Изобретение относится к области биотехнологии и производству антирабических вакцин, а именно к способу определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабических вакцин с применением спектрометрического метода измерения количества молекул РНК, выделенных из полных частиц вируса бешенства, и разработанной линейной модели зависимости числа молекул вирусной РНК от концентрации рибонуклеопротеина вируса бешенства.
Бешенство занимает первоочередное место в ряду вирусных болезней человека и животных, является одним из самых опасных зоонозов, вызывая поражение центральной нервной системы, энцефаломиелиты, параличи с неизбежным летальным исходом [1]. Возбудитель бешенства принадлежит к порядку Mononegavirales, семейству Rhabdoviridae, роду Lyssavirus, виду Rabies lyssavirus [2]. Вирионы вируса бешенства имеют пулевидную форму, длиной ≈180 нм, диаметром ≈75 нм. На наружной поверхности полных вирусных частиц имеются выступы в виде шипов длиной 10 нм, которые прикреплены к двуслойной липидной оболочке [1]. Геном вируса бешенства представлен несегментированной одноцепочечной негативной спиральной РНК длиной около 12000 н.о., который кодирует 5 основных белков: нуклеопротеин (N-белок), фосфопротеин (Р-белок), матриксный белок (М-белок), гликопротеин (G-белок), РНК-зависимую РНК-полимеразу (L-белок). Снаружи РНК покрыта нуклеопротеином, в результате чего формируется нуклеокапсид. Рибонуклеопротеин (РНП) состоит из геномной РНК, N-белка, Р-белка и L-белка. Нуклеокапсид имеет спиральный тип симметрии. Между G- и L-цистронами располагается псевдоген (ψ-фрагмент) [2].
Заболевание приводит к значительным экономическим затратам, которые связаны с гибелью животных, ликвидацией последствий вспышек болезни, проведением профилактических и карантинных мероприятий, регулированием численности диких плотоядных животных, отловом бродячих кошек и собак и осуществлением лабораторных исследований по постановке диагноза [3]. Система мер по борьбе и профилактике заболевания предусматривает иммунизацию восприимчивых животных, а также контроль уровня напряженности поствакцинального иммунитета [4].
В процессе промышленного производства антирабических вакцин особое внимание уделяют концентрации рибонуклеопротеина, который существенно влияет на иммуногенную активность вакцинных препаратов [1]. В связи с этим каждую партию сырья исследуют на определение концентрации рибонуклеопротеина, применяя колориметрический метод Мериона Брэдфорда и/или прямой сэндвич-вариант реакции иммуноферментного связывания (ИФА). Метод Брэдфорда основан на реакции красителя Coomassie Brilliant Blue G-250 с аргинином и гидрофобными аминокислотными остатками. Связанная форма имеет голубую окраску с максимумом поглощения при 595 нм. Значение абсорбции раствора при этой длине волны пропорционально количеству белка в растворе. Однако данный метод имеет ряд ограничений: невысокая аналитическая чувствительность анализа (не менее 2 мкг/мл), невысокая специфичность метода [5]. Метод прямого сэндвич-варианта ИФА хорошо зарекомендовал себя, однако он также имеет ряд ограничений. А именно:
1) возможности увеличения чувствительности метода ограничиваются фоном анализируемого соединения (т.е. его наличием не только в анализируемом образце, но и в используемых реактивах и растворителях), субстратной специфичностью фермента и аффинностью антител;
2) к ограничениям ИФА относится наличие в тестируемых образцах кофакторов, ингибиторов и стимуляторов активности ферментов;
3) ИФА не позволяет разделять нативные белки и их биологические неактивные фрагменты, сохранившие антигенные детерминанты;
4) возможное изменение каталитической активности фермента при его конъюгировании с антигеном;
5) применимость ИФА лишь к хорошо изученным системам, где используются очищенные антигены и высокоспецифические антитела [1].
В связи с этим целесообразно провести поиск способа определения концентрации рибонуклеопротеина вируса бешенства в вируссодержащем сырье для вакцины на основе альтернативного метода.
Задачей настоящего изобретения являлась разработка высокочувствительного, специфичного, быстрого способа определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины с целью устранения вышеуказанных недостатков.
Данная задача была решена благодаря созданию нового способа определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины с применением спектрометрического метода анализа РНК. С помощью данного метода предложена модель определения количества молекул РНК, выделенных из полных частиц вируса бешенства, вида:
NРНК bб=4,3734109×K×(OD260пробы-OD260К--OD320пробы),
где: K - кратность разведения элюата,
OD260 - значение экстинкции элюата РНК вируса бешенства при длине волны 260 нм,
О260К _ значение экстинкции отрицательного контроля при длине волны 260 нм,
OD320 пробы - значение экстинкции элюата РНК вируса бешенства при длине волны 320 нм.
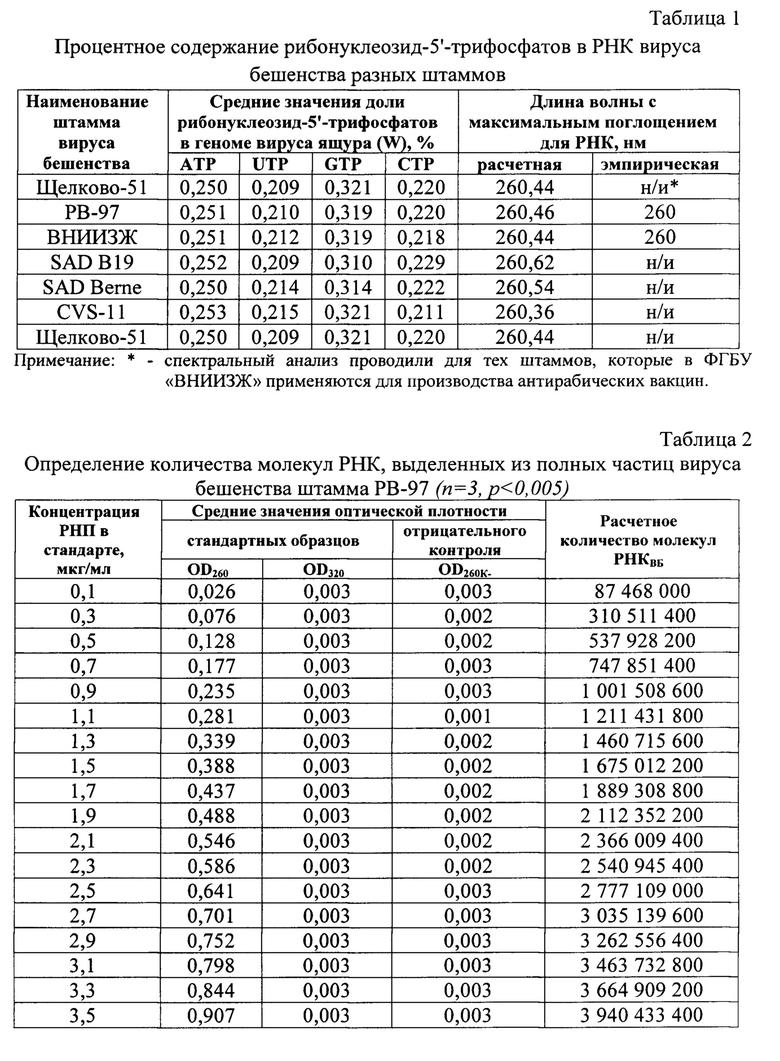
С применением разработанного способа между значениями концентрации рибонуклеопротеина вируса бешенства (СРНП Вб) и количеством молекул РНК, экстрагированных из полных вирусных частиц (NPHКИБ), выявлена зависимость, отраженная в виде линейной функции СРНП ВБ=9×10-10×NРНКВБ+0,022 с высокой достоверностью аппроксимации R2=0,9998. Предложенная модель позволяет оценивать концентрацию рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины.
Сущность изобретения отражена на графических изображениях.
Фиг. 1 - Зависимость концентрации рибонуклеопротеина вируса бешенства от количества молекул РНК, экстрагированной из полных частиц после иммунного захвата (количество молекул выражено в экспоненциальном формате);
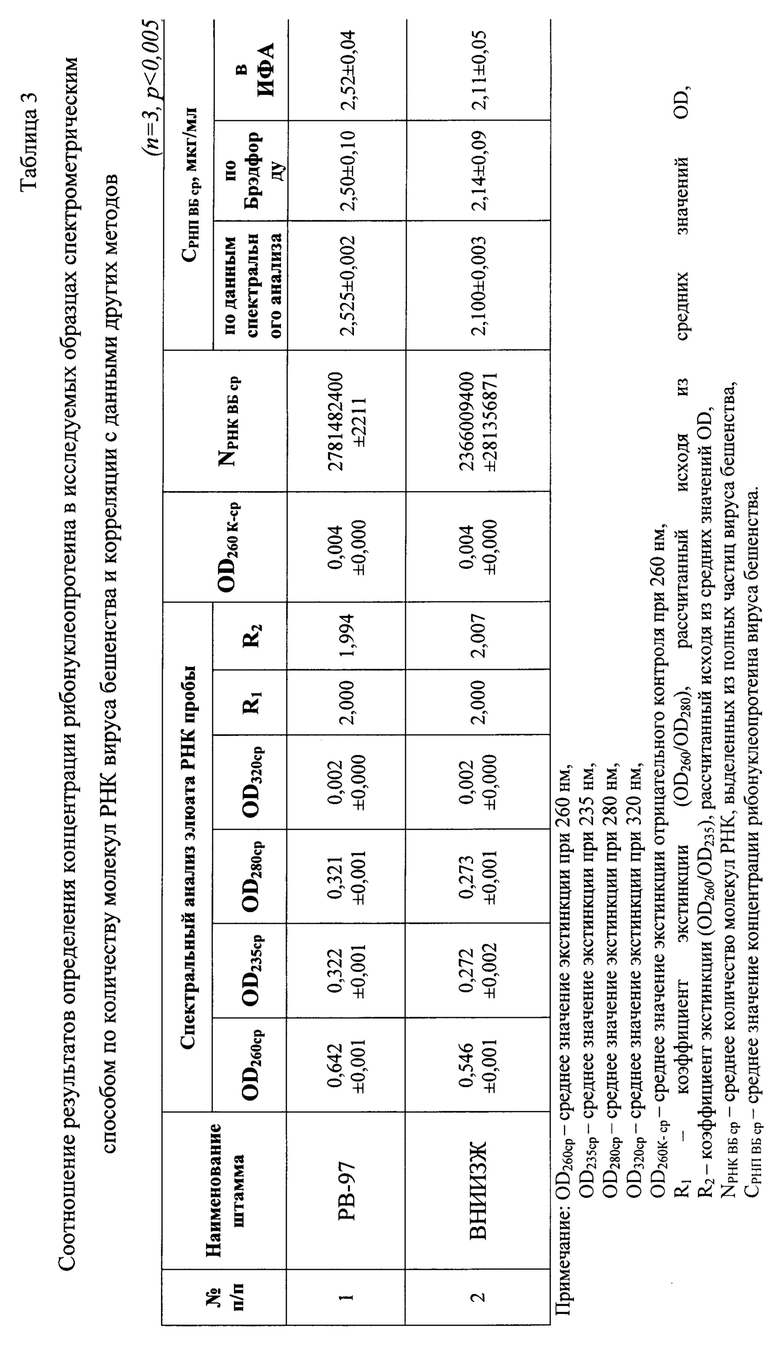
Фиг. 2 - Спектрограммы разведений стандартного 30х элюата РНК вируса бешенства (сверху вниз отражены графики для некоторых разведений элюатов, соответствующих концентрациям полных вирусных частиц: 3,5, 3,0, 2,5, 2,0, 1,5, 1,0, 0,5, 0,1 мкг/мл);
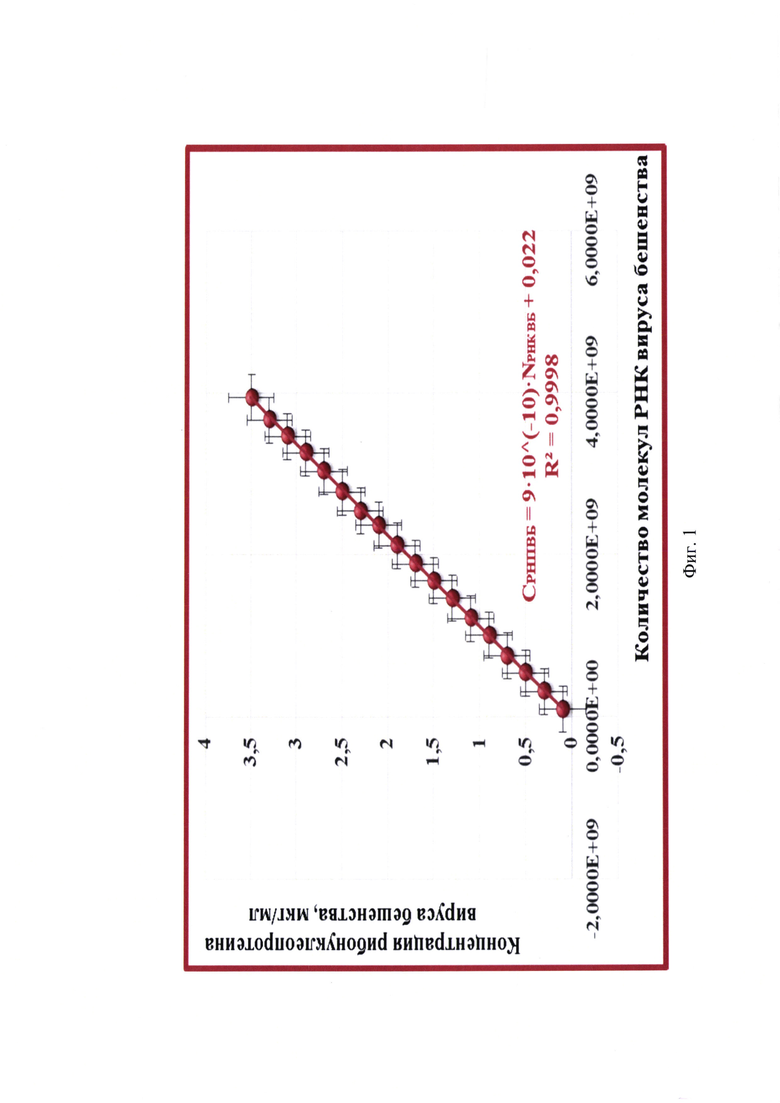
Фиг. 3 - Спектр поглощения элюатов РНК вируса бешенства штамма РВ-97 для оценки чистоты и определения концентрации рибонуклеопротеина в исходных суспензиях;
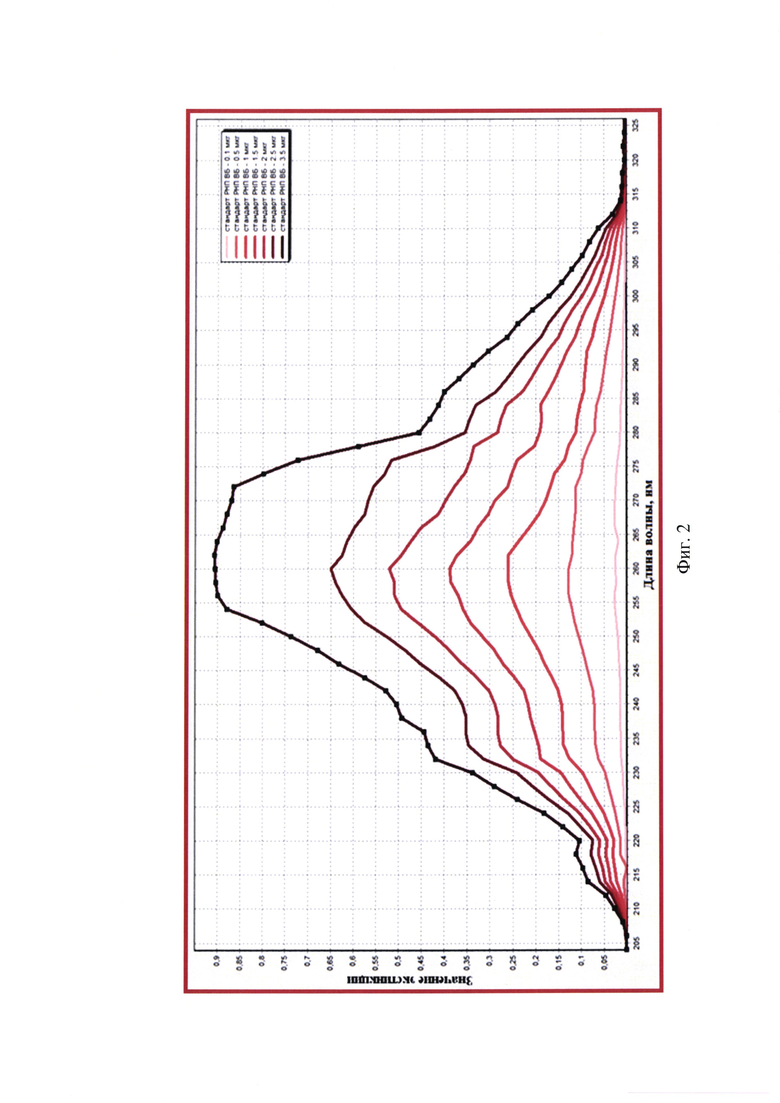
Фиг. 4 - Спектр поглощения элюатов РНК вируса бешенства штамма ВНИИЗЖ для оценки чистоты и определения концентрации рибонуклеопротеина в исходных суспензиях.
Сущность изобретения заключается в новом подходе по определению концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины с помощью спектрометрического метода и разработанной степенной модели.
Заявляемый способ основан на проведении реакции штаммоспецифического связывания вируса бешенства, выделении РНК из образующихся иммунных комплексов, спектрометрическом анализе количества молекул вирусной РНК и расчете концентрации полных вирусных частиц с помощью предложенной модели. По итогам анализа оценивают степень чистоты полученного экстракта РНК вируса бешенства с помощью спектрального анализа, определяют количество молекул вирусной РНК по расчетной формуле на основе значений экстинкции. На основании выборки известных значений концентрации рибонуклеопротеина вируса бешенства и установленных спектрометрическим методом количеств молекул вирусной РНК разработана линейная модель, позволяющая определять содержание рибонуклеопротеина в сырье для антирабической вакцины по результатам спектрометрического анализа.
По итогам анализа научных сведений предложенный способ оценки концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины до настоящего времени не применялся. Для определения концентрации рибонуклеопротеина вируса бешенства используют метод М. Брэдфорда и прямой сэндвич-вариант ИФА [1]. По сравнению с методом М. Брэдфорда разработанный способ характеризуется более высокой аналитической чувствительностью и специфичностью. В сравнении с прямым сэндвич-вариантом ИФА предложенный способ является более экономичным, не предполагает использования ферментов и изменения их каталитической активности, позволяет проводить исследование проб за 2-3 часа. Исходя их этого, предложенный способ актуально применять для определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины. Ключевым элементом заявляемого способа является выявление зависимости количества молекул РНК вируса бешенства от значения экстинкции при длине волны 260 нм, а также установление линейной зависимости между содержанием рибонуклеопротеина вируса бешенства в сырье для вакцины и количеством молекул РНК, экстрагированных из полных вирусных частиц.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении спектрометрического метода оценки количества молекул РНК, выделенных из полных частиц вируса бешенства, и разработанной линейной модели для определения концентрации рибонуклеопротеина вируса бешенства в неинактивированном сырье для вакцины.
Сведений о разработке предлагаемого способа определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины авторами не обнаружено.
Сущность предлагаемого изобретения пояснена на графическом материале - графике зависимости концентрации рибонуклеопротеина вируса бешенства от количества молекул РНК, экстрагированных из полных частиц после штаммспецифического связывания (Фиг. 1).
Сенсибилизируют иммунологические 24-луночные планшеты, поверхность которого свободна от ДНК и РНК, DNase и RNase, очищенными штаммоспецифическими поликлональными антителами против вируса бешенства в объеме 3,0 мл суспензии с концентрацией иммуноглобулинов G 10 мкг/мл при температуре 4±2°С в течение 18 часов. Дно лунок планшетов подвергают трехкратному промыванию 1/15 М фосфатно-солевым буферным раствором (ФСБР), открытые сайты связывания блокируют 0,5% суспензией бычьего сывороточного альбумина (BSA) при температуре 37±1°С в течение 1 часа и лунки промывают 1/15 М ФСБР 5 раз. Данные операции по подготовке планшета осуществляют заранее, до начала проведения исследования.
Для анализа проб вируса бешенства с неизвестной концентрацией рибонуклеопротеина применяют положительный контроль (суспензия вируса бешенства штамма РВ-97, репродуцированного в суспензионной культуре клеток почки новорожденного сирийского хомячка (ВНК-21)); отрицательный контроль (не инфицированная вирусом бешенства суспензия клеток ВНК-21 с концентрацией 2,8⋅106-3,0⋅106 клеток/мл). В лунки с сенсибилизированными поликлональными штаммоспецифическими антителами против вируса бешенства вносят по 1,0 мл контрольных образцов и проб, инкубируют при температуре 37±1°С в течение 1 часа. В результате взаимодействия антител и антигенов вируса бешенства на поверхности лунок формируются иммунные комплексы. Проводят отмывание иммунных комплексов от балластных компонентов с использованием 1/15 М ФСБР и их ресуспендируют в 0,5 мл 1/15 М ФСБР.
Выделяют связанную иммунным комплексом РНК из рибонуклеопротеина вируса бешенства за счет лизиса полипептидных и нуклеопротеидных структур с последующим фракционированием РНК от ДНК, денатурированных белков и липопротеинов. Лизис проводят с использованием 10 мл раствора, содержащего 50% карболовой кислоты (рН 6,5) и 50% 3М раствора гуанидинизотиоцианата (ГТЦ), который смешивают с 0,5 мл суспензии иммунного комплекса. Инкубацию смеси проводят при температуре 22-25°С в течение 15 минут для полного диссоциирования полипептидных и нуклеопротеидных комплексов. Полученный лизат очищают от денатурированных белков и конгломератов с помощью центрифугирования при 9000 g в течение 5 минут. Супернатант переносят в центрифужную пробирку, добавляют 2,5 мл хладона-20 и инкубируют содержимое в течение 5 минут с периодическим перемешиванием. После инкубирования смесь подвергают центрифугированию при 9000g в течение 10 минут при температуре 4±2°С. В результате осаждения происходит разделение содержимого пробирки на три фазы: 1) нижнюю фазу соломенного цвета, содержащую комплекс карболовой кислоты, ГТЦ и хладона-20 со связанными белками и липопротеинами; 2) среднюю фазу белого цвета, включающую в свой состав пептидные составляющие и клеточные ДНК; 3) верхнюю фазу, представляющую собой прозрачный экстракт вирусной РНК [6, 7]. Полностью отбирают верхнюю фазу, не затрагивая остальные фракции, и очищают от балластных компонентов с помощью 5 мл 100%-ного пропанола-2. Смесь инкубируют в течение 10 минут при температуре 22-25°С, после этого содержимое пробирки центрифугируют при 9000g в течение 6 минут при температуре 23-25°С. Супернатант удаляют, оставляя осадок РНК вируса бешенства, к которому добавляют 3,0 мл 80%-ого этанола. Содержимое перемешивают и центрифугируют при 10000 g в течение 6 минут при температуре 23-25°С. Супернатант удаляют, осадок РНК высушивают в потоке воздуха комнатной температуры в течение 3 минут. К высушенному осадку добавляют 100 мкл буфера ТЕ (10 мМ трис(оксиметил)аминометан, 1 мМ ЭДТА, рН 7,3), свободного от РНКаз и Mg, прогревают содержимое пробирки при температуре 58-60°С в течение 2-3 минут для полного растворения РНК вируса бешенства. Буфер ТЕ с указанным показателем кислотности защищает РНК вируса бешенства от деградации [8]. В результате получают по 100 мкл 30-кратных (30х) экстрактов РНК вируса бешенства относительно исходного объема вирусной суспензии (3000 мкл), вносимой для инкубации в планшет на стадии штаммоспецифического связывания.
Для определения количества молекул вирусной РНК и оценки степени чистоты элюатов проводят спектрометрический анализ, оценивая поглощение исследуемым образцом монохроматического ультрафиолетового света. Измерения спектральной поглощающей способности образцов проводят в кварцевых кюветах (длина оптического пути (1) составляет 10 мм, минимальный объем аналита - 50 мкл) при длинах волны в диапазоне 205-325 нм и температуре 23-25°С. Оценивают содержание в препарате остатков фосфолипидов, полисахаридов и ГТЦ, карболовой кислоты, полипептидов и крупных конгломератов, определяя значения оптической плотности (OD, optical density) при длинах волны 205, 235, 270, 280 и 320 нм, соответственно [9]. Экстракт РНК считают свободным от примесей полипептидов, если коэффициент экстинкции R1 (OD260/OD280) находится в пределах 1,8-2,2 и оптимально составляет примерно 2,0. Более низкие значения R1 указывают на наличие ДНК, белковых составляющих и остатков карболовой кислоты в элюате. Более высокие значения коэффициента R1 свидетельствуют о деградации РНК и наличии свободных рибонуклеотидов. Экстракт РНК вируса бешенства считают незагрязненным полисахаридами, если коэффициент экстинкции R2 (OD260/OD235) соответствует значению 2,0 [10]. При замещении 1% РНК на полисахаридные составляющие R2 уменьшается на 0,002. Значения коэффициента R2 большие 2,000 могут указывать на деградацию РНК. Отсутствие крупных частиц в полученном экстракте подтверждается, если OD320 приближено к нулевому значению [10, 11]. Если OD205-259 и/или OD261-325 превышают OD260, а также наблюдаются выраженные двойные (и более) пики на спектрограмме, то в элюате содержатся балластные компоненты и достоверность определения количества молекул РНК и, как следствие, концентрации рибонуклеопротеина вируса бешенства значительно снижается. При низкой степени очистки экстракта РНК вируса бешенства требуется повторно провести этап экстрагирования и спектральный анализ вновь полученного элюата.
Из экстракта РНК, выделенного из рибонуклеопротеина вируса бешенства, получают серию разведений стандарта, соответствующих концентрациям рибонуклеопротеина от 0,1 до 3,5 мкг/мл с шагом 0,2 мкг/мл. Измеряют экстинкцию полученных стандартных образцов при длине волны 260 нм (OD260) и устанавливают зависимость между количеством молекул РНК вируса бешенства (NPHК вб) и OD260 для получения математической модели расчета NPHК вб. Проводят установление зависимости концентрации рибонуклеопротеина в сырье для антирабической вакцины от числа молекул РНК, экстрагированных из вируса бешенства, в процессе построения положительной линейной модели. Определяют степень достоверности аппроксимации (R2) и рассчитывают значение концентрации рибонуклеопротеина вируса бешенства в сырье для антирабической вакцины.
Пример 1. Получение элюата РНК вируса бешенства штамма РВ-97 и оценка степени чистоты полученного экстракта.
На первом этапе исследования проводили штаммоспецифическое связывание вируса бешенства штамма РВ-97 с применением иммунологического планшета, который сенсибилизирован очищенными штаммоспецифическими поликлональными антителами. В качестве стандарта используют неинактивированную суспензию вируса бешенства штамма РВ-97, репродуцированного в суспензионной клеточной линии ВНК-21, с концентрацией рибонуклеопротеина 3,5 мкг/мл. Отрицательным контрольным образцом служит суспензия клеточной линии ВНК-21 с концентрацией клеток 2,8⋅106-3,0⋅106 клеток/мл, не инфицированных вирусом бешенства.
В лунки иммунологического планшета с сенсибилизированными антителами против вируса бешенства штамма РВ-97 вносят стандарт и отрицательный контроль по 2,0 мл в 3 лунки и инкубируют при температуре 37±1°С в течение 1 часа. Исследование проводят в 3 повторениях. В результате формируются иммунные комплексы. Из лунок планшета удаляют суспензию, проводят трехкратное промывание поверхности лунок с использованием 1/15 М ФСБР. Полученные иммунные комплексы ресуспендируют в 0,5 мл 1/15 М ФСБР.
Смешивают 0,5 мл полученных суспензий и 10,0 мл раствора, содержащего 50% фенола (рН 6,5) и 50% 3М раствора ГТЦ. Смесь инкубируют при температуре 22-25°С в течение 15 минут. Полученные лизаты подвергают центрифугированию при 9000 g в течение 5 минут. Надосадок после осаждения переносят в центрифужные пробирки, добавляют по 2,5 мл хладона-20, и инкубируют содержимое в течение 5 минут с периодическим перемешиванием. Суспензию подвергают центрифугированию при 9000g в течение 10 минут при температуре 4-5°С. В результате этого происходит разделение содержимого пробирки на три фазы: нижнюю, среднюю и верхнюю. В новые пробирки аккуратно без остатка отбирают верхнюю фракцию и добавляют по 3,0 мл 100%-ного пропанола-2. Полученные смеси инкубируют в течение 10 минут при температуре 23-25°С. Содержимое пробирок осаждают при 10000g в течение 6 минут при температуре 23-25°С. Надосадок удаляют, оставляя РНК вируса бешенства штамма РВ-97. К осадку РНК добавляют по 3,0 мл 80%-ного этанола, перемешивают содержимое каждой пробирки и центрифугируют при 10000 g в течение 6 минут при температуре 23-25°С. Супернатанты удаляют, осадок РНК высушивают в потоке воздуха комнатной температуры в течение 3 минут. РНК растворяют в 100 мкл буфера ТЕ (10 мМ трис(оксиметил)аминометан, 1 мМ ЭДТА, рН 7,0-7,2), содержимое прогревают при 60±2°С в течение 2-3 минут, тем самым получая 20х элюат РНК вируса бешенства штамма РВ-97 (n=3).
Проводят оценку степени чистоты элюата РНК вируса бешенства штамма РВ-97 стандарта и его разведений с помощью спектрального анализа. Образец РНК сканируют в кварцевой кюветы (1=10 мм) при температуре 22-25°С и регистрируют значения экстинкции в диапазоне от 205 до 325 нм каждые 2 нм, осуществляя полную запись спектра поглощения РНК с помощью компьютерной программы Specrtrum v. 5.0 (Фиг. 2).
В результате исследования 30-ти кратного экстракта РНК вируса бешенства штамма РВ-97 определили, что значения OD205-259 и OD261-325 не превышают OD260 (0,001-0,904<0,905 и 0,903-0,002<0,905), что является признаком высокой степени чистоты полученного элюата РНК (n=3). Из данных спектрального анализа стандарта видно отсутствие выраженных пиков на диаграмме при длинах волны 205, 235, 270, 280 и 320 нм, что свидетельствует о практически полном отсутствии загрязнения препарата примесями фосфолипидов, полисахаридов и ГТЦ, карболовой кислоты, полипептидов и крупных конгломератов, соответственно. Коэффициент экстинкции R1 составляет 2,002 (OD260/OD280=0,905/0,0,452) (близко к норме 2,000), что подтверждает отсутствие клеточной ДНК и наличие лишь следовых количеств примесей белка и остатков карболовой кислоты. Деградации нуклеиновой кислоты и наличия свободных нуклеотидов в растворе не наблюдается, поскольку коэффициент R1 не превышает 2,000. Экстракт РНК вируса бешенства представленного стандарта не загрязнен полисахаридами и остатками ГТЦ, поскольку коэффициент экстинкции R2 (OD260/OD235=0,905/0,453) соответствует значению 1,998. Учитывая, что при замещении 1% РНК на полисахаридные составляющие, значение R2 уменьшается на 0,002, в полученном элюате наличие примесей углеводов составляет не более 1%. Полученные разведения стандарта также соответствовали требованиям (Фиг. 2).
Пример 2. Установление зависимости между количеством молекул РНК вируса бешенства (NPHК вб) и OD260 для получения математической модели расчета NРНК ВБ.
На следующем этапе исследования выявляют зависимость концентрации рибонуклеопротеина культурального вируса бешенства от количества молекул вирусной РНК. Максимальный уровень поглощения одноцепочечных рибонуклеиновых кислот отмечается при длинах волн 252-271 нм, что объясняется высокой спектральной поглощающей способностью рибонуклеозид-5'-трифосфатов в данном диапазоне, а именно, для аденозин-5'-трифосфата (λАТР) - 259 нм, уридин-5'-трифосфата (λUTP) - 262 нм, гуанозин-5'-трифосфата (λGTP) - 252 нм, цитидин-5'-трифосфата (λСТР) - 271 нм [12, 13]. Спектральные исследования доказывают, что поглощения мономеров цельных рибонуклеиновых кислот сливаются, обеспечивая максимальную экстинкцию в указанном диапазоне длин волн [12, 14, 15]. Проведя нуклеотидный анализ РНК штаммов Щелково-51, РВ-97, ВНИИЗЖ, SAD В19, SAD Berne, CVS-11 вируса бешенства, определили средние процентные соотношения рибонуклеозид-5'-трифосфатов (WATP, Wutp, WGTP, Wctp) в составе нуклеиновой кислоты. Результаты исследования представлены в таблице 1.
Полученные данные позволили рассчитать средние значения длин волн (λmax) с максимальным уровнем поглощения РНК для каждого из представленных штаммов возбудителя бешенства, пользуясь формулой:
λmax=λАТР×WATP+λUTP×WUTP+λGТР×WGTP+λСТР×WCTP.
Так, определено, что для РНК вируса бешенства штамма Щелково-51 λmax составляет 261,64 нм (259×0,250+264×0,209+252×0,321+271×0,220), для штамма РВ-97-261,51 нм (259×0,251+264×0,210+252×0,319+271×0,220), для штамма ВНИИЗЖ - 261,68 нм (259×0,251+264×0,212+252×0,319+271⋅0,218), для штамма SAD В19 - 261,63 нм (259×0,252+264×0,209+252×0,310+271×0,229), для штамма SAD Berne - 261,74 нм (259×0,250+264×0,214+252×0,314+271×0,222), для штамма CVS-11 - 261,70 нм (259×0,253+264×0,215+252×0,321+271×0,211).
В работе при производстве антирабических вакцин в ФГБУ «ВНИИЗЖ» применяют штаммы РВ-97 и ВНИИЗЖ, для которых спектрометрическим способом определили значения λmax. Полученные данные составили 260 нм, что подтверждает истинность расчетных значений.
Содержание молекул РНК в чистом препарате рассчитывают, пользуясь законом Бугера-Ламберта-Бера, в соответствии с которым отмечается ослабление параллельного монохроматического пучка света при прохождении через поглощающую среду. На основе данного закона возможно найти соотношение концентрации молекул, поглощающих излучение, и количества поглощенного света [16]. При длине волны 260 нм средний коэффициент экстинкции для одноцепочечной РНК составляет 0,024 (мкг/мл)-1 см-1. Исходя из этого, раствор с содержанием нуклеиновой кислоты 1,00 мг/мл при прохождении ультрафиолетового света с длиной волны 260 нм имеет значение OD260, соответствующее 24,000 [15], и, как следствие, оптическая плотность, равная 1,000, соответствует элюату, содержащему 41,67 мкг РНК/мл.
Применение закона Бугера-Ламберта-Бера справедливо для определения числа молекул вирусной РНК вируса бешенства в разбавленных элюатах, поскольку при высоких концентрациях аналита (более 1,00 мг/мл) расстояние между молекулами поглощающего УФ-свет вещества значительно снижается. По этой причине увеличивается влияние каждой молекулы на распределение поверхностного заряда между соседними молекулами, что может изменить способность РНК поглощать излучение при данной длине волны. Иными словами, для определения количества молекул РНК вируса бешенства в элюатах с высокой концентрацией требуется получать его разведения с использованием буфера ТЕ. До измерения абсорбции разведения экстракта необходимо проводить вычитание фоновых значений указанного буфера. Для расчета содержания РНК в элюате требуется учитывать фактор пересчета для нуклеиновой кислоты возбудителя бешенства (FPHКвб=41,67), кратность разведения (К), а также вычитать фоновое значение исследуемого образца (OD320) и отрицательного контроля (OD260 K-) из OD260 пробы. При определении количества молекул РНК вируса бешенства (NPHКвя) учитывают константу Авогадро (6,02214076×1023 моль-1), средний молекулярный вес рибонуклеозида (Мwрибонуклеозида=340,5 Да), длину РНК вируса бешенства (L=12000 н.о.) [1, 2], а также производят перевод массы нуклеиновой кислоты из [мкг] в [г] в соответствии с требованиями Международной системы единиц Le Systeme International d'Unites [15, 16] (коэффициент 1/106) и пересчет количества молекул из 30х элюата в 1х материал (коэффициент 1/101,48) (общий коэффициент пересчета составляет 1/107,48).
В процессе культивирования вируса бешенства в чувствительных клеточных линиях происходит формирование частиц рибонуклеопротеина, каждая из которых включает в свой состав одну молекулу РНК. Считается, что в суспензиях вируса бешенства РНК находится не только в составе рибонуклеопротеина (97-99%), но и в незначительном количестве (1-3%) в свободном состоянии [1]. Иными словами, при расчете количества РНК, выделенных из рибонуклеопротеина вируса бешенства, следует учитывать коэффициент 0,97.
Вычисляют количество молекул РНК, экстрагированных из рибонуклеопротеина вируса бешенства (NPHК вб), с использованием расчетной формулы: NPHК вб=n×Na [16], где n - количество вещества РНК вируса бешенства. При этом n=m/М, где m - масса вещества, М - молярная масса вещества. Массу вещества оценивают спектрометрическим методом с учетом кратности разведения, фактора пересчета для одноцепочечной РНК, коэффициентов пересчета единиц измерения массы и оптической плотности исследуемого элюата РНК вируса бешенства с учетом вычитания OD320 пробы и OD260 K-: m=1/107,48×FPHКвб×K×(OD260пробы - О260к- - OD320 пробы). Молекулярная масса среднестатистической РНК вируса бешенства (на примере штаммов РВ-97 и ВНИИЗЖ) находим с учетом длины РНК вируса бешенства (L=12000 н.о.) и среднего молекулярного веса рибонуклеозида (Мwрибонуклеозида=340,5 Да): М=LРНК ВБ×Мwрибонуклеозида.
Отсюда следует, что количество вещества РНК вируса бешенства можно определить по формуле: n=(FPHквб×K×(OD260пробы - OD260к- - OD320пробы)/(107,48×LРНК ВБ ×Mwрибонуклеозида). Иными словами, формула для расчета числа структурных частиц (молекул РНК вируса бешенства) представляет собой следующее отношение:
NРНК ВБ=(0,97×Na×FРНКвб×K×(OD260пробы - OD260k- - OD320пробы))/(107,48×L×Мwрибонуклеозида). Проведя ряд расчетов и математических упрощений, получаем итоговую формулу расчета количества молекул РНК вируса бешенства в элюате:
NРНК ВБ=4,3734×109×K×(ОВ260пробы- OD260K-- OD320 пробы),
где: K - кратность разведения элюата,
OD260 - значение экстинкции элюата РНК вируса бешенства при длине волны 260 нм,
О260К- - значение экстинкции отрицательного контроля при длине волны 260 нм,
OD320 пробы - значение экстинкции элюата РНК вируса бешенства при длине волны 320 нм.
Пример 3. Установление зависимости концентрации рибонуклеопротеина в сырье для антирабической вакцины от числа молекул РНК, экстрагированных из вируса бешенства.
Для выявления зависимости концентрации рибонуклеопротеина вируса бешенства от количества молекул вирусной РНК получают серию разведений 30х стандарта РНК, выделенной из суспензии вируса бешенства штамма РВ-97 с содержанием рибонуклеопротеина 3,5 мкг/мл, соответствующих концентрации РНП от 0,1 до 3,5 мкг/мл с шагом 0,2 мкг/мл. До определения экстинкции полученных стандартных образцов осуществляют автоматическое вычитание фоновых значений буфера ТЕ. Проводят спектральный анализ приготовленных положительных стандартов, определяя значения экстинкции при длинах волны 260 и 320 нм. Значение экстинкции отрицательного контроля измеряют для выявления наличия неспецифических молекул РНК, максимальное поглощение для которых отмечается также при длине волны 260 нм. Результаты измерений и вычисления количества молекул РНК, выделенных из полных частиц вируса бешенства, отражены в таблице 2, из которой видно, что значения NPHК вб для всех разведений 30х стандарта с концентрацией рибонуклеопротеина вируса бешенства от 0,1 до 3,5 мкг/мл находятся в диапазоне от 87468000 до 3940433400, соответственно.
На основе полученных данных о количестве молекул вирусной РНК в разведениях стандарта и концентрации рибонуклеопротеина им соответствующих построен калибровочный график зависимости СРНП вб и NРНК вб, который представлен в виде линейной функции СРНП вб=9×10-10×NРНКВБ+0,022 с достоверностью аппроксимации R2=0,9991 (Фиг. 1). В отрицательном контроле вирус бешенства не обнаружен.
Полученная модель на основе количества молекул РНК вируса бешенства, выделенной из полных частиц после их иммунного связывания, позволяет определять концентрацию рибонуклеопротеина вируса бешенства в культуральных вирусных суспензиях.
Пример 4. Определение концентрации рибонуклеопротеина вируса бешенства штамма РВ-97 в суспензии для аттенуированной антирабической вакцины.
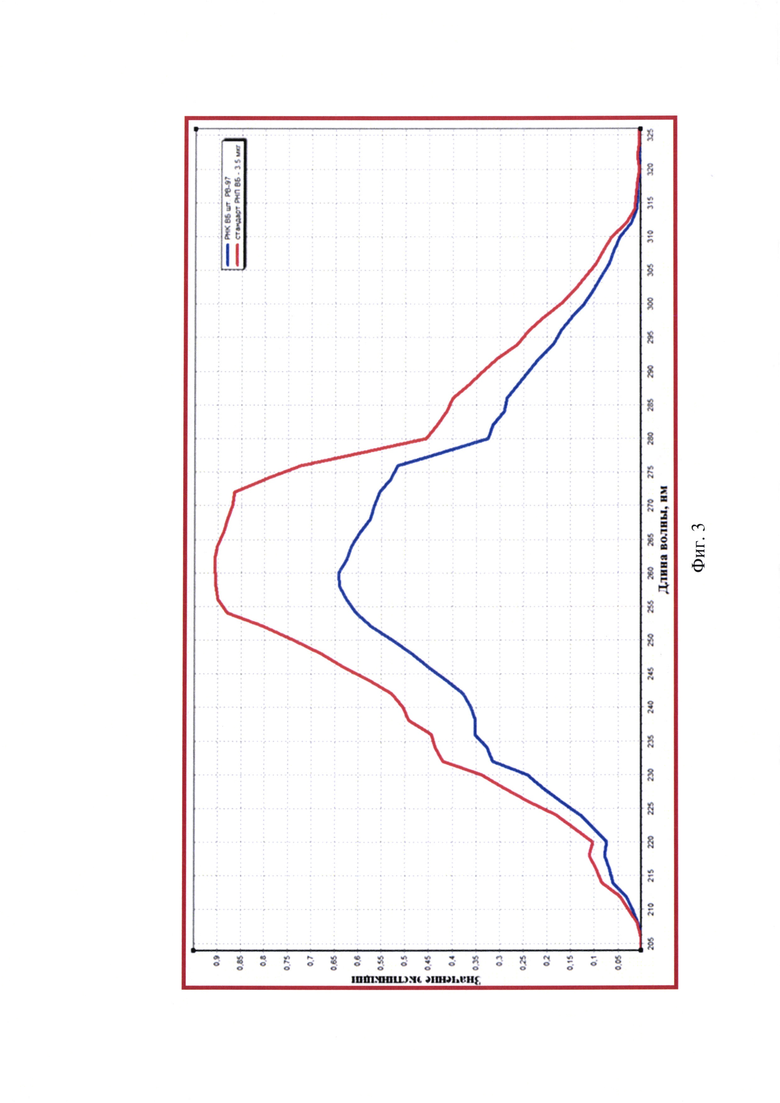
На данном этапе работы проводили тестирование суспензии вируса бешенства штамма РВ-97 для аттенуированной вакцины. Манипуляции по штаммоспецифическому связыванию полных частиц вируса бешенства и выделению из них РНК, а также оценку степени чистоты полученного 30х элюата РНК с помощью спектрального анализа для пробы, положительного и отрицательного контроля проводили так, как описано в примере 1. Результаты исследования представлены на фиг. 3.
Из данных спектрограммы видно, что средние значения OD205-259 и OD261-325 не превышают OD260 (0,001-0,640<0,642 и 0,641-0,002<0,642), следовательно, элюат выделенной РНК вируса бешенства характеризуется высокой степенью чистоты. Полученный препарат не загрязнен примесями фосфолипидов, полисахаридов и остатками ГТЦ, карболовой кислоты, полипептидов и крупных конгломератов, поскольку на графике отсутствуют выраженные пики при длинах волны 205, 235, 270, 280 и 320 нм, соответственно. Коэффициент экстинкции R1 составляет 2,000 (OD260/OD280=0,642/0,321), что соответствует норме и означает высокую степень чистоты элюата РНК вируса бешенства штамма РВ-97 и практически полное отсутствие белка и остатков карболовой кислоты после этапа выделения РНК. Коэффициент экстинкции R2 составляет 1,994 (OD260/OD235=0,642/0,322), что близко к 2,000 и обуславливает высокую степень чистоты элюата. При замещении 1% РНК на полисахаридные составляющие значение R2 уменьшается на 0,002, следовательно, в полученном элюате наличие полисахаридных примесей составляет не более 3%, что допустимо.
По итогам спектрометрического исследования средние значения OD260 составляют 0,642±0,001, OD320 - 0,002±0,000, OD260к-- 0,004±0,000. Исходя из полученных данных, рассчитывают число молекул РНК в элюате, пользуясь формулой:
NРНК ВБ=4,3734×109×K×(OD260пробы-OD260K--OD320 пробы)
Значение кратности разведения (К) принимаем за 1, поскольку цельный 30х элюат не разбавляли. Среднеарифметическое количество молекул РНК, выделенных из полных частиц вируса бешенства штамма РВ-97, составляет 2781482400±2211. Пользуясь разработанной линейной моделью связи СРНП ВБ и NРНКВБ, отраженной функцией СРНП вб=9×10-10×NPНКВБ+0,022, определили значение концентрации рибонуклеопротеина вируса бешенства штамма РВ-97, которое составило 2,525±0,002 мкг/мл, что коррелирует с результатами метода Брэдфорда (2,50±0,10 мкг/мл) и ИФА (2,52±0,04 мкг/мл) (Табл. 3). Отсюда следует, что разработанная с помощью данных спектрального анализа линейная модель позволяет оценивать концентрацию рибонуклеопротеина вируса бешенства штамма РВ-97 в сырье для аттенуированной антирабической вакцины.
Пример 5. Определение концентрации рибонуклеопротеина вируса бешенства штамма ВНИИЗЖ в суспензии для инактивированной антирабической вакцины.
Исследовали суспензию вируса бешенства штамма ВНИИЗЖ для инактивированной антирабической вакцины. Манипуляции по штаммоспецифическому связыванию полных частиц вируса бешенства и выделению из них РНК, а также оценку степени чистоты полученного 30х элюата РНК с помощью спектрального анализа для пробы, положительного и отрицательного контроля проводили так, как описано в примере 1. Результаты исследования представлены на фиг. 4.
Из данных спектрограммы видно, что средние значения OD205-259 и OD261-325 не превышают OD260 (0,001-0,544<0,546 и 0,545-0,002<0,546), следовательно, элюат выделенной РНК вируса бешенства штамма ВНИИЗЖ характеризуется высокой степенью чистоты. Полученный препарат не загрязнен примесями фосфолипидов, полисахаридов и остатками ГТЦ, карболовой кислоты, полипептидов и крупных конгломератов, поскольку на графике отсутствуют выраженные пики при длинах волны 205, 235, 270, 280 и 320 нм, соответственно. Коэффициент экстинкции R1 составляет 2,000 (OD260/OD280=0,546/0,273), что соответствует норме и означает высокую степень чистоты элюата РНК вируса бешенства штамма ВНИИЗЖ и полное отсутствие белка и остатков карболовой кислоты после этапа выделения РНК. Коэффициент экстинкции R2 составляет 2,007 (OD260/OD235=0,546/0,272), что близко к 2,000 и обуславливает высокую степень чистоты элюата. При замещении 1% РНК на полисахаридные составляющие значение R2 уменьшается на 0,002, следовательно, в полученном элюате наличие полисахаридных примесей не обнаружено.
По итогам спектрометрического исследования средние значения OD26o составляют 0,546±0,002, OD320 - 0,002±0,000, OD260к-- 0,004±0,000. Исходя из полученных данных, рассчитывают число молекул РНК в элюате, пользуясь формулой:
NРНК ВБ=4,3734×109×K×(ОВ260пробы-OD260к--OD320 пробы) Значение кратности разведения (К) принимаем за 1, поскольку цельный 30х элюат не разбавляли. Среднеарифметическое количество молекул РНК, выделенных из полных частиц вируса бешенства штамма ВНИИЗЖ, составляет 2366009400±2829. Пользуясь разработанной линейной моделью связи СРНП вб и NРНКВБ, отраженной функцией СРНКП ВБ=9⋅10-10.NPHКВБ+0,022, определили значение концентрации рибонуклеопротеина вируса бешенства штамма ВНИИЗЖ, которое составило 2,100±0,003 мкг/мл, что коррелирует с результатами метода Брэдфорда (2,14±0,09 мкг/мл) и ИФА (2,11±0,05 мкг/мл) (Табл. 3). Отсюда следует, что разработанная с помощью данных спектрального анализа линейная модель позволяет оценивать концентрацию рибонуклеопротеина вируса бешенства штамма ВНИИЗЖ в сырье для инактивированной антирабической вакцины.
Пример 6. Выявление корреляции результатов определения концентрации рибонуклеопротеина вируса бешенства с помощью разработанного способа и ранее применяемых методов.
Проводили тестирование предложенного способа для определения концентрации рибонуклеопротеина вируса бешенства вакцинных штаммов РВ-97 и ВНИИЗЖ в 355 вируссодержащих суспензиях. Исследования параллельно проводили методом Брэдфорда и в прямом сэндвич-варианте ИФА в трех повторениях. Совпадение фактических результатов, полученных с помощью метода Брэдфорда, и ожидаемых результатов по определению концентрации РНП вируса бешенства спектрометрическим методом по количеству молекул РНК составило 95,4-99,0%. При сравнении с данными метода ИФА совпадение фактических и ожидаемых результатов соответствовало значениям 96,0-99,7%. Для положительного контроля совпадение фактических и ожидаемых результатов составило 99,2-99,7%. В отрицательном контрольном образце РНК и полные частицы рибонуклеопротеина вируса бешенства не обнаружены, что также соответствовало ожиданиям. Таким образом, по результатам проведенного исследования корреляция способа спектрометрического определения концентрации рибонуклеопротеина вируса бешенства по оценке количества молекул вирусной РНК в сырье для антирабических вакцин с методами по Брэдфорду и прямого сэндвич-варианта ИФА составила 95,4-99,7%.
Пример 7. Оценка специфичности способа спектрометрического определения концентрации рибонуклеопротеина вируса бешенства по оценке количества молекул вирусной РНК в сырье для антирабических вакцин.
Для оценки специфичности предложенного способа опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабических вакцин спектрометрическим способом по оценке количества вирусной РНК, исследовали суспензии вируса бешенства штамма РВ-97 (СРНП ВБ=2,70 мкг/мл), вируса бешенства штамма ВНИИЗЖ (СРНП ВБ=3,30 мкг/мл), вируса ящура штамма О/Забайкальский/2016, инфекционного некроза гемопоэтической ткани лососевых, весенней виремии карповых, инфекционного некроза поджелудочной железы. Количество инфекционных доз вирусов в суспензиях составлял 106,0 ТЦД50/см3. Исследования проводили в трех повторностях.
Этапы выделения РНК, оценки степени их чистоты и расчеты количества молекул вирусной нуклеиновой кислоты, концентрации рибонуклеопротеина вируса бешенства проводили, как описано в примере 4. Выделенные экстракты РНК характеризовались высокими показателями степени чистоты, поскольку коэффициенты экстинкции R1 и R2 находились в диапазоне 1,995-2,001, что соответствовало требованиям [6].
Для элюатов РНК вируса бешенства штаммов РВ-97 и ВНИИЗЖ, выделенных после штаммоспецифического связывания полных частиц вируса, при длине волны 260 нм получили значения оптических плотностей проб 0,700 и 0,845, что соответствовало заявленным концентрациям 2,70 и 3,30 мкг/мл. Для проб, содержащих другие вирусы, не наблюдалось формирования пиков на спектрограммах и не выявили наличия вируса бешенства, что соответствовало действительности. Иными словами, предложенный способ является специфичными по отношению к вирусу бешенства штаммов РВ-97 и ВНИИЗЖ при использовании специфических поликлональных антител на этапе штаммоспецифического связывания.
Основным преимуществом предлагаемого изобретения является возможность исследования суспензий вируса бешенства для определения концентрации рибонуклеопротеина в сырье для антирабических вакцин в течение 2-3 часов. В предлагаемом изобретении между концентрацией рибонуклеопротеина и количеством молекул РНК, выделенных из полных вирусных частиц после штаммоспецифического связывания и определенных спектрометрическим методом по формуле:
NРНК вб=4,3734×109×K×(OD260пробы-OD260к--OD320 пробы),
установлена зависимость, которая представлена в виде линейной функции с высокой достоверностью аппроксимации R2=0,9998. Разработанная модель имеет вид СРНП ВБ=9⋅10-10⋅NРНКВБ+0,022 и дает возможность оценивать значение концентрации рибонуклеопротеина вируса бешенства в сырье для производства антирабических вакцин.
Предлагаемое изобретение позволяет быстро и с высокой степенью достоверности определять значение концентрации рибонуклеопротеина вируса бешенства в сырье для антирабических вакцин на основе спектрометрического метода с последующим применением предложенной линейной модели.
Источники информации
1. Груздев К.Н., Метлин А.Е. Бешенство животных. - Владимир: ФГБУ «ВНИИЗЖ», 2019. - 394 с.
2. OIE. Manual of diagnostic tests and vaccines for terrestrial animals. - Paris, 2018. - Chapter 3.1.17. - P. 578-612.
3. Безопасность и иммуногенность оральных антирабических вакцин / С.С. Рыбаков, Е.В. Велик, А.Е. Метлин // Ветеринария. - 2010. - №8. - С. 18-22.
4. Вакцины против бешенства: современное состояние и перспективы развития / Е.С.Стародубова, О.В. Преображенская, Ю.В. Кузьменко // Молекулярная биология. - 2015. - Т. 49, №4. - С. 577-584.
5. Bradford, М. М. (1976) A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. - V. 72. - 1976. - P. 248-254.
6. Chomczynski P., Sacchi N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nat Protoc. 1 (2). - 2006. - P. 581-585.
7. Peirson S.N. RNA extraction from mammalian tissues / S.N. Peirson, J.N. Butler // Methods in Molecular Biology. - 2007. - Vol. 362. - P. 315-327.
8. Ross K.S. Repeated freezing and thawing of peripheral blood and DNA in suspension: effects on DNA yield and integrity / K.S. Ross, N.E. Haites, K.F. Kelly // Journal of Medical Genetics. - 1990. - Vol. 27 (9). - P. 569-570.
9. Владимиров Ю.А., Потапенко А.Я. Физико-химические основы фотобиологических процессов. - М.: Высшая школа, 1989. - С. 20-22.
10. Glasel J. Validity of nucleic acid purities monitored by 260nm/280nm absorbance ratios // BioTechniques. - 1995. - Vol. 18 (1). - P. 62-63.
11. The Analysis of DNA or RNA using Its Wavelengths: 230 nm, 260 nm, 280 nm. Bioteachnology.com (13 января 2010). Проверено 15 мая 2020 г. http://bioteachnology.com/dna/analysis-dna-rna-wavelengths-230-260-280-nm.
12. Voet D. Absorption spectra of nucleotides, polynucleotides, and nucleic acids / D. Voet, W.B. Gratzer, R.A. Cox, P. Doty // Biopolymers. - 1963. - Vol. 1(3). - P. 193-208.
13. Zhang R. Carbon black induced DNA damage and conformational changes to mouse hepatocytes and DNA molecule: A combined study using comet assay and multi-spectra methods / R. Zhang, X. Zhang, C. Jia, J. Pan, R. Liu // Ecotoxicol Environ Saf. - 2019. - Vol. 170. - P. 732-738.
14. Tataurov A.V. Predicting ultraviolet spectrum of single stranded and double stranded deoxyribonucleic acids / A.V. Tataurov, Y. You, R. Owczarzy // Biophys. Chem. - 2008. - Vol. 133 (1-3). - P. 66-70.
15. Досон P. Справочник биохимика: Пер. с англ. - М.: Мир. - 1991. -С. 467.
16. Крешков А.П. Основы аналитической химии. Физико-химические (инструментальные) методы анализа. - Изд. «Химия». - 1970. - 472 с.
17. Капорский Л. Н. Оптическая плотность // Физическая энциклопедия / Гл. ред. А.М. Прохоров. - М.: Большая Российская энциклопедия. - 1992. - Т. 3. - С. 441.

Изобретение относится к области биотехнологии. Описан способ спектрометрического определения концентрации рибонуклеопротеина вируса бешенства по оценке количества молекул вирусной РНК, выделенных из полных частиц вируса бешенства после их штаммоспецифического связывания (NРНК ВБ), на основании линейной функции зависимости NРНК ВБ от содержания рибонуклеопротеина вируса бешенства в вирусной суспензии. Количество молекул РНК, выделенных из полных частиц вируса бешенства, определяют спектрометрическим методом при оценке экстинкции при длине волны 260 нм. На основании установленной величины оптической плотности (OD260) рассчитывают число молекул выделенной РНК вируса бешенства и определяют концентрацию рибонуклеопротеина вируса бешенства (Срнп вб) с применением разработанной линейной функции Срнп вб=9×10-10×NРНКВБ+0,022. Изобретение расширяет арсенал спектрометрических способов определения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабических вакцин. 3 з.п. ф-лы, 3 табл., 7 пр., 4 ил.
1. Способ спектрометрического определения концентрации рибонуклеопротеина вируса бешенства по оценке количества молекул вирусной РНК в сырье для антирабических вакцин, включающий следующие стадии:
- проведение штаммоспецифического связывания полных частиц вируса бешенства из сырья для антирабической вакцины;
- выделение РНК из полученных связанных частиц;
- оценка степени чистоты экстрактов РНК с помощью спектрального анализа;
- подготовка серии разведений положительного стандарта РНК вируса бешенства с количествами молекул, соответствующих концентрациям рибонуклеопротеина от 0,1 до 3,5 мкг/мл с шагом разведения 0,2 мкг/мл;
- расчет количества молекул РНК, выделенных из полных частиц вируса бешенства, с помощью данных спектра поглощения;
- установление зависимости концентрации рибонуклеопротеина и количества молекул РНК, экстрагированных из полных частиц вируса бешенства, в виде линейной функции;
- определение степени достоверности аппроксимации (R2) разработанной модели;
- расчет значения концентрации рибонуклеопротеина вируса бешенства в сырье для антирабических вакцин на основе разработанной модели.
2. Способ по п. 1, отличающийся тем, что проводится совмещение процессов штаммоспецифического связывания полных частиц вируса бешенства из сырья для антирабических вакцин с последующим экстрагированием высокоочищенной вирусной РНК и оценкой ее чистоты методом спектрального анализа.
3. Способ по п. 1, отличающийся тем, что на основании спектрометрического исследования элюата РНК вируса бешенства определяют значение экстинкции при длине волны 260 нм (OD260) и рассчитывают количество молекул РНК, экстрагированных из полных частиц вируса бешенства (NРНК ВБ), с применением предложенной функции
NРНК ВБ=4,3734×109×K×(ОD260пробы-OD260k--OD320 пробы).
4. Способ по п. 1, отличающийся тем, что на основании данных о количестве молекул РНК, выделенных из полных частиц вируса бешенства, и определенных в спектрометрическом исследовании, рассчитывают концентрацию рибонуклеопротеина вируса бешенства в сырье для антирабических вакцин с применением разработанной линейной функции Срнп вб=9×10-10×NРНКВБ+0,022.
| Способ определения концентрации 146S-компонента вируса ящура в вируссодержащем сырье для вакцины с применением метода обратной транскрипции-полимеразной цепной реакции в режиме реального времени | 2016 |
|
RU2619878C1 |
| Трансформатор для дуговой сварки | 1928 |
|
SU17526A1 |
| Приспособление для поперечной устойчивости аэропланов | 1921 |
|
SU1414A1 |
| СПОСОБ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ ФИКСИРОВАННОГО ВИРУСА БЕШЕНСТВА ШТАММА "МОСКВА 3253" | 2012 |
|
RU2511440C2 |

