Изобретение относится к оценке степени загрязнения окружающей среды гептилом, а также может быть использовано в исследованиях биологических объектов.
Известен способ определения гептила в окружающей среде химическими средствами [1-3]. Существо этого способа заключается в проведении фотометрического анализа продуктов распада гептила или высокоразрешающего хроматографического (газовая хроматография) разделения химических соединений с масс-спектрометрическим детектированием [1], и идентификации гептила и его продуктов распада с помощью методов ЯМР и инфракрасной спектроскопии [2]. Недостатком химических методов является необходимость использования дорогостоящей аппаратуры, а также сложность проведения анализа в связи с высокой нестабильностью гептила [3].
В настоящее время широкое распространение для определения загрязнения окружающей среды токсичными веществами получили методы биотестирования с использованием lux-биосенсоров. Известны методы, основанные на тушении биолюминесценции токсикантами, в которых используется механизм ингибирующего действия ядовитых веществ на метаболизм клетки, в основном, на дыхательную цепь, что опосредованно влияет на люциферазную реакцию, вызывая ослабление интенсивности биолюминесценции клеток. В этой серии методик, используемых в качестве экспресс-контроля токсичности природных сред, наибольшее распространение в странах Европы и в США получил т.н. "Микротокс" ("Microtox 5TM), в котором в качестве биосенсора используются лиофилизированные морские бактерии Photobacterium phosphoreum. Недостатком метода является неспецифичность реакции и невысокая чувствительность. Для идентификации химического соединения, вызвавшего уменьшение интенсивности свечения клеток, требуется дополнительный анализ.
Известно также обнаружение токсикантов с использованием lux-биосенсоров на основе бактерий Escherichia coli, содержащих плазмиды с lux-генами под контролем индуцибельных стрессовых промоторов [4]. Эти lux-биосенсоры, как указано в приведенном патенте США, были использованы для определения нитратов, фенолов, бензина и других токсикантов, но не использовались для обнаружения в среде гептила.
Техническим результатом предложенного изобретения является возможность осуществления быстрого, не требующего дорогой и сложной аппаратуры, как в стационарных, так и в полевых условиях, контроля за содержанием гептила в окружающей среде.
Для достижения этого результата предложено использовать комплекс lux-биосенсоров на основе бактерий Escherichia coli, содержащих гибридные плазмиды с lux-генами под контролем индуцибельных стрессовых промоторов РkatG, PsoxS, PrecA, PgrpE. В предложенном изобретении для определения гептила использована группа биосенсоров, обладающих свойством индукции (усиления) сигнала (биолюминесценции) клеток при воздействии токсикантом, а также высокой чувствительностью и специфичностью, так как определяется особенностью взаимодействия белка-рецептора (репрессора или активатора) с химическим соединением.
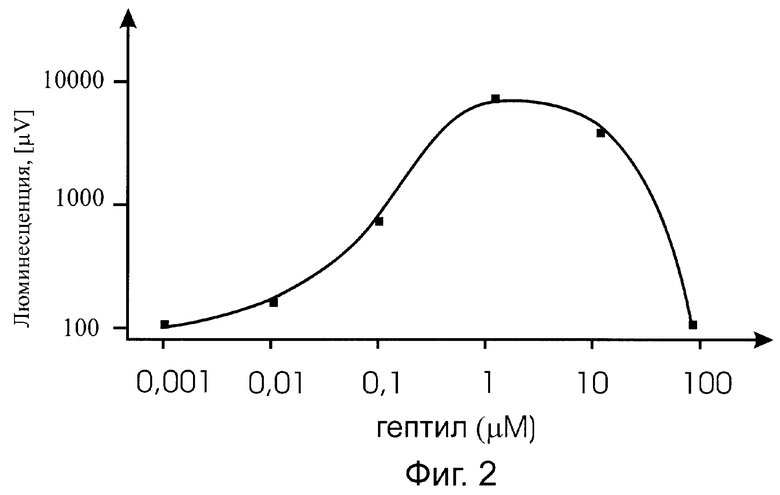
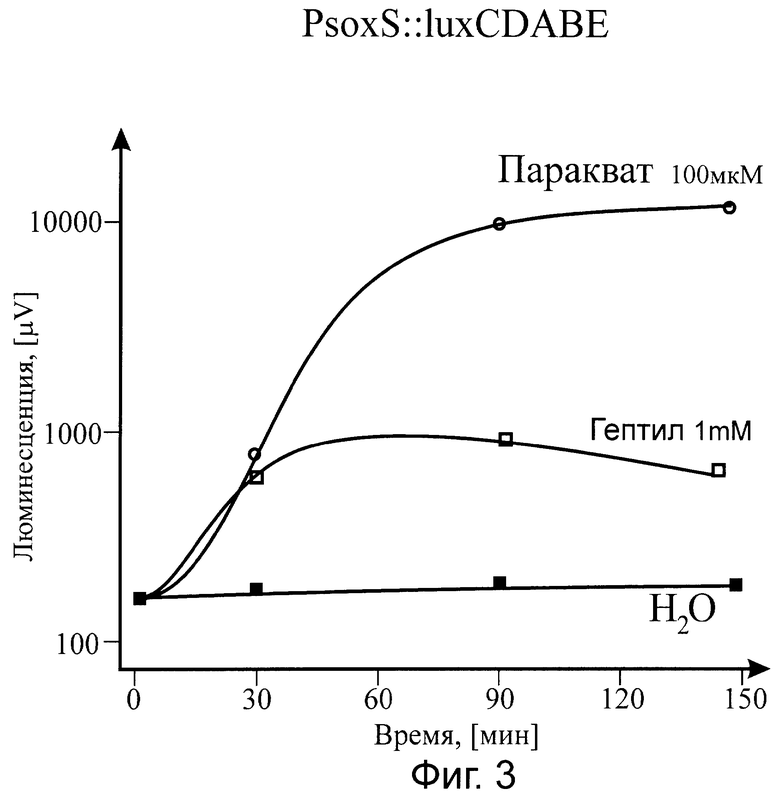
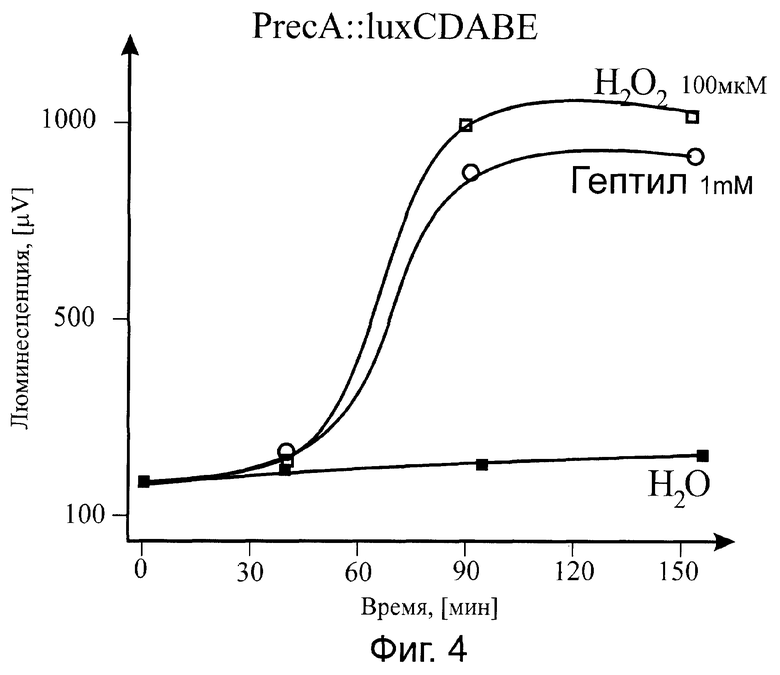
Существо изобретения поясняется прилагаемыми графиками, где на фиг.1 показана зависимость от времени действия гептила (при фиксированной концентрации) на биосенсор PkatG::lux; на фиг.2 - приведена зависимость величины эффекта (при оптимальном времени выдерживания препарата) от концентрации гептила в образце, также для биосенсора PkatG::lux; на фиг.3 показана реакция на гептил (в зависимости от времени действия) на биосенсор PsoxS::lux; на фиг.4 приведена временная зависимость действия гептила на биосенсор PrecA::lux.
В процессе действия гептила на клетку происходит индукция биолюминесценции у биосенсоров PkatG::lux, PsoxS::lux, PrecA::lux, но не PgrpE::lux (поэтому данные для биосенсора PgrpE::lux не приводятся). Как видно из графиков, представленных на фиг.1-4, при выдерживании проб с фиксированной концентрацией гептила наблюдается со временем значительное усиление (примерно в 100 раз) интенсивности биолюминесценции клеток - биосенсоров. На фиг.2 приведена зависимость эффективности действия гептила от его количества в пробе для биосенсора с промотором PkatG. В кривой можно выделить три характерных области. При количестве гептила в пробе более 20 микромолей наблюдается значительный токсический эффект гептила на клетку, что проявляется в гашении биолюминесценции. Этот результат соответствует данным по летальному действию гептила на бактерии Е. coli: гибель бактерий фиксировалась при количестве гептила в пробе выше 20 микромолей [5]. При количествах гептила примерно от 10 до 0,1 микромоля наблюдается значительное усиление биолюминесценции, превышающее исходное значение I0 более чем в 10 раз. И, наконец, при количестве гептила в пробе менее 0,01 микромоля влияние гептила снижается практически до фона, т.е. пороговая концентрация гептила, фиксируемая lux-биосенсором с промотором PkatG, равна 0,001 мкМ на пробу.
У бактерий можно выделить регуляторные системы, специфически реагирующие на токсиканты, действующие на: 1) клеточные мембраны; 2) белки; 3) хромосому (ДНК), а также 4) индуцирующие в клетке окислительный стресс. В качестве биосенсора на токсиканты, действующие на клеточные белки, предложено использовать промотор PgrpE. Этот промотор в бактериальном геноме расположен перед генами "теплового шока" и открывается лишь при появлении в клетке модифицированных, денатурированных белков. В роли биосенсора на ДНК-тропные агенты используется SOS-промотор РrecA. Промотор РrecA открывается лишь при индукции повреждений в геноме, т.е. в молекулах ДНК. Для детекции веществ, индуцирующих в клетке окислительный стресс (образующих в клетке гидроксильный радикал, супероксид-ион-радикал, перекись водорода) были использованы промоторы PkatG и PsoxS. Промотор PkatG (активатор OxyR) специфически реагирует на перекись водорода, органические пероксиды. Промотор PsoxS открывается при появлении в среде супероксид-ион-радикала.
В данном изобретении сконструированы основанные на этих индуцируемых промоторах специфические lux-биосенсоры. Все используемые промоторы с соответствующими регуляторными участками были получены из генома бактерий Escherichia coli K12 MG1655 с помощью метода ПЦР с использованием специальных, синтезированных праймеров. В качестве вектора использовали беспромоторный вектор с репликоном ColE1 и геном bla, определяющим резистентность к ампициллину (селективный маркер). Встраивание промоторной области в плазмиду проводили по сайтам EcoRI-BamHI. В качестве lux-кассеты был выбран lux-оперон Photorhabdus luminescens, состоящий из пяти генов, luxCDABE. Данная кассета имеет два преимущества: во-первых, люцифераза Ph. luminescens (гены luxAB) отличается сравнительно высокой термостабильностью, а, во-вторых, к суспензии клеток не надо добавлять субстрат люциферазной реакции - алифатический альдегид, так как он синтезируется в клетке при помощи белков, кодируемых генами luxCDE [6].
Методика измерения влияния токсиканта на биолюминесценцию биосенсора была примерно одинаковой для всех lux-биосенсоров.
Определение гептила в среде с помощью lux-биосенсоров требует проведения следующих операций.
1. Приготовление проб, содержащих соответствующие биосенсоры;
- выращивание клеток Е. coli, являющихся биосенсорами, до экспоненциальной фазы (OD=0,2-0,4);
- отбор проб по 200 мкл в виалы (6-8 виал для каждого биосенсора).
2. Добавление к пробам гептила или жидкой среды с гептилом в различных концентрациях;
- приготовление для каждого биосенсора положительного и отрицательного контролей. В качестве положительного контроля используют гептил (разведенный в воде) в концентрации 20 мкг/мл (отрицательный контроль - дистиллированная вода);
- добавление по 20 мкл исследуемой среды к каждой пробе каждого биосенсора.
(Все пробы следует приготовить в двойном или тройном экземплярах и затем использовать средние значения полученных экспериментальных величин).
3. Измерение интенсивностей биолюминесценции проб в течение 1-2 часов для биосенсоров PkatG::lux, PsoxS::lux, PgrpE:lux. Для биосенсора PrecA::lux измерение следует проводить 2,5-3 часа.
4. Обработка полученных результатов. Полученные графики на контрольных образцах сравнивают с исследуемым образцом.
Культуру клеток, содержащих гибридную плазмиду с соответствующим промотором и lux-кассетой, растили при 28°С или 37°С на качалке до ранней или средней экспоненциальной фазы (OD=0,2-0,4). Аликвоты этой культуры (по 200 мкл) переносили в стерильные виалы и добавляли в них по 10 мкл тестируемого вещества требуемой концентрации. В контрольную пробирку добавляли 10 мкл дистиллированной воды. Затем пробы инкубировали без перемешивания при 30°С и через каждые 15 мин измеряли интенсивность биолюминесценции при комнатной температуре. Усиление сигнала фиксировалось уже через 15-20 мин - время, необходимое для синтеза люциферазы. Максимальный ответ биосенсора, как правило, наблюдался через 40-60 мин (в случае промотора PrecA максимальный сигнал фиксировался через 90 мин) и при оптимальной концентрации токсиканта превышал начальный сигнал в 100-1000 раз.
В результате наших исследований было показано, что основными промоторами, открывающимися при воздействии на клетки гептила, являются промоторы PkatG, PsoxS, PrecA. Биосенсор С промотором PgrpE не изменял интенсивность биолюминесценции. Поэтому данные по данному промотору не приведены на фиг.1-4.
В табл.1 приведены данные о минимальных (пороговых) концентрациях стандартных, специфических для данного промотора веществ, индуцирующих заметный (в 2-3 раза) эффект усиления биолюминесценции, в конструированных lux-биосенсорах (митомицин С индуцирует повреждения в ДНК, этанол - в белках, перекись водорода и паракват как генератор супероксид-ион-радикалов индуцируют окислительный стресс).
Гептил, по всей видимости, в отличие от представленных в табл.1 соединений, не обладает высокой специфичностью к тому или другому биосенсору, так как в связи с высокой реакционной активностью (очень сильный восстановитель) способен повреждать макромолекулы. Поэтому оптимальным решением проблемы фиксации гептила в среде является использование всех вышеуказанных lux-биосенсоров.
Так как промотор PkatG открывается лишь при воздействии перекиси водорода на белок-регулятор OxyR (окисление активного центра [Fe-S]), a промотор PsoxS открывается лишь при воздействии супероксид-ион-радикала, то можно считать доказанным, что действие гептила на бактериальную клетку в основном определяется формированием активных форм кислорода, в частности, гидроперекиси и супероксид-ион-радикала (в результате восстановления кислорода воздуха до перекиси водорода).
Гидроперекись, образованная в результате химических реакций гептила H2NN(CH3)2 в основном с кислородом O2, открывает промотор PkatG, причем столь же эффективно, как и перекись водорода, используемая в качестве стандарта:
O2+H2NN(CH3)2=O2H (супероксид-радикал) +HNN(CH3)2, причем супероксид-радикал быстро переходит в перекись водорода:
2O2Н=Н2O2+O2
Превращение супероксид-ион-радикала в перекись водорода происходит как спонтанно, так и в результате действия клеточного фермента супероксид-дисмутазы.
Действие гептила на ДНК и, соответственно, на промотор РrecA, носит косвенный характер и определяется в основном формирующимися активными формами кислорода. Этот вывод следует из данных о влиянии перекиси водорода на биосенсор PrecA::lux, а также из данных по практическому совпадению концентрационных зависимостей эффектов гептила и перекиси водорода на биосенсоры PkatG::lux и PrecA::lux до концентраций 10-7-10-6 М/л.
Влияние гептила на биосенсор PgrpE::lux практически отсутствует, поэтому данные о воздействии гептила на этот биосенсор не приводятся. Гептил не модифицирует клеточные белки, так как не проникает в цитоплазму клетки, а весь расходуется (окисляется) вне клетки.
Изобретение позволяет создать высокочувствительный, дешевый и быстрый тест-метод на основе измерения интенсивности биолюминесценции.
Источники информации
1. Дмитриев О.Ю., Иваненко С.И., Овсянников Д.А., Смирнова С.С., Чистова Ж.А. В Сб. "Труды Российской инженерной академии, Секция "Инженерные проблемы стабильности и конверсии", Выпуск 11. "Экологические проблемы разработки и эксплуатации ракетно-космической техники", Москва, СИП РИА, 2004 г., стр.40-43.
2. Лопырев В.А., Долгушин Г.В., Ласкин Б.М. (2001) Журнал Рос. хим. об-ва им. Д.И.Менделеева, т.XLV, №5-6, стр.149-156.
3. Кузнецова Л.В. В Сб. "Труды Российской инженерной академии, Секция "Инженерные проблемы стабильности и конверсии", Выпуск 12. "Экологические проблемы разработки и эксплуатации ракетно-космической техники", Москва, СИП РИА, 2004 г., стр.24-25.
4. Патент США №5683868, C12Q1/68 - прототип.
5. Anti Poso, Atte von Wright, Jukka Gynther (1995) Mutation Research. V.332. PP.63-71.
6. Завильгельский Г.Б., Зарубина А.П., Манухов И.В. (2002) Молекулярная биология. т.36, стр.792-804.
| название | год | авторы | номер документа |
|---|---|---|---|
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ГЕНОТОКСИЧНЫХ ПРОДУКТОВ НЕПОЛНОГО ОКИСЛЕНИЯ НЕСИММЕТРИЧНОГО ДИМЕТИЛГИДРАЗИНА В СРЕДЕ | 2014 |
|
RU2569156C1 |
| Набор lux-биосенсоров для детекции токсичных продуктов неполного окисления несимметричного диметилгидразина в среде | 2015 |
|
RU2626569C2 |
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ДЕТЕРГЕНТОВ ГИДРОФОБНОЙ ПРИРОДЫ В СРЕДЕ | 2007 |
|
RU2355760C1 |
| Штамм бактерии Escherichia coli/pTdcR-TurboYFP, обладающий чувствительностью к терагерцовому излучению | 2018 |
|
RU2691308C1 |
| Бактериальный lux-биосенсор на основе промотора гена бактериальной ДНК-полимеразы II | 2023 |
|
RU2832445C1 |
| Бактериальный lux-биосенсор с повышенной чувствительностью для детекции ацильных производных гомосерин лактона | 2020 |
|
RU2777196C2 |
| Способ определения токсичности химических веществ, генерирующих активные формы кислорода | 2016 |
|
RU2614267C1 |
| Набор стрессиндуцируемых lux-биосенсоров на основе клеток Bacillus subtilis для исследования механизмов токсичности | 2022 |
|
RU2811895C2 |
| Способ выявления из естественных сред перспективных пробиотических штаммов | 2021 |
|
RU2772351C1 |
| ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PHIF И СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ | 2000 |
|
RU2180687C1 |
Изобретение относится к биотехнологии и генной инженерии и может быть использовано при оценке степени загрязнения окружающей среды гептилом. Методом рекомбинантных ДНК получены клетки Escherichia coli, содержащие плазмиды с бактериальными luxCDABE-генами под контролем индуцибельных стрессовых промоторов PkatG, PsoxS, PrecA, PgrpE. На их основе разработан набор проб бактериальных клеток Escherichia coli для определения гептила в окружающей среде. Применение изобретения позволяет быстро в стационарных или полевых условиях определять содержание гептила в окружающей среде. 4 ил., 1 табл.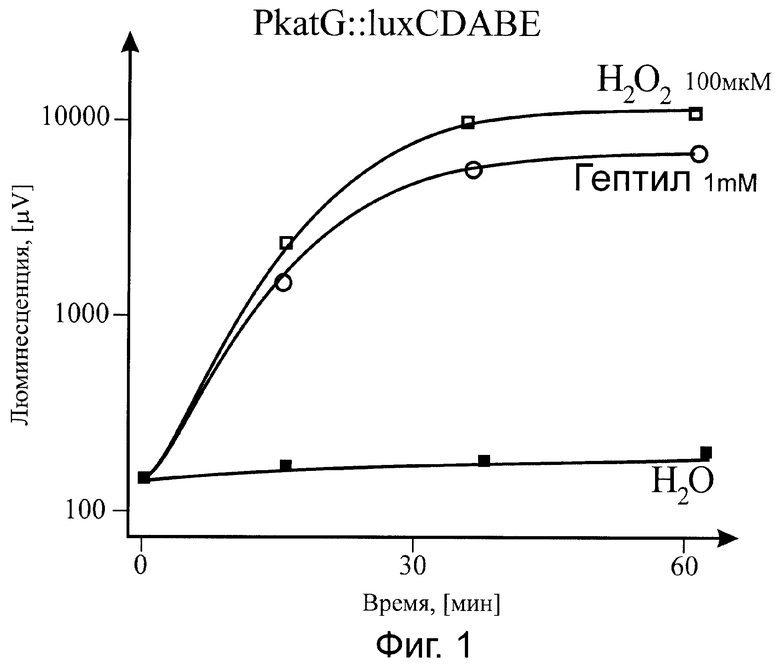
Набор lux-биосенсоров для определения гептила в среде, состоящий из пробы клеток Escherichia coli, трансформированных плазмидой, содержащей бактериальные luxCDABE-гены под контролем промотора PkatG, пробы клеток Escherichia coli, трансформированных плазмидой, содержащей бактериальные luxCDABE-гены под контролем промотора PsoxS, пробы клеток Escherichia coli, трансформированных плазмидой, содержащей бактериальные luxCDABE-гены под контролем промотора PгесА, пробы клеток Escherichia coli, трансформированных плазмидой, содержащей бактериальные luxCDABE-гены под контролем промотора PgrpE.
| СПОСОБ ОПРЕДЕЛЕНИЯ ГЕНОТОКСИЧНОСТИ ХИМИЧЕСКИХ ВЕЩЕСТВ | 2000 |
|
RU2179581C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МАССОВОЙ КОНЦЕНТРАЦИИ НЕСИММЕТРИЧНОГО ДИМЕТИЛГИДРАЗИНА В ВОДЕ | 1996 |
|
RU2090863C1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ПТИЦИН Л.Р | |||
| Биохимический анализ SOS-ответа клеток Escherichia coli | |||
| Генетика | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| Cancer Res | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| Biochem.y | |||
| Приспособление к индикатору для определения момента вспышки в двигателях | 1925 |
|
SU1969A1 |

