Настоящее изобретение относится к кристаллам белка Гремлин-1 человека и белку Гремлин-1 человека в комплексе с ингибирующим антителом. Изобретение также относится к структуре Гремлин-1 человека (отдельно или в комплексе с антителом) и к применению этих структур при скрининге агентов, модулирующих активность Гремлин-1. Изобретение также относится к антителам, которые связываются с аллостерическим ингибирующим сайтом на Гремлин-1, а также к фармацевтическим композициям и применению в медицине таких антител и агентов, идентифицированных методами скрининга.
УРОВЕНЬ ТЕХНИКИ
Гремлин-1 (также известный как Drm и CKTSF1B1) представляет собой гликопротеин, состоящий из 184 аминокислот, который является членом семейства DAN секретируемых белков, содержащих цистиновые узлы (помимо Cerberus и Dan). Помимо подтвержденной проангиогенной функции, Гремлин связывает и подавляет способность BMP-2, 4 и 7 к передаче сигналов, возможно, через агонизм к VEGFR2. Гремлин-1 играет важную роль в процессе развития, в котором он жизненно необходим, в процессе формирования почек и зачатков конечностей. В связи с этими жизненно важными функциями, гомозиготный нокаут Гремлин является летальным для эмбрионов мышей.
В зрелом возрасте повышенные уровни Гремлин связаны с идиопатическим легочным фиброзом и легочной артериальной гипертензией, при которой передача сигналов BMP-2, 4 и 7 снижается при повышении уровней TGF-β. Как при диабетической, так и при хронической нефропатии аллотрансплантата экспрессия Гремлин-1 коррелировала с показателем фиброза.
Повышенные уровни Гремлин также связаны со склеродермией, диабетической нефропатией и колоректальным раком. Было показано, что Гремлин-1 активирует инвазию и пролиферацию раковых клеток, и считается, что он играет роль в развитии саркомы и карциномы шейки матки, легких, яичников, почек, молочной железы, толстой кишки и поджелудочной железы.
На сегодняшний день имеется целый ряд проблем, связанных с изучением Гремлин-1, при этом отсутствует общее понимание природы Гремлин-1 (и его партнера Гремлин-2). Биология BMP сложна, и между видами наблюдается высокая гомология. Гремлин-1 является сложным для работы белком, и для изучения его биологии отсутствуют подходящие инструменты и реагенты. Получение Гремлин-1 также не является простым процессом; известно, что белки с цистеиновыми узлами трудно продуцировать, и свободный цистеин в Гремлин-1 усугубляет эту проблему. Гремлин-1 сложно экспрессировать, не говоря уже о его очистке. До настоящего времени отсутствовала информация о структуре этого белка, и в литературе очень мало информации об этом белке.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Термин Гремлин-1, используемый в настоящем изобретении, как правило, относится к последовательности, указанной в базе данных UniProt под номером O60565 (SEQ ID NO:1). Термин Гремлин-1 также может относиться к полипептиду Гремлин-1, который:
(а) содержит или состоит из аминокислотной последовательности SEQ ID NO:1 с или без N-концевого сигнального пептида, т.е. может содержать или состоять из последовательности зрелого пептида SEQ ID NO:21; или
(b) представляет собой производное, имеющее одну или более аминокислотных замен, модификаций, делеций или вставок относительно аминокислотной последовательности SEQ ID NO:1 с или без N-концевого сигнального пептида (SEQ ID NO:21), сохраняя при этом активность Гремлин-1, такую как аминокислотная последовательность SEQ ID NO:20.
(c) является его вариантом, такие варианты обычно сохраняют по меньшей мере примерно 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94% или 95% идентичности с SEQ ID NO:1 (или SEQ ID NO:20 или 21) (или даже примерно 96%, 97%, 98% или 99% идентичности). Другими словами, такие варианты могут сохранять от примерно 60% до примерно 99% идентичность с SEQ ID NO:1, предпочтительно от примерно 80% до примерно 99% идентичность с SEQ ID NO:1, более предпочтительно от примерно 90% до примерно 99% идентичность с SEQ ID NO:1 и наиболее предпочтительно от примерно 95% до примерно 99% идентичность с SEQ ID NO:1. Варианты описаны ниже.
Как обсуждается ниже, номера остатков обычно указаны на основе последовательности SEQ ID NO:1. Однако специалист может легко экстраполировать нумерацию остатков на производную последовательность или вариант, указанные выше. Если указаны номера остатков, изобретение также охватывает эти остатки в варианте или в производной последовательности.
Авторы настоящего изобретения получили кристаллы Гремлин-1 человека в кристаллическом виде отдельно и в комплексе с антителом, названным Ab 7326 (Fab фрагменты). Кристаллизация Гремлин-1 позволила определить предполагаемые остатки в BMP-связывающем сайте. Кроме того, кристаллизация с антителом Ab 7326, которое является аллостерическим ингибирующим антителом, позволила определить остатки в эпитопе антитела. Антитела, связывающие этот эпитоп, могут быть эффективны в качестве терапевтических средств для лечения заболеваний, связанных с Гремлин-1.
Соответственно, настоящее изобретение относится к кристаллу Гремлин-1.
Настоящее изобретение также относится к структуре Гремлин-1 человека, определенной координатами в Таблице 1.
Кроме того, изобретение относится к:
Машиночитаемому носителю данных, который содержит материал хранения данных, закодированный машиночитаемыми данными, определенными координатами структуры Гремлин-1 в таблице 1 или координатами, определяющими гомологов структуры;
Применению структуры Гремлин-1, определенной координатами в Таблице 1, в качестве структурной модели;
Агентам, идентифицированным с использованием структурной модели;
Способу скрининга агентов, модулирующих активность Гремлин-1, включающему следующие этапы:
(а) идентификацию лиганд-связывающего сайта из структурных координат в таблице 1;
(b) идентификацию агентов-кандидатов, которые взаимодействуют с по меньшей мере частью лиганд-связывающего сайта; и
(с) получение или синтез указанного агента;
Агенту, модулирующему активность Гремлин-1, идентифицированному методом скрининга;
Антителу, которое связывается с эпитопом на Гремлин-1, содержащему по меньшей мере один остаток, выбранный из Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175, где нумерация остатков основана на SEQ ID NO:1;
Антителу к Гремлин-1, которое содержит последовательности определяющих комплементарность областей тяжелой цепи (HCDR), содержащиеся в вариабельной области тяжелой цепи (HCVR) SEQ ID NO:10 или 12, и/или последовательности определяющих комплементарность областей легкой цепи (LCDR), содержащиеся в вариабельной области легкой цепи (LCVR) SEQ ID NO:11 или 13;
Антителу к Гремлин-1, которое содержит по меньшей мере одну последовательность HCDR, выбранную из SEQ ID NO:3, 4, 5 и 6, и/или по меньшей мере одну последовательность LCDR, выбранную из SEQ ID NO:7, 8 и 9;
Выделенному полинуклеотиду, кодирующему антитела;
Вектору экспрессии, несущему этот полинуклеотид;
Клетке-хозяину, содержащей этот вектор;
Способу получения антитела, включающему культивирование клетки-хозяина в условиях, позволяющих продуцировать антитело, и извлекать продуцированное антитело;
Фармацевтической композиции, содержащей антитело;
Антителу или фармацевтической композиции для применения в способе терапевтического лечения организма человека или животного;
Способу лечения или профилактики почечного фиброза, такого как диабетическая нефропатия, идиопатического легочного фиброза, легочной артериальной гипертензии, ангиогенеза и/или рака, включающему введение терапевтически эффективного количества антитела или фармацевтической композиции нуждающемуся в этом пациенту.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 (таблица 1) представлены структурные данные для кристаллографии Гремлин-1.
На фиг. 2 показана структура Гремлин-1 человека. Ленты, представляющие каждый мономер, показаны различными оттенками серого, пальцы 1 и 2 (F1 и F2) отмечены вместе с областями «запястье» (wrist) (w) и цистиновыми узлами (CK). Цистеины, образующие дисульфидные связи, показаны в виде черных стержней. Верхнее изображение - вид сбоку, показывающий дугообразную структуру. Нижнее изображение - вид сверху, показывающий границу раздела в димере.
На фиг. 3 показано выравнивание последовательностей Гремлин-1 человека и мышиного Гремлин-2 (PRDC). Остатки, отмеченные звездочкой, важны для связывания BMP, а остатки, образующие ключевые контакты на границе раздела в димере, отмечены квадратами.
На фиг. 4 показана визуализация поверхности с выделением гидрофобных BMP-связывающих остатков. Мономеры показаны двумя оттенками серого, а шесть ключевых остатков, участвующих в связывании BMP, показаны белым.
На фиг. 5 представлено наложение Гремлин-1 человека и мышиного Гремлин-2. Верхнее изображение представлено в виде ленты с двумя выровненными белками. Показан вид сбоку, демонстрирующий схожесть полной укладки молекул при наложении Гремлина-1 человека и Гремлина-2 мыши. На нижнем изображении подробно показаны аминокислоты, участвующие в связывании BMP в виде стержней (мышиный Гремлин-2 показан белым, и Гремлин-1 человека - черным). Показан вид сбоку, демонстрирующий остатки, участвующие в связывании ВМР.
На фиг. 6(A) представлены ингибирующие эффекты полноразмерного Гремлин-1 в анализе активности репортерного гена Id1 в клетках Hek.
На фиг. 6(B) представлены ингибирующие эффекты укороченного Гремлин-1 в анализе активности репортерного гена Id1 в клетках Hek.
На фиг. 7 показан процент восстановления сигнала для антител, полученных в результате иммунизации, в анализе активности репортерного гена Hek-Id1.
На фиг. 8 показан процент восстановления сигнала для антител, полученных из библиотеки, в анализе активности репортерного гена Hek-Id1.
На фиг. 9 показаны результаты анализа активности репортерного гена Hek-Id1 с титрами человеческого Гремлин (фиг. 9А) и мышиного Гремлин (фиг. 9В) и влияния антитела 7326 (показанного в виде антитела РВ376) на восстановление передачи сигналов BMP.
На фиг. 10 показана структурная модель комплекса Гремлин-Fab с указанием возможных BMP-связывающих областей и эпитопа Fab.
На фиг. 11 показана визуализация поверхности, изображающая каждый мономер Гремлин-1 двумя оттенками серого, шесть ключевых остатков, идентифицированных мутагенезом, которые участвуют в связывании BMP (показаны черным цветом), и все остатки на поверхности в пределах 6 от этих шести ключевых остатков.
от этих шести ключевых остатков.
На фиг. 12 приведены результаты оценки систолического давления правого желудочка (СДПЖ). Влияние антител к Гремлин 1 на СДПЖ оценивали у мышей C57Bl/6 с нормоксией и гипоксией/получавших SU5416. Влияние анти-Гремлин 1 (n=8), контрольного антитела IgG1 (n=6), PBS (n=2), иматиниба (n=8) на развитие легочной артериальной гипертензии (ЛАГ) определяли у самок мышей C57Bl/6, которым подкожно вводили SU5416 (20 мг/кг) каждый день в течение трех дней после индуцирования у них хронической нормобарической гипоксии (10% О2) или нормоксии в течение 21 дней. Определяли СДПЖ и вычисляли среднее значение СДПЖ ± SEM.
*Р<0,05; **P<0,01; ***Р<0,005; ****P<0,001 согласно однофакторному ANOVA.
На фиг. 13 приведены результаты оценки среднего системного артериального давления (САД). Влияние антител к Гремлин 1 на САД оценивали у C57Bl/6 мышей с гипоксией/SU5416, которым вводили анти-Гремлин 1 (n=4), контрольное антитело IgG1 (n=4), иматиниб (n=4), и у C57Bl/6 мышей с нормоксией/SU5416, которым вводили анти-Гремлин 1 (n=4), контрольное антитело IgG1 (n=4), и САД ± SEM наносили на график через 21 день.
*Р<0,05; **P<0,01; ***Р<0,005; ****P<0,001 согласно однофакторному ANOVA.
На фиг. 14 приведены результаты оценки правожелудочковой гипертрофии. Влияние антител к Гремлин 1 на правожелудочковую гипертрофию (RV/LV+S) оценивали на C57Bl/6 мышах с гипоксией/SU5416. Влияние анти-Гремлин 1 (n=8), контрольного антитела IgG (n=6), PBS (n=2), иматиниба (n=8) на развитие легочной артериальной гипертензии (ЛАГ) определяли у самок мышей C57Bl/6, которым подкожно вводили SU5416 (20 мг/кг) каждый день в течение трех дней после индуцирования у них хронической нормобарической гипоксии (10% О2) или нормоксии в течение 21 дней. Определяли СДПЖ и вычисляли среднее значение СДПЖ ± SEM.
*Р<0,05; **P<0,01; ***Р<0,005; ****P<0,001 согласно однофакторному ANOVA.
На фиг. 15 представлена гистологическая оценка мускуляризации сосудов легких. Оценивали влияние антител к Гремлин 1. Залитые парафином срезы легких, полученные от мышей, получавших анти-Гремлин 1 (n=6), контрольное антитело IgG (n=6), или иматиниб (n=6), окрашивали на актин гладких мышц (αSMA) для оценки степени мышечной активности и фактором фон Виллебранда (vWF) для идентификации эндотелиальных клеток помощью иммуногистохимии. Срезы легких оцифровывали, используя систему виртуальной микроскопии Nanozoomer (Hamamatsu, Welwyn Garden City, Великобритания), и более 40 сосудов на группу были оценены независимыми слепыми наблюдателями как немускуляризованные, частично или полностью мускуляризованные. Средние показатели ± SEM для каждой группы модального показателя каждого сосуда наносили на график. Репрезентативные изображения окрашенных на αSMA и vWF срезов легких в парафине из групп IgG1 нормоксии; IgG1 гипоксии/SU5416; иматиниба гипоксии/SU5416; анти-Гремлин 1 гипоксии/SU5416.
*Р<0,05; **P<0,01; ***Р<0,005; ****P<0,001 согласно однофакторному ANOVA.
Краткое описание списка последовательностей
SEQ ID NO:1 показывает последовательность Гремлин-1 человека, включающую N-концевую сигнальную последовательность из 24 аминокислот (Uniprot ID O60565).
SEQ ID NO:2 показывает последовательность укороченного Гремлин-1 человека, использованного в кристаллографии, включая N-концевую метку.
SEQ ID NO:3 показывает HCDR1 Ab 7326 (Chothia).
SEQ ID NO:4 показывает HCDR1 Ab 7326 (Kabat).
SEQ ID NO:5 показывает HCDR2 Ab 7326 (Kabat).
SEQ ID NO:6 показывает HCDR3 Ab 7326 (Kabat).
SEQ ID NO:7 показывает LCDR1 Ab 7326 (Kabat).
SEQ ID NO:8 показывает LCDR2 Ab 7326 (Kabat).
SEQ ID NO:9 показывает LCDR3 Ab 7326 (Kabat).
SEQ ID NO:10 показывает вариабельную область тяжелой цепи Ab 7326 (вариант 1).
SEQ ID NO:11 показывает вариабельную область легкой цепи Ab 7326 (вариант 1).
SEQ ID NO:12 показывает вариабельную область тяжелой цепи Ab 7326 (вариант 2).
SEQ ID NO:13 показывает вариабельную область легкой цепи Ab 7326 (вариант 2).
SEQ ID NO:14 показывает полноразмерную тяжелую цепь IgG1 мышиного Ab 7326 (вариант 1).
SEQ ID NO:15 показывает полноразмерную легкую цепь IgG1 мышиного Ab 7326 (вариант 1).
SEQ ID NO:16 показывает полноразмерную тяжелую цепь человеческого IgG1 Ab 7326 (вариант 2).
SEQ ID NO:17 показывает полноразмерную легкую цепь человеческого IgG1 Ab 7326 (вариант 2).
SEQ ID NO:18 показывает тяжелую цепь Fab мышиного Ab 7326 (вариант 1).
SEQ ID NO:19 показывает легкую цепь Fab мышиного Ab 7326 (вариант 1).
SEQ ID NO:20 показывает последовательность укороченного Гремлин-1 человека, использованного в кристаллографии, без N-концевой метки.
SEQ ID NO:21 показывает последовательность зрелого Гремлин-1 (SEQ ID NO:1 без сигнального пептида).
SEQ ID NO:22 показывает тяжелую цепь человеческого IgG4P (вариант 1).
SEQ ID NO:23 показывает легкую цепь человеческого IgG4P (вариант 1).
SEQ ID NO:24 показывает ДНК тяжелой цепи человеческого IgG1 (вариант 1).
SEQ ID NO:25 показывает ДНК легкой цепи человеческого IgG1 (вариант 1).
SEQ ID NO:26 показывает ДНК тяжелой цепи человеческого IgG4P (вариант 1).
SEQ ID NO:27 показывает ДНК легкой цепи человеческого IgG4P (вариант 1).
SEQ ID NO:28 показывает полноразмерную тяжелую цепь мышиного IgG1 (вариант 2).
SEQ ID NO:29 показывает полноразмерную легкую цепь мышиного IgG1 (вариант 2).
SEQ ID NO:30 показывает полноразмерную тяжелую цепь человеческого IgG1 (вариант 1).
SEQ ID NO:31 показывает полноразмерную легкую цепь человеческого IgG1 (вариант 1).
SEQ ID NO:32 показывает тяжелую цепь Fab (вариант 2).
SEQ ID NO:33 показывает легкую цепь Fab (вариант 2).
SEQ ID NO:34 показывает тяжелую цепь человеческого IgG4P (вариант 2).
SEQ ID NO:35 показывает легкую цепь человеческого IgG4P (вариант 2).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Кристаллическая структура Гремлин-1
Настоящее изобретение относится к структурным координатам Гремлин-1 человека. Полные координаты приведены на фиг. 1 (таблица 1).
В настоящем изобретении также предлагается кристалл Гремлин-1 человека, состоящий из пространственной группы C2 с размерами элементарной ячейки a=84,55 , b=107,22 и с=77,09 .
В настоящем изобретении также предлагается кристалл Гремлин-1 в комплексе с антителом, более конкретно, Fab с тяжелой цепью SEQ ID NO:18 и легкой цепью SEQ ID NO:19.
В настоящем изобретении также предлагается машиночитаемый носитель данных, который содержит материал хранения данных, закодированный с машиночитаемыми данными, определенными координатами структуры Гремлин-1 в таблице 1 или координатами, определяющими гомологи структуры.
Изобретение предусматривает применение структурных данных в таблице 1 и машиночитаемого носителя данных, в качестве структурной модели для Гремлин-1. Такая структурная модель может использоваться для скрининга агентов, взаимодействующих с Гремлин-1. Скрининг может быть высокопроизводительным скринингом.
Агент, который взаимодействует с Гремлин-1, обычно является агентом, который связывается с Гремлин-1. Агенты, взаимодействующие с Гремлин-1, могут модулировать активность Гремлин-1. Ингибирующий модулирующий агент может оказывать влияние на любую из функций Гремлин-1, но обычно уменьшает связывание Гремлин-1 с BMP (BMP 2/4/7). Гремлин-1 является негативным регулятором BMP, поэтому пониженное связывание увеличивает передачу сигналов через BMP. Активирующий модулирующий агент может усиливать связывание Гремлин-1 с BMP.
Связывание BMP и передачу сигналов можно детектировать любым способом, известным в данной области. Например, примеры в настоящей заявки описывают анализ фосфорилирования SMAD. Фосфорилирование SMAD1, 5 и 8 происходит при передаче сигналов BMP. Следовательно, увеличение степени фосфорилирования SMAD можно использовать в качестве показателя повышенной передачи сигналов BMP, что может отражать уменьшение связывания с Гремлин-1.
В примерах также описан анализ активности репортерного гена Id1, причем ген Id1 является геном-мишенью сигнального пути BMP. Поэтому ускорение восстановления сигнала в этом анализе также можно использовать для определения, подавляет ли агент связывание Гремлин-1 с BMP.
Упомянутый в настоящем описании агент может представлять собой любую молекулу, которая потенциально может взаимодействовать с Гремлин-1, но предпочтительно представляет собой малую молекулу или антитело.
Изобретение также относится к методу скрининга агентов, модулирующих активность Гремлин-1, включающий следующие этапы:
(а) идентификацию лиганд-связывающего сайта из структурных координат в таблице 1;
(b) идентификацию агентов-кандидатов, которые взаимодействуют с по меньшей мере частью лиганд-связывающего сайта; и
(с) получение или синтез указанного агента.
Лиганд-связывающий сайт может быть любым предполагаемым сайтом на Гремлин-1, который взаимодействует с белком (лигандом). Лиганд-связывающий сайт обычно представляет собой BMP-связывающий сайт. Как показано в примерах, авторы настоящего изобретения идентифицировали предполагаемый BMP-связывающий сайт на основе кристаллической структуры Гремлин-1. Этот связывающий сайт содержит следующие аминокислоты: Trp93, Phe117, Tyr119, Phe125, Tyr126 и Phe138, где нумерация остатков основана на SEQ ID NO:1.
Следовательно, способ скрининга по изобретению может включать идентификацию агентов, которые взаимодействуют с одним или более из этих остатков, предпочтительно, по меньшей мере, с 2, 3, 4 или всеми 6 из этих остатков.
Взаимодействие агента с остатками белка может быть определено любым подходящим способом, известным в данной области, таким как определение расстояния между остатком и агентом методом рентгеноструктурного анализа (обычно менее 6 или менее 4 ). Как обсуждается в примерах ниже, область Гремлин-1, на которую может воздействовать терапевтическое средство, может включать аминокислоты Asp92-Leu99, Arg116-His130, Ser137-Ser142, Cys176-Cys178. Они находятся в пределах 6 от аминокислот, мутированных на поверхности Гремлин-1.
Этапы (а) и (b) способа скрининга обычно выполняют in silico, и агент может быть получен и синтезирован любым способом, известным в данной области.
В одном из вариантов осуществления настоящее изобретение относится к антителу, которое связывается с эпитопом на Гремлин-1, содержащим по меньшей мере один остаток, выбранный из: Trp93, Phe117, Tyr119, Phe125, Tyr126 и Phe138, где нумерация остатков соответствует SEQ ID NO:1. Настоящее изобретение также относится к антителу, которое связывается с эпитопом, включающим все остатки: Trp93, Phe117, Tyr119, Phe125, Tyr126 и Phe138.
Изобретение также относится к применению этой BMP-связывающей области Гремлин-1 для генерации (потенциально ингибирующих) антител. Например, изобретение относится к антигену, содержащему, по меньшей мере, один (предпочтительно все) из перечисленных выше остатков, который можно использовать для генерации антител.
Вместо взаимодействия с BMP-связывающим сайтом Гремлин-1 агент может оказывать аллостерическое действе. В настоящем описании агент связывается в стороне от нормального связывающего сайта, и при этом он все еще способен модулировать активность Гремлин-1, например, через индуцированные конформационные изменения в белке. Следовательно, структурная модель и способ скрининга по изобретению также можно использовать для идентификации аллостерических модуляторов Гремлин-1.
Было обнаружено, что антитело Ab 7326 по изобретению оказывает аллостерическое действие. Эпитоп этого антитела содержит следующие остатки: Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175, причем нумерация остатков основана на SEQ ID NO:1. Соответственно, способ скрининга по изобретению может включать идентификацию агентов, которые взаимодействуют по меньшей мере с 1, 2, 3, 4, 5 или со всеми 9 из этих остатков. Затем такие агенты могут быть протестированы, например, с помощью анализов, описанных в примерах, на подавление связывания BMP. Предпочтительно Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175 расположены на одном мономере Гремлин-1, а Ile131 расположен на другом мономере Гремлин-1 (димеры Гремлин-1 связываются с димерами BMP).
Еще раз следует отметить, что изобретение также включает антиген, содержащий по меньшей мере один (предпочтительно все) из этих остатков для продуцирования антител к Гремлин-1.
Как уже указано выше, агенты могут быть идентифицированы как взаимодействующие с этими остатками любым подходящим способом, известным в данной области. Агент предпочтительно представляет собой малую молекулу или антитело.
Антитела
Настоящее изобретение относится к антителам, которые связываются с Гремлин-1.
Термин «антитело» в контексте настоящего описания включает полноразмерные антитела и любой антигенсвязывающий фрагмент (т.е. «антигенсвязывающую часть») или их отдельные цепи. Антитело относится к гликопротеину, содержащему по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями, или его антигенсвязывающей части. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначенной в настоящем описании как HCVR или VH) и константной области тяжелой цепи. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначенной в настоящем описании как LCVR или VL) и константной области легкой цепи. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые чередуются с более консервативными областями, называемыми каркасными областями (FR).
Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) системы комплемента классического пути.
Антитело по изобретению может представлять собой моноклональное антитело или поликлональное антитело и обычно является моноклональным антителом. Антитело по изобретению может представлять собой химерное антитело, CDR-привитое антитело, нанотело, человеческое или гуманизированное антитело или антигенсвязывающую часть любого из них. Для получения как моноклональных, так и поликлональных антител обычно используют экспериментальное животное, которое является млекопитающим, не являющимся человеком, такое как коза, кролик, крыса или мышь, но антитело также может быть получено с использованием других видов.
Поликлональные антитела могут быть получены обычными методами, такими как иммунизация подходящего животного представляющим интерес антигеном. Впоследствии можно провести забор крови животного, и очистить фракцию IgG.
Антитела к Гремлин-1 могут быть получены, когда необходима иммунизация животного, путем введения полипептидов животному, например, животному, не являющемуся человеком, в соответствии с хорошо известными и традиционными протоколами, см., например, Handbook of Experimental Immunology, D. M. Weir (ed.), Vol 4, Blackwell Scientific Publishers, Oxford, England, 1986). Многие теплокровные животные могут быть иммунизированы, например, кролики, мыши, крысы, овцы, коровы, верблюды или свиньи. Тем не менее, мыши, кролики, свиньи и крысы, как правило, являются наиболее подходящими.
Моноклональные антитела могут быть получены любым способом, известным в данной области, например, методом гибридомы (Kohler & Milstein, 1975, Nature, 256:495-497), методом триомы, методом гибридомы В-клеток человека (Kozbor et al., 1983, Immunology Today, 4:72) и методом EBV-гибридомы (Cole et al., Monoclonal Antibodies and Cancer Therapy, pp77-96, Alan R Liss, Inc., 1985).
Антитела по изобретению также могут быть получены методами генерации антител отдельными лимфоцитми путем клонирования и экспрессии молекул кДНК вариабельной области иммуноглобулина, полученных из отдельных лимфоцитов, отобранных для продуцирования специфических антител, например, способами, описанными у Babcook, J. et al., 1996, Proc., Natl. Акад. Sci. USA 93 (15):7843-7848l; WO92/02551; WO 2004/051268 и WO 2004/106377.
Антитела по настоящему изобретению также могут быть получены различными способами фагового дисплея, известными в данной области, включая способы, описанные у Brinkman et al. (J. Immunol. Methods, 1995, 182:41-50), Ames et al. (J. Immunol. Methods, 1995, 184:177-186), Kettleborough et al. (Eur. J. Immunol. 1994, 24:952-958), Persic et al. (Gene, 1997, 187, 9-18), Burton et al. (Advances in Immunology, 1994, 57:191-280) и WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401; и US 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743 и 5969108.
Полностью человеческими антителами являются антитела, в которых вариабельные области и константные области (если присутствуют) как тяжелой, так и легкой цепей имеют человеческое происхождение или по существу идентичны последовательностям человеческого происхождения, но необязательно происходящие из одного и того же антитела. Примеры полностью человеческих антител могут включать антитела, продуцируемые, например, способами фагового дисплея, описанными выше, и антитела, продуцируемые мышами, у которых мышиные гены вариабельных и, необязательно, константных областей иммуноглобулина заменены человеческими аналогами, например, как описано в общих чертах в ЕР 0546073, US 5545806, US 5569825, US 5625126, US 5633425, US 5661016, US 5770429, EP 0438474 и EP 0463151.
Альтернативно, антитело по изобретению может быть получено способом, включающим иммунизацию млекопитающего, не являющегося человеком, иммуногеном Гремлин-1; получение препарата антител из указанного млекопитающего; выделение из него моноклональных антител, которые распознают Гремлин-1.
Молекулы антител по настоящему изобретению могут содержать полную молекулу антитела, имеющую полноразмерные тяжелые и легкие цепи, или фрагмент или антигенсвязывающую часть антитела. Термин «антигенсвязывающая часть» антитела относится к одному или более фрагментам антитела, которые сохраняют способность селективно связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может осуществляться фрагментами полноразмерного антитела. Антитела и их фрагменты и их антигенсвязывающие части могут представлять собой, без ограничения, Fab, модифицированный Fab, Fab', модифицированный Fab', F(ab')2, Fv, однодоменные антитела (например, VH или VL или VHH), scFv, би-, три- или тетравалентные антитела, Bis-scFv, диатела, триатела, тетратела и эпитоп-связывающие фрагменты любого из вышеперечисленного (см., например, Holliger and Hudson, 2005, Nature Biotech. 23 (9):1126- 1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2 (3), 209-217). Способы создания и производства этих фрагментов антител хорошо известны в данной области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Другие фрагменты антител, используемые в настоящем изобретении, включают фрагменты Fab и Fab', описанные в международных заявках на патент WO 2005/003169, WO 2005/003170 и WO 2005/003171, и фрагменты Fab-dAb, описанные в международной заявке на патент WO2009/040562. Поливалентные антитела могут содержать множество специфичностей или могут быть моноспецифичными (см., например, WO 92/22853 и WO 05/113605). Эти фрагменты антител могут быть получены традиционными методами, известными специалистам в данной области, затем фрагменты могут быть подвергнуты скринингу в отношении их применимости таким же образом, что и интактные антитела.
Домены константной области молекулы антитела по настоящему изобретению, если имеются, могут быть выбраны с учетом предполагаемой функции молекулы антитела и, в частности, эффекторных функций, которые могут потребоваться. Например, домены константной области могут быть доменами человеческого IgA, IgD, IgE, IgG или IgM. В частности, в тех случаях, когда молекула антитела предназначена для терапевтического применения и требуются эффекторные функции антитела, можно использовать домены константной области человеческого IgG, особенно изотипов IgG1 и IgG3. Альтернативно, изотипы IgG2 и IgG4 могут использоваться, когда молекула антитела предназначена для терапевтических целей, и эффекторные функции антитела не требуются.
Антитело по изобретению может быть получено, экспрессировано, создано или выделено с помощью рекомбинантных средств, таких как (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным в отношении представляющих интерес генов иммуноглобулина, или полученной из него гибридомы, (b) антитела, выделенные из клетки-хозяина, трансформированной для экспрессии представляющего интерес антитела, например, трансфектомы, (c) антитела, выделенные из рекомбинантной библиотеки комбинаторных антител, и (d) антитела, полученные, экспрессированные, созданные или выделенные любыми другими средствами, которые включают сплайсинг последовательностей генов иммуноглобулина с другими последовательностями ДНК.
Антитело по изобретению может представлять собой человеческое антитело или гуманизированное антитело. Предполагается, что термин «человеческое антитело» в контексте настоящего описания включает антитела, имеющие вариабельные области, в которых как каркасные области, так и CDR области получены из иммуноглобулиновых последовательностей зародышевой линии человека. Кроме того, если антитело содержит константную область, эта константная область также происходит из иммуноглобулиновых последовательностей зародышевой линии человека. Человеческие антитела по изобретению могут включать аминокислотные остатки, не кодируемые последовательностями человеческого иммуноглобулина зародышевой линии (например, мутации, введенные путем случайного или сайт-специфического мутагенеза in vitro, или соматические мутации in vivo). Однако термин «человеческое антитело», как используется в настоящем описании, не включает антитела, в которых последовательности CDR, полученные из зародышевой линии другого вида млекопитающих, таких как мышь, были привиты на каркасные последовательности человека.
Такое человеческое антитело может быть человеческим моноклональным антителом. Такое человеческое моноклональное антитело может быть продуцировано гибридомой, которая включает В-клетку, полученную от трансгенного нечеловеческого животного, например, трансгенной мыши, имеющей геном, содержащий человеческий трансген тяжелой цепи и трансген легкой цепи, слитую с иммортализованной клеткой.
Человеческие антитела могут быть получены путем иммунизации человеческих лимфоцитов in vitro с последующей трансформацией лимфоцитов вирусом Эпштейна-Барра.
Термин «производные человеческого антитела» относится к любой модифицированной форме человеческого антитела, например, конъюгату антитела с другим агентом или антителом.
Термин «гуманизированное антитело» предназначен для обозначения молекул CDR-привитых антител, в которых последовательности CDR, полученные из зародышевой линии других видов млекопитающих, таких как мышь, привиты на каркасные последовательности человека. В человеческие каркасные последовательности могут быть введены дополнительные модификации каркасной области.
Используемый здесь термин «молекула CDR-привитого антитела» относится к молекуле антитела, в которой тяжелая и/или легкая цепь содержит одну или более CDR (включая, при необходимости, одну или более модифицированных CDR), полученных из донорского антитела (например, мышиного или крысиного моноклонального антитела), привитых в каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, человеческого антитела). Для обзора, см. Vaughan et al., Nature Biotechnology, 16, 535-539, 1998. В одном из вариантов осуществления, вместо переноса всей CDR, в каркас человеческого антитела переносят только один или более определяющих специфичность остатков из любой CDR, описанной выше в настоящей заявке (см., например, Kashmiri et al., 2005, Methods, 36, 25-34). В одном из вариантов осуществления в каркас человеческого антитела переносят только определяющие специфичность остатки из одной или более CDR, описанных выше в настоящей заявке. В другом варианте осуществления в каркас человеческого антитела переносят только определяющие специфичность остатки из каждой из описанных выше в настоящей заявке CDR.
В случае прививки CDR или определяющих специфичность остатков, можно использовать любую подходящую акцепторную каркасную последовательность вариабельной области с учетом класса/типа донорского антитела, из которого получены CDR, включая каркасные области мыши, приматов и человека. Соответственно, CDR-привитое антитело по настоящему изобретению имеет вариабельный домен, содержащий акцепторные каркасные области человека, а также одну или более CDR или определяющих специфичность остатков, описанных выше. Таким образом, в одном из вариантов осуществления представляется нейтрализующее CDR-привитое антитело, в котором вариабельный домен содержит человеческие акцепторные каркасные области и нечеловеческие донорские CDR.
Примерами человеческих каркасов, которые можно использовать в настоящем изобретении, являются KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (Kabat et al., см. выше). Например, KOL и NEWM можно использовать для тяжелой цепи, REI можно использовать для легкой цепи, а EU, LAY и POM можно использовать как для тяжелой, так и для легкой цепей. Альтернативно, можно использовать последовательности зародышевой линии человека; они доступны, например, по адресу: http://www.vbase2.org/ (см. Retter et al., Nucl. Acids Res. (2005) 33 (приложение 1), D671-D674).
В CDR-привитом антителе по настоящему изобретению акцепторные тяжелая и легкая цепи необязательно должны быть получены из одного и того же антитела и, при необходимости, могут содержать композитные цепи, имеющие каркасные области, полученные из разных цепей.
Кроме того, в CDR-привитом антителе по настоящему изобретению каркасные области не обязательно должны иметь точно такую же последовательность, как у акцепторного антитела. Например, необычные остатки могут быть заменены на более часто встречающиеся остатки для этого класса или типа акцепторной цепи. Альтернативно, выбранные остатки в акцепторных каркасных областях могут быть изменены таким образом, чтобы они соответствовали остатку, находящемуся в том же положении в донорском антителе (см. Reichmann et al., 1998, Nature, 332, 323-324). Такие изменения должны быть сведены к минимуму, необходимому для восстановления сродства донорского антитела. Протокол для выбора остатков в акцепторных каркасных областях, которые, возможно, придется изменить, изложен в WO 91/09967.
Специалисту в данной области также будет понятно, что антитела могут подвергаться различным посттрансляционным модификациям. Тип и степень этих модификаций часто зависят от линии клеток-хозяев, используемых для экспрессии антитела, а также от условий культивирования. Такие модификации могут включать варианты гликозилирования, окисления метионина, образования дикетопиперазина, изомеризации аспартата и дезамидирования аспарагина. Частой модификацией является потеря карбоксиконцевого основного остатка (такого как лизин или аргинин) в результате воздействия карбоксипептидаз (как описано в Harris, RJ. Journal of Chromatography 705:129-134, 1995).
В одном из вариантов осуществления тяжелая цепь антитела содержит домен CH1, а легкая цепь антитела содержит домен CL, либо каппа, либо лямбда.
Биологические молекулы, такие как антитела или фрагменты, содержат кислотные и/или основные функциональные группы, что дет суммарный положительный или отрицательный заряд молекулы. Количество общего «наблюдаемого» заряда будет зависеть от абсолютной аминокислотной последовательности объекта, локального окружения заряженных групп в трехмерной структуре и условий окружающей молекулу среды. Изоэлектрическая точка (pI) - это pH, при котором конкретная молекула или ее поверхность не несет суммарного электрического заряда. В одном из вариантов осуществления антитело или фрагмент по настоящему изобретению имеет изоэлектрическую точку (pI), равную по меньшей мере 7. В одном из вариантов осуществления антитело или фрагмент имеет изоэлектрическую точку, равную по меньшей мере 8, например 8,5, 8,6, 8,7, 8,8 или 9. В одном из вариантов осуществления pI антитела равно 8. Для прогнозирования изоэлектрической точки антитела или фрагмента можно использовать программы, такие как **ExPASY http://www.expasy.ch/tools/pi_tool.html (см. Walker, The Proteomics Protocols Handbook, Humana Press (2005), 571-607).
Антитела, которые связываются с эпитопом, раскрытым в настоящем описании, могут включать по меньшей мере одну, по меньшей мере две или все три последовательности CDR тяжелой цепи SEQ ID NO:4-6 (HCDR1/HCDR2/HCDR3, соответственно). Они представляют собой последовательности HCDR1/HCDR2/HCDR3 антитела Ab 7326 из примеров, определенные по методу Kabat.
Методы Kabat и Chothia для определения последовательностей CDR хорошо известны в данной области (так же, как и другие методы). Последовательности CDR могут быть определены любым подходящим методом, и в настоящем изобретении, несмотря на то, что обычно используется метод Kabat, также могут использоваться другие методы. В данном случае SEQ ID NO:3 представляет последовательность HCDR1 Ab 7326, определенную путем комбинированного метода Chothia & Kabat.
Антитела по изобретению могут содержать по меньшей мере одну, по меньшей мере две или все три последовательности CDR легкой цепи SEQ ID NO:7-9 (LCDR1/LCDR2/LCDR3, соответственно). Они представляют собой последовательности LCDR1/LCDR2/LCDR3 Ab 7326 определенные методом Kabat.
Антитело предпочтительно содержит по меньшей мере последовательность HCDR3 SEQ ID NO:6.
Как правило, антитело содержит по меньшей мере одну последовательность CDR тяжелой цепи, выбранную из SEQ ID NO:3-5, и по меньшей мере одну последовательность CDR легкой цепи, выбранную из SEQ ID NOS 7-9. Антитело может содержать по меньшей мере две последовательности CDR тяжелой цепи, выбранные из SEQ ID NO:3-5, и по меньшей мере две последовательности CDR легкой цепи, выбранные из SEQ ID NO:7-9. Антитело обычно содержит все три последовательности CDR тяжелой цепи SEQ ID NO:3-5 (HCDR1/HCDR2/HCDR3, соответственно) и все три последовательности CDR легкой цепи SEQ ID NO:7-9 (LCDR1/LCDR2/LCDR3, соответственно). Антитела могут быть химерными, человеческими или гуманизированными антителами.
Антитело может содержать последовательность вариабельной области тяжелой цепи (HCVR) SEQ ID NO:10 или 12 (HCVR вариантов 1 и 2 Ab 7326). Антитело может содержать последовательность вариабельной области легкой цепи (LCVR) SEQ ID NO:11 или 13 (LCVR вариантов 1 и 2 Ab 7326). Антитело предпочтительно содержит последовательность вариабельной области тяжелой цепи SEQ ID NO:10 или 12 и последовательность вариабельной области легкой цепи SEQ ID NO:11 или 13 (особенно пары HCVR/LVCR SEQ ID NO:10/11 или 12/13).
Антитело может содержать последовательность тяжелой цепи (Н-цепи):
вариант 1 тяжелой цепи полноразмерного мышиного IgG1 с SEQ ID NO:14 или
вариант 2 тяжелой цепи полноразмерного мышиного IgG1 с SEQ ID NO:28, или
вариант 1 тяжелой цепи полноразмерного человеческого IgG1 с SEQ ID NO:30, или
вариант 2 тяжелой цепи полноразмерного человеческого IgG1 с SEQ ID NO:16, или
вариант 1 тяжелой цепи полноразмерного человеческого IgG4P с SEQ ID NO:22, или
вариант 2 тяжелой цепи полноразмерного человеческого IgG4P с SEQ ID NO:34, или
вариант 1 тяжелой цепи Fab с SEQ ID NO:18, или
вариант 2 тяжелой цепи Fab с SEQ ID NO:32.
Антитело может содержать последовательность легкой цепи (L-цепи):
вариант 1 легкой цепи полноразмерного мышиного IgG1 с SEQ ID NO:15 или
вариант 2 легкой цепи полноразмерного мышиного IgG1 с SEQ ID NO:29, или
вариант 1 легкой цепи полноразмерного человеческого IgG1 с SEQ ID NO:31, или
вариант 2 легкой цепи полноразмерного человеческого IgG1 с SEQ ID NO:17, или
вариант 1 легкой цепи полноразмерного человеческого IgG4P с SEQ ID NO:23, или
вариант 2 легкой цепи полноразмерного человеческого IgG4P с SEQ ID NO:35, или
вариант 1 легкой цепи Fab с SEQ ID NO:19, или
вариант 2 легкой цепи Fab с SEQ ID NO:33.
В одном из примеров антитело содержит пару последовательностей тяжелая/легкая цепи:
вариант 1 полноразмерного мышиного IgG15 с SEQ ID NO:14/15 или
вариант 2 полноразмерного мышиного IgG1 с SEQ ID NO:28/29, или
вариант 1 полноразмерного человеческого IgG1 с SEQ ID NO:30/31, или
вариант 2 полноразмерного человеческого IgG1 с SEQ ID NO:16/17, или
вариант 1 полноразмерного человеческого IgG4P с SEQ ID NO:22/23, или
вариант 2 полноразмерного человеческого IgG4P с SEQ ID NO:34/35, или
вариант 1 легкой цепи Fab с SEQ ID NO:18/19, или
вариант 2 легкой цепи Fab с SEQ ID NO:32/33.
Вариантные формы соответствующих последовательностей могут быть взаимозаменяемыми. Например, антитело может содержать пару последовательностей тяжелая/легкая цепи:
вариант 1 тяжелой цепи/вариант 2 легкой цепи полноразмерного мышиного IgG1 с SEQ ID NO:14/29 или
вариант 2 тяжелой цепи/вариант 1 легкой цепи полноразмерного мышиного IgG1 с SEQ ID NO:28/15, или
вариант 1 тяжелой цепи/вариант 2 легкой цепи полноразмерного человеческого IgG1 с SEQ ID NO:30/17, или
вариант 2 тяжелой цепи/вариант 1 легкой цепи полноразмерного человеческого IgG1 с SEQ ID NO:16/31, или
вариант 1 тяжелой цепи/вариант 2 легкой цепи полноразмерного человеческого IgG4P с SEQ ID NO:22/35, или
вариант 2 тяжелой цепи/вариант 1 легкой цепи полноразмерного человеческого IgG4P с SEQ ID NO:34/23, или
вариант 1 тяжелой цепи/вариант 2 легкой цепи Fab с SEQ ID NO:18/33, или
вариант 2 тяжелой цепи/вариант 1 легкой цепи Fab с SEQ ID NO:32/19.
Антитела могут быть химерными, человеческими или гуманизированными антителами.
Антитело альтернативно может представлять собой или может содержать вариант одной из конкретных указанных выше последовательностей. Например, вариант может представлять собой вариант замены, делеции или добавления любой из указанных выше аминокислотных последовательностей.
Вариантное антитело может содержать 1, 2, 3, 4, 5, до 10, до 20 или более (обычно максимум до 50) аминокислотных замен и/или делеций в специфических последовательностях, обсуждаемых выше. Варианты «делеции» могут содержать делецию отдельных аминокислот, делецию небольших групп аминокислот, например 2, 3, 4 или 5 аминокислот, или делецию более крупных аминокислотных областей, таких как делеция определенных аминокислотных доменов, или других признаков. Варианты «замены» обычно включают замену одной или более аминокислот таким же количеством аминокислот и являются консервативными аминокислотными заменами. Например, аминокислота может быть заменена альтернативной аминокислотой с аналогичными свойствами, например, другой основной аминокислотой, другой кислотной аминокислотой, другой нейтральной аминокислотой, другой заряженной аминокислотой, другой гидрофильной аминокислотой, другой гидрофобной аминокислотой, другой полярной аминокислотой, другой ароматической аминокислотой или другой алифатической аминокислотой. Некоторые свойства 20 основных аминокислот, которые можно использовать для выбора подходящих заместителей, являются следующими:
«Производные» или «варианты» обычно включают последовательности, в которых вместо встречающейся в природе аминокислоты присутствует аминокислота, которая является ее структурным аналогом. Аминокислоты, используемые в последовательностях, также могут быть дериватизированными или модифицированными, например, меченными, при условии сохранения функции антитела.
Производные и варианты, описанные выше, могут быть получены во время синтеза антитела или путем его модификации после продуцирования, или, в случае рекомбинантного антитела, с помощью известных методов сайт-направленного мутагенеза, случайного мутагенеза или ферментативного расщепления и/или лигирования нуклеиновых кислот.
Варианты антитела могут иметь аминокислотную последовательность, которая имеет более примерно 60% или более примерно 70%, например 75 или 80%, обычно более примерно 85%, например, более примерно 90 или 95% аминокислотной идентичности с аминокислотными последовательностями, раскрытыми в настоящей заявке (в частности, с последовательностями HCVR/LCVR и последовательностями H- и L-цепей). Кроме того, антитело может представлять собой вариант, который имеет более примерно 60% или более примерно 70%, например, 75 или 80%, обычно более примерно 85%, например, более примерно 90 или 95% аминокислотной идентичности с последовательностями HCVR/LCVR и последовательностями H- и L-цепей, раскрытыми в настоящей заявке, с сохранением точных CDR, раскрытых для этих последовательностей. Варианты могут иметь по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с последовательностями HCVR/LCVR и с последовательностями H- и L-цепей, раскрытыми в настоящей заявке (при некоторых обстоятельствах, при условии сохранения точных CDR).
Варианты обычно имеют от примерно 60% до примерно 99% идентичности, от примерно 80% до примерно 99% идентичности, от примерно 90% до примерно 99% идентичности или от примерно 95% до примерно 99% идентичности. Этот процент идентичности аминокислот можно наблюдать по всей длине соответствующей последовательности SEQ ID NO или части последовательности, состоящей, например из примерно 20, 30, 50, 75, 100, 150, 200 или более аминокислот, в зависимости от размера полноразмерного полипептида.
Что касается аминокислотных последовательностей, «идентичность последовательности» относится к последовательностям, которые имеют заявленное значение при оценке с помощью программы ClustalW (Thompson et al., 1994, выше) со следующими параметрами:
Параметры для попарного выравнивания - Метод: точный; Матрица: PAM; Штраф за открытие пропуска: 10,00; Штраф за удлинение пропуска: 0,10;
Параметры для выравнивания нескольких последовательностей -Матрица: PAM; Штраф за открытие пропуска: 10,00; % идентичности за счет задержки: 30; штраф за внесение концевого пропуска: вкл; расстояние между пропусками: 0; отрицательная матрица: нет; штраф за удлинение пропуска: 0,20; штраф за пропуск конкретного остатка: вкл.; штрафы за пропуск гидрофильного остатка: вкл.; гидрофильные остатки: GPSNDQEKR. Предполагается, что идентичность последовательностей для конкретного остатка включает идентичные остатки, которые были просто изменены.
Поэтому предложены антитела, имеющие специфические последовательности и варианты с сохранением функции или активности этих цепей.
Антитела могут конкурировать за связывание с Гремлин-1 или связываться с тем же эпитопом, что и антитела, которые определены выше в отношении последовательностей H-цепи/L-цепи, HCVR/LCVR или CDR. В частности, антитело может конкурировать за связывание Гремлин-1 или связываться с тем же эпитопом, что и антитело, которое содержит комбинацию последовательности HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3 с SEQ ID NO:4/5/6/7/8/9. Антитело может конкурировать за связывание с Гремлин-1 или связываться с тем же эпитопом, что и антитело, которое содержит пару последовательностей HCVR и LCVR с SEQ ID NO:10/11 или 12/13 или полноразмерные цепи с SEQ ID NO:14/15 или 16/17.
Термин «эпитоп» представляет собой область антигена, которая связывается с антителом. Эпитопы могут быть определены как структурные или функциональные. Функциональные эпитопы, как правило, представляют собой подмножество структурных эпитопов и имеют остатки, которые непосредственно участвуют в обеспечении сродства взаимодействия. Эпитопы также могут быть конформационными, т.е. состоять из нелинейных аминокислот. В некоторых вариантах осуществления эпитопы могут включать детерминанты, которые представляют собой химически активные поверхностные группы молекул, такие как аминокислоты, боковые цепи сахара, фосфорильные группы или сульфонильные группы, и, в некоторых вариантах осуществления, могут иметь конкретные трехмерные структурные характеристики и/или характеристики удельного заряда.
Используя обычные способы, известные в данной области, можно легко определить, связывается ли антитело с тем же эпитопом, или конкурирует за связывание с эталонным антителом. Например, для определения, связывается ли тестируемое антитело с тем же эпитопом, что и эталонное антитело по изобретению, связывание эталонного антитела с белком или пептидом выполняют в условиях насыщения. Затем оценивают способность тестируемого антитела связываться с белком или пептидом. Если тестируемое антитело способно связываться с белком или пептидом после его связывания эталонным антителом в условиях насыщения, можно сделать вывод, что тестируемое антитело связывается с другим эпитопом, а не с тем, с которым связано эталонное антитело. С другой стороны, если тестируемое антитело не способно связываться с белком или пептидом после его связывания эталонным антителом в условиях насыщения, тогда тестируемое антитело может связываться с тем же эпитопом, что и эпитоп, с которым связано эталонное антитело по изобретению.
Для определения, конкурирует ли антитело за связывание с эталонным антителом, описанный выше метод связывания выполняется в двух вариантах ориентации. В первом варианте ориентации выполняют связывание эталонного антитела с белком/пептидом в условиях насыщения с последующей оценкой связывания тестируемого антитела с той же молекулой белка/пептида. Во втором варианте ориентации связывание тестируемого антитела с белком/пептидом выполняют в условиях насыщения с последующей оценкой связывания эталонного антитела с тем же белком/пептидом. Если в обоих вариантах ориентации только первое (насыщающее) антитело способно связываться с белком/пептидом, то делается вывод, что тестируемое антитело и эталонное антитело конкурируют за связывание с белком/пептидом. Как будет понятно специалисту в данной области, антитело, которое конкурирует за связывание с эталонным антителом, необязательно может связываться с эпитопом, идентичным тому, с которым связывается эталонное антитело, оно может пространственно блокировать связывание эталонного антитела за счет связывания с перекрывающим или смежным эпитопом.
Два антитела связываются с одним и тем же или перекрывающим эпитопом, если каждое конкурентно ингибирует (блокирует) связывание другого с антигеном. Иными словами, 1-, 5-, 10-, 20- или 100-кратный избыток одного антитела ингибирует связывание другого по меньшей мере на 50, 75, 90 или даже 99%, согласно результатам анализа конкурентного связывания (см., например, Junghans et al., Cancer Res, 1990: 50:1495-1502). Альтернативно, два антитела имеют один и тот же эпитоп, если по существу все аминокислотные мутации в антигене, которые уменьшают или устраняют связывание одного антитела, уменьшают или устраняют связывание другого. Два антитела имеют перекрывающиеся эпитопы, если некоторые аминокислотные мутации, которые уменьшают или устраняют связывание одного антитела, уменьшают или устраняют связывание другого.
Затем можно провести дополнительные рутинные эксперименты (например, анализы пептидных мутаций и связывания), чтобы подтвердить, действительно ли наблюдаемое отсутствие связывания тестируемого антитела является результатом связывания эталонного антитела с тем же самым эпитопом, или наблюдаемое отсутствие связывания является результатом пространственного блокирования (или другого явления). Эксперименты такого рода могут быть выполнены методами ELISA, RIA, поверхностного плазмонного резонанса, проточной цитометрии или с помощью любого другого количественного или качественного анализа связывания антител, доступного в данной области.
Антитела можно тестировать в отношении связывания с Гремлин-1, например, стандартным ELISA или Вестерн-блоттингом. ELISA-анализ также можно использовать для скрининга гибридом, которые проявляют положительную реактивность с целевым белком. Селективность связывания антитела также может быть определена путем мониторинга связывания антитела с клетками, экспрессирующими целевой белок, например, методом проточной цитометрии. Таким образом, метод скрининга может включать стадию идентификации антитела, которое способно связывать Гремлин-1, путем проведения ELISA или вестерн-блоттинга или проточной цитометрии.
Антитела могут селективно (или специфически) распознавать Гремлин-1. Антитело или другое соединение «селективно связывается» или «селективно распознает» белок, если оно связывается с преимущественным или высоким сродством с белком, в отношении которого оно является селективным, но по существу не связывается или связывается с низким сродством с другими белками. Селективность антитела может быть дополнительно изучена путем определения, связывается ли это антитело с другими родственными белками, как обсуждалось выше, или же оно способно различать их. Антитела по изобретению обычно распознают Гремлин-1 человека.
Антитела также могут обладать перекрестной реактивностью с родственными белками или Гремлин-1 человека и Гремлин-1 других видов.
Под термином специфический (или селективный) следует понимать, что антитело связывается с представляющим интерес белком без существенной перекрестной реактивности с какой-либо другой молекулой. Перекрестную реактивность можно оценить любым подходящим способом, описанным в настоящей заявке. Перекрестная реактивность антитела считается существенной, если антитело связывается с другой молекулой, по меньшей мере, примерно на 5, 10, 15, 20, 25, 30, 35, 40, 45, 50%. 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 100%, настолько же эффективно, насколько оно связывается с представляющим интерес белком. Антитело, которое является специфическим (или селективным), может связываться с другой молекулой менее чем на 90%, 85%, 80%, 75%, 70%, 65%, 60%, 55%, 50%, 45%, 40% 35%, 30%, 25% или 20% эффективности связывания, с которой оно связывается с представляющим интерес белком. Антитело может связываться с другой молекулой менее чем примерно на 20%, менее чем примерно на 15%, менее чем примерно на 10% или менее чем примерно на 5%, менее чем примерно на 2% или менее чем примерно на 1% эффективности связывания, с которой оно связывается с представляющим интерес белком.
Антитела против Гремлин были описаны ранее. Например, в WO2014/159010A1 (Regeneron) описаны антитела к Гремлин, которые ингибируют активность Гремлин-1, со значениями сродства связывания KD в диапазоне от 625 пМ до 270 нМ при 25°C. Ciuclan et al. (2013) описывают моноклональное антитело к Гремлин-1 со сродством связывания KD, равным 5,6×10-10 М.
Антитела против Гремлин-1 по изобретению являются аллостерическими ингибиторами активности Гремлин-1 и связываются с новым эпитопом, удаленным от сайта связывания BMP. Антитела связываются с Гремлин-1 с исключительно высоким сродством со значениями Kd <100 пМ. Следовательно, антитела по изобретению демонстрируют значительное улучшение по сравнению с доступными в настоящее время антителами и, как ожидается, будут особенно полезными для лечения заболеваний, опосредованных Гремлин-1.
Таким образом, антитела, подходящие для применения по настоящему изобретению, могут иметь высокое сродство связывания с Гремлин-1 (человека). Антитело может иметь константу диссоциации (KD) менее чем <1 нМ и предпочтительно <500 пМ. В одном из примеров антитело имеет константу диссоциации (KD) менее 200 пМ. В одном из примеров антитело имеет константу диссоциации (KD) менее 100 пМ. Для определения сродства связывания антитела с его антигеном-мишенью можно использовать различные методы, такие как анализ поверхностного плазмонного резонанса, анализ насыщения или иммуноанализ, такой как ELISA или RIA, которые хорошо известны специалистам в данной области. Примером метода определения сродства связывания является анализ поверхностно-плазмонного резонанса на приборе BIAcore™ 2000 (Biacore AB, Freiburg, Германия) с использованием сенсорных чипов CM5, который описан у Krinner et al., (2007) (Mol. Immunol. February; 44 (5):916-25. (Epub 2006 May 11)).
Антитела по изобретению обычно являются ингибирующими антителами. Гремлин-1 ингибирует BMP-2, 4 и 7, поэтому ингибирование Гремлин-1 приводит к усилению передачи сигналов через BMP.
Как упомянуто выше, в примерах настоящей заявки описаны два функциональных анализа для скрининга антитела, способного ингибировать Гремлин 1, а именно анализ фосфорилирования SMAD и анализ активности репортерного гена Id1 в клетках HEK. Как правило, ингибирующее антитело восстанавливает фосфорилирование SMAD и/или восстанавливает передачу сигналов BMP в анализе активности репортерного гена Id1 в клетках Hek. Фосфорилирование SMAD может быть восстановлено по меньшей мере на 80%, 90% или 100% по сравнению с контрольным BMP. В анализе активности репортерного гена Id1 в клетках Hek ингибирующее антитело может иметь IC50 менее 10 нМ, предпочтительно менее 5 нМ.
После идентификации и выбора подходящего антитела можно определить аминокислотную последовательность антитела способами, известными в данной области. Гены, кодирующие антитело, можно клонировать с помощью вырожденных праймеров. Может быть получено рекомбинантное антитело с помощью обычных способов.
Настоящее изобретение также относится к выделенной последовательности ДНК, кодирующей вариабельные области (участки) тяжелой и/или легкой цепи (или полноразмерные Н- и L-цепи) молекулы антитела по настоящему изобретению.
Вариант полинуклеотида может содержать 1, 2, 3, 4, 5, до 10, до 20, до 30, до 40, до 50, до 75 или более замен нуклеиновых кислот и/или делеций в последовательностях, приведенных в списке последовательностей. Обычно вариант имеет 1-20, 1-50, 1-75 или 1-100 замен и/или делеций.
Подходящие варианты могут быть по меньшей мере примерно на 70% гомологичны полинуклеотиду любой из последовательностей нуклеиновых кислот, раскрытых в данном документе, обычно они гомологичны по меньшей мере примерно на 80 или 90% и более предпочтительно по меньшей мере примерно на 95, 97 или 99%. Варианты могут иметь по меньшей мере примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности. Обычно идентичность вариантов находится в диапазоне от примерно 60% до примерно 99%, от примерно 80% до примерно 99%, от примерно 90% до примерно 99% или от примерно 95% до примерно 99%. Эти уровни гомологии и идентичности обычно имеют по меньшей мере кодирующие области полинуклеотидов. Методы измерения гомологии хорошо известны в данной области, и специалистам в данной области будет понятно, что в настоящем контексте гомология рассчитывается на основе идентичности нуклеиновых кислот. Такую гомологию могут иметь области, состоящие по меньшей мере из примерно 15, по меньшей мере из примерно 30, например, по меньшей мере из примерно 40, 60, 100, 200 или более смежных нуклеотидов (в зависимости от длины). Такая гомология может быть по всей длине немодифицированной полинуклеотидной последовательности.
Способы измерения гомологии или идентичности полинуклеотидов известны в данной области. Например, пакет UWGCG предоставляет программу BESTFIT, которую можно использовать для расчета гомологии (например, используя настройки по умолчанию) (Devereux et al (1984) Nucleic Acids Research 12, p387-395).
Алгоритмы PILEUP и BLAST также могут быть использованы для вычисления гомологии или для выравнивания последовательностей (обычно, используя настройки по умолчанию), например, как описано в Altschul S.F. (1993) J Mol Evol 36:290-300; Altschul S., F et al. (1990) J. Mol. Biol. 215:403-10.
Программное обеспечение для выполнения анализа BLAST доступно для общественного пользования через Национальный центр биотехнологической информации (http://www.ncbi.nlm.nih.gov/). Этот алгоритм включает сначала идентификацию пар последовательностей с высокой степенью сходства (HSP) путем определения коротких слов длиной W в запрашиваемой последовательности, которые либо соответствуют, либо удовлетворяют некоторому положительному пороговому значению T при сопоставлении со словом той же длины в последовательности базы данных. Параметр T означает пороговую величину, используемую для оценки соседства слов (Altschul et al., см. выше). Указанная исходная величина степени сходства слов служит в качестве затравки для первоначального поиска с целью нахождения более длинных HSP, содержащих их. Попадания в слова расширяют в обоих направлениях вдоль каждой последовательности до тех пор, пока кумулятивная оценка выравнивания может увеличиваться. Расширение совпадения в слово в каждом направлении прекращают, когда: кумулятивная оценка выравнивания опускается до нуля или ниже из-за накопления одного или более результатов сопоставления в виде остатков с негативным показателем; или при достижении конца любой последовательности. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость сопоставления. В программе BLAST по умолчанию используются следующие параметры: длина слова (W)=11, оценочная матрица BLOSUM62 (см. Henikoff and Henikoff (1992) Proc. Natl. Acad. Sci. USA 89:10915-10919), выравнивание (B)=50, ожидание (E)=10, M=5, N=4, и сравнение обеих нитей.
Алгоритм BLAST выполняет статистический анализ сходства между двумя последовательностями; см., например, Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5787. Одним из критериев сходства, оцениваемых в рамках алгоритма BLAST, является наименьшая сумма вероятностей (P (N)), которая указывает на вероятность случайного спаривания между двумя нуклеотидными или аминокислотными последовательностями. Например, последовательность считается аналогичной другой последовательности, если наименьшая сумма вероятностей при сравнении первой последовательности со второй последовательностью меньше чем примерно 1, обычно меньше чем примерно 0,1, предпочтительно меньше чем примерно 0,01 и наиболее предпочтительно меньше чем примерно 0,001. Например, наименьшая сумма вероятностей может находиться в диапазоне от 1 до 0,001, чаще от 0,01 до 0,001.
Гомолог может отличаться от последовательности соответствующего полинуклеотида менее чем на 3, 5, 10, 15, 20 или более мутаций (каждая из которых может представлять собой замену, делецию или вставку). Например, гомолог может отличаться на 3-50 мутаций, чаще на 3-20 мутаций. Эти мутации могут находиться в пределах по меньшей мере 30, например, по меньшей мере примерно 40, 60 или 100 или более смежных нуклеотидов гомолога.
В одном из вариантов осуществления вариантная последовательность может отличаться от конкретных последовательностей, приведенных в списке последовательностей, из-за вырожденности генетического кода. Код ДНК содержит 4 основных остатка нуклеиновой кислоты (A, T, C и G), которые используются для «записи» трехбуквенных кодонов, представляющих аминокислоты белков, кодируемых в генах организма. Линейная последовательность кодонов транслируется вдоль молекулы ДНК в линейную последовательность аминокислот в белке (белках), кодируемом этими генами. Код является сильно вырожденным, поскольку 61 кодон кодирует 20 природных аминокислот, и 3 кодона представляют собой «стоп»-сигналы. Таким образом, большинство аминокислот кодируется более чем одним кодоном - фактически, некоторые из них кодируются четырьмя или более различными кодонами. Следовательно, вариант полинуклеотида по изобретению может кодировать такую же полипептидную последовательность, что и другой полинуклеотид по изобретению, но может иметь другую последовательность нуклеиновой кислоты в связи с использованием разных кодонов для кодирования одних и тех же аминокислот.
Последовательность ДНК по настоящему изобретению может содержать синтетическую ДНК, например, полученную путем химической обработки, кДНК, геномную ДНК или любую их комбинацию.
Последовательности ДНК, которые кодируют молекулу антитела по настоящему изобретению, могут быть получены способами, хорошо известными специалистам в данной области. Например, последовательности ДНК, кодирующие часть или все тяжелые и легкие цепи антитела, могут быть синтезированы, при необходимости, из определенных последовательностей ДНК или на основе соответствующих аминокислотных последовательностей.
Общие способы, позволяющие создавать векторы, способы трансфекции и способы культивирования хорошо известны специалистам в данной области. Более подробно см. «Current Protocols in Molecular Biology», 1999, F. M. Ausubel (ed), Wiley Interscience, New York and the Maniatis Manual produced by Cold Spring Harbor Publishing.
Также предоставляется клетка-хозяин, содержащая один или более векторов клонирования или экспрессии, содержащих одну или более последовательностей ДНК, кодирующих антитело по настоящему изобретению. Для экспрессии последовательностей ДНК, кодирующих молекулу антитела по настоящему изобретению может быть использована любая подходящая система клетка-хозяин/вектор. Можно использовать бактериальные, например, E.coli, и другие микробные системы, или можно использовать эукариотические системы экспрессии клеток-хозяев, например, системы млекопитающих. Подходящие клетки-хозяева млекопитающих включают клетки СНО, миеломы или гибридомы.
Настоящее изобретение также относится к способу получения молекулы антитела по настоящему изобретению, включающему культивирование клетки-хозяина, содержащей вектор по настоящему изобретению, в условиях, подходящих для экспрессии белка из ДНК, кодирующей молекулу антитела по настоящему изобретению, и выделение молекулы антитела.
Антитело Ab 7326 по изобретению идентифицировано для связывания следующих остатков Гремлин-1: Ile110 (131), Lys126 (147), Lys127 (148), Phe128 (149), Thr129 (150), Thr130 (151), Arg148 (169), Lys153 (174) и Gln154 (175), где Lys126 (147), Lys127 (148), Phe128 (149), Thr129 (150), Thr130 (151), Arg148 (169), Lys153 (174) и Gln154 (175) присутствуют на одном мономере Гремлин-1, а Ile110 (131) присутствует на втором мономере Гремлин-1. Нумерация, указанная не в скобках, дана на основе структурного файла (и соответствует нумерации мышиного Гремлин-2, полученного в результате структурного выравнивания). Числа в скобках указывают остатки согласно записи O60565 последовательности SEQ ID NO:1 в базе данных UniProt. Как обсуждается в разделе «Примеры», эти остатки эпитопа идентифицированы с помощью анализа NCONT в пределах 4 в комплексе Гремлин-1-Fab Ab 7326.
Следовательно, антитела по изобретению могут связываться с эпитопом, который содержит по меньшей мере один остаток, выбранный из Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175 (с нумерацией остатков согласно SEQ ID NO:1). Антитела по изобретению могут связываться с эпитопом, который содержит 2, 3, 4, 5, 6, 7, 8 или все 9 из этих остатков (предпочтительно, по меньшей мере, 5 остатков).
Антитела по изобретению также могут распознавать эпитоп, в котором Ile131 присутствует на другом мономере Гремлин-1, который не содержит другие указанные остатки.
Хотя эти остатки предоставлены для конкретной последовательности Гремлин-1 человека, специалист в данной области, используя рутинные методы, может легко экстраполировать положения этих остатков на другие соответствующие последовательности Гремлин (например, мыши). Поэтому антитела, связывающиеся с эпитопами, содержащими соответствующие остатки в таких других последовательностях Гремлин, также входят в объем настоящего изобретения.
Для скрининга антител, которые связываются с конкретным эпитопом, может быть проведен обычный анализ перекрестного блокирования, такой как описано в Antibodies, Harlow and Lane (Cold Spring Harbor Press, Cold Spring Harb., NY). Другие методы включают аланин-сканирующие мутанты, пептидные блоты (Reineke (2004) Methods Mol Biol 248: 443-63) или анализ расщепления пептидов. Кроме того, могут быть использованы такие методы, как вырезание эпитопов, экстракция эпитопа и химическая модификация антигенов (Tomer (2000) Protein Science 9:487-496). Эти способы хорошо известны в данной области.
Эпитопы антител также могут быть определены с помощью рентгеноструктурного анализа. Поэтому антитела по настоящему изобретению можно оценивать с помощью рентгеноструктурного анализа антитела, связанного с Гремлин-1. Таким образом, эпитопы могут быть идентифицированы, в частности, путем определения остатков на Гремлин-1 в пределах 4 от остатка паратопа антитела.
Фармацевтические композиции, дозировки и режимы дозирования
Антитело по изобретению или агент, который модулирует Гремлин-1, идентифицированный методами скрининга, может быть предоставлено в фармацевтической композиции. Фармацевтическая композиция обычно является стерильной и обычно включает фармацевтически приемлемый носитель и/или адъювант. Фармацевтическая композиция по настоящему изобретению может дополнительно содержать фармацевтически приемлемый адъювант и/или носитель.
Используемый в настоящей заявке термин «фармацевтически приемлемый носитель» включает любые растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и задерживающие абсорбцию агенты и т.п., которые являются физиологически совместимыми. Носитель может быть подходящим для парентерального введения, например внутривенного, внутримышечного, внутрикожного, внутриглазного, внутрибрюшинного, подкожного, спинального или другого парентерального пути введения, например, путем инъекции или инфузии. Альтернативно, носитель может быть подходящим для непарентерального введения, например, местного, эпидермального или мукозального пути введения. Носитель может быть пригодным для перорального введения. В зависимости от пути введения модулятор может быть покрыт материалом, защищающим соединение от действия кислот и других природных условий, которые могут инактивировать соединение.
Фармацевтические композиции по изобретению могут включать одну или более фармацевтически приемлемых солей. «Фармацевтически приемлемая соль» относится к соли, которая сохраняет необходимую биологическую активность исходного соединения и не оказывает каких-либо нежелательных токсикологических эффектов. Примеры таких солей включают аддукты кислот и аддукты оснований.
Фармацевтически приемлемые носители содержат водные носители или разбавители. Примеры подходящих водных носителей, которые могут быть использованы в фармацевтических композициях по изобретению, включают воду, забуференную воду и солевой раствор. Примеры других носителей включают этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Во многих случаях желательно включить в композицию изотонические агенты, например сахара, полиспирты, такие как маннит, сорбит или хлорид натрия.
Терапевтические композиции обычно должны быть стерильными и стабильными в условиях производства и хранения. Композиция может быть составлена в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного вещества.
Фармацевтические композиции по изобретению могут содержать дополнительные активные ингредиенты.
Также в объем настоящего изобретения входят наборы, содержащие антитела или модулирующие агенты по изобретению и инструкции по применению. Набор может дополнительно содержать один или более дополнительных реагентов, таких как дополнительный терапевтический или профилактический агент, как обсуждалось выше.
Модуляторы и/или антитела по изобретению или их составы, или композиции можно вводить для профилактического и/или терапевтического лечения.
В случаях терапевтического применения соединения вводят субъекту, уже страдающему от расстройства или состояния, описанного выше, в количестве, достаточном для излечения, облегчения или частичного купирования состояния или одного или более его симптомов. Такое терапевтическое лечение может приводить к уменьшению проявления симптомов заболевания или увеличению частоты или продолжительности бессимптомных периодов. Количество, достаточное для достижения этого, определяется как «терапевтически эффективное количество».
В случаях профилактического применения составы вводят субъекту с риском развития расстройства или состояния, описанного выше, в количестве, достаточном для предотвращения или уменьшения вероятности проявления эффектов состояния или одного или более его симптомов. Количество, достаточное для достижения этого, определяется как «профилактически эффективное количество». Эффективные количества для каждой цели будут зависеть от тяжести заболевания или травмы, а также от веса и общего состояния субъекта.
Субъектом для введения может быть человек или животное, не являющееся человеком. Термин «животное, не являющееся человеком» включает всех позвоночных животных, например млекопитающих и не млекопитающих, таких как приматы, не являющиеся человеком, овцы, собаки, кошки, лошади, коровы, цыплята, земноводные, рептилии и т.д. Обычно вводят людям.
Антитело/модулятор или фармацевтическую композицию по изобретению можно вводить одним или более способами введения, используя один или более из множества способов, известных в данной области. Как будет понятно специалисту в данной области, путь и/или способ введения будет меняться в зависимости от необходимых результатов. Примеры путей введения соединений или фармацевтических композиций по изобретению включают внутривенный, внутримышечный, внутрикожный, внутриглазной, внутрибрюшинный, подкожный, спинальный или другие парентеральные пути введения, например, путем инъекции или инфузии. Используемая в настоящем описании фраза «парентеральное введение» означает способы введения, отличные от энтерального и местного введения, и обычно осуществляется путем инъекции. Альтернативно, антитело/модулирующий агент или фармацевтическую композицию по изобретению можно вводить непарентеральным путем, таким как местный, эпидермальный или мукозальный путь введения. Антитело/модулирующий агент или фармацевтическая композиция по изобретению могут быть составлены для перорального введения.
Подходящая доза антитела/модулирующего агента или фармацевтической композиции по изобретению может быть определена квалифицированным врачом. Фактические уровни дозирования активных ингредиентов в фармацевтических композициях по настоящему изобретению могут меняться для получения количества активного ингредиента, которое является эффективным для достижения нужного терапевтического ответа у конкретного пациента, для конкретной композиции и способа введения, при этом эти уровни не должны быть токсичными для пациента. Выбранный уровень дозирования будет зависеть от множества фармакокинетических факторов, включая активность используемых конкретных композиций по настоящему изобретению, способ введения, время введения, скорость выведения конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в комбинации с конкретными используемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и предыдущую историю болезни пациента, получающего лечение, и аналогичные факторы, хорошо известные в области медицины.
Подходящая доза может находиться, например, в диапазоне от примерно 0,01 мкг/кг до примерно 1000 мг/кг массы тела, обычно от примерно 0,1 мкг/кг до примерно 100 мг/кг массы тела пациента, подлежащего лечению. Например, подходящая доза может находиться в интервале от примерно 1 мкг/кг до примерно 10 мг/кг массы тела в сутки или от примерно 10 мкг/кг до примерно 5 мг/кг массы тела в сутки.
Режим дозирования может быть скорректирован для обеспечения оптимального требуемого ответа (например, терапевтического ответа). Например, может быть введена одна доза, несколько разделенных доз в течение определенного времени, или доза может быть пропорционально уменьшена или увеличена, при необходимости, в зависимости от терапевтической ситуации. Используемая здесь единичная лекарственная форма относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; каждая единица содержит заданное количество активного соединения, рассчитанное для получения нужного терапевтического эффекта в сочетании с необходимым фармацевтическим носителем.
Введение может осуществляться в виде одной или более доз. Многократные дозы могут вводиться одним и тем же или разными способами в одни и те же или разные места. Альтернативно, дозы могут быть введены в виде состава с замедленным высвобождением, и в этом случае требуется менее частое введение. Дозирование и частота могут варьировать в зависимости от периода полувыведения антагониста из организма пациента и требуемой продолжительности лечения.
Как упомянуто выше, модуляторы/антитела или фармацевтические композиции по изобретению могут вводиться вместе с одним или более терапевтическими агентами.
Комбинированное введение двух или более агентов может быть достигнуто несколькими различными способами. Оба могут быть введены вместе в одной композиции или могут быть введены в отдельных композициях как часть комбинированной терапии. Например, один может быть введен до, после или одновременно с другим.
Терапевтические показания
Антитела по настоящему изобретению или модулирующие агенты, идентифицированные способами скрининга по изобретению, можно использовать для лечения, профилактики или улучшения любого состояния, ассоциированного с активностью Гремлин-1. Например, любое состояние, которое полностью или частично является результатом передачи сигналов через Гремлин-1. Другими словами, изобретение относится к лечению, профилактике или улучшению состояний, опосредованных или находящихся под влиянием Гремлин. Такие состояния включают фиброзные заболевания, включая фиброз почек (например, диабетическую нефропатию и хроническую нефропатию аллотрансплантата) и идиопатический фиброз легких, легочную артериальную гипертензию, ангиогенез и рак (например, колоректальный рак).
Приведенные ниже примеры предназначены для иллюстрации изобретения.
Пример 1. Экспрессия, очистка, рефолдинг и определение структуры белка.
Экспрессия белка и подготовка телец включения
Укороченную последовательность, кодирующую Гремлин-1 человека (SEQ ID NO:20), оптимизированную для экспрессии в E.coli, клонировали в модифицированный вектор pET32a (Merck Millipore) с помощью BamHI/XhoI, создавая вектор, кодирующий последовательность Гремлин с N-терминальной меткой 6His-TEV (pET-hГремлин1).
Экспрессируемая последовательность:
MGSSHHHHHHSSGENLYFQGSAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD; SEQ ID NO:2 (не относящиеся к Гремлин остатки метки 6His-TEV выделены курсивом). Нумерация последовательности дана на основе UniProt O60565 и SEQ ID NO:1.
Плазмидную ДНК pET-hГремлин1 использовали для трансформации клеток BL21 (DE3). Единичную колонию, устойчивую к ампициллину, отбирали из чашки с агаром LB/Amp и использовали для инокуляции 100 мл стартовой культуры LB/Amp. После встряхивания (200 об/мин) в течение 16 часов при 37°C 25 мл стартовой культуры использовали для инокуляции 500 мл среды 2xTY/Amp. Культуру встряхивали (250 об/мин) при 37°С до достижения величины OD600, равной 3. Затем к культуре добавляли 20 мл питательной смеси MOPS+глицерин (1M MOPS pH 7,4, 40% глицерин, 0,5% MgSO4, 0,42% MgCl2), индуцированной 300 мкМ IPTG, и дополнительно инкубировали при 17°C, 180 об/мин в течение 16 часов. Клетки собирали на центрифуге (4000 g в течение 20 минут при 4°С).
Клеточные осадки ресуспендировали в буфере для лизиса (PBS pH 7,4, 0,35 мг/мл лизоцима, 10 мкг/мл ДНКазы и 3 мМ MgCl2) при 4°C, и нерастворимую фракцию собирали центрифугированием при 3500 g в течение 30 минут при 4°C. Гранулированные тельца включения трижды промывали путем ресуспендирования в промывочном буфере (50 мМ Трис, 500 мМ NaCl, 0,5% Тритон Х-100, рН 8,0) с последующим центрифугированием при 21000 g в течение 15 минут. Две дополнительные промывки выполняли промывочным буфером без Triton X-100.
Солюбилизация
Тельца включения ресуспендировали в денатурирующем буфере (8М мочевина, 100 мМ Трис, 1 мМ ЭДТА, 10 мМ Na2S4O6 и 100 мМ Na2SO3, рН 8,5), перемешивали в течение 16 часов при комнатной температуре и осветляли центрифугированием при 21000 g в течение 15 минут.
Очистка перед рефолдингом
Солюбилизированные тельца включения наносили на колонку с сефакрилом Sephacryl S-200 26/60 (120 мл), уравновешенную 8 М мочевиной, 50 мМ MES, 200 мМ NaCl, 1 мМ ЭДТА, рН 6,0. Фракции, содержащие белок Гремлин-1, разбавляли 6 М мочевиной, 20 мМ MES, рН 6,0 и загружали в катионообменные колонки HiTrap SP HP, и элюировали градиентом 1М NaCl в объеме, равном 10 объемам колонки (10 CV). Фракции, содержащие очищенный денатурированный белок hГремлин-1, объединяли.
Рефолдинг
Денатурированный очищенный белок Гремлин-1 добавляли по каплям в буфер для рефолдинга (50 мМ Трис, рН 8,5, 150 мМ NaCl, 5 мМ GSH и 5 мМ GSSG, 0,5 мМ цистеина, 5 мМ ЭДТА, 0,5 М аргинина) до получения конечной концентрации 0,1 мг/мл и выдерживали при 4°С при постоянном перемешивании в течение 5 дней. Через 5 дней белок Гремлин-1 подвергали диализу против 20 мМ HEPES, 100 мМ NaCl, рН 7,5.
После диализа белок наносили на колонку с гепарином HiTrap и элюировали, используя градиент 0-100% буфера для элюирования гепарина (20 мМ HEPES, 1 М NaCl, pH 7,5) объемом 20 CV. Правильно свернутый белок элюировался при 1 М NaCl, тогда как любой неправильно свернутый белок элюировался при более низких концентрациях соли.
Белок, элюированный при 1 М NaCl, концентрировали и дополнительно очищали на колонке S75 26/60, уравновешенной 20 мМ Hepes, pH 7,5, 1 М NaCl.
Характеристики белка получали с помощью SDS PAGE (сдвиг в геле), с помощью жидкостной хроматографии и масс-спектрометрии (ЖХ-МС) было продемонстрировано, что белок имеет требуемую молекулярную массу и правильное расположение дисульфидных связей, а активность белка оценивали с помощью клеточного анализа (анализ активности репортера ID1).
Определение структуры Гремлин 1
Кристаллы белка Гремлин 1 выращивали, используя метод висячих капель, смешивая раствор Гремлин 1 с концентрацией 6,6 мг/мл в 0,1 М лимонной кислоты при pH 4,1 М с хлоридом лития и 27% полиэтиленгликоля (PEG) 6000 в соотношении 1:1. Перед сбором данных кристаллы подвергали криозащите, добавляя 20% глицерина в буфер для кристаллизации. Данные дифракции собирали на алмазном источнике света и обрабатывали с помощью программы XDS (Kabsch, Wolfgang (2010) Acta Crystallographica Section D 66, 125-132). Статистические данные по дифракции суммированы в приведенной ниже таблице:
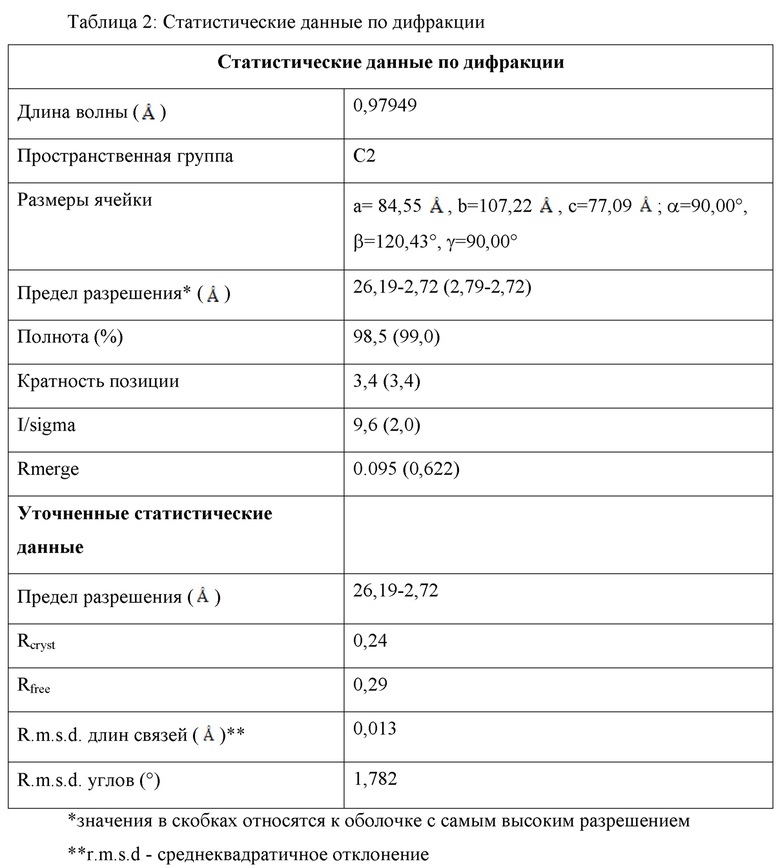
Структура Гремлин-1 была определена путем молекулярного замещения с использованием программы Phaser (McCoy et al., J Appl Cryst (2007), 40, 658-674) и модели Гремлин-1, доступной из запатентованных координат комплекса Гремлин-1/Fab. Полученная модель Гремлин-1 содержала четыре копии мономера Гремлин-1, организованные в виде двух димеров. Модель корректировали методом Coot (Emsley et al. Acta Crystallographica Section D: Biological Crystallography 66 (4), 486-501), а координаты уточняли методом Refmac (Murshudov et al REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallographica Section D: Biological Crystallography. 2011;67(Pt 4):355-367). Окончательные координаты подтверждали методом Molprobity (Chen et al. (2010) MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallographica D66:12-21). Результаты уточненных статистических данных модели приведена в таблице 2 выше.
Пример 2 - BMP-связывающие остатки на Гремлин-1
Как обсуждалось выше, Гремлин-1 принадлежит к семейству белков-антагонистов костного морфогенного белка (BMP) в подгруппе, известной как семейство DAN. В семействе DAN Гремлин-1 имеет наибольшую гомологию с Гремлин-2 (PRDC).
Структура Гремлин-1 человека 2,7 , описанная в примере 1, имеет много общих признаков с опубликованной структурой мышиного Гремлин-2 (Nolan et al (2013), Structure, 21, 1417-1429). Они имеют схожую полную укладку молекул с двумя копиями Гремлин-1, образующими антипараллельный нековалентный димер, расположенный в виде дуги. Каждый мономер имеет характерное расположение «палец-запястье-палец» с мотивом цистинового узла в направлении к концу запястья, напротив пальцев (фиг. 2). Идентичность последовательности между белками составляет 52% и возрастает до 67% в последовательности, видимой в двух структурах. Наиболее высоко консервативная область находится на обширной границе раздела в димере, на которой все ключевые контактные остатки сохранены на 100% (фиг. 3).
Остатки, участвующие в связывании BMP 2, 4 и 7 с мышиным Гремлин-2 (PRDC) и DAN (NBL1), идентифицировали с помощью мутагенеза (Nolan et al (2013), Structure, 21, 1417-1429 и Nolan et al (2014) J. Biol. Chem. 290, 4759-4771). Прогнозируемый BMP-связывающий эпитоп включает гидрофобный участок, охватывающий оба мономера на выпуклой поверхности димера (фиг. 4 и 5). Шесть остатков идентифицированы с помощью мутагенеза: Trp72, Phe96, Tyr98, Phe104, Tyr105 и Phe117, и являются консервативными на 100% в человеческом Гремлин-1 (нумерация основана на последовательности мышиного Гремлин-2). Степень гомологии распространяется на расположение боковых цепей, имеющих одинаковую конформацию в обоих белках (фиг. 5).
Нумерация аминокислот, используемая в файле Гремлин PDB, соответствует нумерации в опубликованной структуре мышиного Гремлин-2, полученной в результате структурного выравнивания. Это позволяет проводить сопоставимое сравнение аминокислот при описании структур. Однако для ясности, ключевые остатки, идентифицированные как участвующие в связывании BMP, приведены ниже с нумерацией на основе файла PDB и файла UniProt с SEQ ID NO:1 в скобках:
Trp72 (93), Phe96 (117), Tyr98 (119), Phe104 (125), Tyr105 (126) и Phe117 (138).
Как в мышином Гремлин-2, так и в человеческом Гремлин-1 гидрофобный BMP-связывающий эпитоп частично закрыт альфа-спиралью, образованной N-концевыми остатками каждого белка. Была предложена модель связывания BMP, при которой N-конец может изгибаться, полностью обнажая BMP-связывающую поверхность (Nolan et al (2013), Structure, 21, 1417-1429). На фиг. 4 N-концевые остатки были удалены из структур Гремлин-1 человека и мышиного Гремлин-2 перед визуализацией поверхности, чтобы выявить сходство BMP-связывающих поверхностей на каждом белке.
В литературе описан только мутагенез шести остатков, которые влияют на связывание BMP. Возможно, что фактический эпитоп BMP охватывает более обширную площадь поверхности, в которую попадают соседние аминокислоты. Отметив на поверхности Гремлин-1 все остатки, расположенные в пределах 6 от мутированных остатков, обнаруживается более обширная область Гремлин-1, на которую потенциально может быть направлено терапевтическое средство (фиг. 11). Этот более обширный регион охватывает следующие аминокислоты Гремлин-1 человека:
Asp92-Leu99
Arg116-His130
Ser137-Ser142
Cys176-Cys178
(нумерация дана на основе SEQ ID NO:1).
Суммируя опубликованную информацию и информацию о кристаллической структуре Гремлин-1 человека, были идентифицированы области Гремлин-1 человека, которые можно использовать для терапевтического вмешательства, блокирующего его взаимодействие с BMP.
Пример 3 - Анализ активности репортерного гена Id1 в клетках Hek
Общие сведения
В анализе активности репортерного гена Id1 в клетках Hek используются клетки клона 12 Hek293 с репортерным геном Id1. Эту клеточную линию стабильно трансфицировали транскрипционным фактором Id1. Id1 является транскрипционным фактором в сигнальном пути BMP. Гремлин, как известно, связывается с BMP, предотвращая связывание BMP с рецепторами, уменьшая сигнал люциферазы от репортерного гена. Следовательно, используя этот анализ активности репортерного гена, можно проводить скрининг антител к Гремлин и определять наличие антител, которые блокируют взаимодействие Гремлин с BMP. В случае блокирования этого взаимодействия в этих клетках наблюдается восстановление сигнала люциферазы.
Способ
Клетки клона 12 культивировали в среде DMEM, содержащей 10% FCS, 1x L-глутамин и 1x NEAA. Клетки также выращивают в присутствии гигромицина B (200 мкг/мл) для гарантии того, что клетки все еще способны экспрессировать ген Id1. Клетки анализировали в DMEM, содержащей 0,5% FCS, 1x L-глутамин и 1x NEAA. В течение короткого времени, необходимого для анализа клеток, гигромицин В не нужен.
Клетки промывали в PBS, извлекали с помощью буфера для диссоциации клеток, центрифугировали и подсчитывали перед посевом в количестве 5×104/лунку в 70 мкл (плотность 7,14×105/мл). Используемые чашки представляли собой 96-луночные стерильные, планшеты белого цвета, покрытые непрозрачным поли-D-лизином. Клетки помещали в инкубатор примерно на 3-4 часа, чтобы они осели. Гетеродимеры BMP восстанавливали до 200 мкг/мл в 4 мМ HCL. BMP разбавляли до 10 мкг/мл в среде для анализа, используя стеклянный флакон с получением нового маточного раствора.
В полипропиленовой чашке разводили Гремлин-1 1:2 для 8-точечной кривой доза-ответ с максимальной конечной дозой 1 мкг/мл.
Добавляли дополнительные 20 мкл среды на лунку, и планшеты инкубировали при 37°С в течение 45 минут.
Приготовленный 100х BMP добавляли во все лунки, кроме лунок, содержащих только клетки. Объем во всех лунках доводили до 60 мкл аналитической средой и инкубировали в течение еще 45 минут при 37°С.
После инкубации 30 мкл образца переносили в лунку аналитического планшета и инкубировали в течение 20-24 часов перед измерением люминесцентного сигнала.
Реагент Cell Steady Glo оттаивали заранее при комнатной температуре. Планшеты для анализа охлаждали до комнатной температуры в течение примерно 10-15 минут перед добавлением реагента. Сигнал люциферазы детектировали путем добавления реагента Cell Steady Glo (100 мкл) в течение 20 минут на шейкере при комнатной температуре и измерения люминесценции выполняли согласно протоколу Cell-Titre Glo на фотометре Synergy 2.
Максимальный сигнал генерировался в лунках, содержащих BMP, а минимальный сигнал генерировался в лунках, содержащих только клетки.
Результаты
Полноразмерные и укороченные формы Гремлин-1 тестировали в анализе активности репортерного гена Hek-Id1 для подтверждения активности, блокирующей BMP4/7. Результаты для полноразмерного белка показаны на фиг. 6А, а результаты для укороченного белка показаны на фиг. 6В.
Процент ингибирования, полученный в результате анализа доза-ответ, рассчитывали на основе максимальных и минимальных сигналов, полученных в анализе, и данных, полученных путем подгонки кривой к 4-параметрической логистической модели. IC50 вычисляли на основе точки перегиба кривой.
Таблица 3: Результаты эффективности для полноразмерного Гремлин-1 и укороченного Гремлин-1 в анализе активности репортерного гена Hek-Id1.
Заключение
Гремлин 1 способен ингибировать передачу сигналов BMP 4/7 в анализе активности репортерного гена Hek-Id1.
Пример 4 - Получение антител к Гремлин-1
Антитела к Гремлин-1 получали иммунизацией и пэннингом библиотеки. Библиотеку создавали в лаборатории в виде наивной человеческой библиотеки с V-областями, амплифицированными из крови доноров.
В результате первого раунда иммунизации получали 26 различных антител, связывающих Гремлин-1. Эти антитела масштабировали и очищали для скрининг-тестирования.
25 человеческих и мышиных перекрестно-реактивных антител из библиотеки подвергали пэннингу с использованием рекомбинантного человеческого Гремлин из R & D Systems. 10 антител отбирали для масштабирования и очищали в виде scFv для скрининг-тестирования.
Пример 5 - Скрининг анти-Гремлин-1 антителтела
Антитела подвергали скринингу, используя анализ активности репортерного гена Hek-Id1, описанного в примере 3, и путем измерения фосфорилирования SMAD. Фосфорилирование SMAD1, 5 и 8 происходит при передаче сигналов BMP. Следовательно, ингибиторы Гремлин-1 увеличивают уровень фосфорилирования SMAD.
Анализ фосфорилирования SMAD выполняли на клетках A549 или на фибробластах легких человека. Уровни фосфорилирования определяли, используя MSD.
Результаты
В анализе активности репортерного гена Hek-Id1 не было видимых эффектов, обеспечиваемых антителами, полученными в результате иммунизации (при 10-кратном избытке антител, протестированных по отношению к гетеродимеру BMP4/7). Результаты показаны на фиг. 7.
Напротив, несколько антител, полученных из библиотеки, способны восстанавливать сигнал в анализе активности репортерного гена Hek-Id1 (50-кратный избыток антител с 50%-ной дозой Гремлин) (фиг. 8). Из них Ab2416 и Ab2417 содержали высокие уровни эндотоксина. Ab7326 сохранял блокирующую способность при 10-кратном избытке и концентрации Гремлин-1, обеспечивающей 80% ингибирования.
Дополнительные результаты представлены на фиг. 9А (человеческий Гремлин) и 9В (мышиный Гремлин). На этих чертежах показаны титры Ab7326 (обозначено как PB376) вплоть до 15 нМ. Было показано, что Ab7326 восстанавливает передачу сигналов BMP, блокированного человеческим (IC50 1,3 нМ) или мышиным (IC50 0,2 нМ) Гремлин. Это антитело функционирует как человеческий и мышиный IgG1.
Последовательности полноразмерного человеческого и мышиного IgG1 представлены ниже. Для синтеза полноразмерных белков человеческого и мышиного IgG1, вариабельные области Ab7326, полученные из библиотеки, были повторно клонированы в векторы, содержащие соответствующие константные домены антител.
Поскольку Ab7326 был получен из наивной человеческой библиотеки, в которой Ab клонировали как scFvs, для повторного клонирования вариабельных областей 7326 в виде полноразмерных Ab или Fabs было необходимо провести ПЦР-амплификацию VH и VK с использованием пулов праймеров/вырожденных праймеров. Продукты амплифицированной ПЦР затем расщепляли и клонировали одновременно в мышиные и человеческие векторы. Поскольку VH и VK амплифицировали, используя пулы праймеров/вырожденных праймеров, были получены две вариантные формы продуктов, отличающиеся одним аминокислотным остатком из-за небольшого различия праймеров, использованных при отжиге во время процесса ПЦР.
Две вариантные формы вариабельной области тяжелой цепи отличались одной аминокислотой в положении 6, а две вариантные формы вариабельной области легкой цепи отличались одной аминокислотой в положении 7, как показано ниже:
Вариант 1 вариабельной области тяжелой цепи содержит глутаминовую кислоту (E) в положении 6.
Вариант 2 вариабельной области тяжелой цепи содержит глутамин (Q) в положении 6.
Вариант 1 вариабельной области легкой цепи содержит серин (S) в положении 7.
Вариант 2 вариабельной области легкой цепи содержит треонин (T) в положении 7.
Мышиный полноразмерный IgG1 - вариант 1 тяжелой цепи (SEQ ID NO:14)
QVQLVESGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA RGSGSYYPNH FDYWGQGTLV TVSSAKTTPP SVYPLAPGSA AQTNSMVTLG CLVKGYFPEP VTVTWNSGSL SSGVHTFPAV LQSDLYTLSS SVTVPSSTWP SETVTCNVAH PASSTKVDKK IVPRDCGCKP CICTVPEVSS VFIFPPKPKD VLTITLTPKV TCVVVDISKD DPEVQFSWFV DDVEVHTAQT QPREEQFNST FRSVSELPIM HQDWLNGKEF KCRVNSAAFP APIEKTISKT KGRPKAPQVY TIPPPKEQMA KDKVSLTCMI TDFFPEDITV EWQWNGQPAE NYKNTQPIMD TDGSYFVYSK LNVQKSNWEA GNTFTCSVLH EGLHNHHTEK SLSHSPGK
Мышиный полноразмерный IgG1 - вариант 1 легкой цепи (SEQ ID NO:15)
DIVMTQSPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT PTFGQGTRLE IKRTDAAPTV SIFPPSSEQL TSGGASVVCF LNNFYPKDIN VKWKIDGSER QNGVLNSWTD QDSKDSTYSM SSTLTLTKDE YERHNSYTCE ATHKTSTSPI VKSFNRNEC
Мышиный полноразмерный IgG1 - вариант 2 тяжелой цепи (SEQ ID NO:28)
QVQLVQSGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA RGSGSYYPNH FDYWGQGTLV TVSSAKTTPP SVYPLAPGSA AQTNSMVTLG CLVKGYFPEP VTVTWNSGSL SSGVHTFPAV LQSDLYTLSS SVTVPSSTWP SETVTCNVAH PASSTKVDKK IVPRDCGCKP CICTVPEVSS VFIFPPKPKD VLTITLTPKV TCVVVDISKD DPEVQFSWFV DDVEVHTAQT QPREEQFNST FRSVSELPIM HQDWLNGKEF KCRVNSAAFP APIEKTISKT KGRPKAPQVY TIPPPKEQMA KDKVSLTCMI TDFFPEDITV EWQWNGQPAE NYKNTQPIMD TDGSYFVYSK LNVQKSNWEA GNTFTCSVLH EGLHNHHTEK SLSHSPGK
Мышиный полноразмерный IgG1 - вариант 2 легкой цепи (SEQ ID NO:29)
DIVMTQTPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT PTFGQGTRLE IKRTDAAPTV SIFPPSSEQL TSGGASVVCF LNNFYPKDIN VKWKIDGSER QNGVLNSWTD QDSKDSTYSM SSTLTLTKDE YERHNSYTCE ATHKTSTSPI VKSFNRNEC
Человеческий полноразмерный IgG1 - вариант 1 тяжелой цепи (SEQ ID NO:30)
QVQLVESGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL
VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA
RGSGSYYPNH FDYWGQGTLV TVSSASTKGP SVFPLAPSSK STSGGTAALG
CLVKDYFPEP VTVSWNSGAL TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL
GTQTYICNVN HKPSNTKVDK KVEPKSCDKT HTCPPCPAPE LLGGPSVFLF
PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE
EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP
REPQVYTLPP SRDELTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL
SPGK
Человеческий полноразмерный IgG1 - вариант 1 легкой цепи (SEQ ID NO:31)
DIVMTQSPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP
KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT
PTFGQGTRLE IKRTVAAPSV FIFPPSDEQL KSGTASVVCL LNNFYPREAK
VQWKVDNALQ SGNSQESVTE QDSKDSTYSL SSTLTLSKAD YEKHKVYACE
VTHQGLSSPV TKSFNRGEC
Человеческий полноразмерный IgG1 - вариант 2 тяжелой цепи (SEQ ID NO:16)
QVQLVQSGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL
VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA
RGSGSYYPNH FDYWGQGTLV TVSSASTKGP SVFPLAPSSK STSGGTAALG
CLVKDYFPEP VTVSWNSGAL TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL
GTQTYICNVN HKPSNTKVDK KVEPKSCDKT HTCPPCPAPE LLGGPSVFLF
PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE
EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP
REPQVYTLPP SRDELTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL
SPGK
Человеческий полноразмерный IgG1 - вариант 2 легкой цепи (SEQ ID NO:17)
DIVMTQTPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP
KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT
PTFGQGTRLE IKRTVAAPSV FIFPPSDEQL KSGTASVVCL LNNFYPREAK
VQWKVDNALQ SGNSQESVTE QDSKDSTYSL SSTLTLSKAD YEKHKVYACE
VTHQGLSSPV TKSFNRGEC
CDR антител определяли методом Kabat (выделено жирным шрифтом в приведенных выше последовательностях). Дополнительные остатки HCDR1, определенные по Chothia, выделены курсивом. Последовательности константных областей также подчеркнуты.
Восстановление передачи сигналов p-SMAD с помощью антител к Гремлин 1 показано в приведенной ниже таблице 4.
Таблица 4: Восстановление передачи сигналов p-SMAD
50 нг/мл
+/-2,8%
+/-1,9%
+/-1,4%
+/-0,6%
+/-0,8%
+/-2,1%
+/-2,7%
+/-3,5%
+/-1,4%
25 нг/мл
+/-3,0%
+/-3,1%
+/-1,2%
+/-2,2%
+/-1,3%
+/-2,1%
+/-3,3%
+/-3,8%
+/-2,5%
200
нг/мл
+/-3,8%
+/-3,2%
+/-3,4%
+/-1,3%
+/-1,1%
+/-1,2%
+/-4,3%
+/-3,2%
+/-2,5%
50 нг/мл
+/-2,6%
+/-3,6%
+/-2,7%
+/-3,1%
+/-1,5%
+/-2,5%
+/-4,4%
+/-2,9%
+/-2,5%
50 нг/мл
+/-3,1%
+/-4,0%
+/-4,0%
+/-2,1%
+/-1,7%
+/-1,7%
+/-2,5%
+/-3,1%
+/-2,1%
Результаты представлены в виде процента от уровня фосфорилирования SMAD одним BMP (контрольный BMP). Эксперименты выполняли в фибробластах легких, полученных от пациентов с идиопатическим легочным фиброзом. Антитела к rhГремлин-1 и Гремлин-1 предварительно инкубировали в течение 45 минут при комнатной температуре. Затем антитела к rhГремлин-1 и Гремлин-1 добавляли к клеткам с BMP в течение 30 минут.
В таблице 5 показаны дополнительные результаты, полученные в анализе фосфорилирования SMAD, в котором изучали замещение BMP-2 или BMP4/7 из комплексов Гремлин 1-BMP антителами к Гремлин-1. Эксперименты снова выполняли в фибробластах легких, полученных от пациентов с идиопатическим легочным фиброзом. rhBMP-2 или rhBMP 4/7 предварительно инкубировали с rhГремлин-1 в течение 1 часа при комнатной температуре. Комплексы BMP-2- или BMP4/7-Гремлин-1 инкубировали с различными концентрациями антител к Гремлин-1 в течение ночи при 4°С. Концентрации антител представляют собой конечную концентрацию в планшете.
Таблица 5: Замещение BMP-2 или BMP4/7 из комплексов Гремлин 1-BMP антителами к Гремлин-1
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
50 нг/мл
+/-3,5%
+/-2,7%
+/-2,9%
+/-2,6%
+/-2.0%
+/-1,9%
+/-2,8%
+/-0,3%
50 нг/мл
+/-4,2%
+/-1.0%
+/-1,4%
+/-4,9%
+/-4,4%
+/-2,2%
+/-0,6%
+/-1,5%
нг/мл
+/-1,1%
+/-2,4%
+/-3,8%
+/-2,4%
+/-2,2%
+/-4,3%
+/-2,8%
+/-2,9%
50
нг/мл
+/-0,8%
+/-1,8%
+/-4,2%
+/-10.0%
+/-4,2%
+/-3,3%
+/-2,3%
+/-4,5%
Результаты, представленные в таблице 5, показывают, что Ab7326 может вытеснять уже находящийся в комплексе BMP-2 или BMP4/7 из комплексов Гремлин 1-BMP. Ab7326 осуществляет это замещение при гораздо более низких концентрациях, чем антитело сравнения 2484. Это свидетельствует о том, что Ab7326 является аллостерическим ингибитором, что согласуется с открытием авторов изобретения, согласно которому Ab7326-связывающий сайт на Гремлин-1 удален от известных BMP-связывающих областей. Таким образом, Ab7326 может получать доступ к аллостерическому связывающему сайту, даже когда BMP образует комплекс с Гремлин-1, что приводит к значительному улучшению ингибирования активности Гремлин.
Пример 6 - Получение кристаллической структуры Гремлин-1 в комплексе с Fab 7326
Кристаллическую структуру Гремлин-1 человека в комплексе с Fab Ab 7326 получали с разрешением 2,1 . Последовательности Fab показаны ниже:
Тяжелая цепь: SEQ ID NO:18
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
Легкая цепь: SEQ ID NO:19
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Затем с помощью программного обеспечения CCP4 NCONT идентифицировали все контакты в пределах 4 между Гремлин-1 и Fab. Были идентифицированы следующие остатки: Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175 (нумерация основана на последовательности SEQ ID NO:1 в базе данных UniProt (пронумерованных как Ile110, Lys126, Lys127, Phe128, Phe128, Arg148, Lys153 и Gln154 в структурном файле, который соответствует нумерации мышиного Гремлин-2)).
На фиг. 10 показаны структурные модели комплекса Гремлин-Fab, причем остатки эпитопа Fab показаны относительно BMP-связывающих областей.
Ab 7326 представляет собой ингибирующее антитело, которое действует аллостерически, т.е. связывается вне BMP-связывающих областей.
Пример 7. Измерение сродства связывания антитела к Гремлин-1 Ab7326 с Гремлин-1.
Способ
Сродство анти-Гремлин mIgG к человеческому Гремлин 1 определяли с помощью анализа биамолекулярного взаимодействия методом поверхностного плазмонного резонанса (SPR) на системе Biacore T200, GE Healthcare Bio-Sciences AB. Анти-Гремлин mIgG связывали, используя поверхность с иммобилизованным на ней анти-мышиным Fc, и Гремлин 1 титровали относительно связанного mIgG. Выполняли иммобилизацию захватывающего лиганда (Affinipure F(ab')2 фрагмент козьего антимышиного IgG, специфический в отношении Fc-фрагмента, 115-006-071, Jackson ImmunoResearch Inc.) при концентрации 50 мкг/мл в 10 мМ NaAc, pH 5,0, на проточную ячейку 2 с сенсорным чипом CM4 с помощью реакции присоединения амина, используя 600-секундные инъекции активации и деактивации, до уровня ~1600 единиц ответа (RU). В качестве рабочего буфера использовали буфер HBS-EP+ (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 3 мМ EDTA, 0,05% поверхностно-активное вещество P20) со скоростью потока 10 мкл/мин. Эталонную поверхность готовили на проточной ячейке 1 путем активации и деактивации поверхности, как для проточной ячейки 2, но без захватывающего лиганда.
Буфер для анализа представлял собой HBS-EP+ плюс 150 мМ NaCl с получением конечной концентрации NaCl 300 мМ плюс 1% CMD40. 60-секундную инъекцию анти-Гремлин mIgG (при 5 мкг/мл в рабочем буфере) пропускали через проточные ячейки 1 и 2, чтобы получить уровень захвата приблизительно 100 RU на поверхности Fc с иммобилизованным анти-мышиным IgG. Рекомбинантный человеческий Гремлин 1 титровали в 5 нМ рабочем буфере (с использованием 2-кратных разведений) и инъецировали через проточные ячейки 1 и 2 со скоростью потока 30 мкл/мин в течение 3 минут с последующей 5-минутной фазой диссоциации. Также использовали контроль, представляющий собой буфер. Поверхность восстанавливали со скоростью потока 10 л/мин с помощью 60-секундной инъекции 50 мМ HCl, 30-секундной инъекции 5 мМ NaOH и 30-секундной инъекции 50 мМ HCl.
Кинетические характеристики определяли с помощью программного обеспечения Biacore T200.
Измерение сродства связывания выполняли при 25°C.
Результаты
Было установлено, что сродство связывания, выраженное в виде среднего значения KD для 5 определений, находится ниже 100 пМ.
Пример 8. Оценка влияния in vivo антител к Гремлин-1 на хроническую гипоксию/SU5416-индуцированную легочную артериальную гипертензию у мышей.
Краткий обзор
Дисбаланс в суперсемействе TGFβ является одним из важных факторов, связанных с развитием ряда патологий легких, включая легочную артериальную гипертензию (ЛАГ) (Budd & Holmes Pharm Ther 2012). Гремлин-1 участвует в развитии и прогрессировании ЛАГ (Thomas et al AJP 2009; Ciuclan et al AJP 2013). Недавние исследования показали, что антитело к Гремлин-1 может ингибировать развитие легочной артериальной гипертензии в преклинической модели ЛАГ, индуцированной гипоксией/SU5416 (Ciuclan et al AJP 2013). В настоящем примере выполняли оценку влияния антител к Гремлин1 на гемодинамику и ремоделирование сосудов в преклинической мышиной модели ЛАГ, индуцированной гипоксией/SU5416.
Гипоксия/SU5416 привела к значительному увеличению систолического давления в правом желудочке (СДПЖ) и правожелудочковой гипертрофии (фиг. 12 и 15). Введение антитела к Гремлин-1 привело к значимому (P<0,005) снижению СДПЖ по сравнению с контрольными группами IgG1 и PBS. У животных в нормоксии не наблюдали какого-либо значимого влияния антитела к Гремлин-1 на СДПЖ (фиг. 12). Также не наблюдали влияния на системное давление (среднее артериальное давление или САД; фиг. 13) или на правожелудочковую гипертрофию (фиг. 14). В совокупности это исследование подтверждает роль Гремлин-1 в развитии ремоделирования легочных сосудов и повышении СДПЖ в модели ЛАГ, индуцированной хронической гипоксией/SU5416, и применение антител к Гремлин-1 для ее лечения.
Общие сведения
Легочная гипертензия (ЛГ) - это гемодинамическое состояние, при котором давление, измеренное в легочной артерии, является повышенным. Клинически это определяется средним легочным артериальным давлением в покое (сЛАД) >25 мм рт. ст., легочным сосудистым сопротивлением (ЛСС) ≥ 3 единиц Вуда (Wood) и легочным давлением клина ≤ 15 мм рт. ст. (Badesch et al 2009). Патологические характеристики ЛГ являются многофакторными и включают легочное артериальное давление, ремоделирование сосудов мелких и средних артерий, правожелудочковую гипертрофию и, в конечном итоге, правожелудочковую сердечную недостаточность (Faber & Loscalzo 2004). ВОЗ классифицирует ЛГ по пяти основным категориям, включая группу I. Группа I представляет легочную артериальную гипертензию, которая включает идиопатическую ЛАГ и наследственную ЛАГ, а также ассоциированную ЛАГ, которая возникает в сочетании с другими осложнениями, такими как системный склероз (Simonneau et al 2009). Ряд предрасполагающих мутаций связан с развитием ЛАГ у пациентов с наследственной и идиопатической ЛАГ, наиболее преобладающими из которых являются мутации в рецепторе морфогенетического белка кости, BMPR2 (Budd & Holmes Pharm Ther 2012). Кроме того, у пациентов с развивающейся ЛАГ также были зарегистрированы мутации в BMP-активированных компонентах нисходящих сигнальных путей Smad (Budd & Holmes Pharm Ther 2012). В недавно проведенных исследованиях у пациентов с ЛАГ были выявлены повышенные уровни ингибитора BMP, Гремлин (Гремлин-1 и 2) (Cahill et al Circ 2012). В соответствии с функциональной ролью Гремлин-1 в развитии ЛАГ, гапло-дефицит Гремлин-1 усиливал BMP-сигнализацию в гипоксическом легком мыши и приводил к снижению легочного сосудистого сопротивления путем ослабления ремоделирования сосудов (Cahill et al Circ 2012). Эти наблюдения также подтвердили Ciuclan и коллеги (AJP 2013), которые продемонстрировали, что антитело к Гремлин-1 ослабляет легочную артериальную гипертензию у мышей, индуцированную хронической гипоксией/SU5416. Недавние исследования показали, что негенетические механизмы могут способствовать снижению экспрессии BMPR2 у пациентов с системным склерозом и развитию ЛАГ (Gilbane et al AJRCCM). В совокупности, дисбаланс в оси суперсемейства BMP может привести к развитию легочных патологий, таких как ЛАГ.
Целью настоящего исследования была оценка in vivo влияния антитела к Гремлин-1 на развитие ЛАГ в мышиной модели хронической гипоксии/SU5416.
Материалы и методы
Реагенты
Иматиниб: Science Warehouse 1625-1000.
SU5416: R & D Systems 3037.
αSMA антитело: Dako M085129-2.
VWF антитело: Dako A008202-2.
Биотинилированное козье анти-кроличье IgG антитело: Vector Labs BA-1000.
Биотинилированное лошадиное антимышиное IgG антитело, адсорбированное крысное: Vector Labs BA-2001.
Карбоксиметилцеллюлоза: Sigma C5678 419273.
Tween 80: Sigma P1754.
Бензиловый спирт: Sigma 305197; 402834.
Хлорид натрия: Sigma S7653; VWR 10241.
Набор VECTASTAIN ABC-AP: Vector Labs AK-5000.
Набор VECTASTAIN Elite ABC: Vector Labs PK-6100.
Нормальная лошадиная сыворотка: Vector Labs, регистрационный номер S-2000.
Полисорбат: Sigma 59924.
Экспериментальные протоколы
Животные
Мышей C57Bl/6 содержали в специальном помещении, свободном от патогенов, со свободным доступом к пище и воде с 12-часовым циклом свет/темнота. Настоящее исследование на животных лицензировано в соответствии с законом Великобритании о домашних животных (научные процедуры) 1986 года.
Модель легочной артериальной гипертензии у мышей с гипоксией/SU5416
Самок мышей C57Bl/6 в возрасте 8-10 недель распределяли по группам (таблица 6). Животных во всех группах взвешивали, и им вводили подкожно (s.c.) 20 мг/кг SU5416 в 100 мкл носителя (0,5% карбоксиметилцеллюлозы (CMC); 0,9% хлорида натрия (NaCl); 0,4% полисорбата 80; 0,9% бензилового спирта в деионизированной воде), как описано у Ciuclan и др. AJRCCM 2013. При необходимости, вводили вторую s.c. инъекцию, как указано в таблице 6, содержащую: PBS, 30 мг/кг IgG1 или 30 мг/кг антитела к Гремлин-1. При этом, мыши из группы Гипоксия+Иматиниб получали пищу с инъецированным в нее иметинибом для получения 100 мг/кг/день. Затем группы мышей с гипоксией помещали в камеру, в условия нормобарической гипоксии (10% O2), тогда как мышей в группах с нормоксией содержали в нормоксических (21% O2) условиях в той же комнате, в которой находилась камера.
Таблица 6: Группы обработки:
Мышей C57Bl/6 распределяли на группы обработки, указанные в таблице.
В дни 7 и 14 всех мышей взвешивали и давали еще одну s.c. дозу 20 мг/кг SU5416. При необходимости, мышам вводили еще одну s.c. инъекцию PBS, 30 мг/кг IgG1 или 30 мг/кг антитела к Гремлин-1 (как указано в таблице 6). Антитело к Гремлин-1, используемое в этих исследованиях, представляло собой Ab7326, полноразмерный мышиный IgG1, вариант 1, как описано в примере 5. В день 21 измеряли систолическое давления в правом желудочке (СДПЖ) и среднее артериальное давление (САД), и собирали ткани.
Правожелудочковое систолическое давление и сбор тканей
Измерения гемодинамики СДПЖ и САД проводили на животных после трех недельного воздействия гипоксии и соответствующей лекарственной терапии, как указано в таблице 6. Животных анестезировали 1,5% изофлуораном и клали на спину на греющее одеяло, температуру которого поддерживали при 37°С. Во-первых, изолировали правую яремную вену и вводили катетер для измерения давления (катетер для измерения давления Millar mouse SPR-671NR диаметром 1,4F, Millar Instruments, Великобритания) и продвигали в правый желудочек для определения СДПЖ. Во-вторых, САД измеряли путем изоляции левой общей сонной артерии и введения катетера для измерения давления. Как СДПЖ, так и САД регистрировали в предварительно откалиброванной системе PowerLab (ADInstruments, Австралия).
Животных умерщвляли с помощью передозировки изофлуоранового анестетика, и собирали цельную кровь. Цельную кровь центрифугировали (220 × g; 2 мин), а сыворотку извлекали и хранили при -80°C. Сердце извлекали, и записывали вес правого и левого желудочка. Легкие перфузировали 2,5 мл физиологического раствора через правый желудочек. Левое легкое фиксировали путем заполнения 10% формалином перед заливкой парафином и изготовления срезов. Правое легкое подвергали быстрой заморозке в жидком азоте и хранили при -80°С.
Гистология
Предметные стекла депарафинизировали и повторно гидратировали, используя ксилол и градиент концентрации этанола. Предметные стекла погружали в 0,3% H2O2 в метаноле на 30 минут для извлечения антигенов, 3 раза промывали в PBS и блокировали в течение 1 часа в 1:30 смеси нормальной лошадиной сыворотки и PBS. Первичное анти-αSMA антитело добавляли на каждое предметное стекло в соотношениитрации 1:100 и инкубировали при 4°С в течение ночи, затем промывали в PBS в течение 5 минут три раза. Биотинилированное лошадиное антимышиное IgG антитело, адсорбированное крысиное вторичное антитело, разводили в соотношении 1:200 и пипетировали на каждое предметное стекло, затем инкубировали в течение 45 минут; после чего промывали в PBS 3 раза в течение 5 минут. Согласно инструкции производителя, щелочную фосфотазу с комплексом биотина и авидина (ABC-AP) готовили за 30 минут и помещали на каждое предметное стекло на 30 минут; затем промывали 3 раза в течение 5 минут. Субстрат АР готовили в соответствии с прилагаемыми к набору инструкциями и пипетировали на каждое предметное стекло, и давали окраске проявиться; затем промывали 3 раза в течение 5 минут. На каждое предметное стекло добавляли первичное анти-vWF антитело в соотношении 1:100 и инкубировали при 4°С в течение ночи, затем промывали в PBS в течение 5 минут три раза. Биотинилированное козье антикроличье IgG вторичное антитело разводили в соотношении 1:200 и добавляли на каждое предметное стекло, оставляя там на 45 минут; затем промывали в PBS 3 раза в течение 5 минут. Согласно предоставленным в наборе инструкциям, готовили ABC за 30 минут и наносили на каждое предметное стекло на 30 минут; затем промывали 3 раза в течение 5 минут. Субстрат DAB готовили в соответствии с предоставленными в наборе инструкциями и пипетировали на каждое предметное стекло, и давали окраске проявиться в течение ~5-10 минут; затем промывали 3 раза в течение 5 минут. Предметные стекла подвергали контрастной окраске гематоксилином в течение ~40 секунд, обезвоживали и накрывали покровным стеклом, используя Pertex. Все предметные стекла сканировали в цифровом виде с помощью сканера для предметных стекол Hamamatsu NanoZoomer 2.0-HT (Hamamatsu, Welwyn Garden City, Великобритания).
Данные и статистический анализ
Более 40 сосудов, взятых случайным образом из каждой группы с гипоксией, оценивали по степени мускуляризации. Сосуды оценивали как минимум четырьмя независимыми наблюдателями: 0=немускуляризованный; 1=частично мускуляризованный; 2=полностью мускуляризованный; и определяли модальное значение для каждого сосуда. Определяли процент сосудов, полностью, частично или немускуляризованных, и вычисляли среднее значение ± SEM. Для определения значимости *P<0,05; **P<0,01; ***Р<0,005; ****P<0,001 выполняли однофакторный ANOVA.
Результаты
Введение SU5416 (20 мг/кг) после воздействия хронической нормобарической гипоксии (10% О2) привело к значимому (P<0,01) увеличению СДПЖ по сравнению с воздействием нормоксии/SU5416 в течение 21 дня как в группе, получавшей только PBS, так и в контрольной группе IgG1 (фиг. 12). Между группами, получавшими IgG1 и PBS, не было выявлено никаких существенных различий. Влияние медикаментозного лечения на повышенный СДПЖ у мышей, которым вводили SU5416, сохранялось при гипоксии: иматиниб приводил к значимому (P<0,001) снижению СДПЖ по сравнению с контрольными группами. Антитело к Гремлин-1 приводило к значимому (P<0,005) снижению СДПЖ по сравнению с контрольными группами IgG1 и PBS. У животных, содержавшихся в условиях нормоксии не наблюдалось никакого значимого эффекта на СДПЖ, оказываемого антителом к Гремлин-1 (фиг. 12).
Влияние антитела к Гремлин-1 (n=4) или контрольного носителя IgG1 (n=4) на СДПЖ оценивали в условиях гипоксии и нормоксии (фиг. 13). Никаких существенных лечебных эффектов на СДПЖ не наблюдалось.
Определяли правожелудочковую гипертрофию (масса правый желудочек/левый желудочек+перегородка) (фиг. 14). Введение SU5416 (20 мг/кг) после воздействия хронической нормобарической гипоксии (10% О2) привело к значимому (P<0,01) увеличению правожелудочковой гипертрофии (RV/LV+S) по сравнению с нормоксией/SU5416 через 21 день как в группе, получавшей только PBS, так и в группе контрольного IgG1 (фиг. 14). Значимого влияния лекарственных веществ (иматиниба или антитела к Гремлин 1) на повышенный СДПЖ у мышей, содержавшихся в условиях гипоксии, которым вводили SU5416, не наблюдалось.
Степень ремоделирования сосудов оценивали путем окрашивания парафиновых срезов легких. Кровеносные сосуды окрашивали на фактор Виллебранда (vWF) для выявления эндотелиальных клеток и актина гладких мышц (αSMA) для оценки степени мускуляризации. Изображения оцифровывали с помощью слайд-сканера Hamamatsu NanoZoomer 2.0-HT, и по меньшей мере 40 сосудов, взятых случайным образом из IgG1 группы, содержащейся в условиях нормоксии, и каждой группы, содержащейся в условиях с гипоксии, оценивали на степень мускуляризации. Сосуды оценивали как минимум четырьмя независимыми наблюдателями: 0=немускуляризованный; 1=частично мускуляризованный; 2=полностью мускуляризованный; и на график наносили модальное значение для каждого определенного сосуда и процент немускуляризованных, частично и полностью мускуляризованных сосудов (график 15). Введение SU5416 (20 мг/кг) после воздействия хронической нормобарической гипоксии (10% О2) привело к значимому увеличению полностью мускуляризованных (P<0,001) и частично мускуляризованных сосудов (P<0,01) со значимым совокупным уменьшением немускуляризованных сосудов (Р<0,001). Влияние медикаментозного лечения на повышенный СДПЖ у мышей, которым вводили SU5416, сохранялось при гипоксии: иматиниб приводил к значимому (P<0,001) уменьшению полностью мускулизированных (P<0,01) сосудов. Кроме того, антитело к Гремлин 1 приводило к значимому (P<0,001 и P<0,05, соответственно) по сравнению с IgG1 контрольный группой гипоксия/SU5416 (фиг. 15). Значимое влияние медикаментозного лечения (иматиниб или антител к Гремлин-1) на процент частично мускуляризованных сосудов по сравнению с IgG1 контрольной группой гипоксия/SU5416, не наблюдалось. Медикаментозное лечение (иматиниб или антитело к Гремлин-1) привело к увеличению процента немускуляризованных сосудов по сравнению с IgG1 контрольной группой гипоксия/SU5416, получавшей, однако это увеличение не было значимым.
Обсуждение
В этом примере оценивали влияние антитела к Гремлин-1 на развитие ЛАГ на модели хронической гипоксии/SU5416. Антитело к Гремлин-1 приводило к достоверному ингибированию СДПЖ (фиг. 12) и ремоделированию сосудов (фиг. 15), при этом не оказывало влияния на системное давление (САД) (фиг. 13) в мышиной модели ЛАГ, индуцированной гипоксией/SU5416. Значимого влияния антитела к Гремлин-1 на СДПЖ у мышей, содержавшихся в условиях нормоксии, получавших SU5416, не наблюдалось. Эффект антитела к Гремлин-1 согласуется с наблюдениями, описанными Ciuclan и коллегами (AJP 2013), в которых показано, что антагонизм с антителом к Гремлин-1 снижает повышенный СДПЖ и ремоделирование сосудов в мышиной модели ЛАГ, индуцированной хронической гипоксией/SU5416 (фиг. 12 и 15). В отличие от Ciuclan и его коллег в этом исследовании не наблюдалось значимого влияния антитела к Гремлин1 на правожелудочковую гипертрофию (фиг. 14). Однако в этом исследовании не наблюдалось значительное влияние контрольного препарата иматиниба на развитие правожелудочковой гипертрофии у мышей с гипоксией/SU5416. В совокупности это исследование подтверждает роль Гремлин-1 в развитии ремоделирования легочных сосудов и повышенного СДПЖ в модели ЛАГ, индуцированной хронической гипоксией/SU5416, и применение антител к Гремлин-1 при ее лечении.
Литература:
Badesch DB, Champion HC, Sanchez MA, Hoeper MM, Loyd JE, Manes A, McGoon M, Naeije R, Olschewski H, Oudiz RJ, Torbicki A. Diagnosis and assessment of pulmonary arterial hypertension. J Am Coll Cardiol. 2009 Jun 30; 54(1 Suppl):S55-66. doi: 10.1016/j.jacc.2009.04.011. Review. PMID:19555859.
Budd DC, Holmes AM. Targeting TGFβ superfamily ligand accessory proteins as novel therapeutics for chronic lung disorders. Pharmacol Ther. 2012 Sep; 135(3):279-91. doi: 10.1016/j.pharmthera.2012.06.001. Epub 2012 Jun 18.
Cahill E, Costello CM, Rowan SC, Harkin S, Howell K, Leonard MO, Southwood M, Cummins EP, Fitzpatrick SF, Taylor CT, Morrell NW, Martin F, McLoughlin P. Гремлин plays a key role in the pathogenesis of pulmonary hypertension. Circulation. 2012 Feb 21; 125(7):920-30. doi: 10.1161/CIRCULATIONAHA.111.038125. PMID:22247494.
Ciuclan L, Sheppard K, Dong L, Sutton D, Duggan N, Hussey M, Simmons J, Morrell NW, Jarai G, Edwards M, Dubois G, Thomas M, Van Heeke G, England K. Treatment with anti-Гремлин 1 antibody ameliorates chronic hypoxia/SU5416-induced pulmonary arterial hypertension in mice. Am J Pathol. 2013 Nov; 183(5):1461-73. doi: 10.1016/j.ajpath.2013.07.017. PMID:24160323
Ciuclan L, Hussey MJ, Burton V, Good R, Duggan N, Beach S, Jones P, Fox R, Clay I, Bonneau O, Konstantinova I, Pearce A, Rowlands DJ, Jarai G, Westwick J, MacLean MR, Thomas M. Imatinib attenuates hypoxia-induced pulmonary arterial hypertension pathology via reduction in 5-hydroxytryptamine through inhibition of tryptophan hydroxylase 1 expression. Am J Respir Crit Care Med. 2013 Jan 1; 187(1):78-89. doi: 10.1164/rccm.201206-1028OC. PMID:23087024
Farber HW, Loscalzo J. Pulmonary arterial hypertension. N Engl J Med. 2004 Oct 14; 351(16):1655-65. Review. PMID:15483284.
Gilbane AJ, Derrett-Smith E, Trinder SL, Good RB, Pearce A, Denton CP, Holmes AM. Impaired bone morphogenetic protein receptor II signaling in a transforming growth factor-β-dependent mouse model of pulmonary hypertension and in systemic sclerosis. Am J Respir Crit Care Med. 2015 Mar 15;191(6):665-77. doi: 10.1164/rccm.201408-1464OC.
Simonneau G, Robbins IM, Beghetti M, Channick RN, Delcroix M, Denton CP, Elliott CG, Gaine SP, Gladwin MT, Jing ZC, Krowka MJ, Langleben D, Nakanishi N, Souza R. Updated clinical classification of pulmonary hypertension. J Am Coll Cardiol. 2009 Jun 30; 54(1 Suppl):S43-54. doi: 10.1016/j.jacc.2009.04.012. Review. PMID:19555858.
Thomas M, Docx C, Holmes AM, Beach S, Duggan N, England K, Leblanc C, Lebret C, Schindler F, Raza F, Walker C, Crosby A, Davies RJ, Morrell NW, Budd DC. Activin-like kinase 5 (ALK5) mediates abnormal proliferation of vascular smooth muscle cells from patients with familial pulmonary arterial hypertension and is involved in the progression of experimental pulmonary arterial hypertension induced by monocrotaline. Am J Pathol. 2009 Feb; 174(2):380-9. doi: 10.2353/ajpath.2009.080565. Epub 2008 Dec 30.
Списки последовательностей
SEQ ID NO:1 (Гремлин-1 человека; Uniprot ID: O60565)
MSRTAYTVGALLLLLGTLLPAAEGKKKGSQGAIPPPDKAQHNDSEQTQSPQQPGSRNRGRGQGRGTAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO:2 (человеческий укороченный Гремлин-1, использованный в кристаллографии, включая N-концевую метку)
MGSSHHHHHHSSGENLYFQGSAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO:3 (Ab 7326 HCDR1 комбинированный по Chothia & Kabat)
GYTFTDYYMH
SEQ ID NO:4 (Ab 7326 HCDR1 Kabat)
DYYMH
SEQ ID NO:5 (Ab 7326 HCDR2 Kabat)
LVDPEDGETIYAEKFQG
SEQ ID NO:6 (Ab 7326 HCDR3 Kabat)
DARGSGSYYPNHFDY
SEQ ID NO:7 (Ab 7326 LCDR1 Kabat)
KSSQSVLYSSNNKNYLA
SEQ ID NO:8 (Ab 7326 LCDR2 Kabat)
WASTRES
SEQ ID NO:9 (Ab 7326 LCDR3 Kabat)
QQYYDTPT
SEQ ID NO:10 (вариант 1 вариабельной области тяжелой цепи Ab 7326)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSS
SEQ ID NO:11 (вариант 1 вариабельной область легкой цепи Ab 7326 1)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIK
SEQ ID NO:12 (вариант 2 вариабельной области тяжелой цепи Ab 7326)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSS
SEQ ID NO:13 (вариант 2 вариабельной область легкой цепи Ab 7326)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIK
SEQ ID NO:14 (вариант 1 тяжелой цепи мышиного полноразмерного IgG1)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSAKTTPPSVYPLAPGSAAQTNSMVTLGCLVKGYFPEPVTVTWNSGSLSSGVHTFPAVLQSDLYTLSSSVTVPSSTWPSETVTCNVAHPASSTKVDKKIVPRDCGCKPCICTVPEVSSVFIFPPKPKDVLTITLTPKVTCVVVDISKDDPEVQFSWFVDDVEVHTAQTQPREEQFNSTFRSVSELPIMHQDWLNGKEFKCRVNSAAFPAPIEKTISKTKGRPKAPQVYTIPPPKEQMAKDKVSLTCMITDFFPEDITVEWQWNGQPAENYKNTQPIMDTDGSYFVYSKLNVQKSNWEAGNTFTCSVLHEGLHNHHTEKSLSHSPGK
SEQ ID NO:15 (вариант 1 легкой цепи мышиного полноразмерного IgG1)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTDAAPTVSIFPPSSEQLTSGGASVVCFLNNFYPKDINVKWKIDGSERQNGVLNSWTDQDSKDSTYSMSSTLTLTKDEYERHNSYTCEATHKTSTSPIVKSFNRNEC
SEQ ID NO:16 (вариант 2 тяжелой цепи человеческого полноразмерного IgG1)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO:17 (вариант 2 легкой цепи человеческого полноразмерного IgG1)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:18 (вариант 1 тяжелой цепи Fab)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
SEQ ID NO:19 (вариант 1 легкой цепи Fab)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:20 (укороченный Гремлин-1 человека, использованный в кристаллографии, без N-концевой метки)
AMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO:21 (последовательность SEQ ID NO:1 зрелого Гремлин-1 без сигнального пептида аминокислот 1-21)
KKKGSQGAIPPPDKAQHNDSEQTQSPQQPGSRNRGRGQGRGTAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO:22 (вариант 1 тяжелой цепи человеческого IgG4P)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
SEQ ID NO:23 (вариант 1 легкой цепи человеческого IgG4P)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:24 (вариант 1 ДНК тяжелой цепи человеческого IgG1)
caagtgcaactggtggaatccggggccgaagtgaaaaagcccggagccactgtgaagatctcttgcaaagtgtccggctacaccttcaccgactattacatgcactgggtccagcaggcacctgggaagggccttgagtggatgggtctggtcgatcccgaggacggcgaaactatctacgccgagaagttccagggtcgcgtcaccatcaccgccgacacttccaccgacaccgcgtacatggagctgtccagcttgaggtccgaggacacagccgtgtactactgcgccacggatgctcggggaagcggcagctactacccgaaccacttcgactactggggacagggcactctcgtgactgtctcgagcgcttctacaaagggcccctccgtgttcccgctcgctccatcatcgaagtctaccagcggaggcactgcggctctcggttgcctcgtgaaggactacttcccggagccggtgaccgtgtcgtggaacagcggagccctgaccagcggggtgcacacctttccggccgtcttgcagtcaagcggcctttactccctgtcatcagtggtgactgtcccgtccagctcattgggaacccaaacctacatctgcaatgtgaatcacaaacctagcaacaccaaggttgacaagaaagtcgagcccaaatcgtgtgacaagactcacacttgtccgccgtgcccggcacccgaactgctgggaggtcccagcgtctttctgttccctccaaagccgaaagacacgctgatgatctcccgcaccccggaggtcacttgcgtggtcgtggacgtgtcacatgaggacccagaggtgaagttcaattggtacgtggatggcgtcgaagtccacaatgccaaaactaagcccagagaagaacagtacaattcgacctaccgcgtcgtgtccgtgctcacggtgttgcatcaggattggctgaacgggaaggaatacaagtgcaaagtgtccaacaaggcgctgccggcaccgatcgagaaaactatctccaaagcgaagggacagcctagggaacctcaagtctacacgctgccaccatcacgggatgaactgactaagaatcaagtctcactgacttgtctggtgaaggggttttaccctagcgacattgccgtggagtgggaatccaacggccagccagagaacaactacaagactacccctccagtgctcgactcggatggatcgttcttcctttactcgaagctcaccgtggataagtcccggtggcagcagggaaacgtgttctcctgctcggtgatgcatgaagccctccataaccactatacccaaaagtcgctgtccctgtcgccgggaaag
SEQ ID NO:25 (вариант 1 ДНК легкой цепи человеческого IgG1)
gacattgtgatgacccagtcccccgattcgcttgcggtgtccctgggagaacgggccaccattaactgcaagagctcacagtccgtcctgtattcatcgaacaacaagaattacctcgcatggtatcagcagaagcctggacagcctcccaagctgctcatctactgggctagcacccgcgaatccggggtgccggatagattctccggatcgggttcgggcactgacttcactctgactatcaactcactgcaagccgaggatgtcgcggtgtacttctgtcagcagtactacgacaccccgacctttggacaaggcaccagactggagattaagcgtacggtggccgctccctccgtgttcatcttcccaccctccgacgagcagctgaagtccggcaccgcctccgtcgtgtgcctgctgaacaacttctacccccgcgaggccaaggtgcagtggaaggtggacaacgccctgcagtccggcaactcccaggaatccgtcaccgagcaggactccaaggacagcacctactccctgtcctccaccctgaccctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaagtgacccaccagggcctgtccagccccgtgaccaagtccttcaaccggggcgagtgc
SEQ ID NO:26 (вариант 1 ДНК тяжелой цепи человеческого IgG4P)
caagtgcaactggtggaatccggggccgaagtgaaaaagcccggagccactgtgaagatctcttgcaaagtgtccggctacaccttcaccgactattacatgcactgggtccagcaggcacctgggaagggccttgagtggatgggtctggtcgatcccgaggacggcgaaactatctacgccgagaagttccagggtcgcgtcaccatcaccgccgacacttccaccgacaccgcgtacatggagctgtccagcttgaggtccgaggacacagccgtgtactactgcgccacggatgctcggggaagcggcagctactacccgaaccacttcgactactggggacagggcactctcgtgactgtctcgagcgcttctacaaagggcccctccgtgttccctctggccccttgctcccggtccacctccgagtctaccgccgctctgggctgcctggtcaaggactacttccccgagcccgtgacagtgtcctggaactctggcgccctgacctccggcgtgcacaccttccctgccgtgctgcagtcctccggcctgtactccctgtcctccgtcgtgaccgtgccctcctccagcctgggcaccaagacctacacctgtaacgtggaccacaagccctccaacaccaaggtggacaagcgggtggaatctaagtacggccctccctgccccccctgccctgcccctgaatttctgggcggaccttccgtgttcctgttccccccaaagcccaaggacaccctgatgatctcccggacccccgaagtgacctgcgtggtggtggacgtgtcccaggaagatcccgaggtccagttcaattggtacgtggacggcgtggaagtgcacaatgccaagaccaagcccagagaggaacagttcaactccacctaccgggtggtgtccgtgctgaccgtgctgcaccaggactggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgccctccagcatcgaaaagaccatctccaaggccaagggccagccccgcgagccccaggtgtacaccctgccccctagccaggaagagatgaccaagaaccaggtgtccctgacctgtctggtcaagggcttctacccctccgacattgccgtggaatgggagtccaacggccagcccgagaacaactacaagaccaccccccctgtgctggacagcgacggctccttcttcctgtactctcggctgaccgtggacaagtcccggtggcaggaaggcaacgtcttctcctgctccgtgatgcacgaggccctgcacaaccactacacccagaagtccctgtccctgagcctgggcaag
SEQ ID NO:27 (вариант 1 ДНК легкой цепи человеческого IgG4P)
gacattgtgatgacccagtcccccgattcgcttgcggtgtccctgggagaacgggccaccattaactgcaagagctcacagtccgtcctgtattcatcgaacaacaagaattacctcgcatggtatcagcagaagcctggacagcctcccaagctgctcatctactgggctagcacccgcgaatccggggtgccggatagattctccggatcgggttcgggcactgacttcactctgactatcaactcactgcaagccgaggatgtcgcggtgtacttctgtcagcagtactacgacaccccgacctttggacaaggcaccagactggagattaagcgtacggtggccgctccctccgtgttcatcttcccaccctccgacgagcagctgaagtccggcaccgcctccgtcgtgtgcctgctgaacaacttctacccccgcgaggccaaggtgcagtggaaggtggacaacgccctgcagtccggcaactcccaggaatccgtcaccgagcaggactccaaggacagcacctactccctgtcctccaccctgaccctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaagtgacccaccagggcctgtccagccccgtgaccaagtccttcaaccggggcgagtgc
SEQ ID NO:28 (вариант 2 тяжелой цепи мышиного полноразмерного IgG1)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSAKTTPPSVYPLAPGSAAQTNSMVTLGCLVKGYFPEPVTVTWNSGSLSSGVHTFPAVLQSDLYTLSSSVTVPSSTWPSETVTCNVAHPASSTKVDKKIVPRDCGCKPCICTVPEVSSVFIFPPKPKDVLTITLTPKVTCVVVDISKDDPEVQFSWFVDDVEVHTAQTQPREEQFNSTFRSVSELPIMHQDWLNGKEFKCRVNSAAFPAPIEKTISKTKGRPKAPQVYTIPPPKEQMAKDKVSLTCMITDFFPEDITVEWQWNGQPAENYKNTQPIMDTDGSYFVYSKLNVQKSNWEAGNTFTCSVLHEGLHNHHTEKSLSHSPGK
SEQ ID NO:29 (вариант 2 легкой цепи мышиного полноразмерного IgG1)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTDAAPTVSIFPPSSEQLTSGGASVVCFLNNFYPKDINVKWKIDGSERQNGVLNSWTDQDSKDSTYSMSSTLTLTKDEYERHNSYTCEATHKTSTSPIVKSFNRNEC
SEQ ID NO:30 (вариант 1 тяжелой цепи человеческого полноразмерного IgG1)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO:31 (вариант 1 легкой цепи человеческого полноразмерного IgG1)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:32 (вариант 2 тяжелой цепи Fab)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
SEQ ID NO:33 (вариант 2 легкой цепи Fab)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:34 (вариант 2 тяжелой цепи человеческого IgG4P)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
SEQ ID NO:35 (вариант 2 легкой цепи человеческого IgG4P)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
--->
СПИСКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЮСБ Биофарма СПРЛ
<120> Кристаллическая структура Gremlin-1 и ингибирующее антитело
<130> PF0045-WO-PCT
<160> 35
<170> PatentIn version 3.5
<210> 1
<211> 184
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 1
Met Ser Arg Thr Ala Tyr Thr Val Gly Ala Leu Leu Leu Leu Leu Gly
1 5 10 15
Thr Leu Leu Pro Ala Ala Glu Gly Lys Lys Lys Gly Ser Gln Gly Ala
20 25 30
Ile Pro Pro Pro Asp Lys Ala Gln His Asn Asp Ser Glu Gln Thr Gln
35 40 45
Ser Pro Gln Gln Pro Gly Ser Arg Asn Arg Gly Arg Gly Gln Gly Arg
50 55 60
Gly Thr Ala Met Pro Gly Glu Glu Val Leu Glu Ser Ser Gln Glu Ala
65 70 75 80
Leu His Val Thr Glu Arg Lys Tyr Leu Lys Arg Asp Trp Cys Lys Thr
85 90 95
Gln Pro Leu Lys Gln Thr Ile His Glu Glu Gly Cys Asn Ser Arg Thr
100 105 110
Ile Ile Asn Arg Phe Cys Tyr Gly Gln Cys Asn Ser Phe Tyr Ile Pro
115 120 125
Arg His Ile Arg Lys Glu Glu Gly Ser Phe Gln Ser Cys Ser Phe Cys
130 135 140
Lys Pro Lys Lys Phe Thr Thr Met Met Val Thr Leu Asn Cys Pro Glu
145 150 155 160
Leu Gln Pro Pro Thr Lys Lys Lys Arg Val Thr Arg Val Lys Gln Cys
165 170 175
Arg Cys Ile Ser Ile Asp Leu Asp
180
<210> 2
<211> 139
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 2
Met Gly Ser Ser His His His His His His Ser Ser Gly Glu Asn Leu
1 5 10 15
Tyr Phe Gln Gly Ser Ala Met Pro Gly Glu Glu Val Leu Glu Ser Ser
20 25 30
Gln Glu Ala Leu His Val Thr Glu Arg Lys Tyr Leu Lys Arg Asp Trp
35 40 45
Cys Lys Thr Gln Pro Leu Lys Gln Thr Ile His Glu Glu Gly Cys Asn
50 55 60
Ser Arg Thr Ile Ile Asn Arg Phe Cys Tyr Gly Gln Cys Asn Ser Phe
65 70 75 80
Tyr Ile Pro Arg His Ile Arg Lys Glu Glu Gly Ser Phe Gln Ser Cys
85 90 95
Ser Phe Cys Lys Pro Lys Lys Phe Thr Thr Met Met Val Thr Leu Asn
100 105 110
Cys Pro Glu Leu Gln Pro Pro Thr Lys Lys Lys Arg Val Thr Arg Val
115 120 125
Lys Gln Cys Arg Cys Ile Ser Ile Asp Leu Asp
130 135
<210> 3
<211> 10
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 3
Gly Tyr Thr Phe Thr Asp Tyr Tyr Met His
1 5 10
<210> 4
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 4
Asp Tyr Tyr Met His
1 5
<210> 5
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 5
Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe Gln
1 5 10 15
Gly
<210> 6
<211> 15
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 6
Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp Tyr
1 5 10 15
<210> 7
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 7
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 8
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 8
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 9
<211> 8
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 9
Gln Gln Tyr Tyr Asp Thr Pro Thr
1 5
<210> 10
<211> 124
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 10
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 11
<211> 112
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 11
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
<210> 12
<211> 124
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 13
<211> 112
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 13
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
<210> 14
<211> 448
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 14
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr
115 120 125
Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn
130 135 140
Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val
180 185 190
Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val
195 200 205
Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg
210 215 220
Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu
245 250 255
Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro
260 265 270
Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala
275 280 285
Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val
290 295 300
Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile
340 345 350
Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys
355 360 365
Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp
370 375 380
Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp
385 390 395 400
Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser
405 410 415
Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly
420 425 430
Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 15
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 15
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 16
<211> 454
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 17
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 17
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 18
<211> 227
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 18
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys
225
<210> 19
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 19
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 20
<211> 118
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 20
Ala Met Pro Gly Glu Glu Val Leu Glu Ser Ser Gln Glu Ala Leu His
1 5 10 15
Val Thr Glu Arg Lys Tyr Leu Lys Arg Asp Trp Cys Lys Thr Gln Pro
20 25 30
Leu Lys Gln Thr Ile His Glu Glu Gly Cys Asn Ser Arg Thr Ile Ile
35 40 45
Asn Arg Phe Cys Tyr Gly Gln Cys Asn Ser Phe Tyr Ile Pro Arg His
50 55 60
Ile Arg Lys Glu Glu Gly Ser Phe Gln Ser Cys Ser Phe Cys Lys Pro
65 70 75 80
Lys Lys Phe Thr Thr Met Met Val Thr Leu Asn Cys Pro Glu Leu Gln
85 90 95
Pro Pro Thr Lys Lys Lys Arg Val Thr Arg Val Lys Gln Cys Arg Cys
100 105 110
Ile Ser Ile Asp Leu Asp
115
<210> 21
<211> 160
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 21
Lys Lys Lys Gly Ser Gln Gly Ala Ile Pro Pro Pro Asp Lys Ala Gln
1 5 10 15
His Asn Asp Ser Glu Gln Thr Gln Ser Pro Gln Gln Pro Gly Ser Arg
20 25 30
Asn Arg Gly Arg Gly Gln Gly Arg Gly Thr Ala Met Pro Gly Glu Glu
35 40 45
Val Leu Glu Ser Ser Gln Glu Ala Leu His Val Thr Glu Arg Lys Tyr
50 55 60
Leu Lys Arg Asp Trp Cys Lys Thr Gln Pro Leu Lys Gln Thr Ile His
65 70 75 80
Glu Glu Gly Cys Asn Ser Arg Thr Ile Ile Asn Arg Phe Cys Tyr Gly
85 90 95
Gln Cys Asn Ser Phe Tyr Ile Pro Arg His Ile Arg Lys Glu Glu Gly
100 105 110
Ser Phe Gln Ser Cys Ser Phe Cys Lys Pro Lys Lys Phe Thr Thr Met
115 120 125
Met Val Thr Leu Asn Cys Pro Glu Leu Gln Pro Pro Thr Lys Lys Lys
130 135 140
Arg Val Thr Arg Val Lys Gln Cys Arg Cys Ile Ser Ile Asp Leu Asp
145 150 155 160
<210> 22
<211> 451
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 22
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly Lys
450
<210> 23
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 23
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 24
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 24
caagtgcaac tggtggaatc cggggccgaa gtgaaaaagc ccggagccac tgtgaagatc 60
tcttgcaaag tgtccggcta caccttcacc gactattaca tgcactgggt ccagcaggca 120
cctgggaagg gccttgagtg gatgggtctg gtcgatcccg aggacggcga aactatctac 180
gccgagaagt tccagggtcg cgtcaccatc accgccgaca cttccaccga caccgcgtac 240
atggagctgt ccagcttgag gtccgaggac acagccgtgt actactgcgc cacggatgct 300
cggggaagcg gcagctacta cccgaaccac ttcgactact ggggacaggg cactctcgtg 360
actgtctcga gcgcttctac aaagggcccc tccgtgttcc cgctcgctcc atcatcgaag 420
tctaccagcg gaggcactgc ggctctcggt tgcctcgtga aggactactt cccggagccg 480
gtgaccgtgt cgtggaacag cggagccctg accagcgggg tgcacacctt tccggccgtc 540
ttgcagtcaa gcggccttta ctccctgtca tcagtggtga ctgtcccgtc cagctcattg 600
ggaacccaaa cctacatctg caatgtgaat cacaaaccta gcaacaccaa ggttgacaag 660
aaagtcgagc ccaaatcgtg tgacaagact cacacttgtc cgccgtgccc ggcacccgaa 720
ctgctgggag gtcccagcgt ctttctgttc cctccaaagc cgaaagacac gctgatgatc 780
tcccgcaccc cggaggtcac ttgcgtggtc gtggacgtgt cacatgagga cccagaggtg 840
aagttcaatt ggtacgtgga tggcgtcgaa gtccacaatg ccaaaactaa gcccagagaa 900
gaacagtaca attcgaccta ccgcgtcgtg tccgtgctca cggtgttgca tcaggattgg 960
ctgaacggga aggaatacaa gtgcaaagtg tccaacaagg cgctgccggc accgatcgag 1020
aaaactatct ccaaagcgaa gggacagcct agggaacctc aagtctacac gctgccacca 1080
tcacgggatg aactgactaa gaatcaagtc tcactgactt gtctggtgaa ggggttttac 1140
cctagcgaca ttgccgtgga gtgggaatcc aacggccagc cagagaacaa ctacaagact 1200
acccctccag tgctcgactc ggatggatcg ttcttccttt actcgaagct caccgtggat 1260
aagtcccggt ggcagcaggg aaacgtgttc tcctgctcgg tgatgcatga agccctccat 1320
aaccactata cccaaaagtc gctgtccctg tcgccgggaa ag 1362
<210> 25
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 25
gacattgtga tgacccagtc ccccgattcg cttgcggtgt ccctgggaga acgggccacc 60
attaactgca agagctcaca gtccgtcctg tattcatcga acaacaagaa ttacctcgca 120
tggtatcagc agaagcctgg acagcctccc aagctgctca tctactgggc tagcacccgc 180
gaatccgggg tgccggatag attctccgga tcgggttcgg gcactgactt cactctgact 240
atcaactcac tgcaagccga ggatgtcgcg gtgtacttct gtcagcagta ctacgacacc 300
ccgacctttg gacaaggcac cagactggag attaagcgta cggtggccgc tccctccgtg 360
ttcatcttcc caccctccga cgagcagctg aagtccggca ccgcctccgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
tccggcaact cccaggaatc cgtcaccgag caggactcca aggacagcac ctactccctg 540
tcctccaccc tgaccctgtc caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc cagccccgtg accaagtcct tcaaccgggg cgagtgc 657
<210> 26
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 26
caagtgcaac tggtggaatc cggggccgaa gtgaaaaagc ccggagccac tgtgaagatc 60
tcttgcaaag tgtccggcta caccttcacc gactattaca tgcactgggt ccagcaggca 120
cctgggaagg gccttgagtg gatgggtctg gtcgatcccg aggacggcga aactatctac 180
gccgagaagt tccagggtcg cgtcaccatc accgccgaca cttccaccga caccgcgtac 240
atggagctgt ccagcttgag gtccgaggac acagccgtgt actactgcgc cacggatgct 300
cggggaagcg gcagctacta cccgaaccac ttcgactact ggggacaggg cactctcgtg 360
actgtctcga gcgcttctac aaagggcccc tccgtgttcc ctctggcccc ttgctcccgg 420
tccacctccg agtctaccgc cgctctgggc tgcctggtca aggactactt ccccgagccc 480
gtgacagtgt cctggaactc tggcgccctg acctccggcg tgcacacctt ccctgccgtg 540
ctgcagtcct ccggcctgta ctccctgtcc tccgtcgtga ccgtgccctc ctccagcctg 600
ggcaccaaga cctacacctg taacgtggac cacaagccct ccaacaccaa ggtggacaag 660
cgggtggaat ctaagtacgg ccctccctgc cccccctgcc ctgcccctga atttctgggc 720
ggaccttccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat ctcccggacc 780
cccgaagtga cctgcgtggt ggtggacgtg tcccaggaag atcccgaggt ccagttcaat 840
tggtacgtgg acggcgtgga agtgcacaat gccaagacca agcccagaga ggaacagttc 900
aactccacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagagtaca agtgcaaggt gtccaacaag ggcctgccct ccagcatcga aaagaccatc 1020
tccaaggcca agggccagcc ccgcgagccc caggtgtaca ccctgccccc tagccaggaa 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtca agggcttcta cccctccgac 1140
attgccgtgg aatgggagtc caacggccag cccgagaaca actacaagac caccccccct 1200
gtgctggaca gcgacggctc cttcttcctg tactctcggc tgaccgtgga caagtcccgg 1260
tggcaggaag gcaacgtctt ctcctgctcc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgtccct gagcctgggc aag 1353
<210> 27
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 27
gacattgtga tgacccagtc ccccgattcg cttgcggtgt ccctgggaga acgggccacc 60
attaactgca agagctcaca gtccgtcctg tattcatcga acaacaagaa ttacctcgca 120
tggtatcagc agaagcctgg acagcctccc aagctgctca tctactgggc tagcacccgc 180
gaatccgggg tgccggatag attctccgga tcgggttcgg gcactgactt cactctgact 240
atcaactcac tgcaagccga ggatgtcgcg gtgtacttct gtcagcagta ctacgacacc 300
ccgacctttg gacaaggcac cagactggag attaagcgta cggtggccgc tccctccgtg 360
ttcatcttcc caccctccga cgagcagctg aagtccggca ccgcctccgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
tccggcaact cccaggaatc cgtcaccgag caggactcca aggacagcac ctactccctg 540
tcctccaccc tgaccctgtc caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc cagccccgtg accaagtcct tcaaccgggg cgagtgc 657
<210> 28
<211> 448
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 28
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr
115 120 125
Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn
130 135 140
Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val
180 185 190
Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val
195 200 205
Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg
210 215 220
Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu
245 250 255
Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro
260 265 270
Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala
275 280 285
Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val
290 295 300
Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile
340 345 350
Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys
355 360 365
Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp
370 375 380
Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp
385 390 395 400
Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser
405 410 415
Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly
420 425 430
Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 29
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 29
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 30
<211> 454
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 30
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 31
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 31
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 32
<211> 227
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys
225
<210> 33
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 33
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 34
<211> 451
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 34
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly Lys
450
<210> 35
<211> 219
<212> белок
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 35
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОР ГРЕМЛИНА-1 ДЛЯ ЛЕЧЕНИЯ ПЕРЕЛОМА КОСТИ ИЛИ ДЕФЕКТА КОСТИ | 2019 |
|
RU2790992C2 |
| Прогнозирование риска развития нежелательной реакции, связанной с введением антитела к ALK2, и способности отвечать на лечение антителом к ALK2 | 2019 |
|
RU2783762C2 |
| СКОНСТРУИРОВАННЫЕ PH-ЗАВИСИМЫЕ АНТИТЕЛА К CD3, А ТАКЖЕ СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2832079C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| ПОЛИПЕПТИД, СОДЕРЖАЩИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН И ТРАНСПОРТИРУЮЩИЙ СЕГМЕНТ | 2017 |
|
RU2827545C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |





















































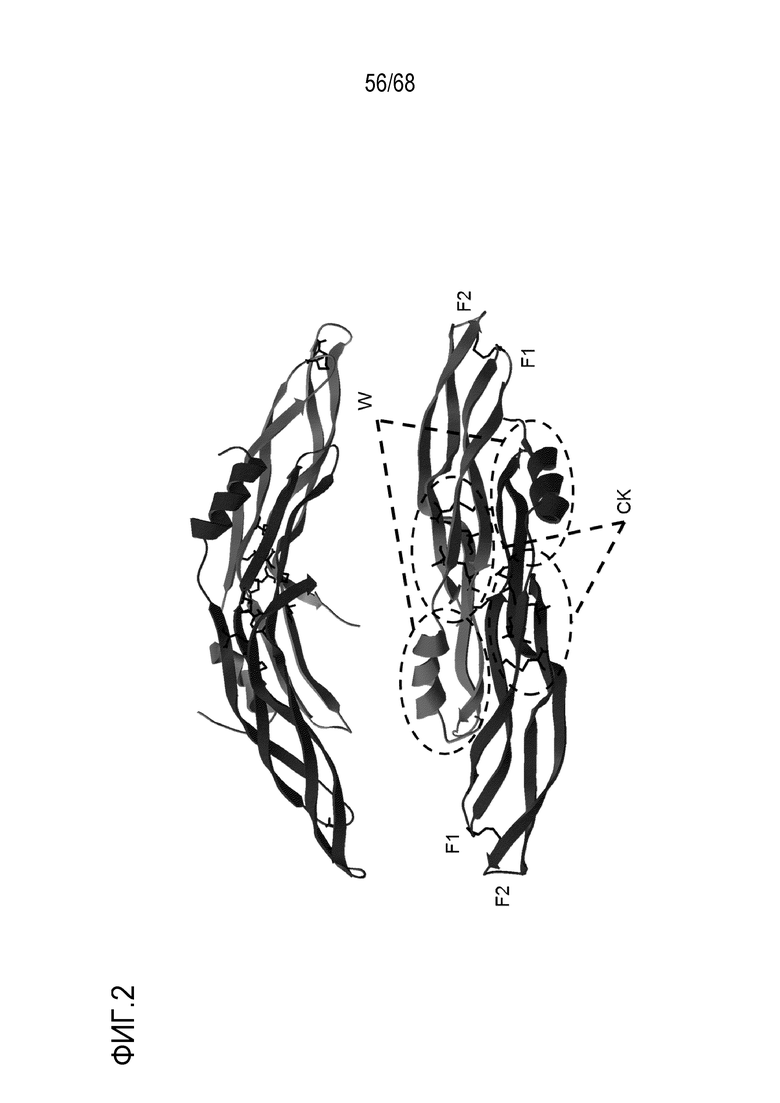
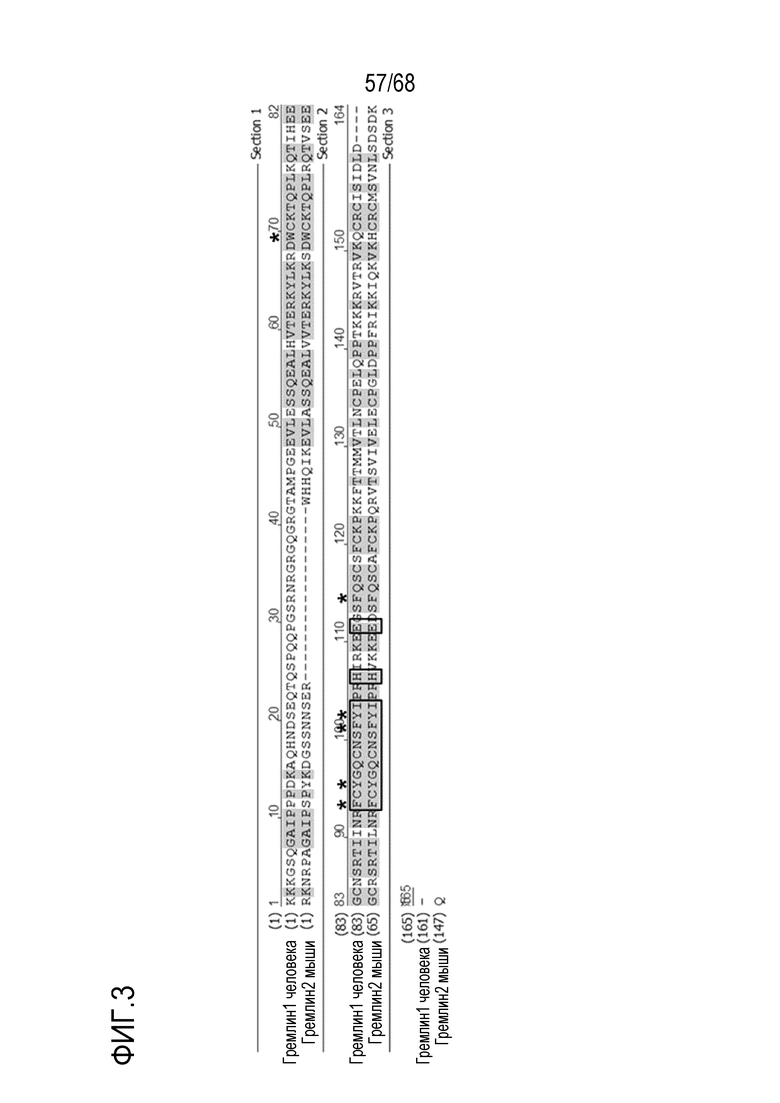
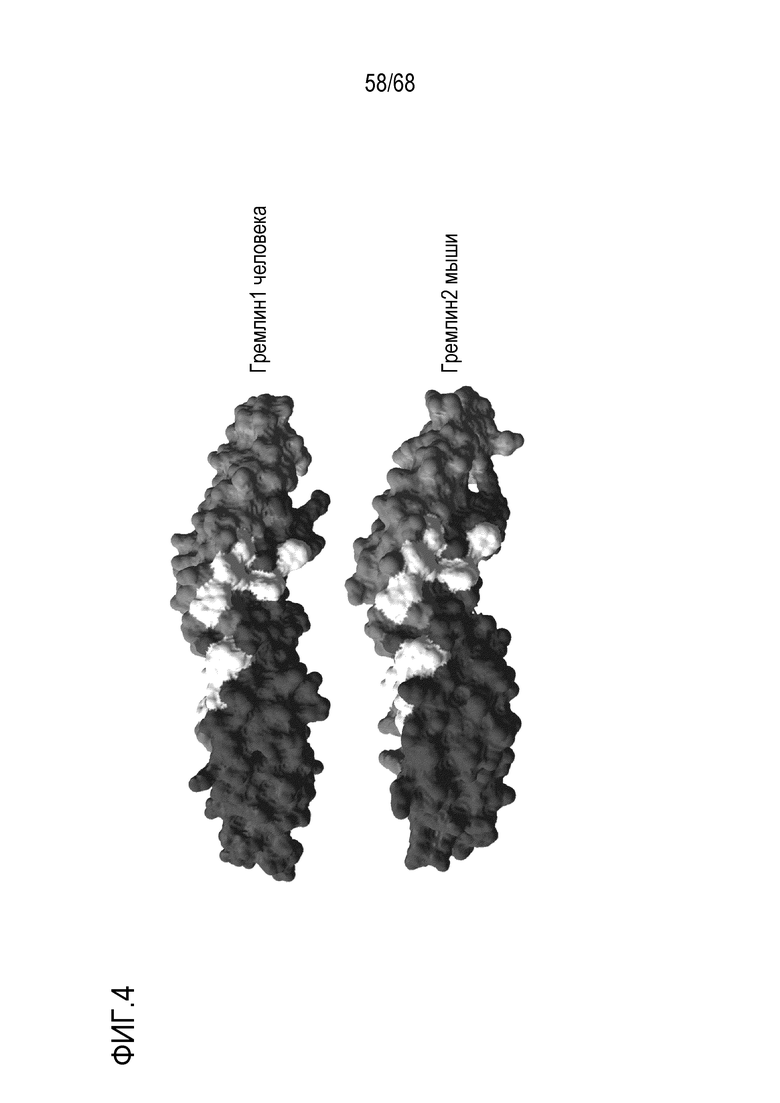

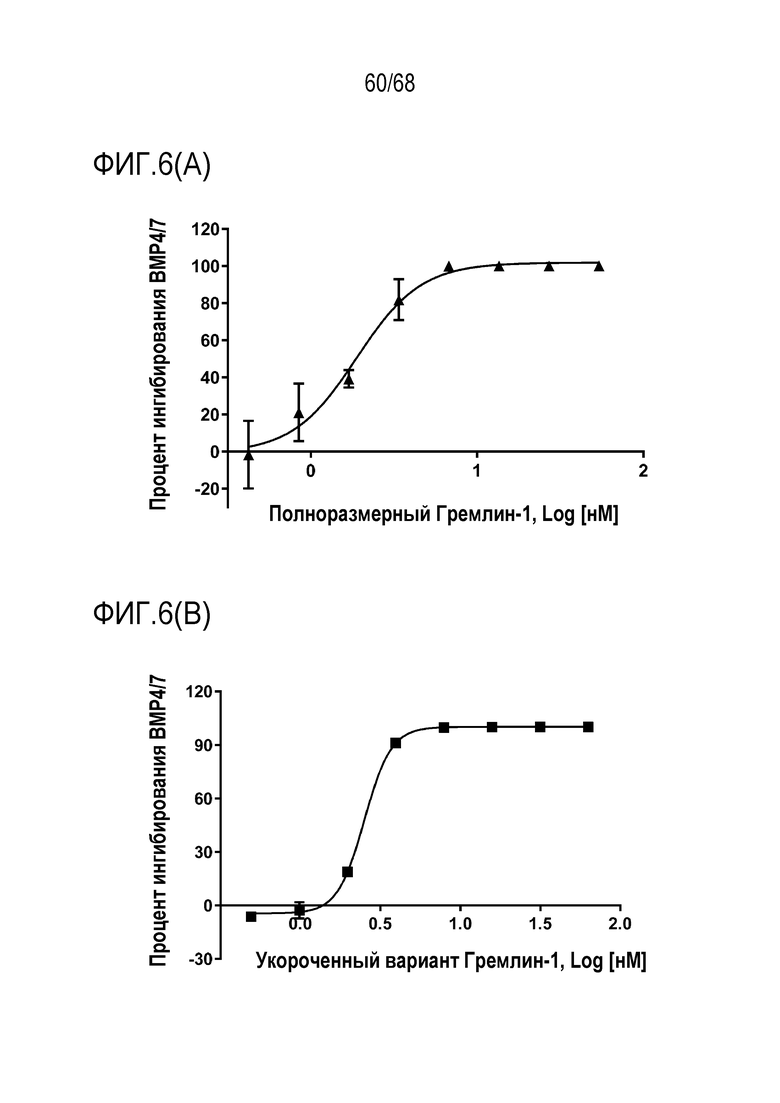
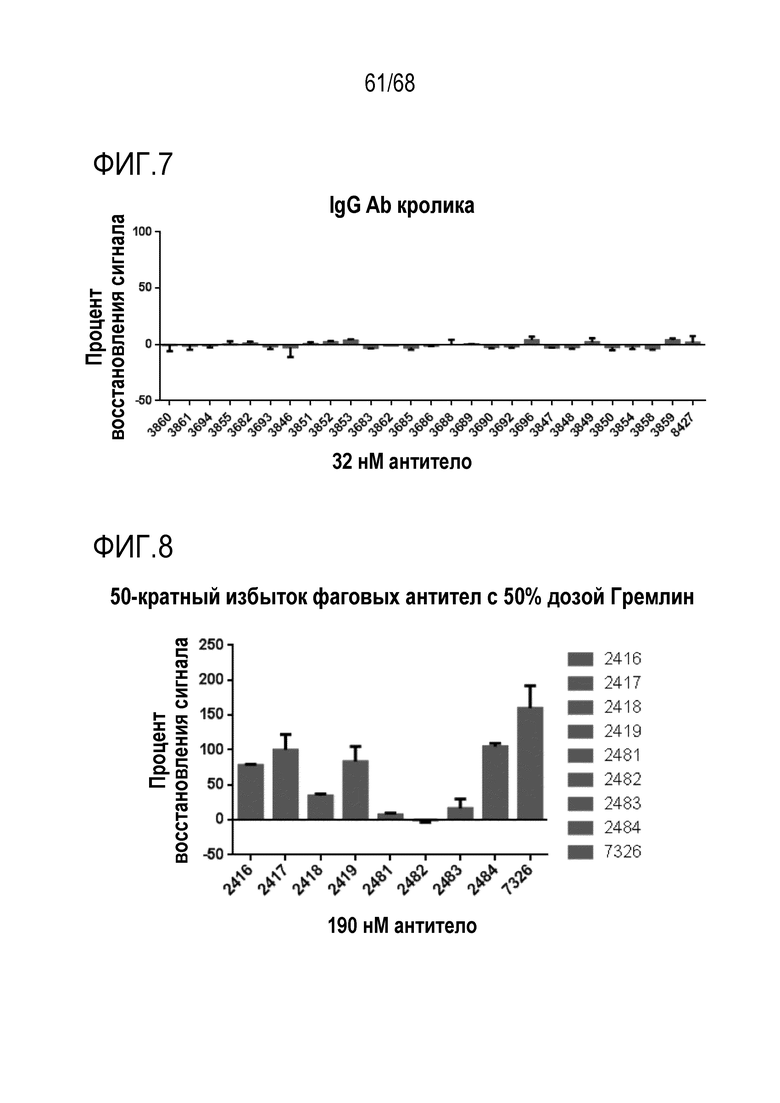
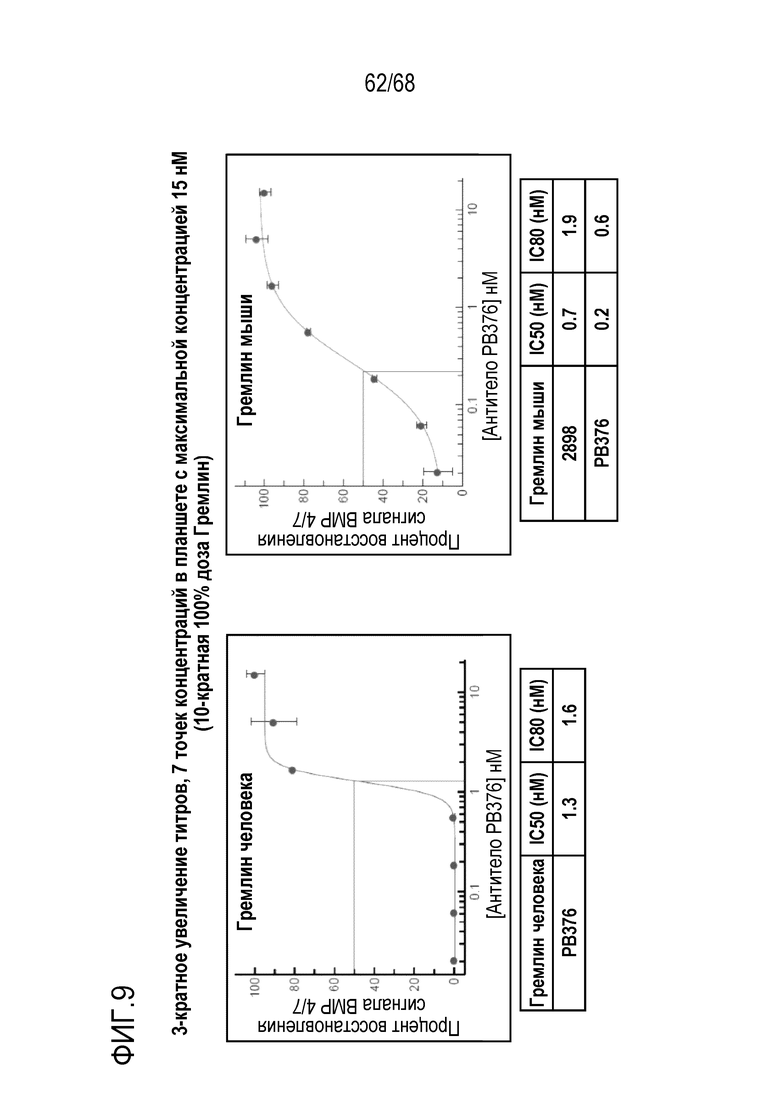
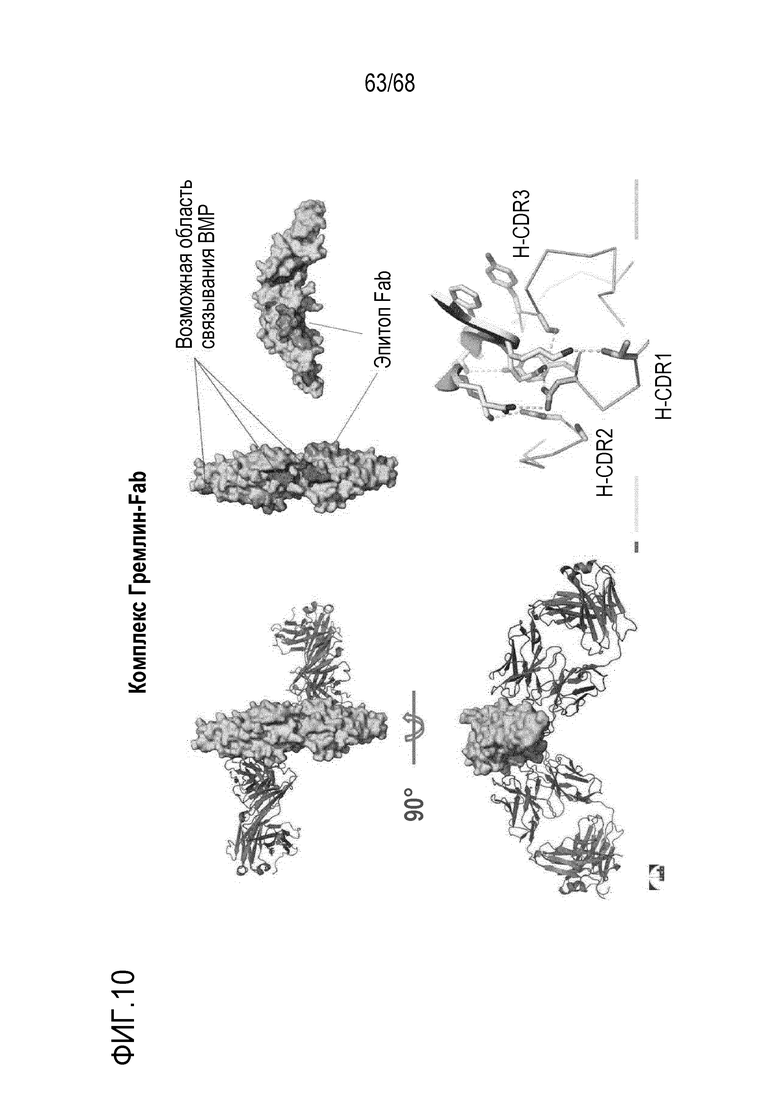

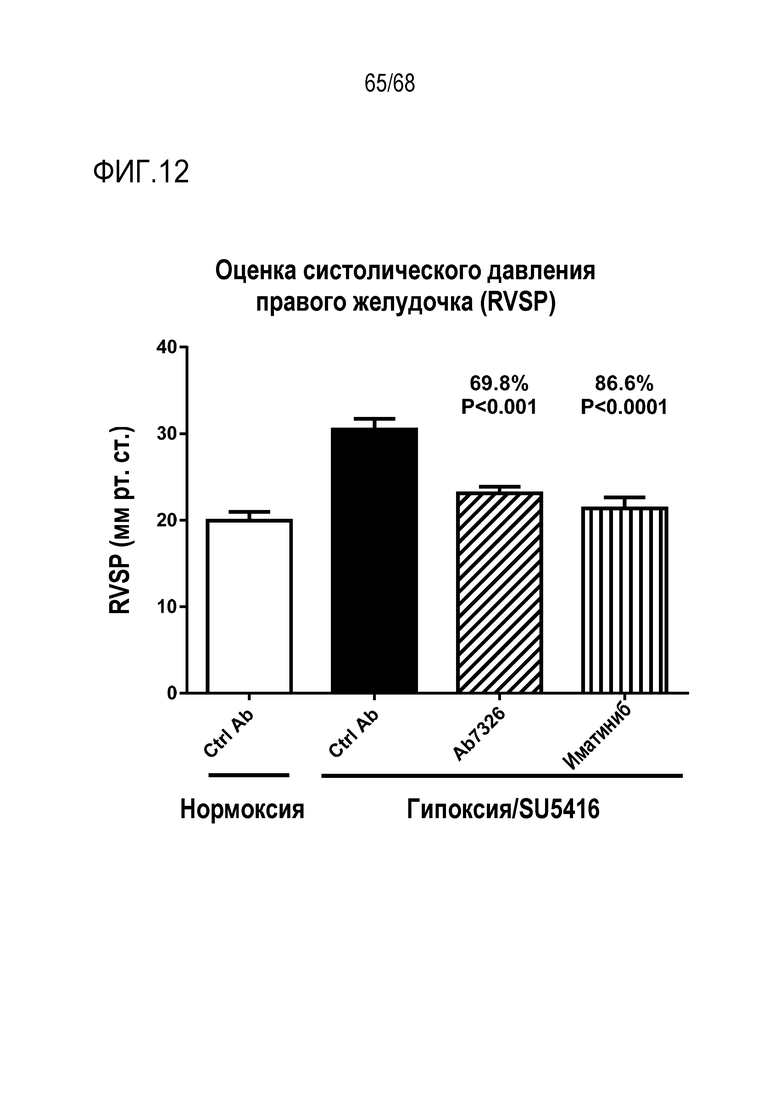
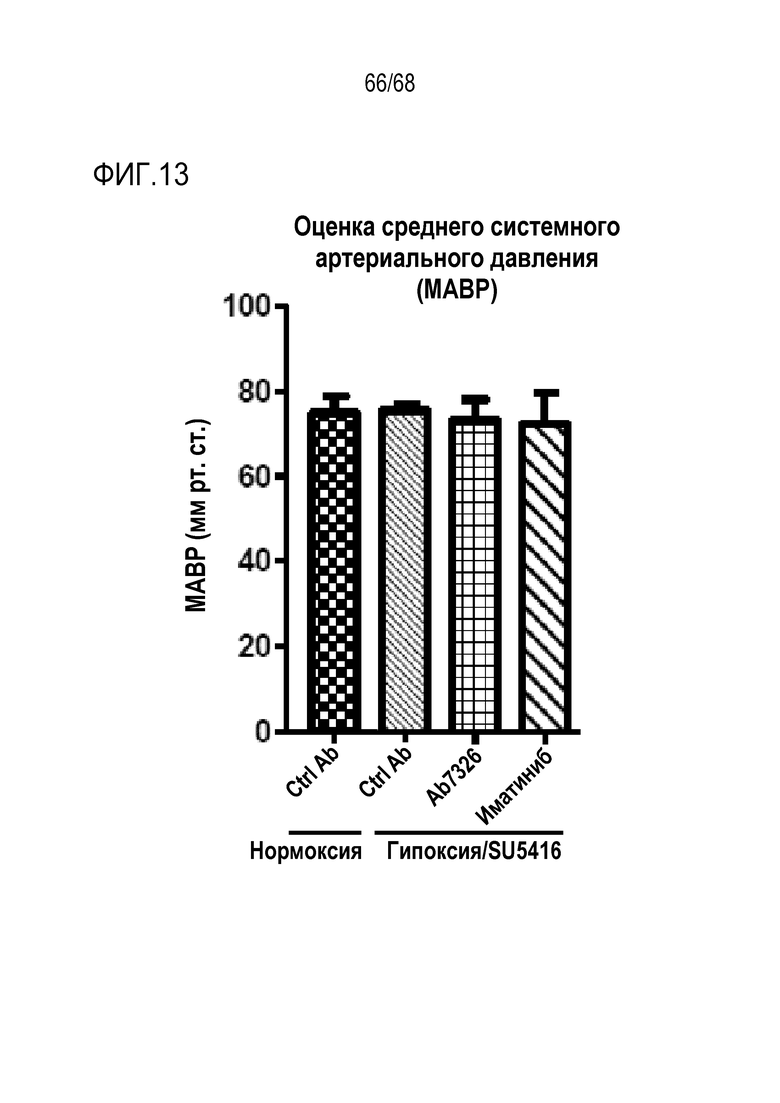
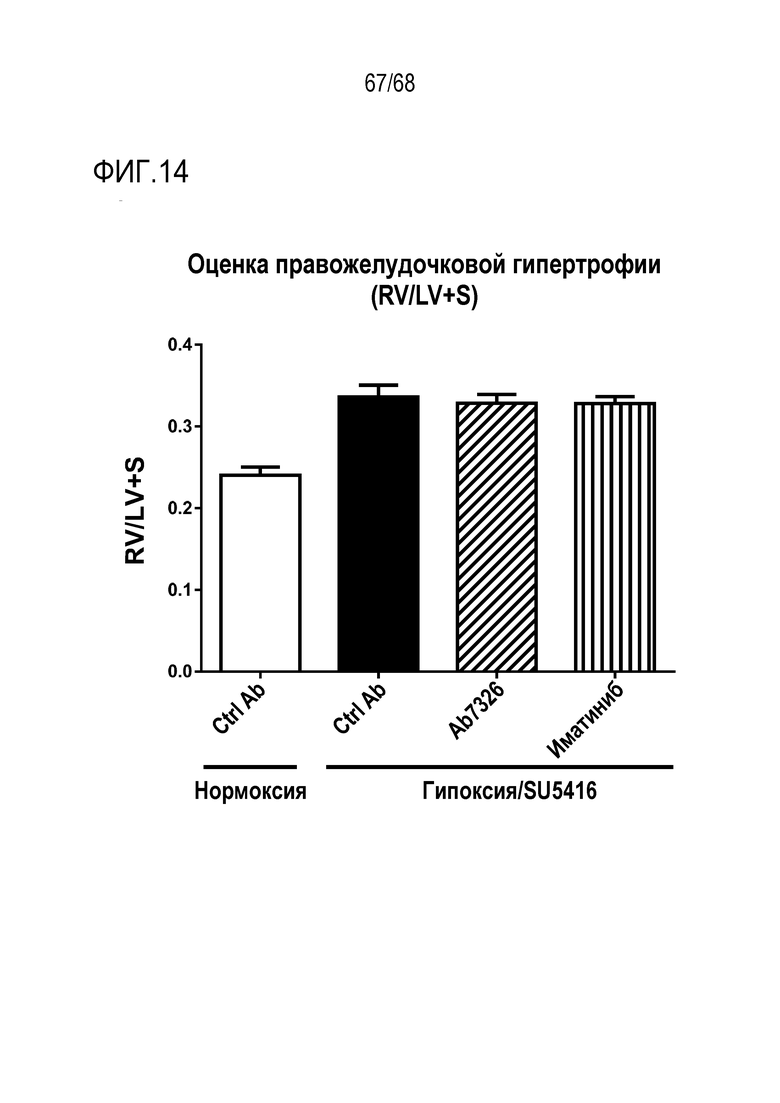
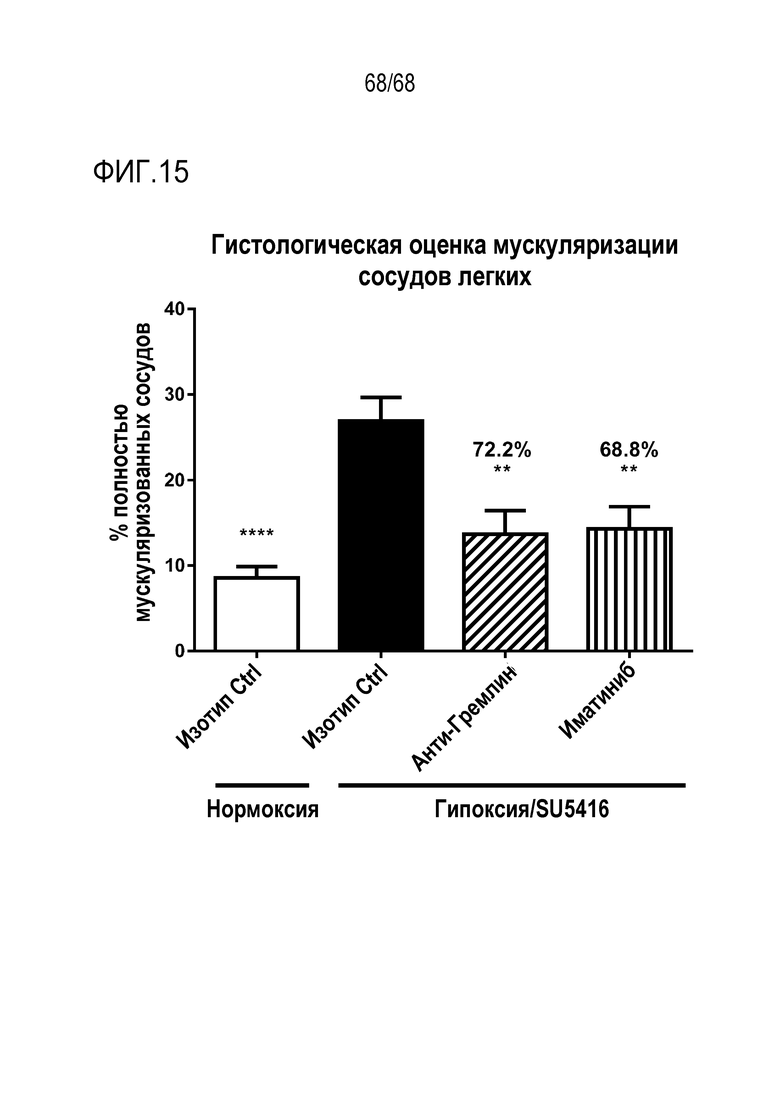
Изобретение относится к области биотехнологии. Предложено антитело, которое специфически связывается с Гремлин-1. Также предложены выделенный полинуклеотид, кодирующий анти-гремлин-1 антитело, включающие указанный полинуклеотид экспрессионный вектор и клетка-хозяин. Также предложен способ получения анти-гремлин-1 антитела, включающий культивирование указанной клетки-хозяина. Кроме того, изобретение относится к применению указанного антитела для лечения или профилактики легочной артериальной гипертензии. Изобретение обеспечивает специфичное связывание с высоким сродством и ингибирующую активность антитела в отношении Гремлин-1. 8 н. и 8 з.п. ф-лы, 16 ил., 6 табл., 8 пр.
1. Анти-гремлин-1 антитело, которое содержит комбинацию последовательностей HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3
SEQ ID NO:4/5/6/7/8/9 или SEQ ID NO:3/5/6/7/8/9.
2. Антитело по п.1, которое содержит
последовательность вариабельной области тяжелой цепи (HCVR) SEQ ID NO:10 или 12 и/или
последовательность вариабельной области легкой цепи (LCVR) SEQ ID NO:11 или 13.
3. Антитело по п.2, которое содержит пару последовательностей HCVR и LCVR SEQ ID NO:10/11 или 12/13.
4. Антитело по п.3, где
последовательности HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3 состоят из SEQ ID NO:4/5/6/7/8/9 или SEQ ID NO:3/5/6/7/8/9 и
оставшиеся HCVR и LCVR по меньшей мере на 95% идентичны SEQ ID NO:10, 11, 12 и/или 13 соответственно.
5. Антитело по любому из пп.1-4, которое содержит
тяжелую цепь с SEQ ID NO:14, 16, 18, 22, 28, 30, 32 или 34 и/или
легкую цепь с SEQ ID NO:15, 17, 19, 23, 29, 31, 33 или 35.
6. Антитело по п.5, которое содержит пару тяжелых и легких цепей SEQ ID NO:14/15, 16/17, 18/19, 22/23, 28/29 или 30/31, 32/33, 34/35.
7. Антитело по п.6, где
последовательности HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3 состоят из SEQ ID NO:4/5/6/7/8/9 или SEQ ID NO:3/5/6/7/8/9 и
оставшиеся тяжелая и легкая цепи по меньшей мере на 95% идентичны SEQ ID NO:14, 15, 16 и/или 17 соответственно.
8. Антитело по любому из пп.1-7, которое представляет собой химерное антитело, антитело человека или гуманизированное антитело.
9. Антитело по любому из пп.1-8, которое представляет собой Fab, модифицированный Fab, Fab', модифицированный Fab', F(ab')2, Fv, однодоменное антитело или scFv.
10. Выделенный полинуклеотид, кодирующий антитело по любому из пп.1-9.
11. Вектор экспрессии, несущий полинуклеотид по п.10.
12. Клетка-хозяин для получения антитела по любому из пп.1-9, где клетка содержит вектор по п.11.
13. Способ получения антитела по любому из пп.1-9, включающий:
культивирование клетки-хозяина по п.12 в условиях, обеспечивающих получение антитела, и
выделение полученного антитела.
14. Фармацевтическая композиция для лечения или профилактики легочной артериальной гипертензии, где композиция содержит эффективное количество антитела по любому из пп.1-9 и фармацевтически приемлемый адъювант и/или носитель.
15. Применение антитела по любому из пп.1-9 или фармацевтическая композиция по п.14 для лечения человека или животного, где заболеванием или состоянием, на которое направлено лечение, является легочная артериальная гипертензия.
16. Способ лечения или профилактики легочной артериальной гипертензии, включающий введение пациенту терапевтически эффективного количества антитела по пп.1-9 или фармацевтической композиции по п.14.
| US 2016024195 A1, 28.01.2016 | |||
| Инкубатор для новорожденных с изменяемой прозрачностью стенок рабочей камеры | 2023 |
|
RU2826790C1 |
| CAHILL E | |||
| et al | |||
| Gremlin plays a key role in the pathogenesis of pulmonary hypertension, Circulation, 2012, vol | |||
| Плуг с фрезерным барабаном для рыхления пласта | 1922 |
|
SU125A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| ЧАЗОВА И.Е | |||
| и др | |||
| Клинические рекомендации по диагностике и лечению легочной гипертонии, ТЕРАПЕВТИЧЕСКИЙ АРХИВ, 2014, том 86, номер 9, с.4-23. | |||

