Изобретение относится к биотехнологии, биосинтезу рекомбинантных белков в клетках Escherichia coli. Разработан экспрессионный вектор на основе элементов LuxR-LuxI системы чувства кворума («quorum sensing», QS) морских психрофильных бактерий вида Aliivibrio logei. Экспрессия целевого гена, клонированного в векторе pBMS10, возможна в двух режимах индукции: автоиндукция при температуре культивирования 22°C и индукция посредством экзогенного добавления 3OxoC6-HSL, при этом температура может как оставаться 22°C, так и повышаться вплоть до 30°C.
Экспрессионные векторы повсеместно используются биотехнологами и молекулярными биологами для создания бактериальных штаммов-продуцентов и дальнейшего синтеза рекомбинантных белков. Для получения штамма-продуцента в такие векторы встраивается ген, кодирующий целевой белок, после чего результирующей плазмидой трансформируется штамм-реципиент, во многих случаях используются бактерии E. coli. В зависимости от регуляторных элементов экспрессионного вектора, индукция экспрессии целевого гена в клетке может происходить в ответ на различные факторы: как химические (в том числе различные сахара), так и физические. Так, например, существуют системы, в которых индукция происходит за счет изменения температуры [1][2]. Индукция понижением температуры удобна для синтеза термолабильных белков в E. coli, т.к. повышается вероятность получения их в нативном состоянии. Большинство экспрессионных систем, индуцируемых понижением температуры, используют промотор холодового шока и обладают температурой индукции 15-18°С [3].
В основе настоящего изобретения лежит использование регуляторных элементов QS-системы психрофильных люминесцентных морских бактерий вида A. logei. Данные бактерии используют LuxI/LuxR систему “quorum sensing” для регуляции экспрессии lux-оперона, отвечающего за люминесценцию. Белок LuxI синтезирует ацильные производные гомосерин лактона - сигнальные молекулы QS системы, называемые аутоиндукторами (АИ), основным из которых в A. logei является 3OxoC6-HSL. Данные молекулы не токсичны и не включены в основные метаболические пути клеток E. coli, что позволяет использовать их в биотехнологии в качестве индуктора экспрессионных систем. Регуляторные белки LuxR1 и LuxR2 A. logei связываются с молекулой АИ, присоединяются к ДНК и активируют транскрипцию с регулируемого ими промотора за счет прямого взаимодействия с РНК-полимеразой [4]. Ранее было показано, что LuxR2 в гетерологичной системе E. coli обладает значительно большей чувствительностью к 3OxoC6-HSL, чем LuxR1: ответ биосенсора, содержащего LuxR2 в качестве белка-рецептора наблюдается при добавлении 1 нМ АИ, когда как LuxR1 способен детектировать в среде присутствие АИ в концентрациях от 1 мкМ [5][6]. Под контролем LuxR-регулируемых промоторов в клетках A. logei расположены гены люминесценции luxCDABEG и ген аутоиндуктор-синтетазы luxI, за счет чего реализуется положительная обратная связь: в присутствии АИ повышается скорость синтеза АИ.
Наиболее близкие экспрессионные системы описаны в работах [7-9]. В работе [7] описано конструирование экспрессионного вектора для клеток E. coli на основе luxR-luxI генов Aliivibrio fischeri, однако у него отсутствует возможность осуществлять температурно-зависимую регуляцию. В патентной заявке [8] и статье [9] описан способ использования элементов LuxI/LuxR QS систем психрофильных бактерий рода Aliivibrio для управления экспрессией целевого гена в клетках E. coli. Патентная заявка [8] является прототипом настоящего изобретения. В ней описано, что присутствие в клетках E. coli гена luxR2 под контролем собственного промотора и целевого гена под контролем LuxR2-регулируемого промотора обеспечивает возможность регуляции экспрессии целевого гена при помощи варьирования температуры культивирования и концентрации 3OxoC6-HSL. Понижение температуры до 2 °C индуцирует экспрессию в присутствии АИ, при повышении температуры культивирования до 3 °C экспрессия целевого белка прекращается. Однако в перечисленных работах экспрессионные векторы не показывали продукции целевого белка более 30% от общего клеточного белка, к тому же для них не была описана возможность использования индукции экзогенным АИ при температурах выше 2 °C.
Техническим результатом предложенного изобретения является экспрессионный вектор pBMS10, предназначенный для клонирования и экспрессии белок-кодирующих генов в клетках E. coli. Плазмида pBMS10 содержит ориджин репликации pBR322 и ген резистентности к канамицину. Для температуро- и АИ-зависимой экспрессии рекомбинантных белков в векторе содержатся luxR2 и luxI гены под контролем собственных промоторов, изолированные из хромосомальной ДНК A. logei K18-44. Ниже гена luxI находится участок SEQ ID NO 1, обеспечивающий инициацию транскрипции и трансляции встраиваемого в экспрессионный вектор целевого гена. SEQ ID NO 1 включает в себя промотор PBMS (нуклеотиды 1-54 SEQ ID NO 1) с полностью палиндромным lux-боксом (нуклеотиды 1-20) и консенсусной последовательностью -10 (нуклеотиды 42-47). За промотором PBMS в плазмиде расположены консенсусная последовательность Шайна-Дальгарно (нуклеотиды 109-115), старт-кодон (нуклеотиды 121-123). На фигуре 1 приведена карта экспрессионного вектора pBMS10.
За счет присутствия гена luxI под контролем LuxR2-регулируемого промотора, а также за счет свойств LuxR2, LuxI и PBMS продуценты на основе данного вектора обладают двумя режимами индукции: автоиндукция при температуре культивирования 22°C и индукция посредством экзогенного добавления 3OxoC6-HSL при температурах не выше 30°C. Данный вектор также обладает низким базовым уровнем экспрессии в условиях отсутствия индукции и способностью к отключению уже запущенной индукции при повышении температуры до 37°C, за счет чего появляется возможность зафиксировать определенное количество целевого белка в клетке.
Пример:
Для апробации изобретения в вектор pBMS10 был встроен ген, кодирующий GFP с His-tag меткой, в качестве целевого. Клетки E. coli MC1061 были трансформированы полученной плазмидой pBMS10-gfpHis. Культуры клеток E. coli MC1061 pBMS10-gfpHis выращивали до OD≈0,5 при 37°C, 200 об/мин в жидкой питательной среде LB в объеме 24 мл и делили на 2 части. К половине культуры добавляли 3OxoC6-HSL до концентрации 1 мкМ. Затем клетки с АИ и без него делили на 4 части по 3 мл и инкубировали при 22, 30, 33, 37°C, 200 об/мин. Через 1 час и через 20 часов были отобраны пробы для оценки экспрессии GFP с помощью флюорометрии и денатурирующего гель-электрофореза.
В таблице 1 приведены коэффициенты индукции флуоресценции MC1061 pBMS10-gfpHis в зависимости от времени инкубации и экзогенного добавления АИ, посчитанные как отношение флуоресценции (в расчете на 1 OD) индуцированных клеток к флуоресценции клеток E. coli без экспрессионной плазмиды, инкубированных то же время при 37°C. Следует отметить, что базовый уровень флуоресценции обусловлен флуоресценцией прочих клеточных компонентов и компонентов питательной среды, на спектре флуоресценции MC1061 pBMS10-gfpHis, выращенных при 37°C, пик GFP полностью отсутствовал. При 22°C не наблюдается значимых различий флуоресценции между образцами с и без экзогенного 3OxoC6-HSL. При этом добавление 1 мкМ 3OxoC6-HSL позволяет использовать pBMS10 для экспрессии целевого белка при температуре как 22°C, так и 30°. При этом при 22°C скорость роста клеток меньше, что отражается на значениях в таблице 1, и для достижения того же абсолютного выхода белка может потребоваться большее время инкубации.
Таблица 1.
индукции
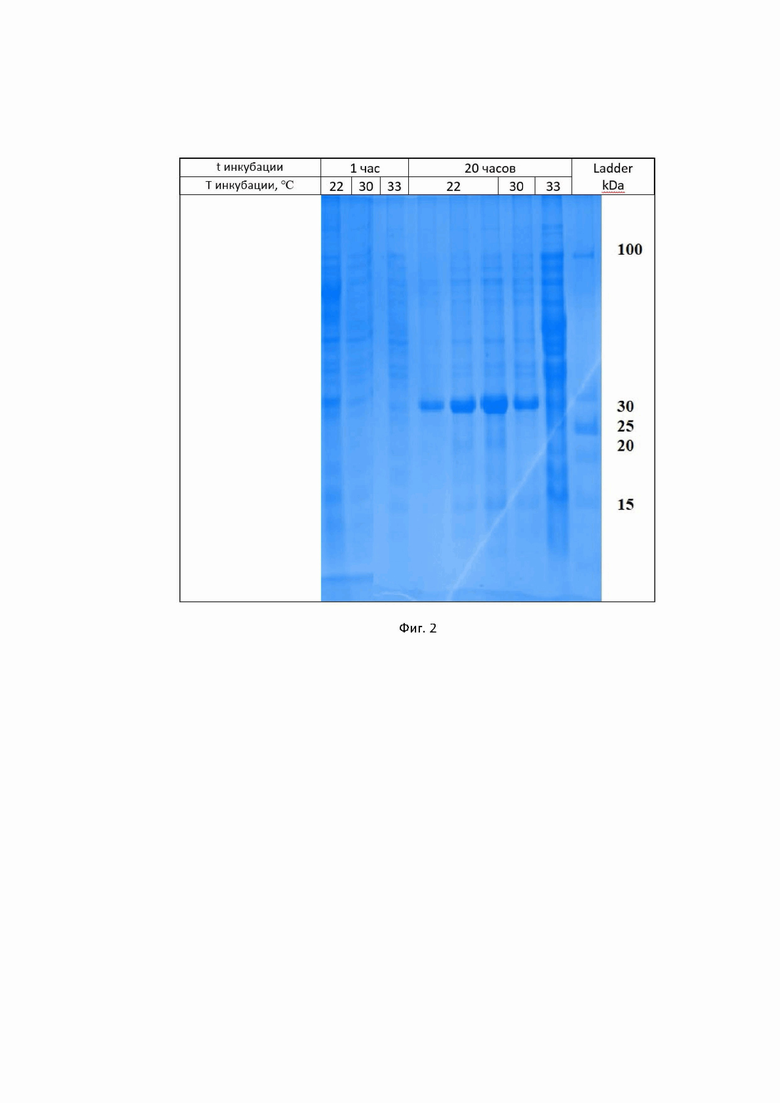
На фигуре 2 приведены результаты гель-электрофореза лизата клеток MC1061 pBMS10-gfpHis, индуцируемых в течение различных промежутков времени при различных температурах после добавления 1 мкМ 3OxoC6-HSL.
Анализ белкового фореза с последующей обработкой результатов при помощи программы TotalLab показал, что экспрессия gfp при клонировании в вектор pBMS10 пропорциональна времени инкубации и зависит от температуры: максимальна при 22°C, существенна при 30°C и незначительна при 33°C. Содержание GFP в лизатах клеток MC1061 pBMS10-gfpHis, культивируемых при 22°C в течение 20 часов после добавления 1 мкМ АИ, максимально и достигает 50 % от общего белка.
Список источников
[1] Menart V.,  and Pavko A., “Constitutive versus thermoinducible expression of heterologous proteins in Escherichia coli based on strong PR,PL promoters from phage lambda,” Biotechnol. Bioeng., vol. 83, no. 2, pp. 181-190, Jul. 2003, doi: 10.1002/BIT.10660.
and Pavko A., “Constitutive versus thermoinducible expression of heterologous proteins in Escherichia coli based on strong PR,PL promoters from phage lambda,” Biotechnol. Bioeng., vol. 83, no. 2, pp. 181-190, Jul. 2003, doi: 10.1002/BIT.10660.
[2] Rosano G. L. and Ceccarelli E. A., “Recombinant protein expression in Escherichia coli: Advances and challenges,” Front. Microbiol., vol. 5, no. APR, p. 79503, Apr. 2014, doi: 10.3389/FMICB.2014.00172.
[3] Vera A.,  Arís A., and Villaverde A., “The conformational quality of insoluble recombinant proteins is enhanced at low growth temperatures,” Biotechnol. Bioeng., vol. 96, no. 6, pp. 1101-1106, Apr. 2007, doi: 10.1002/BIT.21218.
Arís A., and Villaverde A., “The conformational quality of insoluble recombinant proteins is enhanced at low growth temperatures,” Biotechnol. Bioeng., vol. 96, no. 6, pp. 1101-1106, Apr. 2007, doi: 10.1002/BIT.21218.
[4] Stevens A. M., Dolan K. M., and Greenberg E. P., “Synergistic binding of the Vibrio fischeri LuxR transcriptional activator domain and RNA polymerase to the lux promoter region (autoinduction/DNA binding/luminescence/quorum sensing),” Biochemistry, vol. 91, pp. 12619-12623, 1994/
[5] Khrulnova S. A. et al., “Lux-operon of the marine psychrophilic bacterium aliivibrio logei: A comparative analysis of the LuxR1/LuxR2 regulatory activity in escherichia coli cells,” Microbiol. (United Kingdom), vol. 162, no. 4, pp. 717-724, Apr. 2016, doi: 10.1099/mic.0.000253.
[6] Bazhenov S. et al., “Influence of the luxR Regulatory Gene Dosage and Expression Level on the Sensitivity of the Whole-Cell Biosensor to Acyl-Homoserine Lactone,” Biosens. 2021, Vol. 11, Page 166, vol. 11, no. 6, p. 166, May 2021, doi: 10.3390/BIOS11060166.
[7] Nocadello, S.; Swennen, E.F. The new pLAI (lux regulon based auto-inducible) expression system for recombinant protein production in Escherichia coli. Microb. Cell Fact. 2012, 11, 3, doi:10.1186/1475-2859-11-3.
[8] Баженов С.В., Щеглова Е.С., Михайлов А.Э., Манухов И.В. Способ управляемой экспрессии рекомбинантных генов в клетках Escherichia coli с использованием элементов LuxR-LuxI «quorum sensing» системы психрофильных бактерий рода Aliivibrio. Заявка на патент РФ №2022110580/10(022319) от 19.04.2022
[9] Bazhenov, S. V.; Scheglova, E.S.; Utkina, A.A.; Kudryavtseva, A.A.; Al Ebrahim, R.; Manukhov, I. V. New temperature-switchable acyl homoserine lactone-regulated expression vector. Appl. Microbiol. Biotechnol. 2023, 107, 807–818, doi:10.1007/s00253-022-12341-y.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="pBMS10.xml"
softwareName="WIPO Sequence" softwareVersion="2.2.0"
productionDate="2023-11-24">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate></FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>Description file</ApplicantFileReference>
<ApplicantName languageCode="ru">МФТИ, Физтех</ApplicantName>
<ApplicantNameLatin>MIPT</ApplicantNameLatin>
<InventorName languageCode="ru">Баженов Сергей
Владимирович</InventorName>
<InventorNameLatin>Bazhenov Sergey Vladimirovich</InventorNameLatin>
<InventionTitle languageCode="ru">Экспрессионный вектор pBMS10 на
основе элементов LuxR-LuxI «quorum sensing» системы Aliivibrio
logei</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>123</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..123</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Escherichia coli</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acctgtaagatcttacaggtttacctaaataattaccctgctataattt
tctaaaattataaattagtcatttaataaaaatttaacattattttaaatatattaataaggaggtatca
tatg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ управляемой экспрессии рекомбинантных генов в клетках Escherichia coli с использованием элементов LuxR-LuxI "quorum sensing" системы психрофильных бактерий рода Aliivibrio | 2022 |
|
RU2810597C2 |
| Бактериальный lux-биосенсор с повышенной чувствительностью для детекции ацильных производных гомосерин лактона | 2020 |
|
RU2777196C2 |
| ГЛОБАЛЬНЫЕ РЕГУЛЯТОРЫ БАКТЕРИАЛЬНЫХ ПАТОГЕННЫХ ГЕНОВ, ПРОТЕИН ИНАКТИВАЦИИ БАКТЕРИАЛЬНЫХ АУТОИНДУКТОРОВ, ЯВЛЯЮЩИХСЯ МИШЕНЯМИ ДЛЯ СОЗДАНИЯ УСТОЙЧИВОСТИ К БОЛЕЗНЯМ | 1999 |
|
RU2236462C2 |
| Композиция на основе ципрофлоксацина и 7-гидроксикумарина, подавляющая систему чувства кворума LuxI/LuxR-типа у бактерий | 2024 |
|
RU2833847C1 |
| Бактериальный lux-биосенсор на основе промотора гена бактериальной ДНК-полимеразы II | 2023 |
|
RU2832445C1 |
| СПОСОБ ПРИМЕНЕНИЯ КУМАРИНА ДЛЯ ИНГИБИРОВАНИЯ РАЗЛИЧНЫХ СИСТЕМ "КВОРУМ СЕНСИНГА" LUX/LUXR ТИПА У БАКТЕРИЙ | 2019 |
|
RU2744456C1 |
| РЕГУЛЯТОР КОЛЛЕКТИВНОГО ПОВЕДЕНИЯ ("ЧУВСТВО КВОРУМА") У БАКТЕРИЙ | 2012 |
|
RU2534617C2 |
| СПОСОБ ПРИМЕНЕНИЯ ГАММА-ОКТАЛАКТОНА В КАЧЕСТВЕ ИНГИБИТОРА СИСТЕМЫ "КВОРУМ СЕНСИНГА" LuxI/LuxR ТИПА У БАКТЕРИЙ | 2017 |
|
RU2691634C2 |
| Применение кумарина и его производных в качестве ингибиторов системы "кворум сенсинга" LuxI/LuxR типа у бактерий | 2015 |
|
RU2616237C1 |
| ПРИМЕНЕНИЕ ПРОИЗВОДНЫХ 1,3-БЕНЗОДИОКСОЛА В КАЧЕСТВЕ РЕГУЛЯТОРОВ КОЛЛЕКТИВНОГО ПОВЕДЕНИЯ (ЧУВСТВА КВОРУМА) У БАКТЕРИЙ | 2012 |
|
RU2514001C2 |
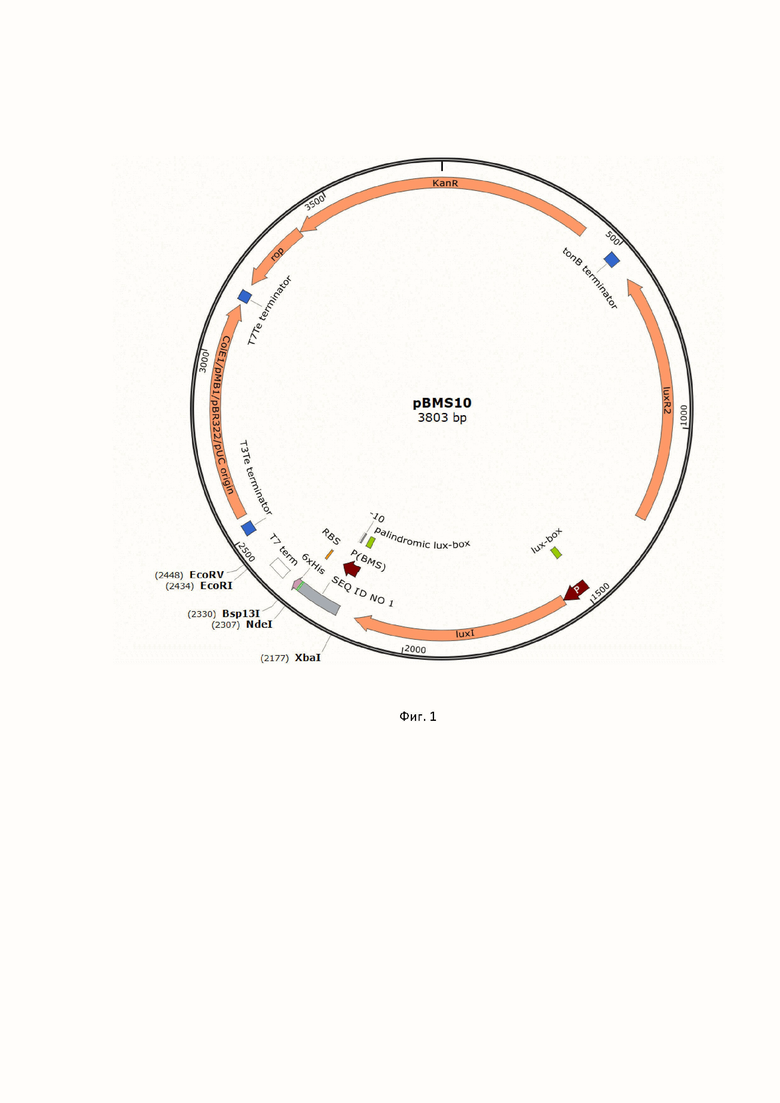
Изобретение относится к области биотехнологии, в частности к экспрессионному вектору pBMS10 на основе элементов LuxR-LuxI системы чувства кворума («quorum sensing», QS) морских психрофильных бактерий вида Aliivibrio logei для клонирования и экспрессии целевых генов в клетках Escherichia coli. Изобретение позволяет осуществлять индуцибельную экспрессию, реализуемую двумя способами: автоиндукцией при температуре культивирования 22°C и индукцией посредством экзогенного добавления 3OxoC6-HSL. Изобретение эффективно для биосинтеза рекомбинантных белков в клетках E. coli с возможностью получения количеств целевого белка, составляющих до 50% от общего белка в клетке, и культивированием в температурном диапазоне 22°C–30°C. 2 ил., 1 табл., 1 пр.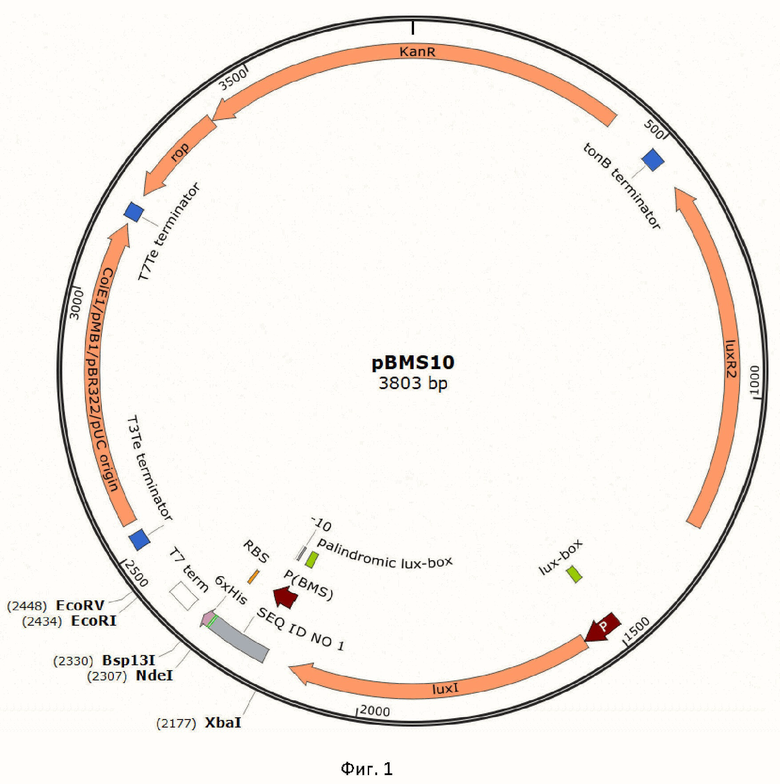
Экспрессионный вектор на основе регуляторных элементов системы чувства кворума бактерий A. logei, позволяющий проводить экспрессию целевого гена в клетках E. coli, состоящий из: ориджина репликации pBR322, гена резистентности к канамицину, luxR2-luxI генов A. logei под собственными промоторами, а также промоторного участка по последовательности SEQ ID NO: 1, включающего LuxR2-регулируемый промотор PBMS с полностью палиндромным lux-боксом и консенсусной последовательностью -10 и консенсусный сайт посадки рибосомы перед мультиклональным сайтом рестрикции для клонирования целевого гена под контроль LuxR2-регулируемого промотора.
| RU2022110580 A1, 19.10.2023 | |||
| The autoinducer synthases LuxI and AinS are responsible for temperature-dependent AHL production in the fish pathogen Aliivibrio salmonicida, BMC Microbiol., 2015, vol | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

