Изобретение относится к области биотехнологии и производству антирабических вакцин, а именно к способу опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени.
Бешенство по своей природе является острым вирусным энцефалитом, одно из немногих заболеваний со 100%-ной летальностью [1]. Возбудителем данной болезни является вирус бешенства порядка Mononegavirales семейства Rhabdoviridae рода Lyssavirus вида Rabies lyssavirus [2]. Геном вируса представлен несегментированной одноцепочечной негативной молекулой РНК длиной около 12000 н. о. Нуклеиновая кислота кодирует 5 основных белков, расположенных в консервативном линейном порядке: нуклеопротеин (N-белок), фосфопротеин (Р-белок), матриксный белок (М-белок), гликопротеин (G-белок), РНК-зависимую РНК-полимеразу (L-белок) [3]. Вирион вируса бешенства имеет пулевидную форму длиной 130-250 нм и диаметром 60-100 нм. Он состоит из следующих функциональных структур: рибонуклеопротеин (РНП), расположенный внутри вириона, и внешняя белково-липидная составляющая. РНП представляет собой внутренний нуклеокапсид, который включает в себя геномную РНК, прочно связанную с нуклеопротеидом и фосфопротеином, а также вирусную полимеразу. Рибонуклеопротеин является важным иммуногенным компонентом вакцинных препаратов против бешенства. В РНП содержится 4% РНК к 96% белка [2]. Снаружи вириона располагается двухслойная липидная оболочка с «шипами» гликопротеина. Матричный белок находится между РНП и внешней составляющей вириона, конденсирует нуклеопротеин и взаимодействует с эндо доменом гликопротеином [3, 4]. Единицы транскрипции разделены короткими не транслируемыми межгенными областями. Исключение составляет межгенная область G-L, которая состоит из 400-700 нуклеотидов и может представлять собой остаток гена, который потерял свою функциональность во время эволюции лиссавирусов [5].
Белки вируса бешенства полифункциональны. Нуклеопротеин защищает вирусный геном от активности клеточной РНКазы и взаимодействует с L- и Р-белками во время транскрипции и репликации. Фосфопротеин отвечает за правильное позиционирование L-белка и действует как шаперон во время синтеза нуклеопротеина, образуя N-P-комплекс, который предотвращают самоагрегацию N-белка и связывание с клеточной РНК [5, 6]. Матриксный белок является структурным элементом, который связывается с рибонуклеопротеином и цитоплазматическим доменом гликопротеина для облегчения процесса сборки вирусных частиц. Гликопротеин - единственный вирусный белок, расположенный на поверхности вириона. Он обеспечивает связывание с рецепторами клетки-хозяина, индуцирует эндоцитоз, а также взаимодействуют с М-белком, облегчая морфогенез вирионов. L-белок выполняет функции, необходимые для транскрипции и репликации генома [7].
Постэкспозиционная профилактика может обеспечить практически полное излечение от болезни при своевременном и правильном применении и доступна уже более века. Тем не менее, данное заболевание по-прежнему ежегодно приводит к гибели десятков тысяч людей и животных из-за плохой доступности соответствующих биологических препаратов, особенно в развивающихся странах тропической Азии и Африки [5, 8]. Как и при любом инфекционном заболевании эффективные программы борьбы с бешенством опираются на строгий лабораторный надзор, надежные диагностические инструменты и высокоэффективные биологические препараты, в том числе антирабические вакцины [5].
В процессе производства антирабических вакцин вируссодержащее сырье исследуют для определения концентрации такого иммуногенного компонента, как рибонуклеопротеина вируса бешенства [5, 7, 8]. В настоящее время для этих целей применяют прямой сэндвич-вариант иммуноферментного анализа (ИФА) (прототип) [9]. Однако, он может иметь ряд недостатков: возможности увеличения чувствительности метода ограничиваются фоном анализируемого соединения, субстратной специфичностью фермента и аффинностью антител; наличие в тестируемых образцах кофакторов, ингибиторов и стимуляторов активности ферментов; ИФА не позволяет разделять нативные белки и их биологические неактивные фрагменты, сохранившие антигенные детерминанты; возможное изменение каталитической активности фермента при его конъюгировании с антигеном [10].
В связи с этим целесообразно разработать способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцин с применением метода оценки максимальных экстремумов графиков второй производной для кривых реакции амплификации в режиме реального времени.
Данный метод позволяет в течение 4 часов с высокой степенью достоверности определять концентрацию рибонуклеопротеина вируса бешенства в суспензиях с высоким содержанием клеточного белка и липопротеинов и не предполагает использование сорбента на этапе экстрагирования нуклеиновой кислоты. Исходя из этого целесообразно предложить новый способ определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины на основе альтернативного метода.
Задачей настоящего изобретения является разработка высокочувствительного и высокоспецифичного способа быстрого определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины с целью устранения вышеуказанных недостатков.
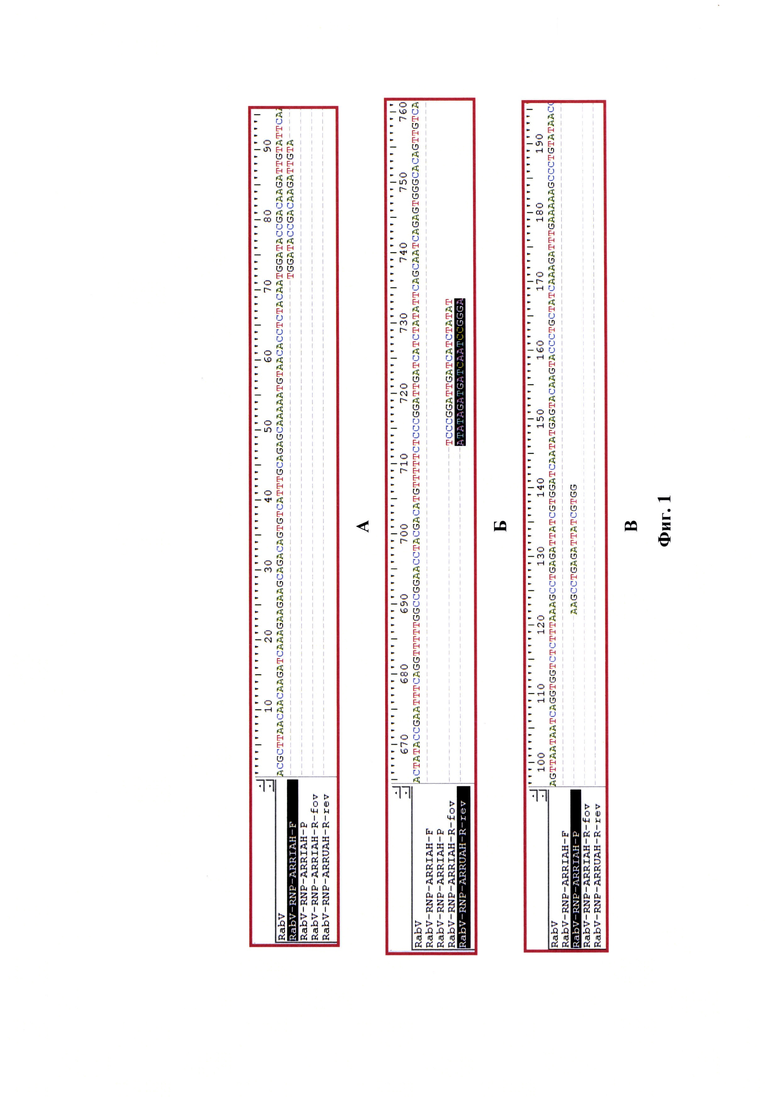
Данная задача решена благодаря созданию нового способа опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени. Предложенный способ позволяет: 1) повысить специфичность анализа проб за счет иммунного связывания рибонуклеопротеина вируса бешенства; 2) исключить возможность увеличения фоновых значений флуоресценции благодарю использованию методики разделения фракций проб с помощью смеси гуанидинизотиоцианата (ГТЦ), трихлорметана, карболовой кислоты и пропанола-2; 3) увеличить чувствительность и специфичность реакции за счет модификации компонентного состава смеси для проведения реакции амплификации; 4) повысить достоверность проводимого анализа благодаря установлению зависимости между значениями концентрации рибонуклеопротеина вируса бешенства (СРНП ВБ) и максимальными экстремумами графиков второй производной для кривых реакции амплификации (Ср), представленной в виде функции второй степени СРНП ВБ = 0,0123(Ср)2-0,0725Ср+8,0129 с высокой достоверностью аппроксимации (R2=0,9952). Предложенная модель позволяет определять концентрацию РНП вируса бешенства в сырье для антирабической вакцины. Сущность изобретения отражена на графических изображениях: Фиг. 1 - Дизайн олигонуклеотидов, гомологичных участку N-гена вируса бешенства, используемых при определении концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины при постановке реакции амплификации.
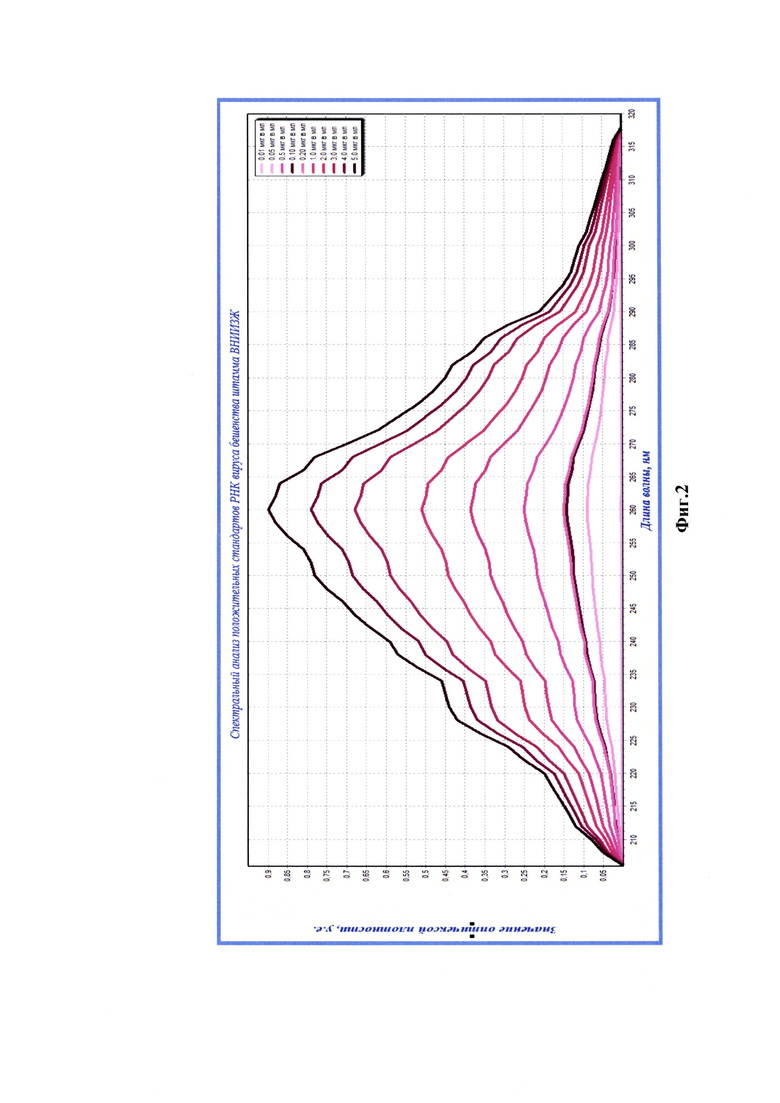
Фиг.2 - Спектрограммы разведений элюатов РНК вируса бешенства штамма ВНИИЗЖ.
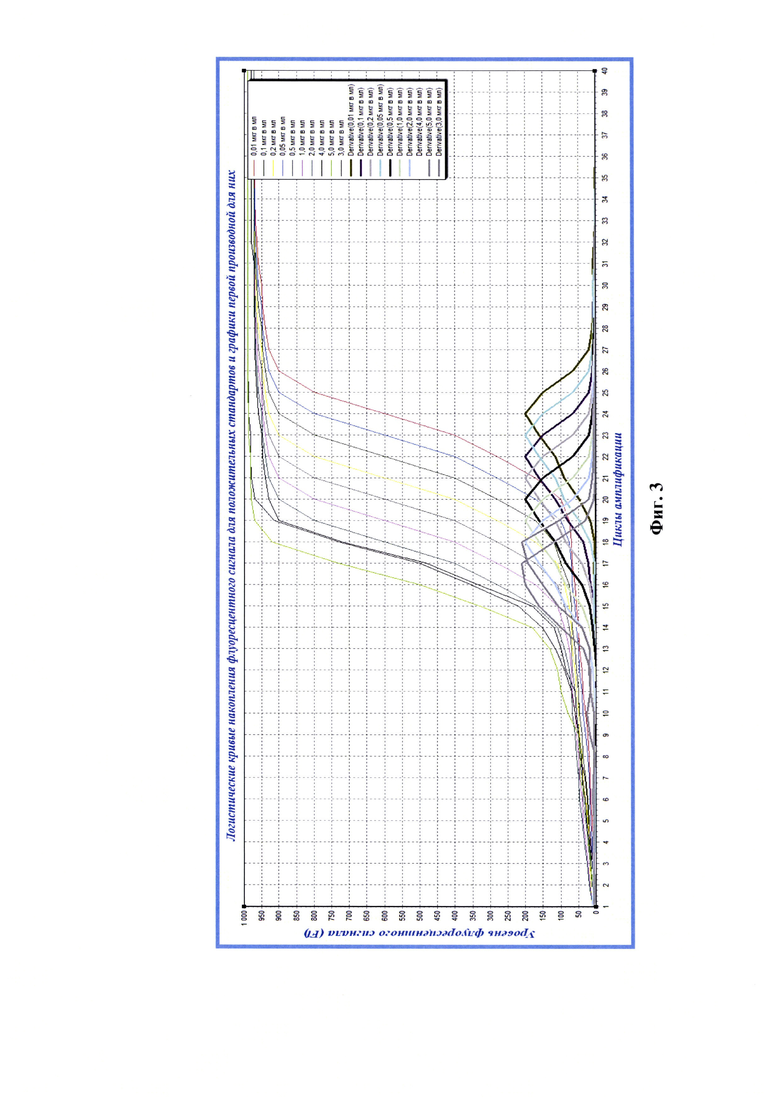
Фиг. 3 - Логистические графики накопления флуоресцентного сигнала при реакции амплификации для положительных стандартов вируса бешенства штамма ВНИИЗЖ с концентрациями вирионов 0,01, 0,05, 0,10, 0,20, 0,50, 1,00, 2,00, 3,00, 4,00, 5,00 мкг/мл (n=3) и графики первой производной (derivative) для данных кривых.
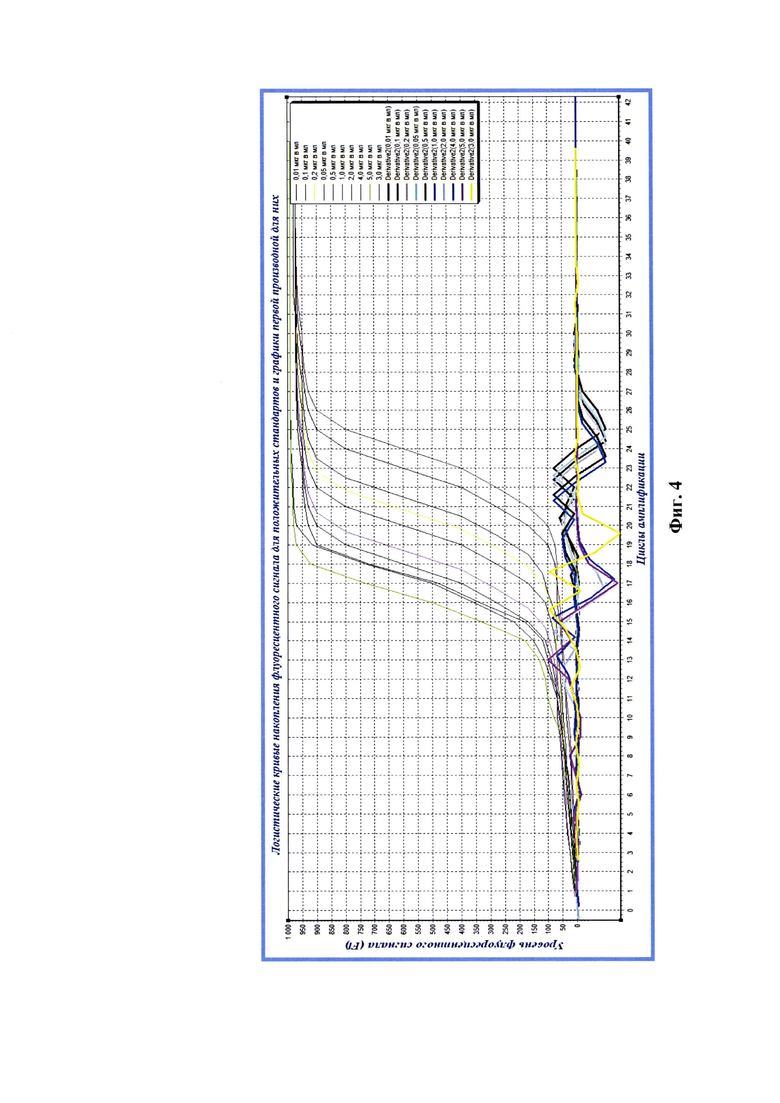
Фиг. 4 - Средние значения критических точек Ср, рассчитанные с помощью второй производной (derivative 2) для графиков реакции амплификации в реальном времени при анализе РНК положительного стандарта вируса бешенства штамма ВНИИЗЖ с концентрациями вирионов 0,01, 0,05, 0,10, 0,20, 0,50, 1,00, 2,00, 3,00, 4,00, 5,00 мкг/мл (n=3).
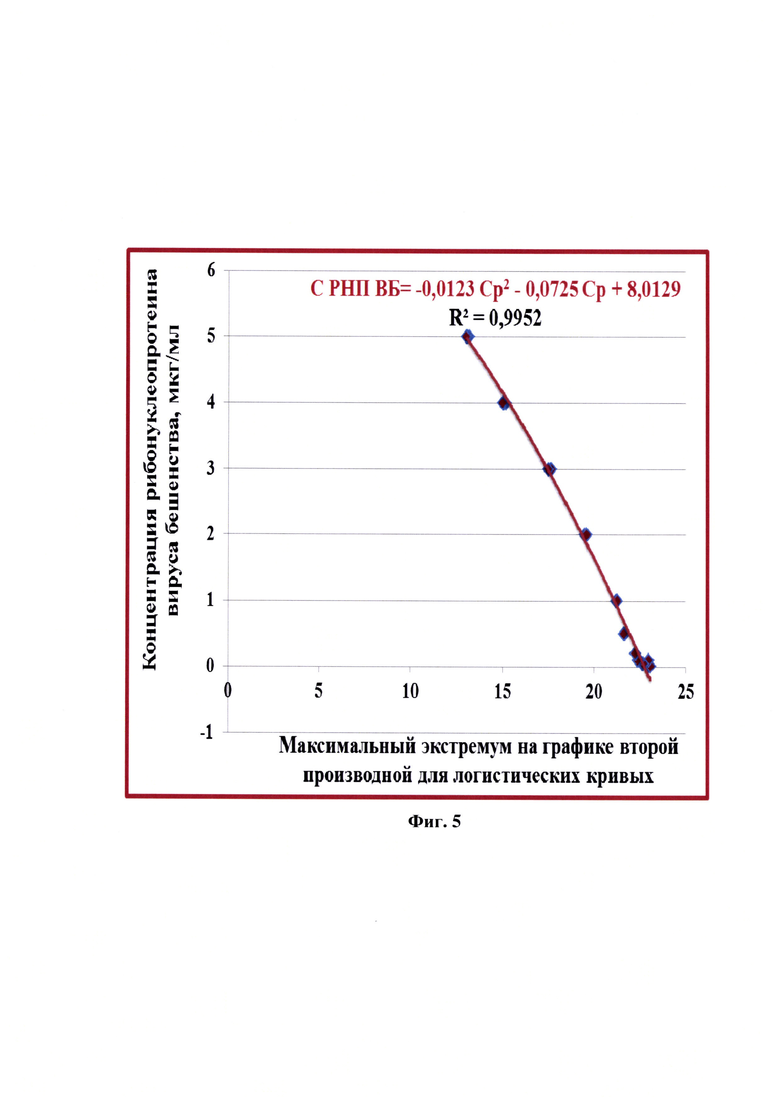
Фиг. 5 - Зависимость критической точки Ср, детектируемой с помощью второй производной для кривой амплификации в режиме реального времени, от концентрации рибонуклеопротеина вируса бешенства (с указанием стандартной погрешности) (n=3).
Сущность изобретения заключается в новом подходе по определению концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины при исследовании на экстремумы графиков второй производной для логистических кривых сигнала флуоресценции относительно количества циклов реакции амплификации. Заявляемый способ основан на проведении серологического связывания рибонуклеопротеина вируса бешенства с использованием очищенных поликлональных антител, получении очищенного элюата вирусной РНК без применения сорбента с использованием метода экстракции смесью карболовой кислоты, ГТЦ, трихлорметана и пропанола-2, постановке реакции амплификации в режиме реального времени с последующим детектированием максимальных экстремумов логистических кривых реакции амплификации с помощью второй производной, определении концентрации рибонуклеопротеина вируса бешенства с использованием разработанной модели зависимости критической точки графика второй производной для кривой накопления флуоресценции и концентрации РНП.
В настоящее время реакцию амплификации в режиме реального времени применяют для детекции генома вируса бешенства в культуральных жидкостях и патологическом материале, преимущественно в тканях головного мозга и слюнных желез (Warner et al., 1997, Fooks et al., 2009) [7, 8]. Разработанный способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при оценке максимальных экстремумов графиков второй производной для логистических кривых реакции амплификации в реальном времени по сравнению с прототипом отличается более высокой чувствительностью и специфичностью при анализе вирусных суспензий с высоким содержанием клеточного белка и липопротеинов.
В отличие от прототипа разработанный способ включает этап серологического связывания рибонуклеопротеина вируса бешенства с применением очищенных поликлональных антител сывороток крови морских свинок против рибонуклеопротеина вируса бешенства; модифицированный этап выделения РНК, исключающий использование сорбента; усовершенствованный этап проведения реверсии и реакции амплификации в режиме реального времени; новый подход к методике определения концентрации рибонуклеопротеина вируса бешенства с применением квадратичной модели зависимости максимального экстремума графика второй производной для логистической кривой флуоресценции и концентрации РНП. Применение предложенного способа повышает достоверность анализа концентрации рибонуклеопротеина вируса бешенства в пробах. При этом также как и прототип разработанный способ позволяет одновременно исследовать несколько десятков проб сырья для антирабической вакцины за довольно короткий промежуток времени (в течение 4 часов). Исходя их этого, актуально применять данный способ для определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины.
Ключевым элементом заявляемого способа является детектирование максимальных экстремумов графиков второй производной для логистических кривых реакции амплификации в режиме реального времени для исследуемых проб и определение концентрации рибонуклеопротеина вируса бешенства с использованием разработанной квадратичной модели зависимости критической точки графика второй производной для кривой накопления флуоресценции и концентрации РНП.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении метода сравнения максимальных экстремумов графиков второй производной для кривых амплификации в режиме реального времени с использованием разработанной квадратичной модели для определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины.
Сведений о разработке предлагаемого способа определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины авторами не обнаружено.
Сущность предлагаемого изобретения пояснена на графическом материале - графике зависимости критической точки Ср, детектируемой с помощью второй производной для кривой амплификации в режиме реального времени, от концентрации рибонуклеопротеина вируса бешенства (фиг.5).
Сенсибилизируют иммунологический 6-луночный планшет, очищенными поликлональными антителами против рибонуклеопротеина вируса бешенства в объеме 1,00 мл суспензии с концентрацией иммуноглобулинов G (Ig G) 5 мкг/мл. После иммобилизации антител при температуре 4±1°С в течение 18-20 часов лунки планшета подвергают трехкратному промыванию 1/15 М фосфатным буферным раствором (ФБР), открытые сайты связывания блокируют 1,0%-ной суспензией бычьего сывороточного альбумина (BSA) при температуре 37±1°С в течение 30 минут и вновь лунки промывают 1/15 М ФБР 5 раз. Процедуру подготовки планшета к работе с пробами осуществляют заранее, до проведения основного анализа. Иммобилизованные планшеты хранят в течение 6 месяцев при температуре 6±2°С
На первом этапе исследования проводят серологическую реакцию связывания рибонуклеопротеина вируса бешенства из исследуемых образцов (разведения положительного стандарта, отрицательный контроль, пробы). В качестве положительного стандарта используют суспензию рибонуклеопротеина вируса бешенства, который репродуцирован в суспензионной культуре клеток почки новорожденного сирийского хомячка (ВНК-21) с известной концентрацией РНП. Отрицательным контролем служит не инфицированная вирусом бешенства суспензия клеток ВНК-21 с концентрацией 3,0±0,2 млн клеток/мл. Применяют контрольную панель готовых разведений положительного стандарта с количествами вирусной РНК, эквивалентными следующим концентрациям рибонуклеопротеина вируса бешенства: 0,01, 0,05, 0,10, 0,20, 0,50, 1,00, 2,00, 3,00, 4,00, 5,00 мкг/мл. Данный диапазон разведений от 0,01 до 5,00 мкг/мл используют, исходя из особенностей процесса производства антирабических культуральных вакцин на этапе получения антигена. В лунки с сенсибилизированными специфическими антителами против рибонуклеопротеина вируса бешенства вносят по 2,0 мл суспензий исследуемых образцов и инкубируют при температуре 37±1°С в течение 30 минут. В результате серологической реакции на поверхности лунок формируется комплексы «рибонуклеопротеин вируса бешенства - специфичные антитела». Лунки отмывают от балластных компонентов с использованием 1/15 М ФБР 3 раза. Образовавшиеся иммунные комплексы ресуспендируют в 1,0 см3 1/15 М ФБР.
На следующем этапе анализа осуществляют выделение связанной иммунным комплексом РНК рибонуклеопротеина вируса бешенства в процессе лизиса белков и липопротеинов, очистки нуклеиновой кислоты от примесей и ее концентрирования. Данный метод выделения РНК вируса бешенства позволяет достичь высокой стандартизации в процедуре экстракции и получить из сырья для вакцины очищенные препараты нуклеиновой кислоты. Представленная ниже модифицированная методика адаптирована к процессу получения очищенного элюата РНК вируса бешенства и имеет отличия по количеству используемых компонентов от прописи Chomczynski Р. [11]. Лизис проводят с применением 5,0 мл раствора, содержащего 50% карболовой кислоты (рН<7,0) и 50% 6М ГТЦ, который смешивают с 1,0 см3 суспензии комплекса «рибонуклеопротеин вируса бешенства - специфичные антитела». Смесь инкубируют при температуре 20-22°С в течение 15-20 минут для полного диссоциирования полипептидных и нуклеопротеидных комплексов. При этом вирусная РНК сохраняет свою целостность благодаря высокой ингибирующей активности карболовой кислоты и ГТЦ в отношении РНКаз. Полученный лизат очищают от конгломератов с помощью центрифугирования при 12000 об/мин в течение 10 минут. Супернатант переносят в центрифужную пробирку с 2,0 мл трихлорметана и инкубируют содержимое в течение 5 минут с перемешиванием на вортексе. Смесь после инкубирования фракционируют при 13 500 об/мин в течение 10 минут при температуре 4-8°С. В результате центрифугирования происходит разделение содержимого пробирки на три фракции: 1) нижняя фракция, содержащая комплекс карболовой кислоты и трихлорметана со связанными липопротеинами и денатурированными полипептидами; 2) средняя фракция, включающая в свой состав белки и клеточную ДНК; 3) верхняя фракция, представляющая собой водный элюат РНК [12, 13]. Верхнюю фракцию полностью отбирают, не затрагивая остальные, и переносят элюат в новую центрифужную пробирку с 3,0 мл 100%-ого пропанола-2 для преципитации одноатомным спиртом с целью концентрирования РНК вируса бешенства. Смесь инкубируют в течение 10 минут при температуре 20-22°С, затем центрифугируют при 13 500 об/мин в течение 6 минут при температуре 20-22°С. Супернатант удаляют, оставляя осадок РНК вируса бешенства, к которому добавляют 2,0 мл 80%-ого пропанола-2. Содержимое перемешивают, инкубируют 3 минуты и осаждают при 14000 об/мин в течение 6 минут при температуре 20-22°С. Надосадок удаляют, осадок РНК высушивают в потоке воздуха комнатной температуры в течение 5 минут. К высушенному осадку добавляют 150 мкл деионизированной для получения элюата РНК вируса бешенства. Таким образом, получают по 150 мкл элюатов вирусной РНК.
На следующем этапе проводят спектральный анализ экстрактов РНК вируса бешенства, определяя поглощение аналитом монохроматического ультрафиолетового света, что позволяет оценить степень чистоты экстракта, который в последующем тестируют в реакции амплификации. Измерения спектральной поглощающей способности образцов проводят при длинах волны в диапазоне 205-325 нм и температуре 20-22°С. В выделенных экстрактах оценивают содержание остатков фосфолипидов, полисахаридов и ГТЦ, карболовой кислоты, полипептидов и крупных взвешенных частиц, определяя значения оптической плотности (OD, optical density) при 205, 235, 270, 280 и 320 нм, соответственно [14, 15]. Наибольшая адсорбция для нуклеиновой кислоты должна отмечаться при длине волны 260 нм. Элюат РНК считают свободным от примесей белка и карболовой кислоты, если OD260/OD280 (коэффициент экстинкции R1) находится в пределах 1,8-2,2 и оптимально составляет примерно 2,0. Более низкие значения R1 указывают на наличие ДНК, протеина и остатков карболовой кислоты в экстракте. Более высокие значения коэффициента R1 свидетельствуют о деградации РНК и наличии свободных рибонуклеотидов. Экстракт нуклеиновой кислоты вируса бешенства считают незагрязненным полисахаридами, если OD260/OD235 (коэффициент экстинкции R2) приближен к значению 2,000 [5]. При замещении 1% РНК на полисахаридные составляющие R2 снижается на 0,002. Значения коэффициента R2 большие 2,000 могут указывать на деградацию молекул РНК. Отсутствие взвеси крупных частиц в элюате подтверждается, если OD320 приближено к нулевому значению [16]. При несоответствии требованиям чистоты повторно проводят этапы серологического связывания и выделения РНК вируса бешенства из исходных материалов.
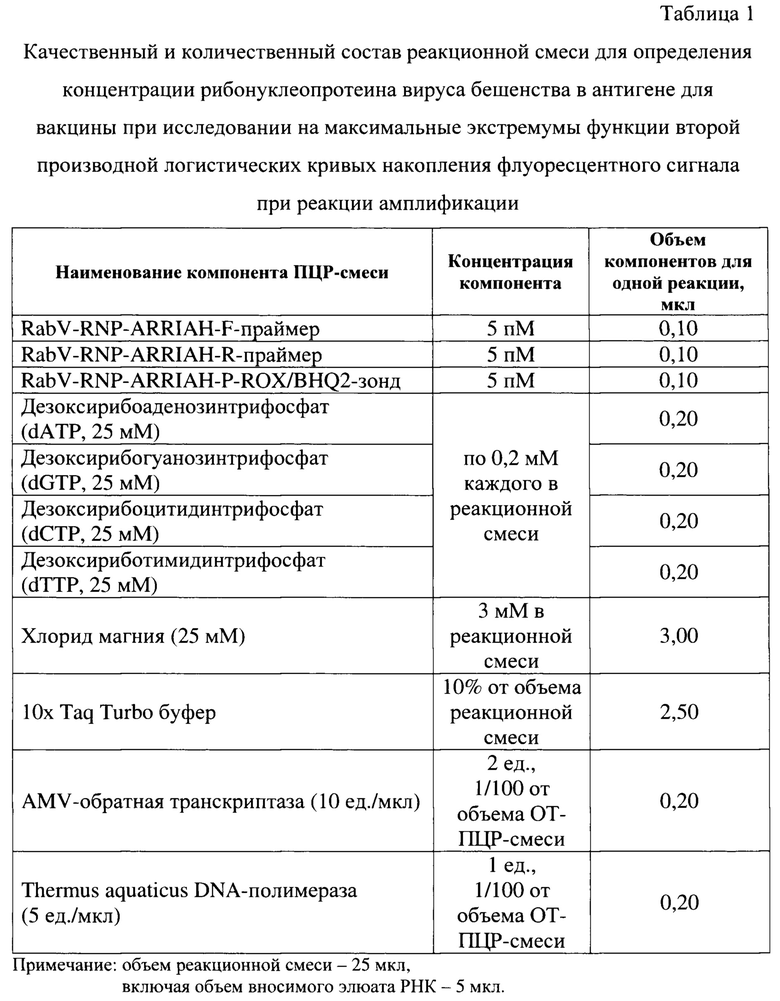
С полученными экстрактами РНК вируса бешенства проводят реверсию и реакцию амплификации. Для постановки реакции готовят реакционную смесь, рецептура которой представлена в таблице 1 и на фиг.1. На данном изображении представлены: А - прямой праймер, Б -обратный праймер - 2 формы - прямая (fov) и реверсивная комплементарная (rev)., В - зонд. В качестве олигонуклеотидов, гомологичных N-гену вируса бешенства, используют RabV-RNP-ARRIAH-F- (5'-TGGATACCGACAAGATTGTA-3'), RabV-RNP-ARRIAH-R-праймеры (5'-ATATAGATGATCAATCCGGGA-3') и RabV-RNP-ARRIAH-Р-ROX/BHQ2-зонд (5'-ROX-AAGCCTGAGATTATCGTGG-BHQ2-3') с концентрациями по 5 пМ на реакцию с внесением в реакционную смесь по 0,1 мкл. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с концентрацией каждого в реакционной смеси по 0,2 мМ. В качестве основы используют 10х Tag Turbo буфер («Евроген»), содержание которого составляет 10% от общего объема реакционной смеси [17], а также добавляют 3 мМ хлорида магния от объема реакционной смеси. В качестве катализаторов обратной транскрипции и реакции амплификации применяют AMV-ревертазу (10 е.a. («Promega») и Thermus aquaticus DNA-полимеразу (10 е.а.) («Promega»). Объем реакционной смеси компонентов для проведения одной реакции составляет 25 мкл, из них 5 мкл - объем элюата РНК вируса бешенства.
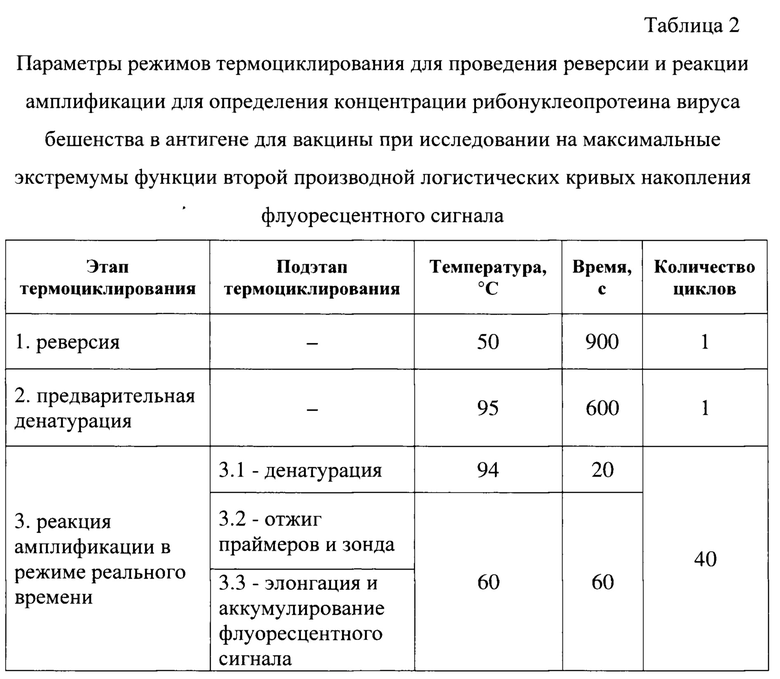
Постановку реакции осуществляют при температурных и временных параметрах, сведения о которых представлены в таблице 2. Реверсию РНК в комплементарную ДНК вируса бешенства проводят при температуре 50°С в течение 900 с за 1 цикл, предварительную денатурацию комплементарной ДНК - при температуре 95°С за 600 с в течение 1 цикла. Реакцию амплификации в режиме реального времени осуществляют в течение 40 циклов, каждый из которых складывается из 3 под этапов: «денатурации», проводимой при температуре 95°С в течение 20 с, а также объединенных под этапов «отжига праймеров и зонда» и «элонгации и аккумулирования флуоресцентного сигнала», осуществляемых при температуре 60°С за 60 с.
Процесс основан на использовании 5'-экзонуклеазной активности Thermus aquticus ДНК-полимеразы [17]. В отсутствии мишени флуорофор ROX и гаситель флуоресценции BHQ2 в составе RabV-RNP-ARRIAH-зонда сближены за счет максимального использования водородных связей между атомами Н, О и N олигонуклеотидов. Благодаря механизму флуоресцентно-резонансного переноса энергии свечение подавлено. За счет 5'-экзонуклеазной активности Thermus aquticus ДНК-полимеразы после отжига специфичных олигонуклеотидов происходит разрушение гибридизованного зонда и ампликона, наблюдается их пространственное разделение, что приводит к росту детектируемого сигнала [18, 19]. Увеличение уровня флуоресценции (Fl) пропорционально количеству образующихся продуктов реакции. Мониторинг сигнала в течение 40 циклов (С) ПЦР в режиме реального времени позволяет построить кинетическую кривую флуоресценции, которая задана функцией Fl = f (С).
Результаты реакции амплификации в режиме реального времени анализируют, оценивая и сравнивая графики накопления флуоресцентного сигнала по значениям максимальных критических точек Ср, определенных с помощью построения графика второй производной функции Fl = f (Ср). Величина Ср является важной характеристикой реакции, прямо пропорциональна количеству копий исходной матрицы РНК и соответственно концентрации рибонуклеопротеина вируса бешенства. Учитывая, что вторая производная функции f (Ср) (f ''(Ср)) непрерывна в некоторой окрестности точки Ср = Cp1 и задана на отрезке циклов амплификации [0; 40], существует определенный участок около точки Ср, для которого во всех координатах на оси О-Ср вторая производная функции f (Ср) будет отрицательна. Поскольку f ''(Ср) является первой производной от функции f '(Ср), то из условия (f '(Ср))'<0, следует, что f '(Ср) на некотором малом отрезке, содержащем точку Ср=Ср1 будет убывающей. Учитывая, что f '(Ср)=0, на участке при Ср<Cp1 первая производная функции f (Ср)>0, а при Ср>Ср1 получаем, что f '(Ср)<0. Иными словами, первая производная функции f (Ср) при переходе через точку Ср=Cp1 изменяет знак с «+» на «-», следовательно, в точке Cp1 функция, отражающая процесс накопления флуоресцентного сигнала, имеет максимальный экстремум. Таким образом, если график реакции амплификации в режиме реального времени представлен функцией Fl=f (Ср), f '(Ср)=0 и f ''(Ср)<0, то при условии, что Ср=Cp1 полученная функция имеет максимум в точке с аргументом Ср1, значение которой учитывают для установления зависимости между концентрацией вирионов вируса ящура и величиной Ср [20].
Преимущество использования второй производной в данном случае заключается в том, что при умножении функции кривой амплификации Fl=f (Ср) на любые множители, в том числе коэффициент пропорциональности а [19, 20], положение максимумов производных не меняется. Максимальный экстремум второй производной находится внутри экспоненциального участка графика накопления флуоресценции, то есть в зоне экспоненты, при анализе которой эффективность реакции амплификации не изменяется [20].
Математически определив значения Ср графиков второй производной для кривых, отражающих накопление флуоресцентного сигнала образцов с разной концентрацией рибонуклеопротеина, устанавливают зависимость между количеством РНП вируса бешенства в сырье для вакцины и максимальным экстремумом Ср, а также оценивают степень достоверности аппроксимации (R2). На основе разработанной модели рассчитывают значение концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины.
Пример 1. Получение положительного стандартного образца рибонуклеопротеина вируса бешенства штамма ВНИИЗЖ для разработки способа опосредованного определения концентрации вирусного рибонуклеопротеина в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени
На первом этапе исследования получали охарактеризованный препарат рибонуклеопротеина вируса бешенства, репродукция которого проводили в течение 48 ч в перевиваемой суспензионной клеточной линии почки сирийского хомячка ВНК-21. Полученную суспензию вируса бешенства после культивирования в объеме 10000 мл подвергали центрифугированию при 1500 g в течение 10 мин. Супернатант удаляли, оставляя осадок клеток объемом 50 мл, в которых содержится РНП. Инфицированные клетки подвергали лизированию, добавляя 50,0 мл ледяной стерильной деионизированной воды, содержащей 25 мкл/мл апротинина (Sigma-Aldrich).
Полученную суспензию инкубировали в течение 1 ч при температуре 4°С и осветляли центрифугированием при 1000 g при той же температуре в течение 20 минут. Супернатант собирали, и процедуру повторяли дважды.
Полученные надосадки из каждого раунда смешивали и центрифугировали при 12000 g и температуре 4,0±0,5°С в течение 8 минут. Супернатант в объеме 50 мл собирали в другую пробирку. Таким образом, получали концентрированный антиген, содержащий рибонуклеопротеин вируса бешенства. Полученный РНП очищали ультрацентрифугированием в ступенчатом градиенте хлорида цезия (CsCl) с концентрациями 17, 18, 19, 20, 22, 24, 26% в соответствии с протоколом, описанным Dietzschold [16]. Центрифужные пробирки разделяли на 7 зон по 4 мл. Вносили 4 мл 26%-ного раствора хлористого цезия в 6 пробирок. Остальные растворы наслаивали с помощью капилляра. Суспензию РНП вируса бешенства разделяли на 6 пробирок и вносили на поверхность градиента. Пробирки уравновешивали. Центрифугирование проводили при 65000 g в течение 2 ч и температуре 15°С. Опалесцирующий слой рибонуклеопротеина вируса бешенства в области 19-21% хлористого цезия (соответствуют плавучей плотности РНП) отбирали в отдельный флакон.
Полученный раствор вносили в центрифужные пробирки, доливали объем пробирок с помощью фосфатно-солевого буферного раствора (ФСБ). Для очищения от остатков хлористого цезия суспензию РНП осаждали с помощью центрифугирования при 60000 g в течение 1 ч при 15°С. Полученный осадок ресуспендировали в 5 мл ФСБ. Иными словами, получали очищенный концентрат рибонуклеопротеина.
В готовой суспензии концентрата определяли содержание рибонуклеопротеина вируса бешенства в количественном сэндвич-варианте ИФА [9]. Проводили вертикальный белковый электрофорез в полиакриламидном геле, содержащем додецилсульфат натрия, для оценки степени чистоты положительного контрольного препарата. По данным количественного сэндвич-варианта ИФА концентрация РНП составляла 2540 мкг/мл. Исходя из результатов вертикального электрофореза, следовало, что получен чистый препарат РНП, включающий в свой состав нуклеопротеин, фосфопротеин и L-белок вируса бешенства с молекулярным весом 57, 260 и 190 kDa, соответственно. Из данного препарата с помощью 1/15 М раствора ФСБ приготовили положительный контрольный образец с концентрацией РНП 5,0 мкг/мл. Иными словами, получили охарактеризованный положительный контрольный образец, который вместе с исследуемыми пробами использовали в дальнейшей работе.
Пример 2. Определение математической зависимости между концентрацией рибонуклеопротеина вируса бешейства штамма ВНИИЗЖ в сырье для антирабической вакцины и значением максимального экстремума графиков второй производной для логистических кривых реакции амплификации в режиме реального времени
Готовили панель положительных контролей со следующими концентрациями РНП: 0,01, 0,05, 0,10, 0,20, 0,50, 1,00, 2,00, 3,00, 4,00, 5,00 мкг/мл. Отрицательным контролем служила не инфицированная вирусом бешенства суспензия клеток ВНК-21 с концентрацией 3,0±0,2 млн клеток/мл. В лунки с сенсибилизированными специфическими антителами против рибонуклеопротеина вируса бешенства вносили по 2,0 мл суспензий исследуемых образцов и инкубировали при температуре 37±1°С в течение 30 минут. В результате серологической реакции на поверхности лунок формировались комплексы «рибонуклеопротеин вируса бешенства - специфичные антитела». Лунки отмывали от балластных компонентов с использованием 1/15 М ФБР 3 раза. Образовавшиеся иммунные комплексы ресуспендировали в 1,0 см3 1/15 М ФБР.
После этапа серологического связывания РНП вируса бешенства штамма ВНИИЗЖ из полученных комплексов выделяли РНК, как описано выше. В результате получали по 150 мкл экстрактов вирусной РНК каждого разведения положительного стандарта и отрицательного контроля. На следующем этапе исследования с помощью спектрального анализа в излучении ультрафиолетового света проводили оценку степени чистоты полученных элюатов РНК вируса бешенства штамма ВНИИЗЖ из разведений положительного стандарта, соответствующих следующим концентрациям рибонуклеопротеина: 0,01, 0,10, 0,50, 1,0, 2,0, 3,0, 4,0, 5,0 мкг/см3. Анализ проводили, как описано выше, с помощью спектрофотометра и записывали данные с помощью компьютерной программы Specrtrum v. 5.0 (Фиг. 2).
По результатам анализа положительных стандартов в указанных выше разведениях выявили, что значения OD205-259 и OD261-325 не превышают OD260, что является признаком высокой степени чистоты полученных элюатов РНК (n=3). Из данных спектрального исследования стандартов отмечали отсутствие выраженных пиков на графиках (фиг.2) при длинах волны 205, 235, 270, 280 и 320 нм, что свидетельствует о практически полном отсутствии загрязнения экстрактов РНК примесями фосфолипидов, полисахаридов и остатков ГТЦ, карболовой кислоты, полипептидов и крупных конгломератов, соответственно. Значения коэффициентов экстинкции (R1) для стандартов приближены к норме 2,000 (R1=1,998-2,000), что подтверждает отсутствие ДНК и наличие лишь следовых количеств примесей белка и остатков карболовой кислоты. Деградации нуклеиновой кислоты и наличия свободных нуклеотидов в элюатах не наблюдалось, так как R1 не превышал 2,000. Экстракты вирусной РНК разведений положительного стандарта не загрязнены полисахаридами и ГТЦ, поскольку значения коэффициента экстинкции (R2) приближены к норме 2,000 и соответствовали 2,000-2,002. Учитывая, что при замещении 1% РНК на углеводы значение R2 уменьшается на 0,002, в полученных экстрактах наличие полисахаридных примесей не было выявлено. Степень разрушения РНК в экстрактах составляла 1,0% ((2,002-2,000)/0,002), что является допустимым. Таким образом, экстракты РНК вируса бешенства, выделенные из положительных стандартных образцов, характеризовались высокой степенью чистоты [14].
После оценки степени чистоты экстрактов выделенных РНК вируса бешенства, полученных из разведений положительного стандарта проводили реверсию РНК вируса бешенства в комплементарную ей ДНК, как описано выше. Полученные данные анализировали с помощью программного обеспечения «Rotor-Gene FRT-Manager», которое позволяет строить логистические кривые накопления флуоресцентного сигнала в режиме реального времени на протяжении заданного количества циклов амплификации (С=40). Применяя технологии компьютерной программы «Maxima» (или аналога), проводили построение графиков первой и второй производной для полученных элюатов РНК вируса бешенства каждого разведения положительного стандарта с определенными концентрациями рибонуклеопротеина вируса бешенства и рассчитывали средние значения максимальных экстремумов графиков второй производной логистических кривых реакции амплификации с проекцией на ось абсцисс О-С (n=3) (Фиг. 3, 4). На изображении снизу вверх отражены графики для разведений экстрактов, соответствующих следующим концентрациям вирионов: 0,01, 0,10, 0,50, 1,00, 2,00, 3,00, 4,00, 5,00 мкг/мл).
Результаты эксперимента по представлению системы параллельной оценки величины максимальных экстремумов графиков второй производной для кривых реакции амплификации в режиме реального времени (Ср) и концентрации рибонуклеопротеина вируса бешенства (СРНП ВБ) в контрольных образцах представлены в таблице 3. Значения Ср для всех разведений положительного стандарта с концентрациями РНП вируса бешенства от 0,01 до 5,00 мкг/мл находились в диапазоне от 23,01±0,02 до 13,02±0,03, соответственно. При исследовании отрицательного контроля накопления флуоресцентного сигнала не наблюдалось, что подтверждало отсутствие РНК вируса бешенства в данном образце. В представленных исследованиях р-уровень значимости меньше 0,01 для контрольных образцов положительного стандарта с концентрациями РНП вируса бешенства от 0,01 до 0,10 мкг/мл и р<0,005 для образцов с концентрациями рибонуклеопротеина вируса бешенства от 0,20 до 5,00 мкг/мл. Зависимость концентрации РНП вируса бешенства и значений максимальных экстремумов графиков второй производной для логистических графиков реакции амплификации в реальном времени представлена на фиг.5 и отражена в виде квадратичной функции СРНП ВБ = 0,0123(СР)2 - 0,0725СР+8,0129 с высокой достоверностью аппроксимации (R2=0,9952). Таким образом, выявлено существование зависимости концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины и максимального экстремума графика второй производной для логистической кривой реакции амплификации в режиме реального времени.
Контрольные образцы параллельно исследовали с помощью прототипного метода ИФА [10, 19]. Полученные данные отражены в таблице 3, из которой следует, что степень различий результатов анализа положительных стандартов разработанным способом и истинными значениями положительных стандартов составляет 0,0-4,8%. Различия в определении содержания рибонуклеопротеина вируса бешенства положительных стандартов прототипным способом по сравнению с истинными значениями для образцов с концентрациями РНП от 0,01 до 5,00 мкг/мл составляли 0,0-4,7%. Степень различия в определении концентрации РНП вируса бешенства разработанным способом и в ИФА для образцов с концентрациями рибонуклеопротеина от 0,01 до 0,20 мкг/мл составляли 0,00-4,8%, с содержанием от 0,50 до 5,00 мкг/мл - 0,00-2,00%. В отрицательном контроле РНК вируса бешенства по N-гену не была выявлена с помощью разработанного способа и не обнаружено наличие антигена вируса бешенства в образце по данным ИФА. Таким образом, разработанный способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени позволяет с высокой степенью достоверности исследовать антиген вируса бешенства, как сырья для вакцины с концентрациями от 0,01 до 5,00 мкг/мл.
Пример 3. Определение концентрации рибонуклеопротеина вируса бешенства штамма ВНИИЗЖ в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени
Для анализа использовали четыре референтные суспензии культурального вируса бешенства штамма ВНИИЗЖ, содержащие рибонуклеопротеин по данным ИФА 0,08; 0,65; 1,25 и 2,45 мкг/мл и высокое содержание клеточного белка 7,21; 8,52; 8,66 и 9,11 мг/мл, соответственно (по данным определения концентрации общего белка по формуле Калькара:
содержание белка = 1,45 ⋅ А280 - 0,74 ⋅ А260 (мг/мл),
где А280 и А260- значения экстинкции при длинах волны 280 и 260 нм, соответственно) [18].
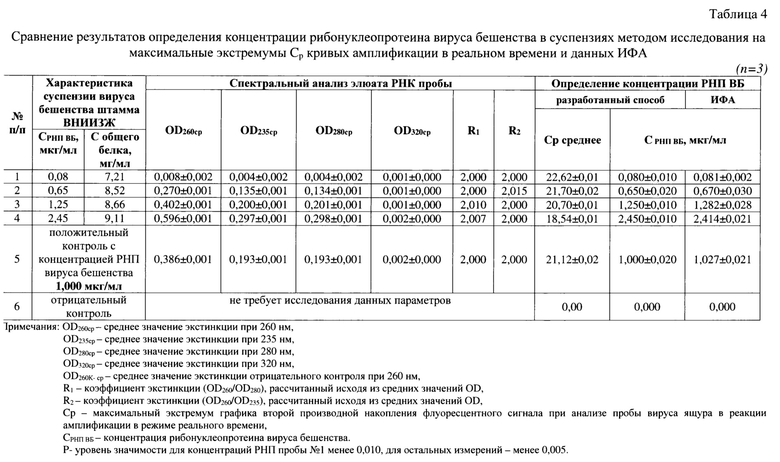
В качестве положительного контроля (К+) применяли суспензию рибонуклеопротеина вируса бешенства с концентрацией РНП 1,00 мкг/мл. Отрицательным контролем (К-) служила не инфицированная вирусом бешенства суспензия клеток ВНК-21 с концентрацией 3,00±0,2 млн клеток/мл. Испытуемую пробу и контрольные образцы исследовали в трех повторностях. Этап серологического специфического связывания вируса бешенства, выделения из них РНК, оценки степени чистоты элюатов, постановку реверсии и реакции амплификации в режиме реального времени проводили, как описано выше. Результаты исследования представлены в таблице 4.
Из данных спектрограмм элюатов РНК четырех исследуемых проб вируса бешенства штамма ВНИИЗЖ (табл.4) видно, что значения экстинкции при длинах волны 205-259 и 261-325 нм не превышали OD260: проба 1 - 0,001-0,007<0,008 и 0,007-0,001<0,008, проба 2 - 0,001-0,269<0,270 и 0,268-0,001<0,270, проба 3 - 0,001-0,401<0,402 и 0,399-0,001<0,402, проба 4 - 0,001-0,594<0,596 и 0,595-0,002<0,596, иными словами, элюат РНК характеризуется высокой степенью чистоты. Для положительного контроля спектральный анализ показал, что значения экстинкции при длинах волны 205-259 и 261-325 нм также не превышали OD260: 0,002-0,385<0,386 и 0,384-0,002<0,386. Экстракт не контаминирован примесями фосфолипидов, полисахаридов и остатков ГТЦ, карболовой кислоты, полипептидов и крупных конгломератов, поскольку отсутствуют выраженные пики при λ 205, 235, 270, 280 и 320 нм, соответственно. Коэффициенты экстинкции (R1) для проб №1, 2, 3, 4 и контроля положительного составляют 2,000; 2,000; 2,010; 2,007; 2,000 что приближено к значению нормы 2,000 и означает высокую степень чистоты элюата РНК вируса бешенства, практически полное отсутствие белка и остатков карболовой кислоты после этапа выделения РНК. Коэффициенты экстинкции (R2) для проб №1, 2, 3, 4 и контроля положительного соответствовали значениям 2,000; 2,015; 2,000; 2,000 и 2,000 (норма), что обуславливало высокую чистоту препарата. Таким образом, полученные экстракты РНК удовлетворяли требованиям чистоты [5, 17].
Полученные элюаты РНК вируса бешенства исследовали с помощью разработанного метода сравнения максимальных экстремумов графиков второй производной для кривых реакции амплификации в режиме реального времени. Результаты исследования представлены в таблице 4, из которой следует, что для проб №1, 2, 3, 4 Ср среднее составляет 22,62±0,01, 21,70±0,02, 20,70±0,01, 18,54±0,01, Ср среднее К+ - 21,12±0,02. Для отрицательного контроля график не формируется. Пользуясь разработанной моделью:
СРНП ВБ = 0,0123(СР)2- 0,725СР + 8,129,
где, СРНП ВБ - концентрация рибонуклеопротеина вируса бешенства,
Ср - значение максимального экстремума графика второй производной для кривой реакции амплификации в режиме реального времени, определили значения концентрации РНП вируса бешенства. Для положительного контроля концентрация РНП составила 1,00±0,01 мкг/мл, что сочетается с эталонным значением (1,00 мкг/мл). В отрицательном контроле вирус бешенства не обнаружен. Значения концентраций РНП вируса бешенства в пробах №1, 2, 3, 4 составили по данным разработанного способа 0,08±0,01, 0,65±0,02, 1,24±0,01, 2,44±0,01 мкг/мл, соответственно. Полученные значения коррелируются с данными ИФА и составляли для проб №1, 2, 3, 4 - 0,081±0,002; 0,670±0,003; 1,282±0,028; 2,414±0,021 мкг/мл, соответственно. Таким образом, разработанный способ исследования максимальных экстремумов графиков второй производной для кривых реакции амплификации в режиме реального времени позволяет опосредованного определять концентрацию РНП вируса бешенства в сырье для антирабической вакцины с высоким количеством балластного клеточного белка.
Пример 4. Определение степени достоверности метода опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени
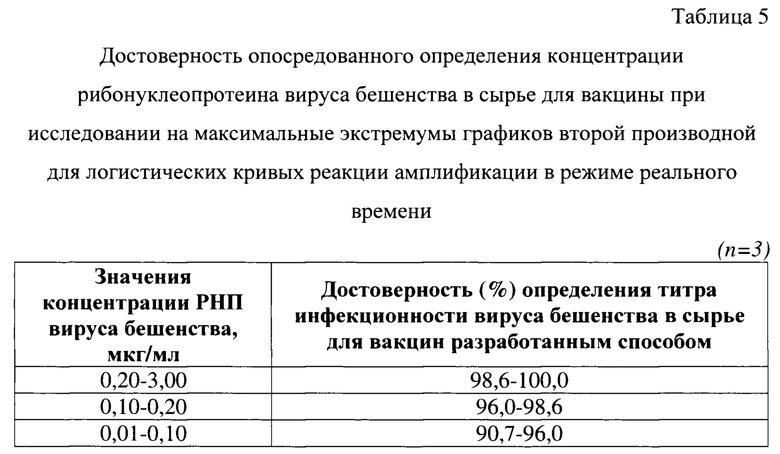
Для исследования использовали 344 суспензии культурального вируса бешенства производственного штамма ВНИИЗЖ с концентрациями рибонуклеопротеина от 0,01 до 5,00 мкг/мл (20 проб с концентрациями 0,01-0,10 мкг/мл, 20 проб с концентрациями 0,10-0,20 и 304 пробы с концентрациями 0,20-5,00 мкг/мл). В качестве положительного контроля применяли суспензию культурального вируса бешенства штамма ВНИИЗЖ с концентрацией РНП 1,00 мкг/мл. В качестве отрицательного контроля применяли суспензию клеток ВНК-21, не зараженную вирусом бешенства. Испытуемые пробы и контрольные образцы исследовали в трех повторностях по методике, представленной выше. Результаты исследования представлены в таблице 5.
Спектральный анализ 344 элюатов РНК проб вируса бешенства показал, что средние значения экстинкции при длинах волны 205-259 и 261-325 нм не превышали OD260, иными словами, элюаты РНК характеризовались высокой степенью чистоты. Экстракты не были контаминированы примесями фосфолипидов, полисахаридов и остатков гуанидинизотиоцианата, полипептидов и крупных конгломератов, поскольку на спектрограммах отсутствовали выраженные пики при λ 205, 235, 270, 280 и 320 нм, соответственно. Коэффициент экстинкции R1 для проб вируса бешенства составлял 1,993-2,000, что приближено к значению нормы 2,000 и означало высокую степень чистоты элюатов вирусной РНК, практически полное отсутствие белка. Коэффициент экстинкции R2 для проб вируса бешенства соответствовал значениям 1,997-2,002, что близко к норме 2,000 и обуславливало высокую чистоту элюатов. Таким образом, полученные препараты удовлетворяли требованиям чистоты [11, 14, 17], и их можно использовать для дальнейших исследований.
С полученными экстрактами РНК вируса бешенства проводили реверсию и реакцию амплификации в режиме реального времени. С помощью разработанного способа опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени получили данные, результат обработки которых отражен в таблице 5. Интерпретацию результатов проводили, пользуясь разработанной функцией второй степени, представленной выше. Для положительного контроля значение концентрации РНП вируса бешенства по данным разработанного способа полностью соответствовало истинному 1,00 мкг/мл. Для отрицательного контроля логистические кривые не были сформированы, что означало отсутствие вируса бешенства в данном образце.
Выявили, что данные, полученные с помощью разработанного способа, коррелировали с методом ИФА (исходные данные) на 90,7-96,0% для концентраций 0,01-0,10 мкг/мл (n=20), на 96,0-98,6% для 0,10-0,20 мкг/мл (n=20), на 98,6-100,0% для концентраций 0,20-5,00 мкг/мл (n=304). Полученные результаты свидетельствовали о высокой степени точности разработанного способа. Таким образом, разработанный способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени характеризуется высокой степенью достоверности результатов.
Пример 5. Определение аналитической чувствительности разработанного способа опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени
Для определения аналитической чувствительности разработанного способа опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени использовали суспензию культурального вируса бешенства штамма ВНИИЗЖ с концентрацией рибонуклеопротеина 1,00 мкг/мл. Подготавливали серию разведений данного стандарта с концентрациями РНП: 1,00; 0,20; 0,05; 0,01; 0,005 мкг/мл. Испытуемые контрольные образцы исследовали в пяти повторностях. Анализ проводили по методике, представленной выше. Результаты исследования представлены в таблице 6, в которой отражен сравнительный анализ определения концентрации РНП вируса бешенства разработанным способом в сравнении прототипным способом.
В результате исследования было установлено, что аналитическая чувствительность способа составляла 0,01 мкг/мл с достоверностью 90,9-100%. При концентрации 0,005 мкг/мл значения Ср определяются, однако достоверность снижается до 20-80%.
Таким образом, аналитическая чувствительность разработанного способа опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени составляла 0,01 мкг/мл.
Основным преимуществом предлагаемого изобретения является возможность определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины, содержащем высокое количество клеточного белка и количеством РНП от 0,01 мкг/мл. Предлагаемое изобретение сочетает осуществление специфического связывания антигена с последующим проведением молекулярно-биологического исследования суспензии культурального вируса бешенства, что повышает чувствительность метода. В предлагаемом изобретении между концентрацией рибонуклеопротеина вируса бешенства и значением максимального экстремума графика второй производной кривой накопления флуоресцентного сигнала реакции амплификации в режиме реального времени установлена зависимость, которая представлена в виде квадратичной функции СРНП ВБ = 0,0123(СР)2 - 0,0725СР + 8,0129 с высокой достоверностью аппроксимации R2=0,9952. Разработанная модель дает возможность определить значение концентрации рибонуклеопротеина вируса бешенства в сырье для производства вакцины с высокой достоверностью.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени»
1. European Pharmacopoeia 7.0. (2012а). Monograph 0451: Rabies vaccine (live, oral) for foxes. European Directorate for the Quality of Medicines and HealthCare (EDQM), Council of Europe, Strasbourg, France.
2. ICTV (International Committee on Taxonomy of Viruses). URL: http://www.ictv online. org/virusTaxonomy. asp ?bhcp=1 (Дата обращения: 12.12.2020).
3. Badrane H., Bahloul C, Perrin P. & Tordo N. (2001). Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J. Virol., 75, 3268-3276.
4. Rupprecht C.E., Barrett J., Briggs D., Cliquet F. (2008). Can rabies be eradicated? Dev. Biol. (Basel), 131, 95-121.
5. OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 7th ed. Paris. - 2018. - Ch. 2.1.17.
6. World Health Organization (2007). Oral Vaccination of Dogs against Rabies. WHO, Geneva, Switzerland. WU X., SMITH T.G. & RUPPRECHT C.E.
(2011). From brain passage to cell adaptation: the road of human rabies vaccine development. Exp.Rev. Vaccines, 10, 1597-1608.
7. Warner C.K., Whitfield S.G., Fekadu M. & Ho H. (1997). Procedures for reproducible detection of rabies virus antigen mRNA and genome in situ in formalin-fixed tissues. J. Virol. Methods, 67, 5-12.
8. Fooks A.R., Johnson N., Freuling CM. (2009). Emerging technologies for the detection of rabies virus: challenges and hopes in the 21st century. PLoS Negl. Trop.Dis., 3, e530.
9. Сухарьков Андрей Юрьевич. Разработка методов оценки оральной антирабической вакцинации животных: диссертация... кандидата биологических наук: 03.02.02 / Сухарьков Андрей Юрьевич. [Место защиты: Федеральное государственное бюджетное учреждение «Федеральный центр охраны здоровья животных»]. - Владимир, 2014. - 141 с.
10. Иммунохимический анализ в лабораторной медицине / В. В. Долгов. - М.-Тверь: ООО «Издательство «Триада», 2015. - С. 34-38. - ISBN 978-5-94789-695-4.
11. Chomczynski P., Sacchi N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nat Protoc. 1 (2). - 2006. - P. 581-585.
12. Peirson S.N. RNA extraction from mammalian tissues / S.N. Peirson, J.N. Butler// Methods in Molecular Biology. - 2007. - Vol.362. - P. 315-327.
13. Peirson S.N., Butler J.M., Foster R.G. Experimantal validation of novel and conventional approaches to quantitative real-time PCR data analysis // Nucleic Acids Res. - 2003. - V. 31:e.73.
14. Glasel J. Validity of nucleic acid purities monitored by 260nm/280nm absorbance ratios // BioTechniques. - 1995. - Vol.18 (1). - P. 62-63.
15. The Analysis of DNA or RNA using Its Wavelengths: 230 nm, 260 nm, 280 nm. Bioteachnology.com (дата обращения 02.06.2019). http://bioteachnology.com/dna/analysis-dna-rna-wavelengths-230-260-280-nm
16. Биохимия белков. URL: http://www.bio.bsu.by/biohirn/files/proteins.pdf (Дата обращения: 14.01.2021).
17. Livak K.J. SNP genotyping by the 5'-nuclease reaction // Methods Mol. Biol., 2003. - V. 212. - P. 129-147.
18. Holland P.M. Detection of specific PCR product by utilizing the 5'-3' exonuclease activity of Thermus aquaticus DNA polymerase / P.M. Holland, R.D. Abramson, R.Watson, D.H. Gelfand // Proc. Natl Acad Sci USA, 1991. - V. 88. -P. 7276-7280.
19. Liu W., Saint D.A. Validation of a quantitative method for real-time PCR kinetics // Biochem. Biophys. Res. Commun. - 2002. - V. 294. - P. 347-353.
20. Rutledge R.G. Mathematics of quantitative kinetic PCR and the application of standard curves / R.G. Rutledge, C. Cote // Nucleic Acide Res. -2004. - V. 31, N. 16. - e.93. P. 1-6.


Изобретение относится к области биотехнологии и производству противоящурных вакцин. Описан способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени. На основании рассчитанного значения максимального экстремума графика второй производной для кривой накопления флуоресцентного сигнала (Ср) определяют концентрацию рибонуклеопротеина вируса бешенства (СРНП ВБ) с применением разработанной математической модели второй степени вида: СРНП ВБ = 0,0123(Ср)2-0,0725Ср+8,0129. Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности определять концентрацию рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины на основе сочетания серологического специфического связывания вируса бешенства и молекулярно-биологического исследования с последующим расчетом значений максимальных экстремумов графиков второй производной для кривых накопления флуоресцентного сигнала в реальном времени и расчетом концентрации аналита с использованием разработанной квадратичной модели 2 з.п. ф-лы, 5 ил., 6 табл., 5 пр.
1. Способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени, отличающийся тем, что в качестве гомологичных для генома вируса бешенства оригинальных олигонуклеотидов используют RabV-RNP-ARRIAH-F - праймер с нуклеотидной последовательностью 5'-TGGATACCGACAAGATTGTA-3', RabV-RNP-ARRIAH-R - праймер с нуклеотидной последовательностью 5'-ATATAGATGATCAATCCGGGA-3' и RabV-RNP-ARRIAH-P-ROX/BHQ2 - зонд с нуклеотидной последовательностью 5'-ROX-AAGCCTGAGATTATCGTGG- BHQ2-3'; включающий следующие стадии:
- внесение в лунки с сенсибилизированными специфическими антителами против рибонуклеопротеина вируса бешенства суспензий образцов и их инкубирование;
- специфическое связывание рибонуклеопротеина вируса бешенства с применением очищенных поликлональных антител сывороток крови морских свинок против рибонуклеопротеина вируса бешенства с формированием иммунных комплексов рибонуклеопротеин вируса бешенства - специфичные антитела;
- выделение вирусной РНК из иммунных комплексов с использованием метода экстракции смесью карболовой кислоты, гуанидинизотиоцианата (ГТЦ), трихлорметана и пропанола-2;
- спектральное исследование степени чистоты элюатов РНК вируса бешенства;
- проведение реверсии РНК в комплементарную ДНК вируса бешенства и реакции амплификации в режиме реального времени;
- исследование логистических кривых накопления флуоресцентного сигнала для проб с построением графиков первой и второй производных и определение значений максимальных экстремумов (Ср) графиков второй производной для кривых реакции амплификации в режиме реального времени;
- расчет концентрации рибонуклеопротеина вируса бешенства в пробах сырья для антирабической вакцины с применением математической модели:
СРНП ВБ = 0,0123(Ср)2 - 0,0725Ср + 8,0129,
где СРНП ВБ означает концентрацию рибонуклеопротеина вируса бешенства, Ср означает значение максимального экстремума графика второй производной для логарифмической кривой накопления флуоресцентного сигнала, с достоверностью аппроксимации R2- 0,9952.
2. Способ по п. 1, отличающийся тем, что смесь компонентов для проведения реверсии и реакции амплификации в режиме реального времени включает в свой состав следующие реагенты: RabV-RNP-ARRIAH-F-, RabV-RNP-ARRIAH-R-праймеры и RabV-RNP-ARRIAH-P-ROX/BHQ2-зонд с концентрациями по 5 пМ на реакцию, дезоксирибонуклеозидтрифосфаты - по 0,2 мМ, 10х Tag Turbo буфер, хлорид магния - 3 мМ, AMV-ревертаза - 10 е.а., Thermus aquaticus DNA-полимераза - 10 е.а.
3. Способ по п. 1, отличающийся тем, что молекулярно-биологический анализ проводят с соблюдением следующих температурных и временных режимов:
- реверсия: 50°С в течение 900 с,
- предварительная денатурация: 95°С в течение 600 с,
- реакция амплификации: денатурация - 95°С в течение 20 с, отжиг праймеров и зонда, элонгация и аккумулирование флуоресцентного сигнала -60°С в течение 60 с.
| Способ спектрометрического определения концентрации рибонуклеопротеина вируса бешенства по оценке количества молекул вирусной РНК в сырье для антирабических вакцин | 2020 |
|
RU2748475C1 |
| СПОСОБ ВЫЯВЛЕНИЯ АНТИГЕНА ВИРУСА БЕШЕНСТВА В ФОРМАЛИНИЗИРОВАННЫХ ПРОБАХ ГОЛОВНОГО МОЗГА ЖИВОТНЫХ | 2004 |
|
RU2266755C1 |
| CN 111560466 A, 21.08.2020. | |||

