Область техники, к которой относится изобретение
Настоящее изобретение относится к области холестеринсодержащих липидов и применению указанных холестеринсодержащих липидов для индуцирования иммунной реакции против B. burgdorferi.
Уровень техники
Болезнь Лима является зоонозом, вызываемым передаваемой клещом спирохетой B.burgdorferi (Steere et al., N. Engl. J. Med., 308:733-740, 1983). B. burgdorferi проникает в кожу восприимчивого хозяина при укусе клеща Ixodes. Спирохеты B. burgdorferi представляют собой подвижные клетки спиральной формы с наружной клеточной мембраной, окружающей протоплазматический цилиндрический комплекс, состоящие из цитоплазмы, клеточной оболочки, внутренней клеточной мембраны и жгутиков, которые расположены не на поверхности клеток, а в периплазматическом пространстве между наружной клеточной мембраной и протоплазматическим цилиндром. Предполагается, что наружная клеточная мембрана и жгутики играют важную роль во взаимодействии хозяин-паразит во время болезни, поэтому было выполнено несколько исследований с целью идентификации основных поверхностных белков в качестве важных иммуногенов (Barbour et al., J. Clin. Invest. 72:504-515, 1983).
Первичным кожным проявлением данного заболевания у человека является хроническая ползучая эритема (erythema chronicum migrans) (ECM), вследствие которой длительное инфекционное поражение кожи вызывает хронический трофический акродерматит (acrodermatitis chronica atrophicans) (Asbrink et al., Acta Derm. Venereol. 64: 506-512, 1984). Кроме того, организмы Borrelia проникают в кровеносную систему хозяина и попадают в различные органы, включая головной мозг и суставы (Barbour et al., Microbiol. Rev. 50:381-400, 1986). Вторичное распространение патогенов вызывает различные клинические синдромы, в том числе лимфоцитарный менингорадикулит (Pfister et al., J. Neurol. 118:1-4, 1984), миокардит (Steere et al., Ann. Intern. Med. 93:8-10, 1980) и хронический артрит (Steere et al., Ann. Intern. Med. 90:286-291, 1979). Инфекционное поражение некоторых тканей, в частности головного мозга и суставов, может сохраняться у многих субъектов в течение многих лет и вызывать серьезные нарушения трудоспособности. Вышеуказанные формы хронической болезни Лима являются следствием неспособности хозяина самому избавиться от инфекционного агента и, возможно, вызвать аутоиммунную реакцию (Steere et al., Ann. Intern. Med. 99: 76-82, 1983).
Установлено, что самые первые антитела IgM против антигенов штамма В31 B. burgdorferi, депонированного в Американскую коллекцию типовых культур в 1983 г. под номером доступа АТСС 35210, воздействуют на специфический для данного рода полипептид патогенного жгутика, именуемый флагеллином и имеющий молекулярную массу, равную 41 кДа (Craft et al., J. Clin. Invest. 78:934-939, 1986), который взаимодействует с моноклональным антителом Н9724 (Barbour et al., Infect. Immun. 52:549-554, 1986). Ранее антитела IgG также вырабатывались против флагеллина с молекулярной массой 41 кДа, но, по мере развития болезни, антитела IgG образуются против других иммуногенов, в частности против двух широко распространенных белков с молекулярной массой 31 кДа и 34 кДа. Установлено, что два указанных белка, именуемых OspA (31 кДа) и OspB (34 кДа), расположены на поверхности B. burgdorferi и входят в состав наружной жидкой клеточной мембраны (Barbour et al., J. Clin. Invest. 72:504-515, 1983).
В патенте США № 4721617 описано применение инактивированных цельных спирохет B. burgdorferi в качестве вакцины против болезни Лима. Кроме того, в патенте США № 6203798 описано применение белковых антигенов, OsрA и OsрB, в качестве кандидатов в вакцины. Однако 25 февраля 2002 г. производитель объявил о прекращении производства вакцины против болезни Лима LIMErix™, содержащей OspA.
Таким образом, по-прежнему существует потребность в реагенте, который может быть использован для индуцирования иммунной реакции против B. burgdorferi, в частности защитной иммунной реакции, с целью создания эффективной вакцины против болезни Лима.
Сущность изобретения
В данном изобретении описаны уникальные соединения и их аналоги, в частности терапевтически приемлемые соли. Одним объектом настоящего изобретения являются новые соединения следующей формулы А, их фармацевтически приемлемые соли или комплексы:
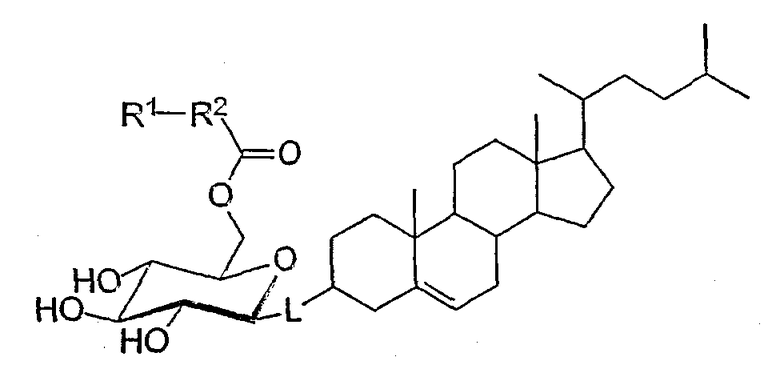
где R1 выбирают из группы, включающей азидо, амино, замещенную аминогруппу, гидразино (например, -NHNH2), гидразид (например, -С(O)NHNH2), семикарбазид (например, -NHC(O)NHNH2) или карбогидразид (например, -NHNHC(O)NHNH2);
R2 выбирают из группы, включающей насыщенную или ненасыщенную углеродную цепь, содержащую 1-25 атомов углерода, в частности 5-20 атомов углерода, либо насыщенную или ненасыщенную замещенную углеродную цепь, содержащую 1-25 атомов углерода, в частности 5-20 атомов углерода; и
L выбирают из О, N, S, P или алкиленового радикала (например, метилена (-СН2-).
Другим объектом настоящего изобретения является очищенное соединение формулы В:
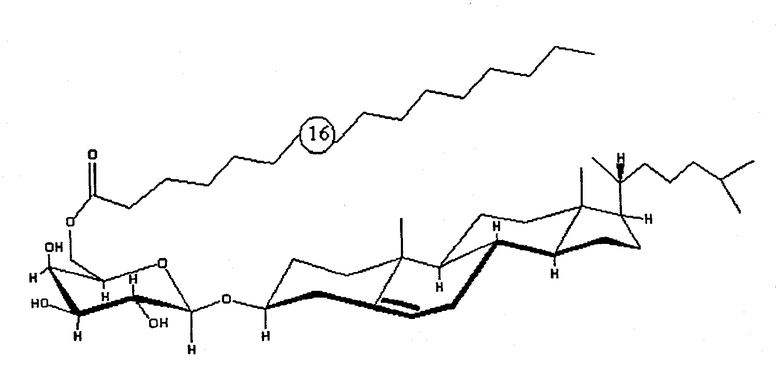
где “16” означает число атомов углерода в алкильном заместителе с прямой цепью. Данное конкретное очищенное соединение исключительно для удобства именуется в данном описании “BBGL-II” (то есть гликолипид-II B. burgdorferi).
Другим объектом настоящего изобретения являются конъюгаты, которые включают (i) носитель и (ii) соединение формулы А или В, связанное с носителем.
Описанные соединения и конъюгаты (формул А и В) могут быть использованы для индуцирования иммунной реакции против B. burgdorferi при введении субъекту терапевтически эффективного количества вышеуказанного соединения или конъюгата. Такое введение позволяет эффективно предотвратить или лечить болезнь Лима у субъекта. Вышеуказанные соединения и их терапевтически приемлемые соли могут быть использованы для получения фармацевтических или иммуногенных композиций.
Вышеуказанные и другие особенности и преимущества данного изобретения станут более очевидными из нижеследующего подробного описания нескольких вариантов осуществления изобретения со ссылкой на прилагаемые чертежи.
Краткое описание чертежей
На фиг.1 показано цифровое изображение анализа методом ТСХ липидных экстрактов из B. burgdorferi. Все липидные экстракты, полученные из штаммов В31 (полоса А), N40 (полоса В) и BL303 (полоса С), обрабатывали парами иода, опыляли антроном (полоса D) и подвергали иммунному окрашиванию антисывороткой мыши против BBGL-II (полоса Е). Альтернативно обработанные ультразвуком клетки В31 вводили в колонку с детоксикирующим гелем, связанное вещество элюировали дезоксихолатом и его липиды подвергали воздействию паров иода (полоса F).
На фиг.2 показано цифровое изображение части масс-спектра MALDI-TOF натриевого аддукта BBGL-II и BBGL-III. Два главных пика псевдомолекулярных ионов [M+Na+] при m/z 809,87 и 835,87 отображают холестерилгалактозу с производными пальмитиновой кислоты (МI) и олеиновой кислоты (МII) BBGL-II (A), и два пика псевдомолекулярных ионов [M+Na]+ при m/z 780,1 и 806,1 отображают моногалактозилдиацилглицерин (BBGL-III), замещенный дипальмитоильным (МIII) и пальмитоилолеильным (МIV) производными (В).
На фиг.3а-в показаны цифровые изображения редактирования 13С ЯМР-спектра DEPT гликолипида BBGL-II в CDCl3 при 126 МГц. На фиг.3а показано цифровое изображение только сигналов СН, полученное при выполнении эксперимента DEPT-90. На фиг.3в показано цифровое изображение положительного резонанса СН3 и отрицательного резонанса СН2, полученное в результате компьютерного вычисления из DEPT-150 - ƒ × DEPT-90, где ƒ означает экспериментально установленный коэффициент.
На фиг.4 показан неполный двумерный 1Н/13С ЯМР-спектр НМВС гликолипида BBGL-II-Ac3 в CDCl3 при 500/126 МГц, где изображен промежуточный остаток, 1Н/13С связности для 1Н Gal/13C ацетилкарбонильной группы, 1H Gal/13С карбонильной группы сложного эфира жирной кислоты и комбинации холестерин Н-3/Gal С-1'. На данной фигуре показаны также связности двух промежуточных остатков.
На фиг.5 показан двумерный 1H/1H ЯМР-спектр COSY-30 BBGL-II-Ac3 в CDCl3 при 500 МГц, где изображены 1Н/1Н связности вицина и гермина.
На фиг.6 показан двумерный 1Н/1Н ЯМР-спектр TOCSY гликолипида BBGL-III в CDCl3 при 500 МГц. На данной фигуре показаны также характеристические подспектры Gal, Gro и жирной кислоты 18:1.
На фиг.7 показан неполный двумерный 1Н/13С ЯМР-спектр НМВС гликолипида BBGL-III-Ac4 в CDCl3 при 500/126 МГц, где изображены 1Н/13С связности промежуточных остатков для четырех протонов Gal с ацетилкарбонильными ядрами 13С, трех глицерильных протонов с карбонильными ядрами 13С сложного эфира жирной кислоты и двух глицерильных протонов с С-1' Gal.
На фиг.8 показано схематическое изображение структуры основной фракции природного BBGL-II, холестерил-6-О-пальмитоил-b-d-галактопиранозида.
На фиг.9 показано схематическое изображение структуры основной фракции природного BBGL-III, 1-олеил, 2-пальмитоил, 3-[О-α-D-галактопиранозил]-sn-глицерина.
На фиг.10 изображен график, показывающий титры антител сывороток кроликов, иммунизированных гликолипидом формулы В. Титры антител, измеренные описанным ниже методом ELISA, представляют собой среднее значение, полученное для четырех разных сывороток кроликов, разведенных в отношении 1:100, каждая из которых была подвергнута трем независимым анализам. Незаштрихованные столбцы, титры IgG; серые столбцы, титры IgM; линии над столбцами показывают стандартные отклонения.
На фиг.11 изображен график, показывающий титры антител сывороток мышей, иммунизированных гликолипидом формулы В. Мышей (10 мышей в одной группе) иммунизировали описанным ниже способом гликолипидом формулы В в адъюванте Фрейнда (CFA); PBS; ДМСО или сквалене. Титры антител, измеренные методом ELISA в сыворотках, разведенных в отношении 1:100, представляют собой среднее значение трех независимых анализов. Линии над столбцами показывают стандартные отклонения.
На фиг.12-15 показана схема синтеза синтетического липида, представленного в данном описании изобретения.
Подробное описание изобретения
I. Термины
За исключением особо оговоренных случаев технические термины использованы в обычно употребляемых значениях. Определения общепринятых терминов в молекулярной биологии можно найти в публикациях Benjamin Lewin, Genes V, published by Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew et al. (eds.), The Еncyclopedia of Molecular Biology, published by Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Robert A. Meyers (ed.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, published by VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Ниже приведены разъяснения некоторых терминов, облегчающие понимание различных вариантов осуществления изобретения.
Термин “введение” соединения означает доставку в организм субъекта соединения, пролекарства соединения, конъюгата соединения или фармацевтической композиции, содержащей описанное соединение.
Аналог: молекула, которая отличается по своей химической структуре от исходного соединения, например гомолог (отличающийся приращением химической структуры, например длины алкильной цепи), молекулярный фрагмент, структура, отличающаяся одной или несколькими функциональными группами либо изменением ионизации. Структурные аналоги часто обнаруживают путем определения количественных соотношений структурной активности (QSAR) методами, описанными в публикации Remington: The Science and Practice of Pharmacology, 19th Edition (1995), chapter 28.
Антиген: соединение, композиция или вещество, которое может стимулировать иммунную реакцию, например продуцирование антител или Т-клеточную иммунную реакцию у животного, в том числе композиции, которые инъецируют или вводят внутрь животному. Антиген взаимодействует с продуктами специфического гуморального или клеточного иммунитета, включая продукты, индуцированные гетерологичными иммуногенами. Термин “антиген” включает все родственные антигенные детерминанты.
Антитело: молекулы иммуноглобулина и иммунологически активные части молекул иммуноглобулина, то есть молекулы, содержащие антигенсвязывающий сайт, который специфически связывает антиген (иммунологически взаимодействует с антигеном).
Природное антитело (например, IgG, IgM, IgD) включает четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные дисульфидными связями. Однако установлено, что антигенсвязывающую функцию антитела могут выполнять фрагменты природного антитела. Поэтому такие антигенсвязывающие фрагменты также входят в определение термина “антитело”. Конкретные, не ограничивающие примеры связывающих фрагментов, определяемых термином “антитело”, включают (i) Fab-фрагмент, содержащий VL, VH, CL и СН1 домены; (ii) Fd-фрагмент, содержащий VH и СН1 домены; (iii) Fv-фрагмент, содержащий VL и VH домены одного плеча антитела; (iv) dAb-фрагмент (Ward et al., Nature 341:544-546, 1989), содержащий VH домен; (v) выделенный гипервариабельный участок (CDR) и (vi) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области.
Животное: живые многоклеточные позвоночные организмы, относящиеся к категории, в которую входят, например, млекопитающие и птицы. Термин “млекопитающее” означает человека и млекопитающих, отличных от человека. Аналогичным образом термин “субъект” означает как человека, так и животных.
Bоrrelia (B.) burgdorferi : спирохета, которая впервые была описана Джонсоном и др. в 1984 г. B. burgdorferi можно культивировать из векторов на основе членистоногих или позвоночных хозяев в модифицированной среде Келли, именуемой BSK (Barbour-Stoenner-Kelly). В среде BSK можно успешно культивировать Borrelia, выделенную из клещей и крови, кожи и цереброспинальной жидкости субъектов, страдающих болезнью Лима. Среда BSK, отвержденная 1,3% агарозой, позволяет получать колонии из отдельных организмов. B. burgdorferi растет медленно по сравнению с большинством бактерий. Каждая спирохета делится на две клетки через 12-24 часов элонгации. Тип Borrelia, инфицирующий людей в США, получил название B. burgdorferi sensu stricto. В Европе болезнь Лима также вызывают B. burgdorferi sensu stricto и два родственных вида Borrelia, B. garinii и B. afzelii. В Азии болезнь Лима вызывают только B. garinii и B. afzelii.
Borrelia, в том числе B. burgdorferi, представляют собой гибкие спиральные клетки, состоящие из протоплазматического цилиндра, окруженного клеточной мембраной, 7-11 периплазматических жгутиков и наружной мембраны, которая свободно связана с нижними структурами. Последовательность ДНК штамма В31 B. burgdorferi, обнародованная в 1997 г., содержит линейную хромосому длиной 950 т.п.о., 9 линейных плазмид и 12 кольцевых плазмид. Наружная мембрана B. burgdorferi и других бактерий Borrelia уникальна тем, что гены, кодирующие ее белки, расположены на линейных плазмидах; указанные внехромосомные гены определяют антигенную идентичность данных организмов и, предположительно, помогают указанным бактериям адаптироваться и выживать в клещах и других млекопитающих-хозяевах.
Конъюгат: термин “конъюгат” означает любую структуру, которая включает описанное здесь иммуногенное соединение, связанное с фармацевтически приемлемым носителем. Таким образом, определение термина “конъюгат” не ограничивается конъюгатом иммуногенного соединения, ковалентно связанного с белком-носителем (данный конкретный тип конъюгата часто определяется в данной области как “конъюгатная вакцина”).
Производное: производное является биологически активной молекулой, выделенной из основной молекулярной структуры.
Иммунная реакция: реакция клетки иммунной системы, такой как В-клетка, Т-клетка или моноцит, на определенный раздражитель. В одном варианте осуществления изобретения данная реакция является специфической для определенного антигена (“антигенспецифическая реакция”). В другом варианте осуществления изобретения иммунная реакция является Т-клеточной иммунной реакцией, такой как CD4+ реакция или CD8+ реакция. В другом варианте осуществления изобретения данная реакция является В-клеточной реакцией, в результате которой образуются специфические антитела. “Иммуногенная композиция” является любой композицией, вызывающей иммунную реакцию у млекопитающего-хозяина при инъецировании или другом способе введения иммуногенной композиции. Иммунная реакция может быть гуморальной, клеточной или обеими вместе. Термин “бустер” означает повышенную иммунную реакцию на иммуногенную композицию, возникающую при повторном введении млекопитающему-хозяину такой же иммуногенной композиции.
Выделенный: “выделенный” биологический элемент (такой как липид) по существу полностью отделен или очищен от других биологических элементов в клетке организма, в которой данный элемент обычно существует, то есть от других хромосомных и внехромосомных ДНК и РНК, белков, липидов и органелл. “Выделенными” липидами являются липиды, очищенные стандартными методами очистки. В определение данного термина входят также химически синтезированные липиды и другие компоненты.
Метка: детектируемое соединение или композиция, которые прямо или непрямо конъюгированы с другой молекулой для облегчения обнаружения данной молекулы. Конкретные неограничивающие примеры меток включают флуоресцентные метки, конъюгацию с ферментом и радиоактивные изотопы.
Болезнь Лима: болезнь Лима возникает вследствие инфицирования бактериями Borrelia burgdorferi. Указанные бактерии передаются человеку при укусе оленьим клещом и ежегодно становятся причиной заражения более чем 16000 человек.
Болезнь Лима наиболее часто проявляется в виде характерного покраснения глаз, появления ползучей эритемы, сопровождающейся неспецифическими симптомами, такими как лихорадка, недомогание, утомление, головная боль, мышечные боли (миалгия) и боли в суставах (артралгия).
Инкубационный период от инфицирования до появления ползучей эритемы обычно составляет 7-14 дней, но может быть равен всего 3 дням или продолжаться в течение 30 дней. У некоторых инфицированных больных отсутствуют распознаваемые признаки (бессимптомное инфицирование, определяемое при помощи серологческой реакции) или проявляются только неспецифические симптомы, такие как лихорадка, головная боль, утомление и миалгия. Спирохеты, вызывающие болезнь Лима, распространяются с места укуса клеща через кожу, лимфатическую и кровеносную системы. Признаки распространенного инфицирования на начальной стадии обычно проявляются через несколько дней или недель после появления отдельного поражения ползучей эритемой. Помимо множественных (вторичных) поражений ползучей эритемой распространенная инфекция на начальной стадии может проявляться в виде заболевания нервной системы, костно-мышечной системы или сердца. Ранние неврологические проявления включают лимфоцитарный менингит, невропатию (в частности, паралич лицевого нерва) и радикулоневрит. Костно-мышечные проявления могут включать перемещающиеся боли в суставах и мышцах при наличии или отсутствии объективных признаков опухания суставов. Проявления заболевания сердца встречаются редко, но могут включать миокардит и временную атриовентрикулярную блокаду различной степени тяжести. Заражение B. burgdorferi у больного, не прошедшего курс лечения или прошедшего недостаточный курс лечения, прогрессирует до поздней стадии в течение недель или месяцев после инфицирования. Наиболее обычным объективным проявлением болезни Лима на поздней стадии является перемежающееся опухание и боль в одном или нескольких суставах, обычно в крупных, испытывающих нагрузку суставах, таких как коленный сустав. У некоторых больных развивается хроническая полиневропатия аксонов или энцефалопатия, причем последнее заболевание обычно проявляется в виде нарушения познавательной способности, расстройства сна, утомления и изменений личности. Болезнь Лима нечасто протекает в тяжелой хронической форме с нарушением трудоспособности. У некоторых больных после лечения болезни Лима возникает постлечебный синдром, вызываемый плохим состоянием здоровья.
Лимфоциты: тип белых клеток крови, которые участвуют в иммунной защите организма. Существуют два основных типа лимфоцитов: В-клетки и Т-клетки.
Фармацевтически приемлемые носители: “носитель” является физиологически приемлемым веществом, с которым связано терапевтически или биологически активное соединение. Носитель может облегчать введение терапевтически или биологически активного соединения определенного типа и/или усиливать иммунную реакцию, вызываемую терапевтически или биологически активным соединением. Тип носителя обычно зависит от определенного способа введения. Например, препараты для парентерального введения обычно представляют собой инъекционные жидкости, которые включают в качестве наполнителя фармацевтически и физиологически приемлемые жидкости, такие как вода, физиологический раствор, сбалансированные растворы солей, водный раствор декстрозы, глицерин или тому подобные. В случае твердых композиций (таких, как порошки, пилюли, таблетки или капсулы) обычно применяемые нетоксичные твердые носители могут включать, например, маннит, лактозу, крахмал или стеарат магния фармацевтических сортов. Помимо биологически нейтральных носителей фармацевтические композиции могут содержать небольшие количества нетоксичных вспомогательных веществ, таких как смачивающие вещества или эмульгаторы, консерванты, буферные агенты, регулирующие рН, и тому подобные, например ацетат натрия или монолаурат сорбита. В публикации Remington's Pharmaceutical Sciences, by E.W. Martin, Mack Publishing Co., Easton, PA, 15th Edition (1975) описаны композиции и препараты, пригодные для фармацевтической доставки BBGL-II, представленного в настоящем описании изобретения.
“Фармацевтически приемлемые соли” описанных соединений включают соли, образуемые из катионов, таких как натрий, калий, алюминий, кальций, литий, магний, цинк, и оснований, таких как аммиак, этилендиамин, N-метилглутамин, лизин, аргинин, орнитин, холин, N,N'-дибензилэтилендиамин, хлорпрокаин, диэтаноламин, прокаин, N-бензилфенетиламин, диэтиламин, пиперазин, трис(гидроксиметил)аминометан и гидроксид тетраметиламмония. Указанные соли могут быть получены стандартными методами, например, в результате взаимодействия свободной кислоты с приемлемым органическим или неорганическим основанием. Любое химическое соединение, приведенное в данном описании изобретения, можно альтернативно вводить в виде его фармацевтически приемлемой соли. “Фармацевтически приемлемые соли” включают также свободные кислоты, основания и цвиттерионные формы. Описание приемлемых фармацевтически приемлемых солей можно найти в публикации Handbook of Pharmaceutical Salts, Properties, Selection and Use, Wiley VCH (2002).
“Фармацевтически приемлемые комплексы” представленных в данном описании изобретении эстрогенных агентов включают комплексы или координационные соединения, образуемые из ионов металлов. Такие комплексы могут включать лиганд или хелатообразователь, используемый для связывания с эстрогенным агентом.
Полипептид: любая цепь аминокислот независимо от длины или посттрансляционной модификации (например, гликозилирования или фосфорилирования).
Очищенный: термин “очищенный” не подразумевает достижения абсолютной чистоты и, скорее, имеет относительное значение. Так, например, очищенный пептидный препарат является препаратом, в котором пептид или белок более обогащены по сравнению с пептидом или белком, присутствующим в естественной среде внутри клетки. В одном варианте осуществления изобретения препарат очищают в такой степени, что содержание описанного здесь липида составляет по крайней мере 50%, более предпочтительно по крайней мере около 90% и наиболее предпочтительно по крайней мере около 98% от общего количества липида в препарате, содержащем соединение по настоящему изобретению.
“Насыщенная или ненасыщенная углеродная цепь” включает алкильные, алкенильные, алкинильные, алкадиенильные или циклоалкильные группы.
“Замещенная аминогруппа” означает группу -NZ1Z2, в которой по крайней мере один из элементов Z1 или Z2 независимо выбирают из алкильной, циклоалкильной, гетероциклоалкильной, арильной, гетероарильной, алкоксильной, арилоксигруппы и их комбинаций.
“Замещенная углеродная цепь” означает углеродную цепь, в которой один или несколько атомов водорода углеродной цепи заменены другой группой, такой как галоген, арил, циклоалкил, аминогруппа или их комбинацией.
Т-клетка: белая клетка крови, имеющая важное значение для иммунной реакции. Т-клетки включают, не ограничиваясь ими, CD4+ Т-клетки и CD8+ Т-клетки. CD4+ Т-лимфоцит является иммунокомпетентной клеткой, на поверхности которой имеется маркер, известный как “дифференцировочный кластер 4” (CD4). Указанные клетки, известные также как Т-клетки-хелперы, помогают сформировать иммунную реакцию, включающую образование антител и Т-клеток-киллеров. CD8+ Т-клетки несут маркер “дифференцировочного кластера 8” (CD8). В одном варианте осуществления изобретения CD8 Т-клетки являются цитотоксическими Т-лимфоцитами. В другом варианте осуществления изобретения CD8-клетки являются Т-клетками-супрессорами.
Терапевтически активный агент: агент, индуцирующий иммунную реакцию, измеряемую при помощи клинической реакции (например, увеличение популяции иммунокомпетентных клеток, образование специфически связывающегося антитела или измеряемая устойчивость к инфицированию B. burgdorferi). Терапевтически активные агенты могут также включать органические или другие химические соединения, имитирующие действие BBGL-II.
Описания вышеуказанных терминов приведены только для облегчения понимания изобретения и не должны ограничивать объем значений, известных специалисту в данной области, или объем прилагаемой формулы изобретения. Если не оговорено особо, все технические и научные термины, используемые в данном описании изобретения, имеют значения, известные специалистам в данной области. Термины, используемые в единственном числе, включают также множественное число данных терминов, за исключением тех случаев, когда из контекста следует обратное. Аналогичным образом союз “или” означает также “и” за исключением тех случаев, когда из контекста следует обратное. Кроме того, должно быть понятно, что все размеры оснований или аминокислот и все значения молекулярной массы, указанные для соединений, являются приблизительными и приведены только для описания. Хотя при осуществлении или проверке данного изобретения могут быть использованы способы и вещества, подобные или эквивалентные представленным в данном описании изобретения, ниже приведены приемлемые способы и вещества. Термин “содержит” означает “включает”. Кроме того, вещества, способы и примеры имеют лишь иллюстративное значение и не ограничивают объем изобретения. Все химические соединения, рассмотренные в данном описании изобретения, включают как (+), так и (-) стереоизомеры (а также либо (+), либо (-) стереоизомеры) и любые их таутомеры.
Как было указано выше, в данном описании изобретения приведены все биологически активные (например, иммуногенные) гликолипиды формулы А. Указанные гликолипиды являются синтетическими аналогами BBGL-II в том смысле, что их получают химическим синтезом в отличие от экстракции и очистки из B. burgdorferi. Соединение формулы В, с другой стороны, можно получить экстракцией из B. burgdorferi (другими словами, данное соединение является природным BBGL-II).
Природный BBGL-II определяется в данном описании изобретения как “холестерил-6-О-пальмитоил-β-D-галактопиранозид”. Экстрагированный природный BBGL-II выделяют и очищают. Например, очищенный природный изолят BBGL-II по существу не содержит других липидов, соединений или молекулярных фрагментов из B. burgdorferi. В одном примере очищенный изолят природного BBGL-II содержит менее одного массового процента нуклеиновых кислот и менее одного массового процента белков.
Аналоги BBGL-II также имеют терапевтическое значение (см. формулу А). Например, в таком аналоге ацильная группа (например, пальмитоильная), показанная в формуле BBGL-II, может быть заменена другими насыщенными или ненасыщенными углеродсодержащими группами или цепями, содержащими 1-25 атомов углерода, в частности другими ацильными группами, выделенными из органических жирных кислот. Холестерильные и галактопиранозидные кольцевые структуры могут включать в любом положении кольца замещающие группы, такие как алкил (в частности, ацильную группу, выделенную из жирной кислоты), карбоксил, замещенный карбоксил (-СOR, где R означает алкил, карбоновую кислоту или сложный эфир), арил, алкокси, гетероцикл, галоген или аминогруппы. Гетероатом О, соединяющий мостиковой связью холестерильные и галактопиранозидные кольца, может быть заменен алкильным радикалом (например, -CH2-) или таким гетероатомом, как N, S или Р.
В соединениях формулы А R2 (вместе с группой -С(О)О-) может быть ацильная группа, выделенная из органической жирной кислоты. Такие ацильные группы включают пальмитоил, лауроил, стеароил, миристоил, олеил и линолеил. Получение производного ацильной группы позволяет получить сайт (например, ацилазид) для ковалентного связывания с белком-носителем.
В некоторых вариантах группа R1 может образовывать ковалентную связь с белком-носителем (другими словами, конъюгировать с белком-носителем), как более подробно описано ниже. Конъюгацию с белком-носителем можно также осуществлять в результате образования производного по крайней мере одной из алкильных групп или цепей холестерильной части. Производное можно также получить описанным ниже способом. Альтернативно можно использовать мелкую молекулу, такую как дигидразид адипиновой кислоты или 1,6-диаминогексан.
BBGL-II и/или его аналоги можно выделить или очистить из культуры B. burgdorferi описанным ниже способом, или они могут быть синтезированы химическим способом. Например, соединение формулы А можно синтезировать, получив сначала галактозилгалогенид, в котором гидроксильные группы галактозного кольца имеют защитные группы. Галактозилгалогенид затем подвергают взаимодействию с холестерином с образованием галактозилхолестерина. Защитные группы в галактозном кольце удаляют и заменяют замещающими защитными группами. Отдельно получают промежуточную азидоацильную кислоту. Промежуточную азидоацильную кислоту конденсируют с галактозилхолестерином, получая при этом азидоацилированный холестерин-β-D-галактопиранозид.
В одном приведенном ниже примере терапевтически эффективное количество очищенного природного BBGL-II (формула В) или химически синтезированного аналога BBGL-II (формула А) означает количество, используемое для индуцирования иммунной реакции либо для лечения или профилактики инфицирования субъекта B. burgdorferi. Лечение означает терапевтическое вмешательство, которое позволяет ослабить признаки или симптомы инфицирования B. burgdorferi либо признаки или симптомы болезни Лима.
Иммуногенные композиции и их применение
Один вариант осуществления изобретения относится к способу лечения субъекта, инфицированного B. burgdorferi, профилактике или ингибированию инфекции или возникновения клинических проявлений болезни Лима. Альтернативно данный способ можно использовать для торможения развития уже существующей инфекции. Указанный способ включает введение субъекту терапевтически эффективного количества очищенного природного BBGL-II или соединения формулы А, таким образом излечивая или предотвращая инфицирование, замедляя развитие или излечивая болезнь Лима. Для получения композиции, предназначенной для индуцирования иммунной реакции у субъекта или вакцинации субъекта, используют BBGL-II или соединение формулы А.
Иммуногенные композиции по настоящему изобретению вызывают иммунную реакцию против B. burgdorferi у млекопитающего-хозяина, в том числе у человека и других животных. Иммунная реакции может быть клеточной или гуморальной реакцией либо обеими вместе, кроме того, данная реакция может вызывать иммунологическую память или бустер-эффект либо то и другое вместе у млекопитающего-хозяина. Антигены, вызывающие образование антител при помощи Т-клеток, известны как Т-зависимые антигены. Антигены, которые не нуждаются в помощи Т-клеток для образования антител и могут непосредственно активировать В-клетки, известны как Т-независимые антигены. Указанные иммуногенные композиции являются полезными вакцинами и могут вызывать защитную реакцию у млекопитающего или хозяина на инфицирование патогенным микроорганизмом.
В частности, описанные здесь вакцины можно получить в виде Т-клетконезависимых вакцин и/или Т-клеткозависимых вакцин. В зависимости от конкретного варианта осуществления изобретения активное соединение формулы А или В первоначально является Т-клетконезависимым, но затем его превращают в Т-клеткозависимую вакцину в результате конъюгации с носителем (в частности, с белком-носителем).
Соединения формулы А или В по данному изобретению могут быть использованы вместе с носителем. Носитель может находиться в смеси с одним или несколькими иммуногенными соединениями или может быть конъюгирован с одним или несколькими иммуногенными соединениями при помощи химического взаимодействия или связи. Например, одно или несколько иммуногенных соединений могут быть конъюгированы с макромолекулярным носителем. Носителем может быть полимер, с которым иммуногенное соединение связано гидрофобной нековалентной связью, в частности пластик, такой как полистирол, или полимер, с которым иммуногенное соединение связано ковалентной связью, в частности полисахарид или полипептид, например бычий сывороточный альбумин, овальбумин или гемоцианин лимфы улитки. Носитель предпочтительно должен быть нетоксичным и неаллергенным. Иммуногенное соединение может быть многовалентно связано с макромолекулярным носителем, так как подобная связь способствует усилению иммуногенности вакцинного препарата.
В одном варианте осуществления изобретения носителем является цепь аминокислот (например, полипептид или белок) или других частей молекулы. В другом варианте осуществления изобретения носителем является димер, олигомер или полимер с более высокой молекулярной массой последовательности аминокислот полипептида B. burgdorferi. Примеры приемлемых иммуногенных носителей включают гемоцианин лимфы улитки (KLH), альбумины, такие как бычий сывороточный альбумин (BSA) и овальбумин, PPD (очищенное белковое производное туберкулина), эритроциты, столбнячный токсоид, холерный токсоид, агарозные гранулы, активированный уголь или бентонит.
Липиды, рассмотренные в данном описании изобретения, могут быть присоединены к любому представляющему интерес белку, включающему, не ограничиваясь ими, rARU, рекомбинантный белок, содержащий повторяющиеся звенья токсина А Clostridium difficile. Носители выбирают с возможностью увеличения иммуногенности полисахарида и/или образования антител против носителя, которые являются предпочтительными в медицинском отношении. В данной области описаны носители, удовлетворяющие указанным критериям. Полимерный носитель может быть природным или синтетическим веществом, содержащим одну или несколько функциональных групп, используемых для конъюгации, таких как первичные и/или вторичные аминогруппы, азидогруппы, альдегиды, гидразиды, эпоксиды, тиолы или карбоксильные группы. Носитель может растворяться или не растворяться в воде.
Водорастворимые пептидные носители являются предпочтительными и включают, не ограничиваясь ими, природные или синтетические полипептиды или белки, такие как бычий сывороточный альбумин, бактериальные или вирусные белки, нетоксичные мутанты или их полипептидные фрагменты, например столбнячный токсин или токсоид, дифтерийный токсин или токсоид, экзотоксин или токсоид Pseudomonas aeruginosa, рекомбинантный экзобелок А Pseudomonas aeruginosa, коклюшный токсин или токсоид, экзотоксины или токсоиды Clostridium perfringens и Clostridium welchii, мутантный нетоксичный голотоксин токсина Шига, токсины Шига 1 и 2, субъединица В токсинов Шига 1 и 2, поверхностный антиген и ядерный антиген гепатита В.
Альтернативными носителями являются некоторые вещества животного, растительного или минерального происхождения, которые являются физиологически приемлемыми и служат для представления липида BBGL-II иммунной системе. Таким образом, многие носители являются приемлемыми и включают инертные вещества или вещества, обладающие биологической активностью и/или стимулирующие иммунную реакцию. В частности, примеры белка-носителя включают, не ограничиваясь ими, белок и гемоцианин лимфы улитки. В качестве носителей можно также использовать полисахариды с молекулярной массой от 10000 до 1000000, такие как крахмалы, декстран, агароза, фиколл или его карбоксилметильное производное и карбоксиметилцеллюлоза.
Полиаминокислоты могут быть также использованы в качестве носителей, при этом они включают наряду с прочими веществами полилизин, полиаланилполилизин, полиглутаминовую кислоту, полиаспарагиновую кислоту и поли(С2-С10)аминокислоты.
В качестве носителей могут быть использованы органические полимеры, которые включают, например, полимеры и сополимеры аминов, амидов, олефинов, винилов, сложных эфиров, ацеталей, полиамидов, карбонатов, простых эфиров и тому подобных. Молекулярная масса указанных полимеров может изменяться в широких пределах. Такие полимеры могут иметь от двух до нескольких тысяч повторяющихся звеньев, например две тысячи повторяющихся звеньев. Число повторяющихся звеньев должно соответствовать применению такой композиции для иммунизации животного-хозяина. Такие полимеры должны иметь более низкую молекулярную массу, в частности, от 10000 до 100000 (молекулярную массу определяют ультрацентрифугированием).
Можно также использовать неорганические полимеры. Указанными неорганическими полимерами могут быть неорганические полимеры, содержащие органические части. В частности, в качестве носителя можно использовать силикаты и гидроксид алюминия. Носитель предпочтительно является иммунологическим адъювантом. В таких случаях особенно желательно, чтобы адъювантом был мурамилдипептид или его аналоги.
Носителем может быть также остаток перекрестно-сшивающего агента, используемого для связывания нескольких синтетических пептидсодержащих цепей. Перекрестно-сшивающие агенты содержат такие функциональные группы, как альдегид (такой, как глутаральдегид), карбоксил, амин, амидо, имидо или азидофенильная группа. В частности, в качестве перекрестно-сшивающего агента желательно использовать бутиральдегид, двухвалентный сложный имидоэфир или карбодиимид.
Настоящее изобретение далее относится к способам получения иммуногенной композиции, которая включает конъюгирование липидов, рассмотренных в данном описании изобретения, с носителем. Примеры таких носителей включают приведенные выше, а также полипептидную или непептидную часть, которая может служить в качестве носителя или адъюванта или обладать другой биологической активностью в комбинации с липидами. Соединения формулы А или В могут быть конъюгированы с белком-носителем путем смешивания указанных соединений с белком-носителем в присутствии реагента, способствующего образованию ковалентной связи между соединением и белком-носителем. Иллюстративные связывающие реагенты включают глутаральдегид, гидроксисукцинимиды и карбодиимиды. Альтернативно мелкую химическую молекулу можно присоединить к белку-носителю, и данная молекула благодаря своей реакционной способности служит в качестве линкерной молекулы между соединением и белком-носителем. Иллюстративные линкеры включают дигидразид адипиновой кислоты, аминогексаноевую кислоту, хлоргексанолдиметилацеталь, D-глюкуроно-лактон и п-нитрофениламин.
Например, соединения формулы А или В могут быть конъюгированы с полипептидом различными способами, один из которых, например, включает получение производного полипептида сукцинилированием и последующее конъюгирование липидного компонента с полипептидом в результате осуществления взаимодействия данного полипептида и соединения с гидрохлоридом 1,этил-3-(3-диметиламинопропил)карбодиимида. Кроме того, липидный компонент можно активировать при помощи любого из нескольких реагентов, предпочтительно бромида цианогена.
Соединения формулы А или В можно также вводить субъекту при помощи липосомной системы доставки с целью усиления их устойчивости и/или иммуногенности. Доставка соединений липосомами может быть особенно предпочтительной, так как липосома может быть интернализована фагоцитами в организме подвергаемого лечению субъекта. Такие клетки после приема липосомы внутрь гидролизуют липосомную мембрану и затем представляют полипептиды иммунной системе вместе с другими молекулами, необходимыми для индуцирования сильной иммунной реакции. Липосомная система может представлять собой любую разновидность однослойных пузырьков, многослойных пузырьков или устойчивых многослойных пузырьков, может быть получена и введена способами, хорошо известными специалистам в данной области, например способами, описанными в патентах США № 5169637, 4762915, 5000958 или 5185154.
Настоящее изобретение относится к введению субъекту терапевтически эффективной дозы фармацевтической композиции, содержащей по крайней мере одно соединение формулы А или В. Фармацевтическая композиция может включать фармацевтически приемлемый носитель. Фармацевтическую композицию по настоящему изобретению можно вводить любым способом, известным квалифицированному специалисту. Субъектом является любое млекопитающее, включая человека.
Фармацевтические композиции предпочтительно получают и вводят в виде дозированных лекарственных форм. Твердыми дозированными лекарственными формами являются таблетки, капсулы и суппозитории. Для лечения субъекта необходимы разные суточные дозы в зависимости от активности соединения, способа введения, характера и тяжести заболевания, возраста и массы тела субъекта. Однако в определенных условиях могут быть приемлемы более высокие или более низкие суточные дозы. Суточную дозу можно вводить однократно в виде отдельной дозированной лекарственной формы или нескольких более мелких дозированных лекарственных форм, а также многократно в виде разделенных доз через определенные периоды времени. Введение можно производить в виде постоянной дозы в течение более продолжительного периода времени, такого как, например, 1-10 недель.
Фармацевтические композиции обычно вводят перорально или парентерально. Способы парентерального введения включают, не ограничиваясь ими, подкожные инъекции (SQ и депо-SQ), внутривенные (IV), внутримышечные (IM и депо-IM), внутригрудинные инъекции или вливания, подоболочечное, чрескожное и местное применение, введение в нос (ингалация) и глаза. Данное соединение можно также вводить из имплантированных резервуаров или насосов. Приемлемые твердые или жидкие фармацевтические препараты включают, например, гранулы, порошки, таблетки, таблетки с покрытием, (микро)капсулы, суппозитории, сиропы, эмульсии, суспензии, кремы, аэрозоли, капли или инъекционные растворы в ампулах, а также препараты с пролонгированным высвобождением активных соединений, при получении которых используют вышеописанные наполнители и добавки и/или вспомогательные вещества, такие как дезинтеграторы, связывающие вещества, вещества для нанесения покрытия, агенты, вызывающие набухание, смазывающие вещества, ароматизаторы, подсластители или солюбилизаторы. Указанные фармацевтические композиции пригодны для использования в разных системах доставки лекарственных средств. Для краткого ознакомления с существующими методами доставки лекарственных средств см. публикацию Langer, Science 249:1527-1533, которая включена в данное описание изобретения в качестве ссылки. Инокуляты обычно получают в виде растворов в физиологически переносимых (приемлемых) разбавителях, таких как вода, физиологический раствор, физиологический раствор с фосфатным буфером или тому подобные, с целью приготовления водной фармацевтической композиции. В состав композиций могут также входить адъюванты, такие как гидроксид алюминия.
Фармацевтические композиции можно вводить местно или системно. Количества, оказывающие эффективное действие при терапевтическом применении, несомненно, зависят от тяжести заболевания, массы тела и общего состояния субъекта. Дозы, используемые in vitro, позволяют определить количества, необходимые для введения фармацевтической композиции in situ, и, кроме того, эффективные дозы для лечения конкретных заболеваний можно определить с помощью животных моделей. Различные факторы, определяющие выбор доз, описаны, например, в публикациях Gilman et al., eds., Goodman and Gilman: The Pharmacological Bases of Therapeutics, 8th ed., Pergamon Press, 1990; и Remington's Pharmaceutical Sciences, 17th ed., Mack Publishing Co., Easton, Pa., 1990, которые включены в данное описание изобретения в качестве ссылки.
Эффективные дозы соединений по данному изобретению могут изменяться в зависимости от характера и тяжести подлежащего лечению заболевания, возраста и состояния здоровья субъекта и других клинических факторов. Таким образом, соответствующая схема лечения должна быть окончательно определена лечащим врачом. Доза соединения по настоящему изобретению обычно находится в пределах от около 0,1 мкг/кг массы тела до около 100 мг/кг массы тела. Другие приемлемые пределы включают дозы от около 1 мкг/кг до 10 мг/кг массы тела. Схема применения лекарственного средства может изменяться один раз в неделю и даже ежедневно в зависимости от ряда клинических факторов, таких как чувствительность субъекта к липиду. В случае более тяжелого заболевания желательно вводить вышеуказанные дозы разными способами, включающими внутривенное или подоболочечное введение. Приемлемым способом введения может быть также непрерывная инфузия.
Очищенные терапевтически активные молекулы, предназначенные для введения животным, обычно объединяют с фармацевтически приемлемым носителем. Фармацевтические препараты могут содержать терапевтическую молекулу только одного типа или комбинацию из терапевтических молекул нескольких типов. Тип носителя обычно зависит от конкретного выбранного способа введения. Например, препараты для парентерального введения обычно содержат в качестве растворителя инъекционные жидкости, которыми являются фармацевтически и физиологически приемлемые жидкости, такие как вода, физиологический раствор, сбалансированные солевые растворы, водный раствор декстрозы, глицерин или тому подобные. Для получения твердых композиций (таких, как порошки, пилюли, таблетки или капсулы) обычно используют нетоксичные твердые носители, которые могут включать, например, маннит, лактозу, крахмал или стеарат магния фармацевтических сортов. Помимо биологически нейтральных носителей, предназначенные для введения фармацевтические композиции могут содержать небольшие количества нетоксичных вспомогательных веществ, таких как смачивающие вещества или эмульгаторы, консерванты, буферные агенты, регулирующие рН, и тому подобные, например ацетат натрия или сорбитан монолаурат.
Как известно в данной области, фармацевтические препараты на основе белка не могут быть эффективно доставлены в результате приема внутрь. Однако фармацевтические белки в виде пилюль можно вводить подкожно, особенно тогда, когда они входят в состав композиции пролонгированного действия. Препараты пролонгированного действия могут быть получены путем объединения целевого белка с биологически совместимой матрицей, такой как холестерин. Другим возможным способом введения белковых препаратов является использование миниосмотических насосов. Как было указано выше, биологически совместимый носитель должен также соответствовать способу доставки.
Фармацевтические композиции можно вводить любым способом, обеспечивающим достижение поставленной цели. Количества и схемы введения терапевтических молекул могут быть легко определены специалистами в области лечения болезни Лима и любых других заболеваний, обусловленных инфицированием B. burgdorferi. Для лечения указанных заболеваний молекулы вводят в количестве, достаточном для ингибирования репликации В. burgdorferi. Указанные лекарственные средства первоначально вводят в количествах, достаточных для достижения таких концентраций в ткани на месте действия, которые, как считают, обеспечивают требуемое действие in vitro. Соединения формулы А или В можно вводить хозяину in vivo системно, например, внутривенно или внутрибрюшинно. Кроме того, соединения формулы А или В можно вводить в место поражения, то есть пептид или белок инъецируют непосредственно в пораженный участок. Чтобы повысить иммунную реакцию, примерно через 4-6 недель после первой инъекции можно ввести повторную или бустер-дозу. Последующие дозы можно вводить в соответствии с настоящим изобретением или по желанию лечащего врача.
Настоящее изобретение проиллюстрировано нижеследующими неограничивающими примерами.
ПРИМЕРЫ
Пример 1
Материалы и методы
Организм и условия роста. Штаммы В31 B. burgdorferi (АТСС 35210), BL303 и N40 культивировали в среде BSK-H (Sigma). Среды инокулировали 2%(об.) замороженной культуры и инкубировали в статичных условиях при 37°С в течение 72 часов до полулогарифмической фазы роста (рН 7,0). Клетки собирали центрифугированием с ускорением 12000 g в течение 30 минут, трижды промывали холодным физиологическим раствором с фосфатным буфером (PBS) и хранили при -20°С до использования.
Экстракция липидов и анализы. Липиды экстрагировали из промытых клеток методом Брига и Дайера (14). Фазу хлороформа выпаривали в роторном испарителе и обрабатывали потоком азота, после чего высушенные липиды (0,1-0,2 мг/мл клеточного белка) вторично растворяли в 1-2 мл хлороформа. Количественное разделение BBGL производили хроматографией на колонке с силикагелем. Общий липидный экстракт (10 мг) в 5 мл хлороформа вводили в колонку с силикагелем (20 × 3 см, кизельгель 60, Merk, 230-400 меш) и элюировали в объеме, равном 10 слоям, хлороформом (фракция 1); 2,5% смесью метанол:хлороформ (об.:об., фракция 2); 5% смесью метанол:хлороформ (об.:об., фракция 3), 10% смесью метанол:хлороформ (об.:об., фракция 4) и метанолом (фракция 5). Фракции упаривали досуха в роторном испарителе и обрабатывали потоком азота, вторично растворяли в 1 мл хлороформа и хранили при -20°С. Фракция 3 почти полностью содержала BBGL-II, в то время как фракция 4 содержала BBGL-III. Для выполнения количественного анализа липидов общую липидную фракцию (200 мкг в 20 мкл) хроматографировали на алюминиевых пластинках, покрытых силикагелем НR (Merck, Darmstadt, Germany), и проявляли, используя смесь хлороформ:метанол (9:1 по объему). Пятна липида обнаруживали при помощи паров иода и гликолипиды обнаруживали путем распыления антрона.
Мечение радиоактивным изотопом. Мечение B. burgdorferi холестерином выполняли, добавляя [4-14C]холестерин (Amersham Pharmacia, GB, courtesy of W.Prinz, NIDDK, NIH) в 100 мл среды во время инокуляции. Меченый холестерин (удельная активность 58,0 мCi/ммоль) добавляли в количестве 0,1 мкCi/мл в смесь твин-80:этанол (1:1 по объему) до конечного объема, равного 50 мкл. Липид экстрагировали указанным методом. Для определения радиоактивности в пятнах липида все липиды разделяли на пластинках для тонкослойной хроматографии, пластинки в течение короткого времени подвергали воздействию паров иода, пятна соскабливали в сцинтилляционную камеру, содержащую 1 мл сцинтилляционной жидкости. Радиоактивность измеряли в сцинтилляционном счетчике Perkin Elmer (модель I450 микробета) и выражали в виде числа распадов в минуту (dpm).
Аффинная хроматография. BBGL-II очищали также аффинной хроматографией, используя эндотоксин-удаляющий гель, так называемый детоксикирующий гель (Pierce, Rockford, IL). Клетки Borrelia 5 раз промывали PBS, вторично суспендировали в том же буфере и разрушали ультразвуком в течение 30 секунд в ультразвуковом дезинтеграторе Брансона, выполняющем 50% рабочих циклов при 100 Вт. Мембраны отделяли от растворимой фракции центрифугированием с ускорением 37000 g в течение 30 минут и супернатант (1 мг общего белка в 1 мл) вводили в колонку с детоксикирующем гелем (иммобилизованный анарозой полимиксин В) (0,9 × 10 см). Колонку промывали PBS в объеме, равном 10 слоям. Связанное вещество элюировали, выполняя промывку PBS, содержащим 1% дезоксихолиновой кислоты (мас./об.), в объеме, равном 10 слоям, в соответствии с инструкциями производителя. Связанную фракцию диализовали против дистиллированной воды, лиофилизовали и липиды экстрагировали вышеуказанным методом.
Иммунизация и иммунное окрашивание. Группы, состоящие из десяти самок белых швейцарских мышей NIH (в возрасте 5-6 недель, общего назначения), иммунизировали внутрибрюшинно 2 дозами BBGL-II, эмульгированного в полном адъюванте Фрейнда (первая доза) или в неполном адъюванте Фрейнда (вторая доза), с интервалом в два недели. Указанные дозы, содержащие 10 мкг BBGL-II на 0,1 мл, вводили внутрибрюшинно в количестве 0,1 мл. Альтернативно мышей иммунизировали три раза с интервалом в 2 недели, используя 10 мкг BBGL-II, эмульгированного в 0,1 мл ДМСО, сквалена или PBS. У мышей брали кровь через 2 недели после введения второй дозы и полученную таким образом сыворотку хранили при -20°С до использования. Кроликов (самки, новозеландские белые, в возрасте 8 недель) иммунизировали двумя дозами BBGL-II, по 75 мкг каждая, в адъюванте Фрейнда, которые вводили внутримышечно и внутрикожно в количестве 20 × 0,05 мл.
Иммунное окрашивание мембранных липидов выполняли ранее описанным методом (15). Пластинки с проявленными хроматограммами, содержащие пятна липидов, покрывали раствором полиизобутилметакрилата (0,05% в гексане) и сушили. Затем пластинки блокировали PBS, содержащим 1% BSA и 0,05% твина-20, в течение 15 минут и инкубировали с антисывороткой против BBGL-II, разведенной в буфере с PBS-BSA в отношении 1:100, в течение 1 часа при 22°С. Пластинки 5 раз промывали буфером с PBS-BSA и инкубировали с IgG крысы против мыши, конъюгированным с щелочной фосфатазой (KPL, MD), разведенным в отношении 1:25000 в PBS-BSA, в течение 1 часа при 22°С. Затем пластинки промывали и проявляли с использованием BCIP/NBT (KPL, MD).
Аналитические методы. Анализ на сахар выполняли методом Sawardeker et al. (16). 0,5 мг BBGL-II или BBGL-III гидролизовали в 1 М растворе HCl в течение 4 часов при 100°С, затем восстанавливали, перацетилировали и анализировали методом ГЖХ-МС в аппарате компании Hewlett-Packard (модель НР 6890) с использованием стеклянной капиллярной колонки НР-5 (0,32 мм в течение 30 минут), программируя изменение температуры со скоростью 8°С/мин со 125°С до 250°С в режиме электронной ионизации (106 эВ). Жирные кислоты анализировали после метанолиза сухих гликолипидов 1 М раствором HCl/MeOH в течение 5 часов при 80°С. Растворитель удаляли в потоке азота и сложные метиловые эфиры свободных жирных кислот экстрагировали хлороформом. Анализ выполняли методом ГЖХ-МС в описанных выше условиях.
Локализация двойной связи. Положение двойной связи в ненасыщенных жирных кислотах определяли методом ГЖХ-МС после получения производного 4,4-диметилоксазолина (17). Фазу хлороформа сушили в потоке азота, смешивали с 500 мкл 2-амино-2-метилпропанола и нагревали в течение ночи при 150°С. Реакционную смесь охлаждали, растворяли в 3 мл дихлорметана и дважды промывали 2 мл дистиллированной воды.
Анализ метилирования. Природный и О-деацилированный гликолипид метилировали методом, описанным в публикации (18). Метилированные продукты гидролизовали 1 М раствором НСl в течение 4 часов при 100°С, превращали в ацетаты альдитов и анализировали методом ГЖХ-МС.
О-Деацилирование. Гликолипиды (2 мг) О-деацилировали 0,33 мл 0,25 М раствора NaОCН3 в метаноле при 37°С. Ход реакции контролировали при помощи ТСХ. Через 2 часа не было обнаружено ни одного пятна, соответствующего исходным, немодифицированным гликолипидам. Растворители выпаривали в потоке азота и продукты экстрагировали смесью хлороформ:вода (1:1 по объему). Органические и неорганические фазы анализировали в отношении сахара и жирных кислот методом ГЖХ-МС.
Определение абсолютной конфигурации. BBGL-II гидролизовали 1 М раствором HCl при 100°С в течение 4 часов. Количественное определение D-галактозы выполняли в высушенном нейтрализованном образце ферментативным методом, используя галактозооксидазу (19). Абсолютную конфигурацию глицерина определяли методом, описанным в публикации (1), производя предварительное О-деацилирование гликолипида (20). При выполнении данного метода первичную гидроксильную группу глицерина, полученного омылением гликолипида, окисляли при помощи TEMPO и превращали в остаток глицериновой кислоты. После выполнения кислотного гидролиза глицериновую кислоту этерифицировали (R)-(-)-2-бутанолом, ацетилировали и анализировали методом ГЖХ-МС. Время удерживания сравнивали с аналогичными образцами, полученными из D- и L-глицериновой кислоты.
Масс-спектрометрия с бомбардировкой ускоренными атомами (FAB). Масс-спектры регистрировали с использованием атомов 6 кэВ, предназначенных для ионизации образцов, полученных из 3-нитробензилового спирта или глицерина, служащего в качестве матрицы. Образцы перацетилировали методом, описанным Деллом (21).
Лазерная десорбция на матрице/ионизационная времяпролетная масс-спектрометрия (MALDI-TOF). Масс-спектры регистрировали в приборе для MALDI-TOF PerSeptive BioSystems Voyager Elite DE-STR (PE-Biosystems, Framingham, MA). Масс-спектры были получены для 100 лазерных циклов при затухании, равном 2600. Прибор действовал в линейном режиме с ускоряющим напряжением 20 кВ и временем запаздывания ионной экстракции, равным 150 нс. Образец и матрицу получали вышеописанным методом (22). BBGL-II и BBGL-III растворяли в смеси хлороформ:метанол (1:1 по объему) до концентрации 4 мкг/мкл и наносили в виде 0,5 мкл капель в разные места в центре пластинки для нескольких образцов. Матрицу такого же объема, 2,5-дигидроксибензойную кислоту, 10 мг/мл воды наносили на каждый образец, сушили и помещали в масс-спектрометр.
ЯМР-спектроскопия. Моногалактозилдиглицерид (MGDG, в основном 1,2-ди-О-стеароил-3-О-β-D-галактопиранозилглицерин) был приобретен в компании Matreya, State College, PA. 1,2-ди-О-пальмитоилглицерин (1,2-дипальмитин) был приобретен в компании NuChek, New Elysian, MN. Метил-α-D-галактопиранозид был получен в собственной лаборатории, и метил-β-D-галактопиранозид был приобретен в компании Aldrich, Milwaukee, WI. Дейтерированные растворители были приобретены в компании Cambridge Isotope Laboratories, Andover, MA.
ЯМР-спектры были получены при 300К без вращения в спектрометре DRX-500 компании Bruker, оснащенном 5 мм широкополосным (ВВО) зондом. Для липидов использовали растворы 5-10 мг соединения в CDCl3 (0,5 мл, плотность 99,96 атом.%) или его смесей с (CD3)2CO (плотность 99,9 атом.%) или CD3OD (плотность 99,8 атом.%) с тетраметилсиланом в качестве эталона химического сдвига для 1Н и 13С ЯМР-спектров. Аномерные метил-D-галактопиранозиды (11 мг) исследовали в виде их растворов в D2O (0,4 мл, плотность 99,96 атом.%) с использованием 4,4-диметил-4-силапентаноат-2,2,3,3-d4 (TSP) натрия в качестве внутреннего эталона для 1Н и 13С ЯМР-спектров. Данные были получены и обработаны с помощью программы XWINNMR компании Bruker, версия 3.0, в процессорах SGI O2 или Octane 2. Для получения одномерных спектров использовали 32768 точек данных, в некоторых случаях с заполнением нулями до 32768 или 65536 точек. Одномерные 1Н ЯМР-спектры регистрировали при частоте 500 МГц, ширине спектра 4,25 кГц, импульсе 30° (3,2 мксек) и времени рецикла 6 с. Одномерные 13С ЯМР-спектры были получены при частоте 126 МГц, ширине спектра 25,1 кГц, импульсе 45° (3 мкс) и времени рецикла 1 с, за исключением исследований с высоким разрешением близко расположенных 13С резонансов холестерина, когда использовали 65536 точек данных с заполнением нулями до 131072 точек. Одномерный 13С ЯМР-спектр редактировали методом DEPT, используя комбинации спектров, полученных с помощью импульсов считывания 30°, 90° и 150° при частоте 1Н. В других случаях использовали импульс считывания 1Н 135° для получения 13С ЯМР-спектров, имеющих резонансы СН2 с отрицательной фазой наряду с резонансами СН и СН3 с положительной фазой. 1Н-связанные 13С ЯМР-спектры были получены с ядерным эффектом Оверхаузера в результате стробированного облучения при частоте 1Н с задержкой релаксации 3,42 с.
Большая часть данных двумерных ЯМР-спектров была получена при помощи последовательностей импульсов, включающих выбор z-градиентной когерентности.
Двумерные 1Н ЯМР-спектры COSY получали с использованием 2048 × 512 точек данных при заполнении нулями до 2048 × 2048 точек и импульсах считывания 30° или 45°. Синусоидальные квадратичные прямоугольные функции без сдвига применяли в обоих направлениях с последующим выполнением преобразования Фурье, после чего данные частоты отображали в виде абсолютных значений. Двумерные 1Н ЯМР-спектры TOCSY были получены с использованием 16384 × 256 точек данных при заполнении нулями до 16384 × 2048 точек с помощью чувствительного к фазе режима эхо/антиэхо с выбранным градиентом. В обоих направлениях применяли синусоидальные квадратичные прямоугольные функции со сдвигом на π/2 радиуса. Одномерные 1Н ЯМР-подспектры отдельных остатков были получены в результате извлечения срезов F2 из двумерных спектров TOCSY. Для некоторых 1Н ЯМР-спектров полученные данные были также подтверждены цифровой, избирательной гомонуклеарной развязкой 1Н.
Двумерные 1Н/13С ЯМР-спектры HSQC и НМВС регистрировали с использованием 2038 × 512 точек данных при заполнении нулями до 2048 × 2048 точек с помощью чувствительного к фазе режима эхо/антиэхо с выбранным градиентом и повышенной чувствительностью для HSQC и последовательности импульсов с выбранным градиентом, фильтрацией низких частот, большим радиусом действия без развязки для НМВС, после чего полученные данные отображали в виде абсолютных значений. Двумерные ЯМР-спектры НМВС были получены с запаздыванием выделения, равным 83 мс, то есть были оптимизированы для 2,3JСН 6,0 Гц. Оптимальная чувствительность была достигнута для спектров HSQC и НМВС в результате использования синусоидальных квадратичных прямоугольных функций со сдвигом на π/2 радиуса в обоих направлениях. Гребни в направлении t1 двумерных спектров при необходимости удаляли путем вычитания среднего ряда в программе AURELIA компании Bruker, версия 2.8.12. Измерения 31Р ЯМР при частоте 202 МГц свидетельствовали об отсутствии фосфора в исследуемых образцах.
Пример 2
Выделение BBGL-II и BBGL-III
Гликолипиды B. burgdorferi (штамм В31) были получены из 2,17 г сухих клеток после экстракции методом Блига и Дайера. Полученные гликолипиды очищали до однородного состояния в колонке с силикагелем (20 × 3 см) и поэтапно элюировали смесями хлороформа и метанола с возрастающей полярностью. Выход BBGL-II из 0,7 г общих липидов был равен 163 г (23,2%), и выход BBGL-III был равен 87 мг (12,4%), из чего следует, что BBGL-II и BBGL-III являются основными липидами в B. burgdorferi. Аналогичные выходы были получены из клинически выделенных штаммов BL303 и N40, в которых состав липидов был таким же, как в штамме В31 (фиг.1). Поскольку выходы биомассы из штамма В31 были намного больше, все последующие эксперименты выполняли с использованием данного штамма.
Пример 3
Аффинная хроматография
BBGL-II связывали с эндотоксин-удаляющим гелем, так называемым детоксикирующим гелем (Pierce). Указанная смола состоит из иммобилизованного полимиксина В на агарозе и используется для удаления эндотоксинов путем связывания с липидом А в липополисахариде (LPS). При введении в указанную колонку обработанных ультразвуком клеток B. burgdorferi наличие BBGL-II в связанном веществе, элюированном из колонки 1% дезоксихолиновой кислотой, можно было продемонстрировать методом ТСХ и иммунным мечением (фиг.1). В связанной фракции не было обнаружено присутствия BBGL-III.
Пример 4
Мечение BBGL-II радиоактивным изотопом
При культивировании в присутствии 14С-холестерина 80% радиоактивность, обнаруженная в общем липидном экстракте, могла быть объяснена наличием BBGL-II (таблица 1). Радиоактивность не была обнаружена в полосах липида, соответствующих BBGL-III, свободному холестерину или сложным эфирам холестерина.
Химический состав гликолипидов B. burgdorferi
bЛокализация двойной связи
сОтносительное количество радиоактивности, обнаруженное в общей липидной фракции (см. пример 1 для более подробного ознакомления)
Пример 5
Композиционный анализ
Анализ на сахар обоих гликолипидов выявил наличие галактозы в качестве единственного моносахарида. Глицерин был обнаружен только в BBGL-III. В результате выполнения метанолиза было обнаружено наличие двух основных жирных кислот, связанных сложноэфирной связью: С16:0 и С18:1, и несколько второстепенных жирных кислот: С14:0, С18:0, С18:2 (таблица 1). Двойная связь была локализована в положении Δ9 и Δ9,12 соответственно в С18:1 и С18:2, из чего следует, что жирными кислотами являются олеиновая и линолевая кислоты.
Ферментативный анализ с использованием галактозооксидазы показал, что галактозная часть имеет D-конфигурацию. Было установлено, что абсолютная конфигурация атома углерода в положении С-2 глицериновой части BBGL-III с остатком сахара в положении С-3 и жирной кислотой в положениях С-1 и С-2 представляет собой L-конфигурацию. Полученный результат совместим с sn-конфигурацией, когда атомы углерода имеют упорядоченную пространственную структуру.
Пример 6
Анализ методом метилирования
Анализ методом метилирования природного BBGL-II выявил наличие 1,5-ди-о-ацетил-2,3,4,6-тетра-О-метилгалацитола и 1,5,6-три-О-ацетил-2,3,4-тетраметилгалацитола в молярном отношении 1,0:0,9, что свидетельствует о присутствии концевого и 6-замещенных остатков галактозы. Предшествующее де-О-ацилирование образца вызвало исчезновение 6-замещенной галактозы, при этом концевая галактоза была единственным обнаруженным элементом. Полученный результат позволяет предположить, что цепь жирной кислоты, которая не была полностью удалена во время метилирования, выполненного методом, описанным в публикации (23), находится в положении С6 галактозы. Анализ методом метилирования BBGL-III позволил обнаружить 1,5-ди-о-ацетил-2,3,4,6-тетра-О-метилгалацитол, что указывает на присутствие концевой галактозы.
Пример 7
Масс-спектроскопия
Спектры MALDI-TOF BBGL-II, полученные в режиме положительных ионов, позволили обнаружить две молекулярные массы, равные 810,1 и 836,1. Разница между указанными молекулярными массами (Δm/z = 26) свидетельствует о вариабельности липидного компонента, вследствие которой натриевый аддукт холестерилгалактозы может быть замещен пальмитиновой или олеиновой жирной кислотой (фиг.2). Данный результат согласуется с результатами, полученными методом ГЖХ-МС, которые показали, что указанные жирные кислоты являются преобладающими частями. Анализ природного BBGL-II методом FAB-MS позволил обнаружить молекулярные массы, равные 809,7 и 835,7, при этом анализ перацетилированного BBGL-II позволил обнаружить молекулярные массы, равные 935,7 и 961,7. Увеличение молекулярной массы Δm/z=126 указывает на включение трех ацетильных групп и подтверждает существование трех свободных гидроксильных групп в BBGL-II. Для подтверждения состава BBGL-II выполняли FAB-MS с высоким разрешением. Теоретическое значение m/z цезиевого аддукта соединения, состоящего из гексозы, холестерина и олеиновой кислоты (С51Н86О7Сs), было вычислено равным 943,5428. Обнаруженная масса m/z была равна 943,5469. Полученный результат подтверждает теоретический состав С51Н86О7Cs (ошибка [м.д./м.м.е.] = +4,3/+4,1.).
Спектры MALDI-TOF BBGL-III позволили обнаружить два иона с массами 780,1 и 806,1, которые соответствовали натриевым аддуктам моногалактозилдиацилглицерина с двумя жирными кислотами 16:0 или с жирными кислотами соответственно 16:0 и 18:1 (фиг.2).
Пример 8
ЯМР-спектроскопия
Структуры BBGL-II и BBGL-III и их перацетильных производных далее исследовали при помощи одномерной (1D) и двумерной (2D) ЯМР-спектроскопии при 500 МГц. Показатели 1Н ЯМР (таблица II) были подтверждены двумерной корреляционной спектроскопией (COSY), общей корреляционной спектроскопией (TOCSY) или экспериментами по избирательной спиновой развязке, и показатели 13С ЯМР (таблица III) были получены методом двумерной гетеронуклеарной одноквантовой корреляции (HSQC) на основании ранее определенных показателей 1Н ЯМР. Связности между остатками и другие данные для показателей 13С были определены в результате двумерной гетеронуклеарной корреляции кратной связи (НМВС).
Таблица II
Химические сдвиги 1 Н (м.д.) остатков галактозы и
глицерина гликолипидов и родственных соединений
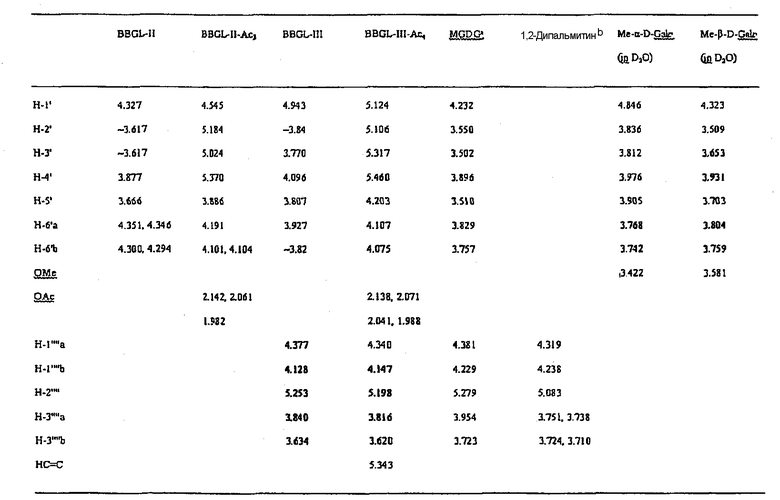
аПромышленно доступный “моногалактозилдиглицерин” является главным образом 1,2-ди-О-стеароил-3-О-β-D-галактопиранозилглицерином
b1,2-ди-О-пальмитоилглицерин
Таблица III
Химические сдвиги 13 С (м.д.) гликолипидов и родственных соединений

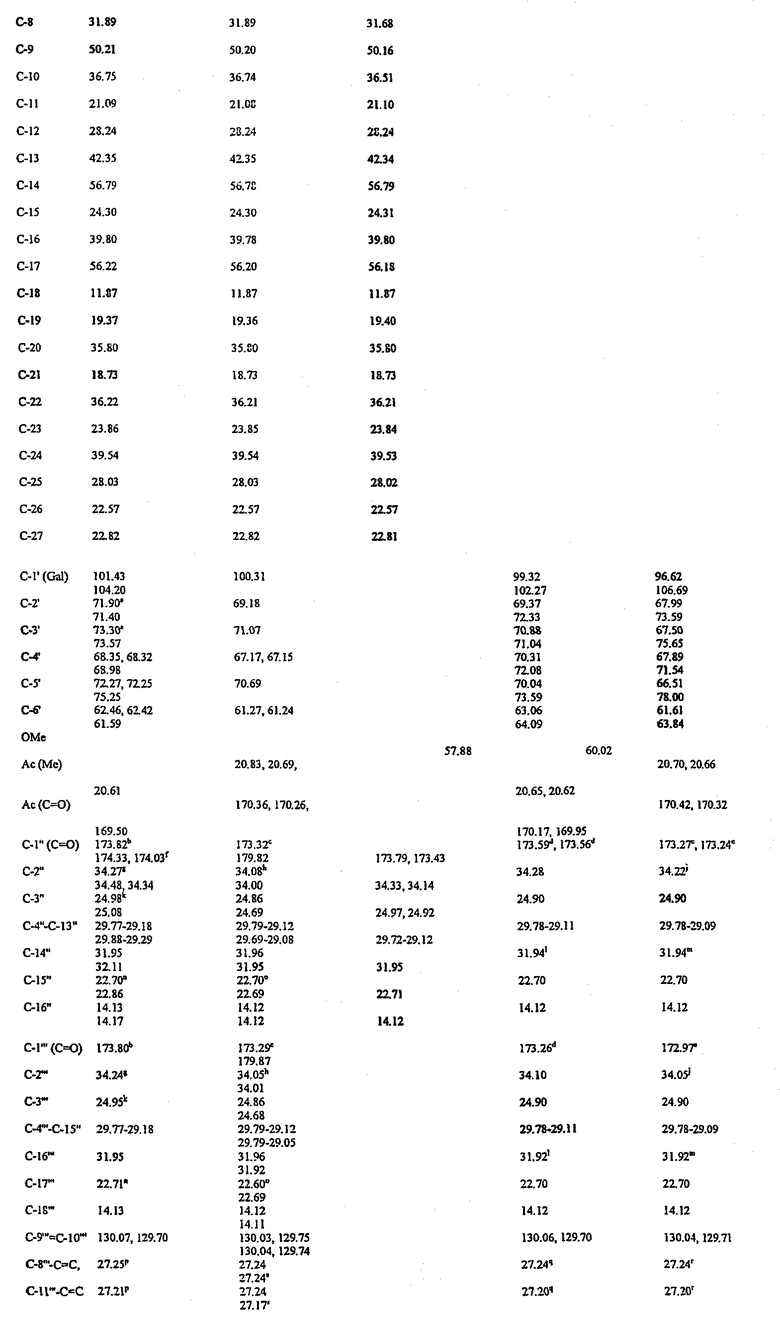

а-еПоказатели являются взаимозаменяемыми.
1Сдвиги в данной группе относятся к стеароильным заместителям, а не к пальмитоилу.
gПоказатели являются взаимозаменяемыми.
Значения некоторых основных констант гомо- и гетеронуклеарного взаимодействия для гликолипидов приведены в таблице IV. Данные ЯМР для нескольких эталонных соединений или структурных компонентов гликолипидов также приведены в таблицах II-IV. Резонансы 13С ЯМР далее определяли, выполняя эксперименты по редактированию спектров ЯМР методом одномерного недеформирующего усиления путем переноса поляризации (DEPT), в результате которых были выявлены углеродные ядра с разным числом присоединенных атомов водорода.
Таблица IV
Константы взаимодействия (J, Гц) остатков галактопиранозы
и глицерина производных гликолипидов и родственных
эталонных соединений
аДанные не определены; bВ растворе (CD3)2CO:CDCl3
(7:3, об./об.)
Пример 9
BBGL-II
Одномерный 1Н ЯМР-спектр BBGL-II в растворе d в хлороформе был не полностью диспергирован при 500 МГц, при этом сигналы Н-2' и Н-3' остатка сахара значительно перекрывали друг друга при ~3,62 м.д. Остатки холестерина, галактозы, пальмитиновой кислоты, олеиновой кислоты и глицерина были отмечены как второстепенные, а синглет, дублет, триплет и квадруполь - как главные. Наблюдалось также наложение мультиплетов (для протонов, отмеченных как a и b, символ “a” относится к протону с резонансом в слабом поле и символ “b” относится к протону с резонансом в сильном поле) Н-1', H-6'a и Н-6'b, и одномерный срез указанных сигналов в двумерном спектре TOCSY позволил получить подспектр, который содержал все семь протонных сигналов сахарной цепи (Н-1'-H-6'b), главным образом в виде двух серий, включающих подгруппы Н-1'-H-4' и Н-5'-H-6'b. Деэкранирование метиленовых протонов сахара в гликолипиде на 0,55 м.д. относительно протонов метил-β-D-галактопиранозида (таблица II) позволяет предположить, что сахар ацилирован в положении О-6'. Результаты эксперимента методом TOCSY также подтвердили, что триплет 1:2:1 триплетов 1:2:1 при δ 5,553 не является частью системы спинов протонов сахара, поэтому данный мультиплет был соотнесен с Н-3 холестериновой части, в частности, потому, что он был также обнаружен в одномерном срезе TOCSY, полученном по сигналам олефиновых протонов BBGL-II. Путем интегрирования было установлено, что последние сигналы соответствовали ~2,5 протона в одномерном 1Н ЯМР-спектре вследствие совмещения одного сигнала олефинового протона (Н-6) холестериновой части с сигналами олефиновых протонов части ненасыщенных жирных кислот в выделенном гликолипиде. В подтверждение данного положения срез TOCSY по сигналам олефиновых протонов также содержал большое число мультиплетов в области алифатических протонов (δ 2,8-0,9) благодаря связности с алифатическими протонами как холестерина, так и ненасыщенных жирных кислот.
В 13С ЯМР-спектре BBGL-II было обнаружено 59 основных разрешенных резонансов, из которых пять относились к ядрам четвертичного 13С, то есть не были обнаружены при выполнении 13С спектроскопии методом DEPT. В направлении от слабого поля к сильному полю (таблица III) пять резонансов четвертичного 13С были определены как два карбонильных резонанса сложного эфира и С-5 (олефиновый), С-13 и С-10 резонансы холестериновой части. Из общего числа резонансов 13С 15 можно было идентифицировать как СН в результате редактирования спектра методом DEPT (фиг.3а), вместе с 27 группами СН2 и шестью группами СН3 (фиг.3b). Пять резонансов СН3 были соотнесены с остатком холестерина (таблица III), и шестой более слабый резонанс при δС 14,13 был соотнесен с комбинацией сигналов атомов углерода ω-метила сложноэфирных групп жирных кислот, которые обычно не могут быть отделены друг от друга. Два основных резонанса олефиновой группы СН при ~130 м.д. были соотнесены с преобладающей сложноэфирной группой ненасыщенной жирной кислоты (олеиновая кислота, 18:1), хотя дополнительные более слабые резонансы в данной области указывали на наличие небольшого числа других сложноэфирных групп ненасыщенных жирных кислот. Резонансы СН при 122,12 и 76,46 м.д. были соотнесены с С-6 и С-3 холестериновой части. Хорошее соответствие было достигнуто между химическими сдвигами 13С BBGL-II и аналогичными сдвигами его липидных компонентов, холестерина, пальмитиновой кислоты и олеиновой кислоты за исключением ядер, расположенных рядом с местами присоединения остатков (таблица III). Показатели 13С для холестерина в растворе d в хлороформе основаны на результатах, приведенных в публикации (24) для раствора d5 в пиридине, причем данные показатели были совместимы с результатами экспериментов методом DEPT.
Остальные пять резонансов СН в диапазоне 101,43-68,32 м.д. были соотнесены с С-1'-C-5' галактозы, которая также давала сигнал СН2 при 61,3 м.д. (таблица III). Было обнаружено удвоение сигналов С-4', C-5' и С-6' галактозы, которое можно было объяснить наличием двух гликолипидов, имеющих разные сложноэфирные группы жирных кислот (пальмитоильная и олеоильная) в положении О-6' галактозы. Наблюдаемое удвоение сигнала С-3 холестерина труднее объяснить подобным образом, так как С-3 более удален от заместителя в положении О-6' галактозы, чем С-4', C-5' и С-6'. Подобное удвоение может быть следствием поворотной изомерии относительно связи С-1'-O-1' (25).
Локализация холестерильной группы в положении О-1' остатка галактозы (Gal) в BBGL-II была установлена по обнаружению перекрестного пика Н-3/C-1' в двумерном спектре НМВС при 3,556/101,49 м.д. Аналогичным образом локализация ацилоксигрупп(ы) в положении С-6' галактозы была установлена по обнаружению перекрестных пиков Н-6'a/C=O и Н-6'b/C=O в спектре НМВС соответственно при 4,351/173,86 м.д. и 4,297/173,86 м.д. В результате анализа мультиплета Н-3 холестерина в 1Н ЯМР-спектре BBGL-II были получены константы взаимодействия J2экв,3 = J4экв,3 = 4,7 Гц и J2акс,3 = J4акс,3 = 11,4 Гц. Полученные величины определяют ориентацию Н-3 как аксиальную, поэтому атом кислорода (О-1'), присоединенный к С-3, занимает экваториальное положение. Таким образом, С-3 имеет обычную стереохимическую конфигурацию, характерную для холестерина. Данные ЯМР для BBGL-II сопоставимы со смесью двух структур, а именно 3-О-(6-О-пальмитоил-β-D-галактопиранозил)холестерин и 3-О-(6-О-олеоил-β-D-галактопиранозил)холестерин. Определение размера кольцевой структуры сахара было основано на сходстве констант взаимодействия кольцевой структуры сахара BBGL-II с константами взаимодействия метил-β-D-галактопиранозида (таблица IV). Ниже рассмотрена аномерная конфигурация BBGL-II.
Пример 10
Гликолипид BBGL-II-Ac 3
Данные ЯМР, подтверждающие наличие галактозы в BBGL-II, были получены в результате его перацетилирования, позволившего получить триацетильное производное, 1Н ЯМР-спектр которого был лучше диспергирован и разделен по сравнению со спектром BBGL-II. В данном спектре был обнаружен узкий квартет Н-4' (J3',4' 3,5 Гц, H4',5' 1,0 Гц), характерный для галактоконфигурации, наряду с широкими квартетами, ожидаемыми для Н-2' и Н-3'. Все три указанных сигнала были в значительной степени деэкранированы (+1,41-1,57 м.д.) относительно своих положений в неацетилированном BBGL-II (таблица II), из чего следует, что ацетилирование произошло в положениях НО-2', HO-3' и НО-4'. Положения трех ацетильных групп были подтверждены в результате обнаружения перекрестных пиков в двумерных спектрах НМВС между Н-2', H-3', H-4' и трех разных групп 13С=О (фиг.4). В данном спектре имеются также перекрестные пики между С-1' Gal и Н-3 холестерина, а также между Н-6'a/H-6'b Gal и С=О сложноэфирной группы жирной кислоты, что подтверждает наличие холестерильной группы в положении О-1' Gal и ацилокси-остатков жирной кислоты в положении С-6'. Показатели сигналов 1Н были подтверждены двумерной спектроскопией COSY (см. фиг.5).
Большие значения J1',2' 7,5 Гц и 8,0 Гц, обнаруженные соответственно для BBGL-II и BBGL-II-Ac3 (таблица IV), указывают на то, что Н-1' и Н-2' имеют трансориентацию в указанных гликолипидах, и поэтому они имеют β-аномерную конфигурацию. Подобный вывод был подтвержден измерением значений 1JС-1',H-1' 158,7 Гц и 157,4 Гц соответственно для BBGL-II и BBGL-II-Ac3 (таблица IV), которые попадают в соответствующий диапазон для β-аномерной конфигурации (26), о чем свидетельствует значение 1JС-1',H-1 160,6 Гц, обнаруженное для метил-β-D-галактопиранозида (Ме-β-D-Galp, таблица IV).
Размер кольцевой структуры сахара для BBGL-II и BBGL-II-Ac3 был определен на основании сходства констант взаимодействия кольцевых структур сахара с константами взаимодействия метил-β-D-галактопиранозида (таблица IV). Различия в значениях J5',6'a и J5',6'b для BBGL-II и BBGL-II-Ac3, с одной стороны, и метил-β-D-галактопиранозида, с другой стороны, могут быть отнесены за счет разных ротамерных распределений относительно связи С-5'-C6', вызванных наличием большого заместителя в положении О-6' гликолипидов. Данные ЯМР для BBGL-II-Ac3 совместимы с результатами его исследования в виде смеси 3-О-(2,3,4-три-О-ацетил-6-О-пальмитоил-β-D-галактопиранозил)холестерина и 3-О-(2,3,4-три-О-ацетил-6-О-олеоил-β-D-галактопиранозил)холестерина.
Пример 11
BBGL-III
1Н ЯМР-спектр BBGL-III в растворе d в хлороформе был хорошо диспергирован; в частности, протоны глицерина полностью диспергированы при 500 МГц, двумерный спектр COSY и двумерный спектр TOCSY (фиг.6) содержат серии из пяти мультиплетов, которые представляют (от слабого поля в сторону сильного поля) протоны глицерина Н-2'''', H-1''''a, H-1''''b, H-3''''a и Н-3''''b. Указанные двумерные спектры имеют одинаковый вид, так как Н-2'''' находится в спиновом взаимодействии со всеми другими протонами в углеродной цепи глицерина и поэтому образует серию из перекрестных пиков в спектре COSY, которая напоминает аналогичную серию в спектре TOCSY. Последний спектр (фиг.5) имеет также серию мультиплетов Н-1'-Y-4', характерную для галактоконфигурации, причем перенос намагниченности от Н-4' к Н-5', H-6'a и Н-6'b обычно подавляется небольшим значением J4',5' (см. таблицу IV). В результате этого мультиплет Н-4' остатка Gal является характерно узким (J3',4' 3,2 Гц, J4',5' 1,1 Гц). Серия из семи мультиплетов в двумерном спектре TOCSY представляет взаимный обмен намагниченности между олефиновыми протонами (δ 5,344) и алифатическими протонами в ненасыщенной жирной кислоте (18:1).
В 13С ЯМР-спектре DEPT-135 BBGL-III были обнаружены шесть резонансов СН и три сигнала CН3 в области сахара, которые сопоставимы с наличием одного остатка глицерина и одного остатка альдозы, а также 25 не полностью разделенных сигналов СН2 и один резонанс СН3 в области алифатического углерода, которые были соотнесены с жирными кислотами. Два сильных резонанса при δС 130,06 и 129,70 позволили предположить наличие одного преобладающего остатка ненасыщенной жирной кислоты. Однако обнаружение двух более слабых пар сигналов в области данного олефинового углерода свидетельствовало о наличии незначительных количеств других ненасыщенных жирных кислот. Три сильных резонанса 13С были обнаружены в области С=О (таблица II) наряду с одним более слабым резонансом. Значительное деэкранирование (соответственно +0,724, +0,567 и +1,470 м.д.) протонов Н-1''''a, H-1''''b и Н-2'''' глицерильного остатка BBGL-III относительно соответствующих протонов исходного глицерина (δН в D2O, 3,653, Н-1''''a; 3,561, Н-1''''b; 3,783, H-2'''') указывает на ацилирование О-1'''' и О-2'''' глицерильного остатка (таблица II). Положения ацильных групп были подтверждены в результате обнаружения перекрестных пиков Н-1''''a/C=O, H-1''''b/C=O и Н-2''''/C=O в двумерном спектре НМВС BBGL-III. В данном спектре был также обнаружен перекрестный пик Н-3''''b/C-1', который указывает на наличие галактозильного остатка в положении О-3'''' глицериновой части.
Пример 12
BBGL-III-Ac 4
В результате перацетилирования BBGL-III был получен продукт, в 1Н и 13С ЯМР-спектрах которого были обнаружены четыре интенсивных сигнала ацетилметильных протонов и четыре сигнала углерода ацетилметильной группы и углерода карбонильной группы, что указывает на образование тетра-О-ацетильного производного. Значительное деэкранирование (от +1,266 до +1,547 м.д.) Н-2', H-3' и Н-4' Gal и более ограниченное деэкранирование (от +0,180 до +0,255 м.д., таблица II) Н-6'a и Н-6'b указывает на ацетилирование О-2', O-3', O-4' и О-6' остатка Gal. Подобный вывод был подтвержден обнаружением перекрестных пиков Н-2'/C=O, H-3'/C=O, H-4'/C=О, H-6'a/C=O и Н-6'b/C=O в двумерном спектре НМВС BBGL-III-Ac4 (фиг.6) в том случае, когда сигналы углерода карбонильной группы в ацетильной группе и сложноэфирной группе жирной кислоты можно легко отличить по химическим сдвигам 13С в областях, соответствующих 170 м.д. и 173 м.д. (таблица III). В области, соответствующей 173 м.д., в двумерном спектре НМВС BBGL-III-Ac4 были обнаружены также перекрестные пики Н-1''''a, H-1''''b и Н-2'''' с атомами углерода карбонильной группы сложного эфира жирной кислоты наряду с перекрестным пиком Н-3''''a/C-1', что подтверждает предполагаемое расположение остатка Gal в положении О-3'''' глицеринового звена.
Низкие значения J1',2' 3,8 Гц и 3,7 Гц соответственно для BBGL-III и BBGL-III-Ac4 (см. таблицу IV) свидетельствуют о наклонной ориентации Н-1' и Н-2' в указанных производных, из чего следует, что их остатки Gal имеют α-аномерную конфигурацию. Подобный вывод был подтвержден высокими значениями JС-1',H-1' 170,5 Гц и 172,4 Гц соответственно для BBGL-III и BBGL-III-Ac4 (таблица IV), которые соответствуют диапазону, ожидаемому для α-аномеров (26), как это имело место для метил-α-D-галактопиранозида (Ме-α-D-Galp, таблица IV).
Данные ЯМР для BBGL-III совместимы со структурой 3-О-α-D-галактопиранозил-1(2)-О-олеоил-2(1)-О-пальмитоилглицерина, и данные ЯМР для BBGL-III-Ac4 подтверждают наличие структуры 3-О-(2,3,4,6-тетра-О-ацетил-α-D-галактопиранозил)-1(2)-О-олеоил-2(1)-О-пальмитоилглицерина. Определение величины кольцевой структуры сахара было основано на сходстве констант взаимодействия кольцевой структуры сахара с константами взаимодействия метил-α-D-галактопиранозида (таблица IV).
Пример 13
Моногалактозилдиглицерид
Авторы настоящего изобретения установили, что промышленно доступный моногалактозилдиглицерид (MGDG) является близким структурным аналогом BBGL-III, поэтому данный продукт был исследован в качестве эталонного соединения для проверки аналитических методов, представленных в данном описании изобретения. 1Н ЯМР-спектр MGDG был хорошо диспергирован при 500 МГц, поэтому были легко обнаружены семь спиновых мультиплетов, относящихся к остатку Gal, включая узкий квартет Н-4' (J3',4' 3,3 Гц, J4',5' 1,1 Гц), который характерен для галактоконфигурации. Кроме того, константы взаимодействия кольцевого протона (таблица IV) указывают на наличие остатка галактозы в виде пиранозильного кольца наряду с вышеуказанными производными гликолипида. Для MGDG не были обнаружены сигналы олефинового протона.
Двумерные 1Н ЯМР-спектры COSY и TOCSY моногалактозилдиглицерида (MGDG) содержали такие же хорошо диспергированные серии из пяти мультиплетов, которые характерны для остатка глицерина, содержащего ацилоксигруппы в положениях С-1'''' и С-2'''', и имеют схему замещения, подтвержденную сдвигами в сторону слабого поля Н-1''''a, H-1''''b и Н-2'''' (таблица II). 13С ЯМР-спектры DEPT-135 характеризовались наличием шести основных резонансов СН и трех преобладающих сигналов СН2 в области сахара (Gal + Gro), а также 14 разрешенных сигналов СН2 и одним резонансом СН3 в алифатической области, которые соответствуют двум сложноэфирным остаткам жирной кислоты, что было подтверждено обнаружением двух сигналов С=О при δС 174,31 и 174,01 в нормальном 13С ЯМР-спектре. Наличие ацилоксигрупп в положениях С-1'''' и С-2'''' было подтверждено обнаружением перекрестных пиков Н-1''''a/C-O, Н-1''''b/C=O и Н-2''''/C=O в двумерном спектре НМВС моногалактозилдиглицерида (MGDG). В данном спектре были также обнаружены перекрестные пики Н-3''''a/C-1' и Н-3''''b/C-1', которые подтверждают связь С-1' остатка Gal с О-3'''' глицериновой части. Хорошее согласование химических сдвигов 13С всех трех атомов углерода глицерильной группы было получено для MGDG, BBGL-III и BBGL-III-Ac4 (см. таблицу III), в отличие от 1,2-дипальмитина, сдвиг С-3'''' которого по существу направлен в сторону сильного поля от соответствующих сдвигов для других соединений, что связано с отсутствием деэкранирующего гликозильного заместителя в данном диглицериде.
Высокое значение J1',2' 7,3 Гц и низкое значение JC-1',H-1' 160,2 Гц (таблица IV) подтвердили, что MGDG имеет β-аномерную конфигурацию, то есть конфигурацию, противоположную конфигурации гликолипида BBGL-III. Объединенные данные ЯМР и ЖХ-МC для MGDG показывают, что данное соединение является по существу 1,2-ди-О-стеароил-3-О-β-D-галактопиранозилглицерином, хотя более слабые пики в спектрах ЯМР и ЖХ-МС свидетельствовали о наличии небольших количеств других элементов. Диастереомерная взаимосвязь MGDG и BBGL-III и отсутствие остатка ненасыщенной жирной кислоты в MGDG являются достаточным фактором, определяющим разную растворимость двух указанных гликолипидов. В отличие от всех других исследованных липидов для растворения MGDG в CDCl3 необходимо добавить примерно 20% CD3OD.
Таким образом, BBGL-II и BBGL-III выделяли из штаммов В31, N40 и BL303 (15) B. burgdorferi и очищали почти до полной однородности хроматографией на силикагеле. При помощи разных аналитических методов (ГЖХ, MALDI-TOF, FAB и ЯМР-спектроскопия) структура основных полярных мембранных гликолипидов B. burgdorferi была определена как холестерил-6-О-ацил-β-D-галактопиранозид (BBGL-II, фиг.8) и 1,2-диацил-3-[O-α-D-галактопиранозил]-sn-глицерин (BBGL-III, фиг.9).
BBGL-III характеризуется высокой структурной гомологией с BOLIP-7, который является ранее описанным моногалактозилдиацилглицерином. Однако полученные результаты ясно показывают, что концевая часть галактозы в BBGL-II действительно связана с холестерином в отличие от BBGL-III, в котором галактоза связана с глицерином.
Дополнительные данные, относящиеся к холестериновой части в BBGL-II, были получены путем метаболического мечения данного гликолипида при культивировании клеток с 14С-холестерином. Результаты указанных экспериментов показали, что 80% радиоактивности в общей липидной фракции может принадлежать BBGL-II. При выполнении указанных экспериментов не было обнаружено значительного количества свободного холестерина, что свидетельствует об отсутствии свободного холестерина в поле мембранных липидов и о его быстром включении в BBGL-II. Установлено, что свободный холестерин или сложные эфиры холестерина входят в состав бактериальных мембран у многих видов, включающих микоплазму, Helicobacter pуlori, Micrococcus lysodeikticus, Bacillus megaterium и Proteus mirabilis. Однако вновь синтезированные холестерилглюкозиды, в которых холестерин, полученный из питательной среды, связан с сахаром и/или липидными частями, до сих пор были обнаружены только в микоплазмах, Helicobacter pylori и B. hermsi, для которых углеводом является глюкоза. Ранее выполненные исследования показали, что холестерин является сильной иммуногенной молекулой. Поэтому большое количество холестеринсодержащего BBGL-III в мембранах B. burgdorferi может привести к повышенным титрам антител против холестерина в процессе патогенеза болезни Лима.
Полимиксин В, поликатионный циклический пептид, был использован в качестве аффинного сорбента для удаления эндотоксинов, главным образом LPS и липида А. При введении обработанных ультразвуком клеток B. burgdorferi в колонку, содержащую иммобилизованный полимиксин В, BBGL-II можно было обнаружить в связанном веществе после элюирования дезоксихолатом. Интересно отметить, что BBGL-III не связывался с колонкой в аналогичных условиях. Побудительной причиной связывания эндотоксинов с полимиксином В является гидрофобное взаимодействие между двумя указанными структурами. Можно признать, что некоторые структурные элементы BBGL-II обладают свойствами, подобными липиду А.
Молекулярная имитация липида А гликолипидом BBGL-II была также обнаружена в результате инициации секреции провоспалительных факторов, таких как интерлейкин-1, IL-6, TNF-альфа и PGE2, при стимуляции культур клеток разными препаратами B. burgdorferi, содержащими BBGL-II. Безотносительно к какой-либо теории вполне вероятно, что BBGL-II действует в качестве “функционально активного липополисахарида (LPS)” в B. burgdorferi.
Пример 14
Результаты иммунологического исследования
Специфические антитела были индуцированы у мышей и кроликов, которым инъецировали разные препараты, содержащие соединение формулы В (BBGL-II) (см. фиг.10 и 11). BBGL-III индуцировал антитела, главным образом IgM, которые взаимодействовали с обоими гликолипидами (данные не приведены). BBGL-II в физиологическом растворе с фосфатным буфером (PBS) (по-видимому, в виде мицелл) был более иммуногенным по сравнению с раствором в ДМСО или сквалене. Самые высокие титры были получены при использовании адъюванта Фрейнда.
Удивительно, что, несмотря на низкую молекулярную массу, BBGL-II вызывал образование антител у мышей и кроликов, главным образом изотипа IgM. Указанные антитела были специфичными к гомологичному гликолипиду. В отличие от этого BBGL-III вызывал образование антител, которые взаимодействовали с обоими гликолипидами.
Пример 15
Синтез аналога BBGL-II (“азидо-BBGL-II”)
Целевое соединение (“азидо-BBGL-II”, соединение 19, см. фиг.15) отличается от природного гликолипида BBGL-II тем, что оно содержит азидогруппу в конце пальмитоильной группы вместо протона. Соединение 19 синтезировали из главных промежуточных соединений, таких как галактозилбромид 4, холестерин 5 и ω-азидопальмитиновая кислота 16.
Соединение 4 (см. фиг.12) получали из галактозы 1, которую обрабатывали пивалоилхлоридом в пиридине. Вместо ожидаемого пентапивалоильного производного было получено частично пивалоилированное соединение 2 с хорошим выходом и высокой чистотой, не требующее хроматографической очистки. Свободная гидроксильная группа в соединении 2 была защищена ацетильной группой (→ 3). Полученный продукт обрабатывали гидробромидом в уксусной кислоте, получая при этом галактозилбромид 4. Бромид 4 связывали с холестерином 5, промотируя реакцию трифторметансульфонатом серебра в дихлорметане, в результате чего происходило стереоселективное образование требуемого β-галактозилхолестерина 6 (см. фиг.13). При выполнении 1Н ЯМР-спектроскопии была обнаружена β-аномерная конфигурация. О-защитные группы затем удаляли, производя обработку метоксидом натрия в смеси безводного дихлорметана и метанола, и получали холестерил-β-D-галактопиранозид 7 в виде аморфного твердого вещества.
Были испробованы многочисленные комбинации защитных групп, позволяющие ввести пивалоильную часть в положение НО-6 тетраола 7. Опыт авторов данного изобретения показал, что использование защитных групп ацильного типа, включающих ацетильные, левулиноильные и хлорацетильные группы, оказалось в основном безуспешным из-за неконтролируемой миграции указанных групп в галактозной части. В конце концов данная проблема была решена благодаря использованию комбинации аллильных, изопропилиденовых и силильных защитных групп. Так первичная гидроксильная группа в соединении 7 была защищена трет-бутилдифенилсилильной группой с образованием соединения 8 без какого-либо влияния на другие гидроксильные группы. Затем триол 8 обрабатывали диметоксипропаном в присутствии камфорсульфоновой кислоты для введения изопропилиденовой группы в положение НО-3 и НО-4 с образованием соединения 9. Промежуточное соединение 9 обрабатывали аллилбромидом/NaH в диметилформамиде с образованием полностью защищенного производного 10, из которого силильную защитную группу избирательно удаляли без какого-либо влияния на другие части молекулы с образованием соединения 11. Данное производное содержит свободную гидроксильную группу в положении НО-6, которая является местом присоединения пальмитоильной части.
Синтез промежуточного соединения 16, представляющего собой 16-азидопальмитиновую кислоту, показан на фиг.14. Промышленно доступную 16-гидроксипальмитиновую кислоту 12 обрабатывали диазометаном с образованием сложного метилового эфира 13 с количественным выходом. Спирт 13 обрабатывали п-толуолсульфонилхлоридом в пиридине с образованием сульфоната 14, который затем обрабатывали азидом натрия в диметилформамиде с образованием сложного азидоэфира 15. Сложный метиловый эфир расщепляли гидроксидом калия в смеси диоксана и воды с образованием кислоты 16.
Холестерилпальмитоилгалактопиранозид 19 получали, конденсируя соединение 11 с азидопальмитиновой кислотой 16 в присутствии дициклогексилкарбодиимида в этилацетете, с количественным выходом (→ 17) (см. фиг. 15). Защитные группы удаляли в две стадии. Сначала соединение 17 обрабатывали уксусной кислотой/водой для удаления изопропилиденовой группы с образованием диола 18. Затем соединение 18 обрабатывали комплексом иридия (MePh2)2(C8H12)IrPF6 в тетрагидрофуране и окисляли для удаления полученной пропенильной группы, используя комбинацию тетроксида осмия и N-оксида триметиламина, с образованием азида 19 в виде аморфного твердого вещества.
Анализы верхнего слоя методами ELISA и ТСХ показали, что антитела против синтетического соединения 19 были образованы в такой же степени, что и при использовании очищенного природного BBGL-II.
Очевидно, что отдельные элементы вышеописанных способов или композиций могут быть изменены или модифицированы без выхода за пределы объема описанного изобретения. Авторы настоящего изобретения заявляют, что все такие модификации и изменения соответствуют объему и духу нижеследующей формулы изобретения.
Ссылки
Список справочной литературы
1. Orloski, К. A., Hayes, E. В., Campbell, G. L., and Dennis, D. T. (2000) MMWR CDC Surveill Summ. 49, 1-11.
2. Steere, A. C. (2001) N.Engl.J.Med. 345,115-125.
3. Sadziene, A. and Barbour, A. G. (1996) Infection 24, 195-202.
4. Rahn, D. W. (2001) Infect.Dis.Clin.North Am. 15, 171-187.
5. Lathrop, S. L., Ball., R., Haber, P., Mootrey, G. Т., Braun, M. M., Shadomy, S. V., Ellenberg, S. S., Chen, R. Т., and Hayes, E. B. (2002) Vaccine 20, 1603-1608.
6. Vinh, T. U., Shi, M. H., Adler, В., and Faine, S. (1989) J.Gen.Microbiol 135 (Pt 10), 2663-2673.
7. Halter, M. R. and Joens, L. A. (1988) Infect.Immun. 56, 3152-3156.
8. Takayama, K., Rothenberg, R. J., and Barbour, A. G. (1987) Infect.Immun. 55, 2311-2313.
9. Cinco, M., Banfi, E., Balanzin, D., Godeas, C., and Panfili, E. (1991) FEMS Microbiol.Immunol. 3, 33-38.
10. Eiffert, H., Letter, H., Jarecki-Khan, K., and Thomssen, R. (1991) Med.Microbiol.Immunol.(Berl) 180, 229-237.
11. Wheeler, C. M., Garcia Monco, J. C., Benach, J. L., Golightly, M. G., Habicht, G. S., and Steere, A. C. (1993) J.Infect.Dis. 167, 665-674.
12. Radolf, J. D., Goldberg, M. S., Bourell, K., Baker, S. I., Jones, J. D., and Norgard, M. V. (1995) Infect.Immun. 63, 2154-2163.
13. Hossain, H., Wellensiek, H. J., Geyer, R., and Lochnit, G. (2001) Biochimie 83, 683-692.
14. Bligh, E. G. and Dyer, W. J. (1969) CanJ.Biochem.Physiol. 37, 911-917.
15. Ben Menachem, G., Wagner, F., Zahringer, U., Rietschel, E. Т., and Rottem, S. (1997) FEMS Microbiol.Lett. 154, 363-369.
16. Sawardeker, J. S., Sloneker, J. H., and Jeanes, A. (1965) Biochem.J. 37, 1602-1604.
17. Fay, L. and Richli, U. (1991) J.Chromatography 541, 89-98.
18. Rasley, A., Anguita, J., and Marriott, I. (2002) J.Neuroimmunol. 130, 22-31.
19. Roth, h., Segal, S., and Bertoli, D. (1965) Anal.Biochem. 10, 32-52.
20. Pasciak, M., Ekiel, I., Grzegorzewicz, A., Mordarska, H., and Gamian, A. (2002 Biochim.Biophys.Acta 1594, 199-205.
21. Dell, A. (1987) Adv.Carbohydr.Chem.Biochem. 45, 19-72.
22. Zahringer, U., Wagner, F., Rietschel, E. Т., Ben Menachem, G., Deutsch, J., and Rottem, S. (1997) J.Biol.Chem. 272, 26262-26270.
23. Ciucanu, I. and Kerek, F. (1984) Carbohydr.Res 131,209-217.
24. Reich, H. J., Jautelat, M., Messe, M. Т., Weigert, F. J., and Roberts, J. D. (1969) Journal of the American chemical society 91, 7445-7453.
25. Hirai, Y., Haque, M., Yoshida, Т., Yokota, K., Yasuda, Т., and Oguma, K. (1995) J.Bacteriol. 177, 5327-5333.
26. Bock, K. and Pedersen, C. (1975) Acta Chemica Scandinavica В 29,258-264.
27. Honarvar, N.. Schaible, U. E., Galanos, C., Wallich, R., and Simon, M. M. (1994) Immunology 82, 389-396.
28. Beck, G., Benach, J. L., and Habicht, G. S. (1990) Biochem.Biophys.Res Commun, 167, 89-95.
29. Beck, G., Habicht, G. S., Benach, J. L., and Coleman, J. L. (1985) J.Infect.Dis. 152, 108-117.
30. Habicht, G. S., Beck, G., Benach, J. L., and Coleman, J. L. (1986) Zentralbl.Bakteriol.Mikrobiol.Hyg.[A] 263, 137-141.
31. Rottem, S. (2002) Biochem.Biophys.Res.Commun. 292, 1289-1292.
32. Razin, S. (1975) J.Bacteriol 124, 570-572.
33. Mayberry, W. R. and Smith, P. F. (1983) Biochim.Biophys.Acta 752, 434-443.
34. Livermore, B. P., Bey, R. F., and Johnson, R. C. (1978) Infect.Immun. 20, 215-220.
35. Alving, C. R. and Swartz, G. M., Jr. (1991) Crit Rev.Immunol, 10,441-453.
36. Anspach, F. B. (2001) J.Biochem.Biophys.Methods 49, 665-681.
37. Srimal, S., Surolia, N., Balasubramanian, S., and Surolia, A. (1996) Biochem.J. 315 (Pt 2), 679-686.
38. Habicht, G. S, Beck, G., Benach, J. L., Coleman, J. L, and Leichtling, K. D. (1985) J.Immunol. 134, 3147-3154.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОВЕРХНОСТНЫЕ ГЛИКАНЫ CLOSTRIDIUM PERFRINGENS И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2798419C2 |
| СФИНГОИДНЫЕ ПОЛИАЛКИЛАМИНОВЫЕ КОНЪЮГАТЫ ДЛЯ ВАКЦИНАЦИИ | 2004 |
|
RU2361577C2 |
| СИСТЕМА ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ | 2002 |
|
RU2294192C2 |
| ХИМЕРНЫЕ ГЕНЫ OSPA, БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2583289C2 |
| ПРИМЕНЕНИЕ ИНГИБИТОРОВ NKG2D ДЛЯ ЛЕЧЕНИЯ СЕРДЕЧНО-СОСУДИСТЫХ И МЕТАБОЛИЧЕСКИХ ЗАБОЛЕВАНИЙ, ТАКИХ КАК ДИАБЕТ 2 ТИПА | 2010 |
|
RU2566264C2 |
| НОСИТЕЛЬ ЛЕКАРСТВА-АНТИГЕНА, ДАЮЩИЙ ВОЗМОЖНОСТЬ ТРАНСМУКОЗАЛЬНОГО И ТРАНСДЕРМАЛЬНОГО ВВЕДЕНИЯ, СПОСОБ ИНДУЦИРОВАНИЯ МУКОЗАЛЬНОГО ИММУНИТЕТА, МУКОЗАЛЬНАЯ ВАКЦИНА И СИСТЕМА ДОСТАВКИ ЛЕКАРСТВА, ИСПОЛЬЗУЮЩИЕ ЕГО | 2005 |
|
RU2396090C2 |
| ПРОИЗВОДНЫЕ ПИПЕРИДИНА, СПОСОБ ИХ ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ИХ ОСНОВЕ | 2002 |
|
RU2279425C2 |
| ХИМЕРНЫЕ ГЕНЫ OSPA, БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2773402C2 |
| ИММУНОМОДУЛИРУЮЩИЙ АГЕНТ | 1989 |
|
RU2120302C1 |
| СУЛЬФОНИЛМОЧЕВИНЫ И РОДСТВЕННЫЕ СОЕДИНЕНИЯ И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2739356C2 |
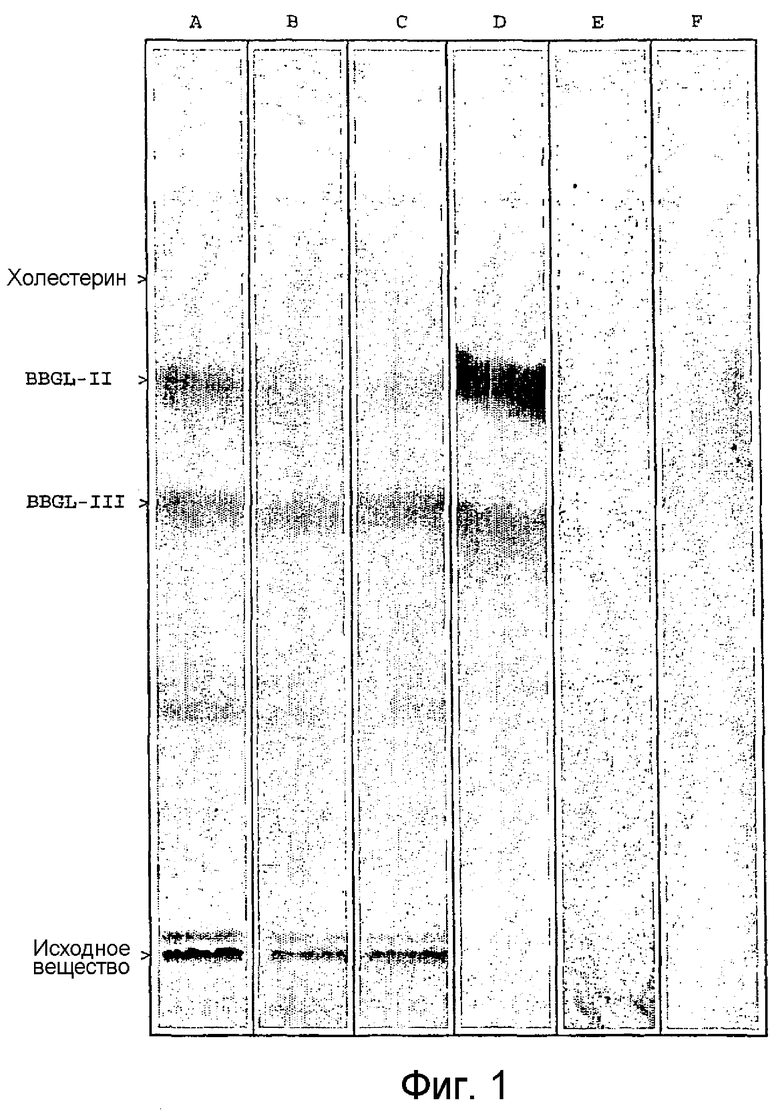
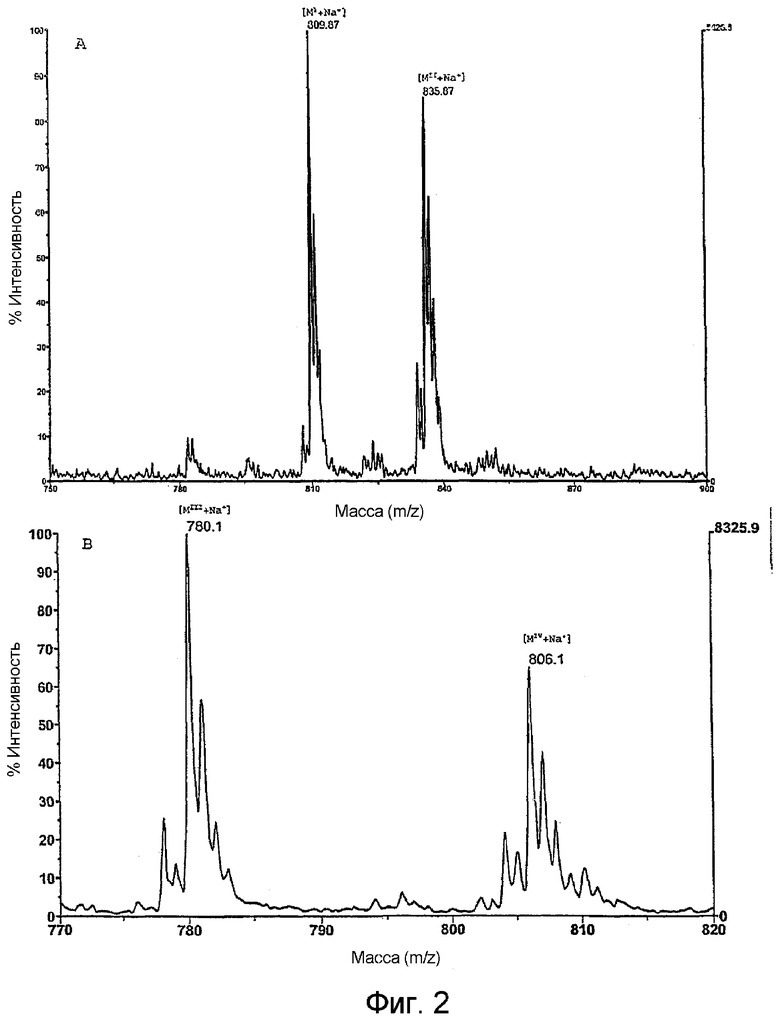
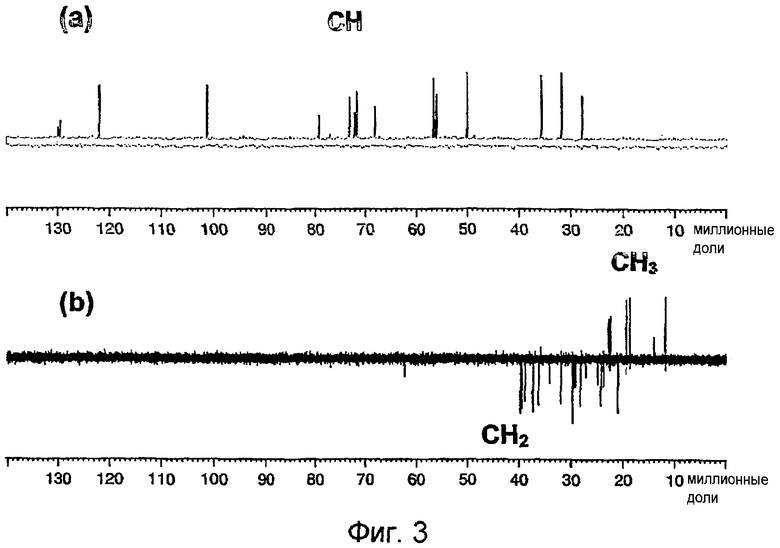
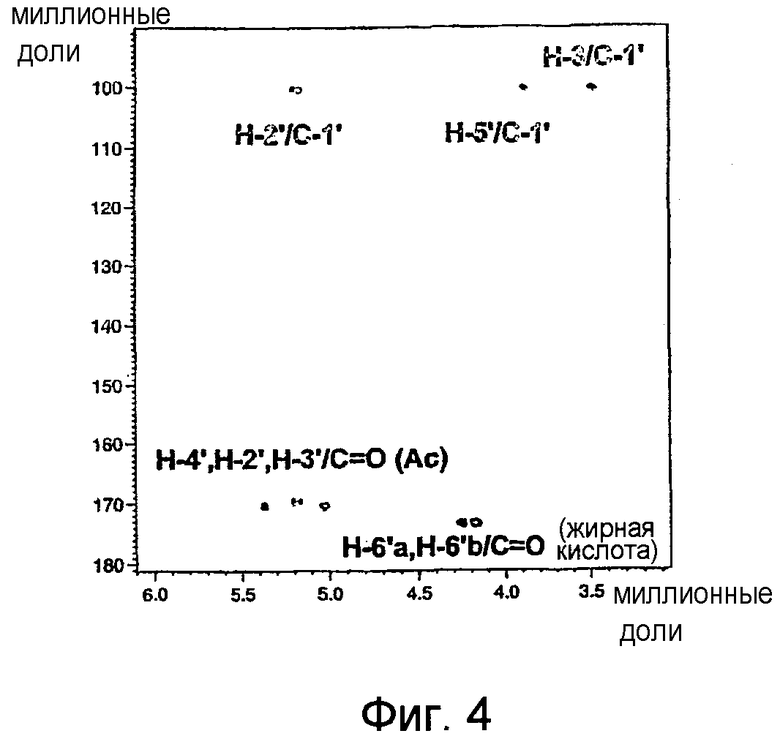
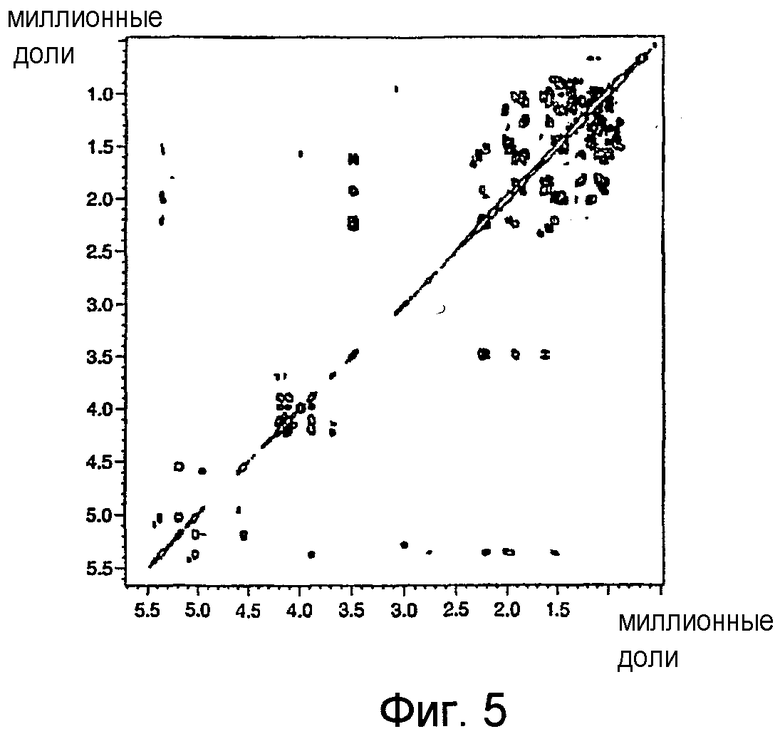
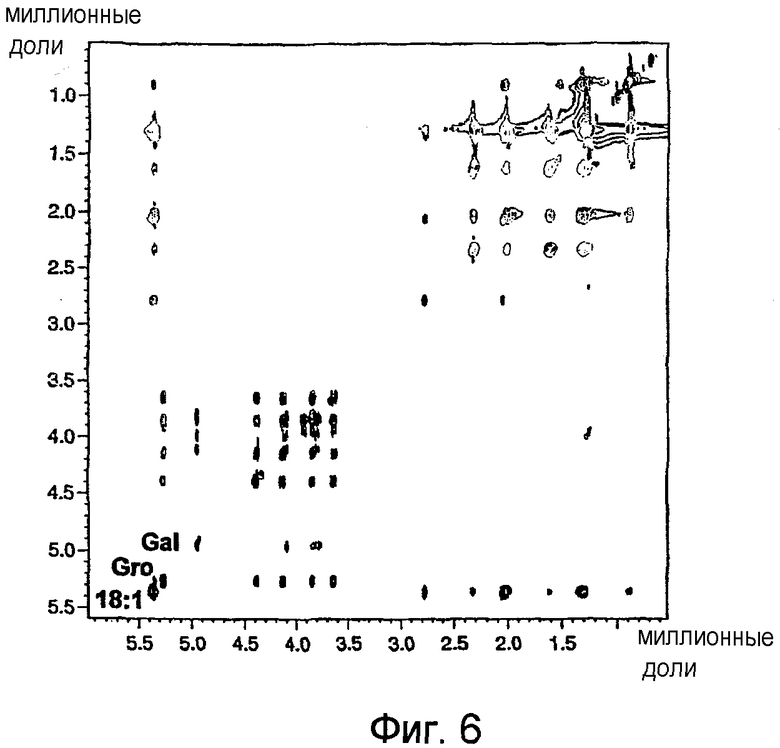
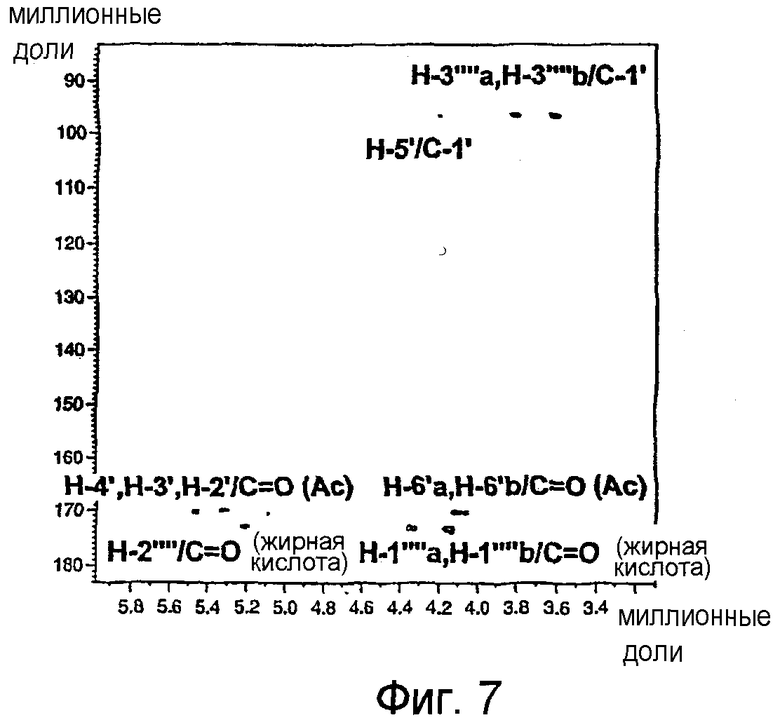
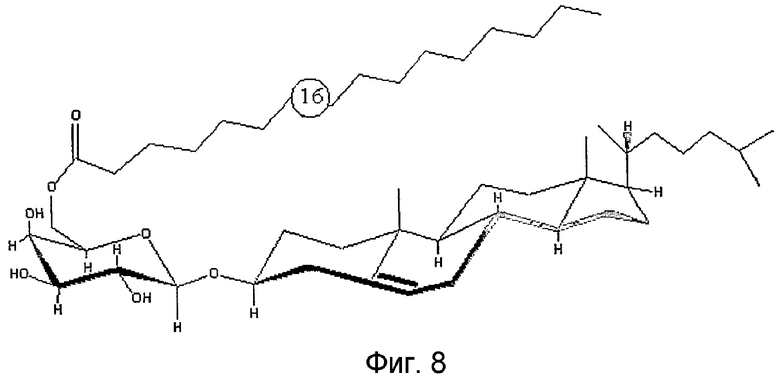
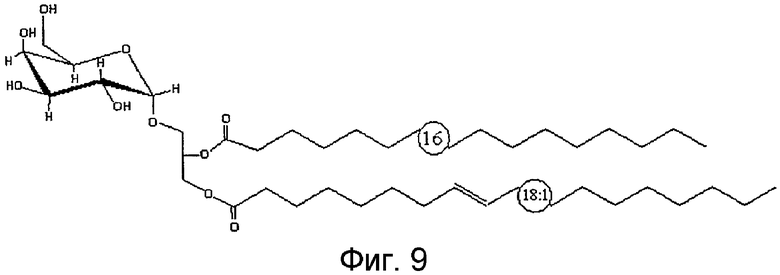
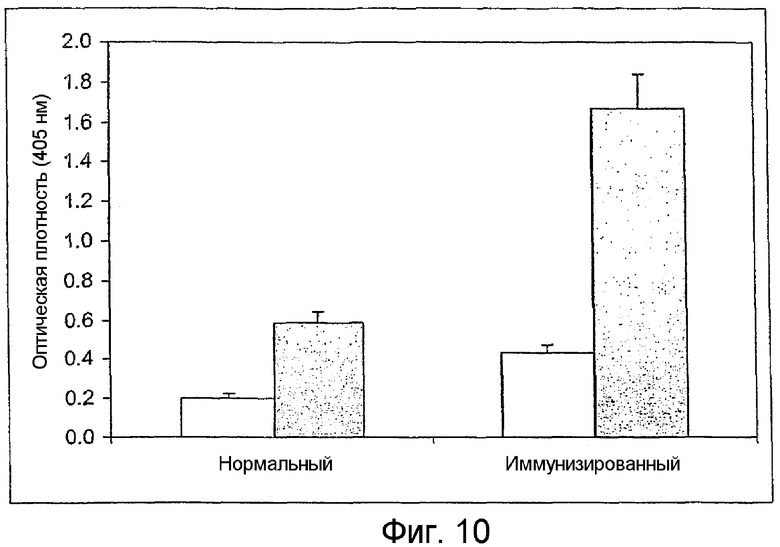
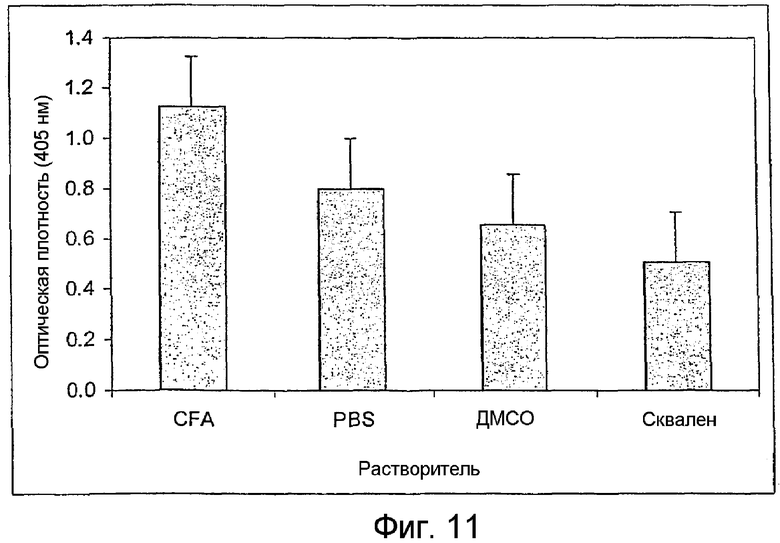
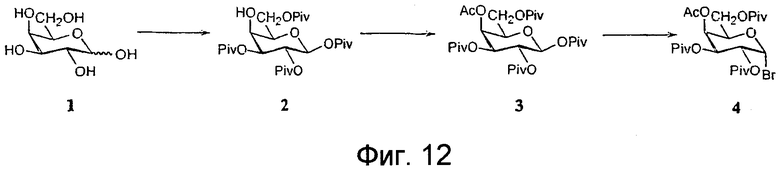
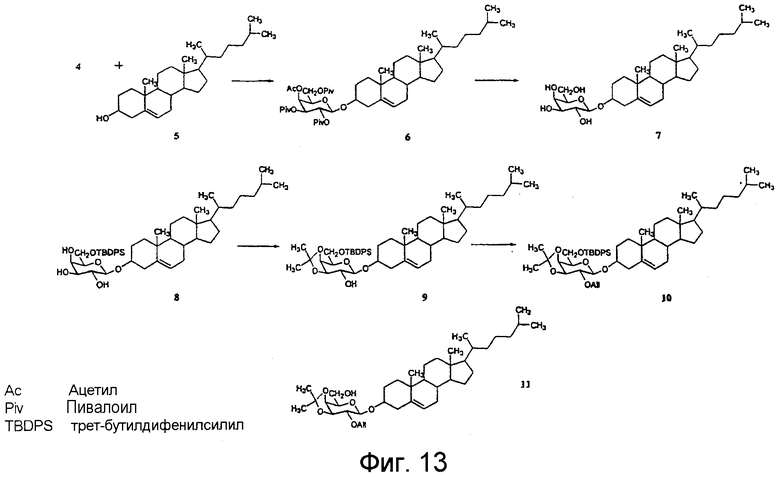
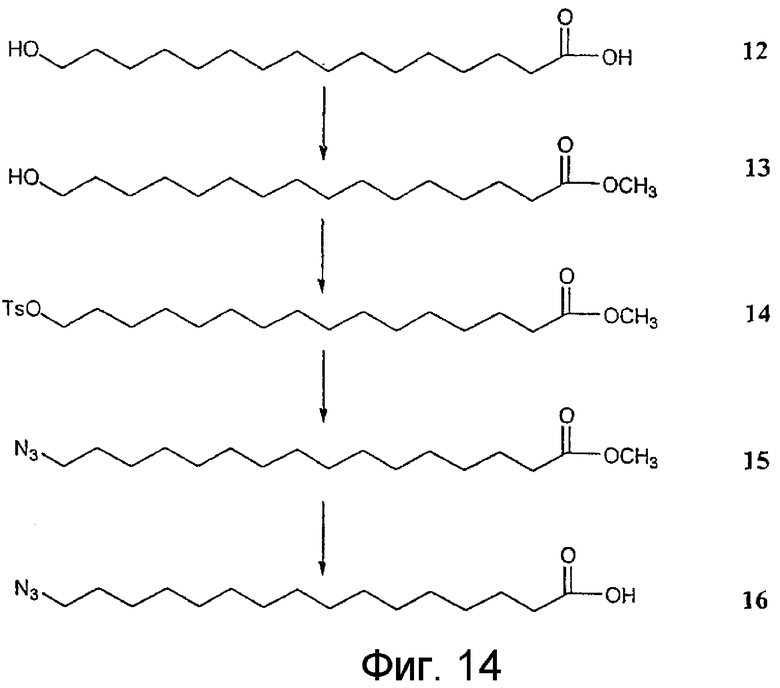
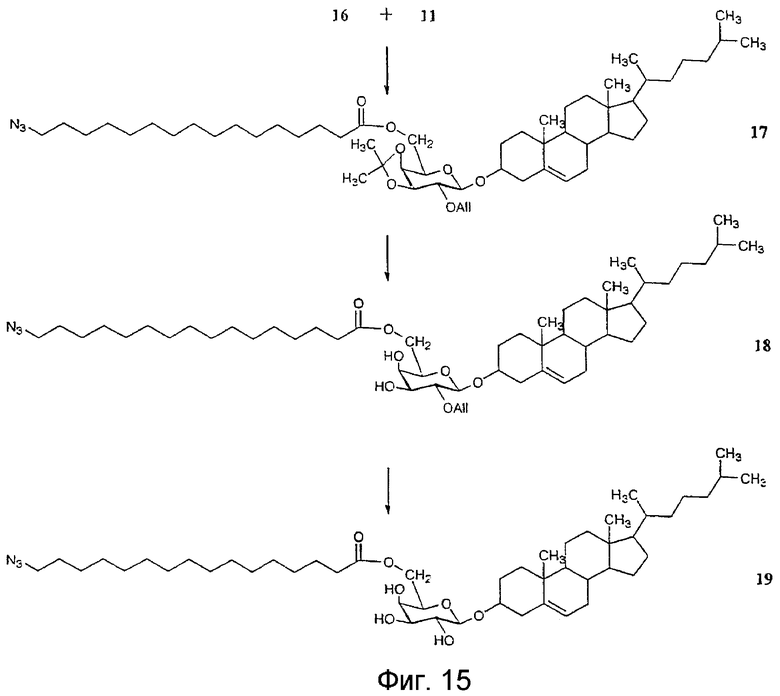
Изобретение относится к холестерил-6-O-ацил-β-D-галактопиранозиду, предназначенному для индуцирования у млекопитающего-хозяина иммунной реакции против В.burgdorferi, где (i) ацильный остаток представляет собой группу, выделенную из органических жирных кислот, содержащих 1-25 атомов углерода; (ii) галактопиранозидная кольцевая структура необязательно включает в любом положении кольца ацильную группу, выделенную из жирной кислоты. Изобретение относится также к смеси 3-O-(6-О-пальмитоил-β-D-галактопиранозил)-
холестерина и 3-O-(6-O-олеоил-β-D-галактопиранозил)холестерина, предназначенной для индуцирования у млекопитающего-хозяина иммунной реакции против В.Burgdorferi. Названные соединения особенно полезны для профилактики или лечения болезни Лима. 2 н. и 5 з. п. ф-лы, 4 табл., 15 ил.
1. Холестерил-6-О-ацил-β-D-галактопиранозид, предназначенный для индуцирования у млекопитающего-хозяина иммунной реакции против B.burgdorferi, где
(i) ацильный остаток представляет собой группу, выделенную из органических жирных кислот, содержащих 1-25 атомов углерода;
(ii) галактопиранозидная кольцевая структура необязательно включает в любом положении кольца ацильную группу, выделенную из жирной кислоты.
2. Соединение по п.1, не модифицированное в соответствии с позицией (ii).
3. Соединение по п.1, представляющее собой холестерил-6-O-пальмитоил-β-D-галактопиранозид.
4. Соединение по п.1, представляющее собой холестерил-6-О-олеоил-β-D-галактопиранозид.
5. Соединение по п.1, предназначенное для лечения субъекта, инфицированного B.Burgdorferi, или для профилактики или ингибирования B.Burgdorferi инфекции или развития клинических симптомов болезни Лима.
6. Смесь 3-O-(6-O-пальмитоил-β-D-галактопиранозил)-холестерина и 3-O-(6-O-олеоил-β-D-галактопиранозил)холестерина, предназначенная для индуцирования у млекопитающего-хозяина иммунной реакции против B.Burgdorferi.
7. Смесь по п.6, предназначенная для лечения субъекта, инфицированного B.Burgdorferi, или для профилактики или ингибирования B.Burgdorferi инфекции или развития клинических симптомов болезни Лима.
| BEN-MENACHEM G; ET AL // PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF USA, NATIONAL ACADEMY OF SCIENCE, WASHINGTON, DC, US | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| Облицовка комнатных печей | 1918 |
|
SU100A1 |
| Насос | 1917 |
|
SU13A1 |
| SCHRÖDER N W J; ET AL // JOURNAL OF BIOLOGICAL CHEMISTRY, AMERICAN SOCIETY OF BIOLOCHEMICAL BIOLOGISTS, BIRMINGHAM, US | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ПАРОВАЯ ИЛИ ГАЗОВАЯ ТУРБИНА | 1914 |
|
SU278A1 |
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |

