Изобретение относится к ветеринарной вирусологии, к средствам молекулярной диагностики, а именно к дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
В области молекулярно-генетических исследований вирусов в настоящее время особое внимание уделяют возможности выявлять нуклеотидные вариации между вакцинными штаммами и полевыми изолятами, что имеет важное прикладное значение [1], в том числе, определять, не вызывает ли аттенуированный штамм заболевание у животных, особенно при использовании живых культуральных вакцин против бешенства.
Бешенство является одним из крайне опасных, смертельных зооантропонозов, который приводит к поражению центральной нервной системы, энцефаломиелитам и параличам людей и животных. Возбудителем данного заболевания является нейротропный вирус, который имеет следующую таксономию: семейство Rhabdoviridae, род Lyssavirus, вид Rabies lyssavirus [2].
Геном возбудителя бешенства представлен несегментированной одноцепочечной негативной молекулой РНК длиной около 11 800-12 000 н. о., для вакцинного штамма «РВ-97» - 11 932 н.о. [3]. Вирусная РНК несет информацию о пяти основных белках, расположенных в строго консервативном линейном порядке: нуклеопротеин (N-белок), фосфопротеин (Р-белок), матриксный белок (М-белок), гликопротеин (G-белок), РНК-зависимую РНК-полимеразу (L-белок) [4, 5]. Пять генов в нуклеиновой кислоте вируса бешенства окружены двумя короткими участками нуклеотидов на 3'- и 5'-концах: лидерная область (leader) и трейлерная область (trailer), соответственно, которые инициируют и завершают транскрипцию и репликацию генома [6].
Бешенство как заболевание приводит к огромным экономическим потерям, которые связаны с гибелью животных, ликвидацией последствий вспышек болезни, введением строгих ограничений, налагаемых на внутреннюю и международную торговлю продукцией животноводства, проведением профилактических и карантинных мероприятий и др. [6].
В соответствии с Кодексом наземных животных и Руководством МЭБ (OIE) по диагностическим тестам и вакцинам для борьбы с бешенством должен применяться следующий комплекс мер по вакцинопрофилактике диких плотоядных животных: программа по оральной вакцинации; вакцинопрофилактика домашних животных; современная лечебно-профилактическая вакцинация людей, обратившихся за антирабической помощью, профилактическая вакцинация людей групп риска, прежде всего, профессионального риска; контроль проводимых антирабических мероприятий, включающих в себя блок задач и методик [6, 7].
Живые вакцины, изготовленные на основе аттенуированного вируса бешенства, широко применяются для профилактики данного заболевания [8]. В Российской Федерации применяют живую культуральную вакцину против бешенства, разработанную с применением производственного штамма «РВ-97» [9]. Учитывая, что не инактивированный вирус бешенства, возможно, после пассирования в организме животных в полевых условиях может восстановить вирулентность и вызвать побочные реакции у вакцинированных животных, крайне важно использовать диагностические инструменты, которые могут быстро и специфически дифференцировать ослабленные вакцинные штаммы и вирулентные полевые изоляты вируса бешенства. Это помогло бы выяснить происхождение вируса в случае легких или системных поствакцинальных реакций у вакцинированных животных [6, 9].
В связи с этим, в настоящее время важно использовать диагностические инструменты, которые могут быстро и специфически дифференцировать вакцинный штамм «РВ-97» и вирулентные полевые изоляты вируса бешенства, подтверждая, что именно данный вакцинный штамм не вызвал инфекционный процесс у животного в случае его заболевания. Исходя из этого, возникает потребность в разработке анализа, позволяющего проводить дифференциацию генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства.
В связи с этим целесообразно провести разработку способа дифференциации генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства. Для решения поставленной задачи требуется провести молекулярно-биологический анализ генома вируса бешенства производственного штамма «РВ-97» и полевых изолятов, представленных в базе данных GenBank, что даст возможность разработать уникальный способ для исследования образцов тканей биологического материала при вспышке заболевания для дифференциации генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
За последние десятилетия методы молекулярно-генетического анализа находят широкое применение в диагностике различных инфекционных заболеваний [1]. Благодаря созданию нового поколения флуоресцирующих красителей, в частности, SYTO 16, и возможности по данным максимума графика производной сигмоидной функции при амплификации специфического участка вирусной нуклеиновой кислоты стал важным и перспективным методом генотипирования микроорганизмов, который возможно применять для дифференциации вакцинных штаммов от полевых изолятов вирусов [10-23].
Анализ максимума графика производной кривых реакции амплификации позволяет косвенно выявлять мутационные изменения и на этой основе проводить генотипирование [14, 16, 18].
Сущность анализа заключается в следующем: после проведения реакции амплификации в режиме реального времени с интеркалирующим красителем смесь ампликонов постепенно нагревают, и при достижении определенной температуры двухцепочечные молекулы кДНК разделяются, флуорофор постепенно высвобождается, и интенсивность флуоресценции снижается. Изменения степени свечения детектируются оптической системой термоциклера. Ампликоны, имеющие мутации, «плавятся» при разной температуре. Для повышения степени достоверности исследования проводят построение графика производной для выявления максимальной точки, основываясь на значениях которой можно дифференцировать вакцинные штаммы и полевые изоляты, в частности, производственный штамм «РВ-97» от полевых изолятов вируса бешенства.
Преимуществом данного метода является то, что чувствительность метода достигает одного нуклеотида, благодаря чему с помощью данного теста можно проводить дифференциацию штаммов и полевых изолятов возбудителя заболеваний [10, 14, 15].
Прототипным вариантом проведения дифференциального анализа является постановка полимеразной цепной реакции с горизонтальным гель-электрофорезом с оригинальными праймерами и с последующим секвенированием по Сенгеру [24]. Однако, данный метод является очень дорогостоящим, продолжительным во времени, поскольку предполагает проведение множества этапов работы с последующим расшифровыванием нуклеотидной последовательности и анализом их с биоинформатических программ. В условиях, при которых требуется осуществлять исследования полевых образцов биоматериала, экономически нецелесообразно использовать данный метод по указанным выше причинам. Таким образом, требуется разработать альтернативный способ дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Задачей настоящего изобретения является разработка чувствительного и специфичного, быстрого способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16 с целью устранения вышеуказанных недостатков.
Данная задача решена благодаря разработки способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
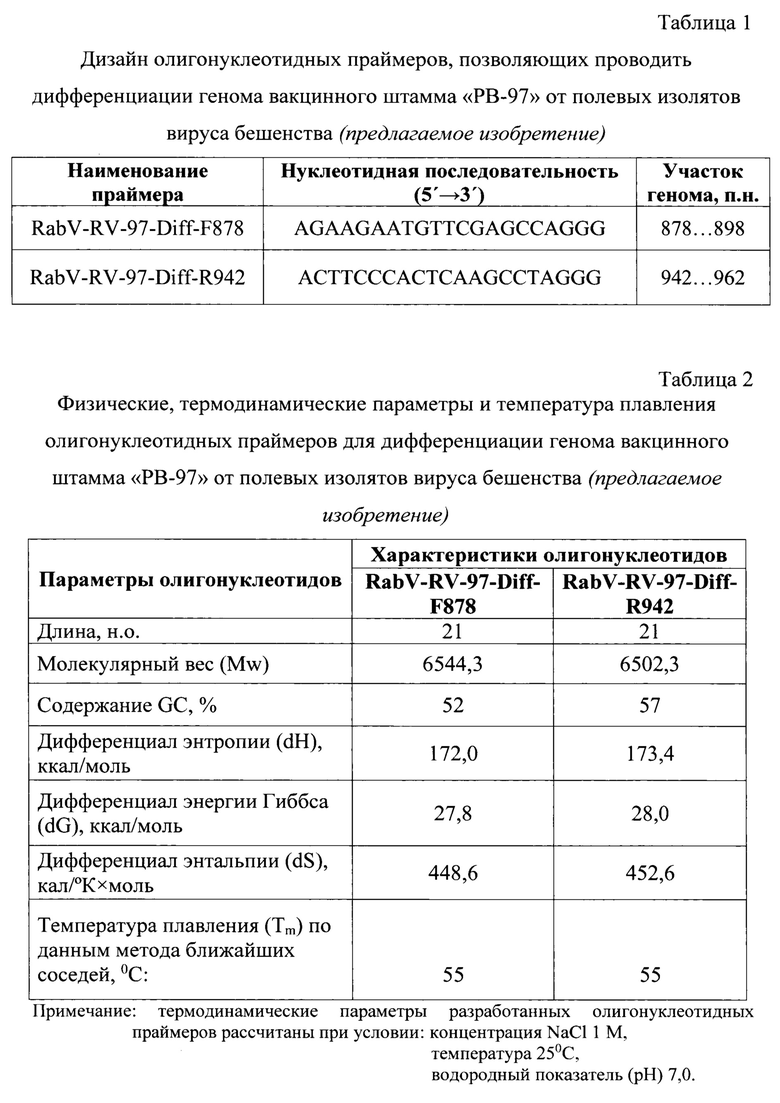
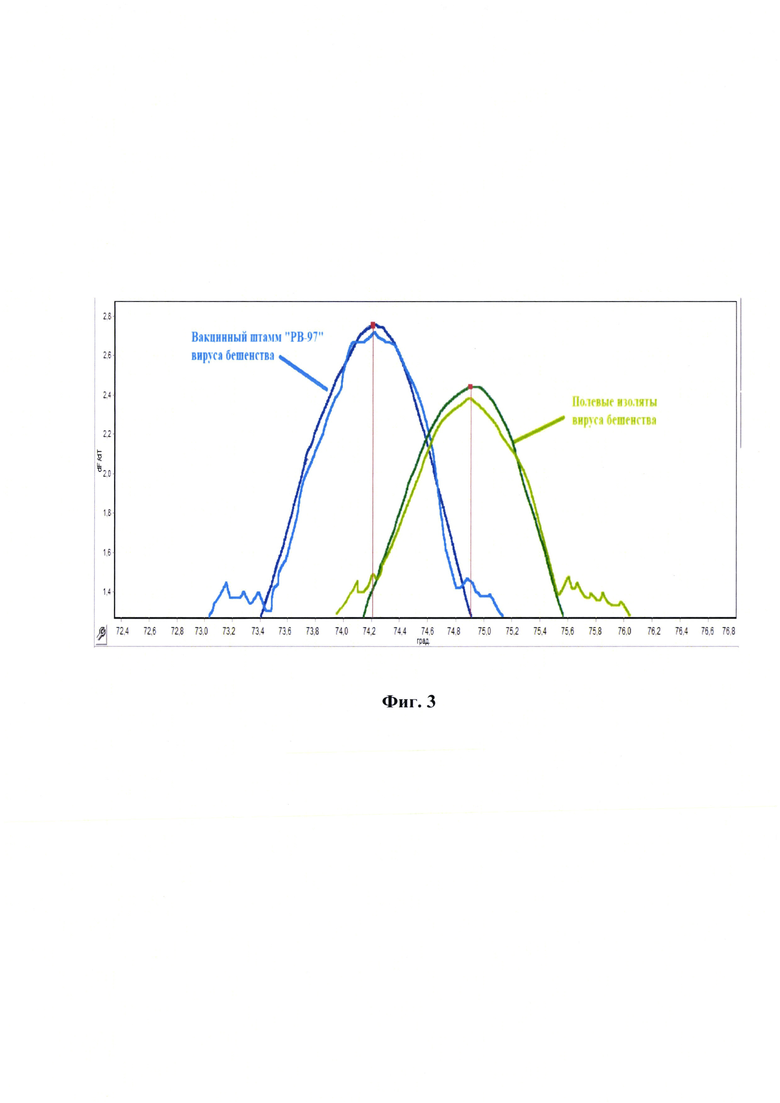
Разработанный способ дает возможность: 1) сократить время проведения анализа до 2,5 ч; 2) применять эффективный флуоресцентный краситель SYTO 16; 3) увеличить чувствительность и специфичность анализа за счет использования высокоспецифичных оригинальных олигонуклеотидных праймеров RabV-RV-97-Diff-F878 и RabV-RV-97-Diff-R942, рассчитанных для участка N-гена кДНК вируса бешенства в диапазоне 878…962 п. н. (размер ампликона составляет 85 п. н.), поскольку именно данная область содержит уникальные мутации для вакцинного штамма «РВ-97» вируса бешенства для однозначной дифференциации данного штамма от полевых изолятов вируса бешенства.
Сущность изобретения отражена на графических изображениях:
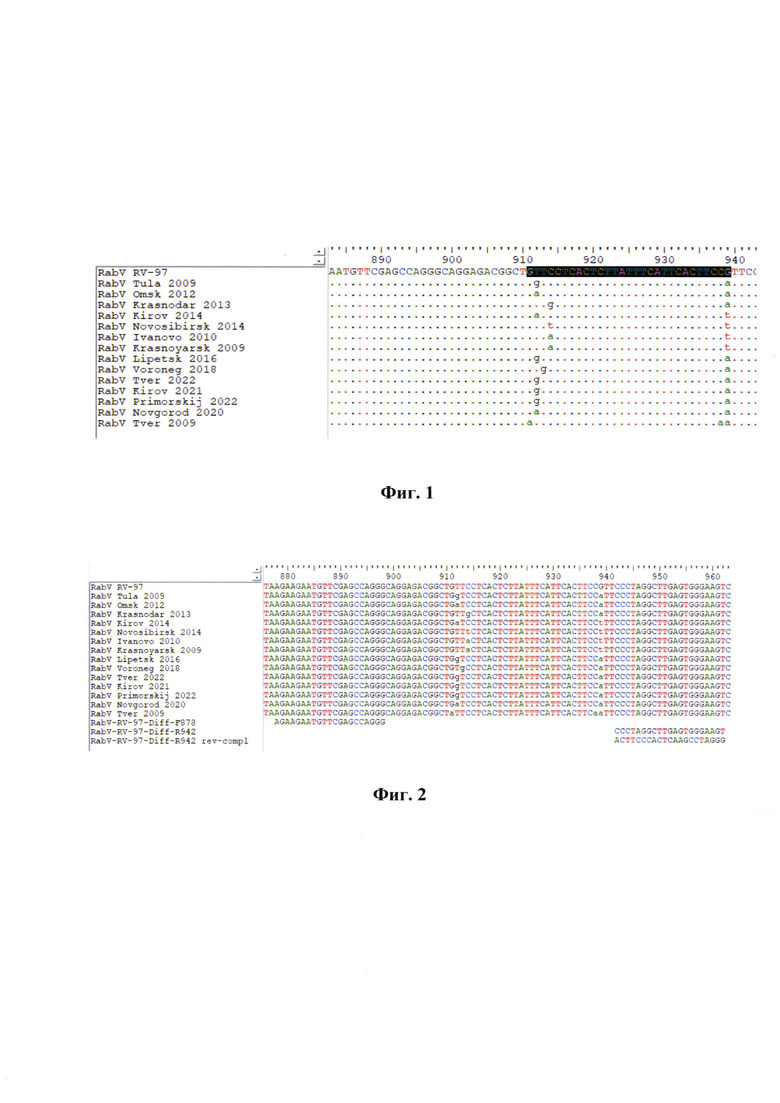
Фиг. 1 - Множественные выравнивания последовательностей полевых изолятов «RabV Tver 2009», «RabV Tula 2009», «RabV Omsk 2012», «RabV Krasnodar 2013», «RabV Kirov 2014», «RabV Novosibirsk 2014», «RabV Ivanovo 2010», «RabV Krasnoyarsk 2009», «RabV Lipetsk 2016», «RabV Voroneg 2018», «RabV Tver 2022», «RabV Kirov 2021», «RabV Primorskij 2022», «RabV Novgorod 2020» и вакцинного штамма «РВ-97» (обозначение - «RV-97») вируса бешенства. Примечание: указаны 6 точечных мутаций в позициях №№911, 912, 913, 914, 938 и 939 п. н.
Фиг. 2 - Дизайн оригинальных олигонуклеотидных праймеров для разработки способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16. Примечание для обратного праймера показана прямая направленность и «revers/compl» - обратная комплементарная.
Фиг. 3 - Максимумы графиков производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16 для дифференциации генома вакцинного штамма «РВ-97» от полевых изолятов разработанным способом. Представлен график для штамма «РВ-97» вируса бешенства и один график для полевых изолятов (для всех исследуемых полевых изолятов вид и топология графика одинакова, они наложены друг на друга, по этой причине продемонстрирован один эталонный график).
Сущность изобретения пояснена в перечне последовательностей, в котором:
SEQ ID N0:1 представляет последовательность нуклеотидов N-гена кДНК вируса бешенства штамма «РВ-97»;
SEQ ID NO:2 представляет последовательность аминокислот, которые кодируются N-гена кДНК вируса бешенства штамма «РВ-97»
SEQ ID NO:3 представляет последовательность праймера RabV-RV-97-Diff-F878;
SEQ ID NO:4 представляет последовательность праймера RabV-RV-97-Diff-R942.
Сущность изобретения заключается в альтернативном подходе по дифференциации генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Заявляемый способ основан на: 1) выделение РНК вируса бешенства из исследуемого биологического материала; 2) проведении обратной транскрипции и реакции амплификации специфического фрагмента кДНК вируса бешенства в диапазоне 878…962 п. н. с применением разработанных оригинальных олигонуклеотидных специфических праймеров RabV-RV-97-Diff-F878 и RabV-RV-97-Diff-R942 (размер ампликона составляет 85 п. н.); 3) проведении плавления полученных ампликонов в разработанном режиме с использованием красителя SYTO 16; 4) построении графика производной для сигмоид и детекции результатов анализа с определением максимального значения температуры плавления ампликонов на данном графике, проведении инструментального дифференциального анализа генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства.
В научных публикациях показана необходимость проведения дифференциации вакцинных штаммов и полевых изолятов вируса бешенства [21-23]. В настоящее время доступно несколько анализов для индикации вируса бешенства с помощью молекулярно-биологических методов исследования [1, 14, 16], но сведений об аналогах предлагаемого способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16 авторами не обнаружено.
Разработанный способ дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16 по сравнению с прототипом отличается не меньшей точностью, при этом имеет явные преимущества - экономическая целесообразность, высокая скорость и простота выполнения анализа.
В отличие от прототипа разработанный способ позволяет провести анализ выравнивания множественных нуклеотидных последовательностей геномов вакцинного штамма «РВ-97» и полевых изолятов вируса бешенства и идентифицировать следующие области с уникальными заменами нуклеотидов в вакцинном штамме «РВ-97»: 911 п. н. - замена А на G, 912 п. н. - G и А на Т, 913 п. н. - G на Т, 938 п. н. - А на С, 939 п. н. - А, Т на G.
Разработанный способ предусматривает проведение реакции амплификации специфического фрагмента кДНК вируса бешенства в диапазоне 878…962 п. н. с применением разработанных оригинальных олигонуклеотидных специфических праймеров RabV-RV-97-Diff-F878 и RabV-RV-97-Diff-R942 для амплификации короткого фрагмента размером 85 п. н., содержащего указанные выше замены.
Разработанный способ предусматривает проведение плавления ампликонов в разработанном режиме с использованием красителя SYTO 16 и осуществление с высокой степенью достоверности дифференциации заявленного вакцинного штамма и полевых изолятов по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной к ДНК.
Ключевым элементом заявляемого способа является определение максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК после реакции амплификации с разработанными оригинальными олигонуклеотидными праймерами и использованием красителя SYTO 16 для дифференциация генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении разработанного способа с использованием оригинальных специфичных олигонуклеотидных праймеров RabV-RV-97-Diff-F878 и RabV-RV-97-Diff-R942, рассчитанных для амплификации участка кДНК вируса бешенства в диапазоне 878…962 п. н., красителя SYTO 16 и технологии анализа максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной к ДНК.
Технический результат изобретения заключается в том, что разработанный способ дает возможность: 1) повысить чувствительность и специфичность за счет применения высокоспецифичных оригинальных праймеров, рассчитанных для целевого участка N-гена вируса бешенства, содержащего в диапазоне 878…962 п. н. шесть уникальных для вакцинного штамма «РВ-97» вируса бешенства мутаций среди представленных изолятов; 2) увеличить достоверность проводимого анализа благодаря применению эффективного SYTO 16; 3) проводить дифференциацию генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК. Характеристики праймеров представлены в таблицах 1 и 2.
Сущность предлагаемого изобретения пояснена на графическом материале - Максимумы графиков производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16 для дифференциации генома вакцинного штамма «РВ-97» от полевых изолятов разработанным способом (фиг. 3).
На первом этапе исследования проводят выделение нуклеиновой кислоты из анализируемых образцов, содержащих вирус бешенства, спомощью набора «РНК-сорб» в соответствии с инструкцией производителя («АтрН Sens»).
Проводят обратную транскрипцию с применением следующих реагентов: деионизированная вода - 45 мкл, буфер 5-кратный - 20 мкл, дезоксирибонуклеозидтрифосфаты (25 мМ) - 10 мкл, олигонуклеотидные праймеры - по 5 мкл, MMLV-ревертаза - 5 мкл, элюат РНК - 10 мкл.
Для постановки ПЦР в режиме реального времени готовят реакционную смесь, которая включает в свой состав следующие компоненты: PCR buffer 10х - 5 мкл, хлорид магния 25 мМ - 5 мкл, олигонуклеотидыне праймеры - по 1 мкл, краситель SYTO 16-2 мкл, Taq ДНК-полимераза - 2 мкл, кДНК - 5 мкл, деионизированная вода - 4 мкл. Итоговый объем смеси для проведения одной реакции составляет 25 мкл.
При проведении реакции применяют положительный контроль (культуральная лиофильно высушенная суспензия вируса бешенства штамма «РВ-97» с титром инфекционной активности не ниже 7,0 lg ККИД50/СМ3) и отрицательный контроль (деионизированная вода без нуклеаз MilliQ вместо к ДНК-матрицы).
Постановку реакции осуществляют в детектирующем термоциклере CFX 96, или аналоге, при температурных и временных параметрах, сведения о которых представлены в таблице 3. Обратную транскрипцию проводят при температуре 42°С в течение 15 минут. Предварительную денатурацию осуществляют при температуре 95°С в течение 4 минут. ПЦР в режиме реального времени включает в себя 3 подэтапа: денатурацию, отжиг олигонуклеотидов, элонгацию. Денатурацию проводят при температуре 95°С в течение 10 секунд, отжиг олигонуклеотидов и элонгацию - при температуре 59°С в течение 45 секунд. Количество циклов реакции амплификации составляет 35. Далее следует важный этап для осуществления дифференциального анализа - плавление, который проводят начиная с 65 до 90°С. Увеличение температуры составляет - 0,1°С за каждый шаг. При этом после первого шага плавления требуется ждать 90 секунд при температуре первого шага. Для каждого последующего шага время ожидания составляет 2 секунды.
Для детекции сигнала устанавливают канал детекции на амплификаторе HRM, для которого длина волны источника составляет 460 нм, детектора - 610 нм. В качестве красителя зеленого цвета применяется SYTO 16 с длиной волны излучения 518-525 нм.
Исследование графика детекции температуры плавления ампликонов для исследуемых образцов выполняют с использованием программного обеспечения для сканирования генов на амплификаторе марки CFX 96, или аналоге. Инструментально осуществляется построение кривых плавления в виде графиков параболы, проводится построение графика производной и детектируются максимумы на нем для полученных ампликонов. Выявляют, что для производственного штамма «РВ-97» вируса бешенства в отличии от полевых изолятов характерен очень узкий индивидуальный диапазон температуры плавления, по данным которого можно судить о принадлежности исследуемого образца к вакцинному штамму «РВ-97» или полевым изолятам вируса бешенства.
Сущность предлагаемого изобретения пояснена примерами его использования, которые не ограничивают объем изобретения.
Пример 1. Определение максимумов на графике производной температур плавления ампликонов для разработки способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Для определения показателей, позволяющих проводить дифференциацию генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16, осуществляли этапы работы, как указано выше.
Исследование кривой детекции температуры плавления для исследуемых образцов выполняли с использованием программного обеспечения для сканирования генов на амплификаторе марки CFX 96, или аналоге. Инструментально осуществляли построение кривых плавления в виде графиков параболы, графика производной и детектировали пики температур плавления для полученных ампликонов размером 85 п. н.
Тестировали кДНК вируса бешенства вакцинного штамма «РВ-97» и 14 следующих полевых изолятов: «RabV Tver 2009», «RabV Tula 2009», «RabV Omsk 2012», «RabV Krasnodar 2013», «RabV Kirov 2014», «RabV Novosibirsk 2014», «RabV Ivanovo 2010», «RabV Krasnoyarsk 2009», «RabV Lipetsk 2016», «RabV Voroneg 2018», «RabV Tver 2022», «RabV Kirov 2021», «RabV Primorskij 2022», «RabV Novgorod 2020».
Выявили, что для вакцинного штамма «РВ-97» вируса бешенства характерен очень узкий индивидуальный за счет уникальных мутаций диапазон максимума на графике производной температуры плавления ампликонов, полученных в реакции амплификации с использованием оригинальных олигонуклеотидных праймеров RabV-RV-97-Diff-F878 и RabV-RV-97-Diff-R942, рассчитанных для амплификации участка кДНК вируса бешенства в диапазоне 878…962 п. н., красителя SYTO 16, равный 74,20±0,02°С (n=30, р<0,005). Для полевых изолятов вируса бешенства «RabV Tver 2009», «RabV Tula 2009», «RabV Omsk 2012», «RabV Krasnodar 2013», «RabV Kirov 2014», «RabV Novosibirsk 2014», «RabV Ivanovo 2010», «RabV Krasnoyarsk 2009», «RabV Lipetsk 2016», «RabV Voroneg 2018», «RabV Tver 2022», «RabV Kirov 2021», «RabV Primorskij 2022», «RabV Novgorod 2020» - 74,91±0,04°C (n=30, p<0,005) (фиг. 3).
Таким образом, для вакцинного штамма «РВ-97» и 14 исследуемых полевых изолятов вируса бешенства, взятых из разных регионов РФ, характерны индивидуальные отличающиеся пики температур плавления ампликонов, что дало возможность проводить дифференциацию генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Пример 2. Исследование аналитической специфичности разработанного способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Специфичность анализа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16 оценивали путем тестирования следующих образцов: культуральных суспензий вируса бешенства вакцинного штамма «CVS», вируса инфекционного некроза гемопоэтической ткани лососевых, вируса инфекционного некроза поджелудочной железы, вируса ящура серотипа О, вируса чумы мелких жвачных животных изолята «PPRV/Israel-2536/Hebron/1997», суспензии Mycoplasma capricolum ssp.
В качестве положительных контролей использовали культуральную лиофильно высушенную суспензию вакцинного штамма «РВ-97» вируса бешенства и патологический материал мозга собаки, содержащей полевой изолят «RabV Primorskij 2022». В качестве отрицательного контроля использовали деионизированную воду. Для проведения исследования использовали детектирующий термоциклер марки CFX-96. В результате исследования амплификация с неспецифичными нуклеиновыми кислотами других инфекционных агентов не была обнаружена (табл.4).
В результате исследования обнаружили, что для проб, не содержащих нуклеиновую кислоту вируса бешенства, и для отрицательного контроля не были сформированы графики плавления и не были обнаружены пиковые значения температур плавления ампликонов. При исследовании положительных контролей - культуральной лиофильно высушенной суспензии вируса бешенства вакцинного штамма «РВ-97» и патологического материла мозга собаки, содержащей полевой изолят «RabV Primorskij 2022» (лиофилизат), получили следующие пиковые значения температур плавления: 74,20±0,01 и 74,90±0,01°С (n=14, р<0,005), соответственно.
Таким образом, полученные результаты свидетельствовали о 100%-ной специфичности разработанного способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Пример 3. Определение диагностических показателей разработанного способа дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Для определения диагностической чувствительности разработанного способа анализировали 455 культуральных суспензий вакцинного штамма «РВ-97» вируса бешенства с разными титрами инфекционной активности (7,00-9,25 lg ККИД50/СМ3), полученных в течение 2017-2023 гг. в рамках производственного процесса изготовления антирабической вакцины. Данные пробы являлись заведомо положительными. Постановку анализа проводили, как отражено выше. С помощью разработанного способа (предлагаемое изобретение) определили, что из 455 исследуемых культуральных суспензий вируса бешенства вакцинного штамма «РВ-97» все определены в качестве положительных (максимумы на графике производной сигмоидной функции при амплификации участка N-гена вирусной кДНК составили 74,20±0,02°С) и подтверждено наличие именно того штамма, который содержался в анализируемых суспензиях. Данные результаты были подтверждены с помощью секвенирования по Сенгеру [24].
Для исследования специфичности метода тестировали 234 суспензии полевых изолятов вируса бешенства, полученных из суспензий ткани головного мозга больных животных. В результате исследования с помощью разработанного способа (предлагаемое изобретение) определили, что все 234 проб содержали геном вируса бешенства (максимумы на графике производной сигмоидной функции при амплификации участка N-гена вирусной кДНК составили 71,91±0,04°С), при этом не содержали вакцинный штамм «РВ-97», что было подтверждено с помощью секвенирования по Сенгеру. Пользуясь представленными выше статистическими методами анализа определили, что в 95%-ном доверительном интервале диагностическая чувствительность (DSe) составила 99,19-100,00%, диагностическая специфичность (DSp) - 98,44-100,0%, k-критерий - 1,000; общая точность (DAc) - 99,47-100,00% (табл. 5).
Основными преимуществами предлагаемого изобретения является возможность проводить за короткий промежуток времени (2,5 ч) дифференциацию генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16.
Разработанный способ впервые применили для решения данной задачи в отношении вакцинного штамма «РВ-97» вируса бешенства. Выявили, что в анализируемом участке кДНК вируса бешенства штамма «РВ-97» в диапазоне 878…962 п. н. амплифицируемого ампликона имеются следующие нуклеотидные замены: 911 п. н. - замена А на G, 912 п. н. - G и А на Т, 913 п. н. - G на Т, 938 п. н. - А на С, 939 п. н. - А, Т на G., которые характерны для вакцинного штамма «РВ-97» и отсутствует в полевых изолятах данного вируса, что позволило разработать следующие специфические олигонуклеотидные праймеры: RabV-RV-97-Diff-F878 и RabV-RV-97-Diff-R942, рассчитанных для амплификации участка кДНК вируса бешенства в диапазоне 878…962 п. н. Разработанный способ характеризуется аналитической специфичностью, равной 100%. В 95%-ном доверительном интервале в 95%-ном доверительном интервале диагностическая чувствительность (DSe) составила 99,19-100,00%, диагностическая специфичность (DSp) - 98,44-100,0%, k-критерий - 1,000; общая точность (DAc)-99,47-100,00%.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16»:
1. Aitman TJ, Petretto E, Behmoaras J. Genetic mapping and positional cloning. Methods Mol Biol. 2010;597:13-32.
2. OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. - 7th ed. - Paris, 2022. - Vol. 1, Chap. 2.1.17.
3. GenBank. Rabies virus. Strain «RV-97» URL: https://www.ncbi.nlm.nih.gov/nuccore/EF542830.1 (Дата обращения: 25.05.2023).
4. Finke S., Conzelmann К.K. (August 2005). «Replication strategies of rabies virus». Virus Res. 111(2): 120-131.
5. Hooper DC, Roy A, Barkhouse DA, Li J, Kean RB. Rabies virus clearance from the central nervous system. Adv Virus Res. 2011;79:55-71.
6. Gomme EA, Wanjalla CN, Wirblich C, Schnell MJ. Rabies virus as a research tool and viral vaccine vector. Adv Virus Res. 2011;79:139-64.
7.  Genetic characterisation of the rabies virus vaccine strains used for oral immunization of foxes in Poland to estimate the effectiveness of vaccination. Arch Virol. 2015 Feb;160(2):509-15.
Genetic characterisation of the rabies virus vaccine strains used for oral immunization of foxes in Poland to estimate the effectiveness of vaccination. Arch Virol. 2015 Feb;160(2):509-15.
8. Вирусвакцина для оральной иммунизации диких плотоядных животных против бешенства Синраб URL: https://www.vettorg.ru/catalog/item-588.html. (Дата обращения: 10.07.2023).
9. Вакцина антирабическая аттенуированная для плотоядных животных производства ФГБУ «ВНИИЗЖ» URL: https://shop.arriah.ru/catalog/search/?q=бешенство (Дата обращения: 17.07.2023).
10. Er ТК, Chang JG. High-resolution melting: applications in genetic disorders. Clin Chim Acta. 2012 Dec 24;414:197-201.
11. Hussmann D, Hansen LL. Methylation-Sensitive High Resolution Melting (MS-HRM). Methods Mol Biol. 2018;1708:551-571.
12. Nikodem D, Clapa T, Narożna D. Technologia HRM-PCR w diagnostyce medycznej [HRM-PCR in medical diagnostic]. Postepy Biochem. 2021 Mar4;67(l):59-63. Polish.
13. Pakbin B, Basti AA, Khanjari A, Brück WM, Azimi L, Karimi A. Development of high-resolution melting (HRM) assay to differentiate the species of Shigella isolates from stool and food samples. Sci Rep.2022 Jan 10;12(1):473.
14. Robinson CV, Uren Webster TM, Consuegra S. Data on optimisation of a multiplex HRM-qPCR assay for native and invasive crayfish as well as the crayfish plague in four river catchments. Data Brief. 2018 May 29; 19:1092-1109.
15. Rojas A, Rojas D, Montenegro VM, Baneth G. Detection of Dirofilaria immitis and other arthropod-borne filarioids by an HRM real-time qPCR, blood-concentrating techniques and a serological assay in dogs from Costa Rica. Parasit Vectors. 2015 Mar 23;8:170.
16. Schiwek S, Beule L, Vinas M, Pfordt A, von Tiedemann A, Karlovsky P. High-Resolution Melting (HRM) Curve Assay for the Identification of Eight Fusarium Species Causing Ear Rot in Maize. Pathogens. 2020 Apr 7;9(4):270.
17. Crossley BM, Bai J, Glaser A, Maes R, Porter E, Killian ML, Clement T, Toohey-Kurth K. Guidelines for Sanger sequencing and molecular assay monitoring. J Vet Diagn Invest. 2020 Nov;32(6):767-775.
18. Davis BM, Rail GF, Schnell MJ. Everything You Always Wanted to Know About Rabies Virus (But Were Afraid to Ask). Annu Rev Virol. 2015 Nov;2(l):451-71.
19. Takahashi T, Inukai M, Sasaki M, Potratz M, Jarusombuti S, Fujii Y, Nishiyama S, Finke S, Yamada K, Sakai H, Sawa H, Nishizono A, Sugiyama M, Ito N. Genetic and Phenotypic Characterization of a Rabies Virus Strain Isolated from a Dog in Tokyo, Japan in the 1940s. Viruses. 2020 Aug 20;12(9):914.
20. Li Y, Zhao H, Wilkins K, Hughes C, Damon IK. Real-time PCR assays for the specific detection of monkeypox virus West African and Congo Basin strain DNA. J Virol Methods. 2010 Oct;169(l):223-7.
21. Yang DK, Kim HH, Lee S, Yoo JY. Establishment of multiplex RT-PCR for differentiation between rabies virus with and that without mutation at position 333 of glycoprotein. J Vet Sci. 2020 Mar;21(2):e22.
22. Wadhwa A, Wilkins K, Gao J, Condori Condori RE, Gigante CM, Zhao H, Ma X, Ellison JA, Greenberg L, Velasco-Villa A, Orciari L, Li Y. A Pan-Lyssavirus Taqman Real-Time RT-PCR Assay for the Detection of Highly Variable Rabies virus and Other Lyssaviruses. PLoS Negl Trop Dis. 2017 Jan 12;ll(l):e0005258.
23. David D, Yakobson B, Rotenberg D, Dveres N, Davidson I, Stram Y. Rabies virus detection by RT-PCR in decomposed naturally infected brains. Vet Microbiol. 2002 Jun 20;87(2):111-8.
24. Estrada-Rivadeneyra D. Sanger sequencing. FEBS J. 2017 Dec;284(24):4174.

--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="RabV RV-97 Diff
xml.xml" softwareName="WIPO Sequence" softwareVersion="2.1.2"
productionDate="2024-04-01">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>0</ApplicationNumberText>
<FilingDate>2023-07-27</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>529</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>0</ApplicationNumberText>
<FilingDate>2023-07-27</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">ФГБУ "Федеральный центр охраны
здоровья животных" (ФГБУ "ВНИИЗЖ")</ApplicantName>
<ApplicantNameLatin> Federal State-Financed Institution Federal
Centre for Animal Health (FGBI ARRIAH)</ApplicantNameLatin>
<InventorName languageCode="ru">Доронин Максим
Игоревич</InventorName>
<InventorNameLatin>Doronin Maksim Igorevich </InventorNameLatin>
<InventionTitle languageCode="ru">Способ дифференциации вакцинного
штамма «РВ-97» от полевых изолятов вируса бешенства по данным
максимума графика производной сигмоидной функции при ампликации
участка N-гена вирусной кДНК с использованием красителя SYTO
16</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>1350</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1350</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q1">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>RabV</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggataccgacaagattgtattcaaagttaataatcaggtggtctctt
taaagcctgagattatcgtggatcaatatgagtacaagtaccctgctatcaaagatttgaaaaagccctg
tataaccctagggaaagcccccgacttgaacaaagcatacaaatcagttttatcaggcatgaatgctgcc
aaacttgatcccgatgatgtatgttcctacttggcagcagcaatgcagttctttgaggggacgtgtccgg
aagactggaccagctacgggatcttgattgcacggaaaggagacaagatcaccccaaattctctagtgga
aataaagcgtactgatgtagaagggaattgggctctgacaggaggcatggaactgacaagggaccccact
gtccctgagcatgcatctttggtcggtcttctcctgagtctgtataggttgagcaaaatatcaggacaaa
acaccggtaactataaaacaaacattgcagataggatagagcaggttttcgagacagccccttttattaa
aatcgtggagcaccatactctaatgacaactcacaaaatgtgtgctaattggagtactataccgaatttc
aggtttttggccggaacctacgacatgtttttctcccggattgatcatctatattcagcaatcagagtgg
gcacagttgtcactgcttatgaagactgttcagggctggtatcgtttactgggtttgtaaagcagatcaa
tctcactgcaagagaagcaatactctacttcttccataagaactttgaggaagaaataagaagaatgttc
gagccagggcaggagacggctgttcctcactcttatttcattcacttccgttccctaggcttgagtggga
agtctccttattcatcgaatgccgttggtcatgtgttcaatctcattcactttgttggatgttatatggg
tcaagtcagatccctaaatgcaacagttattgccgcatgtgctcctcatgagatgtctgttctagggggc
tatctaggggaggaattctttgggaaaggaacatttgagagaagattcttcagagatgagagagaacttc
aagaatacgaggcagctgaactgacaaagaccgacgtggcattggcagatgacggaactgtcaactctga
tgacgaggactacttctccggtgaaaccagaagtccagaagctgtttatgctcgaatcatgatgaatgga
ggtcgactaaaaagatcgcatatacggagatatgtctcagtcagttccaatcatcaagcccgtccaaact
cattcgccgagtttctaaacaagacatattcgagtgactct</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>450</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..450</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>RabV</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MDTDKIVFKVNNQVVSLKPEIIVDQYEYKYPAIKDLKKPCITLGKAPDL
NKAYKSVLSGMNAAKLDPDDVCSYLAAAMQFFEGTCPEDWTSYGILIARKGDKITPNSLVEIKRTDVEGN
WALTGGMELTRDPTVPEHASLVGLLLSLYRLSKISGQNTGNYKTNIADRIEQVFETAPFIKIVEHHTLMT
THKMCANWSTIPNFRFLAGTYDMFFSRIDHLYSAIRVGTVVTAYEDCSGLVSFTGFVKQINLTAREAILY
FFHKNFEEEIRRMFEPGQETAVPHSYFIHFRSLGLSGKSPYSSNAVGHVFNLIHFVGCYMGQVRSLNATV
IAACAPHEMSVLGGYLGEEFFGKGTFERRFFRDERELQEYEAAELTKTDVALADDGTVNSDDEDYFSGET
RSPEAVYARIMMNGGRLKRSHIRRYVSVSSNHQARPNSFAEFLNKTYSSDS</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>RabV</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>agaagaatgttcgagccaggg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q5">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>RabV</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acttcccactcaagcctaggg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к ветеринарной вирусологии. Описан способ дифференциации вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства по данным максимума графика производной сигмоидной функции при ампликации участка N-гена вирусной кДНК с использованием красителя SYTO 16. Основным преимуществом предлагаемого изобретения является возможность проводить за короткий промежуток времени - 2,5 ч дифференциации генома вакцинного штамма «РВ-97» от полевых изолятов вируса бешенства с помощью разработанного способа. Способ характеризуется аналитической специфичностью, равной 100%, диагностической чувствительностью в 95%-ном доверительном интервале, равной 99,19-100,00%, диагностической специфичностью - 98,44-100,0%, общей точностью - 99,47-100,00%. 1 з.п. ф-лы, 3 ил., 5 табл., 3 пр.
1. Способ дифференциации вакцинного штамма «РВ-97» вируса бешенства от полевых изолятов по данным максимума графика производной сигмоидной функции при амплификации участка N-гена вирусной кДНК с использованием красителя SYTO 16, включающий применение высокоспецифичных олигонуклеотидных праймеров:
RabV-RV-97-Diff-F878 с дизайном 5’-AGAAGAATGTTCGAGCCAGGG-3’, RabV-RV-97-Diff-R942 с дизайном 5’-ACTTCCCACTCAAGCCTAGGG-3’, рассчитанных для участка N-гена вирусной кДНК в диапазоне 878…962 п. н., красителя SYTO 16, и при этом максимум температуры плавления фрагмента кДНК размером 85 п. н. для вакцинного штамма «РВ-97» составляет значение 74,20±0,02°С, а для полевых изолятов вируса бешенства данный параметр имеет значение 74,91±0,04°С.
2. Способ по п. 1, где аналитическая специфичность составляет 100%, диагностическая чувствительность в 95%-ном доверительном интервале - 99,19-100,00%, диагностическая специфичность - 98,44-100,0%, общая точность - 99,47-100,00%.
| Способ опосредованного определения количества инфекционных доз вируса бешенства штамма PB-97 в сырье для аттенуированной антирабической вакцины методом обратной транскрипции и полимеразной цепной реакции в режиме реального времени | 2020 |
|
RU2761535C1 |
| Способ дифференциации генома вакцинного штамма НИСХИ от полевых изолятов вируса оспы овец методом полимеразной цепной реакции в режиме реального времени с анализом пиков высокого разрешения | 2022 |
|
RU2786213C1 |
| KR 19950000887 B1, 03.02.1995. | |||

