Изобретение относится к области биотехнологии и производству антирабических вакцин, а именно к способу определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии.
Бешенство - острое вирусное заболевание, поражающее почти все виды млекопитающих, включая человека [1, 2]. Смертность от данного заболевания составляет почти 100%. По оценкам ВОЗ, ежегодно в мире в результате укусов зараженными животными вирусом бешенства регистрируется более 55000 смертей людей. Возбудителем бешенства является нейротропный вирус, принадлежащий к роду Lyssavirus семейства Rhabdoviridae. За подавляющее большинство случаев ответственен вирус бешенства генетической группы RABV [1].
Цикл репродукции вируса включает адсорбцию на поверхности клеток, проникновение в клетку, «раздевание», транскрипцию, трансляцию и посттрансляционную модификацию, репликацию, сборку и почкование вируса [1, 2].
Для первичной транскрипции нет необходимости в синтезе структурных белков вируса. Для возбудителя бешенства характерно 2 типа процесса транскрипции: 1) с геномной РНК сначала транскрибируется лидерная РНК, затем последовательно 5 моноцистронных мРНК, с которых в цитоплазме транслируются N-, Р-, М-, L-белки, а мРНК G-гена транслируется в полисомах, связанных с мембраной комплекса Гольджи; 2) транскрипция с геномной РНК антигеномных РНК (+), которые служат матрицами для синтеза новых генераций геномных РНК (-). В отличие от первичной транскрипции, репликация геномной РНК по описанному механизму может происходить только одновременно с трансляцией и синтезом структурных белков, в особенности нуклеопротеина и фосфопротеина, необходимых для инкапсидации РНК в нуклеокапсидные структуры. Если L-, N- и Р-белки контролируют, главным образом, транскрипцию и репликацию РНК, то М-белок занимает промежуточное положение между нуклеокапсидом и оболочкой вириона, участвуя в конденсации РНК. М-белок играет ключевую роль в регуляции синтеза РНК и почковании вируса. На финальной стадии сборки и почкования нуклеокапсиды «одевают» вирусную оболочку, представленную гликопротеином, который интегрирован в клеточную мембрану. Гликопротеин детерминирует нейровирулентность и нейроинвазивность вируса бешенства [3, 4].
Место формирования вирусных частиц зависит от клеток хозяина. Синтез и сборка нуклеокапсидов происходят в цитоплазме, а почкование - через плазматическую мембрану клетки, от которой вирион получает дополнительный липидный слой.
Вирус бешенства не вызывает в клетках характерные цитопатические изменения. Все события транскрипции и репликации происходят в цитоплазме внутри телец Бабеша-Негри, диаметр которых составляет 2-10 мкм. Данные структуры являются типичными при заражении возбудителем бешенства и используются при диагностике в качестве патогномического признака наличия смертельно опасной инфекции [1].
Для борьбы с бешенством требуется осуществлять вакцинацию восприимчивых животных. Ослабление вирулентности вируса является необходимым условием для разработки антирабических вакцин. Для глобальной профилактики и борьбы с этим заболеванием недостаточно и не хватает доступных высококачественных вакцин. Стабильные, аттенуированные производственные штаммы вируса бешенства с широкой перекрестной активностью против различных вариантов возбудителя являются идеальными кандидатами для создания надежных, безопасных и эффективных препаратов [1, 2, 3]. Следует отметить, что необходимо прилагать значительные усилия для разработки безопасных, эффективных и относительно недорогих вакцин для глобальной борьбы с бешенством в будущем [1].
С момента первой разработки антирабической вакцины Пастером в конце 19 века препараты значительно усовершенствовались, а иммунизация стала проводиться как для домашних и сельскохозяйственных животных, так и для видов-резервуаров. Долгое время для изготовления вакцин применяли ткани головного мозга, пораженные вирусом. Сообщалось о серьезных побочных реакциях при использовании антирабических вакцин из нервных тканей, либо из птичьих эмбрионов. Появление современных технологий промышленного культивирования и ферментации клеток значительно расширило возможности производства вакцинных препаратов с высоким качеством и количеством иммуногенных компонентов, в частности рибонуклеопротеина вируса бешенства [1, 2].
В процессе репродукции аттенуированного штамма возбудителя данного заболевания в клеточных линиях, в частности, в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, который представляет собой важнейший этап производства антирабических вакцин, требуется проводить контроль состояния ДНК клеток для определения оптимальных показателей культивирования вируса с целью повышения концентрации иммуногенных компонентов, что непосредственно будет влиять на иммуногенность получаемого вакцинного препарата.
Традиционно для определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин применяют метод визуального анализа окрашенных клеток с определением титра инфекционной активности и концентрации рибонуклеопротеина вируса бешенства с помощью метода ОТ-ПЦР-РВ (прототип). Этот способ позволяет достоверно определять количественные показатели репродукции вируса бешенства, однако на это требуется не менее 4-5 ч, что критично для процесса измерения концентрации иммуногенного компонента во время культивирования возбудителя данного заболевания в реакторах объемами не менее 2000 л [5, 6, 7]. Требуется быстро в режиме on-line получать сведения о том, продолжать репродукцию вируса, либо останавливать процесс. Прототипный способ является простым в применении, однако он не дает возможности детально определять трансформационные изменения ДНК в клетках и, тем самым, оценивать состояние клеток в суспензии, при которых следует останавливать процесс репродукции вируса. Дело в том, что при затягивании процесса культивирования высокими являются риски разрушения вирусных частиц, а при неполной репродукции и частичной сборке вирионов суспензия будет содержать недостаточное количество иммуногенных компонентов. Оба этих явления негативны с биологической и экономической точек зрения и не позволяют получить максимальный выход продукта для изготовления вакцин, в частности, иммуногенного компонента - рибонуклеопротеина вируса бешенства.
В связи с этим целесообразно провести поиск способа определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии для оптимизации процесса производства вакцинных препаратов.
Проточная цитометрия представляет собой метод, обеспечивающий быстрый, качественный и мультипараметрический анализ клеток, который получил широкое распространение в ветеринарной и медицинской иммунологии, фармакологии, цитологии, онкологии, гематологии, генетики и для исследования инфекционных болезней человека и животных [8, 9].
Проточные питометры сконструированы таким образом, что за счет гидродинамического фокусирования через луч лазера единовременно проходит одна клетка. Как только клетка пересекает луч источника света, детектор начинает регистрацию сигнала, который достигает максимума при попадании клетки в центр луча. Когда клетка покидает луч, сигнал исчезает. Проходящие через луч лазера клетки рассеивают свет под разными углами. Рассеяние света зависит от диаметра клетки и сложности ее внутренней структуры. Регистрация рассеянного света под малыми углами (до 19°, так называемое прямое светорассеяние) используется для определения относительного диаметра клетки [8]. Это сигнал, определяемый как свет того же цвета, что и освещающий поток, т.е. показывающий способность отражать свет по сравнению с окружающей клетку жидкостью. Для регистрации внутриклеточных структур используется боковое светорассеяние, при котором значительное количество света рассеивается под углом 90°. Использование этого параметра позволяет косвенно судить о внутриклеточных изменениях, в том числе трансформационных изменениях ДНК [10, 11].
Предложенный метод является высокочувствительным, объективным и позволяет в течение не более 30 мин определять трансформационные изменения ДНК клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства методом проточной цитометрии для контроля качества антирабических вакцин на разных этапах репродукции возбудителя. Данный метод характеризуется высокой скоростью анализа (до 100000 эпизодов в секунду), обладает возможностью определять клеточные линии, способен проводить определение различных параметров любых клеток и клеточных структур (в том числе и редко встречающихся). Следует отметить, что метод отличается высокой степенью объективности в измерении интенсивности флуоресценции, а, следовательно, дает возможность получить результаты с высокими показателями достоверности.
Разработанный способ дает возможность детально исследовать фазы клеточного цикла клетки по трансформационным изменениям клеточной ДНК.
Эукариотический клеточный цикл состоит из четырех различных фаз: G1-фаза, S-фаза (синтез), G2-фаза (в совокупности известная как интерфаза) и М-фаза (митоз и цитокинез) [8]. Так же существует фаза G0 - состояние покоя, когда клетка вышла из цикла и перестала делиться. Клеточный цикл начинается с этой фазы.
Не пролиферативные клетки у многоклеточных эукариот обычно переходят из G1-фазы в состояние покоя G0-фазу и могут оставаться в состоянии покоя в течение длительных периодов времени, возможно, на неопределенный срок (часто бывает с нейронами). Это очень характерно для клеток, которые полностью дифференцированы. Многие клетки не попадают в G0-фазу и продолжают делиться на протяжении всей жизни организма, например, эпителиальные клетки. Термин «постмитотический» иногда используется для обозначения как покоящихся, так и стареющих клеток. Клеточное старение происходит в ответ на повреждение ДНК и внешний стресс и обычно представляет собой остановку в фазе G1. Клеточное старение может сделать потомство клетки нежизнеспособным. Зачастую это есть биохимическая альтернатива саморазрушению такой поврежденной клетки путем апоптоза.
Интерфаза представляет собой фазу между двумя последовательными М-фазами. Это серия изменений, которые происходят во вновь образованной клетке и ее ядре, прежде чем она снова становится способной к делению. Это также называется подготовительной фазой или промежуточным митозом. Обычно интерфаза длится не менее 91% от общего времени, необходимого для клеточного цикла. Интерфаза протекает в три стадии: G1-, S- и G2-фазы, за которыми следует цикл митоза и цитокинеза. Содержимое ядерной ДНК клетки дублируется во время S-фазы.
Первая фаза внутри интерфазы, от конца предыдущей М-фазы до начала синтеза ДНК, называется G1-фазой, или фазой роста. В это время биосинтетическая активность клетки, которая значительно замедляется во время М-фазы, возобновляется с высокой скоростью. Продолжительность фазы G1 сильно варьируется даже среди разных клеток одного и того же вида. На этой фазе клетка увеличивает запас белков, увеличивает количество органелл (таких как митохондрии, рибосомы) и увеличивается в размерах. Решающая точка называется контрольной и регулируется циклинами G1/S, которые вызывают переход от фазы G1 к фазе S. Прохождение контрольной точки G1 приводит клетку к процессу деления.
Последующая S-фаза начинается, когда осуществляется синтез молекул ДНК. В это время количество ДНК в клетке удваивается, хотя плоидность и количество хромосом остаются неизменными. Скорости транскрипции РНК и синтеза белка на этой фазе очень низкие.
Фаза G2 наступает после репликации ДНК и представляет собой период синтеза белка и быстрого роста клеток для подготовки клетки к митозу. Микротрубочки начинают реорганизовываться, образуя веретено деления (препрофазу). Прежде чем перейти к митотической фазе, клетки должны быть проверены на контрольной точке G2 на наличие любых повреждений ДНК в хромосомах. Эта точка в основном регулируется опухолевым белком р53. Если ДНК повреждена, то фактор р53 либо восстанавливает ДНК (осуществляется процесс репарации), либо запускает апоптоз клетки. Если р53 дисфункционален или мутировал, то клетки с поврежденной ДНК могут продолжать свой клеточный цикл, что приводит к развитию рака. Далее осуществляется митоз [8, 9, 10, 11]. Для исследования клеточного цикла по данным трансформации ДНК клеток важны те фазы, которые представлены выше.
Задачей настоящего изобретения является разработка высокочувствительного, быстрого и объективного (с помощью инструментального анализа) способа определения трансформационных изменений ДНК клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства методом проточной цитометрии для контроля качества антирабических вакцин с целью устранения вышеуказанных недостатков.
Данная задача решена благодаря созданию нового способа определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии. Разработанный способ дает возможность получать данные о фазах репродуктивного цикла клеток, а также результаты, позволяющие оценить продолжительность и доминирование таких фаз клеточного цикла, как G1, S, G2, на разных этапах культивирования. Следует отметить, что характер изменений клеточного цикла при репликации в клетке вируса является важным для понимания процесса его репродукции. Это, позволяет вовремя останавливать процесс культивирования вируса бешенства для получения максимальной концентрации его иммуногенных компонентов в конечной вируссодержащей суспензии.
Сущность изобретения отражена на графических изображениях:
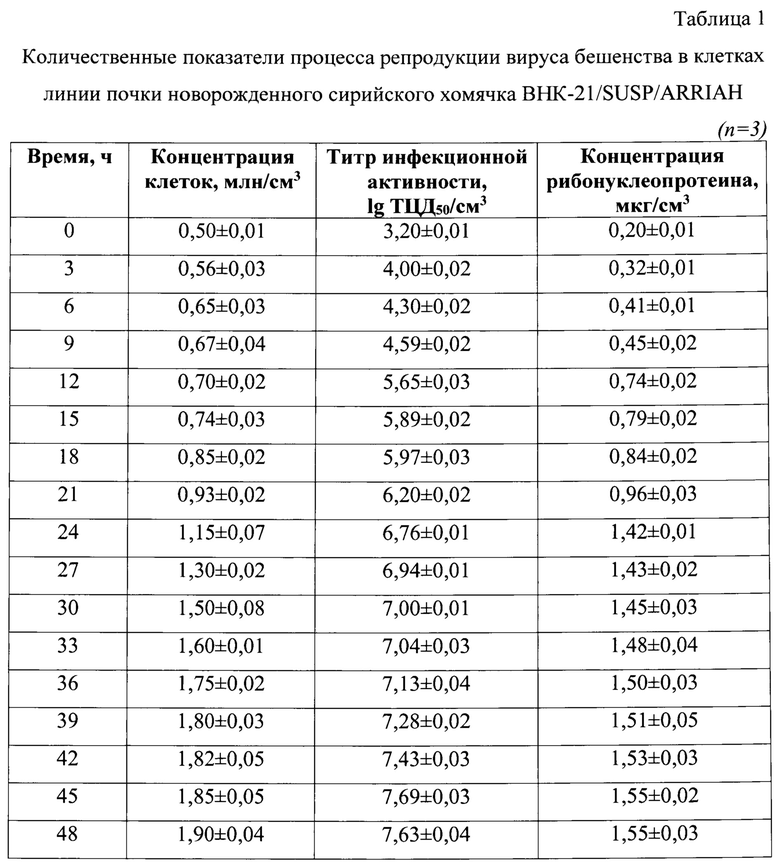
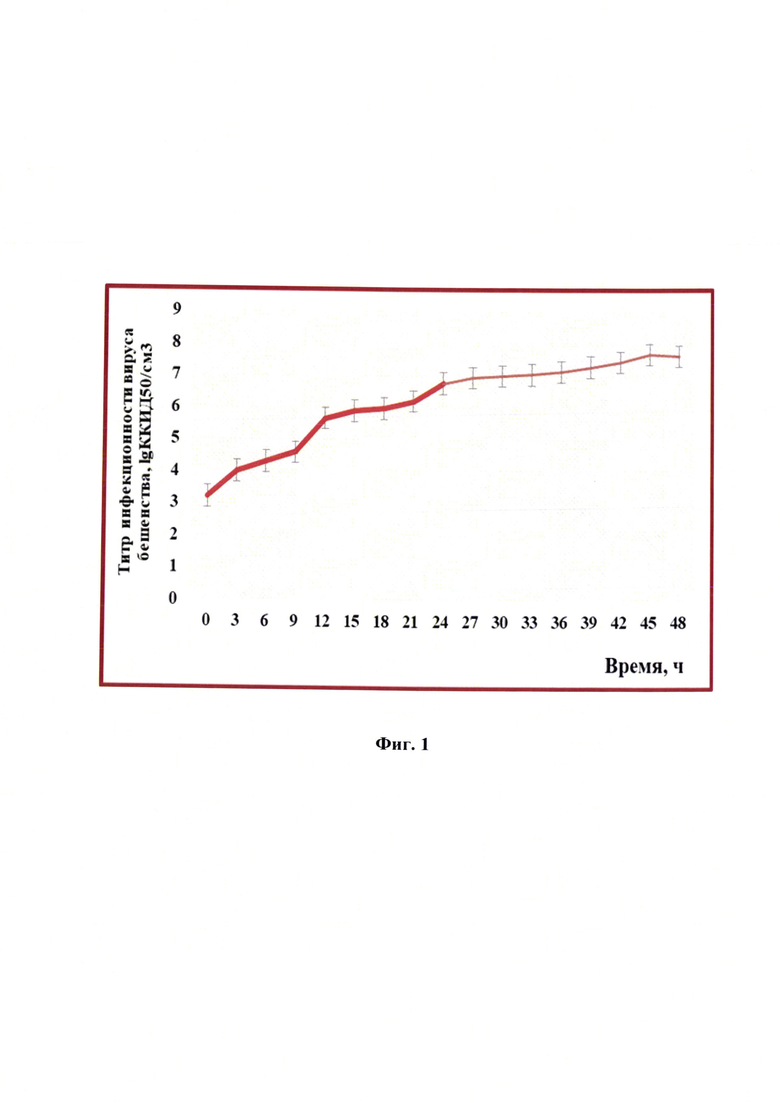
Фиг. 1 - Динамика изменения титра инфекционной активности при репродукции вируса бешенства (n=3, р<0,05).
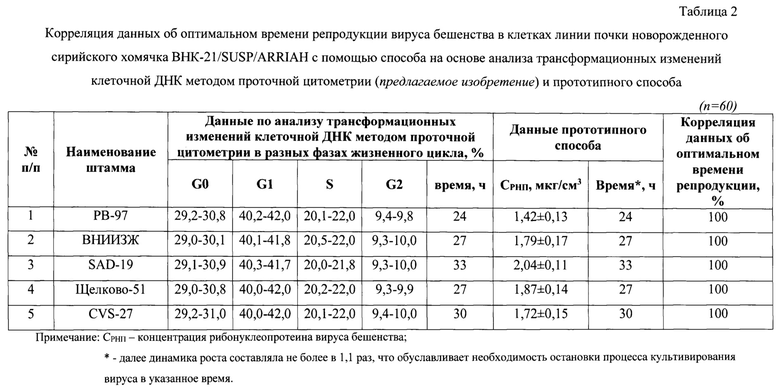
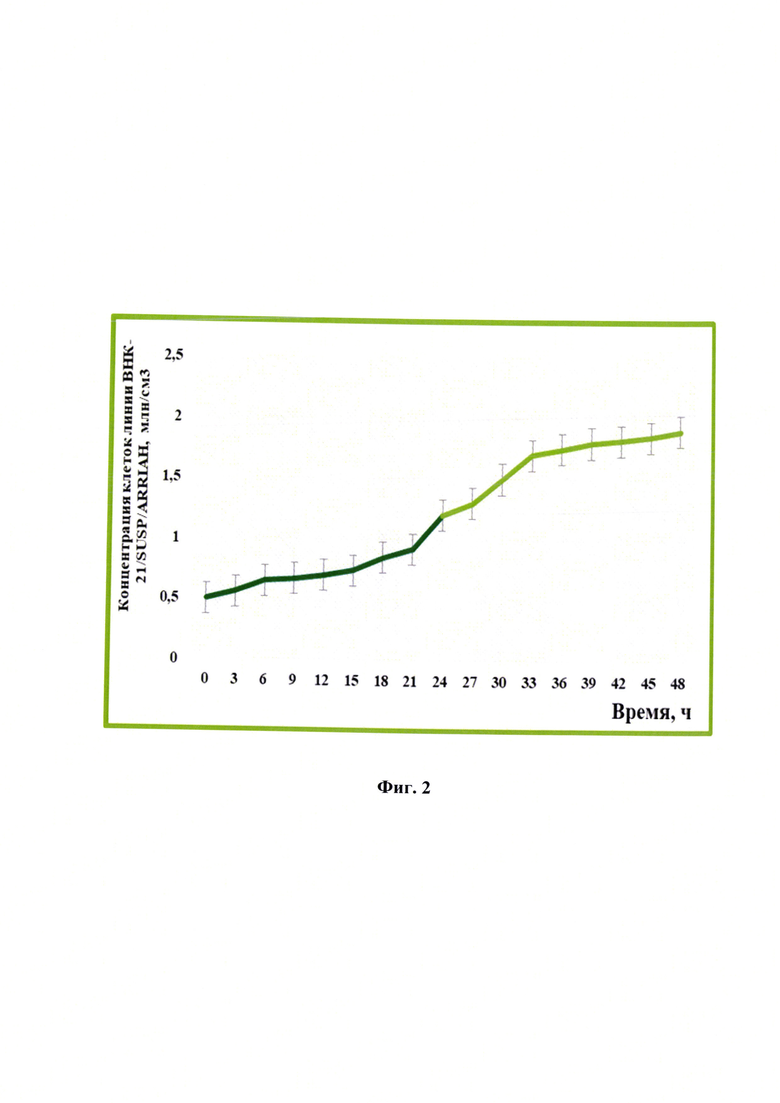
Фиг. 2 - Динамика изменения концентрации клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции вируса бешенства (n=3, р<0,05).
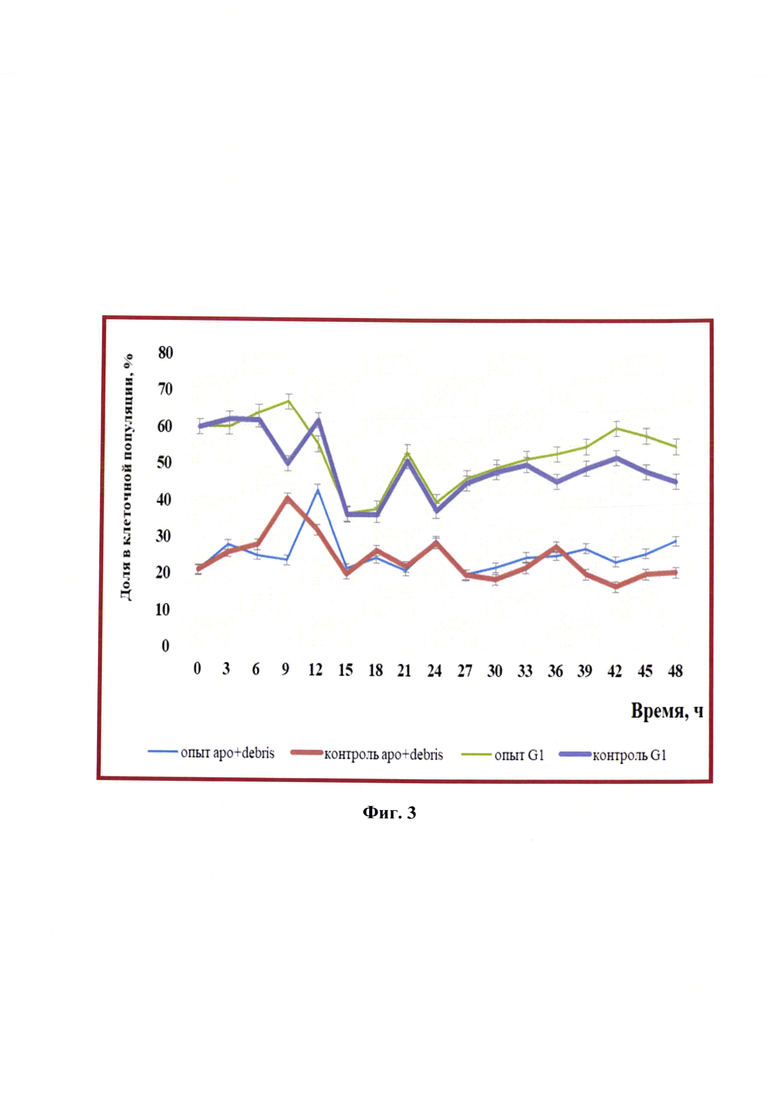
Фиг. 3 - Изменение процентного соотношения клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции вируса бешенства, находящихся на стадиях апоптоза/дебриса и фазы G1, (n=3, р<0,05).
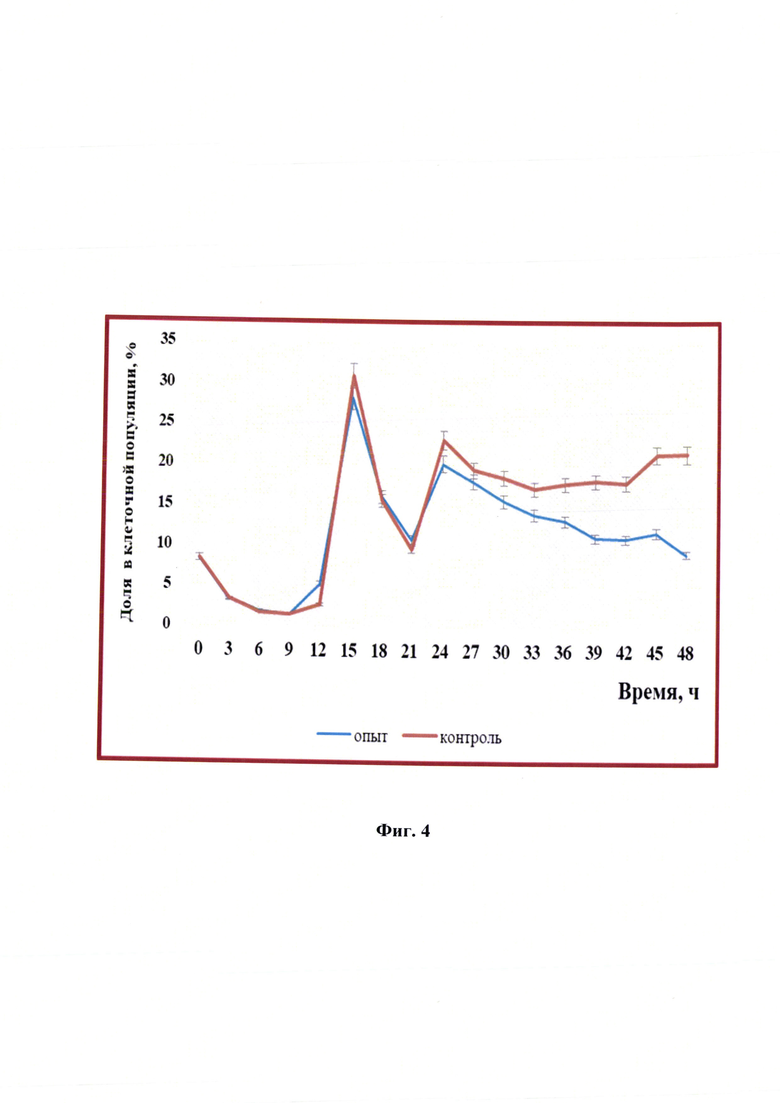
Фиг. 4 - Изменение процентного соотношения клеток, находящихся в S-фазе в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, при репродукции вируса бешенства (n=3, р<0,05).
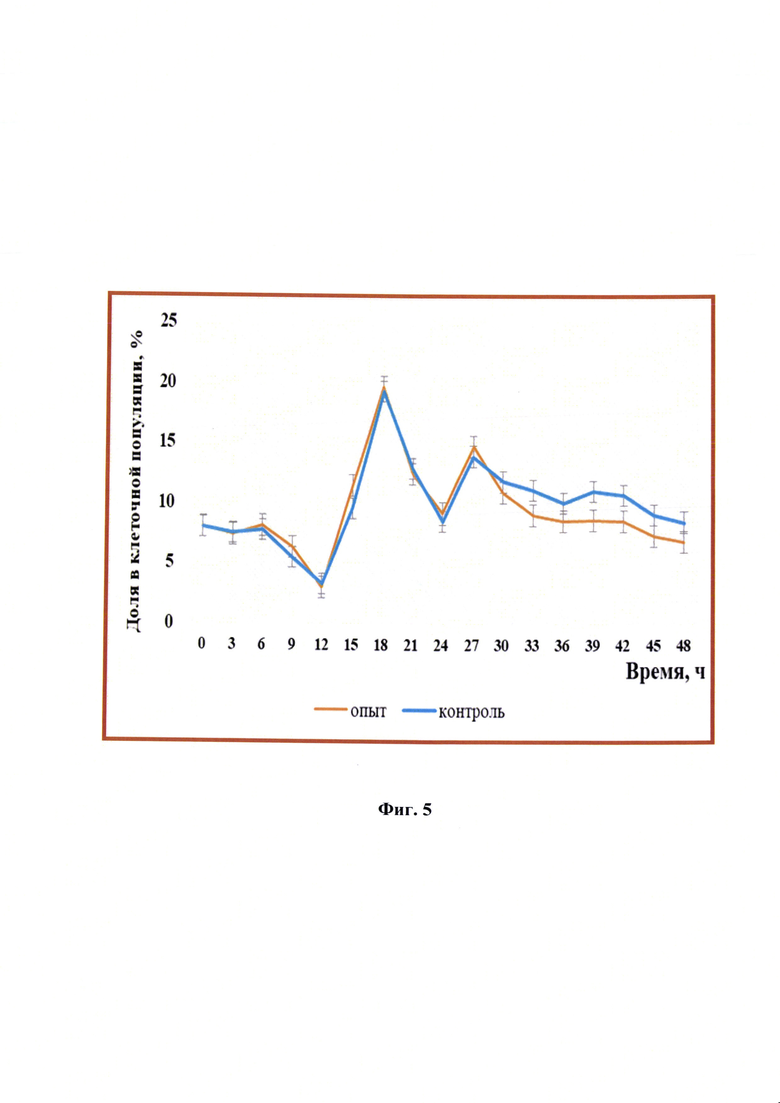
Фиг. 5 - Изменение процентного соотношения клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции вируса бешенства, находящихся на стадии G2+M (n=3, р<0,05).
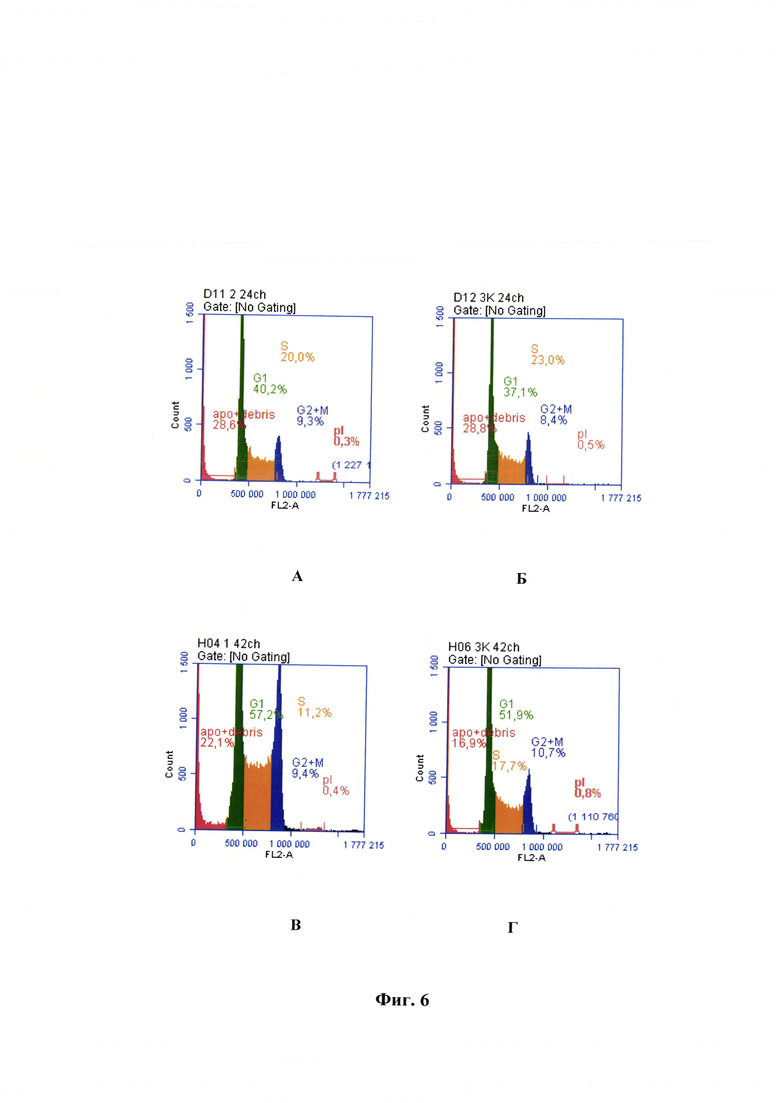
Фиг. 6 - Сравнение ДНК-гистограмм клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства и контрольных образцах после 24 ч репродукции (А - опыт, Б - контроль), через 48 ч (В - опыт, Г - контроль).
Сущность изобретения заключается в новом подходе по определению оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии.
Заявляемый способ основан на: 1) определении стадии клеточного цикла с помощью проточного цитометра «Accuri С6» и концентрации клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH; 2) исследовании трансформационных изменений клеточной ДНК с применением дисульфидсодержащего детергента группы DCD и флуоресцирующего красителя SYTOX™ Green Ready Flow™; 3) заключении по исследованию трансформационных изменений ДНК в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, позволяющем определять оптимальное время завершения процесса репродукции различных штаммов возбудителя бешенства. Подтверждение результатов, получаемых с помощью разработанного способа, осуществляют, проводя следующие количественные исследования: 1) определение титра инфекционной активности вируса бешенства в сырье для вакцин с помощью обратной транскрипции и полимеразной цепной реакции в режиме реального времени (ОТ-ПЦР-РВ) [5]; 2) определение концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени [7].
В настоящее время методом проточной цитометрии проводят количественное определение содержания ДНК в клетке, специфических белковых молекул, узнаваемых моноклональными антителами, а также исследуют клеточный метаболизм, изучают транспортировку катионов кальция и кинетику ферментативных реакций [9]. ДНК-проточная цитометрия используется в прогнозировании течения опухолевого процесса при колоректальных опухолях человека, в мониторинге эндопротезирования коленного и тазобедренного суставов, в биохимическом анализе, при анализе вируснейтрализующей активности антирабических сывороток и иммуноглобулинов и др. [12, 13, 14, 15, 16, 17].
Для определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин метод проточной цитометрии не использовался. Прототипным вариантом для проведения исследования является метод визуального анализа окрашенных клеток, о недостатках которого информация представлена выше. Таким образом, сведений об аналогах использования предлагаемого способа определения трансформационных изменений ДНК в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства методом проточной цитометрии для контроля качества антирабических вакцин авторами не обнаружено.
Разработанный способ определения трансформационных изменений ДНК в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства методом проточной цитометрии для контроля качества антирабических вакцин по сравнению с прототипом отличается высокой чувствительностью, объективностью и возможностью получать количественные показатели процесса репродукции возбудителя бешенства в указанной культуре клеток.
В отличие от прототипа разработанный способ включает этапы оценки стадии клеточного цикла и состояния ДНК пораженных клеток. Применение предложенного способа дает возможность сделать заключение о точном времени завершения процесса репродукции различных штаммов вируса бешенства по состоянию трансформированной ДНК и стадии клеточного цикла для того, чтобы получить сырье с высокими значениями концентрации рибонуклеопротеина. Таким образом, актуально применять предложенный способ для определения трансформационных изменений ДНК в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства методом проточной цитометрии для контроля качества антирабических вакцин.
Ключевым элементом заявляемого способа является количественное определение доли клеток, находящихся в разных фазах клеточного цикла по трансформированной ДНК и заключении об оптимальном времени завершения процесса репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/APvRIAH для изготовления антирабической вакцины.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключается в применении метода проточной цитометрии для исследования стадий клеточного цикла, использовании дисульфидсодержащиего детергента группы DCD и флуоресцирующего красителя SYTOX™ Green Ready Flow™ для детекции трансформационных процессов клеточной ДНК. Полученные данные дают возможность определить оптимальное временя остановки культивирования различных штаммов вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при производстве антирабических вакцин.
Сущность предлагаемого изобретения пояснена на графическом материале - Сравнение ДНК-гистограмм клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства и контрольных образцах после 24 ч репродукции (А - опыт, Б -контроль), через 48 ч (В - опыт, Г - контроль) (фиг. 6).
На предварительном этапе исследования проводят заражение клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH с концентрацией 0,5 млн клеток/см3. Доза заражения вирусом бешенства составляет 0,005 ТЦД50/клетка. Общее время репродукции возбудителя данного заболевания для разработки способа составляет 48 ч. До заражения, а также через каждые 3 ч после внесения вирусной суспензии к клеткам проводят отбор проб. В данных пробах определяют концентрацию клеток, значение титра инфекционной активности и содержание рибонуклеопротеина вируса бешенства. Следует отметить, что в анализе применяют контрольные клетки линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH без заражения и контаминации какими-либо вирусами, бактериями, микоплазмами и грибами.
На основном этапе работы осуществляют повышение проницаемости мембраны клеток для повышения доступа к ДНК клетки с помощью детергента группы DCD, который содержит дисульфидную связь в гидрофобном хвосте. DCD образует мицеллы меньшего размера, чем соответствующие детергенты с линейными углеводородными цепями, обеспечивая при этом хорошую солюбилизацию и восстановление мембранных белков.
Для осуществления процесса повышения проницаемости клеточной мембраны к 100 мкл суспензии клеток, инфицированных вирусом бешенства, добавляют 450 мкл детергирующего соединения, проводят инкубирование в течение 3 мин при комнатной температуре. Клетки осаждают с помощью центрифуги при скорости 2000 об/мин в течение 0,5 мин. В пробирку вносят 40%-ный раствор изопропилового спирта в количестве 500 мкл. Проводят осаждение клеток при 2000 об/мин в течение 0,5 мин. Далее процедуру повторяют с 70% и 80%-ным изопропанолом-2. Пробирки вновь подвергают центрифугированию при скорости 2000 об/мин. К осадку вносят деионизированную воду в количестве 50 мкл, инкубируют их при температуре 60±0,5°С в течение 2 мин, затем проводят элюирование с помощью центрифугирования при 13500 об/мин в течение 1 мин. Полученную суспензию переносят в чистую пробирку и применяют в дальнейшей работе.
Суспензию клеток, пораженных вирусом бешенства, смешивают с флуоресцентным красителем SYTOX™ Green Ready Flow™ в количестве 1:50. Спектральные характеристики данного флуорофора идеально подходят для применения в проточной цитометрии. Концентрация красителя составляет 50 мкг/см3. Смесь инкубируется при температуре 20-25°С в течение 15 мин.
Реагент SYTOX™ Green Ready Flow™ позволяет отличать мертвые клетки от живых в анализах проточной цитометрии. Данный флуорофор возбудим при 488 нм с профилем излучения, подобным флуоресцеинизотиоцианату. Он был выбран из множества флуоресцентных красителей по причине своей высокой стабильности при комнатной температуре, а также дает исключительно ярко-зеленую флуоресценцию при связывании с ДНК и не требует проводить этап отмывания. Краситель SYTOX™ Green Ready Flow™ проникает в клетку после потери целостности мембраны и связывается с ДНК, тем самым, его можно анализировать при возбуждении при 488 нм и детекции при 523 нм.
Смесь помещают в ячейку цитометра для анализа и построения гистограммы. Гидродинамическое фокусирование струи исследуемого материала в струе дисперсионной среды приводит к тому, что клетки выстраиваются поодиночке и в таком порядке пересекают пучок сфокусированных лазерных лучей. Под воздействием определенных световых волн происходит одновременное возбуждение молекулы флуоресцирующего красителя. Свет, исходящий от флуорохрома, фокусируют при помощи оптической системы, состоящей из нескольких зеркал и линз, а затем раскладывают на определенные компоненты. Полученные световые сигналы прибор подвергает анализу и преобразованию в электрические импульсы.
Результаты анализа учитывают инструментальным путем с помощью гистограммы, на которой отражены относительные количественные показатели клеток (в %), находящихся на разных стадиях жизненного цикла с разными вариантами трансформации ДНК. Определяют относительное количество клеток, находящихся в разных фазах клеточного цикла и, тем самым, судят об оптимальном времени прекращения процесса репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH.
Пример 1. Определение динамики изменения концентрации клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них штамма РВ-97 вируса бешенства.
В процессе репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH через каждые 3 ч проводили определение концентрации клеток и оценивали общую динамику ее изменения. Как видно на фиг. 2, количество клеток увеличивалось с 0,5 до 1,9 млн/см3 (в 3,8 раз). Количественные данные для каждых 3 ч детекции отражены в таблице 1. Увеличение концентрации клеток связано с тем, что в процессе культивирования были оптимальные условия. Отмечено, что через 24 ч кратность прироста клеток замедлялась за счет накопления метаболитов. Благодаря увеличению концентрации клеток появилась возможность инфицировать большее их количество и, тем самым, получить более высокие значения титра инфекционной активности и концентрации рибонуклеопротеина вируса бешенства, который является важнейшим иммуногенным компонентом для будущей вакцины.
Пример 2. Определение динамики изменения титра инфекционной активности штамма РВ-97 вируса бешенства в процессе репродукции в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH.
Проводили определение динамики изменения титра инфекционной активности штамма РВ-97 вируса бешенства в процессе репродукции в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH. Пробы отбирали до начала культивирования вируса бешенства, а также через каждые 3 ч в течение 48 ч (фиг. 1). На фиг. 1 отображена динамика накопления титра инфекционной активности при репродукции вируса бешенства. Данные для каждых 3 ч детекции продемонстрированы в таблице 1. Отмечено, что титр инфекционной активности увеличивался от 3,2±0,2 lg ККИД50/см3 при инокуляции вируса до 7,63±0,3 lg ККИД50/см3 через 48 ч репродукции, но более интенсивно он рос в первые 24 часа. Эти данные соотносятся с результатами определения концентрации клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH.
Пример 3. Определение динамики изменения концентрации рибонуклеопротеина штамма РВ-97 вируса бешенства в процессе репродукции в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH.
Проводили определение динамики изменения концентрации рибонуклеопротеина вируса бешенства в процессе репродукции в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH. Пробы отбирали до начала культивирования вируса бешенства, а также через каждые 3 ч в течение 48 ч. Результаты исследования отражены в таблице 1, из которой следует, что за весь период наблюдения содержание иммуногенного компонента увеличилось с 0,20 до 1,55 мкг/см3, при этом за первые 24 ч концентрация рибонуклеопротеина возросла с 0,20 до 1,42 мкг/см3 (в 7 раз), а за вторые 24 ч - с 1,42 до 1,55 мкг/см3 (только в 1,09 раз). Исходя из этого следует, что культивирование штамма РВ-97 вируса бешенства достаточно проводить в течение 24 ч, что обусловлено биологической природой процесса и экономической целесообразностью. Однако, определять данный показатель каждый раз для штаммов вируса бешенства трудоемко, кроме того, существует много факторов, влияющих на течение процесса репродукции вируса бешенства в клеточной линии. Исходя из этого, актуально провести количественный анализ методом ДНК-проточной цитометрии для подтверждения полученных данных.
Пример 4. Определение оптимального времени репродукции штамма РВ-97 вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии.
Определение оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии проводили в соответствии с методикой, представленной выше. Осуществляли пробоподготовку по повышению проницаемости клеток, пораженных вирусом бешенства, а также окрашивание их предложенным флуорофором, детектирующим клеточную ДНК. Результаты исследования, в том числе определения количества клеток, находящихся в разных фазах жизненного цикла, отражены на фиг. 3-6.
Из данных, представленных на фиг. 3 видно, что динамика изменений популяции клеток, находящихся в апоптозе, в контроле и опыте одинакова до 36 ч культивирования (отмечен небольшой временной сдвиг в 3 ч между опытом и контролем через 9 и 12 ч репродукции). Через 36 ч опытные клетки на 28-42% в большей степени переходили в состояние апоптоза по сравнению с контролем.
Содержание опытных клеток, находящихся в стадии фазы G1, через 9 ч увеличивалось на 11,7%, в контрольных отмечали уменьшение данной фазы на 16,6%. В дальнейших исследованиях фазы G1, клетки в контроле и в опыте, изменялись одинаково: через 15-18 ч было уменьшение на 40%, далее ростовой скачок на 45-46%), потом опять снижение на 39-40%, затем снова наблюдалось увеличение. После 33 ч репродукции и до окончания количество опытных клеток, находящихся в фазе G1, было значительно больше, чем контрольных (на 12-21%).
При сравнении количества клеток, находящихся в фазах/состояниях «апоптоз-дебрис» и G1, установили некоторую синхронность между этими фазами: через 9 ч культивирования вируса фаза G1 увеличилась на 10-12%, на 17-19% в это время увеличивался апоптоз. Более выраженную «зеркальность» данных фаз клеточного цикла наблюдали после 24 ч культивирования.
В клеточном цикле интактной популяции клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH также преобладала диплоидная фаза клеточного цикла (G1), во время которой начинался синтез мРНК структурных белков, других клеточных компонентов. На эту фазу приходилось от 30 до 75% клеток в зависимости от условий культивирования.
Известно, что в период G1-фазы клетки начинают увеличиваться в размерах, активируется мРНК и ферменты, необходимые для будущей репликации ДНК [8]. Клетки, находящиеся в фазе G1, в суспензионной культуре поддерживались на уровне 37-70% на протяжении всего времени репродукции вируса, т.е. инфицированные вирусом бешенства клетки служили источником энергии и материалом для синтеза вирусных компонентов, а также для сборки цельных вирионов.
Переход от диплоидной фазы G1 в синтетическую (S-фазу) является одной из контрольных точек клеточного цикла. В зависимости от количества питательных веществ и энергии, а также от внешних факторов культивирования клетка «принимает решение» - вступать в клеточный цикл или же перейти в неделящееся состояние покоя, известное как G0-фаза, которая трансформируется в апоптоз.
Основным событием S-фазы является репликация ДНК, которая имеет свои особенности. Процент клеток в данной фазе в опыте и контроле в первые сутки репродукции вируса бешенства был одинаковым, при этом наблюдали резкое увеличение в виде скачков через 15 и 24 часа в 3,4 и 2,4 раз, соответственно (фиг. 4). Через 24 ч и до окончания репродукции различия между опытными клетками и контрольными начинали возрастать с 8 до 137%. Можно предположить, что клетки этой фазы также участвовали в репродукции вируса.
G2-фаза - последняя из трех последовательных фаз интерфазной стадии клеточного цикла вместе с митотической фазой образовывала тетраплоидную популяцию. В интактных клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, и в инфицированных вирусом бешенства, на фазу подготовки к митозу и сам митоз приходилось от 2 до 20% всей популяции. Через 30 ч репродукции количество опытных клеток в данных фазах начинало уменьшаться на 17-28% по сравнению с контрольными (фиг. 5). Вероятно, это было связано с тем, что ингибировался общий биосинтез и функции клетки-хозяина в результате формирования иммуногенных компонентов вируса бешенства, в том числе рибонуклеопротеина, поэтому уменьшалось содержание клеток, находящихся в фазах G2+M.
Количество полиплоидных клеток в опыте было примерно равно на протяжении 45 часов, только в последние часы репродукции вируса наблюдали их резких скачок в 2,0-2,5 раз, в то время, как в контрольных клетках плоидность популяции имела скачкообразный характер: увеличение через 9, 21, 42 и 48 ч в 2,06; 2,7; 1,56 и 3,18 раз, соответственно. Можно предположить, что имелась частичная резистентность полиплоидных клеток к вирусу.
Информативны ДНК-гистограммы клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства через 24 после начала заражения и через 42 часа культивирования (фиг. 6). Они обобщают графики, представленные на фиг. 3-5. При их анализе можно сделать те же выводы, которые были обозначены выше в данном примере при анализе фаз клеточного цикла. При 29%-ном содержании клеток в фазе G0, 40%-ном для G1-фазы, 20%-ном для S-фазы, 9,3%-ном для G2-фазы достигаются оптимальные показатели культивирования вируса бешенства. При этом концентрация рибонуклеопротеина штамма РВ-97 вируса бешенства составляет 1,42 мкг/см3 (достаточное содержание для составления антирабической вакцины с высокой иммуногенностью и формированием защитного титра антител - ≥0,5 МЕ/см3 [18]. Спустя 25 ч (дополнительный отбор проб) показатели клеток относительно фаз жизненного цикла были следующими: 31% - для фазы G0, 42% - для G1-фазы, 20% - для S-фазы, 10% -для G2-фазы. Далее наблюдалась следующая динамика: количество клеток в состоянии апоптоза и дебриса уменьшалось (на 9% до 48 ч), в G1-фазе увеличивалось (на 17% до 48 ч), в S-фазе уменьшалось (на 9% до 48 ч), в G2-фазе сохранялось на уровне 9-10%. Однако после 24 ч динамика роста концентрации рибонуклеопротеина была очень слабой (в 1,09 раз увеличилась за период 24-48 ч).
Таким образом, по всем показателям (концентрация рибонуклеопротеина как важнейшего иммуногенного компонента и состоянию трансформированной ДНК клеток, инфицированных возбудителем данного заболевания) можно сделать вывод о том, что с биологической и экономической точек зрения 24 ч вполне достаточно для репродукции вируса бешенства штамма РВ-97. По результатам исследования пришли к выводу, что анализируемые параметры при этом должны иметь следующие показатели: относительное количество клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH составляет 29-31% - для фазы G0, 40-42% - для G1-фазы, 20-22% - для S-фазы, 9,3-10,0% - для G2-фазы.
Пример 5. Определение оптимального времени репродукции различных штаммов вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии (предлагаемое изобретение).
Изучены фазы клеточного цикла при репродукции в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH штаммов ВНИИЗЖ, SAD-19, Щелково-51, CVS-27 вируса бешенства и получены результаты, позволяющие оценить продолжительность и доминирование соответствующих фаз клеточного цикла G1, S, G2+M на разных этапах культивирования возбудителя данного заболевания.
При изучении динамики, происходящей в ДНК инфицированных штаммом ВНИИЗЖ вируса бешенства клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, замечено, что через 27 ч культивирования количество клеток в фазе G1 составляло 40,9%, S-фазе - 20,2%, G2-фазе - 9,9%. Полученные результаты были подтверждены данными по концентрациям рибонуклеопротеина вируса бешенства: 0 ч - 0,2; 24 ч - 1,58; 27 ч - 1,79; 30 ч - 1,81; 33 ч - 1,84; 36 ч - 1,84; 39 ч - 1,87; 42 ч - 1,87; 45 ч - 1,89; 48 ч - 1,89 мкг/см3. Из приведенного ряда концентраций видно, что до 27 ч наблюдался рост концентрации иммуногенного компонента, однако, далее до 48 ч культивирования динамика накопления рибонуклеопротеина была очень слабой (только в 1,06 раз). Таким образом, оптимальное время для культивирования штамма ВНИИЗЖ вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH - 27 ч.
При изучении динамики, происходящей в ДНК инфицированных штаммом SAD-19 вируса бешенства клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, замечено, что через 33 ч культивирования вируса бешенства количество клеток в фазе G1 составляло 40,5%, S-фазе - 21,0%, G2-фазе - 10,0%. Полученные результаты были подтверждены данными по концентрациям рибонуклеопротеина вируса бешенства: 0 ч - 0,2; 24 ч - 1,29; 27 ч - 1,39 ч; 30 ч - 1,71; 33 ч - 2,04; 36 ч - 2,04; 39 ч - 2,06; 42 ч - 2,06; 45 ч - 2,07; 48 ч - 2,08 мкг/см3. Так, до 33 ч наблюдался рост концентрации иммуногенного компонента, однако, далее до 48 ч культивирования динамика накопления рибонуклеопротеина была очень слабой (в 1,02 раз). Таким образом, оптимальное время для культивирования штамма SAD-19 вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH - 33 ч.
При изучении динамики, происходящей в ДНК инфицированных штаммом Щелково-51 вируса бешенства клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, замечено, что через 27 ч культивирования вируса бешенства количество клеток в фазе G1 составляло 40,9%, S-фазе - 21,8%, G2-фазе - 9,5%. Полученные результаты были подтверждены данными по концентрациям рибонуклеопротеина вируса бешенства: 0 ч - 0,2; 24 ч - 1,41; 27 ч - 1,72 ч; 30 ч - 1,75; 33 ч - 1,75; 36 ч - 1,76; 39 ч - 1,78; 42 ч - 1,79; 45 ч - 1,80; 48 ч - 1,80 мкг/см3. Видно, что до 27 ч наблюдался рост концентрации иммуногенного компонента, однако, далее до 48 ч культивирования динамика накопления рибонуклеопротеина была очень слабой (в 1,05 раз). Таким образом, оптимальное время для культивирования штамма Щелково-51 вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH - 27 ч.
При изучении динамики, происходящей в ДНК инфицированных штаммом CVS-27 вируса бешенства клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH, отмечено, что через 30 ч культивирования вируса бешенства количество клеток в фазе G1 составляло 42,0%, S-фазе - 22,0%, G2-фазе - 9,2%. Полученные результаты были подтверждены данными по концентрациям рибонуклеопротеина вируса бешенства: 0 ч - 0,2; 24 ч - 1,28; 27 ч - 1,48 ч; 30 ч - 1,87; 33 ч - 1,88; 36 ч -1,89; 39 ч - 1,89; 42 ч - 1,90; 45 ч - 1,92; 48 ч - 1,93 мкг/см3. Рост концентрации иммуногенного компонента наблюдался до 27 ч, однако, далее до 48 ч культивирования динамика накопления рибонуклеопротеина была очень слабой (в 1,03 раз). Таким образом, оптимальное время для культивирования штамма CVS-27 вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH - 30 ч.
Пример 6. Установление корреляции данных по определению оптимального времени репродукции различных штаммов вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии (предлагаемое изобретение) и прототипным способом.
Для установления корреляции данных по определению оптимального времени репродукции различных штаммов вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии (предлагаемое изобретение) и прототипным способом проводили опыт по 240 культивациям возбудителя данного заболевания. Применяли штаммы РВ-97 (n=60), ВНИИЗЖ (n=60), Щелково-51 (n=60), SAD-19 (n=60) и CVS-27 (n=60). По результатам анализа получили данные, которые представлены в таблице 2.
Обнаружено, что для штамма РВ-97 клетки с трансформированной ДНК находились в G0-фазе в количестве 29,2-30,8%, в фазе G1 - 40,2-42,0%, в фазе S - 20,1-22,0%, в фазе G2 - 9,4-9,8% по итогам 24 ч репродукции вируса бешенства. Данное время подтвердилось прототипным способом (24 ч с концентрацией рибонуклеопротеина - 1,42±0,13 мкг/см3). Далее динамика роста составляла не более, чем в 1,1 раз, что обуславливает необходимость остановки процесса культивирования вируса в указанное время.
Для штамма ВНИИЗЖ клетки с трансформированной ДНК находились в G0-фазе в количестве 29,0-30,1%, в фазе G1 - 40,1-41,8%, в фазе S - 20,5-22,0%, в фазе G2 - 9,3-10,0% по итогам 27 ч репродукции вируса бешенства. Данное время подтвердилось прототипным способом (27 ч с концентрацией рибонуклеопротеина - 1,79±0,17 мкг/см3). Далее динамика роста составляла не более, чем в 1,1 раз, что обуславливает необходимость остановки процесса культивирования вируса в указанное время.
Для штамма SAD-19 клетки с трансформированной ДНК находились в G0-фазе в количестве 29,1-30,9%, в фазе G1 - 40,3-41,7%, в фазе S - 20,0-21,8%, в фазе G2 - 9,3-10,0% по итогам 33 ч репродукции вируса бешенства. Данное время подтвердилось прототипным способом (33 ч с концентрацией рибонуклеопротеина - 2,04±0,11 мкг/см3). Далее динамика роста составляла не более, чем в 1,1 раз, что обуславливает необходимость остановки процесса культивирования вируса в указанное время.
Для штамма Щелково-51 клетки с трансформированной ДНК находились в G0-фазе в количестве 29,0-30,8%, в фазе G1 - 40,0-42,0%, в фазе S - 20,2-22,0%, в фазе G2 - 9,3-9,9% по итогам 27 ч репродукции вируса бешенства. Данное время подтвердилось прототипным способом (27 ч с концентрацией рибонуклеопротеина - 1,87±0,14 мкг/см3). Далее динамика роста составляла не более, чем в 1,1 раз, что обуславливает необходимость остановки процесса культивирования вируса в указанное время.
Для штамма CVS-27 клетки с трансформированной ДНК находились в G0-фазе в количестве 29,2-31,0%, в фазе G1 - 40,0-42,0%, в фазе S - 20,1-22,0%, в фазе G2 - 9,4-10,0% по итогам 30 ч репродукции вируса бешенства. Данное время подтвердилось прототипным способом (30 ч с концентрацией рибонуклеопротеина - 1,72±0,15 мкг/см3). Далее динамика роста составляла не более, чем в 1,1 раз, что обуславливает необходимость остановки процесса культивирования вируса в указанное время.
Основными преимуществами предлагаемого изобретения является возможность быстро, в течение 30 мин, определить, на какой стадии репродукции вируса бешенства находится процесс и, тем самым, остановить его с высоким накоплением рибонуклеопротеина, являющегося важнейшим иммуногенным компонентов при производстве антирабических вакцин. Разработанный способ высокочувствительный, объективный и позволяет определять трансформационные изменения ДНК клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH при репродукции в них вируса бешенства методом проточной цитометрии для контроля качества антирабических вакцин на разных этапах репродукции возбудителя. Данный метод характеризуется высокой скоростью анализа, обладает возможностью определять клеточные линии, способен проводить детектирование ДНК клетки в разных фазах жизненного цикла.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии»:
1. Груздев, К.Н. Бешенство животных / К.Н. Груздев, В.В. Недосеков. - Москва: ООО "Аквариум ЛТД", 2001. - 303 с. - ISBN 5-85684-622-2. - EDN PvFDAGX.
2. Анисина, О.В. Оценка методов контроля и профилактики бешенства собак / О.В. Анисина, В.И. Клюкина, М.Н. Романенко // Вопросы нормативно-правового регулирования в ветеринарии. - 2015. - №2. - С. 21-23. - EDN TWNFVJ.
3. Осипова, Н.И. Изучение напряженности иммунитета у лис, вакцинированных против бешенства вирус-вакцинами из штаммов "РВ-97" и "SAD-Bern", и их потомства / Н.И. Осипова // Ветеринария. Реферативный журнал. - 2011. - №2. - С. 432. - EDN NTNCYN.
4. Генетическая характеристика изолятов вируса бешенства, выявленных на территории Владимирской области / С.А. Чупин, М.И. Доронин, Е.В. Чернышова, М.И. Шульпин // Ветеринария сегодня. - 2015. - №4(15). - С. 46-48. - EDN VODNYT.
5. Патент РФ №2755925, 03.08.2020 Способ опосредованного определения титра инфекционной активности вируса бешенства штамма ВНИИЗЖ в сырье для инактивированной антирабической вакцины методом ПЦР в режиме реального времени // Заявка 2020126182 / Доронин М.И., Михалишин Д.В., Борисов А.В., Луговская Н.Н., Оковытая Т.В.
6. Патент РФ №2761535, 02.11.2020 Способ опосредованного определения количества инфекционных доз вируса бешенства штамма РВ-97 в сырье для аттенуированной вакцины методом обратной транскрипции и полимеразной цепной реакции в режиме реального времени // Заявка 2020136135 / Доронин М.И., Михалишин Д.В., Борисов А.В.
7. Патент РФ №2775745, 07.07.2022 Способ опосредованного определения концентрации рибонуклеопротеина вируса бешенства в сырье для вакцины при исследовании на максимальные экстремумы графиков второй производной для логистических кривых реакции амплификации в режиме реального времени // Заявка №2021121290 / Доронин М.И., Михалишин Д.В., Борисов А.В., Оковытая Т.В.
8. Ищенко, И.О. Проточная цитометрия как современный метод биохимического анализа / И.О. Ищенко, М.Д.Л. Оппедизано // Forcipe. - 2020. - Т. 3. - №S1. - С. 121-122. - EDN OFQELM.
9. Битанова, Э.Ж. Проточная цитометрия - преимущества метода и области применения / Э.Ж. Битанова, А.С. Тарабаева // Вестник Казахского национального медицинского университета. - 2017. - №4. - С. 463-465. - EDN YOSNZD.
10. Хайдуков, С.В. Проточная цитометрия как современный метод анализа в биологии и медицине / С.В. Хайдуков, А.В. Зурочка // Медицинская иммунология. - 2007. - Т. 9. - №4-5. - С. 373-378. - EDN RRWKOJ.
11. Волотовский, И.Д. Проточная цитометрия. Основы технологии и ее применение в биологии / И.Д. Волотовский, С.В. Пинчук // Известия Национальной академии наук Беларуси. Серия биологических наук. - 2022. - Т. 67. - №2. - С. 229-242. - DOI 10.29235/1029-8940-2022-67-2-229-242. - EDN PDHKJC.
12. Стахнева, Е.М. Современные методы исследования атеросклероза и ишемической болезни сердца: проточная цитометрия / Е.М. Стахнева, Ю.И. Рагино // Бюллетень сибирской медицины. - 2021. - Т. 20. - №2. - С. 184-190. - DOI 10.20538/1682-0363-2021-2-184-190. - EDN UGOZQY.
13. Калашникова, А.А. Проточная цитометрия в диагностикеи оценке эффективности терапии системных бактериальных инфекций и сепсиса / А.А. Калашникова, Н.В. Макарова, Н.М. Калинина // Медицинская иммунология. - 2015. - Т. 17. - №S. - С. 364. - EDN UQDDFH.
14. Набиева, У.П. Проточная цитометрия: принципы метода и применение в биомедицинских исследованиях / У.П. Набиева, Т.Р. Хегай // Журнал теоретической и клинической медицины. - 2018. - №4. - С. 46-49. - EDN JJGAYE.
15. Проточная цитометрия при анализе вируснейтрализующей активности антирабических сывороток и иммуноглобулина / С.В. Генералов, А.Л. Кравцов, В.А. Кожевников [и др.] // Инфекция и иммунитет. - 2019. - Т. 9. - №1. - С. 107-114. - DOI 10.15789/2220-7619-2019-1-107-114. - EDN ZGXQRF.
16. ДНК-проточная цитометрия в прогнозировании течения опухолевого процесса при колоректальных опухолях человека / Т.Г. Николаева, С.В. Крутий, В.В. Пророков, Я.В. Добрынин // Российский биотерапевтический журнал. - 2004. - Т. 3. - №2. - С. 71. - EDN HZAATN.
17. Нурмуканов, А.С. Микробиологический экспресс-анализ: проточная цитометрия / А.С. Нурмуканов, Н.В. Башкова, Е.П. Гаврюнькина // Молочная промышленность. - 2011. - №4. - С. 37-38. - EDN NULSJD.
18. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019 (OIE). Rabies. - 2019. - Ch. 3.1.17. - P. 578-612.
Изобретение относится к области биотехнологии и производству антирабических вакцин. Предложен способ определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка BHK-21/SUSP/ARRIAH для изготовления антирабических вакцин. Способ осуществляют с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии. Основными преимуществами предлагаемого изобретения является возможность в течение 30 мин определять, на какой стадии репродукции вируса бешенства находится процесс, и тем самым останавливать его с высоким накоплением рибонуклеопротеина, являющегося важнейшим иммуногенным компонентом при производстве антирабических вакцин. Данный метод характеризуется высокой скоростью анализа и отличается высокой степенью объективности в измерении интенсивности флуоресценции. 1 з.п. ф-лы, 6 ил., 2 табл., 6 пр.
1. Способ определения оптимального времени репродукции вируса бешенства в клетках линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH для изготовления антирабических вакцин с помощью анализа трансформационных изменений клеточной ДНК методом проточной цитометрии, включающий определение относительного содержания клеток на G0-, G1-, S-, G2-фазах клеточного цикла с помощью проточного цитометра «Accuri С6» по трансформациям ДНК и основанный на следующих показателях фаз жизненного цикла клеток, при которых требуется завершать процесс репродукции вируса бешенства в указанной культуре клеток: относительное количество клеток линии почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH составляет 29-31% - для фазы G0, 40-42% - для G1-фазы, 20-22% - для S-фазы, 9,3-10,0% - для G2-фазы.
2. Способ по п. 1, отличающийся тем, что для детекции трансформационных процессов клеточной ДНК используют дисульфидсодержащий детергент группы DCD и флуоресцирующий краситель SYTOX™ Green Ready Flow™.
| Способ спектрометрического определения концентрации рибонуклеопротеина вируса бешенства по оценке количества молекул вирусной РНК в сырье для антирабических вакцин | 2020 |
|
RU2748475C1 |
| М | |||
| И | |||
| ДОРОНИН и др., Исследование биологических свойств перевиваемой суспензионной линии клеток ВНК-21/SUSP/ARRIAH, Ветеринария сегодня | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| CN111560466 A, 21 | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |

