Область техники, к которой относится изобретение
Настоящее изобретение относится к искусственно измененному фактору, управляющему функцией ШК, для управления функцией ШК и/или лечения или облегчения заболевания, вызванного нарушением функции ШК, и к его применению. Более конкретно, настоящее изобретение относится к системе, способной выполнять искусственное управление функцией ШК и/или лечить или облегчать заболевание, вызванное нарушением функции ШК, причем система содержит: искусственно измененный фактор, управляющий функцией ШК, для управления функцией ШК и/или лечения или облегчения заболевания, вызванного нарушением функции ШК; и/или композицию для лечения или облегчения заболевания, вызванного нарушением функции ШК.
Уровень техники
Шванновские клетки (далее также сокращенно обозначаемые как ШК) или нейролеммоциты являются основными глиальными клетками периферической нервной системы. Функция глиальных клеток состоит в поддержке нейронов и в периферической нервной системе они представляют собой клетки-сателлиты, обкладочные нейроэпителиальные клетки, глиальные клетки кишечника, глиальные клетки, которые находятся на чувствительных нервных окончаниях, таких как тельце Пачини и тому подобное.
В миелиновых аксонах шванновские клетки образуют миелиновую оболочку. Оболочка не является непрерывной, и отдельные шванновские клетки охватывают около 100 мкм аксона. Разрывы между смежными шванновскими клетками называют узлами Ранвье. Нервная система позвоночных изолирована миелиновой оболочкой для поддержания емкости мембраны аксона. Потенциал действия в узлах Ранвье переходит от узла к узлу. Посредством этого процесса скорость проводимости увеличивается до 10 раз без увеличения диаметра аксона, и энергия может быть сохранена. Шванновские клетки могут быть аналогами олигодендроцитов, которые играют ту же роль в центральной нервной системе. Однако в отличие от олигодендроцитов шванновские клетки образуют миелиновую оболочку только в одном аксоне.
Болезнь Шарко-Мари-Тута (CMT), которая является наследственным заболеванием, представляет собой заболевание, при котором мышцы конечностей постепенно уменьшаются из-за аномалий шванновских клеток, образующих миелиновую оболочку в периферической нервной системе. Среди своих типов болезнь Шарко-Мари-Тута типа 1А (CMT1A) является одним из генетических заболеваний, наиболее часто встречающихся в периферической нервной системе, и существует очень большая разница в генетической информации между моделями человека и животных для выявления генетических причин CMT1A и нахождения решения для CMT1A с помощью информации, полученной из моделей животных, что связано с проблемами в сопоставлении.
CMT1A составляет более половины всех случаев CMT и около 70% случаев CMT1, и частота CMT составляет 1/2500. CMT1A проявляет патологические характеристики, такие как мышечная слабость и потеря, снижение рефлексов, акропарестезия, дисмелия, снижение скорости нервной проводимости (NCV), гипертрофическая сегментарная демиелинизация и ремиелинизация, которая выглядит как луковицеобразные утолщения.
В настоящее время не существует необходимого терапевтического средства или терапевтического способа лечения заболеваний, вызванных дисфункцией шванновских клеток, такой как CMT1A. Существует только способ частичного уменьшения обострения симптомов с использованием физиотерапии, ортопедических средств и ортопедической хирургии. Следовательно, существует большая потребность в разработке необходимого терапевтического способа лечения заболевания, вызванного дисфункцией шванновских клеток.
Сущность изобретения
Техническая проблема
Для решения этой проблемы настоящее изобретение относится к искусственно измененной системе, управляющей функцией ШК, имеющей улучшенный эффект управления функцией ШК. Более конкретно, настоящее изобретение относится к искусственно измененному фактору, управляющему функцией ШК, и системе, управляющей функцией ШК (также обозначаемой как система модификации управления функцией ШК), которая искусственно управляет функцией ШК и/или модифицирует экспрессию фактора, управляющего функцией ШК.
В настоящем изобретении предоставлен генетически измененный или модифицированный фактор, управляющий функцией ШК для конкретной цели.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления искусственно измененной системы, управляющей функцией ШК.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления искусственно измененной системы модификации фактора, управляющего функцией ШК.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления искусственно измененного фактора, управляющего функцией ШК, и продукта его экспрессии.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления композиции для манипуляции с геном для манипуляции фактором, управляющим функцией ШК, и способа его применения.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления способа управления функцией ШК.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления терапевтической или фармацевтической композиции для лечения заболевания, связанного с дисфункцией ШК, и разных вариантов ее применения.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления факторов искусственно измененного PMP22, управляющих функцией ШК, и/или продукта их экспрессии.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления композиции для генной манипуляции для искусственной манипуляции с факторами PMP22, управляющими функцией ШК.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления терапевтического применения факторов искусственно измененного PMP22, управляющих функцией ШК, и/или композиции для генной манипуляции для искусственной манипуляции.
В качестве варианта осуществления настоящее изобретение предназначено для предоставления дополнительного применения факторов искусственно измененного PMP22, управляющих функцией ШК, и/или композиции для генной манипуляции для искусственной манипуляции.
Техническое решение
Чтобы решить эту проблему, настоящее изобретение относится к системе, способной искусственно управлять функцией ШК, причем система содержит: искусственно измененный фактор, управляющий функцией ШК, для управления функцией ШК и/или лечения или облегчения заболевания, связанного с дисфункцией ШК, и/или композицию, способную искусственно изменять фактор, управляющий функцией ШК.
В настоящем изобретении предоставлен искусственно измененный фактор, управляющий функцией ШК, для конкретной цели.
«Фактор, управляющий функцией ШК» относится ко всем элементам, непосредственно участвующим или косвенно влияющим на управление функцией ШК. В этом случае элемент может быть ДНК, РНК, геном, пептидом, полипептидом или белком. Кроме того, фактор, управляющий функцией ШК, включает в себя все элементы, непосредственно участвующие или косвенно влияющие на управление функцией шванновских клеток, а также глиальных клеток и/или фибробластов.
В вариантах осуществления элемент включает в себя все различные материалы, включенные в неестественное, то есть искусственно измененное управление функцией ШК. Например, элемент может быть генетически измененным или модифицированным геном или белком, экспрессируемым в шванновских клетках.
Фактор, управляющий функцией ШК, может подавлять или ингибировать рост шванновских клеток, глиальных клеток и/или фибробластов или стимулировать или увеличивать рост шванновских клеток.
Фактор, управляющий функцией ШК, может прерывать или останавливать прохождение клеточного цикла шванновских клеток, глиальных клеток и/или фибробластов или стимулировать прохождение клеточного цикла шванновских клеток.
Фактор, управляющий функцией ШК, может стимулировать или подавлять дифференцировку шванновских клеток, глиальных клеток и/или фибробластов.
Фактор, управляющий функцией ШК, может стимулировать или подавлять гибель шванновских клеток, глиальных клеток и/или фибробластов.
Фактор, управляющий функцией ШК, может способствовать или нарушать выживаемость периферических нервных клеток.
Фактор, управляющий функцией ШК, может способствовать или нарушать поддержание и передачу сигналов периферических нервных клеток.
Фактор, управляющий функцией ШК, может управлять образованием миелиновой оболочки аксонов нервных клеток.
В этом случае образование миелиновой оболочки включает в себя весь процесс образования миелиновой оболочки, такой как образование миелиновой оболочки, дегенерация миелиновой оболочки, регенерация миелиновой оболочки, поддержание миелиновой оболочки и компактного миелина миелиновой оболочки, и все механизмы, связанные с дегенерацией миелиновой оболочки или функцией миелиновой оболочки.
Фактор, управляющий функцией ШК, может быть использован для облегчения и лечения заболевания, вызванного дисфункциональными или дефектными шванновскими клетками.
В варианте осуществления настоящего изобретения фактором, управляющим функцией ШК, может быть, например, искусственно измененный ген PMP22.
В варианте осуществления настоящего изобретения фактор, управляющий функцией ШК, может включать один или более искусственно измененных генов. Например, ген PMP22 может быть искусственно изменен.
Следовательно, в варианте осуществления настоящего изобретения представлен ген PMP22, который имеет модификацию в последовательности нуклеиновой кислоты в качестве искусственно измененного фактора, управляющего функцией ШК.
Модификация в последовательности нуклеиновой кислоты может быть неограниченно искусственно изменена с помощью комплекса гидовая нуклеиновая кислота-белок-редактор.
Термин «комплекс гидовая нуклеиновая кислота-белок-редактор» относится к комплексу, образованному в результате взаимодействия между гидовой нуклеиновой кислотой и белком-редактором, и комплекс нуклеиновая кислота-белок содержит гидовую нуклеиновую кислоту и белок-редактор.
Комплекс гидовая нуклеиновая кислота-белок-редактор может служить для модификации объекта. Объектом может быть мишень в виде нуклеиновой кислоты, гена, хромосомы или белка.
Например, ген может быть фактором, управляющим функцией ШК, искусственно измененным с помощью комплекса гидовая нуклеиновая кислота-белок-редактор,
при этом искусственно измененный фактор, управляющий функцией ШК, содержит одну или более модификаций нуклеиновых кислот, которые являются
по меньшей мере одним из делеции или вставки одного или более нуклеотидов, замещения одним или более нуклеотидами, отличными от гена дикого типа, и вставки одного или более чужеродных нуклеотидов в последовательность мотива, примыкающего к протоспейсеру (PAM), в последовательности нуклеиновой кислоты, составляющей фактор, управляющий функцией ШК, или в непрерывный участок последовательности оснований от 1 до 50 п.н., примыкающий к ее 5'-концу и/или 3'-концу,
или
химической модификацией одного или более нуклеотидов в последовательности нуклеиновой кислоты, составляющей фактор, управляющий функцией ШК.
Модификация нуклеиновых кислот может происходить в промоторном участке гена.
Модификация нуклеиновых кислот может происходить в экзонном участке гена.
Модификация нуклеиновых кислот может происходить в интронном участке гена.
Модификация нуклеиновых кислот может происходить в энхансерном участке гена.
PAM-последовательность может быть, например, одной или более из следующих последовательностей (описаны в направлении от 5' к 3'):
NGG (N представляет собой A, T, C или G);
NNNNRYAC (каждый из N независимо представляет собой A, T, C или G, R представляет собой A или G и Y представляет собой C или T);
NNAGAAW (каждый из N независимо представляет собой A, T, C или G, а W представляет собой A или T);
NNNNGATT (каждый из N независимо представляет собой A, T, C или G);
NNGRR(T) (каждый из N независимо представляет собой A, T, C или G, R представляет собой A или G); и
TTN (N представляет собой A, T, C или G).
Белок-редактор может быть получен из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptosporangium roseum, AlicyclobacHlus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor bescii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsonii, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus или Acaryochloris marina.
В одном примере варианта осуществления белок-редактор может быть одним или более, выбранными из группы, состоящей из белка Cas9, полученного из Streptococcus pyogenes, белка Cas9, полученного из Campylobacter jejuni, белка Cas9, полученного из Streptococcus thermophilus, белка Cas9, полученного из Streptococcus aureus, белка Cas9, полученного из Neisseria meningitidis, и белка Cpf1. В качестве примера, белок-редактор может представлять собой белок Cas9, полученный из Streptococcus pyogenes, или белок Cas9, полученный из Campylobacter jejuni.
Кроме того, в другом варианте осуществления
в настоящем изобретении предоставлена гидовая нуклеиновая кислота, нацеленная на часть хромосомы 17, содержащей участок от 15,267,977 до 15,273,977 (регуляторный участок); хромосомы 17 от 15,229,777 до 15,265,326 (кодирующий участок); или хромосомы 17 от 15,268,191 до 15,437,045, хромосомы 17 от 15,239,833 до 15,258,667 или хромосомы 17 от 15,342,770 до 15,435,639 (некодирующий участок), в последовательности нуклеиновой кислоты гена PMP22.
В примере варианта осуществления в настоящем изобретении предоставлена гидовая нуклеиновая кислота, которая способна образовывать комплементарную связь по отношению к последовательностям-мишеням SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 в последовательностях нуклеиновых кислот гена PMP22, соответственно.
Гидовая нуклеиновая кислота может образовывать комплементарную связь с частью последовательностей нуклеиновых кислот гена PMP22. Она может содержать от 0 до 5, от 0 до 4, от 0 до 3 или от 0 до 2 ошибочно спаренных оснований. В качестве иллюстративного примера, гидовая нуклеиновая кислота может представлять собой нуклеотиды, образующие комплементарную связь с одной или более последовательностями-мишенями SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53, соответственно.
Например, в настоящем изобретении может быть представлена одна или более гидовых нуклеиновых кислот, выбранных из группы, как описано ниже:
гидовая нуклеиновая кислота, способная образовывать комплементарную связь по отношению к последовательностям-мишеням SEQ ID NO: 1, 3, 25, 27, 28, 41, 44, 45 и 53 в последовательности нуклеиновой кислоты гена PMP22, соответственно;
гидовая нуклеиновая кислота может иметь неограниченное количество нуклеотидов от 18 до 25 п.н., от 18 до 24 п.н., от 18 до 23 п.н., от 19 до 23 п.н. или от 20 до 23 п.н. Это может привести к ошибочно спаренным основаниям в количестве от 0 до 5, от 0 до 4, от 0 до 3 или от 0 до 2.
Кроме того, в настоящем изобретении предоставлена композиция для генной манипуляции, которая может быть использована для искусственной манипуляции фактором, управляющим функцией ШК, для конкретной цели.
В другом варианте осуществления композиция для генной манипуляции может содержать комплекс гидовая нуклеиновая кислота-белок-редактор или кодирующую его последовательность нуклеиновой кислоты.
В качестве примера варианта осуществления композиция для генной манипуляции может содержать:
(a) гидовую нуклеиновую кислоту, нацеленную на часть хромосомы 17, содержащей участок от 15,267,977 до 15,273,977 (регуляторный участок); хромосомы 17 от 15,229,777 до 15,265,326 (кодирующий участок); или хромосомы 17 от 15,268,191 до 15,437,045, хромосомы 17 от 15,239,833 до 15,258,667 или хромосомы 17 от 15,342,770 до 15,435,639 (некодирующий участок) в последовательности нуклеиновой кислоты гена PMP22, или кодирующую ее последовательность нуклеиновой кислоты; и
(b) белок-редактор, образующий комплекс с гидовой нуклеиновой кислотой для расщепления или модификации сайта-мишени в последовательности нуклеиновой кислоты гена PMP22, или кодирующую его последовательность нуклеиновой кислоты.
В примере варианта осуществления композиция для генной манипуляции может содержать:
(а) гидовую нуклеиновую кислоту, которая способна образовывать комплементарную связь по отношению к последовательностям-мишеням SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 в последовательностях нуклеиновых кислот гена PMP22, соответственно, или кодирующую ее последовательность нуклеиновой кислоты; и
(b) белок-редактор, представляющий собой один или более белков, выбранных из группы, состоящей из белка Cas9, полученного из Streptococcus pyogenes, белка Cas9, полученного из Campylobacter jejuni, белка Cas9, полученного из Streptococcus thermophilus, белка Cas9, полученного из Streptococcus aureus, белка Cas9, полученного из Neisseria meningitidis, и белка Cpf1, соответственно, или кодирующую его последовательность нуклеиновой кислоты.
В одном примере варианта осуществления гидовая нуклеиновая кислота может представлять собой последовательность нуклеиновой кислоты, которая образует комплементарную связь по отношению к одной или более последовательностям-мишеням SEQ ID NO: 1, 3, 25, 27, 28, 41, 44, 45 и 53 в последовательности нуклеиновой кислоты гена PMP22, соответственно.
В одном примере варианта осуществления композиция для генной манипуляции может представлять собой вирусную векторную систему.
Вирусный вектор может содержать один или более элементов, выбранных из группы, состоящей из ретровируса, лентивируса, аденовируса, аденоассоциированного вируса (AAV), вируса коровьей оспы, поксвируса и вируса простого герпеса.
В еще одном примере варианта осуществления в настоящем изобретении может быть представлен способ искусственной манипуляции с клетками, в котором имеется:
(a) гидовая нуклеиновая кислота, нацеленная на часть хромосомы 17, содержащей участок от 15,267,977 до 15,273,977 (регуляторный участок); хромосомы 17 от 15,229,777 до 15,265,326 (кодирующий участок); или хромосомы 17 от 15,268,191 до 15,437,045, хромосомы 17 от 15,239,833 до 15,258,667 или хромосомы 17 от 15,342,770 до 15,435,639 (некодирующий участок) в последовательности нуклеиновой кислоты гена PMP22, или кодирующая ее последовательность нуклеиновой кислоты; и
(b) белок-редактор, образующий комплекс с гидовой нуклеиновой кислотой для расщепления или модификации сайта-мишени в последовательности нуклеиновой кислоты гена PMP22, или кодирующая его последовательности нуклеиновой кислоты.
В примере варианта осуществления в настоящем изобретении предоставлен способ искусственной манипуляции с клетками, который включает: введение (а) гидовой нуклеиновой кислоты, которая способна образовывать комплементарную связь по отношению к последовательностям-мишеням SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 в последовательностях нуклеиновых кислот гена PMP22, соответственно, или кодирующей ее последовательности нуклеиновой кислоты; и
(b) белка-редактора, содержащего один или более белков, выбранных из группы, состоящей из белка Cas9, полученного из Streptococcus pyogenes, белка Cas9, полученного из Campylobacter jejuni, белка Cas9, полученного из Streptococcus thermophilus, белка Cas9, полученного из Streptococcus aureus, белка Cas9, полученного из Neisseria meningitidis, и белка Cpf1, соответственно, или кодирующей его последовательности нуклеиновой кислоты в клетки.
Гидовая нуклеиновая кислота и белок-редактор могут присутствовать в одном или более векторах в виде последовательности нуклеиновой кислоты или могут присутствовать в комплексе, образованном путем соединения гидовой нуклеиновой кислоты с белком-редактором.
Введение может быть выполнено in vivo или ex vivo.
Введение может быть выполнено одним или более способами, выбранными из электропорации, липосом, плазмид, вирусных векторов, наночастиц и метода белка слияния с доменом транслокации белка (PTD).
Вирусный вектор может быть одним или более, выбранными из группы, состоящей из ретровируса, лентивируса, аденовируса, аденоассоциированного вируса (AAV), вируса коровьей оспы, поксвируса и вируса простого герпеса.
В другом примере варианта осуществления настоящее изобретение относится к фармацевтической композиции для лечения заболевания, связанного с дисфункцией ШК, и/или заболевания, связанного с фактором, управляющим функцией ШК.
Фармацевтическая композиция может представлять собой композицию для генной манипуляции, которая может быть использована для искусственной манипуляции фактором, управляющим функцией ШК.
Состав композиции для генной манипуляции такой же, как описано выше.
В варианте осуществления предоставлен способ предоставления информации о последовательности положения-мишени, которое можно искусственно изменять в объекте посредством секвенирования гена PMP22.
Кроме того, в варианте осуществления предоставлен способ создания библиотеки с помощью использования информации, предоставленной этим способом.
В варианте осуществления предоставлен набор для генной манипуляции, содержащий следующее:
(a) гидовую нуклеиновую кислоту, нацеленную на часть хромосомы 17, содержащей участок от 15,267,977 до 15,273,977 (регуляторный участок); хромосомы 17 от 15,229,777 до 15,265,326 (кодирующий участок); или хромосомы 17 от 15,268,191 до 15,437,045, хромосомы 17 от 15,239,833 до 15,258,667 или хромосомы 17 от 15,342,770 до 15,435,639 (некодирующий участок) в последовательности нуклеиновой кислоты гена PMP22, или кодирующую ее последовательность нуклеиновой кислоты; и
(b) белок-редактор, образующий комплекс с гидовой нуклеиновой кислотой для расщепления или модификации сайта-мишени в последовательности нуклеиновой кислоты гена PMP22, или кодирующую его последовательность нуклеиновой кислоты.
В примере варианта осуществления в настоящем изобретении предоставлен набор для генной манипуляции, который содержит следующие компоненты:
(а) гидовую нуклеиновую кислоту, способную образовывать комплементарную связь по отношению к последовательностям-мишеням SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 в последовательностях нуклеиновых кислот гена PMP22, соответственно, или кодирующую ее последовательность нуклеиновой кислоты; и
(b) белок-редактор, представляющий собой один или более белков, выбранных из группы, состоящей из белка Cas9, полученного из Streptococcus pyogenes, белка Cas9, полученного из Campylobacter jejuni, белка Cas9, полученного из Streptococcus thermophilus, белка Cas9, полученного из Streptococcus aureus, белка Cas9, полученного из Neisseria meningitidis, и белка Cpf1, соответственно, или кодирующую его последовательность нуклеиновой кислоты.
Интересующий ген может быть искусственно изменен с использованием такого набора.
В одном примере варианта осуществления в настоящем изобретении может быть представлена композиция для лечения нарушения из-за дисфункции ШК, которая содержит:
гидовую нуклеиновую кислоту, способную образовывать комплементарную связь с одной или более последовательностями-мишенями в последовательностях нуклеиновых кислот гена PMP22, соответственно, или кодирующую ее последовательность нуклеиновой кислоты; и
белок-редактор или кодирующую его последовательность нуклеиновой кислоты.
Последовательность-мишень может быть частью хромосомы 17, содержащей участок от 15,267,977 до 15,273,977 (регуляторный участок); хромосомы 17 от 15,229,777 до 15,265,326 (кодирующий участок); или хромосомы 17 от 15,268,191 до 15,437,045, хромосомы 17 от 15,239,833 до 15,258,667 или хромосомы 17 от 15,342,770 до 15,435,639 (некодирующий участок), в последовательности нуклеиновой кислоты гена PMP22.
Последовательности-мишени могут представлять собой одну или более последовательностей SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53.
В одном примере варианта осуществления изобретения в качестве белка-редактора может быть использован белок Cas9, полученный из Campylobacter jejuni, Streptococcus pyogenes.
В одном примере нарушение из-за дисфункции ШК может быть заболеванием, связанным с фактором, управляющим функцией ШК.
В одном примере заболевание, связанное с фактором, управляющим функцией ШК, представляет собой болезнь Шарко-Мари-Тута типа 1А (CMT1A), болезнь Дежерина-Сотта (DSS), врожденную гипомиелинизирующую нейропатию (CHN) или синдром Русси-Леви (RLS).
В одном примере варианта осуществления в настоящем изобретении предоставлены все аспекты применения искусственно измененного фактора, управляющего функцией ШК, или композиции для генной манипуляции, которую используют для искусственной манипуляции фактором, управляющим функцией ШК, для лечения заболевания у объекта.
Объектами лечения могут быть млекопитающие, включая приматов, таких как люди, обезьяны и т.д., грызунов, таких как мыши, крысы и т.д., и тому подобное.
Предпочтительные результаты
Искусственно измененный фактор, управляющий функцией ШК, и система, управляющая функцией ШК, функции которой искусственно модифицированы посредством него, могут быть использованы для эффективного терапевтического применения для нарушения из-за дисфункции ШК, например, заболевания, связанного с фактором, управляющим функцией ШК, такого как болезнь Шарко-Мари-Тута типа 1А (CMT1A), болезнь Дежерина-Сотта (DSS), врожденная гипомиелинизирующая нейропатия (CHN) или синдром Русси-Леви (RLS). Посредством различных механизмов in vivo, в которых участвуют факторы, управляющие функцией ШК, эффективность системы, управляющей функцией ШК, может быть улучшена.
Например, может быть использован ген PMP22.
Краткое описание чертежей
ФИГ.1 представляет собой набор результатов, иллюстрирующих частоту инделей из-за SpCas9-огРНК-опосредованной генной манипуляции, и иллюстрирует частоту инделей каждого из (a) CDS, (b) ТАТА-бокса и (c) Энхансера, на которые разделен сайт-мишень огРНК.
ФИГ.2 представляет собой набор результатов, иллюстрирующих частоту инделей из-за CjCas9-огРНК-опосредованной генной манипуляции, и иллюстрирует частоту инделей каждого из (a) CDS, (b) ТАТА-бокса и (c) Энхансера, на которые разделен сайт-мишень огРНК.
ФИГ.3 иллюстрирует результаты генной манипуляции с помощью SpCas9-огРНК, нацеленной на управляющие факторы (регуляторные элементы) человеческого гена PMP22 в шванноподобных клетках.
ФИГ.4 иллюстрирует коэффициенты мутации со сдвигом рамки, индуцированные с помощью SpCas9-огРНК, нацеленных на CDS человеческого PMP22.
ФИГ.5 иллюстрирует делеции критических сайтов для регуляторных элементов человеческого PMP22 при обработке двойными огРНК.
ФИГ.6 представляет собой график, иллюстрирующий снижение экспрессии мРНК человеческого PMP22 с помощью SpCas9-огРНК в человеческих шванноподобных клетках.
ФИГ.7 представляет собой график, иллюстрирующий эффективное и специфическое снижение экспрессии PMP22 с помощью SpCas9-огРНК в каждом сайте-мишени человеческого гена PMP22 в человеческих первичных шванновских клетках, и (а) иллюстрирует результаты измерения частоты инделей с помощью SpCas9-огРНК в каждом сайте-мишени, (b) иллюстрирует результаты сравнения относительной экспрессии мРНК для PMP22, которые измеряют с помощью количественного анализа ОТ-ПЦР с обработкой или без обработки сигнального фактора миелинизации и комплекса RNP для каждого сайта-мишени (n=3, однофакторный дисперсионный анализ и апостериорные тесты Тьюки: * p < 0,05) и (c) иллюстрирует результаты измерений частоты инделей с помощью SpCas9-огРНК, нацеленных на сайты дистального энхансера (участки дистального энхансера) B и C.
ФИГ.8 представляет собой график, иллюстрирующий эффективное и специфическое снижение экспрессии PMP22 с помощью CRISPR-Cas9, нацеленной на сайт ТАТА-бокса человеческого гена PMP22 in vitro, и (а) иллюстрирует последовательность-мишень, нацеленную на промоторный сайт положения человеческого PMP22, и крайний левый график, средний график и крайний правый график на (b) иллюстрируют результаты измерений частоты инделей с использованием направленного глубокого секвенирования в человеческих первичных шванновских клетках, результаты измерения частоты мутации ТАТА-бокса 1 (n=3) среди общих частот инделей, и результаты сравнения относительной экспрессии мРНК для PMP22, которые измерены с помощью количественного анализа ОТ-ПЦР с обработкой или без обработки сигнального фактора миелинизации и комплекса RNP в человеческих первичных шванновских клетках (n=3, однофакторный дисперсионный анализ и апостериорные тесты Тьюки: * р < 0,05) соответственно.
ФИГ.9 иллюстрирует частоты инделей с помощью PMP22-TATA RNP в нецелевых участках и целевых участках, обнаруженные с помощью анализа нецелевых участков in silico с помощью направленного глубокого секвенирования в человеческих первичных шванновских клетках, и (a) представляет собой график, иллюстрирующий частоты инделей, (b) иллюстрирует примеры инделей с высокой частотой, и (c) показывает нецелевые положения, обнаруженные посредством анализа нецелевых участков in silico.
ФИГ.10 представляет собой набор результатов, иллюстрирующих положения, расщепленные с помощью PMP22-TATA RNP во всем геноме человека, и (a) иллюстрирует график Circos по всему геному, (b) иллюстрирует нецелевые положения, полученные с помощью Digenome-seq среди нецелевых положений, обнаруженных в результате анализа нецелевых участков in silico, и (c) представляет собой график, иллюстрирующий частоты инделей в нецелевых положениях.
ФИГ.11 схематически иллюстрирует терапевтический подход с использованием терапии PMP22-TATA RNP у мышей С22.
ФИГ.12 представляет собой набор результатов, иллюстрирующих ослабление фенотипа заболевания посредством ингибирования экспрессии PMP22 с помощью CRISPR/Cas9 у мышей с CMT1A, и (а) представляет собой график, иллюстрирующий частоты инделей с использованием направленного глубокого секвенирования в седалищном нерве, обработанном mRosa26 или комплексом PMP22-TATA RNP (n=3), (b) представляет собой результат измерения частоты мутации ТАТА-бокса 1 (n=3) среди всех частот инделей, и (c) представляет собой график, сравнивающий относительные количества мРНК, экспрессируемой из PMP22, с использованием количественного анализа ОТ-ПЦР из седалищного нерва, обработанного mRosa26 или комплексом PMP22-TATA RNP.
ФИГ.13 представляет собой набор результатов, иллюстрирующих нецелевые положения и частоты инделей PMP22-TATA огРНК в геноме мыши с помощью анализа in silico, и (а) иллюстрирует нецелевые положения, а (b) представляет собой график, иллюстрирующий частоту инделей в каждом нецелевом положении.
ФИГ.14 представляет собой набор результатов, иллюстрирующих ослабление фенотипа заболевания посредством ингибирования экспрессии PMP22 с помощью CRISPR/Cas9 у мышей с CMT1A, и (a) представляет собой набор изображений полутонкого среза ткани седалищного нерва, обработанного mRosa26 или комплексом PMP22-TATA RNP, а верхний график и нижний график на (b) представляют собой диаграмму рассеяния, иллюстрирующую увеличение коэффициента g у мышей, которых лечили PMP22-TATA RNP, и график, иллюстрирующий увеличение диаметра миелинового аксона у мышей, получавших PMP22-TATA RNP, соответственно.
ФИГ.15 представляет собой набор результатов, иллюстрирующих электрофизиологические изменения посредством ингибирования экспрессии PMP22 с помощью CRISPR/Cas9 у мышей с CMT1A, и (a) представляет собой график, иллюстрирующий изменение в дистальной латентности (DL), (b) представляет собой график, иллюстрирующий изменение в скорости проводимости двигательных нервов (NCV), и (c) представляет собой график, иллюстрирующий изменение электрического вызванного ответа мышц (CMAP) (n=7 для mRosa26 RNP; n=10 для PMP22-TATA).
ФИГ.16 представляет собой набор результатов анализа локомоторного поведения вследствие ингибирования экспрессии PMP22 с помощью CRISPR/Cas9 у мышей с CMT1A, а верхний график и нижний график в (a) представляют собой результат теста вращающегося стержня (n=7 для mRosa26 RNP, n=11 для PMP22-TATA) и результаты теста вращающегося стержня, измеряемые еженедельно до тех пор, пока мышам не станет от 8 недель до 16 недель (n=7 для mRosa26 RNP, n=11 для PMP22-TATA) соответственно, а верхний график и нижнее изображение на (b) представляют собой график, иллюстрирующий соотношение массы икроножной мышцы к массе тела мыши C22, обработанной mRosa26 или комплексом PMP22-TATA RNP, и набор изображений икроножной мышцы мыши C22, обработанной mRosa26 или комплексом PMP22-TATA RNP, соответственно.
Способы осуществления изобретения
Если не указано иное, все технические и научные термины, используемые в настоящем документе, имеют то же значение, которое обычно понимают специалисты в области техники, к которой относится настоящее изобретение. Хотя способы и материалы, подобные или идентичные тем, что описаны в настоящем документе, можно использовать на практике или при тестировании настоящего изобретения, подходящие способы и материалы описаны ниже. Все публикации, заявки на патенты, патенты и другие ссылки, упомянутые в настоящем документе, включены посредством ссылки в своем полном объеме. Кроме того, материалы, способы и примеры являются просто иллюстративными и не предназначены для ограничения.
Аспект настоящего изобретения относится к
искусственно измененной системе, управляющей функцией ШК, которая влияет на улучшение или устранение дисфункции шванновских клеток.
В частности, аспект настоящего изобретения относится к конфигурации различных аспектов, способных облегчать или лечить нарушение экспрессии фактора, управляющего функцией ШК, или заболевание, связанное с дисфункцией шванновских клеток, с помощью искусственного изменения фактора, управляющего функцией ШК, для регуляции экспрессии фактора, управляющего функцией ШК, или восстановления после аномальной функции шванновских клеток. Аспект настоящего изобретения включает в себя искусственно экспрессируемый фактор, управляющий функцией ШК, и/или фактор, управляющий функцией ШК, функции которых модифицированы, способ его получения, композицию, содержащую его, его терапевтическое применение и тому подобное.
Другой аспект настоящего изобретения относится к
дополнительной системе для управления третьим механизмом in vivo, сопутствующим различным функциям искусственно экспрессируемого фактора, управляющего функцией ШК, и/или фактора, управляющего функцией ШК, функции которых модифицированы.
В частности, другой аспект настоящего изобретения может управлять соответствующим механизмом с помощью нацеливания не только на функцию миелинизации, в которой участвует искусственно измененный фактор, управляющий функцией ШК, но также на третью функцию in vivo. Другой аспект настоящего изобретения включает в себя искусственно экспрессируемый фактор, управляющий функцией ШК, и/или фактор, управляющий функцией ШК, функции которых модифицированы, способ его получения, композицию, содержащую его, его терапевтическое применение, способное облегчать или лечить заболевание, связанное с третьей функцией, и тому подобное.
Функция ШК
Вариант осуществления настоящего изобретения относится к усовершенствованию и модификации системы, управляющей функцией ШК.
«Управление функцией ШК» относится к общему функциональному управлению фактора функции ШК, например, шванновских клеток, глиальных клеток и/или фибробластов, на которые влияет функция гена PMP22, и в настоящем документе функция включает в себя весь процесс развития и роста из роста, дифференцировки и гибели шванновских клеток, глиальных клеток и/или фибробластов, а также включает все функции, такие как выживаемость и поддержание периферических нервных клеток шванновских клеток и образование миелиновой оболочки (миелинизации) на аксоне. Кроме того, управление функцией ШК включает общую функцию шванновских клеток, глиальных клеток и/или фибробластов, например, управление всем процессом развития и роста, начиная от клеточного роста и дифференцировки до гибели клеток.
Управление функцией ШК включает управление всеми механизмами, которые подавляют или ингибируют рост шванновских клеток, глиальных клеток и/или фибробластов, или стимулируют или увеличивают рост шванновских клеток, глиальных клеток и/или фибробластов.
Управление функцией ШК включает управление всеми механизмами, которые прерывают или останавливают прохождение клеточного цикла шванновских клеток, глиальных клеток и/или фибробластов, или стимулируют прохождение клеточного цикла шванновских клеток, глиальных клеток и/или фибробластов.
Управление функцией ШК включает управление всеми механизмами, которые стимулируют или подавляют дифференцировку шванновских клеток, глиальных клеток и/или фибробластов.
Управление функцией ШК включает управление всеми механизмами, такое как управление механизмом, который стимулирует или подавляет гибель шванновских клеток, глиальных клеток и/или фибробластов, например, апоптоз.
Кроме того, управление функцией ШК включает управление всеми механизмами, которые способствуют или нарушают выживаемость периферических нервных клеток.
Управление функцией ШК включает в себя все механизмы, которые способствуют или нарушают поддержание и передачу сигналов периферических нервных клеток.
Управление функцией ШК включает управление всеми механизмами, которые участвуют в образовании миелиновой оболочки аксона нервных клеток.
В этом случае образование миелиновой оболочки включает в себя весь процесс образования миелиновой оболочки, такой как образование миелиновой оболочки, дегенерация миелиновой оболочки, регенерация миелиновой оболочки, поддержание миелиновой оболочки и компактного миелина миелиновой оболочки, и все механизмы, связанные с дегенерацией миелиновой оболочки или функцией миелиновой оболочки.
Кроме того, управление функцией ШК включает управление всеми механизмами, которые подавляют или ингибируют рост фибробластов или глиальных клеток, или стимулируют или увеличивают рост фибробластов или глиальных клеток.
Управление функцией ШК включает управление всеми механизмами, которые участвуют в активности фибробластов или глиальных клеток.
В одном варианте осуществления управление функцией ШК может участвовать в лечении или облегчении заболевания, вызванного дупликацией PMP22.
Например, фактор, управляющий функцией ШК, может участвовать в лечении или облегчении заболевания, вызванного, например, модификацией экспрессии (включая мутацию) PMP22.
В варианте осуществления управление функцией ШК может управлять образованием миелиновой оболочки.
Например, поскольку, например, фактор, управляющий функцией ШК, например, сверхэкспрессия PMP22, имеет характеристику, вызывающую нейропатию путем дестабилизации миелиновой оболочки, которая приводит к вмешательству в поддержание и образование миелиновой оболочки, управление функцией ШК посредством управления экспрессией фактора, управляющего функцией ШК, может управлять образованием миелиновой оболочки.
В другом варианте осуществления управление функцией ШК может управлять дифференцировкой шванновских клеток.
Например, поскольку фактор, управляющий функцией ШК, например, PMP22, имеет характеристику увеличения экспрессии во время дифференцировки шванновских клеток, управление функцией ШК посредством управления экспрессией фактора, управляющего функцией ШК, может управлять дифференцировкой шванновских клеток.
В варианте осуществления управление функцией ШК может управлять передачей сигналов нервных клеток.
Например, поскольку фактор, управляющий функцией ШК, например, сверхэкспрессия PMP22 имеет характеристику уменьшения передачи сигналов нервных клеток, то есть скорости проводимости, путем дестабилизации миелиновой оболочки, которая приводит к вмешательству в поддержание и образование миелиновой оболочки, то управление функцией ШК посредством управления экспрессией фактора, управляющего функцией ШК, может управлять образованием миелиновой оболочки.
Фактор, управляющий функцией ШК
Вариантом осуществления настоящего изобретения является искусственно измененный или модифицированный фактор, управляющий функцией ШК.
«Фактор, управляющий функцией ШК» относится ко всем элементам, непосредственно участвующим или косвенно влияющим на управление функцией ШК. В этом случае элемент может быть ДНК, РНК, геном, пептидом, полипептидом или белком.
В вариантах осуществления элемент включает в себя все различные материалы, участвующие в неестественном, то есть искусственно измененном управлении функцией ШК. Например, элемент может быть генетически измененным или модифицированным геном или белком, экспрессируемым в шванновских клетках.
Термин «искусственно измененный» относится к искусственно модифицированному состоянию, а не к состоянию, которое возникает в естественном состоянии и присутствует в нем.
Термин «генетически измененный» относится к случаю, когда выполняют манипуляцию с помощью подвергания биологически или небиологически полученного материала, упомянутого в настоящем изобретении, искусственной генетической модификации, и это может быть, например, ген и/или продукт гена (полипептид, белок и тому подобное), в котором геном искусственно модифицирован для конкретной цели.
В качестве предпочтительного примера в настоящем изобретении предоставлен генетически измененный или модифицированный фактор, управляющий функцией ШК, для конкретной цели.
Далее, ген или белок, имеющий перечисленную функцию, может иметь не только один тип функции, связанной с управлением функцией ШК, но также множество функций. Кроме того, в настоящем изобретении может быть предоставлено две или более функции и фактора, связанных с управлением функцией ШК, при необходимости.
Фактор, управляющий функцией ШК, может подавлять или ингибировать рост шванновских клеток, или стимулировать или увеличивать рост шванновских клеток.
Фактор, управляющий функцией ШК, может прерывать или останавливать прохождение клеточного цикла шванновских клеток или стимулировать прохождение клеточного цикла шванновских клеток.
Фактор, управляющий функцией ШК, может стимулировать или подавлять дифференцировку шванновских клеток.
Фактор, управляющий функцией ШК, может стимулировать или подавлять гибель шванновских клеток.
Фактор, управляющий функцией ШК, может способствовать или нарушать выживаемость периферических нервных клеток.
Фактор, управляющий функцией ШК, может способствовать или нарушать поддержание и передачу сигналов периферических нервных клеток.
Фактор, управляющий функцией ШК, может управлять образованием миелиновой оболочки аксонов нервных клеток.
В этом случае образование миелиновой оболочки включает в себя весь процесс образования миелиновой оболочки, такой как образование миелиновой оболочки, дегенерация миелиновой оболочки, регенерация миелиновой оболочки, поддержания миелиновой оболочки и компактного миелина миелиновой оболочки и все механизмы, связанные с дегенерацией миелиновой оболочки или функцией миелиновой оболочки.
Фактор, управляющий функцией ШК, может быть использован для облегчения и лечения заболевания, вызванного дисфункциональными или дефектными шванновскими клетками.
Кроме того, фактор, управляющий функцией ШК, может управлять всеми механизмами, которые подавляют или ингибируют рост фибробластов или глиальных клеток, или стимулируют или увеличивают рост фибробластов или глиальных клеток.
Фактор, управляющий функцией ШК, может управлять всеми механизмами, которые участвуют в активности фибробластов или глиальных клеток.
В вариантах осуществления фактор, управляющий функцией ШК, может быть PMP22.
В любом варианте осуществления фактор, управляющий функцией ШК, может быть PMP22.
Ген периферического миелинового белка 22 (PMP22) относится к гену (полноразмерной ДНК, кДНК или мРНК), кодирующему белок PMP22, также обозначаемому как GAS3 или GAS-3. В одном примере ген PMP22 может быть одним или более, выбранными из группы, состоящей из следующего, но без ограничения: ген, кодирующий человеческий PMP22 (например, идентификационный номер NCBI NP_000295.1, NP_001268384.1, NP_001268385. 1, NP_001317072.1, NP_696996.1, NP_696997.1 и т.п.), например, ген PMP22, представленный идентификационным номером NCBI NM_000304.3, NM_001281455.1, NM_001281456.1, NM_001330143.1, NM_153321.2 NM_153322.2 и тому подобное.
Периферический миелиновый белок 22 (PMP22) представляет собой трансмембранный гликопротеин миелина массой 22 кДа, и PMP22 экспрессируется в шванновских клетках, образующих миелин периферической нервной системы, и играет важную роль в образовании и поддержании компактного миелина. Ген PMP22 отображен в человеческой хромосоме 17p11.2-p12 и кодирует продукцию гликопротеина PMP22. Модификация экспрессии гена PMP22 связана с наследственной демиелинизирующей нейропатией периферических нервов и вызывает аномальный синтез и функцию миелиновых оболочек. Повышенная экспрессия PMP22 из-за дупликации PMP22 является наиболее вероятным механизмом, вызывающим заболевание.
Точечная мутация или мутация со сдвигом рамки PMP22 вызывает наследственную нейропатию со склонностью к параличу от сдавления (HNPP), а миссенс-мутация вызывает различные типы CMT, называемые синдромом Дежерина-Сотта (СДС) и врожденной гипомиелинизирующей нейропатией (CHN). Кроме того, синдром Русси-Леви (RLS) вызван дупликацией PMP22 и обычно рассматривается как вариант фенотипа CMT1A.
Фактор, управляющий функцией ШК, может быть получен от млекопитающего, включая примата, такого как человек и обезьяна, грызуна, такого как крыса и мышь, и тому подобных.
Информация о генах может быть получена из общедоступной базы данных, такой как GenBank Национального центра биотехнологической информации (NCBI).
В качестве варианта осуществления настоящего изобретения фактор, управляющий функцией ШК, например, PMP22, может быть искусственно измененным фактором, управляющим функцией ШК.
В любом варианте осуществления искусственно измененный фактор, управляющий функцией ШК, может быть генетически изменен.
Генная манипуляция или их модификация могут быть получены с помощью искусственно вызванных вставок, делеций, замещений или инверсионных мутаций в частичном или целом участке геномной последовательности гена дикого типа. Кроме того, манипуляция или модификация гена также могут быть получены с помощью слияния двух или более измененных или модифицированных генов.
Например, ген инактивируют с помощью генной манипуляции или их модификации, так что белок, кодируемый геном, не может экспрессироваться в форме белка, имеющего изначально присущую ему функцию.
Например, ген может быть дополнительно активирован с помощью генной манипуляции или их модификации, так что белок, кодируемый геном, должен экспрессироваться в форме белка, имеющего улучшенную функцию по сравнению с изначально присущей ему функцией. В качестве примера, когда функция белка, кодируемого конкретным геном, представляет собой A, функция белка, экспрессируемого измененным геном, может полностью отличаться от A или может иметь дополнительную функцию (A+B), включая A.
Например, слияние двух или более белков может экспрессироваться с использованием двух или более генов, имеющих разные или комплементарные функции из-за генной манипуляции или их модификации.
Например, два или более белка могут экспрессироваться отдельно или независимо в клетках с использованием двух или более генов, имеющих разные или комплементарные функции из-за генной манипуляции или их модификации.
Измененный фактор, управляющий функцией ШК, может подавлять или ингибировать рост шванновских клеток, или стимулировать или увеличивать рост шванновских клеток.
Измененный фактор, управляющий функцией ШК, может прерывать или останавливать прохождение клеточного цикла шванновских клеток, или стимулировать прохождение клеточного цикла шванновских клеток.
Измененный фактор, управляющий функцией ШК, может стимулировать или подавлять дифференцировку шванновских клеток.
Измененный фактор, управляющий функцией ШК, может стимулировать или подавлять гибель шванновских клеток.
Измененный фактор, управляющий функцией ШК, может способствовать или нарушать выживаемость периферических нервных клеток.
Измененный фактор, управляющий функцией ШК, может способствовать или нарушать поддержание и передачу сигналов периферических нервных клеток.
Измененный фактор, управляющий функцией ШК, может управлять образованием миелиновой оболочки аксонов нервных клеток.
В этом случае образование миелиновой оболочки включает в себя весь процесс образования миелиновой оболочки, такой как образование миелиновой оболочки, дегенерация миелиновой оболочки, регенерация миелиновой оболочки, поддержание миелиновой оболочки и компактного миелина миелиновой оболочки, и все механизмы, связанные с дегенерацией миелиновой оболочки или функцией миелиновой оболочки.
Измененный фактор, управляющий функцией ШК, может быть использован для облегчения и лечения заболевания из-за дисфункциональных или дефектных шванновских клеток.
Кроме того, измененный фактор, управляющий функцией ШК, может управлять всеми механизмами, которые подавляют или ингибируют рост фибробластов или глиальных клеток, или стимулируют или увеличивают рост фибробластов или глиальных клеток.
Измененный фактор, управляющий функцией ШК, может управлять всеми механизмами, которые участвуют в активности фибробластов или глиальных клеток.
Манипуляция включает в себя все структурные или функциональные модификации фактора, управляющего функцией ШК.
Структурная модификация фактора, управляющего функцией ШК, включает в себя все модификации, которые не совпадают с диким типом, присутствующим в естественном состоянии.
Например, когда фактором, управляющим функцией ШК, является ген ДНК или РНК,
структурная модификация может быть структурной модификацией, в которой один или более нуклеотидов потеряны.
Структурная модификация может быть структурной модификацией, в которой вставлен один или более нуклеотидов.
В этом случае вставленный нуклеотид включает все нуклеотиды, введенные от объекта, включая фактор, управляющий функцией ШК, или вне объекта.
Структурная модификация может быть структурной модификацией, в которой один или более нуклеотидов замещены.
Структурная модификация может включать химическую модификацию одного или более нуклеотидов.
В этом случае химическая модификация включает все добавления, удаления или замещения функциональных групп.
В качестве другого примера, когда фактор, управляющий функцией ШК, представляет собой пептид, полипептид или белок,
структурная модификация может быть структурной модификацией, в которой одна или более аминокислот потеряны.
Структурная модификация может быть структурной модификацией, в которой вставлена одна или более аминокислот.
В этом случае вставленная аминокислота включает все аминокислоты, введенные от объекта, включая фактор, управляющий функцией ШК, или вне объекта.
Структурная модификация может быть структурной модификацией, в которой одна или более аминокислот замещены.
Структурная модификация может включать химическую модификацию одной или более аминокислот.
В этом случае химическая модификация включает все добавления, удаления или замещения функциональных групп.
Структурная модификация может быть структурной модификацией, в которой присоединена часть или весь другой пептид, полипептид или белок.
В этом случае другой пептид, полипептид или белок может быть фактором, управляющим функцией ШК, или может быть пептидом, полипептидом или белком, который выполняет другую функцию.
Функциональная модификация фактора, управляющего функцией ШК, включает в себя все модификации, имеющие улучшенную функцию или ухудшенную функцию по сравнению с диким типом, присутствующим в естественном состоянии, или имеющие третью другую функцию.
Например, когда фактор, управляющий функцией ШК, представляет собой пептид, полипептид или белок,
функциональная модификация может быть мутацией фактора, управляющего функцией ШК.
В этом случае мутация может быть мутацией, при которой функция фактора, управляющего функцией ШК, улучшена или ингибирована.
Функциональная модификация может быть функциональной модификацией, в которой добавлена функция фактора, управляющего функцией ШК.
В этом случае добавленная функция может быть одной и той же функцией или разными функциями. Кроме того, фактор, управляющий функцией ШК, к которому добавлена функция, может быть слит с другими пептидами, полипептидами или белками.
Функциональной модификацией может быть увеличение функции из-за увеличения экспрессии фактора, управляющего функцией ШК.
Функциональной модификацией может быть снижение функции из-за снижения экспрессии фактора, управляющего функцией ШК.
Функциональная модификация может представлять собой восстановление функции из-за снижения экспрессии фактора, управляющего функцией ШК.
В качестве варианта осуществления фактор, управляющий функцией ШК, может быть индуцирован одним или более из следующего:
полные или частичные делеции фактора, управляющего функцией ШК, то есть гена, который должен быть изменен (далее называемого геном-мишенью), например, делеция нуклеотидов из 1-30 п.н., 1-27 п.н., 1-25 п.н., 1-23 п.н., 1-20 п.н., 1-15 п.н., 1-10 п.н., 1-5 п.н., 1-3 п.н. или 1 п.н. гена-мишени,
замещение нуклеотидов длиной от 1 п.н. или более, например, 1-30 п.н., 1-27 п.н., 1-25 п.н., 1-23 п.н, 1-20 п.н., 1-15 п.н., 1-10 п.н., 1-5 п.н., 1-3 п.н. или 1 п.н. гена-мишени нуклеотидом, отличным от дикого типа, и
вставка нуклеотидов длиной от 1 п.н. или более, например, 1-30 п.н., 1-27 п.н., 1-25 п.н., 1-23 п.н, 1-20 п.н., 1-15 п.н., 1-10 п.н., 1-5 п.н., 1-3 п.н. или 1 п.н. (каждый независимо выбран из A, T, C и G) в любое положение гена-мишени.
Модифицированная часть гена-мишени («участок-мишень») может представлять собой сайт непрерывной последовательности оснований, составляющий 1 п.н. или более, 3 п.н. или более, 5 п.н. или более, 7 п.н. или более, 10 п.н. или более, 12 п.н. или более 15 п.н. или более, 17 п.н. или более, или 20 п.н. или более, например, 1-30 п.н., 3-30 п.н., 5-30 п.н., 7-30 п.н., 10-30 п.н., 12-30 п.н., 15-30 п.н., 17-30 п.н., 20-30 п.н., 1-27 п.н., 3-27 п.н., 5-27 п.н., 7-27 п.н., 10-27 п.н., 12-27 п.н., 15-27 п.н., 17-27 п.н., 20-27 п.н., 1-25 п.н., 3-25 п.н., 5-25 п.н., 7-25 п.н. 10-25 п.н., 12-25 п.н., 15-25 п.н., 17-25 п.н., 20-25 п.н., 1-23 п.н., 3-23 п.н., 5-23 п.н., 7-23 п.н., 10-23 п.н., 12-23 п.н., 15-23 п.н., 17-23 п.н., 20-23 п.н., 1-20 п.н., 3-20 п.н., 5-20 п.н., 7-20 п.н., 10-20 п.н., 12-20 п.н., 15-20 п.н., 17-20 п.н., 21-25 п.н., 18-22 п.н. или 21-23 п.н. гена.
Между тем, другой вариант осуществления настоящего изобретения относится к
дополнительной системе для управления третьим механизмом in vivo, сопутствующим различным функциям вышеописанных факторов, управляющих функцией ШК, функции которых искусственно модифицированы.
В качестве варианта осуществления PMP22 может участвовать в управлении третьим механизмом in vivo.
Поскольку многие исследования подтвердили, что экспрессия PMP22 увеличена в опухолевой клетке, что означает, что экспрессия PMP22 влияет на связанный с пролиферацией механизм опухолевых клеток при различных раковых заболеваниях, искусственно измененный PMP22 может подавлять или ингибировать пролиферацию клеток различных опухолей (например, рака молочной железы, рака желудка, рака поджелудочной железы и тому подобное), например, с помощью ингибирования экспрессии PMP22 или манипуляции для инактивации PMP22. Кроме того, можно подавлять или ингибировать прогрессирование или метастазирование различных раковых заболеваний или обеспечивать эффект облегчения или лечения раковых заболеваний с помощью искусственно измененного PMP22.
Как описано выше, искусственно измененные факторы в примерах настоящего изобретения могут управлять функцией ШК и могут управлять соответствующим механизмом с помощью нацеливания на третью функцию in vivo. Варианты осуществления настоящего изобретения включают в себя фактор, управляющий функцией ШК, функции которого искусственно модифицированы, способ его получения, содержащую его композицию, его терапевтическое применение, способное облегчать или лечить заболевание, связанное с третьей функцией, и тому подобное.
Система, управляющая функцией ШК
Вариант осуществления настоящего изобретения представляет собой систему, управляющую функцией ШК, которая управляет функцией ШК с помощью искусственного изменения фактора, управляющего функцией ШК.
«Система, управляющая функцией ШК» настоящего изобретения представляет собой термин, включающий все явления, влияющие на стимулирование, увеличение, ингибирование, подавление и/или восстановление нормального функционирования функций ШК с помощью изменения экспрессии и/или изменения функции искусственно измененного фактора, управляющего функцией ШК, и включает в себя все материалы, композиции, способы и применения, которые прямо или косвенно участвуют в системе, управляющей функцией ШК.
Соответствующие элементы, составляющие систему, управляющую функцией ШК, также совокупно обозначают как «элементы, управляющие функцией ШК».
Кроме того, система, управляющая функцией ШК, включает в себя систему модификации, управляющей функцией ШК, экспрессии которой модифицированы и/или функции модифицированы с помощью искусственного изменения фактора, управляющего функцией ШК.
«Система модификации фактора, управляющего функцией ШК» в совокупности относится к транскрипции фактора, управляющего функцией ШК, присутствующего в геноме объекта или введенного извне, модификации экспрессии всего фактора, управляющего функцией ШК, экспрессированного из транскрибированной генетической информации (например, мРНК) в виде белка, то есть такому процессу, как снижение экспрессии, увеличение экспрессии и поддержание экспрессии, а также ее продукту.
Кроме того, «система модификации фактора, управляющего функцией ШК» также включает весь процесс вмешательства в экспрессию и/или функцию нормального фактора, управляющего функцией ШК, с помощью мутации фактора, управляющего функцией ШК, или экспрессии нормального фактора, управляющего функцией ШК, такой как экспрессия мутации нормального фактора, управляющего функцией ШК, аномального фактора, управляющего функцией ШК, и искусственно модифицированного фактора, управляющего функцией ШК, и ее продукт.
В качестве варианта осуществления система настоящего изобретения включает в себя фактор, управляющий функцией ШК, то есть систему модификации PMP22, в качестве элемента, управляющего функцией ШК, и в этом случае фактор, управляющий функцией ШК, то есть PMP22, включает в себя искусственно измененный фактор, управляющий функцией ШК.
«Система модификации PMP22» в целом относится к транскрипции гена PMP22, присутствующего в геноме объекта или введенного извне, модификации экспрессии общего PMP22, экспрессируемого из транскрибированной генетической информации (например, мРНК) в виде белка, то есть процессу, такому как уменьшение экспрессии, увеличение экспрессии и поддержание экспрессии, и ее продукту.
Кроме того, «система модификации PMP22» также включает весь процесс вмешательства в экспрессию и/или функцию нормального PMP22 с помощью мутации PMP22 или экспрессии нормального PMP22, такой как экспрессия мутации PMP22, аномального PMP22 и искусственно модифицированного PMP22, и ее продукт.
В системе модификации PMP22 с помощью искусственно измененного PMP22,
в любом варианте осуществления экспрессия PMP22 может быть увеличена или простимулирована.
В любом варианте осуществления экспрессия PMP22 может быть ингибирована или подавлена.
Например, когда ген PMP22 дуплицирован и сверхэкспрессирован, экспрессия может быть подавлена или снижена до нормального уровня экспрессии с помощью искусственно измененного PMP22.
В любом варианте осуществления функция PMP22 может быть усилена или простимулирована.
В любом варианте осуществления функция PMP22 может быть ослаблена или подавлена.
Кроме того, в системе модификации PMP22 посредством искусственно измененного PMP22
в любом варианте осуществления рост шванновских клеток может быть подавлен или ингибирован, или рост шванновских клеток может быть простимулирован или увеличен.
В любом варианте осуществления прохождение клеточного цикла шванновских клеток может быть прервано или остановлено, или прохождение клеточного цикла шванновских клеток может быть простимулировано.
В любом варианте осуществления дифференцировка шванновских клеток может быть простимулирована или подавлена.
В любом варианте осуществления гибель шванновских клеток может быть простимулирована или подавлена.
В любом варианте осуществления выживаемость периферических нервных клеток может быть улучшена или нарушена.
В любом варианте осуществления поддержание или передача сигналов периферических нервных клеток могут быть улучшены или нарушены.
В любом варианте осуществления образованием миелиновой оболочки аксонов нервных клеток можно управлять.
В этом случае образование миелиновой оболочки включает в себя весь процесс образования миелиновой оболочки, такой как образование миелиновой оболочки, дегенерация миелиновой оболочки, регенерация миелиновой оболочки, поддержание миелиновой оболочки и компактного миелина миелиновой оболочки, и все механизмы, связанные с дегенерацией миелиновой оболочки или функцией миелиновой оболочки.
В любом варианте осуществления его можно использовать для облегчения и лечения заболевания, вызванного дисфункциональными или дефектными шванновскими клетками.
В одном варианте осуществления можно управлять всеми механизмами, которые подавляют или ингибируют рост фибробластов или глиальных клеток, или стимулируют или увеличивают рост фибробластов или глиальных клеток.
В любом варианте осуществления можно управлять всеми механизмами, которые участвуют в активности фибробластов или глиальных клеток.
В любом варианте осуществления соответствующим механизмом можно управлять с помощью нацеливания на третью функцию in vivo, в которой участвует PMP22.
В вариантах осуществления система, управляющая функцией ШК, настоящего изобретения включает в себя композицию для изменения фактора, управляющего функцией ШК, в качестве элемента, управляющего функцией ШК.
В вариантах осуществления система модификации фактора, управляющего функцией ШК, настоящего изобретения включает в себя композицию для изменения фактора, управляющего функцией ШК.
Композиция для манипуляции может представлять собой композицию, способную искусственно изменять фактор, управляющий функцией ШК, предпочтительно композицию для генной манипуляции.
Далее будет описана композиция для генной манипуляции.
Композиция для изменения фактора, управляющего функцией ШК
Манипуляция или модификация материала, который участвует в факторе, управляющем функцией ШК, и в системе, управляющей функцией ШК (включая систему модификации фактора, управляющего функцией ШК) настоящего изобретения, могут быть достигнуты предпочтительно с помощью генной манипуляции.
В одном аспекте композиция и способ, которые изменяют ген, могут быть обеспечены с помощью нацеливания на часть или весь регуляторный участок, некодирующий участок или кодирующий участок фактора, управляющего функцией ШК.
Для композиции и способа
в вариантах осуществления один или более из генов, управляющих функцией ШК, которые участвуют в образовании целевой системы, управляющей функцией ШК (включая систему модификации фактора, управляющего функцией ШК), можно изменять или модифицировать для образования целевой системы, управляющей функцией ШК. Манипуляция или модификация могут быть достигнуты посредством модификации нуклеиновой кислоты, составляющей ген. В результат манипуляции включены все формы нокдауна, нокаута и нокина.
В вариантах осуществления часть или весь регуляторный участок, некодирующий участок или кодирующий участок могут быть использованы в качестве мишени.
В любом варианте регуляторный участок среди нуклеиновых кислот, составляющих ген, управляющий функцией ШК, может быть использован в качестве мишени для манипуляции.
Например, частичная или полная последовательность, такая как проксимальный промотор, энхансер, TATA-бокс и регуляторный элемент инициатора, может быть использована в качестве мишени. В качестве конкретного примера, частичная или полная последовательность Промотора, ТАТА-бокса, CAAT-бокса, Сайта Инициации, Сайта Терминации, Донорного сайта сплайсинга, Акцепторного сайта сплайсинга, Поли(А)-Сайта, Энхансера, 3'-Нетранслируемого участка (НТУ), 5'-НТУ, Аттенюатора и GC-бокса могут быть использованы в качестве мишени.
В любом варианте осуществления энхансерный сайт (например, EGR2-, SOX10- или TEAD1-связывающий сайт) или сайт дистального энхансера (участок дистального энхансера) могут быть выделены среди последовательностей нуклеиновых кислот, составляющих ген, управляющий функцией ШК.
В любом варианте осуществления кодирующий участок среди нуклеиновых кислот, составляющих ген, управляющий функцией ШК, может быть использован в качестве мишени для манипуляции.
Например, интронная или экзонная последовательность может быть использована в качестве мишени. Кроме того, кодирующая последовательность, например кодирующий участок, начальный кодирующий участок, может быть выделена для изменения экспрессии и нокаута.
В вариантах осуществления модификация нуклеиновых кислот может представлять собой замещения, делеции и/или вставки нуклеотидов от 1 п.н. или более, например, 1-30 п.н., 1-27 п.н., 1-25 п.н., 1-23 п.н., 1-20 п.н., 1-15 п.н., 1-10 п.н., 1-5 п.н., 1-3 п.н. или 1 п.н.
В вариантах осуществления для нокаута одного или более генов, управляющих функцией ШК, подавления экспрессии одного или более генов или нокаутов одного, двух или трех аллелей, вышеописанный участок может быть выделен так, что один или более генов, управляющих функцией ШК, будут содержать делецию или мутацию.
В примерах вариантов осуществления нокдаун гена может быть использован для снижения экспрессии нежелательных аллелей или транскриптомов.
В вариантах осуществления нокдаун гена может быть использован для изменения гена, управляющего функцией ШК, влияющего на функции шванновских клеток, с помощью нацеливания на часть или весь регуляторный участок, некодирующий участок или кодирующий участок.
В вариантах осуществления активностью гена, управляющего функцией ШК, можно управлять, например, активировать или инактивировать с помощью изменения нуклеиновых кислот гена. Кроме того, функции шванновских клеток могут быть активированы или инактивированы с помощью изменения нуклеиновых кислот гена.
В вариантах осуществления модификация нуклеиновых кислот гена может катализировать расщепление одиночной цепи или двойных цепей, то есть разрывы цепей нуклеиновых кислот в определенном сайте в целевом гене с помощью комплекса гидовая нуклеиновая кислота-белок-редактор, что приводит к инактивации целевого гена.
В вариантах осуществления разрывы цепи нуклеиновой кислоты могут быть репарированы посредством механизма, такого как гомологичная рекомбинация или негомологичное соединение концов (NHEJ).
В этом случае, когда имеет место механизм NHEJ, может быть индуцировано изменение в последовательности ДНК в сайте расщепления, что приведет к инактивации гена. Репарация с помощью NHEJ может приводить к замещениям, вставкам или делециям короткого фрагмента гена и может быть использована для индукции соответствующих нокаутов гена или нокдауна. Кроме того, на функции шванновских клеток и/или их механизм действия может влиять индукция нокаутов или нокдауна гена, управляющего функцией ШК.
В другом аспекте в настоящем изобретении предоставлена композиция для изменения фактора, управляющего функцией ШК.
Композиция для манипуляции может представлять собой композицию, способную искусственно изменять фактор, управляющий функцией ШК, предпочтительно композицию для генной манипуляции.
Композиция может быть использована для генной манипуляции для одного или более генов, управляющих функцией ШК, которые участвуют в образовании целевой системы, управляющей функцией ШК (включая систему модификации фактора, управляющего функцией ШК) для образования целевой системы, управляющей функцией ШК.
Манипуляция c генами может быть выполнена с учетом процесса управления экспрессией генов.
В примерах вариантов осуществления генная манипуляция может быть выполнена с помощью выбора подходящих средств манипуляции для каждого этапа из этапов управления транскрипцией, процессингом РНК, транспортировкой РНК, деградацией РНК, трансляцией и модификацией белка.
В вариантах осуществления малая РНК (тРНК) разрушает мРНК или уменьшает ее стабильность, используя РНК-интерференцию (РНКи) или РНК-сайленсинг, и в некоторых случаях разрушает мРНК, чтобы препятствовать доставке информации о синтезе белка, что приводит к контролю экспрессии генетической информации.
Генная манипуляция может быть выполнена с помощью модификации нуклеиновых кислот, составляющих фактор, управляющий функцией ШК. В качестве результата манипуляции, включены все формы нокдауна, нокаута и нокина.
В любом варианте осуществления модификация нуклеиновых кислот может представлять собой замещения, делеции и/или вставки нуклеотидов от 1 п.н. или более, например, 1-30 п.н., 1-27 п.н., 1-25 п.н., 1-23 п.н., 1 до 20 п.н., 1-15 п.н., 1-10 п.н., 1-5 п.н., 1-3 п.н. или 1 п.н.
В любом варианте осуществления для нокаута одного или более факторов, управляющих функцией ШК, подавления экспрессии одного или более факторов или нокаутов одного, двух или трех аллелей ген может быть изменен так, чтобы один или более факторов, управляющих функцией ШК, содержали делецию или мутацию.
В любом варианте осуществления нокдаун фактора, управляющего функцией ШК, может быть использован для уменьшения экспрессии нежелательных аллелей или транскриптомов.
В любом варианте осуществления модификация нуклеиновых кислот может представлять собой вставку одного или более фрагментов нуклеиновой кислоты или генов. В этом случае фрагмент нуклеиновой кислоты может представлять собой последовательность нуклеиновой кислоты, состоящую из одного или более нуклеотидов, и длина фрагмента нуклеиновой кислоты может составлять 1-40 п.н., 1-50 п.н., от 1 до 60 п.н., от 1 до 70 п.н., 1-80 п.н., от 1 до 90 п.н., 1-100 п.н., 1-500 п.н. или 1-1000 п.н. В этом случае вставленный ген может быть одним из факторов, управляющих функцией ШК, или геном, имеющим другую функцию.
В вариантах осуществления для модификации нуклеиновых кислот может быть использован фермент дикого типа или вариантный фермент, который способен катализировать гидролиз (расщепление) связей между нуклеиновыми кислотами в молекуле ДНК или РНК, предпочтительно молекуле ДНК. Модификация нуклеиновых кислот может использовать комплекс гидовая нуклеиновая кислота-белок-редактор.
Например, ген может быть изменен с помощью использования одной или более нуклеаз, выбранных из группы, состоящей из мегануклеазы, «цинковопальцевой» нуклеазы, CRISPR/Cas9 (белок Cas9), CRISPR-Cpf1 (белок Cpf1) и TALE-нуклеазы, тем самым управляя экспрессией генетической информации.
В любом варианте осуществления генная манипуляция может быть опосредована негомологичным соединением концов (NHEJ) или репарацией, направленной гомологией (HDR) с использованием комплекса гидовая нуклеиновая кислота-белок-редактор, например, системы CRISPR/Cas, но без ограничения.
В этом случае, когда имеет место механизм NHEJ, может быть индуцировано изменение в последовательности ДНК в сайте расщепления, что приводит к инактивации гена или ингибированию его экспрессии. Репарация с помощью NHEJ может приводить к замещениям, вставкам или делециям короткого фрагмента гена и может быть использована для индукции соответствующего нокаута гена или нокдауна.
В другом аспекте в настоящем изобретении может быть представлен сайт генной манипуляции.
В вариантах осуществления, когда ген изменяют посредством NHEJ-опосредованного изменения, сайт генной манипуляции относится к сайту в гене, вызывая уменьшение или подавление экспрессии продукта гена, управляющего функцией ШК.
Например,
сайт может быть в начальном кодирующем участке.
Сайт может быть в промоторной последовательности.
Сайт может быть в энхансерной последовательности.
Сайт может быть в определенной интронной последовательности.
Сайт может быть в определенной экзонной последовательности.
В качестве варианта осуществления композиция для изменения фактора, управляющего функцией ШК
может быть нацелена на ген PMP22, который влияет на функции шванновских клеток в качестве объекта манипуляции.
Примеры сайтов-мишеней, то есть последовательностей-мишеней для сайтов, в которых происходит генная манипуляция или которые обозначены для генной манипуляции, приведены в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7.
Последовательность-мишень может быть нацелена на один или более генов.
Последовательность-мишень может одновременно быть нацелена на два или более генов. В этом случае два или более генов могут быть гомологичными генами или гетерологичными генами.
Ген может содержать одну или более последовательностей-мишеней.
Ген может быть одновременно нацелен на две или более последовательности-мишени.
Ген может быть изменен в положении и количестве генных манипуляций в зависимости от количества последовательностей-мишеней.
Генная манипуляция может быть разработана различными способами в зависимости от количества и положений последовательностей-мишеней.
Генная манипуляция может происходить одновременно в двух или более последовательностях-мишенях. В этом случае две или более последовательности-мишени могут присутствовать в гомологичном гене или гетерологичном гене.
Генная манипуляция может быть выполнена одновременно в отношении двух или более генов. В этом случае два или более генов могут быть гомологичными генами или гетерологичными генами.
Далее примеры последовательностей-мишеней, которые могут быть использованы в варианте осуществления настоящего изобретения, показаны в следующих таблицах:
Таблица 1. Последовательность-мишень для SpCas9 относительно кодирующей последовательности гена PMP22
Таблица 2. Последовательность-мишень для CjCas9 относительно кодирующей последовательности гена PMP22
Таблица 3. Последовательность-мишень для SpCas9 относительно промоторной последовательности гена PMP22
Таблица 4. Последовательность-мишень для CjCas9 относительно промоторной последовательности гена PMP22
Таблица 5. Последовательность-мишен для SpCas9 относительно энхансерной последовательности гена PMP22
Таблица 6. Последовательность-мишень для CjCas9 относительно энхансерной последовательности гена PMP22
Таблица 7. Последовательность-мишень для SpCas9 относительно дистальных энхансерных участков B и C гена PMP22
[Таблица 1]
(SEQ ID NO: 1)
(SEQ ID NO: 2)
(SEQ ID NO: 3)
(SEQ ID NO: 4)
(SEQ ID NO: 5)
(SEQ ID NO: 6)
(SEQ ID NO: 7)
(SEQ ID NO: 8)
[Таблица 2]
(SEQ ID NO: 9)
(SEQ ID NO: 10)
(SEQ ID NO: 11)
(SEQ ID NO: 12)
(SEQ ID NO: 13)
[Таблица 3]
(SEQ ID NO: 14)
(SEQ ID NO: 15)
(SEQ ID NO: 16)
(SEQ ID NO: 17)
(SEQ ID NO: 18)
(SEQ ID NO: 19)
(SEQ ID NO: 20)
(SEQ ID NO: 21)
(SEQ ID NO: 22)
(SEQ ID NO: 23)
(SEQ ID NO: 24)
(SEQ ID NO: 25)
(SEQ ID NO: 26)
(SEQ ID NO: 27)
(SEQ ID NO: 28)
(SEQ ID NO: 29)
[Таблица 4]
(SEQ ID NO: 30)
(SEQ ID NO: 31)
(SEQ ID NO: 32)
(SEQ ID NO: 33)
(SEQ ID NO: 34)
(SEQ ID NO: 35)
(SEQ ID NO: 36)
(SEQ ID NO: 37)
(SEQ ID NO: 38)
(SEQ ID NO: 39)
(SEQ ID NO: 40)
[Таблица 5]
(SEQ ID NO: 41)
(SEQ ID NO: 42)
(SEQ ID NO: 43)
(SEQ ID NO: 44)
(SEQ ID NO: 45)
(SEQ ID NO: 46)
(SEQ ID NO: 47)
(SEQ ID NO: 48)
(SEQ ID NO: 49)
(SEQ ID NO: 50)
(SEQ ID NO: 51)
(SEQ ID NO: 52)
(SEQ ID NO: 53)
[Таблица 6]
(SEQ ID NO: 54)
(SEQ ID NO: 55)
(SEQ ID NO: 56)
(SEQ ID NO: 57)
(SEQ ID NO: 58)
(SEQ ID NO: 59)
(SEQ ID NO: 60)
(SEQ ID NO: 61)
(SEQ ID NO: 62)
(SEQ ID NO: 63)
(SEQ ID NO: 64)
(SEQ ID NO: 65)
(SEQ ID NO: 66)
[Таблица 7]
(SEQ ID NO: 143)
(SEQ ID NO: 144)
(SEQ ID NO: 145)
(SEQ ID NO: 146)
(SEQ ID NO: 147)
(SEQ ID NO: 148)
(SEQ ID NO: 149)
(SEQ ID NO: 150)
Композиция для системы ножниц для генной манипуляции
Система, управляющая функцией ШК (включая систему модификации, управляющей функцией ШК), настоящего изобретения может включать в себя комплекс гидовая нуклеиновая кислота-белок-редактор в качестве композиции для изменения фактора, управляющего функцией ШК, то есть гена PMP22.
Комплекс гидовая нуклеиновая кислота-белок-редактор
Термин «комплекс гидовая нуклеиновая кислота-белок-редактор» относится к комплексу, образованному в результате взаимодействия между гидовой нуклеиновой кислотой и белком-редактором, и комплекс нуклеиновая кислота-белок содержит гидовую нуклеиновую кислоту и белок-редактор.
Термин «гидовая нуклеиновая кислота» относится к нуклеиновой кислоте, способной распознавать мишени в виде нуклеиновой кислоты, гена, хромосомы или белка.
Гидовая нуклеиновая кислота может присутствовать в форме ДНК, РНК или гибрида ДНК/РНК и может иметь последовательность нуклеиновой кислоты от 5 до 150 оснований.
Гидовая нуклеиновая кислота может содержать один или более доменов.
Домены могут быть, но без ограничения, гидовым доменом, первым комплементарным доменом, линкерным доменом, вторым комплементарным доменом, проксимальным доменом или хвостовым доменом.
Гидовая нуклеиновая кислота может включать два или более доменов, которые могут представлять собой одинаковые доменные повторы, или разные домены.
Гидовая нуклеиновая кислота может иметь одну непрерывную последовательность нуклеиновой кислоты.
Например, одна непрерывная последовательность нуклеиновой кислоты может быть (N)m, где N представляет собой A, T, C или G или A, U, C или G, и m представляет собой целое число 1-150.
Гидовая нуклеиновая кислота может иметь две или более непрерывных последовательностей нуклеиновых кислот.
Например, две или более непрерывных последовательностей нуклеиновых кислот могут быть (N)m и (N)o, где N представляет собой A, T, C или G, или A, U, C или G, m и o представляют собой целое число 1-150, и m и o могут быть одинаковыми или отличаться друг от друга.
Термин «белок-редактор» относится к пептиду, полипептиду или белку, который способен напрямую связываться или взаимодействовать с нуклеиновой кислотой без прямого связывания.
Белок-редактор может представлять собой фермент.
Белок-редактор может представлять собой гибридный белок.
В настоящем документе «гибридный белок» относится к белку, который продуцируется с помощью слияния фермента с дополнительным доменом, пептидом, полипептидом или белком.
Термин «фермент» относится к белку, который содержит домен, способный расщеплять нуклеиновую кислоту, ген, хромосому или белок.
Дополнительный домен, пептид, полипептид или белок может представлять собой функциональный домен, пептид, полипептид или белок, который имеет такую же функцию или отличную от фермента.
Гибридный белок может включать дополнительный домен, пептид, полипептид или белок в одном или более участках аминоконца (N-конца) фермента или его пределах; карбоксильного конца (С-конца) или его пределах; средней части фермента; и их комбинацию.
Гибридный белок может включать функциональный домен, пептид, полипептид или белок в одном или более участках N-конца фермента или его пределах; С-конца или его пределах; средней части фермента; и их комбинацию.
Гидовая нуклеиновая кислота-белок-редактор может служить для модификации объекта.
Объектом может быть мишень в виде нуклеиновой кислоты, гена, хромосомы или белка.
Например, комплекс гидовая нуклеиновая кислота-белок-редактор может привести к конечной регуляции (например, ингибированию, подавлению, уменьшению, увеличению или стимулированию) экспрессии представляющего интерес белка, удалению белка или экспрессии нового белка.
В настоящем документе комплекс гидовая нуклеиновая кислота-белок-редактор может действовать на уровне ДНК, РНК, гена или хромосомы.
Комплекс гидовая нуклеиновая кислота-белок-редактор может действовать на стадиях транскрипции и трансляции генов.
Комплекс гидовая нуклеиновая кислота-белок-редактор может действовать на уровне белка.
1.Гидовые нуклеиновые кислоты
Гидовая нуклеиновая кислота представляет собой нуклеиновую кислоту, которая способна распознавать мишень в виде нуклеиновой кислоты, гена, хромосомы или белка и образует комплекс гидовая нуклеиновая кислота-белок.
В настоящем документе гидовая нуклеиновая кислота выполнена с возможностью распознавания или нацеливания на нуклеиновую кислоту, ген, хромосому или белок, на которые направлен комплекс гидовая нуклеиновая кислота-белок.
Гидовая нуклеиновая кислота может присутствовать в форме ДНК, РНК или смеси ДНК/РНК и иметь последовательность нуклеиновой кислоты от 5 до 150.
Гидовая нуклеиновая кислота может присутствовать в линейной или кольцевой форме.
Гидовая нуклеиновая кислота может представлять собой одну непрерывную последовательность нуклеиновой кислоты.
Например, одна непрерывная последовательность нуклеиновой кислоты может быть (N)m, где N представляет собой A, T, C или G или A, U, C или G, а m представляет собой целое число 1-150.
Гидовая нуклеиновая кислота может представлять собой две или более непрерывных последовательностей нуклеиновых кислот.
Например, две или более непрерывных последовательностей нуклеиновых кислот могут быть (N)m и (N)o, где N представляет собой A, T, C или G, или A, U, C или G, m и o представляют собой целое число 1-150, и могут быть одинаковыми или отличаться друг от друга.
Гидовая нуклеиновая кислота может включать один или более доменов.
В настоящем документе доменами могут быть, но без ограничения, гидовый домен, первый комплементарный домен, линкерный домен, второй комплементарный домен, проксимальный домен или хвостовой домен.
Гидовая нуклеиновая кислота может включать два или более доменов, которые могут представлять собой одинаковые доменные повторы или разные домены.
Домены будут описаны ниже.
i) Гидовый домен
Термин «гидовый домен» представляет собой домен, имеющий комплементарную гидовую последовательность, которая способна образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени и служит для специфического взаимодействия с геном-мишенью или нуклеиновой кислотой-мишенью.
Гидовая последовательность представляет собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени в гене-мишени или нуклеиновой кислоты-мишени, которая имеет, например, по меньшей мере 50% или более, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% комплементарность или полную комплементарность.
Гидовый домен может представлять собой последовательность из 5-50 оснований.
В одном примере, гидовый домен может представлять собой последовательность из 5-50, от 10 до 50, от 15 до 50, от 20 до 50, от 25 до 50, 35-50, от 35 до 50, от 40 до 50 или 45-50 оснований.
В другом примере гидовым доменом может быть последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 оснований.
Гидовый домен может иметь гидовую последовательность.
Гидовая последовательность может представлять собой комплементарную последовательность оснований, которая способна образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени.
Гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени в гене-мишени или нуклеиновой кислоте-мишени, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарности или полную комплементарность.
Гидовая последовательность может быть последовательностью из 5-50 оснований.
В одном примере, гидовым доменом может быть последовательность из 5-50, 10-50, 15-50, 20-50, 25-50, 30-50, 35-50, 40-50 или 45-50 оснований.
В другом примере гидовая последовательность может быть последовательностью из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 оснований.
Кроме того, гидовый домен может включать в себя гидовую последовательность и дополнительную последовательность оснований.
Дополнительная последовательность оснований может быть использована для улучшения или ухудшения функции гидового домена.
Дополнительная последовательность оснований может быть использована для улучшения или ухудшения функции гидовой последовательности.
Дополнительная последовательность оснований может представлять собой последовательность из 1-35 оснований.
В одном примере дополнительная последовательность оснований может представлять собой последовательность из 5-35, от 10 до 35, от 15 до 35, от 20 до 35, от 25 до 35 или 30-35 оснований.
В другом примере дополнительная последовательность оснований может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30 или 30-35 оснований.
Дополнительная последовательность оснований может быть расположена на 5'-конце гидовой последовательности.
Дополнительная последовательность оснований может быть расположена на 3'-конце гидовой последовательности.
ii) Первый комплементарный домен
Термин «первый комплементарный домен» представляет собой последовательность нуклеиновой кислоты, включающую последовательность нуклеиновой кислоты, комплементарную второму комплементарному домену, и обладает достаточной комплементарностью так, чтобы образовывать двойную цепь со вторым комплементарным доменом.
Первый комплементарный домен может представлять собой последовательность из 5-35 оснований.
В одном примере первый комплементарный домен может представлять собой последовательность из 5-35, от 10 до 35, от 15 до 35, от 20 до 35, от 25 до 35 или 30-35 оснований.
В другом примере первый комплементарный домен может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30 или 30-35 оснований.
iii) Линкерный домен
Термин «линкерный домен» представляет собой последовательность нуклеиновой кислоты, соединяющую два или более доменов, которые представляют собой два или более идентичных или разных доменов. Линкерный домен может быть соединен с двумя или более доменами с помощью ковалентной связи или нековалентной связи или может соединять два или более домена с помощью ковалентной связи или нековалентной связи.
Линкерный домен может представлять собой последовательность из 1-30 оснований.
В одном примере линкерный домен может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25 или 25-30 оснований.
В другом примере линкерный домен может представлять собой последовательность из 1-30, 5-30, 10-30, 15-30, 20-30 или 25-30 оснований.
iv) Второй комплементарный домен
Термин «второй комплементарный домен» представляет собой последовательность нуклеиновой кислоты, включающую последовательность нуклеиновой кислоты, комплементарную первому комплементарному домену, и обладает достаточной комплементарностью, чтобы образовывать двойную цепь с первым комплементарным доменом.
Второй комплементарный домен может иметь последовательность оснований, комплементарную первому комплементарному домену, и последовательность оснований, не имеющую комплементарности к первому комплементарному домену, например, последовательность оснований, не образующую двойную цепь с первым комплементарным доменом, и может иметь более длинную последовательность оснований, чем у первого комплементарного домена.
Второй комплементарный домен может иметь последовательность из 5-35 оснований.
В одном примере второй комплементарный домен может представлять собой последовательность из 1-35, от 5 до 35, от 10 до 35, от 15 до 35, от 20 до 35, от 25 до 35 или 30-35 оснований.
В другом примере второй комплементарный домен может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30 или 30-35 оснований.
v) Проксимальный домен
Термин «проксимальный домен» представляет собой последовательность нуклеиновой кислоты, расположенную рядом со вторым комплементарным доменом.
Проксимальный домен может иметь в нем комплементарную последовательность оснований и может быть образован в виде двойной цепи из-за комплементарной последовательности оснований.
Проксимальный домен может представлять собой последовательность из 1-20 оснований.
В одном примере проксимальный домен может представлять собой последовательность из 1-20, 5-20, 10-20 или 15-20 оснований.
В другом примере проксимальный домен может представлять собой последовательность из 1-5, 5-10, 10-15 или 15-20 оснований.
vi) Хвостовой домен
Термин «хвостовой домен» представляет собой последовательность нуклеиновой кислоты, расположенную на одном или более концах обоих концов гидовой нуклеиновой кислоты.
Хвостовой домен может иметь в себе комплементарную последовательность оснований и может быть образован в виде двойной цепи из-за комплементарной последовательности оснований.
Хвостовой домен может представлять собой последовательность из 1-50 оснований.
В одном примере, хвостовой домен может представлять собой последовательность из 5-50, 10-50, 15-50, 20-50, 25-50, 30-50, 35-50, 40-50 или 45-50 оснований.
В другом примере хвостовой домен может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 оснований.
Между тем, часть или все последовательности нуклеиновой кислоты, включенные в домены, то есть гидовый домен, первый комплементарный домен, линкерный домен, второй комплементарный домен, проксимальный домен и хвостовой домен, могут выборочно или дополнительно включать химическую модификацию.
Химическая модификация может представлять собой, но без ограничения, метилирование, ацетилирование, фосфорилирование, фосфоротиоатную связь, закрытую нуклеиновую кислоту (LNA), 2'-O-метил-3'-фосфоротиоат (MS) или 2'-O-метил-3'-тиоPACE (MSP).
Гидовая нуклеиновая кислота включает один или более доменов.
Гидовая нуклеиновая кислота может включать гидовый домен.
Гидовая нуклеиновая кислота может включать первый комплементарный домен.
Гидовая нуклеиновая кислота может включать линкерный домен.
Гидовая нуклеиновая кислота может включать второй комплементарный домен.
Гидовая нуклеиновая кислота может включать проксимальный домен.
Гидовая нуклеиновая кислота может включать хвостовой домен.
В настоящем документе может быть 1, 2, 3, 4, 5, 6 или более доменов.
Гидовая нуклеиновая кислота может включать 1, 2, 3, 4, 5, 6 или более гидовых доменов.
Гидовая нуклеиновая кислота может включать 1, 2, 3, 4, 5, 6 или более первых комплементарных доменов.
Гидовая нуклеиновая кислота может включать 1, 2, 3, 4, 5, 6 или более линкерных доменов.
Гидовая нуклеиновая кислота может включать 1, 2, 3, 4, 5, 6 или более вторых комплементарных доменов.
Гидовая нуклеиновая кислота может включать 1, 2, 3, 4, 5, 6 или более проксимальных доменов.
Гидовая нуклеиновая кислота может включать 1, 2, 3, 4, 5, 6 или более хвостовых доменов.
В настоящем документе в гидовой нуклеиновой кислоте, один тип домена может быть дуплицирован.
Гидовая нуклеиновая кислота может включать несколько доменов с дупликацией или без нее.
Гидовая нуклеиновая кислота может включать один и тот же тип домена. В настоящем документе один и тот же тип домена может иметь одну и ту же последовательность нуклеиновой кислоты или разные последовательности нуклеиновой кислоты.
Гидовая нуклеиновая кислота может включать два типа доменов. В настоящем документе два разных типа доменов могут иметь разные последовательности нуклеиновой кислоты или одну и ту же последовательность нуклеиновой кислоты.
Гидовая нуклеиновая кислота может включать три типа доменов. В настоящем документе три разных типа доменов могут иметь разные последовательности нуклеиновой кислоты или одну и ту же последовательность нуклеиновой кислоты.
Гидовая нуклеиновая кислота может включать четыре типа доменов. В настоящем документе четыре разных типа доменов могут иметь разные последовательности нуклеиновой кислоты или одну и ту же последовательность нуклеиновой кислоты.
Гидовая нуклеиновая кислота может включать пять типов доменов. В настоящем документе пять разных типов доменов могут иметь разные последовательности нуклеиновой кислоты или одну и ту же последовательность нуклеиновой кислоты.
Гидовая нуклеиновая кислота может включать шесть типов доменов. В настоящем документе шесть разных типов доменов могут иметь разные последовательности нуклеиновой кислоты или одну и ту же последовательность нуклеиновой кислоты.
Например, гидовая нуклеиновая кислота может состоять из: [гидовый домен]-[первый комплементарный домен]-[линкерный домен]-[второй комплементарный домен]-[линкерный домен]-[гидовый домен]-[первый комплементарный домен]-[линкерный домен]-[второй комплементарный домен]. В настоящем документе два гидовых домена могут включать гидовые последовательности для разных или одинаковых мишеней, два первых комплементарных домена и два вторых комплементарных домена могут иметь одинаковые или разные последовательности нуклеиновой кислоты. Когда гидовые домены включают в себя гидовые последовательности для разных мишеней, гидовые нуклеиновые кислоты могут специфически связываться с двумя разными мишенями, и в настоящем документе специфические связи могут быть выполнены одновременно или последовательно. Кроме того, линкерные домены могут быть расщеплены с помощью специфических ферментов, и гидовые нуклеиновые кислоты могут быть разделены на две или три при наличии специфических ферментов.
В качестве конкретного примера гидовой нуклеиновой кислоты настоящего изобретения, ниже будет описана гРНК.
гРНК
Термин «гРНК» относится к нуклеиновой кислоте, способной специфически нацеливаться на ферментный комплекс гРНК-CRISPR, то есть комплекс CRISPR, по отношению к гену-мишени или нуклеиновой кислоте-мишени. Кроме того, гРНК представляет собой РНК, специфичную по отношению к нуклеиновой кислоте, которая может связываться с ферментом CRISPR и направлять фермент CRISPR к гену-мишени или нуклеиновой кислоте-мишени.
гРНК может включать несколько доменов. Из-за каждого домена взаимодействия могут происходить в трехмерной структуре или активной форме цепи гРНК или между этими цепями.
гРНК может называться одноцепочечная гРНК (одна молекула РНК); или двухцепочечная гРНК (содержащая более одной, как правило, две отдельные молекулы РНК).
В одном примере варианта осуществления одноцепочечная гРНК может включать гидовый домен, то есть домен, включающий гидовую последовательность, способную образовывать комплементарную связь с геном-мишенью или нуклеиновой кислотой-мишенью; первый комплементарный домен; линкерный домен; второй комплементарный домен, то есть домен, имеющий последовательность, комплементарную последовательности первого комплементарного домена, тем самым образуя двухцепочечную нуклеиновую кислоту с первым комплементарным доменом; проксимальный домен; и, необязательно, хвостовой домен в направлении от 5' к 3'.
В другом варианте осуществления двухцепочечная гРНК может включать в себя первую цепь, которая включает в себя гидовый домен, то есть домен, включающий гидовую последовательность, способную образовывать комплементарную связь с геном-мишенью или нуклеиновой кислотой-мишенью и первым комплементарным доменом; и вторую цепь, которая включает второй комплементарный домен, то есть домен, имеющий последовательность, комплементарную последовательности первого комплементарного домена, тем самым образуя двухцепочечную нуклеиновую кислоту с первым комплементарным доменом, проксимальный домен; и, необязательно, хвостовой домен в направлении от 5' к 3'.
В настоящем документе первая цепь может быть обозначена как crРНК, а вторая цепь может быть обозначена как tracrРНК. crРНК может включать в себя гидовый домен и первый комплементарный домен, а tracrРНК может включать второй комплементарный домен, проксимальный домен и, необязательно, хвостовой домен.
В еще одном варианте осуществления одноцепочечная гРНК может включать гидовый домен, то есть домен, включающий гидовую последовательность, способную образовывать комплементарную связь с геном-мишенью или нуклеиновой кислотой-мишенью; первый комплементарный домен; второй комплементарный домен и домен, имеющий последовательность, комплементарную последовательности первого комплементарного домена, тем самым образуя двухцепочечную нуклеиновую кислоту с первым комплементарным доменом в направлении от 5' к 3'.
i) Гидовый домен
Гидовый домен включает комплементарную гидовую последовательность, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени. Гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, имеющую комплементарность к последовательности-мишени в гене-мишени или нуклеиновой кислоте-мишени, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарности или полную комплементарность. Считается, что гидовый домен обеспечивает специфическое взаимодействие комплекса гРНК-Cas, то есть комплекса CRISPR, с геном-мишенью или нуклеиновой кислотой-мишенью.
Гидовый домен может представлять собой последовательность из 5-50 оснований.
В качестве примера варианта осуществления гидовый домен может представлять собой последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В качестве примерного варианта осуществления, гидовый домен может включать в себя последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В настоящем документе гидовый домен может включать в себя гидовую последовательность.
Гидовая последовательность может представлять собой комплементарную последовательность оснований, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени.
Гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени в гене-мишени или нуклеиновой кислоте-мишени, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарности или полную комплементарность.
В одном примере варианта осуществления гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную гену-мишени, то есть последовательности-мишени гена PMP22, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85% 90% или 95% или более комплементарности или полную комплементарность.
Гидовая последовательность может быть последовательностью из 5-50 оснований.
В примере варианта осуществления гидовая последовательность может представлять собой последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В одном примере варианта осуществления гидовая последовательность может включать в себя последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В одном примере варианта осуществления гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени гена PMP22, которая представляет собой последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В настоящем документе последовательности-мишени генов-мишеней, то есть гена PMP22 для гидовой последовательности, перечислены выше в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7, соответственно, но настоящее изобретение этим не ограничено.
В настоящем документе гидовый домен может включать в себя гидовую последовательность и дополнительную последовательность оснований.
Дополнительная последовательность оснований может представлять собой последовательность из 1-35 оснований.
В одном примере варианта осуществления дополнительная последовательность оснований может быть последовательностью из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 оснований.
Например, дополнительная последовательность оснований может представлять собой последовательность одного основания, гуанина (G) или последовательность двух оснований, GG.
Дополнительная последовательность оснований может быть расположена на 5'-конце гидовой последовательности.
Дополнительная последовательность оснований может быть расположена на 3'-конце гидовой последовательности.
Выборочно, часть или вся последовательность оснований гидового домена может включать химическую модификацию. Химическая модификация может быть метилированием, ацетилированием, фосфорилированием, фосфоротиоатной связью, закрытой нуклеиновой кислотой (LNA), 2'-O-метил-3'-фосфоротиоатом (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
ii) Первый комплементарный домен
Первый комплементарный домен включает последовательность нуклеиновой кислоты, комплементарную второму комплементарному домену, и обладает достаточной комплементарностью, так что он способен образовывать двойную цепь со вторым комплементарным доменом.
В настоящем документе первый комплементарный домен может представлять собой последовательность из 5-35 оснований. Первый комплементарный домен может включать последовательность из 5-35 оснований.
В одном примере варианта осуществления первый комплементарный домен может представлять собой последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В другом варианте осуществления первый комплементарный домен может включать в себя последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Первый комплементарный домен может иметь гомологию с естественным первым комплементарным доменом или может быть получен из естественного первого комплементарного домена. Кроме того, первый комплементарный домен может иметь различие в последовательности оснований первого комплементарного домена в зависимости от существующих в природе видов, может быть получен из первого комплементарного домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию с первым комплементарным доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления первый комплементарный домен может иметь частичную, то есть, по меньшей мере 50%, или более или полную гомологию с первым комплементарным доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides или первым комплементарным доменом, полученным из них.
Например, когда первый комплементарный домен представляет собой первый комплементарный домен из Streptococcus pyogenes или первый комплементарный домен, полученный из него, первый комплементарный домен может представлять собой 5'-GUUUUAGAGCUA-3' или последовательность оснований, имеющую частичную, то есть по меньшей мере 50% или более, или полную гомологию с 5'-GUUUUAGAGCUA-3'. В настоящем документе первый комплементарный домен может дополнительно включать в себя (X)n, что приводит к 5'-GUUUUAGAGCUA (X)n-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом от 5 до 15. В настоящем документе (X)n может быть n повторами одного и того же основания или смесью из n оснований A, T, U и G.
В другом варианте осуществления, когда первый комплементарный домен представляет собой первый комплементарный домен из Campylobacter jejuni или первый комплементарный домен, полученный из него, первый комплементарный домен может представлять собой 5'-GUUUUAGUCCCUUUUUUAAUUUCUU-3' или последовательность оснований, имеющую частичную, то есть по меньшей мере 50% или более, или полную гомологию с 5'-GUUUUAGUCCCUUUUUAAAUUUCUU-3'. В настоящем документе первый комплементарный домен может дополнительно включать в себя (X)n, что приводит к 5'-GUUUUAGUCCCUUUUUAAAUUUCUU (X)n-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом от 5 до 15. В настоящем документе (X)n может представлять n повторов одного и то же основания или смесь из n оснований A, T, U и G.
В другом варианте осуществления первый комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию с первым комплементарным доменом из Parcubacteria bacterium (GWC2011_GWC2_44_17), Lachnospiraceae bacterium (MC2017), Butyrivibrio proteoclasiicus, Peregrinibacteria bacterium (GW2011_GWA_33_10), Acidaminococcus sp. (BV3L6), Porphyromonas macacae, Lachnospiraceae bacterium (ND2006), Porphyromonas crevioricanis, Prevotella disiens, Moraxella bovoculi (237), Smiihella sp. (SC_KO8D17), Leptospira inadai, Lachnospiraceae bacterium (MA2020), Francisella novicida (U112), Candidatus Methanoplasma termitum или Eubacterium eligens, или первым комплементарным доменом, полученным из них.
Например, когда первый комплементарный домен представляет собой первый комплементарный домен из Parcubacteria bacterium или первый комплементарный домен, полученный из нее, первый комплементарный домен может представлять собой 5'-UUUGUAGAU-3' или последовательность оснований, имеющую частичную, то есть, по меньшей мере 50% или более гомологию с 5'-UUUGUAGAU-3'. В настоящем документе первый комплементарный домен может дополнительно включать в себя (X)n, что приводит к 5'-(X) nUUUGUAGAU-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом 1-5. В настоящем документе (X)n может представлять n повторов одного и того же основания или смесь из n оснований A, T, U и G.
Выборочно, часть или вся последовательность оснований первого комплементарного домена может иметь химическую модификацию. Химическая модификация может быть метилированием, ацетилированием, фосфорилированием, фосфоротиоатной связью, закрытой нуклеиновой кислотой (LNA), 2'-O-метил 3'-фосфоротиоатом (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
iii) Линкерный домен
Линкерный домен представляет собой последовательность нуклеиновой кислоты, соединяющую два или более доменов, и соединяющую два или более идентичных или разных доменов. Линкерный домен может быть соединен с двумя или более доменами, или может соединять два или более доменов с помощью ковалентной или нековалентной связи.
Линкерный домен может представлять собой последовательность нуклеиновой кислоты, соединяющую первый комплементарный домен со вторым комплементарным доменом для продуцирования одноцепочечной гРНК.
Линкерный домен может быть соединен с первым комплементарным доменом и вторым комплементарным доменом с помощью ковалентной или нековалентной связи.
Линкерный домен может соединять первый комплементарный домен со вторым комплементарным доменом с помощью ковалентной или нековалентной связи.
Линкерный домен может представлять собой последовательность из 1-30 оснований. Линкерный домен может включать последовательность из 1-30 оснований.
В примере варианта осуществления линкерный домен может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25 или 25-30 оснований.
В примере варианта осуществления линкерный домен может включать последовательность из 1-5, 5-10, 10-15, 15-20, 20-25 или 25-30 оснований.
Линкерный домен подходит для использования в одноцепочечной молекуле гРНК и может быть использован для продуцирования одноцепочечной гРНК с помощью соединения с первой цепью и второй цепью двухцепочечной гРНК или соединения первой цепи со второй цепью с помощью ковалентной или нековалентной связи. Линкерный домен может быть использован для продуцирования одноцепочечной гРНК с помощью соединения с crРНК и tracrРНК двухцепочечной гРНК или соединения crРНК с tracrРНК с помощью ковалентной или нековалентной связи.
Линкерный домен может иметь гомологию с естественной последовательностью, например частичной последовательностью tracrРНК, или может быть получен из нее.
Выборочно, часть или вся последовательность оснований линкерного домена может иметь химическую модификацию. Химическая модификация может быть метилированием, ацетилированием, фосфорилированием, фосфоротиоатной связью, закрытой нуклеиновой кислотой (LNA), 2'-O-метил-3'-фосфоротиоатом (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
iv) Второй комплементарный домен
Второй комплементарный домен включает последовательность нуклеиновой кислоты, комплементарную первому комплементарному домену, и обладает достаточной комплементарностью так, чтобы образовывать двойную цепь с первым комплементарным доменом. Второй комплементарный домен может включать в себя последовательность оснований, комплементарную первому комплементарному домену, и последовательность оснований, не имеющую комплементарности с первым комплементарным доменом, например, последовательность оснований, не образующую двойную цепь с первым комплементарным доменом, и может иметь более длинную последовательность оснований, чем у первого комплементарного домена.
В настоящем документе второй комплементарный домен может представлять собой последовательность из 5-35 оснований. Первый комплементарный домен может включать последовательность из 5-35 оснований.
В примере варианта осуществления второй комплементарный домен может представлять собой последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В примере варианта осуществления второй комплементарный домен может представлять собой последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Кроме того, второй комплементарный домен может иметь гомологию с естественным вторым комплементарным доменом или может быть получен из естественного второго комплементарного домена. Кроме того, второй комплементарный домен может иметь различие в последовательности оснований второго комплементарного домена в соответствии с видами, существующими в природе, и может быть получен из второго комплементарного домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию со вторым комплементарным доменом, содержащимся в существующих в природе видах.
В примере варианта осуществления второй комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию со вторым комплементарным доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides или вторым комплементарным доменом, полученным из них.
Например, когда второй комплементарный домен представляет собой второй комплементарный домен из Streptococcus pyogenes или второй комплементарный домен, полученный из нее, второй комплементарный домен может представлять собой 5'-UAGCAAGUUAAAAU-3' или последовательность оснований, имеющую частичную, то есть, по меньшей мере 50% или более гомологию с 5'-UAGCAAGUUAAAAU-3' (последовательность оснований, образующая двойную цепь с первым комплементарным доменом, подчеркнута). В настоящем документе второй комплементарный домен может дополнительно включать в себя (X)n и/или (X)m, что приводит к 5'-(X)nUAGCAAGUUAAAAU(X)m-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а каждый из n и m может представлять количество оснований, в которых n может быть целым числом от 1 до 15, и m может быть целым числом от 1 до 6. В настоящем документе (X)n может представлять n повторов одного и того же основания или смесь n оснований A, T, U и G. Кроме того, (X)m может представлять m повторов одного и того же основания или смесь m оснований A, T, U и G.
В другом примере, когда второй комплементарный домен является вторым комплементарным доменом из Campylobacter jejuni или вторым комплементарным доменом, полученным из него, второй комплементарный домен может представлять собой 5'-AAGAAAUUUAAAAAGGGACUAAAAU-3' или последовательность оснований, имеющую частичную, то есть, не менее 50% или более гомологию с 5'-AAGAAAUUUAAAAAGGGACUAAAAU-3' (последовательность оснований, образующая двойную цепь с первым комплементарным доменом, подчеркнута). В настоящем документе второй комплементарный домен может дополнительно включать (X)n и/или (X)m, что приводит к 5'-(X)nAAGAAAUUUAAAAAGGGACUAAAAU(X)m-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а каждый из n и m может представлять количество оснований, в которых n может быть целым числом от 1 до 15, и m может быть целым числом от 1 до 6. В настоящем документе (X)n может представлять собой n повторов одного и того же основания или смесь n оснований A, T, U и G. Кроме того, (X)m может представлять m повторов того же самого основания или смесь m оснований A, T, U и G.
В другом варианте осуществления второй комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более или полную гомологию с первым комплементарным доменом из Parcubacteria bacterium (GWC2011_GWC2_44_17), Lachnospiraceae bacterium (MC2017), Butyrivibrio proteoclasiicus, Peregrinibacteria bacterium (GW2011_GWA_33_10), Acidaminococcus sp. (BV3L6), Porphyromonas macacae, Lachnospiraceae bacterium (ND2006), Porphyromonas crevioricanis, Prevotella disiens, Moraxella bovoculi (237), Smiihella sp. (SC_KO8D17), Leptospira inadai, Lachnospiraceae bacterium (MA2020), Francisella novicida (U112), Candidatus Methanoplasma termitum или Eubacterium eligens или вторым комплементарным доменом, полученным из них.
Например, когда второй комплементарный домен представляет собой второй комплементарный домен из бактерии Parcubacteria bacterium или второй комплементарный домен, полученный из нее, второй комплементарный домен может представлять собой 5'-AAAUUUCUACU-3' или последовательность оснований, имеющую частичную, то есть, по меньшей мере 50% или более гомологию с 5'-AAAUUUCUACU-3' (последовательность оснований, образующая двойную цепь с первым комплементарным доменом, подчеркнута). В настоящем документе второй комплементарный домен может дополнительно включать в себя (X)n и/или (X)m, что приводит к 5'-(XnAAAUUUCUACU(X)m-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а каждый из n и m может представлять количество оснований, в которых n может быть целым числом 1-10, и m может быть целым числом от 1 до 6. В настоящем документе (X)n может представлять n повторов одного и того же основания или смесь n оснований A, T, U и G. Кроме того, (X)m может представлять собой m повторов одного и то же основания или смесь m оснований A, T, U и G.
Выборочно, часть или вся последовательность оснований второго комплементарного домена может иметь химическую модификацию. Химическая модификация может быть метилированием, ацетилированием, фосфорилированием, фосфоротиоатной связью, закрытой нуклеиновой кислотой (LNA), 2'-O-метил-3'-фосфоротиоатом (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
v) Проксимальный домен
Проксимальный домен представляет собой последовательность из 1-20 оснований, расположенную рядом со вторым комплементарным доменом, и домен, расположенный в направлении 3'-конца второго комплементарном домена. В настоящем документе проксимальный домен может быть использован для образования двойной цепи между комплементарными последовательностями оснований в нем.
В одном примере варианта осуществления проксимальный домен может представлять собой последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 оснований.
В другом варианте осуществления проксимальный домен может представлять собой последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 оснований.
Кроме того, проксимальный домен может иметь гомологию с естественным проксимальным доменом или может быть получен из естественного проксимального домена. Кроме того, проксимальный домен может иметь различие в последовательности оснований в соответствии с видами, существующими в природе, может быть получен из проксимального домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию с проксимальным доменом, содержащимся в существующих в природе видах.
В примере варианта осуществления проксимальный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию с проксимальным доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides или проксимальным доменом, полученным из них.
Например, когда проксимальный домен представляет собой проксимальный домен из Streptococcus pyogenes или проксимальный домен, полученный из него, проксимальный домен может представлять собой 5'-AAGGCUAGUCCG-3' или последовательность оснований, имеющую частичную, то есть, по меньшей мере 50% или более гомологию с 5'-AAGGCUAGUCCG-3'. В настоящем документе проксимальный домен может дополнительно включать (X)n, что приводит к 5'-AAGGCUAGUCCG(X)n-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом от 1 до 15. Здесь (X)n может представлять n повторов одного и того же основания или смесь из n оснований A, T, U и G.
В еще одном примере, когда проксимальный домен представляет собой проксимальный домен из Campylobacter jejuni или проксимальный домен, полученный из него, проксимальный домен может представлять собой 5'-AAAGAGUUUGC-3' или последовательность оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAAGAGUUUGC-3'. В настоящем документе проксимальный домен может дополнительно включать (X)n, что приводит к 5'-AAAGAGUUUGC(X)n-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом 1-40. Здесь (X)n может представлять n повторов одного и того же основания или смесь из n оснований A, T, U и G.
Выборочно, часть или вся последовательность оснований проксимального домена может иметь химическую модификацию. Химическая модификация может представлять собой метилирование, ацетилирование, фосфорилирование, фосфоротиоатную связь, закрытую нуклеиновую кислоту (LNA), 2'-O-метил-3'-фосфоротиоат (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
vi) Хвостовой домен
Хвостовой домен представляет собой домен, который может быть выборочно добавлен к 3'-концу одноцепочечной гРНК или двухцепочечной гРНК. Хвостовой домен может представлять собой последовательность из 1-50 оснований или включать последовательность из 1-50 оснований. В настоящем документе хвостовой домен может быть использован для образования двойной цепи между комплементарными последовательностями оснований в нем.
В примере варианта осуществления хвостовой домен может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 оснований.
В примере варианта осуществления хвостовой домен может включать в себя последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 оснований.
Кроме того, хвостовой домен может иметь гомологию с естественным хвостовым доменом или может быть получен из естественного хвостового домена. Кроме того, хвостовой домен может иметь различие в последовательности оснований в зависимости от существующих в природе видов, может быть получен из хвостового домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию с хвостовым доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления хвостовой домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию с хвостовым доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides или хвостовым доменом, полученным из них.
Например, когда хвостовой домен представляет собой хвостовой домен из Streptococcus pyogenes или хвостовой домен, полученный из него, хвостовой домен может представлять собой 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC-3' или последовательность оснований, имеющую частичную, то есть, по меньшей мере 50% или более гомологию с 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC-3'. В настоящем документе хвостовой домен может дополнительно включать (X)n, что приводит к 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC(X)n-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом от 1 до 15. В настоящем документе (X)n может представлять n повторов того же самого основания или смесь из n оснований, таких как A, T, U и G.
В другом примере, когда хвостовой домен представляет собой хвостовой домен из Campylobacter jejuni или хвостовой домен, полученный из него, хвостовой домен может представлять собой 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3' или последовательность оснований, имеющую частичную, то есть, по меньшей мере 50% или больше гомологию с 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3'. В настоящем документе хвостовой домен может дополнительно включать в себя (X)n, что приводит к 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU(X)n-3'. X может быть выбран из группы, состоящей из оснований A, T, U и G, а n может представлять количество оснований, которое является целым числом от 1 до 15. Здесь (X)n может представлять n повторов одного и того же основания или смесь из n оснований A, T, U и G.
В другом варианте осуществления хвостовой домен может включать последовательность из 1-10 оснований на 3'-конце, вовлеченных в способ транскрипции in vitro или in vivo.
Например, когда промотор T7 используют при транскрипции in vitro гРНК, хвостовой домен может представлять собой произвольную последовательность оснований, присутствующую на 3'-конце матрицы ДНК. Кроме того, когда промотор U6 используют в транскрипции in vivo, хвостовой домен может быть UUUUUU, когда промотор H1 используют в транскрипции, хвостовой домен может быть UUUU, а когда используют промотор pol-III, хвостовой домен может включать несколько оснований урацила или альтернативных оснований.
Выборочно, часть или вся последовательность оснований хвостового домена может иметь химическую модификацию. Химическая модификация может представлять собой метилирование, ацетилирование, фосфорилирование, фосфоротиоатную связь, закрытую нуклеиновую кислоту (LNA), 2'-O-метил-3'-фосфоротиоат (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
гРНК может включать множество доменов, как описано выше, и, следовательно, длина последовательности нуклеиновой кислоты может быть отрегулирована в соответствии с доменом, содержащимся в гРНК, и взаимодействия могут происходить в цепях в трехмерной структуре или в активной форме гРНК или между этими цепями из-за каждого домена.
гРНК может быть обозначена как одноцепочечная гРНК (одна молекула РНК); или двухцепочечная гРНК (включая более чем одну, обычно две отдельные молекулы РНК).
Двухцепочечная гРНК
Двухцепочечная гРНК состоит из первой цепи и второй цепи.
В настоящем документе первая цепь может состоять из:
5'-[гидовый домен]-[первый комплементарный домен]-3', и
вторая цепь может состоять из:
5'-[второй комплементарный домен]-[проксимальный домен]-3' или
5'-[второй комплементарный домен]-[проксимальный домен]-[хвостовой домен]-3'.
В настоящем документе первая цепь может быть обозначена как crРНК, а вторая цепь может быть обозначен как tracrРНК.
Первая цепь
Гидовый домен
В первой цепи гидовый домен включает комплементарную гидовую последовательность, которая способна образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени. Гидовая последовательность представляет собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени в гене-мишени или нуклеиновой кислоте-мишени, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарности или полную комплементарность. Считается, что гидовый домен обеспечивает специфическое взаимодействие комплекса гРНК-Cas, то есть комплекса CRISPR, с геном-мишенью или нуклеиновой кислотой-мишенью.
В настоящем документе гидовый домен может представлять собой последовательность из 5-50 оснований или включать последовательность из 5-50 оснований. Например, гидовый домен может быть или включать последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Кроме того, гидовый домен может включать в себя гидовую последовательность.
В настоящем документе гидовая последовательность может представлять собой комплементарную последовательность оснований, которая способна образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85% 90% или 95% или более комплементарности или полную комплементарность.
В примере варианта осуществления гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную гену-мишени, то есть последовательности-мишени гена PMP22, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85% 90% или 95% или более комплементарности или полную комплементарность.
В настоящем документе гидовая последовательность может быть последовательностью из 5-50 оснований или включать последовательность из 5-50 оснований. Например, гидовая последовательность может быть или включать последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В одном примере варианта осуществления гидовая последовательность представляет собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени гена PMP22. Гидовая последовательность может быть или включать последовательность из 5-50 оснований. Например, гидовая последовательность может быть или включать последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В настоящем документе для гидовой последовательности, гены-мишени, то есть последовательности-мишени гена PMP22, перечислены выше в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7, но настоящее изобретение этим не ограничено.
Выборочно, гидовый домен может включать в себя гидовую последовательность и дополнительную последовательность оснований.
В настоящем документе дополнительная последовательность оснований может представлять собой последовательность из 1-35 оснований. Например, дополнительная последовательность оснований может быть последовательностью из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 оснований.
В одном примере варианта осуществления дополнительная последовательность оснований может включать одно основание, гуанина (G) или два основания, GG.
В настоящем документе дополнительная последовательность оснований может быть расположена на 5'-конце гидового домена или на 5'-конце гидовой последовательности.
Дополнительная последовательность оснований может быть расположена на 3'-конце гидового домена или на 3'-конце гидовой последовательности.
Первый комплементарный домен
Первый комплементарный домен включает последовательность нуклеиновой кислоты, комплементарную второму комплементарному домену второй цепи, и представляет собой домен, обладающий достаточной комплементарностью для образования двойной цепи со вторым комплементарным доменом.
В настоящем документе первый комплементарный домен может быть или включать последовательность из 5-35 оснований. Например, первый комплементарный домен может быть или включать последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Первый комплементарный домен может иметь гомологию с естественным первым комплементарным доменом или может быть получен из естественного первого комплементарного домена. Кроме того, первый комплементарный домен может иметь различие в последовательности оснований в соответствии с видами, существующими в природе, может быть получен из первого комплементарного домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию с первым комплементарным доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления первый комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию с первым комплементарным доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides, или первым комплементарным доменом, полученным из них.
Выборочно, первый комплементарный домен может включать в себя дополнительную последовательность оснований, которая не подвергается комплементарной связи со вторым комплементарным доменом второй цепи.
В настоящем документе дополнительная последовательность оснований может представлять собой последовательность из 1-15 оснований. Например, дополнительная последовательность оснований может представлять собой последовательность из 1-5, 5-10 или 10-15 оснований.
Выборочно, часть или вся последовательность оснований гидового домена и/или первого комплементарного домена может иметь химическую модификацию. Химическая модификация может быть метилированием, ацетилированием, фосфорилированием, фосфоротиоатной связью, закрытой нуклеиновой кислотой (LNA), 2'-O-метил-3'-фосфоротиоатом (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
Следовательно, первая цепь может состоять из: 5'-[гидовый домен]-[первый комплементарный домен]-3', как описано выше.
Кроме того, первая цепь может необязательно включать дополнительную последовательность оснований.
В одном примере первая цепь может быть
5'-(Nмишень)-(Q)m-3'; или
5'-(Х)a-(Nмишень)-(Х)b- (Q)m-(Х)c-3'.
В настоящем документе Nмишень представляет собой последовательность оснований, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени, и участком последовательности оснований, который может быть изменен в соответствии с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени.
В одном примере варианта осуществления Nмишень может представлять собой последовательность оснований, способную образовывать комплементарную связь с геном-мишенью, то есть последовательностью-мишенью гена PMP22.
В настоящем документе (Q)m является последовательностью оснований, включающей в себя первый комплементарный домен, который способен образовывать комплементарную связь со вторым комплементарным доменом второй цепи. (Q)m может быть последовательностью, имеющей частичную или полную гомологию с первым комплементарным доменом из видов, существующих в природе, и последовательность оснований первого комплементарного домена может быть изменена в соответствии с видами происхождения. Каждый Q может быть независимо выбран из группы, состоящей из A, U, C и G, и m может быть количеством оснований, которое является целым числом от 5 до 35.
Например, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Streptococcus pyogenes или первым комплементарным доменом, полученным из Streptococcus pyogenes, (Q)m может быть 5'-GUUUUAGAGCUA-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GUUUUAGAGCUA-3'.
В другом примере, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Campylobacter jejuni или первым комплементарным доменом, полученным из Campylobacter jejuni, (Q)m может быть 5'-GUUUUAGUCCCUUUUUAAAUUUCUU-3', или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GUUUUAGUCCCUUUUUAAAUUUCUU-3'.
В еще одном примере, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Streptococcus thermophilus или первым комплементарным доменом, полученным из Streptococcus thermophilus, (Q)m может быть 5'-GUUUUAGAGCUGUGUUGUUUCG-3', или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GUUUUAGAGCUGUGUUGUUUCG-3'.
Кроме того, каждый из (X)a, (X)b и (X)c является выборочно дополнительной последовательностью оснований, где каждый из X может быть независимо выбран из группы, состоящей из A, U, C и G, а каждый из a, b и c может быть количеством оснований, которое равно 0 или целому числу 1-20.
Вторая цепь
Вторая цепь может состоять из второго комплементарного домена и проксимального домена и выборочно включать хвостовой домен.
Второй комплементарный домен
Во второй цепи второй комплементарный домен включает последовательность нуклеиновой кислоты, комплементарную первому комплементарному домену первой цепи, и обладает достаточной комплементарностью, чтобы образовать двойную цепь с первым комплементарным доменом. Второй комплементарный домен может включать в себя последовательность оснований, комплементарную первому комплементарному домену, и последовательность оснований, не комплементарную первому комплементарному домену, например, последовательность оснований, не образующую двойную цепь с первым комплементарным доменом, и может иметь более длинную последовательность оснований, чем у первого комплементарного домена.
В настоящем документе второй комплементарный домен может представлять собой последовательность из 5-35 оснований или включать последовательность из 5-35 оснований. Например, второй комплементарный домен может быть или включать последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований, но настоящее изобретение этим не ограничено.
Второй комплементарный домен может иметь гомологию с естественным вторым комплементарным доменом или может быть получен из естественного второго комплементарного домена. Кроме того, второй комплементарный домен может иметь различие в его последовательности оснований в соответствии с видами, существующими в природе, может быть получен из второго комплементарного домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию со вторым комплементарным доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления второй комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию со вторым комплементарным доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides, или вторым комплементарным доменом, полученным из них.
Выборочно, второй комплементарный домен может дополнительно включать дополнительную последовательность оснований, которая не подвергается комплементарной связи с первым комплементарным доменом первой цепи.
В настоящем документе дополнительная последовательность оснований может представлять собой последовательность из 1-25 оснований. Например, дополнительная последовательность оснований может представлять собой последовательность из 1-5, 5-10, 10-15, 15-20 или 20-25 оснований.
Проксимальный домен
Во второй цепи проксимальный домен представляет собой последовательность из 1-20 оснований, и домен, расположенный в направлении 3'-конца второго комплементарного домена. Например, проксимальный домен может представлять собой или включать последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 оснований.
В настоящем документе проксимальный домен может иметь двухцепочечную связь между комплементарными последовательностями оснований в нем.
Кроме того, проксимальный домен может иметь гомологию с естественным проксимальным доменом или может быть получен из естественного проксимального домена. Кроме того, проксимальный домен может иметь различие в последовательности оснований в соответствии с видами, существующими в природе, может быть получен из проксимального домена из видов, существующих в природе, или может иметь частичную или полную гомологию с проксимальным доменом из видов, существующих в природе.
В одном примере варианта осуществления проксимальный домен может иметь частичную, то есть, по меньшей мере 50% или более или полную гомологию с проксимальным доменом Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides или проксимальным доменом, полученным из них.
Хвостовой домен
Выборочно, во второй цепи хвостовой домен может быть доменом, выборочно добавленным к 3'-концу второй цепи, а хвостовой домен может представлять собой или включать последовательность из 1-50 оснований. Например, хвостовой домен может быть или включать последовательность из 1-5, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 оснований.
В настоящем документе хвостовой домен может иметь двухцепочечную связь между комплементарными последовательностями оснований в нем.
Кроме того, хвостовой домен может иметь гомологию с естественным хвостовым доменом или может быть получен из естественного хвостового домена. Кроме того, хвостовой домен может иметь различие в последовательности оснований в соответствии с видами, существующими в природе, может быть получен из хвостового домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию с хвостовым доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления хвостовой домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию с хвостовым доменом из Streptococcus pyogenes, Campylobacter jejuni, Streptococcus thermophilus, Streptococcus aureus или Neisseria meningitides или хвостовым доменом, полученным из них.
В другом варианте осуществления хвостовой домен может включать последовательность из 1-10 оснований на 3'-конце, вовлеченных в способ транскрипции in vitro или in vivo.
Например, когда промотор T7 используют при транскрипции in vitro гРНК, хвостовой домен может представлять собой произвольную последовательность оснований, присутствующую на 3'-конце матрицы ДНК. Кроме того, когда промотор U6 используют в транскрипции in vivo, хвостовой домен может быть UUUUUU, когда промотор H1 используют в транскрипции, хвостовой домен может быть UUUU, а когда используют промотор pol-III, хвостовой домен может включать несколько оснований урацила или альтернативных оснований.
Выборочно, часть или все из каждой последовательности оснований второго комплементарного домена, проксимального домена и/или хвостового домена могут иметь химическую модификацию. Химическая модификация может представлять собой метилирование, ацетилирование, фосфорилирование, фосфоротиоатную связь, закрытую нуклеиновую кислоту (LNA), 2'-O-метил-3'-фосфоротиоат (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
Следовательно, вторая цепь может состоять из: 5'-[второй комплементарный домен]-[проксимальный домен]-3' или 5'-[второй комплементарный домен]-[проксимальный домен]-[хвостовой домен]-3', как описано выше.
Кроме того, вторая цепь может выборочно включать дополнительную последовательность оснований.
В одном примере варианта осуществления вторая цепь может быть 5'-(Z)h-(P)k-3'; или 5'-(X)d-(Z)h-(X)e-(P)k-(X)f-3'.
В другом варианте осуществления вторая цепь может быть 5'-(Z)h-(P)k-(F)i-3'; или 5'-(X)d-(Z)h-(X)e-(P)k-(X)f-(F)i-3'.
В настоящем документе (Z)h является последовательностью оснований, включающей второй комплементарный домен, который способен образовывать комплементарную связь с первым комплементарным доменом первой цепи. (Z)h может быть последовательностью, имеющей частичную или полную гомологию со вторым комплементарным доменом из видов, существующих в природе, и последовательность оснований второго комплементарного домена может быть модифицирована в соответствии с видами происхождения. Каждый из Z может быть независимо выбран из группы, состоящей из A, U, C и G, а h может быть количеством оснований, которое является целым числом от 5 до 50.
Например, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Streptococcus pyogenes или вторым комплементарным доменом, полученным из него, (Z)h может представлять собой 5'-UAGCAAGUUAAAAU-3' или последовательность оснований, имеющую по меньшей мере 50% или более гомологию с 5'-UAGCAAGUUAAAAU-3'.
В другом примере, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Campylobacter jejuni или вторым комплементарным доменом, полученным из него, (Z)h может быть 5'-AAGAAAUUUAAAAAGGGACUAAAAU-3', или последовательностью оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAGAAAUUUAAAAAGGGACUAAAAU-3'.
В еще одном примере, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Streptococcus thermophilus или вторым комплементарным доменом, полученным из него, (Z)h может быть 5'-CGAAACAACACAGCGAGUUAAAAU-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-CGAAACAACACAGCGAGUUAAAAU-3'.
(P)k представляет собой последовательность оснований, включающую в себя проксимальный домен, который может иметь частичную или полную гомологию с проксимальным доменом из видов, существующих в природе, и последовательность оснований проксимального домена может быть модифицирована в соответствии с видами происхождения. Каждый из P может быть независимо выбран из группы, состоящей из A, U, C и G, и k может быть количеством оснований, которое является целым числом от 1 до 20.
Например, когда проксимальный домен имеет частичную или полную гомологию с проксимальным доменом из Streptococcus pyogenes или проксимальным доменом, полученным из него, (P)k может представлять собой 5'-AAGGCUAGUCCG-3' или последовательность оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAGGCUAGUCCG-3'.
В другом примере, когда проксимальный домен имеет частичную или полную гомологию с проксимальным доменом из Campylobacter jejuni или проксимальным доменом, полученным из него, (P)k может быть 5'-AAAGAGUUUGC-3' или последовательностью оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAAGAGUUUGC-3'.
В еще одном примере, когда проксимальный домен имеет частичную или полную гомологию с проксимальным доменом из Streptococcus thermophilus или проксимальным доменом, полученным из него, (P)k может представлять собой 5'-AAGGCUUAGUCCG-3' или последовательность оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAGGCUUAGUCCG-3'.
(F)i может представлять собой последовательность оснований, включающую хвостовой домен, и имеющую частичную или полную гомологию с хвостовым доменом из видов, существующих в природе, и последовательность оснований хвостового домена может быть модифицирована в соответствии с видами происхождения. Каждый из F может быть независимо выбран из группы, состоящей из A, U, C и G, а i может быть количеством оснований, которое является целым числом от 1 до 50.
Например, когда хвостовой домен имеет частичную или полную гомологию с хвостовым доменом из Streptococcus pyogenes или хвостовым доменом, полученным из него, (F)i может быть 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC-3', или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC-3'.
В другом примере, когда хвостовой домен имеет частичную или полную гомологию с хвостовым доменом из Campylobacter jejuni или хвостовым доменом, полученным из него, (F)i может быть 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3'.
В еще одном примере, когда хвостовой домен имеет частичную или полную гомологию с хвостовым доменом из Streptococcus thermophilus или хвостовым доменом, полученным из него, (F)i может быть 5'-UACUCAACUUGAAAAGGUGGCACCGAUUCGGUGUUUUUU-3', или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-UACUCAACUUGAAAAGGUGGCACCGAUUCGGUGUUUUUU-3'.
Кроме того, (F)i может включать последовательность из 1-10 оснований на 3'-конце, вовлеченных в способ транскрипции in vitro или in vivo.
Например, когда промотор T7 используют при транскрипции in vitro гРНК, хвостовой домен может представлять собой произвольную последовательность оснований, присутствующую на 3'-конце матрицы ДНК. Кроме того, когда промотор U6 используют в транскрипции in vivo, хвостовой домен может быть UUUUUU, когда промотор H1 используют в транскрипции, хвостовой домен может быть UUUU, а когда используют промотор pol-III, хвостовой домен может включать несколько оснований урацила или альтернативных оснований.
Кроме того, (X)d, (X)e и (X)f могут быть выборочно добавленными последовательностями оснований, где каждый из X может быть независимо выбран из группы, состоящей из A, U, C и G, а каждый из d, e и f могут быть количеством оснований, которое равно 0 или целому числу из 1-20.
Одноцепочечная гРНК
Одноцепочечная гРНК может быть классифицирована на два типа.
i) одноцепочечная гРНК
Во-первых, существует одноцепочечная гРНК, в которой первая цепь или вторая цепь двухцепочечной гРНК связана с помощью линкерного домена, и в настоящем документе одноцепочечная гРНК состоит из: 5'-[первая цепь]-[линкерный домен]-[вторая цепь]-3'.
В частности, одноцепочечная гРНК может состоять из:
5'-[гидовый домен]-[первый комплементарный домен]-[линкерный домен]-[второй комплементарный домен]-[проксимальный домен]-3' или
5'-[гидовый домен]-[первый комплементарный домен]-[линкерный домен]-[второй комплементарный домен]-[проксимальный домен]-[хвостовой домен]-3'.
Каждый домен, кроме линкерного домена, аналогичен описанию каждого домена первой и второй цепей двухцепочечной гРНК.
Линкерный домен
В одноцепочечной гРНК линкерный домен представляет собой домен, соединяющий первую цепь и вторую цепь, и, в частности, представляет собой последовательность нуклеиновой кислоты, которая соединяет первый комплементарный домен со вторым комплементарным доменом для получения одноцепочечной гРНК. В настоящем документе линкерный домен может быть соединен с первым комплементарным доменом и вторым комплементарным доменом или соединять первый комплементарный домен со вторым комплементарным доменом с помощью ковалентной или нековалентной связи.
Линкерный домен может быть или включать последовательность из 1-30 оснований. Например, линкерный домен может быть или включать последовательность из 1-5, 5-10, 10-15, 15-20, 20-25 или 25-30 оснований.
Линкерный домен подходит для использования в одноцепочечной молекуле гРНК и может быть соединен с первой цепью и второй цепью двухцепочечной гРНК или соединять первую цепь со второй цепью с помощью ковалентной или нековалентной связи для использования при получении одноцепочечной гРНК. Линкерный домен может быть соединен с crРНК и tracrРНК двухцепочечной гРНК или соединять crРНК с tracrРНК с помощью ковалентной или нековалентной связи для использования при получении одноцепочечной гРНК.
Линкерный домен может иметь гомологию с естественной последовательностью, например частичной последовательностью tracrРНК, или может быть получен из нее.
Выборочно, часть или вся последовательность оснований линкерного домена может иметь химическую модификацию. Химическая модификация может представлять собой метилирование, ацетилирование, фосфорилирование, фосфоротиоатную связь, закрытую нуклеиновую кислоту (LNA), 2'-O-метил-3'-фосфоротиоат (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение не ограничено этим.
Следовательно, одноцепочечная рРНК может состоять из: 5'- [гидовый домен]-[первый комплементарный домен]-[линкерный домен]-[второй комплементарный домен]-[проксимальный домен]-3' или 5'-[гидовый домен]-[первый комплементарный домен]-[линкерный домен]-[второй комплементарный домен]-[проксимальный домен]-[хвостовой домен]-3', как описано выше.
Кроме того, одноцепочечная гРНК может выборочно включать дополнительную последовательность оснований.
В одном примере варианта осуществления одноцепочечная гРНК может представлять собой:
5'-(Nмишень)-(Q)m-(L)j-(Z)h-(P)k-3'; или
5'-(Nмишень)-(Q)m-(L)j-(Z)h-(P)k-(F)i-3'.
В другом варианте осуществления одноцепочечная гРНК может представлять собой:
5'-(X)a-(Nмишень)-(X)b-(Q)m-(X)c-(L)j-(X)d-(Z)h-(X)e-(P)k-(X)f-3'; or
5'-(X)a-(Nмишень)-(X)b-(Q)m-(X)c-(L)j-(X)d-(Z)h-(X)e-(P)k-(X)f-(F)i-3'.
В настоящем документе Nмишень представляет собой последовательность оснований, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени, и участок последовательности оснований, который может быть изменен в соответствии с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени.
В одном примере варианта осуществления Nмишень представляет собой последовательность оснований, способную образовывать комплементарную связь с геном-мишенью, то есть последовательностью-мишенью гена PMP22.
(Q)m включает в себя последовательность оснований, включающую в себя первый комплементарный домен, который способен образовывать комплементарную связь со вторым комплементарным доменом. (Q)m может быть последовательностью, имеющей частичную или полную гомологию с первым комплементарным доменом из видов, существующих в природе, и последовательность оснований первого комплементарного домена может быть изменена в соответствии с видами происхождения. Каждый из Q может быть независимо выбран из группы, состоящей из A, U, C и G, а m может быть количеством оснований, которое является целым числом от 5 до 35.
Например, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Streptococcuspyogenes или первым комплементарным доменом, полученным из него, (Q)m может быть 5'-GUUUUAGAGCUA-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GUUUUAGAGCUA-3'.
В другом примере, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Campylobacterjejuni или первым комплементарным доменом, полученным из него, (Q)m может быть 5'-GUUUUAGUCCCUUUUUAAAUUUCUU-3' или последовательностью оснований, имеющей не менее 50% или более гомологию с 5'-GUUUUAGUCCCUUUUUAAAUUUCUU-3'.
В еще одном примере, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Streptococcus thermophilus или первым комплементарным доменом, полученным из него, (Q)m может быть 5'-GUUUUAGAGCUGUGUUGUUUCG-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GUUUUAGAGCUGUGUUGUUUCG-3'.
Кроме того, (L)j представляет собой последовательность оснований, включающую линкерный домен и соединяющую первый комплементарный домен со вторым комплементарным доменом, тем самым получая одноцепочечную гРНК. В настоящем документе каждый из Lмодет быть независимо выбран из группы, состоящей из A, U, C и G, а j может быть количеством оснований, которое является целым числом от 1 до 30.
(Z)h является последовательностью оснований, включающей второй комплементарный домен, который способен иметь комплементарную связь с первым комплементарным доменом. (Z)h может быть последовательностью, имеющей частичную или полную гомологию со вторым комплементарным доменом из видов, существующих в природе, и последовательность оснований второго комплементарного домена может быть изменена в соответствии с видами происхождения. Каждый из Z может быть независимо выбран из группы, состоящей из A, U, C и G, а h представляет собой количество оснований, которое может быть целым числом от 5 до 50.
Например, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Streptococcus pyogenes или вторым комплементарным доменом, полученным из него, (Z)h может представлять собой 5'-UAGCAAGUUAAAAU-3' или последовательность оснований, имеющую по меньшей мере 50% гомологию с 5'-UAGCAAGUUAAAAU-3'.
В другом примере, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Campylobacter jejuni или вторым комплементарным доменом, полученным из него, (Z)h может быть 5'-AAGAAAUUUAAAAAGGGACUAAAAU-3' или последовательностью оснований, имеющей не менее 50% или более гомологию с 5'-AAGAAAUUUAAAAAGGGACUAAAAU-3'.
В еще одном примере, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Streptococcus thermophilus или вторым комплементарным доменом, полученным из него, (Z)h может быть 5'-CGAAACAACACAGCGAGUUAAAAU-3'или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-CGAAACAACACAGCGAGUUAAAAU-3'.
(P)k представляет собой последовательность оснований, включающую в себя проксимальный домен, который может иметь частичную или полную гомологию с проксимальным доменом из видов, существующих в природе, и последовательность оснований проксимального домена может быть модифицирована в соответствии с видами происхождения. Каждый из P может быть независимо выбран из группы, состоящей из A, U, C и G, и k может быть количеством оснований, которое является целым числом от 1 до 20.
Например, когда проксимальный домен имеет частичную или полную гомологию с проксимальным доменом из Streptococcuspyogenes или проксимальным доменом, полученным из него, (P)k может представлять собой 5'-AAGGCUAGUCCG-3'илипоследовательность оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAGGCUAGUCCG-3'.
В другом примере, когда проксимальный домен имеет частичную или полную гомологию с проксимальным доменом из Campylobacterjejuni или проксимальным доменом, полученным из него, (P)k может быть 5'-AAAGAGUUUGC-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-AAAGAGUUUGC-3'.
В еще одном примере, когда проксимальный домен имеет частичную или полную гомологию с проксимальным доменом из Streptococcus thermophilus или проксимальным доменом, полученным из него, (P)k может представлять собой 5'-AAGGCUUAGUCCG-3' или последовательность оснований, имеющую по меньшей мере 50% или более гомологию с 5'-AAGGCUUAGUCCG-3'.
(F)i может представлять собой последовательность оснований, включающую хвостовой домен, и имеющую частичную или полную гомологию с хвостовым доменом из видов, существующих в природе, и последовательность оснований хвостового домена может быть модифицирована в соответствии с видами происхождения. Каждый из F может быть независимо выбран из группы, состоящей из A, U, C и G, а i может быть количеством оснований, которое является целым числом от 1 до 50.
Например, когда хвостовой домен имеет частичную или полную гомологию с хвостовым доменом из Streptococcuspyogenes или хвостовым доменом, полученным из него, (F)i может быть 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-UUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC-3'.
В другом примере, когда хвостовой домен имеет частичную или полную гомологию с хвостовым доменом из Campylobacterjejuni или хвостовым доменом, полученным из него, (F)i может быть 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-GGGACUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3'.
В еще одном примере, когда хвостовой домен имеет частичную или полную гомологию с хвостовым доменом из Streptococcus thermophilus или хвостовым доменом, полученным из него, (F)i может быть 5'-UACUCAACUUGAAAAGGUGGCACCGAUUCGGUGUUUUU-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-UACUCAACUUGAAAAGGUGGCACCGAUUCGGUGUUUUU-3'.
Кроме того, (F)i может включать последовательность из 1-10 оснований на 3'-конце, вовлеченных в способ транскрипции in vitro или in vivo.
Например, когда промотор Т7 используют в транскрипции гРНК in vitro, хвостовой домен может представлять собой произвольную последовательность оснований, присутствующую на 3'-конце матрицы ДНК. Кроме того, когда промотор U6 используют в транскрипции in vivo, хвостовой домен может представлять собой UUUUUU, когда промотор H1 используют в транскрипции, хвостовой домен может представлять собой UUUU, а когда используют промотор pol-III, хвостовой домен может включать несколько оснований урацила или альтернативных оснований.
Кроме того, (X)a, (X)b, (X)c, (X)d, (X)e и (X)f могут быть выборочно добавленными последовательностями оснований, где каждый из X может быть независимо выбран из группы, состоящей из A, U, C и G, а каждый из a, b, c, d, e и f может быть количеством оснований, которое равно 0 или целому числу от 1 до 20.
ii) одноцепочечная гРНК
Во-вторых, одноцепочечная гРНК может быть одноцепочечной гРНК, состоящей из гидового домена, первого комплементарного домена и второго комплементарного домена, и в настоящем документе одноцепочечная гРНК может состоять из:
5'-[второй комплементарный домен]-[первый комплементарный домен]-[гидовый домен]-3'; или
5'-[второй комплементарный домен]-[линкерный домен]-[первый комплементарный домен]-[гидовый домен]-3'.
Гидовый домен
В одноцепочечной гРНК гидовый домен включает комплементарную гидовую последовательность, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени. Гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, имеющую комплементарность к последовательности-мишени в гене-мишени или нуклеиновой кислоте-мишени, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарность или полную комплементарность. Считается, что гидовый домен обеспечивает специфическое взаимодействие комплекса гРНК-Cas, то есть комплекса CRISPR, с геном-мишенью или нуклеиновой кислотой-мишенью.
В настоящем документе гидовый домен может представлять собой или включать последовательность из 5-50 оснований. Например, гидовый домен может быть или включать последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Кроме того, гидовый домен может включать в себя гидовую последовательность.
В настоящем документе гидовая последовательность может представлять собой комплементарную последовательность оснований, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарность или полную комплементарность.
В одном примере варианта осуществления гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную гену-мишени, то есть последовательности-мишени гена PMP22, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85% 90% или 95% или более комплементарность или полную комплементарность.
В настоящем документе гидовая последовательность может быть или включать последовательность из 5-50 оснований. Например, гидовая последовательность может быть или включать последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В одном примере варианта осуществления гидовая последовательность может представлять собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени гена PMP22. Гидовая последовательность может быть или включать последовательность из 5-50 оснований. Например, гидовая последовательность может быть или включать последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
В настоящем документе последовательности-мишени генов-мишеней, то есть гена PMP22 для гидовой последовательности, перечислены выше в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7, соответственно, но настоящее изобретение этим не ограничено.
Выборочно, гидовый домен может включать в себя гидовую последовательность и дополнительную последовательность оснований.
В настоящем документе дополнительная последовательность оснований может представлять собой последовательность из 1-35 оснований. Например, дополнительная последовательность оснований может быть последовательностью из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 оснований.
В одном примере варианта осуществления дополнительная последовательность оснований может представлять собой одну последовательность оснований, гуанина (G) или последовательность двух оснований, GG.
В настоящем документе дополнительная последовательность оснований может быть расположена на 5'-конце гидового домена или на 5'-конце гидовой последовательности.
Дополнительная последовательность оснований может быть расположена на 3'-конце гидового домена или на 3'-конце гидовой последовательности.
Первый комплементарный домен
Первый комплементарный домен представляет собой домен, включающий последовательность нуклеиновой кислоты, комплементарную второму комплементарному домену, и обладающую достаточной комплементарностью, чтобы образовывать двойную цепь со вторым комплементарным доменом.
В настоящем документе первый комплементарный домен может быть или включать последовательность из 5-35 оснований. Например, первый комплементарный домен может быть или включать последовательность из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Первый комплементарный домен может иметь гомологию с естественным первым комплементарным доменом или может быть получен из естественного первого комплементарного домена. Кроме того, первый комплементарный домен может иметь различие в последовательности оснований первого комплементарного домена в зависимости от существующих в природе видов, может быть получен из первого комплементарного домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию с первым комплементарным доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления первый комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию с первым комплементарным доменом из Parcubacteria bacterium (GWC2011_GWC2_44_17), Lachnospiraceae bacterium (MC2017), Butyrivibrio proteoclasiicus, Peregrinibacteria bacterium (GW2011_GWA_33_10), Acidaminococcus sp. (BV3L6), Porphyromonas macacae, Lachnospiraceae bacterium (ND2006), Porphyromonas crevioricanis, Prevotella disiens, Moraxella bovoculi (237), Smiihella sp. (SC_KO8D17), Leptospira inadai, Lachnospiraceae bacterium (MA2020), Francisella novicida (U112), Candidatus Methanoplasma termitum или Eubacterium eligens или первым комплементарным доменом, полученным из них.
Выборочно, первый комплементарный домен может включать в себя дополнительную последовательность оснований, которая не подвергается комплементарной связи со вторым комплементарным доменом.
В настоящем документе дополнительная последовательность оснований может представлять собой последовательность из 1-15 оснований. Например, дополнительная последовательность оснований может представлять собой последовательность из 1-5, 5-10 или 10-15 оснований.
Второй комплементарный домен
Второй комплементарный домен включает последовательность нуклеиновой кислоты, комплементарную первому комплементарному домену, и обладает достаточной комплементарностью, чтобы образовывать двойную цепь с первым комплементарным доменом. Второй комплементарный домен может включать в себя последовательность оснований, комплементарную первому комплементарному домену, и последовательность оснований, не имеющую комплементарности с первым комплементарным доменом, например, последовательность оснований, не образующую двойную цепь с первым комплементарным доменом, и может иметь более длинную последовательность оснований, чем у первого комплементарного домена.
В настоящем документе второй комплементарный домен может представлять собой или включать последовательность из 5-35 оснований. Например, второй комплементарный домен может быть последовательностью из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Второй комплементарный домен может иметь гомологию с естественным вторым комплементарным доменом или может быть получен из естественного второго комплементарного домена. Кроме того, второй комплементарный домен может иметь различие в последовательности оснований второго комплементарного домена в соответствии с видами, существующими в природе, и может быть получен из второго комплементарного домена, содержащегося в существующих в природе видах, или может иметь частичную или полную гомологию со вторым комплементарным доменом, содержащимся в существующих в природе видах.
В одном примере варианта осуществления второй комплементарный домен может иметь частичную, то есть, по меньшей мере 50% или более, или полную гомологию со вторым комплементарным доменом из Parcubacteria bacterium (GWC2011_GWC2_44_17), Lachnospiraceae bacterium (MC2017), Butyrivibrio proteoclasiicus, Peregrinibacteria bacterium (GW2011_GWA_33_10), Acidaminococcus sp. (BV3L6), Porphyromonas macacae, Lachnospiraceae bacterium (ND2006), Porphyromonas crevioricanis, Prevotella disiens, Moraxella bovoculi (237), Smiihella sp. (SC_KO8D17), Leptospira inadai, Lachnospiraceae bacterium (MA2020), Francisella novicida (U112), Candidatus Methanoplasma termitum или Eubacterium eligens или вторым комплементарным доменом, полученным из них.
Выборочно, второй комплементарный домен может включать в себя дополнительную последовательность оснований, которая не подвергается комплементарной связи с первым комплементарным доменом.
В настоящем документе дополнительная последовательность оснований может представлять собой последовательность из 1-15 оснований. Например, дополнительная последовательность оснований может представлять собой последовательность из 1-5, 5-10 или 10-15 оснований.
Линкерный домен
Выборочно, линкерный домен представляет собой последовательность нуклеиновой кислоты, соединяющую первый комплементарный домен со вторым комплементарным доменом для получения одноцепочечной гРНК. В настоящем документе линкерный домен может быть соединен с первым комплементарным доменом и вторым комплементарным доменом, или может соединять первый и второй комплементарные домены с помощью ковалентной или нековалентной связи.
Линкерный домен может быть или включать последовательность из 1-30 оснований. Например, линкерный домен может быть или включать последовательность из 1-5, 5-10, 10-15, 15-20, 20-25 или 25-30 оснований.
Выборочно, часть или вся последовательность оснований гидового домена, первого комплементарного домена, второго комплементарного домена и линкерного домена может иметь химическую модификацию. Химическая модификация может представлять собой метилирование, ацетилирование, фосфорилирование, фосфоротиоатную связь, закрытую нуклеиновую кислоту (LNA), 2'-O-метил-3'-фосфоротиоат (MS) или 2'-O-метил-3'-тиоPACE (MSP), но настоящее изобретение этим не ограничено.
Следовательно, одноцепочечная гРНК может состоять из: 5'-[второй комплементарный домен]-[первый комплементарный домен]-[гидовый домен]-3' или 5'-[второй комплементарной домен]-[линкерный домен]-[первый комплементарный домен]-[гидовый домен]-3', как описано выше.
Кроме того, одноцепочечная гРНК может выборочно включать дополнительную последовательность оснований.
В одном примере варианта осуществления одноцепочечная гРНК может быть:
5'-(Z)h-(Q)m-(Nмишень)-3'; или
5'-(X)a-(Z)h-(X)b-(Q)m-(X)c-(Nмишень)-3'.
В другом варианте осуществления одноцепочечная гРНК может быть
5'-(Z)h-(L)j-(Q)m-(Nмишень)-3'; или
5'-(X)a-(Z)h-(L)j-(Q)m-(X)c-(Nмишень)-3'.
В настоящем документе Nмишень представляет собой последовательность оснований, способную образовывать комплементарную связь с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени, и участком последовательности оснований, который может быть изменен в соответствии с последовательностью-мишенью в гене-мишени или нуклеиновой кислоте-мишени.
В одном примере варианта осуществления Nмишень может представлять собой последовательность оснований, способную образовывать комплементарную связь с геном-мишенью, то есть последовательностью-мишенью гена PMP22.
(Q)m является последовательностью оснований, включающей в себя первый комплементарный домен, который способен образовывать комплементарную связь со вторым комплементарным доменом второй цепи. (Q)m может быть последовательностью, имеющей частичную или полную гомологию с первым комплементарным доменом из видов, существующих в природе, и последовательность оснований первого комплементарного домена может быть изменена в соответствии с видами происхождения. Каждый из Q может быть независимо выбран из группы, состоящей из A, U, C и G, и m может быть количеством оснований, которое является целым числом от 5 до 35.
Например, когда первый комплементарный домен имеет частичную или полную гомологию с первым комплементарным доменом из Parcubacteria bacterium или первым комплементарным доменом, полученным из него, (Q)m может быть 5'-UUUGUAGAU-3', или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-UUUGUAGAU-3'.
(Z)h является последовательностью оснований, включающей в себя второй комплементарный домен, который способен образовывать комплементарную связь с первым комплементарным доменом первой цепи. (Z)h может быть последовательностью, имеющей частичную или полную гомологию со вторым комплементарным доменом из видов, существующих в природе, и последовательность оснований второго комплементарного домена может быть модифицирована в соответствии с видами происхождения. Каждый Z может быть независимо выбран из группы, состоящей из A, U, C и G, а h может быть количеством оснований, которое является целым числом от 5 до 50.
Например, когда второй комплементарный домен имеет частичную или полную гомологию со вторым комплементарным доменом из Parcubacteria bacterium или вторым комплементарным доменом, полученным из Parcubacteria bacterium, (Z)h может быть 5'-AAAUUUCUACU-3' или последовательностью оснований, имеющей по меньшей мере 50% или более гомологию с 5'-AAAUUUCUACU-3'.
Кроме того, (L)j является последовательностью оснований, включающей в себя линкерный домен, который соединяет первый комплементарный домен со вторым комплементарным доменом. В настоящем документе каждый из L может быть независимо выбран из группы, состоящей из A, U, C и G, а j может быть количеством оснований, которое является целым числом от 1 до 30.
Кроме того, каждый из (X)a, (X)b и (X)c является выборочно дополнительной последовательностью оснований, где каждый из X может быть независимо выбран из группы, состоящей из A, U, C и G, и a, b и c могут быть количеством оснований, которое равно 0 или целому числу 1-20.
2. Белок-редактор
Белок-редактор относится к пептиду, полипептиду или белку, который способен непосредственно связываться или взаимодействовать с нуклеиновой кислотой без прямого связывания.
Нуклеиновая кислота может представлять собой нуклеиновую кислоту, содержащуюся в мишени в виде нуклеиновой кислоты, гена или хромосомы.
Нуклеиновая кислота может быть гидовой нуклеиновой кислотой.
Белок-редактор может представлять собой фермент.
Белок-редактор может представлять собой гибридный белок.
В настоящем документе гибридный белок относится к белку, полученному с помощью слияния фермента с дополнительным доменом, пептидом, полипептидом или белком.
Фермент относится к белку, включающему домен, который способен расщеплять нуклеиновую кислоту, ген, хромосому или белок.
Фермент может представлять собой нуклеазу, протеазу или рестрикционный фермент.
Дополнительный домен, пептид, полипептид или белок может представлять собой функциональный домен, пептид, полипептид или белок, который имеет такую же функцию или отличную от фермента.
Гибридный белок может включать в себя дополнительный домен, пептид, полипептид или белок на одном или более N-концах фермента или его пределах; С-конце фермента или его пределах; средней области фермента; и их комбинацию.
Гибридный белок может включать функциональный домен, пептид, полипептид или белок на одном или более N-концах фермента или его пределах; С-конце фермента или его пределах; средней области фермента; и их комбинацию.
В настоящем документе функциональный домен, пептид, полипептид или белок может представлять собой домен, пептид, полипептид или белок, обладающий метилазной активностью, деметилазной активностью, активностью активации транскрипции, активностью репрессии транскрипции, активностью высвобождения фактора транскрипции, активностью модификации гистонов, активностью расщепления РНК или активностью связывания нуклеиновых кислот, или метку или репортерный ген для выделения и очистки белка (включая пептид), но настоящее изобретение этим не ограничено.
Функциональный домен, пептид, полипептид или белок могут представлять собой дезаминазу.
Метка включает в себя гистидиновую (His) метку, V5-метку, FLAG-метку, метку гемагглютинина (HA) вируса гриппа, Myc-метку, VSV-G-метку и метку тиоредоксина (Trx), и репортерный ген включает глутатион-S-трансферазу (GST), пероксидазу хрена (HRP), хлорамфеникол-ацетилтрансферазу (CAT), β-галактозидазу, β-глюкоронидазу, люциферазу, аутофлуоресцентные белки, включая зеленый флуоресцентный белок (GFP), HcRed, DsRed, голубой флуоресцентный белок (CFP) желтый флуоресцентный белок (YFP) и синий флуоресцентный белок (BFP), но настоящее изобретение этим не ограничено.
Кроме того, функциональный домен, пептид, полипептид или белок могут представлять собой последовательность или сигнал ядерной локализации (NLS), или последовательность или сигнал ядерного экспорта (NES).
NLS может представлять собой NLS большого T-антигена вируса SV40 с аминокислотной последовательностью PKKKRKV; NLS, полученный из нуклеоплазмина (например, двухчастичный NLS нуклеоплазмина с последовательностью KRPAATKKAGQAKKKK); NLS c-myc с аминокислотной последовательностью PAAKRVKLD или RQRRNELKRSP; NLS hLSNF1 M9 с последовательностью NQSSNFGPMKGGNFGGRSSGPYGGGGQYFAKPRNQGGY; последовательность полученного из импортина-α домена IBB RMRIZFKNKGKDTAELRRRRVEVSVELRKAKKDEQILKRRNV; последовательности Т-белка миомы VSRKRPRP и PPKKARED; последовательность человеческого р53 POPKKKPL; последовательность c-abl IV мыши SALIKKKKKMAP; последовательности вируса гриппа NS1 DRLRR и PKQKKRK; последовательность антигена вируса гепатита D RKLKKKIKKL; последовательность белка Mx1 мыши REKKKFLKRR; последовательность человеческой поли(АДФ-рибозы)-полимеразы KRKGDEVDGVDEVAKKKSKK; или глюкокортикоидную последовательность рецептора стероидных гормонов (человека) RKCLQAGMNLEARKTKK, но настоящее изобретение этим не ограничено.
Белок-редактор может включать полный активный фермент.
В настоящем документе «полный активный фермент» относится к ферменту, имеющему ту же функцию, что и функция фермента дикого типа, и, например, фермент дикого типа, расщепляющий двойную цепь ДНК, обладает полной ферментативной активностью полного расщепления двойной цепи ДНК.
Кроме того, полный активный фермент включает фермент, имеющий улучшенную функцию по сравнению с функцией фермента дикого типа, и, например, специфический тип модификации или манипуляции с ферментом дикого типа, расщепляющим двойную цепь ДНК, имеет полную ферментативную активность, которая улучшена по сравнению с ферментом дикого типа, то есть активность расщепления двойной цепи ДНК.
Белок-редактор может включать в себя неполный или частично активный фермент.
В настоящем документе «неполный или частично активный фермент» относится к ферменту, имеющему некоторые функции фермента дикого типа, и, например, специфический тип модификации или манипуляции с ферментом дикого типа, расщепляющим двойную цепь ДНК, имеет неполную или частичную ферментативную активность расщепления части двойной цепи, то есть, одной цепи ДНК.
Белок-редактор может включать неактивный фермент.
В настоящем документе «неактивный фермент» относится к ферменту, в котором функция фермента дикого типа полностью инактивирована. Например, специфический тип модификации или манипуляции с ферментом дикого типа, расщепляющим двойную цепь ДНК, инактивирован так, чтобы не расщеплять полностью двойную цепь ДНК.
Белок-редактор может представлять собой натуральный фермент или гибридный белок.
Белок-редактор может присутствовать в форме частично модифицированного натурального фермента или гибридного белка.
Белок-редактор может представлять собой искусственно полученный фермент или гибридный белок, который не существует в природе.
Белок-редактор может присутствовать в форме частично модифицированного искусственного фермента или гибридного белка, которого не существует в природе.
В настоящем документе модификация может быть замещением, удалением, добавлением аминокислот, содержащихся в белке-редакторе, или их комбинацией.
Кроме того, модификация может представлять собой замещение, удаление, добавление некоторых оснований в последовательность оснований, кодирующую белок-редактор, или их комбинацию.
В качестве одного примера варианта осуществления белка-редактора настоящего изобретения фермент CRISPR будет описан ниже.
Фермент CRISPR
Термин «фермент CRISPR» является основным белковым компонентом системы CRISPR-Cas и образует комплекс с гРНК, что приводит к системе CRISPR-Cas.
Фермент CRISPR представляет собой нуклеиновую кислоту или полипептид (или белок), имеющий последовательность, кодирующую фермент CRISPR, и, в качестве примера, фермент CRISPR Типа II или фермент CRISPR Типа V широко используют.
Фермент CRISPR Типа II представляет собой Cas9, который может быть получен из различных микроорганизмов, таких как Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptosporangium roseum, AlicyclobacHlus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor bescii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus и Acaryochloris marina.
Термин «Cas9» представляет собой фермент, который связывается с гРНК для расщепления или модификации последовательности-мишени или положения в гене-мишени или нуклеиновой кислоте-мишени и может состоять из домена HNH, способного расщеплять цепь нуклеиновой кислоты, образующую комплементарную связь с гРНК, домена RuvC, способного расщеплять цепь нуклеиновой кислоты, образующую комплементарную связь с гРНК, домена REC, распознающего мишень, и домена PI, распознающего PAM. Для специфических структурных характеристик Cas9 может быть сделана ссылка на Hiroshi Nishimasu et al. (2014) Cell 156:935-949.
Кроме того, фермент CRISPR Типа V может представлять собой Cpf1, который может быть получен из Streptococcus, Campylobacter, Nitratifractor, Staphylococcus, Parvibaculum, Roseburia, Neisseria, Gluconacetobacter, Azospirillum, Sphaerochaeta, Lactobacillus, Eubacterium, Corynebacter, Carnobacterium, Rhodobacter, Listeria, Paludibacter, Clostridium, Lachnospiraceae, Clostridiaridium, Leptotrichia, Francisella, Legionella, Alicyclobacillus, Methanomethyophilus, Porphyromonas, Prevotella, Bacteroidetes, Helcococcus, Letospira, Desulfovibrio, Desulfonatronum, Opitutaceae, Tuberibacillus, Bacillus, Brevibacillus, Methylobacterium или Acidaminococcus.
Cpf1 может состоять из домена RuvC, подобного и соответствующего домену RuvC в Cas9, домена Nuc без домена HNH в Cas9, домена REC, распознающего мишень, домена WED и домена PI, распознающего PAM. Для специфических структурных характеристик Cpf1, может быть сделана ссылка на Takashi Yamano et al. (2016) Cell 165:949-962.
Фермент CRISPR белка Cas9 или Cpf1 может быть выделен из микроорганизма, существующего в природе, или получен искусственно с помощью рекомбинантного или синтетического способа.
Фермент CRISPR Типа II
Кристаллическая структура фермента CRISPR Типа II была определена в соответствии с исследованиями двух или более типов природных микробных молекул фермента CRISPR Типа II (Jinek et al., Science, 343 (6176): 1247997, 2014) и исследованиями Cas9 (SpCas9) из Streptococcus pyogenes в комплексе с гРНК (Nishimasu et al., Cell, 156: 935-949, 2014; и Anders et al., Nature, 2014, doi: 10.1038/nature13579).
Фермент CRISPR Типа II включает в себя две части, то есть части распознавания (REC) и нуклеазы (NUC), и каждая часть включает несколько доменов.
Часть REC включает в себя домен богатой аргинином спирали (BH), домен REC1 и домен REC2.
В настоящем документе домен BH представляет собой длинный участок α-спирали, богатый аргинином, и домены REC1 и REC2 играют важную роль в распознавании двойной цепи, образованной в гРНК, например одноцепочечной гРНК, двухцепочечной гРНК или tracrРНК.
Часть NUC включает в себя домен RuvC, домен HNH и домен PAM-взаимодействия (PI). В настоящем документе домен RuvC охватывает RuvC-подобные домены, или домен HNH используют для включения HNH-подобных доменов.
В настоящем документе домен RuvC имеет структурное сходство с членами семейства микроорганизмов, существующих в природе, имеющих фермент CRISPR Типа II, и расщепляет одиночную цепь, например, некомплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, не образующую комплементарную связь с гРНК. В данной области домен RuvC иногда обозначают как домен RuvCI, домен RuvCII или домен RuvCIII, и обычно называют RuvC I, RuvCII или RuvCIII. Например, в случае SpCas9 домен RuvC собирают из каждого из трех разделенных доменов RuvC (RuvC I, RuvCII и RuvCIII), расположенных в последовательностях аминокислот 1-59, от 718 до 769 и от 909 до 1098 SpCas9, соответственно.
Домен HNH имеет структурное сходство с эндонуклеазой HNH и расщепляет одиночную цепь, например, комплементарную цепь молекулы нуклеиновой кислоты-мишени, то есть цепь, образующую комплементарную связь с гРНК. Домен HNH расположен между мотивами RuvC II и III. Например, в случае SpCas9 домен HNH расположен в аминокислотной последовательности от 775 до 908 SpCas9.
Домен PI распознает специфическую последовательность оснований в гене-мишени или нуклеиновой кислоте-мишени, то есть мотив, примыкающий к протоспейсеру (PAM), или взаимодействует с PAM. Например, в случае SpCas9 домен PI расположен в последовательности аминокислот от 1099 до 1368 SpCas9.
В настоящем документе PAM может варьироваться в зависимости от происхождения фермента CRISPR Типа II. Например, когда фермент CRISPR представляет собой SpCas9, PAM может представлять собой 5'-NGG-3', когда фермент CRISPR представляет собой Cas9 (StCas9) из Streptococcus thermophilus, PAM может представлять собой 5'-NNAGAAW-3' (W=A или T), когда фермент CRISPR представляет собой Cas9 (NmCas9) из Neisseria meningitides, PAM может представлять собой 5'-NNNNGATT-3', и когда фермент CRISPR представляет собой Cas9 (CjCas9) из Campylobacter jejuni, PAM может представлять собой 5'-NNNVRYAC-3' (V=G или C или A, R=A или G, Y=C или T), где N может быть A, T, G или C; или A, U, G или C.
Фермент CRISPR Типа V
Фермент CRISPR Типа V включает сходные домены RuvC, соответствующие доменам RuvC фермента CRISPR Типа II, и может состоять из домена Nuc вместо домена HNH фермента CRISPR Типа II, доменов REC и WED, которые распознают мишень, и домена PI, распознающего PAM. Для специфических структурных характеристик фермента CRISPR Типа V, можно сделать ссылку на Takashi Yamano et al. (2016) Cell 165: 949-962.
Фермент CRISPR Типа V может взаимодействовать с гРНК, тем самым образуя ферментный комплекс гРНК-CRISPR, то есть комплекс CRISPR, и может позволить гидовой последовательности приблизиться к последовательности-мишени, включая PAM-последовательность совместно с гРНК. В настоящем документе способность фермента CRISPR Типа V взаимодействовать с геном-мишенью или нуклеиновой кислотой-мишенью зависит от PAM-последовательности.
PAM-последовательность представляет собой последовательность, присутствующую в гене-мишени или нуклеиновой кислоте-мишени, и может быть распознана доменом PI фермента CRISPR Типа V. PAM-последовательность может варьироваться в зависимости от происхождения фермента CRISPR Типа V. То есть существуют разные PAM-последовательности, которые могут быть специфически распознаны в зависимости от видов.
В одном примере PAM-последовательность, распознаваемая с помощью Cpf1, может быть 5'-TTN-3'(N представляет собой A, T, C или G).
Активность фермента CRISPR
Фермент CRISPR расщепляет двойную или одиночную цепь гена-мишени или нуклеиновой кислоты-мишени и обладает нуклеазной активностью, вызывающей разрыв или делецию двойной или одиночной цепи. Обычно фермент CRISPR типа II или фермент CRISPR типа V дикого типа расщепляет двойную цепь гена-мишени или нуклеиновой кислоты-мишени.
Чтобы изменить или модифицировать вышеописанную нуклеазную активность фермента CRISPR, фермент CRISPR может быть изменен или модифицирован, такой измененный или модифицированный фермент CRISPR можеть быть модифицирован в не полностью или частично активный или неактивный фермент.
Не полностью или частично активный фермент
Фермент CRISPR, модифицированный для изменения ферментативной активности, проявляя тем самым неполную или частичную активность, называют никазой.
Термин «никаза» относится к ферменту CRISPR, который изменяют или модифицируют, чтобы расщеплять только одну цепь двойной цепи гена-мишени или нуклеиновой кислоты-мишени, и никаза обладает нуклеазной активностью по отношению к расщеплению одиночной цепи, например, цепи, которая не является комплементарной, или комплементарной для гРНК гена-мишени или нуклеиновой кислоты-мишени. Следовательно, для расщепления двойной цепи необходима нуклеазная активность двух никаз.
Например, никаза может обладать нуклеазной активностью домена RuvC. То есть никаза может включать нуклеазную активность домена HNH, и с этой целью домен HNH может быть изменен или модифицирован.
В одном примере, при условии, что фермент CRISPR является ферментом CRISPR Типа II, в одном примере варианта осуществления, когда остаток 840 в аминокислотной последовательности SpCas9 мутировал от гистидина до аланина, нуклеазная активность домена HNH инактивирована для использования в качестве никазы. Поскольку полученная таким образом никаза обладает нуклеазной активностью домена RuvC, она способна расщеплять некомплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая не образует комплементарную связь с гРНК.
В другом примере варианта осуществления, когда остаток 559 в аминокислотной последовательности CjCas9 мутировал от гистидина до аланина, нуклеазная активность домена HNH инактивирована для использования в качестве никазы. Полученная таким образом никаза обладает нуклеазной активностью домена RuvC и, таким образом, способна расщеплять некомплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая не образует комплементарную связь с гРНК.
Например, никаза может обладать нуклеазной активностью домена HNH. То есть никаза может включать нуклеазную активность домена RuvC, и с этой целью домен RuvC может быть изменен или модифицирован.
В одном примере при условии, что фермент CRISPR является ферментом CRISPR Типа II, в одном примере варианта осуществления, когда остаток 10 в аминокислотной последовательности SpCas9 мутировал от аспарагиновой кислоты до аланина, нуклеазная активность домена RuvC инактивирована для использования в качестве никазы. Полученная таким образом никаза обладает нуклеазной активностью домена HNH и, таким образом, способна расщеплять комплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая образует комплементарную связь с гРНК.
В другом примере варианта осуществления, когда остаток 8 в аминокислотной последовательности CjCas9 мутировал от аспарагиновой кислоты до аланина, нуклеазная активность домена RuvC инактивирована для использования в качестве никазы. Полученная таким образом никаза обладает нуклеазной активностью домена HNH и, таким образом, способна расщеплять комплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая образует комплементарную связь с гРНК.
Неактивный фермент
Фермент CRISPR, который модифицируют, чтобы сделать ферментативную активность полностью неактивной, называют неактивным ферментом CRISPR.
Термин «неактивный фермент CRISPR» относится к ферменту CRISPR, который модифицирован так, чтобы не расщеплять полностью двойную цепь гена-мишени или нуклеиновой кислоты-мишени, и неактивный фермент CRISPR обладает нуклеазной инактивностью вследствие мутации в домене с нуклеазной активностью фермента CRISPR дикого типа. Неактивный фермент CRISPR может представлять собой фермент, в котором нуклеазная активность домена RuvC и домена HNH инактивирована.
Например, неактивный фермент CRISPR может быть изменен или модифицирован в домене RuvC и домене HNH так, чтобы инактивировать нуклеазную активность.
В одном примере при условии, что фермент CRISPR является ферментом CRISPR Типа II, в одном примере варианта осуществления, когда остатки 10 и 840 в аминокислотной последовательности SpCas9 мутировали от аспарагиновой кислоты и гистидина до аланина, соответственно, нуклеазные активности домена RuvC и домена HNH инактивированы, так что двойная цепь может не расщеплять полностью двойную цепь гена-мишени или нуклеиновой кислоты-мишени.
В другом примере варианта осуществления, когда остатки 8 и 559 в аминокислотной последовательности CjCas9 мутировали от аспарагиновой кислоты и гистидина до аланина, нуклеазные активности домена RuvC и домена HNH инактивированы, так что двойная цепь может не расщеплять полностью двойную цепь гена-мишени или нуклеиновой кислоты-мишени.
Другие активности
Фермент CRISPR может обладать эндонуклеазной активностью, экзонуклеазной активностью или геликазной активностью, то есть способностью отжигать спиральную структуру двухцепочечной нуклеиновой кислоты в дополнение к вышеописанной нуклеазной активности.
Кроме того, фермент CRISPR может быть модифицирован для полной, неполной или частичной активации эндонуклеазной активности, экзонуклеазной активности или геликазной активности.
Направленность фермента CRISPR
Фермент CRISPR может взаимодействовать с гРНК, тем самым образуя ферментный комплекс гРНК-CRISPR, то есть комплекс CRISPR, и приводить гидовую последовательность к приближению к последовательности-мишени, включая PAM-последовательность совместно с гРНК. В настоящем документе способность фермента CRISPR взаимодействовать с геном-мишенью или нуклеиновой кислотой-мишенью зависит от PAM-последовательности.
PAM-последовательность представляет собой последовательность, присутствующую в гене-мишени или нуклеиновой кислоте-мишени, которая может быть распознана доменом PI фермента CRISPR. PAM-последовательность может варьироваться в зависимости от происхождения фермента CRISPR. То есть существуют различные PAM-последовательности, которые могут быть специфически распознаны в зависимости от видов.
В одном примере при условии, что фермент CRISPR представляет собой фермент CRISPR Типа II,
в случае SpCas9 PAM-последовательность может быть 5'-NGG-3', 5'-NAG-3' и/или 5'-NGA-3',
в случае StCas9 PAM-последовательность может быть 5'-NGGNG-3'и/или 5'-NNAGAAW-3' (W=A или T),
в случае NmCas9 PAM-последовательность может быть 5'-NNNNGATT-3'и/или 5'-NNNGCTT-3',
в случае CjCas9 PAM-последовательность может быть 5'-NNNVRYAC-3' (V=G, C или A; R=A или G; Y=C или T),
в случае Cas9 (SmCas9) из Streptococcus mutans PAM-последовательность может быть 5'-NGG-3'и/или 5'-NAAR-3' (R=A или G), и
в случае Cas9 (SaCas9) из Staphylococcus aureus PAM-последовательность может быть 5'-NNGRR-3', 5'-NNGRRT-3' и/или 5'-NNGRRV-3' (R=A или G; V=G, C или A).
В другом примере при условии, что фермент CRISPR представляет собой фермент CRISPR Типа V,
в случае Cpf1 PAM-последовательность может быть 5'-TTN-3'.
В настоящем документе N может быть A, T, G или C; или A, U, G или C.
Фермент CRISPR, способный распознавать специфическую PAM-последовательность, может быть изменен или модифицирован, используя PAM-последовательность, способную быть специфически распознаваемой в зависимости от видов. Например, домен PI SpCas9 может быть заменен доменом PI CjCas9 так, чтобы обладать нуклеазной активностью SpCas9 и распознавать PAM-последовательность, специфичную по отношению к CjCas9, тем самым образуя SpCas9, распознающий PAM-последовательность, специфичную по отношению к CjCas9. Специфически распознаваемая PAM-последовательность может быть изменена путем замещения или замены домена PI.
Мутантный фермент CRISPR
Фермент CRISPR может быть модифицирован для улучшения или ингибирования различных характеристик, таких как нуклеазная активность, геликазная активность, способность взаимодействовать с гРНК и способность приближаться к гену-мишени или нуклеиновой кислоте-мишени, например, PAM-распознающая способность фермента CRISPR.
Кроме того, мутантный фермент CRISPR может представлять собой фермент CRISPR, который взаимодействует с гРНК с образованием ферментного комплекса гРНК-CRISPR, то есть комплекса CRISPR, и модифицирован или изменен для улучшения специфичности к мишени при приближении или локализации к гену-мишени или нуклеиновой кислоте-мишени, так что только двойная или одиночная цепь гена-мишени или нуклеиновой кислоты-мишени расщепляется без расщепления двойной или одиночной цепи нецелевых гена или нуклеиновой кислоты, которая частично образует комплементарную связь с гРНК и нецелевыми геном или нуклеиновой кислотой, которая не образует комплементарную связь с ней.
В настоящем документе эффект расщепления двойной или одиночной цепи нецелевых гена или нуклеиновой кислоты, частично образующей комплементарную связь с гРНК, и нецелевых гена или нуклеиновой кислоты, не образующей комплементарную связь с нею, обозначают как нецелевой эффект, положение или последовательность оснований нецелевых гена или нуклеиновой кислоты, частично образующей комплементарную связь с гРНК, и нецелевых гена или нуклеиновой кислоты, не образующей с нею комплементарную связь, обозначают как нецелевой участок. В настоящем документе может быть одна или более нецелевых участков. С другой стороны, эффект расщепления двойной или одиночной цепи гена-мишени или нуклеиновой кислоты-мишени обозначают как целевой эффект, а местоположение или последовательность-мишень гена-мишени или нуклеиновой кислоты-мишени обозначают как целевой участок.
Мутантный фермент CRISPR модифицирован по меньшей мере в одной из аминокислот встречающегося в природе фермента CRISPR и может быть модифицирован, например, улучшен или ингибирован по одной или более различным характеристикам, таким как нуклеазная активность, геликазная активность, способность взаимодействовать с гРНК, способность приблизиться к гену-мишени или нуклеиновой кислоте-мишени и специфичность к мишени по сравнению с немодифицированным ферментом CRISPR. В настоящем документе модификацией может быть замещение, удаление, добавление аминокислоты или их смесь.
В мутантном ферменте CRISPR,
модификация может представлять собой модификацию одной или двух или более аминокислот, расположенных в участке, состоящем из аминокислот, имеющих положительные заряды, присутствующих во встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой модификацию одной или двух или более аминокислот из положительно заряженных аминокислот, таких как лизин (K), аргинин (R) и гистидин (H), присутствующих во встречающемся в природе ферменте CRISPR.
Модификация может представлять собой модификацию одной или двух или более аминокислот, расположенных в участке, состоящем из отрицательно заряженных аминокислот, присутствующих во встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой модификацию одной или двух или более аминокислот из отрицательно заряженных аминокислот, то есть аспарагиновой кислоты (D), глутаминовой кислоты (E), серина (S), треонина (T), аспарагина (N), глутамина (Q), цистеина (C), пролина (P), глицина (G), аланина (A), валина (V), изолейцина (I), лейцина (L), метионина (M)), фенилаланина (F), тирозина (Y) и триптофана (W), присутствующих во встречающемся в природе ферменте CRISPR.
В другом примере модификация может представлять собой модификацию одной или двух или более аминокислот из незаряженных аминокислот, то есть серина (S), треонина (T), аспарагина (N), глютамина (Q), цистеина (C), пролина (P), глицина (G), аланина (A), валина (V), изолейцина (I), лейцина (L), метионина (M), фенилаланина (F), тирозина (Y) и триптофана (W), присутствующих во встречающемся в природе ферменте CRISPR.
Кроме того, модификация может представлять собой модификацию одной или двух или более аминокислот, имеющих гидрофобные остатки, присутствующих в встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой модификацию одной или двух или более аминокислот из глицина (G), аланина (A), валина (V), изолейцина (I), лейцина (L), метионина (M), фенилаланина (F), тирозина (Y) и триптофана (W), присутствующих во встречающемся в природе ферменте CRISPR.
Модификация может представлять собой модификацию одной или двух или более аминокислот, имеющих полярные остатки, присутствующих во встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой модификацию одной или двух или более аминокислот из серина (S), треонина (T), аспарагина (N), глютамина (Q), цистеина (C), пролина (P), лизина (K), аргинина (R), гистидина (H), аспарагиновой кислоты (D) и глутаминовой кислоты (E), присутствующих во встречающемся в природе ферменте CRISPR.
Кроме того, модификация может представлять собой модификацию одной или двух или более аминокислот, включая лизин (K), аргинин (R) и гистидин (H), присутствующих во встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой замещение одной или двух или более аминокислот, включая лизин (K), аргинин (R) и гистидин (H), присутствующих во встречающемся в природе ферменте CRISPR.
Модификация может представлять собой модификацию одной или двух или более аминокислот, включая аспарагиновую кислоту (D) и глутаминовую кислоту (E), присутствующих во встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой замещение одной или двух или более аминокислот, включая аспарагиновую кислоту (D) и глутаминовую кислоту (E), присутствующих во встречающемся в природе ферменте CRISPR.
Модификация может представлять собой модификацию одной или двух или более аминокислот, включая серин (S), треонин (T), аспарагин (N), глутамин (Q), цистеин (C), пролин (P), глицин (G), аланин (A), валин (V), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), тирозин (Y) и триптофан (W), присутствующих во встречающемся в природе ферменте CRISPR.
Например, модификация может представлять собой замещение одной или двух или более аминокислот, включая серин (S), треонин (T), аспарагин (N), глутамин (Q), цистеин (C), пролин (P), глицин (G), аланин (A), валин (V), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), тирозин (Y) и триптофан (W), присутствующих во встречающемся в природе ферменте CRISPR.
Кроме того, модификация может представлять собой модификацию одной, двух, трех, четырех, пяти, шести, семи или более аминокислот, присутствующих во встречающемся в природе ферменте CRISPR.
Кроме того, в мутантном ферменте CRISPR,
модификация может представлять собой модификацию одной или двух или более аминокислот, присутствующих в домене RuvC фермента CRISPR. В настоящем документе домен RuvC может быть доменом RuvCI, RuvCII или RuvCIII.
Модификация может представлять собой модификацию одной или двух или более аминокислот, присутствующих в домене HNH фермента CRISPR.
Модификация может представлять собой модификацию одной или двух или более аминокислот, присутствующих в домене REC фермента CRISPR.
Модификация может представлять собой модификацию одной, двух или более аминокислот, присутствующих в домене PI фермента CRISPR.
Модификация может представлять собой модификацию двух или более аминокислот, содержащихся по меньшей мере в двух или более доменах из доменов REC, RuvC, HNH и PI фермента CRISPR.
В одном примере модификация может представлять собой модификацию двух или более аминокислот, содержащихся в доменах REC и RuvC фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере двух или более аминокислот из A203, H277, G366, F539, I601, M763, D965 и F1038, содержащихся в доменах REC и RuvC SpCas9.
В другом примере модификация может представлять собой модификацию двух или более аминокислот, содержащихся в доменах REC и HNH фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере двух или более аминокислот из A203, H277, G366, F539, I601 и K890, содержащихся в доменах REC и HNH SpCas9.
В одном примере модификация может представлять собой модификацию двух или более аминокислот, содержащихся в доменах REC и PI фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере двух или более аминокислот из A203, H277, G366, F539, I601, T1102 и D1127, содержащихся в доменах REC и PI SpCas9.
В другом примере модификация может представлять собой модификацию трех или более аминокислот, содержащихся в доменах REC, RuvC и HNH фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере трех или более аминокислот из A203, H277, G366, F539, I601, M763, K890, D965 и F1038, содержащихся в доменах REC, RuvC и HNH SpCas9.
В одном примере модификация может представлять собой модификацию трех или более аминокислот, содержащихся в доменах REC, RuvC и PI, содержащихся в ферменте CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере трех или более аминокислот из A203, H277, G366, F539, I601, M763, D965, F1038, T1102 и D1127, содержащихся в доменах REC, RuvC и PI SpCas9.
В другом примере модификация может представлять собой модификацию трех или более аминокислот, содержащихся в доменах REC, HNH и PI фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере трех или более аминокислот из A203, H277, G366, F539, I601, K890, T1102 и D1127, содержащихся в доменах REC, HNH и PI SpCas9.
В одном примере модификация может представлять собой модификацию трех или более аминокислот, содержащихся в доменах RuvC, HNH и PI фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере трех или более аминокислот из M763, K890, D965, F1038, T1102 и D1127, содержащихся в доменах RuvC, HNH и PI SpCas9.
В другом примере модификация может представлять собой модификацию четырех или более аминокислот, содержащихся в доменах REC, RuvC, HNH и PI фермента CRISPR.
В одном примере варианта осуществления в мутанте SpCas9 модификация может представлять собой модификацию по меньшей мере четырех или более аминокислот из A203, H277, G366, F539, I601, M763, K890, D965, F1038, T1102 и D1127, содержащихся в доменах REC, RuvC, HNH и PI SpCas9.
Кроме того, в мутантном фермента CRISPR,
модификация может представлять собой модификацию одной или двух или более аминокислот, участвующих в нуклеазной активности фермента CRISPR.
Например, в мутанте SpCas9 модификация может представлять собой модификацию одной или двух или более из группы, состоящей из аминокислот D10, E762, H840, N854, N863 и D986, или одной или двух или более из группы, состоящей из аминокислот, соответствующих другим ортологам Cas9.
Модификация может быть модификацией для частичной инактивации нуклеазной активности фермента CRISPR, и такой мутантный фермент CRISPR может представлять собой никазу.
В настоящем документе модификация может представлять собой модификацию для инактивации нуклеазной активности домена RuvC фермента CRISPR, и такой мутантный фермент CRISPR может не расщеплять некомплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая не образует комплементарную связь с гРНК.
В одном примере варианта осуществления, в случае SpCas9, когда остаток 10 аминокислотной последовательности SpCas9 мутировал от аспарагиновой кислоты до аланина, то есть, когда мутировал до D10A, нуклеазная активность домена RuvC инактивирована, и, таким образом, SpCas9 может быть использован в качестве никазы. Полученная таким образом никаза может не расщеплять некомплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая не образует комплементарную связь с гРНК.
В другом примере варианта осуществления в случае CjCas9, когда остаток 8 аминокислотной последовательности CjCas9 мутировал от аспарагиновой кислоты до аланина, то есть, когда мутировал до D8A, нуклеазная активность домена RuvC инактивирована, и, таким образом, CjCas9 может быть использован в качестве никазы. Полученная таким образом никаза может не расщеплять некомплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая не образует комплементарную связь с гРНК.
Кроме того, в настоящем документе модификация может представлять собой модификацию для инактивации нуклеазной активности домена HNH фермента CRISPR, и такой мутантный фермент CRISPR может не расщеплять комплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, образующую комплементарную связь с гРНК.
В одном примере варианта осуществления, в случае SpCas9, когда остаток 840 аминокислотной последовательности SpCas9 мутировал от гистидина до аланина, то есть, когда мутировал до H840A, нуклеазная активность домена HNH инактивирована, и, таким образом, SpCas9 может быть использован в качестве никазы. Полученная таким образом никаза может не расщеплять комплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая образует комплементарную связь с гРНК.
В другом примере варианта осуществления, в случае CjCas9, когда остаток 559 аминокислотной последовательности CjCas9 мутировал от гистидина до аланина, то есть, когда мутировал до H559A, нуклеазная активность домена HNH инактивирована и, следовательно, CjCas9 может быть использован в качестве никазы. Полученная таким образом никаза может не расщеплять комплементарную цепь гена-мишени или нуклеиновой кислоты-мишени, то есть цепь, которая образует комплементарную связь с гРНК.
Кроме того, модификация может представлять собой модификацию для полной инактивации нуклеазной активности фермента CRISPR, и такой мутантный фермент CRISPR может представлять собой неактивный фермент CRISPR.
В настоящем документе модификация может представлять собой модификацию для инактивации нуклеазной активности доменов RuvC и HNH фермента CRISPR, и такой мутантный фермент CRISPR может не расщеплять двойную цепь гена-мишени или нуклеиновой кислоты-мишени.
В одном примере варианта осуществления, в случае SpCas9, когда остатки 10 и 840 в аминокислотной последовательности SpCas9 мутировали от аспарагиновой кислоты и гистидина до аланина, то есть мутировали до D10A и H840A, соответственно, нуклеазные активности домена RuvC и домена HNH инактивированы, двойная цепь гена-мишени или нуклеиновой кислоты-мишени может быть не расщеплена полностью.
В другом примере варианта осуществления, в случае CjCas9, когда остатки 8 и 559 аминокислотной последовательности CjCas9 мутировали от аспарагиновой кислоты и гистидина до аланина, то есть мутировали до D8A и H559A, соответственно, нуклеазные активности доменов RuvC и HNH инактивированы, и, таким образом, двойная цепь гена-мишени или нуклеиновой кислоты-мишени может быть не расщеплена полностью.
Кроме того, мутантный фермент CRISPR может дополнительно включать необязательно функциональный домен в дополнение к изначально присущим характеристикам фермента CRISPR, и такой мутантный фермент CRISPR может иметь дополнительную характеристику в дополнение к изначально присущим характеристикам.
В настоящем документе функциональный домен может быть доменом, обладающим метилазной активностью, деметилазной активностью, активностью активации транскрипции, активностью репрессии транскрипции, активностью высвобождения фактора транскрипции, активностью модификации гистонов, активностью расщепления РНК или активностью связывания нуклеиновых кислот, или меткой или репортерным геном для выделения и очистки белка (включая пептид), но настоящее изобретение этим не ограничено.
Функциональный домен, пептид, полипептид или белок могут представлять собой дезаминазу.
Например, неполный или частичный фермент CRISPR может дополнительно включать цитидиндезаминазу в качестве функционального домена. В одном примере варианта осуществления цитидиндезаминаза, например, редактирующий комплекс 1 аполипопротеина В (APOBEC1), может быть добавлен к никазе SpCas9, тем самым образуя гибридный белок. Образованный таким образом [никаза SpCas9]-[APOBEC1] может быть использован для репарации оснований или редактирования C в T или U, или G в A.
Метка включает в себя гистидиновую (His) метку, V5-метку, FLAG-метку, метку гемагглютинина (HA) вируса гриппа, Myc-метку, VSV-G-метку и метку тиоредоксина (Trx), а репортерный ген включает глутатион-S-трансферазу (GST), пероксидазу хрена (HRP), хлорамфеникол-ацетилтрансферазу (CAT), β-галактозидазу, β-глюкоронидазу, люциферазу, аутофлуоресцентные белки, включая зеленый флуоресцентный белок (GFP), HcRed, DsRed, голубой флуоресцентный белок (CFP), желтый флуоресцентный белок (YFP) и синий флуоресцентный белок (BFP), но настоящее изобретение этим не ограничено.
Кроме того, функциональный домен может представлять собой последовательность или сигнал ядерной локализации (NLS) или последовательность или сигнал ядерного экспорта (NES).
В одном примере фермент CRISPR может включать один или более NLS. В настоящем документе один или более NLS могут быть включены в N-конце фермента CRISPR или в его пределах; С-конце фермента или его пределах; или их комбинацию. NLS может быть последовательностью NLS, полученной из следующих NLS, но настоящее изобретение этим не ограничено: NLS большого T-антигена вируса SV40, имеющего аминокислотную последовательность PKKKRKV; NLS из нуклеоплазмина (например, двухчастичный NLS нуклеоплазмина, имеющий последовательность KRPAATKKAGQAKKKK); NLS c-myc, имеющий аминокислотную последовательность PAAKRVKLD или RQRRNELKRSP; NLS hRNPA1 M9, имеющий последовательность NQSSNFGPMKGGNFGGRSSGPYGGGGQYFAKPRNQGGY; последовательность RMRIZFKNKGKDTAELRRRRVEVSVELRKAKKDEQILKRRNV домена IBB из импортина-α; последовательности VSRKRPRP и PPKKARED Т-белка миомы; последовательность POPKKKPL человеческого p53; последовательность SALIKKKKKMAP c-abl IV мыши; последовательности DRLRR и PKQKKRK вируса гриппа NS1; последовательность RKLKKKIKKL антигена вируса гепатита D; последовательность REKKKFLKRR белка Mx1 мыши; последовательность KRKGDEVDGVDEVAKKKSKK человеческой поли(АДФ-рибозы)-полимеразы; или последовательность NLS RKCLQAGMNLEARKTKK, полученную из последовательности рецептора глюкокортикоидного стероидного гормона (человека).
Кроме того, мутантный фермент CRISPR может включать фермент CRISPR разделенного типа, полученный с помощью разделения фермента CRISPR на две или более частей. Термин «разделенный» относится к функциональному или структурному разделению белка или случайному разделению белка на две или более частей.
В настоящем документе фермент CRISPR разделенного типа может представлять собой полностью, не полностью или частично активный фермент или неактивный фермент.
Например, SpCas9 может быть разделен на две части между остатком 656, тирозином, и остатком 657, треонином, в результате чего образуется разделенный SpCas9.
Кроме того, фермент CRISPR разделенного типа может выборочно включать дополнительный домен, пептид, полипептид или белок для реконструкции.
В настоящем документе «реконструкция» относится к образованию фермента CRISPR разделенного типа, который должен быть структурно таким же или аналогичным ферменту CRISPR дикого типа.
Дополнительный домен, пептид, полипептид или белок для реконструкции могут быть доменами димеризации FRB и FKBP; интеином; доменами ERT и VPR; или доменами, которые образуют гетеродимер в определенных условиях.
Например, SpCas9 может быть разделен на две части между остатком 713, серином, и остатком 714, глицином, тем самым образуя разделенный SpCas9. Домен FRB может быть соединен с одной из двух частей, а домен FKBP может быть соединен с другой. В полученном таким образом разделенном SpCas9 домен FRB и домен FKBP могут быть образованы в димере в среде, в которой присутствует рапамицин, в результате чего образуется реконструированный фермент CRISPR.
Фермент CRISPR или мутантный фермент CRISPR, описанный в настоящем изобретении, может представлять собой полипептид, белок или нуклеиновую кислоту, имеющие последовательность, кодирующую их, и может быть кодон-оптимизирован для объекта, чтобы ввести фермент CRISPR или мутантный фермент CRISPR.
Термин «оптимизация кодонов» относится к процессу модификации последовательности нуклеиновой кислоты с помощью поддержания нативной аминокислотной последовательности при замене по меньшей мере одного кодона нативной последовательности на более часто или наиболее часто используемый в клетках-хозяевах кодон так, чтобы улучшить экспрессию в клетках-хозяевах. Разные виды имеют специфическое смещение к определенному кодону определенной аминокислоты, и смещение кодонов (различие в использовании кодонов между организмами) часто коррелирует с эффективностью трансляции мРНК, которая считается зависимой от характеристики транслированного кодона и наличия специфической молекулы тРНК. Доминирование тРНК, отобранной в клетках, обычно отражает кодоны, наиболее часто используемые в синтезе пептидов. Следовательно, ген может быть изменен с помощью оптимальной экспрессии гена в данном организме на основе оптимизации кодонов.
3. Последовательность-мишень
Термин «последовательность-мишень» представляет собой последовательность оснований, присутствующую в гене-мишени или нуклеиновой кислоте-мишени, и обладает комплементарностью по отношению к гидовой последовательности, содержащейся в гидовом домене гидовой нуклеиновой кислоты. Последовательность-мишень представляет собой последовательность оснований, которая может варьироваться в зависимости от гена-мишени или нуклеиновой кислоты-мишени, то есть объекта для манипуляции или коррекции гена, которая может быть разработана в различных формах в соответствии с геном-мишенью или нуклеиновой кислотой-мишенью.
Последовательность-мишень может образовывать комплементарную связь с гидовой последовательностью, содержащейся в гидовом домене гидовой нуклеиновой кислоты, и длина последовательности-мишени может быть такой же, как у гидовой последовательности.
Последовательность-мишень может представлять собой последовательность из 5-50 оснований.
В одном варианте осуществления последовательность-мишень может представлять собой последовательность из 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 оснований.
Последовательность-мишень может представлять собой последовательность нуклеиновой кислоты, комплементарную гидовой последовательности, содержащейся в гидовом домене гидовой нуклеиновой кислоты, которая имеет, например, по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% или более комплементарность или полную комплементарность.
В одном примере последовательность-мишень может представлять собой или включать последовательность из 1-8 оснований, которая не является комплементарной гидовой последовательности, содержащейся в гидовом домене гидовой нуклеиновой кислоты.
Кроме того, последовательность-мишень может представлять собой последовательность оснований, примыкающую к последовательности нуклеиновой кислоты, которая может быть распознана с помощью белка-редактора.
В одном примере последовательность-мишень может быть непрерывной последовательностью из 5-50 оснований, примыкающей к 5'-концу и/или 3'-концу последовательности нуклеиновой кислоты, которая может быть распознана с помощью белка-редактора.
В одном примере варианта осуществления последовательности-мишени для ферментного комплекса гРНК-CRISPR будут описаны ниже.
Когда ген-мишень или нуклеиновая кислота-мишень являются мишенью ферментного комплекса гРНК-CRISPR,
последовательность-мишень обладает комплементарностью к гидовой последовательности, содержащейся в гидовом домене гРНК. Последовательность-мишень представляет собой последовательность оснований, которая варьируется в зависимости от гена-мишени или нуклеиновой кислоты-мишени, то есть объекта для манипуляции или коррекции гена, которая может быть разработана в различных формах в соответствии с геном-мишенью или нуклеиновой кислотой-мишенью.
Кроме того, последовательность-мишень может представлять собой последовательность оснований, примыкающую к PAM-последовательности, которая способна быть распознана с помощью фермента CRISPR, то есть Cas9 или Cpf1.
В одном примере последовательность-мишень может представлять собой непрерывную последовательность из 5-50 оснований, примыкающую к 5'-концу и/или 3'-концу PAM-последовательности, которую распознает фермент CRISPR.
В одном примере варианта осуществления, когда фермент CRISPR представляет собой SpCas9, последовательность-мишень может представлять собой непрерывную последовательность из 16-25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-NGG-3', 5'- NAG-3' и/или 5'-NGA-3' (N=A, T, G или C; или A, U, G или C).
В другом примере варианта осуществления, когда фермент CRISPR представляет собой StCas9, последовательность-мишень может представлять собой непрерывную последовательность из 16-25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-NGGNG-3' и/или 5'-NNAGAAW-3' (W=A или T и N=A, T, G или C; или A, U, G или C).
В еще одном примере варианта осуществления, когда фермент CRISPR представляет собой NmCas9, последовательность-мишень может быть непрерывной последовательностью из 16-25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-NNNNGATT-3' и/или 5'-NNNGCTT-3' (N=A, T, G или C; или A, U, G или C).
В одном примере варианта осуществления, когда фермент CRISPR представляет собой CjCas9, последовательность-мишень может представлять собой непрерывную последовательность из 16-25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-NNNVRYAC-3' (V=G, C или A; R=A или G, Y=C или T, N=A, T, G или C; или A, U, G или C).
В другом примере варианта осуществления, когда фермент CRISPR представляет собой SmCas9, последовательность-мишень может представлять собой непрерывную последовательность из 16-25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-NGG-3' и/или 5'-NAAR-3' (R=A или G, N=A, T, G или C; или A, U, G или C).
В еще одном примере варианта осуществления, когда фермент CRISPR представляет собой SaCas9, последовательность-мишень может представлять собой непрерывную последовательность из 16 до 25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-NNGRR-3', 5'-NNGRRT-3' и/или 5'-NNGRRV-3' (R=A или G, V=G, C или A, N=A, T, G или C; или A, U, G или C).
В одном примере варианта осуществления, когда фермент CRISPR представляет собой Cpf1, последовательность-мишень может представлять собой непрерывную последовательность из 16-25 оснований, примыкающую к 5'-концу и/или 3'-концу последовательности 5'-TTN-3' (N=A, T, G или C; или A, U, G или C).
В одном примере варианта осуществления настоящего изобретения
последовательность-мишень может быть последовательностью нуклеиновой кислоты, содержащейся в гене PMP22.
Альтернативно,
последовательность-мишень может быть частичной последовательностью нуклеиновой кислоты гена PMP22.
Альтернативно, последовательность-мишень может быть последовательностью нуклеиновой кислоты кодирующего или некодирующего участка или их смесью гена PMP22.
Альтернативно,
последовательность-мишень может представлять собой последовательность нуклеиновой кислоты Промотора, TATA-бокса, CAAT-бокса, Сайта Инициации, Сайта Терминации, Сайта Донорного Сплайсинга, Акцепторного Сайта Сплайсинга, Поли(А)-Сайта, Энхансера, 3'-НТУ (Нетранслируемого участка), 5'-НТУ, Аттенюатора и GC-бокса или их смеси гена PMP22.
Альтернативно,
последовательность-мишень может быть последовательностью нуклеиновой кислоты экзона, интрона или их смесью гена PMP22.
Альтернативно,
последовательность-мишень может быть последовательностью нуклеиновой кислоты, включающей или примыкающей к мутированному участку (например, участку, отличному от гена дикого типа) гена PMP22.
Альтернативно,
последовательность-мишень может представлять собой непрерывную последовательность нуклеиновой кислоты из 5-50 оснований гена PMP22.
В качестве одного примера варианта осуществления настоящего изобретения вышеуказанные последовательности-мишени гена PMP22 приведены в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7.
Продукт для манипуляции фактором, управляющим функцией ШК
4. Комплекс гидовая нуклеиновая кислота-белок-редактор и его применение
Комплекс гидовая нуклеиновая кислота-белок-редактор может модифицировать мишень.
Мишенью может быть мишень в виде нуклеиновой кислоты, гена, хромосомы или белка.
Например, комплекс гидовая нуклеиновая кислота-белок-редактор может быть использован для окончательной регуляции (например, ингибирования, подавления, уменьшения, увеличения или стимулирования) экспрессии представляющего интерес белка, удаления белка, регуляции (например, ингибирования, подавления, уменьшения, увеличения или стимулирования) активности белка или экспрессии нового белка.
В настоящем документе комплекс гидовая нуклеиновая кислота-белок-редактор может действовать на уровне ДНК, РНК, гена или хромосомы.
Например, комплекс гидовая нуклеиновая кислота-белок-редактор может регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) экспрессию белка, кодируемого ДНК-мишенью, удалять белок, регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) активность белка или экспрессировать модифицированный белок посредством манипуляции или модификации ДНК-мишени.
В другом примере комплекс гидовая нуклеиновая кислота-белок-редактор может регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) экспрессию белка, кодируемого ДНК-мишенью, удалять белок, регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) активность белка или экспрессировать модифицированный белок посредством манипуляции или модификации РНК-мишени.
В одном примере комплекс гидовая нуклеиновая кислота-белок-редактор может регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) экспрессию белка, кодируемого ДНК-мишенью, удалять белок, регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) активность белка или экспрессировать модифицированный белок посредством манипуляции или модификации гена-мишени.
В другом примере комплекс гидовая нуклеиновая кислота-белок-редактор может регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) экспрессию белка, кодируемого ДНК-мишенью, удалять белок, регулировать (например, ингибировать, подавлять, уменьшать, увеличивать или стимулировать) активность белка или экспрессировать модифицированный белок посредством манипуляции или модификации хромосомы-мишени.
Комплекс гидовая нуклеиновая кислота-белок-редактор может действовать на стадиях транскрипции и трансляции генов.
В одном примере комплекс гидовая нуклеиновая кислота-белок-редактор может стимулировать или подавлять транскрипцию гена-мишени, тем самым регулируя (например, ингибируя, подавляя, уменьшая, увеличивая или стимулируя) экспрессию белка, кодируемого геном-мишенью.
В другом примере комплекс гидовая нуклеиновая кислота-белок-редактор может стимулировать или подавлять трансляцию гена-мишени, тем самым регулируя (например, ингибируя, подавляя, уменьшая, увеличивая или стимулируя) экспрессию белка, кодируемого геном-мишенью.
Комплекс гидовая нуклеиновая кислота-белок-редактор может действовать на уровне белка.
В одном примере комплекс гидовая нуклеиновая кислота-белок-редактор может изменять или модифицировать белок-мишень, тем самым удаляя белок-мишень или регулируя (например, ингибируя, подавляя, уменьшая, увеличивая или стимулируя) активность белка.
В одном примере варианта осуществления настоящее изобретение относится к комплексу гидовая нуклеиновая кислота-белок-редактор, используемому для изменения фактора, управляющего функцией ШК, например гена PMP22. Предпочтительно предоставляется ферментный комплекс гРНК-CRISPR.
В частности, в настоящем изобретении может быть представлен гРНК, включающую в себя гидовый домен, способный образовывать комплементарную связь с последовательностью-мишенью из гена, например, выделенной или не встречающейся в природе гРНК и ДНК, кодирующей ее. гРНК и последовательность ДНК, кодирующая ее, могут быть устроены так, чтобы они могли комплементарно связываться с последовательностью-мишенью, перечисленной в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7.
Кроме того, участок-мишень гРНК предназначен для обеспечения третьего гена, который имеет модификацию нуклеиновой кислоты, например, с двухцепочечными или одноцепочечными разрывами; или специфическую функцию в сайте-мишени в гене PMP22.
Кроме того, когда две или более гРНК используют для индукции двух или более случаев расщепления в гене-мишени, например, двух- или одноцепочечного разрыва, два или более случая расщепления могут происходить из-за одинаковых или разных белков Cas9.
гРНК может нацеливаться, например, на два или более участка гена PMP22, и
может независимо индуцировать расщепление двойной цепи и/или одиночной цепи гена PMP22, или
может индуцировать вставку одного чужеродного нуклеотида в сайт расщепления гена PMP22.
Кроме того, в другом примере варианта осуществления настоящего изобретения нуклеиновая кислота, составляющая комплекс гидовая нуклеиновая кислота-белок-редактор, может включать:
(а) последовательность, кодирующую гидовую нуклеиновую кислоту, включающую в себя гидовый домен, который является комплементарным последовательности-мишени гена PMP22, как описано в настоящем документе; и
(б) последовательность, кодирующую белок-редактор.
В настоящем документе может быть два или более из (a) в соответствии с участком-мишенью, и (b) может использовать один и тот же или два или более белков-редакторов.
В одном варианте осуществления нуклеиновая кислота может быть предназначена для нацеливания на ее ферментативно активный или частично активный белок-редактор, чтобы разместить его в достаточной степени рядом с сайтом-мишенью для нокаута или нокдауна для того, чтобы уменьшить, снизить или ингибировать экспрессию гена PMP22.
В другом варианте осуществления нуклеиновая кислота может быть предназначена для нацеливания на свой ферментативно неактивный белок-редактор или гибридный белок (например, слияние домена репрессора транскрипции), чтобы разместить его в достаточной степени рядом с сайтом-мишенью для нокдауна для того, чтобы уменьшить, снизить или ингибировать экспрессию гена PMP22.
Кроме того, должно быть очевидно, что вышеописанная структура, функция и все применения комплекса гидовая нуклеиновая кислота-белок-редактор будут использованы для манипуляции фактором, управляющим функцией ШК, то есть геном PMP22.
Использование комплекса гидовая нуклеиновая кислота-белок-редактор
В варианте осуществления для использования комплекса гидовая нуклеиновая кислота-белок-редактор настоящего изобретения манипуляция или модификация мишени в виде ДНК, РНК, генов или хромосом с использованием ферментного комплекса гРНК-CRISPR будет описана ниже.
Генная манипуляция
Ген-мишень или нуклеиновая кислота-мишень могут быть изменены или откорректированы с использованием вышеописанного ферментного комплекса гРНК-CRISPR, то есть комплекса CRISPR. В настоящем документе манипуляция или коррекция гена-мишени или нуклеиновой кислоты-мишени включает все стадии из i) расщепления или повреждения гена-мишени или нуклеиновой кислоты-мишени и ii) репарации поврежденного гена-мишени или нуклеиновой кислоты-мишени.
i) Расщепление или повреждение гена-мишени или нуклеиновой кислоты-мишени
i) Расщепление или повреждение гена-мишени или нуклеиновой кислоты-мишени может представлять собой расщепление или повреждение гена-мишени или нуклеиновой кислоты-мишени с использованием комплекса CRISPR и, в частности, расщепление или повреждение последовательности-мишени в гене-мишени или нуклеиновой кислоте-мишени.
В одном примере расщепление или повреждение гена-мишени или нуклеиновой кислоты-мишени с использованием комплекса CRISPR может представлять собой полное расщепление или повреждение двойной цепи последовательности-мишени.
В одном примере варианта осуществления, когда используют SpCas9 дикого типа, двойная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может быть полностью расщеплена.
В другом примере варианта осуществления, когда используют никазу SpCas9 (D10A) и никазу SpCas9 (H840A), комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), и некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может быть расщеплена с помощью никазы SpCas9 (H840A), и расщепление может происходить последовательно или одновременно.
В еще одном примере варианта осуществления, когда используют никазу SpCas9 (D10A) и никазу SpCas9 (H840A) и две гРНК, имеющие разные последовательности-мишени, комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с первой гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную совязь со второй гНК, может быть расщеплена с помощью никазы SpCas9 (H840A), и расщепление может происходить последовательно или одновременно.
В другом примере расщепление или повреждение гена-мишени или нуклеиновой кислоты-мишени с использованием комплекса CRISPR может быть расщеплением или повреждением только одиночной цепи последовательности-мишени. В настоящем документе одиночная цепь может быть комплементарной одиночной цепью последовательности-мишени, образующей комплементарную связь с гРНК, или некомплементарной одиночной цепью последовательности-мишени, образующей комплементарную связь с гРНК.
В одном примере варианта осуществления, когда используют никазу SpCas9 (D10A), комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), но некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может не быть расщеплена.
В другом примере варианта осуществления, когда используют никазу SpCas9 (H840A), некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может быть расщеплена с помощью никаы SpCas9 (H840A), но комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с гРНК, может не быть расщеплена.
В еще одном примере расщепление или повреждение гена-мишени или нуклеиновой кислоты-мишени с использованием комплекса CRISPR может быть частичным удалением фрагмента нуклеиновой кислоты.
В одном примере варианта осуществления, когда используют две гРНК, имеющие разные последовательности-мишени и SpCas9 дикого типа, двойная цепь последовательности-мишени, образующая комплементарную связь с первой гРНК, может быть расщеплена, а двойная цепь последовательности-мишени, образующая комплементарную связь со второй гРНК, может быть расщеплена, что приводит к удалению фрагментов нуклеиновой кислоты с помощью первой и второй гРНК и SpCas9.
В другом примере варианта осуществления, когда используют две гРНК, имеющие разные последовательности-мишени, SpCas9 дикого типа, никазу SpCas9 (D10A) и никазу SpCas9 (H840A), двойная цепь последовательности-мишени, образующая комплементарную связь с первой гРНК, может быть расщеплена с помощью SpCas9 дикого типа, комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь со второй гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), а некомплементарная одиночная цепь может быть расщеплена с помощью никазы SpCas9 (H840A)), что приводит к удалению фрагментов нуклеиновой кислоты с помощью первой и второй гРНК, SpCas9 дикого типа, никазы SpCas9 (D10A) и никазы SpCas9 (H840A).
В еще одном примере варианта осуществления, когда используют две гРНК, имеющие разные последовательности-мишени, никазу SpCas9 (D10A) и никазу SpCas9 (H840A), комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с первой гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), некомплементарная одиночная цепь может быть расщеплена с помощью никазы SpCas9 (H840A), комплементарная двойная цепь последовательности-мишени, образующая комплементарную связь со второй гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), и некомплементарная одиночная цепь может быть расщеплена с помощью никазы SpCas9 (H840A), что приводит к удалению фрагментов нуклеиновой кислоты с помощью первой и второй гРНК, никазы SpCas9 (D10A) и никазы SpCas9 (H840A).
В еще одном примере варианта осуществления, когда используют три гРНК, имеющие разные последовательности-мишени, никазу SpCas9 дикого типа, никазу SpCas9 (D10A) и никазу SpCas9 (H840A), двойная цепь последовательности-мишени, образующая комплементарную связь с первой гРНК, может быть расщеплена с помощью SpCas9 дикого типа, комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь со второй гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), а некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с третьей гРНК, может быть расщеплена с помощью никазы SpCas9 (H840A), что приводит к удалению фрагментов нуклеиновой кислоты с помощью первой гРНК, второй гРНК, третьей гРНК, SpCas9 дикого типа, никазы SpCas9 (D10A) и никазы SpCas9 (H840A).
В еще одном примере варианта осуществления, когда используют четыре гРНК, имеющие разные последовательности-мишени, никазу SpCas9 (D10A) и никазу SpCas9 (H840A), комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связб с первой гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь со второй гРНК, может быть расщеплена с помощью никазы SpCas9 (H840A), комплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с третьей гРНК, может быть расщеплена с помощью никазы SpCas9 (D10A), а некомплементарная одиночная цепь последовательности-мишени, образующая комплементарную связь с четвертой гРНК, может быть расщеплена с помощью никазы SpCas9 (H840A), что приводит к удалению фрагментов нуклеиновой кислоты с помощью первой гРНК, второй гРНК, третьей гРНК, четвертой гРНК, никазы SpCas9 (D10A) и никазы SpCas9 (H840A).
ii) Репарация или восстановление поврежденного гена-мишени или нуклеиновой кислоты-мишени
Ген-мишень или нуклеиновая кислота-мишень, расщепленные или поврежденные с помощью комплекса CRISPR, могут быть репарированы или восстановлены посредством NHEJ и репарации, направленной гомологией (HDR).
Негомологичное соединение концов (NHEJ)
NHEJ - это способ восстановления или репарации двухцепочечных разрывов в ДНК с помощью соединения обоих концов расщепленной двойной или одиночной цепи и, как правило, когда два совместимых конца, образованных разрывом двойной цепи (например, расщеплением), часто находятся в контакте друг с другом, чтобы полностью соединить два конца, разорванная двойная цепь восстанавливается. NHEJ - это способ восстановления, который может быть использован во всем клеточном цикле, и обычно происходит, когда нет гомологичного генома, который можно использовать в качестве матрицы в клетках, как при G1-фазе.
В процессе репарации поврежденного гена или нуклеиновой кислоты с использованием NHEJ некоторые вставки и/или делеции (индели) в последовательности нуклеиновой кислоты происходят в участке, репарированном NHEJ, такие вставки и/или делеции вызывают сдвиг рамки считывания, приводя к транскриптому мРНК со сдвигом рамки. В результате изначально присущие функции теряются из-за нонсенс-опосредованного распада или неспособности синтезировать нормальные белки. Кроме того, хотя рамка считывания сохраняется, мутации, в которых может быть вызвана вставка или удаление значительного количества последовательности, разрушают функциональность белков. Мутация является локус-зависимой, поскольку мутации в значительном функциональном домене, вероятно, менее допустимы, чем мутации в незначительном участке белка.
В то время как невозможно ожидать, что индель-мутации вырабатываются с помощью NHEJ в естественном состоянии, специфическая индель-последовательность предпочтительна в данном разорванном участке и может происходить из небольшого участка микрогомологии. Обычно длина делеции составляет 1-50 п.н., вставки имеют тенденцию быть короче и часто включают короткую повторяющуюся последовательность, непосредственно окружающую разорванный участок.
Кроме того, NHEJ представляет собой процесс, вызывающий мутацию, и когда нет необходимости создавать специфическую конечную последовательность, может быть использовано для удаления мотива небольшой последовательности.
Специфический нокаут гена, на который нацелен комплекс CRISPR, может быть выполнен с использованием такого NHEJ. Двойная цепь или две одиночные цепи гена-мишени или нуклеиновой кислоты-мишени могут быть расщеплены с использованием фермента CRISPR, такого как Cas9 или Cpf1, и разорванная двойная цепь или две одиночные цепи гена-мишени или нуклеиновой кислоты-мишени могут иметь индели посредством NHEJ, тем самым индуцируя специфический нокаут гена-мишени или нуклеиновой кислоты-мишени. В настоящем документе сайт гена-мишени или нуклеиновой кислоты-мишени, расщепленных с помощью фермента CRISPR, может быть некодирующим или кодирующим участком, и, кроме того, сайт гена-мишени или нуклеиновой кислоты-мишени, восстановленных с помощью NHEJ, может быть некодирующим или кодирующим учатском.
Репарация, направляемая гомологией (HDR)
HDR - это способ коррекции без ошибок, который использует гомологичную последовательность в качестве матрицы для репарации или восстановления поврежденного гена или нуклеиновой кислоты и, как правило, для репарации или восстановления разорванной ДНК, то есть для восстановления изначально присущей информации клеток, разорванную ДНК репарируют с использованием информации комплементарной последовательности оснований, которая не модифицирована, или информации сестринской хроматиды. Наиболее распространенным типом HDR является гомологичная рекомбинация (ГР). HDR - это способ репарации или восстановления, обычно встречающийся в S или G2/M-фазе активно делящихся клеток.
Для репарации или восстановления поврежденной ДНК, используя HDR, вместо использования комплементарной последовательности оснований или сестринского хроматина клеток, матрица ДНК, искусственно синтезированная с использованием информации комплементарной последовательности оснований или гомологичной последовательности оснований, то есть матрица нуклеиновой кислоты, включающая комплементарную последовательность оснований или гомологичную последовательность оснований, может быть обеспечена клеткам, тем самым репарируя разорванную ДНК. В настоящем документе когда последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты дополнительно добавляют к матрице нуклеиновой кислоты для репарации разорванной ДНК, последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, дополнительно добавленные к разорванной ДНК, могут быть подвергнуты нокину. Дополнительно добавленная последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты может представлять собой последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты для коррекции гена-мишени или нуклеиновой кислоты-мишени, модифицированных мутацией в нормальный ген или нуклеиновую кислоту, или ген или нуклеиновую кислоту, которые должны быть экспрессированы в клетках, но настоящее изобретение этим не ограничено.
В одном примере двойная или одиночная цепь гена-мишени или нуклеиновой кислоты-мишени может быть расщеплена с использованием комплекса CRISPR, матрица нуклеиновой кислоты, включающая последовательность оснований, комплементарную последовательности оснований, примыкающей к сайту расщепления, может быть обеспечена клеткам, и расщепленная последовательность оснований гена-мишени или нуклеиновой кислоты-мишени может быть репарирована или восстановлена посредством HDR.
В настоящем документе матрица нуклеиновой кислоты, включающая в себя комплементарную последовательность оснований, может иметь разорванную ДНК, то есть расщепленную двойную или одиночную цепь комплементарной последовательности оснований, и дополнительно включать последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, которые должны быть вставлены в разорванную ДНК. Дополнительная последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты могут быть вставлены в расщепленный сайт разорванной ДНК, то есть ген-мишень или нуклеиновую кислоту-мишень, используя матрицу нуклеиновой кислоты, включающую последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, которые должны быть вставлены в комплементарную последовательность оснований. В настоящем документе последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, которые должны быть вставлены, и дополнительная последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты могут представлять собой последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты для коррекции гена-мишени или нуклеиновой кислоты-мишени, модифицированных мутацией, в нормальный ген или нуклеиновую кислоту, или ген или нуклеиновую кислоту для экспрессии в клетках. Комплементарная последовательность оснований может представлять собой последовательность оснований, имеющую комплементарные связи с разорванной ДНК, то есть правую и левую последовательности оснований расщепленной двойной или одиночной цепи гена-мишени или нуклеиновой кислоты-мишени. Альтернативно, комплементарная последовательность оснований может быть последовательностью оснований, имеющей комплементарные связи с разорванной ДНК, то есть 3'- и 5'-концами расщепленной двойной или одиночной цепи гена-мишени или нуклеиновой кислоты-мишени. Комплементарная последовательность оснований может представлять собой последовательность из 15-3000 оснований, длина или размер комплементарной последовательности оснований могут быть подходящим образом созданы в соответствии с размером матрицы нуклеиновой кислоты или гена-мишени. В настоящем документе в качестве матрицы нуклеиновой кислоты, может быть использована двух- или одноцепочечная нуклеиновая кислота, или она может быть линейной или кольцевой, но настоящее изобретение этим не ограничено.
В другом примере двух- или одноцепочечный ген-мишень или нуклеиновую кислоту-мишень расщепляют с использованием комплекса CRISPR, матрицу нуклеиновой кислоты, включающую гомологичную последовательность оснований с последовательностью оснований, примыкающей к сайту расщепления, обеспечивают клеткам, и расщепленная последовательность оснований гена-мишени или нуклеиновой кислоты-мишени может быть репарирована или восстановлена с помощью HDR.
В настоящем документе матрица нуклеиновой кислоты, включающая гомологичную последовательность оснований, может представлять собой разорванную ДНК, то есть расщепленную двух- или одноцепочечную гомологичную последовательность оснований, и дополнительно включать последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, которые должны быть вставлены в разорванную ДНК. Дополнительная последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты могут быть вставлены в разорванную ДНК, то есть расщепленный сайт гена-мишени или нуклеиновой кислоты-мишени, с использованием матрицы нуклеиновой кислоты, включающей гомологичную последовательность оснований и последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, которые должны быть вставлены. В настоящем документе последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты, которые должны быть вставлены, и дополнительная последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты могут представлять собой последовательность нуклеиновой кислоты или фрагмент нуклеиновой кислоты для коррекции гена-мишени или нуклеиновой кислоты-мишени, модифицированных мутацией, в нормальный ген или нуклеиновую кислоту, или ген или нуклеиновую кислоту для экспрессии в клетках. Гомологичная последовательность оснований может представлять собой разорванную ДНК, то есть последовательность оснований, имеющую гомологию с расщепленной двухцепочечной последовательностью оснований или правой и левой одноцепочечными последовательностями оснований гена-мишени или нуклеиновой кислоты-мишени. Альтернативно, комплементарная последовательность оснований может представлять собой последовательность оснований, имеющую гомологию с разорванной ДНК, то есть 3'- и 5'-концами расщепленной двойной или одиночной цепи гена-мишени или нуклеиновой кислоты-мишени. Гомологичная последовательность оснований может представлять собой последовательность из 15-3000 оснований, и длина или размер гомологичной последовательности оснований могут быть подходящим образом созданы в соответствии с размером матрицы нуклеиновой кислоты или гена-мишени или нуклеиновой кислоты-мишени. В настоящем документе в качестве матрицы нуклеиновой кислоты можно использовать двух- или одноцепочечную нуклеиновую кислоту, которая может быть линейной или кольцевой, но настоящее изобретение этим не ограничено.
Помимо NHEJ и HDR, существуют способы репарации или восстановления разорванной ДНК.
Одноцепочечный отжиг (SSA)
SSA представляет собой метод репарации двухцепочечных разрывов между двумя повторяющимися последовательностями, присутствующими в нуклеиновой кислоте-мишени, и обычно использует повторяющуюся последовательность из более чем 30 оснований. Повторяющуюся последовательность расщепляют (чтобы иметь липкие концы), чтобы иметь одиночную цепь по отношению к двойной цепи нуклеиновой кислоты-мишени на каждом из разорванных концов, и после расщепления одноцепочечный выступ, содержащий повторяющуюся последовательность, покрывают белком RPA, так что предотвращается неправильный отжиг повторяющихся последовательностей друг с другом. RAD52 связывается с каждой повторяющейся последовательностью на выступе, и выстраивается последовательность, способная к отжигу комплементарной повторяющейся последовательности. После отжига одноцепочечный лоскут выступа расщепляют, и синтез новой ДНК заполняет определенный промежуток для восстановления двойной цепи ДНК. В результате этой репарации последовательность ДНК между двумя повторами удаляется, и длина делеции может зависеть от различных факторов, включая расположение двух повторов, используемых в настоящем документе, а также путь или степень процесса расщепления.
SSA, подобно HDR, использует комплементарную последовательность, то есть комплементарную повторяющуюся последовательность, и, напротив, не требует матрицы нуклеиновой кислоты для модификации или коррекции последовательности-мишени нуклеиновой кислоты.
Репарация одноцепочечного разрыва (РОЦР)
Одноцепочечные разрывы в геноме репарируют с помощью отдельного механизма, РОЦР, из вышеописанных механизмов репарации. В случае одноцепочечных разрывов ДНК PARP1 и/или PARP2 распознает разрывы и запускает механизм репарации. Связывание PARP1 и активность в отношении разрывов ДНК являются временными, и РОЦР стимулируют с помощью поддержания стабильности белкового комплекса РОЦР в поврежденных участках. Наиболее важным белком в комплексе РОЦР является XRCC1, который взаимодействует с белком, стимулируя процессинг 3'- и 5'-концов ДНК для стабилизации ДНК. Процессинг концов обычно участвует в репарации поврежденного 3'-конца до гидроксилированного состояния и/или поврежденного 5'-конца до фосфатной группы, и после процессинга концов происходит заполнение промежутка ДНК. Существует два способа заполнения промежутка ДНК, а именно: репарация короткими последовательностями и репарация длинными последовательностями, и репарация короткими последовательностями включает вставку одного основания. После заполнения промежутка ДНК, ДНК-лигаза стимулирует соединение концов.
Репарация ошибочно спаренных оснований (РОО)
РОО работает с ошибочно спаренными основаниями ДНК. Каждый из комплексов MSH2/6 или MSH2/3 обладает активностью АТФазы и, таким образом, играет важную роль в распознавании ошибочного спаривания и инициации репарации, а MSH2/6 в первую очередь распознает ошибочно спаренные основания и идентифицирует одно или два ошибочных спаривания оснований, но MSH2/3 в первую очередь распознает наибольшее ошибочное спаривание.
Эксцизионная репарация оснований (ЭРО)
ЭРО - это способ репарации, который активен на протяжении всего клеточного цикла и используют для удаления небольшого поврежденного участка оснований, без искажения структуры спирали, из генома. В поврежденной ДНК поврежденные основания удаляют с помощью расщепления N-гликозидной связи, соединяющей основание с фосфат-дезоксирибозным остовом, а затем происходит расщепление фосфодиэфирного остова, что приводит к разрывам в одноцепочечной ДНК. Концы разованной одиночной цепи, образованные таким образом, были удалены, промежуток, образованный из-за удаленной одиночной цепи, заполняют новым комплементарным основанием, а затем конец вновь заполненного комплементарного основания лигируется с остовом с помощью ДНК-лигазы, что приводит к репарации поврежденной ДНК.
Эксцизионная репарация нуклеотидов (ЭРН)
ЭРН является эксцизионным механизмом, важным для удаления большого повреждения, искажающего структуру спирали, из ДНК, и когда повреждение распознано, короткий сегмент одноцепочечной ДНК, содержащий поврежденный участок, удаляют, что приводит к одноцепочечному промежутку из 22-30 оснований. Созданный промежуток заполняют новым комплементарным основанием, а конец вновь заполненного комплементарного основания лигируется с остовом ДНК-лигазой, что приводит к репарации поврежденной ДНК.
Эффекты генной манипуляции
Манипуляция или коррекция гена-мишени или нуклеиновой кислоты-мишени могут в значительной степени привести к эффектам нокаута, нокдауна и нокина.
Нокаут
Термин «нокаут» относится к инактивации гена-мишени или нуклеиновой кислоты-мишени, а «инактивация гена-мишени или нуклеиновой кислоты-мишени» относится к состоянию, в котором не происходит транскрипция и/или трансляция гена-мишени или нуклеиновой кислоты-мишени. Транскрипция и трансляция гена, вызывающего заболевание, или гена, имеющего аномальную функцию, может быть ингибирована посредством нокаута, что приводит к предотвращению экспрессии белка.
Например, когда ген-мишень или нуклеиновая кислота-мишень редактируют или корректируют с использованием ферментного комплекса гРНК-CRISPR, то есть комплекса CRISPR, ген-мишень или нуклеиновая кислота-мишень могут быть расщеплены с использованием комплекса CRISPR. Поврежденный ген-мишень или нуклеиновая кислота могут быть восстановлены посредством NHEJ с использованием комплекса CRISPR. Поврежденный ген-мишень или нуклеиновая кислота-мишень могут иметь индели из-за NHEJ, и, таким образом, может быть индуцирован специфический нокаут для гена-мишени или нуклеиновой кислоты-мишени.
Нокдаун
Термин «нокдаун» относится к уменьшению транскрипции и/или трансляции гена-мишени или нуклеиновой кислоты-мишени или экспрессии белка-мишени. Начало заболевания может быть предотвращено, или заболевание можно лечить, регулируя сверхэкспрессию гена или белка посредством нокдауна.
Например, когда ген-мишень или нуклеиновая кислота-мишень редактируют или корректируют с использованием неактивного комплекса домена ингибирующей активности фермент-транскрипции гРНК-CRISPR, то есть неактивного комплекса CRISPR, включающего домен ингибирующей активности транскрипции, неактивный комплекс CRISPR может специфически связываться с геном-мишенью или нуклеиновой кислотой-мишенью, транскрипция гена-мишени или нуклеиновой кислоты-мишени могут быть ингибированы доменом ингибирующей активности транскрипции, включенным в неактивный комплекс CRISPR, тем самым индуцируя нокдаун, при котором ингибируется экспрессия соответствующего гена или нуклеиновой кислоты.
Нокин
Термин «нокин» относится к вставке специфической нуклеиновой кислоты или гена в ген-мишень или нуклеиновую кислоту-мишень, и в настоящем документе «специфическая нуклеиновая кислота» относится к гену или нуклеиновой кислоте, представляющим интерес, которые должны быть вставлены или экспрессированы. Мутантный ген, запускающий заболевание, может быть использован при лечении заболевания с помощью коррекции до нормального или вставки нормального гена для индукции экспрессии нормального гена посредством нокина.
Кроме того, для нокина может дополнительно понадобиться донор.
Например, когда ген-мишень или нуклеиновая кислота-мишень редактируют или корректируютс с использованием ферментного комплекса гРНК-CRISPR, то есть комплекса CRISPR, ген-мишень или нуклеиновая кислота-мишень могут быть расщеплены с использованием комплекса CRISPR. Ген-мишень или нуклеиновая кислота-мишень, поврежденные с использованием комплекса CRISPR, могут быть репарированы посредством HDR. В настоящем документе специфическая нуклеиновая кислота может быть вставлена в поврежденный ген или нуклеиновую кислоту с использованием донора.
Термин «донор» относится к последовательности нуклеиновой кислоты, которая помогает репарации поврежденного гена или нуклеиновой кислоты на основе HDR, и в настоящем документе донор может включать специфическую нуклеиновую кислоту.
Донором может быть двух- или одноцепочечная нуклеиновая кислота.
Донор может присутствовать в линейной или кольцевой форме.
Донор может включать последовательность нуклеиновой кислоты, имеющую гомологию с геном-мишенью или нуклеиновой кислотой-мишенью.
Например, донор может включать последовательность нуклеиновой кислоты, имеющую гомологию с каждой из последовательностей оснований в местоположении, в которое должна быть вставлена специфическая нуклеиновая кислота, например, до (слева) и после (справа) поврежденной нуклеиновой кислоты. В настоящем документе специфическая нуклеиновая кислота, которая должна быть вставлена, может быть расположена между последовательностью нуклеиновой кислоты, имеющей гомологию с последовательностью оснований после поврежденной нуклеиновой кислоты, и последовательностью нуклеиновой кислоты, имеющей гомологию с последовательностью оснований до поврежденной нуклеиновой кислоты. В настоящем документе гомологичная последовательность нуклеиновой кислоты может иметь по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более гомологию или полную гомологию.
Донор может необязательно включать дополнительную последовательность нуклеиновой кислоты. В настоящем документе дополнительная последовательность нуклеиновой кислоты может служить для повышения стабильности донора, эффективности нокина или эффективности HDR.
Например, дополнительная последовательность нуклеиновой кислоты может представлять собой A, T-обогащенную последовательность нуклеиновой кислоты, то есть A-T-обогащенный домен. Кроме того, дополнительная последовательность нуклеиновой кислоты может представлять собой Скаффолд/матрикс-связывающий участок (SMAR).
В одном примере варианта осуществления, относящегося к эффекту генной манипуляции настоящего изобретения, измененный ген-мишень, полученный с использованием ферментного комплекса гРНК-CRISPR, то есть измененный фактор, управляющий функцией ШК, предпочтительно, измененный PMP22, может иметь следующую конструкцию.
В одном примере варианта осуществления, когда PMP22 представляет собой ген,
строение искусственно измененного PMP22 с помощью ферментного комплекса гРНК-CRISPR может включать модификацию одной или более нуклеиновых кислот из:
делеции или вставки одного или более нуклеотидов;
замещения одним или более нуклеотидами, отличными от гена дикого типа; и
вставки одного или более чужеродных нуклеотидов
в непрерывном участке из 1-50 п.н., 1-40 п.н. или 1-30 п.н., предпочтительно 3-25 п.н. в последовательности оснований, который находится в PAM-последовательности в последовательности нуклеиновой кислоты, составляющей ген PMP22 или примыкающей к его 5'-концу и/или 3'-концу.
Кроме того, химическая модификация одного или более нуклеотидов может быть включена в последовательность нуклеиновой кислоты, составляющую ген PMP22.
В настоящем документе «чужеродный нуклеотид» представляет собой концепцию, включающую все экзогенные, например, гетерологичные или искусственно синтезированные нуклеотиды, за исключением нуклеотидов, изначально включенных в ген PMP22. Чужеродный нуклеотид также включает в себя нуклеотид размером в несколько сотен, тысяч или десятков тысяч п.н. для экспрессии белка, обладающего специфической функцией, а также небольшой олигонуклеотид размером в 50 п.н. или менее. Такой чужеродный нуклеотид может быть донором.
Химическая модификация может включать метилирование, ацетилирование, фосфорилирование, убиквитинирование, АДФ-рибозилирование, миристоилирование и гликозилирование, например, замещение некоторых функциональных групп, содержащихся в нуклеотиде, на любой из атома водорода, атома фтора, -O-алкильной группы, -O-ацильной группы и аминогруппы, но настоящее изобретение не ограничено ими. Кроме того, для увеличения трансферабельности молекулы нуклеиновой кислоты функциональные группы также могут быть замещены любым из -Br, -Cl, -R, -R'OR, -SH, -SR, -N3 и -CN (R=алкил, арил, алкилен). Кроме того, фосфатный остов по меньшей мере одного нуклеотида может быть замещен любой из алкилфосфонатной формы, фосфороамидатной формы и боранофосфатной формы. Кроме того, химическая модификация может представлять собой замещение по меньшей мере одного типа нуклеотида, содержащегося в молекуле нуклеиновой кислоты, любой из закрытой нуклеиновой кислоты (LNA), открытой нуклеиновой кислоты (UNA), морфолино и пептидной нуклеиновой кислоты (PNA), и химическая модификация может представлять собой соединение молекулы нуклеиновой кислоты с одним или более, выбранными из группы, состоящей из липида, проникающего в клетку пептида и лиганда клетки-мишени.
Чтобы сформировать необходимую систему, управляющую функцией ШК (включая систему модификации, управляющей функцией ШК), искусственная модификация с использованием ферментного комплекса гРНК-CRISPR может быть применена к нуклеиновой кислоте, составляющей фактор, управляющий функцией ШК, то есть ген PMP22.
Участок, включающий модификацию нуклеиновой кислоты гена PMP22, может представлять собой участок-мишень или последовательность-мишень.
Такая последовательность-мишень может быть мишенью для ферментного комплекса гРНК-CRISPR, и последовательность-мишень может включать или не включать PAM-последовательность, распознаваемую с помощью фермента CRISPR. Такая последовательность-мишень может предоставить допустимую норму на стадии конструирования гРНК специалистам в данной области техники.
Такая модификация нуклеиновой кислоты включает «расщепление» нуклеиновой кислоты.
Термин «расщепление» в участке-мишени относится к разрыву ковалентного остова полинуклеотидов. Расщепление включает ферментативный или химический гидролиз фосфодиэфирной связи, но настоящее изобретение этим не ограничено, а также включает различные другие способы. Расщепление может быть выполнено как с одиночной, так и с двойной цепью, и расщепление двойной цепи может быть результатом четкого расщепления одиночной цепи. Двухцепочечное расщепление может привести к тупым концам или ступенчатым концам.
Когда используют инактивированный фермент CRISPR, он может индуцировать фактор, обладающий специфической функцией, для приближения к определенному участку участка-мишени или гена PMP22 без процесса расщепления. Химическая модификация одного или более нуклеотидов в последовательности нуклеиновой кислоты гена PMP22 может быть включена в соответствии с такой специфической функцией.
В одном примере могут происходить различные индели из-за целевых и нецелевых активностей посредством расщепления нуклеиновой кислоты, образованного с помощью ферментного комплекса гРНК-CRISPR.
Термин «индель» является общим термином для инсерционной или делеционной мутации, возникающей между некоторыми основаниями в последовательности оснований ДНК. Индель может быть введен в последовательность-мишень во время репарации с помощью механизма HDR или NHEJ, когда ферментный комплекс гРНК-CRISPR расщепляет нуклеиновую кислоту (ДНК или РНК) фактора, связанного с неоваскуляризацией, как описано выше.
Искусственно измененный ген PMP22 настоящего изобретения относится к модификации последовательности нуклеиновой кислоты нативного гена с помощью расщепления, инделей или вставки с использованием донора такой нуклеиновой кислоты и участвует в необходимой системе, управляющей функцией ШК (включая систему модификации фактора, управляющего ШК), например, проявляет эффект управления функциями шванновских клеток посредством экспрессии и/или модификации функции PMP22.
Например,
экспрессия и активность специфического белка могут быть ингибированы с помощью искусственно измененного гена PMP22.
Специфический белок может быть инактивирован с помощью искусственно измененого гена PMP22.
В качестве примера, специфический сайт-мишень гена PMP22 в геноме может быть расщеплен для нокдауна или нокаута гена.
В качестве другого примера, направленный нокдаун может быть опосредован с использованием ферментативно инактивированного фермента CRISPR, слитого с доменом репрессора транскрипции или белком модификации хроматина для того, чтобы изменить транскрипцию, например, блокируя, уменьшая или снижая транскрипцию гена PMP22.
Рост шванновских клеток может быть подавлен или ингибирован, или может быть простимулирован или увеличен с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Прохождение клеточного цикла шванновских клеток может быть прервано или остановлено, или может быть простимулировано с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Дифференцировка шванновских клеток может быть простимулирована или подавлена с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Гибель шванновских клеток может быть простимулирована или подавлена с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Выживаемости периферических нервных клеток может способствовать или мешать с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Поддержанию и передаче сигналов периферических нервных клеток может способствовать или мешать с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Образованием миелиновой оболочки аксонов нервных клеток можно управлять с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
В этом случае образование миелиновой оболочки включает в себя весь процесс образования миелиновой оболочки, такой как образование миелиновой оболочки, дегенерация миелиновой оболочки, регенерация миелиновой оболочки, поддержание миелиновой оболочки и компактного миелина миелиновой оболочки, и все механизмы, связанные с дегенерацией миелиновой оболочки или функцией миелиновой оболочки.
Облегчение и лечение из-за дисфункциональных или дефектных шванновских клеток может быть выполнено с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Кроме того, можно управлять всеми механизмами, которые подавляют или ингибируют рост фибробластов или глиальных клеток, или стимулируют или увеличивают рост фибробластов или глиальных клеток с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
Можно управлять всеми механизмами, которые участвуют в активности фибробластов или глиальных клеток, с помощью искусственно измененного фактора, управляющего функцией ШК, то есть искусственно измененного гена PMP22.
В одном примере варианта осуществления настоящего изобретения искусственно измененный фактор, управляющий функцией ШК, то есть искусственно измененный ген PMP22, может обеспечивать различный искусственно измененный ген PMP22 в соответствии со структурной характеристикой ферментного комплекса гРНК-CRISPR (например, включенного в участок-мишень гена PMP22 или в другой в смежной основной PAM-последовательности).
Далее, хотя были проиллюстрированы репрезентативные примеры ферментов CRISPR, они являются просто конкретными примерами, и, таким образом, настоящее изобретение не ограничено ими.
Например, когда фермент CRISPR представляет собой белок SpCas9, PAM-последовательность представляет собой 5'-NGG-3' (N представляет собой A, T, G или C), и участок расщепленной последовательности оснований (участок-мишень) может быть непрерывным участком из 1-25 п.н., например, 17-23 п.н. или 21-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NGG-3' в гене-мишени.
Настоящая заявка может предоставить искусственно измененный ген PMP22, который получают с помощью
а) делеции одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NGG-3' (N представляет собой A, T, C или G).
б) замещения одного или более нуклеотидов непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NGG-3' нуклеотидами, отличными от нуклеотидов гена дикого типа,
c) вставки одного или более нуклеотидов в непрерывный участок из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NGG- 3', или
d) комбинации двух или более, выбранных из а) до с)
в последовательности нуклеиновой кислоты гена PMP22.
Например, когда фермент CRISPR представляет собой белок CjCas9, PAM-последовательность представляет собой 5'-NNNNRYAC-3' (каждый из N независимо представляет собой A, T, C или G, R представляет собой A или G, а Y представляет собой C или T), и участок расщепленной последовательности оснований (участок-мишень) может представлять собой непрерывный участок из 1-2 п.н., например, 17-23 п.н. или 21-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNNNRYAC-3' в гене-мишени.
В настоящем изобретении может быть представлен искусственно измененный ген PMP22, который получают с помощью:
а') делеции одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу 5'-NNNNRYAC-3' (каждый из N независимо представляет собой A, T, C или G, R представляет собой A или G, а Y представляет собой C или T),
b') замещения одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNNNRYAC-3' нуклеотидами, отличными от нуклеотидов гена дикого типа,
c') вставки одного или более нуклеотидов в непрерывный участок из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNNNRYAC-3', или
d') комбинации двух или более, выбранных из а') до с') в последовательности нуклеиновой кислоты гена PMP22.
Например, когда фермент CRISPR представляет собой белок StCas9, PAM-последовательность представляет собой 5'-NNAGAAW-3' (каждый из N независимо представляет собой A, T, C или G, а W представляет собой A или T), а участок расщепленной последовательности оснований (участок-мишень) может быть непрерывным участком из 1-25 п.н., например, 17-23 п.н. или 21-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNAGAAW-3' в гене-мишени.
Настоящее изобретение может предоставить искусственно измененный ген PMP22, который получают с помощью:
a'') делеции одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNAGAAW-3' (каждый из N независимо представляет собой A, T, C или G, а W представляет собой A или T),
b'') замены одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNAGAAW-3' нуклеотидами, отличными от нуклеотидов гена дикого типа,
c'') вставки одного или более нуклеотидов в непрерывный участок из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к последовательности 5'-NNAGAAW-3', или
d'') комбинации двух или более, выбранных из a'') до c'') в последовательности нуклеиновой кислоты гена PMP22.
Например, когда фермент CRISPR представляет собой белок NmCas9, PAM-последовательность представляет собой 5'-NNNNGATT-3' (каждый из N независимо представляет собой A, T, C или G), и участок расщепленной последовательности оснований (участок-мишень) может представлять собой непрерывный участок из 1-25 п.н., например, 17-23 п.н. или 21-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNNNGATT-3' в гене-мишени.
Настоящее изобретение может предоставить искусственно измененный ген PMP22, который получают с помощью:
a''') делеции одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу 5'-NNNNGATT-3' (каждый из N независимо представляет собой A, T, C или G),
b''') замещения одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNNNGATT-3' нуклеотидами, отличными от нуклеотидов гена дикого типа,
c''') вставки одного или более нуклеотидов в непрерывный участок из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNNNGATT-3', или
d''') комбинации двух или более, выбранных из a''') до c''') в последовательности нуклеиновой кислоты гена PMP22.
Например, когда фермент CRISPR представляет собой белок SaCas9, PAM-последовательность представляет собой 5'-NNGRR(T)-3' (каждый из N независимо представляет собой A, T, C или G, R представляет собой A или G, и (T) представляет собой произвольно добавляемую последовательность), и участок расщепленной последовательности оснований (участок-мишень) может представлять собой непрерывный участок из 1-25 п.н., например, 17-23 п.н. или 21-23 п.н., в последовательности оснований, примыкающей к 5'-концу/или 3'-концу последовательности 5'-NNGRR(T)-3' в гене-мишени.
Настоящее изобретение может предоставить искусственно измененный ген PMP22, который получают с помощью:
a'''') делеции одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNGRR(T)-3' (каждый из N независимо представляет собой A, T, C или G, R представляет собой A или G, и (T) представляет собой произвольно добавляемую последовательность),
b'''') замещения одного или более нуклеотидов из непрерывного участка из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-NNGRR(T)-3' нуклеотидами, отличными от нуклеотидов гена дикого типа,
c'''') вставки одного или более нуклеотидов в непрерывный участок из 1-25 п.н., например, 17-23 п.н., в последовательности оснований, примыкающей к последовательности 5'-NNGRR(T)-3', или
d'''') комбинации двух или более, выбранных из a'''') до c'''') в последовательности нуклеиновой кислоты гена PMP22.
Например, когда фермент CRISPR представляет собой белок Cpf1, PAM-последовательность представляет собой 5'-TTN-3' (N представляет собой A, T, C или G), и участок расщепленной последовательности оснований (участок-мишень) может быть непрерывным участком из 10-30 п.н., например, 15-26 п.н., 17-30 п.н. или 17-26 п.н., в последовательности оснований, примыкающей к 5'-концу или 3'-концу последовательности 5'-TTN-3'.
Белок Cpf1 может быть получен из микроорганизма, такого как Parcubacteria bacterium (GWC2011_GWC2_44_17), Lachnospiraceae bacterium (MC2017), Butyrivibrio proteoclasiicus, Peregrinibacteria bacterium (GW2011_GWA_33_10), Acidaminococcus sp. (BV3L6), Porphyromonas macacae, Lachnospiraceae bacterium (ND2006), Porphyromonas crevioricanis, Prevotella disiens, Moraxella bovoculi (237), Smiihella sp. (SC_KO8D17), Leptospira inadai, Lachnospiraceae bacterium (MA2020), Francisella novicida (U112), Candidatus Methanoplasma termitum, или Eubacterium eligens, например, Parcubacteria bacterium (GWC2011_GWC2_44_17), Peregrinibacteria bacterium (GW2011_GWA_33_10), Acidaminococcus sp. (BV3L6), Porphyromonas macacae, Lachnospiraceae bacterium (ND2006), Porphyromonas crevioricanis, Prevotella disiens, Moraxella bovoculi (237), Leptospira inadai, Lachnospiraceae bacterium (MA2020), Francisella novicida (U112), Candidatus Methanoplasma termitum, или Eubacterium eligens, но настоящее изобретение этим не ограничено.
Настоящее изобретение может предоставить искусственно измененный ген PMP22, который получают с помощью:
a''''') делеции одного или более нуклеотидов из непрерывного участка из 10-30 п.н., например 15-26 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-TTN-3' (N представляет собой A, T, C или G),
b''''') замещения одного или более нуклеотидов из непрерывного участка из 10-30 п.н., например, 15-26 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-TTN-3' нуклеотидами, отличными от нуклеотидов гена дикого типа,
c''''') вставки одного или более нуклеотидов из непрерывного участка из 10-30 п.н., например, 15-26 п.н., в последовательности оснований, примыкающей к 5'-концу и/или 3'-концу последовательности 5'-TTN-3', или
d''''') комбинации двух или более элементов, выбранных из a''''') до c'''') в последовательности нуклеиновой кислоты гена PMP22.
В другом примере варианта осуществления, когда PMP22 представляет собой белок,
искусственно измененный белок включает все белки, участвующие в образовании новых или модифицированных кровеносных сосудов с помощью прямого или косвенного действия ферментного комплекса гРНК-CRISPR.
Например, искусственно измененный белок может быть белком, экспрессируемым с помощью гена PMP22, искусственно измененного с помощью ферментного комплекса гРНК-CRISPR, или другим белком, увеличенным или уменьшенным под воздействием такой белковой активности, но настоящее изобретение этим не ограничено.
Искусственно измененный белок PMP22 может иметь аминокислотную композицию и активность, соответствующую композиции искусственно измененного гена PMP22.
В качестве варианта осуществления,
может быть обеспечен (i) искусственно измененный белок, который изменяют по характеристикам экспрессии.
Например, модификация белка может иметь одну или более характеристик:
уменьшение или увеличение уровня экспрессии в соответствии с делецией или вставкой одного или более нуклеотидов в непрерывном участке из 1-50 п.н., 1-40 п.н., 1-30 п.н., и предпочтительно 3-25 п.н. в последовательности оснований PAM-последовательности в последовательности нуклеиновой кислоты гена PMP22 или примыкающей к ее 5'-концу и/или 3'-концу;
уменьшение или увеличение уровня экспрессии в соответствии с замещением одним или более нуклеотидами, отличными от нуклеотидов гена дикого типа;
уменьшение или увеличение уровня экспрессии, экспрессии гибридного белка или независимой экспрессии специфического белка в соответствии со вставкой одного или более чужеродных нуклеотидов; и
уменьшение или увеличение уровня экспрессии третьего белка под влиянием характеристик экспрессии вышеописанных белков.
Может быть обеспечен (ii) искусственно измененный белок, который изменяют по структурным характеристикам.
Например, модификация белка может иметь одну или более характеристик:
изменение кодонов, аминокислот и трехмерной структуры в соответствии с делецией или вставкой одного или более нуклеотидов в непрерывном участке из 1-50 п.н., 1-40 п.н., 1-30 п.н., и предпочтительно 3-25 п.н. в последовательности оснований PAM-последовательности в последовательности нуклеиновой кислоты гена PMP22 или примыкающей к ее 5'-концу и/или 3'-концу;
изменение кодонов, аминокислот и трехмерной структуры таким образом, в соответствии с замещением одним или более нуклеотидами, отличными от гена дикого типа;
изменение кодонов, аминокислот и трехмерной структуры или структуры слияния со специфическим белком или независимой структурой, из которой выделен конкретный белок в соответствии со вставкой одного или более чужеродных нуклеотидов; и
изменение кодонов, аминокислот и трехмерной структуры третьего белка под влиянием вышеуказанного белка, измененного по структурной характеристике.
Может быть обеспечен (iii) искусственно измененный белок, измененный по функциональным характеристикам.
Например, модификация белка может иметь одну или более характеристик:
активация или инактивация специфической функции или введение новой функции с помощью модификации белка, вызванной делецией или вставкой одного или более нуклеотидов в непрерывном участке из 1-50 п.н., 1-40 п.н., 1-30 п.н. и предпочтительно 3-25 п.н. в последовательности оснований PAM-последовательности в последовательности нуклеиновой кислоты гена PMP22 или примыкающей к ее с 5'-концуи или 3'-концу;
активация или инактивация специфической функции или введение новой функции с помощью модификации белка, вызванной замещением одним или более нуклеотидами, отличными от нуклеотидов гена дикого типа;
активация или инактивация специфической функции или введение новой функции с помощью модификации белка, вызванной вставкой одного или более чужеродных нуклеотидов, в частности, введение третьей функции в существующую функцию вследствие слияния или независимой экспрессии специфического белка; и
изменение функции третьего белка под влиянием вышеописанного белка, измененного по функциональным характеристикам.
Кроме того, может быть включен белок, искусственно измененный с помощью химической модификации одного или более нуклеотидов в последовательности нуклеиновой кислоты, составляющей ген PMP22.
Например, могут быть изменены одно или более из экспрессии, структурных и функциональных характеристик белка, вызванных метилированием, ацетилированием, фосфорилированием, убиквитинированием, АДФ-рибозилированием, миристоилированием и гликозилированием.
Например, третья структура и функция могут быть получены с помощью связывания третьего белка с последовательностью нуклеиновой кислоты гена вследствие химической модификации нуклеотидов.
5. Другие дополнительные компоненты
Дополнительный компонент может быть выборочно добавлен для повышения эффективности комплекса гидовая нуклеиновая кислота-белок-редактор или повышения эффективности репарации поврежденного гена или нуклеиновой кислоты.
Дополнительный компонент может быть выборочно использован для улучшения эффективности комплекса гидовая нуклеиновая кислота-белок-редактор.
Активатор
Дополнительный компонент может быть использован в качестве активатора для повышения эффективности расщепления мишени в виде нуклеиновой кислоты, гена или хромосомы комплекса гидовая нуклеиновая кислота-белок-редактор.
Термин «активатор» относится к нуклеиновой кислоте, служащей для стабилизации связи между комплексом гидовая нуклеиновая кислота-белок-редактор и мишенью в виде нуклеиновой кислоты, гена или хромосомы, или чтобы позволить комплексу гидовая нуклеиновая кислота-белок-редактор легче приблизиться к мишени в виде нуклеиной кислоты, гена или хромосомы.
Активатор может представлять собой двухцепочечную нуклеиновую кислоту или одноцепочечную нуклеиновую кислоту.
Активатор может быть линейным или кольцевым.
Активатор может быть разделен на «хелпер», который стабилизирует связь между комплексом гидовая нуклеиновая кислота-белок-редактор и мишенью в виде нуклеиновой кислоты, гена или хромосомы, и «эскортер», который служит для того, чтобы позволить комплексу гидовая нуклеиновая кислота-белок-редактор легче приблизиться к мишени в виде нуклеиновой кислоты, гена или хромосомы.
Хелпер может повысить эффективность расщепления комплекса гидовая нуклеиновая кислота-белок-редактор по отношению к мишени в виде нуклеиновой кислоты, гена или хромосомы.
Например, хелпер включает последовательность нуклеиновой кислоты, имеющую гомологию с мишенью в виде нуклеиновой кислоты, гена или хромосомы. Следовательно, когда комплекс гидовая нуклеиновая кислота-белок-редактор связан с мишенью в виде нуклеиновой кислоты, гена или хромосомы, гомологичная последовательность нуклеиновой кислоты, включенная в хелпер, может образовывать дополнительную комплементарную связь с мишенью в виде нуклеиновой кислоты, гена или хромосомы для стабилизации связи между комплексом гидовая нуклеиновая кислота-белок-редактор и мишенью в виде нуклеиновой кислоты, гена или хромосомы.
Эскортер может повысить эффективность расщепления комплекса гидовая нуклеиновая кислота-белок-редактор по отношению к мишени в виде нуклеиновой кислоты, гена или хромосомы.
Например, эскортер включает последовательность нуклеиновой кислоты, имеющую гомологию с мишенью в виде нуклеиновой кислоты, гена или хромосомы. В настоящем документе гомологичная последовательность нуклеиновой кислоты, включенная в эскортер, может частично образовывать комплементарную связь с комплексом гидовая нуклеиновая кислота-белок-редактор. Следовательно, эскортер, частично образующий комплементарную связь с комплексом гидовая нуклеиновая кислота-белок-редактор, может частично образовывать комплементарную связь с мишенью в виде нуклеиновой кислоты, гена или хромосомы и, как следствие, может позволить комплексу гидовая нуклеиновая кислота-белок-редактор точно приблизиться к положению мишени в виде нуклеиновой кислоты, гена или хромосомы.
Гомологичная последовательность нуклеиновой кислоты может иметь, по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более гомологию, или полную гомологию.
Кроме того, дополнительный компонент может быть выборочно использован для улучшения эффективности репарации поврежденного гена или нуклеиновой кислоты.
Ассистент
Дополнительный компонент может быть использован в качестве ассистента для повышения эффективности репарации поврежденного гена или нуклеиновой кислоты.
Термин «ассистент» относится к нуклеиновой кислоте, которая служит для участия в процессе репарации или повышения эффективности репарации поврежденного гена или нуклеиновой кислоты, например гена или нуклеиновой кислоты, расщепленных с помощью комплекса гидовая нуклеиновая кислота-белок-редактор.
Ассистентом может быть двухцепочечная нуклеиновая кислота или одноцепочечная нуклеиновая кислота.
Ассистент может присутствовать в линейной или кольцевой форме.
Ассистент может быть разделен на «ассистента NHEJ», который участвует в процессе репарации с использованием NHEJ или повышает эффективность репарации, и «ассистента HDR», который участвует в процессе репарации с использованием HDR или повышает эффективность репарации в соответствии со способом репарации.
Ассистент NHEJ может участвовать в процессе репарации или улучшить эффективность репарации поврежденного гена или нуклеиновой кислоты с использованием NHEJ.
Например, ассистент NHEJ может включать последовательность нуклеиновой кислоты, имеющую гомологию с частью поврежденной последовательности нуклеиновой кислоты. В настоящем документе гомологичная последовательность нуклеиновой кислоты может включать последовательность нуклеиновой кислоты, имеющую гомологию с последовательностью нуклеиновой кислоты на одном конце (например, 3'-конце) поврежденной последовательности нуклеиновой кислоты, и включать последовательность нуклеиновой кислоты, имеющую гомологию с последовательностью нуклеиновой кислоты на другом конце (например, 5'-конце) поврежденной последовательности нуклеиновой кислоты. Кроме того, может быть включена последовательность нуклеиновой кислоты, имеющая гомологию с каждой из последовательностей оснований до и после поврежденной последовательности нуклеиновой кислоты. Последовательность нуклеиновой кислоты, имеющая такую гомологию, может помочь расположить две части поврежденной последовательности нуклеиновой кислоты в непосредственной близости, тем самым увеличивая эффективность репарации поврежденной нуклеиновой кислоты с помощью NHEJ.
Помощник HDR может участвовать в процессе репарации или улучшить эффективность репарации поврежденного гена или нуклеиновой кислоты с использованием HDR.
Например, ассистент HDR может включать последовательность нуклеиновой кислоты, имеющую гомологию с частью поврежденной последовательности нуклеиновой кислоты. В настоящем документе гомологичная последовательность нуклеиновой кислоты может включать последовательность нуклеиновой кислоты, имеющую гомологию с последовательностью нуклеиновой кислоты на одном конце (например, 3'-конце) поврежденной последовательности нуклеиновой кислоты, и последовательность нуклеиновой кислоты, имеющую гомологию с последовательностью нуклеиновой кислоты на другом конце (например, 5'-конце) поврежденной последовательности нуклеиновой кислоты. Альтернативно, может быть включена последовательность нуклеиновой кислоты, имеющая гомологию с каждой из последовательностей оснований до и после поврежденной последовательности нуклеиновой кислоты. Последовательность нуклеиновой кислоты, имеющая такую гомологию, может служить в качестве матрицы поврежденной последовательности нуклеиновой кислоты для повышения эффективности репарации поврежденной нуклеиновой кислоты с помощью HDR.
В другом примере ассистент HDR может включать последовательность нуклеиновой кислоты, имеющую гомологию с частью поврежденной последовательности нуклеиновой кислоты и специфической нуклеиновой кислотой, например, нуклеиновой кислотой или геном, которые должны быть вставлены. В настоящем документе гомологичная последовательность нуклеиновой кислоты может включать последовательность нуклеиновой кислоты, имеющую гомологию с каждой из последовательностей оснований до и после поврежденной последовательности нуклеиновой кислоты. Специфическая нуклеиновая кислота может быть расположена между последовательностью нуклеиновой кислоты, имеющей гомологию с последовательностью оснований после поврежденной нуклеиновой кислоты, и последовательностью нуклеиновой кислоты, имеющей гомологию с последовательностью оснований до поврежденной нуклеиновой кислоты. Последовательность нуклеиновой кислоты, имеющая такую гомологию и специфическую нуклеиновую кислоту, может служить в качестве донора для вставки специфической нуклеиновой кислоты в поврежденную нуклеиновую кислоту, тем самым повышая эффективность HDR для нокина.
Гомологичная последовательность нуклеиновой кислоты может иметь по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более гомологию или полную гомологию.
6. Объект
Термин «объект» относится к организму, в который вводят гидовую нуклеиновую кислоту, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор, организму, в котором действует гидовая нуклеиновая кислота, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор, или пробе или образцу, полученным из организма.
Объектом может быть организм, включающий мишень в виде нуклеиновой кислоты, гена, хромосомы или белка комплекса гидовая нуклеиновая кислота-белок-редактор.
Организм может быть клетками, тканью, растением, животным или человеком.
Клетки могут быть прокариотическими клетками или эукариотическими клетками.
Эукариотические клетки могут быть растительными клетками, клетками животных или клетками человека, но настоящее изобретение не ограничено ими.
Ткань может быть тканью тела животного или человека, такой как кожа, печень, почка, сердце, легкое, мозг или мышечная ткань.
Объектом может быть проба или образец, включающие мишень в виде нуклеиновой кислоты, гена, хромосомы или белка комплекса гидовая нуклеиновая кислота-белок-редактор.
Проба или образец могут быть получены из организма, включающего мишень в виде нуклеиновой кислоты, гена, хромосомы или белка, и могут представлять собой слюну, кровь, кожную ткань, шванновские клетки, раковые клетки или стволовые клетки.
В настоящем изобретении, в качестве конкретного примера, объект может включать ген-мишень или нуклеиновую кислоту-мишень из комплекса гидовая нуклеиновая кислота-белок-редактор.
В настоящем документе ген-мишень может быть фактором, управляющим функцией ШК.
Фактором, управляющим функцией ШК, может быть ген PMP22.
Ген-мишень может быть дикого типа или модифицированной формой дикого типа.
В одном примере варианта осуществления настоящего изобретения объект может включать ген или нуклеиновую кислоту, измененные с помощью комплекса гидовая нуклеиновая кислота-белок-редактор.
В настоящем документе измененный ген может быть фактором, управляющим функцией ШК.
Фактором, управляющим функцией ШК, может быть ген PMP22.
В настоящем документе гидовая нуклеиновая кислота может нацеливаться на ген PMP22.
Гидовая нуклеиновая кислота может представлять собой последовательность нуклеиновой кислоты, комплементарную последовательности-мишени гена PMP22.
Гидовая нуклеиновая кислота может нацеливаться на один или более генов.
Гидовая нуклеиновая кислота может одновременно нацеливаться на два или более генов. В настоящем документе два или более генов могут быть гомологичными или гетерологичными генами.
Гидовая нуклеиновая кислота может нацеливаться на одну или более последовательностей-мишеней.
Гидовая нуклеиновая кислота может быть разработана в различных формах в зависимости от количества или расположений последовательностей-мишеней.
В одном примере варианта осуществления настоящего изобретения гидовая нуклеиновая кислота может представлять собой последовательность нуклеиновой кислоты, комплементарную одной или более последовательностям-мишеням из последовательностей, перечисленных в Таблице 1, Таблице 2, Таблице 3, Таблице 4, Таблице 5, Таблице 6 и Таблице 7.
В конкретном варианте осуществления для искусственной манипуляции с геном PMP22 последовательность гидовой нуклеиновой кислоты соответствует любой из последовательностей-мишеней из SEQ ID NO: 1-66.
В конкретном варианте осуществления для искусственной манипуляции с PMP22, обеспечен белок-редактор, который взаимодействует с последовательностью гидовой нуклеиновой кислоты, соответствующей, например, образующей комплекс с любой одной из последовательностей-мишеней из SEQ ID NO: 1-66, например SEQ ID NO: 1-8, 14-29 или 41-53.
В конкретном варианте осуществления обеспечены продукт модификации нуклеиновой кислоты каждого гена, в котором происходит искусственная манипуляция в участке последовательности-мишени любой из SEQ ID NO: 1-66, например, SEQ ID NO: 1-79, 14-29 или 41-53, и продукт ее экспрессии.
7. Доставка
Гидовая нуклеиновая кислота, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор могут быть доставлены или введены объекту с помощью различных способов доставки и различных форм.
Гидовая нуклеиновая кислота может быть доставлена или введена объекту в форме ДНК, РНК или в смешанной форме.
Белок-редактор может быть доставлен или введен объекту в форме ДНК, РНК, смеси ДНК/РНК, пептида, полипептида, который кодирует белок-редактор, или белка.
Комплекс гидовая нуклеиновая кислота-белок-редактор может быть доставлен или введен в мишень в форме ДНК, РНК или их смеси, которая кодирует каждый компонент, то есть гидовую нуклеиновую кислоту или белок-редактор.
Комплекс гидовая нуклеиновая кислота-белок-редактор может быть доставлен или введен объекту в виде комплекса гидовой нуклеиновой кислоты, имеющей форму ДНК, РНК или их смеси, и белка-редактора, имеющего форму пептида, полипептида или белка.
Кроме того, дополнительный компонент, способный увеличивать или ингибировать эффективность комплекса гидовая нуклеиновая кислота-белок-редактор, может быть доставлен или введен объекту с помощью различных способов доставки и в различных формах.
Дополнительный компонент может быть доставлен или введен объекту в форме ДНК, РНК, смеси ДНК/РНК, пептида, полипептида или белка.
i) Доставка в форме ДНК, РНК или их смеси
Форма из ДНК, РНК или их смеси, которая кодирует гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена объекту с помощью способа, известного в данной области техники.
Или форма из ДНК, РНК или их смеси, которая кодирует гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена объекту с помощью вектора, не вектора или их комбинации.
Вектор может представлять собой вирусный или невирусный вектор (например, плазмиду).
Не вектором может быть «голая» ДНК, комплекс ДНК или мРНК.
Введение, основанное на векторах
Последовательность нуклеиновой кислоты, кодирующая гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена объекту с помощью вектора.
Вектор может включать последовательность нуклеиновой кислоты, кодирующую гидовую нуклеиновую кислоту и/или белок-редактор.
Например, вектор может одновременно включать последовательности нуклеиновой кислоты, которые кодируют гидовую нуклеиновую кислоту и белок-редактор, соответственно.
Например, вектор может включать последовательность нуклеиновой кислоты, кодирующую гидовую нуклеиновую кислоту.
В качестве примера, домены, включенные в гидовую нуклеиновую кислоту, могут содержаться все в одном векторе или могут быть разделены и затем содержаться в разных векторах.
Например, вектор может включать последовательность нуклеиновой кислоты, кодирующую белок-редактор.
В одном примере, в случае белка-редактора, последовательность нуклеиновой кислоты, кодирующая белок-редактор, может содержаться в одном векторе или может быть разделена и затем содержаться в нескольких векторах.
Вектор может включать в себя один или более регуляторных/контрольных компонентов.
В настоящем документе регуляторные/контрольные компоненты могут включать в себя промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козак, сайт внутренней посадки рибосомы (IRES), акцепторный сайт сплайсинга и/или последовательность 2А.
Промотор может быть промотором, распознаваемым с помощью РНК-полимеразы II.
Промотор может быть промотором, распознваемым с помощью РНК-полимеразы III.
Промотор может быть индуцируемым промотором.
Промотор может быть промотором, специфичным к объекту.
Промотор может быть вирусным или невирусным промотором.
В качестве промотора может использоваться подходящий промотор в соответствии с контрольным участком (то есть последовательностью нуклеиновой кислоты, кодирующей гидовую нуклеиновую кислоту или белок-редактор).
Например, промотор, используемый для гидовой нуклеиновой кислоты, может представлять собой промотор H1, EF-1a, тРНК или U6. Например, промотор, используемый для белка-редактора, может представлять собой промотор CMV, EF-1a, EFS, MSCV, PGK или CAG.
Вектор может представлять собой вирусный вектор или рекомбинантный вирусный вектор.
Вирус может быть ДНК-вирусом или РНК-вирусом.
В настоящем документе ДНК-вирус может быть вирусом двухцепочечной ДНК (дцДНК) или вирусом одноцепочечной ДНК (оцДНК).
В настоящем документе РНК-вирус может быть вирусом одноцепочечной РНК (оцРНК).
Вирус может быть ретровирусом, лентивирусом, аденовирусом, аденоассоциированным вирусом (AAV), вирусом коровьей оспы, поксвирусом или вирусом простого герпеса, но настоящее изобретение этим не ограничено.
Обычно вирус может инфицировать хозяина (например, клетки), тем самым вводя нуклеиновую кислоту, кодирующую генетическую информацию вируса, в хозяина или вставляя нуклеиновую кислоту, кодирующую генетическую информацию, в геном хозяина. Гидовая нуклеиновая кислота и/или белок-редактор могут быть введены объекту с использованием вируса, имеющего такую характеристику. Гидовая нуклеиновая кислота и/или белок-редактор, введенные с использованием вируса, могут временно быть экспрессированы в объекте (например, в клетках). Альтернативно, гидовая нуклеиновая кислота и/или белок-редактор, введенные с использованием вируса, могут непрерывно быть экспрессированы в объекте (например, в клетках) в течение длительного времени (например, 1, 2 или 3 недели, 1, 2, 3, 6 или 9 месяцев, 1 или 2 года или постоянно).
Упаковочная способность вируса может варьироваться от 2 кб до 50 кб в зависимости от типа вируса. В зависимости от такой упаковочной способности может быть разработан вирусный вектор, включающий гидовую нуклеиновую кислоту или белок-редактор, или вирусный вектор, включающий как гидовую нуклеиновую кислоту, так и белок-редактор. Альтернативно, может быть разработан вирусный вектор, включающий гидовую нуклеиновую кислоту, белок-редактор и дополнительные компоненты.
В одном примере последовательность нуклеиновой кислоты, кодирующая гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена с помощью рекомбинантного лентивируса.
В другом примере последовательность нуклеиновой кислоты, кодирующая гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена с помощью рекомбинантного аденовируса.
В еще одном примере последовательность нуклеиновой кислоты, кодирующая гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена с помощью рекомбинантного AAV.
В еще одном примере последовательность нуклеиновой кислоты, кодирующая гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена с помощью гибридного вируса, например, одного или более гибридов вируса, перечисленных в настоящем документе.
Введение, основанное на не векторах
Последовательность нуклеиновой кислоты, кодирующая гидовую нуклеиновую кислоту и/или белок-редактор, может быть доставлена или введена объекту с использованием не вектора.
Не вектор может включать последовательность нуклеиновой кислоты, кодирующую гидовую нуклеиновую кислоту и/или белок-редактор.
Не вектором может быть «голая» ДНК, комплекс ДНК, мРНК или их смесь.
Не вектор может быть доставлен или введен объекту с помощью электропорации, бомбардировки частицами, сонопорации, магнитофекции, транзиентного клеточного сжатия или сдавливания (например, описано в литературе [Lee, et al, (2012) Nano Lett., 12, 6322-6327]), липид-опосредованной трансфекции, дендримера, наночастиц, фосфата кальция, диоксида кремния, силиката (Армосил) или их комбинации.
В качестве примера, доставка посредством электропорации может быть осуществлена с помощью смешивания клеток и последовательности нуклеиновой кислоты, кодирующей гидовую нуклеиновую кислоту и/или белок-редактор, в контейнере, камере или кювете, и применения электрических стимулов с заранее определенной продолжительностью и амплитудой для клеток.
В другом примере не вектор может быть доставлен с использованием наночастиц. Наночастицами могут быть неорганические наночастицы (например, магнитные наночастицы, диоксид кремния и т.д.) или органические наночастицы (например, липид, покрытый полиэтиленгликолем (ПЭГ) и т.д.). Наружная поверхность наночастиц может быть конъюгирована с положительно заряженным полимером, который является присоединяемым (например, полиэтиленимин, полилизин, полисерин и т.д.).
В конкретном варианте осуществления не вектор может быть доставлен с использованием липидной оболочки.
В конкретном варианте осуществления не вектор может быть доставлен с использованием экзосомы. Экзосома является эндогенной нановезикулой для переноса белка и РНК, которая может доставлять РНК в мозг и другой орган-мишень.
В конкретном варианте осуществления не вектор может быть доставлен с использованием липосомы. Липосома представляет собой сферическую везикулярную структуру, которая состоит из одного или более слоистых липидных бислоев, окружающих внутренние жидкостные компартменты, и внешнего липофильного фосфолипидного бислоя, который является относительно непрозрачным. При этом липосома может быть изготовлена из нескольких различных типов липидов; фосфолипиды наиболее широко используют для получения липосомы в качестве носителя лекарственного средства.
Могут быть включены другие добавки.
ii) Доставка в форме пептида, полипептида или белка
Белок-редактор в форме пептида, полипептида или белка может быть доставлен или введен объекту с помощью способа, известного в данной области техники.
Пептидная, полипептидная или белковая форма может быть доставлена или введена объекту с помощью электропорации, микроинъекции, транзиентного клеточного сжатия или сдавливания (например, описано в литературе [Lee, et al, (2012) Nano Lett., 12, 6322-6327]), липид-опосредованной трансфекции, наночастиц, липосомы, пептид-опосредованой доставки или их комбинации.
Пептид, полипептид или белок могут быть доставлены с последовательностью нуклеиновой кислоты, кодирующей гидовую нуклеиновую кислоту.
В одном примере перенос с помощью электропорации может быть осуществлен с помощью смешивания клеток, в которые будет введен белок-редактор с гидовой нуклеиновой кислотой или без нее в контейнере, камере или кювете, и применения электрических стимулов с заранее определенной продолжительностью и амплитудой для клеток.
iii) Доставка в форме смеси нуклеиновой кислоты-белка
Гидовая нуклеиновая кислота и белок-редактор могут быть доставлены или введены объекту в форме комплекса гидовая нуклеиновая кислота-белок-редактор.
Например, гидовой нуклеиновой кислотой может быть ДНК, РНК или их смесь. Белок-редактор может представлять собой пептид, полипептид или белок.
В одном примере гидовая нуклеиновая кислота и белок-редактор могут быть доставлены или введены объекту в форме комплекса гидовая нуклеиновая кислота-белок-редактор, содержащего гидовую нуклеиновую кислоту типа РНК и белок-редактор типа белка, то есть рибонуклеопротеин (RNP).
В настоящем изобретении в качестве варианта осуществления способа доставки гидовой нуклеиновой кислоты и/или белка-редактора объекту доставка гРНК, фермента CRISPR или ферментного комплекса гРНК-CRISPR будет описана ниже.
В одном из вариантов осуществления настоящего изобретения последовательность нуклеиновой кислоты, кодирующая гРНК и/или фермент CRISPR, будет доставлена или введена объекту с использованием вектора.
Вектор может включать последовательность нуклеиновой кислоты, кодирующую гРНК и/или фермент CRISPR.
Например, вектор может одновременно включать последовательности нуклеиновой кислоты, кодирующие гРНК и фермент CRISPR.
Например, вектор может включать последовательность нуклеиновой кислоты, кодирующую гРНК.
В одном примере домены, содержащиеся в гРНК, могут содержаться в одном векторе или могут быть разделены и затем содержаться в разных векторах.
Например, вектор может включать последовательность нуклеиновой кислоты, кодирующую фермент CRISPR.
В одном примере, в случае фермента CRISPR, последовательность нуклеиновой кислоты, кодирующая фермент CRISPR, может содержаться в одном векторе или может быть разделена и затем содержаться в нескольких векторах.
Вектор может включать в себя один или более регуляторных/контрольных компонентов.
В настоящем документе регуляторные/контрольные компоненты могут включать в себя промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козак, сайт внутренней посадки рибосомы (IRES), акцепторный сайт сплайсинга и/или последовательность 2А.
Промотор может быть промотором, распознаваемым с помощью РНК-полимеразы II.
Промотор может быть промотором, распознаваемым с помощью РНК-полимеразы III.
Промотор может быть индуцируемым промотором.
Промотор может быть промотором, специфичным к объекту.
Промотор может быть вирусным или невирусным промотором.
В качестве промотора может использоваться подходящий промотор в соответствии с контрольным участком (то есть последовательностью нуклеиновой кислоты, кодирующей фермент гРНК и/или CRISPR).
Например, промотор, используемый для гРНК, может представлять собой промотор H1, EF-1a, тРНК или U6. Например, промотор, используемый для фермента CRISPR, может представлять собой промотор CMV, EF-1a, EFS, MSCV, PGK или CAG.
Вектор может представлять собой вирусный вектор или рекомбинантный вирусный вектор.
Вирус может быть ДНК-вирусом или РНК-вирусом.
В настоящем документе ДНК-вирус может быть вирусом двухцепочечной ДНК (дцДНК) или вирусом одноцепочечной ДНК (оцДНК).
В настоящем документе РНК-вирус может быть вирусом одноцепочечной РНК (оцРНК).
Вирус может быть ретровирусом, лентивирусом, аденовирусом, аденоассоциированным вирусом (AAV), вирусом коровьей оспы, поксвирусом или вирусом простого герпеса, но настоящее изобретение этим не ограничено.
Обычно вирус может инфицировать хозяина (например, клетки), тем самым вводя нуклеиновую кислоту, кодирующую генетическую информацию вируса, в хозяина или вставляя нуклеиновую кислоту, кодирующую генетическую информацию, в геном хозяина. гРНК и/или фермент CRISPR могут быть введены объекту с использованием вируса, имеющего такую характеристику. гРНК и/или фермент CRISPR, введенные с использованием вируса, могут быть временно экспрессированы в объекте (например, в клетках). Альтернативно, гРНК и/или фермент CRISPR, введенные с использованием вируса, могут быть непрерывно экспрессированы в объекте (например, в клетках) в течение длительного времени (например, 1, 2 или 3 недели, 1, 2, 3, 6 или 9 месяцев, 1 или 2 года или постоянно).
Упаковочная способность вируса может варьироваться от 2 кб до 50 кб в зависимости от типа вируса. В зависимости от такой упаковочной способности может быть разработан вирусный вектор, включающий в себя только гРНК или фермент CRISPR, или вирусный вектор, включающий как гРНК, так и фермент CRISPR. Альтернативно, может быть разработан вирусный вектор, включающий в себя гРНК, фермент CRISPR и дополнительные компоненты.
В одном примере последовательность нуклеиновой кислоты, кодирующая гРНК и/или фермент CRISPR, может быть доставлена или введена с помощью рекомбинантного лентивируса.
В другом примере последовательность нуклеиновой кислоты, кодирующая гРНК и/или фермент CRISPR, может быть доставлена или введена с помощью рекомбинантного аденовируса.
В еще одном примере последовательность нуклеиновой кислоты, кодирующая гРНК и/или фермент CRISPR, может быть доставлена или введена с помощью рекомбинантного AAV.
В еще одном примере последовательность нуклеиновой кислоты, кодирующая гРНК и/или фермент CRISPR, может быть доставлена или введена с помощью одного или более гибридов из гибридных вирусов, например, вирусов, описанных в настоящем документе.
В одном примере варианта осуществления настоящего изобретения ферментный комплекс гРНК-CRISPR может быть доставлен или введен объекту.
Например, гРНК может присутствовать в форме ДНК, РНК или их смеси. Фермент CRISPR может присутствовать в форме пептида, полипептида или белка.
В одном примере фермент гРНК и CRISPR может быть доставлен или введен объекту в форме ферментного комплекса гРНК-CRISPR, включающего в себя РНК типа РНК и CRISPR типа белка, то есть рибонуклеопротеин (RNP).
Ферментный комплекс гРНК-CRISPR может быть доставлен или введен объекту с помощью электропорации, микроинъекции, транзиентного клеточного сжатия или сдавливания (например, описано в литературе [Lee, et al, (2012) Nano Lett., 12, 6322-6327]), липид-опосредованной трансфекции, наночастиц, липосомы, пептид-опосредованной доставки или их комбинации.
8. Трансформант
Термин «трансформант» относится к организму, в который вводят гидовую нуклеиновую кислоту, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор, к организму, в котором экспрессируется гидовая нуклеиновая кислота, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор, или пробе или образцу, полученным из организма.
Трансформант может представлять собой организм, в который вводят гидовую нуклеиновую кислоту, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор в форме ДНК, РНК или их смеси.
Например, трансформант может представлять собой организм, в который вводят вектор, включающий последовательность нуклеиновой кислоты, кодирующую гидовую нуклеиновую кислоту и/или белок-редактор. В настоящем документе вектором может быть невирусный вектор, вирусный вектор или рекомбинантный вирусный вектор.
В другом примере трансформант может представлять собой организм, в который вводят последовательность нуклеиновой кислоты, кодирующую гидовую нуклеиновую кислоту и/или белок-редактор, в форме не вектора. В настоящем документе не вектором может быть «голая» ДНК, комплекс ДНК, мРНК или их смесь.
Трансформант может представлять собой организм, в который вводят гидовую нуклеиновую кислоту, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор в форме пептида, полипептида или белка.
Трансформант может представлять собой организм, в который вводят гидовую нуклеиновую кислоту, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор в форме ДНК, РНК, пептида, полипептида, белка или их смеси.
Например, трансформант может представлять собой организм, в который вводят комплекс гидовая нуклеиновая кислота-белок-редактор, включающий гидовую нуклеиновую кислоту типа РНК и белок-редактор типа белка.
Трансформант может представлять собой организм, включающий мишень в виде нуклеиновой кислоты, гена, хромосомы или белка из комплекса гидовая нуклеиновая кислота-белок-редактор.
Организм может быть клетками, тканью, растением, животным или человеком.
Клетки могут быть прокариотическими клетками или эукариотическими клетками.
Эукариотические клетки могут быть растительными клетками, клетками животного или клетками человека, но настоящее изобретение этим не ограничено.
Ткань может быть ткань тела животного или человека, такая как кожа, печень, почка, сердце, легкое, мозг или мышечная ткань.
Трансформант может представлять собой организм, в который вводят или экспрессируют гидовую нуклеиновую кислоту, белок-редактор или комплекс гидовая нуклеиновая кислота-белок-редактор, или пробу, или образец, полученные из организма.
Пробой или образцом может быть слюна, кровь, кожная ткань, шванновские клетки, раковые клетки или стволовые клетки.
Применение
Вариант осуществления настоящего изобретения представляет собой применение для лечения объекта с заболеванием, связанным с дисфункцией шванновских клеток, с использованием фактора, управляющего функцией ШК, предпочтительно, композиции для искусственно измененного PMP22 или искусственно измененного фактора, управляющего функцией ШК, предпочтительно, искусственно измененного PMP22.
Объектом, подлежащим лечению, может быть млекопитающее, включая примата, такого как человек и обезьяна, грызуна, такого как крыса и мышь и тому подобное.
Заболевания, подлежащие лечению
В вариантах осуществления заболевание, подлежащее лечению, может представлять собой заболевание, связанное с дисфункцией ШК.
«Заболевание, связанное с дисфункцией ШК» относится ко всем состояниям, вызванным аномальными функциями шванновских клеток. «Заболевание, связанное с дисфункцией ШК» включает заболевание, вызванное аномальными или дефектными шванновскими клетками, которые могут возникать в процессе общего развития и роста, начиная от роста и дифференцировки шванновских клеток и заканчивая гибелью шванновских клеток, а также включает все заболевания, вызванные аномалией или дефектом функции, такой как выживаемость и поддержание периферических нервных клеток шванновских клеток и образование миелиновой оболочки (миелинизация) в аксонах.
Кроме того, заболевание, связанное с дисфункцией ШК, также включает все состояния, вызванные аномальными функциями фибробластов и/или глиальных клеток. Заболевание, связанное с дисфункцией ШК, включает заболевание, вызванное аномальными или дефектными клетками, которые могут возникать в процессе общего развития и роста, начиная от роста и дифференцировки фибробластов и/или глиальных клеток и заканчивая гибелью фибробластов и/или глиальных клеток.
Примеры заболеваний, связанных с дисфункцией ШК, включают болезнь Шарко-Мари-Тута (CMT), синдром Гийена-Барре (СГБ), тип острой воспалительной демиелинизирующей полирадикулопатии, шванноматоз и хроническую воспалительную демиелинизирующую полинейропатию (ХВДП), но без ограничения.
В одном примере, заболевание, связанное с дисфункцией ШК, может быть демиелинизирующим заболеванием.
В этом случае демиелинизирующее заболевание включает в себя все заболевания, возникающие вследствие повреждения, дефектов, нестабильности, дегенерации и ремиелинизации миелиновой оболочки.
Кроме того, заболевание, связанное с дисфункцией ШК, включает заболевание, связанное с фактором, управляющим функцией ШК.
«Заболевание, связанное с фактором, управляющим функцией ШК» относится ко всем состояниям, возникающим вследствие мутации фактора, управляющего функцией ШК, дупликации гена, управляющего функцией ШК, аномальной экспрессии фактора, управляющего функцией ШК, и тому подобное.
Примеры заболевания, связанного с фактором, управляющего функцией ШК, включают болезнь Шарко-Мари-Тута (CMT), болезнь Дежерина-Сотта, врожденную гипомиелинизирующую нейропатию (CHN), синдром Русси-Леви (RLS) и наследственную нейропатию со склонностью к параличу от сдавливания (HNPP), но без ограничения.
В одном примере заболевание, связанное с фактором, управляющим функцией ШК, может представлять собой заболевание, связанное с PMP22.
В этом случае заболевание, связанное с PMP22, включает все заболевания, возникающие вследствие мутации PMP22, дупликации гена PMP22, аномальной экспрессии PMP22 и тому подобное.
В одном варианте осуществления заболевание, связанное с фактором, управляющим функцией ШК, может представлять собой заболевание, возникающее вследствие дупликации гена PMP22.
Предпочтительно заболевание, связанное с фактором, управляюшим функцией ШК, может представлять собой заболевание Шарко-Мари-Тута Типа 1А (CMT1А), болезнь Дежерина-Сотта (DSS), врожденную гипомиелинизирующую нейропатию (CHN) или синдром Русси-Леви (RLS).
- Болезнь Шарко-Мари-Тута (CMT)
Болезнь CMT - это наследственное заболевание, вызванное дупликацией генов, которая происходит в хромосомах человека, а гены, участвующие в развитии периферических нервов в руках и ногах, дуплицируются мутациями, тем самым вызывая деформацию, например в форме перевернутой бутылке шампанского. Болезнь CMT является относительно распространенным неврологическим генетическим заболеванием, которое встречается у 36 из 100 000 человек в Соединенных Штатах, а число пациентов во всем мире составляет 2,8 миллиона человек, и, по оценкам, около 17 000 человек даже в Корее. Болезнь CMT в общем классифицируется всего на 5 типов: CMT1, CMT2, CMT3, CMT4 и CMTХ в соответствии с унаследованным аспектом, CMT1, CMT2 и CMT3 являются доминантными и наследуются с вероятностью 50% у детей, а CMT4 является рецессивной и наследуется с вероятностью 25%. CMT1 и CMT2 наследуются в основном у большинства семейных пациентов (80% и 20-40%, соответственно), а CMT3 и CMT4 встречаются крайне редко. CMTХ наследуется по материнской линии по Х-хромосомам, но его частота составляет 10-20%.
CMT1 - это заболевание, вызванное неспособностью осуществлять нормальный процесс экспрессии генов из-за дупликации генов, участвующих в образовании белков миелиновой оболочки, окружающей нейрональный аксон. CMT1A является аутосомно-доминантным генетическим заболеванием, вызванным дупликацией гена PMP22, расположенного на хромосоме № 17, 17p11.2-p12, что приводит к структурным и функциональным аномалиям миелиновой оболочки, вызванным сверхэкспрессией PMP22, который является важным компонентом миелиновой оболочки.
CMT2 связан с аномалиями аксонов и представляет собой нейропатию со значительно сниженным потенциалом действия двигательных и чувствительных нервов, в то время как скорость нервной проводимости близка к нормальному состоянию, а CMT3 встречается в раннем детстве как крайне редкое аутосомно-рецессивное генетическое заболевание и является типом, при котором клинические симптомы и снижение скорости нервной проводимости являются очень тяжелыми. CMT4 также относится к типу, при котором возраст ее возникновения является ранним, а клинические симптомы тяжелыми, наследуется по аутосомно-рецессивному типу, а CMTХ возникает в связи с Х-хромосомами, и ее симптомы у мужчин более тяжелые, чем у женщин.
- Болезнь Дежерина-Сотта, (DSS)
DSS представляет собой демиелинизирующую моторно-сенсорную нейропатию, возникающую в раннем возрасте и представляющую собой заболевание, которое обычно наследуется по аутосомно-доминантному типу, но также является аутосомно-рецессивно наследуемым, проявляется тяжелой демиелинизирующей нейропатией, проявляется аномалиями двигательных нервов с младенчества и характеризуется проявлением очень медленной нервной проводимости и увеличением количества специфических белков в спинномозговой жидкости. Болезнь Дежерина-Сотта имеет очень высокую скорость прогрессирования и характеризуется тем, что нарушение походки начинается с раннего возраста и также наследуется, но также происходит спорадически. Подобно CMT1A, дупликация PMP22 обнаруживается у некоторых пациентов с DSS, и, кроме того, было подтверждено, что присутствовала миссенс-мутация соответствующего гена.
- Врожденная гипомиелинизирующая нейропатия (CHN)
CHN - это заболевание нервной системы, симптомы которого появляются сразу после рождения и в качестве его основных симптомов проявляются дыхательная недостаточность, слабость мышц, нарушение движения мышц, снижение тонуса мышц, арефлексия, нарушение моторики (кинезионевроз; атаксия) и паралич, или дизестезия и влияют на мужчин и женщин с одинаковой скоростью. CHN является генетическим заболеванием, при котором нарушение возникает в двигательных и чувствительных нервах и характеризуется уменьшением образования миелиновой оболочки, тогда как демиелинизация и ремиелинизация миелиновой оболочки повторяются.
- Синдром Русси-Леви (RLS)
RLS является редким типом наследственной моторно-сенсорной нейропатии и впервые был описан Русси и Леви и др. в 1926 году и представляет собой случай, когда тремор конечностей, потеря походки и тому подобное являются более серьезными, чем при других наследственных моторно-сенсорных нейропатиях, но те же симптомы были позже обнаружены в различных подтипах наследственной моторно-сенсорной нейропатии, так что RLS в настоящее время рассматривается как один из симптомов, который появляется при наследственной моторно-сенсорной нейропатии. Что касается RLS, мутация гена MPZ в виде гена основного миелинового белка была обнаружена в генетическом тесте пациентов, у которых впервые сообщалось о наличии RLS, и у других пациентов, в случае, когда сообщалось о дупликации гена PMP22 в качестве гена миелинового белка 22 периферических нервов.
В других вариантах осуществления в настоящем изобретении может быть представлен дополнительную систему для управления третьим механизмом in vivo, сопутствующим различным функциям специфического фактора (например, фактора, управляющего функцией ШК), функции которого искусственно модифицированы.
Например, искусственно модифицированный специфический фактор может представлять собой ген PMP22.
Третий механизм может представлять собой механизм in vivo, в котором участвует ген, отличный от функции шванновских клеток.
Фармацевтическая композиция
Один пример варианта осуществления настоящего изобретения относится к композиции для применения при лечении заболевания с использованием искусственно измененного фактора, управляющего функцией ШК.
Композиция может представлять собой композицию для манипуляции, способную искусственно изменять фактор, управляющий функцией ШК. Композиция может быть обозначена в качестве терапевтической композиции или фармацевтической композиции.
В примере варианта осуществления композиция может представлять собой композицию для манипуляции, способную искусственно изменять фактор, управляющий функцией ШК.
Композиция для манипуляции может включать комплекс гидовая нуклеиновая кислота-белок-редактор.
Композиция для манипуляции может включать гидовую нуклеиновую кислоту и/или белок-редактор.
Композиция для манипуляции может включать нуклеиновую кислоту, кодирующую гидовую нуклеиновую кислоту и/или белок-редактор.
Композиция для манипуляции может включать вирус, содержащий нуклеиновую кислоту, кодирующую гидовую нуклеиновую кислоту и/или белок-редактор.
В другом примере варианта осуществления композиция может дополнительно включать дополнительный элемент.
Дополнительный элемент может включать в себя подходящий носитель для доставки в тело объекта.
В одном примере варианта осуществления будут обеспечены следующие терапевтические композиции:
композиция для лечения нарушения из-за дисфункции ШК и/или заболевания, связанного с фактором, управляющим функцией ШК, которая включает гидовую нуклеиновую кислоту, способную образовывать комплементарную связь с каждой из одной или более последовательностей-мишеней в последовательностях нуклеиновых кислот гена PMP22, или кодирующую их последовательность нуклеиновой кислоты, и
белок-редактор или кодирующую его последовательность нуклеиновой кислоты;
композиция для лечения нарушения из-за дисфункции ШК, и/или заболевания, связанного с фактором, управляющим функцией ШК, которая включает гидовую нуклеиновую кислоту, способную образовывать комплементарную связь с каждой из последовательностей-мишеней из SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 из последовательностей нуклеиновых кислот гена PMP22, или кодирующую их последовательность нуклеиновой кислоты; и
белок-редактор или кодирующую его последовательность нуклеиновой кислоты.
композиция для лечения нарушения из-за дисфункции ШК, и/или заболевания, связанного с фактором, управляющим функцией ШК, которая включает комплекс, образованный из гидовой нуклеиновой кислоты, способной образовывать комплементарную связь с каждой из последовательностей-мишеней из SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 из последовательностей нуклеиновых кислот гена PMP22, или кодирующую их последовательность нуклеиновой кислоты; и белок-редактор.
В настоящем документе гидовая нуклеиновая кислота или последовательность нуклеиновой кислоты, кодирующая ее; и последовательность нуклеиновой кислоты, кодирующая белок-редактор, может присутствовать в форме одного или более векторов. Гидовая нуклеиновая кислота или последовательность нуклеиновой кислоты, кодирующая ее; и последовательность нуклеиновой кислоты, кодирующая белок-редактор, может присутствовать в форме гомологичных или гетерологичных векторов.
Способ лечения
В другом примере варианта осуществления настоящего изобретения обеспечен способ лечения заболевания у пациента, который включает в себя получение вышеописанной композиции и введение эффективного количества композиции пациенту, нуждающемуся в этом.
Лечение с помощью генной манипуляции
Может быть использован способ лечения для регуляции фактора, управляющего функцией ШК, предпочтительно, экспрессии гена PMP22 с помощью изменения гена живого организма. Такой способ лечения может быть получен с помощью непосредственного введения композиции для изменения гена для того, чтобы изменить ген живого организма, в организм.
Композиция для генной манипуляции может содержать в себя комплекс гидовая нуклеиновая кислота-белок-редактор.
Композиция для генной манипуляции может быть введена в конкретное место на теле.
В настоящем документе конкретное место на теле может быть тканью, в которой экспрессия и/или функция фактора, управляющего функцией ШК, предпочтительно PMP22, является аномальной; или место рядом с этой тканью. Например, конкретным местом на теле может быть, например, периферическая нервная ткань.
Объектами для введения композиции могут быть млекопитающие, включая приматов, таких как человек или обезьяна, грызунов, таких как мышь или крыса и т.д.
Композицию можно вводить любым удобным способом, таким как инъекция, переливание, имплантация или трансплантация. Композицию можно вводить подкожно, внутрикожно, внутриглазно, интравитреально, внутриопухолево, интранодально, интрамедуллярно, внутримышечно, внутривенно, внутрилимфатически или внутрибрюшинно.
Доза (фармацевтически эффективное количество для получения заранее определенного, необходимого эффекта) композиции может быть выбрана из всех целых чисел в диапазонах значений из 104-109 клеток, например, от 105 до 106 клеток/кг (масса тела), на кг объекта введения, но настоящее изобретение этим не ограничено. Композиция может быть соответствующим образом назначена с учетом возраста, состояния здоровья и массы тела объекта введения, типов одновременно полученных способов лечения, если они были, частоты комбинированного лечения и характеристик необходимого эффекта.
В одном аспекте настоящее изобретение относится к способу для модификации полинуклеотида-мишени в прокариотических клетках, которая может быть достигнута in vivo, ex vivo или in vitro.
В некоторых вариантах осуществления способ может включать отбор образцов клеток или популяции клеток от человека или животного, не являющегося человеком, и модификацию клетки или клеток. Культивирование может происходить на любом этапе ex vivo. Клетка или клетки также могут быть повторно введены в животное, не являющееся человеком, или растение. Повторно введенные клетки наиболее предпочтительно представляют собой стволовые клетки.
В еще одном примере варианта осуществления в настоящем изобретении может быть представлен способ искусственного изменения клеток, который включает в себя: введение (а) гидовой нуклеиновой кислоты, способной образовывать комплементарную связь с последовательностями-мишенями из SEQ ID NO: 1-66, например, SEQ ID NO: 1-8, 14-29 или 41-53 из последовательностей нуклеиновых кислот гена PMP22, или последовательности нуклеиновой кислоты, кодирующей их; и (b) белка-редактора, включающего один или более белков, выбранных из группы, состоящей из белка Cas9, полученного из Streptococcus pyogenes, белка Cas9, полученного из Campylobacter jejuni, белка Cas9, полученного из Streptococcus thermophilus, белка Cas9, полученного из Streptococcus aureus, белка Cas9, полученного из Neisseria meningitidis, и белка Cpf1, или последовательности нуклеиновой кислоты, кодирующей их в клетках.
Гидовая нуклеиновая кислота и белок-редактор могут присутствовать в одном или более векторах в форме последовательности нуклеиновой кислоты или в комплексе комбинации гидовой нуклеиновой кислоты и белка-редактора.
Этап введения может быть выполнен in vivo или ex vivo.
К методике вышеописанной секции «7. Доставка» можно обратиться до этапа введения.
Например, стадия введения может быть выполнена с помощью одного или более способов, выбранных из электропорации, липосом, плазмид, вирусных векторов, наночастиц и способа белка слияния с доменом транслокации белка (PTD).
Например, вирусный вектор может быть одним или более, выбранными из группы, состоящей из ретровируса, лентивируса, аденовируса, аденоассоциированного вируса (AAV), вируса коровьей оспы, поксвируса и вируса простого герпеса.
Когда фактор, управляющий функцией ШК, предпочтительно PMP22, искусственно изменяют с помощью способа и композиции некоторых вариантов осуществления настоящего изобретения, можно контролировать аномальную экспрессию и/или функцию PMP22, и с помощью контроля аномальной экспрессии/или функция PMP22, экспрессия и/или функция аномального PMP22 обычно улучшается/улучшаются, так что можно получить эффекты восстановления или улучшения аномальных функций шванновских клеток и тому подобное, вызванных аномальной экспрессией PMP22.
Дополнительные применения
В конкретном варианте осуществления в настоящем изобретении может быть представлен набор для приготовления композиции для лечения нарушения из-за дисфункции ШК, и/или заболевания, связанного с фактором, управляющим функцией ШК.
Набор может быть приготовлен с помощью обычного способа приготовления, известного в данной области техники.
Набор может дополнительно включать детектируемую метку. Термин «детектируемая метка» относится к атому или молекуле для специфического детектирования молекулы, содержащей метку, среди молекул того же типа без метки. Детектируемая метка может быть присоединена к антителу, специфически связывающемуся с белком или его фрагментом, белком взаимодействия, лигандом, наночастицами или аптамером. Детектируемая метка может включать радионуклид, флуорофор и фермент.
В конкретном варианте осуществления в настоящем изобретении может быть представлен способ скрининга материала, способного регулировать уровень экспрессии искусственно измененного фактора, управляющего функцией ШК, предпочтительно гена PMP22.
В конкретном варианте осуществления в настоящем изобретении может быть представлен способ предоставления информации о последовательности сайта-мишени, которую можно искусственно изменять в объекте, анализируя последовательности фактора, управляющего функцией ШК, предпочтительно гена PMP22.
Кроме того, обеспечен способ построения библиотеки с использованием информации с помощью такого способа.
В настоящем документе может быть использована известная база данных.
В конкретных вариантах осуществления может быть обеспечено животное или клетки, которые могут быть использованы для исследования с использованием способа настоящего изобретения.
Животное или клетки, которые включают редактирование хромосом в одной или более последовательностях нуклеиновых кислот, связанных с заболеванием, могут быть подготовлены с использованием вышеописанного способа. Такая последовательность нуклеиновой кислоты может быть эталонной последовательностью, которая может кодировать последовательность белка, связанного с заболеванием, или может быть связана с заболеванием.
В одном примере варианта осуществления эффект мутации и возникновения и/или прогрессирования заболевания может быть изучен на животном или клетках с использованием измерений, обычно используемых при исследовании заболевания на животном или клетках, подготовленных с помощью способа настоящего изобретения. Альтернативно, может быть изучен фармацевтический эффект активного соединения при заболевании с использованием такого животного или клеток.
В другом примере варианта осуществления эффект стратегии возможной генной терапии может быть оценен с использованием животного или клеток, подготовленных с помощью способа настоящего изобретения. То есть развитие и/или прогрессирование соответствующего заболевания может быть подавлено или уменьшено с помощью модификации последовательности хромосомы, кодирующей связанный с заболеванием белок. В частности, этот способ включает формирование модифицированного белка с помощью редактирования последовательности хромосомы, кодирующей связанный с заболеванием белок, что приводит к достижению модифицированного ответа животного или клеток. Следовательно, в некоторых вариантах осуществления генетически модифицированное животное может быть сравнено с животным, уязвимым для развития соответствующего заболевания, тем самым оценивая эффект процесса генной терапии.
Применение может включать модель заболевания, фармакологическую модель, экспериментальную модель, функциональную модель клетки и гуманизированную модель. Например, применение может включать модель заболевания, связанного с аномальной функцией, фармакологическую модель, экспериментальную модель, функциональную модель клетки и гуманизированную модель шванновских клеток с помощью PMP22.
Таким образом, искусственно измененный фактор, управляющий функцией ШК, и система, управляющая функцией ШК (система модификации управления функцией ШК), могут быть использованы для эффективного лечения заболевания, связанного с дисфункцией ШК, и/или заболевания, связанного с фактором, управляющим функцией ШК, например, заболевания, связанной с избыточной экспрессией PMP22 или дупликацией гена PMP22. Кроме того, с помощью различных механизмов in vivo, в которых задействован фактор, управляющий функциями ШК, предпочтительно PMP22, можно улучшить эффективность системы, управляющей функцией ШК (включая систему модификации фактора, управляющего функцией ШК).
Далее настоящее изобретение будет описано более подробно со ссылкой на примеры.
Примеры даны просто для того, чтобы описать настоящее изобретение более подробно, и специалистам в данной области техники может быть очевидно, что объем настоящего изобретения не ограничено следующими примерами.
Экспериментальный способ
1. Разработка гРНК
Сайты-мишени CRISPR/Cas9 человеческого гена PMP22 были выбраны с использованием инструментов CRISPR RGEN (www.rgenome.net). Сайты-мишени гена PMP22 могут варьироваться в зависимости от типа фермента CRISPR, последовательностей-мишеней кодирующего сайта (кодирующей последовательности, CDS), сайта TATA-бокса и сайта энхансера (например, EGR2-, SOX10- или TEAD1-связывающего сайта; или сайта дистального энхансера (участка дистального энхансера) B или C) гена PMP22 для SpCas9 приведены в Таблицах 1, 3, 5 и 7, описанных выше, и последовательности-мишени кодирующего сайта (кодирующей последовательности, CDS), сайта TATA-бокса и сайта энхансера (например, EGR2- или SOX10-связывающего сайта) гена PMP22 для CjCas9 приведены в Таблицах 2, 4 и 6.
Все гРНК были получены в форме химерной одноцепочечной РНК (огРНК). Последовательности пептидного остова огРНК, специфичных к Cj и Sp, помимо последовательности-мишени, представляют собой 5'-GUUUUAGUCCCUGAAAAGGGACUAAAAUAAAGAGUUUGCGGGACUCUGCGGGGU UACAAUCCCCUAAAACCGCUUUU-3' и 5'-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGA AAAAGUGGCACCGAGUCGGUGC-3', соответственно.
2. Конструирование и синтез гРНК
огРНК была упакована в вектор AAV или синтезирована с РНК. Для того, чтобы вставить огРНК в вирусный вектор, был разработан и получен с помощью отжига олигонуклеотид ДНК, соответствующий последовательностям оснований от 20 до 22 из огРНК, и лигирован в вектор pRGEN-CAS9 (разработанный собственными силами) с использованием сайта BsmBI. Cas9 и огРНК, включающие вариабельную последовательность-мишень на 5'-конце, были экспрессированы посредством промоторов CMV и U6, соответственно.
Кроме того, для системы доставки с помощью RNP, огРНК была транскрибирована с помощью T7 РНК-полимеразы после того, как матрица была получена с помощью отжина двух частично комплементарных олигонуклеотидов, полученных с помощтю Phusion Taq-опосредованной полимеризации. Транскрибированная огРНК была очищена и количественно оценена с использованием спектрометрии.
3. Очистка белка Cas9
Кодон-оптимизированные последовательности ДНК Cas9, включая эпитопы NLS и HA, были субклонированы в вектор pET28 и экспрессированы в BL21 (DE3) с использованием IPTG при оптимальных условиях культивирования. Экспрессированный белок Cas9 был очищен с использованием агарозных гранул Ni-NTA и диализован с помощью подходящего буфера. Активность Cas9 была подтверждена посредством теста расщепления in vitro с использованием хорошо известной эффективной огРНК.
4. Клеточная культура
Человеческую шваноподобную клеточную линию (АТСС) и человеческие первичные шванновские клетки (ScienCell) были культивированы в соответствии с руководством производителя. Человеческие шваноподобные клетки были культивированы в модифицированной по способу Дульбекко среде Игла (DMEM) (WelGene), содержащей высокую концентрацию глюкозы, с добавлением 1 х пенициллина/стрептомицина (WelGene) и 10% эмбриональной телячьей сыворотки (WelGene).
Человеческие первичные шванновские клетки содержались в растворе для культивирования шванновских клеток (ScienCell), предоставленном поставщиком. Для дифференцировки клетки были культивированы в DMEM (WelGene), содержащей низкую концентрацию глюкозы с добавлением 1% эмбриональной телячьей сыворотки (WelGene), 100 нг/мл Nrg1 (Peprotech) для сигналов образования миелиновой оболочки (миелинизации) и 100 мкМ dbcAMP (Sigma-Aldrich) в течение 7 дней.
5. Трансдукция (трансфекция)
Для трансдукции (трансфекции) комплекс RNP, содержащий 4 мкг белка Cas9 (ToolGen) и 1 мкг огРНК, был инкубирован при комнатной температуре в течение 15 минут. После этого комплекс RNP подвергали электропорации с использованием наконечника электропорации 10 мкл и электропоратора Neon (ThermoFisher) и доставили в 2 × 105 клеток. Для направленного глубокого секвенирования геномную ДНК (гДНК) собрали из трансдуцированных клеток через 72 часа после трансдукции.
6. In vitro ПЦР в реальном времени (количественная ОТ-ПЦР)
мРНК была экстрагирована из человеческих первичных шванновских клеток в соответствии с протоколом производителя с использованием мини-набора RNeasy (Qiagen). После этого 100 нг мРНК подвергали обратной транскрипции с использованием набора для обратной транскрипции кДНК высокой емкости (ThermoFisher). Количественная ОТ-ПЦР была выполнена с 10 нг мастер-микса Taqman Gene expression в соответствии с протоколом производителя с использованием QuantStudio 3 (ThermoFisher). Уровни экспрессии PMP22 были рассчитаны, используя значения Ct, и GAPDH был использован в качестве эндогенного контроля. Taqman-зонды (ThermoFisher), использованные в настоящем исследовании, приведены в Таблице 8 ниже.
[Таблица 8]
7. Направленное глубокое секвенирование
Целевой сайт был амплифицирован с помощью ПЦР из гДНК, экстрагированной из трансдуцированных клеток, с использованием Phusion taq-полимеразы (New England BioLabs). После этого было выполнено глубокое секвенирование спаренных концов с использованием Mi-Seq (Illumina) в качестве продукта амплификации ПЦР. Результаты глубокого секвенирования были проанализированы с использованием онлайн-инструмента Cas-Analyzer (www.rgenome.net). Было подтверждено, произошла ли мутация в 3 п.н до PAM-последовательности в результате инделей с помощью Cas9. Праймеры, использованные в настоящем исследовании, приведены в Таблице 8 ниже.
[Таблица 9]
(SEQ ID NO: 67)
(SEQ ID NO: 68)
(SEQ ID NO: 69)
(SEQ ID NO: 70)
(SEQ ID NO: 71)
(SEQ ID NO: 72)
(In silico, in vitro)
(SEQ ID NO: 73)
(SEQ ID NO: 74)
(SEQ ID NO: 75)
(SEQ ID NO: 76)
(SEQ ID NO: 77)
(SEQ ID NO: 78)
(SEQ ID NO: 79)
(SEQ ID NO: 80)
(SEQ ID NO: 81)
(SEQ ID NO: 82)
(SEQ ID NO: 83)
(SEQ ID NO: 84)
(SEQ ID NO: 85)
(SEQ ID NO: 86)
(SEQ ID NO: 87)
(SEQ ID NO: 88)
(SEQ ID NO: 89)
(SEQ ID NO: 90)
(SEQ ID NO: 91)
(SEQ ID NO: 92)
(SEQ ID NO: 93)
(SEQ ID NO: 94)
(SEQ ID NO: 95)
(SEQ ID NO: 96)
(SEQ ID NO: 97)
(SEQ ID NO: 98)
(SEQ ID NO: 99)
(SEQ ID NO: 100)
(SEQ ID NO: 101)
(SEQ ID NO: 102)
(SEQ ID NO: 103)
(SEQ ID NO: 104)
(SEQ ID NO: 105)
(SEQ ID NO: 106)
(SEQ ID NO: 107)
(SEQ ID NO: 108)
(Digenome-seq, in vitro)
(SEQ ID NO: 109)
(SEQ ID NO: 110)
(SEQ ID NO: 111)
(SEQ ID NO: 112)
(SEQ ID NO: 113)
(SEQ ID NO: 114)
(SEQ ID NO: 115)
(SEQ ID NO: 116)
(SEQ ID NO: 117)
(SEQ ID NO: 118)
(SEQ ID NO: 119)
(SEQ ID NO: 120)
(SEQ ID NO: 121)
(SEQ ID NO: 122)
(SEQ ID NO: 123)
(SEQ ID NO: 124)
(SEQ ID NO: 125)
(SEQ ID NO: 126)
(In silico, In vivo)
(SEQ ID NO: 127)
(SEQ ID NO: 128)
(SEQ ID NO: 129)
(SEQ ID NO: 130)
(SEQ ID NO: 131)
(SEQ ID NO: 132)
(SEQ ID NO: 133)
(SEQ ID NO: 134)
(SEQ ID NO: 135)
(SEQ ID NO: 136)
(SEQ ID NO: 137)
(SEQ ID NO: 138)
(SEQ ID NO: 139)
(SEQ ID NO: 140)
(SEQ ID NO: 141)
(SEQ ID NO: 142)
8. Разработка нецелевого сайта in silico
Нецелевой потенциальный сайт был разработан in silico с использованием онлайн-инструмента (www.rgenome.net). Максимальное количество ошибочно спаренных оснований из 3 п.н. было рассмотрено как нецелевой сайт.
9. Digenome-seq
Геномная ДНК клеток HeLa была очищена в соответствии с протоколом поставщика с использованием набора DNeasy Blood & Tissue (Qiagen). Белок Cas9 (100 нМ) и огРНК (300 нМ), предварительно инкубированные, были смешаны с геномной ДНК (10 мкг) в 1 мл реакционного раствора (100 мМ NaCl, 50 мМ Трис-HCl, 10 мМ MgCl2, 100 мкг/мл БСА, рН 7,9) при 37°С в течение 8 часов. Укороченную геномную ДНК обрабатывали РНКазой А (50 мкг/мл) и снова очищали с использованием набора DNeasy Tissue Kit (Qiagen). 1 мкг укороченной геномной ДНК разделили на фрагменты с использованием системы Covaris, и к фрагментам ДНК был присоединен адаптер для создания библиотеки. После этого библиотека была подвергнута секвенированию всего генома (WGS) с использованием HiSeq x Ten Sequencer (Illumina) на глубине секвенирования от 30 до 40 x (Macrogen). Показатели расщепления in vitro были рассчитаны с помощью системы подсчета расщепления ДНК в положениях каждой последовательности оснований, расщепленной в геноме.
10. Мыши и интраневральная инъекция
Линии мышей C22 (B6; CBACa-Tg (PMP22) C22Clh/H), использованные в настоящем исследовании, были приобретены у MRC Harwell (Оксфордшир, Великобритания). Мышей C22 (4 самца и 7 самок) лечили с помощью PMP22-TATA RNP. Интраневральная инъекция была выполнена так же, как и в предыдущем исследовании (Daisuke I№, J Vis Exp., (2016) 115). Мышей 6-дневного возраста анестезировали, а седалищные нервы мышей подвергали хирургическому воздействию. Для того, чтобы свести к минимуму повреждение нерва, сразу же была произведена интраневральная инъекция в конце седалищной вырезки с использованием вытянутых стеклянных микропипеток, прикрепленных к микроинъектору. Комплекс RNP из 11 мкг белка Cas9 и 2,75 мкг огРНК на мышь был введен мышам вместе с Липофектамином 3000 (In vitrogen, Карлсбад, Калифорния, США). Управление, применение и лечение всех животных, использованных в настоящем исследовании, было выполнено в соответствии с инструкциями, подготовленными Комитетом по управлению и использованию животных Samsung (SMC-20170206001) в соответствии с Международной ассоциацией оценки и аккредитации лабораторных исследований на животных.
11. Эксперимент с вращающимся стержнем (Тест вращающегося стержня)
Координация движений была оценена с использованием устройства вращающегося стержня (B.S. Technolab INC., Корея). Этот эксперимент был выполнен для оценки равновесия и координации движений мышей. До эксперимента мыши прошли 3-дневный тренировочный период. В эксперименте горизонтальный вращающийся стержень (21 об/мин) был использован для эксперимента с вращающимся стержнем. Время удержания на вращающемся стержне мыши было измерено, и мыши позволяли оставаться на стержне до 300 секунд.
12. Электрофизиологический тест
Для того, чтобы оценить электрофизиологическое состояние, тест нервной проводимости (NCS) был выполнен таким же образом, как и в предыдущем исследовании (Jinho Lee., J Biomed sci., (2015) 22, 43). Таким образом, мышей анестезировали углекислым газом, и анестезия поддерживалась с использованием носового конуса для подачи 1,5% изофлурана во время эксперимента. Шерсть была полностью удалена от конца до задней лапы. NCS проведен с использованием устройства Nicolet VikingQuest (Natus Medical). Для теста проводимости двигательных нервов седалищного нерва каждый из ответов от дистальной части и проксимальной части определяли, помещая игольчатый электрод для регистрации активности на икроножную мышцу с контрольным электродом, прикрепленным к сухожилию, и размещая стимулирующий отрицательный электрод в положение рядом с отводящим электродом на расстоянии 6 мм по направлению к центру тела внутрь бедра и центральной линии пост-орбитальной части бедра. Были измерены дистальная латентность (DL), скорость проводимости двигательных нервов (MNCV) и амплитуда электрического вызванного ответа мышц (CMAP). CMAP был измерен при максимальной сверхстимуляции.
13. Гистология нервов и изображения
Седалищные нервы мышей были подвергнуты биопсии, и анатомо-патологическое исследование пораженного образца было выполнено с помощью анализа с микроскопом. Образцы были соответствующим образом зафиксированы с использованием 25 мМ какодилатного буфера, содержащего 2% глутаральдегида. Полутонкие срезы были окрашены толуидиновым синим. После инкубации в 1% OsO4 в течение 1 часа образцы были обезвожены в этаноловом ряду, а затем их возможно было пропустить через пропиленоксид и погрузить в эпоксидную смолу (Epon 812, Oken, Нагано, Япония). Клетки были нарезаны до определенной толщины (1 мкм) с использованием ультрамикротома Leica (Leica Microsystems) и окрашены толуидиновым синим в течение 30-45 секунд. Коэффициент g (диаметр аксона/диаметр волокна) был рассчитан с помощью измерения внутреннего диаметра и наружного диаметра миелина с использованием программы Zeiss Zen 2 (Carl Zeiss, Оберкохен, Германия).
14. Статистический анализ
Статистическую значимость данных, связанных с уровнями экспрессии мРНК, оценивали с помощью однофакторного дисперсионного анализа с использованием множественных сравнений из апостериорных тестов Тьюки. Другие типы представленных данных были рассчитаны с использованием U-критерия Манна-Уитни (http: //www.socscistatistics.com/tests/mannwhitney/Default2.aspx). Данные и графики, полученные из настоящего исследования, были проанализированы с использованием GraphPad Prism. Уровень значимости был установлен на уровне 0,05.
Пример 1. Скрининг огРНК
Для того, чтобы провести скрининг терапевтически эффективных последовательностей огРНК, которые могут снизить экспрессию человеческого PMP22 до нормального диапазона, клеточные линии человека были трансдуцированы с помощью различных огРНК и Cas9, разработанных для нацеливания на кодирующие последовательности (CDS) и TATA-бокс и сайты связывания интронного энхансера гена PMP. Вкратце, человеческие T-клетки Jurkat были использованы для скрининга SpCas9, а клетки HEK293T были использованы для CjCas9. гДНК была собрана из клеток и подвергнута направленному глубокому секвенированию. Различные паттерны мутаций, индуцированных с помощью последовательностей огРНК, были идентифицированы с помощью NHEJ-опосредованного инделя. Несколько SpCas9-огРНК сильно индуцировали индели в CDS и двух регуляторных сайтах (ФИГ.1). Было подтверждено, что в специфической CjCas9-огРНК было индуцировано от 30 до 40% инделей (ФИГ.2).
Пример 2. Манипуляция c генами шванноподобных клеток
Хотя эффективные индель-мутации, вызванные огРНК, были идентифицированы в человеческих клетках, неясно, будет ли эффект также возможен в шванновских клетках. Таким образом, для того, чтобы исследовать эффекты ингибирования экспрессии PMP22 и генной манипуляции в шванновских клетках, эффект SpCas9-огРНК был подтвержден с использованием клеток sNF2.0, которые являются шванноподобными клетками. Эффективная SpCas9-огРНК, идентифицированная в клетках Jurkat, неоднократно тестировалась в клетках sNF02.0. После трансдукции с помощью глубокого секвенирования было подтверждено, что такая же высокая частота инделей была получена с помощью той же самой огРНК. В частности, было подтверждено, что почти 70-80% инделей находились в сдвиге рамки PMP22, происходящем из-за CDS, нацеленной на огРНК (ФИГ.4). Трансдукция одиночной огРНК, нацеленной на промоторный (TATA-бокс) сайт и энхансер-связывающие положения, индуцировала индели на 31% и 59% соответственно (ФИГ.3). Интересно, что небольшая делеция размером 40-50 п.н., содержащая основной управляющий фактор (например, EGR- или SOX10-связывающее положение) гена миелина или важный TATA-бокс, была обнаружена в очень большом количестве клеток из клеток, обработанных двойной огРНК (ФИГ. 5).
Пример 3. Контроль экспрессии PMP22 с помощью генной манипуляции
Для того, чтобы оценить изменение экспрессии PMP22 с помощью эффективной огРНК были дифференцированы шванноподобные клетки, и выполнен количественный ОТ-ПЦР. В результате большинство огРНК, нацеленных на PMP22, эффективно ингибировали экспрессию PMP22 (ФИГ.6). Когда использовали одиночную огРНК, экспрессия PMP22 была снижена примерно на 30% по сравнению с контролем, обработанным только Cas9, а когда была использована двойная огРНК, экспрессия PMP22 была снижена примерно на 50% по сравнению с контролем, обработанным только Cas9.
Пример 4. Генная манипуляция шванновских клеток
После того, как ингибирование экспрессии и эффекты генной манипуляции PMP22 были ранее подтверждены в шванноподобных клетках, было подтверждено, показал ли предыдущий результат аналогичный эффект в человеческих первичных шванновских клетках. Частоту инделей в соответствии с сайтом-мишенью наблюдали с использованием SpCas9-огРНК в каждом сайте-мишени гена PMP22 человека в человеческих первичных шванновских клетках. В результате было подтверждено, что частота инделей была высокой в сайте-мишени в большинстве из огРНК-нацеленных TATA-бокса, энхансера и кодирующих последовательностей гена PMP22 (ФИГ.7A). Кроме того, даже когда использовались двойные огРНК, каждая из которых нацеливалась на TATA-бокс и энхансер, была показана высокая частота инделей. Было подтверждено, что индель появился в сайте-мишени с использованием огРНК-нацеленных последовательностей, кодирующих сайты дистального энхансера B и C (ФИГ. 7C), и в этом случае огРНК-нацеленный APOC3 использовали в качестве контроля.
Кроме того, для того, чтобы подтвердить, вызывает ли SpCas9-огРНК в каждом сайте-мишени уменьшение экспрессии гена PMP22, был выполнен количественный анализ ОТ-ПЦР. Так как PMP22 транскрибируется на заключительной стадии дифференцировки шванновских клеток, человеческие первичные шванновские клетки были обработаны хорошо известным различным сигнальным фактором, включая Нейрегулин-1 (Nrg1) и дибутиральный циклический АМФ (dbcAMP), в течение 7 дней. В результате было подтверждено, что экспрессия PMP22 была увеличена в 9 раз в клетках, обработанных Nrg1 и dbcAMP, по сравнению с клетками, которые не обрабатывались ни Nrg1, ни dbcAMP. Напротив, когда клетки были обработаны SpCas9-огРНК в каждом сайте-мишени, было подтверждено, что экспрессия PMP22 была индуцирована в 4-6 раз. Определено, что это происходит из-за ингибирования экспрессии PMP22 вследствие каждой модификации сайта-мишени PMP22 с помощью SpCas9-огРНК в каждом сайте-мишени (ФИГ.7B).
Пример 5. Эффекты уменьшения эффективной и специфической экспрессии PMP22 с использованием CRISPR/Cas9, нацеленного на сайт ТАТА-бокса человеческого гена PMP22
Эксперимент был выполнен в человеческих первичных клетках шванновских клетках с помощью выбора огРНК_TATA_Sp#1 (далее обозначенного как PMP22-TATA огРНК), который демонстрирует высокую эффективность инделей среди огРНК, нацеленных на сайт TATA-бокса, ранее подвергнутого скринингу, и может нацеливаться на TATA-бокс. Индель был индуцирован с помощью трансдукции человеческих первичных шванновских клеток с помощью комплекса RNP, включающего огРНК и белок Cas9 (ФИГ.8B), и было подтверждено с помощью направленного глубокого секвенирования, что 89,54 ± 1,39% от общего количества инделей были получены в сайте ТАТА-бокса человеческого PMP22 (ФИГ.8C).
Кроме того, чтобы подтвердить, вызывала ли мутация, образованная в TATA-боксе PMP22, снижение экспрессии гена PMP22, был проведен количественный анализ ОТ-ПЦР. Так как PMP22 транскрибируется на заключительной стадии дифференцировки шванновских клеток, человеческие первичные шванновские клетки были обработаны с помощью хорошо известного различного сигнального фактора, включая Нейрегулин-1 (Nrg1) и дибутиральный циклический АМФ (dbcAMP), в течение 7 дней. В результате было подтверждено, что экспрессия PMP22 была увеличена в 9 раз в клетках, обработанных Nrg1 и dbcAMP, по сравнению с клетками, которые не обрабатывались ни Nrg1, ни dbcAMP. Напротив, было подтверждено, что, когда клетки обрабатывали вместе с PMP22-TATA RNP, экспрессия PMP22 была индуцирована в 6 раз. Определено, что это происходит из-за ингибирования экспрессии PMP22 с помощью модификации TATA PMP22 с помощью CRISPR/Cas9 (ФИГ.8D). В контроле, обработанном как сигнальным фактором дифференцировки, так и целевым RNP AAVS1, не было подтверждено никакой разницы в дифференцировке гена PMP22.
Для того, чтобы подтвердить специфичность PMP22-TATA RNP, был проведен нецелевой анализ на основе in silico. Посредством направленного глубокого секвенирования не было подтверждено индель-мутации, превышающей коэффициент ошибки секвенирования (в среднем 0,1%) в нецелевом положении с помощью анализа in silico (ФИГ. 9). Поскольку нецелевой анализ на основе in silico может быть необъективным подходом, Digenome-seq (нецелевой анализ на основе полного секвенирования, который более объективен) также был выполнен. В результате стало возможным подтвердить девять нецелевых положений, расщепленных с помощью PMP22-TATA RNP in vitro (ФИГ.10A, ФИГ.10B). Тем не менее, в результате повторного анализа посредством направленного глубокого секвенирования, аномальная индель-мутация не была обнаружена в нецелевом положении (ФИГ. 10C).
Эти результаты показывают, что эффективная и специфическая модификация TATA-бокса PMP22 с помощью PMP22-TATA RNP может контролировать уровень транскрипции PMP22 в человеческих первичных шванновских клетках.
Пример 6. Эффекты ослабления фенотипа заболевания с помощью ингбирования экспрессии CRISPR/Cas9-опосредованного PMP22 у мышей с CMT1A.
Для того, чтобы протестировать контроль транскрипции PMP22 с помощью PMP22-TATA RNP in vivo, PMP22-TATA RNP, заключенный в липосомы, непосредственно вводили в седалищный нерв мыши C22 (ФИГ. 11). В этом случае, в качестве контроля использовали комплекс RNP, нацеленный на Rosa26 (mRosa26). mRosa26 RNP или PMP22-TATA RNP был введен интранервально и доставлен в левый седалищный нерв (ипсилатеральный) 6-дневной мыши (p6), а правый седалищный нерв использовали в качестве внутреннего контроля (контралатеральный). Через четыре недели после инъекции эффективность интраневральной доставки комплекса RNP была подтверждена с помощью направленного глубокого секвенирования с помощью сбора геномной ДНК из седалищного нерва. В результате все седалищные нервы, обработанные mRosa26 RNP и PMP22-TATA RNP, показали эффективность инделей около 11% (ФИГ.12A). Кроме того, мутация TATA-бокса 98,48 ± 0,15% была подтверждена в общем считывании секвенирования инделей, согласующемся с результатами in vitro (ФИГ.12B).
Кроме того, для того, чтобы подтвердить ингибирование экспрессии PMP22 с помощью мутации TATA-бокса in vivo, на мРНК, экстрагированной из всего седалищного нерва, был проведен количественный анализ ОТ-ПЦР на седалищном нерве, обработанном RNP. Аналогично результатам in vitro было подтверждено, что экспрессия гена PMP22 была снижена на 38% по сравнению с контролем (ФИГ.12C).
Для того, чтобы подтвердить, произошла ли нецелевая мутация в седалищном нерве с помощью PMP22-TATA RNP, был проведен нецелевой анализ на основе in silico. В результате восемь нецелевых участков, включая 3 п.н. или более ошибочно спаренных оснований, были подтверждены в геноме мыши (ФИГ.13A), и в результате выполнения направленного глубокого секвенирования не было подтверждено индель-мутации, превышающей коэффициент ошибки секвенирования от нерва (ипсилатерального), обработанного PMP22-TATA RNP (ФИГ.13B).
Для того, чтобы проверить, может ли уменьшение транскрипции PMP22, вызванное PMP22-TATA РНК, предотвратить демиелинизацию, был получен седалищный нерв мыши C22, обработанный с помощью PMP22-TATA RNP или mRosa26 RNP, и его полутонкие поперечные срезы были окрашены толуидиновым синим (миелиновое окрашивание). Кроме того, для того, чтобы измерить g-коэффициент были измерены диаметр аксона и диаметр волокна (аксона, включая миелин). В результате можно подтвердить, что более толстый миелиновый слой был образован в экспериментальной группе, обработанной PMP22-TATA RNP (ФИГ.14A, ФИГ.14B). Кроме того, когда экспериментальную группу обрабатывали PMP22-TATA RNP по сравнению с контролем, обработанным mRosa26 RNP, было обнаружено, что количество аксонов, имеющих большой диаметр, было увеличено (ФИГ.14A, ФИГ.14B). Результат измерения количества крупных миелинизированных волокон, имеющих диаметр 6 мкм или более, в экспериментальной группе (16,5%), получавшей PMP22-TATA RNP, демонстрирует более четкий терапевтический эффект, чем в контроле (2,6%, р <0,01).
Принимая во внимание значительное улучшение гистологического анализа миелинизации, были изучены электрофизиологические профили двух групп. В результате было подтверждено, что дистальная латентность (DL) была уменьшена, и скорость проводимости двигательного нерва (NCV) была увеличена в седалищном нерве экспериментальной группы, обработанной PMP22-TATA RNP, по сравнению с контролем, обработанным mRosa26 RNP (ФИГ. 15A, ФИГ. 15B), и результаты соответствуют увеличению толщины миелина и диаметра аксона в нерве, обработанном PMP22-TATA RNP. Кроме того, было подтверждено, что амплитуда электрического вызванного ответа мышц (CMAP) была значительно увеличена в нерве, обработанном PMP22-TATA RNP (ФИГ.15C), что соответствует предыдущему результату.
Принимая во внимание гистологически и электрофизиологически улучшенные эффекты с помощью PMP22-TATA RNP, локомоторное поведение мышей было проанализировано с помощью эксперимента с вращающимся стержнем. В результате было подтверждено, что мыши (в возрасте от 11 до 16 недель), обработанные PMP22-TATA RNP, оставались на стержне дольше, чем мыши (в возрасте от 11 до 16 недель), получавшие mRosa26 RNP (ФИГ.16А). Кроме того, было подтверждено, что у мышей, получавших PMP22-TATA RNP, было увеличено количество мышц по сравнению с мышами, получавшими mRosa26 RNP (ФИГ.16B).
Эти результаты показывают терапевтический эффект PMP22-TATA RNP для облегчения или лечения демиелинизации из-за сверхэкспрессии PMP22, такой как CMT1A.
Соответственно, вышеупомянутые результаты показывают эффект ингибирования экспрессии PMP22 с использованием CRISPR/Cas9, нацеленного на сайт промотора PMP22. Кроме того, результаты показывают, что непосредственная невирусная доставка PMP22-TATA RNP к седалищному нерву мыши C22 может улучшить клинические и нейропатологические фенотипы, связанные с демиелинизацией, вызванной сверхэкспрессией PMP22. Следовательно, считается, что CRISPR/Cas9-опосредованная модификация сайта управляющего фактора PMP22, может быть хорошей стратегией для лечения CMT1А и других заболеваний, которые проявляют демиелинизирующие нейропатии.
Промышленная применимость
Искусственно измененный фактор, управляющий функцией ШК, и система, управляющая функцией ШК, чьи функции искусственно модифицированы тем же путем, могут быть использованы для эффективного лечения нарушения из-за дисфункции ШК, например, заболевания, связанного с фактором, управляющим функцией ШК, такого как болезнь Шарко-Мари-Тута Типа 1A (CMT1A), болезнь Дейерина-Сотта (DSS), врожденная гипомиелинизирующая нейропатия (CHN) или синдром Русси-Леви (RLS). Благодаря различным механизмам in vivo, в которых задействованы факторы, управляющие функцией ШК, эффективность системы, управляющей функцией ШК, может быть улучшена.
--->
<110> TOOLGEN INCORPORATED
<120> ИСКУССТВЕННО СОЗДАННАЯ СИСТЕМА УПРАВЛЕНИЯ ФУНКЦИЕЙ ШК
<130> OPP17-045
<150> US 62/421,516
<151> 2016-11-14
<160> 150
<170> KoPatentIn 3.0
<210> 1
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 1
cgatgatact cagcaacagg 20
<210> 2
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 2
ggacgatgat actcagcaac 20
<210> 3
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 3
atggacacgc aactgatctc 20
<210> 4
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 4
gatgatcgac aggatcatgg 20
<210> 5
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 5
gaagatgatc gacaggatca 20
<210> 6
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 6
aactcttcac cctcaccaag 20
<210> 7
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 7
aaaacctgcc ccccttggtg 20
<210> 8
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 8
ggaatcttcc aaattcttgc 20
<210> 9
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 9
gatcgtggag acgaacagca gc 22
<210> 10
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 10
gctgacgatc gtggagacga ac 22
<210> 11
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 11
cgcaactgat ctctggcaga ac 22
<210> 12
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 12
gtgcgtgatg agtgctgcgg cc 22
<210> 13
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 13
gtgtagatgg ccgcagcact ca 22
<210> 14
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 14
ggaccagccc ctgaataaac 20
<210> 15
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 15
ggcgtctttc cagtttattc 20
<210> 16
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 16
gcgtctttcc agtttattca 20
<210> 17
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 17
cgtctttcca gtttattcag 20
<210> 18
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 18
ttcaggggct ggtccaatgc 20
<210> 19
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 19
tcaggggctg gtccaatgct 20
<210> 20
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 20
accatgacat atcccagcat 20
<210> 21
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 21
tttccagttt attcaggggc 20
<210> 22
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 22
cagttacagg gagcaccacc 20
<210> 23
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 23
ctggtctggc ttcagttaca 20
<210> 24
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 24
cctggtctgg cttcagttac 20
<210> 25
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 25
aactggaaag acgcctggtc 20
<210> 26
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 26
gaataaactg gaaagacgcc 20
<210> 27
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 27
tccaatgctg ggatatgtca 20
<210> 28
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 28
aatgctggga tatgtcatgg 20
<210> 29
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 29
atagaggctg agaacctctc 20
<210> 30
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 30
gccctctgaa tctccagtca at 22
<210> 31
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 31
aatctccagt caattccaac ac 22
<210> 32
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 32
aattaggcaa ttcttgtaaa gc 22
<210> 33
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 33
ttaggcaatt cttgtaaagc at 22
<210> 34
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 34
aaagcatagg cacacatcac cc 22
<210> 35
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 35
gcctggtctg gcttcagtta ca 22
<210> 36
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 36
gtgtccaact ttgtttgctt tc 22
<210> 37
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 37
gtattctgga aagcaaacaa ag 22
<210> 38
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 38
cagtcttggc atcacaggct tc 22
<210> 39
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 39
ggacctcttg gctattacac ag 22
<210> 40
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 40
ggagccagtg ggacctcttg gc 22
<210> 41
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 41
ttgggcatgt ttgagctggt 20
<210> 42
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 42
tttgggcatg tttgagctgg 20
<210> 43
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 43
gagctggtgg gcgaagcata 20
<210> 44
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 44
agctggtggg cgaagcatat 20
<210> 45
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 45
tgggcgaagc atatgggcaa 20
<210> 46
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 46
ggcctccatc ctaaacaatg 20
<210> 47
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 47
gggttgggag gtttgggcgt 20
<210> 48
<211> 20
<212> ДНК
<213> Homo sapiens
<400> 48
aggtttgggc gtgggagtcc 20
<210> 49
<211> 19
<212> ДНК
<213> Homo sapiens
<400> 49
ttcagagact cagctattt 19
<210> 50
<211> 19
<212> ДНК
<213> Homo sapiens
<400> 50
ggccacattg tttaggatg 19
<210> 51
<211> 19
<212> ДНК
<213> Homo sapiens
<400> 51
ggctttgggc atgtttgag 19
<210> 52
<211> 19
<212> ДНК
<213> Homo sapiens
<400> 52
aacatgccca aagcccagc 19
<210> 53
<211> 19
<212> ДНК
<213> Homo sapiens
<400> 53
acatgcccaa agcccagcg 19
<210> 54
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 54
ttaaatcaca gaggcaaaga gtt 23
<210> 55
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 55
ttgcatagtg ctagactgtt tt 22
<210> 56
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 56
gggtcatgtg ttttgaaaac ag 22
<210> 57
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 57
cccaaacctc ccaacccaca ac 22
<210> 58
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 58
actcagctat ttctggaatg ac 22
<210> 59
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 59
tcatcgcctt tgtgagctcc at 22
<210> 60
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 60
cagacacagg ctttgctcta gc 22
<210> 61
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 61
caaagcctgt gtctggccac ta 22
<210> 62
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 62
agcagtttgt gcccactagt gg 22
<210> 63
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 63
atgtcaaggt attccagcta ac 22
<210> 64
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 64
gaataactgt atcaaagtta gc 22
<210> 65
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 65
ttcctaatta agaggctttg tg 22
<210> 66
<211> 22
<212> ДНК
<213> Homo sapiens
<400> 66
gagctagttt gtcagggtct ag 22
<210> 67
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 67
cacagggcag tcagagaccc 20
<210> 68
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 68
gcaaacaaag ttggacactg 20
<210> 69
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 69
agactcccgc ccatcttcta gaaa 24
<210> 70
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 70
aagtcgctct gagttgttat cagt 24
<210> 71
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 71
cagtgaaacg caccagacg 19
<210> 72
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 72
aatctgccta acaggaggtg 20
<210> 73
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 73
gagggaatgg ggaccaaagg catt 24
<210> 74
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 74
tcatgtgggg tgatgttcag gaag 24
<210> 75
<211> 23
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 75
agagcagctg acctgaggtc caa 23
<210> 76
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 76
cccaagggta gagtgcaagt aaac 24
<210> 77
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 77
gcatcctagc tcatttggtc tgct 24
<210> 78
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 78
gagaggattc ctcatgaatg ggat 24
<210> 79
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 79
accaaacact acacttggtt actg 24
<210> 80
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 80
ctcccactag caattttaaa gtct 24
<210> 81
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 81
gaatgttcag cacaggtttc cttg 24
<210> 82
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 82
ggtcaaaagg agctccatat ttga 24
<210> 83
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 83
caggacaccc atggccaaat ccag 24
<210> 84
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 84
cagagcctcc tgcagggatg tcaa 24
<210> 85
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 85
gcctgccaag gtgactctca tcta 24
<210> 86
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 86
tgcccaggct gatcttgaac tcct 24
<210> 87
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 87
cccagagtta agaggttctt tcct 24
<210> 88
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 88
gaagctactc cagtgcaact agct 24
<210> 89
<211> 23
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 89
acgcagtctg ttctgtgcag tgt 23
<210> 90
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 90
aggccttccc aaggaagacc ctga 24
<210> 91
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 91
gctgatcact ggccaaatcc agct 24
<210> 92
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 92
gggaaacaat gggatcaagc tgca 24
<210> 93
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 93
gcccctttgt aagttgagga gcat 24
<210> 94
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 94
ccctctacct ctctcaatgg gctt 24
<210> 95
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 95
cagacaagca aatgctgaga gatt 24
<210> 96
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 96
cctgtcatta tgatgttcgc tagt 24
<210> 97
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 97
ccagagttgg cctcctacag agat 24
<210> 98
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 98
gtggatgccc cactactgtt catt 24
<210> 99
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 99
tacccaattt gccagtctgt gtct 24
<210> 100
<211> 23
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 100
accaccaggc ctgccctaca aga 23
<210> 101
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 101
tgtgaatttg atcctggcat tatg 24
<210> 102
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 102
tacagacaag cagatgctga gaga 24
<210> 103
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 103
cagtcaacag agctctaacc tcct 24
<210> 104
<211> 23
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 104
agcacctggt tgcacatcaa ctt 23
<210> 105
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 105
catgtggtcc ctgaacgtga atga 24
<210> 106
<211> 23
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 106
gtctgtcgct tgccctcttc tct 23
<210> 107
<211> 24
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 107
atgcagggcc tctagaccat ttca 24
<210> 108
<211> 23
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 108
ctcagccctt tgtgcactca cct 23
<210> 109
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 109
tgcacatcgc aaacatttcg 20
<210> 110
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 110
tgggtatcgc actgtgtcag 20
<210> 111
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 111
aggttcacat ggcttgtggt 20
<210> 112
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 112
atatctgaaa tgcccgcagg 20
<210> 113
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 113
tgcacatcgc aaacatttcg 20
<210> 114
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 114
tgggtatcgc actgtgtcag 20
<210> 115
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 115
tctttaaagg ccttatctcc 20
<210> 116
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 116
ttctgcttga gaattcatcc 20
<210> 117
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 117
ctcctaatct ttcacttagg 20
<210> 118
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 118
caaagcctgg tataacatag 20
<210> 119
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 119
tcacttcgag catctgtgg 19
<210> 120
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 120
ccaaatgaca ggctgagct 19
<210> 121
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 121
agcaggaagt gaaggctaag 20
<210> 122
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 122
atgtaacgtg gcaactctgg 20
<210> 123
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 123
gtgttgctct cgtcaattag 20
<210> 124
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 124
aggtgttgta catggagaag 20
<210> 125
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 125
tgtgagccac catacccagc 20
<210> 126
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 126
cctgcagtcc tttgcggatc 20
<210> 127
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 127
tcgctgccag tataacatgc 20
<210> 128
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 128
aactccagtc tctagactcg 20
<210> 129
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 129
aatagtttga cgttggagcc 20
<210> 130
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 130
actcccaaca tgttctcctg 20
<210> 131
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 131
atcatcgctc acagagtcc 19
<210> 132
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 132
acgactgcag gatcttaatg 20
<210> 133
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 133
tggatggagg ttgggaatcc 20
<210> 134
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 134
ttgaggcagc agcactctcc 20
<210> 135
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 135
agtctatcct agcagctcc 19
<210> 136
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 136
actgagacca gataatgcag 20
<210> 137
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 137
aagagatgcg agttgttcc 19
<210> 138
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 138
cctcttctac tctgagtgg 19
<210> 139
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 139
acctggttta tcacaagcta 20
<210> 140
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 140
aacgtgaaca gaaggatttc 20
<210> 141
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 141
atcactccat cagagtcagg 20
<210> 142
<211> 20
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> праймер
<400> 142
tggctccttc tattctctcc 20
<210> 143
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 143
tatggaattc ccaagccccc tgg 23
<210> 144
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 144
ggaattccat atgagtcatc tgg 23
<210> 145
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 145
cacagcctac actttgatta tgg 23
<210> 146
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 146
tcaggagcat taagcatata ggg 23
<210> 147
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 147
gaccacggtc catgaattcc tgg 23
<210> 148
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 148
agtatttgca gctgaacaaa agg 23
<210> 149
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 149
atggagtaca gagagacata agg 23
<210> 150
<211> 23
<212> ДНК
<213> Homo sapiens
<400> 150
aaagaatcaa tgcacagcca tgg 23
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИСКУССТВЕННАЯ МОДИФИКАЦИЯ ГЕНОМА ДЛЯ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНА | 2018 |
|
RU2767201C2 |
| МОДИФИЦИРОВАННЫЙ ИММУНОРЕГУЛЯТОРНЫЙ ЭЛЕМЕНТ И ИЗМЕНЕНИЕ ИММУНИТЕТА ПОСРЕДСТВОМ ЭТОГО ЭЛЕМЕНТА | 2017 |
|
RU2767206C2 |
| ОПОСРЕДОВАННАЯ НУКЛЕАЗОЙ СБОРКА ДНК | 2015 |
|
RU2707911C2 |
| СПОСОБ СОЗДАНИЯ НОВОГО ГЕНА В ОРГАНИЗМЕ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2832668C1 |
| ПЛАТФОРМА CRISPR/CAS ДЛЯ ИСКЛЮЧАЮЩЕГО СКРИНИНГА ДЛЯ ВЫЯВЛЕНИЯ ГЕНЕТИЧЕСКИХ УЯЗВИМОСТЕЙ, СВЯЗАННЫХ С АГРЕГАЦИЕЙ ТАУ-БЕЛКА | 2020 |
|
RU2808829C2 |
| СИСТЕМА РЕДАКТИРОВАНИЯ ГЕНОМНОЙ ДНК ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ НА ОСНОВЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩЕЙ БЕЛОК SUCAS9NLS | 2022 |
|
RU2804422C1 |
| Животные, отличные от человека, содержащие гуманизированный локус альбумина | 2020 |
|
RU2815514C2 |
| ЭКСПРЕССИРУЮЩИЕ CAS МЫШИНЫЕ ЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ И МЫШИ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2782358C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДИФИКАЦИИ ЦЕЛЕВОГО ЛОКУСА | 2015 |
|
RU2704283C2 |
| СКРИНИНГОВАЯ ПЛАТФОРМА CRISPR/CAS ДЛЯ ИДЕНТИФИКАЦИИ ГЕНЕТИЧЕСКИХ МОДИФИКАТОРОВ ЗАСЕВА ИЛИ АГРЕГАЦИИ ТАУ-БЕЛКА | 2020 |
|
RU2819350C2 |
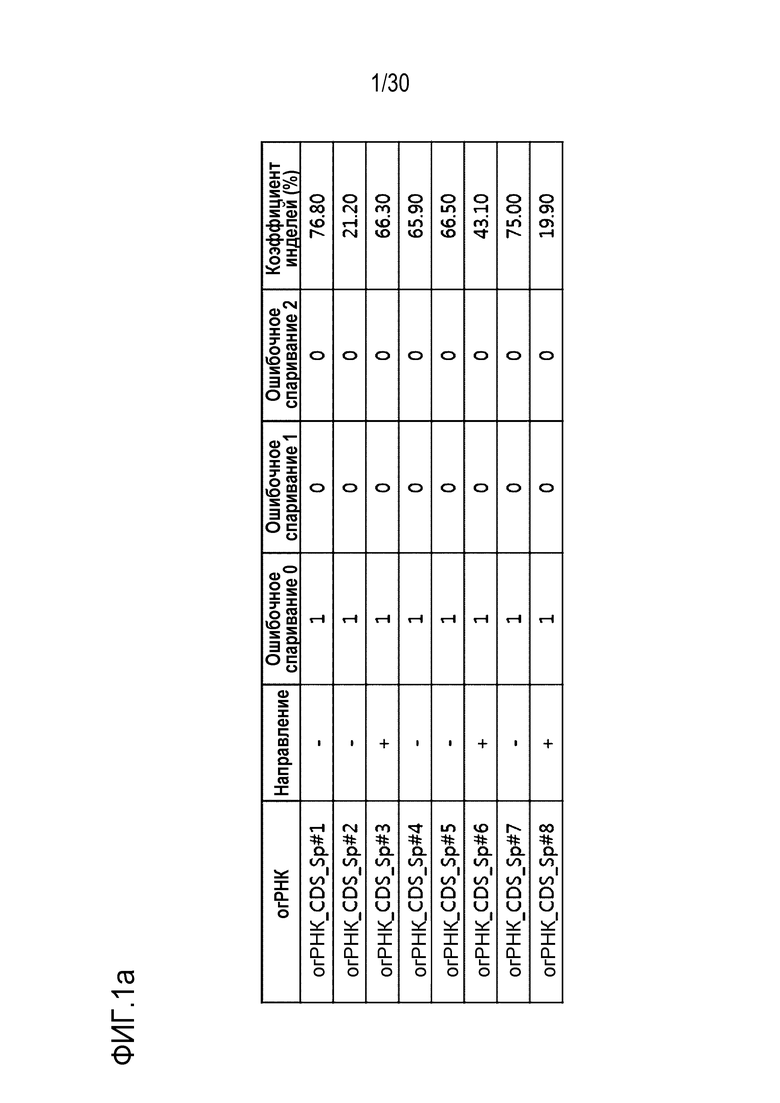
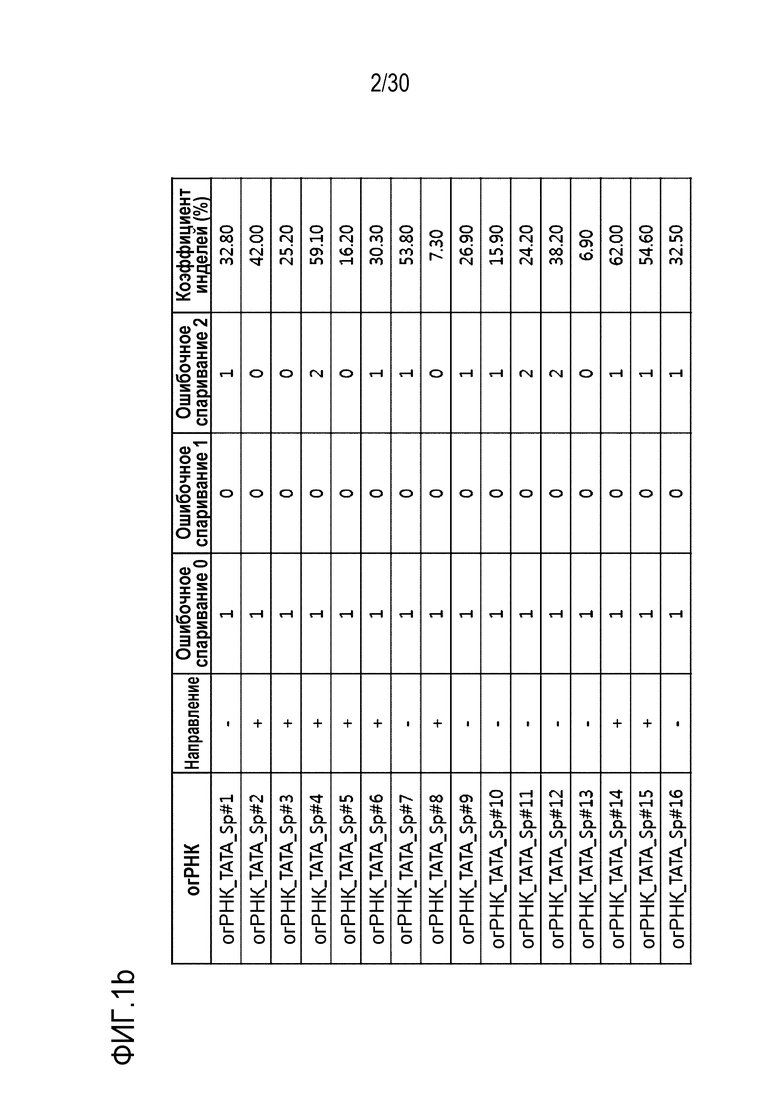
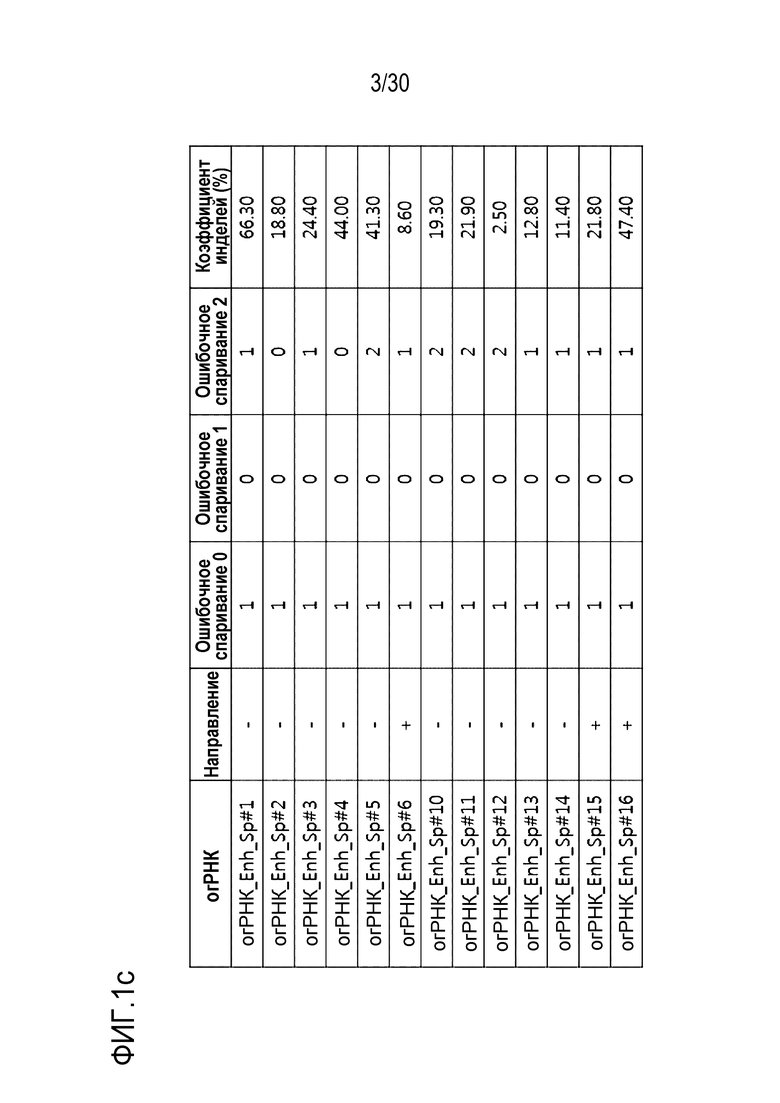
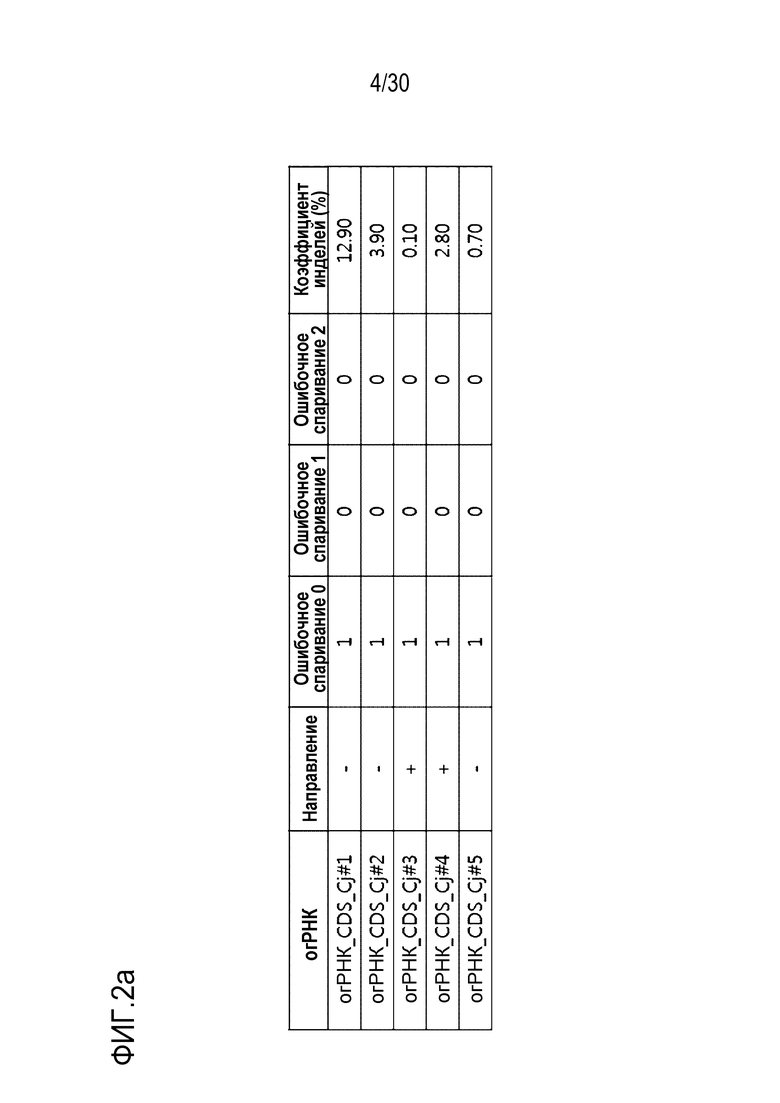
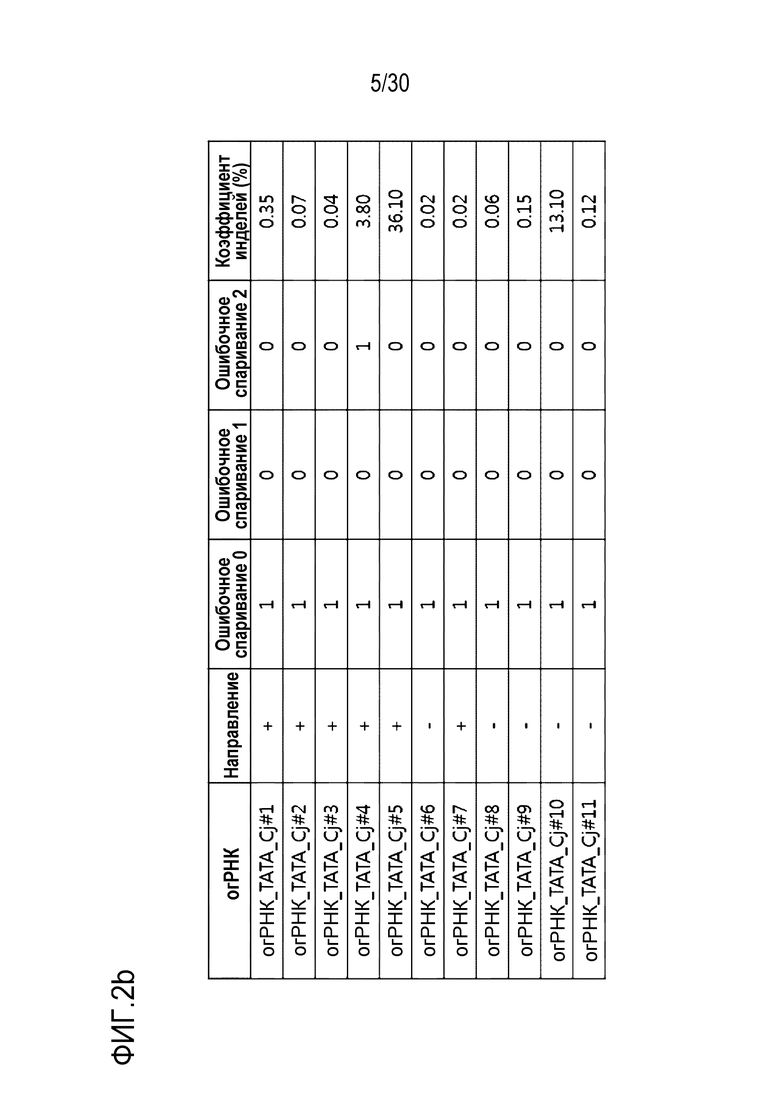
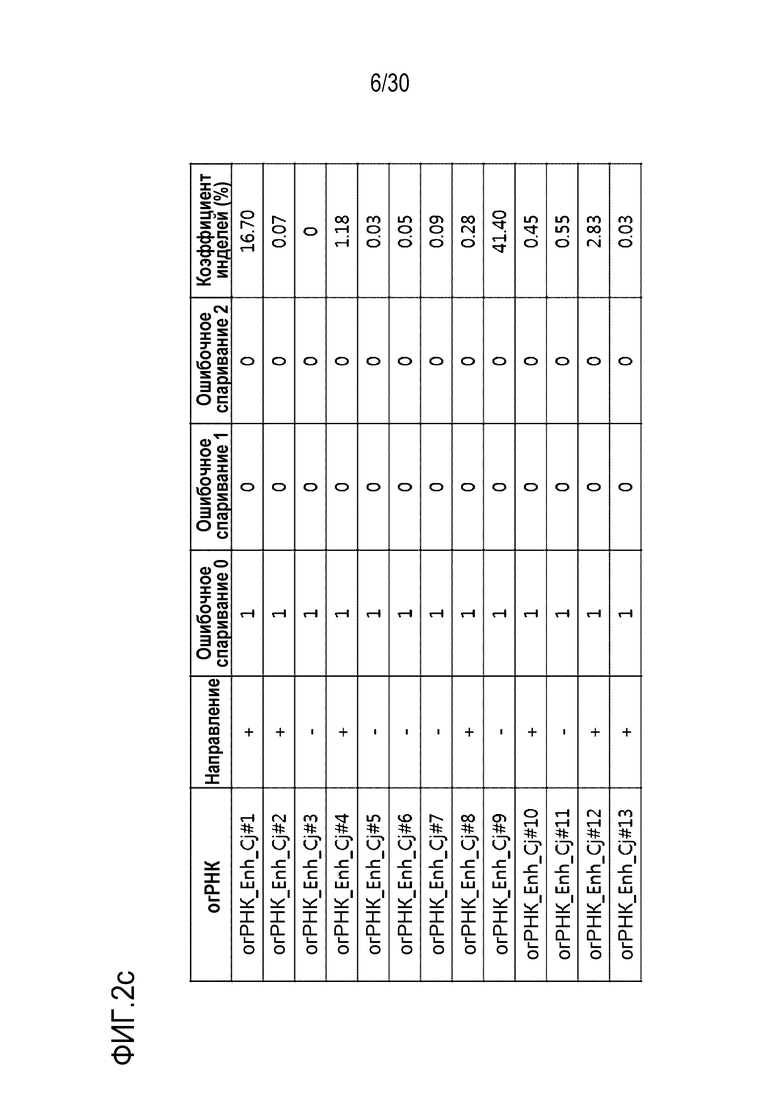
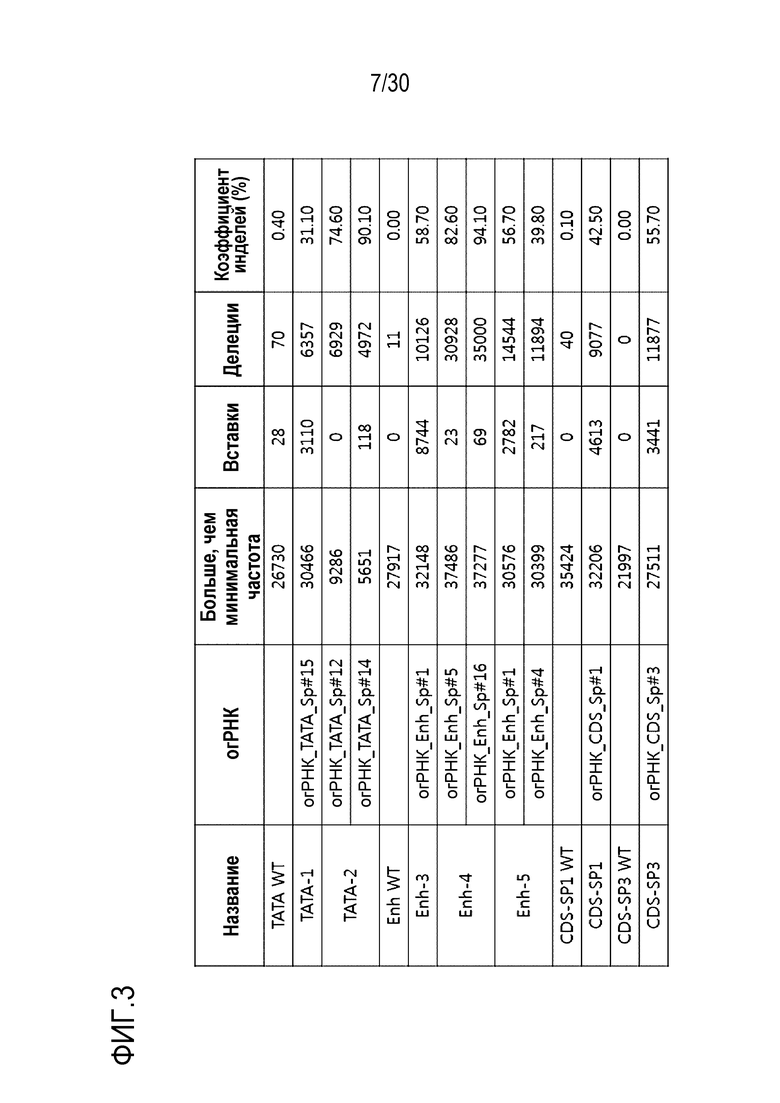
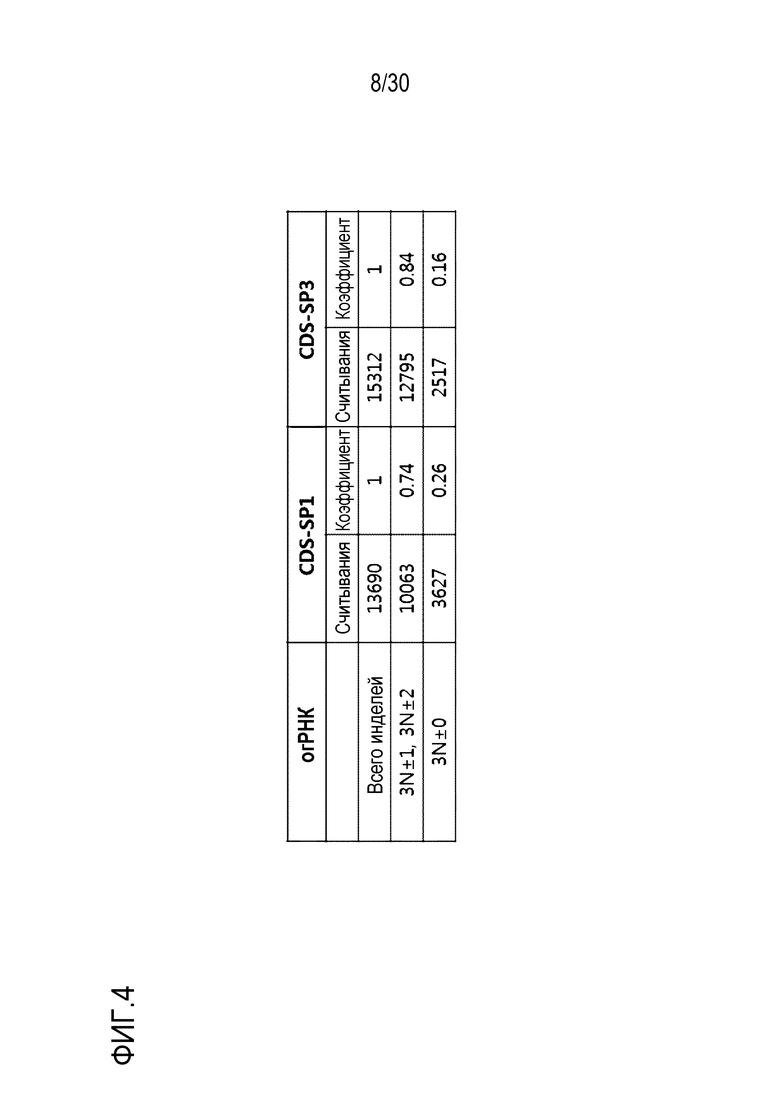
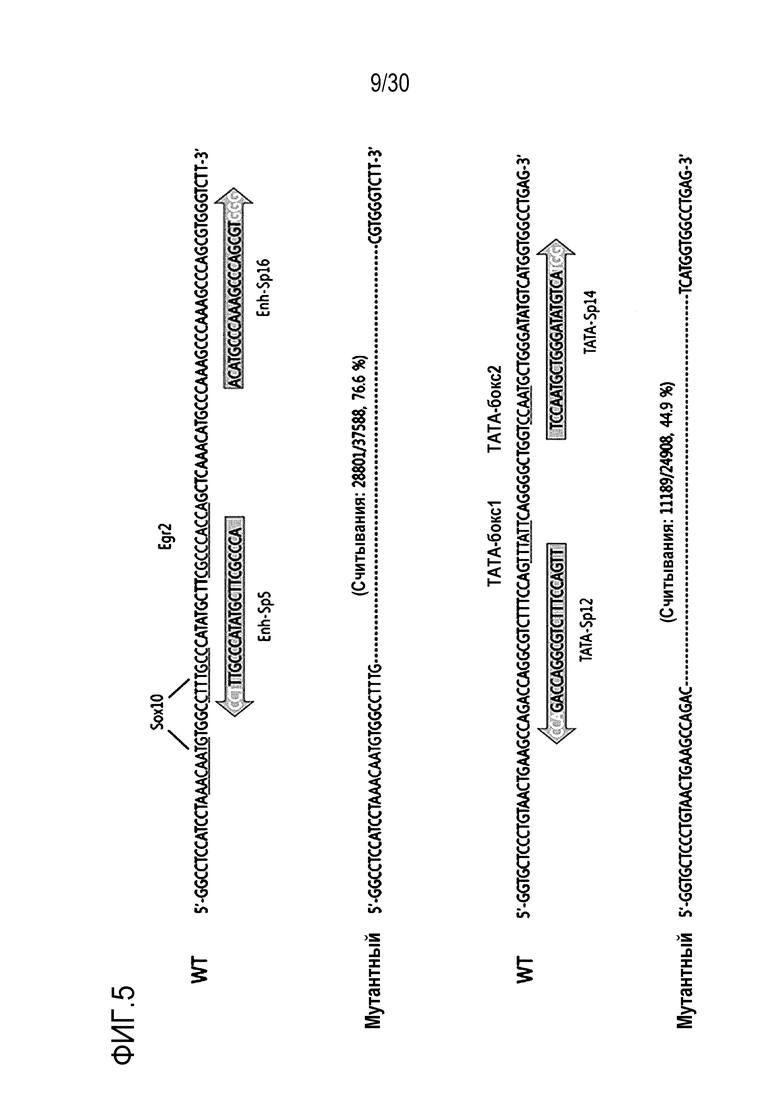
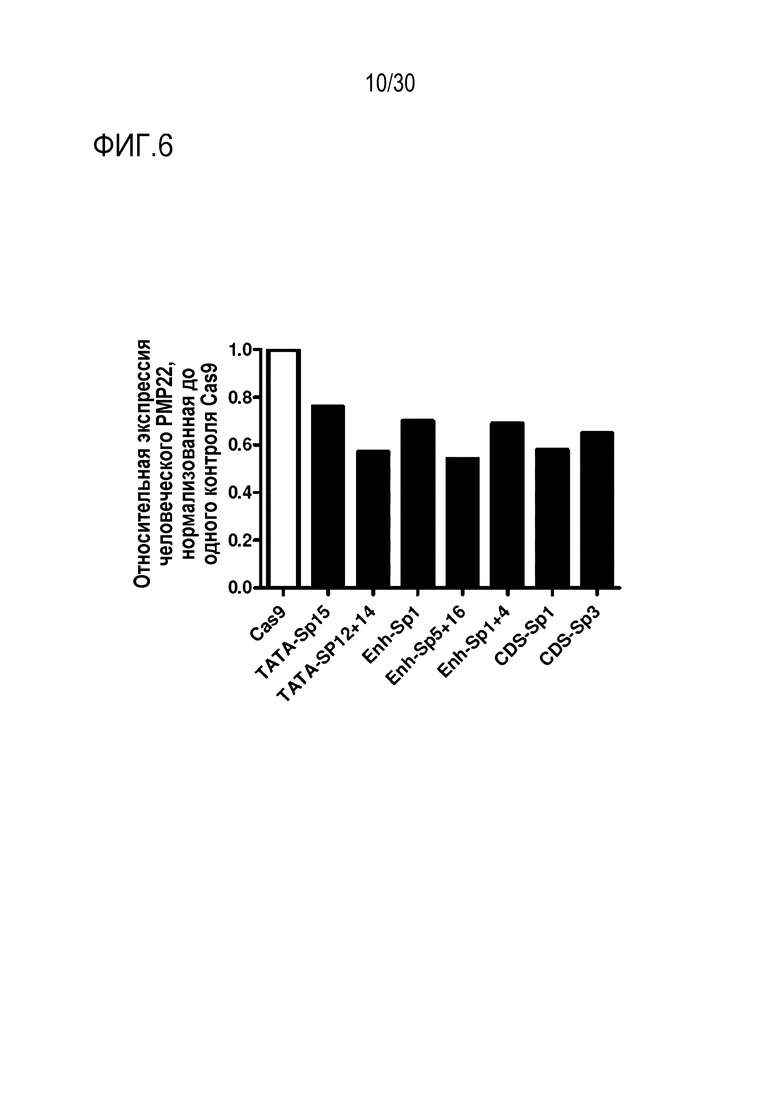
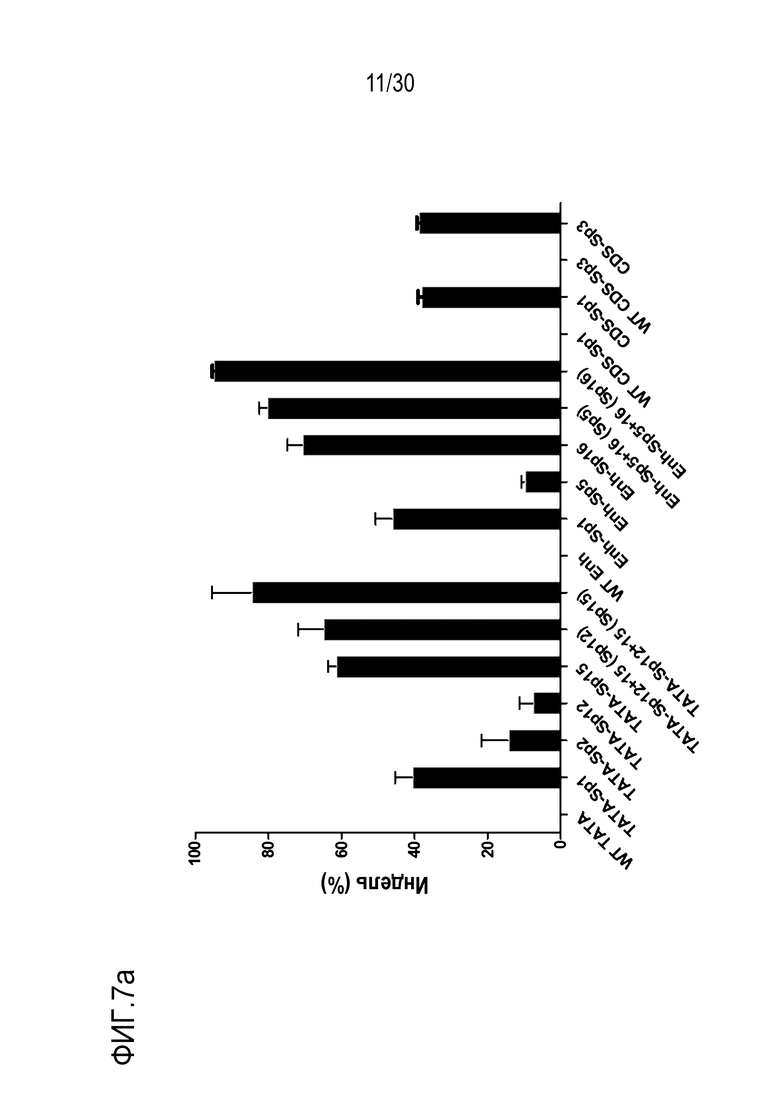
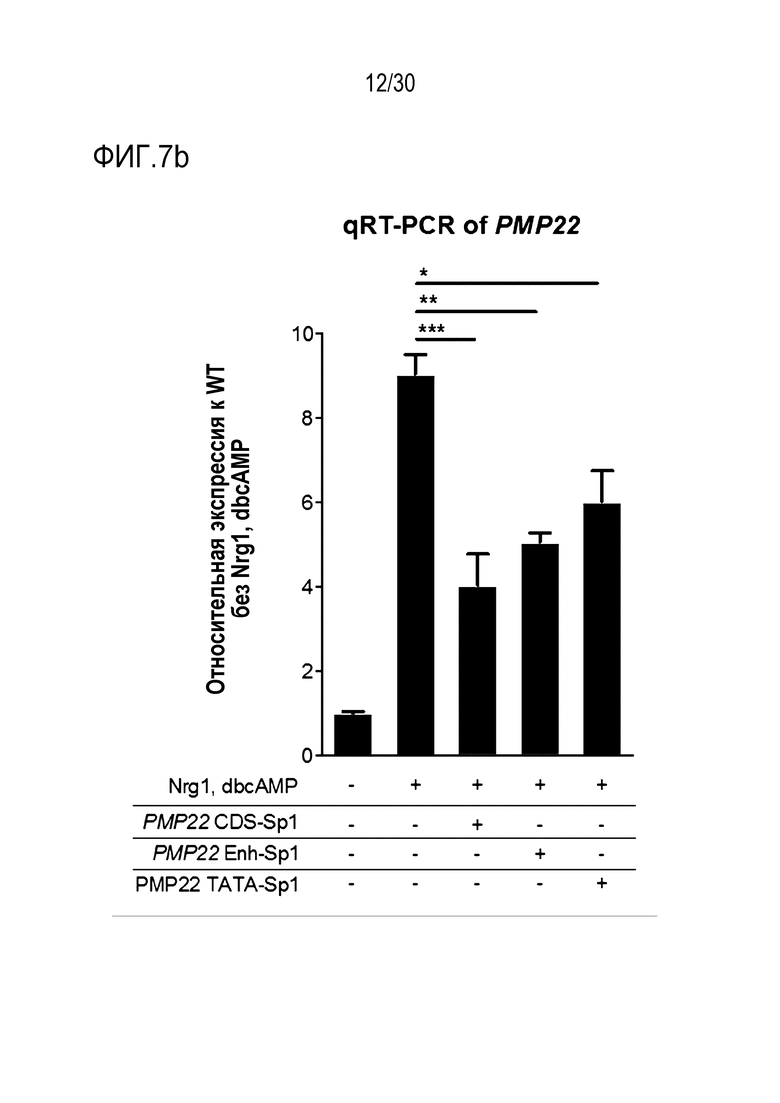
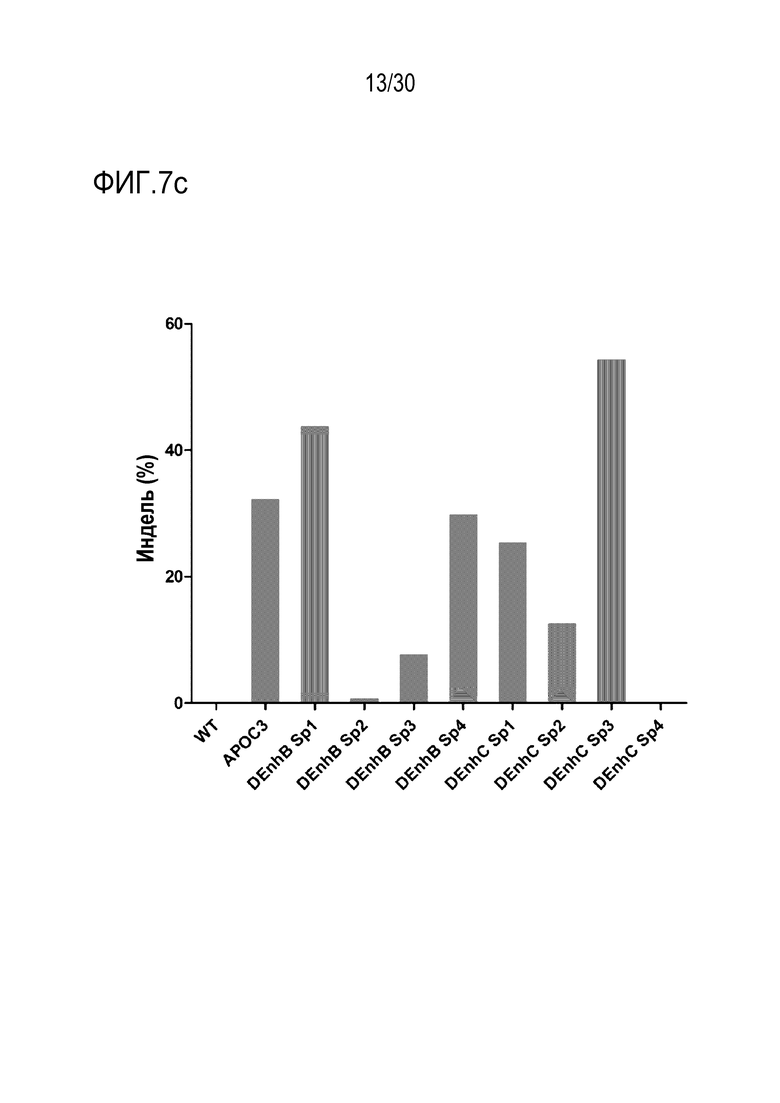
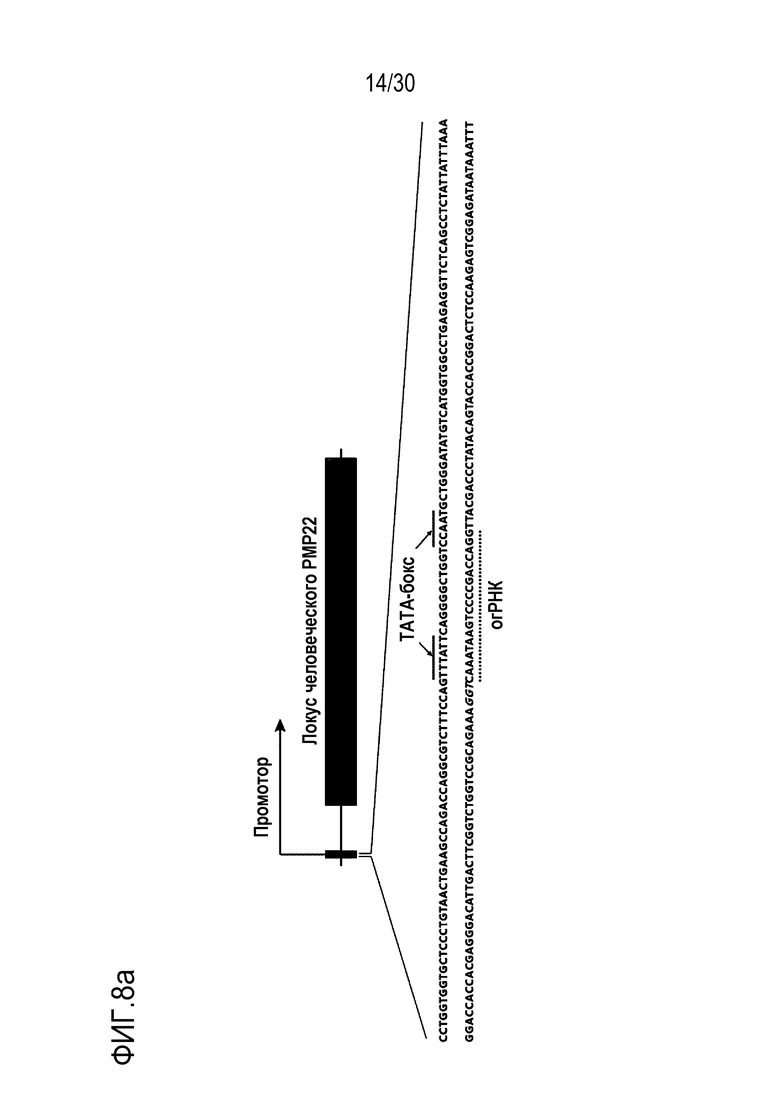
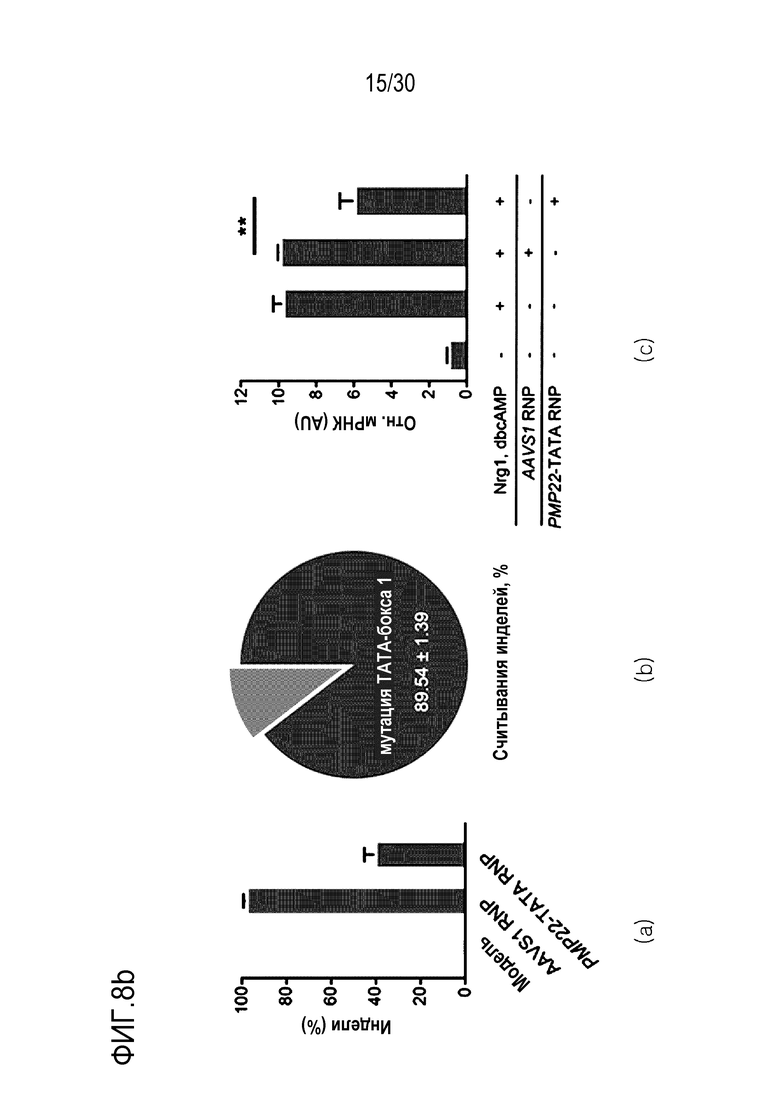
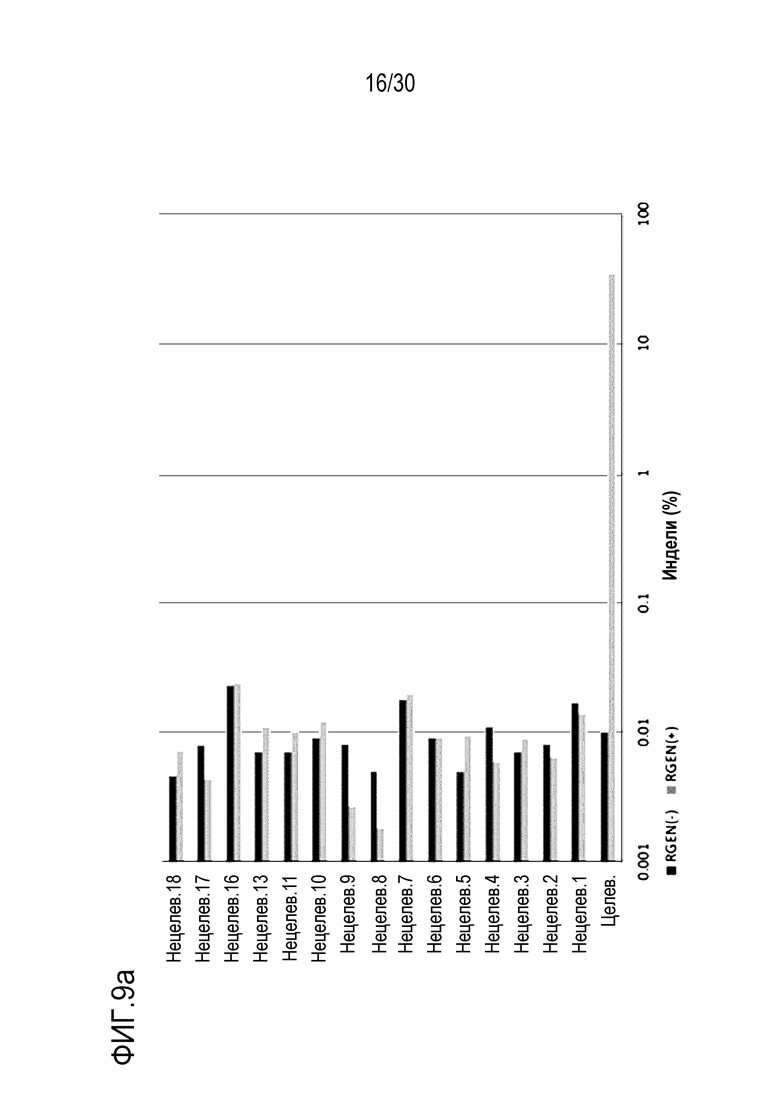
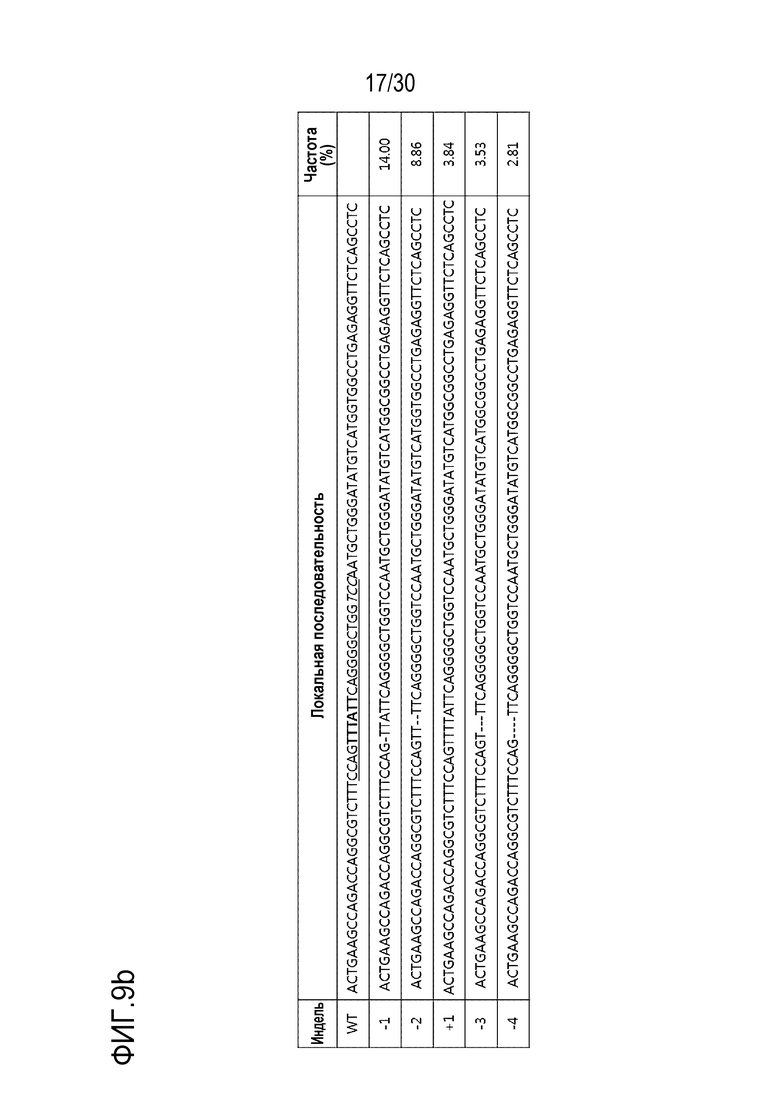
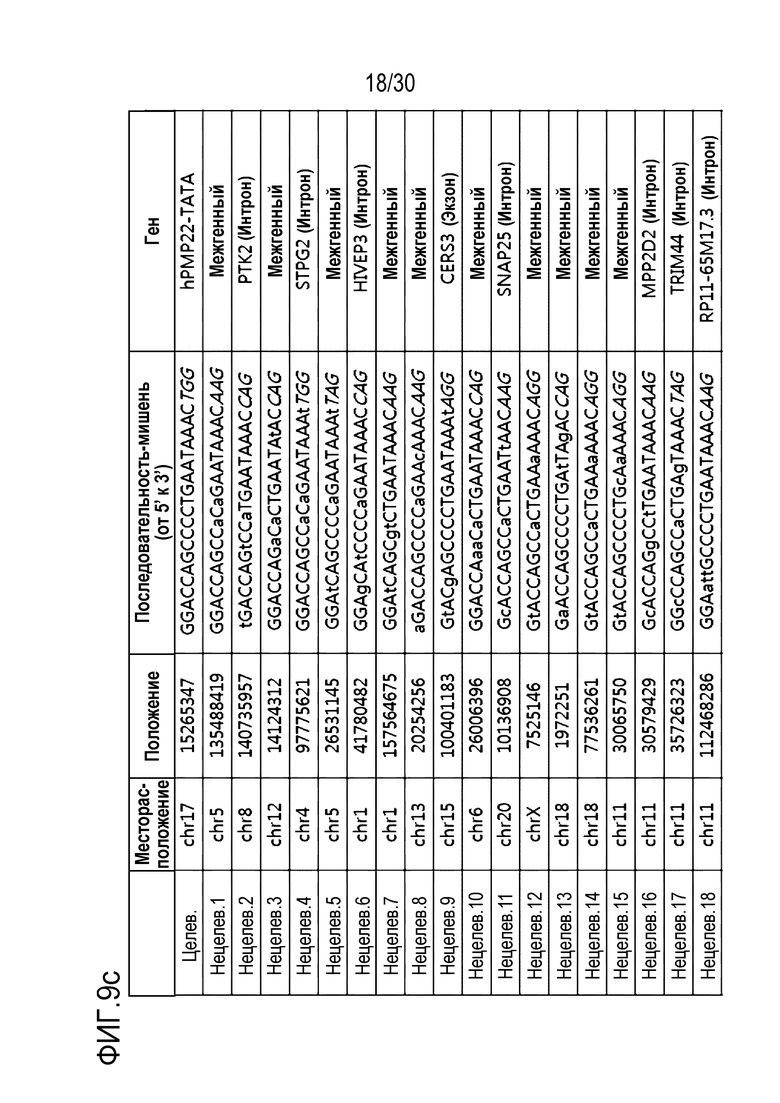
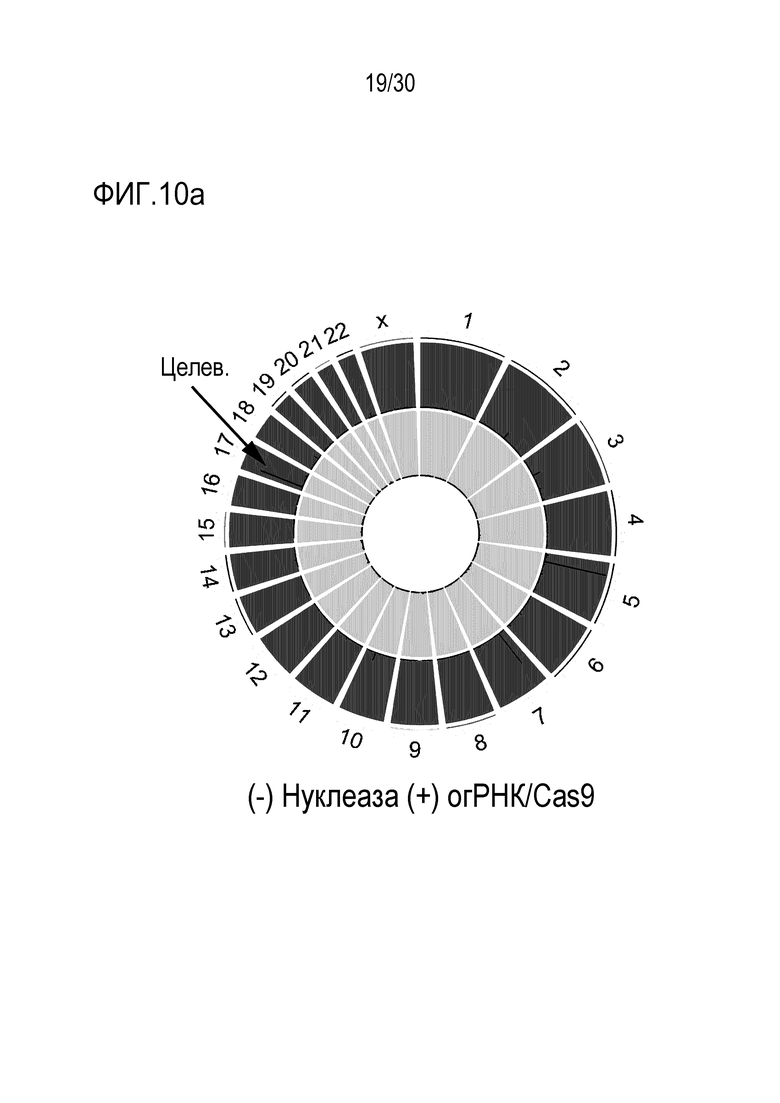
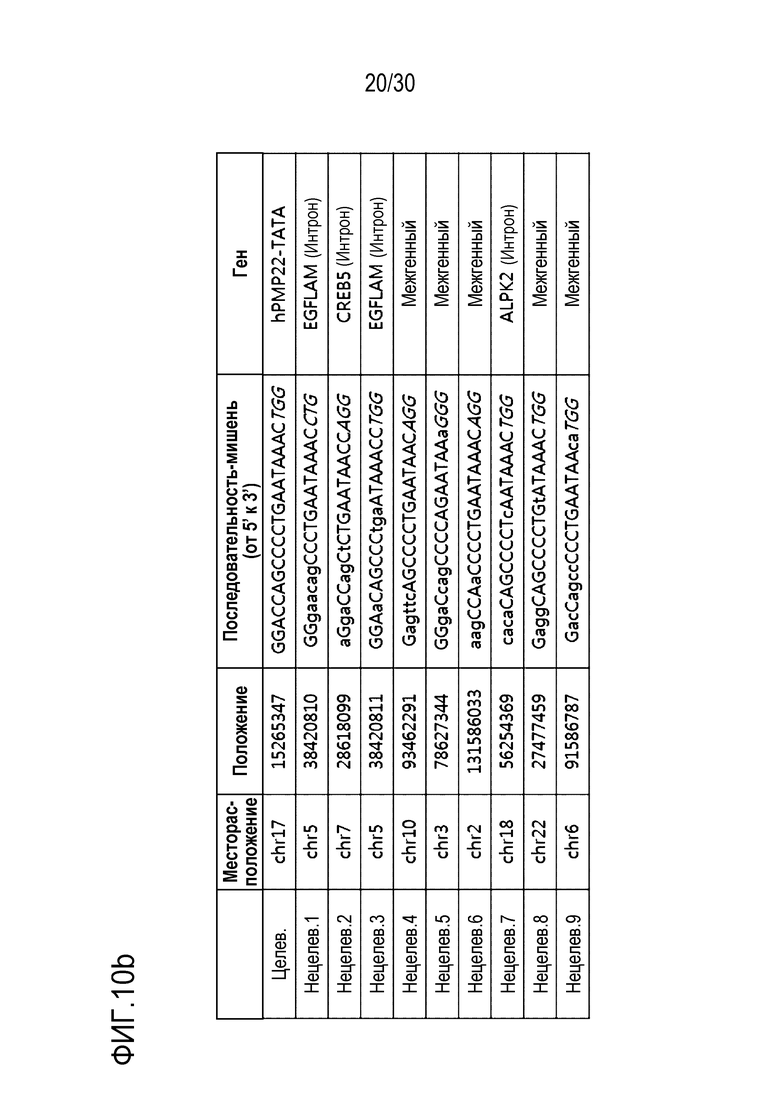
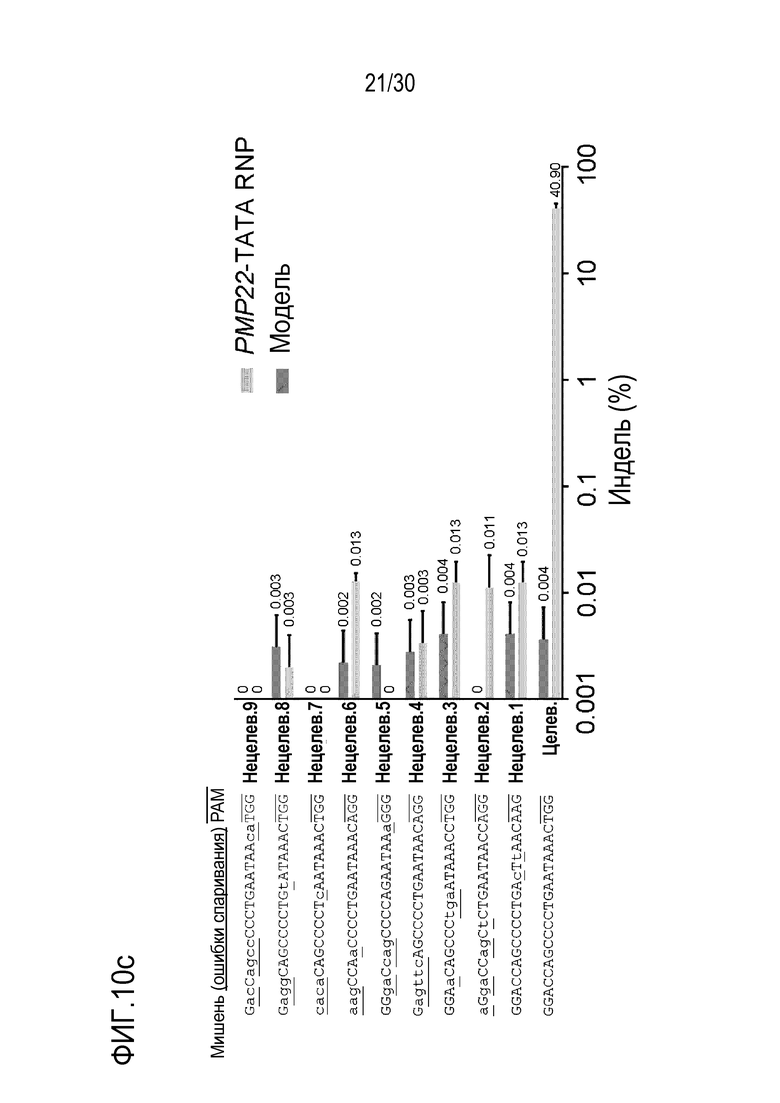
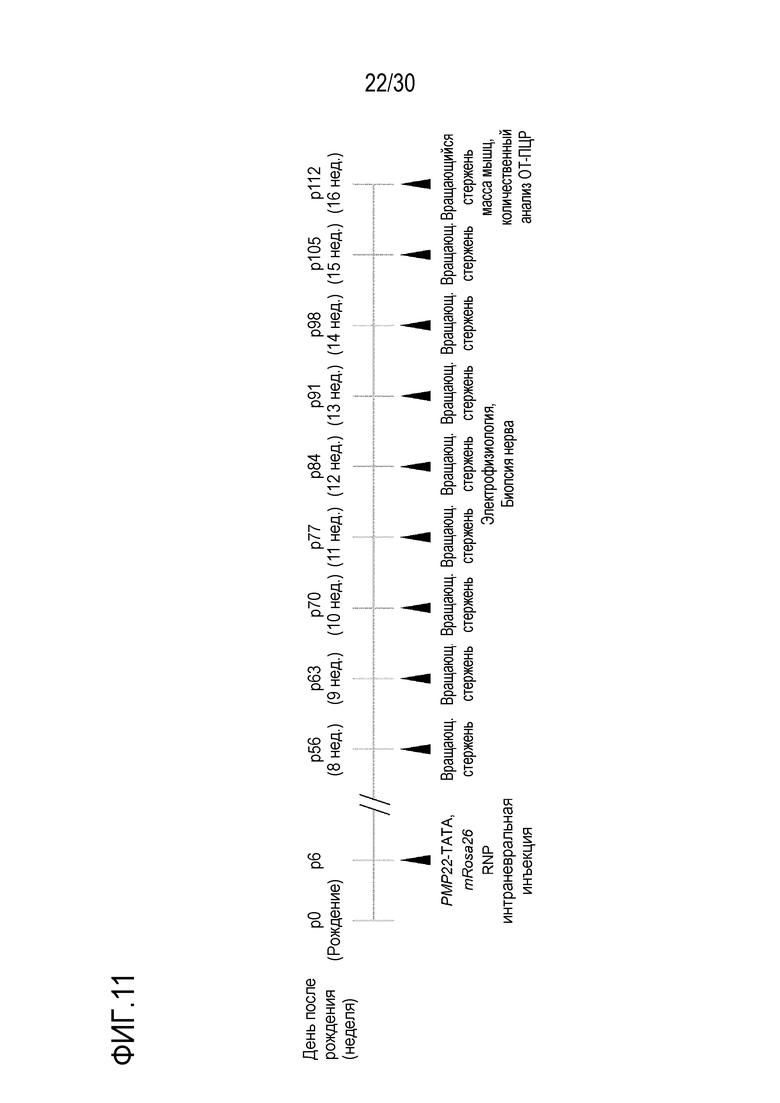
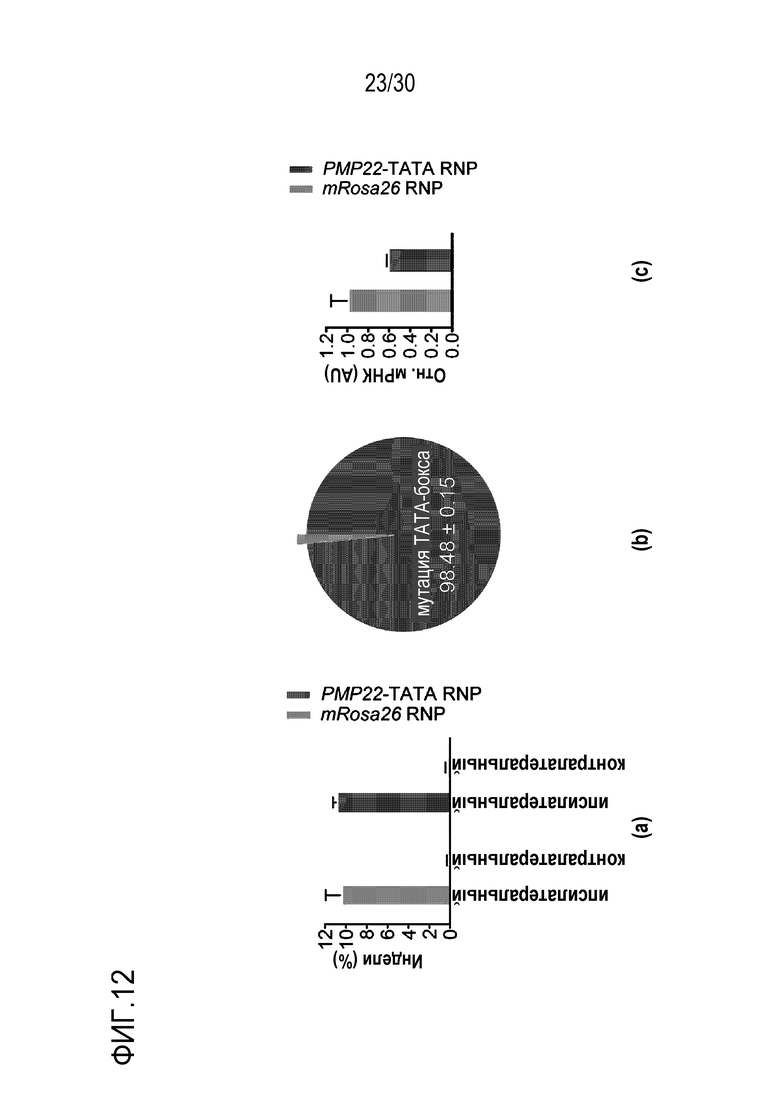
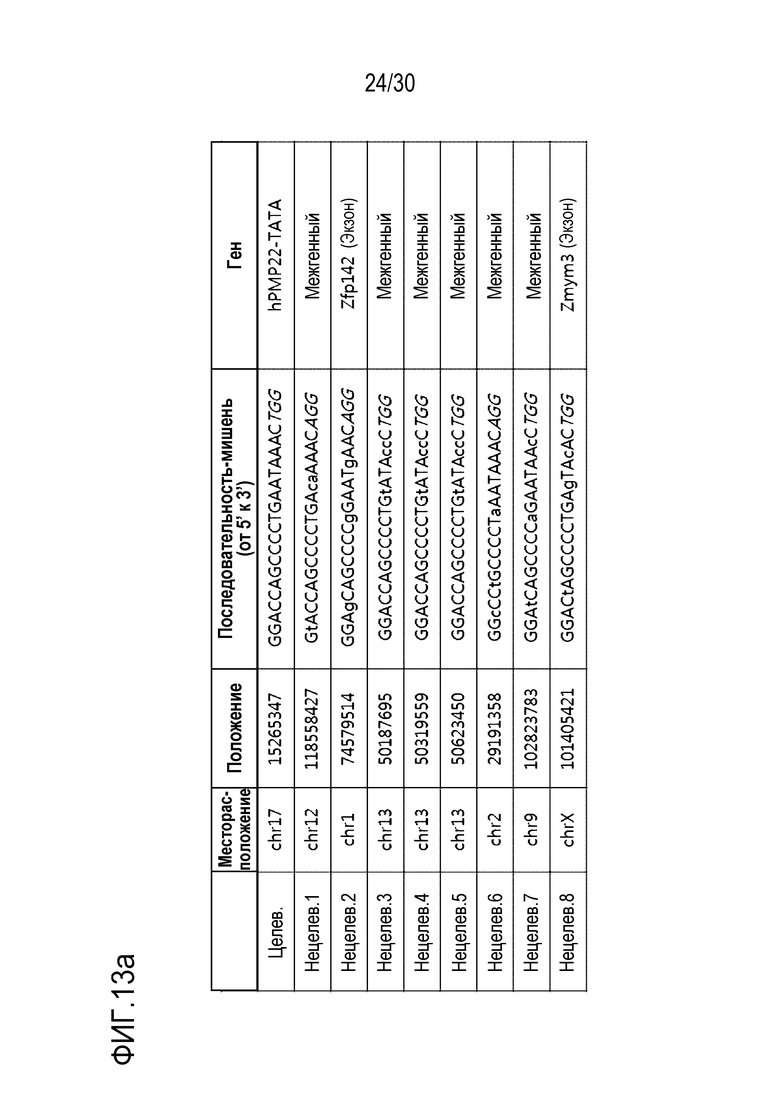
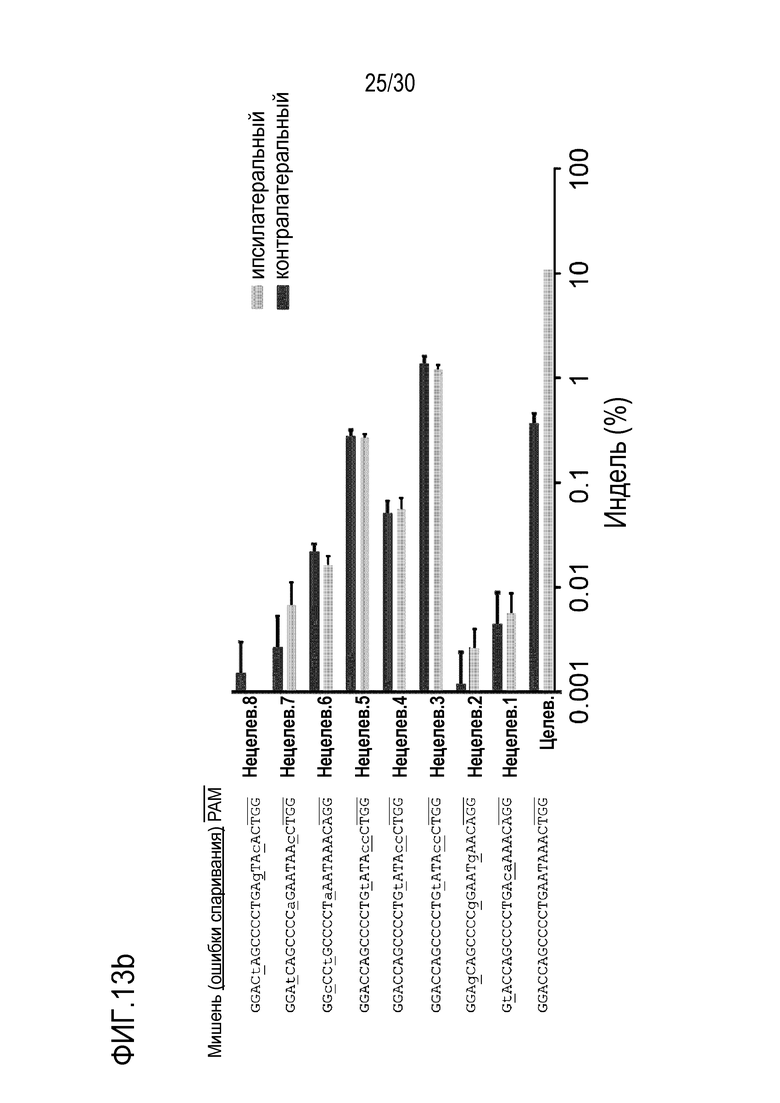

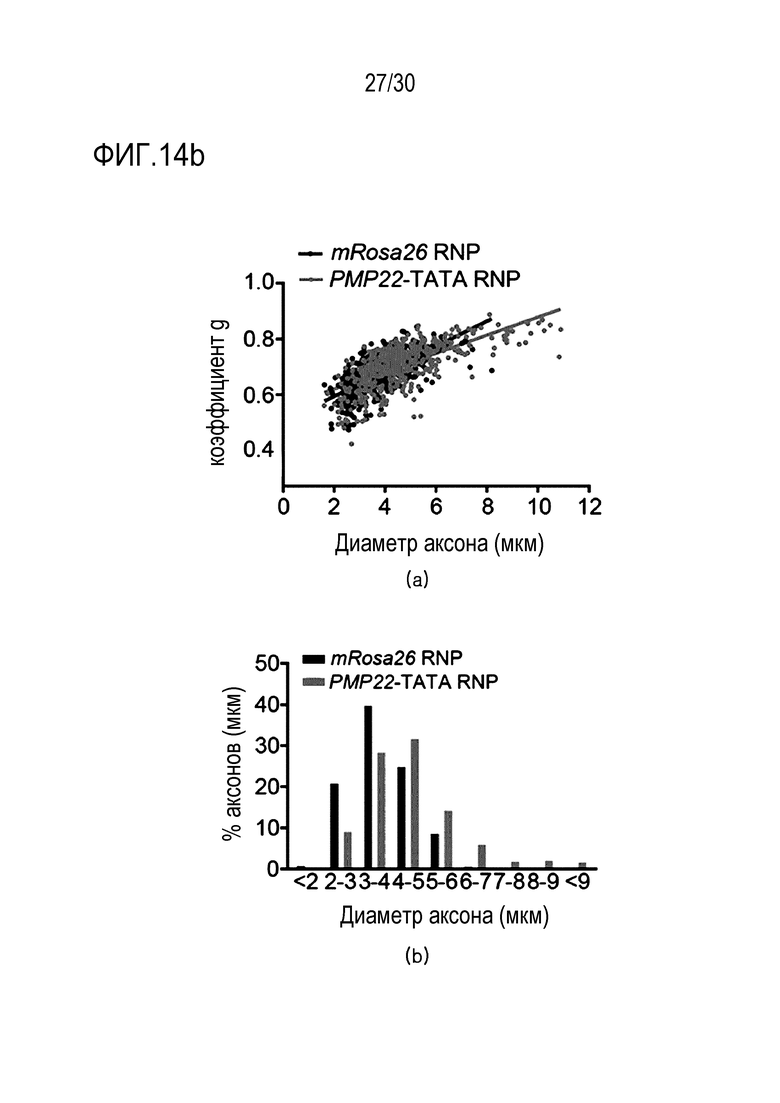
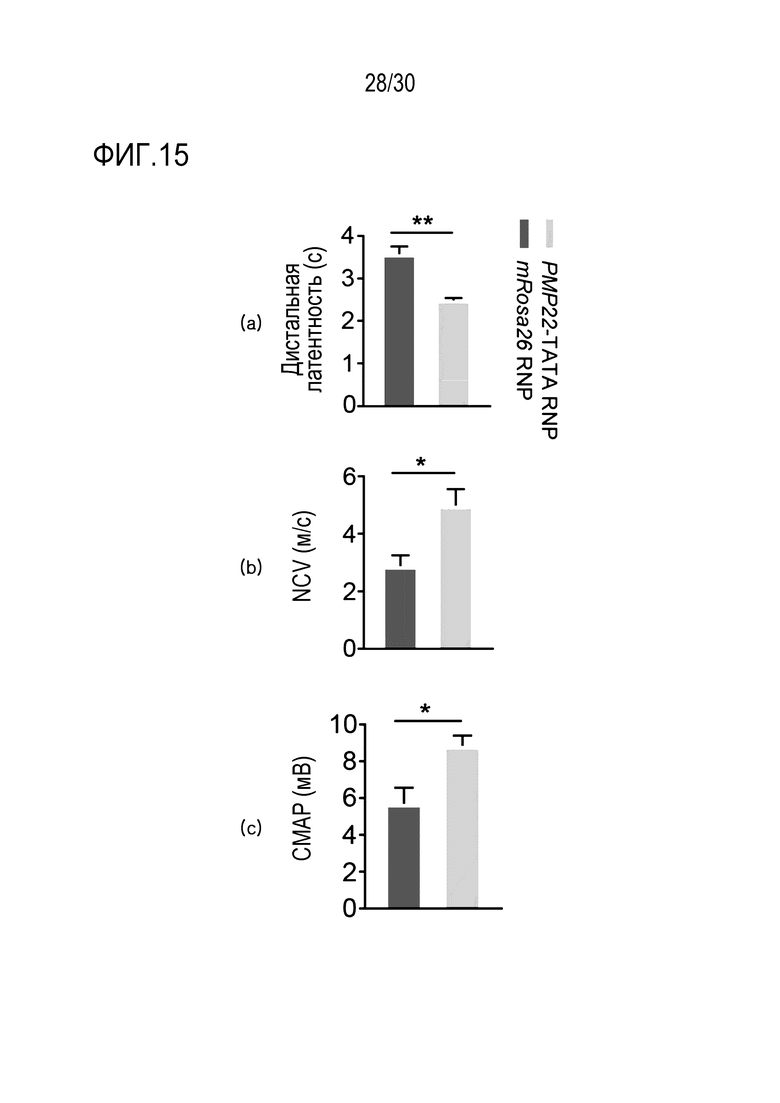
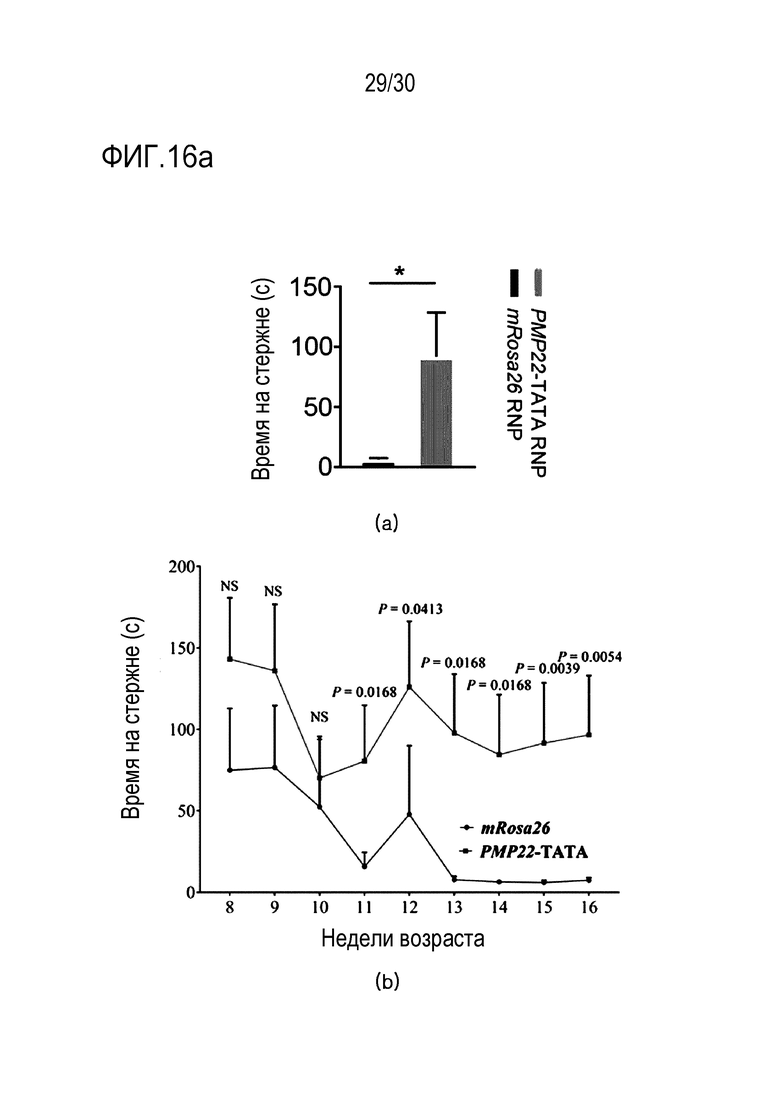
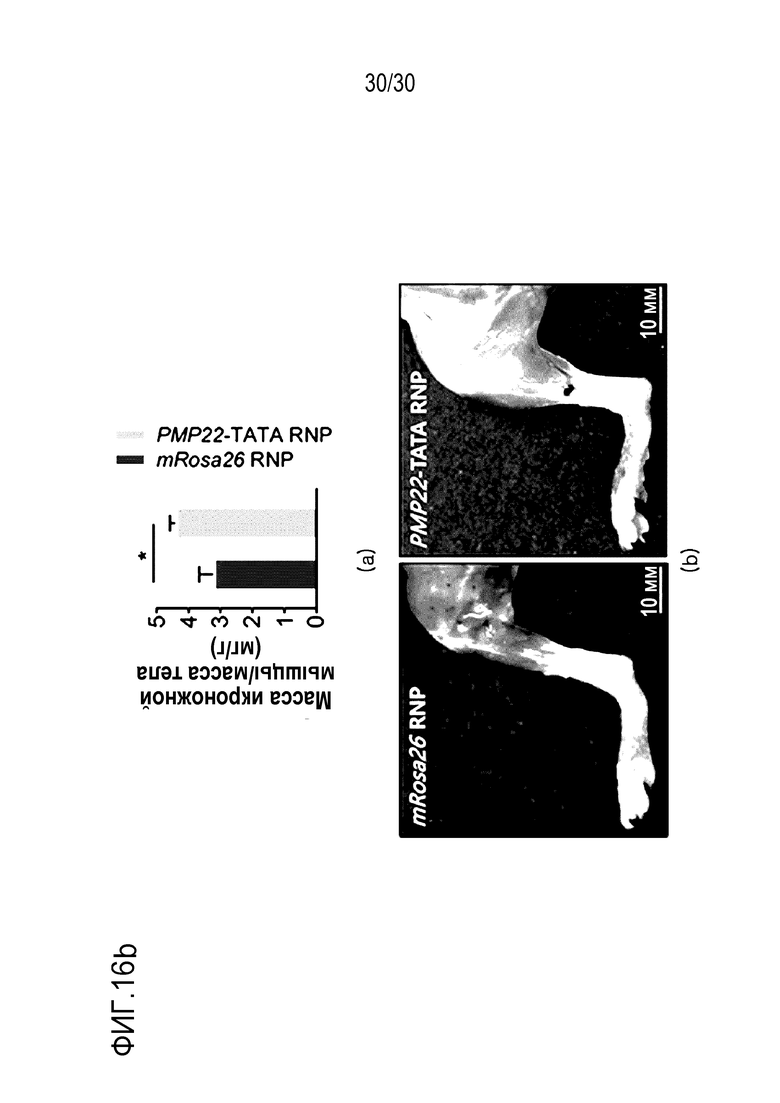
Изобретение относится к области биотехнологии, в частности к композиции, а также к способу редактирования гена PMP22, имеющего последовательность-мишень в клетке, с ее использованием. Указанная композиция содержит белок Cas9 или нуклеиновую кислоту, кодирующую белок Cas9 и гидовую РНК, включающую crРНК и tracrРНК, или нуклеиновую кислоту, кодирующую гидовую РНК, где белок Cas9 происходит из Streptococcus pyogenes или Campylobacter jejuni и где последовательность-мишень расположена в области ТАТА-бокса гена PMP22. Изобретение эффективно для лечения болезни Шарко-Мари-Тута типа 1А. 3 н. и 18 з.п. ф-лы, 16 ил., 9 табл., 6 пр.
1. Композиция, способная редактировать ген PMP22, имеющий последовательность-мишень, в клетке, содержащая:
белок Cas9 или нуклеиновую кислоту, кодирующую белок Cas9; и
гидовую РНК, включающую crРНК и tracrРНК, или нуклеиновую кислоту, кодирующую гидовую РНК,
где белок Cas9 происходит из Streptococcus pyogenes или Campylobacter jejuni,
где последовательность-мишень расположена в области ТАТА-бокса гена PMP22,
где, если белок Cas9 происходит из Streptococcus pyogenes, последовательность-мишень выбрана из SEQ ID NO: 14-29,
где, если белок Cas9 происходит из Campylobacter jejuni, последовательность-мишень выбрана из SEQ ID NO: 30-40,
где crРНК содержит гидовый домен, способный нацеливаться на последовательность-мишень, и первый комплементарный домен,
где гидовый домен и первый комплементарный домен соединены последовательно от 5’- к 3’-концу,
где первый комплементарный домен и tracrРНК способны взаимодействовать с белком Cas9 с образованием комплекса гидовая РНК-Cas.
2. Композиция по п.1,
где гидовая РНК дополнительно содержит линкерный домен,
где первый комплементарный домен crРНК, линкерный домен и tracrРНК соединены последовательно от 5’-конца к 3’-концу с образованием последовательности остова,
где гидовый домен crРНК и остов соединены последовательно от 5’-конца к 3’-концу с образованием одноцепочечной гидовой РНК.
3. Композиция по п.2, где белок Cas9 происходит из Streptococcus pyogenes, где последовательность остова представлена 5’-GUUUUAGUCCCUGAAAAGGGACUAAAAUAAAGAGUUUGCGGGACUC UGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3’.
4. Композиция по п.2, где белок Cas9 происходит из Campylobacter jejuni, где последовательность остова представлена 5’-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAA GUGGCACCGAGUCGGUGC-3’.
5. Композиция по п.1, где композиция содержит белок Cas9 и гидовую РНК, в которой белок Cas9 образует комплекс с гидовой РНК с образованием рибонуклеопротеина (RNP).
6. Композиция по п.1, где композиция представляет собой форму вектора.
7. Композиция по п.6, где вектор выбран из группы, состоящей из плазмидного вектора, ретровируса, лентивируса, аденовируса, аденоассоциированного вируса (AAV), вируса коровьей оспы, поксвируса и вируса простого герпеса.
8. Способ редактирования гена PMP22, имеющего последовательность-мишень, в клетке, включающий:
введение композиции, содержащей белок Cas9 или нуклеиновую кислоту, кодирующую белок Cas9, и гидовую РНК или нуклеиновую кислоту, кодирующую гидовую РНК, субъекту,
где белок Cas9 происходит из Streptococcus pyogenes или Campylobacter jejuni,
где последовательность-мишень расположена в области ТАТА-бокса гена PMP22,
где, если белок Cas9 происходит из Streptococcus pyogenes, последовательность-мишень выбрана из SEQ ID NO: 14-29,
где, если белок Cas9 происходит из Campylobacter jejuni, последовательность-мишень выбрана из SEQ ID NO: 30-40,
где crРНК содержит гидовый домен, способный нацеливаться на последовательность-мишень, и первый комплементарный домен,
где гидовый домен и первый комплементарный домен соединены последовательно от 5’- к 3’-концу,
где первый комплементарный домен и tracrРНК способны взаимодействовать с белком Cas9 с образованием комплекса гидовая РНК-Cas.
9. Способ по п.8,
где гидовая РНК дополнительно содержит линкерный домен,
где первый комплементарный домен crРНК, линкерный домен и tracrРНК соединены последовательно от 5’-конца к 3’-концу с образованием последовательности остова,
где гидовый домен crРНК и остов соединены последовательно от 5’-конца к 3’-концу с образованием одноцепочечной гидовой РНК.
10. Способ по п.9, где белок Cas9 происходит из Streptococcus pyogenes, где последовательность остова представлена 5’-GUUUUAGUCCCUGAAAAGGGACUAAAAUAAAGAGUUUGCGGGACUCUGCGGGGUUACAA UCCCCUAAAACCGCUUUU-3’.
11. Способ по п.9, где белок Cas9 происходит из Campylobacter jejuni, где последовательность остова представлена 5’-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGU GGCACCGAGUCGGUGC-3’.
12. Способ по п.8, где композиция содержит белок Cas9 и гидовую РНК, в которой белок Cas9 образует комплекс с гидовой РНК с образованием рибонуклеопротеина (RNP).
13. Способ по п.8, где композиция представляет собой форму вектора.
14. Способ по п.13, где вектор выбран из группы, состоящей из плазмидного вектора, ретровируса, лентивируса, аденовируса, аденоассоциированного вируса (AAV), вируса коровьей оспы, поксвируса и вируса простого герпеса.
15. Способ лечения болезни Шарко-Мари-Тута типа 1А (CMT1A), включающий:
введение композиции для редактирования гена PMP22 субъекту,
где композиция содержит белок Cas9 или нуклеиновую кислоту, кодирующую белок Cas9, и гидовую РНК или нуклеиновую кислоту, кодирующую гидовую РНК,
где белок Cas9 происходит из Streptococcus pyogenes или Campylobacter jejuni,
где последовательность-мишень расположена в области ТАТА-бокса гена PMP22,
где, если белок Cas9 происходит из Streptococcus pyogenes, последовательность-мишень выбрана из SEQ ID NO: 14-29,
где, если белок Cas9 происходит из Campylobacter jejuni, последовательность-мишень выбрана из SEQ ID NO: 30-40,
где crРНК содержит гидовый домен, способный нацеливаться на последовательность-мишень, и первый комплементарный домен,
где гидовый домен и первый комплементарный домен соединены последовательно от 5’- к 3’-концу,
где первый комплементарный домен и tracrРНК способны взаимодействовать с белком Cas9 с образованием комплекса гидовая РНК-Cas.
16. Способ по п.15, где гидовая РНК дополнительно содержит линкерный домен,
где гидовый домен crРНК, первый комплементарный домен crРНК, линкерный домен и tracrРНК соединены последовательно от 5’-конца к 3’-концу.
17. Способ по п.16, где белок Cas9 происходит из Streptococcus pyogenes, где последовательность остова представлена 5’-GUUUUAGUCCCUGAAAAGGGACUAAAAUAAAGAGUUUGCGGGA CUCUGCGGGGUUACAAUCCCCUAAAACCGCUUUU-3’.
18. Способ по п.16, где белок Cas9 происходит из Campylobacter jejuni, где последовательность остова представлена 5’-GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAA GUGGCACCGAGUCGGUGC-3’.
19. Способ по п.15, где композиция содержит белок Cas9 и гидовую РНК, в которой белок Cas9 образует комплекс с гидовой РНК с образованием рибонуклеопротеина (RNP).
20. Способ по п.15, в котором композиция представляет собой форму вектора.
21. Способ по п.19, где вектор выбран из группы, состоящей из плазмидного вектора, ретровируса, лентивируса, аденовируса, аденоассоциированного вируса (AAV), вируса коровьей оспы, поксвируса и вируса простого герпеса.
| WO2015048577 A2, 02.04.2015 | |||
| KR20060003673 A, 11.01.2006 | |||
| JUN LI et al., The PMP22 Gene and Its Related Diseases, Mol Neurobiol, 2013, Vol.47, pp.673-698 | |||
| WO2016021973 A1 | |||
| АВДЕЕВА Ж.И | |||
| и др., Требования к производству и контролю препаратов на основе моноклональных антител, применяемых для лечения, Биопрепараты, 2010, с.11-14. |

