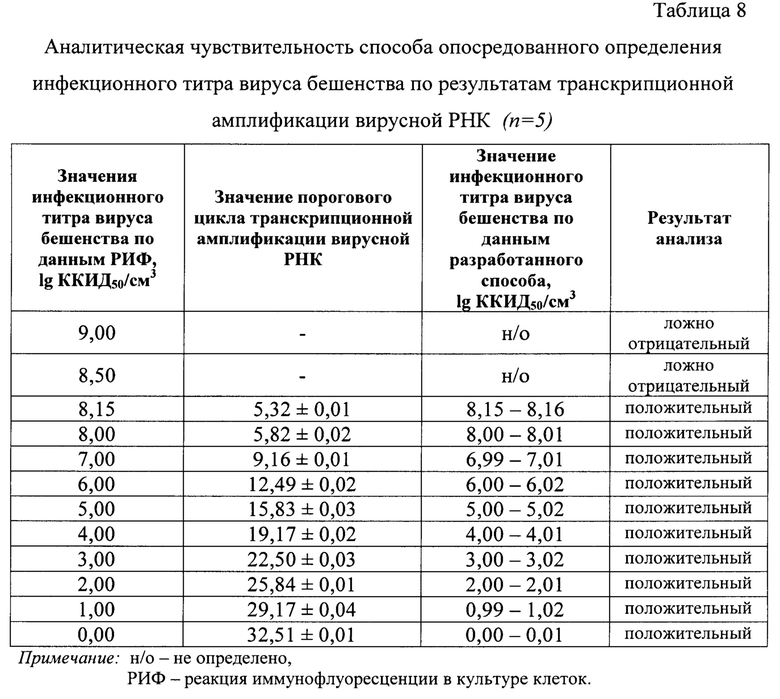
Изобретение относится к области биотехнологии и производству антирабических вакцин, а именно к способу опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для вакцины при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии и разработанной логарифмической функции зависимости величины порогового цикла амплификации РНК и инфекционного титра вируса бешенства.
Бешенство - вирусное заболевание животных и человека, характеризующаяся признаками энцефаломиелита и абсолютной летальностью [1]. Классический возбудитель бешенства Rabies virus (RABV) - нейротропный РНК-(-) содержащий вирус, который принадлежит реалму Riboviria царству Orihornavirae типу Negarnaviricota классу Monjiviricetes порядку Mononegavirales семейству Rhabdoviridae роду Lyssavirus (Rabies lyssa virus) генотипу Rabies virus (RABV) [2, 3].
Вирионы вируса бешенства имеют пулевидную форму длиной 100-300 нм, диаметром 45-100 нм. На наружной поверхности вирусной частицы имеются выступы в виде шипов длиной 10-12 нм, которые прикреплены к двуслойной липидной оболочке [1]. Геном RABV составляет около 12000 нуклеотидных оснований (н.о.) и включает 5 последовательно расположенных генов 3'-N-P-M-G-L-5' (N - нуклеопротеин, G - гликопротеин, Р - фосфопротеин, М - матриксный белок, L - РНК-зависимая РНК-полимераза), кодирующих соответственно 5 белков и лидерную РНК длиной 50 н.о. Гены разделены некодирующими областями. Наиболее консервативным является N-ген, наибольшие изменения, происходящие в изолятах и штаммах вируса бешенства, отмечаются в G-гене [4, 5].
Бешенство приводит к большим экономическим потерям, которые связаны с падежом сельскохозяйственных животных, ликвидацией последствий вспышек заболевания, проведением профилактических и карантинных мероприятий, регулированием численности диких плотоядных животных, отловом бродячих кошек и собак и осуществлением лабораторно-диагностических исследований. Система мер для борьбы с бешенством и его профилактики предусматривает иммунизацию домашних, сельскохозяйственных и диких плотоядных, а также контроль уровня напряженности поствакцинального иммунитета [1, 4].
Иммунизацию домашних и сельскохозяйственных целевых животных проводят с применением инактивированных антирабических вакцин. Для формирования иммунитета у диких плотоядных животных преимущественно используют живые аттенуированные антирабические вакцины [4]. При изготовлении вакцин против бешенства вирусосодержащую суспензию исследуют на определение инфекционного титра вируса бешенства для оценки его активности в клетках. В 1,0 см3 суспензии вируса бешенства определяют количество клеточных культуральных инфекционных доз, вызывающих 50%-ное поражение клеток (ККИД50/см3), что фактически отражает концентрацию полных вирусных частиц, содержащих РНК.
В соответствии с требованиями МЭБ (OIE) для определения инфекционного титра вируса бешенства применяют метод титрования в монослойной клеточной линии почки новорожденного сирийского хомячка ВНК-21 с последующим окрашиванием монослоя с помощью иммуноглобулина G, меченого флуоресцеинизотиоцианатом (ФИТЦ) [1, 4]. Этот метод позволяет рассчитывать минимальную дозу вируса, способную вызвать лизис 50% клеток (прототип). Указанный метод зарекомендовал себя, однако он имеет некоторые недостатки: 1) длительная процедура анализа (не менее 3 суток), связанная с репродукцией вируса в клетках; 2) определенная степень субъективности при оценке результатов исследования; 3) высокая стоимость клеточной линии как тест-системы и затраты на ее поддержание; 4) высокая вероятность риска контаминации клеточной линии; 5) качество приготовленного конъюгата (специфический иммуноглобулин G, меченый ФИТЦ).
В связи с этим целесообразно провести поиск способа определения инфекционного титра вируса бешенства в неинактивированном сырье для вакцины на основе транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Существует метод, моделирующий в условиях in vitro репликацию ретровирусов, где в качестве мишени для амплификации служит одноцепочечная молекула РНК. Методика, названная «Self-Sustained Sequence Replication)) или 3SR, базируется на конкурентном действии трех ферментов, участвующих в ретровирусной репликации - ревертазы, РНКазы Н и ДНК-зависимой РНК-полимеразы. Реакция протекает с участием специфических праймеров при постоянной температуре. В результате реакции амплификации за 60-90 минут образуется до 109-1012 копий РНК [6-17, 30].
Данный экспресс-метод является объективным, высокочувствительным и высокоспецифичным, более дешевым по сравнению с прототипом, характеризуется применением стандартизированных компонентов реакции, не создает ситуаций риска контаминации, поскольку пробирки закрыты на этапе детекции результатов анализа, отличается высокими значениями правильности и позволяет определять инфекционный титр вируса бешенства в неинактивированных вирусосодержащих суспензиях в течение 2-3 часов. Исходя из этого, целесообразно предложить новый способ опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Задачей настоящего изобретения является разработка высокочувствительного и высокоспецифичного экспресс-способа определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии с целью устранения вышеуказанных недостатков.
Данная задача решена благодаря созданию нового способа определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии. Предложенный способ позволяет: 1) сократить время проведения анализа вирусосодержащих суспензий для определения инфекционного титра вируса бешенства до 2-3 ч; 2) исключить вероятность контаминации; 3) повысить объективность анализа; 4) увеличить чистоту РНК вируса бешенства за счет применения магнитных частиц оксида железа (II, III) диаметром 300-320 нм в присутствии 4 М гуанидинизотиоцианата (ГГЦ); 5) увеличить специфичность анализа за счет применения высокоспецифичных оригинальных РНК-праймеров и молекулярного beacon, меченого флуорофором FAM (карбоксифлуоресцеином) (максимальная длина волны поглощения - 520 нм) и гасителем свечения RTQ1 (максимальная длина волны поглощения - 520 нм); 6) увеличить чувствительность анализа за счет амплификации только молекул вирусной РНК; 7) удешевить способ анализа за счет отсутствия использования клеточных культур в качестве тест-систем; 8) применение фермента Т7 ДНК-зависимой РНК-полимеразы позволяет повторно использовать молекулы комплементарной ДНК (кДНК), что приводит к увеличению концентрации РНК-ампликонов не менее, чем на 1 порядок по сравнению с концентрацией РНК-праймеров; 9) повысить достоверность проводимого анализа благодаря установлению зависимости между значениями инфекционного титра вируса бешенства (ТВБ) и пороговыми циклами амплификации РНК (Ct РНК), представленной в виде логарифмической функции: lg ТВБ=-0,2997 × Ct РНК + 9,7524 с высокой достоверностью аппроксимации (R2=0,9997) и эффективностью амплификации 99,41%. Предложенная модель позволяет количественно оценивать инфекционный титр вируса бешенства в неинактивированном сырье для антирабических вакцин.
Сущность изобретения отражена на графических изображениях:
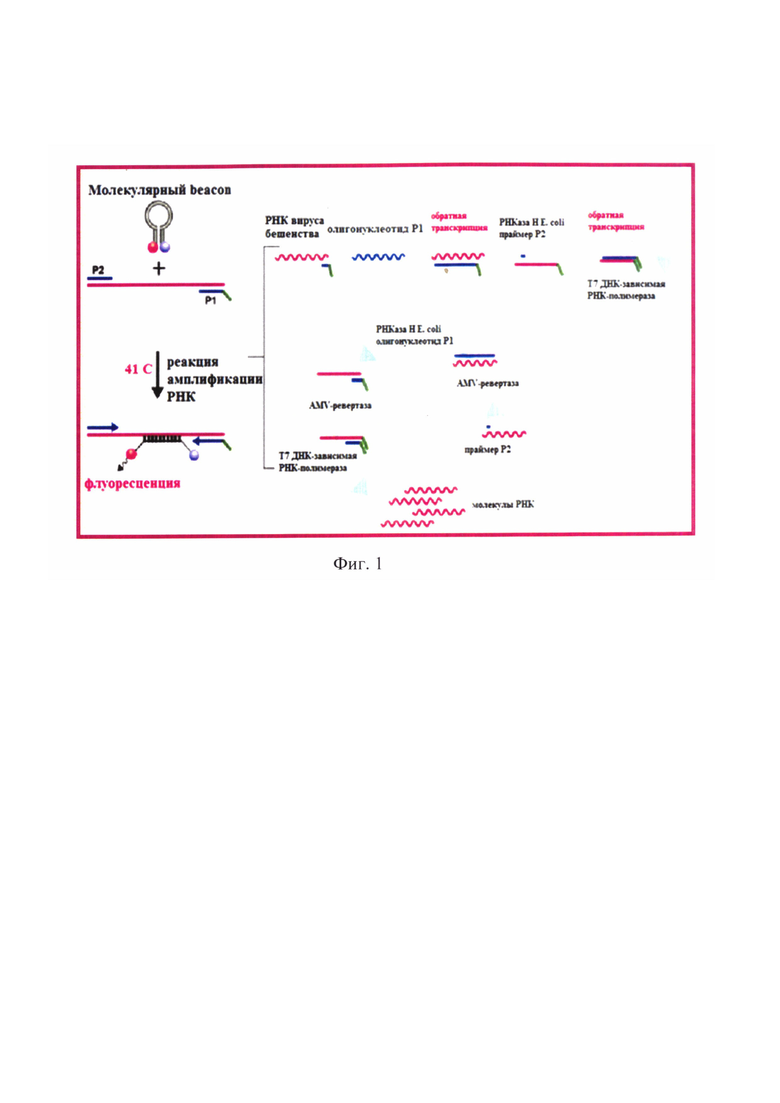
Фиг. 1 - Сущность реакции транскрипционной амплификации нуклеиновой кислоты вируса бешенства.
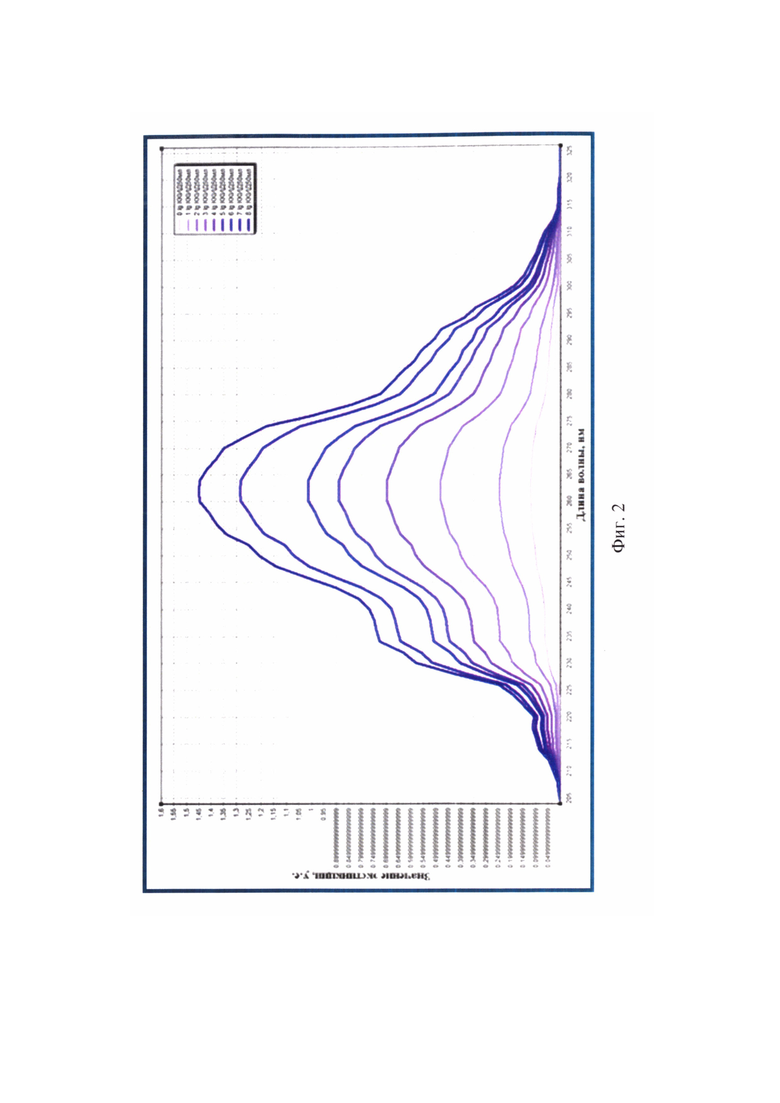
Фиг. 2 - Спектры поглощения экстрактов суммарной РНК вируса бешенства стандартных положительных контролей для оценки степени их чистоты при определении инфекционного титра вируса в исходных суспензиях (максимальные оптические сигналы для РНК вируса бешенства зарегистрированы при длине волны 260 нм) (n=3).
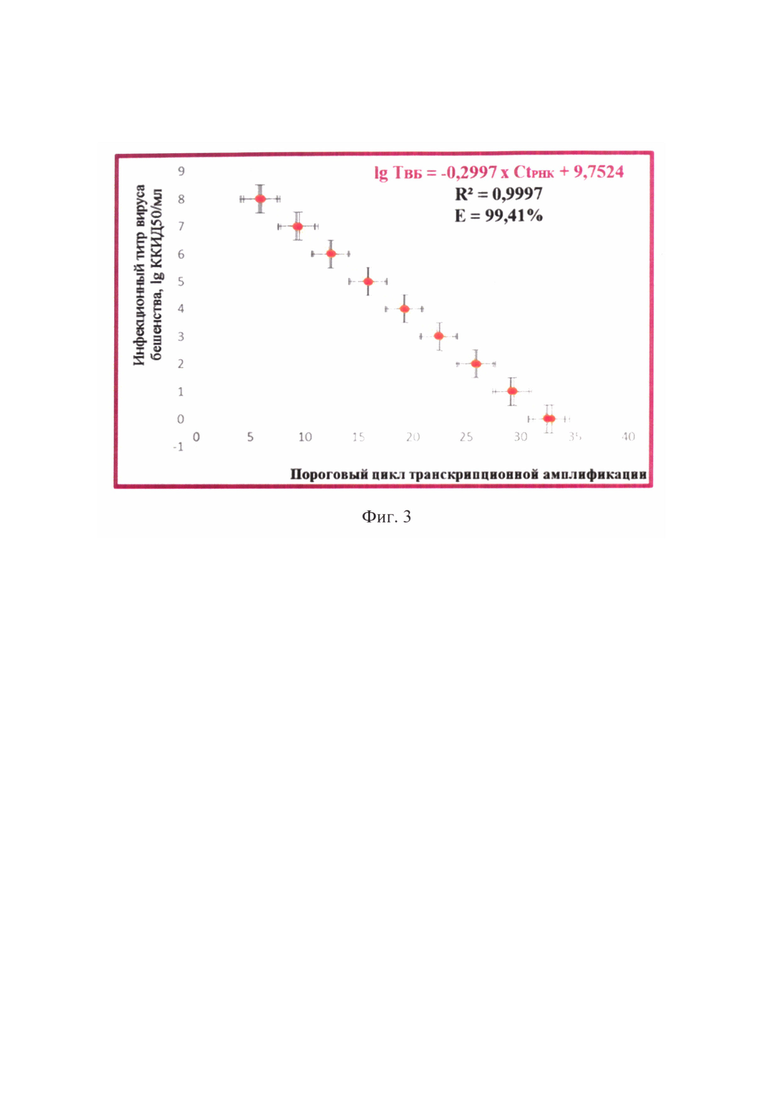
Фиг. 3 - Зависимость порогового цикла амплификации вирусной РНК и инфекционного титра вируса бешенства при транскрипционной амплификации вирусной нуклеиновой кислоты и детекции РНК-ампликонов с применением beacon-технологии (n=3, отмечены точки, отображающие средние значения пороговых циклов амплификации РНК).
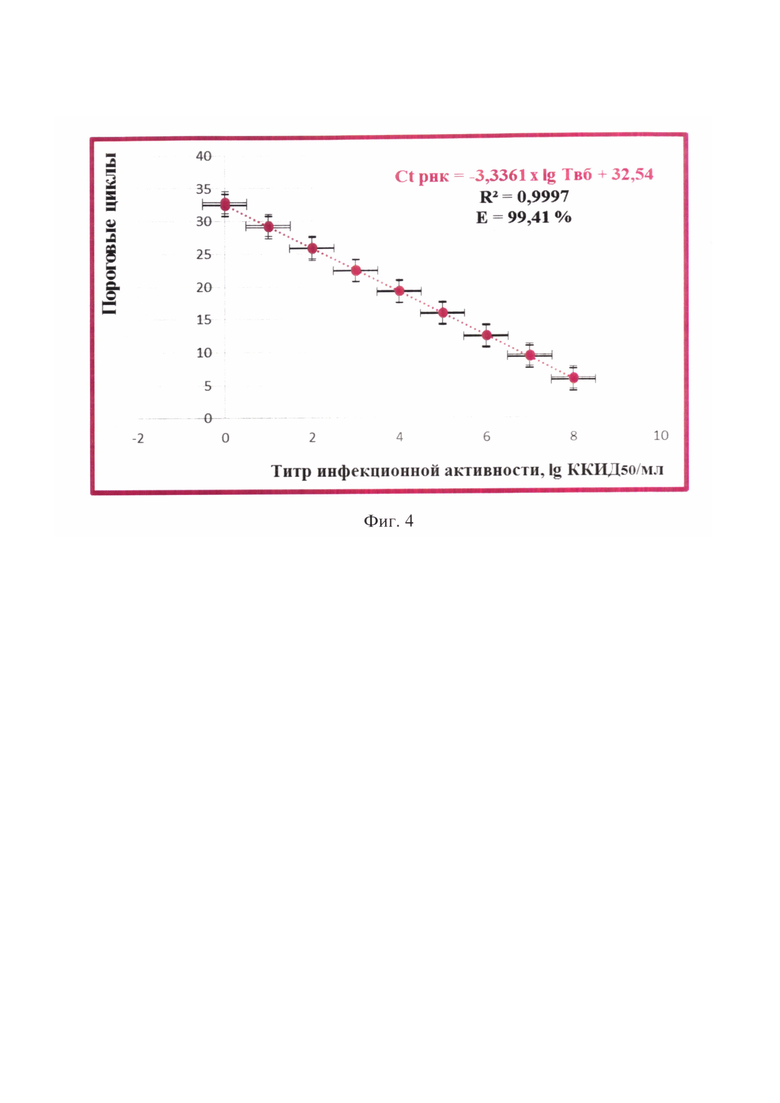
Фиг. 4 - Зависимость инфекционного титра вируса бешенства и порогового цикла амплификации вирусной РНК при детекции РНК-ампликонов с применением beacon-технологии (n=3, отмечены точки, отображающие средние значения пороговых циклов амплификации РНК).
Сущность изобретения заключается в новом подходе к определению инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии. Заявляемый способ основан на: 1) элюировании РНК вируса бешенства с применением магнитных частиц оксида железа (II, III) диаметром 300-320 нм в присутствии 4 М ГТЦ; 2) амплификации специфического фрагмента N-гена РНК вируса бешенства с применение специфического олигонуклеотида Р1 и обратного праймера Р2, а также молекулярного beacon, меченого флуорофором FAM и тушителем свечения RTQ1; 3) обнаружении РНК-ампликонов с помощью флуоресцентного свечения и отображения накопления сигнала в виде логистического графика; 4) расчете инфекционного титра вируса бешенства в сырье неинактивированном сырье для антирабических вакцин с применением логарифмической функции, выраженной в виде уравнения: lg ТВБ=-0,2997 × Ct РНК + 9,7524 при R2=0,9997 и эффективности амплификации (Е)=99,41%.
В настоящее время реакцию транскрипционной амплификации применяют для выявления генома возбудителей различных инфекционных агентов, в частности, возбудителя парагриппа-1, 2, 3, 4, гриппа птиц А, энтеровирусной инфекции, цитомегаловирусной инфекции, аспергиллеза, кандидоза, хламидиоза, микобактериоза КРС, сальмонеллеза животных и др. патогенов [6-17]. При этом для количественной оценки вирусной нагрузки суспензии, в частности, для определения инфекционного титра вируса бешенства ранее данный метод не применялся. Иными словами, сведений об аналогах предлагаемого способа определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин авторами не обнаружено.
Разработанный способ опосредованной оценки инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации нуклеиновой кислоты и детекции продуктов реакции с применением beacon-технологии по сравнению с прототипом отличается быстротой выполнения анализа, его объективностью, более высокой чувствительностью и специфичностью.
В отличие от прототипа разработанный способ включает следующие этапы: 1) элюирование РНК вируса бешенства с применением магнитных частиц оксида железа (II, III) диаметром 300-320 нм в присутствии 4 М ГТЦ; 2) денатурация РНК вируса бешенства; 3) амплификация специфического фрагмента N-гена РНК вируса бешенства с применением специфических оригинальных прямого Р1 и обратного Р2-олигонуклеотидов, а также молекулярного beacon, меченого флуорофором FAM и тушителем свечения RTQ1; 4) детекция РНК-ампликонов с помощью флуоресцентного свечения и отображение накопления сигнала в виде сигмоиды; 5) расчет инфекционного титра вируса бешенства с применением модели зависимости порогового цикла амплификации РНК для кривой флуоресценции и инфекционного титра вируса бешенства в виде логарифмической функции.
Применение разработанного способа позволит сократить время проведения анализа вирусосодержащего сырья для антирабических вакцин для определения инфекционного титра вируса бешенства до 2-3 ч; повысить степень объективности получаемого результата; исключить вероятность контаминации; увеличить чистоту экстракта РНК вируса бешенства; увеличить специфичность и чувствительность анализа; повторно использовать молекулы кДНК, что приводит к увеличению концентрации РНК-ампликонов не менее чем на 1 порядок по сравнению с концентрацией РНК-праймеров; использовать молекулярные beacon, меченые флуорофором FAM и тушителем свечения RTQ1; повысить достоверность и правильность проводимого анализа. Исходя их этого, актуально применять разработанный способ для определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин.
Ключевым элементом заявляемого способа является проведение этапов денатурации, обратной транскрипции, разрушения гетеродуплекса РНК/кДНК и амплификации вирусной РНК с последующим отжигом молекулярного beacon, детектирования пороговых циклов сигмоид для исследуемых проб и опосредованного определения инфекционного титра вируса бешенства с применением разработанной логарифмической модели зависимости порогового цикла амплификации РНК-мишени для логистической кривой накопления флуоресценции и титра вируса бешенства.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении способа амплификации РНК и детекции продуктов реакции с применением beacon-технологии и разработанной логарифмической функции зависимости величины порогового цикла транскрипционной амплификации РНК и инфекционного титра вируса бешенства для опосредованного определения титра вируса бешенства в неинактивированном сырье для антирабических вакцин.
Сущность предлагаемого изобретения пояснена на графическом материале - графике зависимости величины порогового цикла амплификации РНК (Ct РНК) и инфекционного титра вируса бешенства (ТВБ) (фиг. 3).
На первом этапе работы подготавливают панель положительных стандартов вируса бешенства, в качестве которых используют неинактивированные вирусосодержащие суспензии с инфекционными титрами: 0,0; 1,0; 2,0; 3,0; 4,0; 5,0; 6,0; 7,0; 8,0 lg ККИД50/см3. Для получения данных разведений применяли охарактеризованную очищенную с помощью фильтра на 1 мкм суспензию вируса бешенства с титром инфекционной активности 8,0 lg ККИД50/см3. Репродукция вируса проводилась в суспензионной клеточной линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH. В данном анализе взят широкий диапазон возможных значений инфекционного титра вируса бешенства, применяемого при исследовании. Для производства антирабических вакцин используют сырье с титрами ≥ 6,5 lg ККИД50/см3. Отрицательным контролем служила суспензия клеток ВНК-21/SUSP/ARRIAH, не зараженная вирусом бешенства.
Из всех стандартных положительных образцов и отрицательного контроля выделяют РНК вируса бешенства. Для этого взамен частиц кремнезема [18] применяют магнитные частицы оксида железа (II, III) диаметром 300-320 нм в присутствии хаотропного денатурирующего агента 4 М раствора ГТЦ.
К 100 мкл исследуемой суспензии добавляют 500 мкл 4 М раствора ГТЦ, перемешивают на вортексе на низкой скорости в течение 5 мин. К полученному лизату добавляют 30 мкл магнитных частиц оксида железа (II, III) и инкубируют в процессе перемешивания на вортексе на низкой скорости в течение 5 мин. Полученную суспензию осаждают с помощью магнитной подставки, удаляют надосадок и проводят промывание магнитных частиц от ингибиторов реакции амплификации РНК с помощью 4 М раствора ГТЦ, добавляя его к осадку в объеме 400 мкл. Смесь перемешивают на вортексе с высокой амплитудой и осаждают частицы с помощью магнитной подставки, удаляют супернатант. Проводят отмывание магнитных частиц с применением 40 и 70% растворов изопропилового спирта в объеме по 500 мкл с промежуточным осаждением частиц. Полученные частицы промывают 80% раствором диметилкетона, перемешивают на вортексе с высокой амплитудой и осаждают при помощи магнитной подставки. Удаляют супернатант, полученный осадок магнитных частиц с адсорбированными молекулами нуклеиновой кислоты высушивают от остатков диметилкетона с помощью сухого твердотельного термостата при температуре 60±2°С в течение 8-10 мин. К высушенному осадку добавляют 50 мкл буфера ТЕ (10 мМ трис(оксиметил)аминометан, 1 мМ этилендиамин-тетраацетат, рН 7,0-7,3), свободного от РНКаз и ионов Mg2+, прогревают содержимое пробирки при температуре 60±2°С в течение 3-5 минут для получения элюата РНК вируса бешенства. Пробирки помещают на магнитную подставку и отбирают элюат.
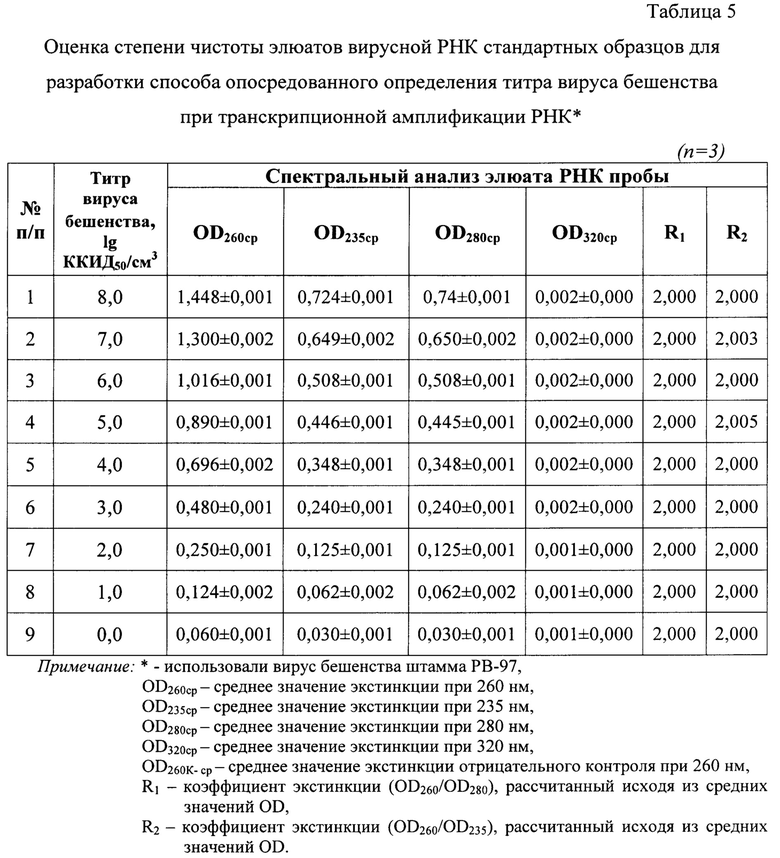
После получения экстракта РНК вируса бешенства осуществляют его спектральное исследование, оценивая поглощение аналитом монохроматического ультрафиолетового света, что позволяет оценить степень чистоты экстракта, который в последующем тестируют в реакции амплификации РНК. Определение спектральной поглощающей способности контролей и проб проводят при длинах волны в диапазоне 205-325 нм и температуре 22-25°С. В выделенных экстрактах оценивают содержание остатков фосфолипидов, полисахаридов и ГТЦ, полипептидов и крупных взвешенных частиц, определяя значения оптической плотности (OD, optical density) при 205, 235, 270, 280 и 320 нм, соответственно [19]. Элюат РНК считают свободным от примесей белка, если коэффициент экстинкции R1 (OD260/OD280) находится в пределах 1,8-2,2 и оптимально составляет примерно 2,0. Более низкие значения R1 указывают на наличие ДНК, полипептидов в экстракте. Более высокие значения коэффициента R1 свидетельствуют о деградации РНК и наличии свободных рибонуклеотидов. Экстракт нуклеиновой кислоты вируса бешенства не содержит полисахариды, если коэффициент экстинкции R2 (OD260/OD235) приближен к значению 2,000 [19]. При замещении 1% РНК на полисахаридные составляющие R2 снижается на 0,002. Значения коэффициента R2  2,000 могут указывать на деградацию молекул РНК. Отсутствие взвеси крупных частиц в элюате подтверждается, если OD320 приближено к нулю [19]. При несоответствии требованиям чистоты повторно проводят этап экстрагирования нуклеиновой кислоты из образца.
2,000 могут указывать на деградацию молекул РНК. Отсутствие взвеси крупных частиц в элюате подтверждается, если OD320 приближено к нулю [19]. При несоответствии требованиям чистоты повторно проводят этап экстрагирования нуклеиновой кислоты из образца.
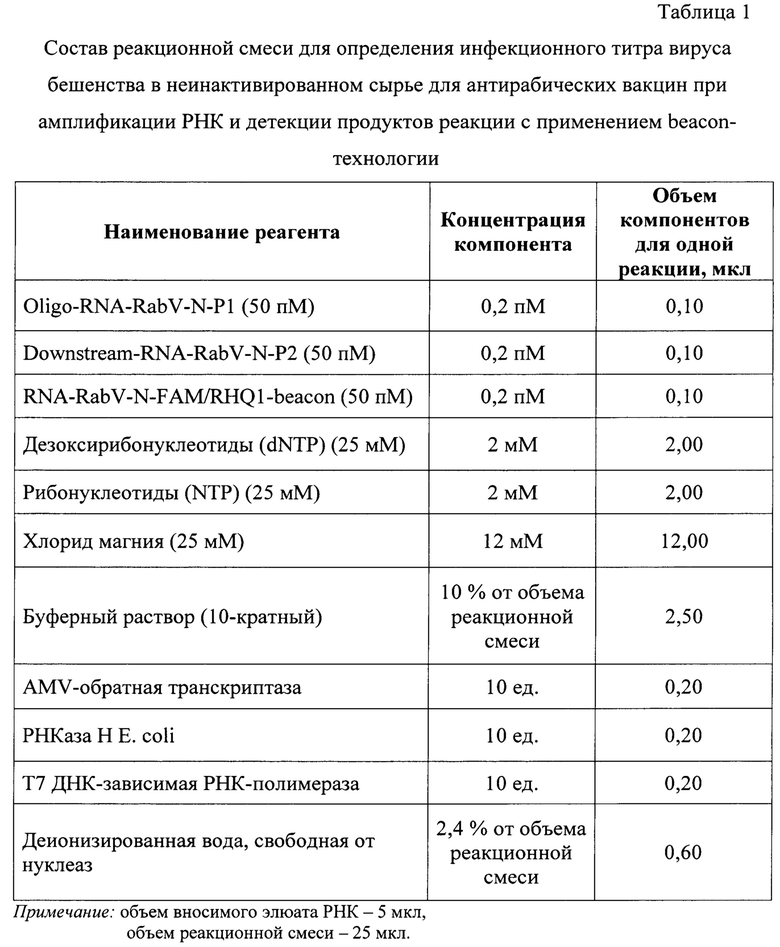
На следующем этапе исследования проводят реакцию амплификации РНК для исследования контрольных образцов и исследуемых проб. Для постановки реакции готовят реакционную смесь, рецептура приготовления которой представлена в таблице 1. Дизайн олигонуклеотидов отражены в таблице 2. Расчет олигонуклеотидных праймеров и beacon осуществляли на основании нуклеотидных последовательностей N-гена вируса бешенства штаммов РВ-97 и ВНИИЗЖ, опубликованных в базах данных GenBank [20] и полученных в рамках исследований в «Федеральном центре охраны здоровья животных» (ФГБУ «ВНИИЗЖ»). Исследовали данные штаммы, поскольку они являются производственными и применяются для изготовления антирабических вакцин в ФГБУ «ВНИИЗЖ».
В качестве гомологичных N-гену вируса бешенства олигонуклеотидов используют Oligo-RNA-RabV-N-P1 (5'-ААТ-ТСТ-ААТ-ACG-ACT-САС-ТАТ-AGG-G-CGU-GGA-GCA-CCA-UAC-UCU-AAG-AA-3') (курсивом обозначена 5'-промоторная часть фермента Т7 ДНК-зависимой РНК-полимеразы), Downstream-RNA-RabV-N-P2 (5'-GGC-UGG-UAU-CGU-UUA-CUG-GGU-AA-3') и RNA-RabV-N-FAM/RHQ1-beacon (5'-FAM-GUG-UGC-UAA-UUG-GAG UAC-UAU-ACC-GAA-RHQ1-3') в концентрации 0,20 пМ на реакцию с внесением в реакционную смесь по 0,10 мкл. Для синтеза нуклеотидных цепей РНК-ампликонов применяют дезоксирибонуклеозидтрифосфаты (dNTP) и рибонуклеозидтрифосфаты (NTP) с их концентрацией в реакционной смеси по 2 мМ. В качестве основы используют буферный раствор (5 х), содержание которого составляет 20% от общего объема реакционной смеси. Буферный раствор включает в свой состав ионы калия (К+) (5⋅10-2М) и диметилсульфооксид (DMSO) (1%). В качестве кофактора применяют добавляют 12 мМ хлорида магния. Содержание деионизированной воды составляет 2,4%. В качестве катализаторов реакции амплификации РНК применяют следующие ферменты: AMV-обратную транскриптазу (10 ед.), РНКаза Н Е. coli (10 ед.) и Т7 ДНК-зависимую РНК-полимеразу (10 ед.). Данные ферменты добавляют в реакционную смесь после прогревания до температуры 65°С и снижения температуры до 41°С, поскольку эти компоненты реакции термолабильны.
Элюаты РНК вируса бешенства каждого образца добавляют к реакционной смеси по 5 мкл. Итоговый объем реакционной смеси для проведения одной реакции составляет 25 мкл.
Праймеры для РНК-матрицы подбирали в соответствии с рядом требований, которые отражены в работах В. Deiman и R. Sooknanan [21, 22]. На 5'-конце Oligo-RNA-RabV-N-P1 расположена 5'-промоторная часть Т7 ДНК-зависимой РНК-полимеразы (5'-AAT-TCT-AAT-ACG-ACT-CAC-TAT-AGG-G-3'). Длины Oligo-RNA-RabV-N-P1 (гибридной части), Downstream-RNA-RabV-N-P2 и RNA-RabV-N-FAM/RHQ1-beacon составляют 23, 23 и 24 н.о., что соответствует требованиям (20-30 н.о.) [21, 22]. Молекулярный вес олигонуклеотидов Oligo-RNA-RabV-N-P1, Downstream-RNA-RabV-N-P2 и RNA-Rab V-N-FAM/RHQ1-beacon равен 7574,5; 7602,4 и 8966,6, соответственно. Процентное содержание G и С в Oligo-RNA-RabV-N-P1 (гибридной части), Downstream-RNA-RabV-N-P2 и RNA-RabV-N-FAM/RHQ1-beacon составляет 48, 48 и 46%, соответственно, что допустимо (40-60%) [21, 22]. На 5'-конце олигонуклеотидных праймеров отмечается преобладание G и С, а на 3'-конце - А и U. На 3'-конце олигонуклеотидных праймеров располагается полиадениловый фрагмент (An). Олигонуклеотиды очищены в полиакриламидном геле и с помощью высокоэффективной жидкостной хроматографии. Гибридная часть молекулярного beacon не комплементарна олигонуклеотидным праймерам. Отсутствуют 4 и более подряд одинаковых нуклеотидов в цепи олигонуклеотидов. В beacon располагается по 6 нуклеотидов с двух сторон, комплементарных друг другу, что требуется для формирования «стеблевой» части. Флуорофор FAM присоединен к 5'-концу, а гаситель флуоресценции RTQ1 - к 3'-концу. Данные условия соответствуют требованиям, предъявляемым к олигонуклеотидам, которые участвуют в реакции амплификации РНК [21, 22]. В качестве флуоресцентного красителя был выбран N-гидроксисукцинимидный эфир 5/6 FAM, SE (5/6 карбоксифлуоресцеин, смесь изомеров) с длиной волны с максимальной флуоресценцией 520 нм, который присоединен к beacon с помощью реакционной аминогруппы концевого нуклеотида. Для тушения свечения использовали гаситель флуоресценции RHQ1 с длиной волны максимального поглощения при 520 нм и возможном диапазоне гашения 470-570 нм. Таким образом, была предложена подходящая пара «флуорофор-гаситель».
При анализе нуклеотидных последовательностей гибридизационной части установили, что для олигонуклеотидов не характерно образование «шпилек» (за исключением «стеблевой» части бикона), а также не выявлено 3'-комплементарности и сайтов, отжигающих сами на себя при условии, когда минимальное количество пар оснований, необходимое для димеризации праймера и минимальное количество пар оснований, необходимое для образования шпильки - 4 [26, 28].
Проведено определение температур плавления (Tm) для гибридизационной части олигонуклеотидов. Точное определение температуры плавления играет очень важную роль в молекулярно-биологических исследованиях, в том числе при подборе РНК-праймеров и бикона для реакции амплификации РНК вируса бешенства. В соответствии с требованиями к реакции амплификации с последующей детекцией РНК-ампликново с помощью beacon-технологии температура плавления олигонуклеотидов (гибридной их части) должна быть выше температуры реакции (41°С) не менее чем на 7-10°С. Для оценки температуры плавления олигонуклеотидов применяли несколько методов. Один из них - метод, учитывающий концентрации ионов К+ и диметилсульфооксида (DMSO) с стандартной применением формулы [25]:
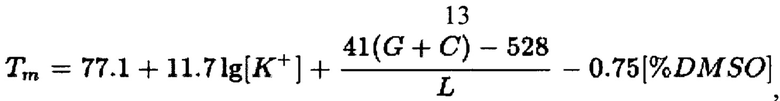
где [К+] - концентрация ионов калия (М), [% DMSO] - количество диметилсульфооксида (%), G - количество остатков рибогуанозинтрифосфата, С - количество остатков рибоцитидинтрифосфата.
Другой метод, который применяли в работе, основан на модели ближайших соседей (nearest neighbour) с применением расчетной формулы [23, 24] с учетом термодинамических показателей олигонуклеотидов:

где dH - дифференциал энтропии (ккал/моль), dG - дифференциал энергии Гиббса (ккал/моль), dS - дифференциал энтальпии (кал/°К⋅моль), С - концентрация олигонуклеотида (М), L - длина олигонуклеотида, [К+] - концентрация ионов калия (М), ln - натуральный логарифм.
В данной формуле учитывается длина олигонуклеотида (L), концентрация ионов калия ([К+]=5⋅10-2 М), газовая постоянная (R=1,987 кал/К⋅моль), концентрация олигонуклеотида (с=2⋅10-7 М), значения энтропии (dH) (в ккал/моль) и энтальпии (dS) (в кал/°К⋅моль). Значения dH и dS вычисляли в соответствии с общеизвестными формулами и термодинамическими параметрами для ближайших соседей для пар нуклеотидов при концентрации NaCl 1М [23-25]. Расчеты термодинамических параметров проводили при условии, что концентрация NaCl 1М, температура 25°С и значение водородного показателя рН 7,0.
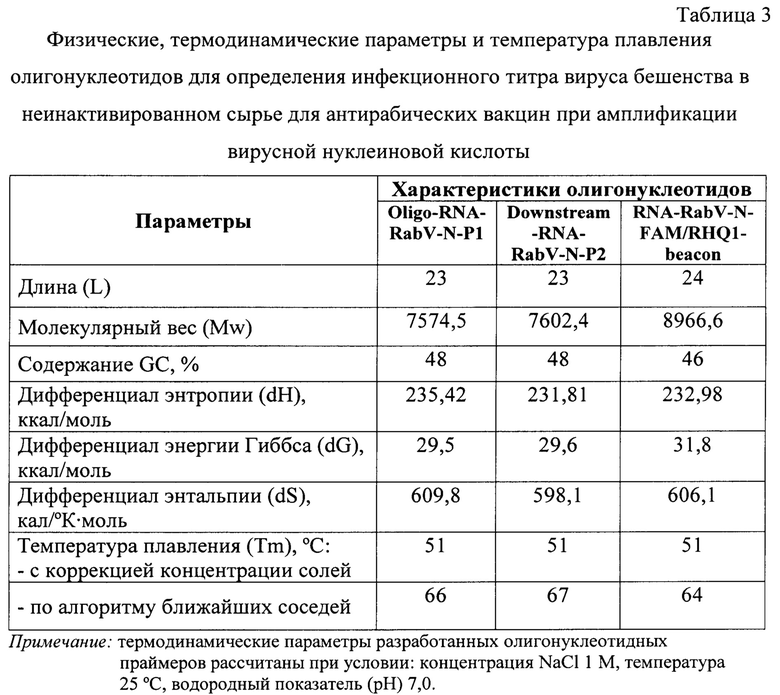
Физические, термодинамические константы и расчет температур плавления [27] разработанных олигонуклеотидных РНК-праймеров и beacon приведены в таблице 3. Из данных таблицы 3 следует, что энтропия, энергия Гиббса и энтальпия для Oligo-RNA-RabV-N-P1 составили 235,42 ккал/моль, 29,5 ккал/моль, 609,8 кал/(°К⋅моль), соответственно. Энтропия, энергия Гиббса и энтальпия для Downstream-RNA-RabV-N-P2 составили 231,81 ккал/моль, 29,6 ккал/моль, 598,1 кал/(°К⋅моль). Энтропия, энергия Гиббса и энтальпия для RNA-RabV-N-FAM/RHQ1-beacon составили 232,98 ккал/моль, 31,8 ккал/моль, 606,1 кал/(°К⋅моль), соответственно. Данные значения были необходимы для расчета температур плавления представленных олигонуклеотидов. Tm при использовании алгоритма ближайших соседей для Oligo-RNA-RabV-N-P1, Downstream-RNA-RabV-N-P2 и RNA-RabV-N-FAM/RHQ1-beacon составили 66, 67 и 64°C, соответственно, что ниже более чем на 7-10°С, чем температура амплификации РНК (41°С) и соответствует общепринятым требованиям, предъявляемым к олигонуклеотидам, используемым для данной реакции [15, 21, 22].
При использовании более простого метода, учитывающего концентрации ионов K+ и диметилсульфооксида (DMSO) [23, 24] Tm для Oligo-RNA-RabV-N-P1, Downstream-RNA-RabV-N-P2 и RNA-RabV-N-FAM/RHQ1-beacon составили 51°C для всех олигонуклеотидов, что на 10°С выше, чем температура амплификации РНК (41°С) и также соответствует общепринятым требованиям, предъявляемым к олигонуклеотидам, используемым для данной реакции [15, 21, 22].
Последовательности разработанных оригинальных олигонуклеотидов были исследованы на наличие нежелательных совпадений с другими последовательностями нуклеиновых кислот с использованием Банка данных последовательности РНК [27]. Последовательности праймеров также проанализировали на наличие внутренних вторичных структур с помощью программы сворачивания нуклеиновых кислот с помощью программы Mfold [28]. Было выявлено, что для разработанных олигонуклеотидов нежелательных совпадений с другими последовательностями нуклеиновых кислот, а также наличия внутренних вторичных структур не обнаружено.
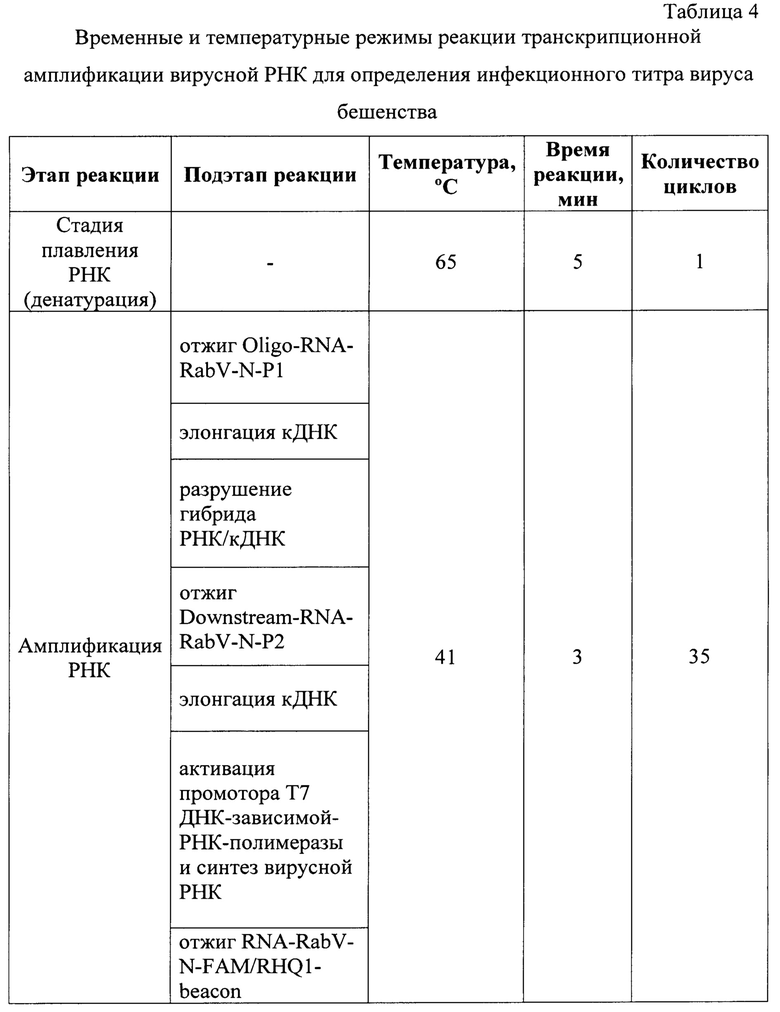
Постановку реакции осуществляют в амплификаторе с наличием флуориметра любой марки при температурных и временных параметрах, сведения о которых отражены в таблице 4. Стадию денатурации РНК проводят при температуре 65°С в течение 5 мин за 1 цикл. Реакцию амплификации РНК осуществляют в течение 35 циклов в изотермических условиях при температуре 41°С в течение 105 минут. Каждый условный цикл длится 3 минуты и складывается из 7 подэтапов: отжиг Oligo-RNA-RabV-N-P1 на вирусной РНК, элонгация комплементарной ДНК (кДНК), разрушение гибрида РНК/к ДНК, отжиг Downstream-RNA-RabV-N-P2, элонгация второй цепи кДНК, активация промотора Т7 ДНК-зависимой РНК-полимеразы, синтез вирусной РНК, отжиг RNA-RabV-N-FAM/RHQ1-beacon. Принципиальная схема процесса представлена на фиг. 1.
Принцип применяемого метода основан на проведении реакции транскрипционной амплификации нуклеиновой кислоты при фиксированной температуре 41°С (изотермические условия) с участием трех ферментов: AMV-ревертазы (ревертаза вируса миелобластоза птиц), РНКазы (RNase) Н Е. coli и Т7 ДНК-зависимой РНК-полимеразы, полученной рекомбинантным способом из бактериофага Т7, специфических прямого и обратного олигонуклеотидных праймеров и молекулярного beacon для амплификации РНК вируса бешенства. В результате этого процесса в ходе реакции происходит накопление миллиардов специфических фрагментов вирусной РНК-мишени. После инкубации РНК вируса бешенства при 65°С в течение 5 мин, при котором осуществляется денатурация нуклеиновой кислоты, начинается линейная стадия реакции амплификации, при которой специфический Oligo-RNA-RabV-N-P1 гибридизируется с участком вирусной РНК. Данный олигонуклеотидный праймер включает в свой состав специфическую гибридизационную часть, а также промоторную последовательность Т7 ДНК-зависимой РНК-полимеразы. При температуре 41°С AMV-ревертаза осуществляет элонгацию, создавая комплементарную ДНК с вирусной РНК-мишени. В результате формируется гетеродуплекс РНК/кДНК. Для РНКазы Н Е. coli данный гибрид выступает в качестве субстрата. Фермент гидролизует РНК, тем самым разрушает гибрид РНК/кДНК, оставляя одноцепочечную к ДНК. С кДНК гибридизуется Downstream-RNA-RabV-N-P2. AMV-ревертаза вновь удлиняет кДНК до 5'-конца с образованием двуцепочечной кДНК (дц кДНК), что приводит промотор Т7 ДНК-зависимой РНК-полимеразы в функциональное состояние. Данный фермент воспринимает дц кДНК с активным промотором в качестве субстрата. В результате производится множество копий фрагмента вирусной РНК вируса бешенства. Молекулярный beacon, комплементарный участку N-гена РНК-мишени вируса бешенства, гибридизутся с ним. В отсутствии мишени флуорофор и гаситель флуоресценции в составе молекулярного beacon сближены за счет максимального использования водородных связей между атомами Н, О и N олигонуклеотида. Благодаря механизму флуоресцентно-резонансного переноса энергии свечение подавлено. После линейной стадии реакция амплификации РНК вступает в циклический процесс.Downstream-RNA-RabV-N-P2 гибридизуется с вновь синтезированной молекулой РНК вируса бешенства. Фермент AMV-ревертаза проводит элонгацию кДНК. В результате образуется гетеродуплекс РНК/кДНК. За счет активности РНКазы Н Е. coli после отжига beacon происходит разрушение гибрида РНК/кДНК за счет гидролиза РНК, наблюдается пространственное разделение флуорофора и гасителя свечения, что приводит к росту детектируемого сигнала при длине волны 520 нм. Oligo-RNA-RabV-N-P1 гибридизируется с кДНК, а AMV-ревертаза удлиняет его с образованием дц кДНК. Фермент Т7 ДНК-зависимая РНК-полимераза синтезирует РНК вируса бешенства. После этого запускается следующий цикл амплификации РНК [13, 15, 16].
Данные реакции транскрипционной амплификации РНК в режиме реального времени с каждого цикла анализируют, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям пороговых циклов амплификации Ct РНК, определенных с помощью пересечения пороговой линии и логарифмическим отображением функции Fl=f (Ct РНК). Флуориметр определяет уровень свечения и строит кинетическую кривую в координатах: уровень флуоресценции - цикл амплификации РНК. В случае присутствия в исследуемой пробе специфической РНК-матрицы кинетическая кривая имеет экспоненциальную зависимость в виде графика сигмоиды. Положительными считаются пробы, которым соответствуют экспонециальные кривые, полученные при анализе флуоресценции красителя, входящего в состав молекулярного beacon, и при значении порогового цикла амплификации менее 32,54. Пробы считаются отрицательными, если при их анализе отсутствует экспонециальная кривая, или график начинает формироваться после 32,54 цикла амплификации. Данное значение порогового цикла амплификации определено экспериментально. При его достижении исследуемый образец имеет значение титра инфекционной активности вируса бешенства 0,0 lg ККИД50/см3 или 1,0 ККИД50/см3.
Выявляют зависимость между Ct РНК и значением десятичного логарифма инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин. Оценивают величину эффективности реакции амплификации (Е) по формуле: Е=1-101/k, где k - угловой коэффициент в зависимости Ct РНК = -k × lg ТВБ + b, а также достоверность аппроксимации (R2). На основе разработанной модели рассчитывают значение инфекционного титра вируса бешенства в неинактивированном сырье для вакцин.
Пример 1. Выражение функции зависимости инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин и порогового цикла реакции транскрипционной амплификации вирусной РНК.
Для определения значения инфекционного титра вируса бешенства штаммов РВ-97 и ВНИИЗЖ подготавливали серию разведений положительных стандартных образцов вируса бешенства, в качестве которых применяли неинактивированные суспензии вируса бешенства с титрами: 0,0; 1,0; 2,0; 3,0; 4,0; 5,0; 6,0; 7,0; 8,0 lg ККИД50/см3. Для получения данных разведений применяли охарактеризованную очищенную с помощью фильтра на 1 мкм суспензию вируса бешенства с титром инфекционной активности 8,0 lg ККИД50/см3. Репродукция вируса проводилась в суспензионной клеточной линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH. Отрицательным контролем служила суспензия клеток ВНК-21/SUSP/ARRIAH, не зараженная вирусом бешенства, с концентрацией 2,5-3,0 млн кл./см3.
Из всех стандартных положительных образцов и отрицательного контроля экстрагировали РНК вируса бешенства. Для этого к 100 мкл исследуемой суспензии добавляли 500 мкл 4 М раствора ГТЦ, перемешивали на вортексе на низкой скорости в течение 5 мин. К полученному лизату добавляли 30 мкл магнитных частиц оксида железа (II, III) и инкубировали в процессе перемешивания на вортексе на низкой скорости в течение 5 мин. Полученную суспензию осаждали с помощью магнитной подставки, удаляли надосадок и проводили промывание магнитных частиц от ингибиторов реакции амплификации РНК с помощью 4 М раствора ГТЦ, добавляя его к осадку в объеме 400 мкл. Смесь перемешивали на вортексе с высокой амплитудой и осаждали частицы с помощью магнитной подставки, удаляли супернатант.Проводили отмывание магнитных частиц с применением 40 и 70% растворов изопропилового спирта в объеме по 500 мкл с промежуточным осаждением частиц. Полученные частицы промывали 80% раствором диметилкетона, перемешивали на вортексе с высокой амплитудой и осаждали при помощи магнитной подставки. Удаляли супернатант, полученный осадок магнитных частиц с адсорбированными молекулами нуклеиновой кислоты высушивали от остатков диметилкетона с помощью сухого твердотельного термостата при температуре 60±2°С в течение 8-10 мин. К высушенному осадку добавляли 50 мкл буфера ТЕ (10 мМ трис(оксиметил)аминометан, 1 мМ этилендиамин-тетраацетат, рН 7,0-7,3), свободного от РНКаз и ионов Mg2+, прогревали содержимое пробирки при температуре 60±2°С в течение 3-5 минут для получения элюата РНК вируса бешенства. Пробирки помещали на магнитную подставку и отбирают элюат.
Полученные элюаты исследовали с помощью спектрального анализа в излучении ультрафиолетового света для оценки степени чистоты полученных образцов. Экстракты РНК сканировали в кварцевых кюветах (l=10 мм) при температуре 23-25°С и регистрировали значения экстинкции в диапазоне от 205 до 325 нм через каждые 2 нм, производя запись спектра поглощения РНК с помощью компьютерной программы Specrtrum v. 5.0 (Фиг. 2, таблица 5).
Обнаружили, что значения OD205-259 и OD261-325 не превышают OD260, что является признаком высокой степени чистоты полученных элюатов РНК (n=3). Из данных спектрального исследования положительных стандартов отмечали отсутствие выраженных пиков на графиках (фиг. 2) при длинах волны 205, 235, 270, 280 и 320 нм, что свидетельствовало о практически полном отсутствии загрязнения элюатов РНК примесями фосфолипидов, полисахаридов и остатков ГТЦ, полипептидов и крупных конгломератов, соответственно. Значения коэффициента экстинкции R1 для положительных стандартов составляли норму 2,000, что подтверждало отсутствие ДНК и наличие лишь следовых количеств примесей полипептидов. Деградации нуклеиновой кислоты и наличия свободных нуклеотидов в элюатах не отмечалось, поскольку R1 не превышал 2,000. Экстракты вирусной РНК не загрязнены полисахаридами и остатками ГТЦ, так как значения коэффициента экстинкции R2 приближены к норме 2,000 и соответствовали 2,000-2,005. Учитывая, что при замещении 1% РНК на полисахариды значение R2 уменьшается на 0,002, в полученных экстрактах наличие углеводных примесей не обнаружено. С помощью спектрального анализа разрушения РНК в экстрактах не обнаружено. Иными словами, элюаты РНК вируса бешенства, выделенные из стандартов, характеризуются высокой степенью чистоты [19] и могут быть использованы для дальнейшей работы.
После оценки степени чистоты полученного экстракта РНК вируса бешенства проводили постановку реакции амплификации РНК для исследования контролей. Состав реакционной смеси представлен в таблице 1. В качестве катализаторов реакции применяли следующие ферменты: AMV-обратную транскриптазу (10 ед.), РНКаза Н Е. coli (10 ед.) и Т7 ДНК-зависимую РНК-полимеразу (10 ед.). Их добавляли в реакционную смесь после прогревания до 65°С и снижения температуры до 41°С. Экстракты нуклеиновых кислот каждого образца добавляли к реакционной смеси по 5 мкл. Итоговый объем смеси для проведения одной реакции составил 25 мкл.
Реакцию денатурации амплификации РНК вируса бешенства осуществляли при температурных и временных параметрах, представленных в таблице 4.
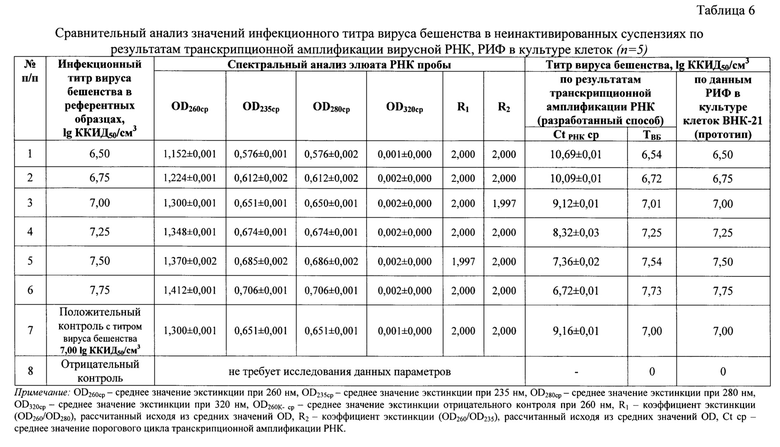
Получены данные реакции амплификации РНК, которые анализировали, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям пороговых циклов амплификации РНК Ct РНК, определенных с помощью пересечения пороговой линии и логарифмическим отображением функции Fl=f (Ct РНК). Устанавливали зависимость между пороговым циклом амплификации РНК и инфекционным титром вируса бешенства в неинактивированном сырье для антирабических вакцин. Полученные результаты отражены на фиг. 3 и выражены в виде логарифмической функции: lg ТВБ = -0,2997 × Ct РНК + 9,7524. Оценивали величину эффективности реакции амплификации (Е) по формуле: Е=1-101/k (k из функции Ct РНК =-3,3361 × lg ТВБ + 32,51) (фиг. 4), а также достоверность аппроксимации (R2). Эффективность реакции транскрипционной амплификации РНК составила 99,41%, достоверность аппроксимации - 0,9997, что соответствовало общепринятым требованиям, предъявляемым к реакциям амплификации [29, 30].
Пример 2. Опосредованное определение инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Для исследования использовали неинактивированные суспензии культурального вируса бешенства штамма ВНИИЗЖ с титрами вируса 6,50, 6,75, 7,00, 7,25, 7,50, 7,75 lg ККИД50/см3 (пробы № 1-6), соответственно. В качестве положительного контроля применяли неинактивированную суспензию культурального вируса бешенства штамма ВНИИЗЖ с титром вируса 7,00 lg ККИД50/см3. Отрицательным контролем служила не инфицированная вирусом бешенства суспензия клеток ВНК-21 с концентрацией 3,00±0,10 млн клеток/см3. Испытуемые пробы и контрольные образцы исследовали в 5 повторностях. Этапы выделения РНК, оценки степени их чистоты и постановку реакции транскрипционной амплификации вирусной РНК проводили, как описано в примере 1. Результаты исследования представлены в таблице 6.
Из данных спектрограмм элюатов РНК проб вируса бешенства следует, что средние значения экстинкции при длинах волны 205-259 и 261-325 нм не превышали OD260, иными словами, элюаты РНК характеризовались высокой степенью чистоты. Экстракты не были контаминированы примесями фосфолипидов, полисахаридов и остатков ГТЦ, полипептидов и крупных конгломератов, поскольку на спектрограммах отсутствовали выраженные пики при А, 205, 235, 270, 280 и 320 нм, соответственно. Коэффициент экстинкции R1 для исследуемых проб вируса бешенства составили 2,000; 2,000; 1,997; 2,000; 2,000; 2,000, что приближено к значению нормы 2,000 и означает высокую степень чистоты элюатов РНК вируса бешенства, практически полное отсутствие белка. Коэффициент экстинкции R2 для анализируемых проб с указанными выше титрами инфекционности соответствовали значению нормы 1,997-2,000, что обуславливало высокую чистоту препаратов. Таким образом, полученные препараты удовлетворяли требованиям чистоты [19] и их можно использовать для дальнейших исследований.
С полученными элюатами неинактивированного вируса бешенства проводили реакцию транскрипционной амплификации РНК. С помощью разработанного способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для вакцины при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии получали данные, отраженные в таблице 6. Из сведений таблицы 6 видно, что средние значения пороговых циклов транскрипционной амплификации РНК для проб вируса бешенства № 1-6 составляли 10,69±0,01, 10,09±0,01, 9,12±0,01, 8,32±0,03, 7,36±0,02, 6,72±0,01, соответственно. Пользуясь разработанной показательной функцией lg ТВБ=-0,2997 × Ct РНК + 9,7524 (R2=0,9997, Е=99,41%), рассчитали средние значения титра вируса бешенства для проб № 1-6, которые составили 6,54, 6,72, 7,01, 7,25, 7,54, 7,73 lg ККИД50/см3, соответственно. Для положительного контроля значение порогового цикла транскрипционной амплификации РНК составило 9,16±0,01, что соответствовало титру вируса бешенства 7,00 lg ККИД50/см3. Для отрицательного контроля график сигмоиды не был сформирован, что означало отсутствие генома вируса бешенства в данном образце.
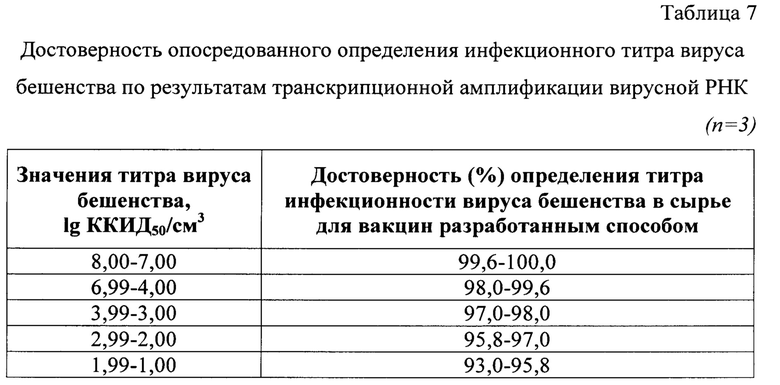
Анализируемые пробы и контроли также тестировали классическим методом РИФ в монослойной клеточной линии ВНК-21 с использованием иммуноглобулина G, меченого ФИТЦ. Выявили, что данные, полученные с помощью разработанного способа, коррелировали с методом РИФ в культуре клеток на 99-100%. Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Иными словами, разработанный способ опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии позволяет оценивать инфекционный титр вируса бешенства в неинактивированном сырье для антирабических вакцин.
Пример 3. Определение степени достоверности опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Для исследования использовали 240 суспензий культурального вируса бешенства производственного штамма РВ-97 с количеством инфекционных доз вируса от 1,00 до 8,00 lg ККИД50/см3. В качестве положительного контроля применяли суспензию культурального вируса бешенства штамма РВ-97 с титром вируса 7,00 lg ККИД50/см3. В качестве отрицательных контролей применяли суспензию клеток ВНК-21/SUSP/ARRIAH, не зараженную вирусом бешенства. Испытуемые пробы и контрольные образцы исследовали в трех повторностях. Этапы выделения РНК, оценки степени их чистоты и постановку транскрипционной амплификации вирусной РНК проводили, как описано в примере 1. Результаты исследования представлены в таблице 7.
Спектральный анализ 240 элюатов РНК проб вируса бешенства показал, что средние значения экстинкции при длинах волны 205-259 и 261-325 нм не превышали OD260, иными словами, элюаты РНК характеризовались высокой степенью чистоты. Экстракты не были контаминированы примесями фосфолипидов, полисахаридов и остатков гуанидинизотиоцианата, полипептидов и крупных конгломератов, поскольку на спектрограммах отсутствовали выраженные пики при λ 205, 235, 270, 280 и 320 нм, соответственно. Коэффициент экстинкции R1 для проб вируса бешенства составлял 1,995-2,000, что приближено к значению нормы 2,000 и означало высокую степень чистоты элюатов вирусной РНК, практически полное отсутствие белка. Коэффициент экстинкции R2 для проб вируса бешенства соответствовал значениям 1,996-2,001, что близко к норме 2,000 и обуславливало высокую чистоту элюатов. Таким образом, полученные препараты удовлетворяли требованиям чистоты [19], и их можно использовать для дальнейших исследований.
С полученными экстрактами РНК вируса бешенства проводили транскрипционную амплификацию. С помощью разработанного способа опосредованного определения инфекционного титра вируса бешенства в сырье для антирабических вакцин получили данные, результат обработки которых отражен в таблице 7. Интерпретацию результатов проводили, пользуясь разработанной показательной функцией lg ТВБ=-0,2997 × Ct РНК + 9,7524 (R2=0,9997, Е=99,41%) с получением значений инфекционного титра для каждой из 240 проб. Для положительного контроля значение порогового цикла амплификации составило 9,16±0,01, что соответствовало инфекционному титру вируса бешенства, равному 7,00 lg ККИД50/см3. Для отрицательного контроля логистические кривые не были сформированы, что означало отсутствие вируса бешенства в данном образце.
Выявили, что данные, полученные с помощью разработанного способа, коррелировали с методом РИФ в культуре клеток (исходные данные) на 99,6-100% для 8,00-7,00 lg ККИД50/см3 (n=30), на 98,0-99,6% для 6,99-4,00 lg ККИД50/см3 (n=30), на 97,0-98,0% для 3,99-3,00 lg ККИД50/см3 (n=30), на 95,8-97,0% для 2,99-2,00 lg ККИД50/см3 (n=30), на 93,0-95,8% для 1,99-1,00 lg ККИД50/см3 (n=30). Полученные результаты свидетельствовали о высокой степени точности разработанного способа опосредованного определения инфекционного титра вируса бешенства в сырье для антирабических вакцин. Таким образом, разработанный способ опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Пример 4. Определение аналитической чувствительности разработанного способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Для определения аналитической чувствительности разработанного способа способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии использовали неинактивированную суспензию культурального вируса бешенства штамма ВНИИЗЖ с инфекционным титром 9,0 lg ККИД50/см3. Подготавливали серию разведений данного стандарта с титрами вируса: 0,0; 1,0; 2,0; 3,0; 4,0; 5,0; 6,0; 7,0; 8,0; 9,0 lg ККИД50/см3. Испытуемые контрольные образцы исследовали в пяти повторностях. Этапы выделения РНК, оценки степени их чистоты и постановку реакции транскрипционной амплификации вирусной РНК проводили, как описано в примере 1. Результаты исследования представлены в таблице 8, в которой отражен сравнительный анализ определения титра вируса бешенства разработанным способом в сравнении прототипным способом.
В результате экспериментов было установлено, что при высоком значении инфекционного титра в положительных образцах (> 8,15 lg ККИД50/см3 при значении Ct РНК - 5,32±0,01) реакция транскрипционной амплификации может приводить к ложно отрицательному результату, поскольку чрезмерное количество исходной матрицы препятствует отжигу праймеров на амплифицированных матрицах, а когда инфекционный титр и соответственно количество молекул РНК снижается предложенный способ позволяет обнаруживать геном вируса бешенства и титр его ифнекционности вплоть до минимальных значений - 0,00 lg ККИД50/см3 (при значении Ct РНК - 32,51±0,01).
Таким образом, аналитическая чувствительность разработанного способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии составляет 0,00 lg ККИД50/см3.
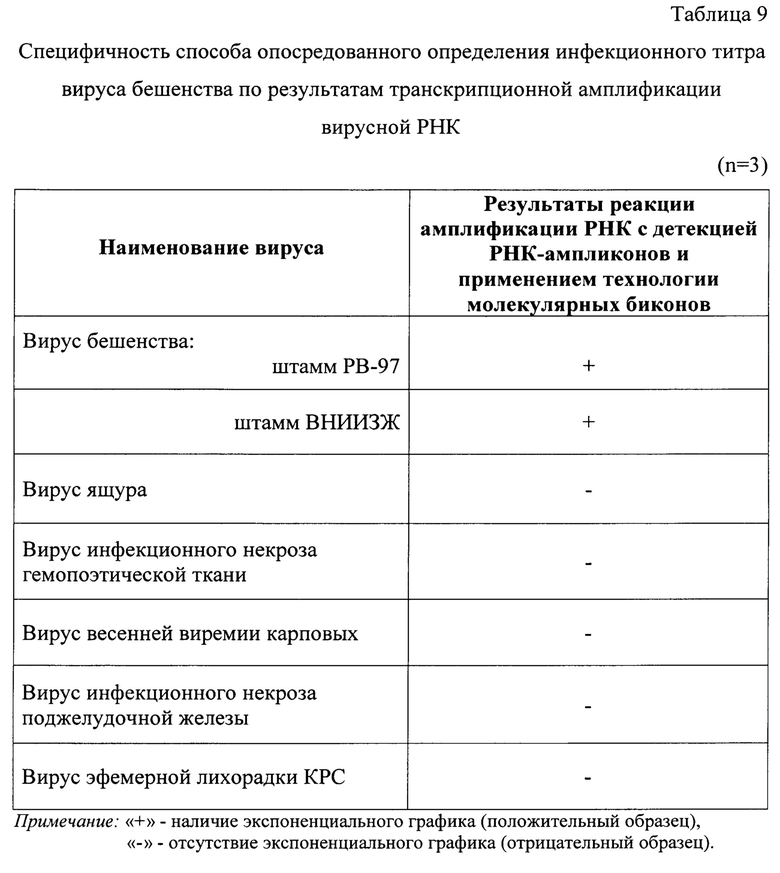
Пример 5. Оценка специфичности разработанного способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин с помощью транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии.
Специфичность предложенного способа определяется олигонуклеотидными праймерами, разработанными для реакции транскрипционной амплификации РНК вируса бешенства. На основе имеющийся информации о структуре генома возбудителя (интересовали производственные штаммы РВ-97 и ВНИИЗЖ, которые являются классическими представителями генетической линии RabV), [20] праймеры были подобраны таким образом, чтобы обеспечить их взаимодействие со строго специфическим участком генома данного вида. Таким образом достигается высокая диагностическая специфичность разработанного способа.
Для оценки специфичности разработанного способа опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии, исследовали неинактивированные суспензии вируса бешенства штаммов РВ-97, ВНИИЗЖ. Также тестировали суспензии вирусов ящура, инфекционного некроза гемопоэтической ткани лососевых, весенней виремии карповых, инфекционного некроза поджелудочной железы, эфемерной лихорадки КРС. Титр вирусов в суспензиях составлял 4,0 lg ККИД (ТЦД)50/см3. Исследования проводили в трех повторностях.
Этапы выделения РНК, оценки степени их чистоты и постановку реакции транскрипционной амплификации вирусной РНК проводили так, как описано в примере 1. Выделенные экстракты РНК характеризовались высокими показателями степени чистоты, поскольку коэффициенты экстинкции R1 и R2 находились в диапазоне 1,994-2,001, что соответствовало требованиям [19].
С полученными элюатами проводили реакцию транскрипционной амплификации РНК с применением указанных выше олигонуклеотидных праймеров и молекулярного beacon, как отражено в примере 1. В ходе реакции транскрипционной амплификации РНК вирусов фиксировали логистические графики накопления флуоресцентного сигнала только для штаммов вируса бешенства. Для проб, содержащих другие вирусы, не наблюдалось формирования графиков экспоненты, и они не выходили за пороговый уровень флуоресцентного сигнала (0,0125 у.е.) (таблица 9). Иными словами, предложенный способ является специфичными по отношению к штамма РВ-97 и ВНИИЗЖ вируса бешенства.
Основными преимуществами предлагаемого изобретения является возможность сократить время проведения анализа неинактивированных вирусосодержащих суспензий для опосредованного определения титра вируса бешенства до 2-3 ч; исключить вероятность контаминации и субъективности при оценке результатов анализа; увеличить чистоту РНК вируса бешенства; увеличить специфичность и чувствительность анализа. В предлагаемом изобретении между инфекционным титром вируса бешенства и пороговым циклом реакции транскрипционной амплификации вирусной РНК с применением разработанных олигонуклеотидов установлена зависимость, отраженная в виде логарифмической функции lg ТВБ=-0,2997 × Ct РНК + 9,7524 высокими достоверностью аппроксимации (R2=0,9997) и эффективностью амплификации 98,41%. Разработанная математическая модель дает возможность оценивать значение инфекционного титра вируса бешенства в неинактивированном сырье для производства антирабических вакцин.
Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности определять значение инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин на основе транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии и с последующим применением разработанной логарифмической модели.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии»:
1. OIE. Manual of diagnostic tests and vaccines for terrestrial animals. - Paris, 2018. - Chapter 3.1.17. - P. 578-612.
2. ICTV Taxonomy history: Rabies lyssavirus URL:
https://talk.ictvonline.org/taxonomy/p/taxonomy
history?taxnode_id=20161105&taxa_name=Rabies%201yssavirus (Дата обращения: 11.04.2020).
3. Baltimore D. Expression of animal virus genomes (англ.) // Microbiology and Molecular Biology Reviews (англ.) русск.: journal. - American Society for Microbiology (англ.). - 1971. - Vol. 35, no. 3. - P. 235-241.
4. Груздев К.Н., Метлин А.Е. Бешенство животных. - Владимир: ФГБУ «ВНИИЗЖ», 2019. - 394 с.
5. Sokol F. Structural Proteins of Rabies Virus / F. Sokol, D. Stancek, H. Koprowski // J. of Virology. - V. 7, №2. - Feb. 1971. - P. 241-249.
6. Baeumner, A. J., Humiston, M. C, Montagna, R. A., and Durst, R. A. (2001) Detection of viable oocysts of Cryptosporidium parvum following nucleic acid sequence based amplification. Anal. Chem. - Vol. 73. - P. 1176-1180.
7. Blok, M. J., Goossens, V. J., van Herle, S. J., et al. (1998a) Diagnostic value of monitoring human cytomegalovirus late pp67 mRNA expression in renalallograft recipients by nucleic acid sequence-based amplification. J. Clin. Microbiol. - Vol. 36. - P. 1341-1346.
8. Mahony, J. В., Song, X., Chong, S., Faught, M., Salonga, Т., and Kapala, J. (2001) Evaluation of the NucliSens Basic Kit for Detection of Chlamydia trachomatis and Neisseria gonorrhoeae in Genital Tract Specimens Using Nucleic Acid Sequence-Based Amplification of 16S rRNA. J. Clin. Microbiol. - Vol. 39. - P. 1429-1435.
9. Morré S. A., Sillekens, P. Т., Jacobs, M. V., et al. (1996) RNA amplification by nucleic acid sequencebased amplification with an internal standard enables reliable detection of Chlamydia trachomatis in cervical scrapings and urine samples. J. Clin. Microbiol. - Vol.34. - P. 3108-3114.
10. Ovyn, C, van Strijp, D., Ieven, M., Ursi, D., van Gemen, В., and Goossens, H. (1996) Typing of Mycoplasma pneumoniae by Nucleic Acid Sequence-Based Amplification, NASBA®. Molecular and Cellular Probes - Vol. 10 - P. 319-324.
11. Song, X., Coombes, В. K., and Mahony, J. B. (2000) Quantitation of Chlamydia trachomatis 16S rRNA using NASBA amplification and a bioluminescent microtiter plate assay. Comb. Chem. High Throughput Screening. - Vol. 3. - P. 303-313.
12. Van der Vliet, G., Cho, S-N., Kampirapap, K., et al. (1996) Use of NASBA® RNA amplification for detection of Mycobacterium leprae in skin biopsies from untreated and treated leprosy patients. Int. J. Lepr. Other Mycobact. Dis. - Vol. 64. - P. 396-403.
13. Van Deursen, P. В. H., Gunther, A. W., Spaargarenvan Riel, С.C, et al. (1999) A novel quantitative multiplex NASBA method: application to measuring tissue factor and CD14 mRNA levels in human monocytes. Nucleic Acids Res. - Vol. 27, e15 (i-vi).
14. Lambrechts, A. C., Bosma, A. J., Klaver, S. G., et al. (1999) Comparison of immunocytochemistry, reverse transcriptase polymerase chain reaction, and Nucleic Acid Sequence-Based Amplification for the detection of circulating breast cancer cells. Breast Cancer Res. Treat. - Vol. 56. - P. 219-231.
15. Loeffler, J., Hebart, H., Cox, P., Flues, N., Schumacher, U., and Einsele, H. (2001) Nucleic acid sequence-based amplification of aspergillus RNA in blood samples. J. Clin. Microbiol. - Vol. 39. - P. 1626-1629.
16. Uyttendaele, M., Bastiaansen, A., and Debevere, J. (1997) Evaluation of the NASBA nucleic acid amplification system for assessment of the viability of Campylobacter jejuni. Int. J. Food Microbiol. - Vol.37. - P. 13-20.
17. Van Strijp D., van Aarle P. NASBA: a method for nucleic acid diagnostics // Methods in molecular medicine-Totowa, NJ: Humana press. - 1995. - P. 331-340.
18. Boom R., Sol C.J.A., Salimans M.M.M., Hansen C.L. et al. Rapid and simple method for the purification of nucleic acids // J. Clin. Microbiol. - 1990. - Vol. 28. - P. 495-503.
19. The Analysis of DNA or RNA using Its Wavelengths: 230 nm, 260 nm, 280 nm. Bioteachnology.com [Электронный ресурс] / URL: http://bioteachnology.com/dna/analysis-dna-rna-wavelengths-230-260-280-nm. (Дата обращения 02.11.2019).
20. GenBank. [Электронный ресурс] / URL: http://www.ncbi.nlm.nih.gov. (Дата обращения: 27.05.2020).
21. Deiman В., van Aarle P., Sillekens P. Characteristics and applications of nucleic acid sequence based amplification // Mol. Biotech. - 2002. - Vol. 20. - P. 163-179.
22. Sooknanan R., van Gemen В., Malek L. Nucleis acid sequence-based amplification // Molecular methods for virus detection-London: Academic press, 1995. - P. 261-285.
23. Nicolas von Ahsen, Carl T. Wittwer, Ekkehard Schütz. Oligonucleotide melting temperatures under PCR conditions: nearest-neighbor corrections for Mg2+, deoxynucleotide triphosphate, and dimethyl sulfoxide concentrations with comparison to alternative empirical formulas (англ.) // Clinical Chemistry: journal. - 2001. - Vol. 47, no. 11. - P. 1956-1961.
24. SantaLucia J. J. A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics (англ.) // Proceedings of the National Academy of Sciences of the United States of America: journal. - 1998. - Vol. 95, no. 4. - P. 1460-1465.
25. SantaLucia J. J., Hicks D. The thermodynamics of DNA structural motifs // Annual Review of Biophysics and Biomolecular Structure: journal. - 2004. - Vol. 33. - P. 11-14.
26. Молекулярный олигокалькулятор. [Электронный ресурс] / URL: http://www.bio.bsu.by/molbiol/oligocalc.html. (Дата обращения: 15.05.2020).
27. The RNA Institute college of arts and science university at Albany. The mfold Web Server. RNA Folding Form. [Электронный ресурс] / URL: htt://bioinfo.math.rpi.tdu/~mfold/dna/form1.cgi (Дата обращения: 01.07.2020).
28. Morre S., Sillekens P., Jacobs M.V., et al. RNA amplification by nucleic acid sequence-based amplification with an internal standard enables reliable detection of Chlamydia trachomatis in cervical scrapings and urine samples // J. Clin. Microbiol. - 1996. - Vol. 34. - P. 3108-3114.
29. Шипицина E.B. Применение метода NUCLEIC ACID SEQUENCE - BASED AMPLIFICATION в реальном времени (NASBA-REAL-TIME) для диагностики урогенитальной хламидийной инфекции // Журнал Акушерства и женских болезней. - Т. LIV выпуск 4/2005. - С. 17-21.
30. Гущин А.Е. Современные методы амплификации нуклеиновых кислот ПЦР и реакция транскрипционной амплификации НАСБА в реальном времени - эффективные инструменты лабораторной диагностики урогенитальной хламидийной инфекции [Электронный ресурс] / URL: https://www.interlabservice.ru/consulting/publications.php?id=8182. (Дата обращения: 22.05.2020).

Изобретение относится к области биотехнологии, а именно к способу опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии и разработанной логарифмической функции зависимости величины порогового цикла транскрипционной амплификации РНК и инфекционного титра вируса бешенства. Значение титра инфекционности вируса бешенства определяют при проведении денатурации и реакции транскрипционной амплификации вирусной РНК с помощью оригинальных специфических олигонуклеотидов по величине порогового цикла амплификации РНК. 6 з.п. ф-лы, 4 ил., 9 табл., 5 пр.
1. Способ опосредованного определения инфекционного титра вируса бешенства в неинактивированном сырье для антирабических вакцин при транскрипционной амплификации и детекции продуктов реакции с применением beacon-технологии, отличающийся тем, что в качестве специфичных олигонуклеотидов для генома вируса бешенства разработаны
- Oligo-RNA-RabV-N-P1 (5'-ААТ-ТСТ-AAT-ACG-ACT-CAC-TAT-AGG-G- CGU-GGA-GCA-CCA-UAC-UCU-AAG-AA-3') (курсивом обозначена 5'-промоторная часть фермента Т7 ДНК-зависимой РНК-полимеразы),
- Downstream-RNA-RabV-N-Р2 (5'-GGC-UGG-UAU-CGU-UUA-CUG-GGU-AA-3'),
- RNA-RabV-N-FAM/RHQ1-beacon (5'-FAM-GUG-UGC-UAA-UUG-GAG UAC-UAU-ACC-GAA-RHQ1-3');
- элюирование РНК вируса бешенства с применением магнитных частиц оксида железа (II, III) диаметром 300-320 нм в присутствии 4 М ГТЦ;
- денатурация РНК вируса бешенства;
- амплификация специфического фрагмента N-гена РНК вируса бешенства с применением специфических оригинальных прямого Р1 и обратного Р2-олигонуклеотидов, а также молекулярного beacon, меченого флуорофором FAM и тушителем свечения RTQ1;
- детекция РНК-ампликонов с помощью флуоресцентного свечения и отображение накопления сигнала в виде сигмоиды;
и расчет инфекционного титра вируса бешенства с применением модели зависимости порогового цикла амплификации (Ct РНК) РНК для кривой флуоресценции и инфекционного титра вируса бешенства (ТВБ) проводят с применением разработанной математической модели: lg ТВБ=-0,2997 × Ct РНК + 9,7524 (R2=0,9997, Е=98,41%), где R2 обозначает достоверность аппроксимации, а E обозначает величину эффективности реакции амплификации.
2. Способ по п. 1, отличающийся тем, что для элюирования РНК вируса бешенства применяют суспензию магнитных частиц оксида железа (II, III) диаметром 300-320 нм.
3. Способ по п. 1, отличающийся тем, что смесь компонентов для проведения реакции транскрипционной амплификации РНК включает в свой состав следующие компоненты: Oligo-RNA-RabV-N-P1, Downstream-RNA-RabV-N-P2, RNA-RabV-N-FAM/RHQ1-beacon (каждый) - по 20 пМ, смесь дезоксирибонуклеозидтрифосфатов (dNTP) и рибонуклеозидтрифосфатов (каждый) - по 2,0 мМ, MgCl2 - 12 мМ, буферный раствор (х10) - 10% от объема реакционной смеси, AMV-ревертаза - 10 ед., РНКаза Н Е. coli - 10 ед., Т7 ДНК-зависимая РНК-полимераза - 10 ед.
4. Способ по п. 1, отличающийся тем, что реакцию амплификации РНК проводят с соблюдением следующих режимов:
- стадия денатурации РНК: 65°С в течение 5 мин (1 цикл);
- реакция амплификации РНК (отжиг Oligo-RNA-Rab V-N-P1, элонгация к ДНК, разрушение гибрида РНК/кДНК, отжиг Downstream-RNA-RabV-N-P2, элонгация кДНК, активация промотора Т7 ДНК-зависимой-РНК-полимеразы и синтез вирусной РНК, отжиг RNA-RabV-N-FAM/RHQ1-beacon): 41°С в течение 3 мин (35 циклов).
5. Способ по п. 1, отличающийся тем, что время проведения анализа вирусосодержащих суспензий для определения инфекционного титра бешенства снижается до 2-3 ч.
6. Способ по п. 1, отличающийся тем, что увеличивается специфичность анализа за счет применения высокоспецифичных оригинальных олигонуклеотидов и увеличивается чувствительность анализа за счет амплификации только РНК.
7. Способ по п. 1, отличающийся тем, что применение фермента Т7 ДНК-зависимой РНК-полимеразы позволяет повторно использовать молекулы комплементарной ДНК, что приводит к увеличению концентрации РНК-ампликонов не менее чем на 1 порядок по сравнению с концентрацией РНК-праймеров.
| Алманиязова С | |||
| Ж | |||
| Бешенство, Медицинский журнал Западного Казахстана, 2012, номер 2 (34), стр | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Khan A | |||
| S | |||
| et al., Report of the international conference on next generation sequencing for adventitious virus detection in biologicals, Biologicals, 2018, Т | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |
| pp | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Khawplod P | |||
| et al., A novel rapid fluorescent focus inhibition test for | |||

