Изобретение относится к области биотехнологии и биохимии, а именно моноклональному антителу, селективно взаимодействующему с RBD фрагментом в составе S белка вируса SARS-CoV-2 и изолированному фрагменту ДНК, кодирующему участки легкой и тяжелой цепи указанного антитела, и антиген-связывающему фрагменту указанного моноклонального антитела. Специфичность моноклонального антитела к RBD фрагменту в составе S белка вируса SARS-CoV-2 подтверждена методами иммуноферментного анализа, иммуноблоттинга, иммуноцитохимического и иммуногистохимического окрашивания. Установлена нуклеотидная последовательность вариабельных доменов моноклонального антитела. Изобретение позволяет получить новые моноклональные антитела, селективно связывающие RBD фрагмент в составе S белка вируса SARS-CoV-2. 8 фиг.
Область техники
Изобретение относится к биотехнологии и биохимии, а именно моноклональному антителу, связывающемуся RBD фрагментом в составе S белка вируса SARS-CoV-2
Уровень техники
Известны различные моноклональные анти-RBD антитела, которые связываются с различными эпитопами RBD домена в составе S белка вируса SARS-CoV-2. Многие, представленные на мировом рынке антитела не открыты по нуклеотидным последовательностям. Как пример можно рассмотреть мышиное моноклональное Компания GeneTex, Inc., клон [1A9] (GTX 632604 https://www.genetex.com/Product/Detail/SARS-CoV-SARS-CoV-2-COVID-19-spike-antibody-1A9/GTX632604) - распознает шиповидные белки SARS-CoV и SARS-CoV-2, но не используется для иммуногистохимического анализа и не охарактеризовано по нейтрализующему потенциалу.
Известна разработка японские исследователей, связанная с получением мышиных моноклональных антител, при этом их последовательности использованных антител неизвестны [https://www.biorxiv.org/content/10.1101/2020.10.01.323220v1.full.pdf]. Недостатком данных моноклональных антител является отсутствие доступа к их вариабельным доменам.
Известна генетическая конструкция для экспрессии рекомбинантного белка RBD вируса SARS-CoV-2, штамм клеток яичника китайского хомячка CHO-S-RBD, продуцирующего рекомбинантный белок - рецептор-связывающий домен (RBD) вируса SARS-CoV-2, который может быть использован для диагностических целей [Патент РФ №2723008, C07K 14/165, опубл. 08.06.2020]. Описан так же способ получения штамма клеток яичника китайского хомячка CHO-S-RBD продуцента рекомбинантного белка RBD вируса SARS-CoV-2, содержащий определенную генетическую конструкцию, введение указанной генетической конструкции в клетки путем липофекции; селекцию клеток на антибиотике Hygromycin B. Раскрыта информация о штамме клеток яичника китайского хомячка CHO-S-RBD, продуценте рекомбинантного белка RBD вируса SARS-CoV-2. Описаны способы получения рекомбинантного белка RBD вируса SARS-CoV-2 включающие: культивирование штамма клеток яичника китайского хомячка CHO-S-RBD; хроматографическую очистку рекомбинантного белка RBD вируса SARS-CoV-2 из культуральной среды штамма клеток яичника китайского хомячка CHO-S-RBD; подтверждение получения рекомбинантного белка RBD вируса SARS-CoV-2. Создан рекомбинантный белок RBD вируса SARS-CoV-2 для выявления антител к SARS-CoV-2. Создана тест-система для иммуноферментного анализа сыворотки или плазмы крови человека. Разработан способ для анализа сыворотки или плазмы крови реконвалесцентов COVID-19 для отбора образцов наиболее перспективных для терапии пациентов, инфицированных SARS-CoV-2. Разработан способ анализа сыворотки или плазмы крови человека для определения титра антител к вирусу SARS-CoV-2. Изобретение предоставляет быструю и эффективную тест-систему для определения наиболее перспективных образцов сыворотки или плазмы крови реконвалесцентов COVID-19 для терапии пациентов, инфицированных SARS-CoV-2. Изобретение позволяет сократить время и упростить процедуру анализа сыворотки или плазмы крови реконвалесцентов COVID-19 для отбора образцов наиболее перспективных для терапии пациентов, инфицированных SARS-CoV-2. Данные технические решения не относятся к моноклональным антителам.
Известен искусственный ген, используемый для создания вакцины против коронавируса SARS-CoV-2, кодирующий искусственный белок-иммуноген, представляющий собой бицистронную структуру, состоящую из последовательностей рецептор-связывающего домена (RBD) гликопротеина S коронавируса SARS-CoV-2, гетерологического сигнального пептида гемагглютинина (HA) вируса гриппа А, линкера, P2A-пептида для расщепления полипротеина во время трансляции и гликопротеина G с мутацией M(1)>P(1), предотвращающей альтернативную инициацию трансляции, длиной 2355 п.н. Описан штамм rVSV-Stbl_RBD_SC2 рекомбинантного вируса везикулярного стоматита, полученный с использованием рекомбинантной плазмиды pStem-rVSV-Stbl_RBD_SC2, обеспечивающий независимый синтез коронавирусного антигена (рецептор-связывающий домен гликопротеина S SARS-CoV-2) и белка G вируса везикулярного стоматита, используемого для создания вакцины против коронавируса SARS-CoV-2 и депонированного в Государственной коллекции возбудителей вирусных инфекций, риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V-984. Техническим результатом является улучшение фолдинга и повышение инфекционного титра рекомбинантного вируса, стабильности целевого трансгена и иммуногенных/антигенных свойств заявляемого рекомбинантного вируса. [Патент РФ №2733834, МПК A61K 39/215 (2006.01), опубл 2020.10.07].
Указанное решение относится к искусственному гену, кодирующему бицистронную структуру, образованную последовательностями рецептор-связывающего домена (RBD) гликопротеина S коронавируса SARS-CoV-2, P2A-пептидом и гликопротеином G вируса везикулярного стоматита, рекомбинантной плазмиде, обеспечивающей экспрессию указанного искусственного гена и рекомбинантному штамму вируса везикулярного стоматита, экспрессирующему антигены коронавируса SARS-CoV-2, индуцирующему специфический иммунный ответ к SARS-CoV-2 и используемому для создания вакцины против коронавируса SARS-CoV-2 и может быть использовано в биотехнологии, молекулярной биологии, генетической инженерии и медицине, но не касается разработки моноклональных антител.
Известно решение по заявке (US, 20190062785 A1, МПК A61K 39/215, A61K 39/205; C12N 15/86, опубл. 28.02.2019 г.), представляющее собой вакцину на основе рекомбинантного вируса бешенства, которая обеспечивает защиту против бешенства и тяжелого острого респираторного синдрома (SARS-CoV). Транскрипционная кассета локализована в межгенном регионе N и P, что гарантирует высокий уровень экспрессии трансгена за счет особенностей организации генома рабдовирусов. В патенте раскрыты основные существующие подходы дизайна целевых коронавирусных иммуногенов/антигенов, такие как полноразмерный гликопротеин S, вариант с усеченным цитоплазматическим доменом (Δ19), а также рецептор-связывающий домен RBD с трансмембранным регионом и цитоплазматическим доменом вируса бешенства. Патент не включает в себя создание моноклональных антител. Патентный поиск нейтрализующих антител показывает ряд патентов - CN 111333722 A (https://patents.google.com/patent/CN111333722A/en), CN 111423508 A (https://patents.google.com/patent/CN111423508A/en), CN 111592595 A (https://patentimages.storage.googleapis.com/ca/4f/d6/8b54080db2f1e3/CN111592595A.pdf), CN 111732655 A (https://patents.google.com/patent/CN111732655A/en), CN 111690059 A (https://patents.google.com/patent/CN111690059A/en), CN 111718411 A (https://patents.google.com/patent/CN111718411A/en), CN 111732654 A (https://patents.google.com/patent/CN111732654A/en). Среди представленных разработок нет мышиных моноклональных антител одновременно обладающих вируснейтрализующей активностью и пригодных для идентификации вируса SARS-CoV-2 всеми методами иммуноанализа.
Сегодня специфические мышиные или кроличьи mAb против S-белка коммерчески доступны во многих странах компании (Abcam- https://www.abcam.com/sars-spike-glycoprotein-antibody-3a2-coronavirus-ab272420.html, GeneTex- https://www.genetex.com/Product/Detail/SARS-CoV-SARS-CoV-2-COVID-19-spike-antibody-1A9/GTX632604, Sino Biological и др.). Их применимость для разных методов описана, но ни одна из этих компаний не предоставляет последовательность антител, хотя эта информация может быть критичной для некоторых экспериментов. Так же для них не описана вируснейтрализующая активность.
Проведенный биоинформатический анализ всех представленных последовательностей по следующим базам данных моноклональных антител к SARS-CoV-2 не выявил совпадающих антител по последовательностям легкой и тяжелой цепи:: по легкой цепи

по тяжелой цепи

В базе данных http://opig.stats.ox.ac.uk/webapps/covabdab/ представлено 407 моноклональных антитела с вируснейтрализующей активностью. Анализ научных публикаций : Wafaa Alsoussi et al., 2020 (https://www.jimmunol.org/content/early/2020/06/23/jimmunol.2000583);
Xiangyang Chi et al., 2020 (https://science.sciencemag.org/content/early/2020/06/19/science.abc6952/tab-pdf); Xiangyu Chen et al., 2020 (https://www.nature.com/articles/s41423-020-0426-7);
Jinkai Wan et al., 2020 (https://www.sciencedirect.com/science/article/pii/S2211124720308998);
Xiaojian Han et al., 2020 https://www.biorxiv.org/content/10.1101/2020.08.19.253369v2.full.pdf+html; Shuo Du et al., 2020 (https://www.biorxiv.org/content/10.1101/2020.07.09.195263v1);
Yunlong Cao et al., 2020 (https://www.sciencedirect.com/science/article/pii/S0092867420306206) Davide Robbiani et al., 2020 (https://www.nature.com/articles/s41586-020-2456-9);
Sarah Clark et al., 2020 (https://www.biorxiv.org/content/10.1101/2020.11.13.381533v1)) показывает отсутствие среди них мышиных моноклональных антител охарактеризованных для исследования клинического материала.
Техническое приемлемой, решаемой настоящим изобретением, является получение моноклонального антитела, способного к связыванию RBD фрагмента в составе S белка вируса SARS-CoV-2 и отличающегося по аминокислотной последовательности вариабельных доменов легкой и тяжелой цепей от известных моноклональных антител к RBD фрагменту в составе S белка вируса SARS-CoV-2, пригодного для исследования клинического материала и обладающего вируснейтрализующей активностью, а также получение изолированных фрагментов ДНК, кодирующих участки легкой и тяжелой цепей указанного антитела и антигенсвязывающего фрагмента указанного моноклонального антитела.
Раскрытие изобретения
Техническим результатом является получение нового моноклонального антитела мыши IgM-изотипа, обладающего вируснейтрализующей активностью и детектирующего до 1 нг рекомбинантного RBD фрагмент S белка вируса SARS-CoV-2 с помощью метода иммуноферментного анализа.
Также техническим результатом является расширение арсенала средств аналогичного назначения, а именно получение нового моноклонального антитела к RBD фрагменту в составе S белка вируса SARS-CoV-2.
Поставленная техническая проблема решается получением моноклонального антитела, селективно
связывающего RBD фрагмент в составе S белка вируса SARS-CoV-2 человека, включающего вариабельный участок тяжелой цепи (VH) указанного антитела, содержащего последовательность аминокислот, SEQ ID NO: 1; а вариабельный участок легкой цепи(VL) указанного антитела содержит последовательность аминокислот, SEQ ID NO: 2. Поставленная техническая проблема решается также тем, что установлен изолированный фрагмент ДНК, кодирующий VH указанного антитела с нуклеотидной последовательностью SEQ IDNO: 3.
Поставленная техническая проблема решается также тем, что получен изолированный фрагмент ДНК, кодирующий VL указанного антитела с нуклеотидной последовательностью SEQ ID NO: 4.
Поставленная техническая проблема решается также тем, что получен антигенсвязывающий фрагмент указанного моноклонального антитела, содержащий вариабельный участок тяжелой цепи (VH) указанного антитела с последовательностью аминокислот SEQ ID NO: 1, и вариабельный участок легкой цепи (VL) указанного антитела с последовательностью аминокислот, SEQ ID NO: 2.
Моноклональное антитело, селективно связывающее RBD фрагмент в составе S белка вируса SARS-CoV-2,человека, настоящего изобретения характеризуется тем, что вариабельный участок тяжелой цепи (VH) указанного антитела содержит:
(i) CDR1 с последовательностью аминокислот QSIVHSNGNTY из SEQ ID NO: 1;
(ii) CDR2 с последовательностью аминокислот KVS из SEQ ID NO: 1;
(iii) CDR3 с последовательностью аминокислот FQGSHVPLT из SEQ ID NO: 1.
При этом вариабельный участок легкой цепи (VL) указанного антитела содержит
(i) CDR1 с последовательностью аминокислот QSLLYSRTRKNY из SEQ ID NO: 2;
(ii) CDR2 с последовательностью аминокислот WAS из SEQ ID NO: 2;
(iii) CDR3 с последовательностью аминокислот KQSYTLYT из SEQ ID NO: 2. Моноклональное антитело получено путем иммунизации мышей Balb/C человеческим рекомбинантным RBD фрагментом S белка вируса SARS-CoV-2, экспрессированным в E.coli (E.coli-RBD) и полноразмерным S белком вируса SARS-CoV-2 в составе клеток линии HEK293, получения и селекции линий гибридом, продуцирующих моноклональные антитела к рекомбинантному RBD фрагменту в составе S белка вируса SARS-CoV-2, анализа аффинности и специфичности отобранного МАт, определения нуклеотидной последовательности его вариабельных доменов. Антитела обычно состоят из двух тяжелых цепей, связанных между собой дисульфидными связями, и легких цепей, ассоциированных с N-концом каждой из тяжелых цепей. Каждая тяжелая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Каждая легкая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий участок. Вариабельные домены легкой и тяжелой цепей обладают похожей общей структурой, и каждый домен включает каркас из четырех участков, последовательности которых являются относительно консервативными, связанных посредством трех участков, определяющих комплементарность (complementarity determining regions, CDRs). Четыре каркасных участка формируют конформацию типа бета-складчатого слоя. Участки CDRs расположены в близком соседстве друг с другом благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDRs и каркасные участки антител могут быть определены путем ссылки на нумерационную систему Кабата (Kabat numbering system, Kabat et al., 1987 "Sequences of Proteins of Immunological Interest", US Dept. of Health and Human Services, US Government Printing Office) в сочетании с данными рентгеноструктурного анализа. Участки CDRs и каркасные участки антител могут также быть определены по номенклатуре Международной информационной системы по иммуногенетике (International Immunogenetics Information System, www.imgt.org). Для получения антитела, которое может связываться с каким-либо специфическим антигеном, обычно используют методику Kohler и Milstein (Kohler et al., (1976) Nature 256: 495-497). Моноклональные антитела получают путем слияния клеток селезенки из иммунизированного животного и клеток миеломы с получением гибридомы. Гибридомы могут быть проверены на способность к продукции нужного антитела, затем гибридомы могут быть выращены, из них могут быть выделены указанные антитела. Термин «выращенные клетки», использованный здесь, означает гибридомы или другие линии клеток, которые производят антитела. Методы получения и проверки таких выращенных клеток описаны Harlow и др. (Antibodies, a Laboratory Manual, Cold Spring Harbor Labs Press, 1988). Получение материала, использующегося в качестве антигена для инъекции животных, включают в себя методики, хорошо известные из уровня техники, например использование полноразмерного белка, использование пептида, выбранного из иммуногенных участков белка, а также любыми другими методами, известными из уровня техники (Harlow и др. Antibodies, a Laboratory Manual, Cold Spring Harbor Labs Press, 1988).
Подходящий способ для выделения антител, эффективных для использования в рамках настоящего изобретения, включает (а) назначение животному эффективного количества белка или пептида с целью получения антител, (b) выделение указанных антител, (с) определение последовательности антител.
Мышей BALB / С в возрасте 5 недель иммунизировали по стандартному протоколу [Methods Mol Biol. 2014; 1131: 33-45. doi: 10.1007/978-1-62703-992-5_3] с несколькими модификациями.
Затем полученные лимфоциты гибридизуют с клетками миеломы X63.
Тестирование супернатантов гибридом проводят методом непрямого иммуноферментного анализа (ИФА) с использованием RBD фрагмента S белка. Все первичные культуры, показавшие активность в ИФА, клонируют методом предельных разведений. Отбирают группы клонов - потомков одной первичной культуры. Клетки каждого из субклонов нарабатывают в культуре в количестве, достаточном для получения асцитных жидкостей.
Моноклональные антитела, выделенные из асцитной жидкости, сравнивают методом непрямого ИФА, по результатам которого было отобрано антитело 11/9.
Затем определили субизотип полученного антитела, оценили специфичность и
иммунореактивность полученного антитела методом непрямого ИФА с сорбированным на планшеты RBD фрагментом S белка с использованием коммерческих типирующих сывороток. Полученные антитела 11/9 определены как IgM.
Мы показали возможность использования МАт 11/9 для анализа клинических образцов, полученных от пациентов с COVID19 и контрольных пациентов без заболевания. В клинической практике чаще всего образцы готовятся в виде парафиновых блоков для облегчения длительного хранения материалов. Однако, длительная фиксация, парафинизация и депарафинизация крайне негативно влияют на структуру и доступность антигена. Таким образом, часто большинство антител способны обнаруживать антиген в экспериментах WB или ELISA, только некоторые из них хорошо работают на клинических образцах.
Например, из более чем 50 антител, доступных на Abcam, GeneTex и Sino Biological только один (Abcam ab275759) был утвержден для работы с клиническими образцами, и это антитело было кроличьим поликлональным.
Ни одно антитело, из перечисленных в Abcam, GeneTex, Sino Biological, не показывает свою эффективность во всех этих методиках одновременно.
Краткое описание чертежей и таблиц, иллюстрирующих изобретение
На Фиг. 1 показаны аминокислотные (SEQ ID NO: 1 и 2) и нуклеотидные (SEQ ID NO: 3 и 4) последовательности МАт 11/9.
На Фиг. 2 показана гель-электрофореграмма рекомбинантного RBD, очищенный из E. Coli используемого для иммунизации
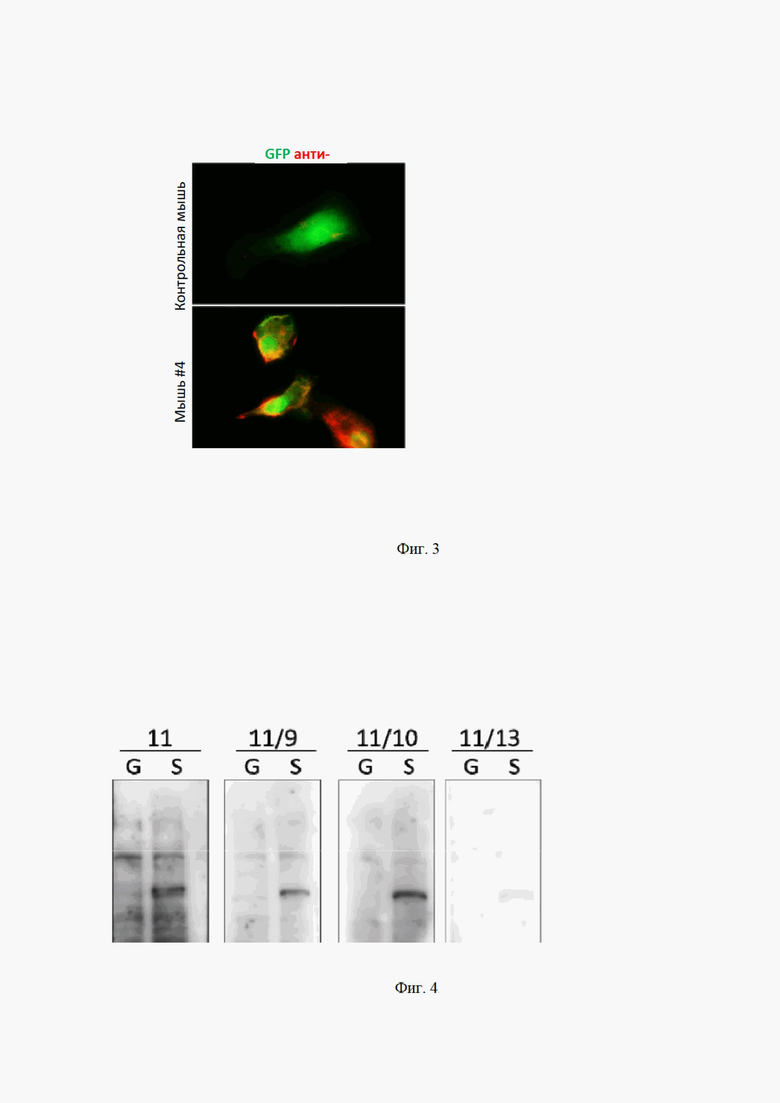
На Фиг. 3 показан иммуноцитохимический анализ клеток, котрансфицированных плазмидами, кодирующими GFP (зеленый) и полноразмерный S белок
окрашенных сывороткой, полученной от мышей (красный). ** р <0,01
На Фиг. 4 показан вестерн-блот анализ лизатов клеток, трансфицированных плазмидой, кодирующей GFP (G) или полноразмерный S-белок (S). Для окрашивания мембран использовали культуральную среду из разных субклонов.
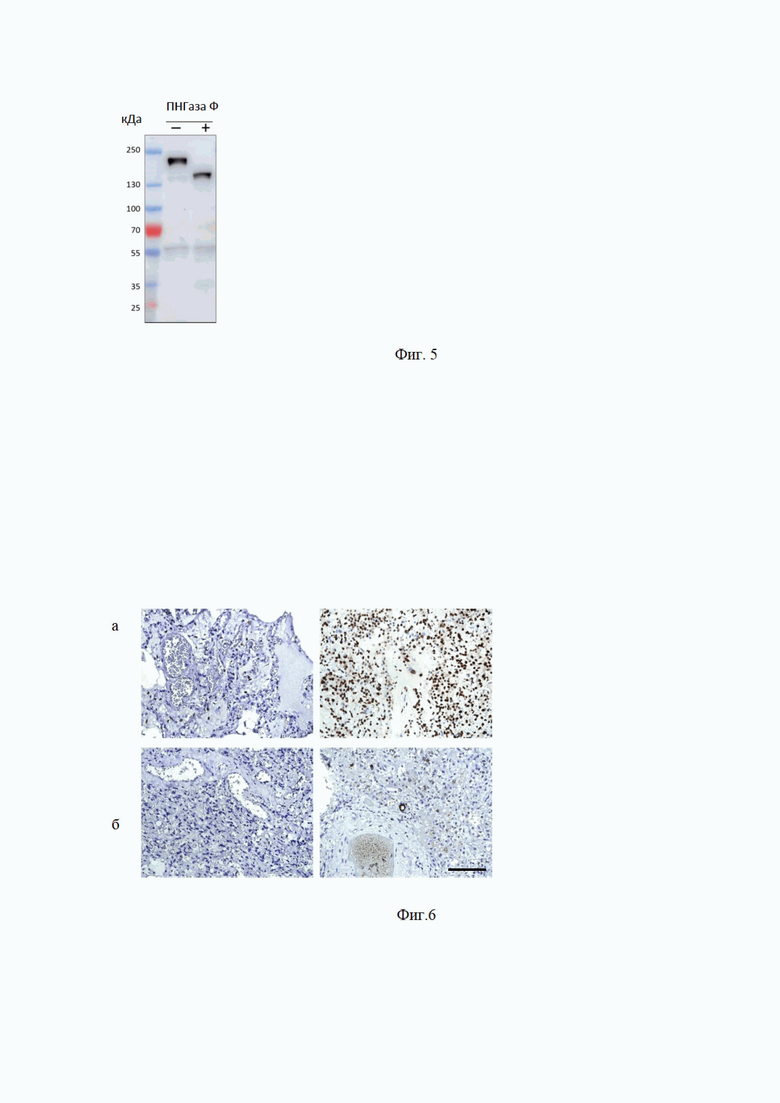
На Фиг. 5 показан вестерн-блот анализ лизата клеток трансфицированных плазмидой, кодирующей полноразмерный S-белок. Лизат инкубировали в присутствии или в отсутствие PNGase F. Мембрану окрашивали МАт 11/9
На Фиг. 6 Иммуногистохимическое окрашивание образцов биопсии легкого, залитых парафином, от контрольных пациентов и пациентов с COVID19. Асцитная жидкость, образованная моноклоном №11 / 9, использовалась в разведениях -а- 1: 1000 и б - 1: 100000. Масштабная линейка 100 мкм.
На Фиг. 7 показаны результаты иммуноферментного анализа с последовательным разведением МАт 11/9 в составе асцитной жидкости, против RBD (красный) или контрольный белок (синий), очищенный от E. coli. А так же результаты иммуноферментного анализа показывающие чувствительность МАт 11/9 в составе асцитной жидкости. Разведение 1:10 000 против различных количеств RBD, очищенного от E. coli и иммобилизованного в лунках планшета.
На Фиг. 8 показаны результаты исследования нейтрализующей активности МАт 11/9 в отношении SARS-CoV-2
Осуществление изобретения
Последующие примеры приведены для целей объяснения и не ограничивают каким-либо образом рамки настоящего изобретения.
Получение мышиного моноклонального антитела против RBD фрагмента в составе S белка вируса SARS-CoV-2.
Первую иммунизацию проводили внутрибрюшинно с помощью 200 мкг рекомбинантного очищенного RBD фрагмента, ресуспендированного в 150 мкл PBS и смешанного с 150 мкл либо полного адъюванта Фрейнда (FCA), либо неполного адъюванта Фрейнда (FIA) (Thermo Fisher). Две недели спустя мышам внутрибрюшинно вводили 200 мкг рекомбинантного очищенного RBD фрагмента, ресуспендированного в 150 мкл PBS, и смешивали с 150 мкл FIA. Через 6 дней кровь из хвостовой вены была собрана для проведения ELISA и IF окрашивания.
Экспрессия рекомбинантного белка и очистка от E.coli
Кодон BL21 (DE3) + RIL Клетки E.coli (Agilent) трансформировали плазмидой pET28-S. Бактерии инкубировали при 37°C на качалке до достижения OD600 0,7. Затем добавляли IPTG до конечной концентрации 1 мМ, и бактерии инкубировали еще 4 часа при 37°C. 200 мл среды с бактериями центрифугировали 15 мин, 5000 g при 4°C и осадок ресуспендировали в 12 мл лизирующего буфера B (200 мМ NaCl, 100 мМ NaH2PO4, 10 мМ Трис-HCl pH 8,0, 8 М мочевина, 0,5 мМ DTT) и инкубировали 1,5 часа при комнатной температуре. Затем раствор центрифугировали в течение 15 мин, 20000 g при 4°C и супернатант инкубировали с 2 мл смолы Ni-NTA в течение 1 часа при постоянном перемешивании. Суспензию переносили в колонку и промывали 20 мл буфера B и 10 мл буфера C (то же, что и буфер B, но с рН 6,3). Связанные белки элюировали буфером D (буфер C с 250 мМ имидазола) и диализовали в течение ночи против PBS с 1 мМ DTT. Чистоту полученного белка оценивали с помощью электрофореза и последующего окрашивания кумасси синим.
На 28, 29 и 30 дни после первой иммунизации мышам подкожно инъецировали 2×106 клеток HEK293, трансфицированных плазмидой pTwist-EF1a-nCoV-2019-S-2xStrep.
Культура клеток
Клетки HEK293 и HT1080 выращивали в среде Игла, модифицированной Дульбекко (DMEM), с добавлением 10% (об. / Об.) Фетальной телячьей сыворотки (FBS), 2 мМ L-глутамина, 1 мМ Na-пирувата и смеси пенициллин-стрептомицин (100 мкг / мл). Трансфекцию проводили реагентом Lipofectamine LTX (Thermo Fisher) в соответствии с протоколом производителя. Трансфицированные клетки окрашивали или вводили мышам через 48 часов после трансфекции. Клетки миеломы и гибридомы X63 выращивали в среде DMEM / F12 с добавлением 15% (об. / Об.) FBS, GlutaMAX (Thermo Fisher), 1 мМ Na-пирувата и смеси пенициллин-стрептомицин (100 мкг / мл). Добавки HAT или HT (Sigma) добавляли в разные моменты времени после слияния, как описано ранее [Methods Mol Biol. 2014;1131:33-45. doi: 10.1007/978-1-62703-992-5_3]. Затем полученные лимфоциты гибридизуют с клетками миеломы SP2/0. Тестирование супернатантов гибридом проводят методом непрямого иммуноферментного анализа (ИФА). Иммуноферментный анализ
Иммуноферментный анализ (ELISA) проводили, как описано ранее [BMC Biotechnol. 2016 Nov 22; 16(1): 83. doi: 10.1186/s12896-016-0314-5]. Вкратце, 0,5 мкг RBD, выделенного из E.coli, RBD, выделенного из клеток или клеток HEK293, или 0,5 мкг контрольного белка, выделенного из E.coli, иммобилизовали в лунках 96-луночного планшета EIA (Corning). После блокирования 1% BSA в TBST (20 мМ Tris pH 7,6, 150 мМ NaCl, 0,1% Tween20) лунки инкубировали с культуральной средой из клеток гибридомы, разведенной 1: 1 в TBST, или с сывороткой мыши, разведенной 1: 500 в TBST. После промывания TBST лунки инкубировали с HRP-конъюгированными вторичными антителами против мыши (Thermo Fisher) (разведение 1: 10000 в TBST) и проявляли с помощью раствора субстрата 1-Step Ultra TMB-ELISA (Thermo Fisher) в соответствии с протоколом производителя.
Мы получили, выделили и очистили рекомбинантный RBD фрагмент S белка из E.coli в денатурирующих условиях (Фиг. 2). Первая иммунизация проводилась белком, смешанным 1:1 с Freund's incomplete adjuvant (FIA). Интересно, что именно использование FIA позволило получить значительно более высокий титр целевых антител у животных. Этот результат говорит в пользу того, что нерастворимый S белок обладает достаточно высокой иммуногенностью для индукции иммунного ответа и не требует дополнительных стимулирующих добавок. Такие данные хорошо согласуются с результатами, полученные ранее, при иммунизации животных поверхностным белком вируса Зика [Biochimie. 2017 Nov; 142: 179-182. doi: 10.1016/j.biochi.2017.09.011]. Иммунофлуоресцентное окрашивание с использованием сыворотки имуннизированных животных показало, что антитела, вырабатываемые в ответ на нерастворимый RBD фрагмент S белка, были способны распознавать нативный S белок, экспрессированный на поверхности человеческих клеток (Фиг. 3). Таким образом, нерастворимый RBD и полноразмерный S белок имеют общие эпитопы и значит могут быть использованы для индукции и селекции антител к SARS-CoV2.
Отбор гибридом, секретируеющих антитела к S белку SARS-CoV2. После иммунизации мыши использовались для выделения спленоцитов, которые сливали с линией миеломы X63. Сразу после слияния полученные клетки были рассеяны на 20 96-луночных планшетов. Через 2 недели было получено 120 моноклонов гибридомных клеток. Антитела, производимые этими гибридомами, проверяли с помощью ELISA на RBD фрагмент S белка, выделенный из клеток HEK293. Использование для первичного скрининга белка, выделенного из другого источника дает возможность отсеять клоны, вырабатывающие антитела на разнообразные примеси, которые неизбежно содержатся в выделенном белке. Кроме того, так как белок из клеток HEK293 предположительно имеет гликозилирование и фолдинг сходные с таковыми у полноразмерного S белка, то антитела, взаимодействующие с RBD из HEK293, со значительной вероятностью, будут взаимодействовать и с нативным S белком.
14 гибридом, показавших наиболее сильную иммунореактивность к RBD, были использованы для дальнейшего анализа. Методом иммуноблотинга мы протестировали взаимодействие соответствующих антител с лизатами человеческих клеток, оверэкспрессирующих полноразмерный S белок или GFP в качестве контроля.
Получение моноклонов, продуцирующих антитела к S белку SARS-CoV2. Ранее многими авторами было показано, что в первые пассажи после слияния клетки гибридомы характеризуются высокой генетической нестабильностью, и поэтому даже потомки одной гибридной клетки, вероятнее всего, будут производить различные антитела [Dev Biol Stand. 1994; 83: 55-64]. По этой причине, гибридома #11, секретирующая наиболее специфичные антитела к RBD по результатам предыдущего анализа, была расклонирована для получения истинных моноклонов. Антитела, продуцируемые 17 субклонами гибридомы #11, были протестированы с помощью ELISA на взаимодействие с RBD, выделенным и E.coli; RBD, выделенным и HEK293; и контрольным белком, выделенным из E.coli. Результаты этого эксперимента позволили выбрать три моноклона для последующего анализа. Иммуноблотинг показал, что все полученные моноклоны обладают значительно большей специфичностью, чем исходная гибридома #11 (Фиг. 4).
Характеризация моноклональных антител к S белку SARS-CoV2.
По результатам предыдущего анализа, наиболее специфичными оказались антитела, производимые моноклоном #11/9. Иммунохцитохимическое окрашивание показывает, что антитела #11/9 окрашивают человеческие клетки, оверэкспрессирующие S белок, и почти не окрашивают соседние не транфицированные клетки. Таким образом, антитела #11/9 взаимодействуют с RBD фрагментом в условиях ELISA; с полноразмерным денатурированным S белком в условиях WB; и с полноразмерным нативным S белком на поверхности клеток.
Ранее было показано, что RBD, как и большинство поверхностных белков, подвергается значительному гликозилированию в клетках [Emerg Microbes Infect. 2020 Mar 17; 9(1): 601-604. doi: 10.1080/22221751.2020.1739565]. Лизат клеток оверэкспрессирующих S белок был обработан дегликозилирующим ферментом PNGase F. Из Фиг. 5 видно, что дегликозилирование уменьшает молекулярную массу S белка как минимум на 30 кДа, однако не оказывает влияние на интенсивность окрашивания антителами #11/9, таким образом, можно утверждать, что участок связывание антител с S белком не экранируется гликозидными группами. Дегликозилирование S-белка из лизата клеток HEK293, трансфицированных плазмидой pTwist-EF1a-nCoV-2019-S-2xStrep, проводили, как описано ранее [Methods Enzymol. 1994; 230: 44-57. doi: 10.1016/0076-6879(94)30006-2], с использованием PNGase F (New England Biolabs).
Иммунофлуоресцентная микроскопия
Клетки HT1080 высевали в лунки камеры Lab-Tek II и котрансфицировали плазмидами pTwist-EF1a-nCoV-2019-S-2xStrep и pTagGFP2-C (Evrogen). Через два дня после трансфекции клетки промывали забуференным фосфатом физиологическим раствором (PBS) и фиксировали 4% PFA в PBS в течение 15 минут при комнатной температуре. Клетки промывали 2 раза PBS и инкубировали с культуральной средой из клеток гибридомы, разведенной 1: 1 в PBS, или с сывороткой мыши, разведенной 1: 500 в PBS. После 5 промываний PBS клетки инкубировали с конъюгированными с AlexaFluor555 вторичными антителами против мыши (Thermo Fisher) (разведение 1: 500 в PBS) и затем окрашивали DAPI. Изображения получали с помощью флуоресцентного микроскопа DIAPHOT 300 (Nikon).
Секвенирование антител
Секвенирование мРНК, кодирующих тяжелую и легкую цепи антител, проводили с использованием метода 5'-SMART RACE, как описано ранее [PLoS One. 2019; 14(6): e0218717] с небольшими модификациями. Первую цепь кДНК синтезировали с использованием набора Mint (Evrogen) с праймерами для k-цепи (TTG TCG TTC ACT GCC ATC AAT C), λ-цепи (GGG GTA CCA TCT ACC TTC CAG) и тяжелой цепи (X) в соответствии с протоколу производителя. Затем кДНК, кодирующая IgG, была амплифицирована с помощью стандартной ПЦР с использованием прямого праймера M1 (AAG CAG TGG TAT CAA CGC AGA GT) и обратных праймеров, специфичных для k-цепи (ACA TTG ATG TCT TTG GGG TAG AAG), λ-цепи (ATC GTA CAC ACC AGT GTG GC) и тяжелой цепи (X). Полученную ДНК очищали и клонировали в вектор pKAN-T (Евроген), который впоследствии секвенировали. Секвенировали по крайней мере 3 разных клона для каждой цепи антител. В частности, антителом согласно настоящему изобретению является моноклональное антитело, обладающее способностью к связыванию RBD фрагмента в составе S белка вируса SARS-CoV-2. Использование праймеров специфическихк цепям k или λ мы продемонстрировали, что Мат 11 / 9 содержит только легкую цепь k. Последовательность действий продуктов ПЦР выявила нуклеотидную и аминокислотную последовательность вариабельных доменов Мат 11 / 9 (Фиг. 1). Биоинформатический анализ показал, что и легкие, и тяжелые цепи не содержат стоп-кодоны. Мы проанализировали сходство Мат 11 / 9 с антителами против SARS-CoV2, которые появляется естественным образом у пациентов с COVID19. С этой целью мы сравнили аминокислотные последовательности определяющая комплементарность область 3 (CDR3) Мат 11 / 9 к ранее опубликованным последовательностям антител, экспрессируемых RBD-связывающими В-клетками, выделенными от 60 пациентов с COVID19 [doi:10.1148/radiol.2020200463]. Важно отметить, что последовательности CDR3, как было показано, играют главную роль в связывании антигена и поэтому В-клетки, которые имеют идентичный CDR3 как для тяжелой, так и для легкой цепей, могут быть отнесены к этому же клонотипу [doi:10.1148/radiol.2020200463]. Сравнение последовательностей Мат 11 / 9 с последовательностями 91 различных клонотипов RBD-связывающего IgG выявили значительное сходство как тяжелых, так и легких цепей Мат 11 / 9 к нейтрализующим антителам, которые появляются у пациентов с COVID19 после заражения. Этот результат указывает на то, что, несмотря на иммунизацию искусственно синтезированным S-белком, мы смогли получить антитела, которые имеют аналогичную последовательность и, скорее всего, связываются с теми же участками белка S, что и естественного происхождения нейтрализующие антитела SARS-CoV2.
Мы использовали МАт 11/9 для окрашивания клинического материала и показали, что они позволяют легко различать контрольные образцы и образцы от пациентов с COVID19 (Фиг. 6). Таким образом, мы продемонстрировали, что МАт 11/9 может идентифицировать S белок с использованием всех иммунохимических методов (WB, ELISA, IHC-P, IF, эксперимент по нейтрализации).
Полученное Мат 11/9 способно выявлять методом иммуноферментного анализа рекомбинантный RBD в концентрации 1 нг (Фиг. 7). В культуральная среда от клеток гибридомы содержит 19,3±0,7 мкг / мл антител.
Мы выполнили анализ нейтрализации вируса с использованием hCoV-19 / Россия / Moscow_PMVL-4 штамм SARS-CoV-2, выделенный из носоглотки / ротоглотки мазок пациента с COVID-19. Вирус пассировали и титровали на клетках Vero E6. Серию пятикратных разведений асцита, образованного моноклоном 11/9, инкубировали с 100 TCID50 SARS-CoV-2 1 час при 37°C. Комплексы антитело-вирус добавляли к монослою культуры клеток Vero E6 в 96-луночные планшеты. Планшеты инкубировали в CO2-инкубаторе при 37°С в течение 72 ч, после чего цитопатический эффект (ЦПЭ) наблюдали микроскопически. Титры нейтрализации определяли как разведения антител, необходимого для 50% нейтрализации вирусной инфекции. Наши данные демонстрируют способность моноклональных антител 11 / 9 эффективно блокировать инфекцию SARS-CoV-2 (Фиг. 8).
Название проекта: Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2
Статус: invalid
Дата создания: 2020-11-19
Общая информация
Текущая заявка
Номер заявки: before
Название изобретения
Заявитель и изобретатель:
Имя заявителя: Федеральное государственное бюджетное учреждение науки Институт биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук
Язык: ru
Название или имя и фамилия латиницей: Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry RAS
Адрес места жительства: 117997, Российская Федеpация, Москва, ГСП-7, улица Миклухо-Маклая, дом 16/10
Адрес для переписки: 117997, Российская Федеpация, Москва, ГСП-7, улица Миклухо-Маклая, дом 16/10
Последовательности
Последовательность 1: " вариабельный участок тяжелой цепи (VH) SEQ ID NO:1"
Характеристики
Последовательность
--->
MRWSCIILFL VATATGVNSQ VQLQQPGAEL VMPGASVKMS CKASGYTFTD YWMHWVKQRP 60
GQGLEWIGAI DTSDSYTSYN QKFKGKATLT VDESSSTAYM QLSSLTSEDS AVYYCARRGY 120
GSSYTWFAYW GQGTLVTVSA 140
<---
Последовательность 2: "вариабельный участок легкой цепи (VL)SEQ ID NO: 2"
Характеристики
Последовательность
--->
MKLPVRLLVL MFWIPASSSD VLMTQTPLSL PVSLGDQASI SCRSSQSIVH SNGNTYLEWY 60
LQKPGQSPKL LIYKVSNRFS GVPDRFSGSG SGTDFTLKIS RVEAEDLGVY YCFQGSHVPL 120
TFGAGTKLEL K 131
<---
Последовательность 3: "Изолированный фрагмент ДНК, кодирующий VH антитела SEQ ID NO: 3"
Характеристики
Последовательность
--->
atgagatgga gctgtatcat cctcttcttg gtagcaacag ctacaggtgt caactcccag 60
gtccaactgc agcagcctgg ggctgagctt gtgatgcctg gggcttcagt gaagatgtcc 120
tgcaaggctt ctggctacac attcactgac tactggatgc actgggtgaa gcagaggcct 180
ggacaaggcc ttgagtggat cggagcgatt gatacttctg atagttatac tagctacaat 240
caaaagttca agggcaaggc cacattgact gtagacgaat cctccagcac agcctacatg 300
cagctcagca gcctgacatc tgaggactct gcggtctatt actgtgcaag aaggggctac 360
ggtagtagct acacctggtt tgcttactgg ggccaaggga ctctggtcac tgtctctgca 420
<---
Последовательность 4: "Изолированный фрагмент ДНК, кодирующий VL антитела SEQ ID NO: 4"
Характеристики
Последовательность
--->
atgaagttgc ctgttaggct gttggtgctg atgttctgga ttcctgcttc cagcagtgat 60
gttttgatga cccaaactcc actctccctg cctgtcagtc ttggagatca agcctccatc 120
tcttgcagat ctagtcagag cattgtacat agtaatggaa acacctattt agaatggtac 180
ctgcagaaac caggccagtc tccaaagctc ctgatctaca aagtttccaa ccgattttct 240
ggggtcccag acaggttcag tggcagtgga tcagggacag atttcacact caagatcagc 300
agagtggagg ctgaggatct gggagtttat tactgctttc aaggttcaca tgttccgctc 360
acgttcggtg ctgggaccaa gctggagctg aaa 393
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817697C1 |
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
| Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2 | 2021 |
|
RU2794141C2 |
| БИСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ SARS-COV-2 | 2022 |
|
RU2791749C1 |
| Гуманизированное моноклональное антитело, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, средство и способ для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2765731C1 |
| Широко нейтрализующее антитело против SARS-CoV-2 | 2022 |
|
RU2810476C1 |
| Однодоменное антитело ламы Н5 и его производное H5-Fc, специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2022 |
|
RU2793967C1 |
| ПОЛИВАЛЕНТНАЯ ВЕКТОРНАЯ ВАКЦИНА HVT | 2020 |
|
RU2832110C1 |
| Универсальный интеграционный вектор SB7G_HIGH и рекомбинантная плазмида SB7G_HIGH_DEV_57, обеспечивающая синтез и секрецию рекомбинантного человеческого антитела к рецептор-связывающему домену (RBD) коронавируса SARS-CoV-2 в клетках млекопитающих и полученная с использованием универсального вектора SB7G_HIGH, и рекомбинантное моноклональное антитело DEV_K57, обладающее вируснейтрализующей активностью в отношении SARS-CoV-2 | 2023 |
|
RU2829359C1 |


Изобретение относится к области биотехнологии и биохимии, а именно моноклональному антителу, селективно взаимодействующему с RBD фрагментом в составе S белка вируса SARS-CoV-2, и изолированному фрагменту ДНК, кодирующему участки легкой и тяжелой цепи указанного антитела, и антигенсвязывающему фрагменту указанного моноклонального антитела. Специфичность моноклонального антитела к RBD фрагменту в составе S белка вируса SARS-CoV-2 подтверждена методами иммуноферментного анализа, иммуноблоттинга, иммуноцитохимического и иммуногистохимического окрашивания. Установлена нуклеотидная последовательность вариабельных доменов моноклонального антитела. Изобретение позволяет получить новые моноклональные антитела, селективно связывающие RBD фрагмент в составе S белка вируса SARS-CoV-2. 4 н.п. ф-лы, 8 ил.
1. Моноклональное антитело, селективно связывающее RBD фрагмент в составе S белка вируса SARS-CoV-2, обладающее вируснейтрализующей активностью, включающее вариабельный участок тяжелой цепи (VH) указанного антитела, содержащий последовательность аминокислот SEQ ID NO: 1, а вариабельный участок легкой цепи (VL) указанного антитела содержит последовательность аминокислот SEQ ID NO: 2.
2. Изолированный фрагмент ДНК, кодирующий VH антитела по п. 1, с нуклеотидной последовательностью SEQ ID NO: 3.
3. Изолированный фрагмент ДНК, кодирующий VL антитела по п. 1, с нуклеотидной последовательностью SEQ ID NO: 4.
4. Антигенсвязывающий фрагмент моноклонального антитела по п. 1, селективно связывающий RBD фрагмент в составе S белка вируса SARS-CoV-2, содержащий вариабельный участок тяжелой цепи (VH) указанного антитела с последовательностью аминокислот SEQ ID NO: 1, и вариабельный участок легкой цепи (VL) указанного антитела с последовательностью аминокислот SEQ ID NO: 2.
| Искусственный ген EctoS_SC2, кодирующий эктодомен гликопротеина S коронавируса SARS-CoV-2 с C-концевым тримеризующим доменом, рекомбинантная плазмида pStem-rVSV-EctoS_SC2, обеспечивающая экспрессию искусственного гена, и рекомбинантный штамм вируса везикулярного стоматита rVSV-EctoS_SC2, используемый для создания вакцины против коронавируса SARS-CoV-2 | 2020 |
|
RU2733834C1 |
| CN 111423508 A, 17.07.2020 | |||
| CN 111620946 A, 04.09.2020 | |||
| TIAN X | |||
| et al., Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody, Emerg Microbes Infect., 2020, Volume 9, Issue 1, pp | |||
| Зеркальный стереовизир | 1922 |
|
SU382A1 |
| HUSSAIN A | |||
| et al., Targeting SARS-CoV2 Spike Protein Receptor Binding | |||

