Изобретение относится к области биотехнологии и производству антирабических вакцин, а именно к способу опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов с применением разработанной линейной функции зависимости величины порогового цикла амплификации ДНК и концентрации RNP-комплекса вируса бешенства (CRNP ВБ)-
Бешенство - вирусное, смертельно опасное заболевание человека и животных, поражает центральную и периферическую нервную систему, вызывает энцефаломиелиты [1]. Возбудитель классического бешенства принадлежит реалму Riboviria царству Orthornavirae типу Negarnaviricota классу Monjiviricetes порядку Mononegavirales семейству Rhabdoviridae роду Lyssavirus {Rabies lyssa virus) генотипу Rabies virus (RABV) [2]. Данный вирус относится к V группе по классификации Балтимора ((-)одноцепочечные РНК-вирусы) [3]. Наибольшее эпидемиологическое значение имеет вирус классического бешенства, так как он является этиологическим агентом большинства случаев бешенства, зарегистрированных у человека и животных [4].
Вирионы вируса бешенства имеют пулеобразную форму длиной 100-300 нм, диаметром 45-100 нм [4]. Геном вируса бешенства представлен несегментированной одноцепочечной негативной спиральной молекулой РНК длиной около 12000 н. о., который кодирует 5 основных белков: нуклеопротеин (N-белок), фосфопротеин (Р-белок), матриксный белок (М-белок), гликопротеин (G-белок), РНК-зависимую РНК-полимеразу (L-белок) [4]. Короткие (58 и 70 нуклеотидов) некодирующие последовательности на 3'- и 5'- концах генома называются лидерной и трейлерной последовательностями, соответственно. Лидерная последовательность является сигнальным участком для узнавания матрицы вирусной РНК-зависимой РНК-полимеразой. Трейлерная последовательность участвует в терминации транскрипции и репликации генома вируса бешенства [4, 5].
Каждый вирион вируса бешенства включает в свой состав 1 молекулу РНК. L- и Р-протеины контролируют процессы репликации и транскрипции генома. Нуклеопротеид вируса бешенства представляет собой спирально свернутую вправо лентообразную нить, построенную из белковых субъединиц, скрепленных РНК. N-белок покрывает молекулу РНК и защищает ее от действия клеточных протеаз. Матриксный белок формирует суперкапсидную оболочку и участвует в конечной фазе сборки вирионов вируса бешенства. G-белок входит в состав суперкапсидной оболочки вириона, образуя на его поверхности шипы. Гликопротеин участвует в адсорбции вириона на клеточной поверхности, проникновении вируса в клетку хозяина. Геномная РНК вируса бешенства, N-белок, Р-белок и РНК-зависимая РНК-полимераза образуют RNP-белковый комплекс (Фиг. 1), который является важным иммуногенным компонентом вакцинных препаратов против бешенства. В RNP-комплексе содержится 4% РНК к 96% белка [5].
По данным ряда исследователей плавучая плотность вириона вируса бешенства 1,16-1,20 г/см33, константа седиментации 600 S, молекулярный вес (Mw) 475 × 106 Да [4, 5]. По данным Sokol F. плавучая плотность RNP-комплекса 1,32 г/см3, константа седиментации 200 S, молекулярная масса 15 Да [5]. Длина РНК вируса 4600 нм, константа седиментации 45 S, плавучая плотность 1,66 г/см3, молекулярный вес 4,4-4,6 × 106 Да [4, 5]. По данным Neurath, количественное соотношение нуклеотидов в РНК разных штаммов вируса бешенства практически одинаково, в среднем в молекуле РНК содержится 25,5-26,3% AMP, 28,1-29,7% UMP, 20,6-24,1% GMP, 21,7-23,8% CMP [5].
При электрофорезе в полиакриламидном геле, содержащем додецилсульфат натрия, в составе вириона обнаруживаются 4 мажорных и 1 минорный белок [4].
Бешенство приводит к значительным экономическим потерям, которые связаны с гибелью животных, ликвидацией последствий вспышек заболевания, проведением профилактических и карантинных мероприятий, регулированием численности диких плотоядных животных, отловом бродячих кошек и собак, осуществлением лабораторных исследований по постановке диагноза. Система мер для борьбы с бешенством и его профилактики предусматривает иммунизацию домашних, сельскохозяйственных и диких плотоядных, а также контроль уровня напряженности поствакцинального иммунитета [1, 4].
В процессе производства антирабических вакцин вируссодержащее сырье исследуют на определение концентрации важного иммуногенного компонента RNP-комплекса вируса бешенства. Традиционно для определения данного компонента применяют колориметрический метод М. Брэдфорда [6] и непрямой количественный сэндвич-вариант иммуноферментного анализа (ИФА) [7] (прототипы). Существенными недостатками метода М. Брэдфорда являются: невысокая аналитическая чувствительность анализа (не менее 2 мкг/мл), невысокая специфичность. Количественный непрямой сэндвич-вариант ИФА зарекомендовал себя, однако он также имеет ряд ограничений: возможности увеличения чувствительности метода ограничиваются фоном анализируемого соединения, субстратной специфичностью фермента и аффинностью антител; наличие в тестируемых образцах кофакторов, ингибиторов и стимуляторов активности ферментов; ИФА не позволяет разделять нативные белки и их биологические неактивные фрагменты, сохранившие антигенные детерминанты; возможное изменение каталитической активности фермента при его конъюгировании с антигеном.
В связи с этим целесообразно провести поиск способа опосредованного определения концентрации RNP-комплекса вируса бешенства в сырье для производства антирабических вакцин с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Данный метод позволяет опосредованно определять концентрацию иммунного RNP-комплекса в сырье для антирабических вакцин в течение 3 часов, не предполагает контаминации исследуемых образцов, поскольку во время анализа пробирки закрыты, предполагает удаление кофакторов и ингибиторов реакции амплификации, предусматривает оценку степени чистоты выделенных элюатов РНК, характеризуется отсутствием изменения каталитической активности фермента, имеет высокую аналитическую чувствительность и специфичность.
Задачей настоящего изобретения является разработка, высокочувствительного и высокоспецифичного экспресс-способа определения концентрации RNP-комплекса вируса бешенства в сырье для производства антирабических вакцин с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов с целью устранения вышеуказанных недостатков.
Данная задача решена благодаря созданию нового опосредованного способа определения концентрации RNP-комплекса вируса бешенства в сырье для производства антирабических вакцин с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов. Предложенный способ позволяет:
1) сократить время проведения анализа сырья для антирабических вакцин по определению концентрации RNP-комплекса вируса бешенства до 3 ч;
2) исключить вероятность контаминации;
3) повысить степень чистоты элюата РНК вируса бешенства;
4) увеличить чувствительность и специфичность анализа за счет применения высокоспецифичных оригинальных олигонуклеотидов;
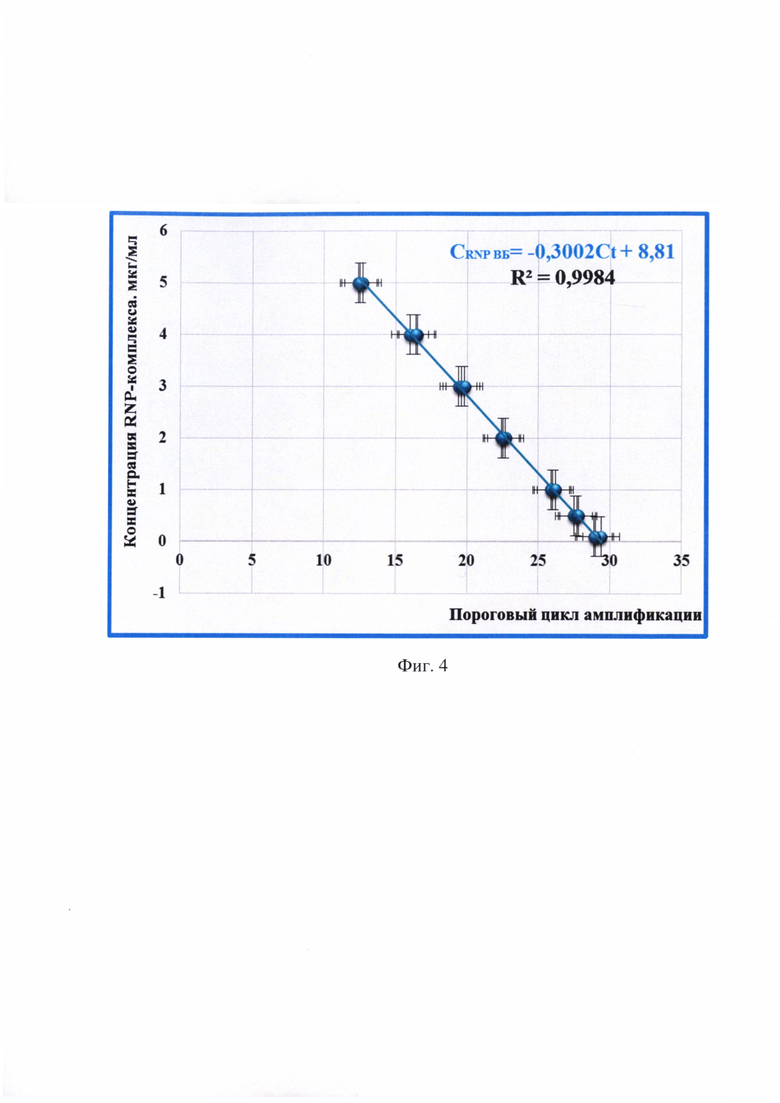
5) повысить достоверность проводимого анализа благодаря установлению зависимости между концентрацией RNP-комплекса вируса бешенства (CRNP ВБ) и пороговым циклом амплификации (Ct), представленной в виде линейной функции: CRNP ВБ=-0,3002 × Ct + 8,81 с высокой достоверностью аппроксимации (R2=0,9984) и эффективностью амплификации (Е) 99,83%. Предложенная модель позволяет опосредованно определять концентрацию RNP-комплекса вируса бешенства в сырье для производства антирабических вакцин.
Сущность изобретения отражена на графических изображениях
Фиг. 1 - Схематическая модель строения RNP-комплекса вируса бешенства.
Фиг. 2 - Электрофореграмма очищенного RNP-комплекса вируса бешенства.
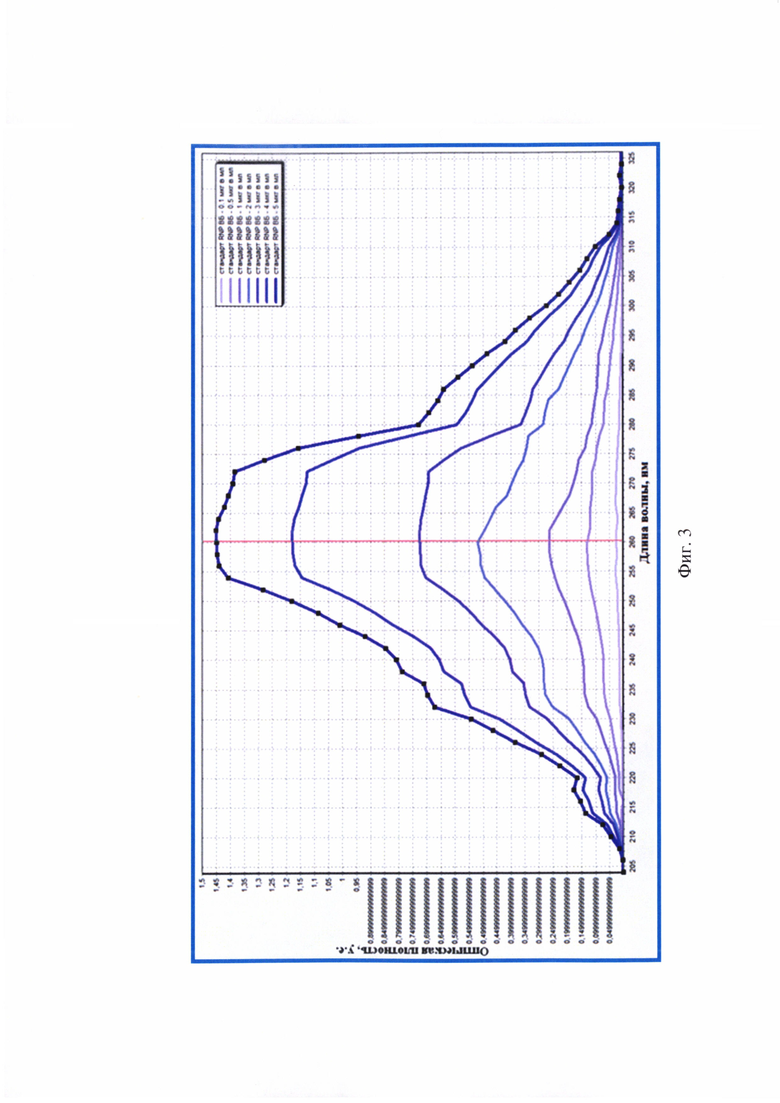
Фиг. 3 - Спектры поглощения экстрактов РНК вируса бешенства положительных стандартов для оценки степени их чистоты (максимальные оптические сигналы для РНК вируса бешенства зарегистрированы при длине волны 260 нм) (n=3).
Фиг. 4 - Зависимость концентрации RNP-комплекса вируса бешенства (CRNP ВБ) и величины порогового цикла амплификации (Ct), определенной с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов (n=3, отмечены точки, отображающие средние значения пороговых циклов реакции амплификации).
Фиг. 5 - Зависимость величины порогового цикла амплификации (Ct) от концентрации RNP-комплекса вируса бешенства (CRNP ВБ), определенной с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов (n=3, отмечены точки, отображающие средние значения пороговых циклов реакции амплификации).
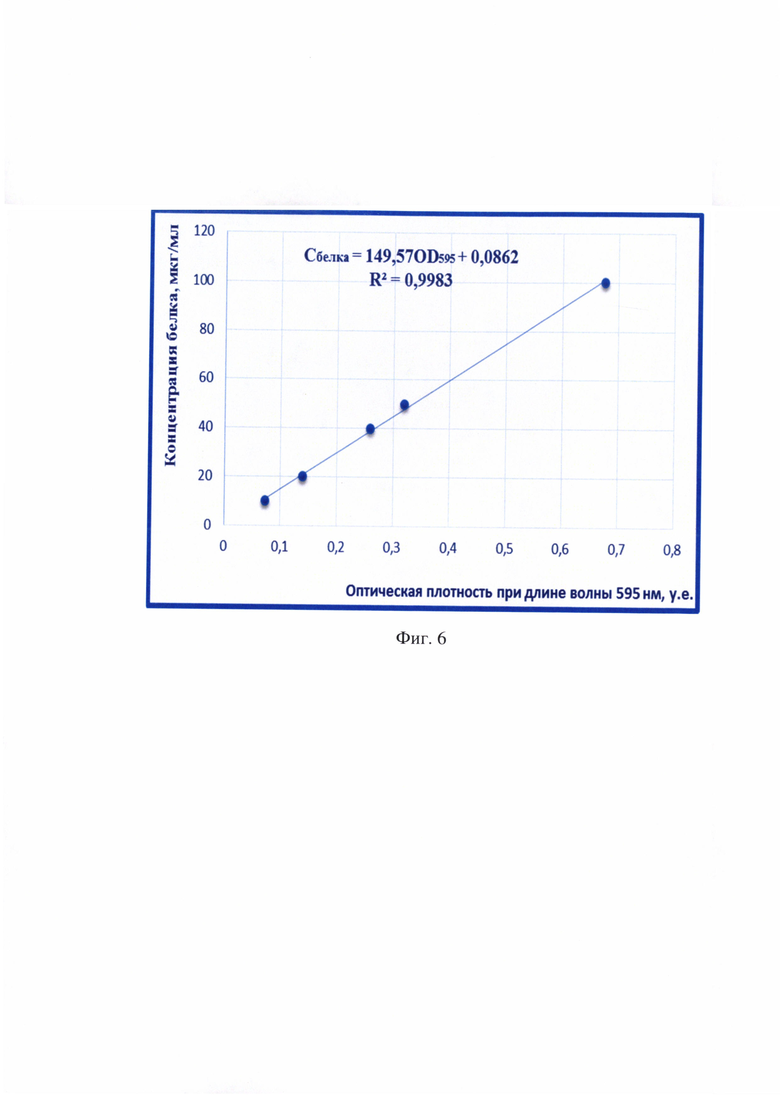
Фиг. 6 - Градуировочная характеристика для определения концентрации белка в суспензии, построенная по стандартным образцам BSA по методу М. Брэдфорда.
Сущность изобретения заключается в новом подходе по опосредованному определению концентрации RNP-комплекса вируса бешенства в сырье для производства антирабических вакцин с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов. Заявляемый способ основан на: 1) осаждении клеток линии почки новорожденного сирийского хомячка ВНК-21, в которых репродуцирован вируса бешенства; 2) твердофазной экстракции РНК вируса бешенства с применением частиц диоксида кремния диаметром 250-300 нм, обработанных 4 М гидроксидом натрия в течение 40 ч; 3) амплификации специфического фрагмента N-гена РНК вируса бешенства с применением оригинальных специфических праймеров и молекулярного зонда, меченого флуоресцентным красителем FAM (λmах флуоресценции = 520 нм) и тушителем свечения RHQ-1 (λmax поглощения = 520 нм); 4) детектировании ампликонов с помощью флуоресцентного свечения и отображения накопления сигнала в виде логистической функции; 5) определении значений пороговых циклов амплификации; 6) расчете концентрации RNP-комплекса вируса бешенства с применением показательной линейной функции: CRNP ВБ=-0,3002 × Ct + 8,81 с высокой достоверностью аппроксимации (R2=0,9984) и эффективностью амплификации 99,83%.
В настоящее время метод амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов применяют для выявления нуклеиновых кислот различных инфекционных агентов, в том числе вируса бешенства. Для количественной оценки вирусной нагрузки суспензии, в частности, для определения концентрации RNP-комплекса вируса бешенства, ранее данный метод с применением разработанной системы оригинальных олигонуклеотидов не применялся. Иными словами, сведений об аналогах предлагаемого способа опосредованного определения концентрации RNP-комплекса вируса бешенства в сырье для антирабических вакцин авторами не обнаружено.
Разработанный способ опосредованного определения концентрации RNP-комплекса в сырье для антирабических вакцин с помощью метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов по сравнению с прототипами отличается более высокой чувствительностью и специфичностью, быстротой выполнения анализа и снижением риска контаминации исследуемых образцов.
В отличие от прототипа разработанный способ включает следующие этапы: осаждение суспензии клеток линии ВНК-21, в которых репродуцирован вируса бешенства; получение суспензии осажденных клеток в фосфатно-солевой буферном растворе (ФСБ); твердофазная экстракция РНК вируса бешенства с применением частиц диоксида кремния диаметром 250-300 нм, обработанных 4 М гидроксидом натрия; обратная транскрипция РНК вируса бешенства; амплификация специфического фрагмента N-гена РНК вируса бешенства с применение оригинальных специфических олигонуклеотидов; детектирование ампликонов с помощью флуоресцентно-гибридизационного метода и отображение накопления сигнала в виде логистической функции; определение значений пороговых циклов амплификации; расчет концентрации RNP-комплекса вируса бешенства с применением показательной линейной функции: CRNP ВБ =-0,3002 × Ct + 8,81 (R2=0,9984, Е=99,83%). Применение предложенного способа позволит повысить чувствительность и специфичность определения концентрации RNP-комплекса; снизить время проведения анализа вируссодержащего сырья до 3 ч; исключить вероятность контаминации; удалить кофакторы и ингибиторы реакции амплификации; увеличить чистоту экстракта РНК вируса бешенства; не вызывать изменения каталитической активности ферментов реакции; повысить достоверность проводимого анализа. Исходя их этого, актуально применять данный способ для опосредованного определения концентрации RNP-комплекса вируса бешенства в сырье для антирабических вакцин.
Ключевым элементом заявляемого способа является определение пороговых циклов логистических кривых реакции амплификации с последующей гибридизационно-флуоресцентной детекции ДНК-ампликонов и расчет концентрации RNP-комплекса вируса бешенства с использованием разработанной линейной функции зависимости данных переменных.
Сопоставительный анализ с прототипами позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении амплификации с последующей гибридизационно-флуоресцентной детекции ДНК-ампликонов, оригинальных специфичных олигонуклеотидов, рассчитанных на участок N-гена вируса бешенства, и разработанной линейной функции зависимости величины порогового цикла реакции амплификации и концентрации RNP-комплекса вируса бешенства для опосредованного определения концентрации последнего в сырье для антирабических вакцин.
Сущность предлагаемого изобретения пояснена на графическом материале - графике зависимости величины порогового цикла амплификации (Ct) и концентрации RNP-комплекса вируса бешенства (CRNP ВБ) (n=3) (фиг. 4).
С целью определения концентрации RNP-комплекса вируса бешенства отдельно подготавливают положительный контрольный образец. Для этого проводят репродукцию вируса бешенства. Суспензионные клетки линии ВНК-21 с концентрацией 3,5-4,0 млн кл./мл инфицируют вирусом бешенства в дозе 0,1-0,5 ККИД50/мл. и инкубируют при температуре 37,0±0,5°С до получения 95-100% поражения клеток в течение 48 ч. После окончания культивирования рН доводят до 7,0 с помощью 7,5%-ного раствора гидрокарбоната натрия.
Полученную суспензию вируса бешенства после культивирования в клеточной линии ВНК-21 подвергают центрифугированию при 1 500 g в течение 10 минут. Супернатант удаляют, оставляя осадок клеток, в которых содержится RNP-комплекс. Инфицированные клетки подвергают лизированию, добавляя ледяную стерильную деионизированную воду, содержащую 25 мкл/мл апротинина (Sigma-Aldrich). Данную смесь инкубируют в течение 1 ч при температуре 4°С и осветляют центрифугированием при 1000 g при той же температуре в течение 20 минут. Супернатант собирают полностью. Указанную процедуру повторяют дважды. Полученные надосадки из каждого раунда смешивают и центрифугируют при 12000 g и температуре 4,0±0,5°С в течение 8 минут. Супернатант собирают в другую пробирку.
На следующем этапе подготовки положительного контрольного образца суспензию RNP-комплекса очищают ультрацентрифугированием в ступенчатом градиенте хлорида цезия (CsCl) (17-24%) в соответствии с протоколом, описанным Dietzschold, 1996 [8]. Центрифугирование проводят при 65000 g в течение 2 ч при температуре 15°С. Опалесцирующий слой отбирают в отдельный пенициллиновый флакон.
Полученный раствор вносят в центрифужные пробирки, доливают объем пробирок с помощью 1/15 М буферного раствора ФСБ. Осаждают RNP-комплекс для очищения от остатков хлористого цезия с помощью центрифугирования при 60000 g в течение 1 ч при 15°С. Полученный осадок растворяют в необходимом количестве буферного 1/15 М раствора ФСБ.
В готовой суспензии RNP-комплекса вируса бешенства определяют концентрацию с помощью метода М. Брэдфорд [6] при длине волны 595 нм по градуировочному графику (фиг. 6) и в непрямом количественном сэндвич-варианте ИФА [7]. Проводят вертикальный белковый электрофорез в полиакриламидном геле, содержащем додецилсульфат натрия, для оценки степени чистоты положительного контрольного препарата. Иными словами, получают охарактеризованный положительный контрольный образец, который вместе с исследуемыми пробами используют в дальнейшей работе.
Составляют контрольную панель положительных стандартов RNP-комплекса вируса бешенства, в качестве которых используют очищенные препараты с концентрациями RNP-комплекса: 0,1; 0,5; 1,0; 2,0; 3,0; 4,0; 5,0 мкг/см3. В качестве отрицательного контроля применяют суспензию клеток ВНК-21, не зараженную вирусом бешенства.
На следующем этапе исследования из всех стандартных положительных и отрицательного контролей, а также тестируемых проб выделяют нуклеиновую кислоту. Для этого применяют метод твердофазной экстракции с применением частиц гидроокиси кремния диаметром 250-300 нм, обработанных 4 М раствором гидроксида натрия.
Принцип очистки РНК вируса бешенства с помощью гидроокиси кремния базируется на высокой степени аффинности отрицательно заряженного остова РНК к положительно заряженным частицам сорбента. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК является полианионом и может взаимодействовать с гидроокисью кремния. Катионы натрия на поверхности частиц сорбента выступают в качестве «мостика», который притягивает анионы кислорода в составе остатка фосфорной кислоты РНК. При высоком значении ионной силы (рН 5,0-5,5) происходит разрушение водородных связей между протоном воды и отрицательно заряженными ионами кислорода в частицах сорбента. В результате молекулы нуклеиновых кислот связываются с носителем, а интенсивное промывание позволяет удалить все ингибиторы реакции амплификации. Очищенные молекулы РНК элюируют при низком значении ионной силы ТЕ-буферного раствора (рН=7,3) [9].
К 100 мкл исследуемого образца добавляют 500 мкл хаотропного денатурирующего агента 4 М гуанидинизотиоцианата (ГТЦ), перемешивают на вортексе в течение 5 минут для денатурирования белковых составляющих. К полученному лизату добавляют 25 мкл суспензии частиц диоксида кремния диаметром 250-300 нм, обработанных 4 М раствором гидроксида натрия. Проводят инкубирование в процессе медленного перемешивания на вортексе в течение 5 мин. Суспензию центрифугируют при 250 g, удаляют супернатант, и проводят промывание частиц от ингибиторов реакции амплификации с помощью 4 М раствора ГТЦ, добавляя его к осадку в объеме 350 мкл. Смесь центрифугируют при 250 g, удаляют надосадочную жидкость. Проводят отмывание частиц сорбента с применением 80%-ного раствора пропанола-2 в объеме по 500 мкл. Осуществляют центрифугирование содержимого пробирки при тех же условиях. Данную процедуру проводят дважды. Осадок частиц сорбента с адсорбированными молекулами нуклеиновой кислоты высушивают от остатков спирта с помощью сухого твердотельного термостата при температуре 60±2°С в течение 8-10 мин. К высушенному осадку добавляют 50 мкл ТЕ-буфера (10 мМ трис(оксиметил)аминометан, 1 мМ этилендиамин-тетраацетат, рН 7,3), свободного от РНКаз и ионов Mg, прогревают содержимое пробирки при температуре 60±2 С в течение 3 минут для получения элюата. Содержимое пробирки центрифугируют при 11000 g в течение 1 минуты, и отбирают экстракт нуклеиновой кислоты.
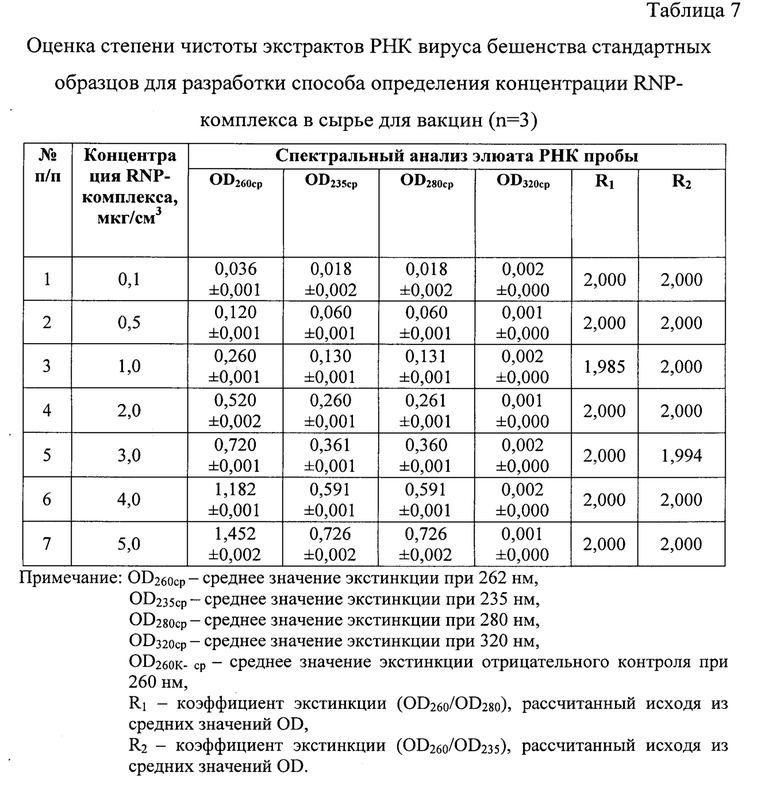
Для полученных элюатов проводят спектральный анализ, определяя поглощение аналитом монохроматического ультрафиолетового света, что позволяет оценить степень чистоты экстракта, который в последующем тестируют в реакции амплификации. Измерения оптической плотности образцов проводят при длинах волны в диапазоне 205-325 нм и температуре 22-25°С. В выделенных экстрактах оценивают содержание остатков фосфолипидов, полисахаридов и ГТЦ, полипептидов и крупных взвешенных частиц сорбента, определяя значения оптической плотности (OD, optical density) при длинах волны 205, 235, 270, 280 и 320 нм, соответственно [10]. Элюат РНК считают свободным от примесей белка, если коэффициент экстинкции R1 (OD260/OD280) находится в пределах 1,8-2,2 и оптимально составляет примерно 2,0. Более низкие значения R1 указывают на наличие молекул ДНК, белковых составляющих в элюате. Более высокие значения коэффициента R1 свидетельствуют о деградации РНК и наличии свободных рибонуклеотидов. Элюат нуклеиновой кислоты вируса бешенства считают незагрязненным полисахаридами, если коэффициент экстинкции R2 (OD260/OD235) приближен к значению 2,0 [9, 10]. При замещении 1% РНК на полисахаридные составляющие R2 снижается на 0,002. Значения коэффициента R2>2,000 могут указывать на деградацию молекул РНК. Отсутствие взвеси крупных частиц в элюате подтверждается, если OD320 приближено к нулевому значению [10]. При несоответствии требованиям чистоты повторно проводят этап выделения РНК вируса бешенства из образцов.
После оценки степени чистоты экстрактов РНК проводят обратную транскрипцию для получения комплементарной ДНК (кДНК) вируса бешенства. Состав реакционной смеси представлен в таблице 1. В качестве основы используют буферный раствор для ПЦР классического состава (10% от объема реакционной смеси), хлорид магния (2 мМ на реакцию), дезоксирибонуклеозидтрифосфаты (dNTP) (по 0,25 мкл на реакцию). В качестве общих праймеров для затравки применяют гексамеры (шестинуклеотидные олигонуклеотиды произвольного состава) (1,25 пМ на реакцию). Для ингибирования РНК-аз в реакционную смесь добавляют РНазин (10 ед./реакцию). В качестве фермента применяют AMV-ревертазу (10 ед./реакцию). Объем вносимого элюата РНК - 2 мкл, общий объем реакционной смеси - 20 мкл.
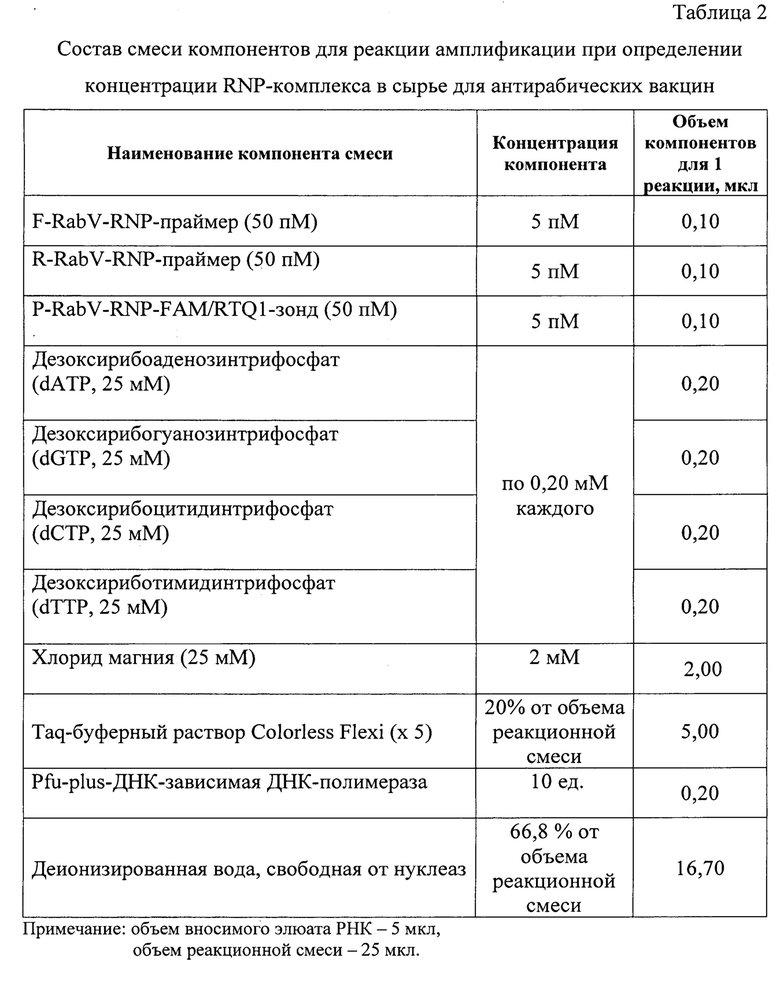
На следующем этапе анализа проводят реакцию амплификации. Для этого готовят реакционную смесь, рецептура которой представлена в таблице 2. Дизайн олигонуклеотидов отражен в таблице 3. Расчет праймеров и зонда осуществляли на основании нуклеотидных последовательностей N-гена вируса бешенства производственных штаммов РВ-97, ВНИИЗЖ и изолятов, опубликованных в базах данных GenBank [11] и полученных в рамках исследований в «Федеральном центре охраны здоровья животных» (ФГБУ «ВНИИЗЖ»).
В качестве гомологичных участку N-гена вакцинных штаммов вируса бешенства используют следующие олигонуклеотиды: F-RabV-RNP-праймер (5' GAA GGT AGA CTG GGA AGG AAA -3'), R-RabV-RNP-праймер (5'-GGA АТС ТАС ТСТ GTC GCA TGA А -3') и P-RabV-RNP-FAM/RTQ1-зонд (5'- FAM-GGC ТТС CAT ТАА ТС A TCG С AG TTG AA-RTQ1-3') в концентрации 5 пМ на реакцию с внесением в реакционную смесь по 0,1 мкл. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с их суммарной концентрацией в реакционной смеси 2 мМ. В качестве основы используют буферный раствор (5-кратный), содержание которого составляет 20% от общего объема реакционной смеси. Буферный раствор включает в свой состав ионы калия (К+) (5⋅10-2 М) и диметилсульфооксид (DMSO) (1%). DMSO включают в реакционную смесь для сведения к минимуму неспецифического связывания олигонуклеотидов. В смесь также добавляют 2 мМ хлорида магния. Содержание деионизированной воды составляет 66,8%). В качестве катализатора реакции амплификации применяют Pyrococcus furiosus P/w-Plus-ДНК-зависимую ДНК-полимеразу (10 ед.). Данная полимераза является термостабильным рекомбинантным ферментом массой приблизительно 92 кДа. Источник происхождения этого фермента - термофильная архебактерия Pyrococcus furiosus. Максимальная активность фермента проявляется при температурах 70-74°С, время полужизни при 94°С составляет не менее 30 мин. Фермент осуществляет синтез ДНК в направлении 5'->3' в присутствии ионов Mg и при соответствующем значении рН, а также обладает 3'->5'-экзонуклеазной активностью («проверочной активностью» - «proofreading activity))). Наличие 3'->5-экзонуклеазной активности делает фермент пригодным для реакции амплификации, где необходимо получение продукта с высокой точностью синтеза. При работе в оптимальных условиях Pfu-plus-ДНК-полимераза допускает минимальное количество ошибок по сравнения с другими термостабильными полимеразами. Особенность данной полимеразы заключается в том, что наличие 3'->5'-экзонуклеазной активности делает этот фермент "медленным", поэтому время проведения элонгации увеличивают до 1-2 минут. Элюаты суммарной РНК вируса бешенства каждого образца добавляют к реакционной смеси по 5 мкл. Итоговый объем смеси для проведения одной реакции составляет 25 мкл.
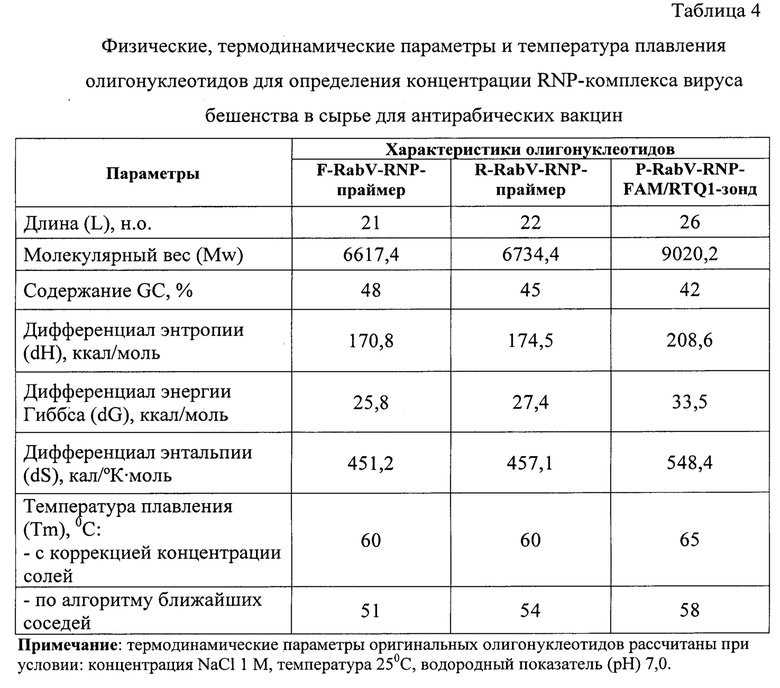
Олигонуклеотиды подбирали в соответствии с рядом общих правил, которые отражены в работах В. Deiman и R. Sooknanan [12, 13]. Длины F-RabV-RNP-праймера, R-RabV-RNP-праймера и P-RabV-RNP-FAM/RTQ1 -зонда составляют 21, 22 и 26 н.о., что соответствует требованиям (20-30 н.о.) [12, 13]. Молекулярный вес олигонуклеотидов F-RabV-RNP-праймера, R-RabV-RNP-праймера и P-RabV-RNP-FAM/RTQ1-зонда равен 6617,4; 6734,4 и 9020,2 (с учетом флуорофора и гасителя свечения), соответственно. Процентное содержание G и С в F-RabV-RNP-праймера, R-RabV-RNP-праймера и P-Rab V-RNP-FAM/RTQ1-зонда составляет 48, 45 и 42%, соответственно, что нормально (допустимо 40-60%) [12, 13]. На 5'-конце праймеров и зонда отмечается преобладание G и С, а на 3'-конце располагается полиадениловый фрагмент (poly(A)). Праймеры и зонд очищены в полиакриламидном геле и с помощью высокоэффективной жидкостной хроматографии, соответственно. Нуклеотидная последовательность зонда не комплементарна олигонуклеотидным праймерам. Отсутствуют 4 и более подряд одинаковых нуклеотидов в цепи праймеров и зонда. Флуорофор FAM присоединен к 5'-концу, а гаситель флуоресценции RHQ1 - к 3'-концу. Данные условия соответствуют требованиям, предъявляемым к олигонуклеотидным праймерам и молекулярному зонду, которые участвуют в реакции амплификации ДНК [12, 13]. В качестве флуоресцентного красителя был выбран FAM с длиной волны максимальной флуоресценцией 520 нм. Для тушения свечения использовали гаситель флуоресценции RHQ1 с длиной волны максимального поглощения при 520 нм и возможном диапазоне гашения 470-570 нм. Иными словами, была выбрана подходящая пара «флуорофор-гаситель».
При анализе нуклеотидных последовательностей олигонуклеотидов установили, что для праймеров и зонда не характерно образование «шпилек», а также не выявлено 3'-комплементарности и сайтов, отжигающих сами на себя при условии, когда минимальное количество пар оснований, необходимое для димеризации праймера и минимальное количество пар оснований, необходимое для образования шпильки - 4 [16].
Проведено определение температур плавления (Тm) для олигонуклеотидов. Для оценки температуры плавления олигонуклеотидных праймеров и зонда применяли 2 метода. Один из них - метод, учитывающий концентрации ионов K+ и диметилсульфооксида (DMSO) с применением формулы [14]:
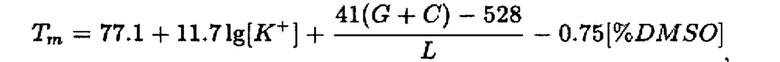
где [K+] - концентрация ионов калия (М),
[% DMSO] - количество диметилсульфооксида (%),
G - количество остатков гуанозинтрифосфата,
С - количество остатков цитидинтрифосфата.
Данный метод позволяет приблизительно оценить температуру плавления олигонуклеотидов.
Другой метод, который применяли в работе, высокоточный и основан на модели ближайших соседей (nearest neighbour, NN) с применением расчетной формулы [17]:
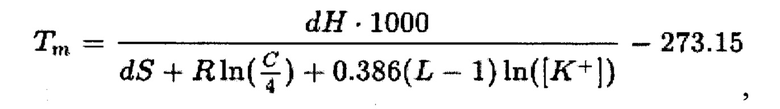
где dH - дифференциал энтропии (ккал/моль),
dG - дифференциал энергии Гиббса (ккал/моль),
dS - дифференциал энтальпии (кал/°К⋅моль),
С - концентрация олигонуклеотида (М),
L - длина олигонуклеотида,
[К+] -концентрация ионов калия (М),
In - натуральный логарифм.
В данной формуле учитывается длина олигонуклеотида (L), концентрация ионов калия ([К+]=5×10-2 М), газовая постоянная (R=1,987 кал/К⋅моль), концентрация олигонуклеотида в (с=2Т(Г7 М), значения энтропии (dH) (в ккал/моль) и энтальпии (dS) (в кал/°К⋅моль). Значения dH и dS вычисляли в соответствии с общеизвестными формулами и термодинамическими параметрами для ближайших соседей пар нуклеотидов при концентрации NaCl 1 М [17, 18]. Расчеты термодинамических параметров проводили при условии, что концентрация NaCl 1 М, температура 25°С и значение водородного показателя рН 7,0.
Физические, термодинамические константы и расчет температур плавления (Тm) [16] разработанных олигонуклеотидных ДНК-праймеров и зонда представлены в таблице 4, из которой следует, что энтропия, энергия Гиббса и энтальпия для F-RabV-RNP-праймера составили 170,8 ккал/моль, 25,8 ккал/моль, 451,2 кал/(°К⋅моль), соответственно. Энтропия, энергия Гиббса и энтальпия для R-RabV-RNP-праймера составили 174,2 ккал/моль, 27,4 ккал/моль, 457,1 кал/(°К⋅моль), соответственно. Энтропия, энергия Гиббса и энтальпия для P-RabV-RNP-зонда составили 208,6 ккал/моль, 33,5 ккал/моль, 548,4 кал/(°К⋅моль), соответственно. Полученные значения использовались для расчета температур плавления представленных олигонуклеотидов. Тm при использовании алгоритма ближайших соседей для F-RabV-RNP-праймера, R-RabV-RNP-праймера и P-RabV-RNP-FAM/RTQ1-зонда составили 51, 54 и 58°С, соответственно. Исходя из полученных данных температура отжига (Та) примерно на 1-5°С ниже Тm (46-50, 49-53, 53-57°С, соответственно).
При использовании более простого метода, учитывающего концентрации ионов К+ и диметилсульфооксида (DMSO), Tm для F-RabV-RNP-праймера, R-RabV-RNP-праймера и P-Rab V-RNP-FAM/RTQ1-зонда составили 60, 60 и 65°С. Иными словами, температура отжига праймеров и зонда должна быть ниже полученных значений на 1-5°С (55-59, 55-59 и 60-64, соответственно).
Экспериментально температуру отжига определяли по кривым плавления гетеродимера олигонуклеотида и матрицы с помощью модели фолдинга с использованием программного обеспечения Hybrid. В результате проведения эксперимента было выявлено, что температура отжига рассматриваемых олигонуклеотидов составляет 52, 53, 55°С. Для проведения реакции амплификации было решено проводить гибридизацию праймеров и зонда с участком N-гена вируса бешенства при температуре 53°С.
Последовательности оригинальных олигонуклеотидов проверили на наличие нежелательных совпадений с другими последовательностями нуклеиновых кислот с использованием Банка данных последовательности нуклеиновых кислот вируса бешенства [11]. Последовательности праймеров и зонда также проанализировали на наличие внутренних вторичных структур с помощью программы сворачивания нуклеиновых кислот с помощью программы Mfold [15]. Выявлено, что для разработанных олигонуклеотидов нежелательных совпадений с другими последовательностями нуклеиновых кислот, а также наличия внутренних вторичных структур не обнаружено.
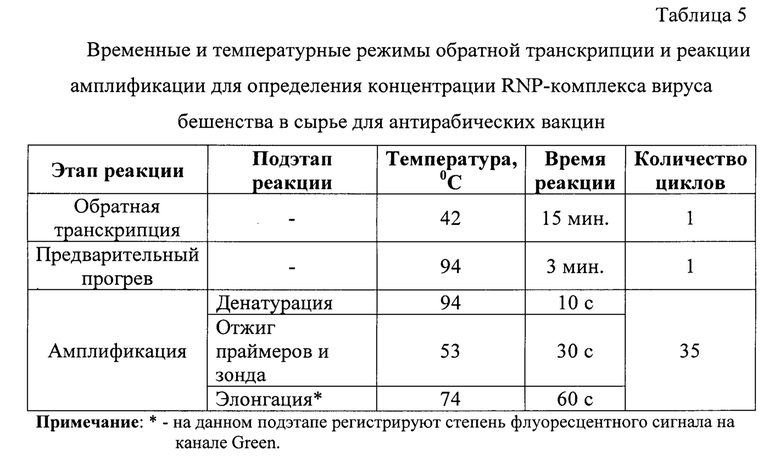
Проведение реакции амплификации осуществляли в детектирующем термоциклере любой марки при температурных и временных параметрах, сведения о которых представлены в таблице 5.
Обратную транскрипцию проводят при температуре 42°С в течение 15 мин. Перед проведением реакции амплификации осуществляют предварительный прогрев смеси при температуре 94°С в течение 3 минут для активации фермента Рfu-ДНК-полимеразы и инактивации AMV-ревертазы. Амплификация включает в себя 3 подэтапа: денатурацию, отжиг олигонуклеотидов, элонгацию. Денатурацию проводят при температуре 94°С в течение 10 с, отжиг праймеров и зонда - при температуре 53°С в течение 30 с, элонгацию - при температуре 74°С в течение 60 с.
Результаты реакции амплификации анализируют, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям пороговых циклов амплификации Ct, определенных с помощью пересечения пороговой линии и логистической функции вида Fl=f(Ct). Иными словами, проводят методом гибридизационно-флуоресцентной детекции ДНК-ампликонов. Учет результатов в реакции происходит на каждом цикле. Прибор определяет уровень флуоресценции и строит кинетическую кривую в координатах: уровень флуоресценции - цикл реакции амплификации. При наличии в исследуемой пробе специфической кДНК-матрицы вируса бешенства сигмоида имеет экспоненциальную зависимость. Положительными считаются пробы, которым соответствуют логистические графики, полученные при анализе флуоресценции красителя FAM, входящего в состав молекулярного зонда. Пробы оцениваются как отрицательные, если при их анализе отсутствует экспоненциальная кривая.
На следующем этапе анализа устанавливают зависимость между пороговым циклом амплификации и концентрацией RNP-комплекса вируса бешенства в сырье для антирабических вакцин. Оценивают величину эффективности реакции амплификации (Е) по формуле: Е=(10-1/k-1), где k - угловой коэффициент в зависимости Ct=k × CRNP ВБ + b, а также достоверность аппроксимации (R2). На основе разработанной модели CRNP ВБ=k × Ct + b рассчитывают значение концентрации RNP-комплекса вируса бешенства в сырье для антирабических вакцин.
Пример 1. Получение положительного контрольного образца для выявления зависимости концентрации RNP-комплекса вируса бешенства и порогового цикла амплификации.
Суспензионные клетки линии ВНК-21 с концентрацией 4,0 млн кл./мл инфицировали вирусом бешенства штамма РВ-97 в дозе 0,1 ККИД50/мл и инкубировали при температуре 37±0,5°С до получения 95-100% поражения клеток в течение 48 ч. После окончания культивирования рН доводили до 7,0 с помощью 7,5%-ного раствора гидрокарбоната натрия.
Полученную суспензию вируса бешенства штамма РВ-97 после культивирования в клеточной линии ВНК-21 в объеме 18000 мл подвергали центрифугированию при 1500g в течение 10 мин. Супернатант удаляли, оставляя осадок клеток объемом 50 мл, в которых содержится RNP-комплекс. Инфицированные клетки подвергали лизированию, добавляя 50,0 мл ледяной стерильной деионизированной воды, содержащей 25 мкл/мл апротинина (Sigma-Aldrich). Полученную суспензию инкубировали в течение 1 ч при температуре 4°С и осветляли центрифугированием при 1000 g при той же температуре в течение 20 минут. Супернатант собирали, и процедуру повторяли дважды. Полученные надосадки из каждого раунда смешивали и центрифугировали при 12000 g и температуре 4,0±0,5°С в течение 8 минут. Супернатант в объеме 50 мл собирали в другую пробирку. Иными словами, получали 360-кратный концентрат, содержащий RNP-комплекс.
На следующем этапе подготовки положительного контрольного образца суспензию RNP-комплекса очищали ультрацентрифугированием в ступенчатом градиенте хлорида цезия (CsCl) с концентрациями 17, 18, 19, 20, 22, 24, 26% в соответствии с протоколом, описанным Dietzschold [8]. Готовили 26%-ный раствора хлористого цезия при растворении 45,092 г хлористого цезия в 140 мл дистиллированной воды. Далее в 6 флаконах составляли растворы хлористого цезия с концентрациями 24, 22, 20, 19, 18, 17% по схеме, представленной в таблице 6.
Центрифужные пробирки разделяли на 7 зон по 4 мл. Вносили 4 мл 26%-ного раствора хлористого цезия в 6 пробирок. Остальные растворы наслаивали с помощью капилляра. Суспензию RNP-комплекса вируса бешенства разделяли на 6 пробирок и вносили на поверхность градиента. Пробирки уравновешивали. Центрифугирование проводили при 65000 g в течение 2 ч и температуре 15°С. Опалесцирующий слой RNP-комплекса вируса бешенства в области 19-21% хлористого цезия (соответствуют плавучей плотности RNP-комплекса) отбирали в отдельный флакон.
Полученный раствор вносили в центрифужные пробирки, доливали объем пробирок с помощью фосфатно-солевого буферного раствора (ФСБ). Для очищения от остатков хлористого цезия суспензию RNP-комплекса осаждали с помощью центрифугирования при 60000 g в течение 1 ч при 15°С. Полученный осадок ресуспендировали в 5 мл ФСБ. Иными словами, получали 3600-кратный концентрат RNP-комплекса относительно исходного объема вируссодержащей суспензии (18000 мл).
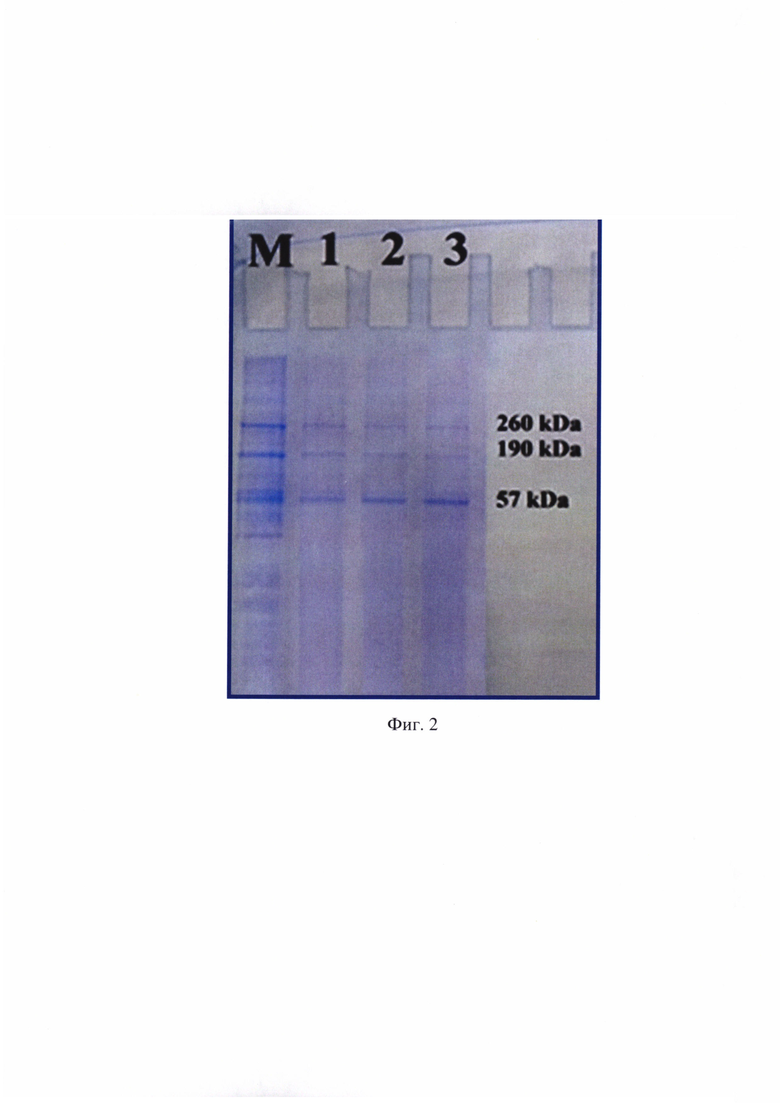
В готовой суспензии определяли концентрацию RNP-комплекса вируса бешенства с помощью метода М. Брэдфорда с применением градуировочного графика (фиг. 6) [6] и в непрямом количественном сэндвич-варианте ИФА [7]. Проводили вертикальный белковый электрофорез в полиакриламидном геле, содержащем додецилсульфат натрия, для оценки степени чистоты положительного контрольного препарата. По данным метода М. Брэдфорда и непрямого количественного сэндвич-варианта ИФА концентрация RNP-комплекса составляла 4900 и 4880 мкг/мл. Результаты вертикального электрофореза отражены на фиг. 2, из которых следует, что получен чистый препарат RNP-комплекса, включающий в свой состав нуклеопротеин, фосфопротеин и L-белок вируса бешенства с молекулярным весом 57, 260 и 190 kDa, соответственно. Из данного препарата с помощью 1/15 М раствора ФСБ приготовили положительный контрольный образец с концентрацией RNP-комплекса 5,0 мкг/мл. Иными словами, получили охарактеризованный положительный контрольный образец, который вместе с исследуемыми пробами использовали в дальнейшей работе.
Пример 2. Выражение функции зависимости концентрации RNP-комплекса вируса бешенства в сырье для антирабических вакцин и порогового цикла реакции амплификации.
Составляли контрольную панель положительных стандартов RNP-комплекса вируса бешенства штамма РВ-97, в качестве которых использовали очищенные препараты с концентрациями RNP-комплекса: 0,1; 0,5; 1,0; 2,0; 3,0; 4,0; 5,0 мкг/см3. В качестве отрицательного контроля применяли суспензию клеток ВНК-21, не зараженную вирусом бешенства.
Из всех стандартных положительных и отрицательного контролен выделяли нуклеиновую кислоту. Для этого применяли метод твердофазной экстракции с применением частиц гидроокиси кремния диаметром 250-300 нм, обработанных 4 М раствором гидроксидом натрия.
К 100 мкл исследуемого образца добавляли 500 мкл хаотропного денатурирующего агента 4 М раствора ГТЦ, перемешивали на вортексе в течение 5 минут для денатурирования белковых составляющих. К полученному лизату добавляли 25 мкл суспензии частиц диоксида кремния диаметром 250-300 нм, обработанных гидроксидом натрия. Проводили инкубирование в процессе медленного перемешивания на вортексе в течение 5 мин. Суспензию центрифугировали при 250 g, удаляли супернатант, и проводили промывание частиц от ингибиторов реакции амплификации с помощью 4 М раствора ГТЦ, добавляя его к осадку в объеме 350 мкл. Смесь осаждали при 250 g, удаляли надосадочную жидкость. Проводили отмывание частиц сорбента с применением 80%-ного раствора пропанола-2 в объеме по 500 мкл. Осуществляли центрифугирование содержимого пробирки при тех же условиях. Данную процедуру проводили дважды. Осадок частиц сорбента с адсорбированными молекулами нуклеиновой кислоты высушивали от остатков спирта с помощью сухого твердотельного термостата при температуре 60±2°С в течение 8-10 мин. К высушенному осадку добавляли 10 мкл ТЕ-буфера (10 мМ трис(оксиметил)аминометан, 1 мМ этилендиамин-тетраацетат, рН 7,3), свободного от РНКаз и ионов Mg, прогревали содержимое пробирки при температуре 60±2°С в течение 3 минут для получения элюата. Содержимое пробирки центрифугировали при 11000 g в течение 1 минуты, и отбирали 10-кратный экстракт нуклеиновой кислоты образцов.
На следующем этапе исследования с помощью спектрального анализа в излучении ультрафиолетового света проводили оценку степени чистоты полученных 10-кратных элюатов. Образцы сканировали в кварцевых кюветах (l=10 мм) при температуре 23-25°С и регистрировали значения экстинкции в диапазоне от 205 до 325 нм через каждые 2 нм, производя запись спектра поглощения РНК с помощью компьютерной программы Specrtrum v. 5.0 (фиг. 3, таблица 7).
По результатам анализа стандартов выявили, что значения OD205-259 и OD261-325 не превышали OD260, что является признаком высокой степени чистоты полученных элюатов РНК (n=3). Из данных спектрального исследования стандартов отмечали отсутствие выраженных пиков на графиках (фиг. 3) при длинах волны 205, 235, 270, 280 и 320 нм, что свидетельствовало о практически полном отсутствии загрязнения экстрактов РНК примесями фосфолипидов, полисахаридов и остатков ГТЦ, полипептидов и крупных конгломератов, соответственно. Значения коэффициентов экстинкции R1 для положительных контролей приближены к норме 2,000 (R1=1,985-2,000), что подтверждало наличие лишь следовых количеств примесей белка. Деградации нуклеиновой кислоты и признаков наличия свободных нуклеотидов в элюатах не отмечалось, поскольку R1 не превышал 2,000. Экстракты вирусной РНК не загрязнены полисахаридами и остатками ГТЦ, поскольку значения коэффициента экстинкции R2 приближены к норме 2,000 и соответствовали 1,994-2,000. Учитывая, что при замещении 1% РНК на углеводы значение R2 уменьшается на 0,002, в полученных экстрактах наличие полисахаридных примесей составило не более 3%, что допустимо. Разрушения РНК в экстрактах данным методом не обнаружено. Следовательно, полученные экстракты, выделенные из контролей, характеризовались высокой степенью чистоты [10].
После оценки степени чистоты полученных контролей проводили обратную транскрипцию и реакцию амплификации. Для постановки реакций готовили смеси компонентов в соответствии с данными таблиц 1, 2. Постановку реакций осуществляли при температурных и временных параметрах, сведения о которых представлены в таблице 5.
Результаты реакции амплификации анализировали, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям пороговых циклов амплификации Ct, определенных с помощью пересечения пороговой линии и логистической функции вида Fl=f(Ct) методом гибридизационно-флуоресцентной детекции ДНК-ампликонов. Учет результатов реакции производили на каждом цикле. Детектирующий амплификатор определял уровень флуоресценции, и строил кинетическую кривую в координатах: уровень флуоресценции - цикл реакции амплификации. Для положительных контрольных образцов были сформированы логистические графики, полученные при анализе флуоресценции красителя FAM, входящего в состав молекулярного зонда. Для отрицательного контрольного образца график не выходил за пределы пороговой линии и не имел экспоненциальной зависимости.
Устанавливали зависимость между пороговым циклом амплификации и концентрацией RNP-комплекса вируса бешенства в сырье для антирабической вакцины в процессе построения графика линейной функции. Полученные данные отражены на фиг. 4 и выражены в виде функции: CRNP ВБ=-0,3002 × Ct + 8,81. Оценивали величину эффективности реакции амплификации (Е) по формуле: Е=(10-1/к-1) (к брали из функции Ct=-3,3263 × CRNP ВБ + 29,38) (Фиг. 5), а также достоверность аппроксимации (R2). Эффективность реакции амплификации составила 99,83%, достоверность аппроксимации - 0,9984, что соответствовало общепринятым требованиям, предъявляемым к молекулярно-биологическим тест-системам [19].
Пример 3. Тестирование способа опосредованного определения концентрации RNP-комплекса вируса бешенства в сырье для антирабических вакцин с применением метода амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Для анализа использовали 6 суспензий культурального вируса бешенства производственных штаммов РВ-97 и ВНИИЗЖ с концентрациями RNP-комплекса 0,60; 0,95; 1,50; 1,75; 2,00; 2,25 мкг/см3, соответственно (пробы №1-6). В качестве положительного контроля применяли суспензию культурального вируса бешенства штамма РВ-97 с концентрацией RNP-комплекса 1,00 мкг/см3. Отрицательным контролем служила суспензия клеток ВНК-21, не зараженная вирусом бешенства. Испытуемые пробы и контрольные образцы исследовали в трех повторностях. Этапы экстрагирования РНК, оценки степени их чистоты и постановку реакции амплификации с последующей гибридизационно-флуоресцентной детекцией проводили, как описано в примере 2.
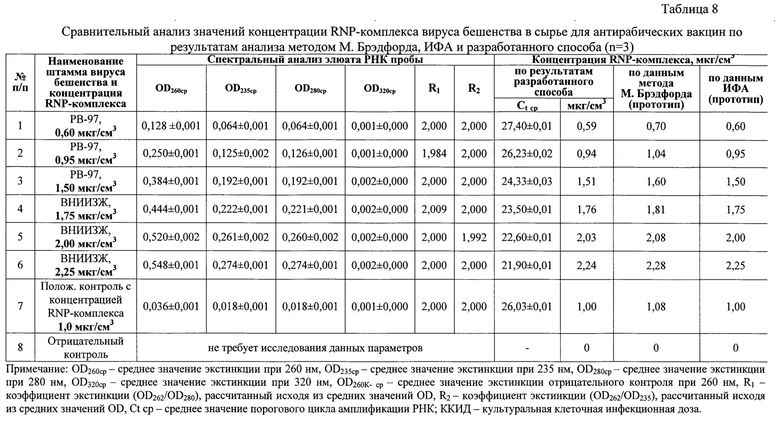
Из данных спектрального анализа (таблица 8) шести элюатов РНК проб вируса бешенства следует, что средние значения экстинкции при длинах волны 205-259 и 261-325 нм не превышали OD260, иными словами, элюаты РНК характеризовались высокой степенью чистоты. Экстракты не были контаминированы примесями фосфолипидов, полисахаридов и остатков ГТЦ, полипептидов и крупных конгломератов, поскольку на спектрограммах отсутствовали выраженные пики при λ 205, 235, 270, 280 и 320 нм, соответственно. Коэффициент экстинкции R1 для проб вируса бешенства находился в диапазоне 1,984-2,009, что приближено к значению нормы 2,000 и означало высокую степень чистоты элюатов вирусной РНК, практически полное отсутствие белка. Коэффициент экстинкции R2 для проб вируса соответствовали значениям 1,992-2,000, что близко к норме 2,000 и обуславливало высокую чистоту экстрактов. Таким образом, полученные элюаты нуклеиновой кислоты удовлетворяли требованиям чистоты [10], и их можно использовать для дальнейшей работы.
С полученными экстрактами РНК вируса бешенства проводят реакцию обратной транскрипции и амплификации кДНК с последующей детекцией ампликонов гибридизационно-флуоресцентным методом. С помощью разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства с помощью амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов получали данные, которые отражены в таблице 8. Из указанной таблицы видно, что средние значения пороговых циклов амплификации для проб №1-6 составляли 27,40±0,01, 26,23±0,02, 24,33±0,03, 23,50±0,01, 22,60±0,01, 21,90±0,01, соответственно. Пользуясь разработанной линейной функцией CRNP ВБ=-0,3002 × Сt + 8,81 (R2=0,9984, Е=99,83%), рассчитали средние значения концентрации RNP-комплекса вируса бешенства для проб №1-6, которые составили 0,59; 0,94; 1,51; 1,76; 2,03; 2,24 мкг/см3, соответственно. Для положительного контроля значение порогового цикла амплификации составило 26,03±0,01, что соответствовало концентрации RNP-комплекса 1,0 мкг/см3. Для отрицательного контроля логистические графики не были сформированы, что означало отсутствие вируса бешенства в данном контроле.
Исследуемые пробы и контроли также тестировали классическим методом М. Брэдфорда и в непрямом количественном варианте ИФА. Выявили, что данные, полученные с помощью разработанного способа, коррелировали с методом М. Брэдфорда на 84,4-97,6% и с методом ИФА на 97,3-99,55% (n=6). Заниженные значения корреляции в первом случае (до 84,4%) объясняются тем, что метод М. Брэдфорда имеет невысокие показатели чувствительности при концентрации белка менее 2 мкг/мл. Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения концентрации RNP-комплекса в сырье для антирабических вакцин. Иными словами, разработанный способ позволяет опосредованно определять концентрацию иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Пример 4. Определение степени достоверности опосредованной оценки концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Для анализа использовали 304 суспензии культурального вируса бешенства производственных штаммов РВ-97 и ВНИИЗЖ с концентрацией RNP-комплекса 0,1-5,0 мкг/см3. В качестве положительного контроля применяли суспензию культурального вируса бешенства штамма РВ-97 с концентрацией RNP-комплекса 1,0 мкг/см3. В качестве отрицательного контроля применяли суспензию клеток ВНК-21, не зараженную вирусом бешенства. Испытуемые пробы и контрольные образцы исследовали в трех повторностях. Этапы элюирования РНК, оценки степени их чистоты полученных экстрактов и постановку обратной транскрипции и реакции амплификации с последующей гибридизационно-флуоресцентной детекцией проводили, как описано в примере 2.
Из данных спектрального анализа 304 элюатов РНК проб вируса бешенства следует, что средние значения оптической плотности при длинах волны 205-259 и 261-325 нм не превышали OD260, иными словами, экстракты РНК характеризовались высокой степенью чистоты. Элюаты не были контаминированы примесями фосфолипидов, полисахаридов и остатков ГТЦ, полипептидов и крупных конгломератов, так как на спектрограммах отсутствовали выраженные пики при длинах волны 205, 235, 270, 280 и 320 нм, соответственно. Коэффициенты экстинкции R1 для проб вируса бешенства составляли 1,995-2,002, что приближено к значению нормы 2,000 и означало высокую степень чистоты элюатов РНК, практически полное отсутствие белка. Коэффициенты экстинкции R2 для проб вируса бешенства соответствовали значениям 1,997-2,000, что близко к норме 2,000 и обуславливало высокую чистоту элюатов. Таким образом, полученные препараты удовлетворяли требованиям чистоты [10], и их можно использовать в дальнейшей работе.
С полученными экстрактами РНК вируса бешенства проводили обратную транскрипцию и реакцию амплификации кДНК вируса бешенства с последующей детекцией продуктов анализа с помощью гибридизационно-флуоресцентного метода. С помощью разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин получили данные, представленные в таблице 9. Интерпретацию результатов проводили, пользуясь разработанной показательной функцией
CRNP ВБ=-0,3002 × Сt + 8,81 (R2=0,9984, Е=99,83%) с получением значений концентрации RNP-комплекса для каждой из 304 исследуемых проб. Для положительного контроля значение порогового цикла амплификации составило 26,03±0,01, что соответствовало концентрации RNP-комплекса вируса бешенства 1,0 мкг/см3. Для отрицательного контроля сигмоиды не были сформированы, что означало отсутствие вируса бешенства в данных контроле.
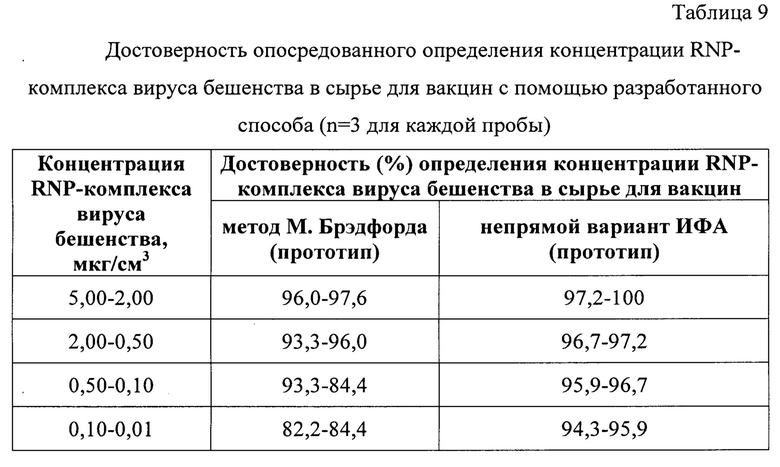
Представленные пробы и контроли также тестировали колориметрическими методами М. Брэдфорда и ИФА. Выявили, что данные, полученные с помощью разработанного способа, коррелировали с методом М. Брэдфорда на 96,0-97,6% для 2,00-5,00 мкг/см3 (n=76), на 93,3-96,0% для 2,00-0,50 мкг/см3 (n=76), на 93,3-84,4% для 0,10-0,50 мкг/см3 (n=76), на 82,2-84,4% для 0,10-0,01 мкг/см3 (n=76). Заниженные значения корреляции (ниже 96,0%) объясняются тем, что метод М. Брэдфорда имеет невысокие показатели чувствительности при концентрации белка менее 2 мкг/мл. Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения концентрации RNP-комплекса в сырье для антирабических вакцин по сравнению с методом М. Брэдфорда.
Данные, полученные с помощью разработанного способа, коррелировали с методом непрямого количественного варианта ИФА на 97,2-100,0% для 2,00-5,00 мкг/см3 (n=76), на 96,7-97,2% для 2,00-0,50 мкг/см3 (n=76), на 95,9-96,7% для 0,10-0,50 мкг/см3 (n=76), на 94,3-95,6% для 0,10-0,01 мкг/см3 (n=76). Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения концентрации RNP-комплекса в сырье для антирабических вакцин по сравнению с методом ИФА.
Иными словами, разработанный способ позволяет с высокой точностью опосредованно определять концентрацию иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Пример 5. Определение аналитической чувствительности разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Для определения аналитической чувствительности разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов подготавливали панель положительных стандартов вируса бешенства со следующими концентрациями RNP-комплекса вируса бешенства: 0,001; 0,005; 0,01; 0,1; 0,5; 1,0; 2,0; 5,0 мкг/см3. Испытуемые контрольные образцы исследовали в 7 повторностях. Все этапы работы осуществляли, как описано в примере 2. Проведен сравнительный анализ концентраций RNP-комплекса вируса бешенства с помощью разработанного способа в сравнении с данными высокоточного прототипного метода (непрямой количественный сэндвич-вариант ИФА). Выявлено, что с достоверностью ≥97,2% разработанным способом определены концентрации RNP-комплекса вируса бешенства со значениями 0,50-5,00 мкг/см3, с достоверностью 94,3-97,2% - с CRNP ВБ 0,01-0,50 мкг/см3, с достоверностью 90,3-94,3% - с CRNP ВБ 0,005-0,010 мкг/см3. Пробы, содержащие 0,001 мкг/см3, характеризовались завышенными значениями пороговых циклов с достоверностью определения 87,7-89,4%. Для производства антирабических вакцин применяют вируссодержащие суспензии с концентрацией RNP-комплекса ≥0,5 мкг/см3. Для этой группы проб степень достоверности составляет ≥94,3%.
По результатам исследования выявлено, что аналитическая чувствительность разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов составляет не менее 0,01 мкг/см3 с достоверностью результатов исследования 94,3% и более.
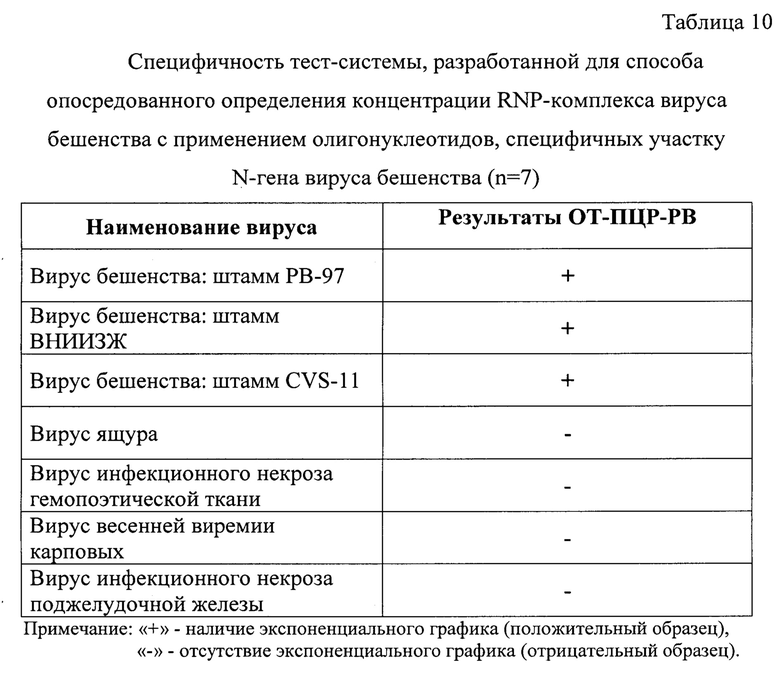
Пример 6. Оценка специфичности разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Для оценки специфичности разработанного способа опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов, исследовали суспензии вируса бешенства штаммов РВ-97, ВНИИЗЖ, CVS-11, вируса ящура штамма Азия-1/Таджикистан/2011, инфекционного некроза гемопоэтической ткани лососевых рыб, весенней виремии карповых, инфекционного некроза поджелудочной железы. Титр инфекционной активности вирусов в суспензиях составлял 7,0 lg ККИД (или ТЦД)50/см3 . Исследования проводили в трех повторностях.
Этапы постановки анализа проводили, как описано в примере 2. Выделенные элюаты нуклеиновой кислоты характеризовались высокими показателями степени чистоты, так как коэффициенты экстинкции R1 и R2 находились в диапазоне 1,997-2,001, что соответствовало требованиям [10].
В ходе проведения реакции амплификации с последующей детекцией ампликонов методом гибридизационно-флуоресцентного анализа с олигонуклеотидами, специфичными для участка N-гена вируса бешенства, наблюдали построение логистических графиков накопления флуоресцентного сигнала только для штаммов вируса бешенства. Для проб, содержащих другие вирусы, не наблюдалось формирования графиков экспоненты, и они не выходили за пороговый уровень флуоресцентного сигнала (0,0125 у.е.). Таким образом, разработанный способ является специфичным, по отношению к вирусу бешенства и может быть использован как высокоспецифичный для опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов.
Основными преимуществами предлагаемого изобретения является сокращение времени проведения анализа сырья для антирабических вакцин по определению концентрации RNP-комплекса вируса бешенства до 3 ч по сравнению с ИФА; исключение вероятности контаминации; повышение степени чистоты элюата РНК вируса бешенства; увеличение чувствительности и специфичности анализа за счет применения высокоспецифичных оригинальных олигонуклеотидов. В предлагаемом изобретении между концентрацией RNP-комплекса вируса бешенства и пороговым циклом амплификации установлена зависимость CRNP ВБ=-0,3002 × Сt + 8,81, отраженная в виде линейной функции с высокими достоверностью аппроксимации (R2=0,9984) и эффективностью амплификации 99,83%. Разработанная математическая модель дает возможность опосредованно определять концентрацию RNP-комплекса вируса бешенства в сырье для производства антирабических вакцин.
Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности опосредованно определять концентрацию RNP-комплекса в сырье для антирабических вакцин на основе реакции амплификации с применением оригинальных специфических олигонуклеотидов с последующей детекцией ампликонов гибридизационно-флуоресцентным методом и применением разработанной математической модели.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов»:
1. OIE. Manual of diagnostic tests and vaccines for terrestrial animals. - Paris, 2018. - Chapter 3.1.17. - P. 578-612.
2. ICTV Taxonomy history: Rabies lyssavirus URL: https://talk.ictvonline.Org/taxonomy/p/taxonomy history?taxnode_id=20161105&taxa_name=Rabies%201yssavirus (Дата обращения: 11.04.2020).
3. Baltimore D. Expression of animal virus genomes (англ.)// Microbiology and Molecular Biology Reviews (англ.) русск.: journal. - American Society for Microbiology (англ.). - 1971. - Vol. 35, no. 3. - P. 235-241.
4. Груздев K.H., Метлин A.E. Бешенство животных. - Владимир: ФГБУ «ВНИИЗЖ», 2019. - 394 с.
5. Sokol F. Structural Proteins of Rabies Virus / F. Sokol, D. Stancek, H. Koprowski // J. of Virology. - V.7, №2. - Feb.1971. - P. 241-249.
6. Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. - 1976. - V. 72. - P. 248-254.
7. Разработка методов оценки оральной антирабической вакцинации животных: автореферат дис.… кандидата биологических наук: 03.02.02 / Сухарьков Андрей Юрьевич. - Владимир, 2014. - 25 с.
8. Dietzschold В. Induction of protective immunity against rabies by immunization with rabies virus ribonucleoprotein / B. Dietzschold, H.H. Wang, C.E. Rupprecht, E. Celis, M. Tollis, E. Heber-Katz, H. Koprowski // Proc. Natl. Acad Sci USA. - 1987. - V. 84. - P. 9165-9169.
9. Основные методы выделения ДНК [Электронный ресурс] / URL: https://genotest.me/osnovnye-metody-vydeleniya-dnk/ (Дата обращения: 30.03.2020).
10. The Analysis of DNA or RNA using Its Wavelengths: 230 nm, 260 nm, 280 nm. Bioteachnology.com [Электронный ресурс] / URL: http://bioteachnology.com/dna/analysis-dna-rna-wavelengths-230-260-280-nm. (Дата обращения 02.11.2019).
11. GenBank. [Электронный ресурс] / URL: http://www.ncbi.nlm.nih.gov. (Дата обращения: 11.12.2019).
12. Deiman В., van Aarle P., Sillekens P. Characteristics and applications of nucleic acid sequence based amplification // Mol. Biotech. - 2002. - Vol.20. - P. 163-179.
13. Sooknanan R., van Gemen В., Malek L. Nucleis acid sequence-based amplification // Molecular methods for virus detection-London: Academic press, 1995. - P. 261-285.
14. SantaLucia J. J., Hicks D. The thermodynamics of DNA structural motifs // Annual Review of Biophysics and Biomolecular Structure: journal. - 2004. - Vol. 33. - P. 11-14.
15. The RNA Institute college of arts and scienceuniversity at Albany. The mfold Web Server. RNA Folding Form. [Электронный ресурс] / URL: http://bioinfo.math.rpi.edu/~mfold/dna/forml.cgi (Дата обращения: 11.01.2020).
16. Молекулярный олигокалькулятор. [Электронный ресурс] / URL: http://www.bio.bsu.by/molbiol/oligocalc.html. (Дата обращения: 19.12.2019).
17. Nicolas von Ahsen, Carl Т. Wittwer, Ekkehard Schiitz. Oligonucleotide melting temperatures under per conditions: nearest-neighbor corrections for Mg, deoxynucleotide triphosphate, and dimethyl sulfoxide concentrations with comparison to alternative empirical formulas (англ.) // Clinical Chemistry: journal. - 2001. - Vol. 47, no. 11.-P. 1956-1961.
18. SantaLucia J. J. A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics (англ.) // Proceedings of the National Academy of Sciences of the United States of America: journal. -1998. - Vol.95, no. 4. - P. 1460-1465.
19. ГОСТ Р 57175-2016 Требования к качеству и безопасности ПЦР-наборов, проведению исследований и испытаний с использованием метода ПЦР при идентификации целевых таксонов микрофлоры, растений и генетически модифицированных организмов.



Изобретение относится к области биотехнологии и производству антирабических вакцин, а именно к способу опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов с применением оригинальных специфических олигонуклеотидов и разработанной линейной функции зависимости величины порогового цикла амплификации и концентрации RNP-комплекса. Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности определять концентрации RNP-комплекса вируса бешенства в сырье для антирабических вакцин. 8 з.п. ф-лы, 6 ил., 10 табл., 6 пр.
1. Способ опосредованного определения концентрации иммуногенного RNP-комплекса вируса бешенства в сырье для антирабических вакцин методом амплификации и гибридизационно-флуоресцентной детекции ДНК-ампликонов, включающий следующие стадии:
- осаждение суспензии клеток линии ВНК-21, в которых репродуцирован вируса бешенства;
- получение вируссодержащей суспензии осажденных клеток в фосфатно-солевом буферном растворе;
- твердофазная экстракция нуклеиновой кислоты вируса бешенства с применением частиц диоксида кремния диаметром 250-300 нм, обработанных 4 М гидроксидом натрия;
- обратная транскрипция РНК вируса бешенства;
- амплификации специфического фрагмента N-гена РНК вируса бешенства с применение оригинальных специфических олигонуклеотидов;
- детектирование ампликонов с помощью флуоресцентного свечения и отображения накопления сигнала в виде логистической функции;
- определение значений пороговых циклов амплификации;
- установление зависимости между пороговым циклом амплификации и концентрацией RNP-комплекса вируса бешенства в сырье для антирабических вакцин в виде линейной функции;
- определение величины эффективности реакции амплификации и достоверность аппроксимации;
- расчет значения концентрации RNP-комплекса вируса бешенства (CRNP ВБ) в образцах сырья для антирабических вакцин на основе разработанной математической модели CRNP ВБ=-0,3002 × Ct + 8,81, где Ct обозначает величину парогового цикла амплификации, R2 озбозначает достоверность аппроксимации, а E обозначает эффективность реакции амплификации;
и положительный контрольный препарат RNP-комплекса вируса бешенства для разработанного способа получают с применением 25 мкг/мл апротинина и ультрацентрифугирования белков вируса бешенства в градиенте плотности хлористого цезия от 17 до 26% при 65000 g в течение 2 ч и переосаждением RNP-комплекса в 1/15 М фосфатно-солевом буферном растворе при 60000 g в течение 1 ч.
2. Способ по п. 1, отличающийся тем, что для элюирования РНК вируса бешенства проводят твердофазную экстракцию нуклеиновой кислоты с применением частиц диоксида кремния диаметром 250-300 нм, обработанных 4 М гидроксидом натрия.
3. Способ по п. 1, отличающийся тем, что определяют степень чистоты элюатов нуклеиновой кислоты с помощью спектрального анализа при длинах волны в диапазоне 205-325 нм.
4. Способ по п. 1, отличающийся тем, что увеличивается чувствительность и специфичность анализа за счет применения высокоспецифичных оригинальных олигонуклеотидных праймеров и молекулярного зонда.
5. Способ по п. 1, отличающийся тем, что смесь компонентов для проведения обратной транскрипции включает в свой состав следующие компоненты: гексамеры (шестинуклеотидные универсальные праймеры) - 5 пМ, смесь дезоксирибонуклеозидтрифосфатов (dNTP) - по 0,25 мМ, MgCl2 - 2 мМ, буферный раствор (10-кратный) - 10% от объема реакционной смеси,
AMV-ревертаза - 10 ед, РНазин - 10 ед, деионизированная вода - 10,0% от объема реакционной смеси, соотношение РНК и реакционной смеси по объему 1/10.
6. Способ по п. 1, отличающийся тем, что смесь компонентов для проведения реакции амплификации с гибридизационно-флуоресцентной детекцией ампликонов включает в свой состав следующие компоненты: F-RabV-RNP-праймер, R-RabV-RNP-праймер и P-RabV-RNP-FAM/RTQ1-зонд (каждый) - по 5 пМ, смесь дезоксирибонуклеозидтрифосфатов (dNTP) - по 0,20 мМ, MgCl2 - 2 мМ, буферный раствор Colorless Flexi (х 5) - 20% от объема реакционной смеси, Pfu-ДНК-зависимая ДНК-полимераза - 10 ед, деионизированная вода - 66,8% от объема реакционной смеси, соотношение ДНК и реакционной смеси по объему 1/5.
7. Способ по п. 1, отличающийся тем, что обратную транскрипцию и реакцию амплификации с гибридизационно-флуоресцентной детекцией ампликонов проводят с соблюдением следующих режимов:
- обратная транскрипция (температура 42°С в течение 15 мин);
- предварительная денатурация (температура 94°С в течение 3 мин);
- реакция амплификации: денатурация (температура 94°С в течение 10 с), отжиг олигонуклеотидов (температура 53°С в течение 30 с), элонгация (температура 74°С в течение 60 с). Гибридизационно-флуоресцентная детекция проводится на подэтапе элонгации ДНК вируса бешенства.
8. Способ по п. 1, отличающийся тем, что время проведения анализа вируссодержащего сырья для производства антирабических вакцин по определению концентрации RNP-комплекса сокращается до 3 ч.
9. Способ по п. 1, отличающийся тем, что достоверность определения концентрации RNP-комплекса ≥97,2% для значений 0,50-5,00 мкг/см3, 94,3-97,2% - для концентраций 0,01-0,50 мкг/см3, 90,3-94,3% - для концентраций 0,005-0,010 мкг/см3.
| Мухамеджанова А | |||
| Г | |||
| и др., Получение антигена вируса бешенства и оценка его активности и специфичности, Ученые записки Казанской государственной академии ветеринарной медицины им | |||
| Н.Э | |||
| Баумана, 2018, Т | |||
| Стеклографический печатный станок с ножной педалью | 1922 |
|
SU236A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Kouznetzoff A., Buckle M., Tordo N., Identification of a region of the rabies virus N protein involved in direct | |||

