Изобретение относится к области биотехнологии, средствам молекулярной диагностики и производству противоящурных вакцин, а именно к способу опосредованного контроля полноты инактивации антигена вируса ящура с применением nested обратно-транскриптазной полимеразной цепной реакции (nested ОТ-ПЦР) с последующим горизонтальным электрофорезом ампликонов в агарозном геле.
Ящур - высококонтагиозное вирусное заболевание парнокопытных животных, которое относится к категории трансграничных инфекций [1]. Возбудитель принадлежит к порядку Picornavirales, семейству Picornaviridae, роду Aphthovirus [2, 3]. Характерной особенностью вируса ящура является наличие 7 типов: А, О, С, Asia-1, SAT-1, SAT-2, SAT-3. В пределах каждого типа существует множество генетических вариантов вируса [3,4].
Вирион вируса ящура имеет диаметр около 23-25 нм, геном представлен одноцепочечной позитивной РНК, состоящей приблизительно из 8500 н.о. с одной большой открытой рамкой считывания длиной около 7000 н.о., которая состоит из 1А-, 1В-, 1С-, 1D-, 2А-, 2В-, 2С-, 3А-, 3В-, 3С-, 3D-генов. Весь геном возбудителя ящура характеризуется высокой степенью вариабельности нуклеотидов, за исключением 2В-гена, а также генов, кодирующих неструктурные белки и 5'-нетранслируемой области (5'-NTR), которые стабильны для всех вакцинных штаммов вируса ящура. Обнаружение генов, кодирующих неструктурные белки или 5'-NTR позволяет детектировать вирус в исследуемом материале [5-11].
Инактивация вируса ящура может достигаться за счет влияния различных физических и химических факторов. Так, при увеличении температуры прогревания до 45°С уменьшается относительное процентное содержание 146S компонента при сохранении титра инфекционной активности вируса ящура на уровне контрольного образца, хранящегося при температуре 4°С, что свидетельствует о разрушении неинфекционных 146S частиц. Температура 55-60°С вызывает спонтанный распад полных вирионов и резкое снижение титра инфекционной активности вируса ящура. Воздействие ультрафиолетовых лучей инактивирует вирус через 5-10 минут. Солнечный свет при температуре 5-6°С обезвреживает вирус через 5-7 дней, при 16-18°С - через 3-4 дня, при 37°С - через 40 часов [1, 2].
Вирус инактивируют различными химическими агентами: 0,5-1,0% раствор фенола при температуре плюс 4°С через 8 недель, при температуре плюс 18-20°С - через 10-14 дней, при температуре плюс 37°С - через 3 суток; 2% раствор формалина при температуре плюс 4°С - через 2 часа; 45-70% раствор этилового спирта при температуре плюс 4°С - через 1-2 ч [2].
При производстве цельновирионных инактивированных вакцин против ящура особое внимание уделяется вопросу безопасности изготовленного препарата. Технология изготовления данных вакцин обязательно предусматривает проведение исследования на отсутствие вирулентного вируса ящура в сырье [4]. В соответствии с требованиями Всемирной организации по охране здоровья животных при производстве вакцин против ящура для получения инактивированного антигена применяют инактиванты первого порядка, бинарный этиленимин/этиленимин (BEI/EI) (Bahnemann, 1990), в частности 1,2-аминоэтилэтиленимин (АЭЭИ) [4, 12].
Данный химический агент представляет собой циклический амин, который вступает в реакцию замещения водорода в N-H-связи вирусной РНК с сохранением цикла инактиванта. Данный процесс приводит к деградации нуклеиновой кислоты вируса, а также формированию соединений, подобных 7-(2-аминоэтил)гуанидину [12], в результате этого утрачивается вирулентность возбудителя, но при этом сохраняется антигенные свойства частицы вируса.
АЭЭИ вызывает потерю инфекционных свойств вируса ящура за счет многочисленных делеций и повреждений вирусной РЕК, присоединения новых функциональных групп к нуклеотидам, и даже разрушения вирусного генома до фрагментов [12, 13]. В результате процессы обратной транскрипции, трансляции и процессинга белков вируса и, как следствие, репродукции и сборки вириона in vivo и in vitro становятся невозможными. Разрывы в 1D-, 1В-, 1С-, 1А-генах приводят к прекращению процесса синтеза важнейших белков вируса ящура VP1, VP2, VP3, VP4, соответственно, которые являются структурными и отвечают за формирование икосаэдрического капсида, состоящего из 60 копий, каждая из которых представлена четырьмя указанными протеинами. Повреждения в 5'-NTR нуклеиновой кислоты вируса ящура препятствуют синтезу регуляторных белков, а в 3'-NTR -структур, отвечающих за терминацию процесса репликации. При разрушениях в области L-гена блокируется синтез первоначального полипептида, следовательно, не происходит посттрансляционный процессинг с образованием L-, Р1-, Р2- и Р3-белков. Кроме того, по причине отсутствия L-белка не протекают протеолитический процесс индуцирования расщепления факторов трансляции вирусной РНК. При повреждениях в 2А-, 2В, 2С-, 3А-, 3В-генах не формируются неструктурные белки, выполняющие ферментативные функции при сборке вирионов. Нуклеотидные разрушения в 3С- и 3D-генах не позволяют синтезировать 3С-протеазу и 3D-белок (РНК-зависимую-РНК-полимеразу). Таким образом, при одном из указанных серьезных повреждений вирус ящура утрачивает свои вирулентные свойства. Этот феномен можно определить с помощью молекулярно-биологического исследования.
Полноту инактивации верифицируют с использованием метода на остаточное содержание вирулентного вируса ящура в чувствительной клеточной линии почки свиньи СП по факту развития цитопатического действия [4, 14]. Существенными недостатками указанного метода являются: длительная процедура анализа, связанная с репродукцией вируса (не менее 3 суток для каждого из трех пассажей); субъективность при оценке результатов исследования; высокая стоимость культуры клеток и затраты на ее поддержание.
Проблемой является отсутствие чувствительного и специфичного вспомогательного экспресс-способа контроля полноты инактивации антигена вируса ящура с целью устранения вышеуказанных недостатков.
Данная проблема была решена благодаря разработке нового способа опосредованного контроля полноты инактивации антигена вируса ящура в сырье для вакцин с применением nested ОТ-ПЦР, предполагающей применением двух систем праймеров, детектирующих протяженные участки в 5'NTR- - 3D-генах вируса с размерами 7289 и 7244 п.о., соответственно, с последующим электрофорезом ампликонов в агарозном геле, с помощью которого возможно выявлять неповрежденный участок нуклеиновой кислоты вируса ящура до процесса инактивации и определять отсутствие вирулентного вируса ящура после воздействия инактивантами. Данная возможность позволит для подтверждения результатов культурального метода проводить контроль полноты инактивации антигена вируса ящура, который используется для изготовления инактивированных вакцин для домашних и сельскохозяйственных животных.
В связи с этим целесообразно разработать способ контроля полноты инактивации антигена вируса ящура с применением молекулярно-биологического исследования, который является более быстрым, менее затратным методом и отличается при этом высокой степенью надежности. Так, для определения остаточной вирулентности вирусов классической чумы свиней, инфекционного бронхита кур, эховирусов и др. в последние годы стали применять обратно-транскриптазную полимеразную цепную реакцию (ОТ-ПЦР) с последующей детекцией продуктов реакции с помощью горизонтального электрофореза, а также биофизический метод с применением рестрикционного и электрофоретического анализа (Закутский Н.И., Луницин А.В., 2016) [15]. Для контроля полноты инактивации антигена вируса ящура в вирионных вакцинах данный метод ранее не применялся. Для работы с РНК-содержащим вирусом ящура было решено взять за основу ОТ-ПЦР с последующим электрофорезом.
В настоящее время данный метод применяют только для проведения диагностики ящура, типирования и молекулярно-эпидемиологических исследований вируса, а также для дифференциации штаммов с применением типоспецифических праймеров (Vangrysperre and De Clerq (1996), Callahan et al., (2002), Reid et al., (2003)) [4, 16-18]. Данный метод отличается высокой чувствительностью и специфичностью, объективностью анализа данных и быстротой проведения исследования. За счет оптимизации условий постановки анализа и разработке штаммоспецифичных оригинальных праймеров, позволяющих детектировать большеразмерные фрагменты, подборе необходимых контролей, можно проводить анализ степени повреждения вирусного генома и, как следствие, исследование полноты инактивации антигена вируса ящура.
Для контроля полноты инактивации антигена вируса ящура в сырье для вакцины используют перевиваемую монослойную клеточную линию почки свиньи СП (прототип). По сравнению с прототипом предлагаемый способ на основе метода nested ОТ-ПЦР с последующей детекцией ампликонов в горизонтальном электрофорезе отличается высокими диагностическими показателями, является более экономичным, позволяет одновременно исследовать несколько десятков проб инактивированного вируссодержащего материала для вакцины, а время проведения анализа сократить до 10 ч. Исходя их этого, актуально применять метод nested ОТ-ПЦР с последующим электрофорезом ампликонов в агарозном геле для контроля полноты инактивации антигена вируса ящура при изготовлении инактивированных вакцин.
Сущность изобретения заключается в новом подходе по контролю полноты инактивации антигена вируса ящура в сырье для инактивированных вакцин с применением nested ОТ-ПЦР с последующим горизонтальным электрофорезом ампликонов в агарозном геле.
Заявляемый способ основан на проведении выделения нуклеиновой кислоты вируса ящура до и после процесса инактивации с помощью хаотропного агента (гуанидинизотиоцианата), денатурирующего белок и протеиназы К; проведении nested ОТ-ПЦР с применением двух систем оригинальных разработанных праймеров, детектирующих протяженный участок 5'NTR- - 3D-генов вируса ящура с размерами 7289 и 7244 п.о., соответственно, с последующей детекцией ампликонов в горизонтальном электрофорезе. В результате проведенного исследования оценивают наличие или отсутствие продуктов ПЦР в виде светящихся полос в потоке ультрафиолетового света при длине волны 312 нм. На основании полученных треков ампликонов делают заключение о целостности участков генома вируса ящура и, как следствие, о полноте инактивации антигена.
Ключевым элементом заявляемого способа является выделение нуклеиновой кислоты до и после процесса инактивации с применением хаотропного агента в сочетании с протеиназой К, а также проведение nested ОТ-ПЦР с использованием двух оригинальных систем праймеров, позволяющих амплифицировать 2 продолжительных участков ДНК с размерами 7289 и 7244 п.о., соответственно, детектирование полученных ампликонов с помощью горизонтального электрофореза и установление зависимости между разрушением продолжительного участка нуклеиновой кислоты вируса ящура и авирулентностью антигена после воздействия применяемых инактивантов.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении экстрагирования РНК вируса ящура с помощью хаотропного агента и протеиназы К до и после инактивации, проведении nested ОТ-ПЦР с использованием двух оригинальных систем праймеров для опосредованного контроля полноты инактивации антигена вируса ящура, применяемого для изготовления инактивированных вакцин.
Сведений об аналогах предлагаемого способа опосредованного контроля полноты инактивации антигена вируса ящура в сырье для инактивированных вакцин авторами не обнаружено. Возможность применения метода ОТ-ПЦР с проведением горизонтального электрофореза в иной модификации и с другими праймерами описана для оценки полноты инактивации вируса классической чумы свиней и бешенства [15, 19]. Сущность изобретения отражена на графических изображениях: Фиг. 1 - Дизайн праймеров для амплификации большеразмерных фрагментов кДНК вируса ящура в nested ОТ-ПЦР (rev/compl - обратный комплементарный праймер, А - прямые праймеры, Б - обратные праймеры).
Фиг. 2 - Электрофореграмма по результатам контроля полноты инактивации антигена вируса ящура в сырье для вакцин (Примечание: 1 - ПКОвыд-е РНК (положительный контрольный образец на этапе выделения РНК - исходный не инактивированный вирус ящура), 2 - ПКОПЦР (положительный контрольный образец на этапе ПЦР - проверенная комплементарная (кДНК) не инактивированного вируса ящура), 3 - ВКОвыд-е РНК (внутренний контрольный образец на этапе выделения РНК - исходный не инактивированный вирус инфекционного некроза гемопоэтической ткани лососевых рыб), 4 - ВКОПЦР (внутренний контрольный образец на этапе ПЦР - проверенная кДНК не инактивированного вируса инфекционного некроза гемопоэтической ткани лососевых рыб), 5 - ОКО (отрицательный контрольный образец, контроль фона - деионизированная вода), 6 - ОИКОвыд-е РНК (отрицательный инактивированный контрольный образец на этапе выделения РНК - инактивированная суспензия вируса ящура), 7 - ОИКОПЦР (отрицательный инактивированный контрольный образец на этапе ПЦР - проверенная кДНК вируса ящура, выделенная из инактивированной суспензии).
Фиг. 3 - Кинетика инактивации вируса ящура штамма О/Забайкальский/16 с помощью 0,025% раствора АЭЭИ при температуре плюс 37±0,5°С.
При проведении контроля полноты инактивации антигена вируса ящура использовали следующие контрольные образцы:
- ПКОвыд-е РНК - положительный контрольный образец на этапе выделения РНК - исходный не инактивированный вирус ящура;
- ПКОПЦР - положительный контрольный образец на этапе ПЦР - проверенная комплементарная (кДНК) не инактивированного вируса ящура;
- ВКОвыд-е РНК - внутренний контрольный образец на этапе выделения РНК - исходный не инактивированный вирус инфекционного некроза гемопоэтической ткани лососевых рыб;
- ВКОПЦР - внутренний контрольный образец на этапе ПЦР - проверенная кДНК не инактивированного вируса инфекционного некроза гемопоэтической ткани лососевых рыб;
- ОКО - отрицательный контрольный образец, контроль фона - деионизированная вода;
- ОИКОвыд-е РНК - отрицательный инактивированный контрольный образец на этапе выделения РНК - инактивированная суспензия вируса ящура;
- ОИКОПЦР - отрицательный инактивированный контрольный образец на этапе ПЦР - проверенная кДНК вируса ящура, выделенная из инактивированной суспензии.
Из суспензии исследуемого образца, а также из ПКОвыд-е РНК, ВКОвыд-е РНК, ОКО и ОИКОвыд-е РНК выделяют нуклеиновую кислоту в процессе лизиса белков и липопротеинов, очистки нуклеиновой кислоты от примесей и ее концентрирования. Данный метод выделения РНК позволяет достичь высокой стандартизации в процедуре экстракции и получить из образцов высокоочищенные препараты нуклеиновой кислоты. Представленная ниже модифицированная методика адаптирована к процессу получения высокоочищенного элюата нуклеиновой кислоты вируса ящура. Лизис проводят с применением раствора, содержащего 50% 6М раствора гуанидинизотиоцианата (ГТЦ) и 50% протеиназы К. Данную смесь добавляют в количестве 0,9 мл к 0,1 мл исследуемой суспензии. Полученную смесь инкубируют при температуре 20±2°С в течение 20 минут для полного диссоциирования полипептидных и нуклеопротеидных комплексов. При этом нуклеиновая кислота сохраняет свою целостность благодаря высокой ингибирующей активности ГТЦ в отношении РНКаз. Полученный лизат очищают от конгломератов с помощью центрифугирования при 14000 об/мин в течение 10 минут. Супернатант переносят в центрифужную пробирку с 0,2 мл хлороформа и инкубируют содержимое в течение 5 минут с перемешиванием на вортексе. Смесь после инкубирования фракционируют при 14000 об/мин в течение 10 минут при температуре 4-8°С. В результате центрифугирования происходит разделение содержимого пробирки на три фракции: 1) нижняя фракция, содержащая комплекс связанных денатурированных полипептидов; 2) средняя фракция, включающая в свой состав белки и клеточную ДНК; 3) верхняя фракция, представляющая собой элюат РНК [13]. Верхнюю фракцию полностью отбирают, не затрагивая остальные, и переносят элюат в новую центрифужную пробирку с 0,3 мл 100%-ого изопропилового спирта для преципитации с целью концентрирования РНК. Смесь инкубируют в течение 10 минут при температуре 20±2°С, затем центрифугируют при 14000 об/мин в течение 5 минут при температуре 20±2°С. Супернатант удаляют, оставляя осадок РНК, к которому добавляют 0,3 мл 80%-ого этилового спирта. Содержимое перемешивают, инкубируют 3 минуты и осаждают при 14000 об/мин в течение 5 минут при температуре 20±2°С. Надосадок удаляют, осадок РНК высушивают в потоке воздуха комнатной температуры в течение 5 минут. К высушенному осадку добавляют 0,1 мл буфера ТЕ (10 мМ трис(оксиметил)аминометан, 1 мМ этилендиамин-тетраацетат, рН 7,0-7,3), свободного от РНКаз и ионов Mg2+, прогревают содержимое пробирки при температуре 60±2°С в течение 3-5 минут и отбирают в отдельную пробирку элюат РНК.
На следующем этапе исследования полученные экстракты РНК исследуют в nested ОТ-ПЦР. Для постановки реакции готовят ОТ-смесь с использованием набора реагентов для обратной транскрипции с гексамерами Applied Biosystems™ GeneAmp™. Рецептура приготовления смеси для обратной транскрипции представлена в таблице 1.
Для проведения обратной транскрипции РНК вирусов ящура и внутреннего контрольного образца (вируса инфекционного некроза гемопоэтической ткани лососевых) применяют хлорид магния с концентрацией 5 мМ на реакцию. В качестве буферного раствора используют Buffer II в количестве 1х на реакцию. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с концентрацией каждого в ОТ-смеси по 1,0 мМ. В качестве универсальных праймеров применяют Random Hexamers с концентрацией 2,5 мкМ. В качестве катализатора обратной транскрипции используют ревертазу Maxima Н Minus (Thermo Scientific) (2,5 ед./мкл). Для защиты ферментативного процесса от РНКаз применяют RNase Inhibitor с активностью 1 ед. на мкл реакционной смеси. Содержание деионизированной воды составляет 1/20 от объема реакционной смеси. Количество элюата РНК составляет 1/10 от объема смеси. Объем реакционной ОТ-смеси компонентов для проведения одной реакции вместе с элюатом вирусной РНК составляет 20 мкл. Обратную транскрипцию проводят при температуре 50°С в течение 25 мин за 1 цикл (таблица 1).
В данной реакции используется указанная ревертаза Maxima Н Minus, которая обладает РНК-зависимой и ДНК-зависимой полимеразной активностью, но не обладает активностью РНКазы Н из-за мутации в РНКазном Н-домене M-MuLV ревертазы. Данный фермент применяли благодаря значительно улучшенной термостабильности, в 50 раз большей технологичности, устойчивости и повышенной скорости синтеза по сравнению с M-MuLV ревертазы дикого типа. Активность РНКазы Н позволяет ферменту вырабатывать очень длинные РНК-транскрипты до 20 т.п.н. Благодаря высокой термостабильности фермент сохраняет полную активность в течение всей реакции обратной транскрипции и генерирует высокие выходы кДНК. Температура реакции при использовании данного фермента может быть увеличена до 65°С для эффективной транскрипции РНК и для улучшения специфичности с использованием ген-специфических праймеров. Чрезвычайно высокая технологичность фермента Maxima Н Minus приводит к повышенной устойчивости к обычным ингибиторам реакции, таким как гуанидин, формамид и этанол [20]. Кроме того, благодаря использованию данного фермента повышается чувствительность реакции.
Перед постановкой nested ПЦР осуществляли предварительный прогрев смеси при температуре 95°С в течение 5 мин для активации ДНК-полимеразы Platinum High Fidelity и инактивации ревертазы Maxima Н Minus (таблица 2).
Nested ПЦР состояла из двух этапов: ПЦР1 и ПЦР2. ПЦР1 включала в себя следующие подэтапы: денатурацию, отжиг праймеров системы №1, элонгацию. Денатурацию проводили при температуре 95°С в течение 10 с, отжиг олигонуклеотидов системы №1 - при температуре 60°С в течение 30 с, элонгацию - при температуре 72°С в течение 7,5 мин, поскольку размер ампликона составлял 7289 п.н. Количество циклов амплификации - 35. ПЦР2 также включала в себя следующие подэтапы: денатурацию, отжиг праймеров системы №2, элонгацию. Денатурацию проводили при температуре 95°С в течение 10 с, отжиг праймеров системы №2 - при температуре 55°С в течение 30 с, элонгацию - при температуре 72°С в течение 7,5 мин, поскольку размер ампликона составлял 7244 п.н. Количество циклов амплификации - 25 (таблица 2). Продолжительность nested ОТ-ПЦР составляла около 8 ч 45 мин.
Рецептура приготовления ПЦР1 и ПЦР2-смеси представлена в таблице 3. Компонентный состав обеих реакций одинаковый, за исключением праймеров. Для ПЦР1 применяли систему оригинальных праймеров №1, для ПЦР2 - систему оригинальных праймеров №2 (дизайн представлен на фиг. 1). Праймеры применяли в концентрации по 5 пМ на реакцию с внесением в смесь по 0,1 мкл. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с концентрацией в смеси по 2,00 мМ. В качестве основы используют Thermo Scientific™ 10Х DreamTaq™ Buffer, содержание которого составляет 10% от общего объема реакционной смеси. DreamTaq™ Buffer специально оптимизирован для ПЦР с использованием ДНК-полимеразы DreamTaq™. Буфер содержит как KCl, так и (NH4)2SO4 для обеспечения высокой специфичности отжига праймеров в широком диапазоне концентраций MgCl2. В реакционную смесь добавляют 4,0 мМ хлорида магния. В качестве фермента применяют ДНК-полимеразу Platinum High Fidelity (Invitrogene) (10 e.a.).
Высокоточная полимераза Platinum® Taq DNA High Fidelity идеально подходит для амплификации большеразмерных фрагментов ДНК (до 15000 п.н.), когда требуются высокая точность и выход ампликонов. Высокая точность обеспечивается смесью ДНК-полимеразы Platinum® Taq и коррекционной полимеразы Pyrococcus species GB-D с 3'→5'-экзонуклеазной активностью. Следует отметить, что данная полимераза проявляется точностью, которая на порядок выше по сравнению с Taq-ДНК-полимеразой [21].
На завершающем этапе исследования проводят детекцию ампликонов ДНК с помощью горизонтального электрофореза в 1,0%-ном агарозном геле, содержащем 0,0005% бромистого этидия. В расплавленный гель агарозы добавляют 0,0005% бромистого этидия. Полученные после реакции образцы в соотношении 5:1 подкрашивают смесью ксиленового голубого и бромфенолового синего с добавлением 15% глицерина для визуализации процесса электрофореза. Разделение молекул ампликонов под влиянием градиента напряжения применяют стандартный ТАЕ-буфер, содержащий ЭДТА, трис (оксиметиламинометан) и ледяную уксусную кислоту. Электрофорез проводят при силе тока 90А в течение 15-20 минут. Движение ПЦР-продуктов осуществляется от отрицательно заряженного катода к положительному аноду. Бромистый этидий интеркалирует между азотистыми основаниями ампликона и в трансиллюминаторе в потоке ультрафиолетового света при длине волны 312 нм флуоресцирует, что обеспечивает детекцию результатов анализа в виде наличия или отсутствия светящихся полос. Результаты электрофореза документируют с помощью видеосистемы «Взгляд» в цветном исполнении.
Проводят интерпретацию полученных данных. Предложены критерии интерпретации данных nested ОТ-ПЦР при контроле полноты инактивации антигена вируса ящура в сырье для вакцин. Важным условием качественно проведенных исследований являются результаты испытания контрольных образцов. Если ПКОвыд-е РНК, ПКОПЦР, ВКОвыд-е РНК, ВКОПЦР после всех стадий исследования в горизонтальном электрофорезе дают положительный результат (наличие ампликонов), а ОКО, ОИКОвыд-е РНК, ОИКОПЦР - отрицательный результат (отсутствие ампликонов), то в таком случае стадии выделения РНК, обратной транскрипции и nested ПЦР проведены верно, и можно учитывать результаты исследуемых проб. Если в треке исследуемой пробы обнаружены светящиеся продукты ПЦР2, то анализируемый участок нуклеиновой кислоты вируса ящура не поврежден, следовательно, данный образец считается не инактивированным или не полностью инактивированным. Если во всех треках свечение ампликона отсутствует, то данный участок нуклеиновой кислоты вируса ящура поврежден, тем самым, репликация вируса невозможна, и антиген считается авирулентным (фиг. 2). Иными словами, на основании отсутствия ПЦР-продуктов исследуемой пробы по сравнению с положительными контролями позволяет сделать заключение о разрушении нуклеиновой кислоты вируса ящура в инактивированных суспензиях и, как следствие, об отсутствии вирулентного вируса. В том случае, если обнаружены несоответствия в результатах анализа контролей, требуется повторно провести исследование.
Пример 1. Расчет олигонуклеотидных праймеров для детекции протяженного участка 5'NTR- - 3D-генов вируса ящура при постановке nested ОТ-ПЦР.
Расчет олигонуклеотидных праймеров проводили на основании нуклеотидных полногеномных последовательностей генома вируса ящура, полученных в рамках исследований в ФГБУ «Федеральный центр охраны здоровья животных» (ФГБУ «ВНИИЗЖ»).
В качестве гомологичных участкам 5'NTR- и 3D-генов вируса ящура олигонуклеотидных праймеров использовали:
первая система праймеров №1:
5'NTR-F1 (5'-GCCTGGTCTTTCCAGGTCTA-3'),
3D-R1 (5'-CGGCGAGTCCTGCCACGGA-3'),
вторая система праймеров №2:
5'NTR-F2 (5'-GAGGGGTGACACTTTGTA-3'),
3D-R2 (5'-CTTCTCCTGTATGGTCCCA-3').
Олигонуклеотиды для кДНК-матрицы подбирали в соответствии с рядом общих правил, которые отражены в работах В. Deiman и R. Sooknanan [22, 23]. Указанная система праймеров позволяла амплифицировать ПЦР-продукты генома вируса ящура в ПЦР1 с размером 7289 п.н., в ПЦР2 (nested) - 7244 п.н. Информация о дизайне оригинальных праймеров представлена в таблице 4 и на фиг. 1.
Длины 5'NTR-F1- и 3D-R1-, 5'NTR-F2- и 3D-R2-праймеров составляют 20 и 19, 18 и 19 и.о., что соответствует требованиям (15-30 н.о.) [22, 23]. Молекулярный вес 5'NTR-F1- и 3D-R1-, 5'NTR-F2- и 3D-R2-праймеров равен 6075,0 и 5814,8, 5594,7 и 5705,7 соответственно. Процентное содержание G и С в праймерах составляет 55 и 74, 50 и 53%, соответственно, что является допустимым. На 5'-конце праймеров и зонда отмечается преобладание G и С, а на 3'-конце - А и Т. Отсутствуют 4 и более подряд одинаковых нуклеотидов в цепи праймеров и зонда. При анализе нуклеотидных последовательностей олигонуклеотидов установили, что для праймеров не характерно образование «шпилек», а также не выявлено 3'-комплементарности и сайтов, отжигающих сами на себя. Расчет вероятности образования «шпилек» и димеров олигонуклеотидов проводили при условии, что минимальное количество пар оснований, необходимых для димеризации - 5, а для образования «шпилек» - 4.
Проведено определение температур плавления (Tm) для олигонуклеотидных праймеров, пользуясь методом ближайших соседей и учитывающим концентрацию солей в буферном растворе. Физические, термодинамические константы и расчет температур плавления разработанных олигонуклеотидных праймеров представлены в таблице 5.
Из нее следует, что энтропия, энергия Гиббса и энтальпия для 5'NTR-F1-праймера составили 164,7 ккал/моль, 25,9 ккал/моль, 431,3 кал/(°K×моль), соответственно. Энтропия, энергия Гиббса и энтальпия для 3D-R1-праймера составили 174,8 ккал/моль, 30,3 ккал/моль, 449,7 кал/(°K×моль), соответственно. Энтропия, энергия Гиббса и энтальпия для 5'NTR-F2-праймера составили 148,3 ккал/моль, 21,7 ккал/моль, 392,2 кал/(°K×моль), соответственно. Энтропия, энергия Гиббса и энтальпия для 3D-R2-праймера составили 153,4 ккал/моль, 23,5 ккал/моль, 402,8 кал/(°K×моль), соответственно. Данные значения необходимы для расчета температур плавления представленных олигонуклеотидов. Tm при использовании алгоритма ближайших соседей для прямого и обратного праймеров для ПЦР1 и ПЦР2 составили 53 и 60, 46 и 49°С, соответственно. Температура отжига олигонуклеотидов (Та) должна быть примерно на 1-5°С ниже Tm. Исходя из полученных данных, она должна находиться в диапазоне 55-65°С для ПЦР1 и 48-59°С для ПЦР2.
При использовании метода, учитывающего концентрации солей Tm для прямого и обратного праймеров для ПЦР1 и ПЦР2 составили 60 и 66, 54 и 57°С, соответственно. Температура отжига олигонуклеотидов (Та) должна быть примерно на 1-5°С ниже Tm и, исходя из полученных данных, она должна находиться в диапазоне 49-56°С для ПЦР1 и 41-48°С для ПЦР2.
Экспериментально было выявлено, что температура отжига рассматриваемых олигонуклеотидов для ПЦР1 составляет 58 и 62°С, для ПЦР2 - 56 и 54. Для проведения ОТ-ПЦР было решено проводить гибридизацию праймеров с участками 5'NTR - и 3D-генов вируса ящура при температурах 60°С для ПЦР1 и 55°С для ПЦР2.
Последовательности разработанных праймеров проверили на наличие нежелательных совпадений с другими последовательностями нуклеиновых кислот с использованием Банка данных последовательности нуклеиновых кислот вируса ящура [4]. Последовательности олигонуклеотидных праймеров также проанализировали на наличие внутренних вторичных структур с помощью программы сворачивания нуклеиновых кислот с помощью программы Mfold. Выявлено, что для разработанных олигонуклеотидов нежелательных совпадений с другими последовательностями нуклеиновых кислот, а также наличия внутренних вторичных структур не обнаружено.
Таким образом, проведен расчет олигонуклеотидных праймеров для детекции протяженного участка 5'NTR - 3D-генов вируса ящура при постановке nested ОТ-ПЦР.
Пример 2. Подбор условий проведения nested ОТ-ПЦР при разработке метода опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцин.
Для разработки высокочувствительного и специфичного метода опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцин с помощью nested ОТ-ПЦР, а также увеличения скорости проведения реакции амплификации для детекции большеразмерного фрагмента 5'NTR-3D-генов вируса ящура требовалось подобрать условия проведения реакции. Исследовали следующие пробы: не инактивированные суспензии вируса ящура штаммов А/Забайкальский/13 и О/Забайкальский/16 с титрами инфекционной активности 7,0 lg ТЦД50/мл (пробы №1 и 2), а также эти суспензии после инактивации с помощью АЭЭИ в концентрации 0,010% в течение 12 ч при температуре 37±0,5°С (пробы №3 и 4). Данные образцы анализировали в монослойной клеточной линии СП, подтвердив, что первые две пробы были не инактивированными, а вторые две получили статус инактивированного антигена. Из данных проб выделили нуклеиновую кислоту с помощью 6М раствора ГТЦ и изопропанола по методике, представленной выше. С полученными элюатами провели ОТ-ПЦР с применением тест-системы, отраженной в работах Vangrysperre W., De Clercq K. (1996) для выявления высоко консервативного 2В-гена вируса ящура [16]. Анализ проводили в трех повторностях. Результаты, полученные при исследовании, представлены в таблице 6, из которой видно, что имеющаяся тест-система не позволяет идентифицировать по небольшому участку 2В-гена вируса статус пробы в отношении полноты инактивации. В итоге предложено использовать две системы олигонуклеотидных праймеров, которые позволяют амплифицировать большеразмерный участок нуклеиновой кислоты вируса ящура с nested ОТ-ПЦР.
С целью достижения высоких диагностических показателей предложенного способа исследования полноты инактивации антигена вируса ящура в сырье для вакцин подбирали оптимальные концентрации компонентов nested ОТ-ПЦР.
Для поиска оптимальной концентрации хлорида магния применяли эмпирический подход. Проводили nested ОТ-ПЦР с последующим горизонтальным электрофорезом со следующими количествами хлорида магния в смеси для реакции: 1,0; 2,0; 3,0; 4,0; 5,0 мМ. Исследования проводили в трех повторностях. Полученные данные отражены в таблице 7.
Из данных проведенного исследования видно, что при анализе с не инактивированным материалом оптимальной является концентрация хлорида магния 4,0 мМ. Для анализа полноты инактивации было решено использовать реакционные смеси с данным количеством катионов Mg2+.
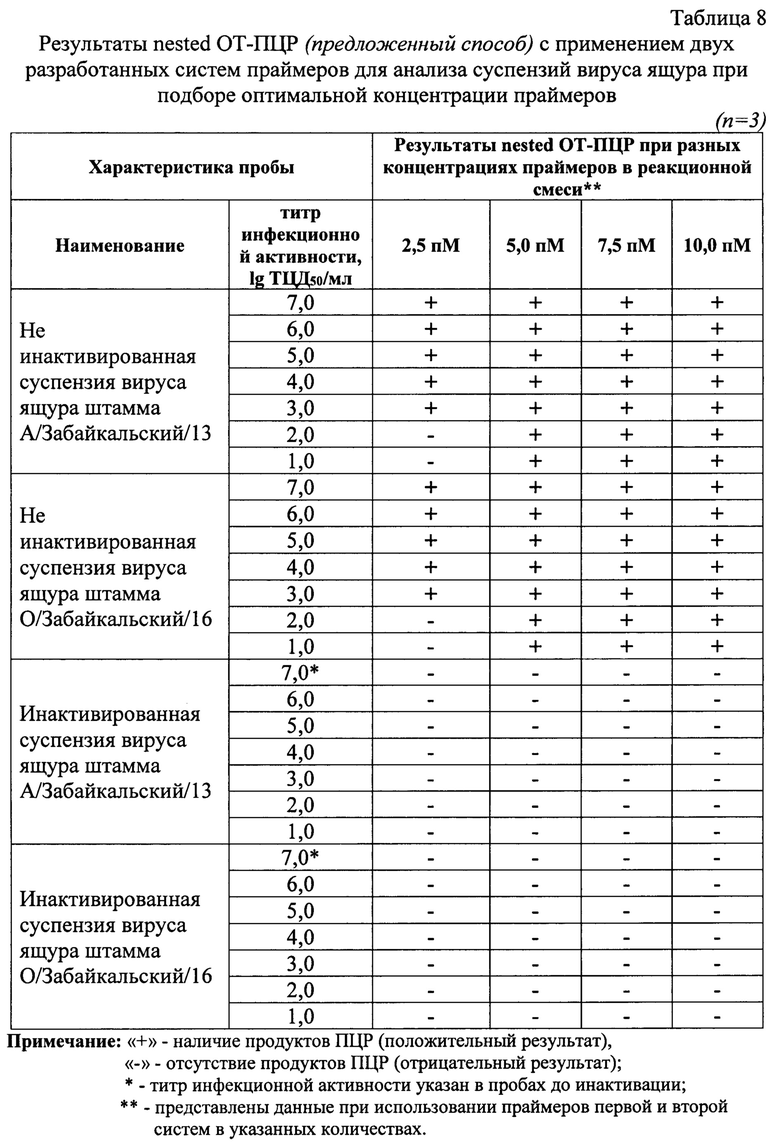
Осуществляли подбор оптимальной концентрации двух разработанных систем олигонуклеотидных праймеров. Для анализа использовали следующие количества праймеров: 2,5; 5,0; 7,5 и 10,0 пМ/реакция. Постановку реакций проводили в трех повторностях. Результаты исследования отражены в таблице 8, из которой видно, что при повышении количеств «затравочных» олигонуклеотидов отмечалось снижение чувствительности реакции. При концентрации каждого из праймеров по 2,5 пМ/реакция снижалась чувствительность реакции и пробы с титром инфекционной активности 2,00 lg ТЦД50/мл и ниже не были идентифицированы. Обнаружено, что оптимальная концентрация олигонуклеотидных праймеров составляла по 5 пМ/реакция.
Проводили исследования по подбору оптимального температурно-временного режима для nested ОТ-ПЦР. Реакцию обратной транскрипции осуществляли с использованием высокоточной ревертазы Maxima Н Minus при температуре 50°С в течение 25 мин. Денатурацию молекулы ДНК проводят, как правило, при 90-96°С, синтез ДНК - при 68-72°С, когда скорость работы высокоточной ДНК-полимеразы Platinum High Fidelity (Thermo FS) составляет более 100 нуклеотидов/с [21]. Для высокоэффективного и быстрого прохождения реакции амплификации требовалось подобрать оптимальную температуру отжига олигонуклеотидных праймеров для ПЦР1 и ПЦР2. Для осуществления ПЦР1 гибридизацию праймеров осуществляли при следующих температурах: 56, 58, 60, 62°С, для ПЦР2 - 53, 55, 57, 59°С. Результаты по подбору оптимальной температуры при проведении реакции для представленных олигонуклеотидов отражены в таблице 9.
По результатам анализа видно, что сопоставимые данные электрофореза отмечались при выборе температурного режима отжига праймеров системы №1 в диапазоне 56-60°С. Идентичные данные электрофореза отмечались при выборе температурного режима отжига праймеров системы №2 в диапазоне 53-55°С. При снижении температуры повышается концентрация гибридизованных праймеров, однако, есть вероятность увеличения количества неспецифических продуктов реакции. Для данного подэтапа для гибридизации олигонуклеотидов системы №1 оптимальной температурой считали 60°С, для системы №2 - 55°С.
Таким образом, nested ОТ-ПЦР для исследования полноты инактивации антигена вируса ящура проводили при температурно-временных показателях, отраженных в таблице 10.
Обратную транскрипцию проводили при температуре 50°С в течение 25 мин, что допустимо при использовании ревертазы Maxima Н Minus (Thermo Scientific). Температура реакции при использовании данного фермента может быть увеличена до 65°С для эффективной транскрипции РНК и для улучшения специфичности с использованием ген-специфических праймеров. Чрезвычайно высокая технологичность фермента Maxima Н Minus приводит к повышенной устойчивости к обычным ингибиторам реакции, таким как гуанидин, формамид и этанол. Кроме того, благодаря использованию данного фермента повышается чувствительность реакции.
Перед постановкой nested ПЦР осуществляли предварительный прогрев смеси при температуре 95°С в течение 5 мин для активации ДНК-полимеразы Platinum High Fidelity и инактивации ревертазы Maxima Н Minus.
Nested ПЦР состояла из двух этапов. ПЦР1 включала в себя следующие подэтапы: денатурацию, отжиг праймеров системы №1, элонгацию. Денатурацию проводили при температуре 95°С в течение 10 с, отжиг олигонуклеотидов системы №1 - при температуре 60°С в течение 30 с, элонгацию - при температуре 72°С в течение 7,5 мин, поскольку размер ампликона составлял 7289 п.н. Количество циклов амплификации - 35. ПЦР2 также включала в себя следующие подэтапы: денатурацию, отжиг праймеров системы №2, элонгацию. Денатурацию проводили при температуре 95°С в течение 10 с, отжиг праймеров системы №2 - при температуре 55°С в течение 30 с, элонгацию - при температуре 72°С в течение 7,5 мин, поскольку размер ампликона составлял 7244 п.н. Количество циклов амплификации - 25. Продолжительность nested ОТ-ПЦР составляла около 8 ч 45 мин.
Для элонгации ДНК применяли высокоточную полимеразу Platinum® Taq DNA High Fidelity, которая идеально подходит для амплификации большеразмерных фрагментов ДНК (до 15000 п.н.), когда требуются высокая точность и выход ампликонов. После реакции проводили горизонтальный электрофорез в 1,0%-ном агарозном геле.
Таким образом, подобраны оптимальные характеристики для проведения nested ОТ-ПЦР для опосредованного контроля полноты инактивации антигена вируса ящура в сырье для вакцины.
Пример 3. Разработка этапов исследования при проведении метода опосредованного контроля полноты инактивации антигена вируса ящура с применением nested ОТ-ПЦР с последующим электрофорезом ампликонов в агарозном геле.
Проводили исследования по разработке метода опосредованного контроля полноты инактивации антигена вируса ящура при производстве вакцин для ветеринарии с применением nested ОТ-ПЦР. Для этого выделяли РНК вируса ящура до и после процесса инактивации, проводили синтез кДНК в реакции обратной транскрипции и nested ПЦР, предполагающую использование системы оригинальных праймеров, детектирующих протяженный участок 5'NTR- - 3D-генов вируса ящура с размерами 7289 и 7244 п.н. в ПЦР1 и ПЦР2, соответственно, с последующей детекцией ампликонов методом горизонтального электрофореза. По итогам анализа оценивали наличие или отсутствие продуктов полимеразной цепной реакции в виде светящихся полос в потоке ультрафиолетового света при длине волны 312 нм. На основании полученных треков ампликонов делали заключение о целостности участков генома вируса ящура и, как следствие, о полноте инактивации антигена. Время проведения всего исследования составляет 10 ч, что в 7,2 раз меньше по сравнению с 72 ч, которые затрачиваются на контроль полноты инактивации в клеточной линии почки свиньи СП.
При проведении контроля полноты инактивации антигена вируса ящура использовали следующие контрольные образцы, представленные выше.
Предложены критерии интерпретации данных nested ОТ-ПЦР при контроле полноты инактивации антигена вируса ящура в сырье для вакцин. Важным условием качественно проведенных исследований являются результаты испытания контрольных образцов. Если ПКОвыд-е РНК, ПКОПЦР, ВКОвыд-е РНК, ВКОПЦР после всех стадий исследования в горизонтальном электрофорезе дают положительный результат - наличие ампликонов, а ОКО, ОИКОвыд-е РНК, ОИКОПЦР - отрицательный результат - отсутствие ампликонов, то в таком случае стадии выделения РНК, обратной транскрипции и nested ПЦР проведены верно, и можно учитывать результаты исследуемых проб. Если в треке исследуемой пробы обнаружены светящиеся продукты ПЦР2, то анализируемый участок нуклеиновой кислоты вируса ящура не поврежден, следовательно, данный образец считается не инактивированным или не полностью инактивированным. Если во всех треках свечение ампликона отсутствует, то данный участок нуклеиновой кислоты вируса ящура поврежден, тем самым, репликация вируса невозможна, и антиген считается авирулентным (фиг. 2). Иными словами, на основании отсутствия ПЦР-продуктов исследуемой пробы по сравнению с положительными контролями позволяет сделать заключение о разрушении нуклеиновой кислоты вируса ящура в инактивированных суспензиях и, как следствие, об отсутствии вирулентного вируса. В том случае, если обнаружены несоответствия в результатах анализа контролей, требуется повторно провести исследование.
Таким образом, проведена разработка этапов исследования при проведении метода опосредованного контроля полноты инактивации антигена вируса ящура с применением nested ОТ-ПЦР и представлена интерпретация данных с помощью горизонтального электрофореза ампликонов в агарозном геле.
Пример 4. Исследование полноты инактивации суспензии вируса ящура штамма О/Забайкальский/16 после воздействия АЭЭИ.
Исследовали процесс инактивации вируса ящура штамма О/Забайкальский/16 в течение 24 ч 1,2-аминоэтилэтиленимином с помощью nested ОТ-ПЦР в сравнении с культуральным методом. Для работы использовали не инактивированную суспензию вируса ящура с титром инфекционной активности в культуре клеток СП 8,25 lg ТЦД50/МЛ. Суспензию вируса сразу после его культивирования подвергали процессу инактивации с применением АЭЭИ (рН 8,2-8,6) с концентрацией 0,025% при температуре 37±0,5°С в течение 24 ч. Водородный показатель вирусной суспензии поддерживали в диапазоне 7,2-7,6. Суспензию подвергали тщательному перемешиванию в течение 3-5 минут через каждый час. Параллельно исследовали вирус ящура того же штамма, не подвергнутый инактивации, экспозиция которого проводилась при той же температуре и в тот период времени. Исследование проводили в 5 повторностях.
Для определения времени полной инактивации после добавления химического агента через 2, 4, 7, 8, 12, 20 и 24 ч в стерильных условиях производили отбор проб. Полученные образцы исследовали на наличие инфекционной активности вируса ящура в монослойной культуре клеток СП, а также с помощью предложенного способа. Результаты исследования отобранных суспензий вируса ящура отражены в таблице 11 и на фиг. 3. Выведена формула для графика инактивации вируса ящура штамма О/Забайкальский/16 в указанных условиях в виде линейного уравнения:
ТВЯ=-1,1662×tинактивации+8,1814,
где: ТВЯ - титр инфекционной активности вируса ящура,
tинактивации - время инактивации. Из полученных данных видно, что по итогам nested ОТ-ПЦР контроль инактивации (вирус ящура, не подвергнутый инактивации, экспозиция при температуре 37±0,5°С в течение 24 ч) сохранил вирусную кДНК без повреждений в участке длиной 7244 п.н. Полная инактивация вируса ящура штамма О/Забайкальский/16 1,2-аминоэтилэтиленимином с концентрацией 0,025% при температуре плюс 37±0,5°С достигалась через 7 ч. Эти данные получены с помощью культурального метода (фиг. 3) и подтверждены в nested ОТ-ПЦР.
Таким образом, способ опосредованного контроля полноты инактивации вируса ящура с помощью nested ОТ-ПЦР не уступает по своей эффективности культуральному методу и позволяет существенно сократить время и материальные затраты на проведение исследования.
Пример 5. Получение вакцины и исследование ее авирулентности на животных.
Полученную инактивированную суспензию вируса ящура штамма О/Забайкальский/16 концентрировали в 14 раз с помощью тангенциальной установки фильтрации и с полученным концентратом проводили получение сорбированной инактивированной противоящурной вакцины с использованием гидроокиси алюминия и сапонина. Количество гидроокиси алюминия составляло 11000-15000 мкг/мл, сапонина - 500-1500 мкг/мл, поддерживающей среды - до 1000000 мкг/мл.
Полученную вакцину исследовали на остаточную инфекционность путем введения крупному рогатому скоту. Для проверки вакцины использовали 2 головы КРС в возрасте 18-24 мес.весом не менее 250 кг. Животные были не ниже средней упитанности, клинически здоровые и не иммунные к ящуру, полученные из благополучных по ящуру зон страны, где иммунизация животных против ящура не проводилась в течение последних двух лет. Препарат вводили каждой из двух голов КРС интрадермолингвально в количестве 2,0 см3 (по 0,1 см3 в 20 точек). Наблюдение за животными вели в течение 14 сут. В течение этого срока у животных ежедневно утром измеряли температуру тела. У КРС через 12-18 ч после введения вакцины наблюдали повышение температуры до 40,1°С, что допустимо.
В период наблюдения у животных не появились клинические признаки ящура, а после окончания контроля на безвредность при патологоанатомическом исследовании не было обнаружено изменений, характерных для ящура. Иными словами, на животных была доказана безвредность вакцины и отсутствие живого вируса ящура в готовом препарате. Таким образом, разработанный способ опосредованного контроля полноты инактивации антигена вируса ящура с помощью nested ОТ-ПЦР с последующей детекцией ампликонов в горизонтальном электрофорезе.
Пример 6. Определение диагностических показателей метода опосредованного контроля полноты инактивации антигена вируса ящура в сырье для вакцин.
Проводили определение следующих диагностических показателей способа опосредованного контроля полноты инактивации антигена вируса ящура в сырье для вакцин с помощью nested ОТ-ПЦР: диагностической чувствительности (DSe), диагностической специфичности (DSp), k-критерия (индекс Каппа Коэна), прогностичности положительного результата (PPV), прогностичности отрицательного результата (NPV), диагностической точности (DAc). Для определения чувствительности предложенного способа исследовали 578 образцов суспензий вируса ящура, которые являлись заведомо положительными (вирулентными) по данным исследованиям культуральным методом в клеточной линии СП (титр инфекционной активности составлял 1,00-8,25 lg ТЦД50/мл). Постановку анализа проводили, как отражено выше в примере. При интерпретации результатов исследования положительными считали пробы, которые содержали живой вирус ящура по данным культурального метода, отрицательными - пробы, в которых живой вирус ящура отсутствовал. В результате проведения nested ОТ-ПЦР определили, что из 578 истинно положительных образцов суспензий 576 определены в качестве положительных, 2 - в качестве ложноотрицательных (титр инфекционной активности 1,00 lg ТЦД50/мл). Для исследования диагностической специфичности метода тестировали 550 образцов суспензий антигена вируса ящура, которые по результатам культурального метода были авирулентными. В результате исследования в nested ОТ-ПЦР определили, что из 550 истинно отрицательных суспензий 549 определены в качестве отрицательных, 1 - в качестве ложноположительной. Иными словами, количество истинно положительных проб (а) - 578, количество ложноотрицательных (b) - 2, количество ложноположительных проб (с) - 1, количество истинно отрицательных (d) - 550.
Пользуясь представленными выше статистическими методами анализа определили, что диагностическая чувствительность (DSe) составила 99,84% (в 95%-ном доверительном интервале: 99,12-100,0%), диагностическая специфичность (DSp) - 99,67% (в 95%-ном доверительном интервале: 98,82-99,96%), k-критерий (индекс Каппа Коэна) - 0,995, прогностичность положительного результата (PPV) - 99,68% (в 95%-ном доверительном интервале: 98,75-99,92%), прогностичность отрицательного результата (NPV) - 99,84% (в 95%-ном доверительном интервале: 98,85-99,98%), диагностическая точность (DAc) - 99,76% (в 95%-ном доверительном интервале: 99,30-99,95%) (таблица 12).
Основным преимуществом предлагаемого изобретения является возможность одновременного исследования большого количества проб (до 89) для опосредованного контроля полноты инактивации антигена вируса ящура в сырье для инактивированных вакцин в течение 10 ч, что в 7,2 раз быстрее по сравнению с классическим культуральным методом. В качестве гомологичных участкам 5'NTR- и 3D-генов вируса ящура олигонуклеотидных праймеров используют две системы праймеров:
система праймеров для ПЦР1:
5'NTR-F1 (5'-GCCTGGTCTTTCCAGGTCTA-3'),
3D-R1 (5'-CGGCGAGTCCTGCCACGGA-3'), которая позволяет амплифицировать фрагмент размером 7289 п.н., и
система праймеров для ПЦР2:
5 NTR-F2 (5'-GAGGGGTGACACTTTGTA -3'),
3D-R2 (5'-CTTCTCCTGTATGGTCCCA-3'), которая позволяет амплифицировать фрагмент размером 7244 п.н.
В данном изобретении предлагается применение nested ОТ-ПЦР с использованием высокоточной ревертазы Maxima Н Minus и ДНК-полимеразы Platinum High Fidelity с подобранными временными и температурными режимами проведения анализа, с последующим горизонтальным электрофорезом ампликонов в агарозном геле, с помощью которого возможно выявлять неповрежденный продолжительный участок генома вируса ящура (по итогам ПЦР2 - 7244 п.н.) до процесса инактивации и определять отсутствие вирулентного вируса ящура после воздействия инактивантами.
Результаты предложенного метода коррелируются с данными в клеточной линии почки свиньи СП. Предлагаемый способ характеризуется высокими диагностическими показателями, в частности, диагностическая чувствительность (DSe) составила 99,84% (в 95%-ном доверительном интервале: 99,12-100,0%), диагностическая специфичность (DSp) - 99,67% (в 95%-ном доверительном интервале: 98,82-99,96%), k-критерий (индекс Каппа Коэна) - 0,995, прогностичность положительного результата (PPV) - 99,68% (в 95%-ном доверительном интервале: 98,75-99,92%), прогностичность отрицательного результата (NPV) - 99,84% (в 95%-ном доверительном интервале: 98,85-99,98%), диагностическая точность (DAc) - 99,76% (в 95%-ном доверительном интервале: 99,30-99,95%).
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ контроля полноты инактивации антигена вируса ящура с применением nested обратно-транскриптазной полимеразной цепной реакции с последующим электрофорезом ампликонов в агарозном геле»:
1. Ящур / Пер. с нем. Г.А. Сурковой; Под ред. и с предисл. канд. вет. наук П.В. Малярца. - Москва: Колос, 1971. - 432 с.
2. Пономарев А.П., Узюмов В.Л., Груздев К.Н. Вирус ящура: структура, биологические и физико-химические свойства. Владимир: Фолиант. - 2006. - 250 с.
3. Таксономия микроорганизмов. [Электронный ресурс] / URL: https://www.ncbi.nlm.nih.gov/guide/taxonomy/ (Дата обращения: 16.04.2021).
4. OIE. Manual of diagnostic tests and vaccines for terrestrial animals. - 7th ed. - Paris, 2018. - Vol. 1, Chap. 2.1.8.
5. Fry E.E., Stuart D.I., Rowlands D.J. The structure of foot-and-mouth disease virus // Curr Top Microbiol Immunol. - 2005. - №288. - P. 1-101. doi: 10.1007/3-540-27109-0_4. PMID: 15648175.
6. Allaire, M., Chernaia, M.M., Malcolm, B.A. and James, M.N. Picornaviral 3C cysteine proteinases have a fold similar to chymotrypsin-like serine proteinases // Nature. - 1994. - №369. - P. 72-76.
7. Beckman, M.T. and Kirkegaard, K. (1998) Site size of cooperative single-stranded RNA binding by poliovirus RNA- dependent RNA polymerase // J. Biol. Chem. - 1998. - №273. - P. 6724-6730.
8. Baranowski E., Sevilla N., Verdaguer N., Ruiz-Jarabo С.M. (1998) Multiple virulence determinants of foot-and-mouth disease virus in cell culture // J. Virol. - 1998. - №72. - P. 6362-6372.
9. Falk M.M. FMDV protease 3C inducing specific proteolitic cleavage of host cell histon H3 / M.M. Falk, P.R. Crigera, J.E. Bergmann // J. Virol. - 1990. - V. 64, N. 2.-P. 748-756.
10. Luz N. A cellular 57 kD protein binds to two region of the internal translation initiation site of FMDV / N. Luz, E. Beck // FEBS. - 1990. - V. 269, N. 2. - P. 311-314.
11. Belsham G.J. Translation and replication of FMDV RNA // Curr Top Microbiol Immunol. - 2005. - V.288. - P. 43-70.
12. Ethyleneimine [Электронный ресурс]. - URL: http://www.cdc.gov/niosh/npg/npgd0274.html (Дата обращения: 21 декабря 2020 г.).
13. Волков А.И., Жарский И.М. Большой химический справочник. Мн.: 2005-608 с.
14. Alexandersen, S. The pathogenesis and diagnosis of foot and mouth disease / S. Alexandersen, Z. Zhang, A.L. Donaldson // J. Compr. Pathol. - 2003 - V. 129. - P. 268-282.
15. Закутский, Н.И. Экспресс-метод технологического контроля полноты инактивации ДНК-содержащих вирусов при изготовлении убитых вакцин. / Н.И. Закутский, С.Ж. Цыбанов, А.В. Луницин, С.Г. Юрков // Ветеринария. - 2016. - №11. - С. 58-61.
16. Vangrysperre W. & De Clerco K. (1996). Rapid and sensitive polymerase chain reaction based detection and typing of foot-and-mouth disease virus in clinical samples and cell culture isolates, combined with a simultaneous differentiation with other genomically and/or symptomatically related viruses // Arch. Virol. - 1996. - №141. - P. 331-344.
17. Callahan J.D., Brown F., Csorio F. Use of a portable real-time reverse transcriptase-polymerase chain reaction assay for rapid detection of foot-and-mouth disease virus / Callahan J.D., Brown F., Csorio F.A. et al. // J. Am. Vet. Med. Assoc. - 2002. - №220. - P. 1636-1642.
18. Reid S. M. Evaluation of automated RTPCR to accelerate the laboratory diagnosis of foot-and-mouth disease virus // J. Virol. Methods. - 2003. - №107. - P. 129-139.
19. Патент РФ №2746588, 02.11.2020 Способ опосредованного определения полноты инактивации антигена вируса бешенства с применением обратно-транскриптазной полимеразной цепной реакции с последующим электрофорезом ампликонов в агарозном геле // Заявка 2020136034, Бюл. №11 / Доронин М.И., Борисов А.В., Михалишин Д.В., Шульпин М.И., Чупин С.А., Назаров Н.А.
20. Maxima H Minus Reverse Transcriptase [Электронный ресурс] / URL: https://www.thermofisher.com/order/catalog/product/EP0752#/EP0752
(Дата обращения: 12.03.2021).
21. Platinum® Taq DNA g/product/11304011#/11304011/ (Дата обращения: 14.03.2021). High Fidelity [Электронный ресурс] / URL: https://www.thermofisher.com/order/catalo
22. Deiman В., van Aarle P., tech. - 2002. - Vol. 20. - P. 163-179.
23. Sooknanan R., van Gemen Sillekens P. Characteristics and applications of nucleic acid sequence based amplification // Mol. BioВ., Malek L. Nucleis acid sequence-based amplification // Molecular methods for virus detection-London: Academic press, 1995. - P. 261-285.



Изобретение относится к области биотехнологии, а именно к способу опосредованного контроля полноты инактивации антигена вируса ящура с применением nested обратно-транскриптазной полимеразной цепной реакции с последующим электрофорезом ампликонов в агарозном геле. Полноту инактивации антигена вируса ящура определяют методом nested ОТ-ПЦР с использованием двух систем оригинальных праймеров: 5'NTR-F1 и 3D-R1 (амплификация участка генома вируса ящура размером 7289 п.н.), 5'NTR-F2 и 3D-R2 (амплификация участка генома вируса ящура размером 7244 п.н.). Детекцию результатов опосредованного контроля полноты инактивации вируса ящура осуществляют в потоке ультрафиолетового света при длине волны 312 нм после проведения горизонтального электрофореза. При наличии флуоресцирующих ампликонов, полученных в ПЦР2 с использованием второй системы оригинальных праймеров, образец вирулентен, при отсутствии светящихся ПЦР2-продуктов образец инактивирован. Предлагаемое изобретение позволяет одновременно исследовать до 89 проб антигена вируса ящура, позволяет в 7,2 раз быстрее по сравнению с культуральным методом и с высокой степенью достоверности опосредованно определять полноту инактивации антигена вируса ящура. 6 з.п. ф-лы, 3 ил., 12 табл., 6 пр.
1. Способ опосредованного контроля полноты инактивации антигена вируса ящура в сырье для вакцины с применением nested обратно-транскриптазной полимеразной цепной реакции с последующим горизонтальным электрофорезом в агарозном геле, отличающийся тем, что применяют праймеры со следующими последовательностями нуклеотидов:
- система праймеров для ПЦР1: 5'NTR-F1 и 3D-R1 - амплификация участка генома вируса ящура размером 7289 п.н.:
5'-GCCTGGTCTTTCCAGGTCTA-3',
5'-CGGCGAGTCCTGCCACGGA-3' соответственно,
- система праймеров для ПЦР2: 5'NTR-F2 и 3D-R2 - амплификация участка генома вируса ящура размером 7244 п.н.:
5'-GAGGGGTGACACTTTGTA-3',
5'-CTTCTCCTGTATGGTCCCA-3' соответственно,
что, тем самым, позволяет устанавливать наличие ампликонов нужного размера - наличие вирулентного вируса ящура либо отсутствие ампликона в агарозном геле - отсутствие вирулентного вируса ящура в сырье при изготовлении противоящурных инактивированных вакцин.
2. Способ по п. 1, отличающийся тем, что основан на проведении элюирования нуклеиновой кислоты вируса ящура до и после процесса инактивации с помощью гуанидинизотиоцианата, протеиназы К и изопропилового спирта.
3. Способ по п. 1, отличающийся тем, что для проведения обратной транскрипции РНК вируса ящура применяют высокоточную ревертазу Maxima Н Minus в количестве 2,5 ед./мкл, для проведения nested ОТ-ПЦР используют высокоточную и эффективную ДНК-полимеразу Platinum High Fidelity в концентрации 2,5 ед./мкл.
4. Способ по п. 1, отличающийся тем, что для проведения реакции nested ОТ-ПЦР применяют хлорид магния с концентрацией 4,0 мМ и олигонуклеотидные праймеры в количестве 5 пМ на реакцию.
5. Способ по п. 1, отличающийся тем, что nested ОТ-ПЦР проводят с соблюдением следующих режимов:
- обратная транскрипция: температура 50°С в течение 25 мин;
- активация ДНК-полимеразы Platinum High Fidelity: температура 95°С в течение 1 мин;
- ПЦР1: денатурация: температура 95°С в течение 30 с, отжиг олигонуклеотидов: температура 60°С в течение 30 с, элонгация: температура 72°С в течение 7,5 мин;
- ПЦР2: денатурация: температура 95°С в течение 30 с, отжиг олигонуклеотидов: температура 55°С в течение 30 с, элонгация: температура 72°С в течение 7,5 мин.
6. Способ по п. 1, отличающийся тем, что является экономичным, позволяет одновременно исследовать до 89 проб вируссодержащего материала для вакцин, а время проведения анализа сократить до 10 часов.
7. Способ по п. 1, отличающийся тем, что его диагностическая чувствительность составляет 99,84% (в 95%-ном доверительном интервале: 99,12-100,0%), диагностическая специфичность - 99,67% (в 95%-ном доверительном интервале: 98,82-99,96%), k-критерий (индекс Каппа Коэна) - 0,995, прогностичность положительного результата - 99,68% (в 95%-ном доверительном интервале: 98,75-99,92%), прогностичность отрицательного результата - 99,84% (в 95%-ном доверительном интервале: 98,85-99,98%), диагностическая точность - 99,76% (в 95%-ном доверительном интервале: 99,30-99,95%).

