Область техники
Изобретение относится к областям молекулярной биологии, вирусологии и клеточной технологии. Изобретение потенциально может быть использовано для лечения ВИЧ.
Работа выполнена при финансовой поддержке Министерства Науки и Высшего образования Российской Федерации в рамках Соглашения №075-15-2019-1661 от 31.10.2019 и при финансовой поддержке РФФИ по соглашению 18-29-07052.
Уровень техники
Ранее нашей лабораторией была разработана технология создания Т-клеточной устойчивости к ВИЧ-1 на основе генетически-кодируемых GPI-заякоренных пептидов из gp41, заявка на патент 2020142800 от 24.12.2020. Технология представляет собой генетическую конструкцию, включающую последовательность GPI-заякоренного белка CD52, между двумя частями которого (сигналом экспорта белка и сигналом GPI-заякоривания) вставлен один из защитных пептидов из оболочечного белка ВИЧ-1 gp41, а также способы доставки генетической конструкции на мембрану клетки с помощью системы геномного редактирования CRISPR/Cas9.
Система геномного редактирования CRISPR/Cas9 является относительно новой технологией, развитием которой занимается большое количество исследовательских групп по всему миру. Технология обладает большим потенциалом и широким спектром возможностей для лечения ВИЧ инфекции. Несмотря на существенные достижения в этой области, увеличение эффективности вносимых с помощью системы CRISPR/Cas9 терапевтических последовательностей является важной задачей, которую необходимо решить для эффективного применения технологии в клинике.
Для достижения этой цели ведутся исследования в области усиления гомологичной рекомбинации (HDR) путем различных модификаций донорной ДНК. Было показано, что удлинение плеч гомологии приводит к увеличению эффективности HDR (Tsunekawa Y, Terhune RK, Fujita I, Shitamukai A, Suetsugu T, Matsuzaki F. Developing a de novo targeted knock-in method based on in utero electroporation into the mammalian brain. Development. 2016;143(17):3216-3222. doi:10.1242/dev.136325; Kurihara T, Kouyama-Suzuki E, Satoga M, et al. DNA repair protein RAD51 enhances the CRISPR/Cas9-mediated knock-in efficiency in brain neurons. Biochem Biophys Res Commun. 2020;524(3):621-628. doi:https://doi.org/10.1016/j.bbrc.2020.01.132) в пирамидальных нейронах мыши. Несколько исследований показали, что использование плазмидной ДНК в качестве донора позволяет увеличить эффективность HDR (Sung JJ, Park C-Y, Leem JW, Cho MS, Kim D-W. Restoration of FVIII expression by targeted gene insertion in the FVIII locus in hemophilia A patient-derived iPSCs. Exp Mol Med. 2019;51(4):1-9. doi:10.1038/s12276-019-0243-1; Liu J-T, Corbett JL, Heslop JA, Duncan SA. Enhanced genome editing in human iPSCs with CRISPR-CAS9 by co-targeting ATP1a1. PeerJ. 2020;8:e9060. doi:10.7717/peerj.9060). Увеличения эффективности HDR удалось добиться путем модификации донорной ДНК введением в нее полноразмерных (Yao X, Wang X, Hu X, et al. Homology-mediated end joining-based targeted integration using CRISPR/Cas9. Cell Res. 2017;27(6):801-814. doi:10.1038/cr.2017.76; Yao X, Wang X, Liu J, Shi L, Huang P, Yang H. CRISPR/Cas9-mediated Targeted Integration In Vivo Using a Homology-mediated End Joining-based Strategy. J Vis Exp. 2018;(133):56844. doi:10.3791/56844) или укороченных сайтов распознавания гидовой РНК (гРНК) (Nguyen DN, Roth TL, Li PJ, et al. Polymer-stabilized Cas9 nanoparticles and modified repair templates increase genome editing efficiency. Nat Biotechnol. 2020;38(1):44-49. doi:10.1038/s41587-019-0325-6).
Другим подходом к решению этой задачи является модификация Cas9. Показано, что увеличение в Cas9 числа сигналов ядерной локализации (NLS) позволяют улучшить эффективность доставки конструкции в ядро, что приводит к повышению уровня редактирования (Torres-Ruiz R, Martinez-Lage M, Martin MC, et al., Efficient Recreation of t(11;22) EWSR1-FLI1(+) in Human Stem Cells Using CRISPR/Cas9. Stem cell reports. 2017;8(5):1408-1420. doi:10.1016/j.stemcr.2017.04.014; Maggio I, Zittersteijn HA, Wang Q, et al., Integrating gene delivery and gene-editing technologies by adenoviral vector transfer of optimized CRISPR-Cas9 components. Gene Ther. 2020;27(5):209-225. doi:10.1038/s41434-019-0119-y).
Эффективным способом доставки донорной ДНК конструкции в ядро является добавление к ней фрагмента энхансера из промотора вируса SV40, так называемого DTS (DNA nuclear transporting sequence). Присоединение фрагмента повышает транспорт плазмид в ядро in vivo (Dean DA, Dean BS, Muller S, Smith LC. Sequence requirements for plasmid nuclear import. Exp Cell Res. 1999;253(2):713-722. doi:10.1006/excr.1999.4716). Это объясняется тем, что DTS SV40 имеет сайты посадки нескольких транскрипционных факторов, экспрессирующихся и работающих практически по всех типах клеток. В качестве эффективной доставки плазмидной ДНК в ядро была отдельно использована последовательность сайта посадки NFkB (Mesika A, Grigoreva I, Zohar M, Reich Z. A Regulated, NFκB-Assisted Import of Plasmid DNA into Mammalian Cell Nuclei. Mol Ther. 2001;3(5):653-657. doi:https://doi.org/10.1006/mthe.2001.0312).
Применение сайта DTS в донорной конструкции упоминается только в одной работе, посвященной получению нокина в зиготах крыс (Remy S, Chenouard V, Tesson L, et al. Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation. Sci Rep. 2017;7(1):16554. doi:10.1038/s41598-017-16328-y). Однако авторам не удалось добиться увеличения уровня нокина, что возможно связано с очень большим (4kb) размером донорной конструкции.
Описанные методы применялись с целью разработки эффективных методов лечения различных генетических заболеваний, однако не применялись для создания клеток, резистентных к ВИЧ.
Патентуемый способ является существенным усовершенствованием созданной ранее системы, позволяющий вносить в геном Т-клеток конструкцию, придающую им устойчивость к ВИЧ с большей эффективностью.
Техническая задача - усовершенствование технологии создания Т-клеточной устойчивости к ВИЧ-1 на основе генетически-кодируемых GPI-заякоренных пептидов из gp41 - решается за счет создания эффективной технологии направленного редактирования гена CXCR4 в CD4-лимфоцитах. Технический результат - повышение эффективности внесения защитной конструкции и повышение устойчивости клеток к ВИЧ.
Внесенные усовершенствования позволили добиться значительного увеличения эффективности внесения защитной конструкции, что является ключевым фактором для потенциальной возможности применения технологии для терапии ВИЧ. При этом преимущества изначальной технологии, а именно, экономичность наработки донорной ДНК и совместимость технологии с разными системами редактирования генома на основе CRISPR/Cas9 и других, позволяющая легко адаптировать ее к последним разработкам в этой области, были сохранены.
Раскрытие сущности изобретения
Настоящее изобретение представляет собой существенное усовершенствование разработанной нами ранее технологии создания Т-клеточной устойчивости к ВИЧ-1 на основе генетически-кодируемых GPI-заякоренных пептидов из gp41, при которой защитный пептид из gp41 доставляется на поверхность плазматической мембраны клетки - мишени за счет внесения кодирующей его нуклеотидной последовательности в геном с помощью CRISPR/Cas9 в составе генетической конструкции, представляющей собой белок CD52, между двумя частями которого (сигналом экспорта белка и сигналом GPI-заякоревания) встроен целевой пептид.
Увеличение эффективности внесения целевой конструкции было достигнуто путем следующих модификаций донорной ДНК: 1. Донорная ДНК вносится в клетку в контексте плазмидной ДНК. 2. Плечи гомологии увеличены приблизительно со 100 до 500 нуклеотидов. 3. Окружающие последовательность целевой конструкции сайты гРНК и DTS увеличивают эффективность доставки донорной ДНК к целевому месту в геноме.
Для осуществления изобретения в клетки вводится два компонента: плазмида, кодирующая нуклеазу Cas9 и гРНК, в последовательности которой есть несколько сайтов NLS (pX330-Cas9-BFP-gRNAX4ex2) и плазмида, кодирующая донорную ДНК. Гидовая РНК направляет Cas9 на определенное место в геноме. Донорная ДНК содержит целевую генетическую конструкцию, фланкированную плечами гомологии, на концах которых расположены сайт посадки Cas9 (протоспейсер X4ex2) и сайт связывания транскрипционных факторов DTS, увеличивающие эффективность доставки донорной ДНК к месту редактирования, и служит матрицей для гомологичной рекомбинации.
В альтернативном варианте осуществления изобретения плазмида, кодирующая нуклеазу Cas9 и гРНК заменена рибонуклеопротеиновым комплексом (RNP), включающим рекомбинантный белок Cas9 в комплексе с гРНК, полученной путем транскрипции in vitro.
Настоящее изобретение предлагает способы эффективной доставки целевой генетической конструкции на мембрану клетки.
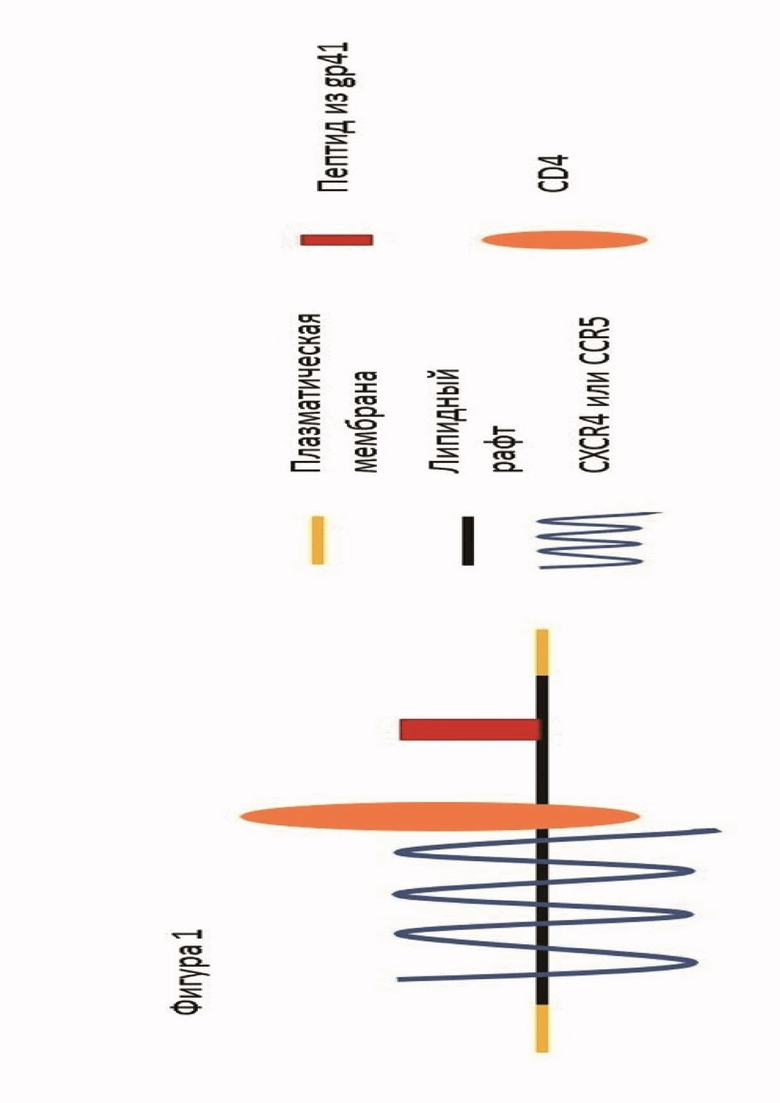
Целевая генетическая конструкция, закодированная в донорной ДНК, представляет собой последовательность GPI-заякоренного белка CD52, между сигналом экспорта и сигналом мембранного заякоревания которого находится последовательность, кодирующая защитный пептид из gp41 - MT-C34. SEQ ID NO: 1 является сигналом экспорта белка. SEQ ID NO:2 служит сигналом для мембранного заякоривания конструкции. Последовательность, кодирующая защитный пептид из gp41 МТ-С34 - SEQ ID NO: 3. Такая конструкция доставляет пептид на поверхность клетки в специфическую зону - липидный рафт, где локализуются рецептор CD4 и корецепторы CCR5 и CXCR4, распознаваемые ВИЧ и используемые им для проникновения в клетку (фиг. 1). Пептид, экспонируемый на поверхности клетки, взаимодействует с N-концевым гептадным повтором gp41 вирусного белка слияния, и препятствует образованию шестиспирального пучка, защищая таким образом клетку от проникновения ВИЧ.
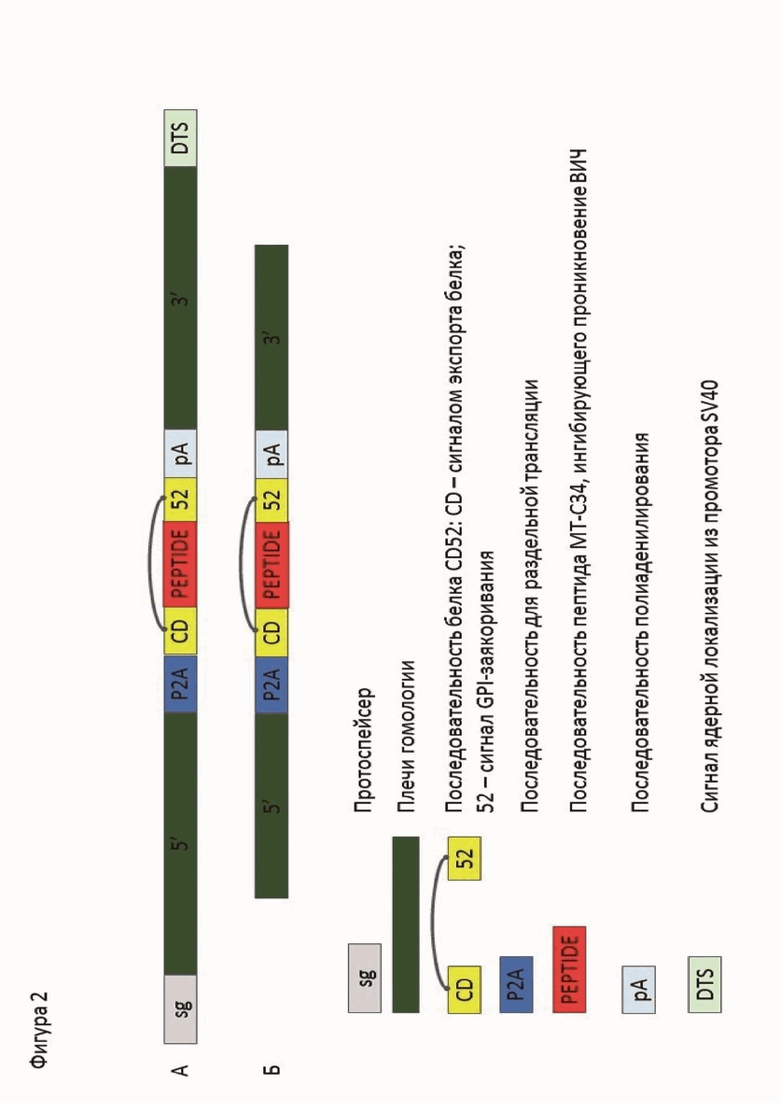
Донорная ДНК представлена следующей последовательностью: протоспейсер X4ex2 (SEQ ID NO: 4), 5’-плечо гомологии (SEQ ID NO: 5), последовательность Р2А, позволяющая рибосоме пропускать образование ковалентной связи между глицином и пролином, иначе говоря, позволяющей получить с одной РНК раздельную трансляцию пептида и белка CXCR4 (SEQ ID NO: 6), сигнал экспорта белка (SEQ ID NO: 1), последовательность, кодирующая защитный пептид из gp41 МТ-С34 (SEQ ID NO: 3), сигнал для мембранного заякоривания (SEQ ID NO: 2), последовательность полиаденилирования (SEQ ID NO: 7), 3’-плечо гомологии (SEQ ID NO: 8), последовательность DTS (SEQ ID NO: 9), как показано на фигуре 2 А. Донорная ДНК вносится в клетку в контексте коммерческого вектора для клонирования pJet (Thermo scientific США).
Наибольшей эффективность обладает гРНК, направляющая Cas9 на второй экзон, CXCR4 (SEQ ID NO: 10).
Краткое описание чертежей
Фигура 1. Схема расположения защитного пептида на поверхности клеточной мембраны.
Фигура 2. Схемы последовательностей, используемых в качестве донорных ДНК для внесения целевых мутаций: А - патентуемая технология; Б - изначальная технология.
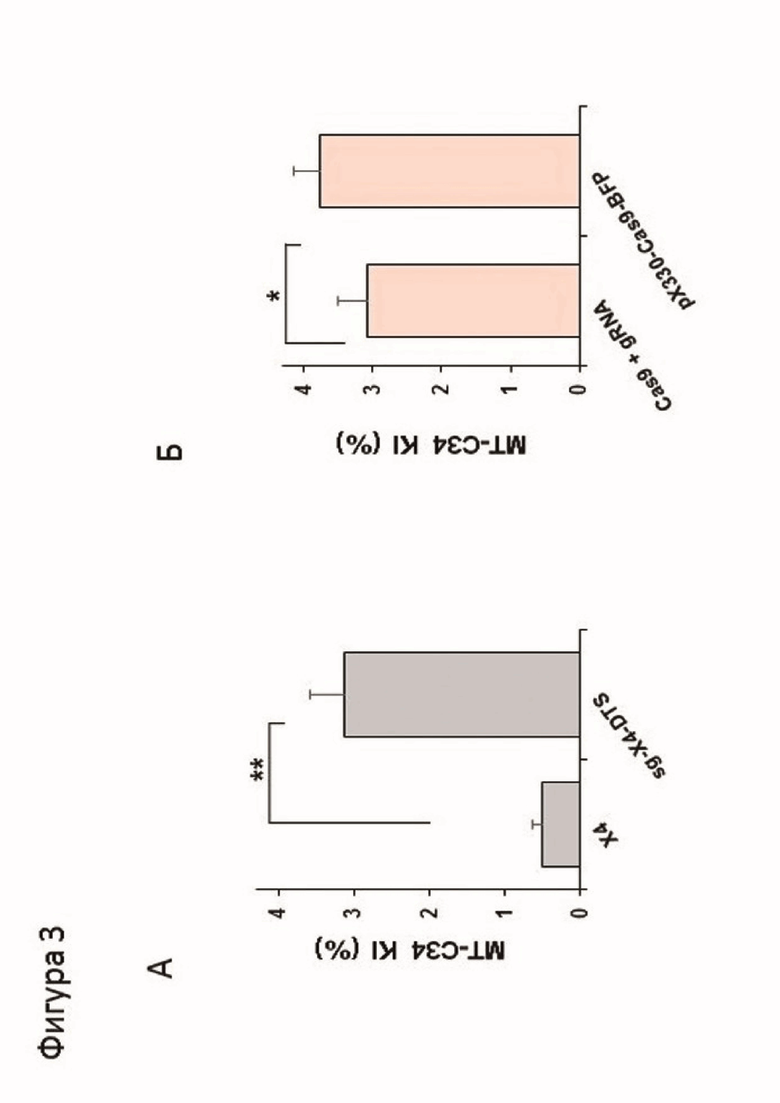
Фигура 3. Сравнение эффективности внесения нокина изначальным и модифицированным методами в клеточную линию CEM/R5: А - Сравнение эффективности внесения нокина конструкции в геном клеток CEM-R5 с использованием изначальной конструкции донорной ДНК (X4) и с использованием модифицированной донорной ДНК (sg-X4-DTS). Б - Увеличение эффективности внесения нокина при использовании плазмиды pX330-Cas9-BFP-gRNAX4ex2 по сравнению с Cas9 дикого типа. * - p <0,05 ** - p <0,01 по t-критерию Стьюдента.
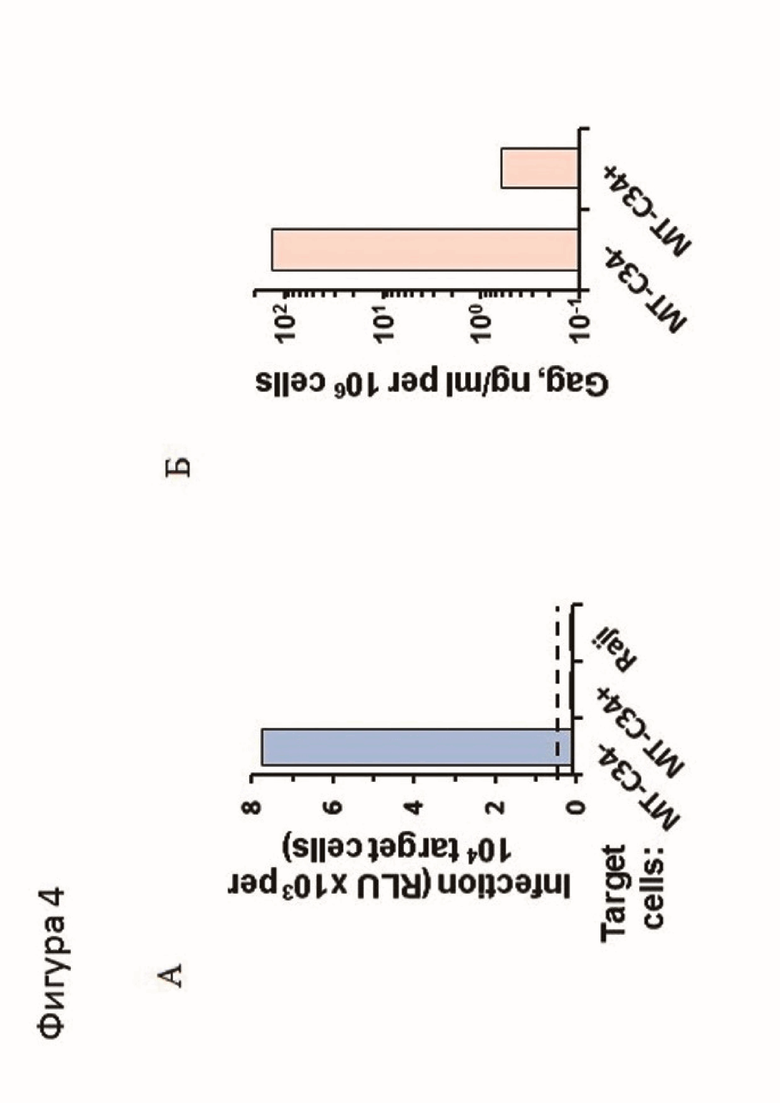
Фигура 4. Инфекционный тест на межклеточную инфекцию от Raji к CD4+MT-С34 А - Люциферазная активность клеток с нокином конструкции (MT-C34+), контрольных клеток (MT-C34-) и клеточной линии Raji. Б - данные содержание вирусного белка Gag в среде через три дня после инфицирования ВИЧ-1 CD4+ лимфоцитов без (MT-C34-) и с (МТ-С34+) нокина конструкции. Данные получены с помощью ИФА на р24.
Осуществление изобретения
Пример 1. Получение клеточной линии, несущей целевую конструкцию с последующей оценкой эффективности метода
На основе клеточной линии CD4+ лимфобластов CEM-CCRF (CEM) была получена сублиния клеток CEM/R5, чувствительных к инфицированию ВИЧ-1 разной тропности за счет экспрессии молекул CD4, CCR5 и CXCR4, одна из которых была привнесена путем лентивирусной трансдукции CCR5-экспрессирующей конструкции.
На основе этой сублинии путем внесения нокина были получены клеточные линии, несущие целевые конструкции с пептидом. Для этого использовали электропорацию с помощью прибора Neon Thermo Fisher в режиме, рекомендуемом производителем.
Смесь для электропорации 1 млн клеток включала следующие компоненты:
- 4 мкг pX330-Cas9-BFP-gRNAX4ex2 - плазмида, кодирующая Cas9 дикого типа, флюоресцентный белок BFP и гРНК;
- 6 мкг матрица-донор в контексте коммерческого вектора pJet (Thermo scientific США), описанная выше (фиг. 2А);
- коммерческий буфер R до 110 мкл.
Описанным способом была получена клеточная линия CEM/R5/MT-С34-new с пептидом в контексте GPI-заякоренного белка CD52. Сортировка клеток с целевой конструкцией проводилась путем иммунофлюоресцентного окрашивания. В качестве первичных антител были использованы антитела к пептиду МТ-С34, полученные в нашей лаборатории; в качестве вторичных антител использовались антитела, коньюгированные с флюоресцентной меткой Alexa546. Выделение клеток с целевой конструкцией проводилось методом FACS - сортировки до достижения чистоты культуры не менее 98%.
С помощью RNP была получена клеточная линия CEM/R5/MT-С34-RNP-new путем электропорации клеток CEM/R5. Смесь для электропорации 1 млн клеток включала:
- комплекс RNP;
- матрица-донор в контексте коммерческого вектора pJet, описанная выше (фиг. 2А);
- коммерческий буфер R до 110 мкл;
Использовался режим, рекомендованный Neon Thermo Fisher.
Для сравнения эффективности внесения нокина модифицируемым и модифицированным методом была проведена электропорация клеток CEM/R5. Смесь для электропорации 1 млн клеток включала следующие компоненты:
- 3 мкг pcDNA3.3-hCas9 (Addgene #41815) - плазмида, кодирующая Cas9 дикого типа;
- 1 мкг вектор pKS gRNA BB, экспрессирующий гРНК, в состав которого включена гРНК SEQ ID NO: 4;
- 1 мкг матрица-донор, полученный путем амплификации ПЦР, и представляющий собой последовательность, описанную на фигуре 2 Б;
- коммерческий буфер R до 110 мкл
Использовался режим, рекомендованный Neon Thermo Fisher.
На третий день после электропорации было проведено окрашивание как описано выше. Установлено, что модифицированный способ значительно эффективнее модифицируемого. Результаты получены путем проведения трех независимых экспериментов и представлены на фигурах 3А и Б.
Пример 2. Внесение целевой конструкции в CD4-лимфоциты человека с последующей оценкой эффективности работы конструкции
Первичные клетки PBMC выделялись из донорской крови. Для этого в стерильных условиях донорская кровь смешивалась с PBS в соотношении 1:1, после чего наслаивалась на раствор фиколла (ПанЭко). Полученная двухфазная смесь центрифугировалась 40 минут с центробежным ускорением 400 g с минимальным ускорением и торможением. В результате эритроцитарная масса оседала на дно пробирки, а целевые клетки образовывали слой на поверхности фиколла. PBMC отбирались и отмывались PBS. Полученная клеточная масса инкубировалась в течении 10 минут в буфере для лизиса эритроцитов (Nh4Cl 155mM, KHCO3 10mM, EDTA 0,1 mM, pH=7.3) и отмывалась в PBS.
Выделение CD4-лимфоцитов проводилась с помощью набора Dynabeads Untouched Kit (Invitrogen, США) следуя инструкции производителя. После выделения клетки активировали CD2/CD3/CD28 магнитными шариками (Miltenyi Biotec, Германия) согласно инструкции производителя и инкубировали с добавлением 100 ед/мл рекомбинантного человеческого интерлейкина-2 (Ронтолейкин, Биотех, Россия). На 3 день после активации клетки электропорировали с помощью прибора Neon Thermo Fisher в режиме 1,600 V, 10 ms x 3.
Смесь для электропорации 2 млн клеток включала следующие компоненты:
- 4 мкг pX330-Cas9-BFP-gRNAX4ex2 - плазмида, кодирующая белок Cas9 дикого типа, флюоресцентный белок BFP и гРНК;
- 6 мкг матрица-донор в контексте коммерческого вектора pJet, описанная выше (фиг. 2)
- коммерческий буфер R до 110 мкл
Описанным способом была получена популяция CD+ лимфоцитов с нокином целевой конструкции (CD4+MT-С34) с пептидом MT-C34 в контексте CD52. Сортировка клеток с целевой конструкцией проводилась путем иммунофлюоресцентного окрашивания. В качестве первичных антител были использованы антитела к пептиду МТ-С34, полученные в нашей лаборатории; в качестве вторичных антител использовались антитела, коньюгированные с флюоресцентной меткой Alexa546. Выделение клеток с целевой конструкцией проводилось методом FACS - сортировки.
Для исследования эффективности работы внесенной в клетки генетической конструкции были проведены инфекционные тесты на межклеточную передачу с вирусом ВИЧ, псевдотипированным белком Env штамма ZM135.
Для этого клетки Raji электропорировали с помощью прибора Neon Thermo Fisher, используя рекомендуемый производителем режим. Смесь для электропорации включала плазмиды pUCHR-inLuc-mR, pCMV-d8.2R и энвелопом ZM135. Через 6 часов электропорированные клетки смешивали с тестируемыми клетками CD4+MT-С34 в соотношении 3:1. Проведенные инфекционные тесты показали, что конструкция эффективно защищает первичные клетки от проникновения ВИЧ-1. Результаты представлены на фигуре 4.
Изобретение потенциально может быть использовано для терапии ВИЧ.
Цитируемая литература
1. Tsunekawa Y, Terhune RK, Fujita I, Shitamukai A, Suetsugu T, Matsuzaki F. Developing a de novo targeted knock-in method based on in utero electroporation into the mammalian brain. Development. 2016;143(17):3216-3222. doi:10.1242/dev.136325
2. Kurihara T, Kouyama-Suzuki E, Satoga M, et al., DNA repair protein RAD51 enhances the CRISPR/Cas9-mediated knock-in efficiency in brain neurons. Biochem Biophys Res Commun. 2020;524(3):621-628. doi:https://doi.org/10.1016/j.bbrc.2020.01.132
3. Sung JJ, Park C-Y, Leem JW, Cho MS, Kim D-W. Restoration of FVIII expression by targeted gene insertion in the FVIII locus in hemophilia A patient-derived iPSCs. Exp Mol Med. 2019;51(4):1-9. doi:10.1038/s12276-019-0243-1
4. Liu J-T, Corbett JL, Heslop JA, Duncan SA. Enhanced genome editing in human iPSCs with CRISPR-CAS9 by co-targeting ATP1a1. PeerJ. 2020;8:e9060. doi:10.7717/peerj.9060
5. Yao X, Wang X, Hu X, et al. Homology-mediated end joining-based targeted integration using CRISPR/Cas9. Cell Res. 2017;27(6):801-814. doi:10.1038/cr.2017.76
6. Yao X, Wang X, Liu J, Shi L, Huang P, Yang H. CRISPR/Cas9-mediated Targeted Integration In Vivo Using a Homology-mediated End Joining-based Strategy. J Vis Exp. 2018;(133):56844. doi:10.3791/56844
7. Nguyen DN, Roth TL, Li PJ, et al. Polymer-stabilized Cas9 nanoparticles and modified repair templates increase genome editing efficiency. Nat Biotechnol. 2020;38(1):44-49. doi:10.1038/s41587-019-0325-6
8. Torres-Ruiz R, Martinez-Lage M, Martin MC, et al., Efficient Recreation of t(11;22) EWSR1-FLI1(+) in Human Stem Cells Using CRISPR/Cas9. Stem cell reports. 2017;8(5):1408-1420. doi:10.1016/j.stemcr.2017.04.014
9. Maggio I, Zittersteijn HA, Wang Q, et al., Integrating gene delivery and gene-editing technologies by adenoviral vector transfer of optimized CRISPR-Cas9 components. Gene Ther. 2020;27(5):209-225. doi:10.1038/s41434-019-0119-y
10. Dean DA, Dean BS, Muller S, Smith LC. Sequence requirements for plasmid nuclear import. Exp Cell Res. 1999;253(2):713-722. doi:10.1006/excr.1999.4716
11. Mesika A, Grigoreva I, Zohar M, Reich Z. A Regulated, NFκB-Assisted Import of Plasmid DNA into Mammalian Cell Nuclei. Mol Ther. 2001;3(5):653-657. doi:https://doi.org/10.1006/mthe.2001.0312
12. Remy S, Chenouard V, Tesson L, et al., Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation. Sci Rep. 2017;7(1):16554. doi:10.1038/s41598-017-16328-y/.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Федеральное государственное бюджетное учреждение науки Институт биологии гена
Российской академии наук (Institute of Gene Biology Russian Academy of Sciences)
<120> Способ усиления направленного редактирования гена CXCR4 в CD4-лимфоцитах
человека в целях создания резистентности клеток к ВИЧ-1
<160> 10
<210> 1
<211> 69
<212> ДНК
<213> Homo sapiens
<400> 1
aag cgc ttc ctc ttc ctc cta ctc acc atc agc ctc ctg gtt atg gta cag ata caa act 60
gga ctc tca 69
<210> 2
<211> 81
<212> ДНК
<213> Homo sapiens
<400> 2
tca gca tcc agc aac ata agc gga ggc att ttc ctt ttc ttc gtg gcc aat gcc ata atc 60
cac ctc ttc tgc ttc agt tga 81
<210> 3
<211> 108
<212> ДНК МТ-С34
<213> Homo sapiens
<400> 3
atg acc tgg atg gag tgg gac cgg gag atc aat aac tac acc tca ctg atc cac agc ctg 60
atc gag gag tcc cag aac cag cag gag aag aac gag cag gag ctc ctg 108
<210> 4
<211> 24
<212> ДНК
<213> Homo sapiens
<400> 4
cacttcagat aactacaccg agg 24
<210> 5
<211> 546
<212> ДНК
<213> Homo sapiens
<400> 5
ttt gcg ctt tag gag aat gag tct ttg caa cgc ccc cgc cct ccc ccc gtg atc ctc cct 60
tct ccc ctc ttc cct ccc tgg gcg aaa aac ttc tta caa aaa gtt aat cac tgc ccc tcc 120
tag cag cac cca ccc cac ccc cca cgc cgc ctg gga gtg gcc tct ttg tgt gta ttt ttt 180
ttt cct cct aag gaa ggt ttt ttt tct tcc ctc tag tgg gcg ggg cag agg agt tag cca 240
aga tgt gac ttt gaa acc ctc agc gtc tca gtg ccc ttt tgt tct aaa caa aga att ttg 300
taa ttg gtt cta cca aag aag gat ata atg aag tca cta tgg gaa aag atg ggg agg aga 360
gtt gta gga ttc tac att aat tct ctt gtg ccc tta gcc cac tac ttc aga att tcc tga 420
aga aag caa gcc tga att ggt ttt tta aat tgc ttt aaa aat ttt ttt taa ctg ggt taa 480
tgc ttg ctg aat tgg aag tga atg tcc att cct ttg cct ctt ttg cag ata tac act tca 540
gat aac 546
<210> 6
<211> 66
<212> ДНК
<213> Homo sapiens
<400> 6
ggatccggcg caacaaactt ctctctgctg aaacaagccg gagatgtcga agagaatcct 60
ggaccg 66
<210> 7
<211> 49
<212> ДНК
<213> Homo sapiens
<400> 7
ataaaatatc tttattttca ttacatctgt gtgttggttt tttgtgtg 49
<210> 8
<211> 472
<212> ДНК
<213> Homo sapiens
<400> 8
agg aaa tgg gct cag ggg act atg act cca tga agg aac cct gtt tcc gtg aag aaa atg 60
cta att taa taa aat ctt cct gcc cac cat cta ctc cat cat ctt ctt aac tgg cat tgt 120
ggg caa tgg att ggt cat cct ggt cat ggg tta cca gaa gaa act gag aag cat gac gga 180
caa gta cag gct gca cct gtc agt ggc cga cct cct ctt tgt cat cac gct tcc ctt ctg 240
ggc agt tga tgc cgt ggc aaa ctg gta ctt tgg gaa ctt cct atg caa ggc agt cca tgt 300
cat cta cac agt caa cct cta cag cag tgt cct cat cct ggc ctt cat cag tct gga ccg 360
cta cct ggc cat cgt cca cgc cac caa cag tca gag gcc aag gaa gct gtt ggc tga aaa 420
ggt ggt cta tgt tgg cgt ctg gat ccc tgc cct cct gct gac tat tcc cga t 472
<210> 9
<211> 71
<212> ДНК
<213> artificial sequence
<220>
<223> последовательность DTS
<400> 9
atgctttgca tacttctgcc tgctggggag cctggggact ttccacaccc taactgacac 60
acattccaca g 71
<210> 10
<211> 71
<212> ДНК
<213> Homo sapiens
<400> 10
cacttcagat aactacaccg 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения линии гуманизированных мышей, содержащих инсерцию 3974insT в гене mGrin2a (mice glutamate [NMDA] receptor subunit epsilon-1), приводящую к преждевременному прекращению трансляции белка grin2a | 2021 |
|
RU2764650C1 |
| Технология создания Т-клеточной устойчивости к ВИЧ-1 на основе генетически кодируемых GPI-заякоренных пептидов из gp41 | 2020 |
|
RU2769567C1 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| Искусственные гены, кодирующие белки-иммуногены EV.CTL и EV.Th, рекомбинантные плазмидные ДНК pEV.CTL и pEV.Th, обеспечивающие экспрессию искусственных генов, и искусственные Т-клеточные полиэпитопные белки-иммуногены EV.CTL и EV.Th, содержащие эпитопы антигенов вируса Эбола, используемые для создания вакцины против вируса Эбола | 2018 |
|
RU2713723C1 |
| УНИВЕРСАЛЬНЫЙ АНТИТЕЛООПОСРЕДОВАННЫЙ БИОСЕНСОР | 2016 |
|
RU2746486C2 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| Способ получения линии гуманизированных мышей, трансгенных по hACE2 | 2020 |
|
RU2757114C1 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| Генетическая конструкция, адаптированная для доставки гена SMN1 человека с помощью аденоассоциированного вируса серотипа 2 для обеспечения нейроспецифичной экспрессии | 2022 |
|
RU2801848C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
Изобретение относится к области биохимии, в частности к донорной ДНК для доставки защитного пептида из gp41 МТ-С34. Также раскрыты способы доставки защитного пептида из gp41 МТ-С34. Изобретение позволяет эффективно редактировать ген CXCR4 в CD4-лимфоцитах. 3 н.п. ф-лы, 4 ил., 2 пр.
1. Донорная ДНК для доставки защитного пептида из gp41 МТ-С34 последовательностью SEQ ID NO: 3 на поверхность плазматической мембраны, при этом донорная ДНК имеет следующую последовательность:
• протоспейсер X4ex2 SEQ ID NO: 4,
• 5’-плечо гомологии SEQ ID NO: 5,
• последовательность Р2А, позволяющая получить с одной РНК раздельную трансляцию пептида и белка CXCR4 SEQ ID NO: 6,
• сигнал экспорта белка SEQ ID NO: 1,
• последовательность, кодирующая защитный пептид из gp41 МТ-С34 SEQ ID NO: 3,
• сигнал для мембранного заякоривания SEQ ID NO: 2,
• последовательность полиаденилирования SEQ ID NO: 7,
• 3’-плечо гомологии SEQ ID NO: 8,
• сайт связывания транскрипционных факторов DTS SEQ ID NO: 9.
2. Способ доставки защитного пептида из gp41 МТ-С34 последовательностью SEQ ID NO: 3 на поверхность плазматической мембраны клетки-мишени с помощью введения в клетку эффективного количества: 1) первой плазмиды pX330-Cas9-BFP-gRNAX4ex2, кодирующей нуклеазу Cas9 и гРНК последовательностью SEQ ID NO: 10, в последовательности которой есть несколько сайтов NLS; и 2) второй плазмиды, кодирующей донорную ДНК по п. 1.
3. Способ доставки защитного пептида из gp41 МТ-С34 SEQ ID NO: 3 на поверхность плазматической мембраны клетки-мишени с помощью введения в клетку эффективного количества: 1) рибонуклеопротеинового комплекса, включающего рекомбинантный белок Cas9 в комплексе с гРНК последовательностью SEQ ID NO: 10; и 2) плазмиды, кодирующей донорную ДНК по п. 1.
| WO 2011119773 A1, 29.09.2011 | |||
| CN 0107893074 A, 10.04.2018 | |||
| CN 0106801056 A, 06.06.2017 | |||
| WO 2015031775 A1, 05.03.2015 | |||
| Рекомбинантный химерный полипептид-иммуноген nTBI, обладающий способностью индуцировать антитела, нейтрализующие вирус иммунодефицита человека 1 типа, и предназначенный для использования в качестве компонента вакцины против ВИЧ-1 | 2016 |
|
RU2642258C1 |

