ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка изъявляет приоритет по предварительной заявке США с серийным № 61/974738, поданной 3 апреля 2014 года, содержание которой включено в данный документ в полном объеме посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к модифицированным рекомбинантным J-цепям и содержащим J-цепь связывающим молекулам, например, антителам.
УРОВЕНЬ ТЕХНИКИ
J-цепь представляет собой кислый полипептид 15 кДа, ассоциированный с пентамером IgM и димером IgA посредством дисульфидных связей, в формировании которых участвует предпоследний остаток цистеина в хвостовой части (х.ч.) секреторного пептида длиной 18 остатков на C-конце тяжелой цепи IgM μ или IgA α. Три дисульфидных мостика образуются между Cys 12 и 100, Cys 71 и 91, а также Cys 108 и 133, соответственно. См., например, Frutiger et al. 1992, Biochemistry 31, 12643-12647. О структурных требованиях для включения J-цепи в состав IgM и IgA человека и для сборки полимерных молекул иммуноглобулинов, а также для ассоциации с J-цепью, сообщалось в источнике Sorensen et al. 2000, Int. Immunol. 12(1): 19-27 и Yoo et al. 1999, J. Biol. Chem. 274(47):33771-33777, соответственно. О получении растворимой формы J-цепи рекомбинантными методами сообщалось в источнике Redwan et al. 2006, Human Antibodies 15:95-102.
Способы получения гибридных антител IgA/IgG и IgM/IgG известны в данной области. Так, например, о получении гибридных антител IgA2/IgG1 и IgM/IgG рекомбинантными методами сообщалось в источнике Chintalacharuvu et al. 2001, Clin Immunol 101(1):21-31. Сообщалось, что присоединение хвостовой частиα или хвостовой части μ на конце тяжелой цепи IgG γ способствует полимеризации и усиливает эффекторную функцию, как, например, активацию комплемента (Smith et al., J. Immunol. J Immunol 1995, 154:2226-2236). Гибридные антитела обладают свойствами как IgA, так и IgG.
Несмотря на достижения, сделанные в области разработки антител, сохраняется необходимость в модифицированных антителах с улучшенными свойствами, как, например, улучшенная аффинность, специфичность и/или авидность.
С прогрессом в данной области с помощью инновационных методов белковой инженерии была усовершенствована функциональность антител, в частности, для достижения более высокой аффинности и продолжительного периода полувыведения и/или лучшего распределения в тканях, а также для обеспечения более высокой точности клеточной деструкции посредством сочетания технологий малых и больших молекул (доставки токсичного молекулярного груза, например, конъюгатов антитела и лекарственного препарата). В другом подходе к улучшению функции антител используется бивалентное связывание структуры иммуноглобулина G (IgG), что позволяет одной молекуле IgG связываться с двумя антигенами. В действительности в некоторых случаях применения асимметричных антител потенциал реализации их полезных функций значителен при одновременном связывании двух различных целевых антигенов. Для удовлетворения этой потребности был создан ряд конструкций с целью получения одной молекулы, способной связываться с двумя разными антигенами, обладающей функциональными свойствами, ранее не встречавшимися в природе. Примером такой биспецифической молекулы является «блинатумомаб» (MT103 или AMG103), который связывается с рецепторами CD3 и CD19 на T- и B-клетках, соответственно. Такая привязка цитотоксической Т-клетки к раковой В-клетке обеспечивает эффективное лечение В-клеточного лейкоза.
Однако сохраняются значительные технические трудности в конструировании, экспрессии и получении биспецифических антител. Благодаря способности одновременно связывать два различных антигена биспецифические антитела рассматриваются в качестве многообещающих терапевтических средств, тем не менее их практическая ценность ограничена ввиду недостаточной стабильности и сложности изготовления.
Для эффективного получения биспецифических IgG были использованы различные методы белковой инженерии с целью ассоциации гетерологичных тяжелых цепей и соответствующей попарной ассоциацией тяжелой и легкой цепей. Кроме того, различные форматы биспецифических антител включают квадромы, гетероконъюгаты, полученные путем химического связывания, рекомбинантные конструкции, полученные путем гетеродимеризации выбранных доменов, и рекомбинантные конструкции минимального размера, состоящие из двух антигенсвязывающих сайтов минимального размера.
Однако все эти усилия были сопряжены с затруднениями.
Таким образом, несмотря на усилия, направленные на разработку генно-инженерных, как, например, биспецифических антител, сохраняется острая необходимость в разработке более эффективных платформ. Это особенно касается терапевтических антител, поскольку разработка и производство новых, модифицированных антител и антителоподобных молекул с множественной специфичностью могут сократить временные рамки между разработкой и клиническим применением таких терапевтических веществ.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение по меньшей мере частично основано на признании того факта, что J-цепь антитела IgM или IgA может быть модифицирована путем введения одного или более связывающих фрагментов в нативную последовательность J-цепи, а модифицированная J-цепь может быть введена в антитела IgM, IgA, IgG/IgM или IgG/IgA без ухудшения функциональных свойств антитела-реципиента или связывания модифицированной J-цепи с ее мишенью. Это обеспечивает взаимодействие модифицированной J-цепи с помощью связывающего фрагмента с одним набором антигенов-мишеней, тогда как антитело IgA, IgM, IgG/IgM или IgG/ IgA может реагировать с другим набором антигенов-мишеней.
Настоящее изобретение дополнительно основано на признании того факта, что путем придания модифицированной J-цепи способности взаимодействовать с эффекторной клеткой, такой как T-клетка, NK-клетка, макрофаг или нейтрофил, может быть активирован иммунный ответ организма, а также может быть улучшена антителозависимая клеточноопосредованная цитотоксичность (ADCC).
В одном аспекте настоящее изобретение касается модифицированной J-цепи, содержащей чужеродный связывающий фрагмент, введенный в нативную последовательность J-цепи.
В одном варианте реализации изобретения нативная последовательность J-цепи представляет собой нативную последовательность J-цепи человека SEQ ID NO.: 1 или ее функциональный фрагмент.
В другом варианте реализации изобретения чужеродный связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1 путем прямого или непрямого слияния.
Еще в одном варианте реализации изобретения связывающий фрагмент вводится путем прямого слияния через пептидный линкер.
В одном варианте реализации изобретения непрямое слияние осуществляется через пептидный линкер, расположенный на C- и/или N-конце связывающего фрагмента или вблизи него.
В другом варианте реализации изобретения чужеродный связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1, расположенную на или вблизи от C- и/или N-конца, как, например, на расстоянии около 10 остатков от C-конца и/или N-конца.
В дополнительном варианте реализации изобретения чужеродный связывающий фрагмент вводится в нативную последовательность J-цепи между цистеиновыми остатками 92 и 101 SEQ ID NO.: 1.
Еще в одном варианте реализации изобретения чужеродный связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1, в или вблизи от сайта гликозилирования.
Длина пептидного линкера, в случае наличия такового, может составлять, например, около 10-20 аминокислот или около 15-20 аминокислот, или 15 аминокислот.
В дополнительном варианте реализации изобретения чужеродный связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1 путем дериватизации с применением химических или химико-ферментативных методов. Химический линкер может представлять собой расщепляемый или нерасщепляемый линкер, причем расщепляемый линкер, например, может представлять собой линкер, неустойчивый к действию химических веществ или ферментов.
В дополнительном варианте реализации изобретения линкер выбран из группы, состоящей из N-сукцинимидил-3-(2-пиридилдитио)пропионата (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилата (SMCC), N-сукцинимидил-4-(2-пиридилдитио)пентаноата (SPP), иминотиолана (IT), бифункциональных производных имидоэфиров, активных эфиров, альдегидов, бис-азидосоединений, производных бис-диазония, диизоцианатов и бис-активных соединений фтора.
В другом варианте реализации изобретения J-цепь модифицирована путем вставки сайта узнавания ферментом и посттрансляционного присоединения чужеродного связывающего фрагмента к сайту узнавания ферментом через пептидный или непептидный линкер.
Во всех вариантах реализации изобретения чужеродный связывающий фрагмент может быть выбран, например, из группы, состоящей из антител, антигенсвязывающих фрагментов антител, конъюгатов антитела и лекарственного препарата, антителоподобных молекул, антигенсвязывающих фрагментов антителоподобных молекул, растворимых и мембраносвязанных белков, лигандов, рецепторов, вирусоподобных частиц, белковых токсинов, ферментов и альтернативных каркасных белков. Примеры альтернативных каркасных белков включают дарпины, домены фибронектина, аднектины и ноттины. Типичные антигенсвязывающие фрагменты включают F(ab')2, F(ab)2, Fab', Fab, Fv, scFv и однодоменные антитела. В предпочтительном варианте реализации изобретения антигенсвязывающий фрагмент представляет собой scFv.
В одном варианте реализации изобретения чужеродный связывающий фрагмент модифицированной J-цепи связывается с эффекторной клеткой, причем эффекторная клетка может быть выбрана, например, из группы, состоящей из T-клеток, натуральных киллеров (NK), макрофагов и нейтрофилов.
В одном варианте реализации изобретения эффекторная клетка представляет собой T-клетку, причем чужеродный связывающий фрагмент может связываться, например, с CD3ε на T-клетке.
В другом варианте реализации изобретения эффекторная клетка представляет собой NK-клетку, причем мишень для чужеродного связывающего фрагмента модифицированной J-цепи может быть выбрана, например, из группы, состоящей из CD16, CD64 и NKG2D на NK-клетке.
В дополнительном варианте реализации изобретения эффекторная клетка представляет собой макрофаг, причем чужеродный связывающий фрагмент модифицированной J-цепи может связываться, например, с CD14 на макрофаге.
В дополнительном варианте реализации изобретения эффекторная клетка представляет собой нейтрофил, причем чужеродный связывающий фрагмент модифицированной J-цепи может связываться, например, с CD16b или CD177 на нейтрофиле.
В одном аспекте настоящее изобретение касается антитела, содержащего модифицированную J-цепь, которая описана выше в данном документе, или антигенсвязывающего фрагмента такого антитела. Антитело может представлять собой антитело IgM, IgA, IgG/IgM или IgG/IgA и включает мультиспецифические, например, биспецифические антитела.
В одном варианте реализации изобретения антитело представляет собой антитело IgM, антитело IgA или антитело IgG, содержащее хвостовую часть или ее антигенсвязывающий фрагмент.
В другом варианте реализации изобретения наблюдалось специфическое связывание антител с одной или более мишенями для связывания, выбранными из группы, состоящей из клеток-мишеней, растворимых форм мишеней для связывания, рецепторов клеточной поверхности, белков матрикса, рецепторов транспортеров.
Еще в одном варианте реализации изобретения антитело связывается с опухолевой клеткой.
В дополнительном варианте реализации изобретения антитело связывается с опухолеассоциированным антигеном-мишенью, который представлен на Фиг. 19, причем J-цепь, например, может быть модифицирована таким образом, что она способна связывать любую из мишеней эффекторных клеток, приведенных на Фиг. 19.
Еще в одних вариантах реализации изобретения модифицированная J-цепь содержится в антителах в ориентации V-линкер-J или в ориентации J-линкер-V.
В одном варианте реализации изобретения опухоль представляет собой гематологическую злокачественную опухоль или солидную опухоль, причем гематологическая злокачественная опухоль может представлять собой, например, лейкоз, лимфому, миелому и миелодиспластический синдром, в частности, включая острый миелоидный лейкоз, острый лимфобластный лейкоз, хронический миелогенный лейкоз или хронический лимфоцитарный лейкоз, лимфому Ходжкина или неходжкинскую лимфому. В таких вариантах реализации изобретения антитело может связываться, например, с одним или более из CDIM, CD19, CD20, CD22, CD33, CD70, CD56, CD138, а модифицированная J-цепь может связываться с CD3ε.
В другом варианте реализации изобретения опухоль представляет собой эпителиальную опухоль.
Еще в одном варианте реализации изобретения антитело связывается с мишенью на углеводной основе на опухоли.
В дополнительном варианте реализации изобретения антитело связывается с вирусным антигеном, таким как антиген HBV или антиген HIV, например, PreS1 или GP120.
В дополнительном аспекте настоящее изобретение касается композиции, содержащей антитела IgM, IgA, IgG/IgM, IgG/IgA, содержащие модифицированную J-цепь, которая описана в данном документе. Композиция может представлять собой, например, фармацевтическую композицию или диагностическую композицию.
Еще в одном аспекте настоящее изобретение касается способа лечения опухоли или вирусного заболевания, включающего введение субъекту, который в этом нуждается, эффективного количества антитела IgM, IgA, IgG/IgM, IgG/IgA с модифицированной J-цепью, что описана в данном документе.
В другом аспекте настоящее изобретение касается применения антитела IgM, IgA, IgG/IgM, IgG/IgA с модифицированной J-цепью, которая описана в данном документе, в получении лекарственного препарата для лечения опухоли или вирусного заболевания.
Еще в одном аспекте настоящее изобретение касается применения антитела IgM, IgA, IgG/IgM, IgG/IgA с модифицированной J-цепью, которая описана в данном документе, в лечении опухоли или вирусного заболевания.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 проиллюстрирована структура пентамера IgM, содержащего J-цепь, причем цепи A и B идентичны нативному IgM.
На ФИГ. 2 представлены структурные схемы IgA, димера IgA и секреторного IgA (sIgA).
На ФИГ. 3 показана аминокислотная последовательность зрелой J-цепи человека (SEQ ID NO.: 1).
На ФИГ. 4 представлено выравнивание зрелой J-цепи человека (SEQ ID NO.: 1) и J-цепи из различных видов животных (SEQ ID NO.: 1-44).
Hum=NP_653247 23-159 (SEQ ID NO.: 1) человека
Chm=XP_001160135 39-175 (SEQ ID NO.: 2) шимпанзе
Gor=XP_004038830 23-159 (SEQ ID NO.: 3) гориллы
Ora=NP_001125381 23-159 (SEQ ID NO.: 4 орангутана
Gib=XP_003265761 39-175 (SEQ ID NO.: 5) гиббона.
Mar=XP_003732538 32-168 (SEQ ID NO.: 6) игрунки обыкновенной
Cyn=NP_001247815 23-159 (SEQ ID NO.: 7) яванского макака
Bab=XP_003898844 23-159 (SEQ ID NO.: 8) бабуина
Squ=XP_003931978 23-159 (SEQ ID NO.: 9) беличьей обезьяны
Shr=XP_006142955 25-160 (SEQ ID NO.: 10) обыкновенной тупайи
Dol=XP_004328961 25-158 (SEQ ID NO.: 11) дельфина
Kil=XP_004283629 25-158 (SEQ ID NO.: 12) дельфина-косатки
Ele=XP_003414110 25-158 (SEQ ID NO.: 13) слона
Sea=XP_006729388 25-158 (SEQ ID NO.: 14) тюленя
Rhi=XP_004419202 25-157 (SEQ ID NO.: 15) носорога
Cat=XP_003985350 25-158 (SEQ ID NO.: 16) домашней кошки
Wol=XP_532398 26-159 (SEQ ID NO.: 17) волка
Pan=EFB23253 1-134 (SEQ ID NO.: 18) большой панды
Fer=XP_004766376 26-158 (SEQ ID NO.: 19) фретки
Hor=NP_001271464 25-158 (SEQ ID NO.: 20) лошади
Gui=NP_001265705 25-160 (SEQ ID NO.: 21) морской свинки
Cam=XP_006188738 25-158 (SEQ ID NO.: 22) верблюда
Goa=XP_005681786 25-158 (SEQ ID NO.: 23) козы
Chn=XP_005392838 94-229 (SEQ ID NO.: 24) шиншиллы
Ham=XP_005068228 24-160 (SEQ ID NO.: 25) хомяка
She=XP_004009937 25-158 (SEQ ID NO.: 26) овцы
BBa=XP_006094475 25-158 (SEQ ID NO.: 27) бурой ночницы
Ant=XP_005983836 25-158 (SEQ ID NO.: 28) антилопы
Cow=NP_786967 25-157 (SEQ ID NO.: 29) коровы
Mou=NP_690052 23-159 (SEQ ID NO.: 30) мыши
Rat=EDL88516 23-159 (SEQ ID NO.: 31) крысы
Hed=XP_004703237 25-157 (SEQ ID NO.: 32) ежа
Rab=P23108 1-136 (SEQ ID NO.: 33) кролика
Opo=XP_003341415 29-162 (SEQ ID NO.: 34) опоссума
All=XP_006270094 26-159 (SEQ ID NO.: 35) аллигатора
Tur=XP_005304167 26-159 (SEQ ID NO.: 36) черепахи
Tas=XP_003772863 27-160 (SEQ ID NO.: 37) сумчатого дьявола
Pla=XP_003430954 36-160 (SEQ ID NO.: 38) утконоса
Par=XP_005142787 28-160 (SEQ ID NO.: 39) волнистого попугайчика
Duc=XP_005031370 28-160 (SEQ ID NO.: 40) утки
Chi=NP_989594 26-158 (SEQ ID NO.: 41) курицы
Tur=XP_003205717 27-159 (SEQ ID NO.: 42) индейки
Fal=XP_005243236 29-160 (SEQ ID NO.: 43) сокола
Spa=XP_005492217 29-158 (SEQ ID NO.: 44) воробья
На ФИГ. 5 показано выравнивание аминокислотных последовательностей J-цепей человека и летучей мыши (маврикийская летучая лисица, Pteropus alecto) (SEQ ID NO.: 1 и 45, соответственно).
На ФИГ. 6: антитело IgM, в котором отсутствует J-цепь (отрицательный контроль), и IgM, содержащее J-цепь, разделяли на ДСН-ПААГ в восстанавливающих условиях и анализировали с помощью вестерн-блота с использованием антитела к J-цепи. Антитело к J-цепи реагировало только с IgM, содержащим J-цепь.
На ФИГ. 7: собранный супернатант из трансфецированных клеток CHO поровну разделяли для иммунопреципитации либо с использованием аффинной матрицы с антителами к мю-цепи или белкам с эпитопной меткой Myc. Клетки CHO экспрессировали так называемые CDIM (молекулы, индуцирующие клеточную гибель) - IgM, специфичный к В-клеткам. Это антитело IgM известно как IGM-55.5. Иммунопреципитированные белки разделяли на ДСН-ПААГ в восстанавливающих условиях и анализировали с помощью вестерн-блота с использованием антител к J-цепи и белкам с эпитопной меткой Myc. Как клетки, трансфецированные плазмидой, содержащей модифицированную специфичную к CD3 J-цепь с модифицирующим одноцепочечным Fv-фрагментом на С-конце, так и клетки, трансфецированные плазмидой, содержащей модифицированную специфичную кCD3 J-цепь с модифицирующим одноцепочечным Fv-фрагментом на N-конце, характеризовались наличием полосы на геле, соответствующей белку около 51 кДа, который реагировал как с антителами к J-цепи, так и с антителами к белкам с эпитопной меткой Myc.
На ФИГ. 8 проиллюстрированы две разные ориентации биспецифического пентамера IgM, содержащего модифицированную J-цепь, со специфичностью связывания по отношению к Т-клеткам (Т-клеточный антиген CD3).
На ФИГ. 9: последовательность (SEQ ID NO.: 46) и структура одноцепочечного Fv-фрагмента, специфичного к CD3, содержащего J-цепь на C-конце (модифицированная анти-CD3 J-цепь с модифицирующим одноцепочечным Fv-фрагментом на С-конце). SP=сигнальный пептид.
На ФИГ. 10: последовательность (SEQ ID NO.: 47) и структура одноцепочечного Fv-фрагмента, специфичного к CD3, содержащего J-цепь на N-конце (модифицированная анти-CD3 J-цепь с модифицирующим одноцепочечным Fv-фрагментом на N-конце). SP=сигнальный пептид.
На ФИГ. 11 представлено схематическое изображение асимметрического биспецифического пентамера IgM со специфичностью связывания по отношению к антигену-мишени, содержащего связывающий домен, ковалентно связанный с J-цепью со специфичностью связывания по отношению к эффекторной клетке.
На ФИГ. 12 представлены вестерн-блоты пентамера IgM, специфичного к CD20, с J-цепью, ковалентно модифицированной с использованием домена, связывающего CD3 scFv. С помощьюПААГ-электрофореза в невосстанавливающих условиях с окраской раствором Кумасси синим удается обнаружить полноразмерный IgM молекулярной массой приблизительно 1 миллион дальтон. Модифицированная J-цепь встраивается в состав пентамеров IgM в обеих ориентациях: CD3-J и J-CD3.
На ФИГ. 13 показаны результаты транзиентных котрансфекций плазмидами, несущими гены тяжелой цепи (HC), легкой цепи (LC) и модифицированной J-цепи биспецифического IgM, специфичного к CD20 (CD20IgM), причем модифицированная J-цепь обеспечивает специфичность связывания по отношению к CD3 (CD3J).
На ФИГ. 14 показано, что встраивание модифицированной J-цепи в состав IgM к CD20, не оказывает никакого отрицательного действия на CDC-активность. В дополнение к этому, полученные данные показывают, что молекула IgM, содержащая модифицированную J-цепь в любой ориентации (V15J или J15V, где «15» обозначает длину линкера), обладает намного большей активностью, чем антитело IgG к CD20.
На ФИГ. 15 показано, что молекула биспецифического антитела IgM к антигену CD20 В-лимфоцитов, содержащая J-цепь к антигену CD3 Т-лимфоцитов, обладает значительной активностью при истощении популяции B-клеток. Это обусловлено высокой Т-клеточной цитотоксичностью в отношении В-лимфоцитов благодаря специфичности модифицированной J-цепи к антигену CD3. Raji, линию CD19+CD20+ B-клеток, кокультивировали с T-ALL, линией цитолитических CD8+ Т-клеток) с CD20 IgM x J-дикого типа или CD20 IgM x CD3-J-цепь в течение 24 часов при 37 градусах, 5% CO2. Клетки собирали и окрашивали флуоресцентными антителами к CD3 и CD19, а также анализировали с помощью проточной цитометрии для оценки жизнеспособности B-клеток.
На ФИГ. 16: биспецифическая молекула IgM, специфичная к CD20, с модифицированной J-цепью, обеспечивающей специфичность связывания по отношению к CD3, обладает значительно улучшенной цитотоксической активностью по сравнению с пентамером IgM, специфичным к CD20, с J-цепью дикого типа. Биспецифический пентамер IgM, вовлекающий Т-клетки, является эффективным в концентрации 1 нг/мл и обладает в 300 раз большей активностью по сравнению с IgM, специфичным к CD20, с J-цепью дикого типа.
На ФИГ. 17 показано истощение популяции B-клеток in vivo при использовании IgG к CD20, IgM и биспецифического IgM. Были приобретены гуманизированные мыши линии NSG, которым были пересажены стволовые клетки CD34+ человека (Jackson Laboratory; номер популяции: 007799), а цельную кровь анализировали в отношении уровней CD19+ B-клеток до введения препарата. Препарат CD20 IgM x CD3-J-цепь был получен для внутривенного введения животным при 0,5 мг/кг. Через 6 часов после введения у животных получали цельную кровь и анализировали с помощью проточной цитометрии в отношении уровней циркулирующих в крови В-клеток человека с использованием флуоресцентно меченого антитела к CD19+.
На ФИГ. 18 представлен вестерн-блот специфичной к CD16 J-цепи, несущей Vhh-домен верблюжьего антитела.
На ФИГ. 19 приведены мишени антитела IgM и мишени эффекторных клеток для модифицированной J-цепи. Любая из мишеней IgM, перечисленных в левом столбце таблицы, может быть комбинирована с любой из мишеней модифицированной J-цепи, перечисленных в правом столбце.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Определения
До того как настоящее изобретение будет описано более подробно, надо понимать, что это изобретение не ограничивается никакими описанными конкретными вариантами реализации изобретения и в силу этого, разумеется, может быть изменено. Также надо понимать, что терминология, использованная в данном документе, предназначена для описания только конкретных вариантов реализации изобретения и не подразумевает ограничения, поскольку объем настоящего изобретения ограничивается только прилагаемой формулой изобретения.
Если представлен диапазон значений, надо понимать, что каждое из промежуточных значений до одной десятой доли нижнего предела, за исключением случаев, когда в контексте четко указано иное, между верхним и нижним пределами этого диапазона и любое другое установленное или промежуточное значения охватываются настоящим изобретением. Верхние и нижние пределы этих меньших диапазонов (кроме пределов, специально исключенных из таких диапазонов) могут быть независимо включены в меньшие диапазоны, охваченные настоящим изобретением.
Если не указано иное, технические и научные термины, используемые в данном документе, имеют такое значение, которое обычно понятно специалисту в данной области техники, к которой относится настоящее изобретение. Источник Singleton et al., Dictionary of Microbiology and Molecular Biology 2nd ed., J. Wiley & Sons (New York, NY 1994) может послужить специалистам в данной области техники в виде общего руководства к пониманию многих из терминов, использованных в настоящей заявке.
Все публикации, упомянутые в данном документе, в явном виде включены в данный документ посредством ссылки для раскрытия и описания способов и/или материалов, в связи с чем цитируются указанные публикации.
Термин «антитело» охватывает моноклональные антитела (включая полноразмерные антитела, которые имеют Fc-область иммуноглобулина), одноцепочечные молекулы, а также фрагменты антител (например, Fab, F(ab')2 и Fv). В данном документе термин «иммуноглобулин» (Ig) используется взаимозаменяемо с термином «антитело». Основным структурным элементом антитела, состоящего из 4 цепей, является гетеротетрамерный гликопротеин, состоящий из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Если не указано иное, термин «антитело» в данном документе используется в самом широком смысле и, в частности, охватывает все изотипы, подклассы и формы антител, включая антитела IgG, IgM, IgA, IgD и IgE и их фрагменты, предпочтительно антигенсвязывающие фрагменты. Предпочтительные антитела по настоящему изобретению включают антитела IgM и IgA и их антигенсвязывающие фрагменты, которые могут быть модифицированы таким образом, что они будут включать последовательности из других изотипов, как, например, IgG, для получения химерных антител.
В случае IgG, молекулярная масса структурного элемента, состоящего из 4 цепей, как правило, составляет 150000 дальтон. Каждая L-цепь связана с H-цепью с помощью одной ковалентной дисульфидной связи, тогда как две H-цепи связаны друг с другом с помощью одной или более дисульфидных связей в зависимости от изотипа H-цепи. В каждой H- и L-цепи также равномерно располагаются внутрицепочечные дисульфидные мостики. Каждая H-цепь имеет N-конец, вариабельный домен (VH), за которым расположены три константных домена (CH) для каждой из цепей α и γ, и четыре CH домена для изотипов μ и ε. Каждая L-цепь имеет N-конец, вариабельный домен (VL), за которым расположен константный домен на другом конце. VL выравнивается с VH, а CL выравнивается с первым константным доменом тяжелой цепи (CH1). Предполагается, что определенные аминокислотные остатки образуют поверхность контакта между вариабельными доменами легкой цепи и тяжелой цепи. Попарное соединение VH и VL приводит к образованию одного антигенсвязывающего сайта.
IgM представляет собой гликопротеин, который формирует полимеры, в которых многие молекулы иммуноглобулина ковалентно связаны вместе с помощью дисульфидных связей. IgM главным образом встречается в виде пентамера, а также в виде гексамера и, таким образом, содержит 10 или 12 антигенсвязывающих сайтов. Пентамерная форма обычно содержит дополнительный полипептид, называемый J-цепью, но также может быть получена в отсутствие J-цепи. Молекула пентамера IgM имеет молекулярную массу приблизительно 970 кДа. Ввиду полимерной природы IgM обладает высокой авидностью и является особенно эффективным в активации комплемента. В отличие от IgG тяжелая цепь в мономерах IgM состоит из одной вариабельной и четырех константных доменов. Константные домены IgM обозначены в данном документе как CM1 или Cμ1, CM2 или Cμ2, CM3 или Cμ3 и CM4 или Cμ4, причем обозначения «CM» и Cμ» используются взаимозаменяемо. Структура пентамера IgM проиллюстрирована на ФИГ. 1.
Термин «IgM» в данном документе используется в самом широком смысле и, в частности, охватывает моно- и мультиспецифические (включая биспецифические) молекулы IgM, такие как, например, мультиспецифические связывающие молекулы IgM, описанные в заявке PCT № PCT/US2014/054079, полное описание которой в явном виде включено в данный документ посредством ссылки.
Термин «связывающий элемент IgM» или «связывающий элемент антитела IgM» используется в самом широком смысле и, в частности, охватывает полипептид константной области тяжелой цепи антитела IgM, содержащий по меньшей мере константный домен CM4, слитый с последовательностью вариабельного домена (VH), связывающейся с мишенью (например, антигеном) в присутствии или в отсутствие соответствующей последовательности вариабельного домена легкой цепи антитела (VL).
Термин «биспецифический связывающий элемент IgM» или «биспецифический связывающий элемент антитела IgM» используется в самом широком смысле и, в частности, охватывает пру полипептидов константной области тяжелой цепи антитела IgM, содержащих по меньшей мере константный домен CM4, слитый с последовательностью вариабельного домена (VH), причем каждая последовательность вариабельного домена связывается с разной мишенью в присутствии или в отсутствие соответствующих последовательностей вариабельного домена легкой цепи антитела (VL). В одном варианте реализации изобретения биспецифическое антитело IgM содержит две антигенсвязывающие области VHVL, причем каждая из них способна к связыванию с разным эпитопом на одном антигене или эпитопами на двух разных антигенах. Биспецифические связывающие элементы антитела IgM могут представлять собой полноразмерные элементы из одного вида или могут быть химеризованными или гуманизированными. Биспецифические антитела IgM по настоящему изобретению имеют пента- или гексамерную кольцевидную структуру, состоящую из пяти или шести связывающих биспецифических элементов антитела IgM.
Термин «мультиспецифический IgM» используется в данном документе в самом широком смысле для обозначения антител IgM, обладающих двумя или более специфичностями связывания. Следовательно, термин «мультиспецифический» охватывает «биспецифический», например, биспецифические антитела или биспецифические связывающие элементы, включая пентамеры IgM, содержащие по меньшей мере две моноспецифические субъединицы, причем каждая субъединица связывается с разным антигеном (AA, BB), или пять, или шесть биспецифических субъединиц, причем каждая субъединица связывается с двумя разными антигенами (AB, AB). Следовательно, биспецифические и мультиспецифические пентамеры IgM могут включать пять идентичных биспецифических связывающих элементов, моноспецифические связывающие элементы, причем по меньшей мере два из них обладают разными специфичностями, или их комбинацию.
«Тяжелая цепь полноразмерного антитела IgM» представляет собой полипептид, состоящий (в направлении от N-конца к C-концу) из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 (CM1 или Cμ1) тяжелой цепи антитела, константного домена 2 (CM2 или Cμ2) тяжелой цепи антитела, константного домена 3 (CM3 или Cμ3) тяжелой цепи антитела и константного домена 4 (CM4 или Cμ4) тяжелой цепи антитела. Биспецифические полноразмерные антитела IgM, как указано в данном документе, состоят из пяти или шести мономеров (связывающих элементов), причем каждый мономер содержит два антигенсвязывающих сайта, которые специфически связываются с двумя разными мишенями для связывания (эпитопами). Для обозначения C-конца тяжелой или легкой цепи полноразмерного антитела служит последняя аминокислота на C-конце тяжелой или легкой цепи. Для обозначения N-конца тяжелой или легкой цепи полноразмерного антитела служит первая аминокислота на N-конце тяжелой или легкой цепи.
Нативный IgA представляет собой тетрамерный белок, содержащий две идентичные легкие цепи (κ или λ) и две идентичные тяжелые цепи (α). Для человека характерны два изотипа IgA, IgA1 и IgA2. IgA по аналогии с IgG содержит три константных домена (A1-CA3 или Cα1-Cα3) с шарнирной областью между доменами Cα1 и Cα2, причем обозначения «CA» и «Cα» используются взаимозаменяемо. Во всех изотипах IgA содержится «хвостовая часть» длиной 18 аминокислот, которая расположена на C-конце по отношению к домену Cα3, что способствует образованию полимерной молекулы Ig (см., например, Garcia-Pardo et al., 1981, J. Biol. Chem. 256, 11734-11738 and Davis et al., 1988, Eur. J. Immunol. 18, 1001-1008). Сывороточный IgA представляет собой мономер, но он также может иметь форму полимеров. В форме секреторной молекулы IgA содержит 2-5 основных состоящих из 4 цепей элемента, связанных J-цепью, которая может включать хвостовую часть, и может быть ассоциирована с секреторным компонентом. Структуры хвостовой части, димерного IgA и секреторного IgA, ассоциированного с секреторным компонентом (sIgA), проиллюстрированы на Фиг. 2. Антитела IgA дополнительно можно поделить на подклассы IgA1 и IgA2. Термин антитело «IgA» используется в данном документе, чтобы, в частности, охватить все подклассы, т.е. антитела IgA1 и IgA2, включая димерные и мультимерные формы с секреторным компонентом и без него, а также фрагменты, предпочтительно антигенсвязывающие фрагменты, таких антител. Предпочтительно для целей настоящего изобретения антитело IgA представляет собой димер, в котором две хвостовые части соединены с помощью J-цепи (см., Фиг. 2).
Термин «IgA» в данном документе используется в самом широком смысле и, в частности, охватывает моно- и мультиспецифические молекулы IgA, такие как, например, мультиспецифические связывающие молекулы IgA, описанные в заявке PCT № PCT/US2015/015268, полное описание которой в явном виде включено в данный документ посредством ссылки.
Термин «мультиспецифический IgA» используется в данном документе в самом широком смысле для обозначения антител IgA, обладающих двумя или более специфичностями связывания. Следовательно, термин «мультиспецифический» охватывает «биспецифический», например, биспецифические антитела или биспецифические связывающие элементы, включая димеры IgA, содержащие две моноспецифические субъединицы, причем каждая субъединица связывается с разным антигеном (AA, BB), или две биспецифические субъединицы, причем каждая субъединица связывается с двумя разными антигенами (AB, AB).
В одном варианте реализации изобретения димерные мультиспецифические молекулы IgA состоят из двух моноспецифических связывающих элементов, причем каждый связывающий элемент обладает специфичностью связывания по отношению к разным мишеням для связывания (AA, BB). В другом варианте реализации изобретения в димерных молекулах IgA один из двух связывающих элементов обладает двумя специфичностями связывания (т.е. является биспецифическим, например, AA, A, B или AA, BC). В другом варианте реализации изобретения каждый из двух связывающих элементов обладает двумя специфичностями, которые могут быть одинаковыми (AB, AB) или разными (например, AC, CD или AB, AC).
Термин «биспецифический связывающий элемент антитела IgA» используется в самом широком смысле и, в частности, охватывает пру полипептидов константной области тяжелой цепи антитела IgA, содержащих по меньшей мере константный домен CA3, связанный с последовательностью вариабельного домена (VH), причем каждая последовательность вариабельного домена связывается с разной мишенью в присутствии или в отсутствие соответствующих последовательностей вариабельного домена легкой цепи антитела (VL). В одном варианте реализации изобретения биспецифическое антитело IgA содержит две антигенсвязывающие области VHVL, причем каждая из них способна к связыванию с разным эпитопом на одном антигене или эпитопами на двух разных антигенах. Биспецифические связывающие элементы антитела IgA могут представлять собой полноразмерные элементы из одного вида или могут быть химеризованными или гуманизированными.
«Тяжелая цепь полноразмерного антитела IgA» представляет собой полипептид, состоящий (в направлении от N-конца к C-концу) из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 (CA1 или Cα1) тяжелой цепи антитела, константного домена 2 (CA2 или Cα2) тяжелой цепи антитела и константного домена 3 (CA3 или Cα3) тяжелой цепи антитела. Би- или мультиспецифические полноразмерные антитела IgA по настоящему изобретению содержат два мономера (связывающие элементы), каждый из которых могут быть моно- или биспецифическим, с секреторным компонентом или без него. Следовательно, мультиспецифические антитела IgA по настоящему изобретению могу включать моноспецифические и биспецифические связывающие элементы, при условии, что полученный в результате IgA обладает по меньшей мере двумя специфичностями связывания. Для обозначения C-конца тяжелой или легкой цепи полноразмерного антитела служит последняя аминокислота на C-конце тяжелой или легкой цепи. Для обозначения N-конца тяжелой или легкой цепи полноразмерного антитела служит первая аминокислота на N-конце тяжелой или легкой цепи.
Дополнительную информацию о структуре и свойствах разных классов антител см., например, Basic and Clinical Immunology, 8th Edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds), Appleton & Lange, Norwalk, Conn., 1994, страница 71 и глава 6.
Используемый в данном документе термин «поверхность контакта» используется для обозначения области, которая содержит «контактные» аминокислотные остатки (или другие неаминокислотные группы, такие как, например, углеводные группы) в первой константной области тяжелой цепи IgM, которые контактируют с одним или более «контактных» аминокислотных остатков (или других неаминокислотных групп) во второй константной области тяжелой цепи IgM.
Термин «асимметричная поверхность контакта» используется для обозначения поверхности контакта (определение которой приведено выше в данном документа), образующейся между двумя цепями антител, как, например, первой и второй константной областью тяжелой цепи IgM и/или между константной областью тяжелой цепи IgM и соответствующей ей легкой цепью, причем контактные остатки в первой и второй цепях различаются по строению, включая комплементарные контактные остатки. Асимметричная поверхность контакта может быть получена вследствие взаимодействий типа выступы/впадина и/или образования соляных мостиков (обращение заряда) и/или других методов, известных в данной области техники, таких как, например, подход CrossMab для присоединения тяжелой μ-цепи к соответствующей ей легкой цепи.
«Впадина» или «углубление» относится по меньшей мере к одной аминокислотной боковой цепи, которая удалена от поверхности контакта со вторым полипептидом и, таким образом, допускает формирование соответствующего выпячивания («выступа») на соседней поверхности контакта с первым полипептидом. Впадина (углубление) может изначально располагаться на поверхности контакта или может быть введена искусственно (например, путем изменения нуклеиновой кислоты, кодирующей белки, формирующие поверхность контакта). Как правило, нуклеиновую кислоту, кодирующую поверхность контакта со вторым полипептидом, изменяют таким образом, чтобы она была способна кодировать впадину. Для достижения этого нуклеиновую кислоту, кодирующую по меньшей мере один «оригинальный» аминокислотный остаток на поверхности контакта со вторым полипептидом, заменяют на ДНК, кодирующую по меньшей мере один «вводимый» аминокислотный остаток, что может иметь боковую цепь меньшего объема, чем оригинальный остаток. Следует понимать, что могут быть использованы несколько оригинальных и соответствующих вводимых остатков. Верхний предел для числа оригинальных остатков, которые подлежат замене, представляет собой число остатков на поверхности контакта со вторым полипептидом. Предпочтительные вводимые остатки для формирования впадины обычно представляют собой природные аминокислотные остатки и предпочтительно выбираются из аланина (A), серина (S), треонина (T), валина (V) и глицина (G). Наиболее предпочтительными аминокислотными остатками являются серин, аланин или треонин, наиболее предпочтительно аланин. В предпочтительном варианте реализации изобретения оригинальный остаток для формирования выступа имеет боковую цепь большого объема, содержащую тирозин (Y), аргинин (R), фенилаланин (F) или триптофан (W).
«Оригинальный» аминокислотный остаток - это остаток, который заменяется на «вводимый» аминокислотный остаток, что может иметь боковую цепь большего или меньшего объема, чем оригинальный остаток. Вводимый аминокислотный остаток может представлять собой встречающийся в природе или не встречающийся в природе аминокислотный остаток, но предпочтительно представляет собой последний.
Под «не встречающимся в природе аминокислотным остатком» подразумевается остаток, который не кодируется с помощью генетического кода, но который способен к ковалентному связыванию со смежным(и) аминокислотным(и) остатком(и) в полипептидной цепи. Примерами не встречающихся в природе аминокислотных остатков являются норлейцин, орнитин, норвалин, гомосерин и другие аналоги аминокислотных остатков, как те, что описаны, например, в следующем источнике Ellman et al., Meth. Enzym. 202:301-336 (1991). Для получения таких не встречающихся в природе аминокислотных остатков могут быть использованы методики Noren et al. Science 244: 182 (1989) и Ellman et al., supra. Вкратце, это включает химическую активацию супрессорной тРНК с использованием не встречающегося в природе аминокислотного остатка с последующей транскрипцией и трансляцией РНК in vitro. Способы по настоящему изобретению в определенных вариантах реализации изобретения включают замену по меньшей мере одного оригинального аминокислотного остатка в тяжелой цепи IgM, но могут быть заменены несколько оригинальных остатков. Как правило, замене подлежат не все оригинальные аминокислотные остатки, расположенные на поверхности контакта первого и второго полипептидов. Предпочтительные оригинальные остатки, используемые для замены, являются «спрятанными». Под термином «спрятанный» подразумевается, что остаток, по существу, недоступен растворителю. Предпочтительно, чтобы вводимый остаток отличался от цистеина для предотвращения окисления или ошибочного попарного соединения при образовании дисульфидных связей.
Выступ «может располагаться во впадине», что означает, что пространственное расположение выступа и впадины на поверхности контакта первого и второго полипептидов, соответственно, а также размеры выступа и впадины таковы, что выступ может располагаться во впадине без значительного нарушения нормальной ассоциации первого и второго полипептидов на поверхности контакта. Поскольку выступы, формируемые, например, Tyr, Phe и Trp, как правило, не выступают перпендикулярно оси поверхности контакта и имеют предпочтительные конформации, выравнивание выступа с соответствующей впадиной основано на моделировании пары выступ/впадина, исходя из трехмерной структуры, как, например, структуры, полученной с помощью рентгеноструктурного анализа или ядерно-магнитного резонанса (ЯМР). Этого можно достичь с использованием методов, широко распространенных в данной области, включая методы молекулярного моделирования.
Под «оригинальной нуклеиновой кислотой» подразумевается нуклеиновая кислота, кодирующая представляющий интерес полипептид, которая может быть «изменена» (т.е. изменена методами генетической инженерии или путем введения мутаций) таким образом, чтобы она была способна кодировать белок с выступом и впадиной. Оригинальная или исходная нуклеиновая кислота может представлять собой встречающуюся в природе нуклеиновую кислоту или может включать нуклеиновую кислоту, которая ранее подвергалась изменению (например, гуманизированный фрагмент антитела). Под «изменением» нуклеиновой кислоты подразумевается, что оригинальная нуклеиновая кислота подвергается мутации путем вставки, делеции или замены по меньшей мере одного кодона, кодирующего представляющий интерес аминокислотный остаток. Как правило, кодон, кодирующий оригинальный остаток, заменяется на кодон, кодирующий вводимый остаток. Методы генетической модификации ДНК таким способом были рассмотрены в Mutagenesis: a Practical Approach, M. J. McPherson, Ed., (IRL Press, Oxford, UK. (1991) и включают, например, сайт-направленный мутагенез, кассетный мутагенез и мутагенез с помощью полимеразной цепной реакции (ПЦР).
Выступ и впадина могут быть «введены» на поверхности контакта первого и второго полипептида с помощью способов синтеза, например, с помощью рекомбинантных методов, синтеза пептидов in vitro, тех методов, которые предназначены для введения не встречающихся в природе ранее описанных аминокислотных остатков путем ферментативного или химического соединения пептидов, или другой комбинации этих методов. В соответствии с этим, выступ или впадина, которые «вводятся», являются «неприродными» или «ненативными», что означает, что они не существуют в природе или в оригинальном полипептиде (например, гуманизированное моноклональное антитело).
Предпочтительно, чтобы вводимый аминокислотный остаток для образования выступа имел относительно небольшое количество «ротамеров» (например, 3-6). «Ротамер» представляет собой энергетически выгодную конформацию боковой цепи аминокислот. Количество ротамеров для различных аминокислотных остатков рассмотрено в источнике Ponders and Richards, J. Mol. Biol. 193: 775-791 (1987).
Если не указано иное, термин «антитело», в частности, охватывает нативные человеческие и отличные от человеческих антитела IgG1, IgG2, IgG3, IgG4, IgE, IgA, IgD и IgM, включая встречающиеся в природе варианты. Следовательно, например, последовательность IgM человека имеет идентификационный номер в базе данных GenBank X14940.1, тогда как согласно сообщениям варианты имеют номера CAB37838.1, CAC20458.1, AFM37312.1, X57331.1 и J00260.1. в базе данных GenBank.
Термин «нативный» в отношении полипептида (например, антитело или J-цепь) используется в данном документе для обозначения полипептида, имеющего последовательность, которая встречается в природе независимо от способа получения. Следовательно, термины «нативный» и «нативная последовательность» используются в данном документе взаимозаменяемо и в явном виде охватывают рекомбинантные полипептиды с последовательностью, которая встречается в природе.
Используемые в данном документе термины «нативная последовательность J-цепи» или «нативная J-цепь» относится к J-цепи антител IgM или IgA с нативной последовательностью любого вида животных, включая зрелую J-цепь человека, аминокислотная последовательность которой показана на Фиг. 3 (SEQ ID NO.: 1), и J-цепи антител видов животных, отличных от человека, включая, в том числе, полипептиды с нативной последовательностью J-цепей SEQ ID NO.: 2-44, показанные на Фиг. 2, или содержащий J-цепь полипептид летучей мыши (SEQ ID NO.: 45), показанный на Фиг. 5.
Термин «модифицированная J-цепь» используется в данном документе для обозначения вариантов нативной последовательности содержащих J-цепь полипептидов, содержащих чужеродный связывающий фрагмент, введенный в нативную последовательность. Введение может быть достигнуто любым способом, включая прямое или непрямое слияние чужеродного связывающего фрагмента или путем химического присоединения через линкер. Термин «модифицированная J-цепь», в частности, охватывает, в том числе, J-цепь человека с нативной аминокислотной последовательностью SEQ ID NO.: 1, модифицированную путем введения связывающего фрагмента. Указанный термин, в частности, охватывает, в том числе, J-цепь человека с нативной аминокислотной последовательностью человека SEQ ID NO.: 1, модифицированную путем введения чужеродного связывающего фрагмента, который не уменьшает эффективность полимеризации (димеризации) IgM или IgA и связывание таких полимеров (димеров) с мишенью.
Термин «связывающий фрагмент» используется в данном документе в самом широком смысле для обозначения любого химического средства, способного к специфическому связыванию с мишенью, такой как антиген. Примеры связывающих фрагментов включают, в том числе, антитела, антигенсвязывающие фрагменты антител, конъюгаты антитела и лекарственного препарата, антителоподобные молекулы, антигенсвязывающие фрагменты антителоподобных молекул, растворимые и мембраносвязанные белки, лиганды, рецепторы, вирусоподобные частицы, белковые токсины, ферменты и альтернативные каркасные белки. К предпочтительным связывающим фрагментам относятся полипептиды (включая пептиды), предпочтительно с биологической функцией. Примером биологической функции может послужить способность связывающего фрагмента к связыванию и активации эффекторной клетки, такой как B-клетка, T-клетка или натуральный киллер (NK).
Термин «полипептид» используется в данном документе в самом широком смысле охватывает пептидные последовательности. Термин «пептид», в целом, служит для описания линейных молекулярных цепей аминокислот, содержащих до около 60, предпочтительно до около 30 аминокислот, ковалентно связанных с помощью пептидных связей.
Термин «чужеродный» применительно к «связывающему фрагменту» используется в данном документе для обозначения связывающего фрагмента, который отсутствует в эталонной нативной полипептидной последовательности в том же самом положении. Следовательно, чужеродная полипептидная последовательность (включая пептидные последовательности) может содержаться в пределах соответствующей нативной последовательности, но в другом положении. В предпочтительном варианте реализации изобретения «чужеродная» последовательность отсутствует в соответствующей нативной последовательности в любом положении.
Используемый в данном документе термин «моноклональное антитело» относится к антителу, полученному из популяции, по существу, гомогенных антител, т.е. отдельные антитела, входящие в состав такой популяции, являются идентичными, за исключением возможных естественных мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высокоспецифическими, при этом они направлены против одного антигенного сайта. Кроме того, в отличие от препаратов стандартных (поликлональных) антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение "моноклональное" означает, что антитело получено из по существу гомогенной популяции антител и не требует применения способа получения, обычно применяемого для получения антител. Например, моноклональные антитела, которые используются по настоящему изобретению, могут быть получены гибридомным методом, описанным Kohler et al. (1975) Nature 256:495, или могут быть получены методами рекомбинантной ДНК (см., например, патент США № 4816567). «Моноклональные антитела» также могут быть получены из фаговых библиотек антител с использованием методов, описанных, например, в источнике Clackson et al. (1991) Nature 352:624-628 и Marks et al. (1991) J. Mol. Biol. Biol. 222:581-597.
Моноклональные антитела, описанные в данном документе, в частности, включают «химерные» антитела (иммуноглобулины), при этом часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, полученных у определенного вида, тогда как оставшаяся(оставшиеся) цепь(цепи) идентична(идентичны) или гомологична(гомологичны) соответствующим последовательностям антител, полученных у другого вида, а также фрагменты таких антител, при условии, что они проявляют желаемую биологическую активность (патент США № 4816567; и Morrison et al. (1984) Proc. Natl. Acad. Sci. USA 81:6851-6855).
«Гуманизированные» формы антител, отличных от человеческих (например, мышиных), относятся к антителам, которые содержат последовательность минимальной длины, полученную из иммуноглобулина, отличного от человеческого. В большинстве случаев гуманизированные антитела представляют собой иммуноглобулины (антитело-реципиент), в которых остатки из гипервариабельной области антитела-реципиента заменены на остатки из гипервариабельной области антитела, принадлежащего виду, отличному от человека (антитело-донор), как, например, мышь, крыса, кролик или нечеловекообразная обезьяна, обладающего желаемой специфичностью, аффинностью и иммунологическим потенциалом. В некоторых случаях остатки Fv каркасного участка (FR) иммуноглобулина человека также заменяют на соответствующие остатки антитела, отличного от человеческого. Кроме того, гуманизированные антитела могут содержать остатки, которые не встречаются в антителе-реципиенте или в антителе-доноре. Эти изменения выполнены для дальнейшего усовершенствования параметров антител. В целом, гуманизированное антитело содержит по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют петлям иммуноглобулина, отличного от человеческого, и все или по существу все каркасные участки представляют собой каркасные участки последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также содержит по меньшей мере часть константной области (Fc) иммуноглобулина, обычно иммуноглобулина человека. Дополнительную информацию см. в источнике Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-329; и Presta (1992) Curr. Op. Struct. Biol. 2:593-596.
«Очищенное» антитело, описанное в данном документе, представляет собой антитело, которое было идентифицировано и выделено и/или отделено от компонента своей естественной среды в рекомбинантной клетке-хозяине. Примеси в естественной среде относятся к веществам, которые создают препятствия для применения антител в диагностических и терапевтических целях, и могут включать ферменты, гормоны и другие растворенные вещества белковой и небелковой природы, а также нежелательные побочные продукты производства. В предпочтительном варианте реализации изобретения антитела, описанные в данном документе, очищены (1) более чем на 95% по массе или более чем на 98% по массе, или более чем на 99% по массе, что установлено с помощью методов ДСН-ПААГ-электрофореза и эксклюзионной ВЭЖХ, (2) до степени, достаточной для получения по меньшей мере 15 аминокислотных остатков, расположенных на N-конце или внутри аминокислотной последовательности, с использованием секвенатора аминокислот или (3) до достижения гомогенности, оцененной с помощью ДСН-ПААГ-электрофореза в восстанавливающих или невосстанавливающих условиях с использованием окрашивания Кумасси синим или предпочтительно серебром. Как правило, очищенное антитело получают с использованием по меньшей мере одного этапа очистки.
Термин «специфическое связывание» или «специфически связывается с», или «специфичный по отношению к» относится к связыванию связывающего фрагмента с мишенью для связывания, как, например, связыванию антитела с антигеном-мишенью, например, эпитопом на определенном полипептиде, пептиде, или другой мишенью (например, гликопротеиновой мишенью), и означает связывание, которое отличается от неспецифического взаимодействия при измерении (например, примером неспецифического взаимодействия может послужить связывание с бычьим сывороточным альбумином или казеином). Специфическое связывание может быть измерено, например, путем определения уровня связывания связывающего фрагмента или антитела, или антитела, модифицированного посредством введения связывающего фрагмента, с молекулой-мишенью по сравнению с уровнем связывания с контрольной молекулой. Например, специфическое связывание можно определить по конкурентному связыванию с контрольной молекулой, которая похожа на мишень, например, по связыванию со взятой в избытке немеченой мишенью. В этом случае специфическое связывание выявляется, если связывание меченой мишени с образцом конкурентно подавляется взятой в избытке немеченой мишенью. Используемый в данном документе термин «специфическое связывание» или «специфически связывается с», или «специфичный по отношению к» определенному эпитопу на определенной полипептидной мишени может относиться к молекуле, имеющей Kd по отношению к мишени, равную по меньшей мере около 200 нМ, альтернативно по меньшей мере около 150 нМ, альтернативно по меньшей мере около 100 нМ, альтернативно по меньшей мере около 60 нМ, альтернативно по меньшей мере около 50 нМ, альтернативно по меньшей мере около 40 нМ, альтернативно по меньшей мере около 30 нМ, альтернативно по меньшей мере около 20 нМ, альтернативно по меньшей мере около 10 нМ, альтернативно по меньшей мере около 8 нМ, альтернативно по меньшей мере около 6 нМ, альтернативно по меньшей мере около 4 нМ, альтернативно по меньшей мере около 2 нМ, альтернативно по меньшей мере около 1 нМ или больше. В определенных случаях термин «специфическое связывание» относится к связыванию, при котором молекула связывается с определенным полипептидом или эпитопом на определенном полипептиде, по существу, не связываясь ни с каким другим полипептидом или полипептидным эпитопом.
«Аффинность связывания» относится к силе суммарных нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иное, используемый в данном документе термин «аффинность связывания» означает характерную аффинность связывания, которая отражает соотношение 1:1 между компонентами пары, участвующей в связывании (например, антитело и антиген). Аффинность молекулы X по отношению к ее партнеру Y в целом может быть представлена с помощью константы диссоциации (Kd). Например, Kd может составлять около 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или больше. Аффинность может быть измерена с помощью методов, известных в данной области техники, включая те, что описаны в данном документе. Низкоаффинные антитела, в целом, связывают антиген медленно и имеют тенденцию к быстрой диссоциации, тогда как высокоаффинные антитела, в целом, связывают антиген быстрее и имеют тенденцию к тому, чтобы дольше оставаться связанными. В данной области известны различные методы изменения аффинности связывания.
Используемый в данном документе термин «Kd» или «значение Kd» относится к константе диссоциации, измеряемой по методу, который подходит для антитела и целевой мишени, например, с использованием анализов поверхностного плазмонного резонанса, например, с использованием BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Пискатауэй, Нью-Джерси) при 25°C с помощью чипов CM5, содержащих иммобилизованные антигены, при около 10 единицах ответа (RU).
Термины «конъюгат», «конъюгированный» и «конъюгация» относятся ко всем возможным формам ковалентной или нековалентной связи и включают, в том числе, прямое слияние на уровне генов или с использованием химических веществ, присоединение через линкер или перекрестносшивающие средство и нековалентную связь.
Термин «слияние» используется в данном документе для обозначения объединения аминокислотных последовательностей разного происхождения в одну полипептидную цепь путем соединения кодирующих их нуклеотидных последовательностей с сохранением рамки считывания. Термин «слияние» в явной форме охватывает типы внутреннего слияния, т.е. вставку последовательностей разного происхождения в полипептидную цепь, а также слияние с одним из ее концов. Термин «слияние» используется в данном документе для обозначения объединения аминокислотных последовательностей разного происхождения.
Используемый в данном документе термин «валентный» обозначает наличие определенного числа сайтов связывания на антителе. В силу этого термины «бивалентный», «тетравалентный» и «гексавалентный» обозначает наличие двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания, соответственно. Следовательно, если в биспецифическом антителе IgA в соответствии с настоящим изобретением каждый связывающий элемент является бивалентным, биспецифическое антитело IgA будет иметь 4 валентности.
Термин «эпитоп» охватывает любую молекулярную детерминанту, способную к специфическому связыванию антитела. В определенных вариантах реализации изобретения антигенная детерминанта включает в себя химически активные поверхностные группы, такие как аминокислоты, боковые цепи сахаров, фосфорильные или сульфонильные группы и, в определенных вариантах реализации изобретения, может обладать специфическими характеристиками трехмерной структуры и/или специфическими характеристиками заряда. Эпитоп представляет собой область антигена, с которой связывается антитело. «Область связывания» представляет собой область на мишени для связывания, с которой связывается связывающая молекула.
«Полиэпитопная специфичность» относится к способности к специфичному связыванию с двумя или более разными эпитопами на одной и той же или другой(их) мишени(ях). «Моноспецифический» относится к способности к связыванию только с одним эпитопом. В соответствии с одним вариантом реализации изобретения биспецифическое антитело IgM связывается с каждым эпитопом с аффинностью по меньшей мере 10-7M или 10-8 M или более высокой аффинностью.
Термин «мишень» или «мишень для связывания» используется в самом широком смысле и, в частности, охватывает полипептиды, в том числе, нуклеиновые кислоты, углеводы, липиды, клетки и другие молекулы, обладающие биологической функцией или не обладающие таковой в той форме, в которой они существуют в природе.
Термин «антиген» относится к структурной единице или ее фрагменту, которая может связываться с антителом или запускать клеточный иммунный ответ. Иммуноген относится к антигену, который может вызывать иммунный ответ в организме, в частности, животного, более конкретно млекопитающего, включая человека. Термин антиген охватывает области, известные как антигенные детерминанты или эпитопы согласно вышеприведенному определению.
Используемый в данном документе термин «иммуногенный» относится к веществам, которые вызываю выработку антител и/или активируют T-клетки и/или другие реактивные иммунокомпетентные клетки, направленные против антигена иммуногена.
«антигенсвязывающий сайт» или «антигенсвязывающий участок» антитела по настоящему изобретению обычно содержит шесть областей, определяющая комплементарность (CDR), которые в разной степени влияют на аффинность сайта связывания по отношению к антигену. Существует три вариабельных домена CDR тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три вариабельных домена CDR легкой цепи (CDRL1, CDRL2 и CDRL3). Размер CDR и каркасных участков (FR) определяется путем сравнения с составленной базой данных аминокислотных последовательностей, в которых эти участки были определены в соответствии с вариабельностью последовательностей и/или информацией по структуре комплексов антитело/антиген. Также в объем настоящего изобретения включены функциональные антигенсвязывающие сайты, состоящие из меньшего количества CDR (т.е. если специфичность связывания определяется тремя, четырьмя или пятью CDR). Для связывания с некоторыми мишенями может быть достаточно менее полного набора из 6 CDR. Следовательно, в некоторых случаях достаточно только наличия CDR VH- или VL-доменов. Кроме того, определенные антитела могут иметь не ассоциированные с CDR сайты связывания антигена. Такие сайты связывания, в частности, охватываются настоящим определением.
Используемый в данном документе термин «клетка-хозяин», обозначает любой вид клеточной системы, которая может быть получена методами генной инженерии для получения антител по настоящему изобретению. В одном варианте реализации изобретения в качестве клеток-хозяев используются клетки яичника китайского хомячка (CHO).
Используемые в данном документе выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо и такие обозначения охватывают клетки-потомки. Следовательно, слова «трансформанты» и «трансформированные клетки» охватывают первичные клетки субъекта или культуры, полученные из них, не учитывая количество переносов. Также надо понимать, что все потомки могут быть не полностью идентичными по содержанию ДНК вследствие случайных или искусственных мутаций. Также включены варианты потомков, которые обладают такой же функцией или биологической активностью, что были выявлены в первоначально трансформированных клетках.
Нуклеиновая кислота «функционально связана», когда она находится в функциональной зависимости от другой последовательности нуклеиновой кислоты. Например, ДНК, кодирующая препоследовательность или лидерную последовательности секретируемых пептидов, функционально связана с ДНК, кодирующей полипептид, если она экспрессируется в виде белка-предшественника, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосом функционально связан с кодирующей последовательностью, если он расположен таким образом, что способствует трансляции. Как правило, «функционально связанный» означает, что последовательности ДНК, которые необходимо соединить, являются смежными и в случае лидерной последовательности секретируемых пептидов - смежными и расположенными в рамке считывания. Однако энхансеры не обязательно должны быть смежными. Соединение осуществляется путем легирования в удобно расположенных сайтах рестрикции. В случае отсутствия таких сайтов синтетические олигонуклеотидные адаптерные последовательности или линкеры используются в соответствии с уровнем техники.
ПОДРОБНОЕ ОПИСАНИЕ
Конструирование и получение антител с модифицированной J-цепью
IgM представляет собой иммуноглобулин, вырабатываемый ранее всего В-клетками в ответ на стимуляцию антигеном, и содержится в сыворотке крови в концентрации около 1,5 мг/мл с периодом полувыведения 5 суток. IgM представляет собой пентамерную или гексамерную молекулу. Точно так же, как и в случае с IgG мономеры IgM состоят из двух легких и двух тяжелых цепей. Однако несмотря на то, что IgG содержит три константных домена тяжелой цепи (CH1, CH2 и CH3), тяжелая (μ)-цепь IgM дополнительно содержит четвертый константный домен (CH4) по аналогии с тяжелыми ε-цепями в IgE. Этот дополнительный константный домен расположен в богатой пролином шарнирной области IgG и IgA, которая отвечает за вращательную гибкость антигенсвязывающих Fab-доменов по сравнению с Fc-доменом антител IgG и IgA.
Пять мономеров IgM формируют комплекс с дополнительной небольшой полипептидной цепью (J-цепь), что приводит к образованию нативной молекулы IgM. Считается, что J-цепь способствует полимеризации μ-цепей до секреции IgM антителопродуцирующими клетками. Тогда как общеизвестно, что получение IgM в кристаллическом виде оказалось чрезвычайно трудным, Czajkowsky и Shao (PNAS 106(35):14960-14965, 2009) недавно опубликовали статью со структурной моделью IgM на основе гомологии, полученную исходя из структуры Fc-домена IgE и известного образования пар дисульфидных связей. Авторы сообщают о том, что пентамер IgM человека представляет собой молекулу в форме гриба со смещенным наклоном. Тяжелая цепь IgM (μ) содержит пять N-связанных сайтов гликозилирования: Asn-171, Asn-332, Asn-395, Asn-402 и Asn-563.
Иммуноглобулин A (IgA), как основной класс антител, присутствующих в секретах слизистых оболочек большинства млекопитающих, выполняет функцию первой линии защиты против инвазии патогенов, попадающих в организм с вдыхаемым воздухом и пищей. IgA также встречается в значительных концентрациях в сыворотке крови многих видов, где он выполняет функцию второй линии защиты, опосредуя элиминацию патогенов, которые проникли через поверхность слизистой оболочки. Рецепторы, специфичные к Fc-области IgA, FcαR, являются ключевыми медиаторами эффекторной функции IgA. Для IgA известно два разных гена константных областей (Cα) тяжелой цепи IgA, которые обуславливают наличие двух подклассов, IgA1 и IgA2. Основные различия между IgA1 и IgA2 заключаются в шарнирной области, которая располагается между двумя плечами Fab и Fc-областью. В IgA1 содержится шарнирная область, выступающая вследствие вставки двойной копии цепочки аминокислот, который отсутствует в IgA2. IgA обладает способностью к образованию димеров, причем считается, что две мономерные единицы, каждая из которых содержит две тяжелые цепи и легкие цепи, располагаются в конфигурации «конец-в-конец», стабилизированной дисульфидными мостиками и включением J-цепи. Димерный IgA, вырабатываемый локально в областях слизистых оболочек, транспортируется через эпителиальный барьер во внешний секрет за счет взаимодействия с полимерным рецептором иммуноглобулина (pIgR). На протяжении этого процесса расщепляется pIgR и происходит ковалентное присоединение основного фрагмента, называемого секреторным компонентом (СК), к димеру IgA.
Как в IgA, так и в IgM содержится выступ длиной 18 аминокислот на C-конце, называемый «хвостовая часть» (х.ч.). Хвостовые части IgM (х.ч. μ) и IgA (х.ч. α) различаются по семи аминокислотным положениям. Хвостовая часть IgM и IgA является высококонсервативной у различных видов животных. Было показано, что консервативный предпоследний цистеиновый остаток в хвостовых частях IgA и IgM вовлечен в полимеризацию. Обе хвостовые части содержат сайты присоединения N-связанных углеводов, наличие которых необходимо для образования димеров в IgA и включения J-цепи, а также для и образования пентамеров в IgM. Однако структура и состав N-связанных углеводов в хвостовых частях различается, свидетельствуя о наличии различий в доступности гликанов для процессинга гликозилтрансферазами.
Сообщалось о нуклеотидных и/или белковых последовательностях J-цепей человека и различных видов позвоночных животных, таких как корова, мышь, птица, амфибия и кролик. J-цепь человека содержит восемь цистеиновых остатков, причем два остатка (Cys13 и Cys69) вовлечены в образование дисульфидных мостиков с α- или μ-цепями (в IgA и IgM, соответственно), а шесть остатков вовлечены в образование внутрицепочечных дисульфидных мостиков (Cys13: Cys101, Cys72: Cys92, Cys109: Cys134). Сообщения о трехмерной кристаллической структуре J-цепи отсутствуют.
Настоящее изобретение по меньшей мере частично основано на признании того факта, что J-цепь антитела IgM или IgA может быть модифицирована путем придания ей специфичности связывании (за счет связывающего фрагмента) без ухудшения способности антитела IgM или IgA к связыванию с его мишенью(мишенями) для связывания. В соответствии с этим, настоящее изобретение касается модифицированных J-цепей, содержащих связывающий фрагмент, введенный в J-цепь с нативной последовательностью, как, например, J-цепь человека с нативной последовательностью SEQ ID NO.: 1. Настоящее изобретение дополнительно касается связывающих молекул, содержащих модифицированную J-цепь. Связывающая молекула может представлять собой, например, антитело IgM, антитело IgA или гибридное антитело IgG/IgM или IgG/IgA, которое может содержать хвостовую часть IgM или IgA в тяжелой цепи IgG и, следовательно, обладает сочетанными свойствами IgG и IgA или IgA, включая способность к включению и образованию полимеров с модифицированной J-цепью по настоящему изобретению. Дополнительную информацию о гибридных антителах IgG/IgM and IgG/IgA см., например, Koteswara et al., Clinical Immunology 2001, 101(1):21-31.
Модифицированная J-цепь содержит чужеродный связывающий фрагмент, который включает, в том числе, полипептид (включая пептиды), способный к специфическому связыванию с мишенью для связывания, или каталитические компоненты, такие как ферментоподобные протеазы. Как обсуждалось выше, связывающие фрагменты включают, в том числе, антигенсвязывающие фрагменты антител, конъюгаты антитела и лекарственного препарата, антителоподобные молекулы, антигенсвязывающие фрагменты антителоподобных молекул, растворимые и мембраносвязанные белки, лиганды, рецепторы, вирусоподобные частицы, белковые токсины, каталитические компоненты, такие как ферменты и ферментоподобные протеазы и альтернативные каркасные белки. Следует подчеркнуть, что согласно идее настоящего раскрытия в J-цепь может быть введен связывающий фрагмент любого типа путем соответствующего подбора положения, типа присоединения (например, прямое или непрямое слияние, химическое присоединение и т.д.).
В предпочтительном варианте реализации изобретения связывающий фрагмент представляет собой антитело или антигенсвязывающий фрагмент антитела (также называемый «фрагментом антитела»), включая моноспецифические, биспецифические и мультиспецифические антитела и фрагменты антител. Термин «фрагмент антитела» используется в самом широком смысле и охватывает, в том числе, Fab, Fab', F(ab')2, scFv и (scFv)2 -фрагменты, фрагменты областей, определяющих комплементарность, линейные антитела, молекулы одноцепочечных антител, миниантитела и мультиспецифические антитела, образующиеся из фрагментов антител. В предпочтительном варианте реализации изобретения фрагмент антитела представляет собой scFv.
В другом предпочтительном варианте реализации связывающий фрагмент представляет собой антителоподобную молекулу, такую как, например, доменное антитело человека (dAb), молекулу переориентирующегося антитела с двойной аффинностью (DART), ди-диатело, антитело с двойным вариабельным доменом, антитело с многоярусными вариабельными доменами (Stacked Variable Domain antibody), иммунопрепарат на основе модульного белка с малым размером молекул (SMIP), суррогатное антитело, антителос доменами, сконструированными посредством замены цепей (SEED) (strand-exchange engineered domain (SEED)-body), или TandAb.
Связывающий фрагмент может представлять собой лиганд, такой как нейротрофин, интерлейкин, растворимый молекулярный фактор или ростовой фактор.
Рецепторы в качестве связывающих молекул включают рецепторы клеточной поверхности, сопряженные с ионными каналами, с G--белком и с ферментом. Конкретные примеры включают, в том числе, ErbB1, ErbB2, ErbB3, ErbB4, ФНОR, PDL1, и CTLA-4.
В дополнительном предпочтительном варианте реализации изобретения связывающий фрагмент представляет собой альтернативный каркасный белок. В этом контексте термин «каркас» предназначен для описания белкового каркаса, в котором содержатся измененные аминокислоты или вставки последовательности, что придают вариантам белка способность к связыванию специфических мишеней. Альтернативные каркасные белки включают, в том числе, CTLA-4, тендамистат, фибронектин, неокрациностатин, липокалины, T-клеточный рецептор, CBM4-2, домен белка A, Im9, сконструированные белки с анкириновым повтором, сконструированные TPR-белки, цинковые пальцы, pVIII, птичий панкреатический полипептид, GCN4, WW-домен, гомологичные Src-домены 2 и 3, PDZ-домены, TEM-I, β-лактамазу, GFP, тиоредоксин, стафилококковую нуклеазу, PHD-палец, cl-2, BPT1, APPI, HPSTI, экотин, LACI-D1, LDTI, MTI-II, токсины скорпиона, ноттины, пептид дефенсин A насекомых, EETI-II, Min-23, CBD, PBP, цитохром b562, домен A рецептора LDL, γ-кристаллин, убиквитин, трансферрин и лектиноподобный домен C-типа. К предпочтительным альтернативным каркасным белкам относятся дарпины, домены фибронектина, аднектины. Дополнительную информацию см. в источнике Binz HK et al, 2005 Nature Biotechnology 23(10):1257-1268.
Связывающий фрагмент может быть введен в нативную последовательность J-цепи в любом положении, которое способствует связыванию связывающего фрагмента с его мишенью для связывания, не нарушая связывания молекулы-реципиента IgM, IgA, IgG/IgM или IgG/IgA с ее мишенью для связывания или мишенями для связывания. Предпочтительные положения включают положения, находящиеся на C-конце или вблизи от него, на N-конце или вблизи от него или внутреннее положение, которое становится доступным при образовании трехмерной структуры J-цепи. В предпочтительных вариантах реализации изобретения связывающий фрагмент вводится в нативную последовательность J-цепи на расстоянии около 10 аминокислотных остатков от C-конца или на расстоянии около 10 аминокислотных остатков от N-конца, причем нативная последовательность J-цепи предпочтительно представляет собой J-цепь антитела человека SEQ ID NO.: 1. В другом варианте реализации изобретения чужеродный связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1 между цистеиновыми остатками 92 и 101 SEQ ID NO.: 1 или в аналогичное положение другой нативной последовательности J-цепи. В дополнительном варианте реализации изобретения чужеродный связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1 в или вблизи от сайта гликозилирования. Наиболее предпочтительно связывающий фрагмент вводится в J-цепь человека с нативной последовательностью SEQ ID NO.: 1 на расстоянии около 10 аминокислотных остатков от N-конца.
Связывающий фрагмент можно вводить посредством прямого или непрямого слияния, сочетая аминокислотные последовательности связывающего фрагмента и J-цепи одной полипептидной цепи, что достигается сочетанием кодирующих их нуклеотидных последовательностей с сохранением рамки считывания с использованием пептидного линкера или без него. Длина пептидного линкера (непрямое слияние) при использовании может составлять, например, около 1-50 или около 1-40, или около 1-30, или около 1-20, или около 1-10, или около 10-20 аминокислотных остатков, и он может быть расположен на одном или обоих концах связывающего фрагмента, который вводится в последовательность J-цепи. В предпочтительном варианте реализации длина пептидного линкера составляет около 10-20 аминокислот. В другом предпочтительном варианте реализации длина пептидного линкера составляет 10-15 аминокислот.
Связывающий фрагмент также может быть присоединен к нативной последовательности J-цепи посредством химической связи с использованием гетеробифункциональных кросслинкеров белков, содержащих две разные функциональные группы, которые обладают их собственной реакционной способностью и избирательностью. Эти кросслинкеры могут быть использованы в одностадийном процессе или могут быть использованы для создания активированных белков, которые зачастую хранят и используют для реакции со второй биомолекулой на отдельном этапе. Следовательно, для формирования конъюгатов между J-цепью и связывающим фрагментом может быть использован, например, гетеробифункциональный сшивающий реагент. Реакционноспособные группы включают, в том числе, иминные реакционноспособные группы (такие как NHS или сульфо-NHS), малеимидные группы и тому подобное. Такие кросслинкеры, которые могут быть расщепляемыми и нерасщепляемыми, были использованы, например, при формировании белков-носителей гаптенов и при получении конъюгатов фермента с антителом. С химической точки зрения, расщепляемые кросслинкеры, в частности, включают, в том числе, линкеры на основе дисульфидной связи, гидразоновые и пептидные линкеры. Широко известным и хорошо изученным линкером, неустойчивым к действию ферментов, является валин-цитруллиновый линкер, но другие пептидные линкеры также известны и являются подходящими. Типичные образцы нерасщепляемых линкеров включают тиоэфиры, такие как SMCC (N-сукцинимидил-4-(N-малеимидометил)-циклогексан-1-карбоксилат). Дополнительную информацию см., например, в источнике Ducry L and Stump B, Bioconjugate Chem. 2010, 21:5-13, полное описание которого в явном виде включено в данный документ посредством ссылки. В отношении перечня дополнительных подходящих линкеров см., например, Klein et al., Protein Engineering, Design & Selection; 2014, 27(10): 325-330, полное описание которого в явном виде включено в данный документ посредством ссылки.
Несмотря на то, что модифицированная J-цепь обычно содержит один чужеродный связывающий фрагмент, также существует возможность для введения нескольких связывающих фрагментов в J-цепь.
Модифицированная J-цепь может быть получена с помощью широко известных методов технологии рекомбинантной ДНК путем экспрессии нуклеиновой кислоты, кодирующей модифицированную J-цепь, в подходящих прокариотических или эукариотических клетках-хозяевах, как, например, клетках CHO или E. coli. Следовательно, модифицированную J-цепь можно экспрессировать, например, в клетках E. coli, как описано Symersky et al., Mol Immunol 2000, 37:133-140.
В одном варианте реализации изобретения J-цепь первоначально может быть модифицирована путем вставки сайта узнавания ферментом и может быть модифицирована посттрансляционно путем вставки пептидного или непептидного линкера, с помощью которого можно присоединить любой чужеродный связывающий фрагмент к J-цепи, такой как, например, цитотоксическая малая молекула, для получения конъюгата антитела и лекарственного препарата (ADC).
Модифицированная J-цепь также может быть коэкспрессирована с тяжелой и легкой цепями антител-реципиентов IgM, IgA, IgG/IgM или IgG/IgA. Несмотря на то, что в силу сложности структуры массовое производство рекомбинантного IgM было сопряжено с трудностями, имеются сообщения о нескольких системах для получения рекомбинантного IgM с использованием клеток нелимфоидного ряда, включая коэкспрессию тяжелой (H) и легкой (L) цепей IgM в клетках глиомы линии C6, клетках CHO и клетках HeLa (см., например, W089/01975 и Wood et al., J. Immunol. 145, 3011-3016 (1990) в отношении экспрессии в клетках CHO). Экспрессия моноклонального антитела IgM в клетках E. coli с J-цепью и без нее описана, например, в источнике Azuma et al., Clin Cancer Res 2007, 13(9):2745-2750. Получение IgM в линии иммортализованных клеток сетчатки глаза человека, экспрессирующих белки E1A и E1B аденовируса, описано в заявке на патент США № 20060063234.
Антитело-реципиент может быть моноспецифическим, биспецифическим или мультиспецифическим. Биспецифические и мультиспецифические связывающие молекулы IgM и IgA, включая антитела, описаны, например, в заявке патент США с серийными № 61/874277 и 61/937984, полные содержания которых в явном виде включены в данный документ посредством ссылки.
Применения антител с модифицированной J-цепью
Антитела, содержащие модифицированную J-цепь по настоящему изобретению, находят широкое применение в терапевтических и диагностических целях.
В одном варианте реализации изобретения антитела, содержащие модифицированную J-цепь, связываются с двумя или более сайтами на одной и той же растворимой форме мишени, такой как, например, VEGF, ФНОα или IL6. Цель может заключаться, например, в антагонистическом связывании многих сайтов на белке и/или повышении авидности к конкретной мишени.
В другом варианте реализации изобретения антитела, содержащие модифицированную J-цепь, описанную в данном документе, связывают два или более сайта на одной и той же мишени (рецепторе) клеточной поверхности, такой как EGFR или HER2 (ErbB2). Следовательно, мишенями таких антител могут быть, например, как эпитопы 4D5, так и 2C4 на молекуле HER2. Этот подход может увеличить биологическую активность и/или авидность к конкретной мишени.
Еще в одном варианте реализации изобретения антитела, содержащие модифицированную J-цепь по настоящему изобретению, связывают две или более разные растворимые формы мишеней (глобулярные белки или пептиды), например, ФНОα и ИЛ6, VEGFα и Ang2 или два цитокина. Этот подход может привести, например, к более полной блокировке специфического сигнального пути; блокировке так называемой «цитокиновой бури», т.е. нежелательной активации Т-клеток, обусловленной действием определенных мультивалентных биспецифических антител, таких как биспецифические антитела к антигену CD3, или координации фермента и его субстрата, например, Фактора IXa и Фактора X. Конкретные примеры включают, в том числе, биспецифические антитела с модифицированной J-цепью, причем первая специфичность направлена по отношению к VEGF, а вторая специфичность направлена по отношению к Ang2 или DLL4 (противоангиогенные свойства), или первая специфичность направлена по отношению к ФНО, а вторая специфичность направлена по отношению к Ang2 или ИЛ-17 (противовоспалительные свойства), причем J-цепи можно придать любую специфичность, характерную для антител IgM, IgA, IgG/IgM или IgG/IgA или их антигенсвязывающего фрагмента.
В дополнительном варианте реализации изобретения антитела, содержащие модифицированную J-цепь, могут связывать одну или более растворимых мишеней и одну или более мишеней - рецепторов клеточной поверхности, таких как ангиогенный фактор и рецептор, ассоциированный с неоваскуляризацией. Цель этого подхода также может заключаться в улучшении доставки и блокировки в определенных участках и тканях.
Еще в одном варианте реализации изобретения конструируют антитела, содержащие модифицированную J-цепь, для связывания двух или более мишеней - рецепторов клеточной поверхности, таких как, например, HER1, HER2 (ErbB2) и HER3 (ErbB3), подавляя функцию многих мишеней через одни и те же или разные сигнальные пути. Это может привести к повышению специфичности и избирательности, и/или к более полной блокировке конкретного сигнального пути. Конкретные примеры таких антител включают, в том числе, биспецифические антитела с модифицированной J-цепью, причем одна специфичность направлена по отношению к HER2, а другая специфичность направлена по отношению к HER3; или одна специфичность направлена по отношению к EGFR (HER1), а другая специфичность направлена по отношению к HER2. Другие биспецифические антитела IgM, IgA, IgG/IgM или IgG/IgA с модифицированной J-цепью могут связываться, например, с EGFR и HER3, ИЛ-1α и ИЛ-1β, ИЛ-4 и ИЛ-13, Ang-2 и VEGF-A, фактором IXA и фактором X или ИЛ-17A и ИЛ-17F.
Антитела, содержащие модифицированную J-цепь по настоящему изобретению, также могут быть предназначены для связывания одной или более растворимых мишеней, или мишеней - рецепторов клеточной поверхности и одной или более мишеней с продолжительным периодом удержания, например, ФНОα и/или VEGF и сывороточный альбумин. Предполагается, что эти молекулы имеют более продолжительный период полувыведения из кровяного русла, чем связывающие молекулы, не обладающие специфичностью к альбумину.
В дополнительном варианте реализации изобретения антитела, содержащие модифицированную J-цепь, описанную в данном документе, могут связывать одну или более растворимых мишеней и один или более белков матрикса и/или субстратов, таких как, например, VEGFα и гиалуроновая кислота. Полученные в результате мультиспецифические связывающие молекулы могут найти применение, например, в антиангиогенной терапии заболеваний глаз, таких как возрастная макулярная дегенерация (ВМД), благодаря их увеличенному периоду удержания во внутриглазном пространстве.
Антитела, содержащие модифицированную J-цепь и связывающие одну или более растворимых, или рецепторных мишеней в дополнение к одному или более транспортных рецепторов (т.е. трансферриновый рецептор), например, антитело к EGFRvIII (мутантная форма с делецией экзона III), который обнаруживается при глиобластоме, в сочетании со специфичностью по отношению к трансферрину, могут найти применение в доставке антител через гематоэнцефалитический барьер.
В предпочтительном варианте реализации изобретения антитела IgM, IgA, IgG/IgM и IgG/IgA, описанные в данном документе, содержат модифицированную J-цепь со специфичностью связывания по отношению к иммунокомпетентной клетке, такой как T-клетка, NK-клетка, макрофаг или нейтрофил, и связываются с антигеном, экспрессированным на пораженной клетке или патогене. Поскольку молекула IgM содержит 5 связывающих элементов, а молекула IgA представляет собой димер, содержащий два связывающих элемента, такие молекулы намного более активны благодаря их большей авидности, чем у биспецифических антител IgG. В дополнение к этому, путем активации и перенацеливания эффекторных клеток, например, эффекторных Т-клеток, на пораженные клетки-мишени или патогены антитела IgM, IgA, IgG/IgM и IgG/IgA, описанные в данном документе, индуцируют иммунный ответ против мишени, тем самым дополнительно увеличивается их активность и эффективность. Благодаря этим полезным свойствам антитела IgM, IgA, IgG/IgM и IgG/IgA, описанные в данном документе, содержащие модифицированную J-цепь, обладают особыми преимуществами в ситуациях, когда антитела IgG связываются с их мишенью с низкой аффинностью. Следовательно, в одном варианте реализации изобретения антитела IgM, IgA, IgG/IgM и IgG/IgA, описанные в данном документе, могут содержать связывающий домен терапевтического антитела IgG.
В определенных вариантах реализации изобретения антитела IgM, IgA, IgG/IgM и IgG/IgA, описанные в данном документе, содержащие модифицированную J-цепь, могут быть использованы для лечения злокачественной опухоли. Предполагается, что воздействие может быть направлено на любой тип опухоли и любой тип опухолеассоциированного антигена. Примеры типов злокачественных опухолей включают, в том числе, острый лимфобластный лейкоз, острый миелоидный лейкоз, рак желчевыводящей системы, рак молочной железы, рак шейки матки, хронический лимфоцитарный лейкоз, хронический миелоидный лейкоз, колоректальный рак, рак эндометрия, рак пищевода, рак желудочно-кишечного тракта, рак головы и шеи, лимфому Ходжкина, рак легких, медуллярный рак щитовидной железы, неходжкинскую лимфому, множественную миелому, рак почек, рак яичников, рак поджелудочной железы, глиому, меланому, рак печени, рак предстательной железы и рак мочевого пузыря. Однако специалисту в данной области понятно, что в данной области известны опухолеассоциированные антигены практически для любого типа злокачественной опухоли.
Опухолеассоциированные антигены, на которые могут быть направлены антитела IgM, IgA, IgG/IgM или IgG/IgA по настоящему изобретению, включают, в том числе, альфа-фетопротеин (АФП), ассоциированные антигены, Ba 733, BAGE, BrE3-антиген, CA125, CAMEL, CAP-1, карбоангидразу IX, CASP-8/m, CCCL19, CCCL21, CD1, CD1a, CD2, CD3, CD4, CD5, CD8, CD11A, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD29, CD30, CD32b, CD33, CD37, CD38, CD40, CD40L, CD44, CD45, CD46, CD52, CD54, CD55, CD59, CD64, CD66a-e, CD67, CD70, CD70L, CD74, CD79a, CD80, CD83, CD95, CD126, CD132, CD133, CD138, CD147, CD154, CDC27, CDK-4/m, CDKN2A, CTLA-4, CXCR4, CXCR7, CXCL12, HIF-1a, p-антиген колоректального рака (CSAp), CEA (CEACAM5), CEACAM6, c-Met, DAM, EGFR (HER1, ErbB1), ErbB2 (HER2), ErbB4 (HER3), EGFRvIII, EGP-1 (TROP-2), EGP-2, ELF2-M, Ep-CAM, фактор роста фибробластов (ФРФ), Flt-1, Flt-3, рецептор фолиевой кислоты, антиген G250, GAGE, gp100, GRO-бета, HLA-DR, HM1.24, хорионический гонадотропин человека (ХГЧ) и его субъединицы, HER2/neu, HMGB-1, фактор, индуцируемый гипоксией (HIF-1), HSP70-2M, HST-2, Ia, IGF-1R, ИФН-γ ИФН -α, ИФН -β, ИФН -λ, ИЛ-4R, ИЛ -6R, ИЛ -13R, ИЛ -15R, ИЛ -17R, ИЛ -18R, ИЛ -2, ИЛ -6, ИЛ -8, ИЛ -12, ИЛ -15, ИЛ -17, ИЛ -18, ИЛ -23, ИЛ -25, инсулиноподобный ростовой фактор-1 (IGF-1), KC4-антиген, KS-1-антиген, KS1-4, Le-Y, LDR/FUT, фактор ингибирования миграции макрофагов (MIF), MAGE, MAGE-3, MART-1, MART-2, NY-ESO-1, TRAG-3, mCRP, MCP-1, MIP-1A, MIP-1B, MIF, MUC1, MUC2, MUC3, MUC4, MUC5ac, MUC13, MUC16, MUM-1/2, MUM-3, NCA66, NCA95, NCA90, антиген PAM4, муцин, вырабатываемый при раке поджелудочной железы, рецептор PD-1, плацентарный фактор роста, p53, PLAGL2, простатическую кислую фосфатазу, PSA, PRAME, PSMA, PlGF, ILGF, ILGF-1R, ИЛ -6, ИЛ -25, RS5, RANTES, T101, SAGE, S100, сурвивин, сурвивин-2B, TAC, TAG-72, тенасцин, рецепторы TRAIL, ФНО-альфа, антиген Tn, антигены Томсена-Фриденрайха, антигены некроза опухоли, VEGFR, фибронектин ED-B, WT-1, 17-1A-антиген, факторы комплемента C3, C3a, C3b, C5a, C5, маркер ангиогенеза, bcl-2, bcl-6, Kras.
Как отмечалось выше, для применения в онкологической практике антитела, содержащие модифицированную J-цепь, могут быть сконструированы для активации функции уничтожения злокачественных клеток-мишеней иммунокомпетентными клетками. Следовательно, J-цепь, например, антител IgM, IgA, IgG/IgM или IgG/IgA, или их антигенсвязывающих фрагментов может быть модифицирована путем придания нативной J-цепи специфичности связывания по отношению к иммунокомпетентной клетке, такой как T-клетка, натуральный киллер (NK), причем антитело IgA, IgG/IgM или IgG/IgA обладает специфичностью связывания по отношению к клетке-мишени, например, онкомаркеру, такому как онкоген на поверхности опухолевой клетки, включая, например, один или более опухолеассоциированных антигенов, перечисленных выше.
В случае T-клеток кластер дифференцировки 3 (CD3) представляет собой мультимерный белковый комплекс, исторически известный как комплекс T3, и состоит из четырех различных полипептидных цепей (ε, γ, δ, ζ), которые собираются и функционируют как три пары димеров (εγ, εδ, ζζ). Комплекс CD3 служит в качестве корецептора T-клеток, который нековалентно связывается с T-клеточным рецептором (TCR). Компоненты этого комплекса CD3, особенно CD3ε, являются мишенями для модифицированной J-цепи антитела IgM, специфически связывающегося с опухолеассоциированным антигеном. Несмотря на то, что модифицированная J-цепь, специфическая по отношению к эффекторным Т-клеткам предпочтительно связывается с антигеном CD3 (CD3ε), известны другие антигены, экспрессированные на эффекторных Т-клетках, и могут послужить мишенью для модифицированной J-цепи. Примеры Т-клеточных антигенов включают, в том числе, CD2, CD3, CD4, CD5, CD6, CD8, CD25, CD28, CD30, CD40, CD40L, CD44, CD45, CD69 и CD90.
Примеры антител IgM, IgA, IgG/IgM или IgG/IgA, содержащих модифицированную J-цепь со специфичностью связывания по отношению к CD3, могут включать области связывания известных антител IgG к опухолеассоциированным антигенам, такие как, например, блинатумомаб (также известный как MT103) (антитело к CD19), CD19hA19 (антитело к CD19, (патент США № 7109304), hPAM4 (антитело к муцину, (патент США № 7282567), hA20 (антитело к CD20, (патент США № 7251164), hIMMU31 (антитело к AFP, (патент США № 7300655), hLL1 (антитело к CD74, (патент США № 7312318), hLL2 (антитело к CD22, (патент США № 7074403), hMu-9 (антитело к CSAp, (патент США № 7387773), hL243 (антитело к HLA-DR, (патент США № 7612180), hMN-14 (антитело к CEACAM5, (Патент США № 6676924), hMN-15 (антитело к CEACAM6, (патент США № 7541440), hRS7 (антитело к EGP-1, (патент США № 7238785), hMN-3 (антитело к CEACAM6, (патент США № 7541440), Ab124 и Ab125 (антитела к CXCR4, (патент США № 7138496), описания которых в явном виде включено в данный документ посредством ссылки.
В конкретном варианте реализации изобретения антитело IgM, IgA, IgG/IgM или IgG/IgA, содержащее CD19-связывающий участок блинатумомаба, содержит модифицированную J-цепь, содержащую CD3-связывающий участок блинатумомаба. Это антитело может быть использовано, например, для лечения неходжкинской лимфомы или острого лимфобластного лейкоза популяции В-клеток (B-ALL).
В другом конкретном варианте реализации изобретения антитело IgM, IgA, IgG/IgM или IgG/IgA, содержащее CD20-связывающий участок ритуксимаба, содержит модифицированную J-цепь со специфичностью по отношению к CD3, как, например, модифицированную J-цепь, содержащую CD3-связывающий участок блинатумомаба.
Еще в одном конкретном варианте реализации изобретения антитело IgM, IgA, IgG/IgM или IgG/IgA, содержащее EpCAM-связывающий участок MT110, содержит модифицированную J-цепь со специфичностью по отношению к CD3, как, например, модифицированную J-цепь, содержащую CD3-связывающий участок MT110. Такие биспецифические антитела могут быть использованы для лечения рака желудочно-кишечного тракта.
Альтернативные антитела, которые могут обладать связывающими участками для применения в комбинации с модифицированной J-цепью со специфичностью по отношению к CD3, включают, например, абциксимаб (антитело к гликопротеину IIb/IIIa), алемтузумаб (антитело к CD52), бевацизумаб (антитело к VEGF), цетуксимаб (антитело к EGFR), гемтузумаб (антитело (к CD33), ибритумомаб (антитело к CD20), панитумумаб (антитело к EGFR), тоситумомаб (антитело к CD20), трастузумаб (антитело к ErbB2), ламбролизумаб (антитело к рецептору PD-1), ниволумаб (антитело к рецептору PD-1), ипилимумаб (антитело к CTLA-4), абаговомаб (антитело к CA-125), адекатумумаб (антитело к EpCAM), атлизумаб (антитело к рецептору IL-6), бенрализумаб (антитело к CD125), обинутузумаб (GA101, антитело к CD20), CC49 (антитело к TAG-72), AB-PG1-XG1-026 (антитело к PSMA, заявка на патент США с серийным № 11/983372, депонировано как ATCC PTA-4405 и PTA-4406), D2/B (антитело к PSMA, WO 2009/130575), тоцилизумаб (антитело к рецептору IL-6), базиликсимаб (антитело к CD25), даклизумаб (антитело к CD25), эфализумаб (антитело к CD11a), GA101 (антитело к D20; Glycart Roche), натализумаб (антитело к интегрину α4), омализумаб (антитело к IgE); антитела к ФНО-α, такие как CDP571 (Ofei et al., 2011, Diabetes 45:881-85), MTNFAI, M2TNFAI, M3TNFAI, M3TNFABI, M302B, M303 (Thermo Scientific, Rockford, Ill.), инфликсимаб (Centocor, Malvern, Pa.), цертолизумаб пегол (UCB, Брюссель, Бельгия), антитело к CD40L (UCB, Брюссель, Бельгия), адалимумаб (Abbott, Abbott Park, Ill.), BENLYSTA.RTM. (Human Genome Sciences); антитела для терапии болезни Альцгеймера, такие как Alz 50 (Ksiezak-Reding et al., 1987, J Biol Chem 263:7943-47), гантенерумаб, соланезумаб и инфликсимаб; антитела к фибрину, такие как 59D8, T2G1s, MH1; антитела к CD38, такие как MOR03087 (MorphoSys AG), MOR202 (Celgene), HuMax-CD38 (Genmab) или даратумумаб (Johnson & Johnson); антитела к вирусу ВИЧ, такие как P4/D10 (Патент США № 8333971), Ab 75, Ab 76, Ab 77 (Paulik et al., 1999, Biochem Pharmacol 58:1781-90), а также антитела к ВИЧ, описанные в (патент США № 5831034, (патент США № 5911989 и Vcelar et al., AIDS 2007; 21(16):2161-2170, и Joos et al., Antimicrob. Agents Chemother. 2006; 50(5):1773-9.
На ФИГ. 19 перечислены примеры антигенов-мишеней для лечения гематологической злокачественной опухоли, эпителиальной опухоли и опухолей с повышенным уровнем гликозилирования, и примеры антигенов на T-клетках, NK-клетках, макрофагах и нейтрофилах для направленного воздействия с использованием модифицированной J-цепи. Следует понимать, что молекула IgM, связывающаяся с любым из перечисленных опухолевых антигенов, может быть комбинирована с модифицированной J-цепью с любой из перечисленных специфичностей связывания. Следовательно, любая из мишеней, перечисленных в левом столбце таблицы, может быть комбинирована с любой из мишеней модифицированной J-цепи, перечисленных в правом столбце.
В предпочтительном варианте реализации изобретения пентамер IgM обладает специфичностью связывания по отношению к клеткам-мишеням, таким как B-клетки, причем связывающий домен, специфичный по отношению к эффекторной клетке, например, T-клетке, может быть ковалентно присоединен к J-цепи антитела IgM. Следовательно, J-цепь может быть модифицирована путем ковалентного присоединения домена, связывающего CD3 (CD3ε). В этой конфигурации пентамер IgM (содержащий 10 копий тяжелой цепи и 10 копий легкой цепи) обладает специфичностью связывания по отношению к B-клеткам-мишеням, причем связывание J-цепи с Т-клетками оказывает цитотоксическое действие. Другими словами, связывание антитела IgM с опухолеассоциированной мишенью дополнительно приобретает функцию связывания с Т-клетками. CD3-связывающий домен, ковалентно связанный с нативной J-цепью или вариантом нативной J-цепь, может представлять собой, например, одноцепочечный Fv-фрагмент (scFv) антитела к CD3 или встречающееся в природе состоящее только из тяжелой цепи антитело, например, представителей семейства верблюдовых (верблюды, ламы, альпаки), или одноцепочечное антитело хрящевых рыб (акулы, скаты), каркасный белок, например, фибронектин (например, фибронектин III) со специфичностью связывания по отношению к CD3). Несмотря на то, что в данном документе делается специальная ссылка на предпочтительные варианты реализации изобретения, следует понимать, что антитела IgM, IgA, IgG/IgM и IgG/IgA со специфичностью связывания по отношению к любой мишени, как, например, любому опухолевому антигену, содержащие модифицированную J-цепь, связывающуюся с Т-клеточным маркером, предусмотрены и охватываются объемом настоящего изобретения.
В одном варианте реализации изобретения мультиспецифическое антитело IgM, IgA, IgG/IgM или IgG/IgA связывается с одной или более опухолеассоциированными мишенями, что перечислены выше, тогда как J-цепь модифицирована для связывания с CD3ε. В предпочтительном варианте реализации изобретения мультиспецифическое антитело IgM, IgA, IgG/IgM или IgG/IgA связывается с одной или более опухолеассоциированными мишенями, что перечислены на Фиг. 19, тогда как J-цепь модифицирована для связывания с CD3ε.
Натуральные киллеры (NK) являются важными компонентами врожденного иммунитета и играют ключевую роль в иммунной защите организма в силу их способности высвобождать цитокины и опосредовать цитолитическую активность против опухолевых клеток и клеток, зараженных вирусом. Антигены NK-клеток включают, в том числе, CD16, CD32a, CD56, CD57, CD64, CD117 (или c-kit), молекулы адгезии, включая лимфоцитарный функционально-ассоциированный антиген-2 (LFA-2 или CD2), LFA-3 (CD58) и LFA-1 (CD11a/CD18).
Примеры биспецифических антител с модифицированной J-цепью, вовлекающих NK-клетки, включают антитела IgM, IgA, IgG/IgM и IgG/IgA со специфичностью связывания по отношению к любым опухолевым антигенам, перечисленным выше, содержащие модифицированную J-цепь, связывающуюся с NK-клетками. В конкретном варианте реализации изобретения биспецифическое антитело IgM, IgA, IgG/IgM и IgG/IgA со специфичностью связывания по отношению к HER2 содержит J-цепь, модифицированную таким образом, что она способна связываться с CD16, CD32a, CD56 или CD64. В другом предпочтительном варианте реализации мультиспецифическое антитело IgM, IgA, IgG/IgM или IgG/IgA связывается с любой из опухолевых мишеней, перечисленных в левом столбце таблицы на Фигуре 19, а J-цепь модифицирована таким образом, что она способна связываться с CD16, CD64 или NKG2D на NK-клетках.
Вовлечение макрофагов при использовании антител с модифицированной J-цепью по настоящему изобретению обеспечивается, например, путем придания специфичности J-цепи по отношению к CD14.
В одном варианте реализации изобретения мультиспецифическое антитело IgM, IgA, IgG/IgM или IgG/IgA связывается с одной или более опухолевыми мишенями, что перечислены выше, тогда как J-цепь модифицирована таким образом, что она способна связываться с CD14. В предпочтительном варианте реализации изобретения мультиспецифическое антитело IgM, IgA, IgG/IgM или IgG/IgA связывается с одной или более опухолеассоциированными мишенями, что перечислены на Фиг. 19, тогда как J-цепь модифицирована для связывания с CD3ε.
Антитела IgM, IgA, IgG/IgM и IgG/IgA с модифицированной J-цепью могут быть направлены на антигены на основе углеводов, см., например, обзорную статью Cazet et al., Breast Cancer Research; 2010, 12:204. Было показано, что опухолевые антигены на основе углеводов в значительной степени ассоциированы с опухолями с альтернативной формой гликозилирования, которая позволяет получить антитела IgG, связывающиеся с такими антигенами с достаточной специфичностью, но необязательно с высокой аффинностью. Применение антител IgM, IgA, IgG/IgM и IgG/IgA с соответствующей повышенной авидностью против этого класса антигенов открывает огромные возможности для получения новых терапевтических антител, особенно в сочетании с мобилизацией эффекторных клеток, достигаемой за счет модификации J-цепи. В одном предпочтительном варианте реализации изобретения антитело IgM, IgA, IgG/IgM и IgG/IgA связывается с одним или более опухолевыми антигенами на основе углеводов, при этом J-цепь модифицирована таким образом, что она способна связывать любую из эффекторных клеток, приведенных на Фиг. 19.
В другом предпочтительном варианте реализации антитела IgM, IgA, IgG/IgM и IgG/IgA с модифицированной J-цепью могут быть использованы как часть антител IgM, IgA, IgG/IgM и IgG/IgA, направленных против патогенов. В предпочтительном варианте реализации изобретения патогены выбраны из группы, состоящей из вируса ВИЧ, Mycobacterium tuberculosis, Streptococcus agalactiae, метициллин-резистентного Staphylococcus aureus, Legionella pneumophilia, Streptococcus pyogenes, Escherichia coli, Neisseria gonorrhoeae, Neisseria meningitidis, Pneumococcus, Cryptococcus neoformans, Histoplasma capsulatum, Hemophilis influenzae B, Treponema pallidum, спирохет, вызывающих болезнь Лайма, Pseudomonas aeruginosa, Mycobacterium leprae, Brucella abortus, вируса бешенства, вируса гриппа, цитомегаловируса, вируса простого герпеса I, вируса простого герпеса II, парвоподобного вируса сыворотки крови человека, респираторно-синцитиального вируса, вируса ветряной оспы, вируса гепатита В, вируса гепатита С, вируса кори, аденовируса, вирусов Т-клеточного лейкоза, вируса Эпштейна-Барр, вируса лейкоза мышей, вируса эпидемического паротита, вируса везикулярного стоматита, вируса Синдбис, вируса лимфатического хориоменингита, папилломавируса, вируса болезни синего языка, вируса Сендай, вируса лейкоза кошек, реовируса, полиовируса, вируса обезьян 40, вируса опухоли молочной железы мышей, вируса денге, вируса краснухи, вируса Западного Нила, Plasmodium falciparum, Plasmodium vivax, Toxoplasma gondii, Trypanosoma rangeli, Trypanosoma cruzi, Trypanosoma rhodesiensei, Trypanosoma brucei, Schistosoma mansoni, Schistosoma japonicum, Babesia bovis, Elmeria tenella, Onchocerca volvulus, Leishmania tropica, Trichinella spiralis, Theileria parva, Taenia hydatigena, Taenia ovis, Taenia saginata, Echinococcus granulosus, Mesocestoides corti, Mycoplasma arthritidis, M. hyorhinis, M. orale, M. arginini, Acholeplasma laidlawii, M. salivarium и M. pneumoniae, как описано в патенте США № 6440416.
В этом варианте реализации изобретения в качестве иммунокомпетентной эффекторной клетки может быть использован, например, нейтрофил.
На Фигуре 19, в частности, перечислены вирусные антигены, являющиеся мишенями для антител IgM, IgA, IgG/IgM и IgG/IgA, описанных в данном документе, в комбинации с маркерами нейтрофилов, на которые направлена модифицированная J-цепь таких антител.
Следует отметить, что антитела IgM, IgA, IgG/IgM и IgG/IgA, описанные в данном документе, также могут направленно связываться с углеводными антигенами на злокачественных клетках, которые богаты ими. Такие антигены включают, например, CEA, CA-125, TADG78, Sialyl Lewis-X (CD15).
Во всех вариантах реализации изобретения связывающий фрагмент (связывающий элемент), использованный для модификации нативной J-цепи, может быть введен перед J-цепью или за ней. Следовательно, модифицированная J-цепь со специфичностью связывания по отношению к CD3 может иметь конфигурацию специфичный к CD3scFv-J или J-специфичный к CD3scFv. Последовательность (SEQ ID NO.: 46) и структура специфичного к CD3 одноцепочечного Fv, содержащего J-цепь на N-конце (J-scFv, специфичный к CD3), показаны на Фигуре 8. Последовательность (SEQ ID NO.: 47) и структура специфичного к CD3 одноцепочечного Fv, содержащего J-цепь на N-конце (J-scFv, специфичный к CD3), показаны на Фигуре 9. Благодаря их повышенной авидности такие антитела являются боле эффективными чем биспецифические антитела IgG. Например, благодаря этому они подходят для направленного связывания с низкокопийными мишенями, как, например, содержащимися на резистентных к ритуксану клетках лимфомы Беркитта, характеризующихся низким уровнем экспрессии CD20. В дополнение к этому, описанные в данном документе антитела IgM, IgA, IgG/IgM и IgG/IgA, содержащие модифицированную J-цепь, обладают значительно большей активностью чем биспецифические антитела IgG.
Фармацевтические композиции антител с модифицированной J-цепью
Для терапевтического применения антитела, содержащие модифицированную J-цепь, могут быть введены в состав фармацевтических композиций. Фармацевтическую композицию по настоящему изобретению можно вводить с использованием различных способов, известных в данной области. Специалисту в данной области понятно, что путь и/или способ введения может варьироваться в зависимости от целевого заболевания или состояния и желательных результатов. Для введения соединения по настоящему изобретению с использованием определенных путей введения может возникнуть необходимость в покрытии соединения определенным веществом или совместном введении соединения с определенным веществом для предотвращения инактивации. Например, соединение может быть введено субъекту в соответствующем носителе, например, липосомах, или в разбавителе. Фармацевтически приемлемые разбавители включают физиологический раствор и водные буферные растворы. Фармацевтические носители включают стерильные водные растворы или взвеси и стерильные порошки для приготовления стерильных инъекционных растворов или взвесей непосредственно перед употреблением. Применение таких сред и средств для приготовления фармацевтически активных субстанций известно в данной области.
Композиции также могут содержать вспомогательные вещества, такие как консерванты, смачивающие вещества, эмульгаторы и/или диспергирующие вещества. Предотвращение присутствия микроорганизмов может обеспечиваться как с помощью методов стерилизации, так и путем включения различных противобактериальных и противогрибковых средств, например, парабена, хлорбутанола, фенола, сорбиновой кислоти и тому подобное. В композиции было бы также целесообразно включить изотонические средства, такие как сахара, хлорид натрия и тому подобное. В дополнение к этому, продолжительная абсорбция инъекционной лекарственной формы может быть вызвана путем включения средств, которые задерживают абсорбцию, как, например, моностеарат алюминия и желатин.
Фактические уровни дозировки активных ингредиентов в фармацевтических композициях по настоящему изобретению могут быть различны, чтобы получить количество активного ингредиента, эффективное для достижения желательного терапевтического ответа для конкретного пациента, композицию и способ введения, не будучи токсичными для пациента. Выбранный уровень дозировки будет зависеть от множества фармакокинетических факторов, включая активность конкретных используемых композиций по настоящему изобретению, путь введения, время введения, скорость выделения конкретного используемого соединения, продолжительность лечения, других лекарственных средств, соединений и/или веществ, используемых в комбинации с используемыми композициями, возраста, пола, веса, состояния, общего состояния здоровья и предварительной истории болезни пациента, который получает лечение, и подобных факторов, известных в области медицины.
Композиция должна быть стерильной и достаточно жидкой, чтобы ее можно было ввести с помощью шприца. Помимо воды предпочтительно, чтобы в качестве носителя использовался изотонический забуференный физиологический раствор.
Следующие примеры, перечень последовательностей и графические материалы, представлены с целью облегчения понимания настоящего изобретения, фактический объем которого изложен в прилагаемой формуле изобретения. Надо понимать, что в изложенные методы, могут быть внесены модификации, не отклоняясь от сущности настоящего изобретения
Дополнительные подробности настоящего изобретения проиллюстрированы в следующих неограничивающих примерах.
Пример 1
Получение биспецифического антитела IgM к CDIM, содержащего модифицированную J-цепь, связывающую CD3
1. Создание ДНК-конструкций с заданными мутациями
a. Синтез ДНК-конструкций. Все ДНК-конструкции с заданными мутациями синтезировали с использованием коммерчески доступных наборов (Genescript) с совместимыми сайтами рестрикции на обоих концах для субклонирования в соответствующие экспрессирующие векторы.
b. Конструирование экспрессирующих векторов. Синтезированные ДНК-конструкции ресуспендировали в буфере Трис-ЭДТА при 1 мкг/мл. ДНК (1 мкг) подвергали ферментативному расщеплению, а синтезированный ген отделяли от несущей плазмидной ДНК путем электрофореза. Расщепленную ДНК легировали в предварительно расщепленную плазмидную ДНК (pCAGGS для J-цепи, Gene 108 (1991) 193-200) с помощью стандартных методов молекулярной биологии. Компетентные клетки бактерий трансфецировали легированной ДНК и высевали на чашки Петри со средой Лурия-Бертани, содержащей целый ряд селективных антибиотиков. Выделяли несколько бактериальных колоний, и получали препараты ДНК с помощью стандартных методов молекулярной биологии. Полученную последовательность ДНК верифицировали с помощью секвенирования. Для получения плазмидной ДНК и впоследствии для трансфекции клеток использовали только бактериальные клоны с 100% совпадением их последовательности ДНК с заданной последовательностью ДНК.
c. Разные J-цепи. Для того, чтобы продемонстрировать, что варианты J-цепи способны к связыванию с IgM, были сконструированы два разных варианта J-цепи с различными сайтами слияния, включающими последовательность антитела к CD3 (OKT3 scFv).
i. Эта конструкция состоит из последовательности scFv OKT3 (антитело к CD3), слитой с N-концом J-цепи человека (CD3scFv-состоящий из 15 а.к. линкер-J): QVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKF KDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYSLDYWGQGTTLTVSSGGGGSGGGG SGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVP AHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEIKGGGGSGGGGSGGGGSQ EDERIVLVDNKCKCARITSRIIRSSEDPNEDIVERNIRIIVPLNNRENISDPTSPLRTRFVYHL SDLCKKCDPTEVELDNQIVTATQSNICDEDSATETCYTYDRNKCYTAVVPLVYGGETKMVETAL TPDACYPDGGGSEQKLISEEDLNSAVDHHHHHH-
(SEQ ID NO.: 46).
Эта конструкция имеет молекулярную массу около 45k кДа и способна к связыванию с растворимой эпсилон-цепью CD3 (Sino Biological) или Т-клетками; и способна к связыванию с моноклональным антителом 4A6 к белкам с эпитопной меткой Myc или другими моноклональными антителами к белкам с эпитопной меткой Myc.
ii. Эта конструкция состоит из scFv OKT3 (антитела к CD3), слитого с C-концом J-цепи человека (J-состоящий из 15 а.к. линкер-CD3scFv):
QEDERIVLVDNKCKCARITSRIIRSSEDPNEDIVERNIRIIVPLNNRENISDPTSPLRT RFVYHLSDLCKKCDPTEVELDNQIVTATQSNICDEDSATETCYTYDRNKCYTAVVPLVYGGETK MVETALTPDACYPDGGGGSGGGGSGGGGSQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMH WVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARY YDDHYSLDYWGQGTTLTVSSGGGGSGGGGSGGGGSQIVLTQSPAIMSASPGEKVTMTCSASSSV SYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSN PFTFGSGTKLEIKEQKLISEEDLNSAVDHHHHHH-
(SEQ ID NO.: 47).
Эта конструкция J-CD3scFv имеет молекулярную массу около 45k кДа и способна к связыванию с растворимой эпсилон-цепью CD3 (Sino Biological) или Т-клетками; и способна к связыванию с моноклональным антителом 4A6 или другими антителами к белкам с эпитопной меткой Myc.
d. Тяжелая цепь IgM: Эта конструкция тяжелой цепи имеет полноразмерную μ-цепь IGM-55.5, которая связывает CDIM на поверхности B-клеток. 
(SEQ ID NO.: 3)
Эта конструкция тяжелой цепи имеет молекулярную массу около 64 кДа, а при коэкспрессии с легкой цепью полученный в результате IgM способен к связыванию В-клеток, позитивных по CDIM.
e. Легкая цепь IGM-55.5, известная как IGM-55.5, который связывает CDIM (молекула, индуцирующая клеточную гибель) на поверхности B-клеток:  (SEQ ID NO.: 5)
(SEQ ID NO.: 5)
Конструкция легкой цепи имеет молекулярную массу около 24 кДа, а при коэкспрессии с соответствующей тяжелой цепью (SEQ ID NO. 3) способа к связыванию с В-клетками, позитивными по CDIM.
2. Экспрессия, очистка и характеристика белков
a. Трансфекция. ДНК, кодирующую тяжелую, легкую и модифицированную J-цепь, трансфецировали в клетки CHO. ДНК для экспрессирующих векторов обычно смешивают в соотношении 1:1:1 с ПЭИ и затем добавляют к клеткам CHO-S. Трансфекцию с использование ПЭИ и клеток CHO-S проводят в соответствии с отработанными методами (см. ʺBiotechnology and Bioengineering, Vol 87, 553-545ʺ).
b. Иммунопреципитация
i. Capture Select IgM (BAC, Thermo Fisher). Белки IgM, полученные из супернатантов при культивировании трансфецированных клеток CHO, частично очищают с помощью иммунопреципитации с использованием аффинной матрицы Capture Select IgM в соответствии с протоколом производителей (GE Life Sciences). После инкубации при комнатной температуре в течение 2 часов, аффинную матрицу отделяют от супернатанта путем центрифугирования. Далее матрицу промывают 3 раза PBS, затем его осторожно удаляют. Связанный белок элюируют из матрицы путем инкубации с буфером для нанесения образцов белка NuPage LDS (Life Technology) в течение 5 минут.
ii. Агарозная аффинная матрица с антителами к белкам с эпитопной меткой Myc (Sigma). Белки IgM, полученные из супернатантов при культивировании трансфецированных клеток CHO, частично очищают с помощью иммунопреципитации с использованием агарозной аффинной матрицы с антителами к белкам с эпитопной меткой Myc в соответствии с протоколом производителей. После инкубации при комнатной температуре в течение 2 часов, аффинную матрицу отделяют от супернатанта путем центрифугирования. Далее матрицу промывают 3 раза PBS, затем его осторожно удаляют после последней промывки. Связанный белок элюируют из матрицы путем инкубации с буфером для нанесения образцов белка NuPage LDS (Life Technology) в течение 5 минут.
c. Гель-электрофорез
i. ДСН-ПААГ в невосстанавливающих условиях
ii. ДСН-ПААГ в невосстанавливающих условиях позволяет разделить IgM и его мутантные формы по размеру. Пентамерный IgM, состоящий из гомодимерных тяжелых и легкой цепей, дает полосу белка на геле с молекулярной массой приблизительно 1000000 дальтон. Буфер для нанесения образцов NuPage LDS Sample Buffer (Life Technologies) добавляют к образцам белка IgM при 25 C в течение 30 минут до нанесения на гель. NativePage Novex 3-12% Bis-Tris гель (Life Technologies) используют с разделяющим буфером Novex Tris-Acetate SDS Running Buffer (Life Technologies). Гель разгоняют до тех пор, пока фронт красителя не достигает нижней части геля.
iii. ДСН-ПААГ в восстанавливающих условиях. Буфер для нанесения образцов NuPage LDS sample buffer (Life Technologies) и восстанавливающее средство NuPage дитиотреитол (Life Technologies) добавляют к белку IgM и нагревают до 80°C в течение 10 минут до нанесения на NuPage Novex 4-12% Bis-Tris гель (Life Technologies). Разделяющий буфер NuPage MES SDS Running Buffer используют для гель-электрофореза. Гели разгоняют до тех пор, пока фронт красителя не достигает нижней части геля. После завершения электрофореза гель вынимают из прибора и окрашивают с использованием набора Colloidal Blue Staining (Life Technologies).
iv. Детекция с помощью вестерн-блота. После завершения электрофореза гель вынимают из XCell SureLock Mini-Cell. Переносят на ПВДФ-мембрану при 30 вольтах в течение 1 часа (см. в руководстве пользователя от Life Technologies). Блокируют с использованием 20 мл 3% БСА в PBST при 25 C в течение 1 часа.
Для анализа антитела к J-цепи с помощью вестерн-блота антитело к J-цепи (SP105, Thermo Fisher) добавляют в разведении 1:500 в 3% БСА в PBST и инкубируют на протяжении ночи при 4C. Промывают четыре раза PBST при комнатной температуре. Добавляют меченные пероксидазой хрена антитела козы к IgG кролика (Jackson Immunology) в разведении 1:5000 в 3% БСА в PBST и инкубируют в течение 1 часа при комнатной температуре. Промывают четыре раза PBST при комнатной температуре. Добавляют 10 мл хемилюминесцентного субстрата пероксидазы хрена (Thermo Fisher) на 10 минут до экспозиции блота на пленке. Антитело к J-цепи реагирует только с IgM, который коэкспрессирован либо с немодифицированной J-цепью (Фигура 6), либо модифицированной J-цепью (Фигура 7).
Для анализа антитела к белкам с эпитопной меткой Myc с помощью вестерн-блота добавляют антитело к белкам с эпитопной меткой Myc (4A6, Millipore) в разведении 1:200 в 3% БСА в PBST для инкубации на протяжении ночи при 4C. Промывают четыре раза PBST при комнатной температуре. Добавляют меченные пероксидазой хрена антитела козы к IgG кролика (Jackson Immunology) в разведении 1:5000 в 3% БСА в PBST и инкубируют в течение 1 часа при комнатной температуре. Промывают четыре раза PBST при комнатной температуре. Добавляют 10 мл хемилюминесцентного субстрата пероксидазы хрена (Thermo Fisher) на 10 минут до экспозиции блота на пленке. Антитело к белкам с эпитопной меткой Myc реагирует только с модифицированной J-цепью с эпитопной меткой Myc(Фиг.2).
3. Функциональный анализ биспецифического связывания
a. FACS-анализ адресного связывания
Связывание IGM-55.5 с CD3scFv-J или IGM-55.5 с J-CD3scFv с Т-клетками подтверждается связыванием антитела с линией Т-клеток (Jurkat, линия CD3-положительных клеток) и линией В-клеток (Nalm6, клеточная линия отрицательного контроля). После отмывки к клеточной суспензии добавляют меченный родамином 4A6. Адресное связывание с клетками оценивается по MFI как клеток положительного, так и отрицательного контролей с антигеном CD3 или без него.
Связывание IGM-55.5 с CD3scFv-J или IGM-55.5 с J-CD3scFv с клетками, экспрессирующими CDIM, подтверждается связыванием антитела с клеточной линией положительного контроля (Daudi, клеточная линия положительного контроля) и клеточной линией отрицательного контроля (Peer). После отмывки к клеточной суспензии добавляют меченный родамином 4A6. Адресное связывание с клетками оценивается по MFI как клеток положительного, так и отрицательного контролей с антигеном CDIM или без него.
b. FACS-анализ опосредованного Т-клетками уничтожения B-клеток при кокультивировании
Raji, линию CD19+CD20+ B-клеток, кокультивировали с T-ALL, линией цитолитических CD8+ Т-клеток с CD20 IgM x J-дикого типа или CD20 IgM x CD3-J-цепь в течение 24 часов при 37 градусах, 5% CO2. Клетки собирали и окрашивали флуоресцентными антителами к CD3 (552852/BD Biosciences) и CD19 (555413/BD Biosciences, а также анализировали с помощью проточной цитометрии для оценки жизнеспособности B-клеток.
c. Комплементзависимая цитотоксичность B-клеток
CD20+ клетки линии Ramos высевали в 96-луночные микропланшеты белого цвета с половинным объемом лунок при 25000 клеток/лунку. Антитело и сыворотку крови человека с компонентами комплемента (конечная концентрация 5%, Quidel) добавляли для вызова комплементзависимой цитотоксичности, а число жизнеспособных клеток измеряли с использованием Cell Titer Glo и протокола производителя. Измеряли интенсивность люминесценции на многорежимном планшетном анализаторе Envision (Perkin Elmer) с использованием времени интеграции 0,1 с на лунку. Процент жизнеспособных клеток рассчитывали путем нормализации значений люминесценции (Относительные единицы люминесценции - RLU) в сравнении с лунками, в которые не добавляли лекарственное вещество. Данные анализировали с использованием GraphPad Prism, и четырехпараметрическую аппроксимацию с верхними и нижними значениями устанавливали на 100 и 0% жизнеспособности, соответственно.
Мультиспецифическое связывание и мультиспецифические функциональные анализы можно осуществлять аналогичным образом с использованием методов, известных в данной области, таких как те, что описаны выше.
Пример 2
Молекулярное клонирование, экспрессия и очистка биспецифического антитела IgM к CD20, содержащего модифицированную J-цепь, несущую связывающий scFv, специфичный к CD3
В этом примере описано получение, молекулярное клонирование, экспрессия и очистка дополнительного антитела IgM, направленного на другой B-клеточный антиген (CD20), и модифицированной J-цепи, связывающейся с CD3. ДНК, соответствующая последовательностям тяжелой и легкой цепей, показанным ниже, была получена с использованием способов, которые описаны в Примере 1.
SEQ ID NO.: 48: Последовательность легкой цепи IgM антитела к CD20 (ритуксимаб)
MDMRVPAQLLGLLLLWLRGARCQIVLSQSPAILSASPGEKVTMTCRASSSVSYIHWFQQKPGSSPKPWIYATSNLASGVPVRFSGSGSGTSYSLTISRVEAEDAATYYCQQWTSNPPTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO.: 49: Последовательность тяжелой цепи IgM антитела к CD20 (ритуксимаб)
MGWSYIILFLVATATGVHSQVQLQQPGAELVKPGASVKMSCKASGYTFTSYNMHWVKQTPGRGLEWIGAIYPGNGDTSYNQKFKGKATLTADKSSSTAYMQLSSLTSEDSAVYYCARSTYYGGDWYFNVWGAGTTVTVSSGSASAPTLFPLVSCENSPSDTSSVAVGCLAQDFLPDSITFSWKYKNNSDISSTRGFPSVLRGGKYAATSQVLLPSKDVMQGTDEHVVCKVQHPNGNKEKNVPLPVIAELPPKVSVFVPPRDGFFGNPRKSKLICQATGFSPRQIQVSWLREGKQVGSGVTTDQVQAEAKESGPTTYKVTSTLTIKESDWLSQSMFTCRVDHRGLTFQQNASSMCVPDQDTAIRVFAIPPSFASIFLTKSTKLTCLVTDLTTYDSVTISWTRQNGEAVKTHTNISESHPNATFSAVGEASICEDDWNSGERFTCTVTHTDLPSPLKQTISRPKGVALHRPDVYLLPPAREQLNLRESATITCLVTGFSPADVFVQWMQRGQPLSPEKYVTSAPMPEPQAPGRYFAHSILTVSEEEWNTGETYTCVVAHEALPNRVTERTVDKSTGKPTLYNVSLVMSDTAGTCY
ДНК, соответствующую этим тяжелым и легким цепям, а также ДНК, соответствующую либо J-цепи дикого типа (д.т.), либо последовательностям J-цепи O15J и J15O, описанным в Примере 1, контрансфециовали в клетки HEK293, а белки были экспрессированы и очищены с использованием аффинной смолы, связывающей однодоменные антитела верблюда. Как показано на Фигуре 13, все четыре белка экспрессировались в необходимом количестве. Гексамер IgM, специфичного к CD20, без J-цепи четко отделяется от содержащих J-цепь пентамеров, это касается пентамера IgM с J-цепью дикого типа, а также биспецифических IgM, причем scFv, специфичный к CD3, связан с J-цепью в любой ориентации (O15J или J15O).
Пример 3
Молекулярное клонирование, экспрессия и очистка биспецифического антитела IgM к CD20, содержащего модифицированную J-цепь, несущую другой связывающий scFv, специфичный к CD3
Для установления того, что сборка биспецифического IgM осуществима с модифицированной J-цепью, несущей scFv, специфичный к CD3, с последовательностью, отличной от той, что была использована в Примерах 1 и 2, была получена J-цепь, несущая вариабельные области из антитела висилизумаб (Nuvion). Ниже показаны последовательности для двух J-цепей, причем scFv соответствует висилизумабу (V), слитому с J-цепью через линкер, содержащий 15 а.к. в двух разных ориентациях - V15J и J15V.
SEQ ID NO.: 50: Последовательность J-цепи для V15J
MGWSYIILFLVATATGVHSQVQLVQSGAEVKKPGASVKVSCKASGYTFISYTMHWVRQAPGQGLEWMGYINPRSGYTHYNQKLKDKATLTADKSASTAYMELSSLRSEDTAVYYCARSAYYDYDGFAYWGQGTLVTVSSGGGGSGGGGSGGGGSDIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQKPGKAPKRLIYDTSKLASGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQWSSNPPTFGGGTKLEIKGGGGSGGGGSGGGGSQEDERIVLVDNKCKCARITSRIIRSSEDPNEDIVERNIRIIVPLNNRENISDPTSPLRTRFVYHLSDLCKKCDPTEVELDNQIVTATQSNICDEDSATETCYTYDRNKCYTAVVPLVYGGETKMVETALTPDACYPD
SEQ ID NO.: 51: Последовательность J-цепи для J15V
MKNHLLFWGVLAVFIKAVHVKAQEDERIVLVDNKCKCARITSRIIRSSEDPNEDIVERNIRIIVPLNNRENISDPTSPLRTRFVYHLSDLCKKCDPTEVELDNQIVTATQSNICDEDSATETCYTYDRNKCYTAVVPLVYGGETKMVETALTPDACYPDGGGGSGGGGSGGGGSQVQLVQSGAEVKKPGASVKVSCKASGYTFISYTMHWVRQAPGQGLEWMGYINPRSGYTHYNQKLKDKATLTADKSASTAYMELSSLRSEDTAVYYCARSAYYDYDGFAYWGQGTLVTVSSGGGGSGGGGSGGGGSDIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQKPGKAPKRLIYDTSKLASGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQWSSNPPTFGGGTKLEIK
Как описано в Примере 1, ДНК, соответствующая этим последовательностям, была синтезирована и трансфецирована в клетки HEK293 наряду с ДНК, кодирующей тяжелую и легкую цепи IgM, специфичного к CD20, для получения белка, который был затем очищен с использованием аффинной матрицы с верблюжьими антителами, специфическими к IgM. Как показано на Фигуре 13, J-цепи, слитые с новым scFv. специфичным к CD3, с помощью линкера длиной 15 а.к. способны к встраиванию в IgM, а пентамерная форма биспецифического IgM с соответствующей J-цепью четко отличимая от гексамерной формы без J-цепи.
Пример 4
Молекулярное клонирование, экспрессия и очистка IgM со вторым связывающим доменом для клетки-эффектора, содержащим модифицированную J-цепь антител верблюда и специфичным к протеинам CD16 - верблюжьи антитела, специфичные к CD16
Этот пример демонстрирует, что существует возможность для соединения связывающего фрагмента, отличного от scFv, с J-цепью, который направлен на другую мишень на эффекторных клетках другой популяции. Была использована показанная ниже последовательность Vhh-домена верблюжьего антитела, которая была отобрана для связывания с антигеном CD16 на натуральных киллерах (NK-клетки). Вновь следует отметить, что эта последовательность была связана с J-цепью с использованием гибкого линкера длиной 15 а.к. для получения C15J. Были выполнены экспрессия и очистка биспецифического IgM (согласно описанию в примере 1), а также его анализ с использованием гибридных гелей. Четко просматривается формирование пентамерных молекул. Кроме того, с использованием вестерн-блота было установлено включение J-цепи C15J в пентамер IgM (Фигура 18).
SEQ ID NO.: 52: Последовательность J-цепи для C15J
MGWSYIILFLVATATGVHSEVQLVESGGELVQAGGSLRLSCAASGLTFSSYNMGWFRRAPGKEREFVASITWSGRDTFYADSVKGRFTISRDNAKNTVYLQMSSLKPEDTAVYYCAANPWPVAAPRSGTYWGQGTQVTVSSGGGGSGGGGSGGGGSQEDERIVLVDNKCKCARITSRIIRSSEDPNEDIVERNIRIIVPLNNRENISDPTSPLRTRFVYHLSDLCKKCDPTEVELDNQIVTATQSNICDEDSATETCYTYDRNKCYTAVVPLVYGGETKMVETALTPDACYPD
Пример 5
Анализ комплементзависимой цитотоксичности для семейства IgM со встроенными J-цепями и без них
Комплементзависимая цитотоксичность является ключевым механизмом уничтожения клеток при участии антител. Известно, что антитела IgM обладают повышенной комплементзависимой цитотоксичностью (CDC), обусловленной их мультимерной формой. Ключевой аспект настоящего изобретения заключается в изучении нарушения связывания с C1q - ключевым компонентом системы комплемента и, следовательно, возможности подавлять CDC, что обусловлено встраиванием модифицированных J-цепей, которые либо на их С-, либо N-концах несут специфичные к эффекторным клеткам связывающие фрагменты scFv или Vhh-домен верблюжьего антитела. Авторами было оценена CDC-активность каждой из конструкций на основе IgM и биспецифического IgM. Как показано на Фигуре 14, авторы установили, что встраивание модифицированной J-цепи к удивлению не оказывает никакого отрицательного влияния на CDC-активность биспецифических IgM. Более того, с использование линкеров изученной длины, авторы установили, что биспецифические IgM обладают CDC-активностью, которая в 60-100 раз выше активности соответствующего IgG в молярном соотношении(Фиг. 14).
Пример 6
Анализ опосредованного Т-клетками уничтожения B-клеток при участии биспецифического IgM при кокультивировании in vitro
Предполагается, что вовлечение эффекторных T-клеток биспецифическими антителами IgM с модифицированной J-цепью приведет к значительному увеличению гибели популяций B-клеток-мишеней по сравнению с IgM, не несущим J-цепь, или несущим J-цепь дикого типа. Для изучения уничтожения клеток при кокультивировании был проведен анализ уничтожения клеток, как описано в Примере 1. Антитело в разовой высокой концентрации (100 мл) инкубировали с клетками CD20+ Raji и эффекторными клетками CD3+ T-ALL. Как показано на Фигуре 15, биспецифический IgM, несущий на его J-цепи scFv, связывающий CD3, способен приводить к полному уничтожению B-клеток. В аналогичном эксперименте, который проводился для оценки дозозависимого эффекта, показано, что уровень уничтожения B-клеток повысился в 300 раз по сравнению с IgM, который не несет scFv, способный к вовлечению T-клеток. Полное уничтожение B-клеток при участии биспецифического IgM наблюдается при использовании концентраций всего лишь 10 нг/мл.
Пример 7
Демонстрация опосредованного Т-клетками уничтожения B-клеток in vivo в модели на мышах линии NSG
Для изучения биспецифических IgM, полученных in vivo, были выполнены эксперименты с использованием гуманизированных мышей линии NSG (с мутацией scid, проявляющейся тяжелым комбинированным иммунодефицитом, и аномалией гамма-цепи рецептора) с диабетом и без ожирения. Иммунная функция этих мышей сильно ослаблена, и у них отсутствуют T- и B-лимфоциты. Им были пересажены стволовые клетки CD34+ человека для получения мышей, по существу содержащих популяции лимфоцитов человека. Когда этим животным внутривенно вводили CD20 IgM x CD3-J-цепь в дозе 0,5 мг/кг, а также получали цельную кровь и анализировали с помощью проточной цитометрии в отношении уровней В-клеток человека в циркуляторном русле, наблюдалось полное истощение популяции B-клеток, даже в случае воздействия в течение всего лишь 6 часов (Фигура 17).
Несмотря на то что в настоящем описании было представлено несколько вариантов реализации изобретения, следует понимать, что описанные системы и способы могут быть воплощены во многих других конкретных формах, не отклоняясь от сущности и объема настоящего изобретения. Примеры настоящего изобретения должны рассматриваться с иллюстративной, а не ограничительной целью, а их назначение не должно ограничиваться приведенными в данном документе подробностями. Различные примеры изменений, замен и поправок очевидны специалистам в данной области и могут быть внесены, не отклоняясь от сущности и объема изобретения, описанного в данном документе.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> IGM BIOSCIENCES, INC.
<120> МОДИФИЦИРОВАННАЯ J-ЦЕПЬ
<130> IGM-0004-WO
<140> PCT/US2015/024149
<141> 2015-04-02
<150> 61/974738
<151> 2014-04-03
<160> 57
<170> PatentIn версии 3.5
<210> 1
<211> 137
<212> ПРТ
<213> Homo sapiens
<400> 1
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Glu Thr Ala
115 120 125
Leu Thr Pro Asp Ala Cys Tyr Pro Asp
130 135
<210> 2
<211> 137
<212> ПРТ
<213> Pan troglodytes
<400> 2
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Arg Ala
100 105 110
Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Glu Thr Ala
115 120 125
Leu Thr Pro Asp Ala Cys Tyr Pro Asp
130 135
<210> 3
<211> 574
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 3
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Arg Met Ala Trp Gly Ala Ser Val Asn Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Ser Ala Ser Ala Pro Thr
115 120 125
Leu Phe Pro Leu Val Ser Cys Glu Asn Ser Pro Ser Asp Thr Ser Ser
130 135 140
Val Ala Val Gly Cys Leu Ala Gln Asp Phe Leu Pro Asp Ser Ile Thr
145 150 155 160
Phe Ser Trp Lys Tyr Lys Asn Asn Ser Asp Ile Ser Ser Thr Arg Gly
165 170 175
Phe Pro Ser Val Leu Arg Gly Gly Lys Tyr Ala Ala Thr Ser Gln Val
180 185 190
Leu Leu Pro Ser Lys Asp Val Met Gln Gly Thr Asp Glu His Val Val
195 200 205
Cys Lys Val Gln His Pro Asn Gly Asn Lys Glu Lys Asn Val Pro Leu
210 215 220
Pro Val Ile Ala Glu Leu Pro Pro Lys Val Ser Val Phe Val Pro Pro
225 230 235 240
Arg Asp Gly Phe Phe Gly Asn Pro Arg Lys Ser Lys Leu Ile Cys Gln
245 250 255
Ala Thr Gly Phe Ser Pro Arg Gln Ile Gln Val Ser Trp Leu Arg Glu
260 265 270
Gly Lys Gln Val Gly Ser Gly Val Thr Thr Asp Gln Val Gln Ala Glu
275 280 285
Ala Lys Glu Ser Gly Pro Thr Thr Tyr Lys Val Thr Ser Thr Leu Thr
290 295 300
Ile Lys Glu Ser Asp Trp Leu Ser Gln Ser Met Phe Thr Cys Arg Val
305 310 315 320
Asp His Arg Gly Leu Thr Phe Gln Gln Asn Ala Ser Ser Met Cys Val
325 330 335
Pro Asp Gln Asp Thr Ala Ile Arg Val Phe Ala Ile Pro Pro Ser Phe
340 345 350
Ala Ser Ile Phe Leu Thr Lys Ser Thr Lys Leu Thr Cys Leu Val Thr
355 360 365
Asp Leu Thr Thr Tyr Asp Ser Val Thr Ile Ser Trp Thr Arg Gln Asn
370 375 380
Gly Glu Ala Val Lys Thr His Thr Asn Ile Ser Glu Ser His Pro Asn
385 390 395 400
Ala Thr Phe Ser Ala Val Gly Glu Ala Ser Ile Cys Glu Asp Asp Trp
405 410 415
Asn Ser Gly Glu Arg Phe Thr Cys Thr Val Thr His Thr Asp Leu Pro
420 425 430
Ser Pro Leu Lys Gln Thr Ile Ser Arg Pro Lys Gly Val Ala Leu His
435 440 445
Arg Pro Asp Val Tyr Leu Leu Pro Pro Ala Arg Glu Gln Leu Asn Leu
450 455 460
Arg Glu Ser Ala Thr Ile Thr Cys Leu Val Thr Gly Phe Ser Pro Ala
465 470 475 480
Asp Val Phe Val Gln Trp Met Gln Arg Gly Gln Pro Leu Ser Pro Glu
485 490 495
Lys Tyr Val Thr Ser Ala Pro Met Pro Glu Pro Gln Ala Pro Gly Arg
500 505 510
Tyr Phe Ala His Ser Ile Leu Thr Val Ser Glu Glu Glu Trp Asn Thr
515 520 525
Gly Glu Thr Tyr Thr Cys Val Val Ala His Glu Ala Leu Pro Asn Arg
530 535 540
Val Thr Glu Arg Thr Val Asp Lys Ser Thr Gly Lys Pro Thr Leu Tyr
545 550 555 560
Asn Val Ser Leu Val Met Ser Asp Thr Ala Gly Thr Cys Tyr
565 570
<210> 4
<211> 137
<212> ПРТ
<213> Pongo abelii
<400> 4
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Ile Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Glu Ala Ala
115 120 125
Leu Thr Pro Asp Ala Cys Tyr Pro Asp
130 135
<210> 5
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 6
<211> 137
<212> ПРТ
<213> Callithrix jacchus
<400> 6
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Asn Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Val Phe Gly Gly Glu Thr Lys Met Val Glu Ser Ala
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 7
<211> 137
<212> ПРТ
<213> Macaca mulatta
<400> 7
Gln Glu Gly Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Thr Tyr Gly Gly Glu Thr Lys Met Val Gln Thr Ala
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 8
<211> 137
<212> ПРТ
<213> Papio anubis
<400> 8
Gln Glu Gly Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Leu Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Thr Tyr Arg Gly Glu Thr Lys Met Val Gln Thr Ala
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 9
<211> 137
<212> ПРТ
<213> Saimiri boliviensis
<400> 9
Gln Glu Asp Glu Arg Ile Val Leu Ala Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Asn Asn Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Val Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Thr
100 105 110
Val Val Pro Leu Val Phe Gly Gly Glu Thr Lys Met Val Lys Ala Thr
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 10
<211> 136
<212> ПРТ
<213> Tupaia chinensis
<400> 10
Glu Asp Lys Gly Thr Val Leu Val Asp Asn Lys Cys Lys Cys Ala Gln
1 5 10 15
Val Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Asn Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Gly Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Leu Thr Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser Ala
85 90 95
Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Thr Met
100 105 110
Val Pro Leu Ile Phe Gly Gly Lys Thr Lys Met Val Gln Thr Ala Leu
115 120 125
Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 11
<211> 134
<212> ПРТ
<213> Tursiops truncatus
<400> 11
Glu Asp Glu Arg Thr Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Leu Thr Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ile Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro
100 105 110
Leu Thr Tyr Gly Gly Lys Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 12
<211> 134
<212> ПРТ
<213> Orcinus orca
<400> 12
Glu Asp Glu Arg Thr Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Leu Thr Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ile Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro
100 105 110
Leu Thr Tyr Gly Gly Lys Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 13
<211> 134
<212> ПРТ
<213> Loxodonta africana
<400> 13
Lys Asp Glu Arg Thr Leu Leu Val Asp Asn Lys Cys Lys Cys Val Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Lys Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Thr Val Pro
100 105 110
Leu Ile Tyr Gly Gly Lys Thr Lys Met Val Thr Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Arg Asp
130
<210> 14
<211> 134
<212> ПРТ
<213> Leptonychotes weddellii
<400> 14
Glu Asp Gly Arg Thr Val Leu Val Asp Asn Lys Cys Gln Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Pro Glu Asn Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Ile Ala Ser Gln Ser Asn Leu Cys Asp Glu Asp Ser Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Trp Val Pro
100 105 110
Phe Thr Tyr Gly Gly Gln Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 15
<211> 133
<212> ПРТ
<213> Ceratotherium simum
<400> 15
Glu Asp Glu Arg Ile Leu Val Asp Asn Lys Cys Lys Cys Val Arg Ile
1 5 10 15
Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Asn Glu Asp Ile Val
20 25 30
Glu Arg His Ile Arg Ile Val Val Pro Leu Asn Asn Arg Glu Asn Ile
35 40 45
Ser Asp Pro Thr Ser Pro Leu Arg Thr Gln Phe Val Tyr His Leu Ser
50 55 60
Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp Asn Gln
65 70 75 80
Val Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser Glu Ile
85 90 95
Cys Tyr Ala Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro Leu
100 105 110
Thr Tyr Gly Gly Glu Thr Lys Val Val Glu Thr Ala Leu Thr Pro Asp
115 120 125
Ser Cys Tyr Pro Asp
130
<210> 16
<211> 134
<212> ПРТ
<213> Felis catus
<400> 16
Glu Asp Glu Arg Thr Val Leu Val Asp Asn Lys Cys Lys Cys Val Arg
1 5 10 15
Val Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Asn Phe Val Tyr His Leu
50 55 60
Ser Asp Val Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Ile Val Met Ala Ser Gln Ser Asn Ile Cys Asp Glu Glu Ser Glu
85 90 95
Val Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro
100 105 110
Phe Thr Tyr Gly Gly Val Thr Arg Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 17
<211> 134
<212> ПРТ
<213> Canis familiaris
<400> 17
Glu Glu Glu Arg Thr Val Leu Val Asp Asn Lys Cys Gln Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Pro Asp Asp Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Leu Val Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Ile Ala Ser Gln Ser Asn Leu Cys Asp Glu Asp Thr Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro
100 105 110
Ile Met Tyr Gly Gly Gln Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 18
<211> 134
<212> ПРТ
<213> Ailuropoda melanoleuca
<400> 18
Glu Asp Gly Arg Thr Val Leu Val Asp Asn Lys Cys Gln Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Gly Asn
65 70 75 80
Gln Ile Val Ile Ala Ser Gln Ser Asn Leu Cys Asp Glu Asp Ser Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro
100 105 110
Phe Thr Tyr Gly Gly Gln Thr Arg Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 19
<211> 133
<212> ПРТ
<213> Mustela furo
<400> 19
Asp Gly Met Thr Val Leu Ala Asp Asn Lys Cys Gln Cys Val Arg Ile
1 5 10 15
Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Asn Glu Asp Ile Ile
20 25 30
Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn Ile
35 40 45
Ser Asp Pro Thr Ser Pro Val Arg Thr Asn Phe Val Tyr His Leu Ser
50 55 60
Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asn Gln
65 70 75 80
Ile Val Ile Ala Ser Gln Ser Asn Leu Cys Asp Glu Asp Ser Glu Thr
85 90 95
Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Trp Val Pro Phe
100 105 110
Thr Tyr Gly Gly Gln Thr Lys Met Val Gln Thr Ala Leu Thr Pro Asp
115 120 125
Ser Cys Tyr Pro Asp
130
<210> 20
<211> 134
<212> ПРТ
<213> Equus caballus
<400> 20
Glu Gly Glu Arg Ile Val Leu Ala Asp Asn Lys Cys Lys Cys Val Arg
1 5 10 15
Val Thr Ser Arg Ile Ile Pro Ser Pro Glu Asn Pro Asn Glu Asp Ile
20 25 30
Leu Glu Arg His Ile Arg Ile Ile Ile Pro Val Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Thr Ala Ser Gln Ser Asn Ile Cys Asp Glu Glu Ser Glu
85 90 95
Thr Cys Tyr Ala Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Pro
100 105 110
Leu Thr Tyr Gly Gly Gln Thr Lys Ile Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 21
<211> 136
<212> ПРТ
<213> Cavia porcellus
<400> 21
Glu Glu Glu Arg Thr Val Leu Ala Asp Asn Lys Cys Gln Cys Ala Arg
1 5 10 15
Val Thr Ser Arg Val Ile Arg Asn Pro Asp Asn Pro Val Glu Asp Ile
20 25 30
Val Glu Arg His Ile Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asp
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Leu Cys Glu Asp Asp Arg Glu
85 90 95
Pro Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Val
100 105 110
Val Pro Leu Ser Tyr Gly Gly Lys Thr Lys Leu Val Thr Ala Ala Leu
115 120 125
Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 22
<211> 134
<212> ПРТ
<213> Camelus ferus
<400> 22
Glu Asp Glu Ser Thr Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Glu Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Leu Thr Glu Val Glu Leu Gly Asn
65 70 75 80
Arg Val Val Thr Ala Thr Gln Ser Asn Leu Cys Asp Asp Asp Ile Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Lys Val Pro
100 105 110
Phe Thr Tyr Asp Gly Glu Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 23
<211> 134
<212> ПРТ
<213> Capra hircus
<400> 23
Glu Asn Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Met Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Thr Arg Glu Val Glu Leu Glu Asp
65 70 75 80
Gln Val Val Thr Ala Ser Gln Ser Asn Ile Cys Asp Arg Asp Thr Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Lys
100 105 110
Leu Asn Tyr Arg Gly Gln Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 24
<211> 136
<212> ПРТ
<213> Chinchilla lanigera
<400> 24
Glu Glu Lys Lys Thr Ile Leu Ala Asp Asn Lys Cys Gln Cys Ala Arg
1 5 10 15
Val Ser Ser Arg Val Ile Arg Ser Pro Glu Asn Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Tyr Ile Arg Ile Ile Val Pro Val Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Arg Arg Thr Gln Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asn Asn
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Leu Cys Glu Glu Asp Gly Glu
85 90 95
Pro Glu Thr Cys Tyr Ala Tyr Asn Arg Asn Lys Cys Tyr Thr Ala Val
100 105 110
Val Pro Leu Thr Tyr Gly Gly Glu Thr Lys Pro Val Lys Val Ala Leu
115 120 125
Thr Pro Asp Ser Cys Tyr Ala Asp
130 135
<210> 25
<211> 137
<212> ПРТ
<213> Mesocricetus auratus
<400> 25
Asp Asn Glu Lys Thr Ile Leu Ala Asp Asn Lys Cys Met Cys Thr Arg
1 5 10 15
Val Thr Ser Arg Ile Ile Arg Ser Thr Glu Asn Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Val Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asp
65 70 75 80
His Ile Val Thr Ala Ser Gln Ser Thr Ile Cys Ser Glu Asp Ser Val
85 90 95
Val Pro Glu Thr Cys Tyr Met Tyr Asn Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Met Val Pro Leu Thr Tyr Arg Gly Glu Thr Lys Met Val Pro Ser Ala
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 26
<211> 134
<212> ПРТ
<213> Ovis aries
<400> 26
Glu Asn Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Met Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Thr Arg Glu Val Glu Leu Glu Asp
65 70 75 80
Gln Val Val Thr Ala Ser Gln Ser Asn Ile Cys Asp Arg Asp Thr Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Lys
100 105 110
Leu Asn Tyr Arg Gly Gln Thr Lys Leu Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 27
<211> 134
<212> ПРТ
<213> Myotis lucifugus
<400> 27
Glu Asp Glu Arg Thr Val Leu Ala Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Val Thr Ser Arg Val Ile Pro Ser Pro Asp Asp Pro Thr Gln Glu Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Val Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Glu Met Cys Lys Lys Cys Asp Thr Thr Gln Val Glu Leu Glu Asn
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Asn Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Lys Val Pro
100 105 110
Phe Ser Tyr Gly Gly Lys Thr Val Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Glu Ser Cys Tyr Pro Asp
130
<210> 28
<211> 134
<212> ПРТ
<213> Pantholops hodgsonii
<400> 28
Glu Asn Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Val Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Met Arg Thr Lys Phe Val Tyr His Leu
50 55 60
Ser Asp Leu Cys Lys Arg Cys Asp Thr Arg Glu Val Glu Leu Glu Asp
65 70 75 80
Gln Val Val Thr Ala Ser Gln Ser Asn Ile Cys Asp Arg Asp Thr Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Lys
100 105 110
Leu Asn Tyr Arg Gly Gln Thr Lys Met Val Glu Thr Ala Leu Thr Pro
115 120 125
Asp Ser Cys Tyr Pro Asp
130
<210> 29
<211> 133
<212> ПРТ
<213> Bos taurus
<400> 29
Glu Asn Glu Arg Ile Val Val Asp Asn Lys Cys Lys Cys Ala Arg Ile
1 5 10 15
Thr Ser Arg Ile Ile Pro Ser Ala Glu Asp Pro Ser Gln Asp Ile Val
20 25 30
Glu Arg Asn Val Arg Ile Ile Val Pro Leu Asn Ser Arg Glu Asn Ile
35 40 45
Ser Asp Pro Thr Ser Pro Met Arg Thr Lys Phe Val Tyr His Leu Ser
50 55 60
Asp Leu Cys Lys Lys Cys Asp Thr Thr Glu Val Glu Leu Glu Asp Gln
65 70 75 80
Val Val Thr Ala Ser Gln Ser Asn Ile Cys Asp Ser Asp Ala Glu Thr
85 90 95
Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Arg Val Lys Leu
100 105 110
Ser Tyr Arg Gly Gln Thr Lys Met Val Glu Thr Ala Leu Thr Pro Asp
115 120 125
Ser Cys Tyr Pro Asp
130
<210> 30
<211> 137
<212> ПРТ
<213> Mus musculus
<400> 30
Asp Asp Glu Ala Thr Ile Leu Ala Asp Asn Lys Cys Met Cys Thr Arg
1 5 10 15
Val Thr Ser Arg Ile Ile Pro Ser Thr Glu Asp Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Val Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Arg Asn Phe Val Tyr His Leu
50 55 60
Ser Asp Val Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Glu Asp
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Ile Cys Asn Glu Asp Asp Gly
85 90 95
Val Pro Glu Thr Cys Tyr Met Tyr Asp Arg Asn Lys Cys Tyr Thr Thr
100 105 110
Met Val Pro Leu Arg Tyr His Gly Glu Thr Lys Met Val Gln Ala Ala
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 31
<211> 137
<212> ПРТ
<213> Rattus norvegicus
<400> 31
Asp Asn Lys Gly Thr Ile Leu Ala Asp Asn Lys Cys Met Cys Thr Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Asn Glu Asp Ile
20 25 30
Val Glu Arg Ser Ile Arg Ile Val Val Pro Leu Asn Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Asn Phe Val Tyr His Leu
50 55 60
Ser Asp Val Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Glu Asp
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Ile Cys Ser Gly Asp Ser Gly
85 90 95
Val Pro Glu Thr Cys Tyr Met Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Met Val Pro Leu Arg Tyr His Gly Glu Thr Lys Met Val Gln Thr Ala
115 120 125
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 32
<211> 133
<212> ПРТ
<213> Echinops telfairi
<400> 32
Lys Asp Glu Gly Thr Val Leu Val Asp Asn Lys Cys Lys Cys Val Arg
1 5 10 15
Val Thr Ala Arg Ser Ile Pro Ser Thr Glu Asp Pro Asn Glu Glu Ile
20 25 30
Leu Glu Lys Asn Ile Arg Ile Ile Val Pro Leu Gly Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Asn Ser Pro Val Arg Thr Thr Phe Val Tyr His Met
50 55 60
Thr Asp Leu Tyr Lys Lys Cys Asp Pro Val Glu Val Glu Leu Asp Asn
65 70 75 80
Gln Val Val Thr Ala Thr Gln Ser Asn Gly Cys Asp Glu Asp Glu Thr
85 90 95
Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Asn Val Val Pro Leu
100 105 110
Thr Tyr Gly Gly Lys Thr Tyr Met Val Arg Thr Ala Leu Thr Pro Asp
115 120 125
Ser Cys Tyr Pro Asp
130
<210> 33
<211> 136
<212> ПРТ
<213> Oryctolagus cuniculus
<400> 33
Glu Asp Glu Ser Thr Val Leu Val Asp Asn Lys Cys Gln Cys Val Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Arg Asp Pro Asp Asn Pro Ser Glu Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Thr Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Glu Phe Lys Tyr Asn Leu
50 55 60
Ala Asn Leu Cys Lys Lys Cys Asp Pro Thr Glu Ile Glu Leu Asp Asn
65 70 75 80
Gln Val Phe Thr Ala Ser Gln Ser Asn Ile Cys Pro Asp Asp Asp Tyr
85 90 95
Ser Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Thr Leu
100 105 110
Val Pro Ile Thr His Arg Gly Gly Thr Arg Met Val Lys Ala Thr Leu
115 120 125
Thr Pro Asp Ser Cys Tyr Pro Asp
130 135
<210> 34
<211> 134
<212> ПРТ
<213> Monodelphis domestica
<400> 34
Glu Asp Glu Gly Arg Ile Leu Val Asp Asn Lys Cys Lys Cys Val Arg
1 5 10 15
Val Thr Ser Arg Leu Val Pro Ser Lys Asp Asn Pro Glu Glu Lys Val
20 25 30
Val Glu Arg Asn Ile Arg Leu Ile Val Pro Leu Arg Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Arg Phe Val Tyr Arg Leu
50 55 60
Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Leu Glu Leu Asn Asn
65 70 75 80
Glu Val Val Thr Ala Thr Gln Ser Asn Asn Cys Asp Asp Thr Ser Glu
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ser Thr Ala Ala
100 105 110
Leu Tyr Leu Glu Gly Glu Thr Arg Leu Val Thr Thr Ala Leu Thr Pro
115 120 125
Glu Ser Cys Tyr Asn Asp
130
<210> 35
<211> 134
<212> ПРТ
<213> Alligator mississippiensis
<400> 35
Glu Glu Trp Glu Glu His Val Leu Val Asp Asn Lys Cys Gln Cys Val
1 5 10 15
Arg Val Thr Ser Lys Phe Val Pro Ser Lys Asp Asn Pro Gln Glu Glu
20 25 30
Ile Leu Glu Arg Asn Ile Arg Val Ile Val Pro Leu Lys Ser Arg Met
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Thr Phe Val Tyr Arg
50 55 60
Leu Ser Glu Leu Cys Lys Lys Cys Asp Pro Thr Glu Ile Glu Leu Gly
65 70 75 80
Gly Gln Ile Val Thr Ala Gln Gln Ser Thr Asp Cys Ser Lys Ser Asp
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Thr Thr Phe Pro
100 105 110
Phe Ala Tyr Gly Gly Gln Thr Lys Asn Ile Glu Ala Ala Leu Thr Pro
115 120 125
Ala Ser Cys Tyr Ala Asp
130
<210> 36
<211> 134
<212> ПРТ
<213> Chrysemys picta
<400> 36
Glu Glu Ala Glu Glu His Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Val Thr Ser Lys Phe Val Pro Ser Lys Asp Asn Pro Gln Glu Glu
20 25 30
Val Leu Val Arg Asn Ile Arg Val Ile Val Pro Leu Leu Ser Arg Lys
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Val Arg Thr Thr Phe Val Tyr Arg
50 55 60
Leu Ser Glu Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Gly
65 70 75 80
Asp Arg Val Val Thr Ala Glu Gln Ser Asn Asn Cys Ser Ser Ser Asp
85 90 95
Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Thr Thr Phe Pro
100 105 110
Phe Phe Tyr Gly Gly Lys Ile Asn Thr Val Gln Ala Ala Leu Thr Pro
115 120 125
Glu Ser Cys Tyr Ala Asp
130
<210> 37
<211> 132
<212> ПРТ
<213> Sarcophilus harrisii
<400> 37
Glu Asp Glu Gly Arg Ile Leu Val Asp Asn Lys Cys Gln Cys Val Arg
1 5 10 15
Val Thr Ser Arg Leu Val Pro Ser Lys Asp Asn Pro Glu Val Glu Val
20 25 30
Val Glu Arg Asn Ile Arg Leu Ile Val Pro Leu Lys Asn Arg Glu Asn
35 40 45
Ile Ser Asp Pro Asn Ser Pro Val Arg Thr Asn Phe Val Tyr Arg Leu
50 55 60
Ser Asn Leu Cys Lys Lys Arg Asp Pro Ile Glu Leu Glu Leu Gly Asn
65 70 75 80
Glu Ile Val Thr Ala Thr Gln Ser Asn Asn Trp Ser Cys Gly Thr Cys
85 90 95
Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ser Thr Ala Leu Leu Asp
100 105 110
Leu Gly Gly Lys Ser Thr Met Val Thr Thr Ala Leu Thr Pro Glu Ser
115 120 125
Cys Tyr Ser Asp
130
<210> 38
<211> 125
<212> ПРТ
<213> Ornithorhynchus anatinus
<400> 38
Asn Lys Cys Lys Cys Thr Arg Val Thr Ser Lys Phe Val Pro Ser Lys
1 5 10 15
Glu Asp Pro Ser Val Lys Val Leu Glu Arg Asn Ile Arg Val Ile Ile
20 25 30
Pro Leu Lys Ala Arg Gln Asn Ile Ser Asp Pro Thr Ser Pro Pro Arg
35 40 45
Thr Arg Phe Val Tyr Glu Leu Ser Lys Leu Cys Gln Lys Cys Asp Gln
50 55 60
Thr Glu Val Glu Leu Asp His Glu Val Val Thr Ala Thr Arg Gly Asn
65 70 75 80
Ser Cys Asp Arg Pro Gly Asp Asp Cys Tyr Thr Tyr Asp Arg Asn Lys
85 90 95
Cys Tyr Met Ser Thr Ile Pro Phe Thr Tyr Gly Gly Glu Thr Lys Val
100 105 110
Val Asn Thr Val Leu Thr Pro Glu Ser Cys Tyr Pro Asp
115 120 125
<210> 39
<211> 133
<212> ПРТ
<213> Melopsittacus undulatus
<400> 39
Glu Asp Met Glu Arg Val Leu Val Asn Asn Lys Cys Gln Cys Val Thr
1 5 10 15
Val Thr Ser Lys Leu Val Arg Ser Lys Glu Asn Pro Asn Glu Glu Ile
20 25 30
Leu Gln Arg Asn Ile Arg Ile Leu Val Pro Leu Lys Ala Arg Glu Asn
35 40 45
Ile Ser Asp Pro Leu Ser Pro Leu Arg Thr Thr Phe Val Tyr Arg Met
50 55 60
Thr Asp Leu Cys Lys Lys Cys Asp Pro Val Glu Phe Glu Leu Gly Gly
65 70 75 80
Glu Ile Tyr Lys Ala Gln Gln Ser Asn Ser Cys Asp Glu Pro Glu Thr
85 90 95
Cys Tyr Thr Tyr Asp Arg Asp Lys Cys Tyr Thr Thr Asn Phe Pro Phe
100 105 110
Leu Tyr His Gly Glu Thr Lys Asn Leu Gln Ala Val Leu Thr Pro Ala
115 120 125
Ser Cys Tyr Ala Asp
130
<210> 40
<211> 133
<212> ПРТ
<213> Anas platyrhynchos
<400> 40
Asp Gly Glu Glu Arg Val Leu Val Asn Asn Lys Cys Lys Cys Thr Thr
1 5 10 15
Val Thr Ser Arg Leu Val Pro Ser Lys Glu Asn Pro Asp Glu Glu Ile
20 25 30
Leu Glu Arg Asn Ile Arg Ile Thr Val Pro Leu Arg Ser Arg Gln Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Thr Phe Val Tyr Arg Met
50 55 60
Ala Glu Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Gly Gly
65 70 75 80
Glu Ile Tyr Glu Ala Gln Gln Ser Thr Ser Cys Asn Glu Pro Glu Thr
85 90 95
Cys Tyr Thr Tyr Asn Arg Asp Lys Cys Tyr Thr Thr Thr Phe Pro Phe
100 105 110
Ile Tyr His Gly Glu Thr Lys His Ile Gln Ala Ala Leu Thr Pro Ala
115 120 125
Ser Cys Tyr Ala Asp
130
<210> 41
<211> 133
<212> ПРТ
<213> Gallus gallus
<400> 41
Asp Gly Glu Glu Arg Val Leu Val Asn Asn Lys Cys Lys Cys Val Thr
1 5 10 15
Val Thr Ser Lys Phe Val Pro Ser Lys Asp Asn Pro Glu Glu Glu Val
20 25 30
Leu Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Lys Ser Arg Glu Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Thr Phe Val Tyr Arg Met
50 55 60
Thr Glu Leu Cys Lys Lys Cys Asp Pro Val Glu Ile Glu Leu Gly Gly
65 70 75 80
Glu Thr Tyr Gln Ala Gln Gln Ser Asn Ser Cys Asn Glu Pro Glu Thr
85 90 95
Cys Tyr Thr Tyr Asn Arg Asp Lys Cys Tyr Thr Thr Thr Phe Pro Phe
100 105 110
Val Tyr His Gly Glu Thr Lys His Ile Gln Ala Ala Leu Thr Pro Thr
115 120 125
Ser Cys Tyr Ala Glu
130
<210> 42
<211> 133
<212> ПРТ
<213> Meleagris gallopavo
<400> 42
Asp Gly Glu Glu Arg Val Leu Val Asn Asn Lys Cys Lys Cys Val Thr
1 5 10 15
Val Thr Ser Lys Phe Val Pro Ser Lys Asp Asn Pro Glu Glu Glu Val
20 25 30
Leu Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Lys Ser Arg Gln Asn
35 40 45
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Asn Phe Val Tyr Arg Met
50 55 60
Thr Glu Leu Cys Lys Lys Cys Asp Pro Val Glu Ile Glu Leu Gly Gly
65 70 75 80
Glu Thr Tyr Gln Ala Gln Gln Ser Thr Ser Cys Asn Glu Pro Glu Thr
85 90 95
Cys Tyr Thr Tyr Asn Arg Asp Lys Cys Tyr Thr Thr Thr Ile Pro Phe
100 105 110
Val Tyr Gln Gly Glu Thr Lys Gln Ile Gln Ala Ala Leu Thr Pro Ala
115 120 125
Ser Cys Tyr Ala Asp
130
<210> 43
<211> 132
<212> ПРТ
<213> Falco peregrinus
<400> 43
Asp Asp Glu His Val Leu Val Asn Asn Lys Cys Gln Cys Val Thr Val
1 5 10 15
Thr Ser Lys Phe Val Pro Ser Lys Glu Asp Pro Gly Glu Glu Val Leu
20 25 30
Glu Arg Asn Ile Arg Ile Leu Val Pro Leu Arg Ala Arg Glu Asn Ile
35 40 45
Ser Asp Pro Met Ser Pro Leu Arg Thr Thr Phe Val Tyr Arg Met Thr
50 55 60
Glu Leu Cys Lys Lys Cys Asp Pro Val Glu Val Glu Leu Gly Gly Glu
65 70 75 80
Ile Tyr Lys Ala Gln Gln Ser Thr Ser Cys Asn Glu Pro Glu Thr Cys
85 90 95
Tyr Thr Tyr Asn Arg Asp Lys Cys Tyr Thr Thr Thr Phe Pro Phe Ser
100 105 110
Tyr His Gly Glu Val Lys Lys Val Gln Ala Val Leu Thr Pro Ala Ser
115 120 125
Cys Tyr Ala Asp
130
<210> 44
<211> 130
<212> ПРТ
<213> Zonotrichia albicollis
<400> 44
Asp Glu Glu Tyr Val Leu Val Asn Asn Lys Cys Gln Cys Val Thr Val
1 5 10 15
Thr Ser Lys Phe Val Pro Ser Gln Glu Asn Pro Gln Glu Glu Val Leu
20 25 30
Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Lys Ala Arg Glu Asn Ile
35 40 45
Ser Asp Pro Leu Ser Pro Leu Arg Thr Thr Phe Val Tyr Arg Leu Ser
50 55 60
Glu Leu Cys Lys Asn Cys Glu Pro Ile Glu Ile Asp Leu Gly Gly Ala
65 70 75 80
Ile His Gln Ala Gln Gln Gly Asn Ser Cys Glu Glu Pro Gln Thr Cys
85 90 95
Tyr Thr Tyr Asp Arg Asn Glu Cys Tyr Ser Ser Pro Val Pro Leu Leu
100 105 110
His His Gly Glu Val Met Gln Val Pro Ala Ala Leu Thr Pro Asp Ser
115 120 125
Cys Phe
130
<210> 45
<211> 169
<212> ПРТ
<213> Pteropus alecto
<400> 45
Glu Asp Glu Arg Ile Leu Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
1 5 10 15
Ile Thr Ser Arg Ile Ile Pro Ser Pro Glu Asp Pro Ser Gln Asp Ile
20 25 30
Val Glu Arg Asn Ile Arg Ile Lys Leu Gly Asp Glu Lys Ser Met Val
35 40 45
Asn Leu Trp Asn Cys Lys Leu Leu Cys Thr Ala Gly Leu Gln Ser Lys
50 55 60
Glu Gly Gly Met Val Ala Asp Thr Ala Gly Lys Val Pro Leu Asn Asn
65 70 75 80
Arg Glu Asn Ile Ser Asp Pro Thr Ser Pro Arg Arg Thr Asn Phe Val
85 90 95
Tyr His Leu Ser Asp Leu Cys Lys Asn Cys Asp Pro Thr Glu Val Glu
100 105 110
Leu Asp Asn Gln Val Val Thr Val Thr Gln Ser Asn Ile Cys Asp Glu
115 120 125
Asp Asn Glu Thr Cys Tyr Ala Tyr Asp Arg Asn Lys Cys Tyr Thr Asn
130 135 140
Arg Val Pro Leu Leu Tyr Gly Gly Lys Thr Ile Met Val Glu Thr Ala
145 150 155 160
Leu Thr Pro Asp Ser Cys Tyr Pro Asp
165
<210> 46
<211> 436
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 46
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser
145 150 155 160
Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys
165 170 175
Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser
180 185 190
Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser
195 200 205
Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser
210 215 220
Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
225 230 235 240
Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu
245 250 255
Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
260 265 270
Gly Ser Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys
275 280 285
Cys Ala Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn
290 295 300
Glu Asp Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn
305 310 315 320
Arg Glu Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val
325 330 335
Tyr His Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu
340 345 350
Leu Asp Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu
355 360 365
Asp Ser Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr
370 375 380
Thr Ala Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Glu
385 390 395 400
Thr Ala Leu Thr Pro Asp Ala Cys Tyr Pro Asp Gly Gly Gly Ser Glu
405 410 415
Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Ser Ala Val Asp His His
420 425 430
His His His His
435
<210> 47
<211> 439
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 47
Met Lys Asn His Leu Leu Phe Trp Gly Val Leu Ala Val Phe Ile Lys
1 5 10 15
Ala Val His Val Lys Ala Gln Glu Asp Glu Arg Ile Val Leu Val Asp
20 25 30
Asn Lys Cys Lys Cys Ala Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser
35 40 45
Glu Asp Pro Asn Glu Asp Ile Val Glu Arg Asn Ile Arg Ile Ile Val
50 55 60
Pro Leu Asn Asn Arg Glu Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg
65 70 75 80
Thr Arg Phe Val Tyr His Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro
85 90 95
Thr Glu Val Glu Leu Asp Asn Gln Ile Val Thr Ala Thr Gln Ser Asn
100 105 110
Ile Cys Asp Glu Asp Ser Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg
115 120 125
Asn Lys Cys Tyr Thr Ala Val Val Pro Leu Val Tyr Gly Gly Glu Thr
130 135 140
Lys Met Val Glu Thr Ala Leu Thr Pro Asp Ala Cys Tyr Pro Asp Gly
145 150 155 160
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val
165 170 175
Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val
180 185 190
Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met
195 200 205
His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr
210 215 220
Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp
225 230 235 240
Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln
245 250 255
Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg
260 265 270
Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr
275 280 285
Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
290 295 300
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
305 310 315 320
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser
325 330 335
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
340 345 350
Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His
355 360 365
Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly
370 375 380
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
385 390 395 400
Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Gly Gly
405 410 415
Gly Ser Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Ser Ala Val
420 425 430
Asp His His His His His His
435
<210> 48
<211> 235
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 48
Met Asp Met Arg Val Pro Ala Gln Leu Leu Gly Leu Leu Leu Leu Trp
1 5 10 15
Leu Arg Gly Ala Arg Cys Gln Ile Val Leu Ser Gln Ser Pro Ala Ile
20 25 30
Leu Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser
35 40 45
Ser Ser Val Ser Tyr Ile His Trp Phe Gln Gln Lys Pro Gly Ser Ser
50 55 60
Pro Lys Pro Trp Ile Tyr Ala Thr Ser Asn Leu Ala Ser Gly Val Pro
65 70 75 80
Val Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
85 90 95
Ser Arg Val Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Thr Ser Asn Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
115 120 125
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
130 135 140
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
145 150 155 160
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
165 170 175
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
180 185 190
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
195 200 205
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
210 215 220
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 49
<211> 593
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 49
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Asn Met His Trp Val Lys Gln Thr Pro Gly Arg Gly Leu
50 55 60
Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asn
115 120 125
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Gly Ser Ala Ser
130 135 140
Ala Pro Thr Leu Phe Pro Leu Val Ser Cys Glu Asn Ser Pro Ser Asp
145 150 155 160
Thr Ser Ser Val Ala Val Gly Cys Leu Ala Gln Asp Phe Leu Pro Asp
165 170 175
Ser Ile Thr Phe Ser Trp Lys Tyr Lys Asn Asn Ser Asp Ile Ser Ser
180 185 190
Thr Arg Gly Phe Pro Ser Val Leu Arg Gly Gly Lys Tyr Ala Ala Thr
195 200 205
Ser Gln Val Leu Leu Pro Ser Lys Asp Val Met Gln Gly Thr Asp Glu
210 215 220
His Val Val Cys Lys Val Gln His Pro Asn Gly Asn Lys Glu Lys Asn
225 230 235 240
Val Pro Leu Pro Val Ile Ala Glu Leu Pro Pro Lys Val Ser Val Phe
245 250 255
Val Pro Pro Arg Asp Gly Phe Phe Gly Asn Pro Arg Lys Ser Lys Leu
260 265 270
Ile Cys Gln Ala Thr Gly Phe Ser Pro Arg Gln Ile Gln Val Ser Trp
275 280 285
Leu Arg Glu Gly Lys Gln Val Gly Ser Gly Val Thr Thr Asp Gln Val
290 295 300
Gln Ala Glu Ala Lys Glu Ser Gly Pro Thr Thr Tyr Lys Val Thr Ser
305 310 315 320
Thr Leu Thr Ile Lys Glu Ser Asp Trp Leu Ser Gln Ser Met Phe Thr
325 330 335
Cys Arg Val Asp His Arg Gly Leu Thr Phe Gln Gln Asn Ala Ser Ser
340 345 350
Met Cys Val Pro Asp Gln Asp Thr Ala Ile Arg Val Phe Ala Ile Pro
355 360 365
Pro Ser Phe Ala Ser Ile Phe Leu Thr Lys Ser Thr Lys Leu Thr Cys
370 375 380
Leu Val Thr Asp Leu Thr Thr Tyr Asp Ser Val Thr Ile Ser Trp Thr
385 390 395 400
Arg Gln Asn Gly Glu Ala Val Lys Thr His Thr Asn Ile Ser Glu Ser
405 410 415
His Pro Asn Ala Thr Phe Ser Ala Val Gly Glu Ala Ser Ile Cys Glu
420 425 430
Asp Asp Trp Asn Ser Gly Glu Arg Phe Thr Cys Thr Val Thr His Thr
435 440 445
Asp Leu Pro Ser Pro Leu Lys Gln Thr Ile Ser Arg Pro Lys Gly Val
450 455 460
Ala Leu His Arg Pro Asp Val Tyr Leu Leu Pro Pro Ala Arg Glu Gln
465 470 475 480
Leu Asn Leu Arg Glu Ser Ala Thr Ile Thr Cys Leu Val Thr Gly Phe
485 490 495
Ser Pro Ala Asp Val Phe Val Gln Trp Met Gln Arg Gly Gln Pro Leu
500 505 510
Ser Pro Glu Lys Tyr Val Thr Ser Ala Pro Met Pro Glu Pro Gln Ala
515 520 525
Pro Gly Arg Tyr Phe Ala His Ser Ile Leu Thr Val Ser Glu Glu Glu
530 535 540
Trp Asn Thr Gly Glu Thr Tyr Thr Cys Val Val Ala His Glu Ala Leu
545 550 555 560
Pro Asn Arg Val Thr Glu Arg Thr Val Asp Lys Ser Thr Gly Lys Pro
565 570 575
Thr Leu Tyr Asn Val Ser Leu Val Met Ser Asp Thr Ala Gly Thr Cys
580 585 590
Tyr
<210> 50
<211> 412
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 50
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Ile Ser Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Met Gly Tyr Ile Asn Pro Arg Ser Gly Tyr Thr His Tyr Asn
65 70 75 80
Gln Lys Leu Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ala Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ser Ala Tyr Tyr Asp Tyr Asp Gly Phe Ala Tyr
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln
145 150 155 160
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
165 170 175
Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys
180 185 190
Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala
195 200 205
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe
210 215 220
Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr
225 230 235 240
Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr Phe Gly Gly Gly Thr Lys
245 250 255
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
260 265 270
Gly Gly Ser Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys
275 280 285
Lys Cys Ala Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro
290 295 300
Asn Glu Asp Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn
305 310 315 320
Asn Arg Glu Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe
325 330 335
Val Tyr His Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val
340 345 350
Glu Leu Asp Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp
355 360 365
Glu Asp Ser Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys
370 375 380
Tyr Thr Ala Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val
385 390 395 400
Glu Thr Ala Leu Thr Pro Asp Ala Cys Tyr Pro Asp
405 410
<210> 51
<211> 415
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 51
Met Lys Asn His Leu Leu Phe Trp Gly Val Leu Ala Val Phe Ile Lys
1 5 10 15
Ala Val His Val Lys Ala Gln Glu Asp Glu Arg Ile Val Leu Val Asp
20 25 30
Asn Lys Cys Lys Cys Ala Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser
35 40 45
Glu Asp Pro Asn Glu Asp Ile Val Glu Arg Asn Ile Arg Ile Ile Val
50 55 60
Pro Leu Asn Asn Arg Glu Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg
65 70 75 80
Thr Arg Phe Val Tyr His Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro
85 90 95
Thr Glu Val Glu Leu Asp Asn Gln Ile Val Thr Ala Thr Gln Ser Asn
100 105 110
Ile Cys Asp Glu Asp Ser Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg
115 120 125
Asn Lys Cys Tyr Thr Ala Val Val Pro Leu Val Tyr Gly Gly Glu Thr
130 135 140
Lys Met Val Glu Thr Ala Leu Thr Pro Asp Ala Cys Tyr Pro Asp Gly
145 150 155 160
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val
165 170 175
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala Ser Val
180 185 190
Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ile Ser Tyr Thr Met
195 200 205
His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Tyr
210 215 220
Ile Asn Pro Arg Ser Gly Tyr Thr His Tyr Asn Gln Lys Leu Lys Asp
225 230 235 240
Lys Ala Thr Leu Thr Ala Asp Lys Ser Ala Ser Thr Ala Tyr Met Glu
245 250 255
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
260 265 270
Ser Ala Tyr Tyr Asp Tyr Asp Gly Phe Ala Tyr Trp Gly Gln Gly Thr
275 280 285
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
290 295 300
Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
305 310 315 320
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser
325 330 335
Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
340 345 350
Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser
355 360 365
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
370 375 380
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser
385 390 395 400
Ser Asn Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
405 410 415
<210> 52
<211> 293
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 52
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Val Glu Ser Gly Gly Glu Leu Val Gln
20 25 30
Ala Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe
35 40 45
Ser Ser Tyr Asn Met Gly Trp Phe Arg Arg Ala Pro Gly Lys Glu Arg
50 55 60
Glu Phe Val Ala Ser Ile Thr Trp Ser Gly Arg Asp Thr Phe Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Val Tyr Leu Gln Met Ser Ser Leu Lys Pro Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Ala Asn Pro Trp Pro Val Ala Ala Pro Arg Ser Gly
115 120 125
Thr Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Glu Asp Glu
145 150 155 160
Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg Ile Thr Ser
165 170 175
Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp Ile Val Glu Arg
180 185 190
Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn Ile Ser Asp
195 200 205
Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His Leu Ser Asp Leu
210 215 220
Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp Asn Gln Ile Val
225 230 235 240
Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser Ala Thr Glu Thr
245 250 255
Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala Val Val Pro Leu
260 265 270
Val Tyr Gly Gly Glu Thr Lys Met Val Glu Thr Ala Leu Thr Pro Asp
275 280 285
Ala Cys Tyr Pro Asp
290
<210> 53
<211> 137
<212> ПРТ
<213> Gorilla gorilla
<400> 53
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Lys Thr Ala
115 120 125
Leu Thr Pro Asp Ala Cys Tyr Pro Asp
130 135
<210> 54
<211> 137
<212> ПРТ
<213> Nomascus leucogenys
<400> 54
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile His Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Ile Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Leu Tyr Gly Gly Glu Thr Lys Met Val Glu Thr Ala
115 120 125
Leu Thr Pro Asp Ala Cys Tyr Pro Asp
130 135
<210> 55
<211> 417
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 55
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
130 135 140
Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser
145 150 155 160
Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser
165 170 175
Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
180 185 190
Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
195 200 205
Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
210 215 220
Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
225 230 235 240
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
245 250 255
Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala Arg
260 265 270
Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp Ile
275 280 285
Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu Asn
290 295 300
Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His Leu
305 310 315 320
Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp Asn
325 330 335
Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser Ala
340 345 350
Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala Val
355 360 365
Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Glu Thr Ala Leu
370 375 380
Thr Pro Asp Ala Cys Tyr Pro Asp Gly Gly Gly Ser Glu Gln Lys Leu
385 390 395 400
Ile Ser Glu Glu Asp Leu Asn Ser Ala Val Asp His His His His His
405 410 415
His
<210> 56
<211> 413
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
полипептид"
<400> 56
Gln Glu Asp Glu Arg Ile Val Leu Val Asp Asn Lys Cys Lys Cys Ala
1 5 10 15
Arg Ile Thr Ser Arg Ile Ile Arg Ser Ser Glu Asp Pro Asn Glu Asp
20 25 30
Ile Val Glu Arg Asn Ile Arg Ile Ile Val Pro Leu Asn Asn Arg Glu
35 40 45
Asn Ile Ser Asp Pro Thr Ser Pro Leu Arg Thr Arg Phe Val Tyr His
50 55 60
Leu Ser Asp Leu Cys Lys Lys Cys Asp Pro Thr Glu Val Glu Leu Asp
65 70 75 80
Asn Gln Ile Val Thr Ala Thr Gln Ser Asn Ile Cys Asp Glu Asp Ser
85 90 95
Ala Thr Glu Thr Cys Tyr Thr Tyr Asp Arg Asn Lys Cys Tyr Thr Ala
100 105 110
Val Val Pro Leu Val Tyr Gly Gly Glu Thr Lys Met Val Glu Thr Ala
115 120 125
Leu Thr Pro Asp Ala Cys Tyr Pro Asp Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly
145 150 155 160
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala
165 170 175
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
180 185 190
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
195 200 205
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
210 215 220
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
225 230 235 240
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
245 250 255
Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
260 265 270
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile
275 280 285
Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
290 295 300
Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
305 310 315 320
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
325 330 335
Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser
340 345 350
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala
355 360 365
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly
370 375 380
Ser Gly Thr Lys Leu Glu Ile Lys Glu Gln Lys Leu Ile Ser Glu Glu
385 390 395 400
Asp Leu Asn Ser Ala Val Asp His His His His His His
405 410
<210> 57
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: синтетический
6His tag"
<400> 57
His His His His His His
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Композиции и способы лечения и предупреждения инфекций, вызванных Staphylococcus aureus | 2015 |
|
RU2764981C1 |
| СОДЕРЖАЩИЙ ЛЕКАРСТВЕННОЕ СРЕДСТВО ПЕРЕНОСЧИК В КЛЕТКУ ДЛЯ ФОРМИРОВАНИЯ ИММУННОГО КОМПЛЕКСА | 2012 |
|
RU2739792C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| СРЕДСТВА НА ОСНОВЕ КОНСТАНТНОЙ ОБЛАСТИ ИММУНОГЛОБУЛИНА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР | 2008 |
|
RU2729829C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ МНОГОКРАТНО СВЯЗЫВАТЬСЯ С МНОЖЕСТВОМ АНТИГЕННЫХ МОЛЕКУЛ | 2018 |
|
RU2757786C2 |
| АНТИТЕЛО ПРОТИВ LAG-3 | 2017 |
|
RU2744866C2 |
| АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ C N-КОНЦЕВОЙ ОБЛАСТЬЮ ЛИЗИЛ-тРНК-СИНТЕТАЗЫ, ЭКСПОНИРОВАННОЙ НА КЛЕТОЧНОЙ МЕМБРАНЕ | 2018 |
|
RU2739393C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАРИАНТОВ FcyRIIB-СПЕЦИФИЧЕСКОЙ Fc-ОБЛАСТИ | 2013 |
|
RU2820161C1 |

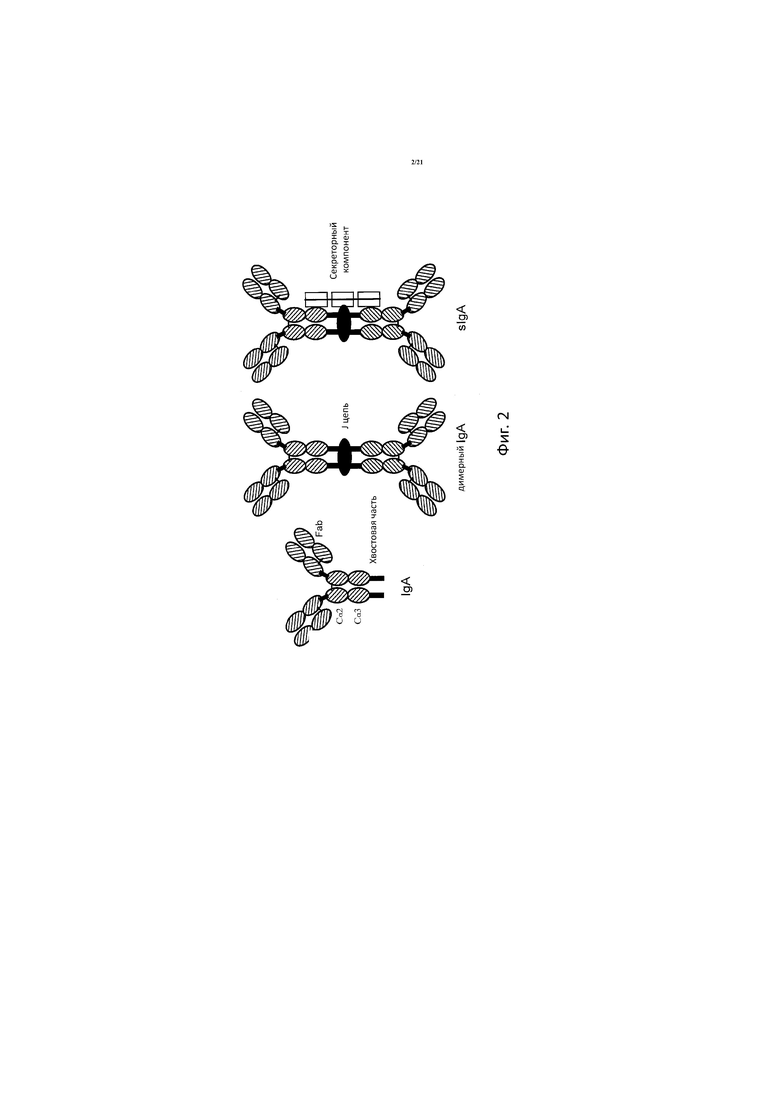













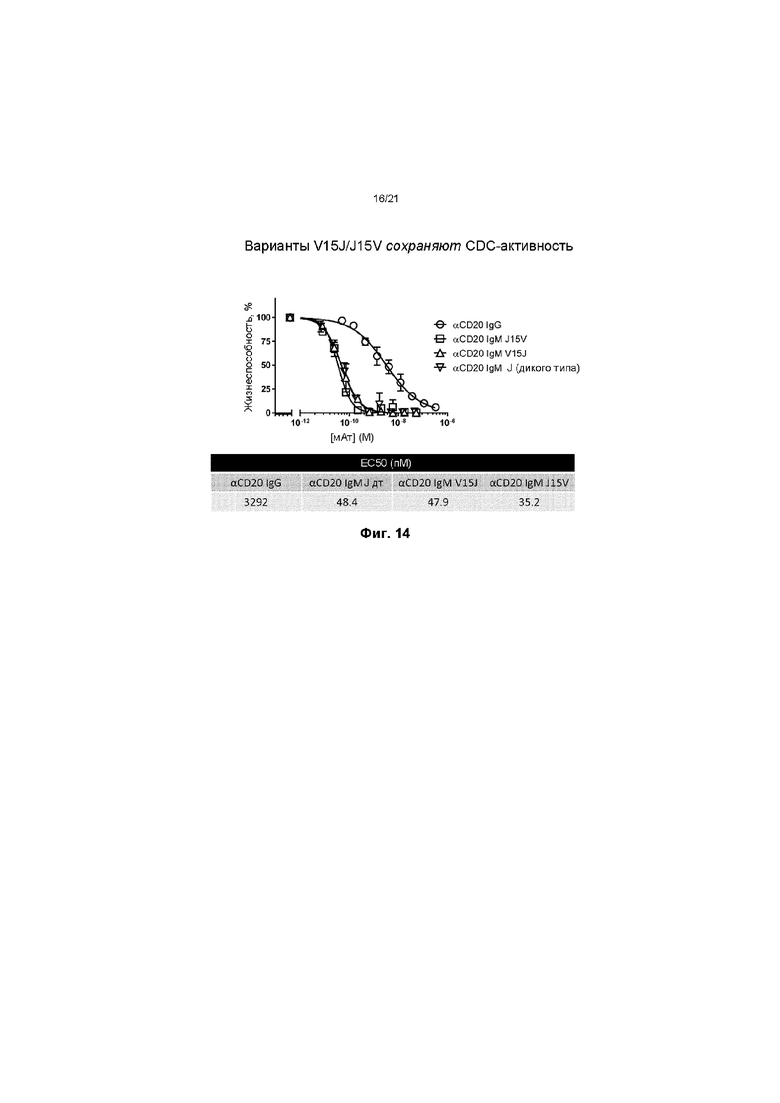
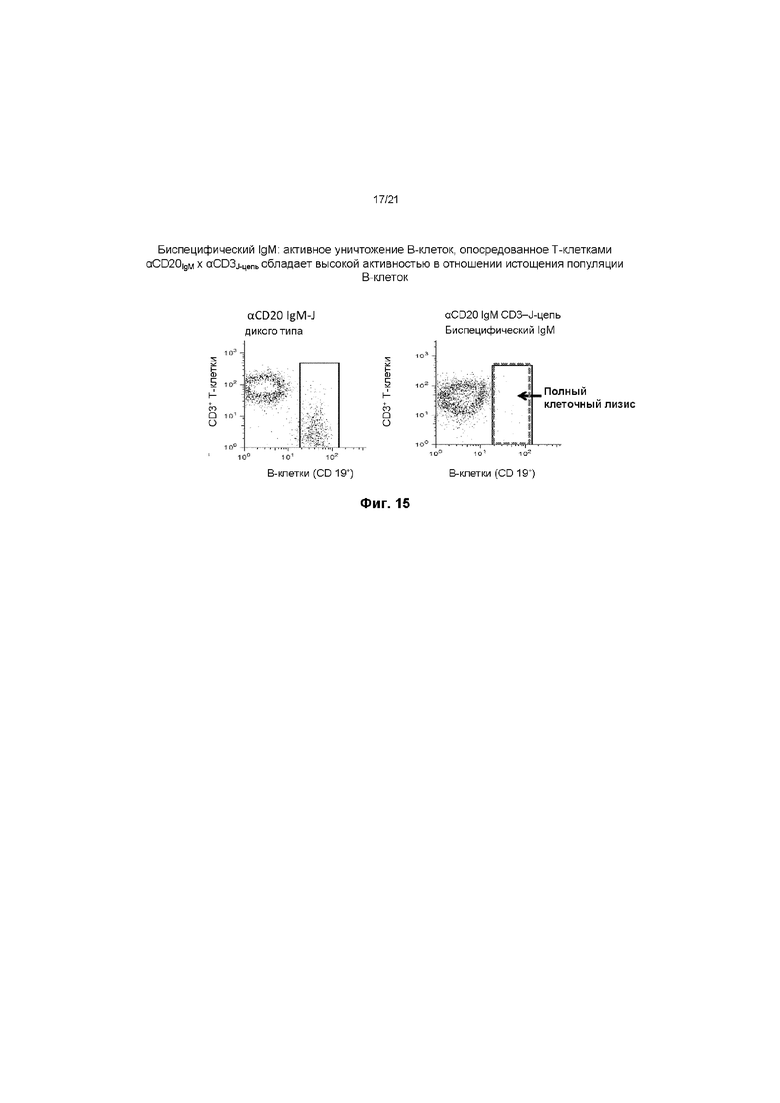
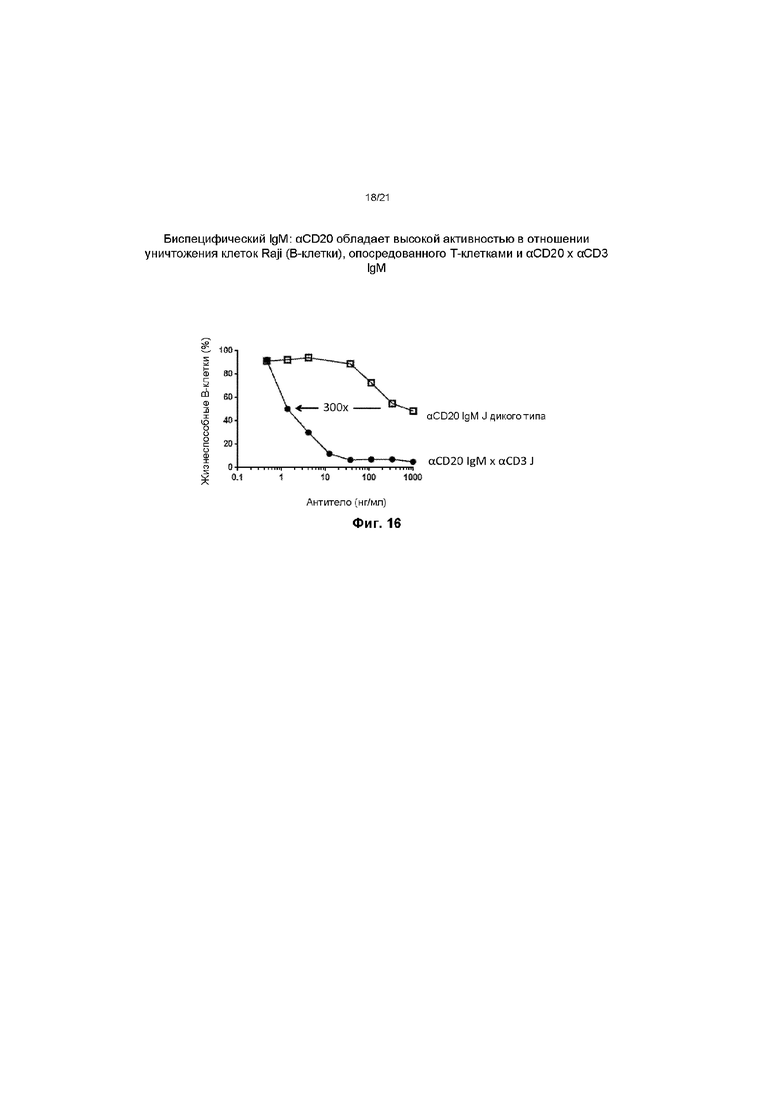
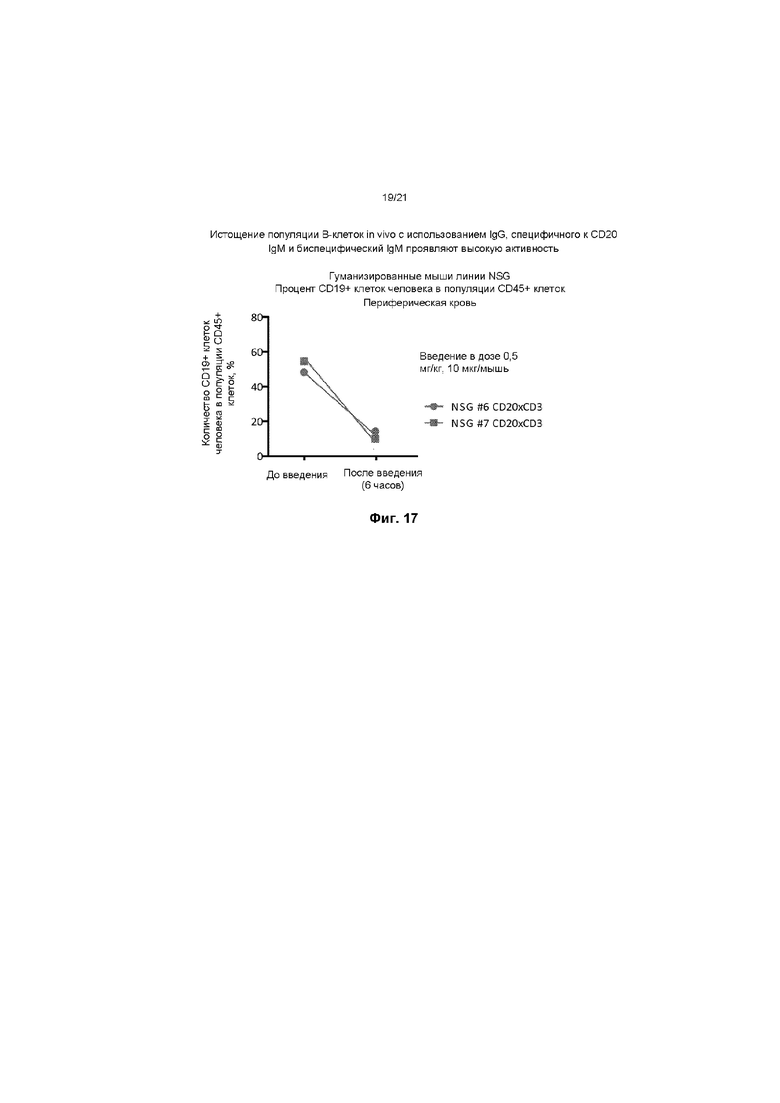


Изобретение относится к области биотехнологии. Предложено антитело IgM, IgA, IgG/IgM или IgG/IgA, которое является биспецифичным или мультиспецифичным и содержит модифицированную J-цепь. Указанная J-цепь содержит чужеродный связывающий фрагмент, введенный в нативную последовательность J-цепи путём непрямого слияния через пептидный линкер, расположенный на C- и/или N-конце указанного чужеродного связывающего фрагмента или в пределах приблизительно 10 остатков от C- и/или N-конца. Также изобретение относится к способу получения указанного антитела и к его применению для лечения злокачественной опухоли. Изобретение обеспечивает правильным образом сконструированные каркасы поливалентных антител, которые содержат модифицированную J-цепь, в последовательность которой введен дополнительный связывающий фрагмент. 4 н. и 11 з.п. ф-лы, 19 ил., 7 пр.
1. Антитело IgM, IgA, IgG/IgM или IgG/IgA, которое является биспецифичным или мультиспецифичным, содержащее тяжелую цепь IgM, IgA, IgG/IgM или IgG/IgA, которая способна связываться с мишенью, и модифицированную J-цепь, причём указанная модифицированная J-цепь содержит чужеродный связывающий фрагмент, введенный в нативную последовательность J-цепи, и при этом указанная тяжелая цепь IgG/IgM или IgG/IgA представляет собой гибридную тяжелую цепь, которая содержит хвостовую часть IgM или IgA и тяжелую цепь IgG и обладает способностью включать и образовывать полимеры с модифицированной J-цепью, при этом нативная последовательность J-цепи представляет собой нативную последовательность J-цепи человека SEQ ID NO: 1, причём указанный чужеродный связывающий фрагмент введён в нативную последовательность J-цепи путём непрямого слияния через пептидный линкер, расположенный на C- и/или N-конце указанного чужеродного связывающего фрагмента или в пределах приблизительно 10 остатков от C- и/или N-конца.
2. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 1, отличающееся тем, что указанный пептидный линкер имеет длину от приблизительно 10 до приблизительно 20 аминокислот.
3. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 1 или 2, отличающееся тем, что чужеродный связывающий фрагмент выбран из группы, состоящей из антител, антигенсвязывающих фрагментов антител, конъюгатов антитела и лекарственного препарата, антителоподобных молекул, антигенсвязывающих фрагментов антителоподобных молекул, растворимых белков, лигандов, рецепторов, вирусоподобных частиц, белковых токсинов, ферментов и альтернативных каркасных белков.
4. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 3, отличающееся тем, что альтернативный каркасный белок выбран из группы, состоящей из дарпинов, доменов фибронектина, аднектинов и ноттинов.
5. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 3, отличающееся тем, что чужеродный связывающий фрагмент представляет собой антигенсвязывающий фрагмент антитела, выбранный из группы, состоящей из F(ab')2, F(ab)2, Fab', Fab, Fv, scFv и однодоменного антитела.
6. Антитело IgM, IgA, IgG/IgM или IgG/IgA по любому из пп. 1-5, отличающееся тем, что чужеродный связывающий фрагмент связывается с эффекторной клеткой, при этом указанная эффекторная клетка выбрана из группы, состоящей из T-клеток, натуральных киллеров (NK), макрофагов и нейтрофилов.
7. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 6, отличающееся тем, что эффекторная клетка представляет собой T-клетку, и при этом указанный чужеродный связывающий фрагмент связывается с полипептидом CD3ε на данной T-клетке.
8. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 6, отличающееся тем, что эффекторная клетка представляет собой NK-клетку, и при этом чужеродный связывающий фрагмент связывается с мишенью, выбранной из группы, состоящей из CD16, CD64 и NKG2D на указанной NK-клетке.
9. Антитело IgM, IgA, IgG/IgM или IgG/IgA по любому из пп. 1-8, отличающееся тем, что модифицированная J-цепь содержит множество чужеродных связывающих фрагментов.
10. Антитело IgM, IgA, IgG/IgM или IgG/IgA по любому из пп. 1-9, отличающееся тем, что указанное антитело связывается с клеткой гематологического рака или клеткой солидной опухоли, при этом указанное антитело связывается с одним или более из следующего: CDIM, CD19, CD20, CD22, CD33, CD70, CD56, CD138.
11. Антитело IgM, IgA, IgG/IgM или IgG/IgA по п. 10, отличающееся тем, что указанное антитело связывается с CD20 и модифицированная J-цепь связывается с CD3ε.
12. Антитело IgM, IgA, IgG/IgM или IgG/IgA по любому из пп. 1-9, отличающееся тем, что указанное антитело связывается с вирусным антигеном, где указанный вирусный антиген представляет собой антиген PreS1 HBV или антиген gp120 ВИЧ.
13. Фармацевтическая композиция, содержащая эффективное количество антитела IgM, IgA, IgG/IgM или IgG/IgA по любому из пп. 1-12 и фармацевтически приемлемый носитель.
14. Способ лечения злокачественной опухоли, причем указанный способ включает введение нуждающемуся в этом субъекту эффективного количества антитела IgM, IgA, IgG/IgM или IgG/IgA по любому из пп. 1-11, где указанное антитело способно специфичным образом связывать антиген опухоли.
15. Способ получения антитела IgM, IgA, IgG/IgM или IgG/IgA, которое является биспецифичным или мультиспецифичным, и содержит тяжелую цепь IgM, IgA, IgG/IgM или IgG/IgA, легкую цепь IgM, IgA, IgG/IgM или IgG/IgA, которая способна связываться с мишенью, и модифицированную J-цепь, причём указанная модифицированная J-цепь содержит чужеродный связывающий фрагмент, введенный в нативную последовательность J-цепи или в её функциональный фрагмент, и при этом указанная тяжелая цепь IgG/IgM или IgG/IgA представляет собой гибридную тяжелую цепь, которая содержит хвостовую часть IgM или IgA и тяжелую цепь IgG и обладает способностью включать и образовывать полимеры с модифицированной J-цепью, включающий:
обеспечение нуклеиновой кислоты, кодирующей нативную последовательность J-цепи, которая содержит чужеродный связывающий фрагмент, введённый через пептидный линкер, расположенный на C- и/или N-конце указанного чужеродного связывающего фрагмента или в пределах приблизительно 10 остатков от C- и/или N-конца, причём указанная нативная последовательность J-цепи человека имеет SEQ ID NO: 1, с получением последовательности модифицированной нативной J-цепи, нуклеиновой кислоты, кодирующей тяжелую цепь IgM, IgA, IgG/IgM или IgG/IgA, которая способна связываться с мишенью, и нуклеиновой кислоты, кодирующей легкую цепь антитела IgM, IgA, IgG/IgM или IgG/IgA, которая способна связываться с мишенью; и
коэкспрессию нуклеиновых кислот, кодирующих модифицированную J-цепь, нуклеиновой кислоты, кодирующей указанную тяжелую цепь антитела-реципиента IgM, IgA, IgG/IgM или IgG/IgA, и указанную легкую цепь антитела IgM, IgA, IgG/IgM или IgG/IgA.
| WO 2013158748 A1, 24.10.2013 | |||
| US 2003224443 A1, 04.12.2003 | |||
| WO 9830592 A1, 16.07.1998 | |||
| JOHANSEN F.E | |||
| et al., Role of J Chain in Secretory Immunoglobulin Formation, Scandinavian Journal of Immunology, 2000, vol | |||
| Устройство для устранения мешающего действия зажигательной электрической системы двигателей внутреннего сгорания на радиоприем | 1922 |
|
SU52A1 |
| Русская печь | 1919 |
|
SU240A1 |
| SYMERSKY J | |||
| et al., Expression of the recombinant human immunoglobulin J chain in Escherichia coli, MOLECULAR | |||

