Изобретение относится к экологии, токсикологии и фармацевтике. Позволяет исследовать механизмы токсичности веществ и смесей, детекцию биологически активных веществ в среде. Изобретение предоставляет возможности для оценки загрязнения почв и водоёмов. Данное изобретение позволяет по увеличению уровня биолюминесценции судить об активации транскрипции стрессовых индуцируемых промоторов в ответ на появление в среде токсичных и биологически активных веществ. Lux-биосенсоры, составляющие набор, представляют собой три штамма Bacillus subtilis с гибридными плазмидами, содержащими lux-гены под контролем стрессовых промоторов. Биосенсорные клетки способны специфично увеличивать уровень биолюминесценции за счёт усиления транскрипции lux-генов при возникновении окислительного стресса, повреждений ДНК под воздействием ДНК-тропных агентов, вызывающих SOS-ответ, и алкилирования ДНК.
На данный момент есть два подхода к исследованиям химических соединений на токсичность с помощью люминесцирующих бактерий: наблюдение за снижением уровня общей люминесценции [1-3] или наблюдение за индукцией люминесценции [4-13]. В экспериментах с индукцией биолюминесценции используются стресс-индуцируемые lux-биосенсоры, полученные методом генной инженерии. Данные клетки содержат в себе генетическую конструкцию с репортерными генами люминесценции под контролем регуляторных элементов. Таким образом, можно получить lux-биосенсоры специфично реагирующие на различные типа клеточного стресса, к примеру на повреждение ДНК, тепловой стресс, окислительный стресс и др. [7] Стресс-индуцируемые lux-биосенсоры на основе Escherichia coli на данный момент активно используются в различных исследованиях по токсичности, например, по исследованию токсичности ракетного топлива и его перспективных компонентов [14]. Также их используют для экологического мониторинга, но для исследования почв биосенсоры на основе грамотрицательных бактерий мало эффективны [15]. Разработка lux-биосенсоров на основе грамположительных бактерий B. subtilis способствует анализу механизмов токсичности различных веществ, в том числе, лекарственных средств, которые специфически взаимодействуют с грамположительными бактериями, а также экологическому мониторингу загрязнения почвы.
Конструирование lux-оперона, который эффективно экспрессировался бы у грамположительных бактерий, затруднено из-за необходимости замены последовательности Шайна-Дальгарно перед каждой рамкой считывания. Ранее были получены люминесцентные бактерии родов Streptococcus, Staphylococcus, Bacillus и др., которые использовались в основном для медицинских целей [2], [16], [17] и в исследованиях термоинактивации и рефолдинга белков в B. subtilis [1]. Люминесценция этих бактерий была конститутивной, и по ней определялось интегральное действие биологически активных веществ на клетки. В предлагаемом изобретении мы представляем набор стресс-индуцируемых lux-биосенсоров на основе B. subtilis.
Техническим результатом изобретения является набор из трех штаммов на основе грамположительной бактерии B. subtilis, позволяющий оценивать токсичность биологически активных веществ и дифференцировать тип воздействия как один из трех типов клеточного стресса: окислительного стресса, SOS-ответа и алкилирования ДНК. В сравнении с аналогичными lux-биосенсорами на основе E. coli, они обладают большей чувствительностью к специфическим для грамположительных бактерий токсикантам и позволяют более эффективно исследовать образцы почв на содержание токсичных агентов. В совокупности с набором lux-биосенсоров на основе E. coli появляется возможность проводить полноценные исследования воздействия различных соединений на бактерии, как грамположительные, так и грамотрицательные. Измерения проводятся на клетках, выращенных в жидкой среде, требуют навыков ведения бактериальной культуры и физического прибора по измерению люминесценции, но не требуют сложной и дорогостоящей аппаратуры, могут выполняться в лаборатории и в полевых условиях.
Задача решена путем конструирования биосенсорных плазмид, содержащих последовательность генов lux-оперона Photorhabdus luminescens с измененным порядком генов - luxABCDE и рибосом-связывающими сайтами, узнающимися рибосомой B. subtilis. Перед кассетой генов люминесценции были клонированы следующие промоторы B. subtilis: PdinC – промотор являющийся частью регулона SOS-ответа, обладающий высокой чувствительностью и высокой амплитудой ответа: 36-175 раз [18], [19], PalkA – промотор ДНК-гликозилазы, активирующийся в ответ на алкилирование ДНК; активация промотора в клетках B. subtilis не высока, а именно - примерно 2,5 раза, что определялось изменением высвобожденного 7-метилгуанина после обработки нитрозогуанидином [20], PmrgA – промотор к гену, кодирующий ферритин-подобный ДНК-связывающий белок, индуцирующийся в ответ на присутствие пероксида водорода [21], [22]. Данные промоторные участки были клонированы в беспомоторный вектор pPL_ABCDExen [23]. Полученные плазмиды были названы pNK-dinC, pNK-alkA и pNK-mrgA, соответственно. Данными плазмидами были трансформированы клетки B. subtilis и получены, соответственно, биосенсоры B. subtilis pNK-dinC, B. subtilis pNK-alkA, B. subtilis pNK-mrgA. Следует отметить, что использованные индуцируемые промоторы будут активироваться в различных штаммах B. subtilis, если только в них не делетировали специально соответствующие регуляторные системы [24-30]. Кроме того, используемый вариант lux-кассеты способен экспрессироваться не только в различных штаммах B. subtilis, но и в других грамположительных бактериях [17, 31]. Таким образом мы утверждаем, что штаммы на основе клеток B. subtilis, полученные описываемым способом будут представлять из себя lux-биосенсоры.
Методика исследования биологически активных веществ с помощью новых lux-биосенсоров в общем виде
Методика исследования биологически активных веществ с помощью новых lux-биосенсоров требует проведения следующих операций:
Приготовление биосенсоров к измерениям;
- выращивание биосенсорных клеток B. subtilis в жидкой среде Brain Heart Infusion (BHI) с добавлением триптофана в концентрации 50 мг/л при температуре 37º C при постоянном перемешивании до экспоненциальной фазы роста (OD = 0,4-0,6);
- отбор аликвот по 180 мкл и распределение их по кюветам или лункам планшета.
Добавление к пробам исследуемых образцов в различных концентрациях;
- приготовление отрицательного и положительных контролей. В качестве отрицательного контроля используется дистиллированная вода. В качестве положительного контроля используются: митомицин С (MitC) в концентрации 10 мкМ (для биосенсора B. subtilis pNK-dinC), пероксид водорода (H2O2) в концентрации 100 мМ (для биосенсора B. subtilis pNK-mrgA) и метилметансульфонат (MMS) в концентрации 10 мМ (для биосенсора B. subtilis pNK-alkA)
- добавление по 20 мкл исследуемых образцов в разных концентрациях к каждой пробе соответственного биосенсора;
(Все пробы следует приготовить в трёх повторах и затем использовать средние значения полученных экспериментальных величин).
Кинетику люминесценции от времени следует измерять на протяжении 4,5-5 часов при комнатной температуре.
Обработка полученных результатов. Полученные графики на исследуемых образцах сравнивают с контрольными образцами.
Технический результат изобретения поясняется фигурами.
На фиг. 1, 2 и 3 приведены примеры контрольной проверки работоспособности каждого из трех биосенсоров с токсическими соединениями, специфически активирующими соответствующие промоторы.
На фиг. 4 и 5 приведены примеры исследования генотоксичности органического вещества.
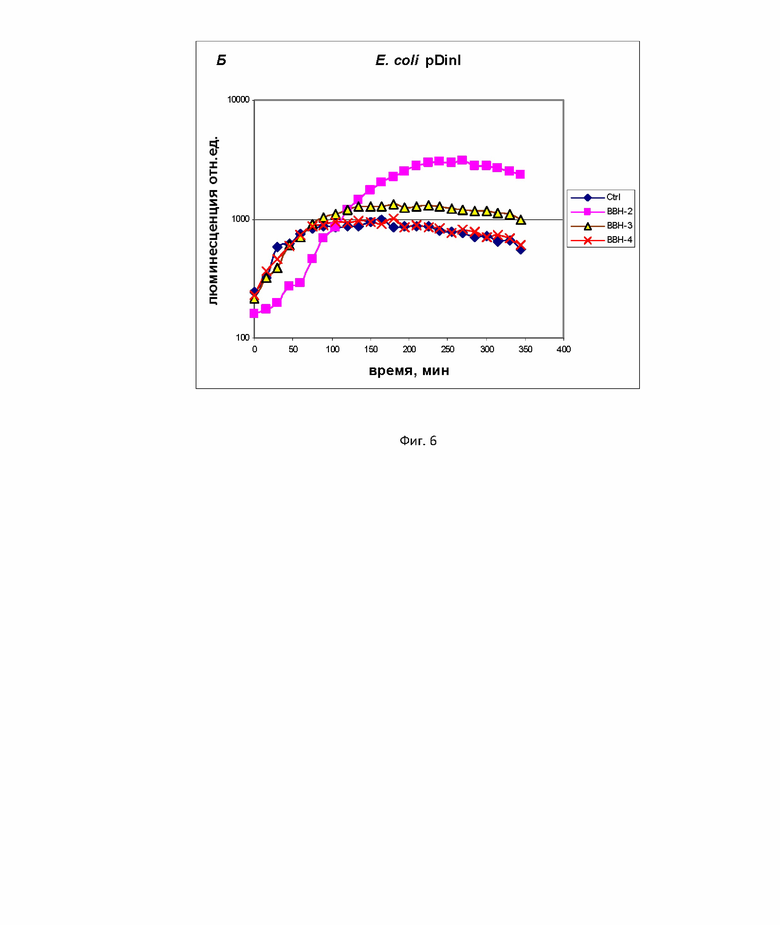
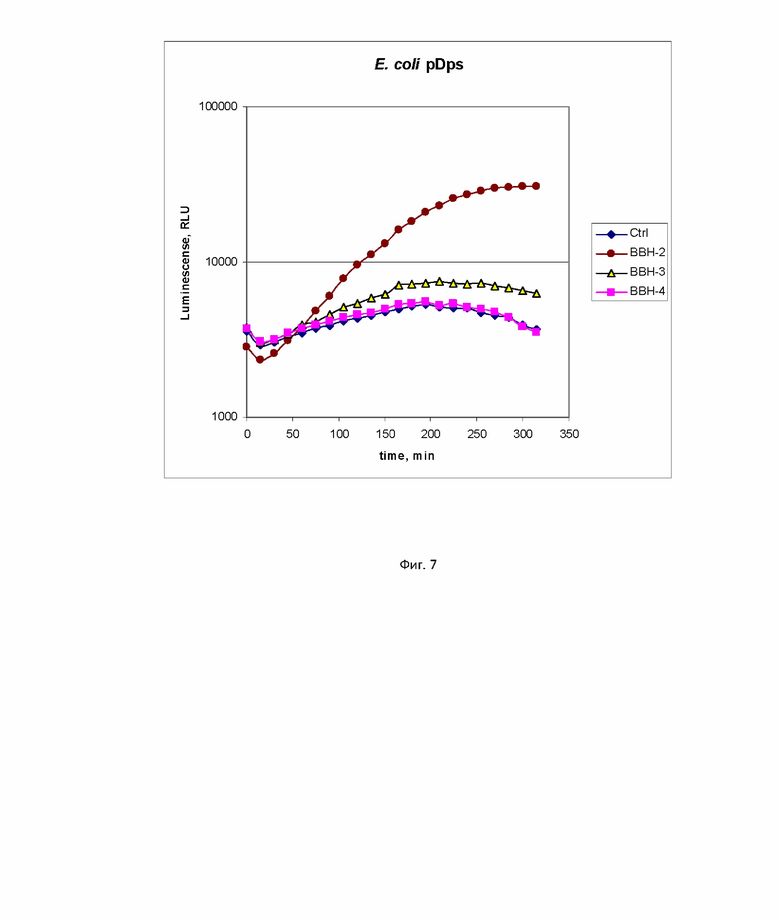
На фиг. 6 и 7 примеры сравнения полученных биосенсоров с существующими аналогами на основе клеток E. coli.
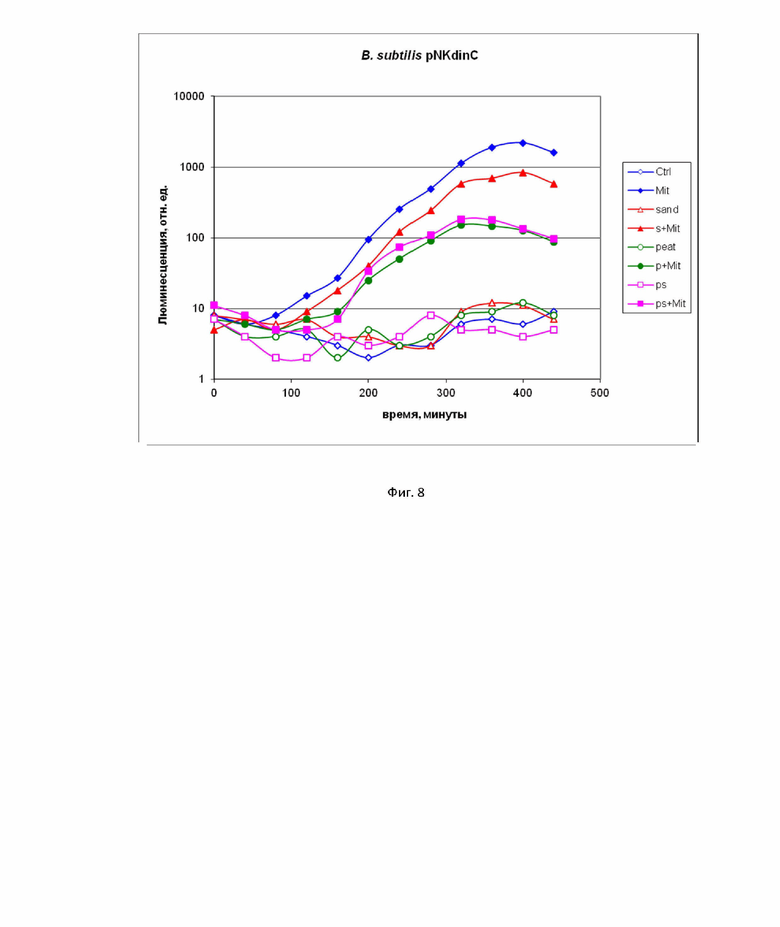
На фиг. 8 пример применения биосенсора типа B. subtilis pNK-DinC для поиска генотоксичных соединений в почве.
Пример 1. Проверка работоспособности биосенсорного штамма B. subtilis 168 pNK-DinC с помощью вещества, останавливающего репликационную вилку (митомицина С).
Данные, представленные на фиг. 1, представляют из себя кривые зависимости люминесценции культуры биосенсорного штамма B. subtilis 168 pNK-DinC от времени. Представлены несколько кинетических кривых: “Ctrl” – отрицательный контроль, а именно биосенсорная культура без добавления токсиканта, “MitC -5”, “MitC -6”, “MitC -7”, “MitC -8” – биосенсорная культура с добавлением митомицина С, вызывающего ковалентные сшивки в ДНК, в конечных концентрациях 10 мкМ, 1 мкМ, 100 Нм, 10 Нм, соответственно. Фиг. 1 показывает, что биосенсорный штамм индуцируется митомицином C и, следовательно, чувствителен к повреждению ДНК. Порог концентрации MitC составляет около 10 нМ, что в целом соответствует чувствительности лучшего из существующих биосенсоров на основе клеток E.coli [7]. Время начала активации биосенсора составляет около 1 часа с максимальной активацией через 3 часа. Это примерно в два раза медленнее, чем для биосенсорных клеток на основе E. coli. Максимальная амплитуда ответа превышает один порядок и составляет 40 раз для концентрации MitC 10 мкМ.
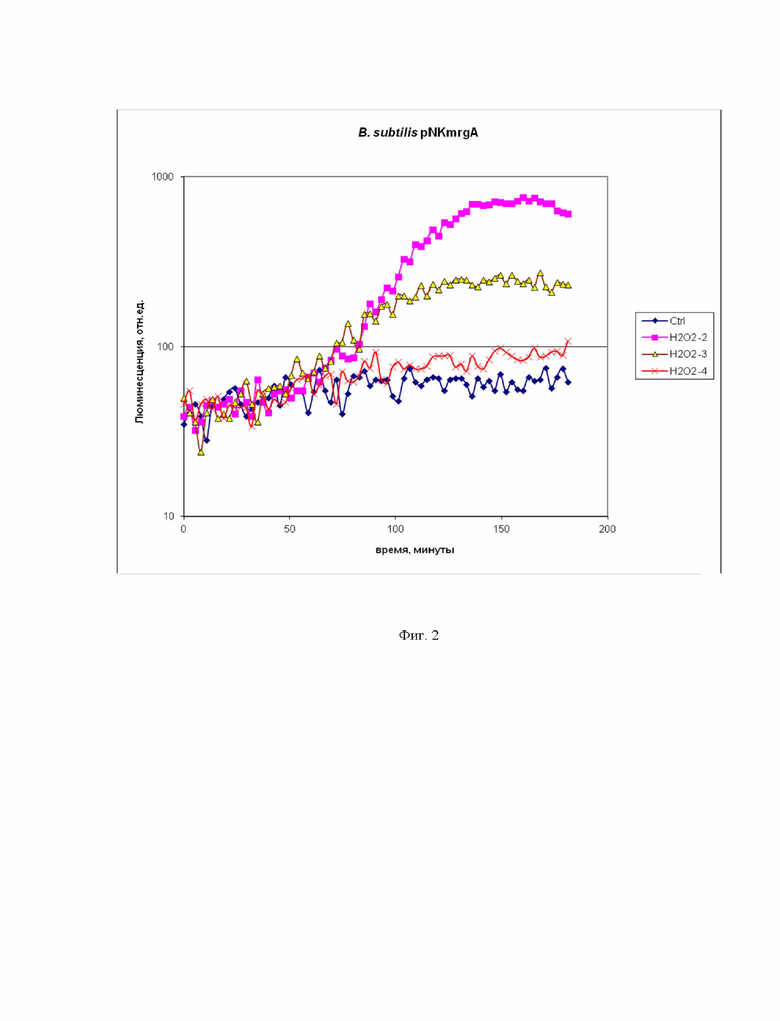
Пример 2. Проверка работоспособности биосенсорного штамма B. subtilis 168 pNK-mrgA, опредляющего окислительные повреждения клетки, с помощью перекиси водорода.
На фиг. 2 приведены кривые зависимости люминесценции культуры биосенсорного штамма B. subtilis 168 pNK-mrgA от времени. Представлены несколько кинетических кривых: “Ctrl” – отрицательный контроль, а именно биосенсорная культура без добавления токсиканта, “H2O2 -2”, “H2O2 -3”, “H2O2 -4” – биосенсорная культура с добавлением пероксида водорода в конечной концентрации 10 мМ, 1 мМ, 100 мкМ, соответственно. Из данных на фиг. 2 видно, что промотор PmrgA в клетках B. subtilis индуцируется пероксидом водорода, т.е. при окислительном стрессе. Индукция биосенсора начинается через 1 час после добавления пероксида водорода и достигает максимальной амплитуды через 2,5 часа. Пороговая концентрация пероксида водорода для этого биосенсора составляет 100 мкМ. Максимальная амплитуда ответа превышает порядок величины и составляет 14 раз для концентрации пероксида водорода 10 мМ.
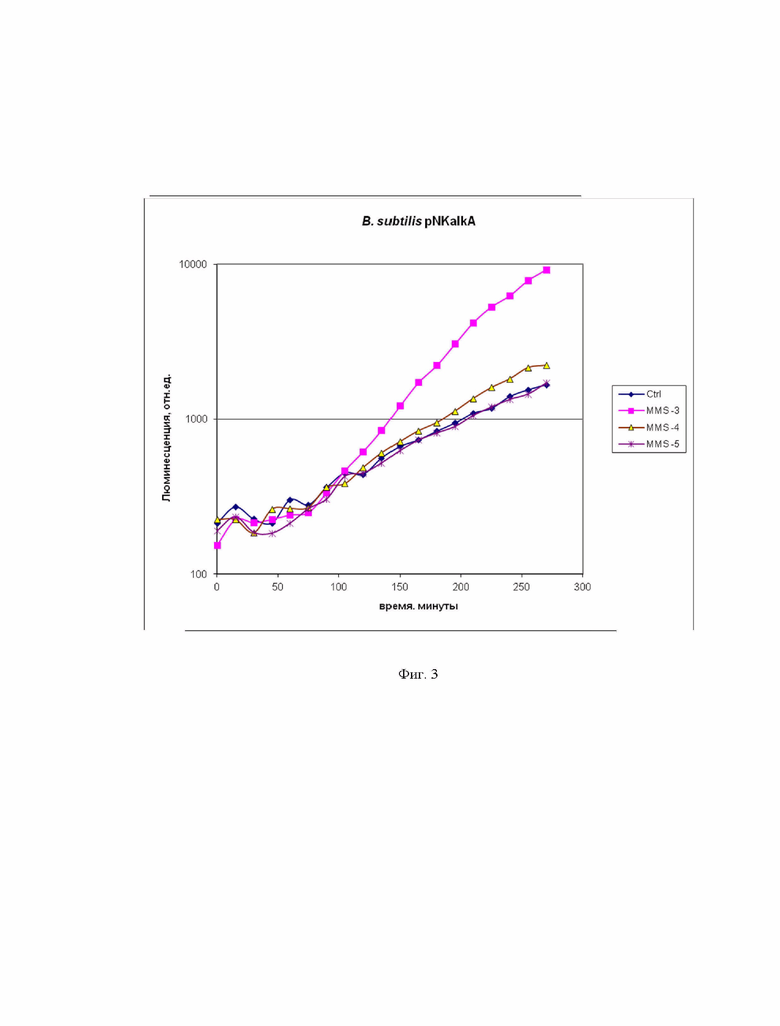
Пример 3. Проверка работоспособности биосенсорного штамма B. subtilis 168 pNK-alkA, с помощью алкилирующего вещества метил-метан-сульфоната.
На фиг. 3 приведены кривые зависимости люминесценции культуры биосенсорного штамма B. subtilis 168 pNK-alkA от времени. Представлены несколько кинетических кривых: “Ctrl” – отрицательный контроль, а именно биосенсорная культура без добавления токсикантов, “MMS -3”,“MMS -4”, “MMS -5” – биосенсорная культура с добавлением метилметансульфоната, в конечных концентрациях 1 мМ, 100 мкМ, 10 мкМ, соответственно. Как видно из данных на фиг. 3, B. subtilis 168 pNK-AlkA индуцируется с помощью метилметансульфоната (известно, что он вызывает алкилирование ДНК). Пороговая концентрация ММS составляет около 100 мкМ. Время отклика биосенсора составляет около 2 часов, максимальная активация - через 4 часа. Максимальная амплитуда ответа не превышает порядка и составляет 6 раз для концентрации MMS 1 мМ. Клетки B. subtilis 168 pNK-AlkA имеют такую же чувствительность, как и биосенсоры на основе клеток E.coli [24].
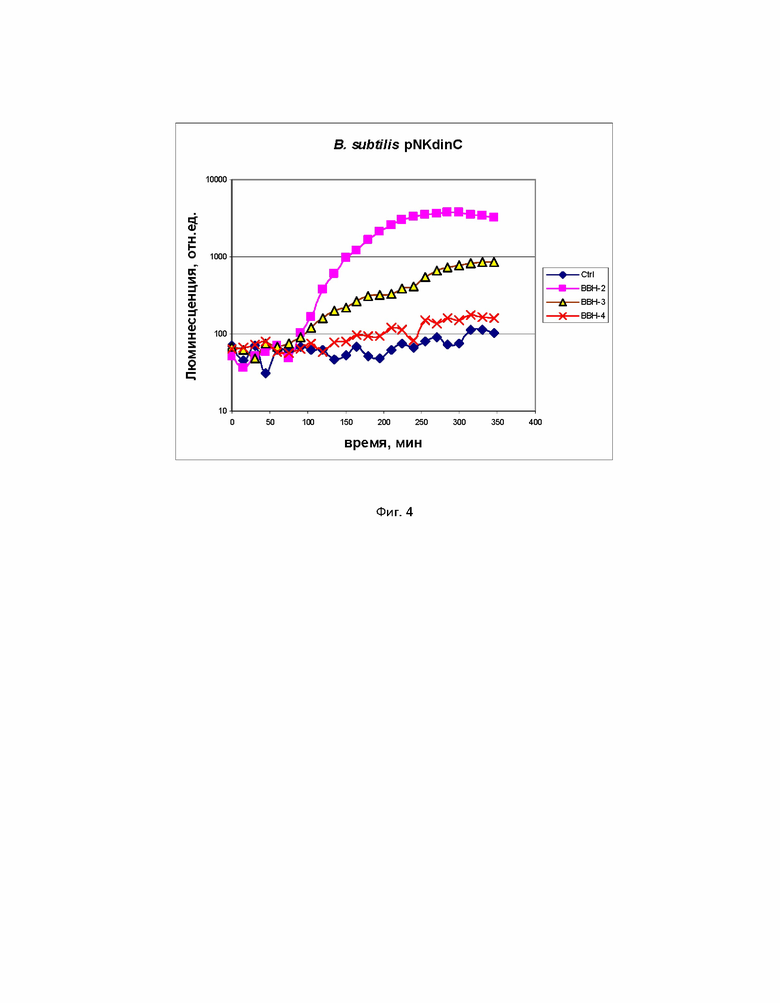
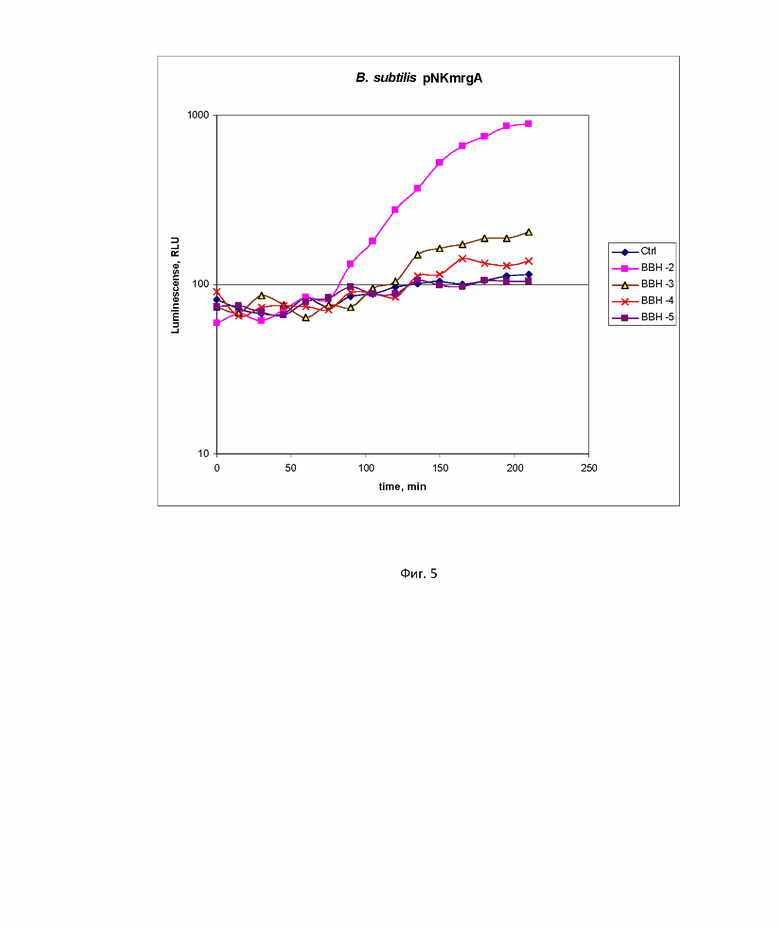
Пример 4. Применение набора биосенсоров B. subtilis 168 pNK-dinC, B. subtilis 168 pNK-alkA и B. subtilis 168 pNK-mrgA для исследования генотоксичности 2,2'-бис (бицикло [2.2.1] гептана).
На фиг. 4 и 5 с помощью биосенсоров B. subtilis 168 pNK-dinC, B. subtilis 168 pNK-alkA и B. subtilis 168 pNK-mrgA было исследовано токсическое действие 2,2'-бис (бицикло [2.2.1] гептана) (БЦГ), перспективного соединения для использования его в качестве компонента ракетного топлива. На фиг. 4 представлены кинетические кривые люминесценции для штамма B. subtilis 168 pNK-dinC. “Ctrl” – отрицательный контроль, а именно биосенсорная культура без добавления токсиканта, “BBH -2 ”, “BBH -3”, “BBH -4”– биосенсорная культура c добавлением БЦГ в концентрациях 10 мМ, 1 мМ, 100 мкМ соответственно. На фиг. 5 представлены кинетические кривые люминесценции для штамма B. subtilis 168 pNK-mrgA. “Ctrl” – отрицательный контроль, а именно биосенсорная культура без добавления токсиканта, “BBH -2 ”, “BBH -3”, “BBH -4”, “BBH -5”– биосенсорная культура c добавлением БЦГ в концентрациях 10 мМ, 1 мМ, 100 мкМ, 10 мкМ соответственно. Кинетические кривые люминесценции для биосенсора B. subtilis 168 pNK-alkA не приведены, так как заметной индукции при добавлении БЦГ нет. Данные были обработаны, были подсчитаны коэффициенты индукции (значение люминесценции индуцированной пробы, деленное на значение люминесценции отрицательного контроля).
Пример 5. Сравнение свойств биосенсоров B. subtilis 168 pNK-DinC и B. subtilis 168 pNK-MrgA с аналогичными биосенсорами на основе E. coli.
Для сравнения свойств lux-биосенсоров на основе B. subtilis и E. coli использовали биосенсоры E. coli MG1655 pDinI (фиг. 6) и E. coli MG1655 pDps (фиг. 7). “Ctrl” – отрицательный контроль, а именно биосенсорная культура без добавления токсиканта, “BBH -2 ”, “BBH -3”, “BBH -4”– биосенсорная культура c добавлением БЦГ в концентрациях 10 мМ, 1 мМ, 100 мкМ соответственно. Согласно полученным данным пороговая концентрация БЦГ, оказывающая генотоксическое воздейчтвие (SOS-ответ), для B. subtilis 168 pNK-DinC составляет 0,1 г/л. Для биосенсора E. coli MG1655 pDinI пороговая концентрация БЦГ, вызывающая SOS-ответ, составляет 1 г/л, как и для биосенсора E. coli MG1655 pColD-lux, согласно работе [14]. Максимальная амплитуда ответа для B. subtilis 168 pNK-DinC была 50-кратной, в то время как для E. coli pDinI она была равна 4. Время начала индукции биосенсоров в ответ на БЦГ примерно одинаково для E. coli и B. subtilis - около 1,5 ч.
Пороговая концентрация БЦГ, вызывающая окислительный стресс, измеренная lux-биосенсором B. subtilis 168 pNK-MrgA составляет от 0,1 г/л до 1 г/л. Это более высокая чувствительность по сравнению с E. coli pDps (1 г/л) и биосенсорами E. coli pOxyR-lux (специфичными для пероксида водорода, согласно [14], его чувствительность к БЦГ составляет 1 г/л). Максимальная амплитуда ответа как для B. subtilis 168 pNK-MrgA, так и для E. coli pDps была примерно равна 10. Время начала индукции биосенсора примерно такое же - около 1 ч.
Пример 6. Применение биосенсорного штамма B. subtilis 168 pNK-DinC для анализа почв на содержание генотоксичных соединений.
На фиг. 8 приведены кинетические кривые при исследовании почв с помощью биосенсоров. B. subtilis 168 pNK-DinC был выбран для проверки эффективности биосенсоров в полевых условиях при тестировании образцов почвы, загрязненных токсичными веществами. В этих экспериментах MitC добавляли в различные почвы арктического региона, собранные на побережье Кандалакшского залива Белого моря: песчаные (образцы подписанные “sand” или “s”), торфяные (образцы подписанные “peat” или “p”) и смешанные(“ps”). 30-миллиграммовые образцы почвы с MitC (образцы подписанные +Mit) или без него добавляли к 200 мкл клеточной культуры биосенсора. Конечная концентрация MitC в среде с культурой клеток биосенсора составляла 100 нМ. В качестве положительного контроля MitC добавляли непосредственно к культуре клеток биосенсора до конечной концентрации 100 нМ (образец “Mit”). “Ctrl” – отрицательный контроль, а именно культура клеток без добавления токсикантов. Затем образцы инкубировали при 16–17 °С с периодическим измерением люминесценции (фиг. 8). Как видно из данных, представленных на фиг. 8, сами исследуемые почвы генотоксическими свойствами не обладают. Добавление песчаной почвы приводит к уменьшению амплитуды ответа биосенсора на MitC примерно в 2 раза. Торфосодержащие почвы уменьшают амплитуду отклика примерно в 10 раз. Такое снижение отклика можно объяснить частичным экранированием люминесценции темными частицами почвы и частичным связыванием токсиканта. Кислотность почвы также играет значительную роль, в то время как торфяники в Карелии характеризуются низким pH [32],что может значительно снизить характеристики биосенсоров на основе E. coli [15].
Таким образом, заявляемые штаммы B. subtilis pNK-alkA, B. subtilis pNK-dinC и B. subtilis pNK-mrgA способны специфически реагировать на различные виды клеточного стресса, их амплитуда ответа коррелирует с концентрациями. При измерении токсичности соединения, перспективного как компонент ракетного топлива БЦГ было показано, что lux-биосенсоры B. subtilis 168 pNK-DinC и B. subtilis 168 pNK-MrgA обладают примерно на порядок большей чувствительностью, чем подобные биосенсоры на основе E. coli. Было также показано, что с помощью биосенсоров на основе B. subtilis можно исследовать почвенные образцы на содержание токсичных веществ.
Изобретение позволяет создать высокочувствительный тест-метод на основе измерения интенсивности биолюминесценции, позволяющий определить класс токсиканта по специфичности его воздействия на клетку, степень его токсичности и исследовать механизмы токсичности новых соединений.
Использованные источники информации
[1] Gnuchikh E.Yu et al. Kinetics of the thermal inactivation and the refolding of bacterial luciferases in Bacillus subtilis and in Escherichia coli differ. PLoS One. 2019. 14(12):1–11. doi: 10.1371/journal.pone.0226576.
[2] Дерябин Д.Г.и др. Сравнение чувствительности люминесцирующих штаммов Photobacterium phosphoreum, Escherichia coli и Bacillus subtilis при оценке биотоксичности углеродных наноматериалов и наночастиц металлов. Микробиология. 2016. 85(2):198-206. DOI: 10.7868/S0026365616020051.
[3] Данилов, В.С; Исмаилов А.Д. Бактериальная люцифераза как биосенсор биологически активных соединений. Биотехнология 1989, 11, 39–78. http://europepmc.org/abstract/MED/2650767.
[4] Завильгельский Г.Б.и др. Наночастицы диоксида титана (TiO2) индуцируют в бактериях стрессовые реакции, фиксируемые специфическими lux-биосенсорами. Российские Нанотехнологии 2011, 6(5–6):75-78. doi: 10.1134/S1995078011030165.
[5] Абилев С.К. и др. Специфические Lux-биосенсоры Escherichia сoli, содержащие плазмиды pReca::Lux, pCold::Lux И pDini::Lux, для детекции генотоксичных агентов. Генетика. 2020. 56(6): 648-655S. doi: 10.1134/S1022795420060022.
[6] G. B. Zavilgelsky, V. Y. Kotova, and I. V Manukhov, “Action of 1 , 1-dimethylhydrazine on bacterial cells is determined by hydrogen peroxide,” vol. 634, pp. 172–176, 2007, doi: 10.1016/j.mrgentox.2007.07.012.
[7] V. Y. Kotova, I. V. Manukhov, and G. B. Zavilgelskii, “Lux-biosensors for detection of SOS-response, heat shock, and oxidative stress,” Appl. Biochem. Microbiol., vol. 46, no. 8, pp. 781–788, 2010, doi: 10.1134/S0003683810080089.
[8] T. K. Van Dyk, W. R. Majarian, K. B. Konstantinov, R. M. Young, P. S. Dhurjati, and R. A. LaRossa, “Rapid and sensitive pollutant detection by induction of heat shock gene-bioluminescence gene fusions,” Appl. Environ. Microbiol., vol. 60, no. 5, pp. 1414–1420, 1994, doi: 10.1128/aem.60.5.1414-1420.1994.
[9] R. A. LaRossa, Bioluminescence Methods and Protocols. 1998.
[10] A. C. Vollmer, S. Belkin, D. R. Smulski, T. K. Van Dyk, and R. A. LaRossa, “Detection of DNA damage by use of Escherichia coli carrying recA’::lux, uvrA’::lux, or alkA’::lux reporter plasmids,” Appl. Environ. Microbiol., vol. 63, no. 7, pp. 2566–2571, 1997, doi: 10.1128/aem.63.7.2566-2571.1997.
[11] J. Kurittu, S. Lönnberg, M. Virta, and M. Karp, “Qualitative detection of tetracycline residues in milk with a luminescence-based microbial method: The effect of milk composition and assay performance in relation to an immunoassay and a microbial inhibition assay,” J. Food Prot., vol. 63, no. 7, pp. 953–957, 2000, doi: 10.4315/0362-028X-63.7.953.
[12] Igonina E. V. и др. “Lux Biosensors: Screening Biologically Active Compounds for Genotoxicity,” Russ. J. Genet. Appl. Res., vol. 8, no. 1, pp. 87–95, 2018, doi: 10.1134/S2079059718010082.
[13] S. A. Khrulnova et al., “Lux-operon of the marine psychrophilic bacterium aliivibrio logei: A comparative analysis of the LuxR1/LuxR2 regulatory activity in Escherichia coli cells,” Microbiol. (United Kingdom), vol. 162, no. 4, pp. 717–724, 2016, doi: 10.1099/mic.0.000253.
[14] Kessenikh A. et al., “Genotoxic effect of 2,2’-bis(bicyclo[2.2.1] heptane) on bacterial cells,” PLoS One, vol. 15, no. 8 August 2020, pp. 1–11, 2020, doi: 10.1371/journal.pone.0228525.
[15] Манухов, И В, и др. “Использование lux-биосенсоров для детекии НДМГ в почве,” Двойные технологии, vol. 3, no. 44, 2008.
[16] D. G. Deryabin, I. F. Karimov, I. V Manukhov, N. A. Tolmacheva, and V. P. Balabanov, “Differential Analysis of Bactericidal Systems of Blood Serum with Recombinant Luminescent Escherichia coli and Bacillus subtilis Strains,” vol. 154, no. 1, pp. 59–63, 2012.
[17] K. P. Francis et al., “Visualizing pneumococcal infections in the lungs of live mice using bioluminescent Streptococcus pneumoniae transformed with a novel gram-positive lux transposon,” Infect. Immun., vol. 69, no. 5, pp. 3350–3358, 2001, doi: 10.1128/IAI.69.5.3350-3358.2001.
[18] N. Au et al., “Genetic composition of the Bacillus subtilis SOS system,” J. Bacteriol., vol. 187, no. 22, pp. 7655–7666, 2005, doi: 10.1128/JB.187.22.7655-7666.2005.
[19] D. L. Cheo, K. W. Bayles, and R. E. Yasbin, “Cloning and characterization of DNA damage-inducible promoter regions from Bacillus subtilis,” J. Bacteriol., vol. 173, no. 5, pp. 1696–1703, 1991, doi: 10.1128/jb.173.5.1696-1703.1991.
[20] F. Morohoshi, K. Hayashi, and N. Munakata, “Bacillus subtilis alkA gene encoding inducible 3-methyladenine DNA glycosylase is adjacent to the ada operon,” J. Bacteriol., vol. 175, no. 18, pp. 6010–6017, 1993, doi: 10.1128/jb.175.18.6010-6017.1993.
[21] J. Mostertz, C. Scharf, M. Hecker, and G. Homuth, “Transcriptome and proteome analysis of Bacillus subtilis gene expression in response to superoxide and peroxide stress,” Microbiology, vol. 150, no. 2, pp. 497–512, 2004, doi: 10.1099/mic.0.26665-0.
[22] J. D. Helmann et al., “The global transcriptional response of Bacillus subtilis to peroxide stress is coordinated by three transcription factors,” J. Bacteriol., vol. 185, no. 1, pp. 243–253, 2003, doi: 10.1128/JB.185.1.243-253.2003.
[23] Гнучих Е.Ю. и др. "Биосенсоры для исследования активности промоторов и шаперонов в клетках Bacillus subtilis," Биотехнология. 2020, 36(6):68–77. doi: 10.21519/0234-2758-2020-36-6-68-77
[24] Y. Nakabeppu, T. Miyata, H. Kondo, S. Iwanaga, and M. Sekiguchi, “Structure and expression of the alkA gene of Escherichia coli involved in adaptive response to alkylating agents.,” J. Biol. Chem., vol. 259, no. 22, pp. 13730–13736, 1984, doi: https://doi.org/10.1016/S0021-9258(18)89806-4.
[25] Au N, Kuester-Schoeck E, Mandava V, Bothwell LE, Canny SP, Chachu K, Colavito SA, Fuller SN, Groban ES, Hensley LA, O'Brien TC, Shah A, Tierney JT, Tomm LL, O'Gara TM, Goranov AI, Grossman AD, Lovett CM. Genetic composition of the Bacillus subtilis SOS system. J Bacteriol. 2005 Nov;187(22):7655-66.
[26] Cheo DL, Bayles KW, Yasbin RE. Cloning and characterization of DNA damage-inducible promoter regions from Bacillus subtilis. J Bacteriol. 1991 Mar;173(5):1696-703.
[27] Helmann JD, Wu MF, Gaballa A, Kobel PA, Morshedi MM, Fawcett P, Paddon C. The global transcriptional response of Bacillus subtilis to peroxide stress is coordinated by three transcription factors. J Bacteriol. 2003 Jan;185(1):243-53.
[28] Haike Antelmann, Susanne Engelmann, Roland Schmid, And Michael Hecker. General and Oxidative Stress Responses in Bacillus subtilis: Cloning, Expression, and Mutation of the Alkyl Hydroperoxide Reductase Operon. JOURNAL OF BACTERIOLOGY, Nov. 1996, p. 6571–6578 Vol. 178, No. 22 0021-9193
[29] Mostertz J, Scharf C, Hecker M, Homuth G. Transcriptome and proteome analysis of Bacillus subtilis gene expression in response to superoxide and peroxide stress. Microbiology. 2004 Feb;150(Pt 2):497-512.
[30] Hayashi K, Ohsawa T, Kobayashi K, Ogasawara N, Ogura M. The H2O2 stress-responsive regulator PerR positively regulates srfA expression in Bacillus subtilis. J Bacteriol. 2005 Oct;187(19):6659-67.
[31] Contag PR. Bioluminescence imaging to evaluate infections and host response in vivo. Methods Mol Biol. 2008;415:101-18. doi: 10.1007/978-1-59745-570-1_6.32Morozova, R.M.; Fedorets, N.G.; Bahmet, O.N Soils and soil cover of Karelian Zaonezhje. Tr. Karel. Nauchnogo Cent. Ross. Akad. Nauk, vol. 6, pp. 69–89, 2004.
| название | год | авторы | номер документа |
|---|---|---|---|
| Бактериальный lux-биосенсор на основе промотора гена бактериальной ДНК-полимеразы II | 2023 |
|
RU2832445C1 |
| Бактериальный lux-биосенсор с повышенной чувствительностью для детекции ацильных производных гомосерин лактона | 2020 |
|
RU2777196C2 |
| Набор lux-биосенсоров для детекции токсичных продуктов неполного окисления несимметричного диметилгидразина в среде | 2015 |
|
RU2626569C2 |
| Экспрессионный вектор pBMS10 на основе элементов LuxR-LuxI "quorum sensing" системы Aliivibrio logei | 2023 |
|
RU2816447C1 |
| Способ управляемой экспрессии рекомбинантных генов в клетках Escherichia coli с использованием элементов LuxR-LuxI "quorum sensing" системы психрофильных бактерий рода Aliivibrio | 2022 |
|
RU2810597C2 |
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ГЕНОТОКСИЧНЫХ ПРОДУКТОВ НЕПОЛНОГО ОКИСЛЕНИЯ НЕСИММЕТРИЧНОГО ДИМЕТИЛГИДРАЗИНА В СРЕДЕ | 2014 |
|
RU2569156C1 |
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ДЕТЕРГЕНТОВ ГИДРОФОБНОЙ ПРИРОДЫ В СРЕДЕ | 2007 |
|
RU2355760C1 |
| НАБОР LUX-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ГЕПТИЛА В СРЕДЕ | 2005 |
|
RU2297450C2 |
| Плазмида для редактирования генома бактерий рода Bacillus и способ внесения модификаций в геном бактерий рода Bacillus | 2022 |
|
RU2815835C1 |
| Рекомбинантная плазмида pBU-LipA, обеспечивающая синтез белка липазы А штамма Bacillus natto IAN | 2023 |
|
RU2808501C1 |

Изобретение относится к набору штаммов B. subtilis pNK-alkA, B. subtilis pNK-dinC и B. subtilis pNK-mrgA, обеспечивающему комплексное исследование токсикологических характеристик ДНК-тропных и вызывающих окислительный стресс соединений, специфично воздействующих на грамположительные бактерии. 3 з.п. ф-лы, 8 ил., 6 пр.
1. Набор штаммов B. subtilis pNK-alkA, B. subtilis pNK-dinC и B. subtilis pNK-mrgA, обеспечивающий комплексное исследование токсикологических характеристик ДНК-тропных и вызывающих окислительный стресс соединений, специфично воздействующих на грамположительные бактерии, при этом штаммы были получены путем трансформации штамма бактерий вида B. subtilis плазмидами pNK-dinC, pNK-alkA и pNK-mrgA, которые были сконструированы путем клонирования промоторов B. subtilis: PdinC, PalkA и PmrgA – перед кассетой генов люминесценции (lux-оперона) Photorhabdus luminescens.
2. Набор по п. 1, в котором штамм B. subtilis pNK-alkA – lux-биосенсор, полученный путем трансформации клеток B. subtilis 168 плазмидой с генами luxABCDE из P. luminescens под контролем стресс-индуцируемого промотора PalkA, специфично реагирующий на алкилирование ДНК различными соединениями.
3. Набор по п. 1, в котором штамм B. subtilis pNK-dinC – lux-биосенсор, полученный путем трансформации клеток B. subtilis 168 плазмидой с генами luxABCDE из P. luminescens под контролем стресс-индуцируемого промотора PdinC, специфично реагирующий на повреждения ДНК, вызывающие SOS-ответ.
4. Набор по п. 1, в котором штамм B. subtilis pNK-mrgA,– lux-биосенсор, полученный путем трансформации клеток B. subtilis 168 плазмидой с генами luxABCDE из P. luminescens под контролем стресс-индуцируемого промотора PmrgA, специфично реагирующий на окислительный стресс.
| НАБОР lux-БИОСЕНСОРОВ ДЛЯ ОПРЕДЕЛЕНИЯ ГЕНОТОКСИЧНЫХ ПРОДУКТОВ НЕПОЛНОГО ОКИСЛЕНИЯ НЕСИММЕТРИЧНОГО ДИМЕТИЛГИДРАЗИНА В СРЕДЕ | 2014 |
|
RU2569156C1 |
| ЗАВИЛЬГЕЛЬСКИЙ Г.Б | |||
| и др., Сенсорные биолюминесцентные системы на основе lux-оперонов для детекции токсичных веществ, Химическая физика, 2012, т.31, н.10, стр.15-20 | |||
| АБИЛЕВ С.К | |||
| и др., Специфические lux-биосенсоры Escherichia coli, содержащие плазмиды pRecA::lux, pColD::lux и pDinI::lux, для детекции генотоксичных | |||

