Настоящее изобретение относится к способам лечения перелома кости или дефекта кости. Изобретение раскрывает эффективное применение анти-гремлин-1-антитела для ускорения заживления и соединения костной ткани при сегментарных дефектах разрыва; и демонстрирует, что ингибиторы активности гремлина-1 могут обеспечить усовершенствованную терапию для лечения или предупреждения несращения перелома.
Уровень техники
Перелом кости представляет собой разрыв или трещину в костной ткани и может быть результатом травматического повреждения, такого как падение или удар, но также может возникнуть в результате заболеваний, которые оказывают отрицательное влияние на целостность кости.
Нестабилизированные переломы костей заживают в процессе эндохондрального окостенения, которое инициируется образованием сгустка крови или гематомы. Это связано с воспалительным ответом, который модулирует иммунные клетки и популяции окружающих скелетных стволовых клеток. Впоследствии гематома заменяется минерализованной хрящевой мозолью благодаря действию различных ростовых факторов, включая трансформирующий фактор роста бета (TGFβ) (Cho et al.; 2002), факторы роста фибробластов (FGF) (Schmid et al.; 2009) и костные морфогенные белки (BMP) (Yu et al.; 2010). Под действием остеокластов и остеобластов минерализованная мозоль заменяется костной тканью. Заключительная стадия ремоделирования включает замену костной ткани пластинчатой костью. Завершение этого процесса может занять много лет в зависимости от возраста и состояния заболевания пациента.
В клинической практике переломы костей обычно лечат фиксацией, с использованием опоры, такой как шина, слепок или скоба. В экстремальных случаях, связанных со сложными переломами, может потребоваться хирургическое вмешательство и использование внутренних и внешних фиксаторов, которые прикрепляются непосредственно к кости. Даже с этими мерами примерно у 10% пациентов процесс восстановления тканей является недостаточным (Einhorn et al.; 2014), что приводит к замедленному сращению кости (неспособности достижения сращения через 6 месяцев после перелома) или отсутствию сращения. Несращение определяется как неполное заживление в течение 9 месяцев, в сочетании с отсутствием рентгенологических показателей, свидетельствующих о заживлении перелома, наблюдаемого в течение трех последовательных месяцев (Buza et al.; 2016). Современные хирургические методы лечения несращенных переломов и критических дефектов кости часто ограничиваются с точки зрения количества и качества доступных материалов. Обычно используемые методы лечения включают аутологичный или аллогенный трансплантат, однако они несут дополнительный риск поражения донорных участков (Goulet et al.; 1997) и развития инфекции (Bostrom et al.; 2005), соответственно.
Костный дефект представляет потерю костной ткани в результате травмы или заболевания.
В настоящее время существует большая неудовлетворенная медицинская потребность в улучшенном лечении переломов костей и дефектов костей. Следовательно, целью настоящего изобретения является обеспечение новых способов лечения перелома кости или дефекта кости.
Настоящее изобретение обеспечивает ингибиторы активности гремлина-1 для применения в лечении перелома кости или дефекта кости. Изобретение раскрывает эффективное применение анти-гремлин-1-антитела для ускорения заживления и соединения костной ткани при дефектах сегментарного зазора; и демонстрирует, что ингибиторы активности гремлина-1 могут обеспечить усовершенствованную терапию для лечения или предупреждения несращения перелома.
Описание изобретения
Если не указано иное, то все научные и технические термины, используемые здесь, имеют то же значение, которое обычно понимается специалистом в данной области. Все публикации, упомянутые здесь, включены посредством ссылки.
Понятно, что любой из описанных здесь вариантов осуществления можно комбинировать.
Настоящее изобретение обеспечивает ингибитор активности гремлина-1 для применения в лечении перелома кости или дефекта кости. Изобретение также обеспечивает применение ингибитора активности гремлина-1 для производства лекарственного средства для лечения перелома кости или дефекта кости. Изобретение также относится к способу лечения перелома кости или дефекта кости, включающему введение терапевтически эффективного количества ингибитора активности гремлина-1.
Гремлин-1 (также известный как Drm и CKTSF1B1) представляет собой гликопротеин из 184 аминокислот, который входит в семейство DAN белков с секретируемым цистеиновым узлом (наряду с Cerberus и Dan, среди прочих). Гремлин связывается с и ингибирует способность BMP-2, 4 и 7 передавать сигналы наряду с установленной проангиогенной функцией, возможно, через агонизм с VEGFR2. Основная роль гремлина-1 проявляется в период развития, где он жизненно необходим при формировании почек и формировании зачатка конечностей.
Известно, что сигнальный путь с участием костного морфогенетического белка (BMP) контролирует образование эндохондральной кости, где гремлин-1 (GREM1) является одним из природных антагонистов этого пути за счет его связывания с BMP2, BMP4 и BMP7 (Hsu et al.; 1998). Условная делеция GREM1 в остеобластах приводит к сенсибилизации сигнального пути/активности BMP и усилению формирования кости in vivo (Gazzerro et al.; 2007), тогда как условная избыточная экспрессия в клетках того же типа вызывает остеопению и спонтанные переломы (Gazzerro et al.; 2005). Кроме того, хотя полный нокаут является эмбриональным летальным на фоне BL6, 49% плодов выживали дольше 24 ч после рождения на смешанном генетическом фоне C57BL/6/FVB, и в то время как дефекты развития скелета присутствовали в большом количестве, повышенные скорости образования кости могли наблюдаться (Canalis et al.; 2012). Несмотря на эту функцию GREM1 в период развития, отсутствуют данные, позволяющие предположить, что ингибирование только этого белка будет способствовать восстановлению перелома кости в постнатальном периоде жизни. Действительно, хотя формирование эндохондральной кости является основным механизмом скелетогенеза на эмбриональных стадиях, механизмы, которые регулируют рекрутинг клеток, являются разными процессами по сравнению с восстановлением перелома в постнатальном периоде (Ferguson et al.; 1999). Роль воспаления была указана в качестве ключевого фактора восстановления кости у взрослых, поэтому факторы развития, контролирующие процессы скелетогенеза, нельзя просто экстраполировать на механизмы восстановления в постнатальном периоде.
Как здесь используется, термин гремлин-1 обычно имеет последовательность с идентификационным номером UniProt O60565 (SEQ ID NO: 1). Термин гремлин-1 может также относиться к полипептиду гремлина-1, который:
(а) содержит или состоит из аминокислотной последовательности SEQ ID NO: 1 с или без N-концевого сигнального пептида, т.е. может содержать или состоять из последовательности зрелого пептида, показанной в SEQ ID NO: 21; или
(b) представляет производное, имеющее одну или более аминокислотных замен, модификаций, делеций или инсерций относительно аминокислотной последовательности SEQ ID NO: 1 с или без N-концевого сигнального пептида (как показано в SEQ ID NO: 21), которое сохраняет активность гремлина-1, такую как аминокислотная последовательность SEQ ID NO: 20;
(c) представляет его варианты, где такие варианты обычно сохраняют, по меньшей мере, примерно 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94% или 95% идентичность с SEQ ID NO: 1 (или SEQ ID NO: 20 или 21) (или даже примерно 96%, 97%, 98% или 99% идентичность). Другими словами, такие варианты могут сохранять примерно 60% - примерно 99% идентичность с SEQ ID NO: 1, предпочтительно примерно 80% - примерно 99% идентичность с SEQ ID NO: 1, более предпочтительно примерно 90% - примерно 99% идентичность с SEQ ID NO: 1 и наиболее предпочтительно примерно 95% - примерно 99% идентичность с SEQ ID NO: 1. Варианты дополнительно описаны ниже.
Как обсуждается далее ниже, номера остатков обычно указываются на основе нумерации в последовательности SEQ ID NO: 1. Однако специалист может легко экстраполировать нумерацию остатков на последовательность производного или варианта, которые обсуждались выше. Там, где указаны номера остатков, изобретение также охватывает эти остатки в последовательности варианта или производного.
Заявители настоящего изобретения кристаллизовали человеческий гремлин-1 один и в комплексе с антителом, названным Ab 7326 (фрагменты Fab). Кристаллизация гремлина-1 позволила определить предполагаемые остатки в сайте связывания с BMP. Кроме того, кристаллизация с Ab 7326, которое является аллостерическим ингибирующим антителом, позволила определить остатки в эпитопе антитела (WO 2018/115017 A2). Антитела, связывающиеся с этим эпитопом, имеют потенциал применения в качестве терапевтических агентов в лечении перелома кости или дефекта кости.
Ингибиторы активности гремлина-1
Ингибитор активности гремлина-1 по настоящему изобретению представляет агент, который снижает или блокирует активность гремлина-1. Ингибиторы по настоящему изобретению могут частично или полностью ингибировать активность гремлина-1. Ингибиторы для применения в настоящем изобретении включают без ограничения ингибиторы, которые способны связываться с гремлином-1 или с молекулой нуклеиновой кислоты, кодирующей гремлин-1, или способны ингибировать экспрессию гремлина-1. Такими ингибиторами могут быть, без ограничения, белки, полипептиды, пептиды, пептидомиметики, нуклеиновые кислоты (например, ДНК, РНК, антисмысловые РНК и миРНК), углеводы, липиды и небольшие молекулы.
В одном варианте осуществления ингибитор активности гремлина-1 представляет анти-гремлин-1-антитело или его функционально активный фрагмент, вариант или производное.
Термин «антитело» в контексте настоящего описания включает целые антитела и любой антигенсвязывающий фрагмент (т.е. «антигенсвязывающий участок») или их отдельные цепи. Антитело или иммуноглобулин обычно относится к гликопротеину, содержащему, по меньшей мере, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные дисульфидными связями, или его антигенсвязывающему участку. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначенной здесь как HCVR или VH) и константной области тяжелой цепи. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначенной здесь как LCVR или VL) и константной области легкой цепи. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с областями, которые являются более консервативными, называемыми каркасными областями (FR).
Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Антитело для применения в настоящем изобретении может представлять моноклональное антитело или поликлональное антитело, и обычно оно представляет моноклональное антитело. Антитело для применения в изобретении может представлять химерное антитело, CDR-привитое антитело, нанотело, человеческое или гуманизированное антитело, или антигенсвязывающий фрагмент любого из них.
Поликлональные антитела можно получить обычными методами, такими как иммунизация подходящего животного антигеном, представляющим интерес. Затем можно отобрать кровь у животного и выделить иммуноглобулиновую фракцию.
Анти-гремлин-1-антитела могут быть получены там, где необходима иммунизация животного, введением полипептидов животному, например, животному, отличному от человека, с использованием хорошо известных и рутинных протоколов, см., например, «Справочник по экспериментальной иммунологии», D.M. Weir (ed.), Vol. 4, Blackwell Scientific Publishers, Oxford, Англия, 1986). Можно иммунизировать многих теплокровных животных, таких как кролики, мыши, крысы, овцы, козы, коровы, верблюды, ламы или свиньи. Однако кролики, мыши и крысы обычно являются наиболее подходящими.
Моноклональные антитела могут быть получены любым способом, известным в данной области, таким как метод гибридомы (Kohler & Milstein, 1975, Nature, 256: 495-497), метод триомы, метод гибридомы В-клеток человека (Kozbor et al., 1983, Immunology Today, 4:72) и метод EBV-гибридомы (Cole et al., Monoclonal Antibodies and Cancer Therapy, pp.77-96, Alan R Liss, Inc., 1985).
Антитела для применения в изобретении также могут быть получены с использованием методов получения антител с одним лимфоцитом посредством клонирования и экспрессии кДНК вариабельной области иммуноглобулина, полученных из отдельных лимфоцитов, отобранных для продукции специфических антител, например, способами, описанными Babcook, J. et al., 1996 , Proc. Natl. Acad. Sci. USA 93 (15): 7843-7848l; WO 92/02551; WO 2004/051268 и WO 2004/106377.
Антитела для применения в настоящем изобретении также можно получить с использованием различных методов фагового дисплея, известных в данной области, и они включают методы, раскрытые Brinkman et al. (J. Immunol. Methods, 1995, 182: 41-50), Ames et al. (J. Immunol. Methods, 1995, 184: 177-186), Kettleborough et al. (Eur. J. Immunol. 1994, 24: 952-958), Persic et al. (Gene, 1997, 187, 9-18), Burton et al. (Advances in Immunology, 1994, 57: 191-280) и в WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401; и в патенте США № 5698426; патенте США № 5223409; патенте США № 5403484; патенте США № 5580717; патенте США № 5427908; патенте США № 5750753; патенте США № 5821047; патенте США № 5571698; патенте США № 5427908; патенте США № 5516637; патенте США № 5780225; патенте США № 5658727; патенте США № 5733743 и патенте США 5969108.
Полностью человеческие антитела представляют собой антитела, в которых вариабельные области и константные области (где они присутствуют) как тяжелой, так и легкой цепей являются человеческими или по существу идентичными человеческим последовательностям, но необязательно из одного и того же антитела. Примеры полностью человеческих антител могут включать антитела, полученные, например, способами фагового дисплея, описанными выше, и антитела, продуцированные мышами, где гены вариабельной области и, необязательно, гены константной области мышиного иммуноглобулина заменены их человеческими аналогами, например, как, в общем, описано в ЕР 0546073, патентах США № 5545806, 5569825, 5625126, 5633425, 5661016, 5770429, EP 0438474 и EP 0463151.
Альтернативно, антитело по изобретению может быть получено способом, включающим иммунизацию млекопитающего, отличного от человека, иммуногеном гремлином-1; получение препарата антител от указанного млекопитающего; получение из них моноклональных антител, которые распознают гремлин-1.
Молекулы антител для применения в настоящем изобретении могут включать полную молекулу антитела, имеющую полноразмерные тяжелые и легкие цепи, или ее фрагмент или антигенсвязывающий участок. Термин «антигенсвязывающий участок» антитела относится к одному или более фрагментам антитела, которые сохраняют способность избирательно связываться с антигеном. Было показано, что антигенсвязывающая функция антитела может выполняться фрагментами полноразмерного антитела. Антитела и их фрагменты и их антигенсвязывающие участки могут представлять, не ограничиваясь этим, Fab, модифицированный Fab, Fab', модифицированный Fab', F(ab')2, Fv, однодоменные антитела (например, VH или VL или VHH), scFv, би-, три- или тетравалентные антитела, Bis-scFv, диатела, триатела, тетратела и эпитоп-связывающие фрагменты любого из вышеуказанного (см., например, Holliger and Hudson, 2005, Nature Biotech. 23 (9): 1126- 1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2 (3), 209-217). Способы создания и получения этих фрагментов антител хорошо известны в данной области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Другие фрагменты антител для применения в настоящем изобретении включают фрагменты Fab и Fab', описанные в международных заявках на патент WO 2005/003169, WO 2005/003170 и WO 2005/003171, и фрагменты Fab-dAb, описанные в международной заявке на патент WO 2009/040562. Поливалентные антитела могут иметь много специфичностей или могут быть моноспецифичными (см., например, WO 92/22853 и WO 2005/113605). Эти фрагменты антител можно получить с использованием обычных методик, известных специалистам в данной области техники, и фрагменты могут быть скринированы на пригодность таким же образом, как и интактные антитела.
В одном примере функционально активный фрагмент антитела для применения в настоящем изобретении представляет Fab, Fab', F(ab')2, Fv или scFv.
Домены константной области молекулы антитела для применения в настоящем изобретении, если они присутствуют, могут быть выбраны с учетом эффекторных функций, которые могут необходимыми. Например, домены константной области могут представлять домены человеческих IgA, IgD, IgE, IgG или IgM. В частности, можно использовать домены константной области человеческого IgG, особенно изотипов IgG1 и IgG3, когда требуются эффекторные функции антител. Альтернативно, могут использоваться изотипы IgG2 и IgG4, когда эффекторные функции антитела не требуются. В одном примере изотипом является IgG4P, как описано в публикации Angal S. et al., Mol Immunol, Vol. 30 (1), p105-108, 1993.
Антитело для применения в изобретении может быть получено, экспрессировано, создано или выделено рекомбинантными способами, например (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным в отношении генов иммуноглобулина, представляющих интерес, или гибридомы полученной из них, (b) антитела, выделенные из клетки-хозяина, трансформированные для экспрессии представляющего интерес антитела, например, из трансфектомы, (c) антитела, выделенные из комбинаторной библиотеки рекомбинантных антител, и (d) антитела, полученные, экспрессированные, созданные или выделенные любым другим способом, который включает сплайсинг последовательностей гена иммуноглобулина с другими последовательностями ДНК.
Антитело для применения в изобретении может представлять собой человеческое антитело или гуманизированное антитело. Предусматривается, что термин «человеческое антитело» в контексте настоящего описания включает антитела, имеющие вариабельные области, в которых как каркасные, так и CDR-области получены из последовательностей иммуноглобулина зародышевой линии человека. Кроме того, если антитело содержит константную область, то константная область также происходит из последовательностей иммуноглобулина зародышевой линии человека. Человеческие антитела для применения в изобретении могут включать аминокислотные остатки, не кодированные последовательностями человеческого иммуноглобулина зародышевой линии (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro, или соматическая мутация in vivo). Однако термин «человеческое антитело», как здесь используется, не предназначен для включения антител, в которых последовательности CDR, полученные из зародышевой линии другого вида млекопитающих, таких как мышь, были привиты на каркасные последовательности человека.
Такое человеческое антитело может представлять человеческое моноклональное антитело. Такое человеческое моноклональное антитело может быть продуцировано гибридомой, которая включает В-клетку, полученную от трансгенного животного, отличного от человека, например, трансгенной мыши, имеющей геном, содержащий трансген тяжелой цепи и трансген легкой цепи человека, слитый с иммортализованной клеткой.
Человеческие антитела можно получить иммунизацией человеческих лимфоцитов in vitro с последующей трансформацией лимфоцитов вирусом Эпштейна-Барра.
Термин «производное» относится к любой модифицированной форме антитела, например, конъюгату антитела и другого агента или эффекторной молекулы.
Эффекторная молекула может включать одну эффекторную молекулу или две или более таких молекул, связанных таким образом, чтобы образовалась единая молекула, которая может быть присоединена к антителам для применения в настоящем изобретении. Когда желательно получить фрагмент антитела, связанный с эффекторной молекулой, то его можно получить стандартными химическими методами или методами рекомбинантной ДНК, в которых фрагмент антитела связывают либо напрямую, либо через связывающий агент с эффекторной молекулой. Способы конъюгирования таких эффекторных молекул с антителами хорошо известны в данной области (см. Hellstrom et al., Controlled Drug Delivery, 2nd Ed., Robinson et al., Eds., 1987, pp. 623-53; Thorpe et al. , 1982, Immunol. Rev., 62: 119-58 и Dubowchik et al., 1999, Pharmacology and Therapeutics, 83, 67-123). Конкретные химические процедуры включают, например, те, которые описаны в WO 93/06231, WO 92/22583, WO 89/00195, WO 89/01476 и WO 2003/031581. Альтернативно, когда эффекторная молекула представляет собой белок или полипептид, то связь может быть достигнута с использованием процедур рекомбинантной ДНК, например, как описано в WO 86/01533 и EP 0392745.
Эффекторная молекула может увеличить период полураспада антитела in vivo и/или уменьшить иммуногенность антитела и/или усилить доставку антитела через эпителиальный барьер к иммунной системе. Примеры подходящих эффекторных молекул такого типа включают полимеры, альбумин, альбумин-связывающие белки или альбумин-связывающие соединения, такие как описаны в WO 2005/117984.
Термин «гуманизированное антитело» относится к молекулам CDR-привитых антител, в которых последовательности CDR, полученные из зародышевой линии других видов млекопитающих, таких как мышь, были привиты к человеческим каркасным последовательностям. Дополнительные модификации каркасной области могут быть сделаны в человеческих каркасных последовательностях.
Как здесь используется, термин «молекула CDR-привитого антитела» относится к молекуле антитела, в которой тяжелая и/или легкая цепь содержит один или более CDR (включая, если желательно, один или более модифицированных CDR) из донорного антитела (например, мышиного или крысиного моноклонального антитела), были привиты на каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, человеческого антитела). Обзор смотри в публикации Vaughan et al., Nature Biotechnology, 16, 535-539, 1998. В одном варианте осуществления, вместо переноса всего CDR, только один или более определяющих специфичность остатков любого из CDR, описанных здесь выше, переносят на каркас человеческого антитела (см., например, Kashmiri et al., 2005, Methods, 36, 25-34). В одном варианте осуществления переносятся только определяющие специфичность остатки из одного или более CDR, описанных здесь выше, на каркас человеческого антитела. В еще одном варианте осуществления только остатки, определяющие специфичность, из каждого из CDR, описанных здесь выше, переносятся на каркас человеческого антитела.
Когда CDR или определяющие специфичность остатки привиты, то может использоваться любая подходящая акцепторная каркасная последовательность вариабельной области с учетом класса/типа донорного антитела, из которого получены CDR, включая каркасные области мыши, приматов и человека. Следовательно, CDR-привитое антитело для применения в настоящем изобретении имеет вариабельный домен, включающий акцепторные каркасные области человека, а также один или более CDR или остатков, определяющих специфичность, описанных выше. Таким образом, в одном варианте осуществления обеспечивается нейтрализующее CDR-привитое антитело, в котором вариабельный домен включает акцепторные каркасные области человека и донорные CDR, отличные от человеческих.
Примерами человеческих каркасов, которые можно использовать в настоящем изобретении, являются KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (Kabat et al., выше). Например, KOL и NEWM можно использовать для тяжелой цепи, REI можно использовать для легкой цепи, и EU, LAY и POM можно использовать как для тяжелой цепи, так и для легкой цепи. Альтернативно, можно использовать последовательности зародышевой линии человека; они доступны, например, на: http://www.vbase2.org/ (см. Retter et al., Nucl. Acids Res. (2005) 33 (supplement 1), D671-D674).
В CDR-привитом антителе для применения в настоящем изобретении акцепторные тяжелая и легкая цепи необязательно должны быть получены из одного и того же антитела и могут, если желательно, включать составные цепи, имеющие каркасные области, полученные из разных цепей.
Также в CDR-привитом антителе для применения в настоящем изобретении каркасные области не должны иметь точно такую же последовательность, как последовательности акцепторного антитела. Например, необычные остатки могут быть заменены на более часто встречающиеся остатки для данного класса или типа акцепторной цепи. Альтернативно выбранные остатки в каркасных областях акцептора могут быть заменены так, чтобы они соответствовали остатку, обнаруженному в таком же положении в донорном антителе (см. Reichmann et al., 1998, Nature, 332, 323-324). Такие изменения должны быть сведены к минимуму, необходимому для восстановления аффинности донорного антитела. Протокол для выбора остатков в акцепторных каркасных областях, которые, возможно, необходимо изменить, описан в WO 91/09967.
Специалисту в данной области также должно быть понятно, что антитела могут подвергаться различным посттрансляционным модификациям. Тип и степень этих модификаций часто зависят от линии клеток-хозяев, используемых для экспрессии антитела, а также от условий культивирования. Такие модификации могут включать вариации гликозилирования, окисления метионина, образования дикетопиперазина, изомеризации аспартата и дезамидирования аспарагина. Частой модификацией является потеря карбоксиконцевого основного остатка (такого как лизин или аргинин) в результате действия карбоксипептидаз (как описано в публикации Harris, RJ. Journal of Chromatography 705: 129-134, 1995).
В одном варианте осуществления тяжелая цепь антитела содержит домен CH1, и легкая цепь антитела содержит домен CL, либо каппа, либо лямбда.
Биологические молекулы, такие как антитела или фрагменты, содержат кислотные и/или основные функциональные группы, что придает молекуле суммарный положительный или отрицательный заряд. Количество суммарного «наблюдаемого» заряда будет зависеть от абсолютной аминокислотной последовательности молекулы, локального окружения заряженных групп в трехмерной структуре и условий окружающей среды молекулы. Изоэлектрическая точка (pI) представляет pH, при котором конкретная молекула или поверхность не несет суммарного электрического заряда. В одном варианте осуществления антитело или фрагмент по настоящему изобретению имеет изоэлектрическую точку (pI), равную, по меньшей мере, 7. В одном варианте осуществления антитело или фрагмент имеет изоэлектрическую точку, равную, по меньшей мере, 8, например, 8,5, 8,6, 8,7, 8,8 или 9. В одном варианте осуществления pI антитела составляет 8. Программы, такие как ** ExPASY http://www.expasy.ch/tools/pi_tool.html (см. Walker, The Proteomics Protocols Handbook, Humana Press (2005), 571 -607) можно использовать для прогнозирования изоэлектрической точки антитела или фрагмента.
Антитела для применения в изобретении могут включать, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR тяжелой цепи SEQ ID NO: 4-6 (HCDR1/HCDR2/ HCDR3 соответственно). Это последовательности HCDR1/HCDR2/HCDR3 антитела Ab7326 из примеров, как определено с использованием методологии Kabat.
Системы нумерации Kabat и Chothia для определения последовательностей CDR хорошо известны в данной области (так же как и другие системы). Последовательности CDR можно определить с использованием любого подходящего метода, и в настоящем изобретении, хотя обычно используется система нумерации по Kabat, можно также использовать другие методы. В данном случае SEQ ID NO: 3 представляет последовательность HCDR1 Ab7326, определенную с использованием комбинированного определения по системе Chothia & Kabat.
Антитела для применения в изобретении могут содержать, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR легкой цепи SEQ ID NO: 7-9 (LCDR1/LCDR2/ LCDR3 соответственно). Это последовательности LCDR1/LCDR2/LCDR3 Ab7326 с использованием методологии Kabat.
В одном варианте осуществления антитело содержит, по меньшей мере, последовательность HCDR3 SEQ ID NO: 6.
Как правило, антитело содержит, по меньшей мере, одну последовательность CDR тяжелой цепи, выбранную из SEQ ID NO: 4-6, и, по меньшей мере, одну последовательность CDR легкой цепи, выбранную из SEQ ID NOS 7-9. Антитело может содержать, по меньшей мере, две последовательности CDR тяжелой цепи, выбранные из SEQ ID NO: 4-6 и, по меньшей мере, две последовательности CDR легкой цепи, выбранные из SEQ ID NO: 7-9. Антитело обычно включает все три последовательности CDR тяжелой цепи SEQ ID NO: 4-6 (HCDR1/HCDR2/HCDR3 соответственно) и все три последовательности CDR легкой цепи SEQ ID NO: 7-9 (LCDR1/LCDR2/LCDR3 соответственно). Антитела могут быть химерными, человеческими или гуманизированными антителами.
Антитело может содержать последовательность вариабельной области тяжелой цепи (HCVR) SEQ ID NO: 10 или 12 (HCVR вариантов 1 и 2 Ab7326). Антитело может содержать последовательность вариабельной области легкой цепи (LCVR) SEQ ID NO: 11 или 13 (LCVR вариантов 1 и 2 Ab7326). Антитело предпочтительно содержит последовательность вариабельной области тяжелой цепи SEQ ID NO: 10 или 12 и последовательность вариабельной области легкой цепи SEQ ID NO: 11 или 13 (в частности, пары HCVR/LVCR с SEQ ID NO: 10/11 или 12/13).
Варианты 1 и 2 Ab7326 различаются одной аминокислотой в вариабельной области тяжелой цепи и одной аминокислотой в вариабельной области легкой цепи следующим образом:
- вариант 1 вариабельной области тяжелой цепи содержит глутаминовую кислоту (E) в положении 6 (SEQ ID NO: 10)
- вариант 2 вариабельной области тяжелой цепи содержит глутамин (Q) в положении 6 (SEQ ID NO: 12)
- вариант 1 вариабельной области легкой цепи содержит серин (S) в положении 7 (SEQ ID NO: 11)
- вариант 2 вариабельной области легкой цепи содержит треонин (T) в положении 7 (SEQ ID NO: 13).
Таким образом, в одном варианте осуществления антитело содержит последовательность вариабельной области тяжелой цепи (HCVR) SEQ ID NO: 10, где остаток глутаминовой кислоты в положении 6 замещен остатком глутамина (E6Q); где нумерация остатков соответствует нумерации в SEQ ID NO: 10.
В одном варианте осуществления антитело содержит последовательность вариабельной области тяжелой цепи (HCVR) SEQ ID NO: 12, где остаток глутамина в положении 6 замещен остатком глутаминовой кислоты (Q6E); где нумерация остатков соответствует нумерации в SEQ ID NO: 12.
В одном варианте осуществления антитело содержит последовательность вариабельной области легкой цепи (LCVR) SEQ ID NO: 11, где остаток серина в положении 7 замещен остатком треонина (S7T); где нумерация остатков соответствует нумерации в SEQ ID NO: 11.
В одном варианте осуществления антитело содержит последовательность вариабельной области легкой цепи (LCVR) SEQ ID NO: 13, где остаток треонина в положении 7 замещен остатком серина (T7S); где нумерация остатков соответствует нумерации в SEQ ID NO: 13.
В одном варианте осуществления антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3, последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3; и где вариабельная область тяжелой цепи включает последовательность, имеющую, по меньшей мере, 95% идентичность (например, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичность), с последовательностью SEQ ID NO: 10, и вариабельная область легкой цепи содержит последовательность, имеющую, по меньшей мере, 95% идентичность (например, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичность) с последовательностью SEQ ID NO: 11.
В одном варианте осуществления антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3, последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3; и где вариабельная область тяжелой цепи содержит последовательность, имеющую, по меньшей мере, 95% идентичность (например, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичность), с последовательностью SEQ ID NO: 12, и вариабельная область цепи содержит последовательность, имеющую, по меньшей мере, 95% идентичность (например, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичность) с последовательностью SEQ ID NO: 13.
Антитело может содержать последовательность тяжелой цепи (Н-цепи):
SEQ ID NO: 14 варианта 1 тяжелой цепи мышиного полноразмерного IgG1, или
SEQ ID NO: 28 варианта 2 тяжелой цепи мышиного полноразмерного IgG1, или
SEQ ID NO: 30 варианта 1 тяжелой цепи человеческого полноразмерного IgG1, или
SEQ ID NO: 16 варианта 2 тяжелой цепи человеческого полноразмерного IgG1, или
SEQ ID NO: 22 вариант 1 тяжелой цепи человеческого полноразмерного IgG4P или
SEQ ID NO: 34 варианта 2 тяжелой цепи человеческого полноразмерного IgG4P, или
SEQ ID NO: 18 варианта 1 тяжелой цепи Fab, или
SEQ ID NO: 32 варианта 2 тяжелой цепи Fab.
Антитело может содержать последовательность легкой цепи (L-цепи):
SEQ ID NO: 15 варианта 1 легкой цепи мышиного полноразмерного IgG1, или
SEQ ID NO: 29 варианта 2 легкой цепи мышиного полноразмерного IgG1, или
SEQ ID NO: 31 варианта 1 легкой цепи человеческого полноразмерного IgG1, или
SEQ ID NO: 17 варианта 2 легкой цепи человеческого полноразмерного IgG1, или
SEQ ID NO: 23 варианта 1 легкой цепи человеческого полноразмерного IgG4P или
SEQ ID NO: 35 варианта 2 легкой цепи человеческого полноразмерного IgG4P, или
SEQ ID NO: 19 варианта 1 легкой цепи Fab, или
SEQ ID NO: 33 варианта 2 легкой цепи Fab.
В одном примере антитело содержит пару последовательностей тяжелой цепи/легкой цепи:
SEQ ID NO: 14/15 варианта 1 мышиного полноразмерного IgG1, или
SEQ ID NO: 28/29 варианта 2 мышиного полноразмерного IgG1, или
SEQ ID NO: 30/31 варианта 1 человеческого полноразмерного IgG1 или
SEQ ID NO: 16/17 варианта 2 человеческого полноразмерного IgG1, или
SEQ ID NO: 22/23 варианта 1 человеческого полноразмерного IgG1Р, или
SEQ ID NO: 34/35 варианта 2 человеческого полноразмерного IgG1Р, или
SEQ ID NO: 18/19 варианта легкой цепи 1 Fab или
SEQ ID NO: 32/33 варианта легкой цепи 2 Fab.
Вариантные формы соответствующих последовательностей могут быть взаимозаменяемыми. Например, антитело может содержать пару последовательностей тяжелой цепи/легкой цепи:
SEQ ID NO: 14/29 варианта 1 тяжелой цепи /варианта 2 легкой цепи мышиного полноразмерного IgG1, или
SEQ ID NO: 28/15 варианта 2 тяжелой цепи/варианта 2 легкой цепи мышиного полноразмерного IgG1, или
SEQ ID NO: 30/17 варианта 1 тяжелой цепи/варианта 2 легкой цепи человеческого полноразмерного IgG1, или
SEQ ID NO: 16/31 варианта 2 тяжелой цепи/варианта 1 легкой цепи человеческого полноразмерного IgG1, или
SEQ ID NO: 22/35 варианта 1 тяжелой цепи/варианта 2 легкой цепи человеческого полноразмерного IgG4Р, или
SEQ ID NO: 34/23 варианта 2 тяжелой цепи/варианта 1 легкой цепи человеческого полноразмерного IgG4Р, или
SEQ ID NO: 18/33 варианта 1 тяжелой цепи/варианта 2 легкой цепи Fab, или
SEQ ID NO: 32/19 варианта 2 тяжелой цепи/варианта 1 легкой цепи Fab.
Антитела могут быть химерными, человеческими или гуманизированными антителами.
Альтернативно антитело может представлять собой или может включать вариант одной из конкретных последовательностей, указанных выше. Например, вариант может представлять вариант замены, делеции или добавления любой из вышеуказанных аминокислотных последовательностей.
Вариантное антитело может содержать 1, 2, 3, 4, 5, до 10, до 20 или более (обычно максимум до 50) аминокислотных замен и/или делеций из специфических последовательностей, указанных выше. «Делеционные» варианты могут включать делецию отдельных аминокислот, делецию небольших групп аминокислот, например, 2, 3, 4 или 5 аминокислот, или делецию более крупных аминокислотных областей, таких как делеция определенных аминокислотных доменов, или другие свойства. Варианты «замены» обычно включают замену одной или более аминокислот одним и тем же числом аминокислот и проведение консервативных аминокислотных замен. Например, аминокислота может быть замещена альтернативной аминокислотой, имеющей сходные свойства, например, другой основной аминокислотой, другой кислой аминокислотой, другой нейтральной аминокислотой, другой заряженной аминокислотой, другой гидрофильной аминокислотой, другой гидрофобной аминокислотой, другой полярной аминокислотой, другой ароматической аминокислотой или другой алифатической аминокислотой. Некоторые свойства 20 основных аминокислот, которые можно использовать для выбора подходящих заместителей, представляют следующие:
Свойства аминокислот
«Производные» или «варианты», как правило, включают такие, в которых вместо встречающейся в природе аминокислоты находится аминокислота, которая представляет ее структурный аналог. Аминокислоты, используемые в последовательностях, также могут быть дериватизированы или модифицированы, например, помечены, обеспечивая тем самым функцию антитела, которая существенно не изменилась.
Производные и варианты, описанные выше, можно получить во время синтеза антитела или посредством модификации после его получения, или когда антитело находится в рекомбинантной форме с использованием известных методик сайт-направленного мутагенеза, случайного мутагенеза или ферментативного расщепления и/или лигирования нуклеиновых кислот.
Вариантные антитела могут иметь аминокислотную последовательность, которая имеет более чем примерно 60% или более чем примерно 70%, например, 75% или 80%, обычно более чем примерно 85%, например, более чем примерно 90% или 95% идентичность аминокислот с аминокислотными последовательностями, раскрытыми здесь (в частности, последовательности HCVR/LCVR и последовательности H- и L-цепей). Кроме того, антитело может представлять вариант, который имеет более чем примерно 60% или более чем примерно 70%, например, 75% или 80%, обычно более чем примерно 85%, например, более чем примерно 90% или 95% идентичность аминокислот с последовательностями HCVR/LCVR и последовательностями H- и L-цепей, раскрытыми здесь, при сохранении точных CDR, раскрытых для этих последовательностей. Варианты могут сохранять, по меньшей мере, примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с последовательностями HCVR/LCVR и с последовательностями H- и L- цепей, раскрытыми здесь (в некоторых обстоятельствах при сохранении точных CDR).
Варианты обычно сохраняют примерно 60% - примерно 99% идентичность, примерно 80% - примерно 99% идентичность, примерно 90% - примерно 99% идентичность или примерно 95% - примерно 99% идентичность. Такой уровень идентичности аминокислот можно наблюдать по всей длине соответствующей последовательности SEQ ID NO или по части последовательности, такой как примерно 20, 30, 50, 75, 100, 150, 200 или более аминокислот, в зависимости от размера полипептида полной длины.
В отношении аминокислотных последовательностей «идентичность последовательности» относится к последовательностям, которые имеют указанное значение при оценке с использованием ClustalW (Thompson et al., 1994, выше) со следующими параметрами:
параметры попарного выравнивания - метод: точный, матрица: PAM, штраф за открытие разрыва: 10,00, штраф за продолжение разрыва: 0,10;
параметры множественного выравнивания - матрица: PAM, штраф за открытие разрыва: 10,00, % идентичности за задержку: 30, штраф разрывы конца: вкл., расстояние разделения разрыва: 0, отрицательная матрица: нет, штраф за продолжение разрыва: 0,20, штраф за разрывы для остатков: включено, штрафы за гидрофильный разрыв: включено, гидрофильные остатки: GPSNDQEKR. Предполагается, что идентичность последовательности в конкретном остатке включает идентичные остатки, которые были просто дериватизированы.
Антитела, имеющие конкретные последовательности и производные и варианты, которые сохраняют функцию или активность этих цепей, следовательно, предназначены для применения в настоящем изобретении.
Предполагается, что «производные» в контексте настоящего описания включают реакционноспособные производные, например, тиол-селективные реакционноспособные группы, такие как малеимиды и т.п. Реакционноспособная группа может быть связана непосредственно или через линкерный сегмент с полимером. Должно быть понятно, что остаток такой группы будет в некоторых случаях образовывать часть продукта в виде связывающей группы между фрагментом антитела и полимером.
Полимер может быть синтетическим или встречающимся в природе полимером, например, необязательно замещенным полиалкиленовым, полиалкениленовым или полиоксиалкиленовым полимером с прямой или разветвленной цепью или разветвленным или неразветвленным полисахаридом, например, гомо- или гетерополисахаридом.
Конкретные необязательные заместители, которые могут присутствовать в синтетическом полимере, включают одну или более гидроксигрупп, метильных групп или метоксигрупп. Конкретные примеры синтетических полимеров включают необязательно замещенный поли(этиленгликоль) с прямой или разветвленной цепью, поли(пропиленгликоль), поли(виниловый спирт) или его производные, особенно необязательно замещенный поли(этиленгликоль), такой как метоксиполи(этиленгликоль) или его производные. Конкретные встречающиеся в природе полимеры включают лактозу, амилозу, декстран, гликоген или их производные.
Размер полимера может варьироваться, если желательно, но обычно он находится в диапазоне средней молекулярной массы от 500 до 50000 Да, например, от 5000 до 40000 Да, например, от 20000 до 40000 Да. Размер полимера, в частности, может быть выбран на основе предполагаемого применения продукта, например, способности локализоваться в определенных тканях или для увеличения периода полураспада в кровотоке (для обзора см. Chapman, 2002, Advanced Drug Delivery Reviews, 54, 531-545). Таким образом, например, когда продукт предназначен для того, чтобы выходить из кровотока и проникать в ткани, то может быть выгодным использовать полимер с небольшой молекулярной массой, например, с молекулярной массой примерно 5000 Да. Для применений, где продукт остается в кровотоке, может быть выгодным использовать полимер с более высокой молекулярной массой, например, имеющий молекулярную массу в диапазоне от 20000 Да до 40000 Да.
Подходящие полимеры включают полиалкиленовый полимер, такой как поли(этиленгликоль) или, особенно, метоксиполи(этиленгликоль) или его производное, и особенно с молекулярной массой в диапазоне примерно от 15000 Да до примерно 40000 Да.
В одном примере антитела для применения в настоящем изобретении присоединены к поли(этиленгликолевым) (ПЭГ) группам. В одном конкретном примере антитело представляет собой фрагмент антитела, и молекулы ПЭГ могут быть присоединены через любую доступную аминокислотную боковую цепь или концевую аминокислотную функциональную группу, расположенную во фрагменте антитела, например, любую свободную аминокислоту, иминогруппу, тиоловую, гидроксильную или карбоксильную группу. Такие аминокислоты могут встречаться в природе во фрагменте антитела или могут быть встроены во фрагмент с использованием методов рекомбинантной ДНК (см., например, патент США 5219996; патент США 5667425; WO98/25971, WO2008/038024). В одном примере молекула антитела представляет модифицированный Fab-фрагмент, где модификация представляет добавление к С-концу его тяжелой цепи одной или более аминокислот для обеспечения возможности присоединения эффекторной молекулы. Соответственно, дополнительные аминокислоты образуют модифицированную шарнирную область, содержащую один или более остатков цистеина, к которым может быть присоединена эффекторная молекула. Для присоединения двух или более молекул ПЭГ можно использовать несколько сайтов.
Антитела могут конкурировать за связывание с гремлином-1 или связываться с тем же эпитопом, что и те, которые определены выше в отношении последовательностей H-цепи/L-цепи, HCVR/LCVR или CDR. В частности, антитело может конкурировать за связывание с гремлином-1 или связываться с тем же эпитопом, что и антитело, которое содержит комбинацию последовательностей HCDR1/HCDR2/ HCDR3/LCDR1/LCDR2/LCDR3 из SEQ ID NO: 4/5/6/7/8/9. Антитело может конкурировать за связывание с гремлином-1 или связываться с тем же эпитопом, что и антитело, которое содержит пару последовательностей HCVR и LCVR SEQ ID NO: 10/11 или 12/13 или полноразмерных цепей SEQ ID NO: 14/15 или 16/17.
«Эпитоп» представляет собой область антигена, которая связывается антителом. Эпитопы можно определить как структурные или функциональные. Функциональные эпитопы, как правило, представляют подмножество структурных эпитопов и имеют остатки, которые непосредственно обеспечивают аффинность взаимодействия. Эпитопы также могут быть конформационными, т.е. состоять из нелинейных аминокислот. В некоторых вариантах осуществления эпитопы могут включать детерминанты, которые представляют химически активные поверхностные группы молекул, такие как аминокислоты, боковые сахарные цепи, фосфорильные группы или сульфонильные группы, и, в некоторых вариантах осуществления, могут иметь конкретные трехмерные структурные характеристики и/или характеристики удельного заряда.
Можно легко определить, связывается ли антитело с тем же эпитопом, что связывается референсное антитело или конкурирует с ним, используя обычные способы, известные в данной области. Например, чтобы определить, связывается ли тестируемое антитело с тем же эпитопом, что и референсное антитело, для применения в изобретении, референсному антителу дают возможность связываться с белком или пептидом в условиях насыщения. Затем оценивают способность тестируемого антитела связываться с белком или пептидом. Если тестируемое антитело связывается с белком или пептидом после насыщаемого связывания с референсным антителом, то можно сделать вывод, что тестируемое антитело связывается с другим эпитопом, чем референсное антитело. С другой стороны, если тестируемое антитело не связывается с белком или пептидом после насыщаемого связывания с референсным антителом, то тогда тестируемое антитело может связываться с тем же эпитопом, что и эпитоп, связанный с референсным антителом по изобретению.
Для того чтобы определить, конкурирует ли антитело за связывание с референсным антителом, описанная выше методика связывания выполняется в двух ориентациях. В первой ориентации референсному антителу дают возможность связываться с белком/пептидом в условиях насыщения с последующей оценкой связывания тестируемого антитела с молекулой белка/пептида. Во второй ориентации тестируемому антителу дают возможность связываться с белком/пептидом в условиях насыщения с последующей оценкой связывания референсного антитела с белком/пептидом. Если в обеих ориентациях только первое (насыщающее) антитело связывается с белком/пептидом, то делается вывод, что тестируемое антитело и референсное антитело конкурируют за связывание с белком/пептидом. Как будет понятно специалисту в данной области, антитело, которое конкурирует за связывание с референсным антителом, необязательно может связываться с идентичным эпитопом, что и референсное антитело, но может стерически блокировать связывание референсного антитела посредством связывания перекрывающегося или смежного эпитопа.
Два антитела связываются с одним и тем же или перекрывающимся эпитопом, если каждое конкурентно ингибирует (блокирует) связывание другого антитела с антигеном. То есть 1-, 5-, 10-, 20- или 100-кратный избыток одного антитела ингибирует связывание другого антитела, по меньшей мере, на 50%, 75%, 90% или даже 99%, как измерено в анализе конкурентного связывания (см., например, Junghans et al., Cancer Res, 1990: 50: 1495-1502). Альтернативно, два антитела имеют один и тот же эпитоп, если по существу все мутации аминокислот в антигене, которые уменьшают или элиминируют связывание одного антитела, уменьшают или элиминируют связывание другого. Два антитела имеют перекрывающиеся эпитопы, если некоторые мутации аминокислот, которые снижают или элиминируют связывание одного антитела, уменьшают или элиминируют связывание другого.
Затем можно провести дополнительное рутинное экспериментирование (например, мутации пептидов и анализы связывания), чтобы подтвердить, действительно ли наблюдаемое отсутствие связывания тестируемого антитела связано со связыванием с тем же эпитопом, что и у референсного антитела, или стерическое блокирование (или другой феномен) ответственено за отсутствие наблюдаемого связывания. Эксперименты такого рода можно выполнить с использованием ELISA, RIA, поверхностного плазмонного резонанса, проточной цитометрии или любого другого количественного или качественного анализа связывания антител, доступного в данной области.
Было обнаружено, что анти-гремлин-1-антитело из примеров, т.е. антитело Ab7326, связывается со следующими остатками гремлина-1: Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175; где Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175, которые присутствуют на одном мономере гремлина-1, и Ile131, который присутствует на втором мономере гремлина-1. Нумерация основана идентификационном номере последовательности SEQ ID NO: 1 UniProt O60565. Как обсуждается в разделе «Примеры», эти остатки эпитопа были идентифицированы с использованием анализа NCONT при 4  из комплекса гремлин-1-Ab7326 Fab.
из комплекса гремлин-1-Ab7326 Fab.
Следовательно, антитела для применения в изобретении могут связываться с эпитопом, который содержит, по меньшей мере, один остаток, выбранный из Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175 (с нумерацией остатков на основе нумерации в SEQ ID NO: 1). Антитела для применения в изобретении могут связываться с эпитопом, который содержит 2, 3, 4, 5, 6, 7, 8 или все 9 из этих остатков (предпочтительно, по меньшей мере, 5 остатков).
Антитела для применения в изобретении могут также распознавать эпитоп, где Ile131 присутствует на другом мономере гремлина-1 с другими остатками.
Несмотря на то, что эти остатки представлены для конкретной последовательности человеческого гремлина-1, специалист в данной области может экстраполировать положения этих остатков на другие соответствующие последовательности гремлина, используя обычные методы. Поэтому антитела, связывающиеся с эпитопами, содержащими соответствующие остатки в этих других последовательностях гремлина, также предусматриваются для применения в настоящем изобретении.
Для скрининга антител, которые связываются с конкретным эпитопом, можно провести обычный анализ перекрестного блокирования, такой как описан в «Antibodies, Harlow and Lane (Cold Spring Harbor Press, Cold Spring Harb., NY). Другие методы включают аланиновое сканирование мутантов, пептидный блоттинг (Reineke (2004) Methods Mol Biol 248:443-63) или анализ расщепления пептидов. Кроме того, могут быть использованы методы, такие как вырезание эпитопа, экстракция эпитопа и химическая модификация антигенов (Tomer (2000) Protein Science: 9: 487-496). Такие методы хорошо известны в данной области.
Эпитопы антител также можно определить с помощью рентгеновской кристаллографии. Следовательно, антитела для применения в настоящем изобретении можно оценить с помощью рентгеновской кристаллографии антитела, связанного с гремлином-1. Эпитопы, в частности, могут быть идентифицированы определением остатков на гремлине-1 в пределах 4 от остатка паратопа антитела.
Антитела можно тестировать на связывание с гремлином-1, например, стандартным анализом ELISA или вестерн-блоттингом. Анализ ELISA также можно использовать для скрининга гибридом, которые показывают положительную реактивность с белком-мишенью. Селективность связывания антитела также можно определить мониторингом связывания антитела с клетками, экспрессирующими белок-мишень, например, проточной цитометрией. Таким образом, способ скрининга может включать стадию идентификации антитела, способного связываться с гремлином-1, проведением ELISA или вестерн-блоттинга или проточной цитометрии.
Антитела могут избирательно (или специфически) распознавать гремлин-1. Антитело или другое соединение «селективно связывает» или «избирательно распознает» белок, когда он связывается с преимущественной или высокой аффинностью к белку, для которого оно селективно, но по существу не связывается или связывается с низкой аффинностью с другими белками. Селективность антитела может быть дополнительно исследована определением того, связывается ли антитело с другими родственными белками, как обсуждалось выше, или насколько оно различает их. Антитела для применения в изобретении обычно распознают человеческий гремлин-1.
Антитела также могут обладать перекрестной реактивностью для родственных белков или человеческого гремлина-1 и гремлина-1 других видов.
Под термином «специфическое (или селективное)» антитело следует понимать, что антитело связывается с белком, представляющим интерес, без существенной перекрестной реактивности с какой-либо другой молекулой. Перекрестная реактивность может быть оценена любым подходящим способом, описанным здесь. Перекрестная реактивность антитела может считаться значительной, если антитело связывается с другой молекулой, по меньшей мере, примерно на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 100% так сильно, как оно связывается с интересующим белком. Антитело, которое является специфическим (или селективным), может связываться с другой молекулой менее чем на 90%, 85%, 80%, 75%, 70%, 65%, 60%, 55%, 50%, 45%, 40% 35%, 30%, 25% или 20% от силы, с которой оно связывается с интересующим белком. Антитело может связываться с другой молекулой с силой менее чем примерно 20%, менее чем примерно 15%, менее чем примерно 10% или менее чем примерно 5%, менее чем примерно 2% или менее чем примерно 1%, с которой оно связывается с представляющим интерес белком.
Таким образом, антитела, подходящие для применения в настоящем изобретении, могут иметь высокую аффинность связывания с (человеческим) гремлином-1. Антитело может иметь константу диссоциации (KD) ниже <1 нМ и предпочтительно <500 пМ. В одном примере антитело имеет константу диссоциации (KD) ниже 200 пМ. В одном примере антитело имеет константу диссоциации (KD) ниже 100 пМ. Для определения аффинности связывания антитела с его антигеном-мишенью можно использовать различные методы, такие как анализ поверхностным плазмонным резонансом, анализ насыщения или иммуноанализы, такие как ELISA или RIA, которые хорошо известны специалистам в данной области. Примерным методом определения аффинности связывания является поверхностный плазмонный резонансный анализ на приборе BIAcore™ 2000 (Biacore AB, Фрайбург, Германия) с использованием сенсорных чипов CM5, как описано Krinner et al., (2007) Mol. Immunol. Feruary; 44 (5): 916-25. (Epub 2006 11 May)).
Анти-гремлин-1-антитело из примеров, т.е. Ab7326, является аллостерическим ингибитором активности гремлина-1, которое связывается с эпитопом, удаленным от сайта связывания с BMP (WO 2018/115017 A2). Ab7326 связывается с гремлином-1 с исключительно высокой аффинностью со значением Kd <100 пМ, и полагается, что оно будет особенно пригодным для применения в настоящем изобретении.
Ингибитор активности гремлина-1 может оказывать влияние на любую из функций гремлина-1, но обычно уменьшает связывание гремлина-1 с BMP (BMP 2, 4 и/или 7). Гремлин-1 является негативным регулятором BMP и, таким образом, пониженное связывание повышает передачу сигналов через BMP.
Связывание и передачу сигналов BMP можно детектировать любым способом, известным в данной области. В примерах настоящей заявки описываются два функциональных анализа для тестирования того, насколько агент уменьшает связывание гремлина-1 с BMP. В примере 3 описан анализ репортерного гена Id1, где ген Id1 является геном-мишенью сигнального пути BMP. Повышение сигнала в данном анализе можно использовать для определения того, насколько агент уменьшает связывание гремлина-1 с BMP. В примере 5 описан анализ фосфорилирования SMAD. SMAD 1, 5 и 8 фосфорилируются на сигнальном пути BMP. Следовательно, повышение фосфорилирования SMAD можно использовать для определения того, насколько агент уменьшает связывание гремлина-1 с BMP.
Как только подходящее антитело идентифицировано и выбрано, то может быть идентифицирована аминокислотная последовательность антитела методами, известными в данной области. Гены, кодирующие антитело, могут быть клонированы с использованием вырожденных праймеров. Антитело можно получить рекомбинантным путем с использованием обычных методов.
Примеры последовательностей ДНК, кодирующих полноразмерные тяжелые цепи и легкие цепи Ab7326, представлены в списке последовательностей:
SEQ ID NO: 24 (вариант 1 тяжелой цепи человеческого IgG1)
SEQ ID NO: 25 (вариант ДНК легкой цепи человеческого IgG1)
SEQ ID NO: 26 (вариант 1 тяжелой цепи ДНК человеческого IgG4P)
SEQ ID NO: 27 (вариант 1 легкой цепи человеческого IgG4P).
Фармацевтические композиции, дозы и режимы дозирования
Ингибитор активности гремлина-1 для применения в настоящем изобретении может находиться в фармацевтической композиции. Фармацевтическая композиция обычно должна быть стерильной и обычно будет включать фармацевтически приемлемый носитель и/или адъювант. Фармацевтическая композиция для применения в изобретении может дополнительно содержать фармацевтически приемлемый адъювант и/или носитель.
Фармацевтические композиции для применения в изобретении могут включать одну или более фармацевтически приемлемых солей. «Фармацевтически приемлемая соль» относится к соли, которая сохраняет желаемую биологическую активность исходной молекулы и не оказывает каких-либо нежелательных токсикологических эффектов. Примеры таких солей включают аддитивные соли кислоты и аддитивные соли основания.
Как здесь используется, термин «фармацевтически приемлемый носитель» включает любые и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие всасывание агенты и тому подобное, которые являются физиологически совместимыми. Носитель может быть подходящим для парентерального введения, например внутривенного, внутримышечного, внутрикожного, внутриглазного, внутрибрюшинного, подкожного, интраспинального или другого парентерального пути введения, например, путем инъекции или инфузии. Альтернативно, носитель может быть подходящим для непарентерального введения, такого как местный, эпидермальный или мукозальный путь введения. Носитель может быть пригоден для перорального введения. В зависимости от пути введения ингибитор может быть покрыт материалом, защищающим его от действия кислот и других естественных условий, которые могут инактивировать ингибитор.
Фармацевтически приемлемые носители включают водные носители или разбавители. Примеры подходящих водных носителей, которые можно использовать в фармацевтических композициях для применения в изобретении, включают воду, забуференную воду и физиологический раствор. Примеры других носителей включают этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Во многих случаях желательно включить в композицию изотонические агенты, например сахара, полиспирты, такие как маннит, сорбит или хлорид натрия.
Фармацевтические композиции обычно должны быть стерильными и стабильными в условиях производства и хранения. Композиция может быть формулирована в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для обеспечения высокой концентрации лекарственного средства.
Фармацевтические композиции для применения по изобретению могут содержать дополнительные активные ингредиенты.
Также обеспечиваются наборы, содержащие ингибитор активности гремлина-1 и инструкции по применению в способе лечения согласно изобретению.
Агенты для применения в изобретении или их составы или композиции можно вводить для терапевтического и/или профилактического лечения.
В терапевтических применениях агенты вводят субъекту, уже страдающему расстройством или патологическим состоянием, в количестве, достаточном для излечения, облегчения или частичного купирования состояния или одного или более его симптомов. Такое терапевтическое лечение может привести к уменьшению выраженности симптомов или увеличению частоты или продолжительности бессимптомных периодов. Количество, достаточное для достижения этого, определяется как «терапевтически эффективное количество».
В профилактических применениях агенты вводят субъекту, подверженному риску развития расстройства или патологического состояния, в количестве, достаточном для предупреждения или ослабления последующих негативных эффектов патологического состояния или одного или более его симптомов. Количество, достаточное для достижения этого, определяется как «профилактически эффективное количество». Эффективные количества для каждой цели будут зависеть от тяжести заболевания или травмы, а также от массы тела и общего состояния субъекта.
Субъектом, которому проводят введение, может представлять человека или животное, отличное от человека. Термин «животное, отличное от человека» включает всех позвоночных животных, например млекопитающих и не млекопитающих, таких как приматы, отличные от человека, собаки, кошки, лошади, овцы, коровы, цыплята, амфибии, рептилии и т. п. Типичным является введение людям.
Агент или фармацевтическую композицию для применения в изобретении можно вводить одним или несколькими путями введения, используя один или более из множества способов, известных в данной области. Как будет понятно специалисту в данной области, путь и/или способ введения будут варьироваться в зависимости от желаемых результатов. Примеры путей введения агентов или фармацевтических композиций для применения в изобретении включают парентеральные пути, такие как внутривенный, внутримышечный, внутрикожный, внутриглазный, внутрибрюшинный, подкожный или интраспинальный пути введения, например, путем инъекции или инфузии. Альтернативно, агент или фармацевтическую композицию можно вводить непарентеральным путем, таким как местный, эпидермальный или мукозальный путь введения. Агент или фармацевтическая композиция могут быть формулированы для перорального введения.
Подходящую дозу ингибирующего агента или фармацевтической композиции для применения в изобретении может определить квалифицированный практикующий врач. Фактические уровни доз активных ингредиентов в фармацевтических композициях для применения в настоящем изобретении могут варьироваться таким образом, чтобы получить количество активного ингредиента, эффективное для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения, не будучи токсичным для пациента. Выбранный уровень доз будет зависеть от многих фармакокинетических факторов, включая активность конкретных используемых композиций, путь введения, время введения, скорость выведения конкретного используемого соединения, продолжительность лечения, возраст пол, массу тела, состояние, общее состояние здоровья и предшествующая история болезни пациента, который подвергается лечению, и подобные факторы, хорошо известные в медицине.
Подходящая доза может находиться, например, в диапазоне примерно от 0,01 мкг/кг до примерно 1000 мг/кг массы тела, обычно примерно от 0,1 мкг/кг до примерно 100 мг/кг массы тела пациента, который подвергается лечению. Например, подходящая дозировка может составлять примерно от 1 мкг/кг до примерно 10 мг/кг массы тела в день или примерно от 10 мкг/кг до примерно 5 мг/кг массы тела в день.
Режимы дозирования можно корректировать для обеспечения получения оптимального желаемого ответа (например, терапевтического ответа). Например, может быть введена одна доза, несколько разделенных доз можно ввести во времени, или дозу можно пропорционально снизить или повысить, что диктуется терапевтической ситуацией. Как здесь используется, разовая лекарственная форма относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; где каждая единица содержит заданное количество активного агента, рассчитанное для получения желаемого терапевтического эффекта в сочетании с необходимым фармацевтическим носителем.
Введение может проводиться в одной или многократных дозах. Многократные дозы можно вводить одним и тем же или разными путями и в одинаковые или разные места введения. Альтернативно, дозы могут быть обеспечены с помощью композиции с замедленным высвобождением, и в этом случае требуется менее частое введение. Дозировка и частота введения могут варьироваться в зависимости от периода полувыведения ингибирующего агента у пациента и желаемой продолжительности лечения.
Агенты, составы или фармацевтические композиции для применения в изобретении можно вводить совместно с одним или более другими терапевтическими агентами. Комбинированное введение двух или более агентов может быть достигнуто различными способами. Оба можно вводить вместе в одной композиции, или они могут быть введены в отдельных композициях в виде части комбинированной терапии. Например, один может быть введен до, после или одновременно с другим.
Терапевтические показания
Ингибиторы активности гремлина-1 по настоящему изобретению обеспечены для лечения перелома кости или дефекта кости. Перелом кости представляет собой разрыв или трещину в костной ткани и может быть результатом травматического повреждения, такого как падение или удар, но также может возникать в результате заболеваний, которые отрицательно влияют на целостность кости. Костный дефект представляет потерю костной ткани вследствие травмы или заболевания.
Перелом может представлять перелом любой кости в организме.
Дефект кости может быть костным дефектом в любой кости тела.
В одном варианте осуществления перелом кости представляет собой перелом с замедленным или несращенным соединением. Перелом с замедленным сращением определяется как перелом, который не достигает сращения в течение 6 месяцев после перелома. Несращенный перелом определяется как неполное заживление в течение 9 месяцев, в сочетании с отсутствием рентгенологических показателей, свидетельствующих о заживлении перелома, наблюдаемого в течение трех последовательных месяцев. (Einhorn et al.; 2014; Buza et al.; 2016). Примеры переломов, которые склонны к развитию с замедленным или несращенным соединением, включают перелом большеберцовой кости, дистального радиуса, шейки бедра и ладьевидной кости.
В одном варианте осуществления перелом кости или дефект кости происходит в результате заболевания, которое оказывает отрицательное влияние на целостность кости. Примеры заболеваний, которые негативно влияют на целостность кости, включают, не ограничиваясь этим, остеопороз, несовершенный остеогенез, диабет, болезнь Педжета, ревматоидный артрит, анкилозирующий спондилит, множественную миелому, первичный рак кости (например, остеосаркома, саркома Юинга и хондросаркома), рак с метастазами в кости (например, рак молочной железы, рак предстательной железы и рак легкого), диффузный идиопатический гиперостоз скелета, остеомиелит, почечную недостаточность, мышечную дистрофию Дюшенна и талассемию.
Краткое описание фигур
На фиг.1 показано процентное восстановление сигнала для антител, полученных иммунизацией, в анализе репортерного гена HEK-ID1.
На фиг.2 показано процентное восстановление сигнала для антител, полученных из библиотеки, в анализе репортерного гена HEK-ID1.
На фиг.3 приведены результаты анализа репортерного гена HEK-ID1 с титрованием человеческого гремлина (фиг.3А) и мышиного гремлина (фиг.3В), и влияние антитела 7326 (показано как антитело РВ376) в восстановлении передачи сигналов с участием BMP.
На фиг.4 показана структурная модель комплекса гремлин-Fab с выделением возможных областей связывания BMP и эпитопа Fab.
На фиг. 5 показано исследование области, лишенной костной мозоли/костной ткани, во время восстановления перелома на полученных рентгеновских снимках. Область внутри дефекта, которая была лишена ткани, определяли количественно с использованием анализа изображений и затем сравнением контрольной группы и группы, обработанной анти-гремлином 1-антителом. Результаты представлены в виде среднего значения ± SD для 10 крыс/группе. * Р <0,05; ** Р <0,01; *** P <0,001, как измерено U-критерием Манна-Уитни.
На фиг. 6 показано исследование LMB (кость с низким содержанием минералов; новообразованная кость) и HMB (кость с высоким содержанием минералов; зрелая кость) в пределах 3 мм дефекта бедренной кости. Панель A: 3D микро-КТ анализ области дефекта бедренной кости для выявления новообразованной или зрелой кости. Измеряли процент объема кости/объема ткани и сравнивали всех субъектов (в целом) в контрольной группе и группе, обработанной анти-гремлин 1-антителом. Также сравнивали контрольную группу с опытной группой, обработанной анти-гремлин 1-антителом, у животных, разделенных на респондеры с низким уровнем ответа (неполное соединение LR) и респондеры с высоким уровнем ответа (полное соединение HR). Результаты представлены в виде среднего значения ± стандартное отклонение. * Р <0,05; ** Р <0,01; *** P <0,001, как измерено U-критерием Манна-Уитни. Панель В: репрезентативная микро-КТ, показывающая 3D изображения объема кости в группах LR и HR в контроле и после обработки анти-гремлин 1-антителом.
На фиг.7 приведены результаты гистоморфометрического анализа дефекта бедренной кости. Процентное соотношение объема кости/объема ткани (BV/TV (%)), трабекулярное число (Tb.N) и трабекулярное разделение (Tb.Sp) сравнивали между контрольной группой и группой, обработанной анти-гремлин 1-антителом. Результаты представлены в виде среднего значения ± SD для 10 крыс / группа. * Р <0,05; ** Р <0,01; *** P <0,001, как измерено U-критерием Манна-Уитни.
На фиг. 8 показана корреляция 3D микро-КТ анализа и 2D гистоморфометрического анализа общего BV/TV%. Корреляции проводили в обеих группах на LMB (кость с низким содержанием минералов; новообразованная кость) и HMB (кость с высоким содержанием минералов; зрелая кость) в пределах 3 мм дефекта бедренной кости и по сравнению с данными оценки 2D гистоморфометрии (n=20). Оценка анализом Пирсона указывает на наличие достоверной корреляции BV/TV% между данными 3D микро-КТ анализа и 2D гистоморфометрического анализа.
Следующие примеры иллюстрируют изобретение.
Примеры
Пример 1. Экспрессия, очистка, рефолдинг и определение структуры белка.
Экспрессия белка и получение телец включения
Усеченную кодирующую последовательность человеческого гремлина-1 (SEQ ID NO: 20), оптимизированную для экспрессии в E.coli, клонировали в модифицированный вектор pET32a (Merck Millipore) с использованием BamHI/XhoI, с получением вектора, кодирующего последовательность гремлина с N-концевой меткой 6His-TEV (pET-hGremlin1).
Экспрессированная последовательность:
MGSSHHHHHHSSGENLYFQGSAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD; SEQ ID NO: 2 (с остатками, не относящимися к остаткам гремлина, меткой 6His-TEV, выделенной курсивом). Нумерация последовательностей основана на UniProt O60565 и SEQ ID NO: 1.
ДНК-плазмиду pET-hGremlin1 использовали для трансформации клеток BL21 (DE3). Единственную колонию, резистентную к ампициллину, собирали с чашки с агаром LB/Amp и использовали для инокуляции 100 мл стартовой культуры LB/Amp. После встряхивания (200 об/мин) в течение 16 ч при 37°С 25 мл стартовой культуры использовали для инокуляции 500 мл среды 2xTY/Amp. Культуру встряхивали (250 об/мин) при 37°С до достижения OD 600, равной 3. Затем к культуре добавляли 20 мл питательной смеси MOPS + глицерин (1M MOPS pH 7,4, 40% глицерин, 0,5% MgSO4, 0,42% MgCl2), индуцировали 300 мкМ IPTG и затем инкубировали при 17°C, 180 об/мин в течение 16 ч. Клетки собирали центрифугированием (4000 g в течение 20 мин при 4°С).
Клеточные осадки после центрифугирования ресуспендировали в буфере для лизиса (PBS pH 7,4, 0,35 мг/мл лизоцима, 10 мкг/мл ДНКазы и 3 мМ MgCl2) при 4°C, и нерастворимую фракцию собирали центрифугированием при 3500 g в течение 30 мин при 4°C. Отцентрифугированные тельца включения трижды промывали ресуспендированием в промывочном буфере (50 мМ Трис, 500 мМ NaCl, 0,5% Тритон Х-100, рН 8,0) с последующим центрифугированием при 21000 g в течение 15 мин. Два дополнительных промывания выполняли с использованием промывочного буфера без тритона X-100.
Солюбилизация
Тельца включения ресуспендировали в денатурирующем буфере (8 М мочевины, 100 мМ Трис, 1 мМ ЭДТА, 10 мМ Na2S4O6 и 100 мМ Na2SO3, рН 8,5), перемешивали в течение 16 ч при комнатной температуре и осветляли центрифугированием при 21000g в течение 15 мин.
Очистка перед рефолдингом
Солюбилизированные тельца включения наносили на колонку с Sephacryl S-200 26/60 (120 мл), уравновешенную буфером из 8 М мочевины, 50 мМ MES, 200 мМ NaCl, 1 мМ ЭДТА, рН 6,0. Фракции, содержащие белок гремлин-1, разбавляли 6 М мочевиной, 20 мМ MES, pH 6,0 и загружали на катионообменные колонки HiTrap SP HP и элюировали в градиенте 1 М NaCl 10 объемами колонки (10 CV). Фракции, содержащие очищенный денатурированный белок hGremlin-1, объединяли.
Рефолдинг
Денатурированный очищенный белок гремлин-1 добавляли по каплям в буфер для рефолдинга (50 мМ Трис, рН 8,5, 150 мМ NaCl, 5 мМ GSH и 5 мМ GSSG, 0,5 мМ цистеина, 5 мМ ЭДТА, 0,5 М аргинина) до конечной концентрации 0,1 мг/мл и инкубировали при 4°С при постоянном перемешивании в течение 5 суток. Через 5 суток белок гремлин-1 подвергали диализу против 20 мМ HEPES, 100 мМ NaCl, рН 7,5.
После диализа белок наносили на колонку с гепарином HiTrap и элюировали, используя градиент 0-100% буфера для элюирования с гепарином (20 мМ HEPES, 1 М NaCl, pH 7,5) с использованием 20 CV. Правильно свернутый белок элюируется при 1 М NaCl, тогда как любой неправильно свернутый белок элюируется при более низких концентрациях соли.
Белок, элюированный при 1 М NaCl, концентрировали и дополнительно очищали на колонке S75 26/60, уравновешенной 20 мМ Hepes, pH 7,5, 1 М NaCl.
Белок характеризовали SDS-PAGE (сдвиг в геле), было показано, что он имеет ожидаемую молекулярную массу и правильное расположение дисульфидных связей с использованием жидкостной хроматографии, масс-спектрометрии (LC-MS) и активен в клеточном анализе (репортерный анализ ID1).
Определение структуры гремлина-1
Кристаллы белка гремлина-1 выращивали, используя метод «висячей капли», смешивая раствор гремлина-1 в концентрации 6,6 мг/мл и 0,1 М лимонной кислоты при pH 4, 1 М хлорида лития и 27% полиэтиленгликоля (PEG) 6000 в соотношении 1:1. Перед сбором данных, кристаллы подвергались криопротекции добавлением 20% глицерина в буфер для кристаллизации. Данные дифракции собирали на алмазном источнике света Diamond Light Source и обрабатывали с использованием XDS (Kabsch, Wolfgang (2010) Acta Crystallographica Section D 66, 125-132). Данные диффракционной статистики обобщены в таблице ниже:
Данные диффракционной статистики ), b=107,22 , c=77,09 ; α=90,00°, β=120,43°, γ=90,00°)))**
**r.m.s.d среднеквадратичное отклонение.
Структура гремлина-1 была разрешена посредством молекулярного замещения с использованием Phaser (McCoy et al., J Appl Cryst (2007), 40, 658-674) и модели гремлина-1, доступной из определенных координат комплекса гремлин-1/Fab. Полученная модель гремлина-1 содержала четыре копии мономера гремлина-1, организованные в виде двух димеров. Модельные корректировки делали с использованием Coot (Emsley et al. Acta Crystallographica Section D: Biological Crystallography 66 (4), 486-501), и координаты уточняли с помощью Refmac (Murshudov et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallographica Section D: Biological Crystallography. 2011;67(Pt4):355-367). Конечные координаты подтверждали с помощью Molprobity (Chen et al. (2010) MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallographica D66:12-21). Обобщение статистики модели уточнения приведено в таблице 2 выше.
Пример 2. Остатки на гремлине-1, связывающиеся с BMP
Как обсуждалось выше, гремлин-1 относится к семейству белков-антагонистов костного морфогенного белка (BMP) в подгруппе, известной как семейство DAN. В семействе DAN гремлин-1 разделяет наибольшую гомологию с гремлином-2 (PRDC).
Структура гремлина-1 человека 2,7 , разрешенная в примере 1, имеет много признаков, общих с опубликованной структурой гремлина-2 мыши (Nolan et al. (2013), Structure, 21, 1417-1429). Общий фолдинг очень похож: две копии гремлина-1 образуют антипараллельный нековалентный димер, расположенный в изгибе. Каждый мономер имеет характерное расположение «палец-запястье-палец» с мотивом цистинового узла в направлении к концу «пальца», противоположном «запястью». Идентичность последовательности между белками составляет от 52% до 67% в последовательности, видимой в двух структурах. Наиболее высоко консервативная область находится на обширной поверхности раздела димера, где все ключевые контактные остатки сохранены на 100%.
Остатки, вовлеченные в связывание BMP 2, 4 и 7 с мышиным гремлином-2 (PRDC) и DAN (NBL1), были идентифицированы с использованием мутагенеза (Nolan et al. (2013), Structure, 21, 1417-1429 и Nolan et al. (2014). J. Biol. Chem. 290, 4759-4771). Предсказанный эпитоп связывания BMP включает гидрофобный участок, охватывающий оба мономера на выпуклой поверхности димера. С использованием мутагенеза было идентифицировано шесть остатков; Trp72, Phe96, Tyr98, Phe104, Tyr105 и Phe117 и являются на 100% консервативными в человеческом гремлине-1 (нумерация основана на нумерации последовательности мышиного гремлина-2). Степень гомологии распространяется на расположение боковых цепей, которые принимают одинаковую конформацию в обоих белках.
Нумерация аминокислот, используемая в файле Gremlin PDB, соответствует нумерации в опубликованной структуре мышиного гремлина-2 на основе структурного выравнивания. Это позволяет проводить одинаковое сравнение аминокислот при описании структур. Однако для ясности ключевые остатки, идентифицированные как играющие важную роль в связывании с BMP, показаны ниже с нумерацией на основе файла PDB и файла UniProt с SEQ ID NO: 1 в скобках:
Trp72 (93), Phe96 (117), Tyr98 (119), Phe104 (125), Tyr105 (126) и Phe117 (138).
Как в мышином гремлине-2, так и в человеческом гремлине-1 гидрофобный связывающий эпитоп BMP частично находится под альфа-спиралью, образованной N-концевыми остатками каждого белка. Была предложена модель связывания BMP, посредством которой N-конец может изгибаться, обнажая полную поверхность связывания BMP (Nolan et al. (2013), Structure, 21, 1417-1429). В настоящем анализе N-концевые остатки удаляли из структур человеческого гремлина-1 и мышиного гремлина-2 перед визуализацией поверхности, для выявления сходства поверхностей, связывающих BMP, на каждым белке.
В литературе описан мутагенез только шести остатков, которые влияют на связывание BMP. Возможно, что фактический эпитоп BMP охватывает большую площадь поверхности, включающую соседние аминокислоты. Выделив все остатки, в пределах 6 от тех, которые мутировали, на поверхности гремлина-1, обнаружена большая область гремлина-1, на которую потенциально может быть нацелено терапевтическое средство. Этот более обширная область охватывает следующие аминокислоты человеческого гремлина-1:
Asp92-Leu99
Arg116-His130
Ser137-Ser142
Cys176-Cys178
(нумерация основана на нумерации SEQ ID NO: 1).
Сочетая опубликованную информацию с информацией о кристаллической структуре человеческого гремлина-1, были идентифицированы области человеческого гремлина-1, которые предположительно можно рассматривать в качестве потенциального пути для терапевтического вмешательства, блокирующего его взаимодействие с BMP.
Пример 3. Анализ репортерного гена Hek Id1.
Основа
В анализе репортерного гена Hek Id1 используются репортерные клетки клона 12 Hek293-Id1. Эту клеточную линию стабильно трансфектировали транскрипционным фактором Id1. Id1 является транскрипционным фактором в сигнальном пути BMP. Гремлин, как известно, связывается с BMP, предотвращая связывание с их рецепторами, уменьшая сигнал люциферазы из репортерного гена. Следовательно, используя этот репортерный анализ, можно проводить скрининг антител против гремлина и выявить антитела, которые блокируют взаимодействие гремлина с BMP. В этих клетках наблюдается восстановление сигнала люциферазы, если происходит блокирование этого взаимодействия.
Метод
Клон 12 клеток культивировали в среде DMEM, содержащей 10% FCS, 1× L-глутамина и 1× NEAA. Клетки также культивировали в присутствии гигромицина B (200 мкг/мл) для гарантии того, что клетки не теряют экспрессию гена Id1. Клетки анализировали в DMEM, содержащей 0,5% FCS, 1× L-глутамин и 1× NEAA. Гигромицин В не требуется в течение короткого периода времени, когда клетки находятся в анализе.
Клетки промывали с использованием PBS, снимали с использованием буфера для диссоциации клеток, центрифугировали и подсчитывали перед посевом из расчета 5×104/лунку в 70 мкл (плотность 7,14×105/мл). Используемые планшеты представляли 96-луночные стерильные, покрытые поли-D-лизином непрозрачные планшеты белого цвета. Клетки помещали в термостат примерно на 3-4 ч для оседания. Гетеродимеры BMP восстанавливали до 200 мкг/мл в 4 мМ HCl. BMP разбавляли до 10 мкг/мл в среде для анализа, используя стеклянный флакон для получения нового рабочего стокового запаса.
В полипропиленовом планшете гремлин-1 разводили 1:2 для получения 8-точечной кривой доза-ответ с максимальной конечной дозой 1 мкг/мл.
Добавляли дополнительный объем 20 мкл среды на лунку и планшеты инкубировали при 37°С в течение 45 мин.
BMP, полученный при 100×, добавляли во все лунки, кроме лунок, содержащих только клетки. Содержимое всех лунок доводили до 60 мкл аналитической средой и инкубировали в течение еще 45 мин при 37°С.
После инкубации 30 мкл образца переносили в лунку планшета для анализа и инкубировали в течение 20-24 ч перед измерением сигнала люминесценции.
Реагент Cell Steady Glo оттаивали заранее при комнатной температуре. Планшеты для анализа охлаждали до комнатной температуры в течение примерно 10-15 мин перед добавлением реагента. Сигнал люциферазы детектировали добавлением реагента Cell Steady Glo (100 мкл) в течение 20 мин на шейкере при комнатной температуре и измерением люминесценции с использованием протокола определения титра клеток на Synergy 2.
Максимальный сигнал генерировался из лунок, содержащих BMP, и минимальный сигнал генерировался из лунок, содержащих только клетки.
Результаты
Полноразмерные и усеченные формы гремлина-1 тестировали в анализе репортерного гена Hek-Id1, чтобы подтвердить блокирующую активность против BMP4/7.
Процент ингибирования в анализе доза-ответ рассчитывали на основе максимальных и минимальных сигналов в анализе и данных, подобранных с использованием 4-параметрической логистической модели. Значение IC50 рассчитывали на точке перегиба кривой.
Результаты эффективности для полноразмерного гремлина-1 и усеченного гремлина-1 в анализе репортерного гена Hek-Id1
Заключение
Гремлин-1 был способен ингибировать сигнальный путь BMP 4/7 в анализе репортерного гена Hek-Id1.
Пример 4. Получение анти-гремлин-1-антитела
Анти-гремлин-1-антитела получали иммунизацией с использованием очищенного гремлина-1, как описано в примере 1, и посредством пэннинга библиотеки. Библиотека была создана на месте в виде наивной человеческой библиотеки с V-областями, амплифицированными из предоставленной крови.
В результате иммунизации было получено 26 различных антител, связывающихся с гремлином-1, из первого раунда иммунизации. Эти антитела были масштабированы и очищены для тестирования в скрининговых анализах.
25 перекрестно-реактивных антител человека и мыши из библиотеки подвергали пэннингу с использованием рекомбинантного человеческого гремлина от R & D Systems. 10 антител были отобраны для масштабирования и выделены в виде scFv для тестирования в скрининговых анализах.
Пример 5. Скрининг анти-гремлин-1 антител
Антитела подвергали скринингу с использованием анализа репортерного гена Hek-Id1, описанного в примере 3, и измерением фосфорилирования SMAD. SMAD 1, 5 и 8 фосфорилируются на сигнальном пути BMP. Следовательно, ингибиторы гремлина-1 повышают фосфорилирование SMAD.
Анализ фосфорилирования SMAD проводили на клетках A549 или на фибробластах легких человека. Уровни фосфорилирования определяли с использованием MSD.
Результаты
В анализе репортерного гена Hek-Id1 отсутствовали видимые попадания с антителами, полученными в результате иммунизации (с 10-кратным избытком антител, тестированных против гетеродимера BMP4/7). Результаты показаны на фиг. 1.
Напротив, ряд антител, полученных из библиотеки, был способен восстанавливать сигнал в анализе репортерного гена Hek-Id1 (50-кратный избыток антител с 50% дозой гремлина) (фиг. 2). Из них Ab2416 и Ab2417 содержали высокие уровни эндотоксина. Ab7326 поддерживало блокирующую способность при 10-кратном избытке и 80% ингибировании концентрации гремлина-1.
Дополнительные результаты представлены на фиг. 3А (человеческий гремлин) и 3В (мышиный гремлин). На этих фигурах показано титрование Ab7326 (обозначено как PB376) до 15 нМ. Было показано, что Ab7326 восстанавливает передачу сигналов BMP при блокировании человеческим (IC50 1,3 нМ) или мышиным (IC50 0,2 нМ гремлина). Антитело функционирует в виде человеческого и мышиного IgG1.
Последовательности полноразмерного мышиного и человеческого IgG1 представлены ниже. Для синтеза полноразмерных IgG1 белков мыши и человека вариабельные области Ab7326, полученные из библиотеки, повторно клонировали в векторы, содержащие соответствующие константные домены антител.
Поскольку Ab7326 было получено из наивной человеческой библиотеки, где Ab клонировали в виде scFvs, для повторного клонирования вариабельных областей 7326 в виде полноразмерных Ab или Fab, было необходимо провести ПЦР-амплификацию VH и VK с использованием пулов праймеров/вырожденных праймеров. Затем продукты, амплифицированные ПЦР, расщепляли и клонировали одновременно в векторы мыши и человека. Поскольку VH и VK были амплифицированы пулами праймеров/вырожденных праймеров, то получали две формы продуктов, отличающихся одним аминокислотным остатком, полученным в результате несколько различного отжига праймеров во время процесса ПЦР.
Две вариантные формы вариабельной области тяжелой цепи отличались одной аминокислотой в положении 6, и две вариантные формы вариабельной области легкой цепи отличались одной аминокислотой в положении 7, как показано ниже:
- вариант 1 вариабельной области тяжелой цепи содержит глутаминовую кислоту (E) в положении 6;
- вариант 2 вариабельной области тяжелой цепи содержит глутамин (Q) в положении 6;
- вариант 1 вариабельной области легкой цепи имеет серин (S) в положении 7;
- вариант 2 вариабельной области легкой цепи содержит треонин (T) в положении 7.
Мышиный полноразмерный IgG1 - вариант 1 тяжелой цепи (SEQ ID NO: 14)
QVQLVESGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA RGSGSYYPNH FDYWGQGTLV TVSSAKTTPP SVYPLAPGSA AQTNSMVTLG CLVKGYFPEP VTVTWNSGSL SSGVHTFPAV LQSDLYTLSS SVTVPSSTWP SETVTCNVAH PASSTKVDKK IVPRDCGCKP CICTVPEVSS VFIFPPKPKD VLTITLTPKV TCVVVDISKD DPEVQFSWFV DDVEVHTAQT QPREEQFNST FRSVSELPIM HQDWLNGKEF KCRVNSAAFP APIEKTISKT KGRPKAPQVY TIPPPKEQMA KDKVSLTCMI TDFFPEDITV EWQWNGQPAE NYKNTQPIMD TDGSYFVYSK LNVQKSNWEA GNTFTCSVLH EGLHNHHTEK SLSHSPGK
Мышиный полноразмерный IgG1 - вариант 1 тяжелой цепи (SEQ ID NO: 15)
DIVMTQSPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT PTFGQGTRLE IKRTDAAPTV SIFPPSSEQL TSGGASVVCF LNNFYPKDIN VKWKIDGSER QNGVLNSWTD QDSKDSTYSM SSTLTLTKDE YERHNSYTCE ATHKTSTSPI VKSFNRNEC
Мышиный полноразмерный IgG1 - вариант 2 тяжелой цепи (SEQ ID NO: 28)
QVQLVQSGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA RGSGSYYPNH FDYWGQGTLV TVSSAKTTPP SVYPLAPGSA AQTNSMVTLG CLVKGYFPEP VTVTWNSGSL SSGVHTFPAV LQSDLYTLSS SVTVPSSTWP SETVTCNVAH PASSTKVDKK IVPRDCGCKP CICTVPEVSS VFIFPPKPKD VLTITLTPKV TCVVVDISKD DPEVQFSWFV DDVEVHTAQT QPREEQFNST FRSVSELPIM HQDWLNGKEF KCRVNSAAFP APIEKTISKT KGRPKAPQVY TIPPPKEQMA KDKVSLTCMI TDFFPEDITV EWQWNGQPAE NYKNTQPIMD TDGSYFVYSK LNVQKSNWEA GNTFTCSVLH EGLHNHHTEK SLSHSPGK
Мышиный полноразмерный IgG1 - вариант 2 тяжелой цепи (SEQ ID NO: 29)
DIVMTQTPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT PTFGQGTRLE IKRTDAAPTV SIFPPSSEQL TSGGASVVCF LNNFYPKDIN VKWKIDGSER QNGVLNSWTD QDSKDSTYSM SSTLTLTKDE YERHNSYTCE ATHKTSTSPI VKSFNRNEC
Человеческий полноразмерный IgG1 - вариант 1 тяжелой цепи (SEQ ID NO: 30)
QVQLVESGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL
VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA
RGSGSYYPNH FDYWGQGTLV TVSSASTKGP SVFPLAPSSK STSGGTAALG
CLVKDYFPEP VTVSWNSGAL TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL
GTQTYICNVN HKPSNTKVDK KVEPKSCDKT HTCPPCPAPE LLGGPSVFLF
PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE
EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP
REPQVYTLPP SRDELTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSLSPGK
Человеческий полноразмерный IgG1 - вариант 1 легкой цепи (SEQ ID NO: 31)
DIVMTQSPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP
KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT
PTFGQGTRLE IKRTVAAPSV FIFPPSDEQL KSGTASVVCL LNNFYPREAK
VQWKVDNALQ SGNSQESVTE QDSKDSTYSL SSTLTLSKAD YEKHKVYACE
VTHQGLSSPV TKSFNRGEC
Человеческий полноразмерный IgG1 - вариант 2 тяжелой цепи (SEQ ID NO: 16)
QVQLVQSGAE VKKPGATVKI SCKVSGYTFT DYYMHWVQQA PGKGLEWMGL
VDPEDGETIY AEKFQGRVTI TADTSTDTAY MELSSLRSED TAVYYCATDA
RGSGSYYPNH FDYWGQGTLV TVSSASTKGP SVFPLAPSSK STSGGTAALG
CLVKDYFPEP VTVSWNSGAL TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL
GTQTYICNVN HKPSNTKVDK KVEPKSCDKT HTCPPCPAPE LLGGPSVFLF
PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE
EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP
REPQVYTLPP SRDELTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL
SPGK
Человеческий полноразмерный IgG1 - вариант 2 легкой цепи (SEQ ID NO: 17)
DIVMTQTPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP
KLLIYWASTR ESGVPDRFSG SGSGTDFTLT INSLQAEDVA VYFCQQYYDT
PTFGQGTRLE IKRTVAAPSV FIFPPSDEQL KSGTASVVCL LNNFYPREAK
VQWKVDNALQ SGNSQESVTE QDSKDSTYSL SSTLTLSKAD YEKHKVYACE
VTHQGLSSPV TKSFNRGEC
CDR антител определяли с использованием метода Kabat (выделены жирным шрифтом в приведенных выше последовательностях). Дополнительные остатки HCDR1 с использованием определения по методу Chothia выделены курсивом. Последовательности константных областей подчеркнуты.
Восстановление передачи сигналов p-SMAD под действием анти-гремлин 1-антител показано в таблице 4 ниже.
Восстановление передачи сигналов p-SMAD
50 нг/мл
+/-2,8%
+/-1,9%
+/-1,4%
+/-0,6%
+/-0,8%
+/-2,1%
+/-2,7%
+/-3,5%
+/-1,4%
25 нг/мл
+/-3,0%
+/-3,1%
+/-1,2%
+/-2,2%
+/-1,3%
+/-2,1%
+/-3,3%
+/-3,8%
+/-2,5%
200
нг/мл
+/-3,8%
+/-3,2%
+/-3,4%
+/-1,3%
+/-1,1%
+/-1,2%
+/-4,3%
+/-3,2%
+/-2,5%
50 нг/мл
+/-2,6%
+/-3,6%
+/-2,7%
+/-3,1%
+/-1,5%
+/-2,5%
+/-4,4%
+/-2,9%
+/-2,5%
50 нг/мл
+/-3,1%
+/-4,0%
+/-4,0%
+/-2,1%
+/-1,7%
+/-1,7%
+/-2,5%
+/-3,1%
+/-2,1%
Результаты представлены в виде процента от фосфорилирования SMAD одним BMP (контроль с BMP). Эксперименты проводили с использованием фибробластов легких от пациентов с идиопатическим легочным фиброзом. Антитела rhGremlin-1 и анти-гремлин-1-антитела предварительно инкубировали в течение 45 мин при комнатной температуре. Затем к клеткам в течение 30 мин добавляли антитела rhGremlin-1 и анти-гремлин-1-антитела с BMP.
Ниже в таблице 5 показаны дополнительные результаты анализа фосфорилирования SMAD, где исследовали вытеснение BMP-2 или BMP4/7 из комплексов гремлин-1-BMP антителами против гремлина-1. Опыты снова проводили с использованием фибробластов легких от пациентов с идиопатическим легочным фиброзом. rhBMP-2 или rhBMP 4/7 предварительно инкубировали с rhGremlin-1 в течение 1 ч при комнатной температуре. Комплексы BMP-2- или BMP4/7-гремлин-1 инкубировали с различными концентрациями анти-гремлин 1-антител в течение ночи при 4°С. Концентрации антител представляют собой конечную концентрацию в планшете.
Вытеснение BMP-2 или BMP4/7 из комплексов гремлин-1-BMP анти-гремлин-1-антителами
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
мкг/мл
50 нг/мл
+/-3,5%
+/-2,7%
+/-2,9%
+/-2,6%
+/-2,0%
+/-1,9%
+/-2,8%
+/-0,3%
50 нг/мл
+/-4,2%
+/-1,0%
+/-1,4%
+/-4,9%
+/-4,4%
+/-2,2%
+/-0,6%
+/-1,5%
+/-1,1%
+/-2,4%
+/-3,8%
+/-2,4%
+/-2,2%
+/-4,3%
+/-2,8%
+/-2,9%
50 нг/мл
+/-0,8%
+/-1,8%
+/-4,2%
+/-10,0%
+/-4,2%
+/-3,3%
+/-2,3%
+/-4,5%
Результаты, представленные в таблице 5, демонстрируют, что Ab7326 может вытеснять уже комплексованный BMP-2 или BMP4/7 из комплексов гремлин-1-BMP. Ab7326 может достигать этого вытеснения в значительно более низких концентрациях, чем антитело сравнения 2484. Это свидетельствует о том, что Ab7326 является аллостерическим ингибитором, что согласуется с открытием заявителей о том, что сайт связывания для Ab7326 находится дистальнее от известных областей связывания BMP на гремлине-1. Таким образом, Ab7326 может получить доступ к аллостерическому сайту связывания, даже когда BMP образует комплекс с гремлином-1, что приводит к значительному повышению ингибирования активности гремлина.
Пример 6. Получение кристаллической структуры гремлина-1 в комплексе с Fab 7326
Кристаллическая структура человеческого гремлина-1 в комплексе с Ab7326 Fab была получена с разрешением 2,1 . Последовательности Fab показаны ниже:
Тяжелая цепь: SEQ ID NO: 18
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
Легкая цепь: SEQ ID NO: 19
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Затем использовали CCP4 программное обеспечение NCONT для идентификации всех контактных остатков при 4 между гремлином-1 и Fab. Были идентифицированы следующие остатки: Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175 (нумерация основана на последовательности UniProt SEQ ID NO: 1 (нумерованы как Ile110, Lys126, Lys127, Phe128, Phe128, Arg148, Lys153 и Gln154 в структурном файле, который соответствует нумерации мышиного гремлина-2).
На фиг. 4 показаны структурные модели комплекса гремлин-Fab, где остатки эпитопа Fab показаны относительно областей связывания BMP.
Ab7326 представляет ингибирующее антитело, которое функционирует аллостерически, т. е. оно связывается с областями связывания BMP.
Пример 7. Измерение аффинности связывания антитела против гремлина-1 Ab7326 с гремлином-1.
Метод
Аффинность анти-гремлин mIgG к человеческому гремлину-1 определяли с помощью анализа бимолекулярного взаимодействия с использованием технологии поверхностного плазмонного резонанса (SPR) на системе Biacore T200, GE Healthcare Bio-Sciences AB. Анти-гремлин mIgG захватывалось иммобилизованным на поверхности анти-мышиным Fc, и гремлин-1 титровали поверх захваченного mIGg. Захватывающий лиганд (Affinipure F(ab')2-фрагмент козьего антимышиного IgG, специфичный для Fc-фрагмента, 115-006-071, Jackson ImmunoResearch Inc.) иммобилизовали при 50 мкг/мл в 10 мМ NaAc, pH 5,0 в проточной ячейке 2 сенсорного чипа CM4 с помощью реакции сочетания аминов с использованием 600-сек инъекций активации и дезактивации до уровня ~ 1600 единиц отклика (RU). В качестве рабочего буфера использовали буфер HBS-EP+ (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 3 мМ ЭДТА, 0,05% поверхностно-активное вещество Р20) со скоростью потока 10 мкл/ мин. Контрольная поверхность была подготовлена в проточной ячейке 1 активацией и деактивацией поверхности, как для проточной ячейки 2, но без захватывающего лиганда.
Буфер для анализа представлял HBS-EP+ плюс 150 мМ NaCl с получением конечной концентрации NaCl 300 мМ плюс 1% CMD40. 60-сек инъекцию анти-гремлин mIgG (при 5 мкг/мл в рабочем буфере) пропускали через проточные ячейки 1 и 2 для получения уровня захвата примерно 100 RU на иммобилизованном на поверхности анти-мышином IgG, Fc. Рекомбинантный человеческий гремлин-1 титровали в рабочем буфере от 5 нМ (с использованием 2-кратных разведений) и инъецировали через проточные ячейки 1 и 2 со скоростью потока 30 мкл/мин в течение 3 мин с последующей 5-мин фазой диссоциации. Также включали контроль, представляющий только буфер. Поверхность регенерировали со скоростью потока 10 мкл/мин путем 60-сек инъецированием 50 мМ HCl, 30-сек инъецированием 5 мМ NaOH и 30-сек инъецированием 50 мМ HCl.
Кинетические данные определяли с использованием оценочного программного обеспечения Biacore T200.
Измерения аффинности проводили при 25°C.
Результаты
Было установлено, что аффинность связывания, представленная в виде среднего значения KD для 5 определений, составляла ниже 100 пМ.
Пример 8. Ингибирование активности гремлина-1 ускоряет заживление и сращение на модели восстановления перелома кости in vivo
8.1. Материалы и методы
Крысиная модель перелома и введение препарата
Модели сегментарного дефекта длинных костей широко использовались для исследования заживления и регенерации кости (Sato et al; 2014). В настоящем исследовании 3-мм дефект бедренной кости воспроизводили у 10-недельных самцов крыс и фиксировали с использованием пластины с 8-отверстиями PEEK (RIS. 602.105, RISystem, Швейцария). Пластину прикрепляли к кости с помощью зажимов в середине диафиза, перед тем как кость была просверлена и зафиксирована винтами. Создавали 3 мм зазор разрушения с использованием пилы Gigly 0,44 мм. Размер/консистенцию/фиксацию дефекта тщательно контролировали с помощью рентгеновского визуализирования с использованием Faxitron (MX-20-DC5, Faxitron Bioptics LLC, США), эту временную точку определяли как сутки 0.
Еженедельное введение начинали на сутки 1 в течение 8 недель, как указано в таблице 1.
Затем получали рентгеновские снимки в течение прижизненной стадии исследования на сутки 11, 25, 39 и 57 для того, чтобы оценить образование костной мозоли и ход заживления. Анализ изображений Definiens использовали для количественной оценки площади дефекта, лишенного костной ткани, на полученных рентгеновских снимках.
Группы обработки
Микро-КТ анализ заживления переломов
Бедренную кость (переломанная сторона) сканировали с разрешением 17,2 мкм с использованием микро-КТ (SkyScan 1076). Была получена область примерно в 15 мм от мозоли с переломом в центре. Сканы реконструировали с использованием программного обеспечения Skyscan NRECON (1.7.10), и затем реконструированные срезы дополнительно сегментировали, чтобы исключить фиксаторные штифты в определенной области 3 мм, рассчитанной по средней точке дефекта перелома бедра. Гистоморфометрический анализ костной мозоли перелома в 3D выполняли с помощью программного обеспечения SkyScan (v. 1.13.1). Определяли среднюю точку в пределах 3 мм дефекта перелома бедренной кости, и срезы на 1,5 мм дистальнее и проксимальнее средней точки были сегментированы для каждой измеренной конечности. Затем проводили бинаризацию реконструированных наборов данных и сегментацию после двух определенных порогов, один для определения мозоли с низкой минерализацией (таким образом, количественно определения новообразованной кости) и другой для определения зрелой кости. Дальнейшую сегментацию этих данных проводили на бедренной кости животных, отнесенных к группе с низким уровнем ответа, на основе удовлетворения критериев неполного преодоления дефекта бедренной кости, или показателей с высоким уровнем ответа, проявляющих соединение в месте перелома.
Гистоморфометрический анализ перелома
Бедренную кость фиксировали в 10% нейтральном забуференном формалине в течение 24 ч, дегидратировали и помещали в метилметакрилат (ММА) при низкой температуре. Срезы толщиной 50 мкм окрашивали толуидиновым синим для количественной оценки костных элементов дефекта заживающего зазора. Гистоморфометрические показатели определяли на губчатой кости в месте дефекта перелома. Измерения выполняли с помощью анализа изображений.
Статистический анализ
Результаты представлены в виде средних значений ± SD. Статистический анализ проводили с использованием двустороннего U-критерия Манна-Уитни с программным обеспечением GraphPad Prism, если не указано иное.
8.2 Результаты
Анализ рентгеновских снимков, полученных на прижизненной стадии исследования, показал, что анти-гремлин-1-антитело ускоряло заживление перелома у животных, где статистически значимые различия между контрольной и обработанной группами проявлялись через 25 суток (P <0,05). Этот эффект был очевиден для оставшейся части исследования (фиг. 5).
Микро-СТ анализ конечных образцов показал, что обработка анти-гремлин 1-антителом (30 мг/кг/раз в неделю) приводила к увеличению новообразованной кости в мозоли на месте перелома (P = 0,06).
Частота возникновения несращенных переломов на этой модели без какого-либо лечебного вмешательства составляет примерно 60% (Sato et al.; 2014). Для проверки того, насколько ингибирование гремлина-1 снижало частоту несращения, животных разделяли на респондеров с низким ответом (LR) и респондеров с высоким ответом (HR). Ингибирование гремлина-1 приводило к достоверному увеличению процентного соотношения объема кости/объема ткани (BV/TV%) в пределах LMB (кость с низким содержанием минералов; новообразованная кость) (P <0,01) и HMB (кость с высоким содержанием минералов; зрелая кость) (P <0,01) в группе респондеров с низким ответом по сравнению с контрольной группой, что свидетельствует о прогрессирующем восстановлении когорты, которая может образовывать несращение.
Кроме того, имела место тенденция (незначительная) к увеличению LMB и HMB BV/TV% в группе с высоким уровнем ответа на обработку анти-гремлин 1-антителом (фиг. 6A, репрезентативные изображения LR и HR показаны на фиг. 6B) ,
Двумерный гистоморфометрический анализ параметров кости выполняли на гистологических срезах места перелома (фиг.7). Обработка анти-гремлин 1-антителом достоверно увеличивала процентное соотношение объема кости/объема ткани (BV/TV%) (P <0,05) по сравнению с контролем. Анти-гремлин 1-антитело достоверно увеличивало трабекулярное число (Tb.N) (P <0,001) и достоверно уменьшало трабекулярное разделение (Tb.Sp) (P <0,01), что указывает на увеличение трабекулярной кости в результате обработки анти-гремлин 1-антителом.
Проводили корреляции между результатами двумерного гистоморфометрического анализа и трехмерного микро-КТ анализа сравнением групп LMB и HMB, сегментированных в анализе микро-КТ, и двумерном гистоморфометрическом анализе срезов перелома (фиг. 8). Сравнения, измеренные по корреляции Пирсона, выявили положительную и значимую корреляцию между гистоморфометрией и микро-КТ анализом в группах LMB (P <0,0001) и HMB (P <0,0001), что подтверждает достоверность данных из каждого ряда данных.
8.3. Вывод
Ингибирование активности гремлина-1 с использованием нейтрализующего анти-гремлин 1-антитела приводило к ускоренному восстановлению перелома, при этом статистически значимые различия между контрольной и обработанной группами проявлялись через 25 суток (3 дозы антитела). Кроме того, конечный анализ места перелома показал усиленное образование костной ткани у животных с низким уровнем ответа, у которых в противном случае могло бы образовываться несращение. Следовательно, ингибирование активности гремлина-1 является многообещающей терапией для профилактики или лечения несращенных переломов и может иметь особое значение для лечения переломов, склонных к несращению, например большеберцовой кости, дистального радиуса, шейки бедра и ладьевидной кости.
Список последовательностей
SEQ ID NO: 1 (человеческий гремлин-1; Uniprot ID: O60565)
MSRTAYTVGALLLLLGTLLPAAEGKKKGSQGAIPPPDKAQHNDSEQTQSPQQPGSRNRGRGQGRGTAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO: 2 (человеческий усеченный гремлин-1, использованный в кристаллографии с N-концевой меткой)
MGSSHHHHHHSSGENLYFQGSAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO: 3 (Ab7326 HCDR1 объединенная система нумерации по Kabat & Chothia)
GYTFTDYYMH
SEQ ID NO: 4 (Ab7326 HCDR1 Kabat)
DYYMH
SEQ ID NO: 5 (Ab7326 HCDR2 Kabat)
LVDPEDGETIYAEKFQG
SEQ ID NO: 6 (Ab7326 HCDR3 Kabat)
DARGSGSYYPNHFDY
SEQ ID NO: 7 (Ab7326 LCDR1 Kabat)
KSSQSVLYSSNNKNYLA
SEQ ID NO: 8 (Ab7326 LCDR2 Kabat)
WASTRES
SEQ ID NO: 9 (Ab7326 LCDR3 Kabat)
QQYYDTPT
SEQ ID NO: 10 (вариант 1 вариабельной области тяжелой цепи Ab7326)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSS
SEQ ID NO: 11 (вариант 1 вариабельной области легкой цепи Ab7326)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIK
SEQ ID NO: 12 (вариант 2 вариабельной области тяжелой цепи Ab7326)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSS
SEQ ID NO: 13 (вариант 2 вариабельной области легкой цепи Ab7326)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIK
SEQ ID NO: 14 (вариант 1 тяжелой цепи мышиного полноразмерного IgG1 1)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSAKTTPPSVYPLAPGSAAQTNSMVTLGCLVKGYFPEPVTVTWNSGSLSSGVHTFPAVLQSDLYTLSSSVTVPSSTWPSETVTCNVAHPASSTKVDKKIVPRDCGCKPCICTVPEVSSVFIFPPKPKDVLTITLTPKVTCVVVDISKDDPEVQFSWFVDDVEVHTAQTQPREEQFNSTFRSVSELPIMHQDWLNGKEFKCRVNSAAFPAPIEKTISKTKGRPKAPQVYTIPPPKEQMAKDKVSLTCMITDFFPEDITVEWQWNGQPAENYKNTQPIMDTDGSYFVYSKLNVQKSNWEAGNTFTCSVLHEGLHNHHTEKSLSHSPGK
SEQ ID NO: 15 (вариант 1 легкой цепи мышиного полноразмерного IgG1 1)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTDAAPTVSIFPPSSEQLTSGGASVVCFLNNFYPKDINVKWKIDGSERQNGVLNSWTDQDSKDSTYSMSSTLTLTKDEYERHNSYTCEATHKTSTSPIVKSFNRNEC
SEQ ID NO: 16 (вариант 2 тяжелой цепи человеческого полноразмерного IgG1)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 17 (вариант 2 легкой цепи человеческого полноразмерного IgG1)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 18 (вариант 1 тяжелой цепи Fab)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
SEQ ID NO: 19 (вариант 1 легкой цепи Fab)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 20 (человеческий усеченный гремлин-1, использованный в кристаллографии без N-концевой метки)
AMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO: 21 (последовательность зрелого гремлина-1 SEQ ID NO: 1 без сигнального пептида из аминокислот 1-21)
KKKGSQGAIPPPDKAQHNDSEQTQSPQQPGSRNRGRGQGRGTAMPGEEVLESSQEALHVTERKYLKRDWCKTQPLKQTIHEEGCNSRTIINRFCYGQCNSFYIPRHIRKEEGSFQSCSFCKPKKFTTMMVTLNCPELQPPTKKKRVTRVKQCRCISIDLD
SEQ ID NO: 22 (вариант 1 тяжелой цепи человеческого IgG4P)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
SEQ ID NO: 23 (вариант 1 легкой цепи человеческого IgG4P)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 24 (вариант 1 ДНК тяжелой цепи человеческого IgG1)
caagtgcaactggtggaatccggggccgaagtgaaaaagcccggagccactgtgaagatctcttgcaaagtgtccggctacaccttcaccgactattacatgcactgggtccagcaggcacctgggaagggccttgagtggatgggtctggtcgatcccgaggacggcgaaactatctacgccgagaagttccagggtcgcgtcaccatcaccgccgacacttccaccgacaccgcgtacatggagctgtccagcttgaggtccgaggacacagccgtgtactactgcgccacggatgctcggggaagcggcagctactacccgaaccacttcgactactggggacagggcactctcgtgactgtctcgagcgcttctacaaagggcccctccgtgttcccgctcgctccatcatcgaagtctaccagcggaggcactgcggctctcggttgcctcgtgaaggactacttcccggagccggtgaccgtgtcgtggaacagcggagccctgaccagcggggtgcacacctttccggccgtcttgcagtcaagcggcctttactccctgtcatcagtggtgactgtcccgtccagctcattgggaacccaaacctacatctgcaatgtgaatcacaaacctagcaacaccaaggttgacaagaaagtcgagcccaaatcgtgtgacaagactcacacttgtccgccgtgcccggcacccgaactgctgggaggtcccagcgtctttctgttccctccaaagccgaaagacacgctgatgatctcccgcaccccggaggtcacttgcgtggtcgtggacgtgtcacatgaggacccagaggtgaagttcaattggtacgtggatggcgtcgaagtccacaatgccaaaactaagcccagagaagaacagtacaattcgacctaccgcgtcgtgtccgtgctcacggtgttgcatcaggattggctgaacgggaaggaatacaagtgcaaagtgtccaacaaggcgctgccggcaccgatcgagaaaactatctccaaagcgaagggacagcctagggaacctcaagtctacacgctgccaccatcacgggatgaactgactaagaatcaagtctcactgacttgtctggtgaaggggttttaccctagcgacattgccgtggagtgggaatccaacggccagccagagaacaactacaagactacccctccagtgctcgactcggatggatcgttcttcctttactcgaagctcaccgtggataagtcccggtggcagcagggaaacgtgttctcctgctcggtgatgcatgaagccctccataaccactatacccaaaagtcgctgtccctgtcgccgggaaag
SEQ ID NO: 25 (вариант 1 ДНК легкой цепи человеческого IgG1)
gacattgtgatgacccagtcccccgattcgcttgcggtgtccctgggagaacgggccaccattaactgcaagagctcacagtccgtcctgtattcatcgaacaacaagaattacctcgcatggtatcagcagaagcctggacagcctcccaagctgctcatctactgggctagcacccgcgaatccggggtgccggatagattctccggatcgggttcgggcactgacttcactctgactatcaactcactgcaagccgaggatgtcgcggtgtacttctgtcagcagtactacgacaccccgacctttggacaaggcaccagactggagattaagcgtacggtggccgctccctccgtgttcatcttcccaccctccgacgagcagctgaagtccggcaccgcctccgtcgtgtgcctgctgaacaacttctacccccgcgaggccaaggtgcagtggaaggtggacaacgccctgcagtccggcaactcccaggaatccgtcaccgagcaggactccaaggacagcacctactccctgtcctccaccctgaccctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaagtgacccaccagggcctgtccagccccgtgaccaagtccttcaaccggggcgagtgc
SEQ ID NO: 26 (вариант 1 ДНК тяжелой цепи человеческого IgG4P)
caagtgcaactggtggaatccggggccgaagtgaaaaagcccggagccactgtgaagatctcttgcaaagtgtccggctacaccttcaccgactattacatgcactgggtccagcaggcacctgggaagggccttgagtggatgggtctggtcgatcccgaggacggcgaaactatctacgccgagaagttccagggtcgcgtcaccatcaccgccgacacttccaccgacaccgcgtacatggagctgtccagcttgaggtccgaggacacagccgtgtactactgcgccacggatgctcggggaagcggcagctactacccgaaccacttcgactactggggacagggcactctcgtgactgtctcgagcgcttctacaaagggcccctccgtgttccctctggccccttgctcccggtccacctccgagtctaccgccgctctgggctgcctggtcaaggactacttccccgagcccgtgacagtgtcctggaactctggcgccctgacctccggcgtgcacaccttccctgccgtgctgcagtcctccggcctgtactccctgtcctccgtcgtgaccgtgccctcctccagcctgggcaccaagacctacacctgtaacgtggaccacaagccctccaacaccaaggtggacaagcgggtggaatctaagtacggccctccctgccccccctgccctgcccctgaatttctgggcggaccttccgtgttcctgttccccccaaagcccaaggacaccctgatgatctcccggacccccgaagtgacctgcgtggtggtggacgtgtcccaggaagatcccgaggtccagttcaattggtacgtggacggcgtggaagtgcacaatgccaagaccaagcccagagaggaacagttcaactccacctaccgggtggtgtccgtgctgaccgtgctgcaccaggactggctgaacggcaaagagtacaagtgcaaggtgtccaacaagggcctgccctccagcatcgaaaagaccatctccaaggccaagggccagccccgcgagccccaggtgtacaccctgccccctagccaggaagagatgaccaagaaccaggtgtccctgacctgtctggtcaagggcttctacccctccgacattgccgtggaatgggagtccaacggccagcccgagaacaactacaagaccaccccccctgtgctggacagcgacggctccttcttcctgtactctcggctgaccgtggacaagtcccggtggcaggaaggcaacgtcttctcctgctccgtgatgcacgaggccctgcacaaccactacacccagaagtccctgtccctgagcctgggcaag
SEQ ID NO: 27 (вариант 1 ДНК легкой цепи человеческого IgG4P)
gacattgtgatgacccagtcccccgattcgcttgcggtgtccctgggagaacgggccaccattaactgcaagagctcacagtccgtcctgtattcatcgaacaacaagaattacctcgcatggtatcagcagaagcctggacagcctcccaagctgctcatctactgggctagcacccgcgaatccggggtgccggatagattctccggatcgggttcgggcactgacttcactctgactatcaactcactgcaagccgaggatgtcgcggtgtacttctgtcagcagtactacgacaccccgacctttggacaaggcaccagactggagattaagcgtacggtggccgctccctccgtgttcatcttcccaccctccgacgagcagctgaagtccggcaccgcctccgtcgtgtgcctgctgaacaacttctacccccgcgaggccaaggtgcagtggaaggtggacaacgccctgcagtccggcaactcccaggaatccgtcaccgagcaggactccaaggacagcacctactccctgtcctccaccctgaccctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaagtgacccaccagggcctgtccagccccgtgaccaagtccttcaaccggggcgagtgc
SEQ ID NO: 28 (вариант 2 тяжелой цепи мышиного полноразмерного IgG1)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSAKTTPPSVYPLAPGSAAQTNSMVTLGCLVKGYFPEPVTVTWNSGSLSSGVHTFPAVLQSDLYTLSSSVTVPSSTWPSETVTCNVAHPASSTKVDKKIVPRDCGCKPCICTVPEVSSVFIFPPKPKDVLTITLTPKVTCVVVDISKDDPEVQFSWFVDDVEVHTAQTQPREEQFNSTFRSVSELPIMHQDWLNGKEFKCRVNSAAFPAPIEKTISKTKGRPKAPQVYTIPPPKEQMAKDKVSLTCMITDFFPEDITVEWQWNGQPAENYKNTQPIMDTDGSYFVYSKLNVQKSNWEAGNTFTCSVLHEGLHNHHTEKSLSHSPGK
SEQ ID NO: 29 (вариант 2 легкой цепи мышиного полноразмерного IgG1)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTDAAPTVSIFPPSSEQLTSGGASVVCFLNNFYPKDINVKWKIDGSERQNGVLNSWTDQDSKDSTYSMSSTLTLTKDEYERHNSYTCEATHKTSTSPIVKSFNRNEC
SEQ ID NO: 30 (вариант 1 тяжелой цепи человеческого полноразмерного IgG1)
QVQLVESGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 31 (вариант 1 легкой цепи человеческого полноразмерного IgG1)
DIVMTQSPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 32 (вариант 2 тяжелой цепи Fab)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
SEQ ID NO: 33 (вариант 2 легкой цепи Fab)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 34 (вариант 2 тяжелой цепи человеческого IgG4P)
QVQLVQSGAEVKKPGATVKISCKVSGYTFTDYYMHWVQQAPGKGLEWMGLVDPEDGETIYAEKFQGRVTITADTSTDTAYMELSSLRSEDTAVYYCATDARGSGSYYPNHFDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
SEQ ID NO: 35 (вариант 2 легкой цепи человеческого IgG4P)
DIVMTQTPDSLAVSLGERATINCKSSQSVLYSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTINSLQAEDVAVYFCQQYYDTPTFGQGTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Ссылки
Bostrom M. P. & Seigerman D. A. (2005), HSS journal: The Musculoskeletal Journal of Hospital for Special Surgery 1, 9-18. The clinical use of allografts, demineralized bone matrices, synthetic bone graft substitutes and osteoinductive growth factors: a survey study.
Buza J. A., 3rd & Einhorn, T. (2016), Clinical cases in mineral and bone metabolism: The Official Journal of the Italian Society of Osteoporosis, Mineral Metabolism, and Skeletal Diseases 13, 101-105. Bone healing in 2016.
Canalis E., Parker K. & Zanotti S. (2012), J.Cell Physiol 227, 269-277. Gremlin1 is required for skeletal development and postnatal skeletal homeostasis.
Cho T. J., Gerstenfeld L. C. & Einhorn T. A. (2002), Journal of Bone and Mineral Research: The Official Journal of the American Society for Bone and Mineral Research 17, 513-520. Differential temporal expression of members of the transforming growth factor beta superfamily during murine fracture healing.
Einhorn T. A. & Gerstenfeld L. C. (2015), Nat. Rev. Rheumatol. 11, 45-54. Fracture healing: mechanisms and interventions.
Ferguson C., Alpern E., Miclau T. & Helms J. A. (1999), Mechanisms of development 87, 57-66. Does adult fracture repair recapitulate embryonic skeletal formation?
Gazzerro E. et al. (2005), Endocrinology 146, 655-665. Skeletal overexpression of gremlin impairs bone formation and causes osteopenia.
Gazzerro E. et al. (2007), J.Biol.Chem. 282, 31549-31557. Conditional deletion of gremlin causes a transient increase in bone formation and bone mass.
Goulet J. A., Senunas L. E., DeSilva G. L. & Greenfield M. L. (1997), Clinical Orthopaedics and Related Research, 76-81. Autogenous iliac crest bone graft. Complications and functional assessment.
Hsu D. R., Economides A. N., Wang X., Eimon P. M. & Harland R. M. (1998), Mol. Cell 1, 673-683. The Xenopus dorsalizing factor Gremlin identifies a novel family of secreted proteins that antagonize BMP activities.
Sato K., Watanabe Y., Harada N., Abe S., Matsushita T., Yamanaka K., Kaneko T., & Sakai Y. (2014) Tissue Eng Part C. Methods 20, 1037-1041.
Schmid G. J., Kobayashi C., Sandell L. J. & Ornitz D. M. (2009), Developmental Dynamics: An Official Publication of the American Association of Anatomists 238, 766-774. Fibroblast growth factor expression during skeletal fracture healing in mice.
Yu Y. Y. et al. (2010), Bone 46, 841-851. Immunolocalization of BMPs, BMP antagonists, receptors, and effectors during fracture repair.
--->
Список последовательностей
<110> UCB Biopharma SPRL
<120> ИНГИБИТОР ГРЕМЛИНА-1 ДЛЯ ЛЕЧЕНИЯ ПЕРЕЛОМА КОСТИ ИЛИ ДЕФЕКТА КОСТИ
<130> PF0117-WO-PCT
<160> 35
<170> PatentIn version 3.5
<210> 1
<211> 184
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 1
Met Ser Arg Thr Ala Tyr Thr Val Gly Ala Leu Leu Leu Leu Leu Gly
1 5 10 15
Thr Leu Leu Pro Ala Ala Glu Gly Lys Lys Lys Gly Ser Gln Gly Ala
20 25 30
Ile Pro Pro Pro Asp Lys Ala Gln His Asn Asp Ser Glu Gln Thr Gln
35 40 45
Ser Pro Gln Gln Pro Gly Ser Arg Asn Arg Gly Arg Gly Gln Gly Arg
50 55 60
Gly Thr Ala Met Pro Gly Glu Glu Val Leu Glu Ser Ser Gln Glu Ala
65 70 75 80
Leu His Val Thr Glu Arg Lys Tyr Leu Lys Arg Asp Trp Cys Lys Thr
85 90 95
Gln Pro Leu Lys Gln Thr Ile His Glu Glu Gly Cys Asn Ser Arg Thr
100 105 110
Ile Ile Asn Arg Phe Cys Tyr Gly Gln Cys Asn Ser Phe Tyr Ile Pro
115 120 125
Arg His Ile Arg Lys Glu Glu Gly Ser Phe Gln Ser Cys Ser Phe Cys
130 135 140
Lys Pro Lys Lys Phe Thr Thr Met Met Val Thr Leu Asn Cys Pro Glu
145 150 155 160
Leu Gln Pro Pro Thr Lys Lys Lys Arg Val Thr Arg Val Lys Gln Cys
165 170 175
Arg Cys Ile Ser Ile Asp Leu Asp
180
<210> 2
<211> 139
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 2
Met Gly Ser Ser His His His His His His Ser Ser Gly Glu Asn Leu
1 5 10 15
Tyr Phe Gln Gly Ser Ala Met Pro Gly Glu Glu Val Leu Glu Ser Ser
20 25 30
Gln Glu Ala Leu His Val Thr Glu Arg Lys Tyr Leu Lys Arg Asp Trp
35 40 45
Cys Lys Thr Gln Pro Leu Lys Gln Thr Ile His Glu Glu Gly Cys Asn
50 55 60
Ser Arg Thr Ile Ile Asn Arg Phe Cys Tyr Gly Gln Cys Asn Ser Phe
65 70 75 80
Tyr Ile Pro Arg His Ile Arg Lys Glu Glu Gly Ser Phe Gln Ser Cys
85 90 95
Ser Phe Cys Lys Pro Lys Lys Phe Thr Thr Met Met Val Thr Leu Asn
100 105 110
Cys Pro Glu Leu Gln Pro Pro Thr Lys Lys Lys Arg Val Thr Arg Val
115 120 125
Lys Gln Cys Arg Cys Ile Ser Ile Asp Leu Asp
130 135
<210> 3
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 3
Gly Tyr Thr Phe Thr Asp Tyr Tyr Met His
1 5 10
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 4
Asp Tyr Tyr Met His
1 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 5
Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe Gln
1 5 10 15
Gly
<210> 6
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 6
Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp Tyr
1 5 10 15
<210> 7
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 7
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 8
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 8
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 9
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 9
Gln Gln Tyr Tyr Asp Thr Pro Thr
1 5
<210> 10
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 10
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 11
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 11
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
<210> 12
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 13
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 13
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
<210> 14
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 14
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr
115 120 125
Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn
130 135 140
Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val
180 185 190
Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val
195 200 205
Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg
210 215 220
Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu
245 250 255
Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro
260 265 270
Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala
275 280 285
Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val
290 295 300
Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile
340 345 350
Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys
355 360 365
Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp
370 375 380
Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp
385 390 395 400
Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser
405 410 415
Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly
420 425 430
Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 15
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 15
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 16
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 17
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 17
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 18
<211> 227
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 18
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys
225
<210> 19
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 19
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 20
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 20
Ala Met Pro Gly Glu Glu Val Leu Glu Ser Ser Gln Glu Ala Leu His
1 5 10 15
Val Thr Glu Arg Lys Tyr Leu Lys Arg Asp Trp Cys Lys Thr Gln Pro
20 25 30
Leu Lys Gln Thr Ile His Glu Glu Gly Cys Asn Ser Arg Thr Ile Ile
35 40 45
Asn Arg Phe Cys Tyr Gly Gln Cys Asn Ser Phe Tyr Ile Pro Arg His
50 55 60
Ile Arg Lys Glu Glu Gly Ser Phe Gln Ser Cys Ser Phe Cys Lys Pro
65 70 75 80
Lys Lys Phe Thr Thr Met Met Val Thr Leu Asn Cys Pro Glu Leu Gln
85 90 95
Pro Pro Thr Lys Lys Lys Arg Val Thr Arg Val Lys Gln Cys Arg Cys
100 105 110
Ile Ser Ile Asp Leu Asp
115
<210> 21
<211> 160
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 21
Lys Lys Lys Gly Ser Gln Gly Ala Ile Pro Pro Pro Asp Lys Ala Gln
1 5 10 15
His Asn Asp Ser Glu Gln Thr Gln Ser Pro Gln Gln Pro Gly Ser Arg
20 25 30
Asn Arg Gly Arg Gly Gln Gly Arg Gly Thr Ala Met Pro Gly Glu Glu
35 40 45
Val Leu Glu Ser Ser Gln Glu Ala Leu His Val Thr Glu Arg Lys Tyr
50 55 60
Leu Lys Arg Asp Trp Cys Lys Thr Gln Pro Leu Lys Gln Thr Ile His
65 70 75 80
Glu Glu Gly Cys Asn Ser Arg Thr Ile Ile Asn Arg Phe Cys Tyr Gly
85 90 95
Gln Cys Asn Ser Phe Tyr Ile Pro Arg His Ile Arg Lys Glu Glu Gly
100 105 110
Ser Phe Gln Ser Cys Ser Phe Cys Lys Pro Lys Lys Phe Thr Thr Met
115 120 125
Met Val Thr Leu Asn Cys Pro Glu Leu Gln Pro Pro Thr Lys Lys Lys
130 135 140
Arg Val Thr Arg Val Lys Gln Cys Arg Cys Ile Ser Ile Asp Leu Asp
145 150 155 160
<210> 22
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 22
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly Lys
450
<210> 23
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 23
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 24
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 24
caagtgcaac tggtggaatc cggggccgaa gtgaaaaagc ccggagccac tgtgaagatc 60
tcttgcaaag tgtccggcta caccttcacc gactattaca tgcactgggt ccagcaggca 120
cctgggaagg gccttgagtg gatgggtctg gtcgatcccg aggacggcga aactatctac 180
gccgagaagt tccagggtcg cgtcaccatc accgccgaca cttccaccga caccgcgtac 240
atggagctgt ccagcttgag gtccgaggac acagccgtgt actactgcgc cacggatgct 300
cggggaagcg gcagctacta cccgaaccac ttcgactact ggggacaggg cactctcgtg 360
actgtctcga gcgcttctac aaagggcccc tccgtgttcc cgctcgctcc atcatcgaag 420
tctaccagcg gaggcactgc ggctctcggt tgcctcgtga aggactactt cccggagccg 480
gtgaccgtgt cgtggaacag cggagccctg accagcgggg tgcacacctt tccggccgtc 540
ttgcagtcaa gcggccttta ctccctgtca tcagtggtga ctgtcccgtc cagctcattg 600
ggaacccaaa cctacatctg caatgtgaat cacaaaccta gcaacaccaa ggttgacaag 660
aaagtcgagc ccaaatcgtg tgacaagact cacacttgtc cgccgtgccc ggcacccgaa 720
ctgctgggag gtcccagcgt ctttctgttc cctccaaagc cgaaagacac gctgatgatc 780
tcccgcaccc cggaggtcac ttgcgtggtc gtggacgtgt cacatgagga cccagaggtg 840
aagttcaatt ggtacgtgga tggcgtcgaa gtccacaatg ccaaaactaa gcccagagaa 900
gaacagtaca attcgaccta ccgcgtcgtg tccgtgctca cggtgttgca tcaggattgg 960
ctgaacggga aggaatacaa gtgcaaagtg tccaacaagg cgctgccggc accgatcgag 1020
aaaactatct ccaaagcgaa gggacagcct agggaacctc aagtctacac gctgccacca 1080
tcacgggatg aactgactaa gaatcaagtc tcactgactt gtctggtgaa ggggttttac 1140
cctagcgaca ttgccgtgga gtgggaatcc aacggccagc cagagaacaa ctacaagact 1200
acccctccag tgctcgactc ggatggatcg ttcttccttt actcgaagct caccgtggat 1260
aagtcccggt ggcagcaggg aaacgtgttc tcctgctcgg tgatgcatga agccctccat 1320
aaccactata cccaaaagtc gctgtccctg tcgccgggaa ag 1362
<210> 25
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 25
gacattgtga tgacccagtc ccccgattcg cttgcggtgt ccctgggaga acgggccacc 60
attaactgca agagctcaca gtccgtcctg tattcatcga acaacaagaa ttacctcgca 120
tggtatcagc agaagcctgg acagcctccc aagctgctca tctactgggc tagcacccgc 180
gaatccgggg tgccggatag attctccgga tcgggttcgg gcactgactt cactctgact 240
atcaactcac tgcaagccga ggatgtcgcg gtgtacttct gtcagcagta ctacgacacc 300
ccgacctttg gacaaggcac cagactggag attaagcgta cggtggccgc tccctccgtg 360
ttcatcttcc caccctccga cgagcagctg aagtccggca ccgcctccgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
tccggcaact cccaggaatc cgtcaccgag caggactcca aggacagcac ctactccctg 540
tcctccaccc tgaccctgtc caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc cagccccgtg accaagtcct tcaaccgggg cgagtgc 657
<210> 26
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 26
caagtgcaac tggtggaatc cggggccgaa gtgaaaaagc ccggagccac tgtgaagatc 60
tcttgcaaag tgtccggcta caccttcacc gactattaca tgcactgggt ccagcaggca 120
cctgggaagg gccttgagtg gatgggtctg gtcgatcccg aggacggcga aactatctac 180
gccgagaagt tccagggtcg cgtcaccatc accgccgaca cttccaccga caccgcgtac 240
atggagctgt ccagcttgag gtccgaggac acagccgtgt actactgcgc cacggatgct 300
cggggaagcg gcagctacta cccgaaccac ttcgactact ggggacaggg cactctcgtg 360
actgtctcga gcgcttctac aaagggcccc tccgtgttcc ctctggcccc ttgctcccgg 420
tccacctccg agtctaccgc cgctctgggc tgcctggtca aggactactt ccccgagccc 480
gtgacagtgt cctggaactc tggcgccctg acctccggcg tgcacacctt ccctgccgtg 540
ctgcagtcct ccggcctgta ctccctgtcc tccgtcgtga ccgtgccctc ctccagcctg 600
ggcaccaaga cctacacctg taacgtggac cacaagccct ccaacaccaa ggtggacaag 660
cgggtggaat ctaagtacgg ccctccctgc cccccctgcc ctgcccctga atttctgggc 720
ggaccttccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat ctcccggacc 780
cccgaagtga cctgcgtggt ggtggacgtg tcccaggaag atcccgaggt ccagttcaat 840
tggtacgtgg acggcgtgga agtgcacaat gccaagacca agcccagaga ggaacagttc 900
aactccacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagagtaca agtgcaaggt gtccaacaag ggcctgccct ccagcatcga aaagaccatc 1020
tccaaggcca agggccagcc ccgcgagccc caggtgtaca ccctgccccc tagccaggaa 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtca agggcttcta cccctccgac 1140
attgccgtgg aatgggagtc caacggccag cccgagaaca actacaagac caccccccct 1200
gtgctggaca gcgacggctc cttcttcctg tactctcggc tgaccgtgga caagtcccgg 1260
tggcaggaag gcaacgtctt ctcctgctcc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgtccct gagcctgggc aag 1353
<210> 27
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 27
gacattgtga tgacccagtc ccccgattcg cttgcggtgt ccctgggaga acgggccacc 60
attaactgca agagctcaca gtccgtcctg tattcatcga acaacaagaa ttacctcgca 120
tggtatcagc agaagcctgg acagcctccc aagctgctca tctactgggc tagcacccgc 180
gaatccgggg tgccggatag attctccgga tcgggttcgg gcactgactt cactctgact 240
atcaactcac tgcaagccga ggatgtcgcg gtgtacttct gtcagcagta ctacgacacc 300
ccgacctttg gacaaggcac cagactggag attaagcgta cggtggccgc tccctccgtg 360
ttcatcttcc caccctccga cgagcagctg aagtccggca ccgcctccgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
tccggcaact cccaggaatc cgtcaccgag caggactcca aggacagcac ctactccctg 540
tcctccaccc tgaccctgtc caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc cagccccgtg accaagtcct tcaaccgggg cgagtgc 657
<210> 28
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 28
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr
115 120 125
Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn
130 135 140
Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val
180 185 190
Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val
195 200 205
Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg
210 215 220
Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu
245 250 255
Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro
260 265 270
Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala
275 280 285
Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val
290 295 300
Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile
340 345 350
Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys
355 360 365
Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp
370 375 380
Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp
385 390 395 400
Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser
405 410 415
Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly
420 425 430
Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 29
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 29
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 30
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 30
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 31
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 31
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 32
<211> 227
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys
225
<210> 33
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 33
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 34
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 34
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asp Ala Arg Gly Ser Gly Ser Tyr Tyr Pro Asn His Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly Lys
450
<210> 35
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> рекомбинантная последовательность
<400> 35
Asp Ile Val Met Thr Gln Thr Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Tyr Tyr Asp Thr Pro Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КРИСТАЛЛИЧЕСКАЯ СТРУКТУРА ГРЕМЛИН-1 И ИНГИБИРУЮЩЕЕ АНТИТЕЛО | 2017 |
|
RU2769285C2 |
| АНТИТЕЛА, НАЦЕЛИВАЮЩИЕСЯ НА КОСТНУЮ ТКАНЬ | 2018 |
|
RU2801206C2 |
| СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ РАКОВОГО МЕТАСТАЗА И РАЗРЕЖЕНИЯ КОСТИ, СВЯЗАННОГО С РАКОВЫМ МЕТАСТАЗОМ | 2007 |
|
RU2470665C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛ К СКЛЕРОСТИНУ ДЛЯ ЛЕЧЕНИЯ НЕСОВЕРШЕННОГО ОСТЕОГЕНЕЗА | 2017 |
|
RU2789033C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
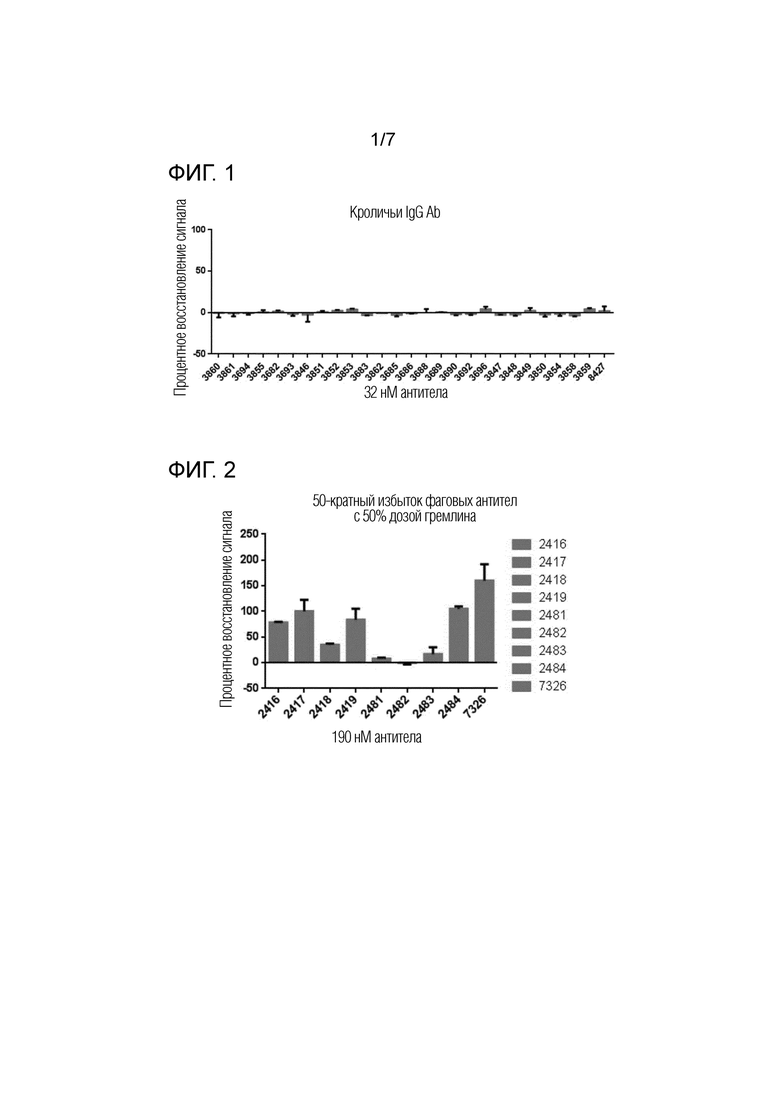
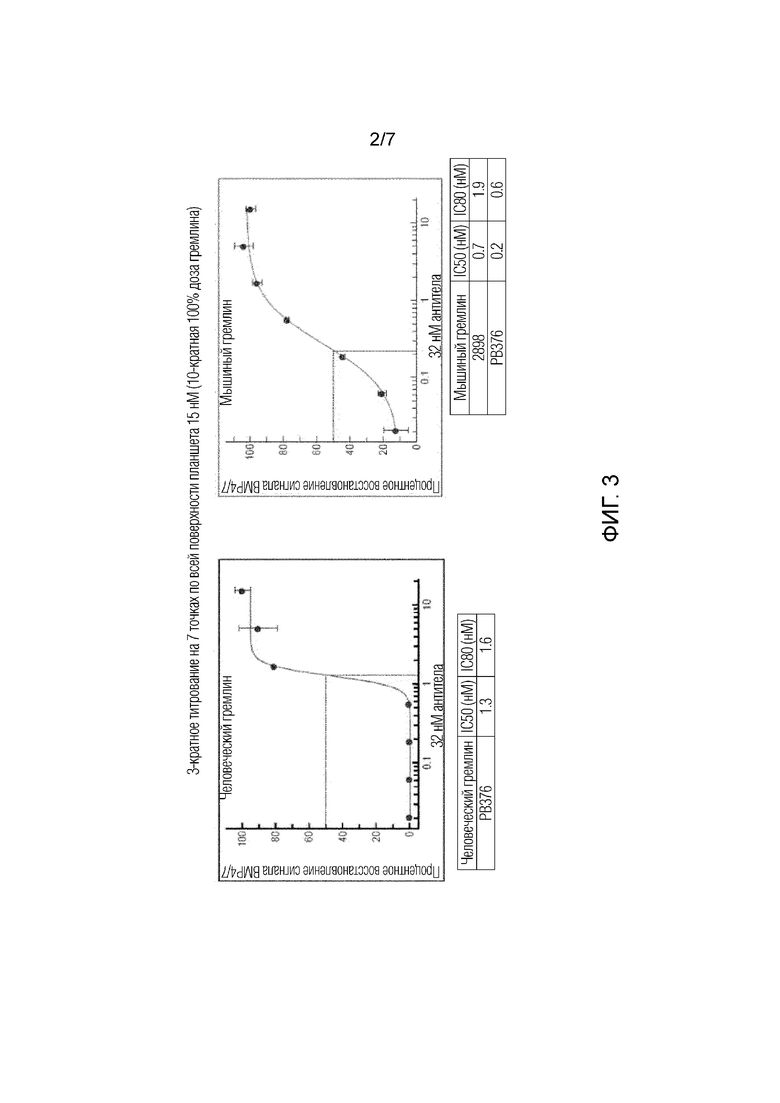
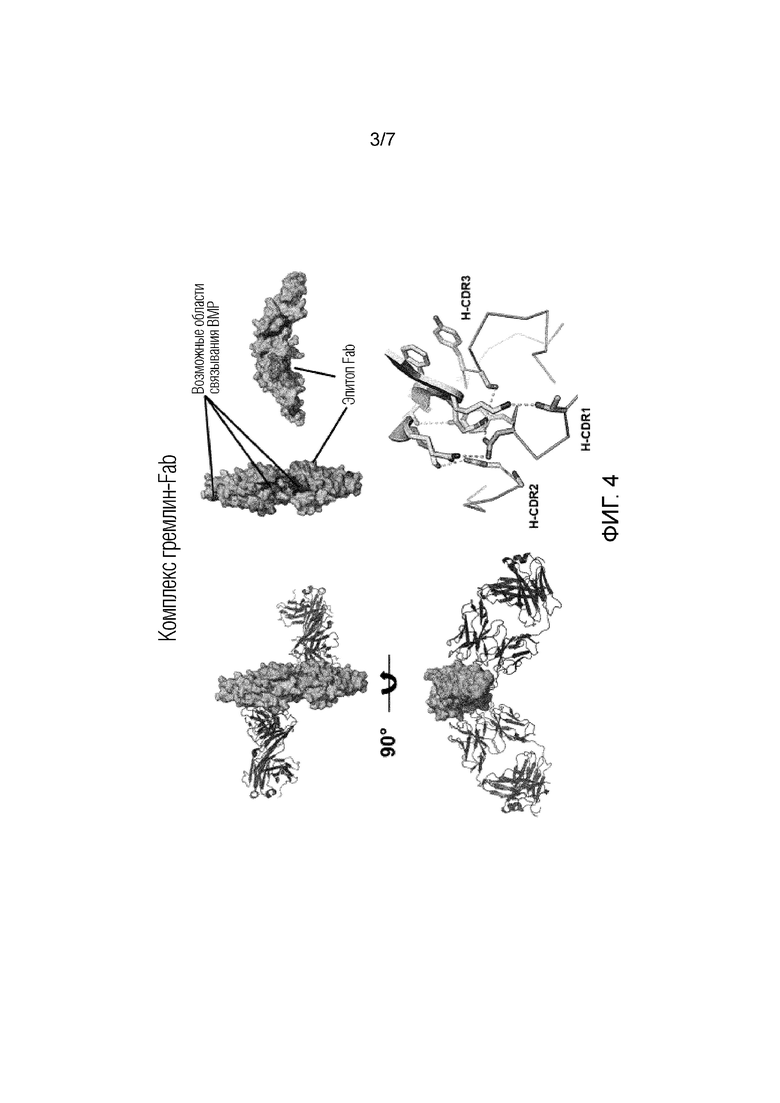
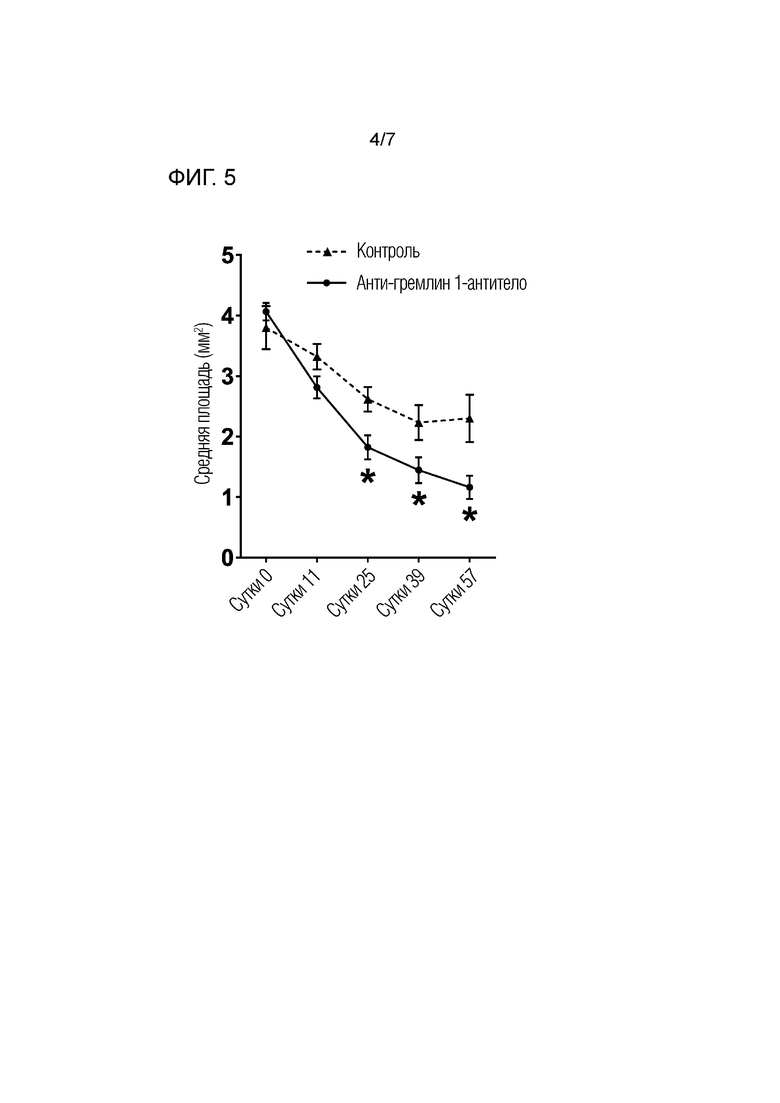
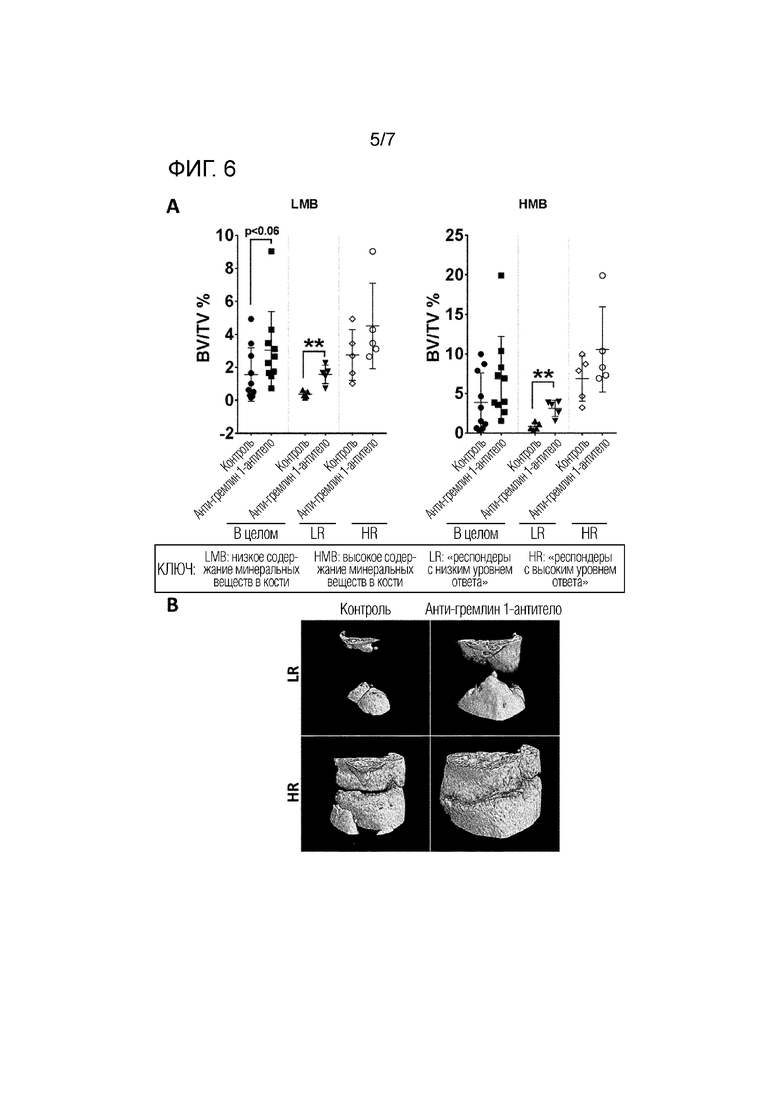
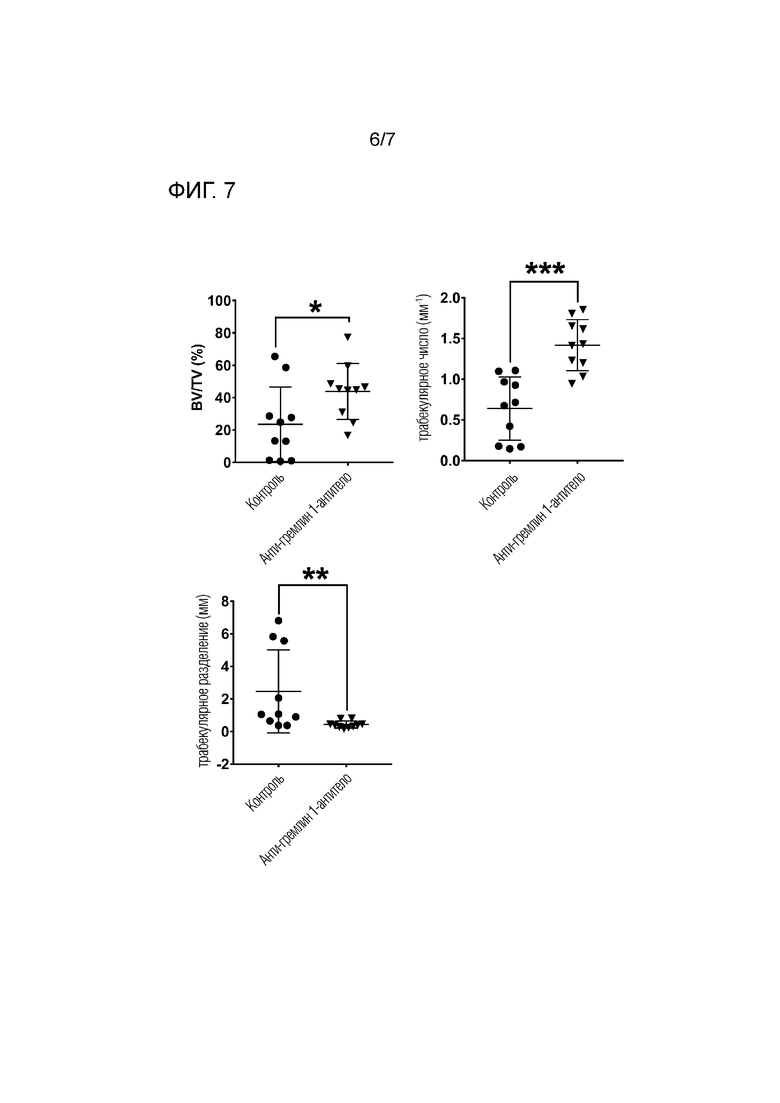
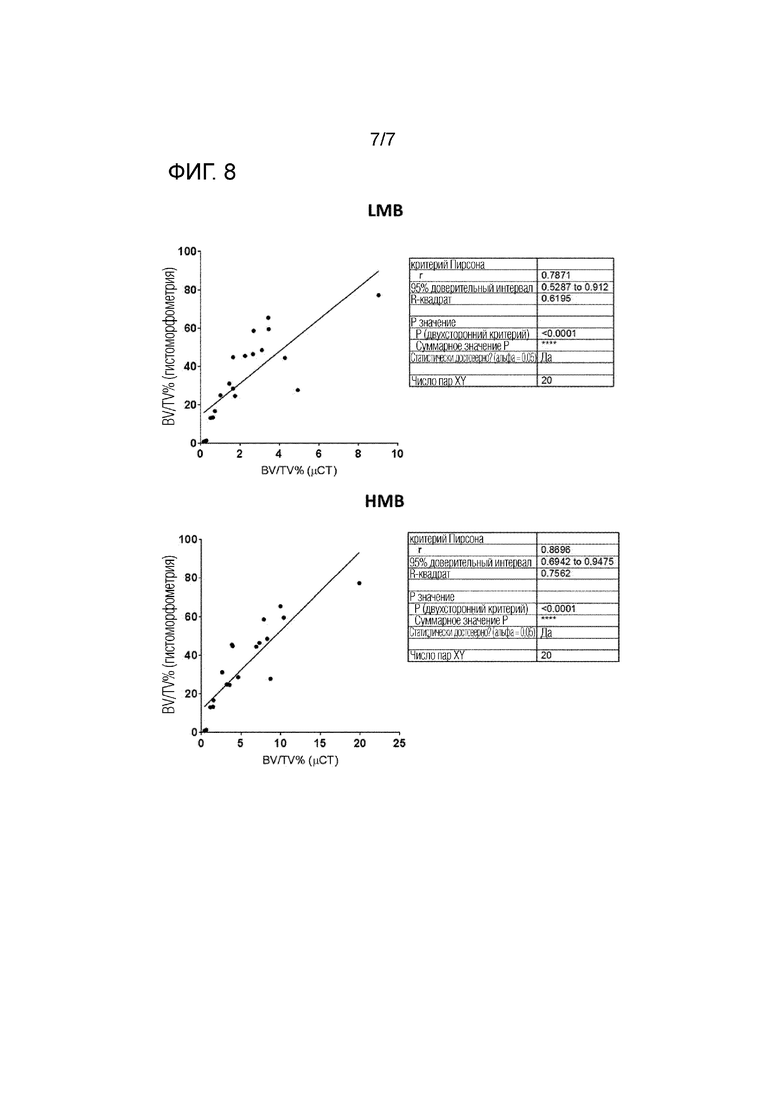
Группа изобретений относится к лечению перелома кости или дефекта кости. Предложены применение ингибитора активности гремлина-1 для производства лекарственного средства для лечения перелома кости или дефекта кости и способ лечения перелома кости или дефекта кости, включающий введение терапевтически эффективного количества ингибитора активности гремлина-1. Ингибитор представляет анти-гремлин 1-антитело или его функционально активный фрагмент. Антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3, последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3. Изобретения обеспечивают ускорение заживления и сращения костной ткани при дефектах сегментарного зазора, и могут обеспечить усовершенствованную терапию для лечения или предупреждения несращения перелома. 2 н. и 14 з.п. ф-лы, 8 ил., 6 табл., 8 пр.
1. Применение ингибитора активности гремлина-1 для производства лекарственного средства для лечения перелома кости или дефекта кости, где ингибитор представляет анти-гремлин 1-антитело или его функционально активный фрагмент, и где антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3, последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3.
2. Применение по п.1, где функционально активный фрагмент антитела представляет Fab, Fab', F(ab')2, Fv или scFv.
3. Применение по п.1 или 2, где антитело содержит вариабельную область тяжелой цепи (HCVR) с SEQ ID NO: 10 и вариабельную область легкой цепи (LCVR) с SEQ ID NO: 11.
4. Применение по п.1 или 2, где антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3 последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3; и где вариабельная область тяжелой цепи содержит последовательность, имеющую, по меньшей мере, 95% идентичность с последовательностью SEQ ID NO: 10, и вариабельная область легкой цепи содержит последовательность, имеющую, по меньшей мере, 95% идентичность с последовательностью SEQ ID NO: 11.
5. Применение по любому из пп.1-4, где антитело связывается с эпитопом на гремлине-1, содержащем по меньшей мере один остаток, выбранный из Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175, где нумерация остатков соответствует нумерации в SEQ ID NO: 1.
6. Применение по любому из предшествующих пунктов, где перелом кости или дефект кости представляет перелом с замедленным сращением или несращенный перелом.
7. Применение по любому из предшествующих пунктов, где перелом кости или дефект кости ассоциирован с заболеванием, которое отрицательно влияет на целостность кости.
8. Применение по п.7, где заболевание, которое оказывает отрицательное влияние на целостность кости, представляет остеопороз, несовершенный остеогенез, диабет, болезнь Педжета, ревматоидный артрит, анкилозирующий спондилит, множественную миелому, первичный рак кости, рак с метастазами в кости, диффузный идиопатический гиперостоз скелета, остеомиелит, почечную недостаточность, мышечную дистрофию Дюшенна и талассемию.
9. Способ лечения перелома кости или дефекта кости, включающий введение терапевтически эффективного количества ингибитора активности гремлина-1, где ингибитор представляет анти-гремлин 1-антитело или его функционально активный фрагмент, и где антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3, последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3.
10. Способ по п.9, где функционально активный фрагмент антитела представляет Fab, Fab', F(ab')2, Fv или scFv.
11. Способ по п.9 или 10, где антитело содержит вариабельную область тяжелой цепи (HCVR) с SEQ ID NO: 10 и вариабельную область легкой цепи (LCVR) с SEQ ID NO: 11.
12. Способ по п.9 или 10, где антитело содержит последовательность SEQ ID NO: 3 или 4 для HCDR1, последовательность SEQ ID NO: 5 для HCDR2, последовательность SEQ ID NO: 6 для HCDR3, последовательность SEQ ID NO: 7 для LCDR1, последовательность SEQ ID NO: 8 для LCDR2 и последовательность SEQ ID NO: 9 для LCDR3; и где вариабельная область тяжелой цепи содержит последовательность, имеющую по меньшей мере 95% идентичность с последовательностью SEQ ID NO: 10, и вариабельная область легкой цепи содержит последовательность, имеющую по меньшей мере, 95% идентичность с последовательностью SEQ ID NO: 11.
13. Способ по любому из пп.9-12, где антитело связывается с эпитопом на гремлине-1, содержащем по меньшей мере один остаток, выбранный из Ile131, Lys147, Lys148, Phe149, Thr150, Thr151, Arg169, Lys174 и Gln175, где нумерация остатков соответствует нумерации в SEQ ID NO: 1.
14. Способ по любому из предшествующих пунктов, где перелом кости или дефект кости представляет перелом с замедленным сращением или несращенный перелом.
15. Способ по любому из предыдущих пунктов, где перелом кости или дефект кости ассоциирован с заболеванием, которое отрицательно влияет на целостность кости.
16. Способ по п.15, где заболевание, которое оказывает отрицательное влияние на целостность кости, представляет остеопороз, несовершенный остеогенез, диабет, болезнь Педжета, ревматоидный артрит, анкилозирующий спондилит, множественную миелому, первичный рак кости, рак с метастазами в кости, диффузный идиопатический гиперостоз скелета, остеомиелит, почечную недостаточность, мышечную дистрофию Дюшенна и талассемию.
| Инкубатор для новорожденных с изменяемой прозрачностью стенок рабочей камеры | 2023 |
|
RU2826790C1 |
| Уплотнительная прокладка-компенсатор температурно-деформационного шва наружной стены здания | 1988 |
|
SU1571159A1 |
| SEBALD H-J et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| GAZZERRO E | |||
| et al | |||
| Skeletal Overexpression of Gremlin Impairs Bone Formation and Causes | |||

