Область техники, к которой относится изобретение
Настоящее изобретение относится к новым вариантам гиалуронидазы человека, обладающим повышенной ферментативной активностью и термической стабильностью по сравнению с гиалуронидазой человека, которая представляет собой фермент, гидролизующий гиалуроновую кислоту, и, более конкретно, к вариантам гиалуронидазы PH20 или их фрагментам, которые содержат одну или несколько замен аминокислотных остатков в области, соответствующей области альфа-спирали и/или ее линкерной области в PH20 дикого типа, имеющем аминокислотную последовательность, представленную в SEQ ID NO: 1, предпочтительно в зрелом PH20 дикого типа, состоящем из аминокислотных остатков L36-S490, и в которых один или несколько N-концевых или C-концевых аминокислотных остатков избирательно удалены, к способу их получения и содержащей их фармацевтической композиции.
Предшествующий уровень техники
Кожа человека состоит из эпидермиса, дермы и подкожного жирового слоя, и в коже имеется шесть типов гликозаминогликанов. Эти гликозаминогликаны включают гиалуроновую кислоту, хондроитинсульфат, дерматансульфат, гепарансульфат, гепарин и кератинсульфат.
Эти гликозаминогликаны состоят из повторяющихся дисахаридных сахарных единиц. Количество дисахаридных сахарных единиц различается среди гликозаминогликанов, но варьирует от нескольких сотен до тысяч. Среди гликозаминогликанов, содержание гиалуроновой кислоты в коже составляет более чем половину ее количества в организме. Гиалуроновая кислота синтезируется гиалуронан-синтазой, присутствующей в клеточной мембране, присутствует сама по себе, не связываясь с протеогликанами, и является единственным гликозаминогликаном, не имеющим сульфатной группы. Другие гликозаминогликаны связываются с протеогликанами и имеют сульфатную группу. Гиалуроновая кислота состоит из глюкуроновой кислоты и N-ацетилглюкозамина, соединенных чередующимися β-1,4 и β-1,3 связями, и состоит из примерно 5000 повторяющихся единиц этих дисахаридов. Известно, что ежедневно в организме человека обновляется около одной трети (5 г) гиалуроновой кислоты.
Гиалуронидазы представляют собой ферменты, которые разрушают гиалуроновую кислоту, присутствующую во внеклеточном матриксе. Известно, что у человека имеется шесть типов гиалуронидаз: это Hyal1, Hyal2, Hyal3, Hyal4, HyalPS1 и PH20/SPAM1. Человеческие Hyal1 и Hyal2 экспрессируются в большинстве тканей. PH20/SPAM1 (в дальнейшем в настоящем документе называемый как PH20) экспрессируется в плазматической мембране сперматозоида и акросомной мембране. Однако HyalPS1 не экспрессируется, так как он представляет собой псевдоген. В соответствии со способом отщепления гиалуроновой кислоты гиалуронидазы делятся на три типа: ферменты (EC 3.2.1.35), которые расщепляют связи β-1,4 между N-ацетилглюкозамином и глюкуроновой кислотой путем использования H2O; ферменты (EC 3.2.1.36), которые расщепляют связи β-1,3 между N-ацетилглюкозамином и глюкуроновой кислотой путем использования H2O; и бактериальные гиалуронидазы (EC 4.2.99.1), которые расщепляют связи β-1,4 без использования H2O.
Каталитическими аминокислотами Hyal1 являются D129 и E131, которые гидролизуют гиалуроновую кислоту путем катализа с использованием субстрата (substrate-assisted catalysis). Hyal1 проявляет оптимальную активность при кислотном pH 3-4 и не обладает ферментативной активностью при pH 4,5 или выше. В отличие от Hyal1, PH20 проявляет ферментативную активность в широком диапазоне pH 3-8.
Arming et al. установили, что каталитическими аминокислотами PH20 являются D111 и E113 (Arming et al., 1997). Arming et al. отметили Leu как первую аминокислоту зрелого белка, и, таким образом, каталитические аминокислоты полноразмерного PH20 с сигнальным пептидом соответствуют D146 и E148, соответственно.
Гиалуронидаза гидролизует гиалуроновую кислоту, тем самым уменьшая вязкость гиалуроновой кислоты во внеклеточном матриксе и увеличивая ее проницаемость в ткани (кожу). Подкожный участок кожи имеет нейтральное значение pH, равное примерно 7,0-7,5. Таким образом, среди различных типов гиалуронидаз PH20 широко применяют в клинической практике (Bookbinder et al., 2006). В примерах, в которых PH20 применяют в клинической практике, PH20 применяют в качестве релаксанта для глаз и анестезирующей добавки в офтальмологической хирургии, а также вводят совместно с терапевтическим агентом на основе антител, который инъецируют подкожно (Bookbinder et al., 2006). Кроме того, на основе свойства гиалуроновой кислоты, которая сверхэкспрессируется в опухолевых клетках, PH20 применяют для гидролиза гиалуроновой кислоты во внеклеточном матриксе опухолевых клеток, тем самым увеличивая доступ противоракового терапевтического агента к опухолевым клеткам. Кроме того, ее также применяют для ускорения резорбции жидкостей организма и крови, которые в избыточном количестве присутствуют в тканях.
PH20 впервые идентифицировали в сперме морских свинок Lathrop et al., и известно также, что PH20 экспрессируется в сперматозоидах разных видов. Ген PH20 человека был клонирован Lin et al. и Gmachl et al. PH20 человека имеет аминокислотную последовательность, представленную в SEQ ID NO: 1, которая состоит из 509 аминокислотных остатков и демонстрирует 60%-ную аминокислотную идентичность с геном PH20 морской свинки. Фермент PH20 человека кодируется геном SPAM1 (молекула адгезии спермы-1), и Ser490 PH20 присутствует в виде связывания с гликозил-фосфатидил-инозитолом (GPI) на поверхности плазматической мембраны сперматозоидов и в акросомной мембране. Сперматозоид гидролизует гиалуроновую кислоту с помощью PH20, когда он проникает в ооциты через богатый гиалуронаном кумулюсный слой ооцитов. PH20 присутствует в количестве, соответствующем 1% или менее количества белков в сперматозоидах, и имеет шесть сайтов N-гликозилирования (N82, N166, N235, N254, N368 и N393).
В настоящее время коммерчески доступный PH20 получают путем экстракции из семенников крупного рогатого скота или овец. Его примеры включают Amphadase® (бычья гиалуронидаза) и Vitrase® (овечья гиалуронидаза).
Бычью тестикулярную гиалуронидазу (BTH) получают путем удаления сигнального пептида и 56 аминокислот на С-конце из бычьего PH20 дикого типа в ходе посттрансляционной модификации. BTH также является гликопротеином и имеет содержание маннозы 5% и содержание глюкозамина 2,2% в расчете на общие компоненты, включая аминокислоты. Когда гиалуронидазу животного происхождения повторно вводят в организм человека в высокой дозе, может образовываться нейтрализующее антитело. Поскольку гиалуронидаза животного происхождения содержит другие биоматериалы в дополнение к PH20, она может вызывать аллергическую реакцию при введении в организм человека (Bookbinder et al., 2006). В частности, получение и применение PH20, выделенного из крупного рогатого скота, может быть ограничено из-за опасений, связанных с коровьим бешенством. Для решения этой проблемы были проведены исследования рекомбинантного белка PH20 человека.
Рекомбинантный белок PH20 человека, согласно сообщениям, экспрессируется в дрожжах (P. pastoris), клетках насекомых DS-2 и клетках животных. Рекомбинантные белки PH20, продуцируемые в клетках насекомых и дрожжах, отличаются от PH20 человека с точки зрения характера N-гликозилирования в ходе посттрансляционной модификации.
Среди гиалуронидаз определены только трехмерные структуры Hyal1 (PDB ID: 2PE4) (Chao et al., 2007) и гиалуронидазы пчелиного яда PDB ID: 1FCQ, 1FCU, 1FCV). Hyal1 состоит из двух доменов, каталитического домена и EGF-подобного домена. Каталитический домен находится в форме (β/α)8, в которой альфа-спираль и бета-цепь, которые характеризуют вторичную структуру белка, повторяются по восемь раз каждая (Chao et al., 2007). EGF-подобный домен является полностью консервативным в вариантах, в которых C-конец Hyal1 сплайсирован по-другому. Аминокислотные последовательности Hyal1 и PH20 являются идентичными на 35,1%, и белковая структура PH20 еще не обнаружена.
Рекомбинантный белок РН20 человека разработан фирмой Halozyme Therapeutic, Inc. и реализуется на рынке под торговым названием Hylenex® (Bookbinder et al., 2006; Frost, 2007).
Когда D146 и E148, которые являются каталитическими аминокислотами PH20, были мутированы в аспарагин (D146N) и глутамин (E148Q), соответственно, ферментативная активность отсутствовала (Arming et al., 1997). Кроме того, когда R246 PH20 заменяли глицином, ферментативная активность снижалась на 90%, а когда E319 заменяли глутамином и R322 заменяли треонином, ферментативная активность исчезала. Вариант, в котором удалены 36 аминокислот на С-конце PH20 (усечение аминокислот 474-509), показал снижение ферментативной активности на 75% по сравнению с PH20 дикого типа. Этот вариант не секретировался внеклеточно и оставался в клетках HeLa. Когда в PH20 удалили C-концевые 134 аминокислоты, PH20 не обладал ферментативной активностью и не секретировался внеклеточно. В соответствии с Frost et al., C-концевая область 477-483 PH20 является важной для достаточной экспрессии (Frost, 2007). Активность варианта PH20, C-конец которого был усечен по одному из положений 477-483 составила лишь 10% от полноразмерного PH20 (1-509) или варианта PH20, C-конец которого был усечен по положению 467 (Frost, 2007).
Между тем, рекомбинантный PH20 все еще обладает недостаточной термической стабильностью или уровнями экспрессии в рекомбинантных клетках. Таким образом, в промышленности существует большая потребность в рекомбинантной гиалуронидазе, имеющей дополнительно улучшенные биологические и физико-химические характеристики.
Раскрытие изобретения
Техническая проблема
Целью настоящего изобретения является обеспечение варианта гиалуронидазы PH20 или его фрагмента с улучшенной термической стабильностью, ферментативной активностью и уровнем экспрессии по сравнению с PH20 дикого типа, предпочтительно зрелым PH20 дикого типа.
Еще одной целью настоящего изобретения является обеспечение композиции для лечения рака, содержащей вышеописанный вариант гиалуронидазы PH20 или его фрагмент, и способ лечения рака с его использованием.
Техническое решение
Для достижения вышеуказанных целей настоящее изобретение обеспечивает вариант гиалуронидазы PH20 или его фрагмент, который содержит одну или несколько замен аминокислотных остатков в области, соответствующей области альфа-спирали и/или ее линкерной области в аминокислотной последовательности PH20 дикого типа, предпочтительно зрелом PH20 дикого типа, и в котором один или несколько N-концевых или C-концевых аминокислотных остатков избирательно удалены.
Настоящее изобретение также обеспечивает композицию для лечения рака, содержащую описанный выше вариант гиалуронидазы PH20 или его фрагмент, и способ лечения рака с его использованием.
Краткое описание чертежей
На фигуре 1 показана модель третичной структуры белка PH20. Третичная структура белка PH20 смоделирована с использованием в качестве матрицы Hyal1 (PDB ID: 2PE4) (Chao et.al., 2007), белковая кристаллическая структура которого была найдена на сервере Swiss-Model (https://swissmodel.expasy.org/).
На фигуре 1А показана модель третичной структуры белка PH20 и указаны D146 и E148, которые представляют собой каталитические аминокислоты. Модель третичной структуры белка PH20 состоит из восьми повторов бета-цепи и альфа-спирали.
На фигуре 1B показана петля eta (η) 8, в которой расположены альфа-спираль 8 PH20 и G340-I344, которые образуют линкерную область в N-концевой области альфа-спирали 8. Показан каждый из остатков G340, T341, L342, S343 и I344.
На фигуре 1C показаны аминокислотные остатки (C351, Y357 и N363), которые взаимодействуют с прилегающей вторичной структурой, из числа аминокислот, расположенных в альфа-спирали 8 PH20. C351 образует дисульфидную связь с C60, расположенным в альфа-спирали 1, Y357 гидрофобно взаимодействует с F315, расположенным между бета-цепью 7 и альфа-спиралью 7, и N363 образует водородную связь с остатком D69, расположенным в альфа-спирали 1.
На фигуре 2 показано сравнение уровней экспрессии белка WT (дикого типа) и вариантов, созданных в соответствии с настоящим изобретением. WT и варианты экспрессировали путем транзиентной трансфекции в клетках ExpiCHO. Уровень экспрессии WT составил 16,1 мг/л. Уровни белковой экспрессии вариантов на основе вариантов HM1 и HM6 были выше, чем у WT, и уровни белковой экспрессии HM4 и HM7 были самыми высокими. Уровень белковой экспрессии HM11, полученного путем введения дополнительных аминокислотных замен (Y365F и I367L) в вариант HM6, снизился до 6,4 мг/мл.
На фигуре 3 показаны экспериментальные результаты для вариантов HM1 и HM6.
На фигуре 3A показаны результаты SDS-PAGE после очистки WT и вариантов HM1 и HM6. Очистку проводили с использованием колонки HisTrap и колонки Q Sepharose. Молекулярная масса WT и вариантов HM1 и HM6 составила ~ 70 кДа (подпись к фигуре: M, маркер молекулярной массы; CS, супернатант; FT, проточный; и Элюирование, элюированные фракции).
На фигуре 3B показаны величины ферментативной активности WT и вариантов HM1 и HM6, измеренные с помощью турбидиметрического анализа при pH 7,0. В настоящем изобретении величина ферментативной активности, измеренная с помощью турбидиметрического анализа, выражена в виде специфической активности.
На фигуре 3C показаны ферментативная активность WT и вариантов HM1 и HM6, измеренная с помощью субстрат-гель-анализа. После удаления SDS с помощью 2,5% Triton X-100 (масса/объем) при 4°С ферментативную реакцию проводили при 37°С в течение 1-4 ч. Вариант HM6 ренатурировал быстрее, чем WT и вариант HM1, и, таким образом, быстрее гидролизовал гиалуроновую кислоту на полиакриламидном геле. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 3D показана ферментативная активность WT и варианта HM1, измеренные с помощью субстрат-гель-анализа при pH 5-8. WT и вариант HM1 продемонстрировали активность в диапазоне pH 5-8, и показали самую высокую ферментативную активность при pH 5,0. Вариант HM1 содержит сигнальный пептид человеческого сывороточного альбумина или Hyal1 человека. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 3E показаны результаты разделения WT и вариантов HM1 и HM6 на фенильной колонке. Варианты элюировались с фенильной колонки быстрее, чем WT.
На фигуре 4 показаны результаты анализа конечного продукта гиалуроновой кислоты, расщепленной под действием WT и варианта HM6, через 10 мин и 1 ч с помощью колонки Amide-80.
На фигуре 5 показаны экспериментальные результаты для мутаций аминокислот G340-I344 PH20.
На фигуре 5A показаны результаты SDS-PAGE после очистки на колонке HisTrap для вариантов HM7, HM8, HM9, HM10 и HM21.
На фигуре 5B показаны результаты измерения ферментативной активности WT и вариантов HM6, HM8, HM9, HM10, HM21 и HM7 с помощью турбидиметрического анализа при pH 7,0.
На фигуре 5C показаны результаты измерения ферментативной активности WT и вариантов HM6, HM8, HM9, HM10, HM21 и HM7 с помощью субстрат-гель-анализа. Столбчатая диаграмма в нижней части фигуры 5С показывает степень ферментативной активности, полученную путем количественной оценки полосы после окрашивания геля альциановым синим. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 5D показаны результаты анализа WT и вариантов HM8, HM9, HM10, HM21 и HM7 с помощью хроматографии на фенильной колонке.
На фигуре 5E показаны результаты разделения WT и вариантов HM6, HM8, HM9, HM10, HM21 и HM7 в зависимости от их изоэлектрических точек при pH 3-7 методом IEF в геле.
На фигуре 6 показаны экспериментальные результаты для варианта HM11.
На фигуре 6A показаны результаты очистки белка хроматографией на колонке HisTrap для варианта HM11.
На фигуре 6B показаны результаты измерения ферментативной активности WT и варианта HM11 при pH 7,0 с помощью турбидиметрического анализа.
На фигуре 7 показаны экспериментальные результаты для вариантов с усеченным N-концом HM40, HM13, HM41, HM24, HM42 и HM25 PH20.
На фигуре 7A показаны результаты очистки белка хроматографией на колонке HisTrap для вариантов HM40, HM13, HM41, HM24, HM42 и HM25 PH20.
На фигуре 7B показаны уровни экспрессии вариантов HM40, HM13, HM41, HM24, HM42, HM25, HP61 и HP62 PH20 в клетках ExpiCHO.
На фигуре 7С показана ферментативная активность вариантов HM40, HM13, HM41, HM24, HM42, HM25, HP61 и HP62 PH20, измеренная при pH 7,0 с помощью турбидиметрического анализа и выраженная в виде специфической активности.
На фигуре 7D показана ферментативная активность вариантов PH20 HM40, HM13, HM41, HM24 и HM42, измеренная с помощью субстрат-гель-анализа. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 7E показаны результаты анализа WT и вариантов HM40, HM13, HM41, HM24 и HM42 с помощью хроматографии на фенильной колонке.
На фигуре 7F показано изменение размера частиц при увеличении температуры для вариантов HM40, HM13, HM41 и HM42 PH20.
На фигуре 8 показаны экспериментальные результаты для вариантов с усеченным С-концом НМ14, НМ15 и НМ16, сконструированных с использованием НМ6 в качестве матрицы.
На фигуре 8A показаны результаты SDS-PAGE после очистки HisTrap для вариантов HM14, HM15 и HM16. В качестве контроля были включены WT и вариант HM6.
На фигуре 8B показаны результаты измерения ферментативной активности WT и вариантов HM6, HM14, HM15 и HM16 при pH 7,0 с помощью турбидиметрического анализа.
На фигуре 8C показаны результаты измерения ферментативной активности WT и вариантов HM6, HM14, HM15 и HM16 для 1, 2 и 4 ч с помощью субстрат-гель-анализа. Правый график на фигуре 8C представляет собой столбчатую диаграмму, показывающую ферментативную активность, измеренную после окрашивания альциановым синим после 1 ч ферментативной реакции. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 8D показаны результаты анализа WT и вариантов HM6, HM14, HM15 и HM16 с помощью хроматографии на фенильной колонке.
На фигуре 8E показаны результаты разделения вариантов HM14, HM15 и HM16 в зависимости от их изоэлектрических точек при pH 3-7 методом IEF в геле.
На фигуре 9 показаны экспериментальные результаты для вариантов HM19 и HM20 PH20, сконструированных с использованием HM10 в качестве матрицы.
На фигуре 9A показаны результаты очищенного белка с помощью хроматографии на колонке HisTrap для вариантов HM19 и HM20 PH20.
На фигуре 9В показаны результаты сравнения ферментативной активности варианта HM19 и HM20 PH20 при pH 7,0 с помощью турбидиметрического анализа.
На фигуре 9C показаны результаты окрашивания геля SDS красителем альциановым синим после ферментативной реакции в течение 1 ч при 37°С с помощью субстрат-гель-анализа для WT и вариантов HM10, HM19 и HM20. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 10 показаны результаты измерения температур агрегации WT и вариантов PH20 с помощью системы динамического рассеяния света (далее в настоящем документе именуемой DLS). Измерения выполняли в трех повторах и выражали в виде среднего значения ± стандартная ошибка(S.E.).
На фигуре 11 показан график Штерна-Фольмера, полученный после измерения изменения флуоресценции остатков триптофана WT и вариантов PH20 при добавлении акриламида (0-0,5 М). Среди аминокислот триптофан возбуждается при 295 нм и излучает максимальную флуоресценцию при 340 нм. Акриламид представляет собой малую молекулу, которая может проникать в структуру белка и гасить флуоресценцию триптофана. Поскольку структура белка более гибкая, тушение флуоресценции акриламидом больше. F0 представляет собой величину флуоресценции в отсутствие акриламида, и F представляет собой величину флуоресценции в присутствии акриламида (0-0,5 М). Изменение измеренной величины флуоресценции выражено в виде отношения F0/F.
На фигуре 11A показан график Штерна-Фольмера для WT и вариантов HM1, HM4, HM6 и HM7.
На фигуре 11B показаны графики Штерна-Фольмера для WT и вариантов HM14, HM15 и HM16.
На фигуре 12 показаны уровни экспрессии вариантов PH20 на основе HM10 в клетках ExpiCHO.
На фигуре 12A графически показаны уровни экспрессии соответствующих вариантов.
На фигуре 12B показаны уровни экспрессии соответствующих вариантов в таблице. WT и варианты PH20 имели 6xHis-метку на С-конце, и уровни экспрессии белка после очистки на колонке HisTrap выражали в мг/л. Варианты HM30-HM33 не экспрессировались в клетках ExpiCHO.
На фигуре 13 показаны результаты Вестерн-блоттинга для клеточных культур вариантов HM29, HM30, HM31, HM32 и HM33. С-конец вариантов HM29, HM30, HM31, HM32 и HM33 на основе HM10 отщепляли после A467, C464, D461, C358 или C455, соответственно. HM29 с отщепленным С-концом экспрессировался в клетках ExpiCHO, но варианты, имеющие C-конец, отщепленный по C464 или короче по длине, не экспрессировались в клетках ExpiCHO. Первичное антитело представляло собой кроличье поликлональное антитело против PH20 (Abcam) в разведении 1:500. Вторичное антитело представляло собой козье анти-кроличье IgG HRP в разведении 1:2000.
На фигуре 14 показаны экспериментальные результаты для усеченных с С-конца вариантов, сконструированных с использованием HM10 в качестве матрицы.
На фигуре 14A показаны результаты измерения ферментативной активности при pH 7,0 с помощью турбидиметрического анализа для вариантов с отщепленным С-концом, сконструированных с использованием HM10 в качестве матрицы.
На фигуре 14B показано сравнение ферментативной активности в зависимости от сайтов C-концевого отщепления 17 вариантов PH20 (HM43, HM44, HM45, HM20, HM19, HM35, HM36, HM37, HM38, HM39, HM47, HM48, HM49, HM50, HM51, HM52 и HM10), сконструированных с использованием HM10 в качестве матрицы.
На фигуре 14C показаны результаты окрашивания в геле сдодецилсульфатом натрий (SDS) красителем альциановым синим после ферментативной реакции в течение 1 ч при 37°С с помощью субстрат-гель-анализа для некоторых (HM29, HM35, HM36, HM37, HM38, HM39, HM43, HM44 и HM45) вариантов PH20, сконструированных с использованием HM10 в качестве матрицы. Белая полоса показывает гиалуроновую кислоту, расщепленную под действием WT и вариантного белка.
На фигуре 15 показан гель SDS после прохождения через последнюю колонку в ходе очистки белка для HP34 (фигура 15A) и HP46 (фигура 15B), экспрессированных в клетках ExpiCHO. HP34 подвергали четырехступенчатой процедуре очистки хроматографией, состоящей из колонок Q Sepharose, Butyl HP, Heparin и Blue Sepharose, и гель SDS является результатом, полученным после хроматографии на колонке Blue Sepharose. HP46 подвергали трехступенчатой процедуре очистки хроматографией, состоящей из колонок Q Sepharose, Butyl HP и Heparin, и гель SDS является результатом, полученным после хроматографии на колонке Heparin.
На фигуре 16 показана ферментативная активность вариантов HP34 и HP46 PH20 без 6xHis-меток, сконструированных с использованием HM21 в качестве матрицы.
На фигуре 16A показаны результаты измерения ферментативной активности WT и вариантов HM21, HP34 и HP46 при pH 7,0 с помощью турбидиметрического анализа.
На фигуре 16B показаны результаты измерения ферментативной активности WT (HW2) и вариантов HM21, HP34 и HP46 при pH 5,3 с помощью анализа Моргана-Элсона (Km: константа Михаэлиса-Ментена, kcat: число оборотов фермента и kcat/Km: каталитическая эффективность).
На фигуре 17 показаны результаты снятия характеристик вариантов PH20 на основе HM21.
На фигуре 17A показаны результаты измерения температур агрегации с помощью DLS для вариантов HP34 и HP46 PH20, не содержащих 6xHis-метки, сконструированных с использованием HM21 в качестве матрицы. В качестве контролей показаны температуры агрегации HW2 и HM21.
На фигуре 17B показаны результаты измерения ферментативной активности в течение 1 ч с помощью субстрат-гель-анализа для HW2 и вариантов PH20 (HP20, HP34 и HP46).
На фигуре 17C показаны результаты выполнения субстрат-гель-анализа после выдерживания образцов вариантов (HW2 и HP46) при pH 3,0 и pH 7,0 в течение 14 ч. После SDS-PAGE SDS удаляли с помощью 2,5% Triton X-100 (масса/объем) и проводили ферментативную реакцию при 37°С в течение 1 ч.
На фигуре 17D показаны уровни экспрессии вариантов HM21, HM53, HM54, HM55, HM56, HP59 и HP60 PH20 в клетках ExpiCHO.
На фигуре 17E показано выражение величины ферментативной активности вариантов HM21, HM53, HM54, HM55, HM56, HP59 и HP60 PH20, измеренной с помощью турбидиметрического анализа, в виде специфической активности при pH 5,3.
На фигуре 18 показаны результаты измерения индекса стимуляции CD4+ Т-клеток при лечении с помощью PH20 и вариантов PH20 в концентрациях 1,5 нг/мл и 15 нг/мл, соответственно.
На фигуре 19 показаны результаты измерения индекса стимуляции CD8+ Т-клеток при лечении с помощью PH20 и вариантов PH20 в концентрациях 1,5 нг/мл и 15 нг/мл, соответственно.
Наилучший режим осуществления изобретения
Если не определено иначе, все технические и научные термины, используемые в настоящем документе, имеют то же значение, что и обычно понимаемое специалистом в области, к которой относится изобретение. Как правило, используемая в настоящем документе номенклатура и экспериментальные способы, которые будут описаны ниже, хорошо известны и традиционно применяются в данной области техники.
Настоящее изобретение обеспечивает вариант гиалуронидазы PH20 или его фрагмент, который содержит одну или несколько замен аминокислотных остатков в области, соответствующей области альфа-спирали и/или ее линкерной области, предпочтительно области альфа-спирали 8 (S347-C381) и/или линкерной области (A333- R346) между альфа-спиралью 7 и альфа-спиралью 8 в аминокислотной последовательности PH20 дикого типа, предпочтительно зрелого PH20 дикого типа, и в котором один или несколько N-концевых или C-концевых аминокислотных остатков избирательно отщепляются и удаляются.
В настоящем изобретении положения аминокислотных остатков в каждом варианте соответствуют аминокислотным положениям PH20 дикого типа, имеющего последовательность, представленную в SEQ ID NO: 1.
Кроме того, в настоящем изобретении «зрелый PH20 дикого типа» означает белок, состоящий из аминокислотных остатков L36-S490 последовательности SEQ ID NO: 1, в котором отсутствуют M1-T35, образующие сигнальный пептид, и A491-L509, которые не связаны с важной ферментативной функцией PH20, в аминокислотной последовательности PH20 дикого типа, имеющего последовательность, представленную в SEQ ID NO: 1.
Таблица 1. Аминокислотная последовательность PH20 дикого типа (SEQ ID NO: 1)
MGVLKFKHIFFRSFVKSSGVSQIVFTFLLIPCCLTLNFRAPPVIPNVPFLWAWNAPSEFCLGKFDEPLDMSLFSFIGSPRINATGQGVTIFYVDRLGYYPYIDSITGVTVNGGIPQKISLQDHLDKAKKDITFYMPVDNLGMAVIDWEEWRPTWARNWKPKDVYKNRSIELVQQQNVQLSLTEATEKAKQEFEKAGKDFLVETIKLGKLLRPNHLWGYYLFPDCYNHHYKKPGYNGSCFNVEIKRNDDLSWLWNESTALYPSIYLNTQQSPVAATLYVRNRVREAIRVSKIPDAKSPLPVFAYTRIVFTDQVLKFLSQDELVYTFGETVALGASGIVIWGTLSIMRSMKSCLLLDNYMETILNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLSATMFIVSILFLIISSVASL
В частности, вариант PH20 или его фрагмент в соответствии с настоящим изобретением может содержать одну или несколько мутаций, предпочтительно замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341G, S343E, M345T, K349E, L353A, L354I, N356E и I361T, более предпочтительно выбранных из группы, состоящей из T341A, T341C, L354I и N356E в PH20 дикого типа, имеющем аминокислотную последовательность, представленную в SEQ ID NO: 1.
В настоящем изобретении термин «вариант PH20» включает мутацию одного или нескольких аминокислотных остатков, предпочтительно замену одного или нескольких аминокислотных остатков в аминокислотной последовательности PH20 дикого типа, а также наличие делеции одного или нескольких аминокислотных остатков на N-конце или C-конце вместе с заменой аминокислотных остатков, и используется в основном в том же значении, что и выражение «вариант PH20 или его фрагмент».
В настоящем изобретении третичную структуру белка PH20, расположенную за пределами активного сайта, изучали путем моделирования структуры белка PH20 человека на основе Hyal1 (SEQ ID NO: 2), который представляет собой гиалуронидазу человека, третичная структура белка которого является известной. В результате аминокислоты, расположенные в области альфа-спирали 8 PH20 отбирали и заменяли аминокислотной последовательностью альфа-спирали 8 Hyal1, тем самым пытаясь повысить термическую стабильность белковой структуры без воздействия на каталитическую активность фермента. В частности, поскольку альфа-спираль 8 расположена во внешней части третичной структуры белка PH20, взаимодействие с соседней альфа-спиралью или бета-цепью меньше, чем с другими альфа-спиралями PH20. В соответствии с настоящим изобретением было обнаружено, что когда аминокислотная последовательность области альфа-спирали 8 человеческого PH20 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8 частично заменена аминокислотной последовательностью области альфа-спирали 8 высокогидрофильного Hyal1 и линкерной областью между альфа-спиралью 7 и альфа-спиралью 8 Hyal1, ферментативная активность при нейтральном pH и температура агрегации белка (Tagg) увеличивается. На основании этих экспериментальных результатов было обнаружено, что может быть обеспечен новый вариант PH20 или его фрагмент, который обладает повышенной ферментативной активностью и термической стабильностью по сравнению с PH20 дикого типа.
Таким образом, вариант PH20 в соответствии с настоящим изобретением содержит одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341G, S343E, M345T, K349E, L353A, L354I, N356E и I361T, предпочтительно выбранных из группы, состоящей из T341A, T341C, L354I и N356E в аминокислотной последовательности PH20 дикого типа (имеющего аминокислотную последовательность, представленную в SEQ ID NO: 1), предпочтительно зрелого PH20 дикого типа (имеющего последовательность, состоящую из аминокислотных остатков L36-S490 в аминокислотной последовательности, представленной в SEQ ID NO: 1).
Вариант PH20 в соответствии с настоящим изобретением также содержит одну или несколько замен аминокислотных остатков в области, соответствующей области альфа-спирали и/или ее линкерной области, предпочтительно области альфа-спирали 8 (S347-C381) и/или линкерной области (A333-R346) между альфа-спиралью 7 и альфа-спиралью 8, более предпочтительно T341-N363, T341-I361, L342-I361, S343-I361, I344-I361, M345-I361 или M345-N363 в PH20 дикого типа, имеющем аминокислотную последовательность, представленную в SEQ ID NO: 1.
В частности, в варианте PH20 в соответствии с настоящим изобретением область альфа-спирали 8 (S347-C381) и/или линкерная область (A333-R346) между альфа-спиралью 7 и альфа-спиралью 8 PH20 дикого типа, предпочтительно зрелого PH20 дикого типа, может быть заменена одним или несколькими аминокислотными остатками соответствующей области (см. таблицы 2 и 3) Hyal1, имеющего последовательность, представленную в SEQ ID NO: 2, но не ограничивается ими.
Таблица 2. Аминокислотная последовательность Hyal1 дикого типа (SEQ ID NO: 2)
MAAHLLPICALFLTLLDMAQGFRGPLLPNRPFTTVWNANTQWCLERHGVDVDVSVFDVVANPGQTFRGPDMTIFYSSQLGTYPYYTPTGEPVFGGLPQNASLIAHLARTFQDILAAIPAPDFSGLAVIDWEAWRPRWAFNWDTKDIYRQRSRALVQAQHPDWPAPQVEAVAQDQFQGAARAWMAGTLQLGRALRPRGLWGFYGFPDCYNYDFLSPNYTGQCPSGIRAQNDQLGWLWGQSRALYPSIYMPAVLEGTGKSQMYVQHRVAEAFRVAVAAGDPNLPVLPYVQIFYDTTNHFLPLDELEHSLGESAAQGAAGVVLWVSWENTRTKESCQAIKEYMDTTLGPFILNVTSGALLCSQALCSGHGRCVRRTSHPKALLLLNPASFSIQLTPGGGPLSLRGALSLEDQAQMAVEFKCRCYPGWQAPWCERKSMW
Более конкретно, новый вариант PH20 или его фрагмент в соответствии с настоящим изобретением предпочтительно содержит замену аминокислотного остатка L354I и/или N356E в аминокислотной последовательности PH20 дикого типа, предпочтительно зрелого PH20 дикого типа,
и дополнительно содержит замену по меньшей мере аминокислотного остатка в одном или нескольких положениях, выбранных из T341-N363, в частности, в одном или нескольких положениях, выбранных из группы, состоящей из T341, L342, S343, I344, M345, S347, M348, K349, L352, L353, D355, E359, I361 и N363, но ими не ограничивается.
Более предпочтительно замена аминокислотного остатка в одном или нескольких положениях, выбранных из группы, состоящей из T341, L342, S343, I344, M345, S347, M348, K349, L352, L353, D355, E359, I361 и N363, может представлять собой одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341D, T341G, T341S, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, D355K, E359D, I361T и N363G, но ими не ограничивается.
Предпочтительно новый вариант PH20 или его фрагмент в соответствии с настоящим изобретением может содержать одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T,
и может дополнительно содержать одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341D, T341G, T341S, L342W, S343E, I344N и N363G, но ими не ограничивается.
Более предпочтительно новый вариант PH20 или его фрагмент в соответствии с настоящим изобретением может содержать любую замену аминокислотного остатка, выбранную из следующих групп замен аминокислотных остатков, но ими не ограничивается:
(a) T341S, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(b) L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(c) M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D, I361T и N363G;
(d) T341G, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(e) T341A, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(f) T341C, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(g) T341D, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(h) I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T; и
(i) S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T.
В настоящем изобретении выражение, описываемое однобуквенным кодом аминокислотного остатка вместе с числами, например, «S347», означает аминокислотный остаток в каждом положении в аминокислотной последовательности, представленной в SEQ ID NO: 1.
Например, «S347» означает, что аминокислотный остаток в положении 347 в аминокислотной последовательности SEQ ID NO: 1 представляет собой серин.
Кроме того, «S347T» означает, что серин в положении 347 SEQ ID NO: 1 заменен треонином.
Вариант PH20 в соответствии с настоящим изобретением интерпретируется как включающий варианты или их фрагменты, в которых аминокислотный остаток в определенном положении аминокислотного остатка является консервативно замененным.
Используемый в настоящем документе термин «консервативная замена» относится к модификациям варианта PH20, которые включают замену одной или нескольких аминокислот на аминокислоты, обладающие аналогичными биохимическими свойствами, которые не приводят к утрате биологической или биохимической функции варианта PH20.
«Консервативная аминокислотная замена» представляет собой замену, при которой аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, определены и хорошо известны в области, к которой относится настоящее изобретение. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин и гистидин), аминокислоты с кислотными боковыми цепями (например, аспарагиновая кислота и глутаминовая кислота), аминокислоты с незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин и цистеин), аминокислоты с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин и триптофан), аминокислоты с бета-разветвленными боковыми цепями (например, треонин, валин и изолейцин) и аминокислоты с ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан и гистидин).
Предполагается, что вариант PH20 или его фрагменты по настоящему изобретению могут все еще сохранять активность, хотя он имеет консервативные аминокислотные замены.
Кроме того, вариант PH20 или его фрагмент в соответствии с настоящим изобретением интерпретируется как включающий варианты PH20 или их фрагменты, обладающие по существу такой же функцией и/или эффектом, что и вариант PH20 или его фрагмент в соответствии с настоящим изобретением, и обладающий гомологией аминокислотной последовательности по меньшей мере 80% или 85%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, наиболее предпочтительно по меньшей мере 99% с вариантом PH20 или его фрагментом в соответствии с настоящим изобретением.
Варианты PH20 в соответствии с настоящим изобретением имеют повышенные уровни экспрессии и скорость рефолдинга белка, тем самым увеличивая высокую термическую стабильность по сравнению со зрелым PH20 дикого типа. Кроме того, ферментативная активность вариантов PH20 была больше или аналогична ферментативной активности зрелого PH20 дикого типа, несмотря на увеличение термической стабильности.
Между тем, даже если варианты зрелого PH20 дикого типа с С-концевой делецией показали уменьшение ферментативной активности, на основании настоящего изобретения, варианты PH20 с С-концевой делецией показали сходную или увеличенную ферментативную активность благодаря более быстрому рефолдингу белка и термической стабильности. Кроме того, варианты PH20 в данном изобретении сохраняли ферментативную активность, когда N-концевые аминокислоты удаляли вплоть до пяти аминокислотных остатков. Это указывает на то, что для экспрессии белка и ферментативной активности важным являлся P41 на N-конце.
Таким образом, вариант PH20 или его фрагмент в соответствии с настоящим изобретением отличается тем, что он содержит одну или несколько замен аминокислотных остатков в области альфа-спирали 8 (S347-C381) и/или в линкерной области (A333-R346) между альфа-спиралью 7 и альфа-спиралью 8 в аминокислотной последовательности PH20 дикого типа, и один или несколько N-концевых и/или C-концевых аминокислотных остатков дополнительно удалены, но ими не ограничиваются.
В одном аспекте вариант PH20 или его фрагмент в соответствии с настоящим изобретением может представлять собой вариант, в котором отщепление происходит перед аминокислотным остатком, выбранным из группы, состоящей из M1-P42 аминокислотной последовательности, представленной в SEQ ID NO: 1, предпочтительно перед аминокислотным остатком L36, N37, F38, R39, A40, P41 или P42 на N-конце, в результате чего один или несколько аминокислотных остатков на N-конце удаляются, и/или отщепление происходит после аминокислотного остатка, выбранного из группы, состоящей из V455-L509, предпочтительно после аминокислотного остатка, выбранного из группы, состоящей из V455-S490, наиболее предпочтительно после аминокислотного остатка V455, C458, D461, C464, I465, D466, A467, F468, K470, P471, P472, M473, E474, T475, E476, P478, I480, Y482, A484, P486, T488 или S490 на C-конце, в результате чего один или несколько аминокислотных остатков на C-конце удаляются.
Выражение «отщепление происходит перед аминокислотным остатком, выбранным из группы, состоящей из M1-P42 на N-конце», означает, что аминокислотный остаток непосредственно перед аминокислотным остатком, выбранным из M1-P42 на N-конце, отщепляется и удаляется.
Например, выражение «отщепление происходит перед аминокислотным остатком L36, N37, F38, R39, A40, P41 или P42», соответственно, означает, что все аминокислотные остатки от M1 до T35 непосредственно перед L36, все аминокислотные остатки от M1 до L36 непосредственно перед N37, все аминокислотные остатки от M1 до N37 непосредственно перед F38, все аминокислотные остатки от M1 до F38 непосредственно перед R39, все аминокислотные остатки от M1 до R39 непосредственно перед A40, все аминокислотные остатки от M1 до A40 непосредственно перед P41 или все аминокислотные остатки от M1 до P41 непосредственно перед P42 в аминокислотной последовательности, представленной в SEQ ID NO: 1, соответственно, отщепляются и удаляются.
Кроме того, выражение «отщепление происходит после аминокислотного остатка, выбранного из группы, состоящей из V455-L509 на С-конце», означает, что аминокислотный остаток непосредственно после аминокислотного остатка, выбранного из V455-L509 на С-конце, отщепляется и удаляется.
Например, выражение «отщепление происходит после аминокислотного остатка V455, C458, D461, C464, I465, D466, A467, F468, K470, P471, P472, M473, E474, T475, E476, P478, I480, Y482, A484, P486, T488 или S490 на С-конце» означает, что аминокислотный остаток после аминокислотного остатка V455, C458, D461, C464, I465, D466, A467, F468, K470, P471, P472, M473, E474, T475, E476, P478, I480, Y482, A484, P486, T488 или S490 в аминокислотной последовательности, представленной в SEQ ID NO: 1, отщепляется и удаляется.
Предпочтительно новый вариант PH20 или его фрагмент в соответствии с настоящим изобретением может быть выбран из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 60-115, но не ограничивается ими.
Наиболее предпочтительно новый вариант PH20 или его фрагмент в соответствии с настоящим изобретением может иметь аминокислотную последовательность, представленную в SEQ ID NO: 99. Новый вариант PH20 или его фрагмент, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 99, может содержат 15 аминокислотных замен T341S, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T, а также делецию перед F38 на N-конце и усечение после F468 на С-конце.
Последовательности замененных или отщепленных аминокислот в вариантах PH20, сконструированных в конкретном варианте осуществления в соответствии с настоящим изобретением, представлены в таблице 11.
Ранее не сообщалось об исследовании, сфокусированном на увеличении ферментативной активности и термической стабильности PH20 за счет замены аминокислот альфа-спирали и ее линкерной области, которые являются вторичными структурами, формирующими третичную структуру белка, как описано в настоящем изобретении. Предыдущие исследования показали, что ферментативная активность PH20 дикого типа изменяется в зависимости от положений отщепления аминокислотных остатков, расположенных в C-концевой области. Однако в настоящем изобретении определенная альфа-спираль, образующая вторичную структуру PH20, была заменена альфа-спиралью другой гиалуронидазы человека, тем самым конструируя варианты PH20, имеющие более высокую стабильность, чем PH20 дикого типа. Эти варианты могут представлять варианты, в которых взаимодействие замененного домена альфа-спирали с частями, образующими другие вторичные структуры PH20, показывает картину, отличную от PH20 дикого типа, что указывает на то, что варианты обладают устойчивой ферментативной активностью независимо от положения С-концевого отщепления.
В определенном варианте осуществления новый вариант PH20 или его фрагмент в соответствии с настоящим изобретением, который обладает увеличенной ферментативной активностью и термической стабильностью по сравнению со зрелым PH20 дикого типа, может представлять собой вариант, который содержит одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341G, S343E, M345T, K349E, L353A, L354I, N356E и I361T, и в котором одна или несколько аминокислот, расположенных в области альфа-спирали 8 (S347-C381) и/или линкерной области (A333-R346) между альфа-спиралью 7 и альфа-спиралью 8 в аминокислотной последовательности PH20 дикого типа, предпочтительно зрелого PH20 дикого типа, заменены другими аминокислотами.
В частности, аминокислотная замена в линкерной области между альфа-спиралью 7 и альфа-спиралью 8 включает замену одного или нескольких аминокислотных остатков в области, состоящей из аминокислотных остатков T341-N363, T341-I361, L342-I361, L342-I361, S343-I361, I344-I361, M345-I361 или M345-N363.
Чтобы изучить эффект С-концевого усечения в вариантах PH20, в которых заменены альфа-спираль 8 и линкерная область между альфа-спиралью 7 и альфа-спиралью 8, в качестве матриц было выбрано три варианта PH20 (HM6, HM10 и HM21).
HM6 представляет собой вариант, в котором аминокислоты в области M345-N363 заменены аминокислотной последовательностью Hyal1 (M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T в SEQ ID NO: 1 заменены). Кроме того, HM6 представляет собой вариант, в котором замена альфа-спирали 8 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8 представляет собой наименее замененный вариант среди вариантов PH20 в соответствии с настоящим изобретением, который не содержит дополнительного С-концевого усечения (то есть форма, в которой С-концевый аминокислотный остаток представляет собой S490, подобно зрелому PH20 дикого типа).
HM10 представляет собой вариант, в котором аминокислоты в области L342-I361 заменены аминокислотной последовательностью Hyal1 (L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T в SEQ ID NO: 1 заменены), и который обладает самой высокой термической стабильностью, при этом обладая ферментативной активностью, сходной с активностью зрелого PH20 дикого типа среди вариантов PH20 в соответствии с настоящим изобретением, которые не содержат дополнительного C-концевого усечения.
HM21 представляет собой вариант, в котором аминокислоты в области T341-I361 заменены аминокислотной последовательностью Hyal1 (T341S, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T в SEQ ID NO: 1 заменены), и который обладает ферментативной активностью, которая примерно в два раза превышает ферментативную активность PH20 дикого типа при pH 7,0 из числа вариантов PH20 в соответствии с настоящим изобретением, которые не содержат дополнительного С-концевого усечения.
Варианты PH20 на основе HM6, сконструированные в настоящем изобретении, имеют N-конец, начинающийся у L36, и С-конец, заканчивающийся у I465, F468 или P471, как показано в таблице 4 ниже.
У вариантов PH20 на основе HM10 обычно N-конец, отщеплен перед остатком F38, и C-конец отщеплен после остатка V455, C458, D461, C464, I465, D466, A467, F468, K470, P472, M473, E474, T475, E476, P478, I480, Y482, A484, P486 или T488, как показано в таблице 5 ниже.
Как показано в примерах вариантов, которые содержат аминокислотные замены в области L342-I361, соответствующей области альфа-спирали 8 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8 HM10 в качестве матрицы, и в которых N-конец был отщеплен перед остатком F38, и С-конец был отщеплен по I465, D466, A467, F468, K470, P472, M473, E474, T475, E476, P478, I480, Y482, A484, P486 или T488, варианты PH20 в соответствии с настоящим изобретением продемонстрировали ферментативную активность, сходную с активностью зрелого PH20 дикого типа, независимо от положения C-концевого отщепления.
Как показано в таблице 6 ниже, два варианта PH20 на основе HM21 обычно характеризуются отщеплением перед остатком F38 с N-конеца, и отщеплением после остатка F468 или K470 на C-конце.
Варианты конструировали с использованием в качестве матрицы HM21, обладающий ферментативной активностью, которая примерно в два раза превышает ферментативную активность зрелого PH20 дикого типа. Эти варианты представляют собой такие варианты, которые содержат аминокислотные замены в области T341-I361, соответствующей области альфа-спирали 8 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8, и в которых N-конец был отщеплен перед остатком F38 и С-конец был отщеплен после F468 или K470. Удивительным образом было обнаружено, что эти варианты сохраняли высокую ферментативную активность HM21 независимо от положения C-концевого отщепления.
В исследовании, проведенном Frost et al., когда длина PH20 является более короткой из-за отщепления перед положением аминокислоты 477, ферментативная активность уменьшалась примерно на 10% у варианта, имеющего отщепление с С-конца после положения 477. Однако в настоящем изобретении, когда одна или несколько аминокислот в альфа-спирали 8 PH20 и его линкерной области были заменены, ферментативная активность сохранялась за счет повышения стабильности белка независимо от положения C-концевого отщепления. Этот результат очень важен, поскольку он решает проблему, заключающуюся в том, что ферментативная активность PH20 дикого типа снижается из-заусечения С-конца PH20 дикого типа.
Кроме того, в настоящем изобретении изучали эффект N-концевых аминокислот PH20, который ранее не был известен.
Для изучения эффектов сайтов N-концевого отщепления в вариантах HM6, в которых один или несколько аминокислотных остатков в области (M345-I361), соответствующей области альфа-спирали 8 PH20 дикого типа и линкерной области между альфа-спиралью 7 и альфа-спиралью 8 (замененные M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D или I361T в SEQ ID NO: 1 и не содержат дополнительного C-концевого отщепления), заменены аминокислотными остатками соответствующей области альфа-спирали 8 Hyal1 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8, варианты, в которых аминокислотные остатки L36-V47 в SEQ ID NO: 1 заменены на FRGPLLPNR, или аминокислотные остатки L36-A52 в SEQ ID NO: 1 заменены на FRGPLLPNRPFTTV, конструировали с использованием HM6 в качестве матрицы. Кроме того, с использованием HM6 в качестве матрицы конструировали варианты HM40, HM13, HM41, HM24, HM42 и HM25, в которых N-конец в аминокислотной последовательности SEQ ID NO: 1 отщеплен перед остатком N37, F38, R39, A40, P41 или P42 (см. таблицу 7).
В результате было показано, что при отщеплении N-конца HM6 перед остатком N37, F38, R39, A40 или P41, на ферментативную активность не оказывалось существенного влияния; однако, в случае отщепления N-конца перед остатком P42, ферментативная активность значительно снижалась, указывая на то, что N-концевая область PH20, расположенная после P41, является важной для экспрессии белка и ферментативной активности. Кроме того, когда одна или несколько аминокислот в N-концевой области L36-V47 или L36-A52 HM6 заменяли аминокислотами Hyal1, вариантный белок не экспрессировался в клетках ExpiCHO, указывая на то, что N-концевая область является важной для экспрессии белка.
Кроме того, в настоящем изобретении была предпринята попытка увеличить экспрессию рекомбинантного белка PH20 в клетках животных путем использования сигнального пептида других белков вместо использования исходного сигнального пептида PH20.
Таким образом, в другом аспекте новый вариант PH20 в соответствии с настоящим изобретением может представлять собой вариант, в котором N-конец дополнительно содержит сигнальный пептид гормона роста человека, имеющий аминокислотную последовательность MATGSRTSLLLAFGLLCLPWLQEGSA SEQ ID NO: 3, сигнальный пептид сывороточного альбумина человека, имеющий аминокислотную последовательность MKWVTFISLLFLFSSAYS SEQ ID NO: 4, или сигнальный пептид Hyal1 человека, имеющий аминокислотную последовательность MAAHLLPICALFLTLLDMAQG SEQ ID NO: 5, как показано в таблице 8 ниже, вместо сигнального пептида PH20 дикого тип, который состоит из M1-T35, но не ограничивается ими.
Выражение «вместо сигнального пептида PH20 дикого типа, который состоит из M1-T35» означает случай, когда сигнальный пептид PH20 дикого типа частично или полностью удален; таким образом, он не выполняет свою функцию. Кроме того, предполагается, что выражение включает случай, когда часть N-конца дополнительно удалена, например, случай, когда отщепление происходит перед остатком N37, F38, R39, A40, P41 или P42, в результате чего происходит дополнительная делеция N-конца вместе с делецией сигнального пептида PH20 дикого типа.
В другом аспекте настоящее изобретение обеспечивает композицию для лечения рака, содержащую новый вариант PH20 в соответствии с настоящим изобретением и способ лечения рака с его использованием.
Раковые заболевания или карциномы, которые можно лечить с помощью нового варианта PH20 в соответствии с настоящим изобретением, конкретно не ограничиваются, но включают как солидные злокачественные опухоли, так и рак крови. Рак может быть выбран из группы, состоящей из рака печени, гепатоцеллюлярной карциномы, рака желудка, рака молочной железы, рака легкого, рака яичника, рака бронхов, рака носоглотки, рака гортани, рака поджелудочной железы, рака мочевого пузыря, рака прямой кишки, рака толстой кишки, рака шейки матки, рак головного мозга, рака предстательной железы, рака костей, рака щитовидной железы, рака паращитовидной железы, рака почек, рак пищевода, рака желчных путей, рака яичка, рака прямой кишки, рака головы и шеи, рака мочеточника, остеосаркомы, нейроцитомы, фибросаркомы, рабдомиосаркомы, астроцитомы, нейробластомы и нейроглиомы, но не ограничиваются ими. Предпочтительно, раковые заболевания, которые можно лечить с помощью композиции в соответствии с настоящим изобретением, могут быть выбраны из группы, состоящей из колоректального рака, рака молочной железы, рака легкого и рака почки, но не ограничиваются ими.
Композиция по настоящему изобретению может представлять собой фармацевтическую композицию. Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый компонент. Компонент, который обычно используется в составе лекарственных средств, может представлять собой один или несколько компонентов, выбранных из группы, состоящей, но без ограничения, из лактозы, декстрозы, сахарозы, сорбита, маннита, крахмала, аравийской камеди, фосфата кальция, альгината, желатина, силиката кальция, микрокристаллической целлюлозы, поливинилпирролидона, целлюлозы, воды, сиропов, метилцеллюлозы, метилгидроксибензоата, пропилгидроксибензоата, талька, стеарата магния и минерального масла. Кроме того, фармацевтическая композиция может дополнительно содержать один или несколько компонентов, выбранных из группы, состоящей из разбавителей, вспомогательных веществ, смазывающих веществ, смачивающих агентов, подсластителей, ароматизирующих веществ, эмульгаторов, суспензий и консервантов.
Фармацевтическую композицию по настоящему изобретению можно вводить перорально или парентерально. Парентеральное введение осуществляется путем внутривенной инъекции, подкожной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, эндотелиального введения, местного введения, интраназального введения, внутрилегочного введения, ректального введения и т.п. Для перорального введения активный ингредиент в композиции должен быть включен в лекарственную форму с покрытием или в лекарственную форму, которая может защитить активный ингредиент от разложения в желудке, учитывая, что пептиды и белки перевариваются в желудке. Альтернативно, настоящую композицию можно вводить через любое устройство, с помощью которого активный ингредиент может перемещаться в представляющую интерес клетку-мишень.
Фармацевтическая композиция может быть составлена в форме растворов, суспензий, сиропов или эмульсий в маслах или водных средах, или в форме экстрактов, частиц, суппозиториев, порошков, гранул, таблеток или капсул, и может дополнительно включать диспергирование. или стабилизирующие агенты для целей составления.
В частности, композицию для лечения рака в соответствии с настоящим изобретением можно применять для комбинированного лечения с другими противораковыми лекарственными средствами.
Противораковое лекарственное средство, которое можно применять для комбинированного лечения с новым вариантом PH20 в соответствии с настоящим изобретением, предпочтительно представляет собой химическое противораковое лекарственное средство, противораковое лекарственное средство на основе антител, биологическое противоопухолевое лекарственное средство, РНКи или клеточный терапевтический агент, но не ограничивается ими.
Предпочтительно противораковое лекарственное средство, которое можно применять для комбинированного лечения с новым вариантом PH20 в соответствии с настоящим изобретением, предпочтительно представляет собой иммуноонкологический агент, более предпочтительно ингибитор иммунных контрольных точек, но не ограничивается ими.
В другом аспекте настоящее изобретение направлено на нуклеиновую кислоту, кодирующую вариант PH20 или его фрагмент.
Нуклеиновые кислоты, используемые в настоящем документе, могут присутствовать в клетках, в клеточном лизате или в частично очищенной или по существу чистой форме. Термин «выделенные» или «по существу чистые» применительно к нуклеиновым кислотам относится к тем нуклеиновым кислотам, которые были очищены от других клеточных компонентов или других загрязняющих веществ, например, других клеточных нуклеиновых кислот или белков стандартными методами, включая обработку щелочью/SDS, разделение в CsCl, колоночную хроматографию, электрофорез в агарозном геле и другие, хорошо известные в данной области. Нуклеиновые кислоты по настоящему изобретению могут представлять собой ДНК или РНК.
В еще одном аспекте настоящее изобретение направлено на рекомбинантный вектор экспрессии, содержащий нуклеиновую кислоту. Для экспрессии варианта PH20 или его фрагмента в соответствии с настоящим изобретением ДНК, кодирующая вариант PH20, может быть получена стандартными методами молекулярной биологии (например, амплификацией PCR или клонированием кДНК с использованием гибридомы, которая экспрессирует вариант PH20), и ДНК может быть вставлена в вектор экспрессии таким образом, что он является «функционально связанным» с последовательностями, контролирующими транскрипцию и трансляцию.
Используемый в настоящем документе термин «функционально связанный» предназначен для обозначения того, что ген, кодирующий вариант PH20 или его фрагмент, лигирован в вектор таким образом, что последовательности контроля транскрипции и трансляции выполняют предназначенную для них функцию регулирования транскрипции и трансляции гена, кодирующего вариант PH20 или его фрагмент. Вектор экспрессии и последовательности контроля экспрессии выбирают таким образом, чтобы они были совместимы с используемой клеткой-хозяином для экспрессии. Гены, кодирующие PH20, вставляют в вектор экспрессии стандартными способами (например, лигирование комплементарных сайтов рестрикционных ферментов на фрагменте гена, кодирующего вариант PH20 или его фрагмент и вектор, или лигирование тупых концов, если сайты рестрикционных ферментов отсутствуют).
Кроме того, рекомбинантные векторы экспрессии несут регуляторные последовательности, которые контролируют экспрессию гена, кодирующего вариант PH20 в клетке-хозяине. Термин «регуляторная последовательность» предназначен для включения промоторов, энхансеров и других элементов контроля экспрессии (например, сигналы полиаденилирования), которые контролируют транскрипцию или трансляцию генов, кодирующих вариант PH20 или его фрагмент. Специалистам в данной области будет понятно, что конструкция вектора экспрессии, включая выбор регуляторных последовательностей, может зависеть от таких факторов, как выбор клетки-хозяина, подлежащей трансформации, уровень экспрессии желаемого белка и т.д.
В еще одном аспекте настоящее изобретение направлено на клетку-хозяина, содержащую нуклеиновую кислоту или вектор. Клетка-хозяин в соответствии с настоящим изобретением предпочтительно выбрана из группы, состоящей из клеток животных, клеток растений, дрожжей, E.coli. и клеток насекомых, но не ограничивается ими.
В частности, клетка-хозяин в соответствии с настоящим изобретением включает прокариотические клетки, такие как E. coli, Bacillus subtilis, Streptomyces sp., Pseudomonas sp., Proteus mirabilis или Staphylococcus sp., грибы, такие как Aspergillus sp., дрожжи, такие как Pichia pastoris, Saccharomyces cerevisiae, Schizosaccharomyces sp. и Neurospora crassa, и эукариотические клетки, такие как низшие эукариотические клетки, и другие высшие эукариотические клетки, такие как клетки насекомых.
Кроме того, клетки-хозяева, которые могут быть использованы в настоящем изобретении, могут происходить из растений или млекопитающих. Предпочтительно примеры клеток-хозяев включают, но без ограничения, клетки почки обезьяны (COS7), клетки NSO, SP2/0, клетки яичника китайского хомячка (CHO), W138, клетки почки детенышей хомячка (BHK), MDCK, клетки миеломы, клетки HuT 78 и клетки HEK293. Более предпочтительно могут быть использованы клетки СНО.
Нуклеиновую кислоту или вектор трансфицируют в клетку-хозяина. Трансфекция может быть выполнена с использованием различных методик, которые обычно используют для введения чужеродной нуклеиновой кислоты (ДНК или РНК) в прокариотические или эукариотические клетки, например, электрофорез, осаждение фосфатом кальция, DEAE-декстран трансфекция или липофекция. Для экспрессии варианта PH20 или его фрагмента по настоящему изобретению могут быть использованы различные комбинации рекомбинантных векторов экспрессии и клеток-хозяев. Предпочтительный вектор экспрессии для эукариотических клеток содержит последовательности регуляции экспрессии генов, полученные, но без ограничения, из SV40, вируса папилломы крупного рогатого скота, аденовируса, аденоассоциированного вируса, цитомегаловируса и ретровируса. Вектор экспрессии, который может быть использован для бактериальных хозяев, включает бактериальные плазмиды, полученные из E. Coli, такие как вектор pET, pRSET, pBluescript, pGEX2T, pUC, col E1, pCR1, pBR322, pMB9 и их производные; плазмиду с широким спектром хозяев, такую как RP4; ДНК фага, представленную различными производными фага лямбда, такими как λgt10, λgt11 и NM989; и другие фаги ДНК, такие как M13 и одноцепочечный нитчатый ДНК фаг. Вектор экспрессии, доступный для клеток дрожжей, может представлять собой плазмиду размером 2 мкм и ее производные. Вектор экспрессии для клеток насекомых включает pVL941.
В дополнительном аспекте настоящее изобретение направлено на способ получения варианта PH20 или его фрагмента, при этом способ включает стадию культивирования клетки-хозяина и экспрессии варианта PH20 или его фрагмента в соответствии с настоящим изобретением.
Когда рекомбинантный вектор экспрессии, способный экспрессировать вариант PH20 или его фрагмент, вводят в клетки-хозяева млекопитающего, вариант PH20 или его фрагмент может быть получен путем культивирования клеток-хозяев в течение такого периода времени, чтобы вариант PH20 или его фрагмент экспрессировался в клетках-хозяевах, предпочтительно в течение такого периода времени, чтобы вариант PH20 секретировался в среду во время культивирования клеток-хозяев.
В некоторых случаях экспрессированный вариант PH20 может быть выделен и очищен из клеток-хозяев. Выделение или очистка варианта PH20 может быть выполнена с помощью обычных способов выделения/очистки (например, хроматографией), которые используются для белков. Хроматография может включать одну или несколько комбинаций, выбранных из аффинной хроматографии, ионообменной хроматографии и гидрофобной хроматографии, но не ограничивается ими. В дополнение к хроматографии для выделения и очистки антитела может быть использована комбинация фильтрации, ультрафильтрации, высаливания, диализа и т.п.
ПРИМЕРЫ
В дальнейшем настоящее изобретение будет описано более подробно со ссылкой на примеры. Для специалиста в данной области будет очевидно, что эти примеры служат только для иллюстративных целей и не должны толковаться как ограничение или изменение объема настоящего изобретения.
Пример 1. Конструирование вариантов PH20
Для конструирования вариантов PH20 кДНК (ID клона: hMU002604) PH20 дикого типа приобретали в Корейском банке генов человека (Korean Human Gene Bank). PH20 дикого типа кодирует аминокислоты от L36 до S490. Ген PH20 амплифицировали с помощью полимеразной цепной реакции (далее в настоящем документе называемой как PCR) и вставляли в сайты рестрикционных ферментов XhoI и NotI вектора pcDNA3.4-TOPO. Для экспрессии в клетках ExpiCHO сигнальный пептид гормона роста человека, человеческий сывороточный гормон или Hyal1 человека использовали в качестве сигнального пептида вместо исходного сигнального пептида PH20. Для очистки белка с использованием колонки HisTrap последовательность ДНК 6xHis-метки была расположена на 3’-конце кДНК PH20. Аминокислотную замену вариантов PH20 проводили с использованием метода PCR, и аминокислотную замену подтверждали с помощью секвенирования ДНК.
Перечень праймеров, используемых в клонировании вариантов PH20, приведен ниже в таблице 9, и конкретные последовательности праймеров приведены ниже в таблице 10.
После обнаружения варианта PH20 с повышенной ферментативной активностью и термической стабильностью также конструировали кДНК без 6xHis-метки варианта PH20.
Когда плотность клеток ExpiCHO достигала 6×106/мл, плазмиду, содержащую кДНК PH20 дикого типа или его варианта, вставленную в вектор pcDNA3.4-TOPO, трансфицировали в клетки ExpiCHO с помощью реагента ExpiFectamine CHO. В качестве среды для культивирования клеток использовали экспрессионную среду ExpiCHO (100-500 мл). После трансфекции клетки ExpiCHO культивировали при встряхивании при 130 об/мин в течение в общей сложности 6 дней, во время которых клетки культивировали при 37°С в течение 1 дня и далее культивировали при более низкой температуре 32°С в течение 5 дней. После завершения культивирования клеточный супернатант собирали путем центрифугирования при 10000 об/мин в течение 30 мин.
Рекомбинантные белки PH20 дикого типа и варианта PH20 с присоединенным 6xHis с C-конца, продуцироваи в клетках ExpiCHO, очищали в три стадии (выполняли с использованием колонки HisTrap, колонки Q Sepharose и колонки Phenyl, соответственно) на хроматографической системе AKTA Prime System.
Для очистки белка с использованием колонки HisTrap готовили буфер A (20 мМ фосфат натрия, pH 7,5, 0,5M NaCl) и буфер B (20 мМ фосфат натрия, pH 7,5, 0,5M NaCl, 0,5M имидазол). Белок связывали с колонкой HisTrap, и колонку промывали 5 объемами колонки (CV) буфера A для удаления неспецифически связанных белков. Получали подтверждение того, что проводимость поддерживалась на постоянном уровне, колонку промывали 5 CV 20% буфера B для элюирования белка. Элюированный белок подвергали диализу буфером для диализа (20 мМ фосфат натрия, pH 7,5, 50 мМ NaCl). Для очистки белка на колонке Q Sepharose готовили буфер A (20 мМ фосфат натрия, pH 7,5) и буфер B (20 мМ фосфат натрия, pH 7,5, 0,5M NaCl). Белок связывали с колонкой Q Sepharose, и колонку промывали 5 CV буфера A для удаления неспецифически связанных белков, и затем 5 CV буфера B промывали при градиенте концентрации 0-100% для элюирования белка.
Для очистки белка с использованием фенильной колонки готовили буфер A (20 мМ фосфат натрия, pH 7,0, 1,5M (NH4)2SO4) и буфер B (20 мМ фосфат натрия, pH 7,0). Белок связывали с фенильной колонкой, и колонку промывали 5 CV буфера A для удаления неспецифически связанных белков, и затем 5 CV буфера B была промывали при градиенте концентрации 0-100% для элюирования белка.
Ферментативную активность PH20 дикого типа и варианта PH20 измеряли с помощью турбидиметрического анализа, субстрат-гель-анализа и анализа Моргана-Эльсона.
Турбидиметрический анализ представляет собой способ измерения оптической плотности осадка, образующегося при смешивании гиалуроновой кислоты с альбумином (BSA). Когда гиалуроновая кислота гидролизуется PH20, оптическая плотность осадка, образующегося при смешивании с альбумином, уменьшается. Гиалуронидазу PH20 (Sigma) разводили до 1, 2, 5, 7,5, 10, 15, 20, 30, 50 и 60 единиц/мл и готовили в каждой пробирке. Очищенный образец белка растворяли в буфере для разведения ферментов (20 мМ TrisHCl, pH 7,0, 77 мМ NaCl, 0,01% (масса/объем) бычий сывороточный альбумин) и разбавляли до 100X, 300X, 600X, 1200X и 2400X, и готовили в каждой пробирке. В свежих пробирках раствор гиалуроновой кислоты, имеющий концентрацию 3 мг/мл, разбавляли 10-кратно до концентрации 0,3 мг/мл таким образом, что объем каждой пробирки становился равным 180 мкл. К разбавленному раствору гиалуроновой кислоты добавляли 60 мкл фермента, смешивали с ним и оставляли реагировать при 37°С в течение 45 мин. После завершения реакции 50 мкл прореагировавшего фермента и 250 мкл кислого раствора альбумина добавляли в каждую лунку 96-луночного планшета и встряхивали в течение 10 мин, а затем измеряли оптическую плотность при 600 нм с помощью спектрофотометра.
В субстрат-гель-анализе белок подвергали электрофорезу в 10% полиакриламидном геле в присутствии додецилсульфата натрия (SDS) (включая 0,17 мг/мл гиалуроновой кислоты) в течение 1 ч, и SDS удаляли с помощью 2,5% Triton X-100 (масса/объем) при 4°С в течение 2 ч. После этого проводили ферментативную реакцию в буфере (50 мМ фосфат натрия, pH 7,0, 150 мМ NaCl) при 37°С (которая является оптимальной температурой для PH20) в течение 1-4 ч, и белок окрашивали в 0,5%-ном растворе реагента альцианового синего. Реагент альциановый синий, не связанный с гиалуроновой кислотой, удаляли с помощью обесцвечивающего раствора. Получали изображение геля SDS, окрашенного альциановым синим и затем количественно оценили полосу.
Термическую стабильность белка измеряли с помощью способа измерения температуры агрегации методом динамического светорассеяния (DLS), способа измерения температуры плавления (Tm) в PCR в реальном времени с использованием красителя Sypro-Orange, способа измерения ферментативной активности после выдерживания белка при заданной температуре в течение заданного времени и т.д. В способе измерения температуры агрегации с помощью DLS агрегацию молекул измеряют с использованием светорассеяния, и, таким образом, чувствительность является высокой, и температура агрегации обычно ниже, чем температура плавления белка.
Последовательности замененных или отщепленных аминокислот в вариантах PH20, сконструированных в соответствии с настоящим изобретением, показаны в таблице 11 ниже.
Среди вариантов в соответствии с настоящим изобретением вариант, имеющий 6xHis-метку, прикрепленную к С-концу PH20, назван HM; вариант, свободный от 6xHis-метки, назван HP; зрелый PH20 (L36-S490), имеющий 6xHis-метку, прикрепленную к С-концу, назван WT; и зрелый PH20 дикого типа (L36-Y482), у которого С-конец отщеплен после Y482, но не содержит 6xHis-метку, назван HW2.
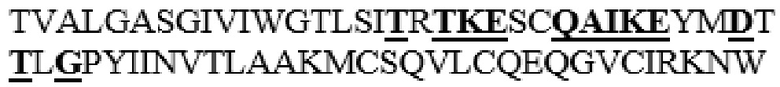
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS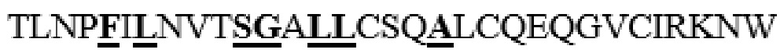
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS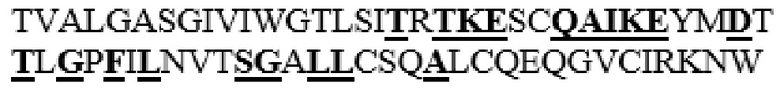
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
NWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS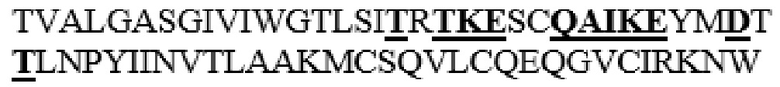
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS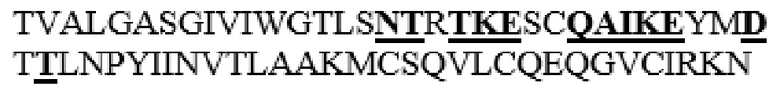
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS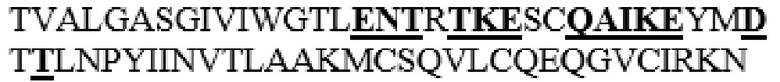
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS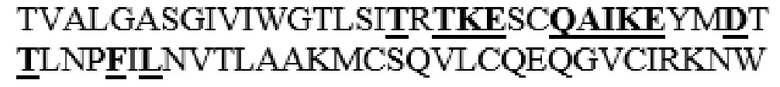
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS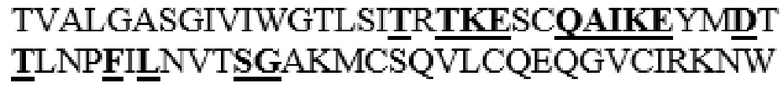
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
DYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS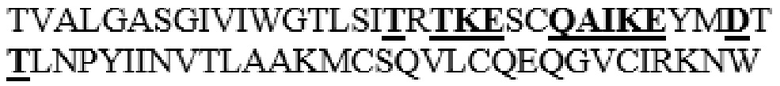
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCI
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF
KSPLPVFAYTRIVFTDQVLKFLSQDELVYTFGE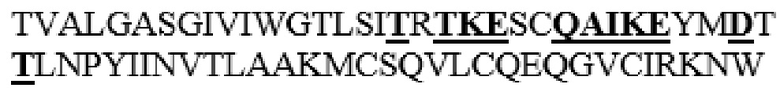
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKP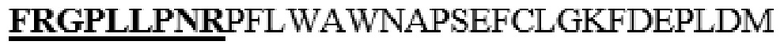
SLFSFIGSPRINATGQGVTIFYVDRLGYYPYIDSITGVTVNGGIPQKISLQDHLDKAKKDITFYMPVDNLGMAVIDWEEWRPTWARNWKPKDVYKNRSIELVQQQNVQLSLTEATEKAKQEFEKAGKDFLVETIKLGKLLRPNHLWGYYLFPDCYNHHYKKPGYNGSCFNVEIKRNDDLSWLWNESTALYPSIYLNTQQSPVAATLYVRNRVREAIRVSKIPDAKSPLPVFAYTRIVFTDQVLKFLSQDELVYTFGETVA
PYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS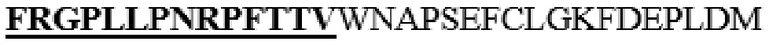
SLFSFIGSPRINATGQGVTIFYVDRLGYYPYIDSITGVTVNGGIPQKISLQDHLDKAKKDITFYMPVDNLGMAVIDWEEWRPTWARNWKPKDVYKNRSIELVQQQNVQLSLTEATEKAKQEFEKAGKDFLVETIKLGKLLRPNHLWGYYLFPDCYNHHYKKPGYNGSCFNVEIKRNDDLSWLWNESTALYPSIYLNTQQSPVAATLYVRNRVREAIRVSKIPDAKSPLPVFAYTRIVFTDQVLKFLSQDELVYTFGETVA
PYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
усечение перед F38 на N-конце, и усечение после K470 на C-конце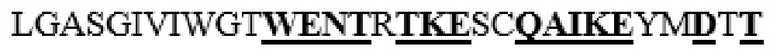
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLK
усечение перед F38 на N-конце, и усечение после F468 на C-конце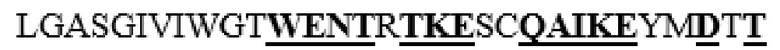
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS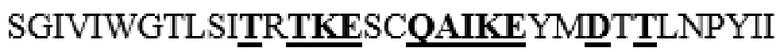
NVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS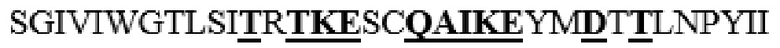
NVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDA
усечение перед L36 на N-конце, и усечение после C464 на C-конце
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVC
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIAD
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVC
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAV
усечение перед F38 на N-конце, и усечение после K470 на C-конце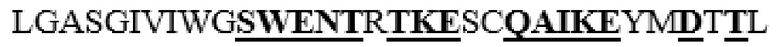
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLK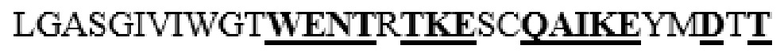
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPP
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPM
усечение перед F38 на N-конце, и усечение перед E474 на C-конце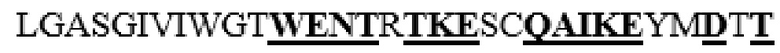
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPME
усечение перед F38 на N-конце, и усечение после T475 на C-конце
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMET
усечение перед F38 на N-конце, и усечение после E476 на C-конце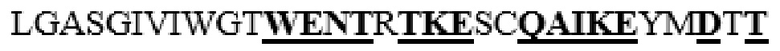
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETE
усечение перед N37 на N-конце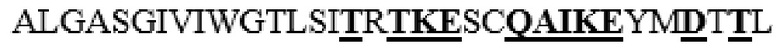
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
усечение перед R39 на N-конце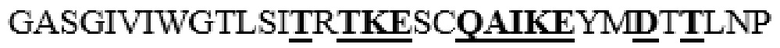
YIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
усечение перед P41 на N-конце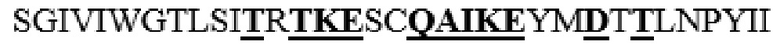
NVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
усечение перед F38 на N-terminus, и усечение после I465 на C-конце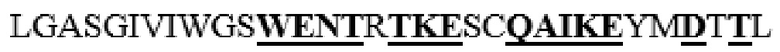
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCI
усечение перед F38 на N-конце, и усечение после D466 на C-конце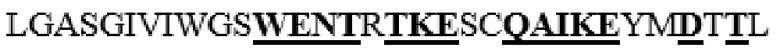
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCID
усечение перед F38 на N-конце, и усечение после A467 на C-конце
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDA
усечение перед F38 на N-конце, и усечение после F468 на C-конце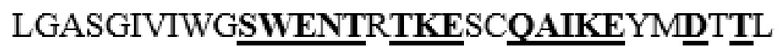
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF
усечение перед F38 на N-конце, и усечение после P478 на C-конце
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEP
усечение перед F38 на N-конце, и усечение после I480 на С-конце
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQI
усечение перед F38 на N-конце, и усечение после Y482 на С-конце
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFY
усечение перед F38 на N-конце, и усечение после A484 на С-конце
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNA
усечение перед F38 на N-конце, и усечение после P486 на С-конце
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASP
усечение перед F38 на N-конце, и усечение после T488 на С-конце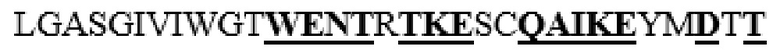
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPST
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQ109IFYNASPSTLS
WNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAFLKPPMETEEPQIFYNASPSTLS
усечение перед F38 на N-конце, и усечение после F468 на С-конце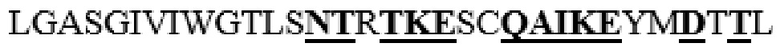
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF
NPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF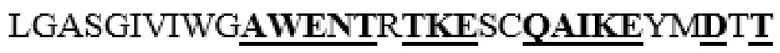
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF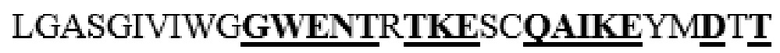
LNPYIINVTLAAKMCSQVLCQEQGVCIRKNWNSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF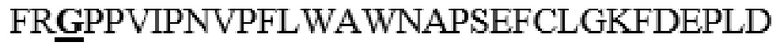
MSLFSFIGSPRINATGQGVTIFYVDRLGYYPYIDSITGVTVNGGIPQKISLQDHLDKAKKDITFYMPVDNLGMAVIDWEEWRPTWARNWKPKDVYKNRSIELVQQQNVQLSLTEATEKAKQEFEKAGKDFLVETIKLGKLLRPNHLWGYYLFPDCYNHHYKKPGYNGSCFNVEIKRNDDLSWLWNESTALYPSIYLNTQQSPVAATLYVRNRVREAIRVSKIPDAKSPLPVFAYTRIVFTDQVLKFLSQDELVYTFGETV
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF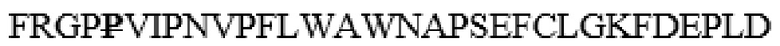
MSLFSFIGSPRINATGQGVTIFYVDRLGYYPYIDSITGVTVNGGIPQKISLQDHLDKAKKDITFYMPVDNLGMAVIDWEEWRPTWARNWKPKDVYKNRSIELVQQQNVQLSLTEATEKAKQEFEKAGKDFLVETIKLGKLLRPNHLWGYYLFPDCYNHHYKKPGYNGSCFNVEIKRNDDLSWLWNESTALYPSIYLNTQQSPVAATLYVRNRVREAIRVSKIPDAKSPLPVFAYTRIVFTDQVLKFLSQDELVYTFGETV
NSSDYLHLNPDNFAIQLEKGGKFTVRGKPTLEDLEQFSEKFYCSCYSTLSCKEKADVKDTDAVDVCIADGVCIDAF
Пример 2. Конструирование вариантов HM1 и HM6 PH20
Как показано на фигуре 1B, аминокислотные остатки M345-N363 PH20 соответствуют области альфа-спирали 8 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8 в модели третичной структуры белка. Среди аминокислот альфа-спирали 8 C351 образует дисульфидную связь с C60 альфа-спирали 1; Y357 образует гидрофобное взаимодействие с F315 альфа-спирали 7; и N363 образует водородную связь с D69 альфа-спирали 1, тем самым стабилизируя вторичные структуры, прилегающие к альфа-спирали 8 (фигура 1C).
Для создания вариантов, обладающих более высокой ферментативной активностью и термической стабильностью по сравнению с WT, путем замены аминокислот, расположенных в области альфа-спирали 8 PH20 и линкерной области между альфа-спиралью 7 и альфа-спиралью 8, конструировали следующие варианты: вариант HM1, в котором были заменены 12 аминокислот в области M345-N363; вариант HM2, в котором были заменены 7 аминокислот в области Y365-V379; и вариант HM3, в котором были заменены 19 аминокислот в области M345-V379. Из числа аминокислот, расположенных в альфа-спирали 8 PH20, C351, которая участвует в связывании дисульфидной связью, и Y357, которая участвует в гидрофобном взаимодействии, не были заменены. Последовательности замененных аминокислот в вариантах HM1, HM2 и HM3 показаны выше в таблице 11. Когда клетки ExpiCHO трансфицировали плазмидой pcDNA3.4-TOPO, содержащей ген варианта HM1, HM2 или HM3, вариант HM1 экспрессировался в клетках ExpiCHO (фигура 3A), а варианты HM2 и HM3 не экспрессировались в клетках. Экспрессию белка подтверждали не только путем измерения ферментативной активности, но также с помощью Вестерн-блоттинга с использованием антитела (Abcam, ab193009) против человеческого PH20. Эпитопом антитела является область Q173-P222. Эти экспериментальные результаты дают основание предположить, что аминокислотная замена в области Y365-V379 аминокислотной последовательности альфа-спирали 8, используемая при конструировании вариантов HM2 и HM3, оказывает значительное влияние на структуру белка; таким образом, белки вариантов HM2 и HM3 не экспрессировались. Уровень экспрессии варианта HM1 был в 3,4 раза выше, чем у дикого типа WT (фигура 2). Вариант HM1 представляет собой вариант, в котором водородная связь между N363 альфа-спирали 8 и D69 альфа-спирали 1 была удалена путем замены N363 глицином (фигура 1C). Для восстановления водородной связи между N363 и D69, G363 в HM1 заменяли аспарагином, тем самым создавая вариант HM6. Замененные аминокислоты в варианте HM6 показаны выше в таблице 11.
Вариант HM6 экспрессировался в клетках ExpiCHO, и уровень его экспрессии был аналогичен уровню экспрессии варианта HM1 (уровень экспрессии был в 3,4 раза выше, чем у WT) (фигура 2). При измерении ферментативной активности с помощью турбидиметрического анализа было показано, что ферментативная активность варианта HM6 в 1,3 раза превышала ферментативную активность WT (фигура 3B). В субстрат-гель-анализе, как правило, SDS удаляли с помощью 2,5% Triton X-100 (масса/объем) после SDS-PAGE, и ферментативную реакцию выполняли при 37°С в течение 1-4 ч, во время которых степень, до которой гиалуронидаза гидролизует гиалуроновую кислоту, измеряли с помощью красителя альцианового синего. Известно, что при удалении SDS из субстратного геля немедленно происходит сворачивание белка, и субстрат не влияет на сворачивание белка. При измерении ферментативной активности WT и вариантов HM1 и HM6 с помощью субстрат-гель-анализа при 37°С в течение 1-4 ч, HM6 продемонстрировал более высокую ферментативную активность по сравнению с WT и вариантом HM1 (фигура 3C). Этот результат дает основание предположить, что сворачивание белков вариантов HM1 и HM6, и результирующая ренатурация были быстрее, чем у WT, и что сконструированные варианты проявляют более высокую ферментативную активность, чем WT. Когда использовали сигнальный пептид самого PH20, уровень экспрессии белка в клетках ExpiCHO был низким, и для решения этой проблемы использовали последовательность сигнального пептида сывороточного альбумина человека или Hyal1 человека. Как показано на фигуре 3C, когда каждый из сигнальных пептидов сывороточного альбумина человека и Hyal1 человека использовали в качестве сигнального пептида варианта HM1, экспрессия белка увеличивалась, и значительное различие между двумя сигнальными пептидами отсутствовало. Когда ферментативную активность измеряли с помощью субстрат-гель-анализа, WT, имеющий сигнальный пептид человеческого сывороточного альбумина, и вариант HM1, имеющий сигнальный пептид человеческого сывороточного альбумина или Hyal1, проявляли ферментативную активность в диапазоне pH от 5 до 8 (фигура 3D). В настоящем изобретении сигнальный пептид сывороточного альбумина человека использовали в качестве сигнального пептида варианта, сконструированного после HM1.
Когда термическую стабильность WT и вариантов HM1 и HM6 сравнивали на основе температур агрегации, температуры агрегации составили 46,5°С, 53,0°С и 50,5°С, соответственно, что указывает на то, что температуры агрегации вариантов HM1 и HM6 были на 6,5°С и 4,0°С выше, чем у WT, соответственно (фигура 10). Результаты измерения температур агрегации согласуются с результатами рефолдинга белка, показанными в субстрат-гель-анализе. Эти результаты впервые дают основание предположить, что водородная связь, образованная остатком N363 альфа-спирали 8, играет важную роль в термической стабильности и ферментативной активности PH20.
Кроме того, когда гидрофильную/гидрофобную природу WT и вариантов HM1 и HM6 сравнивали с использованием хроматографии на фенильной колонке, HM1 и HM6 элюировались раньше, чем WT. Это дает основание предположить, что варианты HM1 и HM6 имеют более гидрофильную природу, чем WT из-за замены аминокислот. Однако варианты HM1 и HM6, элюированные из фенильной колонки, показали два пика, в отличие от WT, и продемонстрировали одинаковую молекулярную массу при обработке PNGase F. Это различие, по-видимому, вызвано N-гликозилированием (фигура 3E).
Пример 3. Конструирование вариантов HM4, HM7, HM8, HM9, HM10, HM11 и HM12 PH20
Считается, что аминокислотная замена областей M345-N363 и M345-I361 путем конструирования вариантов HM1 и HM6 в примере 2, которая привела к увеличению как ферментативной активности, так и термической стабильности PH20, явилась большим достижением в области белковой инженерии. Таким образом, на основе вариантов HM1 и HM6 были дополнительно заменены другие аминокислоты в направлении N-конца и C-конца.
Сначала конструировали варианты HM4 и HM7, которые содержат замененные аминокислоты в вариантах HM1 и HM6, и в которых аминокислоты между G340 и I344 были дополнительно заменены на G340V, T341S, L342W, S343E и I344N. Последовательности замененных аминокислотных последовательностей в вариантах HM4 и HM7 показаны выше в таблице 11. Варианты HM4 и HM7 экспрессировали в клетках ExpiCHO. Результаты очистки белка для HM7 показаны на фигуре 5А. НМ4 и НМ7 показали увеличение в 6,3 раза белковой экспрессии по сравнению с диким животным и показали увеличение температуры агрегации на 10°С и 11,5°С, соответственно, по сравнению с диким типом. Однако ферментативная активность HM4 и HM7, измеренная с помощью турбидиметрического анализа, составила примерно 15% от ферментативной активности WT (фигура 5B). Ферментативная активность и термическая стабильность обычно имеют компромиссное соотношение, но в настоящем изобретении оказалось, что замена, введенная в вариантах HM1 и HM6, увеличивала термическую стабильность вариантов при этом сохраняя ферментативную активность, и ферментативная активность вариантов HM4 и HM7 уменьшилась из-за чрезмерного увеличения термической стабильности. Увеличение температуры агрегации на 11,5°С, которое возникло в вариантах HM4 и HM7, является очень значимым результатом с точки зрения белковой инженерии. Когда структурную гибкость белков анализировали с помощью графика Штерна-Фольмера, было показано, что структурная гибкость HM1, HM6, HM4 и HM7, полученная путем замены альфа-спирали 8 и ее линкерной области, была выше, чем структурная гибкость WT (фигура 11A). Этот результат позволяет предположить, что увеличение локальной термической стабильности привело к увеличению гибкости всей структуры белка.
Различие между вариантами HM6 и HM7 состоит в аминокислотах между G340 и I344. Для идентификации аминокислот, вовлеченных в увеличенную термическую стабильность варианта HM7, конструировали следующие варианты на основе варианта HM6: HM8, в котором заменена I344N; HM9, в котором заменены S343 и I344N; HM10, в котором заменены L342W, S343E и I344N; и HM21, в котором заменены T341S, L342W, S343E и I344N. Последовательности замененных аминокислот в вариантах HM8, HM9, HM10 и HM21 показаны выше в таблице 11. Варианты HM8, HM9, HM10 и HM21 экспрессировались в клетках ExpiCHO (фигура 5A). Поскольку I344N, S343E и L342W вводили в петлю в N-концевом направлении альфа-спирали 8 на основе варианта HM6, температуры агрегации вариантов HM8, HM9 и HM10 повысились до 52,5°С, 53°С и 55,5°С, соответственно (фигура 10). Однако варианты HM8, HM9 и HM10 сохраняли ферментативную активность, аналогичную ферментативной активности WT (фигура 5B). Этот результат дает основание предположить, что аминокислотная замена, введенная в вариантах HM8, HM9 и HM10, оказала локальное влияние на термическую стабильность ферментов, но не имела значительного эффекта на ферментативную активность. Однако HM21 показал пониженную термическую стабильность по сравнению с HM10, но продемонстрировал приблизительно в 2 раза более высокую ферментативную активность по сравнению с WT при pH 7,0. Когда WT и каждый вариант оставляли для взаимодействия с субстратом в течение 1 ч в субстрат-гель-анализе, ферментативная активность уменьшалась в следующем порядке: HM21> HM10> HM9> HM8> HM6> WT (фигура 5C).
Когда физические свойства вариантов HM7, HM8, HM9, HM10 и HM21 исследовали с использованием хроматографии на фенильной колонке, эти варианты PH20 элюировались раньше, чем WT, что позволяет предположить, что все эти варианты имеют гидрофильную природу. Однако картина основного пика в положении аминокислотной замены выглядела иначе (фигура 5D).
Вариант HM7 также показал два пика при хроматографии на фенильной колонке, подобно вариантам HM1 и HM6, что позволяет предположить, что присутствовали два разных типа.
Для изучения картин миграции WT и вариантов в зависимости от их изоэлектрических точек проводили анализ методом изоэлектрического фокусирования (в дальнейшем в настоящем документе именуемый как IEF) (фигура 5E). На геле IEF WT и вариант HM6 и HM8 показали сходные картины миграции, а варианты HM9, HM10, HM21 и HM7, содержащие мутацию S343E, мигрировали в более кислую область. Этот результат дает основание предположить, что изменение изоэлектрической точки белка происходит из-за введения глутаминовой кислоты путем замены S343E аминокислоты между G340 и I344.
Кроме того, на основе варианта HM6 заменяли другие аминокислоты в C-концевых областях альфа-спирали 8, тем самым конструируя варианты HM11 и HM12. Последовательности замененных аминокислот в вариантах HM11 и HM12 показаны выше в таблице 11. Вариант HM11 экспрессировался в клетках ExpiCHO, но уровень его экспрессии был ниже, чем уровень экспрессии WT (фигура 2), и вариант HM12 не экспрессировался в клетках ExpiCHO. Вариант HM11 продемонстрировал активность, соответствующую 32% активности WT (фигура 6B).
Пример 4. Конструирование вариантов PH20 с усечением N-концевых аминокислот на основе HM6
Как уже известно, что С-концевая область PH20 играет важную роль в экспрессии и ферментативной активности PH20, но роль N-концевой области PH20 не очень хорошо известна. Для изучения эффекта отщепления аминокислоты в N-концевой области PH20 на ферментативную активность, варианты HM40, HM13, HM41, HM24, HM42 и HM25, имеющие усечение с N-конца по N37, F38, R39, A40, P41 или P42, конструировали на основе варианта HM6 (таблица 11). Кроме того, дополнительно конструировали HP61 и HP62 с модификациями N-концевых аминокислот.
Варианты HM40, HM13, HM41, HM24, HM42, HP61 и HP62 экспрессировались в клетках ExpiCHO, но HM25 не экспрессировался (фигуры 7A и 7B). Варианты PH20 с усечением по N-концу показали различие в ферментативной активности в зависимости от положения, в котором начинался N-конец. Варианты HM40, HM13 и HM41, в которых были отщеплены одна-три аминокислоты, показали ферментативную активность, сходную с ферментативной активностью матрицы HM6, но HM24 и HM42, в которых было отщеплено четыре-пять аминокислот, показали немного более низкую активность, чем HM6 (фигура 7С). Однако HM25, в котором было отщеплено шесть аминокислот, незначительно экспрессировался в клетках ExpiCHO, и ферментативная активность также была значительно низкой (3,5 Ед/мкг). По-видимому, изменения ферментативной активности HP61 и HP62 с модификациями N-концевых аминокислот являются незначительными.
Что касается ферментативной активности вариантов PH20 с N-концевыми отщеплениями, измеренной с помощью субстрат-гель-анализа (реакция в течение 1 ч), ферментативная активность HM40, HM13 и HM41 была сходной с ферментативной активностью варианта HM6, но ферментативная активность HM24 и HM42 была ниже, чем ферментативная активность HM6 (фигура 7D). НМ25, в котором было отщеплено шесть аминокислот, нельзя было проанализировать, так как количество продуцируемого белка было незначительным.
Когда физические свойства вариантов HM40, HM13, HM41, HM24 и HM42 анализировали с использованием хроматографии на фенильной колонке, эти варианты элюировались из колонки раньше, чем WT, что позволяет предположить, что эти варианты имеют гидрофильную природу (фигура 7E). Этот результат позволяет предположить, что варианты HM40, HM13, HM41, HM24 и HM42, сконструированные на основе варианта HM6, сохранили гидрофильную природу HM6 при рассмотрении характеристик остатков L36-A40.
Температура агрегации вариантов PH20 с усечениями N-конца, измеренная с помощью DLS, различались среди вариантов в зависимости от положения, в котором начиналась аминокислота (фигура 7F). Несмотря на отщепление N-концевых аминокислотных остатков вариантов HM40, HM13, HM41 и HM42, они показали температуру агрегации 50°С или выше, что указывает на то, что характеристики матрицы HM6 остались нетронутыми. Из этих вариантов, варианты HM40 и HM42 продемонстрировали температуру агрегации, которая была на 3-4°С выше, чем температура агрегации HM6, что указывает на то, что термическая стабильность этих вариантов увеличилась. Кроме того, для изучения эффекта замены N-концевых аминокислот PH20 на экспрессию белка и ферментативную активность, конструировали следующие варианты: вариант HM17, в котором N-концевые аминокислотные остатки 36-47 (LNFRAPPVIPNV) PH20 заменены на FRGPLLPNR; и вариант HM18, в котором N-концеваминокислотные остатки 36-52 (LNFRAPPVIPNVPFLWA) PH20 заменены на FRGPLLPNRPFTTV. Последовательности замененных аминокислот в вариантах HM17 и HM18 показаны выше в таблице 11. Варианты HM17 и HM18 не экспрессировались в клетках ExpiCHO. Это дает основание предположить, что даже при отщеплении до пяти аминокислот, расположенных на N-конце, варианты показали экспрессию белка и ферментативную активность; однако замена большего количества аминокислотных остатков, например, 36-47 остатков или 36-52 остатков, оказывала влияние на сворачивание белка.
Пример 5. Конструирование вариантов HM14, HM15 и HM16 PH20 на основе HM6 с усечением С-концевых аминокислот
Известно, что С-концевая область PH20 играет важную роль в экспрессии белка и ферментативной активности. В настоящем изобретении варианты HM14, HM15 и HM16, в которых C-концевые аминокислоты отщеплены по I465, F468 и K471, соответственно, конструировали на основе варианта HM6. Последовательности замененных аминокислот в этих вариантах HM14, HM15 и HM16 показаны выше в таблице 11. Эти варианты HM14, HM15 и HM16 экспрессировались в клетках ExpiCHO (фигура 8A), и уровни их белковой экспрессии находились в порядке HM16 > HM15 > HM14, что указывает на то, что уровни белковой экспрессии снижались по мере увеличения числа отщепленных C-концевых аминокислот (фигура 8A). Однако ферментативная активность вариантов HM14, HM15 и HM16 находилась в порядке HM16 > HM14 (~WT) > HM15 (фигура 8B). Согласно Frost et al., C-концевая область 477-483 PH20 необходима для достаточной экспрессии, и при отщеплении C-конца по 467, ферментативная активность варианта составляла только 10% от варианта PH20, чей C-конец отщеплялся по остаткам 477-483, и при отщеплении С-конца перед остатком 467, вариант не имел ферментативной активности. Однако варианты HM14, HM15 и HM16 с усеченным С-концом, сконструированные на основе варианта HM6 в настоящем изобретении, показали увеличенное сворачивание белка из-за аминокислотной замены области M345-I361 и, таким образом, их термическая стабильность увеличилась. По этой причине, даже при отщеплении C-конца после I465, F468 или P471, вариант показал ферментативную активность, аналогичную ферментативной активности WT, и его ферментативная активность значительно не уменьшалась.
Структурную гибкость WT и вариантов HM14, HM15 и HM16 исследовали методом тушения флуоресценции с использованием акриламида (фигура 11B). Все варианты HM14, HM15 и HM16 были структурно более гибкими, чем WT. Этот результат дает основание предположить, что варианты с С-концевым усечением, сконструированные с использованием варианта HM6, также сохранили свою структурную гибкость.
Ферментативную активность вариантов HM14, HM15 и HM16, измеренную с помощью турбидиметрического анализа, также подтверждали в субстрат-гель-анализе (фигура 8C).
Когда физические свойства вариантов с С-концевым усечением анализировали с использованием хроматографии на фенильной колонке, все варианты HM14, HM15 и HM16 элюировались раньше, чем WT, указывая на то, что они имеют гидрофильную природу. Кроме того, гидрофобность этих вариантов находилась в порядке HM16 > HM14 > HM15 (фигура 8D).
Пример 6. Конструирование вариантов HM19 и HM20 на основе HM10, с отщеплением N-концевых и С-концевых аминокислот
Варианты PH20, сконструированные в настоящем изобретении на основе HM6 и HM8, HM9 и HM10, в которых аминокислотные остатки G340-I344 были дополнительно заменены, показали лучшие характеристики, чем WT, с точки зрения уровней белковой экспрессии, ферментативной активности и термической стабильности. Конструировали варианты HM19 и HM20, которые имели N-конец, отщепленный по F38, и C-конец, отщепленный по F468, на основе HM10, обладающего высокой ферментативной активностью и термической стабильностью среди вариантов HM8, HM9 и HM10. HM19 и HM20, все экспрессировали в клетках ExpiCHO и очищали с использованием хроматографии на колонке HisTrap (фигура 9A). При измерении ферментативной активности этих вариантов с помощью турбидиметрического анализа, HM19 и HM20 показали ферментативную активность, которая на 10% превышала ферментативную активность WT (фигура 9B). В субстрат-гель-анализе HM19 и HM20 также проявили более высокую ферментативную активность, чем WT (фигура 9C).
Пример 7. Характеристика вариантов PH20 на основе HM10
Уровни экспрессии вариантов на основе HM10 с С-концевым усечением в клетках ExpiCHO показали тенденцию к снижению по мере того, как длина C-концевой области становилась короче, и эти варианты не экспрессировались, когда C-конец был усечен по C464 или короче (фигура 12). C464 необходим, так как он образует дисульфидную связь с C437 и является важным для поддержания структуры белка.
Чтобы проверить, не экспрессируется ли вариант в клетках ExpiCHO, когда С-конец отщеплен по остатку 464 или короче, проводили анализ методом Вестерн-блоттинга. Как показано на фигуре 13, варианты HM30, HM31, HM32 и HM33 не были обнаружены при Вестерн-блоттинге.
Ферментативная активность вариантов на основе НМ10 с С-концевым усечением, измеренная с помощью турбидиметрического анализа, показана на фигурах 14A и 14B. Варианты PH20 с С-концевым усечением показали ферментативную активность ± 20% по сравнению с WT. Когда С-конец был отщеплен после I480, активность фермента в целом увеличилась. Кроме того, варианты HP19 и HP20, полученные путем удаления 6xHis-метки из HM19 и HM20, показали снижение ферментативной активности на 23% и 9,6%, соответственно, по сравнению с тем, когда 6xHis-метка присутствовала. Это позволяет предположить, что 6xHis-метка влияет на ферментативную активность.
Когда ферментативную активность вариантов на основе HM10 с С-концевым усечением измеряли с помощью субстрат-гель-анализа, эти варианты показали более высокую ферментативную активность по сравнению с WT, и ферментативную активность, сходную с ферментативной активностью матрицы HM10, что указывает на то, что различие в ферментативной активности в зависимости от длины С-концевой области не было значительным (фигура 14С).
Пример 8. Характеристика вариантов PH20 на основе HM21
Вариант HP34 очищали с помощью четырехступенчатой колоночной хроматографии (фигура 15A), а вариант HP46 очищали с помощью трехступенчатой колоночной хроматографии (фигура 15B). Количество полученных HP34 и HP46 составило 1,73 мг/л и 25,6 мг/л, соответственно. HP34 и HP46 представляют собой варианты, не содержащие 6xHis-метку, и процесс очистки этих вариантов отличается от процесса очистки вариантов, имеющих 6xHis-метку, и поэтому сложно сравнивать уровни экспрессии этих белков.
В турбидиметрическом анализе активность HP34 и HP46 составила 45,6 Ед/мкг и 47,2 Ед/мкг, соответственно, что примерно в 2 раза выше, чем у WT, и примерно на 10% выше, чем у матрицы HM21 (фигура 16А).
Кинетику каждого варианта измеряли с помощью анализа Моргана-Эльсона, и результаты измерений показаны на фигуре 16B. Каталитическая эффективность (kcat/Km) HP34 и HP46 в 1,7-2 раза превышала каталитическую активность у HW2 дикого типа. Этот результат согласуется с результатом, согласно которому специфическая активность была более высокой по сравнению с WT. Константа Михаэлиса (Km) была ниже в этих вариантах, чем у HW2, что указывает на повышение субстратной аффинности этих вариантов. Из этих результатов можно сделать вывод о том, что варианты HM21, HP34 и HP46 прочно связываются с субстратом и обладают свойством превращать субстрат в продукт с высокой эффективностью. Это свойство объясняется эффектом замены Т341 серином. Когда треонин, расположенный в положении 341, заменен аминокислотой, такой как аланин, глицин, аспарагиновая кислота или т.п., эффект замены на ферментативную активность может быть предсказан.
Температура агрегации HP34 и HP46, измеренная с помощью DLS, составила 51,5°С и 51,0°С, соответственно, и была сходной с температурой агрегации матрицы HM21, и примерно на 5°С превышала температуру агрегации HW2, что указывает на то, что эти варианты были термически стабильными (фигура 17A). Ферментативная активность HP20, измеренная с помощью субстрат-гель-анализа, была сходной с ферментативной активностью HP20, тогда как HP46 проявил более высокую ферментативную активность, чем HP20, что указывает на то, что сворачивание белка HM21 в качестве матрицы было лучше, чем сворачивание белка HM10 (фигура 17B).
HW2 дикого типа и вариант HP46 выдерживали в течение ночи при pH 7,0 и pH 3,0, и затем их ферментативную активность сравнивали с помощью субстрат-гель-анализа. В результате было показано, что HP46 проявляет высокую активность не только при pH 7,0, но также и при pH 3,0, что указывает на его превосходную стабильность (фигура 17C).
HM53, HM54, HM55, HM56, HP59 и HP60 представляют собой варианты, имеющие мутацию по аминокислоте в положении 341, среди вариантов на основе HM21. Было получено подтверждение того, что мутация аминокислоты в положении 341 оказывала различные эффекты на уровень экспрессии и активность вариантов (фигуры 17D и 17E).
Пример 9. Анализ иммуногенности in vitro варианта PH20
Биофармацевтические препараты с более высокой молекулярной массой, чем низкомолекулярные синтетические химические соединения, при попадании в организм человека могут вызвать непреднамеренный иммунный ответ. Наружная контактная поверхность, созданная сворачиванием или взаимодействием с прилегающими доменами во вторичной структуре или третичной структуре биоматериала с большой молекулярной массой, может способствовать иммунному ответу на биоматериал путем обеспечения эпитопа для иммунной системы в организме человека. Этот иммунный ответ может продуцировать антитело против лекарственного средства (ADA), и этот ответ может подавлять эффективность лекарственного средства или вызывать гиперчувствительность к лекарственному средству, или способствовать клиренсу лекарственного средства в организме человека. Таким образом, иммунный ответ на лекарственное средство может влиять на результаты клинических испытаний и вызывать серьезные патологические реакции при длительном применении. Этот иммунный ответ может находиться под влиянием различных факторов и запускаться специфическим ответом на лекарственное средство как таковое или заболевание, или факторов, которые зависят от способа введения лекарственного средства или индивидуальных пациентов. Факторы, вызываемые лекарственным средством как таковым, включают сходство или несходство биофармацевтических препаратов с пептидами человека, посттрансляционную модификацию, примеси, образование агрегатов и характеристики состава. Факторы, которые варьируются у разных пациентов, включают пол, реакционную способность в отношении других принимаемых лекарственных средств и генетические факторы в зависимости от типа человеческого лейкоцитарного антигена (HLA).
Этот иммуногенная реакция запускается CD4+ T-клетками или CD8+ T-клетками, которые распознают эпитопы независимо от причины, вызвавшей иммунный ответ. Из-за разнообразия генов HLA класса II у индивидуумов эпитопы CD4+ Т-клеток у разных индивидуумов различаются, и поэтому ответная реакция на биофармацевтические препараты в CD4+ Т-клетках в каждой крови, обеспеченной здоровым донором, может являться очень важным критерием для оценки иммунных ответов, которые могут возникать в клинических процессах. CD4+ Т-клетки активируются антигенпрезентирующими клетками (APC), распознающими антигены, представленные на белках MHC типа II (главного комплекса гистосовместимости). Активированные CD4+ Т-клетки высвобождают цитокины, которые активируют макрофаги, цитотоксические Т-клетки и В-клетки, что приводит к высоким уровням продукции антител. Напротив, CD8+ Т-клетки обладают прямой цитотоксичностью и непосредственно удаляют клетки, инфицированные антигенами, поврежденные клетки или клетки, утратившие свою функцию. CD8+ Т-клетки имеют Т-клеточные рецепторы, которые могут распознавать специфические антигенные пептиды, связанные с молекулами MHC типа I, расположенными на поверхности каждой клетки. CD8+ Т-клетки также могут быть активированы путем распознавания антигенов, представленных антигенпрезентирующими клетками, и эта активация может быть дополнительно усилена цитокинами CD4+ Т-клеток. Следовательно, когда уровень активации CD4+ Т-клеток и CD8+ Т-клеток новыми биоматериалами измеряется in vitro, можно предсказать иммуногенные ответы, которые могут быть индуцированы в клинических процессах. В этом примере, чтобы предсказать иммуногенность варианта PH20 по сравнению с контролем, CD4+ T-клетки и CD8+ T-клетки выделяли из PBMC, а затем обрабатывали 1,5 нг/мл и 15 нг/мл контрольного PH20 и варианта PH20 (HP46), а затем измеряли распределения активированных CD4+ Т-клеток и CD8+ Т-клеток. Уровень активации каждого типа Т-клеток измеряли с использованием индекса стимуляции (Stimulating Index, SI), который определяли следующим образом:
Индекс стимуляции (SI) = (уровень активации Т-клеток после обработки тестируемым образцом) / (уровень активации Т-клеток после обработки носителем)
Если значение SI клеток равно 2 или более, можно подтвердить, что клетки были активированы на значительном уровне.
Иммуногенные ответы могут варьироваться в зависимости от типа HLA. Поэтому эксперименты по измерению ответов в более разнообразных типах HLA выполняли с использованием Т-клеток, выделенных из PBMC, полученных от 10 здоровых доноров. Типы HLA используемых 10 PBMC показаны ниже в таблице 12.
Результаты измерения уровней активации CD4+ и CD8+ Т-клеток, обработанных каждым из PH20 и вариантом PH20, сведены ниже в таблице 13. При рассмотрении результатов измерения уровней активации CD4+ и CD8+ Т-клеток оказалось, что PH20 и вариант PH20, все показали относительно низкие уровни активации. В случае PH20 уровень активации CD4+ Т-клеток по данным измерений был равен 2 или более в двух экспериментах, и в случае варианта PH20 активации CD4+ Т-клеток не было обнаружено. В случае PH20 активация CD8+ Т-клеток была обнаружена в одном эксперименте, и в случае варианта PH20 активация CD8+ Т-клеток также была обнаружена в одном эксперименте. Однако в случае PH20 значение SI по данным измерений составило 2 или более как при 1,5 нг/мл, так и при 15 нг/мл, но в случае варианта PH20 значение SI по данным измерений составило 2 или менее при 1,5 нг/мл, и значение SI составило 2 или более при 15 нг/мл (см. фигуры 18 и 19).
Таким образом, наблюдалось, что при более низкой концентрации уровень активации CD8+ Т-клеток был низким в случае варианта PH20, и было определено, что уровень активации CD8+ Т-клеток под действием PH20 является более высоким, чем уровень активации CD8+ Т-клеток под действием варианта PH20. На основании приведенных выше результатов сделаны следующие выводы:
1) уровни активации CD4+ T-клеток и CD8+ T-клеток под действием PH20 и варианта PH20 являются относительно низкими; и
2) возможность активации CD4+ T-клеток и CD8+ T-клеток вариантом PH20 является более низкой по сравнению с PH20.
На основании этих результатов можно предположить, что вариант PH20 будет иметь более низкую вероятность запуска иммуногенной реакции в клинических процессах, чем PH20.
Полезные эффекты
Варианты PH20 или их фрагменты в соответствии с настоящим изобретением обладают повышенными уровнями экспрессии белка и показывают увеличение температуры агрегации белка на 4-11,5°С или около того при экспрессии в клетках CHO (ExpiCHO), благодаря чему они могут быть эффективно продуцированы, при этом обладая высокой термической стабильностью по сравнению со зрелым PH20 дикого типа.
Кроме того, в результате субстрат-гель-анализа, одного из тестов для измерения активности гиалуронидазы, варианты PH20 или их фрагменты в соответствии с настоящим изобретением улучшили рефолдинг белка, благодаря чему они ренатурировались быстрее по сравнению со зрелым PH20 дикого типа, и исходная ферментативная активность сохранялась независимо от положения C-концевого усечения.
Кроме того, варианты PH20 или их фрагменты в соответствии с настоящим изобретением обладают низкой иммуногенностью, поэтому их можно повторно вводить в организм человека.
Ссылки
Arming, S., Strobl, B., Wechselberger, C., and Kreil, G. (1997). In vitro mutagenesis of PH-20 hyaluronidase from human sperm. Eur J Biochem 247, 810-814.
Bookbinder, L.H., Hofer, A., Haller, M.F., Zepeda, M.L., Keller, G.A., Lim, J.E., Edgington, T.S., Shepard, H.M., Patton, J.S., and Frost, G.I. (2006). A recombinant human enzyme for enhanced interstitial transport of therapeutics. J Control Release 114, 230-241.
Chao, K.L., Muthukumar, L., and Herzberg, O. (2007). Structure of human hyaluronidase-1, a hyaluronan hydrolyzing enzyme involved in tumor growth and angiogenesis. Biochemistry 46, 6911-6920.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> VLP Therapeutics, LLC
<120> ALPHAVIRUS REPLICON PARTICLE
<130> 674559
<150> US62/608,213
<151> 2017-12-20
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 275
<212> PRT
<213> Venezuelan Equine Encephalitis Virus
<400> 1
Met Phe Pro Phe Gln Pro Met Tyr Pro Met Gln Pro Met Pro Tyr Arg
1 5 10 15
Asn Pro Phe Ala Ala Pro Arg Arg Pro Trp Phe Pro Arg Thr Asp Pro
20 25 30
Phe Leu Ala Met Gln Val Gln Glu Leu Thr Arg Ser Met Ala Asn Leu
35 40 45
Thr Phe Lys Gln Arg Arg Asp Ala Pro Pro Glu Gly Pro Ser Ala Lys
50 55 60
Lys Pro Lys Lys Glu Ala Ser Gln Lys Gln Lys Gly Gly Gly Gln Gly
65 70 75 80
Lys Lys Lys Lys Asn Gln Gly Lys Lys Lys Ala Lys Thr Gly Pro Pro
85 90 95
Asn Pro Lys Ala Gln Asn Gly Asn Lys Lys Lys Thr Asn Lys Lys Pro
100 105 110
Gly Lys Arg Gln Arg Met Val Met Lys Leu Glu Ser Asp Lys Thr Phe
115 120 125
Pro Ile Met Leu Glu Gly Lys Ile Asn Gly Tyr Ala Cys Val Val Gly
130 135 140
Gly Lys Leu Phe Arg Pro Met His Val Glu Gly Lys Ile Asp Asn Asp
145 150 155 160
Val Leu Ala Ala Leu Lys Thr Lys Lys Ala Ser Lys Tyr Asp Leu Glu
165 170 175
Tyr Ala Asp Val Pro Gln Asn Met Arg Ala Asp Thr Phe Lys Tyr Thr
180 185 190
His Glu Lys Pro Gln Gly Tyr Tyr Ser Trp His His Gly Ala Val Gln
195 200 205
Tyr Glu Asn Gly Arg Phe Thr Val Pro Lys Gly Val Gly Ala Lys Gly
210 215 220
Asp Ser Gly Arg Pro Ile Leu Asp Asn Gln Gly Arg Val Val Ala Ile
225 230 235 240
Val Leu Gly Gly Val Asn Glu Gly Ser Arg Thr Ala Leu Ser Val Val
245 250 255
Met Trp Asn Glu Lys Gly Val Thr Val Lys Tyr Thr Pro Glu Asn Cys
260 265 270
Glu Gln Trp
275
<210> 2
<211> 275
<212> PRT
<213> Venezuelan Equine Encephalitis Virus
<400> 2
Met Phe Pro Phe Gln Pro Met Tyr Pro Met Gln Pro Met Pro Tyr Arg
1 5 10 15
Asn Pro Phe Ala Ala Pro Arg Arg Pro Trp Phe Pro Arg Thr Asp Pro
20 25 30
Phe Leu Ala Met Gln Val Gln Glu Leu Thr Arg Ser Met Ala Asn Leu
35 40 45
Thr Phe Lys Gln Arg Arg Asp Ala Pro Pro Glu Gly Pro Ser Ala Asn
50 55 60
Lys Pro Lys Lys Glu Ala Ser Gln Lys Gln Lys Gly Gly Gly Gln Gly
65 70 75 80
Lys Lys Lys Lys Asn Gln Gly Lys Lys Lys Ala Lys Thr Gly Pro Pro
85 90 95
Asn Pro Lys Ala Gln Asn Gly Asn Lys Lys Lys Thr Asn Lys Lys Pro
100 105 110
Gly Lys Arg Gln Arg Met Val Met Lys Leu Glu Ser Asp Lys Thr Phe
115 120 125
Pro Ile Met Leu Glu Gly Lys Ile Asn Gly Tyr Ala Cys Val Val Gly
130 135 140
Gly Lys Leu Phe Arg Pro Met His Val Glu Gly Lys Ile Asp Asn Asp
145 150 155 160
Val Leu Ala Ala Leu Lys Thr Lys Lys Ala Ser Lys Tyr Asp Leu Glu
165 170 175
Tyr Ala Asp Val Pro Gln Asn Met Arg Ala Asp Thr Phe Lys Tyr Thr
180 185 190
His Glu Lys Pro Gln Gly Tyr Tyr Ser Trp His His Gly Ala Val Gln
195 200 205
Tyr Glu Asn Gly Arg Phe Thr Val Pro Lys Gly Val Gly Ala Lys Gly
210 215 220
Asp Ser Gly Arg Pro Ile Leu Asp Asn Gln Gly Arg Val Val Ala Ile
225 230 235 240
Val Leu Gly Gly Val Asn Glu Gly Ser Arg Thr Ala Leu Ser Val Val
245 250 255
Met Trp Asn Glu Lys Gly Val Thr Val Lys Tyr Thr Pro Glu Asn Cys
260 265 270
Glu Gln Trp
275
<210> 3
<211> 980
<212> PRT
<213> Venezuelan Equine Encephalitis Virus
<400> 3
Ser Leu Val Thr Thr Met Cys Leu Leu Ala Asn Val Thr Phe Pro Cys
1 5 10 15
Ala Gln Pro Pro Ile Cys Tyr Asp Arg Lys Pro Ala Glu Thr Leu Ala
20 25 30
Met Leu Ser Val Asn Val Asp Asn Pro Gly Tyr Asp Glu Leu Leu Glu
35 40 45
Ala Ala Val Lys Cys Pro Gly Arg Lys Arg Arg Ser Thr Glu Glu Leu
50 55 60
Phe Asn Glu Tyr Lys Leu Thr Arg Pro Tyr Met Ala Arg Cys Ile Arg
65 70 75 80
Cys Ala Val Gly Ser Cys His Ser Pro Ile Ala Ile Glu Ala Val Lys
85 90 95
Ser Asp Gly His Asp Gly Tyr Val Arg Leu Gln Thr Ser Ser Gln Tyr
100 105 110
Gly Leu Asp Ser Ser Gly Asn Leu Lys Gly Arg Thr Met Arg Tyr Asp
115 120 125
Met His Gly Thr Ile Lys Glu Ile Pro Leu His Gln Val Ser Leu Tyr
130 135 140
Thr Ser Arg Pro Cys His Ile Val Asp Gly His Gly Tyr Phe Leu Leu
145 150 155 160
Ala Arg Cys Pro Ala Gly Asp Ser Ile Thr Met Glu Phe Lys Lys Asp
165 170 175
Ser Val Arg His Ser Cys Ser Val Pro Tyr Glu Val Lys Phe Asn Pro
180 185 190
Val Gly Arg Glu Leu Tyr Thr His Pro Pro Glu His Gly Val Glu Gln
195 200 205
Ala Cys Gln Val Tyr Ala His Asp Ala Gln Asn Arg Gly Ala Tyr Val
210 215 220
Glu Met His Leu Pro Gly Ser Glu Val Asp Ser Ser Leu Val Ser Leu
225 230 235 240
Ser Gly Ser Ser Val Thr Val Thr Pro Pro Asp Gly Thr Ser Ala Leu
245 250 255
Val Glu Cys Glu Cys Gly Gly Thr Lys Ile Ser Glu Thr Ile Asn Lys
260 265 270
Thr Lys Gln Phe Ser Gln Cys Thr Lys Lys Glu Gln Cys Arg Ala Tyr
275 280 285
Arg Leu Gln Asn Asp Lys Trp Val Tyr Asn Ser Asp Lys Leu Pro Lys
290 295 300
Ala Ala Gly Ala Thr Leu Lys Gly Lys Leu His Val Pro Phe Leu Leu
305 310 315 320
Ala Asp Gly Lys Cys Thr Val Pro Leu Ala Pro Glu Pro Met Ile Thr
325 330 335
Phe Gly Phe Arg Ser Val Ser Leu Lys Leu His Pro Lys Asn Pro Thr
340 345 350
Tyr Leu Ile Thr Arg Gln Leu Ala Asp Glu Pro His Tyr Thr His Glu
355 360 365
Leu Ile Ser Glu Pro Ala Val Arg Asn Phe Thr Val Thr Glu Lys Gly
370 375 380
Trp Glu Phe Val Trp Gly Asn His Pro Pro Lys Arg Phe Trp Ala Gln
385 390 395 400
Glu Thr Ala Pro Gly Asn Pro His Gly Leu Pro His Glu Val Ile Thr
405 410 415
His Tyr Tyr His Arg Tyr Pro Met Ser Thr Ile Leu Gly Leu Ser Ile
420 425 430
Cys Ala Ala Ile Ala Thr Val Ser Val Ala Ala Ser Thr Trp Leu Phe
435 440 445
Cys Arg Ser Arg Val Ala Cys Leu Thr Pro Tyr Arg Leu Thr Pro Asn
450 455 460
Ala Arg Ile Pro Phe Cys Leu Ala Val Leu Cys Cys Ala Arg Thr Ala
465 470 475 480
Arg Ala Glu Thr Thr Trp Glu Ser Leu Asp His Leu Trp Asn Asn Asn
485 490 495
Gln Gln Met Phe Trp Ile Gln Leu Leu Ile Pro Leu Ala Ala Leu Ile
500 505 510
Val Val Thr Arg Leu Leu Arg Cys Val Cys Cys Val Val Pro Phe Leu
515 520 525
Val Met Ala Gly Ala Ala Gly Ala Gly Ala Tyr Glu His Ala Thr Thr
530 535 540
Met Pro Ser Gln Ala Gly Ile Ser Tyr Asn Thr Ile Val Asn Arg Ala
545 550 555 560
Gly Tyr Ala Pro Leu Pro Ile Ser Ile Thr Pro Thr Lys Ile Lys Leu
565 570 575
Ile Pro Thr Val Asn Leu Glu Tyr Val Thr Cys His Tyr Lys Thr Gly
580 585 590
Met Asp Ser Pro Ala Ile Lys Cys Cys Gly Ser Gln Glu Cys Thr Pro
595 600 605
Thr Tyr Arg Pro Asp Glu Gln Cys Lys Val Phe Thr Gly Val Tyr Pro
610 615 620
Phe Met Trp Gly Gly Ala Tyr Cys Phe Cys Asp Thr Glu Asn Thr Gln
625 630 635 640
Val Ser Lys Ala Tyr Val Met Lys Ser Asp Asp Cys Leu Ala Asp His
645 650 655
Ala Glu Ala Tyr Lys Ala His Thr Ala Ser Val Gln Ala Phe Leu Asn
660 665 670
Ile Thr Val Gly Glu His Ser Ile Val Thr Thr Val Tyr Val Asn Gly
675 680 685
Glu Thr Pro Val Asn Phe Asn Gly Val Lys Ile Thr Ala Gly Pro Leu
690 695 700
Ser Thr Ala Trp Thr Pro Phe Asp Arg Lys Ile Val Gln Tyr Ala Gly
705 710 715 720
Glu Ile Tyr Asn Tyr Asp Phe Pro Glu Tyr Gly Ala Gly Gln Pro Gly
725 730 735
Ala Phe Gly Asp Ile Gln Ser Arg Thr Val Ser Ser Ser Asp Leu Tyr
740 745 750
Ala Asn Thr Asn Leu Val Leu Gln Arg Pro Lys Ala Gly Ala Ile His
755 760 765
Val Pro Tyr Thr Gln Ala Pro Ser Gly Phe Glu Gln Trp Lys Lys Asp
770 775 780
Lys Ala Pro Ser Leu Lys Phe Thr Ala Pro Phe Gly Cys Glu Ile Tyr
785 790 795 800
Thr Asn Pro Ile Arg Ala Glu Asn Cys Ala Val Gly Ser Ile Pro Leu
805 810 815
Ala Phe Asp Ile Pro Asp Ala Leu Phe Thr Arg Val Ser Glu Thr Pro
820 825 830
Thr Leu Ser Ala Ala Glu Cys Thr Leu Asn Glu Cys Val Tyr Ser Ser
835 840 845
Asp Phe Gly Gly Ile Ala Thr Val Lys Tyr Ser Ala Ser Lys Ser Gly
850 855 860
Lys Cys Ala Val His Val Pro Ser Gly Thr Ala Thr Leu Lys Glu Ala
865 870 875 880
Ala Val Glu Leu Thr Glu Gln Gly Ser Ala Thr Ile His Phe Ser Thr
885 890 895
Ala Asn Ile His Pro Glu Phe Arg Leu Gln Ile Cys Thr Ser Tyr Val
900 905 910
Thr Cys Lys Gly Asp Cys His Pro Pro Lys Asp His Ile Val Thr His
915 920 925
Pro Gln Tyr His Ala Gln Thr Phe Thr Ala Ala Val Ser Lys Thr Ala
930 935 940
Trp Thr Trp Leu Thr Ser Leu Leu Gly Gly Ser Ala Val Ile Ile Ile
945 950 955 960
Ile Gly Leu Val Leu Ala Thr Ile Val Ala Met Tyr Val Leu Thr Asn
965 970 975
Gln Lys His Asn
980
<210> 4
<211> 981
<212> PRT
<213> Venezuelan Equine Encephalitis Virus
<400> 4
Ser Leu Val Thr Thr Met Cys Leu Leu Ala Asn Val Thr Phe Pro Cys
1 5 10 15
Ala Gln Pro Pro Ile Cys Tyr Asp Arg Lys Pro Ala Glu Thr Leu Ala
20 25 30
Met Leu Ser Val Asn Val Asp Asn Pro Gly Tyr Asp Glu Leu Leu Glu
35 40 45
Ala Ala Val Lys Cys Pro Gly Ser Gly Gly Gly Ser Ser Thr Glu Glu
50 55 60
Leu Phe Asn Glu Tyr Lys Leu Thr Arg Pro Tyr Met Ala Arg Cys Ile
65 70 75 80
Arg Cys Ala Val Gly Ser Cys His Ser Pro Ile Ala Ile Glu Ala Val
85 90 95
Lys Ser Asp Gly His Asp Gly Tyr Val Arg Leu Gln Thr Ser Ser Gln
100 105 110
Tyr Gly Leu Asp Ser Ser Gly Asn Leu Lys Gly Arg Thr Met Arg Tyr
115 120 125
Asp Met His Gly Thr Ile Lys Glu Ile Pro Leu His Gln Val Ser Leu
130 135 140
Tyr Thr Ser Arg Pro Cys His Ile Val Asp Gly His Gly Tyr Phe Leu
145 150 155 160
Leu Ala Arg Cys Pro Ala Gly Asp Ser Ile Thr Met Glu Phe Lys Lys
165 170 175
Asp Ser Val Arg His Ser Cys Ser Val Pro Tyr Glu Val Lys Phe Asn
180 185 190
Pro Val Gly Arg Glu Leu Tyr Thr His Pro Pro Glu His Gly Val Glu
195 200 205
Gln Ala Cys Gln Val Tyr Ala His Asp Ala Gln Asn Arg Gly Ala Tyr
210 215 220
Val Glu Met His Leu Pro Gly Ser Glu Val Asp Ser Ser Leu Val Ser
225 230 235 240
Leu Ser Gly Ser Ser Val Thr Val Thr Pro Pro Asp Gly Thr Ser Ala
245 250 255
Leu Val Glu Cys Glu Cys Gly Gly Thr Lys Ile Ser Glu Thr Ile Asn
260 265 270
Lys Thr Lys Gln Phe Ser Gln Cys Thr Lys Lys Glu Gln Cys Arg Ala
275 280 285
Tyr Arg Leu Gln Asn Asp Lys Trp Val Tyr Asn Ser Asp Lys Leu Pro
290 295 300
Lys Ala Ala Gly Ala Thr Leu Lys Gly Lys Leu His Val Pro Phe Leu
305 310 315 320
Leu Ala Asp Gly Lys Cys Thr Val Pro Leu Ala Pro Glu Pro Met Ile
325 330 335
Thr Phe Gly Phe Arg Ser Val Ser Leu Lys Leu His Pro Lys Asn Pro
340 345 350
Thr Tyr Leu Ile Thr Arg Gln Leu Ala Asp Glu Pro His Tyr Thr His
355 360 365
Glu Leu Ile Ser Glu Pro Ala Val Arg Asn Phe Thr Val Thr Glu Lys
370 375 380
Gly Trp Glu Phe Val Trp Gly Asn His Pro Pro Lys Arg Phe Trp Ala
385 390 395 400
Gln Glu Thr Ala Pro Gly Asn Pro His Gly Leu Pro His Glu Val Ile
405 410 415
Thr His Tyr Tyr His Arg Tyr Pro Met Ser Thr Ile Leu Gly Leu Ser
420 425 430
Ile Cys Ala Ala Ile Ala Thr Val Ser Val Ala Ala Ser Thr Trp Leu
435 440 445
Phe Cys Arg Ser Arg Val Ala Cys Leu Thr Pro Tyr Arg Leu Thr Pro
450 455 460
Asn Ala Arg Ile Pro Phe Cys Leu Ala Val Leu Cys Cys Ala Arg Thr
465 470 475 480
Ala Arg Ala Glu Thr Thr Trp Glu Ser Leu Asp His Leu Trp Asn Asn
485 490 495
Asn Gln Gln Met Phe Trp Ile Gln Leu Leu Ile Pro Leu Ala Ala Leu
500 505 510
Ile Val Val Thr Arg Leu Leu Arg Cys Val Cys Cys Val Val Pro Phe
515 520 525
Leu Val Met Ala Gly Ala Ala Gly Ala Gly Ala Tyr Glu His Ala Thr
530 535 540
Thr Met Pro Ser Gln Ala Gly Ile Ser Tyr Asn Thr Ile Val Asn Arg
545 550 555 560
Ala Gly Tyr Ala Pro Leu Pro Ile Ser Ile Thr Pro Thr Lys Ile Lys
565 570 575
Leu Ile Pro Thr Val Asn Leu Glu Tyr Val Thr Cys His Tyr Lys Thr
580 585 590
Gly Met Asp Ser Pro Ala Ile Lys Cys Cys Gly Ser Gln Glu Cys Thr
595 600 605
Pro Thr Tyr Arg Pro Asp Glu Gln Cys Lys Val Phe Thr Gly Val Tyr
610 615 620
Pro Phe Met Trp Gly Gly Ala Tyr Cys Phe Cys Asp Thr Glu Asn Thr
625 630 635 640
Gln Val Ser Lys Ala Tyr Val Met Lys Ser Asp Asp Cys Leu Ala Asp
645 650 655
His Ala Glu Ala Tyr Lys Ala His Thr Ala Ser Val Gln Ala Phe Leu
660 665 670
Asn Ile Thr Val Gly Glu His Ser Ile Val Thr Thr Val Tyr Val Asn
675 680 685
Gly Glu Thr Pro Val Asn Phe Asn Gly Val Lys Ile Thr Ala Gly Pro
690 695 700
Leu Ser Thr Ala Trp Thr Pro Phe Asp Arg Lys Ile Val Gln Tyr Ala
705 710 715 720
Gly Glu Ile Tyr Asn Tyr Asp Phe Pro Glu Tyr Gly Ala Gly Gln Pro
725 730 735
Gly Ala Phe Gly Asp Ile Gln Ser Arg Thr Val Ser Ser Ser Asp Leu
740 745 750
Tyr Ala Asn Thr Asn Leu Val Leu Gln Arg Pro Lys Ala Gly Ala Ile
755 760 765
His Val Pro Tyr Thr Gln Ala Pro Ser Gly Phe Glu Gln Trp Lys Lys
770 775 780
Asp Lys Ala Pro Ser Leu Lys Phe Thr Ala Pro Phe Gly Cys Glu Ile
785 790 795 800
Tyr Thr Asn Pro Ile Arg Ala Glu Asn Cys Ala Val Gly Ser Ile Pro
805 810 815
Leu Ala Phe Asp Ile Pro Asp Ala Leu Phe Thr Arg Val Ser Glu Thr
820 825 830
Pro Thr Leu Ser Ala Ala Glu Cys Thr Leu Asn Glu Cys Val Tyr Ser
835 840 845
Ser Asp Phe Gly Gly Ile Ala Thr Val Lys Tyr Ser Ala Ser Lys Ser
850 855 860
Gly Lys Cys Ala Val His Val Pro Ser Gly Thr Ala Thr Leu Lys Glu
865 870 875 880
Ala Ala Val Glu Leu Thr Glu Gln Gly Ser Ala Thr Ile His Phe Ser
885 890 895
Thr Ala Asn Ile His Pro Glu Phe Arg Leu Gln Ile Cys Thr Ser Tyr
900 905 910
Val Thr Cys Lys Gly Asp Cys His Pro Pro Lys Asp His Ile Val Thr
915 920 925
His Pro Gln Tyr His Ala Gln Thr Phe Thr Ala Ala Val Ser Lys Thr
930 935 940
Ala Trp Thr Trp Leu Thr Ser Leu Leu Gly Gly Ser Ala Val Ile Ile
945 950 955 960
Ile Ile Gly Leu Val Leu Ala Thr Ile Val Ala Met Tyr Val Leu Thr
965 970 975
Asn Gln Lys His Asn
980
<210> 5
<211> 11542
<212> DNA
<213> Artificial Sequence
<220>
<223> Genomic RNA expression Vector including Luciferase gene
nucleotide
<400> 5
ttctcatgtt tgacagctta tcatcgatgc accgtgcatg ccgattggtg gaagtaaggt 60
ggtacgatcg tgccttatta ggaaggcaac agacaggtct gacatggatt ggacgaacca 120
ctgaattccg cattgcagag ataattgtat ttaagtgcct agctcgatac aataaacgcc 180
atttgaccat tcaccacatt ggtgtgcacc tccaataggc ggcgcatgag agaagcccag 240
accaattacc tacccaaaat ggagaaagtt cacgttgaca tcgaggaaga cagcccattc 300
ctcagagctt tgcagcggag cttcccgcag tttgaggtag aagccaagca ggtcactgat 360
aatgaccatg ctaatgccag agcgttttcg catctggctt caaaactgat cgaaacggag 420
gtggacccat ccgacacgat ccttgacatt ggaagtgcgc ccgcccgcag aatgtattct 480
aagcacaagt atcattgtat ctgtccgatg agatgtgcgg aagatccgga cagattgtat 540
aagtatgcaa ctaagctgaa gaaaaactgt aaggaaataa ctgataagga attggacaag 600
aaaatgaagg agctcgccgc cgtcatgagc gaccctgacc tggaaactga gactatgtgc 660
ctccacgacg acgagcgtgt cgctacgaag ggcaagtcgc tgtttaccag gatgtatacg 720
cggttgacgg accgacaagt ctctatcacc aagccaataa gggagttaga gtcgcctact 780
ggataggctt tgacaccacc ccttttatgt ttaagaactt ggctggagca tatccatcat 840
actctaccaa ctgggccgac gaaaccgtgt taacggctcg taacataggc ctatgcagct 900
ctgacgttat ggagcggtca cgtagaggga tgtccattct tagaaagaag tatttgaaac 960
catccaacaa tgttctattc tctgttggct cgaccatcta ccacgagaag agggacttac 1020
tgaggagctg gcacctgccg tctgtatttc acttacgtgg caagcaaaat tacacatgtc 1080
ggtgtgagac tatagttagt tgcgacgggt acgtcgttaa aagaatagct atcagtccag 1140
gcctgtatgg gaagccttca ggctatgctg ctacgatgca ccgcgaggga ttcttgtgct 1200
gcaaagtgac agacacattg aacggggaga gggtctcttt tcccgtgtgc acgtatgtgc 1260
cagctacatt gtgtgaccaa atgactggca tactggcaac agatgtcagt gcggacgacg 1320
cgcaaaaact gctggttggg ctcaaccagc gtatagtcgt caacggtcgc acccagagaa 1380
acaccaatac catgaaaaat taccttttgc ccgtagtggc ccaggcattt gctaggtggg 1440
caaaggaata taaggaagat caagaagatg aaaggccact aggactacga gatagacagt 1500
tagtcatggg gtgttgttgg gcttttagaa ggcacaagat aacatctatt tataagcgcc 1560
cggataccca aaccatcatc aaagtgaaca gcgatttcca ctcattcgtg ctgcccagga 1620
taggcagtaa cacattggag atcgggctga gaacaagaat caggaaaatg ttagaggagc 1680
acaaggagcc gtcacctctc attaccgccg aggacgtaca agaagctaag tgcgcagccg 1740
atgaggctaa ggaggtgcgt gaagccgagg agttgcgcgc agctctacca cctttggcag 1800
ctgatgttga ggagcccact ctggaagccg atgtcgactt gatgttacaa gaggctgggg 1860
ccggctcagt ggagacacct cgtggcttga taaaggttac cagctacgat ggcgaggaca 1920
agatcggctc ttacgctgtg ctttctccgc aggctgtact caagagtgaa aaattatctt 1980
gcatccaccc tctcgctgaa caagtcatag tgataacaca ctctggccga aaagggcgtt 2040
atgccgtgga accataccat ggtaaagtag tggtgccaga gggacatgca atacccgtcc 2100
aggactttca agctctgagt gaaagtgcca ccattgtgta caacgaacgt gagttcgtaa 2160
acaggtacct gcaccatatt gccacacatg gaggagcgct gaacactgat gaagaatatt 2220
acaaaactgt caagcccagc gagcacgacg gcgaatacct gtacgacatc gacaggaaac 2280
agtgcgtcaa gaaagaacta gtcactgggc tagggctcac aggcgagctg gtggatcctc 2340
ccttccatga attcgcctac gagagtctga gaacacgacc agccgctcct taccaagtac 2400
caaccatagg ggtgtatggc gtgccaggat caggcaagtc tggcatcatt aaaagcgcag 2460
tcaccaaaaa agatctagtg gtgagcgcca agaaagaaaa ctgtgcagaa attataaggg 2520
acgtcaagaa aatgaaaggg ctggacgtca atgccagaac tgtggactca gtgctcttga 2580
atggatgcaa acaccccgta gagaccctgt atattgacga agcttttgct tgtcatgcag 2640
gtactctcag agcgctcata gccattataa gacctaaaaa ggcagtgctc tgcggggatc 2700
ccaaacagtg cggttttttt aacatgatgt gcctgaaagt gcattttaac cacgagattt 2760
gcacacaagt cttccacaaa agcatctctc gccgttgcac taaatctgtg acttcggtcg 2820
tctcaacctt gttttacgac aaaaaaatga gaacgacgaa tccgaaagag actaagattg 2880
tgattgacac taccggcagt accaaaccta agcaggacga tctcattctc acttgtttca 2940
gagggtgggt gaagcagttg caaatagatt acaaaggcaa cgaaataatg acggcagctg 3000
cctctcaagg gctgacccgt aaaggtgtgt atgccgttcg gtacaaggtg aatgaaaatc 3060
ctctgtacgc acccacctca gaacatgtga acgtcctact gacccgcacg gaggaccgca 3120
tcgtgtggaa aacactagcc ggcgacccat ggataaaaac actgactgcc aagtaccctg 3180
ggaatttcac tgccacgata gaggagtggc aagcagagca tgatgccatc atgaggcaca 3240
tcttggagag accggaccct accgacgtct tccagaataa ggcaaacgtg tgttgggcca 3300
aggctttagt gccggtgctg aagaccgctg gcatagacat gaccactgaa caatggaaca 3360
ctgtggatta ttttgaaacg gacaaagctc actcagcaga gatagtattg aaccaactat 3420
gcgtgaggtt ctttggactc gatctggact ccggtctatt ttctgcaccc actgttccgt 3480
tatccattag gaataatcac tgggataact ccccgtcgcc taacatgtac gggctgaata 3540
aagaagtggt ccgtcagctc tctcgcaggt acccacaact gcctcgggca gttgccactg 3600
gaagagtcta tgacatgaac actggtacac tgcgcaatta tgatccgcgc ataaacctag 3660
tacctgtaaa cagaagactg cctcatgctt tagtcctcca ccataatgaa cacccacaga 3720
gtgacttttc ttcattcgtc agcaaattga agggcagaac tgtcctggtg gtcggggaaa 3780
agttgtccgt cccaggcaaa atggttgact ggttgtcaga ccggcctgag gctaccttca 3840
gagctcggct ggatttaggc atcccaggtg atgtgcccaa atatgacata atatttgtta 3900
atgtgaggac cccatataaa taccatcact atcagcagtg tgaagaccat gccattaagc 3960
ttagcatgtt gaccaagaaa gcttgtctgc atctgaatcc cggcggaacc tgtgtcagca 4020
taggttatgg ttacgctgac agggccagcg aaagcatcat tggtgctata gcgcggcagt 4080
tcaagttttc ccgggtatgc aaaccgaaat cctcacttga agagacggaa gttctgtttg 4140
tattcattgg gtacgatcgc aaggcccgta cgcacaatcc ttacaagctt tcatcaacct 4200
tgaccaacat ttatacaggt tccagactcc acgaagccgg atgtgcaccc tcatatcatg 4260
tggtgcgagg ggatattgcc acggccaccg aaggagtgat tataaatgct gctaacagca 4320
aaggacaacc tggcggaggg gtgtgcggag cgctgtataa gaaattcccg gaaagcttcg 4380
atttacagcc gatcgaagta ggaaaagcgc gactggtcaa aggtgcagct aaacatatca 4440
ttcatgccgt aggaccaaac ttcaacaaag tttcggaggt tgaaggtgac aaacagttgg 4500
cagaggctta tgagtccatc gctaagattg tcaacgataa caattacaag tcagtagcga 4560
ttccactgtt gtccaccggc atcttttccg ggaacaaaga tcgactaacc caatcattga 4620
accatttgct gacagcttta gacaccactg atgcagatgt agccatatac tgcagggaca 4680
agaaatggga aatgactctc aaggaagcag tggctaggag agaagcagtg gaggagatat 4740
gcatatccga cgactcttca gtgacagaac ctgatgcaga gctggtgagg gtgcatccga 4800
agagttcttt ggctggaagg aagggctaca gcacaagcga tggcaaaact ttctcatatt 4860
tggaagggac caagtttcac caggcggcca aggatatagc agaaattaat gccatgtggc 4920
ccgttgcaac ggaggccaat gagcaggtat gcatgtatat cctcggagaa agcatgagca 4980
gtattaggtc gaaatgcccc gtcgaagagt cggaagcctc cacaccacct agcacgctgc 5040
cttgcttgtg catccatgcc atgactccag aaagagtaca gcgcctaaaa gcctcacgtc 5100
cagaacaaat tactgtgtgc tcatcctttc cattgccgaa gtatagaatc actggtgtgc 5160
agaagatcca atgctcccag cctatattgt tctcaccgaa agtgcctgcg tatattcatc 5220
caaggaagta tctcgtggaa acaccaccgg tagacgagac tccggagcca tcggcagaga 5280
accaatccac agaggggaca cctgaacaac caccacttat aaccgaggat gagaccagga 5340
ctagaacgcc tgagccgatc atcatcgaag aggaagaaga ggatagcata agtttgctgt 5400
cagatggccc gacccaccag gtgctgcaag tcgaggcaga cattcacggg ccgccctctg 5460
tatctagctc atcctggtcc attcctcatg catccgactt tgatgtggac agtttatcca 5520
tacttgacac cctggaggga gctagcgtga ccagcggggc aacgtcagcc gagactaact 5580
cttacttcgc aaagagtatg gagtttctgg cgcgaccggt gcctgcgcct cgaacagtat 5640
tcaggaaccc tccacatccc gctccgcgca caagaacacc gtcacttgca cccagcaggg 5700
cctgctcgag aaccagccta gtttccaccc cgccaggcgt gaatagggtg atcactagag 5760
aggagctcga ggcgcttacc ccgtcacgca ctcctagcag gtcggtctcg agaaccagcc 5820
tggtctccaa cccgccaggc gtaaataggg tgattacaag agaggagttt gaggcgttcg 5880
tagcacaaca acaatgacgg tttgatgcgg gtgcatacat cttttcctcc gacaccggtc 5940
aagggcattt acaacaaaaa tcagtaaggc aaacggtgct atccgaagtg gtgttggaga 6000
ggaccgaatt ggagatttcg tatgccccgc gcctcgacca agaaaaagaa gaattactac 6060
gcaagaaatt acagttaaat cccacacctg ctaacagaag cagataccag tccaggaagg 6120
tggagaacat gaaagccata acagctagac gtattctgca aggcctaggg cattatttga 6180
aggcagaagg aaaagtggag tgctaccgaa ccctgcatcc tgttcctttg tattcatcta 6240
gtgtgaaccg tgccttttca agccccaagg tcgcagtgga agcctgtaac gccatgttga 6300
aagagaactt tccgactgtg gcttcttact gtattattcc agagtacgat gcctatttgg 6360
acatggttga cggagcttca tgctgcttag acactgccag tttttgccct gcaaagctgc 6420
gcagctttcc aaagaaacac tcctatttgg aacccacaat acgatcggca gtgccttcag 6480
cgatccagaa cacgctccag aacgtcctgg cagctgccac aaaaagaaat tgcaatgtca 6540
cgcaaatgag agaattgccc gtattggatt cggcggcctt taatgtggaa tgcttcaaga 6600
aatatgcgtg taataatgaa tattgggaaa cgtttaaaga aaaccccatc aggcttactg 6660
aagaaaacgt ggtaaattac attaccaaat taaaaggacc aaaagctgct gctctttttg 6720
cgaagacaca taatttgaat atgttgcagg acataccaat ggacaggttt gtaatggact 6780
taaagagaga cgtgaaagtg actccaggaa caaaacatac tgaagaacgg cccaaggtac 6840
aggtgatcca ggctgccgat ccgctagcaa cagcgtatct gtgcggaatc caccgagagc 6900
tggttaggag attaaatgcg gtcctgcttc cgaacattca tacactgttt gatatgtcgg 6960
ctgaagactt tgacgctatt atagccgagc acttccagcc tggggattgt gttctggaaa 7020
ctgacatcgc gtcgtttgat aaaagtgagg acgacgccat ggctctgacc gcgttaatga 7080
ttctggaaga cttaggtgtg gacgcagagc tgttgacgct gattgaggcg gctttcggcg 7140
aaatttcatc aatacatttg cccactaaaa ctaaatttaa attcggagcc atgatgaaat 7200
ctggaatgtt cctcacactg tttgtgaaca cagtcattaa cattgtaatc gcaagcagag 7260
tgttgagaga acggctaacc ggatcaccat gtgcagcatt cattggagat gacaatatcg 7320
tgaaaggagt caaatcggac aaattaatgg cagacaggtg cgccacctgg ttgaatatgg 7380
aagtcaagat tatagatgct gtggtgggcg agaaagcgcc ttatttctgt ggagggttta 7440
ttttgtgtga ctccgtgacc ggcacagcgt gccgtgtggc agacccccta aaaaggctgt 7500
ttaagcttgg caaacctctg gcagcagacg atgaacatga tgatgacagg agaagggcat 7560
tgcatgaaga gtcaacacgc tggaaccgag tgggtattct ttcagagctg tgcaaggcag 7620
tagaatcaag gtatgaaacc gtaggaactt ccatcatagt tatggccatg actactctag 7680
ctagcagtgt taaatcattc agctacctga gaggggcccc tataactctc tacggctaac 7740
ctgaatggac tacgacatag tctagtccgc caagatggtc ttcacactcg aagatttcgt 7800
tggggactgg cgacagacag ccggctacaa cctggaccaa gtccttgaac agggaggtgt 7860
gtccagtttg tttcagaatc tcggggtgtc cgtaactccg atccaaagga ttgtcctgag 7920
cggtgaaaat gggctgaaga tcgacatcca tgtcatcatc ccgtatgaag gtctgagcgg 7980
cgaccaaatg ggccagatcg aaaaaatttt taaggtggtg taccctgtgg atgatcatca 8040
ctttaaggtg atcctgcact atggcacact ggtaatcgac ggggttacgc cgaacatgat 8100
cgactatttc ggacggccgt atgaaggcat cgccgtgttc gacggcaaaa agatcactgt 8160
aacagggacc ctgtggaacg gcaacaaaat tatcgacgag cgcctgatca accccgacgg 8220
ctccctgctg ttccgagtaa ccatcaacgg agtgaccggc tggcggctgt gcgaacgcat 8280
tctggcgtaa atacagcagc aattggcaag ctgcttacat agaactcgcg gcgattggca 8340
tgccgcctta aaatttttat tttatttttc ttttcttttc cgaatcggat tttgttttta 8400
atatttcaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 8460
aatcgcgaca gacatgataa gatacattga tgagtttgga caaaccacaa ctagaatgca 8520
gtgaaaaaaa tgctttattt gtgaaatttg tgatgctatt gctttatttg taaccattat 8580
aagctgcaat aaacaagtta acaacaacaa ttgcattcat tttatgtttc aggttcaggg 8640
ggaggtgtgg gaggtttttt aggacccggc taggctggcg gggttgcctt actggttagc 8700
agaatgaatc accgatacgc gagcgaacgt gaagcgactg ctgctgcaaa acgtctgcga 8760
cctgagcaac aacatgaatg gtcttcggtt tccgtgtttc gtaaagtctg gaaacgcgga 8820
agtcagcgcc ctgcaccatt atgttccgga tctgcatcgc aggatgctgc tggctaccct 8880
gtggaacacc tacatctgta ttaacgaagc gctggcattg accctgagtg atttttctct 8940
ggtcccgccg catccatacc gccagttgtt taccctcaca acgttccagt aaccgggcat 9000
gttcatcatc agtaacccgt atcgtgagca tcctctctcg tttcatcggt atcattaccc 9060
ccatgaacag aaatccccct tacacggagg catcagtgac caaacaggaa aaaaccgccc 9120
ttaacatggc ccgctttatc agaagccaga cattaacgct tctggagaaa ctcaacgagc 9180
tggacgcgga tgaacaggca gacatctgtg aatcgcttca cgaccacgct gatgagcttt 9240
accgcagctg cctcgcgcgt ttcggtgatg acggtgaaaa cctctgacac atgcagctcc 9300
cggagacggt cacagcttgt ctgtaagcgg atgccgggag cagacaagcc cgtcagggcg 9360
cgtcagcggg tgttggcggg tgtcggggcg cagccatgac ccagtcacgt agcgatagcg 9420
gagtgtatac tggcttaact atgcggcatc agagcagatt gtactgagag tgcaccatat 9480
gcggtgtgaa ataccgcaca gatgcgtaag gagaaaatac cgcatcaggc gctcttccgc 9540
ttcctcgctc actgactcgc tgcgctcggt cgttcggctg cggcgagcgg tatcagctca 9600
ctcaaaggcg gtaatacggt tatccacaga atcaggggat aacgcaggaa agaacatgtg 9660
agcaaaaggc cagcaaaagg ccaggaaccg taaaaaggcc gcgttgctgg cgtttttcca 9720
taggctccgc ccccctgacg agcatcacaa aaatcgacgc tcaagtcaga ggtggcgaaa 9780
cccgacagga ctataaagat accaggcgtt tccccctgga agctccctcg tgcgctctcc 9840
tgttccgacc ctgccgctta ccggatacct gtccgccttt ctcccttcgg gaagcgtggc 9900
gctttctcat agctcacgct gtaggtatct cagttcggtg taggtcgttc gctccaagct 9960
gggctgtgtg cacgaacccc ccgttcagcc cgaccgctgc gccttatccg gtaactatcg 10020
tcttgagtcc aacccggtaa gacacgactt atcgccactg gcagcagcca ctggtaacag 10080
gattagcaga gcgaggtatg taggcggtgc tacagagttc ttgaagtggt ggcctaacta 10140
cggctacact agaaggacag tatttggtat ctgcgctctg ctgaagccag ttaccttcgg 10200
aaaaagagtt ggtagctctt gatccggcaa acaaaccacc gctggtagcg gtggtttttt 10260
tgtttgcaag cagcagatta cgcgcagaaa aaaaggatct caagaagatc ctttgatctt 10320
ttctacgggg tctgacgctc agtggaacga aaactcacgt taagggattt tggtcatgag 10380
attatcaaaa aggatcttca cctagatcct tttaaattaa aaatgaagtt ttaaatcaat 10440
ctaaagtata tatgagtaaa cttggtctga cagttaccaa tgcttaatca gtgaggcacc 10500
tatctcagcg atctgtctat ttcgttcatc catagttgcc tgactccccg tcgtgtagat 10560
aactacgata cgggagggct taccatctgg ccccagtgct gcaatgatac cgcgagaccc 10620
acgctcaccg gctccagatt tatcagcaat aaaccagcca gccggaaggg ccgagcgcag 10680
aagtggtcct gcaactttat ccgcctccat ccagtctatt aattgttgcc gggaagctag 10740
agtaagtagt tcgccagtta atagtttgcg caacgttgtt gccattgctg caggcatcgt 10800
ggtgtcacgc tcgtcgtttg gtatggcttc attcagctcc ggttcccaac gatcaaggcg 10860
agttacatga tcccccatgt tgtgcaaaaa agcggttagc tccttcggtc ctccgatcgt 10920
tgtcagaagt aagttggccg cagtgttatc actcatggtt atggcagcac tgcataattc 10980
tcttactgtc atgccatccg taagatgctt ttctgtgact ggtgagtact caaccaagtc 11040
attctgagaa tagtgtatgc ggcgaccgag ttgctcttgc ccggcgtcaa cacgggataa 11100
taccgcgcca catagcagaa ctttaaaagt gctcatcatt ggaaaacgtt cttcggggcg 11160
aaaactctca aggatcttac cgctgttgag atccagttcg atgtaaccca ctcgtgcacc 11220
caactgatct tcagcatctt ttactttcac cagcgtttct gggtgagcaa aaacaggaag 11280
gcaaaatgcc gcaaaaaagg gaataagggc gacacggaaa tgttgaatac tcatactctt 11340
cctttttcaa tattattgaa gcatttatca gggttattgt ctcatgagcg gatacatatt 11400
tgaatgtatt tagaaaaata aacaaatagg ggttccgcgc acatttcccc gaaaagtgcc 11460
acctgacgtc taagaaacca ttattatcat gacattaacc tataaaaata ggcgtatcac 11520
gaggcccttt cgtcttcaag aa 11542
<210> 6
<211> 11596
<212> DNA
<213> Artificial Sequence
<220>
<223> Genomic RNA expression Vector including Luciferase gene with
Multiple cloning sites
<400> 6
ttctcatgtt tgacagctta tcatcgatgc accgtgcatg ccgattggtg gaagtaaggt 60
ggtacgatcg tgccttatta ggaaggcaac agacaggtct gacatggatt ggacgaacca 120
ctgaattccg cattgcagag ataattgtat ttaagtgcct agctcgatac aataaacgcc 180
atttgaccat tcaccacatt ggtgtgcacc tccaataggc ggcgcatgag agaagcccag 240
accaattacc tacccaaaat ggagaaagtt cacgttgaca tcgaggaaga cagcccattc 300
ctcagagctt tgcagcggag cttcccgcag tttgaggtag aagccaagca ggtcactgat 360
aatgaccatg ctaatgccag agcgttttcg catctggctt caaaactgat cgaaacggag 420
gtggacccat ccgacacgat ccttgacatt ggaagtgcgc ccgcccgcag aatgtattct 480
aagcacaagt atcattgtat ctgtccgatg agatgtgcgg aagatccgga cagattgtat 540
aagtatgcaa ctaagctgaa gaaaaactgt aaggaaataa ctgataagga attggacaag 600
aaaatgaagg agctcgccgc cgtcatgagc gaccctgacc tggaaactga gactatgtgc 660
ctccacgacg acgagcgtgt cgctacgaag ggcaagtcgc tgtttaccag gatgtatacg 720
cggttgacgg accgacaagt ctctatcacc aagccaataa gggagttaga gtcgcctact 780
ggataggctt tgacaccacc ccttttatgt ttaagaactt ggctggagca tatccatcat 840
actctaccaa ctgggccgac gaaaccgtgt taacggctcg taacataggc ctatgcagct 900
ctgacgttat ggagcggtca cgtagaggga tgtccattct tagaaagaag tatttgaaac 960
catccaacaa tgttctattc tctgttggct cgaccatcta ccacgagaag agggacttac 1020
tgaggagctg gcacctgccg tctgtatttc acttacgtgg caagcaaaat tacacatgtc 1080
ggtgtgagac tatagttagt tgcgacgggt acgtcgttaa aagaatagct atcagtccag 1140
gcctgtatgg gaagccttca ggctatgctg ctacgatgca ccgcgaggga ttcttgtgct 1200
gcaaagtgac agacacattg aacggggaga gggtctcttt tcccgtgtgc acgtatgtgc 1260
cagctacatt gtgtgaccaa atgactggca tactggcaac agatgtcagt gcggacgacg 1320
cgcaaaaact gctggttggg ctcaaccagc gtatagtcgt caacggtcgc acccagagaa 1380
acaccaatac catgaaaaat taccttttgc ccgtagtggc ccaggcattt gctaggtggg 1440
caaaggaata taaggaagat caagaagatg aaaggccact aggactacga gatagacagt 1500
tagtcatggg gtgttgttgg gcttttagaa ggcacaagat aacatctatt tataagcgcc 1560
cggataccca aaccatcatc aaagtgaaca gcgatttcca ctcattcgtg ctgcccagga 1620
taggcagtaa cacattggag atcgggctga gaacaagaat caggaaaatg ttagaggagc 1680
acaaggagcc gtcacctctc attaccgccg aggacgtaca agaagctaag tgcgcagccg 1740
atgaggctaa ggaggtgcgt gaagccgagg agttgcgcgc agctctacca cctttggcag 1800
ctgatgttga ggagcccact ctggaagccg atgtcgactt gatgttacaa gaggctgggg 1860
ccggctcagt ggagacacct cgtggcttga taaaggttac cagctacgat ggcgaggaca 1920
agatcggctc ttacgctgtg ctttctccgc aggctgtact caagagtgaa aaattatctt 1980
gcatccaccc tctcgctgaa caagtcatag tgataacaca ctctggccga aaagggcgtt 2040
atgccgtgga accataccat ggtaaagtag tggtgccaga gggacatgca atacccgtcc 2100
aggactttca agctctgagt gaaagtgcca ccattgtgta caacgaacgt gagttcgtaa 2160
acaggtacct gcaccatatt gccacacatg gaggagcgct gaacactgat gaagaatatt 2220
acaaaactgt caagcccagc gagcacgacg gcgaatacct gtacgacatc gacaggaaac 2280
agtgcgtcaa gaaagaacta gtcactgggc tagggctcac aggcgagctg gtggatcctc 2340
ccttccatga attcgcctac gagagtctga gaacacgacc agccgctcct taccaagtac 2400
caaccatagg ggtgtatggc gtgccaggat caggcaagtc tggcatcatt aaaagcgcag 2460
tcaccaaaaa agatctagtg gtgagcgcca agaaagaaaa ctgtgcagaa attataaggg 2520
acgtcaagaa aatgaaaggg ctggacgtca atgccagaac tgtggactca gtgctcttga 2580
atggatgcaa acaccccgta gagaccctgt atattgacga agcttttgct tgtcatgcag 2640
gtactctcag agcgctcata gccattataa gacctaaaaa ggcagtgctc tgcggggatc 2700
ccaaacagtg cggttttttt aacatgatgt gcctgaaagt gcattttaac cacgagattt 2760
gcacacaagt cttccacaaa agcatctctc gccgttgcac taaatctgtg acttcggtcg 2820
tctcaacctt gttttacgac aaaaaaatga gaacgacgaa tccgaaagag actaagattg 2880
tgattgacac taccggcagt accaaaccta agcaggacga tctcattctc acttgtttca 2940
gagggtgggt gaagcagttg caaatagatt acaaaggcaa cgaaataatg acggcagctg 3000
cctctcaagg gctgacccgt aaaggtgtgt atgccgttcg gtacaaggtg aatgaaaatc 3060
ctctgtacgc acccacctca gaacatgtga acgtcctact gacccgcacg gaggaccgca 3120
tcgtgtggaa aacactagcc ggcgacccat ggataaaaac actgactgcc aagtaccctg 3180
ggaatttcac tgccacgata gaggagtggc aagcagagca tgatgccatc atgaggcaca 3240
tcttggagag accggaccct accgacgtct tccagaataa ggcaaacgtg tgttgggcca 3300
aggctttagt gccggtgctg aagaccgctg gcatagacat gaccactgaa caatggaaca 3360
ctgtggatta ttttgaaacg gacaaagctc actcagcaga gatagtattg aaccaactat 3420
gcgtgaggtt ctttggactc gatctggact ccggtctatt ttctgcaccc actgttccgt 3480
tatccattag gaataatcac tgggataact ccccgtcgcc taacatgtac gggctgaata 3540
aagaagtggt ccgtcagctc tctcgcaggt acccacaact gcctcgggca gttgccactg 3600
gaagagtcta tgacatgaac actggtacac tgcgcaatta tgatccgcgc ataaacctag 3660
tacctgtaaa cagaagactg cctcatgctt tagtcctcca ccataatgaa cacccacaga 3720
gtgacttttc ttcattcgtc agcaaattga agggcagaac tgtcctggtg gtcggggaaa 3780
agttgtccgt cccaggcaaa atggttgact ggttgtcaga ccggcctgag gctaccttca 3840
gagctcggct ggatttaggc atcccaggtg atgtgcccaa atatgacata atatttgtta 3900
atgtgaggac cccatataaa taccatcact atcagcagtg tgaagaccat gccattaagc 3960
ttagcatgtt gaccaagaaa gcttgtctgc atctgaatcc cggcggaacc tgtgtcagca 4020
taggttatgg ttacgctgac agggccagcg aaagcatcat tggtgctata gcgcggcagt 4080
tcaagttttc ccgggtatgc aaaccgaaat cctcacttga agagacggaa gttctgtttg 4140
tattcattgg gtacgatcgc aaggcccgta cgcacaatcc ttacaagctt tcatcaacct 4200
tgaccaacat ttatacaggt tccagactcc acgaagccgg atgtgcaccc tcatatcatg 4260
tggtgcgagg ggatattgcc acggccaccg aaggagtgat tataaatgct gctaacagca 4320
aaggacaacc tggcggaggg gtgtgcggag cgctgtataa gaaattcccg gaaagcttcg 4380
atttacagcc gatcgaagta ggaaaagcgc gactggtcaa aggtgcagct aaacatatca 4440
ttcatgccgt aggaccaaac ttcaacaaag tttcggaggt tgaaggtgac aaacagttgg 4500
cagaggctta tgagtccatc gctaagattg tcaacgataa caattacaag tcagtagcga 4560
ttccactgtt gtccaccggc atcttttccg ggaacaaaga tcgactaacc caatcattga 4620
accatttgct gacagcttta gacaccactg atgcagatgt agccatatac tgcagggaca 4680
agaaatggga aatgactctc aaggaagcag tggctaggag agaagcagtg gaggagatat 4740
gcatatccga cgactcttca gtgacagaac ctgatgcaga gctggtgagg gtgcatccga 4800
agagttcttt ggctggaagg aagggctaca gcacaagcga tggcaaaact ttctcatatt 4860
tggaagggac caagtttcac caggcggcca aggatatagc agaaattaat gccatgtggc 4920
ccgttgcaac ggaggccaat gagcaggtat gcatgtatat cctcggagaa agcatgagca 4980
gtattaggtc gaaatgcccc gtcgaagagt cggaagcctc cacaccacct agcacgctgc 5040
cttgcttgtg catccatgcc atgactccag aaagagtaca gcgcctaaaa gcctcacgtc 5100
cagaacaaat tactgtgtgc tcatcctttc cattgccgaa gtatagaatc actggtgtgc 5160
agaagatcca atgctcccag cctatattgt tctcaccgaa agtgcctgcg tatattcatc 5220
caaggaagta tctcgtggaa acaccaccgg tagacgagac tccggagcca tcggcagaga 5280
accaatccac agaggggaca cctgaacaac caccacttat aaccgaggat gagaccagga 5340
ctagaacgcc tgagccgatc atcatcgaag aggaagaaga ggatagcata agtttgctgt 5400
cagatggccc gacccaccag gtgctgcaag tcgaggcaga cattcacggg ccgccctctg 5460
tatctagctc atcctggtcc attcctcatg catccgactt tgatgtggac agtttatcca 5520
tacttgacac cctggaggga gctagcgtga ccagcggggc aacgtcagcc gagactaact 5580
cttacttcgc aaagagtatg gagtttctgg cgcgaccggt gcctgcgcct cgaacagtat 5640
tcaggaaccc tccacatccc gctccgcgca caagaacacc gtcacttgca cccagcaggg 5700
cctgctcgag aaccagccta gtttccaccc cgccaggcgt gaatagggtg atcactagag 5760
aggagctcga ggcgcttacc ccgtcacgca ctcctagcag gtcggtctcg agaaccagcc 5820
tggtctccaa cccgccaggc gtaaataggg tgattacaag agaggagttt gaggcgttcg 5880
tagcacaaca acaatgacgg tttgatgcgg gtgcatacat cttttcctcc gacaccggtc 5940
aagggcattt acaacaaaaa tcagtaaggc aaacggtgct atccgaagtg gtgttggaga 6000
ggaccgaatt ggagatttcg tatgccccgc gcctcgacca agaaaaagaa gaattactac 6060
gcaagaaatt acagttaaat cccacacctg ctaacagaag cagataccag tccaggaagg 6120
tggagaacat gaaagccata acagctagac gtattctgca aggcctaggg cattatttga 6180
aggcagaagg aaaagtggag tgctaccgaa ccctgcatcc tgttcctttg tattcatcta 6240
gtgtgaaccg tgccttttca agccccaagg tcgcagtgga agcctgtaac gccatgttga 6300
aagagaactt tccgactgtg gcttcttact gtattattcc agagtacgat gcctatttgg 6360
acatggttga cggagcttca tgctgcttag acactgccag tttttgccct gcaaagctgc 6420
gcagctttcc aaagaaacac tcctatttgg aacccacaat acgatcggca gtgccttcag 6480
cgatccagaa cacgctccag aacgtcctgg cagctgccac aaaaagaaat tgcaatgtca 6540
cgcaaatgag agaattgccc gtattggatt cggcggcctt taatgtggaa tgcttcaaga 6600
aatatgcgtg taataatgaa tattgggaaa cgtttaaaga aaaccccatc aggcttactg 6660
aagaaaacgt ggtaaattac attaccaaat taaaaggacc aaaagctgct gctctttttg 6720
cgaagacaca taatttgaat atgttgcagg acataccaat ggacaggttt gtaatggact 6780
taaagagaga cgtgaaagtg actccaggaa caaaacatac tgaagaacgg cccaaggtac 6840
aggtgatcca ggctgccgat ccgctagcaa cagcgtatct gtgcggaatc caccgagagc 6900
tggttaggag attaaatgcg gtcctgcttc cgaacattca tacactgttt gatatgtcgg 6960
ctgaagactt tgacgctatt atagccgagc acttccagcc tggggattgt gttctggaaa 7020
ctgacatcgc gtcgtttgat aaaagtgagg acgacgccat ggctctgacc gcgttaatga 7080
ttctggaaga cttaggtgtg gacgcagagc tgttgacgct gattgaggcg gctttcggcg 7140
aaatttcatc aatacatttg cccactaaaa ctaaatttaa attcggagcc atgatgaaat 7200
ctggaatgtt cctcacactg tttgtgaaca cagtcattaa cattgtaatc gcaagcagag 7260
tgttgagaga acggctaacc ggatcaccat gtgcagcatt cattggagat gacaatatcg 7320
tgaaaggagt caaatcggac aaattaatgg cagacaggtg cgccacctgg ttgaatatgg 7380
aagtcaagat tatagatgct gtggtgggcg agaaagcgcc ttatttctgt ggagggttta 7440
ttttgtgtga ctccgtgacc ggcacagcgt gccgtgtggc agacccccta aaaaggctgt 7500
ttaagcttgg caaacctctg gcagcagacg atgaacatga tgatgacagg agaagggcat 7560
tgcatgaaga gtcaacacgc tggaaccgag tgggtattct ttcagagctg tgcaaggcag 7620
tagaatcaag gtatgaaacc gtaggaactt ccatcatagt tatggccatg actactctag 7680
ctagcagtgt taaatcattc agctacctga gaggggcccc tataactctc tacggctaac 7740
ctgaatggac tacgacatag tctagtccgc caagggcgcg ccttcgaagg ccggccgttt 7800
aaacatggtc ttcacactcg aagatttcgt tggggactgg cgacagacag ccggctacaa 7860
cctggaccaa gtccttgaac agggaggtgt gtccagtttg tttcagaatc tcggggtgtc 7920
cgtaactccg atccaaagga ttgtcctgag cggtgaaaat gggctgaaga tcgacatcca 7980
tgtcatcatc ccgtatgaag gtctgagcgg cgaccaaatg ggccagatcg aaaaaatttt 8040
taaggtggtg taccctgtgg atgatcatca ctttaaggtg atcctgcact atggcacact 8100
ggtaatcgac ggggttacgc cgaacatgat cgactatttc ggacggccgt atgaaggcat 8160
cgccgtgttc gacggcaaaa agatcactgt aacagggacc ctgtggaacg gcaacaaaat 8220
tatcgacgag cgcctgatca accccgacgg ctccctgctg ttccgagtaa ccatcaacgg 8280
agtgaccggc tggcggctgt gcgaacgcat tctggcgtaa gcggccgccc tgcagggcga 8340
tcgcatacag cagcaattgg caagctgctt acatagaact cgcggcgatt ggcatgccgc 8400
cttaaaattt ttattttatt tttcttttct tttccgaatc ggattttgtt tttaatattt 8460
caaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaatcgc 8520
gacagacatg ataagataca ttgatgagtt tggacaaacc acaactagaa tgcagtgaaa 8580
aaaatgcttt atttgtgaaa tttgtgatgc tattgcttta tttgtaacca ttataagctg 8640
caataaacaa gttaacaaca acaattgcat tcattttatg tttcaggttc agggggaggt 8700
gtgggaggtt ttttaggacc cggctaggct ggcggggttg ccttactggt tagcagaatg 8760
aatcaccgat acgcgagcga acgtgaagcg actgctgctg caaaacgtct gcgacctgag 8820
caacaacatg aatggtcttc ggtttccgtg tttcgtaaag tctggaaacg cggaagtcag 8880
cgccctgcac cattatgttc cggatctgca tcgcaggatg ctgctggcta ccctgtggaa 8940
cacctacatc tgtattaacg aagcgctggc attgaccctg agtgattttt ctctggtccc 9000
gccgcatcca taccgccagt tgtttaccct cacaacgttc cagtaaccgg gcatgttcat 9060
catcagtaac ccgtatcgtg agcatcctct ctcgtttcat cggtatcatt acccccatga 9120
acagaaatcc cccttacacg gaggcatcag tgaccaaaca ggaaaaaacc gcccttaaca 9180
tggcccgctt tatcagaagc cagacattaa cgcttctgga gaaactcaac gagctggacg 9240
cggatgaaca ggcagacatc tgtgaatcgc ttcacgacca cgctgatgag ctttaccgca 9300
gctgcctcgc gcgtttcggt gatgacggtg aaaacctctg acacatgcag ctcccggaga 9360
cggtcacagc ttgtctgtaa gcggatgccg ggagcagaca agcccgtcag ggcgcgtcag 9420
cgggtgttgg cgggtgtcgg ggcgcagcca tgacccagtc acgtagcgat agcggagtgt 9480
atactggctt aactatgcgg catcagagca gattgtactg agagtgcacc atatgcggtg 9540
tgaaataccg cacagatgcg taaggagaaa ataccgcatc aggcgctctt ccgcttcctc 9600
gctcactgac tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag ctcactcaaa 9660
ggcggtaata cggttatcca cagaatcagg ggataacgca ggaaagaaca tgtgagcaaa 9720
aggccagcaa aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct 9780
ccgcccccct gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac 9840
aggactataa agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc 9900
gaccctgccg cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc 9960
tcatagctca cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg 10020
tgtgcacgaa ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga 10080
gtccaacccg gtaagacacg acttatcgcc actggcagca gccactggta acaggattag 10140
cagagcgagg tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta 10200
cactagaagg acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag 10260
agttggtagc tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg 10320
caagcagcag attacgcgca gaaaaaaagg atctcaagaa gatcctttga tcttttctac 10380
ggggtctgac gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc 10440
aaaaaggatc ttcacctaga tccttttaaa ttaaaaatga agttttaaat caatctaaag 10500
tatatatgag taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc 10560
agcgatctgt ctatttcgtt catccatagt tgcctgactc cccgtcgtgt agataactac 10620
gatacgggag ggcttaccat ctggccccag tgctgcaatg ataccgcgag acccacgctc 10680
accggctcca gatttatcag caataaacca gccagccgga agggccgagc gcagaagtgg 10740
tcctgcaact ttatccgcct ccatccagtc tattaattgt tgccgggaag ctagagtaag 10800
tagttcgcca gttaatagtt tgcgcaacgt tgttgccatt gctgcaggca tcgtggtgtc 10860
acgctcgtcg tttggtatgg cttcattcag ctccggttcc caacgatcaa ggcgagttac 10920
atgatccccc atgttgtgca aaaaagcggt tagctccttc ggtcctccga tcgttgtcag 10980
aagtaagttg gccgcagtgt tatcactcat ggttatggca gcactgcata attctcttac 11040
tgtcatgcca tccgtaagat gcttttctgt gactggtgag tactcaacca agtcattctg 11100
agaatagtgt atgcggcgac cgagttgctc ttgcccggcg tcaacacggg ataataccgc 11160
gccacatagc agaactttaa aagtgctcat cattggaaaa cgttcttcgg ggcgaaaact 11220
ctcaaggatc ttaccgctgt tgagatccag ttcgatgtaa cccactcgtg cacccaactg 11280
atcttcagca tcttttactt tcaccagcgt ttctgggtga gcaaaaacag gaaggcaaaa 11340
tgccgcaaaa aagggaataa gggcgacacg gaaatgttga atactcatac tcttcctttt 11400
tcaatattat tgaagcattt atcagggtta ttgtctcatg agcggataca tatttgaatg 11460
tatttagaaa aataaacaaa taggggttcc gcgcacattt ccccgaaaag tgccacctga 11520
cgtctaagaa accattatta tcatgacatt aacctataaa aataggcgta tcacgaggcc 11580
ctttcgtctt caagaa 11596
<210> 7
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> VEEV_gblock2_fwd primer
<400> 7
tcattcagct acctgagagg g 21
<210> 8
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> VEEV_gblock2_rev primer
<400> 8
aaataaaaat tttaaggcgg catgc 25
<210> 9
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> VEEV_Oligo1_fwd primer
<400> 9
ttaaaatttt tattttattt ttcttttctt ttccgaatcg gattttgttt ttaatatttc 60
<210> 10
<211> 100
<212> DNA
<213> Artificial Sequence
<220>
<223> VEEV_Oligo2_rev primer
<400> 10
catcaatgta tcttatcatg tctgtcgcga tttttttttt tttttttttt tttttttttt 60
tttttttttt tttttttttt tttttgaaat attaaaaaca 100
<210> 11
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Primer
<400> 11
ccggcccgga ccgacaagtc tctatcacc 29
<210> 12
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Primer
<400> 12
ggccgggggc ccctctcagg tagctgaatg 30
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ВАРИАНТЫ ГИАЛУРОНИДАЗЫ С УЛУЧШЕННОЙ СТАБИЛЬНОСТЬЮ И СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2021 |
|
RU2811464C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ВКЛЮЧАЮЩАЯ ВАРИАНТ РН20 ГИАЛУРОНИДАЗЫ ЧЕЛОВЕКА, И ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ПОДКОЖНОГО ВВЕДЕНИЯ | 2020 |
|
RU2810952C2 |
| НОВЫЙ ПОЛИПЕПТИД И СПОСОБ ПОЛУЧЕНИЯ L-ЛЕЙЦИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2811433C1 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| Новый вариант цитратсинтазы и способ получения L-аминокислот c его применением | 2022 |
|
RU2836733C1 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| ВАРИАНТ АЛЬДОЛАЗЫ AROG И СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТЫ С РАЗВЕТВЛЕННОЙ ЦЕПЬЮ С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2826456C1 |
| АНТИТЕЛА ПРОТИВ VSIG4 ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2776638C1 |
| Вариант синтазы ацетогидроксикислот, микроорганизм, содержащий этот вариант, и способ получения L-аминокислоты с разветвленной цепью с использованием данного микроорганизма | 2018 |
|
RU2743964C1 |
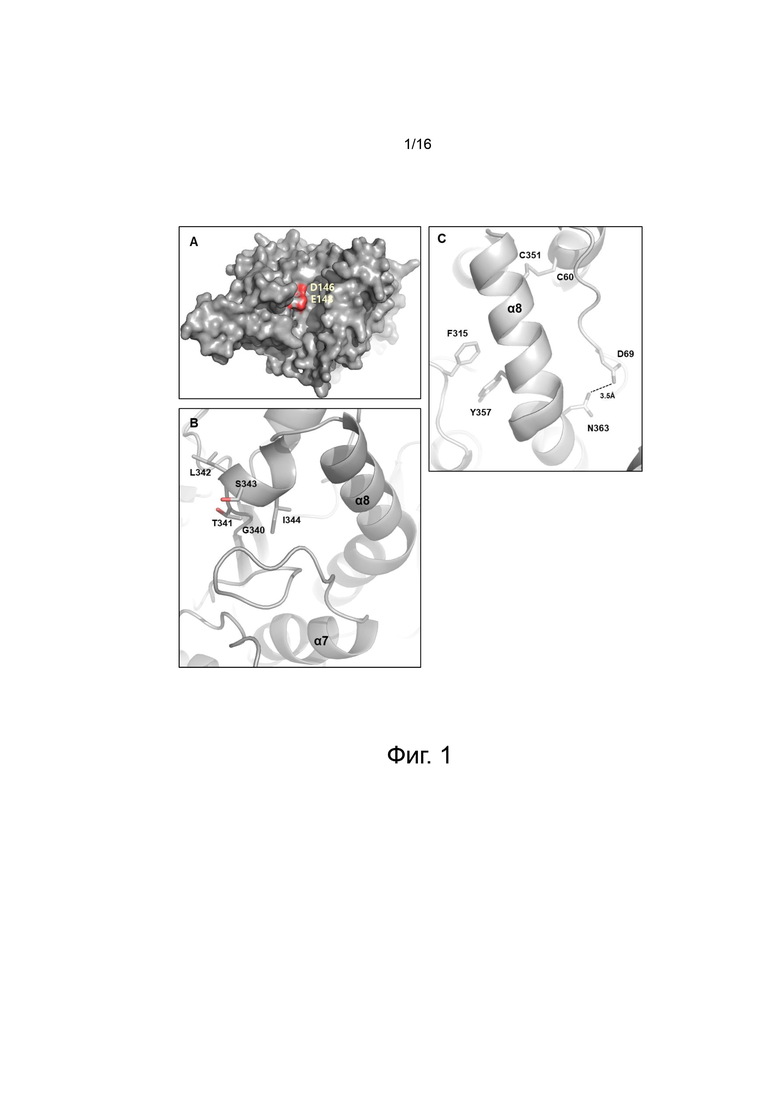
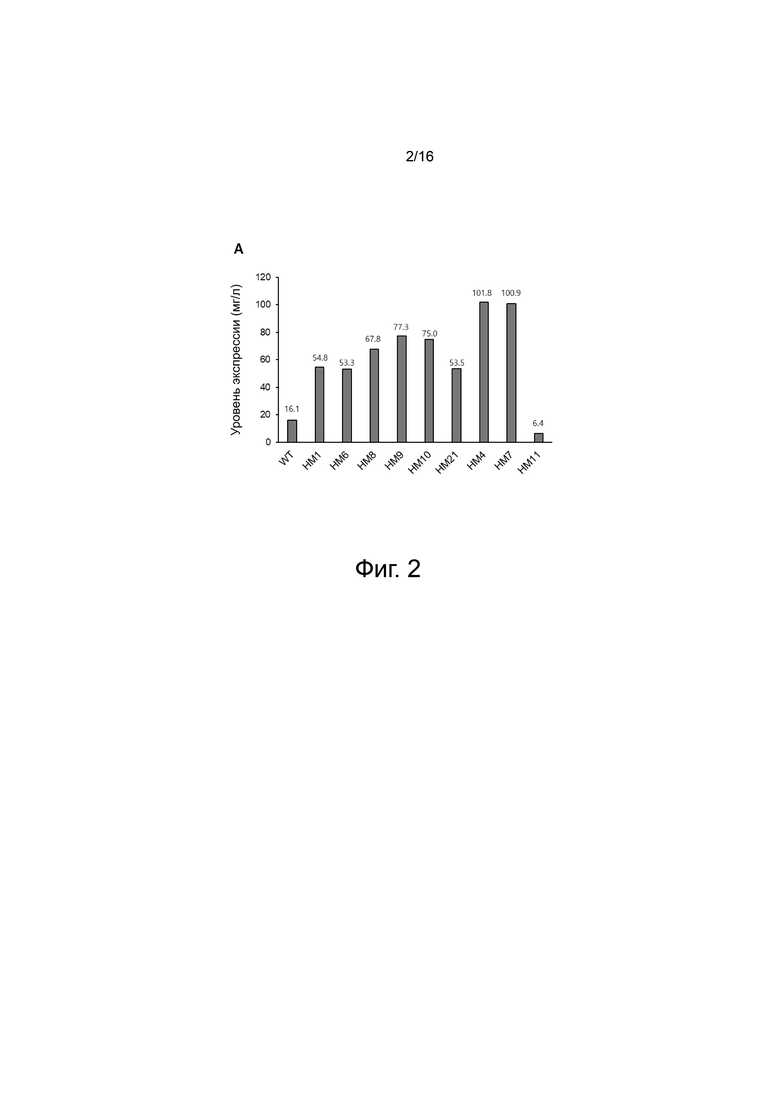
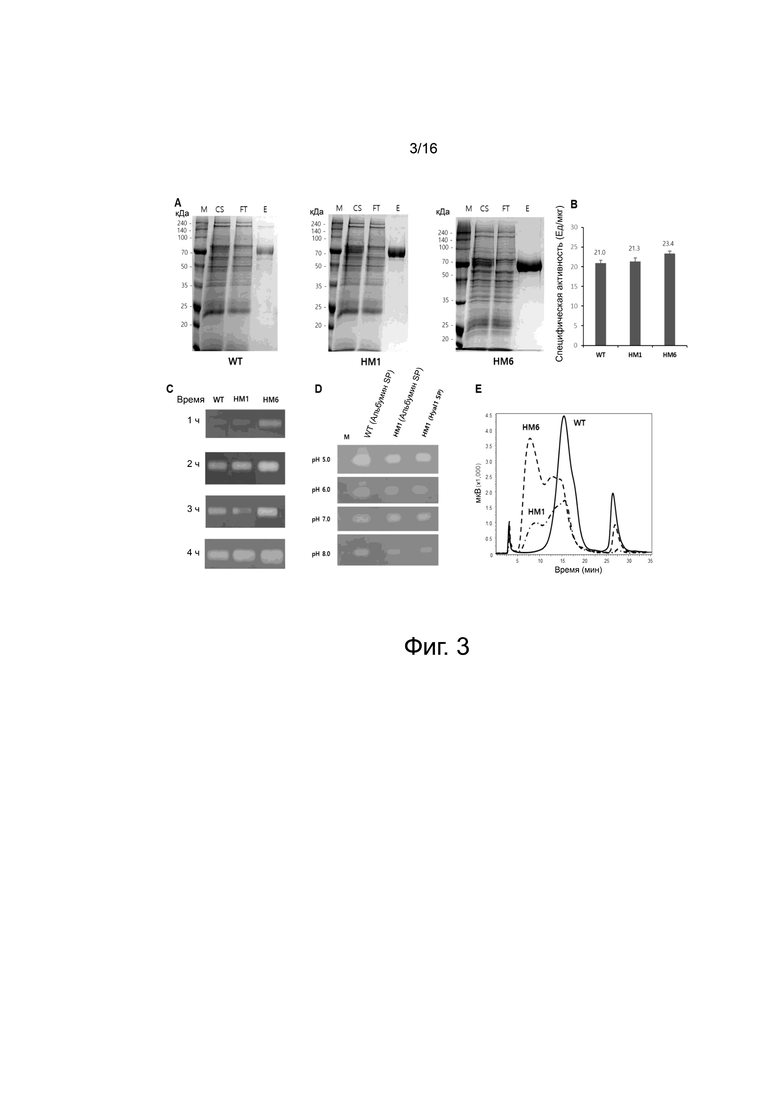
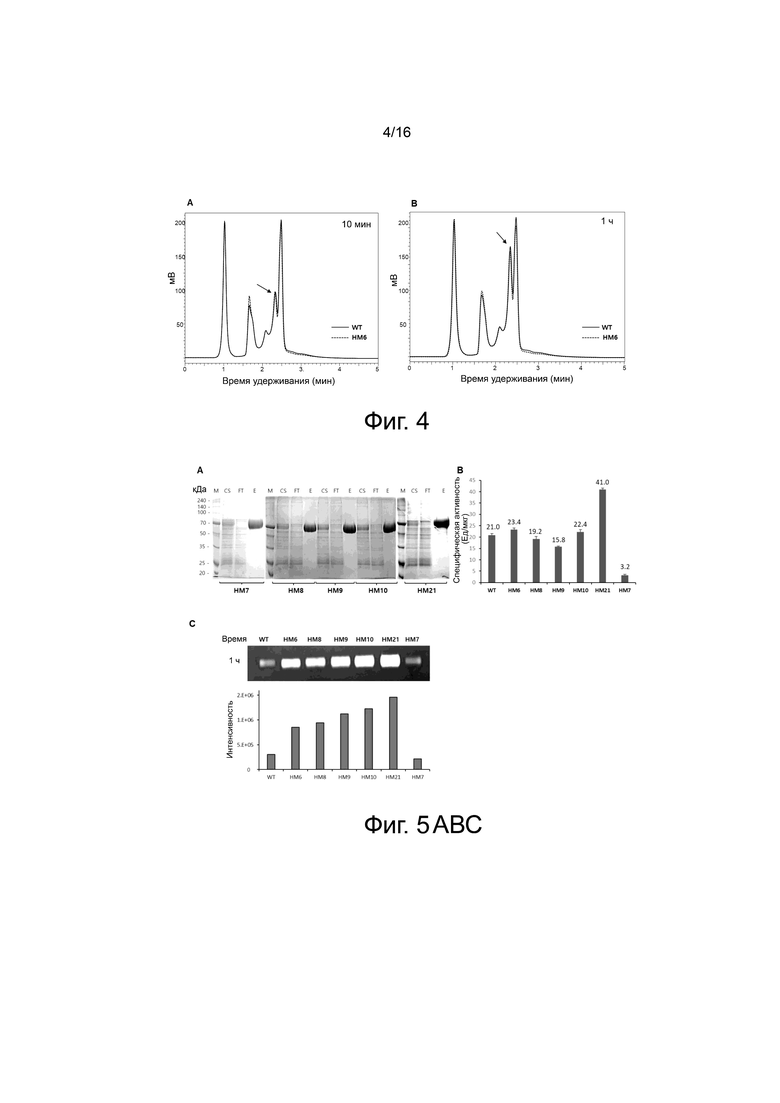
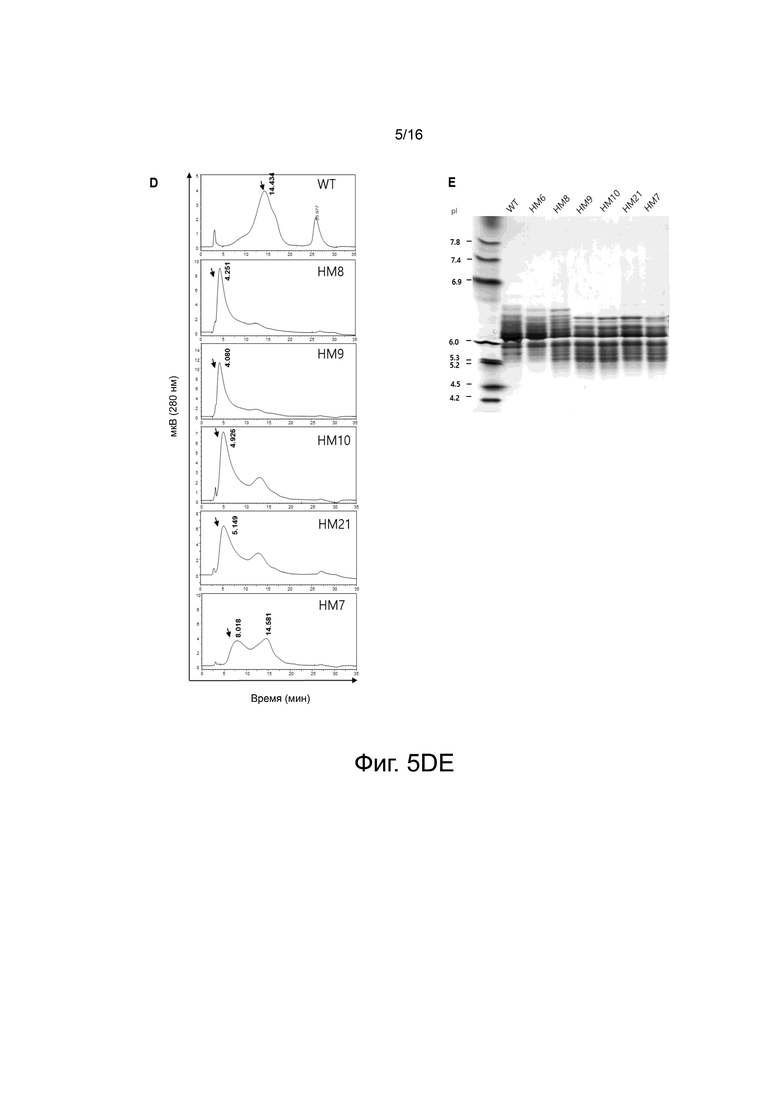
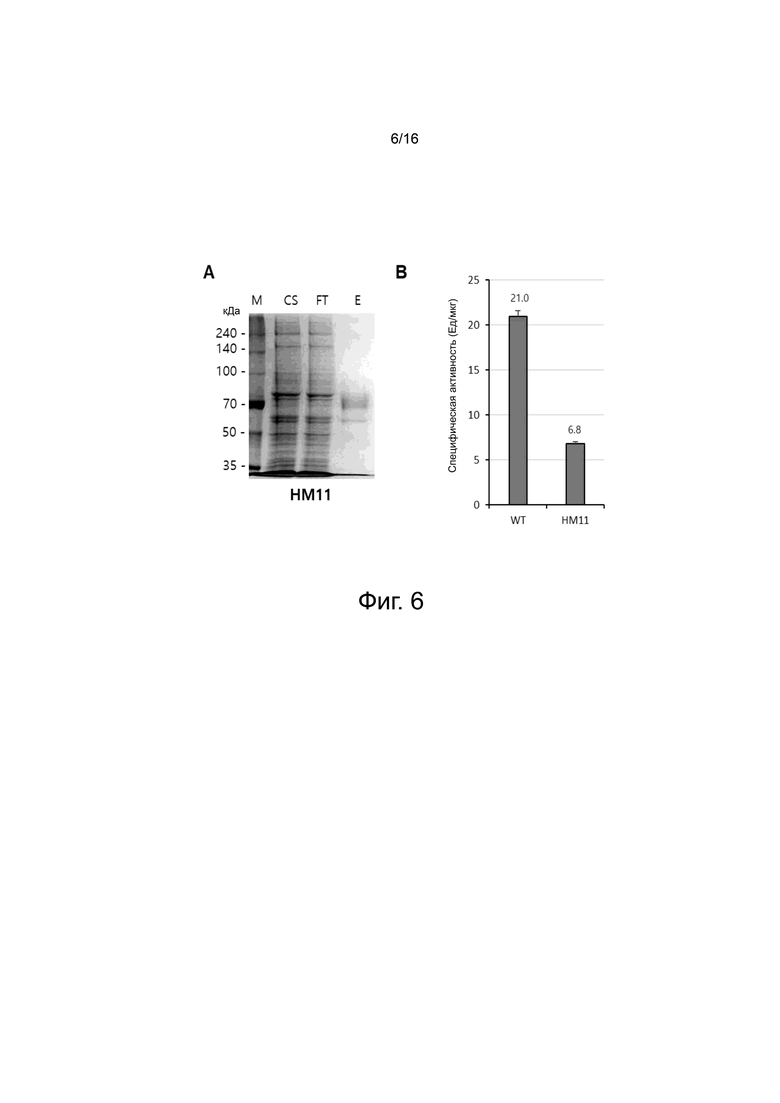
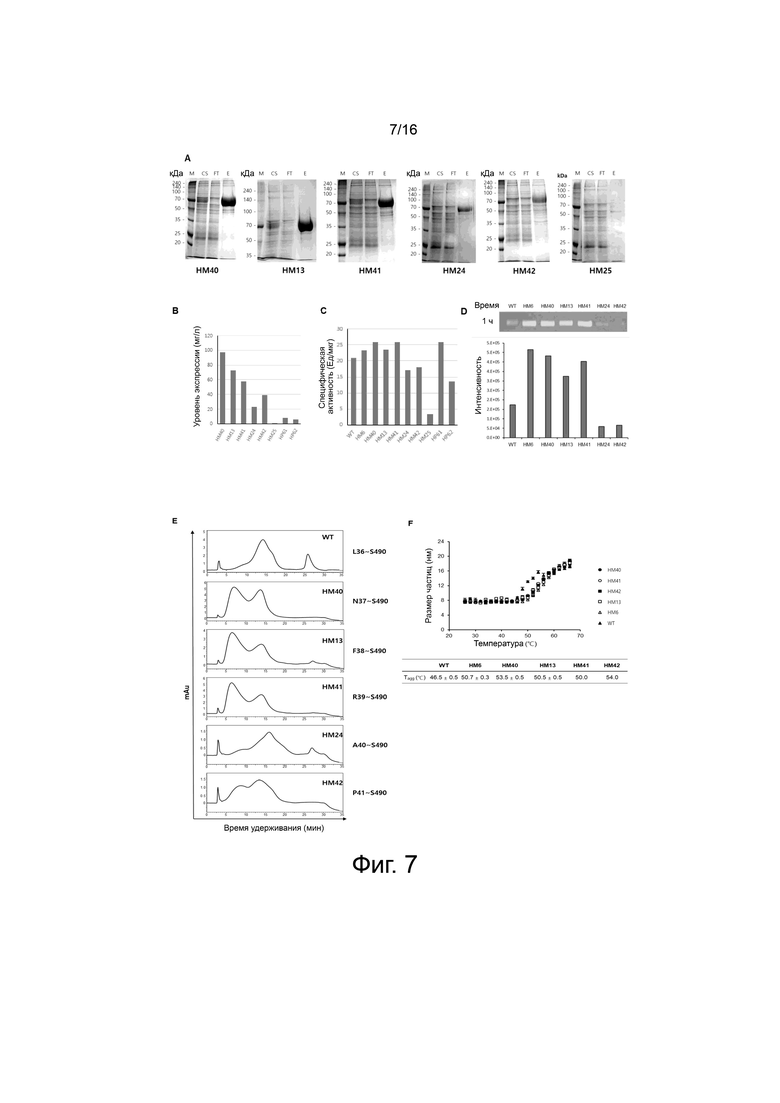
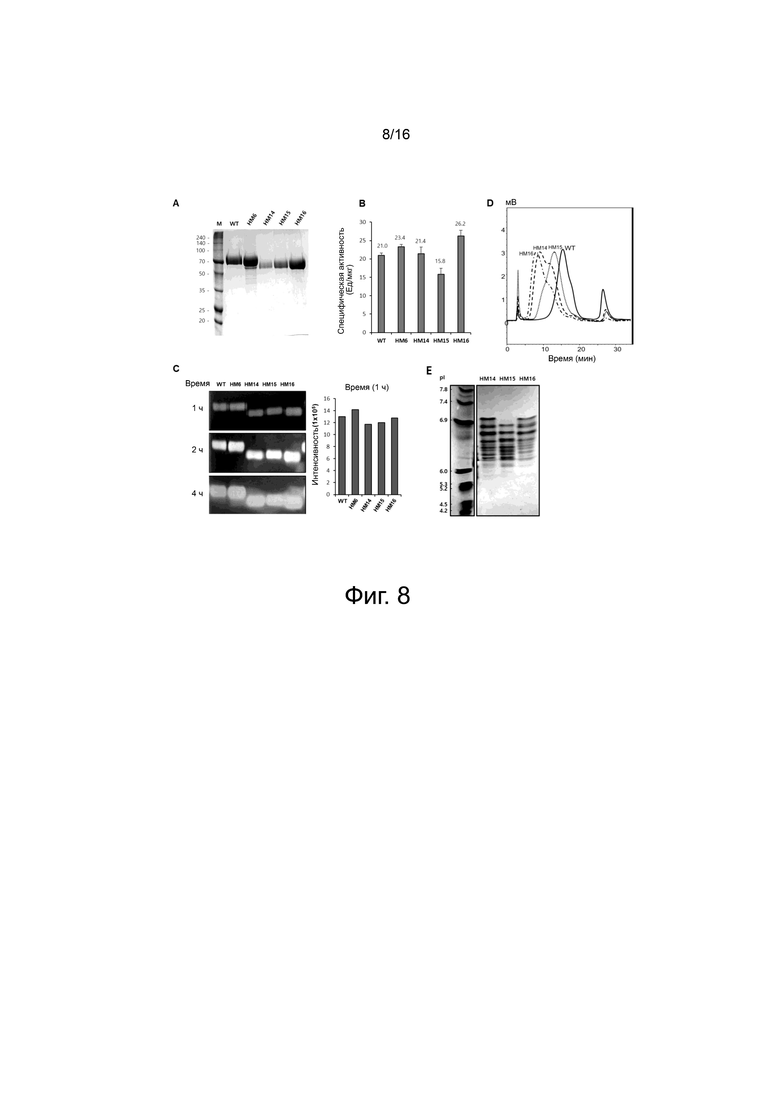
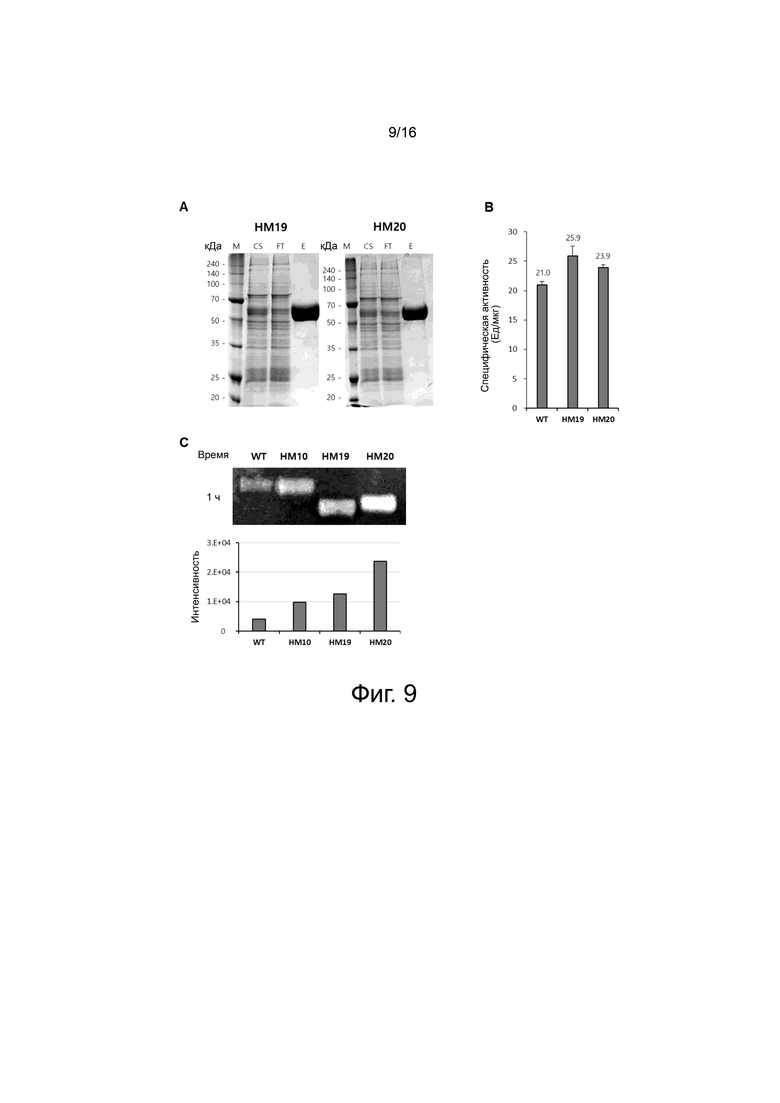

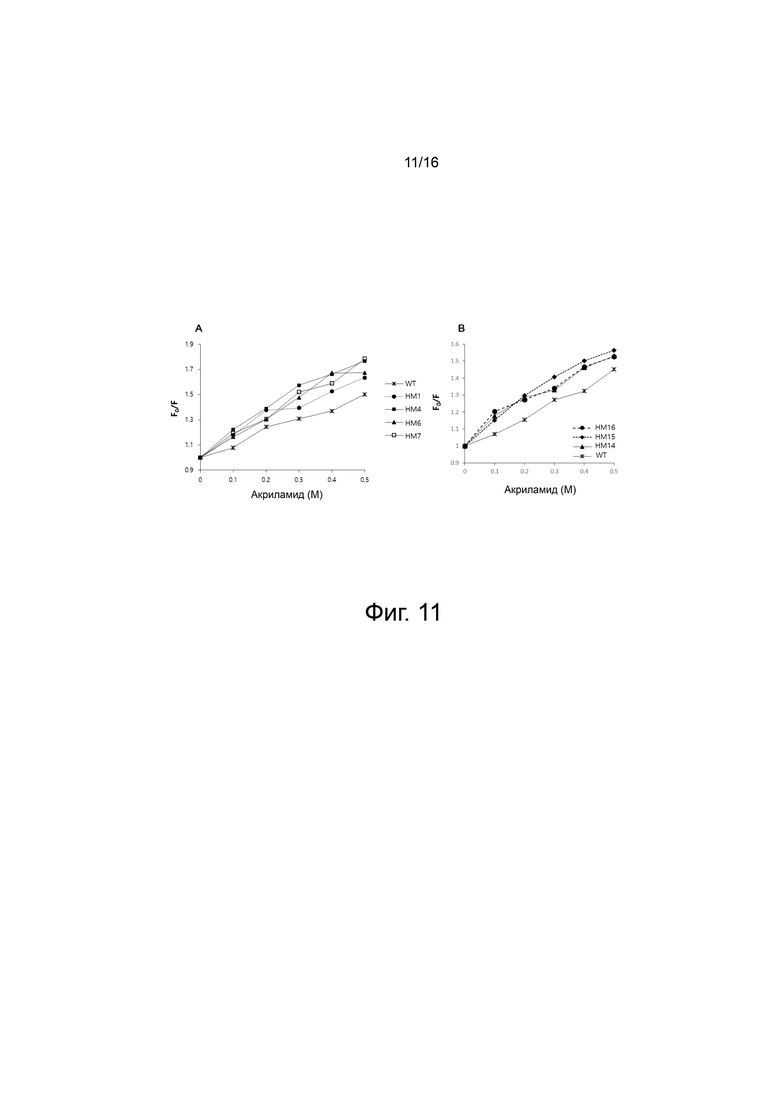

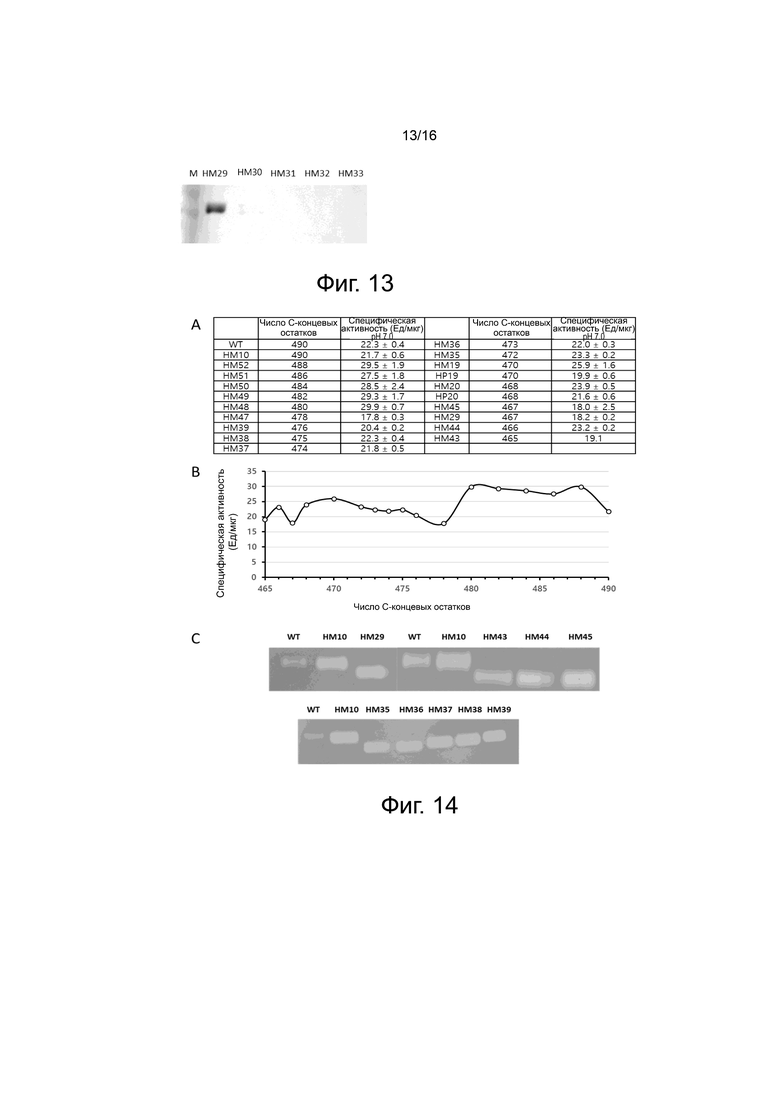
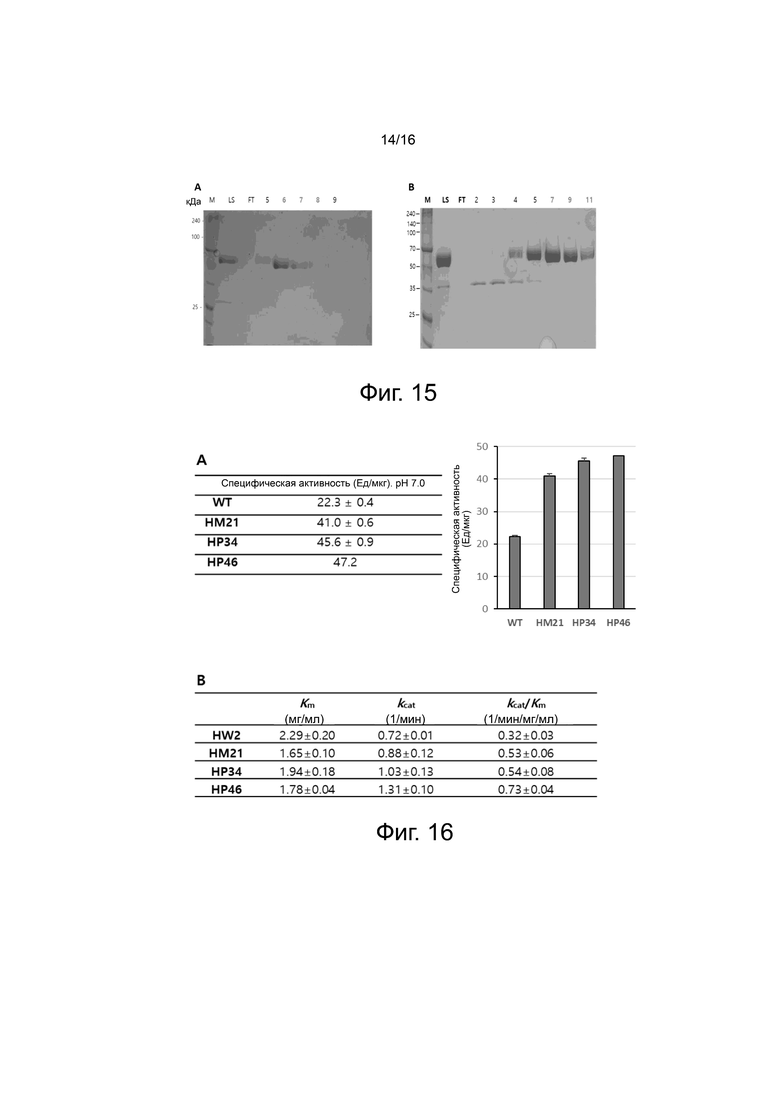
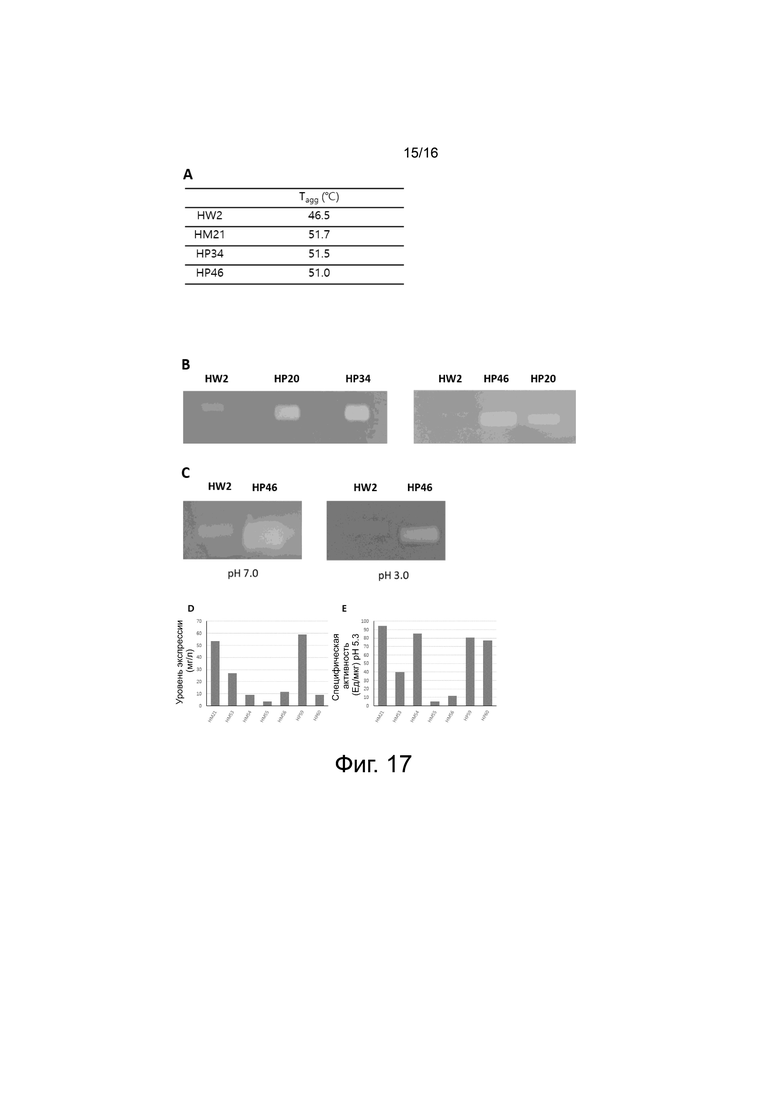
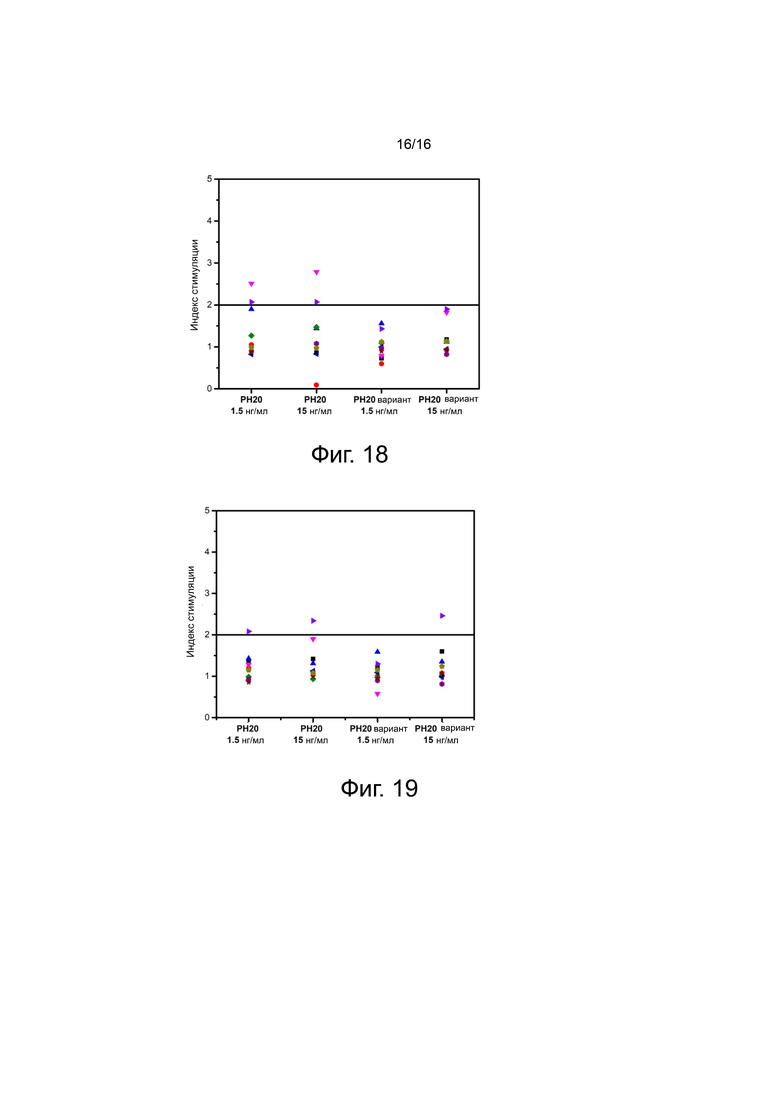
Настоящее изобретение относится к биотехнологии, в частности к области технологии белковой инженерии, которая увеличивает ферментативную активность и термическую стабильность гиалуронидазы человека, которая представляет собой фермент, гидролизующий гиалуроновую кислоту; и более конкретно к вариантам гиалуронидазы PH20 или их фрагментам, которые содержат одну или несколько замен аминокислотных остатков в области, соответствующей области альфа-спирали и ее линкерной области в аминокислотной последовательности PH20 дикого типа из SEQ ID NO: 1, и в которых выборочно один или несколько аминокислотных остатков на N-конце и/или C-конце дополнительно отщеплены. 7 н. и 15 з.п. ф-лы, 19 ил., 13 табл., 9 пр.
1. Вариант PH20 или его фрагмент, где вариант PH20 или фрагмент проявляет активность гиалуронидазы,
содержащие аминокислотные замены L354I и/или N356E; и
необязательно содержащие замену аминокислотного остатка в одном или нескольких положениях, выбранных из группы, состоящей из T341, L342, S343, I344, M345, S347, M348, K349, L352, L353, D355, E359, I361 и N363; и/или
необязательно где один или несколько N-концевых или C-концевых аминокислотных остатков удалены
в PH20 дикого типа, имеющем аминокислотную последовательность, представленную в SEQ ID NO: 1.
2. Вариант PH20 или его фрагмент по п. 1, который содержит замены аминокислотных остатков L354I и/или N356E, и дополнительно содержит одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341D, T341G, T341S, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, D355K, E359D, I361T и N363G.
3. Вариант PH20 или его фрагмент по п. 1, который содержит M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T.
4. Вариант PH20 или его фрагмент по п. 1, который дополнительно содержит одну или несколько замен аминокислотных остатков, выбранных из группы, состоящей из T341A, T341C, T341D, T341G, T341S, L342W, S343E, I344N и N363G.
5. Вариант PH20 или его фрагмент по п. 1, который содержит замену любого аминокислотного остатка, выбранного из следующих групп замен аминокислотных остатков:
(a) T341S, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(b) L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(c) M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D, I361T и N363G;
(d) T341G, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(e) T341A, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(f) T341C, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(g) T341D, L342W, S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T;
(h) I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T; и
(i) S343E, I344N, M345T, S347T, M348K, K349E, L352Q, L353A, L354I, D355K, N356E, E359D и I361T.
6. Вариант PH20 или его фрагмент по п. 1, отличающийся тем, что отщепление происходит перед аминокислотным остатком, выбранным из группы, состоящей из M1-P42 на N-конце, в результате чего один или несколько аминокислотных остатков на N-конце удаляются.
7. Вариант PH20 или его фрагмент по п. 6, отличающийся тем, что отщепление происходит перед аминокислотным остатком L36, N37, F38, R39, A40, P41 или P42 на N-конце, в результате чего один или несколько аминокислотных остатков на N-конце удаляются.
8. Вариант PH20 или его фрагмент по п. 1, отличающийся тем, что отщепление происходит после аминокислотного остатка, выбранного из группы, состоящей из V455-L509 на С-конце, в результате чего один или несколько аминокислотных остатков на С-конце удаляются.
9. Вариант PH20 или его фрагмент по п. 8, отличающийся тем, что отщепление происходит после аминокислотного остатка, выбранного из группы, состоящей из V455-S490 на С-конце, в результате чего один или несколько аминокислотных остатков на С-конце удаляются.
10. Вариант PH20 или его фрагмент по п. 9, отличающийся тем, что отщепление происходит после аминокислотного остатка V455, C458, D461, C464, I465, D466, A467, F468, K470, P471, P472, M473, E474, T475, E476, P478, I480, Y482, A484, P486, T488 или S490 на С-конце, в результате чего один или несколько аминокислотных остатков на С-конце удаляются.
11. Вариант PH20 или его фрагмент по любому из пп. 1-10, отличающийся тем, что вариант PH20 или его фрагмент выбран из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 60-115.
12. Вариант PH20 или его фрагмент по п. 11, который имеет аминокислотную последовательность, представленную в SEQ ID NO: 99.
13. Композиция для гидролиза гиалуроновой кислоты, которая содержит вариант PH20 или его фрагмент по любому из пп. 1-12, и фармацевтически приемлемый компонент.
14. Композиция для гидролиза гиалуроновой кислоты по п. 13, отличающаяся тем, что композицию применяют для комбинированного лечения с другими противораковыми лекарственными средствами.
15. Композиция для гидролиза гиалуроновой кислоты по п. 14, отличающаяся тем, что другие противораковые лекарственные средства представляют собой иммуноонкологические агенты.
16. Композиция для гидролиза гиалуроновой кислоты по п. 15, отличающаяся тем, что иммуноонкологические агенты являются ингибиторами иммунных контрольных точек.
17. Нуклеиновая кислота, кодирующая вариант PH20 или его фрагмент по любому из пп. 1-12.
18. Рекомбинантный вектор экспрессии, содержащий нуклеиновую кислоту по п. 17.
19. Клетка-хозяин для экспрессии варианта PH20 или его фрагмент по любому из пп. 1-12, где клетка трансформирована рекомбинантным вектором экспрессии по п. 18.
20. Клетка-хозяин по п. 19, отличающаяся тем, что клетка-хозяин выбрана из группы, состоящей из клеток животных, клеток растений, дрожжей, E. coli и клеток насекомых.
21. Способ получения варианта PH20 или его фрагмента, проявляющих активность гиалуронидазы, включающий стадию культивирования клетки-хозяина по п. 19.
22. Вариант PH20 или его фрагмент, проявляющие активность гиалуронидазы, содержащие сигнальный пептид, где сигнальный пептид является сигнальным пептидом, произошедшим из: сигнального пептида человеческого гормона роста, имеющего аминокислотную последовательность MATGSRTSLLLAFGLLCLPWLQEGSA согласно SEQ ID NO: 3, произошедшим из сигнального пептида человеческого сывороточного альбумина, имеющего аминокислотную последовательность MKWVTFISLLFLFSSAYS согласно SEQ ID NO: 4, или произошедшим из сигнального пептида Hyal1, имеющего аминокислотную последовательность MAAHLLPICALFLTLLDMAQG согласно SEQ ID NO: 5,
где вариант PH20 или его фрагмент содержат аминокислотные замены L354I и/или N356E; и
необязательно содержат замену аминокислотного остатка в одном или нескольких положениях, выбранных из группы, состоящей из T341, L342, S343, I344, M345, S347, M348, K349, L352, L353, D355, E359, I361 и N363; или
необязательно где один или несколько N-концевых или C-концевых аминокислотных остатков удалены в PH20 дикого типа, имеющем аминокислотную последовательность, представленную в SEQ ID NO: 1.
| US 20150010529 A1 (WEI G), 08.01.2015 | |||
| US 20100143457 A1, 10.06.2010 | |||
| Прибор для записи работы паровоза | 1931 |
|
SU26112A1 |
| WO 2013102144 A2, 04.07.2013. | |||

