Область изобретения
Настоящее изобретение относится к композиции для культивирования естественных клеток-киллеров и способу получения естественных клеток-киллеров с использованием вышеупомянутого.
Предшествующий уровень техники
Разные виды лечения, такие как хирургическое вмешательство, лучевая терапия и химиотерапия, разработаны для лечения рака, но поскольку сообщается о нескольких побочных эффектах, недавно разработана иммунотерапия с использованием иммунной функции пациента. В частности, исследовали иммунотерапию с использованием естественных клеток-киллеров, которые могут подвергаться крупномасштабному производству и замораживанию.
В частности, естественные клетки-киллеры представляют собой тип лимфоцитов, которые распространены в костном мозге, селезенке, периферических лимфоузлах и периферической крови организма. Они составляют примерно 10% лимфоцитов периферической крови и играют важную роль во врожденном иммунном ответе (Ann Rev Immunol., 24: 257-286, 2006)). Корме того, естественные клетки-киллеры являются позитивными в отношении CD56 и CD16, но негативными в отношении CD3. Естественные клетки-киллеры уничтожают клетки посредством высвобождения цитолазматических гранул, содержащих перфорин и гранзим. Естественные клетки-киллеры секретируют разные цитокины, такие как IFN-γ (от англ. Interferon gamma - интерферон гамма), TNF-α (от англ. tumor necrosis factor - фактор некроза опухоли), GM-CSF (от англ. Granulocyte-macrophage colony- stimulating factor - гранулоцитарно-макрофагальный колониестимулирующий фактор) и IL-10 (от англ. Interleukin 10 - интерлейкин 10).
Кроме того, естественные клетки-киллеры экспрессируют несколько рецепторов на клеточной поверхности, и данные рецепторы задействованы в адгезии клеток, активации способности уничтожать клетки или ингибировании способности уничтожать клетки. Однако, наибольшая часть естественных клеток-киллеров в организме нормального субъекта существует в неактивном состоянии. Таким образом, существует необходимость в активированных естественных клетках-киллерах для устранения рака. Кроме того, в случае естественных клеток-киллеров, находящихся в организме пациента с раком, естественные клетки-киллеры имеют функциональные дефекты из-за механизма уклонения раковых клеток от иммунного надзора. Таким образом, очень важно активировать естественные клетки-киллеры для использования естественных клеток-киллеров в качестве терапевтического средства. Кроме того, крайне важно разработать методику масштабной пролиферации и заморозки естественных клеток-киллеров в крови от нормального субъекта или пациента, поскольку число естественных клеток-киллеров, присутствующих в организме, ограничено.
Тем не менее, IL-2, также называемые факторами роста Т-клеток (TCGF - от англ. T-cell growth factor), представляет собой глобулярный гликопротеин, который играет центральную роль в продукции, выживаемости и гомеостазе лимфоцита. IL-2 имеет размер 15,5 кДа - 16 кДа белка и состоит из 133 аминокислот.IL-2 опосредует разные иммунные ответы посредством связывания с рецептором IL-2, который имеет три отличающихся субъединицы.
Корме того, CD80 известен как В7-1 и один из членов семейства В7 среди белков, связанных с мембраной, участвующих в иммунной регуляции посредством связывания с лигандом и, таким образом, передачи костимулирующих ответов и коингибируюших ответов. CD80 представляет собой трансмембранный белок, экспрессируемый на поверхности Т-клеток, В-клеток, дендритных клеток и моноцитов. Известно, что CD80 связывается с CD28, CTLA4 (CD152) и PD-L1.
Таким образом, широко известно, что естественные клетки-киллеры важны для противоракового лечения, но конкретных способов, которые могли бы расширить эффективное использование естественных клеток-киллеров, все еще не достаточно.
Подробное описание изобретения Техническая проблема
Соответственно, авторы настоящего изобретения получили естественные клетки-киллеры посредством использования димера слитого белка, содержащего белок IL-2 и белок CD80, в результате исследования для получения активированных естественных клеток-киллеров в большом количестве. Кроме того, авторы настоящего изобретения идентифицировали, что естественные клетки-киллеры, полученные таким образом, обладают повышенной активностью и демонстрируют превосходное противораковое действие и, таким образом, завершили настоящее изобретение.
Решение проблемы
Для достижения указанной ваше цели, в соответствии с иллюстративным воплощением, предложена композиция для культивирования естественных клеток-киллеров, включающая, в качестве активного ингредиента, димер слитого белка, содержащий IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному иллюстративному воплощению предложен способ культивирования естественных клеток-киллеров, включающий: i) выделение клеток, которые не экспрессируют CD3, из мононуклеарных клеток периферической крови (РВМС); ii) выделение клеток, которые экспрессируют CD56, от клеток, которые не экспрессируют CD3, выделенных на указанной выше стадии; и iii) культивирование выделенных клеток в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному иллюстративному воплощению предложены естественные клетки-киллеры, полученные способом культивирования естественных клеток-киллеров.
Согласно еще одному иллюстративному воплощению предложена фармацевтическая композиция для предупреждения или лечения рака, включающая естественные клетки-киллеры в качестве активного ингредиента.
Согласно еще одному иллюстративному воплощению предложен способ культивирования естественных клеток-киллеров, включающий: i) выделение клеток, которые не экспрессируют CD3, из РВМС; и ii) культивирование выделенных клеток в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному иллюстративному воплощению предложены естественные клетки-киллеры, полученные способом культивирования естественных клеток-киллеров.
Согласно еще одному иллюстративному воплощению предложена фармацевтическая композиция для предупреждения или лечения рака, включающая естественные клетки-киллеры в качестве активного ингредиента.
Согласно еще одному иллюстративному воплощению предложен способ культивирования естественных клеток-киллеров, включающий: i) выделение клеток, которые экспрессируют CD56, из РВМС; и ii) культивирование выделенных клеток в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному иллюстративному воплощению предложены естественные клетки-киллеры, полученные способом культивирования естественных клеток-киллеров.
Согласно еще одному иллюстративному воплощению предложена фармацевтическая композиция для предупреждения или лечения рака, включающая естественные клетки-киллеры в качестве активного ингредиента.
Согласно еще одному иллюстративному воплощению предложен способ стимуляции активности естественных клеток-киллеров в РВМС, включающий культивирование РВМС в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному иллюстративному воплощению предложены РВМС, полученные способом стимуляции активности естественных клеток-киллеров в РВМС.
Согласно еще одному иллюстративному воплощению предложена фармацевтическая композиция для предупреждения или лечения рака, включающая РВМС в качестве активного ингредиента.
Эффект изобретения
Композиция по настоящему изобретению для культивирования естественных клеток-киллеров, включающая в качестве активного ингредиента слитый белок, содержащий IL-2 или его вариант и CD80 или его фрагмент, стимулирует пролиферацию естественных клеток - киллеров, индуцирует экспрессию CD16 и NKp46 и повышает уровень экспрессии и секреции гранзима В и перфорина, и, таким образом, может с пользой использоваться в получении естественных клеток-киллеров, обладающих превосходной противораковой иммунной функцией.
Краткое описание графических материалов
Иллюстративные воплощения могут быть более подробно поняты на основе следующего описания, вместе с прилагаемыми графическими материалами, в которых:
Фиг. 1А представляет собой схематическое изображение димера слитого белка, используемого в настоящем изобретении;
На Фиг. 1В показано изображение SDS-PAGE (от англ. sodium dodecyl sulphate-polyacrylamide gel electrophoresis - электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), подтверждающее полученный димер слитого белка (GI-101);
На Фиг. 1С показано содержание димера слитого белка (GI-101) согласно поглощению;
На Фиг. 1D показан результат анализа полученного димера слитого белка (GI-101) на основе эксклюзионной хроматографии (SEC - от англ. size exclusion chromatography);
На Фиг. 2А показаны изображения SDS-PAGE, подтверждающие полученный димер слитого белка hCD80-Fc;
На Фиг. 2В показаны результаты анализа на основе эксклюзионной хроматографии (SEC) полученного димера слитого белка hCD80-Fc;
На Фиг. 3А показано изображение SDS-PAGE, подтверждающие полученный димер слитого белка Fc-IL2v2;
На Фиг. 3В показаны результаты анализа на основе эксклюзионной хроматографии (SEC) полученного димера слитого белка Fc-IL2v2;
На Фиг. 3С показаны изображения SDS-PAGE, подтверждающие полученный димер слитого белка Fc-IL2wt;
На Фиг. 3D показаны результаты анализа на основе эксклюзионной хроматографии (SEC) полученного димера слитого белка Fc-IL2wt;
На Фиг. 4А показано изображение SDS-PAGE, подтверждающее полученный димер слитого белка hCD80-Fc-IL2wt;
На Фиг. 4В показаны результаты анализа на основе эксклюзионной хроматографии (SEC) полученного димера слитого белка hCD80-Fc-IL2wt;
На Фиг. 5А и 5В показано число NK-клеток при культивировании в условиях AIM-V (5% SR);
На Фиг. 6А и 6В показана жизнеспособность NK-клеток при культивировании в условиях AIM-V (5% SR);
На Фиг. 7А и 7В показано число NK-клеток при культивировании в условиях AIM-V (5% hABS);
На Фиг. 8А и 8В показана жизнеспособность NK-клеток при культивировании в условиях AIM-V (5% hABS);
На Фиг. 9А и 9В показано число NK-клеток при культивировании в условиях X-VIVO (5% hABS);
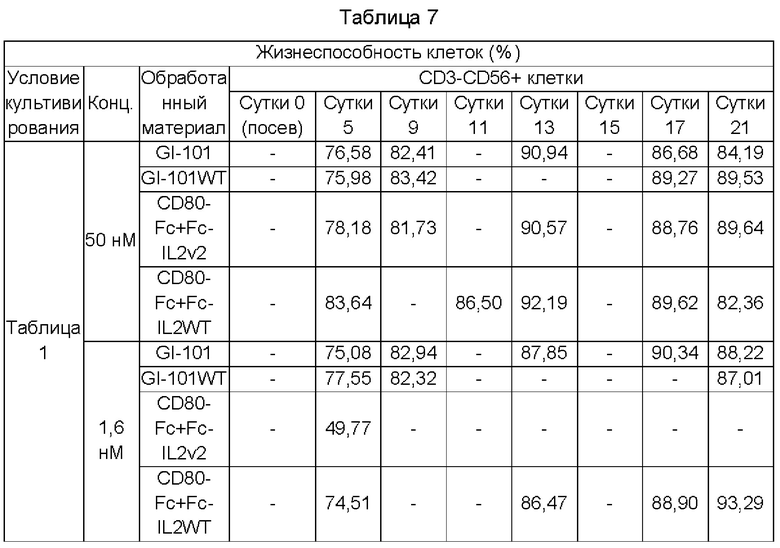
На Фиг. 10А и 10В показана жизнеспособность NK-клеток при культивировании в условиях X-VIVO (5% hABS);
На Фиг. 11А и 11В показано число NK-клеток при культивировании в условиях NK MACS (5% hABS);
На Фиг. 12А и 12В показана жизнеспособность NK-клеток при культивировании в условиях NK MACS (5% hABS);
На Фиг. 13 показан результат анализа FACS (от англ. fluorescence-activated cell sorting - сортировка клеток с активированной флуоресценцией), подтверждающий чистоту естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% SR);
На Фиг. 14 показан результат FACS-анализа, подтверждающий чистоту естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% hABS);
На Фиг. 15 показан результат FACS-анализа, подтверждающий чистоту естественных клеток-киллеров, культивируемых в композиции, содержащей среду X-VIVO15 (5% hABS);
На Фиг. 16 показан результат FACS-анализа, подтверждающий чистоту естественных клеток-киллеров, культивируемых в композиции, содержащей среду NK MACS (5% hABS);
На Фиг. 17 показан результат анализа маркеров активации естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% SR);
На Фиг. 18 показан результат анализа маркеров ингибирования естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% SR);
На Фиг. 19 показан результат анализа маркеров активации естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% hABS);
На Фиг. 20 показан результат анализа маркеров ингибирования естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% hABS);
На Фиг. 21 показан результат анализа маркеров активации естественных клеток-киллеров, культивируемых в композиции, содержащей среду Х-VIVO (5% hABS);
На Фиг. 22 показан результат анализа маркеров ингибирования естественных клеток-киллеров, культивируемых в композиции, содержащей среду X-VIVO (5% hABS);
На Фиг. 23 показан результат анализа маркеров активации естественных клеток-киллеров, культивируемых в композиции, содержащей среду NK MACS (5% hABS);
На Фиг. 24 показан результат анализа маркеров ингибирования естественных клеток-киллеров, культивируемых в композиции, содержащей среду NK MACS (5% hABS);
На Фиг. 25 показан результат анализа маркеров цитотоксичности естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% SR);
На Фиг. 26 показан результат анализа маркеров цитотоксичности естественных клеток-киллеров, культивируемых в композиции, содержащей среду AIM-V (5% hABS);
На Фиг. 27 показан результат анализа маркеров цитотоксичности естественных клеток-киллеров, культивируемых в композиции, содержащей среду X-VIVO15 (5% hABS);
На Фиг. 28 показан результат анализа маркеров цитотоксичности естественных клеток-киллеров, культивируемых в композиции, содержащей среду NK MACS (5% hABS);
На Фиг. 29 показан результат анализа способности к дегрануляции естественных клеток-киллеров, культивируемых в течение 21 суток в композиции, содержащей GI-101 (50 нМ), GI-101_wt (50 нМ), CD80-FC (50 hM)+Fc-IL2v2 (50 нМ) или CD80-Fc (50 HM)+Fc-IL2wt (50 нМ) в среде AIM-V (5% hABS);
На Фиг. 30 показаны результаты анализа эффекта уничтожения раковых клеток естественными клетками-киллерами, культивируемыми в течение 21 суток в композиции, содержащей GI-101 (50 нМ), GI-101_wt (50 нМ), CD80-Fc (50 hM)+Fc-IL2v2 (50 нМ) или CD80-Fc (50 HM)+Fc-IL2wt (50 нМ) в среде AIM-V (5% hABS);
На Фиг. 31 показан результат анализа способности к дегрануляции естественных клеток-киллеров, культивируемых в течение 21 суток в композиции, содержащей GI-101 (50 нМ), GI-101_wt (50 нМ), CD80-Fc (50 hM)+Fc-IL2v2 (50 нМ) или CD80-Fc (50 HM)+Fc-IL2wt (50 нМ) в среде X-VIVO15 (5% hABS);
На Фиг. 32 показаны результаты анализа эффекта уничтожения раковых клеток естественными клетками-киллерами, культивируемыми в течение 21 суток в композиции, содержащей GI-101 (50 нМ), GI-101_wt (50 нМ), CD80-Fc (50 hM)+Fc-IL2v2 (50 нМ) или CD80-Fc (50 HM)+Fc-IL2wt (50 нМ) в среде X-VIVO15 (5% hABS);
На Фиг. 33 показан результат анализа способности к дегрануляции естественных клеток-киллеров, культивируемых в течение 21 суток в композиции, содержащей GI-101 (50 нМ), GI-101_wt (50 нМ), CD80-Fc (50 hM)+Fc-IL2v2 (50 нМ) или CD80-Fc (50 HM)+Fc-IL2wt (50 нМ) в среде NK MACS (5% hABS); и
На Фиг. 34 показаны результаты анализа эффекта уничтожения раковых клеток естественными клетками-киллерами, культивируемыми в течение 21 суток в композиции, содержащей GI-101 (50 нМ), GI-101_wt (50 нМ), CD80-Fc (50 hM)+Fc-IL2v2 (50 нМ) или CD80-Fc (50 HM)+Fc-IL2wt (50 нМ) в среде NK MACS (5% hABS).
Наилучший способ осуществления изобретения Композиция и среда для пролиферации NK-клеток
Согласно аспекту настоящего изобретения предложена композиция для культивирования естественной клетки-киллера (NK-клетки), включающая в качестве активного ингредиента димер слитого белка, содержащий IL-2 или его вариант и CD80 или его фрагмент.Кроме того, предложена среда для культивирования естественных клеток-киллеров, включающая димер слитого белка в качестве активного ингредиента.
Композиция для культивирования естественной клетки-киллера может дополнительно включать любое, выбранное из группы, состоящей из среды, сыворотки, добавки и их комбинации.
Среда для культивирования NK-клеток может представлять собой среду, в которой димер слитого белка, содержащий белок IL-2 и белок CD80, добавляют в среду для культивирования клеток. В данном случае, среда для культивирования клеток может включать любое, выбранное из группы, состоящей из аминокислот, Сахаров, неорганических солей и витаминов. Предпочтительно, среда для культивирования клеток может включать все из аминокислот, Сахаров, неорганических солей и витаминов. В качестве конкретного воплощения среда для культивирования NK-клеток может включать по меньшей мере один из компонентов в указанных ниже Таблице 1 - Таблице 4.
В том виде, в котором он используется в данном документе, термин «среда для культивирования клеток» означает среду, используемую для культивирования клеток, в частности, NK-клеток и более конкретно CD3-CD56+ клеток. Данный термин включает компоненты, которые требуются клеткам для роста и выживаемости клеток in vitro или включает компоненты, которые помогают клеточному росту и выживаемости. Конкретно, компоненты могут представлять собой витамины, незаменимые или заменимые аминокислоты и микроэлементы. Среда может представлять собой среду, используемую для культивирования клеток, предпочтительно эукариотических клеток и более предпочтительно NK-клеток.
Среда для культивирования клеток согласно настоящему изобретению может включать компонент-аминокислоты, компонент-витамины, компонент неорганические соли, другой компонент и чистую воду, где:
а) компонент - аминокислоты представляет собой по меньшей мере одну аминокислоту, выбранную из группы, состоящей из глицина, L-аланина, L-валина, L-лейцина, L-изолейцина, L-треонина, L-серина, L-цистеина, L-метионина, L-аспарагиновой кислоты, L-аспарагина, L-глутаминовой кислоты, L-глутамина, L-лизина, L-аргинина, L-гистидина, L-фенилаланина, L-тирозина, L-триптофана, L-пролина, |3-аланина, у-аминомасляной кислоты, орнитина, цитруллина, гомосерина, трийодтиронина, тироксина и диоксифенилаланина или их комбинации, и предпочтительно по меньшей мере одну аминокислоту, выбранную из группы, состоящей из глицина, L-аланина, L-аргинина, L-цистеина, L-глутамина, L-гистидина, L-лизина, L-метионина, L-пролина, L-серина, L-треонина и L-валина или их комбинации;
b) компонент - витамины представляет собой по меньшей мере один витамин, выбранный из группы, состоящей из биотина, Д-пантотената кальция, фолиевой кислоты, ниацинамида, пиридоксина гидрохлорида, рибофлавина, тиамина гидрохлорида, витамина В12, холина хлорида, и-инозитола и аскорбиновой кислоты или их комбинации, и предпочтительно по меньшей мере один витамин, выбранный из группы, состоящей из и-инозитола, тиамина гидрохлорида, ниацинамида и пиридоксина гидрохлорида или их комбинации;
c) компонент - неорганические соли представляет собой по меньшей мере одну неорганическую соль, выбранную из группы, состоящей из хлорида кальция (CaCl2)(безводный), пентагидрата сульфата меди (CuSO4-5H2O), гептагидрата сульфата железа (II) (FeSO4-7H2O), хлорида магния (безводный), сульфата магния (MgSO4)(безводный), хлорида калия (KCl), хлорида натрия (NaCl), гидрофосфата натрия (Na2HPO4), моногидрата дигидрофосфата натрия (NaH2PO4-H2O), гептагидрата сульфата цинка (ZnSO4-7H2O), нонагидрата нитрата железа (III) (Fe(NO3)3⋅9H2O) и гидрокарбоната натрия (NaHCO3) или их комбинации и предпочтительно по меньшей мере одну неорганическую соль, выбранную из группы, состоящей из хлорида натрия (NaCl), гидрокарбоната натрия (NaHCO3), хлорида калия (KCl), хлорида кальция (CaCl2)(безводный) и моногидрата дигидрофосфата натрия (NaH2PO4-H2O) или их комбинации;
d) другой компонент представляет собой по меньшей мере один другой компонент, выбранный из группы, состоящей из D-глюкозы (декстрозы), пирувата натрия, гипоксантина Na, тимидина, линолевой кислоты, липоевой кислоты, аденозина, цитидина, гуанозина, уридина, 2'-дезоксиаденозина, 2'-дезоксицитидина HCI и 2'-дезоксигуанозина или их комбинации, и он может предпочтительно представлять собой пируват натрия; и
е) очищенная вода используется для растворения аминокислоты, витамина, неорганической соли и другого компонента и может быть получена одним или более способами дистилляции или очищена через фильтр.
Кроме того, среда для культивирования клеток согласно настоящему изобретению может дополнительно включать фактор роста или цитокин. Фактор роста может представлять собой IGF (от англ. insulin-like growth factor - инсулиноподобный фактор роста), bFGF (от англ. basic fibroblast growth factor - основной фактор роста фибробластов), TGF (от англ. transforming growth factor - трансформирующий фактор роста), HGF (от англ. hepatocyte growth factor - фактор роста гепатоцитов), EGF (от англ. epidermal growth factor - эпидермальный фактор роста), VEGF (от англ. vascular endothelial growth factor - фактор роста эндотелия сосудов), PDGF (от англ. platelet derived growth factor - фактор роста тромбоцитов) или т.п., один или по меньшей мере два из них, но не ограничивается ими. Цитокин может представлять собой IL-1, IL-4, IL-6, IFN-γ, IL-10, IL-15, IL-17, IL-21 или т.п., один или по меньшей мере два из них, но не ограничивается ими.
Кроме того, среда для культивирования клеток согласно настоящему изобретению может дополнительно включать антитело для активации естественных клеток-киллеров. Антитело для активации естественных клеток-киллеров может представлять собой антитело против CD3, антитело против CD2, антитело против CD335 или т.п., одно или по меньшей мере два из них, но не ограничивается ими. Кроме того, может быть включен шарик, с которым связывается антитело для активации естественных клеток-киллеров. Кроме того, может быть использован слитый белок, включающий два или более типов антител или их фрагментов вариабельных областей, для активации естественных клеток-киллеров.
В частности, среда для культивирования NK может дополнительно включать одно, выбранное из группы, состоящей из IL-15, IL-21 и их комбинации.
IL-15 и IL-21 могут представлять собой тип интерлейкина (IL) и означают белковые, биоактивные вещества, продуцируемые иммунокомпетентными клетками, такими как лимфоциты или моноциты и макрофаги. IL-15 и IL-21 могут быть использованы при культивировании естественных клеток-киллеров с использованием мононуклеарных клеток в качестве клеток-источников посредством стимуляции пролиферации естественных клеток-киллеров, но существует проблема низкой степени пролиферации и чистоты, когда используются только в отдельности или в комбинации (Biossel L. et al., Biology of Blood and Marrow Transplantation, 14, 1031-1038, 2008).
Конкретно, среда может представлять собой традиционную среду для культивирования животных клеток, такую как DMEM (от англ. Dulbecco's Modified Eagle's Medium - среда Игла, модифицированная по Дульбекко), EDM (от англ. Endothelial differentiation medium - среда для эндотелиальной дифференцировки), MEM (от англ. Minimal Essential Medium - минимальная питательная среда), ВМЕ (от англ. Basal Medium Eagle - базовая питательная среда Игла), RPMI 1640, F-10, F-12, а-МЕМ (от англ. a-Minimal Essential Medium), G-MEM (от англ. Glasgow's Minimal Essential Medium - минимальная питательная среда Глазго), среда Дульбекко в модификации Пскова, среда AIM-V, среда X-VIVO™ 15, среда NK MACS. В одном воплощении настоящего изобретения в качестве среды использовали среду AIM-V, среду X-VIVO™ 15 и среду NK MACS.
Термин «сыворотка», в том виде, в котором он используется в настоящем изобретении, означает прозрачный супернатант, выделенный из крови, после того, как кровь полностью свернулась. Кроме того, требуется добавлять сыворотку к синтетической среде для культивирования животных клеток, и распространено использование бычьей, лошадиной или человеческой сыворотки. В случае сыворотки бычьего происхождения можно использовать фетальную телячью сыворотку (от англ. FBS - fetal bovine serum), сыворотку новорожденных телят, телячью сыворотку, бычью сыворотку или т.п., в зависимости от времени забора крови. В случае сыворотки, происходящей из человека, используют человеческую сыворотку от донора, чей тип крови представляет собой АВ, и можно использовать человеческую сыворотку АВ, которая не содержит антител к антигенам типа крови А и В, таким образом, что может минимизировать иммунную реактивность. Кроме того, в качестве альтернативы «сыворотке» можно использовать CTS Immune Cell SR или т.п. В одном воплощении настоящего изобретения использовали человеческую сыворотку АВ или CTS Immune Cell SR.
GLUTAMAX (GIBCO®), альтернативу L-глутамину, можно использовать в качестве добавки для улучшения стабильности и клеточной активности во время культивирования клеток. Корме того, добавка может представлять собой добавку NK MACS (Miltenyi Biotec, 130-113-102).
Димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент
Термин «IL-2» или «интерлейкин-2» в том виде, как он используется в данном документе, если не утверждается иное, относится к любому IL-2 дикого типа, полученному из любого источника-позвоночного, включая млекопитающих, например, приматов (таких как человек) и грызунов (таких как мыши и крысы). IL-2 может быть получен из животных клеток и также включает IL-2, полученный из рекомбинантных клеток, способных к продукции IL-2. Кроме того, IL-2 может представлять собой IL-2 дикого типа или его вариант.
В настоящем описании изобретения IL-2 или его вариант могут быть совместно выражены термином «белок IL.-2» или «полипептид IL-2». IL-2, белок IL-2, полипептид IL-2 и вариант IL-2 специфично связываются, например, с рецептором IL-2. Это специфичное связывание может быть идентифицировано способами, известными специалистам в данной области.
В одном воплощении IL-2 может иметь аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36. Здесь IL-2 также может находиться в зрелой форме. В частности, зрелый IL-2 может не содержать сигнальную последовательность и может иметь аминокислотную последовательность SEQ ID NO: 10. Здесь IL-2 может использоваться согласно конфигурации, охватывающей фрагмент IL-2 дикого типа, в котором усечена часть N-конца или С-конца IL-2 дикого типа.
Кроме того, данный фрагмент IL-2 может находиться в форме, в которой 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 смежных аминокислот усечены с N-конца белка, имеющего аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36. Кроме того, фрагмент IL-2 может находиться в форме, в которой 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 смежных аминокислот усечены с С-конца белка, имеющего аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36.
Термин «вариант IL-2» в том виде, как он используется в данном документе, относится к форме, в которой часть аминокислот в полноразмерном IL-2 или вышеописанном фрагменте IL-2 заменена. То есть, вариант IL-2 может иметь отличную аминокислотную последоватеьность от IL-2 дикого типа или его фрагмента. Однако вариант IL-2 может иметь эквивалентную или аналогичную активность относительно IL-2 дикого типа. Здесь «активность IL-2» может, например, относиться к специфичному связыванию с рецептором IL-2, причем данное специфичное связывание можно измерять способами, известными специалистам в данной области.
В частности, вариант IL-2 можно получать заменой части аминокислот в IL-2 дикого типа. Одно воплощение варианта IL-2, полученного аминокислотной заменой, может быть получено заменой по меньшей мере одной из 38-ой, 42-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может быть получен заменой по меньшей мере одной из 38-ой, 42-ой, 45-ой, 61-ой или 72-ой аминокислоты в аминокислотной последовательности SEQ ID NO: 10 другой аминокислотой. Кроме того, когда IL-2 находится в форме, в которой часть N-конца аминокислотной последовательности SEQ ID NO: 35 усечена, аминокислота в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10, может быть заменена другой аминокислотой. Например, когда IL-2 имеет аминокислотную последовательность SEQ ID NO: 35, его вариант IL-2 может быть получен заменой по меньшей мере одной из 58-ой, 62-ой, 65-ой, 81-ой или 92-ой аминокислоты в аминокислотной последовательности SEQ ID NO: 35 другой аминокислотой. Данные аминокислотные остатки соответствуют 38-му, 42-му, 45-му, 61-му и 72-му аминокислотному остатку в аминокислотной последовательности SEQ ID NO: 10 соответственно. Согласно одному воплощению могут быть заменены одна, две, три, четыре, пять, шесть, семь, восемь, девять или десять аминокислот, при условии, что такой вариант IL-2 сохраняет активность IL-2. Согласно другому воплощению могут быть заменены от одной до пяти аминокислот.
В одном воплощении вариант IL-2 может находиться в форме, в которой заменены две аминокислоты. В частности, вариант IL-2 может быть получен заменой 38-ой и 42-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой и 45-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой и 45-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может находится в форме, в которой заменяются три аминокислоты. В частности, вариант IL-2 может быть получен заменой 38-ой, 42-ой и 45-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой, 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой, 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может находится в форме, в которой заменяются четыре аминокислоты. В частности, вариант IL-2 может быть получен заменой 38-ой, 42-ой, 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой, 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может находится в форме, в которой заменяются пять аминокислот.В частности, вариант IL-2 может быть получен заменой каждой из 38-ой, 42-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10 другой аминокислотой.
Здесь «другая аминокислота», вводимая заменой, может представлять собой любую аминокислоту, выбранную из группы, состоящей из аланина, аргинина, аспарагина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глутамина, гистидина, изолейцина, лейцина, лизина, метионина, фенилаланина, пролина, серина, треонина, триптофана, тирозина и валина. Однако относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 38-ая аминокислота не может быть заменена аргинином, 42-ая аминокислота не может быть заменена фенилаланином, 45-ая аминокислота не может быть заменена тирозином, 61-ая аминокислота не может быть заменена глутаминовой кислотой, и 72-ая аминокислота не может быть заменена лейцином.
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 38-ая аминокислота - аргинин - может быть заменена аминокислотой, отличной от аргинина. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 38-ая аминокислота - аргинин - может быть заменена аланином (R38A).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 42-ая аминокислота - фенилаланин - может быть заменена другой аминокислотой, чем фенилаланин. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 42-ая аминокислота - фенилаланин - может быть заменена аланином (F42A).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 45-ая аминокислота - тирозин - может быть заменена другой аминокислотой, чем тирозин. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 45-ая аминокислота - тирозин - может быть заменена аланином (Y45A).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 61-ая аминокислота - глутаминовая кислота -может быть заменена другой аминокислотой, чем глутаминовая кислота. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 61-ая аминокислота -глутаминовая кислота - может быть заменена аргинином (E61R).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 72-ая аминокислота - лейцин - может быть заменена другой аминокислотой, чем лейцин. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 72-ая аминокислота - лейцин - может быть заменена глицином (L72G).
В частности, вариант IL-2 может быть получен по меньшей мере одной заменой, выбранной из группы, состоящей из R38A, F42A, Y45A, E61R и L72G в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может быть получен аминокислотными заменами в двух, трех, четырех или пяти положениях среди положений, выбранных в группе, состоящей из R38A, F42A, Y45A, Е61 R и L72G.
Кроме того, вариант IL-2 может находиться в форме, в которой заменены две аминокислоты. В частности, вариант IL-2 может быть получен заменами R38A и F42A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A и Y45A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A и Y45A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами E61R и L72G.
Кроме того, вариант IL-2 может находиться в форме, в которой заменяются три аминокислоты. В частности, вариант IL-2 может быть получен заменами R38A, F42A и Y45A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, Y45A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, E61R и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами Y45A, E61R и L72G.
Кроме того, вариант IL-2 может находиться в форме, в которой заменяются четыре аминокислоты. В частности, вариант IL-2 может быть получен заменами R38A, F42A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A, Y45A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A, E61R и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A, E61R и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, Y45A, E61R и L72G.
Кроме того, вариант IL-2 может быть получен заменами R38A, F42A, Y45A, E61R и L72G.
Предпочтительно одно воплощение варианта IL-2 может содержать замену, которая представляет собой любую замену, выбранную из следующих комбинаций замен (a)-(d) в аминокислотной последовательности SEQ ID NO: 10:
(a) R38A/F42A
(b) R38A/F42A/Y45A
(c) R38A/F42A/E61 R
(d) R38A/F42A/L72G
Здесь, когда IL-2 имеет аминокислотную замену SEQ ID NO: 35, аминокислотная замена может присутствовать в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10. Кроме того, даже когда IL-2 представляет собой фрагмент аминокислотной последовательности SEQ ID NO: 35, аминокислотная замена может присутствовать в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может иметь аминокислотную последовательность SEQ ID NO: 6, 22, 23 или 24.
Кроме того, вариант IL-2 может отличаться наличием низкой токсичности in vivo. Здесь низкая токсичность in vivo может представлять собой побочный эффект, вызванный связыванием IL-2 с альфа цепью рецептора IL-2 (IL-2Rα). Были разработаны разные варианты IL-2 для уменьшения интенсивности побочного эффекта, вызванного связыванием IL-2 с IL-2Rα, и такие варианты IL-2 могут представлять собой варианты, раскрытые в патенте США №5229109 и корейском патенте №1667096. В частности, варианты IL-2, описанные в настоящей заявке, имеют низкую аффинность связывания в отношении альфа цепи рецептора IL-2 (IL-2Rα) и, таким образом, имеют более низкую токсичность in vivo, чем IL-2 дикого типа.
Термин «CD80» в том виде, в котором он используется в данном документе, также именуемый «В7-1», относится к мембранному белку, присутствующему в дендритных клетках, активированных В-клетках и моноцитах. CD80 предоставляет костимулирующие сигналы, важные для активации и выживания Т-клеток. CD80 известен в качестве лиганда двух разных белков - CD28 и CTLA-4, присутствующих на поверхности Т-клеток. CD80 состоит из 288 аминокислот и может, в частности, иметь аминокислотную последовательность SEQ ID NO: 11. Кроме того, термин «белок CD80» в том виде, в котором он используется в данном документе, относится к полноразмерному CD80 или к фрагменту CD80.
Термин «фрагмент CD80» в том виде, в котором он используется в данном документе, относится к усеченной форме CD80. Кроме того, фрагмент CD80 может представлять собой внеклеточный домен CD80. Одно воплощение фрагмента CD80 может быть получено устранением 1-ой-34-ой аминокислот с N-конца, которые представляют собой сигнальную последовательность CD80. В частности, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 288-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 242-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 232-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 139-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 142-ой - 242-ой аминокислот в SEQ ID NO: 11. В одном воплощении фрагмент CD80 может иметь аминокислотную последовательность SEQ ID NO: 2.
Кроме того, белок IL-2 и белок CD80 могут присоединяться друг к другу через линкер или носитель. В частности, IL-2 или его вариант и CD80 (В7-1) или его фрагмент могут присоединяться друг к другу через линкер или носитель. В настоящем описании термины «линкер» и «носитель» могут использоваться взаимозаменяемо.
Линкер связывает два белка. Одно воплощение линкера может содержать от 1 до 50 аминокислот, альбумин или его фрагмент, домен Fc иммуноглобулина или тому подобное. Здесь домен Fc иммуноглобулина относится к белку, который содержит константную область 2 тяжелой цепи (СН2) и константную область 3 тяжелой цепи (СН3) иммуноглобулина, и не содержит вариабельные области тяжелой и легкой цепи, и константную область 1 легкой цепи (CL1) иммуноглобулина. Данный иммуноглобулин может представлять собой IgG, IgA, IgE, IgD или IgM и предпочтительно может представлять собой lgG4. Здесь домен Fc иммуноглобулина G4 дикого типа может иметь аминокислотную последовательность SEQ ID NO: 4.
Кроме того, домен Fc иммуноглобулина может представлять собой вариант домена Fc, а также домен Fc дикого типа. Кроме того, термин «вариант домена Fc» в том виде, как он используется в данном документе, может относиться к форме, которая отличается от домена Fc дикого типа в показателях картины гликозилирования, имеет высокое гликозилирование по сравнению с доменом Fc дикого типа или имеет низкое гликозилирование по сравнению с доменом Fc дикого типа, или представляет собой дегликозилированную форму. Кроме того, в данный документ включается негликозилированный домен Fc. Домен Fc или его вариант может быть адаптирован для наличия скорректированного числа сиаловых кислот, фукозилирований или гликозилирований посредством культуральных условий или генетических манипуляций с хозяином.
Кроме того, гликозилирование домена Fc иммуноглобулина может быть модифицировано традиционными способами, такими как химические способы, ферментативные способы и способы генной инженерии с использованием микроорганизмов. Кроме того, вариант домена Fc может находиться в смешанной форме соответствующих областей Fc иммуноглобулинов - IgG, IgA, IgE, IgD и IgM. Кроме того, вариант домена Fc может находиться в форме, в которой некоторые аминокислоты домена Fc заменяются другими аминокислотами. Одно воплощение варианта домена Fc может иметь аминокислотную последовательность SEQ ID NO: 12.
Данный слитый белок может иметь структуру, в которой с использованием домена Fc в качестве линкера (или носителя) белок CD80 и белок IL-2 или белок IL-2 и белок CD80 связываются с N-концом и С-концом линкера или носителя соответственно (Фиг. 1А). Связь между N-концом или С-концом домена Fc и CD-80 или IL-2 возможно может достигаться посредством линкерного пептида.
В частности, слитый белок может состоять из следующей структурной формулы (I) или (II):
N'-X-[линкер (1)]n-домен Fc-[линкер (2)]m-Y-C' (I)
N'-Y-[линкер (1)]n-домен Fc-[линкер (2)]m-X-C' (II)
Здесь в структурных формулах (I) и (II)
N' представляет собой N-конец слитого белка,
С' представляет собой С-конец слитого белка,
X представляет собой белок CD80,
Y представляет собой белок IL-2,
линкеры (1) и (2) представляют собой пептидные линкеры и n и m каждый независимо равен 0 или 1.
Предпочтительно данный слитый белок может состоять из структурной формулы (I). Белок IL-2 является таким, как описано выше. Кроме того, белок CD80 является таким, как описано выше. Согласно одному воплощению белок IL-2 может представлять собой вариант IL-2 с одной-пятью аминокислотными заменами по сравнению с IL-2 дикого типа. Белок CD80 может представлять собой фрагмент, полученный усечением вплоть до примерно 34 смежных аминокислотных остатков от N-конца или С-конца CD80 дикого типа. В качестве альтернативы, белок CD может представлять собой внеклеточный иммуноглобулиноподобный домен, имеющий активность связывания с рецепторами поверхности Т-клеток CTLA-4 и CD28.
В частности, данный слитый белок может иметь аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30. Согласно другому воплощению данный слитый белок содержит полипептид, имеющий идентичность последовательности 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% с аминокислотной последовательностью SEQ ID NO: 9, 26, 28 или 30. Здесь идентичность представляет собой, например, процент гомологии и может определяться посредством программы для сравнения гомологии, такой как программа BlastN Национального центра биотехнологической информации (NCBI).
Пептидный линкер (1) может содержаться между белком CD80 и доменом Fc. Пептидный линкер (1) может состоять из 5-80 смежных аминокислот, 20-60 смежных аминокислот, 25-50 смежных аминокислот или 30-40 смежных аминокислот.В одном воплощении пептидный линкер (1) может состоять из 30 аминокислот.Кроме того, пептидный линкер (1) может содержать по меньшей мере один цистеин. В частности, пептидный линкер (1) может содержать один, два или три цистеина. Кроме того, пептидный линкер (1) может происходить из шарнира иммуноглобулина. В одном воплощении пептидный линкер (1) может представлять собой пептидный линкер, состоящий из аминокислотной последовательности SEQ ID NO: 3.
Пептидный линкер (2) может состоять из 1-50 смежных аминокислот, 3-30 смежных аминокислот или 5-15 смежных аминокислот.В одном воплощении пептидный линкер (2) может представлять собой (G4S)n (где n представляет собой целое число от 1 до 10). Здесь в (G4S)n n может быть равен 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. В одном воплощении пептидный линкер (2) может представлять собой пептидный линкер, состоящий из аминокислотной последовательности SEQ ID NO: 5.
В другом аспекте настоящего изобретения предложен димер, полученный связыванием двух слитых белков, каждый из которых содержит белок IL-2 и белок CD80. Слитый белок, содержащий IL-2 или его вариант и CD80 или его фрагмент, является таким, как описано выше.
Здесь связывание между слитыми белками, составляющими данный димер, может достигаться посредством дисульфидной связи, образованной цистеинами, присутствующими в линкере, но не ограничиваясь ей. Слитые белки, составляющие димер, могут быть одинаковыми или отличающимися друг от друга слитыми белками. Предпочтительно данный димер может представлять собой гомодимер. Одним воплощением слитого белка, составляющего димер, может быть белок, имеющий аминокислотную последовательность SEQ ID NO: 9.
Способ культивирования NK-клеток 1
Согласно еще одному аспекту настоящего изобретения предложен способ культивирования естественной клетки-киллера, включающий: i) отделение клетки, которая не экспрессирует CD3, из мононуклеарных клеток периферической крови (РВМС); ii) отделение клетки, которая экспрессирует CD56, от клетки, которая не экспрессирует CD3, выделенной на указанной выше стадии; и iii) культивирование выделенных клеток в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Термин «РВМС», в том виде, в котором он используется в настоящем изобретении, означает мононуклеарную клетку периферической крови. РВМС состоят из лимфоцитов (Т-клетки, В-клетки и естественные клетки-киллеры) и моноцитов, и могут быть выделены из цельной крови посредством Ficoll и центрифугирования. РВМС можно выделять из цельной крови, полученной от индивида.
Димер слитого белка представляет собой такой, как подробно описано в случае композиции для культивирования естественных клеток-киллеров. Слитый белок-димер может быть обработан в концентрации 1 нМ - 500 нМ. В частности, слитый белок-димер может быть обработан в концентрации 1 нМ - 500 нМ, 5 нМ -300 нМ или 10 нМ - 150 нМ. В одном воплощении настоящего изобретения димер слитого белка обрабатывали в концентрации 1,6 нМ или 50 нМ.
Способ культивирования выделенных клеток можно осуществлять, используя способ, широко известный в данной области. В частности, температура культивирования на стадии культивирования выделенных клеток может составлять от 27°С до 40°С или от 30°С до 37°С. В одном воплощении настоящего изобретения культивирование можно проводить при температуре 37°С.Кроме того, на стадии культивирования выделенных клеток условие - концентрация CO2 во время культивирования может составлять 1%-10%, и предпочтительно они могут культивироваться в условиях с 5% CO2.
На стадии культивирования выделенных клеток период культивирования может составлять от 5 суток до 25 суток, от 6 суток до 23 суток, или от 7 суток до 21 суток. В одном воплощении настоящего изобретения период культивирования мог составлять 20 суток, и значимое различие в пролиферации появлялось с 5-ых суток.
Полученные NK-клетки и их применение
Согласно еще одному аспекту настоящего изобретения предложены естественные клетки-киллеры, полученные способом культивирования естественных клеток-киллеров.
Естественные клетки-киллеры могут иметь повышенный уровень экспрессии CD16 и NKp46. Естественные клетки-киллеры могут иметь повышенный уровень экспрессии гранзима В и перфорина. Естественные клетки-киллеры, культивируемые согласно способу культивирования естественных клеток-киллеров, можно замораживать, и функция клеток не нарушается, даже при повторном размораживании.
Из-за высокого уровня экспрессии активирующих рецепторов, таких как CD16 и NKp46, естественные клетки-киллеры демонстрируют повышенную способность к уничтожению линии раковых клеток и повышенный уровень секреции гранзима В и перфорина, и, таким образом, можно ожидать превосходный противораковый эффект.Таким образом, может быть получено терапевтическое средство, эффективное в отношении лечения рака, с использованием большого количества активированных естественных клеток-киллеров, которые клинически применимы. Кроме того, естественные клетки-киллеры могут иметь высокий уровень экспрессии NKp30 или DNAM1.
Согласно еще одному аспекту настоящего изобретения предложена фармацевтическая композиция для предупреждения или лечения рака, включающая естественные клетки-киллеры в качестве активного ингредиента.
Корме того, естественные клетки-киллеры, полученные способом культивирования естественных клеток-киллеров, могут быть включены в количестве 10-95 масс. % в расчете на общую массу фармацевтической композиции. Кроме того, фармацевтическая композиция может дополнительно включать, помимо активного ингредиента, по меньшей мере один активный ингредиент, который демонстрирует такие же или похожие функции.
Дозировку фармацевтической композиции можно регулировать в соответствии с разными факторами, включая тип заболевания, тяжесть заболевания, виды и содержание активных ингредиентов и других ингредиентов, включенных в композицию, виды препарата и возраст, массу, общее состояние здоровья, пол и рацион пациента, время введения, путь введения и скорость секреции композиции, продолжительность лечения и одновременно используемые лекарственные средства.
Однако, для желательного эффекта дозировка фармацевтической композиции может составлять 1×102 клеток/кг - 1,0×1013 клеток/кг, и 1×107 клеток/кг - 1,5×1011 клеток/кг в расчете на естественные клетки-киллеры, что является активным ингредиентом. В данном случае доза может быть введена один раз в сутки или может быть разделена на несколько раз.
Кроме того, фармацевтическая композиция может быть введена индивиду разными способами, известными в данной области. Путь введения может быть соответствующим образом выбран специалистом в данной области с учетом способа введения, объема жидкости организма, вязкости или т.п.
Рак может представлять собой любой рак, выбранный из группы, состоящей из рака желудка, рака печени, рака легкого, колоректального рака, рака молочной железы, рака предстательной железы, рака яичника, рака поджелудочной железы, рака шейки матки, рака щитовидной железы, рака гортани, острого лимфобластного лейкоза, опухоли головного мозга, нейробластомы, ретинобластомы, рака головы и шеи, рака слюнных желез и лимфомы.
Способ лечения с использованием полученных NK-клеток
Согласно еще одному аспекту настоящего изобретения предложен способ лечения рака, включающий введение NK-клеток индивиду, имеющему рак. В данном случае NK-клетки и рак представляют собой такие, как описано выше. Согласно еще одному аспекту настоящего изобретения предложено применение NK-клеток для лечения рака.
Способ культивирования NK-клеток 2
Согласно еще одному аспекту настоящего изобретения предложен способ культивирования естественной клетки-киллера, включающий: i) выделение клеток, которые не экспрессируют CD3, из РВМС; и ii) культивирование выделенных клеток в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному аспекту настоящего изобретения предложены естественные клетки-киллеры, полученные способом культивирования естественных клеток-киллеров. Согласно еще одному аспекту настоящего изобретения предложена фармацевтическая композиция для предупреждения или лечения рака, включающая естественные клетки-киллеры в качестве активного ингредиента. Согласно еще одному аспекту настоящего изобретения предложен способ лечения рака, включающий введение NK-клеток индивиду, имеющему рак. В данном случае NK-клетки и рак представляют собой такие, как описано выше. Согласно еще одному аспекту настоящего изобретения предложено применение NK-клеток для лечения рака.
Из-за высокого уровня экспрессии активирующих рецепторов, таких как CD16 и NKp46, естественные клетки-киллеры демонстрируют повышенную способность к уничтожению линии раковых клеток и повышенный уровень секреции гранзима В и перфорина, и, таким образом, можно ожидать превосходный противораковый эффект.Таким образом, может быть получено терапевтическое средство, эффективное в отношении лечения рака, с использованием большого количества активированных естественных клеток-киллеров, которые клинически применимы. Кроме того, естественные клетки-киллеры могут иметь высокий уровень экспрессии NKp30 или DNAM1.
Рак может представлять собой любой рак, выбранный из группы, состоящей из рака желудка, рака печени, рака легкого, коло ректального рака, рака молочной железы, рака предстательной железы, рака яичника, рака поджелудочной железы, рака шейки матки, рака щитовидной железы, рака гортани, острого лимфобластного лейкоза, опухоли головного мозга, нейробластомы, ретинобластомы, рака головы и шеи, рака слюнных желез и лимфомы.
Способ культивирования NK-клеток 3
Согласно еще одному аспекту настоящего изобретения предложен способ культивирования естественных клеток-киллеров, включающий: i) отделение клетки, которая экспрессирует CD56, из РВМС; и ii) культивирование выделенных клеток в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному аспекту настоящего изобретения предложена естественная клетка-киллер, полученная способом культивирования естественных клеток-киллеров. Из-за высокого уровня экспрессии активирующих рецепторов, таких как CD16 и NKp46, естественные клетки-киллеры демонстрируют повышенную способность к уничтожению линии раковых клеток и повышенный уровень секреции гранзима В и перфорина, и, таким образом, можно ожидать превосходный противораковый эффект. Таким образом, может быть получено терапевтическое средство, эффективное в отношении лечения рака, с использованием большого количества активированных естественных клеток-киллеров, которые клинически применимы. Кроме того, естественные клетки-киллеры могут иметь высокий уровень экспрессии NKp30 или DNAM1.
Согласно еще одному аспекту настоящего изобретения предложена фармацевтическая композиция для предупреждения или лечения рака, включающая естественные клетки-киллеры в качестве активного ингредиента. Согласно еще одному аспекту настоящего изобретения предложен способ лечения рака, включающий введение NK-клеток индивиду, имеющему рак. В данном случае NK-клетки и рак представляют собой такие, как описано выше. Согласно еще одному аспекту настоящего изобретения предложено применение NK-клеток для лечения рака.
Рак может представлять собой любой рак, выбранный из группы, состоящей из рака желудка, рака печени, рака легкого, колоректального рака, рака молочной железы, рака предстательной железы, рака яичника, рака поджелудочной железы, рака шейки матки, рака щитовидной железы, рака гортани, острого лимфобластного лейкоза, опухоли головного мозга, нейробластомы, ретинобластомы, рака головы и шеи, рака слюнных желез и лимфомы.
Способ культивирования NK-клеток 4
Согласно еще одному аспекту настоящего изобретения предложен способ стимуляции активности естественных клеток-киллеров в РВМС, включающий культивирование РВМС в присутствии димера слитого белка, содержащего IL-2 или его вариант и CD80 или его фрагмент.
Согласно еще одному аспекту настоящего изобретения предложены РВМС, полученные способом стимуляции активности естественных клеток-киллеров в РВМС. Естественные клетки - киллеры в РВМС имеют высокий уровень экспрессии активирующего рецептора, такого как CD16 и NKp46, таким образом, что повышают способность к уничтожению линий раковых клеток и уровень секреции гранзима В и перфорина, и, таким образом, можно ожидать превосходные противораковые эффекты. Таким образом, терапевтическое средство, эффективное в отношении лечения рака, может быть получено с использованием РВМС, включающее большое количество активированных естественных клеток-киллеров, которые клинически применимы. Кроме того, естественные клетки-киллеры в РВМС могут иметь высокий уровень экспрессии NKp30 или DNAM1.
Согласно еще одному аспекту настоящего изобретения предложена фармацевтическая композиция для предупреждения или лечения рака, включающая РВМС в качестве активного ингредиента. Согласно еще одному аспекту настоящего изобретения предложен способ лечения рака, включающий введение NK-клеток индивиду, имеющему рак. В данном случае NK-клетки и рак представляют собой такие, как описано выше. Согласно еще одному аспекту настоящего изобретения предложено применение NK-клетокдля лечения рака.
Рак может представлять собой любой рак, выбранный из группы, состоящей из рака желудка, рака печени, рака легкого, колоректального рака, рака молочной железы, рака предстательной железы, рака яичника, рака поджелудочной железы, рака шейки матки, рака щитовидной железы, рака гортани, острого лимфобластного лейкоза, опухоли головного мозга, нейробластомы, ретинобластомы, рака головы и шеи, рака слюнных желез и лимфомы.
Способ осуществления изобретения
Ниже в данном документе настоящее изобретение будет описано более подробно посредством следующих примеров. Однако, следующие примеры предназначены лишь для иллюстрации настоящего изобретения, и объем настоящего изобретения не ограничивается ими.
I. Получение GI-101, естественных клеток-киллеров и культуральной композиции естественных клеток-киллеров
Пример получения 1. Получение варианта hCD80-Fc-IL-2 (2М): GI-101
Для получения слитого белка, включающего фрагмент человеческого CD80, домен Fc и вариант IL-2, полинуклеотид, включающий нуклеотидную последовательность (SEQ ID NO: 8), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнир Ig, конъюгированный с линкером (SEQ ID NO: 3), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2M), в котором две аминокислоты заменены (R38A, F42A) (SEQ ID NO: 6), в данном порядке от N-конца синтезировали посредством службы синтеза генов Invitrogen GeneArt ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, вектор вводили в клетки СНО (от англ. Chinese Hamster Ovary - яичник китайского хомяка) (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 9. После введения вектора клетки культивировали в окружающей среде с 37°С, 125 об./мин. и 8% CO2 в течение 7 суток и затем собирали для очистки слитого белка. Очищенный димер слитого белка был назван «GI-101».
Очистку проводили, используя хроматографию, включая смолу с белком А MabSelect SuRe. Слитый белок связывался в условиях 25 мМ Tris, 25 мМ NaCl и рН 7,4. Затем, его элюировали 100 мМ NaCl и 100 мМ уксусной кислотой при рН 3. После помещения 20% 1М Tris-HCl при рН 9 в пробирку для сбора собирали слитый белок. Собранный слитый белок подвергали диализу в буфер PBS (от англ. phosphate buffered saline - фосфатно-солевой буферный раствор) в течение 16 часов для замены.
Затем, поглощение при длине волны 280 нм со временем измеряли посредством использования эксклюзионной хроматографии с помощью колонки TSKgel G3000SWXL (TOSOH Bioscience) с получением высокой концентрации слитого белка. При этом, выделенный и очищенный слитый белок подвергали SDS-PAGE в восстанавливающих (R) или невосстанавливающих (NR) условиях и окрашивали кумасси синим для подтверждения его чистоты (Фиг. 1В). Подтверждали, что слитый белок содержался в концентрации 2,78 мг/мл, как выявлено с использованием NanoDrop (Фиг. 1С). Также, результат, анализируемый с использованием эксклюзионной хроматографии, представлен на Фиг. 1D.
Пример получения 2. Получение димера варианта Fc-IL-2 (2М): Fc-IL-2v2 Для получения слитого белка, содержащего домен Fc и вариант IL-2, полинуклеотид, включающий нуклеотидную последовательность (SEQ ID NO: 45), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), шарнир Ig (SEQ ID NO: 38), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2M), в котором две аминокислоты заменены (R38A, F42A) (SEQ ID NO: 6), в данном порядке от N-конца синтезировали посредством службы синтеза генов Invitrogen GeneArt ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, вектор вводили в клетки СНО (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 44. После введения вектора культуральный раствор культивировали в окружающей среде с 37°С, 125 об./мин. и 8% CO2 в течение 7 суток и затем собирали для очистки димера слитого белка. Очищенный димер слитого белка был назван «Fc-IL2v2».
Очистку и сбор слитого белка осуществляли тем же образом, как в Примере получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в восстанавливающих (R) или невосстанавливающих (NR) условиях и окашивали кумасси синим для подтверждения его чистоты (Фиг. 3А). В результате, подтверждали, что слитый белок образует димер. Также, результат, анализируемый с использованием эксклюзионной хроматографии, представляет собой такой, как показано на Фиг. 3В.
Пример получения 3. Получение димера Fc-IL-2: Fc-IL-2wt
Для получения слитого белка, содержащего домен Fc и IL-2 дикого типа, полинуклеотид, включающий нуклеотидную последовательность (SEQ ID NO: 43), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), шарнир Ig (SEQ ID NO: 38), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и IL-2 дикого типа (SEQ ID NO: 10), в данном порядке от N-конца синтезировали посредством службы синтеза генов Invitrogen GeneArt Gene Synthesis ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, вектор вводили в клетки СНО (EXPI-СНО™) для экспрессии слитого белка SEQ ID NO: 42. После введения вектора клетки культивировали в окружающей среде с 37°С, 125 об./мин. и 8% CO2 в течение 7 суток и затем собирали для очистки димера слитого белка. Очищенный димер слитого белка был назван «Fc-IL2wt».
Очистку и сбор слитого белка осуществляли тем же образом, как в Примере получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в восстанавливающих (R) или невосстанавливающих (NR) условиях и окашивали кумасси синим для подтверждения его чистоты (Фиг. 3С). В результате подтверждали, что слитый белок образует димер. Также, результат, анализируемый с использованием эксклюзионной хроматографии, представляет собой такой, как показано на Фиг. 3D.
Пример получения 4. Получение димера дикого типа hCD80-Fc-IL-2: hCD80-Fc-IL-2wt
Для получения слитого белка, содержащего фрагмент человеческого CD80, домен Fc и белок IL-2 дикого типа, полинуклеотид, включающий нуклеотидную последовательность (SEQ ID NO: 41), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнир Ig, конъюгированный с линкером (SEQ ID NO: 3), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и IL-2 дикого типа (SEQ ID NO: 10), в данном порядке от N-конца, синтезировали посредством службы синтеза генов Invitrogen GeneArt ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, вектор вводили в клетки СНО (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 46. После введения вектора клетки культивировали в окружающей среде с 37°С, 125 об./мин. и 8%-ной концентрацией CO2 в течение 7 суток и затем собирали для очистки димера слитого белка. Очищенный димер слитого белка называли «hCD80-Fc-IL2wt».
Очистку осуществляли с использованием хроматографии, включающей смолу с белком A MabSelect SuRe. Слитый белок связывался в условиях 25 мМ Tris, 25 мМ NaCl и рН 7,4. Затем, его элюировали 100 мМ NaCl и 100 мМ уксусной кислотой при рН 3. После помещения 20% 1М Tris-HCl при рН 9 в пробирку для сбора слитый белок собирали. Собранный слитый белок подвергали диализу в буфер PBS в течение 16 часов для замены.
Затем, поглощение при длине волны 280 нм со временем измеряли посредством использования эксклюзионной хроматографии с помощью колонки TSKgel G3000SWXL (TOSOH Bioscience) с получением высокой концентрации слитого белка. При этом, выделенный и очищенный слитый белок подвергали SDS-PAGE в восстанавливающих (R) или невосстанавливающих (NR) условиях и окашивали кумасси синим для подтверждения его чистоты (Фиг. 4А). В результате, подтверждали, что слитый белок образует димер. Также, результат, анализируемый с использованием эксклюзионной хроматографии, представляет собой такой, как показано на Фиг. 4В.
Пример получения 5. Получение димера hCD80-Fc: hCD80-Fc
Для получения слитого белка, содержащего фрагмент человеческого CD80 и домен Fc, полинуклеотид (SEQ ID NO: 39), включающий нуклеотидную последовательность, кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнир Ig, конъюгированный с линкером (SEQ ID NO: 3), и домен Fc (SEQ ID NO: 4), в данном порядке от N-конца синтезировали посредством службы синтеза генов Invitrogen GeneArt ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, вектор вводили в клетки СНО (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 40. После введения вектора клетки культивировали в окружающей среде с 37°С, 125 об./мин. и 8% CO2 в течение 7 суток и затем собирали для очистки слитого белка - димера. Очищенный димер слитого белка был назван «hCD80-Fc».
Очистку осуществляли с использованием хроматографии, включающей смолу с белком A MabSelect SuRe. Слитый белок связывался в условиях 25 мМ Tris, 25 мМ NaCl и рН 7,4. Затем, его элюировали 100 мМ NaCl и 100 мМ уксусной кислотой при рН 3. После помещения 20% 1М Tris-HCl при рН 9 в пробирку для сбора слитый белок собирали. Собранный слитый белок подвергали диализу в буфер PBS в течение 16 часов для замены.
Затем, поглощение при длине волны 280 нм со временем измеряли посредством использования эксклюзионной хроматографии с помощью колонки TSKgel G3000SWXL (TOSOH Bioscience) с получением высокой концентрации слитого белка. При этом, выделенный и очищенный слитый белок подвергали SDS-PAGE в восстанавливающих (R) или невосстанавливающих (NR) условиях и окашивали кумасси синим для подтверждения его чистоты (Фиг. 2А). В результате, подтверждали, что слитый белок образует димер. Также, результат, анализируемый с использованием эксклюзионной хроматографии, представляет собой такой, как показано на Фиг. 2В.
Пример получения 1. Получение среды для культивирования естественных клеток-киллеров
Среды для культивирования естественных клеток-киллеров получали посредством, соответственно, добавления веществ, в соответствии с условиями добавления 1-4 Таблицы 5, в каждую базовую культуральную среду, имеющую состав Таблицы 1 - Таблицы 4, указанных ниже.







Пример 1. Получение CD3(-)CD56(+) естественных клеток-киллеров, происходящих из мононуклеарных клеток периферической крови (РВМС)
Для получения CD3(-) клеток число РВМС (мононуклеарные клетки периферической крови, Zen-Bio. Inc, NC 27709, США, кат.№: SER-PBMC-200-F) подсчитывали с использованием автоматического счетчика клеток ADAM-MC2 (NanoEnTek, приобретенный у Cosmo Genetech Co., Ltd.). РВМС переносили в новую пробирку и затем центрифугировали при 300×g в течение 5 минут при температуре 4°С. 0,5%-ный (об./об.) бычий сывороточный альбумин (BSA - от англ. bovine serum albumin) и ЭДТА (этилендиаминтетрауксусная кислота) в концентрации 2 мМ включали в PBS для получения буфера MACS (рН 7,2). После завершения центрифугирования осадок клеток обрабатывали 80 мкл буфера MACS и 20 мкл магнитных шариков с CD3 (Miltenyi biotech, 130-050-101) на 1×107 клеток для суспендирования и затем инкубировали при температуре 4°С в течение 15 минут.10 мл буфера MACS добавляли для промывки и центрифугировали при 300×g в течение 10 минут при температуре 4°С, и затем осадок клеток ресуспендировали в 0,5 мл буфера MACS.
2 мл буфера MACS сначала наливали в колонку LD (Miltenyi Biotec, Bergisch Gladbach, Германия, кат. №: 130-042-901), и затем наливали клеточную суспензию.
Затем получали CD3(-) клетки, проходящие через колонку LD. При этом, CD3(-) клетки получали посредством протекания 2 мл буфера MACS три раза таким образом, чтобы можно было достаточно отделить клетки, остающиеся в колонке LD. Полученные CD3(-) клетки подсчитывали с использованием клеточного счетчика и затем помещали в новую пробирку и затем центрифугировали при 300×g в течение 5 минут при температуре 4°С.Затем, супернатант удаляли, и затем 80 мкл буфера MACS и 20 мкл магнитных шариков с CD56 (Miltenyi biotech, кат.№: 130-050-401) добавляли на 1×107 клеток с последующей инкубацией при температуре 4°С в течение 15 минут.10 мл буфера MACS добавляли для промывки и центрифугировали при 300×g в течение 10 минут при температуре 4°С, и затем осадок клеток ресуспендировали в 0,5 мл буфера MACS.
Сначала 3 мл буфера MACS наливали в колонку LS (Miltenyi Biotec, Bergisch Gladbach, Германия, кат. №: 130-042-901), и затем наливали суспензию клеток. При этом, 2 мл буфера MACS три раза наливали таким образом, чтобы клетки, остающиеся в колонке LS, можно было достаточным образом отделить. Затем, после отделения колонки LS от магнитного держателя, добавляли 5 мл буфера MACS, и давление прикладывали посредством поршня с получением CD3(-)CD56(+) естественных клеток-киллеров. Полученные CD3(-)CD56(+) естественные клетки-киллеры помещали в новую пробирку и центрифугировали при 300×g в течение 5 минут при температуре 4°С. После удаления супернатанта клетки суспендировали в базовых культуральных средах, показанных в Таблице 1 - Таблице 4, с учетом условий культивирования. Число суспендированных клеток подсчитывали с использованием счетчика клеток.
Пример 2. Культивирование CD3(-)CD56(+) естественных клеток-киллеров, происходящих из мононуклеарных клеток периферической крови (РВМС)
100 мкл CD335 (NKp46)-биотин и 100 мкл CD2-биотин, включенных в набор для активации/размножения NK-клеток (кат. №: 130-112-968) (Miltenyi Biotec, Bergisch Gladbach, Германия) помещали в микропробирку, объемом 1,5 мл, и смешивали и затем добавляли 500 мкл частиц с антителами против биотина MACSiBead и перемешивали. Затем, добавляли 300 мкл буфера MACS и смешивали при 2°С-8°С в течение 2 часов с использованием ротора для микропробирок. Принимая во внимание число клеток, 5 мкл шариков для активации NK на 1×106 клеток переносили в новую пробирку. 1 мл PBS добавляли и центрифугировали при 300×g в течение 5 минут. После удаления супернатанта среду MACS NK (кат. №: 130-094-483) (Miltenyi Biotec, Bergisch Gladbach, Германия), подлежащую использованию, добавляли из расчета 5 мкл на 106 NK-клеток и высвобождали шарики с последующей инокуляцией в CD3(-)CD56(+) естественные клетки-киллеры, выделенные в Примере 1.
Далее, полученные CD3(-)CD56(+) естественные клетки-киллеры суспендировали в композиции культуральной среды, содержащей добавку, полученную в Примере получения 1, таким образом, что общее число клеток составляло 2,5×105, и высевали в 48-луночный планшет с последующим культивированием в условиях 37°С и 5% CO2. Затем, число клеток определяли каждые 2 суток для субкультивирования в следующем порядке: 48-луночный планшет, 24-луночный планшет, 12-луночный планшет, 6-луночный планшет и колба 25Т, когда клетки сливались на 80% или более сосуда для культивирования (конфлюентность), и, в конечном итоге, все клетки собирали в сутки 21.
Пример 3. Подсчет числа клеток и сравнение жизнеспособности клеток
Общее число клеток и жизнеспособность культивируемых естественных клеток-киллеров подсчитывали, используя счетчик клеток (ADAM-MC2), в сутки 5, 9, 11, 13, 15, 17 и 21. При этом, число клеток подсчитывали в указанные выше даты, когда клетки достигают конфлюентности 80%, которая является критерием для субкультивирования, поскольку степень пролиферации клеток варьирует в зависимости от обрабатываемого материала и типа культуральной среди.
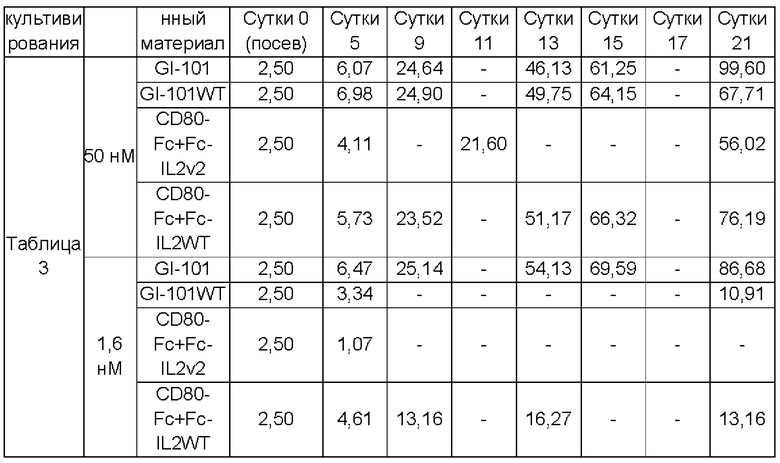
Результаты сравнения общего числа клеток и жизнеспособности CD3-CD56+клеток, культивируемых в условиях композиции культуральной среды, полученной в Примере получения 1, показаны в Таблицах 6-13, и на Фиг. 5А-12В.
В результате, подтвердили, что все композиции культуральных сред, в которые добавляли GI-101, полученный в Примере получения 1, имели общее число естественных клеток-киллеров, больше чем контрольная группа (добавление CD80-Fc+Fc-IL2v2 или CD80-Fc+Fc-IL2WT), несмотря на концентрацию обработки в условиях четырех базовых культуральных сред (Таблицы 1-4) (Фиг. 5А, 5В, 7А, 7В, 9А, 9В, 11А и 11В).
Кроме того, даже в случае жизнеспособности клеток, когда добавляли GI-101, все композиции культуральных сред демонстрировали высокую жизнеспособность, независимо от базовой культуральной среды и концентрации (Фиг. 6А, 6В, 8А, 8В, 10А, 10В, 12А и 12В).
На основе результатов подтвердили, что GI-101 играет важную роль в улучшении способности к пролиферации и жизнеспособности естественных клеток-киллеров, по сравнению с контрольной группой (добавление CD80-Fc+Fc-IL2v2 или CD80-Fc+Fc-IL2WT), независимо от базовой культуральной среды и концентрации.








II. Характеристика естественных клеток-киллеров с использованием композиции для культивирования естественных клеток-киллеров
Пример 4. Измерение чистоты естественных клеток-киллеров
CD3-CD56+ естественные клетки-киллеры, полученные из Примера 2, соответственно, центрифугировали при 300×g в течение 5 минут для удаления супернатанта, и 1 мл буфера FACS добавляли с высвобождением осадка. Затем, 3% (об./об.) FBS, 10 мМ ЭДТА, 20 мМ HEPES, 10 мкг/мл полимиксина В, 100 Ед/мл пенициллина, 100 мкг/мл стрептомицина и 1 мМ пирувата натрия добавляли к PBS с получением буфера FACS, и 1 мл полученного буфера FACS добавляли для ресуспендирования осадка клеток. Далее, его разводили буфером FACS до 2×106 клеток/мл с использованием счетчика клеток.
100 мкл разведенного раствора клеток добавляли в каждую пробирку с 5 мл FACS, и 100 мкл буфера FACS дополнительно добавляли туда с последующей обработкой антителом против человеческого CD3, меченным PerCP (антитело против человеческого CD3, меченное PerCP (клон UCHT1)) и антителом против человеческого CD56, меченным РЕ/су7 (антитело против человеческого CD56, меченное РЕ/су7 (Клон В159)). Затем, после инкубации при 4°С в течение 20 минут, 200 мкл буфера FACS добавляли и центрифугировали при 1500 об./мин. в течение 3 минут.Супернатант удаляли, и 200 мкл буфера FACS добавляли для суспендирования, и затем фенотип клеток определяли с использованием проточного цитометра (CYTEK® Aurora, Cytek, Fremont, СА, США).
Информация об антителах, используемых в эксперименте, показана в Таблице 14. Кроме того, чистоты CD3-CD56+естественных клеток-киллеров, культивируемых в течение 21 суток в условиях композиций культуральных сред, полученных в Примере получения 1, были измерены и показаны на Фиг. 13-16.

Пример 5. Идентификация маркеров активации и ингибирования для естественных клеток-киллеров
CD3-CD56+ естественные клетки-киллеры, полученные в Примере 2, соответственно центрифугировали при 300×g в течение 5 минут для удаления супернатанта, и 1 мл буфера FACS добавляли для высвобождения осадка.
3% (об./об.) FBS, 10 мМ ЭДТА, 20 мМ HEPES, 10 мкг/мл полимиксина В, 100 Ед./мл пенициллина, 100 мкг/мл стрептомицина и 1 мМ пирувата натрия добавляли в PBS с получением буфера FACS, и 1 мл полученного буфера FACS добавляли для ресуспендирования осадка клеток. Затем, его разводили буфером FACS до 2×106 клеток/мл с использованием счетчика клеток. 100 мкл разведенного раствора клеток добавляли в каждую 5 мл пробирку для FACS и подтверждали посредством использования антитела против человеческого CD16, меченного Pe-CF594 (антитело против человеческого CD16, меченное PE-CF594 (клон 3G8)), антитела против человеческого DNAM1, меченного АРС (антитело против человеческого DNAM1, меченное АРС (клон 11А8)), антитела против человеческого NKG2C, меченного BV605 (антитело против человеческого NKG2C, меченное BV605 (клон 134591)), антитела против человеческого NKG2D, меченного BV650 (антитело против человеческого NKG2D, меченное BV650 (Клон 1D11)), антитела против человеческого NKp46, меченного ВВ515 (антитело против человеческого NKp46, меченное ВВ515 (Клон 9Е2)), антитела против человеческого NKp30, меченного BV480 (антитело против человеческого NKp30, меченное BV480 (Клон р30-15)), антитела против человеческого PD-1, меченного РЕ (антитело против человеческого PD-1, меченное РЕ (клон ЕН12.2Н7)), и антитела против человеческого NKG2A, меченного АРС (антитело против человеческого NKG2A, меченное АРС (клон 131411)), с использованием проточного цитометра. Затем, после инкубирования при 4°С в течение 20 минут, 100 мкл буфера FACS добавляли и центрифугировали при 1500 об./мин. в течение 3 минут.
Супернатант удаляли, и 200 мкл буфера FACS добавляли для суспендирования, и затем фенотип клеток определяли с использованием проточного цитометра (CYTEK® Aurora, Cytek, Fremont, СА, США). Информация антител, используемых в эксперименте, показана в Таблице 15. Маркеры активации и ингибирования для CD3-CD56+ естественных клеток-киллеров, культивируемых в течение 21 суток в условиях композиций культуральных сред, полученных в Примере получения 1, идентифицировали и показаны на Фиг. 17-24.

Пример 6. Определение способности естественных клеток-киллеров секретировать гранзим В, перфорин, интерферон гамма
Для определения способности CD3-CD56+ естественных клеток-киллеров, полученных в Примере 2, секретировать гранзим и перфорин, количество экспрессии гранзима В, перфорина и интерферона гамма в естественных клетках-киллерах измеряли посредством внутриклеточного окрашивания. Культивируемые естественные клетки-киллеры центрифугировали в условиях 300×g в течение 5 минут, и супернатант удаляли. Затем, их разводили каждой культуральной композицией до 2×106 клеток/мл с использованием счетчика клеток.
200 мкл полученных клеток дозировали в каждую лунку 96-луночного планшета, и затем добавляли 1%-ную (об./об.) смесь для стимуляции (1х) (Thermo Scientific, Waltham, MA, США) и инкубировали при 37°С, в условиях CO2 в течение 4 часов. Затем, планшеты центрифугировали при 300×g в течение 5 минут, и удаляли супернатант.Затем, 3% (об./об.) FBS, 10 мМ ЭДТА, 20 мМ HEPES, 10 мкг/мл полимиксина В, 100 Ед./мл пенициллина, 100 мкг/мл стрептомицина и 1 мМ пирувата натрия добавляли в PBS с получением FACS, и 100 мкл полученного буфера FACS добавляли для ресуспендирования осадка клеток. Супернатант удаляли, и 100 мкл буфера BD CYTOFIX/CYTOPERM™ (проникающий/фиксирующий буфер, BD science) добавляли для фиксации и проникновения и затем суспендировали с последующей инкубацией при 4°С в течение 30 минут.100 мкл буфера FACS дополнительно добавляли и центрифугировали при 1500 об./мин. в течение 3 минут.
Антителом против человеческого гранзима В, меченным РЕ/су7 (антитело против человеческого гранзима В, меченное РЕ/су7 (Клон NGZB)), антителом против человеческого перфорина, меченным АРС (антитело против человеческого перфорина, меченное АРС (Клон B-D48)), и антителом против человеческого интерферона гамма, меченным BV421 (антитело против человеческого интерферона гамма, меченное BV421 (Клон В27)) осуществляли обработку. Затем, после инкубации при 4°С в течение 20 минут, 100 мкл буфера FACS добавляли и центрифугировали при 1500 об./мин. в течение 3 минут.После удаления супернатанта и добавления 200 мкл буфера FACS (фиксирующий буфер) для суспендирования, количество экспрессии клеток определяли с использованием проточной цитометрии.
Информация об антителах, используемых в эксперименте, показана в Таблице 16. Маркеры для CD3-CD56+ естественных клеток-киллеров, культивируемых в течение 21 суток в условиях композиций культуральных сред, полученных в Примере получения 1, были идентифицированы и показаны на Фиг. 25-28.


III. Анализ способности естественных клеток-киллеров уничтожать раковые клетки в соответствии с культуральной композицией
Пример 8. Подтверждение способности к дегрануляции и эффекта уничтожения е естественных клеток-киллеров в отношении раковых клеток
В частности, линию раковых клеток K562 (Американская коллекция типовых культур, АТСС (от англ. American Type Culture Collection)) разводили в PBS для определения числа клеток, показанного ниже в Таблице 17, и дозировали в каждую лунку.

В частности, линию раковых клеток K562 разводили каждой культуральной композицией до 1×107 клеток/мл и затем дозировали согласно числу клеток, и затем дозировали в соответствии с числом клеток, конкретно определенном в указанной выше таблице для каждой лунки. Затем, естественные клетки-киллеры также разводили каждой культуральной композицией до 5×106 клеток/мл, и затем дозировали в соответствии с числом клеток, конкретно указанным в указанной выше таблице для каждой лунки, и центрифугировали в условиях 30×g в течение 3 минут.Далее, после культивирования при 37°С, в условиях 5% CO2 в течение 4 часов, антителом против человеческого CD3, меченным BV421 (антитело против человеческого CD3, меченное BV421 (клон UCHT1)), антителом против человеческого CD16, меченным РЕ (антитело против человеческого CD16, меченное РЕ (клон 3G8)), антителом против человеческого CD56, меченным РЕ/су7 (антитело против человеческого CD56, меченное РЕ/су7 (Клон В159)), и антителом против человеческого CD107a, меченным FITC (антитело против человеческого CD107a, меченное FITC (клон Н4А3)), осуществляли обработку и инкубацию на льду в течение 20 минут.
Затем, 100 мкл буфера FACS добавляли и центрифугировали при 1300 об./мин., в условиях 4°С в течение 5 минут.После удаления супернатанта осуществляли обработку окрашивающим раствором для определения жизнеспособности 7-AAD, и свет блокировали для осуществления реакции при комнатной температуре в течение 15 минут.Затем, 100 мкл буфера FACS добавляли и центрифугировали при 1300 об./мин., в условиях 4°С в течение 5 минут.Еще раз добавляли 200 мкл буфера FACS, и центрифугировали при 1300 об./мин., в условиях 4°С в течение 5 минут.После повторения указанного выше процесса еще раз супернатант удаляли, и добавляли 400 мкл буфера FACS с последующим анализом с использованием проточного цитометра (CYTEL® Aurora, Cytek, Fremont, СА, США).
Способность к дегрануляции и эффект уничтожения у естественных клеток-киллеров, культивируемых в течение 21 суток, в композиции, в которой 50 нМ добавки в Таблице 5 добавляли к базовой культуральной среде в Таблицах 1-14, соответственно, показаны на Фиг. 29-34.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GI CELL, INC.
<120> КОМПОЗИЦИЯ ДЛЯ КУЛЬТИВИРОВАНИЯ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ
И СПОСОБ ПОЛУЧЕНИЯ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ С ЕЕ ИСПОЛЬЗОВАНИЕМ
<130> PF-B2646
<140> PCT/KR2020/016376
<141> 2020-11-19
<150> 10-2019-0149779
<151> 2019-11-20
<150> 10-2020-0015802
<151> 2020-02-10
<160> 46
<170> PatentIn version 3.5
<210> 1
<211> 25
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> сигнальный пептид (TPA)
<400> 1
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala
20 25
<210> 2
<211> 208
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hB7-1:35-242
<400> 2
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
<210> 3
<211> 30
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> шарнир с линкером
<400> 3
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10 15
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
20 25 30
<210> 4
<211> 216
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> fc иммуноглобулина
<400> 4
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly
210 215
<210> 5
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> линкер
<400> 5
Gly Gly Gly Gly Ser
1 5
<210> 6
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hIL-2M
<400> 6
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 7
<211> 617
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок, содержащий варианты IL-2 и фрагменты CD80
<400> 7
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Ile His Val Thr Lys Glu
20 25 30
Val Lys Glu Val Ala Thr Leu Ser Cys Gly His Asn Val Ser Val Glu
35 40 45
Glu Leu Ala Gln Thr Arg Ile Tyr Trp Gln Lys Glu Lys Lys Met Val
50 55 60
Leu Thr Met Met Ser Gly Asp Met Asn Ile Trp Pro Glu Tyr Lys Asn
65 70 75 80
Arg Thr Ile Phe Asp Ile Thr Asn Asn Leu Ser Ile Val Ile Leu Ala
85 90 95
Leu Arg Pro Ser Asp Glu Gly Thr Tyr Glu Cys Val Val Leu Lys Tyr
100 105 110
Glu Lys Asp Ala Phe Lys Arg Glu His Leu Ala Glu Val Thr Leu Ser
115 120 125
Val Lys Ala Asp Phe Pro Thr Pro Ser Ile Ser Asp Phe Glu Ile Pro
130 135 140
Thr Ser Asn Ile Arg Arg Ile Ile Cys Ser Thr Ser Gly Gly Phe Pro
145 150 155 160
Glu Pro His Leu Ser Trp Leu Glu Asn Gly Glu Glu Leu Asn Ala Ile
165 170 175
Asn Thr Thr Val Ser Gln Asp Pro Glu Thr Glu Leu Tyr Ala Val Ser
180 185 190
Ser Lys Leu Asp Phe Asn Met Thr Thr Asn His Ser Phe Met Cys Leu
195 200 205
Ile Lys Tyr Gly His Leu Arg Val Asn Gln Thr Phe Asn Trp Asn Thr
210 215 220
Thr Lys Gln Glu His Phe Pro Asp Asn Gly Ser Gly Gly Gly Gly Ser
225 230 235 240
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly
245 250 255
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
260 265 270
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg
275 280 285
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
290 295 300
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
305 310 315 320
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
325 330 335
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
340 345 350
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
355 360 365
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
370 375 380
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
385 390 395 400
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
405 410 415
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
420 425 430
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
435 440 445
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala
450 455 460
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly
465 470 475 480
Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
485 490 495
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
500 505 510
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe
515 520 525
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
530 535 540
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
545 550 555 560
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
565 570 575
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
580 585 590
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
595 600 605
Cys Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 8
<211> 1857
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101)
<400> 8
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac ctgatga 1857
<210> 9
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101)
<400> 9
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 10
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hIL-2
<400> 10
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 11
<211> 288
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> CD80
<400> 11
Met Gly His Thr Arg Arg Gln Gly Thr Ser Pro Ser Lys Cys Pro Tyr
1 5 10 15
Leu Asn Phe Phe Gln Leu Leu Val Leu Ala Gly Leu Ser His Phe Cys
20 25 30
Ser Gly Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu
35 40 45
Ser Cys Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile
50 55 60
Tyr Trp Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp
65 70 75 80
Met Asn Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr
85 90 95
Asn Asn Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly
100 105 110
Thr Tyr Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg
115 120 125
Glu His Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr
130 135 140
Pro Ser Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile
145 150 155 160
Ile Cys Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu
165 170 175
Glu Asn Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp
180 185 190
Pro Glu Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met
195 200 205
Thr Thr Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg
210 215 220
Val Asn Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro
225 230 235 240
Asp Asn Leu Leu Pro Ser Trp Ala Ile Thr Leu Ile Ser Val Asn Gly
245 250 255
Ile Phe Val Ile Cys Cys Leu Thr Tyr Cys Phe Ala Pro Arg Cys Arg
260 265 270
Glu Arg Arg Arg Asn Glu Arg Leu Arg Arg Glu Ser Val Arg Pro Val
275 280 285
<210> 12
<211> 215
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> модифицированный Fc
<400> 12
Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys Pro Lys
1 5 10 15
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
20 25 30
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
35 40 45
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
50 55 60
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
65 70 75 80
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
85 90 95
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
100 105 110
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
115 120 125
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
130 135 140
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
145 150 155 160
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
165 170 175
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
180 185 190
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
195 200 205
Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 13
<211> 306
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> mCD80
<400> 13
Met Ala Cys Asn Cys Gln Leu Met Gln Asp Thr Pro Leu Leu Lys Phe
1 5 10 15
Pro Cys Pro Arg Leu Ile Leu Leu Phe Val Leu Leu Ile Arg Leu Ser
20 25 30
Gln Val Ser Ser Asp Val Asp Glu Gln Leu Ser Lys Ser Val Lys Asp
35 40 45
Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His Glu Asp Glu Ser
50 55 60
Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val Val Leu Ser Val
65 70 75 80
Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys Asn Arg Thr Leu
85 90 95
Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly Leu Val Leu Ser
100 105 110
Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys Glu Arg Gly Thr
115 120 125
Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser Ile Lys Ala Asp
130 135 140
Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro Ser Ala Asp Thr
145 150 155 160
Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro Lys Pro Arg Phe
165 170 175
Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile Asn Thr Thr Ile
180 185 190
Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser Ser Gln Leu Asp
195 200 205
Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu Ile Lys Tyr Gly
210 215 220
Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys Pro Pro Glu Asp
225 230 235 240
Pro Pro Asp Ser Lys Asn Thr Leu Val Leu Phe Gly Ala Gly Phe Gly
245 250 255
Ala Val Ile Thr Val Val Val Ile Val Val Ile Ile Lys Cys Phe Cys
260 265 270
Lys His Arg Ser Cys Phe Arg Arg Asn Glu Ala Ser Arg Glu Thr Asn
275 280 285
Asn Ser Leu Thr Phe Gly Pro Glu Glu Ala Leu Ala Glu Gln Thr Val
290 295 300
Phe Leu
305
<210> 14
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI101)
<400> 14
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg 120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa 180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag 240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc 300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag 360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag 420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct 480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt 540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc 600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt 660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc 720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca 780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg 1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt 1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtctct tggaggtggt 1440
ggcggttctg cccctacctc cagctctacc aagaaaaccc agctccagtt ggagcatctg 1500
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc 1560
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacttgcag 1620
tgcctggaag aggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac 1680
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa 1740
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt 1800
ctgaaccggt ggatcacctt ctgccagagc atcatctcca cactgacc 1848
<210> 15
<211> 616
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI101)
<400> 15
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
465 470 475 480
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
485 490 495
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
500 505 510
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
515 520 525
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu
530 535 540
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
545 550 555 560
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
565 570 575
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
580 585 590
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
595 600 605
Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 16
<211> 1437
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101C1)
<400> 16
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctaggtgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc cctgggc 1437
<210> 17
<211> 454
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101C1)
<400> 17
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly
450
<210> 18
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101C2)
<400> 18
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccatctc acgccgctga gtctaagtac ggccctcctt gtcctccatg tcctgctcca 120
gaagctgctg gcggaccctc tgtgttcctg tttcctccaa agcctaagga ccagctcatg 180
atctctcgga cccctgaagt gacctgcgtg gtggtggatg tgtctcaaga ggaccctgag 240
gtgcagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga 300
gaggaacagt tcaactccac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat 360
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca agggcctgcc ttccagcatc 420
gaaaagacca tctccaaggc taagggccag cctagggaac cccaggttta caccctgcct 480
ccaagccaag aggaaatgac caagaaccag gtgtccctga cctgcctggt caagggcttc 540
tacccttccg acattgccgt ggaatgggag tccaatggcc agcctgagaa caactacaag 600
accacacctc ctgtgctgga ctccgacggc tccttctttc tgtactctcg cctgaccgtg 660
gacaagtcta ggtggcaaga gggcaacgtg ttctcctgct ctgtgctgca cgaggccctg 720
cacaatcact acacccagaa gtccctgtct ctgtctcttg gcggaggcgg aggatctgct 780
cctacctcca gctccaccaa gaaaacccag ctccagttgg agcatctgct gctggacctc 840
cagatgatcc tgaatggcat caacaattac aagaacccca agctgaccgc catgctgacc 900
gctaagttct acatgcccaa gaaggccacc gagctgaagc acctccagtg cctggaagag 960
gaactgaagc ccctggaaga agtgctgaat ctggcccagt ccaagaactt ccacctgagg 1020
cctagggacc tgatctccaa catcaacgtg atcgtgctgg aactgaaagg ctccgagaca 1080
accttcatgt gcgagtacgc cgacgagaca gccaccatcg tggaatttct gaaccggtgg 1140
atcaccttct gccagtccat catctccaca ctgacc 1176
<210> 19
<211> 367
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101C2)
<400> 19
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
1 5 10 15
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
20 25 30
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
35 40 45
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
50 55 60
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
85 90 95
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
100 105 110
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
115 120 125
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
130 135 140
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
145 150 155 160
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
165 170 175
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
180 185 190
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
195 200 205
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
210 215 220
Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser
225 230 235 240
Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln
245 250 255
Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala
260 265 270
Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys
275 280 285
His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu
290 295 300
Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile
305 310 315 320
Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr
325 330 335
Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu
340 345 350
Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
355 360 365
<210> 20
<211> 1434
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI101C1)
<400> 20
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg 120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa 180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag 240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc 300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag 360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag 420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct 480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt 540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc 600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt 660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc 720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca 780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg 1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt 1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtccct gggc 1434
<210> 21
<211> 478
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI101C1)
<400> 21
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
465 470 475
<210> 22
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M45)
<400> 22
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Ala Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 23
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M61)
<400> 23
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Arg Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 24
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M72)
<400> 24
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Gly Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 25
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M45)
<400> 25
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttcgccatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 26
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M45)
<400> 26
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Ala Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 27
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M61)
<400> 27
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aaagggaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 28
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M61)
<400> 28
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Arg Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 29
<211> 1857
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M72)
<400> 29
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatggggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac ctgatga 1857
<210> 30
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M72)
<400> 30
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Gly Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 31
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101w)
<400> 31
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
acccgcatgc tgacctttaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 32
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101w)
<400> 32
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 33
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI102-M61)
<400> 33
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg 120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa 180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag 240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc 300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag 360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag 420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct 480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt 540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc 600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt 660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc 720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca 780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg 1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt 1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtctct tggaggtggt 1440
ggcggttctg cccctacctc cagctctacc aagaaaaccc agctccagtt ggagcatctg 1500
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc 1560
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacttgcag 1620
tgcctggaaa gggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac 1680
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa 1740
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt 1800
ctgaaccggt ggatcacctt ctgccagagc atcatctcca cactgacc 1848
<210> 34
<211> 616
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI102-M61)
<400> 34
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
465 470 475 480
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
485 490 495
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
500 505 510
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
515 520 525
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Arg
530 535 540
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
545 550 555 560
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
565 570 575
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
580 585 590
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
595 600 605
Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 35
<211> 153
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hIL-2 дикого типа
<400> 35
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
20 25 30
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
35 40 45
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe
50 55 60
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
65 70 75 80
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
85 90 95
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
100 105 110
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
115 120 125
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
130 135 140
Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150
<210> 36
<211> 158
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> IL-2 с сигнальной последовательностью
<400> 36
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Ala Pro Thr Ser Ser Ser Thr
20 25 30
Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met
35 40 45
Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met
50 55 60
Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His
65 70 75 80
Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn
85 90 95
Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser
100 105 110
Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe
115 120 125
Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn
130 135 140
Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150 155
<210> 37
<211> 474
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеотидов, кодирующая IL-2 с сигнальной последовательностью
<400> 37
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgcccc taccagctcc tctaccaaga aaacccagct ccagttggag 120
catctgctgc tggacctcca gatgattctg aacgggatca acaactataa gaaccccaag 180
ctgacccgca tgctgacctt taagttctac atgcccaaga aggccaccga gctgaagcac 240
ctccagtgcc tggaagaaga actgaagccc ctggaagagg tgctgaatct ggcccagtcc 300
aagaacttcc acctgaggcc acgggacctg atcagcaaca tcaacgtgat cgtgctggaa 360
ctgaagggct ccgagacaac ctttatgtgc gagtacgccg acgagacagc caccatcgtg 420
gaatttctga accggtggat caccttctgc cagagcatca tctccacact gacc 474
<210> 38
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hinge
<400> 38
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 39
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие CD80-Fc белок
<400> 39
ggatccgcca ccatggatgc tatgctgaga ggcctgtgtt gcgtgctgct gctgtgtggc 60
gctgtgttcg tgtctccttc tcacgctgtg atccacgtga ccaaagaagt gaaagaggtc 120
gccacactgt cctgcggcca caacgtttca gtggaagaac tggcccagac caggatctac 180
tggcagaaag aaaagaaaat ggtgctgacc atgatgtccg gcgacatgaa catctggcct 240
gagtacaaga accggaccat cttcgacatc accaacaacc tgtccatcgt gattctggcc 300
ctgaggcctt ctgatgaggg cacctatgag tgcgtggtgc tgaagtacga gaaggacgcc 360
ttcaagcgcg agcacctggc tgaagtgaca ctgtccgtga aggccgactt tcccacacct 420
tccatctccg acttcgagat ccctacctcc aacatccggc ggatcatctg ttctacctct 480
ggcggctttc ctgagcctca cctgtcttgg ctggaaaacg gcgaggaact gaacgccatc 540
aacaccaccg tgtctcagga ccccgaaacc gagctgtacg ctgtgtcctc caagctggac 600
ttcaacatga ccaccaacca cagcttcatg tgcctgatta agtacggcca cctgagagtg 660
aaccagacct tcaactggaa caccaccaag caagagcact tccctgacaa tggatctggc 720
ggcggaggtt ctggcggagg tggaagcgga ggcggaggat ctgctgagtc taagtatggc 780
cctccttgtc ctccatgtcc tgctccagaa gctgctggcg gaccctctgt gttcctgttt 840
cctccaaagc ctaaggacca gctcatgatc tctcggacac ccgaagtgac ctgcgtggtg 900
gtggatgtgt ctcaagagga ccctgaggtg cagttcaatt ggtacgtgga cggcgtggaa 960
gtgcacaacg ccaagaccaa gcctagagag gaacagttca actccaccta cagagtggtg 1020
tccgtgctga ccgtgctgca ccaggattgg ctgaacggca aagagtacaa gtgcaaggtg 1080
tccaacaagg gcctgccttc cagcatcgaa aagaccatct ccaaggctaa gggccagcct 1140
agggaacccc aggtttacac cctgcctcca agccaagagg aaatgaccaa gaaccaggtg 1200
tccctgacct gcctggtcaa gggcttctac ccttccgaca ttgccgtgga atgggagtcc 1260
aatggccagc ctgagaacaa ctacaagacc acacctcctg tgctggactc cgacggctcc 1320
ttctttctgt actctcgcct gaccgtggac aagtctaggt ggcaagaggg caacgtgttc 1380
tcctgctctg tgctgcacga ggccctgcac aatcactaca cccagaagtc cctgtctctg 1440
tccctgggct gatgactcga g 1461
<210> 40
<211> 479
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> CD80-Fc protein
<400> 40
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Ile His Val Thr Lys Glu
20 25 30
Val Lys Glu Val Ala Thr Leu Ser Cys Gly His Asn Val Ser Val Glu
35 40 45
Glu Leu Ala Gln Thr Arg Ile Tyr Trp Gln Lys Glu Lys Lys Met Val
50 55 60
Leu Thr Met Met Ser Gly Asp Met Asn Ile Trp Pro Glu Tyr Lys Asn
65 70 75 80
Arg Thr Ile Phe Asp Ile Thr Asn Asn Leu Ser Ile Val Ile Leu Ala
85 90 95
Leu Arg Pro Ser Asp Glu Gly Thr Tyr Glu Cys Val Val Leu Lys Tyr
100 105 110
Glu Lys Asp Ala Phe Lys Arg Glu His Leu Ala Glu Val Thr Leu Ser
115 120 125
Val Lys Ala Asp Phe Pro Thr Pro Ser Ile Ser Asp Phe Glu Ile Pro
130 135 140
Thr Ser Asn Ile Arg Arg Ile Ile Cys Ser Thr Ser Gly Gly Phe Pro
145 150 155 160
Glu Pro His Leu Ser Trp Leu Glu Asn Gly Glu Glu Leu Asn Ala Ile
165 170 175
Asn Thr Thr Val Ser Gln Asp Pro Glu Thr Glu Leu Tyr Ala Val Ser
180 185 190
Ser Lys Leu Asp Phe Asn Met Thr Thr Asn His Ser Phe Met Cys Leu
195 200 205
Ile Lys Tyr Gly His Leu Arg Val Asn Gln Thr Phe Asn Trp Asn Thr
210 215 220
Thr Lys Gln Glu His Phe Pro Asp Asn Gly Ser Gly Gly Gly Gly Ser
225 230 235 240
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly
245 250 255
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
260 265 270
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg
275 280 285
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
290 295 300
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
305 310 315 320
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
325 330 335
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
340 345 350
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
355 360 365
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
370 375 380
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
385 390 395 400
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
405 410 415
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
420 425 430
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
435 440 445
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala
450 455 460
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
465 470 475
<210> 41
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (hCD80-Fc-IL2wt)
<400> 41
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
acccgcatgc tgacctttaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 42
<211> 367
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Fc-IL2wt
<400> 42
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
1 5 10 15
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
20 25 30
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
35 40 45
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
50 55 60
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
85 90 95
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
100 105 110
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
115 120 125
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
130 135 140
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
145 150 155 160
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
165 170 175
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
180 185 190
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
195 200 205
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
210 215 220
Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser
225 230 235 240
Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln
245 250 255
Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg
260 265 270
Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys
275 280 285
His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu
290 295 300
Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile
305 310 315 320
Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr
325 330 335
Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu
340 345 350
Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
355 360 365
<210> 43
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fc-IL2wt
<400> 43
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgctga gtctaagtat ggccctcctt gtcctccatg tcctgctcca 120
gaagctgctg gcggaccctc tgtgttcctg tttcctccaa agcctaagga ccagctcatg 180
atctctcgga cacccgaagt gacctgcgtg gtggtggatg tgtctcaaga ggaccctgag 240
gtgcagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga 300
gaggaacagt tcaactccac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat 360
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca agggcctgcc ttccagcatc 420
gaaaagacca tctccaaggc taagggccag cctagggaac cccaggttta caccctgcct 480
ccaagccaag aggaaatgac caagaaccag gtgtccctga cctgcctggt caagggcttc 540
tacccttccg acattgccgt ggaatgggag tccaatggcc agcctgagaa caactacaag 600
accacacctc ctgtgctgga ctccgacggc tccttctttc tgtactctcg cctgaccgtg 660
gacaagtcta gatggcaaga gggcaacgtg ttctcctgct ctgtgctgca cgaggccctg 720
cacaatcact acacccagaa gtccctgtct ctgtctcttg gaggtggtgg cggttctgcc 780
cctaccagct cctctaccaa gaaaacccag ctccagttgg agcatctgct gctggacctc 840
cagatgattc tgaacgggat caacaactat aagaacccca agctgacccg catgctgacc 900
tttaagttct acatgcccaa gaaggccacc gagctgaagc acctccagtg cctggaagaa 960
gaactgaagc ccctggaaga ggtgctgaat ctggcccagt ccaagaactt ccacctgagg 1020
ccacgggacc tgatcagcaa catcaacgtg atcgtgctgg aactgaaggg ctccgagaca 1080
acctttatgt gcgagtacgc cgacgagaca gccaccatcg tggaatttct gaaccggtgg 1140
atcaccttct gccagagcat catctccaca ctgacc 1176
<210> 44
<211> 392
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Fc-IL2v2
<400> 44
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Ala Glu Ser Lys Tyr Gly Pro
20 25 30
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
35 40 45
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
50 55 60
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
65 70 75 80
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
85 90 95
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
100 105 110
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
115 120 125
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
130 135 140
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
145 150 155 160
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
165 170 175
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
180 185 190
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
195 200 205
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
210 215 220
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
225 230 235 240
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
245 250 255
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
260 265 270
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
275 280 285
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
290 295 300
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu
305 310 315 320
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
325 330 335
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
340 345 350
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
355 360 365
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
370 375 380
Gln Ser Ile Ile Ser Thr Leu Thr
385 390
<210> 45
<211> 1200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие Fc-IL2v2
<400> 45
ggatccgcca ccatggatgc tatgctgaga ggcctgtgtt gcgtgctgct gctgtgtggc 60
gctgtgttcg tgtctccatc tcacgccgct gagtctaagt acggccctcc ttgtcctcca 120
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 180
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 240
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 300
accaagccta gagaggaaca gttcaactcc acctacagag tggtgtccgt gctgaccgtg 360
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 420
ccttccagca tcgaaaagac catctccaag gctaagggcc agcctaggga accccaggtt 480
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 540
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 600
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 660
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 720
cacgaggccc tgcacaatca ctacacccag aagtccctgt ctctgtctct tggcggaggc 780
ggaggatctg ctcctacctc cagctccacc aagaaaaccc agctccagtt ggagcatctg 840
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc 900
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacctccag 960
tgcctggaag aggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac 1020
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa 1080
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt 1140
ctgaaccggt ggatcacctt ctgccagtcc atcatctcca cactgacctg atgactcgag 1200
<210> 46
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> CD80-Fc-IL2wt
<400> 46
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<---











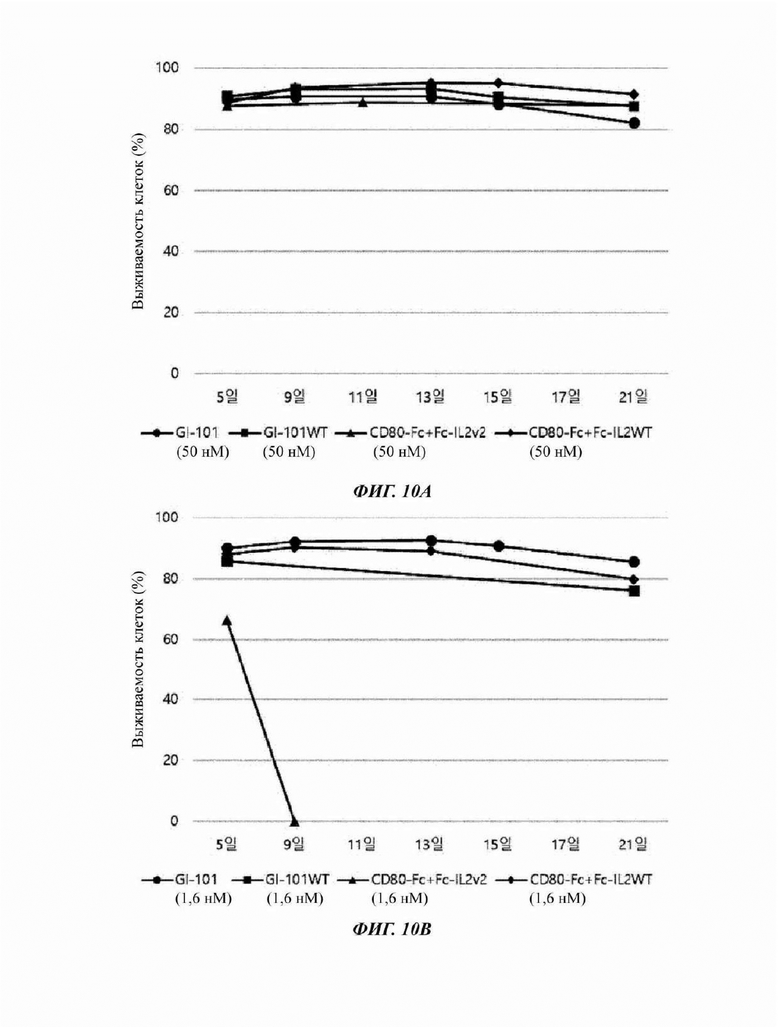



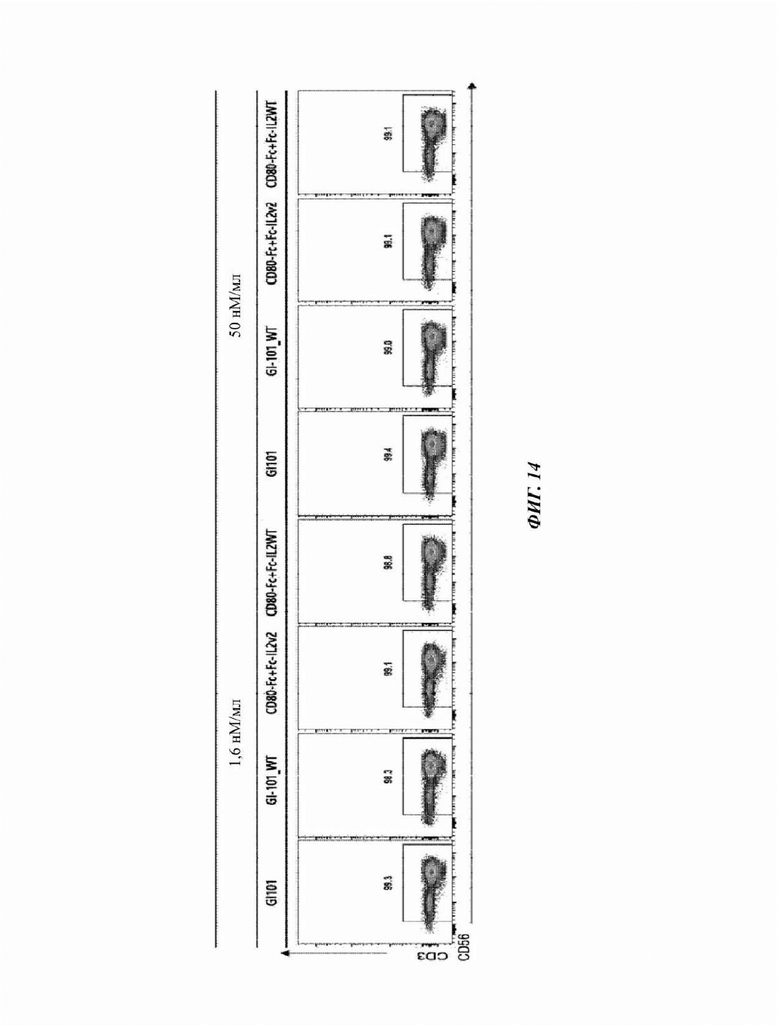



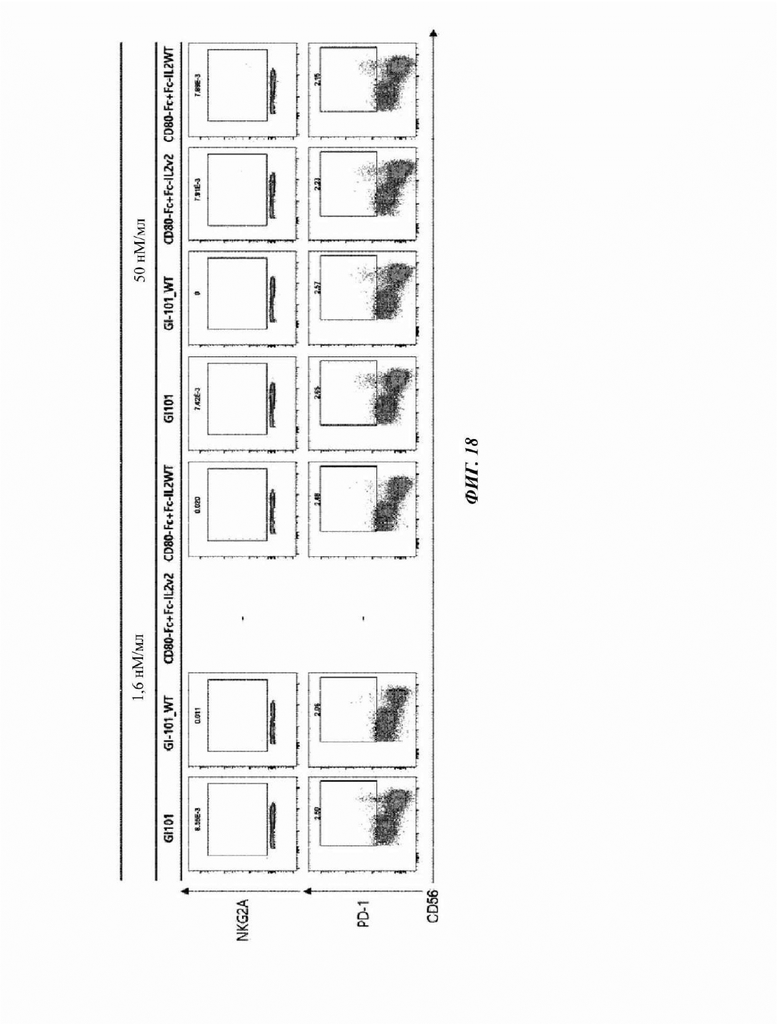
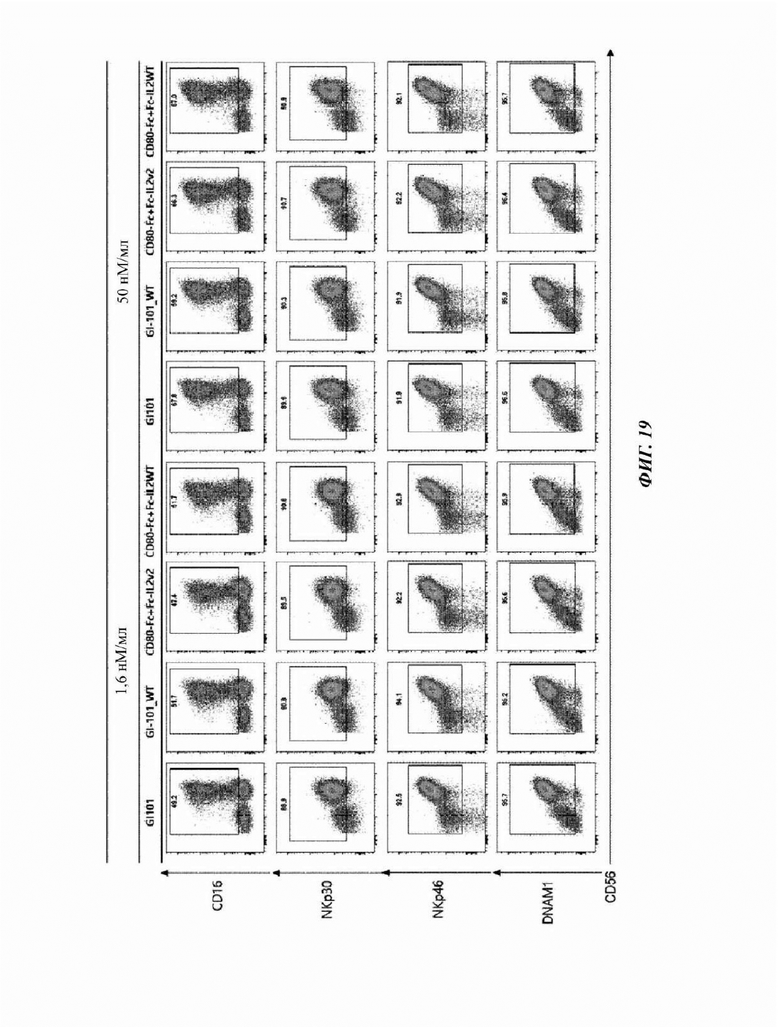
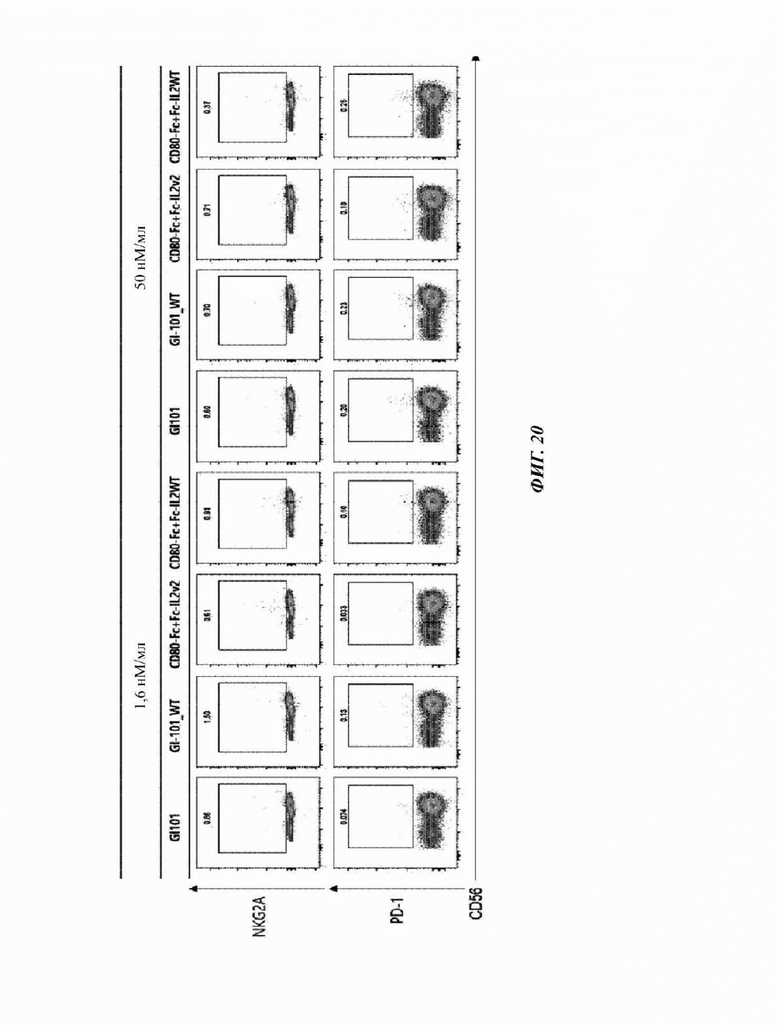











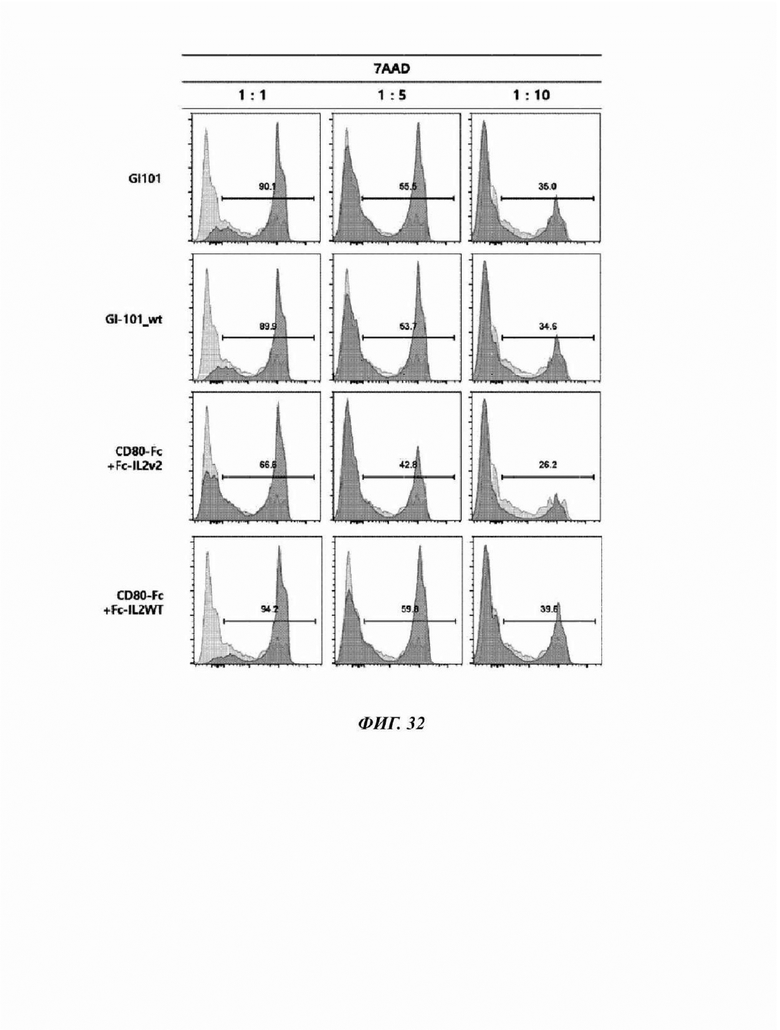
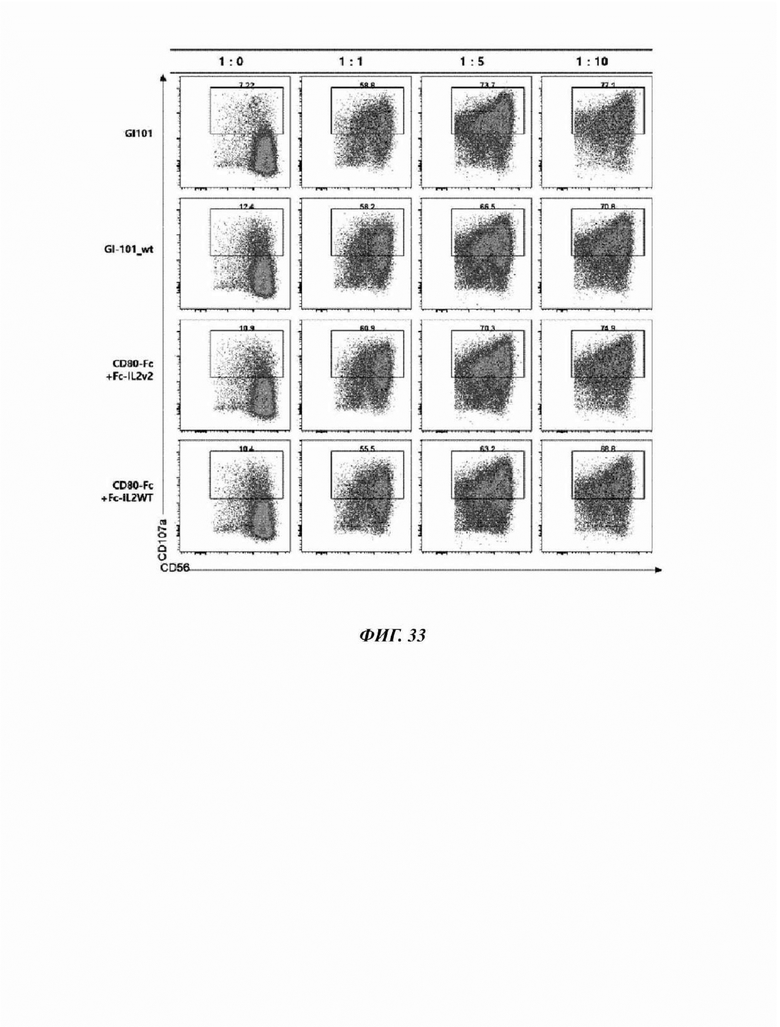

Изобретение относится к биотехнологии. Предложена композиция для культивирования выделенной CD3-CD56+ естественной клетки-киллера, содержащая в качестве активного ингредиента эффективное количество димера слитого белка, где слитый белок содержит аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30. Также предложены варианты способа культивирования выделенной CD3-CD56+ естественной клетки-киллера с использованием указанного димера слитого белка. Изобретение обеспечивает стимулирование пролиферации естественных клеток-киллеров, индуцирование экспрессии CD16 и NKp46 и повышение уровня экспрессии и секреции гранзима В и перфорина. 4 н. и 2 з.п. ф-лы, 34 ил., 17 табл., 8 пр.
1. Композиция для культивирования выделенной CD3-CD56+ естественной клетки-киллера, содержащая в качестве активного ингредиента эффективное количество димера слитого белка, где слитый белок содержит аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30.
2. Способ культивирования выделенной CD3-CD56+ естественной клетки-киллера, включающий стадии, на которых:
i) выделяют клетки, которые не экспрессируют CD3, из мононуклеарных клеток периферической крови (PBMC);
ii) выделяют клетки, которые экспрессируют CD56, из клеток, которые не экспрессируют CD3, выделенных на указанной выше стадии; и
iii) культивируют выделенные клетки в присутствии димера слитого белка, содержащего аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30.
3. Способ по п. 2, в котором димер слитого белка обработан в концентрации 1 нМ - 500 нМ.
4. Способ по п. 2, в котором период культивирования на стадии культивирования составляет от 5 суток до 25 суток.
5. Способ культивирования выделенной CD3-CD56+ естественной клетки-киллера, включающий стадии, на которых:
i) выделяют клетки, которые не экспрессируют CD3, из PBMC; и
ii) культивируют выделенные клетки в присутствии димера слитого белка, содержащего аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30.
6. Способ культивирования выделенной CD3-CD56+ естественной клетки-киллера, включающий стадии, на которых:
i) выделяют клетки, которые экспрессируют CD56, из PBMC; и
ii) культивируют выделенные клетки в присутствии димера слитого белка, содержащего аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30.
| PARK J | |||
| С | |||
| et al., GI101, a novel triple-targeting bispecific CD80-lgG4- IL2 variant fusion protein, elicits synergistic anti-tumor effects in preclinical models | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| US 5229109 A, 20.07.1993 | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| УГНЕТЕНИЕ ОПУХОЛЕЙ С ПОМОЩЬЮ ПЛАЦЕНТАРНОГО ПЕРФУЗАТА ЧЕЛОВЕКА И ВЫДЕЛЕННЫХ ИЗ ПЛАЦЕНТЫ ЧЕЛОВЕКА ВСПОМОГАТЕЛЬНЫХ НАТУРАЛЬНЫХ КЛЕТОК-КИЛЛЕРОВ | 2008 |
|
RU2536242C2 |

