Изобретение относится к биотехнологии, биосинтезу рекомбинантных белков в клетках Escherichia coli и может быть использовано при создании штаммов-продуцентов клеточных метаболитов и белков. Разработан способ плавной регуляции уровня экспрессии целевого гена в клетках E. coli за счет использования элементов системы скоординированного ответа (“quorum sensing”, QS) первого типа психрофильных бактерий рода Aliivibrio. Экспрессия индуцируется при внесении в ростовую среду аутоиндуктора (АИ) (низкомолекулярное вещество - N-(3-оксогексаноил)-лактон L-гомосерина (3-oxo-C6-HSL)) в концентрациях от 1 нМ до 10 мкМ и снижении температуры до 20-22°С. Количество синтезированного белка в расчете на клетку напрямую зависит от концентрации АИ и от продолжительности временного интервала, в течение которого культура клеток экспрессионного штамма инкубировались при температуре 20-22°С.
Экспрессионные системы широко применяются в биотехнологии и молекулярной биологии. Они определяются совокупностью ферментов, участвующих в экспрессии целевых генов и ее регуляции. На сегодняшний день доминирующее положение продолжает занимать биосинтез рекомбинантных белков в клетках E. coli, при этом существует множество различных экспрессионных систем, отличающихся друг от друга по ряду параметров (механизм регуляции экспрессии, используемое вещество-индуктор, базовый уровень экспрессии целевого гена в отсутствие индуктора, амплитуда усиления синтеза целевого продукта в ответ на индукцию и т.п.) [1–6]. В большинстве случаев экспрессионные системы представлены в виде плазмидного вектора с регуляторными элементами, в который необходимо встраивать целевой ген для его последующей экспрессии в клетках E. coli. Наиболее широко используются те, в которых индукция осуществляется с помощью добавления лактозы (или ИПТГ), рамнозы, арабинозы и других сахаров.
В основе настоящего изобретения лежит использование регуляторных элементов QS-системы психрофильных люминесцентных морских бактерий рода Aliivibrio, таких как A. logei и A. salmonicida. Ацильные производные гомосерин лактона, служащие аутоиндуторами QS-систем первого типа (такие системы также называют системами luxR-luxI типа) различных бактерий, являются биологически активными веществами, но не токсичны и не включены в основные метаболические пути клеток E. coli. Регуляторным белком, активатором транскрипции является LuxR или его гомологи [7–9]. Эти белки с высокой специфичностью связываются с молекулой АИ, меняет свою конформацию, после чего связывается с ДНК и активирует транскрипцию с регулируемого им промотора за счет прямого взаимодействия с РНК-полимеразой [10]. В гетерологичной системе клеток E. coli наибольшую активность среди гомологов LuxR из психрофильных бактерий проявляет белок LuxR2 [9]. Белок LuxI отвечает за синтез аутоиндуктора, как правило один вид бактерий может детектировать и синтезировать несколько разных отличающихся по длине ацильной цепи и модификации в третьем положении аутоиндукторов с разной эффективностью. Ген, кодирующий LuxI, расположен под контролем LuxR-регулируемого промотора и его экспрессия возрастает в присутствии АИ, что приводит к повышению уровня синтеза АИ, реализуя таким образом положительную обратную связь. В клетках A. logei по такому механизму регулируется lux-оперон, отвечающий за светимость клеток. Различные элементы lux-оперонов люминесцирующих бактерий применяются для разработки биосенсоров [11–16]. В работах [17,18] промоторы различных QS систем использовались для детекции АИ: PluxICDABEG из A. fischeri, чувствительный к N-(3-оксогексаноил)-лактону L-гомосерина, PlasI из Pseudomonas aeruginosa, чувствительный к N-(3-оксододеканоил)-лактон L-гомосерина, PtraR из Agrobacterium tumefaciens, чувствительный к N-(3-оксооктаноил)-лактону L-гомосерина.
Использование LuxR-регулируемого промотора как такового для создания экспрессионного вектора не является революционной идеей, прототипом настоящего изобретения является экспрессионная система на основе QS системы бактерий Aliivibrio fischeri, описанная в патенте [19] и статье [20]. Однако настоящее изобретение новизной и дает ряд преимуществ по сравнению с ранее созданными образцами. Ключевыми отличиями настоящего изобретения по сравнению с прототипом является низкий базовый уровень экспрессии в условиях отсутствия индукции, а также возможность отключения уже запущенной индукции, за счет чего можно в необходимый момент культивирования клеток кратковременно запускать синтез целевого белка.
Техническим результатом предложенного изобретения является возможность биосинтеза ферментов с положительной и отрицательной регуляцией экспрессии (включение и выключение) посредством внесения АИ в среду и изменения температуры. Активация экспрессии целевого гена происходит в дозозависимой манере при добавлении АИ в широком диапазоне концентраций (от 1 нМ до 10 мкМ) при температурах ниже 25°C и полностью отсутствует при 37°C. При этом экспрессионная система может содержать ген luxI, что дает возможность автоиндукции при достижении оптической плотности более 1,5 единиц при культивировании при температуре 22°C. При 37°C синтез АИ психрофильной АИ-синтетазой LuxI A. logei в клетках E. coli не наблюдается, а промотор, под контролем которого осуществляется клонирование целевого гена остается полностью закрытым. Уровень содержания целевого белка в пробных ферментациях достигал 30% от общего клеточного белка, при этом накопление белка LuxI не наблюдается.
Данные результаты достигнуты путем конструирования плазмиды, содержащей следующие ключевые функциональные элементы:
1. Ген luxR2 A. logei под контролем собственного промотора;
2. Консенсусная последовательность Шайна-Дальгарно, расположенная под контролем LuxR2-регулируемого промотора, в частности промотора гена luxI A. logei;
3. Полилинкер, расположенный сразу за последовательностью Шайна-Дальгарно, для клонирования целевого гена в экспрессионный вектор.
Возможность включения и отключения экспрессии целевого гена обусловлена термолабильностью психрофильного LuxR2 [21]. Основные параметры новой экспрессионной системы были получены путем апробации с использованием гена gfp в качестве целевого. Ниже приведен пример реализации описанного изобретения.
Пример
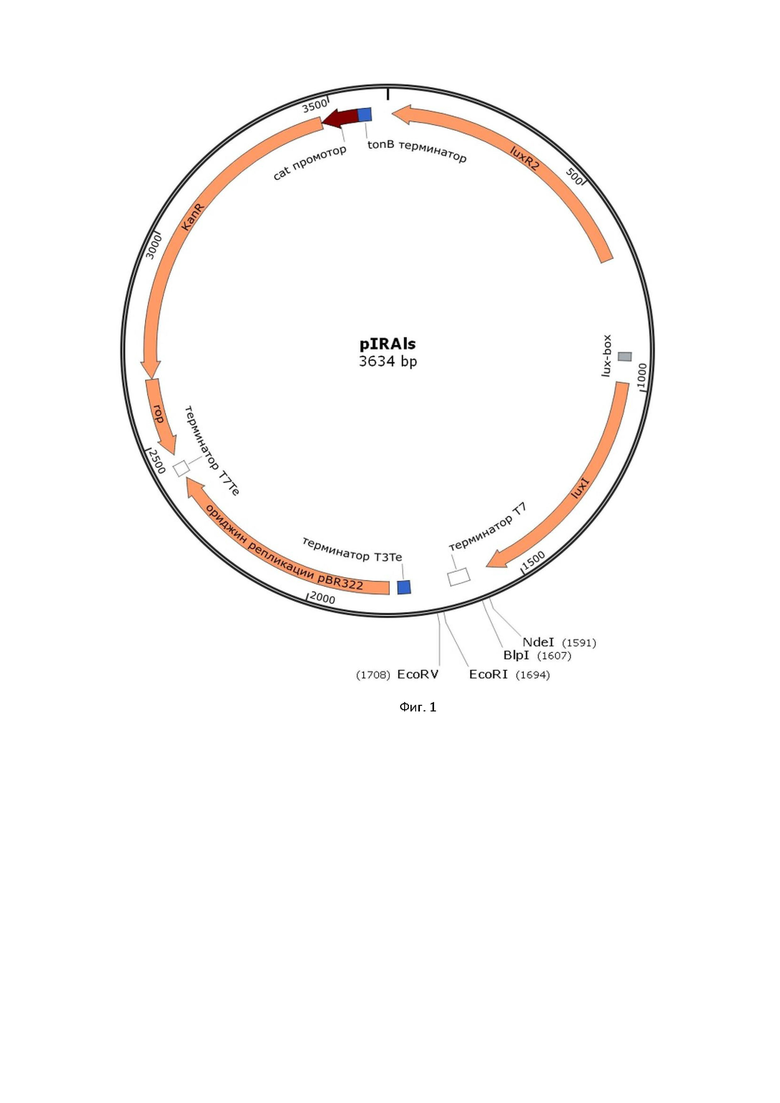
Плазмида pIRAls содержит фрагмент ДНК A. logei K18-44, включающий гены luxR2-luxI (1,5 т.п.н.), консенсусную последовательность Шайна-Дальгарно сразу после рамки считывания гена luxI, сайт рестрикции NdeI для встраивания целевого гена на расстоянии 6 н.п. от последовательности Шайна-Дальгарно, ориджин репликации pBR322 (0,9 т.п.н.) и ген резистентности к канамицину. Общий размер pIRAls составил 3634 п.н.. Для встраивания целевого гена по двум сайтам рестрикции далее за NdeI предусмотрены так же BlpI, EcoRI и EcoRV. Существо изобретения поясняется прилагаемыми иллюстрациями. Карта экспрессионного вектора приведена на фигуре 1.
Методика экспрессии целевого гена в клетках E. coli:
На фиг. 2 показано содержание белка GFP с His-tag меткой в клетках E. coli (белковый форез с окраской кумасси и вестерн-блот с антителами на His-tag) в условиях регуляции его биосинтеза описанной выше экспрессионной системой. Образцы для анализа были получены описанным ниже способом.
Вырастили культуры клеток E. coli MC1061 pIRALs::gfp-his до OD≈0,6 при 37°С, 200 об/мин в жидкой питательной среде LB в объеме 18 мл. Культуру клеток разделили на 6 частей по 3 мл, добавили АИ до итоговой концентрации 1мкМ во все образцы, кроме контрольного. После добавления АИ в среду клетки инкубировали при 22°С в течение различных промежутков времени: от 10 минут до 19 часов для разных образцов, после чего возвращали пробирки в инкубатор на 37°С, где они находились до момента завершения эксперимента (19 часов с момента добавления АИ). В дополнение к полученным культурам была проведена аутоиндукция при комнатной температуре на поверхности твердой среды, АИ в среду не добавляли. На фигуре 2 приведены результаты белкового фореза (а) и вестерн-блота (б) лизата клеток МС1061 pIRALs-gfp-his, индуцируемых в течение различных промежутков времени, отмечены следующие образцы:
• к – контроль без понижения температуры, 19 часов при 37°С
• 10’ – 10 минут при 22°С, затем 19 часов при 37°С;
• 20’ – 20 минут при 22°С, затем 19 часов при 37°С;
• 30’ – 30 минут при 22°С, затем 19 часов при 37°С;
• 40’ – 40 минут при 22°С, затем 19 часов при 37°С;
• с – те же клетки, выращенные на поверхности агаризованной среды LB при комнатной температуре без добавления АИ (пример автоиндукции);
• н – 19 часов при 22°С.
Образцы концентрировали до OD≈40 в буфере PBS и лизировали ультразвуком, на форез наносили 4 мкл надосадочной (240 мкг сырой биомассы) от лизата клеток, на блот наносили 20 мкл надосадочной (1200 мкг сырой биомассы) от лизата клеток.
Как видно, при 37°С экспрессия полностью отсутствует, и образцы содержат целевой белок в количестве пропорциональном времени инкубации при 22°С. При этом содержание целевого белка достигало 27%. Gfp-His имеет массу 27 кДа, что соответствует полосам, видимым на при гель-электрофорезе и вестерн-блоте. При этом мы не наблюдаем накопления LuxI, чья масса равна 22 кДа.
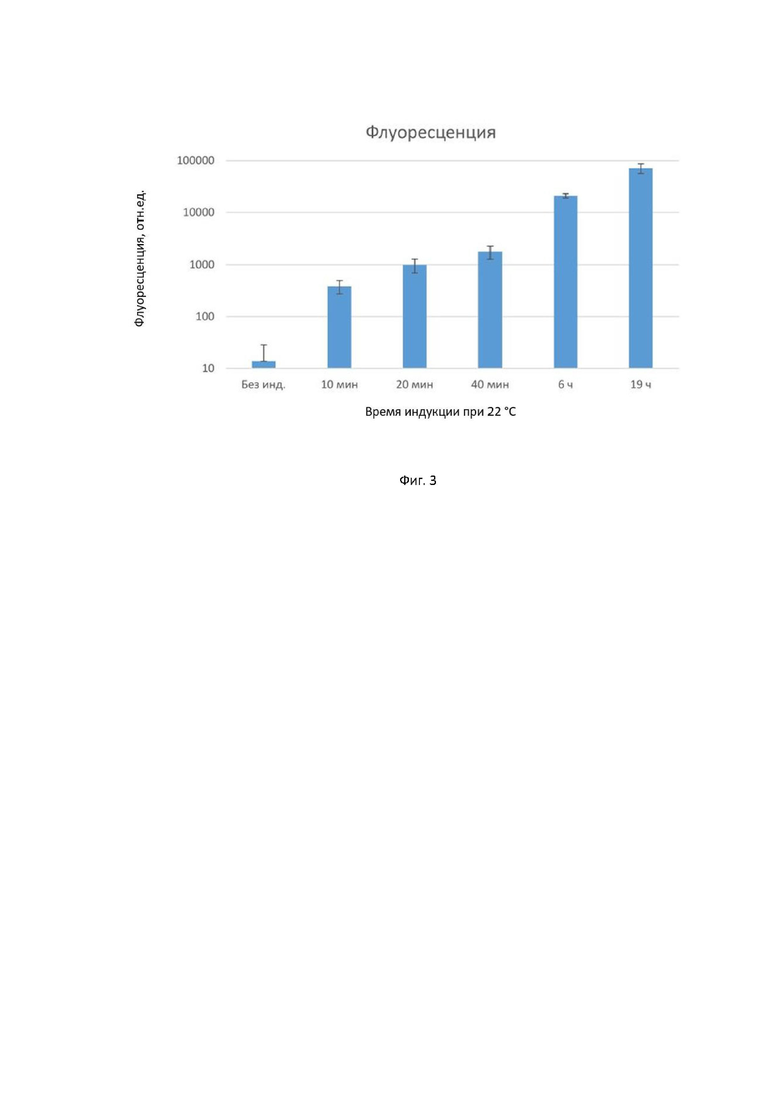
На фигуре 3 приведен уровень флуоресценции культуры клеток, экспрессировавших gfp с разным временем индукции при 22°C. После инкубации на 22°C клетки опять переносились на 37°C и инкубировались далее одинаковое с контрольными образцами время. Общее время инкубации всех образцов составило 19 часов. Из сигнала была вычтена фоновая флуоресценция, измеренная на культуре бесплазмидных клеток.
Изобретение позволяет создавать экспрессионные штаммы с точной регуляцией биосинтеза белков или ферментов, отвечающих за определенные этапы биосинтеза метаболитов.
Использованные источники информации:
1. Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172, doi:10.3389/FMICB.2014.00172/BIBTEX.
2. Chen, R. Bacterial expression systems for recombinant protein production: E. coli and beyond. Biotechnol. Adv. 2012, 30, 1102–1107, doi:10.1016/J.BIOTECHADV.2011.09.013.
3. Choi, Y.J.; Morel, L.; François, T. Le; Bourque, D.; Lucie, B.; Groleau, D.; Massie, B.; Miguez, C.B. Novel, versatile, and tightly regulated expression system for Escherichia coli strains. Appl. Environ. Microbiol. 2010, 76, 5058–5066, doi:10.1128/AEM.00413-10.
4. Hensel, Z. A plasmid-based Escherichia coli gene expression system with cell-to-cell variation below the extrinsic noise limit. 2017, doi:10.1371/journal.pone.0187259.
5. Lozano Terol, G.; Gallego-Jara, J.; Sola Martínez, R.A.; Martínez Vivancos, A.; Cánovas Díaz, M.; de Diego Puente, T. Impact of the expression system on recombinant protein production in Escherichia coli BL21. Front. Microbiol. 2021, 12, 1511, doi:10.3389/FMICB.2021.682001/BIBTEX.
6. Du, F.; Liu, Y.Q.; Xu, Y.S.; Li, Z.J.; Wang, Y.Z.; Zhang, Z.X.; Sun, X.M. Regulating the T7 RNA polymerase expression in E. coli BL21 (DE3) to provide more host options for recombinant protein production. Microb. Cell Fact. 2021, 20, doi:10.1186/S12934-021-01680-6.
7. Sitnikov, D.M.; Schineller, J.B.; Baldwin, T.O. Transcriptional regulation of bioluminesence genes from Vibrio fischeri. Mol. Microbiol. 1995, 17, 801–812.
8. Melkina, O.E.; Goryanin, I.I.; Bazhenov, S. V.; Manukhov, I. V.; Zavilgelsky, G.B. Comparative analysis of Aliivibrio logei luxR1 and luxR2 genes regulation in Escherichia coli cells. Arch. Microbiol. 2019, 201, 1415–1425, doi:10.1007/s00203-019-01691-3.
9. Khrulnova, S.A.; Baranova, A.; Bazhenov, S. V; Goryanin, I.I.; Konopleva, M.N.; Maryshev, I. V; Salykhova, A.I.; Vasilyeva, A. V; Manukhov, I. V; Zavilgelsky, G.B. Lux-operon of the marine psychrophilic bacterium Aliivibrio logei: a comparative analysis of the LuxR1/LuxR2 regulatory activity in Escherichia coli cells. Microbiology 2016, 162, 717–724, doi:10.1099/mic.0.000253.
10. Stevens, A.M.; Dolan, K.M.; Greenberg, E.P. Synergistic binding of the Vibrio fischeri LuxR transcriptional activator domain and RNA polymerase to the lux promoter region. Proc. Natl. Acad. Sci. U. S. A. 1994, 91, 12619–12623, doi:10.1073/pnas.91.26.12619.
11. Bazhenov, S.; Novoyatlova, U.; Scheglova, E.; Fomin, V.; Khrulnova, S.; Melkina, O.; Chistyakov, V.; Manukhov, I. Influence of the luxR regulatory gene dosage and expression level on the sensitivity of the whole-cell biosensor to acyl-homoserine lactone. Biosensors 2021, 11, 166, doi:10.3390/bios11060166.
12. Bazhenov, S. V.; Melkina, O.E.; Fomin, V. V.; Scheglova, E.S.; Krasnik, P. V.; Khrulnova, S.A.; Zavilgelsky, G.B.; Manukhov, I. V. LitR directly upregulates autoinducer synthesis and luminescence in Aliivibrio logei. PeerJ 2021, doi:10.7717/peerj.12030.
13. Kessenikh, A.G.; Novoyatlova, U.S.; Bazhenov, S. V; Stepanova, E.A.; Khrulnova, S.A.; Gnuchikh, E.Y.; Kotova, V.Y.; Kudryavtseva, A.A.; Bermeshev, M. V; Manukhov, I. V Constructing of Bacillus subtilis -Based Lux-Biosensors with the Use of Stress-Inducible Promoters. IJMS 2021, 1–13, doi:https://doi.org/10.3390/ijms22179571.
14. Kessenikh, A.; Gnuchikh, E.; Bazhenov, S.; Bermeshev, M.; Pevgov, V.; Samoilov, V.; Shorunov, S.; Maksimov, A.; Yaguzhinsky, L.; Manukhov, I. Genotoxic effect of 2,2’-bis(bicyclo[2.2.1] heptane) on bacterial cells. PLoS One 2020, 15, doi:10.1371/journal.pone.0228525.
15. Kotova, V.Y.; Manukhov, I. V.; Zavilgelskii, G.B. Lux-biosensors for detection of SOS-response, heat shock, and oxidative stress. Appl. Biochem. Microbiol. 2010, 46, 781–788, doi:10.1134/S0003683810080089.
16. Van Dyk, T.K.; Majarian, W.R.; Konstantinov, K.B.; Young, R.M.; Dhurjati, P.S.; LaRossa, R.A. Rapid and sensitive pollutant detection by induction of heat shock gene- bioluminescence gene fusions. Appl. Environ. Microbiol. 1994, 60, 1414–1420, doi:10.1128/aem.60.5.1414-1420.1994.
17. Winson, M.K.; Swift, S.; Fish, L.; Throup, J.P.; Jorgensen, F.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.S.A.. Construction and analysis of luxCDABE -based plasmid sensors for investigating N -acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett. 1998, 163, 185–192, doi:10.1111/j.1574-6968.1998.tb13044.x.
18. Blackwell, H.E.; Geske, G.D.; O’Neill, J.C. Modulation of bacterial quorum sensing with synthetic ligands. US patent WO/2008/116029, 2008.
19. Swennen, E.; Nocadello, S. Expression of recombinant proteins. US Patent WO2010136897A3, 2011.
20. Nocadello, S.; Swennen, E.F. The new pLAI (lux regulon based auto-inducible) expression system for recombinant protein production in Escherichia coli. Microb. Cell Fact. 2012, 11, 3, doi:10.1186/1475-2859-11-3.
21. Коноплева, М.Н. Механизмы регуляции «quorum sensing» системы первого типа психрофильных люминесцирующих бактерий Aliivibrio logei. Дисс. к.б.н. 2016.
| название | год | авторы | номер документа |
|---|---|---|---|
| Экспрессионный вектор pBMS10 на основе элементов LuxR-LuxI "quorum sensing" системы Aliivibrio logei | 2023 |
|
RU2816447C1 |
| Бактериальный lux-биосенсор с повышенной чувствительностью для детекции ацильных производных гомосерин лактона | 2020 |
|
RU2777196C2 |
| Бактериальный lux-биосенсор на основе промотора гена бактериальной ДНК-полимеразы II | 2023 |
|
RU2832445C1 |
| Набор стрессиндуцируемых lux-биосенсоров на основе клеток Bacillus subtilis для исследования механизмов токсичности | 2022 |
|
RU2811895C2 |
| ГЛОБАЛЬНЫЕ РЕГУЛЯТОРЫ БАКТЕРИАЛЬНЫХ ПАТОГЕННЫХ ГЕНОВ, ПРОТЕИН ИНАКТИВАЦИИ БАКТЕРИАЛЬНЫХ АУТОИНДУКТОРОВ, ЯВЛЯЮЩИХСЯ МИШЕНЯМИ ДЛЯ СОЗДАНИЯ УСТОЙЧИВОСТИ К БОЛЕЗНЯМ | 1999 |
|
RU2236462C2 |
| Композиция на основе ципрофлоксацина и 7-гидроксикумарина, подавляющая систему чувства кворума LuxI/LuxR-типа у бактерий | 2024 |
|
RU2833847C1 |
| СПОСОБ ПРИМЕНЕНИЯ ГАММА-ОКТАЛАКТОНА В КАЧЕСТВЕ ИНГИБИТОРА СИСТЕМЫ "КВОРУМ СЕНСИНГА" LuxI/LuxR ТИПА У БАКТЕРИЙ | 2017 |
|
RU2691634C2 |
| Применение кумарина и его производных в качестве ингибиторов системы "кворум сенсинга" LuxI/LuxR типа у бактерий | 2015 |
|
RU2616237C1 |
| ВЕКТОР НА ОСНОВЕ РЕПЛИКОНА БАКТЕРИОФАГА N15 И РЕКОМБИНАНТНЫЙ ВЕКТОР ДЛЯ РЕГУЛИРУЕМОЙ ЭКСПРЕССИИ ЦЕЛЕВОГО ГЕНА В КЛЕТКАХ ESCHERICHIA COLI, ШТАММ ESCHERICHIA COLI, ОБЕСПЕЧИВАЮЩИЙ ВОЗМОЖНОСТЬ РЕГУЛЯЦИИ ЧИСЛА КОПИЙ ВЕКТОРА, И СИСТЕМА ЭКСПРЕССИИ | 2006 |
|
RU2312146C1 |
| СПОСОБ ПРИМЕНЕНИЯ КУМАРИНА ДЛЯ ИНГИБИРОВАНИЯ РАЗЛИЧНЫХ СИСТЕМ "КВОРУМ СЕНСИНГА" LUX/LUXR ТИПА У БАКТЕРИЙ | 2019 |
|
RU2744456C1 |

Изобретение относится к биотехнологии. Предложен способ управления экспрессией целевого гена в клетках Escherichia coli, заключающийся в клонировании в клетках E. coli целевого гена под контролем LuxR2-регулируемого промотора совместно с геном luxR2 под контролем собственного промотора и варьировании температуры культивации полученных клеток и концентрации аутоиндуктора. При этом ген luxR2, кодирующий регуляторный белок LuxR2, изолирован из генома психрофильных люминесцентных бактерий рода Aliivibrio. При этом при понижении температуры культивации до 22 °C экспрессия целевого гена активируется в дозозависимой манере в присутствии N-(3-оксогексаноил)-лактон L-гомосерина в диапазоне концентраций от 1 нМ до 10 мкМ и прекращается при повышении температуры до 37 °C. Изобретение обеспечивает достижение содержания целевого белка 30% от общего клеточного белка. 1 з.п. ф-лы, 3 ил., 1 пр.
1. Способ управления экспрессией целевого гена в клетках Escherichia coli, заключающийся в клонировании в клетках E. coli целевого гена под контролем LuxR2-регулируемого промотора совместно с геном luxR2 под контролем собственного промотора, где ген luxR2, кодирующий регуляторный белок LuxR2, изолирован из генома психрофильных люминесцентных бактерий рода Aliivibrio, и варьировании температуры культивации полученных клеток и концентрации аутоиндуктора: при понижении температуры культивации до 22 °C экспрессия целевого гена активируется в дозозависимой манере в присутствии N-(3-оксогексаноил)-лактон L-гомосерина в диапазоне концентраций от 1 нМ до 10 мкМ и прекращается при повышении температуры до 37 °C.
2. Способ по п.1, в котором целевой ген клонируется в клетках E. coli под контролем LuxR2-регулируемого промотора совместно с генами luxR2-luxI под контролем собственных промоторов, где гены luxR2, кодирующий регуляторный белок LuxR2, и luxI, кодирующий N-ацил гомосерин лактона синтазу, изолированы из генома психрофильных люминесцентных бактерий рода Aliivibrio, а N-(3-оксогексаноил)-лактон L-гомосерина может быть внесен экзогенно в диапазоне концентраций от 1 нМ до 10 мкМ или синтезирован клетками во время культивации при температуре 22 °C, за счет чего достигается автоиндукция в ходе роста культуры.
| WO 2010136897 A2, 02.12.2010 | |||
| БАЖЕНОВ С | |||
| В | |||
| LuxI/LuxR "Quorum sensing" системы бактерий рода Aliivibrio | |||
| Диссертация на соискание ученой степени кандидата биологических наук | |||
| Москва, 2020 | |||
| Способ получения камфоры | 1921 |
|
SU119A1 |
| КОНОПЛЕВА М | |||
| Н | |||
| Механизмы регуляции "quorum sensing" системы первого типа психрофильных люминесцирующих бактерий Aliivibrio logei | |||
| Диссертация на | |||
