной цитофлуориметрии (FACS)
a) Для каждой опухоли готовили пробирку gentleMACS™ С и 3 мл буферного раствора для расщепления. Буферный раствор для расщепления готовили в соответствии с инструкциями, прилагаемыми к Набору для диссеминации опухоли (miltenyibiotech, 130-096-730), причем свежий раствор готовили непосредственно перед использованием.
b) Мышей умерщвляли, опухоль удаляли с помощью инструментов для очистки, а кровеносные сосуды, жир, фасции и другие ткани, прикрепленные к поверхности опухоли, удаляли после промывания 1×PBS. Чтобы гарантировать полное расщепление опухолевой ткани буфером для расщепления, в каждую пробирку для расщепления следует помещать не более 0,8 г ткани опухоли.
c) Ткань опухоли помещали в лунку 6-луночного планшета, добавляли буфер для расщепления, а ткань опухоли разрезали с помощью чистого пинцета и скальпеля на небольшие кусочки размером примерно 1 мм3.
d) Кусочки ткани помещали в пробирку gentleMACS™ С, лунку промывали остаточным буфером для расщепления, вместе переносили в пробирку для расщепления и помещали на лед.
e) Пробирку для расщепления помещали вверх дном в автоматический процессор тканей gentleMACS, а для расщепления выбирали программу 37_c_m_TDK_1. После выполнения программы пробирку для расщепления удаляли и немедленно центрифугировали при 300×g для сбора клеток и оставшихся тканей.
f) Надосадочную жидкость удаляли, а клетки и ткани повторно суспендировали в 1×PBS. Суспензию клеток добавляли в клеточное сито, которое помещали в пробирку для центрифуги на 50 мл, после чего суспензию просеивали с 10 мл PBS, чтобы получить суспензию отдельных клеток.
g) После центрифугирования при 300×g в течение 5 минут надосадочную жидкость удаляли, а для повторного суспендирования использовали 5 мл PBS. Был проведен подсчет клеток, и концентрация клеток была доведена до 1×106 клеток на образец.
h) Для повторного суспендирования клеток к каждому образцу добавляли 100 мкл раствора PBS с концентрацией 1 мкг/мл Mouse BD Fc Block (№по кат.553141), добавляли 20 мкл реагента для блокировки FcR человека, тщательно перемешивали и инкубировали в течение 10 минут в темноте при комнатной температуре.
i) Добавляли 5 мкг/мл первичного антитела и инкубировали в темноте при температуре 4°С в течение 60 минут.
j) Добавляли 2 мл промывочного буфера FACS, клетки аккуратно повторно суспендировали, центрифугировали при 300×g в течение 5 минут с удалением надосадочной жидкости, что повторяли дважды.
k) Для инкубации клеток в темноте в течение 60 минут добавляли 100 мкл промывочного буфера FACS, содержащего вторичное меченое с помощью РЕ антитело человека IgG Fc и краситель.
l) Добавляли 2 мл промывочного буфера FACS, клетки аккуратно повторно суспендировали, центрифугировали при 300×g в течение 5 минут с удалением надосадочной жидкости, что повторяли дважды.
m) Для повторного суспендирования клеток использовали 200 мкл промывочного буфера FACS и выполняли обнаружение в рабочем положении.
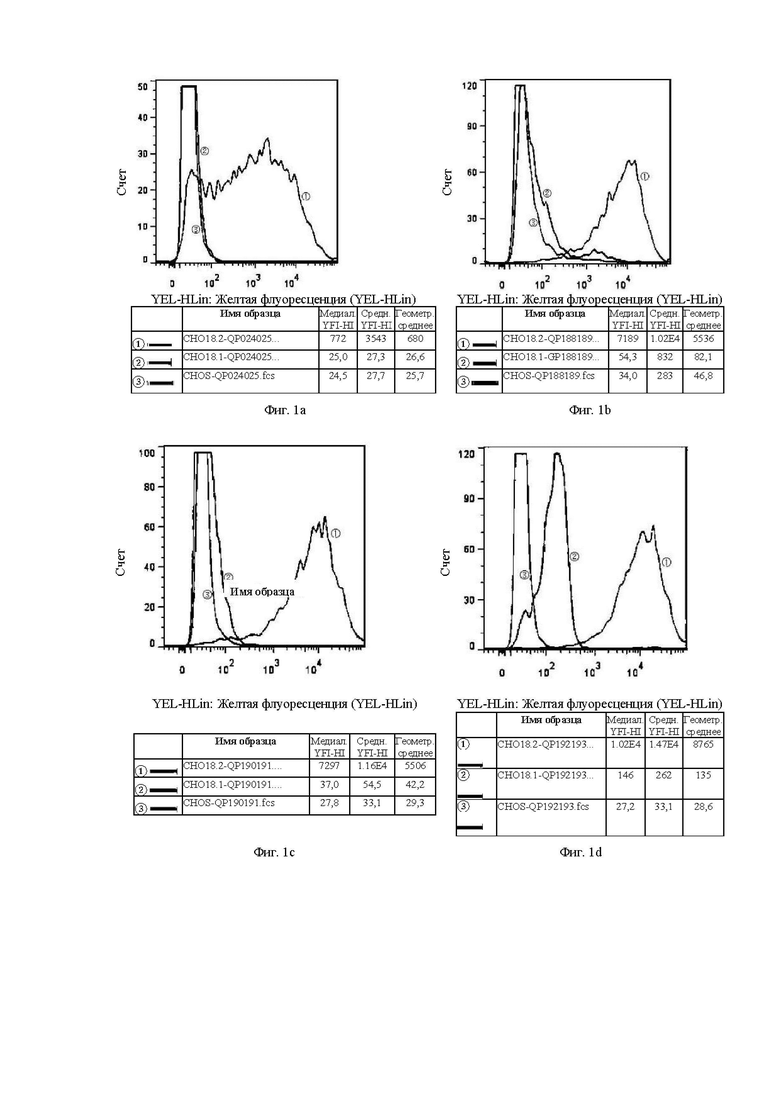
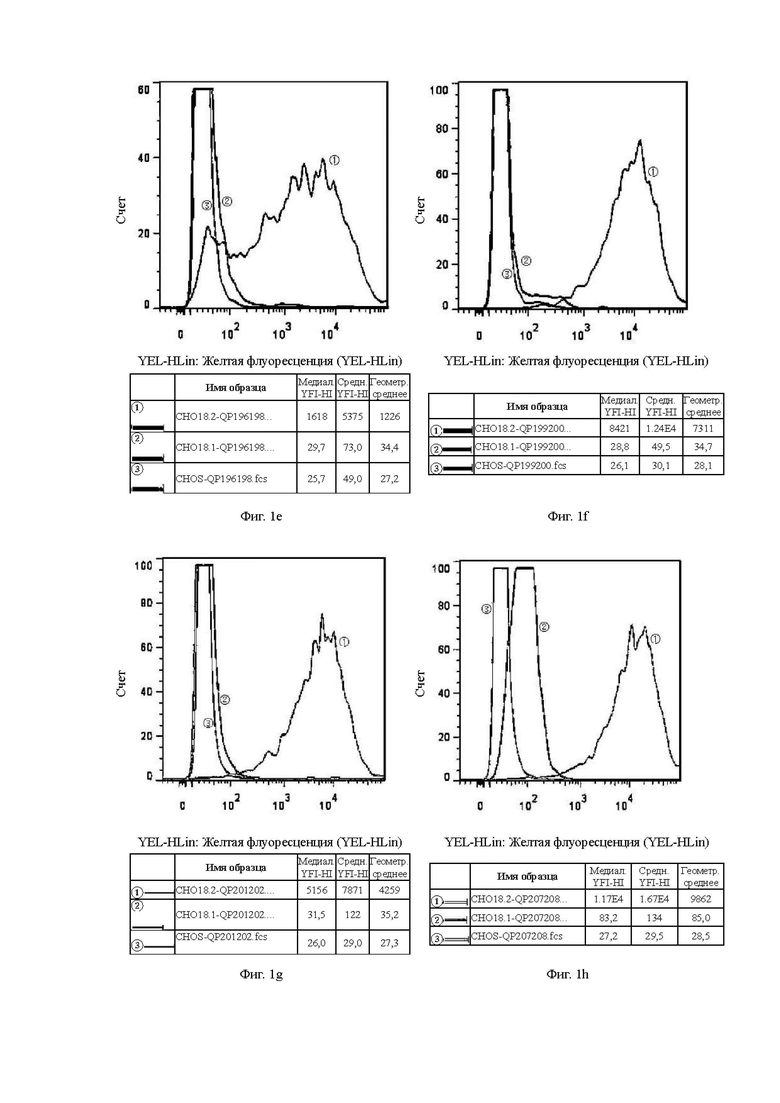
В графических материалах по данному эксперименту клон гибридомы в системе CHOS и клон бактериофага в системе CHOS относятся к способности к связыванию антител, отобранных скринингом из гибридомы и бактериофага, с клетками CHOS, соответственно. В графических материалах СНO18.2: клетки CHOS, трансфицированные геном CLDN18.2 для образования стабильных клеток CHOS-CLDN18.2, СНO18.1: клетки CHOS, трансфицированные геном CLDN18.1 для образования стабильных клеток CHOS-CLDN18.1, CHOS: клетки CHOS, трансфицированные холостым вектором.
CHOS представляет собой клеточную линию, полученную путем иммортализации клеток яичников хомяка. Для определения способности антитела к связыванию с линией клеток использовали методику сортировки способом проточной цитофлуориметрии (FACS).
На Фиг. от 1а до 1h показана способность к связыванию антител, отобранных с помощью скрининга гибридомы, с клетками CHOS.
На Фиг. от 2а до 2h, от 3а до 3h и от 4а до 4d показана способность к связыванию антител, отобранных с помощью скрининга бактериофага, с клетками CHOS.
Цель: Проверить способность антител к связыванию с клаудином 18.1 (неспецифическое связывание) и клаудином 18.2 в клеточных линиях CHOS.
В графических материалах для линий клеток CHOS каждая панель состоит из результатов обнаружения способом FACS трех клеточных линий CHOS с одним тестируемым антителом, при этом:
1. Красная кривая (3) показывает способность к связыванию тестируемого антитела с клеточной линией CHOS (CHOS), трансфицированной холостым вектором, то есть является отрицательным контролем;
2. Синяя кривая (2) показывает способность к связыванию тестируемого антитела с клеточной линией CHOS, трансфицированной клаудином 18.1 (CHO18.1), то есть обнаружение неспецифического связывания.
3. Желтая кривая (1) показывает способность к связыванию тестируемого антитела с клеточной линией CHOS, трансфицированной клаудином 18.2 (CHO18.2), то есть обнаружение способности к связыванию с белком-мишенью.
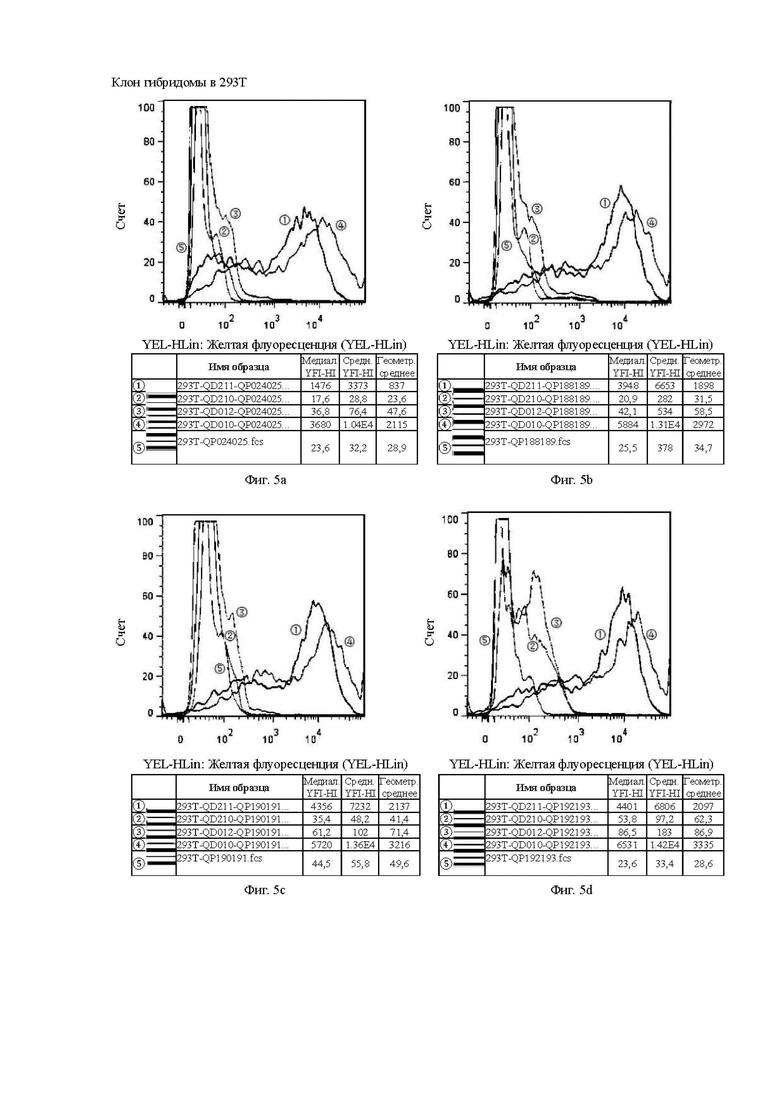
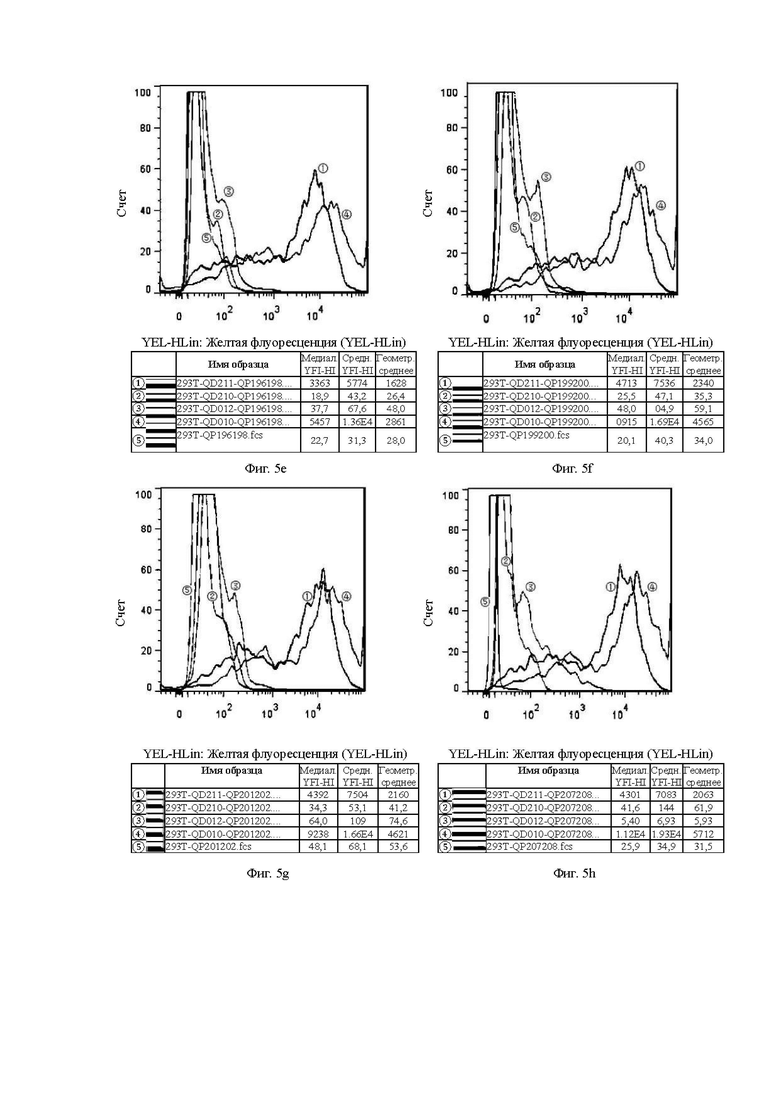
На Фиг. от 5а до 5h показана способность к связыванию антител, отобранных с помощью скрининга гибридомы, с клетками 293Т.
На Фиг. от 6а до 6h и от 7а до 7f показана способность к связыванию антител, отобранных с помощью скрининга бактериофага, с клетками CHOS.
В графических материалах 293T-QD012: 293Т временно трансфицировано CLDN18.1.
293T-QD010: 293Т временно трансфицировано CLDN18.2
Цель: Проверить способность антител к связыванию с клаудином 18.1 (неспецифическое связывание) и клаудином 18.2 в клеточной линии 293-Т.
293T-QD210: Временные клетки 293T-CLDN18.2-18.1 ECD1 (293Т, временно трансфицировали CLDN18.2-18.1 ECD1, первый внеклеточный домен CLDN18.2 был заменен первым внеклеточным доменом CLDN18.1).
293T-QD211: Временные клетки 293T-CLDN18.1-18.2 ECD1 ((293Т, временно трансфицировали CLDN18.1-18.2 ECD1, первый внеклеточный домен CLDN18.1 был заменен первым внеклеточным доменом CLDN18.2).
Цель: Проверить, является ли область связывания антитела доменом, кодируемым экзоном-1 клаудина 18.2 в клеточной линии 293-Т.
Клон гибридомы в 293Т и клон фага в 293Т аналогичным образом относятся к способности к связыванию антител, отобранных скринингом из гибридомы, и на основе бактериофага, с различными временными клеточными линиями 293Т, соответственно.
Цель эксперимента с 293Т такая же, как и с клеточной линией СНО, а значения каждой кривой и ее представление пояснены ниже.
1. Красная кривая (5) показывает способность к связыванию тестируемого антитела с клеточной линией 293Т, временно трансфицированной холостым вектором, то есть является отрицательным контролем.
2. Синяя кривая (4) показывает способность к связыванию тестируемого антитела с клеточной линией 293Т, временно трансфицированной клаудином 18.2 (293T-QD010), для обнаружения способности к связыванию с белком-мишенью.
3. Оранжевая кривая (3) показывает способность к связыванию тестируемого антитела с клеточной линией 293Т, временно трансфицированной клаудином 18.1 (293T-QD012), для обнаружения неспецифического связывания.
4. Темно-зеленая кривая (2) показывает способность к связыванию тестируемого антитела с клеточной линией 293Т, временно трансфицированной клаудином 18.1-18.2 ECD1 (первый внеклеточный домен CLDN18.1 был заменен первым внеклеточным доменом 18.2, который являлся участком, предназначенным для сайта связывания антитела) (293T-QD211), для проверки домена с сайтом связывания антитела.
5. Светло-зеленая кривая (1) показывает способность к связыванию тестируемого антитела с клеточной линией 293Т, временно трансфицированной клаудином 18.2-18.1 ECD1 (первый внеклеточный домен CLDN18.2 был заменен первым внеклеточным доменом 18.1, который ранее являлся участком, предназначенным для сайта связывания антитела) (293T-QD210), для проверки необходимости домена для связывания антитела.
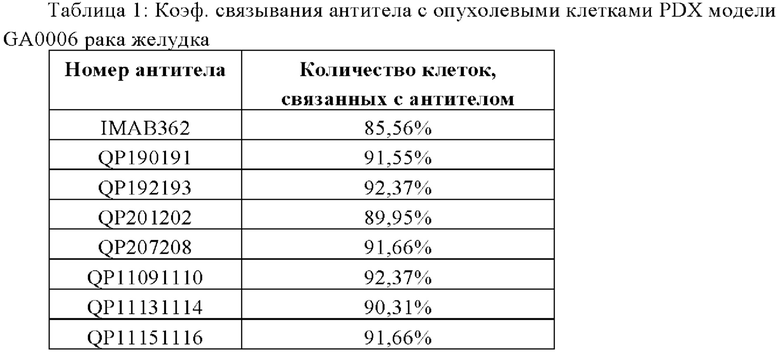
На Фиг. от 8а до 8h показана способность антитела к связыванию с опухолевыми клетками PDX модели GA0006 рака желудка.
Приведенные выше результаты экспериментов показывают, что антитела, предлагаемые в настоящем изобретении, обладают лучшей способностью связывания с белками и более низким неспецифическим связыванием.
В других вариантах осуществления настоящее изобретение также предлагает полинуклеотид, кодирующий антитело, описанное выше в данном документе. Полинуклеотиды, кодирующие антитела, предлагаемые в настоящем изобретении, когда они представлены в форме ДНК, могут содержать некодирующие последовательности, которые будут удалены во время последующей транскрипции и редактирования, или могут содержать только последовательности, кодирующие последовательности, соответствующие антителу, предлагаемому в вариантах осуществления настоящего изобретения, и последовательности, необходимые для экспрессии белка.
Настоящее изобретение также предлагает фармацевтическую композицию, содержащую антитело, описанное в данном документе выше, причем фармацевтическая композиция, предлагаемая в настоящем изобретении, может содержать только одно антитело или комбинацию, по меньшей мере, двух из антител, предложенных в вариантах осуществления.
Специалистам в данной области должно быть понятно, что в фармацевтические композиции также включен фармацевтически приемлемый наполнитель. В качестве ингредиентов, которые следует добавлять в ходе фармацевтического технологического процесса, следует использовать обычные вспомогательные вещества, необходимые для изготовления порошков, таблеток и других лекарственных форм.
Настоящее изобретение также предлагает способ обнаружения присутствия CLDN в каком-либо биологическом образце, причем такой способ содержит:
этап введения антитела в соответствии с любым из вышеперечисленного в биологический образец, причем антитело имеет обнаруживаемую метку, и этап определения наличия обнаруживаемой метки или содержимого обнаруживаемой метки.
Фармакодинамический тест на животных
a) Опухолевые ткани собирали у мышей с опухолью-ксенотрансплантатом рака желудка модели GA0006, и разрезали на опухолевые массы диаметром 2-3 мм, которые инокулировали подкожно в правую переднюю лопатку голых мышей Balb/c.
b) Когда средний объем опухоли у голых мышей Balb/c достигал примерно 100 мм3, мышей случайным образом разделяли на разные группы (по 6 мышей в каждой группе). Всех животных взвешивали, а объем опухоли измеряли с помощью штангенциркуля. Чтобы гарантировать, что объем опухоли в разных группах был одинаковым, был принят метод случайной группировки в соответствии с объемом опухоли. Коэффициент вариации (CV) объема опухоли в каждой группе рассчитывали по формуле CV=SD/MTV×100%, величина которого должна быть меньше 40%. Случайная группировка была выполнена с помощью StudyDirector™. День группировки был принят как Д0, и введение начали в тот же день. Подробные данные о способе введения, дозировке и пути введения приведены в таблице ниже.
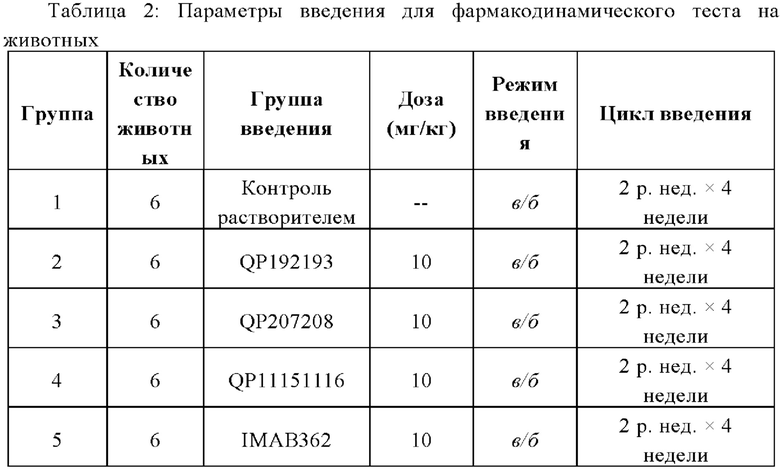
Объем дозировки составлял 10 мкл/г.
Значение номеров антител в Таблицах 1 и 2, таких как QP190191, обозначает комбинацию тяжелой цепи и легкой цепи.
с) После начала введения массу тела и объем опухоли у мышей измеряли дважды в неделю. Формула расчета объема опухоли: Объем опухоли (мм3)=1/2×(а×b2) (где а обозначает длинный диаметр, a b обозначает короткий диаметр). Эксперимент прекращали через неделю после последнего введения, мышей умерщвляли, а опухоли извлекали, взвешивали и фотографировали. Для анализа данных выбраны следующие способы анализа:
Относительная скорость пролиферации опухоли, Т/С (%), то есть процентное значение относительного объема опухоли или массы опухоли между группой лечения и контрольной группой в определенный момент времени. Формула расчета: Т/С %=TRTV/CRTV×100% (TRTV: средний относительный объем опухоли (RTV) в группе лечения; CRTV: средний RTV в контрольной группе; RTV=Vt/V0, где V0 - объем опухоли животного на момент группирования, и Vt - объем опухоли животных после лечения).
Относительная степень ингибирования опухоли, TGI (%), рассчитывают как TGI%=(1-Т/С)×100% (Т и С - относительный объем опухоли (RTV) в группе лечения и контрольной группе в определенный момент времени, соответственно).
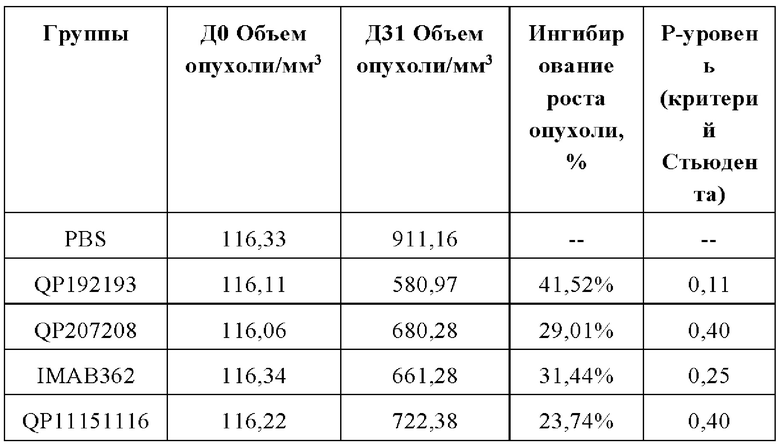
Средний объем опухоли мышей контрольной группы PBS составил 911,16±177,81 мм3 на 31-й день после введения. Средний объем опухоли после введения для антител QP192193 (10 мг/кг), QP207208 (10 мг/кг) и QP11151116 (10 мг/кг) составил на 31-й день 580,97±67,97 мм3, 680,28±193,50 мм3 и 722,38±118,07 мм3, соответственно. Показатель TGI составил 36,34%, 25,34% и 20,72%, соответственно. Средний объем опухоли на 31-й день после введения для контрольной молекулы IMAB362 (10 мг/кг) составил 661,28±104,49 мм3, а показатель TGI составил 27,42%) (см. Таблицу ниже). Все три отобранные скринингом молекулы в определенной степени ингибировали рост опухоли, хотя статистически значимой разницы отмечено не было. QP192193 показало тенденцию к тому, что его эффект был лучше, чем у контрольной молекулы IMAB362, a QP207208 и QP11151116 показали тот же уровень способности ингибировать опухоль, что и IMAB362. Более того, во время процесса введения масса тела мышей существенно не уменьшилась, что указывает на то, что молекулы антител не оказывают на мышей явных токсических и побочных воздействий.
Результаты анализа массы опухоли аналогичны результатам анализа объема опухоли. На 31-й день после введения средняя масса опухоли мышей в контрольной группе PBS составила 722,57±176,32 мг. На 31-й день после завершения введения средняя масса опухоли в группах лечения молекулами антител QP192193, QP207208 и QP11151116 составила 455,5±46,42 мг, 391,93±111,15 мг и 432,03±66,25 мг, соответственно, а показатель ингибирования роста опухоли (TGI) составил 36,96%). 45,76% и 40,21%, соответственно. На 31-й день после введения средний вес опухоли в группе лечения контрольной молекулой IMAB362 составлял 435,78±91 мг, а показатель TGI составлял 39,69% (См. таблицу ниже).
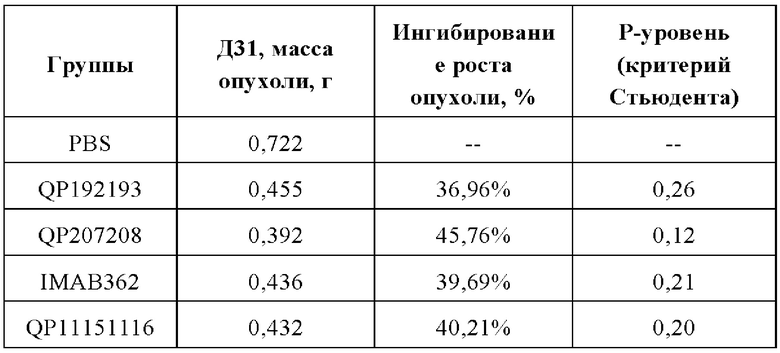
Таким образом, с помощью фармакодинамического теста PDX на животных мы обнаружили, что по сравнению с контрольной молекулой IMAB362, молекула выбранного антитела имеет лучший или такой же уровень способности подавлять опухоль in vivo.
Результаты экспериментов показаны на Фиг. 9-14, а результаты фармакодинамического теста антитела против рака желудка PDX модели GA0006 показаны на Фиг. 9. На Фиг. 10А показана кривая роста опухоли для группы фосфатно-солевого буфера (PBS) (отрицательный контроль) после группировки. На Фиг. 10В показана кривая роста опухоли для группы QP192193 после группировки. На Фиг. 10С показана кривая роста опухоли для группы QP207208 после группировки. На Фиг. 10D показана кривая роста опухоли для группы QP11151116 после группировки. На Фиг. 10Е показана кривая роста опухоли для группы контрольного антитела IMAB362 после группировки. На Фиг. 11 показана масса опухоли на 31-й день в каждой группе мышей с добавлением каждого антитела по настоящему изобретению и контрольного антитела. На Фиг. 12 показана таблица с реальными снимками объема опухоли каждой экспериментальной группы. На Фиг. 13 показана кривая массы тела мышей в каждой группе. На Фиг. 14 показана кривая скорости изменения массы тела мышей в каждой группе. Из результатов экспериментов видно, что в фармакодинамике животных in vivo антитело по данному изобретению демонстрирует лучший эффект в отношении ингибирования роста опухоли, чем IMAB362.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> КЬЮУРЕ БИОТЕХНОЛОДЖИ (ШАНХАЙ) КО., ЛТД.
<120> АНТИ-CLDN АНТИТЕЛО, ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБ ЕГО ОБНАРУЖЕНИЯ
<130> JSP11902614
<150> CN201910410255.8
<151> 2019-05-16
<160> 46
<170> PatentIn version 3.3
<210> 1
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 1
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Asn Trp Val Arg Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Leu Phe Pro Gly Asp Gly Thr Ile Lys Tyr Asn Glu Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Thr Asp Arg Ser Ser Ser Ala Ala Tyr
65 70 75 80
Met Gln Leu Ser Arg Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser
115
<210> 2
<211> 117
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 2
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Ser Ile Ile Ser Gly Gly Arg Thr Tyr Tyr Leu Asp Ser Glu Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Asn Leu Tyr Leu
65 70 75 80
Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys Thr
85 90 95
Arg Ile Tyr Tyr Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 3
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 3
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Ser Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asn Gly Asp Thr Thr Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Gln Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Val Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Phe Val Lys Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 4
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 4
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Ser Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Tyr Pro Gly Asn Gly Asp Thr Thr Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Gln Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Val Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Phe Val Lys Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 5
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 5
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Arg Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Asn Ser Ile Tyr Tyr Val Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Pro Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Asn Ala Tyr Tyr Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 6
<211> 115
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 6
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Phe Val His Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Asp Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Leu Ser Leu Arg Phe Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ala
115
<210> 7
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 7
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Ile Met His Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Asn Asp Gly Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Cys Cys
85 90 95
Ala Arg Leu Gly Phe Thr Thr Arg Asn Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 8
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 8
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Ser Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Phe Cys Gln Asn
85 90 95
Asp Tyr Phe Tyr Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 9
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 9
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Gln Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Asn Met Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 10
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 10
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Asn Pro Gly Gln
35 40 45
Pro Pro Lys Met Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Ile Asp Phe Ser Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Leu Tyr Tyr Cys Gln Asn
85 90 95
Ala Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 11
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 11
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Gly Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Arg Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp His Tyr Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 12
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 12
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Val Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Phe Cys Gln Asn
85 90 95
Asn Tyr Tyr Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 13
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 13
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser His Val Gln Ala Glu Asp Leu Ala Val Tyr Phe Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Asn Leu Glu Leu
100 105 110
Lys
<210> 14
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 14
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Ala Gly
1 5 10 15
Glu Lys Val Thr Met Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Gly Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp His Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 15
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Ala Phe Thr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 16
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ala Tyr Gly Thr Tyr Ser Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 17
<211> 121
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Gly Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Gly Ser Trp Phe Gly Pro Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 18
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Asp Gly Ala Thr Pro Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 19
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ser Tyr Tyr Gly Thr Gly Ala Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 20
<211> 121
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 20
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Gly Tyr Phe Ser Gly Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 21
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Asn Trp Ser Phe Gly Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 22
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Gly Tyr Phe Pro Arg Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 23
<211> 121
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Ser Trp Tyr Trp Leu Phe Gly Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 24
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 24
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Asp Trp Gly Gly Tyr Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 25
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 25
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Tyr Tyr Tyr Phe Trp Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 26
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 26
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Tyr Tyr Tyr Tyr Trp Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 27
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 27
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Tyr Tyr Tyr Tyr Phe Trp Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 28
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 28
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ser Ala Ala Tyr Tyr Tyr Phe Trp Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 29
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 29
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ser Tyr Tyr Tyr Tyr Phe Trp Tyr Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 30
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область тяжелой цепи
<400> 30
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ala Ile Asp Tyr Tyr Thr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 31
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 31
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Met Thr Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 32
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 32
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Asp
85 90 95
Leu Trp Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 33
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 33
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Ala Gln Ser Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 34
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 34
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Asn Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 35
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 35
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Thr
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Met Thr Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 36
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 36
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Arg Gln Phe Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 37
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 37
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Leu Asn Thr Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 38
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 38
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Gln Trp Asp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 39
<211> 111
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 39
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Gln Thr Gly Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 40
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 40
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Thr Thr Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 41
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 41
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 42
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 42
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Tyr Ser Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 43
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 43
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Lys Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 44
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 44
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Ser Tyr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 45
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 45
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 46
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> вариабельная область легкой цепи
<400> 46
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Val Lys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| АНТИТЕЛО К CD3 И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2802272C2 |
| БИСПЕЦИФИЧЕСКИЙ БЕЛОК | 2019 |
|
RU2784486C1 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
| АНТИТЕЛА ПРОТИВ SARS-COV-2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2832025C1 |
| АНТИ-PD-L1 АНТИТЕЛА | 2015 |
|
RU2722212C2 |
| КОНЪЮГАТ АНТИТЕЛА ПРОТИВ CEA И АНАЛОГА ЭКСАТЕКАНА И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2833323C1 |
Группа изобретений относится к биотехнологии. Предложены: анти-CLDN18.2 антитело или его активный фрагмент, способ его обнаружения, полинуклеотид, кодирующий антитело или его активный фрагмент фармацевтическая композиция для лечения опухолей экспрессирующих CLDN18.2, а также применение антитела или его активного фрагмента для получения противоопухолевого лекарственного средства, где опухоль экспрессирует CLDN18.2 и способ лечения опухолей. Изобретение позволяет получить антитело, способность которого к связыванию с клеточными линиями и клетками опухолевой ткани является более выраженной, чем у существующего антитела IMAB362, а противоопухолевый эффект данного антитела также является более мощным, чем у существующего антитела IMAB362. 6 н. и 1 з.п. ф-лы, 14 ил., 4 табл., 2 пр.
1. Анти-CLDN18.2 антитело или его активный фрагмент, которые содержат тяжелую цепь и легкую цепь:
указанная тяжелая цепь содержит три области, определяющие комплементарность (CDR), представленные HCDR1, HCDR2 и HCDR3 соответственно;
указанная легкая цепь содержит три области, определяющие комплементарность (CDR), представленные LCDR1, LCDR2 и LCDR3 соответственно;
а аминокислотная последовательность анти-CLDN18.2 антитела или его активного фрагмента содержат комбинацию областей, определяющих комплементарность (CDR), выбранную из группы, состоящей из:
(i) HCDR1: SYIMH, HCDR2: YINPYNDGTKYNEKFKG, HCDR3: LGFTTRNAMDY, LCDR1: KSSQSLLNSGNQKNYLA, LCDR2: GASTRES, и LCDR3: QNDHSYP;
(ii) HCDR1: NYAMS, HCDR2: SIISGGRTYYLDSEKG, HCDR3: IYYGNSFDY, LCDR1: KSSQSLLNSGNQKNYLT, LCDR2: WASTRES, и LCDR3: QNDYSYPFT;
(iii) HCDR1: SYWMN, HCDR2: QIYPGNGDTTYNGKFKG, HCDR3: FVKGNAMDY, LCDR1: KSSQSLLNSGNQKNYLT, LCDR2: WASTRES, и LCDR3: QNAYSYPFT;
(iv) HCDR1: NYFVH, HCDR2: YINPYNDDTKYNEKFKG, HCDR3: LSLRFFAY, LCDR1: KSSQSLLNSGNQKNYLT, LCDR2: WASTRES, и LCDR3: QNDYSYPLT;
(v) HCDR1: SYAMS, HCDR2: AISGSGGSTYYADSVKG, HCDR3: DYYYYYWFDY, LCDR1: RASQSISSWLA, LCDR2: DASSLES, и LCDR3: QQYSSYSPT;
(vi) HCDR1: SYAMS, HCDR2: AISGSGGSTYYADSVKG, HCDR3: SAAYYYFWFDY, LCDR1: RASQSISSWLA, LCDR2: DASSLES, и LCDR3: QQYLSYPPT; или
(vii) HCDR1: SYAMS, HCDR2: AISGSGGSTYYADSVKG, HCDR3: DSYYYYFWYDY, LCDR1: RASQSISSWLA, LCDR2: DASSLES, и LCDR3: QQYNSYPLT.
2. Анти-CLDN18.2 антитело или его активный фрагмент по п. 1,
в котором тяжелая цепь и легкая цепь антитела образует комбинацию, выбранную из группы, состоящей из:
SEQ ID No: 7 и SEQ ID No: 14,
SEQ ID No: 2 и SEQ ID No: 9;
SEQ ID No: 3 и SEQ ID No: 10;
SEQ ID No: 6 и SEQ ID No: 13;
SEQ ID No: 26 и SEQ ID No: 42;
SEQ ID No: 28 и SEQ ID No: 44;
SEQ ID No: 29 и SEQ ID No: 45.
3. Полинуклеотид, кодирующий антитело или его активный фрагмент по п. 1.
4. Фармацевтическая композиция для лечения опухолей экспрессирующих CLDN18.2, которая содержит терапевтически эффективное количество антитела или его активного фрагмента по п. 1 в качестве активного ингредиента.
5. Применение антитела или его активного фрагмента по п. 1 для получения противоопухолевого лекарственного средства, где опухоль экспрессирует CLDN18.2.
6. Способ обнаружения присутствия CLDN18.2 в биологическом образце, который включает:
этап введения антитела или его активного фрагмента по п. 1 в биологический образец, причем такое антитело имеет обнаруживаемую метку;
и этап обнаружения того, присутствует ли обнаруживаемая метка, или обнаружения содержимого указанной обнаруживаемой метки.
7. Способ лечения опухолей, который включает этап введения антитела или его активного фрагмента по п. 1, где опухоли экспрессируют CLDN18.2.
| WO 2018054973 A1, 29.03.2018 | |||
| US 2018319891 A1, 08.11.2018 | |||
| SINGH P | |||
| et al., Anti-claudin 18.2 antibody as new targeted therapy for advanced gastric cancer, Journal of Hematology & Oncology, vol | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Транспортер для перевозки товарных вагонов по трамвайным путям | 1919 |
|
SU105A1 |
| ЛЕЧЕНИЕ РАКА ПРИ ПОМОЩИ НАПРАВЛЕННЫХ НА МИШЕНЬ АНТИТЕЛ IN VIVO | 2011 |
|
RU2642305C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ К КЛАУДИНУ 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2662066C2 |
