Изобретение относится к оценке обсемененности среды бактериями, имеющими системы скоординированного ответа ("quorum sensing", QS) первого типа, и также может быть использовано в исследованиях систем коммуникации бактерий.
Разработан способ определения аутоиндуктора (АИ) - низкомолекулярного вещества N-(3-оксогексаноил)-лактона L-гомосерина (3-oxo-C6-HSL) QS-систем первого типа с высокой чувствительностью.
Цельноклеточные биосенсоры являются живыми объектами с помощью которых определяется наличие в среде ли внутри клеток биологически активных веществ. Такие биосенсоры востребованы для биотехнологических приложений, в генетической инженерии, в фармацевтике при разработке медицинских препаратов, а также для детекции токсичных агентов при мониторинге окружающей среды [1-5]. В качестве цельноклеточных биосенсоров могут применяться как природные организмы или клеточные культуры, так и полученные с помощью генетической инженерии [6, 7]. Генно-инженерные цельноклеточные бактериальные биосенсоры - это клетка, содержащая генетическую конструкцию в составе плазмидной или хромосомной ДНК, в состав которой встроен индуцируемый (регулируемый) промотор и репортерный ген.
При конструировании lux-биосенсоров на основе клеток Escherichia coli обычно используются luxCDABE гены [8-13], так как в этом случае к клеткам биосенсора не нужно добавлять субстрат люциферазной реакции - алифатический альдегид (luxCDE гены кодируют редуктазу, синтезирующую тетрадеканаль из жирной кислоты [14]. Lux-биосенсоры, в которых luxCDABE кассета транскрипционно слита со стрессовыми промоторами бактерий Е. coli как правило применяются для детекции различных токсикантов и дифференциации их по типу воздействия на клеточные компоненты [10, 17-22]. Использование стрессовых промоторов позволяет специфично детектировать токсиканты действующие избирательно на различные компоненты клетки (белки, мембраны, ДНК), а также детектировать специфично некоторые, встречающиеся в природе токсиканты (активные формы кислорода, тяжелые металлы и т.д.) [8, 13, 16, 20]. В патентах [12, 16, 18] lux-биосенсоры с индуцируемыми промоторами, были использованы для определения нитратов, фенолов, бензина, несимметричного диметил-гидразина и других токсикантов. Ацильные производные гомосерин лактона, зачастую служащие аутоиндуторами QS-систем первого типа (такие системы также называют luxR-luxI типа) различных бактерий, являются биологически активными веществами, но не токсикантами, по этому не могут быть обнаружены с использованием стрессовых промоторов. В работах [21, 22] различные промоторы QS систем использовались для детекции АИ: PluxCDABEG из Aliivibrio fischeri, чувствительный к N-(3-оксогексаноил)-лактону L-гомосерина, PlasI из Pseudomonas aeruginosa, чувствительный к N-(3-оксододеканоил)-лактон L-гомосерина, PtraR из Agrobacterium tumefaciens, чувствительный к N-(3-оксооктаноил)-лактону L-гомосерина.
Прототипом настоящего изобретения является патент [23], в котором для определения АИ применяется промотор PluxCDABEG из A. fischeri. Чувствительность описанных выше биосенсоров как в прототипе, так и в других работах не превышает 1 нМ АИ в среде.
Концентрация АИ 1-10 нМ является пороговой для активации QS системы первого типа. Биосенсоры для детекции ацильных производных гомосерин лактона применяются для определения АИ в среде, для поиска блокаторов QS систем, для изучения внутриклеточных факторов - модуляторов QS систем. Чувствительность биосенсора для определения АИ в природных средах и ряде других работ является важным параметром. Поиск возможностей для детекции малых концентраций АИ в среде оставался до настоящего момента не решенной задачей.
Техническим результатом предложенного изобретения является возможность детекции АИ в окружающей среде и определения АИ-синтезирующих бактериальных штаммов при их культивации на твердых и жидких питательных средах. Чувствительность предложенного биосенсора к 3-охо-С6-HSL по сравнению с прототипами возросла примерно на порядок. Пороговая концентрация активации lux-биосенсора (получение достоверных различий от фоновой люминесценции биосенсорных клеток) составила 30 пМ. Измерения не требуют дорогой и сложной аппаратуры, возможны, как в стационарных, так и в полевых условиях.
Задачи решены путем котрансформации бактериальных клеток Е. coli парой плазмид с репликонами из разных групп несовместимости (ColEI и p15):
1. плазмида pVFR1, несущая гены luxCDABE Photorhabdus luminescens под контролем индуцируемого промотора PluxICDABEG A. fischeri и ген luxR A. fischeri под контролем промотора PluxR;
2. плазмида pSVRAF, несущая дополнительную копию гена luxR A. fischeri под контролем двух промоторов PtetR и PluxR, что обеспечивает повышенное содержание регуляторного белка LuxR в биосенсорной клетке.
Повышенный уровень экспрессии регуляторного белка LuxR, который является АИ-зависимым активатором транскрипции, обусловливает повышение чувствительности биосенсора к АИ.
В предложенном изобретении для определения АИ использован биосенсор, обладающий большей чувствительностью к малым концентрациям, чем биосенсоры применяемые ранее. При попадании ацильных производных гомосерин лактона в среду сигнал (биолюминесценция) клеток индуцируется (увеличивается).
Помимо (pVFR1 + pSVRAF) могут быть использованы следующие комбинации аналогичных плазмид:
1) pSV16 + pIV2 - гены luxCDABE P. luminescens под контролем индуцируемого промотора Pluxl A. logei и ген luxR2 A. logei под контролем промотора PluxR2 + дополнительная копия гена luxR2 A. logei;
2) pIVA + pIV3 - гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxCDABEG A. logei и ген luxR1 A. logei под контролем промотора PluxR1 + дополнительная копия гена luxR1 A. logei;
3) pAS1 + pIVAS1 - гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxCDABEG Aliivibrio salmonicida и ген luxR1 A. salmonicida под контролем промотора PluxR1 + дополнительная копия гена luxR1 A. salmonicida;
4) pAS2 + pIVAS2 - гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxI A. salmonicida и ген luxR2 A. salmonicida под контролем промотора PluxR2 + дополнительная копия гена luxR2 A. salmonicida;
Методика измерения влияния токсиканта на биолюминесценцию биосенсора была примерно одинаковой для всех lux-биосенсоров.
Определение ацильных производных гомосерин-лактонов в среде с помощью lux-биосенсора требует проведения следующих операций:
1. Приготовление проб, содержащих соответствующие биосенсоры;
- выращивание клеток Е. coli, являющихся биосенсорами, при температуре 30-37°С до ранней экспоненциальной фазы (OD=0,05-0,1);
- отбор проб по 200 мкл в виалы (3-4 виал для каждого образца).
2. Добавление к пробам раствора ацил-гомосерина лактона в различных концентрациях или исследуемого образца в различных разведениях;
- приготовление положительного и отрицательного контролей. В качестве положительного контроля используют 3охо-С6-HSL (разведенный в воде) в концентрациях 100 нМ, 10 нМ, 1 нМ и 0,1 нМ. В качестве отрицательного -дистиллированная вода;
- добавление по 20 мкл исследуемой среды к каждой пробе (кроме контрольных);
(Все пробы следует приготовить в двойном или тройном экземплярах и затем использовать средние значения полученных экспериментальных величин).
3. Измерение интенсивностей биолюминесценции проб следует проводить 2,5-3 часа при температуре 20-280°С. Однако наличие высоких концентраций АИ (более 10 нМ) в среде фиксируется уже через 5-10 минут.
4. Обработка полученных результатов. Полученные графики на контрольных образцах сравнивают с графиками исследуемых образцов.
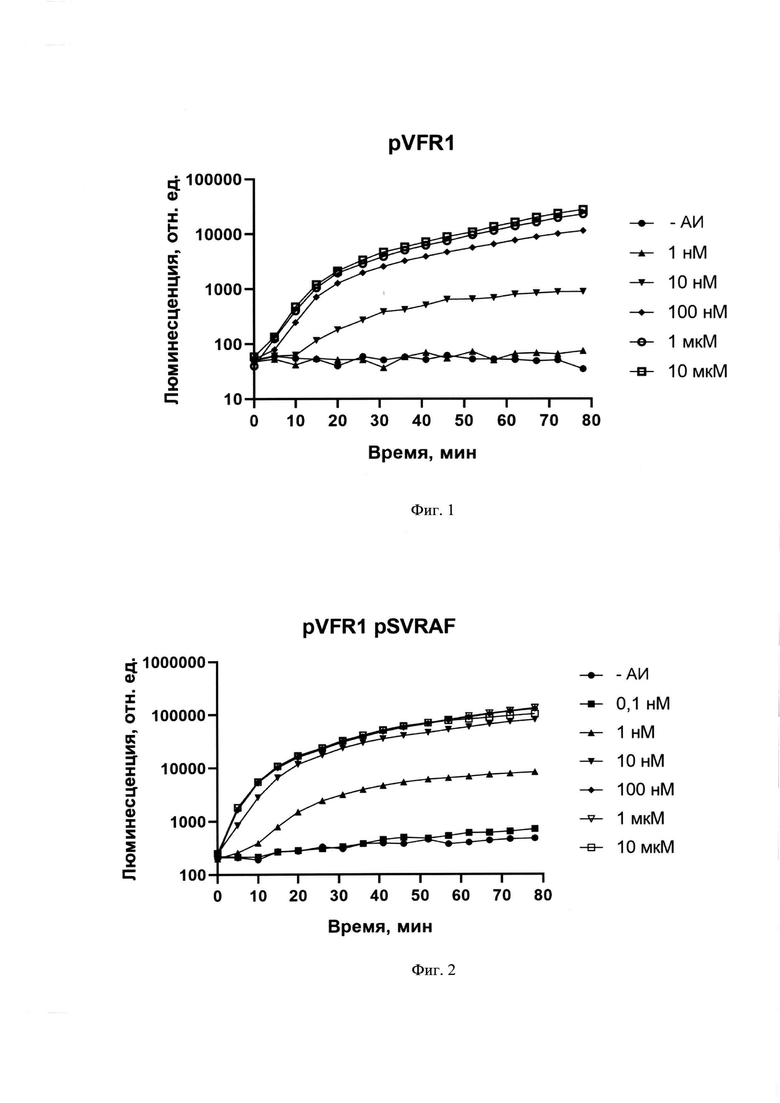
Существо изобретения поясняется прилагаемыми графиками, где на фиг. 1 показаны зависимости люминесценции биосенсоров Е. coli pVFR1 (А) и E.coli pVFR1, pSVRAF (Б) от времени при добавлении 3-охо-С6-HSL в различных концентрациях.
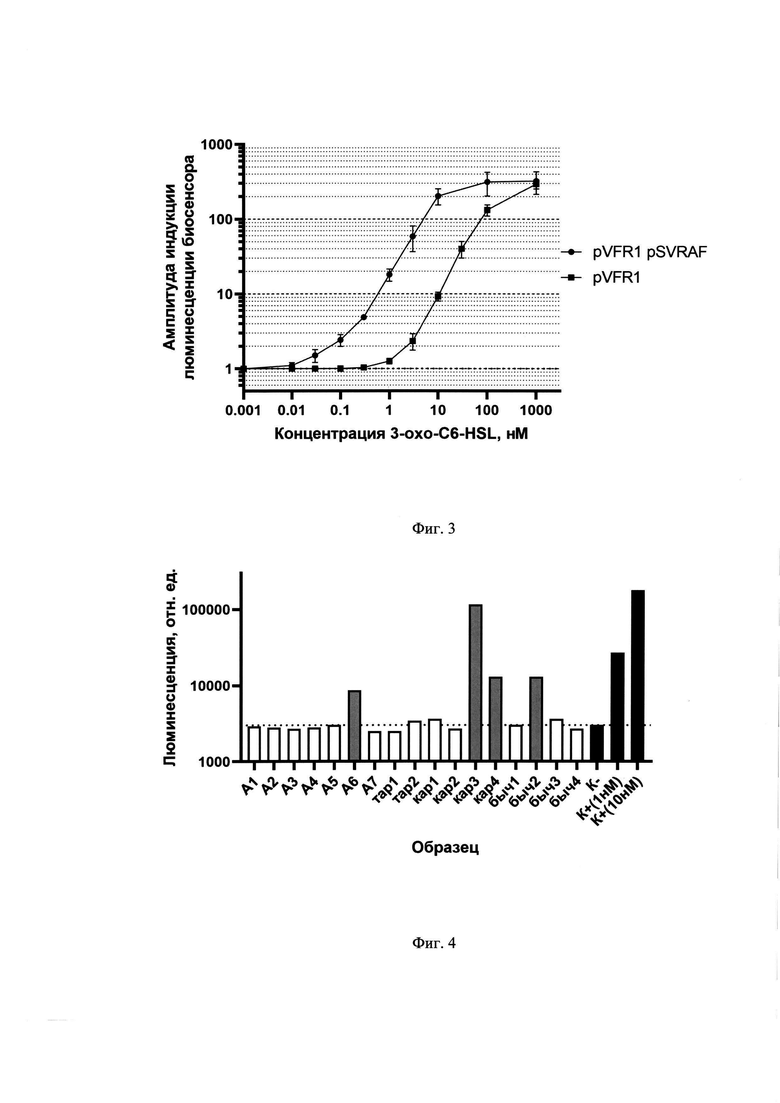
Увеличение дозы регуляторного гена позволило увеличить не только чувствительность биосенсора примерно на порядок, но и скорость отклика. На фиг. 2 приведена амплитудно-концентрационная характеристика биосенсоров с базовым содержанием LuxR (Ε. coli pVFR1) и повышенным содержанием LuxR (Ε. coli pVFR1 pSVRAF). Увеличение содержания регуляторного белка в клетке может быть достигнуто внесением дополнительной копии гена luxR (или его гомологов) не только под собственным промотором, но под более сильным промотором или комбинацией промоторов. На фиг. 3 приведена информация об использовании различных вариантов повышения LuxR белка в клетке, в том числе с использованем более сильного Plac промотора для транскрипции дополнительной копии гена luxR A. fischeri. На графике приводится сравнение АИ-зависимой активации биосенсоров, обладающих одной копией luxR (R), двумя копиями (R+R) и геном luxR под контролем более сильного промотора Plac (Plac-R).
Биосенсор с повышенной чувствительностью на основе LuxR A. fischeri позволяет детектировать присутствие 3охоС6-HSL в растворе в концентрациях от 30 пМ (концентрация вещества в пробе с биосенсорными клетками). Применимость биосенсорного штамма Е. coli MG1655 pVFR1 pSVRAF для поиска ацильных производных гомосерина лактона в окружающей среде продемонстрирована на образцах содержимого кишечника рыб (тар1, тар2, кар1, кар2, кар3, кар4, быч1, быч2, быч3, быч4), пойманных в акватории Таганрогского залива Азовского моря, и образцах воды и илистых донных отложений (Α1, А2…А7), собранных в районе лова рыбы (фиг.4).
Из образцов А6, кар3, кар4 и быч2, содержащих АИ (фиг. 4), были изолированы отдельные бактериальные штаммы и посредством их перекрестного высева на агаризованную среду с биосенсорным штаммом были идентифицированы те из них, которые способны к продукции АИ (фиг.5). Секвенирование генов 16S рРНК отобранных штаммов показало, что они принадлежат видам Aeromonas veronii, Aeromonas hydrophila. Бактерии данного рода - повсеместно распространенные водные бактерии, являющиеся патогенами преимущественно пойкилотермных животных, таких как земноводные, рыбы и амфибии. Данные бактерии обладают QS системой первого типа, ответственной за факторы вирулентности [24].
Полученные результаты подтверждают, что биосенсорный штамм MG1655 pFR pSVRAF с дополнительной копией гена luxR применим для поиска ацильных производных гомосерина лактона в природных образцах и обнаружения бактерий, синтезирующих ацильные производные гомосерина лактона.
Изобретение позволяет создать высокочувствительный, дешевый и быстрый тест-метод на основе измерения интенсивности биолюминесценции.
Использованные источники информации:
1. Su L., Jia W., Hou С.and Lei Y. 'Microbial biosensors: A review'// Biosensors and Bioelectronics. Biosens Bioelectron. 2011. pp. 1788-1799. doi: 10.1016/j.bios.2010.09.005.
2. Woutersen M., Belkin S., Brouwer В., Van Wezel A. P. and Heringa Μ. B. 'Are luminescent bacteria suitable for online detection and monitoring of toxic compounds in drinking water and its sources?'// Analytical and Bioanalytical Chemistry. Springer. 2011. pp. 915-929. doi: 10.1007/s00216-010-4372-6.
3. Chauhan Α., Ripp S., Layton A.C, and Sayler G.S. 'Pseudomonas fluorescens HK44: Lessons learned from a model whole-cellz bioreporter with a broad application history'// Sensors. Multidisciplinary Digital Publishing Institute (MDPI). 2012. pp. 1544-1571. doi: 10.3390/s120201544.
4. Kessenikh A, Gnuchikh E, Bazhenov S, Bermeshev M, Pevgov V, Samoilov V, Shorunov S, Maksimov A, Yaguzhinsky L, Manukhov I. Genotoxic effect of 2,2'-bis(bicyclo[2.2.1] heptane) on bacterial cells. PlosOne 2020 15(8): e0228525. https://doi.org/10.1371/journal.pone.0228525.
5. Гнучих Е.Ю., Манухов И.В., Завильгельский Г.Б. Шаперон DnaK участвует в фолдинге, но не в рефолдинге термоинактивированных белков в Bacillus subtilis. Генетика. 2020. Т. 56. №9. С. 1034-1042.
6. Дерябин Д.Г., Каримов И.Ф. Биохемилюминесцентный способ определения фагоцитарной активности нейтрофилов. Патент РФ 2366953 от
7. Harpaz D., Yeo L., Cecchini F., Koon Т., Kushmaro Α., Tok Α., Marks R. and Eltzov E. 'Measuring artificial sweeteners toxicity using a bioluminescent bacterial panel'// Molecules. 2018. 23(10). p. 2454. doi: 10.3390/molecules23102454.
8. Van Dyk Т.K., Majarian W.R., Konstantinov К.В., Young R.M., Dhurjati P.S. and LaRossa R.A. 'Rapid and sensitive pollutant detection by induction of heat shock gene- bioluminescence gene fusions' // Applied and Environmental Microbiology. 1994. 60(5). pp. 1414-1420. doi: 10.1128/aem.60.5.1414-1420.1994.
9. Van Dyk Т.K., Reed T.R., Vollmer A.C. and LaRossa R.A. 'Synergistic induction of the heat shock response in Escherichia coli by simultaneous treatment with chemical inducers' // Journal of Bacteriology. 1995. 177(20). pp. 6001-6004. doi: 10.1128/jb.177.20.6001 -6004.1995.
10. Van Dyk, Т.К. and Rosson, R.A. (1998) 'Photorhabdus luminescens luxCDABE promoter probe vectors', Methods in molecular biology (Clifton, N.J.), 102, pp. 85-95. doi: 10.1385/0-89603-520-4:85.
11. Bechor O., Smulski D. R., Van Dyk Т.K., LaRossa R.A. and Belkin S. 'Recombinant microorganisms as environmental biosensors: Pollutants detection by Escherichia coli beanngfabA'::lux fusions' // Journal of Biotechnology. 2002. 94(1). pp. 125-132. doi: 10.1016/S0168-1656(01)00423-0.
12. LaRossa R.A., Majarian W.R., Van Dyk Т.K. Highly sensitive method for detecting environmental insults. US patent №5683868 Filed: October 6. 1994
13. Манухов И.В., Котова В.Ю. and Завильгельский Г.Б. 'Lux-биосенсоры для детекции SOS-ответа, теплового шока и окислительного стресса' // Биотехнология. 2009. 6. pp. 16-25
14. Meighen Ε.A. 'Molecular biology of bacterial bioluminescence'// Microbiological Reviews. 1991. 55(1). pp. 123-142. doi: 10.1128/mmbr.55.1.123-142.1991.
15. Завильгельский Г.Б., Котова В.Ю., Манухов И.В., Кондратьев А.Д., Самброс В.В., Шатров Я.Т., Чалкин С.Ф. Набор lux-биосенсоров для определения гептила в среде. Патент РФ №2297450 от 2007.04.20
16. Завильгельский Г.Б., Котова В.Ю., Манухов И.В., Мелькина О.Е. Набор lux-биосенсоров для определения детергентов гидрофобной природы в среде. Патент РФ №2355760 от 2009.05.20.
17. Манухов И.В., Горбунов М.А., Дегтев Д.И., Завильгельский Г. Б., Кессених А.Г., Коноплева М.Н., Котова В.Ю., Краснопеева Е.Д., Мотовилов К.А., Осетрова М.С., Чалкин С.Ф., Шатров Т.Я. Набор lux-биосенсоров для определения генотоксичных продуктов неполного окисления несимметричного диметилгидразина в среде. Патент РФ №2569156 от 18.12.2014.
18. Кессених А.Г., Манухов И.В., Баталова Э.Р., Высоких М.Ю., Коноплева М.Н., Котова В.Ю., Горбунов Μ. Α., Чалкин. С.Ф., Завильгельский Г. Б. Набор lux-биосенсоров для детекции токсичных продуктов неполного окисления несимметричного диметилгидразина в среде. Патент РФ №2626569 от 17.12.2015.
19. Zavilgelsky, G. В.; Kotova, V. Yu.; Manukhov, I. V. (2007) Action of 1,1-dimethylhydrazine on bacterial cells is determined by hydrogen peroxide. // Mutation research-genetic toxicology and environmental mutagenesis. Volume: 634 Issue: 1-2 P. 172-176
20. Горянин И.И., Котова В.Ю., Краснопеева Е.Д., Чубуков П.А., Балабанов В.П., Чалкин С.Ф., Шатров Т.Я., Завильгельский Г.Б., Манухов И.В. Определение генотоксического действия 1,1-диметилгидразина алкилирующими соединениями, возникающими при его окислении, и перекисью водорода. // Труды Московского физико-технического института. 2013. Т. 5. №1-17. С. 103-111.
21. Winson Μ.K., Swift S., Fish L., Throup J. P., rgensen F., Chhabra S.R., Bycroft B.W., Williams P. and Stewart G.S.A. 'Construction and analysis of luxCDABE -based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing' // FEMS Microbiology Letters. 1998. 163(2). pp. 185-192. doi: 10.111 l/j.l574-6968.1998.tb13044.x.
22. Blackwell H.E., Geske G.D., O'Neill J.C. Modulation of bacterial quorum sensing with synthetic ligands. WO/2008/116029 Publication Date: 25.09.2008.
23. Sayler G.S., Ripp S.A., Applegate B. Bioluminescent Biosensor Device. Patent No:US 6544729 Date of Patent: Apr. 08, 2003
24. Natrah F.Μ. I., Alam Μ.I., Pawar S., Harzevili A.S., Nevejan N., Boon N., Sorgeloos P., Bossier P. and Defoirdt T. 'The impact of quorum sensing on the virulence of Aeromonas hydrophila and Aeromonas salmonicida towards burbot (Lota lota L.) larvae'// Veterinary Microbiology. 2012. 159(1-2). pp. 77-82. doi: 10.1016/j.vetmic.2012.03.014.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ управляемой экспрессии рекомбинантных генов в клетках Escherichia coli с использованием элементов LuxR-LuxI "quorum sensing" системы психрофильных бактерий рода Aliivibrio | 2022 |
|
RU2810597C2 |
| Экспрессионный вектор pBMS10 на основе элементов LuxR-LuxI "quorum sensing" системы Aliivibrio logei | 2023 |
|
RU2816447C1 |
| Бактериальный lux-биосенсор на основе промотора гена бактериальной ДНК-полимеразы II | 2023 |
|
RU2832445C1 |
| Набор стрессиндуцируемых lux-биосенсоров на основе клеток Bacillus subtilis для исследования механизмов токсичности | 2022 |
|
RU2811895C2 |
| Набор lux-биосенсоров для детекции токсичных продуктов неполного окисления несимметричного диметилгидразина в среде | 2015 |
|
RU2626569C2 |
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ГЕНОТОКСИЧНЫХ ПРОДУКТОВ НЕПОЛНОГО ОКИСЛЕНИЯ НЕСИММЕТРИЧНОГО ДИМЕТИЛГИДРАЗИНА В СРЕДЕ | 2014 |
|
RU2569156C1 |
| НАБОР LUX-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ГЕПТИЛА В СРЕДЕ | 2005 |
|
RU2297450C2 |
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ДЕТЕРГЕНТОВ ГИДРОФОБНОЙ ПРИРОДЫ В СРЕДЕ | 2007 |
|
RU2355760C1 |
| Применение кумарина и его производных в качестве ингибиторов системы "кворум сенсинга" LuxI/LuxR типа у бактерий | 2015 |
|
RU2616237C1 |
| Способ определения токсичности химических веществ, генерирующих активные формы кислорода | 2016 |
|
RU2614267C1 |

Изобретение относится к бактериальным lux-биосенсорам для детекции субнаномолярных концентраций ацильных производных гомосерин лактона, а также к бактериальным lux-биосенсорам для детекции ацильных производных гомосерин лактона. Биосенсоры состоят из клеток Escherichia coli, трансформированных парой плазмид, которые содержат гены, полученные или из Aliivibrio fischeri, или из Aliivibrio logei, или из Aliivibrio salmonicida, и гены luxCDABE Photorhabdus luminescens. 5 н. и 1 з.п. ф-лы, 5 ил., 1 пр.
1. Бактериальный lux-биосенсор для детекции субнаномолярных концентраций ацильных производных гомосерин лактона, состоящий из клеток Escherichia coli, трансформированных парой плазмид, одна из которых содержит ген luxR из Aliivibrio fischeri под контролем собственного промотора и гены luxCDABE Photorhabdus luminescens под контролем индуцируемого промотора PluxICDABEG из A. fischeri, а вторая содержит дополнительную копию бактериального гена luxR A. fischeri.
2. Бактериальный lux-биосенсор для детекции ацильных производных гомосерин лактона, состоящий из клеток E. coli, трансформированных парой плазмид, одна из которых содержит ген luxR1 из Aliivibrio logei под контролем собственного промотора и гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxCDABEG из A. logei, а вторая содержит дополнительную копию гена luxR1 A. logei.
3. Бактериальный lux-биосенсор для детекции субнаномолярных концентраций ацильных производных гомосерин лактона, состоящий из клеток E. coli, трансформированных парой плазмид, одна из которых содержит ген luxR2 из A. logei под контролем собственного промотора и гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxI из A. logei, а вторая содержит дополнительную копию гена luxR2 A. logei.
4. Бактериальный lux-биосенсор для детекции ацильных производных гомосерин лактона, состоящий из клеток E. coli, трансформированных парой плазмид, одна из которых содержит ген luxR1 из Aliivibrio salmonicida под контролем собственного промотора и гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxCDABEG из A. salmonicida, а вторая содержит дополнительную копию бактериального гена luxR1 A. salmonicida.
5. Бактериальный lux-биосенсор для детекции субнаномолярных концентраций ацильных производных гомосерин лактона, состоящий из клеток E. coli, трансформированных парой плазмид, одна из которых содержит ген luxR2 из A. salmonicida под контролем собственного промотора и гены luxCDABE P. luminescens под контролем индуцируемого промотора PluxI из A. salmonicida, а вторая содержит дополнительную копию бактериального гена luxR2 A. salmonicida.
6. Бактериальный lux-биосенсор по пп.1-5, в котором вторая плазмида содержит дополнительную копию бактериального гена или luxR, или luxR1, или luxR2 под контролем более сильного промотора, чем собственный, или под контролем комбинации промоторов.
| MIN J | |||
| et al., Distinct responses of a recA∷luxCDABE Escherichia coli strain to direct and indirect DNA damaging agents, Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 1999, Vol.442, N2, pp.61-68 | |||
| МАНУХОВ И.В | |||
| Структура lux-оперонов и механизмы регуляции типа "Quorum Sensing" у морских бактерий, автореферат диссертации, 2011, |

