ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
[0001] Данная заявка заявляет приоритет предварительной заявки США № 62/448763, поданной 20 января 2017 года, раскрытие которой включено в данный документ посредством ссылки во всей своей полноте.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка включает перечень последовательностей, который был подан в электронном виде в формате с кодировкой ASCII и, таким образом, включен в данный документ посредством ссылки во всей своей полноте. Указанная ASCII-копия, созданная 11 января 2018 года, имеет название 022548_WO012_SL.txt и имеет размер 75113 байт.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Настоящее изобретение относится к антителам, модифицированным нацеливающими на костную ткань пептидами, и способам их применения для лечения патофизиологической дегенерации кости.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0004] Правильное развитие и поддержание костей в здоровом состоянии являются важными факторами для нормального состояния здоровья. У обычного человека развитие костей происходит до возраста приблизительно 20 лет, когда плотность кости обычно максимальна. После этого плотность кости может уменьшиться без правильного питания и физических нагрузок. Однако для нормального поддержания костей в здоровом состоянии требуется гомеостатическое обновление костной ткани, при котором старая кость удаляется и заменяется новой костью.
[0005] Тем не менее, существуют многочисленные заболевания и состояния, которые могут нарушить развитие и поддержание костей в здоровом состоянии. Например, развитие костей нарушается при таких заболеваниях, как несовершенный остеогенез, при которых прочность костей снижается, приводя к тому, что кости у детей становятся хрупкими и могут легко ломаться. Более того, отсутствие гомеостатического обновления костной ткани может иметь место у здоровых людей с возрастом, что приводит к остеопорозу, когда плотность кости со временем снижается, и в конечном итоге к хрупким костям и переломам костей.
[0006] Кроме того, существуют определенные заболевания, при которых состояние костей нарушается в дополнение к первичному заболеванию и вовлечено в другие сопутствующие осложнения, такие как хроническая болезнь почек (CKD). CKD является прогрессирующим заболеванием, при котором функция почек со временем снижается, что часто приводит к сердечно-сосудистым заболеваниям, связанным с плохим состоянием костей и изменением скорости обновления костной ткани. Было показано, что лечение, которое улучшает состояние костей, одновременно уменьшает тяжесть ассоциированных сердечно-сосудистых заболеваний. Такие сообщения свидетельствуют о том, что нормальная скорость обновления костной ткани может оказывать влияние на другие заболевания, или даже являться их причиной. Следовательно, улучшенные методологии для регуляции развития и/или поддержания костей в здоровом состоянии могут иметь широкое прямое или косвенное влияние на улучшение состояния здоровья индивидуумов, страдающих многочисленными разнообразными заболеваниями и состояниями.
[0007] TGFβ является представителем суперсемейства трансформирующего фактора роста-бета (TGFβ) и играет важную роль в формировании костей в ходе развития млекопитающего (см. Chen et al., Int. J. Biol. Sci. 8(2): 272-88 (2012)). TGFβ, по-видимому, в равной степени важен для гомеостатического поддержания костной ткани в здоровом состоянии. Интересно, что TGFβ, как было показано, экспрессируется на более высоких уровнях у индивидуумов с CKD, позволяя предположить, что он является перспективной мишенью для терапевтического вмешательства. Системная обработка в мышиной модели CKD jck антителами к TGFβ продемонстрировала снижение высокой скорости обновления костной ткани (Liu et al., J. Bone Miner Res. 29(5): 1141-57 (2014)). Однако в этом исследовании не изучали степень, в которой локализация антител к TGFβ в кости может улучшить эффективность лечения. Учитывая, что TGFβ участвует во множестве клеточных процессов, включая ответ на повреждение ДНК, аллергические иммунные ответы и эпителизацию раны, и это лишь некоторые из них, желателен более целенаправленный подход к контролю активности TGFβ для сведения к минимуму потенциальных нежелательных побочных эффектов. Следовательно, необходим более точный подход к регуляции активности TGFβ для обеспечения усовершенствованных способов лечения для регуляции развития и/или поддержания костей в здоровом состоянии.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0008] В данном документе предусмотрены антитела, такие как антитела к TGFβ, которые эффективно нацелены на костную ткань. В первом аспекте в настоящем изобретении предусмотрено антитело или его антигенсвязывающий фрагмент, содержащие тяжелую цепь, легкую цепь и один или несколько пептидов на основе полиаспартата (поли-D). В одном конкретном варианте осуществления антитело или антигенсвязывающий фрагмент содержит тяжелую цепь, легкую цепь и один или несколько пептидов на основе полиаспартата (поли-D), связанных с тяжелой цепью и/или С-концом легкой цепи.
[0009] В одном варианте осуществления антитело или его антигенсвязывающий фрагмент проявляют по меньшей мере 2-кратное увеличение локализации в кости по сравнению с антителом с такой же тяжелой цепью и легкой цепью, но без одного или нескольких поли-D-пептидов.
[0010] В одном варианте осуществления один или несколько поли-D-пептидов связаны с антителом или его антигенсвязывающим фрагментом посредством химического конъюгирования. В другом варианте осуществления один или несколько поли-D-пептидов связаны по шарнирной области тяжелой цепи. В дополнительном варианте осуществления один или несколько поли-D-пептидов связаны с N-концом или C-концом легкой цепи. В еще одном дополнительном варианте осуществления один или несколько поли-D-пептидов связаны с антителом или его антигенсвязывающим фрагментом посредством одного или нескольких спейсеров/линкеров (например, полиэтиленгликолевых (PEG) спейсеров и пептидных линкеров).
[0011] В одном варианте осуществления один или несколько поли-D-пептидов соединены с аминокислотной последовательностью тяжелой цепи и/или один или несколько поли-D-пептидов соединены с аминокислотной последовательностью легкой цепи. Поли-D-пептид, который "соединен" с аминокислотной последовательностью, включен в ту же полипептидную цепь. Например, соединенный поли-D-пептид может транслироваться из той же цепи РНК, что и последовательность тяжелой или легкой цепи, которая может кодироваться из плазмиды с рекомбинантной ДНК. В одном варианте осуществления один или несколько поли-D-пептидов соединены с N-концом и/или один или несколько поли-D-пептидов соединены с С-концом тяжелой цепи. Два или более поли-D-пептида могут быть связаны последовательно, не разделены, разделены одним или несколькими другими аминокислотными остатками (т.e. аминокислотами, отличными от аспартата) или пептидным линкером, связанным с N-концом или C-концом тяжелой цепи. В дополнительном варианте осуществления один или несколько поли-D-пептидов соединены с N-концом, и/или один или несколько поли-D-пептидов соединены с С-концом легкой цепи. Например, два или более поли-D-пептида могут быть связаны последовательно, не разделены, разделены одним или несколькими другими аминокислотными остатками (т.e. аминокислотами, отличными от аспартата) или пептидным линкером, связанным с N-концом или C-концом легкой цепи. В одном варианте осуществления поли-D-пептид соединенный с C-концом тяжелой цепи. В другом варианте осуществления поли-D-пептид соединенный с C-концом тяжелой цепи и поли-D-пептид соединенный с N-концом тяжелой цепи.
[0012] В одном варианте осуществления легкая цепь не содержит поли-D-пептид. В другом варианте осуществления тяжелая цепь не содержит поли-D-пептид.
[0013] В одном варианте осуществления один или несколько поли-D-пептидов, каждый независимо, содержат 2-30 остатков аспарагиновой кислоты. Например, поли-D-пептид может включать 2, или 3, или 4, или 5, или 6, или 7, или 8, или 9, или 10, или 11, или 12, или 13, или 14, или 15, или 16, или 17, или 18, или 19, или 20, или 21, или 22, или 23, или 24, или 25, или 26, или 27, или 28, или 29 или 30 остатков аспарагиновой кислоты. В другом варианте осуществления один или несколько поли-D-пептидов, каждый независимо, содержат 6, 7, 8, 9, 10 или 11 остатков аспарагиновой кислоты. В другом варианте осуществления каждый из одного или нескольких поли-D-пептидов содержит 10 остатков аспарагиновой кислоты; такие пептиды названы "D10" (SEQ ID NO: 1) в данном документе. В некоторых вариантах осуществления антитело или фрагмент могут включать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более 12 поли-D-пептидов.
[0014] В другом варианте осуществления антитело является антителом любого из изотипов IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgM, IgE или IgD. В другом варианте осуществления антитело представляет собой антитело изотипа IgG1 или IgG4. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3, такие как один или несколько из TGFβ1, TGFβ2 и TGFβ3 человека.
[0015] В одном варианте осуществления предполагается, что фрагмент антитела содержит один или несколько пептидов на основе полиаспартата (поли-D). Предполагается, что фрагмент антитела будет демонстрировать по меньшей мере 2-кратное увеличение локализации в кости по сравнению с тем же фрагментом антитела, но без одного или нескольких поли-D-пептидов. Фрагмент антитела может представлять собой, например, любое или комбинацию следующего: Fab, F(ab')2, моноспецифичный Fab2, биспецифичный Fab2, триспецифичный Fab3, моновалентный IgG, scFv, биспецифичное диатело, триспецифичное триотело, scFv-sc, минитело, IgNAR, V-NAR, hcIgG или VhH. В одном варианте осуществления фрагмент антитела связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3, такие как один или несколько из TGFβ1, TGFβ2 и TGFβ3 человека. Антитело или фрагмент антитела в данном документе может быть полностью человеческим, гуманизированным или химерным.
[0016] Во втором аспекте в настоящем изобретении предусмотрен способ получения антитела или его антигенсвязывающего фрагмента, нацеленного на костную ткань, который включает этапы получения тяжелой цепи антитела, получения легкой цепи антитела, получения одного или нескольких поли-D-пептидов, прикрепленных к тяжелой цепи, и/или одного или нескольких поли-D-пептидов, прикрепленных к легкой цепи, и объединения тяжелой цепи и легкой цепи с получением антитела или его антигенсвязывающего фрагмента, нацеленного на костную ткань.
[0017] В одном варианте осуществления один или несколько поли-D-пептидов, прикрепленных к тяжелой цепи, и/или один или несколько поли-D-пептидов, прикрепленных к легкой цепи, присоединяют посредством химического конъюгирования. В другом варианте осуществления один или несколько поли-D-пептидов, прикрепленных к тяжелой цепи, и/или один или несколько поли-D-пептидов, прикрепленных к легкой цепи, присоединяют посредством рекомбинации.
[0018] В третьем аспекте в настоящем изобретении предусмотрено антитело к TGFβ, нацеленное на костную ткань, которое включает тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 2, 3,4 и 5 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 6, 7, 8, 11 и 12, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 2 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 6.
[0019] В четвертом аспекте в настоящем изобретении предусмотрено антитело к TGFβ, нацеленное на костную ткань, которое включает тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14,16 и 17 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 20, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15.
[0020] В пятом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 14 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 15 (например, mAb2 F6). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0021] В шестом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 17 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 15 (например, mAb2 F16). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0022] В седьмом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 16 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 15 (например, mAb2 F11). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0023] В восьмом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 17 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 18 (например, mAb2 F17). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0024] В девятом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 16 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 18 (например, mAb2 F12). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0025] В десятом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 14 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 18 (например, mAb2 F7). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0026] В одиннадцатом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 18 (например, mAb2 F2). Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В одном варианте осуществления антитело специфически связывает TGFβ1.
[0027] В двенадцатом аспекте в настоящем изобретении предусмотрено антитело к TGFβ, нацеленное на костную ткань, включающее тяжелую цепь, содержащую аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, изложенной под любым из SEQ ID NO: 23, 24, 25 и 26 (с кодоном для С-концевого лизина тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, изложенной под любым из SEQ ID NO: 27, 28, 29, 30, 31 и 32, при условии, что аминокислотная последовательность тяжелой цепи не кодируется последовательностью нуклеиновой кислоты, изложенной под SEQ ID NO: 23 (с кодоном для С-концевого лизина тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи кодируется последовательностью нуклеиновой кислоты, изложенной под SEQ ID NO: 27.
[0028] В тринадцатом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, изложенной под SEQ ID NO: 25 (с кодоном для С-концевого лизина тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, изложенной под SEQ ID NO: 27. Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3.
[0029] В четырнадцатом аспекте в настоящем изобретении предусмотрено антитело IgG4 человека, которое включает тяжелую цепь, содержащую аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, изложенной под SEQ ID NO: 26 (с кодоном для С-концевого лизина тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, изложенной под SEQ ID NO: 27. Антитело специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3.
[0030] В пятнадцатом аспекте в настоящем изобретении предусмотрен способ лечения индивидуума от потери костной массы, включающий введение индивидууму эффективного количества антитела к TGFβ или его антигенсвязывающего фрагмента, нацеленного на костную ткань, и выявление по меньшей мере одного из уменьшения уровней TGFβ, снижения активности TGFβ, уменьшения потери костной массы, снижения скорости потери костной массы, увеличения плотности кости, увеличения прочности кости и снижения уровней IL-11.
[0031] В одном варианте осуществления индивидуум является человеком. В другом варианте осуществления антитело к TGFβ или его антигенсвязывающий фрагмент специфически связывает один или несколько из TGFβ1, TGFβ2 и TGFβ3. В дополнительном варианте осуществления антитело к TGFβ включает тяжелую цепь, легкую цепь и один или несколько пептидов на основе полиаспартата (поли-D). Антитело проявляет по меньшей мере 2-кратное увеличение локализации в кости по сравнению с антителом с такой же тяжелой цепью и легкой цепью, но без одного или нескольких поли-D-пептидов. В одном варианте осуществления антитело является антителом любого из изотипов IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgM, IgE и IgD. В другом варианте осуществления антитело представляет собой антитело изотипа IgG1 или IgG4. В одном варианте осуществления индивидуум имеет хроническую болезнь почек и/или заболевание костей, включая метастазирование рака в кость. Заболевание костей может представлять собой несовершенный остеогенез или остеопороз. В одном варианте осуществления эффективное количество антитела к TGFβ или фрагмента антитела, нацеленного на костную ткань, вводят подкожно, внутривенно или внутримышечно.
[0032] В шестнадцатом аспекте в настоящем изобретении предусмотрена фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент по настоящему изобретению и фармацевтически приемлемый носитель. Например, антитело может включать тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 2, 3, 4 и 5 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 6, 7, 8, 11 и 12, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 2 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 6. В другом варианте осуществления антитело может включать тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 29, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15.
[0033] В семнадцатом аспекте в настоящем изобретении предусмотрена выделенная молекула нуклеиновой кислоты, включающая последовательность нуклеиновой кислоты, кодирующую тяжелую цепь, легкую цепь или обе, из антитела к TGFβ, нацеленного на костную ткань, где тяжелая цепь антитела к TGFβ содержит аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17 (с C-концевым лизином тяжелой цепи или без него), и легкая цепь антитела к TGFβ содержит аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 20, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15.
[0034] В восемнадцатом аспекте в настоящем изобретении предусмотрен вектор экспрессии, включающий последовательность нуклеиновой кислоты, кодирующую тяжелую цепь, легкую цепь или обе, из антитела к TGFβ, нацеленного на костную ткань, где тяжелая цепь антитела к TGFβ содержит аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17 (с C-концевым лизином тяжелой цепи или без него); и легкая цепь антитела к TGFβ содержит аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 20, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15.
[0035] В девятнадцатом аспекте в настоящем изобретении предусмотрена клетка-хозяин, содержащая один или несколько векторов экспрессии, включающих последовательности нуклеиновой кислоты, кодирующие антитело к TGFβ, нацеленное на костную ткань, где тяжелая цепь антитела к TGFβ включает тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 29, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15. В одном варианте осуществления клетка-хозяин представляет собой клетку млекопитающего или прокариотическую клетку. В другом варианте осуществления клетка-хозяин представляет собой клетку яичника китайского хомячка (СНО) или клетку Escherichia coli (E. coli).
[0036] В двадцатом аспекте в настоящем изобретении предусмотрен способ получения антитела к TGFβ или его антигенсвязывающего фрагмента, нацеливающихся на костную ткань. Способ включает выращивание клетки-хозяина в условиях, обеспечивающих возможность выработки антитела или его антигенсвязывающего фрагмента. Клетка-хозяин содержит (i) последовательность нуклеиновой кислоты, кодирующую тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17 (с C-концевым лизином тяжелой цепи или без него); и (ii) последовательность нуклеиновой кислоты, кодирующую легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 29, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15. В одном варианте осуществления способ дополнительно включает составление антитела или его антигенсвязывающего фрагмента в виде фармацевтической композиции, содержащей приемлемый носитель.
[0037] В двадцать первом аспекте в настоящем изобретении предусмотрена фармацевтическая композиция, содержащая антитело к TGFβ, нацеленное на костную ткань. Антитело к TGFβ, нацеленное на костную ткань, включает тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 2, 3, 4 и 5 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 6, 7, 8, 11 и 12, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO:2 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 6, или антитело к TGFβ, нацеленное на костную ткань, включает тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17 (с C-концевым лизином тяжелой цепи или без него), и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 29, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13 (с C-концевым лизином тяжелой цепи или без него), когда аминокислотная последовательность легкой цепи представляет собой последовательность под SEQ ID NO: 15. В одном варианте осуществления фармацевтическая композиция составлена в виде жидкого продукта, представляющего собой лекарственное средство. В другом варианте осуществления фармацевтическая композиция составлена в виде лиофилизированного продукта, представляющего собой лекарственное средство.
[0038] В двадцать втором аспекте в настоящем изобретении предусмотрено антитело к TGFβ, нацеленное на костную ткань. Тяжелая цепь антитела содержит: участок 1, определяющий комплементарность, тяжелой цепи (HCDR1), содержащий аминокислотную последовательность под SEQ ID NO: 33, HCDR2, содержащий аминокислотную последовательность под SEQ ID NO: 34, и HCDR3, содержащий аминокислотную последовательность под SEQ ID NO: 35. Легкая цепь антитела содержит: участок 1, определяющий комплементарность, легкой цепи (LCDR1), содержащий аминокислотную последовательность под SEQ ID NO: 36, LCDR2, содержащий аминокислотную последовательность под SEQ ID NO: 37, и LCDR3, содержащий аминокислотную последовательность под SEQ ID NO: 38. Кроме того, антитело дополнительно содержит полипептид D10 на одном или нескольких из N-конца тяжелой цепи, С-конца тяжелой цепи, N-конца легкой цепи и С-конца легкой цепи.
[0039] В двадцать третьем аспекте в настоящем изобретении предусмотрено антитело к TGFβ, нацеленное на костную ткань, где тяжелая цепь антитела содержит определяющие комплементарность участки (CDR) 1-3 тяжелой цепи из последовательности под SEQ ID NO: 39 и CDR1-3 легкой цепи из последовательности под SEQ ID NO: 40, где антитело дополнительно содержит полипептид D10 на одном или нескольких из N-конца тяжелой цепи, С-конца тяжелой цепи, N-конца легкой цепи и С-конца легкой цепи. В некоторых вариантах осуществления антитело содержит вариабельный домен тяжелой цепи (VH или HCVD), содержащий аминокислотную последовательность под SEQ ID NO: 39, и вариабельный домен легкой цепи (VL или LCVD), содержащий аминокислотную последовательность под SEQ ID NO: 40.
[0040] В двадцать четвертом аспекте в настоящем изобретении предусмотрена полинуклеотидная последовательность, кодирующая антитело к TGFβ, нацеленное на костную ткань, где тяжелая цепь антитела содержит HCDR1, содержащий аминокислотную последовательность под SEQ ID NO: 33, HCDR2, содержащий аминокислотную последовательность под SEQ ID NO: 34, и HCDR3, содержащий аминокислотную последовательность под SEQ ID NO: 35, легкая цепь антитела содержит LCDR1, содержащий аминокислотную последовательность под SEQ ID NO: 36, LCDR2, содержащий аминокислотную последовательность под SEQ ID NO: 37, и LCDR3, содержащий аминокислотную последовательность под SEQ ID NO: 38, и антитело дополнительно содержит полипептид D10 на одном или нескольких из N-конца тяжелой цепи, C-конца тяжелой цепи, N-конца легкой цепи и C-конца легкой цепи; или антитело к TGFβ, нацеленное на костную ткань, где тяжелая цепь антитела содержит определяющие комплементарность участки (CDR) 1-3 тяжелой цепи из последовательности под SEQ ID NO: 39 и CDR1-3 легкой цепи из последовательности под SEQ ID NO: 40, где антитело дополнительно содержит полипептид D10 на одном или нескольких из N-конца тяжелой цепи, С-конца тяжелой цепи, N-конца легкой цепи и С-конца легкой цепи.
[0041] В двадцать пятом аспекте в настоящем изобретении предусмотрено нацеливающееся на кости антитело, такое как нацеливающееся на костную ткань антитело к TGFβ или антигенсвязывающий фрагмент по настоящему изобретению, для применения в способе лечения, описанном в данном документе.
[0042] В двадцать шестом аспекте в настоящем изобретении предусмотрено применение нацеливающегося на костную ткань антитела, такого как нацеливающееся на костную ткань антитело к TGFβ или антигенсвязывающий фрагмент по настоящему изобретению, для изготовления лекарственного препарата для описанного в данном документе способа лечения.
[0043] Конкретные варианты осуществления, рассматриваемые в данном документе, дополнительно описаны ниже. Описанные выше и другие признаки и преимущества настоящего изобретения будут более понятны из следующего подробного описания настоящего изобретения, взятого вместе с прилагаемой формулой изобретения. Следует отметить, что объем формулы изобретения определяется приведенными в ней формулировками, а не конкретным обсуждением признаков и преимуществ, изложенных в настоящем описании.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0044] На фиг. 1 изображен пептид D10, химически конъюгированный с антителом к TGFβ (α-TGFβ).
[0045] На фиг. 2 изображен способ химического конъюгирования пептида, несущего линкер (пептидный линкер) и малеимидной функциональной группы с восстановленными дисульфидами шарнирной области IgG1-антитела мыши.
[0046] На фиг. 3A изображен гель SDS-PAGE в восстанавливающих условиях химических конъюгатов пептидного линкера D10 с мышиным к TGFβ, IgG1, mAb1 мыши. Верхняя полоска(-и) представляет(-ют) собой тяжелую цепь, а нижняя полоска - легкую цепь.
[0047] На фиг. 3B изображены значения соотношения пептид-антитело (PAR) относительно соотношения пептид-малеимид:mAb в реакции химического конъюгирования из примера 1, показывающие линейное увеличение PAR с увеличением количества пептидов вплоть до PAR 8 моль:моль.
[0048] На фиг. 4A-4G изображена эксклюзионная хроматография молярных смесей 1:1 TGFβ1 и химических конъюгатов пептида D10 с трастузумабом (Herceptin®) или с антителом к TGFβ mAb1. На фиг. 4A изображен профиль SEC химического конъюгата пептида D10 с mAb1. На фиг. 4B изображен профиль SEC молярной смеси 1:1 химического конъюгата mAb1-D10 с TGFβ1. На фиг. 4C изображен профиль SEC немодифицированного mAb1. На фиг. 4D изображен профиль SEC молярной смеси 1:1 mAb1 с TGFβ1. На фиг. 4E изображен профиль SEC Herceptin®. На фиг. 4F изображен профиль SEC химического конъюгата пептида D10 с Herceptin®. На фиг. 4G изображен профиль SEC химического конъюгата D10 с Herceptin®, смешанного в соотношении 1:1 с TGFβ1.
[0049] На фиг. 5A изображена кривая A280 хроматографии на гидроксиапатите mAb1 и конъюгата D10-mAb1 (ось x=минуты, ось y=поглощение (нормализованное)).
[0050] На фиг. 5B изображен гель SDS-PAGE проточной (FT) фракции и фракции пика 4.
[0051] На фиг. 6A изображена хроматография на гидроксиапатите химических конъюгатов с увеличением количества пептидов. Показано поглощение элюата при 280 нм для каждого конъюгата (ось x=минуты, ось y=поглощение (нормализованное)).
[0052] На фиг. 6B изображена фракция связанного анализируемого вещества (верхняя кривая, кружки, шкала слева) и время удержания (нижняя кривая, треугольники, шкала справа) в зависимости от количества конъюгированных пептидов, как определено при помощи SDS-PAGE.
[0053] На фиг. 7 показан анализ нейтрализации TGFβ in vitro, проведенный в клетках A549 с контрольным конъюгатом (PAR=0) и конъюгатами со средним количеством 4 или 9 пептидов, по сравнению с немодифицированным mAb1.
[0054] На фиг. 8A показано зависящее от времени биораспределение меченого флуорофором mAb1 и химического конъюгата в дозе 1 мг/кг (1 мг/кг массы тела), содержащего приблизительно 4,5 пептида. Моменты времени, в которые животные были запечатлены, указаны на каждой панели. На каждой фотографии левая мышь получала mAb1, а правая мышь получала химический конъюгат. Интенсивность изображения скорректировали, чтобы выявить различия в распределении.
[0055] На фиг. 8B показано соотношение флуоресценции, обнаруженной в представляющем интерес участке, соответствующем дистальному отделу бедренной кости, и представляющем интерес участке, соответствующем сердцу, на изображениях, показанных на фиг. 8А. Кружки соответствуют антителу mAb1, а квадраты соответствуют пептиду D10, конъюгированному с mAb1 (D10-mAb1).
[0056] На фиг. 9A схематически показаны возможные местоположения пептидов D10 на антителе подтипа IgG для создания ряда слитых вариантов антитела mAb1, которые должны быть получены посредством присоединения пептидов D10 к концам тяжелой и/или легкой цепи с использованием рекомбинантных способов. Места добавления пептидов D10 указаны кружками, а применение пептидных линкерных последовательностей обозначено волнистыми линиями (более длинная волнистая линия представляет собой более длинный линкер, чем более короткая волнистая линия).
[0057] На фиг. 9B показан ряд слитых вариантов антитела (слитые варианты), полученных посредством размещения пептидов, как показано на фиг. 9A. Были получены рекомбинантные слитые варианты с различными комбинациями положения присоединения и количества пептидов. Меньшие числа под каждой диаграммой показывают идентичность каждого рекомбинантного слитого варианта, упомянутую в данном документе для ясности. Как указано в данном документе, слитые варианты обозначены либо "слиянием", либо "F", за которым следует предполагаемый номер варианта. Например, "слияние 1" и "F1" оба относятся к антителу, имеющему конфигурацию антитела "1" без пептида D10. Более длинная волнистая линия представляет собой более длинный линкер, чем более короткая волнистая линия.
[0058] На фиг. 10 показан SDS-PAGE указанных очищенных рекомбинантных слитых вариантов mAb1 в восстанавливающих (верхний гель) или невосстанавливающих (нижний гель) условиях.
[0059] На фиг. 11 показана термостабильность рекомбинантных слитых вариантов mAb1, определенная с помощью дифференциальной сканирующей флуориметрии (DSF). Переход в частично денатурированную форму при каждой температуре выявляют по увеличению флуоресценции красителя. Наклон увеличения флуоресценции с температурой (-d(RFU)/dT) рассчитывали и отображали в зависимости от температуры образца. Скорость денатурации максимальна на минимумах кривых, которые представляют среднюю точку тепловых переходов (Tm). Для сравнения структуры каждого из рекомбинантных слитых вариантов mAb1 показаны схематически.
[0060] На фиг. 12A и 12B показана нейтрализация TGFβ при вызванной выработке IL-11 клетками A549 in vitro с помощью восьми рекомбинантных слитых вариантов mAb1, схематически изображенных на фиг. 9B.
[0061] На фиг. 13 показана аффинность рекомбинантных слитых вариантов mAb1 и химических конъюгатов mAb1 к гидроксиапатиту, оцениваемая с помощью колоночной хроматографии на колонке с керамическим гидроксиапатитом.
[0062] На фиг. 14 показано биораспределение выбранных меченых флуорофором рекомбинантных слитых вариантов mAb1 и химических конъюгатов mAb1 у мышей CD-1, полученное посредством визуализации в живом организме в различные моменты времени после введения.
[0063] На фиг. 15 показано количество меченого флуоресцентным красителем антитела, рекомбинантного варианта слияния mAb1 и химического конъюгата mAb1, локализованных в позвоночном столбе после введения мышам CD-1. Флуоресценцию измеряли с помощью прибора IVIS в течение 3 недель. Показан логарифм максимальной флуоресценции в ROI.
[0064] На фиг. 16 показаны флуоресцентные изображения хирургически удаленного позвоночника и бедренных костей мышей, которым вводили рекомбинантные слитые варианты mAb1, mAb1 F6, mAb1 F16 и mAb1 F17 и химический конъюгат mAb1 ("Кон-т") в исследовании, описанном в примере 13 и на фиг. 15, через 10 и 21 день.
[0065] На фиг. 17A и 17В показаны уровни флуоресценции mAb2 F1 и mAb2 F6 в 10 мкл сыворотки и хирургически удаленной поясничной части позвоночника, дистального (трабекулярного) отдела бедренной кости, почки и сердца через 24 и 96 часов, как описано в примере 15.
[0066] На фиг. 18A показано, что нацеливание на костную ткань с помощью mAb1-D10 (mAb1 F6) оказывает глубокое влияние на PK в сыворотке после введения однократной дозы. mAb1 F6 демонстрирует в 13-14 раз более низкую концентрацию в сыворотке (AUC), более быстрый сывороточный клиренс и более короткий период полужизни в сыворотке (t1/2), чем mAb1, как измерено при помощи ELISA. Данные выражены в виде среднего значения ±SD: Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1) наблюдалась при измерении с помощью дисперсионного анализа (AVOVA) и критерия множественного сравнения Даннета. mAb1 представляет собой муринизированное ингибирующее моноклональное антитело к TGFβ, и mAb1 F6 представляет собой рекомбинантное муринизированное ингибирующее моноклональное антитело к TGFβ с полипептидом аспартата D10, присоединенным к С-концу тяжелой цепи mAb1. AUC для визуализации/кости нормализовали до 1,0. Дозы составляли 5 мг/кг для каждого из mAb1 F6 и mAb1.
[0067] На фиг. 18B показано, что mAb1 F6 демонстрирует в кости в 22 раза более высокую концентрацию (AUC) по данным оптической визуализации по сравнению с mAb1. Данные выражены в виде среднего значения ±SD: Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1) наблюдалась при измерении с помощью AVOVA и критерия множественного сравнения Даннета. Дозы составляли 1 мг/кг для каждого из mAb1 F6 и mAb1.
[0068] На фиг. 19 проиллюстрированы PK-профили максимальных-минимальных концентраций при многократном введении доз. Нацеливание на костную ткань (с помощью mAb1 F6) оказывает глубокое влияние на PK в сыворотке максимальных-минимальных концентраций при многократном введении доз. mAb1 F6 демонстрирует более низкие концентрации в сыворотке, чем mAb1, как через 24, так и через 48 часов после введения дозы и после первой дозы и 23-й дозы. Показатели были ниже для mAb1 F6 в 3-4,5 раза через 24 ч. и в 6-9 раз через 48 ч. Накопление также оказалось меньше с mAb1 F6, чем с mAb1. Данные выражены в виде среднего значения ±SD: Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1) наблюдалась при измерении с помощью t-критерия для независимых выборок. Концентрации в сыворотке измеряли с помощью масс-спектрометрии.
[0069] На фиг. 20 показано, что нацеливание на костную ткань (mAb1 F6) и mAb1 увеличивают BV/TV (%) дозозависимым образом у мышей G61°C (OI). Статистически значимые изменения BV/TV (%) по сравнению с мышами G610C, получавшими контрольное антитело 13C4 (мышиное IgG1-антитело), наблюдали в дозах 1 и 5 мг/кг для обоих видов обработки. Мыши G610C, получавшие 13C4 (13C4), демонстрировали статистически значимое снижение BV/TV по сравнению с исходным штаммом WT (WT 13C4). Данные выражены в виде среднего значения ±SD: Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1; #p≤0,05 13C4 по сравнению с WT 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. BV/TV (%) измерено с помощью μCT-визуализации.
[0070] На фиг. 21 показано, что нацеливание на костную ткань (mAb1 F6) и mAb1 увеличивают максимально допустимое усилие до повреждения дозозависимым образом у мышей G61°C (OI). Статистически значимые изменения максимально допустимого усилия до повреждения по сравнению с мышами G610C, получавшими 13C4, наблюдали в дозах 1 и 5 мг/кг для mAb1 F6 и только 5 мг/кг для mAb1. Мыши G610C, получавшие контрольное антитело (13C4), демонстрировали статистически значимое снижение максимально допустимого усилия до повреждения по сравнению с исходным штаммом WT. Данные выражены в виде среднего значения ±SD: Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1; #p≤0,05 13C4 по сравнению с WT 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. Максимально допустимое усилие до повреждения измеряли в биомеханическом испытании на сжатие.
[0071] На фиг. 22 показано влияние mAb1 и mAb1 F6 на BV/TV у мышей G610C. Антитела вводили с различной частотой (3 раза в неделю, 1 раз в неделю, раз в 2 недели или раз в 4 недели) в дозе 5 мг/кг в течение 12 недель. Антитело 13C4 использовали в качестве контроля. Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1; #p≤0,05 13C4 по сравнению с WT 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. BV/TV измеряли с помощью μCT-визуализации.
[0072] На фиг. 23 показано влияние mAb1 и mAb1 F6 на максимально допустимое усилие до повреждения у мышей G610C. Антитела вводили с различной частотой (3 раза в неделю, 1 раз в неделю, раз в 2 недели или раз в 4 недели) в дозе 5 мг/кг в течение 12 недель. Антитело 13C4 использовали в качестве контроля. Статистическая значимость (*p≤0,05 mAb1 или mAb1 F6 по сравнению с 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. Максимально допустимое усилие до повреждения измеряли в биомеханическом испытании на сжатие.
[0073] На фиг. 24 показано влияние mAb1 и mAb1 F6 на BV/TV (%) и средние уровни антител в сыворотке крови мышей G610C. Антитела вводили раз в 2 недели или раз в неделю в дозе 5 мг/кг в течение 12 недель. Антитело 13C4 использовали в качестве контроля. Статистическая значимость (*p≤0,05 mAb1 или mAb1 F6 по сравнению с 13C4; #p≤0,05 13C4 по сравнению с WT 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. BV/TV (%) измеряли с помощью μCT-визуализации.
[0074] На фиг. 25 показано влияние mAb1 и mAb1 F16 на BV/TV (%) у мышей G610C. Антитела вводили 3 раза в неделю в дозе 5 мг/кг в течение 8 недель. Антитело 13C4 использовали в качестве контроля. Статистическая значимость (*p≤0,05 mAb1 или mAb1 F16 по сравнению с 13C4; #p≤0,05 13C4 по сравнению с WT 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. BV/TV (%) измеряли с помощью μCT-визуализации.
[0075] На фиг. 26 показано влияние mAb1 и mAb1 F11 на BV/TV (%) у мышей дикого типа. Антитела вводили 3 раза в неделю в дозе 5 мг/кг в течение 9 недель. Статистическая значимость (*p≤0,05 mAb1 или mAb1 F11 по сравнению с 13C4) наблюдалась при измерении с помощью однофакторного ANOVA. BV/TV (%) измеряли с помощью μCT-визуализации.
[0076] На фиг. 27 показана общая эффективность излучения в поясничном отделе мышей дикого типа после получения однократной внутрибрюшинной дозы носителя или флуоресцентно меченого mAb1 или mAb1 F6. Статистическая значимость (*p≤0,05 mAb1 F6 по сравнению с mAb1) наблюдалась при измерении с помощью однофакторного ANOVA.
[0077] На фиг. 28 показана общая эффективность излучения в сердце мышей дикого типа после получения однократной внутрибрюшинной дозы носителя или флуоресцентно меченого mAb1 или mAb1 F6. Статистическая значимость (*p≤0,05 mAb1 по сравнению с mAb1 F6) наблюдалась при измерении с помощью однофакторного ANOVA.
[0078] На фиг. 29 показана общая эффективность излучения в печени мышей дикого типа после получения однократной внутрибрюшинной дозы носителя или флуоресцентно меченого mAb1 или mAb1 F6. Статистическая значимость (*p≤0,05 mAb1 по сравнению с mAb1 F6) наблюдалась при измерении с помощью однофакторного ANOVA.
[0079] На фиг. 30 показана общая эффективность излучения в кишечнике мышей дикого типа после получения однократной внутрибрюшинной дозы носителя или флуоресцентно меченого mAb1 или mAb1 F6.
[0080] На фиг. 31 показана общая эффективность излучения в указанных тканях мышей дикого типа через 24 ч. или 96 ч. после получения однократной внутрибрюшинной дозы флуоресцентно меченого mAb2 или mAb2 D10. Статистическая значимость (*p≤0,05 mAb1 по сравнению с mAb1 F6) наблюдалась при измерении с помощью t-критерия.
[0081] На фиг. 32 показаны соотношения общей эффективности излучения в поясничном отделе/сыворотке крови мышей дикого типа через 24 ч. или 96 ч. после получения однократной внутрибрюшинной дозы флуоресцентно меченого mAb2 или mAb2 D10.
[0082] На фиг. 33 показаны соотношения общей эффективности излучения в бедренной кости/сыворотке крови мышей дикого типа через 24 ч. или 96 ч. после получения однократной внутрибрюшинной дозы флуоресцентно меченого mAb2 или mAb2 D10. Статистическая значимость (*p≤0,05 mAb2 D10 по сравнению с mAb2) наблюдалась при измерении с помощью t-критерия.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0083] В настоящем изобретении предусмотрены антитела и их антигенсвязывающие фрагменты, которые соединены с одним или несколькими нацеливающими на костную ткань поли-D-пептидами, так что антитела и фрагменты являются преимущественно нацеленными на кости пациента, нуждающегося в этом. Свойство, заключающееся в нацеливании на костную ткань, такого антитела или фрагмента позволяет антителу и фрагменту специфически нацеливаться на костные ткани и снижает системное воздействие антитела или фрагмента на пациента, тем самым повышая эффективность лекарственного средства при сведении к минимуму нежелательных побочных эффектов.
[0084] Используемый в данном документе термин "поли-D-пептид" относится к пептидной последовательности, имеющей множество аминокислот, представляющих собой аспарагиновую кислоту или аспартат или "D", таких как приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30 или более аминокислот (остатков) аспарагиновой кислоты. В одном варианте осуществления поли-D-пептид может включать от приблизительно 2 до приблизительно 30, или от приблизительно 3 до приблизительно 15, или от приблизительно 4 до приблизительно 12, или от приблизительно 5 до приблизительно 10, или от приблизительно 6 до приблизительно 8, или от приблизительно 7 до приблизительно 9, или от приблизительно 8 до приблизительно 10, или от приблизительно 9 до приблизительно 11, или от приблизительно 12 до приблизительно 14 остатков аспарагиновой кислоты. В одном варианте осуществления поли-D-пептиды включают только остатки аспартата. В другом варианте осуществления поли-D-пептиды могут включать одну или несколько других аминокислот или аналогичных соединений. Используемый в данном документе термин "D10" относится к непрерывной последовательности из десяти аминокислот, представляющих собой аспарагиновую кислоту, как видно из последовательности под SEQ ID NO: 1. В некоторых вариантах осуществления антитело или фрагмент антитела по настоящему изобретению могут включать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более 12 поли-D-пептидов.
[0085] Поли-D-пептид может быть соединен с представляющим интерес антителом или антигенсвязывающим фрагментом посредством рекомбинантной технологии или химического конъюгирования. Используемый в данном документе термин "слитый вариант" или "вариант" относится к собранной конструкции антитела (см. фиг. 9B), которая включает по меньшей мере одно из тяжелой цепи или легкой цепи или фрагмента антитела или его части, которая включает или иным образом связана с поли-D-пептидом, таким как последовательность D10. Например, поли-D-пептид может быть соединен с цепью антитела в слитом варианте при помощи рекомбинантной технологии (например, когда последовательность поли-D-пептида является неотъемлемой частью аминокислотной последовательности тяжелой цепи, легкой цепи или фрагмента антитела или его части), химического конъюгирования или обеих.
[0086] Используемый в данном документе термин "химический конъюгат" относится к собранному антителу, которое включает по меньшей мере одно из тяжелой цепи или легкой цепи, или фрагмента антитела, или его части, с которой один или несколько поли-D-пептидов соединены посредством химической реакции, например, с остатками цистеина, присутствующими в аминокислотной последовательности тяжелой цепи, легкой цепи, фрагмента антитела или его части. Типичными остатками цистеина, которые можно использовать для конъюгирования, являются остатки в шарнирной области тяжелой цепи. Остатки цистеина или другие остатки, подходящие для конъюгирования, также можно ввести в цепь антитела посредством мутагенеза. Спейсер/линкер, такой как пептидный линкер или химический фрагмент (например, малеимидная функциональная группа и полиэтиленгликоль (PEG)), можно использовать между поли-D-пептидом и компонентом антитела при конъюгировании. Способы химического конъюгирования требуемых фрагментов с антителами общеизвестны из уровня техники. См., например Behrens and Liu, mAbs 6:1, 46-53 (2014).
[0087] Используемый в данном документе термин "соединенный" относится к соединению поли-D-пептида с цепью антитела посредством рекомбинантной технологии, так что поли-D-пептид транскрибируется с того же РНК-транскрипта, что и цепь антитела, и находится в той же полипептидной последовательности, что и цепь антитела. В таких случаях поли-D-пептид может быть соединен с цепью антитела при помощи какого-либо пептидного линкера или аминокислотного спейсера или без него, на одном или обоих концах цепи антитела, или соединен внутри цепи антитела, не влияя на правильное сворачивание цепи антитела, сборку молекулы антитела или связывание антитела с его антигеном.
[0088] Иллюстративные форматы нацеливающихся на костную ткань антител по настоящему изобретению показаны на фиг. 9B (форматы F2-F20). Нацеливающий на костную ткань пептид (представленный кружками) можно присоединить или слить (например, соединить) с одним или обоими концами тяжелой цепи и/или легкой цепи антитела. В некоторых вариантах осуществления нацеливающийся на костную ткань пептид не присоединен к легкой цепи через N-конец легкой цепи. Присоединение или слияние может быть прямым соединением (т. е. без спейсера или линкера) или через спейсер или линкер (представленный волнистыми линиями; например, пептидный линкер). Конкретные примеры этих форматов приведены в Таблицах 1 и 7 ниже.
[0089] Любой подходящий спейсер или линкер можно использовать в данном документе для присоединения пептида, нацеливающегося на костную ткань, например, с помощью рекомбинантной технологии или химического конъюгирования, к представляющему интерес антителу. Например, можно использовать пептидный линкер, имеющий один, два, три или более повторов пептида G4S (SEQ ID NO: 9). Другие подходящие пептидные линкеры также можно использовать. См., например Chen et al., Adv Drug Deliv Rev 65(10):1357-1369 (2013).
Иллюстративные антитела и их антигенсвязывающие фрагменты, нацеливающиеся на костную ткань
[0090] В настоящем изобретении раскрыты антитела и антигенсвязывающие фрагменты, к которым прикреплен один или несколько поли-D-пептидов (полиаспартат или поли-Asp) (например, последовательность D10). Эти модифицированные антитела и фрагменты имеют улучшенную локализацию в кости. В одном конкретном варианте осуществления эти антитела представляют собой антитела к TGFβ, описываемые в данном документе. Не желая ограничиваться какой-либо теорией, полагают, что эффективное нацеливание антител к TGFβ на костную ткань с помощью одного или нескольких поли-D-пептидов может обеспечить новую терапию для индивидуумов с заболеваниями, характеризующимися патофизиологической дегенерацией кости, связанной с TGFβ.
[0091] Однако, хотя многочисленные варианты осуществления и примеры в данном документе построены в контексте использования антител к TGFβ и последовательностей D10, предполагается, что другие антитела или белки, подходящие для лечения аномального состояния кости или заболевания кости, можно модифицировать нацеливающимися на костную ткань фрагментами, описываемыми в данном документе. Например, терапевтические антитела для лечения потери костной массы, стимуляции роста костей или нацеливания на аномальные клетки (например, раковые клетки) в кости могут быть связаны с одним или несколькими нацеливающимися на костную ткань пептидами, описываемыми в данном документе. Терапевтические антитела могут связываться с белками или пептидами, участвующими в формировании или поддержании кости в здоровом состоянии. Кроме того, можно использовать другие пептиды для локализации в кости или нацеливания на костную ткань.
[0092] Используемые в данном документе термины "антитело α-TGFβ" и "антитело к TGFβ" могут использоваться взаимозаменяемо и относятся к антителу или его антигенсвязывающему фрагменту, которые специфичны в отношении TGFβ1, TGFβ2 и/или TGFβ3. Например, по меньшей мере один антигенсвязывающий сайт (или паратоп) антитела α-TGFβ или его антигенсвязывающего фрагмента связывается с эпитопом, обнаруженным на TGFβ1, TGFβ2 и/или TGFβ3 человека.
[0093] В одном варианте осуществления предполагаемую конструкцию антитело α-TGFβ-D10 можно создать с помощью химического конъюгирования. Например, химическое конъюгирование можно выполнить способами, известными в данной области техники, такими, как раскрытые в патентах США №№ 7763712, 4671958 и 4867973, каждый из которых включен с помощью ссылки. В другом примере пептид или другой линкер можно использовать для присоединения пептида D10 к антителу (см. фиг. 1 и 2). В дополнительном варианте осуществления восстановление тиольных групп в шарнирной области (например, остатков цистеина шарнирной области) антитела позволяет осуществить химическое конъюгирование поли-D-пептидов с использованием спейсера PEG. Подобным образом, другие остатки цистеина рассматриваемых антител и фрагментов антител, либо нативные для антител и фрагментов, либо введенные посредством мутагенеза, можно химически конъюгировать с поли-D-пептидами. Одна такая предполагаемая схема сборки антитела α-TGFβ, химически конъюгированного с пептидом D10, показана на фиг. 2.
[0094] В другом варианте осуществления предполагаемую конструкцию антитело α-TGFβ-D10 можно создать посредством рекомбинантной экспрессии, когда последовательность D10 добавляют к аминокислотной последовательности тяжелой цепи и/или легкой цепи антитела α-TGFβ. Например, последовательности нуклеиновых кислот, кодирующие аминокислотные последовательности тяжелой и/или легкой цепей, можно модифицировать для кодирования последовательности D10, которая будет экспрессироваться на N-конце, на С-конце, либо на обоих N-конце и С-конце тяжелой и/или легкой цепей антитела α-TGFβ. Аналогично, одну или несколько последовательностей D10 можно добавить к аминокислотной последовательности тяжелой цепи антитела в шарнирной области или около нее и/или в аминокислотную последовательность легкой цепи антитела. Каждую последовательность нуклеиновой кислоты для несущей D10 тяжелой и/или легкой цепи можно включить в вектор экспрессии и затем трансфицировать в клетку-хозяина, способную экспрессировать и транслировать последовательность нуклеиновой кислоты в соответствующую аминокислотную последовательность. Кроме того, клетка-хозяин способна собирать экспрессированные аминокислотные последовательности в функциональный белок посредством объединения каждой из тяжелой цепи и легкой цепи с ее комплементарной последовательностью с образованием конструкции антитело α-TGFβ-D10. Примеры предполагаемых рекомбинантных слитых вариантов антитела α-TGFβ-D10 показаны на фиг. 9A и 9B.
[0095] Хотя в данном документе обсуждается поли-D-пептид, другие аналогичные пептиды также можно использовать для обеспечения нацеливания антитела, другого белка или пептида на костную ткань. Например, повторяющиеся последовательности аспарагиновой кислоты могут иметь больше или меньше остатков, чем последовательность D10, например, приблизительно 2, или приблизительно 4, или приблизительно 6, или приблизительно 8, или приблизительно 12, или приблизительно 14, или приблизительно 16, или приблизительно 18, или приблизительно 20, или приблизительно 30, или 6, 7, 8, 9, 10 или 11 остатков и т. п.Кроме того, другие природные аминокислоты с аналогичными химическими свойствами, такие как глутамат или неприродные аминокислоты и/или другие химически эквивалентные соединения также можно использовать в комбинации с аспарагиновой кислотой или заменить ее.
[0096] В одном варианте осуществления предполагается, что антитело с одним или несколькими прикрепленными к нему поли-D-пептидами будет проявлять по меньшей мере приблизительно 2-кратное, или приблизительно 3-кратное, или приблизительно 5-кратное, или приблизительно 10-кратное, или приблизительно 20-кратное увеличение локализации в кости по сравнению с тем же антителом без одного или нескольких поли-D-пептидов.
[0097] Кроме того, хотя в данном документе описано антитело α-TGFβ, любое антитело, которое связывает другие белки, участвующие в формировании кости или поддержании кости в здоровом состоянии, можно при необходимости аналогичным образом модифицировать для нацеливания антитела на костную ткань. Антитела или их антигенсвязывающие фрагменты, рассматриваемые в данном документе, могут быть из любых видов или представлять собой гибридные антитела, объединяющие тяжелые цепи и легкие цепи из разных видов, и могут быть специфичными для любого требуемого эпитопа. Кроме того, антитела, которые можно использовать в данном документе, не ограничены изотипом и могут быть антителами любого из изотипов IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgM, IgE или IgD. Фрагменты антител также можно использовать. Например, последовательности D10 или другие нацеливающиеся на костную ткань соединения можно присоединить к фрагментам Fab и/или Fc или любому другому фрагменту антитела для достижения требуемого результата, как описано в данном документе. Дополнительно, последовательности D10 можно присоединить к фрагментам scFv и другим аналогичным слитым белкам. В другом варианте осуществления последовательности D10 можно присоединить к антителам, имеющим мутацию центральной шарнирной области S228P (пронумерована в соответствии с системой нумерации EU; или, альтернативно, S241P в соответствии с системой Kabat; см. Kabat et al., Sequences of Proteins of Immunological Interest, 4th ed., United States Government Printing Office, 165-492 (1987); и Silva et al. Jour. Biol. Chem. 290:5462-5469 (2015)).
[0098] В дополнительном варианте осуществления антитела и/или другие белки, рассматриваемые в данном документе, можно конъюгировать с дополнительными молекулами. Например, антитела или другие белки, рассматриваемые в данном документе, можно конъюгировать с химическими метками, которые позволяют отслеживать антитела/белки при инъекции или ином введении субъекту. Например, радиоактивные метки, флуоресцентные соединения и тому подобное можно присоединить к антителам/белкам, чтобы помочь их отслеживанию in vivo. Дополнительно, антитела и/или другие белки, рассматриваемые в данном документе, также можно конъюгировать с дополнительными соединениями, обладающими терапевтическим эффектом, такими как малые молекулы, фармацевтические препараты, противоопухолевые средства, гормоны роста, витамины и т.д., так что антитела и/или другие белки могут служить носителем для одного или нескольких таких соединений.
[0099] В некоторых вариантах осуществления нацеливающееся на костную ткань антитело к TGFβ содержит тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 2, 3, 4 и 5, и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 6, 7, 8, 11 и 12, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 2, когда аминокислотная последовательность легкой цепи представляет собой SEQ ID NO: 6. Иллюстративными антителами являются mAb1 F3, mAb1 F4, mAb1 F5, mAb1 F6, mAb1 F8, mAb1 F9, mAb1 F10, mAb1 F11, mAb1 F13, mAb1 F14, mAb1 F15, mAb1 F16, mAb1 F18, mAb1 F19 и mAb1 F20 (таблица 1).
[00100] В других вариантах осуществления нацеливающееся на костную ткань антитело к TGFβ содержит тяжелую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 13, 14, 16 и 17, и легкую цепь, содержащую аминокислотную последовательность, изложенную под любым из SEQ ID NO: 15, 18, 19, 20, 21 и 22, при условии, что аминокислотная последовательность тяжелой цепи не является последовательностью под SEQ ID NO: 13, когда аминокислотная последовательность легкой цепи представляет собой SEQ ID NO: 15. Иллюстративными антителами являются mAb2 F3, mAb2 F4, mAb2 F5, mAb2 F6, mAb2 F8, mAb2 F9, mAb2 F10, mAb2 F11, mAb2 F13, mAb2 F14, mAb2 F15, mAb2 F16, mAb2 F18, mAb2 F19 и mAb2 F20 (таблица 7).
[00101] В некоторых вариантах осуществления антитела по настоящему изобретению, такие как антитела к TGFβ, не имеют С-концевого лизина в тяжелой цепи. С-концевой лизин может быть удален во время изготовления или с помощью рекомбинантной технологии (т.e. кодирующая последовательность тяжелой цепи не включает кодон для концевого лизина на С-конце). Таким образом, в рамках настоящего изобретения также рассматриваются антитела, содержащие аминокислотную последовательность тяжелой цепи под SEQ ID NO: 2 или 13 без С-концевого лизина. Поли-D-пептид может быть присоединен к С-концу тяжелой цепи с С-концевым лизином или без него.
Способы лечения
[00102] В одном конкретном варианте осуществления способ лечения индивидуума, такого как пациент-человек, от потери костной ткани, связанной с TGFβ, включает введение эффективного количества антитела к TGFβ, нацеленного на костную ткань индивидуума. Способ может дополнительно включать этап измерения или выявления снижения уровней или активности TGFβ, снижения потери костной массы или скорости потери костной массы, увеличения плотности кости и/или увеличения прочности кости.
[00103] Используемое в данном документе выражение "эффективное количество" относится к количеству терапевтического средства, такого как антитело α-TGFβ или фрагмент антитела, которое при введении индивидууму, нуждающемуся в этом, улучшает состояние здоровья индивидуума, так, например, посредством снижения уровней или активности TGFβ, связанных с костью, снижения потери костной массы или скорости потери костной массы, увеличения плотности кости и/или увеличения прочности кости.
[00104] Используемый в данном документе термин "индивидуум" относится к животному. Примеры индивидуумов включают людей, одомашненных животных, домашних питомцев и других животных без ограничения. Дополнительные примеры индивидуумов включают животных, имеющих заболевание костей, связанное с TGFβ.
[00105] В другом варианте осуществления рассматриваются фармацевтические составы или композиции антител, включающие водные составы жидких продуктов, представляющих собой лекарственные средства, и составы лиофилизированных продуктов, представляющих собой лекарственные средства, содержащие одно или несколько нацеливающихся на костную ткань антител к TGFβ, таких как химические конъюгаты или рекомбинантные слитые варианты. Фармацевтические композиции, включающие нацеливающееся на костную ткань антитело к TGFβ и/или фрагменты антитела, могут быть составлены, как описано в публикации заявки на патент США № US 2014/0286933 A9, которая включена в данный документ посредством ссылки, или иным образом, как известно в данной области техники.
[00106] В одном конкретном варианте осуществления способ лечения заболевания костей включает введение эффективного количества антитела к TGFβ, нацеленного на костную ткань, индивидууму с заболеванием костей, таким как заболевание костей, связанное с хронической болезнью почек, метастазированием рака в кость или аномальными метаболическими состояниями. В другом конкретном варианте осуществления способ лечения несовершенного остеогенеза включает введение эффективного количества антитела к TGFβ, нацеленного на костную ткань, индивидууму с несовершенным остеогенезом. В дополнительном конкретном варианте осуществления способ лечения остеопороза включает введение эффективного количества антитела к TGFβ, нацеленного на костную ткань, индивидууму с остеопорозом.
[00107] В некоторых вариантах осуществления пациентов лечат комбинацией нацеливающегося на костную ткань антитела или фрагмента антитела по настоящему изобретению и другого терапевтического средства, такого как терапевтическое средство для лечения потери костной массы (например, бисфосфонаты). Антитело или фрагмент антитела и другое терапевтическое средство можно вводить пациенту одновременно или последовательно.
[00108] В некоторых вариантах осуществления нацеливающиеся на костную ткань антитела и другие компоненты, используемые в способах лечения по настоящему изобретению, могут быть предоставлены в наборах или изделиях.
Способы получения антител
[00109] Антитела или фрагменты по настоящему изобретению можно получать способами, хорошо известными в данной области техники. Последовательности ДНК, кодирующие тяжелые и легкие цепи антител, можно вставить в векторы экспрессии, так что гены являются функционально связанными с необходимыми последовательностями для регуляции экспрессии, такими как последовательности для регуляции транскрипции и трансляции. Векторы экспрессии включают плазмиды, ретровирусы, аденовирусы, аденоассоциированные вирусы (AAV), растительные вирусы, такие как вирус мозаики цветной капусты, вирус табачной мозаики, космиды, YAC, полученные из EBV эписомы и т. п.Кодирующая последовательность легкой цепи антитела и кодирующая последовательность тяжелой цепи антитела могут быть встроены в отдельные векторы и могут быть функционально связаны с такими же или разными последовательностями для регуляции экспрессии (например, промоторами). В одном варианте осуществления обе кодирующие последовательности вставлены в один и тот же вектор экспрессии и могут быть функционально связаны с такими же последовательностями для регуляции экспрессии (например, общим промотором), с отдельными идентичными последовательностями для регуляции экспрессии (например, промоторами) или с различными последовательностями для регуляции экспрессии (например, промоторами). Кодирующие последовательности антитела можно вставлять в вектор экспрессии стандартными способами (например, при помощи лигирования комплементарных сайтов рестрикции на фрагменте гена антитела и векторе или лигирования тупых концов, если сайты рестрикции отсутствуют).
[00110] В дополнение к генам цепей антитела рекомбинантные векторы экспрессии могут нести регуляторные последовательности, которые контролируют экспрессию генов цепей антитела в клетке-хозяине. Примеры регуляторных последовательностей для экспрессии в клетках-хозяевах млекопитающих включают вирусные элементы, которые управляют высокими уровнями экспрессии белка в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из ретровирусных LTR, цитомегаловируса (CMV) (такой как промотор/энхансер CMV), вируса обезьян 40 (SV40) (такой как промотор/энхансер SV40), аденовируса (например, главный поздний промотор аденовируса (AdMLP)), полиома, и сильные промоторы млекопитающих, такие как нативные промоторы иммуноглобулина и актина.
[00111] В дополнение к генам цепей антитела и регуляторным последовательностям рекомбинантные векторы экспрессии по настоящему изобретению могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации), и селектируемые маркерные гены. Например, селектируемый маркерный ген придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую был введен вектор. Селектируемые маркерные гены могут включать ген дигидрофолатредуктазы (DHFR) (для применения в dhfr- клетках-хозяевах с селекцией/амплификацией на метотрексате), ген neo (для селекции на G418) и ген глутамат-синтетазы.
[00112] Векторы экспрессии, кодирующие антитела по настоящему изобретению, вводят в клетки-хозяева для экспрессии. Клетки-хозяева культивируют в условиях, подходящих для экспрессии антитела, которое затем собирают и выделяют. Клетки-хозяева включают клетки-хозяева млекопитающих, растений, бактерий или дрожжей. Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, известны в данной области и включают множество иммортализованных клеточных линий, доступных из Американской коллекции типовых культур (АТСС). К ним относятся, inter alia, клетки яичника китайского хомяка (CHO), клетки NS0, клетки SP2, клетки HEK-293T, клетки 293 Freestyle (Invitrogen), клетки NIH-3T3, клетки HeLa, клетки почки новорожденного хомяка (BHK), клетки почки африканской зеленой мартышки (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), клетки A549 и ряд других клеточных линий. Клеточные линии могут быть выбраны на основе их уровней экспрессии. Другими линиями клеток, которые можно использовать, являются линии клеток насекомых, такие как клетки Sf9 или Sf21.
[00113] Дополнительно, экспрессию антител можно усилить с использованием ряда известных методик. Например, система экспрессии гена глутаминсинтетазы (система GS) является общим подходом для усиления экспрессии в определенных условиях.
[00114] Тканевая культуральная среда для клеток-хозяев может включать или не содержать компоненты животного происхождения (ADC), такие как бычий сывороточный альбумин. В некоторых вариантах осуществления для безопасности человека предпочтительны культуральные среды без ADC. Культивирование ткани можно выполнять с использованием способа периодического культивирования с подпиткой, способа непрерывной перфузии или любого другого способа, подходящего для клеток-хозяев и требуемого выхода.
Фармацевтические композиции
[00115] Антитело по настоящему изобретению можно составить для подходящей стабильности при хранении. Например, антитело можно лиофилизировать или сохранить или восстановить для применения с использованием фармацевтически приемлемых вспомогательных веществ. Для комбинированной терапии два или более терапевтических средства, таких как антитела, можно совместно составить, например, смешать и предоставить в одной композиции.
[00116] Термин "вспомогательное вещество" или "носитель" используют в данном документе для описания любого ингредиента, отличного от соединения(-й) по настоящему изобретению. Выбор вспомогательного вещества(-в) будет в значительной степени зависеть от таких факторов, как конкретный способ введения, влияние вспомогательного вещества на растворимость и стабильность и природа лекарственной формы. "Фармацевтически приемлемое вспомогательное вещество" включает все без исключения растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие абсорбцию средства и т. п., которые являются физиологически совместимыми. Некоторыми примерами фармацевтически приемлемых вспомогательных веществ являются вода, солевой раствор, фосфатно-солевой буферный раствор, декстроза, глицерин, этанол и т. п., а также их комбинации. В некоторых случаях в композицию будут включены изотонические средства, например, сахара, многоатомные спирты, такие как маннит, сорбит, или хлорид натрия. Дополнительными примерами фармацевтически приемлемых веществ являются смачивающие средства или незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие средства, консерванты или буферы, которые повышают срок годности или эффективность антитела.
[00117] Фармацевтическую композицию по настоящему изобретению можно получить, упаковать или продать без упаковки в виде единичной стандартной дозы или в виде множества единичных стандартных доз. Используемый в данном документе термин "стандартная доза" представляет собой дискретное количество фармацевтической композиции, содержащее заданное количество активного ингредиента. Количество активного ингредиента обычно равно дозе активного ингредиента, которую вводят субъекту, или удобной доле такой дозы, такой как, например, половина или одна треть такой дозы.
[00118] Фармацевтические композиции по настоящему изобретению обычно подходят для парентерального введения. Используемый в данном документе термин "парентеральное введение" фармацевтической композиции включает любой путь введения, характеризующийся физическим повреждением ткани субъекта и введением фармацевтической композиции через нарушение в ткани, что, как правило, приводит к непосредственному введению в кровоток, в мышцу или во внутренний орган. Парентеральное введение, таким образом, включает, без ограничения, введение фармацевтической композиции посредством инъекции композиции, посредством нанесения композиции через хирургический разрез, посредством нанесения композиции через проникающую в ткань нехирургическую рану и тому подобное. В частности, предполагается, что парентеральное введение включает, без ограничения, подкожное, внутрибрюшинное, внутримышечное, интрастернальное, внутривенное, внутриартериальное, интратекальное, внутрижелудочковое, внутриуретральное, внутричерепное, внутриопухолевое и внутрисиновиальное введение или инфузии; и методы диализной инфузии почек. Регионарная перфузия также рассматривается. Предпочтительные варианты осуществления могут включать внутривенные и подкожные пути.
[00119] Составы фармацевтической композиции, подходящие для парентерального введения, обычно содержат активный ингредиент в комбинации с фармацевтически приемлемым носителем, таким как стерильная вода или стерильный изотонический солевой раствор. Такие составы можно приготовить, упаковать или продать в форме, подходящей для болюсного введения или для непрерывного введения. Инъекционные составы можно приготовить, упаковать или продать в стандартной лекарственной форме, такой как ампулы или многодозовые контейнеры, содержащие консервант. Составы для парентерального введения включают, без ограничения, суспензии, растворы, эмульсии в масляных или водных носителях, пасты и тому подобное. Такие составы могут дополнительно содержать один или несколько дополнительных ингредиентов, включая, без ограничения, суспендирующие, стабилизирующие или диспергирующие средства. В одном варианте осуществления состава для парентерального введения активный ингредиент предоставляют в сухой (т.е. порошковой или гранулированной) форме для восстановления подходящим носителем (например, стерильной апирогенной водой) перед парентеральным введением восстановленной композиции. Парентеральные составы также включают водные растворы, которые могут содержать вспомогательные вещества, такие как соли, углеводы и буферные средства (например, рН от 3 до 9), но для некоторых применений их можно более подходящим образом составить в виде стерильного неводного раствора или в виде высушенной формы для использования в сочетании с подходящим носителем, таким как стерильная апирогенная вода. Иллюстративные формы для парентерального введения включают растворы или суспензии в стерильных водных растворах, например, водных растворах пропиленгликоля или декстрозы. Такие лекарственные формы можно подходящим образом забуферить при необходимости. Другие парентерально вводимые составы, которые являются полезными, включают составы, которые содержат активный ингредиент в микрокристаллической форме или в липосомном препарате. Составы для парентерального введения можно составить для немедленного и/или модифицированного высвобождения. Составы с модифицированным высвобождением включают составы с отсроченным, длительным, импульсным, контролируемым, нацеленным и запрограммированным высвобождением.
[00120] В некоторых вариантах осуществления антитело или антигенсвязывающий фрагмент по настоящему изобретению можно вводить в дозе 40, 20 или 15 мг/кг или меньше (например, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 мг/кг). В некоторых дополнительных вариантах осуществления дозы могут составлять 0,01, 0,02, 0,03, 0,04, 0,05, 0,1, 0,2, 0,3, 0,4 или 0,5 мг/кг. Частота введения доз может быть, например, ежедневной, раз в два, три, четыре или пять дней, еженедельной, раз в две недели или раз в три недели, ежемесячной, раз в два месяца, раз в три месяца, раз в шесть месяцев или раз в двенадцать месяцев или по мере необходимости. Антитело можно вводить внутривенно (например, внутривенной инфузией в течение 0,5-8 часов), подкожно, внутримышечно или любым другим путем введения, который подходит для состояния и лекарственной формы.
Иллюстративные варианты осуществления
[00121] Дополнительные конкретные варианты осуществления настоящего изобретения описываются следующим образом.
1. Антитело или его антигенсвязывающий фрагмент, содержащий тяжелую цепь, легкую цепь и один или несколько пептидов на основе полиаспартата (поли-D), связанных с (i) тяжелой цепью, (ii) С-концом легкой цепи или (iii) с обоими (i) и (ii).
2. Антитело или антигенсвязывающий фрагмент по варианту осуществления 1, где один или несколько поли-D-пептидов связаны с антителом или антигенсвязывающим фрагментом посредством химического конъюгирования.
3. Антитело или антигенсвязывающий фрагмент по варианту осуществления 2, где один или несколько поли-D-пептидов конъюгированы с тяжелой цепью по шарнирной области.
4. Антитело или антигенсвязывающий фрагмент по варианту осуществления 2 или 3, где один или несколько поли-D-пептидов конъюгированы с антителом или антигенсвязывающим фрагментом с помощью полиэтиленгликолевого (PEG) спейсера.
5. Антитело или антигенсвязывающий фрагмент по варианту осуществления 1, содержащие поли-D-пептид, соединенный с аминокислотной последовательностью тяжелой цепи или легкой цепи.
6. Антитело или антигенсвязывающий фрагмент по варианту осуществления 5, содержащие поли-D-пептид, соединенный с N-концом тяжелой цепи.
7. Антитело или антигенсвязывающий фрагмент по варианту осуществления 5, содержащие поли-D-пептид, соединенный с C-концом тяжелой цепи.
8. Антитело или антигенсвязывающий фрагмент по варианту осуществления 5, содержащие первый поли-D-пептид, соединенный с N-концом тяжелой цепи, и второй поли-D-пептид, соединенный с C-концом тяжелой цепи.
9. Антитело или антигенсвязывающий фрагмент по любому из вариантов осуществления 5-8, содержащие поли-D-пептид, соединенный с C-концом легкой цепи.
10. Антитело или антигенсвязывающий фрагмент по любому из вариантов осуществления 5-9, где поли-D полипептид(ы) слит(ы) с тяжелой или легкой цепью посредством пептидного линкера.
11. Антитело или антигенсвязывающий фрагмент по варианту осуществления 10, где пептидный линкер содержит 1-3 повтора аминокислотной последовательности GGGGS (SEQ ID NO: 9).
12. Антитело или антигенсвязывающий фрагмент по любому из предыдущих вариантов осуществления, где один или несколько поли-D-пептидов, каждый независимо, содержат 2-30 остатков аспарагиновой кислоты.
13. Антитело или антигенсвязывающий фрагмент по варианту осуществления 12, где каждый из одного или нескольких поли-D-пептидов содержит 10 остатков аспарагиновой кислоты (SEQ ID NO: 1).
14. Антитело или антигенсвязывающий фрагмент по любому из предыдущих вариантов осуществления, где антитело представляет собой IgG.
15. Антитело или антигенсвязывающий фрагмент по варианту осуществления 15, где антитело представляет собой IgG1 или IgG4.
16. Антитело или антигенсвязывающий фрагмент по любому из предыдущих вариантов осуществления, где антитело или антигенсвязывающий фрагмент специфически связываются с одним или несколькими из TGFβ1, TGFβ2 и TGFβ3.
17. Антитело или антигенсвязывающий фрагмент по варианту осуществления 16, где антитело содержит определяющие комплементарность участки (CDR) 1-3 тяжелой цепи из последовательности под SEQ ID NO: 13 и CDR1-3 легкой цепи из последовательности под SEQ ID NO: 15.
18. Антитело или антигенсвязывающий фрагмент по варианту осуществления 17, где антитело содержит аминокислотную последовательность вариабельного домена тяжелой цепи (VH), соответствующую остаткам 1-120 из последовательности под SED ID NO: 13, и аминокислотную последовательность вариабельного домена легкой цепи (VL), соответствующую остаткам 1-108 из последовательности под SEQ ID NO: 15.
19. Антитело или антигенсвязывающий фрагмент по варианту осуществления 17 или 18, где антитело содержит константный участок IgG4 человека с пролином в положении 228 (нумерация по EU).
20. Антитело или антигенсвязывающий фрагмент по варианту осуществления 19, где тяжелая цепь антитела содержит аминокислотную последовательность под SEQ ID NO: 13 с С-концевым лизином тяжелой цепи или без него и легкая цепь антитела содержит аминокислотную последовательность под SEQ ID NO: 15.
21. Антитело по варианту осуществления 17, где тяжелая цепь содержит аминокислотную последовательность под SEQ ID NO: 13 с С-концевым лизином тяжелой цепи или без него, SEQ ID NO: 14 с лизином, непосредственно предшествующим С-концевой последовательности D10, или без него, SEQ ID NO: 16 с С-концевым лизином тяжелой цепи или без него или SEQ ID NO: 17 с лизином, непосредственно предшествующим С-концевой последовательности D10, или без него, и легкая цепь содержит аминокислотную последовательность под SEQ ID NO: 15, 19, 21 или 22.
22. Антитело или антигенсвязывающий фрагмент по варианту осуществления 16, где антитело представляет собой мышиное антитело 1D11 с аминокислотными последовательностями тяжелой и легкой цепей под SEQ ID NO: 2 с C-концевым лизином или без него и SEQ ID NO: 6 соответственно.
23. Антитело IgG4, которое связывает TGFβ1, TGFβ2 и TGFβ3 человека, где тяжелая цепь антитела содержит аминокислотную последовательность под SEQ ID NO: 14 (с лизином, непосредственно предшествующим С-концевой последовательности D10, или без него) и легкая цепь содержит аминокислотную последовательность под SEQ ID NO: 15.
24. Антитело IgG4, которое связывает TGFβ1, TGFβ2 и TGFβ3 человека, где тяжелая цепь антитела содержит аминокислотную последовательность под SEQ ID NO: 17 (с лизином, непосредственно предшествующим С-концевой последовательности D10, или без него) и легкая цепь содержит аминокислотную последовательность под SEQ ID NO: 15.
25. Антитело или антигенсвязывающий фрагмент по любому из предыдущих вариантов осуществления, где антитело или антигенсвязывающий фрагмент демонстрируют по меньшей мере 2-кратное увеличение локализации в кости по сравнению с антителом с теми же тяжелой цепью и легкой цепью, но не содержащим поли-D-пептида(-ов).
26. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент по любому из предыдущих вариантов осуществления и фармацевтически приемлемое вспомогательное вещество.
27. Способ лечения индивидуума с патологическим состоянием кости, при котором ингибирование TGFβ оказывает благоприятное воздействие, включающий введение индивидууму эффективного количества антитела к TGFβ или антигенсвязывающего фрагмента по любому из пп. 16-25.
28. Способ по варианту осуществления 27, дополнительно включающий выявление по меньшей мере одного из (1) снижения уровней TGFβ, (2) снижения активности TGFβ, (3) уменьшения потери костной массы, (4) уменьшения скорости потери костной массы, (5) увеличения плотности кости, (6) увеличения прочности кости и (7) снижения уровней IL-11.
29. Антитело или антигенсвязывающий фрагмент по любому из вариантов осуществления 16-25 для применения при лечении индивидуума с патологическим состоянием кости, при котором ингибирование TGFβ оказывает благоприятное воздействие.
30. Применение антитела или антигенсвязывающего фрагмента по любому из вариантов осуществления 16-25 для изготовления лекарственного препарата для лечения индивидуума с патологическим состоянием кости, при котором ингибирование TGFβ оказывает благоприятное воздействие.
31. Способ по варианту осуществления 27, антитело или антигенсвязывающий фрагмент для применения по варианту осуществления 29 или применение по варианту осуществления 30, где индивидуумом является человек.
32. Способ, антитело или антигенсвязывающий фрагмент для применения или применение по варианту осуществления 31, где у человека имеется несовершенный остеогенез.
33. Способ, антитело или антигенсвязывающий фрагмент для применения или применение по варианту осуществления 31, где у человека наблюдается потеря костной массы или остеопороз.
34. Способ, антитело или антигенсвязывающий фрагмент для применения или применение по варианту осуществления 31, где у человека имеется хроническая болезнь почек.
35. Способ, антитело или антигенсвязывающий фрагмент для применения или применение по варианту осуществления 31, где человек является пациентом, у которого имеется рак с метастазированием в кость.
36. Выделенная молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую тяжелую цепь, легкую цепь или обе, антитела или антигенсвязывающего фрагмента по любому из вариантов осуществления 1-25.
37. Вектор экспрессии, содержащий выделенную молекулу нуклеиновой кислоты по варианту осуществления 36.
38. Клетка-хозяин, содержащая вектор экспрессии по варианту осуществления 37.
39. Клетка-хозяин по варианту осуществления 38, где клетка-хозяин представляет собой клетку млекопитающего.
40. Способ получения антитела или антигенсвязывающего фрагмента по любому из вариантов осуществления 1-25, включающий:
получение клетки-хозяина, содержащей первую и вторую нуклеотидные последовательности, кодирующие соответственно тяжелую цепь и легкую цепь антитела или антигенсвязывающего фрагмента,
выращивание клетки-хозяина в условиях, обеспечивающих возможность выработки антитела или антигенсвязывающего фрагмента, и
выделение антитела или антигенсвязывающего фрагмента.
41. Способ по варианту осуществления 40, где первая нуклеотидная последовательность предусматривает последовательность под SEQ ID NO: 23, 24, 25 или 26 (с кодоном для С-концевого лизина тяжелой цепи или без него) и вторая нуклеотидная последовательность предусматривает последовательность под SEQ ID NO: 27, 29, 31 или 32.
42. Способ получения нацеливающегося на костную ткань антитела или антигенсвязывающего фрагмента, включающий:
получение антитела или его антигенсвязывающего фрагмента и одного или нескольких поли-D-пептидов и
присоединение поли-D-пептидов к антителу с помощью ковалентной связи посредством химического конъюгирования.
43. Способ по любому из вариантов осуществления 40-42, дополнительно включающий составление антитела или антигенсвязывающего фрагмента в виде фармацевтической композиции, содержащей фармацевтически приемлемый носитель.
44. Способ получения нацеливающейся на костную ткань фармацевтической композиции, включающий:
получение антитела или антигенсвязывающего фрагмента по любому из вариантов осуществления 1-25 и
смешивание антитела или антигенсвязывающего фрагмента с фармацевтически приемлемым носителем.
[00122] Настоящее изобретение будет дополнительно описано в следующих примерах, которые не ограничивают объем настоящего изобретения, описанный в формуле изобретения.
ПРИМЕРЫ
[00123] Приведенные ниже примеры иллюстрируют конкретные варианты осуществления настоящего изобретения и различные их пути применения. Они изложены только в пояснительных целях и не должны рассматриваться в качестве ограничения настоящего изобретения.
Антитело к TGFβ
[00124] Первое антитело к TGFβ, обозначаемое в данном документе как mAb1, представляет собой моноклональное мышиное антитело IgG1, специфичное к TGF-β1, TGF-β2 и TGF-β3 человека ("панспецифичное") и доступное от R&D Systems (клон # 1D11, Миннеаполис, Миннесота). Антитело mAb1 служило в качестве матрицы в примерах.
[00125] Второе антитело к TGFβ, обозначаемое в данном документе как mAb2, использованное в примерах, представляет собой антитело IgG4 к TGFβ человека с мутацией в шарнире S228P (нумерация по EU). Антитело mAb2 аналогично антителам, раскрытым в патенте США № 9090685, который включен посредством ссылки. Антитело mAb2 имеет приблизительную молекулярную массу 144 кДа в негликозилированном виде. Аминокислотные последовательности его тяжелой и легкой цепей представляют собой последовательности под SEQ ID NO: 13 и 15, соответственно. Эти две последовательности показаны ниже. Вариабельные домены выделены курсивом и обозначены в данном документе как вариабельный домен тяжелой цепи (HCVD, SEQ ID NO: 39) и вариабельный домен легкой цепи (LCVD, SEQ ID NO: 40). CDR показаны в рамках и обозначены как определяющий комплементарность участок 1 тяжелой цепи (HCDR1, SEQ ID NO: 33); HCDR2 (SEQ ID NO: 34) и HCDR3 (SEQ ID NO: 35) и определяющий комплементарность участок 1 легкой цепи (LCDR1, SEQ ID NO: 36); LCDR2 (SEQ ID NO: 37) и LCDR3 (SEQ ID NO: 38). Сайт гликозилирования в константном домене тяжелой цепи выделен жирным шрифтом (N297).
QVQLVQSGAE VKKPGSSVKV SCKASGYTFS SNVISWVRQA PGQGLEWMGG VIPIVDIANY
AQRFKGRVTI TADESTSTTY MELSSLRSED TAVYYCASTL GLVLDAMDYW GQGTLVTVSS
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS
GLYSLSSVVT VPSSSLGTKT YTCNVDHKPS NTKVDKRVES KYGPPCPPCP APEFLGGPSV
FLFPPKPKDT LMISRTPEVT CVVVDVSQED PEVQFNWYVD GVEVHNAKTK PREEQFNSTY
RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS SIEKTISKAK GQPREPQVYT LPPSQEEMTK
NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG
NVFSCSVMHE ALHNHYTQKS LSLSLGK (SEQ ID NO: 13)
ETVLTQSPGT LSLSPGERAT LSCRASQSLG SSYLAWYQQK PGQAPRLLIY GASSRAPGIP
DRFSGSGSGT DFTLTISRLE PEDFAVYYCQ QYADSPITFG QGTRLEIKRT VAAPSVFIFP
PSDEQLKSGT ASVVCLLNNF YPREAKVQWK VDNALQSGNS QESVTEQDSK DSTYSLSSTL
TLSKADYEKH KVYACEVTHQ GLSSPVTKSF NRGEC (SEQ ID NO: 15)
[00126] Как описано в данном документе, можно использовать другие антитела, фрагменты антител, белки или пептиды, которые связывают белки, участвующие в формировании кости или поддержании кости в здоровом состоянии.
Пример 1: Получение химических конъюгатов пептида D10 с антителом α-TGFβ mAb1
[00127] Химические конъюгаты пептида D10 и антитела α-TGFβ mAb1 получали способом, изображенным на фиг. 2. mAb1 (2,0 мг) заменяли на дегазированный боратный буфер (25 мМ хлорида натрия, 1 мМ DTPA, 20 мМ бората натрия, рН 8,0) в трех циклах ультрафильтрации через центрифужные фильтры Amicon® Ultra MWCO 50 кДа (EMD Millipore). Дисульфиды шарнирной области затем восстанавливали с помощью 12 моль:моль дитиотреитола (DTT) на mAb в течение 2 ч. при 37°С. Продукт обессоливали на фильтре Amicon® Ultra 4 мл с помощью дегазированного боратного буфера. Аликвоты (1 нмоль, 150 мкг) подвергали реакции при 25°C с возрастающими количествами (1-15 моль:моль) пептида D10-малеимида (Ac-D10-C2-PEG12-C6-малеимид, где PEG12 состоит из PEG определенной длины, содержащего 12 этиленоксидных групп). Через 1,5 ч. оставшиеся непрореагировавшие тиольные группы блокировали добавлением 12 эквивалентов N-этилмалеимида с последующей инкубацией в течение 1,5 ч. Продукты обессоливали посредством ультрафильтрации. На фиг. 3A изображен SDS-PAGE 0,5 мкг каждого продукта на 4-12% геле NuPAGE, окрашенном SimplyBlue™ (Thermo Scientific) и визуализированном с помощью сканера Odyssey® в ближней ИК-области (LiCor). Профили флуоресценции треков объединяли с использованием программного обеспечения AlphaView (ProteinSimple Corp.) и определяли соотношение пептид-антитело (PAR). Предполагалось, что небольшое смещение подвижности в каждой тяжелой или легкой цепи представляет собой добавление одного пептида D10, что соответствует максимуму 5 идентифицируемых полос для тяжелой цепи и 1 для легкой цепи и соответствует числу цистеинов шарнирной области для каждого. Среднее количество пептидов в тяжелой и легкой цепях рассчитывали отдельно из суммы произведений относительной интенсивности каждой второстепенной полосы, умноженной на определенное для нее количество пептидов. PAR рассчитывали по сумме этих чисел для тяжелой и легкой цепей, умноженной на два, поскольку каждая цепь представлена дважды в полном IgG. Значение PAR относительно соотношения пептид-малеимид:mAb в реакции химического конъюгирования изображено на фиг. 3B, которая показывает линейное увеличение PAR с увеличением количества конъюгированных пептидов вплоть до 8 моль:моль.
Пример 2: Связывание химических конъюгатов mAb1-D10 с TGF-β1
[00128] В этом примере набор химических конъюгатов с различным PAR получали таким же образом, как описано в примере 1, за исключением того, что соотношение DTT варьировалось от 8-10 моль:моль, а соотношение малеимид-пептид:mAb составляло 3 или 15 моль:моль. В качестве контроля получали химический конъюгат IgG1 человека (Herceptin®) посредством восстановления его дисульфидов шарнира с помощью 3 моль:моль трис(2-карбоксиэтилфосфина) (ТСЕР) в течение 2 ч. при 37°С в атмосфере аргона с последующей реакцией с 15 моль:моль малеимид-пептида в течение ночи при 25°С и очищения посредством обессоливания с использованием ультрафильтрации.
[00129] Чтобы оценить их способность связывать TGF-β1, химические конъюгаты смешивали с TGF-β1 в молярном соотношении 1:1 с последующей эксклюзионной хроматографией на Superdex 200 (G.E. Healthcare) в фосфатно-солевом буфере (PBS), pH 7,2. Химический конъюгат mAb1-D10 отдельно давал несколько гетерогенный пик, элюирующийся раньше немодифицированного mAb1, как показано на фиг. 4А, что, вероятно, связано с эффектами заряда, создаваемыми конъюгированными пептидами. Добавление TGF-β1 к конъюгату приводило к более раннему пику элюирования, чем у mAb1 или химического конъюгата mAb1-D10, что указывает на образование комплекса с более высокой молекулярной массой (фиг. 4B). Аналогично, добавление TGF-β1 к немодифицированному антителу mAb1 вызывало сдвиг к более раннему времени удержания (фиг. 4C, 4D). Напротив, хотя химическая конъюгация D10 с Herceptin® вызывала сдвиг во времени удержания одного антитела, как и для конъюгата mAb1 (фиг. 4E, 4F), добавление TGF-β1 (1 моль:моль) к конъюгату Herceptin®-D10 не вызывало каких-либо изменений в его времени элюирования или кажущейся молекулярной массе (фиг. 4F, 4G), указывая на то, что не происходило связывания с конъюгатом вследствие конъюгирования.
Пример 3: Зависимое от пептида связывание химического конъюгата mAb1-D10 с гидроксиапатитом
[00130] В этом примере химический конъюгат D10 с mAb1, такой как показанный на фиг. 2, получали посредством восстановления дисульфидов шарнирной области с помощью 12 экв. DTT в 25 мМ NaCl, 1 мМ DTPA, 20 мМ бората натрия, рН 8, в течение 2 ч. при 37°С с последующей реакцией с 2 моль:моль 2,2'-дипиридилдисульфида (Sigma) для превращения части свободных тиолов обратно в дисульфиды. За этим следовала реакция с пептидом D10-малеимидом, описанная в примере 1. Конечный продукт очищали посредством ультрафильтрации. Часть (25 мкг) заменяли на 5 мМ фосфата натрия, рН 7,4, на спин-колонке и подвергали хроматографии на колонке объемом 100 мкл керамического гидроксиапатита (CHT) типа II (BioRad) и элюировали с применением градиента 5-500 мМ фосфата натрия, рН 7,4, при скорости потока 0,5 мл/мин. Немодифицированное mAb1 использовали в качестве контроля. Профиль колонки относительно A280 изображен на фиг. 5A. mAb1 показало только следовое связывание с колонкой, тогда как приблизительно половина конъюгата связывала и элюировала около 0,2 М фосфата. Фракции концентрировали и анализировали с помощью SDS-PAGE (фиг. 5B). В несвязанной (проточной или "FT") фракции в основном присутствовало немодифицированное mAb, тогда как основной пик 4 содержал конъюгат с рассчитанным при помощи SDS-PAGE значением PAR 6.
Пример 4: Влияние диапазона пептидной нагрузки на связывание с гидроксиапатитом
[00131] В этом примере ряд химических конъюгатов пептида D10 с различным количеством пептидов, описанных в примере 1, подвергали хроматографии на колонке CHT типа II, как описано в примере 3. Профили A280 и график фракции связанного конъюгата и времени удержания пика показаны на фиг. 6A. Как видно на фиг. 6B, количество конъюгата, связанного с колонкой, увеличилось до PAR 3,8, выровнялось и начало уменьшаться с увеличением количества пептидов. Напротив, время удержания конъюгата (указывающее на силу взаимодействия) со смолой увеличивалось с увеличением количества пептидов до 9.
Пример 5: Влияние химического конъюгирования пептида D10 с mAb1 на его эффективность нейтрализации TGF-β1 in vitro
[00132] В этом примере набор из трех конъюгатов mAb1-D10, таких как показанные на фиг. 2, получали, как описано в примере 1. Контрольный конъюгат получали посредством исключения пептида D10-малеимида во время конъюгирования. Посредством анализа SDS-PAGE, такого как изображенный на фиг. 3, определили, что пептидная нагрузка (PAR) для этих трех конъюгатов равна 0, 5 или 9. Затем определяли способность этих конъюгатов инактивировать TGF-β1 при совместной инкубации с TGF-β1 в течение 1 ч. Затем серийные разбавления в питательной среде применяли к клеткам A549 человека, экспрессирующим рецептор TGF-β1 человека, с последующей инкубацией в течение ночи. Затем определяли клеточный ответ на активный TGF-β1 по высвобождению IL-11 в питательную среду, которое выявляли в анализе ELISA, специфичном в отношении IL-11. Ответ IL-11 для трех конъюгатов и контрольного mAb1 изображен на фиг. 7. Во всех случаях полумаксимальное ингибирование высвобождения IL-11 (EC50) происходило примерно при 0,1 нМ антитела. Оба конъюгата (PAR=5 и PAR=9) и имитирующий конъюгат без пептида (PAR=0) показали немного лучшее ингибирование, чем mAb1. Количество конъюгированных пептидов не влияло на EC50.
Пример 6: Влияние конъюгированного пептида D10 на биораспределение mAb1, вводимого мышам
[00133] Химический конъюгат D10-mAb1 получали способом, аналогичным описанному в примере 1, как показано на фиг. 2. mAb1 (19,6 мг) заменяли на дегазированный боратный буфер, подвергали реакции с 10,6 экв. (1,06 мкмоль) DTT в течение 1,5 ч. при 37°С с последующей реакцией без замены буфера с 6 экв. пептида D10 (Ac-D10-C2-PEG12-С6-малеимид) в атмосфере аргона в течение 1,5 ч. при 25°С. Неконъюгированные свободные тиолы блокировали добавлением 12 экв. N-этилмалеимида с последующей инкубацией в течение 30 мин. при 25°C. Конечный продукт очищали посредством ультрафильтрации. Соотношение пептид-антитело определяли с использованием SDS-PAGE (PAR ~ 4,8). Легкая цепь показала следовые количества полосы, соответствующей добавлению одного пептида к остатку цистеина легкой цепи.
[00134] Конъюгат и контрольное антитело mAb1 отдельно метили AlexaFluor® 750 (Thermo Fisher), используя условия, описанные производителем. Флуоресцентные тестируемые изделия вводили безволосым мышам SKH-1 и затем визуализировали с помощью прибора IVIS (Perkin Elmer) сразу после введения (0,3-1 ч.) и через 4 ч., а также через 1, 2, 3, 4, 7, 8 и 9 дней. Изображения мышей, которым вводили 1 мг/кг меченого mAb1 и конъюгата, показаны на фиг. 8А. Флуоресценция меченого mAb1 была равномерно распределена по всему организму, и этот паттерн сохранялся, но снижался по интенсивности в течение 9 дней. Напротив, через 1 день конъюгат mAb1-D10 концентрировался по дорсальной средней линии, в конечностях и хвосте или вблизи них, что соответствовало локализации в кости, но был на почти неопределяемых уровнях в других частях тела. В соответствии с изображениями на фиг. 8А, соотношения флуоресценции дистальной части бедренной кости по сравнению с флуоресценцией сердца, измеренные на изображениях фиг. 8А, были статистически значимо повышены (P < 0,05) для конъюгата mAb1-D10 по сравнению с контролем через 24 ч. и оставались статистически значимо повышенными в течение 168 ч. (см. фиг. 8B). Это распределение сохранялось в течение 9 дней с незначительным изменением интенсивности.
Пример 7: Получение рекомбинантных слитых вариантов антитела к TGFβ-D10 мыши
[00135] Набор рекомбинантных слитых вариантов антитела к TGFβ-пептиду D10, показанный на фиг. 9А и 9В, получали, как описано ниже.
Конструкции векторов экспрессии тяжелой и легкой цепи
[00136] Создали панель плазмид для экспрессии тяжелой и легкой цепей mAb1 с пептидами D10 в различных положениях. Кроме того, получили также две плазмиды для тяжелой и легкой цепи mAb1 дикого типа без пептидов D10. Все кодон-оптимизированные последовательности получали синтетически (GeneArt), фланкировали соответствующими сайтами рестриктаз, сконструированными внутри рамки считывания в целях клонирования. Три генных фрагмента, кодирующих константные участки мышиного IgG1 с С-концевыми пептидами D10 или без них и полную легкую цепь mAb1 дикого типа клонировали в пустой эписомный вектор экспрессии млекопитающих pFF, аналог вектора pTT, описанного Durocher et al., (2002, Nucl. Acids Res. 30(2): E9), с получением "mIgG1_CH123_pFF", "mIgG1_CH123D10_pFF" и "mAb1_VLCL_pFF" с использованием рестриктаз ApaLI/HindIII и последующего лигирования. Генные фрагменты, кодирующие вариабельные области с N-концевым пептидом D10 или без него клонировали в векторы с использованием ApaLI/EagI для тяжелых цепей и ApaLI/MfeI для легкой цепи. Генные фрагменты, кодирующие константный участок легкой каппа-цепи мышиного Ig с C-концевым D10 и спейсером G4S (SEQ ID NO: 9) различной длины клонировали в "mAb1_VLCL_pFF" с помощью MfeI/HindIII для замены константного участка дикого типа. Ожидаемую правильную последовательность ДНК каждой конструкции подтверждали секвенированием ДНК (ACGT, Inc.).
Сборка и экспрессия слитых вариантов
[00137] Слитые варианты получали посредством выбора по одному из векторов тяжелой и легкой цепей для котрансфекции, чтобы получить слитые варианты и контроль без пептида, как описано в таблице 1. "PAR" (соотношение пептид-антитело) отражает общее количество пептидов, которые, как ожидается, будут присоединены к конечному экспрессированному антителу как на его тяжелой, так и на легкой цепях. ID слитого варианта "mAb1 F1" представляет собой конструкцию "дикого типа" ("wt"), которая практически идентична последовательности mAb1 без модификации. "HC" обозначает тяжелую цепь mAb1, "LC" обозначает легкую цепь mAb1, "D10" обозначает пептид D10 (SEQ ID NO: 1) и "G4S" обозначает спейсерную последовательность, состоящую из gly-gly-gly-gly-ser (SEQ ID NO: 9), включенную в некоторые конструкции.
Таблица 1 Рекомбинантные слитые варианты mAb1 с D10
[00138] Чтобы оценить способность к экспрессии необходимых рекомбинантных слитых вариантов, шестнадцать вариантов из таблицы 1 оценивали при котрансфекции в клетки Expi293F™ (Life Technologies), используя условия, описанные производителем. Через 4 дня экспрессию определяли с помощью SDS-PAGE кондиционированной среды. Все варианты экспрессировались на уровнях, которые по оценкам составляют 10-30 мкг/мл. Несколько более высокие уровни наблюдали через 5 и 7 дней, но уровни экспрессии зависели от варианта. WT (mAb1 F1), HC-D10:LC-D10 (mAb1 F8) и HC-D10: LC-G4S-D10 (mAb1 F9) показали особенно высокую экспрессию, в то время как D10-HC:D10-LC (mAb1 F12) экспрессировался плохо.
Крупномасштабная экспрессия
[00139] Все двадцать рекомбинантных слитых вариантов mAb1-D10 экспрессировали в клетках Expi293-F в масштабе 30 мл. Кондиционированные среды (CM) собирали на 6-й день и уровни экспрессии оценивали с помощью невосстанавливающего SDS-PAGE. Несколько более высокие уровни экспрессии (30-150 мкг/мл) наблюдали по сравнению с первоначальной оценкой, но относительные уровни экспрессии соответствовали трансфекциям меньшего масштаба.
Пример 8: Характеристика рекомбинантных слитых вариантов mAb1-D10
Уровни экспрессии и связывание TGF- β1
[00140] Количественный анализ уровня экспрессии и способности рекомбинантных слитых вариантов mAb1-D10 связывать TGF-β1 оценивали с использованием Octet® QK384. Биосенсоры Octet® для IgG мыши погружали в кондиционированную среду, разбавленную 1:10 в разбавителе для образца (PBS, pH 7,4, содержащем 0,01% BSA и 0,02% Tween 20), или 2-кратные серийные разбавления очищенного стандарта mAb1 в диапазоне от 1,25 до 100 мкг/мл. Данные по связыванию собирали в течение 2 мин. при встряхивании образцов при 500 об./мин. Титры рассчитывали, используя 4-параметрическое сглаживание начальных скоростей связывания и сравнение с данными, полученными для стандартов. Затем биосенсоры промывали разбавителем в течение 1 мин. для удаления любой среды и восстановления исходного уровня, а затем погружали в лунки, содержащие 40 нМ TGF-β1, в течение 3 мин. при 1000 об./мин. для последующего связывания. За этим следовала стадия диссоциации при переносе сенсоров в разбавитель в течение 3 мин. при 1000 об./мин. Как видно из таблицы 2, концентрации слитых вариантов в среде варьировались от 11 до 178 мкг/мл ("Конц. Octet") и в целом соответствовали тенденциям, наблюдаемым при SDS-PAGE ("Уровень экспрессии"). Кроме того, все варианты слияния были способны связывать TGF-β1, свидетельствуя о том, что присутствие пептида D10 в любом из указанных положений не оказывало серьезного влияния на связывание антигена.
Связывание мышиного FcRn
[00141] Связывание мышиного FcRn рекомбинантными слитыми вариантами mAb1-D10 оценивали с использованием Octet QK384. Все этапы выполняли при 1000 об./мин. Кондиционированную среду, разбавленную до достижения концентрации антител 10 мкг/мл, приводили в контакт с Fv-биосенсорами к IgG мыши в течение 5 минут. Затем биосенсоры погружали в PBSP с pH 6,0 (50 мМ фосфата натрия, pH 6,0, 150 мМ хлорида натрия, 0,005% поверхностно-активного вещества P20), чтобы установить исходный уровень. Затем сенсоры переносили в лунки, содержащие растворимый мышиный FcRn, разбавленный до 1 мкМ в PBSP, рН 6,0, в течение 3 мин. с последующей диссоциацией в течение 3 мин. в лунках, содержащих PBSP, рН 6,0. Как указано в таблице 2, все варианты связывают мышиный FcRn.
Связывание белка G
[00142] Связывание рекомбинантных слитых вариантов mAb1-D10 с белком G оценивали с использованием Octet QK384. Биосенсоры с белком G погружали в лунки, содержащие кондиционированную среду, разбавленную до 10 мкг/мл антитела в разбавителе для образца, и считывали сигнал в течение 3 мин. при 1000 об./мин. Как указано в таблице 2, все варианты связывали белок G.
Таблица 2 Характеристика слитых вариантов mAb1-D10
Octet®
(мкг/
мл)
экспрессии
мышиного
FcRn
белка G
SDS-PAGE
[00143] Очищенную конструкцию WT и рекомбинантные слитые варианты mAb1 F2, mAb1 F6, mAb1 F7, mAb1 F11, mAb1 F12, mAb1 F16 и mAb1 F17 анализировали на 4-20% гелях трис-глицина для SDS-PAGE (Novex, Life Sciences) в восстанавливающих и невосстанавливающих условиях и окрашивали кумасси синим. Изображение в видимом свете, полученное с помощью устройства формирования изображений ProteinSimple®, показано на фиг. 10. Наблюдали небольшое снижение подвижности тяжелой и/или легкой цепи, которое соответствовало ожидаемому присутствию и количеству пептидов D10 на каждой. Примеси и ковалентные агрегаты не обнаружили.
Термическая стабильность, определяемая методом дифференциальной сканирующей флуориметрии (DSF)
[00144] Термическую стабильность нескольких рекомбинантных слитых вариантов mAb1-D10, представляющих все возможные комбинации пептидов D10 на концах тяжелой и легкой цепей, определяли методом дифференциальной сканирующей флуориметрии с использованием SYPRO® оранжевого (Thermo Scientific) в качестве репортерного красителя. Общепринято, что стабильность белков при более высоких температурах предсказывает их стабильность в типичных условиях хранения и, следовательно, может использоваться для оценки их пригодности для изготовления и применения в качестве терапевтических средств. Термическую стабильность можно оценить с использованием красителей, которые демонстрируют увеличение флуоресценции при связывании гидрофобных областей, экспонируемых при развертывании структуры белка на приборах для ПЦР в реальном времени (Lo et al., 2004, Anal. Biochem. 332(1): 153-9). Флуоресценцию образцов (10 мкл белка в концентрации 0,5 мг/мл), содержащих разбавление SYPRO® оранжевого в пропорции 1:1000, отслеживали при повышении температуры в системе детекции ПЦР в реальном времени CFX96. Данные анализировали с использованием CFX Manager 3.0 (Bio-Rad Laboratories). На фиг.11 показана скорость изменения флуоресценции с температурой для нескольких вариантов. Скорость изменения обычно используют вместо абсолютной флуоресценции, чтобы отделить связывание красителя с частями структуры белка, которые не подвержены структурным переходам. Отрицательное смещение отражает увеличение скорости изменения флуоресценции, а минимум представляет среднюю точку перехода в развернутое состояние ("Tm"). Некоторые из протестированных вариантов (mAb1 F6, mAb1 F11, mAb1 F16) показали профили Tm, неотличимые от немодифицированного контрольного антитела ("WT"=mAb1 F1). Два других (mAb1 F2 и mAb1 F7) показали статистически значимое снижение Tm для основного перехода. Два варианта (mAb1 F12 и mAb1 F17) показали самые низкие значения Tm. Примечательно, что все четыре рекомбинантных слитых варианта mAb1-D10, содержащие пептид D10 на N-конце легкой цепи, показали статистически значимое снижение Tm, указывая на то, что размещение пептида в этом положении на антителе mAb1 дестабилизирует его структуру. Наоборот, размещение пептида D10 на любом конце тяжелой цепи не коррелировало с каким-либо изменением Tm. Значения Tm преобладающих переходов для нескольких слитых вариантов, наблюдаемых, как показано на фиг. 11, приведены в таблице 3.
Таблица 3 Дифференциальная сканирующая флуориметрия экспрессированных рекомбинантных слитых вариантов
A см. таблицу 2; B mAb1 F1 представляет собой рекомбинантную версию mAb1.
Пример 9: Эффективность рекомбинантных слитых вариантов mAb1-D10 в отношении нейтрализации TGF-β1 in vitro
[00145] Эффективность рекомбинантных слитых вариантов mAb1-D10 в отношении нейтрализации активности TGF-β1 определяли по ингибированию секреции IL-11 опухолевыми клетками A549 in vitro в ответ на добавление TGF-β1 в среду. Процедуру проводили, как описано в примере 3. Несколько характерных слитых вариантов из примера 7 отбирали на основе их аффинности к белку G (в качестве индикатора легкости их очистки). Профили ингибирования TGF-β1 для восьми вариантов (mAb1 F1, mAb1 F2, mAb1 F6, mAb1 F7, mAb1 F11, mAb1 F12, mAb1 F16 и mAb1 F17) показаны на фиг. 12А и 12В. Все слитые белки показали EC50, аналогичный контрольному mAb1.
Пример 10: Связывание TGF-β1 рекомбинантными слитыми вариантами
[00146] Рекомбинантные слитые варианты mAb1-D10 из примера 7 также количественно оценивали на связывание TGF-β1 с использованием поверхностного плазмонного резонанса (SPR), как выявлено с помощью Biacore®. Образцы, содержащие очищенные варианты, пропускали через сенсорный чип с захваченным аминогруппой TGF-β1 для определения константы равновесия KD. Целевой уровень иммобилизации, составляющий менее 100 единиц ответа (RU) TGF-β1, выбрали для сведения к минимуму вероятности эффектов авидности, возникающих в результате связывания соседних молекул TGF-β1 на чипе. Уровень иммобилизации определяли по изменению единиц ответа после активации поверхности чипа с помощью NHS/EDC, но до гашения этаноламином. Слитые варианты, слитый вариант WT F1 и контрольное mAb1 разбавляли до 30, 10, 3, 1 и 0,37 нМ в буфере HBS-EP и пропускали через чип с TGF-β1 при 30 мкл/мин в течение 3 мин. с последующей диссоциацией в течение 5 мин. в том же буфере. Между прогонами поверхность чипа регенерировали для удаления любого связанного антитела, пропуская две 30-секундные инъекции 40 мМ HCl через чип при скорости потока 75 мкл/мин. Как показано в таблице 4, константы равновесия (KD) для всех слитых вариантов и конструкции WT (F1) находились в пределах 2,4-кратной разницы с контрольным mAb1. Неожиданно, максимальный сигнал (RU), вызванный слитыми слияния, уменьшался с увеличением количества пептидов, причем вариант F17 (D10-HC-D10/D10-LC, PAR=6) демонстрировал самый низкий сигнал. Это противоречит анализу эффективности на клетках A549, описанному в примере 9, который показал, что все протестированные слитые варианты сопоставимы или более эффективны, чем mAb1 или конструкция WT, в отношении нейтрализации TGF-β1. Возможное объяснение этого снижения может отражать электростатическое отталкивание между матрицей чипа (карбоксиметилдекстран) и отрицательно заряженными пептидами D10.
Таблица 4 Константы равновесия для слитых вариантов mAb1-D10, связывающихся с TGF-β1, с применением анализа Biacore
KD (нМ)
A см. таблицу 2
Пример 11: Связывание рекомбинантных слитых вариантов mAb1-D10 с гидроксиапатитом
[00147] Рекомбинантные слитые варианты mAb1-D10 из примера 7 также тестировали на связывание в колонках с керамическим гидроксиапатитом (CHT), как описано в примере 3, для оценки их способности связываться с минеральной структурой кости. Рекомбинантные слитые варианты mAb1-D10 (25 мкг каждый в 5 мМ фосфате натрия, рН 7,4) наносили на колонку объемом 100 мкл с гидроксиапатитом (CHT типа II, BioRad) и элюировали с использованием градиента от 0,005 до 0,5 М Na+ фосфата, рН 7,4. Белок в подвижной фазе, элюируемой из колонки, исследовали на A280. Стандарт (обычно химический конъюгат mAb1-D10) пропускали в начале и в конце каждого набора прогонов для обеспечения согласованности в производительности колонки. На фиг. 13 показаны профили A280 для каждого прогона. Время удержания (RT) и связанная фракция показаны в таблице 5 ниже. Все рекомбинантные слитые варианты, кроме одного, были связаны почти количественно, но демонстрировали различия в аффинности, что отражалось на времени их удержания. Ранговый порядок времени удержания не очень хорошо коррелировал с PAR, свидетельствуя о значительном влиянии местоположения пептида на связывание. В частности, самое сильное связывание (самое высокое RT) наблюдали с вариантом F7 (PAR4), тогда как вариант F17 (PAR6) показал немного более слабое связывание. Один вариант (F6, PAR2) показал более слабое связывание, чем большинство вариантов с PAR4, но статистически значимо лучшее, чем другие варианты с PAR2 (F2 и F11). Один вариант (F2) с пептидом исключительно на N-конце легкой цепи показал только слабое связывание, что отражено в его RT и связанной фракции (23%). Химический конъюгат mAb1-D10 ("CC"; PAR ~ 4) давал два пика с 84%-м элюированием при более раннем RT, чем у всех слитых вариантов с PAR4.
Таблица 5 Связывание гидроксиапатита рекомбинантными слитыми вариантами mAb1-D10 и химическими конъюгатами
B химический конъюгат mAb1-D10 (конъюгированный с цистеинами шарнирной области и цистеинами С-конца легкой цепи, см. фиг. 3A); C не применимо.
Пример 12: Биораспределение рекомбинантных слитых вариантов у мышей
[00148] Биораспределение выбранного подмножества рекомбинантных слитых вариантов mAb1-D10 (F6, F16 и F17) и химического конъюгата mAb1-D10, полученного, как описано в примере 6, определяли после инъекции в хвостовую вену мышам CD-1. Рекомбинантные варианты выбрали на основе нескольких факторов, включая экспрессию и выход после очистки, аффинность связывания TGF-β1, эффективность в клетках и связывание с гидроксиапатитом. Эти варианты включали примеры, содержащие два, четыре или шесть пептидов D10, для оценки того, суммирует ли нацеливание связывание с гидроксиапатитом, или оно зависит от числа пептидов. Белки экспрессировали в клетках HEK293 и очищали белком А. Три рекомбинантных варианта слияния mAb1 детально охарактеризовали in vitro и in vivo. В них пептиды D10 рекомбинантно добавляли либо исключительно к C-концу тяжелой цепи (mAb1 F6), либо к N- и C-концам тяжелой цепи (mAb1 F16), либо к N- и C-концам тяжелой цепи и С-концу легких цепей (mAb1 F17). Рекомбинантные варианты mAb1 F6, F16 и F17 имеют соотношения пептид-антитело (PAR) 2, 4 и 6, соответственно. В качестве положительного контроля выбрали химический конъюгат mAb1-пептид D10 (~ 4,8 PAR), который продемонстрировал нацеливание на костную ткань в исследовании в примере 6.
[00149] Рекомбинантные слитые варианты и химический конъюгат метили в реакции со сложным эфиром NHS на Dylight® 800-4xPEG (Thermo Scientific) в 50 мМ борате натрия, pH 8,65, и при молярном соотношении краситель:белок приблизительно 5:1. Степень введения метки (DOL) поддерживали в пределах 20% (~ 1,2 моль:моль) посредством корректировки соотношения краситель:белок. Затем меченые белки вводили мышам CD-1 в дозе 1 мг/кг посредством инъекции в хвостовую вену. Впоследствии у анестезированных животных проводили визуализацию на устройстве для визуализации в ближней инфракрасной области спектра IVIS для мелких животных (Perkin Elmer) через 24, 48, 168 и 504 часа (3 недели) после введения. Бедренные кости и позвоночник извлекали у животного из каждой группы через 240 и 504 часа для проверки доставки в кость.
[00150] Как показано на изображениях вида сверху на фиг. 14, все слитые варианты mAb1-D10 и химический конъюгат концентрировались по дорсальной средней линии около позвоночного столба и оставались там в течение 3 недель (504 часа), хотя наблюдались статистически значимые различия в интенсивности сигнала для тестируемых изделий. Для количественного анализа использовали представляющий интерес участок (ROI), который включал часть позвоночника между плечами и тазом, как показано на фиг. 14. Максимальную эффективность излучения в пределах ROI, нормализованную по DOL, рассчитывали и наносили на график, как показано на фиг. 15. Рекомбинантный слитый вариант F6, который несет пептид D10 на каждом из С-концов тяжелой цепи, показал самый высокий уровень в позвоночнике среди всех конструкций в течение всего хода исследования (3 недели). Через 48 часов ранговый порядок интенсивности сигнала в пределах ROI представлял собой вариант F6 > вариант F16 > вариант F17 ≈ химический конъюгат, и этот порядок сохранялся на протяжении всего хода исследования (фиг. 15). Площадь под кривой (AUC) для максимальной эффективности излучения в пределах ROI во времени за вычетом фоновой автофлуоресценции (от контрольных животных только с носителем), рассчитанная с использованием программного обеспечения Phoenix® WinNonlin® (Pharsight), показана в таблице 6. Выявлено 8-22-кратное увеличение концентрации в кости (AUC) по сравнению с контрольным mAb1 и небольшие различия (до 2,5-кратных) между содержащими D10 конструкциями. Вариант F6 показал наибольшее увеличение по сравнению с mAb1 (в 21,8 раза, р<0,05). Период полужизни в ткани, рассчитанный с использованием WinNonlin®, также показал >10-кратного увеличения периода полужизни в кости для варианта F6 по сравнению с mAb1 (р<0,05).
[00151] Позвонки и бедренные кости от характерных животных в каждой когорте выделяли через 240 и 504 часов, отделяли от окружающей ткани и визуализировали (фиг. 16). Относительная интенсивность флуоресценции этих образцов соответствовала сигналам изображений вида сверху, наблюдаемым у живых животных, указывая на то, что они отражают присутствие рекомбинантных слитых вариантов и химического конъюгата в кости.
Таблица 6 Концентрация в позвоночнике по данным AUC для флуоресценции
B химический конъюгат mAb1-D10
* P<0,05 по сравнению с mAb1
Пример 13: Получение рекомбинантного слитого варианта антитела к TGFβ-D10 человека
[00152] Вектор экспрессии для получения mAb2 (антитела IgG4 к TGFβ человека с мутацией в шарнире S228P), несущего С-концевую последовательность D10 на тяжелой цепи (т.e. mAb2 HC-D10/mAb2 wt LC (SEQ ID NO: 14/SEQ ID NO: 15), которое имеет соответствующую конфигурацию, как в варианте F6 (см. фиг. 9B), и далее упоминается как mAb2 F6; см. таблицу 7) получали таким же образом, как описано в примере 7. Клетки Expi293F трансфицировали с помощью выделенной и очищенной плазмидной ДНК и через 6 дней собирали кондиционированную среду (60 мл) и полученный белок очищали с помощью белка A HiTrap (G.E. Healthcare).
Таблица 7 Рекомбинантные слитые варианты mAb2 с D10
Пример 14: Биораспределение слитого белка антитела к TGFβ-D10 человека у мышей
[00153] Рекомбинантный вариант mAb2 F6 (mAb2 F6) и контрольные антитела mAb2 метили AlexaFluor® 647 (Thermo Scientific) и вводили внутрибрюшинно мышам C57BL/6 в дозе 1 мг/кг. Через 24 и 96 часов некоторых мышей умерщвляли и позвонки и бедренные кости вырезали и визуализировали на приборе IVIS. Образец сыворотки (10 мкл), полученный при умерщвлении, визуализировали параллельно. Средняя общая эффективность излучения для ROI дистального (трабекулярного) отдела бедренной кости и поясничного отдела позвоночника показана на фиг. 17А и 17В. Относительная интенсивность приведена в таблице 8. Эти результаты показывают значительное увеличение количества антител в поясничном отделе позвоночника и бедренной кости со статистически значимо меньшим содержанием в сыворотке через 96 часов в результате рекомбинантного добавления D10 к mAb2.
Таблица 8 Относительная концентрация в кости слитых белков на основе антитела к TGFβ-D10 человека у мышей C57BL/6
Пример 15: Фармакокинетика в сыворотке и костной ткани однократной дозы слитого белка на основе антитела к TGFβ-D10 мыши у мышей
[00154] В этом примере у мышей измеряли фармакокинетику слитых белков на основе антитела к TGFβ-D10 мыши.
[00155] Однократную дозу mAb1 или рекомбинантного mAb1 F6 (см. таблицу 2) вводили внутрибрюшинно мышам G61°C (модель животных с несовершенным остеогенезом; n=12 на каждый момент времени), и образцы крови собирали через 4 часа или через 2, 7, 15, 22 и 43 дня после приема дозы. Использовали ELISA, оптимизированный для выявления и количественного определения концентраций соответствующих антител в сыворотке.
[00156] Для визуализации костей однократную дозу меченого флуорофором mAb1, рекомбинантного mAb1 F6 или различных других альтернатив D10 вводили внутривенно голым мышам CD-1 (n=3 на каждый момент времени), а оптическую визуализацию in vivo проводили через 4 часа или через 1, 2, 4, 7, 10 и 21 день после введения дозы. Получали флуоресцентные изображения позвоночника мыши, что позволяло провести относительные сравнения тестируемых изделий между mAb1 и mAb1 F6 в кости (не показано).
[00157] Фармакокинетические профили в сыворотке и кости, соответственно, можно видеть на фиг. 18А и 18В, и полученные фармакокинетические параметры приведены в таблице 9 ниже.
[00158] Результаты продемонстрировали фундаментальные различия в фармакокинетике между mAb1 и mAb1 F6 в сыворотке и кости после однократного приема дозы. mAb1 F6 показало в 13 раз меньшую AUC (концентрацию) в сыворотке и в 22 раза более высокую концентрацию в кости по сравнению с mAb1. Кроме того, mAb1 F6 продемонстрировало в 14 раз меньшее t1/2 в сыворотке, чем mAb1, и, соответственно, в 13 раз более быстрый клиренс. И наконец, для кости mAb1 F6 продемонстрировало в 11 раз более длительное t1/2, чем mAb1, и, соответственно, в 17 раз более медленный клиренс. Эти признаки могут быть полезны для человеческой формы mAb1-D10 в клинической области, где периферическое (сывороточное) ингибирование TGFβ может не быть необходимым с точки зрения безопасности, в то время как более высокая концентрация в кости может повысить эффективность.
Таблица 9 Параметры PK после однократной дозы
*p<0,05 по сравнению с mAb1; #AUC нормализовано до 1,0 для mAb1
Пример 16: Фармакокинетика максимальных-минимальных концентраций в сыворотке при многократном введении доз слитого белка на основе антитела к TGFβ-D10 мыши у мышей
[00159] В этом примере проводили фармакокинетическое исследование максимальных-минимальных концентраций при многократном введении доз на животной модели несовершенного остеогенеза.
[00160] mAb1 и mAb1 F6 (см. таблицу 2) вводили внутрибрюшинно в концентрации 0,3 мг/кг и 1 мг/кг 3 раза в неделю в течение 8 недель (24 дозы суммарно) мышам G61°C (n=10) и отбирали образцы крови через 24 и 48 ч. после введения дозы, следующей за 1-й и 23-й дозой (начало и конец исследования). Результаты показаны только для дозы 1 мг/кг (см. фиг. 19). Использовали масс-спектрометрический анализ, оптимизированный для выявления и количественного определения концентрации соответствующих антител в сыворотке.
[00161] Результаты количественно представлены в таблице 10 ниже. Результаты продемонстрировали фундаментальные различия в фармакокинетике между mAb1 и mAb1 F6 в сыворотке как после 1-й, так и после 23-й дозы. Статистически значимо более низкие концентрации в сыворотке наблюдали для mAb1 F6 по сравнению с mAb1 как через 24, так и через 48 ч. после введения 1-й и 23-й дозы. Кроме того, наклон между 24 и 48 ч. после введения дозы для mAb1 F6 был круче по сравнению с mAb1 как при 1-й, так и при 23-й дозе, позволяя предположить, что mAb1 F6 покидает сыворотку (системное кровообращение) с большей скоростью, чем mAb1, вероятно из-за его высокой аффинности в отношении кости (гидроксиапатит). Наконец, как mAb1 F6, так и mAb1, по-видимому, накапливаются в сыворотке с 1-й по 23 дозу, но mAb1 F6, по-видимому, накапливается при сниженной концентрации по сравнению с mAb1 (mAb1 F6: накопление в 2,5-3,5 раза и mAb1: накопление в 4-5,5 раза с 1-й по 23-ю дозу). Эти признаки могут быть полезны для человеческой формы mAb1 F6 в клинической области, где периферическое (сывороточное) ингибирование TGFβ может не быть необходимым с точки зрения безопасности.
Таблица 10 Результаты исследования по введению доз mAb1
(1 мг/кг)
(мкг/мл)
* p≤0,05 по сравнению с mAb1
Пример 17: Исследование эффективности в кости поясничного отдела при многократном введении доз mAb1 и mAb1 F6
[00162] В этом примере проводили исследование эффективности при многократном введении доз на животной модели несовершенного остеогенеза для определения эффективности нацеленного на костную ткань mAb1 F6 в сравнении с не нацеленным mAb1 на плотность и прочность кости.
[00163] mAb1 и mAb1 F6 вводили внутрибрюшинно 3 раза в неделю в дозе 0,3, 1 и 5 мг/кг в течение 8 недель мышам G610C. После получения последней дозы мышей вскрывали и 6-ю кость поясничного отдела визуализировали с помощью μCT для определения отношения объема кости к общему объему (BV/TV) и подвергали биомеханическому тестированию для определения максимально допустимого усилия до повреждения (прочность кости).
[00164] Результаты показаны на фиг. 20 и 21. Статистически значимые изменения BV/TV (%) по сравнению с мышами G610C, получавшими 13C4, наблюдали в дозах 1 и 5 мг/кг для обоих видов обработки. Статистически значимые изменения максимально допустимого усилия до повреждения по сравнению с мышами G610C, получавшими 13C4, наблюдали в дозах 1 и 5 мг/кг для mAb1 F6 и только 1 мг/кг для mAb1. Мыши G610C, получавшие контрольное антитело (13C4), демонстрировали статистически значимое снижение как BV/TV, так и максимально допустимого усилия до повреждения по сравнению с исходным штаммом WT. Эти результаты демонстрируют, что оба вида обработки вызывают аналогичные связанные с дозой изменения BV/TV и максимально допустимого усилия до повреждения у мышей G61°C при данной схеме введения доз 3 раза в неделю. Проявлялась тенденция к повышению эффективности mAb1 F6 в отношении прочности кости, поскольку половина когорты мышей при дозе 5 мг/кг демонстрировала существенно более высокие значения максимально допустимого усилия до повреждения (40 ньютон или выше), чем мыши, получавшие mAb1 в дозе 1 мг/кг или 5 мг/кг.
Пример 18: Исследование частоты введения доз mAb1 и mAb1 F6 в кости поясничного отдела
[00165] В этом примере проводили исследование частоты введения доз на животной модели несовершенного остеогенеза для определения подходящей частоты введения доз mAb1 F6 для достижения оптимальной эффективности антител, нацеленных на костную ткань.
[00166] mAb1 и mAb1 F6 вводили внутрибрюшинно с различной частотой (3 раза в неделю, 1 раз в неделю, 1 раз в 2 недели или 1 раз в 4 недели) в дозе 5 мг/кг в течение 12 недель мышам G610C. Образцы сыворотки для фармакокинетики (PK) отбирали в начале и в конце исследования, чтобы установить максимальные и минимальные значения концентрации как для mAb1, так и для mAb1 F6. После получения последней дозы мышей вскрывали и 6-ю кость поясничного отдела визуализировали с помощью μCT для определения отношения объема кости к общему объему (BV/TV) и подвергали биомеханическому тестированию для определения максимально допустимого усилия до повреждения (прочность кости).
[00167] Результаты показаны на фиг. 22, 23 и 24. Статистически значимые изменения BV/TV (%) по сравнению с мышами G610C, получавшими 13C4, наблюдали для mAb1 при частоте 3 раза в неделю, 1 раз в неделю, 1 раз в 2 недели и 1 раз в 4 недели. Статистически значимые изменения BV/TV (%) по сравнению с мышами G610C, получавшими 13C4, наблюдали для mAb1 F6 при частоте 3 раза в неделю и 1 раз в неделю. При получении mAb1 показано статистически значимо более высокое BV/TV по сравнению с получением mAb1 F6 при частоте введения доз 1 раз в 2 недели и 1 раз в 4 недели. Статистически значимые изменения максимально допустимого усилия до повреждения по сравнению с мышами G610C, получавшими 13C4, наблюдали для mAb1 при частоте 3 раза в неделю, 1 раз в неделю и 1 раз в 2 недели. Статистически значимые изменения максимально допустимого усилия до повреждения по сравнению с мышами G610C, получавшими 13C4, наблюдали для mAb1 F6 при частоте 3 раза в неделю и 1 раз в неделю. Мыши G610C, получавшие контрольное антитело (13C4), демонстрировали статистически значимое снижение BV/TV и тенденцию к более низкому максимально допустимому усилию до повреждения по сравнению с исходным штаммом WT.
[00168] Эти результаты демонстрируют, что как mAb1, так и mAb1 F6 могут вызывать аналогичные максимальные эффекты относительно BV/TV и максимально допустимого усилия до повреждения у мышей G610C. mAb1, по-видимому, имеет преимущество в продолжительности эффективности по сравнению с mAb1 F6, поддерживая статистически значимую эффективность при введении доз один раз в 4 недели в отношении BV/TV и один раз в 2 недели в отношении максимально допустимого усилия до повреждения. Однако средние значения для PK образцов сыворотки при эквивалентно эффективных схемах введения доз (mAb1, 1 раз в 2 недели и mAb1 F6, 1 раз в неделю) давали приблизительно 38 мкг/мл и 8 мкг/мл для mAb1 и mAb1 F6, соответственно. Это говорит о том, что концентрация в сыворотке может быть меньше с mAb1 F6, что может иметь преимущество безопасности для пациентов с OI.
Пример 19: Исследование частоты введения доз mAb1 F16 в кости поясничного отдела
[00169] В этом примере проводили исследование частоты введения доз на животной модели несовершенного остеогенеза, чтобы определить подходящую частоту введения доз mAb1 F16 для достижения оптимального влияния на плотность кости.
[00170] mAb1 и mAb1 F16 вводили внутрибрюшинно 3 раза в неделю в дозе 5 мг/кг в течение 8 недель мышам G610C. После получения последней дозы мышей вскрывали и 6-ю кость поясничного отдела визуализировали с помощью μCT для определения отношения объема кости к общему объему (BV/TV).
[00171] Результаты показаны на фиг. 25. Статистически значимые изменения BV/TV (%) по сравнению с мышами G610C, получавшими 13C4, наблюдали в дозе 5 мг/кг как для mAb1, так и для mAb F16. Мыши G610C, получавшие контрольное антитело (13C4), демонстрировали статистически значимое снижение BV/TV по сравнению с исходным штаммом WT. Эти результаты демонстрируют, что как mAb1, так и mAb F16 вызывают аналогичные связанные с дозой изменения BV/TV у мышей G61°C при этой схеме введения доз.
Пример 20: Исследование частоты введения доз mAb1 F11 в кости
[00172] В этом примере проводили исследование частоты введения доз на мышах дикого типа, чтобы определить подходящую частоту введения доз mAb1 F11 для достижения оптимального влияния на плотность кости.
[00173] mAb1 и mAb1 F11 вводили внутрибрюшинно 3 раза в неделю в дозе 5 мг/кг в течение 11 недель мышам дикого типа. В несколько моментов времени сделали μCT in vivo в ходе прижизненной части исследования. Данные показаны только через 9 недель после введения доз для отношения объема кости к общему объему (BV/TV%).
[00174] Результаты показаны на фиг. 26. Статистически значимые изменения BV/TV (%) по сравнению с мышами дикого типа, получавшими 13C4, наблюдали при 5 мг/кг для обоих видов обработки. Эти результаты демонстрируют, что как mAb1, так и mAb1 F11 вызывают аналогичные связанные с дозой изменения BV/TV у мышей дикого типа при этой схеме введения доз.
Пример 21: Биораспределение нацеливающихся на костную ткань антител к TGFβ mAb1 F6 и mAb2 F6 мышей и человека у мышей
[00175] В этом примере проводили исследование по сравнению биораспределения флуоресцентно меченых mAb1, mAb1 F6, mAb2 и mAb2 D10 (D10 конъюгированы с С-концом тяжелой цепи mAb2; mAb2 F6) у мышей дикого типа. Однократную внутрибрюшинную дозу каждого тестируемого изделия и носителя вводили мышам, которых подвергали эвтаназии в различные моменты времени для сбора ткани. Среди других собранных тканей (данные не показаны), позвонки поясничного отдела, сердце, печень и кишечник собирали через 1, 4, 10, 20, 43 и 98 дней после введения дозы mAb1 и mAb1 F6. Также собирали образцы тканей после введения дозы mAb2 и mAb2 D10 через 24 и 96 часов.
[00176] Результаты показаны на фиг. 27-33. На фиг. 27-30 показана общая эффективность излучения (TRE) в тканях мышей, которым вводили mAb1, mAb1 F6 или носитель, через 1-98 дней после введения дозы. Позвонки поясничного отдела демонстрируют стойкое постоянное присутствие mAb1 F6 по сравнению с mAb1 со статистически значимо более высокой общей эффективностью излучения (TRE) в каждый момент времени. В сердце и печени mAb1 F6 демонстрирует гораздо более низкую TRE по сравнению с mAb1. Наконец, не было отмечено никаких статистически значимых различий между носителем и любым тестируемым изделием в кишечнике.
[00177] Эти результаты демонстрируют, что mAb1 F6 характеризуется высокой аффинностью в отношении кости, что, в противовес этому, приводит к снижению концентрации в других тканях (например, в сердце и печени). Результаты также указывают на безопасность с точки зрения нацеливания на сайт ингибирования TGF-β в кости при одновременном ограничении системного ингибирования TGF-β и снижение побочных эффектов. Отсутствие какой-либо TRE в отношении носителя в кишечнике демонстрирует, что флуорофор сохранил свою метку на соответствующих антителах. Предыдущие данные показали, что если флуорофор не сохраняет свою метку на антителах, он будет выявлен в кишечнике.
[00178] На фиг. 31-33 показаны TRE в тканях мышей, которым вводили mAb2 или mAb2 D10. На фиг. 31 продемонстрирована высокая аффинность в отношении кости mAb2 D10 по сравнению с mAb2. Примечательно, что более высокая TRE наблюдается в бедренной кости для mAb2 D10 по сравнению с mAb2. Такую же общую тенденцию наблюдали в поясничном отделе. Соотношения поясничный отдел/сыворотка и бедренная кость/сыворотка у мышей, получавших дозу mAb2 D10, по сравнению с соотношением у мышей, получавших дозу mAb2, также убедительно поддерживают способность mAb2 D10 нацеливаться на костную ткань (фиг. 32 и 33).
[00179] Описав настоящее изобретение подробно и посредством ссылок на конкретные варианты его осуществления, будет очевидно, что возможны модификации и вариации без отклонения от объема настоящего изобретения, определенного в прилагаемой формуле изобретения. Более конкретно, несмотря на то, что некоторые аспекты настоящего изобретения определены в данном документе как особенно преимущественные, следует иметь в виду, что настоящее изобретение не обязательно ограничено данными конкретными аспектами настоящего изобретения. В некоторых вариантах осуществления значения, раскрытые в данном документе, могут альтернативно изменяться количественно на ±10, 20 или 30% от раскрытых значений и оставаться в рамках объема рассматриваемого изобретения.
[00180] Если не указано иное, научные и технические термины, используемые в связи с настоящим изобретением, должны иметь значения, которые обычно понимаются специалистами в данной области техники. Примерные способы и вещества описаны ниже, хотя способы и вещества, аналогичные или эквивалентные описанным в данном документе, могут быть использованы в практике или при тестировании настоящего изобретения. Все публикации и другие ссылки, упомянутые данном документе, включены посредством ссылки во всей своей полноте. В случае противоречий настоящее описание, в том числе определения, будет иметь преимущественную силу. Хотя ряд документов цитируется в данном документе, цитирование ссылок не является признанием того, что любой из этих документов образует часть общеизвестных знаний в данной области. Кроме того, если иное не требуется по контексту, термины единственного числа включают множественное число, а термины множественного числа включают единственное число. Как правило, номенклатура, используемая в связи с методами и технологиями клеточной и тканевой культуры, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, синтетической органической химии, медицинской и фармацевтической химии, а также химии и гибридизации белков и нуклеиновых кислот, описанными в данном документе, является такой, которая хорошо известна и обычно используется в данной области техники. Ферментативные реакции и методы очистки выполняют в соответствии с описаниями производителя, как обычно осуществляется в данной области техники, или как описано в данном документе. Во всем данном описании и вариантах осуществления слова "иметь" и "содержать" или варианты, такие как "имеет", "имеющий", "содержит" или "содержащий", следует понимать как подразумевающие включение указанного целого числа или группы целых чисел, но не исключение любого другого целого числа или группы целых чисел. Дополнительно, формы существительного единственного числа включают ссылку на формы множественного числа, если контекст четко не определяет иное. Например, ссылка на "антитело" означает одно или несколько антител.
[00181] Для целей описания или определения настоящего изобретения следует отметить, что термин "практически" используется в данном документе для представления характерной степени неопределенности, которая может быть свойственна любому количественному сравнению, значению, измерению или другому представлению. Термин "практически" также используется в данном документе для представления степени, в которой количественное представление может отличаться от принятого эталона, не приводя к изменению основной функции обсуждаемого объекта изобретения.
[00182] Используемый в данном документе термин "приблизительно" относится к ± 10% от данного количества, однако всякий раз, когда рассматриваемое количество относится к неделимому объекту, такому как аминокислота или другой объект, который потерял бы свою идентичность при разделении, "приблизительно" относится к ±1 неделимому объекту. Например, приблизительно 2% воды относится к 1,8-2,2% воды, тогда как приблизительно 6 аминокислотных остатков относится к 5-7 аминокислотным остаткам.
[00183] Используемые в данном документе термины "или" и "и/или" используются для описания множества компонентов в комбинации или исключая друг друга. Например, "x, y и/или z" может относиться только к "x", только к "y", только к "z", "x, y и z", "(x и y) или z", "x или (y и z)" или "x или y или z".
[00184] Последовательности, указанные в настоящем описании, представлены в таблице 11 ниже, а также в списке последовательностей.
Таблица 11 Последовательности
(WT HC)
(SEQ ID NO: 2)
(HC-D10)
(SEQ ID NO: 3)
(D10-HC)
(SEQ ID NO: 4)
(D10-HC-D10)
(SEQ ID NO: 5)
(WT LC)
(SEQ ID NO: 6)
(D10-LC)
(SEQ ID NO: 7)
(LC-D10)
(SEQ ID NO: 8)
1X пептидного линкера (G4S)
(SEQ ID NO: 9)
2X пептидного линкера ((G4S)x2)
(SEQ ID NO: 10)
(LC-G4S-D10)
(SEQ ID NO: 11)
(LC-(G4S)2-D10)
(SEQ ID NO: 12)
(SEQ ID NO: 13)
(SEQ ID NO: 14)
(SEQ ID NO: 15)
(SEQ ID NO: 16)
(SEQ ID NO: 17)
(SEQ ID NO: 18)
(SEQ ID NO: 19)
(SEQ ID NO: 20)
(LC-G4S-D10)
(SEQ ID NO: 21)
(LC-(G4S)2-D10)
(SEQ ID NO: 22)
(SEQ ID NO: 23)
(SEQ ID NO: 24)
(SEQ ID NO: 25)
(SEQ ID NO: 26)
(SEQ ID NO: 27)
(SEQ ID NO: 28)
(SEQ ID NO: 29)
(SEQ ID NO: 30)
(LC-G4S-D10)
(SEQ ID NO: 31)
(LC-(G4S)2-D10)
(SEQ ID NO: 32)
(SEQ ID NO: 33)
(SEQ ID NO: 34)
(SEQ ID NO: 35)
(SEQ ID NO: 36)
(SEQ ID NO: 37)
(SEQ ID NO: 38)
(SEQ ID NO: 39)
(SEQ ID NO: 40)
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GENZYME CORPORATION
<120> АНТИТЕЛА, НАЦЕЛИВАЮЩИЕСЯ НА КОСТНУЮ ТКАНЬ
<130> 022548.WO012
<140>
<141>
<150> 62/448,763
<151> 2017-01-20
<160> 40
<170> PatentIn версия 3.5
<210> 1
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 1
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp
1. 5 10
<210> 2
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 2
His Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Ala
1. 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Thr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Phe Pro Ala Ser Gly Ser Thr Asn Tyr Asn Glu Met Phe
50 55 60
Glu Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Gly Asn Tyr Ala Leu Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser
115 120 125
Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val
130 135 140
Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro
180 185 190
Ser Ser Thr Trp Pro Ser Gln Thr Val Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly
210 215 220
Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys
245 250 255
Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln
260 265 270
Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu
290 295 300
Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg
305 310 315 320
Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro
340 345 350
Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr
355 360 365
Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln
370 375 380
Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly
385 390 395 400
Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu
405 410 415
Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn
420 425 430
His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 3
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 3
His Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Ala
1. 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Thr Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Gln Ile Phe Pro Ala Ser Gly Ser Thr Asn Tyr Asn Glu Met Phe
50 55 60
Glu Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Gly Asn Tyr Ala Leu Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser
115 120 125
Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val
130 135 140
Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro
180 185 190
Ser Ser Thr Trp Pro Ser Gln Thr Val Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly
210 215 220
Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys
245 250 255
Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln
260 265 270
Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu
290 295 300
Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg
305 310 315 320
Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro
340 345 350
Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr
355 360 365
Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln
370 375 380
Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly
385 390 395 400
Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu
405 410 415
Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn
420 425 430
His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys Asp Asp Asp
435 440 445
Asp Asp Asp Asp Asp Asp Asp
450 455
<210> 4
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 4
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp His Val Gln Leu Gln Gln
1. 5 10 15
Ser Gly Pro Glu Leu Val Arg Pro Gly Ala Ser Val Lys Leu Ser Cys
20 25 30
Lys Ala Ser Gly Tyr Ile Phe Ile Thr Tyr Trp Met Asn Trp Val Lys
35 40 45
Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Gln Ile Phe Pro Ala
50 55 60
Ser Gly Ser Thr Asn Tyr Asn Glu Met Phe Glu Gly Lys Ala Thr Leu
65 70 75 80
Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu
85 90 95
Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Gly Asp Gly Asn
100 105 110
Tyr Ala Leu Asp Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
115 120 125
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
130 135 140
Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val
145 150 155 160
Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser
165 170 175
Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu
180 185 190
Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
195 200 205
Gln Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val
210 215 220
Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys
225 230 235 240
Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
245 250 255
Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val
260 265 270
Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp
275 280 285
Asp Val Glu Val His Thr Ala Gln Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys
340 345 350
Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys
355 360 365
Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp
370 375 380
Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys
385 390 395 400
Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser
405 410 415
Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr
420 425 430
Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210> 5
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 5
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp His Val Gln Leu Gln Gln
1. 5 10 15
Ser Gly Pro Glu Leu Val Arg Pro Gly Ala Ser Val Lys Leu Ser Cys
20 25 30
Lys Ala Ser Gly Tyr Ile Phe Ile Thr Tyr Trp Met Asn Trp Val Lys
35 40 45
Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Gln Ile Phe Pro Ala
50 55 60
Ser Gly Ser Thr Asn Tyr Asn Glu Met Phe Glu Gly Lys Ala Thr Leu
65 70 75 80
Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu
85 90 95
Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Gly Asp Gly Asn
100 105 110
Tyr Ala Leu Asp Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
115 120 125
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
130 135 140
Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val
145 150 155 160
Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser
165 170 175
Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu
180 185 190
Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
195 200 205
Gln Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val
210 215 220
Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys
225 230 235 240
Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
245 250 255
Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val
260 265 270
Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp
275 280 285
Asp Val Glu Val His Thr Ala Gln Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys
340 345 350
Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys
355 360 365
Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp
370 375 380
Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys
385 390 395 400
Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser
405 410 415
Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr
420 425 430
Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser
435 440 445
Leu Ser His Ser Pro Gly Lys Asp Asp Asp Asp Asp Asp Asp Asp Asp
450 455 460
Asp
465
<210> 6
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 6
Asn Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105 110
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
115 120 125
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
145 150 155 160
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
180 185 190
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
195 200 205
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 7
<211> 228
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 7
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asn Ile Val Leu Thr Gln
1. 5 10 15
Ser Pro Ala Ser Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser
20 25 30
Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr Gly Asn Ser Phe Met His
35 40 45
Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro Lys Leu Leu Ile Tyr Leu
50 55 60
Ala Ser Asn Leu Glu Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly
65 70 75 80
Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp Pro Val Glu Ala Asp Asp
85 90 95
Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn Glu Asp Pro Leu Thr Phe
100 105 110
Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Ala Asp Ala Ala Pro Thr
115 120 125
Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly Ala
130 135 140
Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn Val
145 150 155 160
Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu Asn Ser
165 170 175
Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser Thr
180 185 190
Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr Cys
195 200 205
Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser Phe Asn
210 215 220
Arg Asn Glu Cys
225
<210> 8
<211> 228
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 8
Asn Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105 110
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
115 120 125
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
145 150 155 160
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
180 185 190
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
195 200 205
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys Asp Asp Asp Asp Asp Asp
210 215 220
Asp Asp Asp Asp
225
<210> 9
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 9
Gly Gly Gly Gly Ser
1. 5
<210> 10
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 10
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10
<210> 11
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 11
Asn Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105 110
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
115 120 125
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
145 150 155 160
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
180 185 190
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
195 200 205
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys Gly Gly Gly Gly Ser Asp
210 215 220
Asp Asp Asp Asp Asp Asp Asp Asp Asp
225 230
<210> 12
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 12
Asn Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Ala Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Asn Asn
85 90 95
Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105 110
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
115 120 125
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
145 150 155 160
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
180 185 190
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
195 200 205
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp
225 230 235
<210> 13
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 13
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn
20 25 30
Val Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Ile Pro Ile Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Thr Leu Gly Leu Val Leu Asp Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 14
<211> 457
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 14
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn
20 25 30
Val Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Ile Pro Ile Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Thr Leu Gly Leu Val Leu Asp Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Asp
435 440 445
Asp Asp Asp Asp Asp Asp Asp Asp Asp
450 455
<210> 15
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 15
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 16
<211> 457
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 16
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Gln Val Gln Leu Val Gln
1. 5 10 15
Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys
20 25 30
Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn Val Ile Ser Trp Val Arg
35 40 45
Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Val Ile Pro Ile
50 55 60
Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe Lys Gly Arg Val Thr Ile
65 70 75 80
Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr Met Glu Leu Ser Ser Leu
85 90 95
Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ser Thr Leu Gly Leu
100 105 110
Val Leu Asp Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
130 135 140
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455
<210> 17
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 17
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Gln Val Gln Leu Val Gln
1. 5 10 15
Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys
20 25 30
Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn Val Ile Ser Trp Val Arg
35 40 45
Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Val Ile Pro Ile
50 55 60
Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe Lys Gly Arg Val Thr Ile
65 70 75 80
Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr Met Glu Leu Ser Ser Leu
85 90 95
Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ser Thr Leu Gly Leu
100 105 110
Val Leu Asp Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
130 135 140
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Leu Gly Lys Asp Asp Asp Asp Asp Asp Asp
450 455 460
Asp Asp Asp
465
<210> 18
<211> 225
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 18
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Glu Thr Val Leu Thr Gln
1. 5 10 15
Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser
20 25 30
Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser Tyr Leu Ala Trp Tyr Gln
35 40 45
Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser
50 55 60
Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
65 70 75 80
Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val
85 90 95
Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro Ile Thr Phe Gly Gln Gly
100 105 110
Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys
225
<210> 19
<211> 225
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 19
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys Asp Asp Asp Asp Asp Asp Asp Asp Asp
210 215 220
Asp
225
<210> 20
<211> 235
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 20
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Glu Thr Val Leu Thr Gln
1. 5 10 15
Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser
20 25 30
Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser Tyr Leu Ala Trp Tyr Gln
35 40 45
Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser
50 55 60
Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
65 70 75 80
Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val
85 90 95
Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro Ile Thr Phe Gly Gln Gly
100 105 110
Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp
225 230 235
<210> 21
<211> 230
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 21
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Asp Asp Asp Asp
210 215 220
Asp Asp Asp Asp Asp Asp
225 230
<210> 22
<211> 235
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 22
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp
225 230 235
<210> 23
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 23
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggata caccttcagt agcaatgtta tcagctgggt gcgccaggcc 120
cctggacaag ggctcgagtg gatggggggg gtcatcccta ttgttgatat tgcgaactac 180
gcacagagat tcaagggcag agtcacgatt accgcggacg aatccactag tacaacttac 240
atggagttga gcagcctgag gtctgaggac acggccgtgt attactgtgc gagcacactt 300
ggtctcgtcc tggatgctat ggactactgg ggtcagggta cgttagtgac ggtctcgagt 360
gcttccacca agggcccatc cgtcttcccc ctggcgccct gctccaggag cacctccgag 420
agcacagccg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacgaagacc 600
tacacctgca acgtagatca caagcccagc aacaccaagg tcgacaagag agttgagtcc 660
aaatatggtc ccccatgccc accatgccca gcacctgagt tcctgggggg accatcagtc 720
ttcctgttcc ccccaaaacc caaggacact ctcatgatct cccggacccc tgaggtcacg 780
tgcgtggtgg tggacgtgag ccaggaagac cccgaggtcc agttcaactg gtacgtggat 840
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagttcaa cagcacgtac 900
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaacggcaa ggagtacaag 960
tgcaaggtct ccaacaaagg cctcccgtcc tccatcgaga aaaccatctc caaagccaaa 1020
gggcagcccc gagagccaca ggtgtacacc ctgcccccat cccaggagga gatgaccaag 1080
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 1140
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 1200
gacggctcct tcttcctcta cagcaggcta accgtggaca agagcaggtg gcaggagggg 1260
aatgtcttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 1320
ctctccctgt ctctggggaa atga 1344
<210> 24
<211> 1374
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 24
gacgacgatg atgacgatga cgacgacgat caggtgcagc tggtgcagtc tggggctgag 60
gtgaagaagc ctgggtcctc ggtgaaggtc tcctgcaagg cttctggata caccttcagt 120
agcaatgtta tcagctgggt gcgccaggcc cctggacaag ggctcgagtg gatggggggg 180
gtcatcccta ttgttgatat tgcgaactac gcacagagat tcaagggcag agtcacgatt 240
accgcggacg aatccactag tacaacttac atggagttga gcagcctgag gtctgaggac 300
acggccgtgt attactgtgc gagcacactt ggtctcgtcc tggatgctat ggactactgg 360
ggtcagggta cgttagtgac ggtctcgagt gcttccacca agggcccatc cgtcttcccc 420
ctggcgccct gctccaggag cacctccgag agcacagccg ccctgggctg cctggtcaag 480
gactacttcc ccgaaccggt gacggtgtcg tggaactcag gcgccctgac cagcggcgtg 540
cacaccttcc cggctgtcct acagtcctca ggactctact ccctcagcag cgtggtgacc 600
gtgccctcca gcagcttggg cacgaagacc tacacctgca acgtagatca caagcccagc 660
aacaccaagg tcgacaagag agttgagtcc aaatatggtc ccccatgccc accatgccca 720
gcacctgagt tcctgggggg accatcagtc ttcctgttcc ccccaaaacc caaggacact 780
ctcatgatct cccggacccc tgaggtcacg tgcgtggtgg tggacgtgag ccaggaagac 840
cccgaggtcc agttcaactg gtacgtggat ggcgtggagg tgcataatgc caagacaaag 900
ccgcgggagg agcagttcaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac 960
caggactggc tgaacggcaa ggagtacaag tgcaaggtct ccaacaaagg cctcccgtcc 1020
tccatcgaga aaaccatctc caaagccaaa gggcagcccc gagagccaca ggtgtacacc 1080
ctgcccccat cccaggagga gatgaccaag aaccaggtca gcctgacctg cctggtcaaa 1140
ggcttctacc ccagcgacat cgccgtggag tgggagagca atgggcagcc ggagaacaac 1200
tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcta cagcaggcta 1260
accgtggaca agagcaggtg gcaggagggg aatgtcttct catgctccgt gatgcatgag 1320
gctctgcaca accactacac acagaagagc ctctccctgt ctctggggaa atga 1374
<210> 25
<211> 1374
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 25
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggata caccttcagt agcaatgtta tcagctgggt gcgccaggcc 120
cctggacaag ggctcgagtg gatggggggg gtcatcccta ttgttgatat tgcgaactac 180
gcacagagat tcaagggcag agtcacgatt accgcggacg aatccactag tacaacttac 240
atggagttga gcagcctgag gtctgaggac acggccgtgt attactgtgc gagcacactt 300
ggtctcgtcc tggatgctat ggactactgg ggtcagggta cgttagtgac ggtctcgagt 360
gcttccacca agggcccatc cgtcttcccc ctggcgccct gctccaggag cacctccgag 420
agcacagccg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacgaagacc 600
tacacctgca acgtagatca caagcccagc aacaccaagg tcgacaagag agttgagtcc 660
aaatatggtc ccccatgccc accatgccca gcacctgagt tcctgggggg accatcagtc 720
ttcctgttcc ccccaaaacc caaggacact ctcatgatct cccggacccc tgaggtcacg 780
tgcgtggtgg tggacgtgag ccaggaagac cccgaggtcc agttcaactg gtacgtggat 840
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagttcaa cagcacgtac 900
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaacggcaa ggagtacaag 960
tgcaaggtct ccaacaaagg cctcccgtcc tccatcgaga aaaccatctc caaagccaaa 1020
gggcagcccc gagagccaca ggtgtacacc ctgcccccat cccaggagga gatgaccaag 1080
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 1140
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 1200
gacggctcct tcttcctcta cagcaggcta accgtggaca agagcaggtg gcaggagggg 1260
aatgtcttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 1320
ctctccctgt ctctggggaa agacgacgat gatgacgatg acgacgacga ttga 1374
<210> 26
<211> 1404
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 26
gacgacgatg atgacgatga cgacgacgat caggtgcagc tggtgcagtc tggggctgag 60
gtgaagaagc ctgggtcctc ggtgaaggtc tcctgcaagg cttctggata caccttcagt 120
agcaatgtta tcagctgggt gcgccaggcc cctggacaag ggctcgagtg gatggggggg 180
gtcatcccta ttgttgatat tgcgaactac gcacagagat tcaagggcag agtcacgatt 240
accgcggacg aatccactag tacaacttac atggagttga gcagcctgag gtctgaggac 300
acggccgtgt attactgtgc gagcacactt ggtctcgtcc tggatgctat ggactactgg 360
ggtcagggta cgttagtgac ggtctcgagt gcttccacca agggcccatc cgtcttcccc 420
ctggcgccct gctccaggag cacctccgag agcacagccg ccctgggctg cctggtcaag 480
gactacttcc ccgaaccggt gacggtgtcg tggaactcag gcgccctgac cagcggcgtg 540
cacaccttcc cggctgtcct acagtcctca ggactctact ccctcagcag cgtggtgacc 600
gtgccctcca gcagcttggg cacgaagacc tacacctgca acgtagatca caagcccagc 660
aacaccaagg tcgacaagag agttgagtcc aaatatggtc ccccatgccc accatgccca 720
gcacctgagt tcctgggggg accatcagtc ttcctgttcc ccccaaaacc caaggacact 780
ctcatgatct cccggacccc tgaggtcacg tgcgtggtgg tggacgtgag ccaggaagac 840
cccgaggtcc agttcaactg gtacgtggat ggcgtggagg tgcataatgc caagacaaag 900
ccgcgggagg agcagttcaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac 960
caggactggc tgaacggcaa ggagtacaag tgcaaggtct ccaacaaagg cctcccgtcc 1020
tccatcgaga aaaccatctc caaagccaaa gggcagcccc gagagccaca ggtgtacacc 1080
ctgcccccat cccaggagga gatgaccaag aaccaggtca gcctgacctg cctggtcaaa 1140
ggcttctacc ccagcgacat cgccgtggag tgggagagca atgggcagcc ggagaacaac 1200
tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcta cagcaggcta 1260
accgtggaca agagcaggtg gcaggagggg aatgtcttct catgctccgt gatgcatgag 1320
gctctgcaca accactacac acagaagagc ctctccctgt ctctggggaa agacgacgat 1380
gatgacgatg acgacgacga ttga 1404
<210> 27
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 27
gaaacggtac tcacgcagtc tccaggtacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtcttggc agcagctact tagcctggta tcagcagaaa 120
cctggtcagg ctcccaggct cctcatctat ggtgcatcca gcagggcacc tggcatccca 180
gacaggttca gtggcagtgg gtctggtacc gacttcactc tcaccatcag ccgactggag 240
cctgaagatt ttgcagttta ttactgtcag cagtatgctg actcaccgat caccttcggc 300
caagggacac gactggagat taaacgtacg gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttag 648
<210> 28
<211> 678
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 28
gacgacgatg atgacgatga cgacgacgat gaaacggtac tcacgcagtc tccaggtacc 60
ctgtctttgt ctccagggga aagagccacc ctctcctgca gggccagtca gagtcttggc 120
agcagctact tagcctggta tcagcagaaa cctggtcagg ctcccaggct cctcatctat 180
ggtgcatcca gcagggcacc tggcatccca gacaggttca gtggcagtgg gtctggtacc 240
gacttcactc tcaccatcag ccgactggag cctgaagatt ttgcagttta ttactgtcag 300
cagtatgctg actcaccgat caccttcggc caagggacac gactggagat taaacgtacg 360
gtggctgcac catctgtctt catcttcccg ccatctgatg agcagttgaa atctggaact 420
gcctctgttg tgtgcctgct gaataacttc tatcccagag aggccaaagt acagtggaag 480
gtggataacg ccctccaatc gggtaactcc caggagagtg tcacagagca ggacagcaag 540
gacagcacct acagcctcag cagcaccctg acgctgagca aagcagacta cgagaaacac 600
aaagtctacg cctgcgaagt cacccatcag ggcctgagct cgcccgtcac aaagagcttc 660
aacaggggag agtgttag 678
<210> 29
<211> 678
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 29
gaaacggtac tcacgcagtc tccaggtacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtcttggc agcagctact tagcctggta tcagcagaaa 120
cctggtcagg ctcccaggct cctcatctat ggtgcatcca gcagggcacc tggcatccca 180
gacaggttca gtggcagtgg gtctggtacc gacttcactc tcaccatcag ccgactggag 240
cctgaagatt ttgcagttta ttactgtcag cagtatgctg actcaccgat caccttcggc 300
caagggacac gactggagat taaacgtacg gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgtgacga cgatgatgac 660
gatgacgacg acgattag 678
<210> 30
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 30
gacgacgatg atgacgatga cgacgacgat gaaacggtac tcacgcagtc tccaggtacc 60
ctgtctttgt ctccagggga aagagccacc ctctcctgca gggccagtca gagtcttggc 120
agcagctact tagcctggta tcagcagaaa cctggtcagg ctcccaggct cctcatctat 180
ggtgcatcca gcagggcacc tggcatccca gacaggttca gtggcagtgg gtctggtacc 240
gacttcactc tcaccatcag ccgactggag cctgaagatt ttgcagttta ttactgtcag 300
cagtatgctg actcaccgat caccttcggc caagggacac gactggagat taaacgtacg 360
gtggctgcac catctgtctt catcttcccg ccatctgatg agcagttgaa atctggaact 420
gcctctgttg tgtgcctgct gaataacttc tatcccagag aggccaaagt acagtggaag 480
gtggataacg ccctccaatc gggtaactcc caggagagtg tcacagagca ggacagcaag 540
gacagcacct acagcctcag cagcaccctg acgctgagca aagcagacta cgagaaacac 600
aaagtctacg cctgcgaagt cacccatcag ggcctgagct cgcccgtcac aaagagcttc 660
aacaggggag agtgtgacga cgatgatgac gatgacgacg acgattag 708
<210> 31
<211> 693
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 31
gaaacggtac tcacgcagtc tccaggtacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtcttggc agcagctact tagcctggta tcagcagaaa 120
cctggtcagg ctcccaggct cctcatctat ggtgcatcca gcagggcacc tggcatccca 180
gacaggttca gtggcagtgg gtctggtacc gacttcactc tcaccatcag ccgactggag 240
cctgaagatt ttgcagttta ttactgtcag cagtatgctg actcaccgat caccttcggc 300
caagggacac gactggagat taaacgtacg gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgtggcgg aggcggcagc 660
gacgacgatg atgacgatga cgacgacgat tag 693
<210> 32
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 32
gaaacggtac tcacgcagtc tccaggtacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtcttggc agcagctact tagcctggta tcagcagaaa 120
cctggtcagg ctcccaggct cctcatctat ggtgcatcca gcagggcacc tggcatccca 180
gacaggttca gtggcagtgg gtctggtacc gacttcactc tcaccatcag ccgactggag 240
cctgaagatt ttgcagttta ttactgtcag cagtatgctg actcaccgat caccttcggc 300
caagggacac gactggagat taaacgtacg gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgtggcgg aggcggcagc 660
ggcggaggcg gcagcgacga cgatgatgac gatgacgacg acgattag 708
<210> 33
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 33
Ser Asn Val Ile Ser
1. 5
<210> 34
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 34
Gly Val Ile Pro Ile Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe Lys
1. 5 10 15
Gly
<210> 35
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 35
Thr Leu Gly Leu Val Leu Asp Ala Met Asp Tyr
1. 5 10
<210> 36
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 36
Arg Ala Ser Gln Ser Leu Gly Ser Ser Tyr Leu Ala
1. 5 10
<210> 37
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 37
Gly Ala Ser Ser Arg Ala Pro
1. 5
<210> 38
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 38
Gln Gln Tyr Ala Asp Ser Pro Ile Thr
1. 5
<210> 39
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 39
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn
20 25 30
Val Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Ile Pro Ile Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Thr Leu Gly Leu Val Leu Asp Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 40
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 40
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ БИСПЕЦИФИЧЕСКИХ АНТИТЕЛ, БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ТАКИХ АНТИТЕЛ | 2018 |
|
RU2795240C2 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| МУТАНТНЫЕ ВАРИАНТЫ Fc IgG1, ЛИШЕННЫЕ ЭФФЕКТОРНЫХ ФУНКЦИЙ | 2018 |
|
RU2764559C2 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| АНТИТЕЛА ПРОТИВ BTN3A И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ РАКА ИЛИ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2800726C2 |
| АНТИТЕЛА ПРОТИВ TFR И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ ПРОЛИФЕРАТИВНЫХ И ВОСПАЛИТЕЛЬНЫХ РАССТРОЙСТВ | 2016 |
|
RU2737637C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ ПРОТИВ TIGIT И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2776714C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ МЕЗОТЕЛИН И CD137 | 2019 |
|
RU2815066C2 |
| КОНЪЮГАТЫ АНТИТЕЛ С ЛЕКАРСТВЕННЫМИ ВЕЩЕСТВАМИ И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ РАКОВЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2799547C2 |
| АНТИТЕЛА К МЕЗОТЕЛИНУ | 2019 |
|
RU2837574C2 |
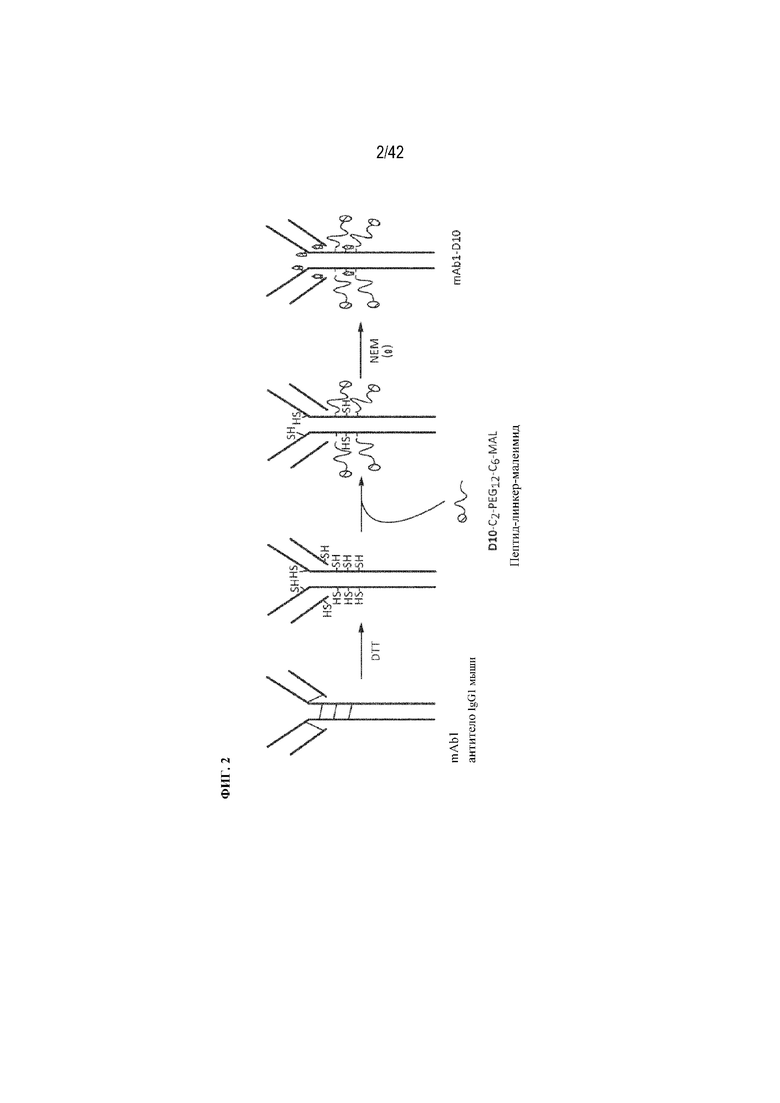

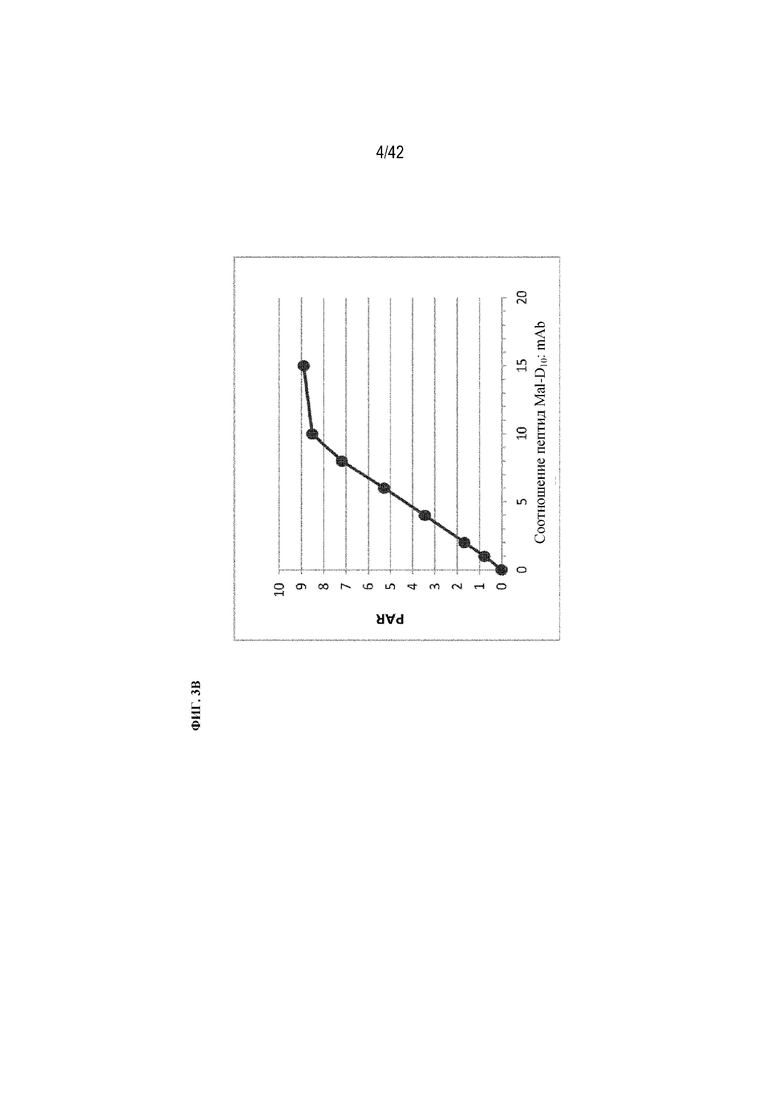
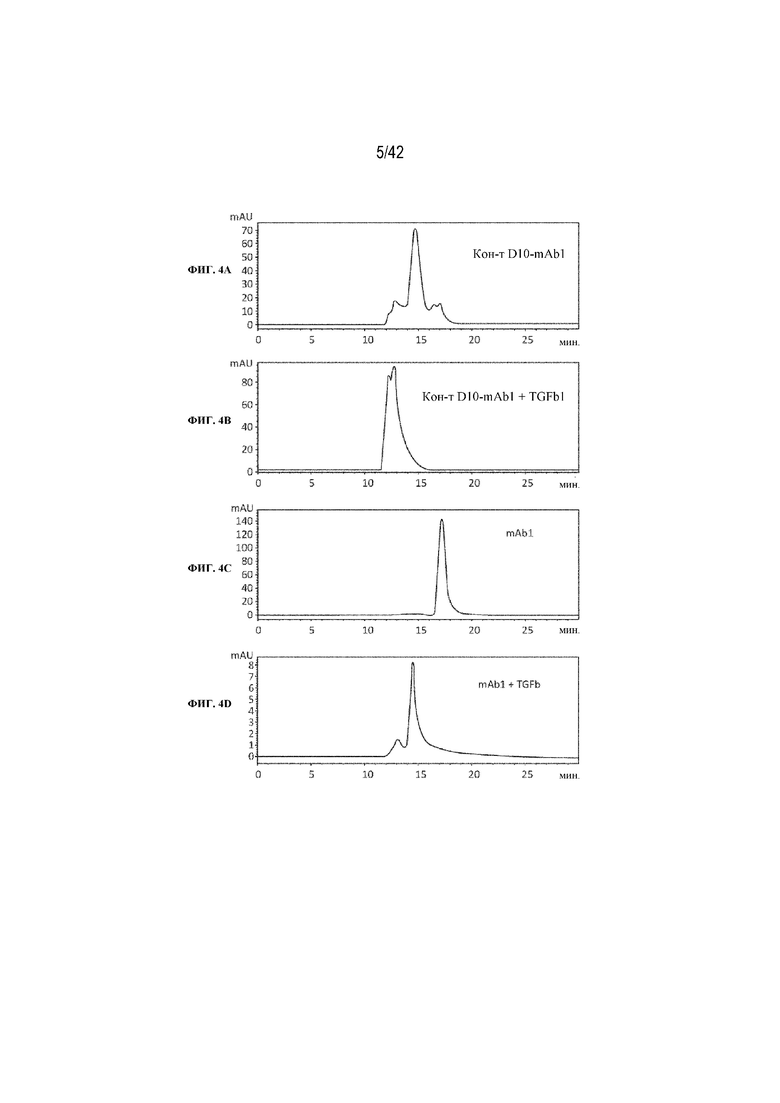
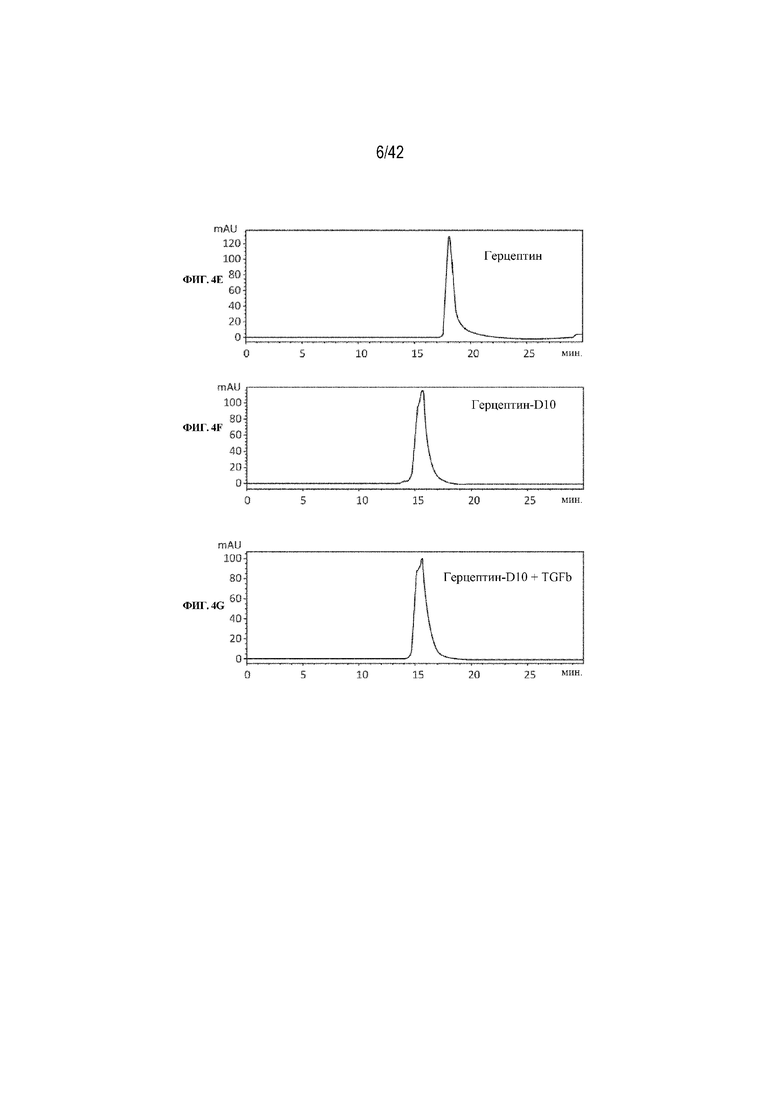
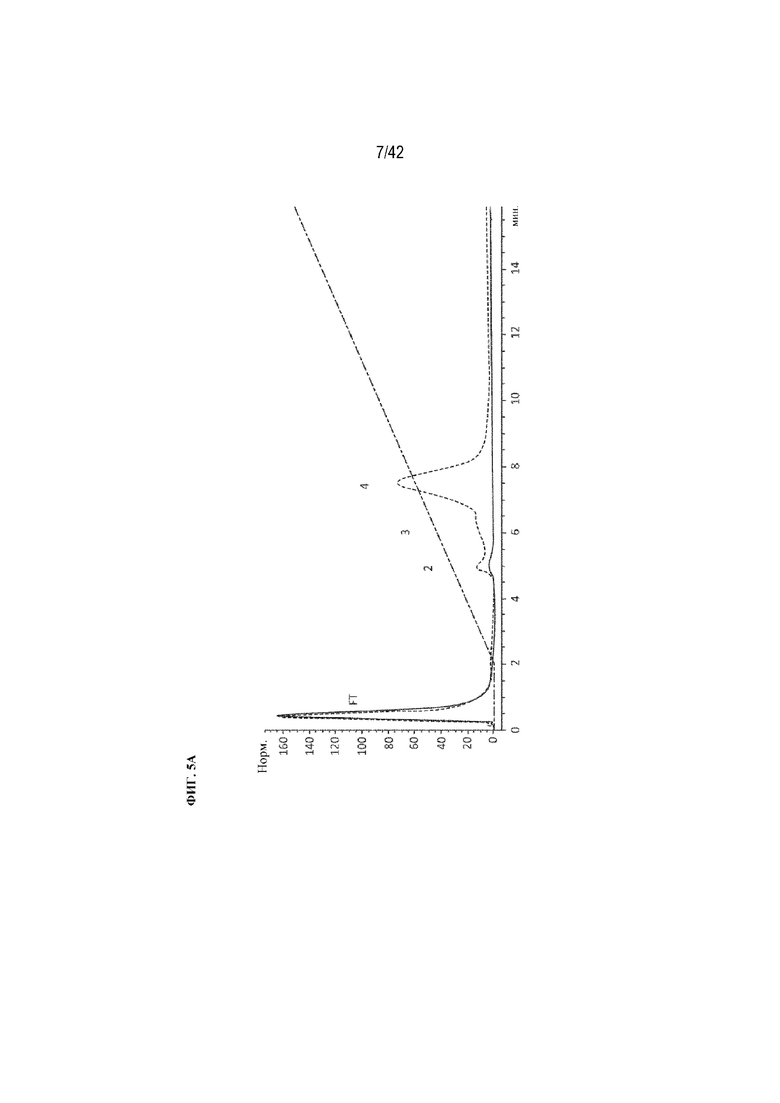
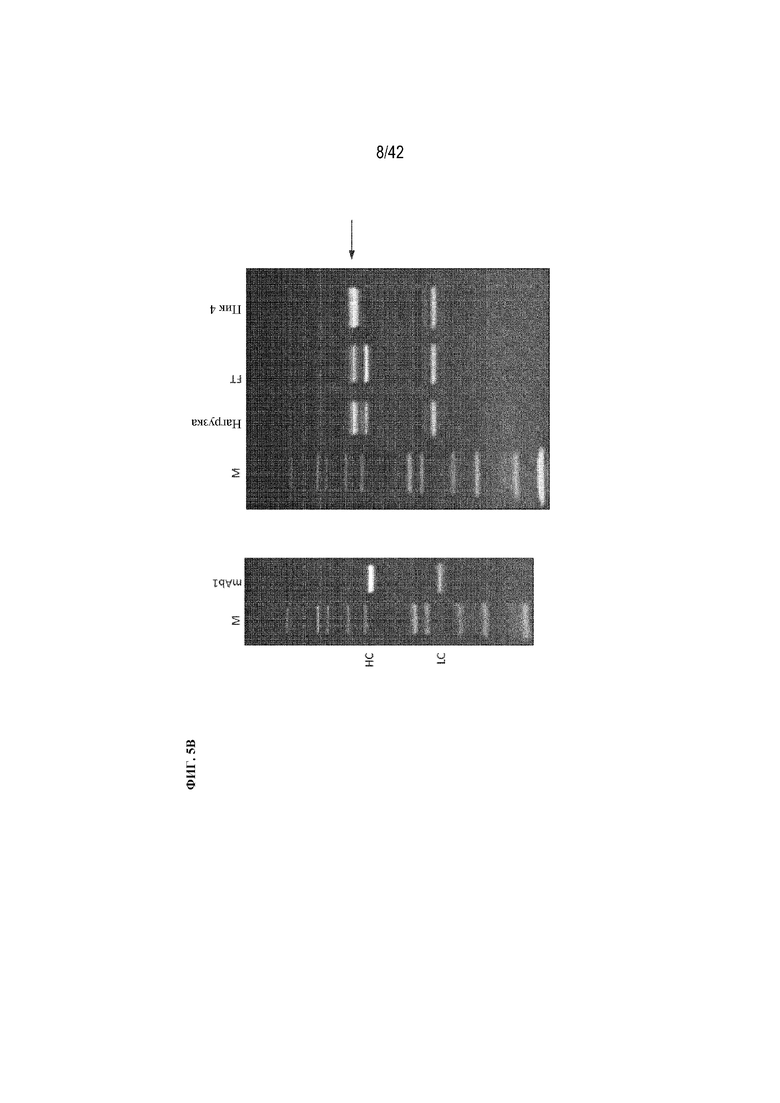
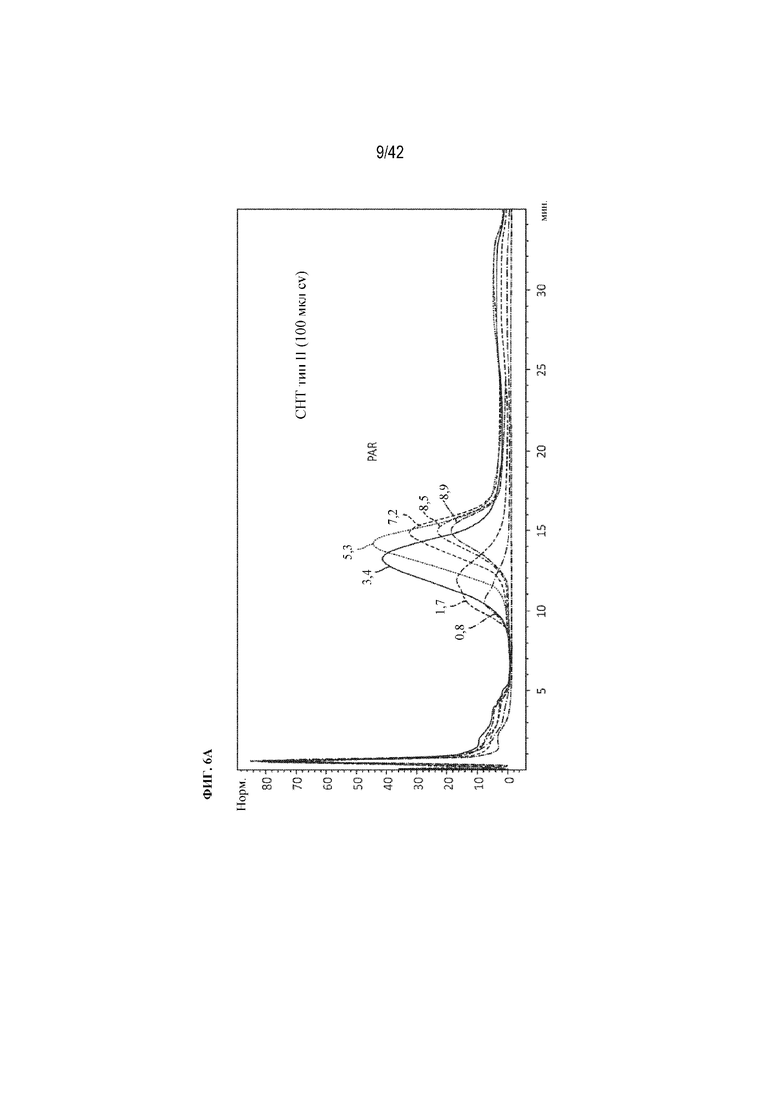
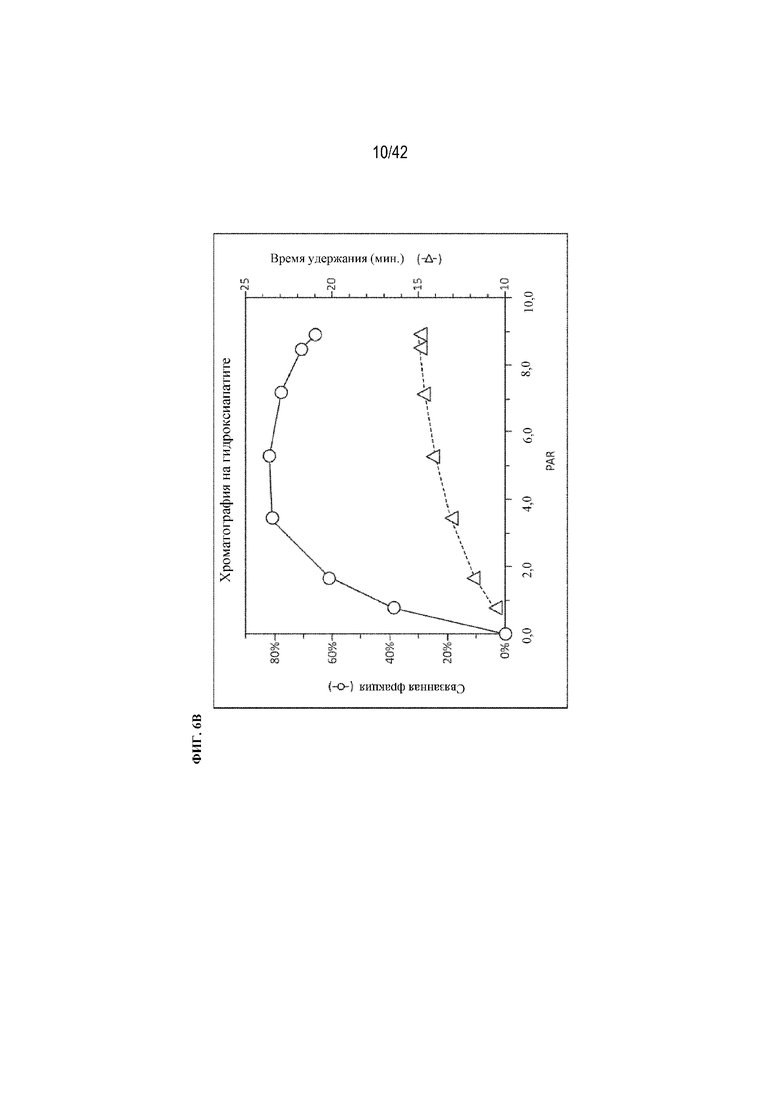
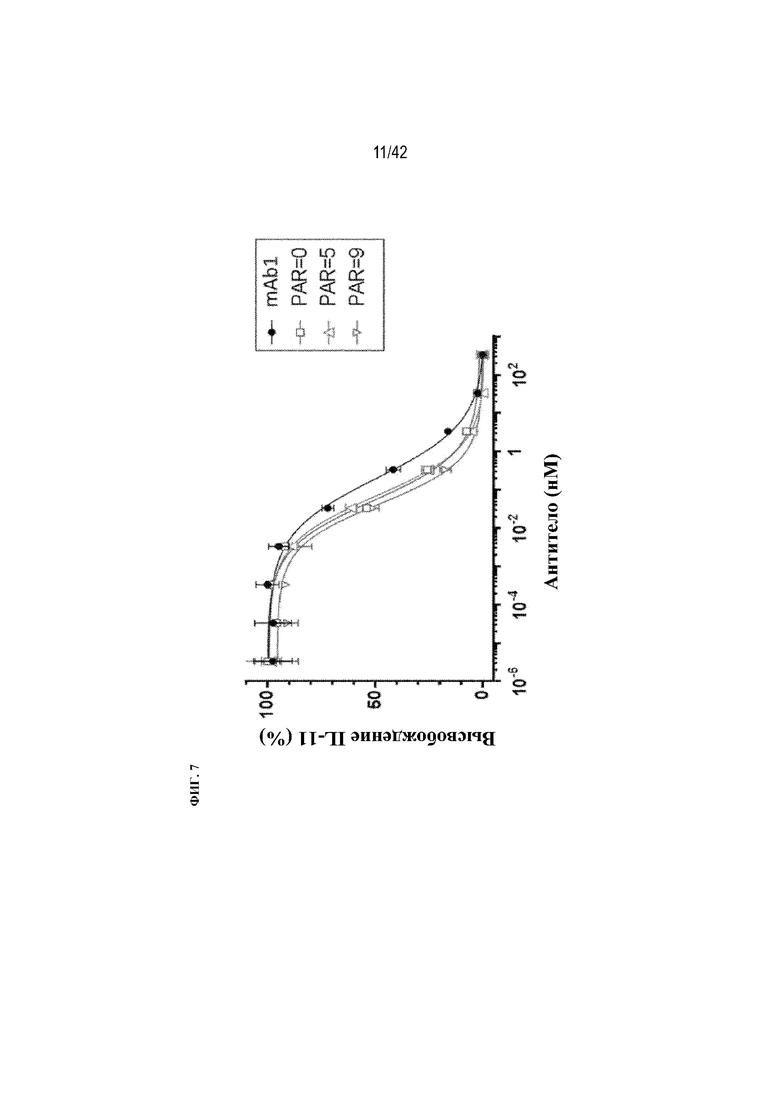
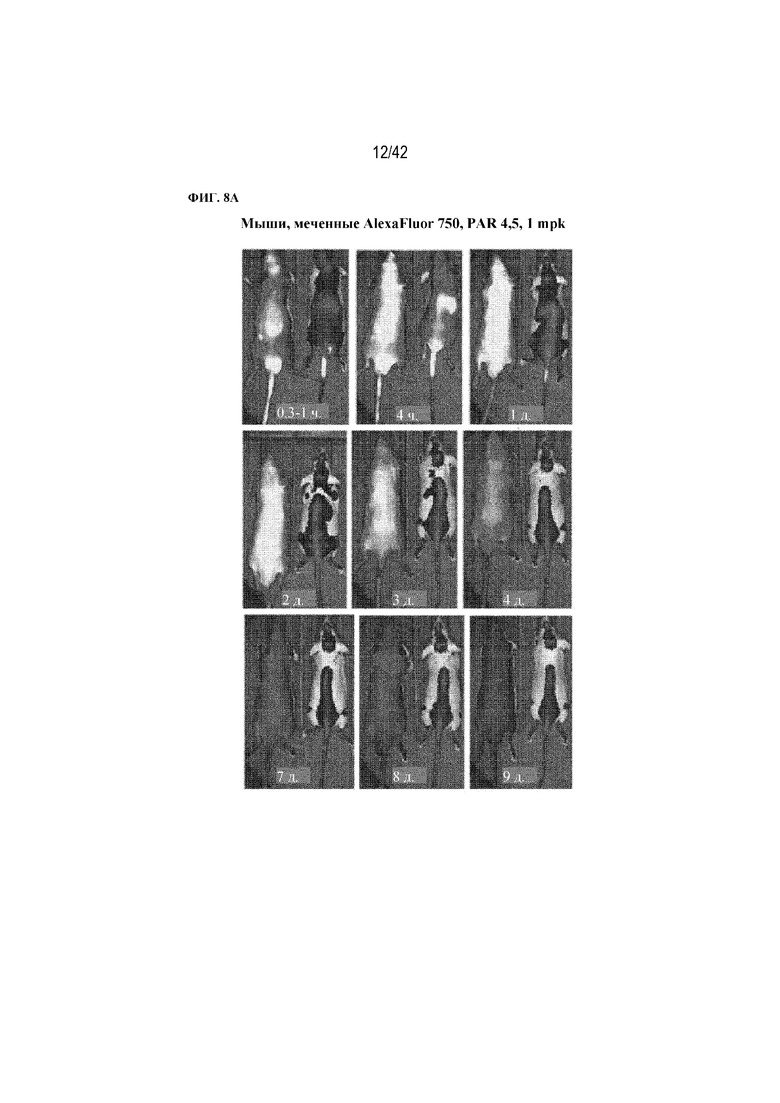
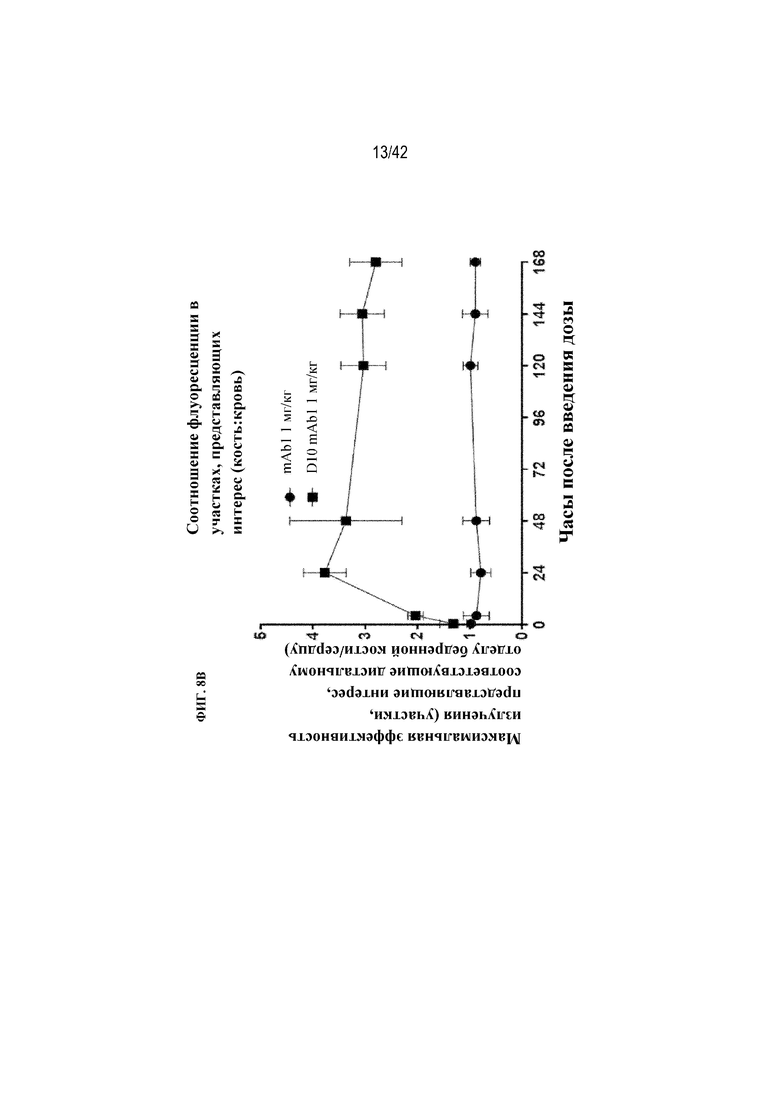
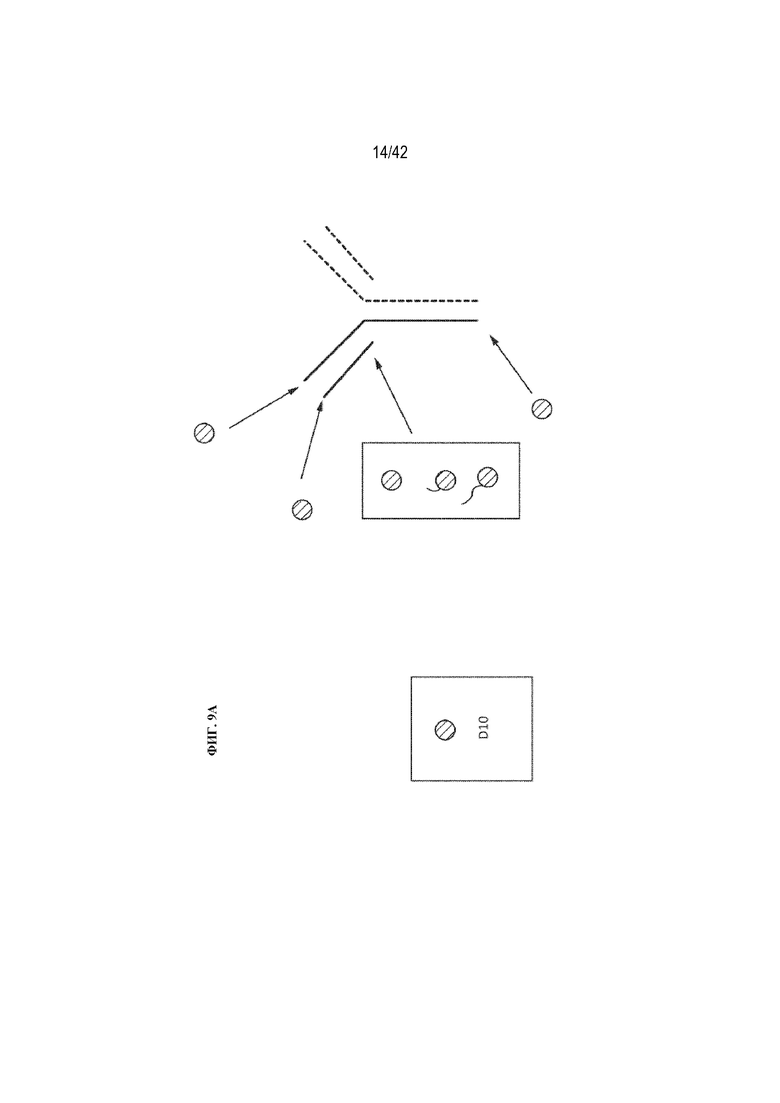
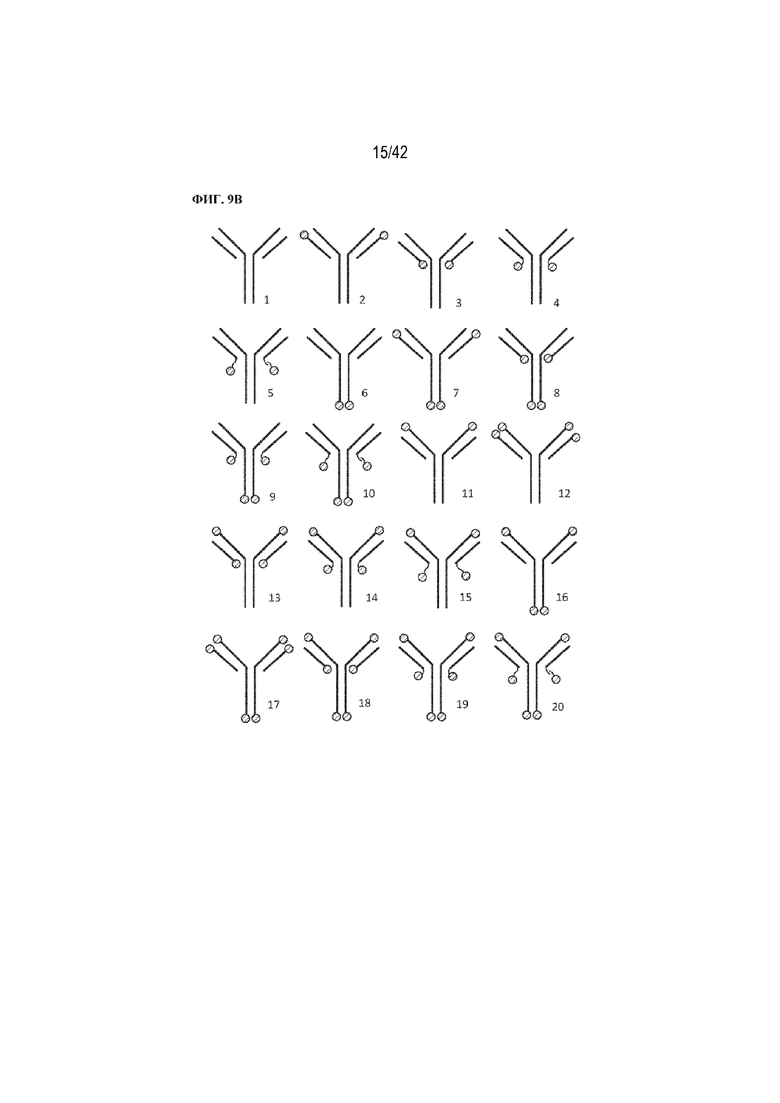

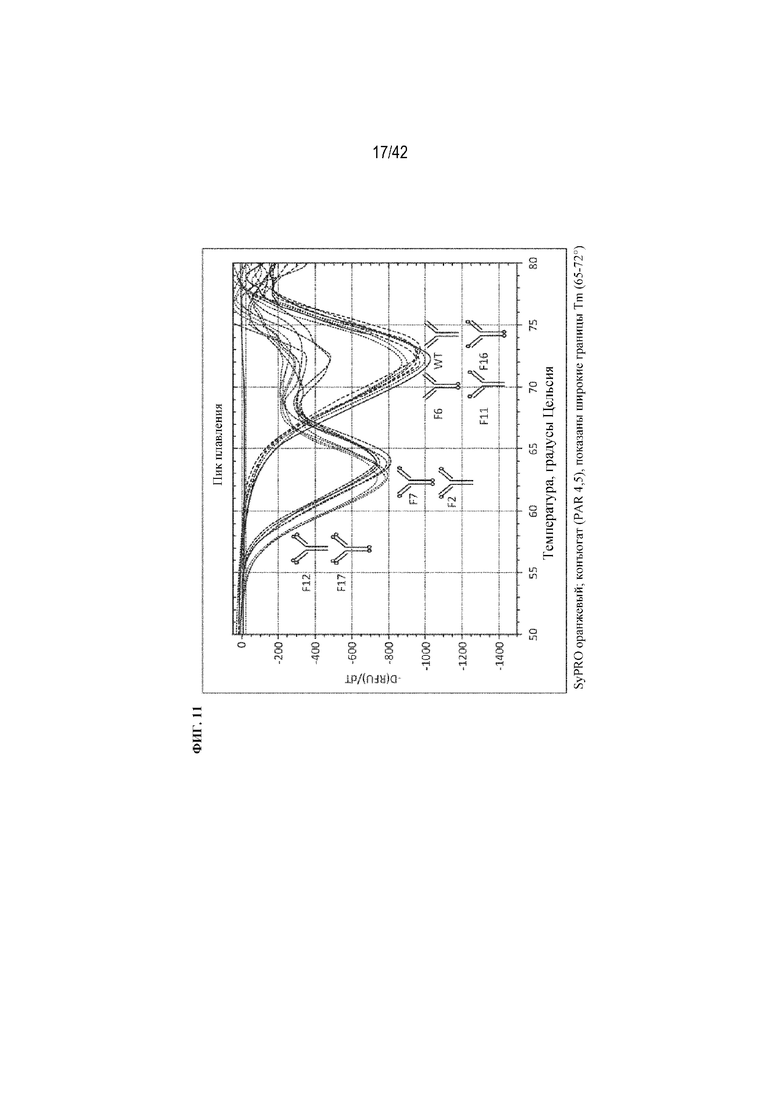
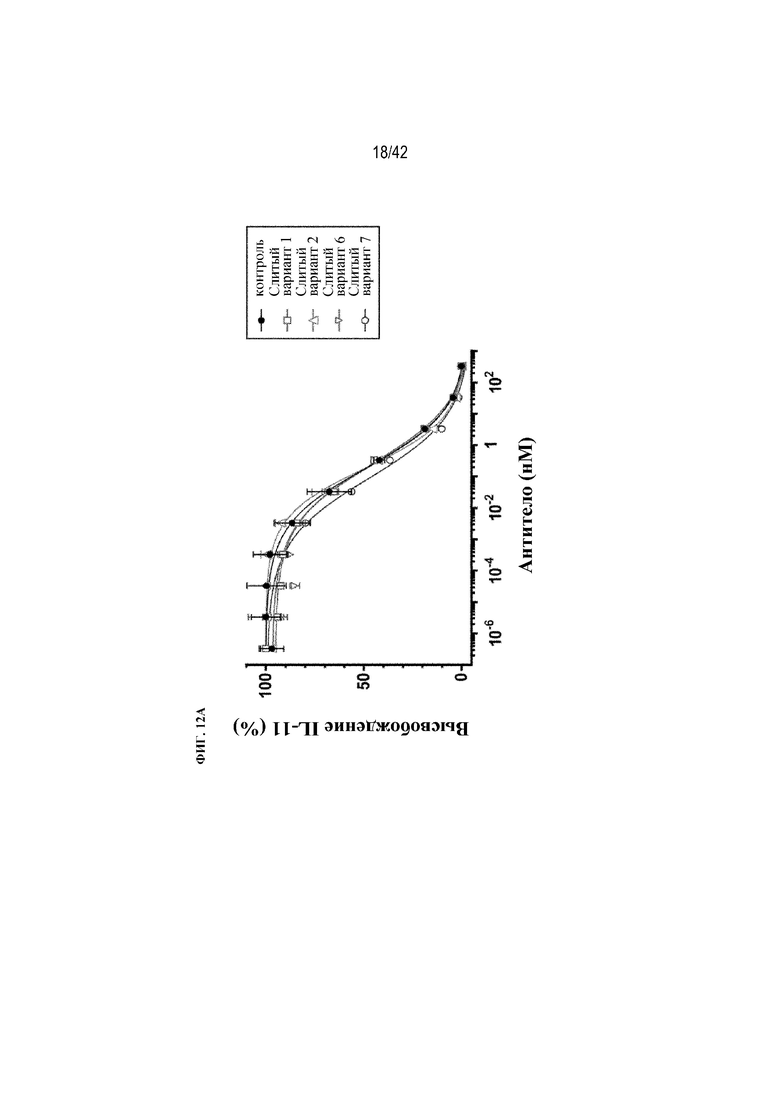
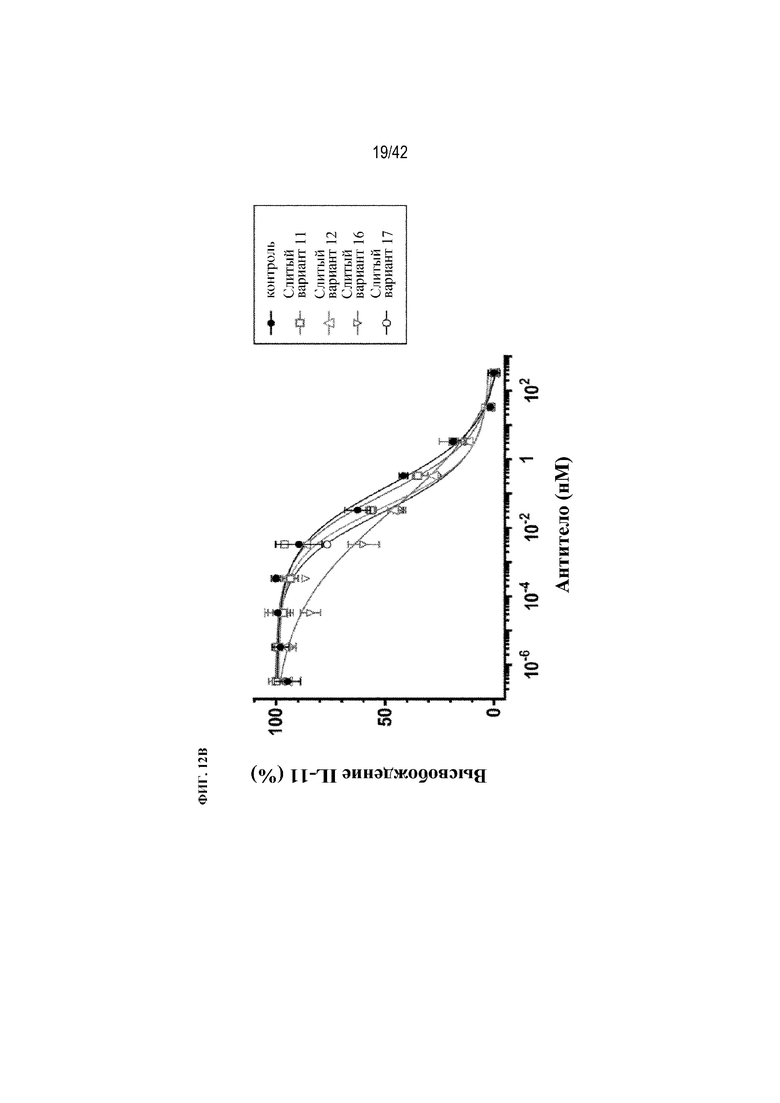
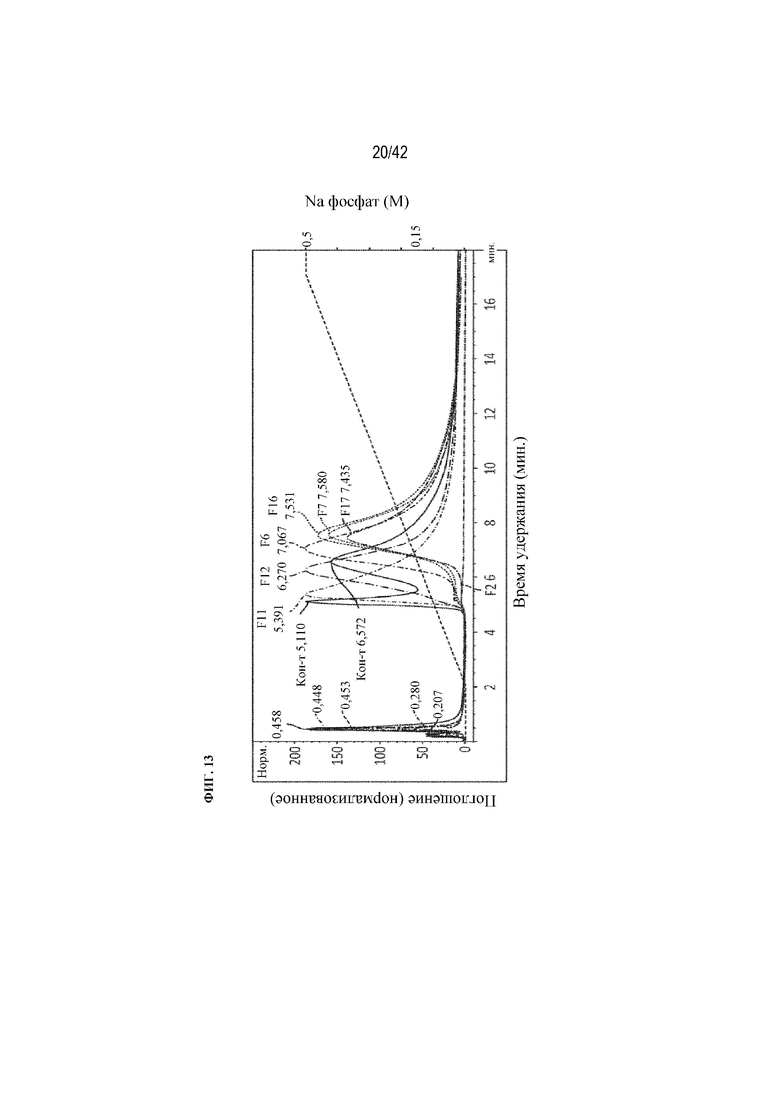
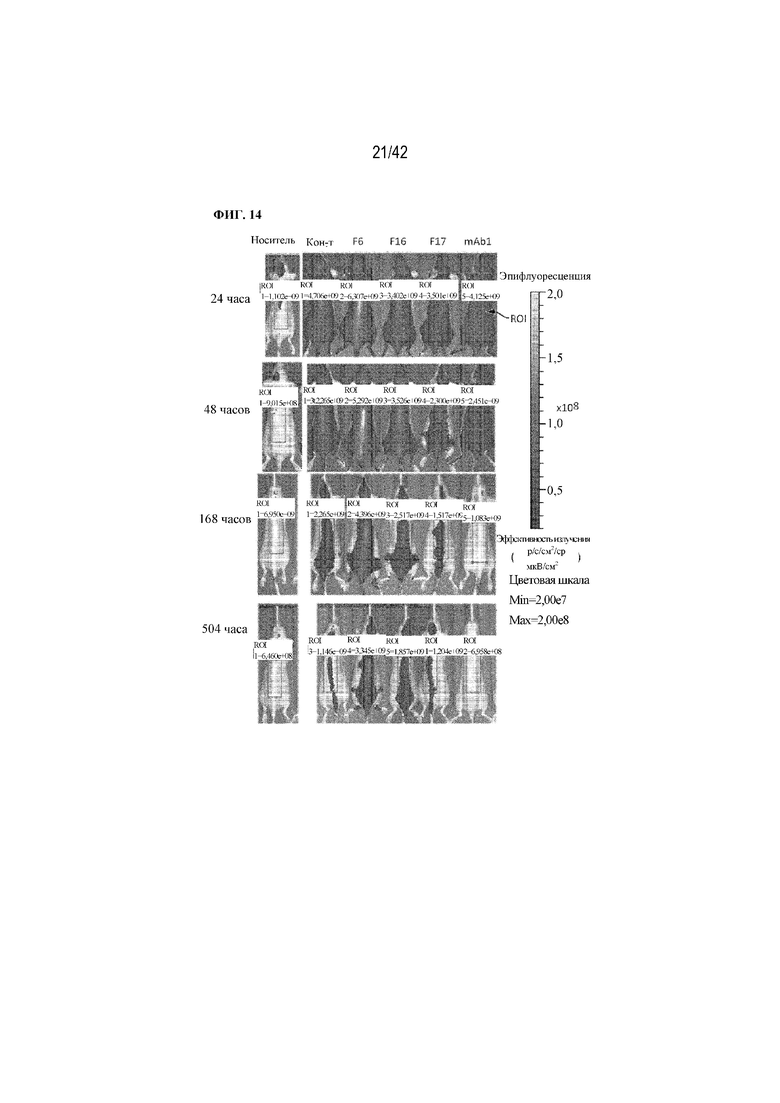


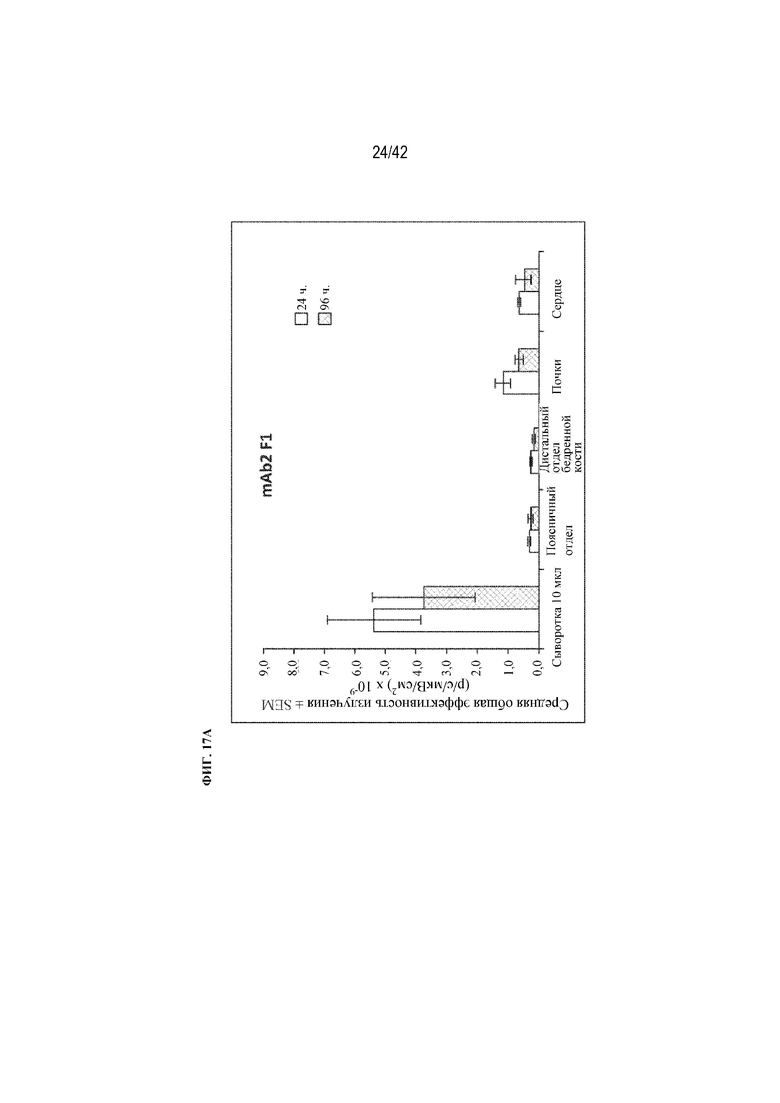
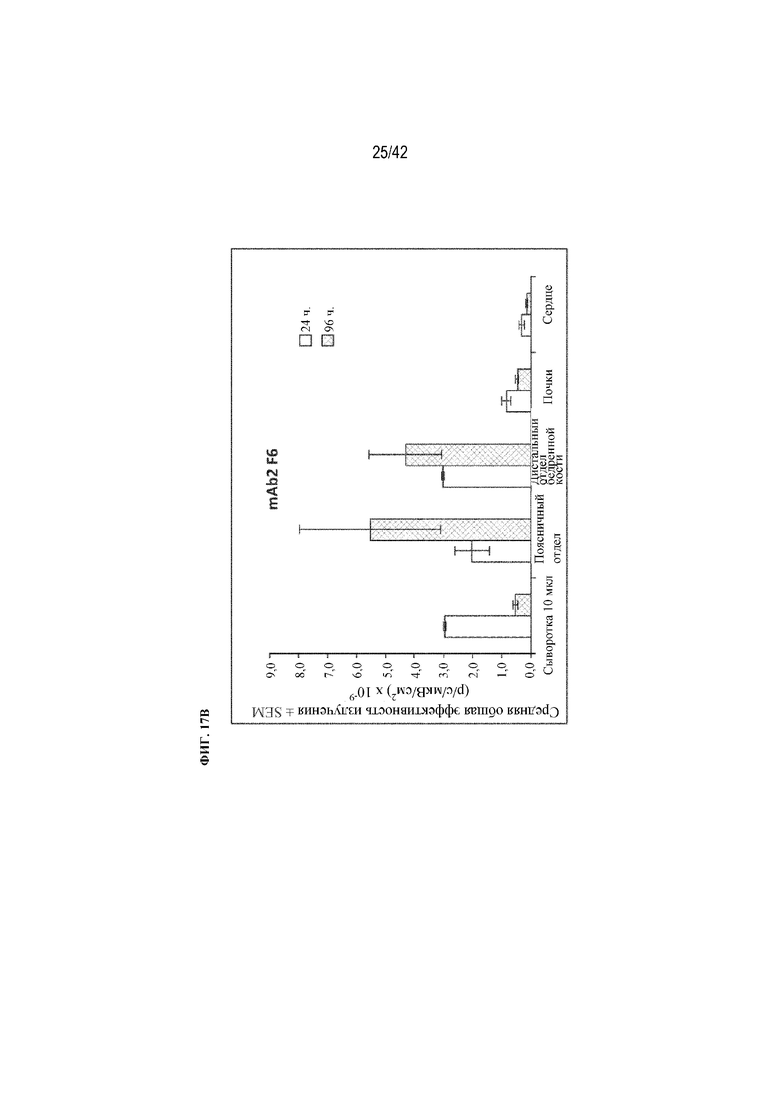
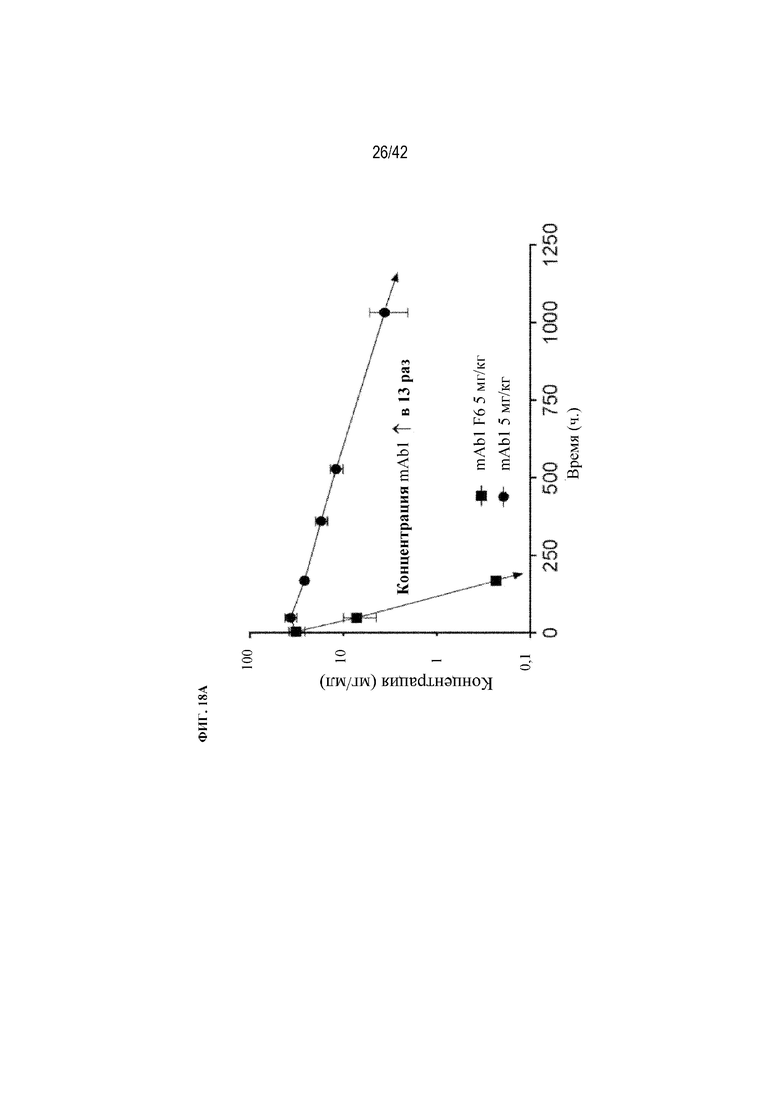
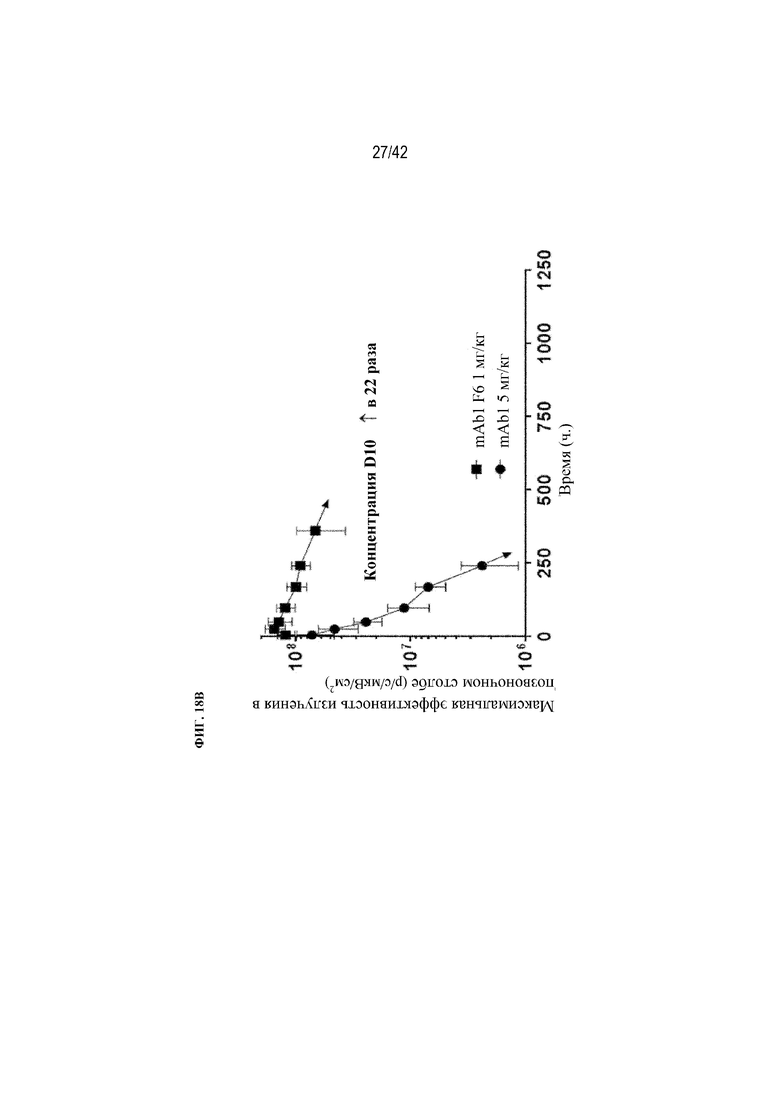
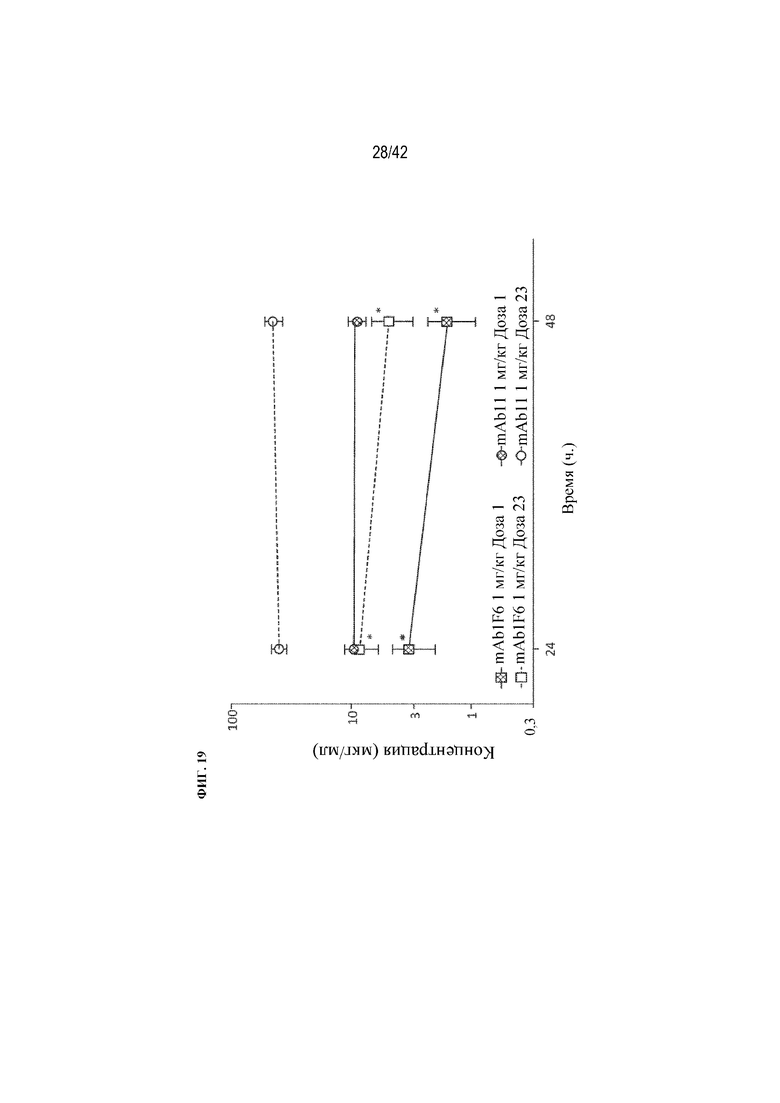
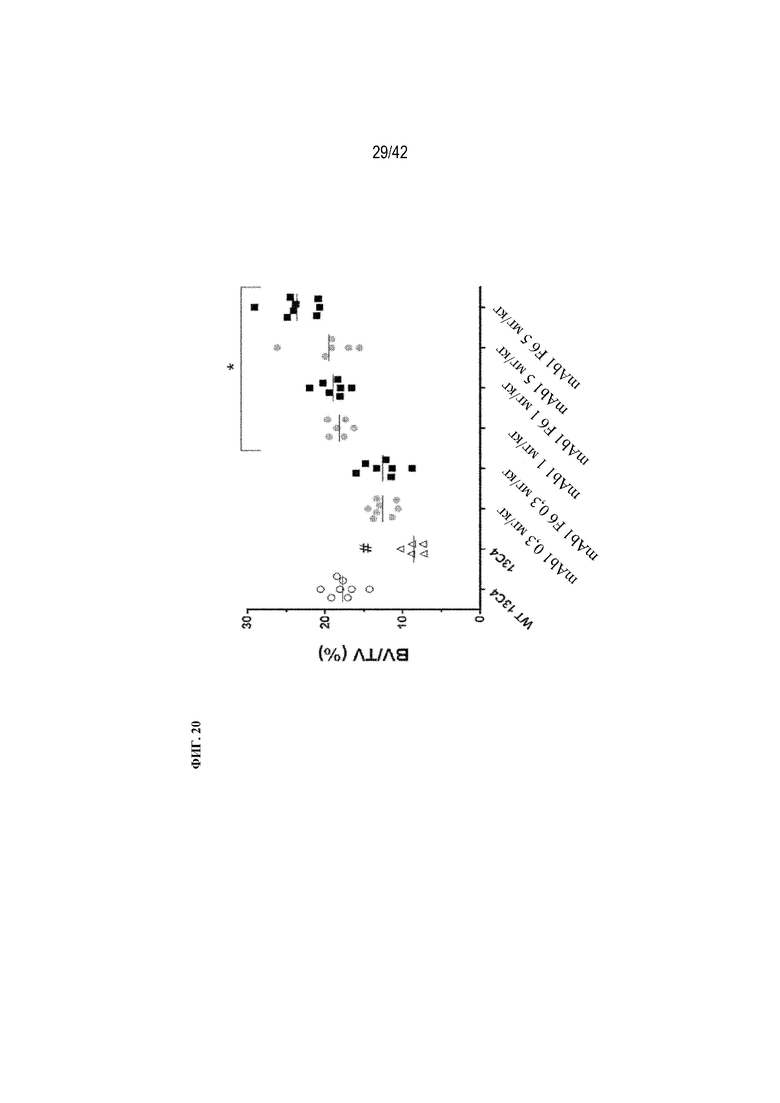
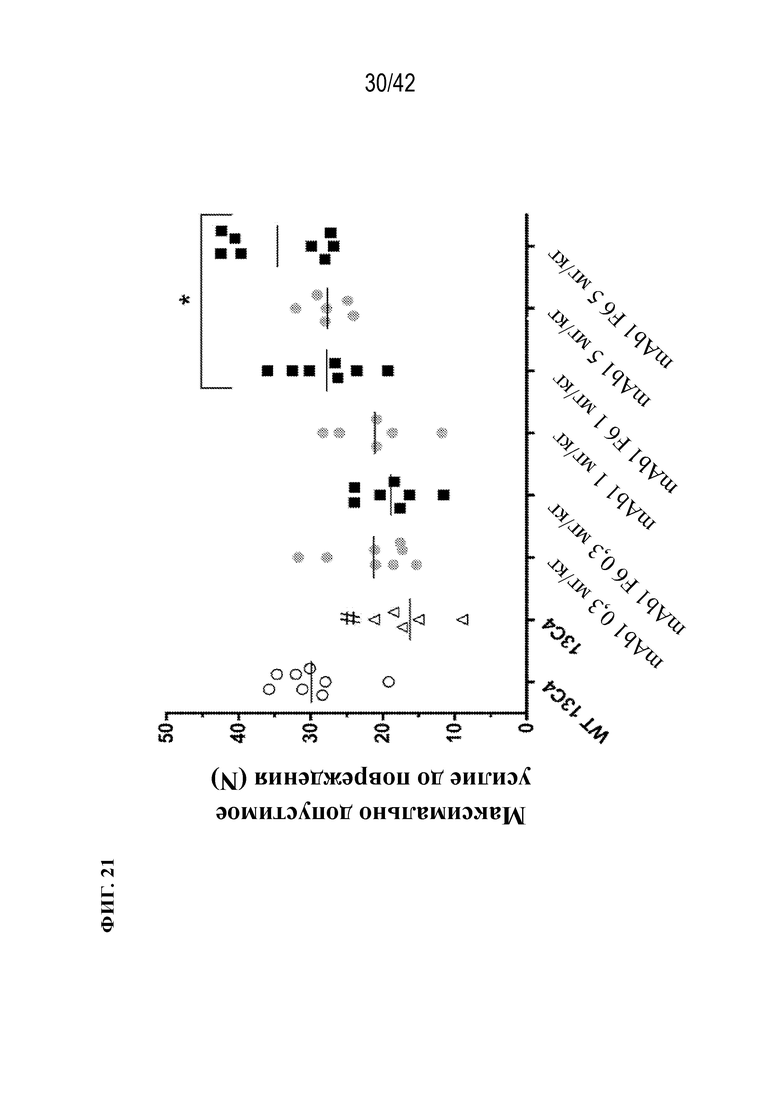
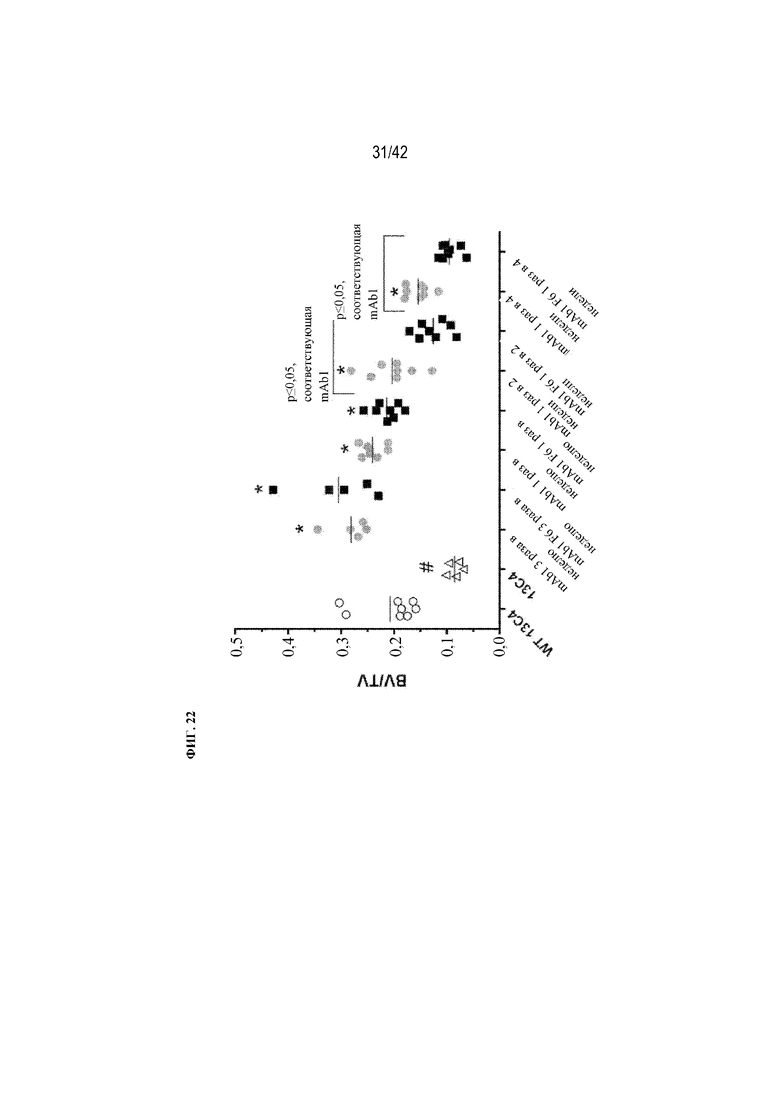
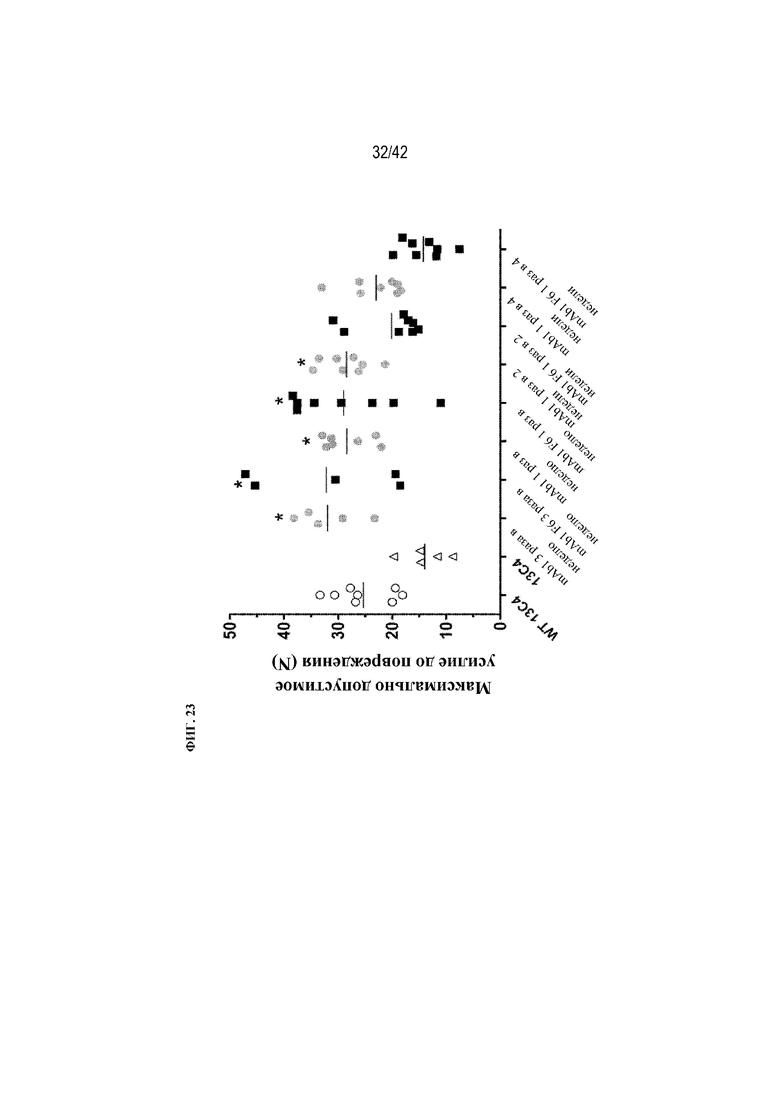
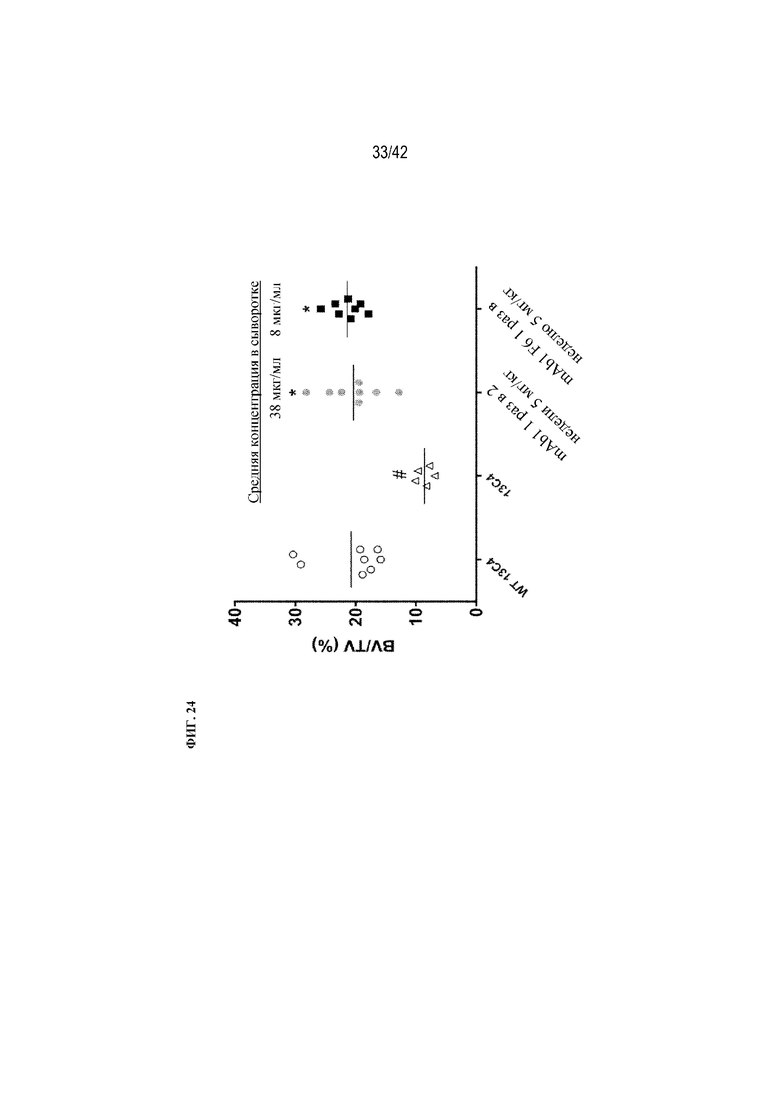
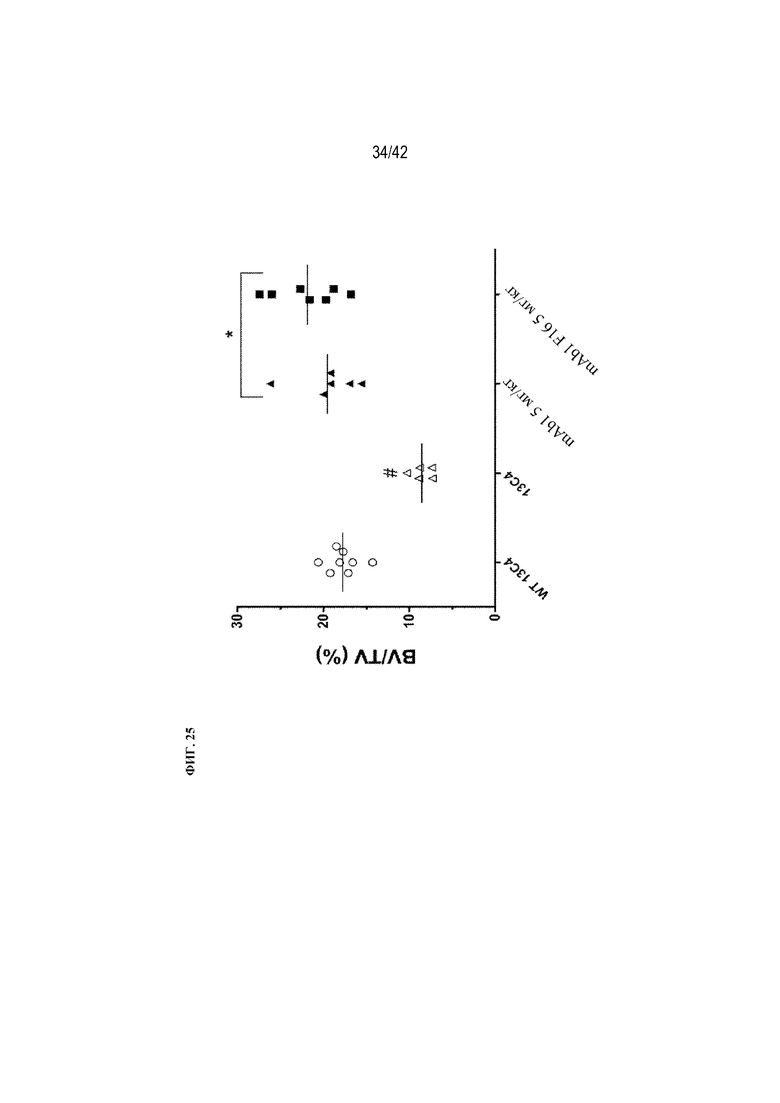
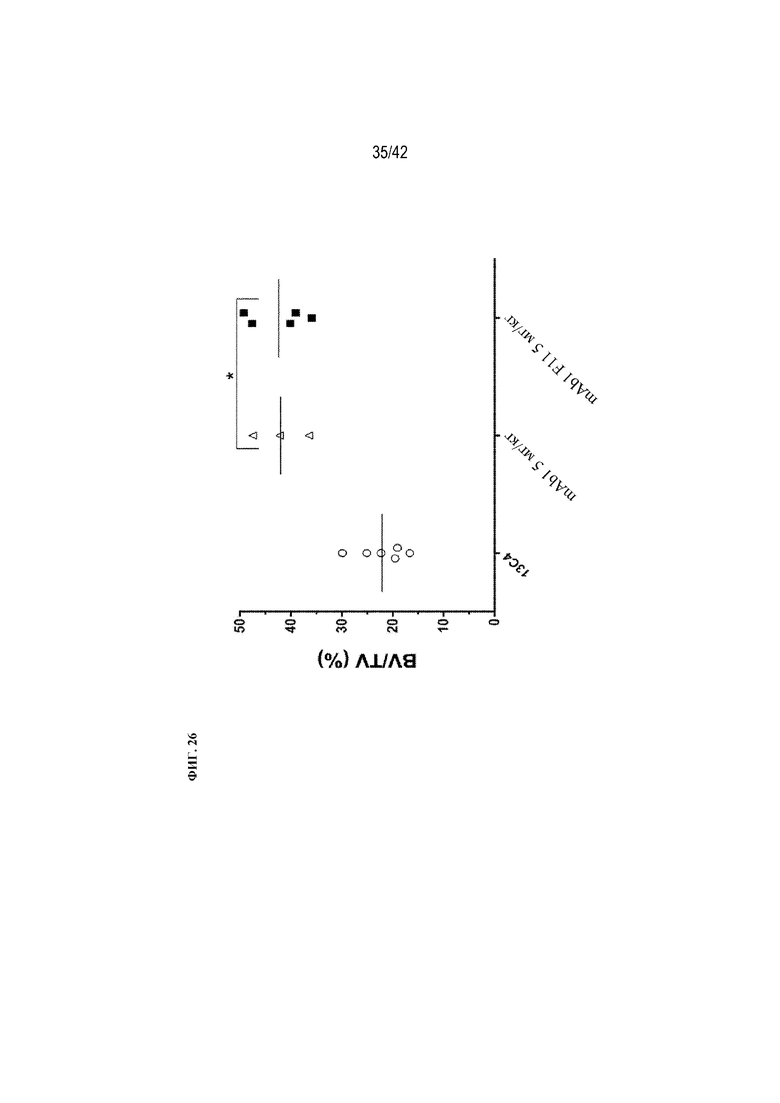
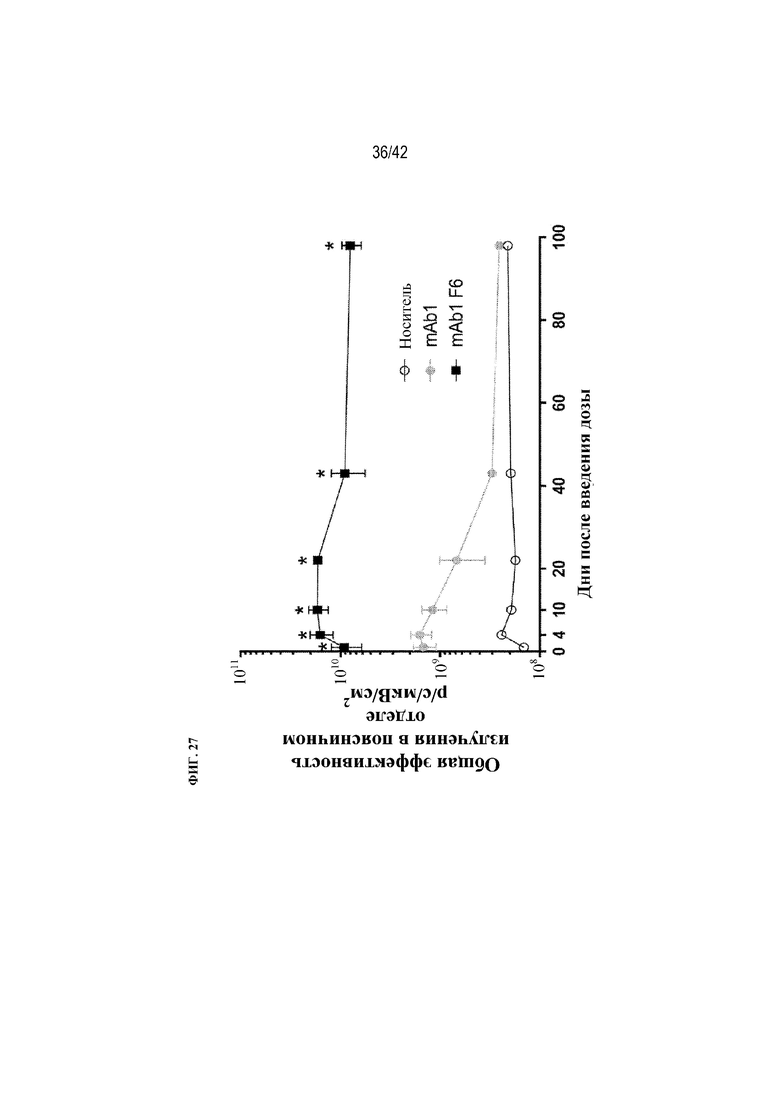
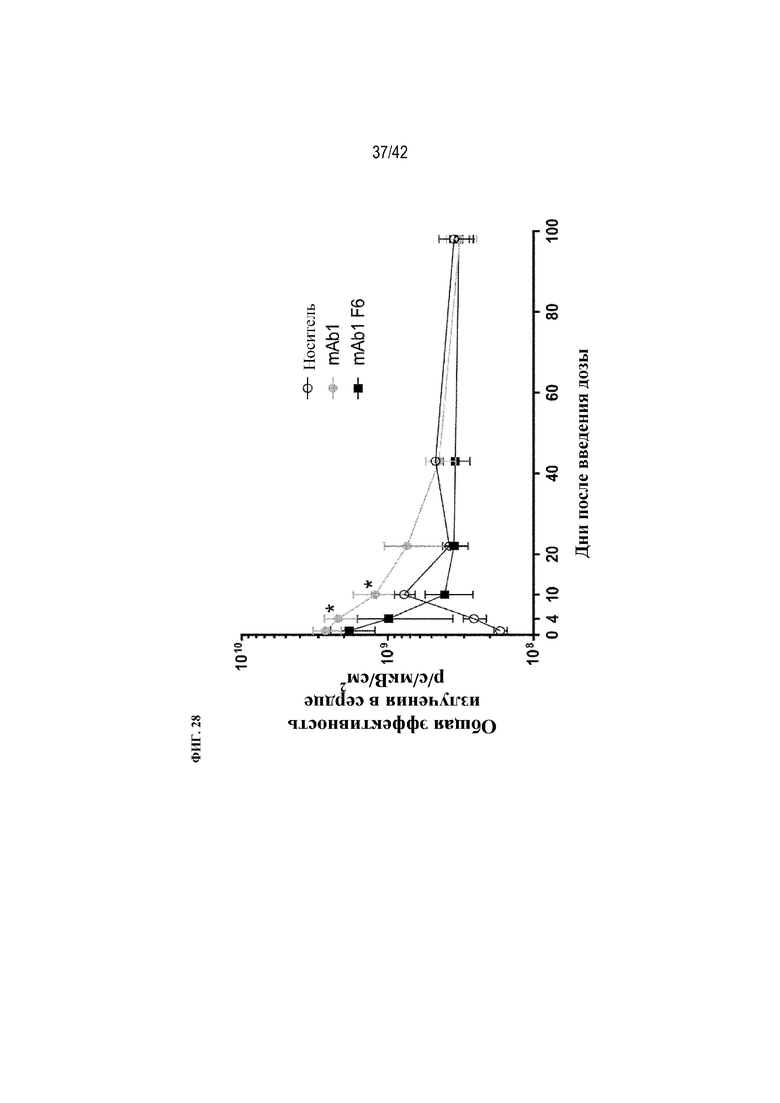

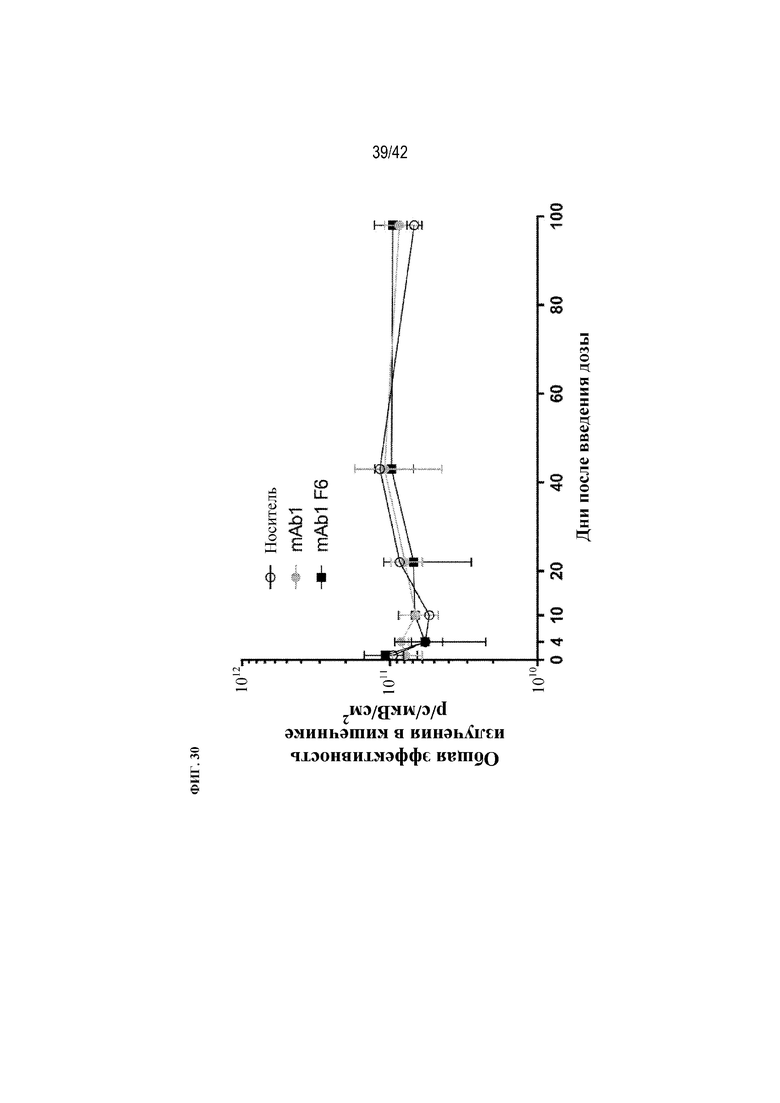

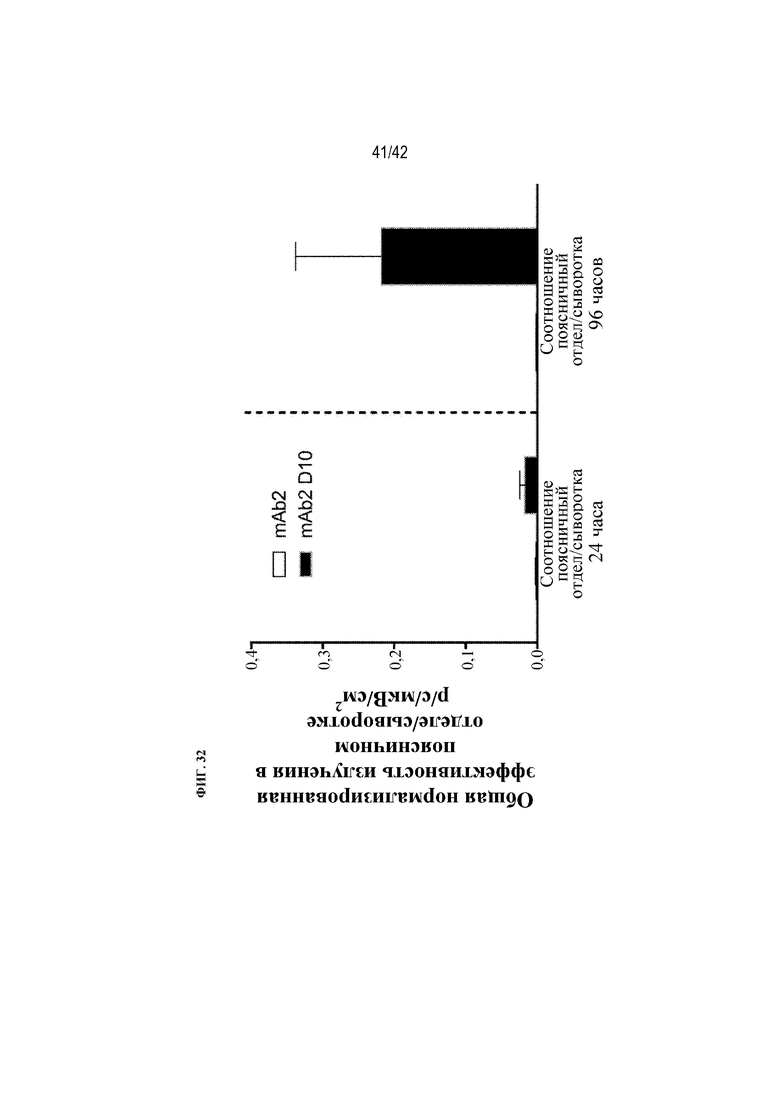
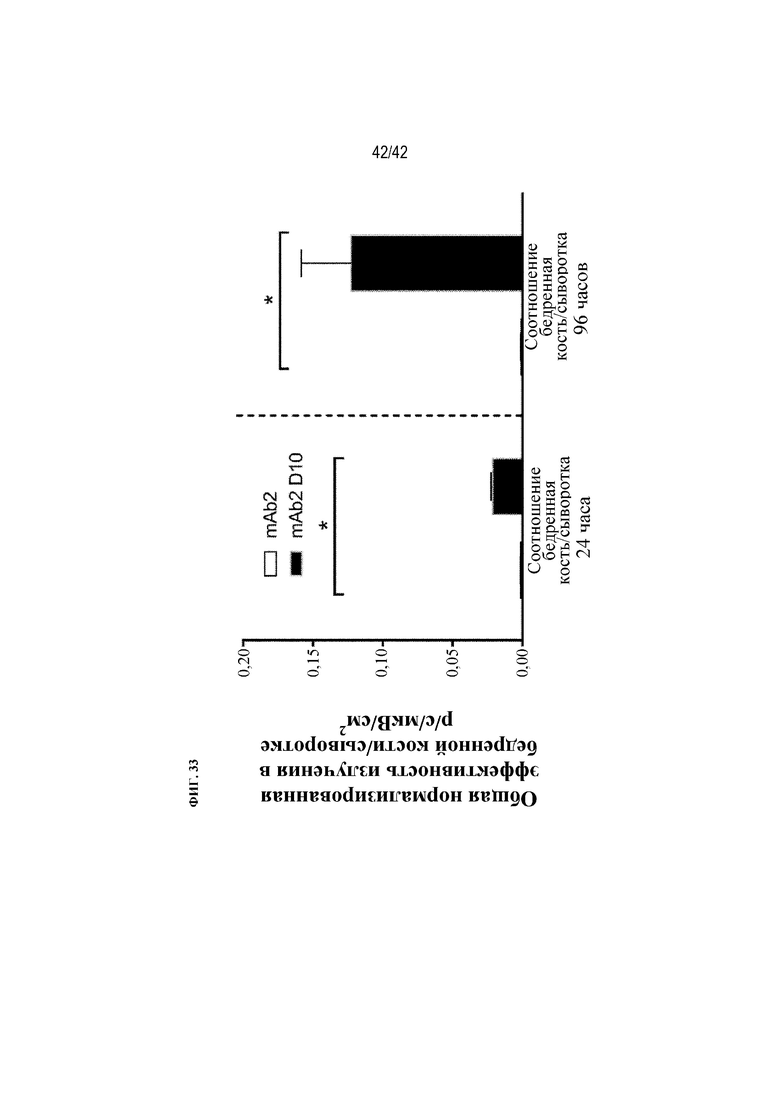
Изобретение относится к области биотехнологии. Предложено нацеленное на костную ткань антитело, содержащее тяжелую цепь и легкую цепь, которое специфически связывается с TGFβ1, TGFβ2 и TGFβ3. Антитело модифицировано одним или несколькими пептидами на основе поли-аспарагиновой кислоты для улучшения локализации антитела в кости. Также раскрыты способы получения и применения указанного антитела для лечения человека с заболеванием костей. Изобретение обеспечивает улучшенную эффективность лечения патофизиологической дегенерации кости при минимизации потенциальных нежелательных побочных эффектов. 8 н. и 14 з.п. ф-лы, 47 ил., 11 табл., 21 пр.
1. Нацеленное на костную ткань антитело, которое специфически связывается с TGFβ1, TGFβ2 и TGFβ3, содержащее тяжелую цепь и легкую цепь, где
а) тяжелая цепь представляет собой слитый полипептид, содержащий пептид на основе поли-аспарагиновой кислоты (поли-D) либо на своем N-конце, либо на своем C-конце, либо обоих концах; и/или
b) легкая цепь представляет собой слитый полипептид, содержащий поли-D-пептид на своем C-конце,
где каждый поли-D-пептид содержит SEQ ID NO: 1.
2. Антитело по п. 1, где тяжелая цепь содержит
(i) поли-D-пептид на своем N-конце;
(ii) поли-D-пептид на своем С-конце, или
(iii) и то, и другое из (i) и (ii).
3. Антитело по п. 1 или 2, где легкая цепь содержит поли-D-пептид на своем C-конце.
4. Антитело по любому из пп. 1-3, где слитый полипептид(ы) содержит пептидный линкер.
5. Антитело по любому из пп. 1-4, где антитело представляет собой IgG.
6. Антитело по п. 5, где антитело содержит определяющие комплементарность участки (CDR) 1-3 тяжелой цепи в последовательности SEQ ID NO: 13 и CDR1-3 легкой цепи в последовательности SEQ ID NO: 15.
7. Антитело по п. 5, где антитело содержит
аминокислотную последовательность вариабельного домена тяжелой цепи (VH), соответствующую остаткам 1-120 последовательности SED ID NO: 13, и
аминокислотную последовательность вариабельного домена легкой цепи (VL), соответствующую остаткам 1-108 последовательности SEQ ID NO: 15.
8. Антитело по п. 6 или 7, где антитело содержит константный участок IgG4 человека с пролином в положении 228 (нумерация по EU).
9. Антитело по п. 1, где
тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 13, и
легкая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 15.
10. Антитело по п. 1, где
тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 13, 14, 16 или 17, и
легкая цепь содержит аминокислотную последовательность SEQ ID NO: 15, 19, 21 или 22.
11. Антитело по п. 1, где антитело содержит
тяжелую цепь, содержащую вариабельный домен с SEQ ID NO: 2, и
легкую цепь, содержащую вариабельный домен с SEQ ID NO: 6.
12. Антитело по п. 1, где
тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 14 или 17, и
легкая цепь содержит аминокислотную последовательность SEQ ID NO: 15.
13. Фармацевтическая композиция для лечения человека с заболеванием костей, при котором состояние здоровья человека улучшается посредством ингибирования TGFβ, содержащая антитело по любому из пп. 1-12 и фармацевтически приемлемое вспомогательное вещество.
14. Применение антитела по любому из пп. 1-12 для изготовления лекарственного препарата для лечения человека с патологическим состоянием кости, при котором ингибирование TGFβ оказывает благоприятное воздействие.
15. Применение по п. 14, где лечение приводит по меньшей мере к одному из следующего:
(1) к снижению уровней TGFβ,
(2) к снижению активности TGFβ,
(3) к уменьшению потери костной массы,
(4) к уменьшению скорости потери костной массы,
(5) к увеличению плотности кости,
(6) к увеличению прочности кости, и
(7) к снижению уровней IL-11 у человека.
16. Применение по п. 14 или 15, где у человека имеется несовершенный остеогенез, потеря костной массы или остеопороз, хроническая болезнь почек или рак с метастазированием в кость.
17. Выделенный полинуклеотид, содержащий кодирующую тяжелую цепь последовательность, содержащую SEQ ID NO: 24, 25 или 26.
18. Выделенный полинуклеотид, содержащий кодирующую легкую цепь последовательность, содержащую SEQ ID NO: 29, 31 или 32.
19. Вектор экспрессии, содержащий
(i) выделенный полинуклеотид, содержащий последовательность, кодирующую тяжелую цепь, содержащую SEQ ID NO: 24, 25 или 26; и
(ii) выделенный полинуклеотид, содержащий последовательность, кодирующую легкую цепь, содержащую SEQ ID NO: 29, 31 или 32.
20. Клетка-хозяин для получения антитела, где клетка содержит вектор экспрессии по п. 19.
21. Клетка-хозяин по п. 20, где клетка-хозяин представляет собой клетку млекопитающего.
22. Способ получения антитела по любому из пп. 1-12, где способ включает:
получение клетки-хозяина, содержащей
(i) выделенный полинуклеотид, содержащий последовательность, кодирующую тяжелую цепь, содержащую SEQ ID NO: 24, 25 или 26; и
(ii) выделенный полинуклеотид, содержащий последовательность, кодирующую легкую цепь, содержащую SEQ ID NO: 29, 31 или 32,
выращивание клетки-хозяина в условиях, обеспечивающих возможность продукции указанного антитела, и
выделение антитела.
| Способ разрушения пены | 1982 |
|
SU1074563A1 |
| WO 2014153435 A1, 25.09.2014 | |||
| EDWARDS J.R | |||
| et al., Inhibition of TGF-β signaling by 1D11 antibody treatment increases bone mass and quality in vivo, Journal of Bone and Mineral Research, 2010, Vol | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| ГУМАНИЗИРОВАННЫЕ АНТИ-ТФР-БЕТА-АНТИТЕЛА | 2005 |
|
RU2386638C2 |
